Предложенный способ относится к области медицины, биологии и биотехнологии и может быть использован при оценке степени пролиферации клеток путем митоза. В частности, этот метод может использоватся для оценки функциональной активности лимфоцитов in vitro с помощью реакции бластной трансформации лимфоцитов путем полимеразной цепной реакции (ПЦР) с регистрацией накопления продуктов реакции «в реальном времени».
Классической моделью оценки функциональной активности лимфоцитов in vitro является реакция бластной транформации лимфоцитов (РБТЛ). Принцип метода основан на митогенном влиянии на лимфоциты некоторых веществ растительного и бактериального происхождения, а также различных веществ, влияющих на функционирование иммунной системы (цитокинов, хемокинов, CpG и т.д.). Фитогемаглютинин (ФГА), получаемый из семян фасоли, оказывает митогенное воздействие в основном на Т-лимфоциты. В классической методике реакции бластной трансформации инкубация монононуклеарных клеток периферической крови в присутствии ФГА осуществляется в течение 72 часов, результаты реакции оценивают по включению меченого тритием 3Н-тимидина через 6-18 часов инкубации после его внесения [1].
Полимеразная цепная реакция (ПЦР) широко используется для амплификации нуклеиновых кислот и позволяет, в частности, выявить присутствие и определить количество нуклеиновой кислоты с определенной последовательностью нуклеотидов в ДНК- или РНК-образце. Результаты полимеразной цепной реакции анализируют либо по окончании реакции, либо непосредственно в ходе реакции. Для регистрации результатов ПЦР в ходе процесса ее проводят в детектирующем амплификаторе в присутствии либо флуоресцентных интеркалирующих красителей, либо флуоресцентно меченых олигонуклеотидных праймеров или проб [2].
Ранее уже предпринимались попытки оценивать степень пролиферативного ответа клеток с помощью маркерных генов, однако для этой оценки применяли другие методы. Например, известен способ оценки пролиферативного ответа путем определения включившегося в ДНК бромодезоксиуридина (BrdU) с помощью метода ELISA [3]. Однако этот метод обладает недостаточно высоким соотношением сигнал-шум (около 10:1) и его чувствительность ограничена пределами определения оптической плотности при 450 нм (от 0.0 до 3.0).
С помощью метода Нозерн-блот было показано, что уровень мРНК гена топоизомеразы II альфа (tpa) и гена-контролера клеточного цикла 2 (cdc2) изменяется в течение клеточного цикла, причем уровень мРНК этих генов возрастает либо непосредственно (cdc2) [5], либо сразу после митоза (tpa) [6]. Таким образом, можно определять степень пролиферативного ответа клеток с помощью метода Нозерн-блот, но этот метод требует гибридизации с радиоактивно меченой пробой. Кроме того, уровень экспрессии мРНК гена топоизомеразы II альфа оценивали в делящихся и неделящихся клетках с помощью метода защиты от рибонуклеаз (ribonuclease protection assay) [4]. С помощью этого метода тоже можно оценивать степень пролиферации клеток, однако он также требует гибридизации с радиоактивно меченой пробой, а чувствительность подобных методов проигрывает чувствительности метода ПЦР «в реальном времени».
С помощью метода Вестерн-блот определяли уровень экспрессии белка топоизомеразы II альфа и бета в процессе клеточного цикла [7], что также позволяет исследовать степень пролиферации клеток, но чувствительность данного метода невысока.
Существует способ оценки степени стимуляции лимфоцитов с помощью проточной цитометрии путем измерения уровня экспрессии специфических маркерных молекул на поверхности клеток. Например, при стимуляции определенных CD-маркеров моноклональными антителами возрастает экспрессия CD38 на поверхности лимфоцитов, и увеличение количества CD38 определяли с помощью проточной цитометрии [3]. Однако в данном методе повышение экспрессии маркерных молекул на поверхности клеток не всегда совпадает с их пролиферацией, и часто зависит от возраста и состояния здоровья организма.
Технический результат, достигаемый при осуществлении предложенного изобретения, заключается в надежном определении степени пролиферации клеток в ответ на исследуемые стимулы либо при спонтанной пролиферации.
Задача, решаемая предложенным изобретением, заключается в использовании метода ПЦР «в реальном времени» для оценки степени пролиферативного ответа клеток с помощью мРНК маркерных генов, активность продуктов которых возрастает при митозе. В частности, для этого могут использоваться гены: топоизомеразы II альфа (topoisomerase II alpha, tpa, OMIM 126430) и гена-контролера клеточного цикла 2 (cell division cycle 2, cdc2, OMIM 116940).
Предложенный способ оценки пролиферативного ответа клеток иммунной системы в крови для достижения указанного технического результата предусматривает:
- взятие венозной крови в вакутейнер с гепарином натрия (BD Vacutainer, sodium heparin) или использование какого-либо другого материала (тканей);
- выделение клеток мононуклеарной фракции на градиенте плотности фиколла (фиколл или фиколл-верографин с плотностью 1,077 г/мл) как описано в стандартной методике [1] в течение 12 часов после взятия крови*;
- подсчет клеток, окрашенных трипановым синим, в камере Горяева на инвертированном микроскопе или в гемацитометре;
- разведение клеток полной питательной ростовой средой до концентрации 1 млн/мл (например, среда RPMI-1640, Sigma #R0883, с добавлением 10% сыворотки Standart Fetal Bovine Serum, HyClone #SH30088.03, 2мМ глутамина ПанЭко #Ф032, 20мМ HEPES-буфера ПанЭко #Ф134);
- раскапывание автоматической пипеткой со стерильным наконечником в лунки 96-луночного планшета по 1-2х105 полученной суспензии клеток в объеме 100-200 мкл среды;
- добавление в лунки митогенов или исследуемых веществ в необходимой концентрации (в случае использования митогена фитогемагглютинина его концентрация составляет 2.5-5.0 мкг/мл среды с клетками; в случае исследования влияния других различных веществ концентрацию подбирает экспериментатор). Под исследуемым веществом следует понимать вещество, влияние которого на пролиферацию клеток исследуется;
- инкубацию планшета в течение исследуемого времени в СО2-инкубаторе при 37°С, 5% СО2, в атмосфере насыщенного водяного пара;
- отбор содержимого лунок после инкубации с периодичностью, определяемой исследователем, исходя из целей и задач эксперимента (в нашей работе отбор содержимого лунок производился с периодичностью раз в 12 часов в течение 7 суток);
- лизис клеток и выделение нуклеиновых кислот с помощью набора «Проба-НК» (ЗАО «НПФ ДНК-Технология») либо другого готового набора для выдения РНК и при необходимости ДНК или иного метода согласно установленной методике;
- постановку реакции обратной транскрипции со специфических праймеров для генов tpa и cdc2 с целью получения ДНК, комплементарной РНК исследуемых генов ;
- постановку ПЦР «в реальном времени» с полученной комплементарной ДНК на специфических праймерах и с использованием меченых олигонуклеотидов на гены tpa и cdc2;
- поставновку ПЦР (при взятии в пробирках разного количества клеток) с выделенной ДНК на специфических праймерах и с использованием меченого олигонуклеотида на ген рецептора гормона роста человека (human growth hormone receptor, ghr) для нормализации результатов;
- выражение степени пролиферации клеток иммунной системы в индексе пролиферации, который представляет собой число, показывающее, во сколько раз увеличивается количество мРНК гена tpa и/или cdc2 в процессе культивирования лимфоцитов с/без добавления ФГА или исследуемого вещества по сравнению с количеством мРНК этих генов в свежевыделенных из крови или другого какого-либо материала здорового донора лимфоцитах.
Таким образом, при анализе результатов ПЦР «в реальном времени» определяют, во сколько раз увеличивается количество мРНК исследуемых генов tpa и cdc2 в процессе культивирования лимфоцитов с добавлением митогенов или исследуемых веществ (или без добавления стимуляторов) по сравнению с количеством мРНК этих генов в свежевыделенных лимфоцитах и в первые часы культивации.
Краткое описание чертежей
На фиг.1 приведены графики для определения эффективности амплификации E (%) (см. формулу 1). Для проведения расчетов определяли эффективность амплификации каждого гена (Е) и значения разности пороговых циклов при разведении образца в 10 раз (slope), для чего сделали разведения образцов в 3,2 и 10 раз (шаг 0,5 lg), построили калибровочные прямые и определили необходимые для расчетов параметры согласно формуле 1 и показаниям прибора. Различия в углах наклона прямых (slope) вызвано неоднородностью уровня регистрируемой интенсивности флуоресценции (например, за счет неоднородности температуры термоблока, неоднородностей оптического тракта прибора, разброса оптических параметров реакционных пробирок, погрешностей при приготовлении реакционной смеси и т.д).
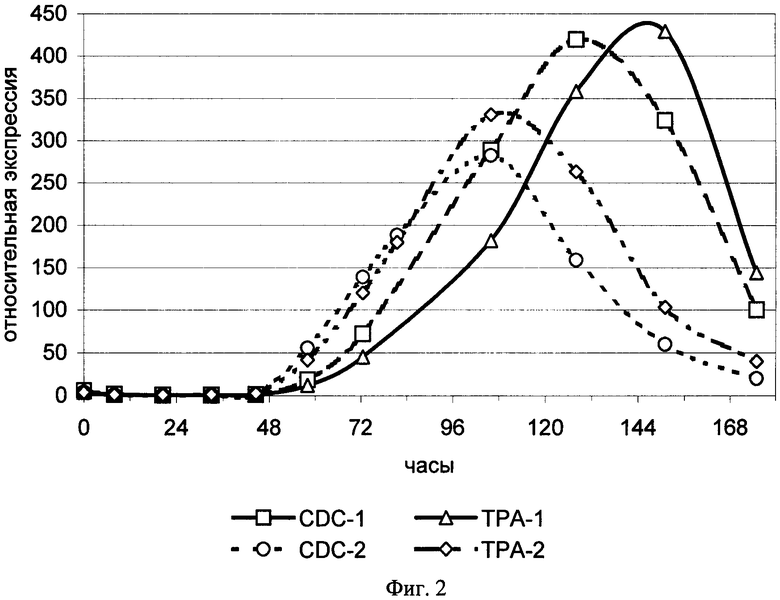
На фиг.2 показана кинетика изменения уровней экспрессии мРНК генов tpa и cdc2 в мононуклеарных клетках периферической крови, взятых у двух здоровых доноров и стимулированных ФГА. CDC-1 - уровень экспрессии мРНК гена cdc2 в популяции клеток донора 1; CDC-2 - уровень экспрессии мРНК гена cdc2 в популяции клеток донора 2; TPA-1 - уровень экспрессии мРНК гена tpa в популяции клеток донора 1; TPA-2 - уровень экспрессии мРНК гена tpa в популяции клеток донора 2.
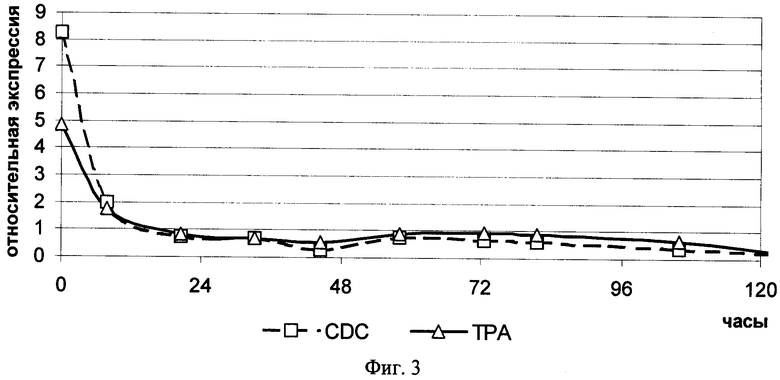
На фиг.3 представлена кинетика изменения уровней экспрессии мРНК генов tpa и cdc2 в мононуклеарных клетках периферической крови, взятых у двух здоровых доноров и культивированных без добавления ФГА. CDC - уровень экспрессии мРНК гена cdc2 в популяции клеток, среднее по двум донорам; TPA - уровень экспрессии мРНК гена tpa в популяции клеток, среднее по двум донорам.
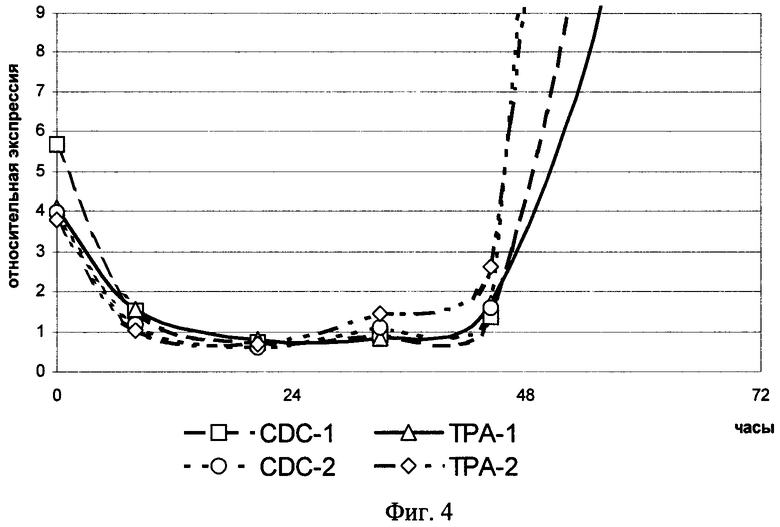
На фиг.4 представлена кинетика изменения уровней экспрессии генов tpa и cdc2 в первые часы культивации мононуклеарных клеток периферической крови, стимулированных ФГА. CDC-1 - уровень экспрессии мРНК гена cdc2 в популяции клеток донора 1; CDC-2 - уровень экспрессии мРНК гена cdc2 в популяции клеток донора 2; TPA-1 - уровень экспрессии мРНК гена tpa в популяции клеток донора 1; TPA-2 - уровень экспрессии мРНК гена tpa в популяции клеток донора 2.
Осуществление изобретения
1. Реакция бластной трансформации
Кровь следует брать натощак в вакутейнеры с гепарином, выделение лимфоцитов из ПК проводить не позднее чем через 24 часа после взятия крови. Выделение фракции мононуклеарных клеток проводят как описано в [1] с небольшими изменениями. Коротко, 7 мл свежей крови, взятые в вакутейнер с гепарином, разбавляют 1:2 по объему средой 199 c солями Хэнкса и глутамином, наслаивают на 3-4 мл раствора фиколла (плотность 1,077 г/мл), центрифугируют при 1600 об/мин, 15°С, 15 мин. Интерфазу аккуратно переносят в чистые пробирки и трижды отмывают 5-8 мл среды 199 в той же центрифуге при 1200 об/мин, 15°С, 10 мин. После отмывок сливают супернатант, полученные клетки ресуспендируют на вортексе в 1 мл полной питательной среды RPMI-1640, определяют число выделенных мононуклеарных клеток в гемоцитометре под микроскопом. Добавляют необходимое количество полной питательной среды RPMI-1640 до концентрации 106 клеток/мл среды.
В стерильный 96-луночный планшет раскапывают по 105 выделенных клеток на лунку в объеме 100 мкл, добавляют по 10 мкл стокового раствора ФГА (итоговая концентрация 2,5 мкг/мл). В лунки с клетками для контроля спонтанной пролиферации стимуляторов не добавляют. Планшет помещают в СО2-инкубатор, насыщенный водяным паром, на 7-8 суток при 37°С, 5% СО2. Через определенное количество часов отбирают пробы клеток.
2. Выделение РНК и ДНК
Для выделения нуклеиновых кислот используется набор “Проба-НК” («ДНК-Технология» #05-017) или любой другой готовый набор или метод выделения РНК и ДНК согласно методике.
3. Реакция обратной транскрипции
Реакцию обратной транскрипции проводят в объеме 20 мкл (в реакцию берут 16,5 мкл полученного препарата рибонуклеиновых кислот). В качестве праймеров для обратной транскрипции используются специфические олигонуклеотиды. Реакцию проводят при температуре 40°С в течение 1 часа, с последующей инактивацией обратной транскриптазы при 95°С в течение 15 минут.
Состав реакции:
- 16,5 мкл препарата РНК;
- 2 мкл 10х буфера* для обратной транскрипции;
- 0,01 мкл праймера для обратной транскрипции в концентрации 100 pM (или каждого праймера по 0,01 мкл, если планируется ставить ПЦР на нескольких системах специфических праймеров);
- 0,24 мкл смеси дезоксирибонуклеотидтрифосфатов четырех типов в концентрации 25 мМ каждого дезоксирибонуклеотидтрифосфата;
- 0,5 мкл фермента обратной транскриптазы (200 ед/мкл);
- вода mQ, обработанная DEPC, до 20 мкл.
*- состав буфера для обратной транскрипции: 500 мМ Tris-HCl pH 8,3 при 25°С; 500 мМ KCl; 40 мМ MgCl2; 100 мМ DTT
4. Проведение ПЦР «в реальном времени»
Для каждого образца проводят ПЦР со специфическими праймерами на гены tpa и cdc2. В качестве нормировочного гена используется ген ghr (ген рецептора гормона роста человека для нормировки по ДНК). Ген hprt1 (ген гипоксантин-фосфорибозилтрансферазы 1, который часто выбирается для нормировки по РНК) в данном случае нельзя использовать, поскольку степень его экспрессии изменяется при стимуляции некоторыми митогенами.
Праймеры и зонды для ПЦР подбираются с учетом структур экзонов и интронов таким образом, чтобы исключить отжиг на матрице геномной ДНК на генах tpa и cdc2. Это позволяет не использовать дополнительный этап обработки нуклеиновых кислот ДНКазой. Отсутствие амплификации на матрице геномной ДНК со специфичных праймеров на гены tpa и cdc2 проверено экспериментально на образцах, не прошедших реакцию обратной транскрипции. Все зонды содержат метку Fam(dT). Соответственно реакции амплификации со специфичных праймеров на tpa, cdc2, и ghr ставят в разных пробирках. Для повышения чувствительности и специфичности ПЦР используют “горячий старт”, который обеспечивается путем разделения компонентов реакции парафином. Амплификацию осуществляют в режиме «реального времени» в объеме 35 мкл по следующей программе:
80°С 30 сек, 94°С 1 мин 1 цикл
94°С 10 сек, 64°С 20 сек 50 циклов.
Для ПЦР «в реальном времени» используют прибор “ДТ-322” (“НПФ ДНК-Технология”) либо любой другой прибор, позволяющий проводить ПЦР «в реальном времени». Измерение уровня флуоресценции проводят на каждом цикле при температуре 64°С.
Дополнительный контроль прохождения реакции осуществляется методом электрофореза продуктов ПЦР в 2% агарозном геле.
Для определения содержания мРНК исследуемых генов были разработаны следующие праймеры и зонды:
мРНК гена tpa: праймер для обратной транскрипции 5'-CATCTTCATCTGACTCTTC-3', прямой праймер 5'-TGTCGTGTCAGACCTTGAAGC-3', обратный праймер 5'-CCTTGGATTTCTTGCTTGTGAC-3', зонд (FAM)-5'-TGTACCACTGTCTTCAAGCCCTCCTGC-3'-(BHQ1)
мРНК гена cdc2: праймер для обратной транскрипции 5'- CTGGAGTTGAGTAACGAG-3', прямой праймер 5'-CTTCACTTGTTAAGAGTTATTTATAC-3', обратный праймер 5'-CCAGAGTGTTACTACCTCATGTG-3', зонд (FAM)-5'-TGCCTTGCCAGAGCTTTTGGAATAC-3'-(BHQ1)
ДНК гена ghr: прямой праймер 5'-CATTCCCATCATTGAGTGTGGAGTGAG-3', обратный праймер 5'-CTGGGGATCAGGTGTTTATGGACCA-3', зонд (BHQ1)-5'-CCTTCTGCCTGGCTTGCTTTCCC-3'-(FAM)
Состав реакционной смеси в пробирке:
- бидистиллированная вода - 25,37 мкл
- реакционный буфер** - 3,5 мкл
- смесь дезоксирибонуклеотидтрифосфатов четырех типов в концентрации 25 мМ каждого дезоксирибонуклеотидтрифосфата - 0,24 мкл
- прямой праймер в концентрации 100 мкМ - 0,125 мкл
- обратный праймер в концентрации 100 мкМ - 0,125 мкл
- детектирующая проба в концентрации 50 мкМ - 0,07 мкл
- Taq - полимераза в концентрации 5 международных единиц активности - 0,5 мкл
- исследуемый образец ДНК - 5 мкл
**- состав реакционного буфера: 100 мМ Tris-HCl (pH 8,8 при 25°С), 500 мM KCl, 0,8% P40, 20 мМ MgCl2
5. Параметры, используемые для описания ПЦР в режиме «реального времени»
1. Ср - значение порогового цикла, автоматически определяемое прибором.
2. Slope - разница в значениях Ср (Δ Ср) при разведении образца в 10 раз.
3. Эффективность амплификации оценивают по формуле:
Е = 10-(1/slope); Е(%)= (Е - 1)х100% (формула 1)
4. Определение уровня экспрессии мРНК tpa относительно ДНК гена ghr.
Определение уровня экспрессии мРНК tpa относительно ДНК гена ghr проводили по формуле:
[tpa]/[ghr] = Eghr Cp1/Eghr Cp2 (формула 2),
где Е - эффективность амплификации,
Сp1 - значение порогового цикла в образце для ghr
Сp2 - значение порогового цикла в образце для tpa.
5. В качестве главной характеристики эксперимента использовали индекс пролиферации - отношение (R) уровня экспрессии маркеров реакции к фоновому значению экспрессии этих маркеров, полученному в первые минуты/часы культивирования лимфоцитов.
Для определения R были использованы два способа:
метод дельта-дельта ЦТ (ΔΔ Ср) с применением нормировочных генов. R считали как соотношение нормированного уровня экспрессии в определенный момент времени [tpai]/[ghri] к фоновому значению [tpaf]/ [ghrf]
R= [tpai]/[ghr1i] х [hprt1f]/[ghrf] (формула 3);
метод без применения нормировочных генов
R = 10 (Cpf-Cpi)/slope (формула 4),
где Cpi - значение порогового цикла в определенный момент времени РБТЛ,
Cpf - фоновое значение порогового цикла в первые часы РБТЛ.
Краткая суть действий при осуществлении способа состоит в следующем.
Взятие клеток, стимулирование их выбранными веществами, затем выделение из них РНК, превращение ее в ДНК (т.к. ПЦР можно ставить с ДНК, а не с РНК).
Проведение ПЦР с мечеными олигонуклеотидами, чтобы определить количество взятой в реакцию ДНК и, следовательно, количество исходной РНК. Чем раньше детектируем сигнал флуоресценции ввиду свечения меченых олигонуклеотидов, тем больше будет ДНК (следовательно, и исходной РНК).
Чтобы исключить ситуацию, когда РНК генов cdc2 и tpa может быть больше ввиду того, что в разных пробирках было взято разное количество клеток, используем нормализацию данных.
Вместе с РНК выделяем из клеток тотальную ДНК. Эта ДНК потом непосредственно используется в ПЦР без обратной транскрипции. В ДНК мы выбрали ген рецептора гормона роста ghr, который существует в некотором определенном числе копий у всех клеток высших эукариот.
Таким образом, в реакции ПЦР одновременно получаем два сигнала: один от меченых олигонуклеотидов на ген cdc2 или tpa, а второй от меченых олигонуклеотидов на ген ghr.
При обработке результатов делим величину полученного сигнала от cdc2 или tpa на величину сигнала от гена ghr. Таким образом, мы получаем нормированные значения для наших генов: величину сигнала от ДНК (следовательно, мРНК) генов cdc2 или tpa в расчете на одну молекулу гена ghr.
Теперь можно напрямую сравнивать значения флуоресценции. Если флуоресценция растет, то это означает, что действительно становится больше молекул мРНК генов cdc2 или tpa, и исключается подозрение, что было взято большее количество клеток в реакцию.
Далее мы делим полученную величину флуоресценции cdc2 или tpa для стимулированных выбранным веществом клеток на величину флуоресценции cdc2 или tpa для нестимулированных клеток, и получаем индекс пролиферации. В этом индексе видно, во сколько раз возрос уровень экспрессии мРНК генов cdc2 или tpa при добавлении исследуемого вещества в момент времени, когда сняли клетки и поместили их в лизирующий раствор.
Если аккуратно подсчитать клетки и брать в реакцию стандартное количество, то можно не определять флуоресценцию от гена ghr, а можно использовать данные флуоресценции от генов cdc2 и tра непосредственно, без нормализации.
Принципиальным отличием предлагаемого метода является то, что степень пролиферации клеток в ответ на исследуемые стимулы или без добавления стимуляторов определяется с помощью ПЦР «в реальном времени» с использованием специфических праймеров и зондов по мРНК генов tpa и cdc2, активность которых возрастает при митозе. Предлагаемый метод является высокоспецифичным и обладает высокой чувствительностью; уровень содержания мРНК выбранных маркерных генов во время пролиферации возрастает в несколько сотен раз по сравнению с уровнем мРНК в неделящихся клетках. Данный метод позволяет непосредственно следить за числом делящихся клеток, оценивая степень экспрессии генов, участвующих в пролиферации и регулирующих клеточное деление, причем вне зависимости от возраста, наличия заболеваний, состояния организма и исследуемых клеток.
Список использованной литературы
1. Current Protocols in Immunology. Ред. Coligan J.E., Kruisbeek A.M., Margulies D.H., Shevach E.M., Strober W. John Wiley & Sons; 2003; ISBN: 0471522767
2. US2002123062, 2002-09-05.
3. Tsehaynesh Messele, Marijke T. L. Roos, Dorte Hamann, Maarten Koot, Arnaud L. Fontanet, Frank Miedema, Peter T. A. Schellekens, and Tobias F. Rinke de Wit. Nonradioactive Techniques for Measurement of In Vitro T-Cell Proliferation: Alternatives to the [3H]Thymidine Incorporation Assay. Clin Diagn Lab Immunol. 2000 July; 7(4): 687-692.
4. Isaacs RJ, Harris AL, Hickson ID. Regulation of the human topoisomerase II alpha gene promoter in confluence-arrested cells. J Biol Chem. 1996 Jul 12; 271(28): 16741-16747.
5. Welch PJ, Wang JY. Coordinated synthesis and degradation of cdc2 in the mammalian cell cycle. Proc Natl Acad Sci USA. 1992 Apr 1; 89(7): 3093-3097.
6. Goswami PC, Roti Roti JL, Hunt CR. The cell cycle-coupled expression of topoisomerase IIalpha during S phase is regulated by mRNA stability and is disrupted by heat shock or ionizing radiation. Mol Cell Biol. 1996 Apr; 16(4): 1500-1508.
7. Christensen MO, Larsen MK, Barthelmes HU, Hock R, Andersen CL, Kjeldsen E, Knudsen BR, Westergaard O, Boege F, Mielke C. Dynamics of human DNA topoisomerases II alpha and II beta in living cells. J Cell Biol. 2002 Apr 1; 157(1): 31-44.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ДИАГНОСТИКИ ВОСПАЛИТЕЛЬНОГО ПРОЦЕССА ПРИ РАННЕМ РЕВМАТОИДНОМ АРТРИТЕ | 2009 |
|
RU2417263C2 |
| Количественный метод определения экспрессии аллелей GNAO1 здоровой формы и с мутацией c.607 G>A | 2021 |
|
RU2777663C1 |
| СПОСОБ ДИАГНОСТИКИ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ ПО УРОВНЮ мРНК TGEβ и TNFα В ПЛАЗМЕ КРОВИ | 2019 |
|
RU2742209C1 |
| Набор реагентов для определения рибонуклеиновых кислот при диагностике лимфопролиферативных заболеваний | 2023 |
|
RU2802064C1 |
| Тест-система и способ выявления A, B, D мутаций гена NPM1 для количественного определения минимальной остаточной болезни | 2023 |
|
RU2830545C1 |
| Набор реагентов для определения малых рибонуклеиновых кислот (микроРНК) при диагностике множественной миеломы | 2024 |
|
RU2836054C1 |
| Набор синтетических олигонуклеотидов для определения уровня экспрессии генов DHN1, DHN7, SGS3, CPK, CYP450, ERD3, CCOAOMT1, LEA и ACT сосны обыкновенной (pinus sylvestris l.) методом ПЦР в реальном времени | 2022 |
|
RU2795627C1 |
| НАБОР ОЛИГОНУКЛЕОТИДНЫХ ПРАЙМЕРОВ И ФЛУОРЕСЦЕНТНОМЕЧЕНОГО ЗОНДА ДЛЯ ВИДОСПЕЦИФИЧНОЙ ЭКСПРЕСС-ИДЕНТИФИКАЦИИ РНК ВИРУСА ХУНИН МЕТОДОМ ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ В РЕАЛЬНОМ ВРЕМЕНИ | 2013 |
|
RU2525938C1 |
| СПОСОБ ДИАГНОСТИКИ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ ПО УРОВНЮ ЭКСПРЕССИИ мРНК IL-10 И/ИЛИ IL-17 В ПЛАЗМЕ КРОВИ | 2020 |
|
RU2752971C1 |
| СПОСОБ ДИАГНОСТИКИ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ И РАКА ЯИЧНИКОВ ПО УРОВНЮ мРНК ММР-9 В ПЛАЗМЕ КРОВИ | 2020 |
|
RU2745424C1 |
Изобретение относится к области биотехнологии, а именно к способу оценки степени пролиферации лимфоцитов с помощью маркерных генов методом полимеразной цепной реакции в реальном времени (ПЦР РВ). Способ включает добавление в часть лунок с лимфоцитами митогенов или исследуемых веществ, инкубацию содержимого лунок, отбор содержимого лунок, лизис клеток и выделение нуклеиновых кислот. Ставят реакцию обратной транскрипции (OT) с праймерами для генов tpa и cdc2: праймер для OT с мРНК гена tpa 5'-CATCTTCATCTGACTCTTC-3', праймер для OT с мРНК гена cdc2 5'-CTGGAGTTGAGTAACGAG-3'. Проводят ПЦР РВ с полученной кДНК на специфических праймерах и с использованием меченых олигонуклеотидов на гены tpa и cdc2: для гена tpa прямой праймер 5'-TGTCGTGTCAGACCTTGAAGC-3', обратный праймер 5'-CCTTGGATTTCTTGCTTGTGAC-3', зонд (FAM)-5'-TGTACCACTGTCTCAAGCCCTCCTGC-3'-(BHQ1), для гена cdc2 прямой праймер 5'-CTTCACTTGTTAAGAGTTATTTATAC-3', обратный праймер 5'-CCAGAGTGTTACTACCTCATGTG-3', зонд (FAM)-5'-TGCCTTGCCAGAGC(FdT)TTTGGAATAC-3'-(BHQ1). Выражают степень пролиферации лимфоцитов в индексе пролиферации, который представляет собой число, показывающее, во сколько раз увеличивается количество мРНК гена tpa и/или cdc2 в процессе культивирования лимфоцитов с/без добавления ФГА или исследуемого вещества по сравнению с количеством мРНК этих генов в свежевыделенных из крови здорового донора лимфоцитах. Предложенное изобретение позволяет непосредственно следить за числом делящихся клеток, оценивая степень экспрессии генов, участвующих и регулирующих клеточное деление. 4 ил., 4 пр.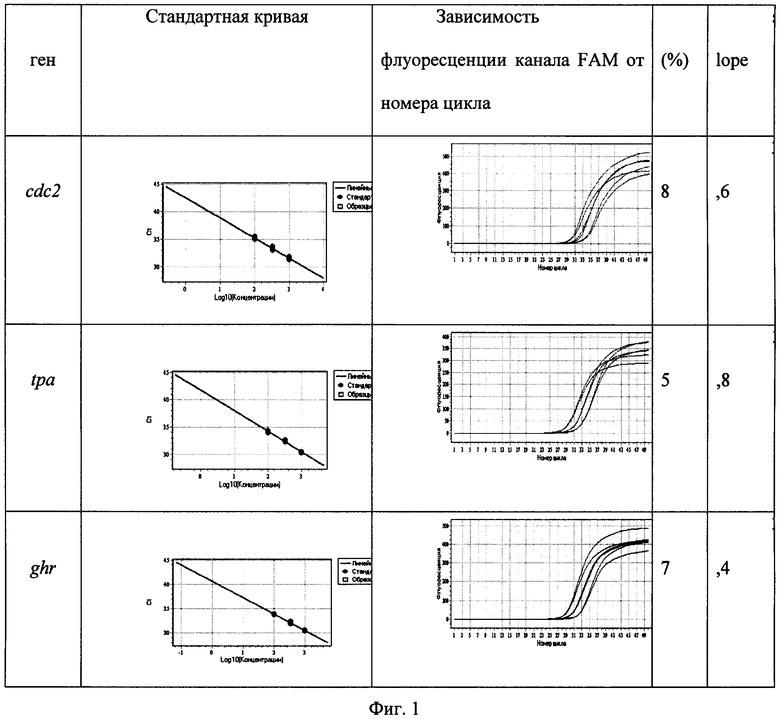
Способ оценки степени пролиферации лимфоцитов с помощью маркерных генов методом полимеразной цепной реакции в реальном времени, включающий процедуру взятия крови,
выделение и культивирование лимфоцитов с последующим их подсчетом,
разведение клеток в полной питательной среде,
раскапывание в лунки планшета полученной суспензии клеток,
добавление в часть лунок митогенов или исследуемых веществ,
инкубацию содержимого лунок планшета,
отбор содержимого лунок после инкубации в течение определенного времени,
лизис клеток и выделение нуклеиновых кислот,
постановку реакции обратной транскрипции с использованием специфических праймеров для генов tpa и cdc2 следующего состава: праймер для обратной транскрипции с мРНК гена tpa 5'-CATCTTCATCTGACTCTTC-3', праймер для обратной транскрипции с мРНК гена cdc2 5'-CTGGAGTTGAGTAACGAG-3',
постановку ПЦР «в реальном времени» с полученной кДНК на специфических праймерах и с использованием меченых олигонуклеотидов на гены tpa и cdc2 следующего состава: для определения количества мРНК гена tpa прямой праймер 5'-TGTCGTGTCAGACCTTGAAGC-3', обратный праймер 5'-CCTTGGATTTCTTGCTTGTGAC-3', зонд (FАМ)-5'-TGTACCACTGTCTCAAGCCCTCCTGC-3'-(BHQ1), для определения количества мРНК гена cdc2 прямой праймер 5'-CTTCACTTGTTAAGAGTTATTTATAC-3', обратный праймер 5'-CCAGAGTGTTACTACCTCATGTG-3', зонд (FAM)-5'-TGCCTTGCCAGAGC(FdT)TTTGGAATAC-3'-(BHQ1),
при необходимости в случае трудностей с выделением РНК из клеток следует провести постановку ПЦР с выделенной ДНК на специфических праймерах и с использованием меченого олигонуклеотида на ген человеческого гормона роста (human growth hormone, ghr) для нормализации результатов, где используют праймеры следующего состава: прямой праймер 5'-CATTCCCATCATTGAGTGTGGAGTGAG-3', обратный праймер 5'-CTGGGGATCAGGTGTTTATGGACCA-3' и зонд (FAM)-5'-CCTTCTGCCTGGCTTGCTTTCCC-3'-(BHQ1),
выражение степени пролиферации лимфоцитов в индексе пролиферации, который представляет собой число, показывающее, во сколько раз увеличивается количество мРНК гена tpa и/или cdc2 в процессе культивирования лимфоцитов с/без добавления ФГА или исследуемого вещества по сравнению с количеством мРНК этих генов в свежевыделенных из крови здорового донора лимфоцитах; в случае с трудностями в выделении РНК индекс пролиферации можно определять путем деления величины сигнала от генов tpa и cdc2 на величину сигнала, полученного от ДНК гена рецептора гормона роста ghr.
| MHSSELE Т | |||
| ЕТ AL., Nonradioactive techniques for measurement of in vitro T-cell proliferation: alternatives to the [(3)H]thymidine incorporation assay, Clin Diagn Lab Immunol., 2000, v.7, no.4, p.687-692 | |||
| ISAACS R.J | |||
| ЕТ AL, Regulation of the human topoisomerase II alpha gene promoter in confluence-arrested cells., Biol Chem., 1996, v.271, no.28, |

