Изобретение относится к антиидиотипическим антителам, которые индуцируют иммунный ответ против опухолей, несущих в качестве антигена рецептор эпидермального фактора роста (EGFR). Предлагаемое изобретение, главным образом, относится к антиидиотипическим антителам, имеющим "внутренний образ" антигена, и способным имитировать внешний домен EGFR человека. В предпочтительном варианте настоящего изобретения указанные антитела происходят от мышиного моноклонального антитела mAb 425, либо от его "очеловеченных" и химерных вариантов. Антитела настоящего изобретения могут быть использованы для иммунотерапии и иммунопрофилактики опухолей.
В описании настоящего изобретения используются некоторые сокращения и специальные термины, которые имеют следующие значения:
"FR" (каркасные участки) представляют собой четыре субрайона вариабельных областей легкой или тяжелой цепи, которые поддерживают при CDR-участка.
"CDR (участки, определяющие комплементарность) представляют собой три субрайона вариабельных областей легкой и тяжелой цепи, которые имеют гипервариабельные последовательности, и образуют петлевые структуры, являющиеся, в основном, ответственными за осуществление непосредственного контакта с антигеном.
"EGF" и "EGFR" означают эпидермальный фактор роста и его рецептор.
"PCR" означает полимеразную цепную реакцию.
"ScFV " означает одноцепочечный FV - фрагмент антитела.
"VL" означает вариабельную область легкой цепи.
"VK" означает вариабельную область легкой каппа-цепи.
"VH" означает вариабельную область тяжелой цепи.
"Химерные" или "частично очеловеченные" антитела представляют собой антитела, содержащие константные области, происходящие от иммуноглобулина человека, и вариабельные области (включая CDR), происходящие от антител, взятых от других источников, не относящихся к человеку, например, от мышиных антител.
"Очеловеченные" или полностью очеловеченные антитела представляют собой антитела, которые содержат константные области и каркасные участки (FR), происходящие от иммуноглобулина человека; и гипервариабельные участки (CDR), происходящие от других источников, не относящихся к человеку.
"Ab1" означает "первое" антитело или исходное антитело, которое индуцирует, при иммунизации, каскад последовательно продуцируемых антител.
"Ab2" означает типичное антиидиотипическое антитело (=антиидиотип), направленное против идиотипов Ab1.
"Ab3" означает антиидиотипическое антитело, направленное против идиотипов Ab2, а поэтому по своей специфичности сравнимое с Ab1.
PBS - означает забуференный фосфатом физиологический раствор.
FCS - означает фетальную телячью сыворотку.
HBSS - означает сбалансированный солевой раствор Хэнкса.
F1TC - означает флуоресцеинизоцианат.
MTC - означает смешанную культуру клеток.
KLH - означает гемоцианин лимфы улитки (хромопротеид, получаемый из моллюска Medathura crenulata, сильный иммуноген, - прим. пер.).
CFA - полный адъювант Фрейнда (ПАФ).
HRPO - означает рекомбинантную пероксидазу человека.
Антиидиотипические антитела (антиидиотипы) представляют собой антитела, направленные против эпитопов (называемых идиотипами), локализованных в антигенсвязывающей области или вариабельной области молекулы другого антитела. Взаимодействия между идиотипами (Ab1) и антиидиотипами (Ab2) играют важную роль в поддержании иммунного гомеостаза. Возникновение теории о сети идиотипов и антиидиотипов привело к объяснению явлений иммунорегуляции (Jerne F. G. 1974, Arn. Immunol. 125C: 373). Антиидиотипические антитела, несущие "внутренний образ" антигена, имитируют трехмерную структуру антигена, распознаваемого антителом Ab1. Введение антитела Ab2 может вызвать продуцирование антител Ab3 против исходного антигена. Идиотипические вакцины были успешно применены для лечения некоторых трансмиссивных болезней, и для лечения рака (см., например,
Uytdehaag, F. G. &., C.M.H.Osterhans, 1986, J.Immunol. 134: 1125; Kennedy, P. C. et al. 1986, Science 232: 220; Hiernaux, J.R., 1988; 56: 1407; Strein K.E. & T.Soder-strom, 1984, J.Exp. Med. 160: 1001).
В опухолевой иммунологии идиотипическая вакцинация была использована для модуляции опухолевого роста в экспериментальных in vivo и in vitro - системах (Smorodinsky, N. I. et al., 1988, Eur J. Immunol. 18: 1713; Viale, G, et al. , 1987, J.Immunal. 139: 1438). Herlyn и др. (1987, Proc. Natl. Acad. Sci. USA, 76: 1438) сообщили о многообещающих результатах, полученных в клинических испытаниях, которые были проведены с участием пациентов, страдающих прогрессирующей карциномой толстой кишки, и в которых наблюдалось прекращение развития метастазов и частичная клиническая ремиссия у пациентов, подвергнутых лечению лишь одними антидиотипами. Антиидиотипические антитела и их использование в противоопухолевой терапии также описаны, например, в патентах США 4918164 и EP 0141783.
Эпидермальный фактор роста (EGF) представляет собой полипептидный гормон, который является митогенным для эпидермальных и эпителиальных клеток. При взаимодействии с чувствительными клетками, EGF связывается с мембранными рецепторами (EGFR). Рецептор для EGF представляет собой трансмембранный гликопротеин размером около 170 кДа, и является генным продуктом протоонкогена c-erb-B.
Mab 425 представляет собой мышиное моноклональное антитело, продуцируемое против хорошо известной клеточной линии карциномы человека A431 (ATCC CRL 1555); причем указанное антитело связывается с внешним доменом EGFR человека, и ингибирует связывание EGF. Было обнаружено, что MAb 425 (ATCC HB 9629) опосредует опухолевую цитотоксичность in vitro, и подавляет in vitro - рост опухолевых клеток эпидермоида и клеточных линий, происходящих от карциномы толстой кишки (Rodeck et al., Cancer Res. 1987, 47: 3692). "Очеловеченные" и химерные варианты MAb 425 описаны в WO 92/15683.
Связывание EGF с рецептором активируют некоторые биохимические процессы, что приводит к репликации ДНК и делению клеток (Cargenter G., 1987, Annu. Rev. Biochem. 56: 881). Недавно было установлено, что система EGFR участвует в онкогенной трансформации клеток (Di Fiore, P.P. et al., 1987, Cell 51: 1063.10), Было высказано предположение, что рост некоторых опухолей, которые экспрессируют EGFR и секретируют EGF или TGF α, происходит за счет пролиферации аутостимулированных клеток (Ennis, B. W.et al., 1989, Mol. Endo. 3: 1830: причем повышенная экспрессия EGFR на клеточной поверхности была обнаружена в опухолях, происходящих от различных тканей, например, в опухолях молочной железы, мочевого пузыря, карциномах, меланомах, и опухолях головного мозга, не происходящих от нервных клеток (Neal D.E. et al., 1985; Lancet, 1, 366; Libermann T.A. et al., 1984, Cancer Res 44: 753; Herlyn M.et al., 1982, J.Clin. Immunol 2: 135). EGFR является терапевтической мишенью, поскольку он непосредственно участвует в клеточной пролиферации некоторых опухолей. Было показано, что чрезмерная экспрессия EGFR имеет плохое прогностическое значение, так как она коррелирует с повышенной инвазивной способностью опухоли (Sainsbury J.R. et al., 1985, Lancet I:364).
Пассивная или активная иммунизация раковых больных против EGFR приводит к индуцированию в кровотоке этих больных специфических антител, которые действуют как антагонисты EGF и TGFα , блокируя их связывание с рецептором, и ингибируя, тем самым, рост опухолей, EGFR является хорошим кандидатом для идиотипической вакцины, поскольку он может быть выделен из опухолевых клеток, хотя и в недостаточном для терапевтических целей количестве.
Несколькими группами исследователей изучались и обсуждались антиидиотипические антитела, и, главным образом, структурное сходство этих антител с их антигенами и "внутренними образами" (см., например, Tsjisaki, M. et.al., 1993, J. Immunol, 150-508; Raychandhuri S., et.al., 1990, J.Immunol 145-760; Bruck C., et al. 1986, Proc. Natl. Acad. Sci. USA, 83: 6578).
Предлагаемое изобретение относится, главным образом, к новым антиидиотипическим антителам (антиидиотипам), которые индуцирует иммунный ответ против рецептора эпидермального фактора роста. В частности, изобретение относится, по крайней мере, к двум таким антиидиотипам (обозначенным 5А6 и 3B6, которые имитируют область антигена на поверхности рецептора человеческого эпидермального фактора роста. Таким образом, антитела настоящего изобретения, а в частности, антитела, которые способны имитировать EGFR, могут быть использованы для индуцирования и усиления иммунного ответа против всех видов опухолей человека, экспрессирующих рецептор эпидермального фактора роста на своей клеточной поверхности, например таких опухолей, как меланома, глиома, и карцинома. Эти антитела могут быть также использованы в диагностических целях для определения локализации опухоли и для ee in vitro и in vivo-оценки.
Таким образом, целью настоящего изобретения является получение нового антитела, которое представляет собой моноклональное антиидиотипическое (Ab2) антитело, индуцирующее иммунный ответ против рецептора эпидермального фактора роста (EGFR).
Более конкретно, настоящее изобретение относится к моноклональному антиидиотипическому антителу, имитирующему "внутренний образ" EGFR - антигена, распознаваемого соответствующим мышиным, "очеловеченным" или химерным моноклональным идиотипическим (Ab1) антителом. Это антитело (Ab1) может не происходить от человека, то есть, оно может быть мышиным, "очеловеченным" или химерным моноклональным антителом, определенным выше. Поскольку указанное антитело является "образно воспроизводимым", т.е., по существу, имеется несколько антител Ab1, направленных против эпитопа EGFR, то настоящее изобретение также относится к антиидиотипическим антителам, которые могут распознавать эту группу антител Ab1 против EGFR.
Кроме того, настоящее изобретение относится к антиидитипическим антителам против EGFR, которые могут быть получены исходя из соответствующего антитела Ab1 против EGFR, например, путем иммунизации.
Таким образом, целью настоящего изобретения является получение моноклонального антиидиотипического антитела, которое может быть продуцировано путем иммунизации животного с использованием соответствующего мышиного, "очеловеченного", или химерного, идиотипического (Ab1) антитела.
Антиидиотипические антитела настоящего изобретения получают, предпочтительно, путем иммунизации животного посредством введения ему моноклонального анти-EGFR антитела 425, либо "очеловеченных" и химерных вариантов этого антитела. MAb 425 продуцируется известной клеточной линией, депонированной под номером допуска ATCC HB 9629.
Таким образом, целью настоящего изобретения является получение моноклонального антиидиотипического антитела, где идиотипическим антителом (Ab1) является mAb 425 (ANCC HB 9629) либо антитело, полученное из mAb 425 путем его "очеловечивания" или химеризации с использованием методики, описанной, например, в WO 92/15683.
Предлагаемое изобретение также относится к специфическим антиидиотипическим антителам, которые имеют хорошо известные аминокислотные последовательности в гипервариабельных (CDR) и вариабельных (FR) областях антител, и которые представлены на фиг. 5A-F.
Другой целью предлагаемого изобретения является получение антиидиотипического моноклонального антитела, где CDR-области и FR-области указанного антитела имеют аминокислотные последовательности, показанные на фиг. 5A-F. Кроме того, целью настоящего изобретения, как предпочтительного варианта его осуществления, является получение антиидиотипического антитела, имеющего аминокислотную или нуклеотидную последовательность, представленную на фиг. 5A-F.
В соответствии с настоящим изобретением, описываемые последовательности могут также иметь модификации и изменения, возникающие в точно определенных последовательностях, изображеных на фиг. 5, вследствие спонтанных, либо химически или физически индуцированных мутаций, инсерций, делеций, и замещений отдельных аминокислотных или нуклеотидных остатков, при условии, что указанные модификации и изменения не оказывают заметного влияния на биологическую активность и свойства конечных антител.
Термин "антиидиотипическое антитело" также относится к частям или фрагментам указанного антитела, например, к одноцепочечным Fv-фрагментам или F(ab)'2 - и Fab' - фрагментам, хорошо известным специалистам (Skerra & Plyckthum, Science, 1988, 240: 1038; Better et al., Science, 1988, 240: 1041). Одноцепочечные Fv - фрагменты (где VL и VH - участки соединены вместе) также описаны в литературе (например, Bird et al., Science 1988, 242: 423; Huston et al., Proc. Natl. Acad. Sci. USA, 1988, 85: 5879).
Кроме того, настоящее изобретение относится к способу получения антиидиотипического антитела, определенного в описании и формуле настоящего изобретения, заключающемуся в иммунизации животного путем введения этому животному соответствующего идиотипического антитела (Ab1), которое связывается с антигеном непосредственно при помощи своих гипервариабельных участков (CDR), с последующим выделением и очисткой синтезированного антиидиотипического антитела (Ab2), которое связывается с идиотипами антитела Ab1, стандартными методами.
Другой целью настоящего изобретения является получение фармацевтической композиции, содержащей антиидиотипическое моноклональное антитело, определенное в настоящем описании изобретения; и необязательно, фармацевтически приемлемый носитель.
И наконец, предлагаемое изобретение также относится к использованию указанного антиидиотипического антитела для изготовления лекарственного средства, обладающего противоопухолевым действием.
В предлагаемой заявке описано получение трех моноклональных антиидиотипических антител (5А6, 3В6 и 15Н8), которые распознают идиотипы, перекрывающиеся с антигенсвязывающим центром (паратопом) антитела mAb 425. Проведенные серологические и иммунологические исследования показали, что антитела 5А6 и 3В6 несут "внутренний образ" эпитопа, распознаваемого антителом mAb 425, индуцируя таким образом гуморальный ответ в сингенной (мыши Ba1b/c) и аллогенной (мыши В6-2F1) системе. Антиидиотипы с истинным внутренним образом должны действовать как суррогаты антигена в организме различных видов животных, однако, эти антиидиотипы были неэффективными в отношении генерирования Ab1-подобного ответа у кроликов и крыс. Клонирование и секвенирование их V - областей выявило наличие аминокислотной гомологии между CDR обеих (легкой и тяжелой) цепей и некоторыми остатками EGFR, локализованными в области связывания с лигандом, что дает основание предположить о имитации антигена. Исследование антигенности гомологичных областей позволило дать приемлемое объяснение отсутствию биологического эффекта в гетерогенной системе, и подтвердило потенциальную эффективность антиидиотипических антител в иммунной терапии.
Фиг. 1 показано ингибирование связывания 425 с EGFR антиидиотипами 5А6, 3В6, 15H8 или неродственными mАb. Процент ингибирования вычисляли как описано в главе "Материалы и методы" с использованием значений ОП, полученных из ELISA.
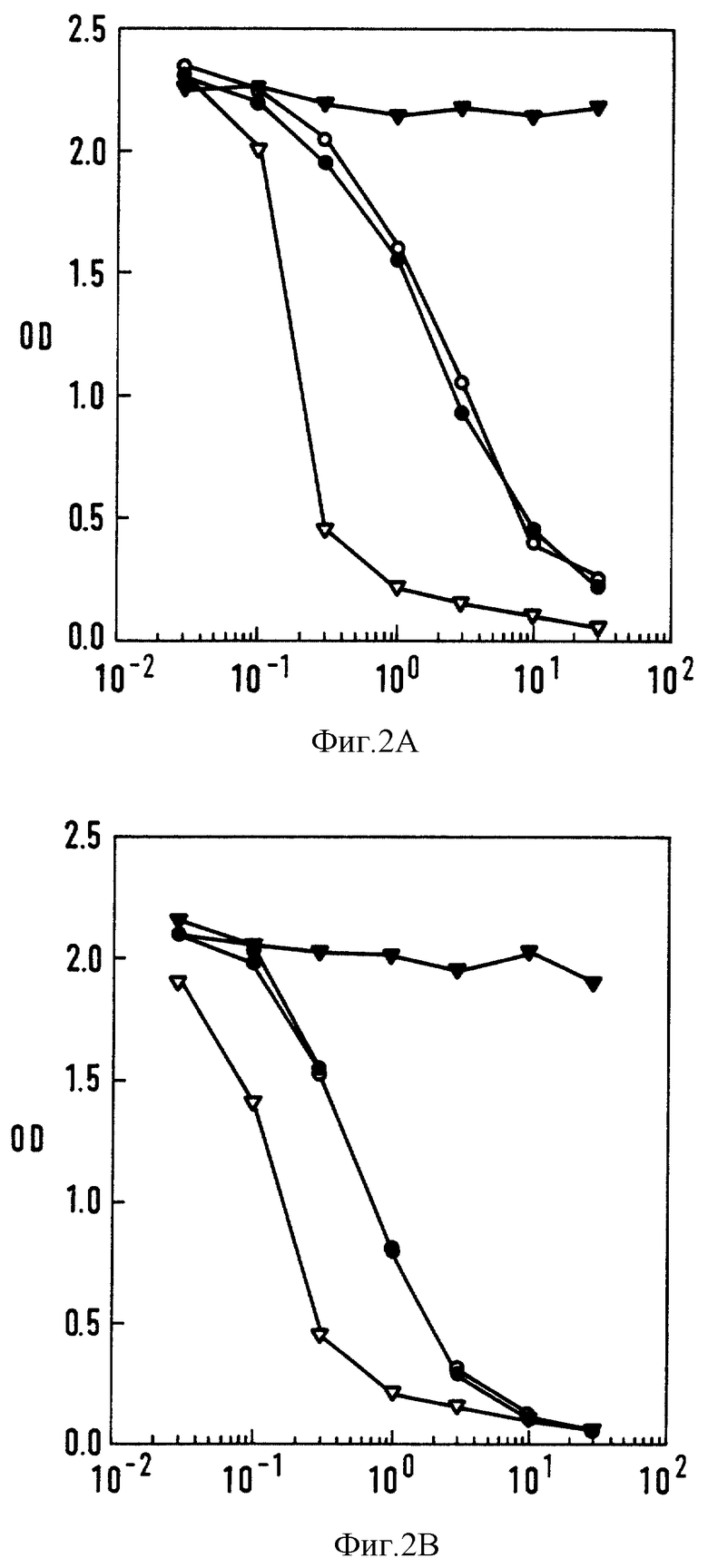
Фиг. 2 - кривые ингибирования антиидиотипами 5А6 (•) 3В6 (°), 15Н8 (▿) и мышиными иммуноглобулинами IgG1 (Sigma)  связывания мышиного (А) и "очеловеченного" варианта (В) антитела 425 с гGFR-экспрессирующими клетками А431. Результаты выражали как среднее значение ОП для двух дубликатных образцов. Ось X: конкурент (мкг/мл); ось y: ОП.
связывания мышиного (А) и "очеловеченного" варианта (В) антитела 425 с гGFR-экспрессирующими клетками А431. Результаты выражали как среднее значение ОП для двух дубликатных образцов. Ось X: конкурент (мкг/мл); ось y: ОП.
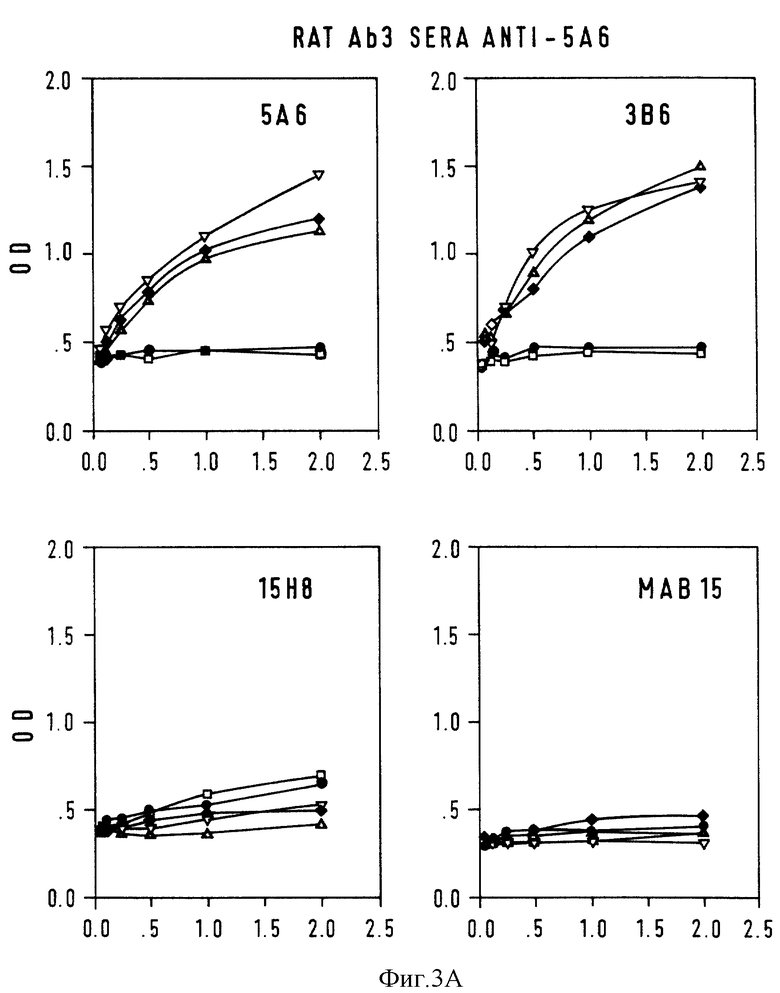
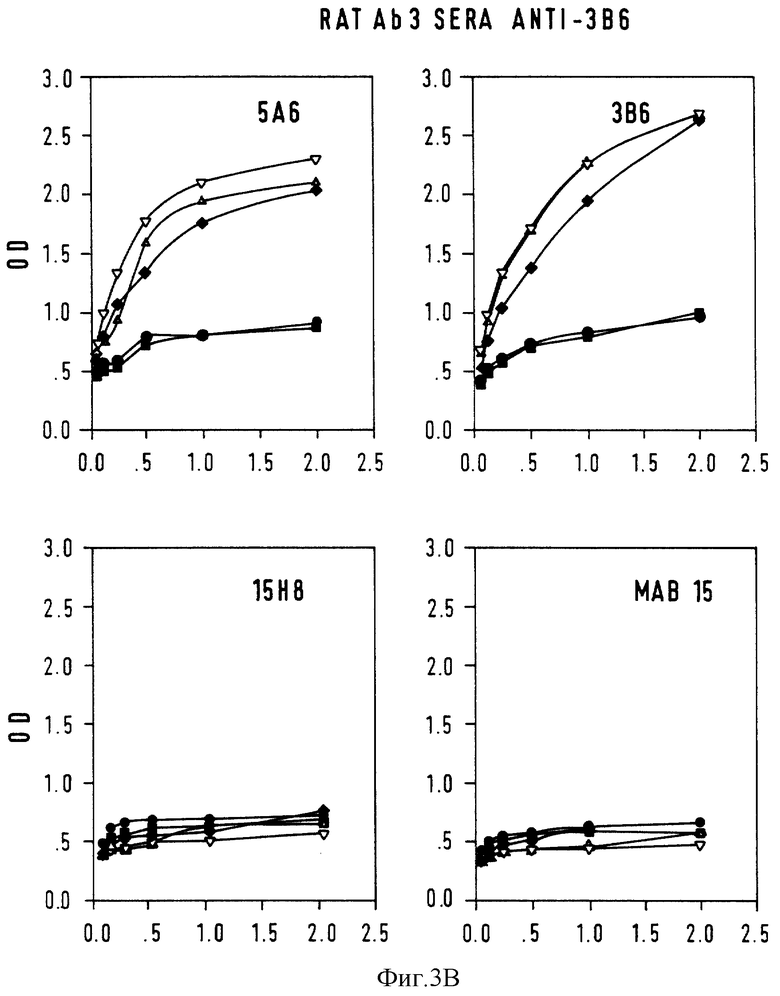
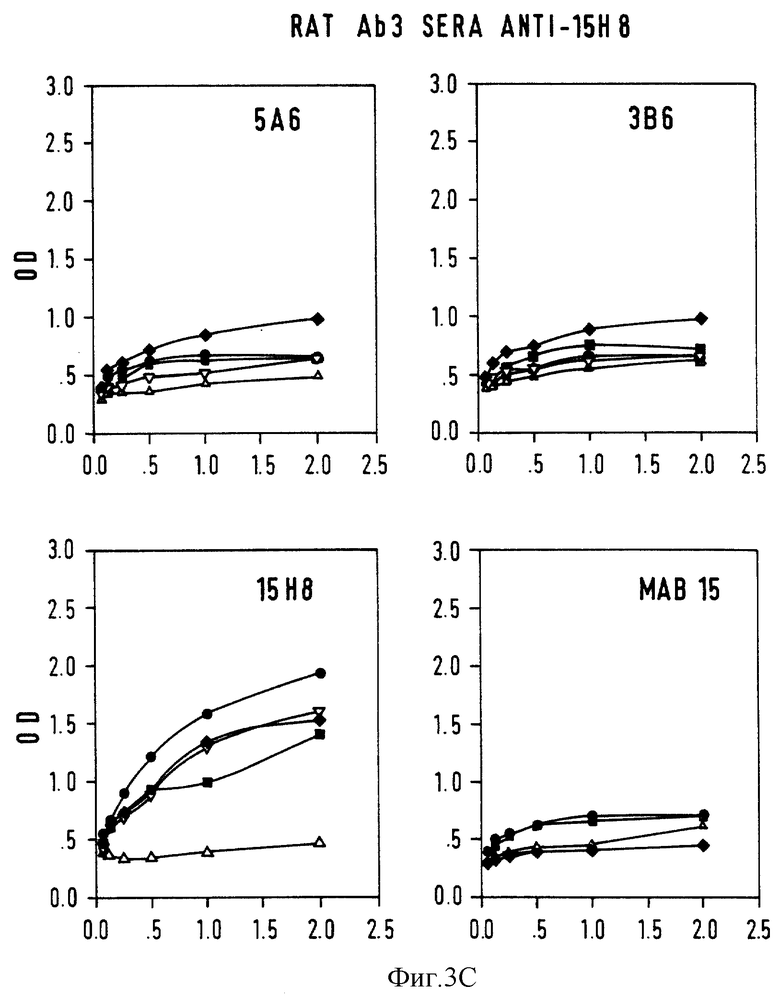
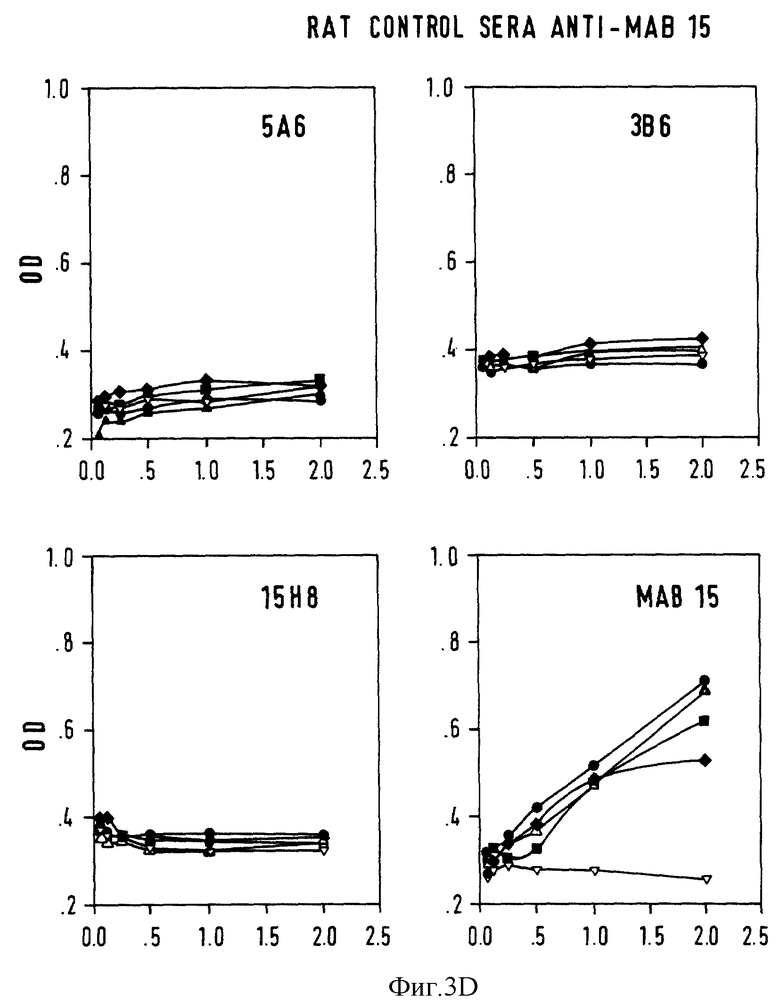
Фиг. 3 - конкурентный ELISA-анализ на Ab3-Ab2-связывание. Серийные разведения крысиной антисыворотки Ab3 анализировали на связывание с нанесенными на твердую фазу Ab2 или неродственными mab 15, предварительно инкубированными с 5А6 (•), 3В6 (▪), и 15H8  В качестве контроля использовали mab 15 (▿) и нормальную мышиную сыворотку.
В качестве контроля использовали mab 15 (▿) и нормальную мышиную сыворотку.  A: крысиная Ab3-содержащая сыворотка против 5A6; B: крысиная сыворотка Ab3-сыворотка против 3B6; C: крысиная AB3-сыворотка против mAb15; D: контрольная крысиная сыворотка против mAb15; ось X: ОП; ось Y: серийные разведения • 10-3.
A: крысиная Ab3-содержащая сыворотка против 5A6; B: крысиная сыворотка Ab3-сыворотка против 3B6; C: крысиная AB3-сыворотка против mAb15; D: контрольная крысиная сыворотка против mAb15; ось X: ОП; ось Y: серийные разведения • 10-3.
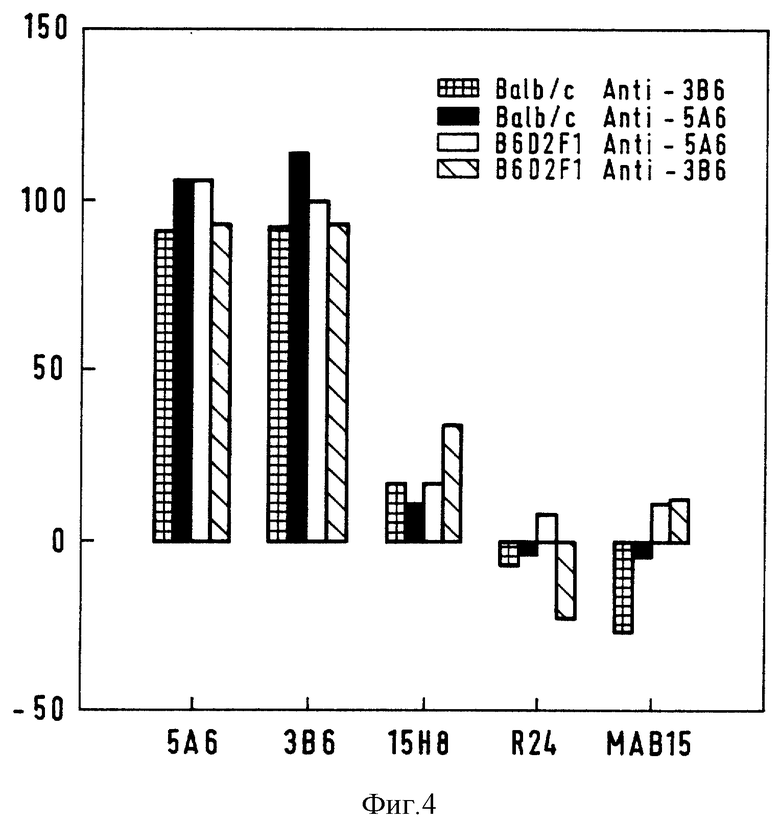
Фиг. 4 - твердофазный ELISA: ингибирование антиидиотопами связывания Ab3 Balb/c и B6D2F1, полученных от 5A6 - и 3B6 - иммунизированных мышей, с очищенным EGF-рецептором.
Фиг. 5 - полная последовательность вариабельной области (включая лидерную последовательность) тяжелой и легкой цепей мышиных моноклональных антител 15H8, 5A6 и 3B6, показаны нуклеотиданая и аминокислотная последовательности. Лидерная последовательность (где имеется надпись) обозначена жирным шрифтом; CDR - последовательности показаны курсивом; а последовательность, используемая для амплификации вариабельных областей, подчеркнута сплошной линией. Тяжелая цепь 15H8, 5A6 и 3B6 имеют характерную структуру группы IIID. Легкая цепь 15H8 и 5A6 имеют характерную структуру каппа-группы Y, тогда как 3B6 имеет структуру, характерную для каппа-группы III согласно классификации Кэбат:
A: тяжелая цепь 15H8
B: легкая цепь 15H8
C: тяжелая цепь 3B6
D: легкая цепь 3B6
E: тяжелая цепь 5A6
F: легкая цепь 5A6
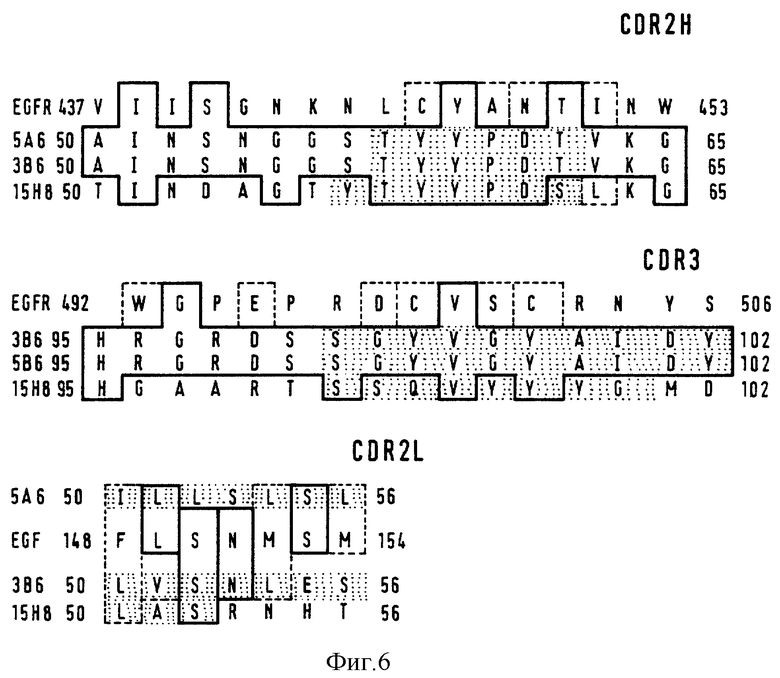
Фиг. 6 - сравнение последовательностей CDR2H, CDR3H и CDR21 антитела Ab2 и внешнего домена EGF-рецептора. Идентичные аминокислоты показаны в сплошных рамках. В пунктирных рамках показаны подобные аминокислоты. Потенциальные антигенные области показаны в заштрихованных рамках.
Таблица I. Константы аффинности 5A6, 3B6 и 15H8 по отношению к 425. Константы диссоциации были вычислены по графикам Скэтчарда, описывающим связывание Ab2 с 425, исходя из величин ОП, полученных путем анализа ELISA, который проводили как описано ниже, в главе "Материалы и методы".
Таблица II. Ингибирование Ab1-Ab2- связывания антителами Ab3 и выявление перекрестно реагирующих идиотипов на Ab2. Двухкратные разведения антисывороток Ab3, полученных от мышей (A), кроликов (B) и крыс (C), иммунизированных антиидиотипами или неродственными mAb, анализировали на ингибирование, связывания иммунизирующих Ab2, других антиидиотипов, и мышиного изотипа и аллотипа, соответствующего mAbs. Указаны титры сыворотки, дающие 100%-ное ингибирование.
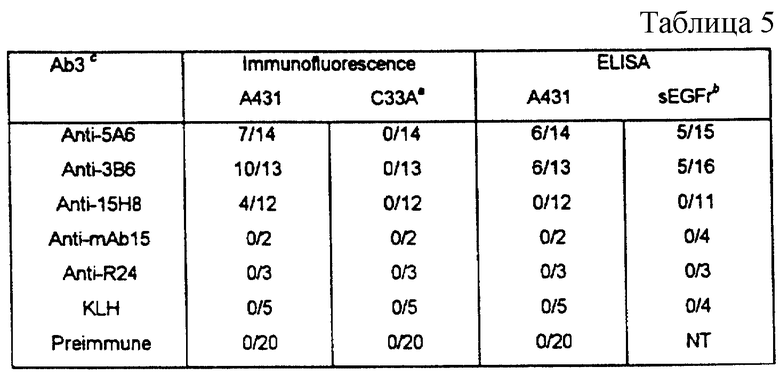
Таблица III. Иммунный ответ против EGFR у мышей Balb/c, определенный путем непрямой иммунофлуоресценции по отношению к EGFR-позитивным (A431) или негативным (С33А) нефиксированным клеткам, и путем ELISA - анализа по отношению к целым клеткам или очищенному внешнему домену.
a) Обнаружение кроличьих антимышиных IgM + IgG с помощью FITC (Флуоресцеин-изотиоцианата).
b) Обнаружение кроличьих антимышиных IgG с помощью FITC.
c) Позитивные животные/иммунизированные животные.
5. Подробное описание изобретения
Биологические материалы и общие методы
Микроорганизмы, клеточные линии, плазмиды, фагмиды, промоторы, маркеры резистентности, сайты инициации репликации, или другие фрагменты векторов, которые упоминаются в настоящей заявке, были закуплены или получены другими доступными путями. При этом следует отметить, что если в настоящей заявке не приводится каких-либо других данных, то все указанные материалы используются лишь в иллюстративных целях, т.е. не имеют решающего значения для настоящего изобретения, и могут быть заменены другими подходящими средствами и биологическими материалами соответственно.
Методы, используемые для осуществления настоящего изобретения, подробно описаны ниже. В предлагаемой заявке не проводится подробного описания стандартных методов, хорошо известных любому специалисту, либо детально описанных в цитируемых источниках, патентных заявках и в общеизвестных работах (например, Harlow E & D. Lane, 1988, Anfibodies, a laborafory mannal. Cold Spring Harbor Laboratory, N.Y.).
Для in vivo-анализов были использованы мыши Balb/c (IFFA CREDO), гибридный штамм B6D2FI, полученный от мышей C57BL/6J и DBA/2 (IFFA CREDO), крысы Wistar (Interfanna Iberica), и белые новозеландские кролики (Biocentre).
A431 представляет собой проскоклеточную (эпидермоидную) карциному (ATCC CRL 1555), экспрессирующую EGFR. C33A (ATCC HTB31) представляет собой карциному шейки матки, и является рецептор-негативной. В качестве "партнера" для слияния при получении гибридомы использовали HL1-Friendly-миелому-653 (Ventrex, bioventures group). Все клеточные линии культивировали в среде RPMI 1640, в которую была добавлена 10%-ная фетальная телячья сыворотка и 2 мМ глутамина.
Антитело mAb 425 (AbI) против EGFR продуцировали по методу Rodeck и др. (см. выше). В различных анализах в качестве контроля использовали несколько мышиных mAb неродственной специфичности, а именно: mab 15 (IgGI, k) против gp85/45 мелкоклеточного рака легких; R24 (IgG3,k) против GD3; 14F9 (IgG3,k) против GD3, и Me36I (IgG2f) против GD2, которые были описаны ранее (Kamma H. et. al. , 1989, Cancer Res. 49(8): 5118; Dippold W.G., 1980, Proc. Natl. Acad. Sci. USA. 77:6114, Masso O.et.al., 1991, Immunologia 10:36; Ihurin I. et.al., 1987, Cancer Res. 47:1229).
FIII (IgGl, k) получали путем ПЭГ-индуцированного слияния спленоцитов, взятых от иммунизированных РНКазой мышей Balb/c и Friengly-миеломы.
Получение идиотипического mAb против mAb 425
Мышиные гибридомы продуцировали с использованием спленоцитов, взятых от мышей Balb/c, иммунизированных антителом mAb 425, и слитых с клетками Friendlu-миеломы 653. Супернатанты тестировали с помощью "сандвич"-анализа ELISA по отношению к mAb 425. Гибридомы 5A6, 3B6 и 15H были получены в результате двух слияний, что давало 2,8%-ную и 10,5%-ную специфическую эффективность, соответственно. После трехкратного клонирования методом серийного разведения, было установлено, их антитела относятся к классу IgGI,k.
Характеризация 5A6, 3B6 и 15HB
Для характеризации Ab2 использовали неочищенные супернатанты или очищенные образцы белка A. Антиидиотипическая природа этого антитела была подтверждена его специфическим связыванием с mAb 425 при тестировании панели с неродственными mAb, имеющими различные антигенные специфичности (FIII, 361, 15 и 14F9).
Для определения локализации 425-идиотопов, распознаваемых антиидиотипами, они были подвергнуты тестированию по отношению к sEGFR посредством ELISA-анализа на ингибирование связывания 425 с EGFR. В результате наблюдалось полное ингибирование реактивности EGFR (фиг. 1), что указывает на то, что антиидиотипы распознают идиотопы внутри антигенсвязывающего сайта.
Для идентификации антиидиотипов 5A6, 3B6 и 15HB как Ab2b были проведены анализы в целях определения совпадают ли идиотопы, распознаваемые на mAb 425, с CDR или антигенсвязывающим сайтом. Для таких анализов путем непрямого ELISA-теста с использованием клеток A431, были построены кривые ингибирования связывания для мышиных (фиг. 2A) и "очеловеченных" (фиг. 2B) вариантов mAb 425. Все три антиидиотипа оказались способными ингибировать связывание обоих вариантов 425; причем каждое из Ab2 имели аналогичные кривые ингибирования связывания, что свидетельствует о том, что идиотопы 425, распознаваемые антителами 5A6, 3B6 и 15H8, непосредственно участвуют в распознавании антигена. В ингибировании связывания клеток 425 с клетками A431 антитело 15H8 обнаруживало эффективность, которая в 10 раз превышала эффективность 5A6 и 3B6. Для определения степени соответствия Ab2 паратопу 425 вычисляли константу диссоциации (KD) антиидиотипов. Константы диссоциации каждого из Ab2 представлены в табл. I. 15H8 обладает более высоким сродством к 425, что коррелирует с его более высокой способностью ингибировать связывание EGFR с 425. Эти результаты позволяют также предположить, что, по крайней мере, два различных идиотопа на mAb 425 распознаются антиидиотипическими антителами.
Индукция сингенных (Balb/c), аллогенных (B6D 2FI) и ксеногенных (крыса, кролик) антител Ab3
Пригодность указанных Ab2 для использования в качестве идиотипических вакцин определяли путем оценки их иммуногенности. Антитела Ab2, несущие истинный "внутренний образ" антигена, должны обладать способностью индуцировать антитела Ab3 с Ab1-подобный специфичностью, выходящей за пределы видового барьера. Ab3 были продуцированы у мышей, крыс и кроликов в соответствии со схемой иммунизации, описанной выше.
Анти-антиидиотипические антитела были обнаружены через 2 недели после введения первой дозы и достигали максимального уровня через 6-8 недель после введения первой дозы. У крыс и у кроликов специфический ответ против иммунизирующего Ab2 поддерживался в течение почти 12 месяцев, тогда, как у мышей этот ответ был более кратковременным, и через 6 месяцев, титр Ab3 снижался. Все кролики продуцировали антимышинные Ig; причем титр антисыворотки Ab3 против иммунизирующего Ab2 был в 5-10 раз выше, чем титры, обнаруженные против изотипических и аллотипических Ig, что свидетельствует о том, что иммунные ответ против константной области Ab2 не нарушает иммунный ответ против v-области.
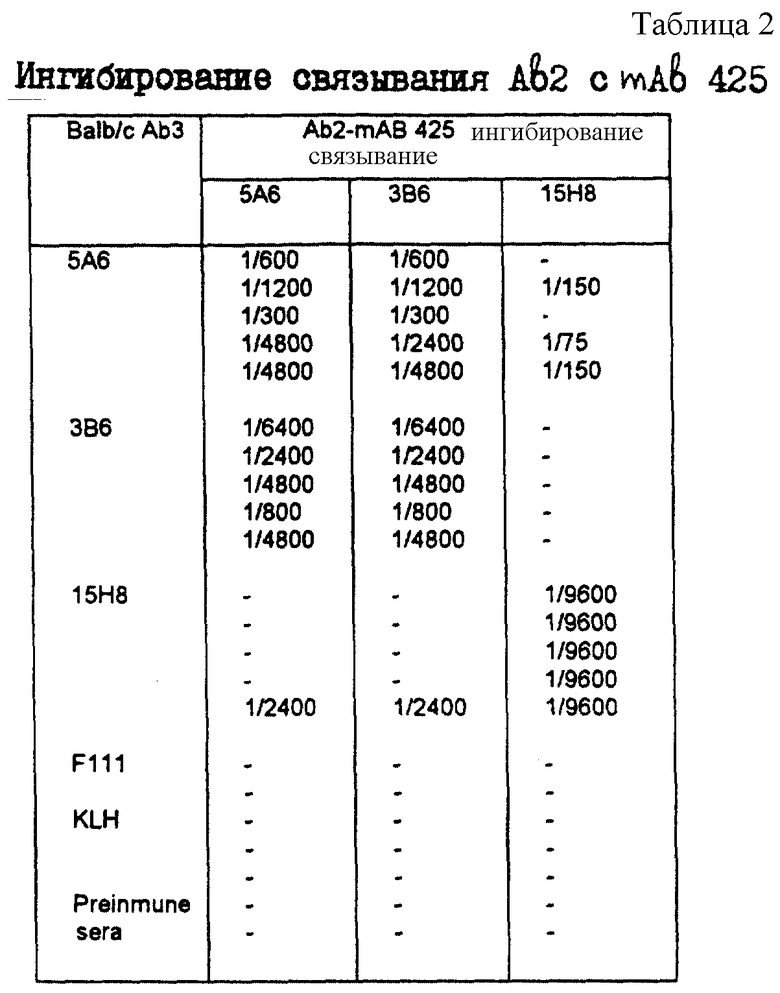
Для определения эффективности иммунизации против тех детерминант Ab2, которые констактируют с паратопом mAb 425 (где должны находиться антиген-имитирующие идиотопы), был проведен анализ на ингибирование связывания, в котором связывание 5A6, 3B6 или 15H8 (антител Ab2) с mAb 425 измеряли в присутствии серийных разведений Ab3-содержащей сыворотки, взятой от животных, иммунизированных антителом Ab2, или неродственными мышиными mAb. В табл. 2-4 разведения Ab3-содержащей сыворотки, которые дают 100%-ное ингибирование связывания Ab2-Ab1, получены от мышей Balb/c (табл. 2), кроликов (табл. 3) и крыс (табл. 4). Как видно из табл. 2-4 все антисыворотки Ab3 против 5A6, продуцированные в этих 3 видах, ингибировали связывание mAb 425 с 5A6 и 3B6 с одинаковой эффективностью. Антисыворотка против 3B6 обнаруживала аналогичный характер ингибирования, что позволяет предположить, что оба эти антиидиотипа могут иметь общие эпитопы. Кроме того, было установлено, что некоторые крысиные и мышиные антисыворотки против 15H8 ингибируют связывание 5A6 и 3B6 с 425, что позволяет сделать вывод о их частичной идиотипический идентичности. Такая реактивность не наблюдалась в случае кроличьих антисывороток против 15H8, что, вероятно, указывает на то, что Ab3 - ответ, продуцируемый у кроликов, может быть направлен, в основном, против других, более иммунодоминантных идиотопов; причем это предположение подтверждается выявлением более высоких ингибирующих титров для кроличьей Ab3 - содержащей сыворотки по сравнению с крысиными и мышиными антисыворотками.
Антигенную идентичность между 5A6 и 3B6 исследовали с помощью конкурентного ELISA-теста, где серийные разведения (10-3-10-5) кроличьих и крысиных антисывороток (Ab3) анализировали на связывание с Ab2 в присутствии равного объема (1 мг/мл) Ab2 или контрольных mAb. Кривые ингибирования связывания для крысиных Ab3-содержащих сывороток показаны на фиг. 3. 5A6 и 3B6 генерируют идентичный спектр Ab3 у крыс и кроликов: антисыворотка против 5A6 с одинаковой эффективностью реагируют с антителами 5A6 и 3B6, нанесенными на ELISA-планшеты, и в равной степени ингибируется обоими Ab2. Кроличья сыворотка Ab3 имеет кривую ингибирования, аналогичную кривым ингибирования, показанным на фиг. 3 для крысиных сывороток.
Идентификация антител Ab3 против EGFR
Антитела к рецептору были обнаружены в Ab3-содержащих сыворотках методом прямой иммунофлуоресценции с использованием клеток A431 и C33A. Полученные результаты представлены в табл. 5.
Антиидиотипическая вакцинация в мышиной системе индуцировала образование антител против EGFR, однако в Ab3-содержащих сыворотках от крыс и кроликов не должны обнаруживаться антитела к EGFR. Продуцирование антител к EGFR у мышей B6D 2FI подтверждает тот факт, что Ab2-вакцинация генетически не ограничивается мышиным штаммом Balb/c. При тестировании против SEGFR с помощью ELISA титры специфических антител достигали значений 1/50 - 1/1600.
Ингибирование связывания Ab3 с EGFR антиидиотипическими антителами 5A6, 3B6 и 15H8
Эти анализы осуществляли для того чтобы определить, имитируют ли антиидиотипы трехмерную структуру EGFR. Для этого различные разведения Ab3-содержащей сыворотки против EGFR, взятой от мышей Balb/c и B6D2F1, смешивали с антиидиотипами и оставляли для реакции на 4 ч при комнатной температуре, после чего их анализировали на антитела к рецептору с использованием SEGFR-покрытых планшетов, и, исходя из величин ОП, вычисляли процент ингибирования, который представлен на фиг. 4. 5П6 и 3B6 давали 100%-ное ингибирование, что свидетельствует о том, что эти антиидиотипы имитируют идентичную трехмерную структуру исходного антигена. Процент ингибирования анти-EGFR Ab3 идиотипом 15H8 составляли 15 - 35% по отношению к 5А6 или 3B6.
Было заявлено, что антиидиотипы, несущие истинный внутренний образ антигена, могут имитировать иммуногенные свойства антигена и индуцировать специфический ответ у различных видов животных, за исключением 5А6 и 3B6, которые не способны индуцировать анти-EGFR антитела у крыс и кроликов; однако, проведенные серологические анализы позволяют предположить, что антитела к рецептору, обнаруженные у мышей, были индуцированы идиотипическими детерминантами 5А6 и 3B6, схожими с EGFR.
Первичные структуры Ab2 и сравнение их аминокислотных последовательностей с EGFR
Для выявления структурообразных особенностей, лежащих в основе иммуногенных свойств Ab2, проводили ДНК-секвенирование. VH и VL - последовательности 5А6, 3B6 и 15H8 представлены на фиг. 5. Аминокислотные последовательности VH-областей были классифицированы в соответствии с критерием Кэбата как относящиеся к подгруппе IIID. Согласно Кэбату VL - области 5А6 и 15H8 принадлежат к группе KV, а 3B6 - к группе kIII. CDR-последовательности сравнивали с аминокислотными последовательностями EGFR человека (GQHUE из банка данных SP1RS). Это сравнение проиллюстрировано на фиг. 6, где идентичные аминокислотные показаны в сплошных рамках, а эквивалентные аминокислоты показаны в пунктирных рамках. Максимальная гомология наблюдалась между остатками 77-84 от CDR2H и остатками 125-129. от CDR3H, и остатками 70-76 от 5А6 и 74-80 от CDR2L, причем области EGFR, имеющие наибольшую гомологию с CDR3H, располагались в гипотетических лиганд-связывающих доменах рецепторного белка, описанного A. Ullrich и др. (30). 15H8 имеет идентичную последовательность CDR2H, но отличающиеся последовательности CDR3H и CDR2L. Для определения какую роль играет каждый гомологичный CDR-участок в формировании "внутреннего образца" EGFR, было проведено сравнение последовательностей 5A6 и 3B6 с известными VH и VL-областями, описанными Кэбатом. В соответствии с настоящим изобретением было установлено, что некоторые VH-цепи обладают высокой степенью гомологии с 5А6 и 3B6, в частности, они имеют аналогичные CDR1H и CDR2H, что дает основание предположить, что последовательности CDR3H играют главную роль в распознавании 425-паратопа и имитации EGFR. Было обнаружено, что CDR3H Ig 63 (в основном мышиного или человеческого происхождения) имеют частичную гомологию (50 - 87%) с CDR3H от 5А6 и 3B6. Гомологичные аминокислоты, очевидно, локализованы в остатках 100H-102, тогда как менее гомологичные аминокислоты были обнаружены в GYVG-сегменте, где наблюдалась максимальная гомология с EGFR. При этом, было установлено, что ни одно антитело мышиного или человеческого происхождения не имело остаток V в положении 100D за исключением 5А6 и 3B6. VL 5А6 и 3B6 обнаруживали гомологию с мышиными каппа-цепями, причем совершенно разные последовательности VL-цепи имели одинаковую специфичность. Очевидно, роль CDR2 не связана со строгой аминокислотной идентичностью. 5А6, 3B6 имеют разные последовательности, образующие идентичную структуру внутреннего образа EGFR, однако, аминокислотные гомологии с EGFR, очевидно, аккумулируются в этой области.
Для оценки иммунологического действия этих антиидиотипов в качестве вакцин, были проанализированы доступность и антигенные свойства CDR2H-, CDR3H- и CDR2D -областей. Антигенный индекс (мера антигенной вероятности) вычисляли с использованием программы ANTIGENIC, а для определения доступности найденных гомологичных последовательностей определяли с использованием программы DSSP.
Анализ вторичной и первичной структуры показал, что указанные гомологичные остатки являются доступными, и, кроме того, совпадают с областями, имеющими положительный антигенный индекс, а поэтому они могут принимать непосредственное участие в продуцировании ответа против "внутреннего образца" антигена. VL5A6- и VL3B6-цепи, представляющие 7 неидентичных антигенных областей и 11024 и 11603 Ангстремов доступного поверхностного белка, соответственно, способы образовывать антигенные изотопы; однако, оба эти антитела генерируют идентичный Ab3-ответ у мышей, кроликов и крыс. Полученные результаты позволяют предположить, что анти-антиидиотипичный ответ, наблюдаемый в этой системе, направлен исключительно на идиотип, определенный VH-цепями 5А6 и 3B6.
Поскольку иммунологический и структурный анализы антиидиотических антител 5А6 и 3B6 показали, что они несут внутренний образ внешнего домена EGFR, что были проведены исследования для того чтобы определить, почему ответ не индуцируется в других видах. Положительную антигенную CDR3H-область GYVGYAIDY, несущую EGFR-эквивалентную сегменты GYVGY, сравнивали с CDR3H-последовательностями мышиного, крысиного и кроличьего антител. Большая группа мышиных антител имеет в предсказанной антигенной области на CDR3H последовательность YAIDY; причем ожидалось, что у мыши, собственный иммуногенный идиотип определяют остатки GYVC. Было установлено, что Ig не имеют последовательность YAIDY, и, по всей вероятности, кроличий анти-антиидиотипический ответ направлен против других идиотипов, определяемых более крупными антигенными областями, что препятствует продуцированию ответа против "внутреннего образа".
Исходя из представленного выше описания изобретения, можно сделать следующий краткий обзор.
Для активной иммунотерапии рака были использованы различные методы. Для получения специфического антиопухолевого ответа, регулируемого взаимодействием "идиотип-антиидиотип", раковым пациентам вводили антиидиотипические антитела, имитирующие иммунологический эффект опухолевых маркеров.
Антиидиотипические mAb против mAb 425 были получены в целях разработки специфических вакцин для EGFR. Для этого были выбраны три антитела (5А6, 3B6 и 15Н8), распознающие сайтассоциированные идиотопы на mAb 425, определяемые их CDR-последовательностями, как было проиллюстрировано посредством ингибирования связывания "очеловеченного" варианта mAb 425 с EGFR.
Иммунологические и серологические анализы показали, что 5А6 и 3B6 действую как "внутренние образцы" антигена, индуцирующие гуморальный специфический ответ у мышей. Ab1-подобные антитела из Ab3-сыворотки связываются с мембраноассоциированным EGF-рецептором на клетках A331, и реагируют против sEGFR. Эта реакция полностью ингибируется обоими антителами 5А6 и 3B6 (независимо от антиидиотипа, используемого для иммуноанализа), что дает основание предположить, что 5A6 и 3B6 несут один и тот же "внутренний образ" EGFR. Иммунологические данные позволяют предположить о частичном идиотипическом сходстве с идиотипом "внутреннего образа" у антител 5А6 и 3B6 и с 425-паратопассоциированными идиотопом от 15H8; и кроме того, серологические анализы свидетельствуют о частичном ингибировании связывания анти-5А6 и анти-3B6 мышиного Ab3 с Ab2 или EGFR в присутствии 15H8. Иммунологические исследования выявили отсутствие биологического эффекта, то есть были индуцированы антитела IgM (реагирующие с клетками А331, но не распознающие sEGFR), которые, однако, не ингибировались антителом 15H8. Полученные результаты позволяют предположить, что в процессе иммунизации были активированы неспецифические клоны.
Для того, чтобы выяснить какая именно структура в антиидиотипах участвует в индуцировании ответа против EGFR, были проведены исследования структурной корреляции между идиотипами и антигеном.
После секвенирования 5А6, 3B6 и 15H8, и сравнительной оценки аминокислотных гомологий по отношению к EGFR была обнаружена серия гомологичных и эквивалентных аминокислотных в антигенных и доступных областях CDR2H, CDR3H и CDR2L. Полученные данные совпадали с несколькими уже имеющимися сведениями, указывающими на присутствие гомологичных последовательностей в CDR-участках (или в смежных участках) VL - VH-цепей, которые участвуют в структурном формировании идиотипов, несущих "внутренний образ" антигена (31-33). Обнаружение того фактора, что различные последовательности в 5A6 и 3B6 образуют два антитела с идентичным иммунологическим действием, послужило поводом для проведения анализа в целях выявления роли гипервариабельных и каркасных областей в индуцировании Ab3-ответа и в формировании структуры "внутреннего образа" EGFR.
Очевидно, что CDR2L, CDR2H и CDR3H от 5А6 и 3B6 участвуют в конструировании "внутреннего образа" антигена, поскольку именно в этих областях была обнаружена максимальная гомология и аминокислотное соответствие с внешним доменом EGFR. Трехмерная имитация, очевидно, играет очень важную роль, что подтверждается следующими полученными данными:
a) CDR21 от 5А6 и 3В6 являются различными, но индуцируют идентичный ответ против EGFR.
b) Гомологичные остатки в CDR2H были обнаружены у неродственных антител, а поэтому, их участие в формировании структуры "внутреннего образа" не зависит от исключительной линейной идентичности.
с) Установленное сродство между CDR -участками и остатками 148-154, 437-452 и 492-502 в EGFR основано, главным образом, на эквивалентности аминокислот.
d) Эквивалентные аминокислоты локализуются именно в CDR, а не в других областях.
Идиотипические анализы нескольких семейств антител (анти-PC, антидекстрановых, антигалактановых, анти-N1P антител), проведенные Davie и др. (34), показали, что, в методе антигенной стимуляции, экспрессия идиотипов предусматривает минимум аминокислот из CDR2H и CDR3H, и объединение со специфическими L-цепями. Данные, полученные авторами настоящей заявки в результате проведения иммунологического анализа и секвенирования, позволяют предположить о продуцировании каскада Ab1-Ab2-Ab3 Id, ограниченного лимитированным набором областей, определенных 425-паратопом
и Ab2-паратопом. Наиболее важным фактом, заслуживающим внимания, является получение двух Ab2, а именно 5А6 и 3В6, обладающих идентичными иммунологическими свойствами, а следовательно, и идиотипическими детерминантами, происходящими от идентичных VH-цепей, соединенных с различными VL-цепями.
Биологическая эффективность антиидиотипических антител коррелирует с их аффинностью в отношении набора идиотипических детерминант в восприимчивых видах. Анализ остатков, участвующих в образовании структуры "внутреннего образа" у антител 5А6 и 3В6, показал, что эти остатки находятся в более крупной антигенной области (остатки 100F 101 в H-цепи), имеющей, как было установлено, гомологию с другими мышиными антителами (из базы данных белков NBPF), но не с кроличьими антителами. Таким образом, структура "внутреннего образа" определяет идиотип, "усматриваемый" мышиной иммунной системой, но замаскированный в более крупной идиотипической области при введении кроликами. Эта гипотеза также подтверждается данными иммунологического анализа; а именно, титры антисыворотки Ab3 против 5A6- и 3B6-паратопов у кроликов выше, чем у мышей. Было также обнаружено, что в антителах человека (из базы данных NBPF) присутствуют гомологичные остатки 100F -101, что дает теоретическую основу для предположения о эффективности 5A6 и 3B6, несущих "внутренний образ" антигена, в человеческой системе.
Эффективность антиидиотипических антител, несущих "внутренний образ" антигена, в качестве вакцин была проиллюстрирована на различных моделях животных и в клинических испытаниях. О получении антиидиотипов, распознающих анти-EGF антитела, недавно сообщил Snarez (Snarez E. et al., 1993, Immunologia 12:122), который получил 4 mAb, не несущих внутренний образ. 5A6 и 3B6 являются первыми антиидиотипическими mAb, несущими "внутренний образ" антигена, о которых стало известно, что они имитируют внешний домен EGP-рецептора человека. Биологические свойства mAb 425 позволили определить, что 5A6 и 3B6 имитируют лиганд-связывающий сайт EGF-рецептора. Серологические исследования, биологическая активность в моделях животных, и структурный анализ этих антиидиотипических антител позволяют сделать вывод о возможности их эффективного использования для вакцинации человека.
6. Терапевтическое и диагностическое использование
Антитела настоящего изобретения могут быть введены человеку в терапевтических целях. Поэтому целью настоящего изобретения является получение фармацевтической композиции, содержащей в качестве активного ингредиента по крайней мере одно антитело или его фрагмент, определенные в описании и формуле изобретения, в комбинации с одним или несколькими фармацевтически приемлемыми носителями, наполнителями или разбавителями.
Антитела настоящего изобретения обычно вводят путем внутривенной инъекции или парентерально. В основном диапазон доз вводимых фрагментов антител широко варьируется так, чтобы данная доза была достаточной для достижения нужного подавления роста опухоли и опухоль-лизирующего эффекта. Эти дозы зависят от возраста, состояния здоровья, пола и тяжести заболевания пациента, и могут варьироваться от 0,1 до 200 мг/кг, а предпочтительно от 0,1 до 100 мг/кг на одну дозу в день, которая может быть введена в виде единой или дробной дозы, в течение одного или нескольких дней.
Препараты для парентерального введения представляют собой стерильный водный или безводный раствор, суспензию или эмульсию. Примерами безводных растворителей, которые могут быть использованы в этих целях, являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло; и инъецируемые органические сложные эфиры, такие как этилолеат; а также другие известные растворители. Антитела настоящего изобретения могут быть использованы в качестве ингредиентов, входящих в состав композиции, содержащей физиологически приемлемый носитель. Примерами подходящих носителей являются физиологический раствор, фосфатно-буферный раствор (PBS), раствор Рингера или лактат-содержащий раствор Ригнера. В фармацевтических композициях могут также присутствовать консерванты, и другие добавки, такие как антибиотики, антиоксиданты, и хелатообразующие агенты.
Антитело (или, необязательно, его фрагмент) может быть также коньюгировано известными методами с цитокинами, такими как 1L-2, в целях поддержания их цитотоксичности.
Фармацевтические композиции настоящего изобретения могут быть использованы для лечения опухолей любых типов, включая меланомы, глиомы, и карциномы, а также опухоли системы кровообращения, и твердые опухоли. В целях диагностики, антитела могут быть коньюгированы, например, с непроницаемым для рентгеновских лучей красителем, либо они могут быть подвергнуты радиоактивному мечению. Предпочтительным способом мечения является метод иодирования. Для диагностических целей, предпочтительно, если антитело будет введено в виде F(ab')2- или scFv-фрагментов. Введение таких антител дает наилучшие результаты, поскольку в этом случае не нужно вводить поправку за фон.
Пример (1). "Очеловеченное" mAb 425
Получение "очеловеченных" mAb 425 было описано в литературе (Kettleborough, C.A. et al., 1991, Protein Eng. 4: 773). Реконструированная форма mAb 425 содержит мышиные CDR, вставленные в человеческие вариабельные области при сохранении структуры антигенсвязывающего центра. Это было сделано для более точного определения локализации идиотипов 425, распознаваемых антителом Ab2.
Пример (2). Продуцирование антиидиотипических mAb против анти-EGFR mAb 425
Мышей Balb/c примировали путем внутрибрюшинной (i.p.) инъекции 75 мг очищенных mAb 425, конъюгированных с KLH (Sigma), фиксированных глутаральдегидом, и смешанных с CFA (Difco). Повторные инъекции животным вводили через 7, 14, 21, 28 и 38 дней после первой инъекции с использованием того же самого иммуногена, эмульгированного в IFA. Слияние клеток проводили через 3 дня после последней инъекции. Слияние клеток селезенки, взятых от иммунизированных мышей, с клетками HL1-Friendly-миеломы 653 с использованием полиэтиленгликоля 1450 (Boehringer Mannheim) осуществляли в соответствии с известной схемой (Carceller A., 1988, Sangre 33 (3):220). Гибридные клетки культивировали в 39% RPMI 1640, 39% гибридомной среде (Gibro), 20% фетальной телячьей сыворотке и 2% среде ГАТ. Положительные гибриды подвергали трехкратному клонированию путем серийного разведения.
Пример (3). Обнаружение и анализ Ab2- активности
Антиидиотипические антитела обнаруживали с помощью "сандвич"-теста ELISA, проводимого с использованием полистироловых планшетов для микротитрования (Nunc, Maxisorb), покрытых 1 мг/мл очищенных mAb 425. Эти планшеты блокировали 2% BSA-PBS и неочищенные супернатанты или сыворотки, разведенные в 0,2% BSA-PBS, инкубировали в течение ночи при 4oC. После промывки PBS - 0,05% Твином 20, добавляли (1 ч, 37oC) биотинилированное mAb 425, полученное методом Harlow и Lane (см. выше), затем планшеты три раза промывали и добавляли HRPO-стрептавидин (Dako), разведенный в отношении 1/1000 в 2% ВSA-PBS. В качестве субстрата использовали раствор 3,3,5,5-тетраметилбензидина (Sigma). Оптическую плотность (ОП) измеряли при 450 нм.
Пример (4). Константа диссоциации (KD) для антиидиотипических Ab2, 5A6, 3B6 и 15H8
Определение констант диссоциации осуществляли по методу Friguet (Friguet B. , et al. , 1985, J. Immunol Methods 77:306), а короче говоря, серийные разведения mAb 425 (2 • 10-8 М - 2 • 10-11) инкубировали с фиксированными концентрациями mAb2 до тех пор, пока не было достигнуто равновесие. Концентрацию свободного Ab2 измеряли посредством непрямого ELISA с использованием планшетов для микротитрования, покрытых 1 мг/мл mAb 425 и коньюгированных с HRPO кроличьих антимышиных иммуноглобулинов IgCI (Zymed).
Пример (5). Очищенный внешний домен EGFR человека (sEGFR)
Очистка внешнего домена EGFR человека и иммуноаффинная хроматография EGFR, секретируемого клетками A431, описаны в литературе (Weber W. et al., 1984, J. Biol. Chem 259 (23):14631). Этот внешний домен EGFR использовали для подтверждения специфичности Ab3 методами непрямого и конкурентного анализов.
Пример (6). Анализ для определения локализации идиотипов, распознаваемых в антителах mAb 425
Для определения локализации идиотипов 425, определенных антителами 5A6, 3B6 и 15H8, был проведен тест на способность этих антител ингибировать связывание mAb 425 с EGF-рецептором. Сначала этот тест осуществляли с использованием очищенных sEGFR. Неочищенные супернатанты и 50 мг/мл очищенных Ab2 смешивали со 125 нг mAb 425. После 4-часового инкубирования при 37oC, 425-активность анализировали с помощью ELISA с использованием планшетов для микротитрования, покрытых 2,5 мг/мл sEGFR. Планшеты три раза промывали смешанным раствором PBS-1% BSA, и добавляли конъюгированные с щелочной фосфатазой (AP) кроличьи антимышиные Ig (Dako). В качестве субстрата использовали 3,3,5,5-тетраметилбензидин, а ОП при 405 нм регистрировали планшет-ридером Tifertek Multiscan.
Эти анализы были дополнены сравнением кривых их ингибирования связывания, построенных для мышиных и реконструированных 425: 100 нг/мл мышиного или реконструированного 425 смешивали с равными объемами возрастающих концентраций (2 • 10-2 М - 2 • 10-1 М) очищенных Ab2, 5A6, 3B6, 15H8 или неродственных мышиных антител IgGI. Остаточную активность оценивали с помощью непрямого клеточного ELISA с использованием клеток A431, предварительно фиксированных 0,1% глутаральдегидом на планшетах для микротитрования (5 • 105 на лунку). После 60-минутного инкубирования при 37oC, клетки 3 раза промывали PBS - 1% BSA - 0,05% Твином 20. Для обнаружения связываемых AbI использовали конъюгированные с HRPO козьи антимышиные IgG2а (Dianova) и конъюгированные с HRPO козьи античеловеческие IgG соответственно, а в качестве субстрата использовали ортофенилдиамид (Sigma).
Пример (7). Продуцирование Ab3 в сингенных (мыши Balb/c), аллогенных (F6D2F1-мыши), и ксеногенных (крысы Wistar и кроликах NZW) системах, и обнаружение Ab3
Очищенные Ab сшивали с KLH (Calbiochem) при помощи глутаральдегида (Sigma), и эмульгировали полным или неполным адъювантом Фрейнда (CFA или IFA), и вводили животным на 0, 15, 30 и 60 день. В некоторых анализах для изучения стабильности Ab3-ответа проводили дополнительные инъекции через 8-12 месяцев. Каждое животное получало дозу 75 мг (мыши), 150 мг (крысы) и 300 мг (кролики) полных иммуноглобулинов путем внутрибрюшинной инокуляции (мыши) или подкожно в несколько различных участков (крысы и кролики). Контрольные группы получали те же самые дозы неспецифических (иррелевантных) KLH-конъюгированных мышиных иммуноглобулинов, либо только KLH.
Пробы крови брали сразу после каждой инъекции и через неделю после инъекции. В планшеты для микротитрования, предварительно покрытые 1 мг/мл Ab2 или другими мышиными иммуноглобулинами, добавляли серийные разведения сыворотки (в смеси PBS и 0,15% сухого молока). Связанные анти-антиидиотопы детектировали с использованием биотинилированных Ab2 (мышиной Ab3-сыворотке); конъюгированных с HRPO кроличьих антикрысиных иммуноглобулинов (Dako), не способных к перекрестной реакции с мышиными Ig; и конъюгированных с HRPO свиных антикроличьих Ig (Dako). Антиидиотипические и аллотипические антитела, вырабатываемые у крыс и кроликов, были элиминированы эктенсивной адсорбцией 1/50-1/100-разведенных сывороточных проб мышиными иммуноглобулинами IgGI после инкубирования в течение ночи при 4oC.
Пример (8). Инкубирование Ab2 с Ab1 антисывороткой Ab3
Этот анализ осуществляли в целях выявления антител, реагирующих с теми участками Ab2, которые контактируют с паратопом mAb 425. Для этого 8-16 нг/мл очищенных 5A6, 3B6, 15H8 и FIII смешивали с серийными разведениями Ab3-содержащей сыворотки, и после инкубирования в течение ночи при 4oC оценивали активность Ab2 с помощью непрямого ELISA с использованием 1 мг/мл 425, нанесенных на полистироловые планшеты. Связанные Ab2 детектировали с использованием конъюгированных со щелочной фосфатазой (PA) кроличьих антимышиных IgGI (Zymed).
Пример (9). Оценка идиотипической идентичности Ab2 посредством взаимной конкуренции за связывание с Ab3
Для определения идиотипической идентичности между всеми антиидиотипическими антителами Ab2 анализировали на ингибирование связывания антисыворотки Ab3 с Ab2-покрытыми планшетами. Серийные разведения крысиной антисыворотки Ab3 (10-3 - 10-5) тестировали на связывание с Ab2 в присутствии равного объема (1 мг/мл) Ab2 или контрольных mAb, предварительно инкубированных в течение ночи при 4oC.
Пример (10). Обнаружение анти-EGFR антител при Ab3-ответе
Мышиные, крысиные и кроличьи антисыворотки Ab3 анализировали на Ab1-подобную активность посредством клеточного ELISA с использованием клеток A431, как описано выше, либо посредством прямого ELISA с использованием sEGFR (2,5 мг/мл). В качестве "второго" антитела были использованы конъюгированные с HRPO кроличьи антимышиные Ig (Dako), кроличьи антикрысиные Ig (Dako) и свиные антикроличьи Ig (Dako) соответственно. Антитела к рецептору были также проанализированы путем прямого иммунофлуоресцентного анализа с использованием нефиксированных живых клеток A 431, культивированных в планшетах Terasaki.
Пример (11). Ингибирование связывания EGFR и Ab3 антиидиотипами 5A6, 3B6 и 15H8
Рецептор и антиидиотипы были исследованы путем конкурентного анализа на наличие общих детерминант. 120 - 240 мг/мл 5A6, 3B6, 15H8 или неродственных IgGI инкубировали в течение ночи при 4oC с 1/25 - 1/50-разведениями антисыворотки Ab3 против EGFR, а затем с помощью ELISA, клеточного ELISA и прямого иммунофлуоресцентного анализа проводили тестирование на противорецепторную активность. Процент ингибирования вычисляли следующим образом: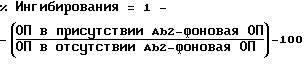
Пример (12). Получение РНК и кДНК
Полную РНК выделяли из 5A6-, 3B6- и 15H8-гибридомных клеточных линий. Клетки, промытые PBS, суспендировали в растворе тиоцианата гуанидиния, после чего выделяли РНК методом ультрацентрифугирования в градиенте хлорида цезия (см. Chirgwin et al., 1979, Biochemistry 18, 5294). Первую цепь кДНК синтезировали с использованием набора Pharmacia. Синтез осуществляли в 15 мкл с 5 мкг РНК в течение 1 ч при 37oC. Первую цепь кДНК использовали непосредственно для амплификации без предварительного клонирования.
Пример (13). PCR-амплификация
Для амплификации копий вариабельных областей легкой цепи мышиного иммуноглобулина и вариабельных областей тяжелой цепи мышиного иммуноглобулина использовали серию PCR - праймеров. 5'- Праймеры были сконструированы для гибридизации с лидерной последовательностью. Эти праймеры представляли собой смесь из 61 олигонуклеотида для вариабельного домена тяжелой цепи и из 405 олигонуклеотидов для вариабельного домена каппа-цепи. 3'-Праймеры были сконструированы для гибридизации с константной областью. Были использованы праймеры, являющиеся специфическими по отношению к IgGI и каппа-цепи мыши.
Для осуществления амплификации с помощью термостабильной ДНК-полимеразы, 25 мкл реакционной смеси, содержащей: 1 мкл гибрида "кДНК-РНК", 250 нМ соответствующей смеси 5'- и 3'-праймеров, 200 мМ каждого dNTP, 1 мМ MgCl2 (для легкой цепи), 2 мМ MgCl2 (для тяжелой цепи), и 1 ед. Tag - полимеразы (Cetus), покрывали сверху минеральным маслом, и подвергали гибридизации при температуре, начиная с 60oC и кончая температурой отжига 55oC. 1/10 PCR - реакции подвергали электрофорезу на 1%-ном агарозном геле TAE, а затем окрашивали этидийбромидом для визуализации полученных PGR - продуктов.
Пример (14). Молекулярное клонирование и секвенирование
1 мкл каждого PCR-продукта лигировали в вектор TA (Invitrogen, San Diego). ДНК-продукты реакции лигирования для областей VH- и VL-цепей каждого Ab2 трансформировали путем теплового шока в компетентные клетки E.coli TGI. В результате было создано шесть ДНК-библиотек: одна для вариабельных областей легкой цепи, а другие для вариабельных областей тяжелой цепи каждого mAb. Колонии отбирали на чашках с LB-средой, содержащей 100 мг/мл карбенициллина, и некоторые из них собирали с помощью зубочистки для последующего анализа. Положительные колонии идентифицировали путем PCR-скрининга или путем плазмидного гидролиза как описано Gussov (Gussov D., Clackson T., 1989. Nucleic Acids Res.,17: 4000). Двухцепочечную плазмидную ДНК для секвенирования получали (Wizarol preps., Promega Corp.) из трансформентов. Затем в соответствии с методикой Сэнгера проводили дидезокси-секвенирование путем обрыва цепи с использованием Tag-полимеразы (Sanger F. et al., 1977, Proc Nat Arad. Sci, USA 74: 5463). Праймеры, используемые в реакциях секвенирования, отжигали в TA-векторе,
Пример (15). Компьютерный анализ и сравнение последовательностей
Для определения вторичной структуры Ab2 и его доступных остатков использовали систему программного обеспечения DSSP (Sybil). Для предварительного определения антигенных участков по методу Kalaskar и Tongaonkar (Kalaskar A. S. et al., 1990, FEBS-Lett 276 (1-2):172) использовали программу ANTIGENIC. Гомологию между Ab2 и EGFR определяли по программе BESTFIT с использованием диска Gencore для таблицы соответствия символов. Сравниваемая последовательность EGFR человека была ранее описана в литературе (Vllrich et al., Nature 309: 418). Ab2 сравнивали с другими известными иммуноглобулинами с использованием банка данных Protein Identification Resourel (PIR ), составленный Национальным фондом по биомедицинским исследованиям National Riomedical Ressarch Fundation (NBRE)) и программы PBPSIS.








Изобретение относится к иммунологии и касается антиидиотипических антител, которые индуцируют иммунный ответ против рецептора эпидермального фактора роста. Сущность изобретения включает моноклональное антитело Ab2, имитирующее внутренний образ EGFR антигена, которое получено из мышиного антитела mAb 425 и продуцируемое линией клеток, депонируемых под номером АТСС НВ 9629, а также способ его получения путем иммунизации животного соответствующим идиотипическим антителом и фармацевтическую композицию на его основе. Технический результат заключается в получении нового продукта, который является хорошим кандидатом для идиотипической вакцины, используемой для ингибирования роста опухоли. 3 с. и 3 з.п.ф-лы, 6 ил., 5 табл.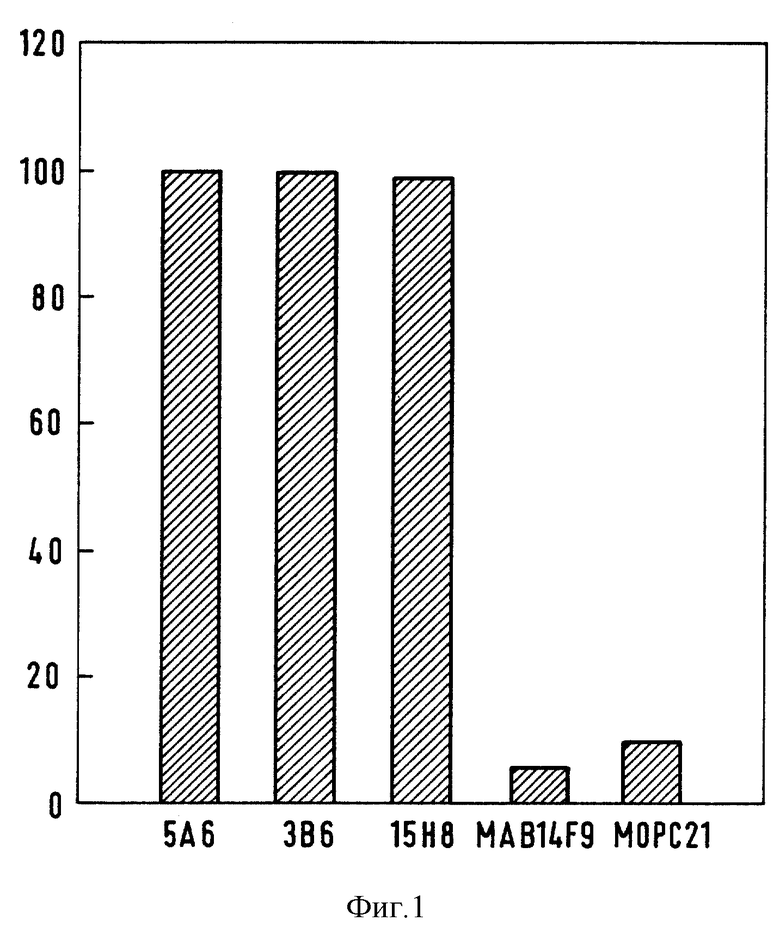
| Огнетушитель | 0 |
|
SU91A1 |
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| ОСТЕРМАН Л.А | |||
| Исследование биологических макромолекул электрофокусированием, иммуноэлектрофорезом и радиоизотопными методами | |||
| - М.: Наука, 1983, с.101-103. | |||
