Изобретение относится к экспериментальной медицине и физиологии и предназначено для проведения физиологических исследований в эксперименте на животных.
Тремор - колебательные движения, вызванные сокращениями реципрокно иннервируемых мышц-антагонистов. Тремор может быть физиологическим и патологическим, вызванным различными заболеваниями нервной системы, связанными с повреждениями мозга различной этиологии. Тремор может наблюдаться в покое (статическая форма тремора) и при осуществлении двигательной активности (активная форма тремора, так называемый интенционный тремор, наблюдаемый при осуществлении целенаправленных движений у человека и животных).
Известен способ моделирования тремора путем электрической стимуляции (Folkerts, Spiegel 1953; Бехтерева с соавт., 1967; Петелин 1970), или сочетанного разрушения различных образований экстрапирамидной системы мозга - стриопаллидума, покрышки среднего мозга, красного ядра (Ward et. al., 1948 - прототип; Peterson et. al., 1949; Critchley 1949; Kaelber 1963; Poirier et. al., 1966; Лагутина и др., 1967).
Недостатками вышеуказанных способов моделирования гиперкинезов являются:
1. Стимуляцией, или разрушением вышеуказанных структур можно получить лишь общую неспецифическую картину тремора отдельных мышечных групп и всего тела, наблюдаемого у испытуемых в состоянии покоя - статический тремор. Активная форма тремора - интенционный тремор, инициируемый целенаправленной реакцией испытуемого, вышеуказанными способами не воспроизводится.
2. Полученная вышеуказанными способами модель статического тремора отличается нестойкостью и, в основном, исчезает через 2-3 недели после проведенной операции.
3. При стимуляции электрическим током, или двусторонних разрушениях различных отделов экстрапирамидной системы могут наблюдаться многочисленные побочные двигательные (гиперкинезы) и поведенческие проявления, включая миоклонические судороги и эпилептические припадки.
4. Вышеуказанные способы моделирования сопровождаются высокой травматичностью подопытных животных и в отдельных случаях могут приводить к летальному исходу животных.
Ранее проведенные исследования показали, что падение уровня стриатного дофамина сопровождается контрактурой мышц-антагонистов, ведущей к ригидности передних конечностей у животных и человека (Andrew 1973; Schultz 1982; Hayashi е.а. 1988; Wickens е.а. 1991; Miklyaeva е.а. 1994). Это дало основание предположить, что нео-стриатум, занимая стратегическую позицию в составе стрио-таламо-кортикальной системы, может играть важную роль в регуляции нормального баланса между антагонистическими группами мышц при осуществлении (неавтоматизированных локомоторных ответов) произвольных движений различной сложности (Hassler 1978; De Long, Wichmann 1993). Однако попытки получить у животных с повреждением неостриатума дискинезию двигательного ответа в виде интенционного тремора долгое время не удавалось. Прежде всего это было связано с отсутствием адекватной поведенческой модели, сенситивной для выявления интенционного тремора у животных. В отличие от эссенциального паркинсонического тремора, выявляемых у паркинсоников в покое, или выполнении любого произвольного движения, наблюдение за каудатэктомированными животными показывают отсутствие дискинезий и интенционного тремора при выполнении произвольных движений передней конечностью (Villablanca е.а. 1976; Denny-Brown, Yanagisawa 1976), осуществлении пищедобывательных движений (Divac е.а. 1978), реакций выжима педали передней конечностью (Benita е.а. 1979), извлечении пищевых шариков из узких трубок-тубусов (Bures е.а. 1991), а также при прослеживании передней лапой движущейся педали или светового пятна (Fabre, Buser 1980).
Цель изобретения: обеспечение условий для моделирования активной (интенционной) формы тремора, возникающего при осуществлении целенаправленной двигательной реакции; снижение травматичности операции и минимизация побочных эффектов.
Для выявления скрытой патологии двигательных нарушений, лежащих в основе интенционного тремора, в наших экспериментах, проведенных на кошках, мы усложнили поведенческую (смысловую) задачу тестируемого инструментального рефлекса. Цель эксперимента заключалась в попытке выявить возможные двигательные нарушения, связанные с локальным повреждением неостриатума, с помощью создания условий повышенной эмоциональной напряженности, требующей мобилизации резервных возможностей всего организма, то есть выявить частный дефект путем активации целого организма. Животным предлагалось в каждой пробе принимать решение о выполнении инструментального ответа нажатия на педаль, или воздерживаться от выполнения этого двигательного ответа в зависимости от порядка предъявления положительного или тормозного условных раздражителей.
Сущность способа моделирования интенционного тремора, включающего стереотаксическое повреждение экстрапирамидной системы мозга и последующее тестирование двигательной активности, заключается в том, что повреждение мозга осуществляют путем избирательного одностороннего электролитического повреждения дорсального отдела головки хвостатого ядра, контрлатерального конечности животного, осуществляющей целенаправленное движение, причем тестирование двигательной активности производят путем подачи зрительных дифференцировочных сигналов, требующих выполнения выработанного двигательного ответа в виде нажатия передней (рабочей) конечностью на педаль, подкрепляемого вознаграждением, или воздержания от его осуществления, а число пусковых зрительных сигналов, требующих двигательного ответа, и тормозных зрительных сигналов, требующих воздержания от выполнения ответа в тестируемой последовательности сигналов является равным, причем дифференцировочные сигналы предъявляют в случайном порядке, при этом интенционный тремор выявляется в виде появления быстрых (5-10 Гц) антагонистических фазических движений рабочей конечностью при выполнении двигательной реакции нажатия на педаль, инициируемой подачей пускового зрительного сигнала.
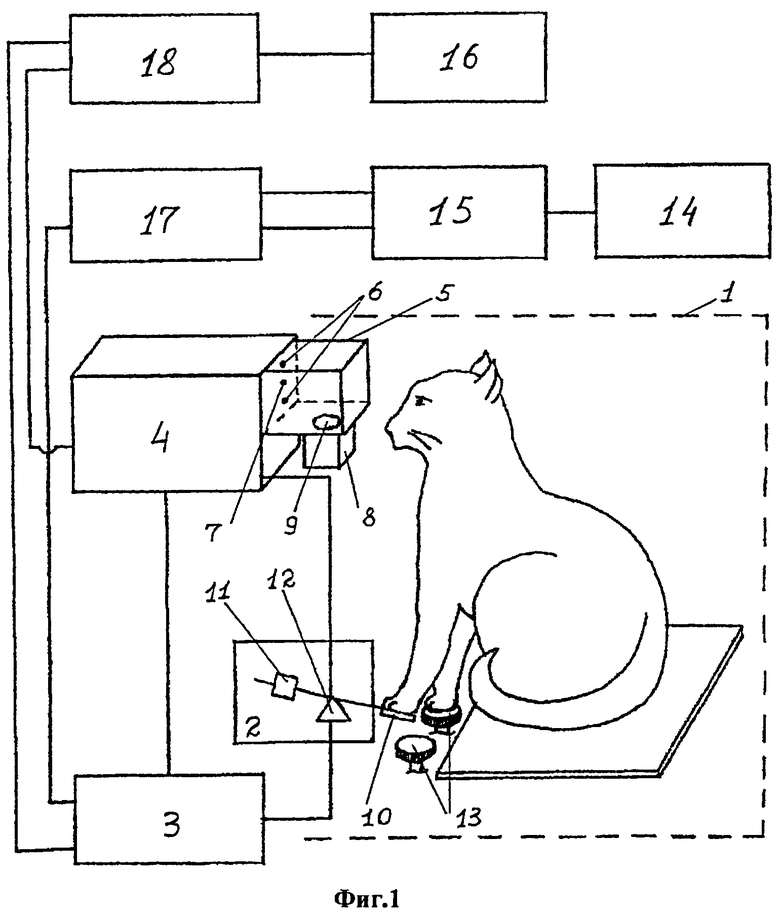
На фиг.1 показана схема устройства для осуществления способа в аксонометрии, где
1 - экспериментальная камера (условно обозначена пунктиром)
2 - пульт управления испытуемого
3 - пульт управления экспериментатора
4 - осциллоскоп
5 - тубус
6 - тормозный зрительный сигнал (два сфокусированных пятна)
7 - пусковой зрительный сигнал (одно сфокусированное пятно)
8 - кормушка
9 - лоток кормушки
10 - педаль
11 - противовес с регулируемым грузом
12 - потенциометр
13 - опорные платформы передних конечностей
14 - тензодатчики опорных платформ, педали и лотка кормушки
15 - усилитель многоканальный
16 - монитор
17 - регистрирующий прибор
18 - компьютер
Устройство для осуществления способа моделирования тремора изображено на фиг 1, 2, 3 и содержит экспериментальную камеру (1), оснащенную опорными платформами (13) для передних конечностей животного, пульт управления у испытуемого (2), пульт управления у экспериментатора (3), кормушку (8), средства экспозиции сигналов, выполненные на основе осциллоскопа (4), на экране которого предъявляются зрительные сигналы (6, 7), тубус для экспозиции сигналов (5). Пульт управления у испытуемого выполнен в виде педали (10), оснащенной противовесом (11) с регулируемым грузом, педаль установлена на оси потенциометра (12), который подключается в электрическую схему развертки лучей осциллоскопа (4), кормушка (8) выполнена автоматизированной и размещена в основании проема тубуса (5). Опорные платформы для конечностей (13), педаль (10) и лоток кормушки (9) оснащены миниатюрными тензодатчиками (на рисунке не показаны), которые включены в электрическую схему, включающую последовательно соединенные многоканальный усилитель (15), регистрирующий прибор (17) и монитор (16). Пульт управления экспериментатора (3) снабжен средствами включения сенсорных сигналов на экране осциллоскопа (19), позволяющим осуществлять подачу зрительных раздражителей. Устройство также оснащено компьютером (18), определяющим изменение исходного местоположения предъявляемых на экране осциллоскопа (4) предъявляемых зрительных сигналов.
Тестирование осуществляют следующим образом (фиг.1, 2, 3):
Подопытным животным за неделю до тестирования под наркозом проводят операцию электролитического повреждения (I=1-2 мА; T=30 с.) дорсального отдела головки хвостатого ядра (см. фиг.2) по стереотаксическим координатам атласа Джаспера и Аймон-Марсана (1954). Каудатэктомированное животное, предварительно обученное тормозному двигательному рефлексу на предъявление на экране осциллоскопа двух световых пятен и положительному рефлексу нажатия передней (рабочей) лапой на педаль при экспозиции одного светового пятна, помещают в экспериментальную камеру (1), оснащенную осциллоскопом (4) с тубусом (5) для экспозиции зрительных сигналов. Пульт управления экспериментатора (3), осциллоскоп (4), усилитель (15), монитор (16), регистрирующий прибор (17) и компьютер (18) подключают к источникам питания. Тензо-датчики, расположенные на опорных платформах (13), педали (10) и лотке кормушки (9) подключают на вход усилителя (15), последовательно соединенного с монитором (16) и регистрирующим прибором (17).
Средства экспозиции сигналов выполнены на основе осциллоскопа (4), на экране которого предъявляются тормозные зрительные сигналы в виде двух сфокусированных световых пятен (6), которые требуют воздержания от выполнения двигательной реакции нажатия на педаль, или предъявление одного светового пятна - пусковой сигнал (7), который означает возможность подкрепления, получаемого выжимом педали (10).
Тестирование двигательной активности производят путем подачи зрительных дифференцировочных сигналов (6, 7), требующих выполнения выработанного двигательного ответа в виде нажатия передней (рабочей) конечностью на педаль (10), подкрепляемого вознаграждением, или воздержания от его осуществления, при этом число пусковых зрительных сигналов (7), требующих двигательного ответа, и тормозных зрительных сигналов (6), требующих воздержания от выполнения ответа в тестируемой последовательности сигналов является равным, причем дифференцировочные сигналы предъявляют в случайном порядке, при этом интенционный тремор выявляется в виде появления быстрых (5-10 Гц) антагонистических фазических движений рабочей конечностью при выполнении двигательной реакции нажатия на педаль (10), инициируемой подачей пускового условного сигнала (фиг.2).
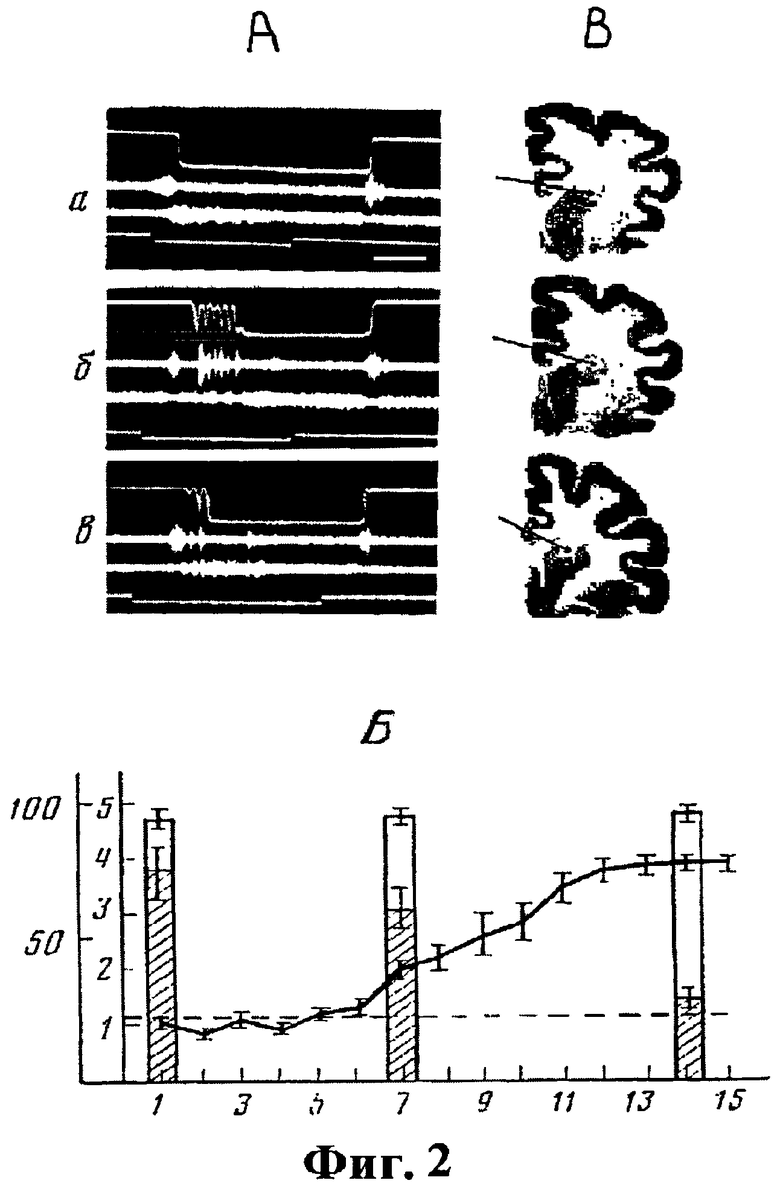
Эксперименты, проведенные на 14 подопытных животных (кошки, самцы) показали, что повреждение дорзального отдела головки хвостатого ядра не нарушало реализацию выработанной двигательной реакции нажатия передней (рабочей) конечностью на педаль при предъявлении положительного сигнала, а также при правильном чередовании положительного и тормозного сигналов (процент правильных решений составлял 95-98%). Также не изменялись по сравнению с интактными животными все регистрируемые параметры двигательного ответа (фиг.2А, а).
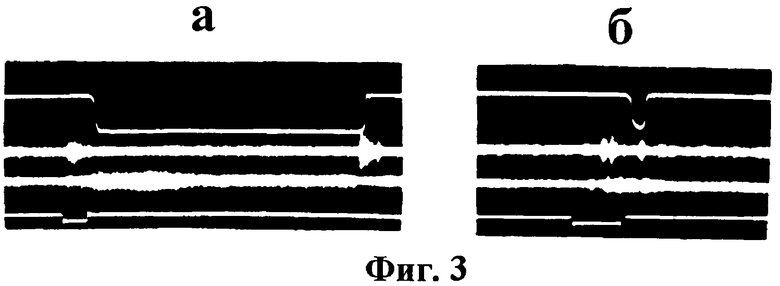
После упрочения динамического стереотипа как у оперированных, так и у интактных животных наблюдалось укорочение латентного периода ИР (до 150±25 мс), а сама двигательная реакция имела характер быстрого устойчивого движения. Экстренная отмена положительного раздражителя в начальной фазе выполнения ответа не прерывала его осуществление - выжим и удержание педали выполнялись в полном объеме (фиг.3а). При выполнении условнорефлекторного ответа в условиях случайного предъявления положительных и тормозных сигналов у интактных и каудатэктомированных животных наблюдалось достоверное увеличение латентных периодов двигательных реакций (до 0,8-1,1 с). Экстренная отмена положительного раздражителя в начальной фазе выполнения ответа, как правило, прерывала его дальнейшее осуществление (фиг.3б).
При действии положительных и тормозных раздражителей в случайном порядке процент правильных решений выработанного двигательного ответа задачи после каудатэктомии в целом не менялся, однако были отмечены изменения, касающиеся реализации программы выученного движения. Они заключались в появлении периодических (5-10 Гц) фазических движений передней рабочей лапы в начальном периоде осуществления двигательного ответа (фиг.2А, б), соответствующих по своим параметрам и условиям возникновения активной форме тремора - интенционному тремору, инициируемому выполнением целенаправленной двигательной реакции. Появление тремора вызывала увеличение (в 4-5 раз) времени реализации двигательного ответа (фиг.2А, б).
В поздние (2-3 месяца) послеоперационные сроки степень выраженности интенционного тремора, а также частота его появления могла снижаться до 25-30% от числа проб в опыте на фоне постепенного увеличения латентного периода ИР (рис.2А, в и 2Б). Условием стойкой демонстрации интенционного тремора в поздние послеоперационные сроки является повышение исходной мотивации путем частичной пищевой депривации, что приводило к попыткам животного к выполнению задачи с меньшим латентным периодом и, как следствие, вновь вызывали стабильный интенционный тремор рабочей лапы (фиг.2А, б). Важно подчеркнуть, что у этих животных, несмотря на повышение пищевой мотиации, при переходе к правильному чередованию сигналов дифференцировочных сигналов появления интенционного тремора рабочей лапы при осуществлении двигательного ответа не наблюдалось.
На фиг.2А показано влияние каудатэктомии на реализацию инструментальных пищевых рефлексов, где
а - правильное чередование пусковых и тормозных сигналов.
б - предъявление дифференцировочных сигналов в случайном порядке;
в - то же, через 2-3 мес. после каудатэктомии.
Сверху вниз: механограмма двигательного ответа нажатия рабочей лапой на педаль, ЭМГ m. biceps brachii, m. triceps brachii рабочей лапы животного, отметка условного сигнала. Калибровка - 1 сек.
На фиг.2Б показано изменение латентного периода двигательных ответов при предъявлении пусковых и тормозных сигналов в случайном порядке в различные послеоперационные сроки, где
Столбики - число правильных ответов (заштриховано - число ответов с двигательными нарушениями), %.
По оси абсцисс - срок после операции, недели.
По оси ординат - величина латентного периода (сек).
Пунктирная линия - средний уровень до операции.
На фиг.2В показаны гистологические срезы мозга оперированных животных.
Стрелки показывают местоположение электролитических повреждений мозга, локализованных в области дорсального отдела головки хвостатого ядра.
На фиг.3. показана реакция животного на экстренную отмену пускового условного раздражителя в начальный период выполнения двигательного ответа при предъявлении положительного и тормозного раздражителей в стереотипном (а), или рандомизированном, случайном (б) порядке. Остальные обозначения, как на рис.2.
Преимущества заявляемого способа:
1. Предлагаемый способ, по сравнению с известными моделями, которые воспроизводят только тремор покоя, позволяет моделировать интенционный тремор действия, инициируемый целенаправленной двигательной реакцией животного.
2. Продуцируемый с помощью предложенного нами способа интенционный тремор воспроизводится после операции в течение длительного времени, не осложнен побочными двигательными и поведенческими проявлениями, характерными для повреждений других отделов экстрапирамидной системы мозга.
3. По сравнению с двусторонними разрушениями бледного шара и других отделов экстрапирамидной системы, операция селективного одностороннего повреждения дорсального отдела хвостатого ядра, минимизирует травматичность животных и исключает возможность летального исхода животных.
Источники информации
Бехтерева Н.П. с сотр. Физиология и патофизиология глубоких структур мозга человека. Л-., М., 1967.
Буреш Я., Бурешова О., Хьюстон Д.П. Методика и основные эксперименты по изусению мозга и поведения. М., Высшая школа, 1991, 398 с.
Лагутина Н.И., с соавт. Сравнительная патология подкорковых образований в свете моделирования гиперкинезов // Журн. невропатол. и психиатр, им. С.С. Корсакова, 1967, т.67, №3, с.368.
Петелин Л.С. Экстрапирамидные гиперкинезы. М., Медицина, 1970, 260 с.
Andrew C.J. Influence of dystonia on the response to long-term L-DOPA therapy in Parkinson's disease // J. of Neurol. Neurosurg. & Psychiatry, 1973, 36: 630-636.
Benita M., et. al Effects of ventrolateral thalamic nucleus cooling on initiationof forelimb ballistic flexion movements by conditioned cats // Exp. Br. Res., 1979, 34: 435-452.
Denny-Brown D., Yanagisawa N. The role of basal ganglia in the initiation of movements // In: the Basal Ganglia (ed. By Yahr M.D.), Raven Press, NY, 1976: 113-159.
Divac I., Markowitch H.J., Pritzel M. Behavioral & anatomical cosequences of small intrastriatal injections of kainic acid 9n the rat // Brain Res., 1978, 151: 523-532.
De Long M.R., Wichmann T. Basal ganglia-thalamocortical circuits in Parkinsonian signs // Clin. Neurosci., 1993, 1: 18-26.
Critchley M. Observation on essential tremor // Brain, 1949, v.72, p.113-139.
Folkerts J., Spiegel E. Tremor on stimulation of the midbrain tegmentum // Confin. Neurologica. 1953, 13.
Fabre M., Buser P. Structures involved in acquisition and performance of visually guided movement in cats // Acta Neurobiol. Exp., 1980, 40: 95-116.
Hassler R. Striatal control of locomotion, intentional actions and of integrations and perceptual activity // J. of Neurol. Sci., 1978, 36: 187-224.
Hayashi A., et. al. Disorder in reciprocal innervation upon initiation of voluntary movement in patients with Parkinson's disease // Exp. Br. Res., 1988, 70: 437-440.
Kaelber W.W. Tremor at rest from tegmental lesion in the cat // J. Neuropath. Exp. Neurol., 1963, v.22, p.695.
Miklayeva E.I., et. al. Differences in the recovery rate of a learned forelimb movement after ablation of the motor cortex in right and left hemisphere in white rats // Behav. Br. Rews., 1993, 56: 145-154.
Peterson E.W., et. al. Production of postural tremor // J. Neurophysiol., 1949, v.61(2): 371-384.
Poirier L.J., et. al. Neuroanatomical study of an experimental postural tremor in monkeys // J. Neurosurg., 1966, v.24, №2, p.191-193.
Schult: Q. Depletion of DA in the striatum as an experimental model of parkinsonism: direct effects and adaptive mechanisms // Progr Neurobiol., 1982, 8: 121-166.
Villablanca J.R., et. al. Effects of caudate nuclei or frontal cortical ablations in cats // Exp. Neurol., 1976, 52: 389-420.
Ward A.A., McCulloch, Magoun H.W. Production of an alternating tremor at rest in monkeys. J. of Neurophysiology, 1948, v.11, №4, p.317-330.
Wickens J.R., et. al. Two dynamic modes of striatal function under dopaminergic-cholinergic control: stimulation and analysis of a model // Synapse, 1991, 8: 1-12.
Изобретение относится к экспериментальной медицине и физиологии и предназначено для моделирования интенционного тремора в эксперименте на кошках. Для этого проводят стереотаксическое повреждение экстрапирамидной системы мозга путем избирательного одностороннего электролитического повреждения дорсального отдела головки хвостатого ядра, контрлатерального конечности животного, осуществляющей целенаправленное движение. Тестирование двигательной активности производят путем подачи зрительных дифференцировочных сигналов, требующих выполнения выработанного двигательного ответа в виде нажатия передней (рабочей) конечностью на педаль, подкрепляемого вознаграждением, или воздержания от его осуществления. Число пусковых зрительных сигналов, требующих двигательного ответа, и тормозных зрительных сигналов, требующих воздержания от выполнения ответа, в тестируемой последовательности сигналов является равным. Дифференцировочные сигналы предъявляют в случайном порядке. Интенционный тремор выявляют в виде появления быстрых (5-10 Гц) антагонистических фазических движений рабочей конечностью при выполнении двигательной реакции нажатия на педаль, инициируемой подачей пускового зрительного сигнала. Способ обеспечивает моделирование интенционного тремора при снижении травматичности и минимизации побочных эффектов. 3 ил.
Способ моделирования интенционного тремора в эксперименте на животных (felis cattus) путем стереотаксического повреждения экстрапирамидной системы мозга и последующим тестированием двигательной активности, отличающийся тем, что повреждение мозга осуществляют путем избирательного одностороннего электролитического повреждения дорсального отдела головки хвостатого ядра, контрлатерального конечности животного, осуществляющей целенаправленное движение, при этом тестирование двигательной активности производят путем подачи зрительных дифференцировочных сигналов, требующих выполнения выработанного двигательного ответа в виде нажатия передней (рабочей) конечностью на педаль, подкрепляемого вознаграждением, или воздержания от его осуществления, а число пусковых зрительных сигналов, требующих двигательного ответа, и тормозных зрительных сигналов, требующих воздержания от выполнения ответа, в тестируемой последовательности сигналов является равным, причем дифференцировочные сигналы предъявляют в случайном порядке, при этом интенционный тремор выявляют в виде появления быстрых (5-10 Гц) антагонистических фазических движений рабочей конечностью при выполнении двигательной реакции нажатия на педаль, инициируемой подачей пускового зрительного сигнала.
| WARD A.A | |||
| et al | |||
| Production of an alternating tremor at rest in monkeys | |||
| J | |||
| of Neurophysioligy, 1948,11,4, pp 317-330 | |||
| СПОСОБ МОДЕЛИРОВАНИЯ ПСИХОТИЧЕСКОГО РАССТРОЙСТВА | 2004 |
|
RU2366464C2 |
| СПОСОБ ЛЕЧЕНИЯ ПАРКИНСОНИЗМА | 2008 |
|
RU2381818C2 |
| CN 101280011 A, 08.10.2008 | |||
| US 7535456 B2, 19.05.2009 | |||
| ЯКИМОВСКИЙ А.Ф | |||
| Структурно-топические основы пикротоксинового хореомиоклонического гиперкинеза | |||
| - Бюл | |||
| эксперим | |||
| биологии | |||

