ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение относится к области рекомбинантных антител, применяемых для терапевтического лечения рака у человека.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Рецептор эпидермального фактора роста (EGFR) играет важную роль в клеточной пролиферации, а также в апоптозе, ангиогенезе и распространении метастазов, процессах, которые имеют ключевое значение для прогрессирования опухолей (Salomon et al., Crit. Rev. Oncology/Haematology, 19:183-232 (1995); Wu et al., J. Clin. Invest., 95:1897-1905 (1995); Karnes et al., Gastroenterology, 114:930-939 (1998); Woodburn et al., Pharmacol. Therap. 82: 241-250 (1999); Price et al., Eur. J. Cancer, 32A:1977-1982 (1996)). Действительно, исследования показали, что EGFR-опосредованный клеточные рост увеличивается при многих солидных опухолях, включая немелкоклеточный рак легкого, рак предстательной железы, рак молочной железы, рак желудка, а также опухоли головы и шеи (Salomon DS et al., Critical Reviews in Oncology/Haematology, 19:183-232 (1995)). Кроме того, сейчас известно, что избыточная активация EGFR на поверхности раковых клеток ассоциирована с поздними стадиями заболевания, развитием метастатического фенотипа и плохим прогнозом у пациентов, страдающих раком (Salomon DS et al., Critical Reviews in Oncology/Haematology 19:183-232 (1995)).
Кроме того, экспрессия EGFR часто сопровождается выработкой клетками, экспрессирующими EGFR, среди прочего, EGFR-лигандов, TGF-альфа и EGF, что позволяет думать об участии аутокринной петли в прогрессировании этих клеток (Baselga, et al. (1994) Pharmac. Therapeut. 64: 127-154; Modjtahedi, et al. (1994) Int. J. Oncology. 4:277-296). Следовательно, блокирование взаимодействия между такими EGFR-лигандами и EGFR способно ингибировать рост и выживание опухолей (Baselga, et al. (1994) Pharmac. Therapeut. 64:127-154).
EGFR представляет собой мембраносвязанный гликопротеин с молекулярным весом приблизительно 170 кДа. EGFR состоит из гликозилированного наружного лигандсвязывающего домена (621 остаток) и цитоплазматического домена (542 остатка), связанных коротким трансмембранным линкером из 23 аминокислот. Внеклеточная часть EGFR содержит 25 дисульфидных мостиков, 12 N-связанных сайтов гликозилирования и обычно считается состоящей из четырех субдоменов. Рентгеновские кристаллические структуры EGFR позволяют полагать, что рецептор может принимать как аутоингибированную связанную конформацию, которая неспособна связывать EGF (Ferguson et al., Mol Cell, 2003, vol 11:507-517), так и активную конформацию, которая может опосредовать связывание лиганда EGF и димеризацию рецептора (Garret et al., Cell 2002, vol 110:763-773; Ogiso et al., Cell, 2002, vol 110:775-787). Было выдвинуто предположение о том, что домен I и домен III, в частности, обеспечивают аддитивный вклад в образование высокоаффинного сайта связывания лиганда. Домены II и IV представляют собой богатые цистеином ламининоподобные области, которые стабилизируют укладку структуры белка и содержат возможную поверхность раздела для димеризации EGFR.
Известно, что EGFR существует на поверхности клетки во множестве различных конформаций, причем наиболее частой из них является связанная или закрытая конформация. Связанная конформация неспособна к димеризации и, следовательно, неактивна. Известно, что терапевтическое антитело Erbitux стабилизирует связанную конформацию за счет связывания с доменом III и создания такой пространственной конфигурации рецептора, которая не позволяет ему достичь несвязанного состояния. Однако некоторые рецепторы могут все еще сохранять способность принять несвязанную конформацию, связать лиганд и димеризироваться. Моноклональное антитело (mAb) в типичном случае способно эффективно связываться только с одной из конформаций, следовательно, не может эффективно нацеливаться на раковые клетки, проявляющие другие конформации, или раковые клетки, проявляющие несколько конформаций.
Моноклональные антитела (mAb), направленные на лигандсвязывающий домен EGFR, могут блокировать взаимодействие с лигандами EGFR и, сопутствующим образом, возникающий в результате этого взаимодействия внутриклеточный сигнальный путь.
Erbitux™ (эрбитукс, цетуксимаб) представляет собой рекомбинантное химерное (человек/мышь) моноклональное антитело, которое специфически связывается с внеклеточным доменом EGFR человека. Эрбитукс состоит из Fv-областей мышиного антитела против EGFR с константными областями тяжелой и каппа-легкой цепи IgG1 человека, имея приблизительный молекулярный вес 152 кДа. Эрбитукс вырабатывается в культуре клеток млекопитающих (мышиная миелома). Эрбитукс официально разрешен для лечения пациентов с метастатическим колоректальным раком, опухоли которых экспрессируют EGFR. В дополнение к этому, эрбитукс применяют в комбинации с лучевой терапией для лечения пациентов с плоскоклеточным раком головы и шеи при невозможности хирургического удаления опухоли или в виде лечения второй очереди, если плоскоклеточный рак головы и шеи не поддается стандартной терапии на основе препаратов платины.
Vectibix™ (вектибикс, панитумумаб) представляет собой рекомбинантное каппа моноклональное антитело IgG2 человека, которое специфически связывается с EGFR человека. Вектибикс имеет приблизительный молекулярный вес 147 кДа. Панитумумаб вырабатывается в генноинженерных клетках млекопитающих (из яичника китайского хомячка). Вектибикс официально разрешен для лечения пациентов с метастатическим колоректальным раком, опухоли которых экспрессируют EGFR, при прогрессировании болезни или после химиотерапии по схемам, содержащим фторпиримидин, оксалиплатин и иринотекан.
На опухолевых клетках человека было идентифицировано множество мутантных рецепторов EGF. Они способны воспроизводить рецепторную активность независимо от связывания лиганда (EGFRvIII), приводя к повышенной онкогенности. Можно генерировать моноклональные антитела против мутантов EGFR, но такие моноклональные антитела не всегда будут эффективными против немутантного EGFR.
У людей, больных раком, были идентифицированы такие мутации EGFR, которые влияют на ответную реакцию на химиотерапию, направленную на EGFR. Документ WO 2006/110478 (Novartis) раскрывает 43 мутации, а также 18 вариантов SNP (однонуклеотидного полиморфизма) в открытой рамке считывания EGFR. В двух или более типах опухолей идентифицированы некоторые бессмысленные мутации. Документ WO 2006/091899 (Amgen) раскрывает еще восемь мутаций, идентифицированных в различных раковых клетках. Одна или несколько таких мутаций могут локализоваться в эпитопе или изменять структуру эпитопа, связываемого с одним из разрешенных терапевтических моноклональных антител. Пациенты, несущие такую мутацию (мутации), будут невосприимчивы к лечению моноклональным антителом.
Кроме того, в литературе имеются сообщения, в которых отмечена гетерогенность по гликозилированию, по меньшей мере, одного из сайтов гликозилирования (Whitson et al., 2005 Biochemistry 44:14920-31; Zhen et al. 2003 Biochemistry 42; 5478-92). Такая гетерогенность может прямым или опосредованным образом приводить к разной экспозиции эпитопов, варьирующих в различных опухолевых клетках.
Антителозависимая клеточноопосредованная цитотоксичность (ADCC) представляет собой альтернативный механизм, посредством которого антитела опосредуют уничтожение опухолевых клеток. Уровень ADCC зависит от нескольких факторов, включая подтип IgG (IgM>IgG1 >IgG2), плотность антител на целевых клетках, характер гликозилирования антитела, а также свойства мишени, как таковой.
Friedmann et al (PNAS 2005, 102:1915-20) показали, что два мышиных моноклональных антитела, отобранных по их способности ингибировать связывание EGF с EGFR за счет связывания различных эпитопов EGFR, проявляют способность к синергическому подавлению экспрессии рецептора в клетках KB и СНО, кратковременно экспрессирующих EGFR. Перекрестно конкурирующие антитела, ингибирующие EGF, не проявляют никакой синергии.
Modjtahedi et al (Cell Biophysics vol 22, 1993, 129-146) протестировали комбинации нескольких крысиных антител против EGFR с неперекрывающимися эпитопами. Антитела принадлежали к разным изотипам. Во всех случаях эффект от применения двух антител был промежуточным между эффектами, наблюдаемыми при применении двух моноклональных антител по отдельности в том же количестве. Это наблюдение было подтверждено как in vivo, так и in vitro.
Документ WO 2004/032960 (патент Merck) раскрывает, что комбинированное применение двух моноклональных антител, Mab 425 и Mab 225 (цетуксимаб), приводит к повышению количества связанных антител на поверхности раковых клеток, экспрессирующих EGFR, по сравнению с применением каждого из моноклональных антител по отдельности в том же количестве. Эта публикация также раскрывает усиление подавления EGFR при использовании комбинации антител по сравнению с двумя моноклональными антителами.
Perera et al (Clin Cancer Res 2005; 11(17):6390-99) раскрыли синергический эффект при лечении мышей, несущих ксенотрансплантаты U8 7MG.de2-7, комбинацией двух мышиных моноклональных антител. Одно из антител (mAb 528) связывает все подтипы EGFR примерно с такой же специфичностью, что и цетуксимаб. Другое антитело (mAB 806) связывает только EGFR подтипа de2-7. Клеточная линия U87MG.de2-7 представляет собой трансфицированную клеточную линию de2-7EGFR. Клеточная линия U87MG.DK экспрессирует вариант EGFR подтипа de2-7 с неактивной киназой. Если для лечения мышей, несущих ксенотрансплантаты U87MG.DK, применяли два антитела, то никаких синергических эффектов не наблюдалось. В модели ксенотрансплантата с клеточной линией A431, экспрессирующей EGFR дикого типа, авторы не нашли никаких подтверждений синергии. EGFR de2-7 представлен только в небольшом числе типов рака, в частности, в глиомах, в некоторой степени в опухолях молочной железы и в опухолях легких.
Хотя эти исследования показали, что в некоторых случаях между двумя мышиными моноклональными антителами может существовать синергия, они также показали, что во многих случаях никакой синергии не наблюдается. Эти исследования также не выявили какой-либо композиции антител против EGFR, которая была бы эффективной против широкого ряда клинически важных линий раковых клеток.
В связи с этим существует потребность в улучшенных терапевтических антителах против EGFR, которые были бы эффективны для лечения и/или профилактики заболеваний, связанных со сверхэкспрессией EGFR, при введении в низких дозах. Также существует потребность в широко применимых терапевтических антителах против рака, которые можно было бы применять, даже не обладая глубокими знаниями о структуре EGFR, экспрессируемого представляющими интерес раковыми клетками.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В одном из аспектов изобретение относится к композиции рекомбинантных антител, содержащей, по меньшей мере, 3 различные молекулы антител против EGFR, где антитела связывают различные первый, второй и третий эпитопы EGFR.
В дополнительном аспекте изобретение относится к композиции рекомбинантных антител, содержащей, по меньшей мере, две различные молекулы антител против EGFR, где одна индивидуальная молекула антитела против EGFR может быть выбрана из группы, состоящей из антител: 992, 1024, 1030, 1042, 1208, 1229, 1254, 1257, 1260, 1261, 1277, 1284, 1308, 1320, 1344 и 1347, или антител, имеющих области CDR указанных антител.
Предпочтительно, чтобы, по меньшей мере, одна индивидуальная молекула антитела против EGFR была выбрана из группы, состоящий из антител 992, 1030, 1024, 1347, 1277, 1254, 1320, 1260, 1261 и 1284, или антител, имеющих области CDR указанных антител. В особенно предпочтительном варианте осуществления изобретения композиция антител содержит антитела 992 и 1024 или два антитела, основанные на их последовательностях CDR3 или на их последовательностях VL и VH, либо содержит два антитела, обладающих, по существу, такой же специфичностью связывания.
Иллюстративные композиции антител по изобретению обладают доказанной эффективностью в ингибировании пролиферации типичных линий раковых клеток, что свидетельствует о пригодности их применения для лечения рака in vivo. Эти результаты были подтверждены в анализах со сфероидами раковых клеток, которые, по-видимому, в большей степени соответствуют ситуации in vivo, когда раковые клетки формируют опухоли. Кроме того, композиция антител по изобретением явно снижает подвижность клеток из раковых сфероидов, то есть уменьшает предрасположенность к метастазированию. Была также продемонстрирована эффективность иллюстративной композиции антител в модели ксенотрансплантата in vivo. Эти результаты были подтверждены при использовании особо предпочтительной композиции антител, состоящей из антител 992 и 1024.
В модели ксенотрансплантата рака человека у мышей иллюстративная композиция антител, предложенная изобретением, приводила к значительно более высокой степени терминальной дифференциации опухолевых клеток по сравнению с имеющимися в продаже антителами вектибикс и эрбитукс. Оказалось, что предпочтительная композиция антител по изобретению реализует свой эффект через другой механизм действия по сравнению с моноклональными антителами, поскольку после прекращения лечения указанной композицией антител возобновления роста опухоли не наблюдалось. Возобновление роста опухоли наблюдается после прекращения лечения моноклональными антителами.
В исследованиях по связыванию авторы настоящего изобретения продемонстрировали, что некоторые из антител, описанных в этой заявке, очевидно, облегчают связывание других антител, благодаря чему увеличивается общее количество антител, связанных с рецептором. Они также продемонстрировали, что связывание трех антител домена III облегчает последующее связывание других антител. Эти наблюдения несомненно поддерживают концепцию применения композиции, содержащей, по меньшей мере, 3 различные молекулы антител против EGFR, где антитела связывают различные первый, второй и третий эпитопы EGFR. Может также наблюдаться эффект при использовании специфической комбинации двух антител по изобретению, за счет выбора антител, обладающих этим специфическим эффектом. Такие антитела являются предпочтительными кандидатами для смешивания с другими антителами.
Композиции по изобретению могут обеспечивать некоторые дополнительные преимущества. Раковые клетки экспрессируют ряд EGFR. Наблюдается вариабельность в их конформации, гликозилировании и первичной структуре (мутации и SNP). Одно моноклональное антитело может быть нацелено на несколько таких вариантов EGFR, но не на все сразу. Мутанты EGFR могут быть невидимыми мутантами для моноклональных антител. Композиция антител, содержащая два антитела по изобретению или три и более различных антител, связывающих разные эпитопы EGFR, менее уязвима для мутантов, SNP, делеционных мутантов и вариаций в гликозилировании. Это доказывается широкой эффективностью смесей антител по настоящему изобретению против целой панели линий раковых клеток, представляющих разнообразные конформации и вариации EGFR.
Введение одного моноклонального антитела может также неполностью выключить киназную активность EGFR. При комбинировании антител можно достичь более эффективного ингибирования передачи сигналов.
Таким образом, может оказаться полезным включение антител, связывающих разные конформации EGFR (например, незакрытые конформации и димеры рецептора) в указанную смесь антител. Такая смесь антител может более эффективно ингибировать активность EGFR, чем моноклональное антитело, связывающее только одну из конформации.
Кроме того, применение подхода с тремя или более антителами против EGFR в композиции создает возможность для увеличения плотности антител на поверхности опухолевой клетки, благодаря чему увеличивается уничтожающее действие ADCC по сравнению с моноклональными антителами.
В дальнейшем аспекте изобретение относится к способу получения композиции антител, включающему
а) трансфицирование первой популяции эукариотических клеток первой экспрессирующей конструкцией, кодирующей первое антитело, содержащее первую когнатную пару цепей VH и VL, способных связывать первый индивидуальный эпитоп EGFR,
b) трансфицирование второй популяции эукариотических клеток второй экспрессирующей конструкцией, кодирующей второе антитело, содержащее вторую когнатную пару цепей VH и VL, способных связывать второй индивидуальный эпитоп EGFR,
c) необязательное повторение стадии b) для третьей или последующих клеточных популяций, экспрессирующих конструкций, когнатных пар и эпитопов EGFR,
d) отбор трансфицированных первой, второй и (необязательно) последующих клеточных популяций,
e) объединение трансфицированных популяций в одном сосуде для получения банка клеток,
f) культивирование клеток из банка клеток в условиях, обеспечивающих экспрессию антител, и
g) восстановление композиции антител из супернатанта и ее очистка.
Для простоты получения, единой технологии производства и выделения целевого продукта, а также для определения параметров все антитела содержат одну и ту же константную область тяжелой цепи.
В дальнейшем аспекте изобретение относится к банку клеток, содержащему, по меньшей мере, две субпопуляции эукариотических клеток, причем каждая субпопуляция трансфицирована или трансдуцирована одной экспрессирующей конструкцией, кодирующей антитело, содержащее когнатную пару цепей VH и VL, способных связывать индивидуальный эпитоп EGFR. Предпочтительно, чтобы клетки были трансфицированы с применением сайт-специфической интеграции.
Кроме того, изобретение относится к способу уменьшения передачи сигнала EGFR, включающему введение в композицию клеток, экспрессирующих EGFR, композиции антител по изобретению, что приводит к уменьшению передачи сигнала EGFR.
Изобретение также относится к способу уничтожения клеток, экспрессирующих EGFR, включающему введение в композицию клеток, экспрессирующих EGFR, композиции антител по изобретению, что приводит к уничтожению клеток, экспрессирующих EGFR.
Также предлагается способ индукции апоптоза в клетках, экспрессирующих EGFR, включающий введение в композицию клеток, экспрессирующих EGFR, композиции антител по изобретению, посредством чего индуцируется апоптоз.
Дальнейший аспект относится к способу ингибирования пролиферации клеток, экспрессирующих EGFR, включающему введение в композицию клеток, экспрессирующих EGFR, композиции антител по изобретению, посредством чего ингибируется пролиферация.
Изобретение относится к способу индуцирования дифференциации опухолевых клеток in vivo, включающему введение индивиду, страдающему раковым заболеванием, композиции антител, по изобретению, посредством чего индуцируется дифференциация опухолевых клеток. Этот аспект основан на наблюдаемых in vivo эффектах терминальной дифференциации раковых клеток при контакте с композицией антител по изобретению.
В дальнейшем аспекте изобретение относится к фармацевтическим изделиям, содержащим композицию антител по изобретению, а также, по меньшей мере, одно соединение, способное индуцировать дифференциацию раковых клеток, в виде комбинации для одновременного, раздельного или последовательного введения по схеме противораковой терапии. Комбинируя композиции антител по изобретению со средствами, о которых известно, что они индуцируют терминальную дифференциацию раковых клеток, можно добиться дополнительного улучшения лечебного эффекта.
Еще в одном аспекте изобретение относится к фармацевтическим изделиям, содержащим композицию антител по изобретению, а также, по меньшей мере, одно химиотерапевтическое или противоопухолевое соединение в виде комбинации для одновременного, раздельного или последовательного введения по схеме противораковой терапии. Вполне вероятно, что композицию антител по изобретению можно использовать для лечения второй очереди, то есть после лечения традиционными химиотерапевтическими или противоопухолевыми средствами или одновременно с таким лечением, после лучевой терапии или одновременно с ней, после хирургической операции или параллельно с ней.
В отдельном аспекте изобретение предлагает полинуклеотид, выбранный из группы, состоящей из нуклеиновой кислоты, имеющей последовательность нуклеиновой кислоты, показанную на фиг.23 (SEQ ID NO 100), нуклеиновой кислоты, кодирующей полипептид, имеющий аминокислотную последовательность, показанную на фиг.23 (SEQ ID NO 101), нуклеиновой кислоты, имеющей последовательность нуклеиновой кислоты, показанную на фиг.34A (SEQ ID NO 102), и нуклеиновой кислоты, кодирующей полипептид, имеющий аминокислотную последовательность, показанную на фиг.34B (SEQ ID NO 103). Кроме того, предлагается полипептид, содержащий аминокислотную последовательность, показанную на фиг.23 (SEQ ID NO 101), и полипептид, содержащий аминокислотную последовательность, показанную на фиг.34B (SEQ ID NO 103), векторы экспрессии, содержащие указанную нуклеиновую кислоту, как определено выше, функционально связанные с промоторной последовательностью, способной управлять экспрессией указанной нуклеиновой кислоты, и клетка, трансфицированная или трансдуцированная указанным вектором экспрессии.
Эти последовательности составляют полинуклеотидные и полипептидные последовательности EGFR у обезьян Cynomolgous, например, у Macaca fascicularis. Указанный вид обезьян широко используется в токсикологических исследованиях на животных. Для того чтобы определенный животный вид имел какую-либо ценность в токсикологических исследованиях с антителами против аутоантигенов человека, необходимо, чтобы антитела также связывали целевой белок у животного, подвергнутого токсическому воздействию, предпочтительно, приблизительно с такой же аффинностью. Благодаря вкладу авторов настоящего изобретения теперь стали доступны тестирующие антитела для связывания с EGFR обезьян cynomolgous. EGFR обезьян cynomolgus и человека являются высоко гомологичными белками, но было обнаружено на удивление много антител с весьма разной аффинностью к EGFR человека и обезьян cynomolgus. Это подчеркивает важность строгого применения для скрининга того белка EGFR Cynomolgus, который был предложен авторами настоящего изобретения.
Кроме того, предлагается способ скрининга антител по связыванию с EGFR cynomolgous, включающий следующие стадии:
- получение, по меньшей мере, одного испытуемого антитела,
- проведение анализа для определения связывания антитела с внеклеточным доменом EGFR cynomolgous (фиг.23, SEQ ID NO 101)) или с полноразмерным EGFR cynomolgous (фиг.34B, SEQ ID NO 103)), либо с поверхностью клетки, экспрессирующей внеклеточный домен EGFR cynomolgous или экспрессирующей полноразмерный EGFR cynomolgous,
- выбор, по меньшей мере, одного антитела, которое связывается с внеклеточным доменом EGFR cynomolgous.
Способ может дополнительно включать скрининг по связыванию с EGFR человека или по связыванию с клетками, экспрессирующими EGFR человека.
В дальнейшем аспекте изобретение относится к способу идентификации антител против EGFR, способных усиливать одновременное связывание другого антитела против EGFR с EGFR, где указанный способ включает:
a. В первом анализе определение максимальной способности связывания первого антитела с фиксированным количеством антигена EGFR,
b. Во втором анализе насыщение фиксированного количества антигена EGFR вторым антителом против EGFR,
c. Контактирование комплекса EGFR-антитело с указанным первым антителом и определение максимальной способности связывания, и
d. Сравнение способностей связывания для определения того, превышает ли максимальная способность связывания на стадии c. максимальную способность связывания на стадии a.
Этот анализ можно использовать для идентификации дополнительных комбинаций антител, обладающих сходными свойствами с антителами 992 и 1024.
Определения
Термин "антитело" описывает функциональный компонент сыворотки и часто относится или к совокупности молекул (антител или иммуноглобулина), или к одной молекуле (антитела или иммуноглобулина). Молекула антитела способна связываться или взаимодействовать со специфическим антигенным детерминантом (антигеном или антигенным эпитопом), который, в свою очередь, может привести к индукции иммунологических эффекторных механизмов. Индивидуальная молекула антитела обычно рассматривается как моноспецифическая, а композиция молекул антитела может быть моноклональной (то есть состоящей из идентичных молекул антитела) или поликлональной (то есть состоящей из двух или более различных молекул антитела, взаимодействующих с одним и тем же или различными эпитопами на одном антигене или даже на отдельных, различных антигенах). Каждая молекула антитела имеет уникальную структуру, которая позволяет ей специфически связываться с соответствующим антигеном, а все природные молекулы антитела имеют одну и ту же общую базовую структуру, состоящую из двух идентичных легких цепей и двух идентичных тяжелых цепей. Антитела в собирательном значении также известны как иммуноглобулины. Термин антитело или антитела при использовании здесь дополнительно включает химерные и одноцепочечные антитела, а также такие связывающие фрагменты антител как Fab, фрагменты Fv или фрагменты scFv, а также такие мультимерные формы, как димерные молекулы IgA или пентавалентные IgM. Антитело может быть человеческим, мышиным, химерным, гуманизированным или реконструированным.
Терминологическое определение "когнатная кодирующая пара VH и VL" описывает кодирующие последовательности исходной пары VH и VL, содержащиеся в одной и той же клетке, продуцирующей антитело, или извлеченные из этой клетки. Таким образом, когнатная пара VH и VL представляет собой пару VH и VL, исходно представленную у донора, от которого получена такая клетка. Терминологическое определение "антитело, экспрессируемое из кодирующей пары VH и VL", указывает, что антитело или фрагмент антитела выработан из вектора, плазмиды или т.п. конструкций, содержащих кодирующую последовательность VH и VL. Когда когнатная кодирующая пара VH и VL экспрессирует либо полное антитело, либо его стабильный фрагмент, эти продукты сохраняют аффинность связывания и специфичность антитела, изначально экспрессируемого из той клетки, откуда они были извлечены. Библиотеку когнатных пар также терминологически обозначают как репертуар или совокупность когнатных пар, которые могут быть представлены по отдельности или в виде общего пула.
Термин "CDR" - определяющая комплементарная область (или гипервариабельный участок) соответствует определению, данному в ссылке Lefranc et al (2003) IMGT unique numbering for immunoglobulin and T cell receptor variable domains and Ig superfamily V-like domains. Dev. Comp Immunol 27, 55-77.
Термин "отдельный член рекомбинантного поликлонального белка" обозначает одну молекулу белка из белковой композиции, содержащей разные, но гомологичные молекулы белка, в которой каждая молекула белка гомологична другим молекулам композиции, но также содержит один или несколько фрагментов вариабельной полипептидной последовательности, который (которые) характеризуется различиями в аминокислотной последовательности между отдельными членами поликлонального белка.
Терминологическое определение "промоторы головка к головке" относится к паре промоторов, расположенных очень близко друг к другу, в результате чего транскрипция двух фрагментов гена, управляемая этими промоторами, идет в противоположных направлениях. Промотор головка к головке можно также сконструировать при помощи фрагмента-вставки, кодирующего посторонние аминокислоты и находящегося между двумя промоторами. Такой вставочный фрагмент вполне может содержать более 500 нуклеотидов. Промоторы головка к головке также могут упоминаться под названием двунаправленные промоторы.
Термин "иммуноглобулин" обычно употребляют как собирательное обозначение смеси антител, обнаруживаемых в крови или сыворотке, но его также используют для обозначения смеси антител, полученной из других источников.
Термин "молекула иммуноглобулина" означает индивидуальную молекулу антитела, например, представляющую собой часть иммуноглобулина или часть любой композиции поликлональных или моноклональных антител.
Терминологическое определение "библиотека вариантных молекул представляющей интерес нуклеиновой кислоты" используется для описания совокупности молекул нуклеиновой кислоты, которые вместе кодируют "представляющий интерес рекомбинантный поликлональный белок". При использовании для трансфекции библиотека вариантных молекул представляющей интерес нуклеиновой кислоты содержится в библиотеке векторов экспрессии. Такая библиотека в типичном случае имеет, по меньшей мере, 2, 3, 5, 10, 20, 50, 1000, 104, 105 или 106 отдельных членов.
Термин "перенос массы" используют для описания переноса последовательностей представляющей интерес нуклеиновой кислоты из одной совокупности векторов в другую совокупность векторов, который осуществляется для каждой ДНК одновременно, не прибегая к выделению представляющей интерес индивидуальной ДНК. Такие совокупности векторов могут представлять собой библиотеки, содержащие, например, те вариабельные области, промоторы, лидерные или энхансерные элементы, которые представляют интерес. Затем эти последовательности можно перемещать без предварительного выделения, например, из фагового вектора в вектор экспрессии млекопитающих. Особенно применительно к библиотекам антител эта методика позволяет, чтобы разнообразие в сцеплении между VH и VL не утрачивалось при перемещении библиотек, например, из вектора отбора (например, вектора фагового дисплея) в вектор экспрессии млекопитающих. Таким образом, сохраняется первоначальное спаривание VH и VL.
При использовании здесь термин "функционально связанный" относится к сегменту, сцепленному с другим сегментом, при том условии, что он попадает в функциональную взаимосвязь с другим сегментом. Например, ДНК, кодирующая сигнальную последовательность, функционально связана с ДНК, кодирующей полипептид, если она экспрессируется как лидер, принимающий участие в переносе полипептида в эндоплазматическую сеть. Таким же образом, промотор или энхансер функционально связан с кодирующей последовательностью, если он стимулирует транскрипцию этой последовательности.
Термин "поликлональное антитело" описывает композицию различных молекул антитела, которая способна связываться или взаимодействовать с несколькими различными специфическими антигенными детерминантами на одном и том же или на различных антигенах. Обычно принято считать, что вариабельность поликлонального антитела локализована в так называемых вариабельных областях поликлонального антитела. Однако в контексте настоящего изобретения термин поликлональность можно также понимать как описание различий между индивидуальными молекулами антитела, локализованных в так называемых константных областях, например, как в случае смесей антител, содержащих два или более изотипов антитела, таких как изотипы человека IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2 или мышиные изотипы IgG1, IgG2a, IgG2b, IgG3 и IgA. В целях настоящего изобретения такое поликлональное антитело также можно называть "композицией антител".
Термин "эпитоп" обычно используют для описания доли более крупной молекулы или части более крупной молекулы (например, антигена или антигенного сайта), проявляющей антигенную или иммуногенную активность у животного, предпочтительно, млекопитающего и, наиболее предпочтительно, человека. Эпитоп, обладающий иммуногенной активностью, представляет собой часть более крупной молекулы, которая вызывает у млекопитающего ответную реакцию антител. Эпитоп, обладающий антигенной активностью, представляет собой часть более крупной молекулы, с которой иммуноспецифически связывается антитело, как это можно определить любым способом, хорошо известным в данной области техники, например, описанными здесь иммунологическими анализами. Антигенные эпитопы необязательно должны быть иммуногенными. Антигены представляют собой вещества, например, токсины, вирусы, бактерии, белки или ДНК, с которыми иммуноспецифически связывается антитело или фрагмент антитела. Антигены или антигенные сайты, за исключением очень маленьких, часто имеют более одного эпитопа и часто способны стимулировать иммунный ответ. Эпитопы могут быть линейными или конформационными. Линейный эпитоп состоит приблизительно из 6-10 прилегающих аминокислот в белковой молекуле и распознается антителом. В отличие от этого конформационный эпитоп состоит из аминокислот, которые не упорядочены последовательно. В данном случае антитело распознает только 3-мерную структуру. Когда белковая молекула сворачивается в трехмерную структуру, аминокислоты, образующие эпитоп, сопоставляются, позволяя антителу распознать последовательность. В денатурированном белке может быть распознан только линейный эпитоп. Конформационный эпитоп по определению должен располагаться на наружной поверхности свернутого белка. Антитело, которое распознает конформационный эпитоп, может связываться с ним только в условиях мягких, не денатурирующих процедур. Антитела, связывающиеся с различными эпитопами на одном и том же антигене, могут разным образом влиять на активность антигена, с которым они связываются, в зависимости от локализации эпитопа. Антитело, связывающееся с эпитопом в активном сайте антигена, может полностью заблокировать функцию антигена, тогда как другое антитело, связывающееся с другим эпитопом, может незначительно влиять или совсем не влиять на активность одиночного антигена. Однако такие антитела все еще способны активировать комплемент, благодаря чему происходит элиминация антигена, кроме того, возможны синергические эффекты при комбинировании одного или нескольких антител, связывающихся с различными эпитопами на одном и том же антигене. В настоящем изобретении эпитоп, предпочтительно, является частью внеклеточного домена EGFR. Антигены, рассматриваемые настоящим изобретением, предпочтительно, являются внеклеточными доменами белков, полипептидов EGFR или их фрагментов, с которыми иммуноспецифически связываются антитела или фрагменты антител. EGFR-ассоциированный антиген также может представлять собой аналог или производное внеклеточного домена полипептида EGFR или его фрагмента, с которым иммуноспецифически связывается антитело или фрагмент антитела.
Антитела, способные конкурировать друг с другом за связывание с одним и тем же антигеном, могут связываться с одними и теми же или с перекрывающимися эпитопами либо могут иметь сайты связывания, расположенные в близком соседстве друг с другом, благодаря чему конкуренция обусловлена, главным образом, стерическим препятствием. Способы определения конкуренции между антителами описаны в примерах.
При использовании здесь термины "поликлональный белок" или "поликлональность" относятся к белковой композиции, содержащей разные, но гомологичные молекулы белка, предпочтительно, выбранные из суперсемейства иммуноглобулинов. Таким образом, каждая молекула белка гомологична другим молекулам композиции, но также содержит один или несколько участков вариабельной последовательности полипептида, которые характеризуются различиями в аминокислотной последовательности между индивидуальными элементами поликлонального белка. Известные примеры таких поликлональных белков включают молекулы антитела или иммуноглобулина, рецепторы T-клеток и рецепторы B-клеток. Поликлональный белок может состоять из определенной подгруппы молекул белка, которые были охарактеризованы таким общим признаком как совпадающая активность связывания с желаемой мишенью, например, в случае поликлонального антитела, направленного против желаемого антигена-мишени.
Под "белком" или "полипептидом" подразумевается любая цепочка аминокислот независимо от ее длины или посттрансляционной модификации. Белки могут существовать в виде мономеров или мультимеров, содержащих две или более собранные полипептидные цепочки, фрагментов белка, полипептидов, олигопептидов или пептидов.
Термин "RFLP" относится к "полиморфизму длины рестрикционных фрагментов", то есть к способу, посредством которого анализируют характер миграции фрагментов нуклеиновой кислоты в геле после ее расщепления рестрикционными ферментами.
Термин "скремблирование" описывает ситуацию, когда два или более различных элемента поликлонального белка, состоящие из двух различных полипептидных цепочек, например, из суперсемейства иммуноглобулинов, экспрессируются из отдельной клетки. Эта ситуация может возникнуть, когда отдельная клетка интегрировала в пределах генома более одной пары сегментов гена, причем каждая пара сегментов гена кодирует отдельный элемент поликлонального белка. В такой ситуации можно получить непреднамеренные комбинации полипептидных цепочек, экспрессируемых из сегментов гена. Такие непреднамеренные комбинации полипептидных цепочек, возможно, не будут давать никакого терапевтического эффекта.
Примером определенного выше скремблирования является термин "скремблирование цепочек VH-VL". В этом примере сегменты гена, кодирующие VH и VL, составляют пару сегментов гена. Скремблирование встречается тогда, когда непреднамеренные комбинации полипептидов VH и VL вырабатываются клеткой, в которую интегрированы две различные пары сегментов гена, кодирующих VH и VL. Такая скремблированная молекула антитела, вероятно, не будет сохранять первоначальную специфичность, то есть, может не проявлять никакого терапевтического эффекта.
Термин "трансфекция" при использовании здесь употребляется в широком смысле, обозначая введение в клетку инородной ДНК. Этот термин также охватывает другие функционально равноценные способы введения инородной ДНК в клетку, например, такие как трансформация, инфекция, трансдукция или слияние донорской клетки с акцепторной клеткой.
Термины "вариабельная последовательность полипептида" и "вариабельная область" могут употребляться взаимозаменяемо.
Термин "отдельные эпитопы" означает, что, когда два различных антитела связывают такие эпитопы, конкуренция за связывание с антигеном составляет менее 100%, предпочтительно, конкуренция за связывание с антигеном составляет менее 50% и, более предпочтительно, пор существу нет конкуренции за связывание с антигеном. Анализ пар антител на "отдельные эпитопы" в типичных случаях проводят посредством экспериментов со связыванием в условиях насыщения антителом либо по методике FACS на клетках, экспрессирующих EGFR, и антителах с индивидуальной флуоресцентной меткой или по методике поверхностного плазмонного резонанса с применением антигена EGFR, захваченного или конъюгированного с поверхностью проточной кюветы, как это описано в примерах.
Терминологическое определение способности "ингибировать связывание EGF" применительно к одной молекуле антитела означает, что молекула антитела демонстрирует значение 1С 50 в отношении связывания EGF с EGFR менее 10 нМ, предпочтительно, менее 8 нМ, более предпочтительно, менее 7 нМ, более предпочтительно, менее 5 нМ, более предпочтительно, менее 4 нМ, более предпочтительно, менее 3 нМ, более предпочтительно, менее 2 нМ, более предпочтительно, менее 1 нМ.
Термины "рецептор эпидермального фактора роста", "EGFR" и "антиген EGFR" употребляются здесь взаимозаменяемо и включают варианты, изоформы и специфические гомологи EGFR человека. В предпочтительном варианте осуществления изобретения связывание антитела по изобретению с антигеном EGFR ингибирует рост клеток, экспрессирующих EGFR (например, опухолевых клеток) за счет ингибирования или блокирования связи лиганда EGFR с EGFR. Термин "лиганд EGFR" охватывает все (например, физиологические) лиганды для EGFR, включая, но не ограничиваясь ими, EGF, альфа-TGF, гепаринсвязывающий EGF (HB-EGF), амфирегулин (AR), херегулин, бета-целлюлин и эпирегулин (EPI). В другом предпочтительном варианте осуществления изобретения связывание антитела по изобретению с антигеном EGFR опосредует фагоцитоз эффекторных клеток и/или уничтожение клеток, экспрессирующих EGFR.
Структура домена EGFR: Внеклеточная часть зрелого EGFR (SwissProt асе. #Р00533) состоит из 621 аминокислоты и четырех рецепторных доменов: домен I охватывает остатки 1-165, домен II остатки 166-312, домен III - остатки 313-481 и домен IV - остатки 482-621 (Cochran et al. 2004 J immunol. Methods 287, 147-158). Было высказано предположение о том, что домены I и III дают вклад в образование высокоаффинных сайтов связывания для лигандов. Домены II и IV представляют собой богатые цистеином ламининоподобные области, которые стабилизируют укладку структуры белка и содержат возможную поверхность раздела для димеризации EGFR.
При использовании здесь термин "ингибирует рост" (например, применительно к клеткам) включает любое поддающееся измерению уменьшение пролиферации (увеличения количества клеток) или клеточного метаболизма при контакте с антителом против EGFR по сравнению с ростом таких же клеток, не контактировавших с антителом против EGFR, например, ингибирование роста клеточной культуры, по меньшей мере, приблизительно на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 99% или 100%.
При использовании здесь термины "ингибирует связывание" и "блокирует связывание" (например, применительно к ингибированию/блокированию связывания лиганда EGFR с EGFR) употребляются взаимозаменяемо и охватывают как частичное, так и полное ингибирование/блокирование. Ингибирование/блокирование связи лиганда EGFR с EGFR, предпочтительно, снижает или изменяет нормальный уровень или тип клеточных сигналов, который имеет место, когда лиганд EGFR связывается с EGFR без ингибирования или блокирования. Ингибирование или блокирование также включают любое поддающееся измерению снижение аффинности связывания лиганда EGFR с EGFR при наличии контакта с антителом против EGFR по сравнению с той ситуацией, когда лиганд не контактирует с антителом против EGFR, например, блокирование связи лиганда EGFR с EGFR, по меньшей мере, приблизительно на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 99% или 100%.
Термин "рекомбинантное антитело" употребляется для описания молекулы или нескольких молекул антитела, которые экспрессируются из клетки или клеточной линии, трансфицированной вектором экспрессии, содержащим кодирующую последовательность антитела, которая в естественных условиях не ассоциирована с клеткой.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1. Сортировка спленоцитов (подробности см. в примере 1). Сделаны следующие каналы (отображены):
• Канал 1: Живые клетки (график FSC/ иодид пропидия) (нижняя левая панель)
• Канал 2: Клетки плазмы пропущены как позиция CD43/позиция CD138 (нижняя правая панель)
• Канал 3: распознавание дублета (верхняя правая панель)
Фиг.2. Мышиная модель - mSymplex™ PCR. Расширение мультиплексного перекрывания RT-PCR для амплификации и когнатного сцепления генов тяжелой и легкой цепи антитела из единичной клетки. Подробности см. в примере 1.
Фиг.3. Клонирование мышиного репертуара. Пул продуктов mSymplex™ PCR, кодирующих пары генов VH/VL из единичных плазматических клеток, был сплайсирован до гена, кодирующего константную легкую каппа-цепь человека, посредством сплайсинга через расширение перекрывания. Пул генов, кодирующих полноразмерные химерные антитела человека-мыши, был вставлен в вектор экспрессии с последующей вставкой двунаправленной промоторной кассеты (2×CMV).
Фиг.4. Схематическое представление вектора экспрессии 00-VP-002 полноразмерного антитела млекопитающих. Amp и Amp pro - ген устойчивости к ампициллину и его промотор, pUC origin - начало репликации pUC, CMV - промотор млекопитающих, управляющий экспрессией легкой цепи и тяжелой цепи, IGHV Leader - геномный лидер тяжелой цепи человека, H stuffer - вставка, которая заменена на кодирующую последовательность вариабельной области тяжелой цепи, IGHG1 - последовательность, кодирующая константную область тяжелой цепи изотипа G1 геномного иммуноглобулина (последовательность показана в приложении 2), Rabbit B-globin A - полиА последовательность бета-глобина кролика, IGKV Leader - мышиный каппа-лидер, L Stuffer - вставка, которая заменена на кодирующую последовательность легкой цепи, SV40 term - терминаторная последовательность вакуолизирующего обезьяньего вируса 40, FRT - целевой сайт распознавания Flp, Neo - ген устойчивости к неомицину, SV40 poly A - полиА сигнальная последовательность вакуолизирующего обезьяньего вируса 40.
Фиг.5. Кластерный анализ различий в поглощении на уровне 450-620 нм. Супернатанты сгруппированы по реактивности, на что указывают цифры (от 1 до 4) после номера клона. Темно-серый цвет указывает на уменьшение количества метаболически активных клеток, тогда как светло-серый цвет указывает на увеличение количества метаболически активных клеток. Черный цвет указывает на супернатанты, не оказывающие влияния на количество метаболически активных клеток.
Фиг.6. Степень ингибирования антител против EGFR перечисленными эталонными антителами, направленными против специфических доменов EGFR, по результатам конкурентного анализа ELISA. А) Расчет ингибирования. В) Расчет ингибирования следующим образом: 25-49%: Умеренная конкуренция (+), 50-74%: сильная конкуренция (++), 75-100%: очень сильная конкуренция (+++). Ячейки, отображающие значительное ингибирование (50-100%) затенены серым цветом. Эрбитукс и вектибикс показаны по два раза (четыре независимых эксперимента), чтобы проиллюстрировать воспроизводимость анализа. Ab2 (225) - мышиный предшественник, который ведет к эрбитуксу.
Фиг.7. Иллюстрация одного цикла картирования эпитопа, проведенного аппаратом Biacore 3000 SPR, когда образец mAb конкурирует за связывание внеклеточного домена EGFR с четырьмя разными эталонными антителами.
Фиг.8. Степень ингибирования антител против EGFR перечисленными эталонными антителами, направленными против специфических доменов EGFR, по результатам конкурентного анализа с применением технологии SPR. А) Расчет ингибирования. В) Расчет ингибирования следующим образом: 25-49%: Умеренная конкуренция (+), 50-74%: сильная конкуренция (++), 75-100%: очень сильная конкуренция (+++). Клетки, отображающие значительное ингибирование (50-100%) затенены серым цветом. Клон 1229, маркированный символом *, не проявлял связывания в анализе Biacore.
Фиг.9. Определение кластеров эпитопа в пределах репертуара антител против EGFR посредством конкурентного анализа пар антитела против EGFR методом SPR. Антитела сгруппированы в соответствии с предполагаемым распознаванием домена EGFR. Клетки, в которых комбинации антител, связанные с перекрывающимися эпитопами, приводили к ингибированию более чем на 50%, затенены серым цветом. Клетки, в которых определения не были получены, закрашены черным цветом. A) Расчет ингибирования. B) Расчет ингибирования следующим образом: 25-49%: Умеренная конкуренция (+), 50-74%: сильная конкуренция (++), 75-100%: очень сильная конкуренция (+++).
Фиг.10. Эпитопные карты эталонных антител и антител против EGFR, направленных против внеклеточного домена EGFR, по результатам анализа Biacore. A) Эпитопная карта антител, направленных против домена I или домена I/II внеклеточного домена (ECD) EGFR. B) Эпитопная карта антител, направленных против домена III ECD EGFR.
Фиг.11. Исследование одновременного связывания олигоклональной смеси антител, направленных против неперекрывающихся эпитопов на EGFR. A) Последовательное добавление антител против домена III, домена I или неизвестной специфичности. Величины ингибирования единственного образца mAb, протестированного по отношению к смесям различных mAb или единственному mAb, показаны в затененных ячейках. Также показаны величины Ru max, использованные для расчета ингибирования. B) Анализ конкуренции шести различных образцов mAb, направленных против неперекрывающихся эпитопов на EGFR, и смеси антител, содержащей шесть протестированных антител. В качестве положительного контроля были использованы смеси антител, куда не были включены протестированные образцы антител. Величины ингибирования единственного образца mAb, протестированного по отношению к смесям различных mAb, показаны в затененных ячейках. Также показаны величины Ru max, использованные для расчета ингибирования. C) Соответствующие сенсограммы по результатам анализа в пункте B, иллюстрирующие блокаду связывания антителами, а в некоторых случаях усиление связывания антителами. D) Испытание дополнительных антител, направленных против домена I, I/II и неизвестной специфичности, по отношению к смеси шести антител mAb.
Фиг.12. Определение опосредованной антителами блокады лиганда EGF титрованием антител на полноразмерном EGFR и выявление связывания биотинилированного лиганда EGF с реактивом стрептавидин-HRP. Эрбитукс, вектибикс и Synagis IgG (паливизумаб) использовали как положительный и отрицательный контроль, соответственно. После блокады распознанного эпитопа антитела испытуемыми антителами визуализировали степень конкуренции лиганда EGF, добавляя 0,1 мкг/мл биотинилированного лиганда EGF и вторичный конъюгат стрептавидин-HRP.
Фиг.13. Влияние предварительной обработки индикаторными антителами на индуцированное EGF (50 нг/мл) фосфорилирование EGFR в клетках HN5. Антитела (10 мкг/мл), указанные на графике, были инкубированы с клетками в течение 30 минут перед добавлением EGF на 7,5 минут. Наборы данных, маркированные символом *, статистически значимо отличались от набора контрольных данных ((-)ctrl) (p<0,05). A. Антитело 1208 проявляло выраженный защитный эффект в отношении фосфорилирования EGFR. B. Антитела 1277 и 1320 статистически значимо защищали от фосфорилирования, индуцированного EGF. Отрезки, изображающие величину ошибки, представляют стандартные отклонения по результатам трех независимых экспериментов.
Фиг.14. Внутриклеточный вестерн-анализ фосфорилированного EGFR (pEGFR) и EGFR в клетках HN5. Образец mix обозначает эквимолярную смесь антител 992, 1030 и 1042 до конечной концентрации 10 мкг/мл, другие антитела были использованы в концентрации 10 мкг/мл каждое. Для того чтобы стимулировать фосфорилированием EGFR, перед фиксацией 7,5 минут добавляли EGF из расчета 50 мкг/мл). Отрезки, изображающие величину ошибки, представляют стандартные отклонения 6 отдельных (ctlr-) или 3 отдельных точек данных (992, 1030, 1042, mix или эрбитукс). Образцы 992, 1030, mix и эрбитукс проявляли статистически значимый (*=p<0,05) защитный эффект в отношении фосфорилирования.
Фиг.15. Влияние инкубации антител на интернализацию EGFR. Данные показаны в виде процентных величин, отражающих удаление рецепторов с клеточной поверхности по сравнению с начальным окрашиванием. Отрезки, изображающие величину ошибки, соответствуют SEM (стандартной ошибке среднего).
Фиг.16. Кривые роста клеток A431-NS в присутствии переменных концентраций антител 992, 1030, 1042 и их смесей, рассчитанные по процентным показателям метаболически активных клеток по сравнению с необработанным контролем. Образец 1001 представлял собой нефункциональное антитело со сходным изотипом, которое было использовано в качестве отрицательного контроля.
Фиг.17. Кривые роста клеток A431-NS в присутствии 10 мкг/мл антител 992, 1030, 1042 и их смесей, а также переменных концентраций EGFR-лиганда EGF, рассчитанные по поглощению на длине волны 450 нм. Образец 1001 представлял собой нефункциональное антитело со сходным изотипом, которое было использовано в качестве отрицательного контроля.
Фиг.18. Кривые роста клеток A431-NS в присутствии переменных концентраций антитела 992 и смесей 992 с антителами, имеющими неперекрывающиеся эпитопы, представленные в домене I, II или III. Образец 1001 представлял собой нефункциональное антитело со сходным изотипом, которое было использовано в качестве отрицательного контроля.
Фиг.19. Апоптоз в клетках A431NS. Образцы смеси EGFR, отдельных моноклональных антител, препаратов эрбитукс и вектибикс были протестированы в 10-кратных разведениях. Комплекс гистон-ДНК из апоптотических клеток измеряли с применением набора ELISA производства компании Roche.
Фиг.20. В четырех группах мышей nude Balb/C Nu/Nu численностью по 10 особей были инокулированы клетки A431NS в количестве 1×106 на животное. После того как опухоли достигали размера приблизительно 100 мм3, начиналось лечение животных. Животным в группах проводили инъекции 1 мг/мл антител пять раз по ходу эксперимента, как это указано стрелками. Диаметр опухолей измеряли нутромерами с цифровой индикацией. Результаты представлены в виде среднего объема опухоли (+/- SEM).
Фиг.21. После того как в эксперименте, показанном на фиг.20, отдельных животных умерщвляли, производили иссечение и взвешивание опухолей. Показаны средние величины +/- SEM. Звездочки указывают статистическую значимость на уровне P<0,05.
Фиг.22. Рост сфероидов A431-NS в присутствии 10 мкг/мл антител 1001, эрбитукса, вектибикса и смеси трех антител с неперекрывающимися эпитопами 992+1030+1042. 1001 представляет собой нефункциональное антитело со сходным изотипом, которое было использовано в качестве отрицательного контроля.
Фиг.23. ДНК (SEQ ID No. 100) и белковая последовательность (SEQ ID NO. 101) внеклеточного домена EGFR Cynomolgus клонированы из кДНК, полученной из эпидермиса обезьяны Cynomolgus.
Фиг.24. Выверка полученной белковой последовательности ECD EGFR Cynomolgus (SEQ ID NO. 101) с ECD EGFR человека (SEQ ID NO 108), полученной из GENBANK (инвентарный номер X00588). Также показана консенсусная последовательность (SEQ ID NO 109).
Фиг.25. Пример аналитического разделения методом ELISA между перекрестно реактивными и видоспецифическими антителами, связывающими ECD EGFR человека, cynomolgus или оба типа доменов.
Фиг.26. Микрофотографии репрезентативных срезов опухолей в каждой из четырех экспериментальных групп мышей с ксенотрансплантатами. При увеличении 200× стрелки указывают на очаги терминальной дифференциации клеток A431 in vivo. Следует обратить внимание на заметно более крупные и более многочисленные очаги терминальной дифференциации в опухолях при лечении смесью трех клонов антител против EGFR (992+1030+1042), две верхние панели.
Фиг.27. A) Изображения сфероидов HN5 при увеличении 40× через 24 часа после добавления 10 мкг/мл контрольного антитела (ритуксимаб, anti CD-20) или смеси антител против EGFR 992 и 1024. B) Количественное определение площади, покрытой клетками, при использовании компьютерной программы Image J (* p<0,01).
Фиг.28. Диаграмма, показывающая уровень инволюкрина в четырех группах лечения в виде процентного показателя в контрольной группе, не получавшей лечения (*#¤p<0,005 по сравнению с эрбитуксом, вектибиксом и группой отрицательного контроля, соответственно).
Фиг.29. A) Изображения клеток HN5 и A431NS, инкубированных с 10 мкг/мл эрбитукса, меченного Alexa-488, или смеси антител 992+1024 в течение 2 часов (при увеличении 60×). B) Изображения с мелким точечным проколом клеток A431NS, инкубированных с 10 мкг/мл эрбитукса, меченного Alexa-488, или смеси антител 992+1024 в течение 2 часов (при увеличении 60×).
Фиг.30. Изображения клеток HN5, инкубированных с 10 мкг/мл эрбитукса, меченного Alexa-488, или смеси антител 992+1024 в течение указанных промежутков времени (при увеличении 60×).
Фиг.31. Определение специфичности представления антигена антителами Fab 992, 1024 и 1030 при серийном титровании антител на клетках A431-NS и очищенном полноразмерном EGFR методом ELISA. Связанные антитела Fab были визуализированы вторичным специфическим конъюгатом HRP с козьим антителом Fab против человека. A) Антитела Fab, протестированные против очищенного полноразмерного EGFR из клеток A431. B) Антитела Fab, протестированные против EGFR, экспрессированного на поверхности клеток A431-NS.
Фиг.32. Определение функциональной аффинности IgG и фрагментов Fab антител 992, 1024, 1030, эрбитукса и вектибикса серийным титрованием на клетках A431-NS, фиксированных параформальдегидом, в анализе по методу ELISA. Связанные антитела Fab и IgG были визуализированы вторичным специфическим конъюгатом HRP с козьим антителом Fab против человека. В качестве отрицательного контрольного антитела было использовано белковое антитело против RSV F Synagis, которое не показало никакого связывания в проведенном анализе ELISA. A) Функциональное связывание антител IgG с клетками A431-NS. B) Функциональное связывание антител Fab с клетками A431-NS.
Фиг.33. Определение усиления связывания IgG с EGFR на клетках A431-NS при предварительном насыщении рецептора фрагментами Fab, связывающими неперекрывающиеся эпитопы. Указанным фрагментам Fab давали возможность насытить распознанный эпитоп EGFR на клетках A431-NS в течение 30 минут, после чего указанные антитела IgG были подвергнуты серийному титрованию, а связанные IgG с добавлением или без добавления Fab визуализированы вторичным конъюгатом HRP с мышиным Fc против человека. A) Характеристики связывания IgG 992 с клетками A431-NS при предварительном насыщении рецептора указанными фрагментами Fab или без такого насыщения. B) Характеристики связывания IgG 1024 с клетками A431-NS при предварительном насыщении рецептора указанными фрагментами Fab или без такого насыщения. С) Характеристики связывания IgG 1030 с клетками A431-NS при предварительном насыщении рецептора указанными фрагментами Fab или без такого насыщения.
Фиг.34. кДНК полноразмерного EGFR cynomolgus (фиг.34A, SEQ ID NO 102) и кодируемый белок (фиг.34В, SEQ ID NO 103).
Фиг.35. Апоптоз, полученный в клетках A431NS при воздействии 1 мкг/мл указанных антител/комбинаций. Комплексы гистон-ДНК определяли с применением набора ELISA производства компании Roche. Уровень апоптоза определяли по отношению к положительному контролю (максимальный апоптоз).
Фиг.36. Мышам Balb/C nu/nu инъецировали 1×106 клеток A431NS. После того как опухоли достигали среднего размера приблизительно 100 мм3, начиналось лечение животных. Мыши получали 17 инъекций антител. Первое лечение начинали в 8-й день, а последнее в 34-й день. Антитела/композиции инъецировали в количестве 0,5 мг на дозу или 0,17 мг на дозу. Показаны средние величины объема опухолей +/- SEM.
Фиг.37. Ингибирование пролиферации клеток A431NS. На оси X показаны разные репрезентативные комбинации 3-х антител по изобретению. На оси Y показана метаболическая активность в виде процентной величины по сравнению с необработанным контролем. Отрезки, изображающие величину ошибки, представляют +/- SEM. Дополнительные подробности см. в примере 6.
Фиг.38. Эффект ингибирования роста при воздействии двух разных доз смеси 992+1024 по сравнению с препаратом эрбитукс в ксенотрансплантатах опухолевых клеток человека A431NS. Мышам Balb/C nu/nu инокулировали 106 клеток A431NS. После того как опухоли достигали среднего размера 100 мм3 (8-й день), мышей рандомизировали на группы численностью по 9 особей, и начинали лечение. Указанные антитела инъецировали в количестве 0,5 мг на дозу или 1 мг на дозу два раза в неделю (в общей сложности 9 инъекций). Светло-серая область на графике показывает период лечения. Начало пунктирной линии обозначает точку времени, в которой первая мышь из данной группы была умерщвлена из-за избыточного размера опухоли. Статистически значимые различия между группами, получавшими смесь 992+1024 (2 мг в неделю) и эрбитукс (2 мг в неделю), а также смесь 992+1024 (1 мг в неделю) и эрбитукс (2 мг в неделю), были получены на 60-й день, когда лечение во всех группах кроме 992+1024 (2 мг в неделю) было прекращено. Размер опухоли у животных, исключенных до 60-го дня, был определен вплоть до конца эксперимента, таким образом график показывает накопленный объем опухоли у всех мышей в данной группе. Показаны средние величины +/- SEM.
Фиг.39. График Каплана-Мейера по данным о выживании мышей, получавших лечение смесью антител 992+1024, препаратом эрбитукс или контрольным антителом (тот же эксперимент, который показан на фиг.38). Результаты представлены в виде процентных показателей выживания мышей, получавших то или иное лечение. При сравнении групп, получавших смесь антител 992+1024 и эрбитукс, наблюдались статистически значимые различия по процентному показателю выживания мышей при высоких дозах (2 мг в неделю, P=0,0008) и низких дозах (1 мг в неделю, P=0,0004). Кроме того, низкая доза смеси 992+1024 давала статистически значимо лучший результат по сравнению с высокой дозой препарата эрбитукс (P = 0,0087). Статистическую значимость различий вычисляли с применением логарифмического рангового критерия (Мантела-Кокса).
Фиг.40. Анализ перекрестной реактивности антител IgG 992, 1024 и 1320 против клеток CHO, трансфицированных полноразмерным EGFR человека и cynomolgus, по методике FACS. Связанные антитела выявляли при помощи козьего F(ab')2 IgG FC против человека. Дискриминацию проводили по однородным клеткам (со свойствами SCC/FCS), экспрессирующим EGFR. Связывание выражено в виде % от максимального связывания антитела в концентрации 1 нМ.
Фиг.41. Выверка аминокислотных последовательностей вариабельных областей мышиных (chi) и гуманизированных (hu) вариабельных областей-кандидатов как в тяжелых, так и в легких цепях антител 992 (A) и 1024 (B) на основе компьютерной программы Clustalw2. Области CDR, определенные по IMGT, подчеркнуты, гэпы представлены символом (-), идентичные аминокислоты символом (*), консервативные мутации символом (:), полуконсервативные мутации символом (.). Выделенные жирным шрифтом аминокислоты указывают те положения, в которых будут проведены обратные мутации к первоначально идентифицированному мышиному остатку, если варианты полностью человеческого каркаса проявят сниженную аффинность связывания. Идентификационные номера последовательностей даны следующим образом: Гуманизированная VH 992 (SEQ ID NO 104). Гуманизированная VL 992 (SEQ ID NO 105). Гуманизированная VH 1024 (SEQ ID NO 106). Гуманизированная VL 1024 (SEQ ID NO 107). Химерная VH 992 (a.к. 3-124 из SEQ ID NO 40). Химерная VL 992 (a.к. 3-109 из SEQ ID NO 72). Химерная VH 1024 (a.к. 3-120 из SEQ ID NO 41). Химерная VL 1024 (a.к. 3-114 из SEQ ID NO 73).
Фиг.42A: Схематическое представление генов, кодирующих двойной вариабельный домен 992L1024. Последовательность IGHV 992L1024 (751 bp) представлена от 5'-сайта рестрикции AscI, сопровождается IGHV 992, линкером ASTKGP, IGHV 1024 и заканчивается на 3'-сайте рестрикции XhoI. Последовательность IGKV 992L1024 (1071 п.о.) представлена от 5'-сайта рестрикции NheI, сопровождается IGKV 992, линкером TVAAP, IGKV 1024, IGKC и заканчивается на 3'-сайте рестрикции NotI.
Фиг.42B: Схематическое представление генов, кодирующих двойной вариабельный домен 1024L992. Последовательность IGHV 1024L992 (751 п.о.) представлена от 5'-сайта рестрикции AscI, сопровождается IGHV 1024, линкером ASTKGP, IGHV 992 и заканчивается на 3'-сайте рестрикции XhoI. Последовательность IGKV 1024L992 (1071 п.о.) представлена от 5'-сайта рестрикции NheI, сопровождается IGKV 1024, линкером TVAAP, IGKV 992, IGKC и заканчивается на 3'-сайте рестрикции NotI.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Смеси антител
В одном из вариантов осуществления изобретение относится к композиции антител, содержащих молекулы антитела, способные связываться, по меньшей мере, с тремя различными эпитопами EGFR, предпочтительно с тремя неперекрывающимися эпитопами EGFR. Неперекрывающуюся природу антител, предпочтительно, определяют, применяя по-разному меченые антитела, в анализе FACS с клетками, экспрессирующими EGFR или, применяя поверхностный плазмонный резонанс с антигеном EGFR, захваченным или конъюгированным на поверхности проточной кюветы. Можно также использовать способы анализа на основе ELISA, как это описано в примерах. Композицию, связывающую три неперекрывающихся эпитопа EGFR, можно использовать против более широкого круга EGFR-зависимых раковых заболеваний, поскольку такая композиция в меньшей степени чувствительна к различиям в конформации EGFR и в меньшей степени зависима от мутаций по сравнению с композицией моноклональных антител, нацеленной на один или два эпитопа. Кроме того, композиция антител, связывающая три неперекрывающихся эпитопа EGFR, может обеспечить лучшую эффективность по сравнению с композицией, нацеленной на меньшее число эпитопов. В особенности, указанная композиция антител может обеспечить лучшую эффективность в отношении терминальной дифференциации раковых клеток in vivo. Фиг.37 приводит многочисленные примеры эффективных композиций антител, связывающих три различных эпитопа EGFR человека, которые иллюстрируют общую приемлемость концепции настоящего изобретения.
При терапии моноклональными антителами против EGFR некоторая часть пациентов не будет адекватно реагировать на проводимое лечение, которое окажется неэффективным. У некоторых пациентов это обусловлено тем, что происходит быстрое выведение антител, или тем, что антитело вызывает у больного иммунную реакцию на себя. У некоторой части пациентов отсутствие лечебной реакции может быть связано с тем, что при той конкретной форме EGFR-зависимого ракового заболевания, которым страдает больной, EGFR экспрессируется в такой конформации, что моноклональной антитело не может связать его эпитоп. Причиной этого могут служить различия в гликозилировании, делеция домена, мутации и/или SNP.
К тому же при некоторых раковых заболеваниях большое значение имеет аутокринная стимуляция EGFR, обусловленная выработкой лиганда раковыми клетками, хотя в других случаях EGFR, экспрессируемый раковыми клетками, не нуждается в стимуляции лигандом. При раковых заболеваниях последнего типа антитело, способное ингибировать связывание лиганда, будет неэффективным.
Композиция антител, в которой антитела способны связывать, по меньшей мере, три различных эпитопа на EGFR, должна найти более широкое применение, поскольку вероятность того, что все три эпитопа изменены по сравнению с эпитопом (эпитопами), распознаваемыми антителами, уменьшается. К тому же, вероятность того, что все антитела будут выведены из организма больного, становится намного меньше. Наконец, примеры показывают, что в функциональных анализах смесь, содержащая три антитела, которые связывают разные эпитопы, превосходит моноклональное антитело и смесь, содержащую два антитела. Превосходство наиболее четко было продемонстрировано в плане индукции терминальной дифференциации раковых клеток при использовании трех антител к домену III с неперекрывающимися эпитопами. Такая эффективная терминальная дифференциация раковых клеток, индуцированная антителами, не была описана ранее и представляет собой значительный шаг вперед в разработке эффективной противораковой терапии на основе антител. Более поздние результаты показали, что похожие или даже лучшие результаты можно получить, применяя особую комбинацию двух антител.
Для того чтобы повысить клиническую эффективность и расширить полезное применение при большем числе EGFR-зависимых типов рака, можно увеличить число антител в композиции. Так, например, композиция может содержать антитела, способные связываться с четырьмя неперекрывающимися эпитопами. Композиция может содержать антитела, способные связываться с пятью неперекрывающимися эпитопами. Композиция может содержать антитела, способные связываться с шестью неперекрывающимися эпитопами. Примеры, приведенные в настоящей заявке, показывают, что одновременно связываться с EGFR могут, по меньшей мере, шесть различных антител (пример 3). Это не исключает возможности и даже предпочтительного варианта, связанного с разработкой композиции, содержащей антитела, способные связываться более чем с шестью, например, с семью или восемью неперекрывающимися эпитопами, путем тщательного подбора антител.
Еще в одном варианте осуществления изобретения композиция содержит более одной молекулы антитела, например, два антитела, связывающие разные, но перекрывающиеся эпитопы. Может оказаться выгодным включать в композицию антитела с перекрывающимися эпитопами, поскольку это увеличивает вероятность связывания эпитопа. Одно логическое обоснование такого подхода заключается в том, что у некоторых пациентов и/или в некоторых раковых клетках эпитоп может измениться вследствие конформационных изменений, мутаций или SNP. Хотя это может повлиять на связывание одного антитела, связывающая способность другого антитела, взаимодействующего с перекрывающимся эпитопом, затронута не будет. Кроме того, существует риск, что одно из антител будет выводиться из организма больного, поскольку оно будет распознано как антиген. Включая в композицию два антитела, связывающие разные, но перекрывающиеся эпитопы, можно уменьшить последствия элиминации одного из двух антител и последствия мутации в эпитопе.
Так, в одном из вариантов осуществления изобретения композиция содержит два антитела, связывающие разные, но перекрывающиеся эпитопы. Еще в одном варианте осуществления изобретения композиция содержит две различные молекулы антитела, связывающие один и тот же эпитоп. Антитела, связывающие один и тот же эпитоп или перекрывающиеся эпитопы, могут иметь одинаковый изотип или разные изотипы.
Таким образом, композиция, содержащая антитела, направленные против трех неперекрывающихся эпитопов, может включать четыре, пять или шесть различных молекул антител, причем два антитела связывают два перекрывающихся эпитопа или один и тот же первый эпитоп, два других антитела связывают два других перекрывающихся эпитопа или один и тот же второй эпитоп, а еще два антитела связывают два других перекрывающихся эпитопа или один и тот же третий эпитоп. Разумеется, композиция может включать более двух, например, три или четыре молекулы антител, способные связывать перекрывающиеся эпитопы или способные связывать один и тот же эпитоп. Таким образом, общее число антител, включенных в композицию, может превышать 6, причем для каждого эпитопа имеется более одного антитела, или имеется несколько антител с перекрывающимися эпитопами. При сохранении постоянной общей дозы антител включение в композицию каждого дополнительного антитела подразумевает снижение концентрации каждого антитела. Следовательно, ожидается, что существует предел по числу антител, которые можно включать в композицию, не снижая ее приемлемой эффективности. Основываясь на наблюдениях, полученных в исследованиях связывания с поверхностным плазмонным резонансом и анализах пролиферации, а также принимая во внимание проблемы производства, следует ожидать, что увеличение числа антител в композиции с 6 до 7, 8, 9, 10 или более принесет совсем небольшие дополнительные преимущества (если вообще принесет их). Разумеется, это не исключает того, что композиция может включать более 10 антител, например, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 антител и более, например, 25 антител или более, например, 30 антител или более, например, 40 антител или более, например, 50 антител или более.
Хотя предпочтительно включать в композицию антител по изобретению, антитела, способные связывать, по меньшей мере, три неперекрывающихся эпитопа EGFR, превосходные результаты также были получены при использовании специфических комбинаций антител, способных связывать два неперекрывающихся эпитопа EGFR. Такие предпочтительные композиции "двух антител" более подробно описаны ниже, наряду с руководящими указаниями по составлению композиций антител в соответствии с настоящим изобретением. Оказалось, что по сравнению с композицией из трех антител, включающей антитела 992, 1030 и 1042, примерно такую же или даже большую эффективность можно получить при использовании композиции, состоящей только из двух антител: 992 и 1024. Поскольку антитела 1024 и 1042 относятся к одной и той же группе и, следовательно, имеют одинаковую специфичность связывания, по сути дела, результаты, наблюдаемые при использовании композиции из трех антител, включая эффект терминальной дифференциации, можно приписать только двум специфичностям связывания в композиции (992 и 1024/1042).
В одном из вариантов осуществления изобретения, по меньшей мере, одно антитело в композиции связывает эпитоп домена III, более предпочтительно, чтобы композиция включала, по меньшей мере, два антитела, связывающие эпитопы домена III, а также композиция может включать три антитела, связывающие эпитопы домена III.
Предпочтительно, чтобы композиция включала, по меньшей мере, одно антитело, связывающее эпитоп домена I, или, по меньшей мере, два антитела, связывающие эпитопы домена I.
Предпочтительно, чтобы композиция включала, по меньшей мере, одно антитело, связывающее эпитоп домена II, или антитела, связывающие два эпитопа домена II.
Композиция также может включать антитело, связывающее эпитоп домена I/II, как это определено здесь.
Композиция может включать антитело, способное связывать эпитоп домена IV.
Предпочтительно, чтобы композиция включала, по меньшей мере, одну молекулу антитела, способную ингибировать связывание EGF.
Еще в одном предпочтительном варианте осуществления изобретения композиция может включать антитело, способное предотвращать фосфорилирование EGFR.
Кроме того, композиция может включать антитело, способное усиливать интернализацию/разрушение EGFR.
В предпочтительном варианте осуществления изобретения композиция содержит, по меньшей мере, одно антитело к домену III и, по меньшей мере, одно антитело к домену I/II. Еще в одном предпочтительном варианте осуществления изобретения композиция содержит, по меньшей мере, два антитела к домену III и одно антитело к домену I.
В следующем предпочтительном варианте осуществления изобретения композиция содержит, по меньшей мере, два антитела к домену III, например, три антитела к домену III.
Антитела, входящие в состав композиции, могут представлять собой химерные антитела с вариабельными цепями происхождения, отличного от человека, и константными цепями человека. Вариабельные цепи происхождения, отличного от человека, могут быть получены от мышей, крыс, овец, свиней, кур, приматов, не являющихся людьми, или других подходящих животных. Для того чтобы получить полностью человеческие антитела, их можно генерировать в организме трансгенного животного с генами антител человека. Антитела также могут представлять собой так называемые гуманизированные антитела, в которых последовательности CDR нечеловеческого происхождения были пересажены в каркасные последовательности человека.
Предпочтительно, чтобы константная цепь человека относилась к изотипу IgG1 или IgG2. С точки зрения простоты получения более предпочтительно, чтобы все антитела в составе композиции относились к одному и тому же изотипу. Однако может быть полезным включение в композицию антител с разным изотипом.
Предпочтительно, чтобы композиции антител по изобретению включали антитела, способные связываться с EGFR, выбранным из группы, состоящей из EGFR человека, мутантного EGFR человека и делеционных вариантов EGFR человека. Предпочтительно, чтобы антитела были способны связывать как EGFR человека, так и EGFR приматов не являющихся людьми, то есть их можно было бы испытать перед клиническими экспериментами в релевантных токсикологических исследованиях. Предпочтительным приматом, не являющимся человеком, является обезьяна cynomolgous (Macaca fascicularis).
Для того чтобы поддержать описанную выше концепцию лечения EGFR-зависимого рака с применением антител, связывающих три или более различных эпитопов, настоящее изобретение создает основу для идентификации, изготовления и описания свойств серии химерных антител (мышь/человек), направленных против EGFR. Эти химерные антитела были сравнены по отдельности и в смесях с современными моноклональными антителами, примерами которых являются эрбитукс (Erbitux™) и вектибикс (Vectibix™).
В таблице 1 приведены сводные данные по отдельным химерным антителам и связанным с ними признакам. Номер антитела представляет собой регистрационный номер, используемый во всем тексте настоящей заявки. В столбце Специфичность указан домен EGFR, с которым связывается антитело, как это показано в примере 3. В столбце EGFR дельта приведены данные о способности антитела связывать мутант (EGFRvIII), как это описано в примере 1. В столбце EGFR cynomolgous указана способность антитела связывать EGFR обезьян cynomolgous (пример 10). В столбце Ингиб. EGF указана способность антитела ингибировать связывание EGF (пример 4). В столбце Пролиферация указана способность антитела ингибировать пролиферацию линий раковых клеток А431 и HN-5 (пример 6).
По данным, полученным при анализе химерных антител, исследованных по отдельности или в комбинации в отношении таких признаков как пролиферация, связывание, разрушение/инактивация рецептора и анализы подвижности, а также в исследованиях на животных моделях можно сделать много выводов.
Результаты, полученные с двумя линиями раковых клеток, HN-5 и A431 (пример 6) были воспроизведены на разных линиях раковых клеток (MDA-MB-468 - линия клеток рака молочной железы, DU145 - линия клеток рака предстательной железы). Из этих экспериментов с очевидностью следует, что комбинации антител, предлагаемые авторами настоящего изобретения, проявляют эффективность против очень широкого диапазона линий раковых клеток, поддерживая эффективность композиций антител против ряда конформаций EGFR.
Также было показано, что превосходство смесей антител в большей степени проявляется в анализах пролиферации при добавлении к питательной среде физиологических концентраций лиганда (EGF) по сравнению с такой же схемой анализа, но без добавления EGF (фиг.17). Согласно литературным данным (Hayashi and Sakamoto 1998, J Pharmacobiodyn 11;146-51) сыворотка содержит приблизительно 1-1,8 нг/мл или 0,2-0,3 нМ EGF, тогда как желудочный сок содержит 0,3 нг/мл (приблизительно 0,05 нМ) (Pessonen et al. 1987 Life Sci. 40; 2489-94). По-видимому, в условиях in vivo представлены EGF и другие лиганды EGFR, следовательно, важным признаком смесей антител по настоящему изобретению является их способность проявлять свою эффективность в присутствии таких лигандов EGFR.
Химерные антитела мышь/человек в соответствии с настоящим изобретением обеспечивают лучшие результаты при использовании в комбинации по сравнению с их применением по отдельности. Этот факт иллюстрируется в нескольких экспериментах (см., например, пример 6), когда антитела, тестируемые по отдельности, проявляют лишь умеренные антипролиферативные воздействия на линию раковых клеток (A431-NS), но при использовании в любой комбинации дают намного лучшие результаты. Эти результаты были подтверждены при исследовании множества комбинаций химерных антител по настоящему изобретению. Особенно превосходные результаты были получены при использовании композиции, содержащей антитела 992 и 1024.
Например, некоторые из антител были протестированы в антипролиферативном анализе с линиями клеток A431-NS и HN-5 вместе с любым из антител 992, 1208, 1254 и 1277.
Исследования по связыванию рецептора показали, что некоторые антитела могут фактически стимулировать связывание добавочных антител таким образом, что конкретное антитело связывается с рецептором в большем количестве после насыщения рецептора одним или несколькими антителами. Связывание антитела 992, направленного против домена III, очевидно, извлекает пользу из этого синергического эффекта, полученного при предварительном насыщении рецептора одним или несколькими антителами, которые связывают неперекрывающиеся эпитопы. Еще один пример такого кооперативного эффекта наблюдается, когда антитело 1396, направленное против неизвестного эпитопа, тестируется против EGFR, насыщенного антителами, которые связывают неперекрывающиеся эпитопы.
Исследования по связыванию рецептора также показали, что одновременно связываться с внеклеточным доменом EGFR могут, по меньшей мере, 6 антител. Эти 6 антител представлены 3 антителами к домену III, одним антителом к домену I, одним антителом к домену I/II и одним антителом, связывающим неизвестный эпитоп. Интересно, что связывание трех антител к домену III, по-видимому, облегчает последующее связывание добавочных антител. Это, очевидно, поддерживает концепцию создания композиций антител с несколькими антителами, связывающими разные эпитопы.
При разработке композиции антител против EGFR предпочтительны антитела с неперекрывающимися эпитопами, поскольку они обеспечивают более высокий синергический эффект.
Также предпочтительно, чтобы, по меньшей мере, одно из антител в смеси (когда оно тестируется изолированно), было способно ингибировать связывание лиганда с EGFR, например, способно ингибировать связывание EGF, и/или способно ингибировать связывание альфа-TGF, и/или способно ингибировать связывание амфирегулина. Предпочтительно, чтобы антитело, способное ингибировать связывание EGF, было выбрано из группы, состоящей из антител 992, 1030, 1024, 1042, 1208, 1254, 1277, 1284, 1320 и 1428, более предпочтительно, из группы, состоящей из антител 1208, 1260, 1277 и 1320.
Также предпочтительно, чтобы, по меньшей мере, одно антитело, входящее в состав смеси антител, было способно уменьшать фосфорилирование EGFR. Примеры антител по изобретению, которые обладают такими свойствами, включают: 992, 1030, 1042, 1208, 1277 и 1320.
Домен III EGFR важен для связывания лиганда с рецептором. Кроме того, антитело, связывающееся с доменом III, может стабилизировать EGFR в связанной мономерной конформации, которая не приводит к передаче сигналов рецептором. По этим соображениям предпочтительно, чтобы композиция антител содержала, по меньшей мере, одно антитело со специфичностью к домену III. Предпочтительные антитела к домену III включают антитела 992, 1024, 1030, 1208, 1254, 1277 и 1320. Более предпочтительно, чтобы, по меньшей мере, антитело к домену III было выбрано из группы, состоящей из антител 992, 1254, 1277, 1208 и 1320. Предпочтительная композиция антител может содержать более одного антитела к домену III, например, по меньшей мере, 3 антитела к домену III, например, по меньшей мере, 4 антитела к домену III, например, по меньшей мере, 5 антител к домену III, например, по меньшей мере, 6 антител к домену III.
Еще в одном предпочтительном варианте осуществления изобретения композиция антител содержит, по меньшей мере, одно антитело к домену I. Предпочтительно, чтобы, по меньшей мере, одно антитело к домену I было выбрано из группы, состоящей из антител 1284, 1308, 1344 и 1347. Более предпочтительно, чтобы, по меньшей мере, одно антитело к домену I было выбрано из группы, состоящей из антител 1284 и 1347.
Еще в одном предпочтительном варианте осуществления изобретения композиция антител содержит, по меньшей мере, одно антитело к домену I/II. Предпочтительно, чтобы, по меньшей мере, одно антитело к домену I/II было выбрано из группы, состоящей из антител 1257, 1260, 1261, 1428 и 1434. Более предпочтительно, чтобы, по меньшей мере, одно антитело к домену I/II было выбрано из группы, состоящей из антител 1261 и 1260.
Эффективные специфические комбинации двух антител по настоящему изобретению включают:
Антитело 1280 вместе с 1024, 1320, 1308, 1284, 1260 или 1030, предпочтительно, с 1320 или 1284.
Антитело 1254 вместе с 1024, 1030, 1260, 1284, 1308 или 1320, предпочтительно, с 1320, 1284 или 1260.
Антитело 1277 вместе с 1024, 1030, 1260, 1284, 1308 или 1320, предпочтительно, с 1320, 1284 или 1260.
Антитело 992 вместе с 1030, 1260, 1284, 1308, 1320 или 1024, предпочтительно, с 1320, 1024 или 1284.
Примеры лучших и предпочтительных смесей двух антител включают 992+1024, 992+1320, 992+1042, 1277+1320, 1208+1320. Особенно предпочтительна смесь 992+1024.
Предпочтительные смеси с тремя антителами включают: Антитела 992+1030+1042, 992+1320+1024, 992+1024+1030, 1320+1284+1261, 1320+1214+1320, 992+1284+1320, 992+1255+1024, 992+1030+1320, 992+1024+1214, 992+1261+1320, 992+1024+1284, 992+1024+1211, 992+1024+1030, 1260+1214+1254, 992+1255+1320, 992+1211+1320, 992+1030+1261, 992+1260+1030, 992+1260+1320, 992+1030+1214.
Предпочтительные смеси с четырьмя антителами включают: Антитела 992+1320+1024+1030, 992+1024+1030+1284, 1277+1320+1260+1347, 1277+1320+1261+1347, 1277+1320+1261+1284, 1254+1320+1260+1347, 1254+1320+1261+1347, 1254+1320+1261+1284, 1254+1024+1260+1347, 1254+1024+1261+1347, 1254+1024+1261+1284, 1277+1024+1260+1347, 1277+1024+1261+1347, 1277+1024+1261+1284.
Предпочтительные смеси с 5 антителами включают: 992+1030+1024+1260+1347, 992+1030+1024+1261+1347, 992+1030+1024+1261+1284, 992+1030+1320+1260+1347, 992+1030+1320+1261+1347, 992+1030+1320+1261+1284.
Одна предпочтительная смесь с 8 антителами включает: 992+1030+1024+1277+1254+1320+1260+1261+1284+1347.
Кроме того, чтобы можно было провести токсикологическое исследование приматов, не являющихся людьми, предпочтителен такой вариант, когда все антитела в композиции связываются с EGFR человека и, по меньшей мере, с одним дополнительным EGFR примата, например, EGFR шимпанзе, macaca mulatta, макака-резус и других макаков или обезьян cynomolgous. Обезьяна cynomolgous представляет собой относительно небольшое животное и очень хорошо подходит для токсикологических исследований. Таким образом, предпочтительным EGFR примата является EGFR обезьяны cynomolgous. Предпочтительно, чтобы антитела связывали EGFR человека и EGFR примата, не являющегося человеком, приблизительно с одинаковой аффинностью.
Настоящее изобретение продемонстрировало лучшие результаты в одном или более функциональных анализов при комбинировании в одной композиции 2, 3, 4, 5, 6, 7 и 8 антител. Хотя эти данные дают руководящую основу для выбора числа антител в композиции, их никоим образом не следует воспринимать как ограничительные. Композиция может содержать более 8 антител, даже несмотря на то, что экспериментальные данные демонстрируют одновременное связывание только для 6 антител. Могут иметь место другие соображения для включения в композицию более 6 антител, например, связанные с различиями в скорости выведения различных антител.
Следующим предпочтительным признаком антител в композициях является их белковая однородность, то есть возможность достаточно простой очистки антител. Применительно к отдельным членам композиции антител предпочтителен профиль ионообменной хроматографии с одним четко выраженным пиком, что облегчает определение параметров. Четкий профиль ионообменной хроматографии также предпочтителен для простоты определения параметров конечной композиции антител. Также предпочтительно комбинировать в композиции такие антитела, которые можно отличить друг от друга при помощи ионообменной хроматографии, чтобы можно было определить параметры композиции со всеми антителами при одном прогоне.
Антитела могут иметь любое происхождение, например, человеческое, мышиное, кроличье, куриное, свиное, овечье или от ламы. Антитела также могут быть химерными, как это описано в примерах, а также могут быть гуманизированными, супергуманизированными или их переформированными версиями, созданными с применением хорошо известных способов, описанных в данной области техники.
Предпочтительная композиция антител
Как показано в прилагаемых примерах, композиция антител против EGFR, основанная на антителах 992 и 1024, имеет уникальные отличительные свойства. Связывание антитела 992 усиливается за счет связывания других антител, включая 1024. В отличие от поступающих в продажу антител, как 992, так и 1024 преимущественно связываются с представленными на клетках конформационными эпитопами (примеры 14 и 15). Эпитопы антител 992 и 1024 перекрываются с эпитопами эрбитукса и вектибикса, но отличаются от них. В отличие от многих других композиций из двух антител, в которых отдельные антитела связываются с неперекрывающимися эпитопами, композиция, основанная на специфичностях связывания антител 992 и 1024 быстро и эффективно запускает интернализацию рецептора. На животной модели после лечения композициями антител, основанными на антителах 992 и 1024, наблюдался новый механизм действия, включающий терминальную дифференциацию, сопровождающуюся повышенной экспрессией инволюкрина и появлением жемчужин кератина. Этот уникальный механизм действия приводит к более эффективному и длительному ингибированию роста раковых клеток как in vitro, так и in vivo. Указанные явления наиболее отчетливо наблюдаются в примерах in vivo, когда опухоли продолжают уменьшаться и после окончания лечения. В контрольной группе, получавшей эрбитукс, опухоли возобновляли рост после окончания лечения. Это явно указывает на разные механизмы действия.
Авторы изобретения полагают, что новый механизм действия достигается при использовании комбинации двух специфичностей связывания, проявляемых антителами 992 и 1024 в одной композиции антител. Такой механизм действия также наблюдается при использовании третьего антитела, не конкурирующего с антителами 992 и 1024, например, в тройной комбинации антител 992, 1024 и 1030.
Эти наблюдения привели к разработке композиции, содержащей, по меньшей мере, 2 разные молекулы антител против EGFR человека, в которой первая индивидуальная молекула антитела против EGFR выбрана из группы, состоящей из антитела 992, антитела, содержащего последовательности антитела 992 VL (аминокислоты 3-109 из SEQ ID NO 72) и VH (аминокислоты 3-124 из SEQ ID NO 40), антитела, имеющего области CDR3 из антитела 992 (SEQ ID NO 116 и 111), антитела, связывающегося с тем же эпитопом, что и антитело 992, и антитела, способного ингибировать связывание антитела 992 с EGFR человека, а вторая индивидуальная молекула антитела против EGFR выбрана из группы, состоящей из антитела 1024, антитела, содержащего последовательности антитела 1024 VL (аминокислоты 3-114 из SEQ ID NO 73) и VH (аминокислоты 3-120 из SEQ ID NO 41), антитела, имеющего области CDR3 из антитела 1024 (SEQ ID NO 120 и 114), антитела, связывающегося с тем же эпитопом, что и антитело 1024, и антитела, способного ингибировать связывание антитела 1024 с EGFR человека.
Предпочтительно, чтобы указанная первая индивидуальная молекула антитела против EGFR была выбрана из группы, состоящей из антитела 992, антитела, содержащего последовательности VL и VH из антитела 992, антитела, имеющего области CDR3 из антитела 992, и антитела, связывающегося с тем же эпитопом, что и антитело 992, а указанная вторая индивидуальная молекула антитела против EGFR была выбрана из группы, состоящей из антитела 1024, антитела, содержащего последовательности VL и VH из антитела 1024, антитела, имеющего области CDR3 из антитела 1024, и антитела, связывающегося с тем же эпитопом, что и антитело 1024.
Настоящее изобретение рассматривает мутации в последовательностях CDR3 антител 992 и 1024, чтобы создавать антитела с такой же специфичностью связывания. Поэтому в одном из вариантов осуществления изобретения антитело, имеющее такую же специфичность связывания, что и антитело 992, содержит CDRH3, имеющий следующую формулу: CTX1X2X3X4X5X6X7X8X9X10X11X12X13X14X15W, где позиции от Х1 до Х15 индивидуально выбраны из перечисленных ниже групп аминокислот:
X1=R или K,
Х2=N, D, Е или Q,
Х3=G, А, V или S,
Х4=D, Е, N или Q,
Х5=Y, F, W или Н,
X6 = Y, F, W или H,
X7 = V, I, L или A,
X8 = S, T, G или A,
X9 = S, T, G или A,
X10 = G, A, V или S,
X11 = D, E, N или Q,
X12 = A, G, V или S,
X13 = M, L, I или V,
X14 = D или E, и
X15 = Y или F,
а CDRL3 описывается следующей формулой: CX1X2X3X4X5X6PPTF, где позиции от X1 до X6 индивидуально выбраны из перечисленных ниже групп аминокислот:
X1 = Q или H,
X2 = H, E или Q,
X3 = Y, F, W или H,
X4 = N, Q или H,
X5 = T, S, G или A, и
X6 = V, I, L или A.
В одном из вариантов осуществления изобретения антитело, имеющее такую же специфичность связывания, что и антитело 1024, содержит CDRH3, имеющий следующую формулу: CVX1X2X3X4X5X6X7X8X9X10X11W, где позиции от X1 до X11 индивидуально выбраны из перечисленных ниже групп аминокислот:
X1 = R или K,
X2 = Y, F W или H,
X3 = Y, F, W или H,
X4 = G, A, V или S,
X5 = Y, F, W или H,
X6 = D, E, N или Q,
X7 = E или D,
X8 = A, G, V или S,
X9 = M, L, I или V,
X10 = D, E, N или Q, и
X11 = Y или F,
а CDRL3 описывается следующей формулой: CX1X2X3X4X5X6PX7TF, где позиции от X1 до X7 индивидуально выбраны из перечисленных ниже групп аминокислот:
X1 = A, G или V,
X2 = Q или H,
X3 = N, Q или H,
X4 = L, I, M или V,
X5 = E, D, N или Q,
X6 = L, I, M или V, и
X7 = Y, F, W или H.
Применяя стандартные методики, можно получить антитела с мутантными областями CDR3, экспрессировать их и протестировать на связывание, используя описанные здесь способы.
В соответствии с этим аспектом изобретения антитела могут быть химерными, человеческими, гуманизированными, переформированными или супергуманизированными. Этих результатов можно достичь, применяя способы, известные в данной области техники. Например, антитела 992 и 1024 можно гуманизировать, применяя способы, описанные в примере 18. Способы "супергуманизации" описаны в патенте США №6881557.
Более предпочтительно, чтобы указанная первая индивидуальная молекула антитела против EGFR была выбрана из группы, состоящей из антитела 992, антитела, содержащего последовательности VL и VH из антитела 992, и антитела, имеющего области CDR3 из антитела 992, а указанная вторая индивидуальная молекула антитела против EGFR была выбрана из группы, состоящей из антитела 1024, антитела, содержащего последовательности VL и VH из антитела 1024, и антитела, имеющего области CDR3 из антитела 1024.
Более предпочтительно, чтобы указанная первая индивидуальная молекула антитела против EGFR была выбрана из группы, состоящей из антитела 992 и антитела, содержащего последовательности VL и VH из антитела 992, а указанная вторая индивидуальная молекула антитела против EGFR была выбрана из группы, состоящей из антитела 1024 и антитела, содержащего последовательности VL и VH из антитела 1024.
Наиболее предпочтительно, чтобы композиция состояла из антител 992 и 1024.
Как описано здесь, предпочтительно, чтобы первое и второе антитела против EGFR не ингибировали связывание друг друга с EGFR человека. Еще более предпочтительно, чтобы, по меньшей мере, одно из антител было способно увеличивать максимальную мощность связывания другого антитела с EGFR человека. Этот эффект наблюдается для антител 992 и 1024 (пример 16).
Соотношение между двумя антителами необязательно должно составлять точную величину 1:1. Следовательно, доля первого антитела в композиции по отношению ко второму антителу может составлять от 5 до 95%, например, от 10 до 90%, предпочтительно, от 20 до 80%, более предпочтительно, от 30 до 70%, более предпочтительно, от 40 до 60%, например, от 45 до 55%, например, приблизительно 50%.
Предпочтительно, чтобы первое и второе антитела относились к изотипу IgG1 или IgG2.
Примерами антител, связывающимися с тем же эпитопом, что и антитело 992, которые были идентифицированы авторами настоящего изобретения, являются антитела из кластера антител, содержащего клоны 1209, 1204, 992, 996, 1033 и 1220.
Примерами антител, связывающимися с тем же эпитопом, что и антитело 1024, которые были идентифицированы авторами настоящего изобретения, являются антитела из кластера антител, содержащего клоны 1031, 1036, 1042, 984, 1024, 1210, 1217, 1221 и 1218.
Специфичность связывания антител определяют последовательности CDR3. В предпочтительных вариантах осуществления изобретения антитело, содержащее область CDR3 из антитела 992, дополнительно содержит CDR1 и CDR2 из VH и VL антитела 992. Таким же образом, предпочтительное антитело, содержащее область CDR3 из антитела 1024, дополнительно содержит CDR1 и CDR2 из VH и VL антитела 1024. Последовательности CDR антител можно найти в таблице 12 (пример 17).
В других вариантах осуществления изобретения антитело, конкурирующее с антителом 992, выбирают из группы, состоящей из антител 1208, 1254 и 1277. Таким же образом, антитело, конкурирующее с антителом 1024, можно выбрать из группы, состоящей из антител 1042 и 1320.
В одном из вариантов осуществления изобретения композиция не содержит добавочных антител в дополнение к указанным первому и второму антителам, более предпочтительно, никаких дополнительных антител против EGFR.
В других вариантах осуществления изобретения композиция дополнительно содержит третье индивидуальное антитело против EGFR, причем молекула указанного третьего индивидуального антитела против EGFR может быть выбрана из группы, состоящей из антитела 1030, антитела, содержащего последовательности VL (аминокислоты 3-113 из SEQ ID NO 74) и VH (аминокислоты 3-120 из SEQ ID NO 42) антитела 1030, антитела, имеющего области CDR3 из антитела 1030 (SEQ ID NO 112 и 119), антитела, связывающегося с тем же эпитопом, что и антитело 1030, и антитела, способного ингибировать связывание антитела 1030 с EGFR человека. Предпочтительно, чтобы указанное третье антитело приводило к усилению связывания указанного первого и/или второго антитела с EGFR человека. В одном из вариантов осуществления изобретения композиция не содержит добавочных антител в дополнение к указанным первому, второму и третьему антителам, более предпочтительно, никаких дополнительных антител против EGFR.
Антитело, связывающееся с тем же эпитопом, что и антитело 1030, может быть выбрано из кластера антител, состоящего из клонов 1195, 1030, 1034, 1194, 980, 981, 1246 и 1223.
Антитело, содержащее область CDR3 из антитела 1030, может дополнительно содержать области CDR1 и CDR2 из VH и VL антитела 1030.
Антитела, входящие в композицию, могут находиться в общем контейнере для введения. Однако их можно производить, очищать и исследовать их свойства по отдельности, составляя композицию в двух или трех разных контейнерах, то есть, как набор частей с одним антителом в каждом контейнере. Эти части набора, как таковые, можно вводить одновременно, последовательно или по отдельности.
В дальнейшем аспекте две специфичности связывания антител 992 и 1024 комбинируются в одной биспецифической связывающей молекуле. Предпочтительно, чтобы биспецифическая связывающая молекула содержала последовательности CDR из антител 992 и 1024, более предпочтительно, последовательности VH и VL антител 992 и 1024. Биспецифическая связывающая молекула может представлять собой антитело с двойным вариабельным доменом, как это описано в примере 19. Биспецифическая связывающая молекула также может быть сконструирована в виде биспецифического фрагмента Fab, биспецифического scFV или диатела, как это описано в литературе.
Предпочтительно, чтобы композиции антител, основанные на специфичностях связывания антител 992 и 1024, приводили к одной или нескольким интернализаций рецептора, к регрессу опухолей A431NS in vivo, к индуцированию терминальной дифференциации клеток A431NS in vivo и к повышающей регуляции экспрессии опухолевого инволюкрина in vivo.
В настоящем документе приведено несколько примеров антител, проявляющих такие же или сходные эффекты, как и комбинация антител 992 и 1024. Примеры такого рода включают антитела, полученные в результате одной и той же иммунизации и относящиеся к одним и тем же кластерам, а также антитела, индивидуально конкурирующие с одним или двумя антителами. Композиции антител с одинаковым или сходным эффектом можно сконструировать на основе последовательностей VL и VH антител 992 и 1024, а также на основе CDR этих антител, особенно, CDR3 двух антител.
Дополнительные композиции антител с одинаковым эффектом или сходными эффектами можно изготовить, проводя иммунизацию и скрининг, по существу, так, как это описано в примерах. Антитела с такой же специфичностью связывания, как и антитела 992 и 1024, можно идентифицировать в двух отдельных анализах на конкуренцию, как это описано здесь. Наконец, композиции антител, в которых одно антитело усиливает связывание другого антитела, можно идентифицировать, проводя эксперименты по связыванию, по существу, таким образом, как это описано в примере 16. Композиции антител, как это описано в примерах, можно дополнительно скринировать на эффекты в отношении интернализации рецептора, на эффективность in vitro и in vivo, на аффинность связывания и т.д.
Применение композиций антител по изобретению
Для применения in vivo в целях лечения и профилактики заболеваний, связанных с экспрессией EGFR (например, свехэкспрессией), антитела по изобретению вводят пациентам (например, человека) в терапевтически эффективных дозах (например, таких дозах, которые приводят к ингибированию роста, фагоцитозу, уменьшению подвижности, к терминальной дифференциации и/или уничтожению опухолевых клеток, экспрессирующих EGFR) с использованием любого подходящего способа введения, например, инъекции и других способов введения, известных в данной области техники для клинических продуктов на основе антител.
Типичные EGFR-зависимые заболевания, которые можно лечить, облегчать и/или предупреждать с применением антител по изобретению, включают, но не ограничиваясь ими, аутоиммунные и раковые заболевания. Например, раковые заболевания, которые можно лечить, облегчать и/или предупреждать, включают рак мочевого пузыря, рак молочной железы, рак матки/шейки матки, рак почки, рак яичника, рак предстательной железы, почечно-клеточный рак, рак поджелудочной железы, рак толстой кишки, рак прямой кишки, рак желудка, плоскоклеточный рак, рак легких (немелкоклеточный), рак пищевода, рак головы и шеи, рак кожи. Аутоиммунные заболевания, которые можно лечить, включают, например, псориаз.
Еще в одном варианте осуществления изобретение относится к способу лечения, облегчения и/или профилактики глиобластомы, включая мультиформную глиобластому; астроцитомы, включая детскую астроцитому, глиомы; нейробластомы; нейроэндокринных опухолей желудочно-кишечного тракта; бронхоальвеолярной карциномы; фолликулярной папиллокарциномы; карциномы слюнных желез; амелобластомы; злокачественной периферийной невриномы; эндокринных опухолей поджелудочной железы или тестикулярных гермином, включая семиному, эмбриональную карциному, опухоль желточного мешка, тератому и хориокарциному.
Выделение и отбор кодирующих пар тяжелых вариабельных цепей и легких вариабельных цепей
Процесс генерирования композиции рекомбинантных антител против EGFR включает выделение из подходящего источника последовательностей, кодирующих вариабельные тяжелые цепи (VH) и вариабельные легкие цепи (VL), то есть генерирование репертуара кодирующих пар для VH и VL. Обычно подходящим источником для получения кодирующих последовательностей VH и VL являются фракции клеток, содержащие лимфоциты, в частности образцы крови, селезенки или костного мозга от животного не являющегося человеком, иммунизированного/вакцинированного полипептидом или пептидом EGFR человека или белками EGFR, полученными из клеток, экспрессирующих EGFR человека, или клетками, экспрессирующими EGFR человека, или фракциями таких клеток. Предпочтительно, чтобы фракции, содержащие лимфоциты, были получены от млекопитающих, не являющихся людьми, или трансгенных животных, с генами иммуноглобулина человека. Собранная фракция клеток, содержащая лимфоциты, может быть дополнительно обогащена для получения специфической популяции лимфоцитов, например, клеток из линий B-лимфоцитов. Предпочтительно, чтобы обогащение проводилось с применением сортировки клеток на магнитных частицах (MACS) и/или сортировки клеток с флуоресцентной активацией (FACS), пользуясь линиеспецифическими маркерными белками на клеточной поверхности, например, для B-клеток, плазмабластов и/или плазмацитов. Предпочтительно, чтобы клеточная фракция, содержащая лимфоциты, была обогащена и отсортирована на B-клетки, плазмабласты и/или плазмациты. Еще более предпочтительно, чтобы клетки с высокой экспрессией CD43 и CD138 были выделены из селезенки или крови. Эти клетки иногда называют циркулирующими плазматическими клетками, ранними плазматическими клетками или плазмабластами. Например, в настоящем изобретении их называют только плазмацитами, хотя взаимозаменяемо могут употребляться и другие термины.
Выделение кодирующих последовательностей VH и VL можно осуществлять классическим способом, когда кодирующие последовательности VH и VL случайно комбинируются в векторе для создания комбинаторной библиотеки пар кодирующих последовательностей VH и VL. Однако в настоящем изобретении предпочтительно отражать различие, аффинность и специфичность антител, выработанных при гуморальном иммунном ответе на иммунизацию EGFR. Это включает сохранение спаривания VH и VL, изначально представленного у донора, то есть создание репертуара пар последовательностей, в котором каждая пара кодирует вариабельную тяжелую цепь (VH) и вариабельную легкую цепь (VL), соответствующие паре цепей VH и VL, изначально представленной в антителе, вырабатываемом тем донором, от которого были выделены эти последовательности. Это также называют когнатной парой кодирующих последовательностей VH и VL, а антитело называют когнатным антителом. Предпочтительно, чтобы кодирующие пары VH и VL, рассматриваемые в настоящем изобретении (комбинаторные или когнатные), были получены от доноров мышей, то есть последовательности были бы мышиными.
Существует несколько разных подходов к генерированию когнатных пар кодирующих последовательностей VH и VL, причем один из подходов включает амплификацию и выделение кодирующих последовательностей VH и VL из единичных клеток, отсортированных из фракции клеток, содержащей лимфоциты. Для того чтобы получить репертуар пар кодирующих последовательностей VH и VL, который отражает разнообразие пар последовательностей VH и VL у донора, предпочтительно применять высокопроизводительный способ по возможности с минимальным скремблированием (случайным комбинированием) пар VH и VL, например, как это описано в документе WO 2005/042774 (включенном сюда в качестве ссылки).
Кодирующие последовательности VH и VL можно амплифицировать по отдельности и спаривать на втором этапе либо можно спаривать их во время амплификации (Coronella et al. 2000. Nucleic Acids Res. 28: E85; Babcook et al 1996. PNAS 93: 7843-7848 и WO 2005/042774). Второй подход включает внутриклеточную амплификацию и спаривание кодирующих последовательностей VH и VL (Embleton et al. 1992. Nucleic Acids Res. 20: 3831-3837; Chapal et al. 1997. BioTechniques 23: 518-524). Третий подход заключается в применении способа избранных лимфоцитарных антител (SLAM), который сочетает в себе локальный гемолиз в геле с клонированием кДНК VH и VL (Babcook et al. 1996. PNAS 93:7843-7848). Еще один способ, который можно использовать на мышах, представляет собой стандартную методику гибридомы, сопровождающуюся скринингом и отбором ведущих кандидатов с последующим клонированием кодируемых антител.
В предпочтительном варианте осуществления настоящего изобретения репертуар кодирующих пар VH и VL, в котором разные пары-члены отражают пары генов, ответственные за гуморальный иммунный ответ на иммунизацию EGFR, создается в соответствии со способом, включающим этапы i) получения фракции клеток, содержащей лимфоциты, от животного донора, иммунизированного EGFR человека, ii) факультативного обогащения указанной клеточной фракции B-клетками или плазмацитами, iii) получения популяции выделенных единичных клеток, содержащей дистрибутивные клетки из указанной клеточной фракции по отдельности во множестве сосудов, iv) амплификации и результирующего сцепления кодирующих пар VH и VL в мультиплексной процедуре RT-PCR с удлинением перекрывания и с применением матрицы, полученной из указанных единичных клеток, v) факультативного проведения вложенной PCR сцепленных кодирующих пар VH и VL. Предпочтительно, чтобы выделенные когнатные кодирующие пары VH и VL были подвергнуты процедуре скрининга, как это описано ниже.
После того как были генерированы пары последовательностей VH и VL, проводят процедуру скрининга для идентификации последовательностей, кодирующих пары VH и VL с реактивностью связывания в отношении EGFR-ассоциированного антигена. Предпочтительно, чтобы EGFR-ассоциированный антиген содержал такую внеклеточную часть EGFR как домен III, II, I и/или IV, фрагменты доменов или полный внеклеточный домен. Другие антигены включают такие мутанты, как делеционные мутанты EGFR или SNP, либо их фрагменты. Если пары последовательностей VH и VL являются комбинаторными, можно применить процедуру фагового дисплея для обогащения пар VH и VL, которые кодируют фрагменты антител, связывающиеся с EGFR, перед проведением скрининга.
Для того чтобы отразить разнообразие, аффинность и специфичность антител, вырабатываемых в ходе гуморального иммунного ответа на иммунизацию EGFR, настоящее изобретение позволило разработать процедуру скрининга на когнатные пары, позволяющую получить по возможности наиболее широкое разнообразие. В целях скрининга репертуар кодирующих пар когнатных VH и VL экспрессируют индивидуально либо как фрагменты антител (например, scFv или Fab), либо как полноразмерные антитела, применяя или бактериальный или происходящий от млекопитающих вектор скрининга, трансфицированный в подходящую клетку-хозяина. Репертуар Fab/антител можно скринировать (без ограничений) на реактивность по отношению к EGFR, на антипролиферативную активность против линий раковых клеток, экспрессирующих EGFR, а также на способность ингибировать связывание лиганда (например, EGF) с EGFR, на ингибирование фосфорилирования, индукцию апоптоза, интернализацию EGFR.
В параллельной серии анализов репертуар Fab/антител скринируют на действие против избранных антигенов, например, пептидов EGFR человека и, факультативно, обезьян cynomolgous или шимпанзе, или макака резус.Антигенные пептиды можно выбирать, например, из внеклеточного домена EGFR человека, мутантного внеклеточного домена EGFR человека и внеклеточного домена EGFR cynomolgous, или из их фрагментов. Пептиды могут быть биотинилированы для облегчения их иммобилизации на микрочастицах или планшетах при скрининге. Можно также использовать альтернативные средства для иммобилизации. Антигены выбирают, основываясь на знании биологии EGFR и на ожидаемом нейтрализующем и/или защитном эффекте, который потенциально могут дать антитела, способные связываться с этими антигенами. Таким же образом, эту процедуру скрининга можно применить к комбинаторной библиотеке фагового дисплея.
Рекомбинантные белки EGFR, используемые для скрининга, могут экспрессироваться в бактериях, клетках насекомых, клетках млекопитающих или другой подходящей системе экспрессии. Для правильного процессинга (включая гликозилирование) белки экспрессируются в клетках млекопитающих. Белок EGFR-ECD может либо экспрессироваться как растворимый протеин (без трансмембранной и внутриклеточной областей), либо гибридизироваться с третьим белковым компонентом для увеличения стабильности. Если белок EGFR экспрессируется со свободным хвостом для гибридизации, перед скринингом гибридный партнер может быть отщеплен. В дополнение к описанному выше первичному скринингу может быть проведен вторичный скрининг с той целью, чтобы убедиться, что ни одна из отобранных последовательностей не кодирует ложноположительные продукты.
Для скрининга, проводимого в соответствии с настоящим изобретением, обычно подходят иммунологические анализы. Такие анализы хорошо известны в данной области техники и к их числу относятся, например, ELISPOT, ELISA, FLISA, мембранные анализы (в том числе вестерн-блоттинг), матрицы на фильтрах и FACS. Анализы можно проводить без каких-либо этапов предварительного обогащения, с применением полипептидов, вырабатываемых из последовательностей, кодирующих пары VH и VL. В том случае когда репертуар кодирующих пар VH и VL представлен когнатными парами, перед скринингом нет никакой необходимости в обогащении, например, посредством фагового дисплея. Однако при скрининге комбинаторных библиотек предпочтительно проводить иммуноанализы в комбинации со способами обогащения или после способов обогащения, таких как фаговый дисплей, рибосомный дисплей, поверхностный бактериальный дисплей, дрожжевой дисплей, эукариотический вирусный дисплей, РНК-дисплей или ковалентный дисплей (см. обзор в ссылке FitzGerald, K., 2000. Drug Discov. Today 5, 253-258).
Кодирующие последовательности пар VH и VL, отобранные по результатам хода скрининга, обычно подвергают секвенированию и анализируют на различия в вариабельных областях. Особый интерес представляют различия в областях CDR, но также интересно исследовать представительство семейства VH и VL. Основываясь на этих анализах, проводят отбор последовательностей, кодирующих пары VH и VL, которые представляют общее разнообразие EGFR-связывающих антител, выделенных от одного или нескольких животных-доноров. Предпочтительно выбирать последовательности с различиями в областях CDR (CDRH1, CDRH2, CDRH3 и CDRL1, CDRL2 и CDRL3). Если встречаются последовательности с одной или несколькими идентичными или очень похожими областями CDR, которые относятся к разным семействам VH или VL, их также отбирают. Предпочтительно, чтобы в отобранных парах последовательностей отличалась, по меньшей мере, область CDR3 вариабельной тяжелой цепи (CDRH3). Потенциально выбор пар последовательностей VH и VL может быть основан исключительно на вариабельности области CDRH3. Во время прайминга и амплификации последовательностей могут встречаться мутации в каркасных участках вариабельной области, особенно в первом каркасном участке. Предпочтительно, чтобы возможные ошибки в первом каркасном участке были скорректированы, давая уверенность в том, что последовательности полностью или, по меньшей мере, на 98% соответствуют последовательностям, происходящим из зародышевой линии, например, последовательности VH и VL полностью были бы мышиными.
Если имеется уверенность в том, что общее разнообразие отобранных последовательностей, кодирующих пары VH и VL, высоко репрезентативно в отношении разнообразия, наблюдаемого на генетическом уровне в гуморальном ответе на иммунизацию EGFR, то можно ожидать, что общая специфичность антител, экспрессируемых из коллекции отобранных кодирующих пар VH и VL также окажется репрезентативной в отношении специфичности антител, вырабатываемых у животных, иммунизированных EGFR. Указание о том, репрезентативна ли специфичность антител, экспрессируемых из коллекции отобранных кодирующих пар VH и VL, в отношении специфичности антител, вырабатываемых донорами, можно получить, сравнивая титры антител из крови донора к избранным антигенам со специфичностью антител, экспрессируемых из коллекции отобранных кодирующих пар VH и VL. В дополнение к этому можно и далее анализировать специфичность антител, экспрессируемых из коллекции отобранных кодирующих пар VH и VL. Степень специфичности коррелируется с числом различных антигенов, по отношению к которым определяют связывающую реактивность. В следующем варианте осуществления настоящего изобретения специфичность отдельных антител, экспрессируемых из коллекции отобранных кодирующих пар VH и VL, анализируют посредством картирования эпитопов.
Картирование эпитопа можно осуществить, применяя множество методик, которые необязательно исключают применение друг друга. Один из способов картирования эпитопной специфичности молекулы антитела заключается в оценке связывания с пептидами разной длины, полученными из первичной структуры целевого антигена. Такие пептиды могут быть как линейными, так и конформационными, причем их можно применять во многих форматах анализа, включая ELISA, FLISA и поверхностный плазмонный резонанс (SPR, Biacore, FACS). Кроме того, можно осуществлять рациональный отбор пептидов, используя доступные данные о последовательности и структуре, чтобы отобразить, например, внеклеточные области или консервативные области целевого антигена, либо можно сконструировать панель перекрывающихся пептидов, представляющую избранную часть антигена или весь антиген (Meloen RH, Puijk WC, Schaaper WMM. Epitope mapping by PEPSCAN. В книге: Immunology Methods Manual. Ed Iwan Lefkovits 1997, Academic Press, pp 982-988). Специфическая реактивность клона антитела с одним или несколькими такими пептидами, в целом, будет указывать на эпитопную специфичность. Однако пептиды во многих случаях являются плохими имитаторами эпитопов, распознаваемых антителами, вырабатываемыми против белковоподобных антигенов, как вследствие утраты природной или специфической конформации, так и вследствие значительно большей скрытой площади поверхности взаимодействия между антителом и белковым антигеном по сравнению с антителом и пептидом. Второй способ эпитопного картирования, который учитывает определение специфичностей непосредственно на белковом антигене, заключается в избирательной маскировке эпитопа с применением существующих, хорошо определенных антител. Ограниченное связывание второго, зондирующего антитела с антигеном после блокирования, в целом, указывает на совмещенные или перекрывающиеся эпитопы. Эпитопное картирование посредством избирательной маскировки можно осуществлять посредством многих иммуноанализов, включая, но не ограничиваясь ими, ELISA и Biacore, хорошо известные в данной области техники (см. например, ссылки Ditzel et al. 1997. J. Mol. Biol. 267:684-695; Aldaz-Carroll et al. 2005. J. Virol. 79: 6260-6271). Еще один потенциальный способ для определения эпитопной специфичности антител против EGFR заключается в отборе "ускользнувших" мутантов в присутствии антитела. Это можно осуществить, например, применяя сканирование аланином. Секвенирование представляющего интерес гена (генов) таких "ускользнувших" мутантов обычно будет показывать, какие аминокислоты в антигене (антигенах) важны для распознавания антителом, то есть составляют эпитоп (часть эпитопа).
Выработка композиции антител против EGFR из отобранных кодирующих пар VH и VL
Композиция антител по настоящему изобретению может быть выработана из поликлональной экспрессирующей клеточной линии в одном или нескольких биореакторах или их эквивалентах. После реализации такого подхода антитела против EGFR могут быть очищены на выходе из реактора как единый препарат без попытки разделения в процессе отдельных членов смеси, составляющих композицию антител против EGFR. Если композиция антител вырабатывается более чем в одном биореакторе, то очищенную композицию антител против EGFR можно получить посредством объединения антител, полученных из индивидуально очищенных супернатантов из каждого реактора.
Один из способов выработки композиции рекомбинантных антител описан в документах WO 2004/061104 и WO 2006/007850 (эти источники включены сюда в качестве ссылки). Описанный там способ основан на сайт-специфической интеграции кодирующей последовательности антитела в геном индивидуальных клеток-хозяев при достаточной уверенности в том, что белковые цепи VH и VL в процессе выработки композиции сохраняют свое первоначальное спаривание. Кроме того, сайт-специфическая интеграция сводит к минимуму эффекты положения, благодаря чему можно ожидать, что рост и свойства экспрессии индивидуальных клеток в поликлональной клеточной линии будут очень сходными. Обычно указанный способ включает следующее: i) клетку-хозяина с одним или несколькими сайтами распознавания рекомбиназы, ii) вектор экспрессии, по меньшей мере, с одним сайтом распознавания рекомбиназы, совместимым с таким же сайтом клетки-хозяина, iii) генерирование коллекции векторов экспрессии посредством переноса отобранных кодирующих пар VH и VL из вектора скрининга в вектор экспрессии, чтобы из вектора могло экспрессироваться полноразмерное антитело или фрагмент антитела (такой перенос необязателен, если вектор скрининга идентичен вектору экспрессии), iv) трансфекция клетки-хозяина коллекцией векторов экспрессии и вектором, кодирующим рекомбиназу, которая способна комбинировать сайты распознавания в геноме клетки-хозяина с такими же сайтами в векторе, v) получение/генерирование поликлональной клеточной линии из трансфицированной клетки-хозяина, vi) экспрессирование и накопление композиции антител из поликлональной клеточной линии.
Если в одной композиции используется небольшое число антител (2-3 или немного более), они могут быть экспрессированы и очищены по отдельности способом, аналогичным производству моноклональных антител, например, как это описано в документе WO 2004/085474. Очищенные антитела можно смешивать после очистки или упаковывать в отдельные флаконы для смешивания перед введением или для раздельного введения.
Предпочтительно использовать такие клетки млекопитающих как клетки CHO, клетки COS, клетки BHK, клетки миеломы (например, клетки Sp2/0 или NS0), такие фибробласты как NIH 3T3, и иммортализованные клетки человека, в частности, клетки HeLa, клетки HEK 293 или PER C6. Однако также можно использовать эукариотические или прокариотические клетки, происходящие не от млекопитающих, например, растительные клетки, клетки насекомых, дрожжевые клетки, грибковые клетки, E. coli и т.д. Подходящая клетка-хозяин содержит в своем геноме один или более соответствующих сайтов распознавания рекомбиназы. Клетка-хозяин также должна содержать способ отбора, который функционально связан с сайтом интеграции, чтобы обладать способностью отбора на интегранты (то есть клетки, имеющие в сайте интеграции встроенную копию вектора экспрессии Ab против EGFR или фрагмент вектора экспрессии). Приготовление клеток, имеющих в своем геноме сайт FRT в предопределенном положении, было описано, например, в патенте США 5677177. Предпочтительно, чтобы клетка-хозяин имела только один сайт интеграции, который расположен в месте, допускающем высокую степень экспрессии интегранта (так называемая горячая точка).
Подходящий вектор экспрессии содержит сайт распознавания рекомбинации, соответствующий сайту (сайтам) распознавания рекомбиназы в клетке-хозяине. Предпочтительно, чтобы сайт распознавания рекомбиназы был сцеплен с подходящим геном, отличающимся от гена отбора, который используется для конструирования клетки-хозяина. Гены отбора хорошо известны в данной области техники и включают ген глутаминсинтетазы (GS), ген дигидрофолатредуктазы (DHFR) и ген устойчивости к неомицину, причем гены GS или DHFR можно использовать для генетической амплификации вставленной последовательности VH и VL. Вектор также может содержать два разных сайта распознавания рекомбиназы, позволяющие осуществлять опосредованный рекомбиназой кассетный обмен (RMCE) последовательности, кодирующей антитело, вместо полной интеграции вектора. Описание RMCE приведено в ссылках (Langer et al 2002; Schlake and Bode 1994). Пригодные сайты распознавания рекомбинации хорошо известны в данной области техники и включают сайты FRT, lox и attP/attB. Предпочтительно, чтобы вектор интеграции представлял собой изотипкодирующий вектор, в котором константные области (предпочтительно, включая интроны) представлены еще до переноса кодирующей пары VH и VL из вектора скрининга (или константные области уже представлены в векторе скрининга, если скрининг проводится на полноразмерных антителах). Константные области, представленные в векторе, могут представлять собой или всю константную область тяжелой цепи (от CH1 до CH3 или до CH4), или константную область, кодирующую фрагмент Fc антитела (от CH2 до CH3 или до CH4). Константная область каппа или лямбда легкой цепи также может быть представлена еще до переноса. Выбор числа представленных константных областей, если это вообще имеет место, зависит от использованной системы скрининга и переноса. Константные области тяжелой цепи можно выбирать из изотипов IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgM, IgD и IgE. Предпочтительны изотипы IgG1, IgG2 и/или IgG3. Далее, вектор экспрессии для сайт-специфической интеграции нуклеиновой кислоты, кодирующей антитело против EGFR, содержит подходящие промоторы или эквивалентные последовательности, управляющие высоким уровнем экспрессии каждой из цепей VH и VL. Фиг.4 иллюстрирует один из возможных способов конструирования вектора экспрессии, хотя существует множество других способов.
Перенос выбранных кодирующих пар VH и VL из вектора скрининга можно произвести общепринятым способом расщепления рестрикционным ферментом и лигирования, чтобы каждая молекула вектора экспрессии содержала одну кодирующую пару VH и VL. Предпочтительно, чтобы перенос кодирующих пар VH и VL осуществлялся в индивидуальном порядке, однако по желанию их перенос можно осуществлять в общей массе. Если в вектор экспрессии переносят все отобранные кодирующие пары VH и VL, то получают коллекцию или библиотеку векторов экспрессии. По желанию можно также использовать альтернативные способы переноса. Если вектор скрининга идентичен вектору экспрессии, то библиотека векторов экспрессии составляется из пар последовательностей VH и VL, отобранных при скрининге, которые располагаются в векторе скрининга/экспрессии.
Способы трансфицирования последовательности нуклеиновой кислоты в клетку-хозяина известны в данной области техники. Для большей уверенности в сайт-специфической интеграции в клетке-хозяине также должна присутствовать подходящая рекомбиназа. В предпочтительном варианте это достигается посредством котрансфекции плазмиды, кодирующей рекомбиназу. Подходящими рекомбиназами являются, например, Flp, Cre или фаговая ЦC31 интеграза, применяемые вместе с системой клетка-хозяин/вектор, где имеются соответствующие сайты распознавания рекомбиназы. Клетка-хозяин также может быть трансфицирована в массе, а это означает, что библиотека векторов экспрессии трансфицируется в клеточную линию в ходе общей единичной реакции, благодаря чему удается получить поликлональную клеточную линию. В альтернативном варианте коллекцию векторов экспрессии можно трансфицировать в клетку-хозяина индивидуально, посредством чего можно генерировать коллекцию индивидуальных клеточных линий (каждая клеточная линия продуцирует антитело с особой специфичностью). Клеточные линии, генерированные посредством трансфекции (индивидуальные или поликлональные), затем подвергаются отбору на сайт-специфические интегранты и адаптируются к выращиванию в суспензии и бессывороточной среде, если перед трансфекцией они уже не обладали такими свойствами. Если трансфекция проводилась индивидуально, то индивидуальные клеточные линии далее анализируют на их ростовые свойства и на выработку антител. Предпочтительно, чтобы для создания поликлональной клеточной линии отбирались клеточные линии со сходными темпами пролиферации и сходным уровнем выработки антител. После этого генерируют поликлональную клеточную линию, смешивая индивидуальные клеточные линии в предопределенном соотношении. Обычно из поликлональной клеточной линии берут свое начало поликлональный маточный банк клеток (pMCB), поликлональный поисковый банк клеток (pRCB) и/или поликлональный рабочий банк клеток (pWCB). Поликлональную клеточную линию генерируют, смешивая индивидуальные клеточные линии в предопределенном соотношении. Поликлональную клеточную линию распределяют по ампулам, создавая таким образом поликлональный поисковый банк клеток (pRCB) или маточный банк клеток (pMCB), причем далее можно генерировать рабочий банк клеток (pWCB) посредством наращивания объемов клеток из поискового или маточного банка клеток. Поисковый банк клеток, прежде всего, предназначен для проведения исследований с целью подтверждения правильности концепции, в которых поликлональная клеточная линия может содержать не столько индивидуальных антител, как поликлональная клеточная линия в маточном банке клеток. Обычно pMCB дополнительно наращивают, чтобы заложить pWCB в целях масштабной выработки антител. При истощении pWCB можно нарастить клеточный материал из новой ампулы pMCB, чтобы заложить новый pWCB.
Один из вариантов осуществления настоящего изобретения заключается в создании поликлональной клеточной линии, способной экспрессировать композицию рекомбинантных антител против EGFR, предлагаемую настоящим изобретением.
Следующий вариант осуществления настоящего изобретения заключается в создании поликлональной клеточной линии, в которой каждая индивидуальная клетка способна экспрессировать единственную кодирующую пару VH и VL, причем поликлональная клеточная линия в целом способна экспрессировать коллекцию кодирующих пар VH и VL, где каждая пара VH и VL кодирует антитело против EGFR. Предпочтительно, чтобы коллекция кодирующих пар VH и VL представляла собой когнатные пары, полученные в соответствии со способами, предлагаемыми настоящим изобретением.
Композицию рекомбинантных антител, предлагаемую настоящим изобретением, можно получить, культивируя одну ампулу из pWCB в соответствующей питательной среде в течение такого промежутка времени, который позволяет осуществить достаточную экспрессию антител, причем поликлональная клеточная линия в это время остается стабильной (такой промежуток времени составляет приблизительно от 15 дней до 50 дней). Можно использовать такие способы культивирования как подпитка или перфузия. Композицию рекомбинантных антител получают из культуральной среды и очищают, применяя традиционные методики очистки. Для очистки IgG часто применяют аффинную хроматографию в сочетании с последующими этапами очистки, такими как ионообменная хроматография, гидрофобные взаимодействия и фильтрация в геле. После очистки оценивают присутствие всех индивидуальных членов в композиции поликлональных антител, например, посредством ионообменной хроматографии. Определение характеристик такой композиции антител подробно описано в документе WO 2006/007853 (включенном сюда в качестве ссылки).
Альтернативный способ экспрессирования смеси антител в рекомбинантном хозяине описан в документе WO 2004/009618. Этот способ позволяет продуцировать антитела с разными тяжелыми цепями, связанными с одной и той же легкой цепью из одной клеточной линии. Такой подход может быть применим при выработке композиции антител против EGFR из комбинаторной библиотеки.
Терапевтические композиции
Еще один из аспектов настоящего изобретения связан с фармацевтической композицией, содержащей в качестве активного ингредиента композицию антител против EGFR либо композицию рекомбинантного Fab или другого фрагмента рекомбинантного антитела против EGFR, либо биспецифическую связывающую молекулу, рассматриваемую в изобретении. Предпочтительно, чтобы активным ингредиентом такой композиции была композиция рекомбинантных антител против EGFR, как это описано в настоящем изобретении. Такая композиция предназначена для облегчения и/или профилактики, и/или для лечения рака. Предпочтительно, чтобы субъектом, которому вводится композиция, был человек, сельскохозяйственное животное или домашнее животное.
Фармацевтическая композиция содержит дополнительно фармацевтически приемлемый наполнитель.
Композицию антител против EGFR или фрагментов этих антител можно вводить в фармацевтически приемлемом разбавителе, носителе или наполнителе в стандартной форме дозирования. Для изготовления походящих лекарственных составов или композиций с целью их введения раковым пациентам можно действовать в соответствии с традиционной фармацевтической практикой. В предпочтительном варианте осуществления изобретения введение лекарства считается терапевтическим, если оно проводится после того как было диагностировано раковое заболевание. Можно использовать любой приемлемый способ введения лекарства, например, способ введения может быть парентеральным, внутривенным, внутриартериальным, подкожным, внутримышечным, внутрибрюшинным, интраназальным, пероральным, а также в виде аэрозолей или суппозиториев. Например, фармацевтические составы могут иметь форму жидких растворов или суспензий. При пероральном введении необходима защита лекарства от разрушения в желудке. При интраназальном введении лекарственных составов антитела можно использовать в виде порошков, назальных капель или аэрозолей.
В соответствии с настоящим изобретением фармацевтические композиции можно изготовлять способом, известным per se, например, посредством таких традиционных процессов как растворение, лиофилизация, смешение, гранулирование или конфекционирование. Фармацевтические композиции можно составлять в соответствии с общепринятой фармацевтической практикой (см., например, ссылки Remington: The Science and Practice of Pharmacy (20th ed.), ed. A.R. Gennaro, 2000, Lippincott Williams & Wilkins, Philadelphia, PA and Encyclopedia of Pharmaceutical Technology, eds. J. Swarbrick and J.C. Boylan, 1988-1999, Marcel Dekker, New York, NY).
Для приготовления фармацевтической композиции в соответствии с настоящим изобретением предпочтительно использовать растворы или суспензии активного ингредиента, особенно изотонические водные растворы или суспензии. В случае применения лиофилизированных композиций, которые содержат активный ингредиент в чистом виде или с носителем, например, маннитом, такие растворы или суспензии, если это возможно, приготавливают перед использованием. Фармацевтические композиции могут быть подвергнуты стерилизации и/или могут содержать наполнители, например, консерванты, стабилизаторы, увлажняющие и/или эмульгирующие агенты, солюбилизаторы, соли для регулирования осмотического давления и/или буферы, и могут быть изготовлены способом, известным per se, например, посредством применения общепринятых процессов растворения и/или лиофилизации. Указанные растворы или суспензии могут содержать вещества, усиливающие вязкость, например, натрий-карбоксиметилцеллюлозу, карбоксиметилцеллюлозу, декстран, поливинилпирролидон или желатин.
Инъекционные композиции готовят традиционным способом в стерильных условиях, то же самое относится и к расфасовке композиций в ампулы или флаконы, а также к герметизации контейнеров.
Содержание активного ингредиента в композиции составляет приблизительно от 1% до 95%, предпочтительно, приблизительно от 20% до 90%. В соответствии с настоящим изобретением фармацевтические композиции могут быть представлены в виде стандартных доз, например, в ампулах, флаконах, суппозиториях, таблетках, пилюлях или капсулах. Лекарственные составы могут вводиться человеку в терапевтически или профилактически эффективном количестве (например, в таком количестве, которое позволяет предупредить, элиминировать или облегчить патологическое состояние) для лечения/профилактики заболевания или патологического состояния. Предпочтительная дозировка вводимого терапевтического агента, по-видимому, зависит от таких переменных как тяжесть рака, общее состояние здоровья конкретного больного, наличие в композиции соединений-наполнителей и способ введения лекарства.
Терапевтическое применение композиций в соответствии с настоящим изобретением
В соответствии с настоящим изобретением предлагаемые фармацевтические композиции могут быть использованы для лечения или облегчения заболевания и млекопитающего. Патологические состояния, которые можно лечить или предупреждать предлагаемыми здесь фармацевтическими композициями, включают раковые заболевания и состояния. Предпочтительно подвергать пациентов терапевтическому лечению фармацевтическими композициями, предлагаемыми настоящим изобретением.
Один из вариантов осуществления настоящего изобретения заключается в способе профилактики, лечения или облегчения у млекопитающего одного или более симптомов, связанных с раком, причем указанный способ включает введение указанному млекопитающему эффективного количества композиции рекомбинантных антител против EGFR, предлагаемой настоящим изобретением.
Следующий вариант осуществления настоящего изобретения заключается в применении композиции рекомбинантных антител против EGFR, предлагаемой настоящим изобретением для приготовления фармацевтической композиции, используемой в целях лечения, облегчения или профилактики у млекопитающего одного или более симптомов, связанных с раком.
Предпочтительно, чтобы млекопитающим, упоминаемым в указанных выше вариантах осуществления изобретения, был человек, сельскохозяйственное или домашнее животное.
В соответствии с настоящим изобретением антитела показаны к применению для лечения некоторых солидных опухолей. Основываясь на ряде факторов, включая среди прочего уровень экспрессии EGFR, следует отметить, что предпочтительным показанием к применению предлагаемых настоящим изобретением композиций являются следующие типы опухолей: рак молочной железы, яичника, толстой кишки, прямой кишки, предстательной железы, мочевого пузыря, поджелудочной железы, головы и шеи, а также немелкоклеточный рак легкого. В связи с каждым из этих показаний четкий потенциал для клинического успеха открывают три клинических пути.
Адъюнктивная терапия: При адъюнктивной терапии пациенты получают лечение антителами, предлагаемыми настоящим изобретением, в сочетании с химиотерапевтическим или противоопухолевым средством и/или лучевой терапией. Целевые типы рака, перечисленные выше, будут подвергаться лечению по протоколу с добавлением антител, предлагаемых изобретением, к стандартной терапии первой и второй очереди. Схемы протоколов рассматривают оценку эффективности по снижению массы опухоли, а также по возможности снизить обычные дозы стандартных химиотерапевтических препаратов. Такое снижение доз создает возможности для дополнительной и/или более длительной терапии за счет снижения связанной с дозой токсичности химиотерапевтического средства. Ранее известные в данной области техники антитела против EGFR были использованы и продолжают находить применение в нескольких клинических испытаниях с адъюнктивной терапией, где они используются в комбинации с указанными ниже химиотерапевтическими или противоопухолевыми средствами: адриамицин (эрбитукс: запущенная карцинома предстательной железы), цисплатин (эрбитукс: запущенные карциномы головы/шеи и легких), таксол (эрбитукс: рак молочной железы), а также доксорубицин (эрбитукс).
Изобретение предлагает фармацевтические продукты, содержащие композицию антител, предлагаемую изобретением, а также, по меньшей мере, одно соединение, способное индуцировать дифференциацию раковых клеток, в виде комбинации для одновременного, раздельного или последовательного введения в схеме противораковой терапии. Комбинируя композиции антител, предлагаемые изобретением, со средствами, заведомо индуцирующими терминальную дифференциацию раковых клеток, можно добиться дополнительного улучшения лечебного эффекта.
По меньшей мере, одно соединение может быть выбрано из группы, состоящей из ретиноевой кислоты, транс-ретиноевых кислот, цис-ретиноевых кислот, фенилбутирата, фактора роста нервов, диметилсульфоксида, активной формы витамина D(3), рецептора активатора пролиферации пероксисом, 12-О- тетрадеканоилфорбол 13-ацетата, гексаметилен-бис-ацетамида, трансформирующего бета-фактора роста, бутировой кислоты, циклического АМФ и веснаринона. Предпочтительно, чтобы соединение было выбрано из группы, состоящей из ретиноевой кислоты, фенилбутирата, полностью транс-ретиноевой кислоты, активной формы витамина D.
Фармацевтические продукты, содержащие композицию антител, предлагаемую изобретением, и, по меньшей мере, одно химиотерапевтическое или противоопухолевое соединение, могут быть использованы в виде комбинации для одновременного, раздельного или последовательного введения при лечении рака. Химиотерапевтическое соединение может быть выбрано из группы, состоящий из адриамицина, цисплатина, таксола, доксорубицина, топотекана, фторпиримидина, оксалиплатина и иринотекана.
Монотерапия: Антитела, предлагаемые настоящим изобретением можно использовать для монотерапии опухолей, то есть вводить их пациентам без химиотерапевтического или противоопухолевого средства. Доклинические результаты, полученные в результате применения антител в соответствии с настоящим изобретением и обсуждаемые здесь, продемонстрировали положительные эффекты при автономной терапии указанными антителами.
Радиофармацевтическое средство: В результате связывания радионуклида (например, иттрия (90Y)) с антителами, предлагаемыми настоящим изобретением, можно ожидать, что рассматриваемые здесь антитела при наличии радиоактивной метки могут быть использованы как диагностическое радиофармацевтическое средство. В такой роли рассматриваемые здесь антитела способны локализовать как солидные опухоли, так и метастатические повреждения, клетки которых экспрессируют EGFR. В связи с применением рассматриваемых здесь антител в качестве радиофармацевтических средств указанные антитела можно использовать при вспомогательном хирургическом лечении солидных опухолей как для дооперационного скрининга, так и для послеоперационного контроля, то есть для определения того, какая опухоль частично осталась и/или рецидивировала. Антитело (111In)-эрбитукс было использовано как радиофармацевтическое средство в клиническом испытании I фазы на пациентах с неоперабельным плоскоклеточным раком легких (Divgi et al. J. Natl. Cancer Inst. 83:97-104 (1991). Пациентов обследовали при помощи стандартной передней и задней гамма-камеры. Предварительные данные показали, что были идентифицированы все первичные и крупные метастатические очаги опухолей, тогда как мелкие метастазы (менее 1 см) были выявлены только в половине случаев.
Ингибиторы тирозинкиназы (TKI) представляют собой синтетические молекулы с низким молекулярным весом, главным образом, производные хиназолина, которые взаимодействуют с внутриклеточным тирозинкиназным доменом рецепторов и ингибируют индуцированное лигандом фосфорилирование рецепторов за счет конкуренции за внутриклеточный сайт связывания Mg-АТР. Некоторые TKI в клиническом аспекте, включая гефитиниб (Iressa, ZD1839), эрлобтиниб (Tarceva, OSI-774), лапатиниб, (Tykerb, GW572016), канертиниб (CI-1033), ЕКВ-569 и PKI-166, нацелены на EGFR. Комбинированное лечебное применение TKI и антител против EGFR продемонстрировало полезные эффекты как in vivo, так и in vitro, направленные против EGFR-зависимых раковых клеток. Фармацевтические продукты, содержащие композицию антител, предлагаемую изобретением, и, по меньшей мере, один TKI, нацеленный на EGFR, могут быть использованы в виде комбинации для одновременного, раздельного или последовательного введения при лечении рака. Кроме того, мелкомолекулярные ингибиторы включают:
сорафениб (raf и множественные RTK), сунитиниб (множественные RTK), темсиролимус (mTOR), RAD001 (mTOR) и AZD217 (VEGFR2).
В других вариантах осуществления изобретения предлагаемые композиции антител применяются в комбинации с другими терапевтическими антителами. Примеры таких антител включают, например, антитела против HER2 (герцептин) и VEGF (авастин). Кроме того, в других вариантах осуществления изобретения предлагаемые композиции антител применяются в комбинации со средством, заведомо стимулирующим клетки иммунной системы, причем такое комбинированное лечение приводит к иммуноопосредованному повышению эффективности предлагаемых изобретением композиций антител. Примеры таких иммуностимулирующих средств включают, но не ограничиваясь ими, рекомбинантные интерлейкины (например, IL-21 и IL-2).
Доза и способ введения
Хотя специфические дозировки антител в соответствии с настоящим изобретением еще не были определены, некоторые соображения по этом поводу можно провести через сравнение со сходным продуктом (ImClone C225 (эрбитукс)), который был разрешен к официальному применению. Антитело C225 типично вводят в диапазоне доз от 5 до 400 мг/м2, причем низшие дозы применяются только в связи с исследованиями по безопасности. В соответствии с этим мы можем ожидать, что дозировка антител, предлагаемых настоящим изобретением, приблизительно соответствует указанной или несколько ниже нее, возможно находясь в диапазоне от 50 до 300 мг/м2, сохраняя при этом эффективность. Дозировка в мг/м2, в отличие от традиционного представления доз в мг/кг, является измерением, основанным на площади поверхности и удобна тем, что ее можно применять для пациентов всех размеров от младенцев до взрослых.
Доступная информация по предписаниям для препарата эрбитукс (Cetuximab) включает первоначальное внутривенное вливание в течение 120 минут в дозе 400 мг/м2, сопровождающееся последующими еженедельными 60-минутными вливаниями в дозе 250 мг/м2. Эти дозировки рекомендованы как для монотерапии, так для комбинированного лечения в сочетании с лучевой терапией. Для препарата вектибикс (panitumumab) рекомендуемая доза составляет 6 мг/кг при 60-минутном введении каждые 14 дней.
Предполагаемая клиническая дозировка антитела HuMaxEGFr (zumutumumab) производства компании Genmab составляет начальную дозу 8 мг/кг HuMax-EGFr, с последующими еженедельными вливаниями поддерживающей дозы до тех пор, пока заболевание не начнет прогрессировать. Поддерживающую дозу подбирают как необходимую, увеличивая ее до тех пор, пока у больного не появится кожная сыпь, но не более 16 мг/кг HuMax-EGFr (дозировки в базовом исследовании III фазы, доступные из описания продукта фирмы Genmab).
Клиническая дозировка композиций антител, предлагаемых настоящим изобретением, по-видимому также будет ограничиваться степенью кожных высыпаний, как это наблюдается при современном использовании в клинической практике моноклональных антител против EGFR (эрбитукс и вектибикс). Данные, полученные в шестинедельном токсикологическом исследовании на обезьянах Cynomolgus, не показали признаков кожных высыпаний, если композицию антител, предлагаемых изобретением, вводили животным в дозе, эквивалентной дозировке, применяемой при лечении одним из моноклональных антител, используемых в клинике (пример 20). Таким образом, предлагаемые изобретением композиции антител можно вводить внутривенно в еженедельной дозе 250 мг/м2, что соответствует 7,5 мг/кг для человека с поверхностью тела 1,8 м2 и массой тела 60 кг. Кроме того, до введения последующих еженедельных доз можно дать начальную загрузочную дозу 400 мг/м2 (соответствует 12 мг/кг для человека с поверхностью тела 1,8 м2 и массой тела 60 кг).
Предполагается, что для доставки в организм субъекта антител, предлагаемых изобретением, могут быть полезными три разных способа. Традиционная внутривенная доставка, предположительно, будет стандартным способом доставки лекарств для лечения большинства опухолей. Однако применительно к опухолям, расположенным в брюшной полости, например, опухолям яичников, желчного протока, других протоков и т.п., предпочтительным может оказаться внутрибрюшинное введение, позволяющее получить высокие дозы антител в опухоли и свести к минимуму клиренс антитела. Примерно так же, некоторые солидные опухоли обладают сосудистой сетью, что создает преимущества при региональном введении. Региональная перфузия позволяет получить высокую дозу антитела в месте расположения опухоли и свести к минимуму краткосрочный клиренс антитела.
Как и при любой терапии, основанной на вливаниях белков или антител, проблемы безопасности связаны, прежде всего, (i) с синдромом выброса цитокинов, то есть падением давления, лихорадкой, тремором, ознобом, (ii) с развитием иммуногенной реакции на вводимый материал (то есть развитием у больного антител человека против лечебного антитела, либо развитием реакций HAHA или HACA), и (iii) с токсичностью по отношению к нормальным клеткам, экспрессирующим рецептор EGF, например, гепатоцитам, которые экспрессируют EGFR. Для отслеживания каждой из этих проблем безопасности можно использовать стандартные тесты и контрольное наблюдение. Особенно большое внимание при клинических испытаниях надо уделять частым проверкам функции печени, чтобы можно было оценить повреждение печени, если таковое происходит.
Диагностическое применение
Еще один вариант осуществления изобретения направлен на диагностические наборы. Применительно к настоящему изобретению такие наборы включают композицию антител против EGFR, изготовленную в соответствии с изобретением, в которой белок может быть снабжен поддающейся выявлению меткой или не иметь метки для выявления в немеченом состоянии. Набор может быть использован для идентификации индивидов, страдающих раком с избыточной экспрессией EGFR.
ПРИМЕРЫ
ПРИМЕР 1: Клонирование антител против EGFR
Иммунизации
Самки мышей BALB/c, линии A или C57B16 (в возрасте 8-10 недель) были использованы для иммунизации посредством инъекций различных очищенных белков в дополнение к клеткам, избыточно экспрессирующим EGFR.
В некоторых случаях для иммунизации были использованы имеющиеся в продаже белки EGFR (R&D systems № по каталогу 1095-ER или Sigma № по каталогу E3641). В других случаях иммунизации были использованы рекомбинантные EGFR и EGFRvIII человека, полученные как гибридные белки, состоящие из ECD EGFR или EGFRvIII и гормона роста человека (hGH), а также включающие сайт расщепления вируса гравировки табака (TEV) в дополнение к His-tag, как это описано в примере 10b. В некоторых случаях ECD EGFR выделяли посредством расщепления TEV-протеазой с последующей очисткой в никелевой колонке.
Для клеточной иммунизации была использована линия клеток рака головы и шеи человека HN5 (Easty DM, Easty GC, Carter RL, Monaghan P, Butler LJ. Br J Cancer. 1981 Jun; 43(6):772-85. Ten human carcinoma cell lines derived from squamous carcinomas of the head and neck.), экспрессирующая приблизительно 107 рецепторов на клетку. Клетки были культивированы в среде DMEM с добавками 10% FBS (эмбриональной телячьей сыворотки), 3мМ глицерина, 5 мМ пирувата натрия и 1% пенициллина/стрептомицина. Перед каждой иммунизацией клетки отмывали в PBS, трипсинизировали ферментом TrypLE и ресуспендировали в питательной среде для выращивания. Затем клеточные суспензии дважды отмывали в PBS при центрифугировании на скорости 250×g в течение 5 минут с выбиванием клеток и ресуспендированием в 15 мл стерильного PBS.
Клетки или антиген разбавляли в PBS, после чего смешивали в соотношении 1:1 с адъювантом Фрейнда. Адъювант применяют для усиления и модуляции иммунного ответа. Для первых иммунизаций использовали полный адъювант Фрейнда (CFA), а для последующих иммунизаций использовали неполный адъювант Фрейнда (IFA). IFA представляет собой эмульсию типа масло-в-воде, состоящую из минеральных масел, а CFA представляет собой IFA, К которому добавлены термически убитые и высушенные микобактерии. Оба адъюванта обладают депонирующим эффектом. CFA дает начало длительной персистенции иммунного ответа, и его применяют при первых иммунизациях для стимуляции иммунного ответа, а IFA применяют для последующих иммунизаций. Проверку эмульсий осуществляли, нанося каплю эмульсии на поверхность стекла с водой. Если капля остается каплей, то эмульсия стабильна, и ее можно вводить посредством инъекций. Мышам вводили только стабильные эмульсии.
В зависимости от плана (см. таблицу 2) для каждой инъекции использовали 25-100 мкг антигена или 107 клеток. В общей сложности мыши получали 4 инъекции. Всем мышам впрыскивали или 300 мкл, или 200 мкл эмульсии. В зависимости от плана инъекции проводили подкожно (s.c.), внутрибрюшинно (i.p.) или внутривенно (i.v.).
В конце эксперимента мышей умерщвляли, сворачивая им шею, после чего извлекали селезенки и переносили их на клеточное сито с размером ячеек 74 мкм (Corning#136350-3479). Клетки мацерировали через фильтр, ресуспендировали в холодной среде RPMI 1640 с 10% FBS и центрифугировали на скорости 300×g в течение 5 минут. Клеточный осадок ресуспендировали в среде RPMI 1640 с 1% FBS, фильтровали через шприцевой фильтр с размером ячеек 50 мкм (BD# 340603) и накапливали центрифугированием. Клеточный осадок криоконсервировали после ресуспендирования в FCS с 10% DMSO, а замороженные клетки хранили при температуре -80°C вплоть до сортировки по методу FACS.
Сортировка мышиных плазматических клеток методом FACS
Флаконы с замороженными спленоцитами оттаивали при 37°C и переносили в 15 мл пробирку со льдом. В пробирку при постоянном покручивании добавляли по каплям 10 мл охлажденной на льду среды RPMI, 10% FBS (эмбриональной телячьей сыворотки). После однократного отмывания в 10 мл FACS PBS добавляли 5 мл FCS PBS перед фильтрованием клеток через Filcon с размером ячеек 50 мкм. Затем клетки осаждали центрифугированием, ресуспендировали в 1 мл PBS с 2% FBS (конечный объем) и окрашивали против CD43-FITC и против CD138-PE в соответствии со специфичным разбавлением до конечной концентрации приблизительно 5 мкг/мл. Клетки инкубировали при 4°C в течение 20 минут в темноте. Затем клетки дважды отмывали в 2 мл буфера FACS. Добавляли до 15 мл FACS PBS. Иодид пропидия (PI) добавляли в соотношении 1:100 (1 часть PI на 100 частей буфера FACS PBS), после чего клетки сортировали в 96-луночных планшетах PCR, содержащих реакционный буфер PCR (см. ниже) и центрифугировали в течение 2 минут на скорости 400×g перед замораживанием планшетов при температуре -80°C. Плазматические клетки сортировали как CD43-позитивные/CD-138-позитивные как показано на фиг.1.
Сцепление когнатных пар VH и VL
Сцепление кодирующих последовательностей VH и VL проводили на одиночных клетках, отсортированных как плазмациты, что облегчало когнатное спаривание кодирующих последовательностей VH и VL. При этом была использована двухэтапная процедура PCR, основанная на одноэтапной мультиплексной RT-PCR с перекрыванием-удлинением с последующим этапом вложенной PCR. Смеси праймеров, использованные в настоящем примере, амплифицируют только легкие каппа-цепи. Однако по желанию к мультиплексной смеси праймеров и к смеси праймеров вложенной PCR можно добавить праймеры, способные амплифицировать легкие лямбда-цепи. Если добавляют праймеры лямбда, то процедуру сортировки следует адаптировать так, чтобы не исключались лямбда-позитивные клетки. Принцип сцепления когнатных последовательностей VH и VL проиллюстрирован на фиг.2.
Полученные 96-луночные планшеты для PCR оттаивали, а отсортированные клетки служили матрицей для мультиплексной RT-PCR с перекрыванием-удлинением. Сортировочный буфер добавляли в каждую лунку перед реакционным буфером для сортировки одиночных клеток (OneStep RT-PCR Buffer; Qiagen), праймерами для RT-PCR (см. таблицу 3) и ингибитором РНК-азы (RNasin, Promega). Это дополняли смесью ферментов для одноэтапной RT-PCR (25-кратное разведение, Qiagen) и смесью dNTP (по 200 мкМ каждой) для получения заданной конечной концентрации в реакционном объеме 20 мкл. Планшеты инкубировали в течение 30 минут при 55°C для получения обратной транскрипции РНК из каждой клетки. После проведения RT планшеты проводили через следующий цикл PCR: 10 минут при 94°C, 35×(40 секунд при 94°C, 40 секунд при 60°C, 5 минут при 72°C), 10 минут при 72°C.
Реакции PCR проводили в термоциклере H20BIT с корзинкой со съемным уплотнением для 24 96-луночных планшетов (ABgene) для повышения пропускной способности. После циклической обработки планшеты для PCR хранили при -20°C.
Для проведения этапа вложенной PCR 96-луночные планшеты для PCR готовили со следующей смесью в каждой лунке (реакции на 20 мкл), чтобы получить заданную конечную концентрацию: 1× буфера FastStart (Roche), смесь dNTP (200 мкМ каждого компонента), смесь вложенных праймеров (см. таблицу 4), ДНК-полимераза Phusion (0,08 U, Finnzymes) и высококачественная смесь ферментов FastStart (0,8 U, Roche). В качестве матрицы для вложенной PCR, переносили 1 мкл материала из реакций мультиплексной PCR с перекрыванием-удлинением. Планшеты для вложенной PCR проводили через следующие термоциклы: 35×(30 секунд при 95°C, 30 секунд при 60°C, 90 секунд при 72°C), 10 минут при 72°C.
Для верификации присутствия фрагментов перекрывания-удлинения длиной приблизительно 890 пар оснований (п.о.) случайно отобранные реакции были проанализированы на 1% агарозном геле.
Планшеты хранили при -20°C до следующего процессинга фрагментов PCR.
Репертуар сцепленных кодирующих пар VH и VL из вложенной PCR накапливали, на смешивая пары от разных доноров, и очищали электрофорезом в препаративном 1% агарозном геле. Кодирующая последовательность константной легкой каппа-цепи человека была сплайсирована посредством перекрывающегося удлинения в кодирующую область VL объединенных продуктов PCR сцепленных кодирующих пар VH и VL (фиг.3). Кодирующая последовательность константной легкой каппа-цепи человека была амплифицирована из плазмиды, содержащей кодирующую последовательность антитела человека с легкой каппа-цепью, в реакции, содержащей связующий фермент (2 U, Finnzymes), 1× связующего буфера, смесь dNTP (200 мкМ каждого компонента), праймер hKCforw-v2 и праймер Kappa3' (таблица 5), а также плазмидную матрицу pLL138 (10 нг/мкл) в общем объеме 50 мкл. Реакцию проводили через следующие термоциклы: 25×(30 секунд при 95°C, 30 секунд при 55°C, 45 секунд при 72°C), 10 минут при 72°C. Полученный в результате PCR фрагмент очищали электрофорезом в препаративном 1% агарозном геле.
Очищенные объединенные фрагменты PCR из каждого репертуара сплайсировали в амплифицированный и очищенный фрагмент PCR константной кодирующей каппа-области человека (приложение 2) при последующем сплайсинге через PCR с перекрыванием-удлинением (общий объем 50 мкл), содержащую фрагмент кодирующей константной каппа-области человека (1,4 нг/мкл), очищенный накопленный фрагмент PCR (1,4 нг/мкл), связующую ДНК-полимеразу (0,5 U, Finnzymes), высококачественную смесь ферментов FastStart (0,2 U, Roche), 1× буфера FastStart (Roche), смесь dNTP (200 мкМ каждого компонента), праймер mhKCrev и набор праймеров mJH (см. таблицу 5). Реакцию проводили через следующие термоциклы: 2 минуты при 95°C, 25×(30 секунд при 95°C, 30 секунд при 55°C, 1 минута при 72°C), 10 минут при 72°C. Полученный в результате PCR фрагмент (длиной приблизительно 1070 п.о.) очищали электрофорезом в препаративном 1% агарозном геле.
Вставление когнатных кодирующих пар VH и VL в вектор скрининга
Для того чтобы идентифицировать антитела, обладающие специфичностью связывания с EGFR, полученные кодирующие последовательности VH и VL были экспрессированы в виде полноразмерных антител. Это включало вставление репертуара кодирующих пар VH и VL в вектор экспрессии и трансфекцию в клетку-хозяина.
Для генерирования репертуара векторов экспрессии, содержащих сцепленные кодирующие пары VH и VL, была использована двухэтапная процедура клонирования. Со статистической точки зрения, если репертуар векторов экспрессии содержит в десять раз больше рекомбинантных плазмид, чем число когнатных спаренных VH и VL, как продуктов PCR, использованных для генерирования репертуара скрининга, то существует 99% вероятность того, что представлены все уникальные пары генов. Таким образом, если было получено 400 фрагментов V-гена с перекрыванием-удлинением, то репертуар, генерированный для скрининга, составит, по меньшей мере 4000 клонов.
Вкратце, очищенный продукт PCR из репертуаров сцепленных кодирующих пар VH и VL, сплайсированный в кодирующую константную каппа-область человека, был расщеплен ДНК-эндонуклеазами XhoI и NotI в сайтах распознавания, введенных в концы продуктов PCR. Расщепленные и очищенные фрагменты были лигированы в вектор экспрессии IgG млекопитающих, переваренный XhoI/NotI, OO-VP-002 (фиг.4) посредством стандартных процедур лигирования. Лигирующая смесь была электропорирована в E. coli, добавлена в 2×YT планшеты, содержащие подходящий антибиотик, и инкубирована при 37°C в течение ночи. Амплифицированный репертуар векторов был очищен из клеток, восстановленных из планшетов с применением стандартных способов очистки ДНК (Qiagen). Плазмиды были подготовлены для вставления фрагментов промотор-лидер посредством расщепления эндонуклеазами AscI и NheI. Сайты рестрикции для этих ферментов были расположены между парами генов, кодирующих VH и VL. После очистки вектора двунаправленный фрагмент промотор-лидер млекопитающих, переваренный AscI-NheI, был вставлен в сайты рестрикции AscI и NheI посредством стандартных процедур лигирования. Лигированный вектор был амплифицирован в E. coli, а плазмида была очищена стандартными способами. Генерированный репертуар векторов скрининга был трансформирован в E. coli посредством общепринятых процедур. Полученные колонии были объединены в 384-луночные мастер-планшеты и заложены на хранение. Число выстроенных колоний превышало число входных продуктов PCR, по меньшей мере, в 3 раза, что давало 95%-ную вероятность присутствия всех полученных уникальных пар V-генов.
Скрининг на связывание с внеклеточным доменом EGFR
В целом, скрининг был проведен как двухэтапная процедура. Библиотеки антител были скринированы на реактивность по отношению к рекомбинантному белку EGFR по методике ELISA, после чего способ FMAT (FLISA) был использован как подход на клеточной основе с клеточной линией NR6wtEGFR для выявления антител против EGFR, связывающихся с EGFR, который экспрессируется на клеточной поверхности. Для библиотек 101 и 108/109 (таблица 2) анализ по методике ELISA был проведен с рекомбинантным EGFR, представляющим внеклеточный домен EGFR.
Вкратце, для проведения ELISA, планшеты Nunc maxisorb (№ по каталогу 464718) были покрыты 1 мкг/мл белка (собственного производства), разбавленного в PBS и выдержанного при 4°C в течение ночи. Перед блокированием в 50 мкл 2%-Milk-PBS-T планшеты были однократно отмыты смесью PBS+0,05% Tween 20 (PBS-T). Затем были добавлены 20 мкл 2%-milk-PBS-T и 5 мкл супернатантов из трансфектантов FreeStyle CHO-S (см. ниже), проведена инкубация материала в течение 1Ѕ часа при комнатной температуре, после чего планшеты были однократно отмыты PBS-T из расчета 20 мкл на лунку. Для выявления антител, связанных с лунками, было добавлено вторичное антитело (HRP-Goat-anti-human IgG, Jackson, № по каталогу 109-035-097), разбавленное в соотношении 1:10000 в 2% milk-PBS-T с последующей инкубацией в течение 1 часа при комнатной температуре. Планшеты были однократно отмыты в PBS-T перед добавлением 25 мкл субстрата (Kem-en-tec Diagnostics, № по каталогу 4390) и инкубацией материала в течение 5 минут. После инкубации добавляли 25 мкл 1M серной кислоты для остановки реакции. Специфический сигнал выявляли считывающим устройством ELISA на длине волны 450 нм.
Для выявления антител против EGFR способом FMAT на клеточной основе клетки SKBR-3 (ATCC #HTB-30) или NR6wtEGFR (Welsh et al., 1991, J Cell Biol, 114, 3, 533-543) хранили в питательной среде для выращивания, как это описано в ссылке. Клетки подсчитывали и разбавляли до концентрации 125000 клеток на миллилитр антителом Alexa-647-конъюгированный козий IgG против человека (H-L) (молекулярный зонд № A21445, партия № 34686A) при разведении 1:40000. Общее количество 20 мкл этой суспензии переносили в 384-луночные планшеты Nunc с прозрачным дном. Впоследствии к клеткам добавляли 10 мкл трансфекционного супернатанта. Сигнал FMAT из реакции измеряли после 6-10 часов инкубации.
Данные скрининга указывают на то, что 221 клон (4,8%) от общего числа клонов оказался позитивным в ELISA. 93 (2,0%) этих клонов также были позитивными в анализе FMAT. В целом, 220 (4,8%) клонов были позитивными в анализе FMAT, а среди них 127 (220-93) уникально позитивными на антиген клеточной поверхности. Библиотека 111 была скринирована сходным образом, но поскольку процедура иммунизации была направлена на выработку антител, специфичных по отношению к делеционному мутанту рецептора EGF (EGFRvIII), скрининговые тесты ELISA включали анализы для выявления как EGFR дикого типа, так и EGFRvIII. Семь клонов были идентифицированы как специфичные на EGFRvIII в анализах ELISA, причем интересно, что именно эти клоны были негативными в отношении окрашивания клеток, экспрессирующих wtEGFR по результатам FMAT. 13 клонов в анализах FMAT и ELISA были идентифицированы как позитивные на wtEGFR, но не на EGFRvIII, что оказалось уникальным для этой библиотеки по сравнению с библиотеками 101 и 108/109. Все позитивные клоны по результатам ELISA были отобраны для дальнейшего анализа.
Анализ последовательности и отбор клона
Клоны, идентифицированные как EGFR-специфичные по результатам ELISA, были извлечены из исходных мастер-планшетов (384-луночного формата) и объединены в новых планшетах. Из клонов была выделена ДНК, которую подвергли секвенированию на V-гены. Была проведена выверка последовательностей и отбор всех уникальных клонов. Множественные выверки полученных последовательностей отразили уникальность каждого конкретного клона и позволили провести идентификацию уникальных антител. В результате анализа 220 клонов были идентифицированы 70 генетически различных кластеров последовательностей антител. Каждый кластер родственных последовательностей, по-видимому, был получен через соматические гипермутации общего клона предшественника. В общем, из каждого кластера были выбраны один или два клона для проверки последовательности и специфичности. Вариабельные последовательности отобранных антител показаны в приложении 1. Нуклеотидные последовательности включают сайты рестрикции на обоих концах. Следовательно, соответствующие транслированные аминокислотные последовательности (с применением третьей рамки считывания последовательности ДНК) включают на N-конце две аминокислоты, которые не образуют части последовательностей VH и VL по определению IMGT (Lefranc et al (2003) IMGT unique numbering for immunoglobulin and T cell receptor variable domains and Ig superfamily V-like domains. Dev. Comp Immunol 27, 55-77). Все показанные последовательности VL включают одну и ту же константную каппа-область человека, которая начинается с аминокислот -TVAAP- и заканчивается на C-конце фрагментом -NRGEC. В целях настоящего изобретения термин «последовательность VL» применительно к специфическому антителу исключает константную каппа-область и две N-концевые аминокислоты (LA-). Термин последовательность применительно к специфическому антителу исключает две N-концевые аминокислоты (RA-).
Проверка достоверности последовательности и специфичности
Для того чтобы проверить достоверность клонов, кодирующих антитела, была приготовлена плазмида ДНК и проведена трансфекция клеток FreeStyle CHO-S (Invitrogen) по шкале 2 мл для экспрессии. Через 96 часов после трансфекции собирали супернатант. Уровень экспрессии оценивали в стандартном анализе ELISA против IgG, а специфичность определяли посредством EGFR-специфичного и EGFRvIII-специфичного анализа ELISA. Было продемонстрировано, что правильную специфичность и последовательность имеют 85% клонов.
Скрининг на антипролиферативные эффекты
Клеточное повреждение неизбежно приводит к утрате способности клетки поддерживать и обеспечивать энергию для метаболической функции и роста клетки. На этой предпосылке основаны анализы метаболической активности. Обычно они направлены на измерение митохондриальной активности. Реактив клеточной пролиферации WST-1 (Roche, № по каталогу 11 644 807 001) представляет собой готовый к употреблению субстрат, который измеряет метаболическую активность жизнеспособных клеток. Далее допускается, что метаболическая активность коррелируется с количеством жизнеспособных клеток. В этом примере анализ с WST-1 был использован для измерения количества метаболически активных клеток после обработки супернатантами клеточных культур, содержащими различные антитела против EGFR.
Перед проведением анализа с WST-1 разные объемы 2-мл супернатантов (0, 10, 25, 50 и 150 мкл) переносили в соответствующие лунки 96-луночного планшета.
Затем клетки HN5 отмывали 1×PBS и разделяли трипсинизацией с применением 3 мл раствора трипсина. Затем добавляли 17 мл полной среды, и клетки центрифугировали на скорости 300×g (1200 rcf) в течение 5 минут. Супернатант удаляли, а клетки ресуспендировали в смеси DMEM+0,5% FBS. Клетки подсчитывали, их концентрацию корректировали, и в лунки с супернатантами добавляли 1500 клеток, так, что каждая лунка содержала в общей сложности 200 мкл среды. Планшеты инкубировали 4 дня в увлажненном инкубаторе при 37°C. Затем в каждую лунку добавляли 20 мкл реактива WST-1, и планшеты инкубировали в течение 1 часа при 37°C. Далее планшеты переносили в орбитальный планшетный шейкер и оставляли еще на один час. Поглощательную способность измеряли на уровне 450 и 620 нм (опорная длина волны) считывающим устройством ELISA. Различие в уровне метаболически активных клеток (MAC) вычисляли как процентный показатель по сравнению с контрольными супернатантами, исходя из следующей формулы:

Затем эти величины были использованы как основа для контролируемого иерархического кластерного анализа (кластеризация основана на реактивности в ELISA), проведенного с применением бесплатного программного обеспечения Cluster и TreeView.
Предпочтительно иметь возможности для скрининга на функциональные антитела на раннем этапе процесса отбора антител. Супернатанты культур из 83 2-мл трансфекций были использованы для скрининга на функции подавления роста в анализе пролиферации, проведенном с применением клеток HN5 в 0,5% FBS. Результаты были визуализированы посредством простого иерархического кластерного анализа. Как можно увидеть из кластерного анализа (фиг.5), было обнаружено, что многие супернатанты снижают количество метаболически активных клеток HN5 (темно-серый цвет) с эффектом зависимости от концентрации
(кластер 2). Сходным образом, некоторые супернатанты увеличивают количество метаболически активных клеток HN5 (светло-серый цвет) с эффектом зависимости от концентрации (кластеры 1, 3 и 4). Интересное наблюдение заключалось в том, что те супернатанты, которые снижали количество метаболически активных клеток HN5, имели реактивность 2 (черные стрелки), тогда как супернатанты, увеличивающие количество метаболически активных клеток HN5, имели реактивность 1 (серые стрелки). Супернатанты с реактивностью 2 были позитивными в анализах ELISA как на wtEGFR, так и на EGFRvIII, тогда как супернатанты с реактивностью 1, были реактивными только в отношении wtEGFR. Таким образом, указанные анализы могут установить связь между реактивностью антител в ELISA и их функциональностью в клеточных анализах.
Репарация клонов
При использовании мультиплексного подхода PCR ожидается некоторая степень перекрестного примирования во внутри- и меж-V-генном семействе вследствие вырожденности праймеров и высокой степени гомологии. Перекрестное примирование внедряет в каркас иммуноглобулина аминокислоты, не встречающиеся в норме, что влечет за собой некоторые потенциальные последствия, например, структурные изменения и увеличение иммуногенности, причем все они приводят к снижению терапевтической активности.
Для того чтобы устранить эти препятствия и быть уверенным в том, что отобранные клоны воспроизводят естественный гуморальный иммунный ответ, такие перекрестно-примирующие мутации были скорректированы в процессе, называемом репарацией клонов.
На первом этапе процедуры репарации клонов последовательность VH была амплифицирована в PCR с набором праймеров, содержащим последовательность, соответствующую последовательности гена VH из представляющего интерес родоначального клона, благодаря чему можно было скорректировать любые мутации, внесенные перекрестным примированием. Фрагмент, полученный в PCR, подвергали перевариванию ферментами XhoI и AscI с последующим обратным лигированием в вектор экспрессии млекопитающих, переваренный ферментами XhoI/AscI (фиг.4) с применением общепринятых приемов лигирования. Лигированный вектор был амплифицирован в E. coli, а плазмида была очищена стандартными способами. Последовательность VH была секвенирована для верификации исправлений, а вектор был подвергнут перевариванию ферментами NheI/NotI для его подготовки к вставлению легкой цепи.
На втором этапе полноразмерная легкая цепь была амплифицирована в PCR с набором праймеров, содержащим последовательность, соответствующую последовательности гена VL из представляющего интерес родоначального клона, благодаря чему можно было скорректировать любые мутации, внесенные перекрестным примированием. Полученный в PCR фрагмент был переварен ферментами NheI/NotI и лигирован в приготовленный ранее вектор, содержащий VH. Продукт лигирования был амплифицирован в E. coli, а плазмида была очищена стандартными способами. Впоследствии легкая цепь была секвенирована для верификации исправлений.
В том случае, если константная каппа-область выбранного клона содержит мутации, внесенные во время амплификации генов, его замещают константной областью без мутаций. Это осуществляют в PCR с перекрыванием, где отремонтированный ген VL (амплифицированный без константной области) гибридизируют с константной областью, имеющей правильную последовательность (его получают в отдельной PCR). Всю последовательность амплифицируют и клонируют в вектор, содержащий VH, как это описано выше, после чего исправленную легкую цепь секвенируют для верификации исправлений.
ПРИМЕР 2: Выработка антител против EGFR млекопитающих
Для кратковременной экспрессии антител против EGFR была использована система экспрессии FreeStyle MAX CHO (Invitrogen). Антитела были экспрессированы в объеме 200-2000 мл.
Приблизительно за 24 часа до трансфекции проводили пересев клеток CHO-S, чтобы довести их концентрацию до 0,5×106 клеток на миллилитр. Плазмиду (1,25 мкг на миллилитр среды для культивирования клеток) разбавляли бессывороточной средой OptiPro и смешивали c раствором реактива для трансфекции FreeStyle MAX в соответствии с рекомендациями поставщика. Реактивы для трансфекции переносили в клеточную культуру, а через 6 дней после этого собирали супернатант.
Экспрессированные антитела очищали из супернатанта культуры с применением этапа аффинной хроматографии в колонке Protein A-Sepharose (MabSelect Sure, GE Health Care) для очистки молекул IgG1. Антитела были элюированы из колонки с применением 0,1M глицина 2,7. Фракции, содержащие антитела, определенные при измерении поглощательной способности на уровне 280 нм, были объединены и подвергнуты диализу 5 мМ ацетатом натрия, 150 мМ NaCl, pH 5. Очищенные образцы антител были протестированы на наличие эндотоксина по аналитической методике LAL.
ПРИМЕР 3: Определение специфичностей эпитопа
Конкурентный анализ ELISA с эталонными антителами
На основе применения эталонных антител, связывающихся с известными доменами EGFR, как это описано в ссылке (J.R. Cochran et. al., JIM 2004: 287; 147-158), был разработан конкурентный анализ ELISA, при помощи которого можно было провести различие между связывающими эпитопами антител против EGFR при инкубации с вторичным реактивом, который был специфичен в отношении области Fc антител против EGFR человека и не проявлял перекрестной реактивности с мышиными или крысиными Fc IgG. Методика ELISA была адаптирована из описаний, опубликованных в ссылке Ditzel et al., 1995, The Journal of Immunology, Vol 154, Issue 2 893-906.
Анализ ELISA с блокированием эпитопа проводили посредством разбавления антигена полноразмерного EGFR до концентрации 0,5 мкг/мл в PBS и покрытия из расчета 50 мкл на лунку ELISA в течение ночи при 4°C. На следующее утро лунки дважды отмывали PBS-T и блокировали в течение одного часа смесью PBS-T-1% BSA при комнатной температуре с последующим двукратным отмыванием в PBS-T. Затем 25 мкл мышиных или крысиных эталонных антител mAb добавляли в независимые лунки ELISA в разведении, известном из предыдущих экспериментов, чтобы увеличить максимальное связывание антигена в 200 раз. Через 15 минут 25 мкл антител против EGFR в концентрации 2 мкг/мл добавляли в лунки, предварительно инкубированные с эталонными антителами, или в лунки, содержащие 25 мкл PBS. После перемешивания компонентов это давало конечную концентрацию антитела против EGFR 1 мкг/мл и 100-кратное максимальное связывание антигена относительно эталонных антител. Антитела были инкубированы в течение 45 минут при комнатной температуре, после чего лунки четыре раза отмывали PBS-T. Вторичный конъюгат козьего IgG против человека с HRP разбавляли в соотношении 1:3000, и 50 мкл этого материала добавляли в каждую лунку с последующей 30-минутной инкубацией при комнатной температуре. Наконец, лунки четыре раза отмывали PBS-T, планшеты разрабатывали, добавляя TMB (50 мкл на лунку) и проводили считывание на волне 620 нм каждые 5-15-30 минут. Степень ингибирования рассчитывали по формуле: % ингибирования = (1-(OD конкуренции/OD без конкуренции (PBS))) × 100.
Реактивы ELISA:
1) Покровный буфер: 1×PBS, Gibco № по каталогу: 20012-019
2) Антигены: Полноразмерный EGFR дикого типа, очищенный от клеток A431, Sigma E3641
3) Планшет ELISA: NUNC Maxisorp, № по каталогу: 442404
4) Буфер блокирования/разбавления: 1% BSA в PBS-T (PBS-T-1% BSA)
5) Отмывочный буфер: 1×PBS/0,05% Tween 20 (PBS-T)
6) Положительный контроль: Эрбитукс (Merck KGaA, 64271 Дармштадт, Германия, № по каталогу: 018964, Cetuximab), Вектибикс (Amgen Inc, One Amgen Center Drive, Саузенд Оукс, штат Калифорния 91320-1799, США, № по каталогу: 3241400, Panitumumab)
7) Эталонные антитела:
• ICR10 (крысиное), Abcam, Ab231
• 199.12 (мышиное), Lab Vision Ab-11, MS-396-PABX
• EGFR.1 (мышиное), Lab Vision Ab-3, MS-311-PABX
• H11 (мышиное), Lab Vision Ab-5, MS-316-PABX
• B1D8 (мышиное), Lab Vision Ab-16, MS-666-PABX
• 111.6 (мышиное), Lab Vision Ab-10, MS-378-PABX
• 225 (мышиное), Lab Vision Ab-2, MS-269-PABX
• 528 (мышиное), Lab Vision Ab-1, MS-268-PABX
8) Конъюгат козьего IgG против человека и HRP, Serotec, Star 106P
9) TMB Plus, KemEnTec, № по каталогу: 4390L
10) 1M H2SO4
Результаты конкурентного анализа ELISA показаны на фиг.6. Конкурентные анализы ELISA были использованы для ранжирования супернатантов с антителами против EGFR в соответствии с доменной специфичностью использованных эталонных антител против внеклеточного домена EGFR. Значения ингибирования в диапазоне 50-100% расценивали как указание на значительную конкуренцию между парами антител, связывающими перекрывающиеся эпитопы или эпитопы, находящиеся в большой близости с антигеном, тогда значения ингибирования ниже 50% указывали на то, что эпитопы, распознанные парами антител, не соседствуют друг с другом, что приводит к снижению стерического несоответствия. Было обнаружено, что антитела против EGFR связывают несколько эпитопов на ECD EGFR, включая домены I, II и III. Для некоторых антител этот анализ не позволяет различить специфическую направленность mAb против домена I или домена II. Такие специфичности получили обозначение домен I/II. Далее оказалось, что некоторые антитела связывают уникальные эпитопы, которые не удается в дальнейшем расшифровать при помощи использованного конкурентного анализа ELISA (например, клоны 1229 и 1320, фиг.6). Вполне возможно, что некоторые из этих антител направлены против домена IV, для которого авторы изобретения не располагают какими-либо эталонными реактивностями антител. Интересно, что антитела к домену III далее можно было подразделить на четыре подгруппы, основываясь на различном характере конкуренции, полученном с проверенными мышиными эталонными антителами, направленными против этого домена. Группа I состояла только из mAb 992, конкурирующего за связывание с эталонными антителами Ab1 и Ab2. Группа II состояла из mAb 1024 и 1042, которые были получены из одной и той же реаранжировки Ig и, следовательно, демонстрировали очень близкую гомологию последовательностей, как на уровне ДНК, так и на уровне аминокислот. Было обнаружено, что эти два антитела конкурируют за связывание только с Ab2. Группа III состояла из mAb 1030, 1208 и 1277, которые конкурировали за связывание с эталонными антителами Ab1, Ab5 и Ab10. И, наконец, группа IV состояла из mAb 1254, конкурирующего за связывание со всеми использованными эталонными антителами, направленными против домена III: Ab1, Ab2, Ab5 и Ab10.
Конкурентный анализ для разных эпитопов с эталонными или одинаковыми видами антител при использовании технологии поверхностного плазмонного резонанса
Анализ SPR проводили в аппарате Biacore 3000 с четырьмя проточными кюветами. Чип CM5 Biacore был конъюгирован с 10000 резонансных единиц (Ru) поликлонального антитела против His в проточных кюветах 1-4 в соответствии с инструкциями производителя. При скорости потока 5 мкл/мин 15 мкл 6×His ECD EGFR в концентрации 20 мкг/мл впрыскивали для захвата во все четыре проточные кюветы, с которыми было конъюгировано поликлональное антитело против His. Сразу же после впрыскивания антигена в каждой проточной кювете при базисном прогоне было установлено максимальное связывание mAb против EGFR без конкуренции. Вкратце, 5 мкл антитела в концентрации 40 мкг/мл впрыскивали во все проточные кюветы с захваченным EGFR при последующем отслаивании комплекса антиген/антитело посредством кислого промывания с низким pH (время контакта с 10 мМ смеси глицин-HCl, pH2, - 10 секунд). После определения максимального связывания антитела против EGFR в каждой проточной кювете проводили конкурентный прогон в таком же цикле аппарата Biacore. Сначала проточные кюветы насыщали антигеном ECD EGFR с последующим впрыскиванием различных эталонных антител или антител против EGFR в разные проточные кюветы при тех же условиях насыщения антигеном, как было описано выше. Этот этап немедленно сопровождался вторым впрыскиванием антитела против EGFR в проточные кюветы, насыщенные антигеном EGFR и конкурентным антителом, для сведения к минимуму диссоциации или антигена, или блокирующего антитела. Затем комплексы антиген/антитело отслаивали посредством кислого промывания с низким pH (время контакта с 10 мМ смеси глицин-HCl, pH2, - 10 секунд), после чего повторяли с новым антителом против EGFR весь цикл, начиная с базисного прогона. Степень ингибирования испытуемых антител против EGFR определяли путем сравнения величины Ru max для индивидуального антитела против EGFR до и после конкуренции при введении контрольных точек, зафиксированных за две секунды до и через две секунды после впрыскивания каждого образца. Пример одного цикла Biacore показан на фиг.7.
Реактивы:
1. Чип CM5, Biacore, № по каталогу: BR-1000-14
2. NHS, Biacore BR-1000-50
3. EDC, Biacore BR-1000-50
4. 10 мМ ацетатный буфер pH 4,5, Biacore, № по каталогу: BR-1003-50
5. Антитело Tetra-His (без BSA), Qiagen, № по каталогу: 34670
6. Этаноламин, 1,0M pH 8,5, Biacore BR-1000-50
7. Подвижный буфер 10×HBS-EP: 0,01M HEPES pH 7,4, 0,15M NaCl, 3 мМ EDTA, 0,005% об./об. сурфактанта P20
8. Антиген: Внеклеточный домен рекомбинантного EGFR человека собственного производства с 6×His.
9. 10 мМ глицин HCl pH 2,0
10. Эталонные антитела:
• ICR10 (крысиное), Abcam, Ab231
• 199.12 (мышиное), Lab Vision Ab-11, MS-396-PABX
• EGFR.1 (мышиное), Lab Vision Ab-3, MS-311-PABX
• H11 (мышиное), Lab Vision Ab-5, MS-316-PABX
• B1D8 (мышиное), Lab Vision Ab-16, MS-666-PABX
• 111.6 (мышиное), Lab Vision Ab-10, MS-378-PABX
• 225 (мышиное), Lab Vision Ab-2, MS-269-PABX
• 528 (мышиное), Lab Vision Ab-1, MS-268-PABX
Для подтверждения результатов эпитопного анализа, полученных по методике конкурентного ELISA, и для проведения дальнейшего эпитопного анализа с конкуренцией между теми же видами пар антител был проведен анализ конкуренции, основанный на связывании антител, с измерениями в режиме реального времени по методике поверхностного плазмонного резонанса. Полученная в результате этого анализа эпитопная карта клонов против EGFR, протестированных в сравнении с панелью эталонных антител, показана ниже на фиг.8. Значения ингибирования в диапазоне 50-100% расценивали как указание на значительную конкуренцию между парами антител, связывающими перекрывающиеся эпитопы или эпитопы, находящиеся в большой близости с антигеном, тогда как значения ингибирования ниже 50% указывали на то, что эпитопы, распознанные парами антител, не соседствуют друг с другом, что приводит к снижению стерического несоответствия. Значения ингибирования ниже 25% не были включены в анализ для перекрывающихся эпитопов, поскольку они были расценены как показатель статистически незначимого ингибирования. Было обнаружено, что все испытуемые антитела, кроме 1320, конкурируют с одним или более использованных эталонных антител, а это указывало на то, что антитело 1320 было направлено против неизвестного эпитопа, для которого еще не располагают какими-либо эталонными активностями антител. Полностью человеческие или гуманизированные антитела вектибикс и эрбитукс были включены в анализ и, в соответствии с полученными результатами, оказалось, что они связывают перекрывающиеся эпитопы. Данные, полученные из конкурентных анализов ELISA и SPR в целом хорошо коррелировались в отношении установленной доменной специфичности антител против EGFR. Однако в двух анализах иногда наблюдались небольшие различия по характеру конкуренции между отдельными эталонными антителами, что, по-видимому, было связано с тем фактом, что в конкурентном анализе ELISA был использован антиген полноразмерного рецептора EGF, а в конкурентном анализе SPR - рекомбинантный внеклеточный домен EGFR.
После того как эпитопное картирование антител против EGFR было подтверждено в двух разных анализах конкуренции, был проведен конкурентный анализ тех же видовых комбинаций пар антител против EGFR с той целью, чтобы выяснить, какие пары антител распознавали разные эпитопы, и можно ли далее разделить на эпитопные кластеры пары антител, распознающих перекрывающиеся эпитопы. Результаты этого анализа показаны на фиг.9. И вновь в этом анализе значения ингибирования в диапазоне 50-100% расценивали как указание на значительную конкуренцию между парами антител, связывающих перекрывающиеся эпитопы. Этот критерий представлялся достоверным, поскольку антитела, протестированные против самих себя и, следовательно, распознающие полностью перекрывающиеся эпитопы, дали значения ингибирования в диапазоне 70%-100%, как это показано на фиг.9. Кроме того, это наблюдение иллюстрирует, что диссоциация пар антигенов или антител в промежутке времени проведенного анализа не оказывает влияния на итоги эксперимента для проходивших тестирование антител. При группировке антител в соответствии с предполагаемой доменной специфичностью ECD EGFR, определенной в предыдущих разделах, было обнаружено, что антитела связывающиеся исключительно с доменом I либо с доменом I или II (I/II), главным образом, формируют кластер с антителами, имеющими такую же специфичность, но не с антителами, распознающими домен III. Сходным образом, антитела к домену III, которые, как было выяснено, конкурируют за связывание только с антителами, распознающими домен III, но не с антителами, распознающими домен EGFR I или I/II. Хотя было обнаружено, что два антитела к домену III (1024 и 1042) полученные из одной и той же реаранжировки Ig, распознают перекрывающиеся эпитопы, важное наблюдение заключалось в том, что попарные комбинации 1024 или 1042 с 992 или 1030 не приводят к значительной конкуренции. Таким образом, был сделан вывод, что антитела 992, 1030 и 1024/1042 распознавали три неперекрывающихся эпитопа домена III антигена ECD EGFR. Наконец, было обнаружено, что mAb 1320 конкурирует за связывание с mAb 1024 и 1449, которые направлены против домена III, но не с другими антителами к домену III, участвовавшими в тестировании (конкуренция 1320 с 1042 не наблюдалась). Таким образом, было предположено, что mAb 1320 осуществляет связывание на периферии домена III на внеклеточном домене EGFR. Обзорное представление об эпитопной специфичности можно получить на фиг.10, где проиллюстрированы эпитопные карты антител, направленных против доменов I, I/II или III EGFR.
После выяснения того факта, что попарные комбинации антител 992, 1030 и 1024/1042 не приводят к значительной конкуренции, как это определяется в анализе SPR, были запланированы новые эксперименты Biacore, чтобы исследовать вопрос о том, как много антител может одновременно связываться с рецепторным антигеном. Прежде всего было исследовано, какое влияние оказывает насыщение домена III тремя антителами 992, 1024 и 1030 на связывание антител, направленных на другие специфичности EGFR, не относящиеся к домену III. Результат этого анализа показан на фиг.11A. Ингибирование отдельных антител выявляли посредством их тестирования в комбинациях или с единичным антителом, или со смесью (до трех антител), полученных при постепенном добавлении одного нового антитела в каждом цикле Biacore. Чтобы обеспечить полную блокаду распознаваемого эпитопа, антитела были протестированы в индивидуальных концентрациях 40 мкг/мл. Как показано на фиг.11A, было обнаружено, что три антитела к домену III (992, 1024 и 1030) связываются с рецептором одновременно без какого-либо взаимного ингибирования. Наблюдаемые отрицательные значения ингибирования, увеличивающиеся при каждом последующем добавлении антитела, позволили предположить синергический эффект связывания для связывания следующих антител. Важно отметить, что, если домен III был инкубирован с тремя антителами, то другие антитела, направленные против неперекрывающихся эпитопов на домене I/II (mAb 1261), домене I (1347) или на неизвестной специфичности (1361), проявляли связывание без блокирования эпитопов смесью трех mAb. Кроме того, эти протестированные антитела демонстрировали небольшие отрицательные величины ингибирования, свидетельствующие о том, что они связывались лучше после насыщения рецептора смесью трех mAb. Следовательно, по результатам этого эксперимента можно было предположить, что шесть протестированных антител могли связываться с антигеном ECD рецептора EGF одновременно. Для того чтобы далее проверить этот наблюдаемый феномен, была приготовлена смесь, состоявшая из всех протестированных антител (1261, 1347, 992, 1024, 1030 и 1361), которую проанализировали на ингибирование каждого индивидуального образца антитела в смеси. В качестве положительного контроля при тестировании были использованы смеси антител, в которые не были включены испытуемые образцы антител. Как представлено на фиг.11B/C, было обнаружено, что при тестировании на связывание с рецептором EGF, который был предварительно инкубирован с полной смесью антител, все шесть испытуемых антител ингибируются на 80-116%. Однако, если из этой смеси удаляли отдельные образцы антител, значительного ингибирования конкретных образцов не наблюдалось, а это доказывало, что отдельные антитела в смеси блокировали связывание с рецептором EGF только для самих себя. Этот эксперимент четко проиллюстрировал, что одновременно связываться с EGFR могут, по меньшей мере, шесть антител, распознающих неперекрывающиеся эпитопы. В заключительном эксперименте было исследовано, могут ли связываться с EGFR другие антитела, направленные против домена I (1284), I/II (1257) или против кластера неизвестной специфичности (1183, 1255), если этот рецептор был предварительно инкубирован со смесью из шести антител. Как представлено на фиг.11D, ни одно из протестированных антител не было способно осуществлять значительное связывание с EGFR при предварительной инкубации этого рецептора со смесью из шести антител. Это могло быть обусловлено тем, что коллекция не включала антител против тех сайтов, которые оставались незанятыми смесью из шести антител. Альтернативное объяснение: фактически, возможно, что все сайты на протестированных доменах были блокированы антителами.
ПРИМЕР 4: Ингибирование активации EGFR
Определение опосредованной антителами блокады связывания лиганда EGF с рецептором EGFR посредством конкурентного анализа ELISA
Для того чтобы верифицировать связывание протестированных антител против EGFR с рецептором EGFR и одновременное блокирование ими связывания биотинилированного лиганда EGF, лунки ELISA покрывали из расчета 80 мкл на лунку полноразмерным EGFR в концентрации 0,5 мкг/мл в PBS на ночь при температуре 4°C. На следующее утро лунки дважды отмывали PBS-T и на один час блокировали 150 мкл PBS-T-1% BSA при комнатной температуре с последующим двукратным отмыванием в PBS-T. Затем в лунки добавляли 80 мкл антител против EGFR в серийном разведении, а также контрольные антитела, после чего планшеты инкубировали 30 минут при комнатной температуре. После инкубации антител 20 мкл биотинилированного лиганда EGF в концентрации 0,5 мкг/мл добавляли во все лунки с разведениями антител против EGFR или в лунки, содержащие только PBS-T 1% BSA, после чего планшеты инкубировали при комнатной температуре в течение 1 часа. Затем лунки пять раз отмывали PBS-T, после чего инкубировали 30 минут при комнатной температуре с вторичным реактивом стрептавидин-HRP, разбавленным в соотношении 1:1000 блокирующим буфером (из расчета 100 мкл на лунку). Наконец, лунки еще пять раз отмывали PBS-T, после чего планшеты разрабатывали, добавляя субстрат ТМВ (100 мкл на лунку), и инкубировали в течение 60 минут. После инкубации реакцию останавливали,, добавляя 1М H2SO4 (100 мкл на лунку), и считывали планшеты на уровне 0D 450 нм.
Реактивы ELISA:
1) Покровный буфер: 1×PBS, Gibco №по каталогу: 20012-019
2) Антиген: Полноразмерный EGFR дикого типа, очищенный от клеток А431, Sigma E2645
3) Планшет ELISA: NUNC Maxisorp, №по каталогу: 442404
4) Буфер блокирования/разбавления: 1% BSA в PBS-T (PBS-T-1% BSA)
5) Отмывочный буфер: 1×PBS/0,05% Tween 20 (PBS-T)
6) Положительный контроль: эрбитукс, вектибикс
7) Отрицательный контроль: Synagis (Medimmune Inc, Palivizumab, № по каталогу: NDC 60574-4111-1)
8) Биотинилированный лиганд EGF, Invitrogen, № по каталогу: Е3477
9) Стрептавидин-HRP, сверхчувствительный: Sigma S 2438
10) ТМВ Plus, KemEnTec, №по каталогу: 4390L
11) 1М H2SO4
Конкурентные анализы ELISA были проведены для ранжирования способности антител против EGFR ингибировать связывание биотинилированного лиганда EGF с полноразмерным рецептором EGFR, внесенным в виде покрытия в лунки ELISA. Как представлено на фиг.12, как эрбитукс, так и вектибикс очень мощно блокировали связывание лиганда EGF, тогда как антитело Synagis в отрицательном контроле, не направленное против EGFR, не ингибировало связывания лиганда EGF. Как показано на фиг.12А, три антитела (992, 1030 и 1042), направленные против домена III и распознающие неперекрывающиеся эпитопы, были протестированы по отдельности или в эквимолярной смеси на их способность ингибировать связывание лиганда EGF. Из трех протестированных антител только антитело mAb 1030 продемонстрировало умеренную ингибиторную активность в отношении лиганда EGF по сравнению с эрбитуксом и вектибиксом. Эквимолярная смесь mAb 992, 1030 и 1042 оказалась более эффективной в ингибировании связывания лиганда EGF по сравнению с теми же антителами при их тестировании по отдельности. При общей концентрации IgG 1 мкг/мл было обнаружено, что эквимолярная смесь ингибирует связывание лиганда EGF приблизительно в два раза эффективнее, чем mAb 1030 и в четыре раза эффективнее, чем mAb 992 и 1042 при их тестировании по отдельности, а это свидетельствовало о синергическом эффекте смешивания трех антител к домену III, распознающих неперекрывающиеся эпитопы. Как показано на фиг.12В, в этом анализе также были протестированы клоны антител против EGFR 1208, 1260, 1277 и 1320. Эти четыре клона были способны ингибировать связывание лиганда EGF с эффектом дозовой зависимости, что оказалось более эффективным, чем результат применения клонов 992, 1030 и 1042 по сравнению с контрольным эрбитуксом. Оказалось, что в концентрациях выше 0,33 мкг/мл клоны антител против EGFR 1208, 1260, 1277 и 1320 были столь же эффективными в блокировании связывания лиганда EGF, как и эрбитукс в соответствующих концентрациях.
Способность ингибировать фосфорилирование EGFR, индуцированное EGF, в клетках HN5
Антитела против EGFR были протестированы на реактивность в отношении фосфорилирования EGFR с применением внутриклеточного вестерн-анализа. Внутриклеточная вестерн-процедура позволяет выявить EGFR и фосфорилированный EGFR (pEGFR) в одном и том же образце, а это, в свою очередь, создает возможность для того, чтобы сравнить соотношение экспрессии EGFR и pEGFR при обработке каждым антителом и для каждого набора данных. Клетки HN5 в соответствии с инструкциями, полученными из АТСС, культивировали в среде DMEM с добавкой 10% FCS и пенициллина/стрептомицина. 43000 клеток HN5 высевали в 96-луночные планшеты Nunc (№ по каталогу 167008) за 24 часа до выращивания на минимальной среде. Перед добавлением антител клетки выращивали 16 часов в условиях голодания на среде DMEM. Антитела добавляли в конечной концентрации 10 мкг/мл в 200 мкл DMEM, а смесь пипетировали вверх и вниз, по меньшей мере, пять раз для лучшего перемешивания компонентов. После 30-минутной обработки антителами добавляли EGF в концентрации 50 мкг/мл в соответствующие лунки и оставляли на 7,5 минут.Внутриклеточные вестерн-анализы проводили, главным образом, по инструкциям, предоставленным производителем набора In-cell western (Odyssey, LI-COR biosciences).
После стимулирования EGF клетки фиксировали в 3,7% формальдегиде (Sigma F-8775, партия 71K500, содержащем ~1% метанола) в течение 20 минут. Перед блокированием в блокирующем буфере LI-COR (927-40000) были проведены пять 5-минутных отмываний в PBS-Triton X-100 (0,1%) для того, чтобы сделать клеточные мембраны проницаемыми. Первичные антитела добавляли в концентрациях, соответствующих предоставленным инструкциям, и инкубировали с осторожным встряхиванием при комнатной температуре в течение 2,5 часов (total EGFR mouse, разведение 1:500, BioSource International, № по каталогу AHR5062 и Phospho-EGFR Tyr1173 Rabbit, разведение 1:100, Biosource, № по каталогу 44-794G).
После инкубации с первичными антителами клетки пятикратно отмывали в течение пяти минут в PBS-T (0,1% tween-20), после чего добавляли вторичные антитела (goat-anti-rabbit IRDye 680, разведение 1:200, LI-COR, № по каталогу 926-32221 и goat-против mouse, IRDye 800CW, разведение 1:800, LI-COR, № по каталогу 926-32210) и инкубировали в течение 1 часа при комнатной температуре при осторожном встряхивании планшетов, покрытых алюминиевой фольгой.
Перед проведением измерений на флуоресцентном считывающем устройстве Tecan планшеты пятикратно отмывали в течение пяти минут в PBS-T. Все отмывания завершали резким движением, переворачивая планшет открытой стороной вниз для сбрасывания отмывающего раствора, с последующим постукиванием шаблоном по бумажному полотенцу. (Применительно к обработке планшетов ELISA важно отметить, что во время этой обработки клетки остаются на планшете, а отмывочный раствор при этой процедуре удаляется быстрее и легче, чем при отсасывании, которое может нарушить целостность монослоя клеток). Остатки отмывочного раствора после заключительного отмывания удаляли легким отсосом со стороны лунок многоканальной пипеткой. Флуоресцентный сигнал измеряли в канале 680 нм (возбуждение 675 нм и эмиссия 705 нм, в обоих случаях полоса пропускания 10 нм) и в канале 800 нм (возбуждение 7 62 нм и эмиссия 798 нм, в обоих случаях полоса пропускания 10 нм).
По результатам внутриклеточного вестерн-анализа становится очевидным, что значительное влияние на статус pEGFR клеток HN5 (р<0,05) оказывают три антитела: 1208, 1277 и 1320 (фиг.13).
Смесь антител против EGFR (992, 1030 и 1042) и отдельные антитела в данном случае были протестированы посредством внутриклеточного вестерн-анализа на эффект ингибирования фосфорилирования EGFR, индуцированного EGF. Как можно видеть на фиг.14, антитела 992 и 1030, а также смесь антител против EGFR значительно ингибировали фосфорилирование EGFR, индуцированное EGF (р<0,05).
ПРИМЕР 5: Интернализация рецепторов E6F в клетках A431NS
Клетки A431NS (АТСС# CRL-2592) трипсинизировали из культурального флакона Т175 со слиянием порядка 80-90% при помощи фермента TrypLE. Обособленные клетки отмывали в PBS и суспендировали в бессывороточной среде DMEM. Клеточный материал разделяли на порции по 1-2 мл и инкубировали 30 минут на льду с испытуемыми антителами. Концентрация антител составляла 10 мкг/мл. Клетки три раза отмывали в среде DMEM (250 г, 4 минуты, 4°С) и ресуспендировали в 1,8 мл среды DMEM. Каждую порцию распределяли в шесть пробирок FACS (по 300 мкл клеточной суспензии в каждую). Три пробирки из каждой порции помещали в водяную баню с температурой 37°С ровно на 40 минут, а другие пробирки сразу же ставили на лед. После инкубации клетки дважды отмывали в среде DMEM (250 г, 4 минуты, 4°С), а клеточный осадок повторно растворяли в 100 мкл кроличьего IgG против человека Fcγ F(ab′)2-FITC в среде DMEM. Клетки инкубировали 30 минут при 4°C, после чего три раза отмывали в среде DMEM при 4°C и анализировали в проточном цитометре FACSCalibur.
Результаты показаны на фиг.15. Инкубация с эрбитуксом и вектибиксом продемонстрировала такой же уровень интернализации рецептора порядка 30% при сохранении первоначального окрашивания поверхности на 70%. Инкубация только с антителом 992 приводила к ингибированию рецептора приблизительно на 45%. Инкубация со смесью антител, содержащей два дополнительных антитела с неперекрывающимися эпитопами, приводила к усилению ингибирования рецептора: 992+1024→74%, 992+1024+1030→83%.
Дальнейшее добавление антител не приводило к дополнительному увеличению интернализации рецептора. Таким образом, для достижения максимального уровня интернализации в клетках А431 нужны, по меньшей мере, три антитела.
ПРИМЕР 6: Анализы пролиферации
Клеточное повреждение неизбежно приводит к утрате способности клетки поддерживать и обеспечивать энергию для метаболической функции и роста клетки. На этой предпосылке основаны анализы метаболической активности. Обычно они направлены на измерение митохондриальной активности. Реактив клеточной пролиферации WST-1 (Roche, № по каталогу 11 644 807 001) представляет собой готовый к употреблению субстрат, который измеряет метаболическую активность жизнеспособных клеток. Далее допускается, что метаболическая активность коррелируется с количеством жизнеспособных клеток. В этом примере анализ с WST-1 был использован для измерения количества метаболически активных клеток после обработки разными антителами в различной концентрации.
Перед проведением анализа с WST-1 соответствующие антитела и смеси антител разбавляли в среде DMEM с добавками 0,5% FBS и 1% P/S до конечной общей концентрации антител 20 мкг/мл, что создавало в лунке с самой высокой концентрацией конечную концентрацию антител 10 мкг/мл. Затем 150 мкл этих растворов добавляли в лунки столбца 2 96-луночного планшета и готовили трехкратные серийные разведения вниз до столбца 9, так, что каждая лунка содержала 100 мкл раствора антител. В столбец 11 добавляли 100 мкл среды. В ряды 1 и 8, а также в столбцы 1 и 12 добавляли 200 мкл среды для уменьшения эффекта испарения среды в экспериментальных лунках.
Затем клетки A431-NS отмывали 1×PBS и разделяли трипсинизацией с применением 3 мл раствора трипсина. Затем добавляли 17 мл полной среды, и клетки центрифугировали на скорости 300×g (1200 rcf) в течение 5 минут. Супернатант удаляли, а клетки ресуспендировали в смеси DMEM+0,5% FBS. Проводили подсчет клеток и подгонку их концентрации до 15000 клеток на миллилитр. Затем 100 мкл клеточной суспензии добавляли в экспериментальные лунки столбцов 2-11 из расчета 1500 клеток на лунку. Планшеты инкубировали 4 дня в увлажненном инкубаторе при 37°C. Затем в каждую лунку добавляли 2 0 мкл реактива WST-1, и планшеты инкубировали в течение 1 часа при 37°C. Далее планшеты переносили в орбитальный планшетный шейкер и оставляли еще на один час. Поглощательную способность измеряли на уровне 4 50 и 620 нм (опорная длина волны) считывающим устройством ELISA. Количество метаболически активных клеток (MAC) рассчитывали как процентный показатель по сравнению с необработанным контролем, исходя из следующей формулы:

Для исследований по титрованию EGF лиганд разбавляли до концентрации 20 нМ/мл в среде DMEM+0,5% FBS, получая в лунке с самой высокой концентрацией EGF уровень 10 нМ/мл. Затем 150 мкл этого раствора добавляли в лунки столбца 2 96-луночного планшета и готовили трехкратные серийные разведения вниз до колонки 9, так, что каждая лунка содержала 100 мкл раствора EGF. В столбец 11 добавляли 100 мкл среды. В ряды 1 и 8, а также в колонки 1 и 12 добавляли 200 мкл среды для уменьшения эффекта испарения среды в экспериментальных лунках. Соответствующие антитела и смеси антител разбавляли до конечной общей концентрации антител 40 мкг/мл в среде DMEM с добавками 0,5% FBS и 1% P/S, что давало конечную концентрацию антител в лунках 10 мкг/мл. Затем 50 мкл этих растворов добавляли в лунки столбцов 2-9 96-луночного планшета.
Затем клетки A431-NS отмывали 1×PBS и разделяли трипсинизацией с применением 3 мл раствора трипсина. Затем добавляли 17 мл полной среды, и клетки центрифугировали на скорости 300×g (1200 rcf) в течение 5 минут. Супернатант удаляли, а клетки ресуспендировали в смеси DMEM+0,5% FBS. Проводили подсчет клеток и подгонку их концентрации до 40000 клеток на миллилитр. Затем 50 мкл клеточной суспензии добавляли в экспериментальные лунки столбцов 2-11 из расчета 2000 клеток на лунку. Планшеты инкубировали 4 дня в увлажненном инкубаторе при 37°C. Затем в каждую лунку добавляли 20 мкл реактива WST-1, и планшеты инкубировали в течение 1 часа при 37°C. Далее планшеты переносили в орбитальный планшетный шейкер и оставляли еще на один час. Поглощательную способность измеряли на уровне 4 50 и 620 нм (опорная длина волны) считывающим устройством ELISA. Количество метаболически активных клеток определяется по результату вычитания поглощательной способности на длине волны 4 50 нм из поглощательной способности на опорной длине волны 620 нм.
Количество метаболически активных клеток (MAC) рассчитывали как процентный показатель по сравнению с необработанным контролем, исходя из следующей формулы:

Результаты
Для того чтобы продемонстрировать превосходство смеси из трех антител против EGFR с неперекрывающимися эпитопами в пределах домена III над одиночными антителами, был проведен эксперимент, в котором исследовали ингибирование роста клеток A431-NS. Как можно видеть на фиг.16А, антитела, сами по себе, являются плохими ингибиторами роста клеток A431-NS, но при их применении в комбинации наблюдается синергический эффект в отношении роста клеток A431-NS. Хотя смесь антитела 992 с антителом 1042 или антителом 1030 достаточно мощна, смесь всех трех антител превосходит обе попарные смеси во всем диапазоне концентраций антител.
Было исследовано влияние отдельных антител и смесей антител на рост клеток A4 31-NS, стимулированных различными концентрациями EGF, а результаты этого исследования показаны на фиг.17. Как можно видеть на фиг.17, концентрации EGF выше 0,1 нМ в отсутствие антител токсичны для клеток. Однако очевидно, что смесь трех антител с неперекрывающимися эпитопами в пределах домена III EGFR (992, 1030 и 1042) действует синергически, ингибируя рост клеток A431-NS в присутствии EGF при его концентрации, по меньшей мере, до 0,3 нМ, причем смесь превосходит все моноклональные антитела.
Далее, авторы изобретения продемонстрировали, что синергический ингибиторный эффект в отношении роста клеток А431-NS можно также получить, комбинируя два антитела с неперекрывающимися эпитопами в домене III EGFR с такими антителами, которые направлены на эпитопы I или II EGFR. Как можно видеть на фиг.18, комбинации антител 992 и 1024, которые направлены на домен III EGFR, с антителом, направленным или на домен I EGFR (1284), или на домен I/II EGFR (1434) обладают такой же мощностью, как и комбинация трех антител, реагирующих с неперекрывающимися эпитопами в пределах домена III EGFR (992+1024+1030). В дополнение к этому следует отметить, что эти смеси антител с большей мощностью ингибируют рост клеток А431-NS, чем терапевтические антитела против EGFR эрбитукс и вектибикс.
Сходные анализы были проведены на двух других линиях раковых клеток, DU145 (АТСС#НТВ-81) и MDA-MB-468 (АТСС#НТВ-132). Результаты этих анализов пролиферации показаны на фиг.16B и 16B. В обоих случаях смесь трех антител (992, 1030 и 1042) превосходила смеси двух антител и одиночные антитела. В отношении клеток DU145 смесь трех антител превосходила вектибикс во всех концентрациях, а в отношении клеток MDA-MB-468 - в высоких концентрациях.
Используя способ, подобный описанному выше, авторы изобретения протестировали разные комбинации трех антител против EGFR.
Результаты
Эффекты различных комбинаций трех антител были исследованы на клеточной линии A431NS. Ингибиторная активность в отношении роста клеток для двадцати наиболее эффективных из проанализированных комбинаций показана на фиг.37. Все комбинации ингибировали пролиферацию клеточной линии A431NS более чем на 60% по сравнению с необработанным контролем. Еще одно интересное наблюдение заключается в том, что за исключением комбинаций (992+1024+1254, 992+1024+1320 и 992+1277+1320) все остальные комбинации содержат антитела с неперекрывающимися эпитопами. Это свидетельствует о том, что из трех антител можно сконструировать несколько комбинаций, связывающих разные эпитопы.
ПРИМЕР 7: Апоптоз
Апоптоз представляет собой клеточный механизм, который ведет к смерти клеток. Ранее этот механизм был описан при использовании таких антител против EGFR как эрбитукс (Baselga J. The EGFR as a target for anticancer therapy - focus on cetuximab. Eur J Cancer. 2001 Sep:37, Suppl 4:S 16-22). В связи с этим авторы провели исследование, чтобы ответить на вопрос, в какой степени отдельные антитела против EGFR 992, 1042 и 1030, а также смесь (992+1042+1030) способны индуцировать апоптоз.
Клетки A431NS в исходном количестве 1×104 были инкубированы в среде DMEM с добавками 0,5% FBS и антибиотиков для тройных определений в 96-луночных культуральных планшетах в присутствии смеси антител против EGFR (равные части 992, 1030, 1042), отдельных антител 992, 1030, 1042, эрбитукса или вектибикса в диапазоне концентраций от 0,01 мкг/мл до 10 мкг/мл. Клетки и антитела были инкубированы в течение 22 часов. Были собраны десять супернатантов, которые проанализировали набором ELISA производства компании Roche, № по каталогу: 11774425001 (Базель, Швейцария) на присутствие комплексов гистон-ДНК.
Эффект смеси был сопоставлен с каждым из моноклональных антител по отдельности, а также с эталонными антителами эрбитукс и вектибикс на клеточной линии A431NS (результаты представлены на фиг.19). Антитела были протестированы в 10-кратном разведении. Смесь оказалась значительно более эффективной (P<0,05) по сравнению с отдельными моноклональными антителами, а также вектибиксом при тестировании в концентрациях 1 мкг/мл и 10 мкг/мл. Смесь статистически значимо (p<0,05) увеличивала апоптоз по сравнению с эрбитуксом в концентрации 1 мкг/мл.
ПРИМЕР 7b:
В дополнение к примеру 7, смесь антител 992+1024, а также смесь антител 992+1024+1030 были исследованы на апоптотическую активность теми же способами, как это описано в примере 7 (фиг.35). Фактический уровень апоптоза был оценен по отношению к максимальному положительному контролю. Обе смеси, отдельные моноклональные антитела 992, 1024 и 1030, а также контрольное антитело в концентрации 1 мкг/мл сравнивали с эрбитуксом на клетках A431NS. Смесь антител 992+1024 проявила себя значительно лучше, чем эрбитукс и отдельные моноклональные антитела (во всех сравнениях P<0,05).
ПРИМЕР 8: Эффективность in vivo
Смесь против EGFR, состоящая из антител 992, 1030 и 1042, была исследована на эффективность in vivo на модели ксенотрансплантатов у мышей nude с применением клеток A431NS. Эта модель широко используется для исследования мощности моноклональных противораковых антител, включая антитела против EGFR. Мыши nude характеризуются иммунной недостаточностью и утратой T-клеток. Это позволяет выращивать в мышах клетки человека.
Две группы мышей nude в возрасте 6-8 недель получили подкожные инъекции 1×106 клеток A431NS. После того как средний размер опухолей достиг 100 мм3, было начато лечение мышей. Животные получили пять внутрибрюшинных инъекций антител в дозе 1 мг с интервалом 2-3 дня. Размер опухолей измеряли по двум диаметрам, применяя штангенциркули с цифровой индикацией и вычисляя объем опухолей по следующей формуле:
Объем опухоли (мм3)=L×W2×0,5,
где L означает наибольший диаметр, a W - наименьший диаметр (Teicher BA, Tumor Models in Cancer Research. Humana Press, NJ, USA 2002, р596). В конце эксперимента опухоли иссекали и взвешивали.
В качестве контрольного антитела был использован препарат Synagis. Эксперимент также включал лечение эрбитуксом и вектибиксом в том же количестве и по той же схеме, как и для смеси против EGFR (антитела 992, 1030 и 1024).
Как можно видеть на фиг.20, смесь антител 992, 1030 и 1042 значительно ингибировала рост опухолевых клеток A431NS (Р<0,05). Показатели среднего веса представлены на фиг.21. Результаты коррелируются с измерениями размеров опухолей. Выявлены статистически значимые различия между группой активной терапии и контрольной группой.
ПРИМЕР 8b: Эффективность in vivo
В дополнение к экспериментам in vivo, описанным в примере 8, смеси антител 992+1024 и 992+1024+1030 были исследованы на описанной выше модели ксенотрансплантатов A431NS (фиг.36). Четыре группы мышей nude в возрасте 6-8 недель (каждая численностью по 9 животных) получили подкожные инъекции 1×106 клеток A431NS. Когда средний размер опухолей достиг 100 мм3, мыши получили первую инъекцию антител. Четыре указанные группы получили смесь 992+1024, смесь 992+1024+1030, эрбитукс или контрольное антитело Synagis. В общей сложности мыши получили 17 инъекций в дозе 0,5 мг (4 раза в неделю). Первую инъекцию проводили в 8-й день, а последнюю в 34-й день эксперимента. Размеры опухолей измеряли в течение 56 дней. По окончании лечения антителами у мышей, получавших эрбитукс, опухоли начали увеличиваться в размерах, тогда как в двух группах мышей, получавших смеси антител 992+1024 или 992+1024+1030, опухоли продолжали уменьшаться в размерах. В группе, получавшей смесь 992+1024, никакого увеличения опухолей не наблюдалось в 91-й день от начала эксперимента (57 дней после окончания исследования). Средний размер опухолей в группе, получавшей комбинацию 992+1024, в 56-й день был значительно меньше, чем в группе, получавшей эрбитукс (P<0,01).
В этом эксперименте также прослеживали выживание мышей. Мышей учитывали как умерших, когда опухоль достигала максимального размера, определенного заранее. В представленной ниже таблице указано число мышей, выживших через 56 дней после инокуляции опухолевых клеток. В обеих группах, получавших комбинации, показатели выживания были лучше по сравнению с эрбитуксом.
Дополнительные эксперименты
Предварительные данные по опухолевым лизатам из экспериментов с ксенотрансплантатами, описанных в примере 8, показывают, что комбинация антител 992+1042+1030 индуцирует сильную понижающую регуляцию выработки VEGF клетками A431NS, где VEGF является важным медиатором ангиогенеза. Усиленное образование кровеносных сосудов, феномен, наблюдаемый во многих солидных опухолях, представляет собой механизм, участвующий в непрерывной доставке питательных веществ и т.п., следовательно, тесно связанный с условиями выживания.
Кроме того, другие предварительные данные показывают, что в опухолевых лизатах из эксперимента с ксенотрансплантатами, описанного в примере 8, может наблюдаться повышенный уровень комбинации антител 992+1042+1030 по сравнению с эрбитуксом и вектибиксом.
ПРИМЕР 8c: Усиленная дифференциация опухолевых клеток in vivo
Терминальная дифференциация клеток представляет собой сложный процесс, который включает активацию специфических программ генной экспрессии клеточного типа, управляющих многоэтапным процессом необратимой утраты пролиферативной способности клеток. При злокачественном заболевании раковые клетки часто находятся в недифференцированном состоянии, характеризующемся высоким темпами пролиферации, в связи с чем было высказано предположение о том, что лекарства, способные индуцировать терминальную дифференциацию раковых клеток, могут оказаться способными элиминировать злокачественные клетки и восстановить нормальный клеточный гомеостаз (Pierce GB, Speers WC: Tumors as caricatures of the process of tissue renewal: prospects for therapy by directing differentiation. Cancer Res 48:1996-2004, 1988). Ранее были опубликованы сообщения о том, что в определенных экспериментальных условиях моноклональные антитела против EGFR способны увеличивать степень терминальной дифференциации опухолевых клеток при плоскоклеточном раке человека, выросших в ксенотрансплантатах опухолей у мышей с недостаточностью иммунной системы (Milas L, Mason K, Hunter N, Petersen S, Yamakawa M, Ang K, Mendelsohn J, Fan Z: In vivo enhancement of tumor radioresponse by C225 antiepidermal growth factor receptor antibody. Clin Cancer Res 6:701-8, 2000; Modjtahedi H, Eccles S, Sandle J, Box G, Titley J, Dean C: Differentiation or immune destruction: two pathways for therapy of squamous cell carcinomas with antibodies to the epidermal growth factor receptor. Cancer Res 54:1695-701, 1994).
Авторы изобретения гистологическими методами исследовали масштабы терминальной дифференциации раковых клеток A431NS в ксенотрансплантатах опухолей у мышей при лечении антителами против EGFR. Гистологическое исследование включало по 3 случайно отобранных привитых опухоли в каждой из четырех экспериментальных групп эксперимента, описанного в примере 8.
Ткани были рассечены и быстро заморожены, затем смонтированы при помощи устройства Tissue-Tek на криомикротом (Leitz, модель 1720), рассечены на срезы по 5 мкм, нанесены на предметные стекла Superfrost Plus и окрашены гематоксилином/эозином. Затем два независимых наблюдателя проводили микроскопическое исследование всех тканевых срезов по "слепому" принципу, подсчитывая участки кератинизации ("раковые жемчужины"), отражающие масштабы терминальной дифференциации (Modjtahedi et al., 1994). Полученные результаты представлены в таблице 7. Мыши, получавшие лечение смесью трех антител против EGFR (992+1024+1030, группа 1), имели заметно более крупные и более многочисленные очаги терминальной дифференциации раковых клеток по сравнению с мышами, получавшими лечение эталонными антителами эрбитукс и вектибикс (группы 2 и 3, соответственно). В контрольной группе (группа 4), получавшей вместо антител PBS, каких-либо признаков терминальной дифференциации не наблюдалось.
Характерные микроскопические изображения были получены при помощи микроскопа, оснащенного цифровой камерой (см. фиг.26).
В результате проведенного исследования комбинация из трех антител против EGFR с неперекрывающимися эпитопами в пределах домена III (клоны 992, 1030 и 1042) продемонстрировала неожиданно сильный эффект индуцирования дифференциации опухолевых клеток in vivo по сравнению с моноклональными антителами эрбитукс и вектибикс. Наблюдаемые эффекты терминальной дифференциации привели нас к выводу о том, что композиции антител, предлагаемые изобретением, могут быть использованы в комбинированной терапии вместе с другими средствами, индуцирующими дифференциацию, например, ретиноевой кислотой, 4-фенилбутиратом.
ПРИМЕР 8d: Длительный эффект композиции антител, предлагаемой изобретением, по ингибированию роста раковых клеток
Эксперимент с ксенотрансплантатами опухолей, описанный в примерах 8 и 8b, был еще раз осуществлен для исследования эффективности смеси антител 992+1024 in vivo. Вкратце, мыши BALB/c nu/nu получили подкожную инъекцию 106 клеток A431NS в бок туловища. Привитым опухолям давали вырасти до среднего размера 100 мм3 (7-й день), после чего мышей рандомизировали на пять групп по девять животных в каждой и начинали лечение антителами. Эти пять групп получали высокую (2 мг в неделю) или низкую (1 мг в неделю) дозу смеси антител 992+1024 или эталонного антитела эрбитукс, либо высокую дозу (2 мг в неделю) контрольного антитела Synagis. В общей сложности все мыши получили по 9 инъекций (0,5 или 1 мг антитела два раза в неделю), начиная с 7-го дня и кончая 33-м днем исследования.
Высокая доза (2 мг в неделю) смеси антител 992+1024 оказалась очень эффективной в сдерживании начального опухолевого роста и в индуцировании длительной регрессии опухолей по сравнению с эрбитуксом (Р=0,0002, фиг.38). Ни одно из животных, получавших 2 мг в неделю смеси антител 992+1024, не скончалось в период исследования (118 дней от начала эксперимента, фиг.38 и 39), что на уровне статистической значимости отличалось в лучшую сторону от исходов в группе с высокой дозой эрбитукса 2 мг в неделю, где на 60-й день осталось в живых только одно из девяти животных (Р=0,0008, фиг.39). Это свидетельствует о длительном эффекте лечения смесью антител 992+1024 в отношении сроков выживания. Хотя и с несколько меньшим эффектом по сравнению с высокой дозой, низкая доза смеси антител 992+1024 (1 мг в неделю) также оказалась способной сдерживать опухолевый рост, причем лучше на уровне статистической значимости, чем эрбитукс в высокой дозе 2 мг в неделю, как по ингибированию опухолевого роста (Р=0,0135, фиг.38), так и по выживанию (Р=0,0087, фиг.39). Эти результаты демонстрируют превосходящую мощность комбинации 992+1024 по сравнению с эрбитуксом, даже при использовании смеси в низкой дозе. Результаты также демонстрируют длительное ингибирование опухолевого роста комбинацией 992+1024 по сравнению с официально разрешенным моноклональным антителом.
ПРИМЕР 9: Рост сфероидов
Для исследования сфероидов в 96-луночный планшет с круглым дном добавляли 35 мкл раствора Poly-HEMA (120 мкг/мл) и оставляли на ночь для выпаривания под вытяжным колпаком. Poly-HEMA предотвращает прикрепление клеток. Клетки A431-NS обрабатывали, как описано выше, подсчитывали и подгоняли концентрацию до 100000 клеток на миллилитр. Затем в экспериментальные лунки столбцов 2-11 добавляли 5 0 мкл клеточной суспензии (5000 клеток на лунку) вместе с 50 мкл 5% раствора матригеля (Matrigel). Для снижения эффекта испарения среды в экспериментальных лунках в ряды 1 и 8, а также в столбцы 1 и 12 добавляли 200 мкл среды. Планшеты центрифугировали на скорости 300×g в течение 5 минут и оставляли на ночь в увлажненном инкубаторе с температурой 37°C для формирования материала. На следующий день соответствующие антитела и смеси антител разбавляли до конечной общей концентрации антител 20 мкг/мл в пустом 96-луночном планшете. Эту процедуру выполняли в среде DMEM с добавками 0,5% FBS и 1% P/S, получая конечную общую концентрацию антител 10 мкг/мл в лунке с наивысшей концентрацией антител. Затем 150 мкл этих растворов добавляли в лунки столбца 2 96-луночного планшета и готовили трехкратные серийные разведения вниз до столбца 9, так, что каждая лунка содержала 100 мкл раствора антител. В столбец 11 добавляли 100 мкл среды. Затем 100 мкл этих растворов переносили в планшет со сфероидами и оставляли для инкубации на 7 дней. Затем в каждую лунку добавляли 20 мкл реактива WST-1, и планшеты инкубировали в течение 1 часа при 37°C. Далее планшеты переносили в орбитальный планшетный шейкер и оставляли еще на один час. Поглощательную способность измеряли на уровне 4 50 и 620 нм /опорная длина волны) считывающим устройством ELISA. Количество метаболически активных клеток (MAC) рассчитывали как процентный показатель по сравнению с необработанным контролем, исходя из следующей формулы:

Смесь трех антител с неперекрывающимися эпитопами в пределах домена III (992+1030+1042) эффективно ингибировала рост сфероидов A431-NS и была более действенной, чем моноклональные терапевтические антитела против EGFR эрбитукс и вектибикс (фиг.22).
ПРИМЕР 10: Связывание с ECD EGFR Cynomolgus
Клонирование внеклеточного домена EGFR Cynomolgus
Внеклеточный домен EGFR Cynomolgus, исключая сигнальный пептид, клонировали из кДНК Cynomolgus, выделенной из эпидермиса при помощи вложенной PCR и сиквенс-специфических праймеров, извлеченных из опубликованной последовательности полноразмерного EGFR человека (GENBANK X00588, Ullrich, A. et. al. Nature 309(5967),418-425 (1984)).
Реактивы для PCR:
кДНК обезьяны Cynomolgous, выделенная из здорового кожного эпидермиса:
CytoMol Unimed, № по каталогу: ccy34218, № партии: A711054.
Реакционный буфер Phusion (5×): Finnzymes, № по каталогу: F-518, № партии: 11.
Фермент Phusion: Finnzymes, F-530S (2 единицы на мкл).
dNTP 25 мМ: Bioline, № По каталогу: BIO-39029, № партии: DM-103F.
Праймеры для амплификации ECD EGFR Cynomolgus, включая частичную сигнальную последовательность и трансмембранный домен:
праймер 5' ATG: 5'-TCTTCGGGAAGCAGCTATGC-3' (SEQ ID NO 135)
праймер 3' Tm 2: 5'-TTCTCCACTGGGCGTAAGAG-3' (SEQ ID NO 136)
Праймеры вложенной PCR для амплификации ECD EGFR Cynomolgus Bp 1-1863 и инкорпорирования XbaI, сайтов рестрикции MluI и стоп-кодона перед трансмембранным доменом:
5' EGFR XbaI: 5'-ATCTGCATTCTAGACTGGAGGAAAAGAAAGTTTGCCAAGGC-3' (SEQ ID NO 137)
3' EGFR MluI: 5'-TACTCGATGACGCGTTTAGGATGGGATCTTAGGCCCGTTCC-3' (SEQ ID NO 138)
Условия PCR:
30 циклов: 98°C/30 секунд плавление, 55°C/30 секунд отжиг, 72°C/60 секунд удлинение. После 30 циклов PCR продуктам реакции давали еще 5 минут для удлинения.
Реакции PCR проводили с 1 мкл матрицы и 2 единицами фермента Phusion в общем объеме 50 мкл реакционного буфера, содержащего 0,2 мМ dNTP, 0,5 мкМ праймера.
Полученная конечная полоска PCR с кажущейся длиной приблизительно 1800-1900 пар оснований была клонирована в векторе экспрессии. ДНК- и белковая последовательность клонированного внеклеточного домена EGFR Cynomolgus показаны на фиг.23, а белковая последовательность ECD EGFR Cynomolgus, выверенная по ECD EGFR человека, показана на фиг.24. Выверка последовательностей ДНК ECD EGFR человека и ECD EGFR Cynomolgus показала идентичность последовательностей на 97,6%, тогда как выверка соответствующих белковых последовательностей показала их идентичность на 98,6%.
Демонстрация перекрестной реактивности антител между внеклеточными доменами EGFR человека и Cynomolgus в анализе ELISA
Для того чтобы подтвердить тот факт, что протестированные антитела против EGFR одинаково хорошо связываются с ECD EGFR, как человека, так и обезьяны Cynomolgus, и в соответствии с этим обосновать проведение токсикологических исследований на обезьянах Cynomolgus, серийные четырехкратные разведения антител, начиная с 1 мкг/мл, были протестированы методом ELISA на связывание с рекомбинантными белками ECD EGFR человека и Cynomolgus. Демонстрация в этом анализе идентичных профилей связывания антител была расценена, как свидетельство хорошей перекрестной реактивности испытуемых антител с EGFR разных биологических видов. Лунки ELISA были покрыты из расчета 50 мкл на лунку полноразмерным EGFR в концентрации 1 мкг/мл в PBS на ночь при 4°C. На следующее утро лунки дважды отмывали PBS-T и на один час блокировали 100 мкл PBS-T-1% BSA при комнатной температуре с последующим двукратным отмыванием в PBS-T. Затем в лунки добавляли 50 мкл серийно разведенных антител против EGFR, а также контрольные антитела и инкубировали материал 1 час при комнатной температуре. После инкубации антител лунки пятикратно отмывали PBS-T, с последующей 30-минутной инкубацией при комнатной температуре с вторичным реактивом стрептавидин-HRP, разбавленным блокирующим буфером в соотношении 1:3000 (из расчета 50 мкл на лунку). Наконец, лунки пятикратно отмывали PBS-T, планшеты разрабатывали, добавляя субстрат TMB (50 мкл на лунку), и инкубировали при комнатной температуре. После инкубации реакцию останавливали, добавляя 1M H2SO4 (100 мкл на лунку), и производили считывание планшетов на уровне OD 450 нм.
Реактивы ELISA:
1. Планшет ELISA, NUNC Maxisorp, № по каталогу: 442404
2. Антиген: ECD rEGFR человека, ECD rEGFR Cynomolgus
3. Покровный буфер: 1×PBS, Gibco № по каталогу: 20012-019
4. Отмывочный буфер: 1×PBS/0,05% Tween 20 (PBS-T)
5. Буфер блокирования/разбавления: 1% BSA в PBS-T
6. Конъюгат козьего IgG против человека и HRP: Serotec, Star 106Р
7. ТМВ Plus (KemEnTec, № по каталогу 4390L)
8. (1М H2SO4)
Как показано на фиг.25, описанный анализ ELISA позволяет провести различие между перекрестной реактивностью антител против EGFR с ECD EGFR человека и Cynomolgus (фиг.25А) и видовой специфичностью антител, распознающих только ECD EGFR человека, которые были использованы для иммунизации мышей (фиг.25В).
ПРИМЕР 11: Ингибирование подвижности
В большинстве случаев смертельный исход рака связан с диссеминацией опухолевых клеток и их последующим ростом в отдаленных участках. Местная инвазия прилегающих здоровых тканей нарушает гомеостатические функции и препятствует хирургическому или радиологическому иссечению опухоли. Недавние исследования пролили свет на центральную роль индуцированной подвижности раковых клеток в этом феномене рассеивания раковых опухолей. Известно, что EGFR способствует клеточной подвижности и рассеиванию опухоли, следовательно, ингибирование опосредованной EGFR подвижности является важным механизмом действия лекарств, нацеленных на EGFR.
Было проведено исследование влияния смеси двух антител 992 и 1024 на подвижность клеток раковой линии карциномы головы и шеи. Сфероиды, состоящие из 10000 клеток, были подготовлены в течение ночи, как это описано в примере 9. Затем сфероиды переносили в культуральные флаконы для клеток NUNC T25 и давали клеткам прикрепиться в течение ночи. Затем добавляли 10 мкг/мл смеси антител 992+1024 или антитело отрицательного контроля, после чего сфероиды инкубировали еще 24 часа. После этого получали изображения с 40-кратным увеличением, а площадь, покрытую клетками, измеряли при помощи компьютерной программы Image J.
Результаты: Как можно видеть на фиг.27А, добавление специфических антител против EGFR 992 и 1024 приводит к значительному снижению площади, покрытой опухолевыми клетками. Количественная оценка подвижности представлена на фиг.27В, которая показывает, что подвижность снизилась приблизительно на 60% по сравнению с применением отрицательного контрольного антитела. Это снижение подвижности высоко значимо статистически (р<0,01).
Таким образом, комбинация антител 992 и 1024 мощно ингибирует подвижность опухолевых клеток, опосредованную EGFR, свидетельствуя о том, что такие комбинации антител против EGFR можно применять для лечения диссеминированного заболевания.
ПРИМЕР 12: Активация инволюкрина композицией антител Sym004
Инволюкрин является маркером ранней дифференциации плоских (чешуйчатых) клеток и представляет собой белок, который участвует в формировании рогового конверта. Поэтому уровень инволюкрина можно использовать, как мерило количества опухолевых клеток, завершивших дифференциацию. Авторы изобретения оценивали уровень инволюкрина в белковых лизатах из ксенотрансплантатов опухолей A431NS, либо не подвергавшихся никаким воздействиям, либо леченных эрбитуксом, вектибиксом или смесью антител 992+1030+1042 с применением поступающего в продажу набора Involucrin ELISA (Biomedical Technologies). Опухолевые лизаты были приготовлены посредством гомогенизации 30-40 мг опухолевых тканей в 1 мл буфера RIPA с применением устройства TissueLyzer компании Qiagen. Концентрацию белка в каждом просветленном лизате определяли, применяя аналитический набор BCA Protein компании Pierce, а уровень инволюкрина оценивали анализом ELISA в 0,4 мкг белка из каждого образца.
Результаты: Как можно видеть на фиг.27, обнаруживается значительно более высокий уровень инволюкрина в группе, получавшей лечение смесью антител 992+1030+1042, по сравнению с отрицательным контролем, а также группами, получавшими эрбитукс и вектибикс. Таким образом, комбинация антител 992, 1030 и 1042 повышает уровень инволюкрина в ксенотрансплантатах опухолей A431NS, то есть предположительно индуцирует высокую степень дифференциации опухолевых клеток A431NS. В этой группе лечения был получен результат, который хорошо коррелируется с большим количеством раковых жемчужин (см. пример 8).
ПРИМЕР 13: Интернализация EGFR композицией антител Sym004
Некоторые антитела функционируют, индуцируя интернализацию своей цели на поверхности клетки. Известно, что EGFR подвергается интернализации при активации таким лигандом, как EGF.
Способность смеси двух антител 992 и 1024 индуцировать интернализацию EGFR была исследована с применением конфокальной микроскопии. Клетки A431NS и HN5 высевали на 8-луночные предметные стекла LabTek и инкубировали в течение ночи в среде DMEM с добавкой 0,5% FBS. Затем к клеткам добавляли 10 мкг/мл смеси антител 992+1024 с меткой Alexa-488 или контрольное антитело эрбитукс с последующей инкубацией в течение разного времени. Затем получали изображения с 60-кратным увеличением при помощи конфокального микроскопа Biorad или с большим, или с маленьким отверстием диафрагмы.
Результаты: Как показано на фиг.29A, добавление специфичных к EGFR антител 992 и 1024 с меткой Alexa-488 на 2 часа приводит к накоплению антител во внутриклеточных везикулах клеток из линий A431NS и HN5. В отличие от этого, эрбитукс обнаруживается, главным образом, на клеточной поверхности. Фиг.29B показывает изображения клеток A431NS с применением меньшего отверстия диафрагмы, которые позволяют получить более тонкие срезы клеток. Из этих изображений явно следует, что антитела 992+1024 локализуются внутри клетки, тогда как эрбитукс находится, главным образом, на клеточной поверхности. Фиг.30 показывает период времени для интернализации, опосредованной смесью антител 992+1024: уже через 30 минут после добавления этих антител их можно обнаружить во внутриклеточных везикулах. Спустя 4 часа почти все антитела 992+1024 обнаруживаются внутри клеток при небольшом или очень слабом окрашивании на клеточной поверхности. В отличие от этого, эрбитукс остается на клеточной поверхности. Также были получены данные, свидетельствующие о том, что интернализация, индуцированная смесью антител 992+1024, приводит к стойкому разрушению EGFR и его исчезновению из клеток.
Таким образом, комбинация антител 992 и 1024 быстро и мощно индуцирует интернализацию EGFR, тогда как эрбитукс не обладает такой способностью.
ПРИМЕР 14: Измерение аффинности антитела поверхностным плазмонным резонансом
Измерение моновалентной аффинности антител IgG Sym004 против рекомбинантного растворимого ECD EGFR
Кинетический анализ полноразмерных антител IgG, предлагаемых изобретением, был проведен в системе BIAcore 2000 по аналитической методике, описанной в ссылке (Canziani, Klakamp, et al. 2004, Anal. Biochem, 325:301-307), которая позволяет измерить моновалентную аффинность целых молекул IgG против растворимого антигена. Вкратце, приблизительно 10000 Ru поликлонального антитела IgG Fc против человека были конъюгированы с поверхностью чипа CM5 согласно инструкциям производителя, что сопровождалось последующим захватом 25 мкг индивидуальных антител против EGFR, предлагаемых изобретением, или отрицательного контрольного антитела Synagis на поверхности чипа против Fc. Плотность захваченного IgG была оптимизирована для каждого клона таким образом, что связывание наивысшей концентрации антигена, задействованной в анализе, не превышало 25 Ru. Затем для генерирования характеристических кривых впрыскивали на скорости потока 25 мкл/мин в серийных двукратных разведениях в буфере HBS-EP 250 мкл растворимого ECD EGFR человека, который по данным вытеснительной гель-хроматографии содержал только моновалентный белок. Поверхность чипа восстанавливали между циклами, снимая с нее комплексы антиген/антитело 10-секундным впрыскиванием 100 мМ H3PO4. Кинетический анализ был проведен посредством первого вычитания реакции проточной кюветы, содержащей отрицательное контрольное антитело Synagis с последующим вычитанием реакции, вызванной изолированным впрыскиванием чистого буфера HBS-EP ("двойная опорная точка"). Константу скорости ассоциации (ka) и константу диссоциации (kd) оценивали глобально по генерированным сенсограммам при помощи ознакомительной компьютерной программы BIA 4.1, предоставленной производителем.
Реактивы:
1. Чип CM5: Biacore, № по каталогу: BR-1000-14
2. NHS: Biacore BR-1000-50
3. EDC: Biacore BR-1000-50
4. 10 мМ ацетатный буфер pH 4,5: Biacore, № по каталогу: BR-1003-50
5. Козий IgG Fc против человека: Caltag, № по каталогу: H10500
6. Этаноламин, 1,0M pH 8,5: Biacore BR-1000-50
7. Подвижный буфер 10×HBS-EP: 0,01M HEPES pH 7,4, 0,15M NaCl, 3 мМ EDTA, 0,005% об./об. сурфактанта P20
8. Антиген: Внеклеточный домен EGFR человека с 6×His.
9. 100 мМ H3PO4
Расчетные показатели моновалентной аффинности полноразмерных IgG, предлагаемых изобретением, против растворимого ECD EGFR человека показаны ниже в таблице 8.
Большинство протестированных антител Sym004 распознавали растворимый ECD EGFR человека с аффинностью в диапазоне 10-20 нМ, за исключением антител 1260, 1277 и 1284, которые имели большую аффинность: 4,2 нМ, 0,4 нМ и 4,6 нМ, соответственно. Наконец, было обнаружено, что антитело 992 связывается с растворимым ECD EGFR намного с меньшей аффинностью, чем другие протестированные антитела. Позднее был проведен кинетический анализ этого антитела по методике Скэтчарда, в результате которого была продемонстрирована аффинность против растворимого ECD EGFR человека 170 нМ.
Измерение аффинности антител Fab Sym004 против иммобилизованного рекомбинантного ECD EGFR
Для того чтобы исследовать возможные различия в презентации антигена между ECD EGFR, представленным в растворимой и иммобилизованной форме, было проведено новое измерение аффинности иммобилизованного химерного антигена рецептора EGF, получившего название EGFR-Fc (R&D Systems, 344-ER), который состоит из ECD EGFR человека, гибридизированного с Fc IgG человека. С этой целью были генерированы фрагменты Fab антител IgG 992, 1024 и 1030, позволяющие провести измерение моновалентной аффинности.
Выработка Fab:
Фрагменты Fab антител 992, 1024 и 1030 были получены посредством переваривания папаином с применением набора для изготовления Fab компании Pierce и в соответствии с инструкциями производителя. Вкратце, 2 мг каждого антитела IgG были подвергнуты буферному обмену в колонках NAP-5 (Amersham Biosciences) со свежеприготовленным буфером переваривания, содержащим 20 мМ цистеина-HCl, pH 7,0, в соответствии с инструкциями производителя. Затем 350 мкл кашицы из папаинового бисера были дважды отмыты в том же буфере для переваривания с последующим осаждением и сбросом супернатанта. Антитела были переварены при добавлении к бисеру 1 мл антитела IgG с буферным обменом и инкубированы в течение ночи при 37°C с встряхиванием на скорости 1000 оборотов в минуту. На следующее утро непереваренный IgG отделяли от чернового материала Fab посредством истощения полноразмерного IgG в колонках HiTrap Protein A (Ge Healthcare). Полученный материал Fab в заключение был подвергнут диализу PBS в течение ночи и проанализирован способом SDS-PAGE в редуцирующих и нередуцирующих условиях. Полоса белка приблизительно 50 кДа в нередуцирующих условиях была расценена как указание на успешную выработку Fab.
Реактивы:
1. Набор ImmunoPure для изготовления Fab, Pierce, № по каталогу: 44885
2. Опресняющая колонка NAP5, Amersham, № по каталогу: 17-0853-02
3. PBS pH 7,2, Gibco; #20012-019
4. HiTrap Protein A HP, колонка 1 мл, GE Healthcare, #17-0402-01
5. NuPAGE 4-12% гель Novex Bis-Tris, Invitrogen, #NP0322BOX
6. Молекулярный маркер, Seeblue Plus 2, Invitrogen, # LC5925
7. Антитела против EGFR - 2,0 мг каждого
Кинетический анализ антител Fab, рассматриваемых в изобретении, был проведен в системе Biacore 2000 с применением рекомбинантного антигена, иммобилизованного на поверхности сенсора с очень низкой плотностью во избежание ограничений в переносе массы. Вкратце, общее количество 285 Ru рекомбинантного химерного ECD-Fc EGFR (R&D Systems, № по каталогу 344-ER) было конъюгировано с поверхностью чипа CM5 в соответствии с инструкциями производителя. Затем фрагменты Fab, полученные из антител, предлагаемых изобретением, были протестированы в серийных двукратных разведениях, начиная с оптимизированной концентрации, которая не приводила к максимальным значениям Ru выше 25 при тестировании на чипе с иммобилизованным EGFR. Кинетический анализ был проведен посредством первого вычитания реакции, вызванной изолированным впрыскиванием чистого буфера HBS-EP. Константу скорости ассоциации (ka) и константу диссоциации (kd) оценивали глобально по генерированным сенсограммам при помощи ознакомительной компьютерной программы BIA 4.1, предоставленной производителем.
Расчетные показатели аффинности протестированных фрагментов Fab антител, предлагаемых изобретением, против растворимого ECD EGFR человека показаны ниже в таблице 9.
Как показано выше в таблице 9, было обнаружено, что фрагменты Fab антител 992 и 1024 имеют аффинность 150 нМ и 26 нМ, соответственно, что соответствует показателям аффинности, представленным в предыдущем примере, что иллюстрирует незначительные различия в распознавании антителами растворимого и иммобилизованного EGFR для этих двух антител. Однако антитело 1030 проявляло в десять раз более высокую аффинность 2,3 нМ против иммобилизованного антигена по сравнению с растворимым рецептором и, вследствие этого, преимущественно распознавало эпитоп, представленный на иммобилизованном антигене.
ПРИМЕР 15: Исследование презентации антигена EGFR и ранжирование функциональной аффинности на клетках A431-NS
Сравнение между презентацией антигена на клетках A431-NS и очищенным полноразмерным рецептором EGFR
Поскольку кинетический анализ показал, что антитело 992 распознавало ECD рекомбинантного EGFR с аффинностью в диапазоне 150-170 нМ, было проведено исследование по вопросу о том, связана ли эта слабая аффинность с тем фактом, что mAb 992 преимущественно связывает нативные конформации EGFR, экспрессируемые на клетках A431-NS, в отличие от конформаций, представленных на ECD рекомбинантного EGFR или от полноразмерного EGFR, очищенного от клеток A431. Для того чтобы исследовать различия в презентациях антигена рецептора EGF, были проведены исследования конкурентным методом ELISA по анализу связывания для субпопуляций антител, предлагаемых изобретением, с применением фрагментов Fab во избежание эффектов авидности на протестированных клетках A431-NS и очищенном полноразмерном EGFR из тех же клеток.
Выработка Fab: Выработку фрагментов Fab осуществляли тем же способом, который был описан в примере 14.
Непрямой анализ ELISA: Для непрямого анализа ELISA, полноразмерный EGFR (Sigma E2645) покрывали из расчета 1 мкг/мл карбонатным буфером (50 мкл на лунку) на ночь при температуре 4°C. На следующее утро лунки дважды отмывали PBS-T и на один час блокировали PBS-T, содержащим 1% BSA при комнатной температуре с последующим двукратным отмыванием в PBS-T. Затем в независимые лунки ELISA добавляли 50 мкл серийных разведений антител Fab в среде DMEM, содержащей 1% BSA и инкубировали материал 1 час при комнатной температуре, после чего лунки четырежды отмывали PBS-T. Затем добавляли 50 мкл конъюгата вторичного козьего антитела против человека (Fab-специфичного) и HRP, разбавленного 1:5000 в среде DMEM, содержащей 1% BSA, и инкубировали материал на льду в течение 30 минут. Наконец, лунки четырежды отмывали PBS-T, планшеты разрабатывали, добавляя субстрат TMB (50 мкл на лунку) и проводили считывание на волне 620 нм каждые 5-15-30 минут. После инкубации с субстратом реакцию останавливали, добавляя 1M H2SO4, и считывали поглощательную способность на волне 450 нм.
Реактивы, непрямой анализ ELISA:
1) Покровный буфер: 50 мМ карбонатный буфер, pH 9,8
2) Антигены: Полноразмерный EGFR дикого типа, очищенный от клеток A431, Sigma E2645
3) Планшет ELISA: NUNC Maxisorp, № по каталогу: 442404
4) Отмывочный буфер: 1×PBS/0,05% Tween 20 (PBS-T)
5) Буфер блокирования/разбавления: 1% BSA в PBS-T (PBS-T-1% BSA)
6) Буфер для разбавления антител: среда DMEM с содержанием 1% BSA
7) Конъюгат козьего антитела против человека (Fab-специфичного) и HRP: Jackson, № по каталогу: 109-035-097
8) Субстрат TMB Plus: KemEnTec, № по каталогу: 4390L
9) 1M H2SO4
Клеточный анализ ELISA: Относительную аффинность связывания, соответствующую молярной концентрации, дающей половину максимальной OD (ED50), определяли посредством титрования антител на клетках A431-NS. Вкратце, 10000 клеток A431-NS выращивали в 96-луночных планшетах ELISA, содержащих среду DMEM с добавками 0,5% FCS и 1% P/S при 37°C, в атмосфере с 5% CO2 в течение ночи. На следующее утро слившиеся клетки (приблизительно 20000 на лунку) дважды отмывали PBS и фиксировали 15-минутной инкубацией с 1% раствором параформальдегида при комнатной температуре с последующим четырехкратным отмыванием PBS. Затем тестируемые антитела EGFR и антитело для отрицательного контроля Synagis серийно разводили в среде DMEM, содержащей 1% BSA, 50 мкл каждого разведения добавляли в лунки и инкубировали 1 час при комнатной температуре, после чего лунки четырежды отмывали PBS. Затем добавляли 50 мкл конъюгата вторичного козьего антитела против человека (Fab-специфичного) и HRP, разбавленного 1:5000 в среде DMEM, содержащей 1% BSA, и инкубировали на льду в течение 30 минут. Наконец, лунки четырежды отмывали PBS, планшеты разрабатывали, добавляя субстрат TMB Plus (50 мкл на лунку) и проводили считывание на волне 620 нм каждые 5-15-30 минут. После инкубации с субстратом реакцию останавливали, добавляя 1M H2SO4, и считывали поглощательную способность на волне 450 нм. Функциональную аффинность, выраженную в величинах ED50, рассчитывали посредством вычитания среднего фонового связывания только с вторичным реактивом и последующей нормализации кривых связывания на построенных графиках процентных величин от максимального связывания применительно к каждому протестированному антителу.
Реактивы, клеточный анализ ELISA:
1) Среда DMEM: Gibco, № по каталогу: 41966-029
2) FCS: Gibco, № по каталогу: 10099-141
3) Пенициллин-стрептомицин (P/S): Gibco, № по каталогу: 15140-122
4) Планшет ELISA: Costar, № по каталогу: 3595
5) Отмывочный буфер (PBS): Gibco, № по каталогу: 20012-019
6) Буфер для разбавления антител: среда DMEM с содержанием 1% BSA
7) Раствор для фиксации клеток: BD Biosciences, № по каталогу: 340181
8) Конъюгат козьего антитела против человека (Fab-специфичного) и HRP: Jackson, № по каталогу: 109-035-097
9) Субстрат TMB Plus: KemEnTec, № по каталогу: 4390L
10) 1M H2SO4
Различия в антигенной презентации рецептора EGF, экспрессируемого на клетках A431-NS и рецептора, очищенного от этих клеток, определяли в исследованиях связывания на основе конкурентного анализа ELISA, с применением такого же реактива вторичного антитела и тех же значений времени инкубации. Результаты показаны на фиг.31. Эксперимент отчетливо показал, что фрагменты Fab антител 992 и 1024 слабо связываются с очищенным полноразмерным EGFR, нанесенным в виде покрытия на лунки ELISA, по сравнению с связыванием Fab 1030 в тех же концентрациях. Однако эта слабая активность связывания антител 992 и 1024 восстанавливалась, когда антитела проходили тестирование на клетках A431-NS, по отношению к которым фрагменты Fab всех трех антител демонстрировали сильную активность связывания. Сравнение двух разных анализов ELISA четко проиллюстрировало преимущественное связывание фрагментов Fab антител 992 и 1024 с нативными конформациями EGFR, экспрессируемыми на клеточной поверхности, в отличие от конформаций, представленных на очищенном антигене в лунках ELISA. Этот результат также позволяет предположить, что очевидно слабая аффинность антитела 992 при измерении методом поверхностного плазмонного резонанса на рекомбинантном и иммобилизованном ECD EGFR была обусловлена неблагоприятной презентацией эпитопа антитела 992 в протестированных системах.
Ранжирование функциональной аффинности на клетках A431-NS.
Клеточные анализы ELISA, проведенные так, как это описано выше, были использованы для ранжирования функциональной аффинности IgG и фрагментов Fab антител 992, 1024, 1030, вектибикс и эрбитукс посредством вычисления половинных значений максимальной OD, выраженных как значения ED50. Результаты этого анализа показаны на фиг.32, а расчетные значения ED50 представлены ниже в таблице 10.
Эксперимент отчетливо показал, что, если принимать во внимание эффекты авидности, то IgG 992 и 1024 связывали клетки A431-NS с более высокой авидностью, чем эрбитукс и вектибикс, тогда как IgG 1030 имел низшую аффинность тестированных антител IgG. Однако при определении моновалентной аффинности к клеткам фрагментов Fab антитело 992 проявляло минимальную аффинность на уровне приблизительно 10 нМ. Однако эта моновалентная функциональная аффинность все-таки была, по меньшей мере, в 15 раз ниже, чем при тестировании в системе BIAcore.
ПРИМЕР 16: Исследование усиления связывания, индуцированного антителами
Конкурентный эксперимент BIAcore, проведенный на парах антител, предлагаемых изобретением, показал, что связывание антител 992 и 1024 усиливалось приблизительно на 55% и 58%, соответственно (фиг.9A), когда эти антитела тестировались против друг друга в обоих направлениях. Для дальнейшего изучения этого феномена был спланирован клеточный анализ ELISA с использованием нефиксированных клеток для исследования эффекта связывания клона антитела IgG при предварительном насыщении рецептора фрагментом Fab другого антитела, связывающего неперекрывающийся эпитоп.
Клеточный анализ ELISA: Анализ ELISA был проведен, по существу, так же, как описано в примере 15, но с некоторыми модификациями. Клетки оставляли нефиксированными, чтобы обеспечить конформационную гибкость EGFR после добавления антитела. Вкратце, 10000 клеток A431-NS выращивали в 96-луночных планшетах ELISA, содержащих среду DMEM с добавками 0,5% FCS и 1% P/S при 37°C, в атмосфере с 5% CO2 в течение ночи. На следующее утро слившиеся клетки (приблизительно 20000 на лунку) дважды отмывали PBS, а лунки для исследования индуцированного антителами усиления связывания преинкубировали с 25 мкл 40 нМ одиночных фрагментов Fab антител 992, 1024 или 1030 либо с 12,5 мкл 80 нМ каждого единичного Fab в описанных выше двойных комбинациях для насыщенного связывания. В лунки, используемые для тестирования антител IgG без добавления фрагментов Fab, добавляли 25 мкл среды DMEM, содержащей 1% BSA. После добавления Fab и среды лунки ELISA в течение 30 минут инкубировали при комнатной температуре, после чего в них добавляли 25 мкл серийных трехкратных разведений антител IgG, предлагаемых изобретением, или антитела Synagis для отрицательного контроля, начиная с концентрации 360 нМ, и инкубировали на льду еще один час. Затем лунки четырежды отмывали PBS, добавляли 50 мкл конъюгата вторичного моноклонального мышиного антитела против человека (Fc-специфичного) и HRP, разбавленного 1:5000 в среде DMEM, содержащей 1% BSA, и инкубировали на льду в течение 30 минут. Наконец, лунки четырежды отмывали PBS, планшеты разрабатывали, добавляя субстрат TMB Plus (50 мкл на лунку) и проводили считывание на волне 620 нм каждые 5-15-30 минут. После инкубации с субстратом реакцию останавливали, добавляя 1M H2SO4, и считывали поглощательную способность на волне 450 нм. Функциональную аффинность, выраженную в величинах ED50, рассчитывали посредством вычитания среднего фонового связывания только с вторичным реактивом и последующей нормализации кривых связывания на построенных графиках процентных величин от максимального связывания применительно к каждому протестированному антителу.
Реактивы, клеточный анализ ELISA:
1) Среда DMEM: Gibco, № по каталогу: 41966-029
2) FCS: Gibco, № по каталогу: 10099-141
3) Пенициллин-стрептомицин (P/S): Gibco, № по каталогу: 15140-122
4) Планшет ELISA: Costar, № по каталогу: 3595
5) Отмывочный буфер (PBS): Gibco, № по каталогу: 20012-019
6) Буфер для разбавления антител: среда DMEM с содержанием 1% BSA
7) Конъюгат мышиного антитела против человека (Fc-специфичного) и HRP: Ab-direct, № по каталогу: MCA647P
8) Субстрат TMB Plus: KemEnTec, № по каталогу: 4390L
9) 1M H2SO4
Исследования усиления связывания, индуцированного антителами, были проведены посредством конкурентного анализа ELISA с применением того же реактива вторичного антитела и тех же временных периодов инкубации. Результаты этого исследования показаны на фиг.33, а расчетные значения ED50 представлены ниже в таблице 11.
Как представлено на фиг.33 и в таблице 11 выше, IgG 992 демонстрировало явное усиление связывания при предварительном насыщении рецептора фрагментами Fab антител 1024, 1030 или комбинацией 1024 и 1030. Инкубация с фрагментами Fab приводила к снижению величин ED50 на 0,4 и 0,5 нМ, соответственно, по сравнению с 0,6 нМ при тестировании одного IgG 992 (без насыщения рецептора фрагментами других антител). Сходным образом, усиленное связывание при предварительном насыщении клеток Fab 992 также демонстрировали IgG 1024 и 1030, а также IgG 1024, если перед ним к клеткам добавляли фрагменты Fab 992 и 1030. Этот результат четко иллюстрирует преимущество обладания более чем одним антителом против неперекрывающихся эпитопов на одном и том же целевом рецепторе.
В этом эксперименте были обнаружены несколько меньшие показатели функциональной аффинности по сравнению с примером 2. Такой результат, вероятно, связан с тем, что в настоящем примере был использован иной вторичный реактив, а также с тем, что протестированные IgG были инкубированы с нефиксированными клетками на льду во избежание интернализации.
ПРИМЕР 16B: Клонирование полноразмерного EGFR Cynomolgus
Полноразмерный EGFR Cynomolgus, включая сигнальный пептид, клонировали из кДНК Cynomolgus, выделенной из эпидермиса при помощи вложенной PCR, и сиквенс-специфических праймеров, извлеченных из опубликованной последовательности полноразмерного EGFR человека (GENBANK X00588, Ullrich, A. et. al. Nature 309(5967), 418-425 (1984)).
Реактивы для PCR:
кДНК обезьяны Cynomolgous выделенная из здорового кожного эпидермиса: CytoMol Unimed, № по каталогу: ccy34218, № партии: A711054.
Реакционный буфер FastStart (10Ч): Roche, № по каталогу: 03 553 361 001
Фермент FastStart: Roche, № по каталогу: 03 553 361 001
Фермент Phusion: Finnzymes, F-530S (2 единицы на мкл).
dNTP 25 мМ: Bioline, № по каталогу: BIO-39029
Праймеры для амплификации полноразмерного EGFR Cynomolgus, включая сигнальную последовательность:
праймер 5' ATG: 5'-TCTTCGGGAAGCAGCTATGC-3' (SEQ ID NO 135)
праймер 3' STOP: 5'-TCATGCTCCAATAAATTCACTG-3' (SEQ ID NO 139)
Условия PCR:
95°C - 2 минуты, 40 циклов: 95°C - 30 секунд, 55°C - 30 секунд, 72°C - 3 минуты 30 секунд с заключительной инкубацией при 72°C в течение 5 минут.
Праймеры для амплификации полноразмерного EGFR Cynomolgus и инкорпорирования сайтов рестрикции Not и Xho во вложенной PCR:
E579 Cyn Not5' 5'-GGAGTCGGCGGCCGCACCATGCGACCCTCCGGGACGG-3' (SEQ ID NO 140)
E580 Cyn Xho5' 5'-GCATGTGACTCGAGTCATGCTCCAATAAATTCACTGC-3' (SEQ ID NO 141)
Условия PCR:
95°C - 2 минуты, затем 30 циклов: 95°C - 30 секунд плавление, 55°C - 30 секунд отжиг, 72°C - 3 минуты удлинение. После 30 циклов PCR продуктам реакции давали еще 10 минут для удлинения.
Реакции PCR проводили с 0,5 мкл шаблона и 0,1 мкл фермента Phusion, 0,4 мкл фермента FastStart в общем объеме реакционного буфера 50 мкл с конечной концентрацией 1Ч буфера FastStart, 0,2 мМ dNTP и 0,2 мкМ каждого праймера.
Фрагмент PCR с кажущейся длиной приблизительно 4000 п.о. был получен и клонирован с применение набора для клонирования TOPO TA (Invitrogen, номер изделия 4506-41), а затем секвенирован. ДНК- и белковая последовательность клонированного EGFR Cynomolgus показана на фиг.34. Выверка белковой последовательности EGFR человека и EGFR Cynomolgus показала идентичность последовательности 99,2%.
Демонстрация перекрестной реактивности антител между полноразмерными EGFR человека и Cynomolgus посредством анализа FACS
Полноразмерные EGFR человека и Cynomolgus были экспрессированы на поверхности клеток CHO посредством стабильной трансфекции, а клетки протестированы на связывание с использованием панели серийно разведенных антител к EGFR методом анализа FACS. Определения были сделаны в KD-зависимых условиях при сохранении молярного избытка антитела на уровне, по меньшей мере, в 5 раз превышающем количество молекул антигена EGFR, экспрессированных на клеточной поверхности фиксированного количества клеток во всех сериях разведений антитела. Такая схема исследования позволяла провести анализ FACS для связывания антител при полном насыщении рецептора во всем диапазоне концентраций тестируемого антитела.
Вкратце, количественный анализ FACS был проведен в матричной системе биоанализатора BD FACS для определения количества молекул EGFR, экспрессированных на поверхности клеток CHO, которые были трансфицированы полноразмерным EGFR человека или обезьяны Cynomolgus. Анализ был проведен посредством титрования IgG эрбитукс с меткой PE на клетках и определения количества молекул эквивалента PE при сравнении со стандартной кривой, полученной по частицам калибровки Rainbow с известной плотностью PE. Количественный анализ показал, что клетки CHO, трансфицированные EGFR, демонстрировали на поверхности каждой клетки приблизительно 350000 молекул. Затем серийные 5-кратные разведения предлагаемых изобретением антител, начиная с 5 нМ, посредством инкубации сопоставляли с 10000 клеток CHO, трансфицированных EGFR, в увеличивающемся объеме, что позволяло создать, по меньшей мере, 5-кратный молярный избыток антитела по сравнению с антигеном EGFR, представленным на поверхности в каждом определении. Антитела инкубировали с клетками в течение 14 часов в шейкере, чтобы стимулировать полное насыщение антигена во всем диапазоне протестированных концентраций антитела, наряду с тем, что добавляли буфер FACS с 0,02% NaN3 и поддерживали температуру на уровне 4°C для профилактики интернализации рецептора. После инкубации клетки осаждали центрифугированием на скорости 1200 об/мин в течение 5 минут при 4°C и ресуспендировали в 200 мкл буфера FACS. Далее клетки окрашивали с вторичным козьим F(ab')2 антителом IgG против человека FcGamma PE в разведении 1:500 и инкубировали в течение 30 минут при 4°C в шейкере. Наконец, клетки дважды отмывали в буфере FACS и анализировали в матричной системе биоанализатора BD FACS с пропусканием клеток CHO, экспрессирующих EGFR, которые демонстрировали однородные рассеивающие свойства вперед/вбок.
Реактивы FACS:
Частицы калибровки Rainbow: BD, № по каталогу: 559123
Буфер FACS: 1ЧPBS + 2% FCS + 0,02% NaN3
Антитело IgG: козий F(ab')2 против человека FcGamma PE: Jackson ImmunoResearch, № по каталогу: 109-116-170
Описанный анализ связывания FACS был применен для определения перекрестной реактивности антител IgG против EGFR 992 и 1024 и их сравнения с контрольным антителом IgG 1320, которое не проявляет перекрестной реактивности с EGFR Cynomolgus. Как показано ниже на фиг.40, описанный анализ FACS позволил четко отделить антитела, проявляющие выраженную перекрестную реактивность между полноразмерными EGFR человека и обезьяны Cynomolgus (фиг.40A - IgG 992 и фиг.40B - IgG 1024), от видоспецифических антител, распознающих только полноразмерный EGFR человека (фиг.40C - IgG 1320). По результатам этого анализа был сделан вывод о том, что антитела IgG 992 и 1024 проявляют блестящую перекрестную реактивность против полноразмерных EGFR человека и обезьяны Cynomolgus, экспрессированных на поверхности стабильно трансфицированных клеток CHO. Различие в связывании между EGFR cynomolgus и EGFR человека удивительно в связи с высокой степенью сходства последовательностей и подчеркивает важность тестирования антител на связывание с точной целевой последовательностью у животных, используемых в доклинических токсикологических исследованиях.
ПРИМЕР 17: Клоны, гомологичные антителам 992, 1024 и 1030
Скрининг клонов антител на связывание с EGFR, основанный на иммуносорбентных анализах (ELISA) и клеточных анализах, привел к идентификации клонов 992, 1024, 1030, как это описано в предыдущих примерах. Были также идентифицированы EGFR-специфичные клоны, гомологичные клонам 992, 1024 и 1030 (таблица 12).
Ожидается, что клоны, принадлежащие к одному и тому же кластеру, будут иметь одну и ту же специфичность связывания, но при этом могут связываться с разной аффинностью. Следовательно, клоны в пределах одного кластера могут замещать друг друга в композициях антител, предлагаемых настоящим изобретением, при том условии, что их аффинность связывания не будет различаться слишком сильно.
Таблица12 IGHV
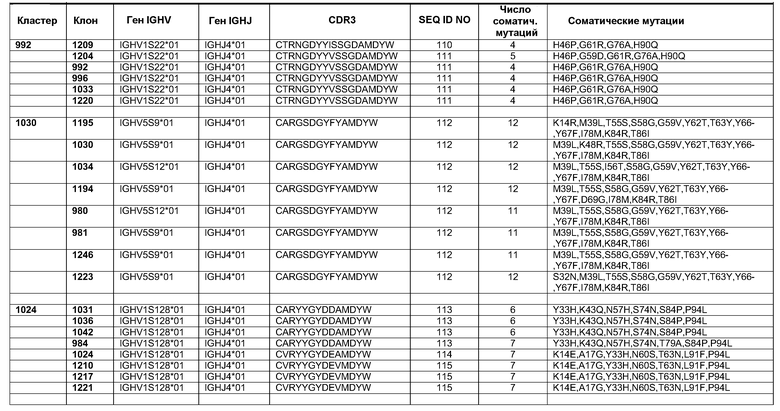
Таблица 12 продолжение IGKV
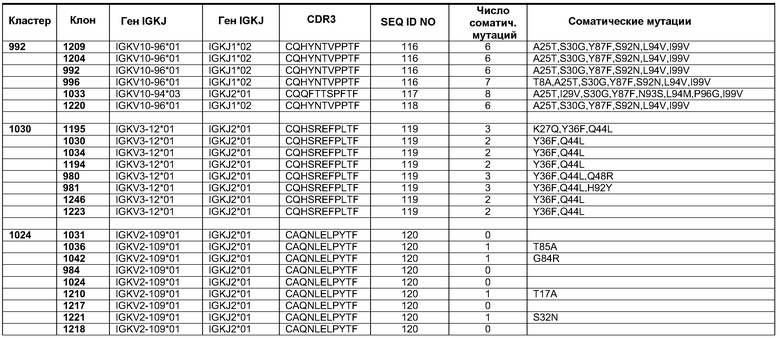
ПРИМЕР 18: Гуманизация антител 922 и 1024
Все антитела содержат для вызова у человека реакции анти-антител. Реакция в какой-то мере коррелируется со степенью "очеловечения" (гуманизации) используемого терапевтического антитела. Предсказать иммуногенность и, следовательно, выработку анти-антител у человека невозможно, но в клинической практике имеется тенденция к предпочтительному использованию антител с высокой степенью гуманизации. Гуманизацию антител, описанных в настоящем изобретении, можно повысить в процессе гуманизации [Reichert JM, Monoclonal antibodies in the clinic. Nature Biotechnol, 2001; 19:819-822; Reichert JM, Rosensweig CJ, Faden LB and Dewitz MC. Monoclonal antibody successes in the clinic. Nature Biotechnol, 2005; 23:1073-1078].
Гуманизация мышиного mAb, в принципе, достигается посредством пересадки участков определения комплементарности (CDR) в каркасные участки (FR) доменов IGHV и IGKV человека с близкородственной последовательностью при помощи процедуры, обычно называемой пересадкой CDR (Jones PT, Dear PH, Foote J, Neuberger MS and Winter G. Replacing the complementarity-determining regions in a human antibody with those from a mouse. Nature, 1986; 321:522-525). Однако простая пересадка CDR только из гипервариабельных участков может привести к снижению аффинности, поскольку некоторые каркасные аминокислоты или участки создают критический контакт с антигеном или поддерживают конформацию петель CDR, связывающих антиген [Queen C, Schneider WP, Selick HE, Payne PW, Landolfi NF, Duncan JF, Avdalovic NM, Levitt M, Junghans RP and Waldmann TA. A humanized antibody that binds to the interleukin 2 receptor. Proc Natl Acad Sci U S A, 1989; 86:10029-10033; Al-Lazikani B, Lesk AM and Chothia C. Standard conformations for the canonical structures of immunoglobulins. J Mol Biol, 1997; 273:927-948]. Поэтому гуманизация антител должна включать как пересадку петель CDR из вариабельных областей мышиного происхождения в высоко гомологичный каркас человека, так и сохранение ключевых остатков мышиного каркаса с документированным влиянием на активность связывания антигена (Winter, G. and W.J. Harris. "Humanized antibodies." Immunol. Today 14. 6 (1993): 243-46). Для достижения гуманизации при сохранении аффинности и функции антител были разработаны и успешно применены несколько способов (см. обзор в ссылке Almagro, J.C. and J. Fransson. "Humanization of antibodies." Front Biosci. 13 (2008): 1619-33). Гуманизации можно достичь рациональными способами, например, пересадкой CDR, перепрофилированием, супергуманизацией, оптимизацией содержимого цепочки человека, причем все эти способы подразумевают конструирование новых кандидатов на роль гуманизированных антител. Аминокислотная последовательность кандидатов основана на понимании и предсказании структуры антитела, а также на вкладе отдельных аминокислот в связывание антигена прямым и опосредованным образом через стабилизацию общей структуры участков, взаимодействующих с антигеном. Как правило, кандидаты должны быть очищены, а некоторые аминокислоты мутированы в обратном направлении к исходным мышиным остаткам, поскольку каждое антитело имеет непредвиденные индивидуальные ограничения. Общим для всех способов является то, что для сохранения аффинности и функциональности может потребоваться несколько успешных этапов конструирования, тестирования и переконструирования. Альтернативой являются более эмпирические подходы, которые подразумевают создание обширных комбинаторных библиотек и извлечение антител с желательными свойствами из пула вариантов посредством отбора такими способами как дрожжевой или фаговый дисплей, либо альтернативные методы скрининга.
Антитела против EGFR, описанные в настоящем изобретении, могут быть гуманизированы посредством пересадки CDR в V-области человека. В предпочтительном сценарии отбор V-области человека основывается на гомологии с исходной мышиной V областью. У человека также можно использовать V-области гена с другими желательными свойствами, например, с низкой иммуногенностью. Настоящий пример описывает способ гуманизации химерных антител против EGFR 992 и 1024. Гуманизированные последовательности, показанные на фиг.41A, были получены посредством пересадки определяемых IMGT участков CDR из IGHV 992 в IGHV1-46/IGHJ4 и из IGKV 992 в IGKV1-27/IGKJ1-01. Аминокислотные последовательности, показанные на фиг.41B, были получены in silico посредством пересадки определяемых IMGT участков CDR из IGHV 1024 в IGHV1-2*02/IGHJ6*02 и из IGKV 1024 в IGKV2-28*01/IGKJ2*01. Были синтезированы искусственные гены, кодирующие указанные гуманизированные антитела, и эти гены были вставлены в вектор экспрессии млекопитающего. Антитела были экспрессированы, очищены и протестированы на активность, как это описано в примере 3. После начального тестирования можно определить кинетику связывания гуманизированных антител методом поверхностного плазмонного резонанса, как это описано в примере 14. Подобным образом, можно определить связывание с hEGFR, экспрессированным на поверхности клеток, как это описано в примере 15.
Если связывающая активность гуманизированных аминокислот оказывается значительно ниже, чем наблюдаемая у исходных антител, то для восстановления аффинности применяют последовательную схему обратных мутаций, начиная с гуманизированных каркасных остатков, расположенных в зоне Vernier, или остатков, предположительно, поддерживающих структуру участков CDR (Foote, J. and G. Winter. "Antibody framework residues affecting the conformation of the hypervariable loops." J Mol. Biol. 224.2 (1992): 487-99; Padlan, E.A. "Anatomy of the antibody molecule." Mol. Immunol 31. 3 (1994): 169-217.). Согласно нумерации IMGT эти остатки для IGHV 992 представляют аминокислоты 13, 45 и 80, для IGKV 992 - аминокислоту 25, для IGHV 1024 - аминокислоты 13, 45, 80 и 82, для IGKL 1024 - аминокислоту 78. Эти мутанты могут быть сконструированы посредством PCR-опосредованного сайт-направленного мутагенеза с применением стандартных методов молекулярной биологии. Сконструированные мутанты подлежат тестированию, как это описано выше. Ожидается, что эти наборы кандидатов приведут к созданию гуманизированных антител с сохраненными свойствами связывания антигенов. Однако нельзя исключить того, что потребуются дополнительные обратные мутации или созревание аффинности посредством введения аминокислотных замен в участках CDR посредством сайт-направленного мутагенеза.
ПРИМЕР 19: Антитело с двойным вариабельным доменом
Белок антитела с двойным вариабельным доменом (DVD) конструируют методами генной инженерии посредством тандемной гибридизации доменов IGHV антител 992 и 1024 за счет линкера из 6 аминокислот (ASTKGP), а также доменов IGKV антител 992 и 1024 за счет линкера из 5 аминокислот (TVAAP) [Wu C, Ying H, Grinnell C, Bryant S, Miller R, Clabbers A, Bose S, McCarthy D, Zhu RR, Santora L, vis-Taber R, Kunes Y, Fung E, Schwartz A, Sakorafas P, Gu J, Tarcsa E, Murtaza A and Ghayur T. Simultaneous targeting of multiple disease mediators by a dual-variable-domain immunoglobulin. Nature Biotechnol, 2007; 25:1290-1297]. Двойные слияния домена IGHV и домена IGKV приводят к созданию доменов IGHC и IGKC, соответственно. В одном полноразмерном антителе DVD (992L1024), IGHV и IGKV 992 составляют N-концевой участок, сопровождающийся линкером, а также, соответственно, IGHV и IGKV 1024. В другом полноразмерном антителе DVD (1024L992), N-концевой участок составляют IGHV и IGKV 1024, после чего идет линкер, а также, соответственно, IGHV и IGKV 992. В качестве матрицы для двухэтапного PCR-опосредованного конструирования генов, кодирующих DVD, используется плазмидная ДНК, кодирующая антитела 992 и 1024. Сначала раздельно амплифицируют два кодирующих участка IGHV and IGKV вариабельного домена, так, чтобы они содержали участки перекрывающегося удлинения в положении участка, кодирующего линкер (комбинации матриц и праймеров см. в таблице 13 и таблице 14). Ген IGKV, кодирующий C-концевой проксимальный вариабельный домен, амплифицируют так, что в кодирующую последовательность включается ген, кодирующий константный домен легкой цепи (IGKC). Кодирующие последовательности и аминокислотные последовательности субъединиц антител с двойным отрицательным доменом показаны в приложении 3.
Первую PCR готовили со следующей смесью в каждой пробирке (реакции по 50 мкл) для получения заданной конечной концентрации: 1Ч буфер FastStart (Roche), смесь dNTP (200 мкм каждого компонента), праймеры (10 пмоль каждого) (см. таблицу 14), смесь ферментов FastStart High Fidelity (2,2 U, Roche) и 100 нг плазмидного шаблона (см. таблицу 14). PCR проводили по следующему термоциклу: 2 минуты при 95°C, 20Ч (30 секунд при 95°C, 30 секунд при 55°C, 1 минута при 72°C), 10 минут при 72°C. Результирующие продукты PCR с правильным размером из первой PCR (см. таблицу 14) очищали препаративным электрофорезом в агарозном геле и использовали на втором этапе, в ходе которого были сплайсированы два вариабельных домена посредством PCR перекрывания-удлинения. Вторую PCR, сплайсинг фрагментов ДНК посредством перекрывания-удлинения, готовили со следующей смесью в каждой пробирке (реакции по 50 мкл) для получения заданной конечной концентрации: 1Ч буфер FastStart (Roche), смесь dNTP (200 мкм каждого компонента), праймеры (10 пмоль каждого) (см. таблицу 15), смесь ферментов FastStart High Fidelity (2,2 U, Roche) и шаблон (100 нг фрагмента PCR, см. таблицу 15). PCR проводили с тем же термоциклом, который был определен выше. Результирующие продукты второго этапа PCR очищали препаративным электрофорезом в агарозном геле и обрабатывали рестрикционными ферментами AscI и XhoI для двойного IGHV, а также NheI и NotI для двойного IGKV (включая IGKC). Фрагменты последовательно вшивали в вектор экспрессии IgG млекопитающего 00-VP-002 (фиг.4) посредством стандартных процедур переваривания рестрикционными ферментами и вшивания. Результирующий плазмидный вектор экспрессии амплифицировали в E. coli, а плазмидный препарат очищали стандартными способами. Антитела DVD экспрессировали и очищали в соответствии с примером 2, а снятие характеристик активности проводили в соответствии с примером 3-13.
Если результирующие антитела проявят слабое связывание или отсутствие связывания в целевым hEGFr, можно протестировать другие линкеры.
(размер п.о.)
(размер п.о.)
(406 п.о.)
(359 п.о.)
(381 п.о.)
(702 п.о.)
(393 п.о.)
(687 п.о.)
1024H_O3'
(392 п.о.)
1024K_O3'
(374 п.о.)
ПРИМЕР 20: Токсикологическое исследование на обезьянах Cynomolgus с 6-недельным внутривенным введением комбинации антител и эрбитукса
Цель исследования: Цель исследования заключалась в том, чтобы определить токсичность испытуемого продукта, смеси антител 992+1024, в результате внутривенного введения обезьянам cynomolgus один раз в неделю на протяжении 6 недель.
Поскольку в клинической практике токсичность является фактором, ограничивающим дозу ингибиторов EGFR наподобие эрбитукса и вектибикса, авторы изобретения сочли важным на раннем этапе оценить переносимость смеси антител 992+1024 в клинически приемлемой дозе. Эта необходимость дополнительно подчеркивается тем фактом, что композиция 992+1024, по-видимому, действует через иной механизм, чем другие продукты, нацеленные на EGFR. Потенциально это чревато новыми неблагоприятными побочными эффектами или утяжелением побочных эффектов, наблюдаемых при лечении другими ингибиторами EGFR.
Группы из трех самок обезьян cynomolgus получали еженедельные внутривенные дозы смеси антител 992+1024 (4/2,7 и 12/8 мг/кг) или эрбитукса (12/8 мг/кг) на протяжении 6 недель. Первые дозы 4 и 12 мг/кг представляли собой загрузочные дозы, а дозы 2,7 и 8 мг/кг поддерживающие дозы, которые были введены 5 раз. Доза 12/8 мг/кг эквивалентна дозе эрбитукса для человека, принятой в клинической практике.
(мг/кг/день)
(мл/кг)
(самки)
По ходу исследования были прослежены указанные ниже параметры: смертность, клинические признаки, вес тела, потребление пищи, гематологические показатели, клиническая химия, вес органов, макроскопические находки.
Результаты
Смертность: На протяжении всего исследования не было ни одного непредусмотренного летального исхода.
Клинические признаки: Неблагоприятных клинических явлений, связанных с лечением, не наблюдалось.
Вес тела: Лечение композицией 992+1024 или эрбитуксом не оказывало какого-либо влияния на вес тела.
Потребление пищи: Заметных влияний на потребление пищи не обнаружено.
Гематология: Не выявлено каких-либо изменений гематологических параметров, которые можно было бы предположительно связать с лечением композицией 992+1024 или эрбитуксом.
Клиническая химия: Не обнаружено никаких изменений в параметрах клинической химии, которые можно было бы предположительно связать с применением любого испытуемого продукта.
На 4-й неделе исследования у одного животного в группе 992+1024 (4,2/2,7 мг/кг) было обнаружено повышение уровня аспарагиновой аминотрансферазы и аланиновой аминотрансферазы по сравнению с исходными показателями до начала лечения. Эти показатели вернулись к норме на 6-й неделе. Учитывая отсутствие подобных эффектов у других животных, получавших лечение, можно говорить о том, что токсикологическое значение увеличения уровня ферментов печени неизвестно.
Вес органов: Не было обнаружено каких-либо различий в весе органов у животных, получавших лечение, и у контрольных животных, которые могли бы иметь токсикологическое значение.
Макроскопические находки: При некропсии не было обнаружено каких-либо единообразных явлений, которые позволили бы предположить влияние композиции 992+1024 или эрбитукса.
Предварительное заключение: Предварительные данные свидетельствуют о том, что смесь антител 992+1024 была хорошо переносима в протестированных дозах, и никаких неблагоприятных эффектов, связанных с лечением, не наблюдалось.
ПРИМЕР 21: Ингибирование роста клеточных линий рака легких
Известно, что клеточные линии рака легких экспрессируют EGFR с мутациями в домене тирозинкиназы (Steiner et al. Clin Cancer Res 13.5 (2007): 1540-51). Способность комбинации двух антител 992 и 1024 ингибировать рост клеточных линий рака легких НСС827 и HI975, несущих разные мутации EGFR, была исследована способом, похожим на тот, что был использован в примере 6.
Результаты
Как можно видеть по данным, представленным в таблице 16 и таблице 17, комбинация антител 992 и 1024 способна ингибировать рост обеих линий раковых клеток. Комбинация превосходит моноклональные антитела 992, 1024 и вектибикс.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ РЕКОМБИНАНТНЫХ АНТИТЕЛ ПРОТИВ РЕЦЕПТОРА ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА | 2009 |
|
RU2540146C2 |
| АНТИТЕЛО ПРОТИВ РЕЦЕПТОРА ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА | 2013 |
|
RU2652880C2 |
| АНТИИДИОТИПИЧЕСКИЕ АНТИТЕЛА, КОТОРЫЕ ИНДУЦИРУЮТ ИММУННЫЙ ОТВЕТ ПРОТИВ РЕЦЕПТОРА ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА | 1996 |
|
RU2172636C2 |
| БЕЛКИ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2549678C2 |
| БЕЛКИ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2673724C2 |
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ АГЕНТЫ С МЕЖВИДОВОЙ СПЕЦИФИЧНОСТЬЮ | 2008 |
|
RU2535992C2 |
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА К РЕЦЕПТОРУ ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА | 2005 |
|
RU2402569C2 |
| АНТИТЕЛА-АНТАГОНИСТЫ ПРОТИВ Notch3 И ИХ ПРИМЕНЕНИЕ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ СВЯЗАННЫХ С Notch3 ЗАБОЛЕВАНИЙ | 2007 |
|
RU2493167C2 |
| БИСПЕЦИФИЧЕСКИЙ АНТИГЕНСВЯЗЫВАЮЩИЙ БЕЛОК EGFR/CD16 | 2019 |
|
RU2792240C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СПЕЦИФИЧНЫЕ ПО ОТНОШЕНИЮ К HER3, И ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2620068C2 |

















































































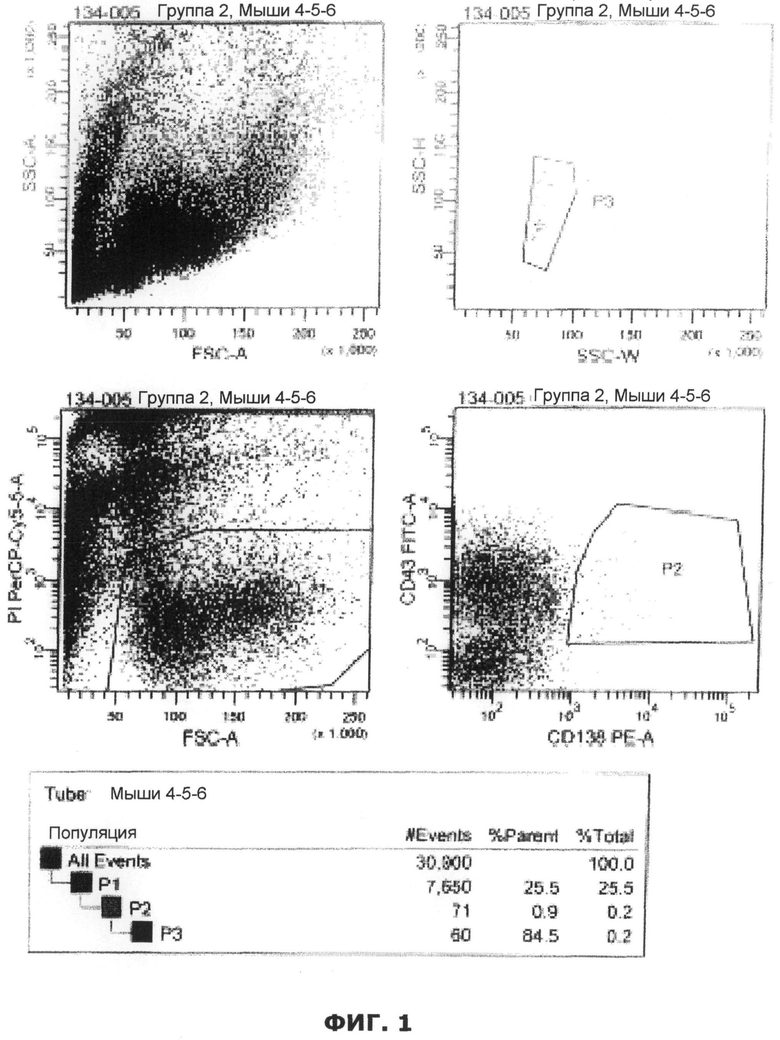
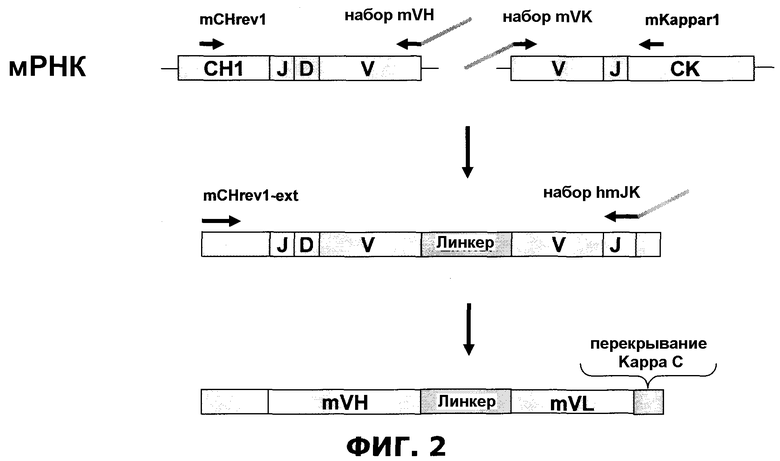
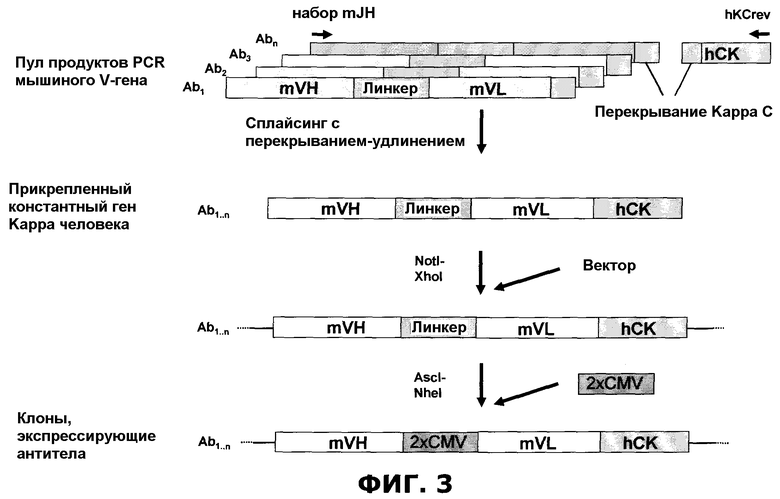
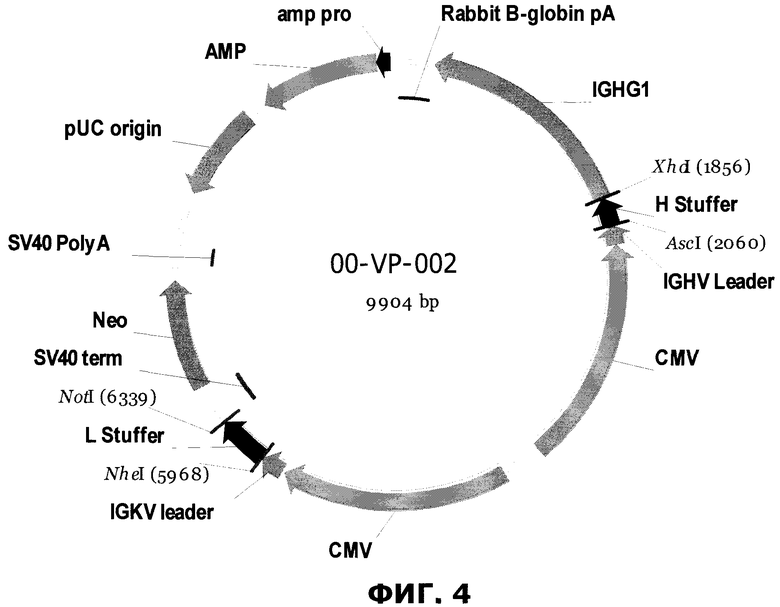
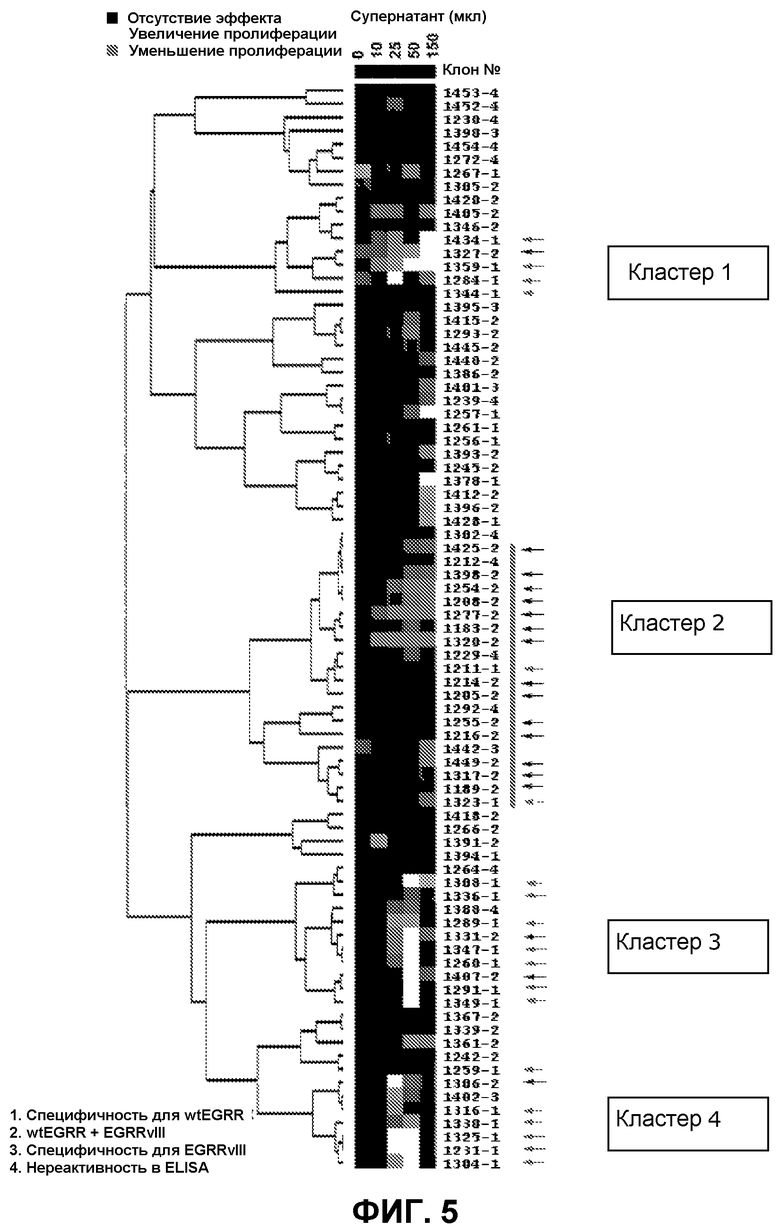
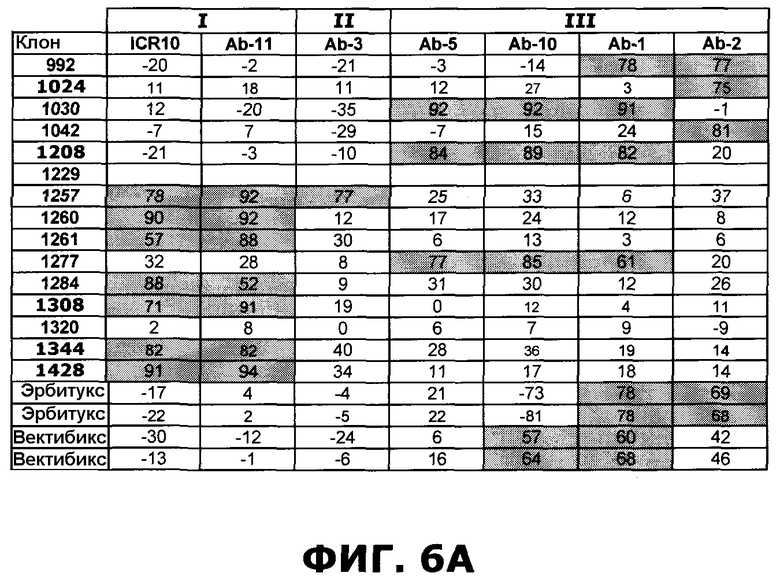
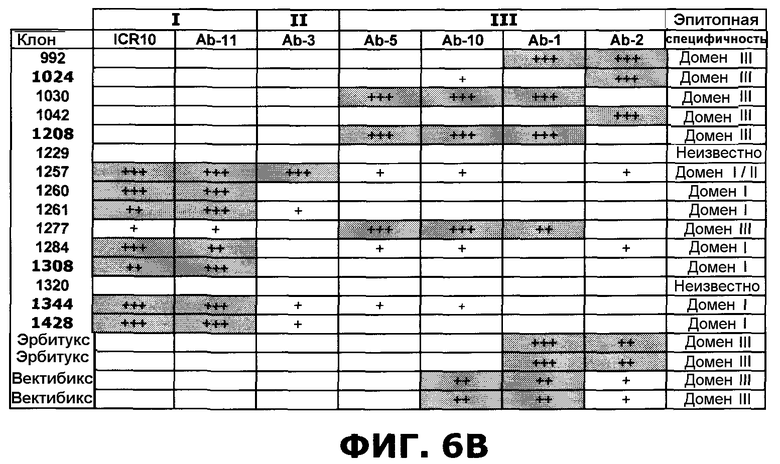
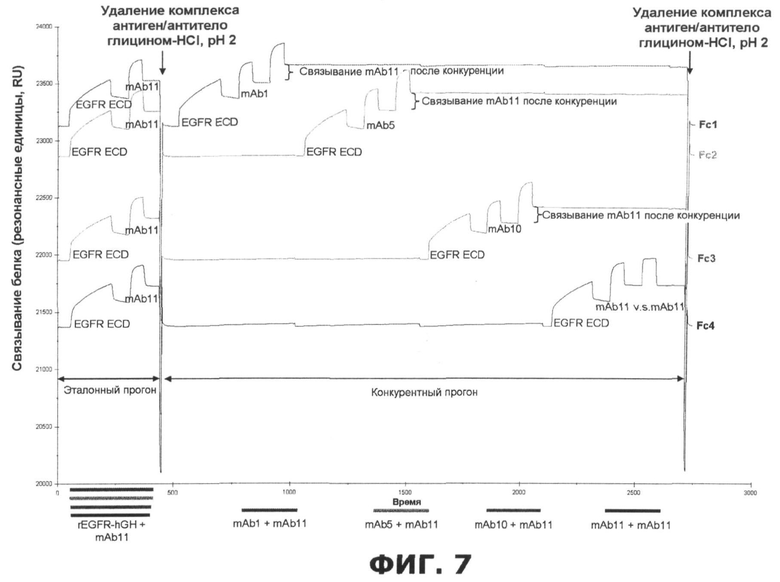
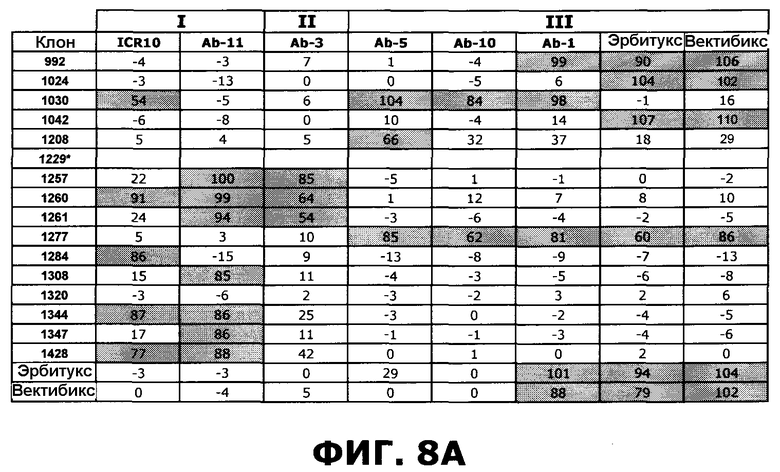
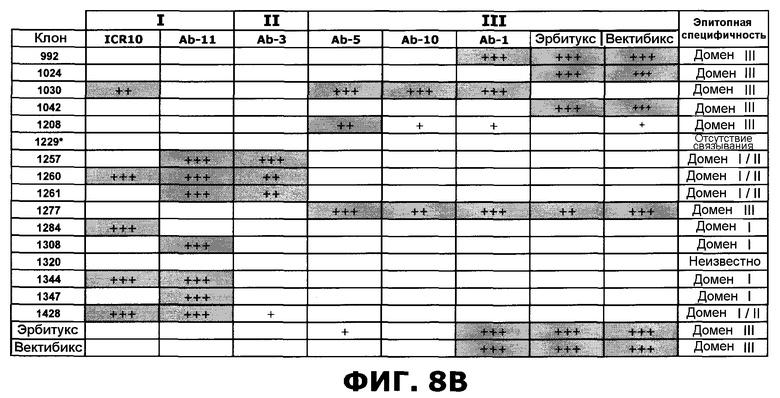
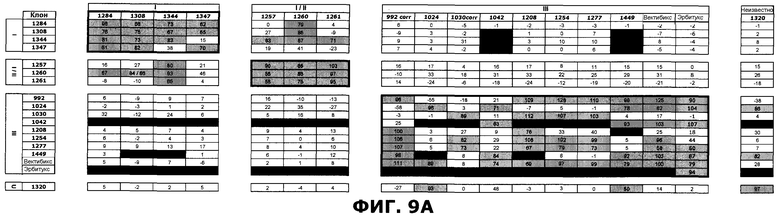
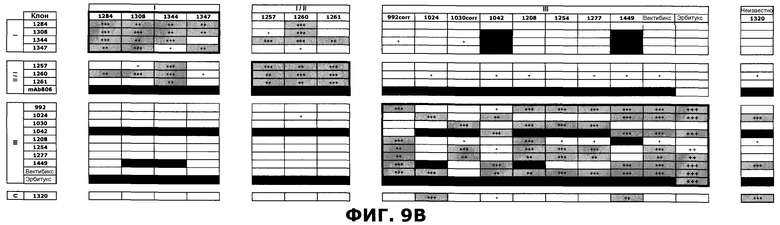
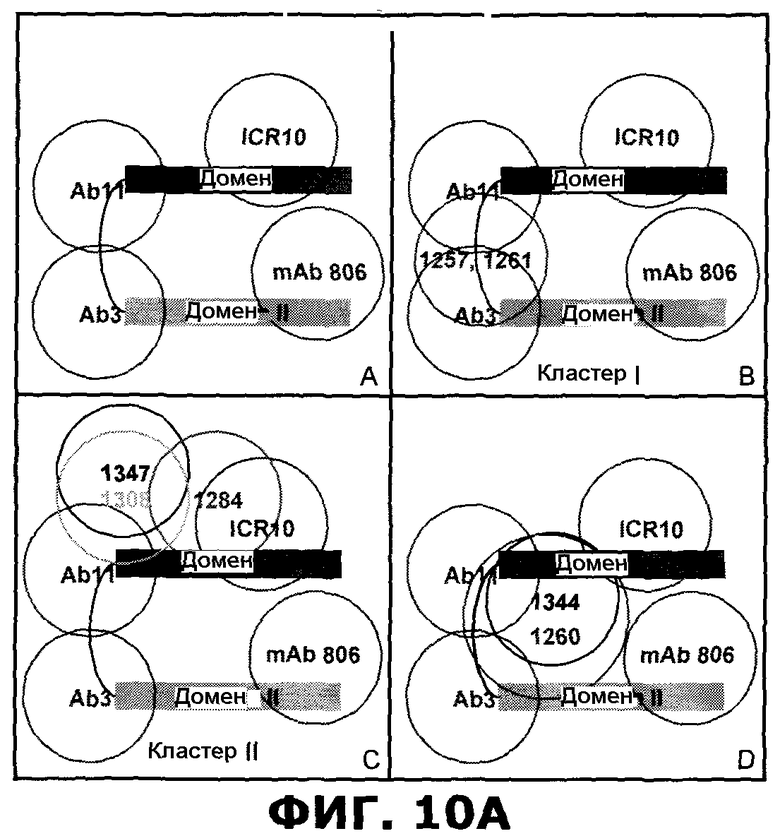
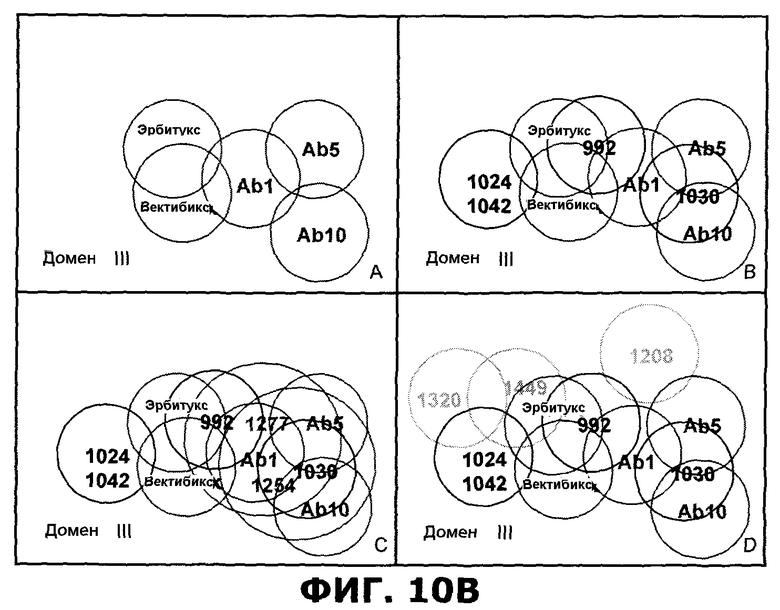

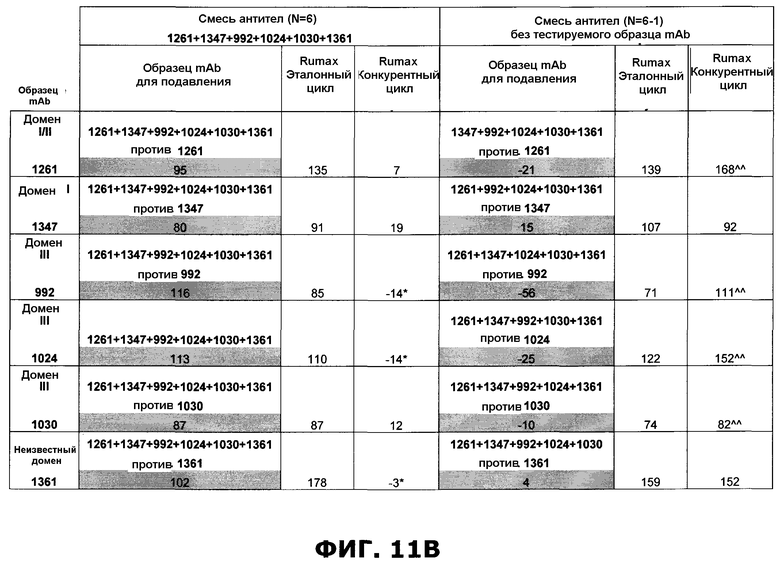
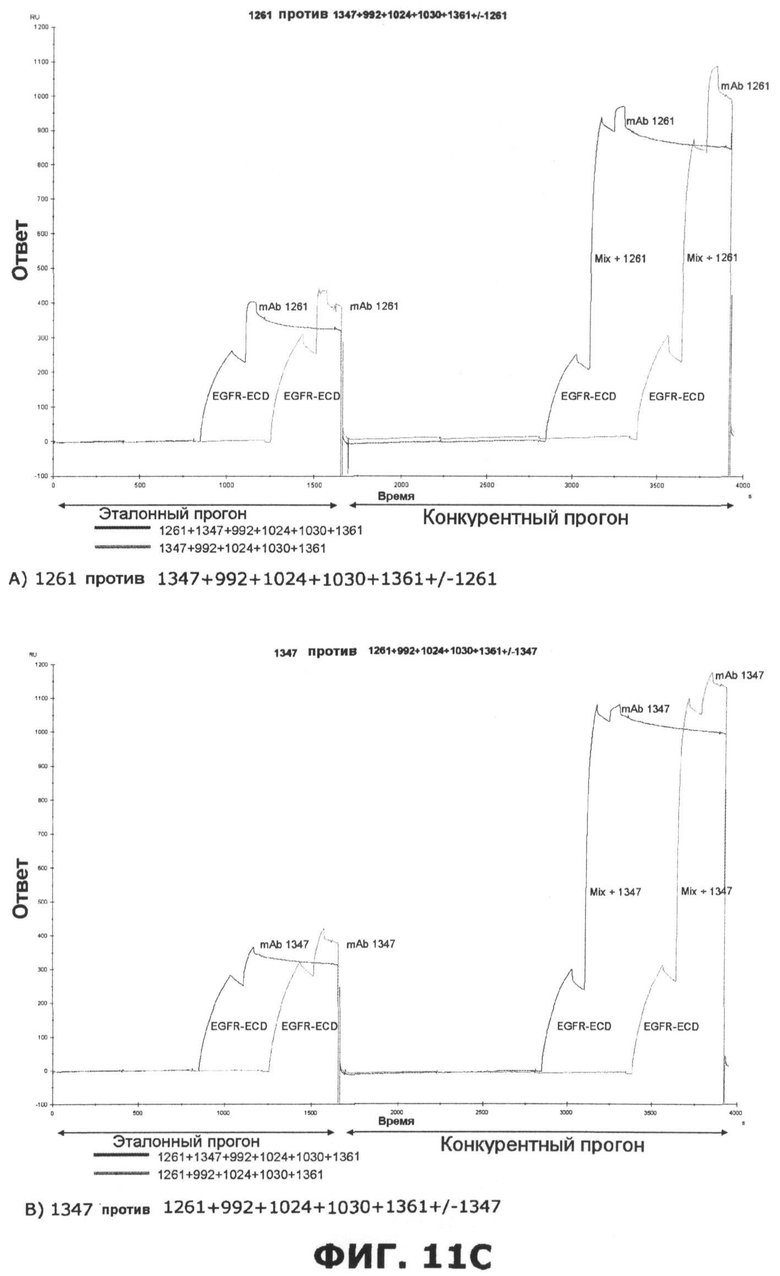
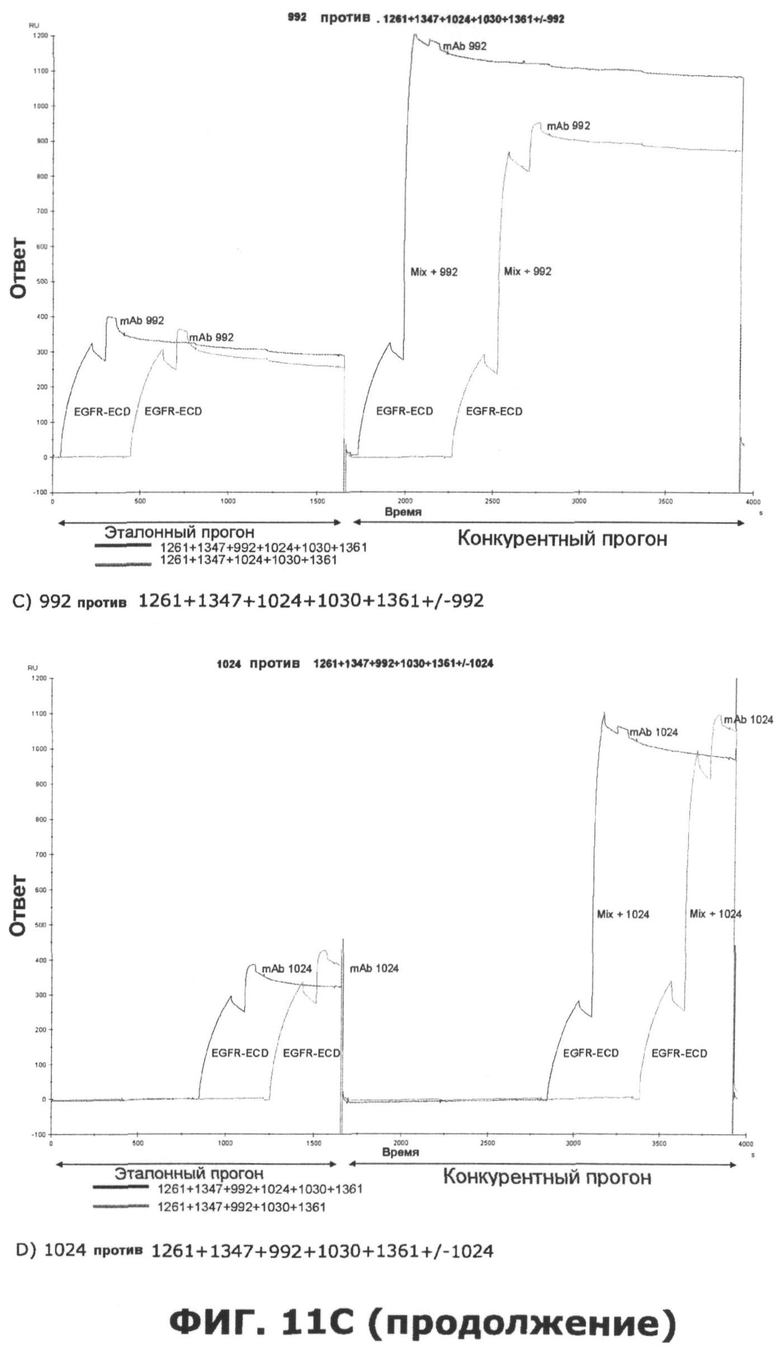
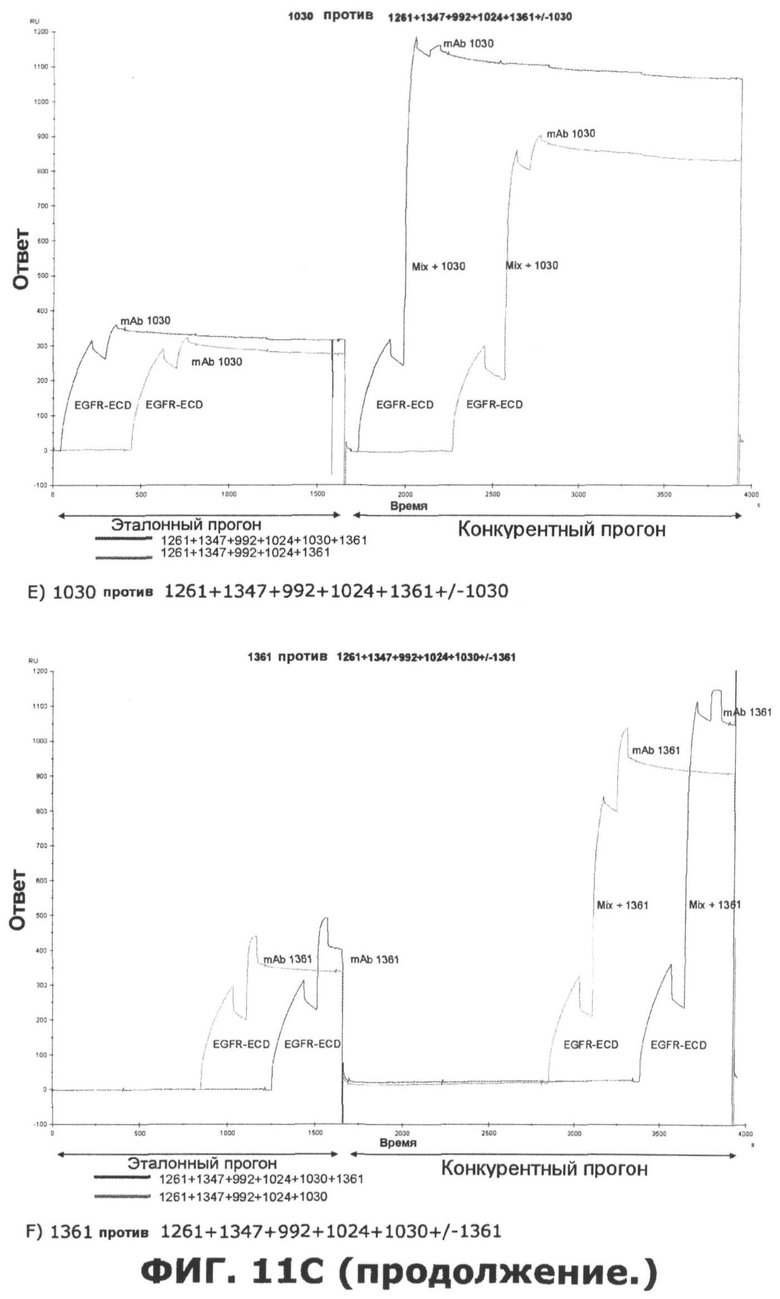
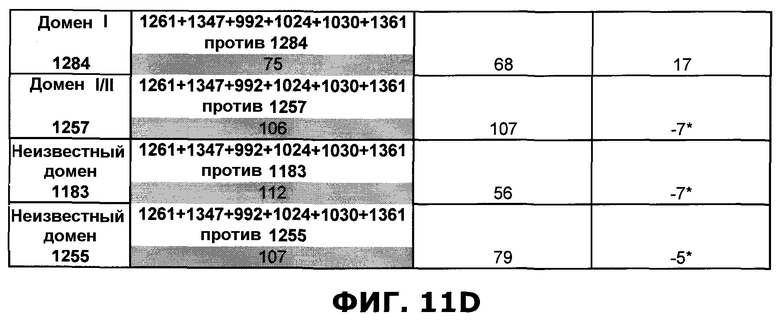
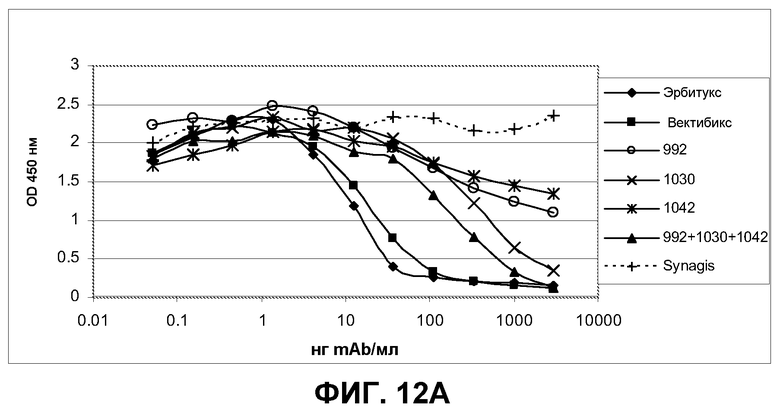
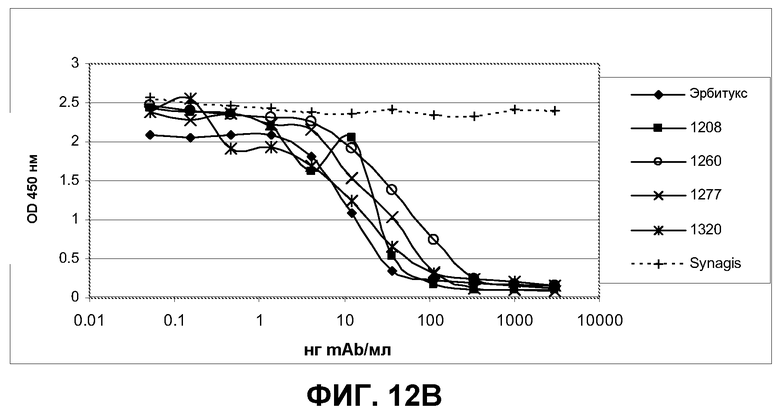
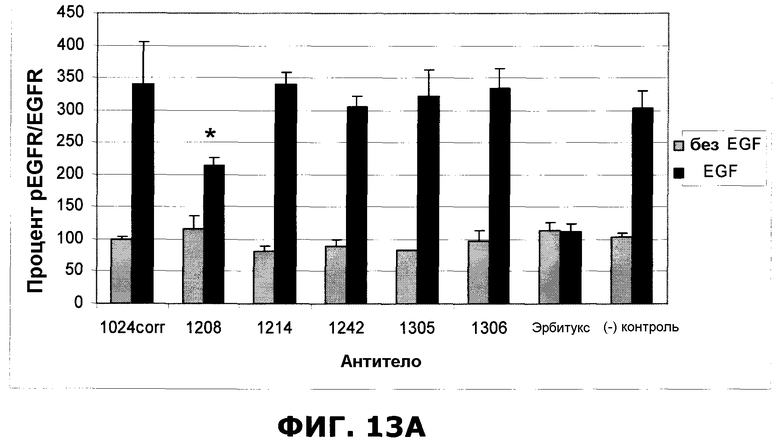
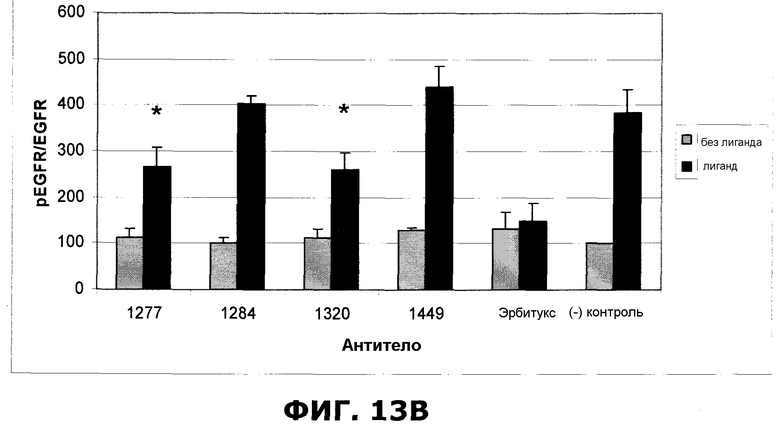
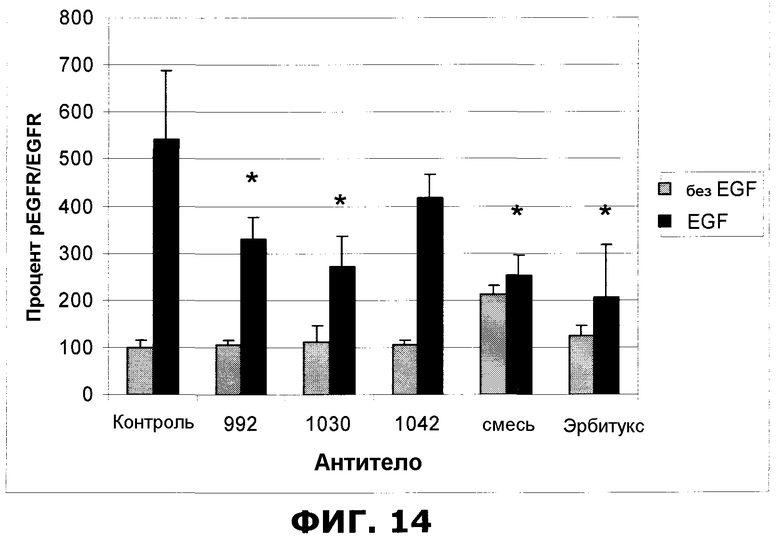
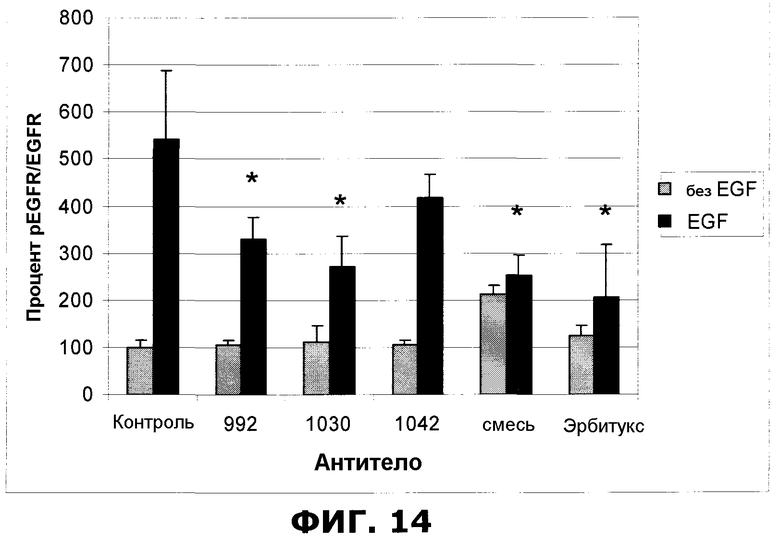
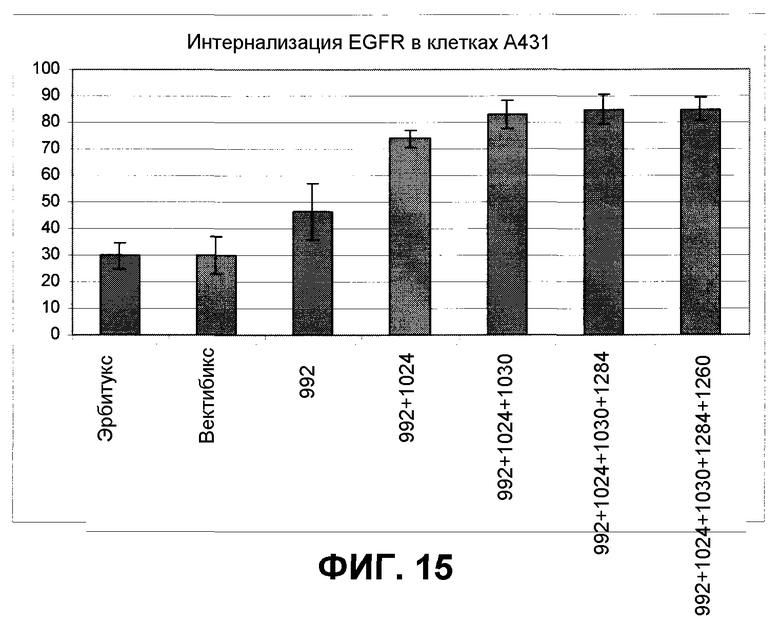
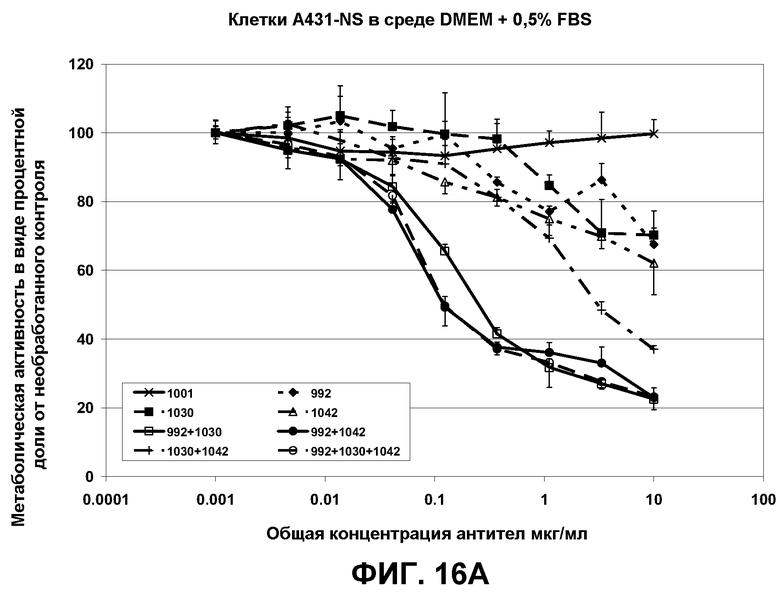
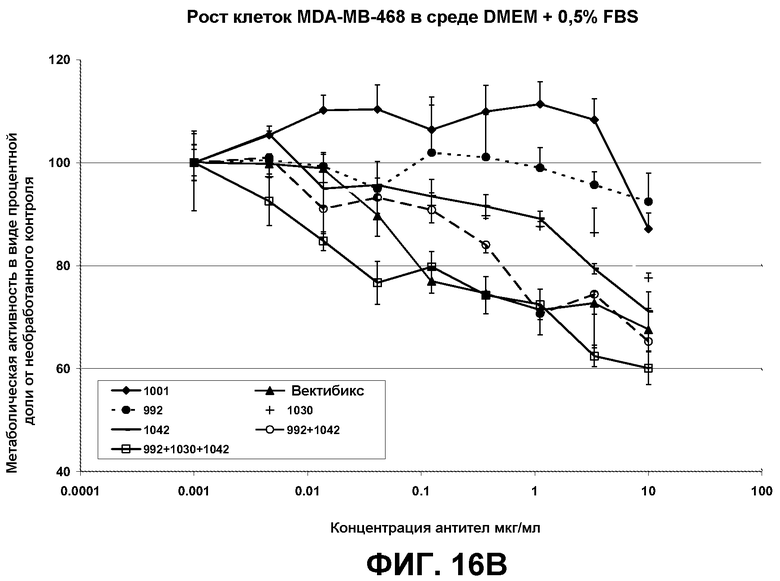
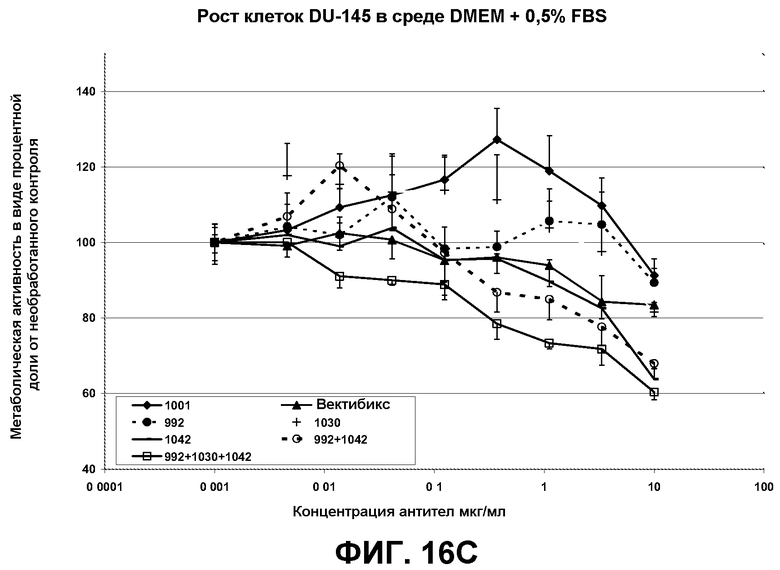
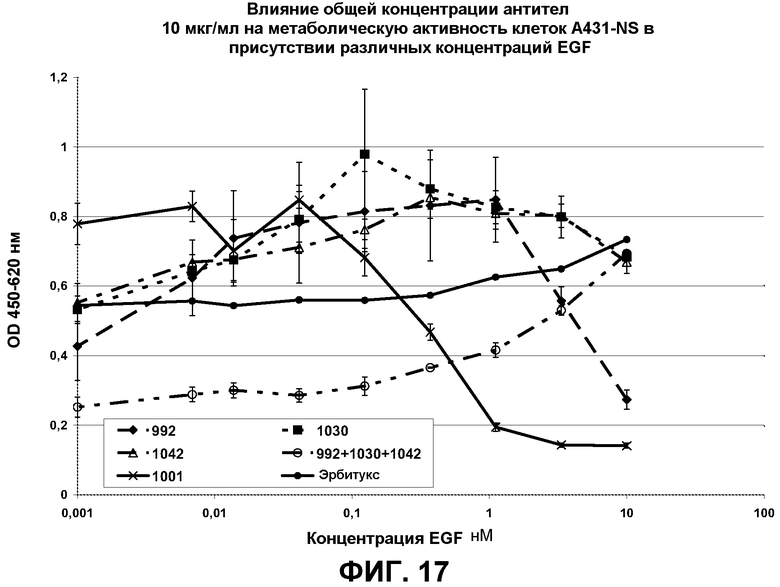
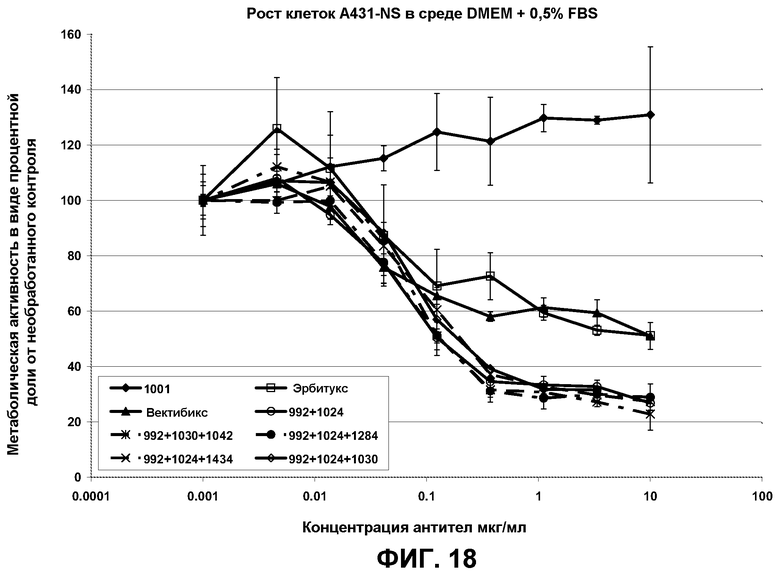
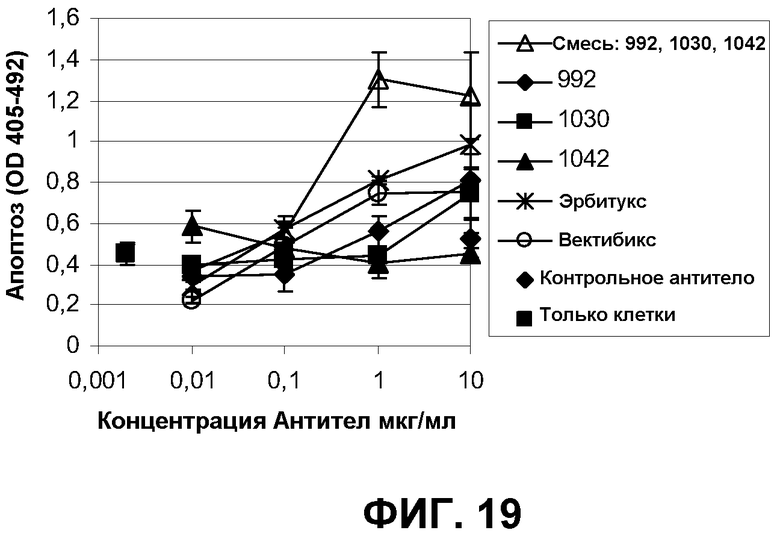
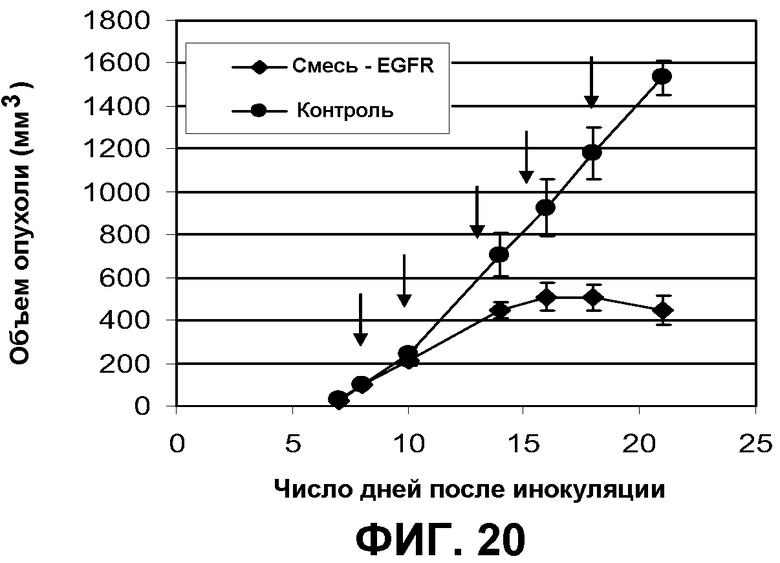
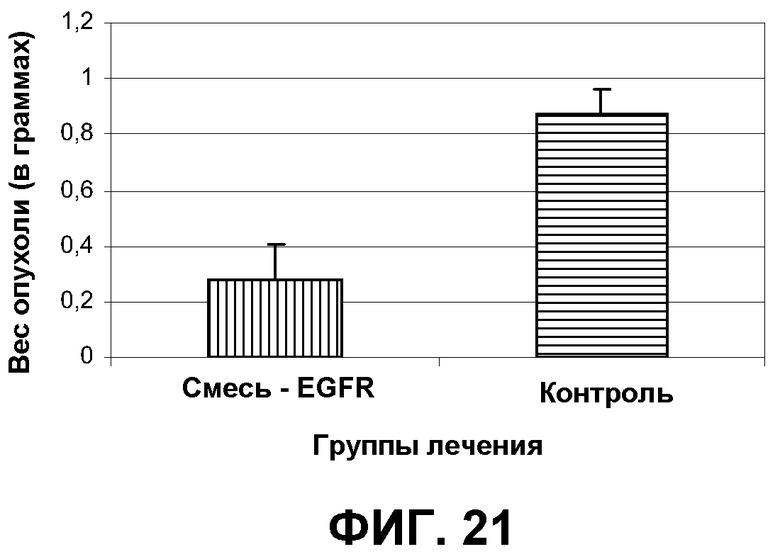
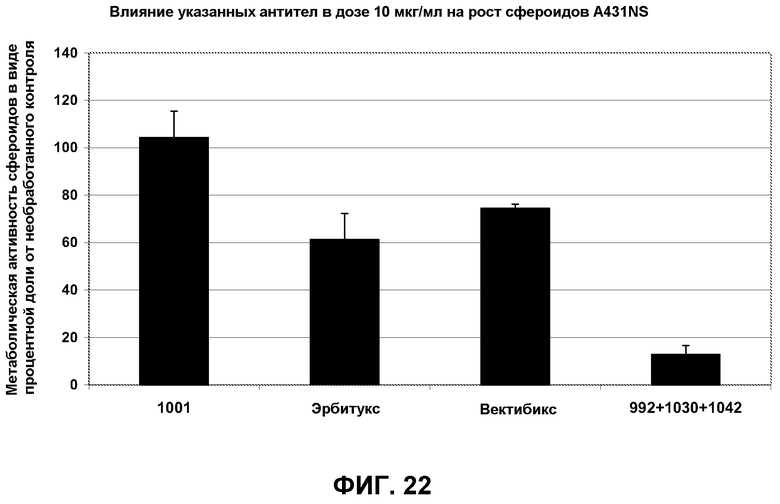
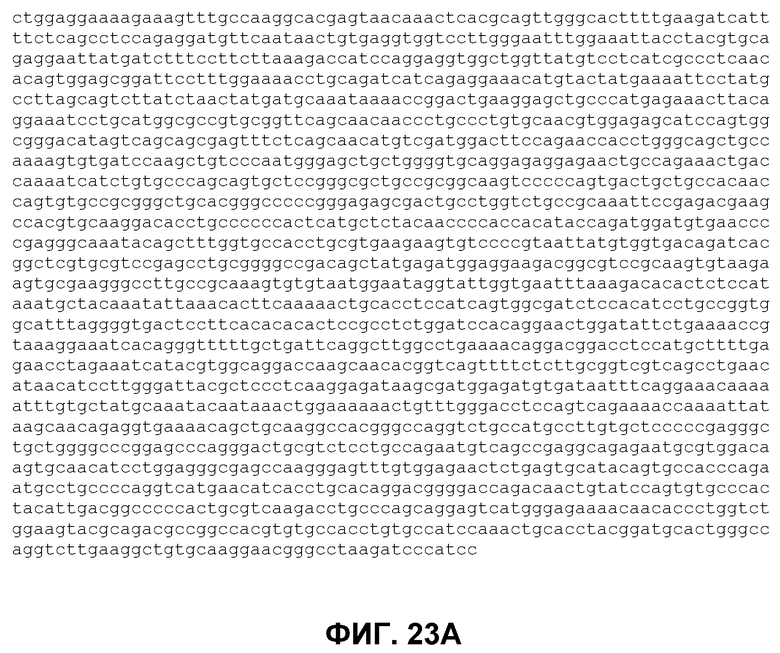


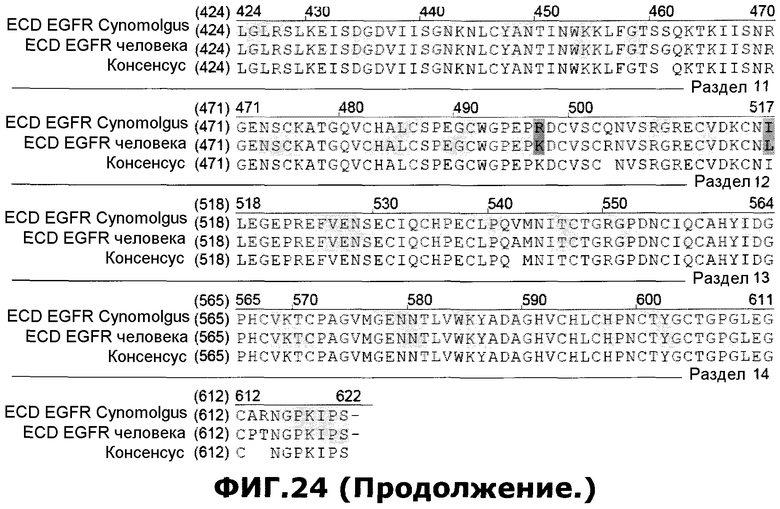
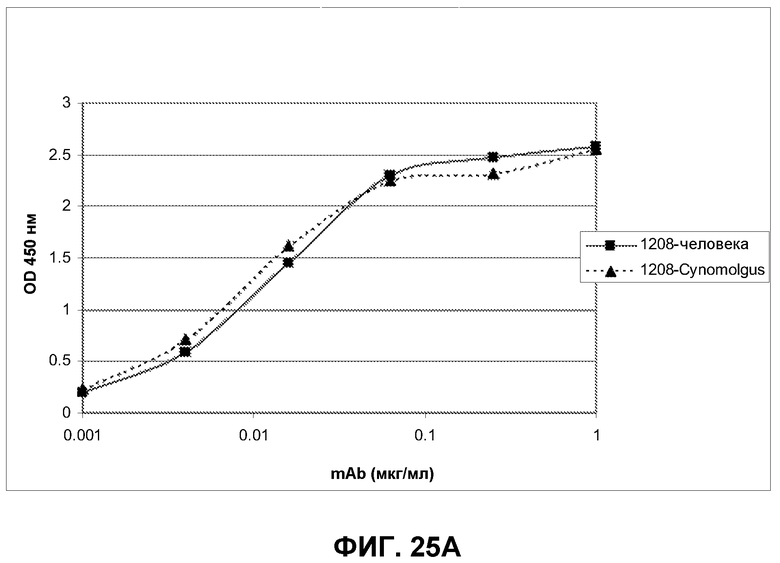
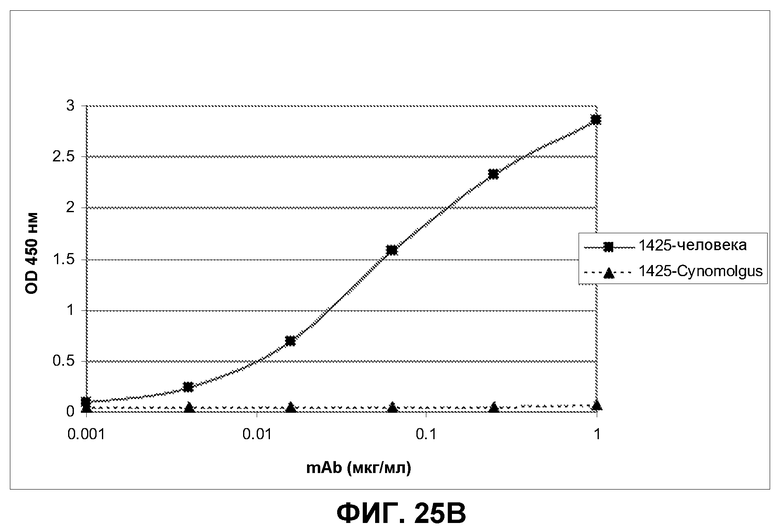

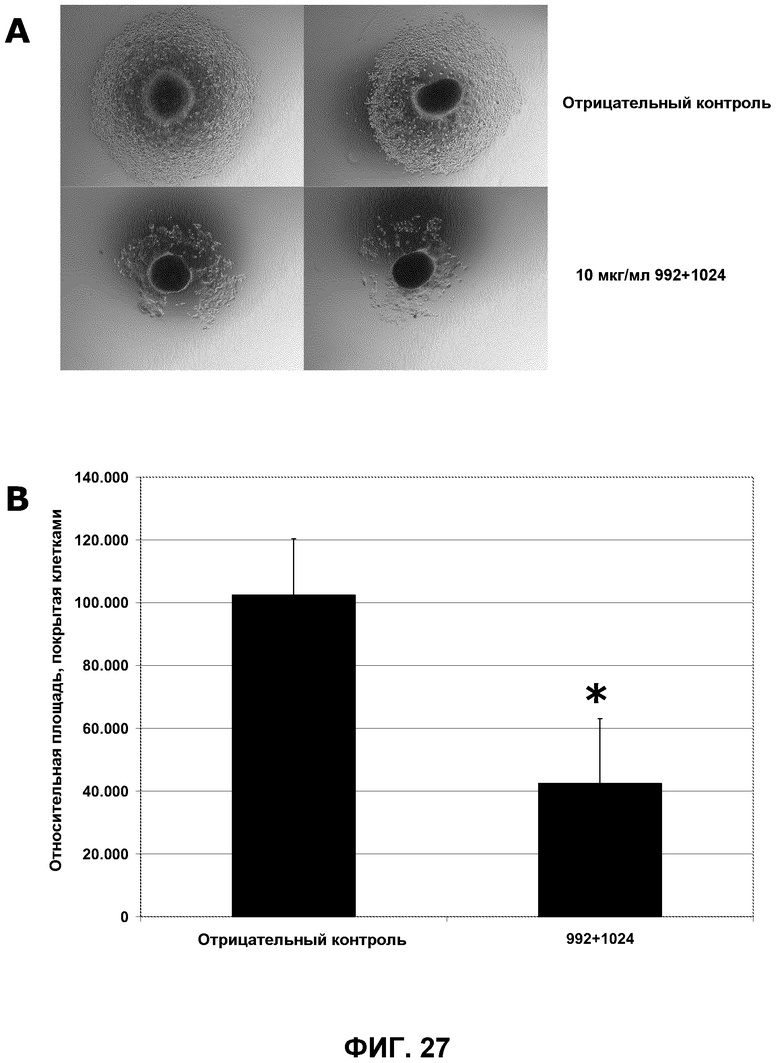
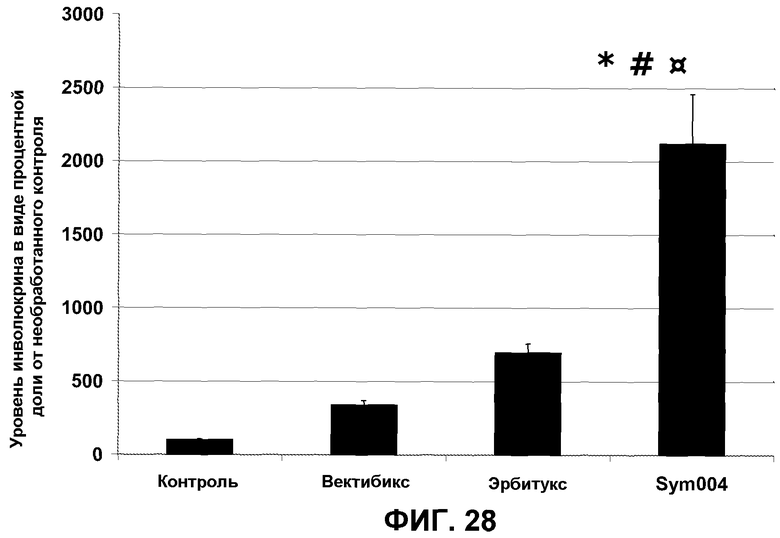
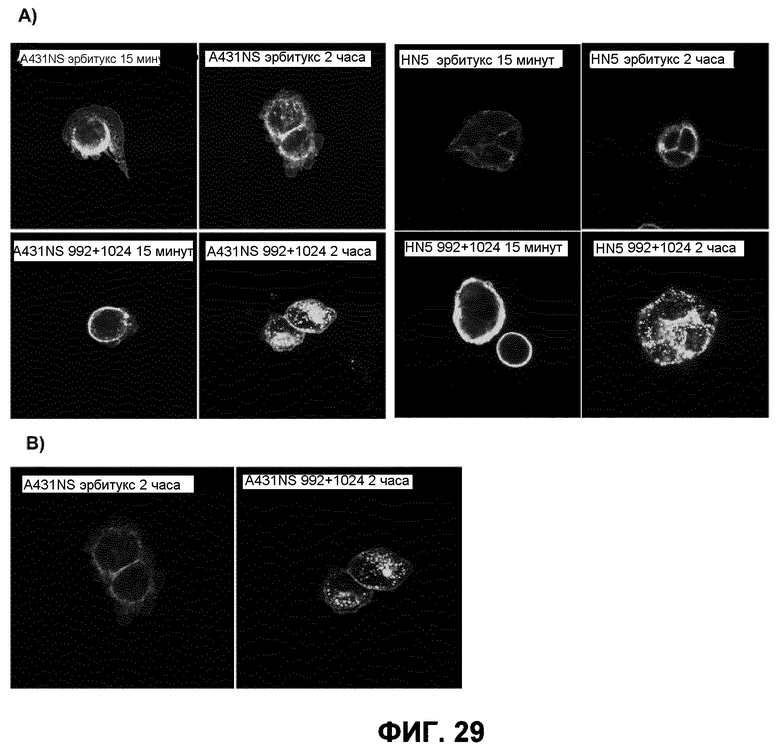
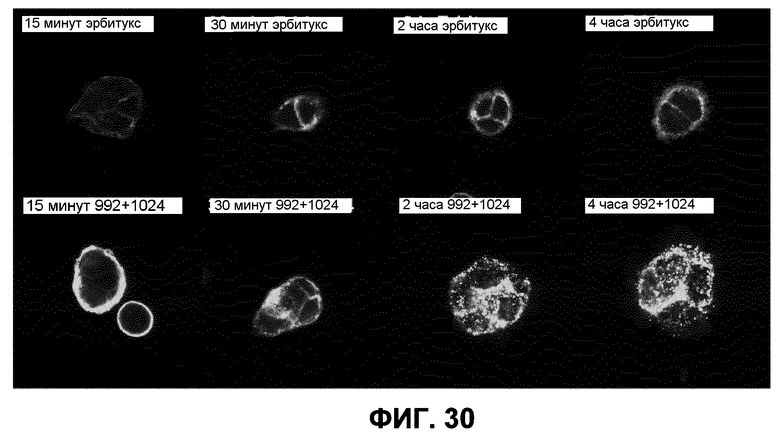
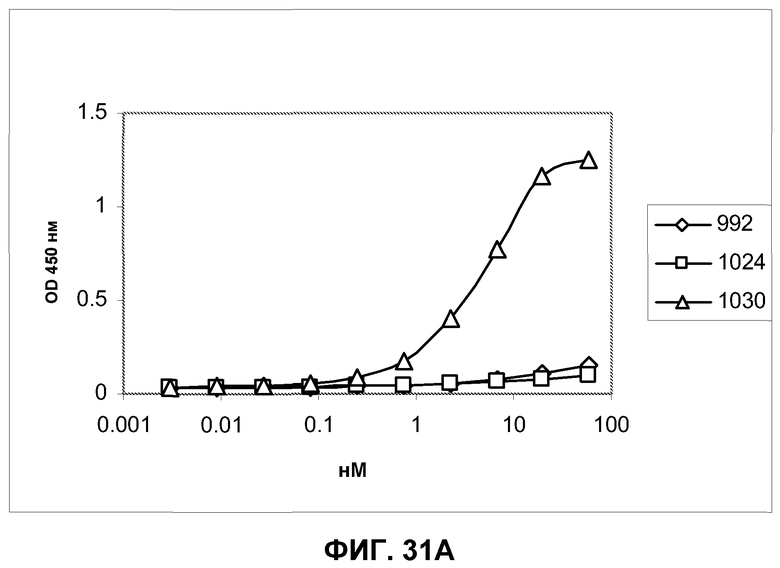
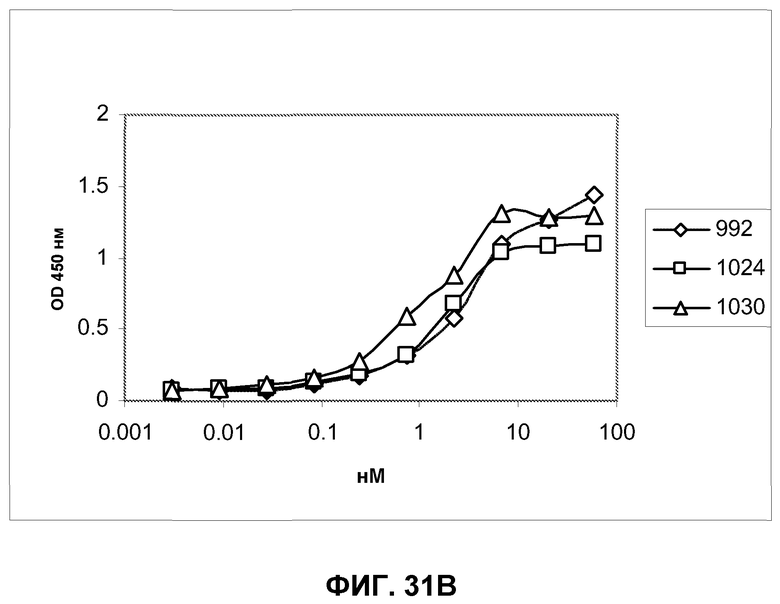
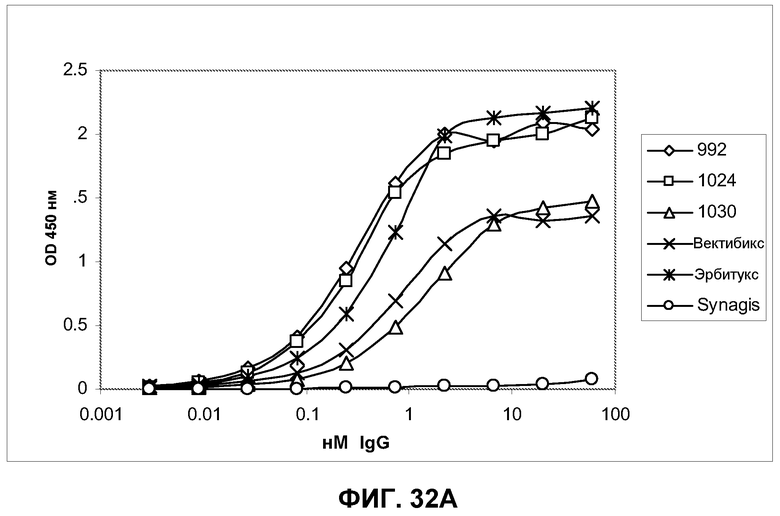
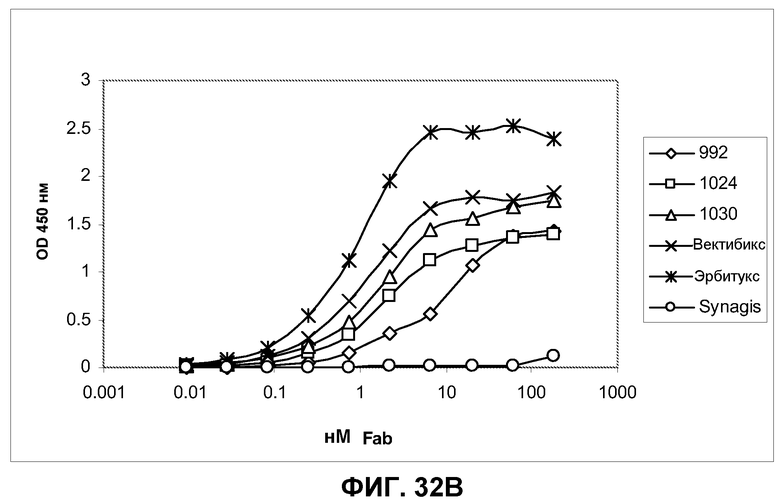
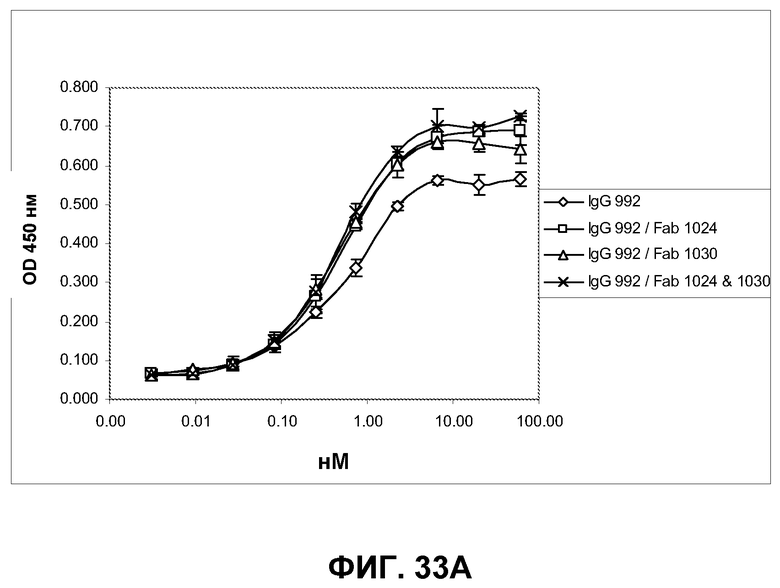
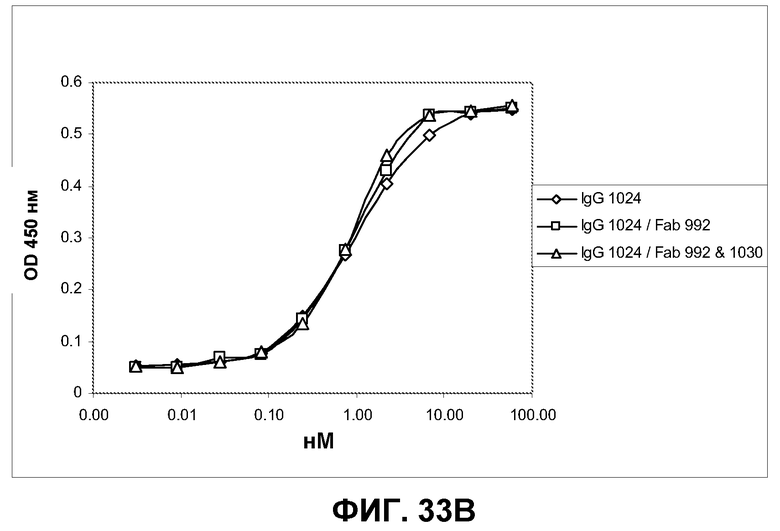
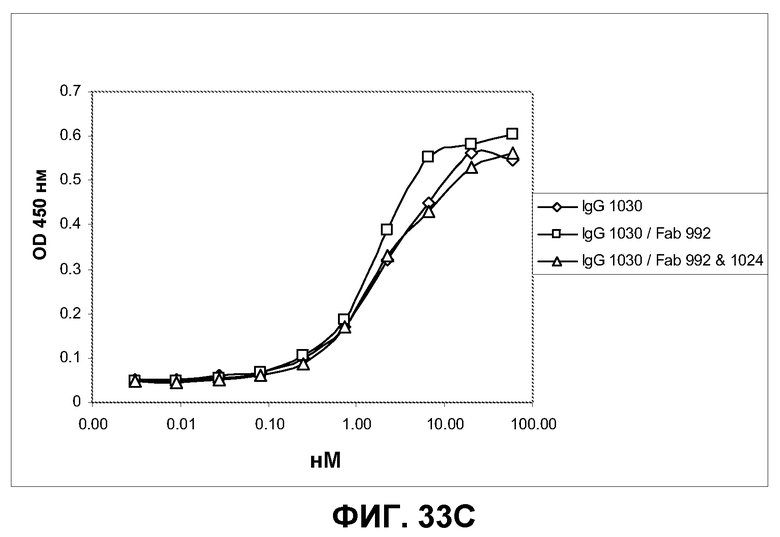


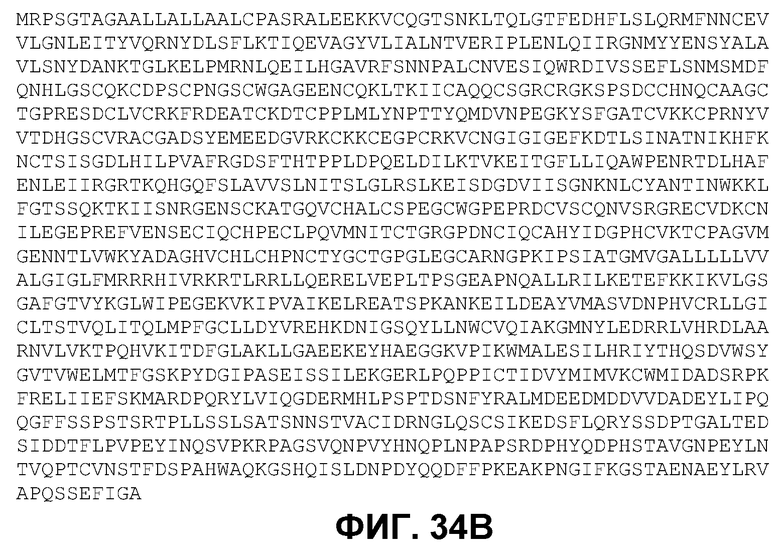
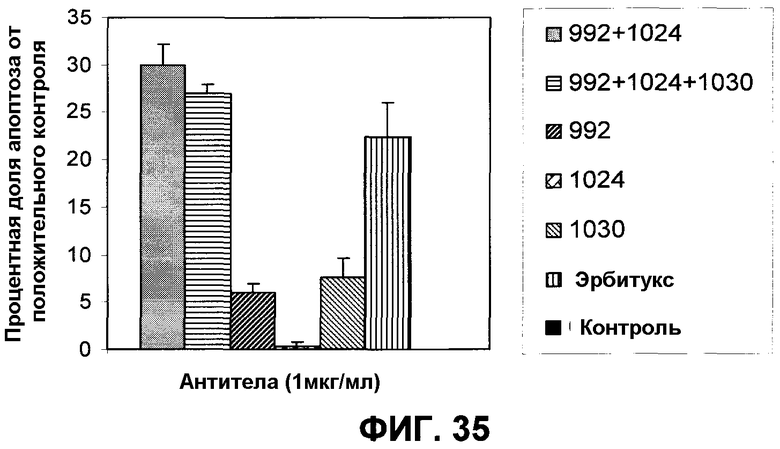
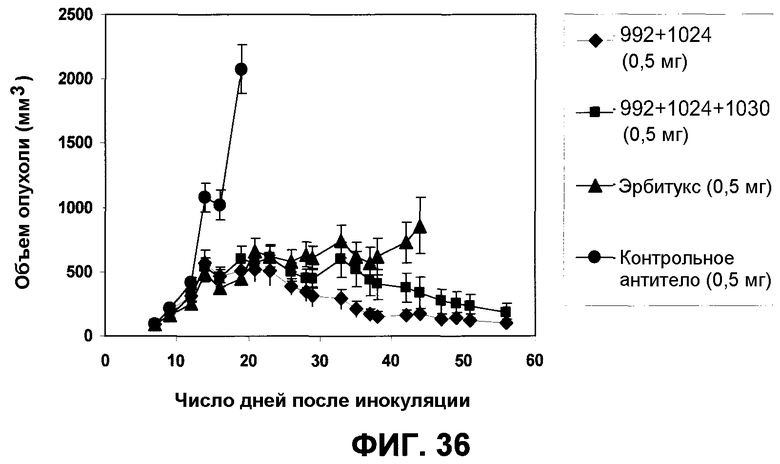
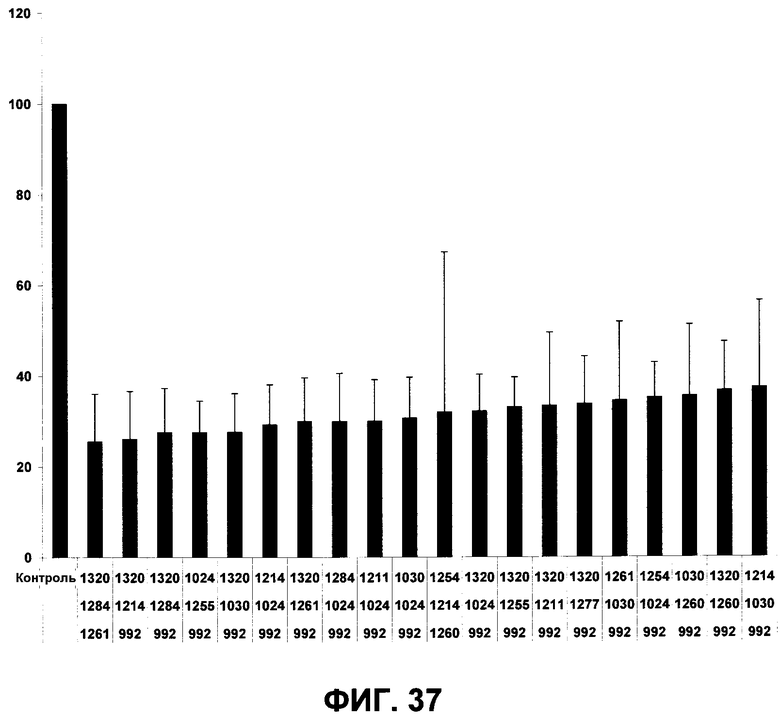
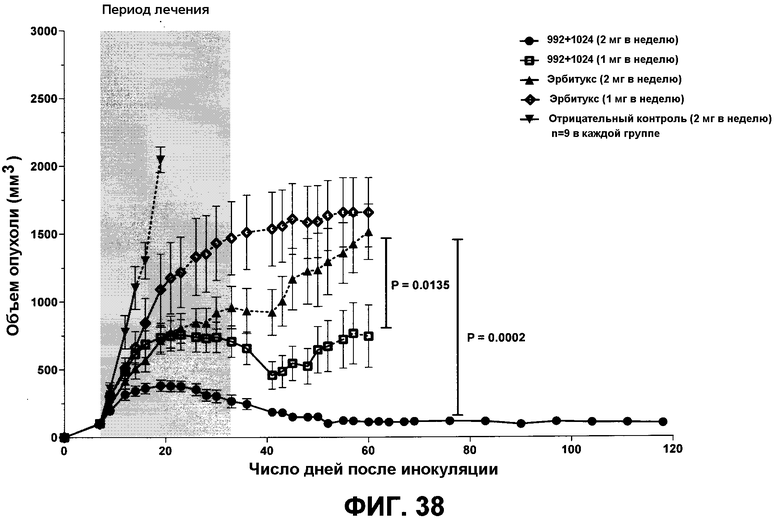
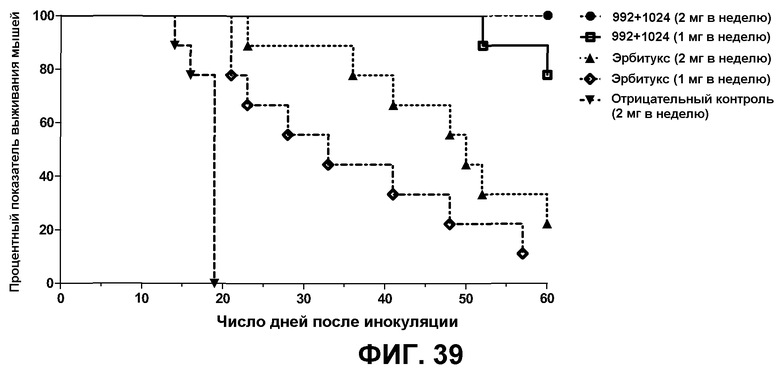
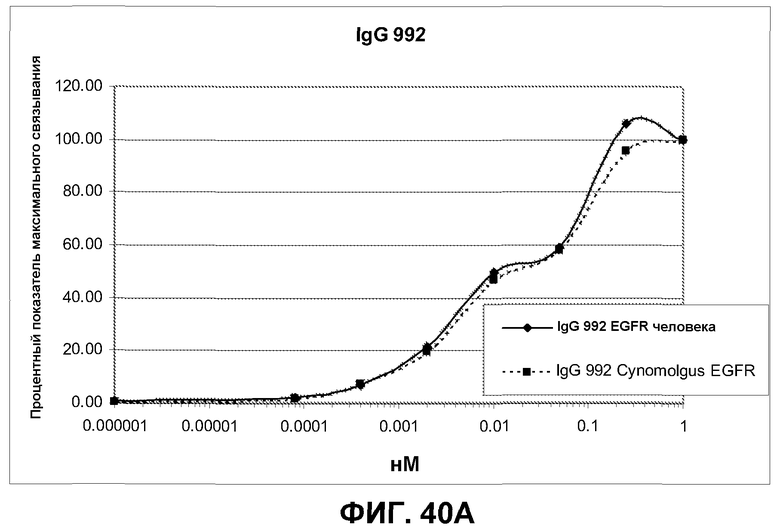
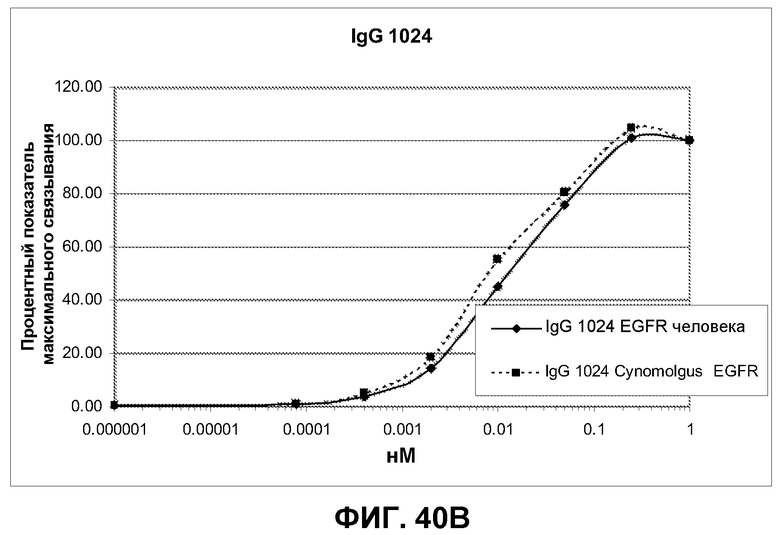
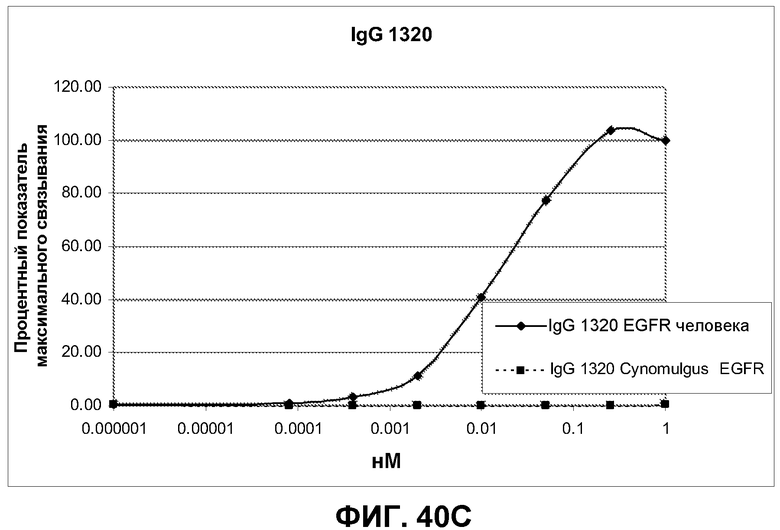
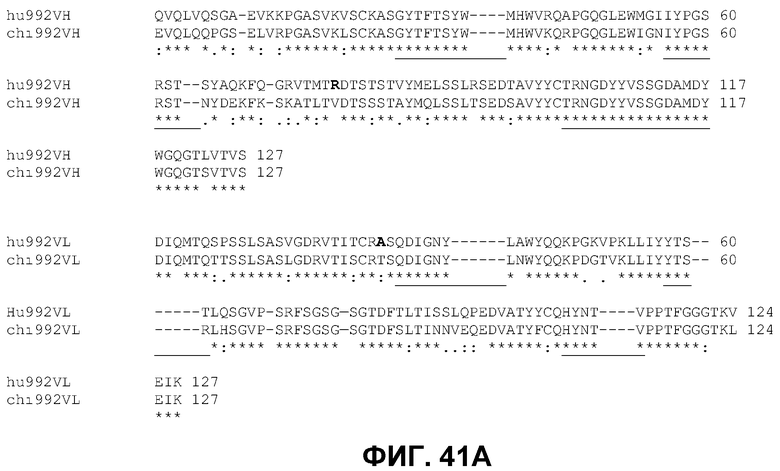

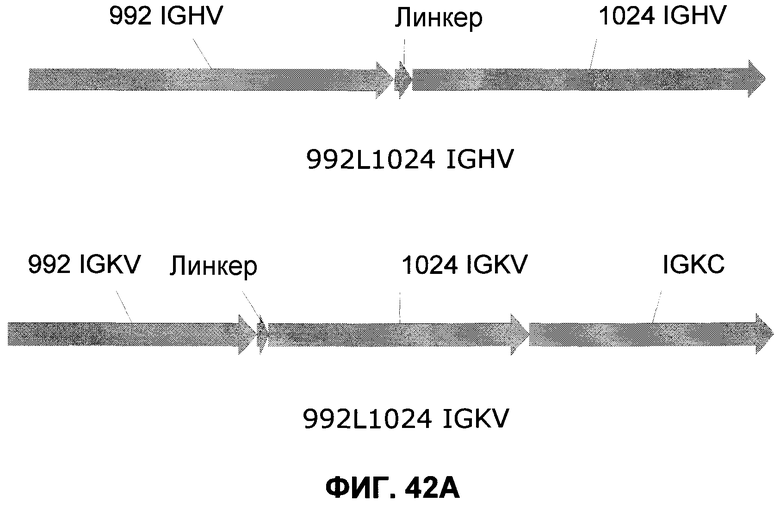
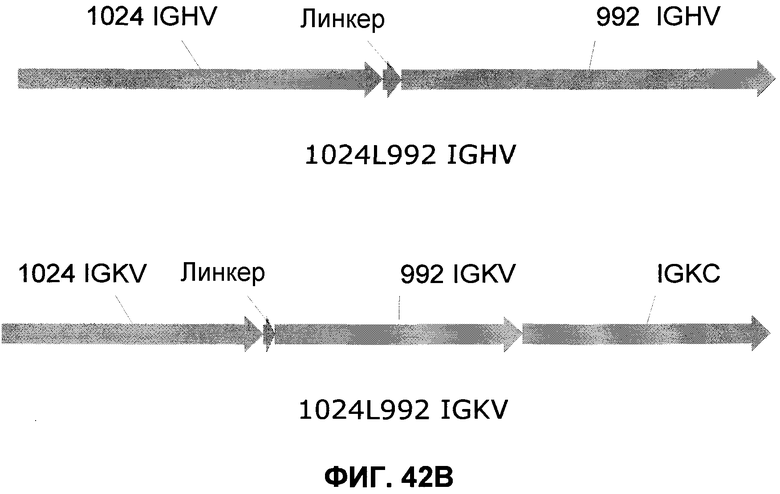
Изобретение относится к области иммунологии. Предложена композиция, содержащая по меньшей мере два антитела, которые связывают различные эпитопы EGFR (ErbB-1) человека, обеспечивая синергетический эффект, а также биспецифическая молекула, представляющая собой антитело или его фрагмент, содержащий последовательности CDR антител в составе композиции по изобретению. Рассмотрено применение композиции по изобретению для изготовления лекарственного средства и фармацевтическая композиция, содержащая композицию по изобретению. Композиция по настоящему изобретению обеспечивает синергетическое действие анти-EGFR антител, что может найти дальнейшее применение в лечении рака. 5 н. и 15 з.п. ф-лы, 42 ил., 17 табл., 21 пр.
1. Композиция антител для лечения и/или профилактики заболеваний, связанных со сверхэкспрессией EGFR, содержащая по меньшей мере 2 различные молекулы антител против EGFR человека, в которой
a. указанную первую молекулу антитела против EGFR выбирают из группы, состоящей из антитела, содержащего CDR1, CDR2 и CDR3 тяжелой цепи (SEQ ID NO: 40) и легкой цепи (SEQ ID NO: 72) антитела 992, и антитела, содержащего последовательности VL (аминокислоты 3-109 из SEQ ID NO: 72) и VH (аминокислоты 3-124 из SEQ ID NO: 40) антитела 992, и
b. указанную вторую молекулу антитела против EGFR выбирают из группы, состоящей из антитела, содержащего CDR1, CDR2 и CDR3 тяжелой цепи (SEQ ID NO: 41) и легкой цепи (SEQ ID NO: 73) антитела 1024, и антитела, содержащего последовательности VL (аминокислоты 3-114 из SEQ ID NO: 73) и VH (аминокислоты 3-120 из SEQ ID NO: 41) антитела 1024.
2. Композиция по п.1, содержащая антитело, содержащее последовательности VL (аминокислоты 3-109 из SEQ ID NO: 72) и VH (аминокислоты 3-124 из SEQ ID NO: 40) антитела 992, и антитело, содержащее последовательности VL (аминокислоты 3-114 из SEQ ID NO: 73) и VH (аминокислоты 3-120 из SEQ ID NO: 41) антитела 1024.
3. Композиция по п.1, в которой первое и второе антитела против EGFR не ингибируют связывание друг друга с EGFR человека.
4. Композиция по п.1, в которой по меньшей мере одно из антител способно увеличивать максимальную способность связывания другого антитела с EGFR человека.
5. Композиция по п.1, в которой доля первого антитела по отношению ко второму антителу в композиции составляет от 5 до 95%, например от 10 до 90%, предпочтительно от 20 до 80%, более предпочтительно от 30 до 70%, более предпочтительно от 40 до 60%, например от 45 до 55%, например приблизительно 50%.
6. Композиция по п.1, в которой первое и второе антитела относятся к изотипу IgG1 или IgG2.
7. Композиция по п.1, которая приводит к интернализации рецептора.
8. Композиция по п.1, которая приводит к регрессии опухолей A431NS in vivo.
9. Композиция по п.1, которая способна индуцировать терминальную дифференциацию клеток A431NS in vivo.
10. Композиция по п.1, которая способна активировать экспрессию опухолевого инволюкрина in vivo.
11. Биспецифическая молекула, связывающая EGFR человека, которая представляет собой антитело или его фрагмент и содержит CDR1, CDR2 и CDR3 тяжелой цепи (SEQ ID NO: 40) и CDR1, CDR2 и CDR3 легкой цепи (SEQ ID NO: 72) антитела 992 и CDR1, CDR2 и CDR3 тяжелой цепи (SEQ ID NO: 41) и CDR1, CDR2 и CDR3 легкой цепи (SEQ ID NO: 73) антитела 1024.
12. Биспецифическая молекула по п.11, представляющая собой антитело с двойным вариабельным доменом.
13. Биспецифическая молекула по п.11, представляющая собой биспецифический фрагмент Fab или биспецифический scFV.
14. Применение композиции антитела по пп.1-10 или биспецифической молекулы по пп.11-13 для изготовления лекарственного средства для лечения, облегчения или профилактики одного или более симптомов, связанных с раком, у млекопитающих.
15. Применение композиции антитела по пп.1-10 или биспецифической молекулы по пп.11-13 для изготовления лекарственного средства для уменьшения передачи сигнала EGFR, уничтожения клеток, экспрессирующих EGFR, индукции апоптоза в клетках, экспрессирующих EGFR, ингибирования пролиферации клеток, экспрессирующих EGFR, индуцирования дифференциации опухолевых клеток in vivo и/или индуцирования интернализации EGFR.
16. Композиция по пп.1-10 для применения в качестве лекарственного средства для лечения, облегчения или профилактики одного или более симптомов, связанных с раком, у млекопитающих.
17. Биспецифическая молекула по пп.11-13 для применения в качестве лекарственного средства для лечения, облегчения или профилактики одного или более симптомов, связанных с раком, у млекопитающих.
18. Композиция по пп.1-10 для применения в качестве лекарственного средства для уменьшения передачи сигнала EGFR, уничтожения клеток, экспрессирующих EGFR, индукции апоптоза в клетках, экспрессирующих EGFR, ингибирования пролиферации клеток, экспрессирующих EGFR, индуцирования дифференциации опухолевых клеток in vivo и/или индуцирования интернализации EGFR.
19. Биспецифическая молекула по пп.11-13 для применения в качестве лекарственного средства для уменьшения передачи сигнала EGFR, уничтожения клеток, экспрессирующих EGFR, индукции апоптоза в клетках, экспрессирующих EGFR, ингибирования пролиферации клеток, экспрессирующих EGFR, индуцирования дифференциации опухолевых клеток in vivo и/или индуцирования интернализации EGFR.
20. Фармацевтическая композиция для лечения и/или профилактики заболеваний, связанных с экспрессией EGFR, содержащая композицию по любому из пп.1-10 или биспецифическую связывающую молекулу по любому из пп.11-13 в терапевтически эффективном количестве.
| FRIEDMAN L.M | |||
| et al., "Synergistic down-regulation of receptor tyrosine kinases by combinations of mAbs: Implications for cancer immunotherapy", PNAS (2005), 102(6):1915-1920 | |||
| SPIRIDON C.I | |||
| et al., "Targeting Multiple Her-2 Epitopes with Monoclonal Antibodies Results in Improved Antigrowth Activity of a Human Breast Cancer Cell Line in Vitro and in |

