Изобретение относится к области иммунологии, энзимологии, микробиологии и генной инженерии и касается нового полинуклеотида, кодирующего функционально активную рекомбинантную IgA1 протеазу Neisseria meningitidis, новой рекомбинантной плазмидной ДНК pBIGAPS1-M и клетки-хозяина, прежде всего нового штамма бактерий Escherichia coli BL21(DE3)/pBIGAPS1-M, который может быть использован для получения высокоочищенной генно-инженерной IgA1 протеазы и в качестве компонента поливалентной вакцины, предназначенной для защиты человека, прежде всего детей, от менингококковой инфекции и других микроорганизмов, патогенность которых обусловлена IgA1 протеазой.
Проблема эффективной вакцинопрофилактики менингококковой инфекции, вызываемой менингококком серогруппы В, до сих пор не решена. В реестре зарегистрированных менингококковых вакцин ВОЗ не зафиксировано ни одного препарата против менинингококка этой серогруппы. Отсутствие эффективной менингококковой группы В вакцины препятствует ликвидации этой инфекции в нашей стране и за ее пределами. Известно, что сериновая IgA протеаза секретируется некоторыми опасными бактериями, прежде всего Neisseria meningitidis, и расщепляет иммуноглобулины класса А на поверхности слизистых оболочек, разрушая первый защитный барьер иммунитета организма-хозяина. Известна менингококковая IgA1 протеаза, которая в опытах на лабораторных животных проявила себя как высокоэффективная менингококковая поливакцина, а следовательно, и вакцина против менингококка серогруппы В (пат. RU 2407792, Аллилуев А.П. и др., 16.08.2010 г). Было установлено, что выделенная из культуральной жидкости или из продуктов очистки вакцины (менингококковой группы А) IgA1 протеаза высокоэффективна ферментативно и иммунологически, но содержится в указанных источниках в микроколичествах, поэтому могут возникнуть сложности при получении протеазы в препаративных количествах.
В предшествующем уровне техники описано множество попыток создания вакцин, предназначенных для защиты от инфекции менингита группы В. В заявках WO 2005/105141 20051110, WO 95/03413, WO 2005/102384 20051103, WO 03/094960 20031120, WO 2006/024954 20060309, WO 2004/014417 20040219, WO 2005/033148 20050414, WO 2004/032958, WO 03/028661 20030410, WO 2004/014419 A1, WO 02/083711 предлагаются композиции и кандидаты в вакцины на основе липополисахарида, капсулярных олигосахаридов и ряда поверхностных белков менингококка для иммунизации пациента против заболевания, вызываемого Neisseria meningitidis различных серогрупп.
Однако указанные композиции не включают IgA1 протеазу и не обеспечивают достаточную эффективную защиту от менингококковой инфекции и, в частности, от менингита группы В.
В заявке WO 2004/102199 А2, LEENHOUTS и др., 25.11.2004, описан способ идентификации, выбора и выделения вакцинного компонента для пассивной и активной иммунизации против микроорганизма Streptococcus pneumonia, который подвергается уничтожению опсонофагоцитарными клетками. Методом компьютерного моделирования (in silico) идентифицированы антигены, индуцирующие опсонофагоцитоз, которые предлагается использовать в качестве компонентов вакцины, предназначенной для иммунизации. Идентифицировано три пневмококковых белка: стрептококклипопротеинароматаза (SlrA), IgA1 протеаза класса металлопротеаз и пневмококковый поверхностный белок адгезии A (PsaA) из Streptococcus pneumonia, получены их рекомбинантные варианты, которые изучены в качестве компонентов вакцины. Установлено, что композиция, содержащая все три рекомбинантных варианта, защищает мышей от указанной инфекции, однако каждый компонент в отдельности не обладает защитными свойствами.
В статье Shishir K. Gupta и др., Vaccine, 2010, т.28, стр.7092-7097, описаны результаты компьютерного моделирования последовательностей белков менингококка серогруппы В и выявлены 6 пептидов-эпитопов Т-хелперных клеток из трех белков: стимулирующий Т-клетки белок А, автотранспортный белок А и IgA1 протеаза. Авторы предполагают, что эти пептиды можно использовать в качестве перспективных агентов против менингококка группы В. Однако экспериментальные данные, подтверждающие данную гипотезу, не приводятся.
Известно получение фрагмента IgA1-протеазы, который используют в качестве пептида-носителя (пат. US 7235242, Achtman и др., 26.06.2007). Описан синтез фрагмента IgA1-протеазы, содержащего от 40 до 200 аминокислотных остатков и включающего 40 аминокислотных остатков последовательности SEQ ID NO:1, с использованием автоматического синтезатора. Однако при конъюгации с полисахаридом IgA1 протеаза обеспечивает защиту только от менингококка серогруппы С и не эффективна против менингококков серогруппы В.
Известно применение бактериальных IgA1 протеаз для лечения заболеваний, связанных с отложениями IgA1 в тканях и органах организма (пат. US7407653, Plaut и др., 05.08.2008). Растворимые IgA1 протеазы, содержащие различные метки, например полигистидиновый фрагмент, получают рекомбинантным способом. Фермент выделяют по известной методике с использованием ультрафильтрации, ионообменной и металл-хелатной хроматографии. Полученные препараты можно использовать в качестве фермента для лечения аутоиммунных заболеваний, связанных с накоплением IgA, но не в качестве вакцины.
Известен также способ получения предшественника IgA1 протеазы. В статье S. Vitovski и J.R. Sayers, Infection and Immunity, 2007, т.75, №6, стр.2875-2885, описано клонирование полноразмерного гена, кодирующего сигнальный пептид, секретируемую форму протеиназы и автотранспортерный домен, и исследование механизма автопроцессинга IgA1 протеазы из ее предшественника. С этой целью был создан ряд мутированных форм белка-предшественника IgA1 протеазы Neisseria meningitidis серогруппы В (NMB), включая мутант с делецией 32-членного пептида в области сайта автопроцессинга, а также мутант, включающий замену остатка Ser в активном центре на остаток Val с целью получения предшественника, не обладающего автопротеолитической активностью. Для получения мутантов использовали штаммы NMB, выделенные из цереброспинальной жидкости больных менингококком. Было установлено, что указанные мутанты не обладали автопротеолитической активностью и не наблюдалось образование секретируемой зрелой протеазы. Но в этой работе получали только предшественник IgA1 протеазы. Протективные и иммуногенные свойства собственно IgA1 протеазы не исследовали, а также не рассматривается возможность создания вакцины на ее основе.
В заявке LONG и др., WO 20101183337 (А2), 14.10.2010, описан способ получения растворимой рекомбинантной IgA1 протеазы с использованием прямого и/или непрямого продуцирования через тельца включения со значительно более высоким выходом по сравнению с известными разработанными ранее способами, причем выход составляет от 20 до 500 мг растворимой IgA1 протеазы/л культуральной среды. Однако полученные указанным способом препараты и композиции IgA1 протеазы являются ферментативно активными и предназначены только для разрушения IgA1 при лечении заболеваний, связанных с отложением этого белка в тканях организма. Кроме того, в работе получали IgA1 протеазу H. influenzae, экспрессированную в Е.coli.
Известен способ получения рекомбинантной IgA1 протеазы Neisseria meningitidis серогруппы В (NMB) (Серова О.В. и др., Биофармацевтический журнал, 2011, т.3, №6), согласно которому получали полноразмерную ферментативно активную IgA1 протеазу, обладающую иммуногенными и протективными свойствами в опытах на мышах в отношении менингококков серогрупп В. Однако указанный препарат обладает ферментативной активностью в отношении IgA1 человека, что может привести к нежелательным последствиям при расщеплении иммуноглобулина А1 на слизистых оболочках организма, например развитие воспалительной реакции. Этот способ как ближайший уровень техники был выбран в качестве прототипа.
Изобретение решает задачу получения рекомбинантной полноразмерной IgA1 протеазы, обладающей повышенными иммуногенными и защитными свойствами в отношении Neisseria meningitidis основных эпидемических серогрупп - А, В и С, но не обладающей ферментативной активностью в отношении IgA1 человека, для создания поливалентной вакцины для защиты человека как от менингококковой инфекции, так и от других патогенов, обусловленных IgA1 протеазой.
Поставленная задача решается за счет полинуклеотида, кодирующего IgA1 протеазу Neisseria meningitidis серогруппы В штамм Н44/76 с мутацией каталитически активного остатка Ser267 на Ala (SEQ ID NO:1);
рекомбинантной плазмидной ДНК, включающей вышеуказанную нуклеотидную последовательность и обеспечивающей экспрессию IgA1 протеазы в клетке-хозяине (pBIGAPS1-M), характеризующейся следующими признаками: длина цепи составляет 8123 п.о.; кодирует секретируемую форму IgA1 протеазы с мутацией каталитически активного остатка Ser267 на Ala, с сигнальной последовательностью на N-конце и гистидиновой меткой на С-конце белка;
нуклеотидную последовательность, кодирующую IgA1 протеазу в плазмиде pBIGAPS1-М, фланкируют уникальные сайты рестрикции эндонуклеазами NdeI и XhoI;
нуклеотидная последовательность, кодирующая IgA1 протеазу в плазмиде pBIGAPS1-M, содержит уникальный сайт рестрикции эндонуклеазой BamHI;
содержит Т7 lac-промотор транскрипции;
генетическим маркером служит нуклеотидная последовательность (kan), определяющая устойчивость трансформированных плазмидой pBIGAPS1-M клеток бактерий к канамицину;
клетки-хозяина, прежде всего штамма-продуцента Е. coli BL21 (DE3)/pBIGAPS1-M, содержащего рекомбинантную плазмидную ДНК, который продуцирует зрелую форму мутантной IgA1 протеазы;
мутантной IgA1 протеазы с молекулярной массой 107,6 кДа и рассчитанной изоэлектрической точкой 7,4, с аминокислотной последовательностью (SEQ ID NO:2), кодируемой вышеуказанной нуклеотидной последовательностью, обладающей иммуногенными и протективными свойствами, предназначенной для получения поливакцины для профилактики менингококковых инфекций, в том числе вызываемых бактериями N. meningitidis серогруппы В;
а также за счет способа получения зрелой формы IgA1 протеазы Neisseria meningitidis серогруппы В, заключающегося в том, что клетку-хозяина трансформируют рекомбинантной плазмидной ДНК, кодирующей мутантную IgA1 протеазу Neisseria meningitidis серогруппы В, с полинуклеотидом (SEQ ID NO:1), культивируют штамм-продуцент, выделяют и очищают целевой продукт из телец включения.
Техническим результатом изобретения является получение высокоочищенной полноразмерной ферментативно неактивной мутантной IgA1 протеазы N. meningitidis серогруппы В (40% фермента от суммарного клеточного белка, 20 мг из 2 г биомассы клеток, чистота препарата - не менее 90%), характеризующейся повышенной иммуногенной и протективной активностью в сравнении с ферментативно активными нативной и рекомбинантной протеазами. Полученный препарат мутантной IgA1 протеазы обладает поливалентным действием в отношении защиты от менингококковой инфекции организма животного независимо от источника ее получения (серогруппы А или В). Отсутствие ферментативной активности в отношении иммуноглобулина А1 человека мутантной IgA1 протеазы позволит избежать дополнительных осложнений при иммунизации человека.
Один объект настоящего изобретения включает рекомбинантную плазмиду, включающую вышеуказанную нуклеотидную последовательность и обеспечивающую экспрессию мутантной IgA1 протеазы в клетке-хозяине.
Для получения мутантной формы IgA1-протеиназы, в которой каталитически активный остаток Ser267 заменен на Ala, был использован метод SOE (splicing by overlap extension, сплайсинг при расширении частично перекрывающихся последовательностей). Для этого были сконструированы специфические праймеры (SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6), перекрывающиеся в области сайта мутации, кроме этого, в эти праймеры был введен сайт рестрикции эндонуклеазы BamHI. Амплификацию фрагмента гена проводили в два этапа, используя в качестве матрицы плазмидную ДНК pBIGAPSI, описанную ранее в заявке RU 2453599, Румш Л.Д. и др., от 18.02.2011 г. Был получен фрагмент ДНК длиной около 2900 п.о. Целевой ПНР-продукт был клонирован в вектор рЕТ-24а(+) (Novagen, кат. №69749-3) при использовании уникальных сайтов рестрикции NdeI и XhoI таким образом, чтобы продуцируемый белок содержал His-метку в C-концевой области для последующей очистки белка с помощью металлохелатной хроматографии. Секвенирование полученной ДНК подтвердило наличие мутации. Полученная плазмидная ДНК была названа pBIGAPS1-M.
Еще один объект настоящего изобретения включает клетку-хозяина, прежде всего штамм Е. coli, содержащий рекомбинантную нуклеиновую последовательность (см. выше), который продуцирует зрелую форму мутантной IgA1 протеазы. В качестве клетки-хозяина использовали, прежде всего, штамм Е. coli BL21(DE3), на основе которого был получен штамм BL21(DE3)/pBIGAPS1-M, описанный ниже.
Другой объект включает способ получения зрелой мутантной формы IgA1 протеазы Neisseria meningitidis серогруппы В, заключающийся в том, что клетку-хозяина Е. coli BL21(DE3) трансформируют описанной выше рекомбинантной плазмидной ДНК, кодирующей IgA1 протеазу Neisseria meningitidis серогруппы В, с полинуклеотидом (SEQ ID NO:1), культивируют клетки-хозяина, как описано ниже, выделяют и очищают целевой продукт из телец включения.
Еще один объект настоящего изобретения включает IgA1 протеазу с молекулярной массой 107,6 кДа и рассчитанной изоэлектрической точкой 7,4 и аминокислотной последовательностью (SEQ ID NO:2), кодируемую описанным выше полинуклеотидом и полученную указанным выше способом, предназначенную для получения поливакцины для профилактики инфекций, в том числе вызываемых бактериями N. meningitidis серогруппы В.
IgA1 протеаза в качестве активного компонента может быть использована для получения поливакцины для профилактики менинкогокковой инфекции серогрупп А, В, С, а также для инфекций, патогенность которых обусловлена IgA1 протеазой, таких как Clostridium ramosum, Bacteroides melaninogenicus, Capnocytophaga ochracea, Streptococcus pneumonia, Streptococcus sanguis, Streptococcus oralis, Streptococcus mitis, Haemophilus influenzae, Haemophilus influenzae, Haemophilus aegyptius, Neisseria gonorrhoeae, Neisseria meningitidis, Haemophilus influenzae, Streptococcus viridans, Streptococcus canis, Streptococcus suis, Staphylococcus aureus, Staphylococcus epidermidis, Moraxella catarrhalis, Helicobacter pylori, Campylobacter jejuni.
Для получения штамма-продуцента IgA1 протеазы трансформируют компетентные клетки Escherichia coli BL21(DE3) (Е. coli В F- dcm ompT hsdS(rB - mB -) gal λ(DE3)) (Stratagene Cat. No 200131) плазмидной ДНК pBIGAPS1-М.
Полученный штамм, названный Е. coli BL21(DE3)/pBIGAPS1-M, характеризуется следующими свойствами.
Культурально-морфологические признаки.
Клетки мелкие, палочковидной формы, грамотрицательные, 1×3,5 мкм, подвижные. Штамм хорошо растет на обычных питательных средах (LB-бульон, LB-агар). При росте на агаризованной среде LB колонии округлые, гладкие, полупрозрачные, блестящие, желтоватого цвета. Край ровный, диаметр колоний 2-3 мм, консистенция пастообразная. Рост в жидкой среде LB характеризуется ровным помутнением, осадок легко седиментирует.
Физиолого-биохимические признаки.
Клетки растут при 4-42°С, оптимум рН 6,8-7,6. В качестве источника азота используют как минеральные соли аммония, так и органические соединения: аминокислоты, пептон, триптон, дрожжевой экстракт.
Генетические признаки.
Клетки проявляют устойчивость к канамицину (до 100 мкг/мл), обусловленную наличием гена устойчивости к антибиотику в рекомбинантной плазмидной ДНК pBIGAPS1-M.
Условия хранения.
Штамм бактерий Е.coli BL21(DE3)/pBIGAPS1-M хранят на чашках и косяках при 4°С. Пересевы на свежие среды проводят один раз в месяц. Может храниться не менее одного года в среде LB, содержащей 15% глицерина, при -70°С.
Устойчивость к антибиотикам.
Клетки штамма-продуцента проявляют устойчивость к канамицину (до 100 мкг/мл), обусловленную наличием гена устойчивости в рекомбинантной плазмидной ДНК pBIGAPS1-M.
В качестве клетки-хозяина можно использовать клетки эукариот (например, дрожжевые клетки, растительные клетки, животные клетки и клетки человека), или клетки прокариот (например, бактериальные клетки и т.п.), или клетки, которые индуцируют иммунный ответ. Специалистам в данной области известно множество указанных клеток и клеточных линий (высших) эукариот, например 293Т (линия эмбриональных клеток почек), HeLa (клетки карциномы шейки матки человека), СНО (клетки яичника китайского хомячка) и другие клеточные линии, включая указанные клетки и клеточные линии, разработанные для лабораторных исследований, такие как, например, клетки hTERT-MSC, HEK293, Sf9 или COS. Пригодные эукариотические клетки, кроме того, включают клетки или клеточные линии, пораженные болезнями или инфекциями, например раковые клетки, прежде всего раковые клетки любого типа рака, упомянутого в данном контексте, клетки, пораженные ВИЧ, и/или клетки иммунной системы или центральной нервной системы (ЦНС). Пригодные клетки можно аналогичным образом получить из эукариотических микроорганизмов, таких как дрожжи, например Saccharomyces cerevisiae (Stinchcomb и др., Nature, т.282, стр.39, (1997)), Schizosaccharomyces pombe, Candida, Pichia и гифомицеты родов Aspergillus, Penicillium и т.п. Пригодные клетки аналогичным образом включают клетки прокариот, таких как, например, клетки бактерий, например Escherichia coli, или бактерий рода Bacillus, Lactococcus, Lactobacillus, Pseudomonas, Streptomyces, Streptococcus, Staphylococcus, предпочтительно Е. coli, и т.п. Клетки человека или клетки животных, например животных, упомянутых в данном контексте, являются прежде всего предпочтительно эукариотическими клетками.
Изобретение иллюстрируют графические материалы.
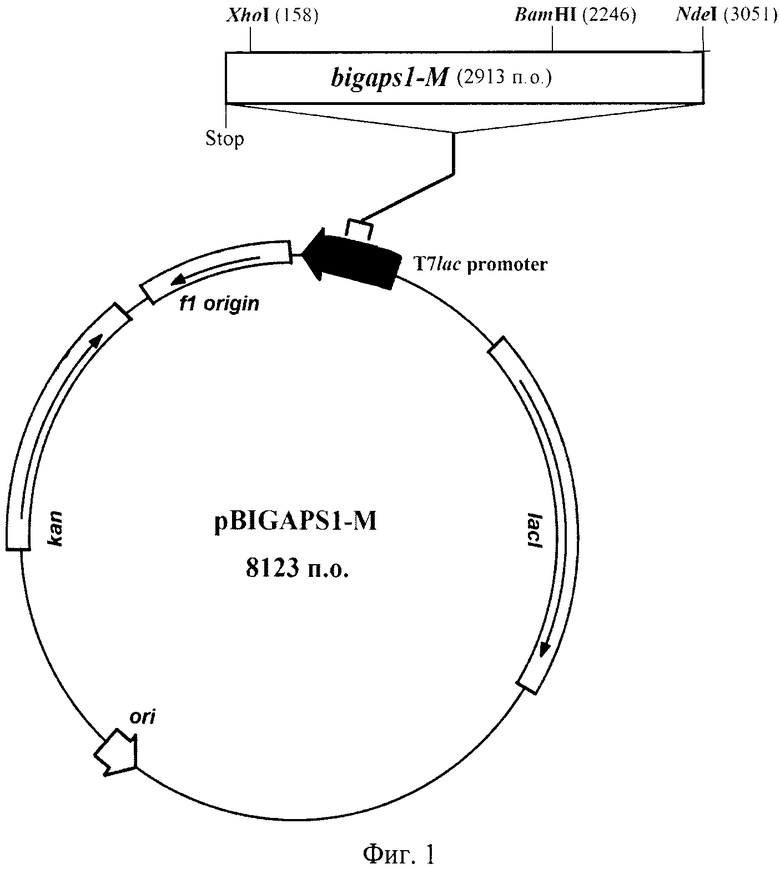
Фиг.1. Физическая карта рекомбинантной плазмидной ДНК pBIGAPS1-M.
Рекомбинантная плазмидная ДНК pBIGAPS1-M, содержащая 8123 п.о., кодирующая секретируемую мутантную форму IgA1-протеиназы, состоящая из:
bigaps1-M - область, кодирующая сигнальный пептид, секретируемую мутантную форму IgA1 протеазы и гистидиновую метку и фланкируемая уникальными сайтами рестрикции эндонуклеазами NdeI (3051) и XhoI (158) (нумерация приведена в соответствии с нумерацией в векторе рЕТ-24а), bigaps1-M содержит также уникальный сайт рестрикции эндонуклеазой BamHI, ori - участок инициации репликации рекомбинантной плазмидной ДНК, T7lac promoter - гибридный промотор транскрипции, Stop - терминатор транскрипции рибосомного оперона Е. coli, kan - ген, определяющий устойчивость микроорганизма к канамицину, f1 origin - участок, позволяющий получать одноцепочечную ДНК, lacI - область, кодирующая lac репрессор.
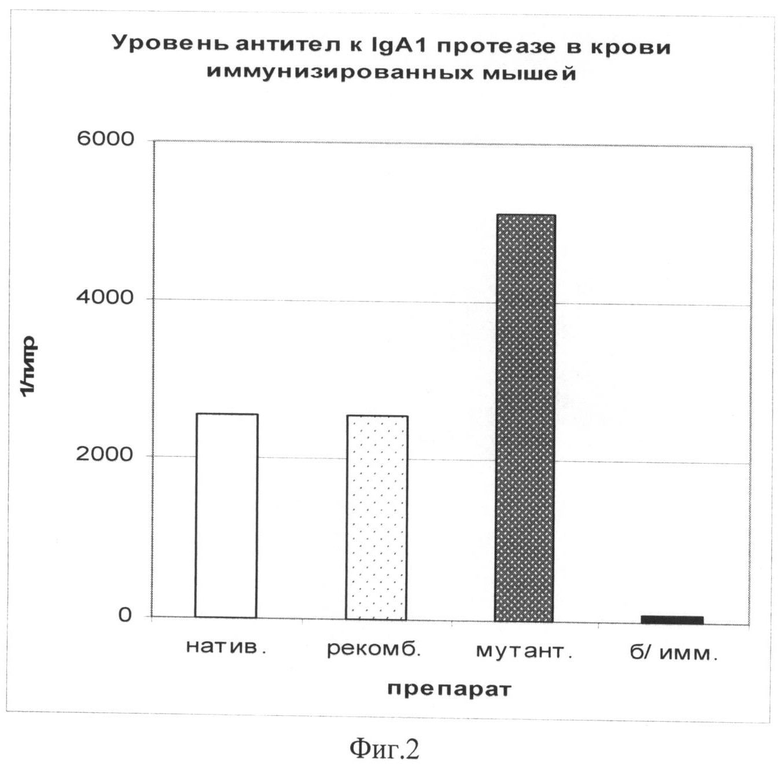
Фиг.2. Уровень специфических антител в крови иммунизированных мышей.
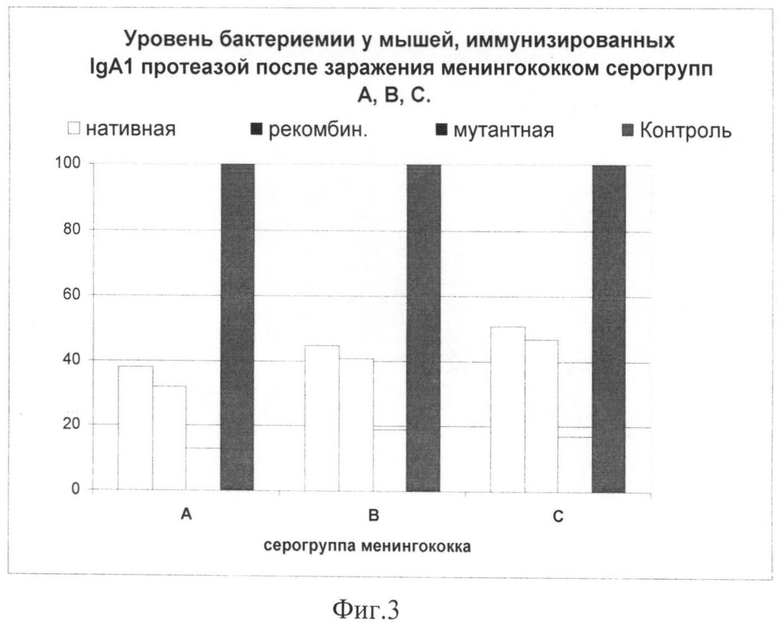
Фиг.3. Защищенность мышей, иммунизированных IgAl-протеазой после заражения менингококком серогруппы В штамм Н44/76.
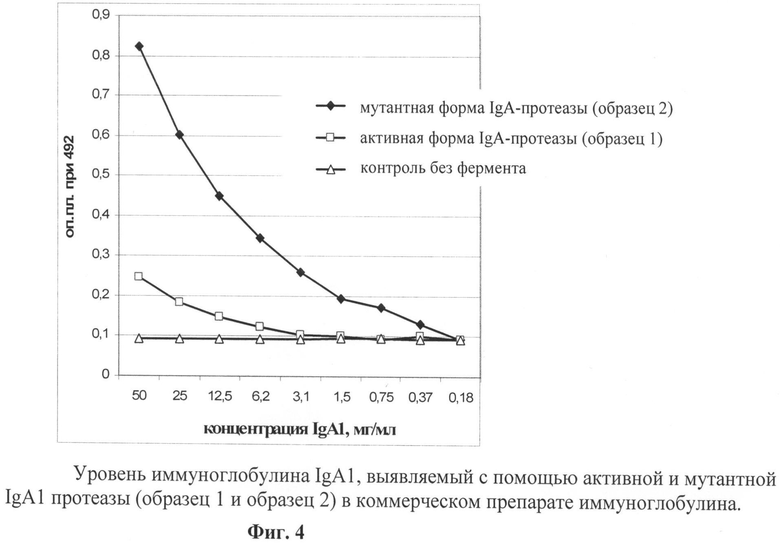
Фиг.4. Уровень иммуноглобулина IgA1, выявляемый в коммерческом препарате.
Фиг.5. Электрофорез в ДСН-ПААГ.
Пример 1
Определение и получение полинуклеотида, кодирующего IgA1 протеазу Neisseria meningitidis серогруппы В штамм Н44/76 с мутацией каталитически активного остатка Ser267 (SEQ ID NO:1)
На основании известной последовательности фрагмента гена iga штамма Н44/76, кодирующего форму протеазы, близкую по размеру к секретируемой (примерно 900-1000 аминокислотных остатков, а.о.), которая описана в дополнительном примере 1, были сконструированы праймеры для замены каталитически активного остатка Ser267 на Ala и последующего клонирования в экспрессионный вектор. Выбранная структура праймеров позволяет амплифицировать продукт полимеразной цепной реакции (ПЦР), пригодный для последующего встраивания в экспрессионные векторы семейства рЕТ (Novagen), используемые для получения белков в штаммах Е.coli, лизогенных по бактериофагу Т7.
Для получения мутантной формы IgA1-протеиназы, в которой каталитически активный остаток Ser267 заменен на Ala, был использован метод SOE (splicing by overlap extension, сплайсинг при расширении частично перекрывающихся последовательностей). Для этого были сконструированы специфические праймеры f3 и f4 (f3 - прямой - 5'-GCTGTGTTAGGCGATGCCGGATCCCCATTATTTGCTTATG-3' SEQ ID №5, f4 - обратный - 5'-CATAAGCAAATAATGGGGATCCGGCATCGCCTAACACAGC-3' SEQ ID №6), перекрывающиеся в области сайта мутации, кроме этого, в эти праймеры был введен сайт рестрикции эндонуклеазы BamHI.
Для последующего клонирования в векторе рЕТ-24(а+) в праймеры (f1 - прямой - 5-CGAGACAGCCATATGAAAACCAAACGTTTTAAAATTAAC-3 SEQ ID №3, f2 - обратный - 5-GGGCTCGAGATTGTACAATCGGGTAATACCG-3 SEQ ID №4), фланкирующие целевой участок гена iga штамма Н44/76, введены последовательности сайтов узнавания рестриктаз NdeI и XhoI (соответственно для f1 и f2), имеющиеся в полилинкере вектора. Строение праймера f2, не содержащего терминирующих кодонов, позволяет получать белок, модифицированный в С-концевой области гексагистидиновой меткой для последующего проведения металлохелатной хроматографии при его выделении.
Амплификацию фрагмента гена проводили в два этапа, используя в качестве матрицы плазмидную ДНК pBIGAPS1, описанную ранее в заявке RU 2011106149.
На первом этапе проводили параллельно две реакции амплификации фрагмента гена, в одной используя праймеры f1 и f4, в другой - f2 и f3. В результате были получены два фрагмента ДНК длиной около 800 и 2100 п.о., соответственно. На втором этапе был получен фрагмент ДНК около 2900 п.о. при использовании в реакции амплификации в качестве матрицы фрагментов ДНК, полученных на первом этапе, а также праймеров f1 и f2.
Амплификацию фрагментов нуклеиновой кислоты проводили по стандартному протоколу за 25 циклов ПЦР. Температуру отжига праймеров подбирали экспериментально. Обработку ПЦР-фрагмента и вектора рестриктазами, лигирование и трансформацию компетентных клеток проводили по стандартным процедурам. Целевой ПЦР-продукт был клонирован в вектор рЕТ-24а(+) (Novagen, кат. №69749-3) при использовании уникальных сайтов рестрикции NdeI и XhoI таким образом, чтобы продуцируемый белок содержал His-метку в C-концевой области для последующей очистки белка с помощью металлохелатной хроматографии. Секвенирование полученной ДНК подтвердило наличие мутации в белке (SEQ ID NO:2). Полученная плазмидная ДНК была названа pBIGAPS1-M.
Пример 2
А) Получение рекомбинантной плазмидной ДНК pBIGAPS1-M
Рекомбинантную плазмидную ДНК получали из клеток ночной культуры стандартным щелочным методом (Maniatis, Т., Fritsch, E.F. and Sambrook, J. (1982) Molecular Cloning: a Laboratory Manual, Cold Sping Harbor Laboratory Press) и использовали для введения в компетентные клетки с помощью теплового шока. Для этого к 50 мкл суспензии компетентных клеток добавляли 100 нг - 1 мкг плазмидной ДНК и инкубировали в течение 40 мин на льду. Полученную смесь инкубировали при 42°С в течение 2 мин, затем инкубировали во льду в течение 2 мин, добавляли 1 мл холодного бульона LB и инкубировали в течение 1 ч при 37°С. После этого 100 мкл суспензии бактериальных клеток высевали на чашки Петри с агаром, содержащим 50 мкг/мл канамицина. Чашки инкубировали в течение ночи при 37°С, бактериальные клетки нескольких выросших колоний использовали для независимого засева микробиологических пробирок с 5 мл жидкой среды LB, содержащей 50 мкг/мл канамицина, с последующим выращиванием бактериальных клеток на качалке при 37°С в течение ночи. После этого в пробирки добавляли глицерин до конечной концентрации 15%, суспензию клеток разливали в пластиковые пробирки и хранили при -70°С.
Физическая карта рекомбинантной плазмидной ДНК приведена на фигуре 1.
Б) Получение штамма
Штамм Е.coli BL21(DE3)/pBIGAPS1-M получали путем введения плазмидной ДНК pBIGAPS1-M в компетентные клетки Е.coli BL21(DE3) с помощью метода теплового шока. Для получения компетентных клеток, бактериальные клетки исходного штамма Escherichia coli BL21(DE3) (E.coli В F- dcm ompT hsdS(rB - mB -) gal λ(DE3)) выращивали в жидкой среде LB в течение ночи при 37°С и использовали для засева 100 мл той же среды. Культуру выращивали до ранней логарифмической фазы на качалке при интенсивной аэрации (до оптической плотности при 600 нм=0.4÷0.5), быстро охлаждали во льду, клетки осаждали центрифугированием в течение 10 мин при 5000 об/мин при +4°С. Осадок дважды промывали в 50 мл 100 мМ CaCl2. Осажденные клетки ресуспендировали в 2 мл 100 мМ CaCl2, с добавлением глицерина до 10%. Разделяли на аликвоты по 50 мкл, быстро замораживали и хранили при -70°С.
Пример 3
Определение продуктивности штамма-продуцента IgA1 протеазы
В 15 мл жидкой среды LB, содержащей 50 мкг/мл канамицина, вносили 1% инокулят ночной культуры клеток и выращивали при 37°С на качалке при 180 об/мин в течение 2 ч до мутности 0,8. Затем добавляли индуктор изопропилтиогалактопиранозид (ИПТГ) до конечной концентрации 0,5 мМ и продолжали инкубацию в тех же условиях в течение 2,0 ч. Отбирали пробу 1 мл и центрифугировали в течение 6 мин при 12000 об/мин, после чего клетки суспендировали в 100 мкл деионизированной воды, добавляли 33 мкл буферного раствора, содержащего 125 мМ Трис-HCl, рН 6,8, 20% глицерин, 3% додецилсульфат натрия, 3% меркаптоэтанол и 0,01% бромфеноловый синий, нагревали в течение 10 мин до 95°С. Отбирали аликвоту и анализировали электрофорезом в 10% полиакриламидном геле, содержащем 0,1% додецилсульфат натрия. Гель окрашивали Кумасси R250 и сканировали на лазерном сканере. Продуктивность штамма Е.coli BL21(DE3)/pBIGAPS1 составляет 40% фермента от суммарного клеточного белка.
Пример 4
Выделение рекомбинантной мутантной IgA1 протеазы
Разрушение клеток и фракционирование лизата
Размороженные после хранения клетки (2 г из 1 л индуцированной культуры) ресуспендировали в семикратном объеме 20 мМ Tris-HCl-буфера, рН 8.5, содержащем лизоцим. Суспензию клеток пятикратно обрабатывали ультразвуком и подвергали центрифугированию (1 час, 20000 g). Разделенные фракции растворимых белков (бесклеточный экстракт, БЭ) и нерастворимая часть клеточного лизата (дебрис и нерастворимые белки) изолировали. Согласно результатам электрофоретического анализа целевой белок формирует нерастворимые тельца включения (ТВ), независимо от температурных условий индукции культуры и типа буфера, использованного при разрушении клеток. Нерастворимую часть лизата клеток, содержащую ТВ после центрифугирования, промывали: а) 60 мл буфера 20 мМ Tris-HCl, 1% Triton X-100, 0.1% SDS, центрифугировали 40 мин 10000 g; б) 60 мл буфера 20 мМ Tris-HCl, I M NaCl, центрифугируют 40 мин, 10000 g. Осадок использовали для проведения хроматографии на Ni-агарозе в денатурирующих условиях.
Металлхелатная хроматография на Ni-агарозе
ТВ, выделенные из 2 г клеток, растворяли в 20 мл 20 мМ Tris-HCl, рН 8.5, содержащего 8 М мочевину и 5 мМ дитиотреитол (ДТТ). Суммарное количество белка, содержащегося в ТВ, составлял около 40 мг при определении по методу Брэдфорд. После центрифугирования и фильтрования раствор инкубировали в течение ночи с 10 мл Ni-агарозы (Qiagen, Cat No 30410), уравновешенной тем же буфером. Затем сорбент промывали 10V колонки буфером, содержащим 20 мМ Tris-HCl рН 8.5, 8 М мочевину, 250 мМ NaCl. Связавшиеся белки элюировали 5 объемами колонки буфером, содержащим 20 мМ Tris-HCl рН 8.5, 8 М мочевину, 250 мМ NaCl, 400 мМ имидазола. Фракции, содержащие очищенный белок, объединяли и использовали для переведения IgA1 протеазы в растворимую форму. Выход частично очищенного препарата целевого белка на этой стадии составляет - 30 мг из 2 г биомассы клеток.
Рефолдинг IgA1 протеазы
Объединенный элюат (30 мл, 1 мг/мл), полученный после проведения хроматографии на Ni-агарозе, подвергали ступенчатому диализу, постепенно снижая концентрацию мочевины до значения 1,5 М (что позволяет получить препарат с высокой концентрацией белка) в 20 мМ Tris-HCl буфере, рН 8,5, 1 мМ DTT. Далее раствор белка подвергали концентрированию, используя метод ультрафильтрации. Далее раствор подвергали центрифугированию (30 мин, 5000 g) и фильтрованию через стерильную мембрану с размером пор 0.2 мкм. Концентрация белка в конечном препарате составляет 1,8 мг/мл, объем 10 мл.
Выход белка на конечной стадии составляет 18 мг из 2 г биомассы клеток. Чистота препарата - не менее 90%.
Пример 5
Определение иммуногенной и протективной активности
Иммуногенную и протективную активность полученной IgA1 протеазы оценивали на мышах линии Balb/C. Для оценки этих показателей животных иммунизировали различными препаратами IgA1 протеазы (нативная IgA1 протеаза, выделенная из культуры Neisseria meningitidis серогруппы А, полученная, как описано в патенте RU 2407792, рекомбинантная IgA1 протеаза серогруппы В, полученная как описано в дополнительном примере 1, мутантная IgAl протеаза серогруппы В, полученная, как описано в настоящем описании, пример 4) однократно в дозе 20 мкг/мышь. По данным иммуноферментного анализа, проведенного, как описано в патенте РФ №2310853, мутантная рекомбинантная IgA1 протеаза в отличие от нативной и рекомбинантной немутантной IgA1 протеазы не обладает специфической ферментативной активностью, т.е. не гидролизует IgA1 человека.
Показателем иммуногенности полученной IgA1 протеазы служит нарастание специфических антител в крови мышей на 30-й день после иммунизации, определяемых методом ИФА.
На фиг.2 показана диаграмма иммуногенной активности указанных выше препаратов IgA1 протеазы. По оси ординат - оптическая плотность сывороток при λ=492 нм. Уже после однократной иммунизации уровень специфических антител в сыворотках мышей, иммунизированных мутантной IgA1 протеазой, возрастает до 1:5000, что превышает этот показатель у мышей, иммунизированных нативной и рекомбинантной IgA1 протеазой (1:2560).
Через 1 месяц после иммунизации мышей подразделяли на три группы и заражали живой вирулентной культурой менингококка серогрупп А штамм А208, В штамм Н44/47 и С штамм 0638.
Протективную активность IgA1 протеазы оценивали по уровню бактериемии у иммунизированных животных через 4 часа после заражения менингококком по сравнению с этим же показателем у контрольных (не иммунизированных) мышей.
На диаграмме фиг.3 представлены уровень бактериемии (в %) у мышей, однократно иммунизированных указанными выше препаратами IgA1 протеазы (заштрихованные столбики), после заражения менингококком серогрупп А, В и С по сравнению с контрольными, не иммунизированными животными (черные столбики). Уровень бактериемии в группе мышей, иммунизированных мутантной протеазой, составлял соответственно для серогрупп А, В и С 13, 19 и 17 КОЕ, в пересчете на 100 КОЕ в контроле (у не иммунизированных мышей).
Дополнительный пример 1
А) Определение и получение нуклеиновой кислоты, кодирующей IgA1 протеазу Neisseria meningitides серогруппы В штамм Н44/76 (SEQ ID NO:7)
Проведение анализа данных геномного банка (http://www.ncbi.nlm.nih.gov/Genbank) выявило, что к настоящему времени для Neisseria meningitidis серогруппы В известен единственный геном штамма МС58. На основании предположения о близости строения генов IgA1 протеаз родственных штаммов, по известной последовательности гена iga МС58, сконструированы праймеры для клонирования фрагмента нуклеиновой кислоты (гена) штамма Н44/76, кодирующего форму протеазы, близкую по размеру к секретируемой (примерно 900-1000 аминокислотных остатков, а.о.). Выбранная структура праймеров позволяет амплифицировать продукт полимеразной цепной реакции (ПЦР), пригодный для последующего встраивания в экспрессионные векторы семейства рЕТ (Novagen), используемые для получения белков в штаммах Е. coli, лизогенных по бактериофагу Т7.
Для последующего клонирования в векторе рЕТ-24(а+) в праймеры (f1 - прямой - 5-CGAGACAGCCATATGAAAACCAAACGTTTTAAAATTAAC-3 SEQ ID №3, f2 - обратный - 5-GGGCTCGAGATTGTACAATCGGGTAATACCG-3 SEQ ID №4), потенциально фланкирующие целевой участок гена iga штамма Н44/76, введены последовательности сайтов узнавания рестриктаз NdeI и XhoI (соответственно для f1 и f2), имеющиеся в полилинкере вектора. Строение праймера f2, не содержащего терминирующих кодонов, позволяет получать белок, модифицированный в С-концевой области гексагистидиновой меткой для последующего проведения металлохелатной хроматографии при его выделении.
Амплификацию фрагмента нуклеиновой кислоты проводят по стандартному протоколу за 25 циклов ПЦР, используя в качестве матрицы геномную ДНК штамма Н44/76. Температуру отжига праймеров подбирают экспериментально. Обработку ПЦР-фрагмента и вектора рестриктазами, лигирование и трансформацию компетентных клеток проводят по стандартным процедурам. Далее проводят секвенирование клонированной ДНК с нескольких первичных клонов, в результате впервые установлена нуклеотидная последовательность фрагмента нуклеиновой кислоты, кодирующего секретируемую форму IgA1 протеазы из Neisseria meningitides Н44/76, на основании которой определена аминокислотная последовательность белка (SEQ ID NO:8). Полученная плазмидная ДНК была названа pBIGAPS1.
Б) Получение рекомбинантной плазмидной ДНК pBIGAPS1
Рекомбинантную плазмидную ДНК получают из клеток ночной культуры стандартным щелочным методом (Maniatis, Т., Fritsch, E.F. and Sambrook, J. (1982) Molecular Cloning: a Laboratory Manual, Cold Sping Harbor Laboratory Press) и используют для введения в компетентные клетки с помощью теплового шока. Для этого к 50 мкл суспензии компетентных клеток добавляют 100 нг - 1 мкг плазмидной ДНК и инкубируют в течение 40 мин на льду. Полученную смесь инкубируют при 42°С в течение 2 мин, затем инкубируют во льду в течение 2 мин, добавляют 1 мл холодного бульона LB и инкубируют в течение 1 ч при 37°С. После этого 100 мкл суспензии бактериальных клеток высевают на чашки Петри с агаром, содержащим 50 мкг/мл канамицина. Чашки инкубируют в течение ночи при 37°С, бактериальные клетки нескольких выросших колоний используют для независимого засева микробиологических пробирок с 5 мл жидкой среды LB, содержащей 50 мкг/мл канамицина, с последующим выращиванием бактериальных клеток на качалке при 37°С в течение ночи. После этого в пробирки добавляют глицерин до конечной концентрации 15%, суспензию клеток разливают в пластиковые пробирки и хранят при -70°С.
Физическая карта рекомбинантной плазмидной ДНК приведена на фигуре 1.
В) Получение штамма
Штамм Е. coli BL21(DE3)/pBIGAPS1 получают путем введения плазмидной ДНК pBIGAPSI в компетентные клетки Е. coli BL21(DE3) с помощью метода теплового шока. Для получения компетентных клеток, бактериальные клетки исходного штамма Escherichia coli BL21(DE3) (E. coli В F- dcm ompT hsdS(rB - mB -) gal λ(DE3)) выращивают в жидкой среде LB в течение ночи при 37°С и используют для засева 100 мл той же среды. Культуру выращивают до ранней логарифмической фазы на качалке при интенсивной аэрации (до оптической плотности при 600 нм=0.4-0.5), быстро охлаждают во льду, клетки осаждают центрифугированием в течение 10 мин при 5000 об/мин при +4°С. Осадок дважды промывают в 50 мл 100 мМ CaCl2. Осажденные клетки ресуспендируют в 2 мл 100 мМ СаС12, с добавлением глицерина до 10%. Разделяют на аликвоты по 50 мкл, быстро замораживают и хранят при -70°С.
Г) Выделение рекомбинантной IgA1 протеазы
Разрушение клеток и фракционирование лизата
Размороженные после хранения клетки (2 г из 1 л индуцированной культуры) ресуспендируют в семикратном объеме 20 мМ Tris-HCl-буфера, рН 8.5, содержащем 1% Тритон-Х100, либо в детергент-содержащем 20 мМ калий-фосфатном буфере, рН 7.5. Суспензия клеток пятикратно обработана ультразвуком и подвергнута центрифугированию (1 час, 20000 g). Разделенные фракции растворимых белков (бесклеточный экстракт, БЭ) и нерастворимая часть клеточного лизата (дебрис и нерастворимые белки) изолируют. Согласно результатам электрофоретического анализа целевой белок формирует нерастворимые тельца включения (ТВ) независимо от температурных условий индукции культуры и типа буфера, использованного при разрушении клеток. Нерастворимую часть лизата клеток, содержащую ТВ, после ресуспендирования в 1% Тритон-Х100 при повторной обработке ультразвуком и последующего центрифугирования используют для проведения хроматографии на Q-сефарозе в денатурирующих условиях.
Анионообменная хроматография на Q-сефарозе
ТВ, выделенные из 2 г клеток, индуцированные при 25°С, растворяют в 20 мл Tris-HCl-буфера, рН 8.5, содержащего 6 М мочевину и 20 мМ дитиотреитол (ДТТ). Суммарное количество белка, содержащегося в ТВ, составляет около 50 мг при определении по методу Брэдфорд. После центрифугирования и фильтрования раствор наносят со скоростью 1 мл/мин на 10 мл анионообменного сорбента (две предупакованные картридж-колонки HiTrap Q Sepharose HP, GE Healthcare, Швеция), уравновешенного тем же буфером. Фракции, содержащие очищенный белок и полученные в результате градиентной элюции хлоридом натрия (100 мл, 0-1 М NaCl) в стартовом буфере, объединяют (в сумме - 20 мл) и используют для переведения IgA1-протеазы в растворимую форму. Выход частично очищенного препарата целевого белка на этой стадии составляет - 40 мг из 2 г биомассы клеток.
Рефолдинг IgA1 протеазы
Объединенный элюат (20 мл, 2 мг/мл), полученный после проведения хроматографии на Q-сефарозе, разбавляют в 10 раз 20 мМ Tris-HCl-буфером, рН 8.5, методом быстрого разбавления (выкапывание раствора белка в буфер). После медленного перемешивания раствора (при концентрации белка 0.2 мг/мл) в течение 20 часов и последующего концентрирования методом ультрафильтрации до исходного объема раствор белка трижды диализуют против 1 л 20 мМ Tris-HCl-буфера, рН 8.5. Далее раствор подвергают центрифугированию (30 мин, 5000 g) и фильтрованию через стерильную мембрану с размером пор 0.2 мкм.
Выход белка на конечной стадии составляет 20 мг из 2 г биомассы клеток. Чистота препарата - не менее 90%.
Дополнительный пример 2
Оценка ферментативной активности образцов рекомбинантой активной и мутантной IgA1 протеазы
В двух образцах рекомбинантной активной и мутантной IgA1 протеазы (образец 1 и образец 2) определяли ферментативную активность методом ИФА с использованием в качестве субстрата миеломного IgA1 человека и конъюгата анти-IgA человека с пероксидазой хрена, как описано в патенте РФ №2310853.
Анализ ферментативной активности полученных образцов рекомбинантной IgA1 протеазы показал (фиг.4) способность активной IgA1 протеазы катализировать гидролиз иммуноглобулина IgA1 и отсутствие протеолитической активности у мутантного образца.
Уровень протеолитической активности IgA1 протеазы определяли также электрофоретическим методом в ДСН-ПААГ. На фиг.5 показан гидролиз IgA1 в присутствии различных концентраций активного фермента и его мутантной формы (образец 1 и образец 2, соответственно).
Видоспецифическую активность полученных препаратов IgA1 протеазы оценивали методом ИФА по уровню IgA1 в сыворотках мышей и человека. Уровень антител к IgA1 протеазе в крови животных, иммунизированных мутантным препаратом, составлял 1:1280 и не зависел от наличия ферментативной активности в исследуемом образце активной протеазы (см. табл.).
Таким образом, на основе полученной плазмидной ДНК pBIGAPS1-M создан экспрессионный штамм Е. coli суперпродуцент IgA1 протеазы серогруппы В, выделен белок и осуществлен его рефолдинг, позволяющий получать IgA1 протеазу в количестве на уровне 10-40 мг из 2 г биомассы клеток.
Впервые получены результаты, свидетельствующие о более высокой иммуногенной и протективной активности мутантной IgA1 протеазы в сравнении с рекомбинантной менингококковой протеазой серогруппы В, как описано в статье Серова О.В. и др., Биофармацевтический журнал, 2012, т.2, №1. Полученный препарат мутантной IgA1 протеазы обладает поливалентным действием в отношении защиты от менингококковой инфекции организма животного независимо от источника ее получения (серогруппы А или В). Отсутствие ферментативной активности в отношении иммуноглобулина А1 человека мутантной IgA1 протеазы позволит избежать дополнительных осложнений при иммунизации человека. Представленные данные по иммуногенности и протективности дают основание считать возможным использование ее в качестве поливакцины для лечения бактерионосительства и профилактики менингококковой инфекции основных эпидемических серогрупп А, В и С, в первую очередь, для профилактики менингококковых инфекций, вызываемых бактериями N. meningitidis серогруппы В, а также других патогенов, активность которых обусловлена наличием IgA1 протеазы.
| название | год | авторы | номер документа |
|---|---|---|---|
| НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ ФУНКЦИОНАЛЬНО АКТИВНУЮ РЕКОМБИНАНТНУЮ IgA1 ПРОТЕАЗУ NEISSERIA MENINGITIDIS СЕРОГРУППЫ B, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, СОДЕРЖАЩАЯ НУКЛЕОТИДНУЮ ПОСЛЕДОВАТЕЛЬНОСТЬ, КОДИРУЮЩУЮ АКТИВНУЮ IgA1 ПРОТЕАЗУ, ШТАММ-ПРОДУЦЕНТ, СОДЕРЖАЩИЙ ПЛАЗМИДНУЮ ДНК, ПРОДУЦИРУЮЩИЙ ЗРЕЛУЮ ФОРМУ IgA1 ПРОТЕАЗЫ, РЕКОМБИНАНТНАЯ Ig ПРОТЕАЗА NEISSERIA MENINGITIDIS СЕРОГРУППЫ В, СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОЙ ФОРМЫ IgA1 ПРОТЕАЗЫ, ОБЛАДАЮЩЕЙ ИММУНОГЕННЫМИ И ПРОТЕКТИВНЫМИ СВОЙСТВАМИ | 2011 |
|
RU2453599C1 |
| Рекомбинантный белок, обладающий протективным действием в отношении менингококков (варианты), полинуклеотид, кодирующий рекомбинантный белок, рекомбинантная плазмидная ДНК, содержащая указанный полинуклеотид, клетка-хозяин, содержащая указанную рекомбинантную плазмидную ДНК, способ получения рекомбинантного белка | 2017 |
|
RU2701964C2 |
| СПОСОБ ПОЛУЧЕНИЯ IgAl-ПРОТЕАЗЫ ИЗ КУЛЬТУРЫ NEISSERIA MENINGITIDIS СЕРОГРУППЫ А И ИММУНОГЕННЫЙ ПРЕПАРАТ НА ЕЕ ОСНОВЕ | 2009 |
|
RU2407792C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ОРЕКСИНА А ЧЕЛОВЕКА, ПЛАЗМИДНАЯ ДНК, ШТАММ-ПРОДУЦЕНТ | 2016 |
|
RU2647771C2 |
| КОМПОЗИЦИИ NEISSERIA MENINGITIDIS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2665841C2 |
| СПОСОБ ВЫСОКОУРОВНЕВОЙ ЭКСПРЕССИИ, СПОСОБ ОЧИСТКИ И СПОСОБ ПЕРЕУКЛАДКИ ПОРИНОВОГО ПРОТЕИНА ВНЕШНЕЙ МЕМБРАНЫ МЕНИНГОКОККА ГРУППЫ В ИЛИ СЛИТОГО С НИМ ПРОТЕИНА, ВЫСОКООЧИЩЕННЫЙ ПОРИНОВЫЙ ПРОТЕИН (ВАРИАНТЫ), ВЫСОКООЧИЩЕННЫЙ ПОРИНОВЫЙ СЛИТЫЙ ПРОТЕИН (ВАРИАНТЫ), ВАКЦИНА ДЛЯ ПРЕДУПРЕЖДЕНИЯ БАКТЕРИАЛЬНОГО МЕНИНГИТА ГРУППЫ В, СПОСОБ ПОЛУЧЕНИЯ КОНЪЮГАТА ПРОТЕИНА МЕНИНГОКОККА ГРУППЫ В И ПОЛИСАХАРИДА, СПОСОБ ПРЕДУПРЕЖДЕНИЯ БАКТЕРИАЛЬНОГО МЕНИНГИТА, ШТАММ Е.COLI (ВАРИАНТЫ) | 1994 |
|
RU2181378C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ ГИБРИДНЫЙ БЕЛОК L-HEP, ШТАММ ESCHERICHIA COLI ПРОДУЦЕНТ УКАЗАННОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА | 2019 |
|
RU2716975C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PTRX-TEVRS-РТН, КОДИРУЮЩАЯ ГИБРИДНЫЙ БЕЛОК, СПОСОБНЫЙ К ПРОТЕОЛИТИЧЕСКОМУ РАСЩЕПЛЕНИЮ С ОБРАЗОВАНИЕМ ФРАГМЕНТА ЭНДОГЕННОГО ЧЕЛОВЕЧЕСКОГО ПАРАТИРЕОИДНОГО ГОРМОНА (1-34), ШТАММ ESCHERICHIA COLI BL21(DE3)/PTRX-TEVRS-РТН - ПРОДУЦЕНТ УКАЗАННОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО PTH (1-34) | 2019 |
|
RU2700452C1 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ТРАНСФЕРРИН-СВЯЗЫВАЮЩИЙ БЕЛОК И HSF ИЗ ГРАМОТРИЦАТЕЛЬНЫХ БАКТЕРИЙ | 2003 |
|
RU2359696C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК ДЛЯ ОБНАРУЖЕНИЯ АГЕНТОВ, ПОВРЕЖДАЮЩИХ ГЕНЕТИЧЕСКИЙ АППАРАТ КЛЕТКИ (ВАРИАНТЫ) | 2005 |
|
RU2311459C2 |

















Изобретение относится к области иммунологии и генной инженерии. Предложена ферментативно неактивная IgA1 протеаза с заменой Ser267Ala для применения в качестве компонента поливалентной вакцины, предназначенной для защиты человека от менингококковой инфекции и других микроорганизмов, патогенность которых обусловлена IgA1 протеазой. Изобретение включает полинуклеотид, кодирующий названную мутантную форму IgA1 протеазы Neisseria meningitidis серогруппы В; содержащую данный полинуклеотид рекомбинантную плазмидную ДНК, штамм Escherichia coli - продуцент мутантной формы IgA1 протеазы по изобретению; способ получения рекомбинантной формы фермента с помощью технологии рекомбинантных ДНК и рекомбинантную инактивированную IgA1 протеазу, проявляющую повышенный в сравнении с ферментом дикого типа уровень иммуногенности. 5 н. и 1 з.п. ф-лы, 1 табл., 5 ил., 7 пр.
1. Полинуклеотид, кодирующий мутантную IgA1 протеазу Neisseria meningitidis с нуклеотидной последовательностью (SEQ ID NO:1).
2. Рекомбинантная плазмидная ДНК, включающая полинуклеотид по п.1 и обеспечивающая экспрессию мутантной IgA1 протеазы в клетке-хозяине.
3. Клетка-хозяин, трансформированная рекомбинантной плазмидной ДНК по п.2, продуцирующая зрелую мутантную форму IgA1 протеазы.
4. Клетка-хозяин по п.3, где прокариотическая клетка-хозяин выбрана из ряда, включающего бактерии рода Escherichia, Bacillus, Lactococcus, Lactobacillus, Pseudomonas, Streptomyces, Streptococcus, Staphylococcus.
5. Способ получения зрелой мутантной формы IgA1 протеазы Neisseria meningitidis серогруппы В, заключающийся в том, что клетку-хозяина трансформируют рекомбинантной плазмидной ДНК по п.2, культивируют трансформированные клетки по п.3 с получением зрелой мутантной формы IgA1 протеазы, а целевой продукт выделяют из телец включения и очищают.
6. Мутантная IgA1 протеаза N.meningitidis серогруппы В с аминокислотной последовательностью SEQ ID NO:2, обладающая повышенным уровнем иммуногенности по сравнению с ферментом дикого типа.
| СЕРОВА О.В | |||
| и др | |||
| - Биофармацевтический журнал, 3 (6), 42-47, 2011 | |||
| ROMANELLO V | |||
| et al | |||
| Protein Expr | |||
| Purif., 45(1), 142-149, 2006 | |||
| VITOVSKI S | |||
| and SAYERS J | |||
| Infect | |||
| Immunol., 75(6), 2875-2885, 2007. |

