ОБЛАСТЬ ТЕХНИКИ
Настоящая заявка заявляет приоритет предварительной заявки на патент США № 60/902949, поданной 21 февраля 2007, полное содержание которой, таким образом, включено сюда посредством ссылки для всех целей.
Настоящее изобретение относится к области биологии, конкретнее, к области терапии рака. В частности, настоящее изобретение относится к новым иммуногенным пептидам, которые служат в качестве очень эффективных вакцин против рака, и содержащим такие пептиды лекарственным средствам для лечения и предупреждения опухолей.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Было показано, что CD8+ цитотоксические T-лимфоциты (CTL) распознают пептидные эпитопы, происходящие из опухолеспецифических антигенов (TAA), представленные на молекулах MHC (главного комплекса гистосовместимости) класса I, и впоследствии лизируют опухолевые клетки. Со времени открытия семейства MAGE в качестве первого примера TAA были открыты многие другие TAA, используя иммунологические подходы (Boon T. (1993) Int. J. Cancer 54: 177-180; Boon T. et al., (1996) J. Exp. Med. 183: 725-729; van der Bruggen P. et al., (1991) Science 254: 1643-1647; Brichard V. et al., (1993) J. Exp. Med. 178: 489-495; Kawakami Y. et al., (1994) J. Exp. Med. 180: 347-352). Некоторые из них в настоящее время клинически разрабатываются в качестве мишеней для иммунотерапии. Открытые на данное число TAA включают MAGE (van der Bruggen P. et al., (1991) Science 254: 1643-1647), gp100 (Kawakami Y. et al., (1994) J. Exp. Med. 180: 347-352), SART (Shichijo S. et al., (1998) J. Exp. Med. 187: 277-288) и NY-ESO-1 (Chen Y.T. et al., (1997) Proc. Natl. Acad. Sci. USA, 94: 1914-1918). С другой стороны, было установлено, что продукты некоторых генов, которые, как показано, до некоторой степени специфическим образом сверхэкспрессированы в опухолевых клетках, распознаются в качестве мишеней для индукции клеточных иммунных ответов. Такие продукты генов включают p53 (Umano Y. et al., (2001) Br. J. Cancer, 84: 1052-1057), HER2/neu (Tanaka H. et al., (2001) Br. J Cancer, 84: 94-99), CEA (Nukaya I. et al., (1999) Int. J. Cancer 80, 92-97) и т.п.
Несмотря на значительный прогресс в основном и клиническом исследовании, касающемся TAA (Rosenberg S.A. et al., (1998) Nature Med, 4: 321-327; Mukherji B. et al., (1995) Proc. Natl. Acad. Sci. USA, 92: 8078-8082; Hu X. et al., (1996) Cancer Res, 56: 2479-2483), в настоящее время имеется только очень ограниченное число TAA-кандидатов, подходящих для лечения раков. TAA, которые очень экспрессированы в раковых клетках и экспрессия которых ограничена раковыми клетками, могли бы быть многообещающими кандидатами на иммунотерапевтические мишени.
И HLA-A24, и HLA-A0201 являются обычными аллелями HLA (главного комплекса гистосовместимости человека) в популяциях японцев и европейцев (Date Y. et al., (1996) Tissue Antigens 47: 93-101; Kondo A. et al., (1995) J. Immunol. 155: 4307-4312; Kubo RT et al., (1994) J. Immunol. 152: 3913-3924; Imanishi et al., Proceeding of the eleventh International Histocompatibility Workshop and Conference Oxford University Press, Oxford, 1065 (1992); Williams F. et al., (1997) Tissue Antigen 49: 129-33.). Таким образом, антигенные пептиды раков, презентируемые этими аллелями HLA, могут найти конкретное применение для лечения раков среди японских и европейских пациентов. Кроме того, известно, что индукция CTL с небольшой аффинностью in vitro обычно является следствием воздействия высоких концентраций пептидов, порождающих высокий уровень комплексов специфический пептид/MHC на антигенпрезентирующих клетках (APC), которые будут эффективно активировать эти CTL (Alexander-Miller et al., (1996) Proc. Natl. Acad. Sci. USA 93: 4102-4107).
В последнее время последовательность пептида, связывающегося с молекулой HLA класса I, можно предположить, используя алгоритмы (Jounal of Immunological Methods, (1995), Vol. 185, p. 181-190, J. Immunol., (1994), Vol.152, p. 163-175, Protein Science, (2000), Vol.9, p. 1838-1846). Однако трудно сказать, что предполагаемый пептидный эпитоп может быть урезан до определенного размера, экспрессирован на поверхности клетки-мишени с молекулой HLA и распознан CTL. Более того, с помощью алгоритма, например BIMAS (http://bimas.dcrt.nih.gov/cgi-bin/molbio/ken_parker_comboform) (Parker K.C. et al., (1994) J. Immunol.; 152(1): 163-175; Kuzushima K. et al., (2001) Blood; 98(6): 1872-1881), можно предположить пептид, связывающийся с молекулой HLA, но предполагаемый пептид не является очень точным (Bachinsky M.M. et. al., Cancer Immun. 2005 Mar 22; 5:6). Следовательно, при отборе TAA все еще остается много проблем и трудностей.
Недавние разработки технологий ранжированных микрорядов кДНК сделали возможным построение полных профилей экспрессии генов в раковых клетках по сравнению с нормальными клетками (Okabe H. et al., (2001) Cancer Res., 61, 2129-2137; Lin Y.M. et al., (2002) Oncogene, 21: 4120-4128; Hasegawa S. et al., (2002) Cancer Res. 62: 7012-7017). Этот подход делает возможным более всестороннее понимание сложной природы раковых клеток и механизмов канцерогенеза и способствует идентификации генов, экспрессия которых дерегулирована в опухолях (Bienz M. et al., (2000) Cell 103, 311-320). Среди транскриптов, экспрессия которых, как установлено, увеличена в раках, в последнее время были открыты CDH3 (входящий № в GenBank NM_001793; SEQ ID NO: 1, 2), EPHA4 (входящий № в GenBank L36645; SEQ ID NO: 3, 4), ECT2 (входящий № в GenBank AY376439; SEQ ID NO: 5, 6), HIG2 (входящий № в GenBank NM_013332; SEQ ID NO: 7, 8) INHBB (входящий № в GenBank NM_002193; SEQ ID NO: 9, 10), KIF20A (входящий № в GenBank NM_005733; SEQ ID NO: 11, 12), KNTC2 (входящий № в GenBank AF017790; SEQ ID NO: 13, 14), TTK (входящий № в GenBank NM_003318; SEQ ID NO: 15, 16) и URLC10 (входящий № в GenBank NM_017527; SEQ ID NO: 17, 18). Полное содержание ссылок включено сюда посредством ссылки. Эти гены, экспрессия которых специфическим образом увеличена в опухолевых клетках различных раковых тканей проанализированных случаев, представляют особый интерес для авторов настоящего изобретения (см. ниже). Таким образом, иммуногенные пептиды, происходящие из CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10, могут найти применение для избирательного уничтожения опухолевых клеток, экспрессирующих такие антигены. Настоящее изобретение обращено к этим и другим потребностям.
Поскольку цитотоксические лекарственные средства, такие как M-VAC, часто вызывают тяжелые побочные реакции, очевидно, что отбор новых молекул мишеней, обдуманный на основе хорошо охарактеризованных механизмов действия, должен быть очень полезен при разработке эффективных противораковых лекарственных средств, имеющих минимизированный риск побочных эффектов. С этой целью были ранее выполнены анализы профилей экспрессии на различных раках и нормальной ткани человека. Такие исследования привели к открытию множества генов, которые специфическим образом сверхэкспрессированы в раках (Lin Y.M. et al., Oncogene. 2002 Jun 13; 21: 4120-4128; Kitahara O. et al., Cancer Res. 2001 May 1; 61: 3544-3549; Suzuki C. et al., Cancer Res. 2003 Nov 1; 63: 7038-7041; Ashida S., Cancer Res. 2004 Sep 1; 64: 5963-5972; Ochi K. et al., Int. J. Oncol. 2004 Mar; 24(3): 647-655; Kaneta Y. et al., Int. J. Oncol. 2003 Sep; 23: 681-691; Obama K., Hepatology. 2005 Jun; 41: 1339-1348; Kato T. et al., Cancer Res. 2005 Jul 1; 65: 5638-5646; Kitahara O. et al., Neoplasia. 2002 Jul-Aug; 4: 295-303; Saito-Hisaminato A. et al., DNA Res. 2002, 9: 35-45). Примеры таких генов, которые, как установлено, являются сверхэкспрессируемыми в различных раках, включают, но без ограничения, CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10. Ранее установлено, что CDH3 сверхэкспрессирован в раке мочевого пузыря, раке шейки матки, холангиоцеллюлярном раке, раке ободочной и прямой кишок, эндометриозе, раке желудка, диффузном раке желудка, немелкоклеточном раке легкого (NSCLC), раке поджелудочной железы, мягкотканевой опухоли и опухоли яичка. EPHA4 был идентифицирован в раке мочевого пузыря, раке шейки матки, холангиоцеллюлярном раке, эндометриозе, диффузном раке желудка, раке яичника, раке поджелудочной железы, раке предстательной железы и мягкотканевой опухоли. ECT2 был идентифицирован в раке мочевого пузыря, раке молочной железы, раке шейки матки, холангиоцеллюлярном раке, хроническом миелоидном лейкозе (CML), раке ободочной и прямой кишок, раке пищевода, NSCLC, лимфоме, раке предстательной железы, раке почки и мелкоклеточном раке легкого (SCLC). HIG2 был идентифицирован в раке почки и SCLC. INHBB был идентифицирован в холангиоцеллюлярном раке, раке пищевода, NSCLC, раке почки, SCLC и мягкотканевой опухоли. KIF20A был идентифицирован в раке мочевого пузыря, раке молочной железы, холангиоцеллюлярном раке, раке пищевода, NSCLC, раке поджелудочной железы, раке предстательной железы, раке почки и SCLC. KNTC2 был идентифицирован в раке мочевого пузыря, раке молочной железы, раке шейки матки, холангиоцеллюлярном раке, CML, раке ободочной и прямой кишок, раке пищевода, NSCLC, лимфоме, остеосаркоме, раке яичника, раке поджелудочной железы, раке предстательной железы, раке почки, SCLC и мягкотканевой опухоли. TTK был идентифицирован в раке мочевого пузыря, раке молочной железы, раке шейки матки, холангиоцеллюлярном раке, CML, раке ободочной и прямой кишок, раке пищевода, раке печени, NSCLC, лимфоме, остеосаркоме, раке предстательной железы, SCLC и мягкотканевой опухоли. URLC10 был идентифицирован в раке мочевого пузыря, раке шейки матки, холангиоцеллюлярном раке, раке пищевода, раке желудка, NSCLC, остеосаркоме, раке поджелудочной железы и SCLC.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение основано отчасти на открытии подходящих для иммунотерапии мишеней. Поскольку TAA часто не обладают иммуногенностью, открытие подходящих мишеней имеет чрезвычайное значение. Как отмечено выше, было установлено, что экспрессия CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10 увеличена в различных раках. Конкретнее, эти гены были идентифицированы, используя анализ профиля экспрессии генов с использованием ранжированного микроряда кДНК всего генома. Как обсуждалось выше, было установлено, что экспрессия CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10 специфическим образом увеличена в различных опухолевых клетках, от клеток рака поджелудочной железы до клеток рака почки. Как изображено в таблице 1, экспрессия CDH3 действительно увеличена в 26 из 34 раков мочевого пузыря, в 17 из 19 раков шейки матки, во всех 19 холангиоцеллюлярных раках, в 30 из 34 раков ободочной и прямой кишок, в 20 из 21 эндометриоза, в 13 из 20 раков желудка, в 7 из 8 диффузных раков желудка, в 36 из 37 NSCLC, во всех 16 раках поджелудочной железы, во всех 21 мягкотканевых опухолях и во всех 10 опухолях яичка.
В таблице 1, кроме того, демонстрируется, что:
экспрессия EPHA4 действительно увеличена в 14 из 34 раков мочевого пузыря, в 8 из 14 раков шейки матки, в 10 из 25 холангиоцеллюлярных раков, в 5 из 15 эндометриозов, в 5 из 8 диффузных раков желудка, во всех 5 раках яичника, во всех 14 раках поджелудочной железы, в 20 из 51 рака предстательной железы и в 14 из 23 мягкотканевых опухолей;
экспрессия ECT2 действительно увеличена в 17 из 19 раков мочевого пузыря, в 5 из 12 раков молочной железы, во всех 14 раках шейки матки, во всех 13 холангиоцеллюлярных раках, во всех 5 CML, в 7 из 8 раков ободочной и прямой кишки, в 12 из 16 раков пищевода, в 6 из 16 NSCLC, в 8 из 10 лимфом, в 1 из 1 рака поджелудочной железы, в 10 из 13 раков предстательной железы, в 3 из 6 раков почки и в 12 из 13 SCLC;
экспрессия HIG2 действительно увеличена в 19 из 20 раков почки и в 7 из 9 мягкотканевых опухолей;
экспрессия INHBB действительно увеличена в 10 из 21 холангиоцеллюлярного рака, во всех 12 раках пищевода, в 10 из 13 NSCLC, в 22 из 24 раков почки, в 8 из 14 SCLC и 45 из 49 мягкотканевых опухолей;
экспрессия KIF20A действительно увеличена во всех 31 раках мочевого пузыря, в 38 из 61 рака молочной железы, в 10 из 11 холангиоцеллюлярных раков, в 7 из 19 раков пищевода, в 21 из 22 NSCLC, во всех 6 раках яичника, в 17 из 36 раков предстательной железы, в 6 из 11 раков почки и во всех 15 SCLC;
экспрессия KNTC2 действительно увеличена в 30 из 32 раков мочевого пузыря, в 47 из 56 раков молочной железы, во всех 10 раках шейки матки, в 16 из 22 холангиоцеллюлярных раков, в 17 из 37 CML, в 3 из 10 раков ободочной и прямой кишок, в 11 из 46 раков пищевода, в 15 из 19 NSCLC, в 7 из 8 лимфом, в 20 из 24 остеосарком, в 3 из 5 раков яичника, во всех 2 раках поджелудочной железы, в 15 из 37 раков предстательной железы, в 14 из 19 раков почки, во всех 15 SCLC и в 40 из 59 мягкотканевых опухолей;
экспрессия TTK действительно увеличена во всех 27 раках мочевого пузыря, в 25 из 30 раков молочной железы, в 15 из 16 раков шейки матки, во всех 10 холангиоцеллюлярных раках, в 5 из 7 CML, в 6 из 10 раков ободочной и прямой кишок, в 24 из 44 раков пищевода, в 8 из 15 раков печени, во всех 12 NSCLC, во всех 6 лимфомах, в 13 из 16 остеобластом, в 12 из 17 раков предстательной железы, во всех 15 SCLC и в 16 из 33 мягкотканевых опухолей;
экспрессия URLC10 действительно увеличена во всех 29 раках мочевого пузыря, в 15 из 16 раков шейки матки, во всех 7 холангиоцеллюлярных раках, в 7 из 19 раков пищевода, во всех 3 раках желудка, в 24 из 27 NSCLC, в 15 из 19 остеосарком, в 4 из 5 раков поджелудочной железы, в 33 из 43 мягкотканевых опухолей.
Настоящее изобретение основано, по крайней мере отчасти, на идентификации специфических пептидных эпитопов продуктов этих генов (CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10), которые обладают способностью индуцировать цитотоксические T-лимфоциты (CTL), специфичные в отношении соответствующих молекул. Как обсуждается подробно ниже, мононуклеарные клетки периферической крови (PBMC) здорового донора были подвергнуты стимуляции, используя связывающиеся с HLA-A*2402 или HLA-A*0201 пептиды-кандидаты, происходящие из CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK или URLC10. Затем были созданы клоны и/или линии CTL со специфической цитотоксичностью в отношении HLA-A24- или HLA-A2-положительных клеток-мишеней, подвергнутых в импульсном режиме воздействию каждого из пептидов-кандидатов. Эти результаты демонстрируют, что эти пептиды являются рестрицированными по HLA-A24 или HLA-A2 пептидными эпитопами, которые могут вызывать сильные и специфические иммунные ответы против клеток, экспрессирующих CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK или URLC10.
Соответственно, настоящим изобретением обеспечиваются способы лечения или предупреждения заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK или URLC10, например рака. Такие способы включают стадию введения нуждающемуся в этом субъекту полипептидов настоящего изобретения CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10. Введение такого пептида(ов) приводит к индукции противоопухолевого иммунитета. Таким образом, настоящим изобретением обеспечиваются способы индукции противоопухолевого иммунитета у субъекта, включающие стадию введения субъекту полипептидов CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, а также фармацевтические композиции для лечения или предупреждения заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/рака URLC10, например рака, которые включают полипептиды CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10. Примеры таких раков включают, но без ограничения, рак мочевого пузыря, рак молочной железы, рак шейки матки, холангиоцеллюлярный рак, CML, рак ободочной и прямой кишок, эндометриоз, рак пищевода, рак желудка, диффузный рак желудка, рак печени, NSCLC,
лимфому, остеосаркому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки, SCLC, мягкотканевую опухоль и опухоль яичка.
Настоящим изобретением, кроме того, обеспечиваются способы предупреждения рецидива после хирургического вмешательства упомянутого выше заболевания.
В отношении конкретных целей и задач, изложенных выше, квалифицированным в данной области техники специалистам будет понятно, что один или несколько аспектов этого изобретения могут соответствовать некоторым целям, в то время как один или несколько других аспектов могут соответствовать некоторым другим целям. Каждая цель может не применяться одинаково, во всех ее отношениях, к каждому аспекту этого изобретения. Как таковые объекты настоящего изобретения можно рассматривать попеременно по отношению к любому аспекту этого изобретения.
Дополнительные объекты и признаки настоящего изобретения проявятся более полно при прочтении следующего подробного описания в соединении с сопроводительными фигурами и примерами. Однако следует понимать, что как предшествующее краткое изложение сущности изобретения, так и следующее подробное описание являются предпочтительными вариантами осуществления и не ограничивают настоящее изобретение или другие альтернативные варианты осуществления настоящего изобретения. В частности, хотя настоящее изобретение описывается здесь относительно ряда конкретных вариантов осуществления, будет понятно, что описание является иллюстрацией настоящего изобретения и не толкуется как ограничение настоящего изобретения. Различные модификации и применения могут приходить на ум квалифицированным в данной области техники специалистам без отступления от сущности и объема настоящего изобретения, характеризуемого прилагаемой формулой изобретения. Подобным образом, другие объекты, признаки, эффекты и преимущества настоящего изобретения будут очевидны из этого краткого изложения сущности изобретения и некоторых вариантов осуществления, описанных ниже, и они будут быстро очевидны квалифицированным в данной области техники специалистам. Такие объекты, признаки, эффекты и преимущества будут очевидны из вышеприведенного описания в соединении с сопроводительными примерами, данными, фигурами и всеми приемлемыми выводами, которые будут сделаны из них, отдельно или с учетом включенных сюда ссылок.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Различные аспекты и применения настоящего изобретения станут очевидными квалифицированному специалисту при учете следующих краткого описания чертежей и подробного описания настоящего изобретения и его предпочтительных вариантов осуществления.
Фиг.1-1. На фиг.1 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для CDH3-A24-10-332 (SEQ ID NO: 34), CDH3-A24-10-470 (SEQ ID NO: 358), CDH3-A24-9-513 (SEQ ID NO: 19), CDH3-A24-9-406 (SEQ ID NO: 22), CDH3-A24-10-807 (SEQ ID NO: 30) и CDH3-A24-10-655 (SEQ ID NO: 344) обнаруживается мощная продукция IFN-гамма. В «a» изображен пример отрицательных пептидов, у которых не могла быть выявлена способность к индукции CTL несмотря на возможную активность связывания с HLA-A*2402. В «b» изображена способность CDH3-A24-10-332 (SEQ ID NO: 34) индуцировать CTL. Для CDH3-A24-10-332 (SEQ ID NO: 34) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #4, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «c» изображена способность CDH3-A24-10-470 (SEQ ID NO: 358) индуцировать CTL. Для CDH3-A24-10-470 (SEQ ID NO: 358) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #4, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «d» изображена способность CDH3-A24-9-513 (SEQ ID NO: 19) индуцировать CTL. Для CDH3-A24-9-513 (SEQ ID NO: 19) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма. В лунке #6, показанной в рамке в левой панели, продемонстрирована специфическая реакция против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. Кроме того, линия CTL, которая была создана из положительной лунки #5, показанной в рамке в срединной панели, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «e» изображена способность CDH3-A24-9-406 (SEQ ID NO: 22) индуцировать CTL. Для CDH3-A24-9-406 (SEQ ID NO: 22) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT
на IFN-гамма, и линия CTL, которая была создана из положительной лунки #2, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа.
Фиг.1-2. На фиг.1 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для CDH3-A24-10-332 (SEQ ID NO: 34), CDH3-A24-10-470 (SEQ ID NO: 358), CDH3-A24-9-513 (SEQ ID NO: 19), CDH3-A24-9-406 (SEQ ID NO: 22), CDH3-A24-10-807 (SEQ ID NO: 30) и CDH3-A24-10-655 (SEQ ID NO: 344) обнаруживается мощная продукция IFN-гамма. В «f» изображена способность CDH3-A24-10-807 (SEQ ID NO: 30) индуцировать CTL. Для CDH3-A24-10-807 (SEQ ID NO: 30) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клон CTL были созданы из положительной лунки #5, показанной в рамке. Созданный клон CTL, индуцированный против этого пептида, продемонстрировал специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном CDH3, и молекулой HLA-A24 (нижний график справа). С другой стороны, COS7, трансфицированные полноразмерным CDH3, но не HLA-A24, и COS7, трансфицированные HLA-A24, но не полноразмерным CDH3, готовили в качестве отрицательного контроля. Клон CTL продемонстрировал высокую специфическую активность CTL в отношении COS7, которые трансфицированы и CDH3, и HLA-A24. В «g» изображена способность CDH3-A24-10-655 (SEQ ID NO: 344) индуцировать CTL. Для CDH3-A24-10-655 (SEQ ID NO: 344) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клон CTL были созданы из положительной лунки #1, показанной в рамке. Созданный клон CTL, индуцированный против этого пептида, продемонстрировал специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном CDH3, и молекулой HLA-A24 (нижний график справа). С другой стороны, COS7, трансфицированные полноразмерным CDH3, но не HLA-A24, и COS7, трансфицированные HLA-A24, но не полноразмерным CDH3, готовили в качестве отрицательного контроля. Клон CTL продемонстрировал высокую специфическую активность CTL в отношении COS7, которые трансфицированы и CDH3, и HLA-A24.
Фиг.2. На фиг.2 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для Epha4-A24-9-453 (SEQ ID NO: 41), Epha4-A24-9-5 (SEQ ID NO: 44), Epha4-A24-9-420 (SEQ ID NO: 48), Epha4-A24-9-869 (SEQ ID NO: 46), Epha4-A24-10-24 (SEQ ID NO: 78), Epha4-A02-9-501 (SEQ ID NO: 376) и Epha4-A02-9-165 (SEQ ID NO: 379) обнаруживается мощная продукция IFN-гамма. В «a» изображен пример отрицательных пептидов, у которых не могла быть выявлена способность к индукции CTL, несмотря на возможную активность связывания с HLA. В «b» изображена способность Epha4-A24-9-453 (SEQ ID NO: 41) индуцировать CTL. Для Epha4-A24-9-453 (SEQ ID NO: 41) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #3, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «c» изображена способность Epha4-A24-9-5 (SEQ ID NO: 44) индуцировать CTL. Для Epha4-A24-9-5 (SEQ ID NO: 44) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #2, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «d» изображена способность Epha4-A24-9-420 (SEQ ID NO: 48) индуцировать CTL. Для Epha4-A24-9-420 (SEQ ID NO: 48) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма. В лунке #6, показанной в рамке в верхней панели, продемонстрирована специфическая реакция против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. Кроме того, линия CTL, которая была создана из положительной лунки #6, показанной в рамке в срединной панели, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «e» изображена способность Epha4-A24-9-869 (SEQ ID NO: 46) индуцировать CTL. Для Epha4-A24-9-869 (SEQ ID NO: 46) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #5, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «f» изображена способность Epha4-A24-10-24 (SEQ ID NO: 78) индуцировать CTL. Для Epha4-A24-10-24 (SEQ ID NO: 78) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #4, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «g» изображена способность Epha4-A02-9-501 (SEQ ID NO: 376) индуцировать CTL. Для Epha4-A02-9-501 (SEQ ID NO: 376) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клон CTL были созданы из положительной лунки #8, показанной в рамке. Цитотоксическую активность созданной линии CTL в отношении клеток-мишеней, подвергнутых в импульсном режиме воздействию пептида, определяли с помощью анализа высвобождения Cr (CRA) (нижний график), и линия CTL обладала очень сильной специфической цитотоксической активностью в отношении клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидов. В «h» изображена способность Epha4-A02-9-165 (SEQ ID NO: 379) индуцировать CTL. Для Epha4-A02-9-165 (SEQ ID NO: 379) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL была создана из положительной лунки #3, показанной в рамке. Цитотоксическую активность созданной линии CTL в отношении клеток-мишеней, подвергнутых в импульсном режиме воздействию пептида, определяли с помощью анализа высвобождения Cr (CRA) (правый график), и линия CTL обладала очень сильной специфической цитотоксической активностью в отношении клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидов.
Фиг.3. На фиг.3 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для ECT2-A24-9-515 (SEQ ID NO: 80), ECT2-A24-10-40 (SEQ ID NO: 100) и ECT2-A24-10-101 (SEQ ID NO: 101) обнаруживается мощная продукция IFN-гамма. В «a» изображен пример отрицательных пептидов, у которых не могла быть выявлена способность к индукции CTL несмотря на возможную активность связывания с HLA. В «b» изображена способность ECT2-A24-9-515 (SEQ ID NO: 80) индуцировать CTL. Для ECT2-A24-9-515 (SEQ ID NO: 80) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT
на IFN-гамма. В лунках #5 и #7, показанных в рамке в левой панели, продемонстрирована специфическая реакция против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. Кроме того, линия CTL, которая была создана из положительной лунки #7, показанной в рамке во второй панели, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. Цитотоксическую активность линии CTL в отношении линии раковых клеток ТЕ6, эндогенно экспрессирующей ECT2 и HLA-A24, определяли с помощью анализа высвобождения Cr (CRA), и клон CTL обладал очень сильной цитотоксической активностью в отношении ТЕ6. С другой стороны, цитотоксическая активность линии CTL в отношении линии раковых клеток TE5, экспрессирующей только ECT2, не была выявлена. В «c» изображена способность ECT2-A24-10-40 (SEQ ID NO: 100) индуцировать CTL. Для ECT2-A24-10-40 (SEQ ID NO: 100) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клон CTL были созданы из положительной лунки #2, показанной в рамке. Созданный клон CTL, индуцированный против этого пептида, продемонстрировал специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном ECT2, и молекулой HLA-A24. С другой стороны, COS7, трансфицированные полноразмерным ECT2, но не HLA-A24, COS7, трансфицированные HLA-A24 и геном URLC10 в качестве замены полноразмерного гена ECT2, и COS7, трансфицированные HLA-A24 и подвергнутые в импульсном режиме воздействию ECT2-10-101, готовили в качестве отрицательного контроля. Клон CTL продемонстрировал высокую специфическую активность CTL в отношении COS7, которые трансфицированы и ECT2, и HLA-A24. В «d» изображена способность ECT2-A24-10-101 (SEQ ID NO: 101) индуцировать CTL. Для ECT2-A24-10-101 (SEQ ID NO: 101) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL была создана из положительной лунки #1, показанной в рамке. Созданная линия CTL, индуцированная против этого пептида, продемонстрировала специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном ECT2, и молекулой HLA-A24. COS7, трансфицированные полноразмерным ECT2,
но не HLA-A24, COS7, трансфицированные HLA-A24 и геном URLC10 в качестве замены полноразмерного гена ECT2, и COS7, трансфицированные HLA-A24 и подвергнутые в импульсном режиме воздействию ECT2-10-40, готовили в качестве отрицательного контроля. Клон CTL продемонстрировал высокую специфическую активность CTL в отношении COS7, которые трансфицированы и ECT2, и HLA-A24.
Фиг.4-1. На фиг.4 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для HIG2-A24-9-19 (SEQ ID NO: 110), HIG2-A24-9-22 (SEQ ID NO: 111), HIG2-A24-9-8 (SEQ ID NO: 387), HIG2-A24-10-7 (SEQ ID NO: 112), HIG2-A24-10-18 (SEQ ID NO: 394), HIG2-A02-9-15 (SEQ ID NO: 116), HIG2-A02-9-4 (SEQ ID NO: 117) и HIG2-A02-10-8 (SEQ ID NO: 121) обнаруживается мощная продукция IFN-гамма. В «a» изображен пример отрицательных пептидов, у которых не могла быть выявлена способность к индукции CTL несмотря на возможную активность связывания с HLA. В «b» изображена способность HIG2-A24-9-19 (SEQ ID NO: 110) индуцировать CTL. Для HIG2-A24-9-19 (SEQ ID NO: 110) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #6, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «c» изображена способность HIG2-A24-9-22 (SEQ ID NO: 111) индуцировать CTL. Для HIG2-A24-9-22 (SEQ ID NO: 111) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клон CTL, которые были созданы из положительной лунки #7, показанной в рамке, продемонстрировали специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «d» изображена способность HIG2-A24-9-8 (SEQ ID NO: 387) индуцировать CTL. Для HIG2-A24-9-8 (SEQ ID NO: 387) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клон CTL, которые были созданы из положительной лунки #5, показанной в рамке, продемонстрировали специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «e» изображена способность HIG2-A02-9-8 (SEQ ID NO: 114) индуцировать CTL. Для HIG2-A02-9-8 (SEQ ID NO: 114) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT
на IFN-гамма, и линия CTL была создана из положительной лунки #10, показанной в рамке. Созданная линия CTL, индуцированная против этого пептида, продемонстрировала специфическую активность CTL в отношении клеток 293Т, трансфицированных и полноразмерным геном HIG2, и молекулой HLA-A02. 293Т, трансфицированные полноразмерным HIG2, но не HLA-A02, 293Т, трансфицированные HLA-A02 и геном FoxP3 в качестве замены полноразмерного гена HIG2, и 293Т, трансфицированные HLA-A02 и подвергнутые в импульсном режиме воздействию HIG2-9-15, готовили в качестве отрицательного контроля. Линия CTL продемонстрировала высокую специфическую активность CTL в отношении 293Т, которые трансфицированы и HIG2, и HLA-A02.
Фиг.4-2. На фиг.4 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для HIG2-A24-9-19 (SEQ ID NO: 110), HIG2-A24-9-22 (SEQ ID NO: 111), HIG2-A24-9-8 (SEQ ID NO: 387), HIG2-A24-10-7 (SEQ ID NO: 112), HIG2-A24-10-18 (SEQ ID NO: 394), HIG2-A02-9-15 (SEQ ID NO: 116), HIG2-A02-9-4 (SEQ ID NO: 117) и HIG2-A02-10-8 (SEQ ID NO: 121) обнаруживается мощная продукция IFN-гамма. В «f» изображена способность HIG2-A24-10-7 (SEQ ID NO: 112) индуцировать CTL. Для HIG2-A24-10-7 (SEQ ID NO: 112) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линии и клон CTL, которые были созданы из положительных лунок #1 и #7, показанных в рамке, продемонстрировали специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «g» изображена способность HIG2-A24-10-18 (SEQ ID NO: 394) индуцировать CTL. Для HIG2-A24-10-18 (SEQ ID NO: 394) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клон CTL, которые были созданы из положительной лунки #7, показанной в рамке, продемонстрировали специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «h» изображена способность HIG2-A02-9-15 (SEQ ID NO: 116) индуцировать CTL. Для HIG2-A02-9-15 (SEQ ID NO: 116) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL была создана из положительной лунки #10, показанной в рамке. Созданная линия CTL, индуцированная против этого пептида, продемонстрировала специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном HIG2, и молекулой HLA-A02. COS7, трансфицированные полноразмерным HIG2, но не HLA-A02, и COS7, трансфицированные HLA-A02 и подвергнутые в импульсном режиме воздействию пептида HIG2-9-8, готовили в качестве отрицательного контроля. Линия CTL продемонстрировала высокую специфическую активность CTL в отношении COS7, которые трансфицированы и HIG2, и HLA-A02.
Фиг.4-3. На фиг.4 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для HIG2-A24-9-19 (SEQ ID NO: 110), HIG2-A24-9-22 (SEQ ID NO: 111), HIG2-A24-9-8 (SEQ ID NO: 387), HIG2-A24-10-7 (SEQ ID NO: 112), HIG2-A24-10-18 (SEQ ID NO: 394), HIG2-A02-9-15 (SEQ ID NO: 116), HIG2-A02-9-4 (SEQ ID NO: 117) и HIG2-A02-10-8 (SEQ ID NO: 121) обнаруживается мощная продукция IFN-гамма. В «i» изображена способность HIG2-A02-9-4 (SEQ ID NO: 117) индуцировать CTL. Для HIG2-A02-9-4 (SEQ ID NO: 117) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клон CTL были созданы из положительной лунки #10, показанной в рамке. Созданная линия CTL, индуцированная против этого пептида, продемонстрировала специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном HIG2, и молекулой HLA-A02 (срединный график). Также готовили COS7, трансфицированные полноразмерным HIG2, но не HLA-A02, COS7, трансфицированные HLA-A02 и геном TTK в качестве замены полноразмерного гена HIG2, и COS7, трансфицированные HLA-A02 и подвергнутые в импульсном режиме воздействию HIG2-9-8, ради отрицательного контроля. Цитотоксическую активность клона CTL в отношении клеток 293Т, трансфицированных и полноразмерным геном HIG2, и молекулой HLA-A02, и линии раковых клеток Caki-1, эндогенно экспрессирующей HIG2 и HLA-A02, определяли с помощью анализа высвобождения Cr (CRA) (нижние графики), и клон CTL обладал очень сильной цитотоксической активностью в отношении клеток, трансфицированных и HIG2, и HLA-A02, и Caki-1. С другой стороны, цитотоксическая активность линии CTL в отношении 293T, трансфицированных только HIG2 или только HLA-A02, и линии раковых клеток A498, экспрессирующей только HIG2, не была выявлена. В «j» изображена способность HIG2-A02-10-8 (SEQ ID NO: 121) индуцировать CTL. Для HIG2-A02-10-8 (SEQ ID NO: 121) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #9, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа.
Фиг.5-1. На фиг.5 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для INHBB-A24-9-180 (SEQ ID NO: 395), INHBB-A24-10-180 (SEQ ID NO: 133), INHBB-A24-10-305 (SEQ ID NO: 135), INHBB-A24-10-7 (SEQ ID NO: 137) и INHBB-A24-10-212 (SEQ ID NO: 426) обнаруживается мощная продукция IFN-гамма. В «a» изображен пример отрицательных пептидов, у которых не могла быть выявлена способность к индукции CTL несмотря на возможную активность связывания с HLA. В «b» изображена способность INHBB-A24-9-180 (SEQ ID NO: 395) индуцировать CTL. Для INHBB-A24-9-180 (SEQ ID NO: 395) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клон CTL были созданы из положительной лунки #7, показанной в рамке. Цитотоксическую активность созданного клона CTL в отношении опухолевых клеток Miapaca2, экспрессирующих и INHBB, и HLA-A02, определяли с помощью анализа высвобождения Cr (CRA), и клетки-эффекторы продемонстрировали высокую специфическую цитотоксическую активность в отношении Miapaca2. С другой стороны, не была продемонстрирована значительная специфическая цитотоксическая активность в отношении Caki-1, экспрессирующих INHBB, но не экспрессирующих HLA-A02. В «c» изображена способность INHBB-A24-10-180 (SEQ ID NO: 133) индуцировать CTL. Для INHBB-A24-10-180 (SEQ ID NO: 133) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL была создана из положительной лунки #3, показанной в рамке. Созданная линия CTL, индуцированная против этого пептида, продемонстрировала высокую специфическую активность CTL в отношении клеток 293Т, трансфицированных и полноразмерным геном INHBB, и молекулой HLA-A24. Также готовили 293Т, трансфицированные полноразмерным INHBB, но не HLA-A24, и 293Т, трансфицированные HLA-A24 и подвергнутые в импульсном режиме воздействию пептида INHBB-10-305, ради отрицательного контроля.
Фиг.5-2. На фиг.5 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для INHBB-A24-9-180 (SEQ ID NO: 395), INHBB-A24-10-180 (SEQ ID NO: 133), INHBB-A24-10-305 (SEQ ID NO: 135), INHBB-A24-10-7 (SEQ ID NO: 137) и INHBB-A24-10-212 (SEQ ID NO: 426) обнаруживается мощная продукция IFN-гамма. В «d» изображена способность INHBB-A24-10-305 (SEQ ID NO: 135) индуцировать CTL. Для INHBB-A24-10-305 (SEQ ID NO: 135) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT
на IFN-гамма, и линия и клон CTL были создана из положительной лунки #2, показанной в рамке. Созданный клон CTL, индуцированный против этого пептида, продемонстрировал высокую специфическую активность CTL в отношении клеток 293Т, трансфицированных и полноразмерным геном INHBB, и молекулой HLA-A24. Также 293Т, трансфицированные полноразмерным INHBB, но не HLA-A24, и 293Т, трансфицированные HLA-A24 и подвергнутые в импульсном режиме воздействию пептида INHBB-10-180, готовили в качестве отрицательного контроля. В «e» изображена способность INHBB-A24-10-7 (SEQ ID NO: 137) индуцировать CTL. Для INHBB-A24-10-7 (SEQ ID NO: 137) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линии CTL были созданы из положительной лунки #8, показанной в рамке в верхней панели, и #2, показанной в рамке в нижней панели. Линия CTL из лунки #8 продемонстрировала специфическую активность CTL в отношении клеток 293T, трансфицированных и полноразмерным геном INHBB, и молекулой HLA-A24. Также готовили 293T, трансфицированные полноразмерным геном INHBB, но не HLA-A24, и 293T, трансфицированные HLA-A24 и подвергнутые в импульсном режиме воздействию пептида INHBB-10-40, ради отрицательного контроля. В «f» изображена способность INHBB-A24-10-212 (SEQ ID NO: 426) индуцировать CTL. Для INHBB-A24-10-212 (SEQ ID NO: 426) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #1, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа.
Фиг.6-1. На фиг.6 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для KIF20A-A24-10-304 (SEQ ID NO: 186), KIF20A-A24-9-383 (SEQ ID NO: 178), KIF20A-A24-10-66 (SEQ ID NO: 194) и KIF20A-A24-9-305 (SEQ ID NO: 174) обнаруживается мощная продукция IFN-гамма. В «a» изображен пример отрицательных пептидов, у которых не могла быть выявлена способность к индукции CTL несмотря на возможную активность связывания с HLA. В «b» изображена способность KIF20A-A24-10-304 (SEQ ID NO: 186) индуцировать CTL. Для KIF20A-A24-10-304 (SEQ ID NO: 186) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма. В лунке #5, показанной в рамке в нижней правой панели, продемонстрирована специфическая реакция против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. Кроме того, линия и клон CTL, которые были созданы из положительной лунки #5, показанной в рамке в верхней левой панели, также продемонстрировали специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. Созданный клон CTL, индуцированный против пептида, продемонстрировал специфическую активность CTL в отношении 24-LCL, трансфицированных полноразмерным геном KIF20A. Также готовили A24-LCL, трансфицированные контрольным вектором, ради отрицательного контроля. Цитотоксическую активность клона CTL в отношении опухолевых клеток Miapaca2, экспрессирующих и KIF20A, и HLA-A24, определяли с помощью анализа высвобождения Cr (CRA), и клон CTL обладал очень сильной специфической цитотоксической активностью в отношении Miapaca2 (нижний график справа). С другой стороны, не была продемонстрирована значительная специфическая цитотоксическая активность в отношении PK59, экспрессирующих KIF20A, но не экспрессирующих HLA-A24. В «c» изображена способность KIF20A-A24-9-383 (SEQ ID NO: 178) индуцировать CTL. Для KIF20A-A24-9-383 (SEQ ID NO: 178) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма. В лунках #3 и #4, показанных в рамках в правой панели, продемонстрирована специфическая реакция против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. Кроме того, линия CTL, которая была создана из положительной лунки #3, показанной в рамке в левой панели, также продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. Созданная линия CTL продемонстрировала высокую специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном KIF20A, и молекулой HLA-A24. Также готовили COS7, трансфицированные полноразмерным KIF20A, но не HLA-A24, и COS7, трансфицированные HLA-A24 и подвергнутые в импульсном режиме воздействию пептида KIF20A-9-621, ради отрицательного контроля.
Фиг.6-2. На фиг.6 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для KIF20A-A24-10-304 (SEQ ID NO: 186), KIF20A-A24-9-383 (SEQ ID NO: 178), KIF20A-A24-10-66 (SEQ ID NO: 194) и KIF20A-A24-9-305 (SEQ ID NO: 174) обнаруживается мощная продукция IFN-гамма. В «d» изображена способность KIF20A-A24-10-66 (SEQ ID NO: 194) индуцировать CTL. Для KIF20A-A24-10-66 (SEQ ID NO: 194) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линии CTL, которые были созданы из положительной лунки #6, показанной в рамке в верхней левой панели, и #3, показанной в рамке в нижней срединной панели, продемонстрировали специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. Кроме того, клон CTL, отобранный из линии CTL из лунки #6 с помощью серийного разведения, продемонстрировал специфическую активность CTL в отношении клеток-мишеней. Созданный клон CTL продемонстрировал специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном KIF20A, и молекулой HLA-A24. Также готовили COS7, трансфицированные полноразмерным KIF20A, но не HLA-A24, COS7, трансфицированные HLA-A24 и геном URLC10 в качестве замены полноразмерного гена KIF20A, и COS7, трансфицированные HLA-A24 и подвергнутые в импульсном режиме воздействию пептида KIF20A-10-308, ради отрицательного контроля. В «e» изображена способность KIF20A-A24-9-305 (SEQ ID NO: 174) индуцировать CTL. Для KIF20A-A24-9-305 (SEQ ID NO: 174) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линии CTL, которые были созданы из положительной лунки #2, показанной в рамке в верхней левой панели, и #6, показанной в рамке в нижней срединной панели, продемонстрировали специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. Кроме того, клон CTL, отобранный из линии CTL из лунки #2 с помощью серийного разведения, продемонстрировал специфическую активность CTL в отношении клеток-мишеней. Цитотоксическую активность клона CTL в отношении опухолевых клеток РК45Р, экспрессирующих и KIF20A, и HLA-A24, определяли с помощью анализа высвобождения Cr (CRA), и клон CTL обладал очень сильной цитотоксической активностью в отношении РК45Р. С другой стороны, не была продемонстрирована значительная специфическая цитотоксическая активность в отношении РК59, экспрессирующих KIF20A, но не экспрессирующих HLA-A24.
Фиг.7-1. На фиг.1 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для KNTC2-А24-9-309 (SEQ ID NO: 196), KNTC2-A24-9-124 (SEQ ID NO: 202), KNTC2-A24-9-154 (SEQ ID NO: 210), KNTC2-A24-9-150 (SEQ ID NO: 213), KNTC2-A24-10-452 (SEQ ID NO: 214), KNTC2-A24-10-227 (SEQ ID NO: 217) и KNTC2-A24-10-273 (SEQ ID NO: 223) обнаруживается мощная продукция IFN-гамма. В «а» изображен пример отрицательных пептидов, у которых не могла быть выявлена способность к индукции CTL, несмотря на возможную активность связывания с HLA. В «b» изображена способность KNTC2-A24-9-309 (SEQ ID NO: 196) индуцировать CTL. Для KNTC2-A24-9-309 (SEQ ID NO: 196) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #8, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «с» изображена способность KNTC2-A24-9-124 (SEQ ID NO: 202) индуцировать CTL. Для KNTC2-A24-9-124 (SEQ ID NO: 202) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #5, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «d» изображена способность KNTC2-A24-9-154 (SEQ ID NO: 210) индуцировать CTL. Для KNTC2-A24-9-154 (SEQ ID NO: 210) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клон CTL, которые были созданы из положительной лунки #5, показанной в рамке, продемонстрировали специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «е» изображена способность KNTC2-A24-9-150 (SEQ ID NO: 213) индуцировать CTL. Для KNTC2-A24-9-150 (SEQ ID NO: 213) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #7, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа.
Фиг.7-2. На фиг.7 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для KNTC2-A24-9-309 (SEQ ID NO: 196), KNTC2-A24-9-124 (SEQ ID NO: 202), KIF20A-A24-9-154 (SEQ ID NO: 210), KNTC2-A24-9-150 (SEQ ID NO: 213), KNTC2-A24-10-452 (SEQ ID NO: 214), KNTC2-A24-10-227 (SEQ ID NO: 217) и KNTC2-A24-10-273 (SEQ ID NO: 223) обнаруживается мощная продукция IFN-гамма. В «f» изображена способность KNTC2-A24-10-452 (SEQ ID NO: 214) индуцировать CTL. Для KNTC2-A24-10-452 (SEQ ID NO: 214) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линии и клон CTL, которые были созданы из положительной лунки #4, показанной в рамке в верхней левой панели, и #5, показанной в рамке в срединной панели, продемонстрировали специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. Кроме того, клон CTL, отобранный из линии CTL из лунки #5 с помощью серийного разведения, продемонстрировал специфическую активность CTL в отношении клеток-мишеней. Созданная из лунки #4 линия CTL продемонстрировала специфическую активность CTL в отношении клеток HEK293, трансфицированных и полноразмерным геном KNTC2, и молекулой HLA-A24. Также готовили HEK293, трансфицированные полноразмерным KNTC2, но не HLA-A24, HEK293, трансфицированные HLA-A24, но не полноразмерным геном KNTC2, и HEK293, трансфицированные HLA-A24 и подвергнутые в импульсном режиме воздействию пептида KNTC-9-309, ради отрицательного контроля. В «g» изображена способность KNTC2-A24-10-227 (SEQ ID NO: 217) индуцировать CTL. Для KNTC2-A24-10-227 (SEQ ID NO: 217) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #1, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «h» изображена способность KNTC2-A24-10-273 (SEQ ID NO: 223) индуцировать CTL. Для KNTC2-A24-10-273 (SEQ ID NO: 223) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT
на IFN-гамма, и линия CTL, которая была создана из положительной лунки #8, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа.
Фиг.8-1. На фиг.8 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для TTK-A02-9-462 (SEQ ID NO: 227), TTK-A02-9-719 (SEQ ID NO: 233), TTK-A02-9-547 (SEQ ID NO: 228) и TTK-A02-10-462 (SEQ ID NO: 254) обнаруживается мощная продукция IFN-гамма. В «a» изображен пример отрицательных пептидов, у которых не могла быть выявлена способность к индукции CTL несмотря на возможную активность связывания с HLA. В «b» изображена способность TTK-A02-9-462 (SEQ ID NO: 227) индуцировать CTL. Для TTK-A02-9-462 (SEQ ID NO: 227) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и два клона CTL, которые были созданы из положительной лунки #4, показанной в рамке, продемонстрировали специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. Созданный клон CTL продемонстрировал высокую специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном TTK, и молекулой HLA-A02. Также готовили COS7, трансфицированные полноразмерным TTK, но не HLA-A02, COS7, трансфицированные HLA-A02, но не полноразмерным геном TTK, и COS7, трансфицированные HLA-A02 и подвергнутые в импульсном режиме воздействию пептида TTK-9-547, ради отрицательного контроля. В «c» изображена способность TTK-A02-9-719 (SEQ ID NO: 233) индуцировать CTL. Для TTK-A02-9-719 (SEQ ID NO: 233) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клоны CTL были созданы из положительной лунки #1, показанной в рамке. Созданная линия CTL продемонстрировала высокую специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном TTK, и молекулой HLA-A02. Также готовили COS7, трансфицированные полноразмерным TTK, но не HLA-A02, и COS7, трансфицированные HLA-A02 и геном HIG2 в качестве замены полноразмерного гена TTK, ради отрицательного контроля.
Фиг.8-2. На фиг.8 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для TTK-A02-9-462 (SEQ ID NO: 227), TTK-A02-9-719 (SEQ ID NO: 233), TTK-A02-9-547 (SEQ ID NO: 228) и TTK-A02-10-462 (SEQ ID NO: 254) обнаруживается мощная продукция IFN-гамма. В «d» изображена способность TTK-A02-9-547 (SEQ ID NO: 228) индуцировать CTL. Для TTK-A02-9-547 (SEQ ID NO: 228) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клоны CTL были созданы из положительной лунки #2, показанной в рамке. Созданная линия CTL продемонстрировала специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном TTK, и молекулой HLA-A02. Также готовили COS7, трансфицированные полноразмерным TTK, но не HLA-A02, COS7, трансфицированные HLA-A02, но не полноразмерным геном TTK, и COS7, трансфицированные HLA-A02 и подвергнутые в импульсном режиме воздействию пептида TTK-10-462, ради отрицательного контроля.
Фиг.8-3. На фиг.8 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для TTK-A02-9-462 (SEQ ID NO: 227), TTK-A02-9-719 (SEQ ID NO: 233), TTK-A02-9-547 (SEQ ID NO: 228) и TTK-A02-10-462 (SEQ ID NO: 254) обнаруживается мощная продукция IFN-гамма. В «e» изображена способность TTK-A02-10-462 (SEQ ID NO: 254) индуцировать CTL. Для TTK-A02-10-462 (SEQ ID NO: 254) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и три клона CTL были созданы из положительной лунки #8, показанной в рамке. Созданный клон CTL продемонстрировал специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном TTK, и молекулой HLA-A02. Также готовили COS7, трансфицированные полноразмерным TTK, но не HLA-A02, COS7, трансфицированные HLA-A02, но не полноразмерным геном TTK, и COS7, трансфицированные HLA-A02 и подвергнутые в импульсном режиме воздействию пептида TTK-9-547, ради отрицательного контроля.
Фиг.9-1. На фиг.9 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для URLC10-A02-9-206 (SEQ ID NO: 271), URLC10-A02-9-212 (SEQ ID NO: 272) и URLC10-A02-10-211 (SEQ ID NO: 288) обнаруживается мощная продукция IFN-гамма. В «a» изображен пример отрицательных пептидов, у которых не могла быть выявлена способность к индукции CTL несмотря на возможную активность связывания с HLA. В «b» изображена способность URLC10-A02-9-206 (SEQ ID NO: 271) индуцировать CTL. Для URLC10-A02-9-206 (SEQ ID NO: 271) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT
на IFN-гамма, и линия CTL, которая была создана из положительной лунки #7, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «c» изображена способность URLC10-A02-9-212 (SEQ ID NO: 272) индуцировать CTL. Для URLC10-A02-9-212 (SEQ ID NO: 272) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия CTL, которая была создана из положительной лунки #3, показанной в рамке, продемонстрировала специфическую реакцию против клеток-мишеней, подвергнутых в импульсном режиме воздействию пептидного эпитопа. В «d» изображена способность URLC10-A02-10-211 (SEQ ID NO: 288) индуцировать CTL. Для URLC10-A02-10-211 (SEQ ID NO: 288) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и линия и клоны CTL были созданы из положительной лунки #5, показанной в рамке.
Фиг.9-2. На фиг.9 изображены результаты отбора пептидных эпитопов, которые, в свою очередь, демонстрируют, что для URLC10-A02-9-206 (SEQ ID NO: 271), URLC10-A02-9-212 (SEQ ID NO: 272) и URLC10-A02-10-211 (SEQ ID NO: 288) обнаруживается мощная продукция IFN-гамма. В «продолжении d» созданный клон CTL продемонстрировал высокую специфическую активность CTL в отношении клеток COS7, Hek293 и 293T, трансфицированных и полноразмерным геном URLC10, и молекулой HLA-A02. Также готовили COS7, Hek293 и 293T, трансфицированные полноразмерным URLC10, но не HLA-A02, и COS7, Hek293 и 293T, трансфицированные HLA-A02 и подвергнутые в импульсном режиме воздействию URLC10-10-64, ради отрицательного контроля.
На этих чертежах «+» означает мишень, подвергнутую в импульсном режиме воздействию пептида, «-» означает мишень, не подвергнутую в импульсном режиме воздействию пептида, «R» означает респондер, «S» означает стимулятор, «E» означает эффектор, а «T» означает мишень.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Используемые здесь термины в единственной числе означают «по крайней мере один», кроме указанных особо случаев.
Кроме случаев, определенных особо, все технические и научные термины, используемые здесь, имеют значение, одинаковое со значением, в котором они обычно понимаются специалистом со средним уровнем компетентности в данной области техники, к которой относится это изобретение.
Настоящее изобретение основано отчасти на открытии подходящих для иммунотерапии мишеней. Идентификация новых TAA, в частности ТАА, которые вызывают сильные и специфические противоопухолевые иммунные ответы, оправдывает дальнейшую разработку клинического применения стратегии вакцинации пептидами при различных типах рака (Boon T. et al., (1996) J. Exp. Med. 183: 725-729; van der Bruggen P. et al., (1991) Science 254: 1643-1647; Brichard V. et al., (1993) J. Exp. Med. 178: 489-495; Kawakami Y. et al., (1994) J. Exp. Med. 180: 347-352; Shichijo S. et al., (1998) J. Exp. Med. 187: 277-288; Chen Y.T. et al., (1997) Proc. Natl. Acad. Sci. USA, 94: 1914-1918; Harris C.C., (1996) J. Natl. Cancer Inst. 88: 1442-1455; Butterfield L.H. et al., (1999) Cancer Res. 59: 3134-3142; Vissers J.L. et al., (1999) Cancer Res. 59: 5554-5559; van der Burg S.H. et al., (1996) J. Immunol. 156: 3308-3314; Tanaka F. et al., (1997) Cancer Res. 57: 4465-4468; Fujie T. et al., (1999) Int. J. Cancer 80: 169-172; Kikuchi M. et al., (1999) Int. J. Cancer 81: 459-466; Oiso M. et al., (1999) Int. J. Cancer 81: 387-394). Поскольку TAA часто не обладают иммуногенностью, открытие подходящих мишеней является чрезвычайно важным результатом.
Как отмечено выше,
CDH3 (входящий № в GenBank NM_001793; SEQ ID NO: 1, 2),
EPHA4 (входящий № в GenBank L36645; SEQ ID NO: 3, 4),
ECT2 (входящий № в GenBank AY376439; SEQ ID NO: 5, 6),
HIG2 (входящий № в GenBank NM_013332; SEQ ID NO: 7, 8),
INHBB (входящий № в GenBank NM_002193; SEQ ID NO: 9, 10),
KIF20A (входящий № в GenBank NM_005733; SEQ ID NO: 11, 12),
KNTC2 (входящий № в GenBank AF017790; SEQ ID NO: 13, 14),
TTK (входящий № в GenBank NM_003318; SEQ ID NO: 15, 16) и
URLC10 (входящий № в GenBank NM_017527; SEQ ID NO: 17, 18) были ранее идентифицированы как сверхэкспрессируемые в различных раках, используя технологии ранжированных микрорядов кДНК.
В настоящем изобретении установлено, что пептиды, происходящие из CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK или URLC10, являются эпитопами TAA, рестрицированными по HLA-A24 и HLA-A2, аллелью HLA, обычно обнаруживаемой в популяциях японцев и европейцев. Конкретно были идентифицированы кандидаты связывающихся с HLA-A24 или HLA-A2 пептидов, происходящих из CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK или URLC10,
с использованием аффинностей их связывания с HLA-A24 или HLA-A2. После in vitro стимуляции T-клеток дендритными клетками (DC), нагруженными этими пептидами, были успешно созданы CTL, используя следующие пептиды:
CDH3-A24-9-513 (SEQ ID NO: 19),
CDH3-A24-9-406 (SEQ ID NO: 22),
CDH3-A24-10-807 (SEQ ID NO: 30),
CDH3-A24-10-332 (SEQ ID NO: 34),
CDH3-A24-10-655 (SEQ ID NO: 344),
CDH3-A24-10-470 (SEQ ID NO: 358),
EphA4-A24-9-453 (SEQ ID NO: 41),
EphA4-A24-9-5 (SEQ ID NO: 44),
EphA4-A24-9-869 (SEQ ID NO: 46),
EphA4-A24-9-420 (SEQ ID NO: 48),
EphA4-A24-10-24 (SEQ ID NO: 78),
EphA4-A02-9-501 (SEQ ID NO: 376),
EphA4-A02-9-165 (SEQ ID NO: 379),
ECT2-A24-9-515 (SEQ ID NO: 80),
ECT2-A24-10-40 (SEQ ID NO: 100),
ECT2-A24-10-101 (SEQ ID NO: 101),
HIG2-A24-9-19 (SEQ ID NO: 110),
HIG2-A24-9-22 (SEQ ID NO: 111),
HIG2-A24-9-8 (SEQ ID NO: 387),
HIG2-A24-10-7 (SEQ ID NO: 112),
HIG2-A24-10-18 (SEQ ID NO: 394),
HIG2-A02-9-8 (SEQ ID NO: 114),
HIG2-A02-9-15 (SEQ ID NO: 116),
HIG2-A02-9-4 (SEQ ID NO: 117),
HIG2-A02-10-8 (SEQ ID NO: 121),
INHBB-A24-9-180 (SEQ ID NO: 395),
INHBB-A24-10-180 (SEQ ID NO: 133),
INHBB-A24-10-305 (SEQ ID NO: 135),
INHBB-A24-10-7 (SEQ ID NO: 137),
INHBB-A24-10-212 (SEQ ID NO: 426),
KIF20A-A24-9-305 (SEQ ID NO: 174),
KIF20A-A24-9-383 (SEQ ID NO: 178),
KIF20A-A24-10-304 (SEQ ID NO: 186),
KIF20A-A24-10-66 (SEQ ID NO: 194),
KNTC2-A24-9-309 (SEQ ID NO: 196),
KNTC2-A24-9-124 (SEQ ID NO: 202),
KNTC2-A24-9-154 (SEQ ID NO: 210),
KNTC2-A24-9-150 (SEQ ID NO: 213),
KNTC2-A24-10-452 (SEQ ID NO: 214),
KNTC2-A24-10-227 (SEQ ID NO: 217),
KNTC2-A24-10-273 (SEQ ID NO: 223),
TTK-A02-9-462 (SEQ ID NO: 227),
TTK-A02-9-547 (SEQ ID NO: 228),
TTK-A02-9-719 (SEQ ID NO: 233),
TTK-A02-10-462 (SEQ ID NO: 254),
URLC-A02-9-206 (SEQ ID NO: 271),
URLC-A02-9-212 (SEQ ID NO: 272) и URLC-A02-10-211 (SEQ ID NO: 288).
Эти пептиды являются пептидными эпитопами каждого TAA, рестрицированными по HLA-A24 или HLA-A2. Поскольку эти антигены сверхэкспрессируются в большинстве раков и тесно связаны с пролиферацией опухолевых клеток, они находят применение в качестве мишеней для иммунотерапии раков. Приводимые в качестве примеров раки включают, но без ограничения, рак мочевого пузыря, рак молочной железы, рак шейки матки, холангиоцеллюлярный рак, CML, рак ободочной и прямой кишок, эндометриоз, рак пищевода, рак желудка, диффузный рак желудка, рак печени, NSCLC, лимфому, остеосаркому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки, SCLC, мягкотканевую опухоль и опухоль яичка.
Соответственно, настоящим изобретением, кроме того, обеспечиваются способы лечения или предупреждения заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, у субъекта, при этом такие способы включают стадии введения субъекту иммуногенного пептида, состоящего из менее приблизительно 40 аминокислот, часто менее приблизительно 20 аминокислот, обычно менее приблизительно 15 аминокислот и имеющего аминокислотную последовательность SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 376, 379, 80, 100, 101, 110, 111, 387, 112, 394, 114, 116, 117, 121, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217, 223, 227, 228, 233, 254, 271, 272 или 288.
В альтернативном случае иммуногенный пептид может иметь аминокислотную последовательность, определенную в SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 376, 379, 80, 100, 101, 110, 111, 387, 112, 394, 114, 116, 117, 121, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217, 223, 227, 228, 233, 254, 271, 272 или 288, в которой заменены, делетированы или в которую добавлены 1, 2 или несколько (например, вплоть до 5) аминокислот, при условии, что результирующий вариант пептида сохраняет иммуногенную активность (т.е. способность индуцировать CTL, специфичные в отношении клеток, экспрессирующих CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков).
Число остатков, подвергаемых замене, делеции или добавлению, составляет, как правило, 5 аминокислот или меньше, предпочтительно 4 аминокислоты или меньше, более предпочтительно 3 аминокислоты или меньше, даже более предпочтительно одну или две аминокислоты. Предполагаемые раки включают, но без ограничения, рак мочевого пузыря, рак молочной железы, рак шейки матки, холангиоцеллюлярный рак, CML, рак ободочной и прямой кишок, эндометриоз, рак пищевода, рак желудка, диффузный рак желудка, рак печени, NSCLC, лимфому, остеосаркому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки, SCLC, мягкотканевую опухоль и опухоль яичка. Кроме того, настоящим изобретением обеспечиваются способы предупреждения рецидива после хирургического вмешательства упомянутых выше заболеваний.
Известно, что варианты пептидов (т.е. пептиды, имеющие аминокислотную последовательность, которая модифицирована путем замены, делеции одного, двух или нескольких аминокислотных остатков в первоначальной аминокислотной последовательности или добавления в нее такое число остатков) сохраняют первоначальную биологическую активность (Mark D.F. et al., (1984) Proc. Natl. Acad. Sci. USA 81: 5662-5666; Zoller M.J. and Smith M., (1982) Nucleic Acids Res. 10: 6487-6500; Dalbadie-McFarland G. et al., (1982) Proc. Natl. Acad. Sci. USA 79: 6409-6413). В контексте настоящего изобретения предпочтительно, чтобы следствием модификации аминокислоты(от) было сохранение свойств боковой цепи первоначальной аминокислоты (процесс, известный как консервативная аминокислотная замена). Примеры свойств боковых цепей аминокислот включают гидрофобные аминокислоты (A, I, L, M, F, P, W, Y, V), гидрофильные аминокислоты (R, D, N, C, E, Q, G, H, K, S, T) и боковые цепи, имеющие совместно следующие функциональные группы или характеристики: алифатическую боковую цепь (G, A, V, L, I, P); содержащую гидроксильную группу боковую цепь (S, T, Y); содержащую атом серы боковую цепь (C, M); боковую цепь, содержащую карбоновую кислоту и амид (D, N, E, Q); содержащую основание боковую цепь (R, K, H) и содержащую ароматическое кольцо боковую цепь (H, F, Y, W). Заметьте, что буквы в скобках означают однобуквенные коды аминокислот.
В предпочтительных вариантах осуществления иммуногенный пептид представляет собой нанопептид (9-мер) или декапептид (10-мер).
Настоящим изобретением, кроме того, обеспечивается способ индукции противоопухолевого иммунитета против заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, у субъекта, при этом такой способ включает стадии введения субъекту иммуногенного пептида настоящего изобретения, а именно пептида, имеющего аминокислотную последовательность SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 376, 379, 80, 100, 101, 110, 111, 387, 112, 394, 114, 116, 117, 121, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217, 223, 227, 228, 233, 254, 271, 272 или 288, или его варианта (т.е. включающего 1, 2 или несколько (например, вплоть до 5) замен, делеций или добавлений аминокислот), нуждающемуся в этом субъекту. Предполагаемые раки включают, но без ограничения, рак мочевого пузыря, рак молочной железы, рак шейки матки, холангиоцеллюлярный рак, CML, рак ободочной и прямой кишок, эндометриоз, рак пищевода, рак желудка, диффузный рак желудка, рак печени, NSCLC, лимфому, остеосаркому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки, SCLC, мягкотканевую опухоль и опухоль яичка.
В контексте настоящего изобретения субъектом предпочтительно является млекопитающее. Приводимые в качестве примеров млекопитающие включают, но без ограничения, например, человека, не являющегося человеком примата, мышь, крысу, собаку, кошку, лошадь или корову.
В настоящем изобретении пептид может вводиться субъекту с использованием in vivo или ex vivo протокола. Кроме того, настоящим изобретением также обеспечивается применение нанопептида или декапептида, выбираемого из пептидов, имеющих аминокислотную последовательность SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 376, 379, 80, 100, 101, 110, 111, 387, 112, 394, 114, 116, 117, 121, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217, 223, 227, 228, 233, 254, 271, 272 или 288 (и их вариантов), для производства иммуногенной композиции для лечения или предупреждения заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков. Предполагаемые раки включают, но без ограничения, рак мочевого пузыря, рак молочной железы, рак шейки матки, холангиоцеллюлярный рак, CML, рак ободочной и прямой кишок, эндометриоз, рак пищевода, рак желудка, диффузный рак желудка, рак печени, NSCLC, лимфому, остеосаркому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки, SCLC, мягкотканевую опухоль и опухоль яичка.
Анализы гомологии следующих пептидов показывают, что они не обладают значительной гомологией с пептидами, происходящими из каких-либо продуктов известных генов человека.
CDH3-A24-9-513 (SEQ ID NO: 19),
CDH3-A24-9-406 (SEQ ID NO: 22),
CDH3-A24-10-807 (SEQ ID NO: 30),
CDH3-A24-10-332 (SEQ ID NO: 34),
CDH3-A24-10-655 (SEQ ID NO: 344),
CDH3-A24-10-470 (SEQ ID NO: 358),
EphA4-A24-9-453 (SEQ ID NO: 41),
EphA4-A24-9-5 (SEQ ID NO: 44),
EphA4-A24-9-869 (SEQ ID NO: 46),
EphA4-A24-9-420 (SEQ ID NO: 48),
EphA4-A24-10-24 (SEQ ID NO: 78),
EphA4-A02-9-501 (SEQ ID NO: 376),
EphA4-A02-9-165 (SEQ ID NO: 379),
ECT2-A24-9-515 (SEQ ID NO: 80),
ECT2-A24-10-40 (SEQ ID NO: 100),
ECT2-A24-10-101 (SEQ ID NO: 101),
HIG2-A24-9-19 (SEQ ID NO: 110),
HIG2-A24-9-22 (SEQ ID NO: 111),
HIG2-A24-9-8 (SEQ ID NO: 387),
HIG2-A24-10-7 (SEQ ID NO: 112),
HIG2-A24-10-18 (SEQ ID NO: 394),
HIG2-A02-9-8 (SEQ ID NO: 114),
HIG2-A02-9-15 (SEQ ID NO: 116),
HIG2-A02-9-4 (SEQ ID NO: 117),
HIG2-A02-10-8 (SEQ ID NO: 121),
INHBB-A24-9-180 (SEQ ID NO: 395),
INHBB-A24-10-180 (SEQ ID NO: 133),
INHBB-A24-10-305 (SEQ ID NO: 135),
INHBB-A24-10-7 (SEQ ID NO: 137),
INHBB-A24-10-212 (SEQ ID NO: 426),
KIF20A-A24-9-305 (SEQ ID NO: 174),
KIF20A-A24-9-383 (SEQ ID NO: 178),
KIF20A-A24-10-304 (SEQ ID NO: 186),
KIF20A-A24-10-66 (SEQ ID NO: 194),
KNTC2-A24-9-309 (SEQ ID NO: 196),
KNTC2-A24-9-124 (SEQ ID NO: 202),
KNTC2-A24-9-154 (SEQ ID NO: 210),
KNTC2-A24-9-150 (SEQ ID NO: 213),
KNTC2-A24-10-452 (SEQ ID NO: 214),
KNTC2-A24-10-227 (SEQ ID NO: 217),
KNTC2-A24-10-273 (SEQ ID NO: 223),
TTK-A02-9-462 (SEQ ID NO: 227),
TTK-A02-9-547 (SEQ ID NO: 228),
TTK-A02-9-719 (SEQ ID NO: 233),
TTK-A02-10-462 (SEQ ID NO: 254),
URLC-A02-9-206 (SEQ ID NO: 271),
URLC-A02-9-212 (SEQ ID NO: 272) и
URLC-A02-10-211 (SEQ ID NO: 288).
Поэтому возможность неизвестных или нежелательных ответов против этих молекул при использовании иммунотерапии значительно снижена.
Что касается антигенов HLA, представленные здесь данные демонстрируют, что использование антигенов типа A-24 или типа A-2 (которые, как указано выше, являются в высокой степени экспрессируемыми среди японцев) благоприятно для получения эффективных результатов. Использование таких подтипов, как A-2402 и A-0201, является даже более предпочтительным. Как правило, в клинике заранее исследуется тип антигена HLA пациента, которому требуется лечение, который, в свою очередь, делает возможным отбор соответствующих пептидов, имеющих высокие степени аффинности связывания с антигеном пациента или обладающих способностью к индукции цитотоксических T-клеток (CTL) при презентации антигена. Кроме того, для получения пептидов, обладающих высокой аффинностью связывания и способностью к индукции CTL, можно произвести замену, делецию или добавление 1, 2 или нескольких (например, вплоть до 5) аминокислот на основе аминокислотной последовательности частичного пептида встречающегося в природе CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10. Здесь термин «несколько» означает 5 или менее, более предпочтительно 3 или менее. Кроме того, помимо пептидов, представляемых в природе, поскольку уже известны соответствующие требования к последовательностям пептидов, представляемым при связывании с антигенами HLA (Kubo R.T. et al., (1994) J. Immunol., 152, 3913-3924; Rammensee H.G. et al., (1995) Immunogenetics. 41:178-228; Kondo A. et al., (1995) J. Immunol. 155: 4307-4312), можно произвести модификации, основанные на таких требованиях, в иммуногенных пептидах настоящего изобретения. Например, можно выгодно использовать пептиды, обладающие высокой аффинностью связывания с HLA-24, в которых вторая с N-конца аминокислота заменена фенилаланином, тирозином, метионином или триптофаном. Подобным образом можно также выгодно использовать пептиды, С-концевая аминокислота которых заменена фенилаланином, лейцином, изолейцином, триптофаном или метионином. С другой стороны, можно выгодно использовать пептиды, обладающие высокой аффинностью связывания с HLA-A2, в которых вторая с N-конца аминокислота заменена лейцином или метионином, и пептиды, C-концевая аминокислота которых заменена валином или лейцином. Замену можно произвести не только в концевых аминокислотах, но также в положении потенциального распознавания TCR пептидов. В нескольких исследованиях продемонстрировано, что замены аминокислот в пептиде могут приводить к пептиду, который равен первоначальному пептиду или лучше него, например CAP1, p53 (264-272), Her-2/neu (369-377) или gp100 (209-217) (Zaremba et al. Cancer Res. 57, 4570-4577, 1997, T. K. Hoffmann et al. J. Immunol. (2002) Feb 1; 168(3): 1338-1347, S. O. Dionne et al. Cancer Immunol. Immunother. (2003) 52: 199-206 и S. O. Dionne et al. Cancer Immunology, Immunotherapy (2004) 53, 307-314). Кроме того, к N-концу и/или C-концу пептида можно добавить 1-2 аминокислоты.
Однако, когда последовательность пептида идентична части аминокислотной последовательности эндогенного или экзогенного белка, имеющего отличную функцию, могут быть вызваны побочные эффекты, такие как аутоиммунные нарушения или симптомы аллергии. Поэтому предпочтительно исключать ситуацию, при которой иммуногенная последовательность соответствует аминокислотной последовательности известного белка. Можно исключить эту ситуацию путем выполнения поиска гомологии, используя имеющиеся базы данных. Если поиски гомологии подтвердят, что пептиды, в которых присутствуют 1, 2 или несколько отличных аминокислот, не существуют в природе, тогда может быть исключена угроза того, что модификации упомянутой выше аминокислотной последовательности, например, увеличат аффинность связывания с антигенами HLA и/или увеличат способность к индукции CTL.
Хотя предполагается, что описанные выше пептиды, обладающие высокой аффинностью связывания с антигенами HLA, являются высокоэффективными в качестве вакцин против рака, пептиды-кандидаты, которые выбраны по наличию высокой аффинности связывания в качестве показателя, необходимо проверить на фактическое наличие способности к индукции CTL. Способность к индукции CTL можно в заведенном порядке подтвердить путем индукции антигенпрезентирующих клеток, несущих антигены MHC человека (например, B-лимфоцитов, макрофагов и дендритных клеток), или конкретнее дендритных клеток, происходящих из мононуклеарных лейкоцитов периферической крови человека, и после стимуляции представляющим интерес пептидом, смешивания с CD8-положительными клетками и определения цитотоксической активности в отношении клеток-мишеней. В качестве реакционной системы можно использовать полученных трансгенных животных, которые экспрессируют антиген HLA человека (например, трансгенных животных, описанных BenMohamed L. и др. (2000) в Hum. Immunol.; 61(8): 764-779, других статьях, книгах на эту же тему, Linkout). Например, в клетки-мишени можно ввести радиоактивную метку 51Cr и т.п., и цитотоксическую активность можно рассчитать на основе радиоактивности, высвободившейся из клеток-мишеней. В альтернативном случае ее можно исследовать путем измерения IFN-гамма, продуцируемого и высвобождаемого CTL в присутствии антигенпрезентирующих клеток, которые несут иммобилизованные пептиды, и визуализации зоны ингибирования на средах, используя моноклональные антитела против IFN-гамма.
В результате проверки способности к индукции CTL описанных выше пептидов было обнаружено, что те пептиды, которые обладают высокой аффинностью связывания с антигеном HLA, не обладают обязательно на высоком уровне способностью к индукции. Однако нанопептиды или декапептиды, выбираемые из группы пептидов, имеющих аминокислотные последовательности, указанные при следующих пептидах, продемонстрировали на особенно высоком уровне способность к индукции CTL.
CDH3-A24-9-513 (SEQ ID NO: 19),
CDH3-A24-9-406 (SEQ ID NO: 22),
CDH3-A24-10-807 (SEQ ID NO: 30),
CDH3-A24-10-332 (SEQ ID NO: 34),
CDH3-A24-10-655 (SEQ ID NO: 344),
CDH3-A24-10-470 (SEQ ID NO: 358),
EphA4-A24-9-453 (SEQ ID NO: 41),
EphA4-A24-9-5 (SEQ ID NO: 44),
EphA4-A24-9-869 (SEQ ID NO: 46),
EphA4-A24-9-420 (SEQ ID NO: 48),
EphA4-A24-10-24 (SEQ ID NO: 78),
EphA4-A02-9-501 (SEQ ID NO: 376),
EphA4-A02-9-165 (SEQ ID NO: 379),
ECT2-A24-9-515 (SEQ ID NO: 80),
ECT2-A24-10-40 (SEQ ID NO: 100),
ECT2-A24-10-101 (SEQ ID NO: 101),
HIG2-A24-9-19 (SEQ ID NO: 110),
HIG2-A24-9-22 (SEQ ID NO: 111),
HIG2-A24-9-8 (SEQ ID NO: 387),
HIG2-A24-10-7 (SEQ ID NO: 112),
HIG2-A24-10-18 (SEQ ID NO: 394),
HIG2-A02-9-8 (SEQ ID NO: 114),
HIG2-A02-9-15 (SEQ ID NO: 116),
HIG2-A02-9-4 (SEQ ID NO: 117),
HIG2-A02-10-8 (SEQ ID NO: 121),
INHBB-A24-9-180 (SEQ ID NO: 395),
INHBB-A24-10-180 (SEQ ID NO: 133),
INHBB-A24-10-305 (SEQ ID NO: 135),
INHBB-A24-10-7 (SEQ ID NO: 137),
INHBB-A24-10-212 (SEQ ID NO: 426),
KIF20A-A24-9-305 (SEQ ID NO: 174),
KIF20A-A24-9-383 (SEQ ID NO: 178),
KIF20A-A24-10-304 (SEQ ID NO: 186),
KIF20A-A24-10-66 (SEQ ID NO: 194),
KNTC2-A24-9-309 (SEQ ID NO: 196),
KNTC2-A24-9-124 (SEQ ID NO: 202),
KNTC2-A24-9-154 (SEQ ID NO: 210),
KNTC2-A24-9-150 (SEQ ID NO: 213),
KNTC2-A24-10-452 (SEQ ID NO: 214),
KNTC2-A24-10-227 (SEQ ID NO: 217),
KNTC2-A24-10-273 (SEQ ID NO: 223),
TTK-A02-9-462 (SEQ ID NO: 227),
TTK-A02-9-547 (SEQ ID NO: 228),
TTK-A02-9-719 (SEQ ID NO: 233),
TTK-A02-10-462 (SEQ ID NO: 254),
URLC-A02-9-206 (SEQ ID NO: 271),
URLC-A02-9-212 (SEQ ID NO: 272) и
URLC-A02-10-211 (SEQ ID NO: 288).
Как отмечено выше, настоящим изобретением обеспечиваются пептиды, обладающие способностью к индукции цитотоксических T-клеток, а именно пептиды, имеющие аминокислотную последовательность SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 376, 379, 80, 100, 101, 110, 111, 387, 112, 394, 114, 116, 117, 121, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217, 223, 227, 228, 233, 254, 271, 272 или 288, или их варианты (т.е. пептиды, в которых заменены, делетированы или в которые добавлены 1, 2 или несколько аминокислот).
Предпочтительно, когда аминокислотные последовательности, состоящие из 9 или 10 аминокислот, показанных в SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 376, 379, 80, 100, 101, 110, 111, 387, 112, 394, 114, 116, 117, 121, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217, 223, 227, 228, 233, 254, 271, 272 или 288, или их варианты не соответствуют аминокислотной последовательности, связанной с другим эндогенным белком.
В частности, примерами предпочтительных вариантов являются замена аминокислоты на лейцин или метионин в положении второй с N-конца аминокислоты, замена С-концевой аминокислоты на валин или лейцин и добавление 1-2 аминокислот к N-концу и/или C-концу.
Квалифицированный в данной области специалист признает, что помимо замен и добавлений аминокислот в способах настоящего изобретения можно также использовать иммунологически активные фрагменты пептидов. Способы определения активных фрагментов широко известны в данной области техники. Клоны CTL, полученные при стимуляции этими модифицированными пептидами, могут распознавать первоначальные пептиды и вызывать повреждение клеток, экспрессирующих первоначальные пептиды.
Пептиды настоящего изобретения можно приготовить, используя широко известные методы. Например, пептиды можно приготовить синтетически, используя или технологию рекомбинантных ДНК, или химический синтез. Пептиды настоящего изобретения можно синтезировать индивидуально или в виде более длинных полипептидов, состоящих из двух или более пептидов. Пептиды настоящего изобретения предпочтительно являются выделенными, т.е. в значительной степени лишены других встречающихся в природе белков и их фрагментов клеток-хозяев.
Пептиды настоящего изобретения могут содержать модификации, такие как гликозилирование, окисление боковой цепи или фосфорилирование; при условии, что модификации не аннулируют биологическую активность описанных здесь пептидов, а именно способность связываться с антигеном HLA и индуцировать CTL. Другие модификации включают введение D-аминокислот или других миметиков аминокислот, которые могут использоваться, например, для увеличения полупериода существования пептидов в сыворотке.
Более того, это изобретение может вмещать способ отбора пептида, в котором 1, 2 или несколько аминокислот заменены, причем первоначальный пептид включает аминокислотную последовательность, выбираемую из группы, состоящей из SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 80, 100, 101, 110, 111, 387, 112, 394, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217 или 223, при этом указанный способ включает стадии:
(a) подтверждения отсутствия значительной гомологии последовательности с полной последовательностью пептида с 1, 2 или несколькими аминокислотными заменами;
(b) определения способности к индукции CTL кандидатов-пептидов с заменами; и
(c) отбора пептида, способность к индукции CTL которого равна или выше способности к индукции первоначального пептида.
Например, в предпочтительных вариантах осуществления настоящим изобретением обеспечивается способ идентификации пептида, обладающего способностью индуцировать CTL против клеток, экспрессирующих по крайней мере один опухолеспецифический антиген, причем опухолеспецифическим антигеном является антиген, выбираемый из группы, состоящей из CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10, при этом указанный способ включает стадии:
(i) обеспечения или создания по крайней мере одной последовательности-кандидата, которая состоит из аминокислотной последовательности, модифицированной путем замены, делеции одного, двух или нескольких аминокислотных остатков в первоначальной аминокислотной последовательности или добавления в нее указанного числа остатков, причем первоначальную аминокислотную последовательность выбирают из группы, состоящей из SEQ ID NO: SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 80, 100, 101, 110, 111, 387, 112, 394, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217 или 223;
(ii) отбора последовательности-кандидата, которая не обладает значительной гомологией с последовательностями пептидов, происходящих из любых продуктов известных генов человека, отличных от указанных опухолеспецифических антигенов;
(iii) приведения пептида, состоящего из последовательности-кандидата, отобранной на стадии (ii), в контакт с антигенпрезентирующими клетками;
(iv) приведения антигенпрезентирующих клеток стадии (iii) в контакт с Т-клетками для оценки способности пептида стимулировать Т-клетки; и
(v) идентификации пептида, способность к индукции CTL которого равна или выше способности к индукции пептида, состоящего из первоначальной аминокислотной последовательности.
Предпочтительно аминокислоту заменяют отличной аминокислотой, в которой свойства боковой цепи аминокислоты сохранены (процесс, известный как консервативная аминокислотная замена). Примеры свойств боковых цепей аминокислот включают гидрофобные аминокислоты (A, I, L, M, F, P, W, Y, V), гидрофильные аминокислоты (R, D, N, С, Е, Q, G, Н, K, S, Т) и боковые цепи, имеющие совместно следующие функциональные группы или характеристики: алифатическую боковую цепь (G, А, V, L, I, P); содержащую гидроксильную группу боковую цепь (S, Т, Y); содержащую атом серы боковую цепь (С, M); боковую цепь, содержащую карбоновую кислоту и амид (D, N, Е, Q); содержащую основание боковую цепь (R, K, Н) и содержащую ароматическое кольцо боковую цепь (H, F, Y, W). Заметьте, что буквы в скобках означают однобуквенные коды аминокислот. В настоящем изобретении значительная гомология представляет собой, например, идентичность на более 90%, предпочтительно на 95%, более предпочтительно на 99% или 100% с продуктом известного гена человека, подвергаемым сравнению.
Пептиды этого изобретения можно приготовить в виде комбинации, которая включает два или более пептидов настоящего изобретения, для применения в качестве вакцины против заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, такой как вакцина, индуцирующая CTL in vivo. Предполагаемые раки включают, но без ограничения, рак мочевого пузыря, рак молочной железы, рак шейки матки, холангиоцеллюлярный рак, CML, рак ободочной и прямой кишок, эндометриоз, рак пищевода, рак желудка, диффузный рак желудка, рак печени, NSCLC, лимфому, остеосаркому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки, SCLC, мягкотканевую опухоль и опухоль яичка. Пептиды могут присутствовать в смеси или могут быть конъюгированы друг с другом, используя стандартные методы. Например, пептиды можно экспрессировать в виде одиночной полипептидной последовательности. Пептиды в комбинации могут быть одинаковыми или различными.
При введении пептидов этого изобретения пептиды презентируются с высокой плотностью на антигенах HLA антигенпрезентирующих клеток, которые, в свою очередь, индуцируют CTL, которые специфическим образом реагируют в отношении комплекса, образованного между представленным пептидом и антигеном HLA. В альтернативном случае антигенпрезентирующие клетки с иммобилизованными пептидами этого изобретения на своей клеточной поверхности, получаемые при извлечении дендритных клеток из субъектов, можно стимулировать пептидами этого изобретения. Введение вновь этих клеток соответствующим субъектам индуцирует CTL, и в результате клеточная цитотоксическая реакция в отношении клеток-мишеней может быть увеличена.
Конкретнее, настоящим изобретением обеспечиваются лекарственные средства для лечения и/или предупреждения пролиферации, метастазирования и т.п. заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, которые включают один или несколько пептидов настоящего изобретения или кодирующий пептиды полинуклеотид. Пептиды или полинуклеотиды настоящего изобретения находят конкретное применение для лечения заболевания с тесной связью с CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков. Предполагаемые раки включают, но без ограничения, рак мочевого пузыря, рак молочной железы, рак шейки матки, холангиоцеллюлярный рак, CML, рак ободочной и прямой кишок, эндометриоз, рак пищевода, рак желудка, диффузный рак желудка, рак печени, NSCLC, лимфому, остеосаркому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки, SCLC, мягкотканевую опухоль и опухоль яичка.
Пептиды этого изобретения можно вводить субъекту непосредственно, в виде фармацевтической композиции, приготовленной с помощью общепринятых способов приготовления лекарственных средств. В таких случаях помимо пептидов этого изобретения носители, наполнители и т.п., которые обычно используются для лекарственных средств, могут быть включены соответствующим образом, без особых ограничений. Иммуногенные композиции этого изобретения могут использоваться для лечения и предупреждения заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков. Предполагаемые раки включают, но без ограничения, рак мочевого пузыря, рак молочной железы, рак шейки матки, холангиоцеллюлярный рак, CML, рак ободочной и прямой кишок, эндометриоз, рак пищевода, рак желудка, диффузный рак желудка, рак печени, NSCLC, лимфому, остеосаркому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки, SCLC, мягкотканевую опухоль и опухоль яичка.
Иммуногенные композиции для лечения и/или предупреждения заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, которые включают в качестве активного ингредиента один или несколько пептидов настоящего изобретения, могут дополнительно включать адъювант, вследствие чего будет эффективно создаваться клеточный иммунитет. В альтернативном случае они могут вводиться с другими активными ингредиентами, такими как противораковые агенты.
Предполагаемые раки включают, но без ограничения, рак мочевого пузыря, рак молочной железы, рак шейки матки, холангиоцеллюлярный рак, CML, рак ободочной и прямой кишок, эндометриоз, рак пищевода, рак желудка, диффузный рак желудка, рак печени, NSCLC, лимфому, остеосаркому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки, SCLC, мягкотканевую опухоль и опухоль яичка. Подходящие композиции включают гранулы. Подходящие адъюванты описаны в литературе (Johnson A.G. (1994) C1in. Microbiol. Rev., 7: 277-289).
Приводимые в качестве примеров адъюванты включают, но без ограничения, алюминия фосфат, алюминия гидроокись и квасцы. Кроме того, можно подходящим образом использовать липосомные композиции, гранулированные композиции, в которых лекарственное средство связано с шариками диаметром несколько микрон, и композиции, в которых липид связан с пептидом. Способ введения может быть оральным, интрадермальным, подкожным, внутривенным введением или т.п. и может включать системное введение или местное введение поблизости от прицельной опухоли.
Дозу пептида(ов) этого изобретения можно соответствующим образом регулировать в соответствии с подвергаемым лечению заболеванием, возрастом пациента, весом, способом введения и т.п. Хотя доза обычно составляет от 0,001 мг до 1000 мг, предпочтительно от 0,01 мг до 100 мг, более предпочтительно от 0,1 мг до 10 мг, предпочтительно вводимых один раз в несколько дней-несколько месяцев, квалифицированный в данной области техники специалист может без труда выбрать соответствующую дозу и способ введения, поскольку выбор и оптимизация этих параметров находятся полностью в пределах знаний установившейся практики.
Настоящим изобретением, кроме того, обеспечиваются внутриклеточные везикулы, называемые экзосомами, которые представляют комплексы, образованные между пептидами настоящего изобретения и антигенами HLA, на своей поверхности. Экзосомы можно приготовить, например, при использовании способов, подробно описанных в опубликованном переводе на японский язык международной публикации № Hei 11-510507 и 2000-512161, и предпочтительно готовят, используя антигенпрезентирующие клетки, полученные от субъектов, являющихся объектами для лечения и/или профилактики. Экзосомы этого изобретения можно инокулировать в качестве вакцин против рака аналогично пептидам этого изобретения.
Тип используемых антигенов HLA должен соответствовать такому типу у субъекта, которому необходимо лечение и/или профилактика. Например, для популяции японцев HLA-A24 или HLA-A2, в частности HLA-A2402 или HLA-A0201, часто является соответствующим.
В некоторых вариантах осуществления композиции в виде вакцин настоящего изобретения включают компонент, который примирует цитотоксические T-лимфоциты. В качестве агентов, способных примировать CTL in vivo против вирусных антигенов, были идентифицированы липиды. Например, остатки пальмитиновой кислоты можно присоединить к эпсилон- и альфа-аминогруппам остатка лизина и затем связать с иммуногенным пептидом настоящего изобретения. Липидизированный пептид можно затем вводить или непосредственно, в мицелле или частице, включенной в липосому, или эмульгировать в адъюванте. В качестве другого примера примирования липидом ответов CTL, для примирования CTL можно использовать липопротеины E. coli, такие как трипальмитоил-S-глицерилцистеинилсерил-серин (P3CSS), ковалентно присоединенные к соответствующему пептиду (см., например, Deres K. et al., (1989) Nature 342: 561-564).
Иммуногенные композиции настоящего изобретения могут также включать нуклеиновые кислоты, кодирующие один или несколько описанных здесь иммуногенных пептидов. См., например, Wolff J.A. et al., (1990) Science 247: 1465-1468; патенты США № 5580859, 5589466, 5804566, 5739118, 5736524 и 5679647 и заявку WO 98/04720. Примеры основанных на ДНК методик доставки включают «голую ДНК», облегченную (опосредованную бупивикаином, полимерами, пептидами) доставку, комплексы с катионными липидами и опосредуемую частицами доставку («генное ружье») или доставку с использованием давления (см., например, патент США № 5922687).
Иммуногенные пептиды настоящего изобретения могут также быть экспрессированы с использованием вирусных или бактериальных векторов. Примеры подходящих экспрессионных векторов включают аттенуированные вирусы-хозяева, такие как вирус коровьей оспы или оспы птиц. Этот подход включает использование вируса коровьей оспы, например, в качестве вектора для экспрессии нуклеотидных последовательностей, кодирующих пептид. При введении в хозяина рекомбинантный вирус коровьей оспы экспрессирует иммуногенный пептид и, таким образом, вызывает иммунный ответ. Векторы на основе вируса коровьей оспы и способы, применимые в протоколах иммунизации, описываются, например, в патенте США № 4722848. Другим подходящим вектором является БЦЖ (бацилла Кальметта-Герена). Векторы на основе БЦЖ описываются Stover C.K. и др. (1991) в Nature 351: 456-460. Широкий выбор других векторов, пригодных для терапевтического введения или иммунизации, например векторы на основе аденовирусов и адено-ассоциированных вирусов, ретровирусные векторы, векторы Salmonella typhi, векторы на основе дезактивированного сибиреязвенного токсина и т.п., известен в данной области техники. См., например, Shata M.T. et al., (2000) Mol. Med. Today 6: 66-71; Shedlock D.J. and Weiner D.B. et al., (2000) J. Leukoc. Biol. 68: 793-806; и Hipp J.D. et al., (2000) In Vivo 14: 571-585.
Настоящим изобретением также обеспечиваются способы индукции антигенпрезентирующих клеток, используя один или несколько пептидов этого изобретения. Антигенпрезентирующие клетки могут быть индуцированы путем индукции дендритных клеток из моноцитов периферической крови и затем приведения их в контакт (их стимулирования) с одним или несколькими пептидами этого изобретения in vitro, ex vivo или in vivo. При введении субъектам пептидов настоящего изобретения в организме субъекта индуцируются антигенпрезентирующие клетки с иммобилизованными на них пептидами этого изобретения. В альтернативном случае после иммобилизации пептидов этого изобретения на антигенпрезентирующих клетках клетки можно ввести субъекту в качестве вакцины. Например, введение ex vivo может включать стадии:
a: получения антигенпрезентирующих клеток из субъекта, и
b: приведения антигенпрезентирующих клеток стадии a в контакт с пептидом настоящего изобретения.
Альтернативно, в соответствии с настоящим изобретением обеспечивается применение пептидов этого изобретения для производства фармацевтической композиции, индуцирующей антигенпрезентирующие клетки. Кроме того, настоящим изобретением также обеспечивается пептид настоящего изобретения для индукции антигенпрезентирующих клеток. Антигенпрезентирующие клетки, полученные на стадии b, можно вводить субъекту в качестве вакцины.
Этим изобретением также обеспечивается способ индукции антигенпрезентирующих клеток, обладающих на высоком уровне способностью к индукции цитотоксических T-клеток, включающий стадию переноса генов, составленных из полинуклеотида(ов), кодирующего один или несколько пептидов этого изобретения, в антигенпрезентирующие клетки in vitro. Введенные гены могут быть в форме ДНК или РНК. Для введения могут подходящим образом, без особых ограничений, использоваться различные способы, обычно выполняемые в этой области, такие как липофекция, электропорация и способ с использованием кальция фосфата. Конкретнее, трансфекцию можно выполнить, как описано Reeves M.E. и др. (1996) в Cancer Res., 56: 5672-5677; Butterfield L.H. и др. (1998) в J. Immunol., 161: 5607-5613; Boczkowski D. и др. (1996) в J. Exp. Med., 184: 465-472; в опубликованном переводе на японский язык международной публикации № 2000-509281. При переносе гена в антигенпрезентирующие клетки ген подвергается транскрипции, трансляции и т.п. в клетке, и затем полученный белок подвергается процессингу MHC класса I или класса II и проходит через путь для представления частичных пептидов.
Настоящим изобретением, кроме того, обеспечиваются способы индукции CTL, используя один или несколько пептидов этого изобретения. При введении субъекту пептидов этого изобретения в организме субъекта индуцируются CTL и таким образом увеличивается эффективность иммунной системы против клеток, экспрессирующих CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раковых клеток в опухолевых тканях.
Предполагаемые раки включают, но без ограничения, рак мочевого пузыря, рак молочной железы, рак шейки матки, холангиоцеллюлярный рак, CML, рак ободочной и прямой кишок, эндометриоз, рак пищевода, рак желудка, диффузный рак желудка, рак печени, NSCLC, лимфому, остеосаркому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки, SCLC, мягкотканевую опухоль и опухоль яичка. В альтернативном случае пептиды настоящего изобретения можно использовать в связи с ex vivo терапевтическим способом, в котором полученные от субъекта антигенпрезентирующие клетки и CD8-положительные клетки или мононуклеарные лейкоциты периферической крови приводят в контакт (стимулируют) с одним или несколькими пептидами этого изобретения in vitro, и после индукции CTL клетки возвращаются субъекту. Например, способ может включать стадии:
a: получения антигенпрезентирующих клеток от субъекта,
b: приведения антигенпрезентирующих клеток стадии a в контакт с пептидом настоящего изобретения,
c: смешивания антигенпрезентирующих клеток стадии b с CD8+ T-клетками и сокультивирования для индукции цитотоксических T-клеток, и
d: сбора CD8+ T-клеток из сокультуры стадии c.
В альтернативном случае в соответствии с настоящим изобретением обеспечивается применение пептидов этого изобретения для производства фармацевтической композиции, индуцирующей CTL. Кроме того, настоящим изобретением также обеспечивается пептид настоящего изобретения для индукции CTL. CD8+ T-клетки с цитотоксической активностью, полученные на стадии d, можно вводить субъекту в качестве вакцины.
Настоящим изобретением, кроме того, обеспечиваются выделенные цитоксические T-клетки, индуцированные с использованием пептидов этого изобретения. Цитотоксические T-клетки, индуцированные при стимуляции антигенпрезентирующей клеткой, представляющей один или несколько пептидов этого изобретения, предпочтительно получают от субъектов, являющихся объектами для лечения и/или профилактики, и могут вводиться отдельно или в комбинации с другими лекарственными средствами, в том числе одним или несколькими пептидами этого изобретения или экзосомами, обладающими противоопухолевой активностью. Получаемые цитотоксические T-клетки действуют специфическим образом против клеток-мишеней, представляющих пептиды этого изобретения, или предпочтительно тот же пептид(ы), который был использован для индукции. Клетками-мишенями могут быть клетки, которые экспрессируют эндогенно CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, или клетки, которые трансфицированы генами CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10. Клетки, которые представляют пептиды этого изобретения на своей клеточной поверхности, вследствие стимуляции этими пептидами, могут также стать мишенями для атаки.
Настоящим изобретением также обеспечиваются антигенпрезентирующие клетки, представляющие комплексы, образованные между антигенами HLA и одним или несколькими пептидами этого изобретения. Антигенпрезентирующие клетки, полученные через контактирование с пептидами этого изобретения или нуклеотидами, кодирующими такие пептиды, предпочтительно получают от субъектов, являющихся объектами для лечения и/или профилактики, и могут вводиться в качестве вакцин отдельно или в комбинации с другими лекарственными средствами, в том числе пептидами, экзосомами или цитотоксическими Т-клетками настоящего изобретения.
Настоящим изобретением также обеспечиваются композиция, состоящая из нуклеиновых кислот, кодирующих полипептиды, которые способны образовывать субъединицу рецептора Т-клеток (TCR), и способы их применения. Субъединицы TCR обладают способностью образовывать TCR, которые придают Т-клеткам специфичность в отношении опухолевых клеток, представляющих CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK или URLC10. Используя известный в данной области техники способ, можно идентифицировать нуклеиновые кислоты альфа- и бета-цепи в качестве субъединиц TCR CTL, индуцированных одним или несколькими пептидами этого изобретения (WO 2007/032255 и Morgan et al., J. Immunol, 171, 3288 (2003)). Производные TCR предпочтительно связываются с клетками-мишенями, представляющими пептид CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK или URLC10, с высокой авидностью и необязательно опосредуют эффективное уничтожение клеток-мишеней, представляющих пептид CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK или URLC10, in vivo или in vitro.
Нуклеиновые кислоты, кодирующие субъединицы TCR, можно встроить в подходящие векторы, например ретровирусные векторы. Эти векторы широко известны в данной области техники. Нуклеиновые кислоты или содержащие их векторы подходящим образом можно перенести в T-клетку, которая предпочтительно является T-клеткой пациента. Преимущественно настоящим изобретением обеспечивается готовая к использованию композиция, позволяющая быстро модифицировать собственные T-клетки пациента (или Т-клетки другого млекопитающего) для быстрого и легкого получения модифицированных T-клеток, обладающих отличными свойствами, связанными с уничтожением раковых клеток.
Также настоящим изобретением обеспечиваются CTL, которые готовят с помощью трансдукции нуклеиновыми кислотами, кодирующими полипептиды субъединицы TCR,
связывающиеся с пептидом CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK или URLC10, например, SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 376, 379, 80, 100, 101, 110, 111, 387, 112, 394, 114, 116, 117, 121, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217, 223, 227, 228, 233, 254, 271, 272 или 288, в связи с HLA-A24 или HLA-A2. Трансдуцированные CTL способны к самонаведению на раковые клетки in vivo, и их можно нарастить с помощью широко известного способа культивирования in vitro (например, Kawakami et al., J. Immunol., 142, 3452-3461 (1989)). T-клетки настоящего изобретения можно использовать для формирования иммуногенной композиции, пригодной для лечения или предупреждения рака у нуждающегося в терапии или защите пациента (WO 2006/031221).
В контексте настоящего изобретения термин «вакцина» (также упоминаемый как иммуногенная композиция) относится к материалу, который индуцирует противоопухолевый иммунитет или подавляет раки при инокуляции животным. В соответствии с настоящим изобретением было выдвинуто предположение, что полипептиды, имеющие аминокислотную последовательность SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 80, 100, 101, 110, 111, 387, 112, 394, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217 или 223, являются рестрицированными по HLA-A24 пептидными эпитопами, а полипептиды SEQ ID NO: 376, 379, 114, 116, 117, 121, 227, 228, 233, 254, 271, 272 или 288 являются рестрицированными по HLA-A2 пептидными эпитопами, которые могут индуцировать сильный и специфический иммунный ответ против клеток, экспрессирующих CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раковых клеток, экспрессирующих CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10. Предполагаемые раки включают, но без ограничения, рак мочевого пузыря, рак молочной железы, рак шейки матки, холангиоцеллюлярный рак, CML, рак ободочной и прямой кишок, эндометриоз, рак пищевода, рак желудка, диффузный рак желудка, рак печени, NSCLC, лимфому, остеосаркому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки, SCLC, мягкотканевую опухоль и опухоль яичка.
Таким образом, настоящим изобретением также охватывается способ индукции противоопухолевого иммунитета, используя полипептиды, имеющие аминокислотную последовательность SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 376, 379, 80, 100, 101, 110, 111, 387, 112, 394, 114, 116, 117, 121, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217, 223, 227, 228, 233, 254, 271, 272 или 288 или ее вариант (т.е. включающую 1, 2 или несколько (например, вплоть до 5) замен, делеций или добавлений аминокислот). Как правило, противоопухолевый иммунитет включает такие иммунные ответы, как следующие:
- индукция цитотоксических лимфоцитов против опухолей, содержащих клетки, экспрессирующие CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10,
- индукция антител, которые распознают опухоли, содержащие клетки, экспрессирующие CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, и
- индукция продукции противоопухолевых цитокинов.
Следовательно, когда некий пептид индуцирует какой-либо один из этих иммунных ответов при инокуляции животному, выносят решение, что пептид обладает индуцирующим противоопухолевый иммунитет эффектом. Индукцию противоопухолевого иммунитета пептидом можно выявить наблюдением за in vivo или in vitro ответом иммунной системы в хозяине на пептид.
Например, широко известен способ выявления индукции цитотоксических T-лимфоцитов. Чужеродный материл, проникающий в живой организм, презентируется T-клеткам и B-клеткам путем действия антигенпрезентирующих клеток (APC). T-клетки, которые реагируют на антиген, презентируемый APC, специфичным для антигена образом, подвергаются дифференцировке в цитотоксические T-клетки (также упоминаемые как цитотоксические T-лимфоциты или CTL) вследствие стимуляции антигеном и затем пролиферируют; этот процесс упоминается здесь как «активация» T-клеток. Следовательно, индукцию CTL неким пептидом можно определить по презентации пептида T-клетке с помощью APC и выявлению индукции CTL. Кроме того, APC оказывают активирующий CD4+ T-клетки, CD8+ T-клетки, макрофаги, эозинофилы и NK-клетки эффект. Поскольку для противоопухолевого иммунитета также важны CD4+ T-клетки, индуцирующее противоопухолевый иммунитет действие пептида можно определить, используя эффект активации этих клеток в качестве показателей.
В данной области техники широко известен способ оценки индуцирующего CTL эффекта, используя дендритные клетки (DC) в качестве APC. DC является репрезентативной APC, обладающей самым сильным индуцирующим CTL эффектом среди APC. В этом способе проверяемый полипептид в начальной стадии приводят в контакт с DC, и затем эту DC приводят в контакт с T-клетками. Выявление T-клеток, обладающих цитотоксическими эффектами в отношении представляющих интерес клеток после приведения в контакт с DC, показывает, что проверяемый полипептид обладает индуцирующей цитотоксические T-клетки активностью. Активность CTL против опухолей можно выявить, используя, например, лизис меченных 51Cr опухолевых клеток в качестве показателя. В альтернативном случае широко известна оценка степени повреждения опухолевых клеток, используя активность поглощения 3H-тимидина или высвобождение LDH (лактозодегидрогеназы) в качестве показателя. Кроме того, ее можно также исследовать путем визуального определения IFN-гамма, продуцируемого и высвобождаемого CTL в присутствии антигенпрезентирующих клеток, которые несут иммобилизованные пептиды, используя антитела против IFN-гамма, такого как анализ ELISPOT.
Помимо DC в качестве АРС можно также использовать мононуклеарные клетки периферической крови (PBMC). Имеется сообщение, что индукция CTL усиливается при культивировании PBMC в присутствии GM-CSF и IL-4. Подобным образом установлено, что CTL индуцируются при культивировании PBMC в присутствии гемоцианина лимфы улитки (KLH) и IL-7.
Проверенные полипептиды, которые, как подтверждено с помощью этих способов, обладают индуцирующей CTL активностью, являются полипептидами, обладающими активирующим DC эффектом и последующей индуцирующей CTL активностью. Следовательно, полипептиды, которые индуцируют CTL против клеток, экспрессирующих CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, пригодны в качестве вакцин против заболеваний с тесной связью с CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков. Кроме того, APC, которые приобрели способность индуцировать CTL против заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, путем приведения в контакт с этими полипептидами, пригодны в качестве вакцин против этого заболевания. Кроме того, CTL, которые приобрели цитотоксичность благодаря презентации полипептидных антигенов APC, можно также использовать в качестве вакцин против заболевания с тесной связью с CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK
и/или URLC10, например раков. Такие терапевтические способы для заболевания с тесной связью с CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, в которых используется противоопухолевый иммунитет, обусловленный APC и CTL, упоминаются как клеточная иммунотерапия. Предполагаемые раки включают, но без ограничения, рак мочевого пузыря, рак молочной железы, рак шейки матки, холангиоцеллюлярный рак, CML, рак ободочной и прямой кишок, эндометриоз, рак пищевода, рак желудка, диффузный рак желудка, рак печени, NSCLC, лимфому, остеосаркому, рак яичника, рак поджелудочной железы, рак предстательной железы, рак почки, SCLC, мягкотканевую опухоль и опухоль яичка.
Как правило, при использовании полипептида для клеточной иммунотерапии эффективность индукции CTL можно увеличить объединением множества полипептидов, имеющих различные структуры, и приведением их в контакт с DC. Следовательно, при стимуляции DC фрагментами белков предпочтительно использовать смесь множества типов фрагментов.
Индукцию полипептидом противоопухолевого иммунитета можно, кроме того, подтвердить с помощью наблюдения за индукцией продукции антител против опухолей. Например, когда у лабораторного животного, иммунизированного полипептидом, индуцируются антитела против этого полипептида, а рост, пролиферация и/или метастазирование опухолевых клеток подавляется этими антителами, устанавливают, что полипептид вызывает противоопухолевый иммунитет.
Противоопухолевый иммунитет можно также индуцировать путем введения вакцины этого изобретения, и индукция противоопухолевого иммунитета делает возможным лечение и предупреждение заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков. Терапия против заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, или предупреждение его возникновения может включать ингибирование роста клеток, экспрессирующих CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, сокращение этих клеток и пресечение появления этих клеток, например раковых клеток. Снижение смертности индивидуумов, имеющих заболевание с тесной связью с CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, уменьшение маркеров заболевания в крови, ослабление выявляемых симптомов, сопровождающих заболевание, и т.п. также включены в терапию или профилактику заболевания, например, раков. Такие терапевтические и профилактические эффекты являются предпочтительно статистически значимыми, например наблюдаемыми на уровне значимости, составляющем 5% или меньше, когда терапевтический или профилактический эффект вакцины против заболевания с тесной связью с CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, сравнивают с контролем без введения вакцины. Например, для определения статистической значимости можно использовать проверку по критерию Стьюдента, U-тест Манна-Уитни или ANOVA (дисперсионный анализ).
Поскольку настоящим изобретением обеспечивается способ лечения или предупреждения заболевания, тесно связанного со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, терапевтические соединения или композиции могут вводиться с профилактической или терапевтической целью субъектам, страдающим этим заболеванием или подверженным риску (или чувствительным к) развития этого заболевания. Таких субъектов можно идентифицировать, используя стандартные клинические способы. В контексте настоящего изобретения профилактическое введение имеет место до обнаружения явных клинических симптомов заболевания, так что заболевание или нарушение предотвращается или альтернативно замедляется его развитие. В контексте медицины термином «предотвращать» охватывается любое действие, которое снижает общие показатели смертности от заболевания или заболеваемости. Предупреждение может происходить на первичном, вторичном и третичном уровне предупреждения. В то время как первичное предупреждение аннулирует развитие заболевания, вторичный и третичный уровни предупреждения включают действия, направленные на предотвращение развития заболевания и появление симптомов, а также на уменьшение отрицательного эффекта уже развившегося заболевания путем восстановления функции и уменьшения связанных с заболеванием осложнений.
В контексте лечения рака термин «эффективное» относится к лечению, которое приводит к уменьшению размера, распространенности или метастатического потенциала рака у субъекта. Когда лечение применяется профилактически, «эффективное» означает, что лечение задерживает или предотвращает возникновение не рака или ослабляет клинический симптом рака. Оценку рака можно провести, используя стандартные клинические протоколы. Кроме того, эффективность лечения можно определить в соединении с любым известным способом диагностики или лечения рака. Например, рак можно диагностировать гистопатологически или путем установления симптоматических отклонений от нормы.
Вышеупомянутый пептид, обладающий иммунологической активностью, или полинуклеотид или вектор, кодирующий такой пептид, можно объединить с адъювантом. Адъювант относится к соединению, которое усиливает иммунный ответ против пептида после введения вместе (или последовательно) с пептидом, обладающим иммунологической активностью. Примеры подходящих адъювантов включают, но без ограничения, холерный токсин, токсин сальмонелл, квасцы и т.п. Кроме того, вакцину этого изобретения можно объединить соответствующим образом с фармацевтически приемлемым носителем. Примерами таких носителей являются стерилизованная вода, физиологический солевой раствор, фосфатный буфер, жидкая среда для культивирования и т.п. Кроме того, вакцина может при необходимости содержать стабилизаторы, взвешенные вещества, консерванты, поверхностно-активные вещества и т.п. Вакцину вводят системно или местно. Введение вакцины может осуществляться с помощью однократного введения или быть повторным при многократных введениях.
При использовании APC или CTL в качестве вакцины этого изобретения заболевание, тесно связанное со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раки, можно лечить или предупреждать, например, с использованием ex vivo способа. Конкретнее, PBMC субъекта, получающего лечение и подвергаемого профилактике, получают, приводят ex vivo в контакт с пептидом настоящего изобретения. После индукции APC или CTL клетки можно вводить субъекту. APC можно также индуцировать при введении вектора, кодирующего пептид, в PBMC ex vivo. До введения APC или CTL, индуцированные in vitro, можно клонировать. Путем клонирования и выращивания клеток, обладающих высокой повреждающей клетки-мишени активностью, клеточную иммунотерапию можно провести более эффективно. Кроме того, выделенные таким образом APC и CTL можно использовать для клеточной иммунотерапии не только индивидуумов, из которых клетки получены, но также схожих типов заболеваний у других индивидуумов.
Аспекты настоящего изобретения описываются в следующих примерах, которые представлены только для иллюстрации настоящего изобретения и помощи специалисту со средним уровнем компетентности в его создании и применении. Никоим образом не подразумевается, что примеры в других отношениях ограничивают объем настоящего изобретения.
Хотя при осуществлении на практике или проверке настоящего изобретения могут использоваться способы и материалы, схожие с описанными здесь способами и материалами или эквивалентные им, подходящие способы и материалы описываются ниже.
ПРИМЕРЫ
Ниже настоящее изобретение демонстрируется, но не ограничивается, следующими примерами. Однако материалы, способы и т.п., описанные здесь, только иллюстрируют аспекты настоящего изобретения, и никоим образом не подразумевается, что они ограничивают объем настоящего изобретения. Как таковые материалы, способы и т.п., схожие с материалами, способами и т.п., описанными здесь, или эквивалентные им, можно использовать при осуществлении на практике или проверке настоящего изобретения.
МАТЕРИАЛЫ И МЕТОДЫ
Линии клеток
Клетки A24-LCL (HLA-A24), линия B-лимфобластоидных клеток человека, были созданы с помощью трансформации вирусом Эпштейна-Барр. T2-клетки, COS7, A498, Caki-2 и HEK293 приобретали из ATCC. Caki-1 и MIAPaca-2 приобретали от JCRB. PK-45P, PK-59, TE-5 и TE-6 приобретали от TKG. 293T приобретали от GenHunter.
Отбор пептида-кандидата, происходящего из CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10
9-мерные и 10-мерные пептиды, происходящие из CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK или URLC10, которые связываются с молекулой HLA-A*2402 или HLA-A*0201, предсказывали, используя программное обеспечение для предсказания связывания «BIMAS» (http://bimas.dcrt.nih.gov/cgi-bin/molbio/ken_parker_comboform) (Parker K.C. et al., (1994) J. Immunol.; 152(1): 163-175; Kuzushima K. et al., (2001) Blood.; 98(6): 1872-1881). Эти пептиды были синтезированы Sigma (Sapporo, Japan) согласно стандартному способу твердофазного синтеза и очищены с помощью HPLC с обращенной фазой. Чистоту (>90%) и идентичность пептидов определяли с помощью аналитической HPLC и масс-спектрометрического анализа соответственно. Пептиды растворяли в диметилсульфоксиде (DMSO) при 20 мг/мл и хранили при -80°C.
In vitro индукция CTL
Происходящие из моноцитов дендритные клетки (DC) использовали в качестве антигенпрезентирующих клеток (APC) для индукции реакций CTL против пептидов, представленных на HLA. DC получали in vitro, как описано в другом месте (Nukaya I. et al., (1999) Int. J. Cancer 80, 92-97, Tsai V. et al., (1997) J. Immunol 158: 1796-802). Вкратце, мононуклеарные клетки периферической крови (PBMC), выделенные из нормального добровольца (HLA-A*2402 и/или HLA-A*0201) с помощью раствора фиколл-пак (Pharmacia), отделяли путем адгезии к пластмассовой колбе для культивирования тканей (Becton Dickinson), для того чтобы обогатить их в отношении фракции моноцитов. Обогащенную в отношении моноцитов популяцию культивировали в присутствии 1000 Е/мл GM-CSF (Genzyme) и 1000 Е/мл IL-4 (Genzyme)
в AIM-V (Invitrogen), содержащей 2% инактивированной нагреванием аутологичной сыворотки (AS). Через 7 дней культивирования получаемые с использованием цитокинов DC подвергали в импульсном режиме воздействию 20 мкг/мл синтезированных пептидов в присутствии 3 мкг/мл бета-2-микроглобулина в течение 4 ч при 20°C в AIM-V. Эти подвергнутые в импульсном режиме воздействию пептидов DC затем инактивировали MMC (30 мкг/мл в течение 30 мин) и смешивали в соотношении 1:20 с аутологичными CD8+ T-клетками, полученными при положительном отборе с использованием Dynabeads M-450 CD8 (Dynal) и DETACHa BEADTM (Dynal). Эти культуры размещали в 48-луночных планшетах (Corning); при этом каждая лунка содержала 1,5×104 подвергнутых в импульсном режиме воздействию пептидов DC, 3×105 CD8+ T-клеток и 10 нг/мл IL-7 (Genzyme) в 0,5 мл AIM-V/2% AS. Спустя три дня в эти культуры добавляли IL-2 (CHIRON) до конечной концентрации 20 МЕ/мл. В день 7 и 14 T-клетки далее повторно стимулировали аутологичными, подвергнутыми в импульсном режиме воздействию пептидов DC. DC готовили каждый раз тем же способом, который описан выше. CTL проверяли против подвергнутых в импульсном режиме воздействию пептидов клеток A24-LCL или T2-клеток после третьего раунда стимулирования пептидами в день 21.
Процедура наращивания CTL
CTL наращивали в культуре, используя способ, схожий с тем, который описан Riddell S.R. и др. (Walter E.A. et al., (1995) N. Engl. J. Med. 333: 1038-1044; Riddel S.R. et al., (1996) Nature Med. 2: 216-223). Общее число 5×104 CTL ресуспендировали в 25 мл AIM-V/5% AS с 2 видами линий B-лимфобластоидных клеток человека, инактивированных MMC, в присутствии 40 нг/мл моноклональных антител против CD3 (Pharmingen). Через день после инициации культур к культурам добавляли 120 МЕ/мл IL-2. В культуры подавали свежую AIM-V/5% AS, содержащую 30 МЕ/мл IL-2, в день 5, 8 и 11.
Создание клонов CTL
Делали разведения для получения 0,3, 1 и 3 CTL/лунку в микротитрационном планшете с 96 круглодонными лунками (Nalge Nunc International). CTL культивировали с 7×104 клеток/лунку 2 видов линий B-лимфобластоидных клеток человека, 30 нг/мл антитела против CD3 и 125 Е/мл IL-2 в общем объеме 150 мкл/лунку AIM-V, содержащей 5% AS. Спустя 10 дней в среду добавляли 50 мкл/лунку IL-2, для того чтобы конечная концентрация IL-2 стала 125 Е/мл. Активность CTL проверяли на день 14 и клоны CTL наращивали, используя тот же способ, который описан выше.
Специфическая активность CTL
Для проверки специфической активности CTL выполняли анализ ELISPOT на IFN-гамма и анализ ELISA на IFN-гамма.
Вкратце, в качестве клеток-стимуляторов готовили подвергнутые в импульсном режиме воздействию пептидов клетки A24-LCL или T2 (1×104/лунку). В качестве клеток-респондеров использовали культивируемые в 48-луночном планшете клетки или клоны CTL после серийного разведения. Анализ ELISPOT и анализ ELISA на IFN-гамма выполняли согласно процедуре производителя.
Создание клеток, принудительно экспрессирующих один из двух или оба целевых гена и HLA-A02 или HLA-A24
кДНК, кодирующую открытую рамку считывания целевых генов или HLA-A02, или HLA-A24, амплифицировали с помощью ПЦР. Амплифицированный с помощью ПЦР продукт клонировали в вектор pcDNA3.1 myc-His (Invitrogen). Плазмиды трансфицировали в нулевую по целевым генам, HLA-A02 и HLA-A24 линию нормальных клеток человека COS7 или 293T, используя липофектамин (Invitrogen), в соответствии с рекомендованными производителем процедурами. В альтернативном случае плазмиду, содержащую целевые гены, трансфицировали в A24-LCL с помощью электропорации, используя GenePulserII (Biorad). Вкратце, 2,5×106 клеток A24-LCL подвергали в импульсном режиме воздействию 10 мкг плазмиды при 140 В и 1000 мкФ. Через 2 дня после трансфекции трансфицированные клетки обрабатывали раствором для диссоциации клеток и использовали в качестве клеток-мишеней для анализа активности CTL.
Анализ цитотоксичности
Цитотоксическую активность определяли с помощью четырехчасового анализа высвобождения 51Cr. Клетки-мишени подвергали в импульсном режиме воздействию пептида в концентрации 20 мкг/мл в течение ночи. Клетки-мишени метили 100 мкКи Na2 51CrO4 при 37°C в течение одного часа и затем промывали три раза RPMI1640. Клетки-мишени (1×104/100 мкл) и 100 мкл различных чисел клеток-эффекторов в общем объеме 200 мкл помещали в микротитрационный планшет с 96 круглодонными лунками (Corning) и культивировали при 37°C в термостате с CO2 в течение четырех часов. После культивирования из каждой лунки отбирали 100 мкл супернатанта и измеряли радиоактивность, используя гамма-счетчик. Самопроизвольным высвобождением была радиоактивность от клеток-мишеней при использовании среды в отсутствие клеток-эффекторов, а максимальным высвобождением была радиоактивность от клеток-мишеней при использовании 1 M HCl.
Процент специфической цитотоксичности определяли путем расчета по следующей формуле:
Специфический лизис в % = [(экспериментальное высвобождение - самопроизвольное высвобождение)/(максимальное высвобождение - самопроизвольное высвобождение)]×100.
РЕЗУЛЬТАТЫ
Увеличенная экспрессия CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10 в раках
Касающиеся общего профиля экспрессии генов данные, полученные из различных раков, используя ранжированный микроряд кДНК, выявили, что была увеличена экспрессия следующих генов:
CDH3 (входящий № в GenBank NM_001793; SEQ ID NO: 1, 2),
EPHA4 (входящий № в GenBank L36645; SEQ ID NO: 3, 4),
ECT2 (входящий № в GenBank AY376439; SEQ ID NO: 5, 6),
HIG2 (входящий № в GenBank NM_013332; SEQ ID NO: 7, 8),
INHBB (входящий № в GenBank NM_002193; SEQ ID NO: 9, 10),
KIF20A (входящий № в GenBank NM_005733; SEQ ID NO: 11, 12),
KNTC2 (входящий № в GenBank AF017790; SEQ ID NO: 13, 14),
TTK (входящий № в GenBank NM_003318; SEQ ID NO: 15, 16) и
URLC10 (входящий № в GenBank NM_017527; SEQ ID NO: 7, 18).
По сравнению с соответствующей нормальной тканью экспрессия CDH3 была действительно увеличена в следующих раках:
в 26 из 34 раков мочевого пузыря,
в 17 из 19 раков шейки матки,
в 19 из 19 холангиоцеллюлярных раков,
в 30 из 34 раков ободочной и прямой кишок,
в 20 из 21 эндометриоза,
в 13 из 20 раков желудка,
в 7 из 8 диффузных раков желудка,
в 36 из 37 NSCLC,
в 16 из 16 раков поджелудочной железы,
в 21 из 21 мягкотканевой опухоли и
в 10 из 10 опухолей яичка.
По сравнению с соответствующей нормальной тканью экспрессия EPHA4 была действительно увеличена в следующих раках:
в 14 из 34 раков мочевого пузыря,
в 8 из 14 раков шейки матки,
в 10 из 25 холангиоцеллюлярных раков,
в 5 из 15 эндометриозов,
в 5 из 8 диффузных раков желудка,
в 5 из 5 раков яичника,
в 14 из 14 раков поджелудочной железы,
в 20 из 51 рака предстательной железы и
в 14 из 23 мягкотканевых опухолей.
По сравнению с соответствующей нормальной тканью экспрессия ECT2 была действительно увеличена в следующих раках:
17 из 19 раков мочевого пузыря,
в 5 из 12 раков молочной железы,
в 14 из 14 раков шейки матки,
в 13 из 13 холангиоцеллюлярных раков,
в 5 из 5 CML,
в 7 из 8 раков ободочной и прямой кишки,
в 12 из 16 раков пищевода,
в 6 из 16 NSCLC,
в 8 из 10 лимфом,
в 1 из 1 рака поджелудочной железы,
в 10 из 13 раков предстательной железы,
в 3 из 6 раков почки и
в 2 из 13 SCLC.
По сравнению с соответствующей нормальной тканью экспрессия HIG2 была действительно увеличена в 19 из 20 раков почки и в 7 из 9 мягкотканевых опухолей.
По сравнению с соответствующей нормальной тканью экспрессия INHBB была действительно увеличена в следующих раках:
в 10 из 21 холангиоцеллюлярного рака,
в 12 из 12 раков пищевода,
в 10 из 13 NSCLC,
в 22 из 24 раков почки,
в 8 из 14 SCLC и
45 из 49 мягкотканевых опухолей.
По сравнению с соответствующей нормальной тканью экспрессия KIF20A была действительно увеличена в следующих раках:
В 31 из 31 рака мочевого пузыря,
в 38 из 61 рака молочной железы,
в 10 из 11 холангиоцеллюлярных раков,
в 7 из 19 раков пищевода,
в 21 из 22 NSCLC,
в 6 из 6 раков яичника,
в 17 из 36 раков предстательной железы,
в 6 из 11 раков почки и
в 15 из 15 SCLC.
По сравнению с соответствующей нормальной тканью экспрессия KNTC2 была действительно увеличена в следующих раках:
в 30 из 32 раков мочевого пузыря,
в 47 из 56 раков молочной железы,
в 10 из 10 раков шейки матки,
в 16 из 22 холангиоцеллюлярных раков,
в 17 из 37 CML,
в 3 из 10 раков ободочной и прямой кишок,
в 11 из 46 раков пищевода,
в 15 из 19 NSCLC,
в 7 из 8 лимфом,
в 20 из 24 остеосарком,
в 3 из 5 раков яичника,
в 2 из 2 раков поджелудочной железы,
в 15 из 37 раков предстательной железы,
в 14 из 19 раков почки,
в 15 из 15 SCLC и
в 40 из 59 мягкотканевых опухолей.
По сравнению с соответствующей нормальной тканью экспрессия TTK была действительно увеличена в следующих раках:
В 27 из 27 раков мочевого пузыря,
в 25 из 30 раков молочной железы,
в 15 из 16 раков шейки матки,
в 10 из 10 холангиоцеллюлярных раков,
в 5 из 7 CML,
в 6 из 10 раков ободочной и прямой кишок,
в 24 из 44 раков пищевода,
в 8 из 15 раков печени,
в 12 из 12 NSCLC,
в 6 из 6 лимфом,
в 13 из 16 остеобластом,
в 12 из 17 раков предстательной железы,
в 15 из 15 SCLC и
в 16 из 33 мягкотканевых опухолей.
По сравнению с соответствующей нормальной тканью экспрессия URLC10 была действительно увеличена в следующих раках:
в 29 из 29 раков мочевого пузыря,
в 15 из 16 раков шейки матки,
в 7 из 7 холангиоцеллюлярных раков,
в 7 из 19 раков пищевода,
в 3 из 3 раков желудка,
в 24 из 27 NSCLC,
в 15 из 19 остеосарком,
в 4 из 5 раков поджелудочной железы,
в 33 из 43 мягкотканевых опухолей.
Предсказание связывающихся с HLA-A24 или HLA-A2 пептидов, происходящих из CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK или URLC10
В таблице 2 представлены связывающиеся с HLA-A*2402 пептиды, происходящие из CDH3, в порядке аффинности связывания. В таблице 2A представлены 9-мерные пептиды, происходящие из CDH3, а в таблице 2B представлены 10-мерные пептиды, происходящие из CDH3.
В таблице 3 представлены связывающиеся с HLA-A*2402 и HLA-A*0201 пептиды, происходящие из EPHA4, в порядке аффинности связывания. В таблице 3A представлены связывающиеся с HLA-A*2402 9-мерные пептиды, происходящие из EPHA4, в таблице 3B представлены связывающиеся с HLA-A*2402 10-мерные пептиды, происходящие из EPHA4, а в таблице 3C представлены связывающиеся с HLA-A*0201 9-мерные пептиды, происходящие из EPHA4.
В таблице 4 представлены связывающиеся с HLA-A*2402 пептиды, происходящие из ECT2, в порядке аффинности связывания. В таблице 4A представлены 9-мерные пептиды, происходящие из ECT2, а в таблице 4B представлены 10-мерные пептиды, происходящие из ECT2.
В таблице 5 представлены связывающиеся с HLA-A*2402 и HLA-A*0201 пептиды, происходящие из HIG2, в таблице 5A представлены связывающиеся с HLA-A*2402 9-мерные пептиды, происходящие из HIG2, в таблице 5B представлены связывающиеся с HLA-A*2402 10-мерные пептиды, происходящие из HIG2, в таблице 5C представлены связывающиеся с HLA-A*0201 9-мерные пептиды, происходящие из HIG2, а в таблице 5D представлены связывающиеся с HLA-A*0201 10-мерные пептиды, происходящие из HIG2.
В таблице 6 представлены связывающиеся с HLA-A*2402 и HLA-A*0201 пептиды, происходящие из INHBB, в таблице 6A представлены связывающиеся с HLA-A*2402 9-мерные пептиды, происходящие из INHBB, в таблице 6В представлены связывающиеся HLA-A*2402 10-мерные пептиды, происходящие из INHBB, в таблице 6С представлены связывающиеся с HLA-A*0201 9-мерные пептиды, происходящие из INHBB, а в таблице 6D представлены связывающиеся с HLA-A*0201 10-мерные пептиды, происходящие из INHBB.
В таблице 7 представлены связывающиеся с HLA-A*2402 пептиды, происходящие из KIF20A, в порядке аффинности связывания. В таблице 7A представлены 9-мерные пептиды, происходящие из KIF20A, а в таблице 7B представлены 10-мерные пептиды, происходящие из KIF20A.
В таблице 8 представлены связывающиеся с HLA-A*2402 пептиды, происходящие из KNTC2, в порядке аффинности связывания. В таблице 8A представлены 9-мерные пептиды, происходящие из KNTC2, а в таблице 8B представлены 10-мерные пептиды, происходящие из KNTC2.
В таблице 9 представлены связывающиеся с HLA-A*0201 пептиды, происходящие из TTK, в порядке аффинности связывания. В таблице 9A представлены 9-мерные пептиды, происходящие из TTK, а в таблице 9B представлены 10-мерные пептиды, происходящие из TTK.
В таблице 10 представлены связывающиеся с HLA-A*0201 пептиды, происходящие из URLC10, в порядке аффинности связывания. В таблице 10A представлены 9-мерные пептиды, происходящие из URLC10, а в таблице 10B представлены 10-мерные пептиды, происходящие из URLC10.
Объяснение и определение терминов в таблицах
Начальное положение означает число аминокислот с N-конца.
Показатель связывания получают из «BIMAS», описанного в разделе «Материалы и методы».
Число положительных доноров означает число доноров, CD8+-T-клетки которых можно индуцировать до специфических CTL с помощью ex vivo стимуляции антигенпрезентирующими клетками. Это представлено как (число положительных доноров/число всех доноров).
Число положительных лунок означает число лунок, в которых можно выявить специфическую продукцию IFN-гамма с помощью анализа ELISPOT на IFN-гамма. Для одного донора можно приготовить от 4 до 8 лунок. Это представлено как (число положительных лунок/число всех лунок, проверенных с помощью анализа ELISPOT на IFN-гамма).
Положительная линия CTL означает число линий CTL, созданных из положительных лунок. Создание линии CTL определяют с помощью ELISA. Это представлено как (число созданных линий CTL/число положительных лунок, проверенных с помощью анализа ELISPOT на IFN-гамма).
Отсутствие положительно донора определяется не по отсутствию положительных лунок, а по отсутствию созданной линии CTL.
Пептиды, показанные полужирным шрифтом в таблицах, обладают стимулирующей T-клетки активностью.
Отсутствие данных в столбцах «число положительных доноров», «число положительных лунок» и «положительная линия CTL», показанное «-», означает, что пептиды нельзя синтезировать по какой-либо причине.
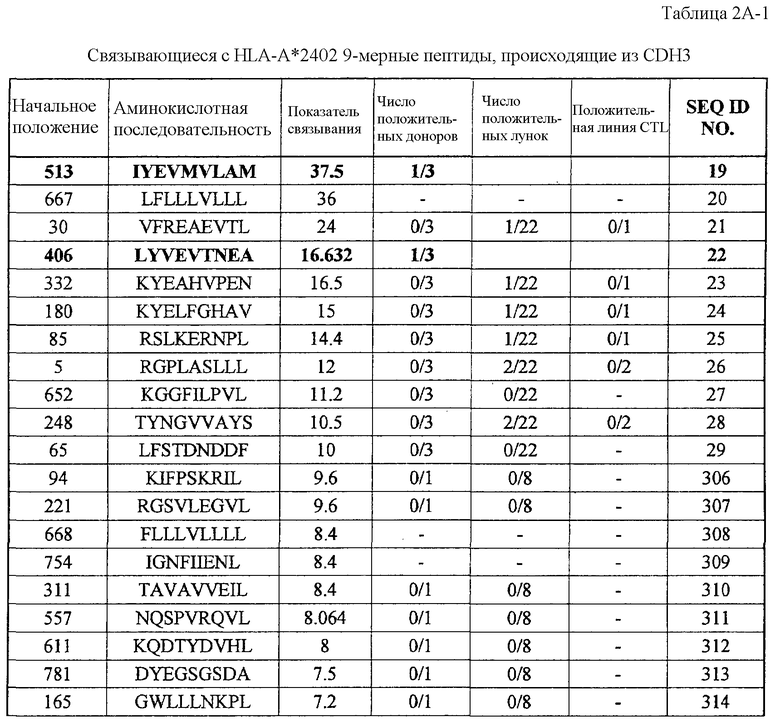
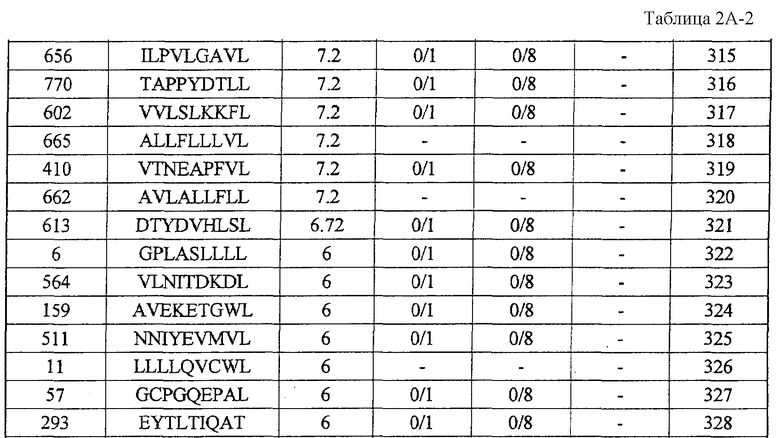
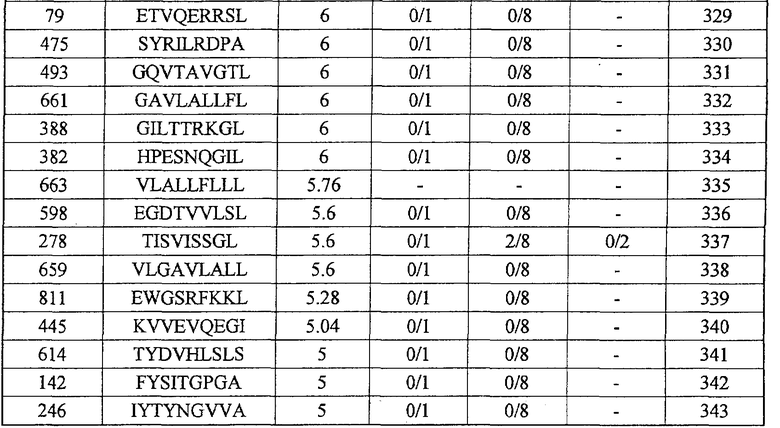
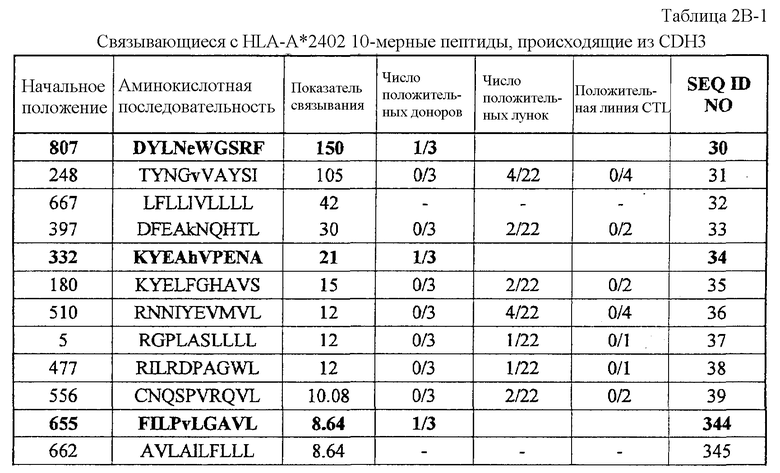
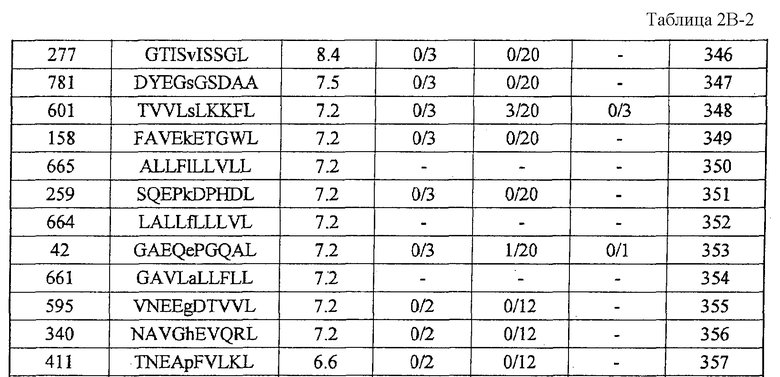
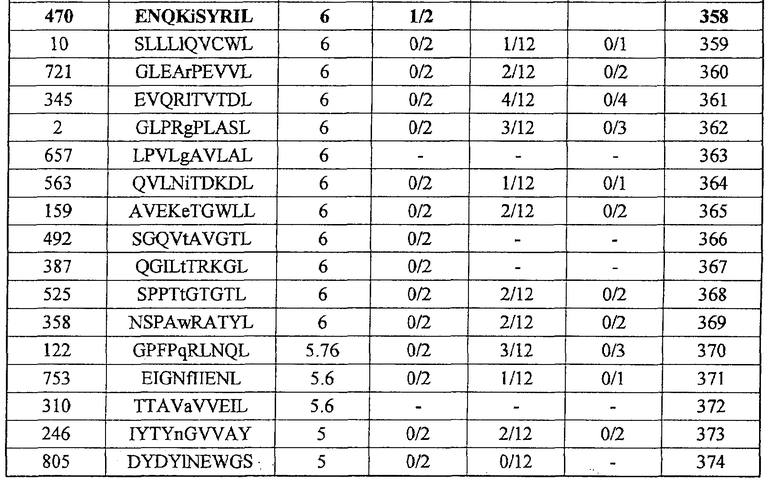
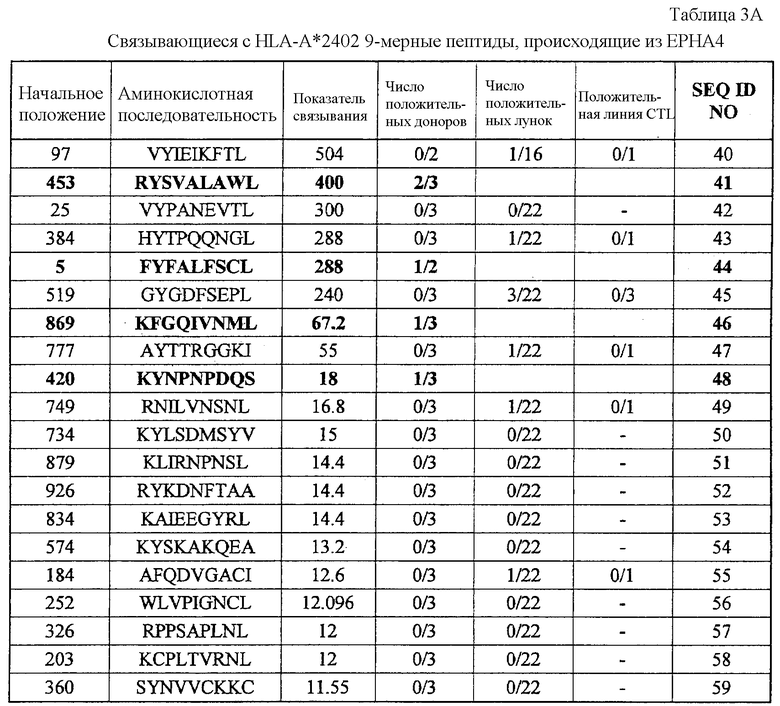
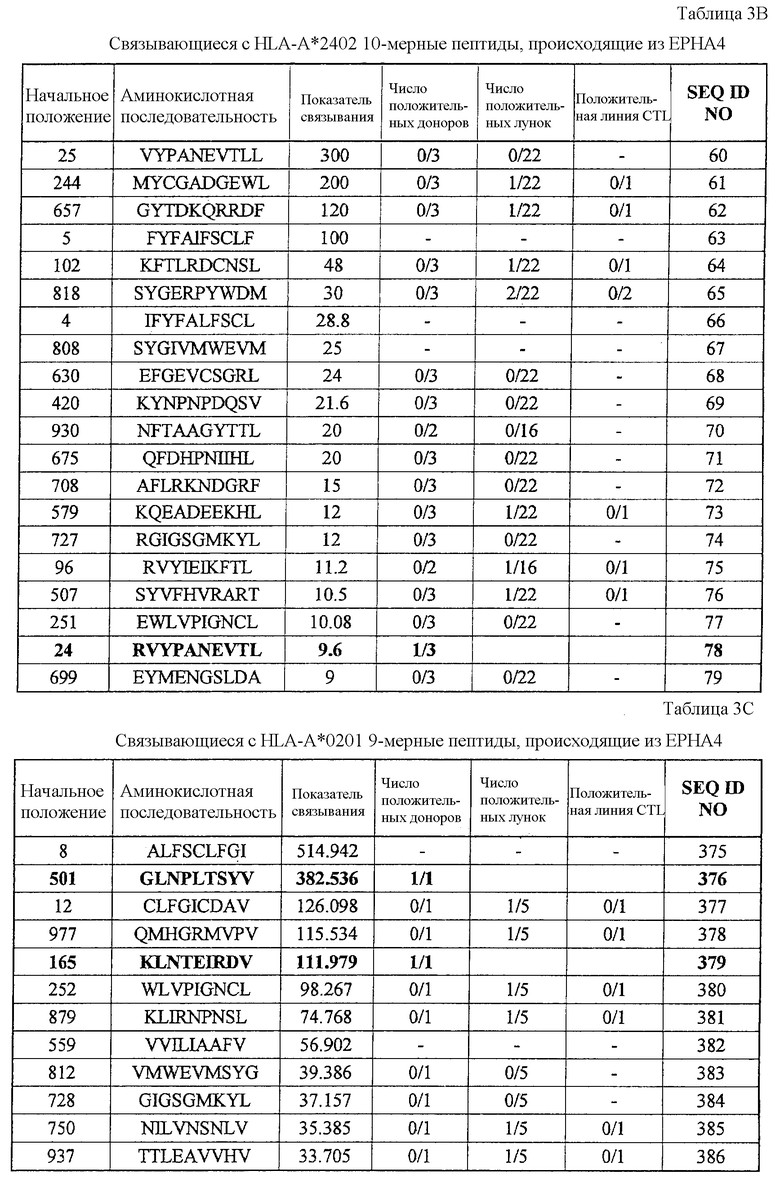
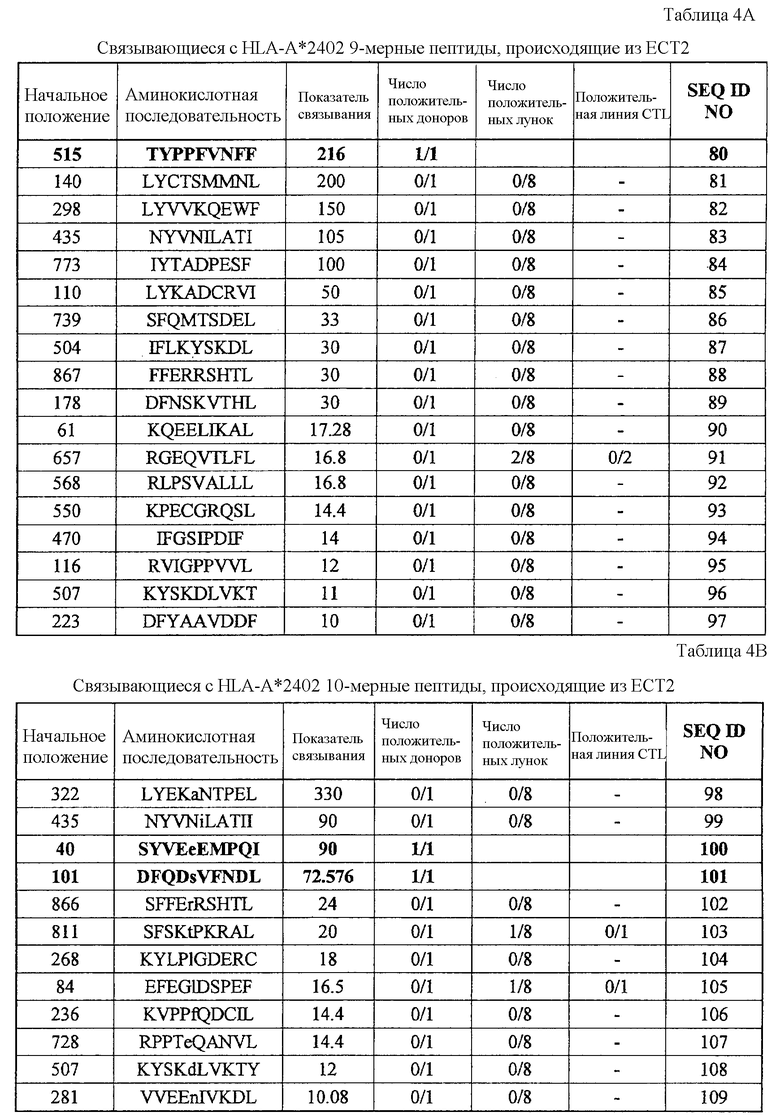
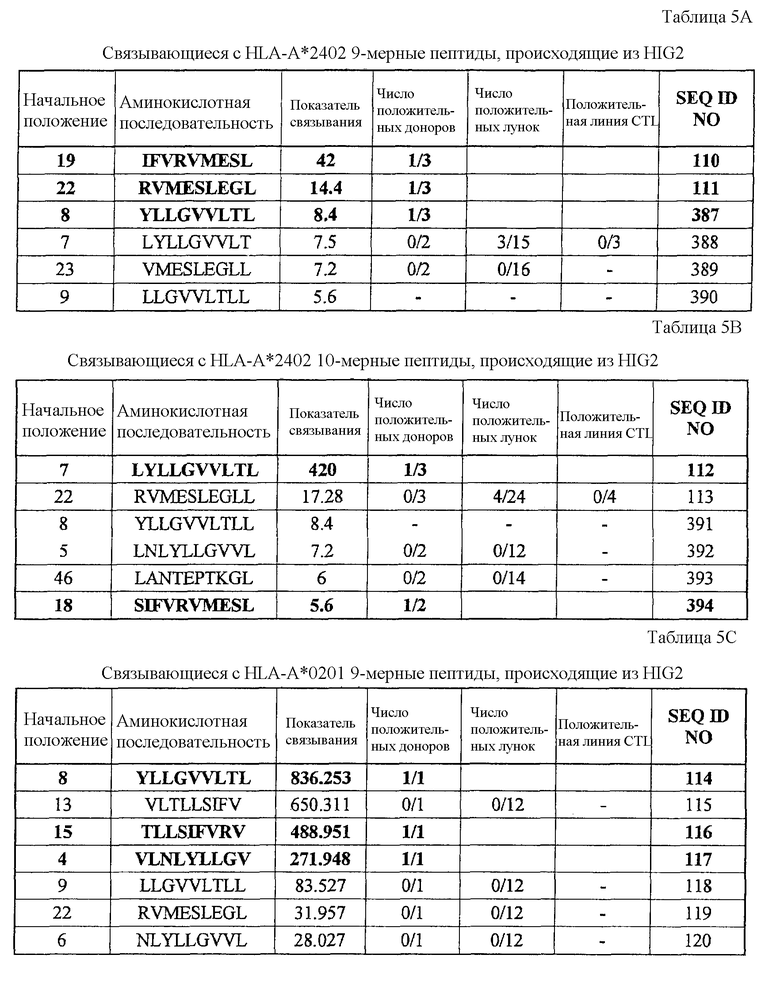
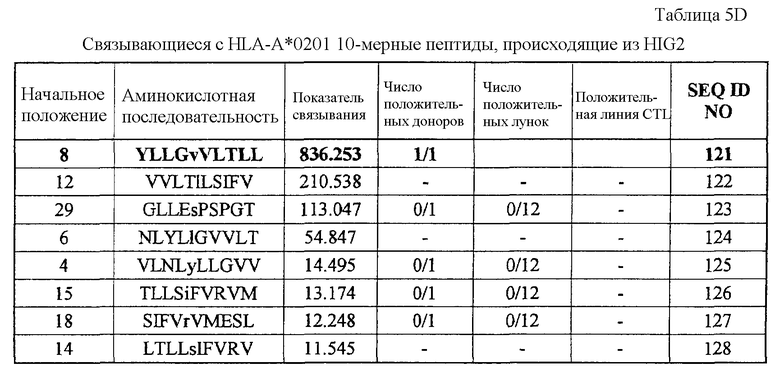
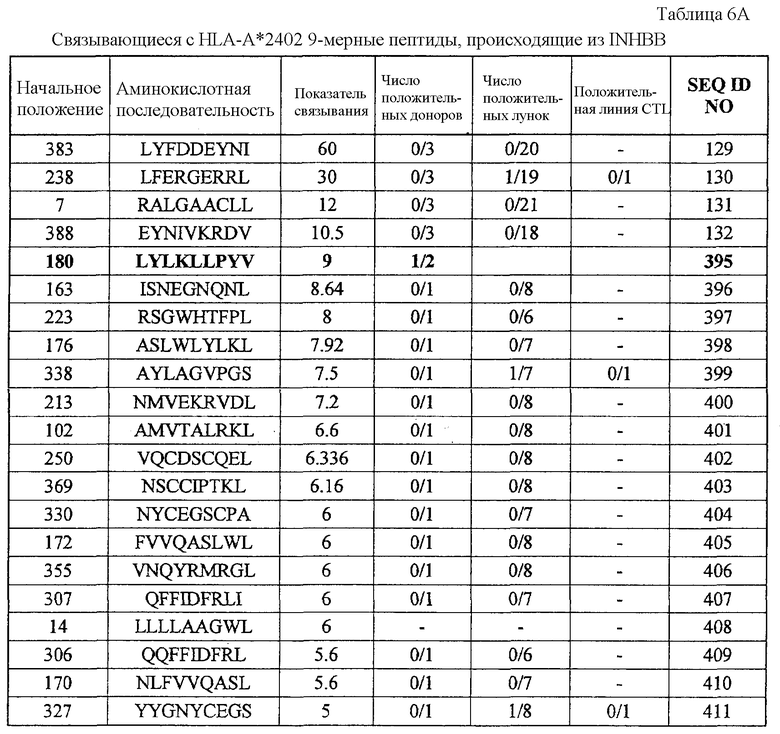

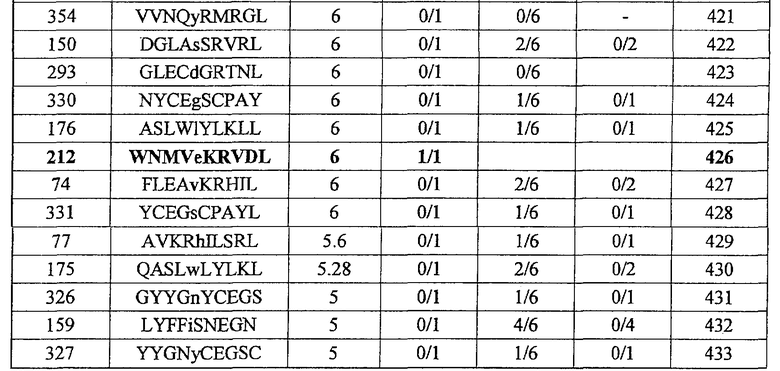
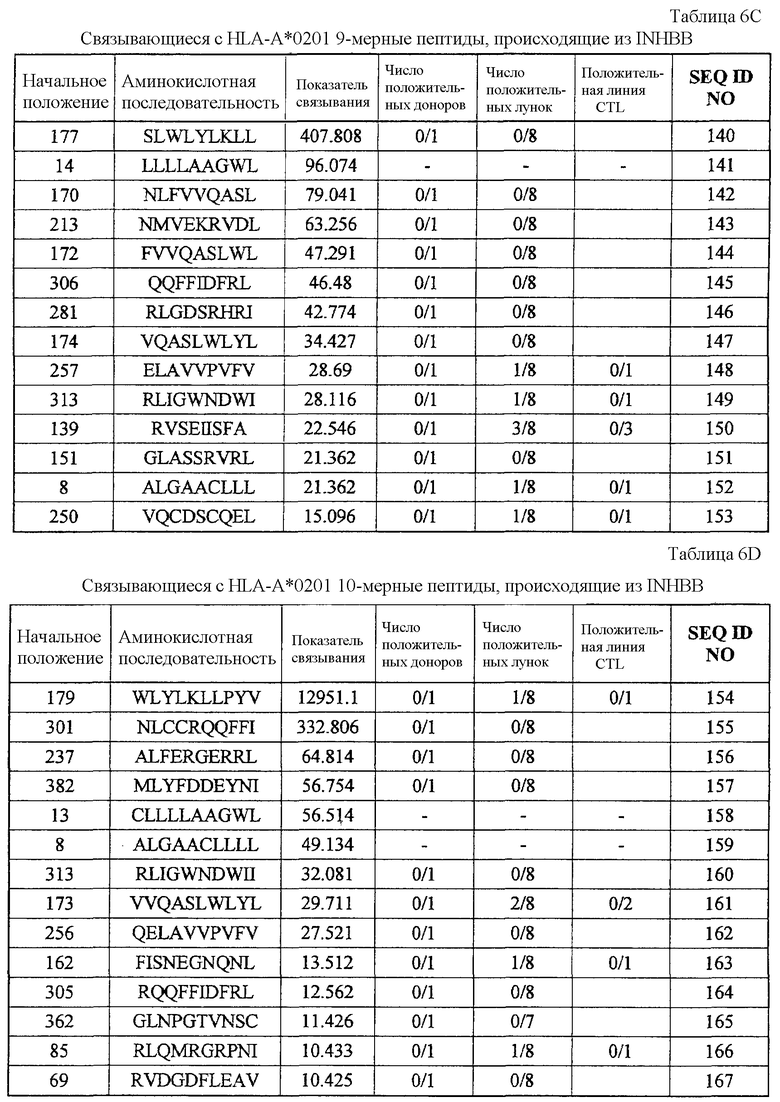
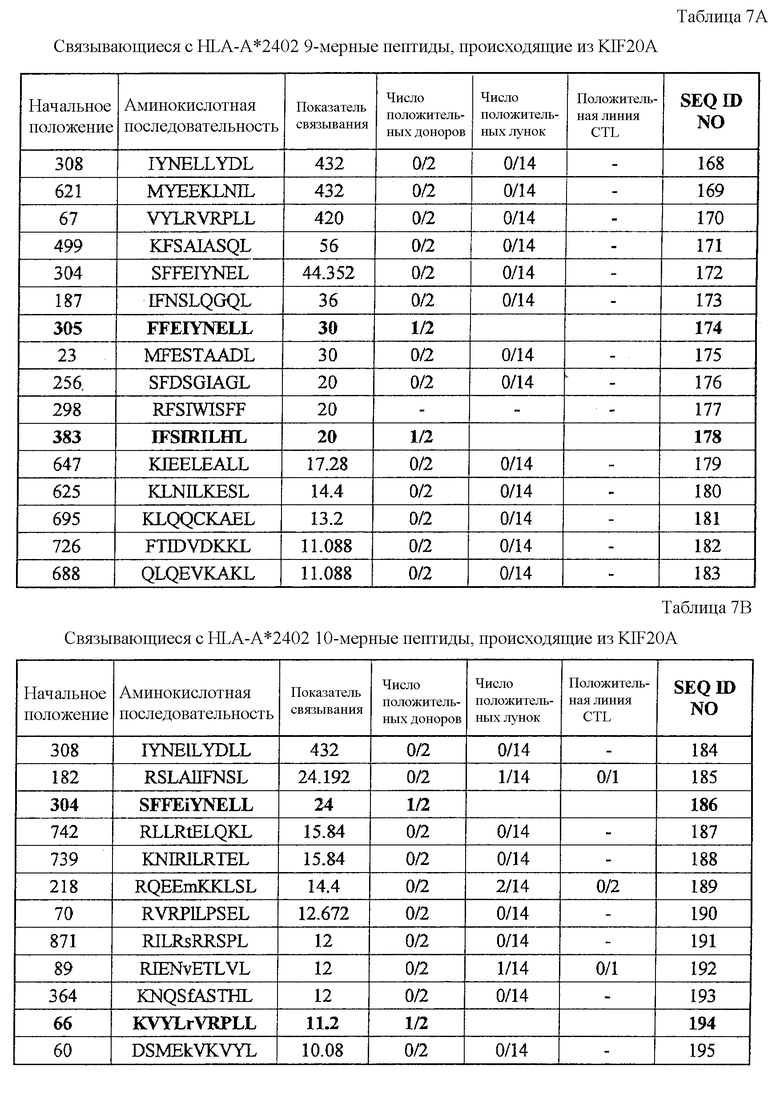
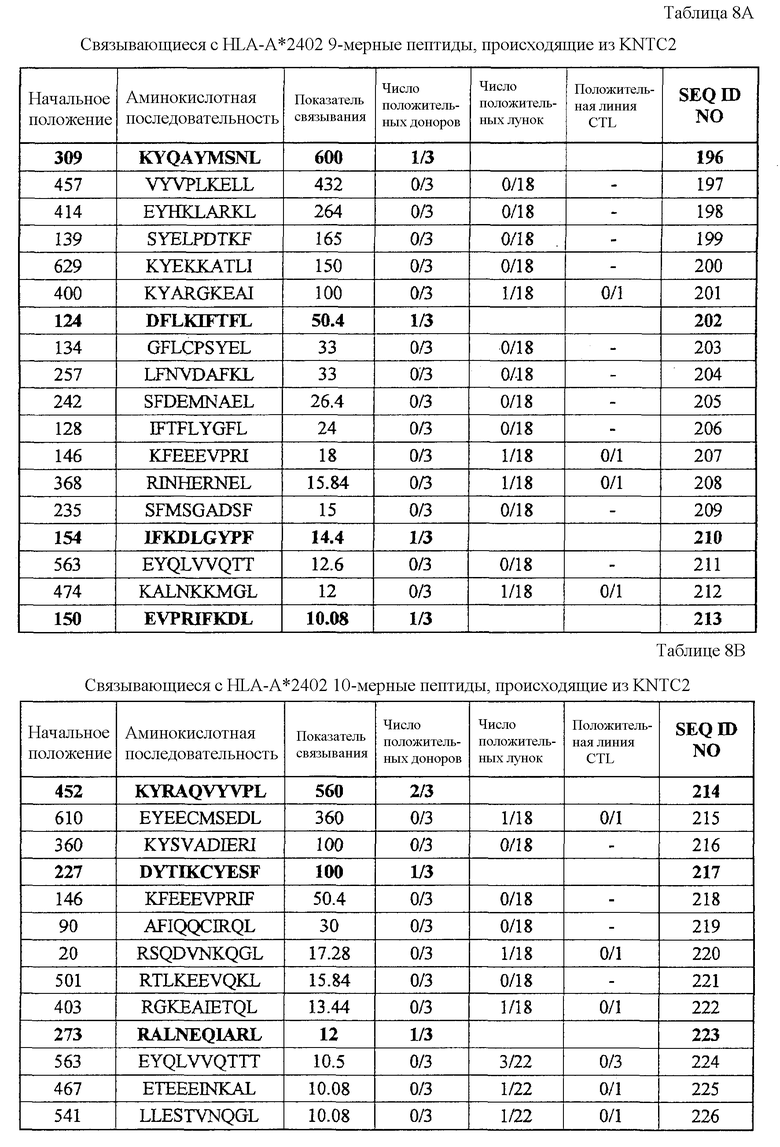
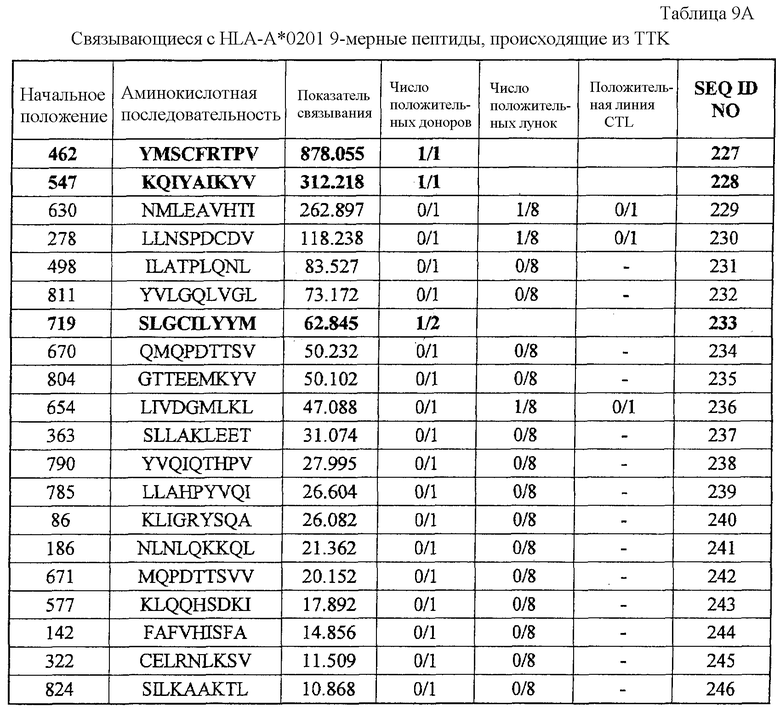
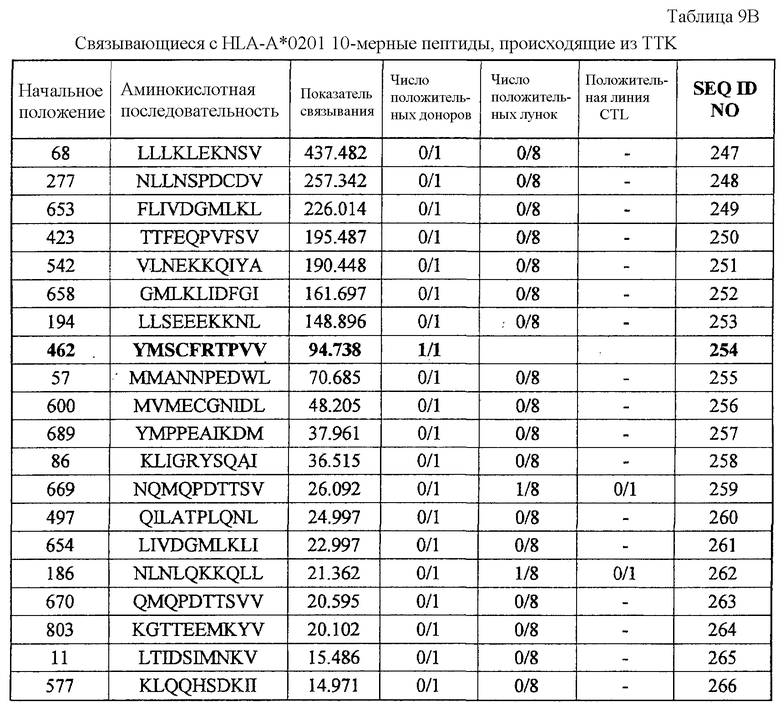
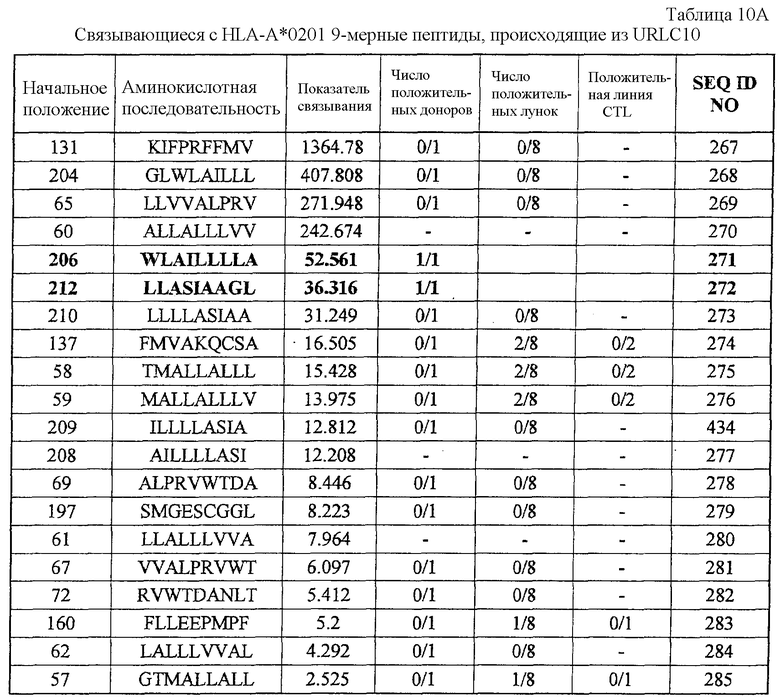
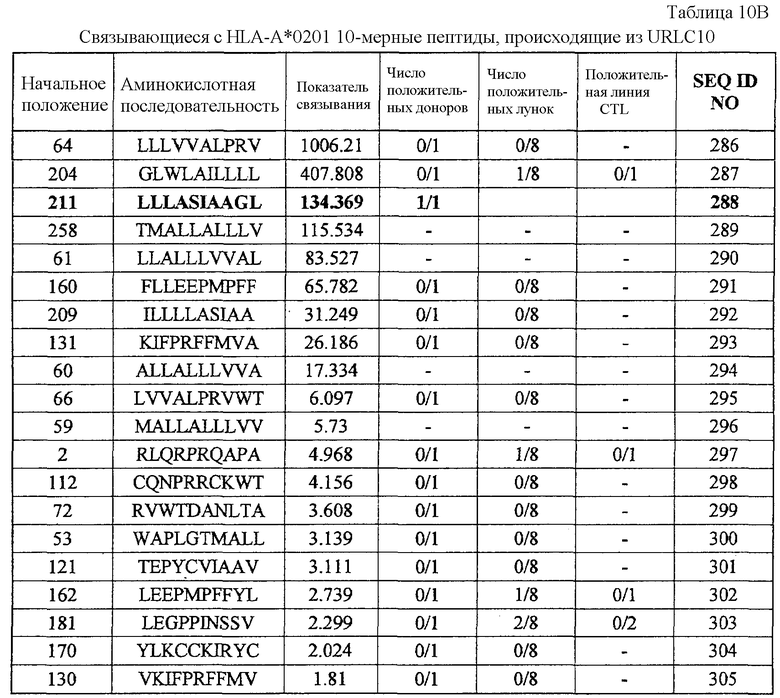
Стимуляция T-клеток с использованием предсказанных пептидов из CDH3, рестрицированных по HLA-A*2402, и создание линий CTL, подвергнутых стимуляции происходящими из CDH3 пептидами
CTL, специфичные для тех пептидов, которые происходят из CDH3, создавали в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Получаемые в результате CTL, обладающие обнаруживаемой специфической активностью CTL, определяемой с помощью анализа ELISPOT на IFN-гамма, продемонстрированы на фиг.1. В частности, для CDH3-A24-9-513 (SEQ ID NO: 19), CDH3-A24-9-406 (SEQ ID NO: 22), CDH3-A24-10-807 (SEQ ID NO: 30), CDH3-A24-10-332 (SEQ ID NO: 34), CDH3-A24-10-655 (SEQ ID NO: 344) и CDH3-A24-10-470 (SEQ ID NO: 358) продемонстрирована мощная продукция IFN-гамма по сравнению с контролем с помощью анализа ELISPOT на IFN-гамма, и клетки в положительной лунке номер #5, подвергнутые стимуляции SEQ ID NO: 19, #2, подвергнутые стимуляции SEQ ID NO: 22, #5, подвергнутые стимуляции SEQ ID NO: 30, #4, подвергнутые стимуляции SEQ ID NO: 34, #1, подвергнутые стимуляции SEQ ID NO: 344, и #4, подвергнутые стимуляции SEQ ID NO: 358, наращивали и создавали линии CTL. Линии CTL, обладающие более высокими специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида, определяли с помощью ELISA. Результаты продемонстрированы на фиг.1. Тогда как с использованием других пептидов, представленных в таблице 2, линии CTL не могли быть созданы несмотря на возможную активность связывания с HLA-A*2402. Например, типичный отрицательный пептид (CDH3-A24-10-248) продемонстрирован на фиг.1a. В этом изобретении пептиды, с использованием которых могла быть создана линия CTL, отбирали в качестве сильно стимулирующего CTL пептида.
Создание клонов CTL, подвергнутых стимуляции происходящими из CDH3 пептидами
Кроме того, осуществляли серийное разведение этих линий CTL в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Создание клонов CTL из линии CTL из лунки #5, подвергнутой стимуляции CDH3-A24-10-807 (SEQ ID NO: 30), и #1, подвергнутой стимуляции CDH3-A24-10-655 (SEQ ID NO: 344), продемонстрировано на фиг.1f и g. Клоны CTL обладали сильными и специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида.
Специфическая активность CTL в отношении клеток-мишеней, экспрессирующих CDH3 и HLA-A*2402
Созданные линии CTL, индуцированные против этих пептидов, проверяли на их способность распознавать клетки-мишени, экспрессирующие CDH3 и HLA-A*2402. Специфическую активность CTL в отношении клеток COS7, трансфицированных полноразмерным геном CDH3 и молекулой HLA-A*2402, которые служат в качестве специфической модели клеток-мишеней, эндогенно экспрессирующих CDH3 и HLA-A*2402, проверяли, используя в качестве клеток-эффекторов линии CTL, индуцированные CDH3-A24-10-807 (SEQ ID NO: 30) и CDH3-A24-10-655 (SEQ ID NO: 344). В качестве контролей готовили COS7, трансфицированные полноразмерным геном CDH3, но не молекулой HLA-A*2402, и COS7, трансфицированные HLA-A*2402, но не полноразмерным геном CDH3. Клонами CTL, демонстрирующими наивысшую специфическую активность CTL в отношении COS7, был клон CTL, трансфицированный и CDH3, и HLA-A2402 (фиг.1f и g).
Эти результаты ясно демонстрируют, что CDH3-A24-10-807 (SEQ ID NO: 30) и CDH3-A24-10-655 (SEQ ID NO: 344) естественно экспрессируются на поверхности клетки-мишени вместе с молекулой HLA-A2402 и распознаются CTL. Кроме того, эти пептиды являются пептидными эпитопами, которые могут служить в качестве противораковых вакцин, направленных на экспрессирующие CDH3 опухоли.
Стимуляция T-клеток с использованием предсказанных пептидов из EPHA4, рестрицированных по HLA-A*2402 или HLA-A*0201, и создание линий CTL, подвергнутых стимуляции происходящими из EPHA4 пептидами
CTL, специфичные для тех пептидов, которые происходят из EphA4, создавали с использованием анализа ELISPOT на IFN-гамма. Получаемые в результате CTL, обладающие обнаруживаемой специфической активностью CTL, определяемой с помощью анализа ELISPOT на IFN-гамма, продемонстрированы на фиг.2. В частности, для EphA4-A24-9-453 (SEQ ID NO: 41), EphA4-A24-9-5 (SEQ ID NO: 44), EphA4-A24-9-869 (SEQ ID NO: 46), EphA4-A24-9-420 (SEQ ID NO: 48), EphA4-A24-10-24 (SEQ ID NO: 78), EphA4-A02-9-501 (SEQ ID NO: 376) и EphA4-A02-9-165 (SEQ ID NO: 379)
продемонстрирована мощная продукция IFN-гамма с помощью анализа ELISPOT на IFN-гамма, и клетки в положительной лунке номер #3, подвергнутые стимуляции EphA4-A24-9-453 (SEQ ID NO: 41), #2, подвергнутые стимуляции EphA4-A24-9-5 (SEQ ID NO: 44), #5, подвергнутые стимуляции EphA4-A24-9-869 (SEQ ID NO: 46), #6,
подвергнутые стимуляции EphA4-A24-9-420 (SEQ ID NO: 48), #4, подвергнутые стимуляции EphA4-A24-10-24 (SEQ ID NO: 78), #8, подвергнутые стимуляции EphA4-A02-9-501 (SEQ ID NO: 376), и #3, подвергнутые стимуляции EphA4-A02-9-165 (SEQ ID NO: 379), наращивали и создавали линии CTL. Линии CTL, обладающие более высокими специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида, определяли с помощью ELISA. Главным образом, линии CTL, подвергнутые стимуляции EphA4-A02-9-501 (SEQ ID NO: 376) и EphA4-A02-9-165 (SEQ ID NO: 379), проверяли с помощью анализа высвобождения 51Cr в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Результаты продемонстрированы на фиг.2a-h. Тогда как с использованием других пептидов, представленных в таблице 3, линии CTL не могли быть созданы несмотря на возможную активность связывания с HLA-A*2402 или HLA-A*0201. Например, типичный отрицательный пептид (EphA4-A24-9-384) продемонстрирован на фиг.2a. В этом изобретении пептиды, с использованием которых могла быть создана линия CTL, отбирали в качестве сильно стимулирующих CTL пептидов.
Стимуляция T-клеток с использованием предсказанных пептидов из ECT2, рестрицированных по HLA-A*2402, и создание линий CTL, подвергнутых стимуляции происходящими из ECT2 пептидами
CTL, специфичные для тех пептидов, которые происходят из ЕСТ2, создавали в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Получаемые в результате CTL, обладающие обнаруживаемой специфической активностью CTL, определяемой с помощью анализа ELISPOT на IFN-гамма, продемонстрированы на фиг.3. В частности, для ЕСТ2-А24-9-515 (SEQ ID NO: 80), ЕСТ2-А24-10-40 (SEQ ID NO: 100) и ECT2-A24-10-101 (SEQ ID NO: 101) продемонстрирована мощная продукция IFN-гамма, и клетки в положительной лунке номер #7, подвергнутые стимуляции ЕСТ2-А24-9-515 (SEQ ID NO: 80), #2, подвергнутые стимуляции ЕСТ2-А24-10-40 (SEQ ID NO: 100), и #1, подвергнутые стимуляции ЕСТ2-А24-10-101 (SEQ ID NO: 101), наращивали и создавали линии CTL. Линии CTL, обладающие более высокими специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида, определяли с помощью ELISA. Результаты продемонстрированы на фиг.3а-d. Тогда как с использованием других пептидов, представленных в таблице 4, линии CTL не могли быть созданы несмотря на возможную активность связывания с HLA-A*2402. Например, типичные отрицательные пептиды (ЕСТ2-А24-10-322, ЕСТ2-А24-9-657 и ЕСТ2-А24-10-811) продемонстрированы на фиг.3а. В этом изобретении пептиды, с использованием которых могла быть создана линия CTL, отбирали в качестве сильно стимулирующего CTL пептида.
Создание клонов CTL, подвергнутых стимуляции происходящими из ЕСТ2 пептидами
Кроме того, осуществляли серийное разведение этих линий CTL в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Создание клонов CTL из линии CTL из лунки #2, подвергнутой стимуляции ECT2-A24-10-40 (SEQ ID NO: 100), продемонстрировано на фиг.3с. Клоны CTL обладали сильными и специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида.
Специфическая активность CTL в отношении клеток-мишеней, экспрессирующих ECT2 и HLA-A*2402
Созданные линии CTL, индуцированные против этих пептидов, проверяли на их способность распознавать клетки-мишени, экспрессирующие ECT2 и HLA-A*2402. Специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном ECT2, и молекулой HLA-A*2402, которые служат в качестве специфической модели клеток-мишеней, эндогенно экспрессирующих ECT2 и HLA-A*2402, проверяли, используя в качестве клеток-эффекторов клон CTL, индуцированный ECT2-A24-10-40 (SEQ ID NO: 100), и линию CTL, индуцированную ECT2-A24-10-101 (SEQ ID NO: 101). В качестве контролей готовили COS7, трансфицированные полноразмерным геном ECT2, но не молекулой HLA-A*2402, и COS7, трансфицированные HLA-A*2402, но не полноразмерным геном ECT2. Линией CTL, демонстрирующей наивысшую специфическую активность CTL в отношении COS7, была линия CTL, трансфицированная и ECT2, и HLA-A2402 (фиг.3c и d).
Эти результаты ясно демонстрируют, что ECT2-A24-10-40 (SEQ ID NO: 100) и ECT2-A24-10-101 (SEQ ID NO: 101) естественно экспрессируются на поверхности клетки-мишени вместе с молекулой HLA-A2402 и распознаются CTL. Кроме того, эти пептиды являются пептидными эпитопами, которые могут служить в качестве противораковых вакцин, направленных на экспрессирующие ECT2 опухоли.
Цитотоксическая активность в отношении линии раковых клеток, эндогенно экспрессирующей HLA-A*2402 и ECT2
Кроме того, цитотоксическую активность проверяли с помощью анализа цитотоксичности в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». В результате, как показано на фиг.3b, клон CTL, подвергнутый стимуляции ECT2-A24-9-515 (SEQ ID NO: 80), продемонстрировал замечательно высокую цитотоксическую активность в отношении HLA-A24-положительных и ECT-положительных линий раковых клеток TE6, по сравнению с активностями в отношении HLA-A24-отрицательных и ECT-положительных линий раковых клеток TE5.
Стимуляция T-клеток с использованием предсказанных пептидов из HIG2, рестрицированных по HLA-A*2402 или HLA-A*0201, и создание линий CTL, подвергнутых стимуляции происходящими из HIG2 пептидами
CTL, специфичные для тех пептидов, которые происходят из HIG2, создавали в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Получаемые в результате CTL, обладающие обнаруживаемой специфической активностью CTL, определяемой с помощью анализа ELISPOT на IFN-гамма, продемонстрированы на фиг.4. В частности, для HIG2-A24-9-19 (SEQ ID NO: 110), HIG2-A24-9-22 (SEQ ID NO: 111), HIG2-A24-9-8 (SEQ ID NO: 387), HIG2-A24-10-7 (SEQ ID NO: 112), HIG2-A24-10-18 (SEQ ID NO: 394), HIG2-A02-9-8 (SEQ ID NO: 114), HIG2-A02-9-15 (SEQ ID NO: 116), HIG2-A02-9-4 (SEQ ID NO: 117) и HIG2-A02-10-8 (SEQ ID NO: 121) продемонстрирована мощная продукция IFN-гамма с помощью анализа ELISPOT на IFN-гамма, и клетки в положительной лунке номер #6, подвергнутые стимуляции HIG2-A24-9-19 (SEQ ID NO: 110), #7, подвергнутые стимуляции HIG2-A24-9-22 (SEQ ID NO: 111), #5, подвергнутые стимуляции HIG2-A24-9-8 (SEQ ID NO: 387), #1, подвергнутые стимуляции HIG2-A24-10-7 (SEQ ID NO: 112), #7, подвергнутые стимуляции HIG2-A24-10-18 (SEQ ID NO: 394), #10, подвергнутые стимуляции HIG2-A02-9-8 (SEQ ID NO: 114), #10, подвергнутые стимуляции HIG2-A02-9-15 (SEQ ID NO: 116), #10, подвергнутые стимуляции HIG2-A02-9-4 (SEQ ID NO: 117), и #9, подвергнутые стимуляции HIG2-A02-10-8 (SEQ ID NO: 121), наращивали и создавали линии CTL. Линии CTL, обладающие более высокими специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида, определяли с помощью ELISA. Результаты продемонстрированы на фиг.4a-j. Тогда как с использованием других пептидов, представленных в таблице 5, линии CTL не могли быть созданы несмотря на возможную активность связывания с HLA-A*2402. Например, типичный отрицательный пептид (HIG2-A24-9-7) продемонстрирован на фиг.4a. В этом изобретении пептиды, с использованием которых могла быть создана линия CTL, отбирали в качестве сильно стимулирующего CTL пептида.
Создание клонов CTL, подвергнутых стимуляции происходящими из HIG2 пептидами
Кроме того, осуществляли серийное разведение этих линий CTL в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Создание клонов CTL из линии CTL из лунки #7, подвергнутой стимуляции HIG2-A24-9-22 (SEQ ID NO: 111), линии CTL из лунки #5, подвергнутой стимуляции HIG2-A24-9-8 (SEQ ID NO: 387), линии CTL из лунки #1, подвергнутой стимуляции HIG2-A24-10-7 (SEQ ID NO: 112), линии CTL из лунки #7, подвергнутой стимуляции HIG2-A24-10-18 (SEQ ID NO: 394), и линии CTL из лунки #10, подвергнутой стимуляции HIG2-A02-9-4 (SEQ ID NO: 117), продемонстрировано на фиг.4c, e, f, g и i. Клоны CTL обладали сильными и специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида.
Специфическая активность CTL в отношении клеток-мишеней, экспрессирующих HIG2 и HLA-A*0201
Созданные линии CTL, индуцированные против этих пептидов, проверяли на их способность распознавать клетки-мишени, экспрессирующие HIG2 и HLA-A*0201. Специфическую активность CTL в отношении клеток 293Т или COS7, трансфицированных и полноразмерным геном HIG2, и молекулой HLA-A*0201, которые служат в качестве специфической модели клеток-мишеней, эндогенно экспрессирующих HIG2 и HLA-A*0201, проверяли, используя в качестве клеток-эффекторов линии CTL, индуцированные HIG2-A02-9-8 (SEQ ID NO: 114), HIG2-A02-9-15 (SEQ ID NO: 116), и клон CTL, индуцированный HIG2-A02-9-4 (SEQ ID NO: 117). В качестве контролей готовили 293Т или COS7, трансфицированные полноразмерным геном ЕСТ2, но не молекулой HLA-A*0201, и 293Т или COS7, трансфицированные HLA-A*0201, но не полноразмерным геном ЕСТ2 (или трансфицированные другим замещающим геном, например FoxP3 или ТТК). Линией CTL, демонстрирующей наивысшую специфическую активность CTL в отношении 293Т или COS7, была линия CTL, трансфицированная и ЕСТ2, и HLA-A*0201 (фиг.4е, h и i).
Эти результаты ясно демонстрируют, что HIG2-A02-9-8 (SEQ ID NO: 114), HIG2-A02-9-15 (SEQ ID NO: 116) и HIG2-A02-9-4 (SEQ ID NO: 117) естественно экспрессируются на поверхности клетки-мишени вместе с молекулой HLA-A2402 или HLA-A0201 и распознаются CTL. Кроме того, эти пептиды являются пептидными эпитопами, которые могут служить в качестве противораковых вакцин, направленных на экспрессирующие HIG2 опухоли.
Цитотоксическая активность в отношении линии раковых клеток, эндогенно экспрессирующей HLA-A*0201 и HIG2
Кроме того, цитотоксическую активность проверяли с помощью анализа цитотоксичности в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». В результате, как показано на фиг.4i, клон CTL, подвергнутый стимуляции HIG2-A02-9-4 (SEQ ID NO: 117), продемонстрировал замечательно высокую цитотоксическую активность в отношении HLA-A02-положительных и HIG2-положительных линий раковых клеток CAki-1 по сравнению с активностями в отношении HLA-A02-отрицательных и HIG2-положительных линий раковых клеток А498.
Стимуляция T-клеток с использованием предсказанных пептидов из INHBB, рестрицированных по HLA-A*2402 или HLA-A*0201, и создание линий CTL, подвергнутых стимуляции происходящими из INHBB пептидами
CTL, специфичные для тех пептидов, которые происходят из INHBB, создавали в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Получаемые в результате CTL, обладающие обнаруживаемой специфической активностью CTL, определяемой с помощью анализа ELISPOT на IFN-гамма, продемонстрированы на фиг.5. В частности, для INHBB-A24-9-180 (SEQ ID NO: 395), INHBB-A24-10-180 (SEQ ID NO: 133), INHBB-A24-10-305 (SEQ ID NO: 135), INHBB-A24-10-7 (SEQ ID NO: 137) и INHBB-A24-10-212 (SEQ ID NO: 426) продемонстрирована мощная продукция IFN-гамма с помощью анализа ELISPOT на IFN-гамма, и клетки в положительной лунке номер #7, подвергнутые стимуляции INHBB-A24-9-180 (SEQ ID NO: 395), #3, подвергнутые стимуляции INHBB-A24-10-180 (SEQ ID NO: 133), #2, подвергнутые стимуляции INHBB-A24-10-305 (SEQ ID NO: 135), #8 и #2, подвергнутые стимуляции INHBB-A24-10-7 (SEQ ID NO: 137), и #1, подвергнутые стимуляции INHBB-A24-10-212 (SEQ ID NO: 426), наращивали и создавали линии CTL. Линии CTL, обладающие более высокими специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида, определяли с помощью ELISA. Результаты продемонстрированы на фиг.5b-e. Тогда как с использованием других пептидов, представленных в таблице 6, линии CTL не могли быть созданы несмотря на возможную активность связывания с HLA-A*2402 и HLA*0201. Например, типичный отрицательный пептид (INHBB-A24-9-238) продемонстрирован на фиг.5a. В этом изобретении пептиды, с использованием которых могла быть создана линия CTL, отбирали в качестве сильно стимулирующего CTL пептида.
Создание клонов CTL, подвергнутых стимуляции происходящими из INHBB пептидами
Кроме того, осуществляли серийное разведение этих линий CTL в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Создание клонов CTL из линии CTL из лунки #7, подвергнутой стимуляции INHBB-A24-9-180 (SEQ ID NO: 395), и линии CTL из лунки #2, подвергнутой стимуляции INHBB-A24-10-305 (SEQ ID NO: 135), продемонстрировано на фиг.5b и d. Клоны CTL обладали сильными и специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида.
Специфическая активность CTL в отношении клеток-мишеней, экспрессирующих INHBB и HLA-A*2402
Созданные линии CTL, индуцированные против этих пептидов, проверяли на их способность распознавать клетки-мишени, экспрессирующие INHBB и HLA-A*2402. Специфическую активность CTL в отношении клеток 293Т, трансфицированных и полноразмерным геном INHBB, и молекулой HLA-A*2402, которые служат в качестве специфической модели клеток-мишеней, эндогенно экспрессирующих INHBB и HLA-A*2402, проверяли, используя в качестве клеток-эффекторов линии CTL, индуцированные INHBB-A24-10-180 (SEQ ID NO: 133) и INHBB-A24-10-7 (SEQ ID NO: 137), и клон CTL, индуцированный INHBB-A24-10-305 (SEQ ID NO: 135). В качестве контролей готовили 293Т, трансфицированные полноразмерным геном INHBB, но не молекулой HLA-A*2402, и 293Т, трансфицированные HLA-A*2402, но не полноразмерным геном INHBB. Линией CTL, демонстрирующей наивысшую специфическую активность CTL в отношении 293Т, была линия CTL, трансфицированная и INHBB, и HLA-A*2402 (фиг.5c, d и e).
Эти результаты ясно демонстрируют, что INHBB-A24-10-305 (SEQ ID NO: 135), INHBB-A24-10-180 (SEQ ID NO: 133) и INHBB-A24-10-7 (SEQ ID NO: 137) естественно экспрессируются на поверхности клетки-мишени вместе с молекулой HLA-A2402 и распознаются CTL. Кроме того, эти пептиды являются пептидными эпитопами, которые могут служить в качестве противораковых вакцин, направленных на экспрессирующие INHBB опухоли.
Цитотоксическая активность в отношении линии раковых клеток, эндогенно экспрессирующей HLA-A*2402 и INHBB
Кроме того, цитотоксическую активность проверяли с помощью анализа цитотоксичности в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». В результате, как показано на фиг.5b, клон CTL, подвергнутый стимуляции INHBB-A24-9-180 (SEQ ID NO: 395), продемонстрировал замечательно высокую цитотоксическую активность в отношении HLA-A24-положительных и INHBB-положительных линий раковых клеток MIAPaca2 по сравнению с активностями в отношении HLA-A24-отрицательных и INHBB-положительных линий раковых клеток CAki-2.
Стимуляция T-клеток с использованием предсказанных пептидов из KIF20A, рестрицированных по HLA-A*2402, и создание линий CTL, подвергнутых стимуляции происходящими из KIF20A пептидами
CTL, специфичные для тех пептидов, которые происходят из KIF20A, создавали в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Получаемые в результате CTL, обладающие обнаруживаемой специфической активностью CTL, определяемой с помощью анализа ELISPOT на IFN-гамма, продемонстрированы на фиг.6. В частности, для KIF20A-A24-9-305 (SEQ ID NO: 174), KIF20A-A24-9-383 (SEQ ID NO: 178), KIF20A-A24-10-304 (SEQ ID NO: 186) и KIF20A-A24-10-66 (SEQ ID NO: 194) продемонстрирована мощная продукция IFN-гамма с помощью анализа ELISPOT на IFN-гамма, и клетки в положительной лунке номер #2, подвергнутые стимуляции KIF20A-A24-9-305 (SEQ ID NO: 174), #3, подвергнутые стимуляции KIF20A-A24-9-383 (SEQ ID NO: 178), #5, подвергнутые стимуляции KIF20A-A24-10-304 (SEQ ID NO: 186), и #6, подвергнутые стимуляции KIF20A-A24-10-66 (SEQ ID NO: 194), наращивали и создавали линии CTL. Линии CTL, обладающие более высокими специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида, определяли с помощью ELISA. Результаты продемонстрированы на фиг.6a-e. Тогда как с использованием других пептидов, представленных в таблице 7, линии CTL не могли быть созданы несмотря на возможную активность связывания с HLA-A*2402. Например, типичные отрицательные пептиды (KIF20A-A24-9-647 и KIF20A-A24-10-182) продемонстрированы на фиг.6a. В этом изобретении пептиды, с использованием которых могла быть создана линия CTL, отбирали в качестве сильно стимулирующего CTL пептида.
Создание клонов CTL, подвергнутых стимуляции происходящими из KIF20A пептидами
Кроме того, осуществляли серийное разведение этих линий CTL в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Создание клонов CTL из линии CTL из лунки #2, подвергнутой стимуляции KIF20A-A24-9-305 (SEQ ID NO: 174), линии CTL из лунки #5, подвергнутой стимуляции KIF20A-A24-10-304 (SEQ ID NO: 186), и линии CTL из лунки #6, подвергнутой стимуляции KIF20A-A24-10-66 (SEQ ID NO: 194), продемонстрировано на фиг.6b, d и e. Клоны CTL обладали сильными и специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида.
Специфическая активность CTL в отношении клеток-мишеней, экспрессирующих KIF20A и HLA-A*2402
Созданные линии CTL, индуцированные против этих пептидов, проверяли на их способность распознавать клетки-мишени, экспрессирующие KIF20A и HLA-A*2402. Специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном KIF20A, и молекулой HLA-A*2402, и A24-LCL, трансфицированных с помощью электропорации полноразмерным геном KIF20A, которые служат в качестве специфической модели клеток-мишеней, эндогенно экспрессирующих KIF20A и HLA-A*2402, проверяли, используя в качестве клеток-эффекторов линии CTL, индуцированные KIF20A-A24-9-383 (SEQ ID NO: 178) и KIF20A-A24-10-304 (SEQ ID NO: 186), и клон CTL, индуцированный KIF20A-A24-10-66 (SEQ ID NO: 194). В качестве контролей готовили COS7, трансфицированные полноразмерным геном KIF20A, но не молекулой HLA-A*2402, и COS7, трансфицированные HLA-A*2402, но не полноразмерным геном KIF20A (или трансфицированные замещающим полноразмерным геном URLC10), COS7, трансфицированные HLA-A*2402 и подвергнутые в импульсном режиме воздействию KIF20A-10-308, и A24-LCL, трансфицированные контрольным вектором. Линией CTL, демонстрирующей наивысшую специфическую активность CTL в отношении COS7, была линия CTL, трансфицированная и KIF20A, и HLA-A2402 (фиг.6b, c и d). Альтернативно линия CTL, подвергнутая стимуляции KIF20A-A24-10-304 (SEQ ID NO: 186), продемонстрировала активность в отношении A24-LCL, трансфицированных KIF20A.
Эти результаты ясно демонстрируют, что KIF20A-A24-9-383 (SEQ ID NO: 178), KIF20A-A24-10-304 (SEQ ID NO: 186) и KIF20A-A24-10-66 (SEQ ID NO: 194) естественно экспрессируются на поверхности клетки-мишени вместе с молекулой HLA-A2402 и распознаются CTL. Кроме того, эти пептиды являются пептидными эпитопами, которые могут служить в качестве противораковых вакцин, направленных на экспрессирующие KIF20A опухоли.
Цитотоксическая активность в отношении линии раковых клеток, эндогенно экспрессирующей HLA-A*2402 и KIF20A
Кроме того, цитотоксическую активность проверяли с помощью анализа цитотоксичности в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». В результате, как показано на фиг.6b и е, клон CTL, подвергнутый стимуляции KIF20A-A24-9-305 (SEQ ID NO: 174) или KIF20A-A24-10-304 (SEQ ID NO: 186), продемонстрировал замечательно высокую цитотоксическую активность в отношении HLA-A24-положительных и KIF20A-положительных линий раковых клеток PK45P или MIAPaca2 соответственно по сравнению с активностями в отношении HLA-A24-отрицательных и KIF20A-положительных линий раковых клеток PK59.
Стимуляция T-клеток с использованием предсказанных пептидов из KNTC2, рестрицированных по HLA-A*2402, и создание линий CTL, подвергнутых стимуляции происходящими из KNTC2 пептидами
CTL, специфичные для тех пептидов, которые происходят из KNTC2, создавали в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Получаемые в результате CTL, обладающие обнаруживаемой специфической активностью CTL, определяемой с помощью анализа ELISPOT на IFN-гамма, продемонстрированы на фиг.7. В частности, для KNTC2-A24-9-309 (SEQ ID NO: 196), KNTC2-A24-9-124 (SEQ ID NO: 202), KNTC2-A24-9-154 (SEQ ID NO: 210), KNTC2-A24-9-150 (SEQ ID NO: 213), KNTC2-A24-10-452 (SEQ ID NO: 214), KNTC2-A24-10-227 (SEQ ID NO: 217) и KNTC2-A24-10-273 (SEQ ID NO: 223) продемонстрирована мощная продукция IFN-гамма с помощью анализа ELISPOT на IFN-гамма, и клетки в положительной лунке номер #8, подвергнутые стимуляции KNTC2-A24-9-309 (SEQ ID NO: 196), #5, подвергнутые стимуляции KNTC2-A24-9-124 (SEQ ID NO: 202), #5, подвергнутые стимуляции KNTC2-A24-9-154 (SEQ ID NO: 210), #7, подвергнутые стимуляции KNTC2-A24-9-150 (SEQ ID NO: 213), #4 и #5, подвергнутые стимуляции KNTC2-A24-10-452 (SEQ ID NO: 214), #1, подвергнутые стимуляции KNTC2-A24-10-227 (SEQ ID NO: 217), и #8, подвергнутые стимуляции KNTC2-A24-10-273 (SEQ ID NO: 223), наращивали и создавали линии CTL. Линии CTL, обладающие более высокими специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида, определяли с помощью ELISA. Результаты продемонстрированы на фиг.7a-h. Тогда как с использованием других пептидов, представленных в таблице 8, линии CTL не могли быть созданы несмотря на возможную активность связывания с HLA-A*2402. Например, типичный отрицательный пептид (KNTC2-A24-10-610) продемонстрирован на фиг.7a. В этом изобретении пептиды, с использованием которых могла быть создана линия CTL, отбирали в качестве сильно стимулирующего CTL пептида.
Создание клонов CTL, подвергнутых стимуляции происходящими из KNTC2 пептидами
Кроме того, осуществляли серийное разведение этих линий CTL в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Создание клонов CTL из линии CTL из лунки #5, подвергнутой стимуляции KNTC2-A24-9-154 (SEQ ID NO: 210), и линии CTL из лунки #5, подвергнутой стимуляции KNTC2-A24-10-452 (SEQ ID NO: 214), продемонстрировано на фиг.7d и f. Клоны CTL обладали сильными и специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида.
Специфическая активность CTL в отношении клеток-мишеней, экспрессирующих KNTC2 и HLA-A*2402
Созданные линии CTL, индуцированные против этих пептидов, проверяли на их способность распознавать клетки-мишени, экспрессирующие KNTC2 и HLA-A*2402. Специфическую активность CTL в отношении клеток НЕК293, трансфицированных и полноразмерным геном KNTC2, и молекулой HLA-A*2402, которые служат в качестве специфической модели клеток-мишеней, эндогенно экспрессирующих KNTC2 и HLA-A*2402, проверяли, используя в качестве клеток-эффекторов клоны CTL, индуцированные KNTC2-A24-10-452 (SEQ ID NO: 214). В качестве контролей готовили НЕК293, трансфицированные полноразмерным геном KNTC2, но не молекулой HLA-A*2402, НЕК293, трансфицированные HLA-A*2402, но не полноразмерным геном KNTC2, и НЕК293, трансфицированные HLA-A*2402 и подвергнутые в импульсном режиме воздействию KNTC2-9-309. Линией CTL, демонстрирующей наивысшую специфическую активность CTL в отношении НЕК293, была линия CTL, трансфицированная и KNTC2, и HLA-A*2402 (фиг.7f).
Эти результаты ясно демонстрируют, что KNTC2-A24-10-452 (SEQ ID NO: 214) естественно экспрессируется на поверхности клетки-мишени вместе с молекулой HLA-A2402 и распознается CTL. Кроме того, этот пептид является пептидным эпитопом, который может служить в качестве противораковых вакцин, направленных на экспрессирующие KNTC2 опухоли.
Стимуляция T-клеток с использованием предсказанных пептидов из TTK, рестрицированных по HLA-A*0201, и создание линий CTL, подвергнутых стимуляции происходящими из TTK пептидами
CTL, специфичные для тех пептидов, которые происходят из TTK, создавали в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Получаемые в результате CTL, обладающие обнаруживаемой специфической активностью CTL, определяемой с помощью анализа ELISPOT на IFN-гамма, продемонстрированы на фиг.8. Как изображено на фиг.8b-d, для TTK-A2-9-462 (SEQ ID NO: 227), TTK-A2-9-547 (SEQ ID NO: 228), TTK-A2-9-719 (SEQ ID NO: 233) и TTK-A2-10-462 (SEQ ID NO: 254) продемонстрирована мощная продукция IFN-гамма с помощью анализа ELISPOT на IFN-гамма, и наращивали клетки в положительной лунке номер #4, подвергнутые стимуляции TTK-A2-9-462 (SEQ ID NO: 227), #2, подвергнутые стимуляции TTK-A2-9-547 (SEQ ID NO: 228), #1, подвергнутые стимуляции TTK-A2-9-719 (SEQ ID NO: 233), и #8, подвергнутые стимуляции TTK-A2-10-462 (SEQ ID NO: 254). Линии CTL, обладающие более высокими специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида, определяли с помощью ELISA. Тогда как с использованием других пептидов, представленных в таблице 9, линии CTL не могли быть созданы несмотря на возможную активность связывания с HLA-A*0201. Например, типичный отрицательный пептид (TTK-A2-9-278) продемонстрирован на фиг.8a. В этом изобретении пептиды, с использованием которых могла быть создана линия CTL, отбирали в качестве сильно стимулирующего CTL пептида.
Создание клонов CTL, подвергнутых стимуляции происходящими из TTK пептидами
Кроме того, осуществляли серийное разведение этих линий CTL в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Создание клонов CTL из линии CTL из лунки #4, подвергнутой стимуляции TTK-A2-9-462 (SEQ ID NO: 227), линии CTL из лунки #2, подвергнутой стимуляции TTK-A2-9-547 (SEQ ID NO: 228), линии CTL из лунки #1, подвергнутой стимуляции TTK-A2-9-719 (SEQ ID NO: 233), и линии CTL из лунки #8, подвергнутой стимуляции TTK-A2-10-462 (SEQ ID NO: 254), продемонстрировано на фиг.8b, c, d и e. Клоны CTL обладали сильными и специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида.
Специфическая активность CTL в отношении клеток-мишеней, экспрессирующих TTK и HLA-A*0201
Созданные клоны CTL, индуцированные против этих пептидов, проверяли на их способность распознавать клетки-мишени, экспрессирующие TTK и HLA-A*0201. Специфическую активность CTL в отношении клеток COS7, трансфицированных и полноразмерным геном TTK, и молекулой HLA-A*0201, которые служат в качестве специфической модели клеток-мишеней, эндогенно экспрессирующих TTK и HLA-A*0201, проверяли, используя в качестве клеток-эффекторов клоны CTL, индуцированные TTK-A2-9-462 (SEQ ID NO: 227), TTK-A02-9-547 (SEQ ID NO: 228), TTK-A2-9-719 (SEQ ID NO: 233) и TTK-A2-10-462 (SEQ ID NO: 254). В качестве контролей готовили COS7, трансфицированные полноразмерным геном ТТК, но не молекулой HLA-A*0201, COS7, трансфицированные HLA-A*0201, но не полноразмерным геном ТТК (или трансфицированные замещающим полноразмерным геном HIG2), и COS7, трансфицированные HLA-A*0201 и подвергнутые в импульсном режиме воздействию различных целевых пептидных эпитопов. Клоном CTL, демонстрирующим наивысшую специфическую активность CTL в отношении COS7, был клон CTL, трансфицированный и ТТК, и HLA-A*0201 (фиг.8b, c, d и e).
Эти результаты ясно демонстрируют, что TTK-A2-9-462 (SEQ ID NO: 227), TTK-A02-9-547 (SEQ ID NO: 228), TTK-A2-9-719 (SEQ ID NO: 233) и TTK-A02-10-462 (SEQ ID NO: 254) естественно экспрессируются на поверхности клетки-мишени вместе с молекулой HLA-A2 и распознаются CTL. Кроме того, эти пептиды являются пептидными эпитопами, которые могут служить в качестве противораковых вакцин, направленных на экспрессирующие ТТК опухоли.
Стимуляция T-клеток с использованием предсказанных пептидов из URLC10, рестрицированных по HLA-A*0201, и создание линий CTL, подвергнутых стимуляции происходящими из URLC10 пептидами
CTL, специфичные для тех пептидов, которые происходят из URLC10, создавали в соответствии с протоколами, изложенными выше в разделе «Материалы и методы». Получаемые в результате CTL, обладающие обнаруживаемой специфической активностью CTL, определяемой с помощью анализа ELISPOT на IFN-гамма, продемонстрированы на фиг.9. Как изображено на фиг.9b-d, для URLC-A2-9-206 (SEQ ID NO: 271), URLC-A2-9-212 (SEQ ID NO: 272) и URLC-A2-10-211 (SEQ ID NO: 288) продемонстрирована мощная продукция IFN-гамма с помощью анализа ELISPOT на IFN-гамма, и наращивали клетки в положительной лунке номер #7, подвергнутые стимуляции URLC-A2-9-206 (SEQ ID NO: 271), #3, подвергнутые стимуляции URLC-A2-9-212 (SEQ ID NO: 272), и #5, подвергнутые стимуляции (SEQ ID NO: 288). Линии CTL, обладающие более высокими специфическими активностями CTL в отношении мишени, подвергнутой в импульсном режиме воздействию пептида, по сравнению с активностями в отношении мишени, не подвергнутой в импульсном режиме воздействию пептида, определяли с помощью ELISA. Тогда как с использованием других пептидов, представленных в таблице 10, линии CTL не могли быть созданы несмотря на возможную активность связывания с HLA-A*0201. Например, типичный отрицательный пептид (URLC-A2-9-58) продемонстрирован на фиг.9a. В этом изобретении пептиды, с использованием которых могла быть создана линия CTL, отбирали в качестве сильно стимулирующего CTL пептида.
Специфическая активность CTL в отношении клеток-мишеней, экспрессирующих URLC10 и HLA-A*0201
Созданные линии CTL, индуцированные против этих пептидов, проверяли на их способность распознавать клетки-мишени, эндогенно экспрессирующие URLC10 и HLA-A*0201. Специфическую активность CTL в отношении COS7, Hek293 и 293T, трансфицированных и полноразмерным геном URLC10, и молекулой HLA-A*0201, которые служат в качестве специфической модели клеток-мишеней, эндогенно экспрессирующих URLC10 и HLA-A*0201, проверяли, используя в качестве клеток-эффекторов линию CTL, индуцированную URLC10-A02-10-211. В качестве контролей готовили COS7, Hek293 или 293T, трансфицированные полноразмерным геном URLC10, но не молекулой HLA-A*0201 (трансфицированные замещающей HLA-A*2402), COS7, Hek293 или 293T, трансфицированные HLA-A*0201, но не полноразмерным геном URLC10, и COS7, трансфицированные HLA-A*0201 и подвергнутые в импульсном режиме воздействию целевого пептидного эпитопа (URLC10-A02-10-64). Линией CTL, демонстрирующей наивысшую специфическую активность CTL в отношении COS7, Hek293 или 293T, была линия CTL, трансфицированная и URLC10, и HLA-A*0201 (фиг.9-2).
Эти результаты ясно демонстрируют, что URLC10-A02-10-211 естественно экспрессируется на поверхности клетки-мишени вместе с молекулой HLA-A*0201 и распознается CTL. Кроме того, этот пептид является пептидным эпитопом, который может служить в качестве противораковых вакцин, направленных на экспрессирующие URLC10 опухоли.
Анализ гомологии являющихся антигенами пептидов
Клоны CTL, созданные против следующих пептидов, продемонстрировали сильную специфическую активность CTL.
CDH3-A24-9-513 (SEQ ID NO: 19),
CDH3-A24-9-406 (SEQ ID NO: 22),
CDH3-A24-10-807 (SEQ ID NO: 30),
CDH3-A24-10-332 (SEQ ID NO: 34),
CDH3-A24-10-655 (SEQ ID NO: 344),
CDH3-A24-10-470 (SEQ ID NO: 358),
EphA4-A24-9-453 (SEQ ID NO: 41),
EphA4-A24-9-5 (SEQ ID NO: 44),
EphA4-A24-9-869 (SEQ ID NO: 46),
EphA4-A24-9-420 (SEQ ID NO: 48),
EphA4-A24-10-24 (SEQ ID NO: 78),
EphA4-A02-9-501 (SEQ ID NO: 376),
EphA4-A02-9-165 (SEQ ID NO: 379),
ECT2-A24-9-515 (SEQ ID NO: 80),
ECT2-A24-10-40 (SEQ ID NO: 100),
ECT2-A24-10-101 (SEQ ID NO: 101),
HIG2-A24-9-19 (SEQ ID NO: 110),
HIG2-A24-9-22 (SEQ ID NO: 111),
HIG2-A24-9-8 (SEQ ID NO: 387),
HIG2-A24-10-7 (SEQ ID NO: 112),
HIG2-A24-10-18 (SEQ ID NO: 394),
HIG2-A02-9-8 (SEQ ID NO: 114),
HIG2-A02-9-15 (SEQ ID NO: 116),
HIG2-A02-9-4 (SEQ ID NO: 117),
HIG2-A02-10-8 (SEQ ID NO: 121),
INHBB-A24-9-180 (SEQ ID NO: 395),
INHBB-A24-10-180 (SEQ ID NO: 133),
INHBB-A24-10-305 (SEQ ID NO: 135),
INHBB-A24-10-7 (SEQ ID NO: 137),
INHBB-A24-10-212 (SEQ ID NO: 426),
KIF20A-A24-9-305 (SEQ ID NO: 174),
KIF20A-A24-9-383 (SEQ ID NO: 178),
KIF20A-A24-10-304 (SEQ ID NO: 186),
KIF20A-A24-10-66 (SEQ ID NO: 194),
KNTC2-A24-9-309 (SEQ ID NO: 196),
KNTC2-A24-9-124 (SEQ ID NO: 202),
KNTC2-A24-9-154 (SEQ ID NO: 210),
KNTC2-A24-9-150 (SEQ ID NO: 213),
KNTC2-A24-10-452 (SEQ ID NO: 214),
KNTC2-A24-10-227 (SEQ ID NO: 217),
KNTC2-A24-10-273 (SEQ ID NO: 223),
TTK-A02-9-462 (SEQ ID NO: 227),
TTK-A02-9-547 (SEQ ID NO: 228),
TTK-A02-9-719 (SEQ ID NO: 233),
TTK-A02-10-462 (SEQ ID NO: 254),
URLC-A02-9-206 (SEQ ID NO: 271),
URLC-A02-9-212 (SEQ ID NO: 272) и
URLC-A02-10-211 (SEQ ID NO: 288).
Это наводит на мысль, что последовательности SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 376, 379, 80, 100, 101, 110, 111, 387, 112, 394, 114, 116, 117, 121, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217, 223, 227, 228, 233, 254, 271, 272 или 288 гомологичны пептидам, происходящим из других молекул, о которых известно, что они сенсибилизируют иммунную систему человека.
Для исключения этой возможности был проведен анализ гомологии с пептидными последовательностями в виде запрашиваемых последовательностей, используя алгоритм BLAST (http://www.ncbi.nlm.nih.gov/blast/blast.cgi). Значительной гомологии последовательностей обнаружено не было.
Эти результаты говорят о том, что последовательности SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 376, 379, 80, 100, 101, 110, 111, 387, 112, 394, 114, 116, 117, 121, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217, 223, 227, 228, 233, 254, 271, 272 или 288 являются уникальными и, следовательно, характеризуются небольшим риском индуцировать не намеченный иммунологический ответ на любые неродственные молекулы.
ОБСУЖДЕНИЕ
Идентификация новых TAA, в частности ТАА, которые индуцируют сильные и специфические противоопухолевые иммунные ответы, оправдывает дальнейшую разработку клинического применения стратегии вакцинации пептидами при различных типах рака (Boon T. et al., (1996) J. Exp. Med. 183: 725-729; van der Bruggen P. et al., (1991) Science 254: 1643-1647; Brichard V. et al., (1993) J. Exp. Med. 178: 489-495; Kawakami Y. et al., (1994) J. Exp. Med. 180: 347-352; Shichijo S. et al., (1998) J. Exp. Med. 187: 277-288; Chen Y.T. et al., (1997) Proc. Natl. Acad. Sci. USA, 94: 1914-1918; Harris C.C., (1996) J. Natl. Cancer Inst. 88: 1442-1445; Butterfield L.H. et al., (1999) Cancer Res. 59: 3134-3142; Vissers J.L. et al., (1999) Cancer Res. 59: 5554-5559; van der Burg S.H. et al., (1996) J. Immunol. 156: 3308-3314; Tanaka F. et al., (1997) Cancer Res. 57:4465-4468; Fujie T. et al., (1999) Int. J. Cancer 80: 169-172; Kikuchi M. et al., (1999) Int. J. Cancer 81: 459-466; Oiso M. et al., (1999) Int. J. Cancer 81: 387-394).
Технологии ранжированных микрорядов кДНК могут выявить полные профили экспрессии генов в раковых клетках (Lin Y.M. et al., Oncogene. 2002 Jun. 13; 21: 4120-4128; Kitahara O. et al., Cancer Res. 2001 May 1; 61: 3544-3549; Suzuki C. et al., Cancer Res. 2003 Nov 1; 63: 7038-7041; Ashida S., Cancer Res. 2004 Sep 1; 64: 5963-5972; Ochi K. et al., Int. J. Oncol. 2004 Mar; 24(3): 647-655; Kaneta Y. et al., Int. J. Oncol. 2003 Sep; 23: 681-691; Obama K., Hepatology. 2005 Jun; 41: 1339-1348; Kato T. et al., Cancer Res. 2005 Jul 1; 65: 5638-5646; Kitahara O. et al., Neoplasia. 2002 Jul-Aug; 4: 295-303; Saito-Hisaminato A. et al., DNA Res. 2002, 9: 35-45.) и находят применение для идентификации потенциальных TAA. Среди транскриптов, экспрессия которых увеличена в различных раках, новые гены человека, названные CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10, были идентифицированы, используя эти технологии.
Как продемонстрировано выше, CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10 сверхэкспрессируются в различных раках, но демонстрируют минимальную экспрессию в нормальных тканях. Кроме того, было установлено, что эти гены имеют значительную функцию, связанную с пролиферацией клеток. Таким образом, пептиды, происходящие из CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10, могут служить в качестве эпитопов TAA, которые, в свою очередь, могут использоваться для вызова значительных и специфических иммунных ответов против раковых клеток.
Таким образом, поскольку CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и URLC10 являются новыми TAA, вакцины, в которых используются эти пептидные эпитопы, находят применение в качестве иммунотерапевтических средств против различных раков или другого заболевания, при которых экспрессируются эти молекулы.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
В настоящем изобретении идентифицированы новые TAA, в частности ТАА, которые вызывают сильные и специфические противоопухолевые иммунные ответы. Такие TAA оправдывают дальнейшую разработку в качестве пептидных вакцин против заболеваний, тесно связанных со сверхэкспрессией CDH3, EPHA4, ECT2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков.
Все патенты, заявки на патенты и публикации, на которые здесь ссылаются, включены посредством ссылки.
Хотя настоящее изобретение описано подробно и относительно его конкретных вариантов осуществления, должно быть понятно, что вышеприведенное описание является примерным и объяснительным по природе и, как подразумевается, иллюстрирует настоящее изобретение и его предпочтительные варианты осуществления. Благодаря обычному экспериментированию квалифицированный в данной области техники специалист быстро поймет, что в нем могут быть осуществлены различные изменения и модификации без отступления от сущности и объема настоящего изобретения. Таким образом, подразумевается, что настоящее изобретение определяется не вышеприведенным описанием, а следующей формулой изобретения и ее эквивалентами.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПЕПТИДНЫЕ ВАКЦИНЫ ПРОТИВ РАКА С ЭКСПРЕССИЕЙ ПОЛИПЕПТИДОВ MPHOSPH1 ИЛИ DEPDC1 | 2007 |
|
RU2469044C2 |
| ПЕПТИДЫ ТТК И ВАКЦИНЫ, ИХ СОДЕРЖАЩИЕ | 2010 |
|
RU2531348C2 |
| ПЕПТИДЫ UBE2T И СОДЕРЖАЩИЕ ИХ ВАКЦИНЫ | 2013 |
|
RU2663350C2 |
| ФАРМАЦЕВТИЧЕСКОЕ СРЕДСТВО, СОДЕРЖАЩЕЕ ЭПИТОПНЫЕ ПЕПТИДЫ HIG2 И URLC10, ДЛЯ ЛЕЧЕНИЯ РАКА, СПОСОБЫ И СРЕДСТВА ДЛЯ ИНДУКЦИИ АНТИГЕНПРЕЗЕНТИРУЮЩЕЙ КЛЕТКИ И ЦИТОТОКСИЧЕСКОГО Т-ЛИМФОЦИТА (ЦТЛ), АНТИГЕНПРЕЗЕНТИРУЮЩАЯ КЛЕТКА И ЦТЛ, ПОЛУЧЕННЫЕ ТАКИМ СПОСОБОМ, СПОСОБ И СРЕДСТВО ИНДУКЦИИ ИММУННОГО ПРОТИВООПУХОЛЕВОГО ОТВЕТА | 2009 |
|
RU2529373C2 |
| ПЕПТИДЫ HJURP И СОДЕРЖАЩИЕ ИХ ВАКЦИНЫ | 2011 |
|
RU2570560C2 |
| ПЕПТИДЫ ТОРК И СОДЕРЖАЩИЕ ИХ ВАКЦИНЫ | 2012 |
|
RU2633503C2 |
| ПЕПТИДЫ ТЕМ8 И СОДЕРЖАЩИЕ ИХ ВАКЦИНЫ | 2008 |
|
RU2498993C2 |
| ПЕПТИДЫ ЭПИТОПА WDRPUH И ВАКЦИНЫ, СОДЕРЖАЩИЕ ИХ | 2009 |
|
RU2514386C2 |
| ПЕПТИДЫ KNTC2 И СОДЕРЖАЩИЕ ИХ ВАКЦИНЫ | 2014 |
|
RU2671395C2 |
| ВАКЦИНЫ НА ОСНОВЕ ПЕПТИДА FOXP3 | 2007 |
|
RU2473557C2 |
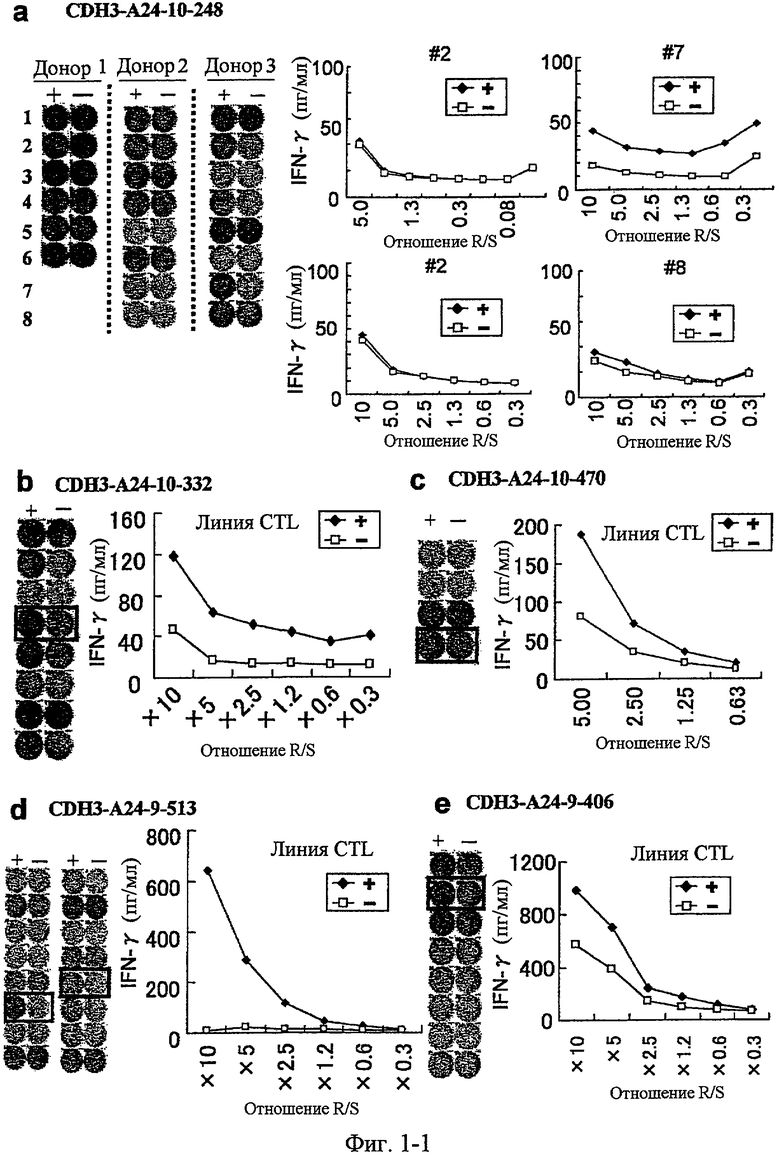
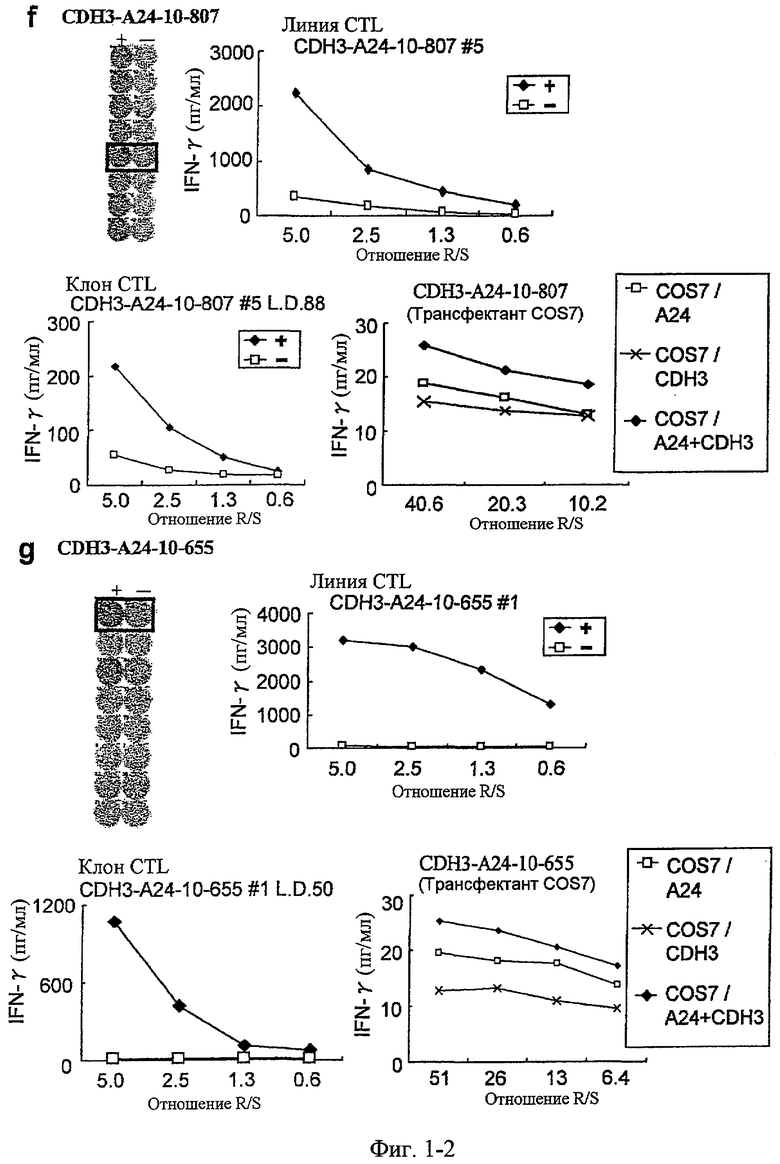
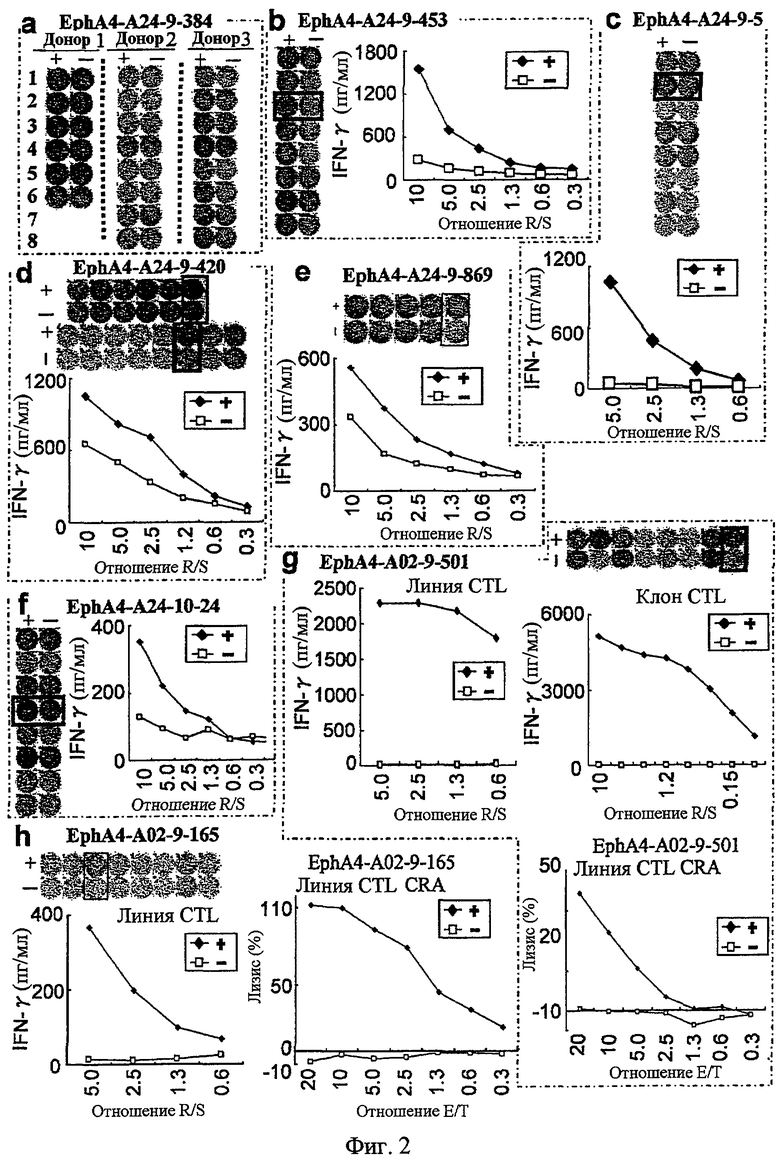
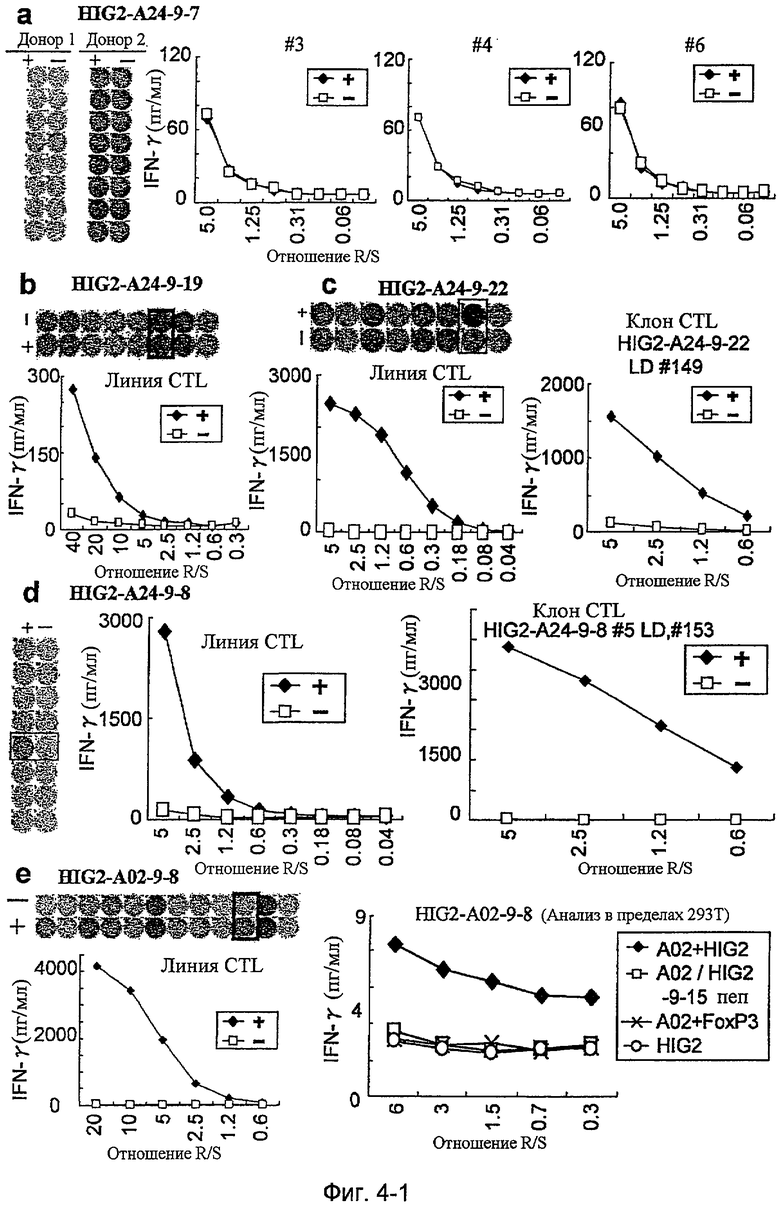
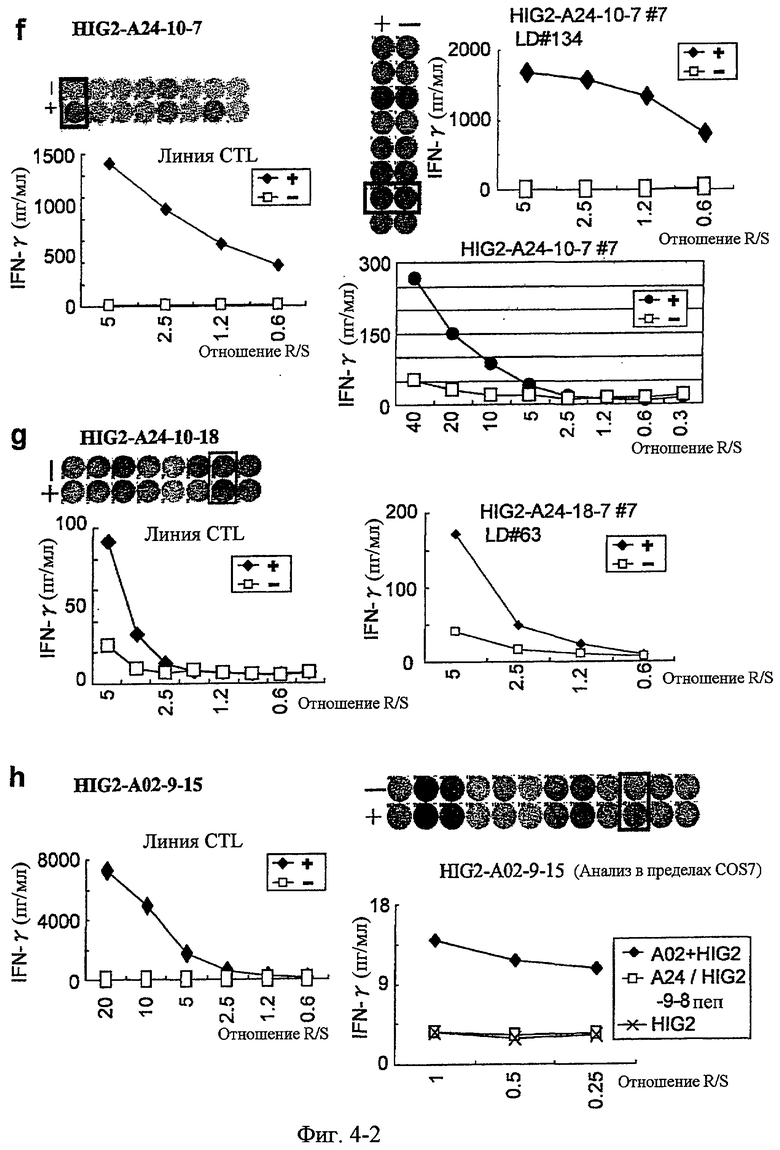
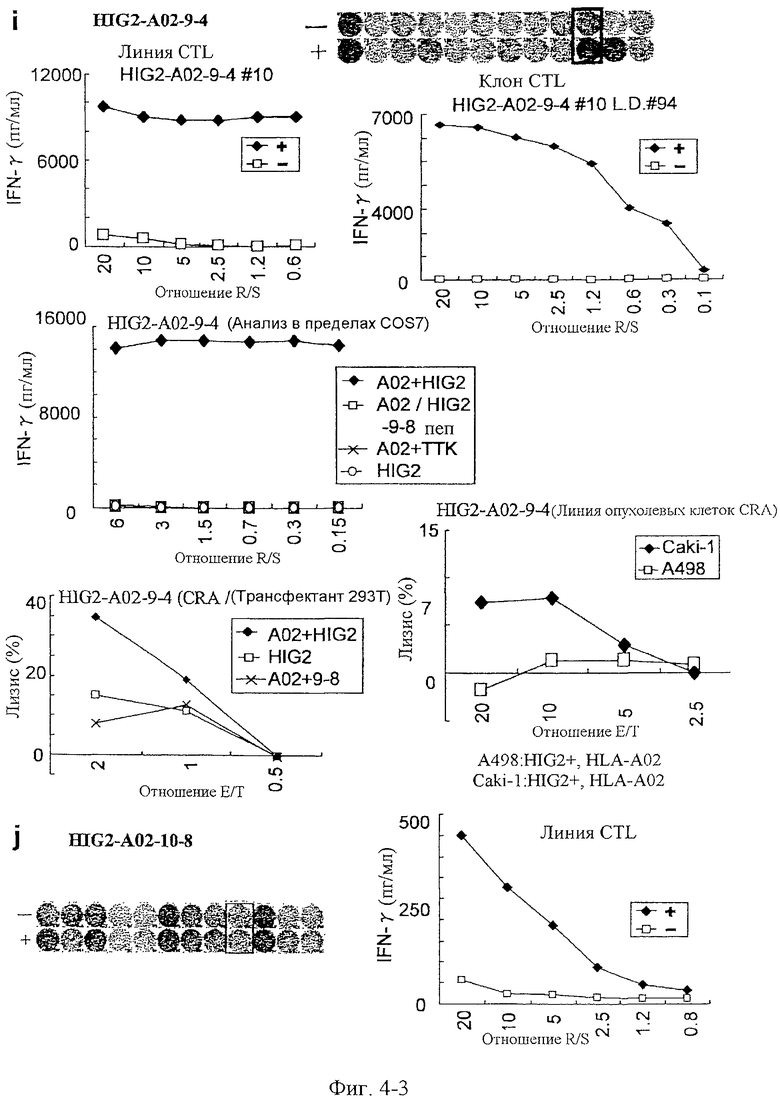
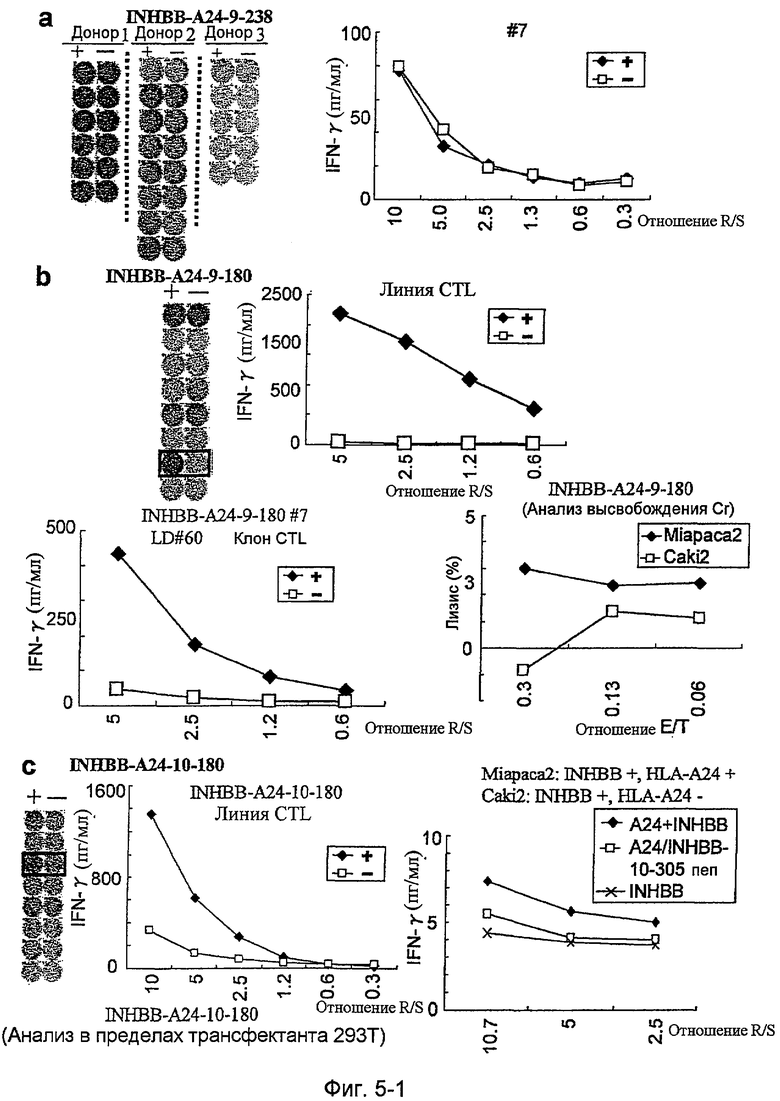
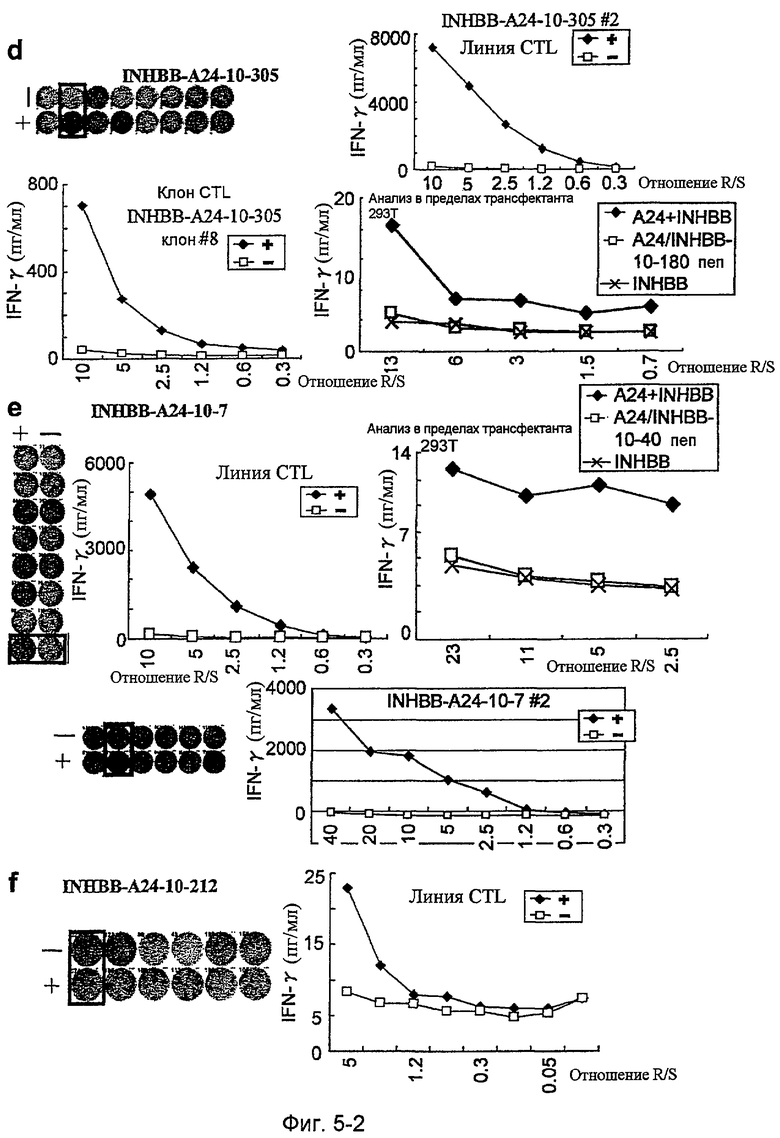
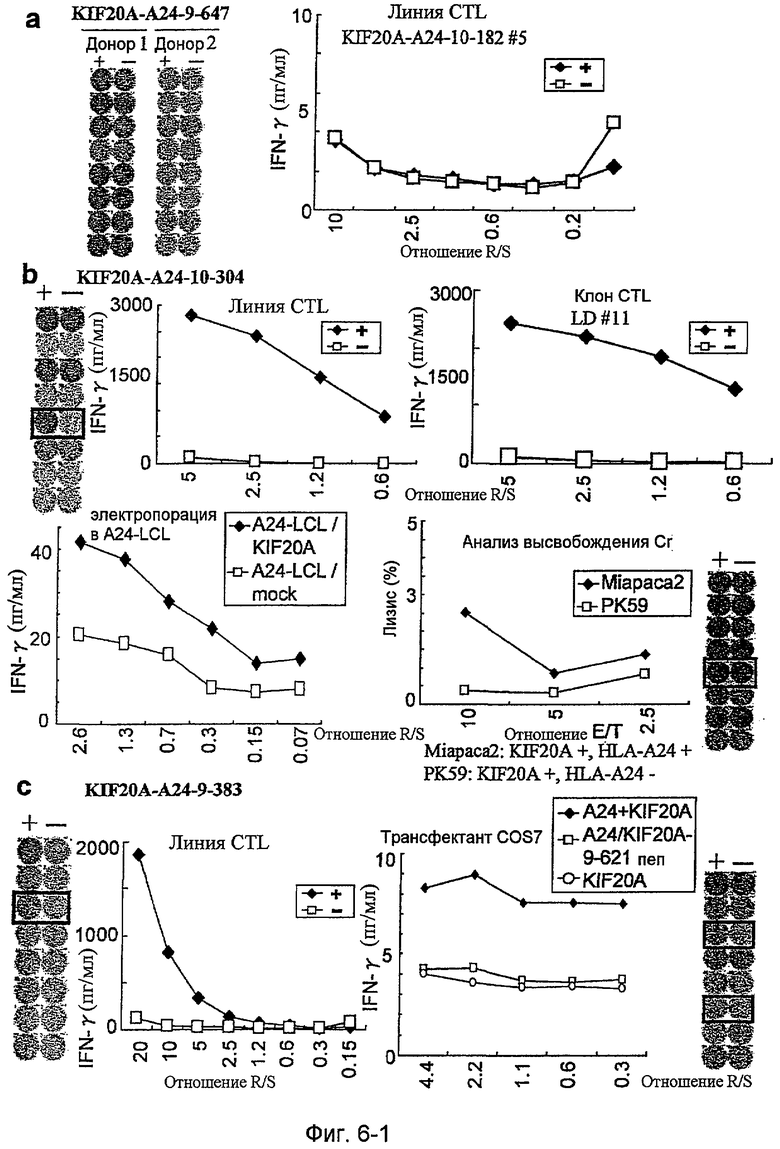
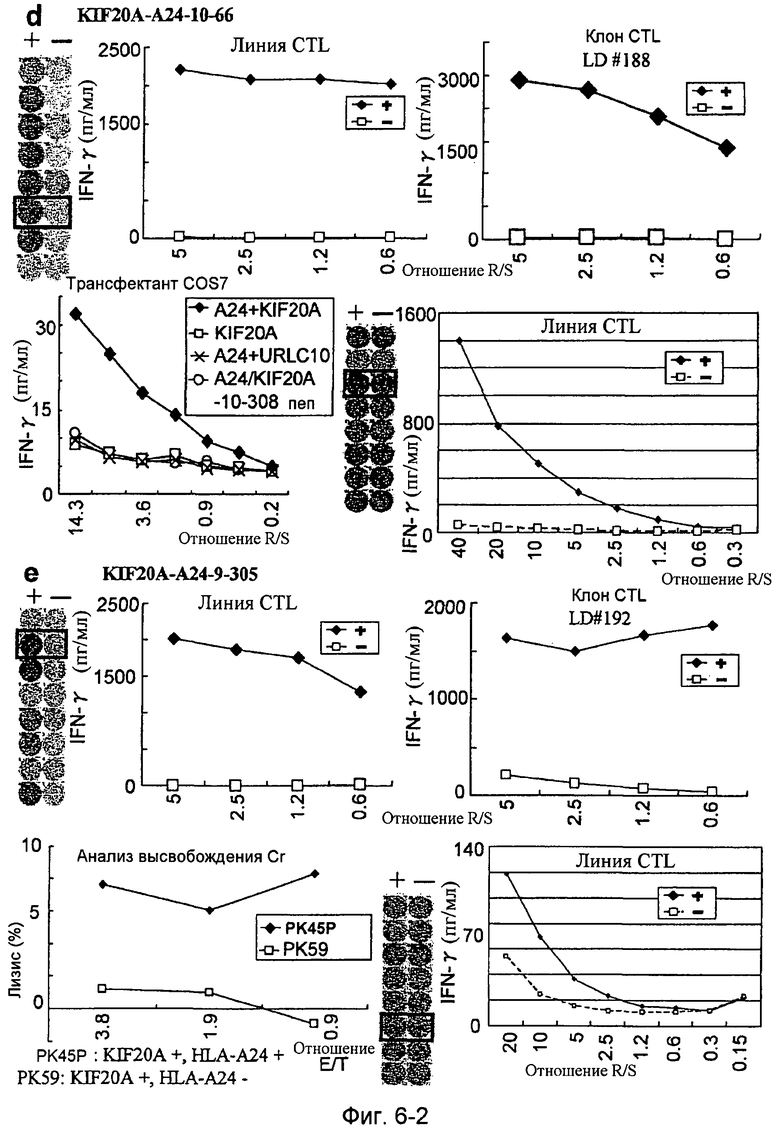
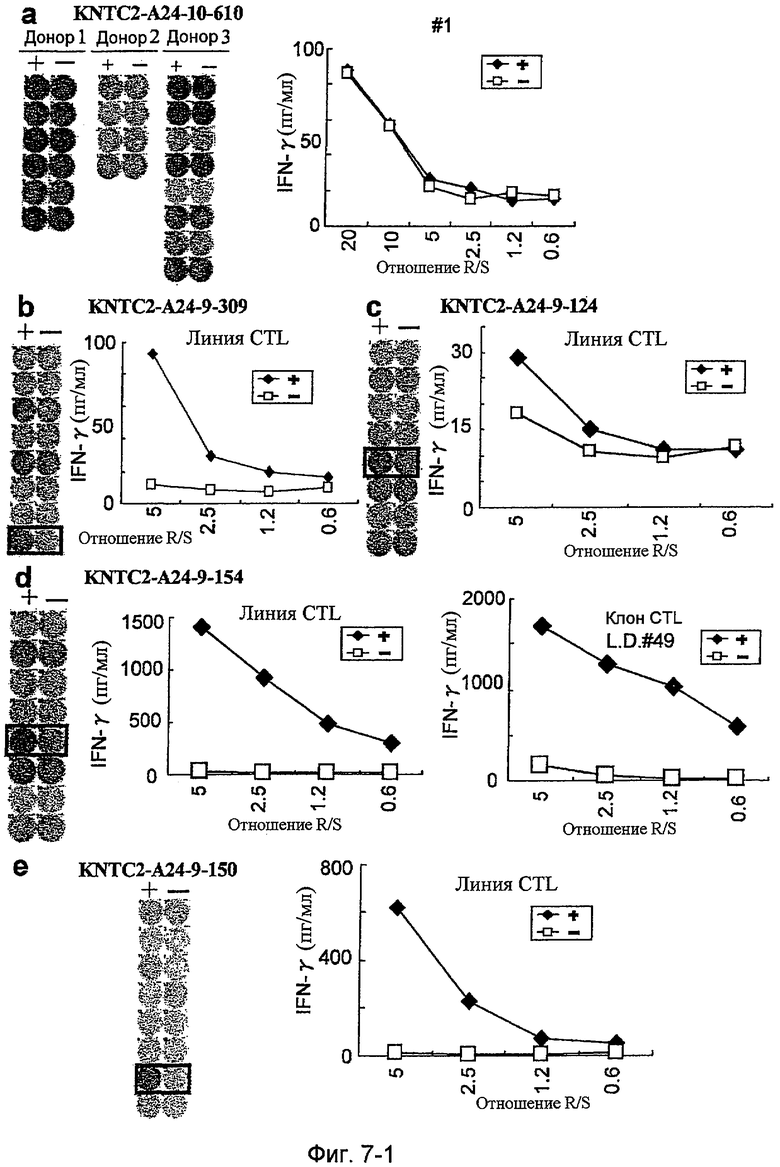
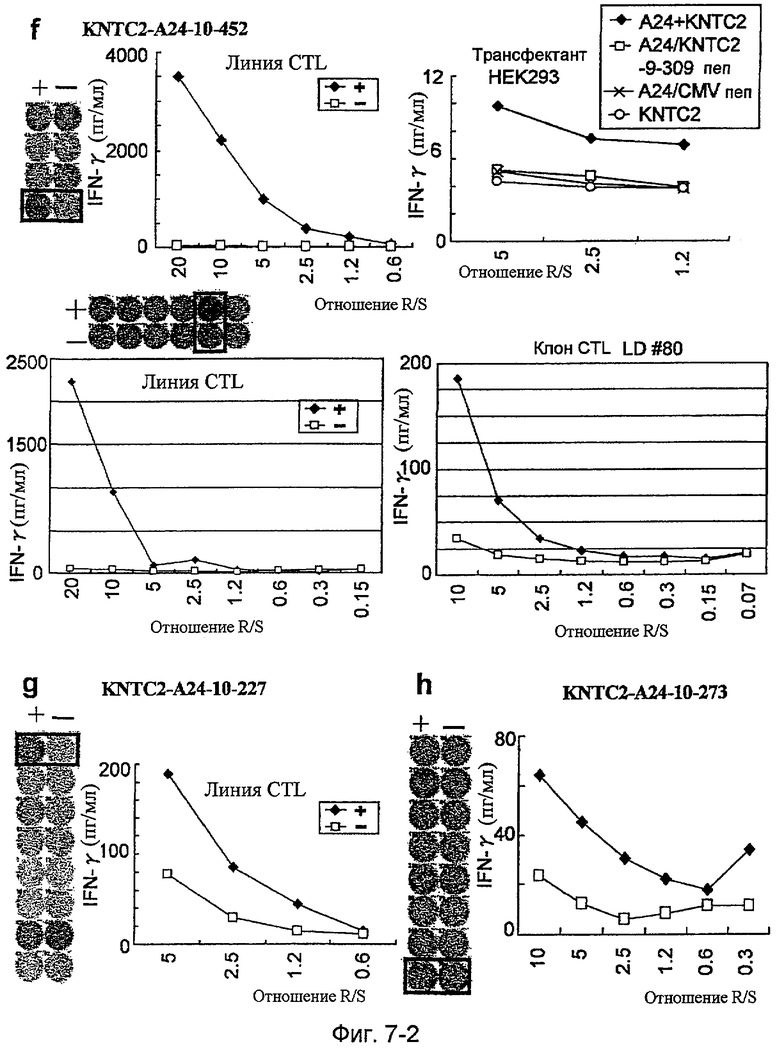

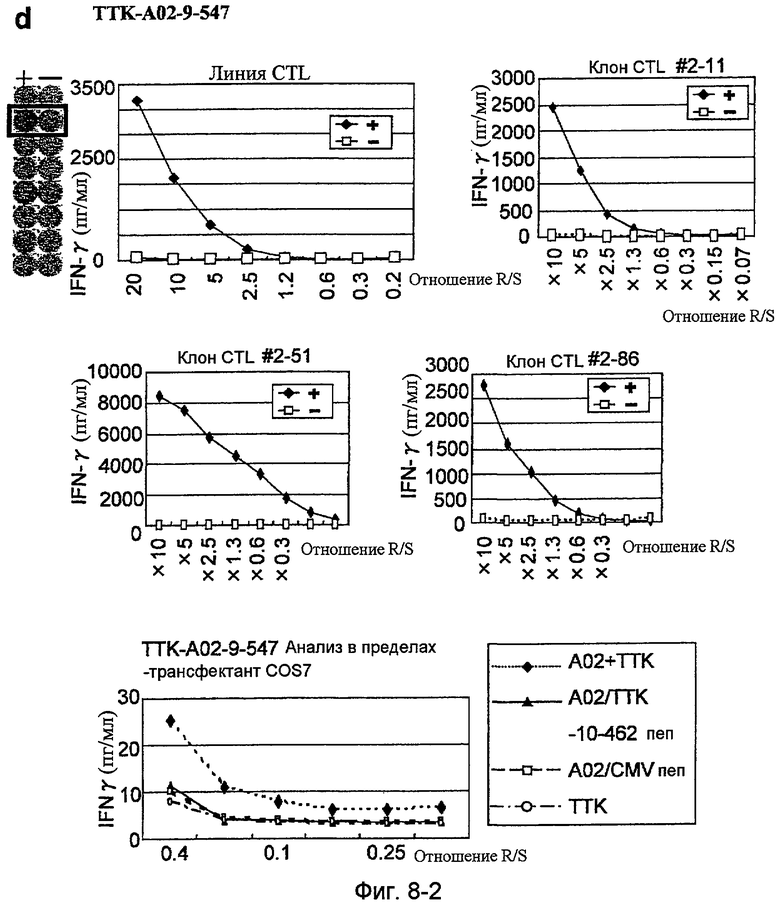
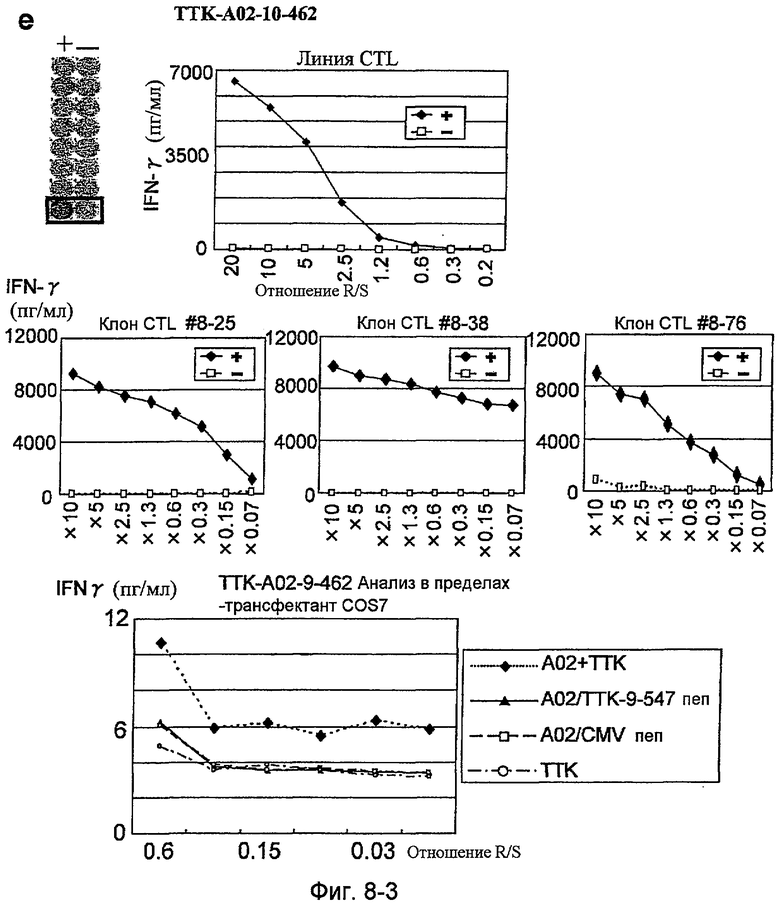
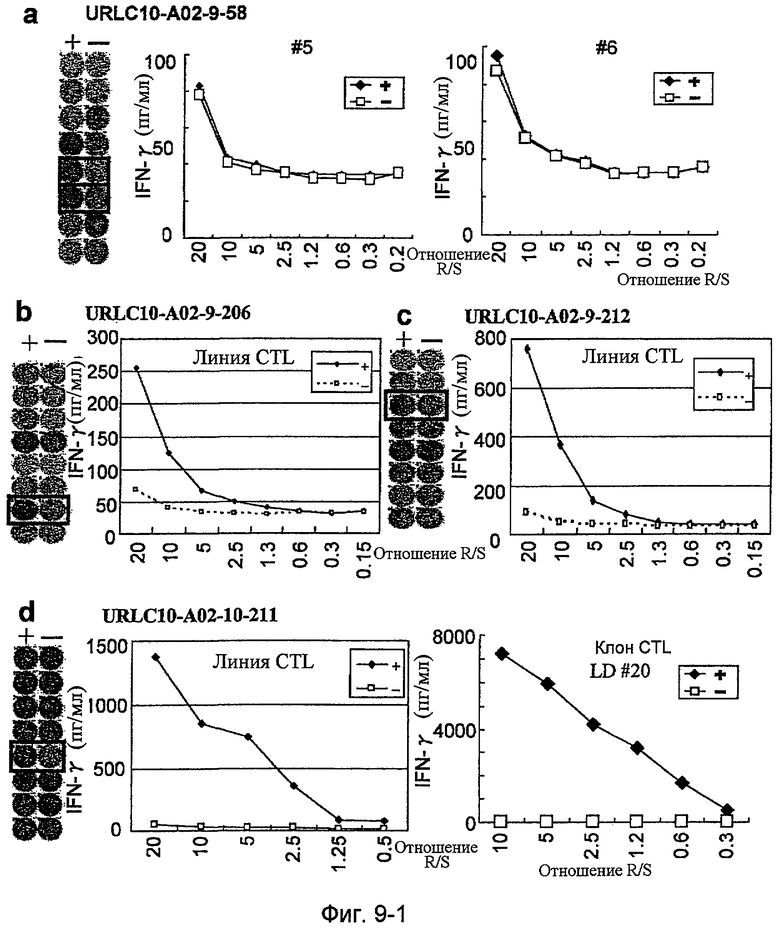
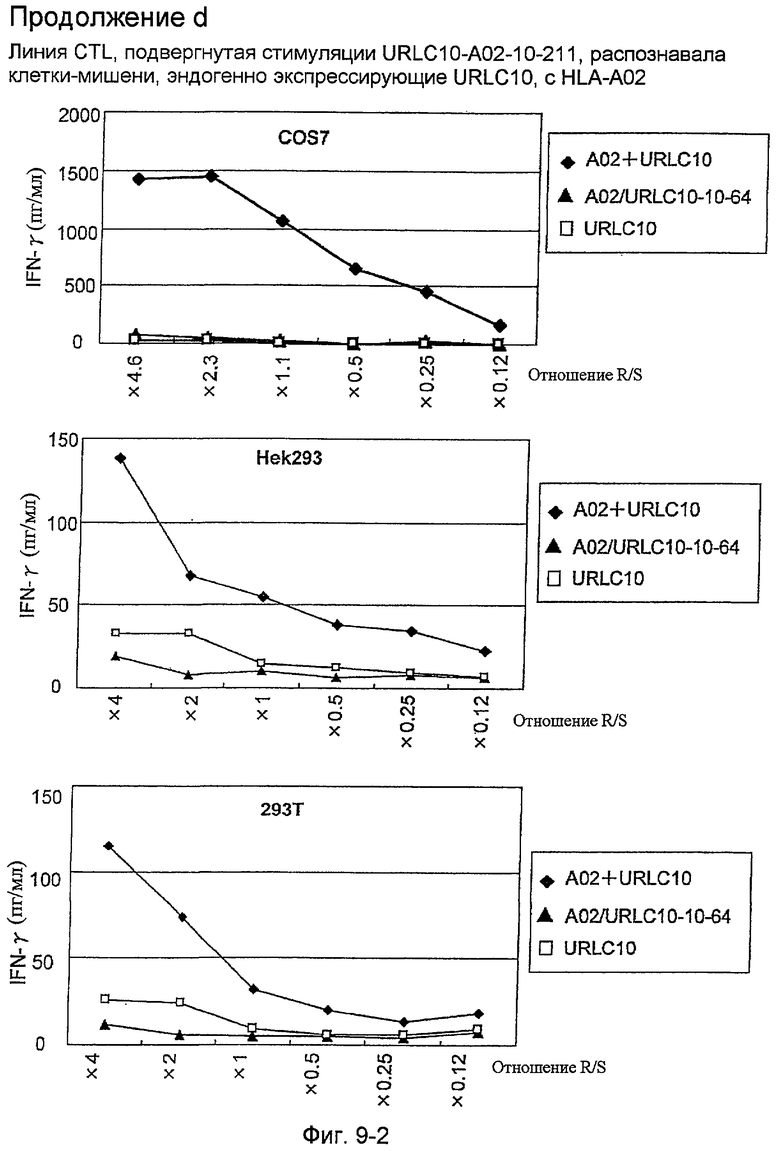
Изобретение относится к биохимии. Описаны выделенные пептиды, обладающие способностью индуцировать цитотоксическую Т-клетку, имеющие аминокислотные последовательности, показанные в SEQ ID NO: SEQ ID NO: SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 80, 100, 101, 110, 111, 387, 112, 394, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217, 223, 376, 379, 114, 116, 117, 121, 227, 228, 233, 254, 271, 272 или 288, а также пептиды, имеющие вышеупомянутые аминокислотные последовательности, в которых заменены, делегированы или в которые добавлены 1 или 2 аминокислоты при условии, что пептиды обладают способностью к индукции цитотоксических Т-клеток, и способ отбора таких пептидов. Настоящим изобретением также обеспечиваются лекарственные средства для лечения или предупреждения заболевания, тесно связанного со сверхэкспрессией CDH3, ЕРНА4, ЕСТ2, HIG2, INHBB, KIF20A, KNTC2, TTK и/или URLC10, например раков, содержащие в качестве активного ингредиента один или несколько этих пептидов. Представлены способы индукции цитотоксической Т-клетки, антигенпрезентирующей клетки, ингибирования пролиферации клетки. Также представлены цитотоксическая Т-клетка и антигенпрезентирующая клетка. 14 н. и 4 з.п. ф-лы, 9 ил., 10 табл.
1. Выделенный пептид, обладающий способностью индуцировать цитотоксическую Т-клетку, где указанный пептид выбран из группы, состоящей из пептидов, состоящих из аминокислотных последовательностей SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 80, 100, 101, 110, 111, 387, 112, 394, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217 или 223.
2. Пептид, обладающий способностью индуцировать цитотоксическую Т-клетку, который состоит из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 19, 22, 30, 34, 358, 41, 44, 46, 48, 78, 80, 100, 101, 110, 111, 387, 112, 394, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217 и 223, в которой заменены 1 или 2 аминокислоты, где замещения 1 или 2 аминокислот представляют собой одно или оба из следующих замещений:
(a) вторая с N-конца аминокислота в аминокислотной последовательности SEQ ID NO: 19, 22, 30, 34, 358, 41, 44, 46, 48, 78, 80, 100, 101, 110, 111, 387, 112, 394, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217 или 223 замещена на фенилаланин, тирозин, метионин или триптофан, и
(b) С-концевая аминокислота аминокислотной последовательности SEQ ID NO: 19, 22, 30, 34, 358, 41, 44, 46, 48, 78, 80, 100, 101, 110, 111, 387, 112, 394, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217 или 223 замещена на фенилаланин, лейцин, изолейцин, триптофан или метионин.
3. Выделенный пептид, обладающий способностью индуцировать цитотоксическую Т-клетку, где указанный пептид выбран из группы, состоящей из пептидов, состоящих из аминокислотных последовательностей SEQ ID NO: 376, 379, 114, 116, 117, 121, 227, 228, 233, 254, 271, 272 или 288.
4. Пептид, обладающий способностью индуцировать цитотоксическую Т-клетку, где указанный пептид состоит из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 376, 379, 114, 116, 117, 121, 227, 228, 233, 254, 271, 272 и 288, в которой заменены 1 или 2 аминокислоты, где замещения 1 или 2 аминокислот представляют собой одно или оба из следующих замещений:
(а) вторая с N-конца аминокислота аминокислотной последовательности SEQ ID NO: 376, 379, 114, 116, 117, 121, 227, 228, 233, 254, 271, 272 или 288 замещена на лейцин или метионин, и (b) С-концевая аминокислота аминокислотной последовательности SEQ ID NO: 376, 379, 114, 116, 117, 121, 227, 228, 233, 254, 271, 272 или 288 замещена на валин или лейцин.
5. Фармацевтическая композиция, ингибирующая пролиферацию клетки, экспрессирующей гены, имеющие последовательности SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15 и/или 17, где указанная композиция содержит эффективное количество по меньшей мере одного пептида по любому из пп.1-4 или полинуклеотида, кодирующего эти пептиды, и носители и/или эксципиенты.
6. Фармацевтическая композиция по п.6, где клетка, экспрессирующая гены, имеющие последовательности SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15 и/или 17, представляет собой раковую клетку.
7. Фармацевтическая композиция по п.6, где рак выбран из группы, состоящей из рака мочевого пузыря, рака молочной железы, рака шейки матки, холангиоцеллюлярного рака, CML, рака ободочной и прямой кишок, эндометриоза, рака пищевода, рака желудка, диффузного рака желудка, рака печени, NSCLC, лимфомы, остеосаркомы, рака яичника, рака поджелудочной железы, рака предстательной железы, рака почки, SCLC, мягкотканевой опухоли и опухоли яичка.
8. Способ индукции антигенпрезентирующей клетки in vitro, обладающей высокой способностью индуцировать цитотоксическую Т-клетку, где способ включает стадию приведения антигенпрезентирующей клетки в контакт с пептидом по любому из пп.1-4.
9. Способ индукции цитотоксической Т-клетки in vitro путем приведения Т-клетки в контакт с пептидом по любому из пп.1-4.
10. Способ индукции антигенпрезентирующей клетки in vitro, обладающей высокой способностью индуцировать цитотоксическую Т-клетку, где указанный способ включает стадию переноса гена, включающего полинуклеотид, кодирующий пептид по любому из пп.1-4, в антигенпрезентирующую клетку.
11. Выделенная цитотоксическая Т-клетка, которая индуцирована in vitro смешиванием антигенпрезентирующих клеток, которые были приведены в контакт с пептидом по любому из пп.1-4, с СВ8+Т-клетками, с последующим сокультивированием.
12. Антигенпрезентирующая клетка, индуцирующая CTL, которая содержит иммобилизованный пептид по любому из пп.1-4, на поверхности указанной клетки.
13. Антигенпрезентирующая клетка, индуцирующая CTL, индуцированная способом по п.9.
14. Способ ингибирования пролиферации клетки, экспрессирующей гены, имеющие последовательности SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15 и/или 17, у субъекта, где способ включает введение указанному субъекту.
15. Способ ингибирования пролиферации клетки, экспрессирующей гены, имеющие последовательности SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15 и/или 17, у субъекта, где способ включает введение указанному субъекту фармацевтической композиции, включающей эффективное количество полинуклеотида, кодирующего пептид по любому из пп.1-4.
16. Способ по п.14 или 15, где клетка, экспрессирующая гены, имеющие последовательности SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15 и/или 17, представляет собой раковую клетку.
17. Способ по п.16, где рак выбран из группы, состоящей из рака мочевого пузыря, рака молочной железы, рака шейки матки, холангиоцеллюлярного рака, CML, рака ободочной и прямой кишок, эндометриоза, рака пищевода, рака желудка, диффузного рака желудка, рака печени, NSCLC, лимфомы, остеосаркомы, рака яичника, рака поджелудочной железы, рака предстательной железы, рака почки, SCLC, мягкотканевой опухоли и опухоли яичка.
18. Способ отбора пептида, в котором 1, 2 или несколько аминокислот заменены, причем первоначальный пептид включает аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 19, 22, 30, 34, 344, 358, 41, 44, 46, 48, 78, 80, 100, 101, 110, 111, 387, 112, 394, 395, 133, 135, 137, 426, 174, 178, 186, 194, 196, 202, 210, 213, 214, 217 и 223, при этом указанный способ включает стадии;
(a) подтверждения отсутствия значительной гомологии последовательности пептида-кандидата с 1, 2 или несколькими аминокислотными заменами, с полной последовательностью указанного первоначального пептида;
(b) определения способности к индукции CTL пептида-кандидата с заменами; и
(c) отбора пептида, у которого способность к индукции CTL равна или выше способности к индукции первоначального пептида.
| Конусное соединение съемных буровых коронок со штангами | 1949 |
|
SU83239A1 |
| ДНК, КОДИРУЮЩАЯ НОВЫЙ ПЕПТИД RG1 | 2000 |
|
RU2283130C2 |

