Область изобретения
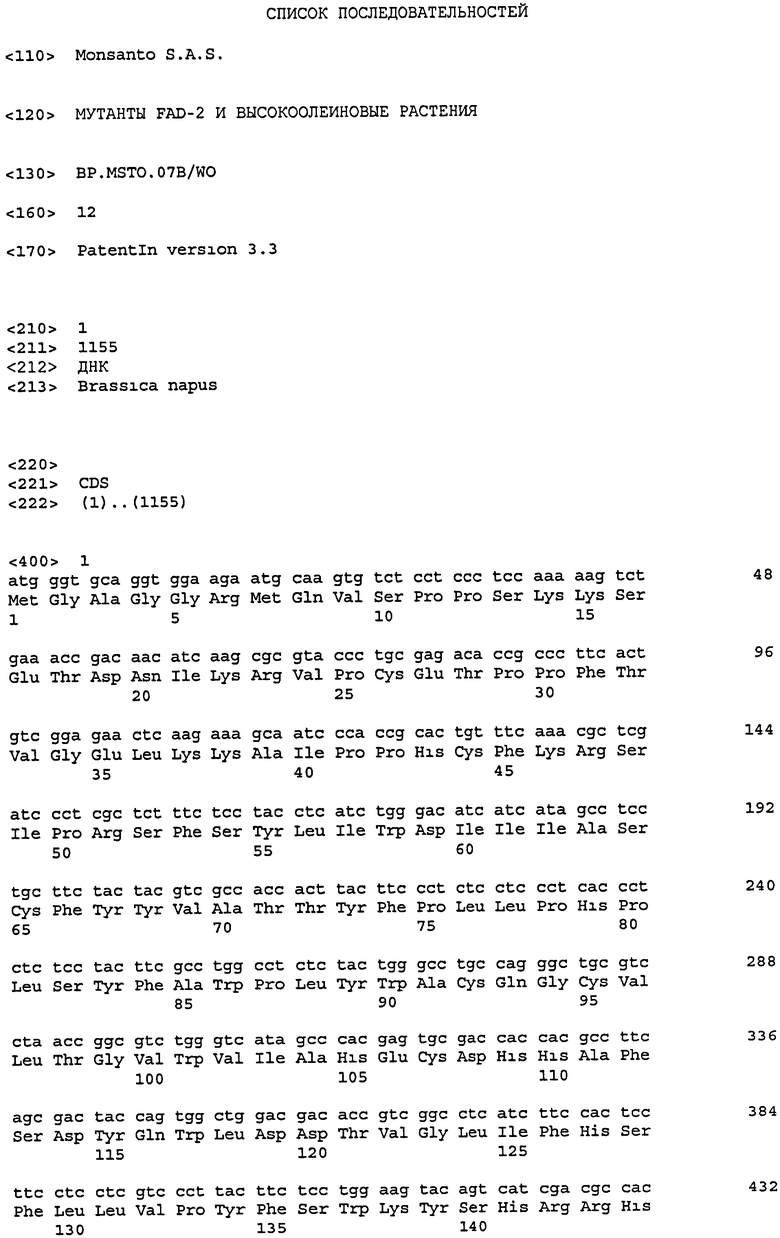
Настоящее изобретение относится к растениям, семенам и продуктам, полученным из них, в особенности к растениям Brassica, семенным продуктам, полученным из них, имеющим мутантные последовательности, создающие высокий профиль олеиновых кислот в масле семян.
Более конкретно, изобретение относится к мутантным последовательностям дельта-12-десатуразы жирных кислот, также упоминаемым здесь как последовательности FAD2, в растениях, которые наделяют масло семян высоким профилем олеиновых кислот.
Уровень техники изобретения
Дельта-12 десатураза жирных кислот (также известная как олеиновая десатураза или олеатдесатураза) вовлечена в энзиматическое превращение олеиновой кислоты в линолевую кислоту.
Разновидности с высоким уровнем олеиновой кислоты (возможно в комбинации с низкими уровнями линоленовой кислоты) требуются для множества различных применений (пищевого использования, медицинского использования, применения в биодизелях и многих других).
Мутантные семена, дающие масло, демонстрирующее высокое содержание олеиновой кислоты (олеиновая кислота выше чем 70 весовых % по отношению к общему весу жирных кислот, присутствующих в масле), ранее описанные в литературе, имели очень малое агрономическое значение и/или плохие характеристики корней, и/или очень низкую урожайность.
Все еще существует потребность в материале, имеющем стабильное, высокое содержание олеиновой кислоты (возможно в сочетании со стабильно низким содержанием линоленовой кислоты) в разных местоположениях на протяжении лет, также с хорошей агрономической производительностью и нормальной морфологией масличного рапса. В особенности растения не должны иметь фасциации и должны иметь нормальное развитие корня.
Краткое описание изобретения
Настоящее изобретение относится к молекуле нуклеиновой кислоты, включающей (или состоящей из) последовательность нуклеиновой кислоты, кодирующей белок дельта-12-олеатдесатуразу (FAD2), причем указанный белок FAD2 имеет замену аминокислоты в положении 108 относительно белка FAD2 дикого типа.
Другим объектом является молекула нуклеиновой кислоты, включающая (или состоящая из) последовательность нуклеиновой кислоты, кодирующая белок FAD2, причем указанный белок FAD2 имеет замену аминокислоты в положении 118 относительно белка FAD2 дикого типа.
Еще одним объектом является молекула нуклеиновой кислоты, включающая (или состоящая из) последовательность нуклеиновой кислоты, кодирующая белок FAD2, причем указанный белок FAD2 имеет замены аминокислот в положении 108 и в положении 118 относительно белка FAD2 дикого типа.
Предпочтительно указанный белок FAD2 представляет собой белок FAD2 Brassica, более конкретно - белок FAD2 Brassica napus.
Предпочтительно указанная замещенная аминокислота в положении 108 представляет собой аспарагиновую кислоту (замещающую глицин в белке FAD2 дикого типа).
Предпочтительно указанная замещенная аминокислота в положении 118 представляет собой фенилаланиновую кислоту (замещающую лейцин в белке FAD2 дикого типа).
Предпочтительная молекула нуклеиновой кислоты по изобретению включает (или состоит из) нуклеиновую кислоту с последовательностью SEQ ID NO: 1, 5, 11 или 12, комплементарной им формы или их РНК-формы.
Молекула нуклеиновой кислоты по изобретению может включать или состоять из нуклеотидной последовательности, имеющей по крайней мере 80%, предпочтительно по крайней мере 85%, более предпочтительно по крайней мере 90% и еще более предпочтительно по крайней мере 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 1 или 11, или с их комплементарной формой или их РНК-формой, кодирующей белок FAD2, имеющий аминокислотную замену в положении 108 относительно белка FAD2 дикого типа.
Молекула нуклеиновой кислоты по изобретению может включать или состоять из нуклеотидной последовательности, имеющей по крайней мере 80%, предпочтительно по крайней мере 85%, более предпочтительно по крайней мере 90% и еще более предпочтительно по крайней мере 95%, 96%, 97%, 98% или 99% идентичности с SEQ ID NO: 5 или 12, или с их комплементарной формой или их РНК-формой, кодирующей белок FAD2, имеющий аминокислотную замену в положении 118 относительно белка FAD2 дикого типа.
Более конкретно, указанный белок FAD2 дикого типа включает (или состоит из) аминокислотной последовательности с SEQ ID NO: 4 или 8.
Также объект настоящего изобретения представляет собой фрагмент по крайней мере из 10, 15, 20, 25, 30, 40, 50, 100 или более нуклеотидов молекулы нуклеиновой кислоты в соответствии с изобретением, указанный фрагмент включает мутантный кодон, соответствующий указанной аминокислотной замене в положении 108 и/или мутантный кодон, соответствующий указанной аминокислотной замене в положении 118.
Указанные фрагменты могут быть использованы в качестве праймеров, зондов и/или селективных маркеров.
Любая из молекул нуклеиновых кислот по изобретению может быть использована в способе селекции растений с помощью маркера, предпочтительно видов Brassica, более предпочтительно - разновидностей Brassica napus, также являются объектом настоящего изобретения.
Другой объект настоящего изобретения представляет собой набор для анализа, который может содержать первый контейнер, содержащий любую из молекул нуклеиновых кислот по изобретению.
Другой объект настоящего изобретения представляет собой белок FAD2, имеющий аминокислотную замену в положении 108, или соответствующем положении 108, относительно белка FAD2 дикого типа, представленному аминокислотной последовательностью SEQ ID NO: 4 или 8.
Другой объект настоящего изобретения представляет собой белок FAD2, имеющий аминокислотную замену в положении 118, или соответствующем положении 118, относительно белка FAD2 дикого типа, представленному аминокислотной последовательностью SEQ ID NO: 4 или 8.
Другой объект настоящего изобретения представляет собой белок FAD2, имеющий аминокислотную замену в положении 108, или соответствующем положении 108, и аминокислотную замену в положении 118, или соответствующем положении 118, относительно белка FAD2 дикого типа, представленному аминокислотной последовательностью SEQ ID NO: 4 или 8.
Предпочтительный белок FAD2 по изобретению включает (или состоит из) аминокислотную последовательность SEQ ID NO: 2 или 6.
Другой объект настоящего изобретения представляет собой вектор, включающий молекулу нуклеиновой кислоты, кодирующую мутантный белок FAD2, в соответствии с изобретением.
Другой объект настоящего изобретения представляет собой клетку-хозяин, включающую вектор по изобретению и/или последовательность нуклеиновой кислоты, кодирующую мутантный белок FAD2 в соответствии с изобретением.
Другой объект настоящего изобретения представляет собой растение, стабильно трансформированное вектором по изобретению.
Растение для трансформации может быть выбрано из группы, состоящей из масличных культур, более конкретно из подсолнечников, соевых бобов, хлопков, кукурузы и/или рапсовых семян.
Другой объект настоящего изобретения представляет собой растение или часть растения, или семя, содержащие нуклеотидную последовательность, кодирующую белок FAD-2, имеющий аминокислотную замену в положении 108 относительно белка FAD-2 дикого типа или положении, соответствующем ему.
Более конкретно, растение, или часть растения, или семя в соответствии с изобретением содержат (или экспрессируют) белок FAD2, имеющий аминокислотную замену, соответствующую положению 108 относительно белка FAD-2 дикого типа или положению, соответствующему ему.
Другой объект настоящего изобретения представляет собой растение или часть растения, или семя, содержащие нуклеотидную последовательность, кодирующую белок FAD-2, имеющий аминокислотную замену в положении 118 относительно белка FAD-2 дикого типа или положении, соответствующем ему.
Более конкретно, растение, или часть растения, или семя в соответствии с изобретением содержат (или экспрессируют) белок FAD2, имеющий аминокислотную замену, соответствующую положению 118 относительно белка FAD-2 дикого типа или положению, соответствующему ему.
Другой объект настоящего изобретения представляет собой растение или часть растения, или семя, содержащие нуклеотидную последовательность, кодирующую белок FAD-2, имеющий аминокислотную замену в положениях 108 и 118 относительно белка FAD-2 дикого типа или положениях, соответствующих им.
Более конкретно, растение или часть растения, или семя в соответствии с изобретением содержат (или экспрессируют) белок FAD2, имеющий аминокислотную замену, соответствующую положениям 108 и 118 относительно белка FAD-2 дикого типа или положениям, соответствующим им.
Другой объект настоящего изобретения представляет собой растение или часть растения, или семя, содержащие первую нуклеотидную последовательность, кодирующую белок FAD2, имеющий аминокислотную замену в положении 108 или положении, соответствующем ему, и вторую нуклеотидную последовательность, кодирующую белок FAD2, имеющий аминокислотную замену в положении 118 или положении, соответствующем ему, относительно белка FAD2 дикого типа.
Более конкретно, растение или часть растения, или семя в соответствии с изобретением содержат (или экспрессируют) два белка FAD2, один из которых имеет аминокислотную замену, соответствующую положению 108 или положению, соответствующему ему, а другой имеет аминокислотную замену, соответствующую положению 118 относительно белка FAD-2 дикого типа или положению, соответствующему ему.
Предпочтительно указанная замещенная аминокислота в положении 108 или положении, соответствующем ему, представляет собой аспарагиновую кислоту (замещающую глицин в белке FAD2 дикого типа).
Предпочтительно указанная замещенная аминокислота в положении 118 или в положении, соответствующем ему, представляет собой фенилаланин (замещающий лейцин в белке FAD2 дикого типа).
Растение, или часть растения, или семя согласно изобретению могут быть получены мутагенной обработкой, более конкретно EMS обработкой.
Потомство, происходящее от указанного растения, или части растения, или семени, также является частью изобретения.
Другой объект настоящего изобретения представляет собой растительное масло, полученное из семян по изобретению, причем указанное масло содержит более чем (приблизительно) 72%, 75%, 80% или 85% олеиновой кислоты по отношению к общему весу жирных кислот, присутствующих в рапсовом масле.
Предпочтительно, указанное масло также включает менее чем (приблизительно) 4%, 3,5% 3%, 2%, 1% или 0,5% линоленовой кислоты.
Изобретение также относится к пищевым или кормовым продуктам, содержащим и/или приготовленным из растения, части растения, семени и/или растительного масла в соответствии с изобретением.
Способ улучшения содержания олеиновой кислоты в растении может включать этап трансформации растения вектором по изобретению.
Альтернативно, способ получения линий растения с высоким содержанием олеиновой кислоты может включать:
(а) скрещивание первого растения по изобретению со вторым растением,
(b) получение семян от скрещивания на этапе (а),
(c) выращивание фертильных растений из таких семян,
(d) получение семян потомства от растений этапа (с),
(e) идентификацию среди потомства тех семян, которые имеют высокое содержание олеиновой кислоты.
Альтернативно, способ получения линий растения с высоким содержанием олеиновой кислоты может включать:
(a) индукцию мутагенеза в, по крайней мере, некоторых клетках из растения, более конкретно растения Brassica, и предпочтительно разновидности Brassica napus, которое имеет содержание олеиновой кислоты в клетках менее чем 70%;
(b) регенерацию растений из, по крайней мере, одной из указанных мутагенезированных клеток;
(c) выбор регенерированных растений, которые имеют любую из последовательностей нуклеиновой кислоты по изобретению и/или которые экспрессируют белок FAD2 в соответствии с изобретением; и
(d) получение дальнейших поколений растений из указанных регенерированных растений.
Краткое описание фигуры
Фиг.1 соответствует списку последовательностей по представленному изобретению.
Подробное описание изобретения
Настоящее изобретение относится к растениям, более конкретно растениям Brassica, и предпочтительно разновидности Brassica napus, которые были получены для получения масла, имеющего содержание олеиновой кислоты выше чем 70%, относительно общего веса жирных кислот, присутствующих в масле.
Более конкретно, растение по изобретению имело по крайней мере один мутантный ген FAD2 по изобретению.
Предпочтительно, указанный мутантный ген FAD2 приводит к высокому содержанию олеиновой кислоты (то есть содержание олеиновой кислоты выше чем 70% вес., относительно общего веса жирных кислот, присутствующих в масле) в семенах указанных растений и в масле, полученном из указанных семян.
Настоящее изобретение относится также к любой части или любому продукту указанного растения, несущему не менее одного указанного мутантного гена FAD2.
В контексте представленного изобретения, часть или продукт растения подразумевается включающим лист, семядолю, стебель, черешок, стебелек, семя или любую другую ткань или фрагмент ткани указанного растения.
Настоящее изобретение относится также к любому потомству указанного растения, несущему по крайней мере один мутантный ген FAD2 по изобретению.
В контексте представленного изобретения термин «потомство» относится к прямым и непрямым наследникам, потомкам и производным от растения или растений по изобретению и включает первое, второе, третье и/или последующие поколения, которые могут быть получены самоопылением, скрещиванием с растениями с одинаковым или другими генотипами, и могут быть модифицированы различными подходящими методами генной инженерии.
Настоящее изобретение также относится к указанным мутированным генам FAD2, которые обеспечивают высокое содержание олеиновой кислоты в семенах при присутствии в растениях.
Конкретно, изобретение относится к новым изолированным молекулам нуклеиновой кислоты, которые кодируют новые варианты форм белка FAD2, имеющего замещенную аминокислоту в положении 108 (или соответствующем положении 108) и/или замещенную аминокислоту в положении 118 (или соответствующем положении 118) относительно белка FAD2 дикого типа, такого как белок FAD2 дикого типа, представленный SEQ ID NO: 4 и/или SEQ ID NO:8.
Выделенная молекула нуклеиновой кислоты по изобретению содержит по крайней мере одну мутацию, приводящую к замещению, предпочтительно к замещению аспарагиновой кислоты глицином в положении 108 (или соответствующем ему) и/или приводящему к замене, предпочтительно замене фенилаланина лейцином в положении 118 (или соответствующей ему) относительно белка FAD2 дикого типа, такого как белок FAD2 дикого типа, представленный SEQ ID NO: 4 и/или SEQ ID NO: 8.
Указанная мутация(и) изменяет(ют) функциональность получаемого продукта гена FAD2, причем уровень олеиновой кислоты изменяется, предпочтительно увеличивается, у растения, экспрессирующего мутантную последовательность(ти), по сравнению с соответствующим уровнем у растений, экспрессирующих последовательность(ти) дикого типа.
В рамках настоящего изобретения, если не указано иначе, термин «в положении 108» должен пониматься как обозначающий аминокислоту в положении 108 белка FAD2 дикого типа, представленного SEQ ID NO: 4 и/или SEQ ID NO: 8, но также относящийся к аминокислоте, соответствующей указанному положению в белке FAD2 дикого типа, который может иметь иную аминокислотную последовательность в связи с делециями или дополнительными аминокислотами в полипептиде.
Сходным образом, термин «в положении 118» должен пониматься как обозначающий аминокислоту в положении 118 белка FAD2 дикого типа, представленного SEQ ID NO: 4 и/или SEQ ID NO: 8, но также относящийся к аминокислоте, соответствующей указанному положению в белке FAD2 дикого типа, который может иметь иную аминокислотную последовательность в связи с делециями или дополнительными аминокислотами в полипептиде.
Термин «соответствующий положению» как он использован здесь, означает, что положение не только определяется числом предшествующих аминокислот. Положение выбранной аминокислоты в соответствии с представленным изобретением может изменяться из-за делеций или дополнительных аминокислот в полипептиде. Таким образом, под «соответствующим положением» в соответствии с представленным изобретением надо понимать, что описываемая аминокислота(ы), может отличаться от указанного номера, но все еще иметь одинаковые соседние аминокислоты в линейной последовательности.
В одном аспекте молекула нуклеиновой кислоты по изобретению кодирует белок FAD2, в котором замена указанной аминокислоты, соответствующей положению 108, представляет собой глутамат, и предпочтительно, аспарагиновую кислоту.
Более конкретно, молекула нуклеиновой кислоты по изобретению кодирует белок FAD2, имеющий замещенную глицином аспарагиновую кислоту в положении 108 относительно белка FAD2 дикого типа, представленного аминокислотной последовательностью SEQ ID NO: 4 или 8.
Молекула нуклеиновой кислоты по изобретению может включать (или состоять из) последовательности нуклеиновой кислоты по SEQ ID NO: 3, 5, 7, 9, 10 или 12, причем кодон, кодирующий аминокислоту в положении 108 имел не менее одной мутации (или был мутирован) так, чтобы он кодировал аминокислоту, отличную от глицина, и предпочтительно, чтобы он кодировал аспарагиновую кислоту в положении 108 в соответствии с белком FAD2 по изобретению.
Предпочтительная молекула нуклеиновой кислоты по изобретению включает (или состоит из) последовательности нуклеиновой кислоты по SEQ ID NO: 1 или 11.
В другом аспекте молекула нуклеиновой кислоты по изобретению кодирует белок FAD2, в котором замена указанной аминокислоты, соответствующей положению 118, представляет собой фенилаланин.
Более конкретно, молекула нуклеиновой кислоты по изобретению кодирует белок FAD2, имеющий замещенный фенилаланином лейцин в положении 118 относительно белка FAD2 дикого типа, представленного аминокислотной последовательностью SEQ ID NO: 4 или 8.
Молекула нуклеиновой кислоты по изобретению может включать (или состоять из) последовательность нуклеиновой кислоты по SEQ ID NO: 1, 3, 7, 9, 10 или 11, причем кодон, кодирующий аминокислоту в положении 118, имел не менее одной мутации (или был мутирован) так, чтобы он кодировал аминокислоту, отличную от лейцина, и предпочтительно, чтобы он кодировал фенилаланин в положении 118 в соответствии с белком FAD2 по изобретению.
Предпочтительная молекула нуклеиновой кислоты по изобретению включает (или состоит из) последовательность нуклеиновой кислоты SEQ ID NO: 5 или 12.
В другом аспекте молекула нуклеиновой кислоты по изобретению может кодировать белок FAD2, имеющий делецию в положении 118 относительно белка FAD2 дикого типа, представленного аминокислотной последовательностью SEQ ID NO: 4 или 8.
Более конкретно, молекула нуклеиновой кислоты по изобретению кодирует белок FAD2, имеющий делецию лейцина в положении 118 относительно белка FAD2 дикого типа, представленного аминокислотной последовательностью SEQ ID NO: 4 или 8.
Молекула нуклеиновой кислоты по изобретению может включать (или состоять из) последовательность нуклеиновой кислоты по SEQ ID NO: 1, 3, 5, 7, 9, 10 или 11, причем кодон, кодирующий аминокислоту в положении 118, был удален.
Специалистом будет признано, что последовательности нуклеиновой кислоты SEQ ID NO: от 1 до 12 (т.е. SEQ ID NO: 1, 3, 5, 7, 9, 10, 11 и 12) не единственные последовательности, которые могут быть использованы для получения белка FAD2 по изобретению. Также предполагается, что существуют другие молекулы нуклеиновой кислоты, имеющие другие последовательности, но которые, в силу вырожденности генетического кода, кодируют белок FAD2, включающий замену аминокислоты в положении 108 (или соответствующей положению 108) и/или замену аминокислоты в положении 118 (или соответствующей положению 118) по отношению к аминокислотной последовательности дикого типа, такой как последовательность белка FAD2 дикого типа, представленная SEQ ID NO: 4 или 8.
В частности, молекула нуклеиновой кислоты по изобретению может включать (или состоять из) нуклеотидной последовательности, имеющей по крайней мере 80%, предпочтительно по крайней мере 85%, более предпочтительно, по крайней мере 90%, и еще более предпочтительно, по крайней мере 95%, 96%, 97%, 98% или 99% идентичности с любым из SEQ ID NO: от 1 до 12 (т.е. SEQ ID NO: 1, 3, 5, 7, 9, 10, 11 и 12), или с их комплементарной формой или их РНК-формой, кодирующей белок FAD2, имеющий замену аминокислоты в положении 108 и/или 118 относительно белка FAD2 дикого типа, такого как белок FAD2 дикого типа, представленный SEQ ID NO: 4 или 8.
Более конкретно, молекула нуклеиновой кислоты по изобретению демонстрирует нуклеотидную последовательность, имеющую по крайней мере 80%, предпочтительно по крайней мере 85%, более предпочтительно, по крайней мере 90%, и еще более предпочтительно, по крайней мере 95%, 96%, 97%, 98% или 99% идентичности с любым из SEQ ID NO: 1, 3, 5, 7, 9, 10, 11 и 12, или с их комплементарной формой или их РНК формой, и кодирующие белок FAD2, имеющий замену глицина аспарагиновой кислотой в положении 108 (или соответствующем положении 108) и/или замену фенилаланина лейцином в положении 118 (или соответствующем положению 118) относительно белка FAD2 дикого типа, представленного аминокислотной последовательностью SEQ ID NO: 4 или 8.
Молекула нуклеиновой кислоты по изобретению может быть получена из разновидностей Brassica napus, таких как MSP05, MSP06, MSP07, MSP11 и/или 28DHS.059.
Более конкретно, молекула нуклеиновой кислоты по изобретению, имеющая мутацию в положении 1540 (также упоминаемую как SNP1540) с последовательностью нуклеиновой кислоты SEQ ID NO: 11, которая вызывает изменение генетического кодона GGC на GAC, приводя к замене аминокислоты в положении 108 (или соответствующем положении 108) относительно белка FAD2 дикого типа, представленного аминокислотной последовательностью SEQ ID NO: 4 или 8.
Выделенная молекула нуклеиновой кислоты по изобретению, содержащая указанную мутацию SNP1540, приводящую к замене аспарагиновой кислоты глицином в положении 108, изменяет функциональность получающегося продукта гена FAD2, причем уровень олеиновой кислоты увеличивается в растении, экспрессирующем мутантную последовательность, по сравнению с соответствующим уровнем в растении, экспрессирующем последовательность дикого типа.
В рамках изобретения термин «SNP1540» относится к полиморфизму единичного нуклеотида, соответствующему указанной мутации в положении 1540 нуклеиновой кислоты SEQ ID NO: 11, и может указывать также на соответствующую мутацию в любой молекуле нуклеиновой кислоты, кодирующей белок FAD2 по изобретению, имеющий замещенную аминокислоту в положении 108 (или соответствующей положению 108) относительно белка FAD2 дикого типа, представленного SEQ ID NO: 4 или 8.
Предполагается любой фрагмент молекулы нуклеиновой кислоты по изобретению из по крайней мере 10, 15, 20, 25, 50, 100 или более нуклеотидов, включающий указанную SNP1540.
В другом аспекте новая молекула нуклеиновой кислоты по изобретению получена из разновидностей Brassica napus, таких как MSP05, MSP11 и/или 28DHS.059, имеющих мутацию (SNP1590), приводящую к замене аминокислоты в положении 118 последовательности FAD2 дикого типа, такой как представлена SEQ ID NO: 8.
Более конкретно, молекула нуклеиновой кислоты по изобретению, имеющая мутацию в положении 1590 (также упоминаемая как SNP1590) с последовательностью нуклеиновой кислоты SEQ ID NO: 12, которая вызывает изменение генетического кодона CTT на TTT, приводя к замене аминокислоты в положении 118 (или соответствующей положению 118) относительно аминокислотной последовательности дикого типа, такой как последовательность белка FAD2 дикого типа, представленного аминокислотной последовательностью SEQ ID NO: 4 или 8.
Выделенная молекула нуклеиновой кислоты по изобретению, содержащая указанную мутацию SNP1590, приводящую к замене фенилаланина лейцином в положении 118, изменяет функциональность получающегося продукта гена FAD2, причем уровень олеиновой кислоты увеличивается в растении, экспрессирующем мутантную последовательность по сравнению с соответствующим уровнем в растении, экспрессирующем последовательность дикого типа.
В рамках изобретения термин «SNP1590» относится к полиморфизму единичного нуклеотида, соответствующему указанной мутации в положении 1590 нуклеиновой кислоты SEQ ID NO: 12, и может указывать также на соответствующую мутацию в любой молекуле нуклеиновой кислоты, кодирующей белок FAD2 по изобретению, имеющий замещенную аминокислоту в положении 118 (или соответствующей положению 118) относительно белка FAD2 дикого типа, представленного SEQ ID NO: 4 или 8.
Предполагается любой фрагмент молекулы нуклеиновой кислоты по изобретению из по крайней мере 10, 15, 20, 25, 50, 100 или более нуклеотидов, включающий указанную SNP1590.
Также предполагается любой фрагмент молекулы нуклеиновой кислоты по изобретению из по крайней мере 10, 15, 20, 25, 50, 100 или более нуклеотидов, включающий указанную SNP1540 и указанную SNP1590.
Предполагается любой фрагмент молекулы нуклеиновой кислоты по изобретению из по крайней мере 10, 15, 20, 25, 50, 100 или более нуклеотидов, включающий по крайней мере одну мутацию, приводящую к белку FAD2 по изобретению.
Другими словами, также предполагается любой фрагмент молекулы нуклеиновой кислоты по изобретению из по крайней мере 10, 15, 20, 25, 30, 40, 50, 100, 500 или более нуклеотидов, включающий по крайней мере одну мутацию в кодоне, кодирующем указанную аминокислоту в положении 108 (или соответствующей положению 108) и/или в кодоне, кодирующем указанную аминокислоту в положении 118 (или соответствующей положению 118) относительно белка FAD2 дикого типа, представленного SEQ ID NO: 4 или 8.
Подобные фрагменты могут быть использованы как праймеры, как зонды и/или как маркеры.
Фрагменты нуклеиновой кислоты по изобретению могут быть использованы в качестве маркеров при генетическом картировании растения и в программах выведения растения.
Подобные маркеры могут включать, например, исследования полиморфизма длины рестрикционных фрагментов (RFLP), детектирование амплификацией со случайными праймерами (RAPD), полимеразную цепную реакцию (ПЦР) или самоподдерживающуюся репликацию последовательности (3SR).
Методы размножения с применением маркера могут быть использованы для идентификации и отслеживания растения в соответствии с изобретением или его потомства, также объекта изобретения, в процессе размножения.
Методы размножения с применением маркера могут быть использованы в дополнение к или как альтернатива другим видам способов идентификации.
Примером размножения с применением маркера является применение праймеров ПЦР, которые специфически амплифицируют молекулу нуклеиновой кислоты по изобретению.
Изобретение предоставляет методы сегрегации и отбора анализов генетических скрещиваний определенных растений, имеющих последовательности нуклеиновой кислоты согласно изобретению.
Способ по изобретению может, например, вовлекать определение присутствия в геноме конкретных аллелей FAD2, содержащих не менее одной мутации, приводящей к замещению (предпочтительно замещению аспарагиновой кислоты глицином) в положении 108 (или соответствующем) и/или приводящей к замещению (предпочтительно замещению фенилаланина лейцином) в положении 118 (или соответствующем) относительно белка FAD2 дикого типа, представленного SEQ ID NO: 4 или 8.
Такое определение может быть, например, достигнуто набором способов, таких как амплификация ПЦР, методом «отпечатков пальцев» ДНК, методом «отпечатков пальцев» РНК, гель-блоттингом и RFLP анализами, анализом защиты от нуклеаз, секвенированием соответствующего фрагмента нуклеиновой кислоты, получением антител (моноклональных и поликлональных), или альтернативными методами, адаптированными для различения белков, продуцируемых сходными аллелями из других вариантов форм этого белка или от дикого типа.
Более конкретно, такие фрагменты могут быть использованы в способе селекции с помощью маркера черт высокой масличности в растениях, предпочтительно видах Brassica, более предпочтительно в разновидностях Brassica napus.
Другой аспект представленного изобретения связан с рекомбинантной нуклеотидной последовательностью, включающей одну или более прилегающих регуляторных нуклеотидных последовательностей, оперативно связанных с нуклеотидной последовательностью в соответствии с изобретением. Указанная прилегающая регуляторная последовательность(ти) предпочтительно происходит из гомологичного организма.
Однако указанные прилегающие регуляторные последовательности могут также происходить из гетерологичных организмов.
Указанные прилегающие регуляторные последовательности представляют собой специфические последовательности, такие как промоторы, энхансеры, секреторные сигнальные последовательности и/или терминаторы.
Другой аспект изобретения относится к вектору, содержащему молекулу нуклеиновой кислоты по изобретению, возможно оперативно связанную с одной или более прилегающей регуляторной последовательностью(ями), происходящими из гомологичных или гетерологичных организмов.
В настоящем контексте слово «вектор» определено как любая биохимическая конструкция, которая может быть использована для введения нуклеотидной последовательности (путем трансдукции, трансфекции, трансформации, инфекции, конъюгации и т.д.) в клетку.
Преимущественно, вектор в соответствии с изобретением выбирается из группы, состоящей из плазмид (включая репликативные и интегративные плазмиды), вирусов, фагмид, хромосом, транспозонов, липосом, катионных везикул, или их смеси. Указанный вектор может уже содержать одну или более прилегающих регуляторных последовательностей, позволяя экспрессию указанной молекулы нуклеиновой кислоты и ее транскрипцию в полипептид по изобретению.
Изобретение также относится к полипептиду FAD2, имеющему аминокислотную замену в положении 108 (или соответствующем ему) относительно белка FAD2 дикого типа, представленного SEQ ID NO: 4 или 8.
Более конкретно, полипептид FAD2 по изобретению включает (или состоит из) аминокислотной последовательности SEQ ID NO: 6, кроме того включающей замену аспарагиновой кислоты глицином в положении 108.
Предпочтительный полипептид FAD2 по изобретению включает (или состоит из) аминокислотной последовательности SEQ ID NO: 2.
Изобретение также относится к полипептиду FAD2, имеющему аминокислотную замену в положении 118 (или соответствующем ему) относительно белка FAD2 дикого типа, представленного SEQ ID NO: 4 или 8.
Более конкретно, полипептид FAD2 по изобретению включает (или состоит из) аминокислотной последовательности SEQ ID NO: 2, кроме того включающей замену фенилаланина лейцином в положении 118.
Предпочтительный полипептид FAD2 по изобретению включает (или состоит из) аминокислотную последовательность SEQ ID NO: 6.
Настоящее изобретение также включает любые фрагменты белка FAD2 по изобретению, имеющие дельта-12-олеатдесатуразную активность и включающие указанную(ые) замену(ы) в положении 108 и/или 118.
Молекулы нуклеиновой кислоты, рекомбинантные молекулы нуклеиновой кислоты и/или векторы по представленному изобретению пригодны для трансформирования растений-мишеней, и таким образом предоставления продукта измененного гена FAD2, причем уровень олеиновой кислоты изменяется, предпочтительно - увеличивается, в растении, экспрессирующем мутантный FDA2 согласно изобретению, по сравнению с соответствующим уровнем в растении, экспрессирующем последовательность дикого типа.
Настоящее изобретение также относится к трансформированной клетке-хозяину, или рекомбинантной клетке-хозяину, содержащей (или имеющей встроенную) одну или более нуклеотидных последовательностей и/или векторов в соответствии с изобретением.
В представленном контексте, «трансформированная клетка-хозяин» или «рекомбинантная клетка», также упоминаемая как «трансформант», представляет собой клетку, имеющую включенными одну или более нуклеотидных последовательностей и/или векторов в соответствии с изобретением. Трансформированная клетка-хозяин может быть клеткой с указанным(и) вектором(ами) и/или указанной(ыми) нуклеотидной(ыми) последовательностью(ями), включенной(ыми) средствами генетической трансформации, предпочтительно средствами гомологической рекомбинации, или другими хорошо известными способами, используемыми для получения рекомбинантного организма.
Любой способ, с помощью которого новая последовательность может быть включена в геном хозяина, предполагается представленным изобретением.
Более предпочтительно, любой способ, с помощью которого новая последовательность может быть включена в геном хозяина, и стабильно наследует его потомство, предполагается представленным изобретением.
В настоящее время существует широкий спектр известных способов достижения прямой или непрямой трансформации высших растений экзогенной ДНК.
Трансформация растительных клеток может быть опосредована использованием векторов. Обычный способ достижения трансформации представляет собой использование Agrobacterium tumefaciens для включения чужеродного гена в целевую клетку растения.
Вирусы растения также предлагают возможные средства для передачи экзогенной ДНК.
Также может быть применено прямое введение в растительные клетки. Обычно, протопласты целевого растения помещаются в культуру в присутствии переносимых молекул нуклеиновой кислоты и фактора, который увеличивает поглощение указанных молекул нуклеиновой кислоты протопластом. Полезными факторами в этом отношении являются полиэтиленгликоль и фосфат кальция.
Альтернативно, поглощение молекул нуклеиновой кислоты может быть стимулировано электропорацией. В этом способе, электрический импульс используется для открытия временных пор в клеточной мембране протопласта, и указанные молекулы нуклеиновой кислоты из окружающего раствора проникают в клетку через поры. Сходным образом, для доставки молекул указанной нуклеиновой кислоты непосредственно в клетку и предпочтительно непосредственно в ядро клетки может применяться микроинъекция.
В этих способах трансформация происходит в растительной клетке в культуре. После события трансформации растительные клетки могут быть регенерированы в целые растения.
Способы для регенерации взрослых растений из каллуса или протопластов хорошо известны.
Также доступны альтернативные способы, которые не требуют применения изолированных клеток и, таким образом, способов регенерации растений для достижения трансформации. Они обычно упоминаются как «баллистические» или способы с «ускорением частиц», в которых молекулы нуклеиновой кислоты, покрывающие металлические частицы, вталкиваются в клетки растения пороховым зарядом или электрическим разрядом. Этим способом растительные клетки в культуре или репродуктивные органы или клетки растений, например пыльца, могут быть стабильно трансформированы интересующими молекулами нуклеиновой кислоты.
Настоящее изобретение может быть применено к трансформации практически любого типа растения, однодольных или двудольных.
Подходящие растения для трансформации представляют собой предпочтительно масличные культуры, такие как подсолнечник, соевый боб, хлопок, кукуруза, и т.п., предпочтительно виды Brassica, более предпочтительно разновидности Brassica napus.
В одном аспекте изобретения, растения включают не менее одной последовательности, кодирующей FAD2 по изобретению.
Растение по изобретению может включать последовательность нуклеиновой кислоты SEQ ID NO: 5 или последовательность нуклеиновой кислоты SEQ ID NO: 12.
Предпочтительно, растение по изобретению содержит последовательность нуклеиновой кислоты SEQ ID NO: 11 или последовательность нуклеиновой кислоты SEQ ID NO: 1, как MSP06 или MSP07.
В другом аспекте изобретения растение содержит две последовательности, кодирующие FAD2 по изобретению.
В частности, растение по изобретению содержит последовательность нуклеиновой кислоты SEQ ID NO: 11 и последовательность нуклеиновой кислоты SEQ ID NO: 12, такую как MSP05, MSP11 или 28DHS.059.
Предпочтительно, растение по изобретению содержит последовательность нуклеиновой кислоты SEQ ID NO: 1 и последовательность нуклеиновой кислоты SEQ ID NO: 5, такую как MSP05, MSP11 или 28DHS.059.
Разновидность MSP06 поддерживается в качестве патентного депонента по Будапештскому Соглашению в NCIMB под номером доступа NCIMB 41367, помещено 22 декабря 2005 г.
Разновидность MSP07 поддерживается в качестве патентного депонента по Будапештскому Соглашению в NCIMB под номером доступа NCIMB 41368, помещено 22 декабря 2005 г.
Разновидность 28DHS.059 поддерживается в качестве патентного депонента по Будапештскому Соглашению в NCIMB под номером доступа NCIMB 41364, помещено 22 декабря 2005 г.
Разновидность MSP05 поддерживается в качестве патентного депонента по Будапештскому Соглашению в NCIMB под номером доступа NCIMB 41233, помещено 9 июля 2004 г.
Разновидность MSP11 поддерживается в качестве патентного депонента по Будапештскому Соглашению в NCIMB под номером доступа NCIMB 41234, помещено 9 июля 2004 г.
Другой объект изобретения представляет собой способ производства высокоолеиновых линий растения, включающий: (a) скрещивание первого растения со вторым растением, имеющим по крайней мере один ген мутантного FAD2 в соответствии с изобретением, (b) получение семян от скрещивания по этапу (a); (c) выращивание фертильных растений из подобных семян; (d) получение семян потомства от растений этапа (с) и (e) идентификацию среди потомства семян, имеющих высокое содержание олеиновой кислоты.
В другом аспекте, изобретение предлагает способ увеличения содержания олеиновой кислоты в растениях, более конкретно в растениях Brassica, и предпочтительно в растениях Brassica napus, включающий этапы:
(a) индукцию мутагенеза в, по крайней мере, некоторых клетках растения, более конкретно растения Brassica, и предпочтительно растения Brassica napus, которое имеет содержание олеиновой кислоты менее чем 70%;
(b) регенерацию растений из по крайней мере одной мутагенизированной клетки;
(c) отбора регенерированных растений, которые имеют последовательность нуклеиновой кислоты по изобретению и/или которые экспрессируют белок FAD2 по изобретению; и
(d) получения дальнейших поколений растений из указанных регенерированных растений.
Предпочтительно семена, полученные от указанных растений, дают масло, имеющее содержание олеиновой кислоты более чем 70% вес., более предпочтительно - более чем 75% вес., по отношению к общему весу жирных кислот, присутствующих в масле.
Другой объект изобретения представляет собой растительное масло, полученное из по крайней мере одного растения в соответствии с изобретением, которое содержит более чем (приблизительно) 70%, 72%, 75%, 80% или 85% олеиновой кислоты.
Более конкретно, растительное масло по изобретению, полученное предпочтительно из, по крайней мере, одного вида Brassica по изобретению, более предпочтительно из одной из разновидностей Brassica napus в соответствии с изобретением, содержит более чем (приблизительно) 70%, 72%, 75%, 80% или 85% олеиновой кислоты. Указанное масло может, кроме того, содержать менее чем (приблизительно) 4%, 3,5%, 3%, 2%, 1% или 0,5% линоленовой кислоты по отношению к общему весу жирных кислот, присутствующих в масле.
Предпочтительно указанное масло содержит более чем (приблизительно) 70%, 72%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89% или 90%, предпочтительно между (приблизительно) 70% и (приблизительно) 90%, более предпочтительно между (приблизительно) 72% и (приблизительно) 89%, олеиновой кислоты. Указанное масло может также включать менее чем (приблизительно) 4%, 3,5%, 3%, 2%, 1% или 0,5%, предпочтительно между (приблизительно) 4% и (приблизительно) 0,4% линоленовой кислоты по отношению к общему весу жирных кислот, присутствующих в масле.
В соответствии с предпочтительным вариантом осуществления две двойные низкие озимые разновидности рапса (ENVOL и LIBERATOR) были отправлены на обработку этилметансульфонатом (EMS) в 1992 г. Обработка EMS проводилась при 2,5% и 5% в течение 4 ч или 8 ч.
Поколение М1 было выращено в оранжерее после 8 недель яровизации в ростовой камере и затем собрано в июле 93 г.
Семена М1 были высажены в поле в сентябре 93 г., закрыты мешками в начале цветения и семена М2 были собраны в июле 94 г.
Семена М2 были высажены в сентябре 94 г., закрыты мешками в начале цветения и семена М3 были собраны в июле 95 г.
Затем состав жирных кислот потомства был проанализирован с применением аналитического метода, основанного на газовой хроматографии, который хорошо известен в этой области технологии.
Все потомки, показывающие содержание олеиновой кислоты выше 68%, были сохранены.
Выбранное потомство было снова высажено в поле в сентябре 1995 г., закрыто мешками в апреле и собрано в июле 1996 г.
На этой стадии потомки были исследованы на хорошие агрономические и морфологические характеристики, такие как хорошая способность к прорастанию, хорошая сила осенью, хорошая устойчивость зимой, хорошая корневая система, хорошая устойчивость к черной ножке и светлой пятнистости (light leaf spot), так же как и превосходная устойчивость к полеганию.
Материал, который был слишком высоким или слишком поздним, был удален, так же как и материал, демонстрирующий высокую фасциацию.
Был снова проведен анализ оставшегося потомства при помощи газовой хроматографии для отбора индивидуумов с уровнями олеиновой кислоты выше чем 68%. Все эти индивидуумы были высажены в поле в сентябре 1996-1997 гг.
Потомство, названное MUT152-96, выглядело особо интересно в отношении агрономических и морфологических характеристик, так же как и в отношении содержания олеиновой кислоты в нем. Оно было культивировано в изоляции во время посевного сезона в сентябре 1996-1997 гг. Наиболее интересное потомство в отношении агрономических и морфологических характеристик было выбрано для изоляции мешками и скрещивания.
Скрещивание проводилось с разновидностями двойного нижнего озимого масличного рапса, имеющими обычный профиль жирной кислоты (т.е. олеиновую кислоту ниже 70%) или с низким содержанием линоленовой кислоты (т.е. менее чем приблизительно 3,5%) для получения линий с высоким содержанием олеиновой кислоты, связанным с низким содержанием линоленовой кислоты (HOLL).
Материал был проведен через линейное разведение, самоопыление проводилось по крайней мере до поколения F7.
На всех поколениях применялось сильное селективное давление против фасциации и в пользу нормального развития растения и нормальной корневой системы.
Состав жирной кислоты отслеживался в каждом поколении и сохранялся только материал с содержанием олеиновой кислоты выше чем 75% и линоленовой кислоты ниже чем 3,5%.
В результате этого процесса были получены следующие разновидности HOLL: MSP05, MSP06, MSP07, MSP11, 28 DHS 059.
Двойными низкими разновидностями с обычными профилями жирной кислоты, использованными в данной работе, были BRISTOL, CAPITOL, CAPVERT, VIVOL и CAIMAN и размножение или поддержание их проводилось по такой же схеме ухода, что и описанная здесь выше для линий HOLL.
Основные семена были использованы для определения содержания жирной кислоты в испытаниях - небольших исследовательских испытаниях (от 6 до 12 м2) или испытаниях при разработке (500 м2) и для работ по секвенированию.
ПРИМЕРЫ
Пример 1
Семена были измельчены в первом растворе, состоящем из метанола (800 мл), триметилпентана (200 мл) и 5 г NaOH. Приблизительно 3 мл раствора было использовано для приблизительно 10 г семян (другими словами, приблизительно от 10 до 50 семян на 1 мл раствора).
Экстракция проводилась в течение 20 минут, после чего был добавлен второй раствор, состоящий из триметиламина (900 мл) и пропанола-2 (100 мл) в том же объеме, что и первый раствор.
Полученный раствор был взболтан и оставлен в покое до формирования верхней фазы.
Верхняя фаза была отобрана и перенесена в сосуды.
Один микролитр ее был впрыснут в газовый хроматограф (Fisons thermo-electron с колонкой DB3 - 30 метров с диаметром 0,25 мм и толщиной 25 мкм). Время пробега было приблизительно 4 мин.
Содержание олеиновой кислоты было обобщено в таблице 1.
Содержание олеиновой кислоты по отношению к общему весу жирной кислоты в выделенном масле.
Пример 2
Растительные материалы, использованные для секвенирования:
- мутантные линии с повышенным содержанием олеиновой кислоты MSP05, MSP06, MSP07, MSP11, 28DHS.059; и
- разновидности дикого типа с нормальным содержанием олеиновой кислоты: Bristol, Capitol, Vivol, Capvert и Caiman.
Все эти линии были выращены в ростовой камере, и у 7-дневных растений были собраны стебли.
Растительные ткани были лиофилизированы и использованы для выделения ДНК.
ДНК была выделена с использованием наборов для выделения растительной ДНК Qiagen (Qiagen INC. - USA, Valencia CA).
ПЦР проводился по протоколу TaqGold (AB Biosystem, Inc).
Реакционная смесь включает 2,5 мкл 10× буфера, 0,2 мкл TaqGold, 0,2 мкл dNTP (25 мМ), 2 мкл праймеров (5 мкМ) и 10 мкл ДНК-матрицы (2 нг/мкл) и 10,1 мкл Н2О.
Циклы ПЦР были следующими: 94°С 5 мин; 8 циклов 94°С 40 сек, 62°С 40 сек, 72°С 1 мин, 94°С 40 сек, 60°С 40 сек, 72°С 1 мин, 94°С 40 сек, 58°С 40 сек, 72°С 1 мин, 94°С 40 сек, 56°С 40 сек, 72°С 1 мин, 3 цикла 94°С 40 сек, 55°С 40 сек, 72°С 1 мин; держать при 72°С 7 мин.
Продукты ПЦР были проанализированы в 1% агарозном геле.
Для секвенирования 5 мкл продуктов ПЦР было отделено в новую пробирку и туда был добавлен 1 мкл Экзонуклеазы I (разведение 1:50) и 1 мкл щелочной фосфатазы из креветок (разведение 1:5).
Смесь была инкубирована при 37°С в течение 20 мин и затем при 80°С в течение 15 мин для инактивации ферментов.
Было добавлено 40 мкл Н2О и 6 мкл было использовано в качестве матрицы с 1 мкл праймера для секвенирования.
Секвенирование проводили на ДНК-анализаторе 3730 (Applied Biosystems).
Последовательности были собраны и сравнены с использованием программы SeqMan II от LAserGene (DNASTAR, INC, Madison. WI).
Пример 3
Четыре последовательности генов дельта-12-олеатдесатуразы (FAD2) Brassica napus, 4684997, 46399190, 8705228 и 4092878, были загружены из Генбанка (NCBI). Эти последовательности были использованы в качестве запросов BLAST против базы данных последовательностей Monsanto.
С использованием программы «blastn» (NCBI), было получено несколько высококомплементарных результатов. Все эти последовательности были загружены и собраны заново программой SeqmanII (DNASTAR,INC, Madison, Wisconsin, USA).
Два различных транскрипта были идентифицированы и обозначены как Fad2-1 (SEQ ID NO: 9) и Fad2-2 (SEQ ID NO: 10). Fad2-1 и Fad2-2 имеют высокую гомологию последовательности, с 97% идентичности последовательности.
Для идентификации значащих мутаций, связанных с высоким содержанием олеиновой кислоты в мутантных линиях и их потомстве, были созданы гнездовые локус-специфичные праймеры для покрытия целых последовательностей.
3' конец праймера всегда находился на нуклеотиде, который отличал Fad2-1 от Fad2-2 за исключением тех, которые расположены на 5' и 3' концах консенсусных последовательностей, в которых нет различающихся у двух генов нуклеотидов.
Праймеры были также созданы таким образом, что один ампликон будет перекрывать другой для обеспечения полного покрытия всей последовательности. Эти праймеры были составлены и использованы для получения локус-специфичных ампликонов мутантного и дикого типа. Результаты секвенирования показали, что все локус-специфические ПЦР-праймеры вели себя как ожидалось.
Последовательности, принадлежащие к одному гену, были собраны вместе с использованием программы SeqManII.
Консенсусные геномные последовательности мутантных генов Fad2-1 и Fad2-2 представлены соответственно SEQ ID NO: 11 и 12.
Таблица 2 обобщает свойства последовательностей обоих генов Fad2-1 и Fad2-2.
Элементы основаны на консенсусных геномных последовательностях из различных прочтений разных генотипов.
И ген Fad2-1 и ген Fad2-2 имеют по одному интрону каждый.
Размеры интронов слегка различаются у двух генов. Для Fad2-1 размер интрона составляет 1105 п.о., начиная с положения 109 до положения 1213, в то время как для Fad2-2 интрон состоит из 1123 п.о., начиная с положения 112 до положения 1234 на консенсусных последовательностях.
Интрон расположен на участке 5'UTR.
Предполагаемые кодоны инициации трансляции расположены в положениях 1218 и 1239 соответственно для Fad2-1 и Fad2-2.
Кодоны терминации трансляции расположены в положениях 2370-2372 и 2391-2393 соответственно для Fad2-1 и Fad2-2.
Последовательности 3'UTR представляют собой 247 пар оснований в случае гена Fad2-1 и 273 пары оснований в случае гена Fad2-2.
Была обнаружена мутация трансляции в положении 1540 (называемая SNP1540) гена FAD2-1 (как представлено SEQ ID NO: 11), которая вызывала изменение генетического кодона GGC на GAC, приводя к изменению аминокислотного остатка с глицина на аспарагиновую кислоту.
Поскольку глицин и аспарагиновая кислота имеют весьма различные свойства в отношении гидрофобности, зарядов и полярности и т.д., мутация вызывает радикальные изменения в работе фермента в мутантных линиях.
Также были описаны высококонсервативные аминокислотные последовательности среди растительных дельта-12-десатураз жирной кислоты и растительных дельта-15-десатураз жирной кислоты (патент США US6872872B1). Среди других, один из указанных консервативных участков аминокислотной последовательности представляет собой AHECGH. Оказалось, что SNP1540 находится в том же участке. «G» этого участка была мутирована в «D». Поскольку консервативность участков обычно означает их функциональную или структурную значимость, мутация в этом консервативном участке вызвала побочные эффекты в ферменте Fad2-1, приводя к высокому содержанию олеиновой кислоты в мутантных линиях MSP11, MSP05, MSP06, MSP07 и 28DHS.059.
Точечная мутация в положении 1590 (названная SNP1590) гена FAD2-2 (как представлено SEQ ID NO: 12) вызывала изменение аминокислотного остатка с лейцина (CTT) на фенилаланин (TTT).
И лейцин и фенилаланин имеют гидрофобную природу и имеют некоторые сходные аминокислотные свойства, но фенилаланин содержит большую жесткую ароматическую группу на боковой цепи, которая вызывает некоторые изменения в работе фермента.
Более того, в комбинации с мутацией SNP1540, мутация вызывает более заметное влияние на фенотип.
Сочетание различных аллелей при таких мутациях создавало градиент на содержание олеиновой кислоты, как отмечено в разных мутантных линиях (см. таблицу 1).
Три мутантные линии, MSP11, MSP05 и 28DHS.059, несли двойные мутации в SNP1540 и SNP1590. Поскольку обе мутации были миссенс-мутациями, функции гена FAD2 были серьезно затронуты, приводя к наибольшему содержанию олеиновой кислоты среди мутантных линий.
Содержание олеиновой кислоты для MSP05 было ниже, чем для других двух мутантов. Это произошло по причине того, что содержание олеиновой кислоты было получено по данным только одного года, которые могут быть объектом вариаций в связи с воздействием окружающей среды.
Две мутантные линии, MSP06 и MSP07, несли единичную точечную мутацию в SNP1540. Поскольку они менее серьезны, чем у двойных мутантов, содержание олеиновой кислоты для этих двух линий было слегка ниже, чем для двойных мутантов.
В общем, данные последовательности четко показывали, что эти мутации в Fad2-1 и Fad2-2 сильно связаны с содержанием олеиновой кислоты в разных мутантных линиях.
Сочетание различных аллелей объясняет все фенотипические вариации содержания олеиновой кислоты в полученных растительных материалах.
Идентификация количественных вариаций последовательности критически важна для создания диагностических анализов, специфичных для каждой мутантной аллели.
Знание о связи между вариациями последовательности и фенотипом могут позволить создать маркерные анализы для точного предсказания содержания олеиновой кислоты в растениях без необходимости химического анализа содержания жирной кислоты.
| название | год | авторы | номер документа |
|---|---|---|---|
| FAD-2-МУТАНТЫ И РАСТЕНИЯ С ВЫСОКИМ СОДЕРЖАНИЕМ ОЛЕИНОВОЙ КИСЛОТЫ | 2007 |
|
RU2461624C2 |
| Brassica juncea КАЧЕСТВА ОМЕГА-9 | 2009 |
|
RU2557316C2 |
| КАНОЛА HO/LL С УСТОЙЧИВОСТЬЮ К ЗАБОЛЕВАНИЮ КИЛОЙ КРЕСТОЦВЕТНЫХ | 2012 |
|
RU2711934C2 |
| КАНОЛА HO/LL С УСТОЙЧИВОСТЬЮ К ЗАБОЛЕВАНИЮ КИЛОЙ КРЕСТОЦВЕТНЫХ | 2012 |
|
RU2618846C2 |
| ЛОКУСЫ ФУНКЦИОНАЛЬНОСТИ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ К УЧАСТКУ-МИШЕНИ СВЯЗЫВАЮЩИЕ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656159C2 |
| МУТАНТНЫЙ КАЛЬРЕТИКУЛИН ДЛЯ ДИАГНОСТИКИ МИЕЛОИДНЫХ НОВООБРАЗОВАНИЙ | 2014 |
|
RU2668808C2 |
| ФУНКЦИОНАЛЬНЫЕ ЛОКУСЫ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ ДЛЯ САЙТА-МИШЕНИ СВЯЗЫВАЮЩИЕСЯ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656158C2 |
| МУТАНТНЫЕ ЛЕНТИВИРУСНЫЕ БЕЛКИ ENV И ИХ ПРИМЕНЕНИЕ В КАЧЕСТВЕ ЛЕКАРСТВЕННЫХ СРЕДСТВ | 2012 |
|
RU2654673C2 |
| НУКЛЕИНОВЫЕ КИСЛОТЫ И СПОСОБЫ ПОЛУЧЕНИЯ СЕМЯН, ИМЕЮЩИХ В ЗАРОДЫШЕ ПОЛНЫЙ ДИПЛОИДНЫЙ ГЕНОМ, КОМПЛЕМЕНТАРНЫЙ МАТЕРИНСКОМУ | 2006 |
|
RU2438297C2 |
| МУТАНТЫ ИНДУЦИРУЕМОЙ АКТИВАЦИЕЙ ЦИТИДИНДЕАМИНАЗЫ (AID) И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2537264C2 |






















Изобретение относятся к области биохимии. Представлены варианты белков дельта-12-олеатдесатуразы (FAD2), где один вариант имеет аминокислотную замену в положении, соответствующем положению 108 на аспарагиновую кислоту, а другой - замены в положениях, соответствующих положениям 108 на аспарагиновую кислоту и 118 на фенилаланин, в белке FAD2 дикого типа с последовательностью SEQ ID NO:4 или SEQ ID NO:8, приведенными в описании. Описаны: нуклеиновые кислоты, кодирующие указанные белки; векторы экспрессии, содержащие указанные нуклеиновые кислоты; клетки-хозяева, содержащие указанные векторы. Предложены фрагменты, состоящие из по крайней мере 20 нуклеотидов выделенной молекулы нуклеиновой кислоты, содержащие указанные замены, для использования в качестве праймера, зонда и/или маркера. Представлено растение, создающее высокий профиль олеиновых кислот в масле семян, трансформированное нуклеиновой кислотой с последовательностью SEQ ID NO:1, или SEQ ID NO:1 и SEQ ID NO:5. Изобретение позволяет увеличить содержание олеиновой кислоты в семенах масличных растений. 17 н. и 2 з.п. ф-лы, 2 табл., 1 ил., 3 пр.
1. Выделенная молекула нуклеиновой кислоты, кодирующая белок дельта-12-олеатдесатуразу (FAD2), причем указанный белок FAD2 имеет аминокислотную замену в положении, соответствующем положению 108 в белке FAD2 дикого типа, представленном SEQ ID NO:4 или SEQ ID NO:8, где указанная аминокислота в положении 108 заменена на аспарагиновую кислоту.
2. Выделенная молекула нуклеиновой кислоты, кодирующая белок FAD2, причем указанный белок FAD2 имеет аминокислотную замену в положении, соответствующем положению 108, и в положении, соответствующем положению 118 в белке FAD2 дикого типа, представленном SEQ ID NO:4 или SEQ ID NO:8, где указанная аминокислота в положении 108 заменена на аспарагиновую кислоту, и указанная аминокислота в положении 118 заменена на фенилаланин.
3. Выделенная молекула нуклеиновой кислоты по п.1, где указанный белок FAD2 является белком FAD2 Brassica, более конкретно - белком FAD2 Brassica napus.
4. Выделенная молекула нуклеиновой кислоты по п.2, где указанный белок FAD2 является белком FAD2 Brassica, более конкретно - белком FAD2 Brassica napus.
5. Выделенная молекула нуклеиновой кислоты, включающая нуклеотидную последовательность, имеющую, по крайней мере, 80% идентичности с SEQ ID NO:1 или 11, или с их комплементарной формой или РНК-формой, и кодирующая белок FAD2, имеющий аминокислотную замену в положении, соответствующем положению 108 в белке FAD2 дикого типа, представленном SEQ ID NO:4 или SEQ ID NO:8, где указанная аминокислота в положении 108 заменена на аспарагиновую кислоту.
6. Фрагмент из, по крайней мере, 20 нуклеотидов выделенной молекулы нуклеиновой кислоты по п.1, причем указанный фрагмент включает мутированный кодон, соответствующий указанной аминокислотной замене в положении 108, где указанный фрагмент предназначен для применения в качестве праймера, зонда и/или маркера по отношению к молекуле нуклеиновой кислоты по п.1.
7. Белок FAD2, имеющий аминокислотную замену в положении 108, или положении, соответствующем положению 108, относительно белка FAD2 дикого типа, такого как белок FAD2 дикого типа, представленный аминокислотной последовательностью SEQ ID NO:4 или 8, где указанная аминокислота в положении 108 заменена на аспарагиновую кислоту, что повышает уровень олеиновой кислоты.
8. Белок FAD2, имеющий аминокислотную замену в положении 108, или положении, соответствующем положению 108, и аминокислотную замену в положении 118, или положении, соответствующем положению 118 относительно белка FAD2 дикого типа, такого как представленный аминокислотной последовательностью SEQ ID NO:4 или 8, где указанная аминокислота в положении 108 заменена на аспарагиновую кислоту, и указанная аминокислота в положении 118 заменена на фенилаланин, что повышает уровень олеиновой кислоты.
9. Белок олеатдесатураза дельта-12 (FAD2), включающий аминокислотную последовательность SEQ ID NO:2.
10. Вектор экспрессии, включающий молекулу нуклеиновой кислоты по п.1.
11. Вектор экспрессии, включающий молекулу нуклеиновой кислоты по п.2.
12. Клетка-хозяин для экспрессии экзогенного генетического материала, включающая вектор по п.10.
13. Клетка-хозяин для экспрессии экзогенного генетического материала, включающая вектор по п.11.
14. Растение, стабильно трансформированное вектором по п.10, в силу чего уровень олеиновой кислоты его семян повышен, причем указанное растение для трансформации выбрано из группы, состоящей из подсолнечника, соевых бобов, хлопка, кукурузы и семени рапса.
15. Растение, стабильно трансформированное вектором по п.11, в силу чего уровень олеиновой кислоты его семян повышен, причем указанное растение для трансформации выбрано из группы, состоящей из подсолнечника, соевых бобов, хлопка, кукурузы и семени рапса.
16. Способ увеличения содержания олеиновой кислоты в растении, включающий трансформирование растения вектором по п.10.
17. Способ увеличения содержания олеиновой кислоты в растении, включающий трансформирование растения вектором по п.11.
18. Растение Brassica napus, создающее высокий профиль олеиновых кислот в масле семян, содержащее последовательность нуклеиновой кислоты SEQ ID NO:1.
19. Растение Brassica napus, создающее высокий профиль олеиновых кислот в масле семян, содержащее последовательность нуклеиновой кислоты SEQ ID NO:1 и последовательность нуклеиновой кислоты SEQ ID NO:5.
| Приспособление для очистки дна бассейнов с закругленными углами от твердых осадков | 1928 |
|
SU30206A1 |
| US 6063947 А, 16.05.2000 | |||
| WO 9721340 A1, 19.06.1997 | |||
| Электронно-лучевая усилительная лампа | 1941 |
|
SU61880A1 |
| ФРАГМЕНТ ДНК (ВАРИАНТЫ), ВЕКТОР, СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ, СПОСОБ ИНДУЦИРОВАНИЯ ПРОДУКЦИИ ГАММА-ЛИНОЛЕНОВОЙ КИСЛОТЫ (ВАРИАНТЫ), СПОСОБ ИНДУЦИРОВАНИЯ ПРОДУКЦИИ ОКТАДЕКАТЕТРАЕНОВОЙ КИСЛОТЫ И ДЕЛЬТА-6-ДЕСАТУРАЗА ЦИАНОБАКТЕРИЙ | 1992 |
|
RU2152996C2 |
| РЕМОНТНО-БУРОВОЙ АГРЕГАТ | 2006 |
|
RU2320842C1 |

