Изобретение относится к области рекомбинантной генетики, экспрессии протеина и созданию вакцин. Изобретение также включает методы выделения поринового протеина наружной мембраны менингококка группы В в рекомбинантной клетке-хозяине. Изобретение включает и методы очистки и переукладки рекомбинантного протеина.
Наружные мембраны вида Neisseria в большей степени, чем другие грам-отрицательные бактерии, обладают полупроницаемой мембраной, позволяющей свободно проникать в обоих направлениях - как в клетку, так и из клетки субстанциям с низкомолекулярной массой, но задерживать молекулы большего размера (Heasley, F.A., et al., "Reconstitution and characterization of the N. gonnorrhoeae outer membrane permeability barrier", в Genetics and Immunobiology of Neisseria gonorrhoeae, Daniellson and Normark, eds., University of Umea, Umea, pp12-15 (1980); Douglas J.T. et al., FEMS Microbiol. Lett. 12: 305-309 (1981). Одним из механизмов, позволяющих осуществлять транспорт молекул из и в клетку, является включение в мембрану протеинов, называемых пориновыми. Порины состоят из трех идентичных полипептидных цепей (Jones R. B. et al. , Infect. Immun. 30:773-780 (1980); Mc Dade Jr. and Jhonston, J. Bacteriol. 141:1183-1191 (1980)), имеют нативную трехмерную конформацию, формируют насыщенные водой вольтаж-зависимые каналы, находящиеся в наружной мембране или других мембранах бактерии, в которые и включены порины (Linch, E. C., et al., Biophys. J. 41:62 (1983); Linch, E.C. et al., Biophys. J. 45: 104-107 (1984); Young J.D.E., et al., Proc. Natl. Acad. Sci USA 80:3831-3835 (1983); Mauro. A. , et al., Proc. Natl.Acad. Sci USA 85:1071-1075 (1988); Young J.D.E., et al., Proc. Natl. Acad. Sci USA 83:150-154 (1986)). В связи с большим количеством протеинов, находящихся в составе наружной мембраны, в зависимости от протеиновых антигенов бактерии подразделены на подгруппы Neisseria meningitidis и Neisseria gonorrhoeae, которые в свою очередь подразделены на серотипы в эпидемиологических целях (Frasch, С. Е. et al., Rev. Infect. Dis. 7:504-510 (1985); Knapp, J.C., et al., "Overview of epidemiological and clinical applications of auxtore/serovar classification N. gonorrhoeae", The Pathogenic Neisseriae, Schoolinck G.K.,ed., American Society of Microbiology, Washington, pp 6-12 (1985)). Многие из этих протеинов, принадлежащих как гонококку, так и менингококку, были очищены (Heckels, J.E. , J. Gen. Microbiol. 99:333-341 (1977); James and Heckels, J. Immunol. Meth. 42: 223-228 (1981); Jadd R.C., Anal. Biochem. 173:307-316 (1988); Blake and Gotschlich, Infect. Immun. 36:277-283 (1982); Wetzler L.M. et al., J. Rxp. Med/168:1883-1897(1988)), клонированы и упорядочены (Gotschlich E.C., et al. , Proc. Natl. Acad. Sci. USA 84: 8135- 8139 (1987); McGuinness, В., et al., J. Exp. Med. 171: 1871-1882 (1990); Carbonetti and Sparling, Proc. Natl. Acad. Sci. USA 84: 9084- 9088 (1987); Feavers, I.M., et al., Infect. Immun. 60: 3620-3629 (1992); Murakami, К. , et al, Infect. Immun. 57:2318-2323 (1989); Wolff and Stern, FEMS Microbiol. Lett. 83: 179-186 (1991); Ward M.J. , et al., FEMS Microbiol., Lett. 73:283-289 (1992)).
Пориновые протеины вначале были изолированы вместе с липополисахаридами. Они были названы "эндотоксин- ассоциированные протеины". Изучения поринов природных штаммов показали, что полная агрегация и олигомеризация не возможны до тех пор, пока в окружающей клетку среде присутствует липополисахарид (ЛПС) соответствующего бактериального штамма (Holcenburg et al., Biochemistry 28:4187-4193 (1989); Sen and Nicaido, J. Biol. Chem. 266:11295-11300 (1991).
Менингококковые порины были подразделены на три большие классификации, которые в предшествующей номенклатуре были известны как Класс 1, 2 и 3 (Frasch, C.E., et al., Rev. Infect. Dis. 7:504-510 (1985)). Каждый изученный менингококк содержит один из аллелей поринового гена Класса 2 или 3, но не оба аллеля (Feavers, I.M. et al., Infect. Immun. 60:3620-3629 (1992); Murakami, К. , et al. , Infect. Immun. 57:2318-2323 (1989)). Наличие или отсутствие гена Класса 1 не является обязательным. Так же все изученные гонококки содержат только один ген порина, имеющий сходство с аллелями 2 или 3 класса (Gotschlich, E. C. et al., Proc. Natl. Acad. Sci. USA 84: 8135-8139 (1987); Carbonetti and Sparling, Proc. Natl. Acad. Sci. USA 84: 9084-9088 (1987)). Обнаружено, что у N. gonorrhoeae полностью отсутствует аллель класса 1. Полученные данные о генах позволяют предположить, что пориновые протеины нейссерии имеют по крайней мере 70% гомологичности друг с другом с различными вариантами основного предмета (Feavers, I.M. et al., Infect. Immun. 60: 3620-3629 (1992)). Было предположено, что различия между пориновыми протеинами нейссерий обусловлены иммунологическим влиянием, вызванным во время инвазии патогенного микроорганизма в человеческий организм. Вместе с тем, очень мало известно, как изменения последовательностей пориновых протеинов влияют на функциональную активность этих белков.
Как было сказано выше, изолированные гонококковые порины аллельного типа Класса 2 по электрофизиологическим свойствам отличаются от изолированных поринов гонококка класса 3, это было показано при изучении их липидной мембраны в отношении ионной селективности и вольтаж-зависимости (Lynch,E.C. et al. , Biophys. J. 41:62 (1983); (Lynch, E.C. et al., Biophys. J, 45:104-107 (1984)). Способность различных пориновых протеинов проникать в двойной липидный слой интактных бактерий коррелирует не только с типом порина, но и с видом нейссерии, из которой они были получены (Linch, E.C., et al., Biophys. J. 45:104-107 (1984)). Как оказалось, некоторые функциональные признаки могли быть отнесены к различным областям последовательности порина. Одна такая функциональная зона, ранее идентифицированная у всех гонококковых класс 2-подобных протеинов - участок расщепления химотрипсином. При расщеплении химотрипсином этот класс поринов теряет способность к ответу на потенциал напряжения и к закрытию каналов мембраны. Класс 3-подобные порины гонококка, так же как и менингококковые порины, не содержат последовательности, чувствительной к воздействию химотрипсина, однако и они не способны отвечать при воздействии потенциального напряжения закрытием мембранных каналов (Greco, F. "The formation of channels in lipid bilayers by gonococcal major outer membraine protein" тезисы, The Rockfeller University, New York (1981); Greco, F. et al., Fed. Proc. 39:1813 (1980)).
Трудности во время исследований возникли вследствие необходимости возможности для легкого манипулирования генами порина с помощью новых молекулярных технологий и при получении достаточно очищенного протеина для биофизической характеристики измененных пориновых протеинов. Как было ранее изучено, клонированные и экспрессированные в Е. coli гены порина нейссерии оказались смертельными для самой Е. coli (Carbonetti and Sparling, Proc. Natl. Acad. Sci. USA 84: 9084-9088 (1987); Carbonetti, N.H. et al., Proc. Natl. Acad. Sci. USA, 85:6841- 6845 (1988); Barlow, A.K., et al., Infect. Immun. 55:2734-2740 (1987)). Таким образом, многие из этих генов были клонированы и упорядочены, как части целого гена, или помещены в плазмиды с небольшим числом копий под контролем экспрессии (Carbonetti N.H., et al., Proc. Natl. Acad Sci. USA 85: 6841-6845 (1988)). В таких условиях, даже если весь ген порина экспрессирован, лишь небольшое количество накопленных протеинов может быть получено и очищено для характеристики этих протеинов.
Другая сторона проблемы - клонирование генов порина в низкокопийный плазмид с контролируемой экспрессией, введение модификаций в ген порина и затем вновь введение модифицированной последовательности назад в Neisseria (Carbonetti N. H. et al., Proc. Natl. Acad. Sci. USA 85:6841-6845. (1988)). Вместе с тем, возникла проблема ограничения эндонуклеазной системы, присутствующей в Neisseria, особенно в гонококках (Davies, J.K., Clin. Microbiol. Rev. 2:S78-S82 (1989)).
Настоящее изобретение направлено на преодоление описанных выше трудностей. Последовательность ДНК зрелых протеинов класса 2, 3 и связанных с ними протеинов может быть амплифицирована с использованием хромосомы менингококковой бактерии в качестве шаблона для реакции полимерразных цепей (РПЦ-реакция. Амплифицированная последовательность порина была связана и клонирована в экспрессионный вектор, содержащий Т7 промотор. Штамм BL21 Е. coli, являющийся лизогенным для DE3 лямбда фага (Studier and Moffatt, J. Mol. Biol. 189: 113-130 (1986)), модифицированный для элиминации ompА гена, был отобран в качестве экспрессионного хозяина для плазмида рЕТ-17b, содержащего ген порина. Во время индукции большое количество поринового протеина скопилось в Е. coli без какого-либо летального воздействия на бактерию-хозяина. Экспрессированные порин-протеины менингококка были экстрагированы и подвергнуты стандартной процедуре, а затем очищены методом хроматографии молекулярного сита и ионообменной хроматографии. При оценке протеинового профиля методом молекулярного сита, пориновые протеины были элюированы из колонки и определены как тримеры. Для уверенности в том, что никаких РПЦ артефактов не было внесено в менингококковый ген порина, учитывая высокий уровень экспрессии, была определена последовательность внесенного Por В гена. Были использованы исследования ELISA ингибиции для того, чтобы убедиться, что полученный рекомбинантный пориновый протеин имел природную антигенную структуру и тримерную конформацию.
Порины различных штаммов и видов Нейссерии различаются по первичной аминокислотной последовательности и по биофизическим характеристикам, как было замечено во время функциональных исследований. При изучении того, как изменения в первичной аминокислотной последовательности поринов Neisseria коррелируют с изменением биофизических свойств, были встречены определенные трудности, связанные с возможностью легко манипулировать клонированными генами порина при помощи новых молекулярных технологий и получить достаточное количество экспрессированного модифицированного поринового протеина для очистки его и применения в функциональных биофизических исследованиях. В данном изобретении ген, кодирующий зрелый Por В протеин, отсутствующий промотор нейссерии и сигнальная последовательность были клонированы в экспрессионный плазмид рЕТ-17b и трансформированы в Е. coli. При индукции было продуцировано большое количество Por В протеина.
Экспрессированный пориновый протеин был восстановлен с получением его нативной тримерной структуры, а затем очищен. Достаточно очищенный рекомбинантный пориновый протеин был получен как в качестве антигена, так и для получения биофизических характеристик. Таким образом, исследование ставит цели, где биофизические характеристики могут быть изучены более детально.
Общим объектом изобретения является обеспечение методом выделения поринового протеина менингококка группы В, в особенности относящегося к классу 2 и 3.
Специальным объектом изобретения является обеспечение методом выделения поринового протеина менингококка группы В класса 2 и 3 в Е. coli, включающим:
(a) трансформация Е. coli вектором, включающим избранный маркер и ген, кодирующий протеин, выбранный из группы, насчитывающей:
(i) зрелый пориновый протеин;
(ii) связанный протеин, включающий зрелый пориновый протеин, включенный в аминокислоты от 1 до 20 гена Т7 10 капсидного протеина;
указанный ген связан с Т7 промотором;
(b) выращивание трансформированной Е. coli в среде, содержащей слективный агент, и
(c) индуцирование экспрессии указанного протеина;
протеин составляет более чем 2% от общего протеина, экспрессированного в Е. coli.
Другим специальным объектом изобретения является обеспечение методом очистки и переукладывания поринового протеина менингококка группы В и связанного с ним протеина, полученных методами, описанными выше.
Дальнейшим объектом изобретения является получение вакцины, содержащей пориновый протеин и связанный с ним протеин менингококка группы В, полученных описанными выше методами, являющейся достаточно эффективно вызывающей выработку защитных антител у животных к N. meningitidis; а также получение фармацевтически приемлемого растворителя, носителя и эксципиента для вакцины.
Другим специфическим объектом исследования является обеспечение методом эффективной профилактики бактериального менингита у животных, включающего назначение животным вакцины, содержащей пориновый протеин и связанный с ним протеин менингококка группы В, полученные в соответствии с методами, описанными выше.
Другой специальный объект изобретения - получение обеспечения методом, позволяющим приготовить полисахаридный конъюгат, включающий: получение вышеуказанным методом поринового и связанного с ним протеина менингококка группы В; получение полисахарида из N. meningitidis; конъюгацию поринового и связанного с ним протеина менингококка группы В с полисахаридом.
Следующий специальный объект изобретения - получение метода очистки поринового и связанного с ним протеина менингококка группы В, включающего: лизирование трансформированной Е. coli для высвобождения поринового и связанного с ним протеина менингококка группы В, как части нерастворимых включений в организм-хозяин; отмывание включений буфером для удаления загрязняющих клеточных протеинов, принадлежащих Е. coli; взвешивание и растворение включений в водном растворе денатуранта; дилюция полученного раствора детергентом и очистка растворенного порина и связанного с ним протеина гель-фильтрацией и ионообменной хроматографией.
Следующий специальный объект изобретения - получение метода переукладки порина и связанного с ним протеина менингококка группы В, описанных выше, включающего: лизирование трансформированной Е. coli для высвобождения поринового и связанного с ним протеина менингококка группы В, как части нерастворимых включений в организм-хозяин; отмывание включений буфером для удаления загрязняющих клеточных протеинов, принадлежащих Е. coli; взвешивание и растворение включений в водном растворе денатуранта; дилюция полученного раствора детергентом и очистка растворенного порина и связанного с ним протеина гель-фильтрацией для получения переуложенного протеина в элюанте.
Следующий специфический объект изобретения - обеспечение штаммом BL21 Е. coli (DE3) дельтаompА клетки-хозяина, включающего вектор, содержащий ДНК молекулу, кодирующую пориновый протеин или связанный с ним протеин менингококка группы В, молекула ДНК присоединена к Т7 промотору вектора.
Следующий специфический объект изобретения - обеспечение штаммом BL21 (DE3) дельта omp E. coli.
Следующие объекты и цели изобретения будут ясны из дальнейшего описания.
Фиг. 1: Диаграмма, показывающая последующую стратегию Por В гена. РПЦ- продукт, описанный в примере 1 (Materials and Methods section), были соединены в BamHI-Xhol сайт экспрессионного плазмида рЕТ-17. Начальная протяженность двойной закрученной спирали была получена с использованием олигонуклеотидной последовательности, направленной противоположно BamHI сайту и перпендикулярно Xhol сайту рЕТ-17b плазмида. Добавочная последовательность была получена путем делеций 3 конца гена, используя экзонуклеаза III/фасолевая нуклеаза реакцию. После повторного связывания и трансформации обратно в Е. coli, несколько клонов было отобрано по размеру вставки и упорядочены.
Упорядочение происходило всегда с 3' конца гена с использованием олигонуклеотидного начала перпендикулярно Bpu11021 сайту.
Фиг.2: Гель-электрофорез, показывающий продукты РПЦ-реакции (электрофорез происходит в 1% агарозе с использованием ТАЕ буфера). Фиг.3 (а) и (б): рис (а): SDS-PAGE анализы лизатов клетки Е. coli, содержащей контрольный рЕТ-17b плазмид без вставки и клона Е. coli, содержащего рЕТ-17b плазмид, включающий вставку из полученного путем РПЦ-реакции продукта, описанного в материалах и методах секции выше. Обе культуры растут в O.D. of 0,6 to 600 nm, добавлен IPTG и инкубированы при t 37oС в течение 2 часов. 1,5 mls каждой культуры отцентрифугировано и бактериальная пилюля растворена в 100 (I) SDS-PAGE приготовленного буфера. Ряд А показывает профиль протеинов, полученный при использовании 10 (I) контрольного образца, ряды В (5(I) и С (10 (I) показывают протеиновый профиль Е. coli как клетки-хозяина, экспрессирующей Por В протеин.
рис (б): Western блот анализ лизатов всей клетки Е. coli, содержащей контрольный плазмид без вставки после 2-часовой индукции с IPTG- ряд А 20 (I); и соответствующий клон E.coli, содержащий porВ-рЕТ-17b плазмид - ряд В (5(I); ряд С 10 (I) и ряд D 20 (I). Моноклональное антитело 4D11 было использовано, как первичное антитело, и метод вестерн блот был проведен, как и описано выше.
Фиг. 4: Нуклеотидная последовательность (SEQ ID NO.1) и транслированная аминокислотная пocлeдoвaтeльнocть (SEQ ID NO.1) зрелого PorВ гена клонированы в экспрессионный плазмид рЕТ-17b. Два нуклеотида, которые отличны от описанного выше серотипа 15 PorВ, подчеркнуты.
Фиг. 5: График, демонстрирующий профиль элюции Сефакрил S-300 колонки дикого типа протеинов класса 3, изолированных из менингококкового штамма 8765 и рекомбинантного протеина класса 3, продуцированного BL21(DE3) - дельта ompА штаммом Е. coli, содержащим r3рЕТ-17b (является ли это копией - это в примерах не указано) плазмид, и как показано при поглощении 280 nm и при помощи SDS-PAGE анализа. Пустой объем колонки показан стрелкой. Фракции, содержащие менингококковый порин и рекомбинантный порин, как определено SDS-PAGE, обозначены линией.
Фиг.6: График, изображающий результат ингибиции ELISA исследований, показывает способность гомологичного дикого типа (wt) PorВ вызывать выработку реактивных антител в 6 человеческих иммунных сыворотках. Арифметическое значение показано рельефной линией.
Фиг. 7: График, изображающий результаты ингибиции ELISA исследований, показывает способность очищенного рекомбинантного PorВ протеина вызывать выработку реактивных антител в 6 человеческих иммунных сыворотках. Арифметическое значение ингибиции показано рельефной линией.
Фиг. 8: График, показывающий сравнение двух значений ингибиции, полученных у wt рекомбинантного Por В протеина.
Фиг. 9А и 9В: Нуклеотидная последовательность (SEQ ID NO.3) и транслированная аминокислотная последовательность (SEQ ID NO.4) зрелого гена порина класса II клонированы в экспрессионный плазмид рЕТ-17b.
Фиг. 10А и 10В: Нуклеотидная последовательность (SEQ ID NO.5) и транслированная аминокислотная последовательность (SEQ ID NO.6), связанных с геном порина класса II, клонированы в экспрессионный плазмид рЕТ-17b.
Фиг.11 ((а) и (б)): Фиг (а) изображает рестрикционную карту плазмида рЕТ-17b. Фиг.(б) показывает нуклеотидную последовательность (SEQ ID NO.7 и SEQ ID NO. 9) между BgIII b Xhol сайтами рЕТ-17b плазмида. Последовательность, обеспеченная плазмидом, находится на нормальной линии, в то время как внесенная из РПЦ - продукта изображена на рельефной линии. Аминокислоты (SEQ ID NO. 8 и SEQ ID NO.10), которые происходят из плазмида, находятся на нормальной линии, в то время как аминокислоты из вставки - на рельефной. Стрелки отграничивают, где последовательность начинает соответствовать последовательности фиг.4 и где заканчивает.
В отличие от пориновых протеинов Е. coli и некоторых других грам-негативных бактерий, сравнительно мало известно поринах Neisseria, в частности о том, как влияет изменение первичной последовательности в порине на его ионоселективность, вольтаж- зависимость и другие биофизические функции. Кристаллические структуры двух поринов Е. coli: OmpF и PhoЕ были растворены в 2,4 А и 3,0 А, соответственно (Cowan, S.W. et al., Nature 358: 727-733 (1992)). Оба порина были хорошо изучены благодаря их необычайной стабильности, поэтому было возможно проведение молекулярно-генетических исследований. Данные, полученные генетиками, хорошо коррелировали с кристаллической структурой обоих поринов. Хотя, как было показано в различных экспериментах с моноклональными антителами для отбора поринов нейссерии, топология поверхности клетки Neisseria имеет близкое сходство с обоими указанными поринами Е. coli (van der Ley, P., et al., Infect. Immun. 59: 2963-2971 (1991)). Практически отсутствует информация о том, как изменения в аминокислотной последовательности в специфических областях поринов нейссерии влияют на их биофизические характеристики, в то время как о поринах Е. coli такие данные есть (Cowan, S.W. et al., Nature 358: 727-733 (1992)).
Для проведения таких исследований существуют две большие проблемы: (1) невозможность проводить с легкостью генетические манипуляции с Neisseria с помощью молекулярных технологий и (2) неспособность экспрессировать достаточное количество поринов в Е. coli для их дальнейшей очистки с целью получения данных для биофизической и биохимической характеристики. Большинство данных о последовательности ДНК менингококковых и гонококковых поринов были получены клонированием частично совпадающих участков гена порина и затем восстановлением информации для выявления всей последовательности гена (Gotschlich, E. C. et al., Proc. Natl. Acad. Sci. USA 84: 8135-8139 (1987); Murakami, К. et al., Infect. Immun. 57: 2318-2323 (1989)). Carbonetti et al. были первыми, кто клонировал гонококковый ген порина в Е. coli с использованием контролируемого рТ7-5 экспрессионного плазмида. Результаты этих исследований показали, что когда произошла индукция гена порина, накопилось небольшое количество пориновго протеина. Экспрессия этого протеина оказалась летальной для Е. coli (Carbonetti and Sparling, Proc. Natl. Acad. Sci. USA 84: 9084-9088 (1987)). Дополнительные эксперименты показали, что изменения в гене порина гонококка могут быть получены в системе Е. coli и затем вновь перенесены в гонококк (Carbonetti et al., Proc. Natl. Acad. Sci. USA 85: 6841-6845 (1988)). Тем не менее оказалось, что возможности легкого проведения данных манипуляций для получения достаточного количества порина с целью биохимической и биофизической характеристики ограничены.
Feavers и соавт. описали метод, при помощи которого амплификация происходит методом РПЦ-реакции поринового гена нейссерии из различных источников, используя два синтезированных олигонуклеотида к общим доменам 5 - конца и 3 - конца поринового гена соответственно (Feavers, I.M. et al., Infect. Immun. 60: 3620-3629 (1992)). Олигонуклеотиды были созданы так, что амплифицированная ДНК могла быть принудительно клонирована в плазмиды, используя рестрикцию эндонуклеаз Bgl II и Xho I.
Используя описанную Feavers ПЦР- систему ДНК, последовательность зрелого PorВ протеина менингококкового штамма 8765 серотипа 15 была амплифицирована и связана в BamHI-Xhol сайт Т7 экспрессионного плазмида рЕТ-17b. Это разместило PorВ протеиновую последовательность в рамках позади Т7 промотора и 20 аминокислот 10 протеин, включая первую последовательность. При добавлении PTG к культуре Е. coli, содержащей этот плазмид, большое количество PorВ протеина накапливается внутри бактерии. Полное объяснение, почему эта конструкция не явилась летальной для Е. coli и произошла экспрессия большого количества протеинов, ожидается при проведении дальнейших изучений. Тем не менее существует одно возможное объяснение замены промотора нейссерии и сигнальной последовательности - на Т7 и 10 соответственно приводит к тому, что продуцированный порин направляется в цитоплазму, а не к наружной мембране. Henning и соавт. обнаружили, что когда E.coli OmpA протеин и его фрагменты экспрессированы, то эти продукты, обнаруженные в цитоплазме, гораздо менее токсичны, чем те, которые направлены в периплазматическое пространство (Klose M., et al., J. Biol. Chem. 263: 13291-13296 (1988); Klose M., et al., J. Biol. Chem. 263: 13297-13302 (1988); FreudI, R., et al., J. Mol. Biol. 205: 771-775 (1989)). Все же каким бы ни было объяснение, но экспрессированный PorВ протеин легче изолировался, подвергался очистке и реформировался в тримерную конформацию, чем нативный протеин. Результаты по исследованию ингибиции ELISA с использованием человеческой иммунной сыворотки дают основание предполагать, что PorВ протеин, полученный способом, описанным выше, обладает почти всеми (если не всеми) антигенными характеристиками протеина PorВ дикого типа, очищенного из менингококка. Эта система дает возможность легкого манипулирования геном порина нейссерии с помощью новых молекулярных технологий. Эта система позволяет получать большое количество чистого поринового протеина для характеристики его. Эта система так же позволяет изучить и охарактеризовать гены из большого числа штаммов как гонококков, так и менингококков.
Таким образом, настоящее изобретение относится к методу выделения поринового протеина наружной мембраны менингококка группы В, в особенности поринов, относящихся к классу 2 и 3.
В одном из вариантов, настоящее изобретение относится к методу выделения поринового протеина менингококка группы В в Е. coli, включающему:
(a) трансформацию Е. coli вектором, включающим избранный маркер и ген, кодирующий протеин, выбранный из группы насчитывающей:
(i) зрелый пориновый протеин,
(ii) связанный протеин, включающий зрелый пориновый протеин, включенный в аминокислоты от 1 до 20 или 22 гена Т7 10 капсидного протеина;
указанный ген связан с Т7 промотором;
(b) выращивание трансформированной Е. coli в среде, содержащей селективный агент, и
(c) индуцирование экспрессии указанного протеина;
протеин составляет более чем 2% от общего протеина, экспрессированного в Е. coli.
В предпочтительном варианте экспрессированный пориновый протеин менингококка группы В и связанный с ним протеин составляли около 5% от общего белка, экспрессированного в Е. coli. В другом исследовании оба экспрессированных протеина составили около 10% от общего количества протеинов, экспрессированных в Е. coli. И, наконец, в другом эксперименте пориновый протеин менингококка группы В и связанный с ним протеин составляют 30% от общих протеинов, экспрессированных в Е. coli.
Примеры плазмидов, которые содержат Т7 индуцирующий промотор, включают экспрессионные плазмиды рЕТ-17b, pET-11a, pET-24a-d(+) и рЕТ-9а, все они являются коммерчески доступными из Novagen (565 Science Drive, Madison, WI 53711). Эти плазмиды включают в последовательности Т7 промотор, необязательно как оператор, рибосомо связывающий сайт, рестрикционные сайты, позволяющие вставить структуральный ген и Т7 терминальную последовательность (см. Novagen каталог, стр 36-43 (1993)).
В предпочтительном варианте, использовался штамм BL21 (DE3) дельта ompA E. coli. Как был отмечено выше, плазмиды могут трансформироваться в этот штамм или в "дикий" штамм BL21 (DE3). Штамм BL (DE3) дельта ompA E. coli предпочтительнее, так как другие OmpA протеины, продуцируемые штаммом, могут загрязнить очищенный порин и создать нежелательные побочные иммуногенные эффекты.
Трансформированная E. coli растет в среде, содержащей селективный агент, например какой-либо бетта-лактам, к которому чувствительная E. coli, такой как ампициллин, рЕТ экспрессионный вектор обеспечивает селективным маркером, который передает резистентность к антибиотикам трансформированному организму.
Высокий уровень экспрессии поринового протеина менигококка группы В может быть токсичным для E. coli. Тем не менее, настоящее изобретение позволяет E. coli экспрессировать протеин на уровне 30% и даже 50% от общего числа клеточных протеинов.
В другом предпочтительном варианте, настоящее изобретение относится к вакцине, включающей пориновый протеин наружной мембраны менингококка группы В и связанный с ним протеин, полученные в соответствии с методами, описанными выше, а также фармакологически приемлемые дилюэнт, носитель и эксципиент, в совокупности с которыми вакцина может быть назначена в количестве, эффективном для вызывания выработки защитных антител у животных к Neisseria meningitidis. В предпочтительном варианте живые существа были выбраны из группы, включающей человека, крупный рогатый скот, овец, свиней и цыплят. В другом предпочтительном варианте был выбран человек.
Другой предпочтительный вариант изобретения - описанная выше вакцина, в которой указанный порин наружной мембраны менингококка и связанный с ним протеин конъюгированы с капсульным полисахаридом (СР), принадлежащим так же менингококку группы В. Этот капсульный полисахарид может быть приготовлен, как описано у Ashton, F.E. et al., Microbiоl. Pathog. 6: 455-458 (1989); Jennings, H.J. et al., J. Immunol. 134: 2651 (1985); Jennings, H.J. et al., J. Immunol. 142: 3585-3591 (1989); Jennings, H.J. "Capsular Polysaccharides as Vaccine Candidates" в Carrent Topics in Microbiology and Immunology, 150: 105-107 (1990).
Предпочтительно, чтобы капсульный полисахарид изолирован методом, предложенным Frasch, C.E. et al., "Production and Control of Neisseria meningitidis Vaccines" in Bacterial Vaccines, Alan R. Liss, Inc., стр. 123-145 (1990), содержание чего полностью представлено ниже:
- выращивание организмов на модифицированной среде Франца от 10 до 20 ч, затем;
- в течение 10 мин содержание культуры при 55oС, убивающей микроорганизм, затем;
- удаление инактивированных клеток путем центрифугирования, затем;
- добавление Цетавлона до 0,1%;
- преципитация капсульного полисахарида из культурального бульона;
- добавление кальция хлорида 1М;
- растворение капсульного полисахарида, затем центрифугирование для удаления клеточных остатков;
- добавление этилалкоголя 25%;
- удаление преципитированной нуклеиновой кислоты центрифугированием;
- добавление этилалкоголя 80%;
- преципитация неочищенного капсульного полисахарида и удаление алкоголя.
Неочищенный капсульный полисахарид в дальнейшем подвергается очистке методом гель-фильтрационной хроматографии, после частичной деполимеризации разведенной кислотой, например ацетиловой кислотой, муравьиной кислотой и трифлюроацетиловой кислотой (0,01-0,05 N), для получения смеси полисахаридов, имеющих среднюю величину молекулярного веса от 12,000 до 16,000. Затем капсульный полисахарид подвергается N-деацилированию с помощью борогидрида и N-пропионилируется с получением N-Pr-GBMP. Таким образом, капсульный полисахарид, используемый для конъюгата вакцины настоящего изобретения, может являться фрагментом капсульного полисахарида(СР), N-деацилированным капсульным полисахаридом или его фрагментом, а также N-Pr капсульным полисахаридом и его фрагментом. При использовании этих фрагментов как части СР- порин-протеинового конъюгата, капсульный полисахарид может индуцировать иммунную активность (см. примеры).
В дополнительном предпочтительном варианте, настоящее изобретение относится к методу приготовления полисахаридного конъюгата, включающего: получение вышеописанным методом поринового и связанного с ним протеина наружной мембраны менингококка группы В; получение капсульного полисахарида из менингококка; конъюгирование протеина и капсульного полисахарида.
Конъюгаты настоящего изобретения могут быть получены путем реакции сведения концевых групп капсульного полисахарида к первичным аминогруппам порина с помощью редуктивного аминирования. Восстановленные группы могут быть получены с помощью селективного гидролиза или специфического окислительного расщепления или комбинации обоих способов. Предпочтительно получение конъюгата капсульного полисахарида с порином методом Jennings et al., U.S. Patent 4, 356, 170, содержание которого находится в сноске, который включает контролируемое окисление капсульного полисахарида с использованием периодата, следующее за редуктивным аминированием поринового протеина.
Вакцина настоящего изобретения представляет собой пориновый протеин и связанный с ним протеин менингококка группы В или конъюгированную вакцину, в количестве, зависящем от способа назначения. Хотя подкожный и внутримышечный способы введения являются предпочтительными, пориновый протеин и связанный с ним протеин менингококка группы В, находящиеся в вакцине в настоящем изобретении, могут быть назначены интраперитонеально и внутримышечно. Лица, сведущие в данной области, будут признательны, если дозы, назначаемые для определенных протоколов лечения, могут быть окончательно определены без каких-либо незаконных, нежелательных экспериментов. Подходящие дозы вакцин могут быть в пределах от 2 микрограмм протеина на кг веса тела до 100 микрограмм на кг вес тела.
Вакцина настоящего изобретения может применяться в таких формах, как капсулы, жидкие растворы, суспензии или эликсиры для орального применения, либо стерильные жидкие формы, такие как растворы или суспензии. Инертными носителями, которые являются предпочтительными для вакцины, являются солевые растворы, фосфатно-буферные солевые растворы, или другие носители, в которых пориновый протеин и связанный с ним протеин менингококка группы В растворим. Вакцина может быть в виде препарата с однократной дозировкой или во флаконах с несколькими дозами, которые могут быть использованы для программ по осуществлению массовых вакцинаций. Ссылка на Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, PA, Osol (ed) (1980) и New Trends and Developments in Vaccines, Voller et al., (eds), University Park Press, Baltimore, MD (1978) о методах приготовления и использования вакцин.
Пориновый и связанный с ним протеин менингококка группы В или конъюгированная вакцина настоящего изобретения в дальнейшем могут использоваться с включением адъюванта, который усиливает продукцию порин-специфических антител. Такими адъювантами могут являться, например, Freund s комплит-адъювант (CFA), стеарил-тирозин (ST, см. U.S. Patent 4.258,029), дипептид, известный как MDP, сапонин, алюминия гидрохлорид и лимфатические цитокины.
Freund s адъювант это эмульсия минерального масла и воды, которая смешана с иммуногенной субстанцией. Хотя данный адъювант обладает сильным действием, он, как правило, не назначался людям. Напротив, адъювант алюм (алюминия гидрохлорид) или ST мог быть использован для назначения людям. Пориновый протеин менингококка группы В или конъюгированная вакцина могут быть абсорбированы алюминием гидрохлоридом, из которого вакцина медленно высвобождается после инъекции. Порин менигококка группы В или конъюгированная вакцина могут также быть инкапсулированы с липосомами в соответствии с Fullerton, U.S. Patent 4235877.
В другом предпочтительном варианте изобретения, настоящее изобретение относится к методам профилактики бактериальных менингитов у живых существ, включая назначение живым существам поринового и связанного с ним протеина менигококка группы В или конъюгированной вакцины, произведенной в соответствии с методами, описанными выше в дозах, эффективных для предупреждения заболевания бактериальным менингитом.
Дополнительный вариант изобретения относится к методам очистки описанного выше поринового протеина и связанного с ним протеина наружной мембраны менингококка группы В с использованием: лизиса трансформированной Е. coli для высвобождения порина и связанного с ним протеина менингококка группы В как части нерастворимых включений; отмывания включений буфером для удаления загрязняющих Е. coli клеточных протеинов; суспензирование и растворение включений в водном растворе денатуранта; дилюции полученного раствора в детергенте и очистки растворенного порина менингококка группы В методом гель-фильтрации.
Первый шаг (лизис) может быть выполнен в соответствии с методами, известными лицам, сведущим в данной области науки, например: разрушение ультразвуком, разрушение ферментами или осмотический шок и др.
Включения могут быть отмыты буфером, который способен растворять клеточные протеины Е. coli без растворения включений, к которым относятся и пориновые протеины менингококка группы В. К данным буферам относятся TEN - буфер (50 mM Tris HCl, 1 mM EDTA, 100 mM NaCl, рН 8,0), Tricine, Bicine и HEPES и др.
Денатуранты, которые могут быть использованы в практике изобретения, включают 2М-8М мочевину или 6М гуанидин HCl, более предпочтительно 4-8 М мочевина или 4-6 М гуанидин HCl, еще более предпочтительны около 8М мочевины и около 6М гуанидина HCl.
Примерами детергентов, которые могут быть использованы для разведения растворимых пориновых протеинов менингококка группы В, являются ион-детергенты, такие как - SDS и цетавлон (Calbiochem); неионные детергенты: Твин, Тритон X, Бридж 35 и октил глюкозид; цвиттерионные детергенты: 3,14 -Цвиттергент, эмпиген ВВ и Чэмпс.
Наконец, растворимый пориновый протеин наружной мембраны менингококка группы В может быть очищен методом гель-фильтрации для разделения высоко- и низкомолекулярных субстанций. Типы фильтрующих гелей включают: Сефакрил-300, Сефароз CL-6B, Био-Гель А-1.5m. Колонка элюирована буфером, использованным для разведения растворенного протеина. Фракции, содержащие порин и связанный с ним протеин, могут быть идентифицированы методом гель-электрофореза. Фракции объединяются, подвергаются диализу и концентрированию.
Наконец, очищенный более чем на 95% пориновый протеин может быть получен пропусканием его концентрированной фракции через колонку с Q сефарозой.
В другом варианте, изобретение относится к экспрессии гена поринового протеина менингококка группы В, который является частью вектора, содержащего Т7 промотор, способный к индукции. Если промотор способен к индукции, то уровень транскрипции увеличивается в ответ на индуцирующий агент. Т7 промотор является способным к индукции при добавлении изопропил бетта-D тиогалактопиранозида (IPTG) в культуральную среду. Альтеранативно могут быть использованы Тас- промотор или промотор теплового шока.
Предпочтительно, чтобы пориновый протеин менингококка группы В был бы экспрессирован из pET-17d экспрессионного вектора или из рЕТ-11а экспрессионного вектора, каждый из которых содержат Т7 промотор.
Клонирование гена поринового протеина менингококка группы В или гена связанного с ним протеина в экспрессионный вектор может быть выполнено в соответствии с традиционными технологиями, включающими связывание окончаний, разрушение рестриктивными ферментами для обеспечения подходящими окончаниями, заполнение связующих окончаний, использование алкалинфосфатазы для того, чтобы избежать нежелательное присоединение и, наконец, связывание соответствующими лигазами. Ссылки сделаны на Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor, New York, Cold Spring Harbor Laboratory Press (1989) по основным методикам клонирования.
Для того чтобы получить структуру, которая по иммунологическим характеристикам является идентичной структуре нативного протеина, порин и связанный с ним протеин наружной мембраны менингококка группы В должны подвергнуться приданию необходимой конформации. Настоящее изобретение относится к методу преукладки (придание конформации) описанных выше протеинов наружной мембраны и связанных с ними протеинов, включающему: лизис трансформированной Е. coli для высвобождения порина и связанного с ним протеина менингококка группы В как части нерастворимых включений; отмывания включений буфером для удаления загрязняющих Е. coli клеточных протеинов; суспензирование и растворение включений в водном растворе денатуранта; дилюции полученного раствора в детергенте и очистки растворенного порина менингококка группы В методом гель-фильтрации для получения протеина, имеющего определенную конформацию в элюанте. Было обнаружено, что в элюанте при использовании гель-фильтрации получены пориновые протеины и связанные с ними протеины менингококка группы В, сложенные в тримерную структуру.
В другом предпочтительном варианте, настоящее изобретение относится к хорошо очищенному, имеющему определенную конформацию порину и связанному с ним протеину наружной мембраны менигококка группы В, полученного в соответствии с вышеописанными методами. Хорошо очищенный протеин является белком, который в основном отсутствует в других клеточных компонентах N. meningitidis, что может быть доказано, например, методом электрофореза. Хорошо очищенный протеин имеет чистоту более 95%, измеренную с помощью денситометрии во время гель-электрофореза после окраски Coomassie голубой или серебряной краской.
Следующие примеры иллюстрируют, но не ограничиваются методами и композициями настоящего изобретения. Другие приемлемые модификации и адаптации к различным условиям и параметрам, встретившиеся в этой технологии, являются очевидными для лиц, сведущих в этом разделе науки, изложены и рассмотрены в настоящем изобретении.
Примеры
Пример 1: Клонирование класса 3 поринового протеина менингококка группы В.
Материалы и методы
Организмы: штамм 8765 (В: 15: Р 1,3) N. meningitidis группы В были получены Dr. Wendel Zollinger (Wolter Reed Army Institute for Research) и выращены на агаровой среде, как описано ранее (Swanson, J.L, Infect. Immun. 21: 292-302 (1978)) в сосуде с угасанием свечи, находясь в инкубаторе при температуре 30oС. Штаммы DME558 Е. coli (из коллекции Benson S.; Silhavy, T. J. et al. , "Experiments with Gene Fusions" Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., 1984), BRE51 (Bremer,E. et al., FEMS Microbiol. Lett. 33: 173-178 (1986)), BL21(DE3) были выращены на LB агаровых плашках при температуре 37oС.
Р1 трансдукция: Plvir лизат штамма DME558 Е. coli был использован для трансдукции маркера резистентности к тетрациклину в штамм BRE51 (Bremer, Е., et al. , FEMS Microbiol. Lett. 33: 173-178 (1986)), в котором произошла делеция всего ompA гена (Silhavy, T.J. et al., Experiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., 1984). Штамм DME558, содержащий тетрациклин резистентный маркер в непосредственной близости к ompA гену, был выращен на LB среде до тех пор, пока он не достиг плотности приблизительно 0.6 OD при 600 nm. Одна десятая миллилитра 0.5 М CaCl2 была добавлена к 10 мл культуры и к 0.1 мл раствора, содержащего 1•109 PFU PIvir. Культура была инкубирована при температуре 37oС в течение 3 ч. После этой процедуры плотность бактериальных клеток визуально уменьшилась. Затем было добавлено 0.5 мл хлороформа и культура фагов, сохраненная при температуре 4oС. Так как 1-2% хромосом Е. coli могут быть поглощены каждым фагом, потребовалось определенное число фагов, поглощающих все хромосомы клетки-хозяина, включая и маркер тетрациклиновой резистентности, близкий к ompA гену.
Другой штамм BRE51, у которого отсутствует ompA ген, был выращен на LB среде при температуре 37oС. Перед этим культура была разведена 1:50 свежей средой LB и выращивалась 2 ч. Клетки были удалены центрифугированием и взвешены в МС соли. 0.1 мл бактериальных клеток был смешан с 0.05 лизатом фагов, описанных выше, и инкубирован 20 мин при комнатной температуре. Затем был добавлен такой же объем 1М содиума цитрата и бактериальные клетки были помещены в LB плашки, содержащие 12.5 μg/ml тетрациклина. Накануне чашки были инкубированы при температуре 37oС. Тетрациклиноустойчивые (12 μg/ml) трансдуктанты были исследованы на отсутствие экспрессии OmpA протеина с помощью SDS-PAGE и Western блот анализов, как описано ниже. Устойчивость бактерии к антибиотику опосредуется геном резистентности к тетрациклину, интегрированному в хромосому практически в том месте, где располагался ompА ген, подвергнувшийся делеции из этого штамма. Определенный штамм был обозначен, как BRE-TR.
Затем был проведен второй круг продукции фага со штаммом BRE-TR, используя те же методы, что были описаны выше. Представители этой популяции фагов содержат как ген тетрациклиновой резистентности и делецию OmpА гена. Затем эти фаги были отобраны и помещены на хранение. Эти фаги были затем использованы для заражения Е. соli штамма BL21(DE3). После заражения, бактерия содержит маркер тетрациклиновой резистентности. Существует высокая вероятность того, что OmpА делеция была селекционирована в LB плашках, содержащих тетрациклин.
Колонии бактерий, выросшие на плашках, затем были пророщены раздельно на LB среде и проверены на наличие ОmрА протеина. У всех колоний, отобранных для исследования, отсутствовал ОmрА протеин, наличие которого оценивалось реактивностью антител в SDS-PAGE western блоте.
SDS-PAGE и Western Блот. SDS-PAGE представляет собой вариант метода Laemmli (Laemmli, U.K., Nature 227: 680-685 (1970)), как было описано ранее (Blake and Gotschlich, J. Exp. Med. 159: 452-462 (1984)). Электрофоретический перенос на Иммобилон Р (Milipore Corp. Bedford, MA) был выполнен в соответствии с методами Towbin et al. (Towbin,H., et al., Proc. Natl. Acad. Sci. USA 76: 4350-4354 (1979)) за исключением того, что бумага вначале была пропитана метанолом. Исследование Western блот было проведено с реагентами, конъюгированными с фосфатазой (Blacke, M.S. et al., Analyt. Biochem. 136: 175-179 (1984)).
Реакция полимеразных цепей (РПЦ): Метод, описанный Feavers et al. (Feavers, I.M. et al., Infect. Immun. 60: 3620-3629 (1992)), был использован для амплификации гена, кодирующего PorВ. Селекционированные праймеры являлись 33 праймером (SEQ ID NO.11) (GGG GTA GAT CTG CAG GTT ACC TTG TAC GGT АСА ATT AAA GCA GGC GT) и 34 праймером (SEQ ID NO.12) (GGG GGG GTG ACC CTC GAG TTA GAA TTT GTG ACG CAG ACC AAC), как ранее описано (Feavers, I.M. et al., Infect. Immun. 60: 3620-3629(1992)). Кратко говоря, компоненты реакции были следующие: хромосомная ДНК (100 ng/μl) менингококкового штамма 8765 - 1μl; 5 и 3 праймеры (1μM) 2μl каждый; dNTP (10 mM биомассы), 4μl каждый; 10 Х РПЦ буфера (100 mМ Tris HCl, 500 mМ KCl, рН 8,3), 10μl; 25 mМ MgCl2, 6μl; дважды дистиллированная вода 62μl; и Taq полимераза (Cetus Corp., 5u/μl), 1 μl. Реакция была выполнена на GTC-2 Генетическом Термоциклере (Precision Inst. Inc. , Chicago, IL), соединенном с Lauda 4/K метанол/ вода охлаждающей системы (Brinkman Instruments, Inc., Westbury, NY) при температуре 0oС. Термоциклер запрограммирован на 30 циклов (94oС - 2 мин; 40oС - мин; 72oС - 3 мин). По окончании всех 30 циклов реакция была продолжена в течение 3 мин при температуре 72oС и окончательно остановлена при температуре 4oС до готовности для проведения анализа на 1% агарозном геле в ТАЕ буфере, как описано Maniatis (Maniatis, Т. et al.,Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY(1982)).
Субклонирование продукта реакции полимеразных цепей: pET-17b плазмид (Novagen, Inc.) был использован для субклонирования и приготовлен путем двухкратного разрушения плазмида с помощью рестрикционных эндонуклеаз BamHI и Xhol (New England Biolabs, Inc. , Beverly, MA). Разрушенные концы затем подвергаются дефосфорилированию с алкалин-фосфатазой из кишечника теленка. (Boehringer Manheim, Indianapolis, IN). Разрушенный плазмид затем изучен в 1% агарозном геле, отрезанный плазмид удален и очищен с использованием набора для очистки генов Gene Clean kit(Bio 101). Продукт реакции полимеразных цепей был расщеплен с помощью рестриктивных эндонуклеаз Bgl II и Xho I (New England Biolabs, Inc.). ДНК была экстрагирована с помощью фенол-хлороформа и преципитирована добавлением 0.1 объема 3 М содиума ацетата, 5 μl гликогена (20 (g/μl)) и 2.5 объема этанола. После отмывания ДНК в 70% этаноле (объем/ объем) она вновь растворена в ТЕ буфере. Расщепленный РПЦ продукт связан с дважды расщепленным pET-17b плазмидом, описанным выше, используя стандартную Т4 лигазную методику при температуре 16oС. (Current Protocols in Molecular Biology, John Wiley & Sons, New York (1993)). Связанный продукт затем трансформирован в BL21 (DE3)- ΔompA методом, описанным выше, что было сделано в соответствии с требованиями метода Chung et al., (Chung, C.T. et al., Proc. Natl. Acad. Sci. USA 86:2172-2175 (1989)). Трансформанты были селекционированы в LB плашках, содержащих 50 μg/ml карбенициллина и 12 μg/ml тетрациклина. Было отобрано несколько трансформантов, они культивированы в LB бульоне, содержащем карбенициллин и тетрациклин в течение 6 ч при температуре 30oС; экспрессия плазмидного гена была индуцирована добавлением IPTG. Температура была увеличена до 37oС и культивирование продолжено еще в течение 2 ч. Клетки каждой культуры были собраны путем центрифугирования, все клеточные лизаты приготовлены и анализированы методом SDS-PAGE и Western блот, используя моноклональные антитела (4D11), которые взаимодействуют со всеми поринами нейссерии.
Анализ нуклеотидной последовательности: Последовательность нуклеотидов в ДНК клонированного порина класса 3 была определена с помощью дидеокси-метода, используя денатурированную дважды скрученную ДНК плазмида, как описано в Current Protocols in Molecular Biology, Jhon Miley & Sons, New York (1993)). Набор Секвеназа II (Unated States Biochemical Corp., Cleveland, ОН) был использован в соответствии с приложенной инструкцией. Три синтезированных олигонуклеотидных праймера (Operon Technologies, Inc., Alameda, СА) были использованы для этих реакций. Один для 5 - конца (SEQ ID NO.13), который насчитывает 5 TCAAGCTTGGTACCGAGCTC и два для 3 конца, (SEQ ID NO.14) 5 TTTGTTAGCAGCCGGATCTG (SEQ ID NO.15) и 5 CTCAAGACCCGTTTAGAGGCC. Совпадающая делеция была достигнута воздействием рестрикционной эндонуклеазы Bpu11021 на плазмидную ДНК и воздействием на концы Thio-dNTP и полимеразы Klenow (Current Protocols in Molecular Biology, John Wiley & Sons, New York (1993)). Полученный плазмид затем расщеплен рестриктивной эндонуклеазой Xhol. exoll/ фасоль нуклеазная делеция использована для получения 3 -делеций плазмида (Stratagene, Inc., La Jolla, CA), как описано в приложении. Карта данной стратегии показана на фиг.1.
Экспрессия и очистка продукта PorВ гена: Используя кончик стерильной микропипетки, единичная колония BL21 (DE3)-ΔompA, содержащая PorВ-рЕТ-17b плазмид, была отобрана и инкубирована в 10 мл LB бульона с содержанием 50 μg/ml карбенициллина. Культура была накануне инкубирована при температуре 30oС со встряхиванием. 10 мл этой культуры затем стерильно добавлено к 1 л LB бульона с такой же концентрацией карбенициллина. Инкубация культуры продолжена при температуре 37oС в шейкерном инкубаторе до тех пор, пока OD600 не достигнет 0,6-1,0. К культуре добавляется 3 мл раствора IPTG (100 mM) и культура инкубируется еще 30 мин. Затем добавлен рифампицин (5,88 мл раствора биомассы; 34 мг/мл в метаноле) и инкубация культуры продолжается еще в течение 2 ч. Клетки собираются путем центрифугирования со скоростью 10 000 об/мин в GS3 центрифуге и взвешиваются. Клетки тщательно взвешены в 3 мл TEN буфера (50 mM Tris HСl, 1 mM EDTA, 100 mM NaCl, pH 8,0) на грамм влажного веса клеток. К этому добавляется 8 μl биомассы раствора PMSF (50 mM ангидрида этанола) и 80 μl раствора биомассы лизозима (10 мг/мл в воде) на грамм влажного веса клеток. Эта смесь помешивается в течение 20 мин при комнатной температуре. При помешивании добавлено 4 мг на грамм влажного веса клеток деоксиколата. Смесь помещена в водяную баню при температуре 37oС и размешана стеклянной палочкой. Когда смесь начинает загустевать, добавляется 20 μl раствора биомассы DN-азы I (1 мг/мл) на грамм влажного веса клеток. Затем, смесь удаляется из водяной бани и оставляется при комнатной температуре до тех пор, пока раствор не перестанет быть густым. Потом смесь в течение 20 мин центрифугируется со скоростью 15 000 об/мин в SS-34 центрифуге при температуре 4oС. Полученная пилюля тщательно промыта дважды TEN буфером. Затем, она взвешена в свежеприготовленном TEN буфере, содержащем 0.1 mM PMSF и 8 М мочевины, и подвержена разрушению ультразвуком в ультразвуковой ванне (Heat Systems, Inc., Plainview, NY). Концентрация протеинов была определена с помощью набора ВСА (Pierce, Rockville, IL), установленная концентрация протеинов достигает менее 10 мг/мл, используя TEN- мочевина буфер. Затем полученный образец разведен 1:1 10% (вес/объем) Цвиттергентом 3,14 (CalBiochem, La Jolla, CA), разрушен ультразвуком и погружен в колонку молекулярного сита Сефакрил S-300. Сефакрил S-300 колонка (2,5•200 см) предварительно уравновешена 100 mM Tris HCl, 200 mM NaCl, 10 mM EDTA, 0,05% Цвиттергент 3,14 и 0,02% азид, рН 8,0. Уровень потока колонки достигал 8 мл/ч, было получено 10 мл фракций. OD280 каждой фракции был измерен, к протеин-содержащим фракциям был применен SDS-PAGE анализ.
Исследования ингибиции ELISA: Плашки с микротитром (Nunc-Immuno Plate IIF, Nunc Inc. , Naperville, IL) были сенсибилизированы добавлением 0,1 мл porB (2 μg/ml), очищенного из штамма дикого типа 8765 в 0,1 М карбонатном буфере, рН 9,6 с 0,02% азидом. Накануне плашки инкубированы при комнатной температуре. Плашки отмыты 5 раз 0,9% NaCl, 0,05% Бридж 35, 10 mM содиума ацетатом рН 7,0, азидом 0,02%. Человеческая иммунная сыворотка против PorB протеина типа 15, класса 3 была получена в Dr. Phillip О. Livingstone, Memorial-Sloan Kettering Cancer Center, New York, NY. Человеческая иммунная сыворотка разведена PBS с 0,05% Бридж 35, добавлена в плашки и инкубирована 2 ч при комнатной температуре. Плашки снова были отмыты указанным выше способом. Вторичные антитела, конъюгированные с алкалинфосфатазой козлиные анти-человеческие иммуноглобулины G (Tago Inc., Burlingame, CA) были разведены PBS-Бридж и добавлены в плашки. Затем, инкубированы при комнатной температуре в течение 1 ч. Плашки снова отмыты способом, описанным выше, и добавлен р-нирофенил фосфат (Sigma Phosphatase Substrate 104) (1 мг/мл) в 0,1 диэтаноламине, 1 mM MgCl2, 0,1 mM ZnCl2, 0,02% азид, рН 9,8. Плашки были инкубированы при температуре 37oС в течение 1 ч и поглощение 405 nm определено с использованием Elida-5 счетчика для микротитруемых плашек (Physica, New York, NY). Контрольные лунки показали отсутствие первичных и/или вторичных антител. Работа была проделана для получения титра для каждой человеческой сыворотки, которые могли дать полумаксимальное считывание в исследовании ELISA. Титр каждой человеческой сыворотки мог быть использован в ингибиции ELISA. ELISA микротитруемые плашки могли быть сенсибилизированы очищенным PorB протеином дикого типа и отмыты, как указано ранее. BV-96 полипропиленовые микротитрованные плашки (unc, Inc.,) было добавлено различное количество другого очищенного протеина дикого типа PorB или рекомбинантного PorB протеина в общем объеме 75 μl. Человеческая сыворотка была разведена HBS-Бридж раствором до удвоения половины максимального титра и добавлена в объеме 75 μl в каждую лунку, содержащую PorB протеин или рекомбинантный протеин PorB. Плашка инкубируется 2 ч при комнатной температуре и центрифугируется в Sorvall RT6000 замороженной центрифуге, оборудованной подставкой для плашек (Wilmington, DE) со скоростью 3000 об/мин в течение 10 мин. Не дотрагиваясь до дна, из каждой лунки удалено 100 μl и перенесено в сенсибилизированные и отмытые LISA микротитрованные плашки. ELISA плашки инкубируются в течение 2 ч, отмываются и в плашки добавляются конъюгированные вторичные антитела, так как было описано ранее. Плашки обрабатываются и прочитываются способом, описанным выше. Процент ингибиции рассчитывается по формуле: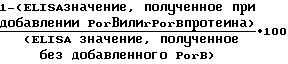
Результаты
Реакция полимеразных цепей и Субклонирование: Разработан метод, позволяющий более легкое клонирование и генетическое манипулирование в конечном счете, приводящее к получению достаточно чистого поринового протеина из различного числа различных генов порина нейссерии для получения в будущем антигенной и биофизической характеристики. Первым шагом к получению этого результата явилось клонирование поринового гена из Neisseria. Используя технологию, описанную Feavers et al. (Feavers, I.M. et al., Infect. Immun. 60: 3620-3629 (1992)), последовательность ДНК зрелого поринового протеина класса 3, порин серотипа 15 был амплифицирован с использованием хромосомы менингококка штамма 8765 как шаблона для реакции полимеразных цепей. Соответствующие сайты синтезированы к концам олигонуклеотидных праймеров так, что когда произошло расщепление, амплифицированная последовательность поринового протеина может быть прямо связана и клонирована в выбранный экспрессионный плазмид. После 30 циклов получен продукт реакции полимеразных цепей, как показано на фиг.2. Большой продукт колеблется между 900 bp и 1000 bp, что соответствует предшествующим исследованиям (Feavers, I. M. et al., Infect. Immun. 60: 3620-3629 (1992)). Тем не менее, продукта с высоким молекулярным весом не было обнаружено, даже если РПЦ проходила в строго соблюденных условиях (40oС, 50 mM KCl).
Способность экспрессионной системы Studier et al. (Studier and Moffatt, J. Mol. Biol. 189: 113-130 (1986)) продуцировать большое количество клонированного поринового протеина была использована и является доступной для коммерческого использования Novagen Inc. Амплифицированный продукт РПЦ был клонирован в BamHI-Xhol сайт рЕТ-17b плазмида. Эти стратегические точки ДНК последовательности поринового протеина находятся за Т7 промотором, ДНК последовательностью, кодирующей лидирующую последовательность из 9 аминокислот и 11 аминокислот зрелого φ10 протеина. Штамм BL21 Studier Е.соli лизогенный для DE3 лямбда производных (Studier and Moffatt, J. Mol. Biol. 189: 113-130 (1986)) был отобран, как экспрессионный хозяин для рЕТ-17b плазмида, содержащего ген порина. Однако, предполагалось, что OmpА протеин, имеющий свое происхождение в экспрессионном хозяине Е.coli, мог иметь тенденцию к очистке вместе с экспрессированным менингококковым пориновым протеином, поэтому была получена модификация этого штамма путем трансдукции Р1, которая удаляет ompА ген из штамма. Таким образом после воздействия рестриктивными эндонуклеазами на оба продукта реакции полимеразных цепей, рЕТ-17b вектор и связывания, продукт был трансформирован в BL21 (DE3)-ΔompA и полученный транформант отобран для ампициллиновой и тетрациклиновой резистентности. Из большого числа колоний, рассмотренных на селекционной чашке, 10 были отобраны для будущей характеристики. Все 10 колоний экспрессировали большое количество протеина, молекулярный вес которого приблизительно равнялся молекулярному весу PorВ протеина, затем пророщены до log-фазы и индуцированы IPTG. Клеточный лизат одной культуры изображен на фиг.3а. Western блот-анализы с 4D11 моноклональным антителом позволили предположить, что экспрессированный протеин является PorВ протеином (фиг.3b). В противоположность другим исследованиям, когда нейссериальный порин был клонирован и экспрессирован в Е. coli, бактериальные клетки-хозяева не выявляли никаких признаков токсического или летального воздействия на них, даже после добавления IPTG. Клетки Е. coli оказались жизнеспособными и могли быть рекультурированы в любое время экспрессионной фазы.
Анализы нуклеотидной последовательности: Количество Por В, экспрессированного в этом эксперименте, было значительно больше по сравнению с предыдущими наблюдениями. Никаких побочных реакций в клетке-хозяине Е. coli в результате этой экспрессии выявлено не было. Для того чтобы быть уверенным, что в ген порина менингококка не внесено никаких артефактов во время реакции полимеразных цепей, весь φ10 порин и связанный с ним протеин были упорядочены путем экстензии двойного скрученного праймера из плазмида. Результаты показаны на фиг. 4. Нуклеотидная последовательность была идентичной другой последовательности менингококкового серотипа 15 PorВ гена, ранее описанной Heckels, et al. (Ward, M. J. , et al., FEMS Microbiol. Lett. 73: 283-289 (1992)) с двумя исключениями, которые показаны. Два различающихся нуклеотида обнаружены в третьей позиции кодона и не меняют аминокислотную последовательность экспрессированного протеина. Таким образом, в нуклеотидной последовательности не обнаружено никаких артефактов РПЦ или мутаций, которые могут являться причиной высокой экспрессии. Так же отсутствует токсичность для Е. coli. Т. е. имеющиеся данные позволяют предполагать, что действительно, продуцирован PorВ протеин.
Очистка продукта экспрессии PorВ гена: PorВ протеин, экспрессированный в Е. coli, был нерастворим в TEN буфере, что позволило предположить, что при экспрессии гена PorВ протеин находился в Е. coli в виде включений. Тем не менее, отмывание PorВ протеина TEN буфером удаляет большинство протеинов, загрязняющих Е. coli. PorВ протеин может быть растворен в свежеприготовленной 8М мочевине и разведен детергентом Цвиттергент 3,14. Окончательная очистка достигается использованием Сефакрил S-300 колонкой молекулярных сит, которые не только удаляют мочевину, но и загрязняющие протеины. Большинство PorВ протеинов, элюированных из колонки, имеют молекулярный вес тримера, гораздо больший, чем у PorВ дикого типа. Результаты сравнительной элюции двух диких типов и PorВ, экспрессированного в Е. coli, показаны на фиг.5. Важно знать, что когда концентрация PorВ протеина в 8М мочевины была более чем 10 мг/мл до того, как произошло разведение детергентом Цвиттергент, относительное количество PorВ протеинов, обнаруженных как тримеры, уменьшилось, и они представляли собой агреганты, элюированные в пустом объеме. Вместе с тем, когда концентрация протеина достигала менее 10 мг/мл в мочевинном буфере, большинство PorВ протеина было элюировано в точно такой же фракции, как и дикий тип PorВ. Так же было установлено с помощью Т7-Таg моноклональных антител и western блот-анализа, что 11 аминокислота зрелого Т7 капсидного протеина поддерживалась как амино-окончание. Общее количество менингококкового поринового протеина, полученного из 1 л Е. coli, составило приблизительно 50 мг.
Исследования ELISA ингибиции. Для того чтобы определить, имеет ли рекомбинантный PorВ протеин сходную антигенную конформацию с PorВ протеином, который продуцирован диким типом менигококкового штамма 8765, для исследований ELISA ингибиции у 6 пациентов, вакцинированных диким типом менигококка тип 15 PorВ протеином, была получена сыворотка. В этом исследовании антитела, реактивные к нативному PorВ протеину, были полностью ингибированы различным количеством очищенного рекомбинантного PorВ протеина или гомологичного очищенного PorВ протеина дикого типа. Результаты ингибиции гомологичных очищенных PorВ каждой из шести сывороток и значения ингибиции представлены на фиг. 6. Соответственно, ингибиция сывороток рекомбинантным очищенным PorВ протеином показана на фиг.6b. Сравнение значений ингибиции фиг.6 и 7 показано на фиг. 8. Эти данные позволяют предположить, что антитела, содержащиеся в сыворотке шести пациентов, имеют сходные эпитопы на гомологичном очищенном PorВ протеине дикого типа и на рекомбинантном очищенном PorВ протеине. Это доказывает, что рекомбинантный PorВ протеин приобрел в большинстве своем, если не весь, нативную конформацию, имеющуюся у дикого типа PorВ протеина.
Пример 2. Клонирование порина класса 2 N. meningitidis группы В штамма BNCV M986.
Геномная ДНК была изолирована из приблизительно 0,5 г менингококка группы В штамма BNCV М 986 (серотип2а) с использованием методов, описанных ранее (Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor, New York, Cold Spring Harbor Laboratory Press (1989)). ДНК в дальнейшем служила шаблоном для двух пориновых специфических олигонуклеотидов класса 2 в стандартной реакции полимеразных цепей. Эти нуклеотиды замышлялись как комплементарные 5 и 3 регионам фланков порина класса 2 и содержащие EcoRI рестрикционные сайты, способствующие клонированию фрагмента. Последовательность нуклеотидов была следующей:
(SEQ [D NO.16)
5 IAGC GGC TTG GAA TTC CCG GCT GGC TTA AAT TTC 3 (SEQ ID NO.17) и
5 CAA ACG AAT GAA TTC AAA TAA AAA AGC CTG 3
Затем была использована реакция полимеразных цепей для того, чтобы получить порин 2 класса. Условия реакции были следующие: BNCV M986 геномная ДНК 200 нг, два олигонуклеотидных праймера, описанных выше по 1 μM каждый, 200 μM каждого dNTP, буфер реакции полимеразных цепей (10 mM Tris HCl, 50 mM KCl, рН 8,3), 1,5 mM MgCl2, 2,5 единицы Taq полимеразы с добавленным до 100 μM дистиллированной водой. Реакционная смесь затем была проведена через 25 циклов при температуре 95oС в течение 1 мин, при температуре 50oС в течение 2 мин и при температуре 72oС в течение 1,5 мин. В конце циклического периода смесь была помещена в 1% агарозный гель и подвергнута электрофорезу в течение 2 ч, после которого полоска при 1.3 kb была удалена и ДНК была приведена в норму при помощи набора по очищению гена Gene Clean kit (Bio 101). ДНК была затем расщеплена с помощью Eco RI, очищена и связана с pUC19, расщепленным так же EcoRI, используя при этом Т4 ДНК лигазу. Связанная смесь затем была использована для перенесения в известную E.coli DH5 альфа. Были отобраны и упорядочены рекомбинантные плазмиды. Была обнаружена вставка, имеющая ДНК последовательность, совместимую с таковой у порина 2 класса. см. Murakami, K. et al., Infect. Immun. 57: 2318-2323 (1989).
Плазмид рЕТ-17b (Novagen) был использован для экспрессии порина 2 класса. Как описано ниже, были созданы два плазмида для того, чтобы получить два различных протеина. Один плазмид был предназначен для продукции зрелого порина 2 класса, в то время как другой был предназначен для получения порина 2 класса, связанного с 20 аминокислотами, принадлежащими φ10 капсидному протеину Т7 гена.
Конструкция зрелого порина класса 2.
Зрелый порин 2 класса был получен путем ампилификации pUC 19- порина 2 класса, используя олигонуклеотиды (SEQ ID NO.18) 5 CCT GTT GCA GCA CAT ATG GAC GTT ACC TTG TAC GGT АСА ATT AAA GC 3 & (SEQ ID NO.19) 5 CGA CAG GCT TTT TCT CGA GAC CAA TCT TTT CAG 3 . Такая стратегия позволила клонирование амплифицированного порина класса 2 в Ndel и Xhol сайты плазмида рЕТ-17b, продуцируя таким образом зрелый протеин класса 2. Стандартная реакция полимеразных цепей была выполнена с использованием pUC 19-класс2 как стандарта и двух олигонуклеотидов, описанных выше. Во время реакции полимеразных цепей был произведен 1.1 kb продукт, изученный затем в 1% агарозном геле. ДНК, полученная во время реакции полимеразных цепей, была гель-очищена и расщеплена с использованием рестрикционных энзимов Ndel и Xhol. 1.1 kb продуцированная ДНК была вновь гель- очищена и связана с рЕТ-17b, расщепленным с помощью тех же рестрикционных энзимов. Связывание происходило с участием Т4 ДНК лигазы. Эта смесь была использована для трансформации в известный штамм Е.coli DH5α. Колонии, содержащие 1.1 kb вставку, были выбраны для последующих анализов. ДНК из DH5α клонов, была проанализирована с помощью рестрикционного картирования и проведено упорядочение выбранных клоновых плазмидов. После этих исследований ДНК, полученная из DH5α клонов была использована для трансформации в Е. coli BL21(DE3)-ΔompA. Трансформанты были отобраны и внесены в LB- агар, содержащий 100 μг/мл карбенициллина. Различные трансформанты были изучены в отношении их способности продуцировать пориновый протеин класса 2. Это осуществлялось путем выращивания клонов на LB- жидкой среде, содержащей 100 μг/мл карбенициллина и 0,4% глюкозу при температуре 30oС до достижения OD600= 0,6 с последующим индуцированием культуры IPTG (0,4 mМ). В дальнейшем клетки были разрушены, и клеточный экстракт был изучен с помощью SDS-PAGE.
Конструкция связанного порина класса 2.
Связанный порин класса 2 был получен путем амплификации pUC19-класс 2 порин с использованием олигонуклеотидов (SEQ ID NO.20) 5 ССТ GTT GCA GCG GAT CCA GAC GTT ACC TTG TAC GGT АСА ATT AAA GC 3 & (SEQ ID NO.21) 5 CGA CAG GCT TTT TCT CGA GAC CAA TCT TTT CAG 3 . Такая стратегия позволила добиться клонирования амлифицированного порина класса 2 в BamHI и Xhol сайты плазмида еРТ-17b, таким образом продуцируя связанный порин класса 2, содержащий в дополнение 22 аминокислоты на N-окончании, производном от Т7 φ10 капсидного протеина, содержащегося в плазмиде. Стандартная реакция полимеразных цепей была проведена с использованием pUC19-класс 2 как стандарта и двух олигонуклеотидов, описанных выше. В ходе реакции полимеразных цепей был произведен 1.1 kb продукт, который затем подвергнут анализу в 1,0% агарозном геле. ДНК, полученная в ходе реакции полимеразных цепей, была гель-очищена и расщеплена энзимами BamHI и Xhol. Полученный 1.1 kb продукт был вновь подвергнут гель-очистке и связан с еРТ-17b, расщепленным теми же энзимами. При связывания использовалась Т4 ДНК лигаза. Полученная смесь была затем трансформирована в известный Е. coli DH5α. Колонии, содержащие 1.1 kb вставку, были отобраны для последующих анализов. ДНК из DH5α клонов была изучена с помощью рестрикционного энзимного картирования, проведено упорядочение клоновых соединений выбранных плазмидов. После этого анализа ДНК, полученная из DH5α колоний, была использована для трансформации в Е. coli BL21(DE3)-ΔompA. Трансформант был селекционирован на LB-агаре, содержащем 100 μг/мл карбенициллина. Различные трансформанты были изучены на наличие способности синтезировать пориновый протеин класса 2. Это было осуществлено путем выращивания колоний на LB жидкой среде, содержащей 100 (μг/мл карбенициллина и 0,4% глюкозу при 30oС до достижения OD600=0.6 и последующей индукции культур с IPTG (0,4 mM). Клетки были затем разрушены и клеточный экстракт проанализирован с помощью SDS-PAGE.
Пример 3. Клонирование и экспрессия зрелого порина класса 3 штамма 8765 N. meningitidis группы В в Е. соli.
Геномная ДНК была изолирована из примерно 0,5 г штамма 8765 менингококка группы В, используя метод, описанный выше (Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd. ed., Cold Spring Harbor, New York, Cold Spring Harbor Laboratory Press (1989)). Эта ДНК была использована как шаблон для специфических олигонуклеотидов поринов 3 класса в стандартной реакции полимеразных цепей.
Зрелый порин 3 класса был создан путем амплификации геномной ДНК 8765 штамма с использованием олигонуклеотидов: (SEQ ID NO.22) 5 GTT GCA GCA CAT ATG GAC GTT ACC CTG TAG GGC ACC 3 (SEQ ID NO.23) & 5 GGG GGG ATG GAT CCA GAT TAG AAT TTG TGG CGC AGA CCG АСА СС 3 . Такая стратегия позволила клонирование амплифицированного порина класса 3 в Ndel и BаmН сайты плазмида рЕТ- 24а (+), таким образом продуцируя зрелый порин класса 3. Стандартная реакция полимеразных цепей была проведена с использованием геномной ДНК, изолированной из 8765 как шаблона, и двух олигонуклеотидов, описанных выше.
Условия реакции были следующие: геномная ДНК штамма 8765 - 200 нг, два олигонуклеотидных праймера, описанных выше по 1 μM каждый, 200 μM каждой dNTP, буфер реакции полимеразных цепей 10 mM Tris HCl, 50 mM KCl, pH 8,3, 1,5 mM MgCl2 и 2,5 единицы Taq полимеразы, дополненных 100 μл дистиллированной воды. Эта смесь была затем была подвергнута 25 циклам, включающим 95oС в течение 1 мин, 50oС в течение 2 мин и 72oС в течение 1,5 мин.
Данная реакция полимеразных цепей позволила получить около 930 bp продукта, что выявлено при анализировании в 1% агарозном геле. ДНК, полученная при помощи реакции полимеразных цепей, была гель-очищена и расщеплена рестрикционными энзимами Ndel и BamHI. 930 bp продукт был вновь затем подвергнут гель- очистке и связан с Ndel и BamHI, расщепленного рЕТ-24а (+), с использованием Т4 лигазы. Полученная смесь была использована для трансформирования известного штамма Е. coli DH5α. Колонии, содержащие 930 bp вставку, были отобраны для дальнейших анализов. Колонии, содержащие 1.1 kb вставку, были отобраны для последующих анализов. ДНК из DH5α клонов была изучена с помощью рестрикционного энзимного картирования, проведено упорядочение клоновых соединений выбранных плазмидов. После этого анализа ДНК, полученная из DH5α колоний, была использована для трансформации в Е. coli BL21(DE3)- ΔompA. Трансформант был селекционирован на LB- агаре, содержащем 50 μг/мл канамицина. Различные трансформанты были изучены на наличие способности синтезировать пориновый протеин класса 3. Это было осуществлено путем выращивания колоний на LB жидкой среде, содержащей 50 μг/мл канамицина и 0,4% глюкозу при 30oС до достижения CD600=0.6 и последующей индукции культур с IPTG (1 mM). Клетки были затем разрушены и клеточный экстракт проанализирован с помощью SDS-PAGE.
Пример 4. Очистка и перескладывание рекомбинантного порина класса 2.
Штамм Е. coli BL21(DE3) ΔompA [pNV-5] выращивается до достижения средней лог- фазы (OD= 0.6 при 600 нм) в бульоне Luria при температуре 30oС. Затем добавляется IPTG (0.4 mM) и клетки выращиваются в течение 2 ч при температуре 37oС. Собранные клетки отмываются различными объемами TEN буфера (50 mM Tris HCl, 0,2 M NaCl, 10 mM EDTA, pH 8,0) и клеточная паста замораживается при температуре -75oС.
Для очистки, предварительно взвешенные клетки размораживаются и взвешиваются в TEN буфере в соотношении 1:15 (g/v). Суспензированные клетки пропускаются через Stansted клеточный разрушитель (Stansted fluid power Ltd.) дважды при 8,000 psi. Полученный раствор затем центрифугируется в условиях 13,000 об/мин в течение 20 мин и полученный супернатант удаляется. Оставшаяся пилюля дважды взвешивается в TEN буфере, содержащем 0,5% деоксиколат и супернатант удаляется. Пилюля затем взвешивается в TEN буфере, содержащем 8М деионизированную мочевину (электрофоретический градиент) и в 0.1 mM PMSF (3 г/10 мл). Суспензия разрушена ультразвуком в течение 10 мин или до тех, пор пока не получена однородная суспензия. Добавлено 10 мл 10% водного раствора Цвиттергента 3,14 (Calbiochem) и раствор тщательно перемешан. Затем раствор вновь подвергается ультразвуковой обработке в течение 10 мин. Оставшиеся нерастворимые частицы удаляются центрифугированием. Определяется протеиновая концентрация. Она достигает 2 мг/мл в 8М мочевине-10% Цвиттергеном буфере (1:1).
Смесь помещается в колонку Сефакрил S-300 размером 2,6•100 см, уравновешенную 100 mM Tris-HCl, 1M NaCl, 10 mM EDTA, 20 mM СаСl2, 0,05% 3,14-цвиттерген, 0,02% содиума азид, рН 8,0. Уровень потока - 1 мл/мин. Собрано 10 мл фракции. Порин укладывается в тримерную конформацию во время гель-фильтрации. Измеренное OD каждой фракции равно 280 нм. Фракции, содержащие протеин, затем подвергнуты SDS гель-электрофорезу для исследования порина. Фракции, содержащие порин, отбираются. Отобранные фракции также диализируюся или разводятся 1: 10 в 50 mM Tris HCl, pH 8.0, 0.05% 3,14-цвиттерген, 5mM EDTA, 0,1 M NaCl. Полученный раствор затем помещен в 2,6•10 см Q сефарозную колонку (Pharmacia), уравновешенную тем же буфером. Порин элюирован с линейным градиентом 0,1-1М NaCl.
Пример 5. Очистка и укладывание рекомбинантного порина класса 3
Штамм Е. coli BL21(DE3) ΔompA, содержащий porВ-рЕТ-17b плазмид, выращивается до достижения средней лог- фазы (OD=0.6 при 600 нм) в бульоне Luria при температуре 30oС. Затем добаляется IPTG (0.4 mM) и клетки выращиваются в течение 2 ч при температуре 37oС. Собранные клетки отмываются различными объемами TEN буфера (50 mM Tris HCl, 0,2 M NaCl, 10 mM EDTA, pH 8,0) и клеточная паста замораживается при температуре -75oС.
Для очистки около 3 г клеток размороживаются и взвешиваются в 9 мл TEN буфера. Добавляется лизозим (Sigma, 0.25 мг/мл), деоксиколат (Sigma, 1.3 мг/мл) плюс PMSF (Sigma, μг/мл) и смесь осторожно встряхивается в течение 1 ч при комнатной температуре. В это время клетки лизируются и высвобожденная ДНК образует раствор, который становится очень вязким. К раствору добавляется ДНК-аза (Sigma, 2 μг/мл) и раствор затем вновь перемешивается в течение 1 ч при комнатной температуре. Смесь центрифугируется в S-600 роторе со скоростью 15К об/мин и супернатант удаляется. Пилюля дважды взвешивается в 10 мл TEN буфера и супернатант удаляется. Затем, пилюля вновь взвешивается в 10 мл 8М мочевины (Pierce) в TEN буфере. Смесь легко перемешивается для удаления каких-либо сгустков. Суспензия разрушена ультразвуком в течение 20 мин или до тех пор, пока не получена однородная суспензия. Добавлено 10 мл 10% водного раствора Цвиттергента 3,14 (Calbiochem) и раствор тщательно перемешан. Затем раствор вновь подвергается ультразвуковой обработке в течение 10 мин. Оставшиеся нерастворимые частицы удаляются центрифугированием. Определяется протеиновая концентрация. Она достигает 2 мг/мл в 8М мочевине-10% Цвиттергеном буфере (1:1).
Смесь помещается в колонку Сефакрил S-300 размером 180•2,5 см, уравновешенную 100 mM Tris HCl, 1M NaCl, 10 mM EDTA, 20 mM CaCl2, 0,05% 3,14-цвиттерген, 0,02% содиума азид, рН 8,0. Уровень потока - 1 мл/мин. Собрано 10 мл фракции. Порин укладывается в тримерную конформацию во время гель-фильтрации. Измеренное OD каждой фракции равно 280 нм. Фракции, содержащие протеин, затем подвергнуты SDS гель-электрофорезу для исследования порина. Фракции, содержащие, порин отбираются.
Отобранные фракции диализируются и концентрируются 4-6 складывание, используя Amicon концентратор с РМ 10 мембраной против буфера, содержащего 100 mМ Tris HCl, 0,1 М NaCl, 10 mM EDTA, 0.05% 3,14-цвиттерген, рН 8,0. Альтернативно-собранные фракции преципитируются с 80% этанолом и взвешиваются в вышеупомянутом буфере. Шесть к 10 мг материала затем применяется в моно 10/10 колонке (Pharmacia), уравновешенной в том же буфере. Порин, элюированный с поверхности 0.1-0.6 М NaCl градиента с 1,2% усилением в мин более 50 мин периода. Уровень потока составляет 1 мл/мин. Максимальное содержание порина собрано и диализировано против TEN буфера и 3,14-цвиттергена. Порин может быть очищен в дальнейшем с помощью другой S-300 гель-хроматографии.
Пример 6. Очистка и химическая модификация полисахаридов.
Капсульный полисахарид из N. meningitidis группы В и Е coli K1 насчитывают α(2-8) полисиаловых кислот (отнесенных как к GBMP, так и к K1 полисахаридам). Высокий молекулярный вес полисахаридов, изолированных из растущей среды путем преципитации (см. Frasch, C.E., "Production and Control N. meningitidis Vaccine" в Bacterial Vaccines, Alan R. Liss, Inc., pages 123-145 (1990)), был очищен и химически модифицирован до того, как соединен с пориновым протеином. Полисахариды с высоким молекулярным весом были частично деполимеризованы 0,1 М ацетиловой кислотой (7 мг полисахаридов/мл), рН 6,0-6,5 (70oС, 3 ч) для получения полисахаридов, имеющих молекулярный вес 12000-16000. После очистки в гель-фильрационной хроматографической колонке (Superdex 200 prep grade, Pharmacia) полисахариды были N-деацетилированы в присутствии NaBH4 и затем N-пропионилированы, методом, описанным Jennings et al. , (J. lmmunol. 137:1808 (1986)) для получения N-Pr-GBMP. Использование NalO4, следующее за гель-фильтрационной очисткой, позволило получить окисленный N-Pr-GBMP, имеющий средний молекулярный вес 12000 Дальтон.
Пример 7. Соединение окисленного N-Pr-GBMP с пориновым протеином (РР) класса 3 менингококка группы В.
Окисленный N-Pr-GBMP (9,5 мг) был добавлен к очищенному пориновому протеину класса 3 (3,4 мг), растворенному в 0,21 мл 0,2 М фосфатном буфере, рН 7.0, который так же содержал 10% октил глюкозид. После растворения полисахарида, был добавлен содиум цианоборогидрид (7 мг) и полученный раствор инкубирован при температуре 37oС в течение 4 дней. После этого смесь была разведена 0,15 М содиумом хлоридом, содержащим 0,01% тимерозаль и разделена гель-фильтрационной хроматографической колонкой, используя Superdex 200 PG. Конъюгат N-Pr-GВМР-РР был получен как единичный пик элюирования около пустого объема. Анализы раствора конъюгата показали наличие 43% полисахарида (относительно веса). В конъюгате было получено 44% поринового протеина и 12% полисахарида. Уровень полученного протеина в различных экспериментах в основном составлял 50-80%, полисахаридов 9-13%.
Пример 8. Исследование иммуногенности.
Иммуногенность N-Pr-GBMP-PP конъюгата и N-Pr-GBMP- Тетанус токсина (N-Pr-GBMP-TT) конъюгата, которые были получены путем сходных процедур, были исследованы на 4-6-недельных мышах женского пола Swiss Webster CFW. Полисахаридный (2 μг) - конъюгат был назначен в солевом растворе, абсорбированном на алюминии гидроксиде или смешанном к стеарил тирозину. ELISA-титры сыворотки против полисахаридного антигена и бактерицидные титры против менингококка группы В показаны в табл.1.
Имея в настоящее время полно описанное изобретение, лицам, сведущим в данной области, понятно, что то же самое может быть выполнено в различных и эквивалентных уровнях условий, формул и других параметров без какого-либо ущерба изобретению или др. его воплощениям. Все патенты и публикации, указанные здесь в полном виде, помещены в ссылках.

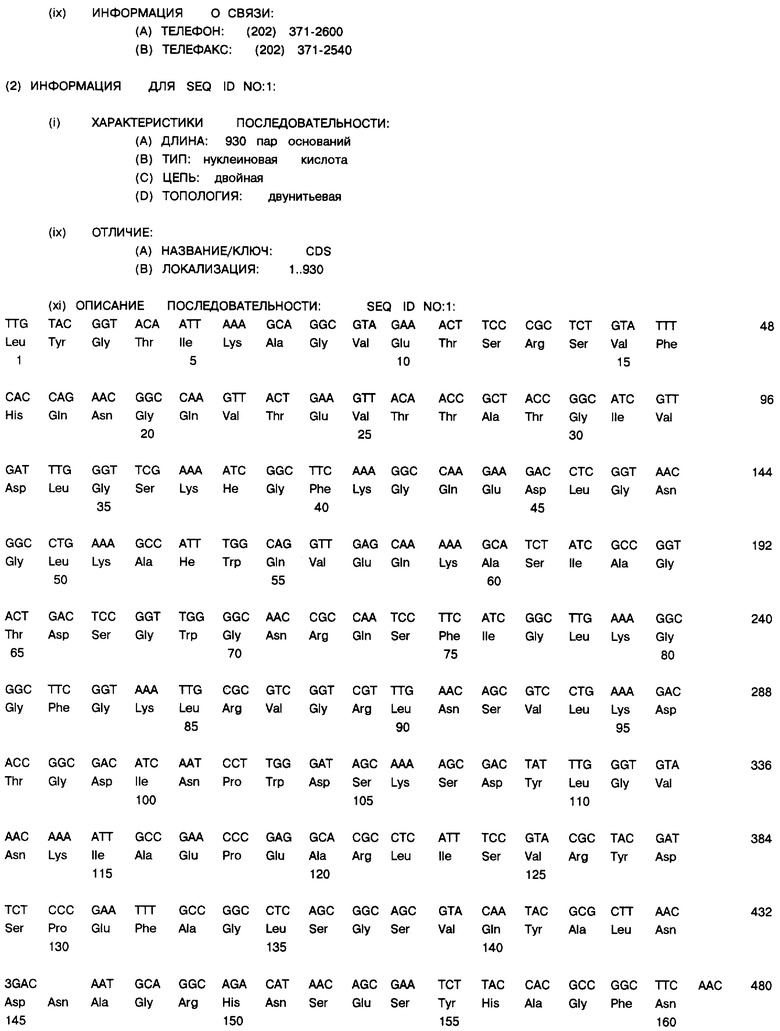

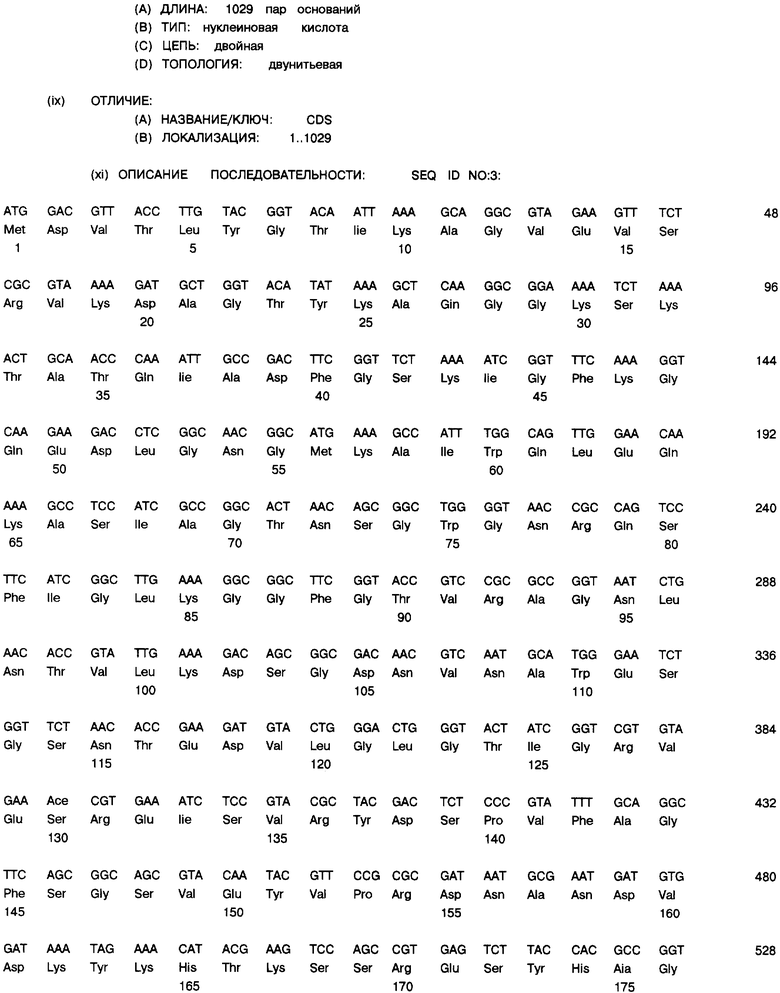

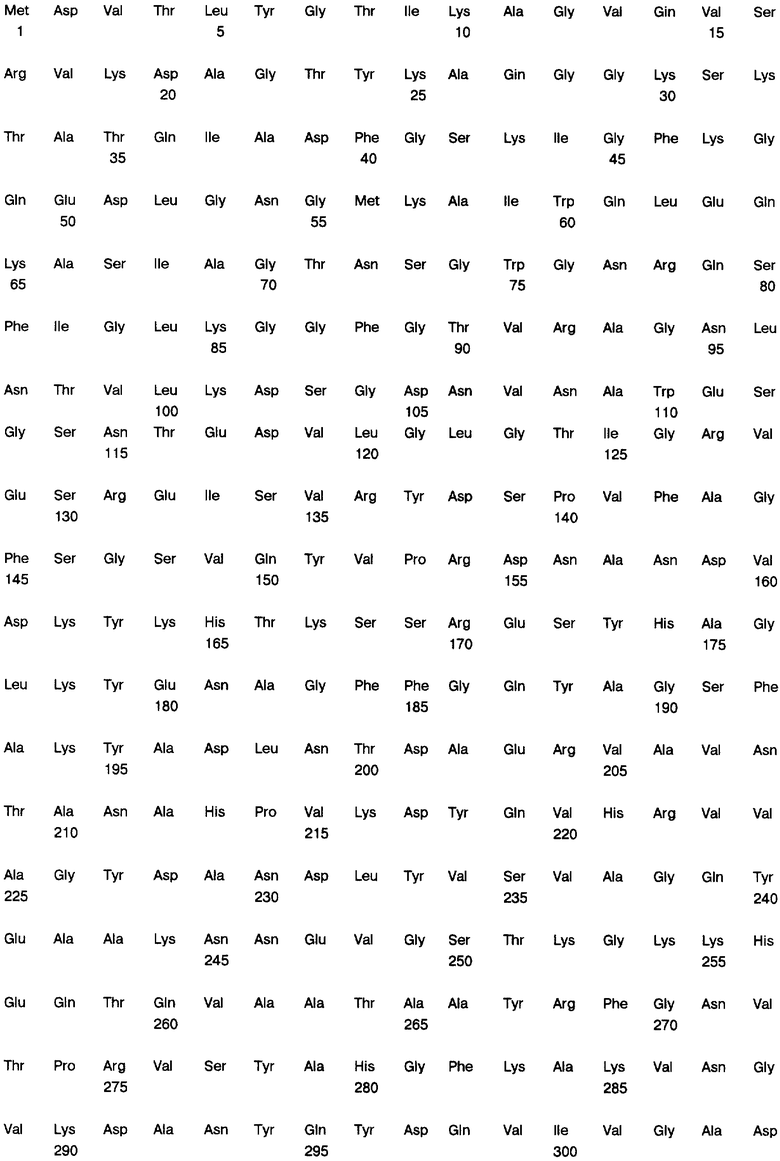
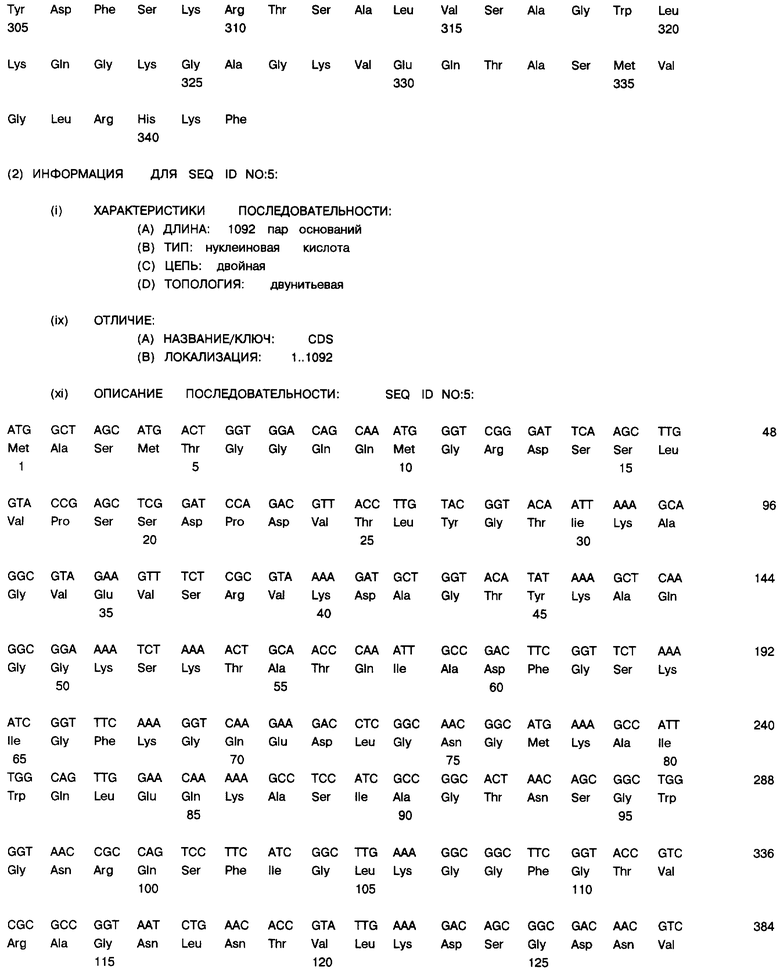
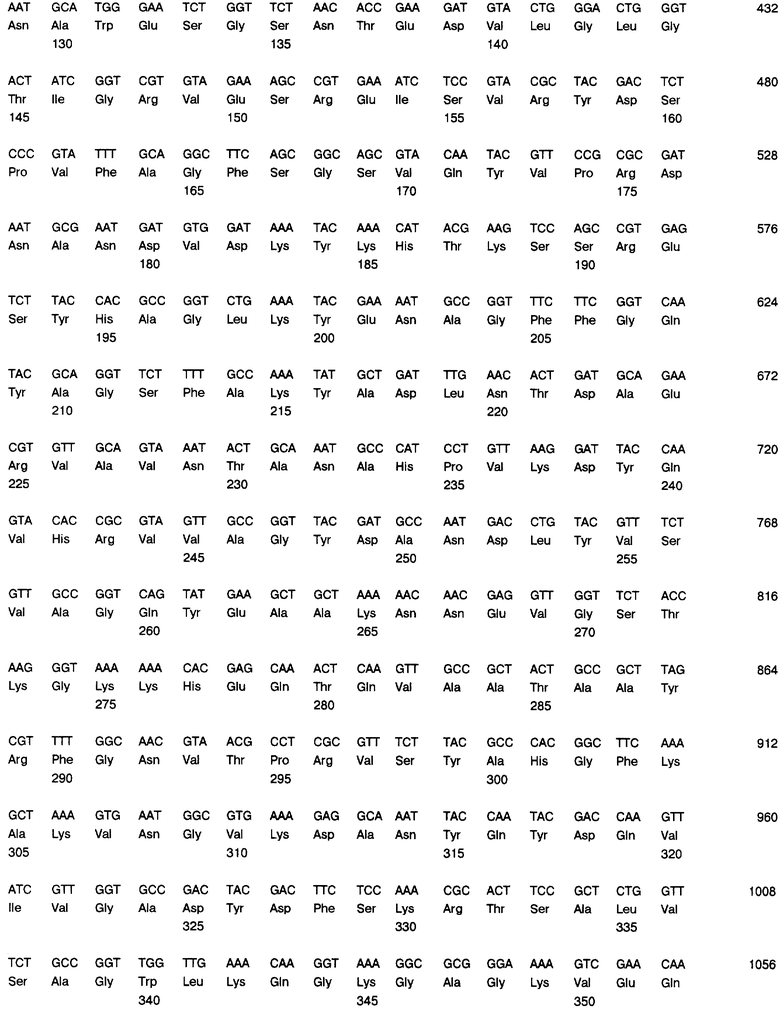






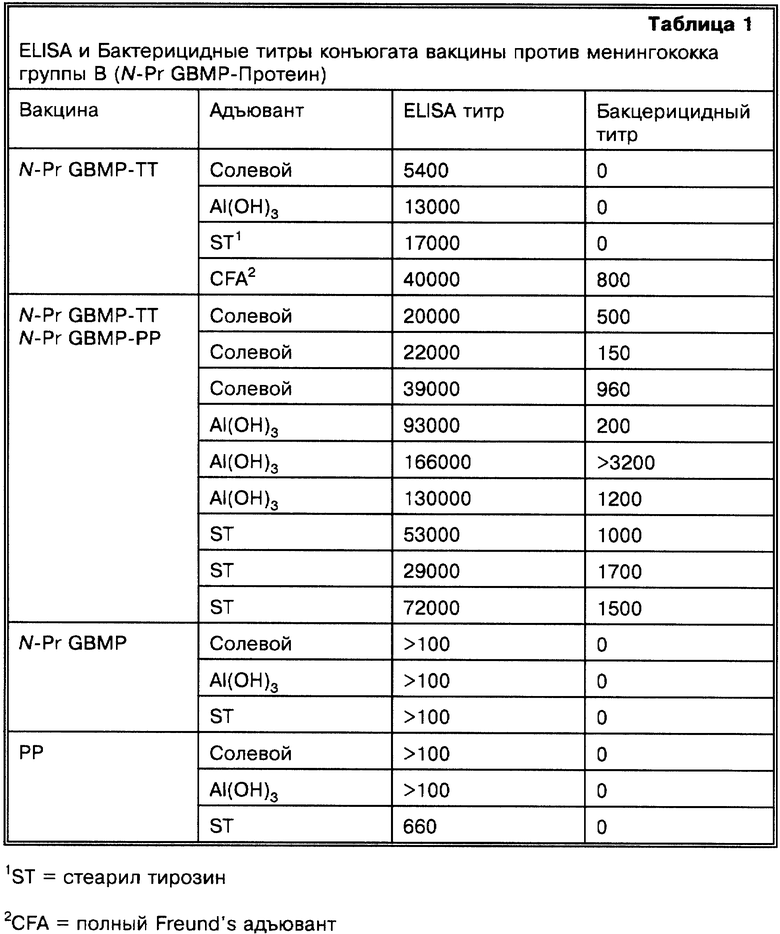
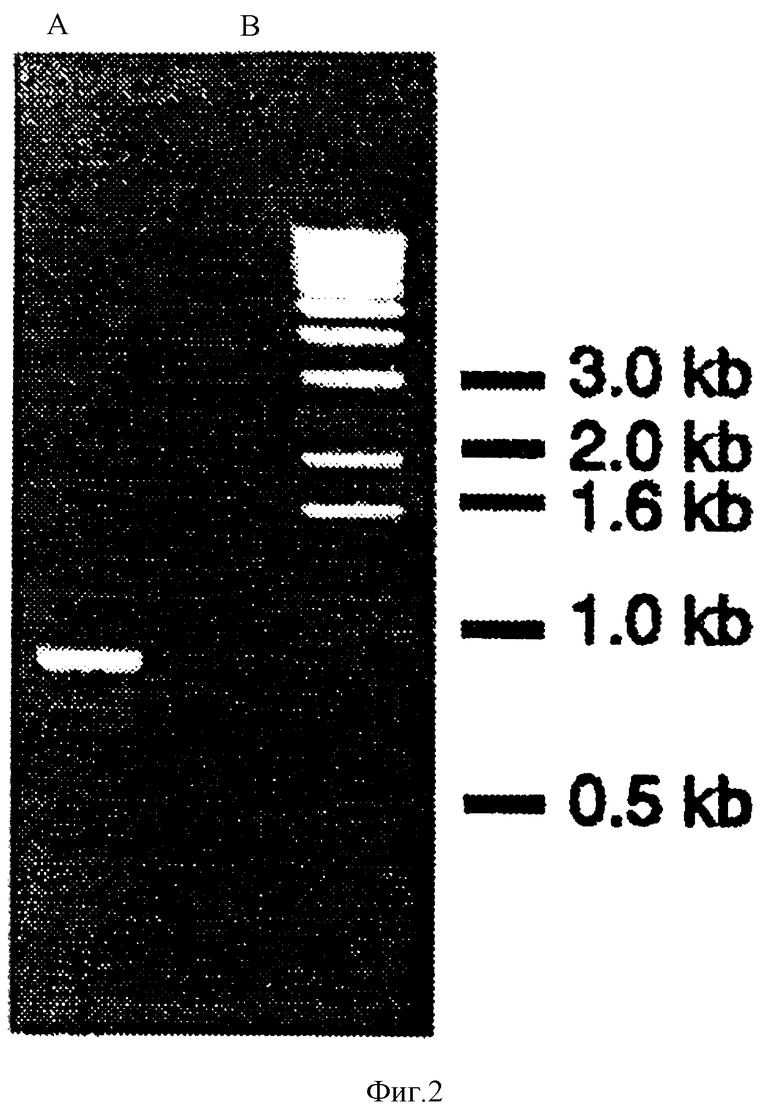
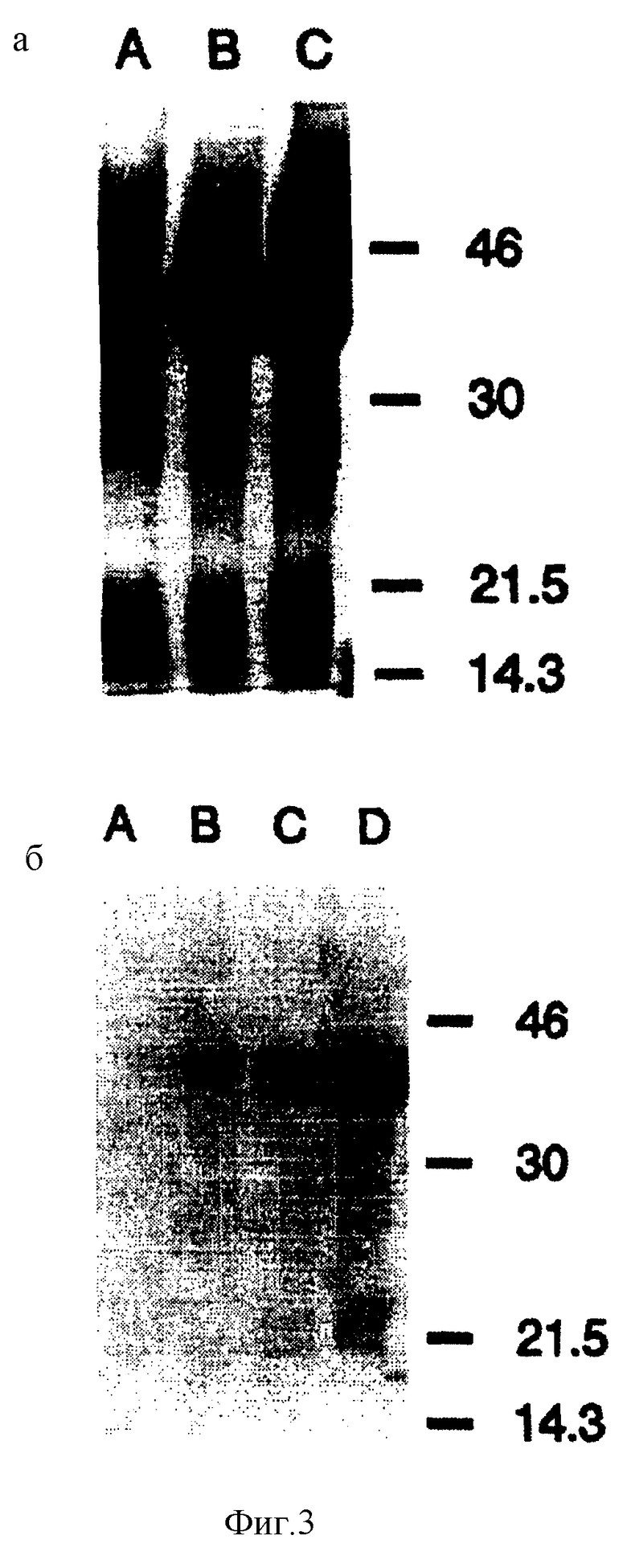


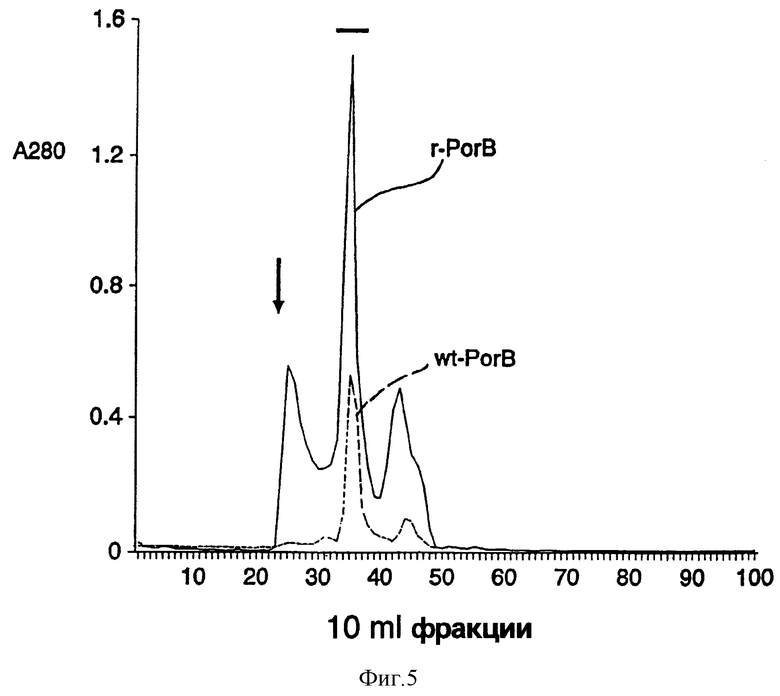
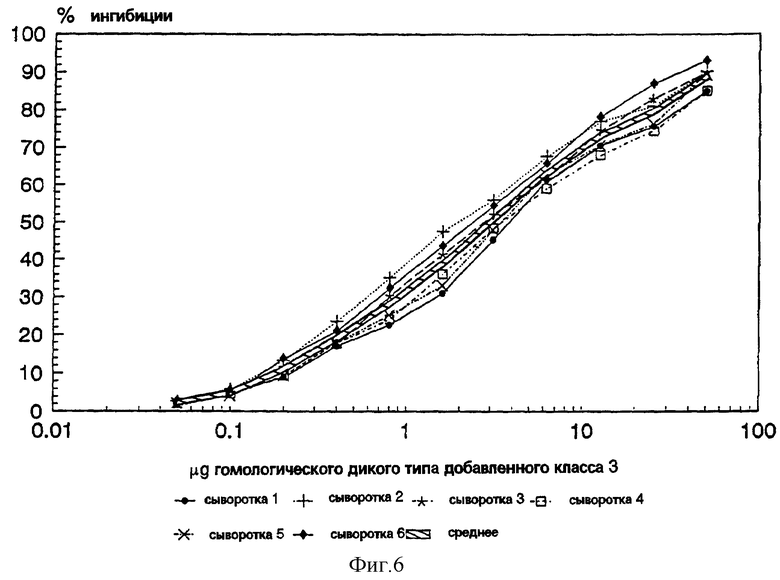
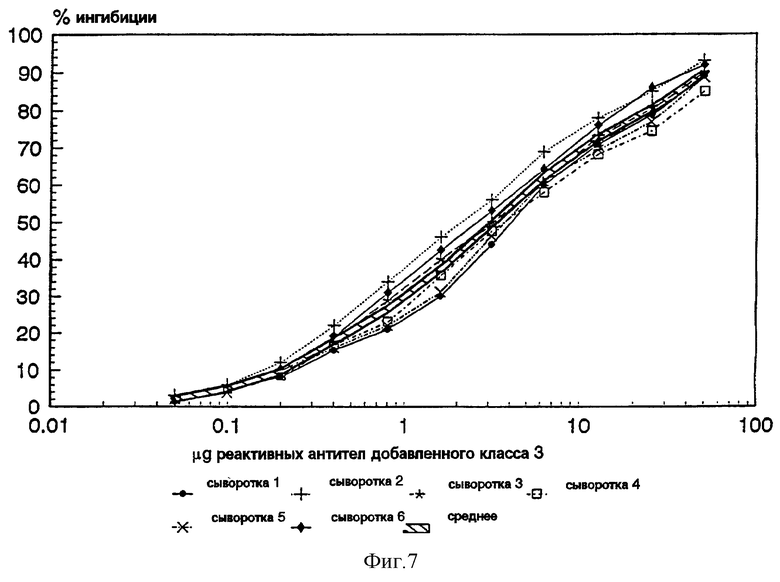
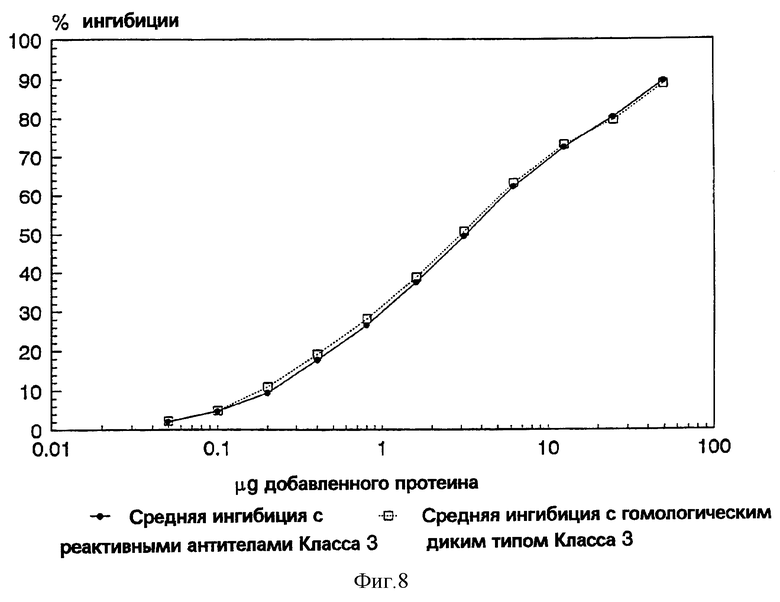







Изобретение относится к генной инженерии. В штамм Е. coli трансформируют вектор. Вектор включает селективный маркер и ген, кодирующий зрелый пориновый протеин или зрелый пориновый протеин, слитый с аминокислотами 1-22 капсидного протеина φ10. Последний кодируется геном Т7. Ген Т7 связан функционально с промотором Т7. Трансформированный штамм Е. coli выращивают в среде. Индуцируют экспрессию указанного протеина. Экспрессированный таким образом протеин составляет более 2% от общего экспрессированного протеина. На стадии индуцирования лизируют Е. coli. Выделенный в виде нерастворимых включений протеин промывают буфером. Суспендируют и растворяют указанные включения в водном растворе денатуранта, разбавляют детергентом и очищают гель-фильтрацией и ионообменной хроматографией. Высокоочищенный пориновый протеин и слитый протеин имеют аминокислотную последовательность, приведенную в описании, и кодируют ДНК с последовательностью, приведенной в описании. Вакцина содержит пориновый протеин внешней мембраны или слитый с ним протеин в эффективном количестве и фармацевтически приемлемый носитель. Полученный переуложенный протеин конъюгируют с полисахаридом Neisseria meningitidis. Особи вводят в эффективном количестве пориновый протеин менингококка группы В или слитый с ним протеин. Штамм Е. coli BL21(DE3) Δ ompA предназначен для получения рекомбинантных штаммов-продуцентов поринового протеина менингококка группы В или слитого с ним протеина. После трансформации данного штамма плазмидным вектором рЕТ, несущим ген, кодирующий пориновый протеин или слитый с порином протеин, он продуцирует пориновый протеин или слитый с ним протеин. Изобретение позволяет на более высоком уровне производить выделение пориновых протеинов наружной мембраны менингококков группы В и связанных с ними протеинов. 16 с. и 8 з.п. ф-лы, 11 ил., 1 табл.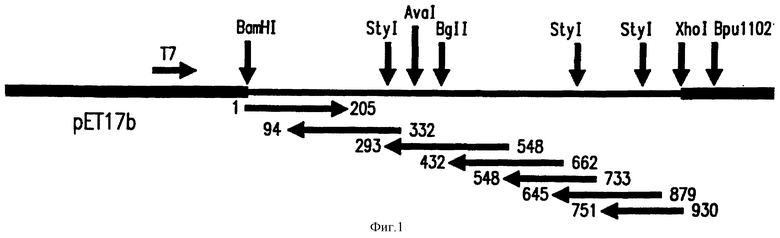
| ЕР 0467714 А, 22.10.1992 | |||
| ВАЛОК ДЛЯ ПРОДОЛЬНОЙ ПРОКАТКИ ТРУ | 0 |
|
SU351604A1 |
| Автомат для мерной резки проводов и зачистки их концов от изоляции | 1972 |
|
SU492964A1 |
| ЕР 0474313 А, 11.03.1992 | |||
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| Штамм бактерий ЕSснеRIснIа coLI | 1990 |
|
SU1761807A1 |

