Изобретение относится к промышленному использованию микроорганизмов и, в частности, относится к способу ферментационного получения L-лизина.
В известной практике получения L-лизина в результате ферментации для повышения продуктивности используют микробный штамм, выделенный из природного окружения, или искусственный мутантный штамм, полученный из этого микробного штамма. Известно большее число искусственных мутантных штаммов, продуцирующих L-лизин. Большинство из них представляют мутантные штаммы, обладающие устойчивостью к S-2-аминоэтилцистеину (АЕС), и принадлежат к роду Brevibacterium, Corynebacterium, Bacillus или Escherichia. Кроме того, были раскрыты различные методики для повышения продуцирования аминокислоты, например, в результате использования трансформантов с применением рекомбинантных ДНК (патент США 4278765).
В отношении тех, которые принадлежат к роду Escherichia, можно указать, например, японскую открытую патентную выкладку 56-18596, патент США 4346170 и Applied Microbiology and Biotechnology 15, 227-231 (1982), в которых раскрыты способы получения L-лизина в результате использования бактериального штамма, в котором усилена активность дигидродипиколинатсинтазы (здесь и далее иногда используют сокращение "DDPS"). Однако DDPS, используемая в этих случаях, является дикого типа синтазой, у которой неприятным побочным эффектом является ингибирование ее по типу обратной связи L-лизином. В результате оказывается невозможным достичь достаточно высокой производительности L-лизина.
Между прочим, в упомянутой статье в Applied Microbiology and Biotechnology, 15, 227-231 (1982) описывается получение L-лизина в количестве 3 г/л гидрохлорида L-лизина из 75 г/л глюкозы, где коэффициент превращения (число г L-лизина, получаемого из 1 г сахара, или его процент) по расчетам оказался 0,04, или 4%.
С другой стороны, в патентной публикации Кореи 92-8382 раскрыт способ получения L-лизина с использованием бактерии, принадлежащей к роду Escherichia, в которую введена DDPS, полученная из бактерии, принадлежащей к роду Corynebacterium, который, как известно, лишен недостатка ингибирования по типу обратной связи с помощью L-лизина (коэффициент превращения 17%). Однако верхний предел температуры для роста бактерий, принадлежащих к роду Соrуnеbacterium, ниже, чем верхний предел температур для роста бактерий, принадлежащих к роду Escherichia, примерно на 10 градусов. Поэтому культивирование следует осуществлять при более низких температурах культивирования, если ДНК, кодирующую DDPS, полученную из бактерий, принадлежащих к роду Corynebacterium, вводят в бактерию, принадлежащую к роду Escherichia с целью использования ее для получения L-лизина. Поэтому слишком рано говорить о том, что трудно продемонстрировать преимущества, которыми обладают бактерии рода Escherichia, то есть что высока температура их культивирования, что они быстро размножаются и что они отличаются высокой скоростью продуцирования L-лизина.
Обычно, если экспрессируется ген, полученный из гетерологичного организма, это иногда вызывает разложение продукта экспрессии с помощью протеазы и образование нерастворимых включений, что приводит к значительно большим трудностям по сравнению со случаем экспрессии гомологичного гена. Далее, если ДНК, кодирующую DDPS, полученную из бактерий, принадлежащих к роду Corynebacterium, вводят в бактерию, принадлежащую к роду Escherichia, для промышленного производства L-лизина, налагаются более строгие ограничения по сравнению со случаем использования рекомбинанта, в который вводят гомологичный ген, в соответствии с требованиями к рекомбинантным ДНК.
Между прочим, дигидродипиколинатсинтаза (DDPS) является энзимом для дегидратации и конденсации аспартополуальдегида и пировиноградной кислоты для синтеза дигидродипиколиновой кислоты. Эта реакция расположена в начале схемы системы биосинтеза L-лизина в биосинтезах аминокислот семейства аспарагиновых кислот. Как известно, этот энзим отвечает за такой важный регуляторный сайт, как аспартокиназа, в бактериях, принадлежащих роду Escherichia.
DDPS кодируется геном под названием dapA в Е. coli [Escherichia coli]. dapA был клонирован, и его нуклеотидная последовательность также была определена (Richaud, F. et al., J. Bacteriol., 297 (1986)).
С другой стороны, аспартокиназа (здесь и далее сокращенно именуемая как "АК") является энзимом для катализирования реакции превращения аспарагиновой кислоты в β-фосфоаспарагиновую кислоту, которая служит основным регуляторным энзимом в системе биосинтеза аминокислот семейства аспарагиновых кислот. АК из Е. coli имеет три типа (АК1, АК11, АК111), два из которых являются комплексными энзимами с гомосериндегидрогеназой (здесь и далее иногда сокращаемой до "HD"). Одним из таких комплексных энзимов является AK1-HD1, кодируемый thrA геном, а другой - AK11-HD11, кодируемый metLM геном. АК1 подвержен супрессии с помощью треонина и изолейцина и ингибируется треонином, тогда как АК1 подавляется метионином.
Напротив, известно, что только АК111 является простым функционирующим энзимом, который является продуктом гена, обозначаемого как lysC, и подвержен супрессии и ингибируется L-лизином. Отношение их внутриклеточных активностей АК1:АК11:АК111 = примерно 5:1:4.
Как было указано ранее, DDPS, полученная из бактерий, принадлежащих роду Corynebacterium, лишена недостатка ингибирования по типу обратной связи с помощью L-лизина. Однако, если ее вводят в бактерию, принадлежащую роду Escherichia, для использования для продуцирования L-лизина, возникает проблема температур, при которых ведут культивирование. Ожидается, что L-лизин может эффективно продуцироваться в результате ферментации с использованием бактерий, принадлежащих роду Escherichia, если удастся получить мутантный энзим DDPS или АК111, полученные из бактерий, принадлежащих роду Escherichia, которые лишены недостатка ингибирования по типу обратной связи с помощью L-лизина. Однако в известной литературе нет материалов, которые раскрывали бы такой мутантный энзим DDPS, и также существует одно сообщение о мутантном энзиме АК111 (Boy, E. et al., J. Bacteriol., 112, 84 (1972)), но в нем нет ни одного примера, который предполагал бы, что такой мутантный энзим может также повысить продуктивность L-лизина.
Настоящее изобретение было выполнено с учетом вышеуказанных точек зрения, его целью было получение DDPS и АК111, полученных из бактерий, принадлежащих к роду Escherichia, с эффективно десенсибилизированным ингибированием по типу обратной связи с помощью L-лизина, и создание способа получения L-лизина в результате ферментации, который был бы усовершенствован по сравнению с известными способами.
В результате длительных повторных исследований для достижения указанных целей авторам удалось получить ДНК, кодирующую DDPS, полученную из бактерий, принадлежащих к роду Escherichia, в которой ингибирование по типу обратной связи с помощью L-лизина было существенно десенсибилизировано. ДНК, кодирующую DDPS, полученную из Е. coli, в которой недостаток, связанный и ингибированием по типу обратной связи с помощью L-лизина, был существенно десенсибилизирован, иногда называют здесь мутантной dapA или dapA*.
Кроме того, авторы создали бактерию, принадлежащую роду Escherichia, несущую мутантную dapA и аспартокиназу, которая лишена недостатка ингибирования по типу обратной связи с помощью L-лизина. ДНК, кодирующая аспартокиназу, полученную из Е. coli, в которой ингибирование по типу обратной связи с помощью L-лизина достаточно десенсибилизировано, иногда здесь называют мутантной lysC или lysC*.
Кроме того, авторы создали бактерию, принадлежащую роду Escherichia, несущую мутантный dapA и мутантный lysC. И было обнаружено, что значительное количество L-лизина можно получить и накопить в культуре в результате культивирования вышеуказанной бактерии, принадлежащей к роду Escherichia в предпочтительной среде.
Кроме того, авторы обнаружили, что производство L-лизина можно далее увеличить, воздействуя на другие гены в системе биосинтеза L-лизина бактерий, принадлежащих к роду Escherichia, несущих мутантный dapA и мутантный lysC.
В настоящем изобретении предложен способ получения L-лизина, включающий стадии культивирования любой из бактерий, принадлежащих роду Escherichia, описанному ранее, в соответствующей среде, продуцирования и накопления L-лизина в этой культуре и выделения L-лизина из этой культуры.
В этом описании ДНК, кодирующая DDPS или АК111, или ДНК, содержащая промотор в дополнении к этому, иногда называется "DDPS геном" или "АК111 геном". Кроме того, мутантный энзим, который десенсибилизирует ингибирование по типу обратной связи с помощью L-лизина, и ДНК, кодирующая его, или ДНК, содержащая промотор в дополнении к нему, иногда просто называются "мутантным энзимом" и "мутантным геном" соответственно. Кроме того, фраза "ингибирование по типу обратной связи с помощью L-лизина десенсибилизировано" означает, что достаточно существенной десенсибилизации ингибирования и нет необходимости в полной десенсибилизации.
Далее настоящее изобретение будет описано более подробно.
1. ДНК, кодирующая мутантную дигидродипиколинатсинтазу (DDPS) настоящего изобретения
ДНК, кодирующая мутантную DDPS настоящего изобретения, содержит мутацию для десенсибилизации ингибирования по типу обратной связи с помощью L-лизина DDPS, кодируемой ДНК, кодирующей дикого типа DDPS. Примерами DDPS могут служить те, которые получены из бактерий, принадлежащих роду Escherichia, особенно DDPS, полученные из Е. coli. Мутации DDPS для десенсибилизации ингибирования по типу обратной связи с помощью L-лизина, представлены следующими мутациями:
(1) мутацией в результате замены 81-го аланинового остатка остатком валина;
(2) мутацией в результате замены 118-го гистидинового остатка тирозиновым остатком; и
(3) мутацией в результате замены 81-го аланинового остатка валиновым остатком, и в результате замены 118-го гистидинового остатка тирозиновым остатком;
считая с N-конца DDPS в аминокислотной последовательности DDPS, представленной в последовательности SEQ ID 4 в описании последовательностей.
ДНК, кодирующая дикого типа DDPS, специально не ограничена при условии, что она кодирует DDPS, полученную из бактерий, принадлежащих роду Escherichia, что конкретно представлено ДНК, кодирующей аминокислотную последовательность, определенную в последовательности SEQ ID 4, и дополнительно конкретно представлено последовательностью, представленной основаниями под номерами 272-1147 в основной последовательности, определенной в последовательности SEQ ID 3. В этих последовательностях те последовательности, которые содержат мутацию в нуклеотидной последовательности для того, чтобы вызвать замену аминокислотных остатков, описанных ранее, находятся ДНК, кодирующие мутантную DDPS настоящего изобретения. Любой кодон, соответствующий замененному аминокислотному остатку, доступен независимо от его типа при условии, что он кодирует идентичный аминокислотный остаток. Кроме того, постулируется, что имеющаяся DDPS слегка отличается в последовательности в зависимости от различий в бактериальных видах и бактериальных штаммах, однако те, у которых есть замена, делеция или вставка аминокислотного остатка (остатков) в положении (положениях), не связанном с энзиматической активностью, также включены в мутантный DDPS ген настоящего изобретения.
Способ получения такого мутантного гена следующий. Вначале ДНК, содержащую дикого типа DDPS ген или DDPS ген, содержащий другую мутацию, подвергают in vitro мутационной обработке и ДНК после мутационной обработки лигируют с векторной ДНК, адаптированной хозяину для получения рекомбинантной ДНК. Рекомбинантную ДНК вводят в микроорганизм хозяина для получения трансформантов. Когда один из них, который экспрессирует мутантную DDPS, выбирают из вышеуказанных трансформантов, такой трансформант содержит мутантный ген. В другом варианте ДНК, содержащую дикого типа DDPS ген или DDPS ген, содержащий другую мутацию, можно лигировать с векторной ДНК, адаптированной хозяину для получения рекомбинантной ДНК. После этого рекомбинантную ДНК подвергают in vitro мутационной обработке и рекомбинантную ДНК после мутационной обработки вводят в микроорганизм хозяина для получения трансформантов. Когда один из них, который экспрессирует мутантную DDPS, выбирают из вышеуказанных трансформантов, такой трансформант содержит также мутантный ген.
Можно также микроорганизм, который продуцирует дикого типа энзим, подвергнуть мутационной обработке для создания мутантного штамма, который продуцирует мутантный энзим, а затем получить мутантный ген из мутантного штамма. В другом варианте трансформант, в который вводят рекомбинантную ДНК, лигированную с дикого типа геном, можно заменить мутантной обработкой для создания мутантного штамма, который продуцирует мутантный энзим. Если после этого выделить рекомбинантную ДНК из мутантного штамма, мутантный ген создается на основе вышеуказанной ДНК.
Агентом для осуществления in vitro мутантной обработки ДНК может быть гидроксиламин и т. п. Гидроксиламин является химическим мутагеном, который вызывает мутацию замены цитозина на тимин в результате изменения цитозина на N4-гидроксицитозин. В другом варианте, если сам микроорганизм подвергают мутационной обработке, эту обработку проводят, используя УФ-излучение, или мутаген, который обычно используют для осуществления искусственных мутаций, например N-метил-N-нитро-N-нитрозогуанидин (NTG) или азотистую кислоту.
Никаких проблем не возникает, если использовать в качестве донорного микроорганизма для ДНК, содержащей дикого типа DDPS ген или DDPS ген, содержащий другую мутацию, описанную ранее, при условии, что этот микроорганизм принадлежит к роду Escherichia. Конкретно можно использовать те, которые описаны в книге, написанной Neidhardt et al. [Neidhardt, F.C. et al., Escherichia coli and Salmonella Typhimurium, American Society for Microbiology; Washington, D.C., 1208, таблица 1]. Так, например, можно использовать штаммы Е. coli JM109 и МС1061. Если в качестве донорного микроорганизма для ДНК, содержащей DDPS ген, используют дикий штамм, получают ДНК, содержащую дикого типа DDPS ген.
(1) Получение дикого типа DDPS гена
Пример получения ДНК, содержащей DDPS ген, будет раскрыт далее. Вначале Е. coli, содержащую дикого типа dapA, например МС1061 штамм, культивируют для получения культуры. Если культивируют описанный ранее микроорганизм, культивирование можно вести в соответствии с обычными способами ведения твердой культуры, однако предпочтительно вести культуру в жидкой среде, учитывая эффективность сбора бактерий. Можно использовать среду, в которую добавляют один или более из источников азота, например дрожжевой экстракт, пептон, мясной экстракт, кукурузный сироп и эксудат сои или пшеницы с одной или более из неорганических солей, таких как первичный кислый фосфат калия, вторичный кислый фосфат калия, сульфат магния, хлорид натрия, хлорид магния, хлорид железа, сульфат железа или сульфат магния, и, кроме того, необязательно добавляют сахара, витамины и т.п. Удобно устанавливать исходное значение рН среды 6-8. Культивирование ведут в течение 4-24 часов при 30-42oС, предпочтительно при около 37oС в культуре с аэрацией и перемешиванием в результате встряхивания, или в стационарной культуре, или т.п.
Полученную таким образом культуру центрифугируют, например, при 3000 об/мин в течение 5 минут для получения клеточного осадка Е. coli MC061 штамма. Хромосомную ДНК можно получить из клеточного осадка, например, с помощью способа Saito and Miura (Biochem. Biophys. Аста., 72, 619 (1963)), или по способу K.S. Kirby (Biochem. J., 64, 405 (1956)).
Для того чтобы выделить DDPS ген из хромосомной ДНК, полученной таким образом, приготавливают библиотеку ДНК. Вначале хромосомную ДНК частично переваривают подходящим рестрикционным энзимом для получения смеси различных фрагментов. Можно использовать широкий круг рестрикционных энзимов, если степень разрезания контролируется временем реакции разрезания и т.п. Так, например, SauЗA1 оставляют реагировать на хромосомной ДНК при температуре не менее 30oС, предпочтительно при 37oС при концентрации энзима 1-10 ед/мл в течение различных промежутков времени (1 минута, до 2 часов) для ее расщепления.
Затем полученные ДНК фрагменты лигируют с векторной ДНК, автономно реплицируемой в клетках бактерий, принадлежащих роду Escherichia, для получения рекомбинантной ДНК. Более конкретно рестрикционный энзим, который создает терминальную нуклеотидную последовательность, которая комплементарна полученной с помощью рестрикционного энзима SauЗA1, использованного для нарезания хромосомной ДНК, например BamHI, оставляют реагировать с векторной ДНК в температурных условиях не менее 30oС и при концентрации энзима 1-100 ед/мл в течение не менее 1 часа, предпочтительно в течение 1-3 часов для полного расщепления, нарезания и отщепления. Далее смесь фрагментов хромосомной ДНК, полученную, как указано ранее, смешивают с расщепленной и нарезанной векторной ДНК, на которую ДНК лигаза, предпочтительно Т4 ДНК лигаза, воздействовала в температурных условиях 4-16oС при концентрации энзима 1-100 ед/мл в течение, по крайней мере, 1 часа, предпочтительно в течение 6-24 часов, для получения рекомбинантной ДНК.
Полученную рекомбинантную ДНК используют для трансформации микроорганизма, принадлежащего к роду Escherichia, например DDPS дефицитного мутантного штамма, такого как Escherichia coli K-12 штамм, предпочтительно JE7627 штамм (ponB704, dacB12, pfv+, tonA2, dapA, lysA, str, malA38, metB1, ilvH611, leuA371, рrоА3, lac-3, tsx-76) для получения библиотеки хромосомных ДНК. Такую трансформацию можно провести, например, по способу D.M. Morrison (Methods in Enzymology 68, 326 (1979)) или способом, в котором реципиентную бактерию обрабатывают хлоридом кальция для повышения проницаемости ДНК (Mandel, M. and Higa, A. , J. Mol. Biol., 53, 159 (1970)). Штамм JE7627 доступен из национального Института Генетики (Mishima-shi, Shizuoka-ken, Japan).
Бактериальный штамм, содержащий рекомбинантную ДНК гена DDPS, получают из штаммов с повышенной активностью DDPS, или из штаммов, в которых ауксотрофия, возникающая в результате дефицита в DDPS гене, компенсирована, среди полученной библиотеки хромосомных ДНК. Так, например, мутантный штамм с дефицитом DDPS требует диаминопимелиновой кислоты. Так, если DDPS дефицитный мутантный штамм используют в качестве хозяина, ДНК фрагмент, содержащий DDPS ген, можно получить, выделяя бактериальный штамм, который становится способным расти на среде, не содержащей диаминопимелиновой кислоты, и выделяя рекомбинантную ДНК из бактериального штамма.
Подтверждением факта, содержит ли штамм-кандидат, содержащий рекомбинантную ДНК, содержащую DDPS ген, действительно рекомбинантную ДНК, в которую DDPS ген клонирован, можно получить в результате получения клеточного экстракта из штамма-кандидата и получения из него неочищенного энзимного раствора для подтверждения того, повысилась ли DDPS активность. Процедуру определения энзиматической активности DDPS можно осуществить по способу Yugari et al. (Yugari, Y. and Gilvard, C., J. Biol. Chem. 240, 4710 (1962)).
Рекомбинантную ДНК, в которой ДНК, содержащая DDPS ген, встроена в векторную ДНК, можно выделить из описанного ранее бактериального штамма, например в результате способа Р. Guerry et al. (J. Bacteriol., 116, 1064 (1973)) или в результате способа D. B. Clewell (J. Bacteriol., 110, 667 (1972)).
Получение дикого типа DDPS гена можно также осуществить в результате получения хромосомной ДНК из штамма, содержащего DDPS ген на хромосоме, по способу Saito and Miura или аналогично, и амплификации DDPS гена с помощью полимеразной цепной реакции (PCR) (см. White, T.J., et al., Trends Genet., 5, 185 (1989)) праймеры ДНК, которые можно использовать в реакции амплификации, должны быть комплементарны с обоими 3'-концами двухцепочечной ДНК, содержащей полный участок или частичный участок DDPS гена. Если амплифицирован только частичный участок DDPS гена, необходимо использовать такие ДНК фрагменты, как праймеры, чтобы осуществить скринирование ДНК фрагмента, содержащего полный участок из библиотеки хромосомных ДНК. Если амплифицирован полный участок DDPS гена, раствор PCR реакции, содержащий фрагменты ДНК, содержащие амплифицированный DDPS ген, подвергают обработке с помощью электрофореза на агарозном геле, а затем экстрагируют целевой ДНК фрагмент. Так, ДНК фрагмент, содержащий DDPS ген, можно выделить.
ДНК праймеры можно соответствующим образом получить на основании, например, последовательности, известной в Е. coli (Richeud F., et al., J. Bacteriol., 297 (1986)).
Более конкретно праймеры, которые позволят амплифицировать участок, содержащий 1150 оснований, кодирующих DDPS ген, предпочтительны, и два вида праймеров, определенные в последовательностях SEQ ID 2 и SEQ ID 1, подходят для этой цели. Синтез праймеров можно осуществить такими обычными способами, как фосфодиамидитный способ (см. Tetrahedron Letters, 22, 1859 (1981)), используя коммерчески доступный ДНК синтезатор (например, ДНК синтезатор Модель 380В от Applied Biosystems Inc.). Кроме того, PCR можно осуществить, используя коммерчески доступный прибор PCR (например, ДНК Thermal Cycler Model PJ2000 от Takara Shuzo Co, Ltd.), используя Taq ДНК полимеразу (поставляемую Takara Shuzo Co., Ltd.) в соответствии с инструкциями изготовителей.
Что касается DDPS гена, амплифицированного в PCR, такая операция, как введение мутаций в DDPS ген, становится легкой, если его лигировать с векторной ДНК, автономно реплицируемой в клетках бактерии, принадлежащей роду Escherichia и введенной в клетки бактерии, принадлежащих роду Escherichia. Векторная ДНК, которую можно использовать, способ трансформации и способ подтверждения присутствия DDPS гена те же, что и в вышеописанной процедуре.
(2) Осуществление мутации в DDPS гене
Способ осуществления такой мутации, как замена, вставка и делеция аминокислотных остатков, можно показать на примере способа рекомбинантной РСР (Higuchi, R. , 61 в PSR Technology (Eriich, H.A., Eds., Stockton press (1989)) и способом сайтспецифического мутагенеза (Kramer, W. and Frits, H.J. , Meth. in Enzymol., 154, 350 (1987); Kunkel Т.A. et al., Meth. in Enzymol., 154, 367 (1987)). Используя эти способы, можно вызвать нужные мутации в нужном сайте.
Кроме того, в соответствии с химическим синтезом нужного гена можно ввести мутацию или получить случайную мутацию в нужном сайте.
Далее, доступен способ, в котором DDPS ген на хромосоме или плазмиде непосредственно обрабатывают гидроксиламином (Hashimoto Т. and Sekiguchi M., J. Bacteriol. , 159, 1039 (1984)). В другом варианте можно использовать способ, в котором бактерию, принадлежащую роду Escherichia, содержащую DDPS ген, облучают УФ-излучением, или способ, основанный на обработке химическим агентом, таким как N-метил-N'-нитрозогуанидин или азотистая кислота. В соответствии с этими способами мутацию можно осуществить статистически.
Что касается выбора способа для мутантного гена, рекомбинантная ДНК, содержащая ДНК фрагмент, содержащий DDPS ген и векторную ДНК, первой подвергают непосредственной мутации с помощью гидроксиламина или т.п., который используют для трансформации, например, Е. coli штамма. Затем трансформированные штаммы культивируют на такой минимальной среде, как М9, содержащей S-2-аминоэтилцистеин (АЕС) в качестве аналога L-лизина. Штаммы, содержащие рекомбинантную ДНК, содержащую дикого типа DDPS ген, не могут синтезировать L-лизин и диаминопимеловую кислоту (ДАР), и их рост подавляется, так как DDPS, экспрессируемая из рекомбинантной ДНК, является ингибитором АЕС. Напротив, штамм, содержащий рекомбинантную ДНК, содержащую DDPS ген, в котором ингибирование L-лизина десенсибилизировано, содержит мутантный энзим, кодируемый DDPS геном в вышеуказанной рекомбинантной ДНК, которая не ингибируется АЕС. Поэтому он способен расти на минимальной среде, в которую добавляем АЕС. Этот факт можно использовать для отбора штамма, который устойчив к АЕС как аналогу L-лизина, то есть штамма, содержащего рекомбинантную ДНК, содержащую мутантный DDPS ген, в котором ингибирование десенсибилизировано.
Полученный таким образом мутантный ген можно ввести как рекомбинантную ДНК в подходящий микроорганизм хозяина и экспрессировать. Так можно получить такой микроорганизм, который содержал бы DDPS, в которой десенсибилизировано ингибирование по типу обратной связи. Хозяином предпочтительно является микроорганизм, принадлежащий роду Escherichia, примером которого может служить Е. coli.
В другом варианте фрагмент мутантного DDPS гена можно взять из рекомбинантной ДНК и встроить в другой вектор для использования. Векторная ДНК, которую можно использовать для целей настоящего изобретения, предпочтительно является плазмидной векторной ДНК, для которой примерами могут служить pUC19, pUC18, pBR322, pHSG299, pHSG298, pHSG399, pHSG398, PSF1010, pMW119, pMW219 и pMW218. Кроме того, можно использовать также векторы фаговой ДНК.
Кроме того, для эффективной экспрессии мутантного DDPS гена другой промотор, который функционирует в микроорганизмах, например, lac, trp и PL, можно лигировать в обратном направлении от ДНК последовательности, кодирующей мутантную DDPS, или промотор, содержащийся в DDPS гене, можно использовать как он есть, или после амплификации промотора.
Далее, как было указано ранее, мутантный ген можно встроить в автономно реплицируемую векторную ДНК, которая введена в хозяина, и оставить ее в хозяине как внехромосомную ДНК, как плазмиду. В другом варианте мутантный ген можно интегрировать в хромосому микроорганизма хозяина способом, в котором используют трансдукцию, транспосон (Berg, D.E. and Berg, C.M. Bio. Technol., 1, 417 (1983)), Mu фаг [японская открытая патентная выкладка 2-109985] или гомологичную рекомбинацию [Experiments in Molecular Genetics, Cold Spring Harbor Lac. (1972)].
2. ДНК, кодирующая мутантную аспартокиназу 111 (ДК111), используемая в настоящем изобретении
ДНК, кодирующая мутантную AK111, используемая в настоящем изобретении, имеет мутацию для десенсибилизации ингибирования по типу обратной связи кодируемой AK111 L-лизином в ДНК, кодирующей дикого типа AK111. Примером мутации для десенсибилизации ингибирования AK111 по типу обратной связи с помощью L-лизина может служить:
(a) мутация для замены 323-го глицинового остатка остатком аспарагиновой кислоты;
(b) мутация для замены 323-го глицинового остатка остатком аспарагиновой кислоты и замены 408-го глицинового остатка остатком аспарагиновой кислоты;
(c) мутация для замены 34-го аргининового остатка цистеиновым остатком и замены 323-го глицинового остатка остатком аспарагиновой кислоты;
(d) мутация для замены 325-го лейцинового остатка фенилаланиновым остатком;
(e) мутация для замены 318-го метионинового остатка изолейциновым остатком;
(f) мутация для замены 318-го метионинового остатка изолейциновым остатком и замены 349-го валинового остатка метиониновым остатком;
(g) мутация для замены 345-го серинового остатка лейциновым остатком;
(h) мутация для замены 347-го валинового остатка метиониновым остатком;
(i) мутация для замены 352-го треонинового остатка изолейциновым остатком;
(j) мутация для замены 352-го треонинового остатка изолейциновым остатком и замены 369-го серинового остатка фенилаланиновым остатком;
(k) мутация для замены 164-го остатка глутаминовой кислоты лизиновым остатком; и
(l) мутация для замены 417-го метионинового остатка изолейциновым остатком и замены 419-го цистеинового остатка тирозиновым остатком;
считая с N-конца АК111 в аминокислотной последовательности АК111, определенной в последовательности SEQ ID 8 в описании последовательностей.
ДНК, кодирующая дикого типа АК111, особенно не ограничена, и примером для ДНК, кодирующей АК111, полученной из бактерии, принадлежащей роду Escherichia, может служить Е. coli. Более конкретно она представлена примерами ДНК, кодирующей аминокислотную последовательность, определенную в последовательности SEQ ID 8, и последовательность, представленную основаниями под 584-1930 в основной последовательности, определенной в последовательности SEQ ID 7. АК111 Е. coli кодируется lysC геном.
В этих последовательностях те, которые содержат мутацию в основной последовательности для того, чтобы вызвать замену аминокислотных остатков, описанных ранее, являются ДНК, кодирующими мутантные АК111 настоящего изобретения. Любой кодон, соответствующий замещенному аминокислотному остатку, доступен независимо от его типа при условии, что он кодирует идентичный аминокислотный остаток. Кроме того, существуют такие, в которых аминокислотные последовательности, содержащие дикого типа АК111, слегка отличаются в зависимости от различия в бактериальных видах и бактериальных штаммах. Те, которые содержат замещения, делении или вставки аминокислотного остатка (остатков) в положении (положениях), не связанном с энзиматической активностью, таким образом также включены в мутантный АК111 ген настоящего изобретения. Так, например, основная последовательность дикого типа lysC гена, полученная в примере 2, описанная далее (SEQ ID 7), отличается от уже опубликованной последовательности lysC Е. coli K-12 JC411 штамма по 6 сайтам (Саssan, M. , Parsot, С., Cohen, G.N., and Patte, J.C., J. Biol Chem., 261, 1052 (1986)). Закодированные аминокислотные остатки отличаются по 2 их сайтам (в lysC JC411 штамма 58-ой глициновый остаток заменен цистеиновым остатком, а 401-ый глициновый остаток заменен аланиновым остатком, считая с N-конца в аминокислотной последовательности lysC, определенной в последовательности SEQ ID 8). Ожидается даже для lysC, имеющей ту же последовательность, что и lysC в Е. coli K-12 JC411 штамма, что lysC с мутацией, в которой ингибирование по типу обратной связи с помощью L-лизина десенсибилизировано, получают, если осуществлена любая из вышеперечисленных мутаций (а) до (l).
Способ получения ДНК, кодирующей мутантные АК111, в которых ингибирование по типу обратной связи с помощью L-лизина десенсибилизировано, следующий. Вначале ДНК, содержащую дикого типа АК111 ген или АК111 ген, содержащий другую мутацию, подвергают in vitro мутационной обработке, а ДНК, полученную в результате такой мутационной обработки, лигируют с векторной ДНК, адаптированной к хозяину для получения рекомбинантной ДНК. Рекомбинантную ДНК вводят в микроорганизм хозяина для получения трансформантов. Когда один из них, который экспрессирует мутантные АК111, выбирают из указанных трансформантов, такой трансформант содержит мутантный ген. В другом варианте ДНК, содержащая дикого типа АК111 ген или АК111 ген, содержащий другую мутацию, может быть лигирована с векторной ДНК, адаптированной к хозяину, для получения рекомбинантной ДНК. После этого рекомбинантную ДНК подвергают in vitro мутационной обработке, а рекомбинантную ДНК после мутационной обработки вводят в микроорганизм хозяина для получения трансформантов. Если один из них, который экспрессирует мутантные АК111, выбирают из вышеуказанных трансформантов, такой трансформант содержит мутантный ген.
В другом варианте можно также микроорганизм, который продуцирует дикого типа энзим, подвергнуть мутационной обработке для создания мутантного штамма, который продуцирует мутантный энзим, а затем из этого мутантного штамма получить мутантный ген. Примером агента для осуществления непосредственной мутационной обработки ДНК может служить гидроксиламин и т.п. Гидроксиламин представляет собой химический мутаген, который вызывает мутацию от цитозина до тимина, в результате замены цитозина на N4-гидроксицитозин. В другом варианте, если сам микроорганизм подвергают мутационной обработке, обработку осуществляют с помощью УФ-излучения, или используя такой мутаген, который обычно используют для искусственной мутации, как N-метил-N'-нитро-N-нитрозогуанидин (NTG).
В качестве донорного микроорганизма для ДНК, содержащей дикого типа АК111 ген или АК111 ген, содержащий любую из других мутаций, указанных выше, можно использовать при условии, что этот микроорганизм принадлежит к роду Escherichia. Более конкретно можно использовать те, которые описаны в книге, написанной Neidhardt et al. (Neidhardt, F.C., et al., Escherichia coli and Salmonella Typhimurium, American Society for Microbiology, Washington D.C., 1208, табл. 1). Так, например, Е. coli M109 штамм и МС1061 штамм могут служить примерами. Если АК111 ген получают из этих штаммов, получение библиотеки хромосомных ДНК и т.п. можно осуществить таким же способом, что и получение DDSR гена, описанное ранее. В качестве хозяина для получения такой библиотеки предпочтительно использовать штамм, полностью лишенный АК1, 11 и 111, такой как, например, Е. coli CT3 штамм (доступный от Е. coli Genetic Stock Center (Connecticut, USA)).
Из полученной библиотеки хромосомных ДНК получают бактериальный штамм, содержащий рекомбинантную ДНК АК111 гена в виде штамма, в котором АК111 активность усилена, или штамм, в котором дополнена ауксотрофия. Клеточные экстракты получают из штаммов-кандидатов, а неочищенные энзимные растворы получают из них для подтверждения АК111 активности. Процедура измерений для АК111 энзиматической активности может быть осуществлена в соответствии со способом Stadtman et al. (Stadtman, E.R., Cohen, G.N. Lebras, G., и Pobichon-Szumajsten H., J. Biol. Chem., 236, 2033 (1961)).
Так, например, если мутантный штамм, полностью лишенный АК, используют в качестве хозяина, ДНК фрагмент, содержащий АК111 ген, можно получить в результате выделения трансформированного штамма, который становится способным к росту на среде, не содержащей L-лизина, L-треонина, L-метионина и диаминопимелиновой кислоты, или на среде, не содержащей гомосерина и диаминопимелиновой кислоты, и выделения рекомбинантной ДНК из бактериального штамма.
Если АК111 ген амплифицирован из хромосомной ДНК в результате PCR, ДНК праймеры, которые следует использовать для PCR реакции, можно соответствующим образом получить на основании, например, последовательности, известной в Е. coli (Cassan, M. , Parsot, С., Cohen, G.N., и Patte, J.C., J. Biol. Chem. , 261, 1052 (1986)). Однако праймеры, которые могут амплифицировать участок, содержащий 1347 оснований, кодирующих lysC ген, также годятся, и, например, подходят два праймера, имеющие последовательность, определенную в последовательности SEQ ID 5 и SEQ ID 6.
Способ осуществления такой мутации, как замещение, вставка и делеция аминокислотного остатка (остатков) на АК111 гене, полученном, как указано ранее, представлен примером рекомбинантной PCR, методом сайтспецифического мутагенеза и т. п. , точно так же, как и мутационной обработкой DDPS гена, описанной ранее. Кроме того, в соответствии с химическим синтезом целевого гена оказывается возможным осуществлять мутацию или получать случайную мутацию в целевом сайте.
Кроме того, доступен способ, в котором ДНК АК111 гена на хромосоме или внехромосомную рекомбинантную ДНК непосредственно обрабатывают гидроксиламином (Hashimoto, Т. and Sekiguchi, M., J. Bacteriol., 159, 1039 (1984)). В другом варианте можно использовать способ, в котором бактерию, принадлежащую к роду Escherichia, содержащую АК111 ген на хромосоме, или внехромосомную рекомбинантную ДНК облучают УФ-светом, или способ, в котором осуществляют обработку таким химическим агентом, как N-метил-N'-нитрозогуанидин или азотистая кислота.
В отношении способа отбора мутантного АК111 гена штамм, полностью лишенный АК, например Е. coli GT3 штамм, вначале трансформируют рекомбинантной ДНК, содержащей АК111 ген, который был подвергнут мутационной обработке. Затем полученные трансформированные штаммы культивируют на минимальной среде, такой как М9, содержащей значительное количество L-лизина. Штаммы, содержащие рекомбинантную ДНК, содержащую дикого типа АК111 ген, не могут синтезировать L-треонин, L-изолейцин, L-метионин и диаминопимелиновую кислоту (ДАР), и их рост подавляется, так как только один АК ингибируется L-лизином. Напротив, штамм, содержащий рекомбинантную ДНК, содержащую мутантный АК111 ген, в котором ингибирование с помощью L-лизина десенсибилизировано, должен быть способен расти на минимальной среде, в которую добавлено значительное количество L-лизина. Этот факт можно использовать для отбора штамма, который устойчив во время роста к L-лизину или АЕС, как аналогу L-лизина, то есть, штамм, содержащий рекомбинантную ДНК, содержащую мутантный АК111 ген, в котором ингибирование десенсибилизировано.
Полученный таким образом мутантный ген можно ввести как рекомбинантную ДНК в подходящий микроорганизм (в хозяина) и экспрессировать. Таким образом можно получить микроорганизм, который содержит АК111, в которой было десенсибилизировано ингибирование в результате обратной связи.
Хозяином предпочтительно является микроорганизм, принадлежащий роду Escherichia, примером которого является Е. coli.
В другом варианте фрагмент мутантного АК111 гена можно получить из рекомбинантной ДНК, которую можно использовать в настоящем изобретении, является, предпочтительно, плазмидной векторной ДНК, примерами которой могут быть pUC19, pUC18, pBP322, pHSG299, pHSG298, pHSG399, pHSG398, PSF1010, pMWll8, pMW219 и pMW218. Кроме того, можно использовать векторы фаговой ДНК.
Далее, для эффективной экспрессии мутантного АК111 гена другой промотор, который функционирует в микроорганизмах, такой как lac, trp и PL, можно лигировать в обратном направлении от ДНК последовательности, кодирующей мутантный АК111, или промотор, содержащий АК111 ген, можно использовать, как он есть, или после его амплификации.
Кроме того, как было указано ранее, мутантный ген может быть встроен в автономно реплицируемую векторную ДНК, встроенную в хозяина, и оставлен в хозяине в качестве такой внехромосомной ДНК, как плазмида. В другом варианте мутантный ген можно интегрировать в хромосому микроорганизма хозяина способом, в котором используют трансдукцию, транспосон (Berg. D.E. и Berg, C.M., Bio. Technol., I, 417 (1983)), Mu фаг [японская открытая патентная выкладка 2-109985] или гомологичную рекомбинацию [Experiments in Molecular Genetics, Cold Spring Harbor Lab. 1972].
3. Получение L-лизина по способу настоящего изобретения
L-лизин можно эффективно получать, культивируя в предпочтительной среде бактерию, трансформированную в результате введения мутантного DDRS гена, полученного, как было указано ранее, и содержащую АК, которая десенсибилизирует ингибирование по типу обратной связи с помощью L-лизина, продуцирования и накопления L-лизина в этой культуре и выделения L-лизина из культуры. А именно, L-лизин можно эффективно получить, представив возможность бактерии, принадлежащей к роду Escherichia, содержать как мутантную DDPS, так и мутантную АК111.
Бактерия, принадлежащая к роду Escherichia, содержащая АК, в которой десенсибилизировано ингибирование по типу обратной связи с помощью L-лизина, может быть представлена бактерией, принадлежащей к роду Escherichia, трансформированной в результате включения в хромосомную ДНК, в ДНК, кодирующую АК111, содержащую мутацию для десенсибилизации ингибирования по типу обратной связи с помощью L-лизина, или в бактерию, принадлежащую роду Escherichia, трансформированную в результате введения в клетки рекомбинантной ДНК, сконструированной в результате лигирования ДНК с векторной ДНК, автономно реплицируемой в клетки бактерии, принадлежащей роду Escherichia. Далее, АК, в которой ингибирование по типу обратной связи с помощью L-лизина, десенсибилизировано, может быть дикого типа АК, без недостатка - ингибирования по типу обратной связи с помощью L-лизина, или АК, в которую такого дикого типа АК ген введен в бактерию, принадлежащую роду Escherichia, таким же способом. Далее, мутантный штамм бактерий, принадлежащих роду Escherichia, который стал продуцировать мутантную АК111 в результате мутационной обработки клеток бактерий, принадлежащих роду Escherichia, также может быть использован.
С другой стороны, для осуществления трансформации в результате введения мутантного DDPS гена в бактерию, принадлежащую роду Escherichia, мутантный DDPS ген можно ввести в хромосомную ДНК для осуществления трансформации или трансформацию можно осуществить в результате лигирования мутантного DDPS гена с векторной ДНК, автономно реплицируемой в клетках бактерии, принадлежащей роду Escherichia.
Если оба мутантных гена и мутантный ген АК111 введены в бактерию, принадлежащую роду Escherichia, оба мутантных гена могут быть интегрированы в хромосомный ДНК бактерий, принадлежащих роду Escherichia, и находится в них, либо они могут находиться в идентичной плазмиде или выделенных плазмидах в клетках в качестве внехромосомной ДНК. Если используют выделенные плазмиды, предпочтительно использовать плазмиды, обладающие стабильным распределительным механизмом, чтобы позволить каждой из них совместимо находиться в клетке. Далее, один из мутантных генов может быть интегрирован в хромосомную ДНК и находиться в ней, а другой мутантный ген может находиться в плазмиде в клетках в качестве внехромосомной ДНК соответственно. Если мутантный DDPS ген и мутантный АК111 ген вводят в бактерию, принадлежащую роду Escherichia, возможен любой порядок введения обоих генов.
Выход L-лизина можно далее увеличить в результате усиления дигидродипиколинатредуктазного гена бактерии, принадлежащей роду Escherichia, в которую были встроены мутантный DDPS ген и мутантный АК111 ген. Выход L-лизина можно еще более повысить в результате введения гена диаминопимелатдегидрогеназы, полученного из coryneform бактерии, в бактерию, принадлежащую роду Escherichia, в которой был усилен ген дигидродипиколинатредуктазы. Этот диаминопимелатдегидрогеназный ген должен быть усилен. В другом варианте выход L-лизина можно повысить в такой же степени в результате усиления гена тетрагидропиколинатсукциминазы и гена сукцинилдиаминопимелатдеацилазы вместо введения диаминопимелатдегидрогеназы.
Под усилением гена здесь подразумевают увеличение активности энзима как продукта экспрессии гена на клетку. Конкретнее, это можно представить как увеличение числа копий гена в клетке, увеличение количества экспрессии на ген в результате использования промотора, обладающего более высокой эффективностью экспрессии, и введения мутации для повышения активности энзима в ген. Для увеличения числа копий гена в клетке ген встраивают в вектор, автономно реплицируемый в бактерии, принадлежащей роду Escherichia, и бактерии, принадлежащие роду Escherichia, могут быть трансформированы этим вектором. Этот вектор является предпочтительно плазмидой мультикопийного типа. В другом варианте число копий можно повысить в результате амплификации ДНК, интегрированной в хромосомную ДНК, в результате использования Мu фага или т. п. Что касается использования плазмиды, если плазмиды используют для введения мутантного DDPS гена и мутантного АК111 гена, такие плазмиды, обладающие стабильным механизмом распределения предпочтительно используют, в которых эти плазмиды стабильно существуют в клетках вместе. Приемлемо любое другое введение генов.
Механизм будет раскрыт далее, причем в этом механизме выход L-лизина можно повысить постадийно в результате последовательного усиления генов системы биосинтеза L-лизина, как было указано ранее. Система биосинтеза, содержащая множество реакций, может быть сравнена с жидкостью, протекающей через множество трубопроводов, соединенных последовательно и имеющих различную толщину. Здесь каждый трубопровод соответствует отдельному энзиму, а толщина трубопровода соответствует скорости энзиматической реакции. Для увеличения количества жидкости, протекающей через трубопровод, необходимо увеличить сечение самого узкого трубопровода. В противном случае нечего ожидать какого-либо эффекта, даже если увеличить сечение толстого трубопровода. Для того чтобы еще увеличить поток на выходе, следует увеличить сечение второго наиболее тонкого трубопровода. С этой точки зрения авторы настоящего изобретения пытались усилить систему биосинтеза L-лизина. Для этой цели, как показано далее в примере 6, был установлен порядок стадий, определяющих скорость системы биосинтеза L-лизина в результате введения генов Е. coli системы биосинтеза L-лизина, полученных из Е. coli, постадийно. В этом исследовании 4 гена dapC (ген тетрагидропиколинатсукцинилазы), dapD (ген тетрагидродиалинатсукцинилазы), dapE (ген сукцинилдиаминопимелатдеацилазы) и dapF (ген диаминопимелатэпимеразы), расположенные в прямом направлении в схеме биосинтеза, были заменены геном DDH, кодирующим DDH (диаминопимелатдегидрогеназу) Brevibacterium lactofermentum, способный катализировать реакции, в которых участвуют сами эти генные продукты. А именно, введение генов для энзимов системы биосинтеза L-лизина и энзимов, кодируемых ими, следующее:
ррс: фосфоенолпируваткарбоксилаза
aspC: аспартатаминотрансфераза
lysC: аспартокиназа 111
lysC*: десенсибилизирующая ингибирование аспартокиназа 111
asd: аспартатполуальдегиддегидрогеназа
dapA: дигидродипиколинатсинтаза
dapA*: ингибирование десенсибилизирующая дигидропиколинатсинтаза
dapB: дигидродипиколинатредуктаза
DDH: диаминопимелатдегидрогеназа (полученная из Brevibacterium lactofermentum)
lysA: диаминопимелатдекарбоксилаза
В результате индивидуального введения каждого из генов в Е. coli было определено получение L-лизина в штаммах, в которые были введены lysC*, dapA или dapA*, и наибольший выход оказался для штамма, в который была введена dapA*. В соответствии с этим результатом было обнаружено, что реакция, катализируемая dapA, является первой, определяющей скорость. Затем, когда каждый из генов системы биосинтеза L-лизина был введен в штамм, содержащий dapA*, оказалось, что lysC* оказывает наибольшее влияние на увеличение выхода L-лизина. Таким образом, было обнаружено, что реакция, которую катализирует lysC, является второй стадией, определяющей скорость. Таким же способом было обнаружено, что реакция, катализируемая dapB, является третьей стадией, определяющей скорость, а реакция, катализируемая DDH, является четвертой. Далее, в результате исследований стадий, определяющих скорость, среди реакций, катализируемых dapC, dapD, dapE и dapF, замененных DDH, было обнаружено, что dapD и dapE связаны с определением скорости.
Способ получения генов системы биосинтеза L-лизина в Е. coli и DDH гена Brevibacterium lactofermentum описывается далее, ррс ген можно получить из плазмиды рS2 [Sabe, Н. et al.. Gene, 31, 279 (1984)] или рТ2, содержащих этот ген. ДНК фрагмент, содержащий ррс ген, получают, разрезая рS2 с помощью Aat11 и Af111. ДНК фрагмент, содержащий ррс ген, также получают, разрезая рТ2 с помощью SmaI и Scal. E. coli F15 штамм (AJ12873), содержащий рТ2, депонирован в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology [postal code: 305, 1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan] под регистрационным номером FERM BP-4732 на основе будапештского соглашения.
Ген аsрС получают из плазмиды pLF4 (Inokuchi, К. et al., Nucleic Acids Res. , 10, 6957 (1982)), содержащей этот ген. ДНК фрагмент, содержащий этот ген, aspC, получают, разрезая pLF4 с помощью Pvu11 и Stu1.
Ген asd получают из плазмиды pAD20 (Haziza, С., et al., ЕМВО, 1, 379 (1982)), содержащей этот ген. ДНК фрагмент, содержащий asd ген, получают, разрезая pAD20 с помощью Ase1 и Cla1.
Ген dapB получают, амплифицируя хромосомную ДНК Е. coli с помощью PCR, используя два вида олигонуклеотидных праймеров (например, последовательность SEQ ID 9 и 10), полученных на основе нуклеотидной последовательности известного dapB гена (Bouver, J. et al., J. Biol. Chem., 259, 14829 (1984)).
DDH ген получают в результате амплификации хромосомной ДНК Brevibacterium lactofermentum в результате PCR, используя два вида олигонуклеотидных праймеров (например, последовательность SEQ ID 11 и 12), полученных на основании известной нуклеотидной последовательности DDH гена Соrуnеbacterium glutamicum (Ishino S. et al., Nucleic Acid Res., 15, 3917 (1987)).
Ген lysA получают в результате амплификации хромосомной ДНК Е. coli с помощью PCR, используя два вида олигонуклеотидных праймеров (например, последовательность SEQ ID 13 и 14), полученных на основании нуклеотидной последовательности, известной как lуsА ген (Stragier, P., et al., J. Mol. Biol., 168, 321 (1983)).
Ген dapD получают в результате амплификации хромосомной ДНК Е. coli W3110 штамма с помощью PCR, используя два вида олигонуклеотидных праймеров (например, последовательность SEQ ID 15 и 16), полученных на основании нуклеотидной последовательности известного dapD гена (Richand, С., et al., J. Biol. Chem., 259, 14824 (1984)).
Ген dapE получают в результате амплификации Е. coli ДНК с помощью PCR, используя два вида олигонуклеотидных праймеров (например, последовательность SEQ ID 17 и 18), полученных на основании нуклеотидной последовательности известного dapE гена (Bouvier, J., et al., J. Bacteriol., 174, 5265 (1992)).
Ген dapF получают в результате амплификации хромосомной ДНК Е. coli с помощью PCR, используя два вида олигонуклеотидных праймеров (например, последовательность SEQ ID 19 и 20), полученных на основании нуклеотидной последовательности известного dapF гена (Richaud, С., et al., Nucleic Acid Res., 16, 10367 (1988)).
В настоящем изобретении любая бактерия, принадлежащая роду Escherichia, может быть использована в качестве хозяина при условии, что промотор мутантного DDPS гена, мутантный АК111 ген или другой ген системы биосинтеза L-лизина, или другой промотор для экспрессии этих генов функционирует в ее клетках, а источник репликации векторной ДНК, которую используют для введения функции в ее клетки, способен к репликации, если мутантный DDPS ген, мутантный АК111 ген или другой ген системы биосинтеза L-лизина вводится в плазмиду в качестве внехромосомной ДНК.
Так, например, они могут быть представлены L-лизинпродуцирующей Е. coli, конкретно мутантным штаммом, обладающим устойчивостью к аналогам L-лизина. Аналогом лизина является такой, который ингибирует пролиферацию бактерий, принадлежащих к роду Escherichia, но подавление полностью или частично десенсибилизируется, если L-лизин параллельно присутствует в среде. Так, например, существуют оксализин, гидроксамат лизина, АЕС, γ-метиллизин, α-хлоркапролактам и т.п. Мутантные штаммы, обладающие устойчивостью к этим аналогам лизина, получают, используя обычные операции искусственного мутагенеза для микроорганизмов, принадлежащих к роду Escherichia. Бактериальный штамм, который используют для получения L-лизина, конкретно можно представить примером Escherichia coli AJ11442 (депонированным как FERM BP-1543 и NRRL В-12185; см. японскую открытую патентную выкладку 56-18596 или патент США 4346170). В аспартокиназе описанных ранее микроорганизмов ингибирование по типу обратной связи с помощью L-лизина десенсибилизировано.
Кроме того, можно привести в качестве примера микроорганизмы, продуцирующие L-треонин, так как ингибирование их аспартокиназ с помощью L-лизина обычно также десенсибилизировано в микроорганизмах, продуцирующих L-треонин. В качестве L-треонин продуцирующих бактерий, принадлежащих к Е. coli, известно, что наивысшей продуктивностью к настоящему времени отличается штамм В-3996. Штамм В-3996 депонирован в Research Institute for Genetic and Industrial Microorganism Breeding под регистрационным RIA 1867.
Среда, которую используют для культивирования трансформантов, содержащих мутантный ген в соответствии с настоящим изобретением, является обычной средой, содержащей источник углерода, источник азота, неорганические ионы и необязательно другие органические компоненты.
В качестве источника углерода можно использовать такие сахара, как глюкоза, лактоза, галактоза, фруктоза или гидролизат крахмала; такие спирты, как глицерин или сорбит; или такие органические кислоты, как фумаровая кислота, лимонная кислота или янтарная кислота.
В качестве источника азота можно использовать неорганические соли аммония, например сульфат аммония, хлорид аммония или фосфат аммония; такие органические источники азота, как соевый гидролизат; аммиак (газообразный) или водный аммиак.
Желательно добавлять такие необходимые вещества, как витамин B1 и L-изолейцин или дрожжевой экстракт для того, чтобы они содержались в соответствующих количествах в качестве органических следовых элементов питания. Кроме того, при необходимости добавляют в небольших количествах фосфат калия, сульфат магния, ионы железа, магния и т.п.
Культивирование ведут предпочтительно в аэробных условиях в течение 16-72 часов. Температуру при культивировании поддерживают в интервале 25-45oС, а рН во время культивирования поддерживают в интервале 5-8. Для поддержания нужного значения рН можно использовать неорганические, органические, кислотные или щелочные вещества, а также газообразный аммиак.
Выделение L-лизина из ферментационного раствора обычно осуществляют, объединяя использование ионообменной смолы и осаждение, а также другие способы.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фиг.1 представляет стадии получения pdapA1 и pdapA2.
Фиг. 2 представляет ингибирование с помощью лизина дикого типа и мутантной DDPS.
Фиг. 3 представляет стадии получения плазмиды pdapAS824, содержащей ген типа dapA* с двойной мутацией.
Фиг.4 представляет стадии получения pLYSC1 и pLYSC2.
Фиг. 5 представляет степень появления и степень мутации трансформантов после обработки гидроксиламином.
Фиг. 6 представляет ингибирование L-лизином дикого типа и мутантной АК111.
Фиг. 7 представляет стадии получения плазмиды RSF24P, полученной из RSF1010, содержащей dapA*24.
Фиг.8 представляет стадии получения плазмиды pLLC*80.
Фиг. 9 представляет стадии получения плазмиды RSFD80, полученной из RSF1010, содержащей dapA*24 и lуsС*80.
Фиг. 10 представляет строение плазмид pdapA и pdapA*, содержащих dapA и dapA*.
Фиг.11 представляет строение плазмид plysC и plysC*, содержащих lysC или lуsС*80.
Фиг.12 представляет строение плазмиды рррс, содержащей ррс.
Фиг.13 представляет строение плазмиды раsрС, содержащей аsрС.
Фиг.14 представляет строение плазмиды pasd, содержащей asd.
Фиг.15 представляет строение плазмиды pdapB, содержащей dapB.
Фиг.16 представляет строение плазмиды pDDH, содержащей DDH.
Фиг.17 представляет строение плазмиды plysA, содержащей lysA.
Фиг. 18 представляет стадии получения плазмиды pCAB1, полученной из RSF1010, содержащей dapA*24, lysC*80 и dapB.
Фиг. 19 представляет стадии получения плазмиды pCABD2, полученной из RSF1010, содержащей dapA*24, lysC*80, dapB и DDH.
Фиг.20 представляет строение плазмиды pdapD, содержащей dapD.
Фиг.21 представляет строение плазмиды pdapE, содержащей dapE.
Фиг.22 представляет строение плазмиды pdapF, содержащей dapF.
Фиг. 23 представляет стадии получения плазмиды pMWdapDEl, содержащей dapE.
Фиг. 24 представляет стадии получения плазмиды pCABDEl, содержащей dapA*24, lуsС*80, dapB, dapD и dapE.
ПРЕДПОЧТИТЕЛЬНЫЙ ВАРИАНТ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Более конкретно настоящее изобретение будет раскрыто со ссылками на примеры.
ПРИМЕР 1: Получение мутантного DDPS гена
1. Клонирование дикого типа dapA гена
Нуклеотидная последовательность dapA гена Е. coli была известна (Richaud, F., et al., J. Bacteriol., 297 (1986)), и было известно, что в ней есть открытая считывающая рамка (ORF), содержащая 876 пар оснований, которая кодирует протеин, содержащий 292 аминокислотных остатка. Так как неизвестно, как регулируется этот dapA ген, участок, содержащий только SD последовательность и ORF за исключением промоторного участка, амплифицируют, используя PCR методику, и клонируют.
Полную геномную ДНК Е. coli K-12 MC1061 штамма экстрагируют в соответствии со способом Saito and Miura (Biochem. Biophys. Acta, 72, 619 (1963)). Два вида праймеров, содержащих последовательности, представленные как SEQ ID 1 и 2, получают и используют для осуществления PCR в соответствии со способом Erlich et al. (PCR Technology, Stockton press (1989)), и мишеневую ДНК амплифицируют. Полученную ДНК встраивают в коммерчески доступный клонирующий вектор pCR1000 для PCR фрагментов (получен от Invitrogen: Ltd., California, USA), как она есть. pCR1000 содержит lacZ промотор (Placz) и продается в состоянии, разрезанном по сайту в прямом направлении от lacZ промотора. Если рекомбинантную ДНК, полученную в результате лигирования PCR фрагмента, между обоими концами разреза pCR1000 вводят в Е. coli, PCR фрагмент транскрибируется под контролем lacZ промотора. После лигирования PCR фрагмента с помощью PCR1000 получают два вида плазмид, которые являются pdapA1 как плазмида, лигированная в нормальной ориентации, и pdapA2 как плазмида, лигированная в обратной ориентации относительно направления транскрипции dapA по отношению к направлению транскрипции с помощью lacZ промотора (фиг.1).
Если эти плазмиды вводят в Е. coli E7627, которая представляет штамм с дефицитом по DDPS, штаммы со встроенными плазмидами дополнены ауксотрофно для диаминопимелиновой кислоты хозяина JE7627. Таким образом, подтверждается, что ДНК фрагменты, встроенные в обе плазмиды, содержат ген dapA, кодирующий активную DDPS.
Трансформированный штамм, полученный в результате введения pdapAl в дикого типа Е. coli W3110 штамм [доступный из National Institute of Genetics (Mishima-shi, Shizuoka-ken, Japan)] , обозначают W3110/pdapAl, а трансформированный штамм, полученный в результате введения pdapA2 в Е. coli W3110 штамм, обозначают W3110/pdapA2 соответственно. Эти два трансформированных штамма культивируют соответственно в минимальной среде М9, содержащей следующий состав и дополненной АЕС в качестве аналога лизина. Штамм W3110 без введенной плазмиды также культивируют в той же самой среде в качестве контроля. Эти два трансформированных штамма и W3110 штамм, не содержащий плазмиды, подавляются в росте с помощью АЕС, однако ингибирование их роста снимается при добавлении L-лизина.
Минимальная среда М9:
А: (20•М9)
Na2HPО4•12H2O - 303 г/л
КН2РО4 - 60 г/л
NaCl - 10 г/л
NH4Cl - 20 г/л
В: 1 М MgSО4;
С: 50% глюкозы;
D: 1 г/л тиамина;
А, В, С и D, указанные выше, отдельно стерилизуют и смешивают в отношении А:В:С:D:вода = 5:0,1:1:0,1:95.
2. Получение мутантного DDPS гена (dapA*)
Предполагается, что штамм, содержащий плазмиду, содержащую dapA*, кодирующую DDPS с десенсибилизированным ингибированием, с помощью L-лизина может расти на минимальной среде М9 с добавлением значительного количества АЕС. Штамм, содержащий плазмиду, содержащую dapA*, отбирают по его устойчивости роста в присутствии АЕС.
Для эффективности получения dapA* dapA на pdapAl и pdapA2, полученные в 1, подвергают мутационной обработке.
(1-2-1) Исследование условий отбора для штамма, содержащего плазмиду, содержащую dapA*
Штамм W3110/pdapAl и штамм W3110/pdapA2, полученные, как указано ранее, культивируют на М9 агарозных пластинах, содержащих различные концентрации АЕС соответственно. Исследуют концентрации АЕС, ингибирующие рост и условия отбора, исследуют для штамма, содержащего плазмиду, содержащую dapA*.
Рост трансформантов на среде М9, содержащей АЕС в различных концентрациях, представлен в таблице 1. В этой таблице знак + означает рост трансформанта, а знак - означает отсутствие роста.
Направление транскрипции dapA гена на pdapA1 совпадает с направлением транскрипции с помощью lacZ промотора (фиг.1). Так было обнаружено, что dapA ген на pdapA1 обеспечивает устойчивость к АЕС в значительных концентрациях, даже если dapA остается как дикий тип, так как его экспрессируемое количество амплифицируется с помощью lacZ промотора, тогда как dapA ген на pdapA2 имеет меньшее экспрессируемое количество и обеспечивает ингибирование роста с помощью АЕС при более низких концентрациях, так как направление транскрипции обратно направлению в отношении lacZ промотора, а промотор самих dapA также в дефиците (рост подавляется в соответствии с добавлением 30 мМ в случае W3110/pdapA1 штамма, и 15 мМ в случае W3110/pdapA2 штамма). Было подтверждено, что ингибирование роста исключается в случае одновременного добавления L-лизина.
Поэтому pdapA используют в качестве объекта для введения мутации. Среду приготавливают, добавляя 60 мМ АЕС к минимальной среде М9, и используют для отбора штамма, содержащего плазмиду, содержащую dapA. Эту среду называют "селекционной средой" в примере 1 далее.
(1-2-2) In vitro мутационная обработка для pdapA2 с помощью гидроксиламина
In vitro способ мутационной обработки, в которой плазмиду непосредственно обрабатывают гидроксиламином, используют для введения мутации в плазмиду pdapA2.
2 мкг ДНК обрабатывают при 75oС в течение 1-4 часов в 0,4 М гидроксиламина [0,1 М KH2PО4 - 1 мМ EDTA (рН 6,0): 100 мкл, 1 М гидроксиламин - 1 мМ EDTA (рН 6,0): 80 мкл, ДНК: 2 мкг, всего: 200 мкл в результате добавления воды] . ДНК после обработки очищают на стеклянном порошке, вводят в Е. coli W3110, и высевают на полную среду [L-бульон: 1% Васто триптон, 0,5% дрожжевой экстракт, 0,5% NaCl, 1,5% агар], и образуются колонии. Их реплицируют на селекционную среду, описанную в разделе (1-2-1), и отбирают те, которые образуют колонии на селекционной среде. Получены предполагаемые мутантные плазмиды, всего 36 штаммов после двукратных экспериментов.
Предполагаемые штаммы из 36 штаммов, полученных таким образом, снова помещают на селекционную среду и подтверждают устойчивость к АЕС.
(1-2-3) Выделение dapA* гена и исследования на dapA* продукте
Выделяют мутантную pdapA2 из указанных ранее 36 штаммов. dapA-дефицитный штамм, JE7627 трансформируют этими плазмидами и дикого типа pdapA2 соответственно. Не содержащий клеток экстракт приготавливают из каждого из трансформированных штаммов и определяют энзиматическую активность DDPS.
Не содержащий клеток экстракт (неочищенный энзимный раствор) приготавливают следующим образом. Трансформированный штамм культивируют в 2•ТУ среде (1,6 Васто триптон, 1% дрожжевой экстракт, 0,5% NaCl) и собирают при оптической плотности на 660 нм (OD660) примерно 0,8. Клеточный осадок промывают 0,85% NaCl при 0oС и суспендируют в 20 мМ калийфосфатного буфера (рН 7,5), содержащего 400 мМ КСl. Клетки разрушают ультразвуком (0oС, 200 Вт, 10 мин). Раствор разрушенных клеток центрифугируют со скоростью 33000 об/мин в течение 1 часа при 0oС до получения надосадочной жидкости, к которой добавляют сульфат аммония до получения 80% насыщения, чтобы хранить при 0oС в течение ночи после центрифугирования. Осадок растворяют в 20 мМ калийфосфатного буфера (рН 7,5) - 400 мМ КСl.
Энзиматическую активность DDPS определяют в соответствии со способом Yugari et al. (Yugari, Y. and Gilvard, C., J. Biol. Chem., 240, 4710 (1962)). Поглощение реакционного раствора со следующим составом определяют при 37oС на спектрофотометре на длине волны 270 нм в зависимости от времени и определяют образующийся дигидродипиколинат. Пируват натрия удаляют из реакционной системы, чтобы использовать в качестве бланкового эксперимента.
Состав реакционного раствора:
50 мМ имидазол-НСl рН 7,4
20 мМ L-аспартатполуальдегид
20 мМ пируват натрия
энзимный раствор
вода до баланса: всего 1,0 мл.
Различные концентрации L-лизина добавляют к энзимному реакционному раствору во время измерений энзиматической активности DDPS и степень ингибирования с помощью L-лизина определяют. Как видно из фиг. 2, дикого типа DDPS страдает из-за ингибирования с помощью L-лизина. Мутантными плазмидами, полученными из трансформированных штаммов, с DDPS, которые не страдают ингибированием с помощью L-лизина по сравнению с дикого типа, оказались три вида из 36 предполагаемых плазмид. Их обозначили как pdapAS8, pdapAS9 и pdapAS24 соответственно. В соответствии со следующим определением нуклеотидных последовательностей было выявлено, что pdapAS8 и pdapAS9 имеют одинаковую мутацию.
Степени десенсибилизации ингибирования с помощью L-лизина различны для трех образцов мутантной DDPS, кодируемой pdapAS8, pdapAS9 и pdapAS24, однако ингибирование с помощью L-лизина было десенсибилизировано во всех трех образцах. Хотя на специфическую активность энзима могут влиять условия роста клеток и получения образцов, было обнаружено, что это в любом случае меньше по сравнению с результатами для дикого типа. Однако признано, что нет существенных проблем при использовании их в качестве материала для культивирования.
(1-2-4) Определение нуклеотидной последовательности мутантного dapA гена
Нуклеотидные последовательности мутантных dapA генов определяют в соответствии с обычной методикой, используя ДНК секвенатор АВ1 Модель 373А (Applied Biosistems Inc). В результате оказывается, что 487-й С изменен на Т в pdapAS8 и pdapA и 597-й С изменен на Т в pdapAS24 на последовательности SEQ ID 3. Поэтому оказалось, что 81-й аланиновый остаток был изменен на валиновый остаток в DDPS, кодируемой pdapAS8 и pdapAS9, и 118-й гистидиновый остаток изменен на тирозиновый остаток в DDPS, кодируемой pdapAS24 в аминокислотной последовательности, представленной в последовательности SEQ ID 4.
(1-2-5) Получение dapA с двойной мутацией
Два образца мутантных dapA генов получают, как указано ранее. Для того чтобы проверить, является ли десенсибилизирование ингибирования аддитивным для этих мутантов, получают плазмиду, содержащую мутантную dapA, содержащую обе эти мутации. Процедура получения представлена на фиг.3. Полученная плазмида с двумя мутациями обозначена как pdapAS824.
ПРИМЕР 2: Получение мутантного АК111 гена
1. Клонирование дикого типа lysC гена
Нуклеотидная последовательность АК111 гена (lysC) E. coli была известна (Cassan, М. , Parsot, С., Cohen, G.N., и Patte, J.C., J. Biol. Chem., 261, 1052 (1986)), и известно, что ее открытая считывающая рамка содержит 1347 пар оснований и кодирует протеин, содержащий 449 аминокислотных остатков. В этом гене присутствует оператор, и он подвергается супрессии с помощью L-лизина. Таким образом, для удаления операторного участка участок, содержащий только SD последовательность и ORF, амплифицируют, используя PCR способ, и клонируют.
Полную геномную ДНК E. coli K-12 MC1061 штамма получают в соответствии со способом Saito and Miura (Biochem. Biophys. Acta., 72, 619 (1963)). Два типа праймеров, имеющих последовательности, представленные в последовательностях SEQ ID 5 и 6, приготавливают, причем их используют для проведения PCR реакции в соответствии со способом Еrlich et al. (PCR Technology, Stockton press (1989)) и lysC ген амплифицируют. Полученную ДНК расщепляют с помощью BamHI и AseI, затем затупляют концы и встраивают в сайт SmaI мультикопийного вектора, рuС18. Этот SmaI сайт расположен в прямом направлении от lacZ промотора, существующего в векторе, и если рекомбинантную ДНК, полученную в результате встраивания ДНК фрагмента в SmaI сайт риС18, вводят в Е. coli, встроенный ДНК фрагмент транскрибируется в результате сквозного прочитывания транскрипции под контролем lacZ промотора. После встраивания PCR фрагмента в SmaI сайт puC18 получают два типа плазмид, одна плазмида - pLYSC1 - плазмида, встроенная в обратной ориентации, и другая плазмида - pLYSC2 - плазмида, встроенная в нормальной ориентации по отношению к направлению транскрипции lysC по отношению к направлению транскрипции lacZ промотора (фиг.4).
Если эти плазмиды используют для трансформации Е. coli GT3 (thrA1016, metM1005, lуsС1004) как полностью дефицитного штамма для АК1, 11 и 111, ауксотрофию GT3 для гомосерина и диаминопимелиновой кислоты дополняют. Таким образом, подтверждают, что ДНК фрагменты, встроенные в обе плазмиды, содержат ген lysC, кодирующий активную АК111.
Трансформированный штамм, полученный в результате введения pLYSCI в АК полностью дефицитный штамм, Е. coli GT3 обозначают как GT3/pLYSC1, а трансформированный штамм, полученный в результате введения pLYSC2 в Е. coli GT3, обозначают как GT3/pLYSC2. Значительное количество L-лизина добавляют к минимальной среде М9 и GT3/pLYSC1 штамм и GT3/pLYSC2 штамм культивируют соответственно. Оба штамма, GT3/pLYSC1 и GT3/pLYSC2, содержат плазмиды, содержащие дикого типа lysC, в которой АК111, кодируемая lysC, на плазмиде является одной АК. Дикого типа АК111 в качестве единственной АК ингибируется с помощью L-лизина. Так, оба штамма не могут синтезировать L-треонин, L-изолейцин, L-метионин и диаминопимелиновую кислоту (DAP), и их рост подавляется.
2. Получение мутантного АК111 гена (lysC*)
Предполагается, что штамм, содержащий плазмиду, содержащую lysC*, кодирующую АК с десенсибилизированным ингибированием с помощью L-лизина, может расти на минимальной среде М9, дополненной значительным количеством L-лизина. Штамм, содержащий плазмиду, содержащую lysC*, отбирают, селектируя штаммы по их устойчивости роста к L-лизину или АЕС, в качестве аналога L-лизина.
Для того чтобы эффективно получать lysC*, lysC на pLYSC1 и pLYSC2, полученные в (1), подвергают мутационной обработке.
(2-2-1) Исследование условий селекции для штамма, содержащего плазмиду, содержащую lysC*
Штаммы GT3/pLYSC1 и GT3/pLYSC2 культивируют на М9 агарных пластинах, содержащих различные концентрации L-лизина или АЕС соответственно. Ингибирующие рост концентрации с помощью L-лизина или АЕС исследуют и исследуют условия селекции для штамма, содержащего плазмиду, содержащую lysC*.
Рост трансформантов на среде М9, содержащей L-лизин или АЕС в различных концентрациях, представлен в таблице 2. В этой таблице знак + означает рост трансформантов, а знак ± означает слабый рост, а знак - соответствует отсутствию роста.
Направление транскрипции lysC гена на pLYSC2 совпадает с направлением транскрипции с помощью lacZ промотора (фиг.4). Так было обнаружено, что lysC ген на pLYSC2 обеспечивает устойчивость к L-лизину и АЕС в значительно более высоких концентрациях, даже если lysC остается как дикого типа, так как количество его экспрессии амплифицируется с помощью lacZ промотора, тогда как lysC ген на pLYSC1 имеет меньшую экспрессию и обеспечивает ингибирование роста с помощью L-лизина и АЕС при более низких концентрациях, так как направление транскрипции оказывается обратным по отношению к lacZ промотору, а сам промотор также в дефиците (рост не подавляется вплоть до добавления 100 мМ для L-лизина и вплоть до добавления 3 мМ АЕС в случае штамма GT3/pLYSC2, тогда как рост полностью подавляется в случае добавления 0,2 мМ как для L-лизина, так и для АЕС в случае штамма GT3/pLYSC1). Подтверждено, что ингибирование роста снимается в результате одновременного добавления гомосерина и диаминопимелиновой кислоты.
Поэтому pLYSC1 используют для экспериментов с введением мутаций. Среду получают, добавляя 10 мМ L-лизина или 0,2 мМ АЕС к минимальной среде М9, которую используют для отбора содержащих плазмиду штаммов, содержащих lysC*. Эту среду называют "селекционной средой" в примере 2 далее.
(2-2-2) In vitro мутационная обработка для pLYSC1 гидроксиламином
Для введения мутации в плазмиду pLYSC1 используют два типа способов, которые представляют собой способ мутационной обработки, в котором плазмиды непосредственно обрабатывают гидроксиламином, дополнительный способ in vivo мутационной обработки, в котором клетки, содержащие плазмиду, обрабатывают нитрозогуанидином (NTG) с последующим экстрагированием плазмиды для обеспечения разнообразия мутаций, а именно, ожидая мутацию, отличную от мутации от цитозина до тиамина под действием гидроксиламина.
(in vitro мутационная обработка гидроксиламином)
2 мкг ДНК обрабатывают в условиях 75oС в течение 1-4 часов в 0,4 М гидроксиламине (0,1 М КН2РО4 - 1 мМ EDTA (рН 6,0): 100 мкл, 1 М гидроксиламин - 1 мМ EDTA (рН 6,0): 80 мкл, ДНК: 2 мкг, всего 200 мкл, дополнено водой). ДНК после обработки очищают на стеклянном порошке, вводят в АК полностью дефицитный штамм, Е. coli GT3 штамм и засевают на полную среду (L-бульон: 1% Васто триптон, 0,5% дрожжевой экстракт, 0,5% NaCl, 1,5% агар) и образуются колонии. Их снова реплицируют на селекционную среду, описанную в разделе (2-2-1), и способные к росту штаммы (на этой селекционной среде) отбирают в качестве штаммов-кандидатов. Степень появления трансформантов и степень мутаций представлены на фиг.5. Мутантные штаммы получены в результате четырехчасовой обработки при достаточно высоком выходе 0,5-0,8%.
(in vivo мутационная обработка с помощью NTG)
В Е. coli MC1061 вводят pLYSC1 и обработку NTG проводят на полных клетках. Клетки после обработки культивируют в течение ночи для фиксации мутаций, а затем плазмиду экстрагируют и вводят в Е. coli GT3. Трансформированный штамм культивируют в 2•ТУ среде (1,6% Васто триптона, 1% дрожжевого экстракта, 0,5% NaCl), собирают при OD660 порядка 0,3, промывают ТМ буфером, описанным далее, затем суспендируют в NTG растворе (полученном в результате растворения NTG в концентрации 0,2 мг/мл в ТМ буфере) и обрабатывают при 37oС в течение 0-90 минут. Клетки промывают ТМ буфером и 2•ТУ средой, а затем мутации фиксируют в результате культивирования в 2•ТУ среде в течение ночи. Затем плазмидную ДНК экстрагируют из клеток и вводят Е. coli GT3 штамм. Скринирование штаммов-кандидатов осуществляют тем же способом, что и in vitro мутации, и получают мутанты, устойчивые к лизину (lysR) и устойчивые к АЕС (АЕСR).
ТМ буфер:
Трис - 50 мМ
Малеиновая кислота - 50 мМ
(NH4)2SO4 - 1 г/л
MgSO4•7Н2О - 0,1 г/л
Ca(NO3)2 - 5 мг/л
FeSO4•7Н2О - 0,25 мг/л
рН доводят до 6,0 с помощью NaOH
Получают всего 180 штаммов-кандидатов (обработка гидроксиламином: 48 штаммов, обработка NTG: 132 штамма); их снова наносят на селекционную среду и подтверждают устойчивость к АЕС и L-лизину для 153 штаммов. Учитывая разницу в характере накопления аминокислот в среде, эти 153 штамма подразделяют на 14 групп и определяют АК активности после отбора представительных штаммов для каждой из групп. Не существует большой разницы в АК активности между мутантными штаммами, полученными в результате обработки гидроксиламином и мутантными штаммами, полученными в результате обработки NTG. Поэтому последующие эксперименты проводят, не делая для них различии.
(2-2-3) Выделение lysC* гена и исследование lysC* продукта
В качестве репрезентативных штаммов вышеуказанных 14 групп выбирают штаммы 24, 43, 48, 60, 80, 117, 126, 149, 150, 156, 158, 167, 169 и 172. Мутантные плазмиды, полученные из pLYSCI, выделяют из каждого из них и обозначают соответственно как pLYSC1*24, pLYSC1*43, pLYSC1*48, pLYSC1*60, pLYSC1*80, pLYSC1*117, pLYSC1*126, pLYSC1*149, pLYSC1*150, pLYSC1*156, pLYSC1*158, pLYSC1*167, pLYSC1*169 и pLYSC1*172. AK полностью дефицитный штамм GT3 трансформируют ими и дикого типа pLYSC1. Не содержащие клеток экстракты получают для каждого из трансформированных штаммов и определяют энзиматическую активность АК111.
Не содержащий клеток экстракт (неочищенный раствор энзима) получают следующим образом. Трансформированный штамм культивируют в 2•ТУ среде и собирают при OD660 около 0,8. Клетки промывают 0,02 М КН2РО4 (рН 6,75) - 0,03 М β-меркаптоэтанола в условиях 0oС, а затем клетки разрушают, обрабатывая ультразвуком (0oС, 100 Вт, 30 мин•4). Раствор разрушенных клеток центрифугируют со скоростью 33000 об/мин в течение 1 часа в условиях 0oС до получения надосадочной жидкости, к которой добавляют сульфат аммония до получения 80% насыщения. После центрифугирования осадок растворяют в 0,02 М КН2РО4 (рН 6,75) - 0,03 М β-меркаптоэтаноле и хранят при 0oС в течение ночи.
Энзиматическую активность АК111 определяют в соответствии со способом Stadtman et al. (Stadtman, E.R., Cohen, G.N., Lebras G., и Pobichon-Szulmajster, H., J. Biol. Chem., 236, 2033 (1961)). А именно: реакционный раствор следующего состава инкубируют при 27oС в течение 45 минут и добавляют раствор FeCl3 (2,8н НСl 0,4 мл + 12% ТСА 0,4 мл + 5% FeCl3•6H2O / 0,1н НСl 0,7 мл) для проявления цвета, что центрифугируют с последующим измерением поглощения надосадочной жидкости на длине волны 540 нм. Активность определяют по количеству гидроксаминовой кислоты, которая образуется в минуту (1 ед = 1 мкмоль/мин). Коэффициент молярного поглощения составляет 600. Аспартат калия удаляют из реакционного раствора для использования в качестве сравнения.
Состав реакционного раствора, мл:
Реакционная смесь*1 - 0,3
Раствор гидроксиламина*2 - 0,2
0,1 М аспартат калия (рН 7,0) - 0,1
Раствор энзима Вода - До баланса
Всего - 1,0 мл
*1: 1 М Трис-НСl (рН 8,1) 9 мл + 0,3 М MgSО4 0,5 мл + 0,2 М АТР (рН 7,0) 5 мл
*2: 8 М раствор гидроксиламина нейтрализуют непосредственно перед использованием с помощью КОН.
Различные концентрации L-лизина добавляют к энзимному реакционному раствору для определения энзиматической активности АК и степень ингибирования с помощью L-лизина исследуют. Результаты представлены на фиг.6 и в таблице 3. Дикого типа и 24, 43, 48, 60, 80, 117 и 126 представлены на фиг.6А. 149, 150, 156, 158, 167, 169 и 172 представлены на фиг.6В.
Как видно из этих результатов, дикого типа АК111 сильно подвержена ингибированию с помощью L-лизина, который ингибирует около 50% при концентрации 0,45 мМ L-лизина и ингибирует около 100% при концентрации 5 мМ. Напротив, мутантные АК111, полученные на этот раз, обладают различными степенями десенсибилизации, однако ингибирование с помощью L-лизина оказалось десенсибилизированным во всех 14 образцах. Особенно в случаях 24, 80, 117, 169 и 172 ингибирование едва наблюдалось даже при концентрации лизина 100 мМ, и они обладали 50% ингибированием при концентрациях, не менее чем в 200 раз превышающих ситуацию, наблюдаемую для дикого типа. Специфическая активность в расчете на полный протеин, на который могут влиять условия роста клеток и приготовления образцов, были равны или были больше, чем для дикого типа почти во всех случаях, в которых было мало проблем снижения активности в результате введения мутаций (таблица 3). В соответствии с этим фактом постулируется, что активный центр АК111 не зависит от регуляторного сайта с помощью L-лизина с каждым из них. В таблице 3 степень десенсибилизации ингибирования (%) рассматривается как АК активность в присутствии 100 мМ L-лизина, что соответствует АК активности в отсутствии L-лизина в реакционном растворе. Термостабильность (%) означает степень сохранения активности после обработки при 55oС в течение 1,5 часа.
Соответственно затем исследуют термостабильность мутантных энзимов. Если нужно повысить активность энзима, важно, чтобы сконструированный энзим стабильно сохранялся в клетках. Измерения in vitro выдвигают ряд проблем из-за различия в активностях внутриклеточной и внеклеточной протеаз и влияния буферов на in vitro хранение энзимов. Однако для удобства термостабильность мутантных АК111 исследуют in vitro как один параметр.
На основании результатов исследований температуры инактивации АК111 в различных условиях определяют степень сохранения активности после обработки при 55oС в течение 90 минут.
Как видно из таблицы 3, половина из образцов энзимов гораздо эффективнее, нежели результаты для дикого типа. Вообще, мутантный энзим часто бывает более нестабильным, нежели энзим дикого типа. Однако некоторые мутантные АК111, полученные на этот раз, превосходят стабильность, характерную для дикого типа, и многие из них, по-видимому, гораздо предпочтительнее для использования при производстве L-лизина.
(2-2-4) Определение основной последовательности дикого типа lysC и мутантного lysC
Нуклеотидную последовательность дикого типа lysC гена, полученного в этих экспериментах, определяют обычным способом, используя ДНК секвенатор АВ1 модель 373А (Applied Biosystems Inc.) (SEQ ID 7). В результате обнаружено различие в шести сайтах (два участка на аминокислотном уровне) по сравнению с уже опубликованной последовательностью 1узС Е. coli K-12 JC411 штамма (Cassan, М. , Parsot, С. Cohen, G.N., и Patte, J.C., J. Biol. Chem., 261, 1052 (1986)). Предполагается, что это различие по шести сайтам связано с различием в использованных бактериальных штаммах.
Таким же образом определяют основные последовательности для каждого из lysC*, существующих в 14 образцах мутантных pLYSC1, и уточняют мутантные участки. Полученные результаты представлены в таблице 4. В этой таблице в скобках указаны мутации аминокислотных остатков на основании мутаций нуклеотидов. Всего имеет место 12 типов мутаций, так как два набора ( 4 и 167, 24 и 80) имеют совершенно одинаковые мутации среди 14 образцов. В отношении типов мутаций мутации 149, 150, 156, 158, 167, 169 и 172 получены в результате обработки гидроксиламином, а 24, 43, 48, 60, 80, 117 и 126 получены в результате NTG обработки. Однако, что касается характера мутаций, любые из них состоят в замене цитозина тимином или гуанина аденином в кодирующей цепочке в результате мутации от цитозина до тимина на некодирующей цепочке.
ПРИМЕР 3: Получение L-лизина в результате ферментации с использованием штамма, в который введена dapA*
Для получения L-лизина с использованием Е. coli по способу, раскрытому в японской открытой патентной выкладке 56-18596, в патенте США 4346170 и в статье в Applied Microbiology and Biotechnology, 15, 227-231 (1982), считается существенным, чтобы хозяин для усиления DDPS имел аспартокиназу, которая изменена таким образом, чтобы не испытывала подавления с помощью L-лизина. Примером такого штамма может служить L-треонин-продуцирующая бактерия. Что касается L-треонин-продуцирующей Е. coli, то штамм В-3996 обладает наиболее высокой продуктивностью среди известных к настоящему времени. Так, штамм В-3996 используют в качестве хозяина для оценки dapA*. Штамм В-3996 содержит в качестве отдельной плазмиды внехромосомно pVIC40. Подробности описаны в японской открытой патентной выкладке 3-501682 (РСТ). Этот микроорганизм депонирован в Research Institute for Genetics and Industrial Microorganism Breeding под регистрационным RIA 1867.
С другой стороны, dapA*, содержащаяся в pdapAS24 (в которой 118-й гистидиновый остаток заменен тирозиновым остатком), выбирают как dapA* для введения в Е. coli на основании степени десенсибилизации ингибирования и специфической активности энзима. Вначале для повышения количества экспрессируемой dapA* и повышения стабильности плазмиды мутантную dapA*, существующую на pdapAS24 (здесь и далее обозначаемую как "dapAS24"), лигируют в прямом направлении от промотора гена устойчивости к тетрациклину pVIC40, получают RSF24P, как показано на фиг.7.
Штамм, полученный в результате введения плазмиды RSF24P в E.coli JM109 штамм, обозначают как AJ12395, который депонирован в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology 28 октября 1993 г. под регистрационным номером FERM P-13935 и передан из исходного депозитария в международный депозитарий на основании будапештского соглашения 1 ноября 1994 г., и был депонирован под регистрационным номером FERM BP-4858. Штаммы, содержащие pdapAS8 и pdapAS9, не были депонированы. Однако все из участков мутаций dapA* на каждой из плазмид были выяснены, как будет раскрыто далее. Это не представляет трудностей для специалистов, и плазмиды выделяют из вышеуказанной депонированной бактерии по способу Maniatis et al. (Sambrook, J., Fritsch, E.F., Maniatis, Т., Molecular Cloning, Cold Spring Harbor Laboratory Press, 1.21 (1989)), и целевой ген получают, используя способ сайтнаправленного мутагенеза (Sambrook, J., et al., см. ранее, 15.63 (1989)).
pVIC40 иссекают из В-3996 штамма в соответствии с обычными способами и штамм В-3996 получают как штамм, не содержащий плазмиду. Плазмиду RSF24P вводят в В-399 штамм в соответствии с обычными способами и получают штамм B-399/RSF24P. Оценивают выход L-лизина с помощью B-399/RSF24P.
С другой стороны, RSFP конструируют в качестве контрольной плазмиды. А именно, большой фрагмент выбирают из расщепления pVIC40 дважды расщепленной BamHI и DraI, как представлено на фиг.7, и затупляют концы фрагментом ДНК полимеразы Кленова. Большой фрагмент с затупленными концами самолигируют для получения плазмидной RSFP. RSFP вводят в штамм В-399 обычными способами и получают штамм B-399/RSFP. Выход L-лизина определяют для B-399/RSFP.
Культивирование ведут при перемешивании со скоростью 114-116 об/мин в условиях культивирования: 37oС в течение 48 часов, используя следующую среду. Полученные результаты представлены в таблице 5.
Среда для получения L-лизина, г/л:
A: (NH4)2SО4 - 16
К2РO4 - 1
MgSО4•7H2O - 1
FeSО4•7H2O - 0,01
MnSО4•5Н2О - 0,01
Дрожжевой экстракт (Difco) - 2
L-метионин - 0,5
L-треонин - 0,1
L-изолейцин - 0,05
рН доводят до 7,0 с помощью КОН в автоклаве при 115oС в течение 10 мин - 16/20 объема
В: 20% глюкоза (автоклав при 115oС в течение 10 минут) - (4/20 объема)
С: Фармацевтический СаСО3 (термически стерилизованный в сухом состоянии при 180oС в течение 2 дней) - 30 г/л
А и В смешивают в отношении А:В=4:1, добавляют 30 г С к 1 л смеси и растворяют, а затем добавляют антибиотик (стрептомицин: 100 мкг/мл, канамицин: 5 мкг/мл).
ПРИМЕР 4: Получение L-лизина в результате ферментации с использованием штамма, в который введены dapA* и lysC* (I)
Влияние мутантной DDPS на производство L-лизина представлено в примере 3. Для осуществления дальнейшего усовершенствования мутантному АК111 гену, полученному в примере 2, позволяют сосуществовать с мутантным DDPS геном. Мутантный АК111 ген для сосуществования с мутантным DDPS геном выбирают из штамма 80 (lysC*80) исходя из энзиматической активности, термостабильности и т.п.
lуsС*80 используют после иссечения его из плазмиды pLLC*80 (фиг.8), полученной чередующимся лигированием lysC*, существующей на pLLSCl*80 (здесь и далее обозначаемой как "lуsС*80" - в прямом направлении от lacZ промотора вектора pHSG399, полученного Takara Shuzo Co., LTD), который содержит сайт со вставкой в обратном направлении по отношению к рuС18 для повышения экспрессии lysC*. pLLC*80 является плазмидой, полученной для создания lуsС*80 для того, чтобы направление транскрипции имело нормальную ориентацию по отношению к lacZ промотору для повышения производительности L-лизина, так как lysC*80 в pLYSCl*80 имеет направление транскрипции в обратной ориентации по отношению к lacZ промотору.
Плазмиду RSFD80, содержащую dapA* и lysC*, получают из pLLC*80 и RSF24P, полученных в примере 3 и представленных на фиг. 9. RSFD80 включает dapA*24 и lуsС*80 в таком порядке, чтобы обеспечить направление транскрипции в нормальной ориентации по отношению к tetP в прямом направлении от промотора (tetP) гена устойчивости к тетрациклину.
Плазмиду RSFD80 вводят в Е. coli JM109 штамм, который обозначают как А 12396, депонирован в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology 28 октября 1993 г. под регистрационным номером FERM P-13936, и передан из исходного депозитария в международный депозитарий на основании Будапештского соглашения 1 ноября 1994 г. под регистрационным номером FERM ВР-4859. Штаммы, содержащие pLYSC1*24, pLYSC1*43, pLYSC1*48, pLYSC1*60, pLYSC1*117, pLYSC1*126, pLYSC1*149, pLYSC1*150, pLYSC1*156, pLYSC1*158, pLYSC1*167, pLYSC1*169 и pLYSC1*172, не были депонированы. Однако все участки мутаций lysC* для каждой из этих плазмид были выяснены, как было указано ранее. Специалисту легко выделить плазмиды из вышеуказанной депонированной бактерии, используя способ Маniatis et al. (Sambrook et al. Molecular Cloning, 1.21 (1989)), и получить целевой ген, используя способ сайтнаправленного мутагенеза (там же 15.63 (1989)). RSFD80 вводят в штамм В-399 в соответствии с обычными способами и получают B-399/RSFD80. Оценивают производительность L-лизина штаммом B-399/RSFD80. В качестве контроля используют B-399/RSFP.
Культивирование осуществляют при перемешивании со скоростью 114-116 об/мин в условиях культивирования: температура 37oС, 48 часов, используя ту же среду для получения L-лизина, что и в примере 3.
Полученные результаты представлены в таблице 6.
ПРИМЕР 5: Получение L-лизина в результате ферментации с использованием штамма, в который введены dapA* и lysC* (II)
В примере 4 было подтверждено, что выход L-лизина может быть увеличен, если бактерия, принадлежащая роду Escherichia, содержит мутантный dapA ген и мутантный lysC ген.
Осуществляют эксперименты, которые подтверждают наличие или отсутствие эффекта при замене хозяина.
В качестве хозяина используют Е. coli W3110(tyrA) штамм. Этот W3110(tyrA) штамм подробно описан в Европейской патентной публикации 488424/92. Способ его получения кратко описан далее. Е. coli W3110 штамм получают из National Institute of Genetics (Mishima-shi, Shiruoka-ken, Japan). Этот штамм высевают на LB пластины, содержащие стрептомицин, и получают штамм, устойчивый к стрептомицину, в результате отбора штаммов, которые образуют колонии. Отобранный устойчивый к стрептомицину штамм смешивают с Е. coli K-12 ME8424 штаммом и стационарно культивируют в полной среде (L-бульон: 1% Васто триптона, 0,5% дрожжевого экстракта, 0,5% NaCl) в условиях 37oС в течение 15 минут для того, чтобы вызвать конъюгацию. Штамм Е. coli K-12 ME8424 имеет генетические характеристики (HfrPО45, thi, relAl, tyrA: Tn10, ung-1, nadB), и доступен от Национального Института Генетики.
Затем культуру засевают на полную среду (L-бульон: 1% Васто триптона, 0,5% дрожжевого экстракта, 0,5% NaCl, 1,5% агара), содержащую стрептомицин, тетрациклин и L-тирозин, и отбирают образующий колонии штамм. Этот штамм обозначают как Е. coli W3110(tyrA) штамм.
Между прочим, в Европейской патентной публикации 488424/92 описывается много штаммов, образованных в результате введения плазмид в W3110(tyrA) штамм. Так, например, штамм, полученный в результате введения плазмиды pHATerm, обозначен как Е. coli W3110(tyrA)/pHATerm штамм, и депонирован в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology, под регистрационным номером FERM BP-3653. Штамм W3110(tyrA)/pHATerm штамма. Выделение плазмиды можно осуществить в соответствии с обычными способами.
Плазмиду RSFD80, содержащую оба dapA* и lysC*, полученную в примере 4, вводят в W3110(tyrA), полученную, как указано ранее, и получают W3110(tyrA)/RSFD80. L-лизина выход оценивают для W3110(tyrA)/RSFD80. В качестве контроля RSFP вводят в W3110(tyrA) штамм в соответствии с обычными способами и получают W3110(tyrA)/RSFP. Выход L-лизина оценивают также и для W3110(tyrA)/RSFP в качестве контроля.
Культивирование осуществляют при перемешивании со скоростью 114-116 об/мин в условиях культивирования: 37oС в течение 48 часов, используя вышеуказанную среду для получения L-лизина. Полученные результаты приведены в таблице 7.
ПРИМЕР 6: Анализы стадий, определяющих скорость системы биосинтеза L-лизина и повышение производительности L-лизина с помощью L-лизинпродуцирующей бактерии, принадлежащей роду Escherichia
Была сделана попытка усовершенствовать выход L-лизина в результате анализа стадий, определяющих скорость системы биосинтеза L-лизина с помощью Е. coli и усиления генов энзимов, которые катализируют эти стадии.
1. Определение первой из стадий, определяющих скорость
(6-1-1) Получение генов системы биосинтеза L-лизина
Стадию, определяющую скорость, определяют, выделяя различные гены системы биосинтеза L-лизина, вводя эти гены в Е. coli, исследуя влияние каждого из генов на выход L-лизина. Вводимые гены для энзимов системы биосинтеза L-лизина и энзимы, кодируемые ими, следующие:
ррс: фосфоенолпируваткарбоксилаза
aspC: аспартатаминотрансфераза
lysC: аспартокиназа 111
lуsС*80: десенсибилизирующая ингибирование аспартокиназа 111
asd: аспартатполуальдегиддегидрогеназа
dapA: дигидродипиколинатсинтаза
dapA*24: десенсибилизирующая ингибирование дигидродипиколинатсинтаза
dapB: дигидродипиколинатредуктаза
DDH: диаминопимелатдегидрогеназа (полученная из Brevibacterium lactofermentum)
lysC: диаминопимелатдекарбоксилаза
Всю систему биосинтеза L-лизина из фосфоенолпировиноградной кислоты можно полностью охватить описанными выше генами. Гены dapC, dapD, dapE и dapF наряду с генами системы биосинтеза L-лизина, которой исходно обладает Е. coli, заменяют на ген DDH, кодирующий DDH (диаминопимелатдегидрогеназу) Brevibacterium lactofermentum, которая катализирует реакции, касающиеся самих этих генных продуктов. Штамм W3110(tyrA) штамм Е. coli K-12 серий используют в качестве хозяина для введения этих генов.
Гены dapA и dapA* соответственно получают в результате иссечения из pdapA2 и pdapA*24 (см. пример 1) с помощью EcoRI и КрnI (фиг.10). Эти гены лигируют с pMW118, которая была расщеплена EcoRI и KpnI до получения pdapA и pdapA*. Гены lysC и 1уsС*80 получают в результате иссечения из pLYSC1 и pLYSC*80 (см. пример 2) с помощью EcoRI и SphI. Эти гены лигируют с pMW119, которая расщеплена EcoRI и SphI до получения plysC и plysC* (фиг.11).
Ген ррс получают из плазмиды рТ2, содержащей этот ген. рТ2 вырезают SmaI и SсaI, и концы затупляют, а затем встраивают в SmaI сайт pMW118 до получения плазмиды рррс (фиг.12). Е. coli F15 (AJ12873), содержащая рТ2, была депонирована в National Institute of Bioscience and Human Technology of Agency of Industrial Science and Technology под регистрационным номером FERM BP-4732.
Ген aspC получают из плазмиды pLF4 (Inokuchi К. et al. Nucleic Acid Res. , 10, 6957 (1982)), содержащей этот ген (фиг.13). pLF4 вырезают с помощью PvuII и StuI и концы затупляют, а затем встраивают в SmaI сайт pMW119 для получения плазмидной paspC.
Ген asd получают из плазмиды pAD20 (Haziza, С., et al., ЕМВО, 1, 379 (1982)), содержащей этот ген. pAD20 вырезают с помощью AseI и ClaI, концы затупляют, а затем встраивают в SmaI сайт pMW118 для получения плазмиды pasd (фиг.14).
Ген dapB получают в результате амплификации dapB гена из хромосомной ДНК Е. coli W3110 штамма с помощью PCR, используя два вида олигонуклеотидных праймеров (последовательности SEQ ID 9 и 10), полученные на основе нуклеотидной последовательности известного dapB гена (Bouvier, J. et al., J. Biol. Chem., 259, 14829 (1984)) (фиг.15). Полученный амплифицированный ДНК фрагмент вырезают с помощью AseI и DraI, концы затупляют, а затем встраивают в SmaI сайт pMW119 для получения плазмиды pdapB.
Ген DDH получают в результате амплификации DDH гена из хромосомной ДНК Brevibacterium lactofermentum ATCC13869 с помощью PCR, используя два вида олигонуклеотидных праймеров (последовательности SEQ ID 11 и 12), полученные на основании известной нуклеотидной последовательности DDH гена Corynebacterium glutamicum (Ishino, S. et al., Nucleic Acids Res., 15, 3917 (1987)). Полученный амплифицированный ДНК фрагмент вырезают с помощью ЕсоТ221 и AvaI, концы затупляют, а затем встраивают в SmaI сайт pMW119 для получения плазмиды pDDH (фиг.16).
Ген lysA получают, амплифицируя lysA ген из хромосомной ДНК Е. coli W3110 штамма с помощью PCR, используя два вида олигонуклеотидных праймеров (последовательности SEQ ID 13 и 14), полученных на основании нуклеотидной последовательности известного lysA гена (Stragier, P. et al., J. Мо1. Biol., 168, 321 (1983)). Полученный амплифицированный фрагмент ДНК вырезают с помощью SplI и BelI, концы затупляют, а затем встраивают в SmaI сайт pMW118 для получения плазмиды plysA (фиг.17).
Подтверждением того факта, что каждый из вышеуказанных генов был клонирован, осуществляют, вырезая их рестрикционными энзимами, представленными на фигурах. Векторы pMW118 и pMW119 (продукция Nippon Gene), использованные для клонирования этих генов, были выбраны благодаря тому, что они способны к сосуществованию в клетках Е. coli вместе с RSF1010 как вектора, используемого для получения плазмид для производства лизина, описанных ранее, и также имеют стабильный механизм распределения.
(6-1-2) Производительность L-лизина с помощью Е. coli с введенными генами системы биосинтеза лизина
Е. coli W3110(tyrA) трансформируют каждой из плазмид, содержащих гены системы биосинтеза L-лизина, описанные ранее, и полученные трансформанты культивируют для осуществления продуцирования L-лизина. Такое культивирование осуществляют в течение 30 часов в условиях культивирования: температура 37oС, перемешивание со скоростью 114-116 об/мин, используя следующую среду. Полученные результаты приведены в таблице 8.
Состав среды, г/л:
Глюкоза - 40
МgSO4•7H2O - 1
(NH4)2SO4 - 16
КН2РО4 - 1
FeSО4•7Н2О - 0,01
MnSО4•5Н2О - 0,01
Дрожжевой экстракт (Difco) - 2
L-тирозин - 0,1
рН устанавливают 7,0 с помощью КОН для того, чтобы обработать в автоклаве при 115oС в течение 10 минут (глюкозу и МgSО4•7H2O стерилизуют отдельно).
Фармацевтический СаСО3 - 25
(термически стерилизованный в сухом состоянии при 180oС в течение 2 дней)
Антибиотики:
стрептомицин - 20
или ампициллин - 50 мг/л
в зависимости от вида плазмиды, подлежащей введению)
E. coli W3110(tyrA) начинает производить L-лизин в результате введения plysC*, pdapA или pdapA*. Так как оба продукта, и продукт lysC и продукт dapA, страдают от ингибирования по типу обратной связи с помощью L-лизина, можно утверждать, что эти энзимы и являются основными регулирующими точками в биоситезе L-лизина. Реакция, катализируемая dapA продуктом, идет в положении разветвления системы биосинтеза L-лизина и является первой стадией системы биосинтеза, присущего L-лизину. Уже сообщалось, что Е. coli также начинает производить L-лизин в результате амплификации дикого типа dapA (Eur. J. Appl. Microbiol. Biotechnol., 15, 227 (1982)), что также подтверждается приведенными ранее результатами. С другой стороны, результат примера 3 был снова подтвержден в том, что выход L-лизина можно повысить еще, если dapA* в качестве десенсибилизирующего ингибирование типа гена вводят в Е. coli.
Неочищенный энзимный раствор приготавливают из W3110(tyrA), W3110(tyrA)/pdapA и W3110(tyrA)/pdapA* по способу примера 1, определяют активность DDPS (дигидродипиколинатсинтазы) и исследуют степень ингибирования DDPS активности с помощью L-лизина. Полученные результаты приведены в таблице 9.
Продукт dapA*, десенсибилизирующий ингибирование, по-видимому, оказывает значительное действие на получение L-лизина, так как отличается высокой степенью десенсибилизации ингибирования, хотя имеет более низкую специфическую активность, нежели энзим дикого типа (около 1/2). Необходимость десенсибилизации ингибирования dapA продукта была показана для производства L-лизина.
Кроме того, тот факт, что lysC* оказывает влияние на продуцирование L-лизина, можно рассматривать следующим образом. Первая стадия, определяющая скорость, является стадией, на которой HD (гомосериндегидрогеназа продукт thrA или metLM) конкурирует с DDPS (продукт dapA) в использовании ASA (аспартат-β-полуальдегид) в качестве субстрата для служения в точке разветвления системы биосинтеза, и если dapA усиливается, как было указано ранее, реакция идет в направлении биосинтеза L-лизина. С другой стороны, можно предположить, что если подаваемое количество ASA повышают в результате усиления lysC, который участвует в реакции далее в обратном направлении от dapA, любые реакции, связанные с HD и DDPS, также облегчаются и, таким образом, продуцируемое количество L-лизина также возрастает. Однако этот эффект едва ли достижим только в результате усиления дикого типа lysC. Это, вероятно, связано с тем, что ингибирование дикого типа АК111 (продукт lys) с помощью L-лизина более жесткое, нежели для дикого типа DDPS (AK111 и DDPS ингибируются примерно на 100% и 80%, соответственно, в присутствии 5 мМ L-лизина).
В соответствии с вышеизложенными фактами можно судить, что реакция с помощью DDPS, обладающего более сильным лизинпродуцирующим эффектом, является первой стадией, определяющей скорость, и при этом постулируется, что реакция с помощью AK111 является второй стадией, определяющей скорость.
2. Определение второй стадии, определяющей скорость
Вторую стадию, определяющую скорость, идентифицируют в результате усиления различных генов системы биосинтеза L-лизина, в штаммах с введенными dapA*. Для того чтобы другие плазмиды стабильно сохранялись, когда их вводят в Е. coli, содержащую плазмиду, содержащую dapA*, dapA* переносят из pdapA в RSF1010 и получают RSF24P (фиг. 7). Е. coli W3110(tyrA) трансформируют плазмидой RSF24P, содержащей dapA*.
Плазмиды, содержащие гены системы биосинтеза L-лизина, вводят в Е. coli W3110(tyrA)/RSF24P. Два вида плазмид, а именно RSF24P и плазмида, содержащая другой ген из системы биосинтеза L-лизина, сосуществуют в каждом из полученных трансформантов. Продуктивность L-лизина исследуют для этих штаммов по способу (6-1-2). Полученные результаты приведены в таблице 10.
В результате оказалось, что заметный эффект усиления продуктивности L-лизина наблюдается только для lysC. Для дикого типа lysC эффект вовсе не наблюдается. Это, вероятно, связано с тем, что ингибирование за счет L-лизина достаточно сильно, как было указано ранее. Таким образом, было подтверждено, что реакция, в которой участвует lysC*, является второй стадией, определяющей скорость.
lysC* встраивают в RSF24P, и получают RSFD80 (фиг.9). Таким же способом lysC встраивают в RSF24P и получают плазмиду, которую обозначают как RSFD1. Эти плазмиды вводят в Е. coli W3110 (ТуrА), получают неочищенный ферментный раствор и определяют АК активность и степень ингибирования АК активностью L-лизином таким же образом, что и в (6-1-3). Полученные результаты представлены в таблице 11.
Специфические активности АК штаммов, содержащих эти плазмиды повышаются в 20-30 раз за счет встраивания lysC и lysC* в RSF24P. E.coli имеет три вида АК, и среди них lysC кодирует АК111. Однако полную активность трех видов АК определяют в описанном ранее эксперименте. Предполагается, что ингибирование с помощью L-лизина также возрастает в штамме, содержащем RSFD1 с встроенным дикого типа lysC, так как степень, занимаемая АК111, выше, нежели для АК1, и АК111 по сравнению с контролем (W3110(tyrA)/RSF24P), что приводит к отсутствию эффекта влияния на увеличение производительности L-лизина. С другой стороны, было выявлено, что ингибирование оказалось десенсибилизированным примерно на 100% АК111 в штамме, содержащем RSFD80, и этот факт вносит вклад в повышение производительности L-лизина.
3. Определение третьей стадии, определяющей скорость
Затем различные плазмиды системы биосинтеза L-лизина вводят в Е. coli W3110 (tyrA)/RSFD80 и культивируют для получения лизина. Результаты такого культивирования приведены в таблице 12.
Эффект увеличения производительности L-лизина наблюдают только для dapB, и было обнаружено, что реакция, в которой участвует dapB, является третьей стадией, определяющей скорость. Так, dapB встраивают в RSFD80 и получают рСАВI (фиг.18). Эту плазмиду вводят в Е. coli W3110(tyrA), получают неочищенный энзимный раствор и определяют энзиматическую активность DDPS (дигидродипиколинатредуктазы) по способу, описанному Tamir, H., and Gilvard, С. , J. Biol. Chem., 249, 3034 (1974). Таким же способом приготавливают неочищенный энзиматический раствор из штамма, содержащего только RSFD80, и штамма, содержащего как RSFD90, так и pdapB, и определяют активность DDPR. Полученные результаты приведены в таблице 13.
DDPR активность возрастает примерно в 3 раза для штамма, содержащего как RSFD80, так и pdapB, и примерно в 6,5 раз для штамма, содержащего pCAB1, в котором dapB был встроен в RSFD80, по сравнению с контролем (штамм, содержащий только RSFD80).
В соответствии с тем фактом, что оба W3110(tyrA)/RSFD80+pdapB и W3110(tyrA)/pCAB1 отличаются одинаковым накоплением L-лизина в 10,8 г/л, был сделан вывод, что dapB определяет достаточное количество производимого L-лизина, и внимание переносится на следующую стадию, определяющую скорость.
4. Определение четвертой стадии, определяющей скорость
Четвертую стадию, определяющую скорость, определяют, используя плазмиду pCABI, содержащую lysC*, dapA* и dapB. Различные плазмиды системы биосинтеза L-лизина вводят в Е. coli W3110(tyrA)/pCAB1 и осуществляют культивирование для получения L-лизина. Результаты культивирования приведены в таблице 14.
Эффект повышения производительности L-лизина наблюдается только для DDH, и было обнаружено, что реакция, катализируемая DDH, является четвертой стадией, определяющей скорость. Кроме того, SDAP (N-сукцинил-L, L-α, ε-диаминопимелиновая кислота) детектируется в культуральном бульоне штамма, в который DDH не был введен. Детектирование SDAP осуществляют с помощью проявления ТСХ (проявляющий растворитель: метанол: вода: 10н. НСl: пиридин 80: 175: 2,5:10) (Bouvier, J., Richard, C., Higgins Bogler, О., and Stragier, P. , J. Bacteriol., 174, 5265 (1992)). Далее цвет бульона был коричневым в случае штамма без DDH, но он изменялся на желтый в случае введения DDH в штамм. Таким образом, DDH вводят в рСАВ1 для получения плазмиды pCABD2 (фиг. 19) и определяют DDH активность Е. coli W3110(tyrA), трансформированного этой плазмидой. DDH энзиматическую активность определяют в соответствии с литературными данными (Azizono, Haruo, Fermentation and Industry, 45, 964 (1987)). Полученные результаты представлены в таблице 15.
DDH активность не определяется в контроле (W3110 (tyrA)/pCAB1), так как DDH исходно в Е. coli не присутствует. Удельная активность DDH штамма, содержащего pCABD2 (W3110(tyrA)/pCABD2) была примерно в 2,5 раза выше, чем для штамма, содержащего pDDH (W3110(tyrA)/pCAB1+pDDH), однако оба штамма дали одинаковое количество накопленного L-лизина (12,23 г/л). Поэтому можно судить, что количество, экспрессируемое в случае DDH pCABD2, является достаточным количеством.
5. Анализ стадий, определяющих скорость среди dapC, dapD, dapE и dapF
Затем для исследования ограничивающего порядка из dapC, dapD, dapE и dapF, замененных DDH в описанном ранее анализе, вначале эти гены клонируют. dapC не был клонирован, так как не была известна его последовательность оснований, однако остальные три образца генов были клонированы в соответствии со способом PCR.
dapD ген получают в результате амплифицирования dapD гена из хромосомной ДНК Е. coli W3110 штамма с помощью PCR, используя два типа олигонуклеотидных праймеров (последовательности SEQ ID 15 и 16), полученных на основании нуклеотидной последовательности известного dapD гена (Richend, С., J. Biol. Chem. , 259, 14824, (1984)). Полученный амплифицированный фрагмент разрезают Есо01091 и SaсI, концы затупляют, а затем встраивают в SmaI сайт pMW118 для получения плазмиды pdapD (фиг.20).
dapE Ген получают в результате амплификации dapE гена из хромосомной ДНК Е. coli W3110 штамма с помощью PCR, используя два вида олигонуклеотидных праймеров (последовательности SEQ ID 17 и 18), полученных на основании нуклеотидной последовательности известного dapE гена (Bouvier, J. et al., J. Bacteriol. , 174, 5265 (1992)). Полученный амплифицированный ДНК фрагмент разрезают MunI и BglII, и концы затупляют, затем встраивают в SmaI сайт pMW118 для получения плазмиды pdapE (фиг.21).
Ген dapF получают в результате амплификации dapF гена из хромосомной ДНК Е. coli W3110 штамма с помощью PCR, используя два вида олигонуклеотидных праймеров (последовательности SEQ ID 19 и 20), полученные на основании нуклеотидной последовательности известного dapF гена (Richand, С., et al., Nucleic Acid Res., 16, 10367 (1988)). Полученный амплифицированный ДНК фрагмент вырезают с помощью PstI и концы затупляют, а затем встраивают в SmaI сайт pMW118 для получения плазмиды pdapF (фиг.22).
Каждую из полученных таким образом плазмид вводят в W3110(tyrA)/pCAB1 и культивируют для получения L-лизина. В ранних экспериментах наблюдалось изменение цвета бульона и в присутствии или в отсутствие накопления промежуточного SDAP в дополнении к получаемому количеству L-лизина между введением DDH, и до этого введения. Таким образом, анализ стадии, определяющей скорость, осуществляют также, используя их в качестве показателей. Полученные результаты приведены в таблице 16.
Получаемое количество L-лизина слегка повышается в результате усиления dapD или dapE, но DDH не превышается. Кроме того, было обнаружено, что изменение цвета бульона и накопление SDAP в качестве промежуточного, наблюдаемое после введения DDH, является независимым друг от друга явлением, причем изменение цвета бульона связано с dapD, а исчезновение SDAP зависит от dapE. Связь между SDAP и dapE можно постулировать на основании характера биосинтеза L-лизина. Усиление dapF не оказывает действия на повышение производительности L-лизина.
dapE иссекают из pdapE и встраивают в pdapD для получения плазмиды pMWdapDE1, содержащей как dapE, так и dapD (фиг.23). Кроме того, фрагмент, содержащий dapE и dapD, иссекают из pMWdapDE1 и встраивают в рСАВ1 для получения pCABDE1 (фиг. 24). Получают штаммы, содержащие pCAB1, pCABDE1 или pCABD2 и штамм, содержащий как pCABDE1, так и pdapF, и культивируют для получения L-лизина. Полученные результаты представлены в таблице 17.
Было обнаружено, что количество продуцируемого L-лизина, цвет бульона и наличие или отсутствие накопления SDAP становится эквивалентным тем, что в случае продуцирования DDH в результате усиления dapD и dapE в сочетании. Кроме того, было обнаружено, что дальнейшее усиление dapF не оказывает влияния на увеличение выхода L-лизина, и реакция, в которой участвует dapF, не представляет ограничения скорости. Полученные результаты можно интерпретировать следующим образом.
После стадии введения рСАВ1 промежуточные накопленные на двух стадиях SKAP (N-сукцинил-ε-кето-L-α-аминопимелиновая кислота) и SDAP. Среди этих промежуточных SDAP детектируется во внеклеточном бульоне. Хотя SKAP не детектируется, можно предположить, что накопление происходит в бактериальных клетках. Причина для такого предположения состоит в цвете бульона. Цвет бульона является желтым в случае дикого типа штамма (W3110(tyrA)) или аналогично не продуцирует L-лизина. Однако бульон становится коричневым, вероятно, из-за бактериолиза или т. п., если штаммы оставляют для роста, Можно предположить, что SDAP оказывает мало влияния на рост, так как он выводится из клеток, и, следовательно, бульон изменяет свой цвет на желтый, хотя накопленное количество SDAP возрастает, если SKAP метаболизирует в результате усиления только dapD. Однако даже если dapD усиливается, накопленное количество L-лизина не возрастает, если только не десенсибилизировать ограничение скорости дальнейшим dapE.
6. Выводы
В соответствии с раскрытыми ранее результатами было обнаружено, что производительность L-лизина возрастает постадийно в результате: (1) введения dapA*, (2) введения lysC*, (3) усиления dapB и (4) усиления DDH или dapD и dapE в бактерии, принадлежащей роду Escherichia. Далее, была получена Е. coli, в которой увеличена продуктивность L-лизина постадийно.
7. Анализ стадии, определяющей скорость системы биосинтеза L-лизина в штамме Е. coli
Для того чтобы определить, можно ли применять полученные выводы к штаммам, отличающимся от штамма Е. coli К-12 серии, стадии, определяющие скорость системы биосинтеза L-лизина с помощью Е. coli, С штамм (IFO 13891) анализируют описанным ранее способом. Условия культивирования были теми же, что и для W3110(tyrA), однако к среде не добавляли L-тирозин.
(1) Определение первой стадии, определяющей скорость
Е. coli С штамм (IFO 13891), трансформированный плазмидами, содержащими гены системы биосинтеза L-лизина, культивируют в среде для получения L-лизина и определяют получаемое количество гидрохлорида L-лизина. Полученные результаты приведены в таблице 18.
Таким же образом, что и в W3110(tyrA), L-лизин накапливается в среде с помощью С штамма при введении дикого типа dapA и дальнейшем десенсибилизировании ингибирования типом dapA*. lysC* не оказывает влияния на продуцирование L-лизина.
(2) Определение второй стадии, определяющей скорость
Плазмиду RSF24P, содержащую dapA*, вводят в Е. coli С штамм и далее вводят плазмиды, содержащие гены системы биосинтеза L-лизина. Полученные трансформанты культивируют в среде для получения L-лизина и определяют количество продуцируемого гидрохлорида L-лизина. Полученные результаты приведены в таблице 19.
Было обнаружено, что lysC* оказывает действие на увеличение продуцирования L-лизина даже в том случае С штамма с трансформированным dapA*, и реакция, в которой принимает участие lysC*, является второй стадией, определяющей скорость.
(3) Определение третьей стадии, определяющей скорость
Плазмиду RSFD90, содержащую dapA* и lysC*, вводят в Е. coli С штамм и затем вводят плазмиды, содержащие гены системы биосинтеза L-лизина. Полученные трансформанты культивируют в среде для получения L-лизина и определяют количество продуцируемого L-лизина. Полученные результаты представлены в таблице 20.
Таким же образом, что и для W3110 штамма, только dapB оказывает влияние на повышение продуцирования L-лизина, и это оказывается третьей стадией, определяющей скорость.
(4) Определение четвертой стадии, определяющей скорость
Плазмиду pCAB1, содержащую dapA*, lysC* и dapB, вводят в Е. coli С штамм, а затем вводят плазмиды, содержащие гены системы биосинтеза L-лизина. Полученные трансформанты культивируют в продуцирующей L-лизин среде и определяют количество продуцируемого гидрохлорид L-лизина. Полученные результаты представлены в таблице 21.
Таким же образом, как и для штамма W3110, только DDH оказывает влияние на повышение выхода L-лизина, и это считают четвертой стадией, определяющей скорость.
(5) Анализ стадий, определяющих скорость, среди dapC, dapD, dapE и dapF
В Е. coli С штамм, содержащий pCAB1, вводят плазмиды, содержащие dapD, dapE или dapF гены, вместо DDH, и осуществляют культивирование для получения L-лизина. Полученные результаты приведены в таблице 22.
Было обнаружено, что две стадии dapD и dapE также имеют отношение к определению скорости для С штамма, таким же образом, что и для W3110 штамма.
Как было указано ранее, штаммы К-12 и С, принадлежащие к различным сериям, имеют один и тот же порядок определения скорости. Таким образом, можно считать, что все виды Е. coli могут быть использованы в концепции, гласящей, что продуцирование L-лизина можно увеличить постадийно в результате введения dapA* и lysC* и усиления dapB и DDH (или (dapD и dapE) в указанном порядке.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
В соответствии с настоящим изобретением был получен DDPS мутантный ген, происходящий из бактерии, принадлежащей роду Escherichia, в котором ингибирование по типу обратной связи с помощью L-лизина существенно десенсибилизировано. Продуцирующие L-лизин бактерии более усовершенствованы, нежели те, которые были получены ранее в результате введения гена в бактерии, принадлежащие роду Escherichia, содержащие аспартокиназу, в которой ингибирование по типу обратной связи с помощью L-лизина было десенсибилизировано.
Кроме того, продуцирование L-лизина можно увеличить постадийно в результате усиления dapB и DDH (или dapD и dapE) в вышеупомянутых продуцирующих L-лизин бактериях в таком порядке.
| название | год | авторы | номер документа |
|---|---|---|---|
| ДНК, КОДИРУЮЩАЯ АСПАРТОКИНАЗУ III БАКТЕРИЙ, И СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 1997 |
|
RU2204604C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА (ВАРИАНТЫ) И РЕКОМБИНАНТНАЯ ДНК, ИСПОЛЬЗУЕМАЯ ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ (ВАРИАНТЫ) | 1996 |
|
RU2197528C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2368659C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2351653C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2250266C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ Methylophilus И СПОСОБ ПОЛУЧЕНИЯ БАКТЕРИАЛЬНЫХ КЛЕТОК Methylophilus | 2000 |
|
RU2334794C2 |
| БАКТЕРИЯ - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2005 |
|
RU2316588C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2009 |
|
RU2518677C2 |
| СПОСОБ ПРОИЗВОДСТВА L-АМИНОКИСЛОТ | 2007 |
|
RU2422530C2 |
| СПОСОБ ПОЛУЧЕНИЯ ОСНОВНОГО ВЕЩЕСТВА | 2005 |
|
RU2350655C2 |












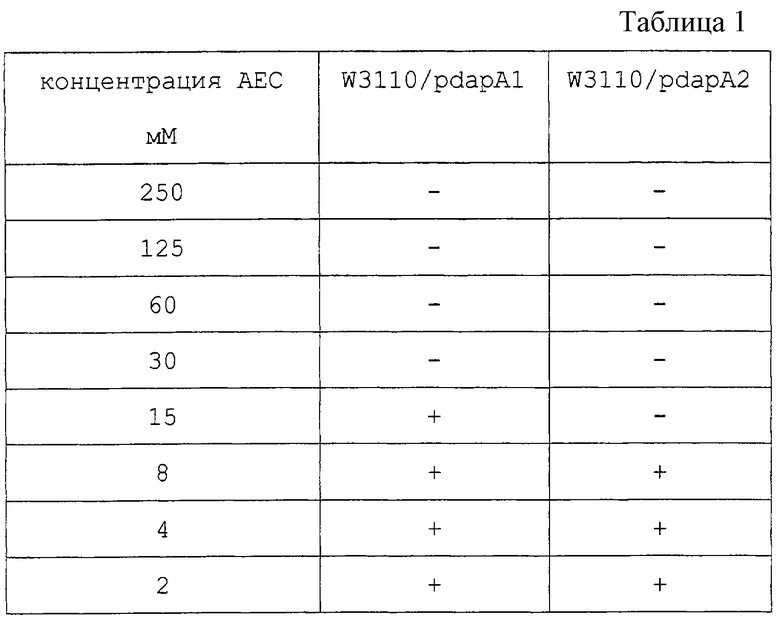

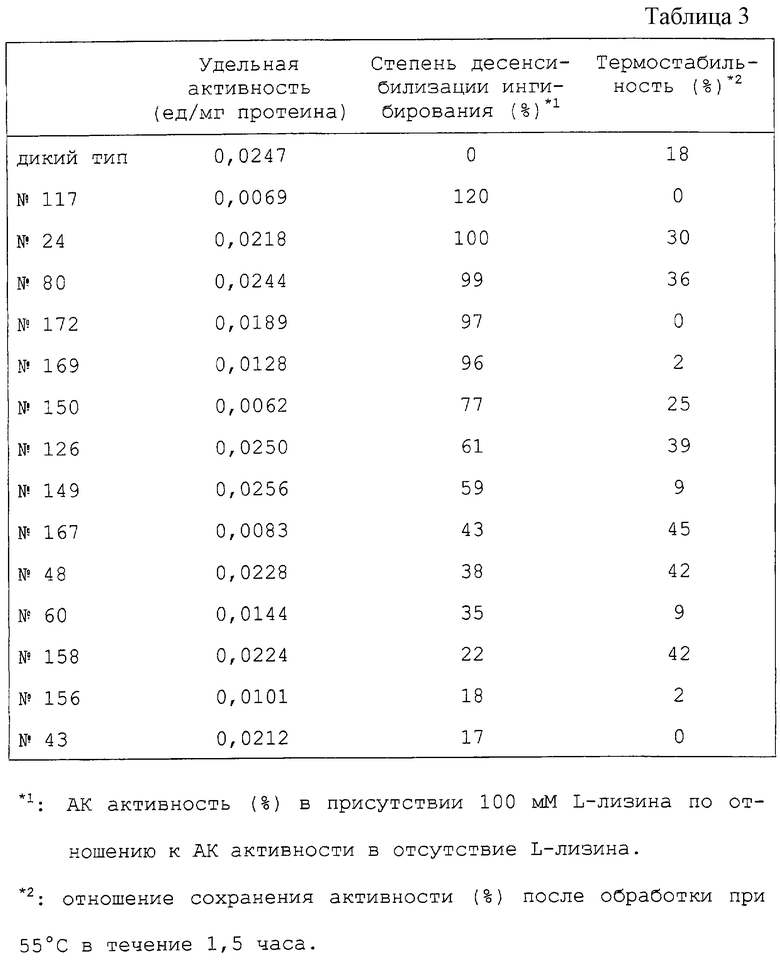



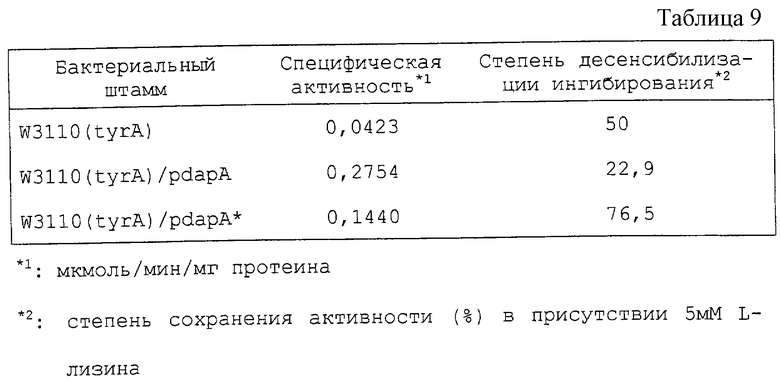
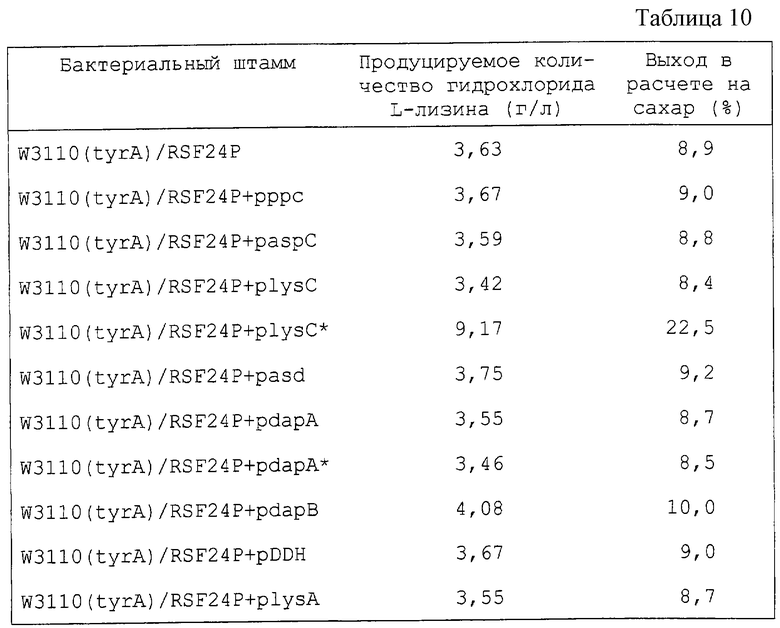
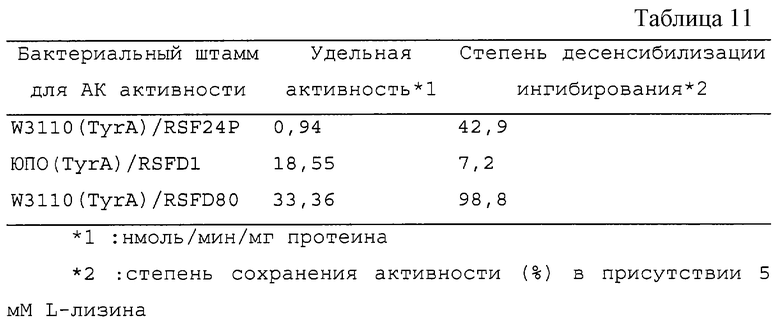
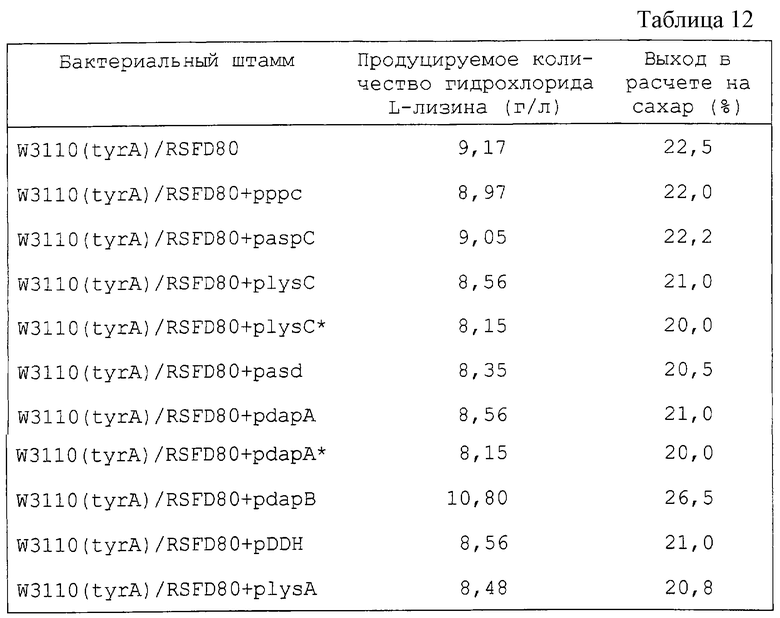
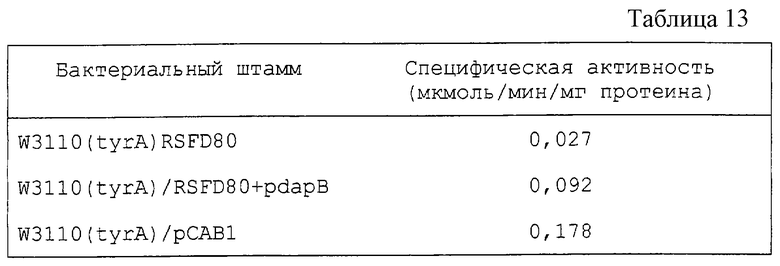
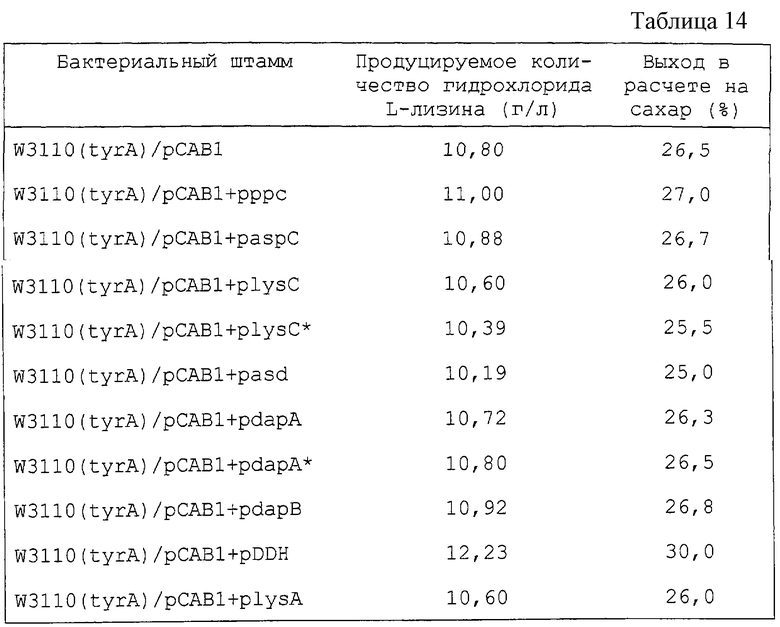
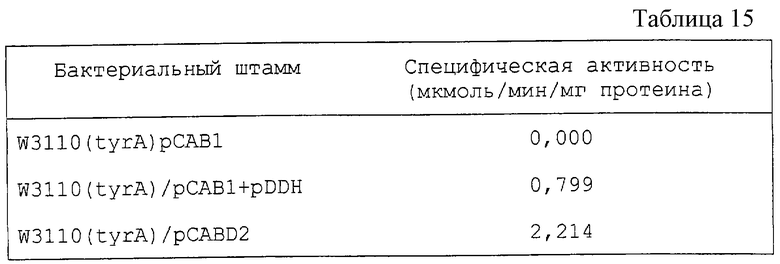
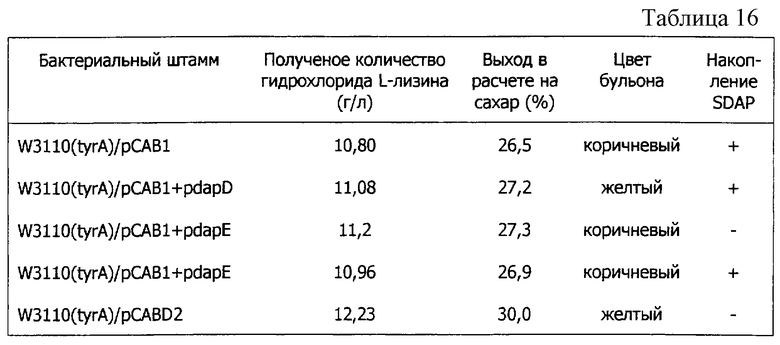
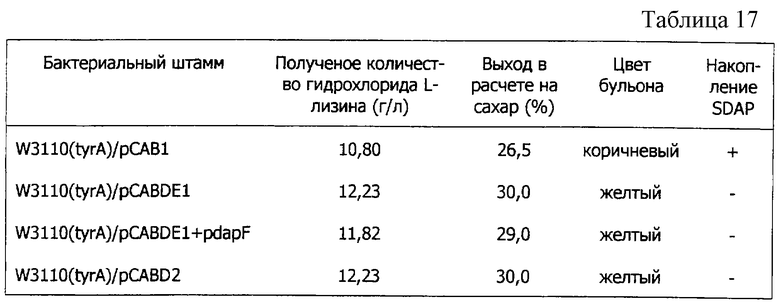
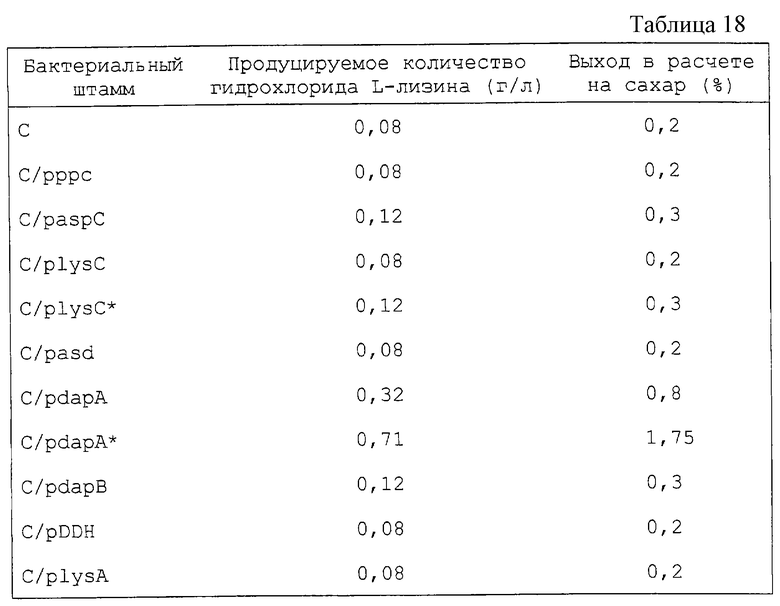
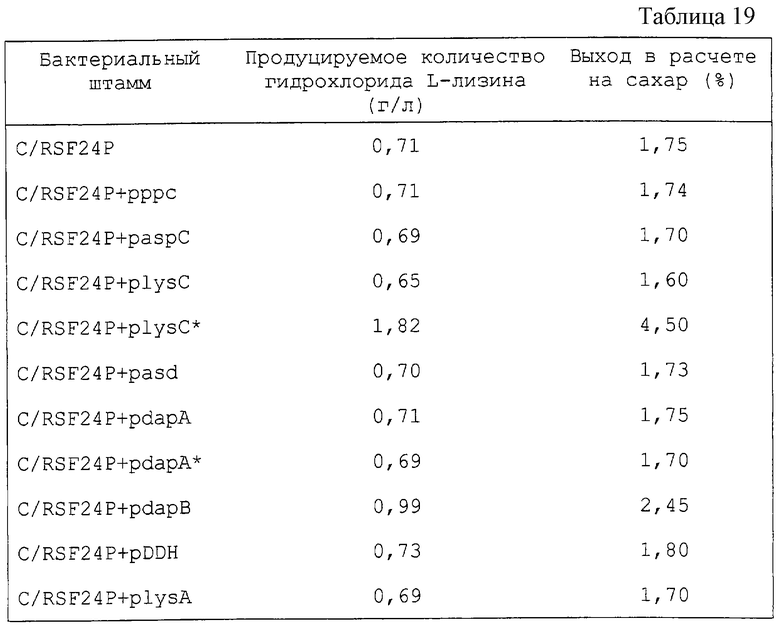
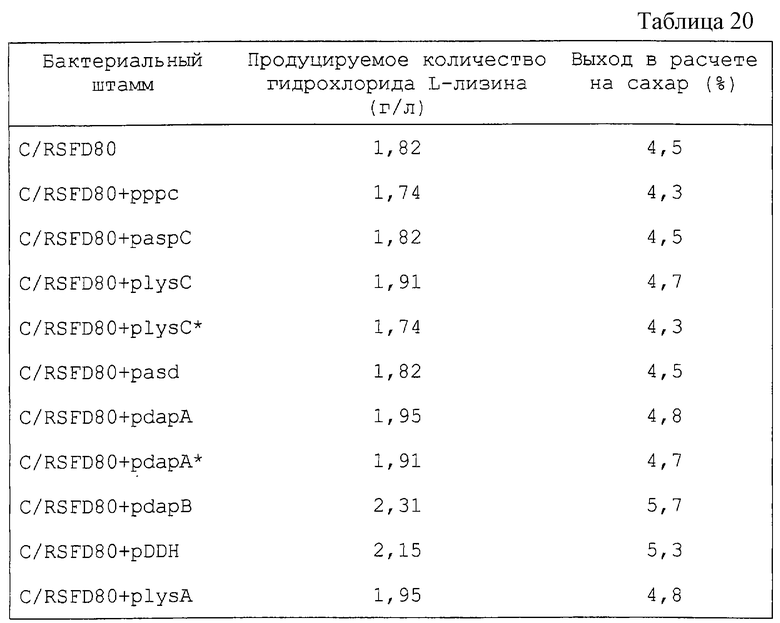
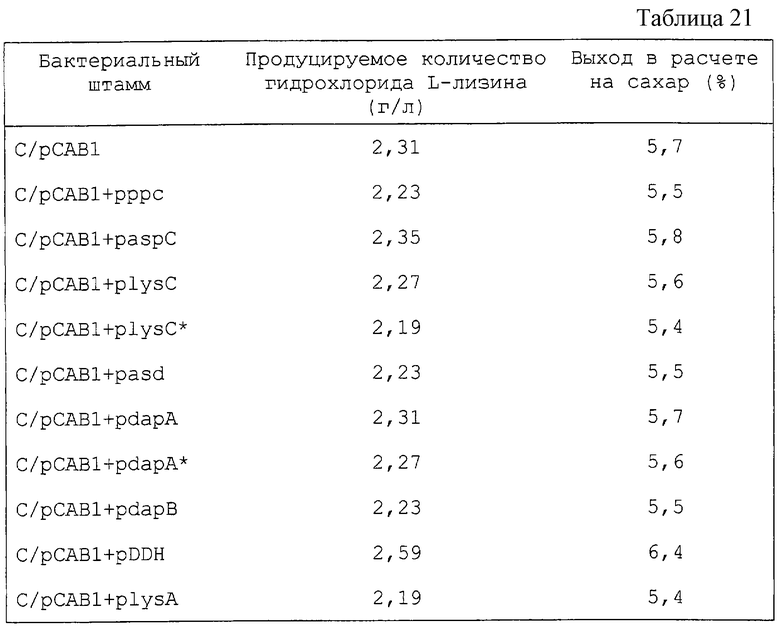
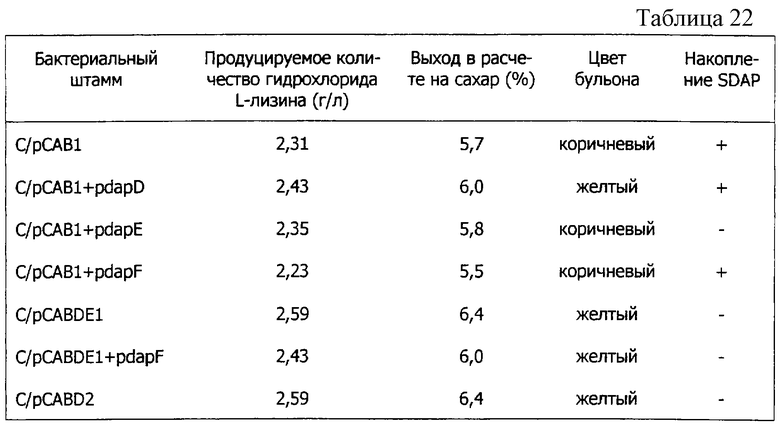
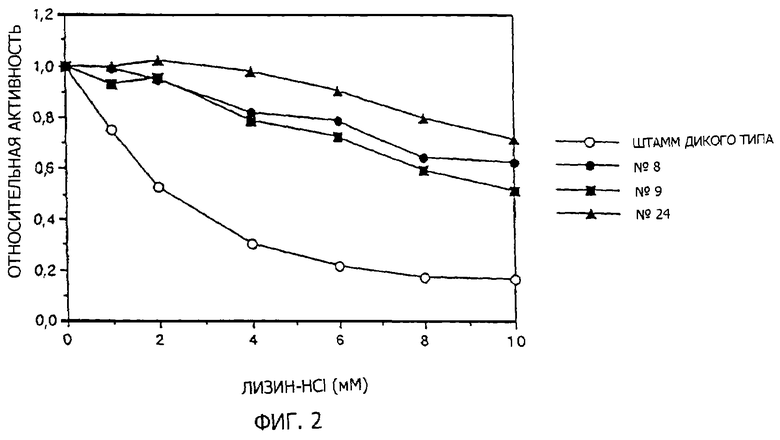
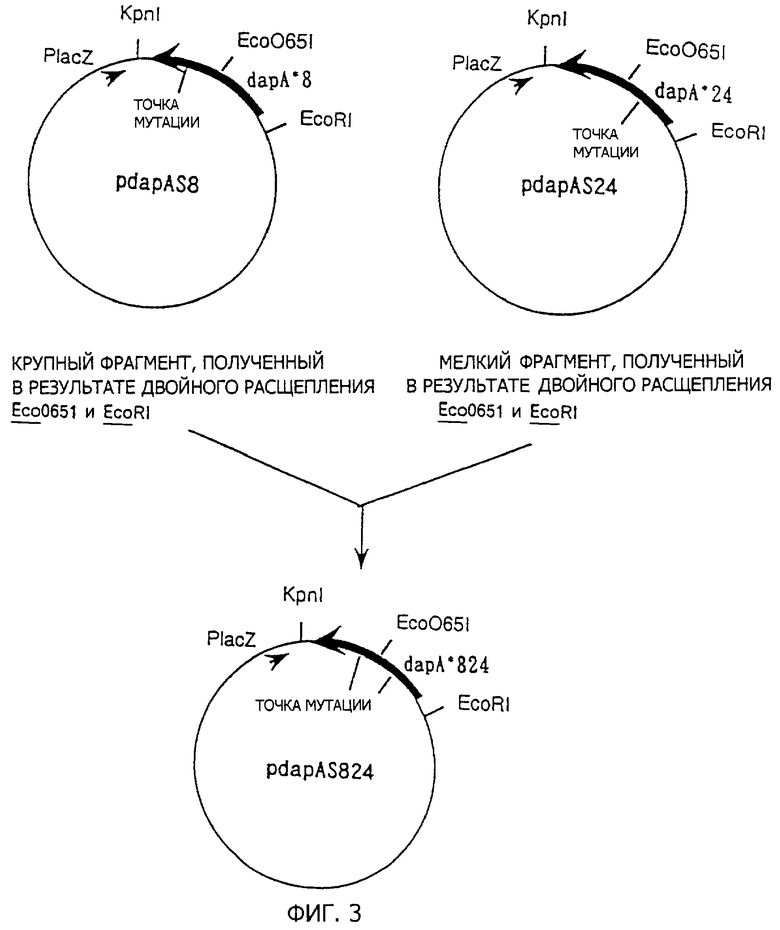
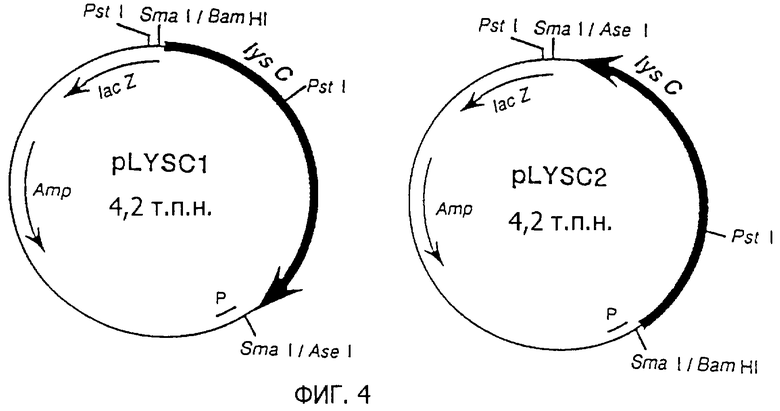
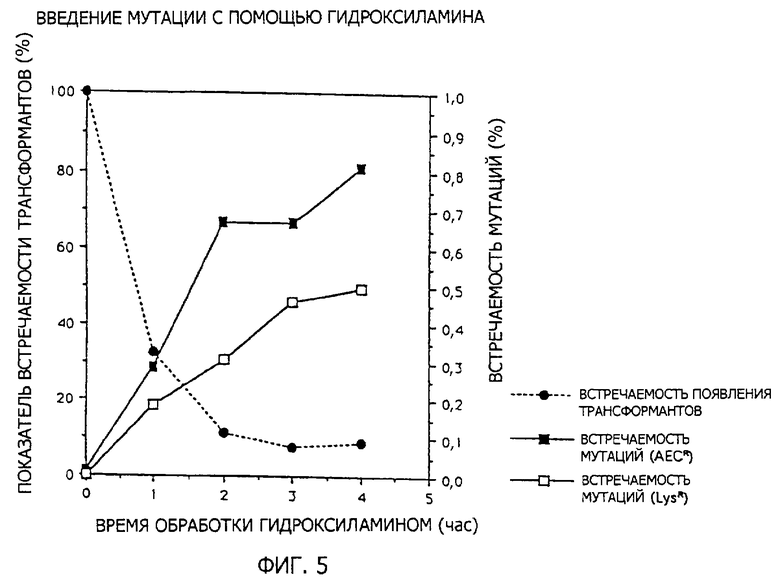

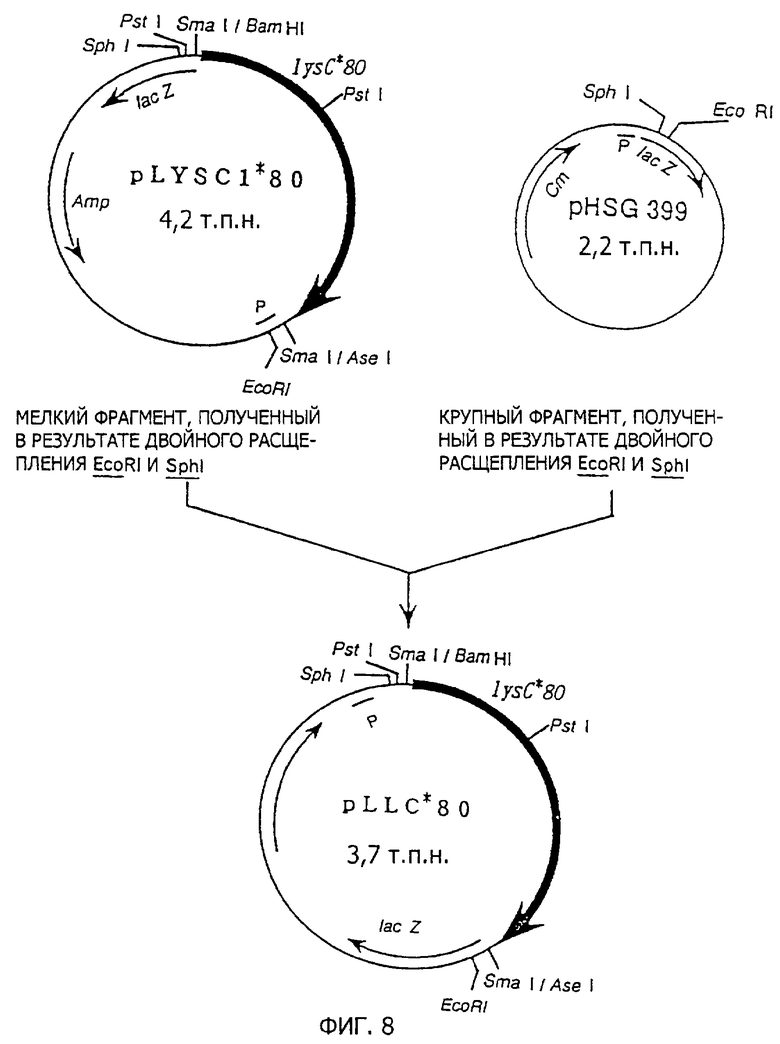

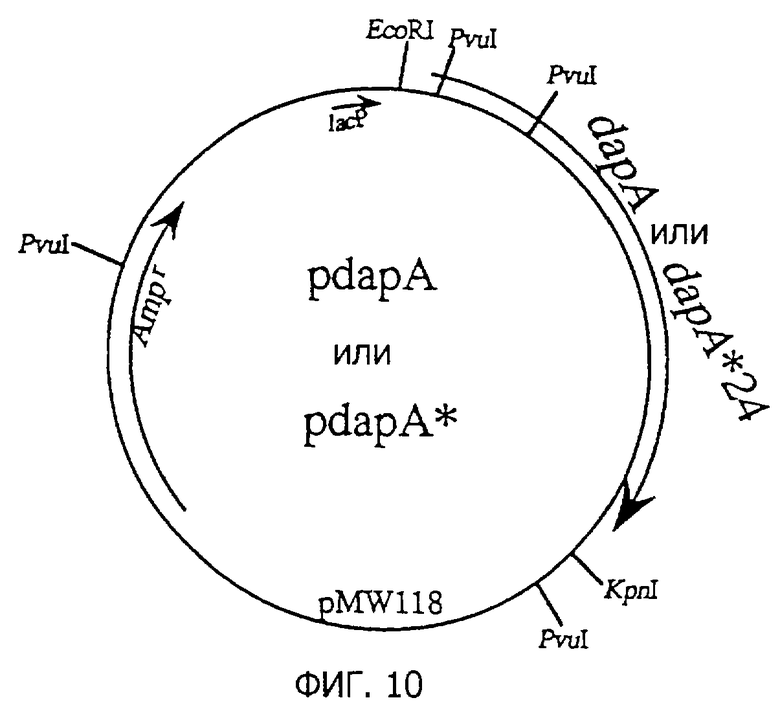
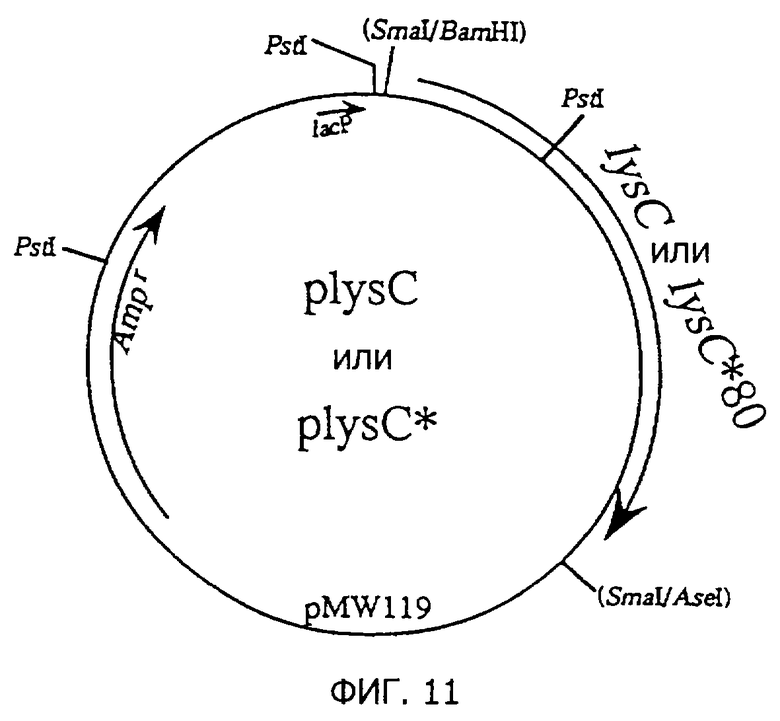
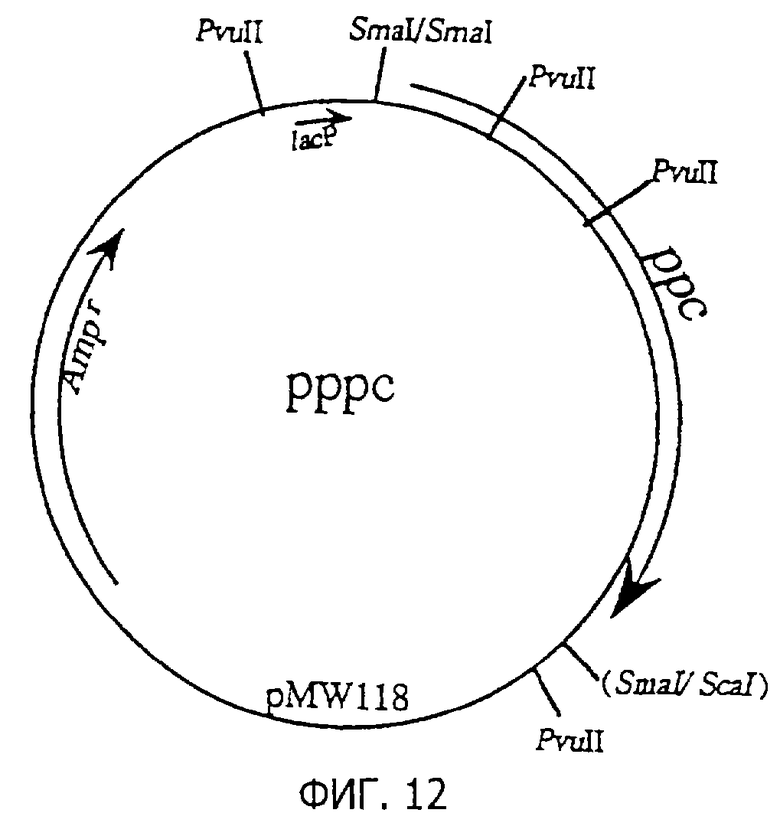


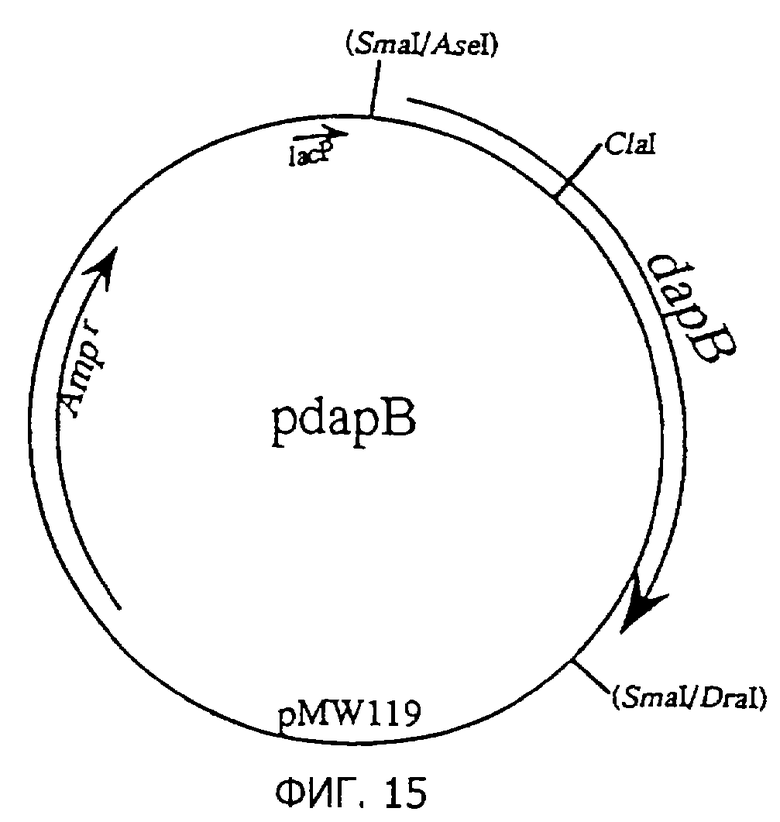
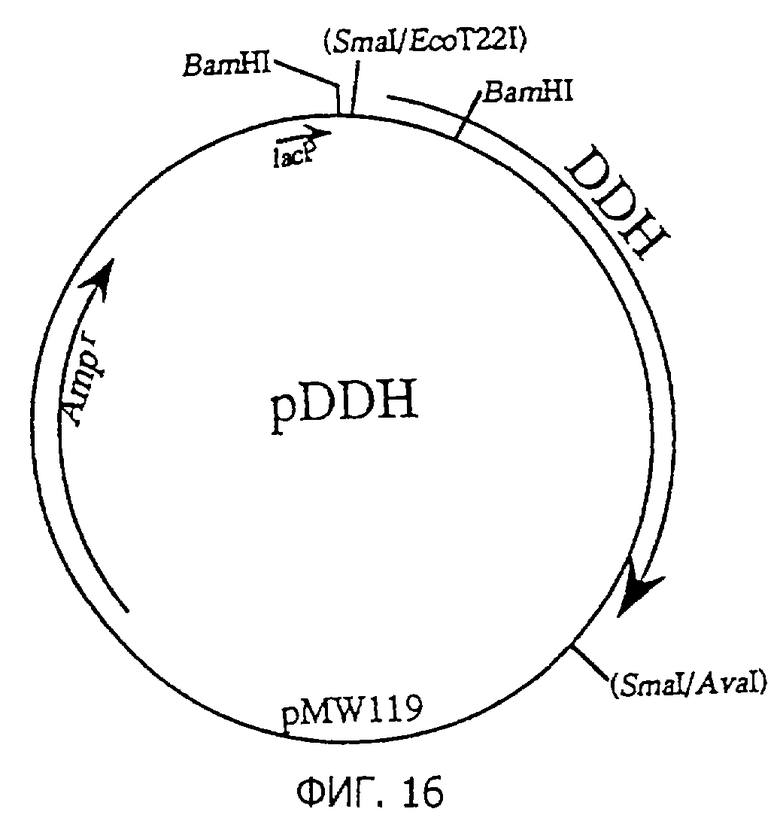
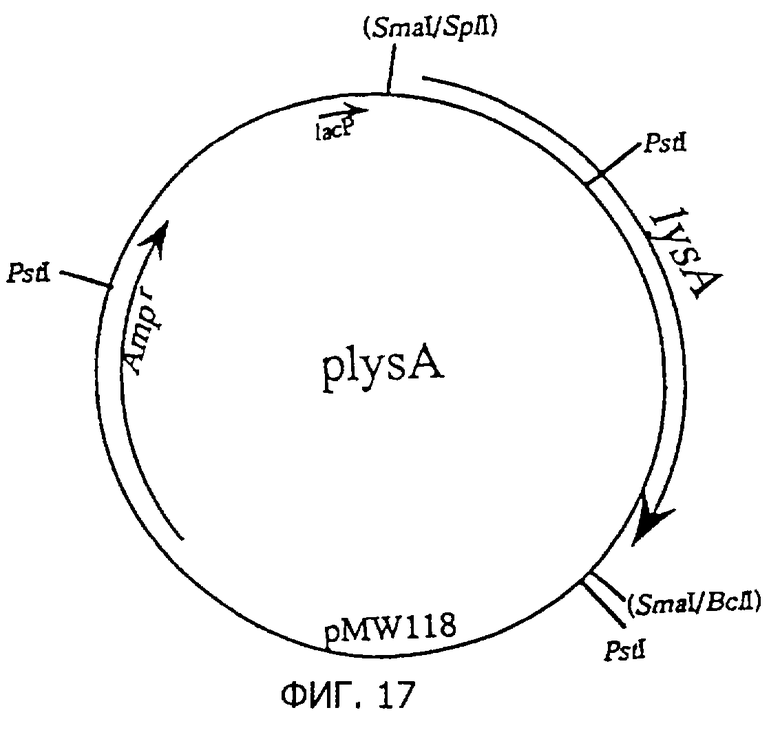
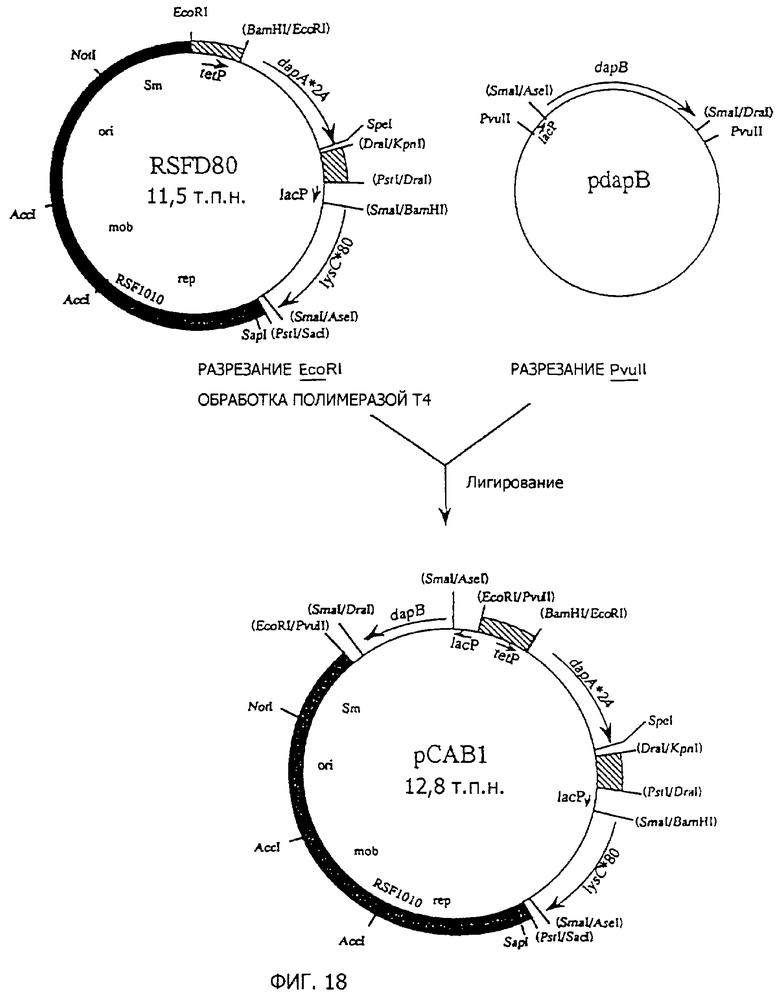
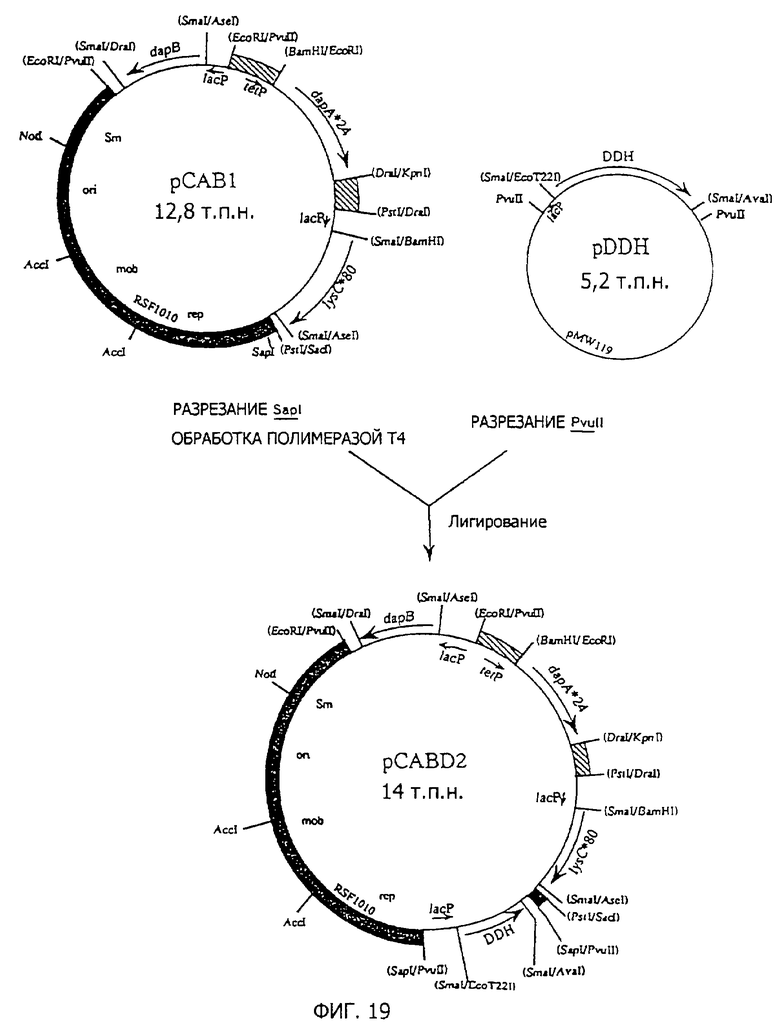
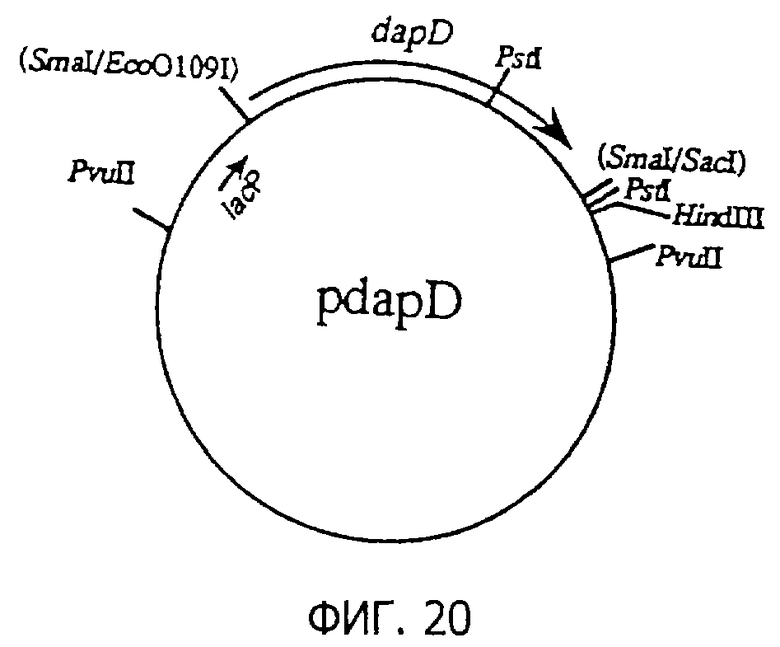

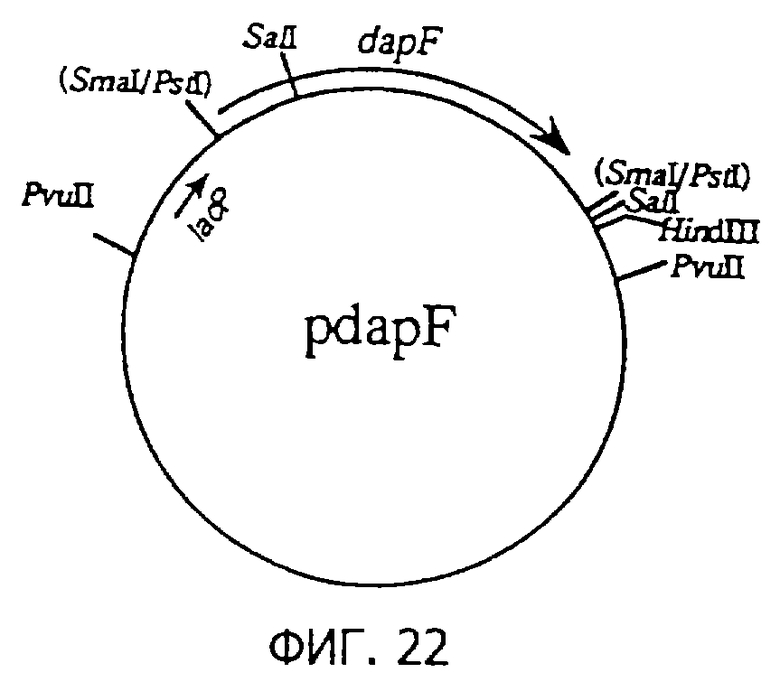

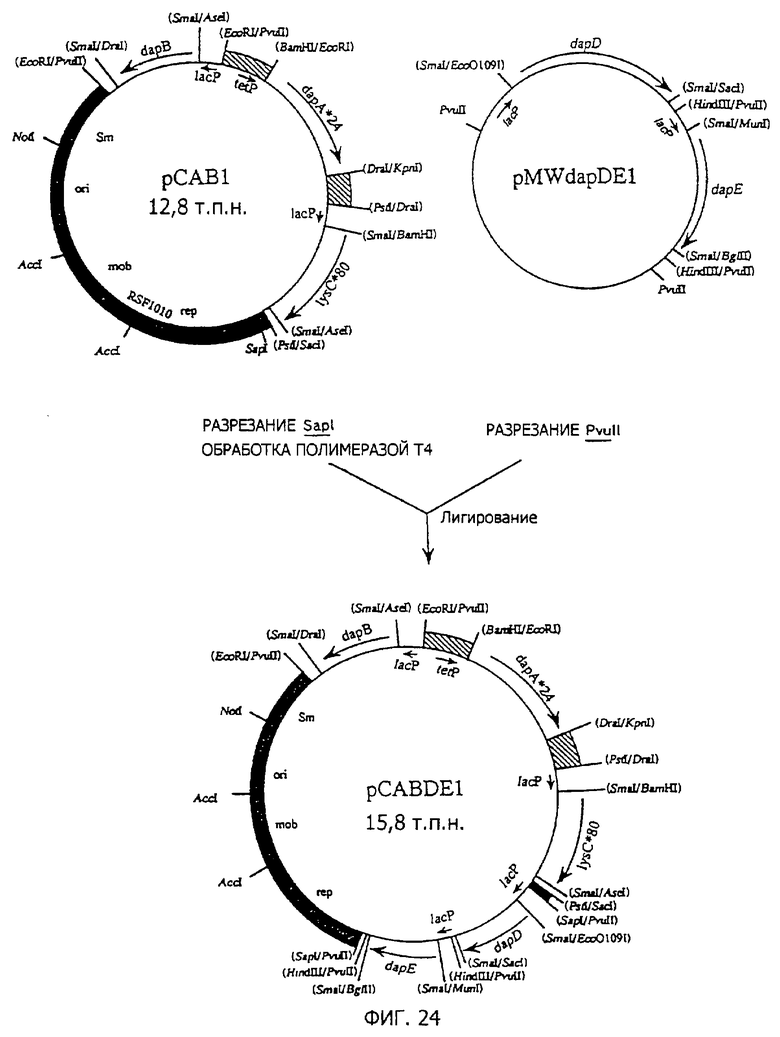
Изобретение относится к промышленному использованию микроорганизмов. Способ включает культивирование бактерий рода Escherichia, накопление L-лизина в ее культуре и сбора L-лизина из культуры, при этом бактерии несут дигидродипиколинатсинтазу, ингибирование которой по типу обратной связи L-лизином десенсибилизировано, и аспартокиназу, ингибирование которой по типу обратной связи L-лизином десенсибилизировано, и экспрессия гена дигидропиколинатредуктазы в бактериях усилена. Способ позволяет увеличить постадийно продукцию L-лизина. 8 з.п.ф-лы, 24 ил, 22 табл.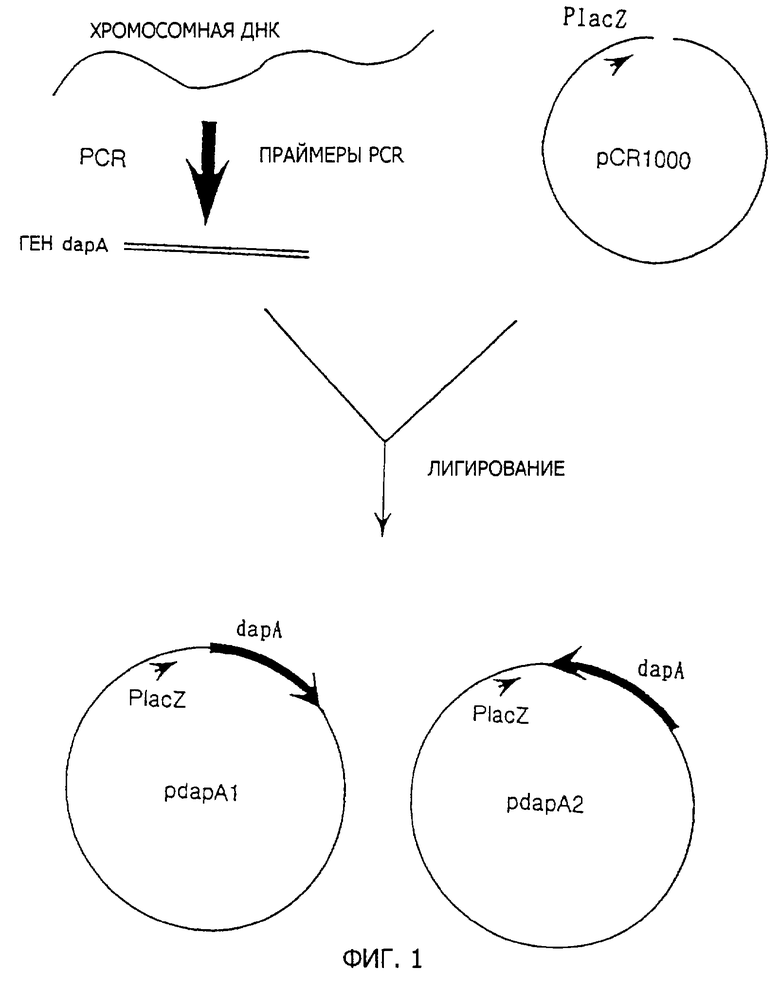
| Способ получения глинозема и щелочи | 1974 |
|
SU485970A1 |
| WО 9319190 A, 30.09.1993 | |||
| Способ получения L-аланина | 1988 |
|
SU1719433A1 |
