ССЫЛКА НА СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ, ПРЕДСТАВЛЕННЫЙ В ЭЛЕКТРОННОМ ВИДЕ
Настоящая заявка включает посредством ссылки список последовательностей, поданный с настоящей заявкой в виде текстового файла, озаглавленного «Her3-100WO1_SL», созданный 12 ноября 2012 и имеющий объем 31,3 килобайт.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к композициям, которые специфично связываются с HER3, и к способам применения таких композиций для лечения злокачественной опухоли.
УРОВЕНЬ ТЕХНИКИ
Рецептор 3 эпидермального фактора роста человека (HER3, также называемый Erbb3) является рецепторной протеинтирозинкиназой и относится к подсемейству EGFR/HER рецепторов эпидермального фактора роста (EGFR) рецепторных протеинтирозинкиназ (RTK), состоящему из EGFR (HER1/Erbb1), HER2/Erbb2, HER3/Erbb3 и HER4/Erbb4. EGFR и HER2 относятся к наиболее хорошо установленным онкогенным RTK, участвующим в образовании опухолей в случае множества типов солидных опухолей, включая основные категории, такие как рак молочной железы, рак прямой и ободочной кишки и рак легкого. Было показано, что тирозинкиназные активности рецепторов EGFR и HER2 важны для их онкогенной активности.
Подобно прототипному EGFR, трансмембранный рецептор HER3 состоит из внеклеточного лиганд-связывающего домена (ECD), домена димеризации в пределах ECD, трансмембранного домена и внутриклеточного протеинтирозинкиназного домена (TKD) и С-концевого домена фосфорилирования (см., например, Kim et al. (1998), Biochem. J. 334, 189-195; Roepstorff et al. (2008), Histochem. Cell Biol. 129, 563-578).
Лиганд херегулин (HRG) связывается с внеклеточным доменом HER3 и активирует опосредованный рецептором сигнальный путь путем стимулирования димеризации с другими членами семейства EGFR (например, другими рецепторами HER) и трансфосфорилирования его внутриклеточного домена. Было показано, что у HER3 отсутствует определяемая тирозинкиназная активность, вероятно вследствие неконсервативной замены некоторых ключевых остатков в тирозинкиназном домене. Поэтому, как следствие киназной недостаточности, для HER3 необходимо образование гетеродимеров с другими RTK, особенно с EGFR и HER2, чтобы претерпеть фосфорилирование и стать функционально активным.
Центральная роль HER3 в онкогенезе заключается в действии в качестве поддерживающего белка для обеспечения возможности максимальной индукции пути PI3K/AKT. Было показано, что HER3 содержит кластер из шести С-концевых тирозин-содержащих мотивов, которые, при фосфорилировании, имитируют консенсусный участок связывания PI3K/p85. Следовательно, при образовании гетеродимеров с HER3 вышерасположенные онкостимуляторы, EGFR, HER2, cMET и FGFR2, могут быть наиболее эффективно сопряжены с путем PI3K/AKT. Поэтому, логично ожидать, что потеря активности HER3 может блокировать прогрессирование злокачественной опухоли в различных системах, индуцируемых дивергентными RTK. Исследования показали, что нокдаун ми-РНК HER3 в злокачественных клетках молочной железы с амплифицированным HER2 приводит к подобным антипролиферативным эффектам, как нокдаун ми-РНК HER2, что также демонстрирует острую необходимость HER3 для злокачественной опухоли.
Было обнаружено, что помимо стимулирования роста опухолей в условиях отсутствия стресса, HER3 в значительной степени вовлечен в придание терапевтической устойчивости ко многим целенаправленным лекарственным средствам, включая ингибиторы тирозинкиназ EGFR, моноклональные антитела против HER2, такие как трастузумаб, а также низкомолекулярные ингибиторы PI3K или AKT или MEK. Полученные данные дополнительно привлекают внимание к HER3 в качестве многообещающей мишени в случае злокачественной опухоли как для циторедукции первичной опухоли, так и для устранения проблем, связанных с резистентностью злокачественной опухоли, которые неизбежно возникают, несмотря на первоначальные клинические ответы.
HER3 имеет два различные пути димеризации со своими партнерами RTK: лиганд-зависимый (в присутствии HRG) или лиганд-независимый. Что касается димеров HER2-HER3, то известно, что в клетках, имеющих экспрессию HER2 от низкого до среднего уровня, HER3 может образовывать комплекс с HER2 только после связывания с лигандом; напротив, в клетках с амплифицированным HER2 (HER2 IHC 3+), они образуют спонтанные димеры без HRG (Junttila et al. (2009) Cancer Cell. 15(5): 429-40). Димеры, образованные в присутствии или отсутствие лиганда, структурно различаются, как было показано в раннем исследовании, которое продемонстрировало, что трастузумаб/герцептин® (моноклональное HER2-антитело Genentech/Roche, одобренное для лечения рака молочной железы HER2 3+) может разрушать только лиганд-независимый димер, но не лиганд-зависимый димер, в то время как пертузумаб/омнитарг® (rhuMAb 2C4, моноклональное HER2-антитело Genentech/Roche в фазе 3 испытаний) может разрушать только лиганд-зависимые димеры.
Образование димеров между членами семейства HER увеличивает потенциал для передачи сигнала от HER3 и представляет собой средства не только для многообразия сигнала, но также для усиления сигнала. Было показано, что HER3 фосфорилируется в контекстах разных клеток. Например, HER3 конститутивно фосфорилируется по остаткам тирозина в подгруппе злокачественных клеток молочной железы человека, сверхэкспрессирующих HER3 (см., например, Kraus et al. (1993) Proc. Natl. Acad. Sci. USA 90, 2900-2904; Kim et al. (1998), Biochem. J. 334, 189-195; Schaefer et al. (2004) Cancer Res. 64, 3395-3405; Schaefer et al. (2006) Neoplasia 8, 612-622). Соответственно, требуется терапия, которая эффективно препятствуют фосфорилированию HER3.
Кроме того, было обнаружено, что HER3 сверхэкспрессируется и/или сверхактивирован в некоторых типах злокачественных опухолей, таких как рак молочной железы, рак яичника, рак простаты, рак печени, рак почки и рак мочевого пузыря, рак поджелудочной железы, рак головного мозга, гемопоэтические неоплазии, ретинобластомы, меланомы, рак прямой и ободочной кишки, рак желудка, рак головы и шеи, рак легкого и т.д. (см., например, Sithanandam and Anderson (2008) Cancer Gene Ther. 15, 413-448). В общем, HER3 часто активирован в случае злокачественных опухолей, экспрессирующих EGFR, HER2, C-Met и FGFRII.
Была показана корреляция между экспрессией HER2/HER3 и прогрессированием от неинвазивной до инвазивной стадии (Alimandi et al., Oncogene 10, 1813-1821; DeFazio et al., Cancer 87, 487-498; Naidu et al., Br. J. Cancer 78, 1385-1390). Таким образом, HER3 может быть использован в качестве диагностического маркера повышенной агрессивности опухолей и низкой выживаемости. Продолжительная активация PI3K/AKT под действие HER3, как было многократно показано, обусловливает резистентность опухолей к ингибиторам EGFR/HER2.
Хотя роль HER3 в развитии и прогрессировании злокачественных опухолей была изучена (см., например, публикацию Horst с соавторами (2005) Int. J. Cancer 115, 519-527; Xue с соавторами (2006) Cancer Res., 66, 1418-1426), HER3 остается в значительной степени недооцененным в качестве мишени для клинического вмешательства. Наиболее современные способы иммунотерапии сфокусированы, в основном, на ингибировании действия HER2 и, в частности, гетеродимеризации комплексов HER2/HER3 (см., например, публикацию Sliwkowski с соавторами (1994), J. Biol. Chem. 269, 14661-14665). Таким образом, целью настоящего изобретения является предоставление улучшенных иммунотерапевтических средств, которые эффективно ингибируют HER3-опосредованную передачу клеточных сигналов, которые можно применять для диагностики, прогнозирования и лечения различных злокачественных опухолей.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к связывающим анти-HER3-молекулам, например, антителам или их антигенсвязывающим фрагментам, например, моноклональным антителам, способным подавлять активность НER3 в лиганд-зависимых и независимых условиях. Напротив, другие моноклональные анти-HER3-антитела, известные в данной области (например, Ат №6 (международная патентная публикация WO 2008/100624) и U1-59 (международная патентная публикация WO 2007077028; также названные в настоящем документе AMG), могут только подавлять лиганд-зависимую активность HER3. Также раскрыты связывающие анти-HER3-молекулы с созревшей аффинностью, обладающие повышенной эффективностью и увеличенным временем полу-жизни, которые последовательно могут быть введены реже с увеличенными интервалами между дозами и в меньших объемах доз. Изобретение также относится к способам лечения заболеваний, таких как злокачественная опухоль, у человека, включающим в себя введение связывающих анти-HER3-молекул. В некоторых конкретных аспектах применяют YTE-мутантное антитело человека, являющееся производным 2C2.
Изобретение относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, который специфично связывается с эпитопом во внеклеточном домене HER3, при этом связывающая молекула специфично связывается с таким же эпитопом HER3, как и антитело или его антигенсвязывающий фрагмент, содержащий вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL) CL16 или 2C2. Также предлагается выделенная связывающая молекула или ее антигенсвязывающий фрагмент, который специфично связывается с HER3 и конкурентно ингибирует связывание HER3 антителом или его антигенсвязывающим фрагментом, содержащим VH и VL CL16 или 2C2.
Изобретение также относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, который специфично связывается с HER3, содержащему VL антитела, при этом VL содержит аминокислотную последовательность:
[FW1]X1GSX2SNIGLNYVS[FW2]RNNQRPS[FW3]AAWDDX3X4X5GEX6[FW4],
где [FW1], [FW2], [FW3] и [FW4] означают каркасные области VL, и где
(a) X1 означает аминокислотные остатки аргинин (R) или серин (S),
(b) X2 означает аминокислотные остатки серин (S) или лейцин (L),
(c) X3 означает аминокислотные остатки серин (S) или глицин (G),
(d) X4 означает аминокислотные остатки лейцин (L) или пролин (P),
(e) X5 означает аминокислотные остатки аргинин (R), изолейцин (I), пролин (P) или серин (S), и
(f) X6 означает аминокислотные остатки валин (V) или аланин (A).
Кроме того, изобретение относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, которая специфически связывается с HER3, содержащей VH антитела, при этом VH содержит аминокислотную последовательность:
[FW5]YYYMQ[FW6]X7IGSSGGVTNYADSVKG[FW7]VGLGDAFDI[FW8], где [FW5], [FW6], [FW7] и [FW8] означают каркасные области VH, и где X7 означает аминокислотные остатки тирозин (Y), изолейцин (I) или валин (V).
Изобретение относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, который специфично связывается с HER3, содержащему VL антитела и VH антитела, при этом VL содержит аминокислотную последовательность:
[FW1]X1GSX2SNIGLNYVS[FW2]RNNQRPS[FW3]AAWDDX3X4X5GEX6[FW4], где [FW1], [FW2], [FW3] и [FW4] означают каркасные области VL, и где
(a) X1 означает аминокислотные остатки аргинин (R) или серин (S),
(b) X2 означает аминокислотные остатки серин (S) или лейцин (L),
(c) X3 означает аминокислотные остатки серин (S) или глицин (G),
(d) X4 означает аминокислотные остатки лейцин (L) или пролин (P),
(e) X5 означает аминокислотные остатки аргинин (R), изолейцин (I), пролин (P) или серин (S), и
(f) X6 означает аминокислотные остатки валин (V) или аланин (A), и где VH содержит аминокислотную последовательность:
[FW5]YYYMQ[FW6]X7IGSSGGVTNYADSVKG[FW7]VGLGDAFDI[FW8], где [FW5], [FW6], [FW7] и [FW8] означают каркасные области VH, и где X7 означает аминокислотные остатки тирозин (Y), изолейцин (I) или валин (V).
Изобретение также относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, который специфично связывается с HER3, содержащему VL антитела, при этом VL содержит аминокислотную последовательность определяющей комплементарность области 1 VL (VL-CDR1), идентичную или идентичную за исключением четырех, трех, двух или одной аминокислотной замены, последовательностям: SEQ ID NO: 18, SEQ ID NO: 19 или SEQ ID NO: 20. Также изобретение относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, которая специфически связывается с HER3, содержащей VL антитела, при этом VL содержит аминокислотную последовательность определяющей комплементарность области 2 VL (VL-CDR2), идентичную или идентичную за исключением четырех, трех, двух или одной аминокислотной замены, последовательности SEQ ID NO: 21.
Кроме того, изобретение относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, который специфично связывается с HER3, содержащей VL антитела, при этом VL содержит аминокислотную последовательность определяющий комплементарность области 3 VL (VL-CDR3), идентичную или идентичную за исключением четырех, трех, двух или одной аминокислотной замены, последовательностям: SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25, SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29 или SEQ ID NO: 30. Также изобретение относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, который специфично связывается с HER3, содержащему VH антитела, при этом VH содержит аминокислотную последовательность определяющей комплементарность области 1 VL (VH-CDR1), идентичную или идентичную, за исключением четырех, трех, двух или одной аминокислотной замены, последовательности SEQ ID NO: 31.
Кроме того, изобретение относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, который специфично связывается с HER3, содержащему VH антитела, при этом VH содержит аминокислотную последовательность определяющий комплементарность области 2 (VH-CDR2), идентичную или идентичную за исключением четырех, трех, двух или одной аминокислотной замены, последовательностям: SEQ ID NO: 32, SEQ ID NO: 33 или SEQ ID NO: 34. Также изобретение относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, который специфично связывается с HER3, содержащему VH антитела, при этом VH содержит аминокислотную последовательность определяющей комплементарность области-3 (VH-CDR3), идентичную или идентичную за исключением четырех, трех, двух или одной аминокислотной замены, последовательности SEQ ID NO: 35.
Изобретение относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, который специфично связывается с HER3, содержащему VL антитела, при этом VL содержит аминокислотные последовательности VL-CDR1, VL-CDR2 и VL-CDR3, идентичные или идентичные за исключением четырех, трех, двух или одной аминокислотной замены в одной или нескольких VL-CDR, последовательностям: SEQ ID NO: 18, 21 и 22, SEQ ID NO: 18, 21 и 26, SEQ ID NO: 18, 21 и 27, SEQ ID NO: 20, 21 и 22, SEQ ID NO: 19, 21 и 22, SEQ ID NO: 18, 21 и 25, SEQ ID NO: 18, 21 и 28, SEQ ID NO: 18, 21 и 29, SEQ ID NO: 18, 21 и 30, SEQ ID NO: 18, 21 и 23, SEQ ID NO: 19, 21 и 23, SEQ ID NO: 20, 21 и 23, SEQ ID NO: 18, 21 и 24 или SEQ ID NO: 18, 21, и 25, соответственно. Изобретение также относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, который специфично связывается с HER3, содержащему VH антитела, при этом VH содержит аминокислотные последовательности VH-CDR1, VH-CDR2 и VH-CDR3, идентичные или идентичные за исключением четырех, трех, двух или одной аминокислотной замены в одной или нескольких VH-CDR, последовательностям: SEQ ID NO: 31, 32 и 35, SEQ ID NO: 32 и 35, SEQ ID NO: 31, 33 и 35 или SEQ ID NO: 31, 34, и 35, соответственно.
Кроме того, изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, который специфично связывается с HER3, содержащему VL и VH, которые включают в себя аминокислотные последовательности VL-CDR1, VL-CDR2, VL-CDR3, VH-CDR1, VH-CDR2 и VH-CDR3, идентичные или идентичные за исключением четырех, трех, двух или одной аминокислотной замены, в одной или нескольких CDR, последовательностям: SEQ ID NO: 18, 21, 22, 31, 32 и 35, SEQ ID NO: 18, 21, 26, 31, 32 и 35, SEQ ID NO: 18, 21, 27, 31, 32 и 35, SEQ ID NO: 20, 21, 22, 31, 32 и 35, SEQ ID NO: 19, 21, 22, 31, 32 и 35, SEQ ID NO: 18, 21, 25, 31, 32 и 35, SEQ ID NO: 18, 21, 28, 31, 32 и 35, SEQ ID NO: 18, 21, 29, 31, 32 и 35, SEQ ID NO: 18, 21, 30, 31, 32 и 35, SEQ ID NO: 18, 21, 23, 31, 32 и 35, SEQ ID NO: 19, 21, 23, 31, 32 и 35, SEQ ID NO: 20, 21, 23, 31, 32 и 35, SEQ ID NO: 18, 21, 24, 31, 32 и 35 или SEQ ID NO: 18, 21, 25, 31, 32 и 35, соответственно. Также предлагается выделенная связывающая молекула или ее антигенсвязывающий фрагмент, который специфично связывается с HER3, содержащий VL антитела и VH антитела, при этом VL содержит аминокислотную последовательность, по меньшей мере, приблизительно на 90-100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16 и SEQ ID NO: 17. Изобретение также относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, который специфично связывается с HER3, содержащему VL антитела и VH антитела, при этом VH содержит аминокислотную последовательность, по меньшей мере, приблизительно на 90-100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 2, SEQ ID NO: 12 и SEQ ID NO: 13. Кроме того, изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, который специфично связывается с HER3, при этом антитело или его антигенсвязывающий фрагмент содержит VL, содержащую последовательность, по меньшей мере, приблизительно на 90-100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16 и SEQ ID NO: 17, и при этом антитело или его антигенсвязывающий фрагмент содержит VH, содержащую последовательность, по меньшей мере, приблизительно на 90-100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 2, SEQ ID NO: 12 и SEQ ID NO: 13.
Изобретение также относится к выделенному антителу или его антигенсвязывающему фрагменту, который содержит VL, содержащую последовательность SEQ ID NO: 49, и VH, содержащую последовательность SEQ ID NO: 50. Кроме того, изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, который содержит VL, содержащую последовательность SEQ ID NO: 3, и VH, содержащую последовательность SEQ ID NO: 2. Также изобретение относится к выделенной связывающей молекуле или ее антигенсвязывающему фрагменту, который специфично связывается с эпитопом во внеклеточном домене HER3, содержащему последовательность VL антитела SEQ ID NO: 3, последовательность VH антитела SEQ ID NO: 2 и последовательность константной области IgG1 SEQ ID NO: 46. Также предлагается выделенная связывающая молекула или ее антигенсвязывающий фрагмент, который специфично связывается с эпитопом во внеклеточном домене HER3, состоящий из последовательности VL антитела SEQ ID NO: 3, последовательности VH антитела SEQ ID NO: 2 и последовательности константной области IgG1 SEQ ID NO: 46.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ/ФИГУР
На фиг. 1 представлена интернализация моноклональных анти-HER3-антител клона 16 в клетки KPL4, показанная в виде истощения поверхностного флуоресцирующего окрашивания. Верхняя панель показывает интернализацию в момент времени = 0. Нижние панели показывают интернализацию после 2,5 часов.
На фиг. 2А представлено множественное выравнивание последовательностей, соответствующих последовательностям VL моноклональных анти-HER3-антител клона 16 (CL16; исходный, родительский клон), клона 16 (GL; клон зародышевой линии), 5H6, 8A3, 4H6, 6E.3, 2B11, 2D1, 3A6 и 4C4. Указано положение CDR1, CDR2 и CDR3. Выделены аминокислотные остатки, которые отличаются по сравнению с антителом CL16 (GL).
На фиг. 2В представлено множественное выравнивание последовательностей, соответствующих последовательностям VН моноклональных анти-HER3-антител клона CL16 (CL16; родительский клон) и клонов 15D12.1 (также называемого 15D12.I) и 15D12.2 (также называемого 15D12.V). Указано положение CDR1, CDR2 и CDR3. Выделены аминокислотные остатки, которые отличаются по сравнению с родительским антителом CL16.
На фиг. 2С представлено множественное выравнивание последовательностей, соответствующих последовательностям VL моноклональных анти-HER3-антител клона 16 (CL16; исходный, родительский клон), CL16 (GL; клон зародышевой линии), 1A4, 2C2, 3E.1, 2F10 и 2B11. Указано положение CDR1, CDR2 и CDR3. Выделены аминокислотные остатки, которые отличаются по сравнению с антителом CL16 (GL).
На фиг. 3 показано подавление фосфорилирования HER3 (pHER3) в зависимых от лиганда клетках MCF-7, при этом HER3 активируется только экзогенным HRG (лигандом). Исследовали моноклональные анти-HER3-антитела 2C2, описанные в литературе моноклональные анти-HER3-антитела AMG и MM и контрольное антитело R347. Представлено максимальное ингибирование pHER3 в процентах и значения IC50.
На фиг. 4 показано подавление роста клеток MDA-MB-175, разработанной модели регуляции HRG по типу аутокринной петли, в которой эндогенный HRG стимулирует активность HER3 и рост клеток. Исследованы моноклональные анти-HER3-антитела 2C2, описанные в литературе моноклональные анти-HER3-антитела AMG и MM и контрольное антитело R347. Представлено максимальное ингибирование роста в процентах и значения IC50.
На фиг. 5 показано подавление роста клеток HMCB, разработанной модели регуляции РКП по типу аутокринной петли, в которой эндогенный HRG стимулирует активность HER3 и рост клеток. Исследованы моноклональные анти-HER3-антитела 2C2, описанные в литературе моноклональные анти-HER3-антитела AMG и MM и контрольное антитело. Представлены значения IC50.
На фиг. 6 показано, что 2C2 не только ингибировало рост клеток HMCB, но также подавляло фосфорилирование HER3 (pHER3) и фосфорилирование AKT (pAKT) в указанной зависимой от лиганда меланоме.
На фиг. 7 показано, что 2C2 подавляло фосфорилирование HER3 (pHER3) и фосфорилирование AKT (pAKT) в зависимых от лиганда клетках NSCLC A549.
На фиг. 8 показано подавление фосфорилирования HER3 (pHER3) в моделях на основе клеток злокачественной опухоли легкого, желудка и молочной железы. На панели А показано подавление pHER3 в линии клеток HCC827, модели NSCLC на основе мутантных EGFR-зависимых клеток, при взаимном влиянии EGFR/HER3. На панели В показано подавление pHER3 в модели NSCLC на основе EGFR-TKI-резистентных HCC827, полученных в результате долговременной обработки EGFR TKI. На панели С показано подавление pHER3 в линии клеток MKN45, модели рака желудка с амплифицированным cMET, при взаимном влиянии cMET-HER3. На панели D показано подавление рHER3 в клеточной линии Kato III, модели рака желудка с FGFR2-амплификацией, при взаимном влиянии FGFR2-HER3. На панели E показано подавление рHER3 в клеточной линии BT-474, не зависимой от лиганда модели рака молочной железы с амплифицированным HER2 (т.е. в клетках отсутствует экспрессия HRG). Исследовали моноклональные анти-HER3-антитела 2C2, описанные в литературе моноклональные анти-HER3-антитела AMG и MM и контрольное антитело R347. Представлено максимальное ингибирование pHER3 в процентах и значения IC50.
На фиг. 9 показано подавление фосфорилирования AKT (pAKT) в клеточной модели рака желудка и молочной железы. На панели A показано подавление pAKT в линии клеток MKN45. На панели B показано подавление pAKT в линии клеток Kato III. На панели С показано подавление pAKT в линии клеток BT-474, не зависимой от лиганда модели рака молочной железы с амплифицированным HER2 (т.е. в клетках отсутствует экспрессия НRG). Исследовали моноклональные анти-HER3-антитела 2C2, описанные в литературе моноклональные анти-HER3-антитела AMG и MM и контрольное антитело R347. Представлено максимальное ингибирование pAKT в процентах и значения IC50.
На фиг. 10 показано, что 2C2 подавляет передачу клеточных сигналов и пролиферацию клеток MDA-MB-361. На панели A показано, что 2C2 подавлял фосфорилирование HER3 (pHER3) в клетках MDA-MB-361 с амплифицированным HER2. На панели В показано, что 2C2 подавляет рост клеток зависимым от дозы образом. Процент ингибирования представлен для 6- и 14-дневных обработок (верхняя и нижняя панели, соответственно).
На фиг. 11 показано, что 2C2 подавляет фосфорилирование HER3 (pHER3) в клетках HARA-B, экспрессирующих высокие уровни HRG.
На фиг. 12 показано, что 2C2 и rhuMab 2C4, но не антагонисты EGFR цетуксимаб и гефитиниб, ингибируют зависимую от лиганда HRG передачу сигналов (нижняя часть панелей А и В). В верхней части панелей А и В показаны базальные клетки, SW620 (панель А, левая часть), SW480 (панель А, средняя часть), Colo205 (панель А, правая часть), LOVO (панель В, левая часть), HCT15 (панель В, средняя часть) и Caco-2 (панель В, правая часть).
На фиг. 13 показан ELISA-анализ связывания HRG-HER3, в котором измеряли прямое блокирование связывания HRG с HER3 клоном 16, описанными в литературе моноклональными анти-HER3-антителами AMG и MM, положительным контрольным блокирующим лиганд моноклональным анти-HER3-антителом и контрольным антителом R347.
На фиг. 14 показано, что 2C2 блокирует димеризацию HER2-HER3. Панель А демонстрирует анализ HRG-индуцируемой димеризации HER2-HER3, который оценивает степень образования комплекса HER2-HER3 в клетках T-47D, зависимой от лиганда модели, показывающей четкую HRG-индуцируемую ассоциацию HER2-HER3, предварительно обработанных моноклональными анти-HER3-антителами 2C2, CL16, AMG и MM. Все анти-HER3 антитела блокировали такую индуцируемую лигандом димеризацию HER2-HER3. Панель В демонстрирует анализ не зависимой от лиганда димеризации HER2-HER3, который оценивает степень образования комплекса HER2-HER3 в клетках BT-47D, предварительно обработанных 2C2 или CL16, которые блокировали такую не зависимую от лиганда димеризацию HER2-HER3.
На фиг. 15 показана интернализация и распад HER3, индуцированные 2C2. На панели А показан основанный на FACS анализ интернализации, который количественно определяет временную зависимость и степень интернализации мишени в ответ на две разные концентрации моноклонального антитела 2C2. На панели B показан распад HER3 в модельных клетках рака прямой и ободочной кишки Lovo, HCT15 и SW620, предварительно обработанных моноклональным анти-HER3-антителом 2C2 или контрольным антителом R347.
На фиг. 16 показан основанный на FACS анализ клеточного цикла, показывающий, что в клетках SkBR3, линии злокачественных клеток молочной железы с аплифицированным HER2, подобной BT-474, как герцептин® (трастузумаб), так и моноклональное антитело CL16 (родительское антитело для моноклонального антитела 2C2), вызывали задержку клеточного цикла в фазе G1. Также показаны результаты, соответствующие клеткам, обработанным контрольным антителом R347м и моноклональным анти-HER2-антителом rhuMAb 2C4 (пертузумаб/омнитарг®).
На фиг. 17 показано ингибирование индуцируемой HRG секреции VEGF анти-HER3-антителами. На панели А показаны изменения секреции VEGF в клетках злокачественной опухоли молочной железы BT-474, предварительно обработанных моноклональными анти-HER3-антителами CL16 и Merrimack MM, моноклональным анти-HER2- антителом герцептином® (трастузумабом) или контрольным антителом R347. На панели В показаны изменения секреции VEGF в модели, основанной на клетках злокачественной опухоли молочной железы MCF-7, предварительно обработанных моноклональными анти-HER3-антителами CL16 и Merrimack MM, моноклональным анти-HER2-антителом герцептином® (трастузумабом) или контрольным антителом R347.
На фиг. 18 показано, что моноклональное анти-HER3-антитело 2C2 связывается на поверхности клеток с HER3 макак-крабоедов (cyno-HER3) на клеточной поверхности, эктопически экспрессированным в клетках Ad293, и модулирует его активность. На панели А показан Вестерн-блот-анализ клеток Ad293, трансфицированных контрольным вектором (левая сторона) или вектором, экспрессирующим cyno-HER3 (правая сторона). Клетки обрабатывали 2C2 или контрольным антителом (R347) с ко-стимуляцией или без ко-стимуляции HRG и исследовали с использованием в качестве зонда антитело против HER3 (средний блот), против pHER3 (верхний блот) и против GAPDH (нижний блот). На панели В показана основанная на денситометрии количественная оценка pHER3 в верхних четырех дорожках, показанных на панели А.
На фиг. 19 показано зависимое от дозы уменьшение объема опухоли после введения моноклонального антитела 2C2 при использовании модели на основе ксенотрансплантата клеток опухоли головы и шеи FADU. На панели А показано, что введение 7 мг/кг 2C2 два раза в неделю было максимально эффективным с 99% dTGI (ингибирование роста опухоли) в данной модели. Панель В демонстрирует сильное уменьшение объема опухоли после комбинированного введения моноклонального антитела 2C2 с моноклональным анти-EGFR-антителом цетуксимабом при использовании модели на основе ксенотрансплантата клеток опухоли головы и шеи FADU. Комбинированная обработка приводила к частичной регрессии в 7 из 10 случаев и к полной регрессии в 2 из 10 случаев.
Фиг. 20 показана нелинейная фармакокинетика для 2C2 после введения однократной дозы и многократных доз 5 мг/кг или 30 мг/кг мышам, несущим опухоли. Полученные данные свидетельствуют, что мышиный HER3 служит в качестве «поглотителя» для связывания с 2C2, вводимого мышам, и что 30 мг/кг в виде однократной дозы является достаточным для насыщения поглотителя.
На фиг. 21 показан полезный противоопухолевый эффект нагрузочной дозы 10 мг/кг моноклонального антитела 2C2 при использовании модели на основе ксенотрансплантата клеток опухоли головы и шеи FADU. Введение нагрузочной дозы 2C2 для насыщения поглотителя HER3 мыши, позволяло 2C2 в дозе 3 мг/кг проявлять высокую противоопухолевую активность, в то время как 2C2 в дозе 3 мг/кг без нагрузочной дозы проявляет только умеренную активность.
На фиг. 22 показано, что обработка 2C2-YTE снижает уровни pHER3 и pAKT в экстрактах ксенотрансплантированных опухолей FADU. В данном эксперименте уровни pHER3 и pAKT были снижены на 59,5% и 51,7%, соответственно. Никакого изменения общих уровней HER3 в данном эксперименте не наблюдали.
На фиг. 23 показано зависимое от дозы уменьшение объема опухоли после введения моноклонального антитела 2C2 при использовании модели на основе ксенотрансплантатов клеток опухоли головы и шеи человека Detroit562. Панель А демонстрирует, что доза 10 мг/кг 2C2, вводимая дважды в неделю, была максимально эффективна со значением dTGI 72%. Панель В демонстрирует уменьшение объема опухоли после комбинированного введения моноклонального антитела 2C2 с моноклональным анти-EGFR-антителом цетуксимабом при использовании модели на основе ксенотрансплантатов клеток опухоли головы и шеи человека Detroit562. Комбинированная обработка приводила к частичным регрессиям в 9 из 10 случаев, в то время как цетуксимаб отдельно приводил к частичным регрессиям в 5/10 случаев. Модель на основе ксенотрансплантатов Detroit562 включает мутацию PIK3CA.
На фиг. 24 показано зависимое от дозы уменьшение объема опухоли после введения моноклонального антитела 2C2-YTE при использовании модели на основе ксенотрансплантов клеток опухоли головы и шеи человека CAL27.
На фиг. 25 показано зависимое от дозы уменьшение объема опухоли после введения моноклонального антитела 2C2 при использовании модели на основе ксенотрансплантатов NSCLC человека A549. Панель А демонстрирует, что 30 мг/кг 2C2, вводимого дважды в неделю, было максимально эффективным с dTGI 91% вплоть до последнего дня фазы обработки (день 33; после этого возобновление роста). Оба антитела 2C2-YTE и 2C2 в дозе 10 мг/кг обладали сравнимой активностью. Панель В демонстрирует уменьшение объема опухоли после комбинированного введения моноклонального антитела 2C2 с моноклональным анти-EGFR-антителом цетуксимабом при использовании модели на основе ксенотрансплантатов NSCLC человека A549. Добавление цетуксимаба к 2C2 увеличивало активность 2C2 в течение фазы обработки и задерживало возобновление роста опухоли в течение фазы возобновления роста опухоли. Модель на основе ксенотрансплантатов A549 содержит мутацию KRAS и делецию LKB-1.
На фиг. 26 показано уменьшение объема опухоли после введения моноклонального антитела 2C2-YTE при использовании модели на основе ксенотрансплантатов плоскоклеточной карциномы человека HARA-B. Доза 30 мг/кг 2C2-YTE, вводимая дважды в неделю, проявила максимальную эффективность с dTGI 64,6%. 2C2-YTE в дозе 10 мг/кг имело сравнимую активность, в то время как 2C2-YTE в дозе 3 мг/кг было неактивным.
На фиг. 27 демонстрирует зависимое от дозы уменьшение объема опухоли после введения моноклонального антитела 2C2 при использовании модели на основе ксенотрансплантатов клеток злокачественной опухоли прямой и ободочной кишки человека HT-29. Доза 30 мг/кг 2C2, вводимая дважды в неделю, проявила максимальную эффективность с dTGI 56% вплоть до последнего дня фазы обработки (день 26; после этого возобновление роста). Оба антитела 2C2-YTE и 2C2 в дозе 30 мг/кг имеют сравнимую активность. Модель на основе ксенотрансплантатов HT-29 содержит мутацию BRAF.
На фиг. 28 показано уменьшение объема опухоли после введения моноклонального антитела 2C2 при использовании модели на основе ксенотрансплантатов клеток злокачественной опухоли прямой и ободочной кишки человека HCT-116. Доза 30 мг/кг 2C2, вводимая дважды в неделю, проявила максимальную эффективность с dTGI. Оба антитела 2C2-YTE и 2C2 в дозе 10 мг/кг обладают сравнимой активностью. Модель на основе ксенотрансплантатов НCT-116 содержит мутацию KRAS.
На фиг. 29 показано уменьшение объема опухоли после введения моноклонального антитела 2C2 при использовании модели на основе ксенотрансплантатов клеток злокачественной опухоли прямой и ободочной кишки человека LOVO. Доза 30 мг/кг 2C2, вводимая дважды в неделю, проявила максимальную эффективность с dTGI 48%. Оба антитела, 2C2-YTE и 2C2, в дозе 10 мг/кг обладают сравнимой активностью. Модель на основе ксенотрансплантатов LOVO содержит мутацию KRAS.
На фиг. 30 показано уменьшение объема опухоли после введения моноклонального антитела 2C2 при использовании модели на основе ксенотрансплантатов клеток злокачественной опухоли простаты человека DU145. Доза 30 мг/кг 2C2, вводимая дважды в неделю, проявила максимальную эффективность с dTGI 77%. Модель на основе ксенотрансплантатов DU145 содержит делецию LKB-1.
На фиг. 31 показано уменьшение объема опухоли после введения моноклонального антитела 2C2 при использовании ортотопической модели на основе ксенотрансплантатов злокачественной опухоли молочной железы человека BT-474. Панель А демонстрирует, что доза 30 мг/кг 2C2, вводимая дважды в неделю, проявляла максимальную эффективность с dTGI 55%. Панель В демонстрирует уменьшение объема опухоли после комбинированного введения моноклонального антитела 2C2 с низкомолекулярным лекарственным средством лапатинибом при использовании ортотопической модели на основе ксенотрансплантатов злокачественной опухоли молочной железы человека BT-474. Добавление 2C2 к лапатинибу увеличивает активность лапатиниба в течение фазы обработки и незначительно замедляет возобновление роста опухоли в течение фазы возобновления роста опухоли. Оба антитела, 2C2-YTE и 2C2, в дозе 30 мг/кг имеют сравнимую активность в течение фазы обработки в виде монотерапевтических средств. Панель С демонстрирует уменьшение объема опухоли после введения моноклонального антитела 2C2 при использовании ортотопической модели на основе ксенотрансплантатов злокачественной опухоли молочной железы человека BT-474. Трастузумаб отдельно был очень активным в данной модели, и незначительное усиление наблюдали при добавлении 2C2 в такой модели. Модель на основе ксенотрансплантатов BT-474 содержит амплифицированный HER2 (3+, HercepTest).
На фиг. 32 показано, что обработка клоном 16 (предшественник 2C2) снижает уровни pHER3 и pAKT в экстрактах ксенотрансплантированных опухолей BT-474. В данном эксперименте уровни pHER3 и pAKT были снижены на 50% и 46,1%, соответственно. Никаких изменений общих уровней HER3 в данном эксперименте не наблюдали.
На фиг. 33 показано уменьшение объема опухоли после введения моноклонального антитела 2C2 при использовании ортотопической модели на основе ксенотрансплантатов злокачественной опухоли молочной железы человека MCF-7. Панель А демонстрирует, что доза 10 мг/кг 2C2, вводимая дважды в неделю, проявляла максимальную эффективность с dTGI 34%. Оба антитела, 2C2-YTE и 2C2, в дозе 10 мг/кг имеют сравнимую активность. Панель В демонстрирует уменьшение объема опухоли после комбинированного введения моноклонального антитела 2C2 с низкомолекулярным лекарственным средством паклитакселом при использовании ортотопической модели на основе ксенотрансплантатов злокачественной опухоли молочной железы человека MCF-7. Добавление 2C2 к паклитакселу увеличивало активность паклитаксела в течение фазы обработки. Модель на основе ксенотрансплантатов MCF-7 содержит низкие уровни HER2 (1+, HercepTest).
На фиг. 34 показано уменьшение объема опухоли после введения моноклонального антитела 2C2-YTE при использовании ортотопической модели на основе ксенотрансплантатов злокачественной опухоли молочной железы человека MDA-MB-361 (панели А-С). Добавление 2C2-YTE к моноклональному антителу трастузумабу повышало активность трастузумаба в течение фазы обработки и замедляло возобновление роста опухоли в течение фазы возобновления роста опухоли (панель А). Добавление 2C2-YTE к моноклональному антителу rhuMAb 2C4 незначительно повышало активность rhuMAb 2C4, но не замедляло возобновление роста опухолей (панель В). Добавление 2C2-YTE к низкомолекулярному лекарственному средству лапатинибу повышало активность лапатиниба, но не замедляло возобновление роста опухолей (панель С).
На фиг. 35 показаны уровни пролонгированной экспозиции моноклонального антитела 2C2-YTE в сыворотке у наивных трансгенных по FcRn человека мышей SCID по сравнению с 2C2 и клоном 16-GL после однократной дозы таких антител 60 мг/кг.
На фиг. 36 показано увеличение уровней белка НER3 в ответ на обработку ингибитором MEK (MEKi) селуметинибом (указан звездочкой). Обработка MEKi в сочетании с 2C2 снижает уровни HER3 обратно к нормальным уровням в моделях злокачественных опухолей с использованием клеток HT-29 (слева), LOVO (середина) и Colo205 (справа). Уровни pHER3 также исследовали в моделях HT-29 и LOVO, и было показано, что они отвечают сходным образом.
На фиг. 37 показано, что сочетание 2C2-YTE и селуметиниба повышает противоопухолевую эффективность любого агента по сравнению с его использованием отдельно в моделях подкожных ксенотрансплантатов злокачественной опухоли и A549 (панель А, верх), HT-29 (панель В, верх,), LOVO (панель СC, верх). Вестерн-блот-анализ лизатов опухолей (модели на основе ксенотрансплантатов A549, HT-29 и LOVO) мышей, обработанных сочетанием препаратов, показал, что фосфо-HER3 и фосфо-ERK были полностью ингибированы (панели А-С, внизу).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к молекулам и их антигенсвязывающим фрагментам, которые связываются с HER3. В некоторых аспектах такими молекулами являются антитела и их антигенсвязывающие фрагменты, которые специфично связываются с HER3. Также предлагаются соответственные полинуклеотиды, композиции, содержащие анти-HER3-антитела или их антигенсвязывающие фрагменты, и способы получения анти-HER3-антител и их антигенсвязывающих фрагментов. Кроме того, предлагаются способы применения новых анти-HER3-антител, такие как способы лечения злокачественной опухоли у субъекта и диагностические применения.
Для того чтобы настоящее изобретение можно было легче понять, вначале определены некоторые термины. Дополнительные определения даны по всему объему подробного описания.
I. ОПРЕДЕЛЕНИЯ
До подробного описания настоящего изобретения следует понимать, что указанное изобретение не ограничено конкретными композициями или стадиями способа, поскольку таковые могут варьировать. Используемые в настоящем описании и прилагаемой формуле изобретения формы единственного числа включают и указание множественного числа, если в контекст явно не диктует иное. Термины в единственном числе, а также термины «один или несколько» и «по меньшей мере, один» могут быть использованы в настоящем описании взаимозаменяемо.
Кроме того, «и/или» при использовании в настоящем описании, следует понимать как конкретное раскрытие каждого из двух конкретных признаков или компонентов вместе с другим или без него. Таким образом, подразумевается, что термин «и/или», употребляемый в такой фразе, как «A и/или B» в настоящем описании, включает «A и B», «A или B», (только) «A» (только) и (только) «B». Подобным образом, подразумевается, что термин «и/или», употребляемый в такой фразе, как «A, B и/или C», охватывает каждый из следующих аспектов: A, B и C; A, B или C; A или C; A или B; B или C; A и C; A и B; B и C; (только) A; (только) B; и (только) C.
Если не указано иное, все технические и научные термины, употребляемые в настоящем описании, имеют такое же значение, которое обычно понимает специалист в области, к которой настоящее изобретение относится. Например, Краткий словарь биомедицины и молекулярной биологии, Juo, Pei-Show, 2nd ed., 2002, CRC Press; Словарь клеточной и молекулярной биологии, 3rd ed., 1999, Academic Press; и Оксфордский словарь биохимии и молекулярной биологии, Revised, 2000, Oxford University Press, предлагают специалисту общий словарь многих из терминов, употребляемых в настоящем изобретении.
Единицы, префиксы и символы указаны в форме, принятой Международной системе единиц (СИ). Диапазоны числовых значений даны с учетом чисел, определяющих границы диапазона. Если не указано иное, аминокислотные последовательности описаны слева направо в направлении от амино- к карбоксильному концу. Заголовки, представленные в настоящем описании, не являются ограничениями различных аспектов, которые могут быть сделаны на основе ссылки на описание в целом. Соответственно, термины, которые определены непосредственно ниже, более полно определены, основываясь на полном описании.
Следует понимать, что повсюду, где аспекты описаны в настоящем описании с использованием выражения «содержащий», также предлагаются другие аналогичные аспекты, описанные с использованием выражения «состоящий из» и/или «состоящий, главным образом, из».
Аминокислоты обозначены в настоящем описании либо посредством их общеизвестных трехбуквенных символов, либо однобуквенных символов, рекомендованных комиссией по биохимической номенклатуре ИЮПАК-ИЮБ. Подобным образом, нуклеотиды обозначены общепринятыми однобуквенными кодами.
Термины «HER3» и «рецептор HER3» используют взаимозаменяемо в настоящем описании, и относят к ErbB3-белку (также называемому в литературе HER3, рецептор ErbB3), который описан в патенте США №5480968 и публикации Plowman с соавторами (1990), Proc. Natl. Acad. Sci. USA 87, 4905-4909; см. также публикацию Kani с соавторами (2005), Biochemistry 44, 15842-15857, и Cho and Leahy (2002) Science 297, 1330-1333. Полноразмерная последовательность зрелого белка HER3 (без лидерной последовательности) соответствует последовательности, представленной на фиг. 4 и последовательности SEQ ID NO: 4, приведенной в патенте США №5480968, минус 19 аминокислот лидерной последовательности, которая отщепляется от зрелого белка.
Термины «ингибирование» и «подавление» использованы в настоящем описании взаимозаменяемо и относятся к любому статистически значимому снижению биологической активности, включая полное блокирование активности. Например, «ингибирование» может относиться к уменьшению биологической активности приблизительно на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100%. Соответственно, когда термины «ингибирование» или «подавление» применяют для описания, например, влияния на опосредованное лигандом фосфорилирование HER3, термин относится к способности антитела или его антигенсвязывающего фрагмента осуществлять статистически значимое уменьшение степени фосфорилирования HER3, индуцируемого EGF-подобным лигандом, относительно фосфорилирования в необработанной (контрольной) клетке. Клетка, которая экспрессирует HER3, может быть встречающейся в природе клеткой или клеточной линией (например, злокачественной клеткой) или может быть рекомбинантно полученной клеткой в результате введения нуклеиновой кислоты, кодирующей HER3, в клетку-хозяина. В одном аспекте, связывающая анти-HER3-молекула, например, антитело или его антигенсвязывающий фрагмент, ингибирует опосредованное лигандом фосфорилирование HER3, по меньшей мере, на 10%, или, по меньшей мере, на 20%, или, по меньшей мере, на 30%, или, по меньшей мере, на 40%, или, по меньшей мере, на 50%, или, по меньшей мере, на 60%, или, по меньшей мере, на 70%, или, по меньшей мере, на 80%, или, по меньшей мере, на 90%, или примерно на 100%, как определено, например, с использованием вестерн-блоттинга с последующим исследованием с использованием в качестве зонда антитела против фосфотирозина, или ELISA, как описано в примерах ниже.
Термин «подавление роста» клетки, экспрессирующей HER3, употребляемый в настоящем описании, относится к способности связывающей анти-HER3-молекулы, например, антитела или его антигенсвязывающего фрагмента, статистически значимо снижать пролиферации клетки, экспрессирующей HER3, относительно пролиферации в отсутствие связывающей анти-HER3-молекулы, например, антитела или его антигенсвязывающего фрагмента. В одном аспекте пролиферация клеток, экспрессирующих HER3 (например, злокачественных клеток), может быть снижена, по меньшей мере, на 10%, или, по меньшей мере, на 20%, или, по меньшей мере, на 30%, или, по меньшей мере, на 40%, или, по меньшей мере, на 50%, или, по меньшей мере, на 60%, или, по меньшей мере, на 70%, или, по меньшей мере, на 80%, или, по меньшей мере, на 90%, или приблизительно на 100%, при контакте клеток со связывающей анти-HER3-молекулой, например, антителом или его антигенсвязывающим фрагментом согласно настоящему изобретению, относительно пролиферации, измеряемой в отсутствие связывающей анти-HER3-молекулы, например, антитела или его антигенсвязывающего фрагмента (контрольные условия). Пролиферацию клеток можно исследовать, используя известные в данной области способы с измерением скорости клеточного деления, фракции клеток в клеточной популяции, претерпевших клеточное деление, и/или скорости потери клеток из клеточной популяции вследствие конечной дифференцировки или гибели клеток (например, по включению тимидина).
Термины «антитело» или «иммуноглобулин», которые используют в настоящем описании взаимозаменяемо, включают целые антитела и их любой антигенсвязывающий фрагмент или одиночные цепи.
Обычное антитело содержит, по меньшей мере, две тяжелые (H) цепи и две легкие (L) цепи, соединенные между собой дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно в настоящем описании VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращено в настоящем описании VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена, CL. Области VH и VL также могут быть дополнительно подразделены на области гипервариабельности, названные определяющими комплементарность областями (CDR), перемежающиеся с областями, которые являются более консервативными, названными каркасными областями (FW). Каждая VH и VL состоит из трех CDR и четырех FW, расположенных от амино-конца к карбоксильному концу в следующем порядке: FW, CDR1, FW2, CDR2, FW3, CDR3, FW4. Вариабельные области тяжелых и легких цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (C1q) классической системы комплемента. Примеры антител согласно настоящему изобретению включают анти-HER3-антитела клона 16 (CL16) (исходные антитела и антитела, приведенные к зародышевой линии), оптимизированные по аффинности клоны, включая, например, анти-HER3-антитело 2C2, и оптимизированные по времени полужизни в сыворотке анти-HER3 антитела, включая, например анти-HER3-антитело 2C2-YTE.
Термин «приведение к зародышевой линии» означает, что аминокислоты в конкретных положениях в антителе подвергают мутациям, приводящим обратно к аминокислотам зародышевой линии. Например, «приведенной к зародышевой линии» антитело CL16 получают из исходного антитела CL16 путем внесения трех точек мутаций, Y2S, E3V и M201, в FW1 областей VL.
Термин «антитело» означает молекулу иммуноглобулина, которая распознает и специфично связывается с мишенью, такой как белок, полипептид, пептид, углевод, полинуклеотид, липид или их сочетание при участии, по меньшей мере, одного распознающего антиген участка в вариабельной области молекулы иммуноглобулина. В используемом в настоящем описании смысле термин «антитело» охватывает интактные поликлональные антитела, интактные моноклональные антитела, фрагменты антител (такие как фрагменты Fab, Fab’, F(ab’)2 и Fv), одноцепочечные мутанты Fv (scFv), полиспецифичные антитела, такие как биспецифичные антитела, полученные, по меньшей мере, из двух интактных антител, химерные антитела, гуманизированные антитела, человеческие антитела, слитые белки, содержащие часть антитела, определяющую антиген, и любую другую модифицированную молекулу иммуноглобулина, содержащую распознающий антиген участок, при условии, что антитела проявляют требуемую биологическую активность. Антителом может быть любое антитело из пяти основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, или их подклассов (изотипов) (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2), на основании идентичности константных доменов их тяжелых цепей, называемых альфа, дельта, эпсилон, гамма и мю, соответственно. Различные классы иммуноглобулинов имеют различные и хорошо известные состоящие из субъединиц структуры и трехмерные конфигурации. Антитела могут быть «голыми» или конъюгированными с другими молекулами, такими как токсины, радиоизотопы и т.д.
«Блокирующее» антитело или «антагонистическое» антитело представляет собой антитело, которое ингибирует или снижает биологическую активность антигена, с которым оно связывается, такого как HER3. В некоторых аспектах блокирующие антитела или антагонистические антитела в значительной степени или полностью ингибируют биологическую активность антигена. Желательно, чтобы биологическая активность была снижена на 10%, 20%, 30%, 50%, 70%, 80%, 90%, 95% или даже 100%.
Термины «антитело к HER3» или «антитело, которое связывается с HER3» или «анти-HER3» относятся к антителу, которое способно связываться с HER3 с достаточной аффинностью, так что антитело оказывается пригодным в качестве терапевтического средства или диагностического реагента для целенаправленного воздействия на HER3. Степень связывания анти-HER3-антитела с неродственным, отличным от HER3 белком, составляет менее чем приблизительно 10% от связывания антитела с HER3, измеренного, например, посредством радиоиммуноанализа (РИА), BIACORE™ (с использованием рекомбинантного НER3 в качестве анализируемого вещества и антитела в качестве лиганда, или наоборот), или других анализов связывания, известных в данной области. В некоторых аспектах антитело, которое связывается с HER3, имеет константу диссоциации (KD), составляющую <1 мкΜ, <100 нM, <10 нM, <1 нM, <0,1 нM, <10 пM, <1 пM или <0,1 пM.
Термин «антигенсвязывающий фрагмент» относится к части интактного антитела и относится к определяющим антиген вариабельным областям интактного антитела. В данной области известно, что антигенсвязывающая функция антитела может быть осуществлена фрагментами, полученными из антитела полной длины. Примеры фрагментов антител включают без ограничения фрагменты Fab, Fab', F(ab')2 и Fv, линейные антитела, одноцепочечные антитела и полиспецифичные антитела, образованные из фрагментов антител.
«Моноклональное антитело» относится к гомогенной популяции антител, участвующих в высокоспецифичном распознавании и связывании отдельной антигенной детерминанты или эпитопа. В противоположность моноклональным антителам, поликлональные антитела обычно включают различные антитела, направленные против различных антигенных детерминант. Термин «моноклональное антитело» охватывает как интактные, так и полноразмерные моноклональные антитела, а также фрагменты антител (такие как Fab, Fab’, F(ab’)2, Fv), одноцепочечные (scFv) мутанты, слитые белки, содержащие часть антитела, и любую другую модифицированную молекулу иммуноглобулина, содержащую участок распознавания антигена. Кроме того, «моноклональное антитело» относится к таким антителам, которые получены любым из ряда способов, включая без ограничения, создание гидридом, селекцию фагов, рекомбинантную экспрессия и трансгенных животных.
Термин «гуманизированное антитело» относится к антителу, полученному из иммуноглобулина животного, отличного от человека (например, мыши), которое было сконструировано так, чтобы оно содержало минимальные последовательности животного, отличного от человека (например, мыши). Обычно, гуманизированными антителами являются человеческие иммуноглобулины, в которых остатки из определяющей комплементарность области (CDR) заменены остатками из CDR вида животного, отличного от человека (например, мыши, крысы, кролика или хомяка), которые обладают требуемой специфичностью, аффинностью и функциональными возможностями (Jones et al., 1986, Nature, 321:522-525; Riechmann et al., 1988, Nature, 332:323-327; Verhoeyen et al., 1988, Science, 239:1534-1536). В некоторых случаях остатки каркасной области Fv (FW) человеческого иммуноглобулина заменяют соответствующими остатками антитела вида животного, отличного от человека, которые имеют требуемую специфичность, аффинность и функциональные возможности.
Гуманизированное антитело также может быть дополнительно модифицировано путем замены дополнительных остатков либо в Fv каркасной области, и/либо в пределах заменяющих остатков иммуноглобулина животного, отличного от человека, чтобы улучшить и оптимизировать специфичность, аффинность и/или функциональные возможности антитела. В общем, гуманизированное антитело будет содержать по существу все из, по меньшей мере, одного и обычно двух или трех вариабельных доменов, содержащих все или по существу все области CDR, которые соответствуют иммуноглобулину животного, отличного от человека, в то время как все или по существу все области FR представляют собой области с консенсусной последовательностью иммуноглобулина человека. Гуманизированное антитело также может содержать, по меньшей мере, часть константной области или домен (Fc) иммуноглобулина, обычно иммуноглобулина человека. Примеры способов, используемых для получения гуманизированных антител, описаны в патентах США №5225539 или 5639641.
«Вариабельная область» антитела относится к вариабельной области легкой цепи антитела или вариабельной области тяжелой цепи антитела, либо отдельно, либо в сочетании. Каждая из вариабельных областей тяжелой и легкой цепи состоит из четырех каркасных областей (FW), соединенных тремя определяющими комплементарность областями (CDR), также известными как гипервариабельные области. CDR в каждой цепи удерживаются вместе в непосредственной близости областями FW, и с CDR из другой цепи вносят вклад в образование антигенсвязывающего участка антител. Существует, по меньшей мере, два способа определения CDR: (1) способ, основанный на межвидовой вариабельности последовательностей (т.е., способ, описанный Кабатом с соавторами, Sequence of Proteins of Immunological Interest, (5th ed., 1991, National Institutes of Health, Bethesda Md.)); и (2) способ, основанный на кристаллографических исследованиях комплексов антиген-антитело (Al-lazikani с соавторами, (1997) J. Molec. Biol., 273: 927-948)). Кроме того, для определения CDR в данной области иногда используют сочетания указанных двух способов.
Номенклатуру Кабата обычно используют в отношении остатка в вариабельном домене (приблизительно остатки 1-107 легкой цепи и остатки 1-113 тяжелой цепи) (например, Kabat et al., Sequences of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)).
Нумерация положения аминокислот согласно Кабату относится к системе нумерации, используемой для вариабельных доменов тяжелой цепи или вариабельных доменов легкой цепи при составлении антител, описанной Kabat с соавторами, Sequences of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991). При использовании указанной системы нумерации действительная линейная аминокислотная последовательность может содержать меньше или может содержать дополнительные аминокислоты, что соответствует укорачиванию, или встраиванию в FW или CDR вариабельного домена. Например, вариабельный домен тяжелой цепи может содержать вставку одной аминокислоты (остаток 52a согласно Кабату) после остатка 52 H2 и встроенные остатки (например, остатки 82a, 82b и 82c, и т.д. согласно Кабату) после остатка 82 FW тяжелой цепи.
Нумерацию остатков по Кабату можно определить для данного антитела путем выравнивания остатков в областях гомологии последовательности антитела со «стандартной» пронумерованной по Кабату последовательностью. Чотиа (Chothia) наоборот основывается на положении структурных петель (Chotia and Lesk, J. Mol. Biol. 196: 901-917 (1987)). Конец петли CDR-H1 по Чотиа при нумерации с использованием правила нумерации Кабата варьирует между H32 и H34 в зависимости от длины петли (это происходит вследствие того, что система нумерации Кабата размещает вставки у H35A и H35B; если ни 35A, ни 35B не присутствует, петля заканчивается у 32; если присутствует только 35A, петля заканчивается у 33; если присутствуют и 35A и 35B, петля заканчивается у 34). Гипервариабельные области AbM представляют собой нечто среднее между CDR по Кабату и структурными петлями по Чотиа, и используются Оксфордскими программными средствами молекулярного моделирования AbM антитела.
IMGT (ImMunoGeneTics) также представляет собой систему нумерации для вариабельных областей иммуноглобулина, включая CDR. См., например, публикацию Lefranc, M.P. et al., Dev. Comp. Immunol. 27: 55-77(2003), которая включена в настоящее описание в виде ссылки. Система нумерации IMGT была основана на выравнивании более чем 5000 последовательностей, структурных данных и характеристики гипервариабельных петель и создает возможность для легкого сравнения вариабельных и CDR-областей для всех видов. Согласно схеме нумерации IMGT, VH-CDR1 расположена в положениях от 26 до 35, область VH-CDR2 расположена в положениях от 51 до 57, VH-CDR3 расположена в положениях от 93 до 102, VL-CDR1 расположена в положениях от 27 до 32, VL-CDR2 расположена в положениях от 50 до 52 и VL-CDR3 расположена в положениях от 89 до 97.
В используемом в настоящем описании смысле, описанные последовательности CDR VH соответствуют классическим положениям в нумерации по Кабату, а именно, VH-CDR1 по Кабату расположена в положениях 31-35, VH-CDR2 расположена в положениях 50-65, и VH-CDR3 расположена в положениях 95-102. VL-CDR2 и VL-CDR3 также соответствуют классическим положениям в нумерации по Кабату, а именно, положениям 50-56 и 89-97, соответственно. В используемом в настоящем описании смысле термины «VL-CDR1» или «CDR1 легкой цепи» соответствуют последовательностям, расположенным по Кабату в положениях 23-34 в VL (напротив, классическое расположение VL-CDR1 согласно системе нумерации Кабата соответствует положениям 24-34).
В используемом в настоящем описании смысле область Fc включает полипептиды, содержащие константную область антитела, за исключением первого домена константной области иммуноглобулина. Таким образом, Fc относится к последним двум доменам константной области иммуноглобулина IgA, IgD и IgG и последним трем доменам константной области иммуноглобулина IgE и IgM и гибкому шарниру, N-концевому по отношению к указанным доменам. В случае IgA и IgM Fc может включать J-цепь. В случае IgG Fc содержит домены иммуноглобулина Cгамма2 и Cгамма3 (Cγ2 и Cγ3) и шарнир между Cгамма1 (Cγ1) и Cгамма2 (Cγ2). Хотя границы Fc-области могут варьировать, Fc-область тяжелой цепи IgG человека обычно определяют как область, которая содержит остатки C226 или P230 на ее карбоксильном конце, при этом нумерация представлена согласно указателю EU, как описано Кабатом (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)). Fc может относиться к такой области отдельно или такой области в контексте антитела, фрагмента антитела или слитого белка Fc. Наблюдали полиморфизм наблюдали в ряде различных положений Fc, включающих без ограничения положения 270, 272, 312, 315, 356 и 358, пронумерованные согласно указателю EU, и, таким образом, могут существовать незначительные различия между представленной последовательностью и последовательностями согласно известному уровню техники.
Термин «человеческое антитело» означает антитело, продуцируемое человеком, или антитело, имеющее аминокислотную последовательность, соответствующую продуцируемому человеком антителу, полученное с использованием любого способа, известного в данной области. Указанное определение человеческого антитела включает интактные антитела или полноразмерные антитела, их фрагменты и/или антитела, содержащие, по меньшей мере, один полипептид тяжелой и/или легкой цепи человека, такие как, например, антитело, содержащее полипептиды легкой цепи мыши и тяжелой цепи человека.
Термин «химерные антитела» относится к антителам, в которых аминокислотная последовательность молекулы иммуноглобулина получена из двух или более видов. Обычно вариабельная область как легких, так и тяжелых цепей соответствует вариабельной области антител, полученных от одного вида млекопитающих (например, мыши, крысы, кролика и т.д.) с требуемой специфичностью, аффинностью и функциональными возможностями, в то время как константные области являются гомологичными последовательностям в антителах, полученных от другого вида (обычно человека), чтобы избежать возникновения иммунного ответа у такого вида.
Термины «YTE» или «YTE-мутант» относятся к мутации в Fc IgG1, которая приводит к увеличению связывания с человеческим FcRn и повышает время полужизни антитела, имеющего мутацию. YTE-мутант содержит сочетание трех мутаций M252Y/S254T/T256E (нумерация EU по Кабату, Kabat et al., (1991) Sequences of Proteins of Immunological Interest, U.S. Public Health Service, National Institutes of Health, Washington, D.C.), введенных в тяжелую цепь IgG1. См. патент США №7658921, который включен в настоящее описание в виде ссылки. Было показано, что YTE-мутации повышают время полужизни антител в сыворотке приблизительно в четыре раза по сравнению с вариантами дикого типа такого же антитела (Dall’Acqua et al., J. Biol. Chem. 281: 23514-24 (2006)). См. также патент США №7083784, который включен в настоящее описание в виде ссылки в полном объеме.
«Аффинность связывания» обычно относится к силе суммарных нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и его партнером по связыванию (например, антигеном). Если не оговорено особо, в используемом в настоящем описании смысле «аффинность связывания» относится к присущей молекулам аффинности связывания, которая отражает взаимодействие 1:1 между членами связывающейся пары (например, антителом и антигеном). Аффинность молекулы X по отношению к ее партнеру Y, как правило, может быть представлена константой диссоциации (KD). Аффинность может быть измерена обычными способами, известными в данной области, включая способы, описанные в настоящей публикации. Низко аффинные антитела обычно связывают антиген медленно и имеют тенденцию легко диссоциировать, тогда как высоко аффинные антитела обычно связывают антиген быстрее и имеют тенденцию дольше оставаться связанными. В данной области известны различные способы измерения аффинности связывания, любой из которых можно применять в целях настоящего изобретения.
«Эффективность» обычно выражают в виде значения IC50 в нМ, если не указано иное. IC50 представляет собой медианную ингибирующую концентрацию молекулы антитела. В функциональных анализах IC50 является концентрацией, которая снижает биологический ответ на 50% от максимального. В исследованиях связывания с лигандом IC50 является концентрацией, которая снижает связывание с рецептором на 50% от максимального уровня специфичного связывания. IC50 можно рассчитать любым из ряда способов, известных в данной области. Повышение эффективности можно определить, проводя измерения, например, по сравнению с родительским моноклональным антителом CL16 (клон 16).
Кратное повышение эффективности антител или полипептидов согласно изобретению по сравнению с антителом клона 16 может быть, по меньшей мере, приблизительно в 2 раза, по меньшей мере, приблизительно в 4 раза, по меньшей мере, приблизительно в 6 раз, по меньшей мере, приблизительно в 8 раз, по меньшей мере, приблизительно в 10 раз, по меньшей мере, приблизительно в 20 раз, по меньшей мере, приблизительно в 30 раз, по меньшей мере, приблизительно в 40 раз, по меньшей мере, приблизительно в 50 раз, по меньшей мере, приблизительно в 60 раз, по меньшей мере, приблизительно в 70 раз, по меньшей мере, приблизительно в 80 раз, по меньшей мере, приблизительно в 90 раз, по меньшей мере, приблизительно в 100 раз, по меньшей мере, приблизительно в 110 раз, по меньшей мере, приблизительно в 120 раз, по меньшей мере, приблизительно в 130 раз, по меньшей мере, приблизительно в 140 раз, по меньшей мере, приблизительно в 150 раз, по меньшей мере, приблизительно в 160 раз, по меньшей мере, приблизительно в 170 раз, или по меньшей мере, приблизительно в 180 раз или больше.
«Зависимая от антител опосредованная клетками цитотоксичность» или «ADCC» относится к форме цитотоксичности, при которой секретированный Ig, связанный с Fc-рецепторами (FcR), присутствующими на некоторых цитотоксических клетках (например, природных клетках-киллерах (NK), нейтрофилах и макрофагах), обеспечивает возможность специфичного связывания таких цитотоксических эффекторных клеток с несущими антиген клетками-мишенями, и затем уничтожения клеток-мишеней цитотоксинами. Специфичные высокоаффинные IgG-антитела, направленные на поверхность клеток-мишеней, «вооружают» цитотоксические клетки и абсолютно необходимы для такого уничтожения. Лизис клетки-мишени является внеклеточным, требует прямого контакта клетки с клеткой и не вовлекает комплемент. Предполагается, что кроме антител, другие белки, содержащие Fc-области, в частности слитые белки Fc, проявляющие способность специфично связываться с несущей антиген клеткой-мишенью, будут способны вызывать опосредованную клетками цитотоксичность. Для простоты опосредованную клетками цитотоксичность, являющуюся результатом активности слитого белка Fc, также называют в настоящем описании ADCC-активностью.
Полипептид, антитело, полинуклеотид, вектор, клетка или композиция, которые являются «выделенными», представляют собой полипептид, антитело, полинуклеотид, вектор, клетку или композицию, которые присутствуют в форме, не встречающейся в природе. Выделенные полипептиды, антитела, полинуклеотиды, векторы, клетки или композиции включают такие, которые были очищены до такой степени, что они больше не существуют в форме, в которой они найдены в природе. В некоторых аспектах, антитело, полинуклеотид, вектор, клетка или композиция, которые являются выделенными, по существу, является очищенными.
Термин «субъект» относится к любому животному (например, млекопитающему), включая без ограничения людей, приматов, отличных от человека, грызунов и тому подобных, которому должен стать реципиентом конкретного лечения. Обычно термины «субъект» и «пациент» используют в настоящем описании взаимозаменяемо по отношению к человеку.
Термин «фармацевтическая композиция» относится к препарату, который существует в такой форме, которая обеспечивает эффективную биологическую активность активного ингредиента, и которая не содержит дополнительных компонентов, которые являются неприемлемо токсичными для субъекта, которому может быть введена композиция. Такая композиция может быть стерильной.
«Эффективное количество» антитела, которое описано в настоящей публикации, представляет собой количество, достаточное для достижения конкретной заявленной цели. «Эффективное количество» можно определить эмпирически и стандартным способом, в зависимости от заявленной цели.
Термин «терапевтически эффективное количество» относится к количеству антитела или другого лекарственного средства, эффективного для «лечения» заболевания или расстройства у субъекта или млекопитающего.
Слово «метка» при использовании в настоящем описании относится к регистрируемому соединению или композиции, которые конъюгированы прямо или опосредованно с антителом таким образом, чтобы создать «меченое» антитело. Метка может быть регистрируемой как таковая (например, радиоизотопные метки или флуоресцирующие метки) или, в случае ферментативной метки, может катализировать химическое изменение субстратного соединения или композиции, которое можно регистрировать.
Такие термины, как «процесс лечения» или «лечение», или «лечить», или «облегчение», или «облегчать» относятся как к (1) терапевтическим мерам, которые позволяют излечивать, замедлять, уменьшать симптомы и/или останавливать прогрессирование диагностированного патологического состояния или расстройства, так и к (2) профилактическим или превентивным мерам, которые предотвращают и/или замедляют развитие целевого патологического состояния или расстройства. Таким образом, субъекты, которые нуждаются в лечении, включают тех субъектов, которые уже имеют расстройство, и тех, у кого необходимо предотвратить расстройство. В некоторых аспектах, у субъекта успешно «лечат» злокачественную опухоль способами согласно настоящему изобретению, если у пациента наблюдают, например, полную, частичную или временную ремиссию определенного типа злокачественной опухоли.
Термины «злокачественная опухоль», «опухоль», «раковое» и «злокачественное» относятся или описывают физиологическое состояние у млекопитающих, которое обычно характеризуется нерегулируемым ростом клеток. Примеры злокачественных опухолей включают без ограничения карциному, включая аденокарциномы, лимфомы, бластомы, меланомы, саркомы и лейкозы. Более конкретные примеры таких злокачественных опухолей включают плоскоклеточный рак, мелкоклеточный рак легкого, немелкоклеточный рак легкого, рак желудочно-кишечного тракта, ходжкинскую и неходжкинскую лимфому, рак поджелудочной железы, глиобластому, глиому, рак шейки матки, рак яичника, рак печени, такой как карцинома печени и гепатома, рак мочевого пузыря, рак молочной железы (включая гормонально опосредованный рак молочной железы, см., например, Innes et al. (2006) Br. J. Cancer 94: 1057-1065), рак ободочной кишки, рак прямой и ободочной кишки, карциному эндометрия, миелому (такую как множественная миелома), карциному слюнных желез, рак почек, атакой как почечноклеточная карцинома и опухоли Вильмса, базальноклеточную карциному, меланому, рак простаты, рак вульвы, рак щитовидной железы, рак семенников, рак пищевода, различные типы рака головы и шеи и злокачественные опухоли, имеющие мукоидное происхождение, такие как мукоидный рак яичника, холангиокарциному (печени) и папиллярную карциному почек.
В используемом в настоящем описании смысле термин «карциномы» относится к злокачественным опухолям эпителиальных клеток, которые представляют собой клетки, которые покрывают поверхность тела, продуцируют гормоны и образуют железы. Примерами карцином являются злокачественные опухоли кожи, легкого, ободочной кишки, желудка, молочной железы, простаты и щитовидной железы.
Термин «мутация KRAS» в используемом в настоящем описании смысле относится к мутациям, обнаруженным в случае некоторых злокачественных опухолей в человеческом гомологе вирусного онкогена саркомы крыс v-Ki-ras2 Kirsten. Не ограничивающие примеры последовательностей мРНК гена KRAS человека включают последовательности с номерами доступа в Genbank NM004985 и NM033360. Сообщалось, что мутации KRAS встречаются в 73% опухолей поджелудочной железе, 35% опухолей прямой и ободочной кишки, 16% опухолей яичника и 17% опухолей легкого. Мутация KRAS обычно встречается в кодонах 12 или 143 гена KRAS человека.
Термины «полинуклеотид» или «нуклеиновая кислота», которые использованы в настоящем описании взаимозаменяемо, относятся к полимеру нуклеотидов любой длины и включают ДНК и РНК. Нуклеотиды могут представлять собой дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания и/или их аналоги или любой субстрат, который может быть включен в полимер ДНК- или РНК-полимеразой. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. Предшествующее описание применимо ко всем полинуклеотидам, упоминаемым в настоящем описании, включая РНК и ДНК.
Термин «вектор» означает конструкцию, которая способна доставлять и, в некоторых аспектах» экспрессировать один или несколько представляющих интерес генов или последовательностей в клетке-хозяине. Примеры векторов включают без ограничения вирусные векторы, экспрессирующие векторы в виде «голой» ДНК или РНК, плазмидные, космидные и фаговые векторы, экспрессирующие ДНК- или РНК-векторы, ассоциированные с катионными конденсирующими агентами, экспрессирующие ДНК- или РНК-векторы, инкапсулированные в липосомы, и некоторые эукариотические клетки, такие как клетки-продуценты.
Термины «полипептид», «пептид, и «белок» используют в настоящем описании взаимозаменяемо по отношению к полимерам аминокислот любой длины. Полимер может быть линейным или разветвленным, он может содержать модифицированные аминокислоты, и он может прерываться другими соединениями, отличными от аминокислот. Термины также охватывают полимер аминокислот, который был модифицирован в природе или в результате вмешательства; например, образования дисульфидной связи, гликозилирования, липидизации, ацетилирования, фосфорилирования или любой другой обработки или модификации, такой как конъюгация с метящим компонентом. Также в определение включены, например, полипептиды, содержащие один или несколько аналогов аминокислот (включая, например, неприродные аминокислоты и т.д.), а также другие модификации, известные в данной области. Понятно, что поскольку полипептиды согласно настоящему изобретению, основаны на антителах, в некоторых аспектах полипептиды могут быть в виде отдельных цепей или ассоциированных цепей.
Термины «идентичный» или «идентичность» в процентах в контексте двух или более нуклеиновых кислот или полипептидов относятся к двум или более последовательностям или подпоследовательностям, которые являются одинаковыми или имеют конкретное процентное содержание нуклеотидов или аминокислотных остатков, которые являются одинаковыми при сравнении и выравнивании (введении, при необходимости, пробелов) для максимального соответствия, без учета каких-либо консервативных аминокислотных замен в качестве части идентичности последовательностей. Идентичность в процентах можно измерить, используя компьютерную программу или алгоритмы сравнения последовательностей, или посредством визуального просмотра. В данной области известны различные алгоритмы и компьютерные программы, которые можно использовать для получения выравнивания аминокислотных или нуклеотидных последовательностей.
Одним таким не ограничивающим примером алгоритма выравнивания последовательностей является алгоритм, описанный в публикации Karlin с соавторами, 1990, Proc. Natl. Acad. Set, 87: 2264-2268, модификация которого описана Karlin с соавторами, 1993, Proc. Natl. Acad. Set, 90: 5873-5877, и который включен в программы NBLAST и XBLAST (Altschul с соавторами, 1991, Nucleic Acid Res., 25: 3389-3402). В некоторых аспектах можно использовать Gapped BLAST, который описан Altschul с соавторами, 1997, Nucleic Acids Res. 25: 3389-3402. BLAST-2, WU-BLAST-2 (Altschul с соавторами, 1996, Methods in Enzymology, 266: 460-480), ALIGN, ALIGN-2 (Genentech, South San Francisco, California) или Megalign (DNASTAR) являются дополнительными общедоступными компьютерными программами, которые можно использовать для выравнивания последовательностей. В некоторых аспектах идентичность в процентах между двумя нуклеотидными последовательностями определяют, используя программу GAP в пакете компьютерных программ GCG (например, используя матрицу NWSgapdna.CMP и вес пробелов 40, 50, 60, 70 или 90, и вес длины 1, 2, 3, 4, 5 или 6). В некоторых альтернативных аспектах можно использовать программу GAP в пакете компьютерных программ GCG, которая включает в себя алгоритм Нидлемана и Вунша (J. Mol. Biol. (48): 444-453 (1970)), для определения идентичности в процентах между двумя аминокислотными последовательностями (например, используя либо матрицу BLOSUM 62, либо матрицу PAM250, и вес пробела 16, 14, 12, 10, 8, 6 или 4 и вес длины 1, 2, 3, 4, 5). Альтернативно в некоторых аспектах идентичность в процентах между нуклеотидными или аминокислотными последовательностями определяют, используя алгоритм Майерса и Миллера (CABIOS, 4: 11-17 (1989)). Например, идентичность в процентах можно определить, используя программу ALIGN (версия 2.0) и используя таблицу остатков PAM120, штраф за длину пробела 12 и штраф за пробел 4. Соответствующие параметры для максимального выравнивания с использованием конкретной компьютерной программы выравнивания может определить специалист в данной области. В некоторых аспектах используют параметры компьютерной программы выравнивания по умолчанию.
В некоторых аспектах идентичность в процентах «X» первой аминокислотной последовательности по отношению ко второй последовательности аминокислот вычисляют как 100×(Y/Z), где Y означает количество аминокислотных остатков, оцениваемых как идентично совпадающие при выравнивании первой и второй последовательностей (которые выравнивают в результате визуального просмотра или с использованием конкретной программы выравнивания последовательностей), и Z означает общее количество остатков во второй последовательности. Если длина первой последовательности больше, чем длина второй последовательности, то идентичность в процентах первой последовательности по отношению ко второй последовательности будет выше, чем идентичность в процентах второй последовательности по отношению к первой последовательности.
«Консервативная аминокислотная замена» представляет собой замену, при которой один аминокислотный остаток заменяют другим аминокислотным остатком, имеющим сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, определены в данной области, включая аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, глицин, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Например, замена тирозина на фенилаланин является консервативной заменой. В некоторых аспектах консервативные замены в последовательностях полипептидов и антител согласно изобретению не отменяет связывание полипептида или антитела, содержащего такую аминокислотную последовательность, с антигеном (антигенами), т.е. HER3, с которым связывается полипептид или антитело. Способы идентификации консервативных нуклеотидных и аминокислотных замен, которые не устраняют связывание антигена, хорошо известны в данной области (см., например, Brummell et al., Biochem. 32: 1180-1 187 (1993); Kobayashi et al., Protein Eng. 12(10): 879-884 (1999); и Burks et al., Proc. Natl. Acad. Sci. USA 94: 412-417 (1997)).
Термин «консенсусная последовательность» в используемом в настоящем описании смысле по отношению к вариабельным областям легкой цепи (VL) и тяжелой цепи (VH) относится к составной или обобщенной последовательности VL или VH, определенной на основе информации о том, какие аминокислотные остатки в цепи VL или VH могут быть подвергнуты модификации без вредя для связывания антигена. Таким образом, в «консенсусной последовательности» цепи VL или VH некоторые положения аминокислот заняты одним из множества аминокислотных остатков, возможных в данном положении. Например, если аргинин (R) или серин (S) находятся в конкретном положении, то такое конкретное положение в консенсусной последовательности может быть занято либо аргинином, либо серином (R или S). Консенсусные последовательности цепи VH и VL могут быть определены, например, с использованием созревания аффинности in vitro (например, рандомизацией каждого положения аминокислоты в определенной CDR с использованием вырожденных кодирующих праймеров), сканирующего мутагенеза (например, мутагенеза на основе сканирования аланином) аминокислотных остатков в CDR антитела или любыми другими способами, известными в данной области, с последующей оценкой связывания мутантов с антигеном, чтобы определить, влияет ли мутантная аминокислота в данном положении на связывание антигена. В некоторых аспектах вводят мутации в области CDR. В других аспектах мутации вводят в каркасные области. В некоторых других аспектах мутации вводят в CDR и каркасные области.
II. Связывающие анти-HER3-молекулы
Настоящее изобретение относится к молекулам, связывающим HER3, например антителам и их антигенсвязывающим фрагментам, который специфично связывает HER3. Полноразмерные аминокислотные (а/к) и нуклеотидные (н.) последовательности HER3 известны в данной области (см., например, номер доступа в UniProt P2186 для HER3 человека или номер доступа в UniProt 088458 для HER3 мыши). В некоторых аспектах связывающие анти-HER3-молекулы представляют собой антитела человека. В некоторых аспектах HER3-связывающие молекулы представляют собой антитела или их антигенсвязывающие фрагменты. В некоторых аспектах HER3-связывающие молекулы, например, антитела или их антигенсвязывающие фрагменты включают Fab, Fab’, F(ab’)2, Fd, одноцепочечный Fv или scFv, связанный дисульфидной связью Fv, домен V-NAR, IgNar, интраантитело, IgGACH2, миниантитело, F(ab’)3, тетраантитело, триантитело, диантитело, однодоменное антитело, DVD-Ig, Fcab, mAb2, (scFv)2 или scFv-Fc. В некоторых аспектах антитело относится к подтипу IgG1 и включает тройной мутант YTE, который описан выше в разделе «Определения».
В некоторых аспектах анти-HER3-антитела или их антигенсвязывающие фрагменты согласно изобретению модифицированы по сравнению с исходным антителом клона 16 (CL16). Модификации могут включать в себя мутации в областях CDR и/или в областях FW по сравнению с CL16. В некоторых аспектах анти-HER3-антитело согласно изобретению содержит модификации в CDR1 и/или CDR3 легкой цепи CL16, включая без ограничения:
1) CDR1 легкой цепи, содержащую консенсусную последовательность X1GSX2SNIGLNYVS, где X1 выбран из m R или S, и X2 выбран из S или L; и
2) CDR3 легкой цепи, содержащую консенсусную последовательность AAWDDX3X4X5GEX6, где X3 выбран из S или G, X4 выбран из L или P, X5 выбран из R, I, P или S, и X6 выбран из V или A.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит модификации в CDR2 тяжелой цепи CL16, включая без ограничения CDR1 тяжелой цепи, содержащую консенсусную последовательность X7IGSSGGVTNYADSVKG, где X7 выбран из Y, I или V.
В одном аспекте анти-HER3-антитело или его антигенсвязывающий фрагмент содержит VL-область, содержащую консенсусную аминокислотную последовательность:
[FW1]X1GSX2SNIGLNYVS[FW2]RNNQRPS[FW3]AAWDDX3X4X5GEX6[FW4],
где [FW1], [FW2], [FW3] и [FW4] означают аминокислотные остатки каркасной области 1 VL (SEQ ID NO: 40 или 44), каркасной области 2 VL (SEQ ID NO: 41), каркасной области 3 VL (SEQ ID NO: 42) и каркасной области 4 VL (SEQ ID NO: 43), и где X1 означает аминокислотные остатки аргинин (R) или серин (S), X2 означает аминокислотные остатки серин (S) или лейцин (L), X3 означает аминокислотные остатки серин (S) или глутаминовую кислоту (E), X4 означает аминокислотные остатки лейцин (L) или пролин (P), X5 означает аминокислотные остатки аргинин (R), изолейцин (I), пролин (P) или серин (S), и X6 означает аминокислотные остатки валин (V) или аргинин (R).
В одном аспекте анти-HER3-антитело или его антигенсвязывающий фрагмент содержит VH-область, содержащую консенсусную аминокислотную последовательность:
[FW5]YYYMQ[FW6]X7IGSSGGVTNYADSVKG[FW7]VGLGDAFDI[FW8],
где [FW5], [FW6], [FW7] и [FW8] означают аминокислотные остатки каркасной области 1 VH (SEQ ID NO: 36), каркасной области 2 VH (SEQ ID NO: 37), каркасной области 3 VH (SEQ ID NO: 38) и каркасной области 4 VH (SEQ ID NO: 39), и где X7 означает аминокислотные остатки тирозин (Y), изолейцин (I) или валин (V).
В одном аспекте анти-HER3-антитело или его антигенсвязывающий фрагмент содержит VL-область, содержащую консенсусную аминокислотная последовательность:
[FW1]X1GSX2SNIGLNYVS[FW2]RNNQRPS[FW3]AAWDDX3X4X5GEX6[FW4],
где [FW1], [FW2], [FW3] и [FW4] означают аминокислотные остатки каркасной области 1 VL 1 (SEQ ID NO: 40 или 44), каркасной области 2 VL (SEQ ID NO: 41), каркасной области 3 VL (SEQ ID NO: 42) и каркасной области 4 VL (SEQ ID NO: 43), и где X1 означает аминокислотные остатки аргинин (R) или серин (S), X2 означает аминокислотные остатки серин (S) или лейцин (L), X3 означает аминокислотные остатки серин (S) или глутаминовую кислоту (E), X4 означает аминокислотные остатки лейцин (L) или пролин (P), X5 означает аминокислотные остатки аргинин (R), изолейцин (I), пролин (P) или серин (S), и X6 означает аминокислотные остатки валин (V) или аргинин (R); и при этом указанное анти-HER3-антитело или его антигенсвязывающий фрагмент дополнительно содержит VH-область, которая содержит консенсусную аминокислотная последовательность:
[FW5]YYYMQ[FW6]X7IGSSGGVTNYADSVKG[FW7]VGLGDAFDI[FW8],
где [FW5], [FW6], [FW7] и [FW8] означают аминокислотные остатки каркасной области 1 VH (SEQ ID NO: 36), каркасной области 2 VH (SEQ ID NO: 37), каркасной области 3 VH (SEQ ID NO: 38) и каркасной области 4 VH (SEQ ID NO: 39), и где X7 означает аминокислотные остатки тирозин (Y), изолейцин (I) или валин (V).
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR1, состоящую из последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 18, 19 и 20. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 18, 19 и 20. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR2, состоящую из последовательности SEQ ID NO: 21. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR2, содержащую последовательность SEQ ID NO: 21. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR3, состоящую из последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 22, 23, 24, 25, 26, 27, 28, 29 и 30. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 22, 23, 24, 25, 26, 27, 28, 29 и 30.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR1, состоящую из последовательности SEQ ID NO: 31. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR1, содержащую последовательность SEQ ID NO: 31. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR2, состоящую из последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 32, 33 и 34. В некоторых аспектах анти- HER3 антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 32, 33 и 34. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR3, состоящую из последовательности SEQ ID NO: 35. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR3, содержащую последовательность SEQ ID NO: 35.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR1, состоящую из последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 18, 19 и 20, за исключением одной, двух, трех или четырех аминокислотных замен. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 18, 19 и 20, за исключением одной, двух, трех или четырех аминокислотных замен. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR2, состоящую из последовательности SEQ ID NO: 21, за исключением одной, двух, трех или четырех аминокислотных замен. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR2, содержащую последовательность SEQ ID NO: 21, за исключением одной, двух, трех или четырех аминокислотных замен. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR3, состоящую из последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 22, 23, 24, 25, 26, 27, 28, 29 и 30, за исключением одной, двух, трех или четырех аминокислотных замен. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 22, 23, 24, 25, 26, 27, 28, 29 и 30, за исключением одной, двух, трех или четырех аминокислотных замен.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR1, состоящую из последовательности SEQ ID NO: 31, за исключением одной, двух, трех или четырех аминокислотных замен. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR1, содержащую последовательность SEQ ID NO: 31, за исключением одной, двух, трех или четырех аминокислотных замен. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR2, состоящую из последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 32, 33 и 34, за исключением одной, двух, трех или четырех аминокислотных замен. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 32, 33 и 34, за исключением одной, двух, трех или четырех аминокислотных замен. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR3, состоящую из последовательности SEQ ID NO: 35, за исключением одной, двух, трех или четырех аминокислотных замен. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR3, содержащую SEQ ID NO: 35, за исключением одной, двух, трех или четырех аминокислотных замен.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR1, состоящую из последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 18, 19 и 20; VL-CDR2, состоящую из последовательности SEQ ID NO: 21; и VL-CDR3, состоящую из последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 22, 23, 24, 25, 26, 27, 28, 29 и 30. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 18, 19 и 20; VL-CDR2, содержащую последовательность SEQ ID NO: 21; и VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 22, 23, 24, 25, 26, 27, 28, 29 и 30.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR1, состоящую из последовательности SEQ ID NO: 31; VH-CDR2, состоящую из последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 32, 33 и 34; и VH-CDR3, состоящую из последовательности SEQ ID NO: 35. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR1, содержащую последовательность SEQ ID NO: 31; VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 32, 33 и 34; VH-CDR3, содержащую последовательность SEQ ID NO: 35.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR1, состоящую из последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 18, 19 и 20, за исключением одной, двух, трех или четырех аминокислотных замен; VL-CDR2, состоящую из последовательности SEQ ID NO: 21, за исключением одной, двух, трех или четырех аминокислотных замен; и VL-CDR3, состоящую из последовательности выбранной из группы, состоящей из последовательностей SEQ ID NO: 22, 23, 24, 25, 26, 27, 28, 29 и 30, за исключением одной, двух, трех или четырех аминокислотных замен. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL-CDR1, содержащую последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 18, 19 и 20, за исключением одной, двух, трех или четырех аминокислотных замен; VL-CDR2, содержащую последовательность SEQ ID NO: 21, за исключением одной, двух, трех или четырех аминокислотных замен; и VL-CDR3, содержащую последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 22, 23, 24, 25, 26, 27, 28, 29 и 30, за исключением одной, двух, трех или четырех аминокислотных замен.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR1, состоящую из последовательности SEQ ID NO: 31, за исключением одной, двух, трех или четырех аминокислотных замен; VH-CDR2, состоящую из последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 32, 33 и 34, за исключением одной, двух, трех или четырех аминокислотных замен; и VH-CDR3, состоящую из последовательности SEQ ID NO: 35, за исключением одной, двух, трех или четырех аминокислотных замен. В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VH-CDR1, содержащую последовательность SEQ ID NO: 31, за исключением одной, двух, трех или четырех аминокислотных замен; VH-CDR2, содержащую последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 32, 33 и 34, за исключением одной, двух, трех или четырех аминокислотных замен; и VH-CDR3, содержащую последовательность SEQ ID NO: 35, за исключением одной, двух, трех или четырех аминокислотных замен.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит модификации в CDR1, CDR2 и/или CDR3 тяжелой и/или легкой цепи, и дополнительно содержит модификации в FW1, FW2, FW3 и/или FW4 тяжелой и/или легкой цепи. В некоторых аспектах FW1 содержит последовательность SEQ ID NO: 40 или 44, FW2 содержит последовательность SEQ ID NO: 41, FW3 содержит последовательность SEQ ID NO: 42, FW4 содержит последовательность SEQ ID NO: 43, FW5 содержит последовательность SEQ ID NO: 36, FW6 содержит последовательность SEQ ID NO: 37, FW7 содержит последовательность SEQ ID NO: 38 и FW8 содержит последовательность SEQ ID NO: 39.
В некоторых аспектах FW1 содержит последовательность SEQ ID NO: 40 или 44, за исключением одной, двух, трех или четырех аминокислотных замен; FW2 содержит последовательность SEQ ID NO: 41, за исключением одной, двух, трех или четырех аминокислотных замен; FW3 содержит последовательность SEQ ID NO: 42, за исключением одной, двух, трех или четырех аминокислотных замен; FW4 содержит последовательность SEQ ID NO: 43, за исключением одной, двух, трех или четырех аминокислотных замен; FW5 содержит последовательность SEQ ID NO: 36, за исключением одной, двух, трех или четырех аминокислотных замен; FW6 содержит последовательность SEQ ID NO: 37, за исключением одной, двух, трех или четырех аминокислотных замен; FW7 содержит последовательность SEQ ID NO: 38, за исключением одной, двух, трех или четырех аминокислотных замен; и FW8 содержит последовательность SEQ ID NO: 39, за исключением одной, двух, трех или четырех аминокислотных замен.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL и VH, содержащие аминокислотные последовательности VL-CDR1, VL-CRD2, VL-CDR3, VH-CDR1, VH-CDR2, и VH-CDR3, идентичные или идентичные, за исключением четырех, трех, двух или одной аминокислотной замены в одной или нескольких CDR последовательностям: SEQ ID NO: 18, 21, 22, 31, 32 и 35, SEQ ID NO: 18, 21, 26, 31, 32 и 35, SEQ ID NO: 18, 21, 27, 31, 32 и 35, SEQ ID NO: 20, 21, 22, 31, 32 и 35, SEQ ID NO: 19, 21, 22, 31, 32 и 35, SEQ ID NO: 18, 21, 25, 31, 32 и 35, SEQ ID NO: 18, 21, 28, 31, 32 и 35, SEQ ID NO: 18, 21, 29, 31, 32 и 35, SEQ ID NO: 18, 21, 30, 31, 32 и 35, SEQ ID NO: 18, 21, 23, 31, 32 и 35, SEQ ID NO: 19, 21, 23, 31, 32 и 35, SEQ ID NO: 20, 21, 23, 31, 32 и 35, SEQ ID NO: 18, 21, 24, 31, 32 и 35, или SEQ ID NO: 18, 21, 25, 31, 32 и 35, соответственно.
Вариабельные домены тяжелой и легкой цепей анти-HER3-антитела или его антигенсвязывающего фрагмента согласно изобретению включают последовательности, перечисленные в таблице 2.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL антитела и VH антитела, при этом VL содержит аминокислотную последовательность, по меньшей мере, примерно на 80%, примерно на 85%, примерно на 90%, примерно на 95%, примерно на 96%, примерно на 97%, примерно на 98%, примерно на 99% или примерно на 100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16 и SEQ ID NO: 17.
В других аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL антитело и VH антитела, при этом VH содержит аминокислотную последовательность, по меньшей мере, примерно на 80%, примерно на 85%, примерно на 90%, примерно на 95%, примерно на 96%, примерно на 97%, примерно на 98%, примерно на 99% или примерно на 100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 2, SEQ ID NO: 12 и SEQ ID NO: 13.
В других аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит VL, содержащую последовательность, по меньшей мере, примерно на 80%, примерно на 85%, примерно на 90%, примерно на 95%, примерно на 96%, примерно на 97%, примерно на 98%, примерно на 99% или примерно на 100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16 и SEQ ID NO: 17, и дополнительно содержит VH, содержащую последовательность, по меньшей мере, примерно на 80%, примерно на 85%, примерно на 90%, примерно на 95%, примерно на 96%, примерно на 97%, примерно на 98%, примерно на 99% или примерно на 100% идентичную эталонной аминокислотной последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO: 2, SEQ ID NO: 12 и SEQ ID NO: 13.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент содержит VH, указанную в таблице 2, и VL, указанную в таблице 2. Антитела на протяжении описания обозначены в соответствии с их VL-цепями. Тяжелые цепи конкретных антител, раскрытых в настоящем описании, соответствуют исходной тяжелой цепи CL16 (SEQ ID NO: 2). Таким образом, «антитело CL16» представляет собой IgG1, содержащий две исходных легких цепи CL16 (SEQ ID NO: 17) и две исходных тяжелых цепи CL16 (SEQ ID NO: 2), тогда как «антитело 2C2» представляет собой IgG1, содержащий две легких цепи 2C2 (VL 2C2 (SEQ ID NO: 3) и две исходных тяжелых цепи CL16 (SEQ ID NO: 2).
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент содержит константную область тяжелой цепи или ее фрагмент. В некоторых конкретных аспектах константная область тяжелой цепи представляет собой константную область IgG. Константная область IgG может содержать константную область легкой цепи, выбранную из группы, состоящей из константной области каппа и константной области лямбда.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению связывает HER3 по существу с такой же или лучшей аффинностью, что и антитело CL16, содержащее исходную тяжелую цепи CL16 (SEQ ID NO: 2) и исходную легкую цепь CL16 (SEQ ID NO: 17). В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению связывает HER3 по существу с такой же или лучшей аффинностью, что и антитело 2C2, содержащее легкую цепь 2C2 (VL 2C2 (SEQ ID NO: 3) и исходную тяжелую цепь CL16 (SEQ ID NO: 2).
В одном аспекте настоящего изобретения анти-HER3-антитело или его антигенсвязывающий фрагмент специфично связывает HER3 и его антигенные фрагменты с константой диссоциации kd (koff/kon) менее чем 10-6 М или менее чем 10-7 М, или менее чем 10-8 М, или менее чем 10-9 М, или менее чем 10-10 М, или менее чем 10-11 М, или менее чем 10-12 М, или менее чем 10-13 М. В конкретном аспекте настоящего изобретения анти-HER3-антитело или его антигенсвязывающий фрагмент специфично связывает HER3 и его антигенные фрагменты с константой диссоциации от 2×10-10 М до 6×10-10 М.
В другом аспекте анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению связывается с HER3 и/или его антигенными фрагментами с Koff менее чем 1×10-3 сек-1, или мене чем 2×10-3 сек-1. В других аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент связывается с HER3 и его антигенными фрагментами с Koff менее чем 10-3 сек-1, менее чем 5×10-3 сек-1, менее чем 10-4 сек-1, менее чем 5×10-4 сек-1, менее чем 10-5 сек-1, менее чем 5×10-5 сек-1, менее чем 10-6 сек-1, менее чем 5×10-6 сек-1, менее чем 5×10-7 сек-1, менее чем 10-8 сек-1, менее чем 5×10-8 сек-1, менее чем 10-9 сек-1, менее чем 5×10-9 сек-1 или менее чем 10-10 сек-1. В конкретном аспекте анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению связывается с HER3 и/или его антигенными фрагментами с Koff от 0,5×10-4 сек-1 до 2,0×10-4 сек-1.
В другом аспекте анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению связывается с HER3 и/или его антигенными фрагментами с константой скорости ассоциации или константой скорости kon, составляющей, по меньшей мере, 105 M-1 сек-1, по меньшей мере, 5×105 М-1 сек-1, по меньшей мере, 106 М-1 сек-1, по меньшей мере, 5×106 М-1 сек-1, по меньшей мере, 107 М-1 сек-1, по меньшей мере, 5×107 М-1 сек-1 или, по меньшей мере, 108 М-1 сек-1 или, по меньшей мере, 109 М-1 сек-1. В другом аспекте анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению связывается с HER3 и/или его антигенными фрагментами с константой скорости ассоциации или константой скорости kon от 1×105 М-1 сек-1 до 6×105 М-1 сек-1.
Последовательности VH и VL, описанные в таблице 1, можно «смешивать и сочетать» для создания других связывающих анти-HER3-молекул согласно изобретению. В некоторых аспектах последовательности VH 15D12.I и 15D12.V смешивают и сочетают. Дополнительно или альтернативно последовательности VL 5H6, 8A3, 4H6, 6E.3, 2B 11, 2D1, 3A6, 4C4, 1A4, 2C2, 3E.1 можно смешивать и сочетать.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит мутации, которые улучшают связывание с FcRn человека и улучшают время полужизни анти-HER3-антитела или его антигенсвязывающего фрагмента. В некоторых аспектах такими мутациями являются мутация замены метионина (M) на тирозин (Y) в положении 252, мутация замены серина (S) на треонин (T) в положении 254 и мутация замены треонина (T) на глутаминовую кислоту (E) в положении 256, пронумерованных в соответствии с индексом EU, как описано Kabat (Kabat, et al. (1991) Sequences of Proteins of Immunological Interest, U.S. Public Health Service, National Institutes of Health, Washington, D.C.), введенные в константный домен IgG1. См. патент США №7658921, который включен в настоящее описание в виде ссылки. Было показано, что такой тип мутанта IgG, называемый «мутантом YTE» имеет примерно в четыре раза увеличенное время полужизни по сравнению с вариантами такого же антитела дикого типа (Dall'Acqua et al., J. Biol. Chem. 281: 23514-24 (2006)). В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент, содержащий константный домен IgG, содержит одну или несколько аминокислотных замен аминокислотных остатков в положениях 251-257, 285-290, 308-314, 385-389, и 428-436, пронумерованных согласно индексу EU, как описано Kabat, при этом такие мутации увеличивают время полужизни в сыворотке анти-HER3-антитела или его антигенсвязывающего фрагмента.
В некоторых аспектах мутант YTE дополнительно содержит замену в положении 434 константного домена IgG, пронумерованного согласно индексу EU, как описано Kabat, аминокислотой, выбранной из группы, состоящей из триптофана (W), метионина (M), тирозина (Y) и серина (S). В других аспектах мутант YTE дополнительно содержит замену в положении 434 константного домена IgG, пронумерованного согласно индексу EU, как описано Kabat, аминокислотой, выбранной из группы, состоящей из триптофана (W), метионина (M), тирозина (Y) и серина (S), и замену в положении 428 константного домена IgG, пронумерованного согласно индексу EU, как описано Kabat, аминокислотой, выбранной из группы, состоящей из треонина (T), лейцина (L), фенилаланина (F) и серина (S).
В еще одном аспекте мутант YTE дополнительно содержит замену в положении 434 константного домена IgG, пронумерованного согласно индексу EU, как описано Kabat, тирозином (Y), и замену в положении 257 константного домена IgG, пронумерованного согласно индексу EU, как описано Kabat, лейцином (L). В некоторых аспектах мутант YTE дополнительно содержит замену в положении 434 константного домена IgG, пронумерованного согласно индексу EU, как описано Kabat, серином (S) и замену в положении 428 константного домена IgG, пронумерованного согласно индексу EU, как описано Kabat, лейцином (L).
В конкретном аспекте анти-HER3-антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи 2C2 (VL 2C2; SEQ ID NO: 3), вариабельную область тяжелой цепи исходного CL16 (SEQ ID NO: 2) и константный домен IgG1, содержащий мутация метионина (M) на тирозин (Y) в положении 252, мутацию серина (S) на треонин (T) в положении 254 и мутацию треонина (T) на глутаминовую кислоту (E) в положении 256 константного домена IgG1, пронумерованного согласно индексу EU, как описано Kabat.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению содержит, по меньшей мере, одну аминокислотную замену в константном домене IgG, выбранную из группы, состоящей из:
(a) замены аминокислоты в положении 252 тирозином (Y), фенилаланином (F), триптофаном (W) или треонином (T),
(b) замены аминокислоты в положении 254 треонином (T),
(c) замены аминокислоты в положении 256 серином (S), аргинином (R), глутамином (Q), глутаминовой кислотой (E), аспарагиновой кислотой (D) или треонином (T),
(d) замены аминокислоты в положении 257 лейцином (L),
(e) замены аминокислоты в положении 309 пролином (P),
(f) замены аминокислоты в положении 311 серином (S),
(g) замены аминокислоты в положении 428 треонином (T), лейцином (L), фенилаланином (F) или серином (S),
(h) замены аминокислоты в положении 433 аргинином (R), серином (S), изолейцином (I), пролином (P) или глутамином (Q),
(i) замены аминокислоты в положении 434 триптофаном (W), метионином (M), серином (S), гистидином (H), фенилаланином (F) или тирозином, и
(j) сочетания двух или более указанных замен, при этом положения пронумерованы согласно индексу EU, как описано Kabat, и при этом модифицированный IgG имеет увеличенное время полужизни в сыворотке по сравнению со временем полужизни в сыворотке IgG, имеющего константный домен IgG дикого типа.
В других аспектах аминокислотные последовательности VH и/или VL могут быть на 85%, 90%, 95%, 96%, 97%, 98% или 99% сходны с последовательностями, указанными выше, и содержать 1, 2, 3, 4, 5 или больше консервативных замен. HER3-антитело, содержащее области VH и VL, имеющие высокое (т.е., 80% или больше) сходство с областями VH, имеющими последовательности SEQ ID NO: 2, 12 или 13, и/или областями VL, имеющими последовательности SEQ ID NO: 1, 3, 4, 5, 6, 7, 8, 9, 10, 11, 14, 15, 16 или 17, соответственно, могут быть получены мутагенезом (например, сайт-специфичным или ПЦР-опосредованным мутагенезом) молекул нуклеиновых кислот, кодирующих последовательности SEQ ID NO: 1-17, с последующим тестированием кодируемого измененного антитела в отношении сохраненной функции с использованием функциональных анализов, описанных в настоящей публикации.
Аффинность или авидность антитела по отношению к антигену может быть определена экспериментально с использованием любого подходящего способа, хорошо известного в данной области, например, проточной цитометрии, твердофазного иммуноферментного анализа (ELISA) или радиоиммуноанализа (РИА) анализа кинетики (например, анализа BIACORE™). Легко можно применять прямые анализы связывания, а также формы анализов конкурентного связывания (см., например, Berzofsky et al., «Antibody - Antigen Interactions», в Fundamental Immunology, Paul, W.E., Ed., Raven Press: New York, N.Y. (1984); Kuby, Immunology, W.H. Freeman and Company: New York, N.Y. (1992); и способы, описанные в настоящей публикации. Измеряемая аффинность конкретного взаимодействия антитело-антиген может варьировать в случае измерения в разных условиях (например, концентрация соли, pH, температура). Поэтому измерения аффинности и других параметров связывания антигена (например, KD или Kd, Kon, Koff) осуществляют с использованием стандартизованных растворов антитела и антигена и стандартизованного буфера, который известен в данной области и такой, как буфер, описанный в настоящей публикации.
В данной области также известно, что аффинности, измеряемые с использованием анализа BIACORE™, могут варьировать в зависимости от того, какой из реагентов связан с чипом. В связи с этим аффинность может быть измерена с использованием такого формата анализа, при котором направленное к мишени антитело (например, моноклональное антитело 2C2) иммобилизуют на чипе (называемого форматом «IgG снизу»), или с использованием формата, в случае которого белок-мишень (например, HER3) иммобилизуют на чипе (называемого, например, форматом «HER3 снизу»).
III. Связывающие молекулы, которые связываются с тем же самым эпитопом, что и анти-HER3-антитела и их антигенсвязывающие фрагменты согласно изобретению
В другом аспекте изобретение содержит HER3-связывающие молекулы, которые связываются с тем же эпитопом, что и различные анти-HER3-антитела, описанные в настоящей публикации. Термин «эпитоп» в используемом в настоящем описании смысле относится к белковой детерминанте, способной связываться с антителом согласно изобретению. Эпитопы обычно состоят из химически активных поверхностных групп молекул, таких как аминокислоты или боковые цепи сахаров, и обычно имеют специфичные характеристики трехмерной структуры, а также специфичные характеристики заряда. Конформационные и неконформационные эпитопы отличаются тем, что связывание с первыми, но не с последними, прекращается в присутствии денатурирующих растворителей. Такие антитела могут быть идентифицированы на основе их способности перекрестно конкурировать (например, конкурентно ингибировать связывание статистически значимым образом) с такими антителами, как антитело CL16, антитело 2C2 или 2C2-мутант YTE, в стандартных анализах связывания HER3. Соответственно, в одном аспекте изобретение относится к анти-HER3-антителам и их антигенсвязывающим фрагментам, например, моноклональным антителам человека, которые конкурируют за связывание с HER3 с другим анти-HER3-антителом или его антигенсвязывающим фрагментом согласно изобретению, таким как антитело CL16 или антитело 2C2. Способность тестируемого антитела ингибировать связывание, например, антитела CL16 или антитела 2C2 свидетельствует о том, что тестируемое антитело может конкурировать с данным антителом за связывание с HER3; согласно не ограничивающей теории такое антитело может связываться с тем же самым или родственным (например, структурно сходным или пространственно близким) эпитопом на HER3, что и анти-HER3-антитело или его антигенсвязывающий фрагмент, с которым оно конкурирует. В одном аспекте анти-HER3-антитело или его антигенсвязывающий фрагмент, который связывается с тем же самым эпитопом на HER3, что и, например, антитело CL16 или антитело 2C2, представляет собой моноклональное антитело человека.
IV. Механизм действия
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3. В других аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование AKT. В следующих аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять образование димеров HER2-HER3. В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять клеточный рост. В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, не обладает ADCC-эффектом. В конкретных аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3, фосфорилирование AKT и/или образование колоний опухолевых клеток в результате не зависимого от лиганда механизма действия.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в HRG-зависимых клетках злокачественной опухоли молочной железы MCF-7, как было измерено в ELISA, с IC50 меньше чем примерно 30 нг/мл, меньше чем примерно 25 нг/мл, меньше чем примерно 20 нг/мл, меньше чем примерно 15 нг/мл или меньше чем примерно 10 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в HRG-зависимых клетках злокачественной опухоли молочной железы MCF-7, как было измерено в ELISA, с IC50 меньше чем примерно 20 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в HRG-зависимых клетках злокачественной опухоли молочной железы MCF-7, как было измерено в ELISA, с IC50 меньше чем примерно 15 нг/мл. В другом конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент может подавлять фосфорилирование HER3 в HRG-зависимых клетках злокачественной опухоли молочной железы MCF-7, как было измерено в ELISA, с IC50 меньше чем примерно 10 нг/мл.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять клеточный рост в клетках злокачественной опухоли молочной железы MDA-MB-175 с IC50 меньше чем примерно 0,90 мкг/мл, меньше чем примерно 0,80 мкг/мл, меньше чем примерно 0,70 мкг/мл, меньше чем примерно 0,60 мкг/мл, меньше чем примерно 0,50 мкг/мл, меньше чем примерно 0,40 мкг/мл, меньше чем примерно 0,30 мкг/мл или меньше чем примерно 0,20 мкг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять клеточный рост в клетках злокачественной опухоли молочной железы MDA-MB-175 с IC50 меньше чем примерно 0,50 мкг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять клеточный рост в клетках злокачественной опухоли молочной железы MDA-MB-175 с IC50 меньше чем примерно 0,40 мкг/мл. В другом конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент может подавлять клеточный рост в клетках злокачественной опухоли молочной железы MDA-MB-175 с IC50 меньше чем примерно 0,30 мкг/мл. В другом конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять клеточный рост в клетках злокачественной опухоли молочной железы MDA-MB-175 с IC50 меньше чем примерно 0,20 мкг/мл.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять клеточный рост в клетках миеломы HMCB с IC50 меньше чем примерно 0,20 мкг/мл, меньше чем примерно 0,15 мкг/мл, меньше чем примерно 0,10 мкг/мл, меньше чем примерно 0,05 мкг/мл, меньше чем примерно 0,04 мкг/мл, или меньше чем примерно 0,03 мкг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять клеточный рост в клетках миеломы HMCB с IC50 меньше чем примерно 0,10 мкг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять клеточный рост в клетках миеломы HMCB с IC50 меньше чем примерно 0,05 мкг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять клеточный рост в клетках миеломы HMCB с IC50 меньше чем примерно 0,04 мкг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять клеточный рост в клетках миеломы HMCB с IC50 меньше чем примерно 0,03 мкг/мл.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в EGFR-зависимых клетках злокачественной опухоли легкого HCC827 с IC50 меньше чем примерно 20 нг/мл, меньше чем примерно 15 нг/мл, меньше чем примерно 10 нг/мл, меньше чем примерно 8 нг/мл, меньше чем примерно 6 нг/мл, меньше чем примерно 4 нг/мл, или меньше чем примерно 2 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в EGFR-зависимых клетках злокачественной опухоли легкого HCC827 c IC50 меньше чем примерно 10 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент может подавлять фосфорилирование HER3 в EGFR-зависимых клетках злокачественной опухоли легкого HCC827 c IC50 меньше чем примерно 8 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в EGFR-зависимых клетках злокачественной опухоли легкого HCC827 с IC50 меньше чем примерно 6 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент может подавлять фосфорилирование HER3 в EGFR-зависимых клетках злокачественной опухоли легкого HCC827 с IC50 меньше чем примерно 4 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в EGFR-зависимых клетках злокачественной опухоли легкого HCC827 с IC50 меньше чем примерно 2 нг/мл.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в EGFR-зависимых клетках злокачественной опухоли легкого HCC827, резистентных к TKI, с IC50 меньше чем примерно 30 нг/мл, меньше чем примерно 25 нг/мл, меньше чем примерно 20 нг/мл, меньше чем примерно 15 нг/мл, меньше чем примерно 10 нг/мл или меньше чем примерно 5 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в EGFR-зависимых клетках злокачественной опухоли легкого HCC827, резистентных к TKI, с IC50 меньше чем примерно 20 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти- HER3 антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в EGFR-зависимых клетках злокачественной опухоли легкого HCC827, резистентных к TKI, с IC50 меньше чем примерно 15 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в EGFR-зависимых клетках злокачественной опухоли легкого HCC827, резистентных к TKI, с IC50 меньше чем примерно 10 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент может подавлять фосфорилирование HER3 в EGFR-зависимых клетках злокачественной опухоли легкого HCC827, резистентных к TKI, с IC50 меньше чем примерно 5 нг/мл.
В некоторых конкретных аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент можно применять для лечения TKI-резистентных злокачественных опухолей.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент может подавлять фосфорилирование HER3 в cMET-зависимых клетках аденокарциномы желудка человека MKN45 с IC50 меньше чем примерно 15 нг/мл, меньше чем примерно 10 нг/мл, меньше чем примерно 9 нг/мл, меньше чем примерно 8 нг/мл, меньше чем примерно 7 нг/мл, меньше чем примерно 6 нг/мл, меньше чем примерно 5 нг/мл или меньше чем примерно 4 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент может подавлять фосфорилирование HER3 в cMET-зависимых клетках аденокарциномы желудка человека MKN45 с IC50 меньше чем примерно 10 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в cMET-зависимых клетках аденокарциномы желудка человека MKN45 с IC50 меньше чем примерно 8 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять фосфорилирование HER3 в cMET-зависимых клетках аденокарциномы желудка человека MKN45 с IC50 меньше чем примерно 6 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент может подавлять фосфорилирование HER3 в cMET-зависимых клетках аденокарциномы желудка человека MKN45 с IC50 меньше чем примерно 4 нг/мл.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению, может подавлять pAKT в cMET-зависимых клетках MKN45 с IC50 меньше чем примерно 15 нг/мл, меньше чем примерно 10 нг/мл, меньше чем примерно 9 нг/мл, меньше чем примерно 8 нг/мл, меньше чем примерно 7 нг/мл, меньше чем примерно 6 нг/мл, меньше чем примерно 5 нг/мл, меньше чем примерно 4 нг/мл или меньше чем примерно 3 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pAKT в cMET-зависимых клетках MKN45 с IC50 меньше чем примерно 8 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pAKT в cMET-зависимых клетках MKN45 с IC50 меньше чем примерно 6 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pAKT в cMET-зависимых клетках MKN45 с IC50 меньше чем примерно 4 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pAKT в cMET-зависимых клетках MKN45 с IC50 меньше чем примерно 3 нг/мл.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению, может подавлять pHER в FGFR2-зависимых клетках перстневидно-клеточной карциномы желудка человека Kato III с IC50 меньше чем примерно 9 нг/мл, меньше чем примерно 8 нг/мл, меньше чем примерно 7 нг/мл, меньше чем примерно 6 нг/мл, меньше чем примерно 5 нг/мл, меньше чем примерно 4 нг/мл, меньше чем примерно 3 нг/мл, меньше чем примерно 2 нг/мл или меньше чем примерно 1 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pHER в FGFR2-зависимых клетках перстневидно-клеточной карциномы желудка человека Kato III с IC50 меньше чем примерно 5 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент может подавлять pHER в FGFR2-зависимых клетках перстневидно-клеточной карциномы желудка человека Kato III с IC50 меньше чем примерно 4 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pHER в FGFR2-зависимых клетках перстневидно-клеточной карциномы желудка человека Kato III с IC50 меньше чем примерно 3 нг/мл. В конкретном аспекте, HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pHER в FGFR2-зависимых клетках перстневидно-клеточной карциномы желудка человека Kato III с IC50 меньше чем примерно 2 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент может подавлять pHER в FGFR2-зависимых клетках перстневидно-клеточной карциномы желудка человека Kato III с IC50 меньше чем примерно 1 нг/мл.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pAKT в FGFR-2-зависимых клетках Kato III с IC50 меньше чем примерно 6 нг/мл, меньше чем примерно 5 нг/мл, меньше чем примерно 4 нг/мл, меньше чем примерно 3 нг/мл, меньше чем примерно 2 нг/мл или меньше чем примерно 1 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pAKT в FGFR-2-зависимых клетках Kato III с IC50 меньше чем примерно 4 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pAKT в FGFR-2-зависимых клетках Kato III с IC50 меньше чем примерно 3 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3 антитело или его антигенсвязывающий фрагмент, может подавлять pAKT в FGFR-2-зависимых клетках Kato III с IC50 меньше чем примерно 2 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pAKT в FGFR-2-зависимых клетках Kato III с IC50 меньше чем примерно 1 нг/мл.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению, может подавлять pHER в не зависимых от лиганда клетках злокачественной опухоли молочной железы BT-474 с IC50 меньше чем примерно 10 нг/мл, меньше чем примерно 9 нг/мл, меньше чем примерно 8 нг/мл, меньше чем примерно 7 нг/мл, меньше чем примерно 6 нг/мл, меньше чем примерно 5 нг/мл, меньше чем примерно 4 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pHER в не зависимых от лиганда клетках злокачественной опухоли молочной железы BT-474 с IC50 меньше чем примерно 8 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pHER в не зависимых от лиганда клетках злокачественной опухоли молочной железы BT-474 с IC50 меньше чем примерно 6 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pHER в не зависимых от лиганда клетках злокачественной опухоли молочной железы BT-474 с IC50 меньше чем примерно 4 нг/мл.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению, может подавлять pAKT в не зависимых от лиганда клетках злокачественной опухоли молочной железы BT-474 с IC50 меньше чем примерно 10 нг/мл, меньше чем примерно 9 нг/мл, меньше чем примерно 8 нг/мл, меньше чем примерно 7 нг/мл, меньше чем примерно 6 нг/мл, меньше чем примерно 5 нг/мл, меньше чем примерно 4 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pAKT в не зависимых от лиганда клетках злокачественной опухоли молочной железы BT-474 с IC50 меньше чем примерно 8 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pAKT в не зависимых от лиганда клетках злокачественной опухоли молочной железы BT-474 с IC50 меньше чем примерно 6 нг/мл. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pAKT в не зависимых от лиганда клетках злокачественной опухоли молочной железы BT-474 с IC50 меньше чем примерно 4 нг/мл. В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может подавлять pHER3, pAKT и образование колоний опухолевых клеток в клетках BT-474, модели не зависимой от лиганда злокачественной опухоли молочной железы.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению, может подавлять HRG-индуцированную секрецию VEGF. В конкретном аспекте HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению, может подавлять HRG-индуцированную секрецию VEGF в не зависимых от лиганда клетках злокачественной опухоли молочной железы BT-474 и/или HRG-зависимых клетка злокачественной опухоли молочной железы MCF-7.
В некоторых аспектах HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению, может вызывать задержку клеточного цикла. В конкретном аспекте, HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению, может вызывать задержку клеточного цикла в злокачественных клетках молочной железы, включая без ограничения клетки SKBR3 или BT474.
V. Получение анти-HER3-антител и антигенсвязывающих фрагментов
Моноклональные анти-HER3-антитела могут быть получены с использованием способов на основе гибридом, таких как способы, описанные в публикации Kohler and Milstein (1975) Nature 256: 495. При использовании способа на основе гибридом, мышь, хомячка или другого соответствующего животного-хозяина иммунизируют, как описано выше, вызывая продуцирование лимфоцитами антител, которые будут специфично связываться с иммунизирующим антигеном. Лимфоциты также могут быть иммунизированы in vitro. После иммунизации лимфоциты выделяют и сливают с подходящей линией клеток миеломы, используя, например, полиэтиленгликоль, чтобы получить клетки гибридомы, которые затем могут быть отобраны из смеси с неслитыми лимфоцитами и клетками миеломы. Затем гибридомы, которые продуцируют моноклональные антитела, специфично направленные против выбранного антигена, которые определяют с использованием иммунопреципитации, иммуноблоттинга или анализа связывания in vitro (например, радиоиммуноанализа (РИА); твердофазного иммуноферментного анализа (ELISA)), могут быть размножены в культуре in vitro с применением стандартных способов (Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, 1986) или в виде асцитных опухолей in vivo у животного. Моноклональные антитела затем могут быть очищены из культуральной среды или асцитной жидкости, как описано для поликлональных антител выше.
Альтернативно моноклональные анти-HER3-антитела также могут быть получены с использованием способов рекомбинации ДНК, которые описаны в патенте США №4816567. Полинуклеотиды, кодирующие моноклональные антитело, выделяют из зрелых B-клеток или клеток гибридомы, например, посредством ОТ-ПЦР с использованием олигонуклеотидных праймеров, которые специфично амплифицируют гены, кодирующие тяжелую и легкую цепи антитела, и их последовательность определяют, используя обычные способы. Выделенные полинуклеотиды, кодирующие легкую и тяжелую цепи, затем клонируют в подходящих экспрессирующих векторах, которые при трансфекции в клетки-хозяева, такие как клетки E. coli, клетки обезьян COS, клетки яичника китайского хомячка (CHO) или клетки миеломы, которые в ином случае не продуцируют иммуноглобулиновый белок, обеспечивают выработку моноклональных антител клетками-хозяевами. Также рекомбинантные моноклональные анти-HER3-антитела или их антигенсвязывающие фрагменты требуемого вида могут быть выделены из библиотек в фаговом дисплее, экспрессирующих CDR требуемого вида, как описано (McCafferty et al., 1990, Nature, 348: 552-554; Clarkson et al., 1991, Nature, 352: 624-628; и Marks et al., 1991, J. Mol. Biol., 222: 581-597).
Полинуклеотид(ды), кодирующий анти-HER3-антитело или его антигенсвязывающие фрагменты, дополнительно могут быть модифицированы различным образом с использованием методики рекомбинантной ДНК с целью создания альтернативных антител. В некоторых аспектах константные домены легкой и тяжелой цепей, например, мышиного моноклонального антитела могут быть заменены (1) например, такими областями антитела человека, чтобы создать химерное антитело или (2) полипептидом, отличным от иммуноглобулина, чтобы создать слитое антитело. В некоторых аспектах константные области укорачивают или удаляют, создавая требуемый фрагмент моноклонального антитела. Сайт-специфичный мутагенез или мутагенез с высокой плотностью вариабельной области можно использовать для оптимизации специфичности, аффинности и т.д. моноклонального антитела.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент представляет собой антитело человека или его антигенсвязывающий фрагмент. Антитела человека можно получать непосредственно, используя различны способы, известные в данной области. Могут быть созданы иммортализованные B-лимфоциты человека, иммунизированные in vitro или выделенные из организма иммунизированного индивидуума, который продуцирует антитело, направленной против антигена-мишени (см., например, Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77 (1985); Boemer et al., 1991, J. Immunol., 147 (1): 86-95; и патент США 5750373).
Также анти-HER3-антитело человека или его антигенсвязывающий фрагмент можно выбрать из фаговой библиотеки, когда такая фаговая библиотека экспрессирует антитела человека, как описано, например, в публикациях Vaughan et al., 1996, Nat. Biotech., 14: 309-314, Sheets et al., 1998, Proc. Nat'l. Acad. Sci., 95: 6157-6162, Hoogenboom and Winter, 1991, J. Mol. Biol., 227: 381 и Marks et al., 1991, J. Mol. Biol., 222: 581). Способы создания и применения фаговых библиотек антител также описаны в патентах США №5969108, 6172197, 5885793, 6521404, 6544731, 6555313, 6582915, 6593081, (6300064, 6653068, 6706484 и 7264963, и в публикации Rothe et al., 2007, J. Mol. Biol., doi: 10.1016/j.jmb.2007.12.018 каждая из публикаций включена в виде ссылки в полном объеме).
Методики созревания аффинности и методики перетасовки цепей (публикация Marks et al., 1992, Bio/Technology 10: 779-783, включенная в виде ссылки в полном объеме) известны в данной области и могут быть применены для создания высоко аффинных антител человека или их антигенсвязывающих фрагментов.
В некоторых аспектах моноклональное анти-HER3-антитело может представлять собой гуманизированное антитело. Способы конструирования, гуманизации или изменения поверхности антител животных, отличных от человека, или антител человека также могут быть использованы и хорошо известны в данной области. Гуманизированное, подвергнутое изменению поверхности или подобным образом сконструированное антитело может иметь один или несколько аминокислотных остатков из источника, который отличен от человека, например, но без ограничения, мыши, крысы, кролика, примата, отличного от человека, или другого млекопитающего. Такие аминокислотные остатки, полученные от животного, отличного от человека, заменяют остатками, которые часто называют «импортируемыми» остатками, которые обычно берут из «импортируемого» вариабельного, константного или другого домена известной последовательности человека. Такие импортируемые последовательности можно использовать для уменьшения иммуногенности или уменьшения, усиления или модификации связывания, аффинности, скорости образования комплекса, скорости распада комплекса, авидности, специфичности, времени полужизни или любой другой подходящей характеристики, известной в данной области. В общем, остатки CDR непосредственно и наиболее значимо вовлечены в осуществление влияния на связывание HER3. Соответственно, часть или все последовательности CDR животного, отличного от человека, или человека сохраняют, в то время как последовательности вариабельных и константных областей животного, отличного от человека, могут быть заменены аминокислотами человека или другими аминокислотами.
Антитела также необязательно могут представлять собой гуманизированные, подвергнутые изменению поверхности, сконструированные или человеческие антитела, сконструированные с сохранением высокой аффинности по отношению к антигену HER3 и других подходящих биологических свойств. Для достижения указанной цели гуманизированные (или человеческие) или сконструированные анти-HER3-антитела и подвергнутые изменению поверхности антитела необязательно могут быть получены способом, основанным на анализе исходных последовательностей и различных концептуальных гуманизированных сконструированных продуктов, с использованием трехмерных моделей исходных, сконструированных и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов общедоступны и известны специалистам в данной области. Доступны компьютерные программы, которые иллюстрируют и представляют трехмерные конформационные структуры выбранных для исследования последовательностей иммуноглобулинов. Просматривание таких представлений позволяет проводить анализ вероятной роли остатков в функционировании выбранной для исследования последовательности иммуноглобулина, т.е. анализ остатков, которые влияют на способность выбранного для исследования иммуноглобулина связываться с антигеном, таким как HER3. Таким образом, могут быть выбраны и объединены каркасные (FW) остатки из консенсусных и импортируемых последовательностей так, чтобы добиться требуемой характеристики антитела, такой как повышенная аффинность по отношению к антигену(ам)-мишени.
Гуманизацию, изменение поверхности или конструирование анти-HER3-антител или их антигенсвязывающих фрагментов согласно настоящему изобретению можно осуществить, используя любой известный способ, такой как без ограничения способы, описанные в публикациях Jones et al., Nature 321:522 (1986); Riechmann et al., Nature 332:323 (1988); Verhoeyen et al., Science 239:1534 (1988)), Sims et al., J. Immunol. 151: 2296 (1993); Chothia and Lesk, J. Mol. Biol. 196:901 (1987), Carter et al., Proc. Natl. Acad. Sci. U.S.A. 89:4285 (1992); Presta et al., J. Immunol. 151:2623 (1993), патентах США №5639641, 5723323, 5976862, 5824514, 5817483, 5814476, 5763192, 5723323, 5766886, 5714352, 6204023, 6180370, 5693762, 5530101, 5585089, 5225539, 4816567, 7557189, 7538195 и 7342110; международных заявках № PCT/US 98/16280, PCT/US 96/18978, PCT/US 91/09630, PCT/US 91/05939, PCT/US 94/01234, PCT/GB 89/01334, PCT/GB 91/01134, PCT/GB 92/01755, международных публикациях заявок на выдачу патента № WO 90/14443, WO 90/14424, WO 90/14430; и публикации европейского патента № EP 229246; каждая из которых включена в настоящее описание в виде ссылки в полном объеме, включая цитированные в таких публикациях ссылки.
Гуманизированные анти-HER3-антитела и их антигенсвязывающие фрагменты также могут быть получены у трансгенных мышей, содержащих локусы иммуноглобулинов человека, которые способны при иммунизации продуцировать полный репертуар антител человека в отсутствие продуцирования эндогенных иммуноглобулинов. Такой способ описан в патентах США №5545807, 5545806, 5569825, 5625126, 5633425 и 5661016.
В некоторых аспектах предлагается фрагмент анти-HER3-антитела. Известны различные способы для получения фрагментов антител. Традиционно такие фрагменты получают в результате протеолитического расщепления интактных антител (например, Morimoto et al., 1993, Journal of Biochemical and Biophysical Methods 24: 107-1 17; Brennan et al., 1985, Science, 229: 81). В некоторых аспектах фрагменты анти-HER3-антитела получают рекомбинантно. Фрагменты антитела Fab, Fv и scFv могут быть экспрессированы и секретированы из E. coli или других клеток-хозяев, с обеспечением таким образом возможности получения больших количеств таких фрагментов. Такие фрагменты анти-HER3-антител также могут быть выделены из фаговых библиотек антител, обсуждаемых выше. Фрагменты анти-HER3-антител также могут представлять собой линейные антитела, которые описаны в патенте США №5641870. Другие способы получения фрагментов антител будут очевидны для специалиста.
Согласно настоящему изобретению способы могут быть адаптированы для получения одноцепочечных антител, специфичных к HER3 (см., например, патент США №4946778). Кроме того, способы могут быть адаптированы для конструирования библиотек, экспрессирующих Fab (см., например, Huse et al., Science 246: 1275-1281 (1989)), чтобы обеспечить быструю и эффективную идентификацию моноклональных Fab-фрагментов с требуемой специфичностью по отношению к HER3 или его производным, фрагментам, аналогам или гомологам. Фрагменты антител могут быть получены способами, известными в данной области, включая без ограничения: (a) F(ab’)2-фрагмент, получаемый расщеплением пепсином молекулы антитела; (b) Fab-фрагмент, образованный восстановлением дисульфидных мостиков F(ab’)2-фрагмента, (c) Fab-фрагмент, образованный в результате обработки молекулы антитела пепсином и восстановителем, и (d) Fv-фрагменты.
Кроме того, может быть желательным, особенно в случае фрагментов антител, модифицировать анти-HER3-антитело или его антигенсвязывающий фрагмент для увеличения его времени полужизни в сыворотке. Это может быть достигнуто, например, включением эпитопа, связывающего рецептор спасения, в антитело или фрагмент антитела в результате мутации соответствующей области в антителе или фрагменте антитела или в результате включения эпитопа в пептидную метку, которую затем сливают с антителом или фрагментом антитела на любом конце или в середине (например, посредством синтеза ДНК или пептида), или в результате мутации YTE. В данной области известны другие способы увеличения времени полужизни антитела или его антигенсвязывающего фрагмента в сыворотке, например, конъюгация с гетерологичной молекулой, такой как ПЭГ.
Гетероконъюгаты анти-HER3-антител и их антигенсвязывающих фрагментов также входят в объем настоящего изобретения. Гетероконъюгаты антител состоят из двух ковалентно связанных антител. Такие антитела были предложены, например, для направления иммунных клеток целенаправленно к нежелательным клеткам (см., например, патент США №4676980). Предполагается, что гетероконъюгированные анти-HER3 антитела и их антигенсвязывающие фрагменты могут быть получены in vitro с использованием способов, известных в области химии синтеза белков, включая способы с использованием поперечно сшивающих агентов. Например, могут быть сконструированы иммунотоксины с использованием реакции дисульфидного обмена или за счет образования тиоэфирной связи. Примеры подходящих для такой цели реагентов включают иминотиолат и метил-4-меркаптобутиримидат.
В некоторых аспектах HER3-связывающие молекулы согласно изобретению, например, антитела или их антигенсвязывающие фрагменты, могут быть объединены с другими терапевтическими средствами или конъюгированы с другими терапевтическими средствами или токсинами с образованием иммуноконъюгатов и/или слитых белков. Примеры таких терапевтических средств и токсинов включают без ограничения цетуксимаб (эрбитукс®), панитумумаб (вектибикс®), лапатиниб (тикерб®/тиверб®) и паклитаксел (таксол®, абраксан®) и производные (например, доцетаксел).
В некоторых аспектах HER3-связывающие молекулы согласно изобретению, например, антитела или их антигенсвязывающие фрагменты, могут быть конъюгированы с антителами или фрагментами антител, мишенью которых является эпидермальный фактор роста (EGFR). В других аспектах HER3-связывающие молекулы согласно изобретению могут быть конъюгированы с ингибиторами тирозинкиназ. В некоторых конкретных аспектах HER3-связывающие молекулы согласно изобретению могут быть конъюгированы с ингибиторами тирозинкиназной активности, ассоциированными с EGFR и/или HER2/neu. В некоторых аспектах HER3-связывающме молекулы согласно изобретению могут быть конъюгированы с антимитотическими средствами. В некоторых конкретных аспектах HER3-связывающие молекулы согласно изобретению могут быть конъюгированы со средствами, которые стабилизируют сборку микротрубочек веретена митотического деления.
В целях настоящего изобретения следует понимать, что модифицированные анти-HER3-антитела или их антигенсвязывающие фрагменты могут содержать любой тип вариабельной области, которая относится к ассоциации антитела или полипептида с HER3. В этой связи вариабельная область может включать вариабельную область или может быть получена от любого типа млекопитающего, которого можно индуцировать к выработке гуморального ответа и образованию иммуноглобулинов против требуемого ассоциированного с опухолью антигена. Источником вариабельной области модифицированных анти-HER3-антител или их антигенсвязывающих фрагментов, как таковой, может быть человек, мышь, примат, отличный от человека (например, макак-крабоед, макаки и т.д.) или люпина. В некоторых аспектах и вариабельные и константные области модифицированных анти-HER3-антитела или их антигенсвязывающих фрагментов являются человеческими. В других аспектах вариабельные области совместимые антитела (обычно полученные от животного, отличного от человека) могут быть сконструированы или специфично оптимизированы для улучшения связывающих свойств или снижения иммуногенности молекулы. В этой связи, вариабельные области, применимые в настоящем изобретении могут быть гуманизированы или иным образом изменены в результате включения импортируемых аминокислотных последовательностей.
В некоторых аспектах вариабельные домены как тяжелой, так и легкой цепей анти-HER3-антитела или его антигенсвязывающего фрагмента изменяют, по меньшей мере, посредством частичной замены одной или нескольких CDR и, при необходимости, посредством частичной замены и изменения последовательности каркасной области. Хотя CDR могут быть получены из антитела такого же класса или даже подкласса, что и антитело, из которого получают каркасные области, предполагается, что CDR могут быть получены из антитела другого класса и в некоторых аспектах из антитела от другого вида. Не обязательно заменять все CDR полными CDR из донорной вариабельной области, чтобы перенести способность связывать антиген одного вариабельного домена к другому. Иногда необходимо перенести только такие остатки, которые нужны для поддержания активности антигенсвязывающего участка. Учитывая объяснения, приведенные в патентах США №5585089, 5693761 и 5693762, специалисты в данной области смогут, либо проводя рутинные эксперименты, либо осуществляя проверку методом проб и ошибок, получить функциональное антитело с уменьшенной иммуногенностью.
Специалистам в данной области будет понятно, что, несмотря на изменения в вариабельной области, модифицированные анти-HER3-антитела или их антигенсвязывающие фрагменты согласно изобретению будут включать антитела (например, полноразмерные антитела или их иммунореактивные фрагменты), в которых, по меньшей мере, часть одного или нескольких доменов константной области было делетировано или иным образом изменено, так чтобы получить требуемые биохимические характеристики, такие как повышенная локализация в опухоли или пониженное время полужизни в сыворотке по сравнению с антителом примерно с такой же иммуногенностью, содержащим нативную или неизмененную константную область. В некоторых аспектах константная область модифицированных антител будет содержать константную область человека. Модификации в константной области, предназначенные для настоящего изобретения, включают добавления, делеции или замены одной или нескольких аминокислот в одном или нескольких доменах. То есть, модифицированные антитела, раскрытые в настоящем описании, могут содержать изменения или модификации одного или нескольких из трех константных доменов тяжелой цепи (CH1, CH2 или CH3) и/или константного домена легкой цепи (CL). В некоторых аспектах предусмотрены модифицированные константные области, в которых один или несколько доменов частично или полностью делетированы. В некоторых аспектах модифицированные антитела будут содержать конструкции или варианты с делетированными доменами, в которых был удален полный домен CH2 (конструкции ΔCH2). В некоторых аспектах исключенный домен константной области будет заменен коротким аминокислотным спейсером (например, 10 остатков), который обеспечивает некоторую гибкость молекулы, обычно нарушаемую придаваемую отсутствующей константной областью.
Кроме конфигурации в данной области известно, что константная область опосредует несколько эффекторных функций. Например, связывание компонента C1 комплемента с антителами активирует систему комплемента. Активация комплемента важна для опсонизации и лизиса клеточных патогенов. Активация комплемента также стимулирует воспалительный ответ и также может быть вовлечена в аутоиммунную гиперчувствительность. Кроме того, антитела связываются с клетками через Fc-область, при этом участок Fc-рецептора в Fc-области антитела связывается с Fc-рецептором (FcR) на клетке. Существует несколько Fc-рецепторов, которые являются специфичными для разных классов антител, включая IgG (гамма-рецепторы), IgE (эта-рецепторы), IgA (альфа-рецепторы) и IgM (мю-рецепторы). Связывание антитела с Fc-рецепторами на поверхности клеток запускает ряд важных и разнообразных биологических ответов, включая поглощение и разрушение покрытых антителами частиц, клиренс иммунных комплексов, лизис покрытых антителами клеток-мишеней клетками-киллерами (так называемая зависимая от антител опосредованная клетками цитотоксичность или ADCC), высвобождение медиаторов воспаления, плацентарный перенос и регуляцию продукции иммуноглобулинов.
В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент обеспечивает измененные эффекторные функции, которые, в свою очередь, влияют на биологический профиль вводимого антитела или его антигенсвязывающего фрагмента. Например, делеция или инактивация (в результате точечных мутаций или другими способами) домена константной области может уменьшить связывание Fc-рецептора циркулирующим модифицированным антителом, тем самым увеличивая локализацию в опухоли. В других случаях вероятно, что модификации константной области согласно настоящему изобретению уменьшают связывание комплемента и таким образом уменьшают время полужизни в сыворотке и неспецифичную ассоциацию конъюгированного цитотоксина. Другие модификации константной области могут быть использованы для устранения дисульфидных связей или олигосахаридных остатков, что обеспечивает возможность для усиленной локализации вследствие повышенной специфичности к антигену или гибкости антитела. Подобным образом, модификации константной области согласно настоящему изобретению легко могут быть осуществлены с использованием хорошо известных биохимических способов или способов молекулярной инженерии, что находится в компетенции специалиста в данной области.
В некоторых аспектах HER3-связывающая молекула, которая представляет собой антитело или его антигенсвязывающий фрагмент, не обладает одной или несколькими эффекторными функциями. Например, в некоторых аспектах антитело или его антигенсвязывающий фрагмент не обладает активностью в зависимой от антител клеточной цитотоксичности (ADCC) и/или активностью в зависимой от комплемента цитотоксичности (CDC). В некоторых аспектах анти-HER3-антитело или его антигенсвязывающий фрагмент не связывается с Fc-рецептор и/или факторы комплемента. В некоторых аспектах антитело или его антигенсвязывающий фрагмент не обладает эффекторной функцией.
Будет понятно, что в некоторых аспектах модифицированные анти-HER3-антитела или их антигенсвязывающие фрагменты могут быть сконструированы так, чтобы они слить домен CH3 непосредственно с шарнирной областью соответствующих модифицированных антител или их фрагментов. В других конструкциях может требоваться наличие пептидного спейсера между шарнирной областью и модифицированными доменами CH2 и/или CH3. Например, совместимые конструкции могут быть экспрессированы в том случае, когда домен CH2 был делетирован и оставшийся домен CH3 (модифицированный или немодифицированный) связан с шарнирной областью с использованием спейсера, состоящего из 5-20 аминокислот. Такой спейсер может быть добавлен, например, для того, чтобы гарантировать, что регуляторные элементы константного домена останутся свободными и доступными или что шарнирная область останется гибкой. Однако следует отметить, что аминокислотные спейсеры в некоторых случаях могут оказаться иммуногенными и вызвать нежелательный иммунный ответ против конструкции. Соответственно, в некоторых аспектах любой спейсер, добавляемый к конструкции будет относительно неиммуногенным, или даже совсем исключен для того, чтобы сохранить требуемые биохимические свойства модифицированных антител.
Кроме делеции целых доменов константных областей, будет понятно, что анти-HER3-антитела и их антигенсвязывающие фрагменты согласно настоящему изобретению могут быть получены с использованием частичной делеции или замены нескольких или даже одной аминокислоты. Например, мутация одной аминокислоты в выбранных областях домена CH2 может быть достаточной для существенного снижения связывания Fc и тем самым увеличения локализации в опухоли. Подобным образом, может требоваться просто делетировать такую часть одного или нескольких доменов константной области, которые контролируют эффекторную функцию (например, связывание комплемента C1Q), подвергаемую модулированию. Такие частичные делеции константных областей могут улучшать выбранные характеристики антитела или его антигенсвязывающего фрагмента (например, время полужизни в сыворотке), оставляя при этом другие требуемые функции, ассоциированные с данным доменом константной области, интактными. Кроме того, как упоминается выше, константные области раскрываемых в настоящем изобретении анти-HER3-антител и их антигенсвязывающих фрагментов могут быть модифицированы в результате мутации или замены одной или нескольких аминокислот, которая усиливает профиль получаемой в результате конструкции. В этой связи можно нарушить активность, обеспечиваемую консервативным участком связывания (например, Fc-связывания), при этом по существу сохранить конфигурацию и иммуногенный профиль модифицированного антитела или его антигенсвязывающего фрагмента. Некоторые аспекты могут включать в себя добавление одной или нескольких аминокислот к константной области, чтобы усилить требуемые характеристики, например, снизить или повысить эффекторную функцию или обеспечить связывание большего количества цитотоксинов или углеводов. В указанных аспектах может требоваться встраивание или дублирование конкретных последовательностей, полученных из выбранных доменов константной области.
Настоящее изобретение, кроме того, охватывает варианты и эквиваленты, которые по существу гомологичны химерным, гуманизированным и человеческим анти-HER3-антителам или их антигенсвязывающим фрагментам, указанным в настоящем описании. Они могут содержать, например, мутации в виде консервативных замен, т.е. замену одной или нескольких аминокислот сходными аминокислотами. Например, консервативная замена относится к замене одной аминокислоты другой в пределах одного и того же общего класса, например, одной кислой аминокислоты другой кислой аминокислотой, одной основной аминокислоты другой основной аминокислотой или одной нейтральной аминокислоты другой нейтральной аминокислотой. Что подразумевается под консервативной аминокислотной заменой хорошо известно в данной области.
Анти-HER3-антитело или его антигенсвязывающий фрагмент может быть дополнительно модифицирован так, чтобы он содержал дополнительные химические остатки, в норме не являющиеся частью белка. Такие дериватизованные остатки могут улучшать растворимость, биологическое время полужизни или абсорбцию белка. Остатки также могут уменьшать или исключать любые требуемые побочные эффекты белков и тому подобное. Обзор, посвященный таким остаткам, можно найти в публикации Remington's Pharmaceutical Sciences, 20th ed., Mack Publishing Co., Easton, PA (2000).
VI. Полинуклеотиды, кодирующие HER3-связывающие молекулы
В некоторых аспектах изобретение охватывает полинуклеотиды, содержащие последовательности нуклеиновых кислот, которые кодируют полипептид, который специфично связывает HER3 или его антигенсвязывающий фрагмент. Например, изобретение относится к полинуклеотиду, содержащему последовательность нуклеиновой кислоты, которая кодирует анти-HER3-антитело, или кодирует антигенсвязывающий фрагмент такого антитела. Полинуклеотиды согласно изобретению могут быть в форме РНК или в форме ДНК. ДНК включает кДНК, геномную ДНК и синтетическую ДНК; и может быть двунитевой или однонитевой, и в том случае, когда она является однонитевой, она может быть кодирующей нитью или некодирующей (антисмысловой) нитью.
В некоторых аспектах полинуклеотиды являются выделенными. В некоторых аспектах полинуклеотиды являются в значительной степени очищенными. В некоторых аспектах полинуклеотиды содержат кодирующую последовательность для зрелого полипептида, слитую в одной и той же рамке считывания с полинуклеотидом, который способствует, например, экспрессии и секреции полипептида из клетки-хозяина (например, лидерная последовательность, которая функционирует в качестве секреторной последовательности для регуляции транспорта полипептида из клетки). Полипептид, имеющий лидерную последовательность, представляет собой пребелок, и может иметь лидерную последовательность, отщепляемую в клетке-хозяине с образованием зрелой формы полипептида. Полинуклеотиды также могут кодировать HER3-связывающий пробелок, который представляет собой зрелый белок плюс дополнительные 5’-аминокислотные остатки.
В некоторых аспектах полинуклеотиды содержат кодирующую последовательность для зрелого HER3-связывающего полипептида, например, анти-HER3-антитела или его антигенсвязывающего фрагмента, слитого в одной рамке считывания с маркерной последовательностью, которая обеспечивает, например, возможность очистки кодируемого полипептида. Например, маркерной последовательностью может быть гексагистидиновая метка, поставляемая в векторе pQE-9, чтобы обеспечить очистку зрелого полипептида, слитого с маркером, в случае бактериального хозяина, или маркерной последовательностью может быть гемагглютининовая (HA) метка, получаемая из белка гемагглютинина вируса гриппа, в том случае, когда используют в качестве хозяина млекопитающего (например, клетки COS-7).
Кроме того, настоящее изобретение относится к вариантам требуемых полинуклеотидов, кодирующих, например, HER3-связывающие фрагменты, аналоги и производные HER3-связывающих молекул согласно изобретению.
Варианты полинуклеотидов могут содержать изменения в кодирующих областях, некодирующих областях или и тех и других. В некоторых аспектах варианты полинуклеотидов содержат изменения, которые дают молчащие замены, добавления или делеции, но не изменяют свойства или активности кодируемого полипептида. В некоторых аспектах нуклеотидные варианты получают в результате молчащих замен вследствие вырожденности генетического кода. Варианты полинуклеотидов могут быть получены по разным причинам, например, чтобы оптимизировать экспрессию кодонов для конкретного хозяина (изменить кодоны в мРНК человека на кодоны, предпочтительные в бактериальном хозяине, таком как E. coli). Также предлагаются векторы и клетки, содержащие полинуклеотиды, описанные в настоящей публикации.
В некоторых аспектах последовательность ДНК, кодирующая HER3-связывающую молекулу, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, может быть сконструирована химическим синтезом с использованием синтезатора олигонуклеотидов. Такие олигонуклеотиды могут быть сконструированы на основе аминокислотной последовательности требуемого полипептида и отбора таких кодонов, которые являются предпочтительными в клетке-хозяине, в которой представляющий интерес рекомбинантный полипептид может быть получен. Можно использовать стандартные способы синтеза выделенной полинуклеотидной последовательности, кодирующей представляющий интерес выделенный полипептид. Например, полная аминокислотная последовательность может быть использована для конструирования обратно транслированного гена. Кроме того, может быть синтезирован ДНК-олигомер, содержащий нуклеотидную последовательность, кодирующую конкретный выделенный полипептид. Например, могут быть синтезированы несколько небольших олигонуклеотидов, кодирующих части требуемого полипептида, и затем лигированы. Отдельные олигонуклеотиды обычно содержат выступающие 5’- или 3’-концы для комплементарной сборки.
После сборки (в результате синтеза, сайт-специфичного мутагенеза или другим способом) полинуклеотидные последовательности, кодирующие конкретный представляющий интерес выделенный полипептид, могут быть встроены в экспрессирующий вектор и оперативно связаны с последовательностью регуляции экспрессии, подходящей для экспрессии белка в требуемом хозяине. Правильная сборка может быть подтверждена секвенированием нуклеотидов, рестрикционным картированием и экспрессией биологически активного полипептида в подходящем хозяине. Как хорошо известно в данной области, чтобы получить высокие уровни экспрессии трансфицируемого гена, ген должен быть оперативно связан с последовательностями регуляции транскрипции и трансляции, которые являются функциональными в выбранном для экспрессии хозяине.
В некоторых аспектах рекомбинантные экспрессирующие используют для амплификации и экспрессии ДНК, кодирующей анти-HER3-антитела или их антигенсвязывающие фрагменты. Рекомбинантные экспрессирующие векторы представляют собой реплицируемые ДНК-конструкции, которые содержат синтетические или полученные из кДНК фрагменты ДНК, кодирующие полипептидную цепь анти-HER3-антитела или его антигенсвязывающий фрагмент, оперативно связанные с подходящими элементами регуляции транскрипции или трансляции, полученными из генов млекопитающих, микроорганизмов, вирусов или насекомых. Транскрипционная единица обычно содержит сборку из (1) генетического элемента или элементов, играющих регуляторную роль в экспрессии генов, например, промоторов или энхансеров транскрипции, (2) структурной или кодирующей последовательности, которая транскрибируется в мРНК и транслируется в белок, и (3) соответствующих последовательностей инициации и терминации транскрипции и трансляции, которые подробно описаны ниже. К таким регуляторным элементам может относиться последовательность оператора для регуляции транскрипции. Дополнительно может быть придана способность к репликации в хозяине, обычно придаваемая началом репликации, и введен селектируемый ген для облегчения распознавания трансформантов. Области ДНК являются оперативно связанными, когда они функционально связаны друг с другом. Например, ДНК для сигнального пептида (лидера секреции) оперативно связана с ДНК для полипептида, если она экспрессируется в виде предшественника, который принимает участие в секреции полипептида; промотор оперативно связан с кодирующей последовательностью, если он контролирует транскрипцию последовательности; или сайт связывания рибосомы оперативно связан с кодирующей последовательностью, если он расположен так, что обеспечивает возможность трансляции. Структурные элементы, предназначенные для применения в системах экспрессии у дрожжей, включают лидерную последовательность, обеспечивающую внеклеточную секрецию транслированного белка клеткой-хозяином. Альтернативно, когда рекомбинантный белок экспрессируется без лидерной или транспортной последовательности, он может содержать N-концевой остаток метионина. Такой остаток необязательно может впоследствии отщепляться от экспрессированного рекомбинантного белка с получением конечного продукта.
Выбор последовательности регуляции экспрессии и экспрессирующего вектора будет зависеть от выбора хозяина. Можно использовать широкое множество сочетаний экспрессирующий хозяин/вектор. Применимые экспрессирующие векторы в случае эукариотических хозяев, включают, например, векторы, содержащие последовательности регуляции экспрессии из SV40, вируса папилломы крупного рогатого скота, аденовируса и цитомегаловируса. Применимые экспрессирующие векторы в случае бактериальных хозяев включают известные бактериальные плазмиды из E. coli, включая pCR1, pBR322, pMB9 и их производные, плазмиды широкого круга хозяев, такие как M13 и нитчатые однонитевые ДНК-фаги.
Подходящие клетки-хозяева для экспрессии HER3-связывающей молекулы, например, анти-HER3-антитела или его антигенсвязывающего фрагмента, включают клетки прокариотов, дрожжей, насекомых или высших эукариот под контролем соответствующих промоторов. Прокариоты включают грамотрицательные или грамположительные организмы, например E. coli или бациллы. Клетки высших прокариот включают постоянные клеточные линии, полученные из клеток млекопитающих, которые описаны ниже. Также можно использовать бесклеточные системы трансляции. Подходящие клонирующие и экспрессирующие векторы для применения в случае хозяев, являющихся бактериями, грибами, дрожжами и млекопитающими, описаны в публикации Pouwels с соавторами (Cloning Vectors: A Laboratory Manual, Elsevier, N.Y., 1985), соответствующее содержание которой включено в настоящее описание в виде ссылки. Дополнительную информацию, имеющую отношение к способам получения белков, включая получение антител, можно найти, например, в публикации патента США №2008/0187954, патентах США №6413746 и 6660501 и международной публикации патента № WO 04009823, при этом каждая публикация включена в настоящее описание в виде ссылки в полном объеме.
Также предпочтительно можно использовать различные системы культур клеток млекопитающих и насекомых, чтобы экспрессировать рекомбинантные HER3-связывающие молекулы, например, анти-HER3-антитела или их антигенсвязывающие фрагменты. Можно осуществлять экспрессию рекомбинантных белков в клетках млекопитающих, так как такие белки обычно подвергаются правильному фолдингу, соответствующим образом модифицированы и полностью функциональны. Примеры подходящих линий клеток-хозяев млекопитающих включают HEK-293 и HEK-293T, линии клеток почки обезьяны COS-7, описанные Gluzman (Cell 23: 175, 1981), и другие клеточные линии, включая, например, L-клетки, линии клеток C127, 3T3, яичника китайского хомячка (CHO), NSO, HeLa и BHK. Экспрессирующие векторы млекопитающих могут содержать нетранскрибируемые элементы, такие как начало репликации, подходящий промотор и энхансер, связанные с геном, который необходимо экспрессировать, и другие 5’- или 3’-фланкирующие нетранскрибируемые последовательности и 5’- или 3’-нетранслируемые последовательности, такие как необходимые сайты связывания с рибосомой, сайт полиаденилирования, донорные и акцепторные участки сплайсинга и последовательности терминации транскрипции. Обзор бакуловирусных систем для получения гетерологичных белков в клетках насекомых приведен в публикации Luckow and Summers, BioTechnology 6: 47 (1988).
HER3-связывающие молекулы, например, анти-HER3-антитела или их антигенсвязывающие фрагменты, продуцируемые трансформированным хозяином, можно очистить согласно любому подходящему способу. Такие стандартные способы включают хроматографию (например, ионообменную, аффинную и хроматографию на колонках по размеру), центрифугирование, определение дифференциальной растворимости или любой другой стандартный способ очистки белков. С белком могут быть связаны аффинные метки, такие как гексагистидин, домен связывания мальтозы, последовательность оболочки вируса гриппа и глутатион-S-трансфераза, чтобы обеспечить легкую очистку в результате пропускания через соответствующую аффинную колонку. Выделенные белки также могут быть охарактеризованы физически с использованием известных способов, таких как протеолиз, ядерный магнитный резонанс и рентгеновская кристаллография.
Например, надосадки, полученные из систем, которые секретируют рекомбинантный белок в культуральную среду, можно сначала сконцентрировать, используя коммерчески доступный фильтр для концентрирования белков, например, установку для ультрафильтрации Amicon или Millipore Pellicon. После стадии концентрирования концентрат можно нанести на подходящий матрикс для очистки. Альтернативно можно использовать анионообменную смолу, например, матрикс или подложку, имеющие боковые группы диэтиламиноэтила (DEAE). Матрикс может быть акриламидным, агарозным, декстрановым, целлюлозным или других типов, обычно используемых при очистке белков. Альтернативно, можно использовать стадию катионного обмена. Подходящие катионообменники включают различные нерастворимые матриксы, содержащие сульфопропильные или карбоксиметильные группы. Наконец, можно использовать одну или несколько стадий обращено-фазовой высокоэффективной жидкостной хроматографии (ОФ-ВЭЖХ) с применением гидрофобной среды для ОФ-ВЭЖХ, например, силикагеля, имеющего боковые метильные или другие алифатические группы, чтобы дополнительно очистить HER3-связывающую молекулу. Также можно использовать некоторые или все указанные выше стадии очистки в различных сочетаниях для получения гомогенного рекомбинантного белка.
Рекомбинантный HER3-связывающий белок, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, получаемый в бактериальной культуре, может быть выделен, например, с использованием сначала экстракции из клеточных осадков с последующей одной или несколькими стадиями концентрирования, высаливания, ионного обмена в воде или эксклюзионной хроматографии по размеру. Высокоэффективную жидкостную хроматографию (ВЭЖХ) можно использовать для конечных стадий очистки. Микробные клетки, используемые для экспрессии рекомбинантного белка, могут быть разрушены любым обычным способом, включая осуществление циклов замораживания-оттаивания, обработку ультразвуком, механическое разрушение или использованием лизирующих агентов.
Способы, известные в данной области для очистки антител и других белков, также включают, например, способы, описанные в публикациях патентов США №2008/0312425, 2008/0177048 и 2009/0187005, каждая из которых включена в настоящее описание в виде ссылки в полном объеме.
В некоторых аспектах HER3-связывающая молекула представляет собой полипептид, который не является антителом. В данной области известно множество способов идентификации и получения не являющихся антителами полипептидов, которые связываются с высокой аффинностью с белком-мишенью. См., например, публикации Skerra, Curr. Opin. Biotechnol., 18:295-304 (2007), Hosse et al., Protein Science, 15:14-27 (2006), Gill et al., Curr. Opin. Biotechnol., 17:653-658 (2006), Nygren, FEBS J., 275:2668-76 (2008) и Skerra, FEBS J., 275: 2677-83 (2008), каждая из которых включена в настоящее описание в виде ссылки в полном объеме. В некоторых аспектах можно использовать методику фагового дисплея для идентификации/получения HER3-связывающего полипептида. В некоторых аспектах полипептид содержит белковый каркас типа, выбранного из группы, состоящей из белка A, липокалина, домена фибронектина, консенсусного повторяющегося домена анкирина и тиоредоксина.
VI. Способы лечения с применением терапевтических анти-HER3-антител
Способы согласно изобретению направлены на применение анти-HER3-связывающих молекул, например, антител, включая их антигенсвязывающие фрагменты, варианты и производные, для лечения пациентов, имеющих заболевание, ассоциированное с экспрессией HER3 или HER3-экспрессирующими клетками. Под «HER3-экспрессирующей клеткой» подразумевают клетку, экспрессирующую HER3. Способы выявления экспрессии HER3 в клетках хорошо известны в данной области и включают без ограничения методику ПЦР, иммуногистохимию, проточную цитометрию, Вестерн-блот, ELISA и тому подобные.
Несмотря на то, что следующее обсуждение относится к способам диагностики и лечению различных заболеваний и расстройств с использованием HER3-связывающей молекулы согласно изобретению, способы, описанные в настоящей публикации, также применимы к анти-HER3-антителам и антигенсвязывающим фрагментам, вариантам и производным таких анти-HER3-антител, которые сохраняют требуемые свойства анти-HER3-антител согласно изобретению, например, способны специфично связывать HER3 и нейтрализовать активность HER3. В некоторых аспектах HER3-связывающие молекулы представляют собой человеческие или гуманизированные антитела, которые не опосредуют ADCC у человека, или антитела, выбранные из известных анти-HER3-антител, которые не опосредуют ADCC, или являются анти-HER3-антителами, которые сконструированы таким образом, что они не опосредуют ADCC. В некоторых аспектах HER3-связывающая молекула представляет собой моноклональное антитело клона 16. В других аспектах HER3-связывающая молекула представляет собой антитело, являющееся YTE-мутантом клона 16. В некоторых аспектах HER3-связывающая молекула представляет собой моноклональное антитело P2B11. В некоторых аспектах HER3-связывающая молекула представляет собой моноклональное антитело 1A4. В некоторых аспектах HER3-связывающая молекула представляет собой моноклональное антитело 2C2. В некоторых аспектах HER3-связывающая молекула представляет собой моноклональное антитело 2F10. В некоторых аспектах HER3-связывающая молекула представляет собой моноклональное антитело 3E1. В некоторых аспектах HER3-связывающая молекула представляет собой моноклональное антитело P2B11, сконструированное для увеличения времени полужизни в сыворотке. В некоторых аспектах HER3-связывающая молекула представляет собой моноклональное антитело 1A4, сконструированное для увеличения времени полужизни в сыворотке. В некоторых аспектах HER3-связывающая молекула представляет собой моноклональное антитело 2C2, сконструированное для увеличения времени полужизни в сыворотке. В некоторых аспектах HER3-связывающая молекула представляет собой моноклональное антитело 2F10, сконструированное для увеличения времени полужизни в сыворотке. В некоторых аспектах HER3-связывающая молекула представляет собой моноклональное антитело 3E1, сконструированное для увеличения времени полужизни в сыворотке. В других аспектах HER3-связывающая молекула представляет собой YTE-мутантное антитело P2B11. В других аспектах HER3-связывающая молекула представляет собой YTE-мутантное антитело 1A4. В других аспектах HER3-связывающая молекула представляет собой 2C2-YTE-мутантное антитело. В других аспектах HER3-связывающая молекула представляет собой YTE-мутантное антитело 2F10. В других аспектах HER3 -связывающая молекула представляет собой YTE-мутантное антитело 3E1.
В одном аспекте лечение включает в себя нанесение или введение анти-HER3-связывающей молекулы, например, антитела или его антигенсвязывающего фрагмента, варианта или производного согласно настоящему изобретению субъекту или пациенту, или нанесение или введение анти-HER3-связывающей молекулы в выделенную ткань или клеточную линию, полученную от субъекта или пациента, при этом субъект или пациент имеет заболевание, симптом заболевания или предрасположенность к заболеванию. В другом аспекте подразумевается, что лечение также включает в себя нанесение или введение фармацевтической композиции, содержащей связывающую анти-HER3-молекулу, например, антитело или его антигенсвязывающий фрагмент, вариант или производное согласно настоящему изобретению субъекту или пациенту, или нанесение или введение фармацевтической композиции, содержащей связывающую анти-HER3-молекулу в выделенную ткань или клеточную линию, полученную от субъекта или пациента, который имеет заболевание, симптом заболевания или предрасположенность к заболеванию.
Анти-HER3-связывающие молекулы, например, антитела или их антигенсвязывающие фрагменты, варианты или производные согласно настоящему изобретению применимы для лечения различных злокачественных опухолей. В одном аспекте изобретение относится к связывающим анти-HER-молекулам, например, антителам или их антигенсвязывающим фрагментам, вариантам или производным для применения в качестве лекарственного средства, в частности для применения с целью лечения или профилактики злокачественной опухоли. Примеры злокачественной опухоли включают без ограничения рак ободочной кишки, рак легкого, рак желудка, плоскоклеточный рак головы и шеи, меланому, рак поджелудочной железы, рак простаты и рак молочной железы.
Согласно способам, предлагаемым в настоящем изобретении, по меньшей мере, одну связывающую анти-HER3-молекулу, например, антитело или его антигенсвязывающий фрагмент, вариант или производное, которые определены в настоящем описании, применяют для того, чтобы вызвать позитивный терапевтический ответ в отношении злокачественной опухоли. Термин «позитивный терапевтический ответ» в отношении лечения злокачественной опухоли относится к улучшению состояния в случае заболевания в связи с активностью таких связывающих анти-HER3-молекул, например, антител или их антигенсвязывающих фрагментов, вариантов или производных, и/или снижению симптомов, ассоциированных с заболеванием. Таким образом, например, улучшение состояния при заболевании можно охарактеризовать как полный ответ. Под «полным ответом» подразумевают отсутствие клинически выявляемого заболевания с нормализацией любых предыдущих результатов тестирования. Альтернативно улучшение состояния при заболевании можно охарактеризовать как частичный ответ. Термин «позитивный терапевтический ответ» охватывает уменьшение или подавление прогрессирования и/или продолжительности существования злокачественной опухоли, уменьшение или ослабление степени тяжести злокачественной опухоли и/или ослабление ее одного или нескольких симптомов в результате введения связывающей анти-HER3-молекулы согласно изобретению. В конкретных аспектах такие термины относятся к одному, двум или трем или большему количеству результатов после введения связывающих анти-HER3-молекул согласно изобретению: (1) стабилизации, уменьшению или удалению популяции злокачественных клеток; (2) стабилизации или снижению роста злокачественной опухоли; (3) нарушению образования злокачественной опухоли; (4) ликвидации, удалению или контролю первичной, регионарной и/или метастатической злокачественной опухоли; (5) снижению смертности; (6) увеличению выживаемости без признаков заболевания, безрецидивной выживаемости, выживаемости без прогрессирования и/или общей выживаемости, продолжительности или частоты выживаемости; (7) увеличению доли ответов, долговременности ответа или количества пациентов, у которых выявляют ответ или которые находятся в состоянии ремиссии; (8) снижению частоты госпитализации, (9) уменьшению продолжительности госпитализации, (10) размер злокачественной опухоли сохраняется и не увеличивается или увеличивается менее чем на 10%, предпочтительно менее чем на 5%, предпочтительно менее чем на 4%, предпочтительно менее чем на 2%, и (12) увеличению количества пациентов в состоянии ремиссии.
Клинический ответ можно оценить, используя способы скрининга, такие как магнитно-резонансная томография (МРТ), рентгенография, компьютерное томографическое (КТ) сканирование, проточная цитометрия или анализ на основе активируемой флуоресценцией сортировки клеток (FACS), гистология, макропатология и химия крови, включая без ограничения изменения, регистрируемые посредством ELISA, РИА, хроматографии и тому подобного. Кроме указанных позитивных терапевтических ответов субъект, подвергаемый терапии связывающей анти-HER3-молекулой, например, антителом или его антигенсвязывающим фрагментом, вариантом или производным, может получать полезное действие в виде ослабления симптомов, ассоциированных с заболеванием.
Связывающие анти-HER3-молекулы, например, антитела или их антигенсвязывающие фрагменты, варианты или производные согласно изобретению можно применять в сочетании с любыми известными средствами терапии злокачественной опухоли, включая любое средство или сочетание средств, которые, как известно, применимы или которые применялись или применяются в настоящее время для лечения злокачественной опухоли, например, рака ободочной кишки, рака легкого, рака желудка, плоскоклеточного рака головы и шеи и рака молочной железы. Второе средство или сочетание средств в фармацевтическом комбинированном препарате или схеме дозирования предпочтительно обладает активностями, дополняющими активность антитела или полипептида согласно изобретению так, что они не оказывают неблагоприятного влияния друг на друга.
Противораковые средства включают лекарственные средства, применяемые для лечения злокачественных образований, злокачественного роста. Лекарственную терапию можно использовать отдельно или в сочетании с другими способами лечения, такими как хирургия или лучевая терапия. Можно использовать несколько классов лекарственных средств для лечения злокачественной опухоли, в зависимости от природы вовлеченного органа. Например, рак молочной железы обычно стимулируется эстрогенами, и его можно лечить лекарственными средствами, которые инактивируют половые гормоны. Подобным образом, рак простаты можно лечить лекарственными средствами, которые инактивируют андрогены, мужские половые гормоны. Противораковые средства для применения в некоторых способах согласно настоящему изобретению включают наряду с прочим антитела (например, антитела, которые связывают IGF-1R, антитела, которые связывают EGFR, антитела, которые связывают HER2, антитела, которые связывают HER3, или антитела, которые связывают cMET), малые молекулы, мишенью которых является IGF1R, малые молекулы, мишенью которых является EGFR, малые молекулы, мишенью которых является HER2, антиметаболиты, алкилирующие агенты, ингибиторы топоизомераз, средства, мишенью которых являются микротрубочки, ингибиторы киназ, ингибиторы синтеза белка, иммунотерапевтические средства, гормональные терапевтические средства, глюкокортикоиды, ингибиторы ароматазы, ингибиторы mTOR, химиотерапевтические средства, ингибиторы протеинкиназы B, ингибиторы фосфатидилинозитол-3-киназы (PI3K), ингибиторы циклинзависимой киназы (CDK), RLr9, CD289, ингибиторы ферментов, анти-TRAIL, ингибиторы MEK и т.д.
В конкретных аспектах HER3-связывающие молекулы согласно изобретению, например, антитела или их антигенсвязывающие фрагменты, можно вводить в сочетании с антителами или фрагментами антител, мишенью которых является рецептор эпидермального фактора роста (EGFR), например, цетуксимабом (эрбитуксом®, Imclone), панитумумабом (вектибиксом®, Amgen), матузумабом/EMD72000 (Merck Serono), олигоклональным MM-151 (Merrimack), нимотузумабом (TheraCIM, InnGene Kalbiotechy), GA201/RG7160 (Roche), Sym004 (Symphogen), MEHD-7945A (с двойной специфичностью к EGFR/HER3, Genentech). В других конкретных аспектах HER3-связывающие молекулы согласно изобретению, например, антитела или их антигенсвязывающие фрагменты, можно вводить в сочетании с антителами или фрагментами антител, мишенью которых является HER2, например, пертузумабом (rhuMAb 2C4/омнитарг®, Genentech), трастузумабом (герцептин®, Genentech/Roche), MM-111 (биспецифичное HER2/HER3-антитело, Merrimack, например, WO 2009/126920). В других конкретных аспектах HER3-связывающие молекулы согласно изобретению, например, антитела или их антигенсвязывающие фрагменты, можно вводить в сочетании с антителами или фрагментами антител, которые также нацелены на HER3, например, MEHD-7945A/RG7597 (EGFR/HER3-двойная специфичность, Genentech, например, WO 2010108127), MM-121 (Merrimack, например, WO 2008/100624), MM-111 (HER2/HER3-биспецифичное антитело, Merrimack, например, WO 2009/126920), AV-203 (Aveo, например, WO 2011/136911), AMG888 (Amgen, WO 2007/077028), HER3-8 (ImmunogGen, например, WO 2012/019024). В следующих конкретных аспектах HER3-связывающие молекулы согласно изобретению, например, антитела или их антигенсвязывающие фрагменты, можно вводить в сочетании с антителами или фрагментами антител, мишенью которых является HER4. В конкретном аспекте HER3-связывающие молекулы согласно изобретению можно вводить в сочетании с антителом, мишенью которого является EGFR или HER2 (например, цетуксимабом или трастузумабом). В следующем конкретном аспекте HER3-связывающие молекулы согласно изобретению можно вводить в сочетании с конъюгатами антител с лекарственными средствами, мишенью которых является HER2 (например, трастузумабом-эмтанзином, Genentech/Roche). Предполагается, что HER3-связывающие молекулы согласно изобретению усиливают интернализацию и распад корецептора, индуцированные связыванием антитела с корецептором, и таким образом будут повышать эффективность антитела и/или конъюгата антитела с лекарственным средством, мишенью которых являются EGFR, HER2 и/или HER4.
В других аспектах HER3-связывающие молекулы согласно изобретению можно вводить в сочетании с ингибиторами тирозинкиназ. В некоторых других конкретных аспектах HER3-связывающие молекулы согласно изобретению можно вводить в сочетании с ингибиторами активности тирозинкиназ, ассоциированных с EGFR и/или HER2/neu, например, лапатинибом. В конкретных аспектах HER3-связывающие молекулы согласно изобретению можно вводить в сочетании с низкомолекулярными ингибиторами рецептора(ов) эпидермального фактора роста (например, EGFR, HER2, HER4) например, гефитинибом (Iressa®, Astrazeneca); канертинибом/CI-1033 (Pfizer); лапатинибом (Tykerb®, GlaxoSmithKline), эрлотинибом (Tarceva®, OSI Pharma), афатинибом (Tovok®/Tomtovok®, Boehringer Ingelheim), нератинибом (HKI-272, Pfizer).
В некоторых аспектах HER3-связывающие молекулы согласно изобретению можно вводить в сочетании с антимитотическими средствами. В некоторых конкретных аспектах HER3-связывающие молекулы согласно изобретению можно вводить в сочетании со средствами, которые стабилизируют сборку микротрубочек веретена митотического деления, например, паклитакселом или доцетакселом.
В некоторых аспектах HER3-связывающие молекулы согласно изобретению можно вводить в сочетании с ингибиторами MEK (киназы активируемых митогенами протеинкиназ (MAPK), также известной как MAPKK), например, селуметинибом (AZD6244, ARRY-142866, AstraZeneca), WX-554 (Wilex), траметинибом (GlaxoSmithKline), рефаметинибом (Ardea Biosciences), E-6201 (Eisai), MEK-162 (Novartis). В конкретном аспекте сочетание ингибитора MEK и HER3-связывающей молекулы согласно изобретению является более эффективным, чем любое средство отдельно. В конкретном аспекте HER3-связывающую молекулу согласно изобретению вводят в сочетании с селуметинибом.
В том случае, когда комбинированная терапия включает в себя введение связывающей анти-HER3-молекулы в сочетании с введением другого терапевтического средства способы согласно изобретению охватывают совместное введение с применением отдельных препаратов или одного фармацевтического препарата и последовательное введение в любом порядке. В некоторых аспектах анти-HER3-антитела, описанные в настоящей публикации, вводят в сочетании с другими лекарственными средствами, при этом антитело или его антигенсвязывающий фрагмент, вариант или производное и терапевтическое средство (средства) можно вводить последовательно в любом порядке или одновременно (т.е. в один момент времени или в одном временной рамке).
Комбинированная терапия может обеспечивать «синергию» и проявлять «синергетический» эффект, т.е. эффект, достигаемый в случае использования активных ингредиентов вместе, который выше, чем сумма эффектов в результате применения соединений по отдельности. Синергетический эффект может быть достигнут, когда активные ингредиенты: (1) приготовлены вместе и введены или доставлены одновременно в комбинированном стандартном дозированном препарате; (2) доставлены поочередно или параллельно в виде отдельных препаратов; или (3) при использовании другой схемы. В случае поочередной доставки в ходе терапии синергетический эффект может быть достигнут, когда соединения вводят или доставляют последовательно, например, используя разные инъекции в отдельных шприцах. В общем, во время терапии с поочередным введением эффективную дозу каждого активного ингредиента вводят последовательно, т.е. периодически, тогда как в случае комбинированной терапии эффективные дозы двух или более активных ингредиентов вводят вместе.
В некоторых аспектах HER3-связывающую молекулу, например, анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению, можно вводить в синергетическом сочетании с ингибитором рецептора эпидермального фактора роста (EGFR). В некоторых аспектах ингибитором EGFR является антитело. В конкретных аспектах антителом, ингибирующим EGFR, является эрбитукс® (цетуксимаб) или панитумумаб (вектибикс®). В конкретных аспектах HER3-связывающие молекулы согласно изобретению, например, антитела или их антигенсвязывающие фрагменты, можно вводить в синергетическом сочетании с ингибиторами тирозинкиназной активности, ассоциированной с EGFR и/или HER2/neu, например, лапатинибом. В некоторых аспектах HER3-связывающую молекулу, например, анти- HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению, можно вводить в синергетическом сочетании с ингибитором HER2. В некоторых аспектах ингибитором HER2 является антитело. В конкретных аспектах антителом, ингибирующим HER2, является пертузумаб (rhuMAb 2C4/омнитарг®, Genentech), трастузумаб (герцептин®, Genentech/Roche) или трастузумаб-эмтанзин (Genentech/Roche). В конкретных аспектах HER3-связывающие молекулы согласно изобретению, например, антитела или их антигенсвязывающие фрагменты, можно вводить в синергетическом сочетании с ингибиторами тирозинкиназной активности, ассоциированной с HER2/neu, например, лапатинибом. В некоторых аспектах HER3-связывающие молекулы согласно изобретению можно вводить в синергетическом сочетании с антимитотическим средством. В некоторых конкретных аспектах антимитотическое средство стабилизирует сборку микротрубочек веретена митотического деления. В некоторых конкретных аспектах антимитотическим средством является паклитаксел или доцетаксел. В некоторых конкретных вариантах антитело 2C2 можно вводить в синергетическом сочетании с ингибитором рецептора фактора роста (EGFR). В некоторых конкретных вариантах ингибитором EGFR является антитело. В конкретных вариантах антителом, ингибирующим EGFR, вводимым синергетически с антителом 2C2, является эрбитукс® (цетуксимаб). В конкретных вариантах антитело 2C2 можно вводить в синергетическом сочетании с ингибиторами тирозинкиназной активности, ассоциированной с EGFR и/или HER2/neu, например, лапатинибом. В некоторых вариантах антитело 2C2 можно вводить в синергетическом сочетании с антимитотическим средством. В некоторых конкретных вариантах антимитотическое средство, вводимое синергетически с антителом 2C2, стабилизирует сборку микротрубочек веретена митотического деления. В некоторых конкретных вариантах антимитотическим средством, вводимым синергетически с антителом 2C2, является паклитаксел.
В одном аспекте злокачественная опухоль содержит мутацию KRAS. В конкретных аспектах мутация KRAS локализована в кодоне 12 гена KRAS человека. Как показано в разделе «Примеры», анти-HER3-антитела, раскрытые в настоящем описании, способны ингибировать рост опухолевых клеток, которые содержат мутацию KRAS, либо в случае применения в виде отдельного средства (монотерапия), либо в сочетании с другим терапевтическим средством.
Следующий аспект относится к применению связывающих анти-HER3-молекул, например, антител или их антигенсвязывающих фрагментов, вариантов или производных, для диагностического мониторинга уровней белка в ткани в качестве части процедуры клинического исследования, например, чтобы определить эффективность данной схемы лечения. Например, выявление можно облегчить, связывая антитело с регистрируемым веществом. Примеры регистрируемых веществ включают различные ферменты, простетические группы, флуоресцирующие вещества, люминесцентные вещества, биолюминесцентные вещества и радиоактивные вещества. Примерами подходящих ферментов являются пероксидаза хрена, щелочная фосфатаза, β-галактозидаза или ацетилхолинэстераза; примерами подходящих комплексов простетических групп являются стрептавидин/биотин и авидин/биотин; примерами подходящих флуоресцирующих веществ являются умбеллиферон, флуоресцеин, изотиоцианат флуоресцеина, родамин, дихлортриазиниламин флуоресцеин, дансилхлорид или фикоэритрин; примером люминесцирующего вещества является люминол; примерами биолюминесцентных вещества являются люцифераза, люциферин и экворин; и примерами подходящих радиоактивных веществ являются 125I, 35S или 3H.
VII. Фармацевтические композиции и способы введения
Способы получения и введения связывающих анти-HER3-молекул, например, антител или их антигенсвязывающих фрагментов, вариантов, или производных согласно изобретению субъекту, нуждающемуся в таком введении, хорошо известны или легко могут быть определены специалистами в данной области. Путь введения связывающей анти-HER3-молекулы, например, антитела или его антигенсвязывающего фрагмента, варианта или производного, может быть, например, пероральным, парентеральным, путем ингаляции или местным. Термин «парентеральное» в используемом в настоящем описании смысле включает, например, внутривенное, внутриартериальное, внутрибрюшинное, внутримышечное, подкожное, ректальное или вагинальное введение. Однако в других способах, совместимых с приведенными в настоящем описании инструкциями, связывающие анти-HER3-молекулы, например, антитела или их антигенсвязывающие фрагменты, варианты или производные согласно изобретению, можно доставлять непосредственно в место с нежелательной клеточной популяцией, таким образом увеличивая экспозицию патологической ткани с терапевтическим средством.
Как обсуждается в настоящем описании, связывающие анти-HER3-молекулы, например, антитела или их антигенсвязывающие фрагменты, варианты или производные согласно изобретению, можно вводить в фармацевтически эффективном количестве для лечения in vivo опосредованных HER3-экспрессирующими клетками заболеваний, таких как некоторые типы злокачественных опухолей.
Фармацевтические композиции, применяемые в настоящем изобретении, могут содержать фармацевтически приемлемые носители, включая, например, воду, ионообменники, белки, буферные вещества и соли. Также могут присутствовать консерванты и другие добавки. Носителем может быть растворитель или дисперсионная среда. Подходящие препараты для применения в терапевтических способах, раскрытых в настоящем описании, описаны в Remington's Pharmaceutical Sciences (Mack Publishing Co.) 16th ed. (1980). В некоторых аспектах HER3-связывающие молекулы согласно изобретению готовят в виде композиции, стабильной в условиях хранения в холодильнике (2-8°C). В конкретном аспекте композиция, стабильная в условиях хранения в холодильнике, содержит 25 мМ гистидин/гистидин HCl, 205 мМ сахарозу, 0,02% полисорбат 80, pH 6,0. В другом конкретном аспекте HER3-связывающие молекулы согласно изобретению готовят в концентрации 25-100 мг/мл в композиции, стабильной в условиях хранения в холодильнике.
В любом случае, может быть приготовлены стерильные инъекционные растворы путем введения активного соединения (например, анти-HER3-антитела или его антигенсвязывающего фрагмента, варианта или производного, как такового или в сочетании с другими активными средствами) в требуемом количестве в соответствующий растворитель с последующей стерилизацией фильтрованием. Кроме того, препараты могут быть упакованы и могут продаваться в форме набора. Такие изделия производства могут иметь ярлыки или вкладыши в упаковку, на которых указано, что вложенные композиции применимы для лечения субъекта, страдающего или предрасположенного к возникновению заболевания или расстройства.
Парентеральные препараты могут быть введены в виде однократной болюсной дозы, инфузии или ударной болюсной дозы с последующим введением поддерживающей дозы. Такие композиции можно вводить с конкретными фиксированными или переменными интервалами, например, один раз в сутки или по мере «необходимости».
Композицию можно вводить в виде одной дозы, множественных доз или на протяжении установленного периода времени при инфузии. Схемы дозирования также можно корректировать, чтобы получить оптимальный требуемый ответ (например, терапевтический или профилактический ответ).
Терапевтически эффективные дозы композиций согласно настоящему изобретению для лечения заболеваний, опосредованных HER3-экспрессирующими клетками, таких как некоторые типы злокачественных опухолей, включая, например, рак ободочной кишки, рак легкого, рак желудка, плоскоклеточный рак головы и шеи, меланому, рак поджелудочной железы, рак простаты и рак молочной железы, варьируют в зависимости от множества разных факторов, включая способы введения, целевое место, физиологическое состояние пациента, является пациент человеком или животным, другие вводимые лекарственные средства и является лечение профилактическим или терапевтическим. Обычно пациентом является человек, но также можно лечить млекопитающих, отличных от человека, включая трансгенных млекопитающих. Доза для лечения можно титровать, применяя обычные способы, известные специалистам в данной области, чтобы оптимизировать безопасность и эффективность.
Количество, по меньшей мере, одной связывающей анти-HER3-молекулы, например, антитела или его связывающего фрагмента, варианта или производного, которое необходимо ввести, легко может определить специалист в данной области без излишнего экспериментирования с учетом описания настоящего изобретения. Факторы, влияющие на способ введения и соответствующее количество, по меньшей мере, одной связывающей анти-HER3-молекулы, например, антитела, его антигенсвязывающего фрагмента, варианта или производного, включают без ограничения тяжесть заболевания, историю заболевания и возраст, массу, состояние здоровья и физическое состояние индивидуума, подвергаемого терапии. Подобным образом, количество связывающей анти-HER3-молекулы, например, антитела или его фрагмента, варианта или производного, которое необходимо ввести, будет зависеть от способа введения и того, будет ли субъекту введена однократная доза или множественные дозы такого средства.
Настоящее изобретение также относится к применению связывающей анти-HER3-молекулы, например, антитела или его антигенсвязывающего фрагмента, варианта или производного, для производства лекарственного средства для лечения определенного типа злокачественной опухоли, включая, например, рак ободочной кишки, рак легкого, рак желудка, плоскоклеточный рак головы и шеи, меланому, рак поджелудочной железы, рак простаты и рак молочной железы.
Изобретение также относится к применению связывающей анти-HER3-молекулы, например, антитела согласно изобретению или его антигенсвязывающего фрагмента, варианта или производного в производстве лекарственного средства для лечения субъекта с определенным типом злокачественной опухоли. В некоторых аспектах лекарственное средство применяют для субъекта, который предварительно подвергался лечению, по меньшей мере, одним другим терапевтическим средством. Термины «предварительно подвергаемый лечению» или «предварительное лечение» подразумевают субъекта, который получал одно или несколько других терапевтических средств (например, подвергался лечению, по меньшей мере, одним другим противораковым терапевтическим средством) до получения лекарственного средства, содержащего связывающую анти-HER3-молекулу, например, антитело или его антигенсвязывающий фрагмент, вариант или производное. Необязательно, чтобы у субъекта наблюдали ответ на предварительное лечение предшествующим терапевтическим средством или терапевтическими средствами. Таким образом, субъект, который получает лекарственное средство, содержащее связывающую анти-HER3-молекулу, например, антитело или его антигенсвязывающий фрагмент, вариант или производное, мог отвечать или мог быть не в состоянии отвечать на предварительное лечение предыдущим средством лечения или одним или несколькими предыдущими средствами лечения, когда предварительное лечение включало в себя применение множества средств лечения.
Изобретение также относится к совместному введению связывающей анти-HER3-молекулы, например, антитела согласно изобретению или его антигенсвязывающего фрагмента, варианта или производного и, по меньшей мере, одного другого терапевтического средства. Анти-HER3-антитело и, по меньшей мере, одно другое терапевтическое средство можно совместно вводить вместе с одной композиции или можно совместно вводить в одно и то же время или перекрывающиеся промежутки времени в виде отдельных композиций.
Изобретение также относится к применению связывающей анти-HER3-молекулы, например, антитела согласно изобретению или его антигенсвязывающего фрагмента, варианта или производного, в производстве лекарственного средства для лечения у субъекта злокачественной опухоли, при этом связывающую анти-HER3-молекулу вводят до лечения субъекта, по меньшей мере, одним другим терапевтическим средством.
VIII. Диагностика
Изобретение, кроме того, относится к диагностическому способу, применимому при диагностике заболеваний, опосредованных HER3-экспрессирующими клетками, таких как некоторые типы злокачественной опухоли, включая, например, рак ободочной кишки, рак легкого, рак желудка, плоскоклеточный рак головы и шеи, меланому, рак поджелудочной железы, рак простаты и рак молочной железы, который включает в себя измерение уровня экспрессии белка или транскрипта HER3 в ткани или других клетках или жидкости организма, полученных от индивидуума, и сравнение измеренного уровня экспрессии со стандартным уровнем экспрессии HER3 в нормальной ткани или жидкости организма, при этом повышение уровня экспрессии по сравнению со стандартом является показателем расстройства.
Анти-HER3-антитела согласно изобретению и их антигенсвязывающие фрагменты, варианты и производные можно применять для анализа уровней белка HER3 в биологическом образце с использованием классических иммуногистологических способов, известных специалистам в данной области (например, см. Jalkanen с соавторами, J. Cell Biol. 707: 976-985 (1985); Jalkanen с соавторами, J. Cell Biol. 105: 3087-3096 (1987)). Другие основанные на антителах способы, применимые для выявления экспрессии белка HER3, включают иммуноанализы, такие как твердофазный иммуноферментный анализ (ELISA), иммунопреципитация или Вестерн-блоттинг. Подходящие анализы описаны более подробно в настоящем описании.
Под «анализом уровня экспрессии полипептида HER3» подразумевают качественное или количественное измерение или оценку уровня полипептида HER3 в первом биологическом образце либо непосредственно (например, путем определения или оценки абсолютного уровня белка), либо относительно (например, путем сравнения с уровнем ассоциированного с заболеванием полипептида во втором биологическом образце). Уровень экспрессии полипептида HER3 в первом биологическом образце можно измерить или оценить и сравнить со стандартным уровнем полипептида HER3, при этом стандарт получают на основе второго биологического образца, получаемого от индивидуума, не имеющего расстройства, или определяют по средним уровням в популяции индивидуумов, не имеющих расстройства. Как будет понятно в данной области, после того «стандартный» уровень полипептида HER3 становится известным, его можно многократно использовать в качестве стандарта для сравнения.
Изобретение, кроме того, относится к диагностическому способу, применимому при диагностике заболеваний, опосредованных HER3-экспрессирующими клетками, таких как некоторые типы злокачественной опухоли, включая, например, рак ободочной кишки, рак легкого, рак желудка, плоскоклеточный рак головы и шеи, меланому, рак поджелудочной железы, рак простаты и рак молочной железы, который включает в себя измерение уровня активности белка HER3 в ткани или других клетках или жидкости организма, полученных от индивидуума, и сравнение измеренного уровня активности со стандартным уровнем активности HER3 в нормальной ткани или жидкости организма, при этом повышение уровня активности по сравнению со стандартом является показателем расстройства.
Изобретение, кроме того, относится к диагностическому способу, применимому при лечении заболеваний, опосредованных HER3-экспрессирующими клетками, таких как некоторые типы злокачественной опухоли, включая, например, рак ободочной кишки, рак легкого, рак желудка, плоскоклеточный рак головы и шеи, меланому, рак поджелудочной железы, рак простаты и рак молочной железы, который включает в себя измерение уровня активности белка HER3 в ткани или других клетках или жидкости организма, полученных от индивидуума во время лечения заболевания, опосредованного HER3-экспрессирующими клетками, и сравнение измеренного уровня активности со стандартным уровнем активности HER3 в нормальной ткани или жидкости организма и/или сравнение измеренного уровня активности со стандартным уровнем активности HER3 в ткани или жидкости организма, полученных от индивидуума до лечения, при этом снижение уровня активности по сравнению со стандартом является показателем ингибирования активности HER3.
Под «анализом уровня активности белка HER3» подразумевают качественное или количественное измерение или оценку активности белка HER3 в первом биологическом образце либо непосредственно (например, путем определения или оценки абсолютного уровня активности) или относительно (например, путем сравнения с уровнем активности во втором биологическом образце). Уровень активности белка HER3 в первом биологическом образце можно измерить или оценить и сравнить со стандартной активностью белка HER3, при этом стандарт получают на основе второго биологического образца, полученного от индивидуума, не имеющего заболевания, или определяют по средним уровням в популяции индивидуумов, не имеющих заболевания, или от индивидуума перед лечением. Как будет понятно в данной области, после того «стандартный» уровень активности белка HER3 становится известным, его можно многократно использовать в качестве стандарта для сравнения. В некоторых аспектах уровень активности HER3 в биологическом образце измеряют или оценивают или сравнивают, выявляя фосфорилированный HER3 в биологическом образце. В конкретном аспекте уровень активности HER3 в биологическом образце измеряют или оценивают или сравнивают, выявляя фосфорилированный HER3 в биопсии кожи, при этом кожу стимулируют HRG до или после биопсии.
Подразумевают, что термин «биологический образец» означает любой биологический образец, полученный от индивидуума, из клеточной линии, культуры ткани или другого источника клеток, потенциально экспрессирующих HER3. Способы получения биопсии ткани и жидкостей организма млекопитающих хорошо известны в данной области.
В некоторых аспектах биологическую активность ингибитора HER3 (например, анти-HER3-антитела согласно изобретению и его антигенсвязывающих фрагментов, вариантов и производных), вводимого субъекту, можно выявить, используя анализ ex-vivo. В конкретных аспектах анализ ex vivo включает в себя определение уровня фосфорилированного HER3 в биопсии кожи, при этом кожу стимулируют HRG до или после биопсии. В конкретном аспекте получают парные образцы биопсии от субъекта, которому был введен ингибитор HER3. В конкретном аспекте HRG инъецируют в первую область кожи, а контрольный буфер инъецируют во вторую область кожи субъекта, которому вводят ингибитор HER3, при этом после требуемого периода времени (например, 10-60 минут) берут биопсию из первой и второй области кожи. В альтернативном аспекте первую биопсию кожи обрабатывают HRG, а вторую биопсию кожи обрабатывают контрольным буфером, при этом первая и вторая биопсии являются парными образцами биопсии кожи, взятыми от субъекта, которому был введен ингибитор HER3. В другом конкретном аспекте в образцах биопсии кожи выявляют уровень фосфорилированного HER3. В некоторых аспектах определяют разницу в уровнях фосфорилированного HER3 между первым (обработанным HRG) и вторым (обработанным контрольным буфером) образцами биопсии. В некоторых аспектах образец биопсии кожи гомогенизируют, и уровень фосфорилированного HER3 выявляют в ELISA. В следующих аспектах уровни фосфорилированного HER3 в образцах биопсии от субъекта, которому был введен ингибитор HER3, сравнивают с уровнями фосфорилированного HER3 в образцах биопсии кожи от контрольного субъекта, которому не вводили ингибитор HER3, при этом снижение уровня фосфорилированного HER3 в образцах биопсии от субъекта, которому был введен ингибитор HER3, является мерой биологической активности ингибитора HER3. В альтернативных аспектах уровни фосфорилированного HER3 в образцах биопсии кожи от субъекта, которому был введен ингибитор HER3, сравнивают с уровнями фосфорилированного HER3 в образцах биопсии кожи от того же субъекта, взятых до введения ингибитора HER3, при этом снижение уровня фосфорилированного HER3 в образцах биопсии кожи от субъекта после введения ингибитора HER3 является мерой биологической активности ингибитора HER3. Другие конкретные аспекты способов подробно описаны в разделе 5.15 «Примеры».
IX. Наборы, содержащие HER3-связывающие молекулы
Настоящее изобретение относится к наборам, которые содержат HER3-связывающую молекулу, например, анти-HER3-антитело или его антигенсвязывающий фрагмент согласно изобретению, описанные в настоящей публикации, и которые можно применять для осуществления способов, описанных в настоящей публикации. В некоторых аспектах набор содержит, по меньшей мере, одно очищенное анти-HER3-антитело или его антигенсвязывающий фрагмент в одной или нескольких емкостях. В некоторых аспектах наборы содержат все компоненты, необходимые и/или достаточные для осуществления анализа для выявления, включая все контроли, инструкции по осуществлению анализов и любую необходимую компьютерную программу для анализа и представления результатов. Специалист в данной области легко поймет, что раскрытая HER3-связывающая молекула, например, анти-HER3-антитело или его антигенсвязывающий фрагмент согласно настоящему изобретению, легко можно включить в одну из разработанных форм наборов, которые хорошо известны в данной области.
X. Иммуноанализы
Связывающие анти-HER3-молекулы, например, антитела или их антигенсвязывающие фрагменты, варианты или производные молекул согласно изобретению можно анализировать в отношении иммноспецифичного связывания любым способом, известным в данной области. Иммуноанализы, которые можно использовать, включают без ограничения системы конкурентного и неконкурентного анализа с использованием методики, такой как Вестерн-блоты, РИА, ELISA (твердофазный иммуноферментный анализ), иммуноанализы типа «сэндвич», анализы иммунопреципитации, реакции преципитации, реакции диффузной преципитации в геле, анализы иммунодиффузии, анализы агглютинации, анализы фиксации комплемента, иммунорадиометрические анализы, флуоресцентные иммуноанализы, иммуноанализы с использованием белка A, при этом названы только некоторые анализы. Такие анализы являются обычными и хорошо известными в данной области (см., например, публикацию Ausubel с соавторами, (1994) Current Protocols in Molecular Biology (John Wiley and Sons, Inc., NY) Vol. 1, которая включена в настоящее описание в виде ссылки в полном объеме).
HER3-связывающие молекулы, например, анти-HER3 антитела или их антигенсвязывающие фрагменты, варианты или производные молекул согласно изобретению можно применять гистологически в анализах на основе иммунофлуоресценции, иммуноэлектронной микроскопии или неиммунологических анализах для выявления in situ рецепторов HER3 или их консервативных вариантов или пептидных фрагментов. Выявление in situ можно осуществлять, извлекая гистологический образец из организма пациента и нанося на него меченую HER3-связывающую молекулу, например, анти-HER3-антитело или его антигенсвязывающий фрагмент, вариант, или производное, предпочтительно нанося наслаиванием меченой HER3-связывающей молекулы (например, антитела или фрагмента) на биологический образец. Применяя такой способ можно определить не только присутствие HER3 или консервативных вариантов или пептидных фрагментов, но также его распределение в исследуемой ткани. Применяя настоящее изобретение, специалисты легко могут понять, что любой из широкого множества гистологических способов (таких как способы окрашивания) можно модифицировать, чтобы добиться такого выявления in situ.
Активность связывания такой партии HER3-связывающих молекул, например, анти-HER3-антител или их антигенсвязывающих фрагментов, вариантов или производных, можно определить согласно хорошо известным способам. Специалисты в данной области смогут определить рабочие и оптимальные условия анализа для каждого определения, используя обычные эксперименты.
Способы и реагенты, подходящие для определения характеристик связывания выделенной HER3-связывающей молекулы, например, анти-HER3-антитела или его антигенсвязывающего фрагмента, варианта или измененного/мутантного производного, известны в данной области и/или коммерчески доступны. Оборудование и компьютерная программа, созданные для таких кинетических анализов, коммерчески доступны (например, BIAcore, компьютерная программа BIAevaluation, GE Healthcare; компьютерная программа KinExa, Sapidyne Instruments).
При практическом осуществлении настоящего изобретения будут использованы, если не указано иное, обычные способы клеточной биологии, культивирования клеток, молекулярной биологии, трансгенной биологии, микробиологии, рекомбинантной ДНК и иммунологии, которые известны специалистам в данной области. Такие способы полностью описаны в литературе. См., например, Sambrook et al., ed. (1989) Molecular Cloning A Laboratory Manual (2nd ed.; Cold Spring Harbor Laboratory Press); Sambrook et al., ed. (1992) Molecular Cloning: A Laboratory Manual, (Cold Springs Harbor Laboratory, NY); D.N. Glover ed., (1985) DNA Cloning, Volumes I and II; Gait, ed. (1984) Oligonucleotide Synthesis; Mullis et al. U.S. Pat. No. 4,683,195; Hames and Higgins, eds. (1984) Nucleic Acid Hybridization; Hames and Higgins, eds. (1984) Transcription And Translation; Freshney (1987) Culture Of Animal Cells (Alan R. Liss, Inc.); Immobilized Cells And Enzymes (IRL Press) (1986); Perbal (1984) A Practical Guide To Molecular Cloning; the treatise, Methods In Enzymology (Academic Press, Inc., N.Y.); Miller and Calos eds. (1987) Gene Transfer Vectors For Mammalian Cells, (Cold Spring Harbor Laboratory); Wu et al., eds., Methods In Enzymology, Vols. 154 and 155; Mayer and Walker, eds. (1987) Immunochemical Methods In Cell And Molecular Biology (Academic Press, London); Weir and Blackwell, eds., (1986) Handbook Of Experimental Immunology, Volumes I-IV; Manipulating the Mouse Embryo, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., (1986); и в Ausubel et al. (1989) Current Protocols in Molecular Biology (John Wiley and Sons, Baltimore, Md.).
Общие принципы конструирования антител приведены в публикации Borrebaeck, ed. (1995) Antibody Engineering (2nd ed.; Oxford Univ. Press). Общие принципы конструирования белков приведены в публикации Rickwood с соавторами, (1995) Protein Engineering, A Practical Approach (IRL Press at Oxford Univ. Press, Oxford, Eng.). Общие принципы антител и связывания антитело-гаптен приведены в публикациях: Nisonoff (1984) Molecular Immunology (2nd ed.; Sinauer Associates, Sunderland, Mass.); и Steward (1984) Antibodies, Their Structure and Function (Chapman and Hall, New York, N.Y.). Кроме того, стандартные способы иммунологии, известные в данной области и не описанные конкретно, обычно осуществляют, следуя описаниям, приведенным в Current Protocols in Immunology, John Wiley & Sons, New York; Stites et al., eds. (1994) Basic and Clinical Immunology (8th ed; Appleton & Lange, Norwalk, Conn.) and Mishell and Shiigi (eds) (1980) Selected Methods in Cellular Immunology (W.H. Freeman and Co., NY).
Стандартные ссылочные работы, описывающие общие принципы иммунологии, включают Current Protocols in Immunology, John Wiley & Sons, New York; Klein (1982) J., Immunology: The Science of Self-Nonself Discrimination (John Wiley & Sons, NY); Kennett et al., eds. (1980) Monoclonal Antibodies, Hybridoma: A New Dimension in Biological Analyses (Plenum Press, NY); Campbell (1984) "Monoclonal Antibody Technology" in Laboratory Techniques in Biochemistry and Molecular Biology, ed. Burden et al., (Elsevere, Amsterdam); Goldsby et al., eds. (2000) Kuby Immunnology (4th ed.; H. Freemand & Co.); Roitt et al. (2001) Immunology (6th ed.; London: Mosby); Abbas et al. (2005) Cellular and Molecular Immunology (5th ed.; Elsevier Health Sciences Division); Kontermann and Dubel (2001) Antibody Engineering (Springer Verlan); Sambrook and Russell (2001) Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Press); Lewin (2003) Genes VIII (Prentice Hall2003); Harlow and Lane (1988) Antibodies: A Laboratory Manual (Cold Spring Harbor Press); Dieffenbach and Dveksler (2003) PCR Primer (Cold Spring Harbor Press).
Все ссылки, цитированные выше, а также все ссылки, цитированные в настоящем описании, включены в настоящее описание в виде ссылки в полном объеме.
Следующие далее примеры предлагаются с целью иллюстрации, но никоим образом не являются ограничивающими.
ПРИМЕРЫ
Аспекты настоящего изобретения могут быть дополнительно определены при обращении к следующим не ограничивающим примерам, в которых подробно описано получение некоторых антител согласно настоящему изобретению и способы применения антител согласно настоящему изобретению. Специалистам в данной области будет понятно, что на практике могут быть осуществлены многочисленные модификации, как материалов, так и способов, не выходя за рамки объема настоящего изобретения.
Пример 1. Способы выделения/оптимизации моноклональных анти-HER3-антител
1.1. Антигены и клеточные линии
Все рекомбинантные химеры человеческий Her1(ECD)/Fc, химеры человеческий HER2(ECD)/Fc, химеры человеческий HER3(ECD)/Fc и человеческий Her4(ECD)/Fc приобретали из R&D Systems (Minneapolis, MN) и сливали с C-концевой 6X-гистидиновой меткой через линкерный пептид. Рекомбинантную химеру мышиный HER3(ECD)/Fc получали своими силами. Клетки злокачественной опухоли молочной железы человека KPL-4 культивировали в DMEM с добавлением 5% фетальной сыворотки теленка (FBS).
1.2. Селекция библиотеки HER3-связывающих агентов - идентификация антитела клона 16 (CL16)
Немеченый и биотинилированный HER3(ECD)/Fc использовали в качестве мишеней для селекции HER3-связывающих агентов из библиотеки Fab человека в фаговом дисплее Dyax Fab 310 (Dyax, Cambridge, MA). Осуществляли два параллельных пэннинга: пэннинг с улавливанием и пэннинг в растворе. В случае пэннинга с улавливанием вводимый фаг сначала инкубировали с поликлональным IgG человека, который предварительно улавливали на иммунологических пробирках посредством иммобилизованного рекомбинантного белка A/G, и затем подвергали селекции, используя немеченую мишень, улавливаемую на иммунологических пробирках посредством иммобилизованного рекомбинантного белка A/G.
В случае пэннинга в растворе вводимому фагу обеспечивали возможность для инкубации с поликлональным IgG человека, магнитными шариками, покрытыми стрептавидином, с погашенным биотином для деселекции и затем подвергали селекции с использованием биотинилированной мишени с последующей инкубацией с магнитными шариками, покрытыми стрептавидином, для улавливания фага, связанного с мишенью. После удаления несвязанного фага в результате тщательной промывки TPBS (1×PBS/0,1% твин-20) связанный фаг элюировали, используя 100 мМ TEA (триэтиламин). Элюированный фаг и фаг, оставшийся на шариках после пэннинга в растворе, затем модифицировали и подвергали следующим раундам селекции. Осуществляли три раунда селекции для каждого из параллельных исследований, проводимых при селекции.
Процент позитивных связывающихся фагов был в диапазоне от менее чем 1% при использованием пэннинга с улавливанием до 68% при использовании трех раундов пэннинга в растворе (таблица 3).
1.3. Скрининг агентов, связывающих HER3 человека и мыши, с использованием ELISA фагов
Фаги, обогащенные в результате второго и третьего раундов селекции, подвергали скринингу в ELISA фагов в отношении связывания HER3 человека и мыши. 96-луночные планшеты с половинным объемом лунок покрывали, используя раствор 5 мкг/мл, по 50 мкл на лунку разных антигенов, разбавленных в 1×PBS, pH 7,4, в течение ночи при 4°C. Покрытые планшеты блокировали 3% (масс./об.) обезжиренным молоком в TPBS в течение 1 часа при комнатной температуре и промывали два раза в TPBS. Затем планшеты инкубировали в течение 1 часа с надосадками фагов, полученных в течение ночи. После промывки десять раз в TPBS планшеты инкубировали с анти-M13-антителом, конъюгированным с пероксидазой хрена (HRP), в течение 1 часа и промывали десять раз. Планшеты проявляли, используя раствор субстрата пероксидазы тетраметилбензидина (TMB), реакции останавливали, добавляя 0,18 М H2SO4, и планшеты регистрировали при 450 нм на устройство для считывания планшетов для ELISA.
Идентифицировали 29 уникальных позитивных связывающих агентов, которые перекрестно взаимодействовали с мышиным HER3 (в виде слияния HER3-Fc). Ни один из идентифицированных связывающих агентов не проявлял перекрестную реактивность по отношению к HER2 или Her4 (данные не показаны).
1.4. Преобразование Fab в целые IgG-антитела и экспрессия
Вариабельную область легкой цепи (VL) и вариабельную область тяжелой цепи (VH) иммуноглобулина из позитивных фаговых клонов создавали, используя ПЦР, и встраивали в вектор экспрессии IgG1 человека, содержащий константную область легкой цепи лямбда и область CH1-шарнир-CH2-CH3 IgG1. Чтобы экспрессировать IgG1-антитела, клетки эмбриональной почки человека 293-F временно трансфицировали векторами с преобразованным IgG, используя реагент 293fectinTM (Invitrogen, Carlsbad, CA). Кондиционированную среду собирали через 10 дней после трансфекции, объединяли и стерильно фильтровали. IgG1 очищали, используя шарики с белком A. Конечный раствор элюированного IgG1 диализовали против PBS, и концентрации IgG1 определяли, используя количественный анализ белка.
Клон 16 (CL16; SEQ ID NO: 1 и 2, аминокислотные последовательности VL и VH, соответственно) преобразовывали в IgG1 человека.
1.5. Определение интернализации антитела клона 16 (CL16) с использованием иммунофлуоресценции
Клетки злокачественной опухоли молочной железы человека KPL-4 метили антителом клона 16 (CL16). Инкубация клеток с CL16 приводила к увеличению эндоцитоза HER3, который предотвращал образование активных сигнальных комплексов рецептора с HER2 на клеточной поверхности.
Связанным с клеточной поверхностью антителам CL16 давали возможность интернализоваться, инкубируя клетки в условиях роста в течение либо нуля (неинтернализованные), либо 2,5 часов (интернализованные) (фиг. 1). Затем все клетки фиксировали 3,7% параформальдегидом, промывали в PBS, делали проницаемыми, обрабатывая 0,5% тритоном X-100 в PBS, и красили, используя 1 мкг/мл козьего антитела против IgG человека, меченого красителем Alexa Fluor® 488 (Invitrogen), перед добавлением антифединговой среды для приготовления препаратов и исследованием с использованием флуоресцентной микроскопии. Было обнаружено, что антитело CL16 интернализуется в клетки KPL-4. В нулевой момент времени наблюдали интенсивное окрашивание поверхности клеток KPL-4 (фиг. 1, 0 часов, верхняя панель), после инкубации в условиях роста в течение 2,5 часов окрашивание клеточной поверхности уменьшалось и сменялось внутриклеточным точечным окрашиванием, являющимся показателем интернализации (фиг. 1, 2,5 часа, нижние панели).
1.6. Конструирование фагового вектора, экспрессирующего Fab клона 16
ДНК, кодирующую антигенсвязывающий фрагмент (Fab) антитела клона 16, клонировали в модифицированном основанном на M13 фаговом экспрессирующем векторе, ранее описанным Dall'Acqua с соавторами (Dall'Acqua с соавторами, 2005, Methods. 36: 43-60). В таком модифицированном векторе была сконструирована ДНК константной области лямбда (λ) человека вместо легкой цепи каппа (κ человека. Экспрессия Fab-фрагмента происходила под контролем промотора LacZ, и секреция Fab-фрагмента была возможна благодаря сигнальным последовательностям фага P3, слитым с N-концами любой VH- и VL-цепи Fab-фрагмента. Клонирование осуществляли, используя основанный на гибридизации мутагенез, как описано Kunkel (Kunkel, T.A., 1985, Proc. Natl. Acad. Sci. USA; 82: 488-492) и Wu (Wu, H., 2003, Methods Mol. Biol. 207: 197-212).
Кратко, вариабельные области IgG клона 16 амплифицировали в полимеразной цепной реакции (ПЦР). Посредством гибридизации с последующей ДНК-полимеразной реакцией вариабельную область легкой цепи клона 16 интегрировали в рамке с константной областью лямбда человека, и вариабельную область тяжелой цепи клонировали в рамке с константной областью 1 тяжелой цепи (CH1) человека, соответственно. Затем фаговый вектор, содержащий Fab-фрагмент клона 16, выращивали в Escherichia coli штамма CJ236, чтобы получить содержащую уридин (U) одноцепочечную ДНК (оцДНК), как описано Wu и An (Wu, H. and An, LL., 2003, Methods Mol. Biol. 207: 213-33). Содержащую уридин оцДНК использовали в качестве матрицы для введения требуемых мутаций с целью повышения аффинности связывания с HER3.
1.7. Приведение клона 16 (CL16) к зародышевой линии
Анализ последовательностей показывает, что каркасы VH клона 16 (CL16) имеют 100% идентичность последовательности с геном VH зародышевой линии 3-23, тогда как каркасы VL отличаются по 6 положениям от своего ближайшего гена зародышевой линии 47*01. Осуществляли сайт-специфичный мутагенез, чтобы изменить каждую и все аминокислоты, которые отличаются от аминокислот, кодируемых геном зародышевой линии 47*01. В частности, вводили шесть точечных мутаций в вариабельные области легкой цепи, а именно: Y2S, E3V, S18R, M21I, H38Q и S50Y, где первая буква означает однобуквенный код аминокислоты из исходного клона 16, цифры означают номер остатка в каркасе (по Кабату), а вторая бука означает однобуквенный код аминокислоты последовательности зародышевой линии. Последовательности, соответствующие VL исходного CL16 и VL CL16 зародышевой линии (GL), см. на фиг. 2A и фиг. 2C, соответственно. Полученные варианты были представлены в виде Fab, и их связывание с рекомбинантным белком HER3 определяли в ELISA.
Результаты связывания показали, что аминокислотная мутация H38Q в каркасе 2 улучшала связывание по сравнению с родительским клоном 16, как было измерено в ELISA. Напротив, мутация S49Y в том же каркасе оказывала негативное влияние на связывание. Другие точечные мутации не оказывали влияния на связывание HER3. В случае полностью приведенного к зародышевой линии мутанта с мутациями во всех 6 аминокислотах, не соответствующих зародышевой линии, наблюдали сходную степень уменьшения связывания, как и в случае точечной мутации S50Y, что свидетельствует о том, что аминокислота S50 принимает участие в связывании. Дополнительное тестирование клона со всеми точечными мутации к зародышевой линии, за исключением S50Y, показало сохранение и/или увеличение связывания с HER3 по сравнению с родительским клоном 16. Такой частично приведенный к зародышевой линии клон, клон 16 (GL) (также называемый в настоящем описании «GL-P6»), использовали в качестве матрицы для дальнейшей оптимизации аффинности.
1.8. Оптимизация аффинности клона 16 (CL16)
Каждую аминокислоту из всех 6 определяющих комплементарность областей (CDR) клона зародышевой линии GL-P6 по отдельности подвергали мутациям, приводящим к другим 20 аминокислотам, используя способ мутагенеза на основе гибридизации (Kunkel, 1985). Использовали два набора праймеров ДНК, один, содержащий кодон NSS, кодирующий 8 аминокислот, и другой, содержащий кодон NWS, кодирующий 12 разных аминокислот, чтобы ввести мутации в каждое целевое положение CDR. Отдельные вырожденные праймеры использовали в реакциях основанного на гибридизации мутагенеза. Кратко, каждый вырожденный праймер фосфорилировали, затем использовали в соотношении 10:1 с оцДНК Fab GL-16, в которую не вводили уридина. Смесь нагревали до 95°C, затем охлаждали до 55°C в течение 1 часа. Затем добавляли лигазу T4 и ДНК-полимеразу T7 и смесь инкубировали в течение 1,5 часа при 37°C. Продукты синтеза, CDR VH и VL CDR, соответственно объединяли; однако библиотеки NSS и NWS хранили по отдельности и подвергали скринингу независимо. Обычно 1 мкл объединенной ДНК библиотеки путем электропорации вводили в XL1-Blue для образования бляшек на бактериальном газоне XL1-Blue или для получения Fab-фрагментов (Wu and An, 2003).
1.9. Первичный скрининг библиотеки Fab
Первичный скрининг заключался в анализе ELISA по одной точке (SPE), который осуществляли, используя надосадок культуры бактерий, выращенной в 96-луночной планшете (с глубокими лунками), и инфицированной отдельными рекомбинантными клонами M13, как описано в других публикация (Wu and An, 2003). Кратко, такой ELISA с улавливанием заключался в покрывании отдельных лунок 96-луночного иммунологического планшета Maxisorp примерно 50 нг антитела овцы против Fd человека (Biodesign International, ME) в карбонатном буфере с pH 8,5 в течение ночи при 4°C. На следующий день планшет блокировали 3% БСА в буфере PBS в течение 1 часа при комнатной температуре. Затем в планшет добавляли Fab-надосадок и инкубировали при комнатной температуре в течение 1 часа. После промывки в лунку добавляли 0,1 мкг биотинилированного белка HER3 и смесь инкубировали в течение 1,5 часа при комнатной температуре. После этого осуществляли инкубацию с конъюгатом нейтравидин-пероксидаза хрена (HRP) (Pierce, IL) приблизительно в течение 40 минут при комнатной температуре. Активность HRP определяли, используя субстрат тетраметилбензидин (TMB), и реакцию гасили, используя 0,2 М H2SO4. Планшеты регистрировали при 450 нм.
Клоны, дающие сигнал оптической плотности (OD) при 450 нм, превышающий сигнал родительского клона Fab GL-P6, собирали и снова выращивали (15 мл) (Wu and An, 2003) и повторно анализировали в ELISA (как описано выше) в двух повторах, чтобы подтвердить положительные результаты. Клоны, которые повторно давали сигнал выше, чем сигнал Fab GL-P6, секвенировали. Затем определяли концентрацию белка Fab каждого клона, который имеет изменение в CDR, в количественном ELISA Fab, при этом Fab с известной концентрацией использовали в качестве эталона сравнения. Концентрацию Fab определяли, сравнивая сигналы ELISA с сигналами, генерируемыми эталонным Fab. Анализ связывания повторяли еще раз для всех позитивных вариантов, используя нормализованные концентрации Fab, чтобы определить относительную аффинность связывания мутантных Fab и родительского Fab GL-P6.
Анализ связывания ELISA показал, что два VH-варианта, обозначаемых как клон 14C7 и клон 15D12, которые содержали точечные мутации Y50I или Y50V, соответственно, в CDR2, демонстрировали приблизительно 5-кратное улучшение связывания HER3 по сравнению с родительским, приведенным к зародышевой линии клоном GL-P6. При проведении мутагенеза VL идентифицировали несколько одиночных мутаций либо в CDR1, например, клон 4H6 (содержащий точечную мутацию S24R), клон 6E3 (содержащий точечную мутацию S27L), либо в CDR3, например, клон 5H6 (содержащий точечную мутацию S94G), клон 8A3 (содержащий точечную мутацию S96aI), клон 4C4 (содержащий точечную мутацию S96aR), клон 2B11 (содержащий точечную мутацию S96aP) и клон 2D1 (содержащий мутацию V97A), демонстрирующие улучшенное связывание.
В частности, замена аминокислоты S96a в VL-CDR3 либо изолейцином (I), либо аргинином (R), либо пролином (P) приводила к 3,5-кратному, 8,6-кратному и 32-кратному улучшению связывания, соответственно.
1.10. Комбинаторный скрининг библиотеки Fab
Точечные мутации в VH и VL, которые, как было определено, полезны для связывания с HER3, дополнительно объединяли, чтобы добиться дополнительной синергии связывания. Комбинаторные мутанты экспрессировали в виде Fab и подвергали скринингу, используя ELISA связывания HER3. При объединении любой одной из точечных мутаций Y50I или Y50V в VH-цепи Fab-фрагмента с мутациями VL, по-видимому, не давало пользы, а снижало связывание с HER3, объединение нескольких мутаций VL дополнительно улучшало связывание. Такие сочетания мутаций VL включают сочетания в клоне 1A4 (содержащем точечные мутации L96P, S97P и V100A), клоне 2C2 (содержащем точечные мутации S26L, L96P, S97P и V100A), клоне 2F10 (содержащем мутации S97P и V100A) и клоне 3E1 (содержащем точечные мутации S23R, L96P, S97P и V100A).
1.11. Превращение оптимизированных по аффинности вариантов Fab в форму IgG и экспрессия антител
Варианты с одиночными мутациями и сочетаниями мутаций, проявляющими улучшенное связывание, превращали в форму IgG для дальнейшей характеристики. Вариабельные области каждого варианта амплифицировали в ПЦРЮ, используя праймеры, которые кодировали сайты рестрикции, чтобы облегчить клонирование в экспрессирующем IgG векторе млекопитающих для экспрессии с использованием клеток HEK 293F. Секретированные растворимые белки IgG1 человека очищали из кондиционированной среды непосредственно на колонках с белком A HiTrap объемом 1 мл (GE Healthcare, NJ) согласно инструкциям производителя. Очищенные образцы IgG1 человека (обычно гомогенность >95%, судя по электрофорезу в полиакриламидном геле с додецилсульфатом натрия) диализовали против PBS, мгновенно замораживали и хранили при -70°C.
Связывание очищенных IgG исследовали, используя ELISA связывания HER3. IgG с сочетанием мутаций проявляли улучшенное связывание, которое определяли по общему сигналу связывания, при этом в случае 2C2 наблюдали наиболее значимое улучшение связывания по сравнению с родительским клоном 16 и другими вариантами с сочетаниями мутаций. Связывание IgG с HER3 мыши и макака-крабоеда также тестировали в ELISA. Результаты показали улучшенное связывание комбинированным мутантов с их паралогичными видами HER3.
Выравнивание аминокислотных последовательностей вариабельных областей легкой и тяжелой цепей для каждой из идентифицированных отдельных мутаций показано на фиг. 2A и фиг. 2B, соответственно, в таблице 4 указаны номера последовательностей SEQ ID NO для каждого клона. Выравнивание вариабельных областей легкой цепи для каждого из клонов с сочетаниями мутаций представлен на фиг. 2C.
[FW1]X1GSX2SNIGLNYVS[FW2]RNNQRPS[FW3]AAWDDX3X4X5GEX6[FW4],
где [FW1], [FW2], [FW3] и [FW4] означают каркасные области VL,
где
(a) X1 означает аминокислотные остатки аргинин (R) или серин (S),
(b) X2 означает аминокислотные остатки серин (S) или лейцин (L),
(c) X3 означает аминокислотные остатки серин (S) или глицин (G),
(d) X4 означает аминокислотные остатки лейцин (L) или пролин (P),
(e) X5 означает аминокислотные остатки аргинин (R), изолейцин (I), пролин (P) или серин (S), и
(f) X6 означает аминокислотные остатки валин (V) или аланин (A).
[FW5]YYYMQ[FW6]X7IGSSGGVTNYADSVKG[FW7]VGLGDAFDI[FW8],
где [FW5], [FW6], [FW7] и [FW8] означают каркасные области VH,
где X7 означает аминокислотные остатки тирозин (Y), изолейцин (I) или валин (V).
1.12. Исследования связывания моноклонального анти-HER3-антитела
Кинетические константы скорости (kon, koff) и равновесные константы диссоциации (KD) для связывания анти-HER3-IgG с внеклеточным доменом белка HER3 человека определяли, используя методику резонанса поверхностного плазмона BIAcore™, путем измерения связывания внеклеточного домена HER3 человека (huHER3(ECD)) с IgG, улавливаемым на поверхности сенсорного чипа. Отдельные константы скорости ассоциации (kon) и диссоциации (koff) затем вычисляли на основании полученных кривых связывания, используя компьютерную программу BIAevaluation, доступную от поставщика. Данные подгоняли к модели связывания 1:1, которая включала член для корректировки ограниченного переносом массы связывания в случае его выявления. Затем на основании таких констант скорости вычисляли кажущуюся константу диссоциации (KD) для взаимодействия IgG с белком внеклеточного домена HER3 человека в виде отношения koff/kon.
На основании графиков BIAcore высокого разрешения константы скорости ассоциации и диссоциации для связывания родительского IgG клона 16 с внеклеточным доменом HER3 человека составляли 5,29×105/М сек и 73,0×10-4/сек, соответственно, что давало значение кажущейся KD 14 нМ. При сравнении константы скорости ассоциации для связывания вариантов IgG с улучшенной аффинностью с внеклеточным доменом HER3 человека были сходны с константами скорости ассоциации, измеренными для родительского IgG, в диапазоне от 3,41×105 до 4,32×105/М сек. Те же графики также использовали для определения соответствующих констант скорости диссоциации для вариантов клона 16, которые были в диапазоне от 1,60×10-4 до 6,21×10-4/сек. Значения кажущейся KD для вариантов клона 16 вычисляли, как описано выше, и они были в диапазоне от 0,429 нМ (вариантный клон 2C2) до 1,44 нМ (вариантный клон P2B11). Индивидуальные ошибки kon и koff были низкими (≤~2% от вычисляемого параметра), и общая подгонка к кинетическим данным показала, что использование модели взаимодействия 1:1 было уместно. Также оценка не показала, что связывание было ограничено переносом массы.
В таблице 5 суммированы биофизические свойства комбинированных моноклональных клонов, представленных на фиг. 2C, включая значения Kon, Koff и KD, а также уровни экспрессии и выходы.
Моноклональные антитело 2C2, содержащее VL 2C2 (SEQ ID NO: 3) и VH исходного C16 (SEQ ID NO: 2), было наилучшим вариантом с наибольшим повышением аффинности, имеющим KD 0,4 нМ, что представляло собой 32-кратное улучшение по сравнению с родительским моноклональным антителом клона 16. Улучшение KD, главным образом, было результатом пониженной скорости распада комплекса. Также оценивали уровень экспрессии и выходы продукции. Все клоны моноклональных антител хорошо экспрессировались в 5-дневном исследовании временной трансфекции, при этом в данном исследовании наблюдали наибольший уровень экспрессии в случае моноклонального антитела 2C2. В случае всех наилучших вариантов с оптимизированной аффинностью наблюдали разные степени улучшения аффинности, но антитело 1A4 выпадало вследствие более низкой эффективности экспрессии.
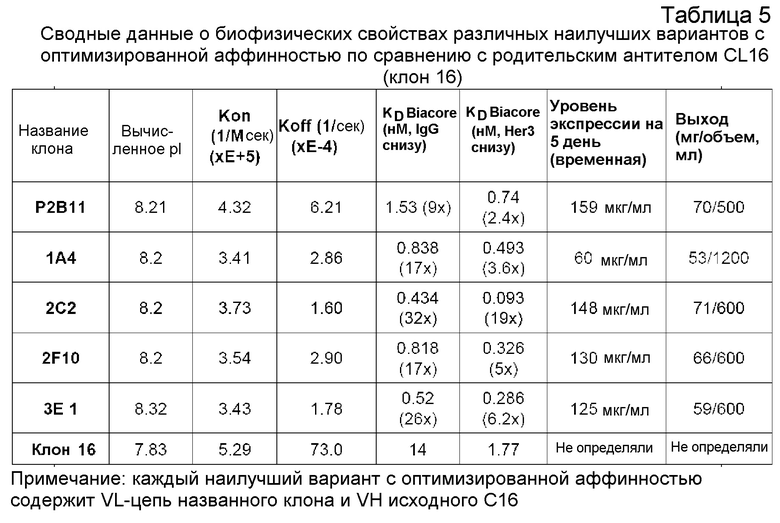
Осуществляли различные основанные на клетках анализы, чтобы оценить функциональное улучшение различных оптимизированных по аффинности наилучших вариантов по сравнению с клоном 16 в не зависимой от лиганда (линия клеток злокачественной опухоли рак молочной железы человека BT-474, ATCC № HTB-20™), а также в зависимой от лиганда (линия клеток злокачественной опухоли молочной железы человека T-47D, ATCC № HTB-133) моделях (обе линии клеток получали из ATCC), включая ингибирование пути передачи сигнала HER3 (pHER3 и pAKT), подавление клеточного роста (кратковременный 6-дневный анализ роста и долговременный анализ клоногенности), и отмена HRG-индуцированного pHER3 в клетках T-47D (дифференцированная эпителиальная подлиния T-47).
Анализы клоногенности осуществляли следующим образом. Клетки BT-474 высевали при плотности 1000 клеток/лунку в 6-луночные планшеты. После прикрепления в течение ночи клетки обрабатывали IgG, используемым в качестве изотипического контроля, или указанными моноклональными HER3-антителами, следуя концентрационной/дозовой кривой. Среду с соответствующими дозами моноклональных антител освежали один раз в неделю в течение трех недель. В конце 21 дня клетки обрабатывали для анализа Cell-titer-Glo (CTG), чтобы оценить ингибирование образования колоний различными моноклональными антителами (используя контрольный IgG as base-line). Значения IC50 получали в анализе Prizm.
Анализ 6-дневного роста BT-474 осуществляли, как указано для фиг. 4 (см. раздел 2.2 в примере 2, ниже). Анализ pAKT в BT-474 осуществляли по существу, как описано для фиг. 10 (см. раздел 2.6.1 в примере 2, ниже). Анализ HRG-индуцируемого pHER3 в T-47D осуществляли по существу, как описано для фиг. 3 (см. раздел 2.1 в примере 2, ниже), и FACS-анализ связывания и интернализации в T-47D осуществляли, используя такой же протокол, который описан для фиг. 16A (см. раздел 3.3.1 в примере 3, ниже).
Получали значения IC50 и максимальные уровни ингибирования в целях сравнения. Как показано в таблице 6, в случае наилучших вариантов с повышенной аффинностью наблюдали соответствующую 2-3-кратно повышенную эффективность в большинстве анализов. Родительский клон 16 и/или соответствующий оптимизированный клон, например, антитело клона 2C2 антитело (также просто называемое 2C2 или моноклональное антитело 2C2) дополнительно характеризовали в нескольких анализах in vitro и in vivo, как описано ниже.
Кроме того, вводили мутации в Fc-область оптимизированного клона 2C2, чтобы продлить время полужизни. В частности, M252Y, S254T, T256E, пронумерованные согласно индексу EU, как описано Kabat. Такая оптимизированная в отношении времени полужизни молекулу названа 2C2-YTE. Будет понятно, что могут быть введены другие мутации вместо или в сочетании с указанными тремя, см., например, патент США №7083784; международную публикацию заявки № WO 2009058492; публикации Dall'Acqua с соавторами, 2002 J. Immunol. 169: 5171-80; Zalevsky с соавторами, 2010, Nature Biotech. 28: 157-9). Показано, что 2C2-YTE ингибирует пролиферацию клеток BT-474 и рост колоний в такой же степени, как и 2C2 (данные не показаны).
Стабильную при хранении в холодильнике (2-8°C) композицию получали, готовя антитела (например, 50 мг/мл) в 25 мМ гистидине/гистин HCl, 205 мМ сахарозе, 0,02% полисорбате 80, pH 6,0.
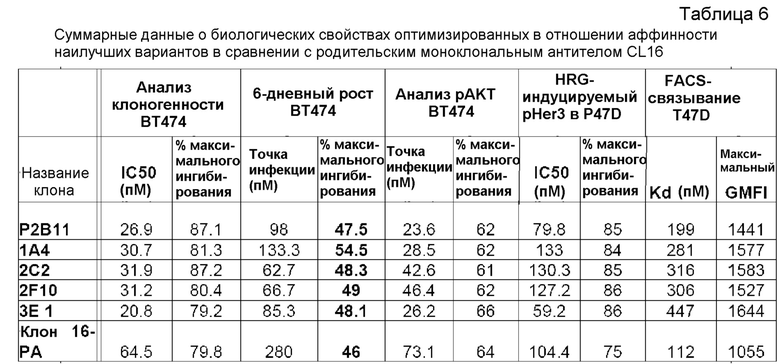
Пример 2. Характеристика моноклональных анти-HER3-антител
2.1. Анализ HRG-индуцируемого фосфорилирования HER3 (pHER3) в клетках MCF-7
MCF-7 (ATCC № HTB-22™) представляет собой линию клеток злокачественной опухоли молочной железы человека с экспрессией HER3, но без эндогенной экспрессии HRG. Клетки MCF-7 высевали при плотности 30000 клеток/лунку в 96-луночный планшет и давали возможность прикрепиться в течение ночи. Затем клетки подвергали голоданию без сыворотки в течение 24 часов перед обработкой. После голодания без сыворотки среду удаляли и заменяли бессывороточной средой, содержащей тестируемые и контрольные антитела, и клетки инкубировали при 37°C в течение 1 часа. Тестируемые антитела, используемые в данном примере и в дополнительных примерах, представленных ниже, включают анти-HER3-антитела, предлагаемые в настоящем описании, такие как клон 16, 2C2, 2C2-YTE; и анти-HER3-антитела, известные в данной области, в частности U1-59 (международная публикация патента WO 2007077028) и Ab#6 (публикация патента WO 2008/100624), обозначаемые в настоящем описании AMG и MM, соответственно. Тем временем готовили исходный раствор херегулина (HRGβ1, R&D Systems, Minneapolis, MN) в 4-кратной концентрации (80 нг/мл) в бессывороточной ростовой среде. В конце 1-часового периода инкубации HRGβ1 инъецировали в лунки (конечная концентрация 20 нг/мл) и инкубировали при 37°C в течение 20 минут. В конце обработки среду удаляли и клетки промывали PBS. Клетки лизировали в 80 мкл лизирующего буфера с тритоном X (Boston Bioproducts, Ashland, MA) с ингибиторами протеаз и фосфатаз (Calbiochem, La Jolla, CA) и хранили при -20°C вплоть до анализа. Затем осуществляли ELISA pHER3, следуя протоколу производителя (R&D Systems, DYC1769), используя 96-луночные планшеты для ELISA с половинным объемом Corning® Costar® 3690 (Corning Life Science, Lowell, MA) и 50 мкл лизата клеток на лунку.
Активацию HER3, отражаемую фосфорилированием HER3 (сокращенно s pHER3), стимулировали обработкой клеток HRGβ1. Предварительная обработка анти-HER3-мАт 2C2 вызывала зависимое от дозы подавление сигнала pHER3 в анализе ELISA pHER3 (фиг. 3, вверху). Описанные в литературе моноклональные анти-HER3-антитела MM и AMG также были активными в данном анализе, однако 2C2 было приблизительно в 5 раз более эффективным, как было определено на основании измерений IC50 (фиг. 3, внизу). Сходные результаты наблюдали для 2C2-YTE (данные не показаны).
2.2. Подавление роста клеток злокачественной опухоли молочной железы MDA-MB-175
MDA-MB-175 (ATCC № HTB-25™) представляет собой стабильную HRG-экспрессирующую (1β-изоформа), которая зависит от пути передачи сигнала HRG-HER3 для роста и жизнеспособности. Клетки высевали с плотностью 2000 клеток/лунку в 96-луночный планшет с белыми стенками и давали возможность прикрепиться в течение ночи. На следующий день среду удаляли и заменяли 100 мкл/лунку свежей полной ростовой среды, содержащей тестируемые и контрольные антитела. Затем планшеты инкубировали всего в течение 6 дней. Чтобы вычислить относительное количество клеток, использовали CellTiter-Glo™ (Promega, Madison, WI) согласно протоколу производителя. После добавления CellTiter-Glo™ планшеты инкубировали при комнатной температуре в течение 10 минут и измеряли люминесценцию, используя устройство для считывания микропланшетов.
Анализ роста осуществляли с использованием моноклональных анти-HER3-антител 2C2, MM или AMG. Как показано на фиг. 4, все три антитела достигали антипролиферативного эффекта в разной степени, при этом 2C2 проявлял наиболее высокую эффективность (IC50=0,14 мкг/мл) (фиг. 4, вверху) и более значительное подавление роста (72%) (фиг. 4, внизу).
2.3. Подавление роста клеток меланомы HMCB
HMCB (ATCC № CRL-9607(TM)) представляет собой стабильную HRG-экспрессирующую модель меланомы (1β-изоформа), зависимую от HRG-индуцируемой гетеродимеризации HER2-HER3. Клетки HMCB высевали с плотностью 750 на лунку в 100 мкл полной среды, содержащей 10% инактивированной нагреванием FBS в 96-луночные планшеты (Costar®). На следующий день антитела для обработки готовили в полной среде. Начальная концентрация всех моноклональных анти-HER3-антител и контрольного IgG составляла 10 мкг/мл, и серийные разведения готовили в полной среде. Среду для посева удаляли и добавляли растворы для обработки по 100 мкл на лунку в трех повторах.
Затем планшеты инкубировали в 5% CO2 при 37°C в течение 6 дней. В каждую лунку добавляли равные объемы реагента CellTiter-Glo™. Планшеты встряхивали на встряхивающем устройстве для планшетов в течение 10 минут при комнатной температуре, чтобы гарантировать полный лизис клеток. Люминесценцию измеряли с использованием устройства для считывания 2104 EnVision® Multilabel (PerkinElmer, Waltham, MA).
Как показано на фиг. 5, 2C2 снова было более эффективным, чем существующие антитела. 2C2 было в 8-30 раз более эффективным, чем описанные в литературе моноклональные анти-HER3-антитела AMG и MM, в ингибировании клеточного роста в линии клеток меланомы HMCB.
2.4. Анализ активности HER3 и AKT в клетках миеломы HMCB и клетках NSCLC A549
Оценивали способность наилучших вариантов HER3-антител подавлять путь передачи сигнала HER3 в моделях с аутокринной регуляцией HRG, HMCB (ATCC № CRL-9607(TM)) и A549 (ATCC № CCL-185). Клетки HMCB высевали по 105 на лунку в 24-луночные планшеты и в среду, содержащую 10% инактивированной нагреванием FBS, и давали возможность достичь слияния на 80% или больше перед обработкой антителами. Среду для посева удаляли и клетки подвергали инкубации с антителами. Моноклональные анти-HER3-антитела и контрольный IgG готовили в полной среде. Начальная концентрация всех анти-HER3-антител составляла 10 мкг/мл, и готовили серийные разведения. Контрольный IgG использовали только в концентрации 10 мкг/мл. Обработки осуществляли после удаления среды для посева. После инкубации в течение 6 часов (клетки HMCB) или 72 часов (клетки A549) в условиях 5% CO2 при 37°C клетки промывали один раз охлажденным на льду PBS и затем лизировали, добавляя восстанавливающий буфер Лэммли (Boston BioProducts, Ashland, MA).
После короткой инкубации собирали клеточные лизаты, равные количества наносили на бис-трис-гели Bis NuPAGE® Novex® (Invitrogen, Carlsbad, CA) и белки переносили на мембраны PVDF (Invitrogen, Carlsbad, CA). Мембраны блокировали 5% обезжиренным сухим молоком и 0,1% твином-20 (Sigma, St. Louis, MO) в TBS (pH 7,4) и инкубировали в течение ночи при 4°C с антителами к HER3 (антитело sc-285, Cell Signaling Technology, Beverly, MA), pHER3 (антитело 4791, Cell Signaling Technology, Beverly, MA), AKT (антитело 9272, Cell Signaling, Technology, Beverly, MA), pAKT (антитело 4060, Cell Signaling Technology, Beverly, MA), нейрегулину-1/HRG (антитело NRG1/HRG) (sc-348, Santa Cruz) и GAPDH (антитело G8795, Sigma, St. Louis, MO).
Мембраны промывали в 0,1% твин-20 в TBS и затем инкубировали в течение 1 часа со вторыми антителами и конъюгированным с перкосидазой хрена стрептавидином (GE Healthcare). После промывки полосы белков выявляют на рентгеновской пленке, используя хемилюминесцентный субстрат SuperSignal® West Femto и хемилюминесцентный субстрат SuperSignal® West Pico (Pierce/Thermo Scientific, Rockford, IL).
Как показано на фиг. 6 и 7, антитело 2C2 отменяло путь передачи сигнала HER3 как в клетках HMCB, так и в клетках A549. 2C2 эффективно подавляло pHER3 и расположенную ниже в каскаде эффекторную молекулу pAKT зависимым от дозы образом и было более эффективным, чем любое из описанных в литературе моноклональных анти-HER3-антител AMG или MM в клетках HMCB. Антитело 2C2 также подавляло pHER3 и расположенную ниже в каскаде эффекторную молекулу pAKT в клетках A549.
2.5. Анализ фосфорилирования HER3 (pHER3) в клеточных моделях рака легкого, желудка и молочной железы
2.5.1. Клеточный анализ pHER3
Клетки (клетки NSCLC HCC827, резистентные к гефитинибу клетки NSCLC HCC827, клетки злокачественной опухоли желудка MKN45, клетки злокачественной опухоли желудка Kato III или клетки злокачественной опухоли молочной железы BT-474 с амплификацией HER2) высевали с плотностью 30000 клеток/лунку в 96-луночный планшет и давали возможность прикрепиться в течение ночи. Затем клетки обрабатывали тестируемым и контрольными антителами согласно указанной дозовой кривой при 37°C в течение 4 часов. В конце обработки среду удаляли и клетки промывали PBS. Клетки лизировали в 80 мкл лизирующего буфера с тритоном X (Boston BioProducts, Ashland, MA) с ингибиторами протеаз и фосфатаз (Calbiochem, La Jolla, CA) и хранили при -20°C вплоть до анализа. Затем осуществляли ELISA pHER3, следуя протоколу производителя (R&D Systems, DYC1769, Minneapolis, MN), используя 96-луночные планшеты для ELISA с половинным объемом (Costar 3690) и 50 мкл клеточного лизата на лунку.
2.5.2. Подавление активности pHER3 в клетках HCC827
Клетки HCC827 (ATCC CRL-2868™), модель мутантного EGFR-зависимого немелкоклеточного рака легкого (NSCLC), обрабатывали тестируемыми или контрольными антителами, как описано выше в разделе примера 2.5.1 (см. выше). Как показано на фиг. 8A, антитело 2C2 способно частично ингибировать сигнал pHER3, тогда как описанные в литературе моноклональные анти-HER3-антитела AMG и MM были менее эффективны, и в 10 раз менее активными, чем 2C2.
2.5.3. Подавление активности pHER3 в резистентных к гефитинибу клетках HCC827
HCC827 несут и зависят от мутантного EGFR, который делает их высокочувствительными к ингибиторам тирозинкиназы EGFR (TKI), таким как гефитиниб. На родительские клетки HCC827 воздействовали постоянной токсической дозой гефитиниба и выделяли резистентные клоны, которые, как было показано, несут амплифицированный cMET, что является известным механизмом, благодаря которому злокачественные опухоли ускользают от терапии TKI. TKI-резистентные клетки HCC827 обрабатывали моноклональными анти-HER3-антителами, как описано выше в разделе примера 2.5.1 (см. выше). Как показано на фиг. 8B, моноклональное анти-HER3-антитело 2C2 подавляло активность HER3 в мутантных HCC827, которые были резистентными к гефитинибу. Подобно результатам, полученным для родительской клеточной линии, 2C2 проявлял более высокую эффективность, чем антитела AMG и MM (примерно в 10 раз более высокая эффективность) в TKI-резистентной клеточной линии HCC827.
2.5.4. Подавление активности pHER3 в клетках MKN45
Даже несмотря на то, что cMET не является членом Her-семейства, было показано, что он способен образовывать димеры с HER3. MKN45, модельную линию клеток злокачественной опухоли желудка с амплификацией cMET, использовали для оценки того, могут ли анти-HER3-антитела антагонизировать зависимую от cMET активацию HER3. Клетки MKN45 обрабатывали моноклональными анти-HER3-антителами, как описано выше в разделе примера 2.5.1. Как показано на фиг. 8C, все три моноклональных анти-HER3-антитела (2C2, AMG и MM) были способны подавлять pHER3 в клетках MKN45, но 2C2 проявляло более высокую эффективность, чем антитела AMG и MM (приблизительно в 5-7 раз более высокую эффективность).
2.5.5. Подавление активности pHER3 в клетках Kato III
Кроме связывания с EGFR, HER2 и cMET, HER3 димеризуется с FGFR2, что способствует его трансформирующему потенциалу. Линию клеток Kato III (ATCC № HTB-103™), модель рака желудка с амплификацией FGFR2, использовали для того, чтобы оценить, могут ли анти-HER3-антитела подавлять зависимую от FGFR2 активацию HER3. Клетки Kato III обрабатывали моноклональными анти-HER3-антителами, как описано выше в разделе примера 2.5.1 (см. выше). В указанной модели все три моноклональных анти-HER3-антитела (2C2, AMG, и MM) достигали сходной максимальной степени подавления pHER3 (~60%), но судя по измерениям IC50, 2C2 было в 15-20 раз более эффективным, чем антитела AMG и MM, соответственно (фиг. 8D).
2.5.6. Подавление активности pHER3 в клетках BT-474
Было показано, что димеры HER2-HER3 являются одним из наиболее трансформирующих онкогенных структур в злокачественной опухоли. Соответственно, авторы исследовали моноклональные анти-HER3-антитела в линии клеток BT-474 (ATCC № HTB-20™), хорошо разработанной модели рака молочной железы с амплификацией HER2, которая не экспрессирует лиганда и предположительно управляется не зависимой от лиганда димеризацией HER2-HER3. Клетки BT-474 обрабатывали тремя моноклональными анти-HER3-антителами, а также 2C2-YTE, как описано выше в разделе примера 2.5.1. В отличие от моделей, в которых все три тестированных моноклональных анти-HER3-антитела были активными, таких как клетки HCC827, резистентные к гефитинибу клетки HCC827, клетки MKN45 и клетки Kato III, 2C2 (как родительское 2C2, так и мутант 2C2-YTE) было единственным среди тестированных моноклональных анти-HER3-антител, проявляющим значимую активность, подавляющую pHER3 (фиг. 8E). Полученные результаты показывают, что 2C2 (как родительская форма 2C2, так и мутант 2C2-YTE) является функциональным в не зависимой от лиганда модели, и демонстрируют бифункциональность 2C2 как в зависимых от лиганда, так и в не зависимых от лиганда условиях.
2.6. Анализ фосфорилирования AKT (pAKT) в клеточных моделях рака желудка и молочной железы
2.6.1. Клеточный анализ pAKT
Клетки (клетки злокачественной опухоли желудка MKN45, клетки злокачественной опухоли желудка Kato III или клетки ИЕ-474 злокачественной опухоли молочной железы с амплификацией HER2) высевали при плотности 30000 клеток/лунку в 96-луночные планшеты и давали возможность прикрепиться в течение ночи. Затем клетки подвергали обрабатывали тестируемыми и контрольными антителами согласно указанной дозовой кривой при 37°C в течение 4 часов. В конце обработки среду удаляли и клетки промывали PBS. Клетки лизировали в 80 мкл лизирующего буфера с тритоном X (Boston Bioproducts, Ashland, MA) с ингибиторами протеаз и фосфатаз (Calbiochem, La Jolla, CA) и хранили при -20°C вплоть до анализа. AKT/pAKT анализировали на основе протокола производителя, включенного в набор для определения в лизатах целых клеток фосфо(Ser473)/суммарного AKT (№ в каталоге K15100D, Meso-Scale Discovery, Gaithersburg, MD), чтобы определить содержание pAKT.
2.6.2. Подавление активности pAKT в клетках MKN45
Чтобы выяснить, может ли 2C2 подавлять нижележащий путь передачи сигнала HER3 в добавление к pHER3, авторы дополнительно оценивали его способность подавлять фосфорилирование AKT в модели рака желудка на клетках MKN45, зависимых от амплифицированного cMET. Клетки MKN45 обрабатывали моноклональными анти-HER3-антителами, как описано выше в разделе примера 2.6.1. В данной модельной системе моноклональное антитело 2C2 вызывало частичное ингибирование pAKT с более высокой эффективностью (приблизительно в 5-7 раз выше), чем моноклональные анти-HER3-антитела AMG и MM (фиг. 9A). Полученный результат демонстрирует, что 2C2 не только ингибирует активность HER3, но также подавляет нижележащие эффекторные молекулы HER3, такие как pAKT.
2.6.3. Подавление активности pAKT в клетках Kato III
Чтобы исследовать, обусловливает ли такая активность более высокую эффективность в подавлении pAKT, эффектора HER3, авторы анализировали ингибирование pAKT различными моноклональными анти-HER3-антителами, используя для такой клеточной линии набор для анализа Meso-Scale Discovery, как описано выше в разделе примера 2.6.1. Как показано на фиг. 9B, в соответствии с данными для pHER3, 2C2 подавляло pAKT в клетках Kato III модели рака желудка, зависимых от амплифицированного FGFR2. 2C2 и в этом случае был более эффективным (судя по измерениям IC50), и вызывал максимальный ответ в виде ингибирования pAKT по сравнению с антителами AMG и MM.
2.6.4. Подавление активности pAKT в клетках BT-474
Было показано, что димеры HER2-HER3 являются одним из наиболее трансформирующих онкогенных структур в злокачественной опухоли. Соответственно, авторы исследовали активность моноклональных анти-HER3-антител в линии клеток BT-474. Клетки BT-474 обрабатывали моноклональными анти-HER3-антителами, как описано выше в разделе примера 2.6.1, выше, а также YTE-мутантной формой 2C2. В отличие от моделей, в которых все три тестированных моноклональных анти-HER3-антитела (2C2, AMG и MM) были активными, таких как клетки MKN45 и Kato III, 2C2 (как родительская форма 2C2, так и мутант 2C2-YTE) было единственным среди тестированных моноклональных анти-HER3-антител, проявляющим значимую активность, подавляющую pAKT (фиг. 9C). Полученные результаты показывают, что 2C2 (как родительская форма 2C2, так и мутант 2C2-YTE) является функциональным в не зависимой от лиганда модели, и демонстрируют бифункциональность 2C2 (как родительской формы 2C2, так и мутанта 2C2-YTE)как в зависимых от лиганда, так и в не зависимых от лиганда условиях.
2.7. Подавление передачи сигнала HER3 клеточной пролиферации в клетках MDA-MB-361.
Чтобы охарактеризовать активность 2C2-YTE в клетках злокачественной опухоли молочной железы с амплификацией HER2, которые не дают высокого ответа на трастузумаб, авторы сфокусировали внимание на MDA-MB-361 (ATCC № HTB-27), модели злокачественной опухоли молочной железы, которая несет активирующую мутацию в PIK3CA (E545K), которая может вносить вклад в резистентность к трастузумабу вследствие свойственной ей активации пути PI3K (Junttila с соавторами, 2009, Cancer Cell. 15: 429-40). Авторы определили влияние 2C2 на передачу сигнала HER3 и пролиферацию клеток в такой модели.
Чтобы оценить передачу сигнала, линию клеток молочной железы человека MDA-MB-361 высевали в 24-луночные планшеты с плотностью 150000 клеток на лунку в RPMI (Invitrogen) с добавлением 10% инактивированной нагреванием фетальной сыворотки теленка (FBS) (Invitrogen). На следующий день среду для посева удаляли и клетки подвергали инкубации с анти-HER3-антителом 2C2 или контрольным антителом в полной среде, используя конечную концентрацию 30 мкг/мл. После инкубации в течение 6 часов в условиях 5% CO2 при 37°C клетки промывали один раз охлажденным на льду PBS и затем лизировали, добавляя восстанавливающий буфер Лэммли (Boston BioProducts, Ashland, MA). После короткой инкубации собирали клеточные лизаты, равные количества наносили на бис-трис-гели Bis NuPAGE® Novex® (Invitrogen, Carlsbad, CA) и белки переносили на мембраны PVDF (Invitrogen, Carlsbad, CA). Мембраны блокировали 5% обезжиренным сухим молоком и 0,1% твином-20 (Sigma, St. Louis, MO) в TBS (pH 7,4) и инкубировали в течение ночи при 4°C с антителами к HER3 (антитело sc-285, Cell Signaling Technology, Beverly, MA) и pHER3 (антитело 4791, Cell Signaling Technology, Beverly, MA). Мембраны промывали в 0,1% твин-20 в TBS и затем инкубировали в течение 1 часа со вторыми антителами и конъюгированным с перкосидазой хрена стрептавидином (GE Healthcare). После промывки полосы белков выявляют на рентгеновской пленке, используя хемилюминесцентный субстрат SuperSignal® West Femto и хемилюминесцентный субстрат SuperSignal® West Pico ((Pierce/Thermo Scientific, Rockford, IL).
Чтобы оценить клеточную пролиферацию, клетки MDA-MB-361 высевали с плотностью 2000 клеток/лунку в 100 мкл среды, содержащей 10% инактивированной нагреванием FBS; использовали 96-луночные планшеты Costar для обработки культуры ткани из белого полистирола с плоским дном (Corning). На следующий день среду, используемую для посева, удаляли и добавляли антибиотики в полной среде до конечного объема 100 мкл на лунку. Затем планшеты инкубировали в условиях 5% CO2 при 37°C в течение 6 или 14 суток. В случае 14-дневного анализа свежие антитела вносили на 7 день. Равные объемы реагента CellTiter-GloR (Promega) добавляли в каждую лунку в конце каждой временной точки. Планшеты встряхивали на встряхивающем устройстве для планшетов в течение 10 минут при комнатной температуре, чтобы гарантировать полный лизис клеток. Люминесценцию измеряли с использованием устройства для считывания 2104 EnVision® Multilabel (PerkinElmer).
2C2 снижало уровни pHER3 (фиг. 10A) и подавляло клеточный рост (фиг. 10B) в данной линии клеток, что свидетельствует о том, что 2C2-YTE является активным не только в чувствительных к трастузумабу злокачественных опухолях с амплификацией HER2, но также активно в злокачественных опухолях с амплификацией HER2, которые менее чувствительны к трастузумабу вследствие мутации в PIK3CA.
2.8. Идентификация новых типов HRG-зависимых злокачественных опухолей
Чтобы идентифицировать дополнительные новые HRG-зависимые типы злокачественные опухоли линии клеток множественной плоскоклеточной карциномы легкого (SCC) подвергали скринингу в отношении активности передачи сигнала HER3 и экспрессии HRG. Линии клеток HARA-B (JCRB № JCRB1080.1) и KNS-62 (JCRB № IFO50358) экспрессировали значимые уровни HER3, HRG, а также pHER3 (данные не показаны). Соответственно, авторы исследовали активность 2C2 в линиях клеток HARA-B и KNS-62. Клетки обрабатывали моноклональными анти-HER3-антителами, по существу, как описано выше в разделе примера 2.4, выше, антитело 2C2 было способно снижать уровни pHER3 в линии клеток HARA-B (фиг. 11) и линии клеток KNS-62 (данные не показаны). Как показано ниже (примеры, раздел 5.4), 2C2-YTE демонстрировал зависимую от дозы противоопухолевую эффективность в модели, основанной на ксенотрансплантатах плоскоклеточной NSCLC человека HARA-B. Таким образом, указанные критерии (т.е. экспрессия HER3, HRG, а также pHER3) могут быть полезными средствами для скрининга с целью идентификации дополнительных типов злокачественных опухолей, отвечающих на анти-HER3-антитела, включая, например, 2C2, AMG, MM, которые описаны в настоящей публикации, и другие антитела, известные в данной области (см., например, международные публикации патентов WO 2011/136911, WO 2012/019024, WO 2010/022814).
2.9. HER2 является основным стимулирующим фактором в некоторых HRG-зависимых типах злокачественных опухолей
В присутствии лиганда HER3 херегулина (HRG) HER3 гетеродимеризуется с EGFR или HER2, что приводит к фосфорилированию HER3 и передаче онкогенного сигнала через фосфоинозитид-3-киназу (PI3K) и протеинкиназу B (PKB), также известную как AKT. Характеризовали набор моделей CRC, чтобы определить, что является основным стимулирующим фактором в передаче сигнала через HER3, рецепторная тирозинкиназа, EGFR или HER2. В частности, шесть разных линий опухолевых клеток CRC, SW620 (ATCC № CCL-227), SW480 (ATCC № CCL-228), Colo205 (ATCC № CCL-222), LOVO (ATCC № CCL-229), HCT15 (ATCC № CCL-225) и Caco-2 (ATCC № HTB-37) обрабатывали антагонистами HER2 или EGFR отдельно или в сочетании с антагонистом HER3, 2C2. Кратко, клетки высевали в 24-луночные планшеты с плотностью 1,5×105 клеток на лунку. На следующий день 2 идентичных набора клеток обрабатывали 10 мкг/мл следующих антител: анти-HER3-антитело 2C2, контрольное IgG-антитело R347, анти-HER2-антитело 2C4 (например, публикация патента WO 2001/00245), анти-EGFR-антитело цетуксимаб или ингибитор тирозинкиназы EGFR гефитиниб в концентрации 5 мкМ. После 5-6-часовой инкубации при 37°C добавляли HRG в концентрации 50 нг/мл в один набор клеток на 15 минут при 37°C. Затем все клетки промывали холодным PBS и лизировали добавлением 60 мкл буфера для образцов с 2x SDS (додецилсульфатом натрия) (Invitrogen). Лизаты переносили в пробирки объемом 1,5 мл и кипятили в течение 5 минут с последующим охлаждением на льду в течение 2 минут. Равные объемы (20 мкл) образцов белка разделяли в бис-трис-гелях NuPAGE Novex (Invitrogen) перед переносом на мембраны из поливинилиденфторида (PVDF) (Invitrogen). Мембраны промывали в забуференном трис физиологическом растворе (KPL), содержащем 0,1% твин-20 (Sigma), и инкубировали в течение ночи при 4°C с антителами к HER3 (Santa Cruz Biotechnology), pHER3-Tyr1289 (Cell Signaling Technology), фосфорилированной AKT (протеинкиназе B (pAKT)) (Cell Signaling Technology), фосфорилированной ERK (активируемая митогенами протеинкиназа/регулируемая внеклеточными сигналами киназа (pERK)) (Cell Signaling Technology) и глицеральдегид-3-фосфатдегидрогеназе (GAPDH) (Sigma). Мембраны промывали в забуференном трис физиологическом растворе (KPL), содержащем 0,1% твин-20 (Sigma) и затем инкубировали в течение 1 часа со вторым антителом, конъюгированным с пероксидазой хрена (GE HealthCare). После промывки полосы белка выявляли на рентгеновской пленке, используя хемилюминесцентный субстрат SuperSignal West Pico (Pierce/Thermo Scientific).
Как показано на фиг. 12, и анти-HER3-антитело и анти-HER2-антитело снижали уровни HER3, pHER3 и pAKT в стимулируемых лигандом клетках, тогда как антагонист EGFR, такой как цетуксимаб и гефитиниб, не оказывали влияния на такие сигнальные молекулы. Полученные данные демонстрируют, что HER2 является основным стимулирующим фактором HRG-индуцируемой передачи сигнала HER3 во всех тестированных моделях злокачественных опухолей.
Пример 3. Исследования механизма действия моноклональные анти-HER3-антител
3.1. Клон 16 частично блокировал связывание лиганда с HER3
Эффективность моноклональных анти-HER3-антител в блокировании индуцированной лигандом активности HER3 может быть следствием их способности непосредственно блокировать связывание лиганда. Чтобы исследовать такой ход событий, авторы разработали анализ связывания HRG-HER3 in vitro, включающий в себя покрывание планшета херегулином (HRG) и связывание с ним меченого рекомбинантного белка HER3.
3.1.1. Анализ связывания HRG-HER3
Лунки микропланшета покрывали 10 нг/мл херегулина (HRGβ1, № в каталоге 377-HB, R&D Systems, Minneapolis, MN) в течение ночи при 4°C. На следующий день планшеты промывали 4 раза PBST (PBS + 0,05% твин-20) и блокировали в PBS + 1 мкг/мл БСА при комнатной температуре в течение 1 часа. Во время блокирования готовили серийные разведения тестируемых антител (клон 16, AMG, MM и позитивное контрольное моноклональное анти-HER3-антитело, блокирующее связывание с лигандом) в отдельном планшете в PBSTB (PBS + 0,05% твин-20 + 0,1% БСА) и объединяли с 5 мкг/мл рекомбинантного HER3 (№ в каталоге 348-RB, R&D Systems, Minneapolis, MN) при комнатной температуре в течение 30 минут. Затем планшеты для ELISA промывали 4 раза PBST перед добавлением смеси антитело-HER3. Планшеты инкубировали при комнатной температуре в течение 1 часа и затем промывали 4 раза PBST. Анти-His-HRP (№ в каталоге 34460, Qiagen, Valencia, CA) добавляли при комнатной температуре на 1 час. Планшеты промывали 4 раза PBST с последующим выявлением с использованием TMB. Планшеты считывали при 450 нм, используя считывающее устройство для микропланшетов. Типичные результаты показаны на фиг. 13.
3.1.2. Результаты
Тестировали способность нескольких моноклональных антител (клон 16, AMG, MM и позитивного контрольного моноклонального анти-HER3-антитела, блокирующего связывание лиганда) мешать связыванию HRG с HER3. Позитивное контрольное моноклональное антитело, блокирующее связывание HER3 с лигандом, эффективно и полностью подавляло связывание HER3 с HRG. Напротив, клон 16 (родительский вариант для 2C2, см. раздел примеров 1.6 «Оптимизация аффинности», выше) только отчасти был эффективным в нарушении такого связывания (приблизительно 30% от максимального ингибирования). Моноклональные антитела AMG и MM оказывали сходное слабое частично блокирующее влияние (фиг. 13). Полученные данные показали, что клон 16 вероятно не функционирует как моноклональное антитело, непосредственно блокирующее лиганд.
3.2. 2C2 нарушает димеризацию HER2-HER3
По своей природе обладающий киназной недостаточностью мономер HER3 является неактивным и нуждается в образовании гетеродимеров с другими RTK, чтобы стать активным. Показано, что димер HER2:HER3 является наиболее онкогенным компонентов в передаче сигналов как в зависимых от лиганда, так и в не зависимых от лиганда условиях (Junttila с соавторами, 2009, Cancer Cell. 15: 429-40). Нарушение димеризации HER2-HER3 под влиянием 2C2 оценивали, используя анализ HRG-индуцированного образования димеров HER2-HER3 в клетках T-47D, модели зависимого от лиганда рака молочной железы, в которой наблюдается HRG-индуцированное образование димеров HER3-HER2, и в не зависимых от лиганда клетках BT-474. Анализ был основан на коиммунопреципитации HER3-HER2.
3.2.1. Анализ индуцированной лигандом димеризации HER2-HER3
Клетки T-47D (ATCC № в каталоге HTB-133(TM)) высевали по 1×106/лунку в 6-луночные планшеты на ночь. На следующее утро клетки обрабатывали моноклональными антителами 2C2, CL16, AMG и MM в концентрации 5 мкг/мл в полной сыворотке в течение 2 часов при 37°C. Контроли не включали обработку антителом или включали обработку контрольным IgG1 R347. После такой обработки следовала обработка 50 нг/мл HRG в течение 10 минут при 37°C (включая контроль, не обрабатываемый HRG). Клетки промывали 3 раза холодным PBS перед добавлением 500 мкл буфера для лизирования клеток, содержащего ингибиторы протеаз и фосфатаз (Sigma, St. Louis, MO). Клетки лизировали на льду в течение, по меньшей мере, 30 минут. Лизаты собирали, используя скребок для клеток. 50 мкл раствора с шариками с белком A, содержащего 25 мкл шариков с белком A, конъюгированных с анти-HER3-мАт (№ в каталоге MAB3481, R&D Systems, Minneapolis, MN), добавляли к 500 мкл клеточного лизата и переносили на колонки для иммунопреципитации (IP). IP-колонки вращали в течение ночи при 4°C. Затем IP-колонки центрифугировали, чтобы удалить лизаты, и шарики промывали холодным буфером для лизиса клеток. 50 мкл 2X буфера для образцов с SDS, содержащего 50 мМ DTT, добавляли в каждую IP-колонку и колонки кипятили в течение 4 минут. Нижний наконечник каждой колонки удаляли и колонки центрифугировали, чтобы собрать элюаты. 20 мкл элюата разделяли, используя SDS-ПААГ. Осуществляли Вестерн-блоттинг для HER2 и HER3 (с использованием анти-HER2-антитела, № в каталоге OP15L, CalBiochem, La Jolla, CA; и анти-HER3-антитела, номер в каталоге No. SC-285, Santa Cruz Biotechnology, Inc., Santa Cruz, CA).
3.2.2. Анализ зависимой от лиганда димеризации HER2-HER3
Клетки BT-474 (ATCC, № в каталоге HTB-20TM) высевали с плотностью 1×106 клеток на лунку в 6-луночной планшет в полную среду для культуры клеток RPMI 1640 с 10% инактивированной нагреванием FBS. На следующий день среду для посева удаляли и заменяли свежей средой RPMI 1640, содержащей насыщающую дозу тестируемых антител. В данном эксперименте тестировали CL16, предшествующий 2C2 вариант с более низкой аффинностью, 2C2 и R347, контрольный IgG, в концентрации 5 мкг/мл. Клетки инкубировали с антителами в течение 2 часов при 37°C. Затем среду удаляли и клетки промывали один раз холодным PBS. Поперечно сшивающий агент 3,3’-дитиобис[сульфосукцинимидилпропионат] (DTSSP) добавляли в концентрации 2 мМ в 1 мл холодного PBS. Клетки инкубировали в течение, по меньшей мере, 1 часа на льду. Затем клетки промывали 3 раза холодным PBS. Добавляли буфер для лизиса клеток (500 мкл), содержащий ингибиторы протеаз и фосфатаз, и клетки помещали на лед, по меньшей мере, на 30 минут, чтобы обеспечить возможность лизиса, затем собирали, используя скребок для клеток. HER2 и HER3 иммунопреципитировали из клеточных лизатов. Клеточные лизаты (500 мкл) объединяли с 50 мкл шариков с белок A-сефарозой (50% взвесь; Invitrogen), предварительно конъюгированных с 1 мкг HER3-мАт (MAB3481, R&D Systems) в центрифужной колонке SigmaPrep (Sigma). Смесь инкубировали с вращением при 4°C в течение ночи. На следующий день шарики отделяли от клеточного лизата центрифугированием. Шарики в колонках промывали четыре раза холодным буфером для лизиса клеток (Cell Signaling Technologies), содержащем ингибиторы протеаз (Sigma) и фосфатаз (EMD Millipore). После процедуры промывки в центрифужные колонки добавляли 50 мкл 2x буфера для образцов с SDS (додецилсульфатом натрия), содержащего 50 мМ дитиотреитола (DTT; EMD Chemicals). Затем колонки кипятили в течение 4 минут. Белки элюировали центрифугированием и использовали сразу же для иммуноблоттинга (как описано в разделе 2.4).
3.2.3. Результаты
Клетки T-47D, обработанные или не обработанные HRG, подвергали лизису. HER3 преципитировали моноклональным анти-HER3-антителом, затем белки в осадке разделяли в SDS-ПААГ и подвергали блоттингу для исследования присутствия HER2 в качестве признака взаимодействия HER2-HER3. Модель была индуцируемой лигандом, так как димер появлялся только после стимуляции лигандом. Предварительная обработка 2C2 эффективно предотвращала образование димера, демонстрируя его способность препятствовать индуцируемому лигандом образованию димера HER2-HER3. Также обнаружено, что другие моноклональные анти-HER3-антитела, включая MM, AMG и родительский клон 16, были эффективными (фиг. 14A). В случае использования поперечно сшивающего агента DTSSP для биохимическлй стабилизации белковых комплексов, улавливали конститутивный гетеродимер HER2:HER3 в отсутствие HRG в клетках BT-474, что свидетельствовало о не зависимом от лиганда образовании гетеродимеров. Предварительная обработка клеток 2C2 или CL16 эффективно нарушала образование гетеродимеров (фиг. 14B).
3.3. Интернализация и разрушение HER3, индуцированные 2C2
Интернализация и разрушение мишени представляют собой два общих механизма, посредством которых моноклональные антитела ингибируют функции мишени. Сначала авторы оценили 2C2-опосредованную интернализацию HER3 в клетках злокачественной опухоли молочной железы BT-474. Затем авторы выяснили, может ли за такой быстрой 2C2-индуцированной интернализацией HER3 следовать разрушение мишени.
3.3.1. Анализ интернализации HER3
Интернализацию HER3 определяли, используя анализ, основанный на активируемой флуоресценцией сортировке клеток (FACS). Клетки BT-474 открепляли ферментом аккутазой и суспендировали клетки в PBS, содержащем 1% БСА (FACS-буфер) до плотности клеток 10×106 клеток/мл. 50 мкл клеток добавляли в каждую лунку 96-луночного планшета с U-образным дном. В каждую лунку добавляли 50 мкл моноклональных анти-HER3-антител плюс контроль изотипа (20 мкг/мл) для достижения конечной концентрации 10 мкг/мл. Планшет инкубировали при 37°C в течение 0,5 часа, 2 часов и 4,5 часа, соответственно. Клетки промывали два раза холодным FACS (клетки осаждали центрифугированием при 1500 об/мин в течение 2 минут). Клетки ресуспендировали в холодном FACS-буфере, содержащем мышиное моноклональное антитело против HER3 человека (« в каталоге MAB3481, R&D Systems, Minneapolis, MN) в концентрации 1 мкг/мл или 10 мкг/мл.
Клетки и антитела против HER3 человека инкубировали на льду в течение 1 часа. Затем клетки два раза промывали холодным FACS-буфером. Затем клетки ресуспендировали в холодном FACS-буфере, содержащем меченое красителем Alexa Fluor® 488 второе антитело (Invitrogen) (1:200 об./об.), и инкубировали на льду в течение 30-45 минут. Затем клетки два раза промывали холодным FACS-буфером и ресуспендировали в 100 мкл холодного FACS-буфера. В данный момент осуществляли FACS. Получали абсолютные геометрические средние интенсивности флуоресценции (GMFI) вычитанием GMFI из контролей, содержащих только второе антитело. Относительный клиренс HER3 с поверхности вычисляли путем сравнения с результатами, полученными с использованием контрольного моноклонального IgG-антитела. Типичные результаты показаны на фиг. 15A.
3.3.2. Анализ разрушения белка HER3
Модельные клетки злокачественной опухоли прямой и ободочной кишки Lovo, HCT15 и SW620 (ATCC, № CCL-229, CCL-225 и CCL-227, соответственно) высевали по 1,5×105/лунку в 24-луночные планшеты. После прикрепления в течение ночи клетки обрабатывали 2C2 и контрольным моноклональным антителом в течение 3-4 часов. Клетки промывали один раз холодным PBS, непосредственно лизировали, используя 50-60 мкл 2X-буфера для образцов с SDS, и кипятили при 100°C в течение 10 минут. По 20 мкл образцов наносили на SDS-ПААГ-гели, разделяли электрофоретически и подвергали Вестерн-блоттингу с использованием антитела против HER3 (Santa Cruz Biotech), чтобы количественно определить общие уровни белка HER3. Антитела против GAPDH (Sigma) также использовали для количественной оценки уровней GAPDH в качестве контроля загрузки общего белка.
3.3.3. Результаты
Как показано на фиг. 15A, обе дозы 2C2 оказывали очень сходное воздействие. 30-минутная обработка вызывала 39% потери популяции поверхностного HER3 (61% оставалось), тогда как 2-часовая обработка вызывала потерю 62% (38% оставалось), что свидетельствует о быстрой интернализации мишени под действием 2C2. Кроме того, когда три разных модели злокачественной опухоли прямой и ободочной кишки инкубировали с 2C2, полное разрушение HER3 наблюдали в клетках SW620, тогда как почти полное разрушение наблюдали в других двух клеточных линиях (фиг. 15B), что свидетельствует о том, что 2C2 обладает высокой способностью вызывать разрушение мишени.
3.4. Эффекторные функции: зависимая от антител опосредованная клетками цитотоксичность (ADCC) и зависимая от комплемента цитотоксичность (CDC)
ADCC является одним из известных путей, посредством которого моноклональное антитело может проявлять свою противоопухолевую эффективность in vivo. Чтобы оценить ADCC-активность клона 16, авторы использовали анализ PBMC-обусловленной ADCC in vitro в двух моделях рака молочной железы с амплификацией HER2: BT-474 и SkBR3. Герцептин/трастузумаб использовали в качестве позитивного контроля, так как было показано, что он оказывает ADCC-эффект в таких типах злокачественных опухолей. В обеих моделях авторы наблюдали значимые эффекты герцептина в уничтожении опухолей, но остальные тестированные моноклональные антитела, клон 16, AMG и MM, были, главным образом, неактивны, что свидетельствует о том, что они не оказывали значимого ADCC-эффекта (данные не показаны). 2C2-YTE тестировали в анализах CDC, используя сыворотку человека в качестве источника комплемента. Кроме того, анти-HER2-антитело трастузумаб и анти-CD20 антитело ритуксимаб включали в качестве контролей. Ни одно из антител, включая и 2C2-YTE, не проявляли какой-либо регистрируемой CDC-активности ни в одной из концентраций (данные не показаны). Клетки SkBR3 не экспрессируют CD20. В качестве позитивного контроля ритуксимаб проявлял значимую активность, приводящую к гибели клеток, в сходном анализе CDC, против клеток Daudi, которые экспрессируют CD20 (данные не показаны).
3.5. Задержка клеточного цикла
3.5.1. Анализ задержки клеточного цикла в клетках злокачественной опухоли молочной железы SkBR3
Клетки BioSante (ATCC № HTB-30) высевали с плотностью 150000 клеток/лунку в 6-луночный планшет и давали возможность прикрепиться в течение ночи. На следующий день среду удаляли и заменяли свежей ростовой средой, содержащей тестируемые и контрольные антитела. Затем клетки инкубировали при 37°C в течение 48 часов. В конце обработки клетки обрабатывали трипсином, объединяли в конической пробирке объемом 15 мл и центрифугировали при 1500 об/мин в течение 5 минут. Затем клетки промывали один раз PBS и фиксировали охлажденным на льду 70% этанолом при -20°C в течение ночи.
После фиксации клетки центрифугировали, как описано выше, промывали один раз в PBS и ресуспендировали в растворе для окрашивания (PBS + 0,1% тритон X-100, 0,2 мг/мл РНК-азы A, не содержащей ДНК-азы, и 20 мкг/мл йодида пропидия). Клетки красили в течение 30 минут при комнатной температуре в темноте и анализировали, используя систему проточной цитометрии LSRII (BD Biosciences). Йодид пропидия выявляли, используя канал техасского красного; данные анализировали с помощью пакета для анализа данных проточной цитометрии FlowJo (Tree Star, Inc., OR), используя модель Dean/Jett/Fox.
3.5.2. Результаты
Основанный на FACS анализ клеточного цикла показал, что в клетках SkBR3, линии клеток злокачественной опухоли молочной железы с амплификацией HER2, сходной с BT-474, и герцептин и клон 16 (родительский вариант для 2C2) вызывали задержку клеточного цикла в G1-фазе (увеличенная популяция G1 при уменьшении популяции S/G2, как показано на фиг. 16).
3.6. Антиангиогенные эффекты за счет блокирования HRG-индуцированной секреции VEGF
Было показано, что HRG стимулирует секрецию VEGF, основного проангиогенного цитокина, в различных моделях злокачественных опухолей. Поэтому авторы оценивали ингибирующие эффекты 2C2 в подавлении HRG-индуцированной секреции VEGF в двух моделях рака молочной железы: MCF-7 и BT-474.
3.6.1. Анализ HRG-индуцированной секреции VEGF
Клетки MCF-7 и клетки BT-474 высевали с плотностью 100000 клеток/лунку в 24-луночный планшет и обеспечивали период покоя в течение 2 суток. Затем среду удаляли и заменяли 500 мкл свежей ростовой среды, содержащей 2% FBS и контрольные и тестируемые антитела. После 24-часовой инкубации среду от культуры клеток собирали и определяли уровни VEGF, используя набор для ELISA VEGF (R&D Systems DY293B). Относительное количество клеток в каждой лунке определяли, добавляя свежую среду к клеткам вместе с Cell Titer Glo (Promega, соотношение 1:1) и инкубируя планшеты в течение 10 минут при комнатной температуре. Люминесценцию регистрировали, используя считывающее устройство для планшетов, и такие значения использовали для нормализации данных.
3.6.2. Результаты
Обработка HRG индуцировала значительное повышение секреции VEGF в обеих модельных линиях клеток злокачественной опухоли молочной железы BT-474 (фиг. 17A) и MCF-7 (фиг. 17B) в диапазоне от 6,5-кратного до 8-кратного. Моноклональные антитела CL16 (клон 16) и MM были способны подавлять большую часть повышения секреции, что свидетельствует о том, что такие моноклональные анти-HER3-антитела могут оказывать дополнительные эффекты в отношении модулирования сосудов.
Пример 4. Перекрестная реактивность с HER3 макак-крабоедов и мышей
4.1. 2C2 связывается с HER3 макака-крабоеда и мыши с аффинностью, сходной с аффинностью по отношению к HER3 человека
Анализы Biacore осуществляли по существу как описано выше, чтобы сравнить аффинность 2C2 по отношению к HER3 человека, макака-крабоеда (cyno) и мыши, чтобы обеспечить возможность отбора соответствующего вида для определения токсичности (верхняя часть таблицы 7). Дополнительные анализы Biacore осуществляли для 2C2-YTE, используя прибор BIAcore более высокого разрешения, альтернативный реагент для улавливания Fc и уточненный протокол инъекций, чтобы скорректировать фоновое связывание. Кратко, реагент для улавливания белок A иммобилизовали на двух соседних проточных ячейках, соединенных последовательно на одном и том же сенсорном чипе CM5, используя стандартный протокол для аминов, который описан производителем оборудования. Одну из таких поверхностей с белком A использовали в качестве эталонной поверхности для данного эксперимента, тогда как другая служила в качестве активной поверхности, используемой для регистрации улавливания IgG и последующего связывания HER3 (ECD). Были зарегистрированы конечные плотности белка A на поверхностях эталонной и активной проточных ячеек 1986 условных единиц и 1979 условных единиц, соответственно. При используемой конфигурации способ был настроен так, что IgG 2C2-YTE сначала улавливали только на активной поверхности с белком A, затем осуществляли инъекцию белка HER3 на поверхности обеих активной и эталонной проточных ячеек. В таком виде в указанной методике корректируют кривую связывания в отношении любого неспецифичного связывания аналита HER3 с поверхностью для улавливания с белком A. Для стадии улавливания IgG такой IgG 2C2-YTE готовили в концентрации 10 нМ в HBS-EP + буфер, поставляемый к оборудованию (0,01 М HEPES, pH 7,4, 0,15 М NaCl, 3 мМ EDTA и 0,05% P20), затем инъецировали на поверхность активной проточной ячейки с белком A в течение 30 секунд со скоростью потока 10 мкл/мин. Белок HER3 человека, макака-крабоеда и мыши сначала готовили в виде маточных растворов с концентрацией 500 нМ в поставляемом к оборудованию буфере, затем делали двукратные серийные разведения каждого, получая конечную концентрацию 0,39 нМ. Затем белок HER3 инъецировали на поверхности обеих активной и эталонной ячеек с белком A в течение 120 секунд со скоростью потока 75 мкл/мин. Данные о диссоциации собирали в течение 15 минут, затем осуществляли два 60-секундных импульсных впрыскиваний 10 мМ Gly-буфера, pH 1,7, между инъекциями, чтобы регенерировать проточные ячейки снова с получением улавливающих поверхностей с белком A. Несколько инъекций буфера также осуществляли в промежутках в серии инъекций. Выбранные инъекции буфера затем использовали вместе с данными, полученными для эталонной ячейки, чтобы скорректировать наборы первичных данных в отношении артефактов инъекции и/или неспецифичных взаимодействий при связывании, такой способ обычно называют «двойным сравнением с эталоном» (Myszka, 1999). Затем осуществляли глобальную подгонку полностью скорректированных данных о связывании к модели связывания 1:1 (компьютерная программа BIAevaluation 4.1), которая включала член для корректировки ограниченного переносом массы связывания в случае его выявления. В указанном анализе определяли кинетические константы скорости (kon, koff), на основании которых затем вычисляли кажущуюся KD в виде отношения koff/kon (нижняя часть таблицы 7). Варьирование значений Kon и Koff между двумя группами экспериментов, вероятно, является следствием различий между двумя протоколами, которые подробно описаны выше, и обычно были в допустимых пределах двукратной ошибки для измерения таких кинетических параметров. Как показано в таблице 7, аффинность 2C2 и 2C2-YTE по отношению к HER3 макака-крабоеда фактически была идентичной аффинности по отношению к HER3 человека. Аффинность по отношению к HER3 мыши была в пределах 3-кратной от аффинности по отношению к HER3 человека.

4.2. Анализ HRG-индуцированного фосфорилирования HER3 макака-крабоеда
Клетки Ad293 (Stratagene № 240085) временно трансфицировали вектором, экспрессирующим полноразмерный cynoHER3, следуя протоколу, прилагаемому к реагенту липофектамину 2000 (Invitrogen). Клеткам обеспечивали возможность для инкубации при 37°C в течение 48 часов перед обработкой. Антитела добавляли в концентрации 10 мкг/мл в полной ростовой среде на 1 час с последующей стимуляцией с использованием 20 нг/мл HRGβ1 (R&D Systems) в течение 10 минут при 37°C. В конце обработки среду удаляли и клетки промывали один раз PBS. Клетки лизировали 2x трис-глициновым буфером для образцов Novex (Invitrogen) и определяли уровни pHER3 и общего HER3, используя иммуноблоттинг (антитело Cell Signaling №4791 и антитело Santa Cruz №285, соответственно). Денситометрию полос осуществляли, используя компьютерную программу ImageJ (IH, imagej.nih.gov/ij/).
4.3. Результаты
Чтобы полно установить и перекрестное модулирование HER3 макака-крабоеда (cyno-HER3) под действием 2C2, получили стабильную линию клеток Ad293, эктопически экспрессирующих полноразмерный cyno-HER3, как показано на Вестерн-блоте (фиг. 18A). При обработке HRG cyno-HER3 подвергался сильной активации, выявляемой по индукции сигнала pHER3 (фиг. 18B). Когда клетки совместно обрабатывали 2C2, но не в том случае, когда их обрабатывали контрольным антителом R347, индукция pHER3 была полностью устранена, что свидетельствует о том, что 2C2 было не только способно связываться с cyno-HER3 на клеточной поверхности, но также способно эффективно устранять его активацию (фиг. 18B). Вместе с приведенными выше данными измерения аффинности в анализе Biacore, показывающими, что 2C2 имело аффинность по отношению к cyno-HER3, идентичную аффинности к HER3 человека, такие результаты подтверждают, что макаки-крабоеды являются соответствующим видом для определения токсичности в случае испытаний 2C2.
Исследования моноклональных анти-HER3-антител in vivo
4.4. Исследования на модели подкожных ксенотрансплантатов опухолевых клеток головы и шеи FADU человека
4.4.1. Способ
Клетки головы и шеи FADU человека (ATCC № HTB-43) поддерживали при 37°C в инкубаторе с 5% CO2 в среде RPMI 1640, содержащей 4,5 г/л глюкозы, L-глутамин, пируват натрия и 10% фетальной сыворотки теленка. Ксенотрансплантаты получали подкожной инъекцией 5×106 клеток на мышь (суспендированных в 50% матригеле) в правый бок бестимусных мышей nu/nu 4-6-недельного возраста. Опухолям давали возможность вырасти до 200 мм3 перед рандомизацией для исследований эффективности. 2C2, 2C2-YTE, цетуксимаб, контрольный IgG2 или сочетание 2C2 с моноклональным антителом цетуксимабом вводили внутрибрюшинно. Для исследований зависимости от дозы 2C2 вводили в дозе 3, 5, 7, и 10 мг на килограмм массы тела (мг/кг), контроль в дозе 10 мг/кг. В случае исследований сочетания 2C2 вводили в дозе 3 мг/кг, цетуксимаб в дозе 30 мг/кг и контрольное антитело в дозе 6 мг/кг. Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
В конце исследований эффективности с использованием 2C2, мышей обрабатывали 2C2 в последний момент времени, который указан, чтобы определить фармакокинетические значения. Осуществляли пункцию сердца, чтобы собрать кровь в пробирки для отделения сыворотки Microtainer (SST). Пробирки с кровью осторожно встряхивали в течение 10 секунд и выдерживали при комнатной температуре в течение 20 минут, чтобы дать возможность сыворотке свернуться. Образцы центрифугировали при 1000×g в течение 10 минут, и образцы сыворотки осторожно переносили в новые пробирки и хранили при -80°C.
Формат непрямого твердофазного иммуноферментного анализа (ELISA) использовали для количественного определения 2C2 в сыворотке мышей. Стандарты, контроли качества и образцы сыворотки мышей инкубировали с антителами козы против IgG человека, которые были иммобилизованы на 96-луночном планшете для микротитрования. После инкубации несвязанные материалы удаляли на стадии промывки и 2C2 определяли, используя антитело козы против IgG человека, конъюгированное с пероксидазой хрена. Добавляли раствор кислоты для остановки и степень ферментативного превращения субстрата определяли, измеряя оптическую плотность при 450 нм. Измеряемая оптическая плотность была прямо пропорциональна концентрации 2C2, присутствующего в сыворотке мышей. Использовали стандартную кривую для анализа 2C2, чтобы интерполировать концентрацию в образцах сыворотки.
4.4.2. Результаты
В случае использования модели, основанной на ксенотрансплантатах клеток опухоли головы и шеи человека FADU, выращенных подкожно у самок мышей nude 2C2 демонстрировало зависимую от дозы противоопухолевую эффективность. Максимальную эффективность с 99% ингибированием роста опухолей (dTGI) наблюдали при использовании дозы 7 мг/кг, вводимой два раза в неделю на протяжении исследования (фиг. 19A).
При комбинированном введении 3 мг/кг 2C2 с 30 мг/кг цетуксимаба, вводимыми два раза в неделю на протяжении фазы обработки (дни 7-18), наблюдали явную синергетическую противоопухолевую эффективность в модели на основе ксенотрансплантатов FADU (фиг. 19B). Такой эффект продолжался долго, и опухоли только начинали расти снова в конце фазы возобновления роста на 40 день. Комбинированная обработка приводили к частичным регрессиям в 7 из 10 случаев и к полным регрессиям в 2 из 10 случаев.
2C2 перекрестно взаимодействовало с HER3 мыши, и было хорошо установлено, что HER3 экспрессируется во многих непатогенных тканях мышей. Таким образом, HER3 хозяина может служить в качестве поглотителя, абсорбируя моноклональные антитело 2C2 до того, как оно достигнет опухолевой ткани. Используя несущих опухоли самок мышей nude, 2C2 в дозе 5 мг/кг вводили таким мышам либо один раз, либо три раза, и с течением времени следили на уровнями экспозиции 2C2. 2C2 можно было выявить только через 1 день после последней дозы 5 мг/кг 2C2, и антитело становилось нерегистрируемым через 3 дня после последней обработки (фиг. 36). С другой стороны, введение дозы 30 мг/кг 2C2 с использованием такой же схемы, как в случае 5 мг/кг приводило к намного более длительному временному интервалу, когда 2C2 можно было измерить в сыворотке. Полученные данные продемонстрировали нелинейную фармакокинетику для 2C2 после введения однократной дозы и многократных доз 5 мг/кг или 30 мг/кг мышам, несущим опухоли. Данные показали, что мышиный HER3 может действовать в качестве поглотителя, связывая 2C2, вводимое мышам, и что 30 мг/кг в виде однократной дозы было достаточно для насыщения поглотителя.
Наличие поглотителя HER3 у мышей в случае 2C2 подтверждали функционально путем введения высокой нагрузочной дозы 2C2 с последующей низкой поддерживающей дозой мышам с ксенотрансплантированными опухолями FADU. В модели опухоли FADU была продемонстрирована противоопухолевая эффективность нагрузочной дозы 2C2 10 мг/кг и поддерживающей дозы 2C2 3 мг/кг. Введение 10 мг/кг 2C2 в виде однократной дозы было только временно эффективным. 2C2, вводимое в дозе 3 мг/кг два раза в неделю, проявляло умеренную, но длительную эффективность. Сочетание нагрузочной дозы 2C2 10 мг/кг с поддерживающей дозой 2C2 3 мг/кг было более эффективным в блокировании роста опухолей по сравнению с любой схемой обработки отдельно (фиг. 21).
Способность 2C2 модулировать фармакокинетические маркеры pHER3 и pAKT тестировали в экстрактах ксентрансплантированных опухолей FADU. 2C2 вводили два раза в дозе 30 мг/кг в течение 48 часов мышам, несущим ксенотрансплантированные опухоли FADU человека, и экстракты анализировали спустя 24 часа. Кратко, бестимусным мышам nude подкожно имплантировали злокачественные клетки головы и шеи FADU. Животным вводили 2C2 в дозе 30 мг/кг два раза в течение 48 часов. Экстракты готовили спустя 24 часа для анализа pHER3, pAKT и общего HER3 (фиг. 22, верхняя, средняя и нижняя панели, соответственно). R347 использовали в качестве контрольного IgG1-антитела. Использовали по 6 животных на группу обработки. Данные представлены в виде среднего ± стандартное отклонение. В данном случае 2C2 ингибировало фосфорилирование и HER3 и AKT на 59,5% и 51,7%, соответственно, по сравнению с опухолями у мышей, обработанных контрольным IgG1 (фиг. 22, верхняя и средняя панели). Модулирования общего HER3 под действием 2C2 не наблюдали (фиг. 22, нижняя панель).
4.5. Исследования на модели подкожных ксенотрансплантатов клеток головы и шеи человека Detroit562
4.5.1. Способ.
Клетки головы и шеи человека Detroit562 (ATCC № CCL-138) поддерживали при 37°C в инкубаторе с 5% CO2 в среде RPMI 1640, содержащей 4,5 г/л глюкозы, L-глутамин, пируват натрия и 10% фетальной сыворотки теленка. Ксенотрансплантаты получали подкожной инъекцией 5×106 клеток на мышь в правый бок бестимусных мышей nu/nu 4-6-недельного возраста. Опухолям давали возможность вырасти до 200 мм3 перед рандомизацией для исследований эффективности. 2C2, 2C2-YTE, цетуксимаб, контрольный IgG2 или сочетание 2C2 с моноклональным антителом цетуксимабом вводили внутрибрюшинно. Для исследований зависимости от дозы 2C2 вводили в дозе 1, 3, 10 и 30 мг на килограмм массы тела (мг/кг). В случае исследований сочетаний 2C2 вводили в дозе 3 мг/кг, цетуксимаб в дозе 30 мг/кг и контрольное антитело в дозе 10 мг/кг. Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
В конце исследований эффективности с использованием 2C2 мышей обрабатывали 2C2 в последний момент времени, который указан, чтобы определить фармакокинетические значения. Осуществляли пункцию сердца, чтобы собрать кровь в SST-пробирки Microtainer. Пробирки с кровью осторожно встряхивали в течение 10 секунд и выдерживали при комнатной температуре в течение 20 минут, чтобы дать возможность сыворотке свернуться. Образцы центрифугировали при 1000×g в течение 10 минут, и образцы сыворотки осторожно переносили в новые пробирки и хранили при -80°C.
Формат непрямого твердофазного иммуноферментного анализа (ELISA) использовали для количественного определения 2C2 в сыворотке мышей. Стандарты, контроли качества и образцы сыворотки мышей инкубировали с антителами козы против IgG человека, которые были иммобилизованы на 96-луночном планшете для микротитрования. После инкубации несвязанные материалы удаляли на стадии промывки и 2C2 определяли, используя антитело козы против IgG человека, конъюгированное с пероксидазой хрена. Добавляли раствор кислоты для остановки и степень ферментативного превращения субстрата определяли, измеряя оптическую плотность при 450 нм. Измеряемая оптическая плотность была прямо пропорциональна концентрации 2C2, присутствующего в сыворотке мышей. Использовали стандартную кривую для анализа 2C2, чтобы интерполировать концентрацию в образцах сыворотки.
4.5.2. Результаты
2C2 проявляло противоопухолевую эффективность в модели, основанной на ксенотрансплантатах клеток головы и шеи человека Detroit562, выращенных подкожно у самок мышей nude. Доза 10 мг/кг 2C2, вводимая два раза в неделю, была максимально эффективной с dTGI 72% (фиг. 23A). Модель Detroit562 содержит мутацию PIK3CA.
Модель опухоли Detroit562 была чувствительной к моноклональному анти-EGFR-антителу цетуксимабу, который вызывал ингибирование роста опухоли в дозе 10 мг/кг, вводимой два раза в неделю. Сочетание 3 мг/кг 2C2 с 10 мг/кг цетуксимаба увеличивало противоопухолевую эффективность цетуксимаба и приводило к частичной регрессии в 9 из 10 случаев, тогда как цетуксимаб отдельно приводил к частичной регрессии в 5 из 10 случаев (фиг. 23B).
4.6. Исследования на модели подкожных ксенотрансплантатов клеток головы и шеи человека CAL27
4.6.1. Способ.
Клетки головы и шеи человека CAL27 (ATCC, № CRL-2095) поддерживали при 37°C в инкубаторе с 5% CO2 в среде RPMI 1640, содержащей 4,5 г/л глюкозы, L-глутамин, пируват натрия и 10% фетальной сыворотки теленка. Ксенотрансплантаты получали подкожной инъекцией 5×106 клеток на мышь в правый бок бестимусных мышей nu/nu 4-6-недельного возраста. Опухолям давали возможность вырасти до 200 мм3 перед рандомизацией для исследований эффективности. 2C2-YTE, цетуксимаб или контрольный IgG1 вводили внутрибрюшинно. Для исследований зависимости от дозы 2C2-YTE вводили в дозе 3, 10 и 30 мг на килограмм массы тела (мг/кг). Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
4.6.2. Результаты.
Зависимая от дозы активность 2C2-YTE была подтверждена в третьей модели опухоли головы и шеи, CAL27, с использованием 2C2-YTE. 2C2-YTE в дозе 3, 10 или 30 мг/кг, вводимое два раза в неделю приводило к TGI 26,4%, 55,2% или 68,8%, соответственно, по сравнению с животными, обработанными контрольным IgG1 (фиг. 24).
Модель опухоли CAL27 была чувствительной к моноклональному анти-EGFR-антителу цетуксимабу, который вызывал ингибирование роста опухоли в дозе 30 мг/кг, вводимой два раза в неделю, с TGI 75,0% (фиг. 24).
4.7. Исследования на модели подкожных ксенотрансплантатов NSCLC человека с мутацией KRAS A549
4.7.1. Способ
Клетки NSCLC A549 (ATCC, № CCL-185), которые содержат мутацию в кодоне 12 гена KRAS, поддерживали при 37°C в инкубаторе с 5% CO2 в среде Хама F12K, содержащей 4,5 г/л глюкозы, L-глутамин, пируват натрия и 10% фетальной сыворотки теленка. Ксенотрансплантаты получали подкожной инъекцией 5×106 клеток на мышь (суспендированных в 50% матригеле) в правый бок бестимусных мышей nu/nu 4-6-недельного возраста. Опухолям давали возможность вырасти до 200 мм3 перед рандомизацией для исследований эффективности. 2C2, 2C2-YTE, цетуксимаб, контрольный IgG2 или сочетание 2C2 с моноклональным антителом цетуксимабом вводили внутрибрюшинно. Для исследований зависимости от дозы 2C2 вводили в дозе 3, 10 и 30 мг на килограмм массы тела (мг/кг) и 2C2-YTE в дозе 10 мг/кг. В случае исследований сочетаний 2C2 и цетуксимаба, каждый из них вводили в дозе 10 мг/кг. Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
4.7.2. Результаты.
2C2 проявляло зависимую от дозы противоопухолевую эффективность в модели на основе ксенотрансплантатов NSCLC человека A549, выращенных подкожно у самок мышей nude. Максимальная эффективность с dTGI 91% была достигнута в случае дозы 2C2 30 мг/кг, вводимой два раза в неделю вплоть до 33 дня (фиг. 25A). 2C2 и 2C2-YTE, вводимые в дозе 10 мг/кг, проявляли сходную противоопухолевую эффективность в такой модели опухоли A549. После прекращения обработки опухоли начинали расти с такой же скоростью, как и опухоли у мышей, обработанных контролем. Модель ксенотрансплантата A549 содержит мутацию KRAS и делецию LKB-1.
Цетуксимаб в дозе 10 мг/кг, вводимый отдельно, был неэффективным в такой модели опухоли A549. Однако добавление цетуксимаба в дозе 10 мг/кг к 2C2, также вводимому в дозе 10 мг/кг, приводило к аддитивной противоопухолевой эффективности во время фазы обработки по сравнению с введением 2C2 отдельно. Кроме того, в группе с комбинированной обработкой наблюдали более медленную скорость возобновления роста опухолей после прекращения обработки (фиг. 25B).
4.8. Исследования на модели подкожных ксенотрансплантатов плоскоклеточной NSCLC человека HARA-B
4.8.1. Способ
Клетки плоскоклеточной NSCLC человека HARA-B, которые экспрессируют ген RAS дикого типа, HRG и pHER3, поддерживали при 37°C в инкубаторе с 5% CO2 в среде RPMI 1640, содержащей 4,5 г/л D-глюкозы, 2,383 г/л HEPES-буфера, L-глутамин, 1,5 г/л бикарбоната натрия, 110 мг/л пирувата натрия и 10% фетальной сыворотки теленка. Ксенотрансплантаты получали подкожной инъекцией 5×106 клеток на мышь (суспендированных в 50% матригеле) в правый бок бестимусных мышей nu/nu 4-6-недельного возраста. Опухолям давали возможность вырасти до 227 мм3 перед рандомизацией для исследований эффективности. 2C2-YTE вводили внутрибрюшинно в дозе 3, 10 и 30 мг на килограмм массы тела (мг/кг), контроль в дозе 30 мг/кг. Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
4.8.2. Результаты.
2C2-YTE проявляло зависимую от дозы противоопухолевую эффективность в модели на основе ксенотрансплантатов плоскоклеточной NSCLC человека HARA-B, выращенных подкожно у самок мышей nude. Максимальная эффективность с dTGI 64,6% была достигнута с использованием дозы 30 мг/кг 2C2-YTE, вводимой два раза в неделю вплоть до 29 дня (фиг. 26). 2C2-YTE, вводимое в дозе 10 мг/кг, проявляло сходную противоопухолевую эффективность в дозе 30 мг/кг; однако 2C2-YTE в дозе 3 мг/кг было неэффективным в такой модели опухоли HARA-B. Модель на основе ксенотрансплантатов HARA-B содержит аллель RAS дикого типа.
4.9. Исследования на модели подкожных ксенотрансплантатов CRC человека HT-29
4.9.1. Способ
Клетки карциномы прямой и ободочной кишки человека HT-29 (ATCC № HTB-38) поддерживали при 37°C в инкубаторе с 5% CO2 в среде RPMI 1640, содержащей 4,5 г/л глюкозы, L-глутамин, пируват натрия и 10% фетальной сыворотки теленка. Ксенотрансплантаты получали подкожной инъекцией 5×106 клеток на мышь в правый бок бестимусных мышей nu/nu 4-6-недельного возраста. Опухолям давали возможность вырасти до 200 мм3 перед рандомизацией для исследований эффективности. Моноклональные антитела 2C2, 2C2-YTE и контрольный IgG1 вводили внутрибрюшинно. 2C2 вводили в дозе 2, 10 и 30 мг на килограмм массы тела (мг/кг), тогда как 2C2-YTE вводили в дозе 30 мг/кг. Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
4.9.2. Результаты.
2C2 проявляло зависимую от дозы противоопухолевую эффективность при использовании модели, основанной на ксенотрансплантатах клеток прямой и ободочной кишки человека HT-29, подкожно инъецированных самкам мышей nude. Доза 2C2 30 мг/кг, вводимая два раза в неделю, была максимально эффективной с dTGI 56% во время фазы обработки (фиг. 27). 2C2-YTE проявляло такую же эффективность, как и 2C2, при введении обоих антител в дозе 30 мг/кг. После прекращения обработки опухоли росли с такой же скоростью, что и контрольные опухоли. Модель на основе ксенотрансплантатов HT-29 содержит мутацию BRAF. Цетуксимаб в дозе 10 мг/кг, вводимый отдельно, не обладал измеряемой противоопухолевой активностью в такой модели. Активность 2C2 в дозе 30 мг/кг в сочетании с цетуксимабом в дозе 10 мг/кг была неотличима от активности 2C2 в дозе 30 мг/кг отдельно в конце фазы обработки (данные не показаны). Результат свидетельствует о том, что такая EGFR-экспрессирующая модель опухоли CRC, которая хорошо отвечает на 2C2, дополнительно не подвергалась ингибированию при добавлении 2C2-YTE к цетуксимабу.
4.10. Исследования на модели подкожных ксенотрансплантатов CRC человека HCT-116
4.10.1. Способ
Клетки карциномы прямой и ободочной кишки человека HCT-116 поддерживали при 37°C в инкубаторе с 5% CO2 в среде RPMI 1640, содержащей 4,5 г/л глюкозы, L-глутамин, пируват натрия и 10% фетальной сыворотки теленка. Ксенотрансплантаты получали подкожной инъекцией 5×106 клеток на мышь в правый бок бестимусных мышей nu/nu 4-6-недельного возраста. Опухолям давали возможность вырасти до 200 мм3 перед рандомизацией для исследований эффективности. Моноклональные антитела 2C2, 2C2-YTE, цетуксимаб и контрольный IgG1 вводили внутрибрюшинно. 2C2 вводили в дозе 3, 10 и 30 мг на килограмм массы тела (мг/кг), тогда как 2C2-YTE вводили в дозе 30 мг/кг. Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
4.10.2. Результаты.
2C2 в нескольких разных концентрациях и 2C2-YTE в дозе 10 мг/кг, вводимые два раза в неделю, проявляли умеренную противоопухолевую эффективность в модели на основе ксенотрансплантатов клеток прямой и ободочной кишки человека HCT-116, инъецируемых подкожно самкам мышей nude (фиг. 28). Максимальная эффективность была отмечена на уровне dTGI 43% в случае 2C2 в дозе 10 мг/кг. Моноклональное анти-EGFR-антитело цетуксимаб не проявляло эффективности в дозе 10 мг/кг. Модель на основе ксенотрансплантатов HCT-116 содержит мутацию KRAS.
4.11. Исследования на модели подкожных ксенотрансплантатов CRC человека LOVO
4.11.1. Способ.
Клетки карциномы прямой и ободочной кишки человека LOVO (ATCC № CCL-229) поддерживали при 37°C в инкубаторе с 5% CO2 в среде Хама F12K, содержащей 4,5 г/л глюкозы, L-глутамин, пируват натрия и 10% фетальной сыворотки теленка. Ксенотрансплантаты получали подкожной инъекцией 5×106 клеток на мышь в правый бок бестимусных мышей nu/nu 4-6-недельного возраста. Опухолям давали возможность вырасти до 200 мм3 перед рандомизацией для исследований эффективности. Моноклональные антитела 2C2, 2C2-YTE, цетуксимаб и контрольный IgG1 вводили внутрибрюшинно. 2C2 вводили в дозе 10 или 30 мг на килограмм массы тела (мг/кг), 2C2-YTE и цетуксимаб вводили в дозе 10 мг/кг и контроль вводили в дозе 30 мг/кг. Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
4.11.2. Результаты
2C2 в дозе 30 мг/кг, вводимое два раза в неделю, достигало противоопухолевой эффективности dTGI 48% в модели на основе ксенотрансплантатов клеток прямой и ободочной кишки человека LOVO, выращенных подкожно у самок мышей nude (фиг. 29). Все антитела 2C2, 2C2-YTE и цетуксимаб в дозе 10 мг/кг обладали сравнимой эффективностью. Модель на основе ксенотрансплантатов LOVO содержит мутацию KRAS.
4.12. Исследования на модели подкожных ксенотрансплантатов карциномы простаты человека DU145
4.12.1. Способ
Клетки карциномы простаты человека DU145 (ATCC № HTB-81) поддерживали при 37°C в инкубаторе с 5% CO2 в среде MEM, содержащей соли Эрла, L-глутамин и 10% фетальной сыворотки теленка. Ксенотрансплантаты получали подкожной инъекцией 5×106 клеток на мышь (суспендированных в 50% матригеле) в правый бок бестимусных мышей nu/nu 4-6-недельного возраста. Опухолям давали возможность вырасти до 200 мм3 перед рандомизацией для исследований эффективности. Моноклональные антитела 2C2, MM и AMG вводили внутрибрюшинно по 30 мг на килограмм массы тела. Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
4.12.2. Результаты
При использовании модели на основе ксенотрансплантатов злокачественных клеток простаты человека DU145, выращенных подкожно у самцов мышей nude 2C2 в дозе 30 мг/кг, вводимое два раза в неделю, проявляло противоопухолевую эффективность с dTGI 77% в такой модели опухоли (фиг. 30). Модель на основе ксенотрансплантатов DU145 содержит делецию LKB-1. Моноклональные анти-HER3- антитела AMG и MM, используемые в дозе 30 мг/кг, проявляли противоопухолевую эффективность, но они были менее эффективны, чем 2C2 в такой же дозе 30 мг/кг.
4.13. Исследования на модели ортотопических ксенотрансплантатов злокачественных клеток молочной железы человека BT-474
4.13.1. Способ
Клетки злокачественной опухоли молочной железы человека BT-474 поддерживали при 37°C в инкубаторе с 5% CO2 в среде RPMI 1640, содержащей 4,5 г/л глюкозы, L-глутамин, пируват натрия и 10% фетальной сыворотки теленка. Ортотопические ксенотрансплантаты получали в результате инъекции 1×107 клеток на мышь (суспендированных в 50% матригеле) в жировую подушку молочной железы на правом боку бестимусных мышей nu/nu 4-6-недельного возраста. Гранулы эстрогена (0,36 мг) помещали под кожу левого бока за 1-2 дня до инъекции клеток. Опухолям давали возможность вырасти до 200 мм3 перед рандомизацией для исследований эффективности. 2C2, 2C2-YTE и/или анти-HER2-антитела, известные в данной области: MM, AMG и трастузумаб (торговое название герцептин®; например, патент США № 5821337) вводили внутрибрюшинно по 30 мг на килограмм массы тела. Лапатиниб вводили через пероральный зонд по 100 мг на килограмм массы тела. Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
4.13.2. Результаты
Используя модель зависимого от HER2 рака молочной железы человека, клетки BT-474, инъецированные ортотопически в жировую подушку молочной железы самок мышей nude, показали, что введение 2C2 в дозе 30 мг/кг, инъецируемой два раза в неделю, приводило к dTGI 55% в ксенотрансплантатах BT-474 (фиг. 31A). BT-474 экспрессирует HER2 на очень высоких уровнях, охарактеризованных 3+ с использованием HercepTest. Оба антитела AMG и MM, вводимые в дозе 30 мг/кг, не проявляли противоопухолевой эффективности в такой HER2-стимулируемой модели.
Лапатиниб представляет собой низкомолекулярной лекарственное средство, ингибирующее EGFR и HER2. Так как опухоли BT-474 стимулируются HER2, в такой модели тестировали лапатиниб и обнаружили, что он вызывает задержку роста опухоли в модели опухоли BT-474. Комбинированная обработка с использованием 30 мг/кг 2C2 и 100 мг/кг лапатиниба приводила к улучшенной противоопухолевой эффективности по сравнению с лапатинибом отдельно, что было наиболее четко видимо по задержке возобновления роста опухолей в отсутствие дополнительных обработок (фиг. 31B). Противоопухолевая активность 2C2-YTE была сходна с активностью 2C2. Анти-HER2-антитело трастузумаб также тестировали в такой модели и показали, что оно очень активно в такой модели HER2-стимулируемого ксенотрансплантата с dTGI 111,6%. Наблюдали небольшое дополнительное усиление активности трастузумаба в дозе 30 мг/кг при добавлении 30 мг/кг 2C2, при этом наблюдали dTGI 118,5% (фиг. 31C).
Способность клона 16 (родительский клон, из которого получено 2C2) модулировать фармакодинамические маркеры pHER3 и pAKT тестировали в экстрактах ксенотрансплантированных опухолей BT-474. Коротко, самкам бестимусных мышей nude ортотопически имплантировали клетки злокачественной опухоли молочной железы BT-474, в высокой степени экспрессирующие HER2. Животным вводили клон 16 в дозе 30 мг/кг два раза в течение 48 часов. Экстракты готовили спустя 24 часа для анализа pHER3, pAKT и общего HER3 (tHER3). Результаты нормализовали по отношению к обработанным PBS контрольным животным. Использовали по три животных на группу. Как показано на фиг. 32, клон 16 ингибировал фосфорилирование как HER3, так и AKT, на 50,0% и 46,1%, соответственно, по сравнению с опухолями от обработанных PBS мышей, и не наблюдали модулирования общего HER3 клоном 16.
4.14. Исследования на модели ортотопических ксенотрансплантатов злокачественных клеток молочной железы человека MCF-7
4.14.1. Способ.
Клетки злокачественной опухоли молочной железы человека MCF-7 поддерживали при 37°C в инкубаторе с 5% CO2 в среде Optimem, содержащей глютамакс, 2,4 г/л бикарбоната натрия, Hepes и 5% фетальной сыворотки теленка. Ортотопические ксенотрансплантаты получали в результате инъекции 5×106 клеток на мышь (суспендированных в 50% матригеле) в жировую подушку молочной железы на правом боку бестимусных мышей nu/nu 4-6-недельного возраста. Гранулы эстрогена (0,36 мг) помещали под кожу на левом боку за 1-2 дня до инъекции клеток. Опухолям давали возможность вырасти до 200 мм3 перед рандомизацией для исследований эффективности. Моноклональные антитела 2C2, 2C2-YTE и трастузумаб вводили внутрибрюшинно. 2C2 вводили в дозе 10 или 30 мг на килограмм массы тела (мг/кг), 2c2-YTE и трастузумаб в дозе 10 мг/кг. Паклитаксел вводили внутривенно в дозе 10 мг на килограмм массы тела. Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
4.14.2. Результаты
2C2 в дозе либо 10 мг/кг, либо 30 мг/кг проявлял умеренную противоопухолевую эффективность с dTGI 34% в модели на основе ксенотрансплантатов клеток злокачественной опухоли молочной железы человека MCF-7, инъецированных ортотопически в жировую подушку молочной железы самок мышей nude. 2C2-YTE в дозе 10 мг/кг имел эффективность, сходную с эффективностью 2C2 в такой же концентрации (фиг. 33A). Трастузумаб не проявлял эффективности в такой HER2-экспрессирующей модели, что свидетельствует о том, что HER2 не достаточен для стимуляции опухолевого роста. Опухоли MCF-7 экспрессировали низкие уровни HER2 (1+), измеренные с использованием HercepTest.
Паклитаксел проявлял явную противоопухолевую эффективность в ортотопической модели рака молочной железы MCF-7 в случае использования в дозе 10 мг/кг каждые два дня в течение десяти дней. Добавление 10 мг/кг 2C2 к обработке паклитакселом повышало противоопухолевую эффективность паклитаксела по сравнению с его применением отдельно в конце фазы обработки (фиг. 33B). Опухоли возобновляли рост с такой же скоростью, как и в случае опухолей, подвергаемых лечению паклитакселом, после прекращения лечения.
4.15. Исследования на модели ортотопических ксенотрансплантатов клеток злокачественной опухоли молочной железы человека MDA-MB-361
4.15.1. Способ.
Клетки злокачественной опухоли молочной железы человека MDA-MB-361 поддерживали при 37°C в инкубаторе с 5% CO2 в среде RPMI 1640, содержащей 4,5 г/л глюкозы, L-глутамин, пируват натрия и 10% фетальной сыворотки теленка. Ортотопические ксенотрансплантаты получали в результате инъекции 5×106 клеток на мышь (суспендированных в 50% матригеле) в жировую подушку молочной железы в правом боку бестимусных мышей nu/nu 4-6-недельного возраста. Гранулы эстрогена (0,36 мг) помещали под кожу левого бока за 1-2 дня до инъекции клеток. Опухолям давали возможность расти до 230 мм3 перед рандомизацией для исследований эффективности. Антитела 2C2-YTE и/или анти-HER2-антитела, известные в данной области, в частности трастузумаб (торговое название герцептин®; например, патент США №5821337) и RhuMAb 2C4 (например, публикация патента WO 2001/00245), называют в настоящем описании трастузумабом и 2C4, соответственно. Моноклональные антитела трастузумаб и 2C4 вводили внутрибрюшинно по 30 мг на килограмм массы тела (2C2-YTE) или по 10 мг на килограмм массы тела (трастузумаб и 2C4). Лапатиниб вводили через пероральный зонд по 100 мг на килограмм массы тела. Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
4.15.2. Результаты
При использовании HER2-стимулируемой модели рака молочной железы человека, клеток MDA-MB-361 (тест Hercept +2), инъецированных ортотопически в жировую подушку молочной железы самок мышей nude, введение 2C2-YTE в дозе 30 мг/кг, инъецируемой два раза в неделю, всего пять доз, приводило к 70,1% dTGI в ксенотрансплантатах MDA-MB-361 (фиг. 34A-C). Клетки MDA-MB-361 экспрессируют HER2 на среднем уровне 2+, судя по характеристике с использованием HercepTest, и оцениваются позитивно в FISH-анализе (флуоресцентный анализ гибридизации in situ). Оба антитела, трастузумаб и rhuMAb 2C4, вводимые в дозе 10 мг/кг, и лапатиниб в дозе 100 мг/кг, вводимый два раза в сутки, также проявляли противоопухолевую эффективность в модели опухоли MDA-MB-361.
Так как опухоли MDA-MB-361 стимулируются HER2, 2C2-YTE сочетали с лекарственными средствами, мишенью которых является HER2, такими как трастузумаб, rhuMAb 2C4 или лапатиниб. Комбинированное лечение с использованием 30 мг/кг 2C2-YTE и 10 мг/кг трастузумаба приводило к аддитивной противоопухолевой эффективности по сравнению с использованием трастузумаба отдельно. Аддитивный эффект также проявлялся в задержке возобновления роста опухолей в отсутствие дополнительного лечения (фиг. 34A). Сочетание 2C2-YTE с трастузумабом было лучше, чем сочетания 2C2-YTE либо с rhuMAb 2C4 (фиг. 34B), либо с лапатинибом (фиг. 34C) в такой модели.
4.16. Трансгенные мыши, экспрессирующие рецептор FcRn человека, для исследования экспозиции антител с YTE-модификацией.
4.16.1. Способ.
Трансгенным самкам мышей SCID, экспрессирующим рецептор FcRn человека, вводили однократную дозу 60 мг/кг антитела клон 16-GL, 2C2 или 2C2-YTE внутривенным путем. От таких мышей собирали сыворотку в нескольких временных точках после введения доз, используя пункцию сердца, и кровь собирали в SST-пробирки Microtainer. Пробирки осторожно встряхивали в течение 10 секунд и выдерживали при комнатной температуре в течение 20 минут, давая возможность крови свернуться. Образцы центрифугировали при 1000×g в течение 10 минут, и образцы сыворотки осторожно переносили в новые пробирки и хранили при -80°C. Формат непрямого твердофазного иммуноферментного анализа (ELISA) использовали для количественного определения 2C2 в сыворотке мышей. Стандарты, контроли качества и образцы сыворотки мышей инкубировали с антителами козы против IgG человека, которые были иммобилизованы на 96-луночном планшете для микротитрования. После инкубации несвязанные материалы удаляли на стадии промывки и 2C2 определяли, используя антитело козы против IgG человека, конъюгированное с пероксидазой хрена. Добавляли раствор кислоты для остановки и степень ферментативного превращения субстрата определяли, измеряя оптическую плотность при 450 нм. Измеряемая оптическая плотность была прямо пропорциональна концентрации 2C2 или 2C2-YTE, присутствующих в сыворотке мышей. Использовали стандартную кривую для анализа 2C2 или 2C2-YTE, чтобы интерполировать концентрацию в образцах сыворотки.
4.16.2. Результаты
В случае антитела 2C2-YTE, которое содержит мутацию YTE в остове 2C2, наблюдали более высокие уровни экспозиции с течением времени по сравнению с 2C2 или клоном 16-GL (фиг. 35). Через четырнадцать дней после однократной дозы антитела, вводимой таким мышам, уровень экспозиции в сыворотке 2C2-YTE был выше 100 мкг/мл, тогда как в случае 2C2 и клона 16-GL уровень был ниже 1 мкг/мл. Полученные данные демонстрируют, что YTE может продлевать время полужизни 2C2-YTE по сравнению с его родительским антителом 2C2.
4.17. Ингибитор MEK индуцирует экспрессию HER3 и в сочетании с анти-HER3-антителом проявляет аддитивную противоопухолевую эффективность.
Мутации KRAS (онкоген вируса саркомы крыс Kirsten V-Ki-ras2) и BRAF (онкоген B1 вируса саркомы мышей v-raf) приводят к конститутивной активации передачи сигнала EGFR через онкогенный путь Ras/Raf/Mek/Erk. Мутация Kras относится к наиболее часто встречающимся мутационным событиям во многих солидных опухолях, особенно при раке прямой и ободочной кишки (CRC, 30-40%) и раке легкого (LC, 20-25%). Мутация Braf также встречается с относительно высокой частотой в случае CRC (~15%). Было показано, что вследствие своей способности конститутивно активировать путь ERK, мутанты Kras и Braf придают опухоли резистентность к терапии RTK, особенно EGFR-мАт, таким как цетуксимаб и панитумумаб. Влияние ингибирования митоген-активируемой протеинкиназы (MEK) на путь HER3 в моделях CRC и LC исследовали, используя ингибитор MEK селуметиниб (AstraZeneca, см., например, WO 03/077914 и WO 2007/076245) отдельно или в сочетании с 2C2 (или 2C2-YTE). Исследовали несколько моделей CRC и LC, включая модели, несущие мутантный Kras (например, A549, LOVO) или мутантный Braf (например, HT-29, Colo205) или RAS дикого типа (например, HARA-B, KNS-62).
4.17.1. Способы
Исследование на культуре клеток: клетки высевали по 105 на лунку в 24-луночные планшеты и в среду, содержащую 10% инактивированной нагреванием FBS, и давали возможность достичь слияния на 80% или больше перед обработкой. 2C2 (10 мкг/мл) или контрольное антитело, ингибитор MEK селуметиниб (1 или 10 мкМ) или сочетание 2C2 (10 мкг/мл) и селуметиниба (10 мкМ) готовили в полной среде. Обработки проводили после удаления среды для посева. После инкубации в течение 24 часов в условиях 5% CO2 при 37°C клетки промывали один раз охлажденным на льду PBS и затем лизировали, добавляя 60 мкл 2x буфера для образцов с додецилсульфатом натрия (SDS) (Invitrogen). Образцы нагревали в течение 5 минут и затем охлаждали на льду в течение 2 минут. Образцы анализировали, используя Вестерн-блоттинг, по существу, как описано выше (см. примеры, раздел 2.4).
Исследования на ксенотрансплантатах: Клетки NSCLC человека A549 (ATCC № CCL-185), которые содержат мутацию в кодоне 12 гена KRAS, поддерживали при 37°C в инкубаторе с 5% CO2 в среде Хама F 12K, содержащей 4,5 г/л глюкозы, L-глутамин, пируват натрия и 10% фетальной сыворотки теленка. Ксенотрансплантаты получали подкожной инъекцией 5×106 клеток на мышь (суспендированных в 50% матригеле) в правый бок бестимусных мышей nu/nu 4-6-недельного возраста. Клетки карциномы прямой и ободочной кишки человека HT-29 (ATCC № HTB- 38) поддерживали при 37°C в инкубаторе с 5% CO2 в среде RPMI 1640, содержащей 4,5 г/л глюкозы, L-глутамин, пируват натрия и 10% фетальной сыворотки теленка. Ксенотрансплантаты получали подкожной инъекцией 5×106 клеток на мышь в правый бок бестимусных мышей nu/nu 4-6-недельного возраста. Клетки карциномы прямой и ободочной кишки человека LOVO (ATCC № CCL-229) поддерживали при 37°C в инкубаторе с 5% CO2 в среде Хама F 12K, содержащей 4,5 г/л глюкозы, L-глутамин, пируват натрия и 10% фетальной сыворотки теленка. Ксенотрансплантаты получали подкожной инъекцией 5×106 клеток на мышь в правый бок бестимусных мышей nu/nu 4-6-недельного возраста. В случае всех трех моделей опухолей опухолям давали возможность вырасти до 200 мм3 перед рандомизацией для исследований эффективности. 2C2-YTE или контрольный IgG1 вводили внутрибрюшинно, селуметиниб вводили перорально. В случае исследований сочетаний 2C2-YTE и селуметиниб вводили в дозе 30 мг/кг или 75 мг/кг, соответственно. Измерения штангенциркулем использовали для вычисления объемов опухолей по формуле:
объем опухоли = π/6(длина×ширина×ширина)
в случае опухолей, выросших у мышей. Противоопухолевые эффекты выражены в виде разницы в ингибировании опухолевого роста в процентах (TGI), которую вычисляли следующим образом:
разница TGI в процентах = 1-(dT/dC)×100,
где dT = изменение среднего объема опухоли в группе обработки по сравнению со значением при стадировании, и dC = изменение среднего объема опухоли в контрольной группе по сравнению со значением при стадировании.
Получение лизатов из замороженных опухолей: Мышей подвергали гуманной эвтаназии путем асфиксии в CO2 согласно протоколу авторов in vivo и опухоли вырезали и переносили в пробирки с матриксом A для лизирования. Добавляли лизирующий буфер RIPA (500 мкл), содержащий смесь ингибиторов протеаз и набор I и II смеси ингибиторов фосфатаз, затем образцы гомогенизировали, используя устройство Fast Prep. Образцы охлаждали на льду в течение 30 минут и подвергали дополнительному циклу гомогенизации перед осветлением центрифугированием при 14000 об./мин в течение 10 минут при 4°C. Осветленные лизаты переносили в свежие пробирки объемом 1,5 мл и измеряли содержание белка. Затем лизаты хранили при -80°C вплоть до анализа. Образцы анализировали, используя Вестерн-блоттинг, по существу, как описано выше (см. примеры, раздел 2.4).
4.17.2. Результаты
Как показано на фиг. 36, уровни и общего и pHER3-белка возрастали после обработки ингибитором MEK селуметинибом в клетках злокачественной опухоли прямой и ободочной кишки HT-29, выращенных в культуре, которые экспрессируют мутантный BRAF, и в клетках LOVO, которые экспрессируют мутантный KRAS (фиг. 36, левый и средний блоты, соответственно). Повышение уровня HER3 также наблюдали в клетках Colo205, которые экспрессируют мутантный BRAF, и в клетках DLD-1 и HCT, которые экспрессируют мутантный KRAS (фиг. 36, правый блот, и не показанные данные), после обработки селуметинибом. Повышения происходили в случае обеих доз 1 мкМ и 10 мкМ селуметиниба. Активность селуметиниба подтверждали на основании снижения pERK во всех клеточных линиях при дозах 1 мкМ и 10 мкМ. Ингибирование MEK приводило к ингибированию фосфорилирования ERK. Анти-HER3-антитело, 2C2, ингибировало как общий HER3, так и pHER3 в клетках HT-29 и LOVO. 2C2 также снижал HER3 в клетках Colo205 и DLD-1. Кроме того, совместная обработка 2C2 и селуметинибом блокировала индукцию общего HER3 и pHER3 селуметинибом в клетках HT-29, LOVO и DLD-1 (фиг. 36 и не показанные данные). Не наблюдали регистрируемых уровней HER3 или pHER3 в клетках злокачественной опухоли прямой и ободочной кишки SW480, которые экспрессируют мутантный KRAS ни в необработанных, ни в обработанных селуметинибом клетках.
Как показано на фиг. 37A, комбинированная обработка 30 мг/кг 2C2-YTE и 75 мг/кг селуметиниба приводила к аддитивной противоопухолевой эффективности в ксенотрансплантатах NSCLC A549 по сравнению с обработкой селуметинибом отдельно. Аддитивный эффект также проявлялся в задержке возобновления роста опухолей в отсутствие дополнительных обработок (верхняя панель). Вестерн-блот-анализ лизатов опухолей от мышей, обработанных сочетанием 30 мг/кг 2C2-YTE и 75 мг/кг селуметиниба в течение 4-дневного периода, показал, что фосфо-HER3 и фосфо- ERK были полностью ингибированы. Оба маркера служили в качестве фармакодинамических показателей действия 2C2-YTE и селуметиниба. Сходные данные получали в случае моделей на основе ксенотрансплантатов CRC HT-29 (фиг. 37B, верхняя и нижняя панели) и LoVo (фиг. 37C, верхняя и нижняя панели). Кроме того, обнаружено, что уровень фосфо-AKT снижается в лизатах опухолей HT-29, обработанных сочетанием 30 мг/кг 2C2-YTE и 75 мг/кг селуметиниба, по сравнению с отдельными обработками (фиг. 37B, нижняя панель). Обработка селуметинибом отдельно в дозе 75 мг/кг приводила к увеличению уровня фосфо-HER3 в экстрактах опухолей LoVo, которое было предотвращено в опухолях, обработанных сочетанием 2C2-YTE и селуметиниба (фиг. 37C, нижняя панель). Сходные результаты наблюдали в HARA-B (данные не показаны).
В культуре клеток наблюдали, что уровни белка HER3 возрастают в ответ на ингибитор MEK в большинстве исследованных моделей, что свидетельствует о том, что путь HER3 может играть роль в резистентности к ингибиторам MEK. В нескольких исследованиях на моделях ортотопических ксенотрансплантатов CRC и LC наблюдали, что сочетание 2C2-YTE и селуметиниба повышает противоопухолевую эффективность любого средства, по сравнению с его использованием отдельно. Полученные данные подтверждают применимость 2C2 в сочетании с ингибитором MEK, подобным селуметинибом, для усиления противоопухолевой активности и предотвращения резистентности.
4.18. Токсикологические исследования на макаках-крабоедах
4.18.1. Способ
Двадцать самцов макак-крабоедов (Macaca fascicularis) распределяли на четыре группы (по 5 животных на группу) и вводили всего пять доз контрольного наполнителя или 2C2-YTE в дозе 10, 30 или 120 мг/кг. Животным вводили дозы один раз в неделю посредством 5-минутной внутривенной инфузии в объеме 5 мл/кг. По три животных на группу умерщвляли на 32 день (через три дня после последнего введения дозы на 29 день фазы дозирования) и по два животных на группу умерщвляли на 43 день фазы восстановления (сорок пятый день после последнего введения дозы на 29 день фазы дозирования). Оценка токсичности была основана на нескольких факторах, включая смертность, клиническое обследование, массу тела, оценку раздражения в месте введения дозы, клинические и патологоанатомические оценки.
Образцы плазмы макак-крабоедов выделяли и анализировали в отношении уровней растворимого HER3 (sHER3), используя форму анализа типа «сэндвич» с анти-HER3-антителами и систему регистрации электрохемилюминесценции (ECL) для количественного определения свободного sHER3. Пустые 96-луночные планшеты Meso Scale Discovery (MSD) (MSD, номер в каталоге L15XA-6/L11XA-6) покрывали 0,5 мкг/мл 2C2-YTE в течение ночи при 2-8°C и затем блокировали MSD-блокатором A (MSD, номер в каталоге R93BA-1). Эталонный стандарт и контроли качества (QC) и неразбавленные тестируемые образцы плазмы макак-крабоедов добавляли в блокированные планшеты на 1 час при комнатной температуре. Добавление биотинилированного анти-hErbB3/HER3-антитела (R&D Systems, номер в каталоге BAM348) с последующим добавлением Sulfo-TAG (MSD, номер в каталоге R32AD-1) приводило к испусканию света при электрохимической стимуляции. Сигнал ECL улавливали и регистрировали на визуализирующем устройстве MSD Sector Imager 2400. Количество непосредственно генерируемого света коррелировало с количеством sHER3 в образцах плазмы макак-крабоедов. Первичные данные (оценка ECL) экспортировали в SOFTmax® PRO. Стандартную кривую, полученную для стандартов рекомбинантного HER3 человека, подгоняли, используя программу подгонки по 5 параметрам. Концентрации HER3 в плазме макак-крабоедов вычисляли на основе стандартной кривой, используя статистическую функцию SOFTmax PRO.
Кроме того, собирали образцы биопсии кожи для биоанализа. Кратко, на коже животного рисовали парные 10-мм-кружки и ~100 мкл PBS или HRG в концентрации 0,1 мг/мл инъецировали интрадермально в центр каждого кружка. Приблизительно через 20 минут образец кожи собирали из каждого места инъекции и мгновенно замораживали. Альтернативно собирали парные образцы биопсии (без предварительной интрадермальной инъекции) и инкубировали приблизительно в течение 30 мин при комнатной температуре в культуральной среде с добавлением или без добавления 100 мкг/мл HRG с последующими двумя промывками охлажденным на льду PBS. Затем промытый образец мгновенно замораживали. Затем ткани гомогенизировали в пробирках с матриксом A для лизирования (MP Biomedicals), содержащих лизирующий буфер RIPA и смесь ингибиторов протеаз (Sigma-Aldrich) и наборы I и II смеси ингибиторов фосфатаз (EMD-Millipore), используя устройство Fast Prep (MP Biomedicals). Затем образцы подвергали циклу замораживания-размораживания и дополнительному циклу гомогенизации перед осветлением центрифугированием при 14000 об./мин в течение 5 минут при 4°C. Осветленные лизаты переносили в свежие пробирки объемом 1,5 мл и измеряли содержание белка. Уровни общего HER3 и pHER3 определяли, используя анализ ELISA типа «сэндвич».
4.18.2. Результаты
Проводимое не в соответствии с GLP 1-месячное исследование токсичности с использованием многократных доз 2C2-YTE с шестинедельной фазой восстановления осуществляли на макаках-крабоедах, чтобы оценить токсичность и активность 2C2-YTE при введении один раз в неделю посредством внутривенной инфузии макакам-крабоедам в течение, по меньшей мере, 1 месяца (всего 5 доз) и чтобы оценить обратимость, стойкость или задержанное появление любых эффектов после 6-недельного периода восстановления. Не отмечали неблагоприятных эффектов после внутривенного введения один раз в неделю (5-минутная инфузия) вплоть до 120 мг/кг/дозу 2C2-YTE в течение 5 недель (всего 5 доз) у самцов макак-крабоедов.
Способность 2C2-YTE блокировать HRG-индуцированный pHER3 в коже макак-крабоедов подтверждали при оценке in vivo и ex vivo. Полное подавление циркулирующего растворимого HER3 наблюдали у всех животных, получающих внутривенный 2C2-YTE. Стимуляция ex-vivo образцов биопсии кожи с использованием HRG приводила к увеличению соотношения pHER3:tHER3, что свидетельствует о том, что HER3, присутствующий в коже макак-крабоедов, может быть активирован под действием HRG, основного лиганда HER3. Полное подавление HRG-индуцированного pHER3 достигали во всех группах, обработанных 2C2-YTE, в конце фазы введения доз (данные не показаны). Таким образом, 2C2-YTE блокировали in vivo и ex vivo HRG-индуцированное фосфорилирование HER3 в образцах биопсии кожи макак-крабоедов.
ВКЛЮЧЕНИЕ В ВИДЕ ССЫЛКИ
Все публикации и патенты, упоминаемые в настоящем описании, включены таким образом в виде ссылки в полном объеме, так же, как в случае, когда указано, что каждая отдельная публикации или патент специально и по отдельно включены в виде ссылки. Кроме того, предварительные заявки на выдачу патента США №: 61/563092, поданная 23 ноября 2011, 61/656670, поданная 7 июня 2012, и 61/722558, поданная 5 ноября 2012, включены в виде ссылки в полном объеме для всех целей.
Предшествующее описание конкретных аспектов настолько полно раскрывает общую природу изобретения, что другие специалисты, применяя знания в данной области техники, легко могут модифицировать и/или адаптировать для различных применений такие конкретные аспекты, без излишнего экспериментирования, не отступая от общей концепции настоящего изобретения. Поэтому полагают, что такие адаптации и модификации входят в общий замысел и диапазон эквивалентов раскрытых аспектов, на основании инструкций и рекомендаций, представленных в настоящем описании. Следует понимать, что фразеология или терминология используется в настоящей публикации в целях описания, а не ограничения, так что специалисту в данной области надлежит интерпретировать терминологию или фразеологию настоящего описания в свете представленных инструкций и руководства.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К HER3 И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2560583C2 |
| ТЕРАПИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ, РЕЗИСТЕНТНОЙ К ПРЕПАРАТАМ НА ОСНОВЕ ПЛАТИНЫ | 2005 |
|
RU2403065C2 |
| БИСПЕЦИФИЧНЫЕ АНТИ-HER2/АНТИ-HER3 АНТИТЕЛА | 2016 |
|
RU2653443C2 |
| КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛО, КОТОРОЕ СВЯЗЫВАЕТСЯ С ДОМЕНОМ II HER2, И ЕГО КИСЛЫЕ ВАРИАНТЫ | 2009 |
|
RU2543664C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СПЕЦИФИЧНЫЕ К ИЛ-21, И ОБЛАСТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2708336C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, АНАЛОГИ АНТИТЕЛ, КОМПОЗИЦИИ И СПОСОБЫ | 2010 |
|
RU2580038C2 |
| АНТИТЕЛА, СПОСОБНЫЕ СПЕЦИФИЧЕСКИ СВЯЗЫВАТЬСЯ С HER2 | 2014 |
|
RU2628094C2 |
| ВВЕДЕНИЕ ФИКСИРОВАННЫХ ДОЗ HER-АНТИТЕЛ | 2005 |
|
RU2438705C2 |
| КОМПОЗИЦИЯ АНТИТЕЛ К HER2 | 2005 |
|
RU2361880C2 |
| ПРИМЕНЕНИЕ КОНЪЮГАТА АНТИ-HER2 АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО ПРИ ЛЕЧЕНИИ УРОТЕЛИАЛЬНОЙ КАРЦИНОМЫ | 2019 |
|
RU2750817C1 |






















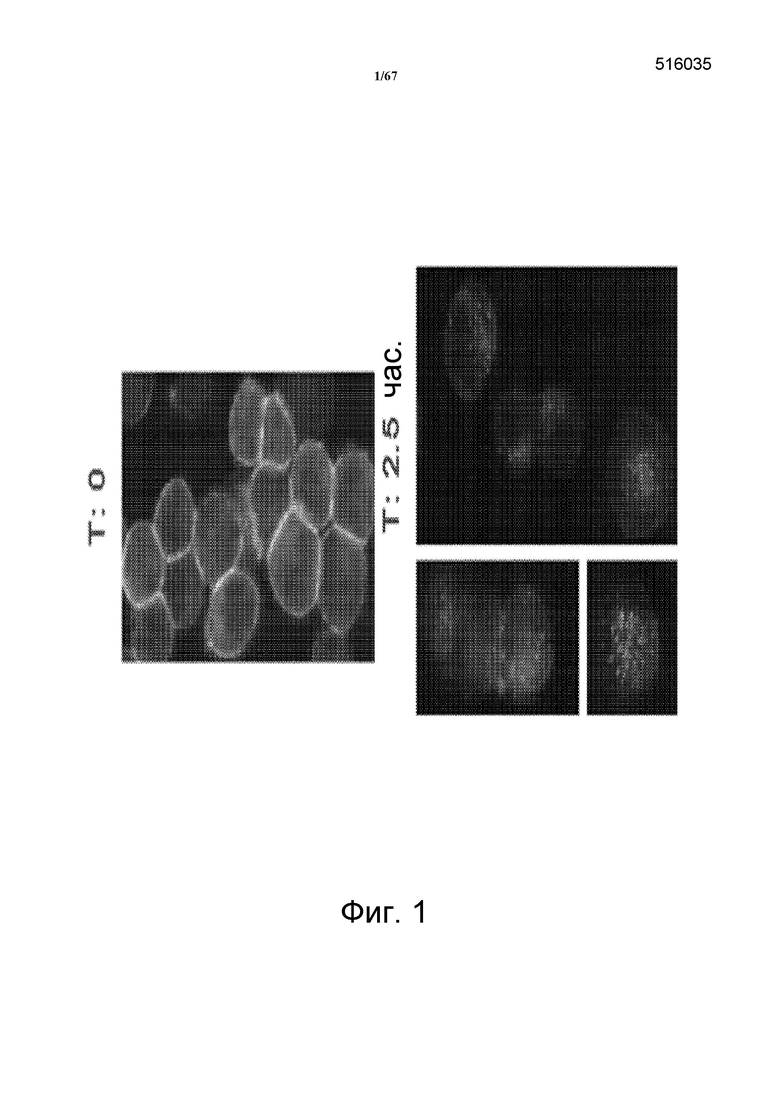


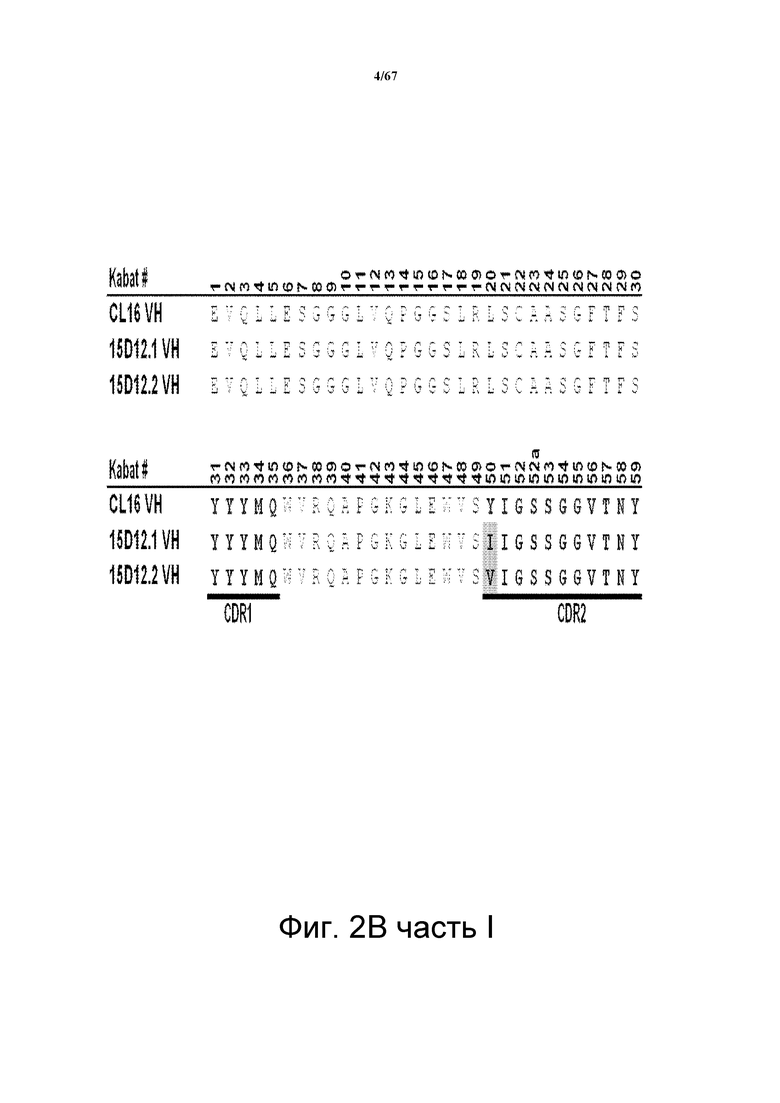

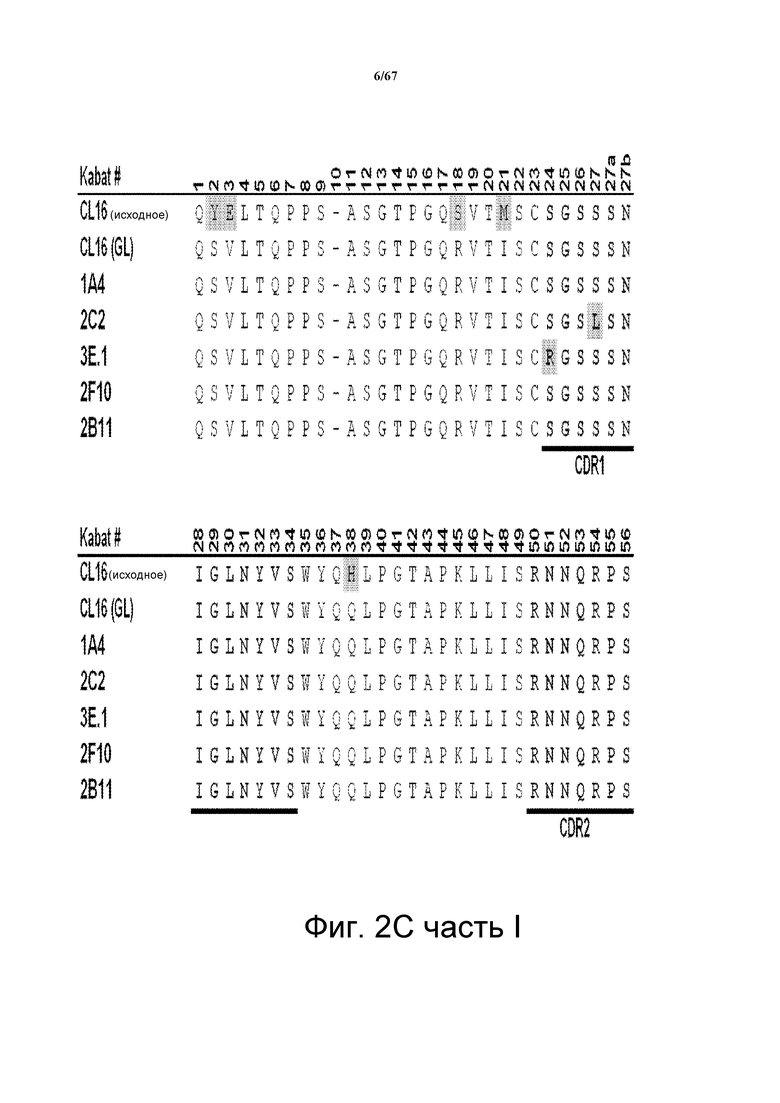
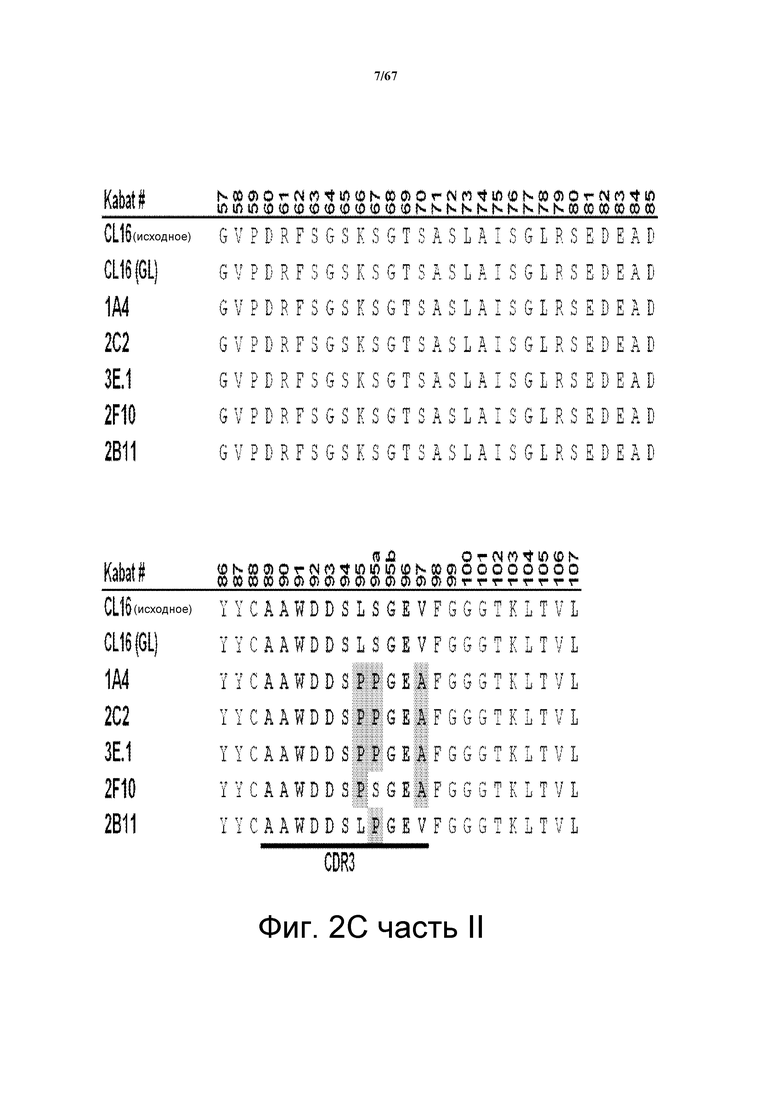
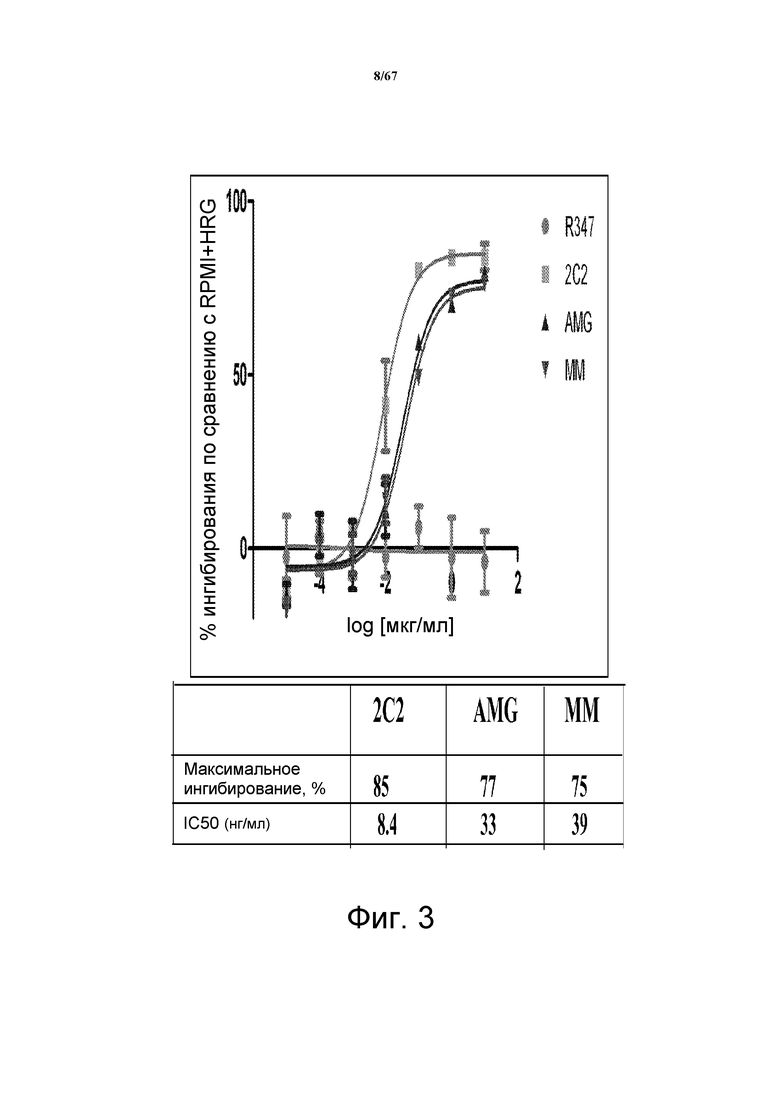
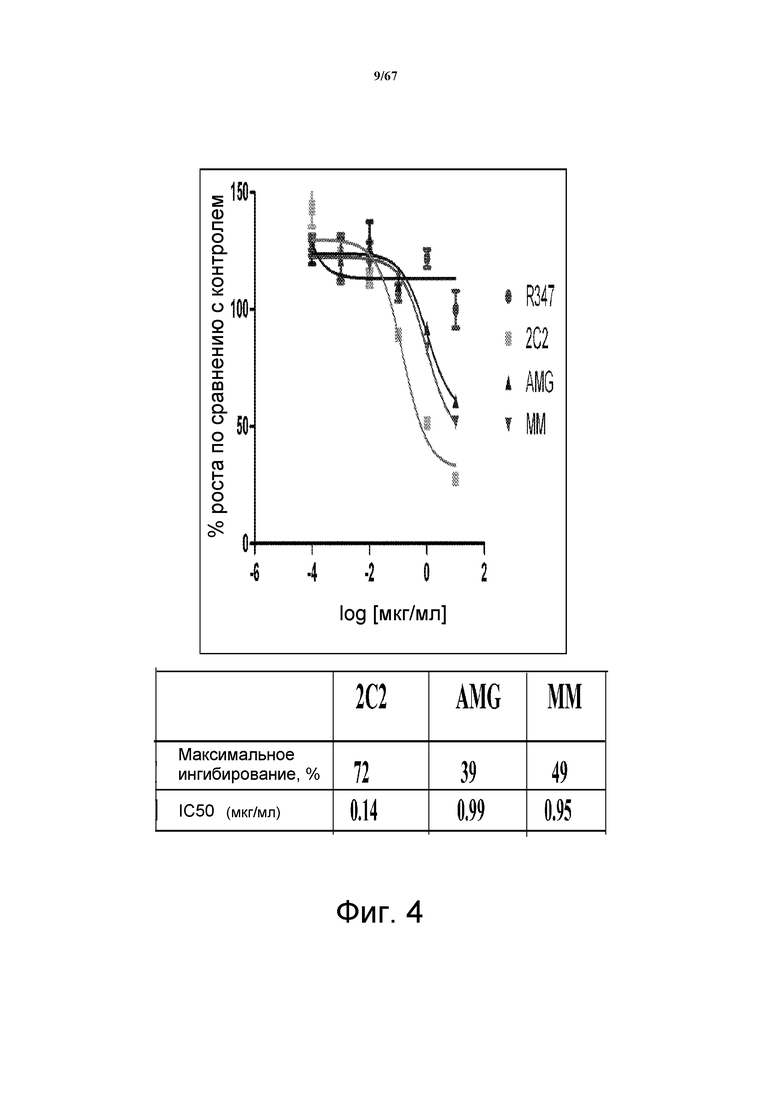
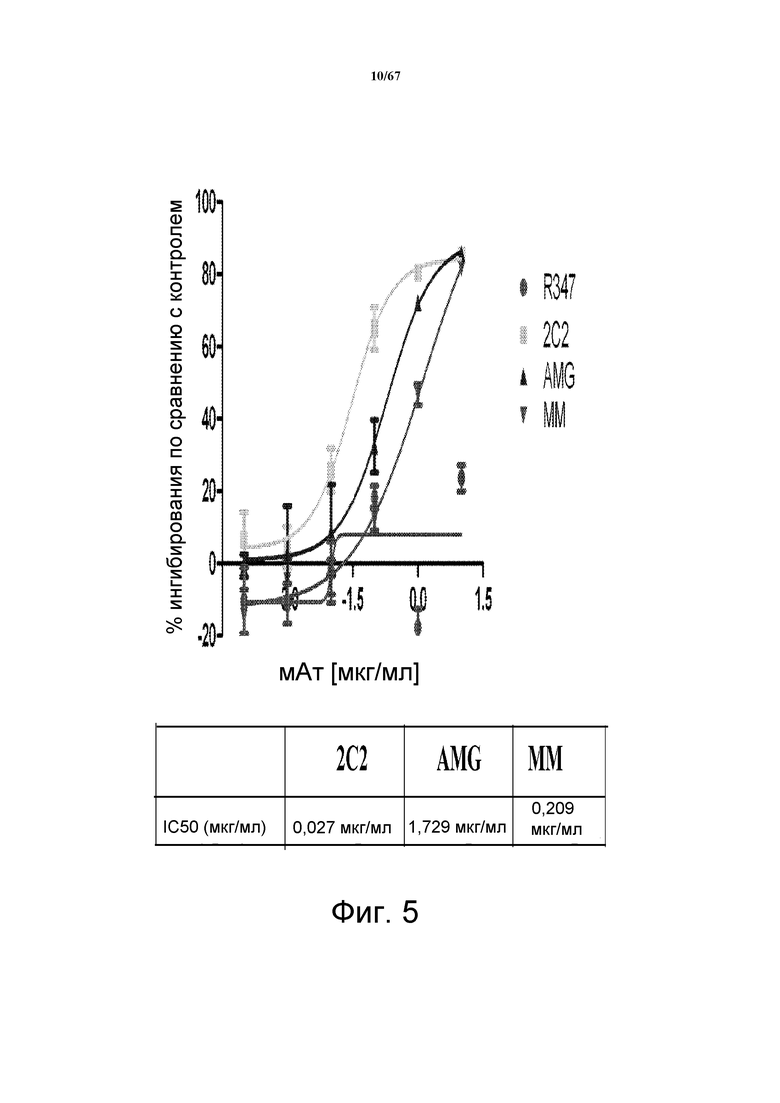
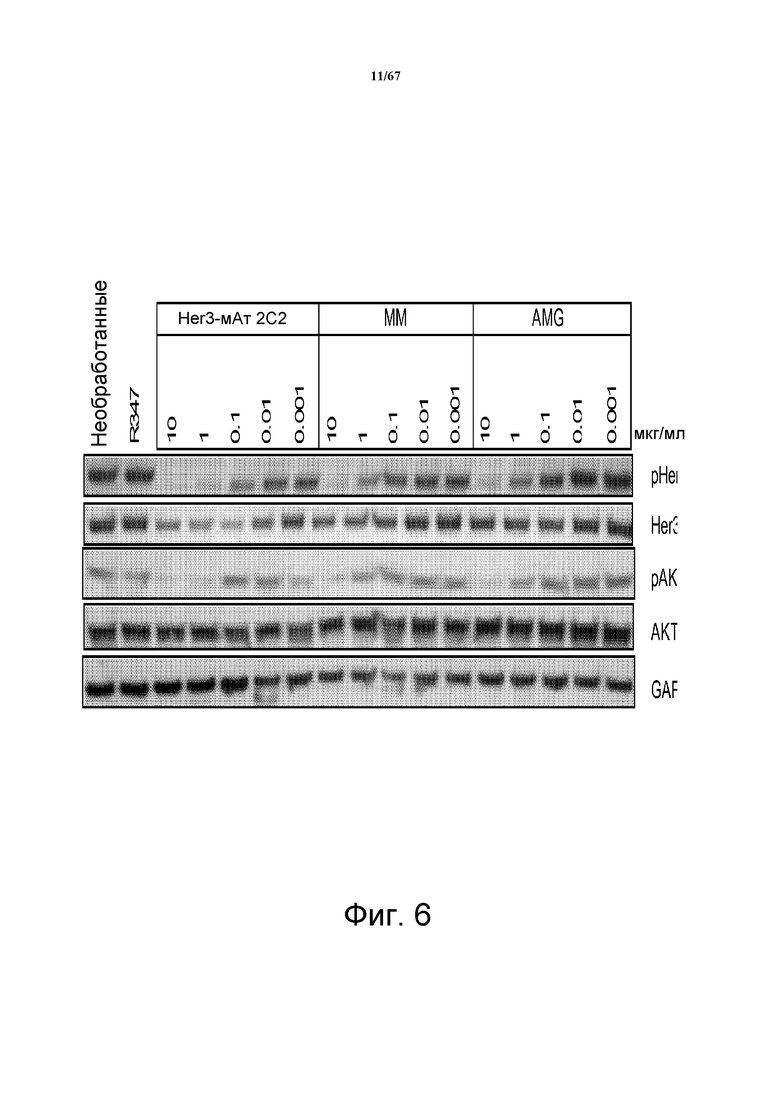
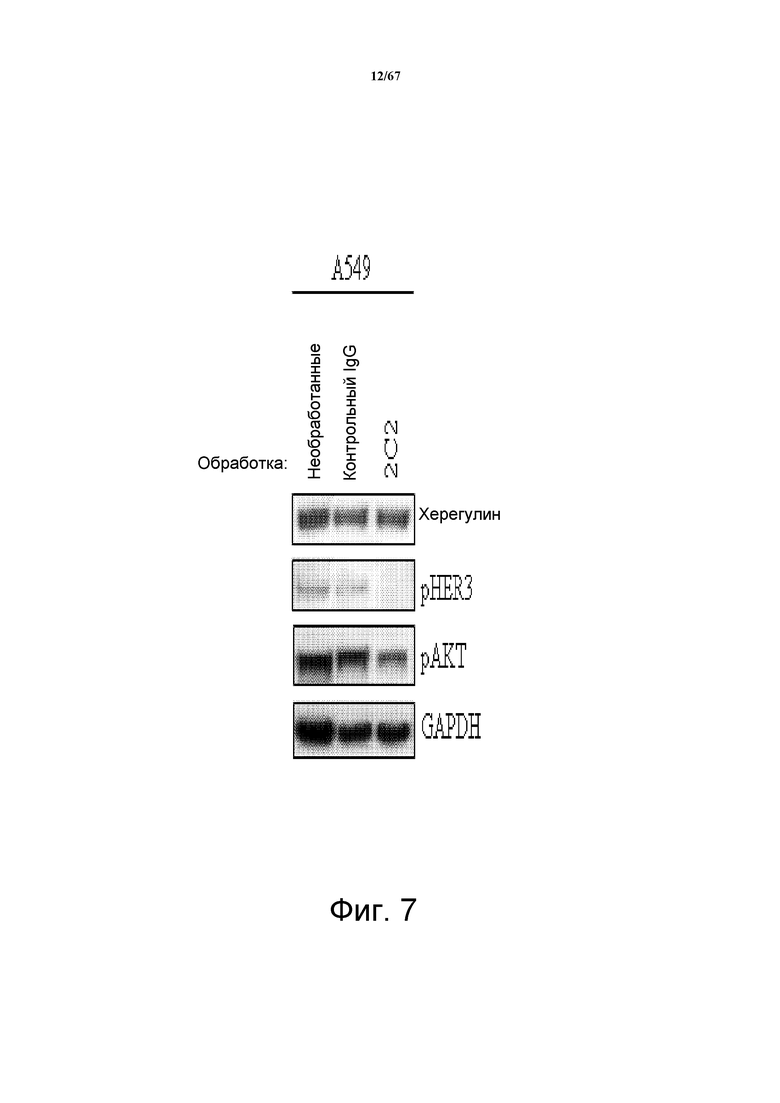
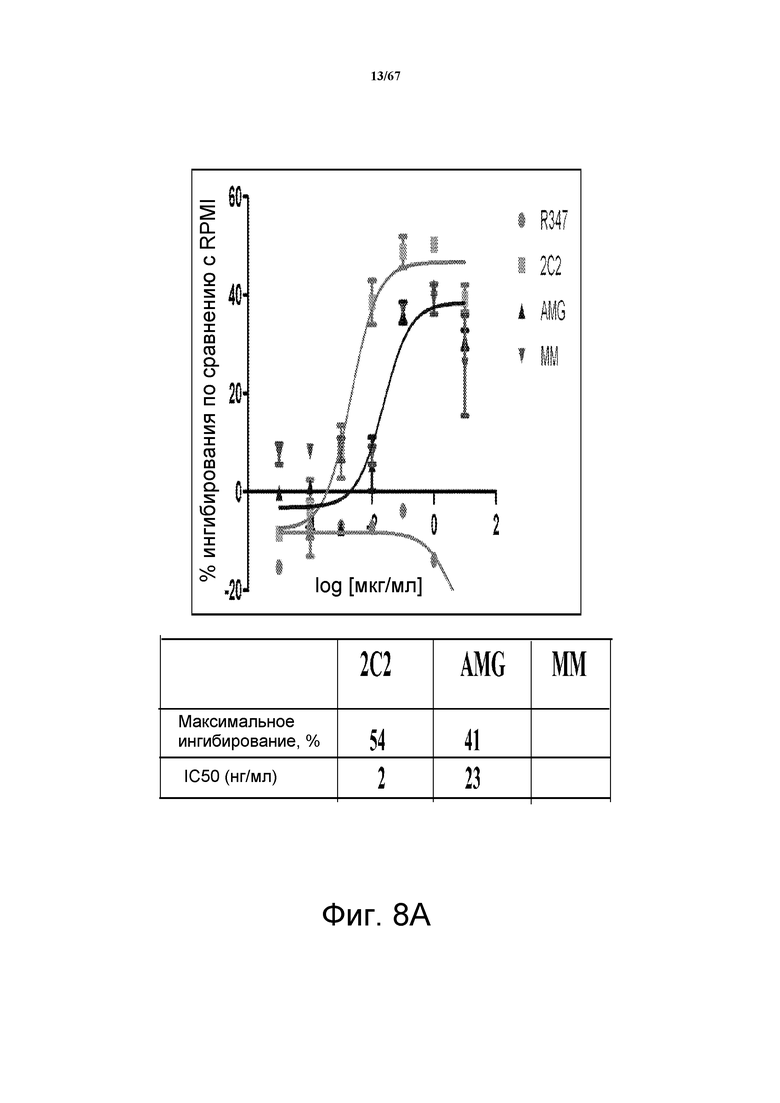
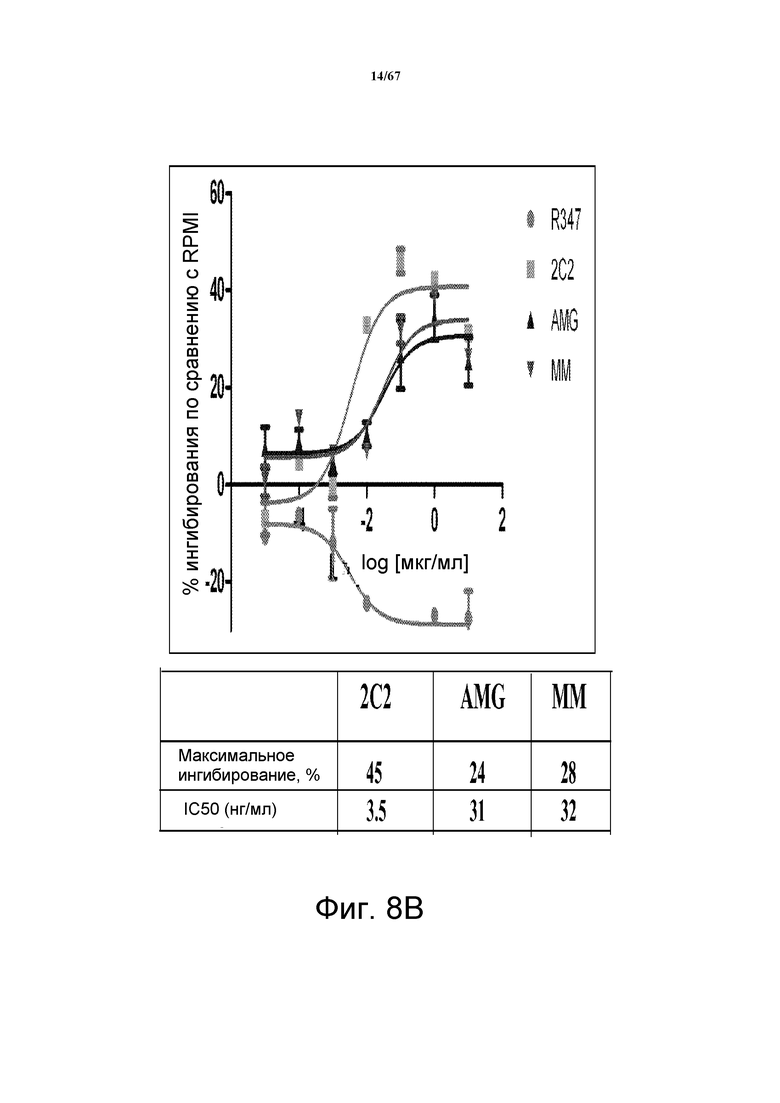
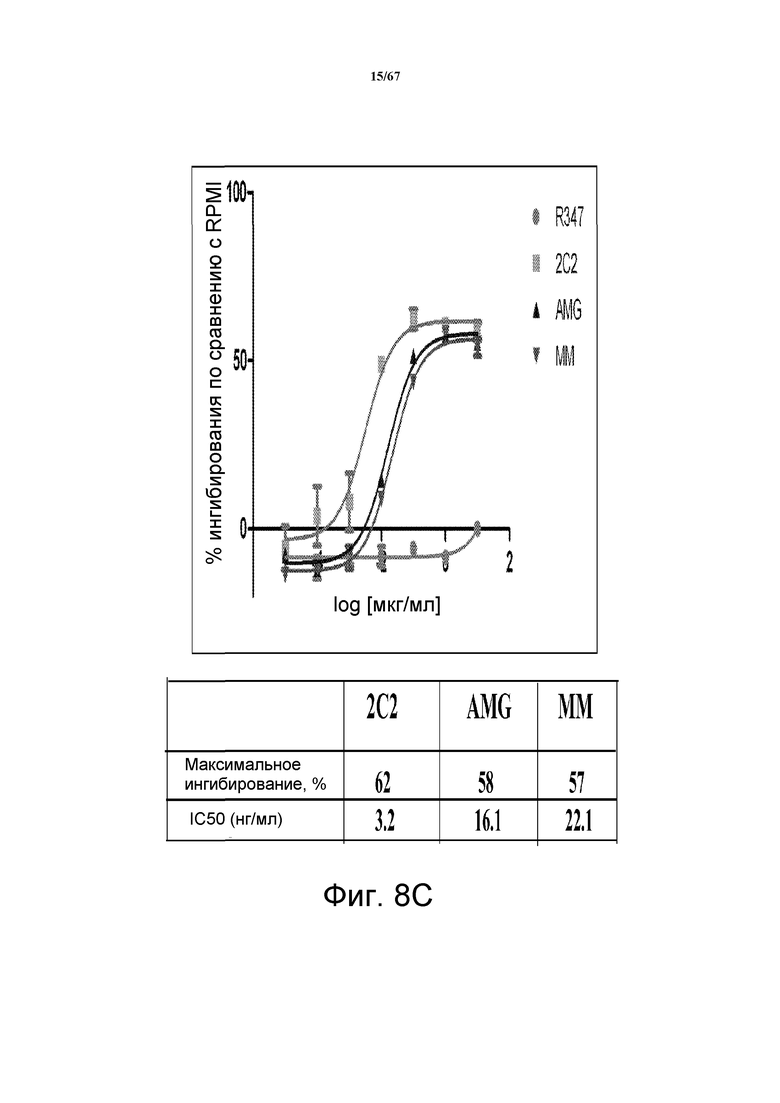
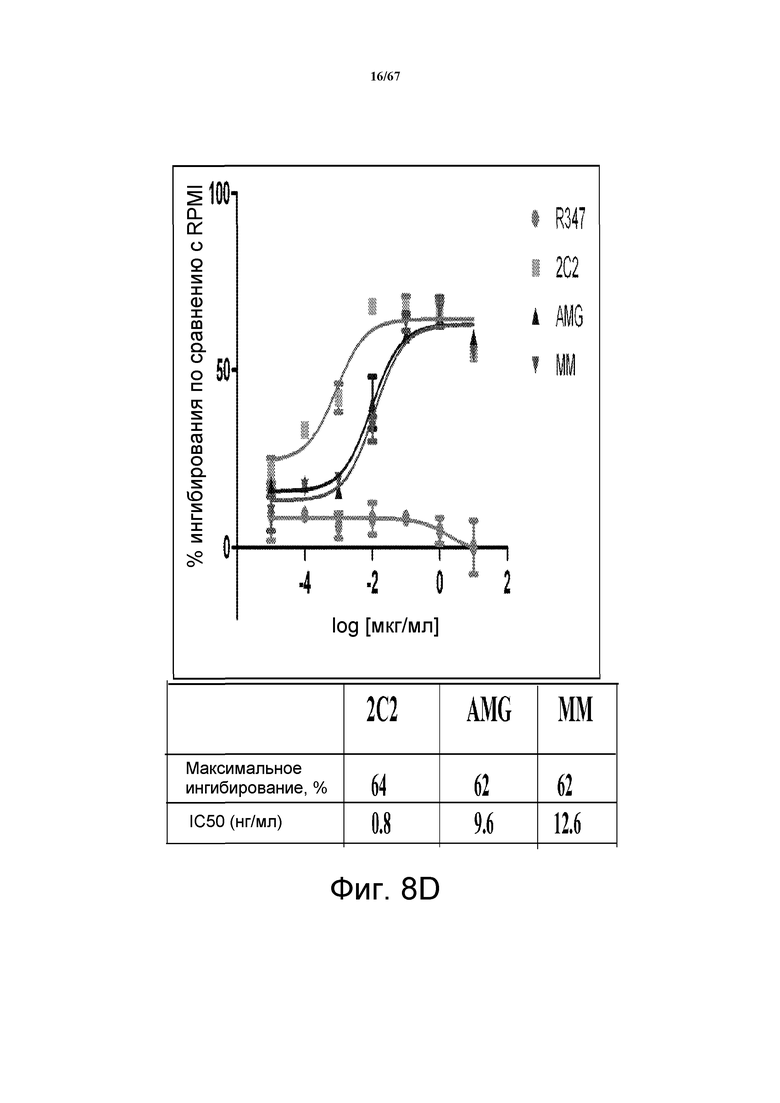
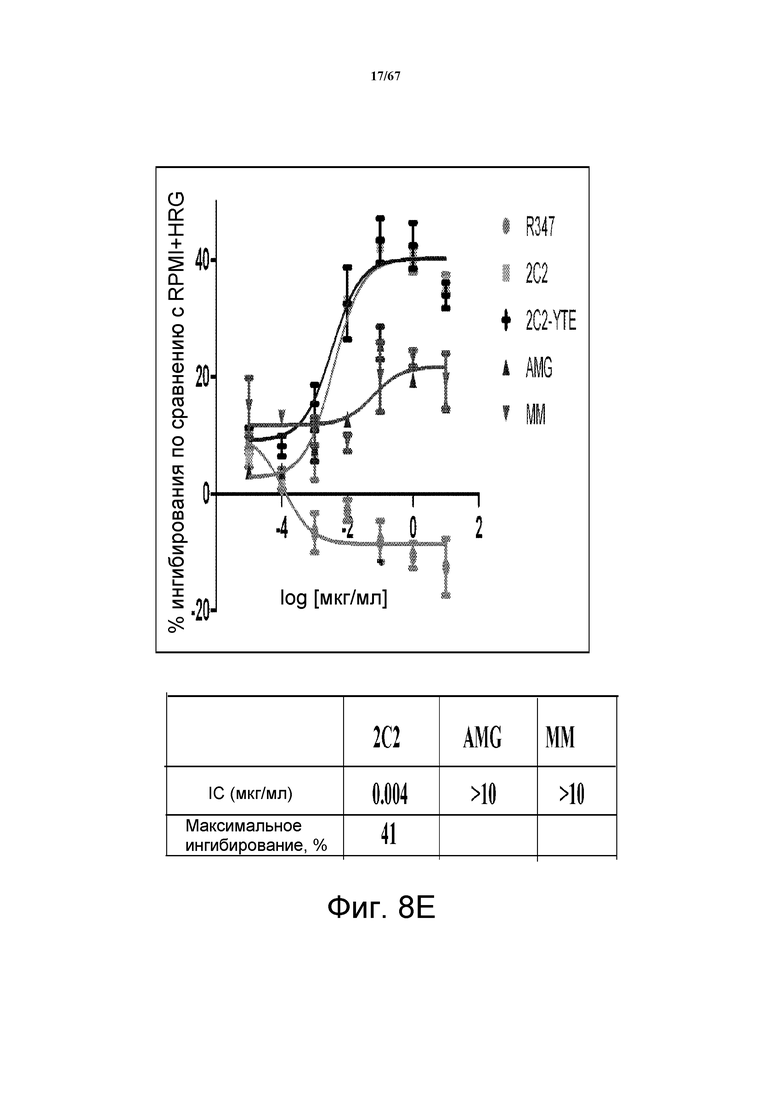
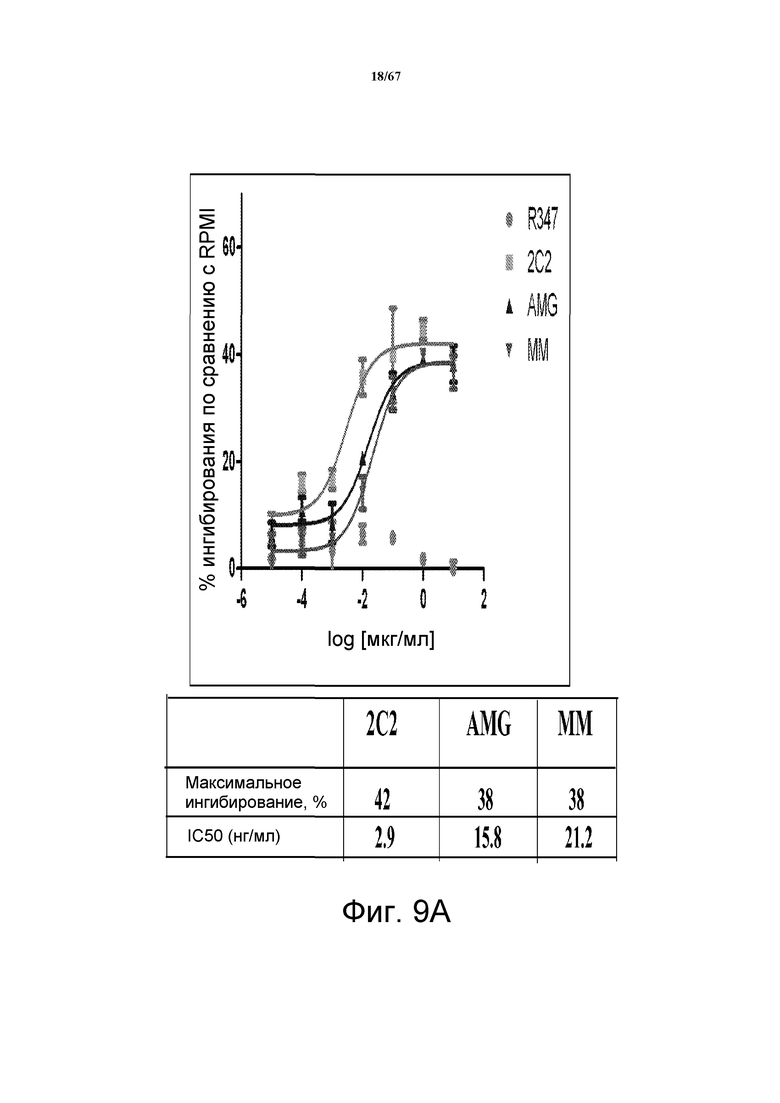
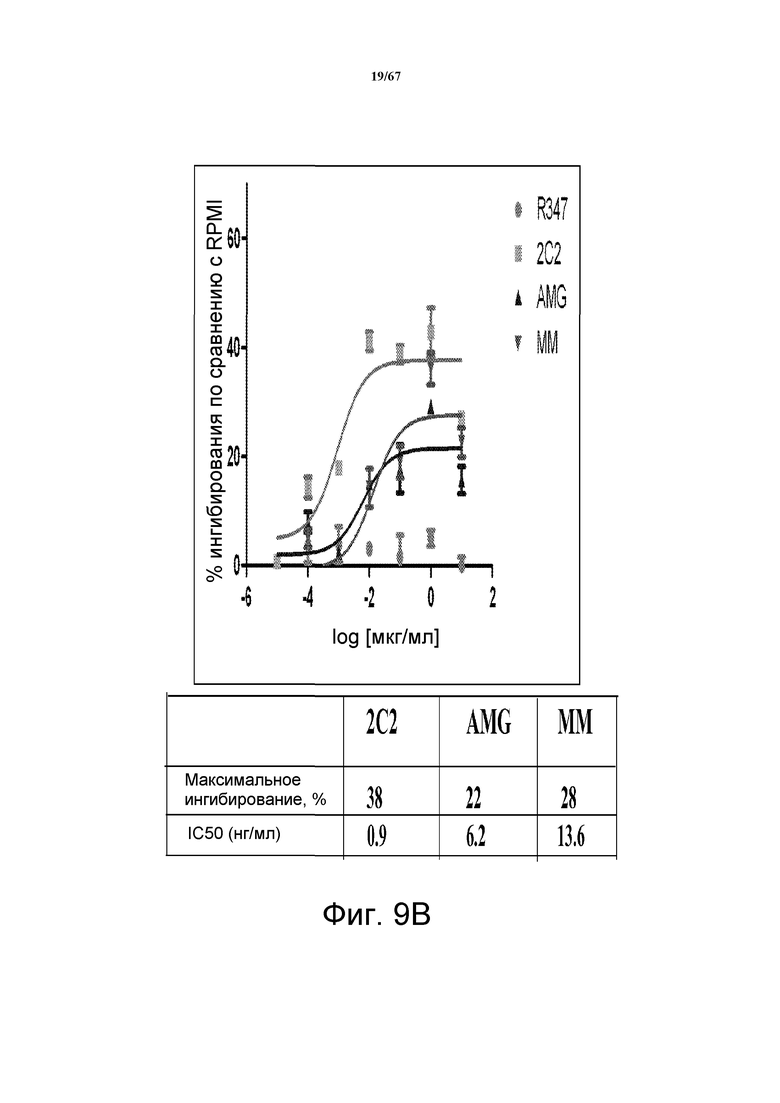
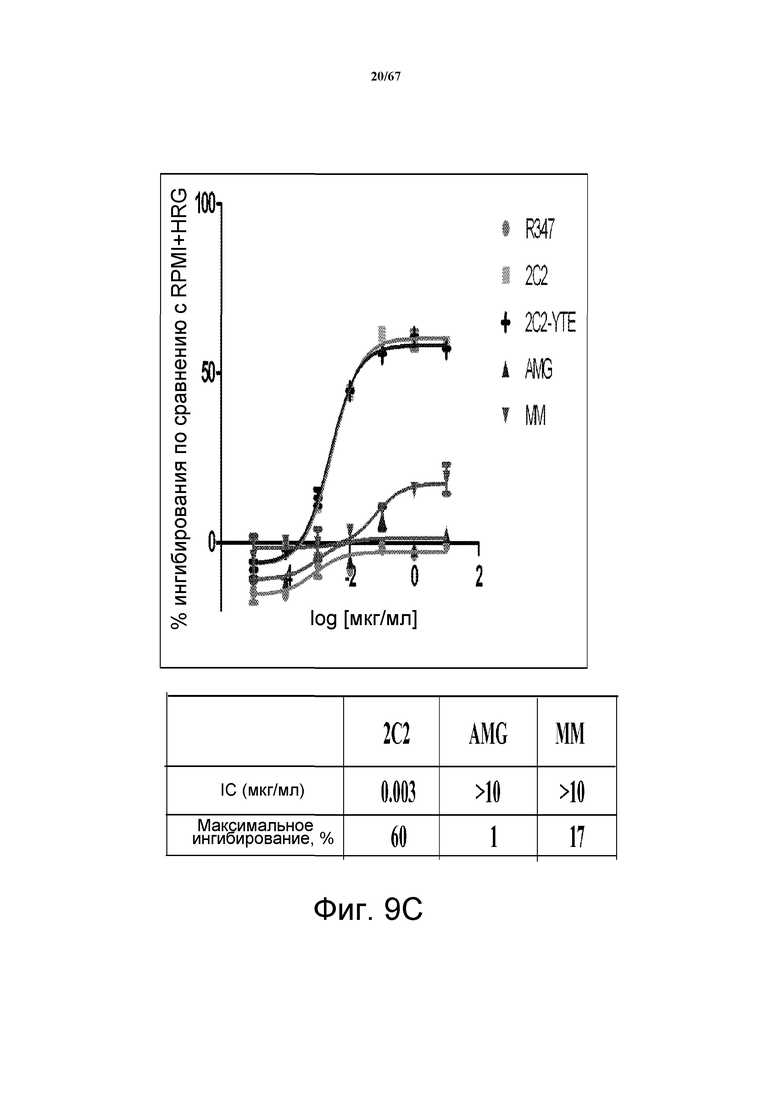
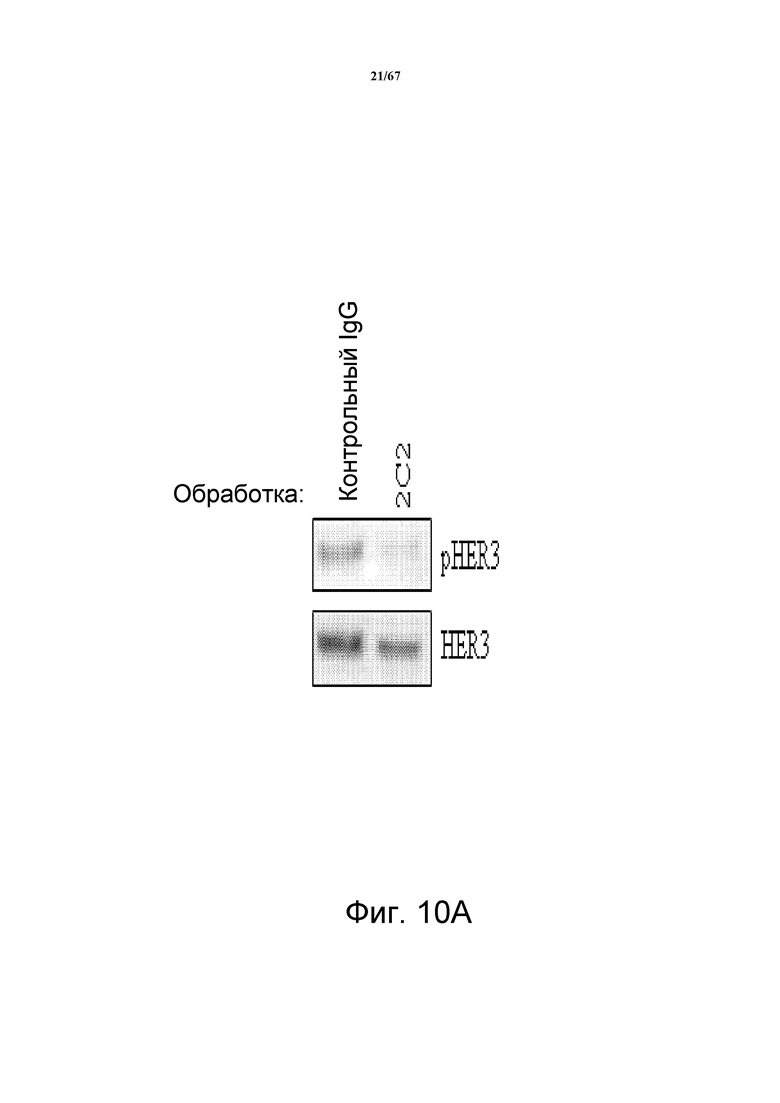
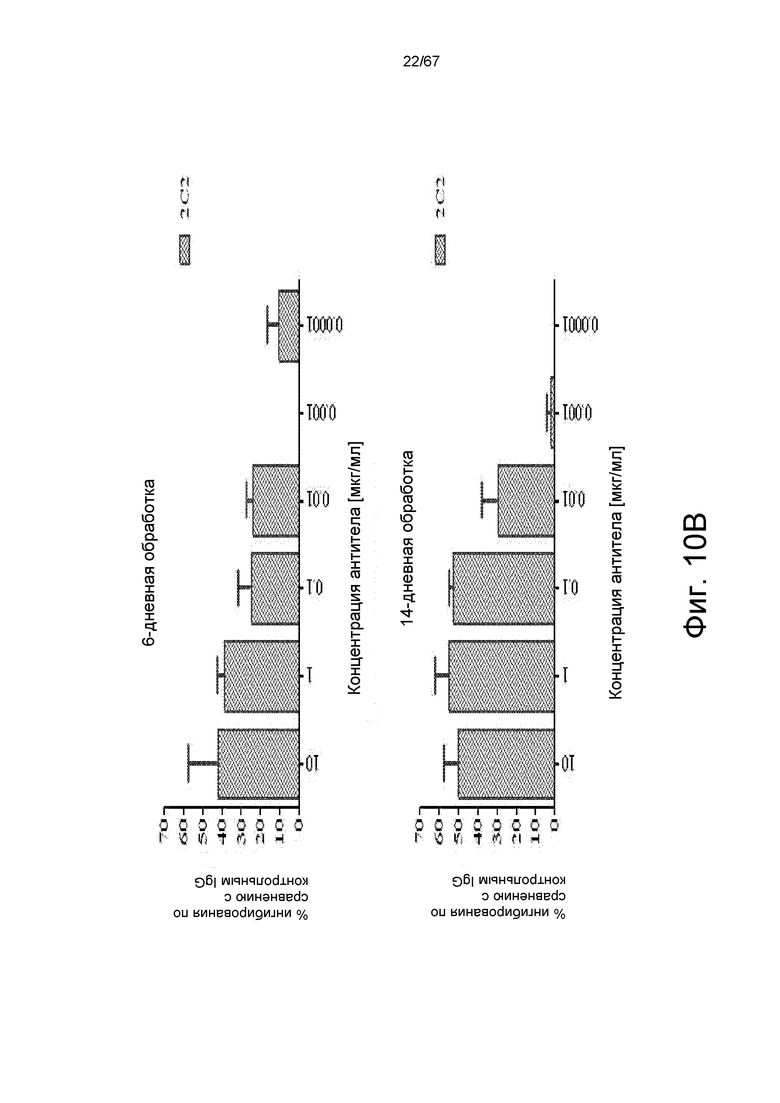
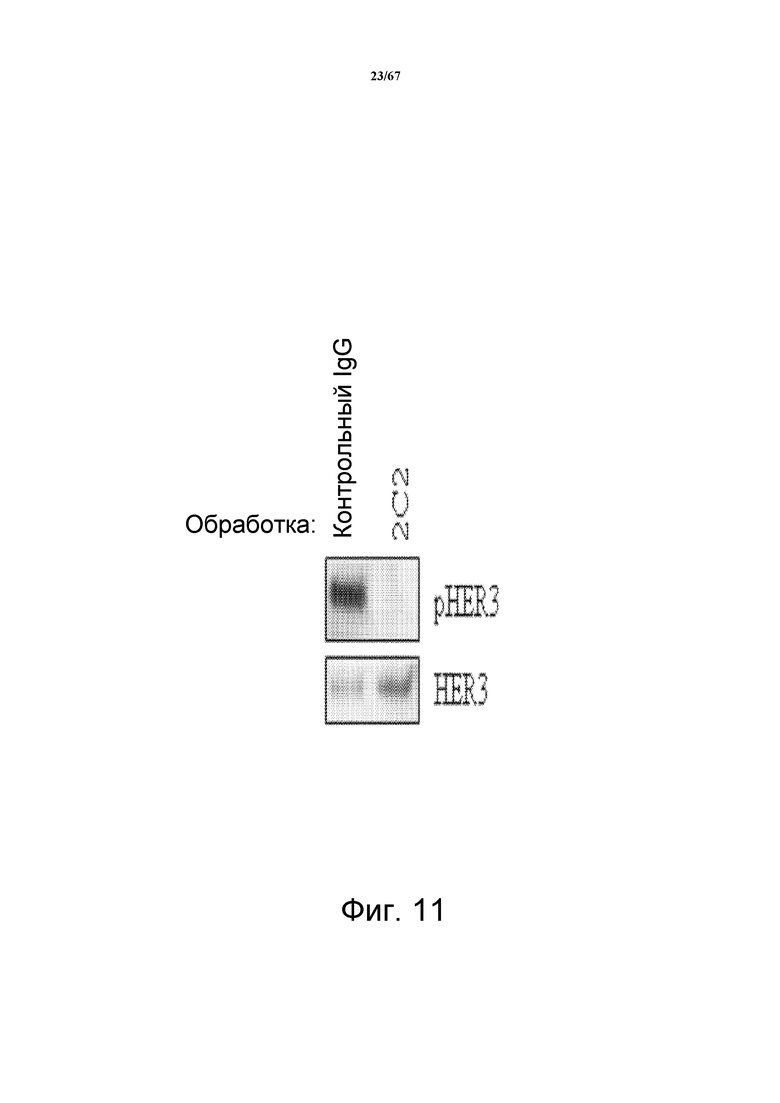
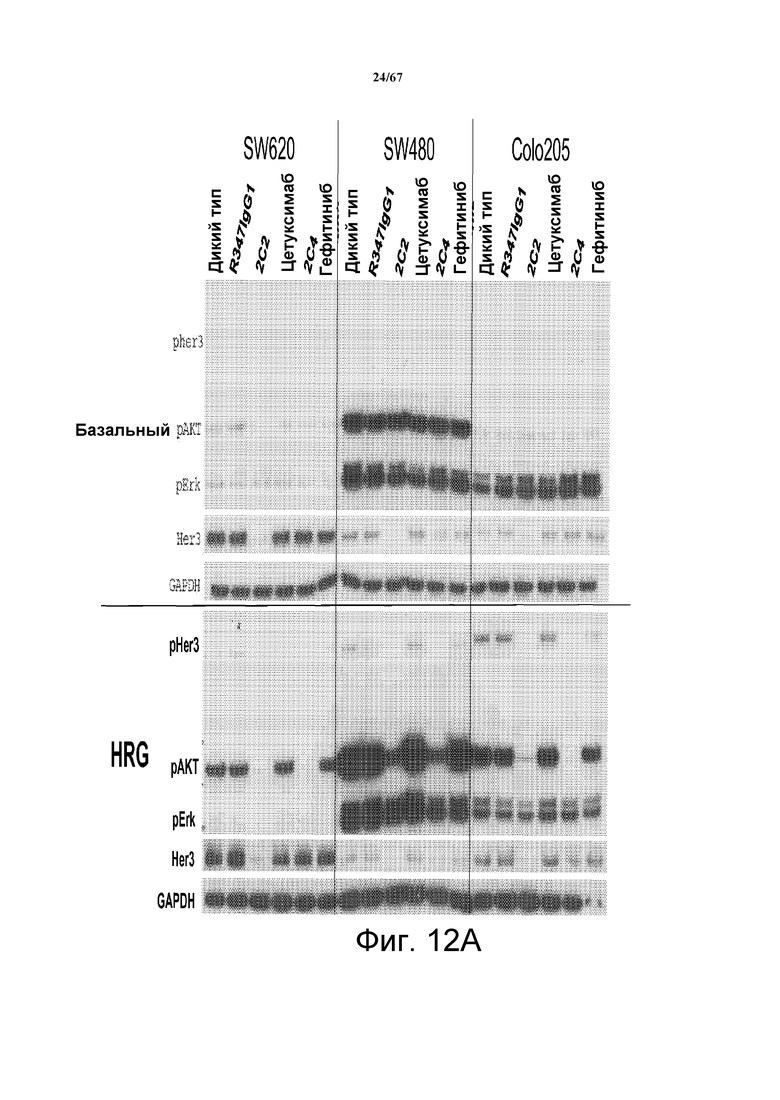
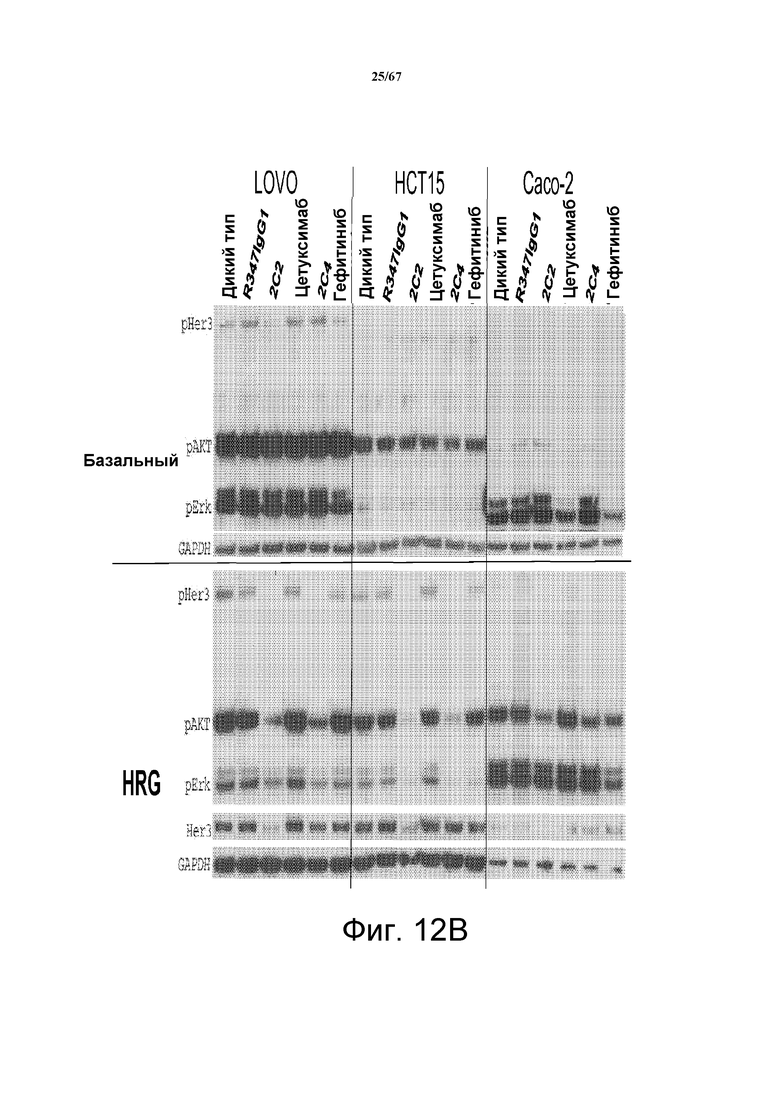
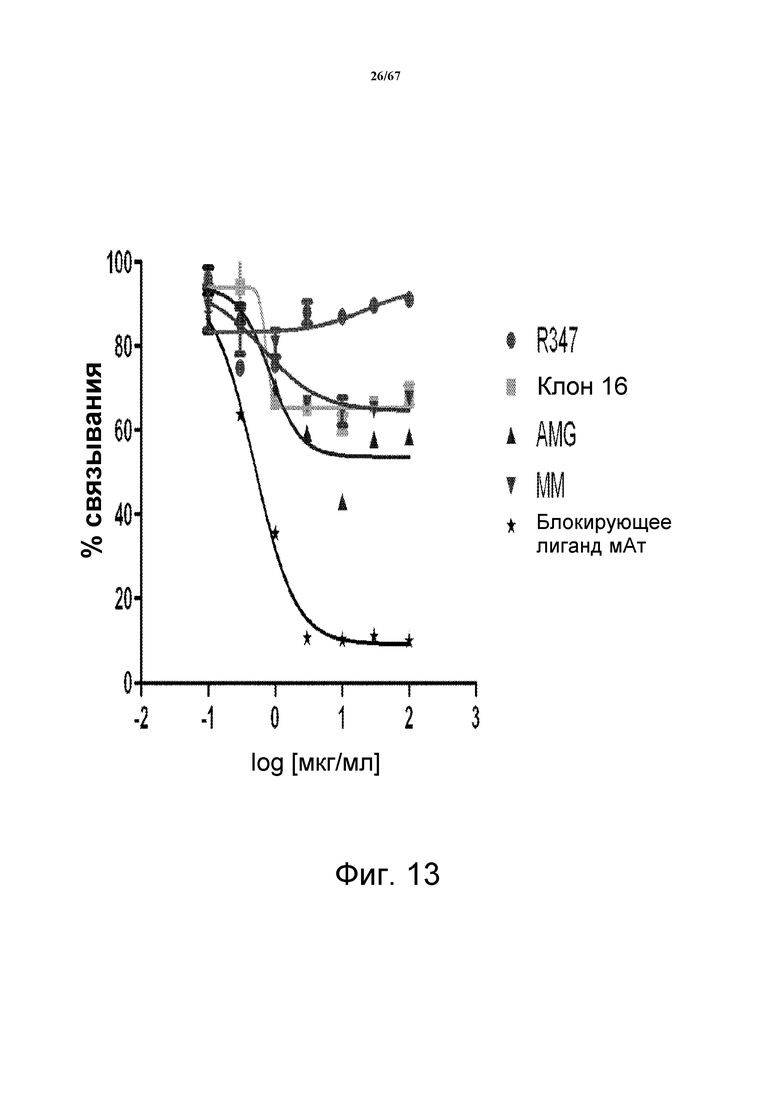
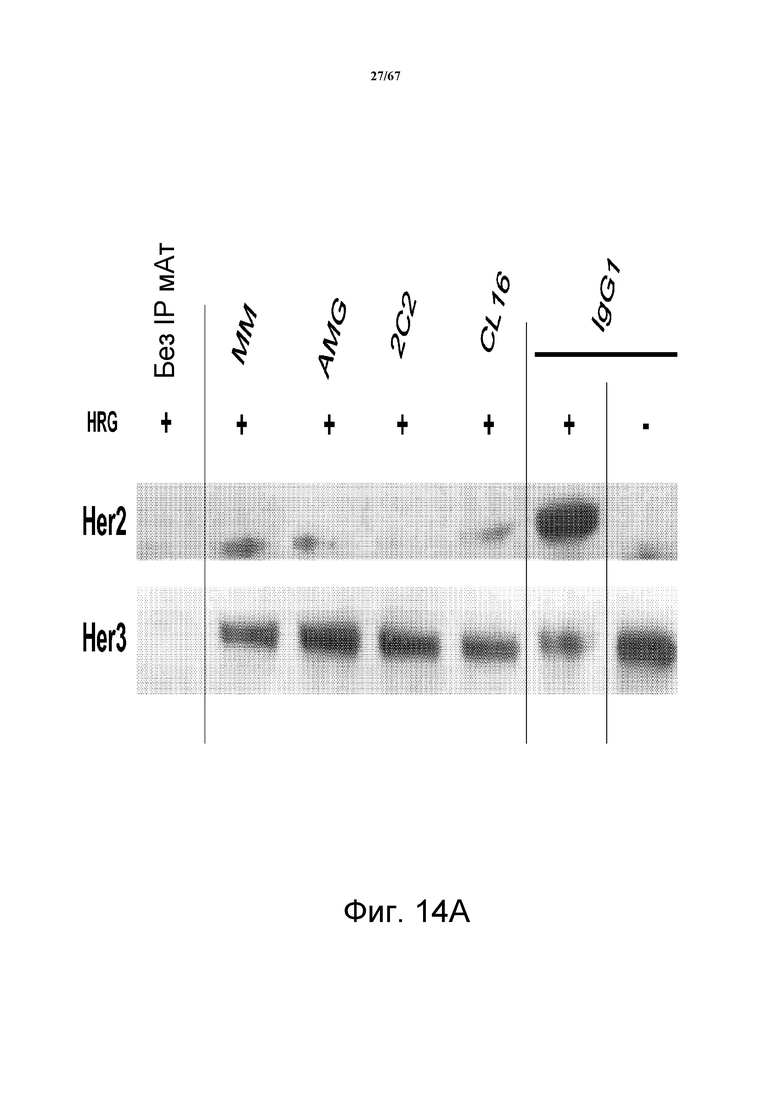
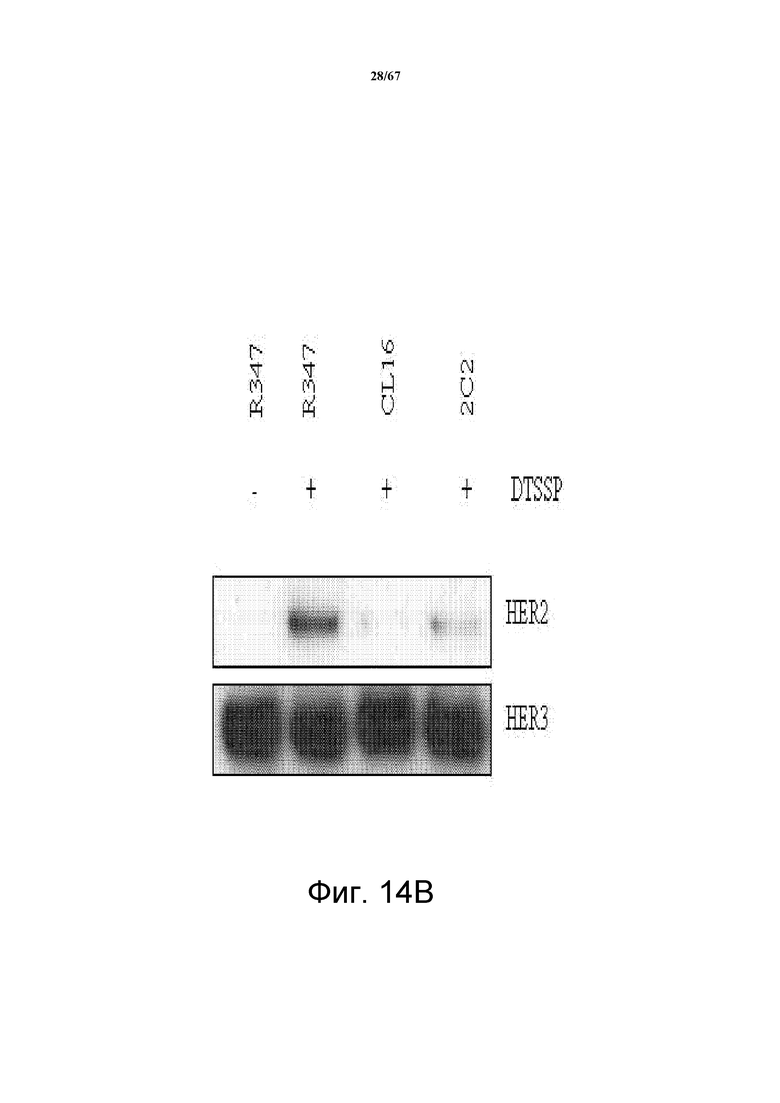
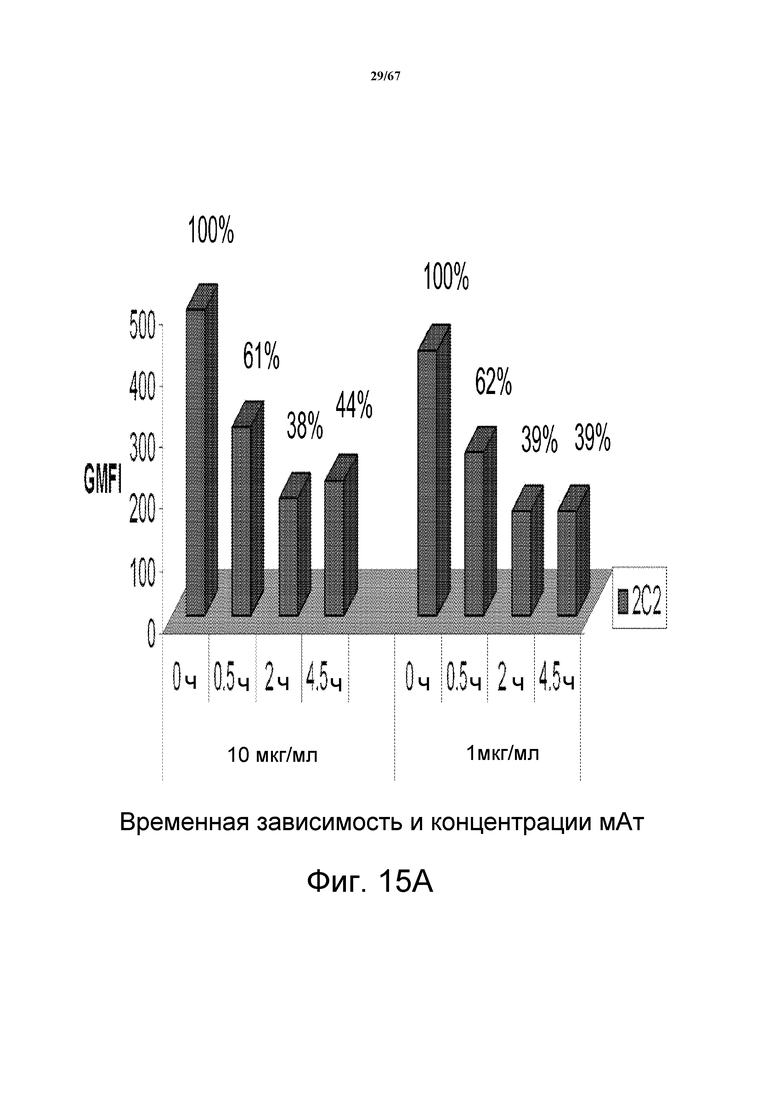
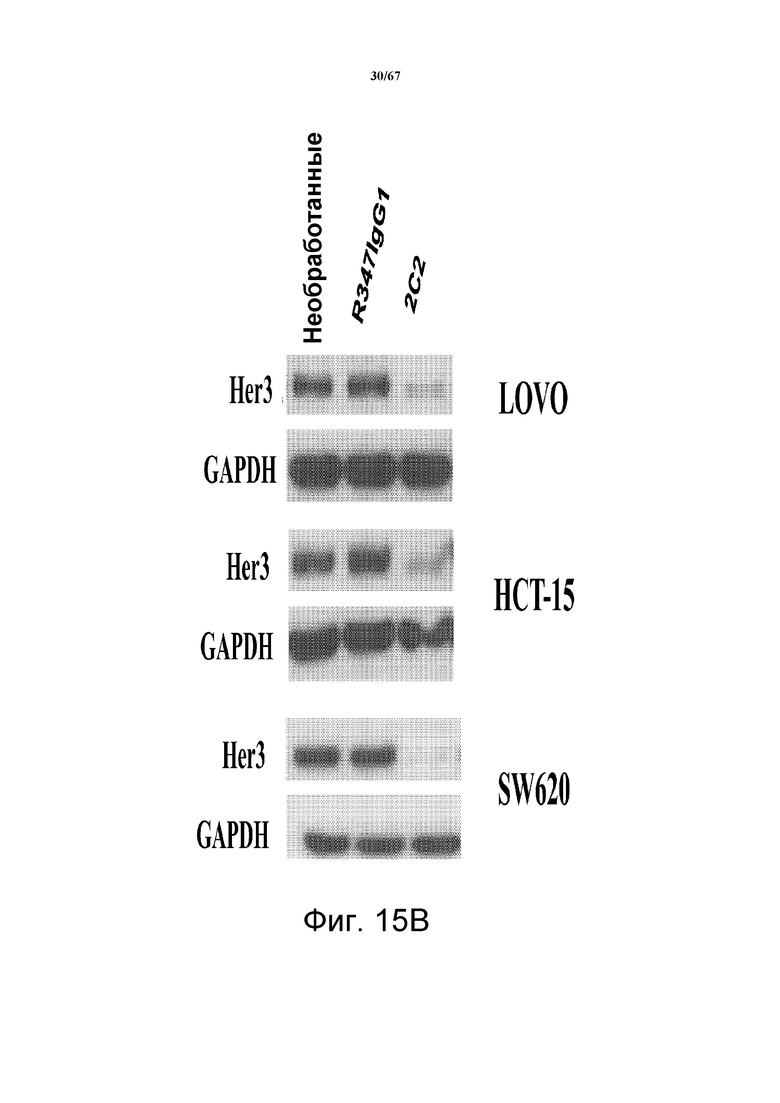
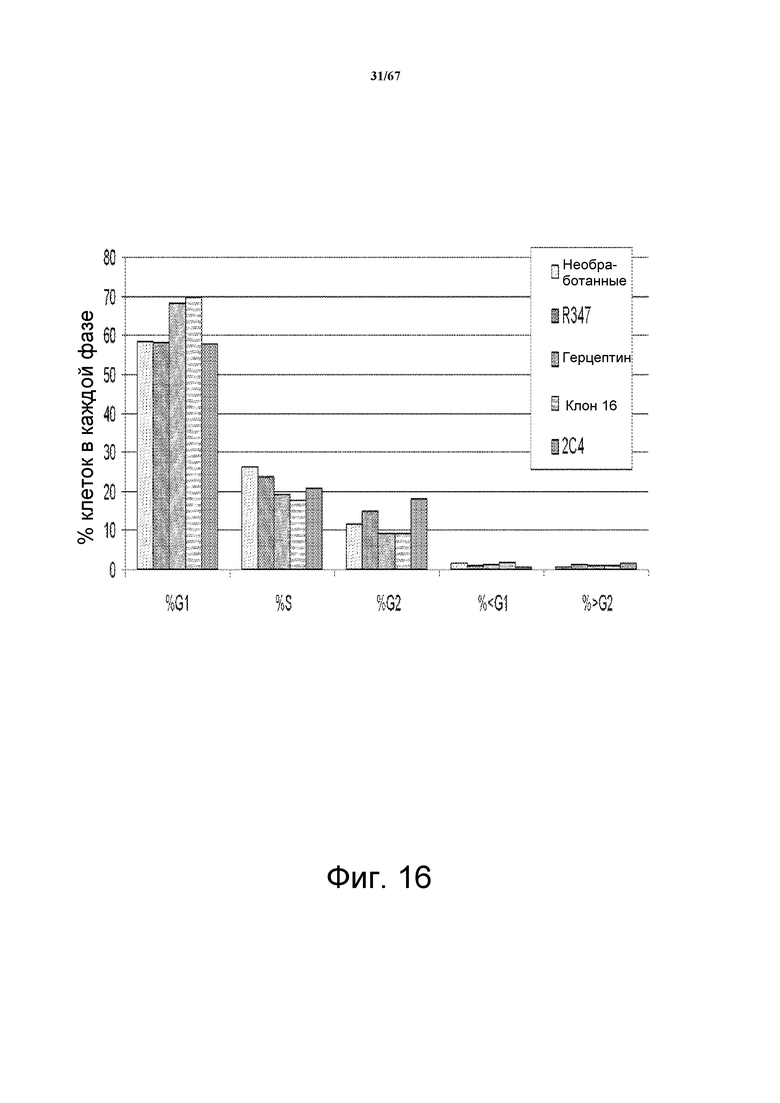
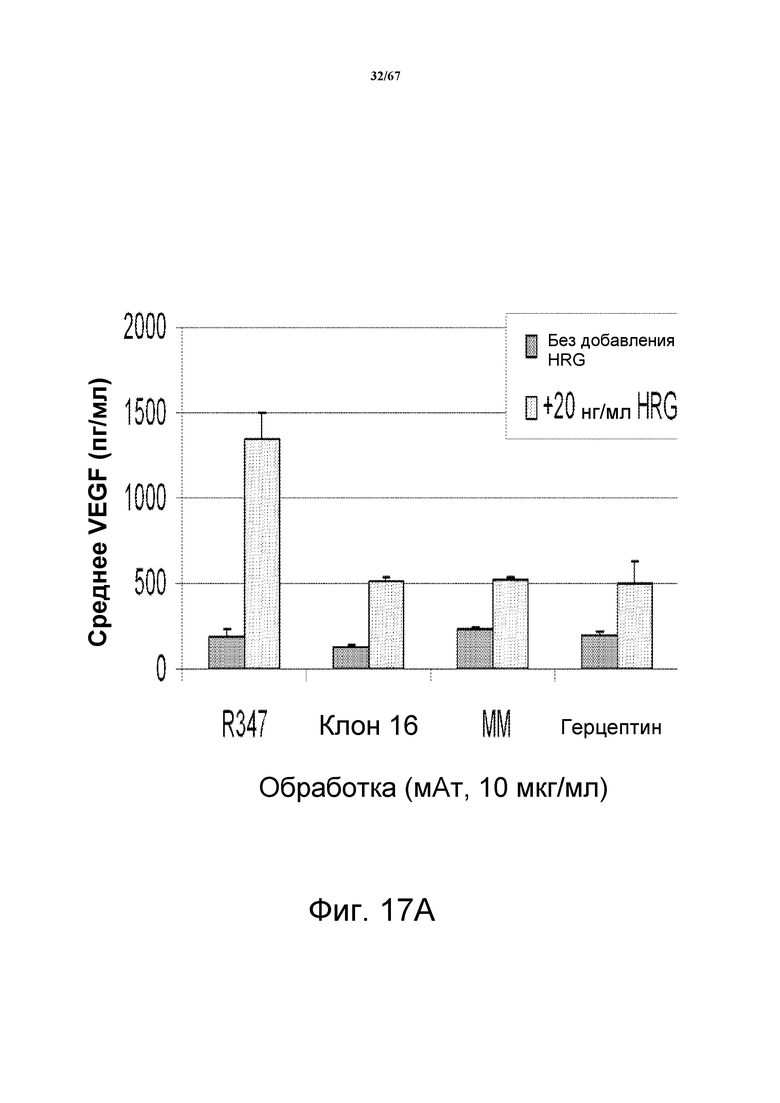
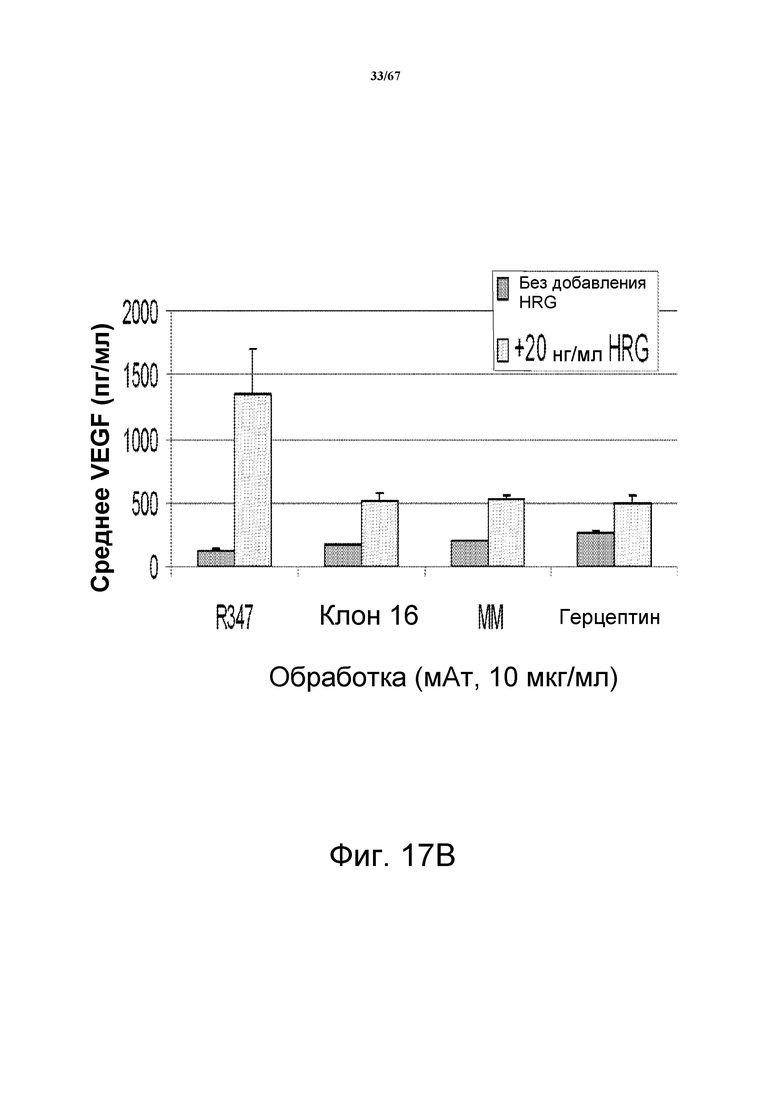
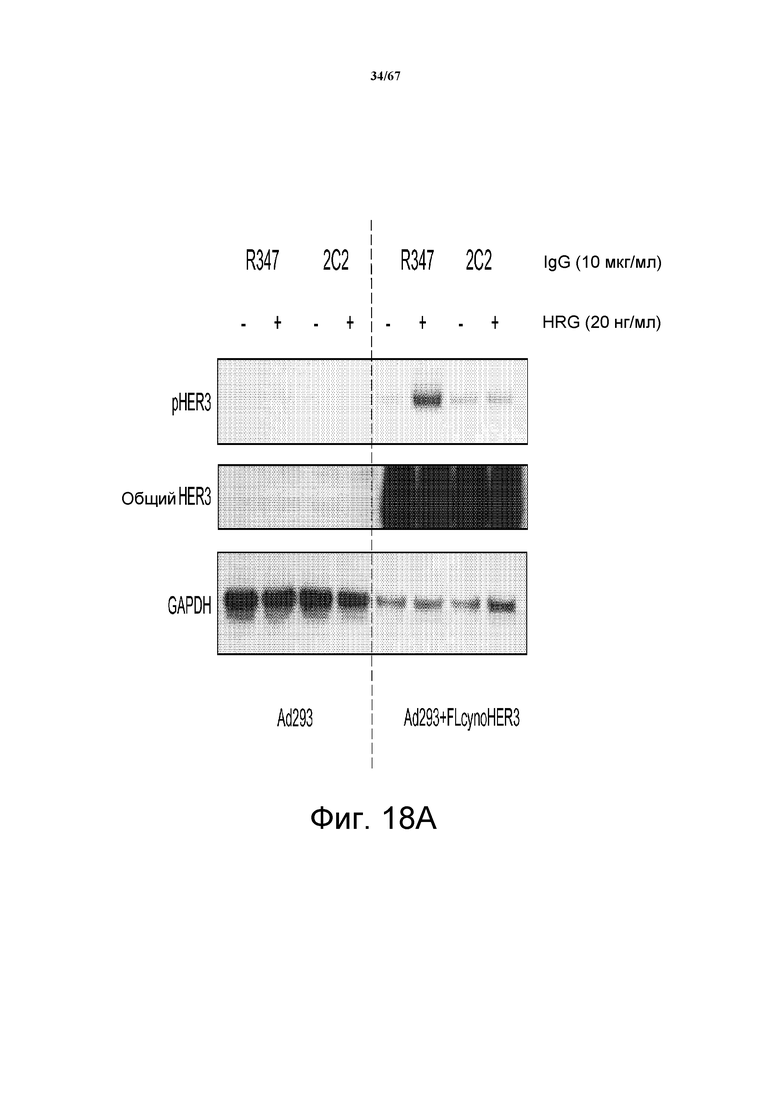
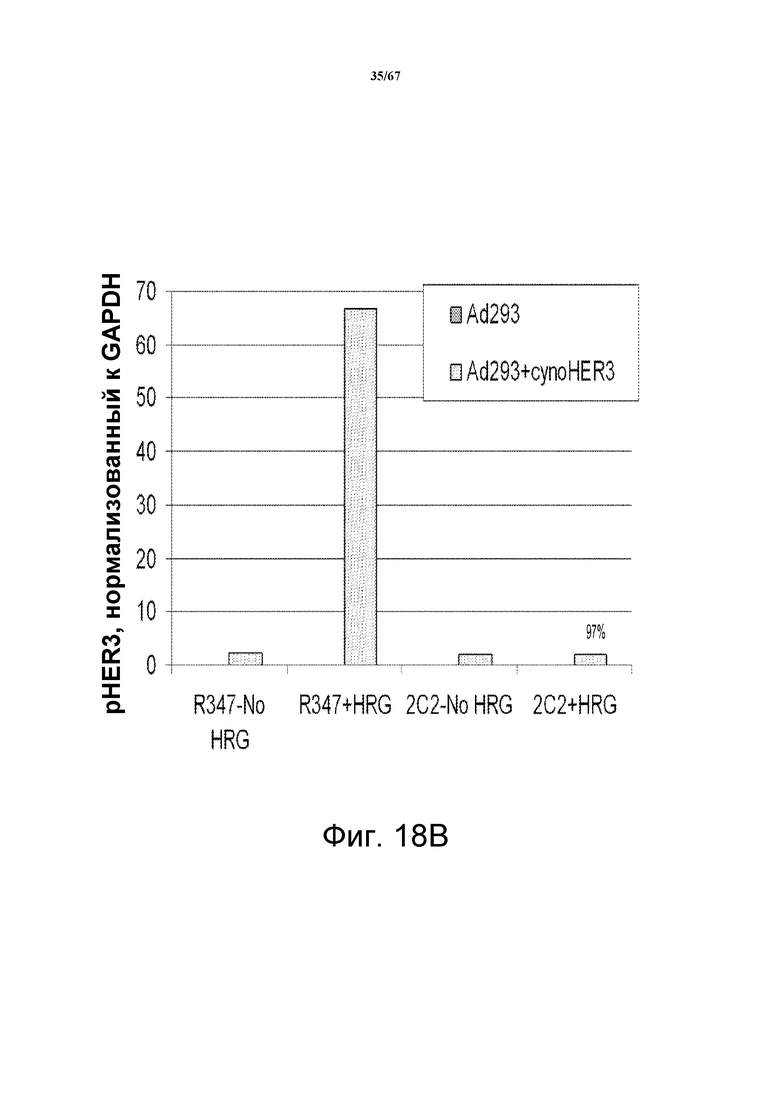
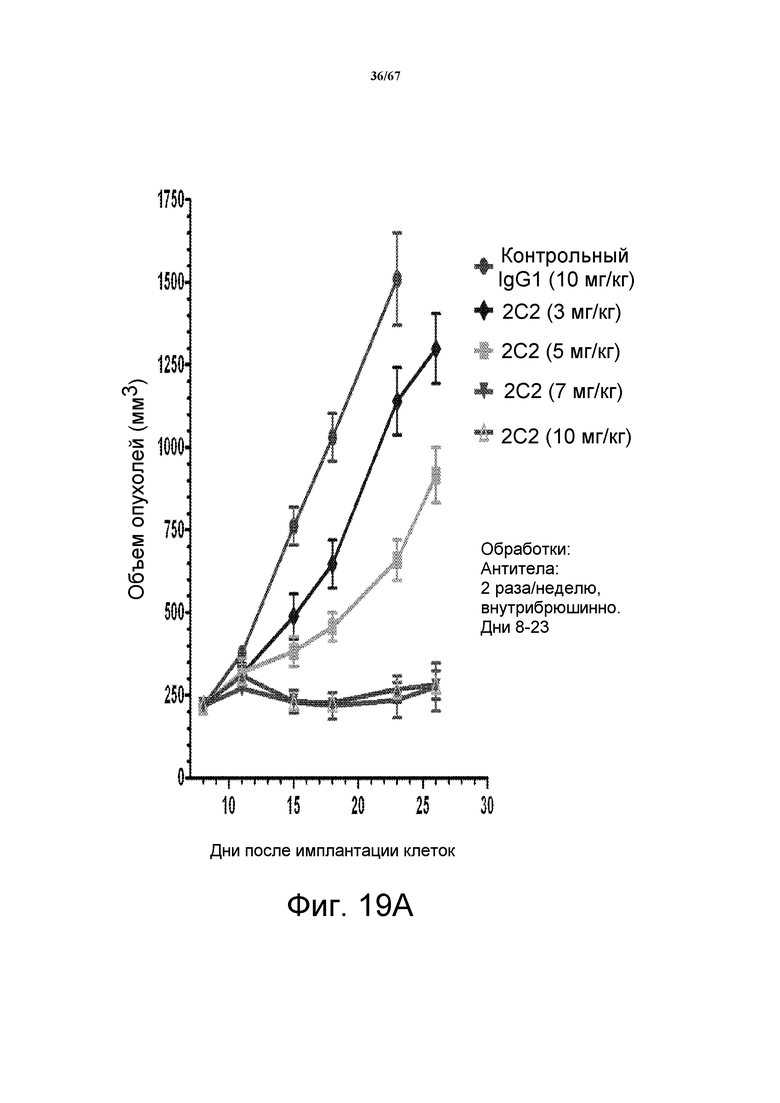
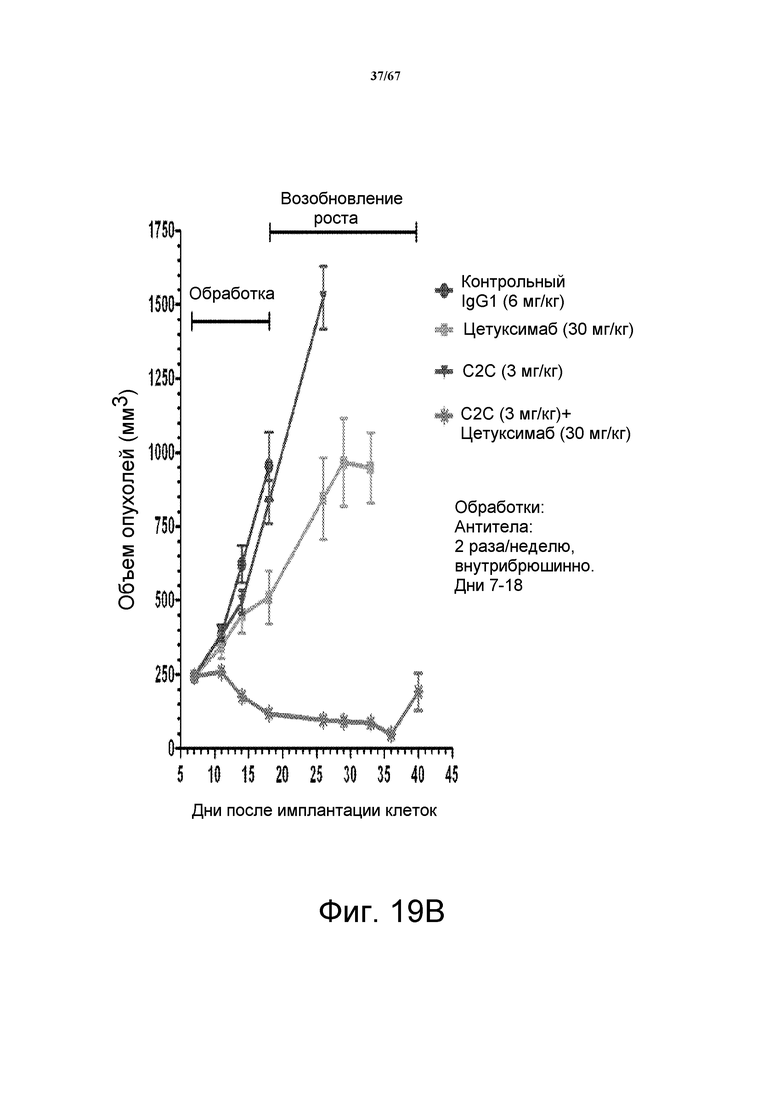
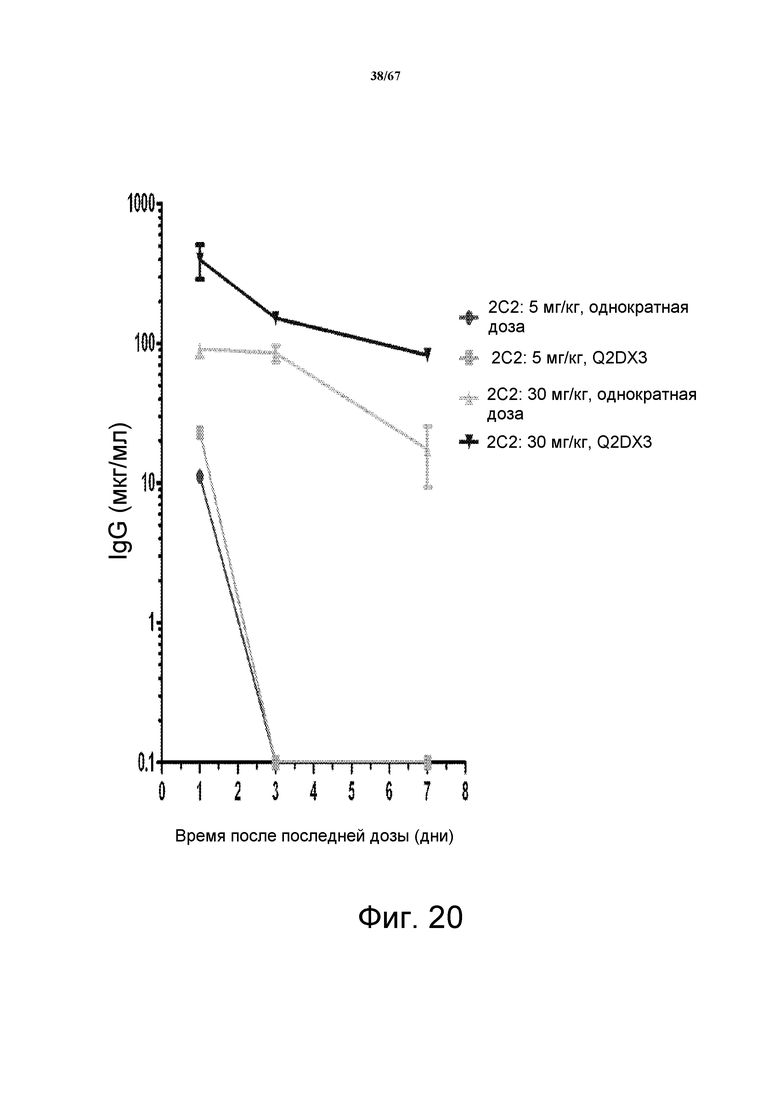
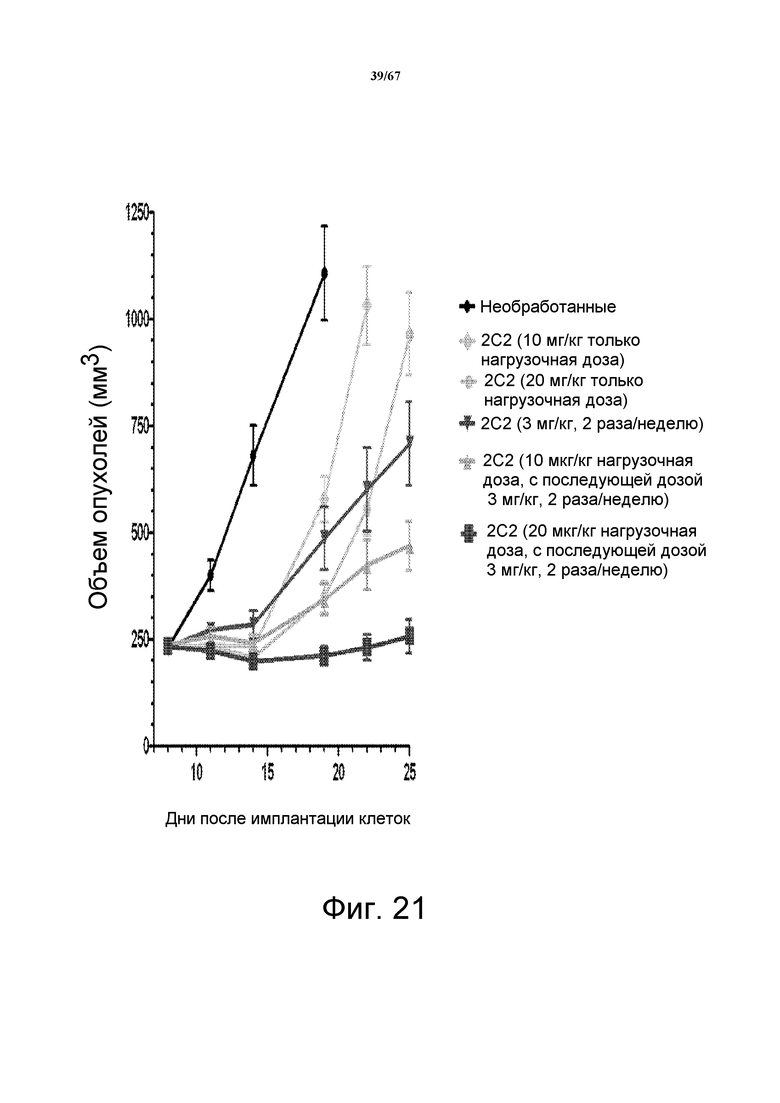

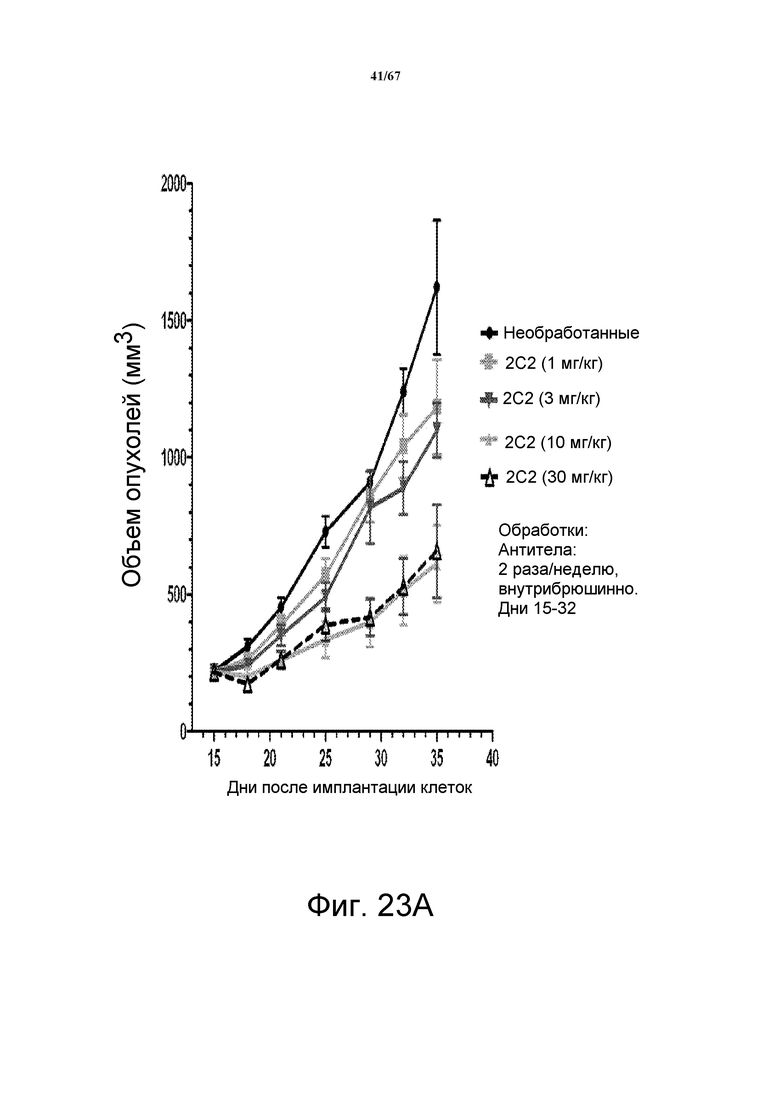
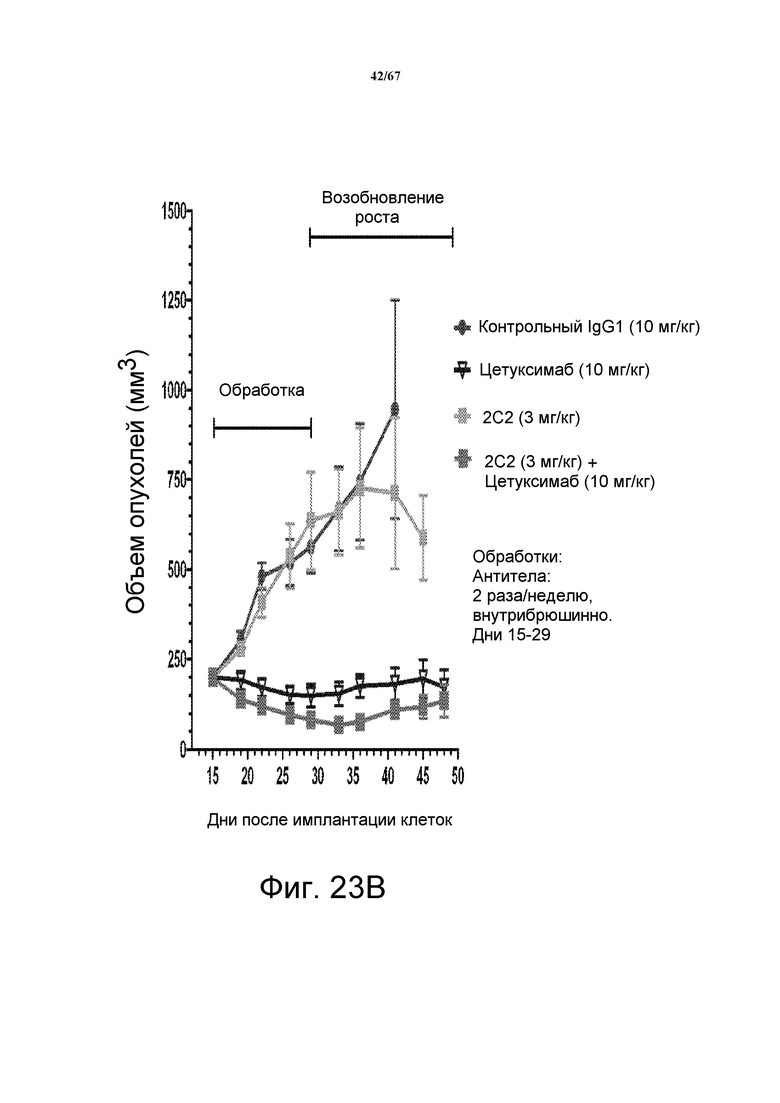
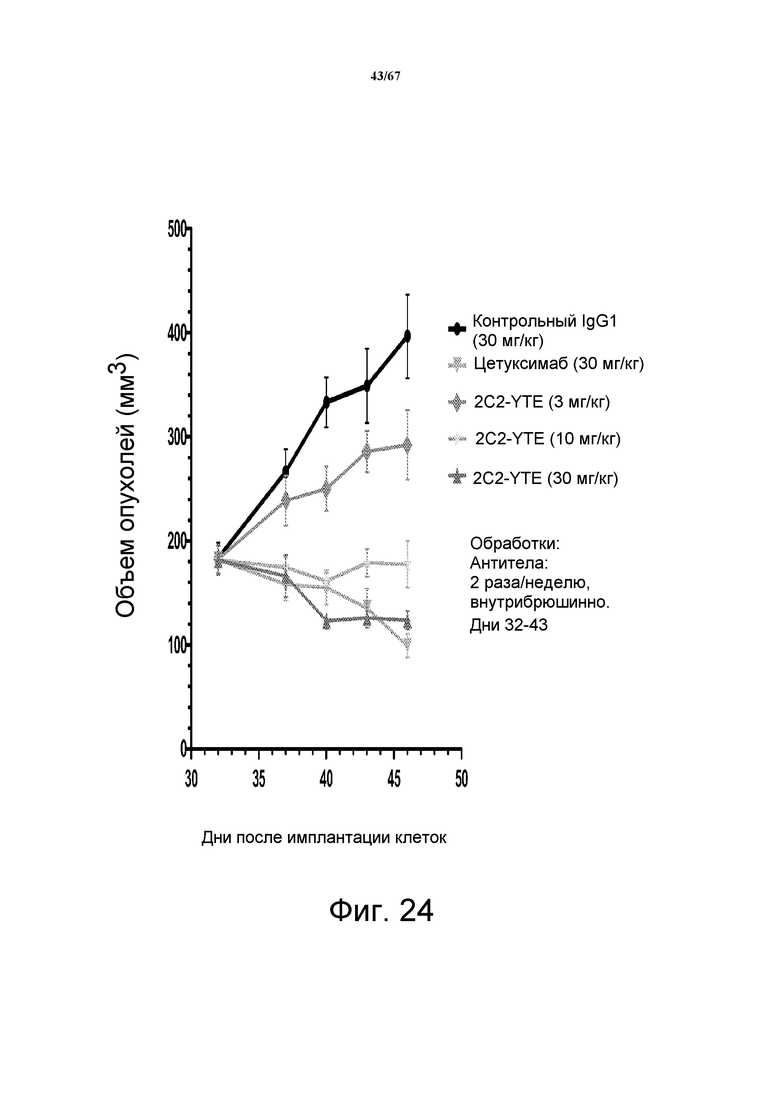
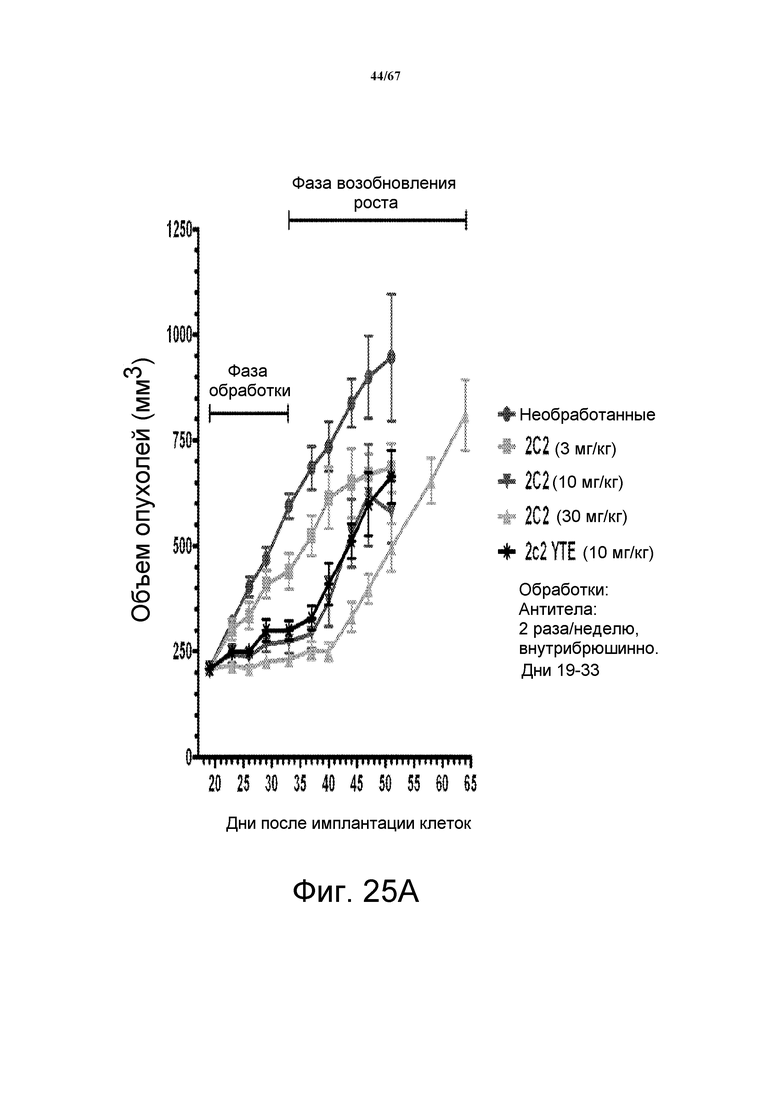
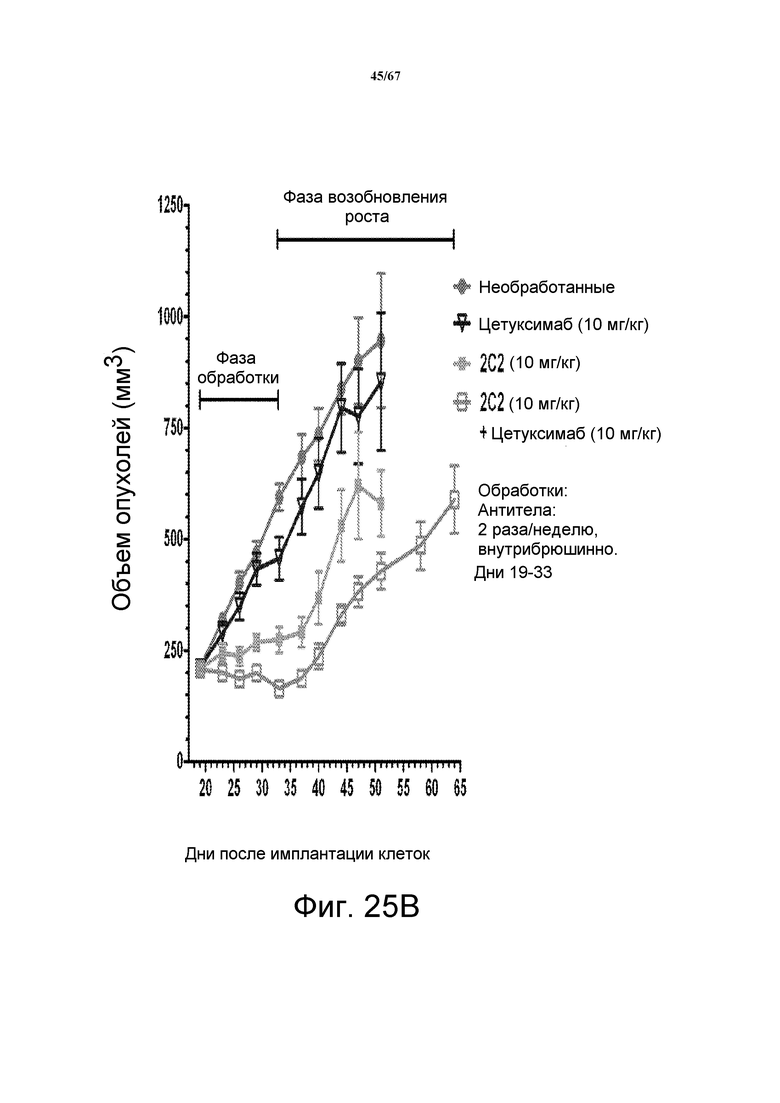
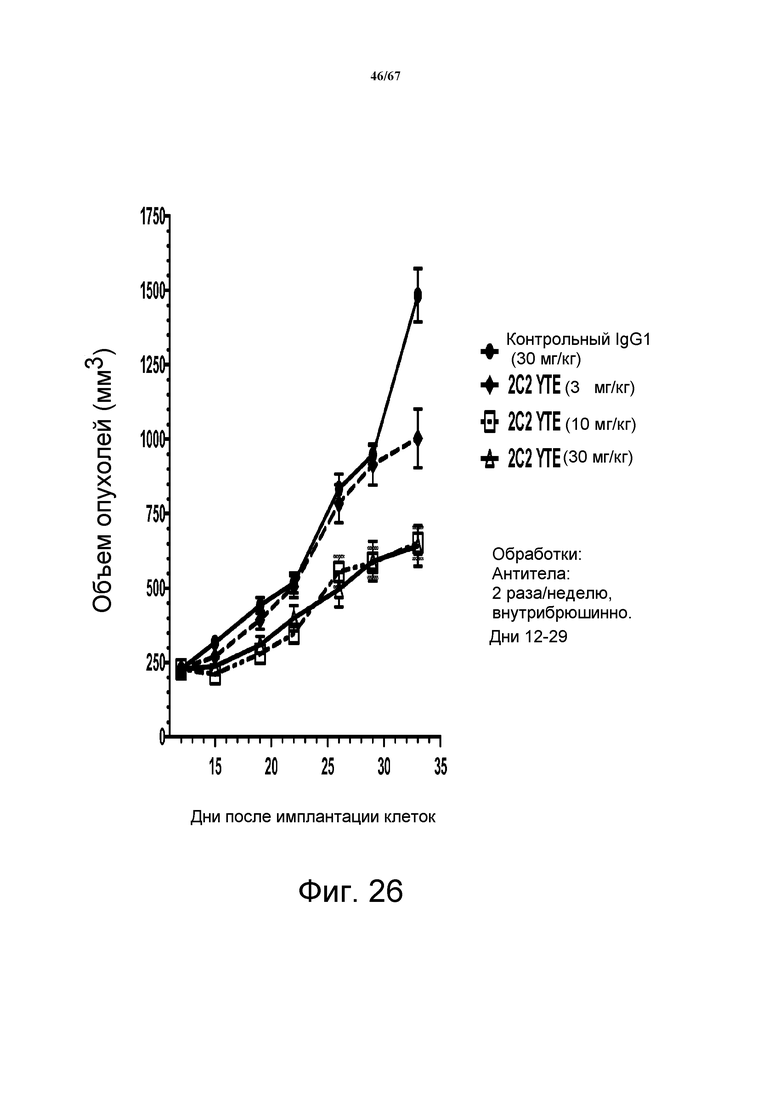
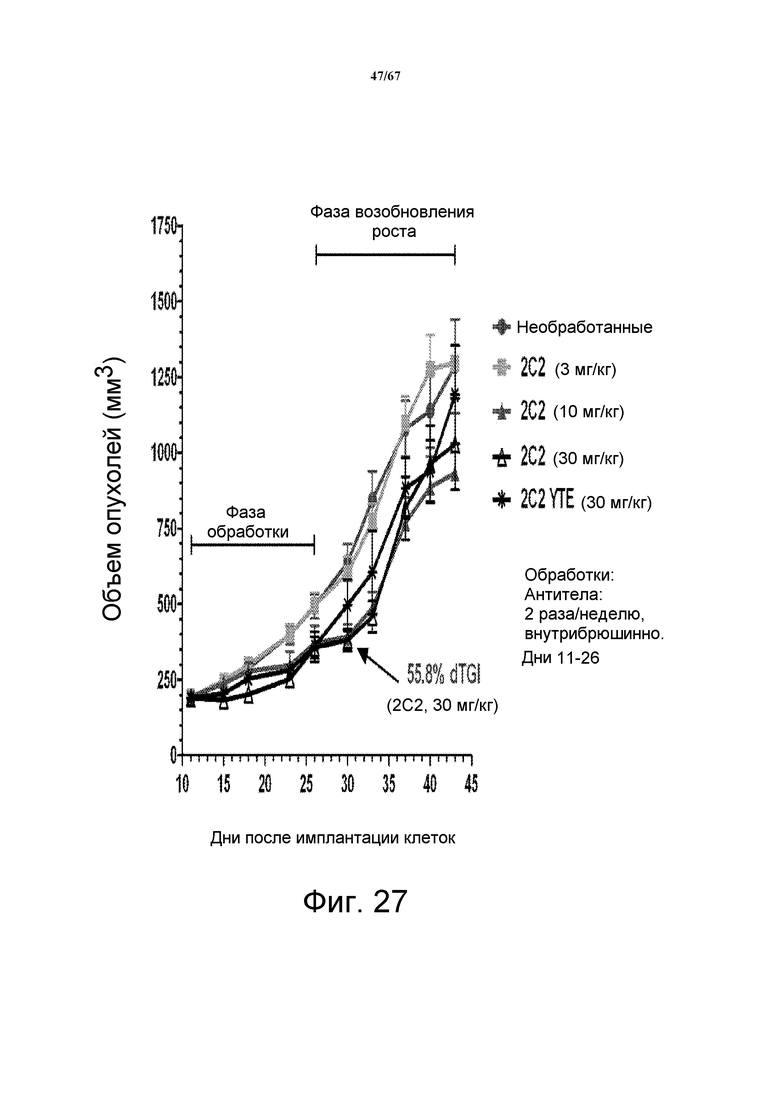
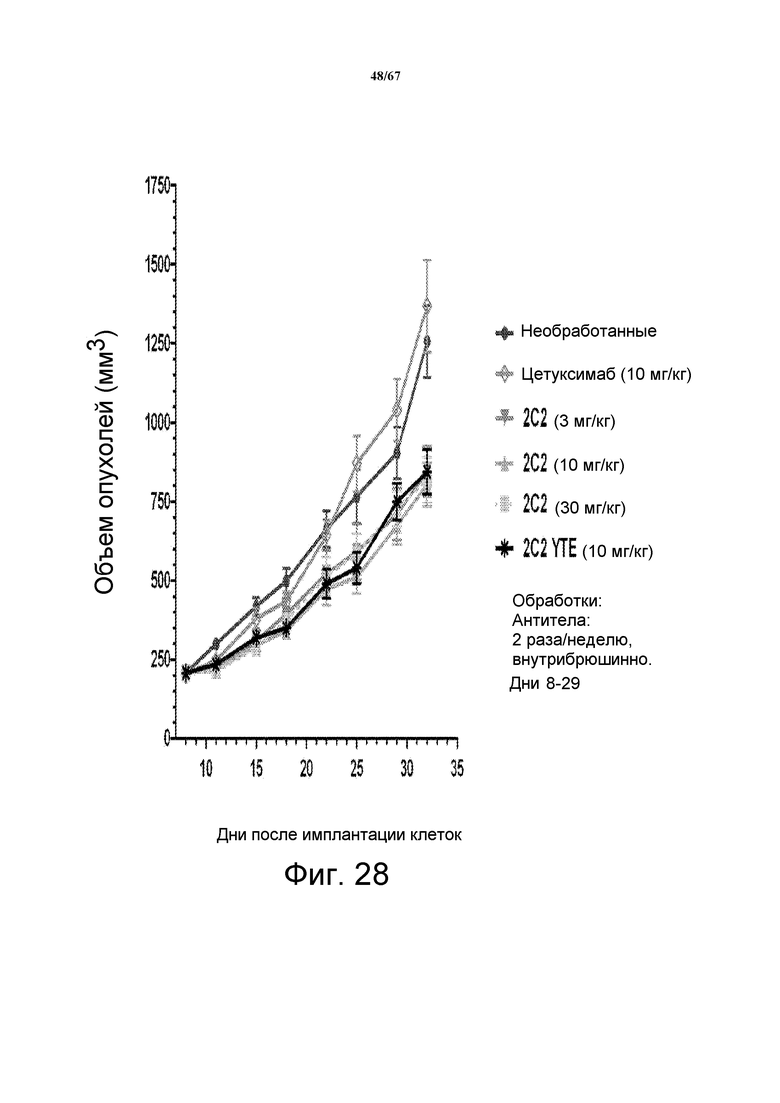
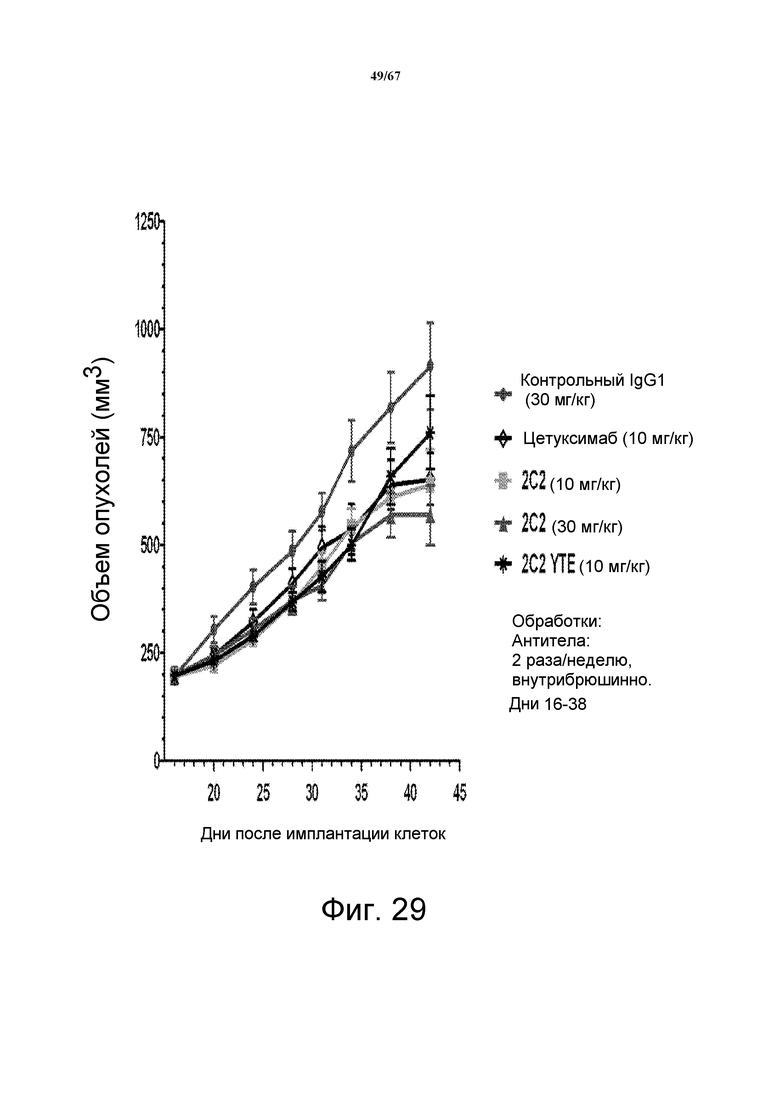
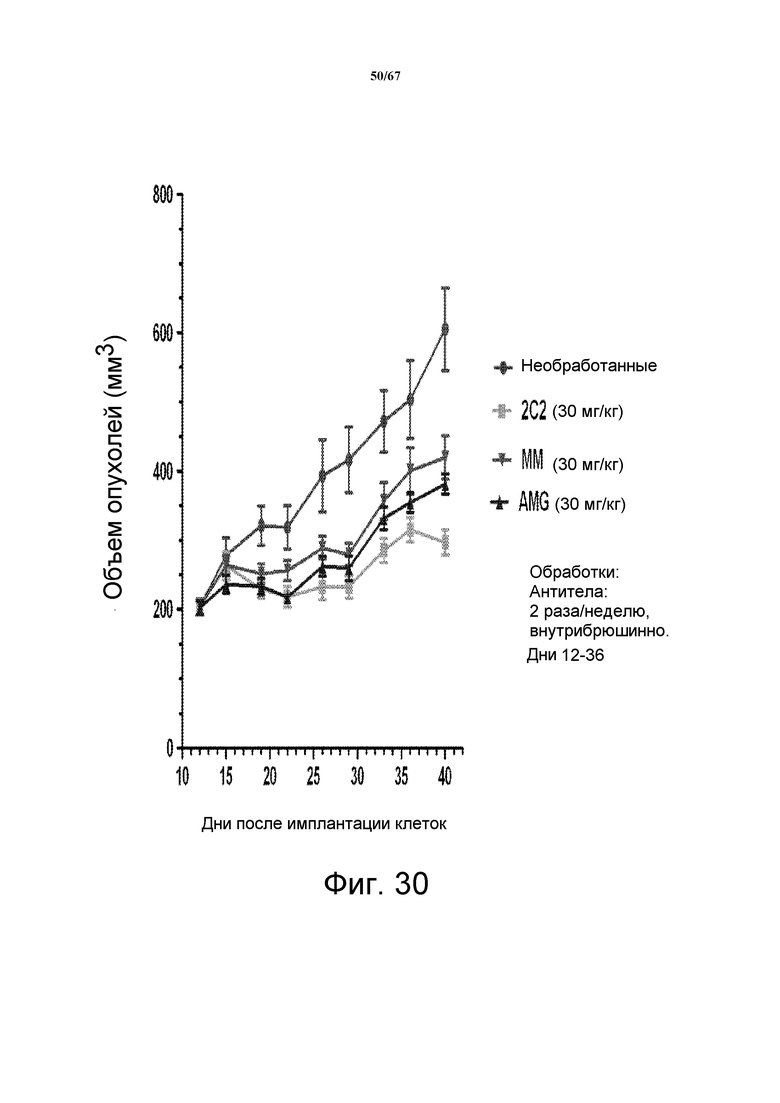

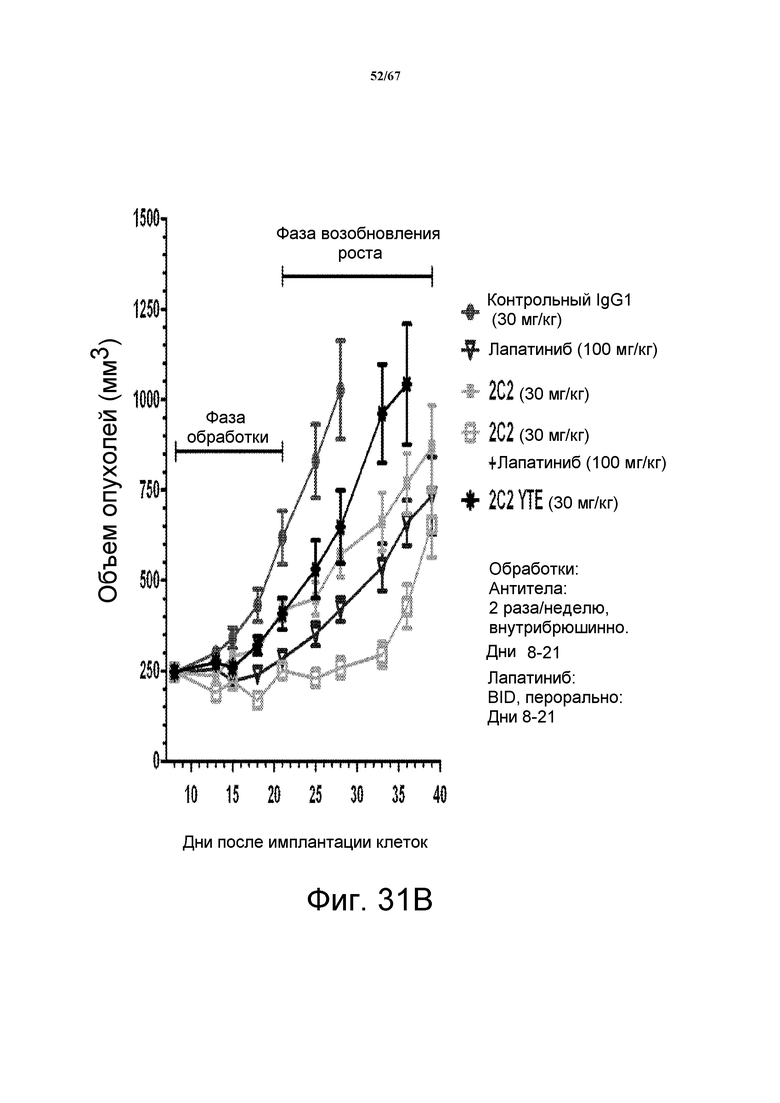
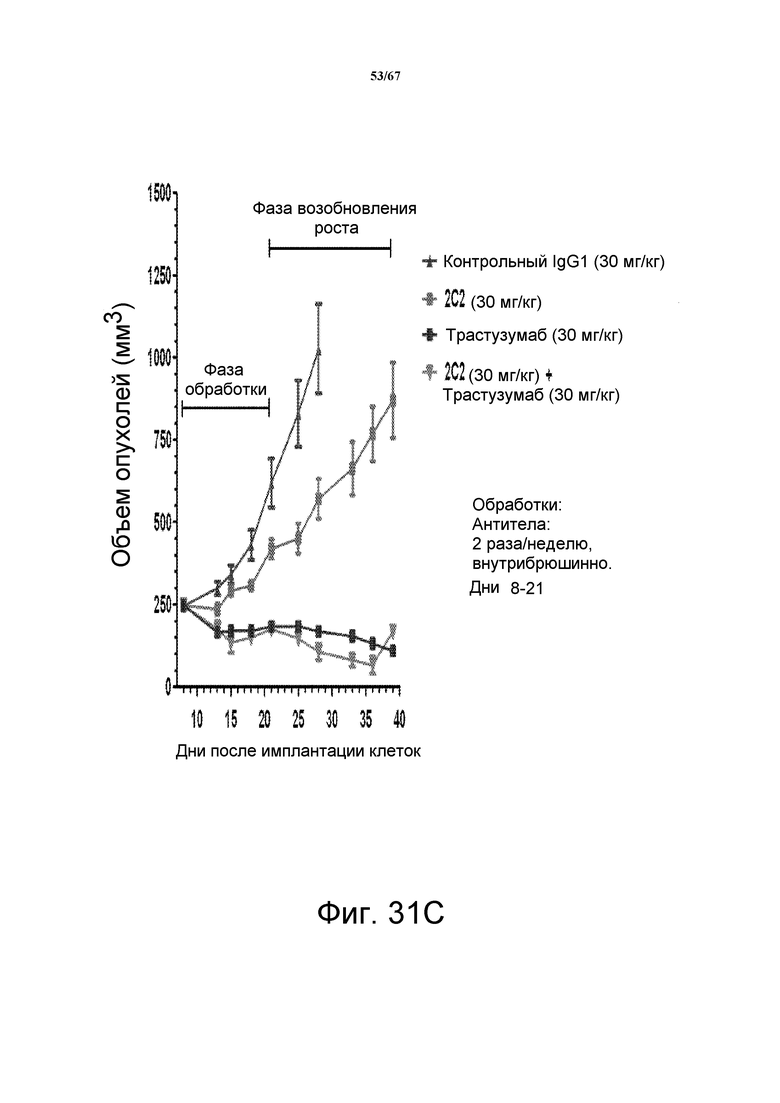
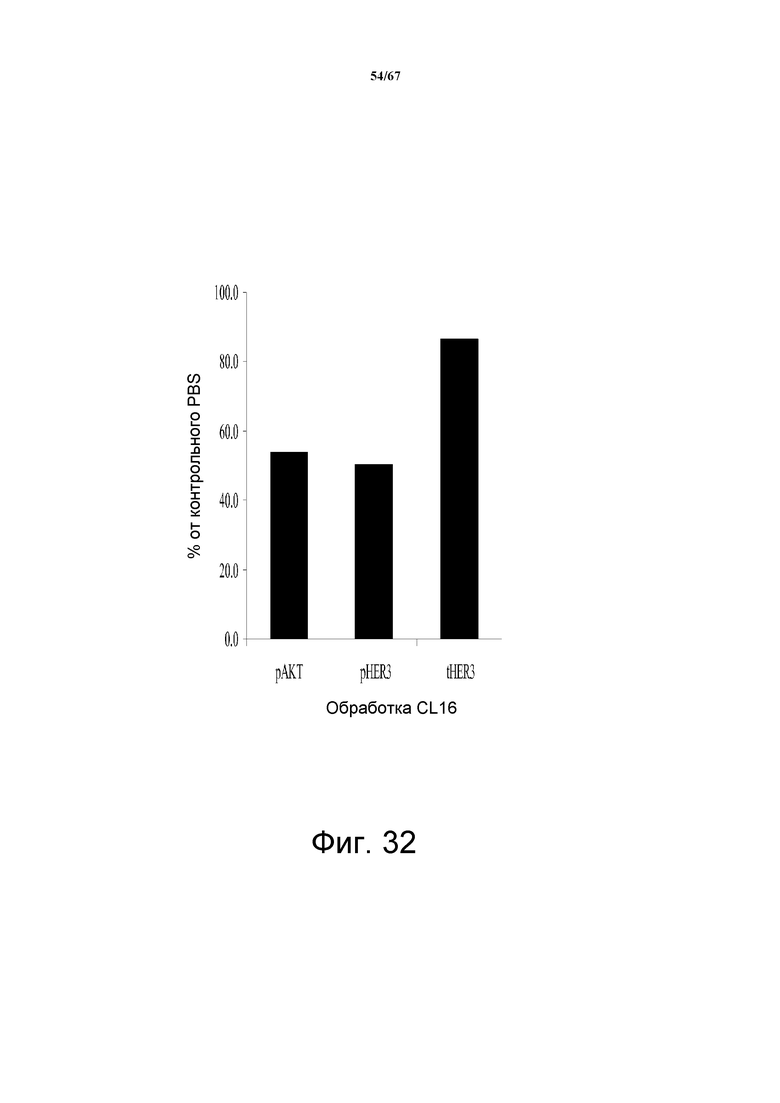
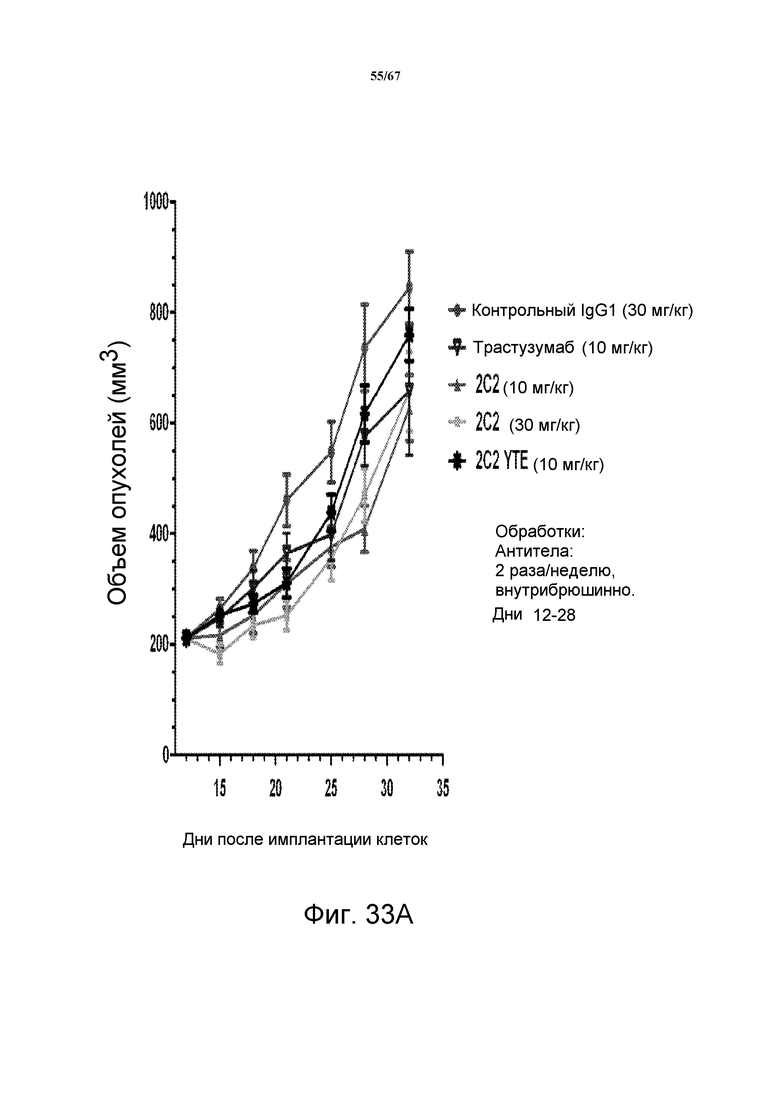

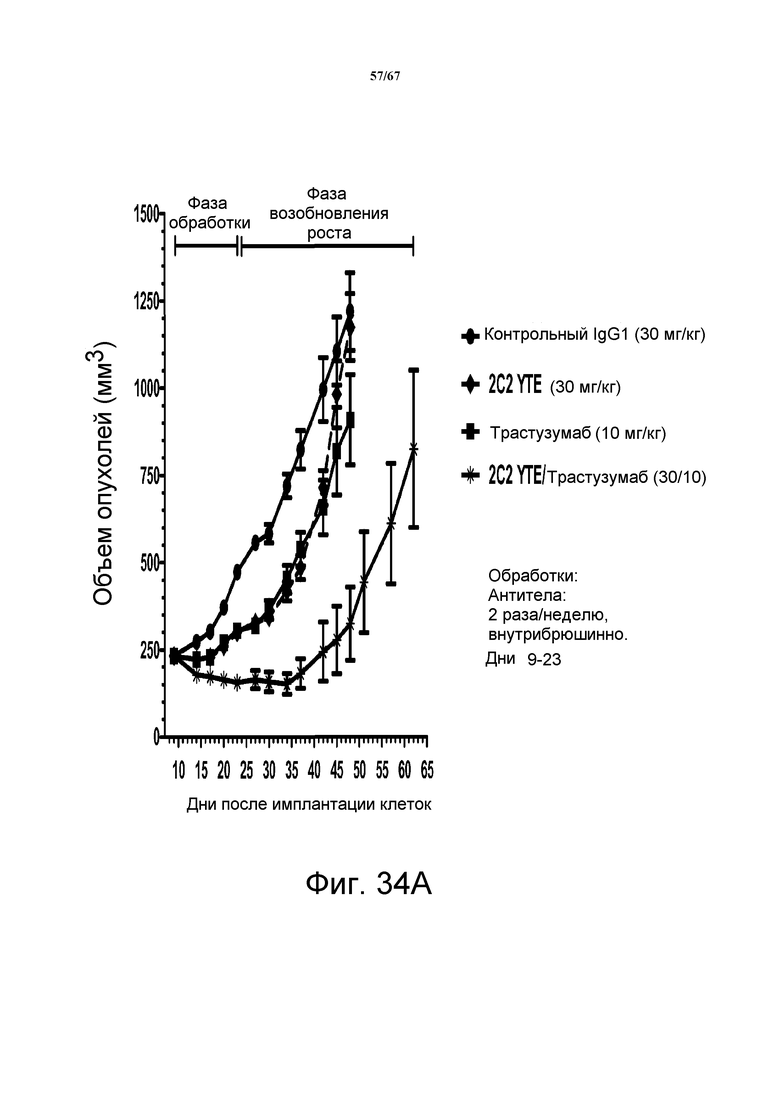
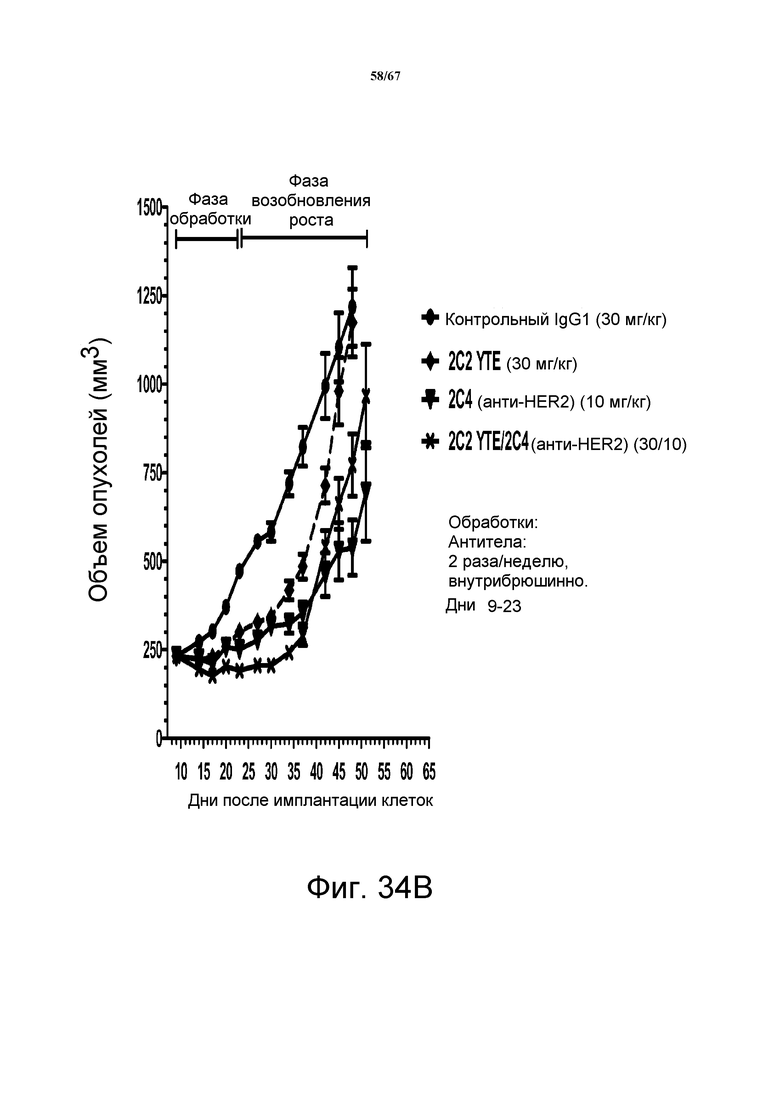
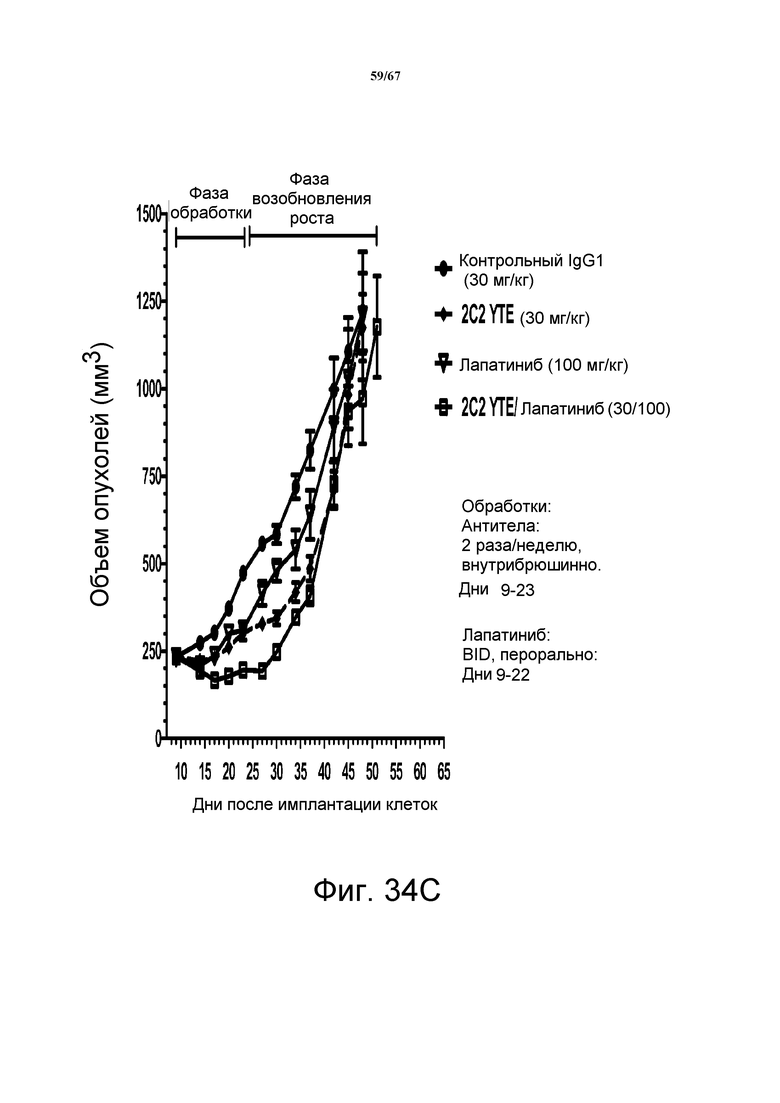
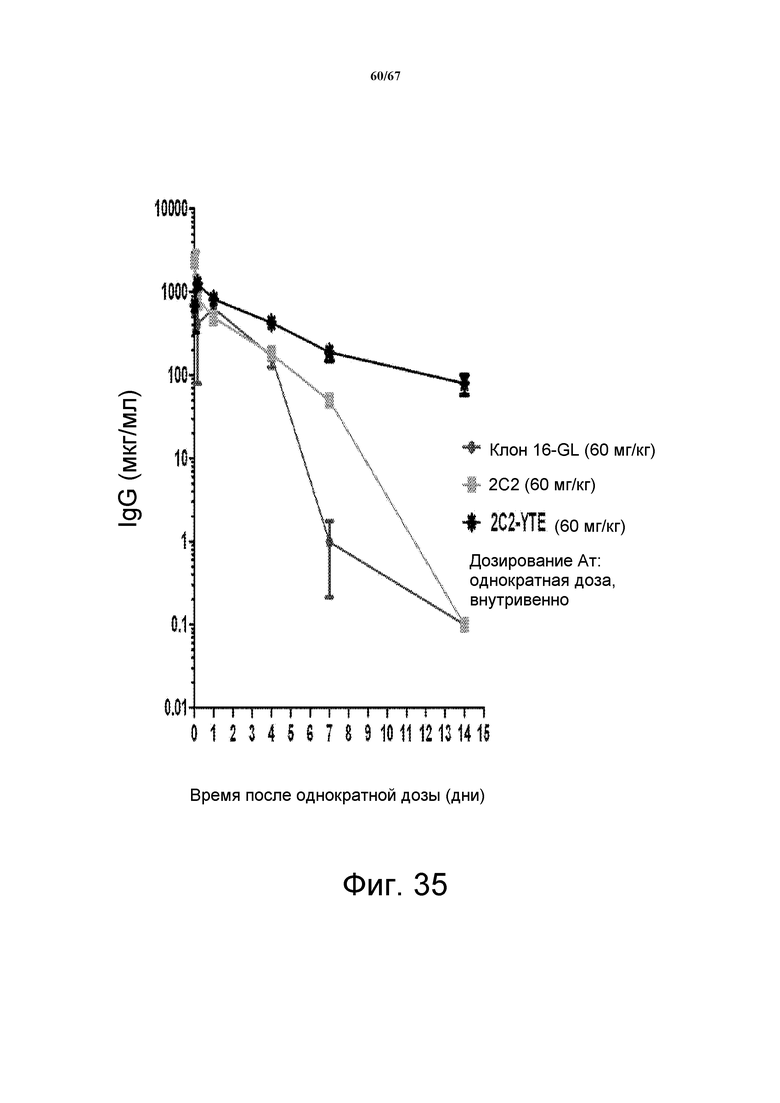
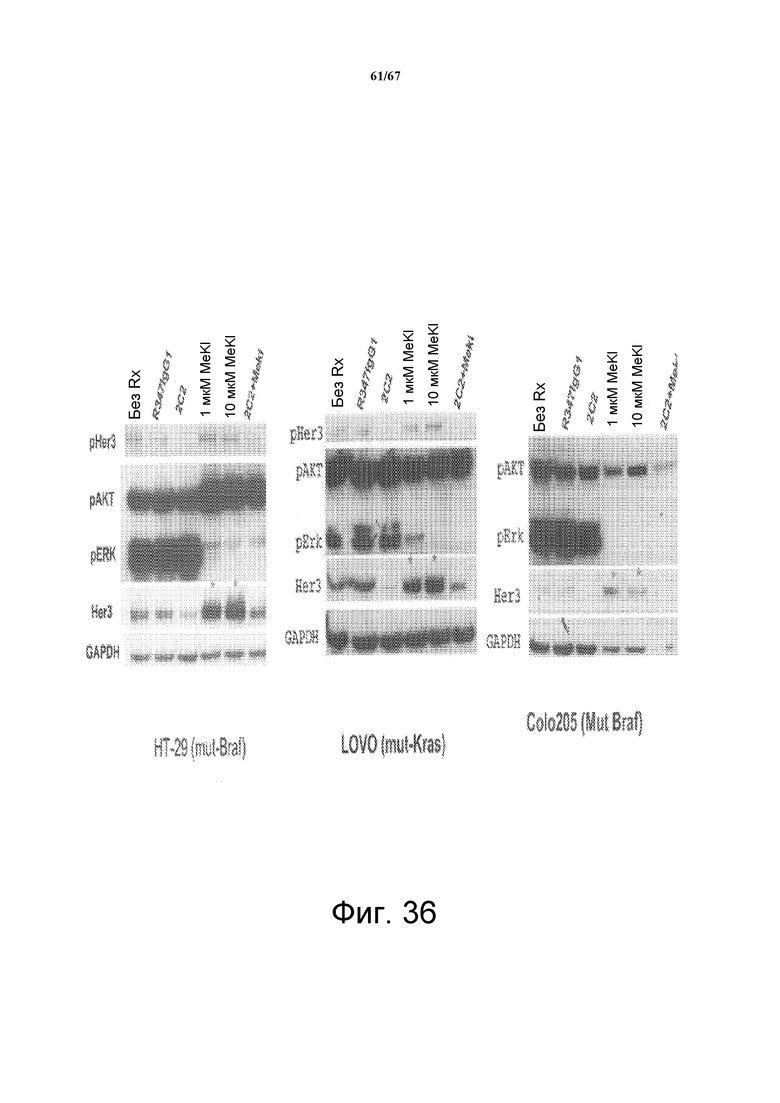
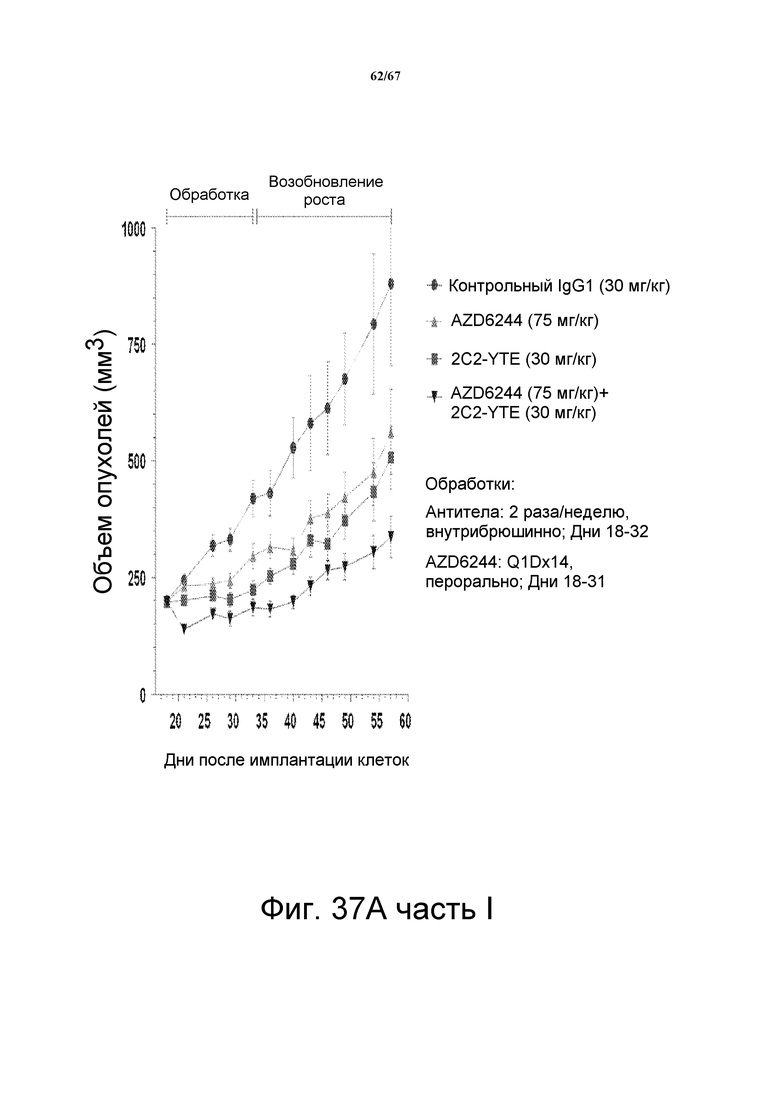
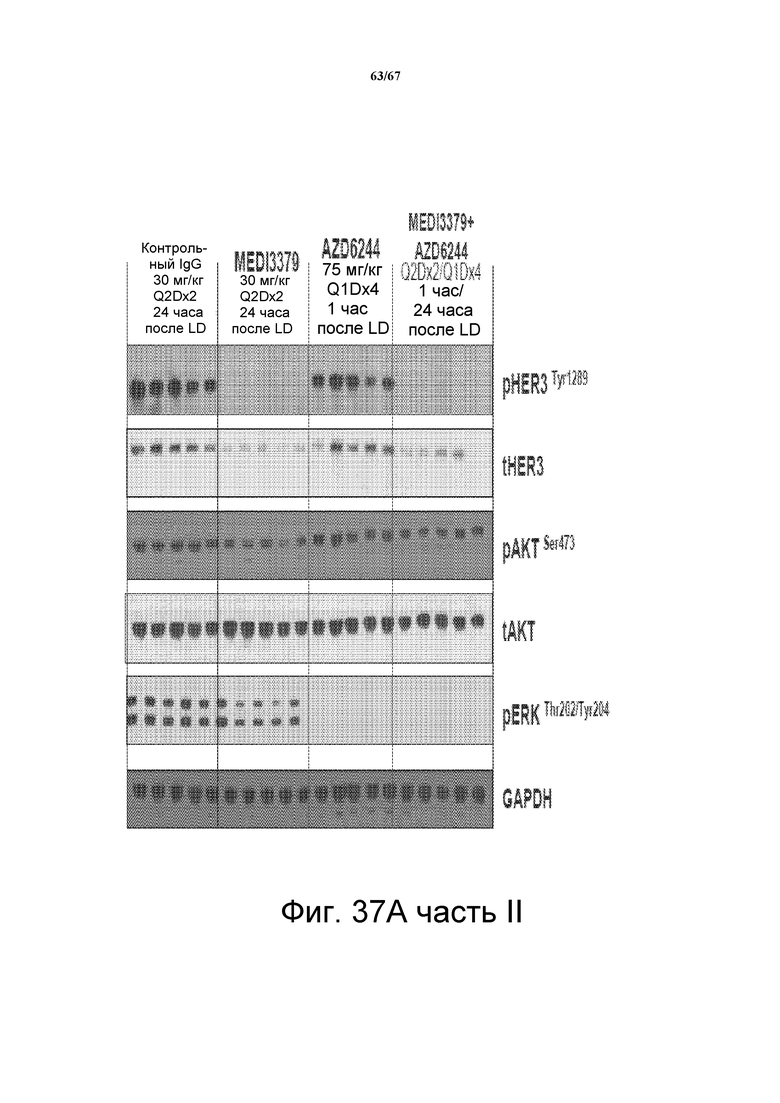
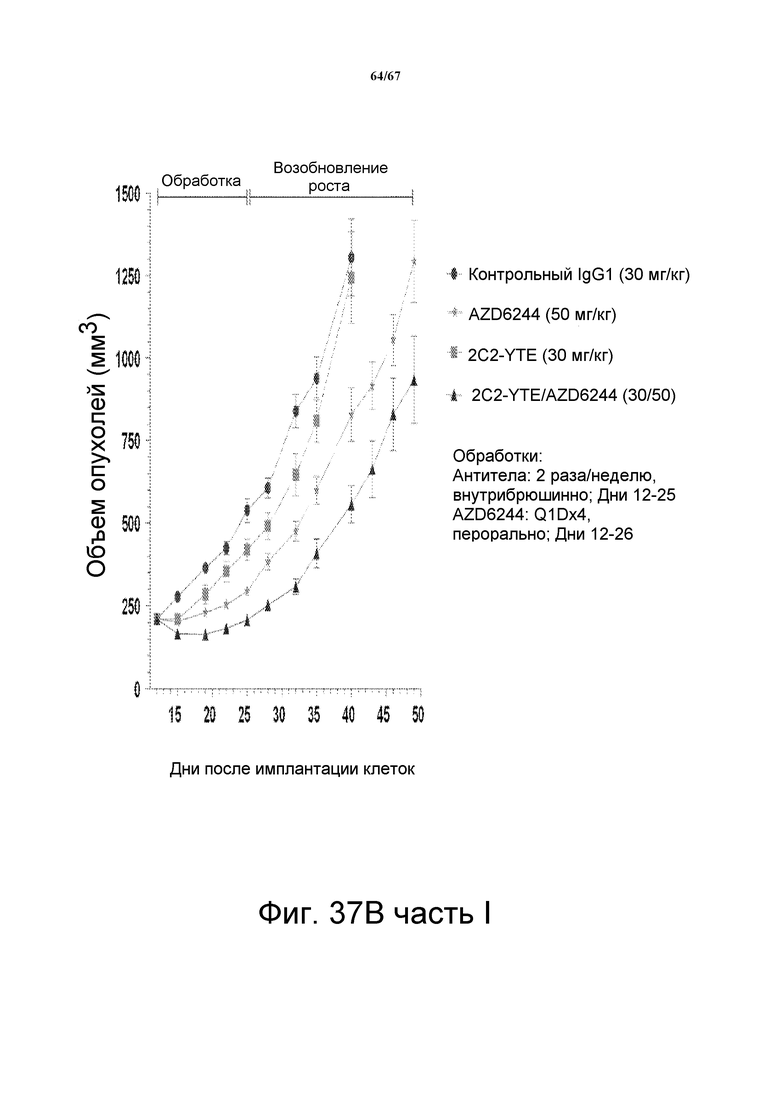
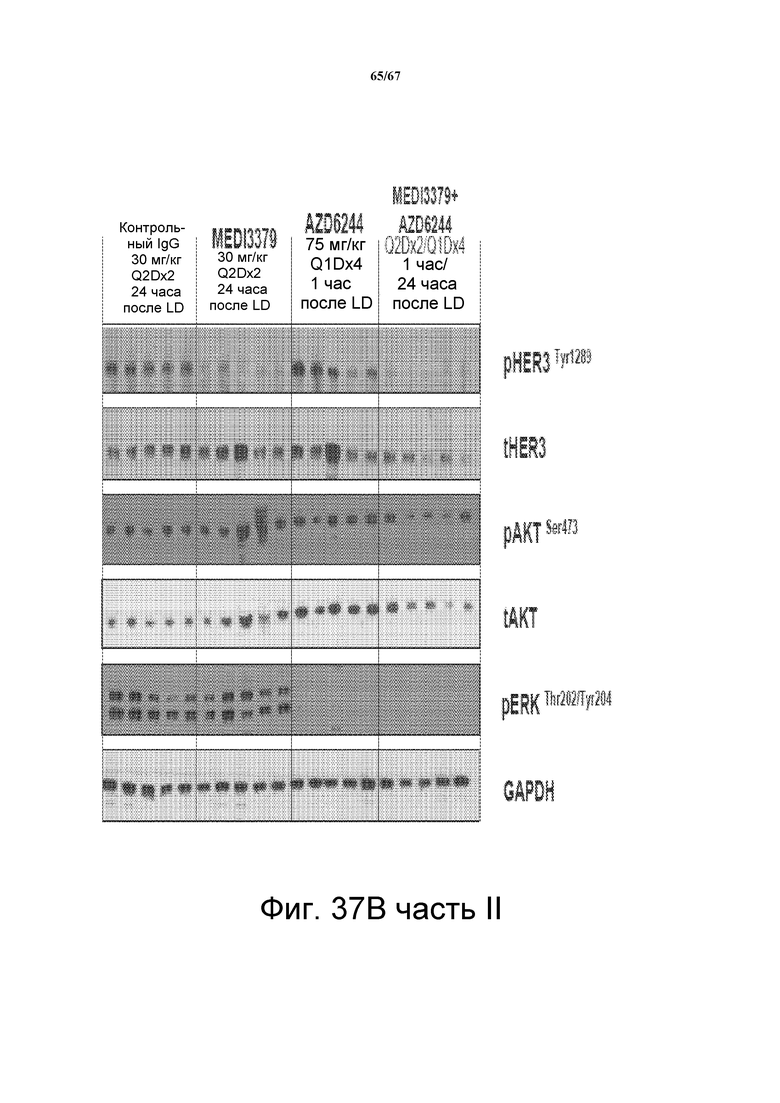
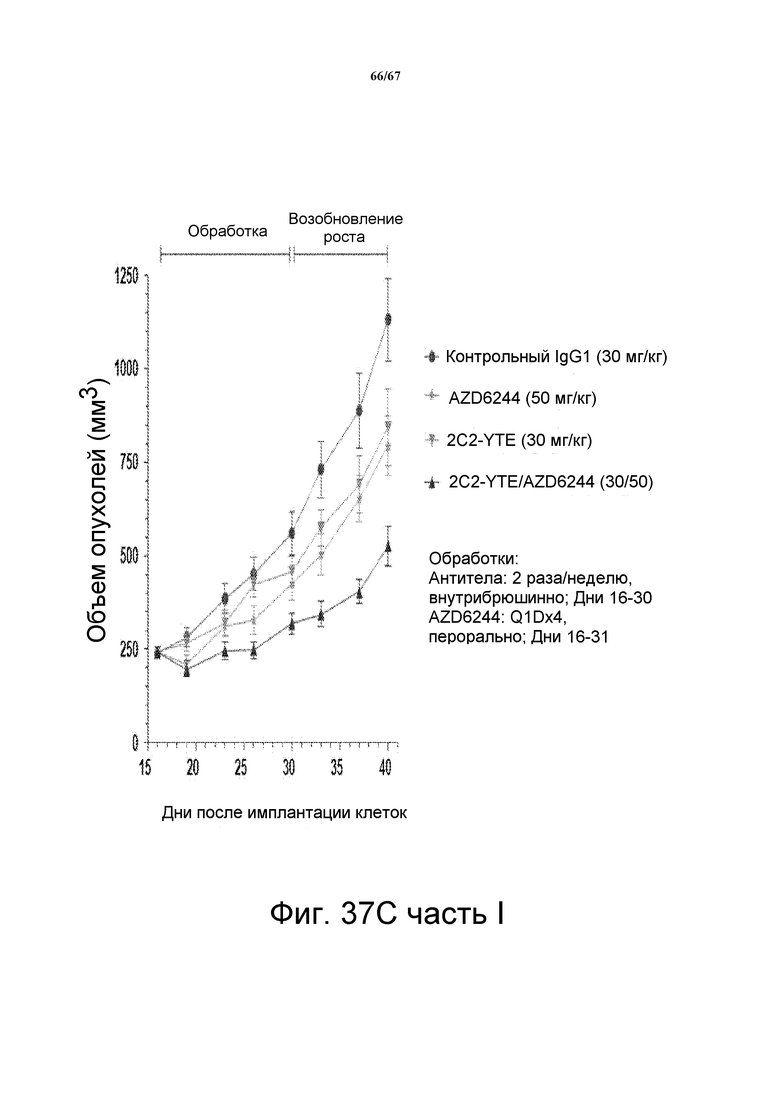

Изобретение относится к биохимии. Описаны антитела и их антигенсвязывающие фрагменты, которые связывают внеклеточный домен рецептора HER3 и ингибируют различные связанные с рецептором HER3 функции посредством зависимого от лиганда и/или независимого от лиганда механизмов. Также представлены композиции с увеличенным временем полужизни. Кроме того, изобретение относится к композициям и способам диагностики и лечения заболеваний, ассоциированных с опосредованной HER3 сигнальной трансдукцией. Изобретение расширяет арсенал средств диагностики и лечения заболеваний, ассоциированных с опосредованной HER3 сигнальной трансдукцией. 10 н. и 13 з.п. ф-лы, 37 ил., 7 табл., 4 пр.
1. Выделенное антитело или его антигенсвязывающий фрагмент, которые специфично связываются с HER3, содержащие вариабельную область легкой цепи антитела (VL) и вариабельную область тяжелой цепи антитела (VH), при этом VL содержит аминокислотную последовательность:
[FW1] X1GSX2SNIGLNYVS [FW2] RNNQRPS [FW3] AAWDDX3X4X5GEX6 [FW4]
где [FW1], [FW2], [FW3] и [FW4] означают каркасные области VL, и где
(a) X1 означает аминокислотные остатки аргинин (R) или серин (S),
(b) X2 означает аминокислотные остатки серин (S) или лейцин (L),
(c) X3 означает аминокислотные остатки серин (S) или глицин (G),
(d) Х4 означает аминокислотные остатки лейцин (L) или пролин (Р),
(e) Х5 означает аминокислотные остатки аргинин (R), изолейцин (I), пролин (Р) или серин (S), и
(f) Х6 означает аминокислотные остатки валин (V) или аланин (А), и
где VH содержит аминокислотную последовательность: [FW5] YYYMQ [FW6] X7IGSSGGVTNYADSVKG [FW7] VGLGDAFDI [FW8], где [FW5], [FW6], [FW7] и [FW8] означают каркасные области VH, и где Х7 означает аминокислотные остатки тирозин (Y), изолейцин (I) или валин (V).
2. Антитело или его антигенсвязывающий фрагмент по п. 1, где VL содержит аминокислотную последовательность определяющей комплементарность области 1 VL (VL-CDR1), выбранную из: SEQ ID NO: 18, SEQ ID NO: 19 или SEQ ID NO: 20, аминокислотную последовательность определяющей комплементарность области 2 VL
(VL-CDR2), выбранную из: SEQ ID NO: 21, аминокислотную последовательность определяющей комплементарность области 3 VL (VL-CDR3), выбранную из: SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 25, SEQ ID NO: 26, SEQ ID NO: 27, SEQ ID NO: 28, SEQ ID NO: 29, или SEQ ID NO: 30, и
где VH содержит аминокислотную последовательность определяющей комплементарность области 1 (VH-CDR1), выбранную из SEQ ID NO: 31, аминокислотную последовательность определяющей комплементарность области 2 (VH-CDR2), выбранную из: SEQ ID NO: 32, SEQ ID NO: 33 или SEQ ID NO: 34, и аминокислотную последовательность определяющей комплементарность области 3 (VH-CDR3), выбранную из: SEQ ID NO: 35.
3. Антитело или его антигенсвязывающий фрагмент по п. 1, где X1 означает серин (S), Х2 означает лейцин (L), Х3 означает серин (S), Х4 означает пролин (Р), X5 означает пролин (Р), Х6 означает аланин (А), и Х7 означает тирозин (Y).
4. Антитело или его фрагмент по п. 1, в котором FW1 содержит SEQ ID NO: 40 или 44, FW2 содержит SEQ ID NO: 41, FW3 содержит SEQ ID NO: 42, FW4 содержит SEQ ID NO: 43, FW5 содержит SEQ ID NO: 36, FW6 содержит SEQ ID NO: 37, FW7 содержит SEQ ID NO: 38, FW8 содержит SEQ ID NO: 39.
5. Антитело или его антигенсвязывающий фрагмент по п. 1, где VL содержит аминокислотную последовательность по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, идентичную аминокислотной последовательности SEQ ID NO: 3, и где VH содержит аминокислотную последовательность по меньшей мере на 80%, по меньшей мере на 90%, по меньшей мере на 95%, идентичную аминокислотной последовательности SEQ ID NO: 2.
6. Антитело или его антигенсвязывающий фрагмент по п. 1, которое включает VL, содержащую SEQ ID NO: 3, и VH, содержащую SEQ ID NO: 2.
7. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6, которая включает константную область тяжелой цепи.
8. Антитело или его антигенсвязывающий фрагмент по п. 7, где константная область тяжелой цепи или ее фрагмент представляет собой константную область IgG, выбранную из константной области IgG1 человека, константной области IgG2 человека, константной области IgG3 человека и константной области IgG4 человека.
9. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6, которое содержит константную область легкой цепи, выбранную из группы, состоящей из константной области каппа человека и константной области лямбда человека.
10. Антитело или его антигенсвязывающий фрагмент по п. 9, которое содержит константную область IgG, и в которой по меньшей мере одна аминокислотная замена в константном домене IgG выбрана из группы, состоящей из:
(a) замены аминокислоты в положении 252 тирозином (Y), фенилаланином (F), триптофаном (W) или треонином (Т),
(b) замены аминокислоты в положении 254 треонином (Т),
(c) замены аминокислоты в положении 256 серином (S), аргинином (R), глутамином (Q), глутаминовой кислотой (Е), аспарагиновой кислотой (D) или треонином (Т),
(d) замены аминокислоты в положении 257 лейцином (L),
(e) замены аминокислоты в положении 309 пролином (Р),
(f) замены аминокислоты в положении 311 серином (S),
(g) замены аминокислоты в положении 428 треонином (Т), лейцином (L), фенилаланином (F) или серином (S),
(h) замены аминокислоты в положении 433 аргинином (R), серином (S), изолейцином (I), пролином (Р) или глутамином (Q),
(i) замены аминокислоты в положении 434 триптофаном (W), метионином (М), серином (S), гистидином (Н), фенилаланином (F) или тирозином, и
(j) сочетания двух или больше указанных замен, при этом нумерация приведена согласно индексу EU, как указано в публикации Kabat.
11. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6, где антитело представляет собой полностью человеческое антитело, гуманизированные антитело, химерное антитело, моноклональные антитело, поликлональное антитело, рекомбинантное антитело, полиспецифичное антитело или его антигенсвязывающий фрагмент, и где антигенсвязывающий фрагмент представляет собой Fv, Fab, F(ab')2, Fab', dsFv, scFv и sc(Fv)2.
12. Антитело или его антигенсвязывающий фрагмент по п. 10, содержащие (i) легкую цепь антитела, содержащую VL антитела, содержащую SEQ ID NO: 3 и константную область лямбда человека, и (ii) тяжелую цепь антитела, содержащую VH, содержащую SEQ ID NO: 2 и константную область IgG1 человека, в котором константный домен IgG1 человека содержит аминокислотные замены по сравнению с константным доменом IgG1 человека дикого типа в положениях 252, 254 и 256, при этом нумерация приведена согласно индексу EU, как указано в публикации Kabat, и в котором
(а) аминокислота в положении 252 (метионин) заменена тирозином (Y),
(b) аминокислота в положении 254 (серин) заменена треонином (Т), и
(c) аминокислота в положении 256 (треонин) заменена глутаминовой кислотой (Е).
13. Композиция для лечения злокачественной опухоли у субъекта, содержащая антитело или его фрагмент по любому из пп. 1-12 в эффективном количестве и фармацевтически приемлемый носитель.
14. Нуклеиновая кислота, кодирующая антитело по любому из пп. 1-12.
15. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 14.
16. Клетка-хозяин для получения антитела или его антигенсвязывающего фрагмента по любому из пп. 1-12, содержащая последовательность нуклеиновой кислоты по п. 14 или вектор по п. 15.
17. Способ получения антитела или антигенсвязывающего фрагмента по любому из пп. 1-12, включающий (a) культивирование клетки по п. 16; и (b) выделение антитела или его антигенсвязывающего фрагмента.
18. Набор для выполнения способа лечения злокачественной опухоли у субъекта, содержащий антитело по любому из пп. 1-12 или композицию по п. 13 и вкладыш в упаковку.
19. Применение антитела или его антигенсвязывающего фрагмента по любому из пп. 1-12 для ингибирования пролиферации клеток, экспрессирующих HER3.
20. Применение антитела или его антигенсвязывающего фрагмента по п. 19, в котором указанная клетка, экспрессирующая HER3, представляет собой опухолевую клетку головы и шеи человека, клетку немелкоклеточного рака легкого человека, колоректальную опухолевую клетку человека, клетку рака простаты человека или клетку рака молочной железы человека.
21. Способ лечения злокачественной опухоли у субъекта, включающий в себя введение субъекту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп. 1-12.
22. Способ по п. 21, в котором злокачественная опухоль выбрана из группы, состоящей из рака ободочной кишки, рака легкого, рака желудка, рака молочной железы, рака головы и шеи, и меланомы, и где рак необязательно включает клетки, содержащие мутацию KRAS.
23. Способ лечения злокачественной опухоли, включающий введение субъекту терапевтически эффективного количества первого средства, которое представляет собой антитело или его антигенсвязывающий фрагмент по любому из пп. 1-12, в сочетании с терапевтически эффективным количеством второго средства, которое представляет собой другое противоопухолевое средство, отличное от первого средства.
| US20110171222 A1, 14.07.2011 | |||
| СПОСОБЫ ИДЕНТИФИКАЦИИ ОПУХОЛЕЙ, ВОСПРИИМЧИВЫХ К ЛЕЧЕНИЮ АНТИТЕЛАМИ ПРОТИВ ErbB2 | 2003 |
|
RU2338751C2 |

