Изобретение относится к области медицинской и молекулярной генетики, преимущественно к области регуляции экспрессии генов и может быть использовано, например, для тестирования веществ, влияющих на процесс старения с использованием способа, основанного на применении специальных нуклеосомных матриц и элонгационного комплекса РНК-полимеразы (ЭКРНКП). При внесении различных агентов на матрицы может происходить модулирование процесса транскрипции, продукты которой детектируются. Способность изменять активность транскрипции позволяет соответственно маркировать тестируемые вещества как влияющие на процесс старения.
Поддержание правильной генетической информации (ДНК последовательность), а также эпигенетических и регуляторных маркеров (модификация ДНК и гистонов) во время осуществления процессов репликации и транскрипции ДНК является необходимым для выживания и нормального функционирования клетки. Нуклеосомная упаковка геномной ДНК вокруг гистоновых белков, формирующая хроматин, играет важную роль в регуляции активности генома [1-2]. Плотно упакованные хроматиновые структуры снижают доступ к ДНК и, соответственно, ограничивают экспрессию генов и повышают геномную стабильность, в то время как частичная декомпактизация хроматина способствует нерегулируемой экспрессии и геномной нестабильности. Таким образом, нуклеосомы структурные единицы хроматина - содержащие октамер гистонов Н2А/Н2В, Н3 и Н4 и накрученную вокруг гистонового октамера ДНК, является ключевым регулятором процесса транскрипции [3-4]. Известно, что гистоновые белки являются объектами специфических пост-трансляционных модификаций (ацетилирование, фосфорилирование и метилирование и др.), которые коррелируют с активностью транскрипции [5-6]. Ряд работ, включая наши исследования, подтвердили, что гистоновое эпигенетическое кодирование критическим образом влияет на процесс корректного формирования элонгационного комплекса РНК-полимеразы на нуклеосомах [7-8]. Полученные нами данные дают основание предполагать, что во время транскрипции через нуклеосомы РНК полимеразой 2 запускается механизм, включающий формирование промежуточного интермедиата (так называемой как ⌀-петли), который облегчает прохождение РНК полимеразы через нуклеосому [9]. Нарушения в системах, контролирующих сохранение хроматина в процессах репликации и транскрипции, в том числе нарушение образования интермедиатов, приводят к развитию различных патологий, включая опухолеобразование и преждевременное старение клетки [10-11]. Таким образом, нуклеосомные поверхности можно использовать в качестве основы для разработки спосособа скрининга агентов, вызывающих изменение эпигенетического состояния и транскрипционной активности клетки. К таким веществам, могут относится противо- и про-опухолевые агенты, а также вещества, влияющие на процесс старения клетки. Что в свою очередь, будет способствовать разработке лекарственных средств для терапии опухолей различного генеза, а также предотвращении преждевременного старения клеток.
Известен способ использования нуклеосом в качестве биосенсоров [12]. Подобный биосенсор состоит, по крайней мере, из одной нуклеосомы состоящей из ДНК-регулирующего транскрипционного элемента, в котором ДНК помечена двумя метками (а гистоновый октамер остается немеченым). Биосенсор имеет две метки и может образовывать донорно-акцепторную пару при сближении. Метки вводят на расстоянии примерно 30 пар оснований друг от друга. Если ДНК сворачивается вокруг гистонов (образуется нуклеосомная структура), то метки сближаются, и происходит передача энергии с донора на акцептор. Если нуклеосомная структура не формируется, метки остаются разделенными на расстоянии, передачи энергии не происходит. Таким образом, метки маркируют, когда элемент имеет нуклеосомную конфигурацию. Эта система может быть использована для высокопроизводительного скрининга определенных веществ. Способ имеет ограниченное применение для определения агонистов и антагонистов ядерных рецепторов. Для этого в качестве ДНК-района используют последовательность, взаимодействующую с лигандом ядерных рецепторов [12]. Однако использование этой системы для детекции изменений в структуре хроматина во время транскрипции или репликации не проводилось и требует ее существенной доработки. Кроме того, эта дорогостоящая система.
Существует также способ диагностики патологических состояний пациентов, например опухоли или аутоиммунные заболевания, основанный на анализе нуклеосом, выделенных из образцов пациентов [13]. Аминокислотные концы белков нуклеосомы могут быть изменены в результате пост-трансляционных модификаций посредством добавления и/или отсоединения химических групп - метиловой, ацетиловой и фосфорной. Подобные модификации играют ключевую роль в поддержании хроматиновой структуры и транскрипционной активности. Было также показано, что профиль эпигенетических маркеров отличается между нормальным и патологическим состоянием, что может служить основой для разработки диагностики различных заболеваний. Выявление модифицированных нуклеосом осуществляется с помощью методов иммунохимического анализа с применением антител, направленных против уникальных эпитопов нуклеосомных белков. Ограничение предложенного способа заключается в том, что с его помощью проводится только диагностика наличия патологических изменений в нуклеосомах. Но с его помощью нельзя проводить скрининг потенциальных лекарственных веществ, обладающих антиопухолевым, антивозрастным и другими потенциалами.
Известен также ряд способов определения агентов, модулирующие активность гистон-модифицирующих ферментов, таких как ацетилазы, деацетилазы, метилтрансферазы, деметилазы, киназы и пр [14-17]. Например, метод [17] состоит из реконструированных, иммобилизованных нуклеосом и флуоресцентных способов анализа продуктов реакции. Способ состоит из внесения тестируемого агента в смесь, содержащую нуклеосомы и один (или более) гистон-модифицирующих ферментов. В случае взаимодействия агента с функциональными группами гистон-модифицирующих белков, их активность меняется, что можно детектировать флуоресцентным функциональным тестом с использованием реконструированных нуклеосом. Подобные способы также имеют ограниченную применимость, поскольку основан на использовании конкретного механизма нуклеосомной модификации: либо ацетилирование/деацетилирование, метилирование или фосфолирировании гистонов.
Наиболее близким техническим решением (прототипом) является метод, который описан нами ранее [18]. Метод позволяет определять способность веществ различного происхождения стабилизировать или дестабилизировать элонгационный комплекс РНК-полимеразы (ЭКРНКП). На основе предложенного подхода нуклеосомы могут быть использованы для определения агентов, влияющих на процесс опухолеобразования посредством выявления веществ, дестабилизирующих комплекс ЭКРНКП. Ранее нами были разработаны и опубликованы специальные нуклеосомные матрицы, позволяющие контролировать процесс формирования данного комплекса и активность транскрипции [18-24]. Ключевым компонентом данных систем являются ДНК-матрицы, в которых молекулярно-генетическими методами были вставлены высоко-аффинные гистон-связывающие последовательности. Ранее нами были отобраны матрицы - 601, 603, 605 (названия соответствуют названиям клонов-продуцентов [25]), последовательность которых не была гомологична, но все они обладали характерным свойством связывать коровые гистоны с высокой аффинностью и в точно определенных позициях на ДНК. Данные системы успешно воспроизводят различные важные аспекты механизма транскрипции хроматина in vivo, такие как котранскрипционное сохранение/восстановление структуры хроматина, вытеснение/обмен гистонов Н2А/Н2В, высокий нуклеосомный транскрипционный барьер и вытеснение всех гистонов при интенсивной транскрипции. Более подробно строение и получение данных матриц описано в статьях [22-25].
Указанный способ включает внесение тестируемого агента в соответствующем буфере в разработанную систему ЭКРНКП, инкубации и заключительного функционального тестирования. Компоненты для формирования ЭКРНКП брали в субоптимальных соотношениях, а сама система ЭКРНКП находилась в состоянии динамического равновесия. Внесение агентов, связывающихся с одним из компонентов системы и влияющих на его функциональную активность, в конечном итоге приводило к изменению транскрипционной активности и детектировалось. Этот способ выбран в качестве прототипа предложенного решения. Недостаток этого способа заключается в ограниченном применении, а именно в том, его помощью предложено тестировать только противоопухолевые агенты.
В то же время есть ряд данных, которые указывают на то, что система ЭКРНКП может быть чувствительна к процессу старения. Известно, что старение организма сопровождается изменениями в структуре хроматина, повышением геномной нестабильности и увеличением ошибок транскрипции [10, 11, 26]. Ярким примером влияния хроматиновой структуры на старение является вовлечение Sir2/Hst2 гистоновых деацетилаз, играющих ключевую роль транскрипционном отключении. В норме при старении уровень Sir2 снижается [27]. Было показано, что инактивация Sir2 или Hst2 сокращает продолжительность жизни [28], в то время как введение дополнительных копий гена, кодирующего с Sir2 увеличивает ее длительность [29]. Есть ряд исследований, указывающий на тесное взаимоотношение процесса укорачивания теломер (признанного механизма старения клетки) и биосинтеза гистонов [30-32]. Таким образом, структура хроматина является, с одной стороны чувствительным детектором процесса старения клетки, а с другой стороны - ключевым регулятором этого процесса.
Технический результат предполагаемого изобретения заключается в расширении функциональных возможностей использования системы ЭКРНКП для тестирования агентов, влияющих на процесс старения клетки. В более широком плане задачей настоящего изобретения является разработка обоснованного и экспериментально подтвержденного способа тестирования эффективности терапевтических препаратов, направленных на лечение различных патологий, связанных со старением клеток.
Указанный технический результат достигается тем, что применяется способ использования специальных нуклеосомных матриц, включающий сборку нуклеосом из донорного хроматина на ДНК-матрицах, сшивку с ЭКРНКП, активацию элонгации, внесение различных агентов, способных стабилизировать или дестабилизировать компоненты матрицы, проведение транскрипции, оценку продуктов транскрипции и отличающийся тем, что способ используют для тестирования веществ, способных влиять на процесс старения клетки. Тестируемые и контрольные вещества вносят согласно описанному ниже протоколу (см. стр.14 - транскрипционный тест). Любые вещества различного происхождения (высокомолекулярные, низкомолекулярные, экзогенные, эндогенные и пр) приводящие в тестируемой системе к изменениям в структуре хроматина, повышению геномной нестабильности и увеличению ошибок транскрипции будут рассматриваться как вещества ускоряющие процесс старения клетки. Вещества с обратным эффектом, то есть способствующие стабилизации генома и сохранению нормальной структуры хроматина, будут рассматриваться как вещества замедляющие процесс старения клетки. Таким образом, в предлагаемой тестируемой системе возможно осуществление скрининга большого количества веществ с целью последующего отбора лекарственных препаратов, направленных против старения.
Возможен вариант, когда транскрипцию проводят различными РНК полимеразами (РНК полимеразы 2 из различных организмов или РНК полимеразу Е. coli).
Можно использовать также вариант, в котором для сборки нуклеосомных матриц используют очищенные препараты гистонов Н2А, Н2В, Н3, Н4 натурального происхождения.
Существует вариант, в котором для сборки гибридных нуклеосомных матриц используют гистоны различных видов животных Возможно применение варианта, в котором для сборки нуклеосомных матриц используют рекомбинантные гистоны эукариотического и прокариотического происхождения, включая использование коммерческих препаратов фирм INVITROGENE, CLONTECH, STRATAGENE, PROMEGA и др. Рекомбинантные гистоны могут содержать различные модификации и быть очищены различными способами. Также могут быть использованы синтетические гистоны, полученные с помощью автоматических белковых синтезаторов.
Существует также вариант, в котором для сборки нуклеосомных матриц помимо гистонов Н2А, Н2В, Н3, Н4 добавляют линкерный гистон Н1.
Возможен вариант, когда для сборки нуклеосомных матриц используют ДНК, несущую определенные мутации, ассоциированные с различными заболеваниями или со старением.
Возможен вариант, в котором сборка нуклеосомных матриц происходит на твердой поверхности, при этом нуклеосомы оказываются ковалентно или нековалетно прикреплены к твердой поверхности. Твердые поверхности могут быть изготовлены из различных материалов и представлять лунки планшетов, различные сорбенты и пр.
Можно использовать вариант, в котором к твердой поверхности прикрепляют ДНК, а не нуклеосомы.
Возможен также вариант, в котором перед внесением агентов в систему проводят предварительное математическое моделирование.
Существует вариант, в котором в систему вместе с тестируемым агентом вносят транскрипционные факторы, например: TFIIS, FACT, NELF и др. [41-42]. Элонгационный фактор TFIIS. TFIIS - транскрипционный фактор, который облегчает расщепление РНК в активном центре РНКП2 в составе ЭК GreB является прокариотическим аналогом TFIIS. Было показано, что TFIIS облегчает транскрипцию и повышает выход транскриптов в 2-2.5 раза [там же]
Можно использовать вариант, в котором в систему вместе с тестируемым агентом вносят факторы, ремоделирующие хроматин, например SWI/SNF, ISW2, ACF и др. Известен ряд факторов, способствующих процессу транскрипции нуклеосомной ДНК (например, белковый комплекс FACT, нуклеолин, другие шапероны гистонов и АТР-зависимые факторы ремоделирования хроматина), которые влияют на свойства нуклеосом и хроматиновые фибрилл [39-40].
Возможен вариант внесения мутаций в гистоновые белки или использования готовых мутантных конструкций, например Sin-мутантов [8]. Все используемые мутации уменьшали аффинность гистонов Н3 или Н4 к нуклеосомной ДНК. Как следствие, Sin-мутации вызывают диссоциацию гистонов во время транскрипции и, таким образом, нарушают процесс транскрипции хроматина, необходимый для нормальной жизнедеятельности клетки.
Кроме того, можно применять модификацию нуклеосомных матриц, связанную с удалением N-концевых доменов коровых гистонов, что также приводит к локализованному и значительному ослаблению нуклеосомного барьера.
Возможен вариант, в котором оценку продуктов транскрипции или интермедиатов проводят электрофоретически.
Возможен вариант, в котором для оценки продуктов или интермедиатов транскрипции используют кристаллографию.
В современной литературе отсутствуют указания на подобный способ скрининга антивозрастных препаратов с одновременным использованием нуклеосомной матрицы и ЭКРНКП2, которая позволяет оценить функциональную активность и механизм действия препарата.
Следовательно, предложение отвечает критериям новизны и изобретательского уровня.
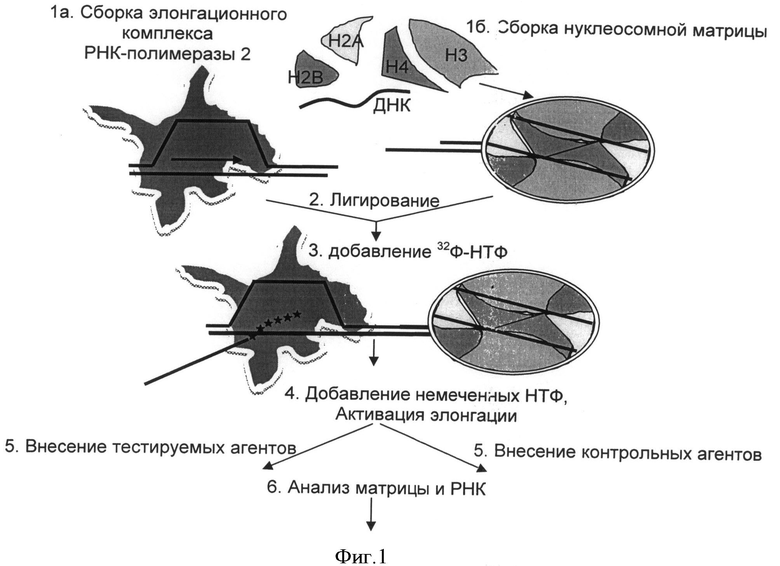
Способ использования специальных функциональных нуклеосомных матриц, для проведения скрининга веществ, способных влиять на процесс старения клетки реализуется следующим образом (фигура 1, на стр.16 заявки приведены методики реализации способа).
Проводят предварительную подготовку всех компонентов смеси. Очистку белков (РНК-полимераз, коровых гистонов, транскрипционных факторов, ремоделеров и пр.) осуществляют согласно опубликованным ранее методам [21-23, 33-35].
Проводят подготовку ДНК-матриц для сборки нуклеосом путем амплификации с заданными праймерами на плазмидах и последующей рестрикцией амплифицированных фрагментов согласно опубликованным ранее методам [21-23, 25]. Для сборки можно использовать различные типы матриц, например - 601, 603, 605. Ранее было показано, что, ДНК-последовательности данных матриц негомологичны, но все они обладали характерным свойством связывать коровые гистоны с высокой аффинностью и в точно определенных позициях на ДНК [21, 24].
В модификациях метода внесение мутаций в матричные последовательности проводят по опубликованным ранее методам [21].
Сборку нуклеосом проводят при субоптимальном соотношении ДНК-матриц и гистоновых белков (в качестве источников гистоновых белков может выступать донорный хроматин, рекомбинантные прокариотические и эукариотические гистоны, синтетические гистоны, а также гистоны, полученные от разных видов животных) с помощью модифицированного метода переноса с использованием продолжительного диализа [21, 25]. Сборку нуклеосом можно выполнять как в растворе, так и на различных сорбентах [21-23, 25].
Оценку качества сборки хроматина осуществляют различными способами, например: электрофорезом в нативных условиях, с помощью определения чувствительности к рестриктазам, с помощью определения чувствительности к рестриктазам и последующего удлинения праймера [22, 23, 34] и др.
Для тестирования действия агентов используют транскрипционный тест. Элонгационные комплексы, содержащие РНК формируют на предсобранных нуклеосомных матрицах согласно описанным ранее протоколам [15-21, 25, 34, 36], инициируют транскрипцию, затем останавливают транскрипцию в момент входа РНК полимеразы в нуклеосому). После этого вносят тестируемые вещества и процесс транскрипции возобновляют. Контролями могут служить следующие образцы: положительный №1 - внесение всех составляющих, без тестируемого агента, положительный №2 - внесение всех составляющих с добавлением фактора FACT, отрицательный №1 - внесение всех составляющих без добавления УТФ. В качестве контролей также можно использовать различные концентраций KCL для изменения условий проведения транскрипции. Известно, что чем выше содержание KCl, тем эффективнее идет процесс транскрипции. Количество транскриптов при 1 М KCl оценивают как 100%.
Для проведения анализа полученных транскриптов и ДНК-гистоновых комплексов используют ряд методов: электрофорез в нативном геле, электрофорез в денатурирующем геле, анализ чувствительности к рестриктазам с последующим проведением электрофореза в нативном геле [21-23, 25], ДНКаза1-футпринтинг [21-23], кристаллографический анализ [37-38] и пр.
Нарушение структуры нуклеосом во время и после транскрипции коррелирует с преждевременным старением, а повышение эффективности этого процесса способно замедлить старение [10-11]. Кроме того, более медленная транскрипция нуклеосомных матрниц (накопление коротких транскриптов) коррелирует с более эффективным сохранением нуклеосом [8, 21]. Поэтому вещества, имеющие анти-возрастную активность, будут приводить к накоплению коротких транскриптов и обеспечивать более эффективное сохранение нуклеосом, которое будет оценено вышеописанными способами. Накопление коротких транскриптов не всегда связано с более эффективным сохранением нуклеосом, поскольку некоторые белковые факторы уменьшают эффективность накопления коротких транскриптов без влияния на эффективность сохранения нуклеосом. Поэтому оценка эффективности сохранения нуклеосом наиболее важна для характеризации веществ, имеющих анти-возрастную активность.
Методики реализации способа
Плазмиды pYP01, pYP02, pYP03, pYP04, pYP06, pYP07, pYP08, pYP09, pGEM-3Z/601, pGEM-SZ/603 and pGEM-3Z/605 и др. [23,24,43] использовали в качестве матрицы для нуклеосомной сборки и транскрипции in vitro. Данные плазмиды содержат последовательность, обладающую высокой аффинностью связывания с гистонами между энхансером и промотором (НПП). Остальная часть плазмиды не содержит какой-либо последовательности ДНК, имеющей высокую аффинность к гистонам, и, соответственно, может служить неспецифическим низкоаффинным конкурентным носителем (своего рода «липучкой») для избытка гистонов и гистоновых октамеров во время хроматиновой сборки. Сборка хроматина на плазмидах может проводиться либо с использованием очищенных гистонов либо донорного хроматина. В обоих случаях присутствие такой «ДНК-липучки» гарантирует полную загрузку НПП и минимальную блокировку промоторов и энхансеров. Фрагменты ДНК из плазмиды амплифицировали помощью праймеров [22, 23, 34] и очищали с помощью набора QIAfilter Plasmid Maxi Kit (Qiagen). Один из праймеров мечен с 5-конца с помощью α-[32]-АТР (Perkin Elmer) и полинуклеотид-киназы (NEB) согласно протоколу фирмы-разработчика. Полученные ДНК фрагменты расщепляли с помощью рестриктазы TspRI.
Гистоновые белки Н2А/Н2В и Н3/Н4 выделяли из крови цыплят с помощью хроматографии на гидрокси-апатите [22-23].
Донорный хроматин из куриных эритроцитов получали согласно методу опубликованному ранее [44]. Кратко: собирали куриные эритроциты с помощью центрифугирования при 1800 g в течение 10 минут при 4°С. Ресуспендировали в стандартном фосфатно-солевом буфере (все буфера были предварительно охлаждены).Собирали при 3000 g в течение 5 минут при 4°С. Повторяли процедуру 3 или 4 раза. Половину осадка ресуспендировали в холодном фосфатно-солевом буфере и затем добавляли буфер A (Buffer А: 15 mM Tris-HCl (pH 7.5), 15 mM NaCl, 60 mM KCl, 0.15 mM spermine, 0.5 mM spermidine, 15 mM β-mercaptoethanol, 0.34 M sucrose, and 0.1 mM PMSF) с добавлением 0.5% NP-40 и смешивали переворачиванием. Собирали ядра центрифугированием при 12,000 g 10 минут при 4°С. Ресуспендировали ядра в буфере А, без NP-40. Снова собирали ядра центрифугированием при 12,000 g 10 минут при 4°С. Повторяли процедуру 3 или 4 раза до исчезновения красного цвета. Ресуспендировали очищенный ядра в небольшом кол-ве буфера А. Затем брали аликвоту (~1 мкл) ядер переносили в 900 мкл НЕ буфера добавляли 100 мкл 10% SDS, и измеряли при А260. Концентрация выделенных ядер должна быть ~200-400 А260/мл.
Из очищенного донорного хроматина удаляли линкерные гистоны Н1.Н5 описанным ранее способом [22-23]. Кратко: разводили хроматин до концентрации 50-100 A260/ml. Промывали сорбент СМ С-25 Sephadex (Pharmacia, 36 mg of resin per mg of chromatin) буфером B (10 mM Tris-HCl (pH 7.5), 350 mM NaCl, 0.5 mM EDTA, and 0.1 mM PMSF.) в течение 1 часа. Медленно добавляли 2М NaCl к хроматину до финальной концентрации 0,35М, затем 1/3 отмытого СМ С-25 Sephadex добавляли к хроматину и медленно помешивали в течении 2 часов при 4°С. Собирали сорбент центрифугированием при 12,000 g 10 минут при 4°С. Удаляли супернатант содержащий растворимый хроматин. Повторяли процедуру 5 или более раз. Проводили анализ полученного -Н1/5 хроматина с помощью 18% полиакриламидного электрофореза в денатурирующих условиях. Затем препарат концентрировали и диализовали в течение ночи против буфера В при 4°С.
Сборку хроматина проводили на полученных фрагментах при их концентрации 10 нМ (50 мкг/ml) и концентрации донорного хроматина 18,75 мкг/мл, (что соответствует соотношению 1:3 хроматин/ДНК) для достижения суб-насыщенного уровня сборки хроматина с помощью модифицированного метода переноса с использованием продолжительного диализа в 1М NaCl. Длинный -Н1-хроматин (лишенный линкерных гистонов) куриных эритроцитов был использован в качестве донора октамеров коровых гистонов. Более подробно: ДНК матрицы и гистоновые белки (или донорный хроматин) смешивали в хроматин-восстанавливающем буфере С (1 М NaCl, 10 мМ Трис-HCl (рН 7.5), 0.2 мМ ЭДТА, 0.1% NP40 and 5 мМ β-меркаптоэтанола) в объеме 120 мкл, и переносили в малые диализные мешки и помещали в стеклянные сосуды, содержащие 100 ml восстанавливающего буфера С.Затем сосуд запечатывали и соединяли с грандиент-мейкером, внутреннюю емкость которого заполняли 500 мл буфера А, а внешнюю - 500 мл буфера D (10 мМ Трис-HCl (рН 7.5), 0.2 мМ ЭДТА, 0.1% NP40 and 5 мМ β-меркаптоэтанола). Диализ выполняли при 4°С при перемешивании в течение ночи со скоростью движения жидкости 1 мл/мин, создаваемой перистальтическим насосом. Присутствие избытка донорного хроматина не приводило к связыванию излишнего количества коровых гистонов. Однако сборку хроматина, используя очищенные гистоны следует проводить очень аккуратно, поскольку даже небольшой избыток очищенных гистонов приводит к полной загрузке промоторов и энхансеров в отсутствие специальной «ДНК-липучки». В таком случае, должна быть добавлена неспецифическая конкурентная ДНК во время сборки.
Оценку качества сборки хроматина проводили несколькими способами: электрофореза в нативных условиях, с помощью определения чувствительности к рестриктазам, с помощью определения чувствительности к рестриктазам и последующего удлинения праймера. Все методы описаны ранее [21-23, 25].
При проведении электрофореза в нативных условиях, комплекс ДНК-белок остается интактным. Подвижность частиц в таком геле определяется зарядом, массой и формой комплексов. Для эксперимента использовали 5'-радиоактивно меченные рестриктные фрагменты плазмид. Состав агарозного геля: 1.2% агарозы, 20 мМ HEPES-Na (рН 8.0), 0.2 мМ ЭДТА, 5% глицерин. Электродный буфер: 20 мМ HEPES-Na (рН 8.0), 0.2 мМ EDTA. Электрофорез проводили на вертикальном аппарате (GibcoBRL V16), используя две стеклянные пластины, разделенные 1 мм спейсером при 120 V в течение 1.5 часов до тех пор, пока бромфеноловый синий не достигнет нижней границы геля. После электрофореза гель переносили на ватманскую бумагу ЗММ, покрывали полиэтиленом и высушивали в течение 20 мин при 50°С и затем в течение 20 мин при 80°С. Высушенный гель экспонировали в течение ночи на PhosphorScreen (Perkin Elmer) и сканировали на Cyclone Phosphorlmager (Perkin Elmer).
Метод оценки сборки хроматина с помощью определения чувствительности к рестриктазам является наиболее простым и быстрым способом оценки «загрузки» ДНК-матрицы нуклеосомами. Метод основан на том, что хроматиновая сборка обеспечивает эффективную защиту нуклеосомной ДНК от действия рестриктаз [45]. Сайты рестрикции должны быть выбраны так, чтобы образовывались ДНК-фрагменты различной длины для облегчения их разделения в агарозном геле и последующего анализа. В методе, представленном ниже, сайты выбирали вне 601-фрагментов плазмиды для оценки степени нежелательной нуклеосомной сборки. Следует отметить, что данная техника может быть использована для любого участка плазмиды [25]. 750 нг ДНК или нуклеосомной матрицы (20 мкг/мл) инкубировали в присутствии избытка рестриктаз (10 ед. акт. каждой) в NEBuffer #2 в течение 2 часов при 37°С. Затем ДНК очищают методом фенол/хлороформной экстракции с последующей преципитацией в этаноле и анализируют с помощью электрофореза в 1% агарозном/ТАЕ геле. Интенсивность полос оценивали количественно с использованием программы OptiQuant (Perkin Elmer). Интенсивность полос, соответствующая финальным продуктам расщепления, уменьшается с возрастанием эффективности хроматиновой сборки. Степень сборки хроматина была прямо пропорциональна уменьшению интенсивности полос. Однако даже в насыщенном хроматине анализируемые сайты рестрикции остаются достаточно доступными, что позволяет предположить, что нуклеосомы плохо формируются на плазмидных участках, не содержащих гистон-связывающие последователи.
Метод оценки сборки хроматина с помощью определения чувствительности к рестриктазам и последующего удлинения праймера также является методом, разработанным нами ранее, и является более поздней модификацией предыдущего метода анализа чувствительности различных участков к рестриктазам.
Данная методика позволяет проводить прямой, количественный анализ высокого разрешения для оценки нуклеосомной загрузки и позиционирования на полинуклеосомных матрицах. Хроматиновые образцы расщепляли в избытке рестриктаз, например AluI, MspI и SeaI, затем проводили фенол/хлороформной экстракцию ДНК с последующей преципитацией в этаноле.
Очищенные ДНК-фрагменты, полученные в результате расщепления AluI, MspI и SeaI, подвергали последующему расщеплению EcoRI рестриктазой для образования 3'-конца. 5'-конец формировали с помощью праймера с последовательностью 5'-gaatttcgagggcatgataacgccttttaggg-3' на EcoRI-расщепленной ДНК. Праймер локализовывался непосредственно перед промотором и был помечен с 5'-конца α-[32]-АТР (Perkin Elmer) с помощью полинуклеотид-киназы согласно инструкции производителя. Затем проводили наращивание цепи с помощью Taq ДНК полимеразы (New England Biolabs) в 1х ThermoPol буфере согласно рекомендациям фирмы-производителя. Полученные продукты очищали методом фенол/хлороформной экстракции с последующей преципитацией в этаноле и анализировали полиакриламидном электрофорезом в денатурирующих условиях. Для этого использовали 8% денатурирующий полиакриламидный гель (19:1) содержащий О.бхТВЕ буфер, 8 М мочевины. Электродный буфер: 0.5х ТВЕ буфер. Электрофорез проводили на вертикальном аппарате (GibcoBRL S2), используя две стеклянные пластины 20×30 см, разделенные 0.4 mm спейсером при 2000 V (мощностью не более 50 ватт) в течение часа до тех пор, пока бромфеноловый синий не достигнет нижней границы геля. После проведения ЭФ оценивали распределение полос в геле, которая отражала чувствительность каждого сайта рестрикции в хроматине к соответствующим ферментам и позволяет проводить количественную оценку загрузки нуклеосомами. Интенсивность полос оценивали с помощью OptiQuant интерфейса (Perkin Elmer). Интенсивность полос соответствовала финальным продуктам расщепления и уменьшалась с увеличением эффективности хроматиновой сборки. Показано, что при 95% хроматиновой упаковке, ДНК становится полностью устойчива к действию рестриктаз, что в свою очередь указывает на полное заполнение ДНК нуклеосомами.
Проведение транскрипционного теста осуществляли следующим образом. Все матрицы линеаризовали по сайтам рестрикции. Комплексы инициации сформированы в 50-мкл аликвотах в ТВ-буфере при концентрации 1 нМ хроматина, 10 нМ коровой RNA полимеразы, 300 нМ σ54, 120 нМ NtrC и 400 нМ NtrB. 5 мкл 40 мМ АТФ в 1х ТВ была добавлена к реакционной смеси, так чтобы финальная концентрация АТФ составляла 4 мМ. Смесь инкубировали при 37°С более 1 минуты для образования открытой формы комплекса инициации. Начинали элонгацию: в реакцию добавляли смесь трех рибонуклеотид-трифосфатов (4 мМ каждого) в 1х ТВ40 (буфер ТВ содержащий 40 мМ KCl) с 2.5 µCi [α-32Ф]-ГТФ (3000 Ci/mmol) и 2 мг/мл гепарина. РНК полимераза останавливается после транскрипции 45 п.н., когда встречает нуклеотид, комплементарный отсутствующему в реакции (УТФ). После этого в реакцию вносят тестируемые вещества вместе с добавлением УТФ. Реакцию продолжали при 37°С в течение 10 минут и останавливали эквивалентным объемом фенол/хлороформа (1:1). Для анализа РНК образцы преципитировали в этаноле, растворяли в формамид-содержащем растворе, денатурировали при 95°С в течение 5 минут, охлаждали на льду и анализировали. Контролями мог служить следующие образцы: положительный №1 - внесение всех составляющих, без тестируемого агента, положительный №2 - внесение всех составляющих с добавлением веществ, заведомо стабилизирующих транскрипционную систему, таких как FACT, транскрипционные факторы и пр., отрицательный №1 - внесение всех составляющих без добавления УТФ. В качестве контролей также можно использовать различные концентраций KCL для изменения условий проведения транскрипции. Известно, что чем выше содержание KCl тем эффективнее идет процесс транскрипции. Количество транскриптов при 1 М KCl оценивают как 100%.
Возможно использование варианта транскрипции, когда собранные ЭКРНКП иммобилизуются на Ni-NTA-агарозе [16]. После интенсивных отмывок комплексы элюировали с Ni-NTA носителя в присутствии 100 мМ имидазола и транскрипцию продолжали в растворе.
Анализ полученных ДНК-гистоновых комплексов можно проводить с помощью анализа чувствительности к рестриктазам с последующим электрофорезом в нативном геле. Нуклеосомы защищают ДНК от расщепления рестриктазами [45]. Сайты рестрикции выбирают таким образом, чтобы продукты полного расщепления имели разную длину и, соответственно, различную подвижность в агарозном геле. Рестрикцию проводили, используя 750 нгДНК или 20 мкг/мл хроматиновой матрицы, которые инкубировали в присутствии избытка рестриктаз (10 единиц каждой). После инкубации образцы анализировали с помощью электрофореза в 4.5% ПААГ. Интенсивность полос оценивали с помощью OptiQuant интерфейса (Perkin Elmer).
Анализ меченных транскриптов проводят с помощью электрофореза в денатурирующих условиях. Высушенный гель экспонировали в течение ночи на PhosphorScreen (Perkin Elmer) и сканировали на Cyclone Phosphorlmager (Perkin Elmer). Данные оценивали с помощи программы OptiQuant. Метод позволяет проводить не только качественную, но и полуколичественную оценку полученных транскриптов.
Анализ полученных ДНК-гистоновых комплексов методом футпринтинга с использованием ДНКазы 1 при конечной концентрации 2.5 мкг/мл меченной с конца матрицы в присутствии 10-кратного избытка -Н1 куриного эритроцитарного хроматина в ТВ-буфере (20 мМ Трис-HCl рН8, 5 мМ MgCl2, 2mM β-меркаптоэтанола, 10 мМ KCl). ДНКаза1 в концентрации 20-50 U/мл была добавлена на 30 сек при 37°С. Реакцию останавливали добавлением ЭДТА. Продукты расщепления были проанализированы в нативном ПААГ и затем экстрагированы из геля, очищены и проанализированы в денатурирующем геле SDS-ПААГ
Моделирование. Для моделирования взаимодействия тестируемого вещества с нуклеосом-РНК-полимераза 2 элонгационным комплексом (НРЭК) можно использовать построенную ранее модель РНКП 2 согласно разработанному протоколу [21] и разрешенным структурам [37, 38]. Электростатическую поверхность белков оценивали и визуализировали с помощью PyMOL скрипта (http://www.pymol.orq) или подобных. Расстояние между отрицательно заряженной поверхностью РНКП2, гистоновым октамером и аминокислотными остатками на поверхности также определяли с помощью PyMOL или подобных программ. Все последовательности белков были взяты из базы данных NCBI (NP_000928.1, NP_010141.1, NP_418415.1, YP_145078.1, and CAB65466.3) (http://www.ncbi.nlm.nih.gov/quide/).
Возможно использование мутантных Н3 и/или Н4 гистонов вместо нормальных аналогов [8]. Sin мутации представляют собой замены одной аминокислоты в Н3 или Н4 гистонах [46]. Нуклеосомы собранные на основе Sin мутантных гистонов демонстрируют ослабление ДНК-гистоновых контактов в области, фланкирующую нуклеосомы. Контакты именно в этой области являются наиболее сильными среди остальных 14 сайтов связывания гистонов с ДНК. Разрушение контактов этой области приводит к разворачиванию матрицы, значительному снижению нуклеосомного барьера для прохождения РНК полимеразы как у дрожжей, так и у людей [8, 46]. Sin-мутантная модельная система удобна не только для тестирования анти-возрастных препаратов, срывающих процесс транскрипции в клетке, но и для изучения самих процессов развития патологических состояний в клетке, которые возникают при нарушении правильности осуществления процесса транскрипции.
Использование факторов, модифицирующих процесс трансрипции [48]. Ремоделирование с помощью ISW2 проводили согласно методу описанному ранее [48]. Кратко: нуклеосомные матрицы (5 нг/мл) инкубировали с 35 нг на 370 нг тотальной ДНК при 300С в течении 30 минут в буфере (25 mM HEPES-KOH (рН 7.6), 5 тМ MgCl2, 40 mM KCl, 0.1 mg/ml BSA, 6 mM Tris-HCl (рН 8.0), 5% glycerol, 30 mM NaCl, 0.3 mM ATP) Затем нуклеосомы анализировали с помощью нативного 4.5% полиакриламидного геля [23].
Таким образом, предложенная заявка предлагает новый способ использования специальных функциональных нуклеосомных матриц, включающий сборку нуклеосом из донорного хроматина на разработанных ранее ДНК-матрицах, активацию элонгации, внесение различных агентов, способных стабилизировать или дестабилизировать компоненты нуклеосомной матрицы, проведение транскрипции, оценку изменения структуры матрицы после транскрипции для скрининга веществ, способных влиять на процесс старения клетки
Выполнение скрининга веществ, влияющих на процесс старения с помощью нуклеосомных матриц, позволит проводить оценку функционального действия препаратов и механизма, лежащего в его основе
Использование комплексной методики в целом, позволит в дальнейшем не только выявлять терапевтические агенты, но и разрабатывать методы, препятствующие и/или замедляющие процессы, связанные с преждевременным старением клетки.
Дополнительные материалы, подтверждающие возможность реализации указанного назначения
Упаковка ДНК в хроматине регулирует все геномные процессы, которые происходят внутри клетки [50]. Плотная упаковка хроматина существенно затрудняет доступ ферментов транскрипции транскрипционных факторов к ДНК. Старение сопровождается изменениями в транскрипционном профиле, хроматиновой плотности и повышенной нестабильности генома [51, 52]. Происходящие изменения в структуре хроматина позволяют объяснить возрастные изменения биологических функций клеток и увеличение заболеваемости состояния с возрастом. Самые ярко выраженные изменения в структуре хроматина заключаются в удалении гистонов с ДНК, которые приводят к формированию более открытой структуры хроматина [53]. Это в свою очередь увеличивает возможность несанкционированного доступа к ДНК ферментов транскрипции и ДНК-повреждающих агентов [54-56]. Как было показано на дрожжах, введение веществ, стабилизирующих количество гистонов на ДНК, увеличивает продолжительность жизни клетки [57]. Подобные результаты получены и в других работах [58, 59]. Трудность использования подобных методических подходов для широкомасштабного скрининга веществ, влияющих на процесс старения заключается в том, что тестирование проводится на живых организмах (дрожжи, нематоды и пр.) или на клетках. Это сопряжено с трудоемкостью и затратностью проведения подобных экспериментов и сложностью последующей интерпретации. Предлагаемая в заявке тест-система представляет собой матрицу, состоящую из гистонов и ферментов транскрипции. Ранее нами было доказано, что данная система воспроизводит все ключевые эффекты транскрипции, проходящей в естественных условиях in vivo [8, 9, 18, 21, 23, 25, 42, 47-48]. При использовании данной системы можно отследить множество эффектов и деталей транскрипции, в частности в контексте предлагаемой заявки важен показатель изменения состояния матрицы (полная или частичная потеря гистонов). Преимущества нашей системы очевидны, поскольку она с одной стороны воссоздает процесс транскрипции в клетке, а с другой - это высокоочищенная система белков и нуклеиновых кислот, которая позволяет проводить количественный и качественный анализ каждого шага транскрипции с помощью использования набора недорогих методов. Что исключает длительную отработку методов интерпретации и анализа продуктов, характерную для более сложных систем. Использование предлагаемой системы позволит проводить широкомасштабный поиск веществ, способных дестабилизировать или наоборот, стабилизировать хроматиновую организацию и процесс транскрипции. Отобранные после тестирования в нашей системе вещества представляют особый интерес, поскольку они с большой вероятностью будут положительно (в случае стабилизации) или отрицательно (в случае дестабилизации) влиять на развитие патологических процессов в клетке, в частности - преждевременного старения.









Изобретение относится к способу скрининга веществ, способных влиять на процесс старения клетки путем использования нуклеосомных матриц. Способ включает в себя сборку нуклеосом из донорного хроматина с использованием очищенных препаратов гистонов Н2А, Н2В, Н3, Н4 натурального происхождения на ДНК-матрицах, где ДНК несет определенные мутации, ассоциированные с различными заболеваниями или со старением, активацию элонгации, внесение различных тестируемых агентов, способных стабилизировать или дестабилизировать компоненты матрицы, проведение транскрипции, оценку продуктов транскрипции. Для оценки продуктов или интермедиатов транскрипции возможно использование различных методов электрофоретического анализа, а также методы кристаллографии. Способность изменять активность транскрипции позволяет соответственно маркировать тестируемые вещества как влияющие на процесс старения. 12 з.п.ф-лы, 1 ил.
1. Способ скрининга веществ, способных влиять на процесс старения клетки путем использования нуклеосомных матриц, включающий:
сборку нуклеосом из донорного хроматина с использованием очищенных препаратов гистонов Н2А, Н2В, Н3, Н4 натурального происхождения на ДНК-матрицах, где ДНК несет определенные мутации, ассоциированные с различными заболеваниями или со старением,
активацию элонгации,
внесение различных тестируемых агентов, способных стабилизировать или дестабилизировать компоненты матрицы,
проведение транскрипции,
оценку продуктов транскрипции.
2. Способ по п.1, отличающийся тем, что транскрипцию проводят различными РНК полимеразами (РНК полимеразы 2 из различных организмов или РНК полимеразу Е. coli).
3. Способ по п.1, отличающийся тем, что для сборки нуклеосомных матриц помимо гистонов Н2А, Н2В, Н3, Н4 добавляют гистон H1.
4. Способ по п.1, отличающийся тем, что для сборки нуклеосомных матриц используют гистоны различных видов животных.
5. Способ по п.1, отличающийся тем, что для сборки нуклеосомных матриц используют рекомбинантные гистоны эукариотического и прокариотического происхождения, а также синтетические гистоны.
6. Способ по п.1, отличающийся тем, что сборка нуклеосомных матриц происходит на твердой поверхности, при этом нуклеосомы оказываются ковалентно или нековалентно прикреплены к твердой поверхности, которая может быть изготовлена из различных материалов, представлять лунки планшетов или различные сорбенты и пр.
7. Способ по п.1, отличающийся тем, что сборка нуклеосомных матриц происходит на твердой поверхности, при этом к твердой поверхности прикрепляют ДНК, а не нуклеосомы.
8. Способ по п.1, отличающийся тем, что перед внесением агентов в систему проводят предварительное математическое моделирование.
9. Способ по п.1, отличающийся тем, что в систему вместе с тестируемым агентом вносят транскрипционные факторы, например TFIIS, FACT, NELF и др.
10. Способ по п.1, отличающийся тем, что в систему вместе с тестируемым агентом вносят факторы, ремоделирующие хроматин, например SWI/SNF, ISW2, ACF и др.
11. Способ по п.1, отличающийся тем, что для сборки нуклеосомных матриц используют мутантные гистоновые белки (Sin-мутанты и др).
12. Способ по п.1, отличающийся тем, что для оценки продуктов или интермедиатов транскрипции проводят электрофоретический анализ.
13. Способ по п.1, отличающийся тем, что для оценки продуктов или интермедиатов транскрипции используют методы кристаллографии.
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| BONDARENKO VA ET AL | |||
| Nucleosomes can form a polar barrier to transcript elongation by RNA polymerase II | |||
| Molecular Cell | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| KIREEVA ML ET AL | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Molecular Cell | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Биохимия: учебник для ВУЗов | |||
| / Под ред | |||
| Северина Е.С | |||
| - М.: ГЭОТАР-МЕД, 2004, с.162-170. | |||

