ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ПАТЕНТНЫЕ ЗАЯВКИ
[0001] Настоящая заявка испрашивает приоритет согласно предварительной заявке на патент USSN 61/406954, поданной 26 октября 2010, и USSN 61/455808, поданной 26 октября 2010, обе указанные предварительные заявки полностью включены в настоящую заявку посредством ссылки для всех целей.
ЗАЯВЛЕНИЕ О ФИНАНСИРУЕМЫХ ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА ИССЛЕДОВАНИЯХ
[Не применимо]
УРОВЕНЬ ТЕХНИКИ
[0002] Взрослые стволовые клетки имеют чрезвычайно большое значение для долговременного поддержания тканевого гомеостаза на протяжении всей жизни. Пролиферативная способность этих клеток, обеспечивающая их самообновление, включает многочисленные тесно связанные процессы, которые поддерживают целостность генома во время деления клетки. Регуляторные механизмы, лежащие в основе старения таких клеток, еще не вполне установлены. Тем не менее, результаты глобальных исследований экспрессии генов стволовых клеток, выделенных у молодых и старых мышей, указывают на участие эпигенетических механизмов в регуляции динамики высокоорганизованного хроматина. Эти исследования показали наличие скоординированной возрастной регуляции активности участков хромосом, перестроек хроматина и генов, определяющих линию дифференцировки (Chambers et al. (2007) PLoS Biol., 5: e201; Rossi et al. (2007) Exp. Gerontol, 42: 385-390; Rossi et al. (2005) Proc. Natl. Acad. Set USA, 102: 9194-9199).
[0003] Все клетки постоянно подвергаются воздействию эндогенных и экзогенных источников повреждения ДНК, в зависимости от характера повреждения они активируют различные механизмы репарации повреждений ДНК (Sinclair et al. (2004) Am. Nat., 164: 396-414). Одновременно клетки активируют механизмы так называемых контрольных точек, которые задерживают прогрессирование клеточного цикла до тех пор, пока целостность генома не будет восстановлена (Shiloh (2001) Curr. Opin. Genet. Dev., 11: 71-77). Согласно одному аспекту гипотезы, о том, что старение организма вызвано старением стволовых клеток, постепенная и скоординированная возрастная потеря способности этих клеток восстанавливать повреждения ДНК приводит к их накоплению с течением времени. Повреждения ДНК будут представлять значительную угрозу для выживания взрослых стволовых клеток из-за изменения паттернов пролиферации и дифференцировки, в конечном итоге приводя к запуску процесса клеточного старения. Таким образом, способность взрослых стволовых клеток отслеживать и эффективно восстанавливать повреждения ДНК имеет ключевое значение для профилактики старения и неопластического перерождения клеток.
[0004] Немного известно о прямой взаимосвязи хроматина и факторов репарации ДНК. Более 50% генома человека состоит из ретротранспозонов (Lander et al. (2001) Nature, 409: 860-921). Из-за высокой степени консервативности последовательности их эпигенетическая структура плохо известна и недостаточно описана на геномном уровне. Фактически многие ретротранспозоны произошли из генов РНК предковых видов и могут представлять генетически активные последовательности, которые кодируют различные типы РНК с еще не известными функциями (McClintock (1956) Cold Spring Harb. Symp. Quant. Biol., 21: 197-216). Однако существуют весомые доказательства того, что ретротранспозонная часть генома оказывает существенное влияние на структуру, целостность и эволюцию генома и транскриптома хозяина (Han et al. (2004) Nature, 429: 268-274; Kazazian (2004) Science, 303: 1626-1632). Растущее число доказательств свидетельствует о том, что во время эволюции млекопитающих большое количество древних ретроэлементов приобрело регуляторные или структурные функции.
[0005] Большая часть ретротранспозонов экспрессируется чрезвычайно сложным образом, который зависит от типа клетки или ткани, и потенциально обеспечивает богатый источник не кодирующих белки РНК, которые направляют клеточную дифференцировку и развитие многоклеточных организмов {Amoral et al. (2008) Science 319: 1787-1789; Birney et al. (2007) Nature, 447: 799-816; Denoeud et al. (2007) Genome Res., 17: 746-759; Dinger et al. (2008) Genome Res., 18: 1433-1445; Dinger et al. (2008) J. Mol. Endocrinol., 40: 151-159; Emanuelsson et al. (2007) Genome Res., 17: 886-897; Faulkner et al. (2009) Nat. Genet., 41: 563-571; Lowe et al. (2007) Proc. Natl. Acad. Sci. USA, 104: 8005-8010; Mattick et al. (2009) Bioessays, 31: 51-59; Mercer et al. (2008) Proc. Natl. Acad. Sci. USA, 105: 716-721; Mikkelsen et al., ed. (2007) Genome-wide maps of chromatin state in pluripotent and lineage-committed cells; Rozowsky et al. (2007) Genome Res., 17: 35 732-745; Trinklein et al. (2007) Genome Res., 17: 720-731). Недавние исследования доказали, что транскрипционная активность ретротранспозонов запускает и управляет такими процессами как: (I) сборка центромерного хроматина, (II) транскрипция генов, (III) компартментализация хроматина, (IV) организация изолирования хроматина при инактивации Х-хромосомы в ядре. Ретротранспозоны также выполняют различные функции при неслучайных хромосомных транслокациях в клетках опухолей (Allen et al. (2004) Nat. Struct. Mol. Biol., 11: 816-821; Chueh et al. (2005) Hum. Mol. Genet., 14: 85-93; Lei and Corces (2006) Cell, 124: 886-888; Lei and Corces (2006) Nat. Genet., 38: 936-941; Lin et al. (2009) Cell, 139: 1069-1083; Lunyak (2008) Curr. Opin. Cell Biol., 20: 281-287; Lunyak et al. (2007) Science, 317: 248-251; Mattick et al. (2009) Bioessays, 31: 51-59; Navarro et al. (2009) Epigenetics Chromatin 2: 8).
[0006] Ретротранспозонная часть генома характеризуется также значительной вариабельностью процесса метилирования ДНК и ковалентных модификаций хроматина, в зависимости от типа ткани, стадии развития или текущего заболевания (Kondo and Issa (2003) J. Biol. Chem. 278: 27658-27662; Estecio et al. (2007) PLoS ONE 2: 15 e399). В полногеномном исследовании, выполненном Martens (Martens et al (2005) EMBO J., 24: 800-812), показано, что длинные рассеянные повторы (LINEs), короткие рассеянные повторы (SINE)/Alu-элементы и другие рассеянные ретротранспозоны имеют переменную степень метилирования гистонов H3K9, H3K27 и H4K20, что обуславливает вероятность того, что посттранскрипционные модификации (РТМ) хроматина ретротранспозонов могут влиять на разнообразные клеточные процессы.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0007] Эффективное восстановление двунитевых разрывов ДНК и поддержание аутентичности генома на уровне хроматина имеет основополагающее значение для корректного самообновления взрослых стволовых клеток человека. Старение стволовых клеток может быть связано с дефектами этих двух процессов. В одном примере мы показываем, что в самообновляющихся взрослых стволовых клетках примерно 65% повреждений естественного происхождения, которые могут быть восстановлены, происходят в мобильных элементах (МЭ). Повышающая регуляция активности транскрипции с ретротранспозонов SINE/Alu препятствует перемещению комплексов конденсина I и когезина в перицентрический хроматин, что приводит к потере эффективности репарации ДНК, что, в свою очередь, приводит к старению. Стабильный общий нокдаун транскриптов ретротранспозонов SINE/Alu в стареющих взрослых стволовых клетках человека восстанавливает способность клеток к самообновлению и, что явилось неожиданным фактом, увеличивает их пластичность, что проявляется в виде повышения экспрессии Nanog и Oct4. Наши результаты демонстрируют функциональное значение ретротранспозонов SINE/Alu и предоставляют возможность механистического понимания их новой роли в качестве промежуточного звена, обеспечивающего взаимодействие между хроматином, репарацией ДНК и старением взрослых стволовых клеток человека.
[0008] Согласно некоторым вариантам реализации в настоящем изобретении предложены способы восстановления нестареющего фенотипа или аспектов нестареющего фенотипа в стареющей клетке (например, стареющая взрослая стволовая клетка). Согласно некоторым вариантам реализации в настоящем изобретении предложены способы поддержания нестареющего фенотипа или аспектов нестареющего фенотипа в клетке (например, стареющей взрослой стволовой клетке). Согласно некоторым вариантам реализации в настоящем изобретении предложены способы индукции и/или восстановления, и/или поддержания нестареющего фенотипа, или его аспектов (например, пролиферативная способность и/или плюрипотентность) в клетке млекопитающего. Способы в общем включают снижение уровня или активности транскриптов ретротранспозонов SINE/Alu в клетке до количества, которое является достаточным для индукции или восстановления способности к пролиферации и/или плюрипотентности указанной клетки млекопитающего.
[0009] Согласно некоторым вариантам реализации в настоящем изобретении предложены способы трансдифференцировки клетки млекопитающего из клетки первого типа или линии в клетку второго типа или линии. Этот способ обычно включает превращение (трансформацию) дифференцированной клетки в плюрипотентную клетку (или восстановление плюрипотентности стволовой клетки или клетки-предшественника) одним из описанных здесь способов (например, путем снижения уровня или активности транскриптов ретротранспозонов SINE/Alu в клетке); культивирование клетки в условиях, которые индуцируют или допускают дифференцировку клетки; отбор клеток, которые дифференцировались в клетки второго типа или линии; и культивирование клеток второго типа или линии. Согласно некоторым вариантам реализации изобретения клетки первого типа представляют собой 30 мезодермальные клетки. Согласно некоторым вариантам реализации изобретения клетки первого типа представляют собой эктодермальные клетки. Согласно некоторым вариантам реализации изобретения клетки первого типа представляют собой энтодермальные клетки. Согласно некоторым вариантам реализации изобретения клетки первого типа или линии представляют собой мезодермальные клетки, и клетки второго типа или линии представляют собой нейроэктодермальные клетки. Согласно некоторым вариантам реализации изобретения клетки первого типа представляют собой эктодермальные клетки, и клетки второй линии представляют собой мезодермальные или энтодермальные клетки. Согласно некоторым вариантам реализации изобретения клетки первого типа представляют собой энтодермальные клетки и клетки второй линии представляют собой эктодермальные или мезодермальные клетки. Согласно некоторым вариантам реализации изобретения клетки первого типа представляют собой адипоциты или клетки костного мозга. Согласно некоторым вариантам реализации изобретения клетки второго типа представляет собой тип клеток, выбранный из группы, состоящей из клетки крови, эмбриональной клетки, эпителиальной клетки, адипоцита, гладкомышечной клетки, нейрональной клетки, бета-клетки поджелудочной железы и кардиомиоцита. Согласно некоторым вариантам реализации изобретения культивирование клетки в условиях, которые индуцируют или допускают дифференцировку указанной клетки, включает культивирование клетки в среде, не содержащей или с пониженным содержанием фактора ингибирования лейкемии (LIF), и/или приведение в контакт (или культивирование) клетки с одним или более реагентами (например, ретиноевой кислотой, тромбоцитарным фактором роста (PDGF), инсулином, арктигенином, транс-ретиноидной кислотой (витамин А), босвеллиевой кислотой, бромелайном и другими протеолитическими ферментами, фенетиловым эфиром кофейной кислоты (САРЕ), флавоноидами (включая апигенин, лютеолин, кверцетин, генистеин и даидзеин), эмодином, эйкозапентаеновой (ЭПК) и докозогексаеновой (ДГК) кислотами, монотерпенами, ресвератролом, 1,25-D3 (витамин D3)), которые индуцируют дифференцировку.
ОПРЕДЕЛЕНИЯ
[0010] МикроРНК представляют собой одноцепочечные РНК, которые обычно имеют в длину 22 нуклеотида и образуются из молекул шпилечных РНК-предшественников, включающих приблизительно ~70-нуклеотидов, под действием нуклеазы семейства РНКазы III известной как Dicer. Так же, как и малые интерферирующие РНК (миРНК), микроРНК могут ингибировать активность генов посредством разрушения гомологичных мРНК у растений или блокировки трансляции мРНК у растений и животных.
[0011] Малая шпилечная РНК (мшРНК или shRNA) - это молекула РНК, которая содержит смысловую нить, антисмысловую нить и короткую петлевую последовательность между смысловым и антисмысловым участками. Благодаря комплементарности последовательностей смыслового и антисмыслового участков такие молекулы РНК, как правило, образуют двухцепочечные РНК, имеющие форму шпильки (дцРНК). Малую шпилечную РНК клонируют в вектор, который обеспечивает экспрессию за счет промотора РНК-полимеразы III (pol III). Экспрессированная мшРНК затем экспортируется в цитоплазму, где она расщепляется под действием Dicer с образованием молекул миРНК, которые далее встраиваются в комплекс РНК-индуцируемого сайленсинга генов (RISC).
[0012] Малые интерферирующие РНК (миРНК) обычно представляют собой 21-23 нуклеотидные двухцепочечные молекулы РНК. После включения в комплекс РНК-индуцируемого сайленсинга генов они облегчают расщепление и деградацию комплементарных им мРНК.
[0013] Piwi-взаимодействующие РНК (пиРНК) - это класс малых некодирующих молекул РНК, которые экспрессируются в клетках животных или могут быть введены в них (см., например Seto et al. (2007) Molecular Cell, 26(5): 603-609; Siomi et al. (2011) Nat. Rev. Mol. Cell Biol., 12:246-258). Взаимодействуя с белками семейства Piwi, молекулы пиРНК образуют РНК-белковые комплексы. Было показано, что пиРНК-содержащие комплексы участвуют в эпигенетическом и посттранскрипционном сайленсинге генов ретротранспозонов и других генетических элементов.
[0014] В настоящей заявке термин «плюрипотентность» и «плюрипотентные стволовые клетки» означает, что такие клетки обладают способностью дифференцироваться во все типы клеток в организме. Плюрипотентные клетки характеризуются экспрессией одного или более маркеров плюрипотентности, известных специалисту в данной области. Указанные маркеры включают, но не ограничиваются ими, щелочную фосфатазу, SSEA3, SSEA4, Sox2, Oct3/4, Nanog, TRA160, TRA181, TDGF 1, Dnmt3b, FoxD3, GDF3, Cyp26al, TERT и zfp42. Согласно некоторым вариантам реализации изобретения плюрипотентные клетки способны формировать или способствовать формированию эктодермальных, мезодермальных и энтодермальных тканей в живом организме.
[0015] В настоящей заявке термины «первичные клетки», «первичные клеточные линии» и «первичные культуры» используют взаимозаменяемо для обозначения клеток и клеточных культур, которые получили от субъекта и выращивали in vitro в течение ограниченного числа пассажей, т.е. разделений культуры. Например, первичные культуры представляют собой культуры, которые, возможно были пассированы 0 раз, 1 раз, 2 раза, 4 раза, 5 раз, 10 раз или 15 раз, но при этом число раз меньше достаточного для перехода критической стадии. Как правило, первичные клетки жировой ткани согласно настоящему изобретению поддерживают в культуре не более 10 пассажей перед использованием.
[0016] В настоящей заявке термины «индивидуум», «субъект», «хозяин» и «пациент» используют взаимозаменяемо и относят к любому млекопитающему, которое нуждается в диагностике, лечении или терапии, в частности, к человеку.
[0017] Под «жировой» понимают любую жировую ткань. Жировая ткань может быть коричневой или белой жировой тканью, полученной из отложений жировой ткани в коже, сальниках/висцеральных отложений, молочной железе, половых органах или других местах. Согласно некоторым вариантам реализации изобретения жировая ткань представляет собой подкожную белую жировую ткань или висцеральную жировую ткань или образец липоаспирата. Жировая ткань может быть получена из любого организма, имеющего жировую ткань. Предпочтительно жировая ткань представляет собой ткань млекопитающего, наиболее предпочтительно, жировую ткань человека. Удобным источником жировой ткани является липосакция, однако источник жировой ткани не ограничивается только этим.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0018] На фигурах 1А-1С приведены показатели, характеризующие старение клеток hADSC (стволовых клеток, полученных из жировой ткани) ex vivo. Фигура 1А: типичная кумулятивная кривая длительного роста. Показаны три различных состояния: SR - самообновление (удвоение популяции <17); preSEN - стадия, предшествующая старению (удвоение популяции 29-38); SEN - старение (удвоение популяции >39). Фигура 1В: иммуногистохимическое определение активности связанной со старением J3-галактозидазы (SA-J3-Gal) и включения 5′-бром-2′дезоксиуридина (BrdU). Примеры морфологических изменений клеток hADSC (10-кратное увеличение) показаны на вставках. Гистограммы соответствуют проценту SA-J3-Gal-положительных клеток (слева) и BrdU-положительных клеток (справа) с прогрессирующим размножением hADSC ex vivo, данные получены на основании трех независимых экспериментов. Планки погрешностей представляют значения стандартного отклонения от среднего. Фигура 1С: ответ на повреждение ДНК (DDR) в стареющих клетках hADSC. Типичный пример окрашивания антителами устойчивых очагов γН2АХ (зеленый)/53ВР1 (красный), формирующихся при старении клеток hADSC. Количественная оценка формирования очагов γН2АХ/53ВР1 при размножении клеток hADSC ex vivo приведена на фиг. 8.
[0019] На фигурах 2A-2G показаны результаты полногеномного анализа локализации γН2АХ. Фигура 2А: схематичное графическое представление анализа нуклеосомной структуры хроматина методом ChIP-seq на платформе SOLiD (ABI). Фигура 2В: относительное хромосомное распределение метки γН2АХ в хромосомах 10 и 2 Самообновляющихся и стареющих клеток. Показаны уровни накопления метки γН2АХ в самообновляющихся и стареющих клетках, которые были рассчитаны как log2 содержания метки в определенном положении, нормированного по неспецифичному фоновому содержанию метки в геноме. Относительная разность уровней обогащения меткой γН2АХ 5 между клетками, рассчитанная как абсолютная величина разности уровней обогащения, специфичных для двух исследуемых типов клеток, показана под дорожками для отдельных клеток. Под дорожками разности показана хромосомная локализация больших кластеров γН2АХ-модифицированных сайтов в самообновляющихся и стареющих клетках. Фигура 2С: различия относительных уровней обогащения γН2АХ в самообновляющихся (SR) клетках по сравнению со стареющими (SEN) для различных функциональных геномных областей. Абсолютные значения нормированной величины разности содержания метки γН2АХ в разных типах клеток показаны на оси ординат. Красные столбики представляют функциональные области генома, которые характеризуются большим процентом включения метки в самообновяющихся клетках (SR), синие столбики представляют функциональные области генома, которые характеризуются высоким процентом включения метки в стареющих клетках. Фигура 2D: диаграммы разброса данных, показывающие зависимость между плотностью генов и плотностью метки γН2АХ в хромосомах человека в самообновляющихся (SR) и стареющих клетках (SEN). На графиках показан наклон и точка пересечения линии линейного тренда с осью ординат (Y-значения), а также коэффициент ранговой корреляции Спирмена (R-значение) и статистический уровень значимости (Р-значение). Фигура 2Е: диаграммы разброса данных, показывающие зависимость между содержанием GC и плотностью метки γН2АХ в хромосомах человека в самообновляющихся (SR) и стареющих клетках (SEN). На графиках показан наклон и точка пересечения линии линейного тренда с осью ординат (Y-значения), а также коэффициент ранговой корреляции Спирмена (R-значение) и статистический уровень значимости (Р-значение). Фигура 2F: уровень обогащения метки γН2АХ в промоторных участках, окружающих сайты инициации транскрипции (TSS), в самообновляющихся (синий) и стареющих (красный) клетках. Уровни обогащения рассчитывают как log2 содержания метки в определенных положениях генома, нормированного к неспецифическому фоновому содержанию метки в геноме. Фигура 2G: показано число генов, содержащих γН2АХ-модифицированные сайты в промоторных участках, определяемых как ±2 тыс. п.о. относительно сайтов, инициации транскрипции, в самообновляющихся (SR) и стареющих клетках (SEN). Число генов с γН2АХ-модифицированными сайтами в промоторных 35 участках, найденное в обоих клеточных фенотипах, указано в области пересечения.
[0020] На фигурах 3А-3Е показано накопление γН2АХ в перителомерных и перицентрических участках хроматина. Фигура 3А: уровни накопления метки γН2АХ в перителомерных участках хромосом самообновляющихся (SR - синий) и стареющих (SEN - красный) клеточных линий. Концы хромосомы (теломеры) показаны в начале оси X, которая затем продлевается на плечи хромосомы. Средний уровень накопления метки γН2АХ вычисляют как log2 содержания метки в определенных положениях генома, нормированного к неспецифическому фоновому содержанию метки в геноме, усредненному по плечам хромосом. Фигура 3В: уровни накопления метки γН2АХ в перицентрических участках самообновляющихся (SR - синий) и стареющих (SEN - красный) клеточных линий. Центромеры показаны как разрыв с центром на оси абсцисс, которая продлевается на плечи хромосом в обоих направлениях. Средние уровни накопления γН2АХ вычисляют как log2 содержания метки в определенных положениях генома, нормированного к неспецифическому фоновому содержанию метки в геноме, усредненному по плечам хромосом. Фигура 3С: статистическая значимость уровней накопления γН2АХ в перителомерных участках хромосом человека, левое и правое плечо, в самообновляющихся (SR - синий) и стареющих (SEN - красный) клетках. Уровни значимости (Р-значения) рассчитывают путем сопоставления уровней накопления метки γН2АХ в перителомерных участках и фонового содержания метки в геноме, и представляют в виде графической зависимости -lnP, где -ln Р=3 (горизонтальная линия) примерно соответствует 95% доверительному интервалу. Фигура 3D: статистическая значимость уровней накопления метки γН2АХ в перицентрических участках хромосом человека в самообновляющихся (SR - синий) и стареющих (SEN - красный) клетках. Уровни значимости (Р-значения) рассчитывают путем сопоставления уровней накопления метки γН2АХ в перицентрических участках и фонового содержания метки в геноме, и представляют в виде графической зависимости -lnP, где -ln Р=3 (горизонтальная линия) примерно соответствует 95% доверительному интервалу. Фигура 3Е: отличие стареющих от самовосстанавливающихся клеток по количеству больших кластеров γН2АХ в перицентрических участках. Абсолютные значения нормированной величины разности содержания кластеров γН2АХ между клеточными фенотипами показаны на оси ординат. Синие столбики соответствуют хромосомам, которые имеют более крупные кластеры в самообновляющихся клетках, красные столбики соответствуют хромосомам, которые имеют более крупные кластеры в стареющих клетках.
[0021] На фигурах 4А-4С показано, что в стареющих клетках hADSC очаги устойчивого повреждения ДНК, связанные с центромерами, являются местом активной транскрипции. Панель А: Иммунофлуоресцентное окрашивание самообновляющихся клеток hADSC. Клетки высевали на покровные стекла и окрашивали антителами к CENP (зеленый) и антителами к 53ВР1 (красный). Окрашивание DAPI показано синим цветом. Изображение типичного интерфазного ядра, полученное методом конфокальной микроскопии, представлено в виде серии снимков с отдельных каналов регистрации и в виде объединенного изображения. Z-срез толщиной 4 мкм анализировали с помощью программного обеспечения Imaris, также показаны изображения для проекций z1, z2 и z3. В самообновляющихся клетках hADSC не обнаружено сайтов, связанных с очагами поражения. Центромерные области отчетливо видны. Панель В: устойчивое повреждение ДНК связано с центромерами. Стареющие клетки hADSC высевали на покровные стекла и проводили иммунное окрашивание, как описано для панели А. Стрелка указывает на совместную локализацию центромерного участки и очагов устойчивого повреждения γН2АХ/53ВР1, связанных со старением. Количественный анализ этих данных и дополнительное комбинированное окрашивание антителами к γН2АХ представлены на фигуре 13. (масштабная полоска 4 мкм). Панель С: связь очагов устойчивого повреждения ДНК при старении с участками с высокой транскрипционной активностью. Комбинированное иммунное окрашивание очагов повреждения ДНК, визуализированных с помощью антител к 53ВР1 (зеленый), с ядерными тельцами PML (синий) и растущей цепью РНК (красный), исследовали методом конфокальной микроскопии. Стареющие клетки hADSC инкубировали с галогенированным предшественником фторуридином в течение 10 мин in vivo, фиксировали и окрашивали антителами. Антитела к BrdU использовали для выявления меченой фторуридином РНК. Показано типичное изображение одиночного ядра. Стрелка указывает на расположение очага повреждения ДНК солокализованного с РНК. На фигуре показано пространственное взаиморасположение сайта включения фторуридина, 53ВР1 и телец PML. Показан один конфокальный срез толщиной 5 мм. Изображение анализировали с помощью программного обеспечения Imaris, показаны плоскости z1, z2 и z3. Картинка демонстрирует ориентацию плоскостей z1, z2 и z3 в пределах одного z-сечения. Конфокальные срезы подтверждают тесную связь растущих транскриптов с сайтами устойчивого повреждения ДНК в стареющих клетках hADSC.
[0022] Фигура 5 демонстрирует, что в стареющих клетках hADSC существует положительная корреляция между устойчивым повреждением ДНК, связанным с центромерами, и активацией транскрипции SINE/Alu ретротранспозонов, а также нарушением привлечения компонентов комплекса когезина и конденсина I. Фигура 5С: Потеря когезина и конденсина I в перицентрическом участке устойчивого повреждения ДНК в стареющих клетках hADSC. ChIP-анализ перицентрических повторов на хромосоме 10 в самообновляющихся (синие столбцы) и стареющих (красные столбцы) клетках hADSC. Повторы оценивали как положения для привлечения TFIIIC, Ecol, а также компонентов комплексов когезина (Rad21) и конденсина I (САР-Н) (n=3, ±SEM). Представлено схематическое изображение субъединиц комплексов когезина и конденсина I, а также рисунок, демонстрирующий установленную ранее функцию Ecol. * р<0,02, ** р<0,2.
[0023] На фигуре 6A-6G показано, что стабильный нокдаун обычного транскрипта ретротранспозонов SINE/Alu в стареющих взрослых стволовых клетках человека восстанавливает их пролиферативные свойства и приводит к развитию ИПК-подобного фенотипа. Фигура 6А: модель ретротранспозона SINE/Alu. Вторичная структура общей РНК SINE/Alu (SEQ ID NO: 1). Участки, формирующие мшРНК, заштрихованы. Фигура 6В: типичный пример эффективности лентивирусной трансдукции hADSCs показан с помощью GFP. Фигура 6С: нозерн-блот гибридизация РНК, выделенной из клеток hADSC, стабильно экспрессирующих мшРНК к SINE/Alu. Стареющие клетки hADSC 1 инфицировали lentiGFP sh-193Alu, lentiGFP sh-132Alu или контрольной конструкцией lentiGFP, не содержащей вставки мшРНК. РНК выделяли через 24 часа после трансдукции, проводили нозерн-гибридизацию с олигонуклеотидами, специфичными в отношении SINE/Alu. Стареющие клетки hADSC, которые стабильно экспрессируют sh-132Alu, продемонстрировали почти полный нокдаун транскриптов ретротранспозонов SINE/Alu. Фигура 6D: после стабильного нокдауна транскриптов ретротранспозонов SINE/Alu в стареющих клетках hADSC пролиферативные свойства клеток восстанавливались. Показан захват [Н3]-тимидина. Стареющие клетки (дикий тип) и стареющие клетки, трансдуцированные lentiGFP (контроль) или lentiGFP sh-132Alu, импульсно метили [Н3]-тимидином с активностью 1 мкКи в течение 24 часов через 24 или 96 ч после инфицирования. Данные представлены как средние значения ± ст.ошиб.ср. для трех измерений. Фигура 6Е: экспрессию маркеров плюрипотентности Nanog и Oct4 измеряли методом кПЦР в стареющих клетках hADSC (дикий тип) и в hADSC с обращением фенотипа старения при стабильном нокдауне транскрипции ретротранспозонов SINE/Alu (lentiGFP sh-132Alu). РНК выделяли из клеток через 96 ч после инфицирования. Результаты представлены как относительное количество (дискретное косинусное преобразование, DCt). Образцы нормировали по β-актину. Данные представлены как среднее значение ± ст.ошиб.ср. (n=3), ***р=6,98е-05, *р=0,03. Фигура 6F: морфологические изменения в клетках hADSC с обращением фенотипа старения при стабильном нокдауне транскрипции ретротранспозонов SINE/Alu. Через 7 дней культивирования без питательной среды клетки образуют GFP-положительные 1 клеточные агрегаты, которые характеризуются положительным окрашиванием на маркер плюрипотентности - щелочную фосфатазу (ЩФ). Фигура 6G: модель, демонстрирующая вмешательство транскриптов ретротранспозонов SINE/Alu в инициирование устойчивого ответа на повреждение ДНК, что приводит к старению взрослых мезенхимальных стволовых клеток человека.
[0024] На фигуре 7А-7В показаны результаты исследования клеток методом проточной цитофлуорометрии и пролиферативные свойства клеток hADSC. Фигура 7А: исследование клеток hADSC методом проточной цитофлуорометрии. Ранние PD hADSC окрашивали конъюгированными с FITC (CD31, CD44 и CD-45) или AlexaFlour-488 (CD105) антителами к поверхностным маркерам клетки и исследовали методом проточной цитофлуорометрии. Клетки имели положительные результаты окрашивания для CD105 и CD 45 и отрицательные для CD 34 и CD 44. Клеточные популяции показаны в виде флуоресцентных сигналов на графиках бокового светорассеяния (вверху) и гистограммах (внизу) распределения окрашенных клеток (синяя линия) по сравнению с неокрашенными клетками (красная линия); также указан процент положительных клеток. Фигура 7В: способность к репликации клеток hADSC снижается со старением ex vivo. Пролиферацию в самообновляющихся (SR), приближающихся к стадии старения (preSEN) и стареющих (SEN) клетках hADSC измеряли по включению [Н3]-тимидина: 1 мкКю добавили к 10000 клеток в 2 мл среды DMEM/F12. После 24-часовой инкубации клетки собирали, выделяли ДНК и измеряли радиоактивность с помощью жидкостного сцинтилляционного счетчика. Результаты представлены как количество [Н3]-тимидина (импульсов в минуту), включившегося в ДНК в процессе синтеза, на 1 мкг выделенной ДНК. ДНК из клеток не обработанных [Н3]-тимидином использовали в качестве контроля фоновой радиоактивности.
[0025] На фигуре 8 показаны результаты количественного анализа накопления очагов устойчивого повреждения ДНК в клетках hADSC, которые перевивали ex vivo. γН2АХ окрашивали аффинно очищенными поликлональными антителами кролика. На гистограмме показан процент клеток, имеющих 1, 2, 3 или более 3 очагов. Типичные примеры приведены ниже. Образование очагов оценивали в самообновляющихся SR (удвоение популяции <17), приближающихся к стадии старения preSEN (удвоение популяции >29-<38) и стареющих SEN (удвоения популяции >39) культурах hADSCs. n=общее число ядер, подсчитанное в 3 независимых экспериментах.
[0026] На фигуре 9 показана активация ответа на повреждение ДНК в стареющих клетках hADSC. Клетки стареющей культуры hADSC окрашивали антителами к фосфорилированной форме Chk1 (S345), Chk2 (Т68) и cdc2 (Tyr 15) (светлее) и DAPI (темнее). Показаны конфокальные срезы толщиной 50 мкм. Chk1 и Chk2 представляют собой трансдукторные киназы, которые действуют ниже каскада киназы ATM/ATR, чтобы обеспечить точку контроля повреждений ДНК. В отличие от повреждений ДНК, индуцированных генотоксическим стрессом или облучением, стареющие клетки hADSC не демонстрируют существенной локализации фосфорилированных форм Chk1 (S345) и Chk2 (Т68) в ядре.
[0027] Фигура 10. Фракции γН2АХ-модифицированных нуклеосом, которые локализованы в различных функциональных геномных областях самообновляющихся (SR, левые столбики) и стареющих (SEN, правые столики) взрослых мезенхимальных стволовых клеток, полученных из жировой ткани (hADSC). Правый хвост распределения увеличен и для ясности показан в виде вставки.
[0028] На фигурах 11А-11С показано сравнение γН2АХ-модифицированных положений мононуклеосомного размера с геномными кластерами γН2АХ-модифицированных нуклеосом. γН2АХ-модифицированные мононуклеосомы и γН2АХ-модифицированные кластеры определяли, как описано в разделе «Дополнительные методы». Фигура НА: распределение частот γН2АХ-модифицированных геномных положений в самообновляющихся (SR) и стареющих (SEN) hADSCs. Доминирующее положение в распределении занимают кластеры мононуклеосомного размера. Соответственно, часть гистограммы, демонстрирующая частоты средних и больших γН2АХ-модифицированных нуклеосомных кластеров, увеличена и показана в виде вставки для удобства наглядного представления. Фигура 11В: процентное отношение функциональных областей генома, занятых γН2АХ-модифицированными мононуклеосомами и большими кластерами γН2АХ-модифицированных нуклеосом. Фигура 11С: показатель относительной энтропии рассчитывали в качестве меры различия 25 распределения частот размеров кластеров γН2АХ между клетками SR и клетками SEN (см. панель рис. 11А.). Соотношение между относительной энтропией и длиной кластера использовали для расчета порогового значения (7400 п.о.), позволяющего разграничить средние и большие кластеры, как описано в разделе «Дополнительные методы».
[0029] На фигуре 12 показано относительное накопление γН2АХ - 30 модифицированных нуклеосом в перителомерных по сравнению с перицентрическими участками генома. Величины накопления рассчитывали как log2 нормированные соотношения содержания метки γН2АХ согласно данным анализа ChIP-seq на одно положение в каждом участке к фоновому содержанию метки в геноме на одно положение. Значения для самообновляющихся (SR) клеток выделены синим цветом, стареющих (SEN) клеток - красным цветом. Содержание γН2АХ в перителомерных участках понижено, тогда как перицентрические участки обогащены γН2АХ.
[0030] На фигуре 13, панели A-D, показаны устойчивые очаги γН2АХ/53ВР1 в стареющих клетках hADSC, связанные с центромерными участками. Панель А: Количественный анализ солокализации CENP-A и 53ВР1 при старении клеток. Стареющие hADSCs окрашивали антителами к CENP-A (зеленый), 53ВР1 (красный) и DAPI (синий). В трех независимых экспериментах провели оценку в общей сложности 200 клеток. Планки погрешностей представляют величину стандартной ошибки среднего. Пример изображения при большем увеличении показан на панели В. Масштабная метка - 1 мкм. Изображения анализировали с помощью программного обеспечения Imaris с представлением оптических сечений, как показано на фигурах 4С и 4D. Панель D: солокализация связанных со старением устойчивых очагов γН2АХ/53ВР1 и кинетохора. Двойное иммунное окрашивание стареющих клетках hADSC антителами к γН2АХ (панель С) или 53ВР1 (панель D) и антителами к внутреннему кинетохору у пациентов с кальцинозом, синдромом Рейно, нарушением моторики пищевода, склеродактилией и телеангиэктазией (далее сокращенно CREST). Масштабные метки показаны для каждого отдельного изображения. Черно-белые изображения, полученные для отдельных каналов регистрации, приведены на панели D для удобства наглядного представления.
[0031] На фигуре 14 показана экспрессия SINE/Alu в клетках hADSC типов SR и SEN. Нозерн-гибридизация РНК самообновляющихся (SR) и стареющих (SEN) клеток hADSC с олигонуклеотидным зондом к SINE/Alu. Указаны полосы, соответствующие SINE/Alu и 7SL. Тотальную РНК массой 2 мкг на полосу загружали, как описано в разделе «Экспериментальные методики». Рибосомные малые РНК можно наблюдать в геле, окрашенном бромистым этидием, для сравнения загрузки. Размеры лесенки маркеров молекулярной массы оцРНК указаны справа.
[0032] Фигура 15А и 15В иллюстрирует нокдаун типичных транскриптов Alu в клетках ADSC, обусловленный лентивирусной мшРНК, и влияние нокдауна на старение клеток и пролиферацию. Фигура 15А схематически иллюстрирует протокол для стабильного нокдауна транскриптов Alu-повторов с использованием мшРНК, которые доставляют в клетку с использованием лентивирусного вектора. Фигура 15В иллюстрирует вектор для доставки, трансфицированные клетки и нозерн-блот с результатами трансфекции.
[0033] На фигуре 16 показано влияние нокдауна обычного транскрипта Alu, вызванного лентивирусной мшРНК, в клетках ADSCs. Пролиферация клеток 15 К заявке №2013119440 представлена как функция времени. Нокдаун транскрипта Alu-повтора восстановил пролиферацию клеток, ранее бывших стареющими.
[0034] На фигуре 17 показана морфология мутантных по Alu мшРНК клеток ADSC в культуре и их начальные характеристики.
[0035] На фигуре 18 показано сравнение «стандартных» протоколов для получения индуцированных плюрипотентных стволовых клеток (ИПСК) и одного из протоколов, описанных в настоящей заявке.
[0036] На фигурах 19А и 19В показана дальнейшая дифференцировка клеток, мутантных по Alu мшРНК. На фигуре 19А показана дифференцировка клеток ADSC в нейробласты. Фигура 19В иллюстрирует применение способов, описанных в настоящей заявке, для трансдифференцировки клеток по другим многочисленным путям дифференцировки или в другие линии.
[0037] На фигурах 20А-20Е показана трансдифференцировка фибробластов почек в нейробласты. На фигуре 20А показаны клетки НЕК 293Т через два. дня после инфицирования sh-132Alu. На фигуре 20В показано образование колонии через 7 дней после выделения одной колонии и разделения на отдельные клетки (среда ES с добавлением LIF). На фигуре 20С показана экспрессия маркеров плюрипотентности (Nanog, oct4 и щелочной фосфатазы). На фигуре 20D показана экспрессия мезодермальных маркеров через два дня после формирования эмбриоидных телец (ЭТ). На фигуре 20Е показана экспрессия остеогенных (остеопонтин), адипогенных (липопротеинлипаза) и глиальных (GAFP) генов через 2 дня после формирования эмбриоидных телец.
[0038] Фигура 21, панели А (слева) и В (справа) иллюстрируют альтернативные модели действия мшРНК Alu-повтора. Панель А: мшРНК к Alu-повтору образует шпилечную мшРНК, которая направляет ядерные белки PIWI, либо эндонуклеазу Dicer к расположению геномного SINE/Alu-повтора, инициируя транскрипционный сайленсинг за счет гетерохроматинизации, которая включает как метилирование ДНК, так и модификацию гистонов. Панель В: мшРНК к Alu-повтору активирует Dicer-зависимый Ago2 путь посттранскрипционного сайленсинга, ведущий к цитоплазматической деградации непроцессированных РНК-транскриптов Alu-повтора.
[0039] На фигуре 22А (вверху) представлен 7SL-консервативный участок в полноразмерной последовательности Alu-повтора. На фигуре 22А (внизу) показана вторичная структура общей полноразмерной вРНК Alu-повтора (SEQ ID NO: 2). Выделенный горизонтально сегмент представляет собой высококонсервативную часть, произошедшую от 7SL, тогда как выделенный вертикально сегмент представляет собой участок, который использовали для аффинного выделения комплексов синтетической РНК и белков-партнеров. На фигуре 22В показана консервативность последовательности мшРНК Alu по сравнению с остальной частью элемента. Средний процент уровня идентичности между рассеянными повторяющимися копиями элементов сравнивают для участков последовательности мшРНК Alu и других участков последовательности Alu четырех подсемейств Alu-повторов. Показатели статистической значимости различий приведены для каждого сравнения. На фигуре 22С показано пошаговое схематическое представление несмещенного аффинного выделения комплексов РНК-белок. На фигуре 22D показан окрашенный серебром денатурирующий 4-12% градиентный бис-трис гель NuPAGE Novex®, загруженный преципитатами после проведения аффинного выделения комплексов РНК-белок, с вырезанными полосами, которые помечены А-Е.
[0040] Фигура 23 представляет численность членов в основных функциональных категориях, схематично показанную в виде изменения интенсивности цвета. Наиболее многочисленные белки в каждой группе отмечены более ярким цветом, который переходит в черный для белков, многочисленность которых равна всего 1.
[0041] На фигуре 24а показаны доверительные уровни для вовлечения выделенных белковых комплексов в различные клеточные процессы, в которых параметры Генной Онтологии (ГО) представлены как -log10 (р-значение) с использованием р-значения, вычисленного с помощью программного обеспечения DAVID. Фигура 24В показывает доверительные уровни для присутствия выделенных белковых комплексов в различных клеточных компонентах, представленное как -log10 (р-значение), вычисленное с помощью программного обеспечения DAVID. Фигура 24С показывает сеть взаимодействий выделенных белков, полученную с помощью программного обеспечения STRING 9.0. Более толстые линии представляют взаимодействия с более высоким доверительным уровнем (экспериментальные данные), тогда как пунктирные линии представляют предполагаемые взаимодействия (более низкий доверительный уровень).
[0042] На фигуре 25 показано схематическое изображение белковых комплексов с использованием параметров генной онтологии (ГО), полученное с помощью программного обеспечения String 9.0. Каждый конкретный клеточный процесс причислен к одной из следующих 5 категорий: транскрипция, репарация ДНК, модификации хроматина, регуляция клеточного цикла и другие.
ПОДРОБНОЕ ОПИСАНИЕ
[0043] Было обнаружено, что в самообновляющихся стволовых клетках значительная часть (например, ~65% в некоторых случаях) образующихся в естественных условиях восстанавливаемых повреждений ДНК происходит в мобильных элементах. Положительная регуляция транскрипционной активности ретротранспозонов SINE/Alu препятствует привлечению комплексов конденсина I и когезина в перицентрический хроматин, что приводит к потере эффективности процесса репарации ДНК и в конечном итоге к старению.
[0044] Было показано, что стабильный нокдаун типичных транскриптов ретротранспозонов SINE/Alu в стареющих взрослых стволовых клетках человека восстанавливает способность клеток к самообновлению и, что явилось неожиданным, увеличивает их пластичность (например, проявляющуюся активацией маркеров плюрипотентности Nanog и Oct4). Результаты, представленные в настоящей заявке, демонстрируют функциональное значение ретротранспозонов SINE/Alu и обеспечивают возможность механистического понимания их новой роли как медиаторов взаимодействия между хроматином, репарацией ДНК и старением взрослых стволовых клеток человека.
[0045] В связи с этим фактом в настоящем изобретении предложены способы индукции, и/или восстановления, и/или поддержания способности к пролиферации и/или плюрипотентности клетки млекопитающего. В целом, способы обеспечивают снижение скорости начала старения или предотвращают старение, или позволяют клеткам избежать старения. Согласно различным вариантам реализации изобретения способы обеспечивают индукцию или восстановление фенотипа дифференцированной и/или стареющей клетки до фенотипа плюрипотентной клетки. Согласно различным вариантам реализации изобретения способы, как правило, включают понижающую регуляцию или ингибирование уровня или активности транскриптов ретротранспозонов SINE/Alu в клетке (см., например, фиг. 18).
[0046] Как показано в настоящей заявке, указанная понижающая регуляция/ингибирование позволяет клеткам (например, взрослым стволовым клеткам) избежать старения, восстанавливая их плюрипотентность и/или способность к пролиферации (см., например, фиг. 6D, 6F 6Е, 15, 16, и 17). Указанная понижающая регуляция имеет несколько способов применения. Например, полагают, что продолжительность существования популяции стволовых клеток в жизнеспособном состоянии (например, линии стволовых клеток) может быть увеличена (возможно, бесконечно) путем понижающей регуляции (например, стабильного снижения активности) транскриптов ретротранспозонов SINE/Alu в этих клетках. При желании стволовые клетки могут быть индуцированы к дифференцировке в эмбриоидные тельца, предшественники или конкретные линии или окончательно дифференцированные клетки в соответствии со стандартными методами, хорошо известными специалистам в данной области техники.
[0047] Соответственно, предложены клетки (включая ранее окончательно дифференцированные клетки млекопитающих, клетки-предшественники, стволовые клетки, линии стволовых клеток и индуцированные плюрипотентные стволовые клетки (ИПСК)), в которых подавлены транскрипты ретротранспозонов SINE/Alu. Согласно различным вариантам реализации изобретения клетки включают нормальные (нестареющие) стволовые клетки, стволовые клетки, которые «избежали» старения за счет понижающей регуляции синтеза или активности транскриптов ретротранспозонов SINE/Alu, индуцированные плюрипотентные стволовые клетки (ИПСК), которые содержат конструкцию, которая отрицательно регулирует уровень или активность транскриптов ретротранспозонов SINE/Alu, эмбриональные стволовые клетки, и т.п.Согласно некоторым вариантам реализации изобретения исключены клетки, которые были обработаны таким образом, чтобы получить ИПСК (например, исключены клетки, в которых активность одного или более из Nanog и/или LIN-28 и/или Oct3/4, и/или Sox2 и/или Klf4 и/или с-myc непосредственно повышена и/или которые содержат гетерологичные конструкции, экспрессирующие Oct3/4, и/или Sox2 и/или Klf4 и/или cmyc).
[0048] Также было показано, что клетки, в которых плюрипотентность была индуцирована и/или восстановлена в соответствии с описанными здесь способами (например, ингибирование ретротранспозонов Alu), могут быть впоследствии индуцированы на дифференцировку с использованием стандартных способов, хорошо известных специалистам в данной области техники (например, удаление LIF из культуральной среды) (см., например, фиг. 19А, 19В и 20А-20Е). Это является еще одним доказательством того, что описанные в настоящей заявке способы обеспечивают индукцию или восстановление плюрипотентности клетки. Соответственно, в некоторые варианты реализации изобретения включены дифференцированные клетки, включающие клетки, которые были индуцированы на дифференцировку из полипотентных стволовых клеток, полученных согласно способам, описанным в настоящей заявке.
[0049] Помимо этого, некоторые варианты реализации изобретения включают способы трансдифференцировки клеток. Способы трансдифференцировки, как правило, включают индукцию или восстановление плюрипотентного фенотипа в клетке согласно описанным в настоящей заявке способам (например, путем ингибирования ретротранспозонов Alu или других компонентов пути) с последующим культивированием полученных плюрипотентных клеток в условиях, обеспечивающих или индуцирующих дифференцировку. Клетки, которые дифференцируются в желаемый тип клеток, (например, бета-клетки поджелудочной железы, мотонейроны, гемопоэтические клетки-предшественники, нервные клетки, дофаминергические нейроны, адипоциты, кардиомиоциты, и т.п.) и/или линии, затем отбирают с возможностью последующего культивирования (например, размножение в культуре) или непосредственного использования.
[0050] Согласно некоторым вариантам реализации способы, описанные в настоящей заявке, могут быть использованы для восстановления плюрипотентности клетки и/или способности клетки к пролиферации (например, клетка, которая детерминирована к дифференцировке по какому-либо пути). Согласно некоторым вариантам реализации способы, описанные в настоящей заявке, могут быть использованы для восстановления клетки, которая демонстрирует один или более признаков старения, до состояния, характеризующегося меньшей степенью старения, или состояния без признаков старения.
[0051] Согласно различным вариантам реализации изобретения стволовая клетка включает клетку, выбранную из группы, состоящей из эмбриональной стволовой клетки, стволовой клетки пуповинной крови, взрослой стволовой клетки и ИПСК. Согласно различным вариантам реализации изобретения стволовая клетка млекопитающего представляет собой стволовую клетку, полученную из ткани, выбранной из группы, состоящей из жировой ткани человека, костного мозга человека, нервной ткани человека, гладких мышц человека, жировой ткани человека, кардиомиоцитов человека, эндотелиальной ткани человека, эпителиальной ткани человека, ткани поджелудочной железы человека, костной или хрящевой ткани человека и других тканей.
[0052] Согласно некоторым вариантам реализации изобретения клетка представляет собой клетку, которая детерминирована к дифференцировке в клетку типа, выбранного из группы, состоящей из эктодермы, мезодермы и энтодермы. Согласно некоторым вариантам реализации изобретения клетка представляет собой клетку, которая детерминирована к дифференцировке в клетку типа, выбранного из группы, состоящей из клеток жировой ткани человека, клеток крови человека, нервных клеток человека, гладкомышечных клеток человека, адипоцитов человека, хондроцитов человека, остеокластов и остеобластов человека, кардиомиоцитов человека, эндотелиальных клеток человека и эпителиальных клеток человека. Согласно некоторым вариантам реализации изобретения клетка включает не способную к самообновлению клетку-предшественника. Согласно некоторым вариантам реализации изобретения клетка млекопитающего включает окончательно дифференцированную клетку.
Понижающая регуляция/ингибирование транскриптов ретротранспозонов SINE/Alu
[0053] Было обнаружено, что ингибирование транскриптов ретротранспозонов SINE/Alu может восстановить пролиферативную способность и/или плюрипотентность в стареющих стволовых клетках или может поддерживать пролиферативную способность и/или плюрипотентность в нестареющих стволовых клетках. Может быть использован любой из различных способов ингибирования транскриптов SINE/Alu ретротранспозонов.
[0054] В различных вариантах реализации изобретения транскрипты ретротранспозонов SINE/Alu можно подавлять/ингибировать с помощью ингибиторной РНК. Ингибиторные РНК, которые можно использовать, включают, но не ограничиваются перечисленными: миРНК, мшРНК, микроРНК, 27-mer дуплексы, являющиеся субстратами Dicer, одноцепочечные интерферирующие РНК и тому подобное.
[0055] Термин миРНК обычно относится к двухцепочечной интерферирующей РНК, если не указано иное. В различных вариантах реализации изобретения молекулы миРНК, которые можно использовать для ингибирования транскриптов ретротранспозонов SINE/Alu, включают молекулы двухцепочечной рибонуклеиновой кислоты, содержащей две нуклеотидные нити, где каждая нить имеет примерно от 19 до примерно 28 нуклеотидов (т.е. примерно 19, 20, 21, 22, 23, 24, 25, 26, 27 или 28 нуклеотидов). Таким образом, выражение «интерферирующая РНК, которая имеет в длину от 19 до 49 нуклеотидов», когда речь идет о двухцепочечной интерферирующей РНК, означает, что антисмысловая и смысловая нити независимо друг от друга имеют в длину от примерно 19 до примерно 49 нуклеотидов, включая молекулы интерферирующих РНК, в которых смысловая и антисмысловая нити соединены линкерной молекулой.
[0056] Помимо молекул миРНК, молекулы других интерферирующих РНК и РНК-подобные молекулы могут ингибировать транскрипты ретротранспозонов SINE/Alu. Примеры молекул других интерферирующих РНК, которые могут ингибировать транскрипты ретротранспозонов SINE/Alu, включают, но не ограничиваются перечисленными: малые шпилечные РНК (мшРНК), одноцепочечные миРНК, микроРНК (миРНК) и 27-mer дуплексы, являющиеся субстратами Dicer. Примеры РНК-подобных молекул, которые могут ингибировать транскрипты ретротранспозонов SINE/Alu, включают, но не ограничиваются ими, молекулы миРНК, одноцепочечные миРНК, микроРНК, piwiPHK и мшРНК, содержащие один или более химически модифицированных нуклеотидов, один или более аналогов нуклеотидов, один или более дезоксирибонуклеотидов, и/или одну или более нефосфодиэфирных связей. Как правило, все РНК или РНК-подобные молекулы, которые могут взаимодействовать с RISC, осуществляющим регуляцию транскриптов ретротранспозонов SINE/Alu, и участвовать в RISC-опосредованных изменениях экспрессии генов, могут называться «интерферирующие РНК» или «интерферирующие РНК молекулы». Таким образом, миРНК, одноцепочечные миРНК, мшРНК, микроРНК и 27-mer дуплексы, являющиеся субстратами Dicer, представляют собой подмножества «интерферирующих РНК» или «интерферирующие РНК молекулы».
[0057] С использованием известные нуклеотидные последовательности для транскрипта(ов) ретротранспозонов SINE/Alu легко могут быть получены соответствующие миРНК. Согласно различным вариантам реализации изобретения миРНК, которая ингибирует транскрипты ретротранспозонов SINE/Alu, может включать частично очищенную РНК, по существу чистую РНК, синтетическую РНК, рекомбинантно полученную РНК, а также модифицированную РНК, которая отличается от природной РНК вставкой, делецией, заменой и/или изменением одного или более нуклеотидов. Такие изменения могут включать, например, добавление ненуклеотидного материала, например, к концу/концам миРНК или к одному или нескольким внутренним нуклеотидам миРНК, включая модификации, которые делают миРНК устойчивой к расщеплению нуклеазами.
[0058] Согласно различным вариантам реализации изобретения одна или обе нити миРНК могут содержать 3′-липкий конец. В настоящей заявке термин «3′-липкий конец» относится, по меньшей мере, к одному непарному нуклеотиду, который выступает на 3′-конца нити РНК. Таким образом, согласно одному варианту реализации изобретения миРНК содержит, по меньшей мере, один 3′-липкий конец от 1 до примерно 6 нуклеотидов (включая рибонуклеотиды или дезоксинуклеотиды) в длину, от 1 до примерно 5 нуклеотидов в длину, от 1 до примерно 4 нуклеотидов в длину или от примерно 2 до примерно 4 нуклеотидов в длину.
[0059] Согласно иллюстративному варианту реализации изобретения, в котором обе нити молекулы миРНК включают 3′-липкий конец, длина липких концов может быть одинаковой или различной для каждой нити. Согласно некоторым вариантам реализации изобретения 3′-липкий конец присутствует на обеих нитях миРНК и имеет в длину один, два или три нуклеотида. Например, каждая нить миРНК может включать 3′-липкие концы из дитимидиловой кислоты (ТТ) или диуридиловой кислоты (УУ).
[0060] Для повышения стабильности миРНК 3′-липкие концы также могут быть стабилизированы в отношении деградации. Согласно одному варианту реализации изобретения липкие концы стабилизируют путем включения в них пуриновых нуклеотидов, например, аденозиновых или гуанозиновых нуклеотидов. Согласно некоторым вариантам реализации изобретения замена пиримидиновых нуклеотидов на модифицированные аналоги, например замещение уридиновых нуклеотидов в 3′-липких концах на 2′-дезокситимидин, является допустимым и не влияет на эффективность деградации, вызванной РНК-интерференцией. В частности, полагают, что отсутствие 2′-гидроксила в 2′-дезокситимидине может значительно повысить устойчивость 3′-липкого конца к действию нуклеазы.
[0061] Согласно некоторым вариантам реализации изобретения миРНК содержит последовательность AA(N19)TT (SEQ ID NO: 1), AA(N21)TT (SEQ ID NO: 2), NA(N21) (SEQ ID NO: 3), и другие последовательности, где N представляет собой любой нуклеотид. Согласно различным вариантам реализации изобретения указанные миРНК содержат примерно 30-70% пар GC, и предпочтительно содержат приблизительно 50% G/C. Последовательность смысловой нити миРНК соответствует (N19)TT (SEQ ID NO: 4) или N21 (SEQ ID NO: 5) (т.е. с 3 до 23-е положения), соответственно. В последнем случае 3′-конец смысловой нити миРНК превращают в ТТ. Цель такого превращения последовательности состоит в создании симметричного дуплекса в отношении состава последовательности 3′-липкого конца смысловой и антисмысловой нитей. Антисмысловую нить РНК затем синтезируют как комплементарную последовательности смысловой нити с 1-го до 21-е положение.
[0062] Поскольку в этих вариантах антисмысловая нить не распознает 1-е положение 23-нуклеотидной смысловой нити последовательность -специфичным образом, 3′-концевые нуклеотидные остатки антисмысловой нити можно выбрать произвольно. Тем не менее, предпоследний нуклеотид антисмысловой нити (комплементарный нуклеотиду в положении 2 23-нуклеотидной смысловой нити в любом варианте), как правило, является комплементарным целевой последовательности.
[0063] Согласно другому иллюстративному варианту реализации изобретения миРНК содержит последовательность NAR(N17)YNN (SEQ ID NO: 6), где R представляет собой пурин (например, А или Г), и Y представляет собой пиримидин (например, Ц или У/Т). Таким образом, согласно этому варианту реализации соответствующие 21-нуклеотидные смысловая и антисмысловая нити РНК обычно начинаются с пуриновых нуклеотидов. Такие миРНК могут быть экспрессированы с Пол III векторов экспрессии без изменения сайта-мишени, поскольку считается, что экспрессия РНК с Пол III промоторов является эффективной, только когда первый транскрибируемый нуклеотид представляет собой пурин.
[0064] В различных вариантах реализации изобретения миРНК согласно настоящему изобретению могут быть нацелены на любой участок, имеющий в длину примерно 10-30 или 15-25, или 19-25 последовательных нуклеотидов в любой из последовательностей мРНК-мишени («последовательность-мишень»). Методики выбора последовательностей-мишеней для миРНК приведены, например, в Tuschl et al., «The siRNA User Guide,» пересмотренной 6 мая 2004 года. С руководством «siRNA User 10 Guide» можно ознакомиться во всемирной паутине на сайте, который поддерживает д-р Thomas Tuschl, также руководство можно найти на сайте Университета Рокфеллера, выполнив поиск по ключевому слову «миРНК». Кроме того, «siRNA User Guide» можно найти, выполнив поиск в Google для «siRNA User Guide» и по ссылке «www.rockefeller.edu/labheads/Tuschl/sirna.html». Методики выбора последовательностей-мишеней для миРНК и микроРНК также могут быть найдены в Sioud (2008) siRNA and miRNA Gene Silencing: From Bench to Bedside (Methods in Molecular Biology), Humana Press.
[0065] В некоторых вариантах реализации изобретения смысловая нить миРНК, согласно настоящему изобретению, содержит нуклеотидную последовательность, идентичную любому непрерывному участку, имеющему в длину от примерно 19 до примерно 25 нуклеотидов в целевом транскрипте(-ах) ретротранспозонов SINE/Alu.
[0066] МиРНК, осуществляющие сайленсинг транскриптов ретротранспозонов SINE/Alu, могут быть получены с использованием ряда способов, известных специалистам в данной области техники. Например, миРНК могут быть синтезированы химическим или рекомбинантным путем с использованием методов, известных в данной области техники, таких как in vitro система Drosophila, описанная в публикации патентной заявке США 2002/0086356.
[0067] Согласно некоторым вариантам реализации изобретения миРНК синтезируют химическим путем с использованием соответствующим образом защищенных фосфорамидитов рибонуклеозидов и обычного ДНК/РНК-синтезатора. миРНК может быть синтезирована в виде двух отдельных, комплементарных молекул РНК или в виде одной молекулы РНК с двумя комплементарными участками. Коммерческие поставщики синтетических молекул РНК или реагентов для синтеза включают: Proligo (Гамбург, Германия), Dharmacon Research (Лафайет, Колорадо, США), Pierce Chemical (подразделение Perbio Science, Рокфорд, Иллинойс, США), Glen Research (Стерлинг, Виржиния, США), ChemGenes (Ашланд, Массачусетс, США) и Cruachem (Глазго, Великобритания). Изготовленные по заказу миРНК могут быть получены от коммерческих поставщиков (см., например, Thermo Fisher Scientific, Лафайет, Колорадо; Qiagen, Валенсия, Калифорния, США; Applied Biosystems, Фостер-сити, Калифорния и т.п.).
[0068] Согласно некоторым вариантам миРНК также могут быть экспрессированы с рекомбинантных кольцевых или линейных ДНК-плазмид с использованием любого подходящего промотора. Подходящие промоторы для экспрессии миРНК с плазмиды включают, например, U6 или H1 РНК-полимеразы III промоторные последовательности и промотор цитомегаловируса. Выбор других подходящих промоторов находится в пределах квалификации специалиста в данной области. Рекомбинантные плазмиды могут также содержать индуцируемые или регулируемые промоторы для экспрессии миРНК в определенной ткани или в определенной внутриклеточной среде.
[0069] МиРНК, экспрессированная с рекомбинантной плазмиды, может быть либо выделена из систем экспрессии на основе культуры клеток с помощью стандартных методик, или может быть экспрессирована внутриклеточно или вблизи целевой области(ей) в живом организме. Применение рекомбинантных плазмид для доставки миРНК в клетки in vivo обсуждается более подробно ниже.
[0070] МиРНК может быть экспрессирована с рекомбинантной плазмиды в виде двух отдельных, комплементарных молекул РНК, либо как одна молекула РНК с двумя комплементарными участками. Выбор подходящей плазмиды для экспрессии миРНК, способы вставки последовательности нуклеиновой кислоты для экспрессии миРНК в плазмиду и способы доставки рекомбинантной плазмиды в клетки-мишени находятся в пределах квалификации специалиста в данной области техники (см., например, Tuschl 25 (2002) Nat. Biotechnol, 20: 446-448; Brummelkamp et al. (2002) Science 296: 550 553; Miyagishi et al (2002) Nat. Biotechnol. 20: 497-500; Paddison et al. (2002) Genes Dev. 16: 948-958; Lee et al. (2002) Nat. Biotechnol. 20: 500-505; Paul et al. (2002) Nat. Biotechnol. 20: 505-508 и т.п.).
[0071] Согласно одному иллюстративному варианту реализации изобретения плазмида, содержащая последовательности нуклеиновых кислот, кодирующие миРНК для ингибирования транскриптов ретротранспозонов SINE/Alu, содержит последовательность ДНК, кодирующую смысловую нить РНК, которая функционально связана с терминаторной последовательностью полиТ под контролем человеческого промотора гена U6-РНК, и последовательность ДНК, кодирующую антисмысловую нить РНК, которая функционально связана с терминаторной последовательностью полиТ под контролем человеческого промотора гена U6-РНК. Эта плазмида, в конечном итоге, предназначена для применения в создании рекомбинантного аденоассоциированного вирусного вектора, содержащего те же самые последовательности нуклеиновой кислоты для экспрессии миРНК.
[0072] В настоящей заявке термин «функционально связанный с терминаторной последовательностью полиТ» означает, что последовательности нуклеиновых кислот, кодирующие смысловую или антисмысловую нити, примыкают к сигнальной терминаторной последовательности полиТ в 5′-направлении или расположены достаточно близко, чтобы в процессе транскрипции смысловой или антисмысловой последовательности с плазмиды сигналы терминации полиТ останавливали транскрипцию после того, как желаемый продукт был транскрибирован.
[0073] В настоящей заявке термин «под контролем» промотора означает, что последовательности нуклеиновых кислот, кодирующие смысловую или антисмысловую нити, расположены в 3′-направлении от промотора, так что промотор может инициировать транскрипцию смысловой или антисмысловой кодирующих последовательностей.
[0074] Согласно некоторым вариантам реализации миРНК могут быть доставлены в виде малых шпилечных РНК или малых шпилечных РНК (мшРНК). мшРНК представляет собой последовательность РНК, которая делает крутой поворот в виде шпильки, который можно использовать для сайленсинга экспрессии генов по механизму РНК-интерференции. В типичных вариантах реализации изобретения для гарантированной экспрессии мшРНК использует вектор, введенный в клетку, и задействует промотор U6. Указанный вектор обычно передается дочерним клеткам, что обеспечивает наследование сайленсинга генов. Структура шпильки мшРНК расщепляется клеточными механизмами до миРНК, которая затем связывается с РНК-индуцированным комплексом сайленсинга (RISC). Этот комплекс связывается и расщепляет мРНК, которая комплементарна миРНК, связанной с ним.
[0075] Согласно некоторым вариантам реализации изобретения смысловая последовательность мшРНК будет составлять в длину от примерно 19 до примерно 30 нуклеотидов (например, приблизительно 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 нуклеотидов), как правило, от примерно 19 до примерно 22 нуклеотидов в длину, антисмысловая последовательность будет составлять в длину от примерно 19 до примерно 30, более предпочтительно от 19 до примерно 22 нуклеотидов (например, приблизительно 19, 20, 21 или 22 нуклеотидов), и участок петли будет составлять от приблизительно 3 до приблизительно 19 нуклеотидов (например, приблизительно 3, 4, 5 и т.д., … вплоть до примерно 19) в длину. Согласно некоторым вариантам реализации изобретения смысловая и антисмысловая последовательности имеют одинаковую длину, то есть мшРНК образует симметричную шпильку, но необязательно. В некоторых случаях смысловая или антисмысловая нить может быть короче, чем ее комплементарная нить и образуется асимметричная шпилька. Кроме того, хотя в некоторых случаях спаривание оснований между смысловой и антисмысловой последовательностями является точным, в других случаях точное спаривание является необязательным. Другими словами, некоторое несоответствие спаривания между последовательностями допустимо, или даже необходимо, например, для уменьшения силы водородных связей между двумя нитями. Тем не менее, в одном иллюстративном варианте реализации изобретения смысловые и антисмысловые последовательности имеют одинаковую длину, а спаривание оснований между двумя нитями является точным и не содержит каких-либо несоответствий. Молекула мшРНК также может содержать 5′-концевую фосфатную группу, которая может быть химически модифицирована. Кроме того, петлевая часть молекулы мшРНК может содержать, например, нуклеотиды, не-нуклеотиды, ликерные молекулы, конъюгированные молекулы и т.д.
[0076] МшРНК/миРНК/пиРНК, описанная в настоящей заявке, нацелена на и вызывает РНКи-опосредованную деградацию транскриптов ретротранспозонов SINE/Alu или их форм, полученных в результате альтернативного сплайсинга, либо участвует в геномном сайленсинге через PIWI РНК пути. Таким образом, в некоторых вариантах реализации изобретения предложены способы ингибирования транскриптов ретротранспозонов SINE/Alu в клетке, включающие введение эффективного количества миРНК/мшРНК/пиРНК, нацеленных на транскрипты ретротранспозонов SINE/Alu, в клетку с тем, чтобы вызвать разрушение мРНК-мишени.
[0077] Согласно различным вариантам реализации изобретения миРНК/мшРНК/пиРНК могут быть экспрессированы с рекомбинантных вирусных векторов, введенных в клетки-мишени. Рекомбинантные вирусные векторы содержат последовательности, кодирующие миРНК/мшРНК, и любой подходящий промотор для экспрессии последовательностей миРНК/мшРНК/пиРНК. Подходящие промоторы включают, но не ограничиваются ими, U6 или H1 РНК-Пол III промоторные последовательности и промотор цитомегаловируса. Выбор других подходящих промоторов находится в пределах квалификации специалиста в данной области. Рекомбинантные вирусные векторы могут также содержать индуцируемые или регулируемые промоторы для экспрессии миРНК/мшРНК в определенной ткани или в определенной внутриклеточной среде.
[0078] МиРНК/мшРНК/пиРНК может быть экспрессирована с рекомбинантного вирусного вектора в виде двух отдельных, комплементарных молекул РНК, либо как одна молекула РНК с двумя комплементарными участками.
[0079] Любой вирусный вектор, способный принимать кодирующие последовательности молекул(-ы) миРНК/мшРНК/пиРНК, которые экспрессируют, может быть использован, например, вектор, полученный из аденовируса (AV); аденоассоциированного вируса (AAV), ретровирусов (например лентивирусов (LV), рабдовирусов, вируса мышиного лейкоза), вируса герпеса и тому подобных вирусов. Тропизм вирусных векторов также может быть модифицирован путем псевдопитирования векторов с помощью белков оболочки или других поверхностных антигенов из других вирусов. Например, аденоассоциированный вирусный вектор может быть псевдотипирован поверхностными белками вируса везикулярного стоматита (VSV), вируса бешенства, вируса лихорадки Эбола, вируса Мокола и подобных вирусов.
[0080] Выбор рекомбинантных вирусных векторов, которые можно использовать в способах введения нуклеиновых кислот для экспрессии миРНК/мшРНК/пиРНК в вектор, и способы доставки вирусного вектора в клетки-мишени известны любому специалисту в данной области (см., например, Domburg (1995) Gene Therap. 2: 301-310; Eglitis (1988) Biotechniques 6: 608-614; Miller (1990) Hum. Gene Therap. 1: 5-14; Anderson (1998) Nature 392: 25-30, и т.д.).
[0081] В некоторых вариантах реализации подходящие вирусные векторы включают, но не ограничиваются ими, лентивирусные векторы. В одном иллюстративном варианте сконструирован лентивирусный мшРНК вектор для нокдауна генетических транскриптов SINE/Alu. Смысловую и антисмысловую нити мшРНК химически синтезируют и ренатурируют в равных количествах, создавая, таким образом, липкие концы, содержащие специфические сайты рестрикции для клонирования, и лигируют в вектор (например, расщепленный по сайтам рестрикции HindIII и BglII и очищенный на геле вектор pENTR/pTER+). Равные количества каждой конструкции смешивают с вектором Plenti-CMV-GFP DEST в реакционной смеси, содержащей фермент LR Clonase®, для рекомбинации клонированных мшРНК-продуцирующих элементов в целевой вектор в соответствии с инструкциями производителя (Invitrogen). Полученную лентивирусную плазмиду трансформируют в клетки Е. coli Stbl3 (Invitrogen) для амплификации.
[0082] Согласно некоторым вариантам реализации подходящие вирусные векторы включают векторы, полученные из AV и AAV. В одном иллюстративном варианте реализации изобретения миРНК/мшРНК/пиРНК экспрессируют в виде двух отдельных, комплементарных одноцепочечных молекул РНК из рекомбинантного AAV вектора, содержащего, например, U6 или H1 РНК промоторы или промотор цитомегаловируса (CMV). Подходящий AV вектор для экспрессии миРНК, способ доставки рекомбинантного AV вектора, а также способ доставки вектора в клетки - мишени описаны в Xia 5 et al. (2002) Nat. Biotech. 20: 1006 1010.
[0083] Подходящие AAV векторы для экспрессии миРНК/мшРНК, способы конструирования рекомбинантного AV вектора и способы доставки векторов в клетки-мишени также описаны в Samulski et al. (1987) J. Virol. 61: 3096-3101; Fisher et al. (1996) J. Virol., 70: 520-532; Samulski et al. (1989) J. Virol. 63: 3822-3826; патенты США 5252479 10 и 5139941, международные патентные заявки WO 1994/013788 и WO 1993/024641 и т.д.
Источники клеток
[0084] Стволовые клетки (например, взрослые стволовые клетки, эмбриональные стволовые клетки, клетки пуповинной крови, ИПСК и т.д.) могут быть получены в соответствии со стандартными методами, хорошо известными специалистам в данной области техники. В некоторых вариантах реализации изобретения стволовые клетки также являются коммерчески доступными.
Дифференциация стволовых клеток
[0085] В некоторых вариантах предложены способы дифференцировки стволовых клеток, в которых уровень или активность транскриптов ретротранспозонов SINE/Alu понижают/ингибируют. Считается, что эти стволовые клетки могут быть дифференцированы в эмбриоидные тельца или окончательно дифференцированные клетки с использованием стандартных способов дифференцировки, которые хорошо известны специалистам в данной области техники.
[0086] Например, в некоторых вариантах реализации изобретения удаление фактора ингибирования лейкемии (LIF) может привести к дифференцировке модифицированных стволовых клеток, описанных в настоящей заявке (например, стволовые клетки, в которых уровень или активность транскриптов ретротранспозонов SINE/Alu понижают/ингибируют) в эмбриоидные тельца.
[0087] Способы дифференцировки стволовых клеток хорошо известны специалистам в данной области техники. Например, экстракт инфаркта миокарда содержит биохимические факторы, которые вызывают дифференциацию мезенхимальных стволовых клеток костного мозга (МСК КМ) в кардиомиоциты (см., например, Ge et al. (2009) Biochem. Biophys. Res. Commun., 381(3): 317-321). Фактор морфогенеза костей (BMP4, BMP7 и BMP8b) может усилить дифференцировку эмбриональных стволовых клеток человека в половые клетки (см., например, Gonsalves et al. (2006) Stem Cells Dev., 15(6): 831-837). Первичные культуры клеток, полученных из жировой ткани или скелетных мышц, дифференцируются в адипоциты при культивировании в среде с высоким содержанием глюкозы (см., например, Aguiari et al. (2008) Proc. Natl. Acad. Sci., USA, 105(4): 1226-1231). ИЛ-27 индуцирует дифференцировку гемопоэтических стволовых клеток (см., например, Seita et al. (2008) Blood, 111(4): 1903-1912).
[0088] В нейрональных клетках белки семейства нейротрофинов являются одним из наиболее важных индуцибельных сигналов для начала дифференцировки. Как известно, член этого семейства фактор роста нервов индуцирует нейрогенез, а нейротрофин 3 вовлечен в развитие олигодендроцитов. Другим важным семейством, участвующем в передаче сигналов к началу дифференцирования, является семейство цитокинов, включающее цилиарный нейротрофический фактор (CNTF) и фактор ингибирования лейкемии (LIF), которые играют ключевую роль в регулировании глиогенеза в развивающейся центральной нервной системе млекопитающих. Кроме того, AICAR (5-аминоимидазол-4-карбоксамид-1-β-D-рибофуранозид;) индуцирует дифференцировку нервных стволовых клеток в астроглию (см., например, Zang et al. (2008) J. Biol. Chem., 283(10): 6201-6208 и ссылки в этом источнике).
[0089] Также известно, что некоторые природные соединения индуцируют дифференцировку стволовых клеток in vitro. Такие соединения включают, но не ограничиваются, перечисленными: ретиноевую кислоту, тромбоцитарный фактор роста PDGF, инсулин, арктигенин, ATRA (витамин А), босвеллиевую кислоту, бромелайн и другие протеолитические ферменты, САРЕ, флавоноиды (в том числе апигенин, лютеолин, кверцетин, генистеин и даидзеин), эмодин, ЭПК и ДГК, монотерпены, ресвератрол, 1,25-D3 (витамин D3) и другие.
[0090] Считается, что плюрипотентные клетки, полученные с использованием способов, описанных в настоящей заявке, могут быть впоследствии индуцированы на дифференцировку с использованием способов, которые, как известно, индуцируют дифференцировку ИПСК. В этой связи следует отметить, что способы дифференцировке индуцированных плюрипотентных клеток в CD34+CD43+ гематопоэтические клетки-предшественники и CD31+CD43- эндотелиальные клетки описаны в Choi et al. (2009) Stem Cells 27: 559. Эти клетки могут быть далее разделены на фенотипически определяемые подмножества примитивных гемопоэтических клеток по способу дифференцировки, который близок к таковому у эмбриональных стволовых клеток. Способы дифференцировки индуцированных плюрипотентных клеток в инсулин-продуцирующие клетки поджелудочной железы описаны в Zhang et al. (2009) Cell Res. 19: 429. После первой стадии дифференцировки в PDX-1-положительные клетки-предшественники, плюрипотентные клетки человека далее дифференцировали в панкреатические клетки, экспрессирующие MafA, GLUT2, инсулин, а в некоторых случаях амилазу и С-пептид. Функциональные кардиомиоциты, демонстрирующие саркомерную структуру и экспрессирующие сердечные маркеры, включая Nkx2.5, сердечный тропонин Т, атриальный натрийуретический фактор и тяжелую и легкую цепи миозин, также были получены из индуцированных плюрипотентных клеток человека и ничем не отличаются от тех, которые были получены из ЭС клеток (см., например, Zhang et al. (2009) Circ. Res. 104:e30). Электрофизиологические эксперименты выявили, что сходно с ЭС клетками, плюрипотентные клетки дифференцируются в клетки, фенотип которых сходен с таковым у клеток узлов, предсердий и желудочков, и реагируют на канонический для кардиомиоцитов бета-адренергический сигнальный путь. Плюрипотентные и ЭС клетки человека эффективно превращаются в нервные клетки при использовании Noggin и препарата SB431542 - двух ингибиторов сигнализации, опосредованной трансформирующим фактором роста бета (TGF-β)/Smad (Chambers et al. (2009) Nat. Biotechnol. 27: 275). Синергитическое действие этих двух ингибиторов привело к образованию Рах6+ примитивных нервных клеток, которые затем удалось дифференцировать в клетки нервного гребня, переднего отдела ЦНС, соматические двигательные нейроны и дофаминергические нейроны.
[0091] Эти подходы предназначены для иллюстративных целей и не являются ограничивающими. Другие способы индукции дифференцировки стволовых клеток хорошо известны специалистам в данной области техники.
[0092] Согласно некоторым вариантам реализации изобретения клетки и композиции, содержащие клетки, которые были дифференцированы в эмбриоидные тельца или дифференцированы далее (например, окончательно дифференцированы), также включены в область изобретения. Такие дифференцированные клетки включают, но не ограничиваются перечисленными: эмбриоидные тельца и/или клетки - предшественники и/или окончательно дифференцированные клетки, которые дифференцируют в линии кардиомиоцитов, клеток крови, эпителиальных клеток, остеобластов, остеокластов, хондроцитов, адипоцитов, гладкомышечные клетки, нервные клетки (нейроны), глиальные клетки, панкреатические β-клетки, мотонейроны и другие клетки.
ПРИМЕРЫ
[0093] Следующие примеры приведены в иллюстративных целях и не ограничивают объем настоящего изобретения.
Пример 1
Изменения хроматина, которые зависят от ретротранспозонов SINE/Alu, индуцируют старение взрослых стволовых клеток ex vivo
[0094] В этом примере представлены механистические аспекты старения взрослых мезенхимальных стволовых клеток ex vivo. С помощью полногеномного сравнительного анализа восстанавливаемых и устойчивых повреждений ДНК во взрослых стволовых клетках человека, переходящих в стадию клеточного старения, раскрыта новая, неожиданная функциональная роль ретротранспозонов в сложных путях клеточного старения. Представлены доказательства того, что большинство восстанавливаемых повреждений ДНК в самообновляющихся взрослых стволовых клетках человека распределяется неслучайным образом и накапливается в ретротранспозонных повторах. Данные показывают, что нарушенная регуляция транскрипционной активности, по меньшей мере, одного класса ретротранспозонов человека (SINE/Alu-повторов) ухудшает сборку когезинового и конденсинового комплексов в перицентрических участках хроматина, обогащенных γН2АХ, тем самым, препятствуя нормальному функционированию клеточного механизма репарации ДНК. Неспособность взрослых стволовых клеток восстановить повреждения ДНК в перицентрических областях приводит к переходу от самообновления к старению. Представлены доказательства того, что это событие является обратимым и предлагается новая механистическая модель, согласно которой потеря надлежащего контроля транскрипционной активности ретротранспозонной части человеческого генома может прямо или косвенно влиять на сложные молекулярные события, что приводит к прекращению самообновления взрослых стволовых клеток и запуску процесса их старения.
Результаты
Репликативное старение взрослых мезенхимальных стволовых клеток человека
[0095] Мезенхимальные стволовые клетки жировой ткани взрослого человека (hADSCs) выделяли из подкожного жира брюшной полости, как описано в разделе «Экспериментальные методики», и характеризовали по поверхностным антигенам клеток (рис. 7А). После четырех недель в культуре клетки становились более однородными, как показано методом проточной цитофлуорометрии. Более 99,3% и 99,9% клеток экспрессировали маркеры CD105 и CD44, специфичные для мезенхимальных стволовых клеток (Dominici et al. (2006) Cytotherapy 8: 315-317), соответственно, однако не экспрессировали маркеры гемопоэтических стволовых клеток (CD45) или эндотелиальных клеток-предшественников (CD31) (рис. 7А). Существует возможность индуцировать дифференцировку клеток hADSC в ряд линий мезенхимальных тканей, включая адипоциты, остеобласты, миоциты и хондроциты (Erickson et al. (2002) Biochem. Biophys. Res. Commun. 290: 763-769; Zuk et al. (2001) Tissue Eng., 7: 211-228). Таким образом, в этих экспериментах клетки hADSC обладали выраженным сходством с мезенхимальными стволовыми клетками (МСК).
[0096] Способность к самообновлению (SR) клеток hADSC оставалась однородной до удвоения популяции (PD17), после этого, с увеличением продолжительности культивирования ex vivo клетки постепенно начинали приобретать фенотип с характерными признаками «старости», как показано на рис. 1А (а также описано в (Bonab et al. (2006) ВМС Cell Biol., 7: 14; Fehrer et al. (2007) Aging Cell, 6: 745-757; Kern et al. (2006) Stem Cells, 24: 1294-1301). Морфологические изменения культуры являются характерными для модели клеточного старения Хейфлика (Juckett (1987) Mech. Ageing Dev., 38: 49-71). К моменту 37 удвоения популяции в культуре hADSC накапливались гигантские неделящиеся клетки, экспрессирующие лизосомальный фермент - связанную с репликативным старением J3-галактозидазу (активна при рН=6) (SA-J3-Gal) (Dimri et al. (1995) Proc. Natl. Acad. Sci. USA, 92: 9363-9367), как показано на рис. 1В. Активность самообновления клеток была недостаточной вследствие уменьшения количества делящихся клеток, как было установлено, по включению бромдезоксиуридина (BrdU) и [Н3]-тимидина в ДНК (рис. 1В и рис. 7В).
[0097]. Чтобы определить, является ли ограниченная способность клеток hADSC к размножению следствием старения, связанного с накоплением устойчивых повреждений ДНК, были проведены эксперименты по окрашиванию антителами ключевых медиаторов ответа на повреждение ДНК (DDR), которые способствуют активации контрольных точек клеточного цикла и систем репарации ДНК. В частности, использовали фосфорилированную форму варианта гистона Н2АХ (γН2АХ) (Shiloh (2003) Nat. Rev. Cancer, 3: 155-168) и связывающего р53 белка-1 (53ВР1) (Aguilera and Gomez-Gonzalez (2008) Nat. Rev. Genet., 9: 204-217; Shiloh (2003) Nat. Rev. Cancer, 3: 155- 168; Stewart (2009) Cell Cycle, 8: 1532-1538), чтобы доказать наличие молекулярных характеристик клеток, имеющих двунитевые разрывы ДНК, в культурах как самообновляющихся, так и стареющих клеток hADSC. С началом старения hADSCs происходит направленная локализация γН2АХ и 53ВР1 (рис. 1В) с образованием характерных очагов, которые были названы очаги устойчивых повреждений ДНК (ранее были описаны в работе Rodier et al. (2009) Nat. Cell Biol., 11: 973-979). Образование устойчивых очагов γH2AX/53BP1 при клеточном старении было связано с наличием невосстановленных двунитевых разрывов ДНК, как определили на основании солокализации очагов с несколькими факторами репарации ДНК (d′Adda di Fagagna et al. (2003) Nature, 426: 194-198). Возникновение очагов накопления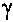 H2AX/53BP1 в самообновляющихся клетках ADSCs регистрировали очень редко, однако их формирование увеличивалось с началом старения клеток hADSC (SEN hADSCs) (рис. 8).
H2AX/53BP1 в самообновляющихся клетках ADSCs регистрировали очень редко, однако их формирование увеличивалось с началом старения клеток hADSC (SEN hADSCs) (рис. 8).
[0098] В совокупности эти результаты показывают, что связанные со старением эффекты в клетках ADSC возникают не только с увеличением продолжительности культивирования, ведущей к старению, но непрерывно приобретаются с самого начала их размножения ех vivo, аналогичный феномен ранее был обнаружен у фибробластов человека и гемопоэтических стволовых клеток человека.
[0099] Было замечено, что стареющие клетки hADSC содержали активированные формы киназ контрольных точек повреждений ДНК - CHK1 и CHK2, фосфорилированных по положениям S345 и Т68, соответственно (рис. 9). Фосфорилирование этих сайтов ATM/ATR киназами необходимо для полной остановки клеточного цикла, индуцированной повреждениями ДНК, в соматических клетках человека (d′Adda di Fagagna et al. (2003) Nature, 426: 194-198; Sedelnikova et al. (2008) Aging Cell 7: 89-100; Tanaka et al. (2006) Cell Prolif., 39: 313-323). Для того, чтобы далее определить состояние этих клеток был проведен дифференциальный (самообновляющиеся клетки hADSC против стареющих) транскрипционный анализ 96 генов, вовлеченных в различные аспекты клеточного цикла (кПЦР-анализ клеточного цикла человека), как подробно описано в разделе «Экспериментальные методики». Гены, активность которых в стареющих клетках hADSC была статистически достоверно подавлена со значением р<0,05, приведены в табл. 1, и включают гены, участвующие в регуляции клеточного цикла, репликации ДНК и митоза, полученные данные позволяют предположить, что в культуре стареющих клеток hADSC реализуется программа ответа на двунитевые разрывы ДНК, которая прямо или косвенно связана с образованием ассоциированных со старением очагов повреждений ДНК. Причинные факторы, которые могут вызывать этот процесс во взрослых стволовых клетках человека, до сих пор неизвестны.
Полногеномный анализ локализации γН2АХ в стареющих клетках hADSC методом ChIP-seq.
[00100] Для выявления геномных локусов, непосредственно вовлеченных в повреждение ДНК в стареющих клетках hADSC, проведено полногеномное профилирование хроматина методом иммунопреципитации хроматина с антителами к модифицированному гистону γH2AX с последующим секвенированием всего пула выделенных фрагментов на приборе массового параллельного секвенирования ДНК ABI SOLiD (ChIP-seq метод) (рис. 2А). Асинхронно делящиеся образцы самообновляющихся клеток hADSC использовали в аналогичных экспериментах, чтобы установить общее распределение γН2АХ-модифицированного хроматина, если он присутствовал, для определения локусов восстанавливаемых повреждений ДНК, которые не оказывают негативного влияния на свойства самообновляющихся клеток. ChIP-seq проводили на четырех повторах образцов каждой из культур самообновляющихся и стареющих клеток hADSC. После геномного картирования полученного распределения меток последовательностей (табл. 2) удаляли выбросы данных, снижали шум и объединяли данные повторных экспериментов. Подробное описание разработанных алгоритмов и подходов, которые использовали для анализа данных ChIP-seq, приведено в разделе «Дополнительные методы». Техническая пригодность использованного экспериментального протокола ChIP-seq и процедуры объединения данных была поддержана хорошей согласованностью результатов картирования по всем повторным экспериментам. После составления карты и обработки данных по распределению метки согласно ChIP-seq, оценивали содержание метки по отношению к эмпирически рассчитанному пороговому значению («Дополнительные методы») для окна в 200 п.н., что примерно соответствует размеру мононуклеосомы, с целью выявления отдельных γН2АХ-модифицированных нуклеосом в геноме человека. Полученное в результате геномное распределение γН2АХ-модифицированных нуклеосом сравнивали для самообновляющихся и стареющих клеток (рис. 2В) для того, чтобы оценить позиционные и количественные изменения в γН2АХ-модифицированном хроматине, связанные с этими фенотипами.
Большая часть γH2AX-модифицированного хроматина локализуется в ретротранспозонной части человеческого генома
[00101] В более ранних исследованиях было показано, что топологические ограничения на структурах хроматина при репликации ДНК и транскрипции представляют наибольшую опасность для целостности ДНК (см. обзор в Shrivastav et al. (2008) Cell Res., 18: 134-147). Всего существует четыре потенциальных источника повреждений ДНК у стареющих взрослых стволовых клеток: кратковременные двунитевые разрывы ДНК, вызванные транскрипцией индуцибельных генов или активностью точки начала репликации ДНК (Ju et al. (2006) Science, 312: 1798-1802; Ju and Rosenfeld (2006) Cell Cycle, 5: 2557-2560; Rampakakis and Zannis-Hadjopoulos (2009) Nucleic Acids Res., 37: 5714-5724), повреждения ДНК, которые образуются в результате препятствий для продвижения репликативных вилок из-за столкновения молекулярных комплексов, обеспечивающих репликацию и транскрипцию, а также трудностей, связанных с особенностями репликации центромерных и теломерных областей генома (Dalai and Bui (2010) Curr. Opin. Cell Biol., 22: 392-402; Dotiwala et al. (2010) Curr. Biol., 20: 328-332; Morris and Moazed (2007) Cell, 128: 647-650; Schoeftner and Blasco (2009) EMBO J. 28: 2323-2336). Недавние исследования распределения γH2AX в геноме клеток Saccharomyces cerevisiae выявили наличие «ломких» локусов генома (Szilard et al. (2010) Nat. Struct. Mol. Biol., 17: 299-305), эти данные позволяют предположить, что картирование сайтов, обогащенных γН2АХ, может быть полезным для точного определения геномных элементов, входящих в группу риска в геномах других видов. Поскольку механистические аспекты «ломких» локусов связаны с регуляцией контрольных точек клеточного цикла и репарации ДНК (Durkin and Glover (2007) Annu. Rev. Genet., 41: 169-192), можно ожидать, что постепенное накопление невосстановленных повреждений ДНК в ломких локусах асинхронно делящихся клеток может привести к запуску процесса клеточного старения.
[00102] Эксперименты по картированию показали, что γН2АХ-модифицированный хроматин распределялся неслучайным образом и, что явилось неожиданным фактом, представлял собой небольшую долю общего хроматина человека в асинхронно делящихся самообновляющихся клетках hADSC: ~1,4% (200,977 γН2АХ-модифицированные нуклеосомы), и еще меньшую долю в стареющих клетках hADSC: ~1,2% от общего хроматина (173,877 γН2АХ-модифицированные нуклеосомы). Большая часть модификаций γН2АХ (SR: 65,4%, SEN: 65,2%) была локализована в мобильных элементах (МЭ), большинство из которых представляли собой SINE/Alu, L1 или LTR ретротранспозоны (рис. 10). Помимо общей распространенности ретротранспозонов, несущих γН2АХ-модифицированные нуклеосомы, для этого геномного элемента величина разности между уровнями относительного накопления метки γН2АХ в SR по сравнению со стареющими клетками hADSC была самой большой (рис. 2С). SINE/Alu элементы содержали большее количество γН2АХ-модифицированных нуклеосом в самообновляющихся клетках по сравнению со стареющими клетками, тогда как L1s элементы имели наибольшее относительное увеличение γН2АХ-модифицированных нуклеосом в стареющих клетках (рис. 2С). Известно, что ретротранспозонная часть генома, за исключением генных последовательностей, препятствует продвижению комплекса репликации (Scheifele et al. (2009) Proc. Natl. Acad. Sci. USA, 106: 13927-13932; Voineagu et al. (2008) Proc. Natl. Acad. Sci. USA, 105: 9936-9941; de la Loza et al. (2009) 20 DNA Repair (Amst), 8: 620-626), это позволяет предположить, что образование части модифицированных гистонов γН2АХ, обнаруженных в асинхронно делящихся клетках (SR образец), может быть вызвано приостановкой или разрушением репликационной вилки, как описано ранее в дрожжах (Admire et al. (2006) Genes Dev., 20: 159-173; Cha and Kleckner (2002) Science, 297: 602-606; de la Loza et al. (2009) DNA Repair (Amst), 8: 620-626; Deshpande and Newlon (1996) Science, 272: 1030-1033).
[00103] Данные указывают на роль ретротранспозонов в клеточном ответе, вызванном повреждениями ДНК, который связан с нормальным прогрессированием клеточного цикла (Curcio et al. (2007) Mol. Cell Biol., 27: 8874-8885; Scheifele et al (2009) Proc. Natl. Acad. Sci. USA, 106: 13927-13932), и свидетельствуют о том, что эти элементы могут способствовать развитию фенотипа старения в hADSCs (Rudin and Thompson (2001) Genes Chromosomes Cancer, 30: 64-71; Wang et al. (1999) Mutat. Res., 433: 147-157). Кроме того, данные дополнительно подтверждают наблюдение, что ретротранспозонная часть генома особенно чувствительна к двунитевым разрывам ДНК - это критический вывод, учитывая, что данный тип разрывов ДНК недавно был непосредственно связан с геномными перестройками при онкологических заболеваниях (Eickbush (2002) Nat. Genet., 31: 126-127; Gosselin et al (2009) Cancer Res., 69: 7917-7925; Lin et al (2009) Cell, 139: 1069-1083; Rudin and Thompson (2001) Genes Chromosomes Cancer, 30: 64-71; Weinstock et al. (2006) DNA Repair (Amst) 5: 1065-1074).
Размер и распределение γH2AX хроматина в самообновляющихся и стареющих клетках hADSC
[00104] Предыдущие исследования показали, что модификация γН2АХ образуется в течение нескольких минут после формирования двунитевого разрыва ДНК и может охватывать большие (от тысяч до миллионов пар оснований) области хроматина (Redon et al. (2003) EMBO Rep., 4: 678-684; Rogakou et al (1999) J. Cell Biol., 146: 905-916). Таким образом, исследовали непрерывность γН2АХ хроматина в асинхронно делящихся самообновляющихся и стареющих клетках hADSC.
[00105] Был разработан алгоритм, который позволяет анализировать сегменты максимального размера для выявления смежных областей генома, которые имеют аномально высокие плотности γН2АХ-модифицированных сайтов («Дополнительные методы»). Указанные области с высокой плотностью γН2АХ принимали за отдельные кластеры γН2АХ хроматина и распределение кластеров оценивали для самообновляющихся и стареющих клеток. Как в самообновляющихся, так и в стареющих клетках был обнаружен один доминирующий пик размера кластеров γН2АХ хроматина, который соответствовал фрагментам, примерный размер которых был характерен для мононуклеосом, которые, вероятно, отмечали одиночные сайты двунитевых разрывов ДНК (рис. ПА). Общее количество γН2АХ мононуклеосомного хроматина существенно не изменилось в процессе клеточного старения взрослых стромальных клеток (рис. 11В): в каждом типе клеток обнаружили ~21 млн.п.о. хроматина мононуклеосомного размера, при этом разность между самообновляющимися и стареющими клетками составила всего ~0,6%. Проведенный компьютерный анализ не выявил конкретной геномной категории, которая существенно обогащена сайтами накопления мононуклеосомных повреждений при остановке клеточного цикла в стареющих клетках hADSC (табл. 3). Тем не менее, большая часть мононуклеосомных γН2АХ сайтов (67,3% в SR и 65,8% в SEN) была связана с ретротранспозонной частью генома, причем 34,9% и 30,0% этих сайтов обнаруживали в элементах SINE, 32,4% и 35,78% в LINEs элементах самообновляющихся и стареющих клеток hADSC, соответственно (табл. 3).
[00106] Характер распределения размеров кластеров γН2АХ в значительной степени совпадал для большинства кластеров, но существенно различался на концах гистограмм, которые занимали большие кластеры (рис. ПА). Эту разницу между самообновляющимися и стареющими клетками в отношении распределения размера γН2АХ кластеров использовали для разграничения средних и больших кластеров. Значения относительной энтропии, рассчитанные как показатель разности между двумя распределениями («Дополнительные методы»), вычисляли в диапазоне пороговых значений для длины кластера. Пороговое значение длины кластера, которое максимально увеличивает относительную энтропию распределения размера γН2АХ кластеров между SR и SEN, принимали как точку отсечения между средними и большими кластерами (рис. 11С).
[00107] В отличие от геномной локализации кластеров, имеющих размер мононуклеосом, были обнаружены существенные различия количества и локализации в геномных категориях больших γН2АХ кластеров. Общее количество больших γН2АХ кластеров снизилось более чем в три раза в образцах стареющих клеток (11243429 п.о.) по сравнению с самообновляющимися (37467892 п.о.) (рис. 11Б). В самообновляющихся клетках 47,9% больших кластеров хроматина было связано с генной частью генома, включая: (I) промоторы генов, (II) интроны, (III) экзоны и (IV) соединения интронов и экзонов (табл. 3). В нереплицирующихся образцах стареющих клеток большие γН2АХ кластеры сдвигались к межгенным участкам с уменьшением генных последовательностей до 35,2%.
[00108] Было предположено, что накопление γН2АХ может быть результатом топологических проблем, связанных со столкновением репликационной вилки и комплекса транскрипции. Полученные данные подтверждают эту гипотезу, так как динамика больших кластеров указывала на сдвиг к межгенным участкам в нереплицирующихся образцах стареющих клеток со значительным снижением экзонных и интронных последовательностей. Это наводит на мысль, что большие генные и межгенные кластеры γН2АХ могут представлять собой горячие точки не только в отношении ломкости хромосом при переходе из G2 в М фазу (SEN образцы), но и в отношении запутывания сестринских хроматид во время фазы S в самообновляющихся клетках hADSC.
γН2АХ хроматин и плотность генов
[00109] Транскрипционная активность может стать препятствием для продвижения репликационных вилок, приводя к их распаду и накоплению двунитевых разрывов ДНК (см. обзор в Zegerman and Diffley (2009) DNA Repair (Amst) 8: 1077-1088). Таким образом, функциональные участки генов, такие как экзоны и промоторы, возможно, относительно обогащены γН2АХ-модифицированными нуклеосомами в SR клетках hADSC, в отличие от стареющих клеток, в которых репликативная активность прекращена. Кроме того, можно ожидать, что хромосомы человека, имеющие высокую плотность генов, будут склонны к накоплению γН2АХ и если это повреждение не будет восстановлено при активации контрольной точки, возможно, вследствие износа комплекса репарации ДНК, может развиться фенотип старения. В этом случае, можно было бы ожидать, что обогащение γН2АХ будет наблюдаться в одних и тех же локусах хроматина самообновляющихся и стареющих клеток и количество γН2АХ хроматина будет положительно коррелировать с плотностью генов в обоих фенотипах.
[00110] Чтобы проверить эти две гипотезы, оценивали перекрытие между сайтами γН2АХ-модифицированного хроматина в самообновляющихся и стареющих клетках и сравнивали плотность генов в хромосомах человека с плотностью метки последовательности γН2АХ самообновляющихся и стареющих клетках. Перекрытие сайтов γН2АХ-модифицированного хроматина для двух типов клеток было небольшим: ~5%. В SR образцах была показана достоверная положительная корреляция плотности генов с плотностью метки γН2АХ, тогда как в образцах стареющих клеток корреляция была отрицательная (рис. 2D и таблица 4).
[00111] Это различие корреляционной зависимости между самообновляющимися и стареющими клетками согласуется с моделью, предполагающей наличие конфликта между комплексами транскрипции и репликации, которая объясняет избыток повреждений, специфичных для генных участков, в самообновляющихся клетках hADSC, и указывает на то, что большинство повреждений в стареющих клетках не связано с накоплением невосстановленных повреждений, вызванных столкновением репликационной вилки и транскрипционного комплекса.
[00112] Учитывая, что около 50% модификации γН2АХ связаны с ретротранспозонной частью генома (рис. 10), а также на основании представленных ранее данных, что две самые распространенные категории ретротранспозонов, такие как SINE и LINE, различаются по степени содержания хроматина с высокой плотностью генов и низкой плотностью генов (межгенные участки), соответственно, с уклоном в сторону GC-богатых ДНК (Lander et al. (2001) Nature, 409: 860-921; Mouse Genome Sequencing Consortium, Waterston et al. (2002) Nature 420: 520-562), далее исследовали корреляционную зависимость между накоплением γН2АХ и содержанием GC-пар в хромосомах человека. GC-богатые области генома также склонны к накоплению повреждений ДНК (Cha and Kleckner (2002) Science, 297: 602-606), однако отсутствует какая-либо безусловная причина ожидать различий в отношении накопления повреждений, связанных с содержанием пар GC, между самообновляющимися и стареющими клетками. Действительно, при сравнении содержания GC пар и плотности метки γН2АХ в хромосомах, как это было сделано для плотности генов, наблюдали достоверную положительную корреляцию как для самообновляющихся, так и стареющих клеток hADSC (рис. 2Е и таблица 4). Положительная корреляция между содержанием GC пар и плотностью метки γН2АХ в хромосомах в стареющих клетках отличалась от отрицательной корреляции между плотностью генов и плотностью метки γН2АХ в тех же клетках. Этот результат является неожиданным, так как известно, что содержание GC пар положительно коррелирует с плотностью генов. Чтобы разделить эти две функциональные области генома, был проведен частичный корреляционный анализ плотности генов и плотности метки γН2АХ в стареющих клетках при контроле содержания GC. После проведения частичного корреляционного анализа в таком варианте значение коэффициента корреляция для плотности генов и плотности метки γН2АХ в стареющих клетках стало более отрицательным и статистически достоверным (табл. 4).
[00113] Полученные данные указывают на перераспределение γН2АХ-модифицированных нуклеосом из генных участков в самообновляющихся клетках в негенные участки в стареющих клетках hADSC. Эти данные согласуются с результатами, которые наблюдали при сравнительном полногеномном анализе относительного обогащения γН2АХ-модифицированных нуклеосом в самообновляющихся и стареющих клетках (рис. 2С). Перераспределение γН2АХ в процессе старения может также объяснить избыток SINE/Alu-связанных γН2АХ-модифицированных нуклеосом в самообновляющихся клетках, так как высокое содержание SINE/Alus-повторов показано как в генах, так и во фланкирующих гены областях, наряду с избытком L1-связанных γН2АХ-модифицированных нуклеосом в стареющих клетках, так как L1s главным образом локализован в межгенных участках (рис. 2С).
Распределение γН2АХ в промоторных областях
[00114] В проведенных ранее исследованиях было установлено, что для активации транскрипции необходимо образование временных двунитевых разрывов ДНК в промоторах индуцируемых генов (Ju et al. (2006) Science, 312: 1798-1802). Было предположено, что нарушение регуляции транскрипции в процессе клеточного старения может быть вызвано невозможностью успешно восстановить двунитевой разрыв ДНК (Tower (2006) Cell Metab., 4: 101-103). В этих условиях временные двунитевые разрывы ДНК в самообновляющихся клетках hADSC могут превратиться в очаги устойчивых повреждений ДНК в стареющих клетках.
[00115] Наблюдали значительные изменения накопления γН2АХ в проксимальных участка промоторов кодирующих белки генов в самообновляющихся клетках по сравнению со стареющими. В соответствии с общим распределением γН2АХ в геноме было зарегистрировано двукратное увеличение накопления γН2АХ в промоторных областях самообновляющихся клеток по сравнению со стареющими клетками; ~3,9% промоторных последовательностей самообновляющихся клеток содержали γН2АХ по сравнению с 2,0% для стареющих клеток (табл. 5). Кроме того, существовало относительно небольшое перекрытие генов, содержащих γН2АХ-модифицированные промоторы, между самообновляющимися и стареющими клетками (рис. 2G). Для обеих культур: самообновляющихся и стареющих клеток hADSC было обнаружено периодические обогащение γН2АХ в определенных положениях от -2 тыс. п.о. до +2 тыс. п.о. относительно точки инициации транскрипции (рис. 2Е). Эта модель обогащения согласуются с фазированием нуклеосом вокруг точки инициации транскрипции самообновляющихся и стареющих клетках, однако распределения были сдвинуты, что указывает на передислокацию γН2АХ-модифицированных нуклеосом по отношению к точке инициации транскрипции в образцах стареющих клеток.
[00116] Были исследованы гены, содержащие γН2АХ-модифицированные промоторные области в стареющих клетках hADSC (табл. 6), для того чтобы оценить их потенциальное воздействие на клеточные функции, связанные с фенотипом старения. Было показано наличие ряда избыточно представленных функциональных категорий генной онтологии (GO) среди этого набора генов, включая связанные категории клеточного цикла (GO:0007049) и пролиферации клеток (GO:0008283) (табл. 7). Гены в этих категориях продемонстрировали воздействие на клеточный цикл и деление клеток (табл. 8). Когда был проведен аналогичный анализ для группы регуляторных генов клеточного цикла (табл. 1, и не представленные данные), не обнаружили какой-либо корреляции между связанной со старением отрицательной регуляцией транскрипционной активности генов и накоплением γН2АХ в их промоторных областях.
[00117] Полученные данные показывают, что понижающая регуляция регуляторных генов клеточного цикла в стареющих клетках hADSC на уровне транскрипции не связана с накоплением двунитевых разрывов ДНК, как это предлагали ранее (Tower (2006) Cell Metab.,4: 101-103).
Распределение γН2АХ хроматина в перителомерном хроматине и старение взрослых стволовых клеток человека
[00118] В проведенных ранее исследованиях было установлено, что возникновение двунитевых разрывов ДНК, приводящих к старению соматических клеток, непосредственно вызвано нарушением функционирования теломер (Celli and de Lange (2005) Nat. Cell Biol., 7: 712-718; d′Adda di Fagagna et al. (2003) Nature, 426: 194-198; Takai et al. (2003) Curr. Biol., 13: 1549-1556). Было предположено, что эрозия теломер может обеспечить резервуар сигналов устойчивых повреждений ДНК, что в результате приводит к устойчивой активации р53 и проявлению фенотипа старения.
[00119] Для того, чтобы установить вовлечены ли непосредственно теломеры с нарушенными функциями в повреждение ДНК в стареющих клетках hADSC, исследовали хромосомное обогащение γН2АХ в теломерных и перителомерных областях человеческого генома.
[00120] Теломерные участки содержат очень большое число повторов и не охарактеризованы полностью на уровне последовательностей, поэтому сложно однозначно картировать распределение метки γН2АХ в теломерных областях генома человека методом ChIP-seq. Чтобы исследовать накопление γН2АХ в теломерах проведен анализ содержания ChIP-seq меток, которые включали специфичный мотив последовательности телемерного повтора (TTAGGG/AATCCC, (SEQ ID NO: 9), по отношению к общему содержанию всех секвенируемых меток (картировались однозначно и не картировались из-за наличия повторов). Образцы самообновляющихся клеток имели в среднем 1,66% теломерных меток, в то время как стареющие клетки имели 1,63%. Эти процентные доли не выявили избыточной представленности теломерных меток, и разница между двумя типами клеток не была значимой.
[00121] Определяли перителомерные области как участки в 100 тыс. п.о. на концах секвенированных хромосом человека и провели исследование участка геномной ДНК в 15 млн.п.о., который продлевается на плечи хромосом в самообновляющихся и стареющих клетках hADSC (рис. 3А). В целом, в SR образцах вся область была обогащена γН2АХ, тогда как образцы SEN были истощены в отношении γН2АХ. Тем не менее, как для асинхронно делящихся взрослых стволовых клеток, так и для стареющих клеток было отмечено снижение плотности γН2АХ в участках, которые прилегали и лежали внутри перителомерных областей (рис. 3А). Фактически общая доля γН2АХ-модифицированного хроматина, найденного в перителомерных участках, была довольно небольшой (SR=0,07%, SEN=0,05%,. фиг. 10), однако это может быть связано с относительно небольшой частью генома, которая включает перителомеры, как определено в настоящей заявке. Чтобы оценить эту возможность, вычислили геномное обогащение перителомерных областей γН2АХ-модифицированным хроматином в самообновляющихся и стареющих клетках как долю модифицированного хроматина, нормированную к доле генома, которая содержит перителомерные последовательности. При оценке перителомерных участков подобным образом было установлено, что содержание в них γН2АХ-модифицированных нуклеосом снизилось в 1,5 раза (SR) и 2,1 раза (SEN) (рис. 12).
[00122] Полученные данные подтверждают мнение, что старение взрослых стволовых клеток человека не может быть непосредственно вызвано интенсивным повреждением перителомерных областей из-за укорочения теломер, хотя это противоречит результатам, которые наблюдали в соматических клетках человека (d′Adda di Fagagna et al. (2003) Nature, 426: 194-198). Это наблюдение дополнительно подтверждает выводы, представленные ранее (Sedelnikova et al. (2004) Nat. Cell Biol., 6: 168-170). Однако при внимательном изучении был выявлен ряд отдельных хромосом со значительным накоплением γН2АХ в теломерах. Распределение γН2АХ в перителомерных областях этих хромосом было сильно асимметричным по отношению к плечам хромосом (рис. 3С). В самообновляющихся клетках теломеры 10, 12, 17 и 18-й хромосомы были обогащены γН2АХ, также как и 4, 7 и 18-й хромосомы в стареющих клетках hADSC. Несмотря на различия между самообновляющимися и стареющими клетками в накоплении γН2АХ в перителомерных областях, асимметрия плеча хромосомы всегда сохранялась. Относительные уровни специфичного в отношении плеча хромосомы накопления γН2АХ в перителомерных областях были идентичны для самообновляющихся и стареющих клеток (рис. 3С).
Распределение γН2АХ в перицентрических областях
[00123] Убедительные доказательства указывают, что перицентрический хроматин имеет очень важное значение для регулирования сборки кинетохора, сцепления сестринских хроматид, присоединения веретена деления и расхождения хромосом (Blower and Karpen (2001) Nat. Cell Biol., 3: 730-739; Bernard et al. (2001) Science 294: 2539-2542; Schueler and Sullivan (2006) Annu. Rev. Genomics Hum. Genet., 7: 301-313). Данные, полученные в экспериментах на дрожжах, также указывают на причинно-следственную связь между остановкой репликационной вилки и хромосомными двунитевыми разрывами ДНК в областях известных как «зоны замедления репликации» (RSZ) (Cha and Kleckner (2002) Science, 297: 602-606), многие из которых связаны с перицентрическим и центромерным хроматином. Кроме того, контрольная точка сборки веретена деления (SAC) оказывает некоторое избыточное действие совместно с контрольной точкой повреждения ДНК, чтобы осуществить долгосрочную остановку клеточного цикла в присутствии невосстановленного двунитевого разрыва ДНК, тем самым, предотвращая расхождение поврежденного хроматина (Dotiwala et al. (2010) Curr. Biol., 20: 328-332; Elledge (1996) Science, 274: 1664-1672; Harrison and Haber (2006) Annu. Rev. Genet., 40: 209-235; Lim et al (2009) Trends Cell Biol., 19: 325-333). В этой связи далее исследовали наличие признаков ломкости перицентрического хроматина человека в процессе старения взрослых стволовых клеток, а также будет ли он устойчив к накоплению γН2АХ в делящихся клетках.
[00124] Согласно данным распределение γН2АХ-модифицированного хроматина в перицентромерных участках существенно отличалось от распределения, которое наблюдали в перителомерных участках. Несмотря на то, что перицентромерные участки также содержали небольшую долю общего γН2АХ-модифицированного хроматина на протяжении всего геном (SR=0,78%, SEN=0,82%;. фиг. 10), в перицентромерных участках обнаружили существенно большую долю γН2АХ-модифицированного хроматина, чем ожидалось на основании размера той части генома, которую они занимают (рис. 3В). Было показано, что ряд отдельных хромосом имеет очень высокие уровни накопления γН2АХ в перицентромерных участках, что согласуется с данными о наличии повреждений в перителомерных участках конкретных хромосом (рис. 3D). В целом, перицентромерные участки демонстрируют увеличение уровня обогащения в 2,0 и 2,1 раза самообновляющихся и стареющих клетках, соответственно (рис. 12), что указывает на внутреннюю чувствительность («ломкость») этих участков хромосом. Перицентромерные участки были не только обогащены γН2АХ, они также представляли собой одну из немногих функциональных областей генома, показавшую относительно высокое содержание γН2АХ в стареющих клетках (рис. 2В, рис. 3D, 3Е). Наиболее высокие уровни обогащения γН2АХ в стареющих клетках обнаружили в 6, 14, 15, 16 и 21-й хромосоме (рис. 3Е).
[00125] Интересным фактом является то, что полученные данные не выявили равномерного и постепенного накопления γН2АХ в перицентрическом хроматине одних и тех же хромосом в процессе старения, это указывает на то, что накопление повреждений в этих участках при старении не обязательно является результатом простой передачи дефекта репарации ДНК, связанного с клеточным циклом.
Солокализация CENP-A и CREST с очагами устойчивых повреждений ДНК в стареющих ADSCs
[00126] Увеличение накопления γН2АХ в перицентрических участках только в определенной части хромосом человека в стареющих клетках hADSC может свидетельствовать о: I) невосстановленных двунитевых разрывах ДНК (возможно, в связи с изменением характеристик их перицентрического гетерохроматина и молекулярных механизмов, отвечающих за его репарацию), и/или II) дефектах сборки кинетохора, что провоцирует образование вызванных веретеном деления повреждений ДНК, которые приводят к активации контрольной точки веретена деления (Guerrero et al. (2010) Proc. Natl. Acad. Sci. USA, 107: 4159-4164). Оба эти события, действуя по отдельности или синергически, могут вызвать старение взрослых стволовых клеток человека.
[00127] Чтобы разграничить два возможных сценария, сначала оценили совместную локализацию устойчивых очагов γН2АХ/53ВР1 и центромер-специфического гистона CENP-A, который является вариантом гистона Н3 и необходим для сборки кинетохора, в стареющих клетках hADSC. Самообновляющиеся и стареющие клетки hADSC окрашивали антителами к 53ВР1 и CENP-A белкам и исследовали с помощью конфокальной флуоресцентной микроскопии. Почти в 75% случаев CENP-A был солокализован с большим очагом 53ВР1/γН2АХ в стареющих клетках hADSC (рис. 4А, В, рис. 13А, В). Сходную картину перекрывания сигналов антительных меток наблюдали, когда оценивали совместную локализацию человеческих антител к кинетохору (α-CREST) и антител к 53ВР1, либо антител к α-CREST и антител к γН2АХ методом конфокальной флуоресцентной микроскопии. Как и ожидалось, не наблюдали (уровень, детектируемый данным методом) ни солокализации центромерных участков с 53ВР1 (рис. 4а) или γН2АХ (данные не показаны) в интерфазе самообновляющихся клеток hADSC, поскольку в этих клетках не была инициирована длительная остановка клеточного цикла (Рис. 1В, рис. 7В), ни образования очагов γН2АХ/53ВР1 (рис. 1С). Данные указывают, что центромерные участки встроены в очаги устойчивых γН2АХ/53ВР1 повреждений ДНК, которые образуются при старении клеток hADSC, также до сих пор не наблюдали дефектов включения CENP-A в центромерный хроматин или сборки внутреннего кинетохора. CENP-А необходим для привлечения многих других белков, ассоциированных с центромерами и кинетохором (McClelland et al. (2007) EMBO J. 26: 5033-5047), за исключением белков, расположенных в соседних доменах гетерохроматина, таких как гетерохроматиновый белок 1 (НР1) (Blower and Karpen (2001) Nat. Cell Biol., 3: 730-739). He исключена возможность того, что дефекты других компонентов кинетохора/веретена деления, которые действуют ниже каскада CENP-A, могут влиять на сборку веретена деления, и что такие дефекты приведут к повышению уровней γН2АХ в перицентромерных участках из-за неправильного расхождения хромосом и остановки клеточного цикла (Dalton et al. (2007) Cancer Res., 67: 11487-11492; Guerrero et al. (2010) Proc. Natl. Acad. Sci. USA, 107: 4159-4164; Quignon et al. (2007) Oncogene, 26: 165-172). Действительно, при старении hADSCs наблюдали значительную понижающую регуляцию транскрипции KNTC1 (hROD), который является компонентом чувствительного к напряженности комплекса кинетохора, известного как RZZ (табл. 1) (Chan et al. (2000) Nat. Cell Biol., 2: 944-947).
Связанные со старением очаги γН2АХ являются сайтами активной Пол-III-зависимой транскрипции
[00128] Существует ряд источников информации, в которых показано, что перицентрический хроматин имеет важное значение для формирования и функционирования центромер (Bernard et al (1994) Exp.Cell Res., 214: 373-380; Dalai and Bui (2010) Curr. Opin. Cell Biol., 22: 392-402). В делящихся дрожжах нативные перицентрические участки хромосом доступны для взаимодействия с РНК Пол-III (Chen et al. (2008) Nature, 451: 734-737; Scott et al. (2007) PLoS One 2: e1099), что свидетельствует о сложной взаимосвязи между сборкой гетерохроматина, созреванием центромерного хроматина и транскрипцией. Ранее было показано, что связанные со старением виды активности, которые оказывают модифицирующее действие на хроматин у млекопитающих, например опосредуемые белками сиртуинами, участвуют не только в формировании факультативного гетерохроматина (Vaquero et al. (2007) Nature 450, 440-444), но также вовлечены в процесс репрессии генов в участках конститутивного гетерохроматина, таких как перицентромерный хроматин. Важно отметить, что образование двунитевых разрывов ДНК, индуцированных окислительным стрессом, приводит к увеличению транскрипции перицентрических сателитных повторов ДНК в модели эмбриональных стволовых клеток млекопитающих (Oberdoerffer et al. (2008) Cell, 135: 907-918).
[00129] С учетом этих фактов исследовали возможность существования транскрипционной активности, связанной с устойчивыми очагами γН2АХ/53ВР1, в стареющих клетках hADSC. При этом объединили метод исследования транскрипционной активности по включению 5-фторуридина (5-FUr) с методом обнаружения 53ВР1 и белка промиелоцитарного лейкоза (PML) при старении клеток hADSC (описано в разделе «Экспериментальные методики» и в Casafont et al. (2006) Neuroscience, 140: 453-462). Включение галогенированного аналога нуклеотида в растущую цепь РНК визуализировали в целом ядре методом иммуноцитохимического окрашивания антителами к BrdU (Boisvert et al. (2000) J. Cell Biol., 148: 283-292). Выбрали специфический ядерный компартмент - ядерные тельца PML, поскольку было показано, что они участвуют в некоторых аспектах регуляции транскрипции и/или сегрегации белка (Carbone et al. (2002) Oncogene 21: 1633-1640; Dellaire et al. (2006) J. Cell Biol., 175: 55-66; Dellaire et al. (2009) Cell Cycle, 8: 3750-3769). Наблюдали близкое взаимное расположение ядерных телец PML и устойчивых очагов 53ВР1, связанных со старением (рис. 4С). Растущие РНК-транскрипты заметно присутствовали в периферической части ядерного тельца PML и демонстрировали очевидную совместную локализацию с сигналом 53ВР1 (рис. 4С). Эти наблюдения согласуются с предыдущими сообщениями о возможном участии РНК в формировании очагов 53ВР1 после ИК-индуцированного повреждения в клетках линий NIH3T3 и Hela (Pryde et al. (2005) J. Cell Sci., 118:2043-2055).
[00130] Было предположено, что транскрипционный контроль посредством ретроэлементов может способствовать транскрипции перицентрических участков (Ugarkovic (2005) EMBO Rep., 6: 1035-1039). В экспериментах большие кластеры γН2АХ хроматина были солокализованы с перицентрическими участками, которые, как известно, обогащены повторами ретротранспозонов SINE/Alu (рис. 3В, 3D, 3Е, таблица 3, см. в Schueler and Sullivan (2006) Annu. Rev. Genomics Hum. Genet., 7: 301-313). С учетом того, что транскрипционная активность SINE/Alu-повторов в значительной степени повышена в стареющих клетках hADSC (рис. 14) и осуществляется Пол-III транскрипционным комплексом (Deininger (1989) Mobile DNA (Washington, DC, American Society for Microbiology)), была изучена динамика формирования устойчивых очагов 53ВР1/γН2АХ и включения 5-FUr в ответ на ингибирование Пол-III-зависимой транскрипции с помощью тагетина (Allen et al. (2004) Nat. Struct. Mol. Biol., 11: 816-821; Wang et al. (2003) Mol. Biol. Cell, 14: 2425-2435).
[00131] Стареющие клетки hADSC культивировали в присутствии 10 мкМ тагетина, ингибитора Пол III-зависимой транскрипционной активности, в течение 2 часов при 37°С (+тагетин) или без обработки ингибитором (-тагетин). Ядерную РНК метили добавлением в культуру 2 мМ FUr на 10 мин. при 37°С. После фиксации клетки окрашивали антителами к BrdU (красный), чтобы обнаружить сайты включения FUr в сочетании с анти-53ВР1. В ходе экспериментов с двойным окрашиванием было установлено, что сайты включения FUr локализованы исключительно возле очагов устойчивых повреждений ДНК на протяжении всей глубины изображений, полученных наложением серии снимков по z-оси. Ингибирование Пол-III-зависимой транскрипции тагетином привело к полному исчезновению сайтов включения FUr и к потере уплотнения сайтов повреждений ДНК, как было обнаружено по более диффузному окрашиванию 53ВР1.
[00132] Полученные данные выявили, что обработка стареющих клеток hADSC ингибитором Пол-III тагетином нарушала их способность к формированию очагов 53ВР1. Кроме того, специфичный в отношении нити сравнительный ОТ-ПЦР анализ ряда последовательностей ретротранспозонов SINE/Alu, связанных с повреждением ДНК в перицентрических областях 10-й (обозначенных MIR и Alu) и 21-й хромосомы (обозначенных AluSx и AluJb), показал, что повышение транскрипционной активности Alu-повторов коррелирует с наличием устойчивых повреждений ДНК в образцах стареющих клеток, как показано для Alu-повтора в непосредственной близости от кластера γН2АХ, имеющего размер 7,6 тыс. п.о. Данные показывают, что, несмотря на то, что транскрипционную активность выбранных повторов (AluJb, AluSx и Alu) регистрируют независимо от состояния hADSC, активность транскрипции Alu повышена в 10-й хромосоме, в которой повреждение наблюдается только при клеточном старении. Транскрипционная активность элемента MIR не была зарегистрирована ни в самообновляющихся, ни в стареющих клетках hADSC. Эти данные также подтверждают наблюдения, что очаги γН2АХ/53РВ1 в стареющих hADSC являются центрами Пол-III-зависимой транскрипционной активности (рис. 4С). С помощью обычного ChIP-seq анализа наблюдали увеличение привлечения Пол-III транскрипционного комплекса (обозначенного как TFIIIC) к MIR повтору при старении клеток даже при отсутствии транскрипции элемента MIR (рис. 5). Это могло быть связано со способностью TFIIIC являться не только критическим фактором для загрузки комплекса Пол-III (Young et al. (1991) Science, 252: 542-546; Kundu et al. (1999) Mol. Cell Biol., 19: 1605-1615; Noma et al. (2006) Cell, 125: 859-872), но и его способностью участвовать в структурной организации когезии хроматина и конденсации внутри эукариотического ядра, как недавно продемонстрировали у дрожжей (Iwasaki et al. (2010) Mol. Biol. Cell, 21: 254-265).
[00133] Наличие больших участков сателлитной ДНК ограничивает выбор специфичных в отношении нити праймеров для обратной транскрипции из-за более высокой степени комплементарности последовательности, препятствуя, тем самым, дальнейшей тщательной оценке множественных, плотно упакованных SINE/Alu повторов в пределах и вокруг этих геномных локусов. Тем не менее, полученные данные в совокупности с результатами нескольких исследований на дрожжах и млекопитающих (Lunyak et al. (2007) Science, 317: 248-251; Noma et al. (2006) Cell, 125: 859-872; Oki and Kamakaka (2005) Mol. Cell, 19: 707-716; Scott et al. (2006) Curr. Biol., 16: 119-129; Willoughby et al (2000) J. Biol. Chem., 275: 759-768) указывают на общую роль Пол-III транскрипции в структуре генома и подтверждают гипотезу о том, что, подобно центромерным генам тРНК у дрожжей (Iwasaki et al. (2010) Mol. Biol. Cell, 21: 254-265), перицентрические SINE/Alu-ретротранспозоны у человека могут играть важную роль в функционировании центромер в самообновляющихся клетках hADSC, а также в старении клеток hADSC.
Нарушенное связывание когезина и конденсина I с перицентрическими очагами γН2АХ в стареющих клетках hADSC
[00134] Несколько линий доказательств указывают на то, что процессы восстановления и сегрегации хромосом напрямую связаны через когезин, который является эволюционно-консервативным белковым комплексом (Heidinger-Pauli et al. (2008) Mol. Cell, 31: 47-56; Kim et al. (2002) Proc. Natl. Acad. Sci. USA, 99: 1241-1246; Sjogren and Nasmyth (2001) Curr. Biol., 11: 991-995; Strom et al. (2004) Mol. Cell, 16: 1003-1015; Unal et al. (2004) Mol. Cell, 16: 991-1002). Также ранее сообщалось о специфическом связывании когезина с SINE/Alu-повторами (Hakimi et al. (2002) Nature, 418L 994-998). Субъединица Scc1/Rad21/Mcd1 этого комплекса играет важную роль в функционировании когезинового комплекса, также было показано, что она необходима для сцепления сестринских хроматид и функционирования кинетохора в клетках позвоночных (Sonoda et al. (2001) Dev. Cell, 1: 759-770), а также контрольных точек повреждения ДНК в фазах G1 и G2-M (Jessberger (2009) EMBO J., 28: 2491-2493). В источниках информации имеются сообщения, что содержание когезинового комплекса увеличивается в участках двунитевых разрывов ДНК, где он способствует репарации ДНК путем гомологичной рекомбинации (HR) (Bekker-Jensen et al (2006) J. Cell Biol., 173: 195-206; Potts et al. (2006) EMBO J., 25: 3377-3388). Неэффективная пострепликативная репарация двунитевых разрывов ДНК может возникнуть в результате дефектов как загрузки когезина, так и превращения когезина в состояние обеспечивающее сцепление под действием фермента Ecol (ctf7) (Strom et al. (2004) Mol. Cell, 16: 1003-1015; Unal et al (2004) Mol. Cell, 16: 991-1002; Unal et al (2007) Science, 317: 245-248). Кроме того, нельзя упускать из виду важность для этих явлений пути, связанного с конденсином. Конденсин загружает когезин на хромосому и, как известно, образует два различных комплекса: конденсин I и II (Samoshkin et al. (2009) PLoS One, 4: e6831). Важно отметить, что комплекс конденсин I был вовлечен в функционирование перицентрического гетерохроматина. Истощение субъединицы Сар-Н комплекса конденсин I приводит к дефектам, связанным с изменениями в структурной целостности проксимального по отношению к центромерам гетерохроматина (Oliveira et al. (2005) Mol. Cell Biol., 25: 8971-8984). Было показано, что истощение Сар-Н не влияет на включение CENP-A в центрический хроматин или сборку кинетохора, однако приводит к серьезному истощению субъединицы когезина Scc1/Rad21/Mcd1, что вызывает дефекты клеточного цикла.
[00135] Было предположено, что транскрипция SINE/Alu в перицентрическом хроматине может влиять на загрузку конденсина и когезина, тем самым блокируя репарацию ДНК путем гомологичной рекомбинации. Невозможность восстановить повреждения ДНК прямо или косвенно приведет к исчезновению способности клеток hADSC самообновляться вследствие запуска процесса клеточного старения. Поэтому исследовали компоненты, обеспечивающие привлечение когезинового комплекса (Scc1/Rad21/Mcd1), комплекса конденсин I (Сар-Н) и ацетилазы гистонов Ecol к SINE/Alu-повторам в непосредственной близости от устойчивых перицентрических кластеров γН2АХ на 10-й хромосоме. Данные, полученные с помощью обычного ChIP анализа, показали статистически достоверную потерю привлечения Scc1/Rad21/Mcd1 и Сар-Н к MIR и Alu-повторам во время старения клеток hADSC (SEN красные полосы на рис. 5С) по сравнению со статусом тех же геномных локусов в самообновляющихся клетках (SR синяя полоса на рис. 5С). Было обнаружено, что привлечение ацетилазы гистонов Ecol в значительной степени коррелировало с формированием устойчивых кластеров γН2АХ при старении, и, по-видимому, не зависело от Scc1/Rad21/Mcd1. Этот результат указывает на другую, еще не идентифицированную, роль белка Ecol в DDR, которая отличается от установленной ранее функции в модулировании сцепленности Scc1/Rad21/Mcd1 (Hakimi et al. (2002) Nature, 418L 994-998; Strom et al. (2004) Mol. Cell, 16: 1003-1015; Unal et al. (2004) Mol. Cell, 16: 991-1002; Unal et al. (2007) Science, 317: 245-248).
[00136] В совокупности эти данные дополнительно поддерживают вывод о наличии корреляции между увеличением транскрипции SINE/Alu-повторов и (I) накоплением повреждений ДНК, (II) дефектами процесса привлечения конденсина I (Сар-Н) и когезина (Scc1/Rad21/Mcd1) в перицентрические области и (III) старением клеток hADSC.
Функциональное значение транскриптов ретротранспозонов SINE/Alu в старении клеток hADSC
[00137] SINE/Alu-элементы занимают 6% генома человека, значительно превосходя по числу копий другие семейства псевдогенов, которые возникли в результате ретротранспозиции (Weiner et al., 1986). Человеческий ретротранспозон SINE/Alu представляет собой тандемный повтор из двух элементов В1, связанных линкером, который богат аденином (рис. 6А), вторичная структура его РНК была установлена ранее (Sinnett et al. (1991) J. Biol. Chem., 266: 8675-8678). Для того, чтобы оценить функциональное значение транскрипции ретротранспозонов SINE/Alu в развертывании и/или индукции процесса клеточного старения при старении ex vivo стареющие клетки hADSC были генетически модифицированы для стабильной экспрессии мшРНК, нацеленной на общий транскрипт ретротранспозонов SINE/Alu. Для этого воспользовались общей областью последовательности ретротранспозонов SINE/Alu для создания ряда конструкций, содержащих мшРНК и несущих GFP для оценки эффективности трансдукции, которые доставляли в клетку в лентивирусных векторах (рис. 6В; «Экспериментальные методы»).
[00138] Нокдаун обычного транскрипта ретротранспозонов SINE/Alu в трансдуцированных клетках hADSC, стабильно экспрессирующих мшРНК, нацеленную на различные части SINE/Alu, был продемонстрирован методом нозерн-блот гибридизации (рис. 6С) с эффективностью трансдукции почти 99% (рис. 6В). Клетки hADSC, трансдуцированные lentiGFP, экспрессирующим sh-132Alu РНК, продемонстрировали почти полный нокдаун обычного транскрипта ретротранспозонов SINE/Alu. hADSCs, трансдуцированные lentiGFP (контроль) или lentiGFP sh-193Alu, не проявили или проявили очень незначительные изменения уровней транскрипции SINE/Alu. Неожиданно было установлено, что линии стареющих клеток hADSC, стабильно экспрессирующие sh-132Alu РНК, имели очень существенные изменения морфологического строения и демонстрировали увеличение пролиферации, как обнаружили в экспериментах по оценке поглощения [Н3]-тимидина при синтезе ДНК. В противоположность этому, в стареющих клетках hADSC, трансдуцированных lentiGFP (контроль) или обработанных полибреном один (РВ), не было выявлено существенных изменений скорости пролиферации по сравнению с диким типом (wt) стареющих клеток hADSC (рис. 6D). Достоверное восстановление синтеза ДНК после нокдауна транскрипта ретротранспозонов SINE/Alu затем регистрировали через 96 часов после трансдукции. Неожиданным явилось то, что при сравнении с wt стареющих hADSC, ранее бывшие стареющими линии клеток hADSC, которые стабильно экспрессировали sh-132Alu РНК, продемонстрировали статистически значимое увеличение активности факторов плюрипотентности Nanog и Oct4, как обнаружено методом количественной ПЦР (рис. 6Е). Уровни Nanog и Oct4 в wt стареющих клетках hADSC были ниже предела обнаружения метода количественной ПЦР.
[00139] В заключение следует отметить, что эти результаты указывают на возможность того, что РНК ретротранспозонов SINE/Alu является неожиданным, но ключевым компонентом механизма, который контролирует плюрипотентность стволовых клеток. В соответствии с полученными результатами наблюдали, что ранее бывшие стареющими hADSC со стабильным нокдауном общей РНК SINE/Alu были жизнеспособны и через 7 дней в культуре формировали клеточные агрегаты (структуры, аналогичные эмбриональным тельцам), которые дали положительный результат при испытании на активность щелочной фосфатазы - маркера индуцированной плюрипотентности (Рис. 6F).
Обсуждение
[00140] Повторяющиеся элементы участвуют в изменении/контроле свойств хроматина во многих системах, однако лишь в немногих исследованиях было изучено их функциональное значение в контексте клеточного старения, в частности, в стволовых клетках взрослого человека. Несколько наблюдений связывали устойчивый ответ на повреждение ДНК с проявлением клеточного фенотипа старения и в конечном итоге со старением клеток. Хотя считается, что клеточное старение представляет собой необратимую в естественных условиях остановку клеточного цикла, индуцированную сигнализацией повреждения ДНК, до сих пор неясно, что определяет неспособность клеток с активированным повреждением ДНК оперативно и правильно исправлять повреждения ДНК и восстанавливать способность к пролиферации. Однако на основании совокупных данных исследований различных моделей клеточного старения стало ясно, что эти детерминанты могут меняться в зависимости от типов клеток и интенсивности, продолжительности и характера повреждения ДНК.
[00141] В этой связи возникает возможность того, что в клетках существуют специфичные участки генома/элементы, где повреждение ДНК не так легко восстанавливается, возможно, по механизму, на который негативно влияют возрастные изменения структуры хроматина и/или организации хромосом. Эти функциональные элементы ранее не были идентифицированы. В этом исследовании была изучена эта проблема.
[00142] Центральное место в данной рабочей гипотезе занимала первая попытка выполнить анализ расположения модификации гистона γН2АХ, которая появляется на нуклеосомах, прилегающих к местам повреждений ДНК. С помощью ChIP-seq анализа, разрешение которого позволяет идентифицировать одну нуклеосому, смогли картировать расположение восстанавливаемых повреждений ДНК, которые не влияли на пролиферативную способность клеток hADSC, и провести сравнительный анализ динамики этих повреждений в начале клеточного старения. На основании данных по распределению модификаций γН2АХ был сделан вывод, что ни восстанавливаемые, ни устойчивые повреждения ДНК не возникают случайным образом в самообновляющихся и стареющих клетках hADSC, и локализация повреждений обоих типов смещена в сторону транспозируемой части генома. Повышенная активность транскрипции ретротранспозонов SINE/Alu в стареющих взрослых стволовых клетках человека достоверно коррелирует с образованием очагов устойчивых повреждений ДНК, что указывает на их роль в препятствовании процессу репарации ДНК. Представляем механистическое объяснение вмешательства, вызванного транскрипционной активностью ретротранспозонов, в реализацию ответа на повреждение ДНК: нарушение привлечения комплексов когезина и конденсина I к перицентрическому хроматину после их активации. Оба эти комплекса имеют решающее значение для организации хроматина и восстановления повреждений этих геномных областей. Предлагается новая модель, согласно которой SINE/Alu транспозоны действуют путем изменения структуры хроматина в непосредственной близости от очага повреждений ДНК, тем самым, блокируя их репарацию (рис. 6G). Это событие является основным для установления/индукции устойчивого ответа на повреждение ДНК при старении взрослых стволовых клеток человека ex vivo.
[00143] Несмотря на то, что до сих пор остается открытым вопрос о том, какой фактор регулирует активность транскрипции ретротранспозонов SINE/Alu, можно предположить, что это может быть связано с потерей эпигенетического контроля метилирования ДНК при старении клеток hADSC ex vivo. Тем не менее, нельзя исключить возможность того, что положительная регуляция транскриптов ретротранспозонов SINE/Alu непосредственно не связана с увеличением их транскрипционной активности, а скорее является результатом вызванных старением этих клеток дефектов посттранскрипционного процессинга, метаболизма или даже деградации. Независимо от их характера выводы представляют собой первый пример установления роли транскрипции ретротранспозонов SINE/Alu в изменении свойств перицентрического хроматина в сочетании с устойчивым ответом на повреждение ДНК и клеточным старением взрослых стволовых клеток человека.
[00144] Следует отметить, что полученные результаты демонстрируют функциональное значение транскриптов ретротранспозонов SINE/Alu для механизма старения hADSC ex vivo и бросают вызов догме, что клеточное старение, и в конечном итоге старость клеток, является необратимым процессом. Поскольку истощение типичных транскриптов SINE/Alu само по себе является достаточным, чтобы восстановить пролиферативную способность клеток hADSC и увеличить пластичность взрослых стволовых клеток человека, ранее ограниченных специфичными тканевыми нишами, вероятно, можно предположить, что SINE/Alu-ретротранспозоны играют важную роль в установлении характеристик мезенхимальной ниши через неидентифицированные пути, которые, тем не менее, взаимосвязаны со старением взрослых стволовых клеток.
Экспериментальные методы
Антитела
[00145] Использованные первичные антитела: b-актин (Abcam #ab62760), 53ВР1 (Bethyl Laboratories #А300-273А), BrdU (BD, 7580), САР-Н (Bethyl Laboratories #A300- 603A), CD31 (Invitrogen #HMCD3101), CD44 (Invitrogen #HMCD4401), CD45 (Invitrogen #HMCD4501), CD105 (Invitrogen #HMCD10520), фосфо-cdc2 (Tyr15) (Cell Signaling Technologies #9111), фосфо-Chk1 (Ser345) (CST #2341), фосфо-Chk2 (Thr68) (CST #2661), CENP-A (Abcam #ab13939), человеческое анти-центромерное аутоантитело, a-CREST (Antibodies Incorporated #15-235), Ecol (Bethyl Laboratories #A300-312A), γH2AX (Millipore #05-636), PML (Santa Cruz Biotechnology #sc-966) и Rad21 (Abeam # ab992). Вторичные антитела: AlexaFlour®, конъюгированные с антителами осла (Invitrogen).
Выделение и размножение стволовых клеток жировой ткани (ADSC) человека
[00146] Стволовые клетки жировой ткани человека выделяли из человеческой подкожной белой жировой ткани, собранной во время процедуры липосакции. Липоаспират ресуспендировали в сбалансированном солевом растворе Хэнка (HBSS), содержащем 3,5% бычьего сывороточного альбумина (BSA), 1% коллагеназы II типа (Sigma) в соотношении 1:3 вес/объем и встряхивали при 37°С в течение 50 мин. Клетки фильтровали через фильтр с размером пор 70 мкм (BD Falcon #352350), обрабатывали буфером для лизиса эритроцитов (150 мМ NH4C1, 10 мМ КНСО3, 0,1 мМ ЭДТА, рН=7,3) и размножали ex vivo в полной среде DMEM/F12 (DMEM/F12, 10% эмбриональной бычьей сыворотки, 100 ед/мл пенициллина, 100 мкг/мл стрептомицина, 2,5 мкг/мл амфотерицина В, Invitrogen) в атмосфере 10% CO2 при 37°С и перевивали при 80% конфлюентности, культуральную среду меняли каждые 72-96 часов. Общее число удвоений популяции рассчитывали путем суммирования удвоений популяции (PD=log(N/N0)×3.33, где N0 - количество клеток, которое помещали в матрас и N - число клеток, собранных при этом пассаже на протяжении нескольких пассажей как функцию количества дней выращивания в культуре.
Характеристика поверхностных маркеров
[00147] Образцы, содержащие 5×105 клеток, инкубировали в течение 30 мин. на льду в темноте с конъюгированными с флуорохромом антителами (CD31, CD44, CD45 и CD 105; Invitrogen) в фосфатно-солевом буфере с 1% БСА (Sigma), отмывали и исследовали с помощью мини-системы Guava EasyCyte (Guava Technologies, Millipore). Анализ данных выполняли на программном обеспечении FlowJo (Tree Star, Ашланд, Орегон, США).
Оценка клеточного старения и фенотипа плюрипотентности
[00148] Количественный анализ активности связанной со старением J3-галактозидазы проводили, как описано в инструкции производителя для набора (BioVision). Изменение клеточного фенотипа на фенотип индуцированных плюрипотентных клеток оценивали с помощью набора для окрашивания щелочной фосфатазы (Stemgent).
Иммунофлюоресценция
[00149] Примерно 1-3×104 клеток/лунку в 4-луночных стеклах фиксировали 4% параформальдегидом и пермеабилизировали 0,5% Тритона Х-100 в фосфатно-солевом буфере. Блокирование и инкубацию антител проводили в 4% нормальной сыворотке осла, приготовленной в фосфатно-солевом буфере (NDS; Jackson Immunochemicals). Ядра окрашивали 100 нг/мл 4′,6-диамидино-2-фенилиндола (DAPI, Sigma), стекла монтировали в водной среде для монтирования ткани ProLong® Gold antifade (Invitrogen).
[00150] Изображения в режиме эпифлуоресценции получали на флуоресцентном микроскопе Olympus ВХ60 с программным обеспечением Spotfire 3.2.4 (Diagnostics Instruments). Конфокальные изображения (толщина среза Z-серии 0,39 мкм) получали на конфокальном двухфотонном микроскопе Carl Zeiss (LSM 510 NLO), снабженном 488 нм аргоновым, 543 нм гелий-неоновым и когерентным титансапфировым Coherent Chameleon лазерами, с использованием 63х Plan Аро объектива и разрешением 0.08×0.08×0.39 мкм вокселей. Деконволюцию изображений для серии срезов проводили с использованием программного обеспечения Huygens Professional 3.4.0 (Scientific Volume Imaging, Нидерланды) и визуализировали в Bitplane Imaris 6.3.1 (Bitplane Inc., Сент-Пол, Миннесота, США).
Включение метки 5-фторуридина для детектирования РНК-транскриптов in situ
[00151] Стареющие клетки hADSC, 1x104 клеток/лунку в 4-луночных стеклах, обрабатывали 2 мМ 5-фторуридина (FUr; Sigma)) в течение 10 минут. Ядра подвергали воздействию ледяного буфера CSK (100 мМ KCl, 300 мМ сахарозы, 10 мМ Pipes, рН=6,8, 3 мМ MgCl2, 1 мМ EGTA, 1,2 мМ фенилметилсульфонилфторида) с 0,5% Тритона Х-100 и фиксировали 4% параформальдегидом. Иммунное окрашивание проводили, как описано выше для иммунофлуоресценции.
Получение профиля экспрессии
[00152] Тотальную РНК от ~1,5×106 самообновляющихся и стареющих клеток ADSC готовили согласно методике с использованием TRIZOL® (Invitrogen). Набор Human Cell Cycle qRCR Array (SABiosciences #PAHS-020C-2) использовали для профилирования (7500 Fast PCR System, Applied Biosystems). Три образца РНК от самообновляющихся клеток hADSC сопоставляли с образцами РНК для демонстрирующих репликативное старение стареющих клеток. Статистическую значимость различий транскрипционной активности рассчитывали с использованием t-теста, как описано в руководстве SABiosciences и на сайте «www.sabiosciences.com/RTPCR». Основанные на использовании модели значения экспрессии рассчитывали и фильтровали для генов, разность в уровне экспрессии которых между образцами самообновляющихся и стареющих клеток была выше или ниже в 2 раза. Категории генов описывали с использованием системы классификации PANTHER (анализ белка через эволюционные связи).
Подготовка лизатов нуклеосом
[00153] Образцы самообновляющихся и стареющих hADSC, содержащие 3×105 клеток, промывали фосфатно-солевым буфером и сшивали 1% формальдегидом. Клетки снимали с чашки в буфере CSK с 0,5% Тритона Х-100, промывали и обрабатывали ультразвуком в буфере с микрококковой нуклеазой (50 мМ Трис-HCl, рН=7,9, 5 мМ CaCl2; NEB) с помощью соникатора Covaris S2 Sonicator (частота развертки на 1% рабочего 35 цикла, 5 интенсивность и 50 циклов в пакете в течение 12 циклов по 10 секунд каждый при 4°С). Соникаты центрифугировали, чтобы избавиться от продуктов распада клеток, супернатанты инкубировали с 25 Ед/мкл микрококковой нуклеазы (NEB).
ChIP-Seq: приготовление нуклеосомного ChIP и библиотеки SOLiD
[00154] Нуклеосомные лизаты разбавляли буфером для связывания с антителом (25 мМ Трис-HCl, рН 7,9, 0,15% ДСН, 1% Тритона Х-100, 150 мМ NaCl) и подвергали иммунопреципитации с 10 мкг антитела к γН2АХ в течение ночи при 4°С. Гранулы белок А-сефароза блокировали 10 мкг/мл гликогена, 10 мкг/мл лизоцима, 5 мкг/мл тРНК и 20 мкг/мл БСА в течение 1 часа при 4°С перед связыванием с иммунными комплексами в течение 1 часа при 4°С. Гранулы промывали буфером для связывания и снова тем же буфером, содержащим 0,5 М NaCl и затем буфером ТЕ (10 мМ Трис-HCl, рН=7,6, 1 мМ ЭДТА). Иммунные комплексы, связанные с гранулами, повторно ресуспендировали в буфере ТЕ, разъединяли межмолекулярные сшивки и очищали путем обработки протеиназой К, экстракцией в смеси фенол:хлороформ и осаждением изопропанолом. Фрагменты ДНК готовили для лигирования адаптера посредством заполнения концов ДНК-полимеразой I (фрагмент Кленова) и фосфорилирования 5′-концов ПЦР-праймеров полинуклеотидкиназой (NEB), лигировали с 30-кратным молярным избытком адаптеров SOLIDTM System 2.0 (Applied Biosystems) для приготовления библиотеки в соответствии с протоколом производителя. Библиотеки ДНК амплифицировали с помощью ПЦР с использованием клонированной ДНК-полимеразы Pfu (Stratagene, Agilent Technologies). Проводили отбор фрагментов ДНК, имеющих мононуклеосомный размер, в соответствии с их размером на 2% агарозном геле, вырезали на уровне примерно 200 п.о. и очищали с помощью QIAQUICK® Gel Extraction Kit (Qiagen). Реакционную смесь для ПЦР в эмульсии (ePCR) для секвенирования препаратов гранул готовили с 0,05 и 0,1 пг/мкл ДНК-библиотеки для каждого образца. Образцы секвенировали на платформе SOLiDTM System 2.0 (Applied Biosystems) в соответствии с протоколом производителя.
Анализ ретротранспозонной транскрипции методами ОТ-ПЦР и кПЦР
[00155] Геномные координаты SINE/Alu-элементов, исследованных методом ОТ-ПЦР, брали из Ассамблеи Браузера генома Калифорнийского университета в Санта-Круз (genome.ucsc.edu) 6 марта 2006 года (NCBI36/hg18); MIR: chr10:41922815-41922906, Alu: chr10:41928992-41929118, AluJb: chr21:10141132-10141429 и AluSx: chr21:10145344-10145644. Общую РНК количеством 100 нг на реакцию использовали для RT2 First Strand Kit (SABiosciences). Праймеры для синтеза первой цепи находятся в локусах, расположенных за пределами последовательностей SINE/Alu-элемента (внешние или обратные праймеры, таблица 9), тогда как прямые праймеры локализованы в последовательности SINE/Alu-элемента (внутренние или прямые праймеры, таблица 9). В качестве положительного контроля использовали RPL13A, глицеральдегид-3-фосфатдегидрогеназу.
[00156] Для количественной ПЦР в режиме реального времени по 1 мг общей РНК каждого образца использовали для синтеза первой нити к ДНК с помощью SUPERSCRIPT® III First-Strand Synthesis System для ОТ-ПЦР (Invitrogen) со случайным праймированием. кПЦР проводили с использованием RT2 SYBR Green qPCR MasterMix (SABiosciences) и выполняли на LightCycler® 480 II (Roche). Праймеры для кПЦР приведены в таблице 9. Анализ данных относительной экспрессии генов выполняли методом дискретного косинусного преобразования 2 типа (2-DCt).
Лентивирусные мшРНК конструкции
[00157] Лентивирусные мшРНК конструкции для нокдауна генетических транскриптов SINE/Alu были сконструированы следующим образом: олигонуклеотиды Ленти sh-132 Alu РНК: 5′-GATCCCCCCACCACGCCCGGCTAATTTTCAAGAGAAATTAGCCGGGCGTGGTGGTTTTTGGAAA-3′ (SEQ ID NO: 7) и Ленти sh-193 Alu РНК 5′-GATCCCCCCCGGGTTCAAGCGATTCTTTCAAGAGAAGAATCGCTTGAACCCGGGTTTTTGGAAA-3′ (SEQ ID NO: 8) ренатурировали с равным количеством их комплементарных нитей, создавая липкие концы, содержащие специфические сайты рестрикции для клонирования, и лигировали в вектор pENTR/PTER+, очищенный на геле после рестрикции HindIII и BglIII (Campeau et al., 2009). Конструкции подтверждали секвенированием (последовательность смысловой нити показана выше). Равные количества каждой конструкции смешивали с вектором pLenti-CMV-GFP DEST (Campeau 25 et al., 2009) в реакции с LR Clonase для того, чтобы рекомбинировать клонированные мшРНК продуцирующие элементы в целевой вектор в соответствии с инструкциями производителя (Invitrogen). Полученную лентивирусную плазмиду трансформировали в клетки E.coli Stbl3 (Invitrogen) для амплификации.
Получение и трансдукция лентивирусов
[00158] Клетки 293Т выращивали в полной среде DMEM (DMEM с высоким содержанием глюкозы, 10% ЭБС, 0,1 М заменимой аминокислоты, 6 мМ L-глутамина, 1 мМ пирувата, 100 Ед/мл пенициллина, 100 мкг/мл стрептомицина) для получения лентивирусов клетки трансфицировали в течение 12 часов pLenti sh-132Alu или pLenti sh-193 Alu и pCgpV, pRSV-Rev, а также вспомогательными плазмидами pCMV-VSV-G (Allele Biotechnology) в молярном соотношении 2:1:1:1 с использованием Липофектамина 2000 (Invitrogen) в соответствии со стандартным протоколом (Campeau et al., 2009). Среду собирали через 48 и 72 часов и фильтровали (фильтр 0,45 мкм). Вирус осаждали ПЭГ и замораживали в аликвотах (-80°С). Лентивирусную трансдукцию проводили в полной среде с 5 мкг/мл Полибрена (Santa Cruz Biotechnology) в течение 12 часов. Титры вирусов определяли путем сравнения содержания GFP-положительных клеток с общей популяцией клеток.
Индекс пролиферации
[00159] Для каждого условия в экспериментах по включению [Н3]-тимидина примерно по 10000 клеток обрабатывали [Н3]-тимидина с активностью 1 мкКи (Perkin-Elmer, Бостон, Массачусетс) в полной среде DMEM/F12 в течение 24 часов. ДНК выделяли из собранных клеток и количественно определяли с помощью NanoDrop (ND-1000; NanoDrop Technologies Inc). Поглощение [Н3]-тимидина в клеточной ДНК измеряли на жидкостном сцинтилляционном счетчике (LS 6500; Beckman Instruments). Лентивирусную трансдукцию проводили, как описано выше, трансдукцию необработанных и обработанных Полибреном контролей проводили параллельно.
Нозерн гибридизация
[00160] РНК выделяли из самообновляющихся и стареющих клеток hADSC (с или без лентивирусной трансдукции) с помощью набора mirVANA (Ambion, Invitrogen), по 2 мг общей РНК/дорожку разделяли на геле, приготовленном в ТЕ буфере с 7 М мочевины и 6% полиакриламида. Гель окрашивали бромистым этидием, фотографировали и подвергали электроблоттингу на HybondTM-N+(Amersham, GE Healthcare). Гибридизацию проводили с использованием последовательных промывок 6×SSC, 4×раствор Денхардта, 0,1% ДСН при 37°С. Олигонуклеотидные зонды метили биотин-16-дУТФ (Roche) с помощью терминальной трансферазы (NEB). Нозерн-блот визуализировали с помощью конъюгата пероксидазы хрена со стрептавидином (Invitrogen) с использованием ECL Plus Western Blotting Detection Reagent (Amersham, GE Healthcare) и Amersham Hyperfilm (GE Healthcare). Использованные в качестве зондов олигонуклеотиды включали: SINE/Alu 132: 5′-CCACCACGCCCGGCTAATTT-3′ (SEQ ID NO: 9) SINE/Alu 90: 5′-CGCGCGCCACCACGCCCGGCTAATTTTTGTATTTTTAGTAGAGACGGGGTTTCACCATGTTGGCC-3′ (SEQ ID NO:10).
Пример 2
Дополнительные методы
Удаление выбросов данных
[00161] Метки γН2АХ ChIP-seq были нанесены на карту референсной последовательности генома человека по состоянию на март 2006 года (NCBI build 36.1, UCSC hg18) с помощью SOLiD™ System Analysis Pipeline Tool (Corona Lite). После картирования расположения меток в геноме согласно данным ChIP-Seq для каждого биологического повторного эксперимента сравнивали содержание меток между повторными экспериментами для идентичных геномных положений во всех хромосомах. Для проведения сравнения определяли содержание меток для каждого положения индивидуального нуклеотида в каждом повторном эксперименте. Затем нуклеотидные положения, характеризующиеся количеством метки, равным 0 во всех повторах, исключали из рассмотрения. Эту процедуру проводили, поскольку положение, имеющее 0 меток в обоих наборах данных, не будет учитываться в последующем анализе из-за отсутствия в ней сигнала. Помимо этого, если положение характеризуется содержанием метки, равным нулю в одном повторе, и ненулевое количество метки в другом повторе эксперимента, то положение считается ненадежным. В отношении нуклеотидных положений, которые имеют ненулевое содержание метки в обоих повторах эксперимента, для сравнения содержания меток по всем нуклеотидным положениям использовали линейный регрессионный анализ. На основании линии регрессии и распределения данных вокруг линии 95% доверительные границы определяли с использованием формулы Воркинга-Хотеллинга следующим образом: для каждого значения х ожидаемое значение у-Е(у) оценивали на основании линейной регрессионной модели Е(у)=mx+b. Затем для каждого приблизительного значения Е(у) вычисляли стандартное отклонение - s(y) - на основании распределения данных вокруг Е(у) для соответствующего значения х. Для каждого значения Е(у) верхние и нижние значения доверительного интервала вычисляли как Е(у)±w·s(y), где константу w определяли на основании F распределения при уровне достоверности а со степенями свободы 2 и n-2: w=√(2·F[(1-α),2, n-2]). Регрессионный анализ и определение доверительных границ проводили для каждой отдельной хромосомы. Определенные таким образом положения нуклеотидов, которые выпадают из доверительных границ, удаляли из дальнейшего рассмотрения. Результаты повторных экспериментов хорошо согласовались между собой.
Снижение шума и объединение данных.
[00162] После удаления выбросов выполняли снижение шума для того, чтобы эмпирически определить пороговое значение содержания метки th, выше которого отдельные положения генома, имеющие размер нуклеосомы (например, мононуклеосомы) рассматривали как модифицированные. Размер окна в 200 б.п.был выбран для этого анализа на основании содержания 147 пар оснований ДНК в одной нуклеосоме плюс линкер. Среднее содержание метки в окне размером 200 пар оснований - λ200 рассчитывали как общее содержание метки в геноме n, разделенное на длину генома 1 и умноженное на размер окна 200:λ200=(n/1)·200. Распределение Пуассона параметризовали значением 200 для моделирования фонового шума и определения порогового значения сигнала содержания метки th для модифицированных положений генома, содержащих нуклеосомы: где τ = количество метки в окне и Р - порог р-значения с поправкой Бонферрони. Для выполнения этой процедуры суммировали соответствующие значения содержания метки, специфичные в отношении положений для повторных экспериментов и сравнивали с распределением Пуассона, параметризованным суммой 200-значений, вычисленных для каждого повтора. Этот подход был использован, поскольку повторы представляют собой независимые биологические повторы, каждый из которых моделируется отдельным распределением Пуассона. Таким образом, суммирование величин содержания метки, специфичных в отношении положений по всему геному, может быть смоделировано распределением Пуассона с параметром, равным сумме отдельных 200-значений каждого повтора. В результате этой процедуры получили классификацию всех геномных положений нуклеосомного размера как модифицированных или немодифицированных на основании данных, которые были объединены между повторами экспериментов и не имели выбросов.
Основанный на максимальном размере сегмента алгоритм для анализа кластеризации смежных модифицированных нуклеосомных положений
[00163] Помимо анализа фрагментов модифицированной геномной ДНК мононуклеосомного размера, описанного выше, также исследовали кластеры смежных солокализованных модифицированных нуклеосом. Кластеры оперативно определяли как непрерывные геномные участки, где количество модифицированных фрагментов мононуклеосомного размера значительно больше, чем средний фоновый геномный уровень модифицированных положений. Для того, чтобы выявить и разграничить такие кластеры мы применили алгоритм максимального размера сегмента. Для выполнения этой процедуры сначала разработали бинарную оценочную схему, которая характеризует фрагменты мононуклеосомного размера (200 тыс. п.о.) как модифицированные или немодифицированные. Эту процедуру используют для определения бинарной полногеномной карты оценок нуклеосом. Затем для определения кластеров к геномной карте бинарных оценок нуклеосом применяли алгоритм максимального сегмента. Подробное описание алгоритма, позволяющего анализировать сегмент максимального размера, представлены в других работах (Ruzzo and Tompa (1999) Proc. Int. Conf. Intell. Syst. Mol. Biol., 234-241). Ниже описана оценочная схема для отдельных положений нуклеосом.
Бинарная схема оценки нуклеосом
[00164] Бинарную оценочную схему реализуют таким образом, чтобы присвоить логарифмическую вероятность того, что отдельные фрагменты мононуклеосомного размера модифицированы или немодифицированы. Оценки присваиваются как: s=ln(q/p), где р - плотность модифицированных нуклеосомных положений по всему геному, вычисленная как количество модифицированных положений, деленное на общее число положений и q - плотность модифицированных нуклеосомных положений в реальных кластерах. Невозможно непосредственно рассчитать q, так как кластеры неизвестны заранее. Для того чтобы оценить q распределение Пуассона параметризовали, используя р, т.е. плотность модифицированных нуклеосомных положений по всему геному. Затем, с учетом доверительного уровня Р, пороговое значение q может быть выбрано с помощью распределения Пуассона, как показано ниже:
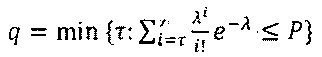
где τ = количество модифицированных нуклеосомных положений. Эта оценочная схема в результате позволила назначить одну положительную оценку всем модифицированным положениям нуклеосом и одну отрицательную оценку всем немодифицированным положениям. Этот подход принят на основании доказательств того, что логарифмические отношения правдоподобия такого рода являются оптимальной оценкой для идентификации смежных геномных сегментов (Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA, 87: 2264-2268). Кроме того, значение q, выбранное для оценочной схемы, обеспечивает контроль выбора кластеров, поскольку плотность модифицированных нуклеосом в кластере будет больше или равна значению q.
Относительная энтропия для определения больших кластеров.
[00165] После получения гистограмм распределения размеров смежных модифицированных нуклеосомных участков (то есть кластеров), классифицировали кластеры на 3 группы: 1) мононуклеосомные кластеры, 2) кластеры среднего размера и 3) большие кластеры. Для того, чтобы установить пороговое значение размера между средними и большими кластерами рассчитали значения относительной энтропии для различных порогов между распределениями размеров в самообновляющихся и стареющих клетках (рис. 11А). В качестве порогового было принято значение, которое обеспечивает максимальную величину относительной энтропии между двумя распределениями (рис. 11С). Таким образом, величина разности долей средних и больших кластеров в двух типах клеток является максимальной.
Сравнение с функциональными геномными областями.
[00166] Картирование распределения меток в геноме и контроль качества процедур, как описано выше (см. разделы Удаление выброса, Снижение шума и объединение данных), позволили получить карты положений нуклеосом в геноме человека, которые можно с уверенностью считать модифицированными гистоном γН2АХ для самообновляющихся (SR) и стареющих (SEN) клеток hADSC.
[00167] Эти карты использовали для сравнения расположения γН2АХ-модифицированных нуклеосом с различными функциональными геномными областями, представляющими интерес, внутри и между самообновляющимися и стареющими клетками. Например, сравнивали расположение модифицированных нуклеосом в геноме с расположением точки инициации транскрипции, экзонов, интронов, последовательностей повторяющихся элементов, перицентрических участков и перителомерных участков. Указанные сравнения выполняли с помощью UCSC Table Browser tool (Karolchik et al. (2004) Nucleic Acids Res., 32: D493-D496), который является частью набора инструментов Браузера Генома человека и аннотаций генома Калифорнийского университета Санта-Круз (Karolchik et al. (2003) Nucleic Acids Res., 31: 30 51-54). Кроме того, пользовательские Perl-скрипты использовали для изменения записей Браузера Генома человека UCSC, таких как записи аннотаций функций генов, чтобы извлечь специфичные области, представляющие интерес, включая расположение точки инициации транскрипции и границы экзон/интрон для дальнейшего анализа.
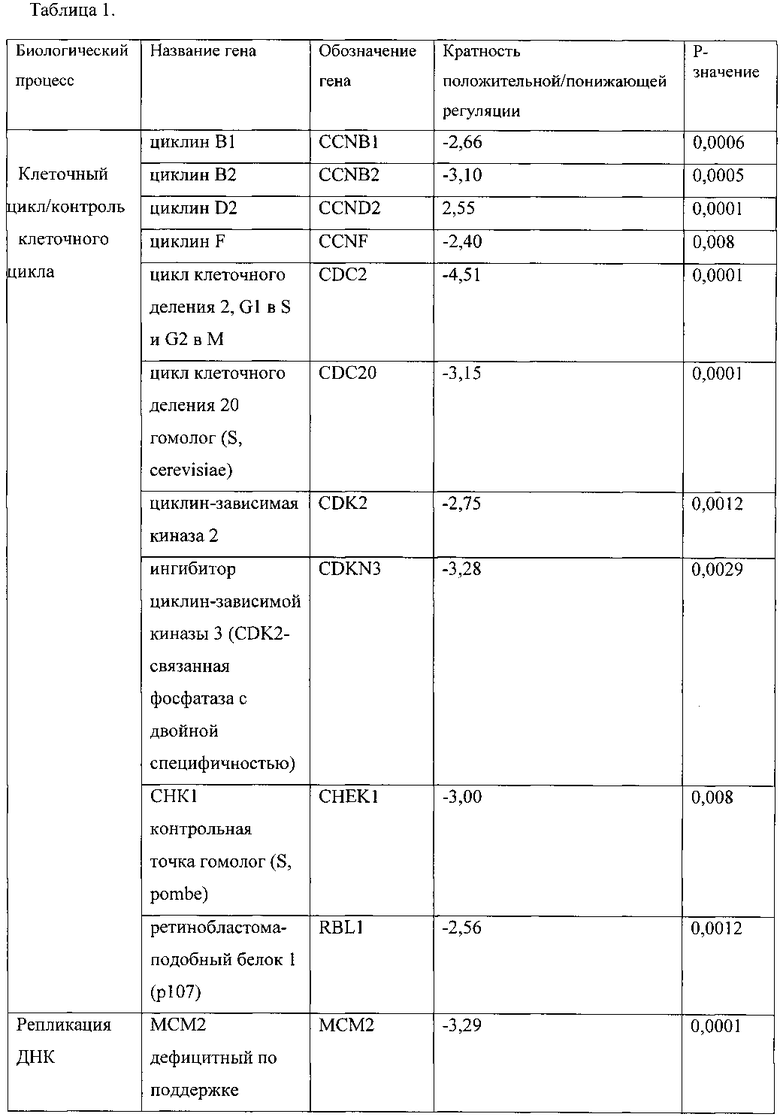
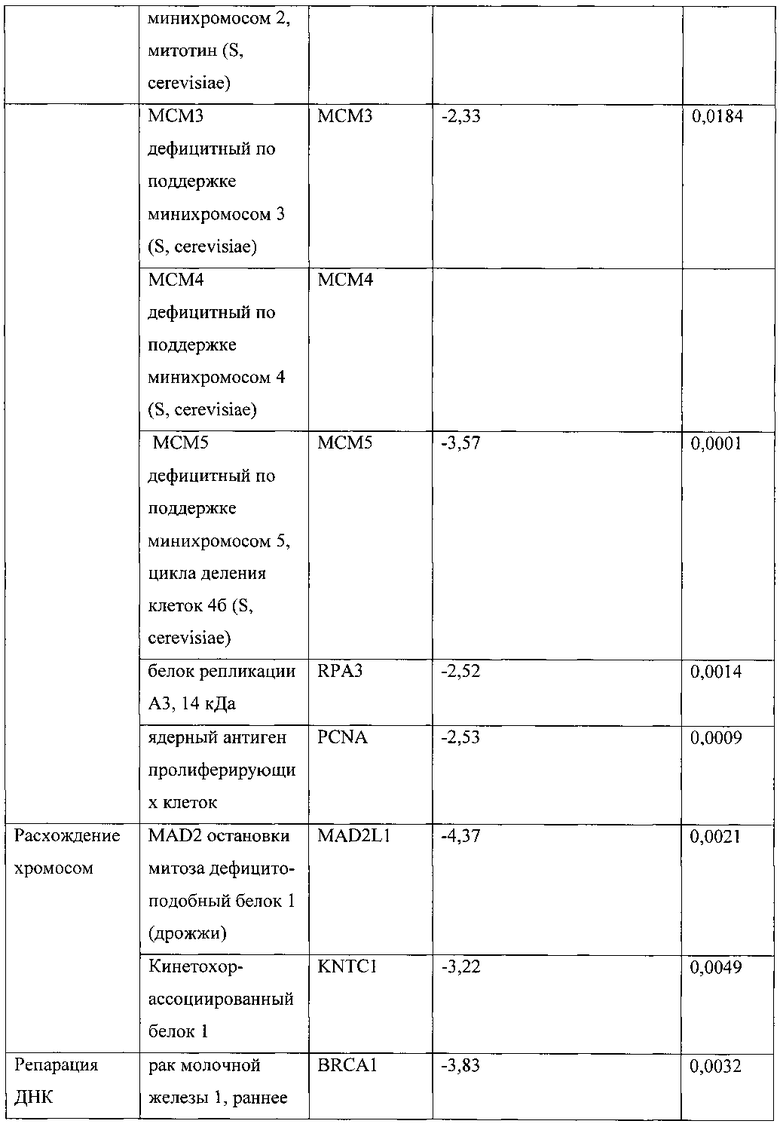
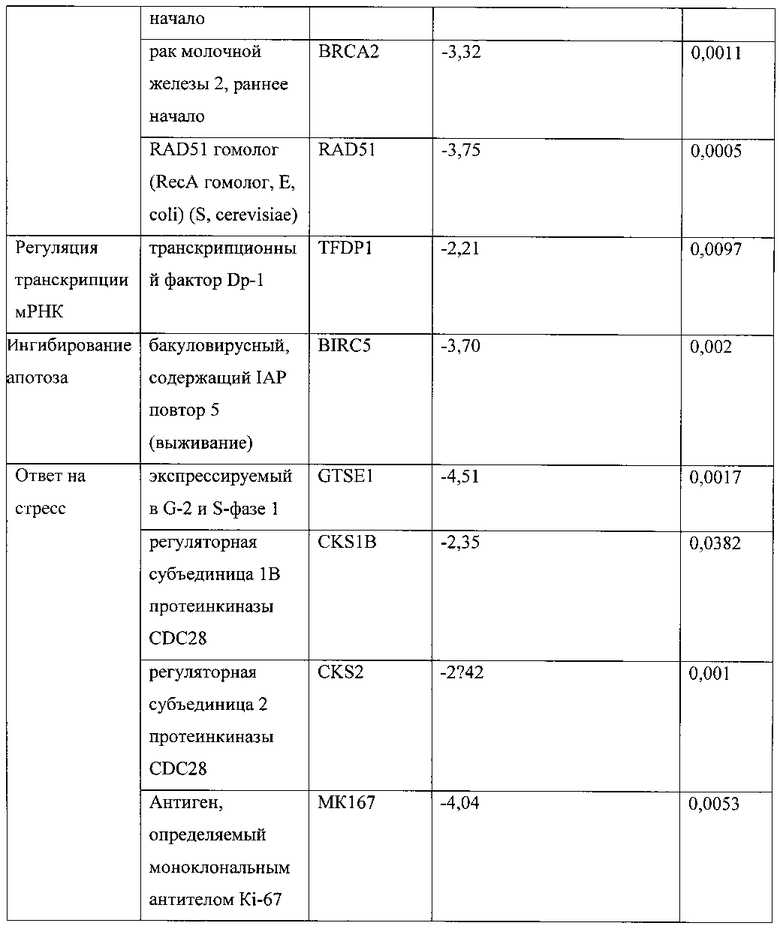
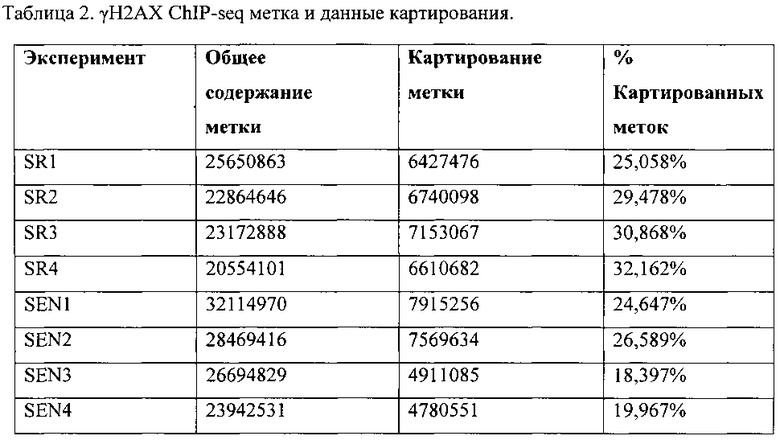
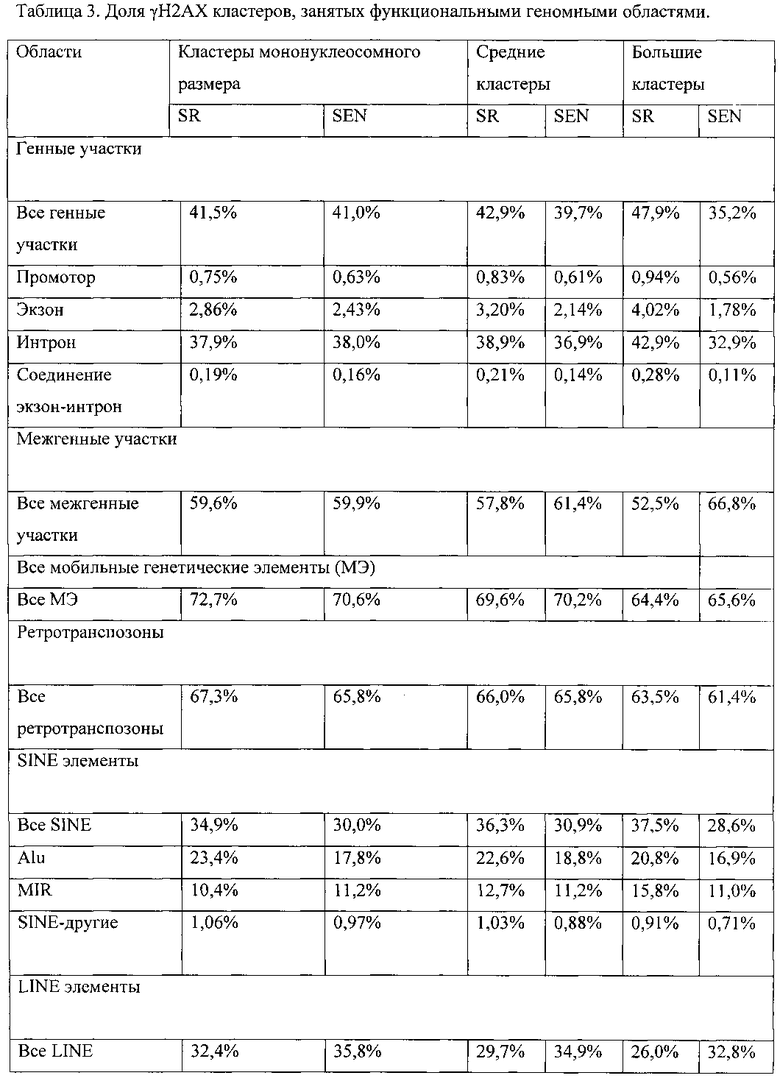
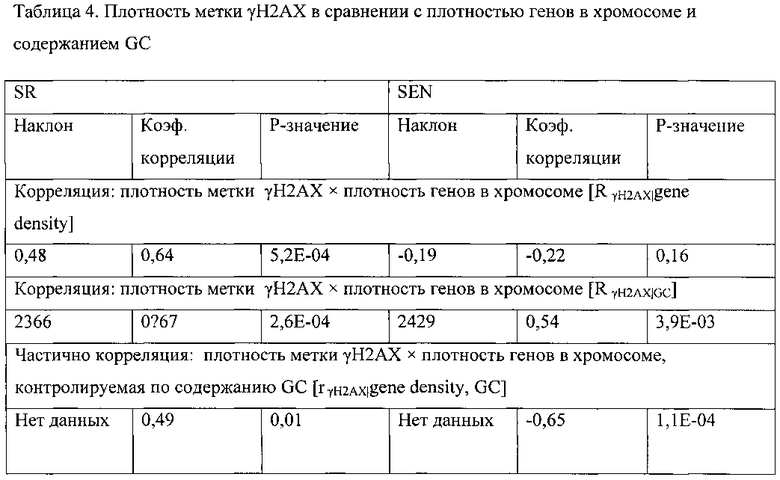
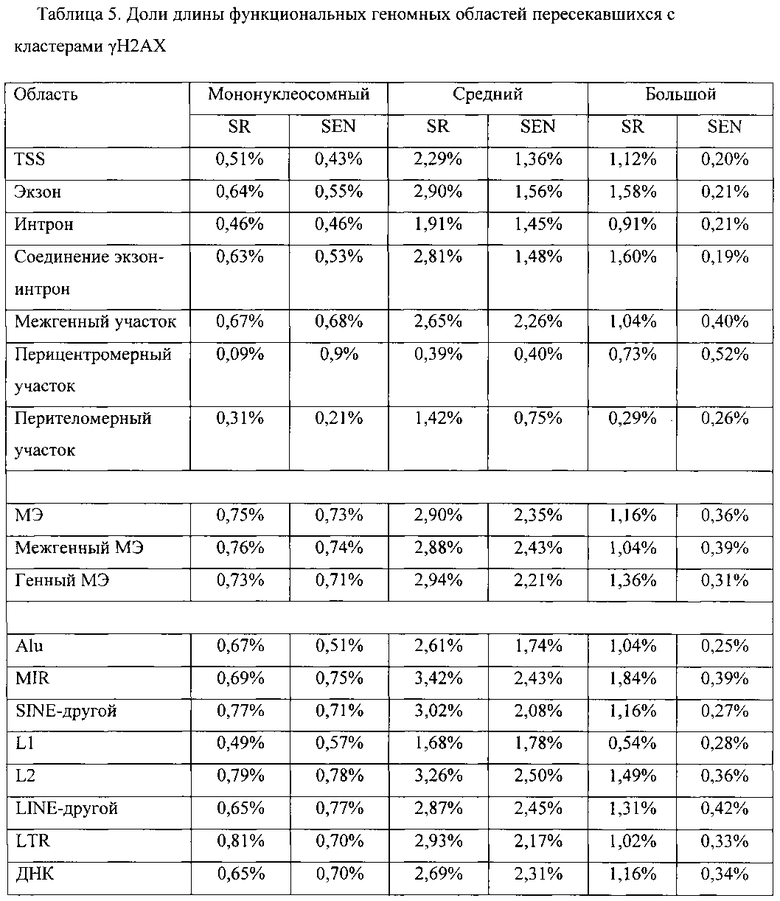
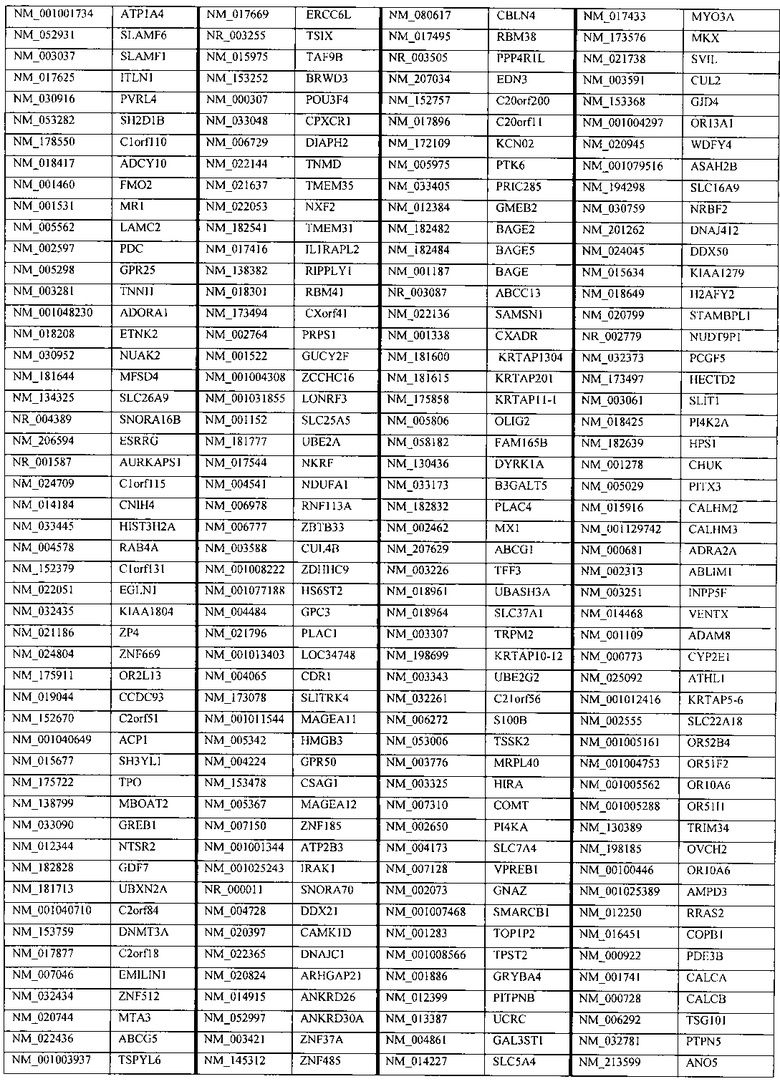
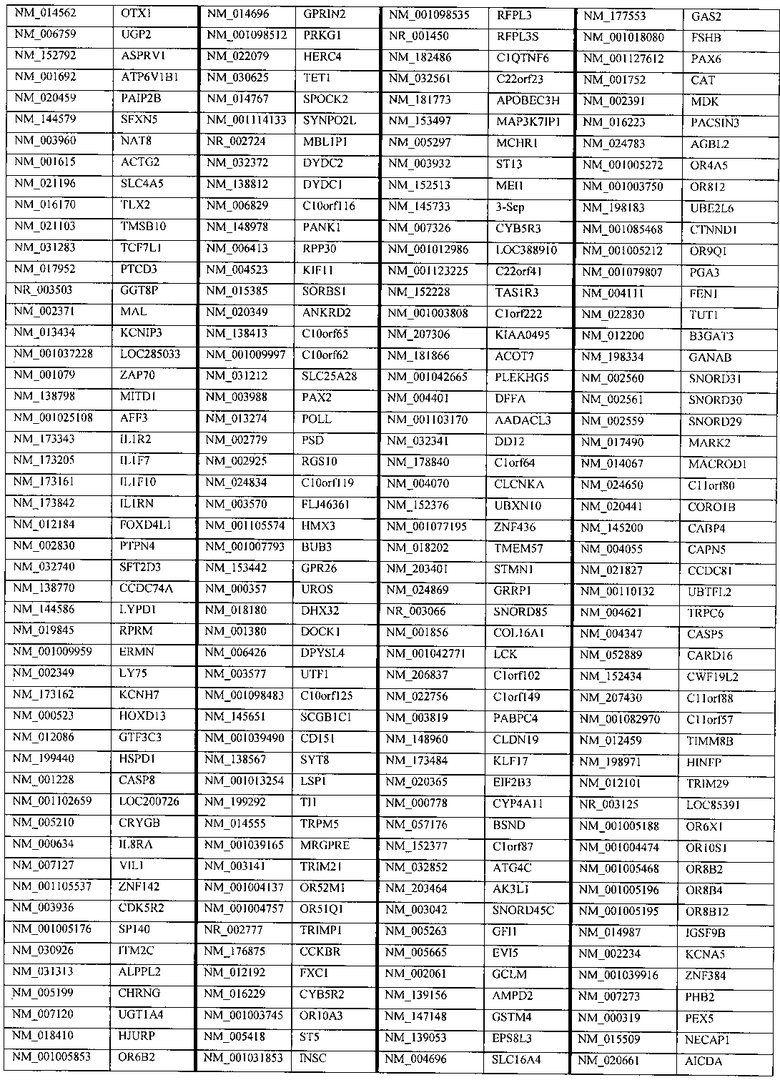
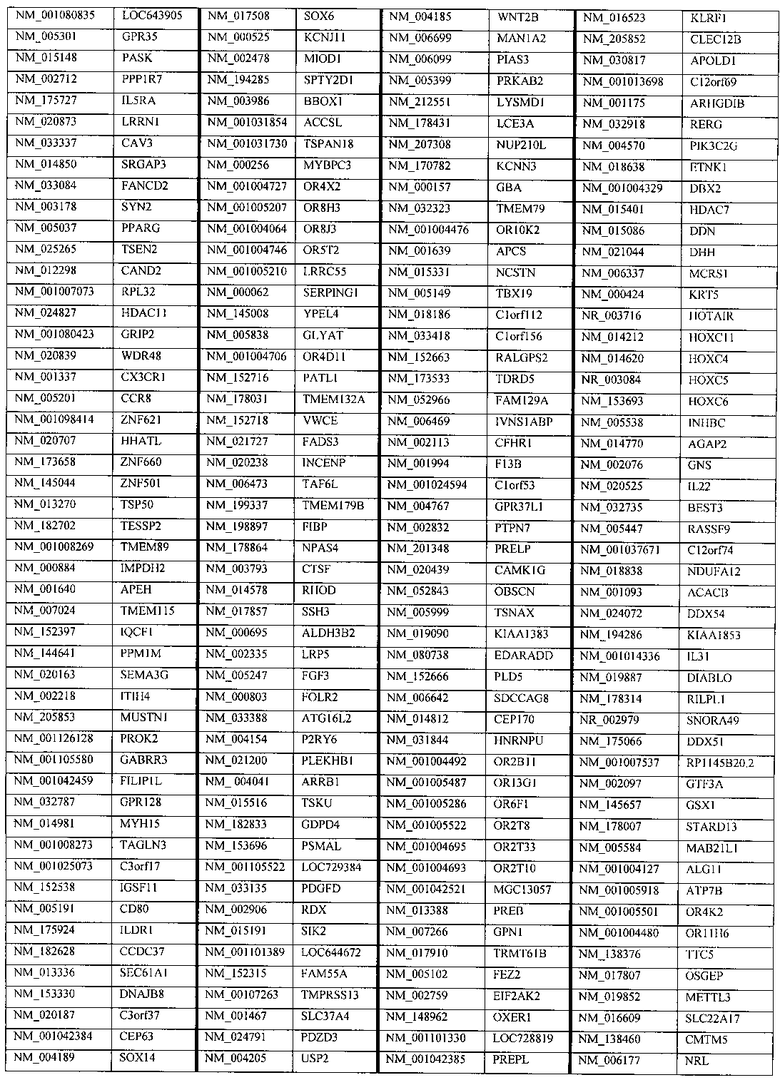
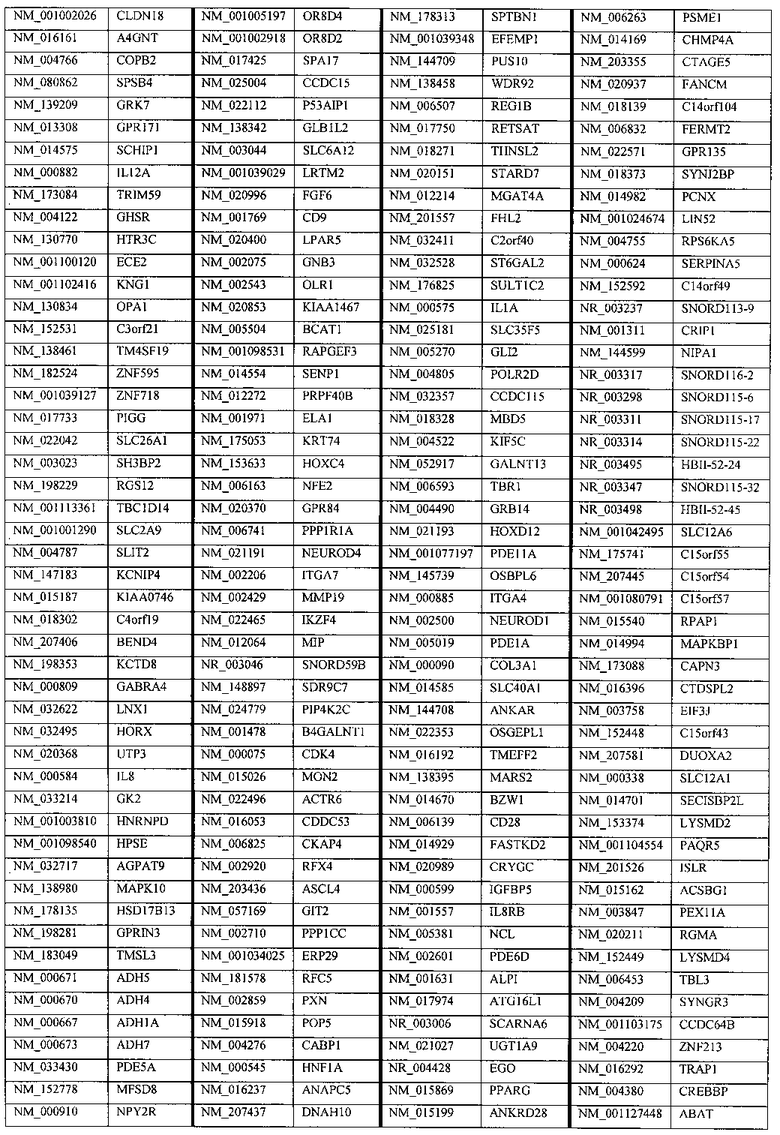
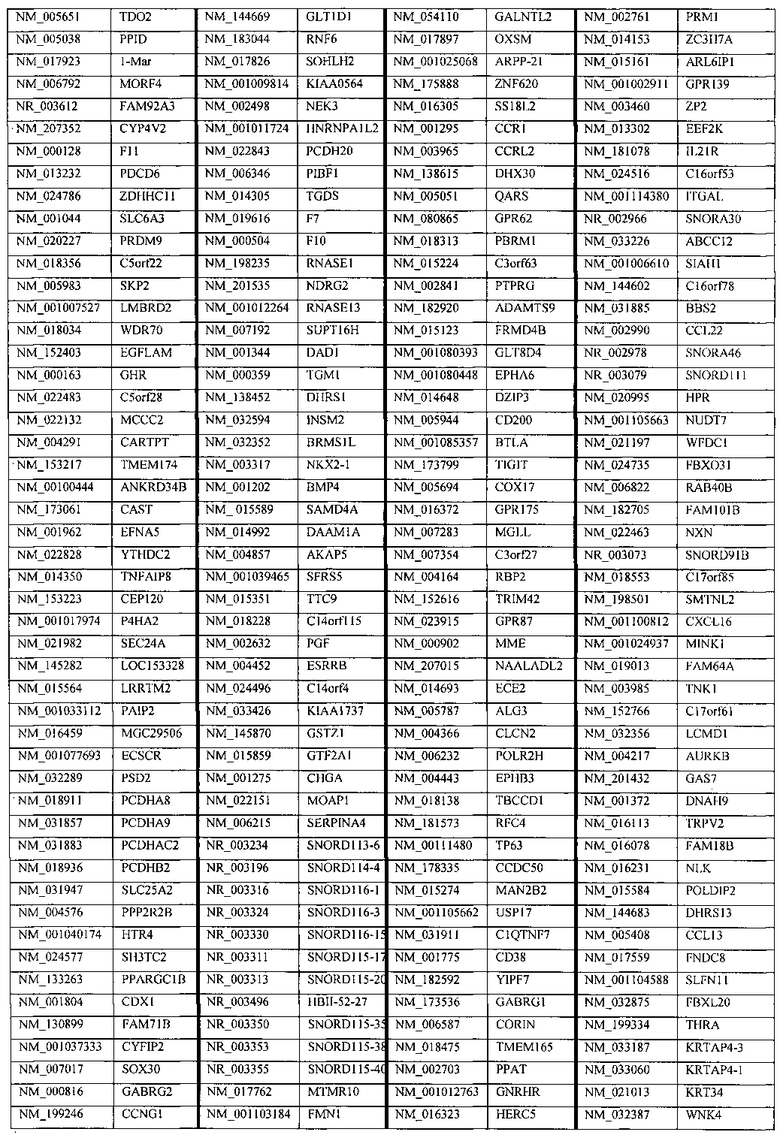
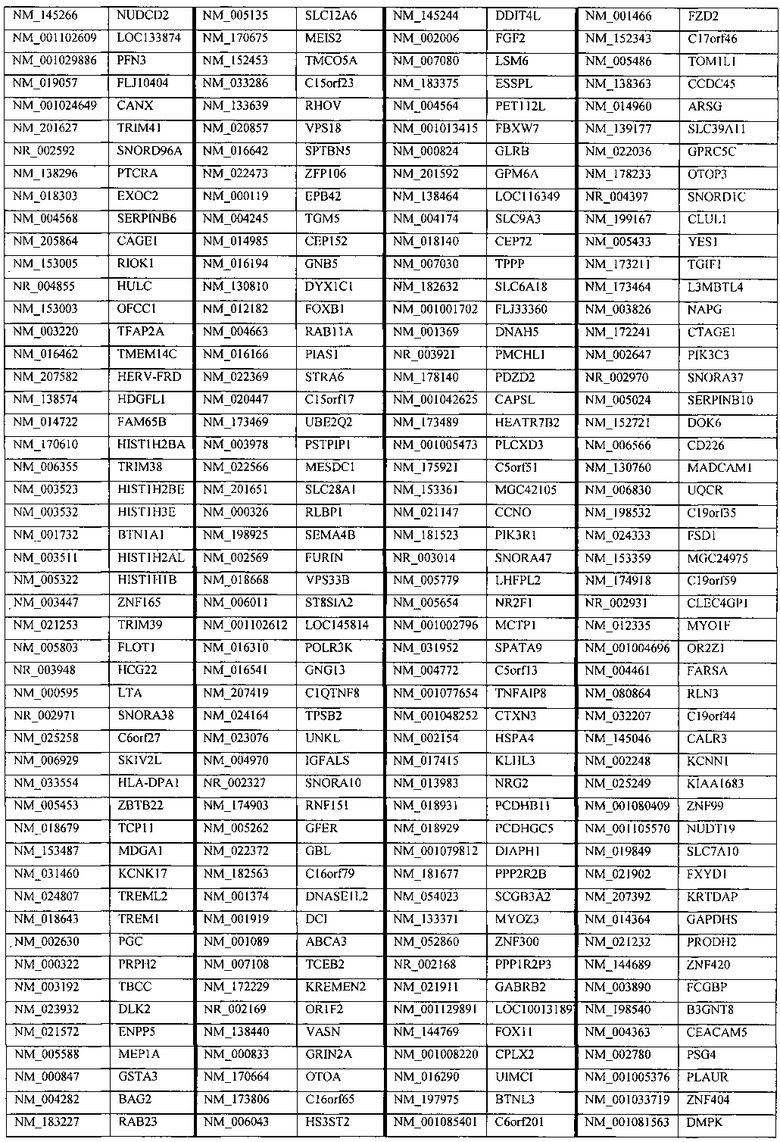
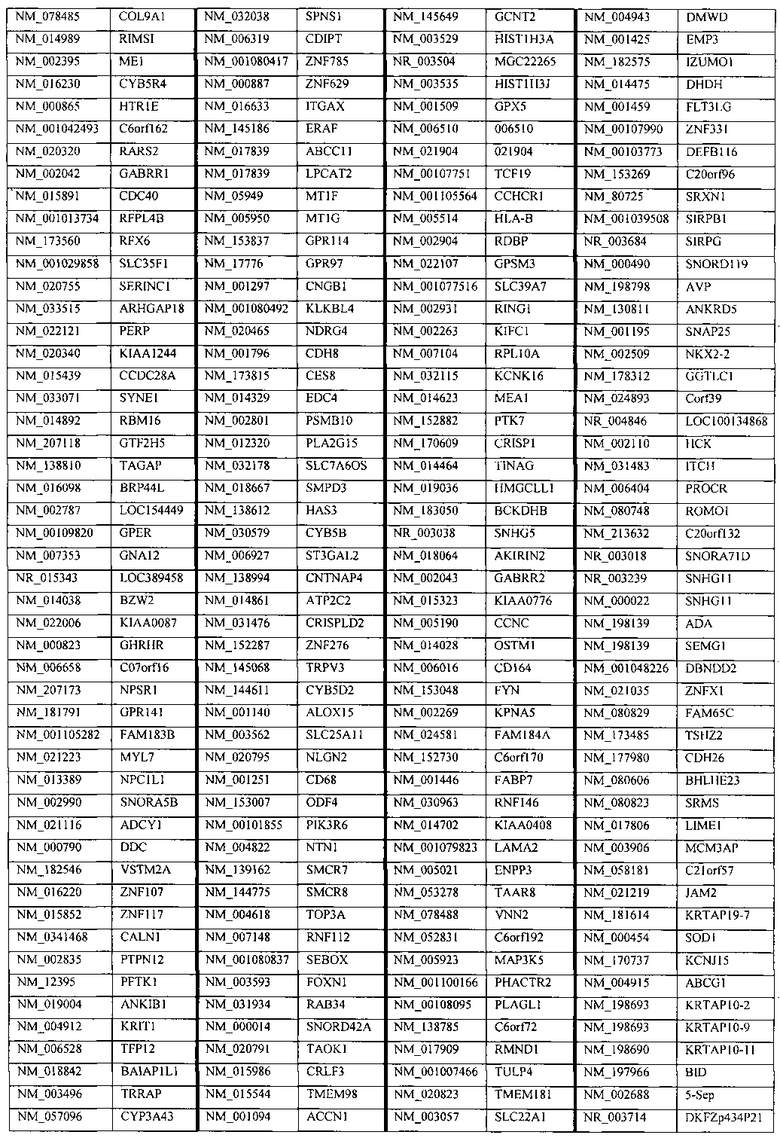
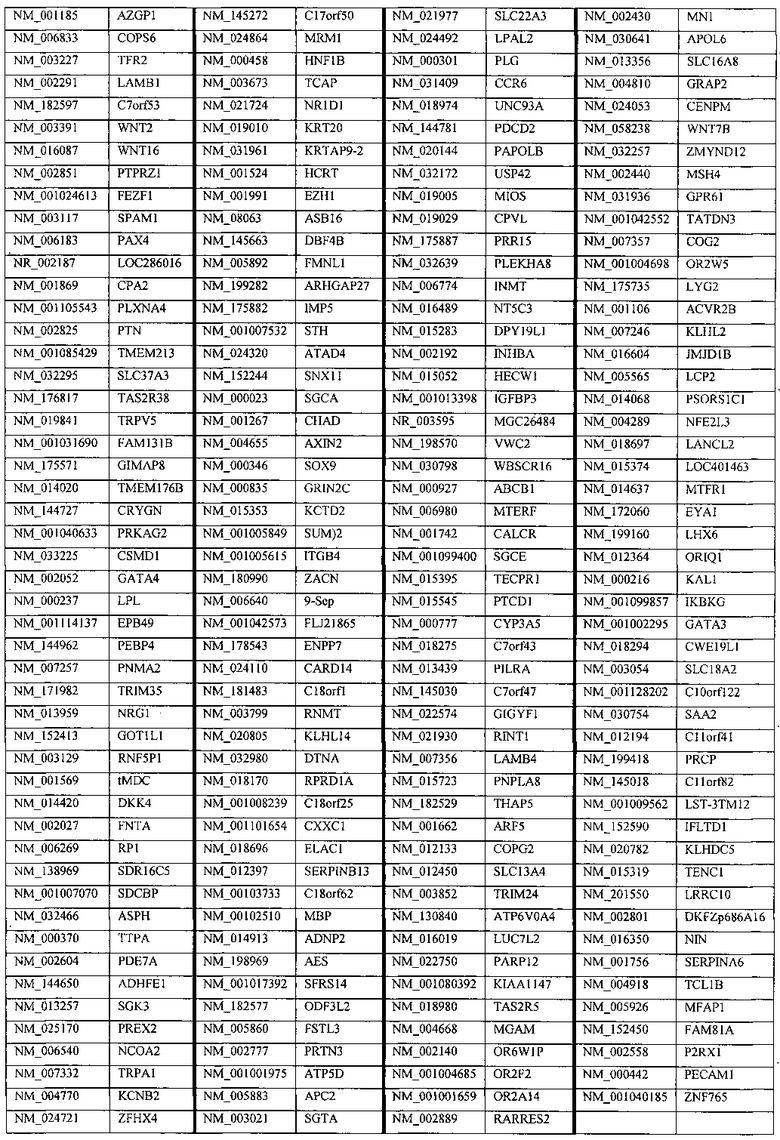

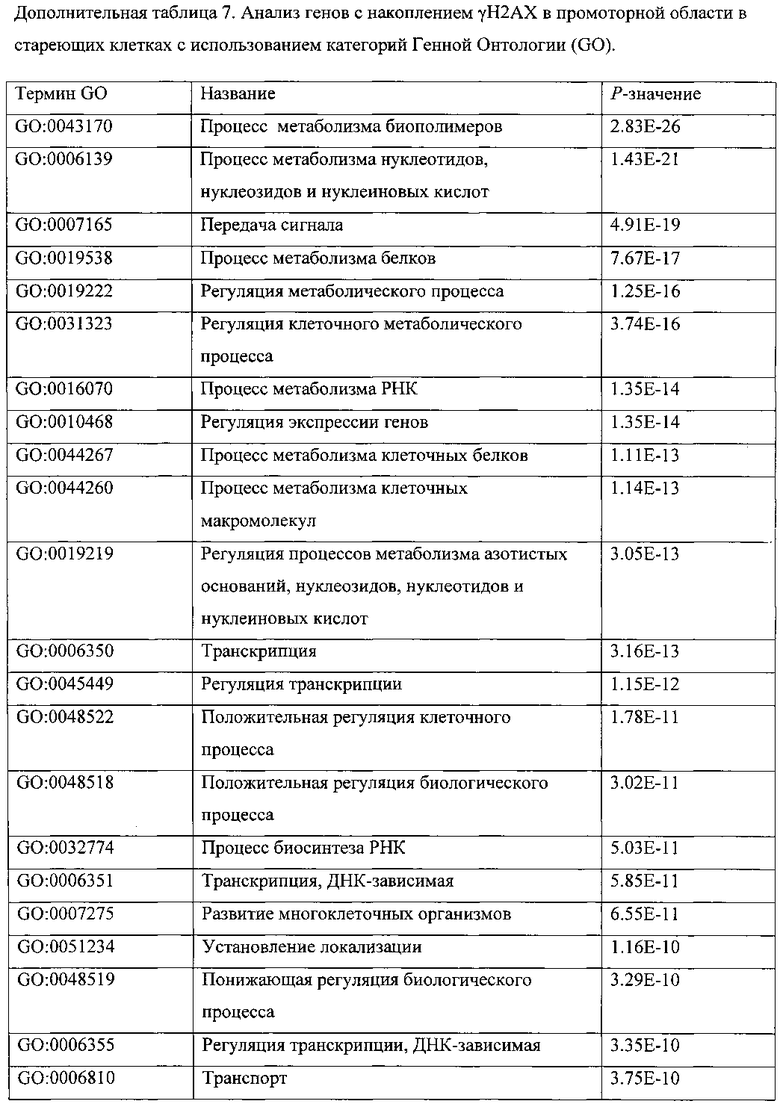
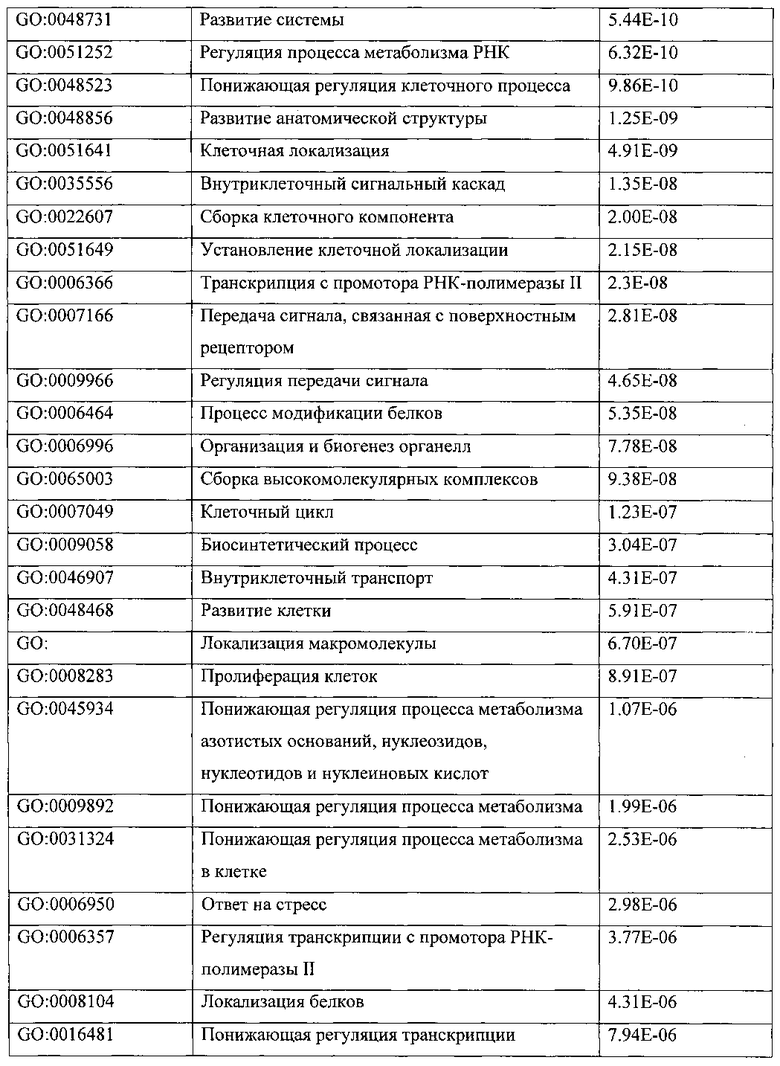
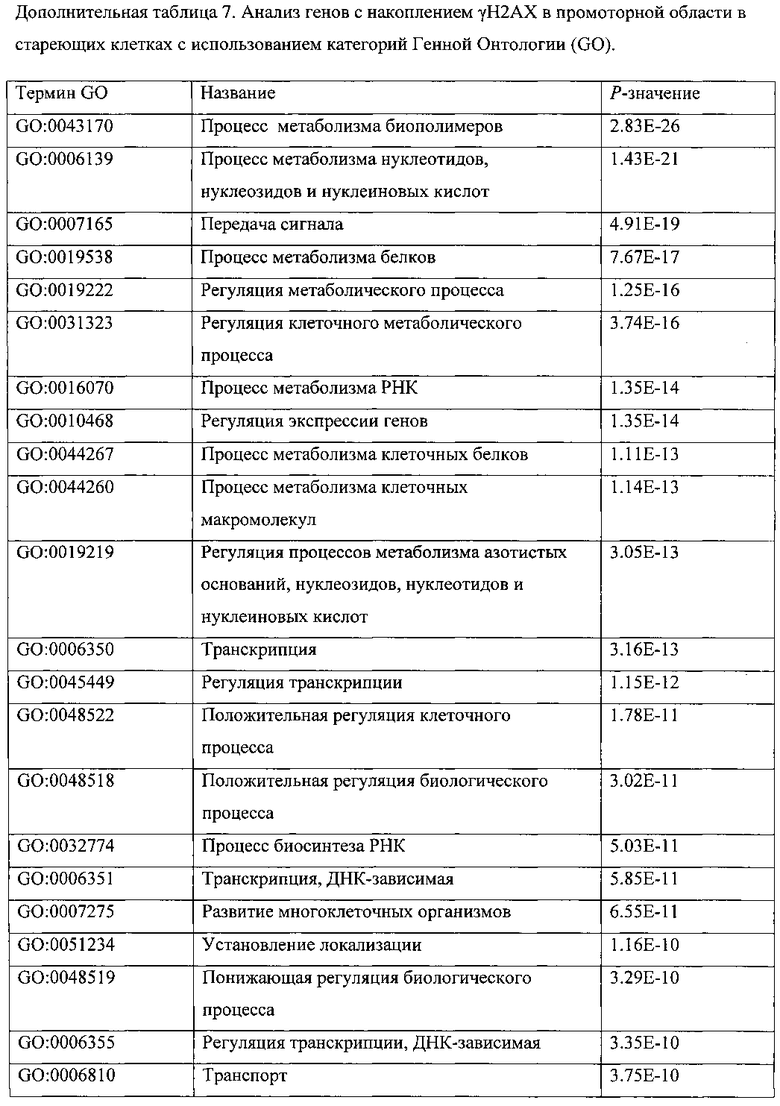
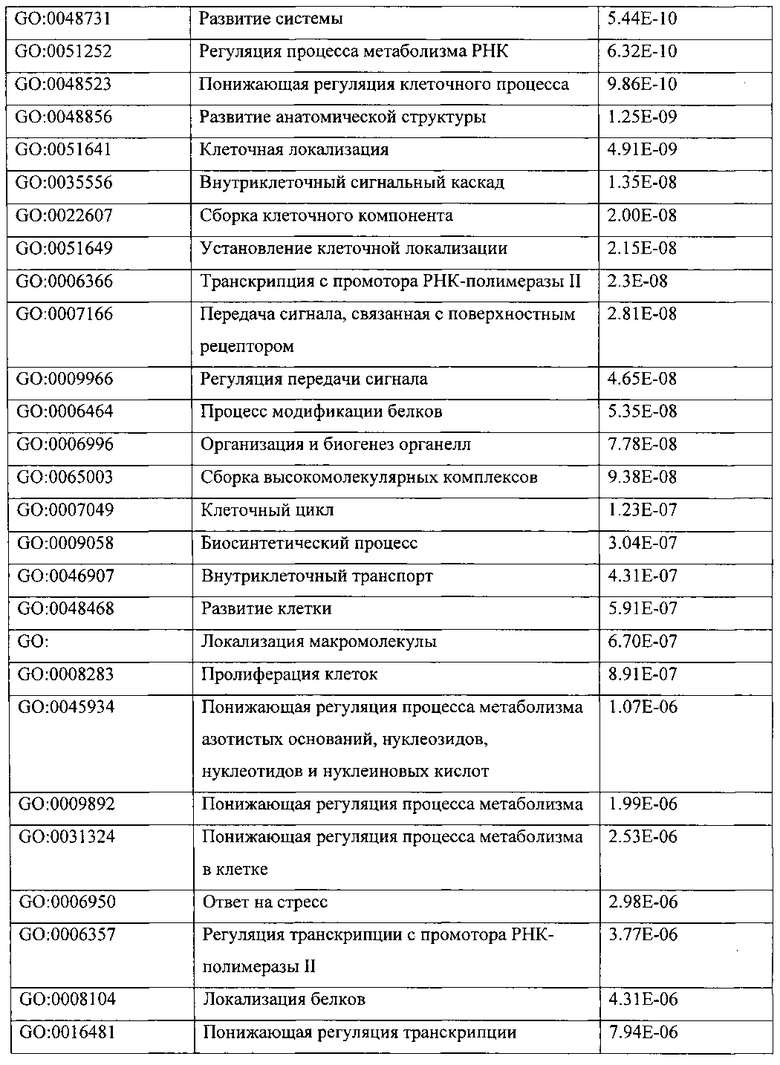

Пример 3
Белковый комплекс, который собирается на пиРНК, полученной из транскрипта Alu-ретротранспозона, указывает на предполагаемое участие ретроРНК в клеточном цикле, репарации ДНК и сборке хроматина.
[00169] Последние данные, полученные в ходе полногеномного анализа, указывают, что тысячи коротких рассеянных ядерных элементов (SINEs) присутствуют в ограниченных элементах, не содержащих экзонов (Lowe and Haussler, PNAS 2007), это позволяет предположить, что указанные повторяющиеся ретротранспозонные элементы могут играть уникальную и еще не установленную роль в формировании и/или специализации геномного ландшафта, а также регуляции генов в эволюции млекопитающих. В данном примере приведены результаты аффинного выделения комплексов РНК-белок с использованием пиРНК транскриптов ретротранспозонов Alu с последующим исследованием методом ЖХ-МС/МС. Данные указывают на роль пиРНК транскриптов ретротранспозонов Alu в регулировании репарации ДНК, клеточного цикла и хроматина.
[00170] Было показано, что большая часть сайтов восстанавливаемого повреждения ДНК в самообновляющихся взрослых стволовых клетках человека связана с ретротранспозонной частью генома, в частности, с Alu-семейством ретротранспозонов (Wang et al. (2011) Cell Cycle 10: 3016-3030). Проведенные исследования показали, что положительная регуляция транскрипционной активности Alu-повторов может быть инициирована генотоксическим повреждением, индуцированным стрессом, и может быть зарегистрирована в пигментном эпителий сетчатки человека (ПЭС) и взрослых стволовых клетках человека (Id.) в процессе старения как in vivo, так и ex vivo, соответственно. Полученные доказательства указывают, что повышенное накопление РНК Alu-повторов в клетках напрямую связано с дефицитом Dicer-1 (Kaneko et al.(2011) Nature 471: 325-330), это позволяет предположить, что накопление транскриптов Alu-повторов не обязательно может быть связано с повышенной транскрипционной активностью ретротранспозонов Alu, а скорее может быть результатом отсутствия сопутствующего процессинга транскриптов ретротранспозонов Alu, ведущего к развитию токсичности для клетки. Последние данные указывают, что накопление непроцессированных транскриптов ретротранспозонов Alu вызывает ухудшение состояния хроматина и потерю способности к репарации ДНК в перицентромерных областях, что вызывает устойчивое повреждение ДНК и в конечном итоге приводит к клеточному старению. Данные также указывают, что взрослые стволовые клетки человека, стабильно экспрессирующие мшРНК против транскриптов ретротранспозонов Alu, преодолевают процесс клеточного старения и восстанавливают способность к репарации (Wang et al. (2011) Cell Cycle 10: 3016-3030), это позволяет предположить, что в этих событиях принимает участие 25 комплекс РНК-интерференции. Данные также указывают на существование двух одинаково возможных механизмов, посредством которых действуют мшРНК против транскриптов ретротранспозонов Alu: 1) путем Dicer-зависимого посттранскрипционного сайленсинга гена через цитоплазматическую деградацию непроцессированных РНК ретротранспозонов Alu или 2) усиливая сайленсинг транскрипции посредством Dicer- или 30 Piwi-зависимого пути РНК-интерференции для непосредственного воздействия на хроматин, как показано на фигуре 21. Каждый их этих путей является возможными и может зависеть от сборки одного или нескольких белковых комплексов, которые способны перекрестно взаимодействовать с путями, отвечающими за обнаружение/репарацию повреждений ДНК и поддержание хроматина и/или центромер. В недавнем исследовании был обнаружен интересный факт, что транскрипты РНК, транскрибируемой с теломерной ДНК (TERRA), взаимодействуют с гетерохроматиновым белком 1 (НР1), триметилированным по 9-му лизину гистоном Н3 (H3K9me3) и основными компонентами комплекса Shelterin, а также членами пути, отвечающего за обнаружение повреждений ДНК (Deng et al. (2009) Mol. Cell, 35: 403-413).
[00171] Также можно предположить, что часть РНК ретротранспозонов Alu, которая является функционально значимой для преодоления фенотипа старения в клетках hADSC (Wang et al. (2011) Cell Cycle 10: 3016-3030), может определять широкий спектр нижележащих эффектов, связанных с транскрипцией ретротранспозонов. Поскольку о цитоплазматической роли РНК ретротранспозонов Alu в сборке частицы узнавания сигнала уже сообщалось в предыдущих работах (Не et al. (1994) J. Cell Sci., 107(Pt 4): 903-912), усилия были направлены именно на установлении ядерных комплексов, которые собираются на РНК ретротранспозонов Alu. Представлены доказательства состава молекулярных комплексов, собирающихся на процессированных РНК-транскриптах ретротранспозонов Alu, установленного с помощью несмещенного аффинного выделения комплексов РНК-белок в сочетании с масс-спектрометрией. Данные указывают на пути и молекулярные процессы, посредством которых процессированные интермедиаты РНК-транскриптов ретротранспозонов Alu могут участвовать во множестве процессов регуляции ядерных событий, происходящих во взрослых стволовых клетках человека.
Результаты
РНК ретротранспозонов Alu и ее гомология с РНК, взаимодействующей с белками группы PIWI (пиРНК) человека
[00172] Ранее сообщалось, что РНК ретротранспозонов Alu участвует в цитоплазматической сборке частицы узнавания сигнала (SRP) в клетках млекопитающих (Не et al. (1994) J. Cell Sci., 107(Pt 4): 903-912). Частица узнавания сигнала млекопитающих состоит из одной 7SL РНК и шести белков (Walter and Blobel (1981) J. Cell Biol., 91: 557-561). SRP9 и SRP14 связываются с 5′-концом РНК (Alu-домен), который участвует в остановке трансляции (Siegel and Walter (1988) Trends Biochem. Sci., 13: 314- 30 316; Siegel and Walter (1986) Nature 320:81-84). Однако функциональная мшРНК, представленная в работе Wang et al. ((2011) Cell Cycle 10: 3016-3030), не комплементарна 7SL РНК. Кроме того, в Alu-повторах консервативность последовательности участка, который соответствует мшРНК, значительно выше, чем остальной части Alu-элементов (рис. 22В). Подобная консервативность согласуется со способностью мшРНК эффективно подавлять транскрипцию копий Alu-повтора, включая членов различных подсемейств. В совокупности эти данные проливают свет на функциональное значение последовательности мшРНК к Alu-повтору, исследование которой было проведено в настоящей заявке, и указывают, что эта последовательность может представлять собой процессированную молекулу РНК Alu-повтора, которая функционирует в ядре. Было подвергнуто сомнению соответствие последовательности мшРНК к Alu-повтору, исследование которой было проведено в настоящей заявке, последовательности РНК, взаимодействующей с piwi (пиРНК), ранее представленной в Girard et al. (2006) Nature 442: 199-202.
[00173] Точная последовательность мшРНК была идентифицирована как взаимодействующая с piwi РНК генов pir-52207 и pir-57460 (Id.) (номер доступа в NCBI # DQ585095 (tgcctgtaat cccagctact caggaggctg, (SEQ ID NO:30)), и обозначается в настоящей заявке piALU. Эти факты свидетельствует о том, что РНК Alu-ретротранспозона процессируется аналогом комплекса PIWI человека, молекулярная функция которого неизвестна, однако указывают, что транскрипт piAlu, полученный из подвергшихся процессингу транскриптов Alu-ретротранспозона, может функционировать в ядре.
Взаимодействие РНК траснкриптов Alu-ретротранспозона с ядерными факторами
[00174] Была предпринята попытка получить полный список белков, которые прямо или косвенно взаимодействуют с piALU. Для этого применили несмещенное аффинное выделение комплексов РНК-белок с использованием синтетических биотинилированных олигонуклеотидов РНК, которые включали последовательность Alu-повтора с 140 по 183 нуклеотид, показанную на рис. 22В, и ядерных экстрактов самообновляющихся взрослых стволовых клеток человека. Указанная часть последовательности Alu-повтора является консервативной в различных типах ретротранспозонов Alu: AluSx, AluJb, AluJo и AluY (рис. 22, В).
[00175] В экспериментах объединили аффинное выделение комплексов piALU РНК-белок с полноценным анализом взаимодействующих белков методом ЖХ-МС/МС (жидкостной хроматографии-тандемной масс-спектрометрии с ионизацией электрораспылением). Поскольку существует путь надзора за ядерной РНК, который защищает от транскрипционного шума, добавили контрольную гексануклеотидную РНК-приманку, описанную в разделе «Материалы и методы», чтобы избежать взаимодействий, которые не специфичны для определенных последовательностей. Методика эксперимента ранее была опубликована в работе Deng et al. ((2009) Mol. Cell, 35: 403-413) и приведена на рис. 22В. Сравнительный анализ связывания РНК-белок методом ПААГ-электрофореза выявил наличие множества полос, специфичных для комплекса зонд-piALU (рис. 22С), которые были вырезаны для идентификации методом ЖХ-МС/МС. Полный перечень идентифицированных белков приведен в дополнительной таблице 1.
Взаимодействия РНК ретротранспозонов Alu с основными модификаторами хроматина, комплексами репарации ДНК, факторами сборки центромерного хроматина/кинетохора и транскрипционными факторами
[00176] Данные масс-спектрометрического анализа преципитатов, полученных в результате несмещенного аффинного выделения комплексов piALU РНК-белок, показал наличие четырех основных категорий белков, таких как модификаторы хроматина, комплексы репарации ДНК, факторы сборки хроматина центромер/кинетохора и транскрипционные факторы (рис. 23). Представленность каждого компонента показана в виде градиента цвета от истинных оттенков до черного для пептидов, представленность которых равна 1. Аналогично белковым взаимодействиям с РНК, транскрибируемой с теломерной ДНК (TERRA) (Id.), связывающиеся с piALU белки включают белки ответа на повреждение ДНК - DNA-PKC, PARP и hnRNPM, эти данные указывают, что РНК piALU может быть неотъемлемой частью комплексов репарации ДНК.
[00177] Чтобы оценить связь РНК piALU с биологическими процессами, применили анализ с использованием терминов Генной Онтологии (GO) (The Gene Ontology Consortium (2010) Nucleic Acids Res., 38(suppl 1): D331-D335; Barrell et al. (2009) Nucleic Acids Res 37:D396-D403) для составления списка взаимодействующих белков. Alu-взаимодействующие белки были достоверно (3,12×10-13≤Р≤1,5×10-3) вовлечены в совокупности в 19 клеточных процессов, включая: репарацию ДНК, организацию органелл, клеточный цикл, ремоделирование хроматина и транскрипцию (рис. 24А). Указанные общие категории включают эксцизионную репарацию поврежденных нуклеотидных участков, репарацию двунитевых разрывов ДНК по механизму гомологичной рекомбинации, V(D)J рекомбинацию, ответ на стресс, расхождение хромосом, контрольные точки клеточного цикла, регулирование остановки клеточного цикла контрольными точками, регулирование интерфазы митотического цикла и митоза (рис. 24А). Alu-взаимодействующие белки также часто включали белки комплексов, связанных с расплетанием ДНК-дуплекса, регуляцией транскрипции и транскрипционным удлинением нити от промоторов РНК-полимеразы II.
[00178] Некоторые ядерные белки были ассоциированы с несколькими терминами GO (n=12), относящимися к клеточным компонентам, включая центросому, нуклеоплазму, кинетохор, веретено деления, микротрубочеки цитоскелета, центромерные и теломерные участки хромосом, а также когезиновому комплексу и ДНК-направленному РНК-полимеразному комплексу (рис. 24А).
[00179] Эти данные указывают на возможную роль РНК Alu-повторов в регуляции организации хроматина в центромерных/перицентромерных участках хромосом, а также процессах репарации ДНК и транскрипционной регуляции генов. Доказано, что все эти клеточные процессы абсолютно необходимы для регуляции прогрессирования клеточного цикла.
Сеть белок-белковых взаимодействий для выделенных аффинными методами piALU РНК-белковых комплексов
[00180] Пониманию клеточной функции(-й) выделенного белкового комплекса(-ов) способствует умение правильно раскрыть и интерпретировать все функциональные взаимодействия между белками, идентифицированными методом ЖХ-МС/МС. Чтобы построить сеть белок-белковых взаимодействий для выделенных аффинными методами piALU РНК-связанных белков, использовали инструмент поиска по сетевой базе данных взаимодействующих генов (STRING), версия 9.0. Этот инструмент обеспечивает уникальный всеобъемлющий охват и легкость доступа к информации об экспериментальных и прогнозируемых взаимодействиях (Szklarczyk et al. (2011) Nucleic Acids Res., 39: D561-D568). Модель сети белок-белковых взаимодействий, построенная согласно базе STRING на основании ядерных взаимодействий белков с РНК piALU по данным ЖХ-МС/МС, показана на рис. 24С. Для построения сети и распределения белков по семействам (более сильные ассоциации представлены более толстыми линиями на фигуре 24С и фиг. 25) использовали только доказательства взаимодействий/связей с высоким доверительным уровнем (полученные экспериментально) в проверенных базах данных в сочетании с алгоритмом кластеризации Маркова (MCL)) (Enright et al. (2002) Nucleic Acids Res., 30: 1575-1584). Был обнаружено, что 9 из 64 РНК Alu-взаимодействующих партнеров участвуют в расхождении хромосом и включают SGOL1, SGOL2, CENPE, CENPF, MRE11A, CDCA5 и еще 4 (PBRM1, CENPL, CDC27 и MAP3K8) напрямую связаны с регуляцией митоза, в то время как добавление PIN4 и KIF15 отображает сложную сеть белков, регулирующих сборку веретена деления. Это наблюдение позволяет предположить, что транскрипты piALU могут формировать молекулярное соединение между когезией сестринских центромер и динамикой микротрубочек на кинетохоре. Показанная ранее потеря белков наружного кинетохора CENP-E и CENP-F при нокдауне Sgo (Salic et al. (2004) Cell 118: 567-578) в совокупности с наблюдением о том, что накопление непроцессированной РНК Alu - повтора блокирует привлечение когезинового комплекса к центромерам и запускает процесс клеточного старения у взрослых стволовых клеток (Wang et al. (2011) Cell Cycle 10: 3016-3030), указывает, что в процессированном виде РНК Alu-повтора может выполнять функцию РНК-адаптора в динамике микротрубочек веретена деления.
[00181] Данные, приведенные на рис. 25, также указывают, что 34 из 64 исследованных piALU РНК взаимодействующих партнеров, так или иначе, участвуют в регуляции клеточного цикла. Наличие белков центросом СЕР192, СЕР110, СЕР152 и СЕР135 указывает на интригующую возможность того, что интермедиаты РНК Alu-повтора участвуют в функционировании этой клеточной органеллы и вместе с CUL4A, GMNN и CTD1 (рис. 25) могут участвовать в регулировании интерфазы и перехода из фазы G2 к М-фазе.
[00182] Чрезвычайно интересным является тот факт, что РНК piALU также преципитировали белковые компоненты системы репарации нуклеотидных повреждений (ERCC3, DDB1, MRE11A), путей репарации ДНК, основанных на негомологичном концевом присоединении (Lig4, XRCC6) и гомологичной рекомбинации (PALB2, RAD50), а также белок BAZ1B, что свидетельствует о предполагаемой роли РНК piALU в репарации двунитевых разрывов. Сборка комплексов репарации повреждений ДНК, описанная в настоящей заявке, может быть критическим событием при старении взрослых стволовых клеток (Kaneko et al. (2011) Nature 471: 325-330; Wang et al. (2011) Cell Cycle 10: 3016-3030). Это приводит к мнению, что отсутствие надлежащей сборки/функции этих комплексов вместе или по отдельности с компонентами ремоделирования хроматина (CHD6, SMARCA2, BAZ1A, ARID1A, INO80, ASXL1 и KDM2A) может объяснить постепенную потерю способности к репарации повреждений ДНК, что инициирует ответ на устойчивые повреждения ДНК и запускает процесс клеточного старения ((Rodier et al. (2009) Nat. Cell Biol., 11: 973-979; Rodier et al (2011). J. Cell Sci., 124: 68-81).
Обсуждение
[00183] Проведенный проект по полногеномному секвенированию РНК методом ChIP-seq выявил, что более трети всех известных крупных консервативных нкРНК могут быть преципитированы всего четырьмя различными модифицирующими хроматин белками (Khalil et al. (2009) Proc. Natl. Acad. Sci. USA, 106: 11667-11672), это указывает на то, что самые длинные нкРНК связываются с модифицирующими хроматин белками. Интерактом РНК piALU, представленный в настоящей заявке, также включает множество модифицирующих хроматин белков, возможно, проливая свет на другой уровень сложности комплексов нкРНК/модифицирующие хроматин белки, которые зависят от наличия более коротких, подвергшихся процессингу форм транскриптов Alu-повтора. Полноразмерная последовательность РНК Alu-повтора может образовывать четыре различные структуры "петля-на-стебле", которые могут действовать в качестве отдельных центров нуклеации для формирования различных комплексов РНК-белок (рис. 22). Только одну из четырех структур «петля-на-стебле» использовали в качестве синтетической РНК-приманки, поскольку она содержит представленную ранее человеческую РНК, взаимодействующую с piwi (Girard et al. (2006) Nature 442: 199-202), и способна вызывать нуклеацию белковых комплексов, которые прямо или косвенно ответственны за восстановление способности стареющих взрослых стволовых клеток самообновляться (Wang et al. (2011) Cell Cycle 10: 3016-3030).
[00184] Действительно, в соответствии с полученными ранее данными наблюдаемые взаимодействия подтверждают значимость указанных ядерных комплексов piALU РНК-белок для контроля клеточного цикла, репарации ДНК, поддержания структуры хроматина и функции центромерных/перицентромерных участков хромосом взрослых стволовых клеток человека. В настоящее время отсутствует функциональная оценка этих данных, однако они все же могут дать ключ к пониманию того, как транскрипция/процессинг транскриптов етротранспозонов регулирует старение взрослых стволовых клеток человека и старение ex vivo.
[00185] Данные, представленные в этом примере, указывают на возможность участия коротких молекул РНК, транскрибированных с ретротранспозонов Alu, в регуляции клеточного цикла, РНК-Пол II транскрипции и поддержании архитектуры хроматина. По-видимому, это участие тесно связано с процессами репарации ДНК.
Материалы и методы
Выделение и размножение ADSC человека
[00186] Стволовые клетки, полученные из жировой ткани человека (hADSCs), выделяли из человеческой подкожной белой жировой ткани, собранной во время процедуры липосакции. Липоаспират суспендировали в сбалансированном солевом растворе Хэнка (HBSS), содержащем 3,5% бычьего сывороточного альбумина (BSA), 1% коллагеназы типа II (Sigma) в соотношении 1:3 вес/объем и встряхивали при 37°С в течение 50 мин. Клетки фильтровали через клеточный фильтр с размером пор 70 мкм (BD Falcon #352350), обрабатывали буфером для лизирования эритроцитов (150 мМ NH4C1, 10 мМ КНСО3, 0,1 мМ ЭДТА, рН 7,3) и размножали ex vivo в полной среде DMEM/F12 (DMEM/F12, 10% FBS, 100 ед/мл пенициллина, 100 мкг/мл стрептомицина, 2,5 мкг/мл амфотерицина В, Invitrogen) в атмосфере 10% CO2 при 37°С, перевивали при 80% конфлюентности, культуральную среду заменяли каждые 72-96 часов.
Выделение ядерного хроматина
[00187] Ядерные и цитоплазматические белки выделяли из самообновляющихся клеток hADSC с использованием набора для выделения цитоплазматических и ядерных белков ProteoJET™ Kit (Fermentas # K0311) в соответствии с протоколом производителя.
Складывание вторичный структуры РНК
[00188] 5′-Биотинилированный одноцепочечный синтетический РНК-зонд, соответствующий участку с 140 по 183 нуклеотид полноразмерной последовательности Alu-повтора, (5′-/Bio/-GCCGGGCGUGAUGGUGGGCGCCUGUAGUCCCAGCUAC15UCGGGAGGC-3′, SEQ ID NO: 27), а также синтетический случайный октамерный повтор 5′-/Bio/-(CACUGA)8 (SEQ ID NO: 28), который использовали в качестве линейного контроля, приобрели в Integrated DNA Technologies (IDT). Указанные олигонуклеотиды очищали с помощью ВЭЖХ, свободной от РНКазы. По 8 мкг каждого олигонуклеотида нагревали в пробирке для ПЦР с 15 мл×2 буфера для восстановления вторичной структуры (40 мМ HEPES, рН 7,9, 0,4 мМ ЭДТА, 2 М NaCl), 9 мл воды DEPC и 50 ед/мл ингибитора РНКазы SUPERase-In (Ambion, #АМ2694). РНК олигонуклеотиды нагревали до 78°С, чтобы индуцировать переход в линейную конформацию, затем температуру реакции уменьшали и поддерживали на уровне 54°С, чтобы обеспечить и поддержать образование желаемый вторичной структуры (имитируя вторичную структуру «петля на стебле» полноразмерной РНК), на основании расчетов, сделанных с помощью программного обеспечения UNAFold (IDT). Контрольный повтор РНК 5′-/Bio/-(CACUGA)8 (SEQ ID NO: 28) не способен формировать вторичную структуру.
Преципитация белков (несмещенное аффинное выделение комплексов РНК-белок)
[00189] По 100 мкл покрытых стрептавидином бусин Dynabeads (Dynabeads М-270 Streptavidin, # 65306) на одну реакцию промывали три раза в 1х буфере D150 (20 мМ HEPES рН=7,9, 20% глицерина, 0,2 мМ ЭДТА, 150 мМ NaCl, 0,05% NP-40, 1 мМ фенилметилсульфонилфторида, 10 мМ β-меркаптоэтанола) перед последней отмывкой в высокосолевом 1х буфере для восстановления вторичной структуры (содержащем 1 М NaCl). В каждую реакционную пробирку вносили по 30 мкл раствора РНК, восстановившей вторичную структуру, и инкубировали при комнатной температуре в течение 15 мин, затем дважды промывали 100 мкл низкосолевого буфера для восстановления вторичной структуры (содержащего 150 мМ NaCl). Связанные с гранулами РНК-зонды инкубировали 5 в течение 30 мин. при комнатной температуре с ядерными экстрактами самообновляющихся клеток hADSC, 1 мкл 0,01 М фенилметилсульфонилфторида и 3 мкл SUPERase-In. После инкубации гранулы Dynabeads пять раз промывали 1х буфером D150, в результате этой процедуры остаются только непосредственно и косвенно взаимодействующие партнеры, прикрепленные к гранулам. В качестве отрицательного контроля использовали инкубацию ядерных экстрактов только с гранулами Dynabeads, что позволило контролировать любое взаимодействие между ядерными молекулами и непосредственно гранулами.
Денатурирующий ПААГ-электрофорез комплексов РНК-белок, выделенных аффинным методом.
[00190] К 10 мкг белка добавляли Буфер образца XT 12 (Sample Buffer 12) (BioRad #161-0791) и восстанавливающий агент XT (XT Reducing Agent) (BioRad #161-0792). После денатурации в течение 5 мин. при 100°С образцы белков загружали в готовый к применению 4-12% акриламидный бис-трис градиентный гель No vex NuPAGE® (Invitrogen, #NP0321BOX) и разделяли при 150 Вольт в течение двух часов. Гели окрашивали ProteoSilver Plus Silver Stain Kit (Sigma, #Prot-SIL1). Предварительно окрашенный стандарт SeeBlue Plus2 (Invitrogen, #LC5925) использовали для визуализации и примерной оценки молекулярной массы белковых образцов. Полосы, соответствующие полосам для зонда к Alu, вырезали и анализировали с помощью ЖХ-МС/МС для определения содержания белка.
Подготовка образцов для масс-спектрометрического анализа
[00191] Образцы геля сначала промывали ацетонитрилом, затем восстанавливали с использованием ДТТ с последующим алкилированием иодацетамидом. Образцы геля трехкратно промывали 50 мМ бикарбоната аммония и ацетонитрила, затем охлаждали и ресуспендировали каждый срез геля в трипсине (5,5 мкг/мл в 50 мМ бикарбоната аммония/10% ацетонитрила), и инкубировали при 37°С в течение 24 часов для протеолитического расщепления белков. Пептиды выделяли однократной промывкой смесью 50 мМ бикарбоната аммония/10% ацетонитрила с последующей однократной промывкой смесью 50% ацетонитрила/0,1% муравьиной кислоты, затем образцы для масс-спектрометрии лиофилизировали и регидратировали в 20 мкл смеси 5% ацетонитрила/0,2% муравьиной кислоты.
Идентификация белковых партнеров методом масс-спектрометрии высокого 5 разрешения ВЖ-МС/МС
[00192] ВЖ-МС/МС высокого разрешения проводили на масс-спектрометре LTQ-Orbitrap XL (Thermo Fisher Scientific), как было описано_ENREF_45 (Lopez et al. (2011) J. Proteome Res., 10: 133-142). Вырезанные и расщепленные трипсином образцы геля загружали в 96-луночные планшеты для масс-спектрометрического анализа на спектрометре LTQ-Orbitrap XL (Thermo Fisher Scientific), как было описано ранее (Lopez et al. (2011) J. Proteome Res., 10: 133-142). С помощью автоматического дозатора Thermo Scientific EASY-nLC вводили по 10 мкл каждого восстановленного образца. Разделение проводили методом обратно-фазовой хроматографии с использованием 20 см колонки, изготовленной из кварцевого стекла с внутренним диаметром 75 мкм, заполненной сорбентом Hypersil GOLD™ С18™ с размером частиц 3 мкм, при скорости потока 250 нл/мин. Градиент создавали с использованием 5-40% ацетонитрила, 0,2% муравьиной кислоты в течение 150 минут. Спектрометр LTQ-Orbitrap запускали в верхней 8 конфигурации с разрешением 60K для полного сканирования, с включенной функцией выбора моноизотопного предшественника и +1, а также отклонением неопределенного зарядового состояния. Анализ на спектрометре LTQ-Orbitrap проводили с использованием CID фрагментации.
Анализ данных ЖХ-МС/МС и идентификация белков
[00193] Анализ данных ЖХ-МС/МС, идентификацию белков и создание списка пиков выполняли с использованием алгоритма Proteome Discoverer (Thermo Fisher Scientific), включающего поисковую систему SEQUEST® и Percolator™ (Kail и др. (2007) Nat, методы, 4.: 923-925), как описано ранее (Lopez et al. (2011) J. Proteome Res., 10: 133-142). Поиск по данным MC/MC проводили используя точность по массе 10 млн.-1 над соотношением масса/заряд предшественников и окно размером 0,5 Да для ионных фрагментов. При анализе допускали поиск по полностью ферментированным трипсином фрагментам, имеющим не более трех пропущенных сайтов расщепления. Окисленные остатки метионина искали в виде переменной модификации, алкилированные остатки цистеина искали в виде фиксированной модификации. Базу данных для человека загружали с сайта NCBI и дополняли общими контаминантами. Пептиды фильтровали для каждого зарядного состояния к 1% уровню ложноположительных результатов (FDR), а затем группировали пептиды в белки с использованием принципа бритвы Оккама.
Анализ сети белковых взаимодействий
[00194] Анализ всех наиболее типичных выделенных членов РНК-белкового комплекса проводили с использованием программного обеспечения STRING 9.0 (Szklarczyk et al. (2011) Nucleic Acids Res., 39: D561-D568). Полученную сеть взаимодействий затем исследовали с использованием плагина BINGO2 для определения статистического обогащения для 14 категорий Генной Онтологии (The Gene Ontology Consortium (2010) Nucleic Acids Res., 38(suppl 1) D331-D335; Barrell et al. (2009) Nucleic Acids Res 37:D396-D403).
[00195] Следует понимать, что примеры и варианты реализации, описанные в настоящей заявке, приведены только для иллюстративных целей и их различные модификации или изменения будут предложены специалистам в данной области техники и должны быть включены в сущность и объем настоящей заявки, а также область прилагаемой формулы изобретения. Все публикации, патенты и патентные заявки, цитируемые в настоящей заявке, полностью включены в заявку посредством ссылки для всех целей.







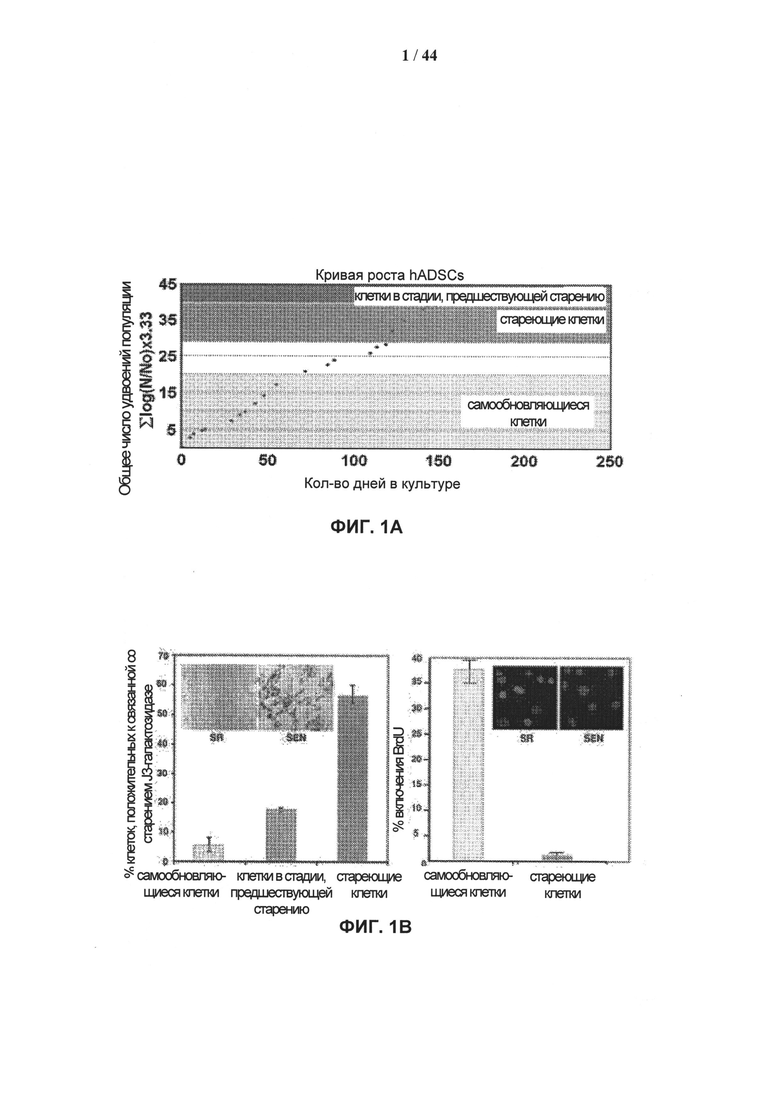
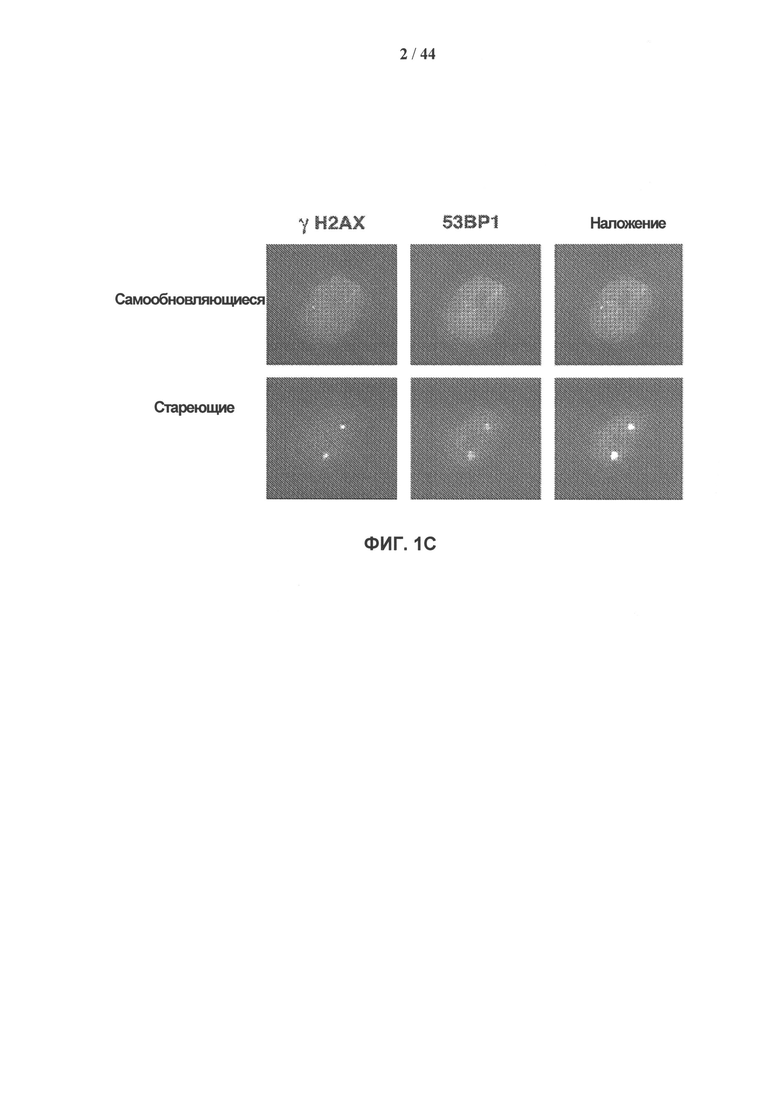
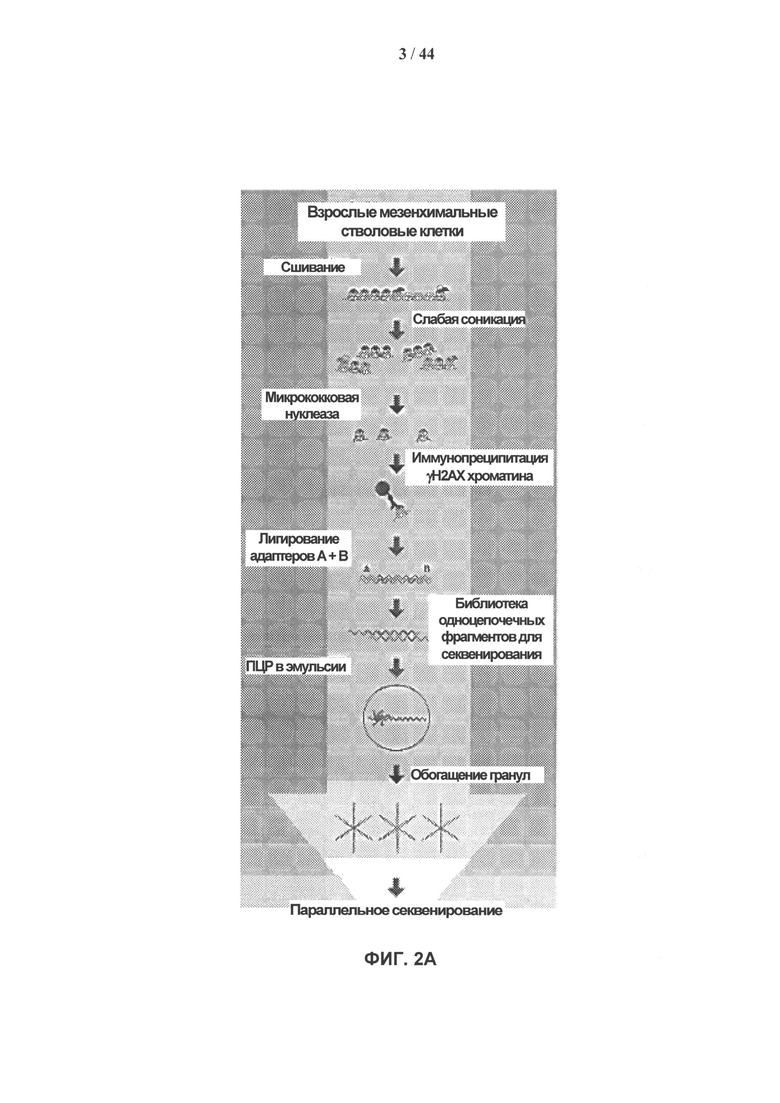
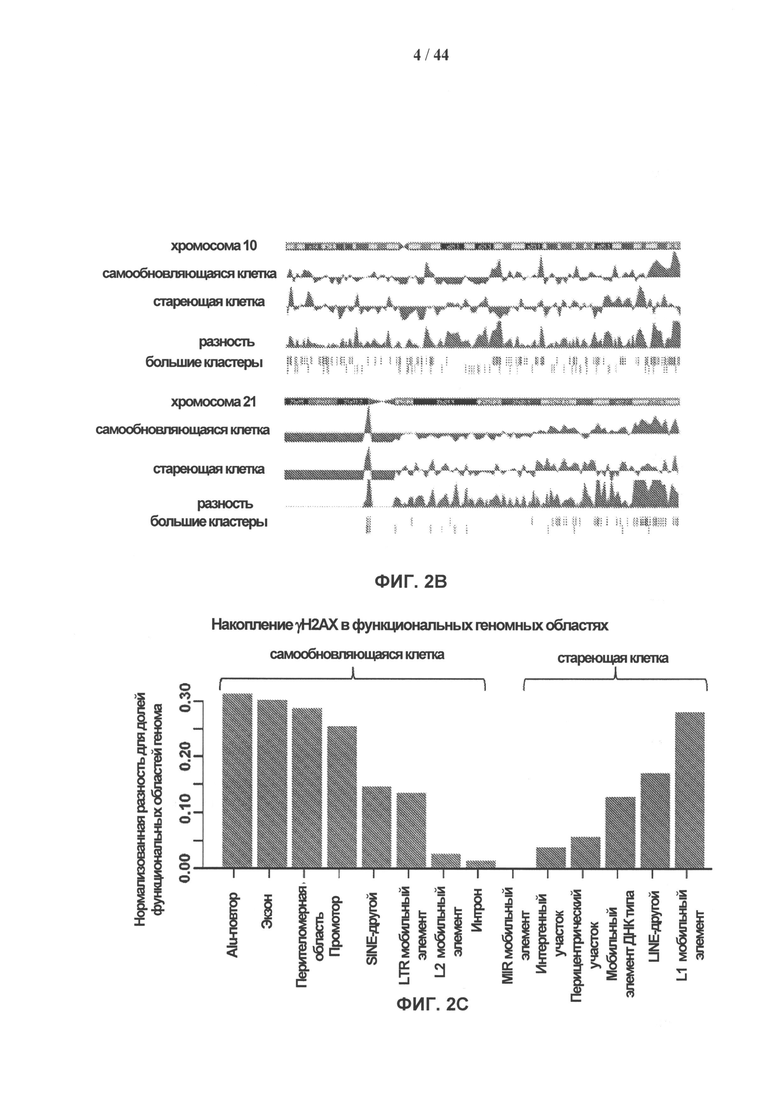
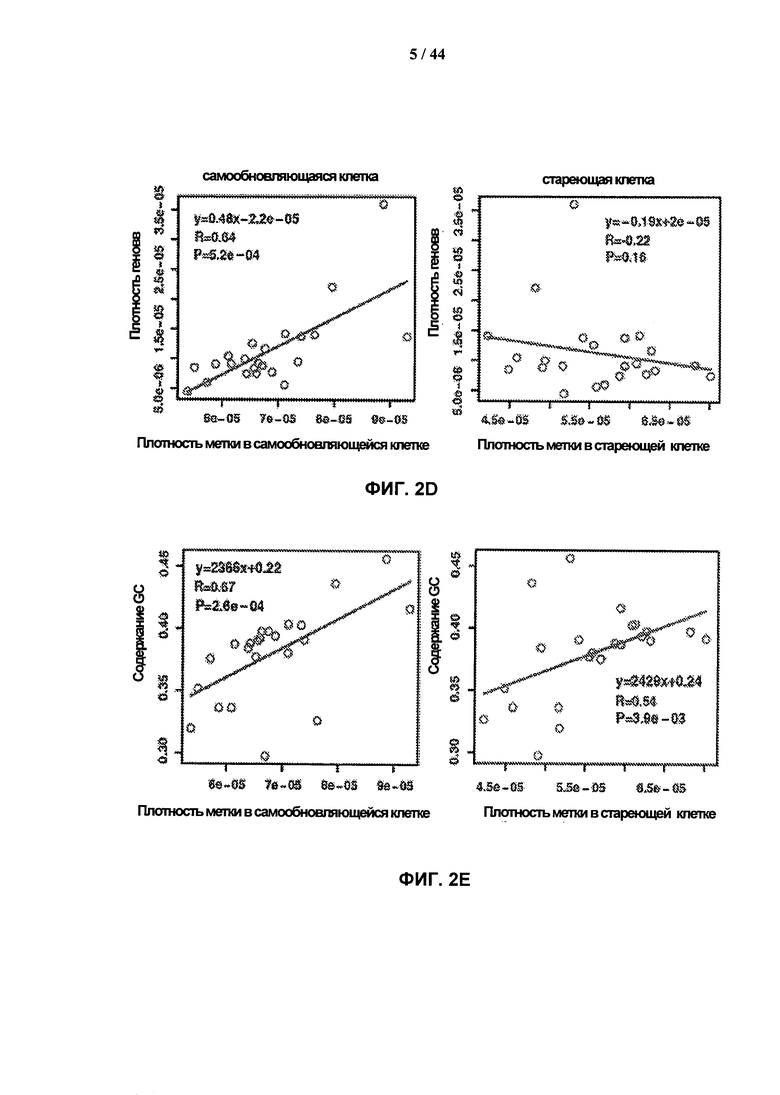
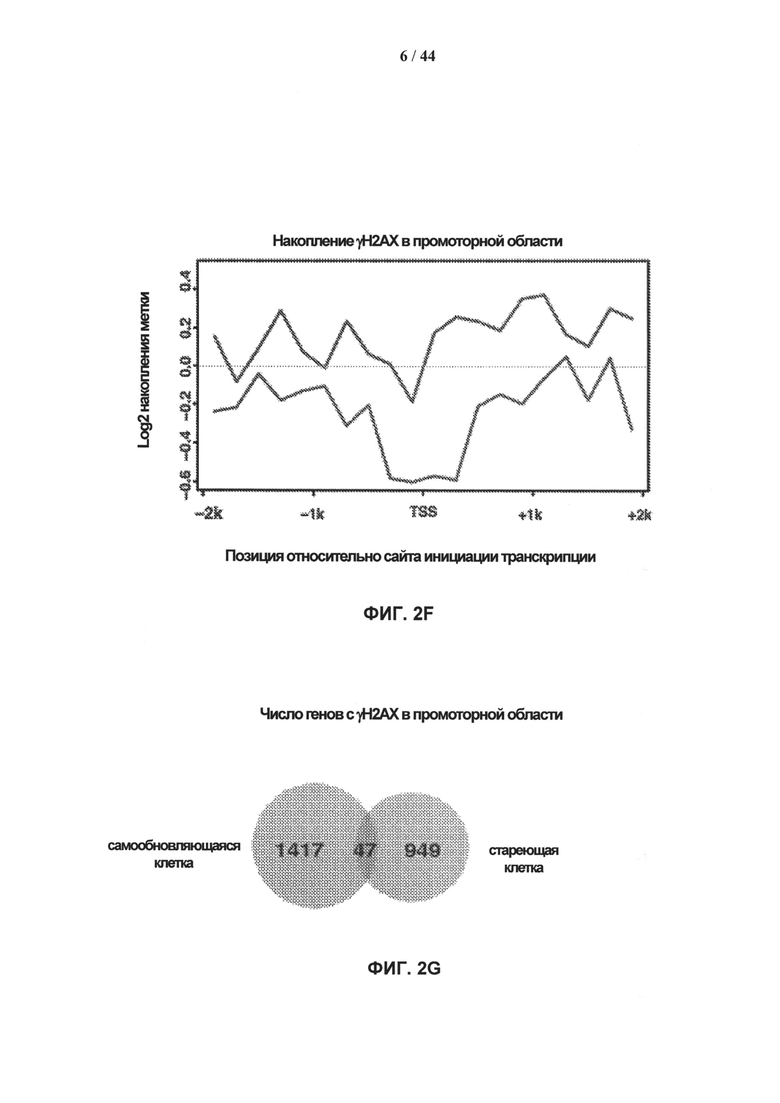
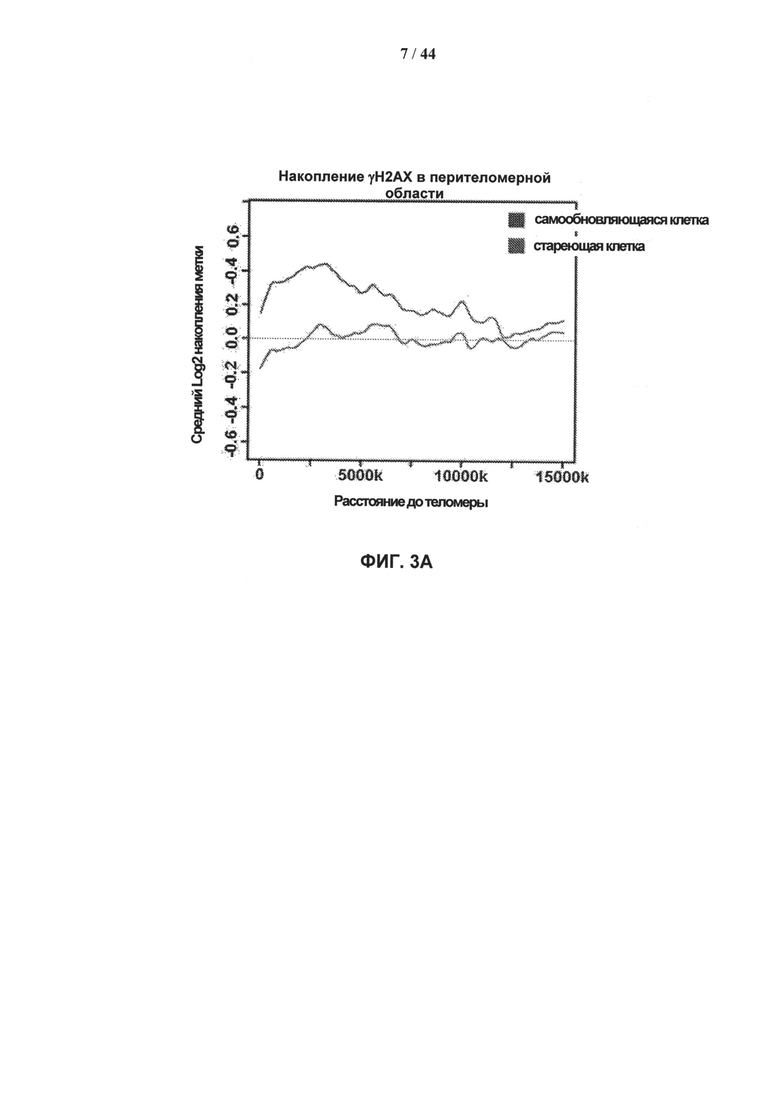
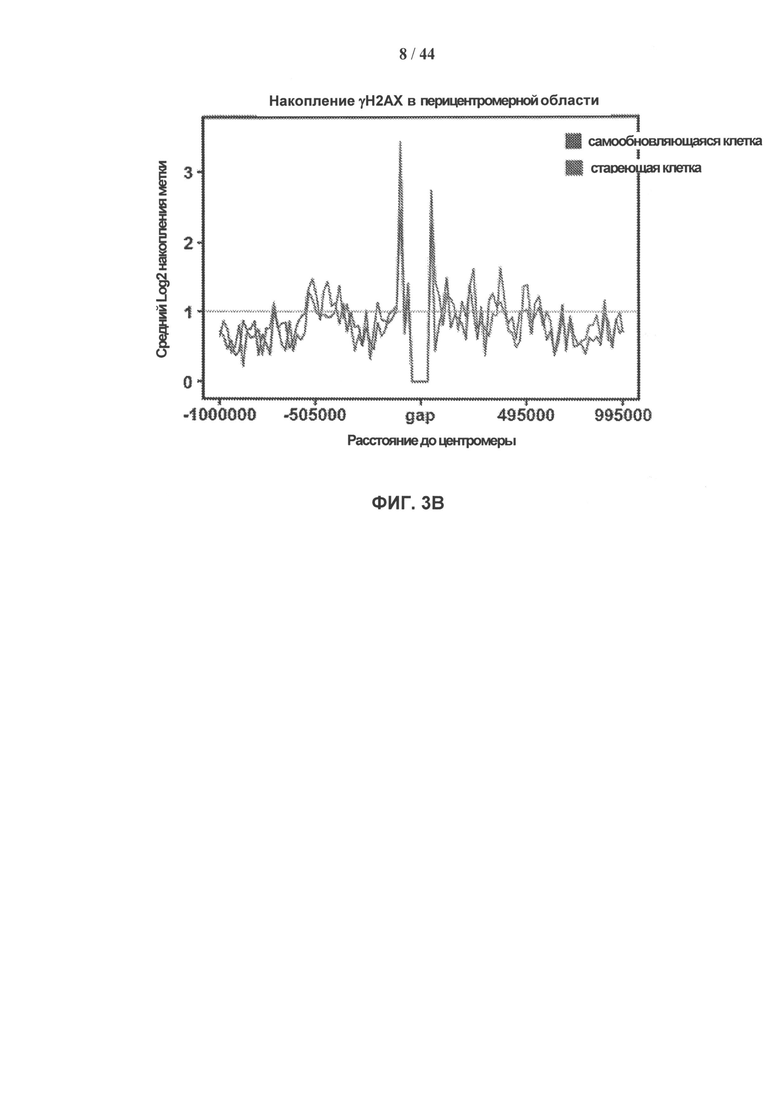
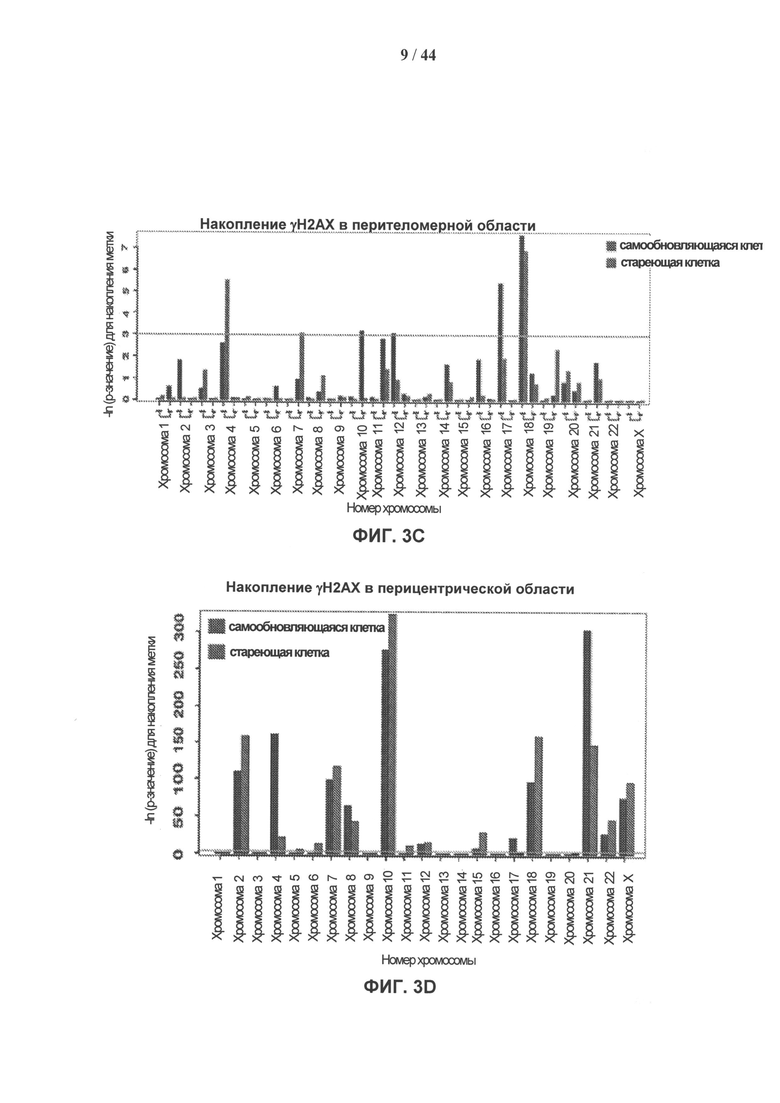
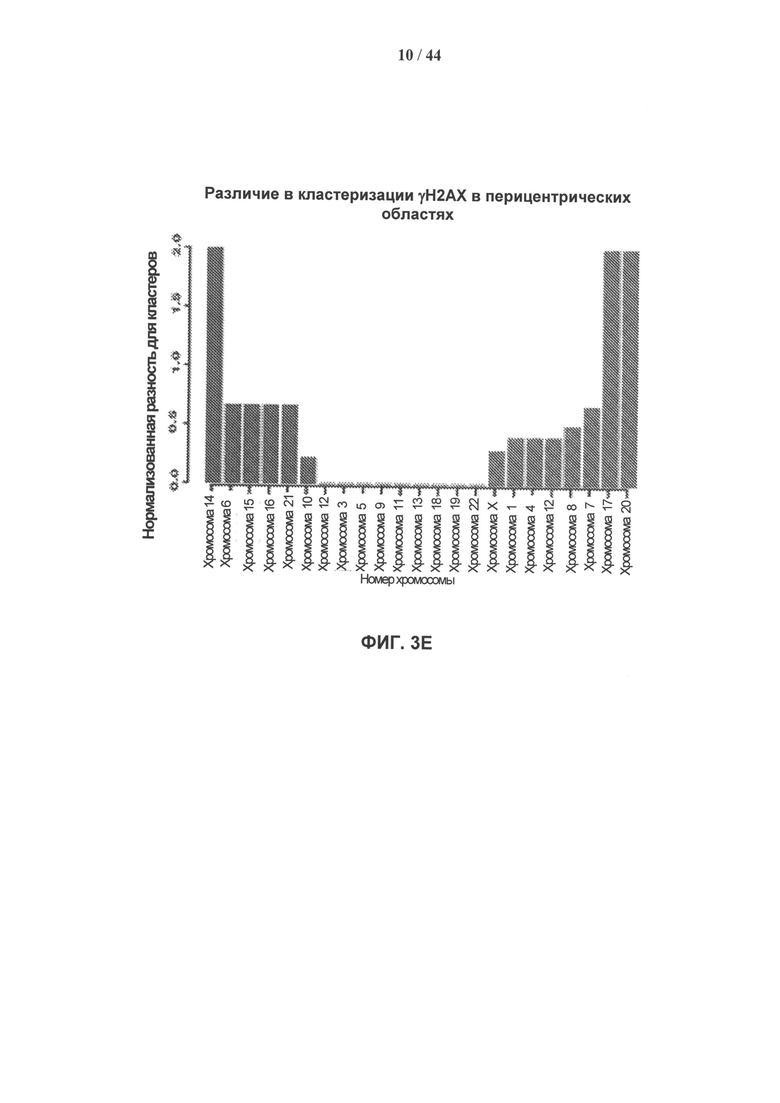
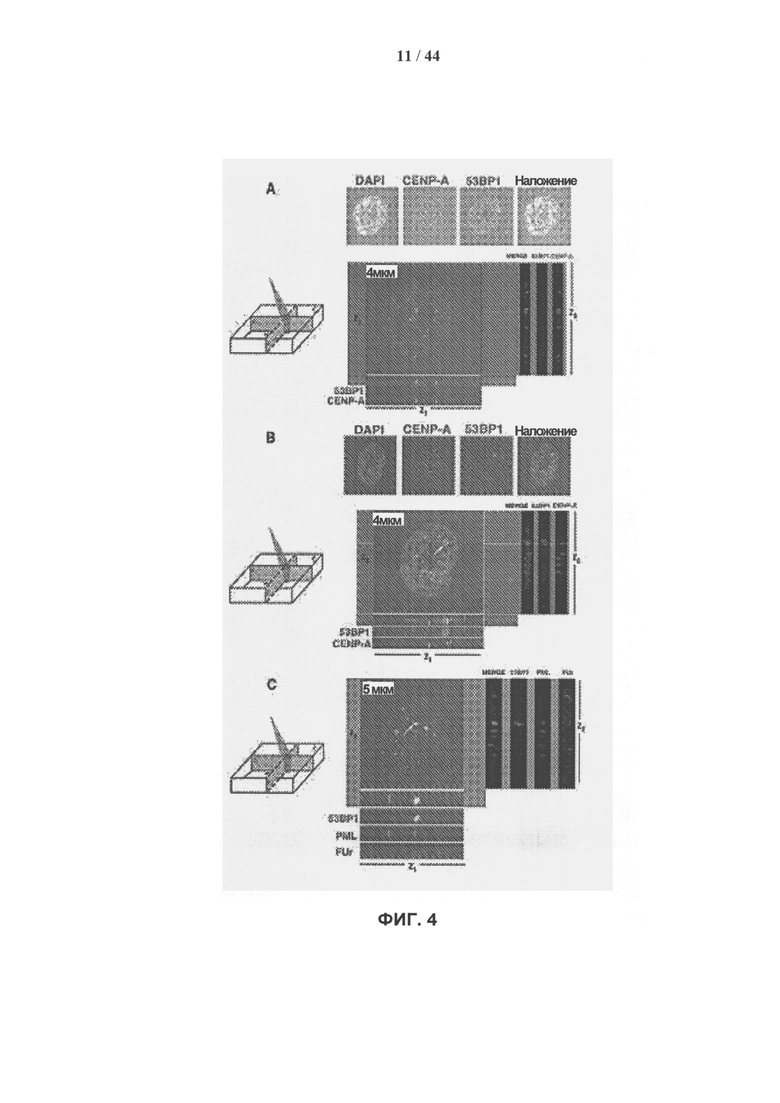
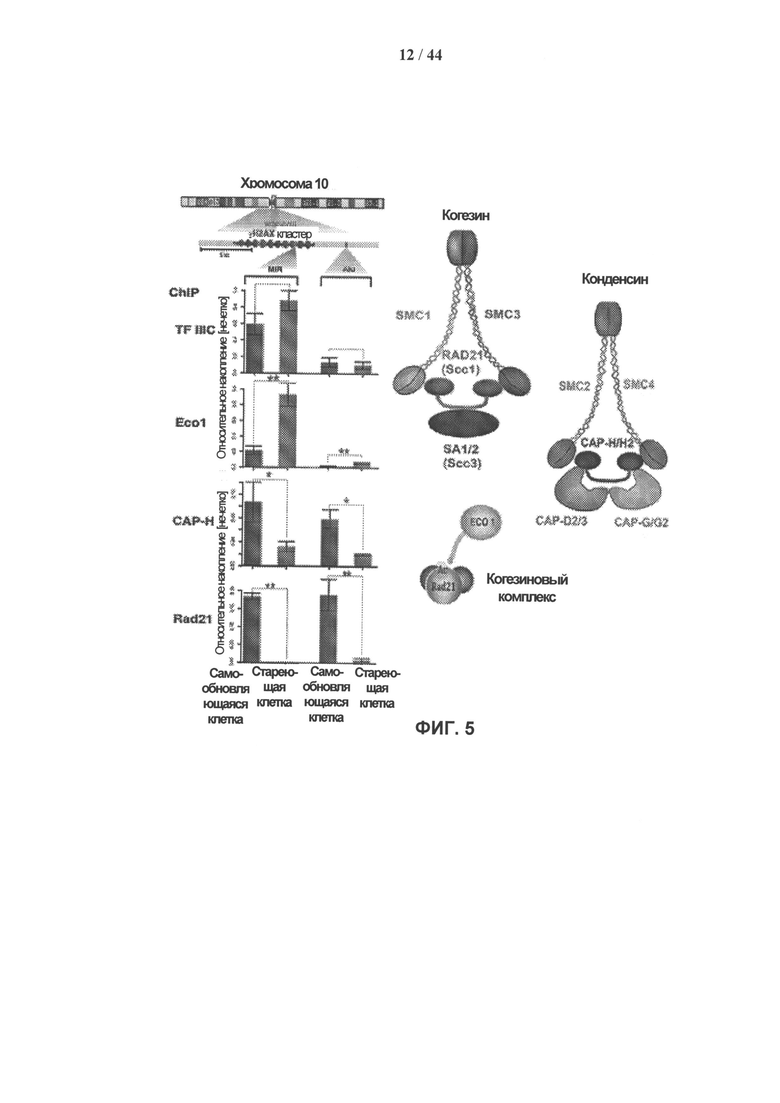
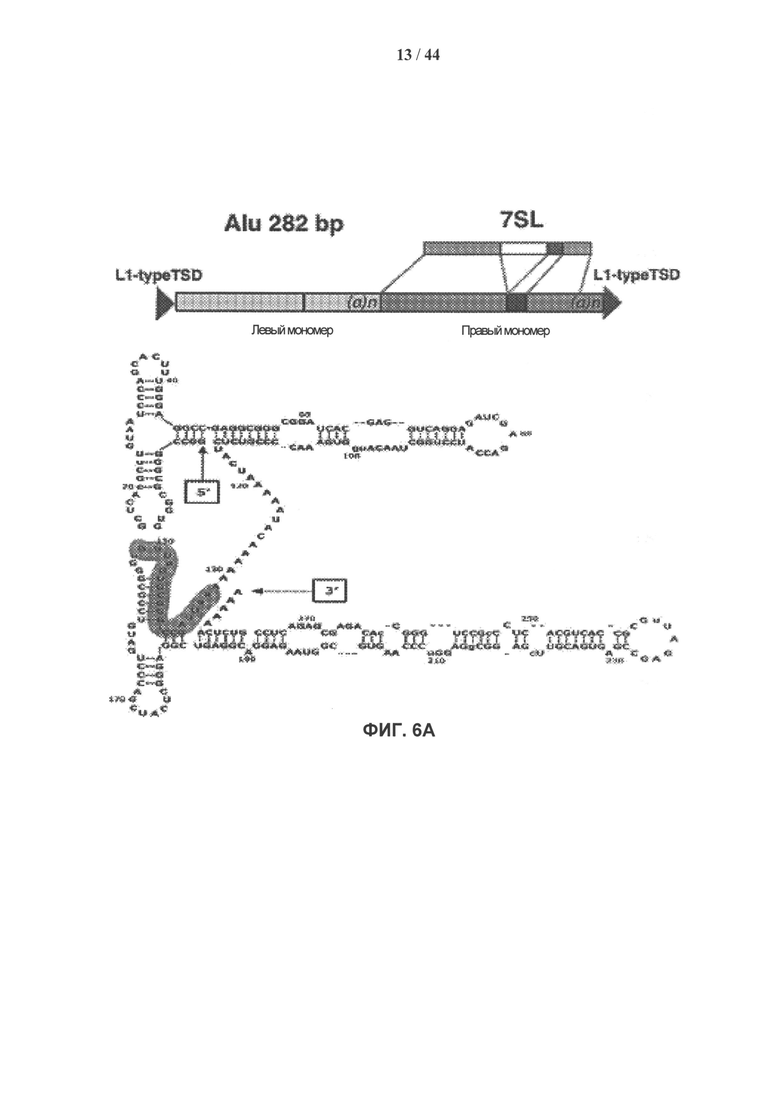
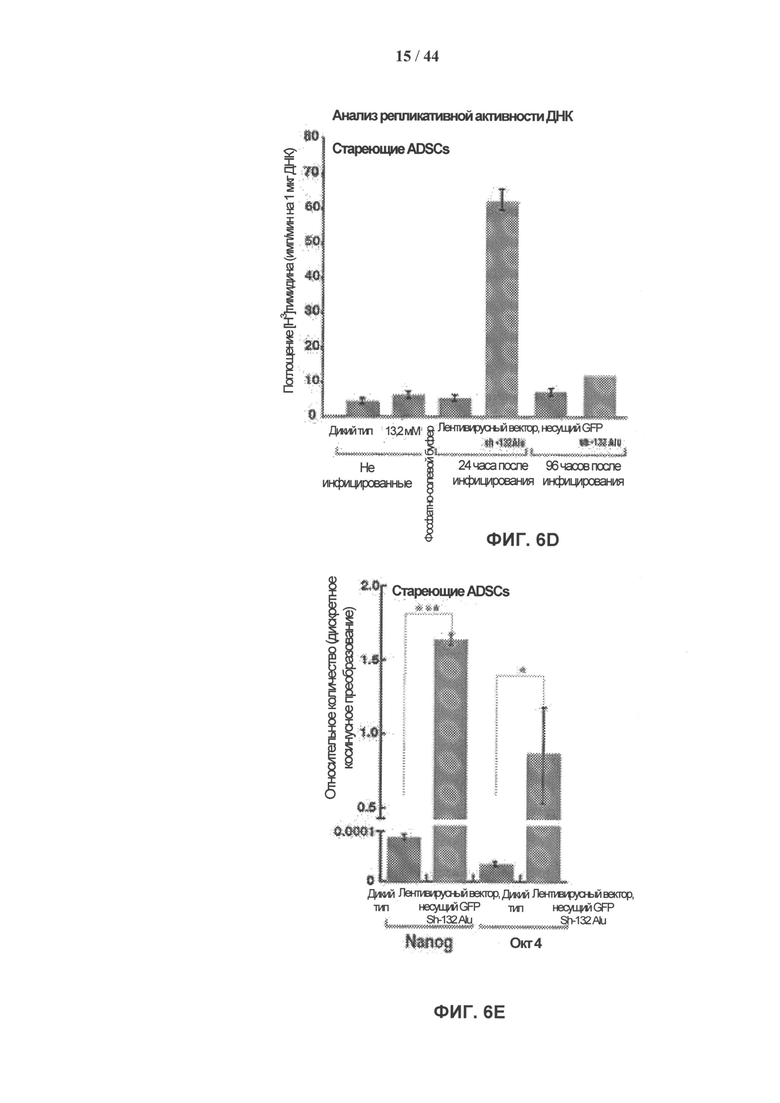
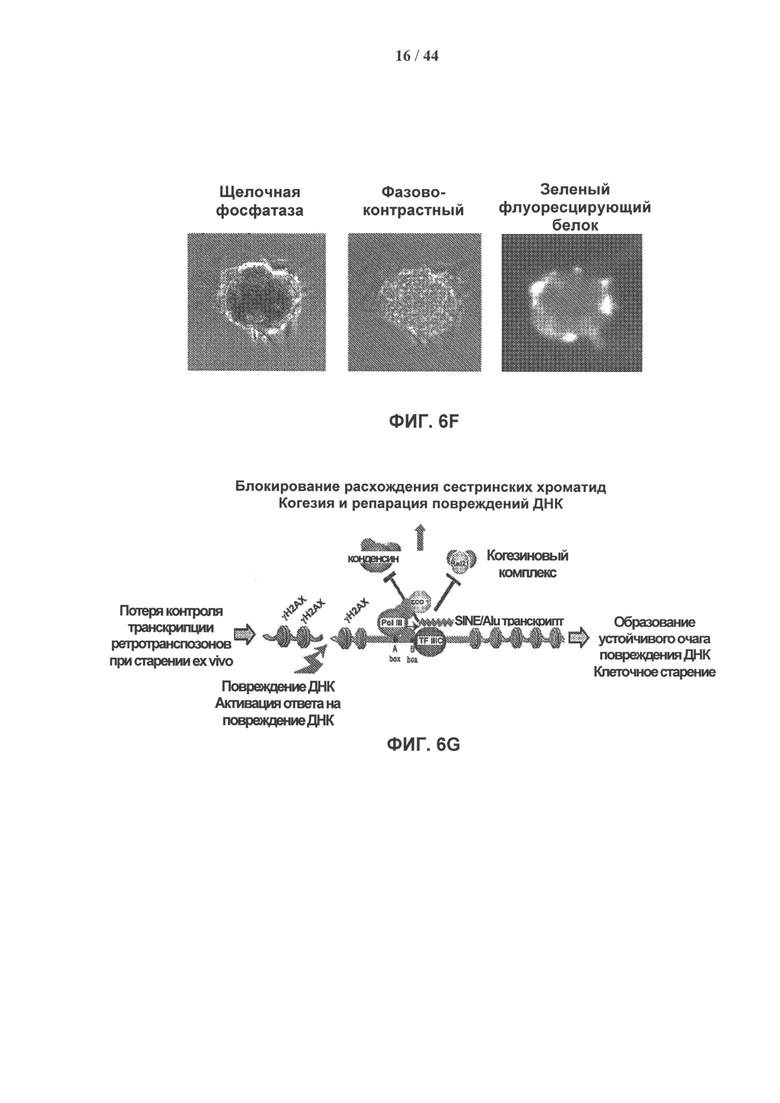


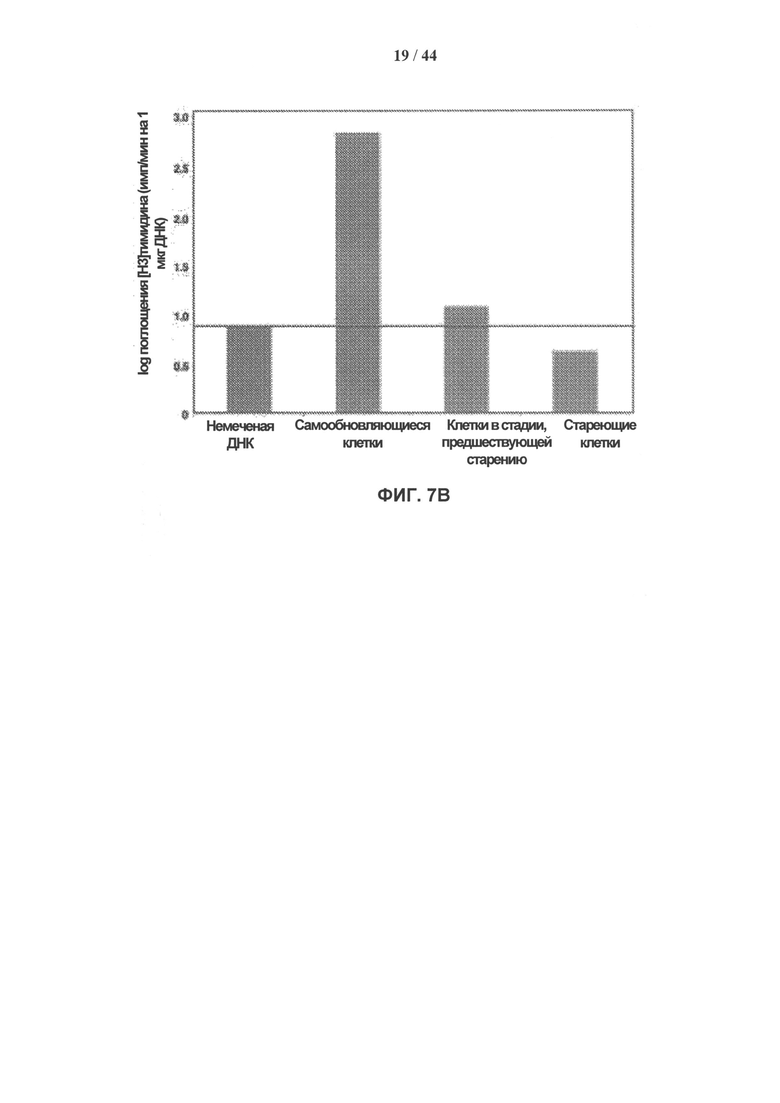
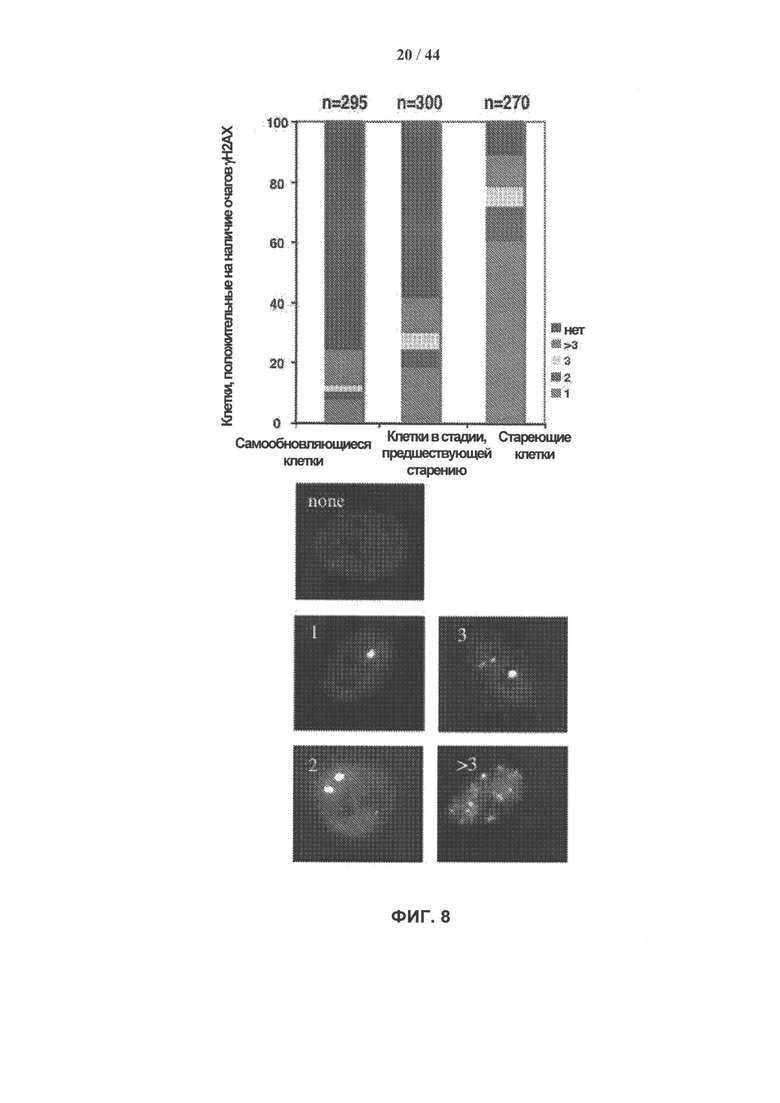
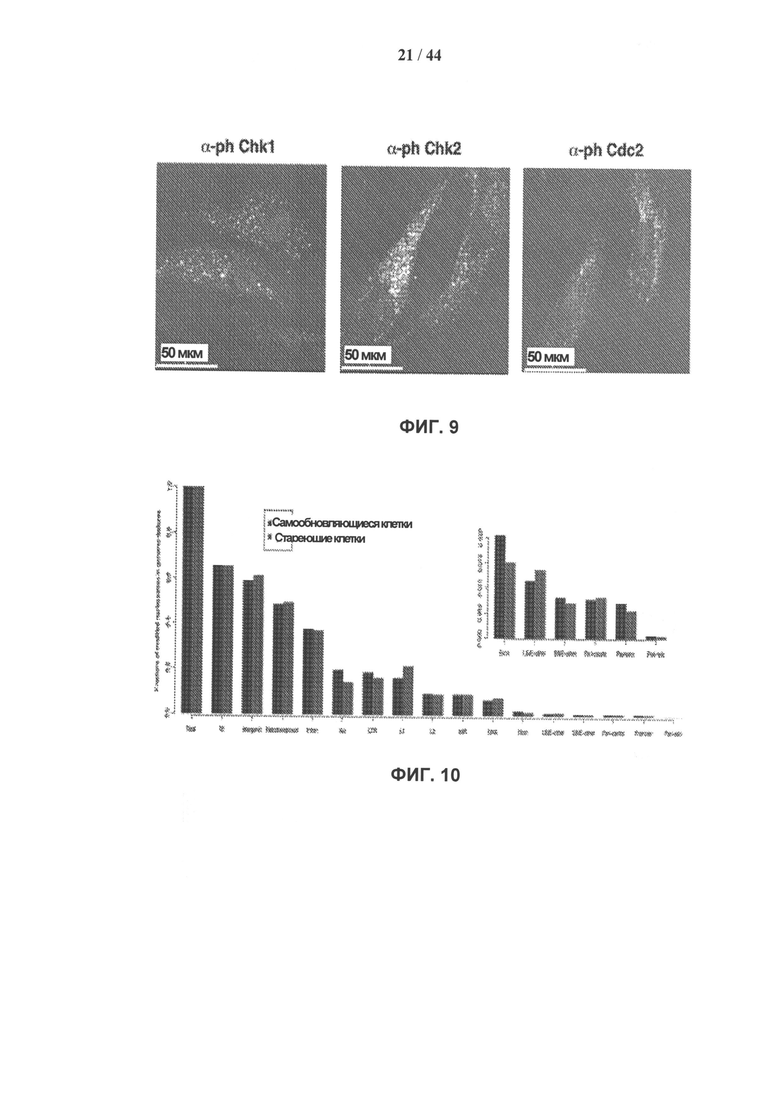
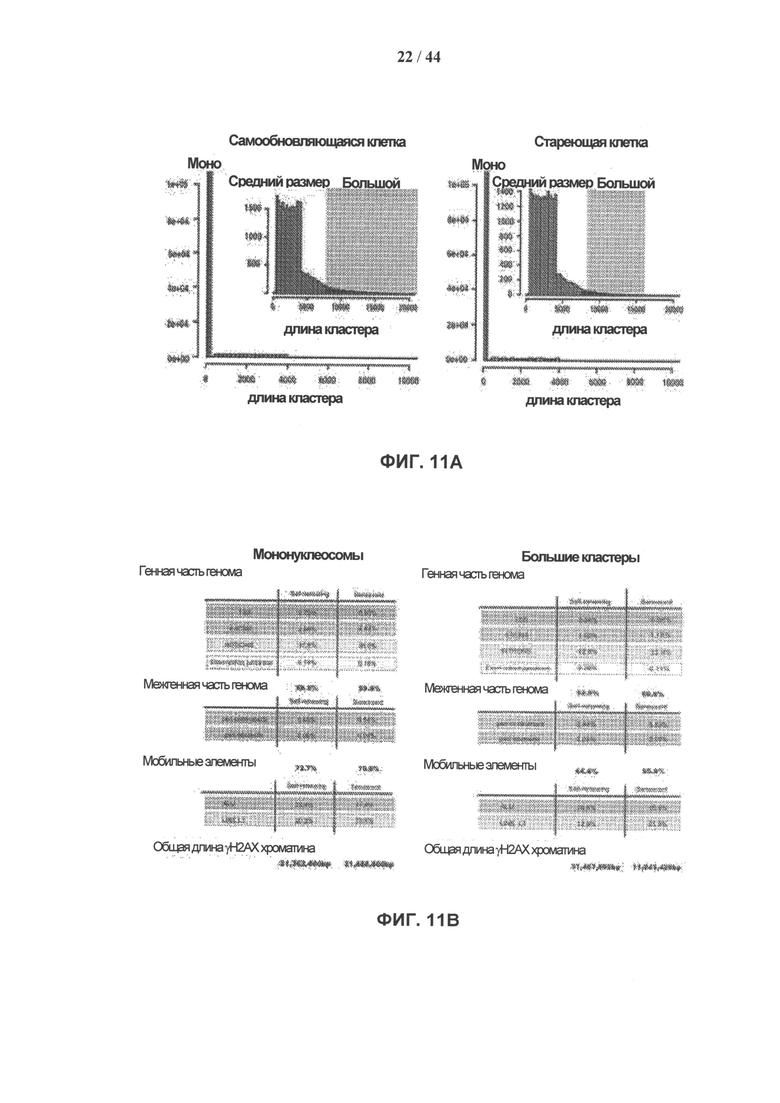
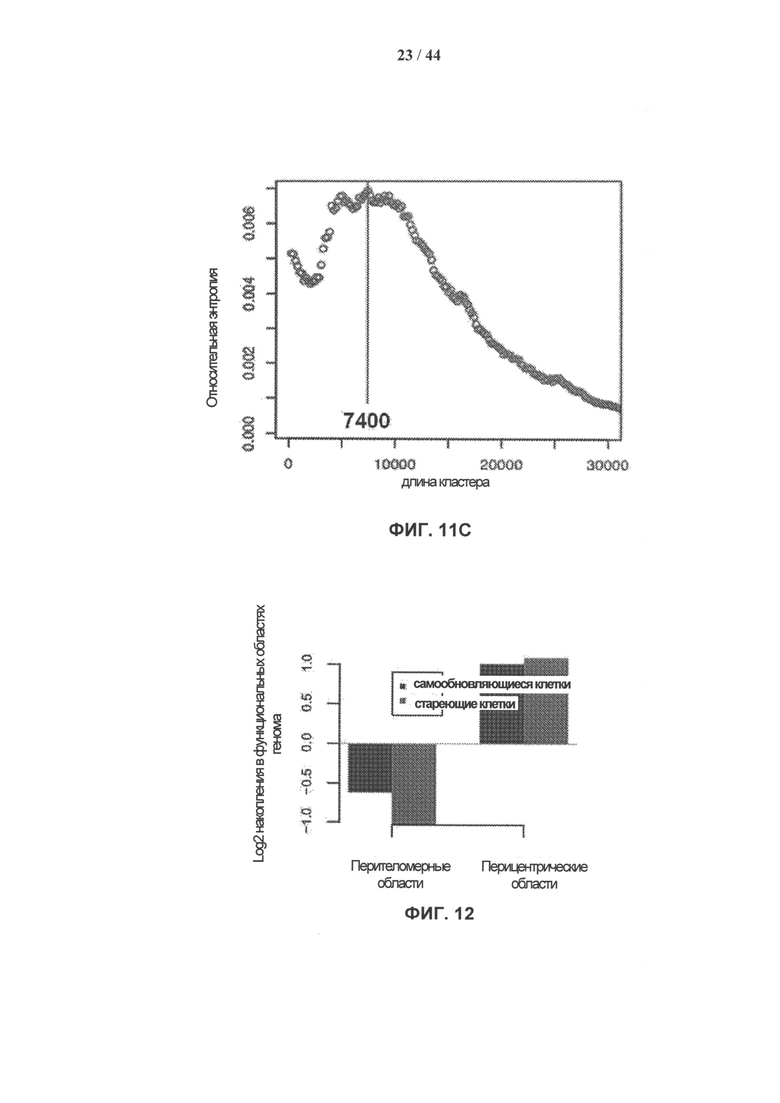
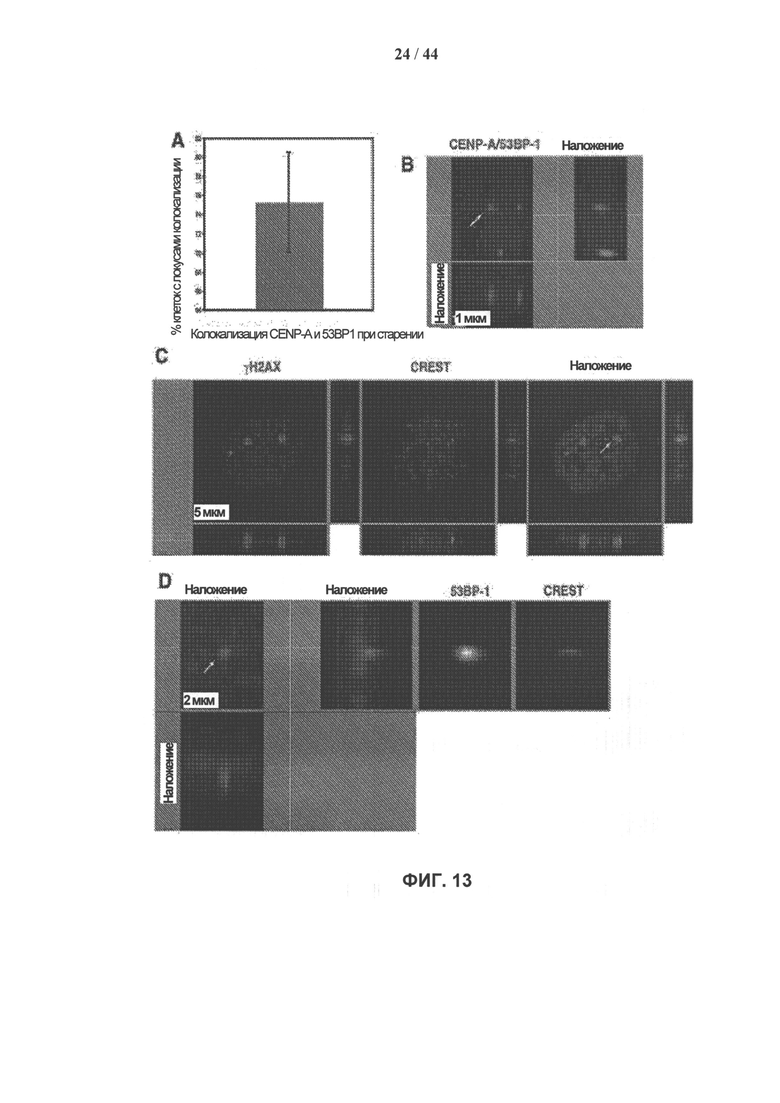
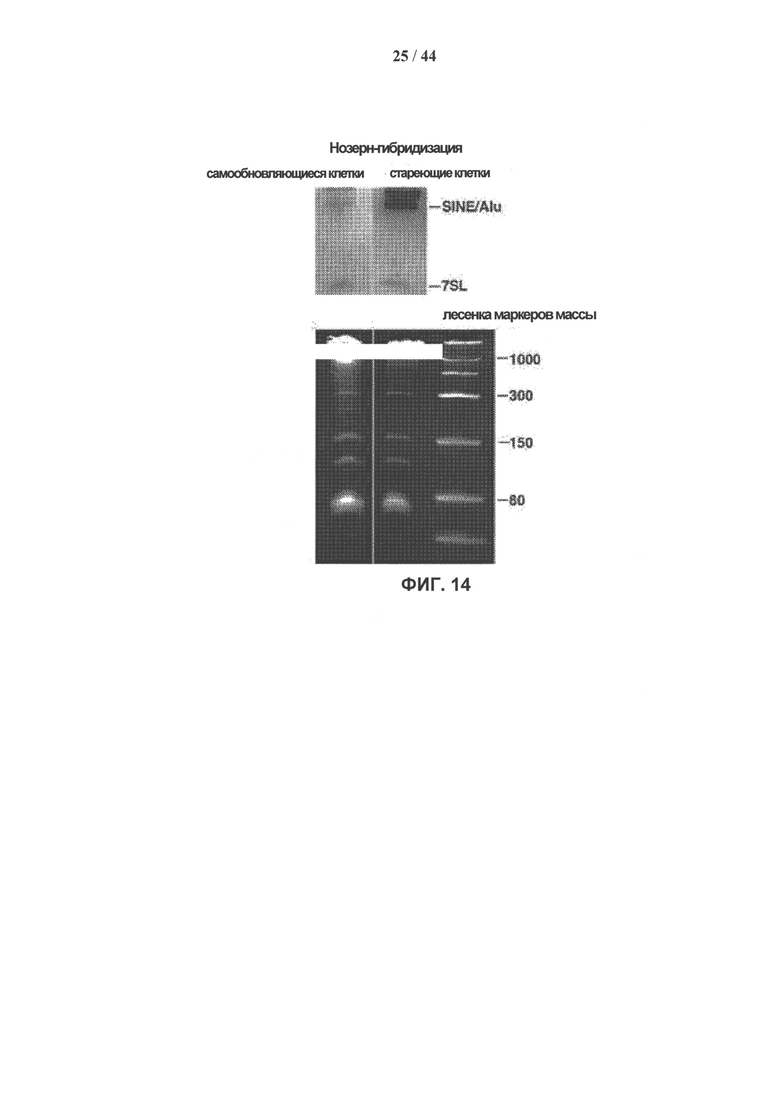
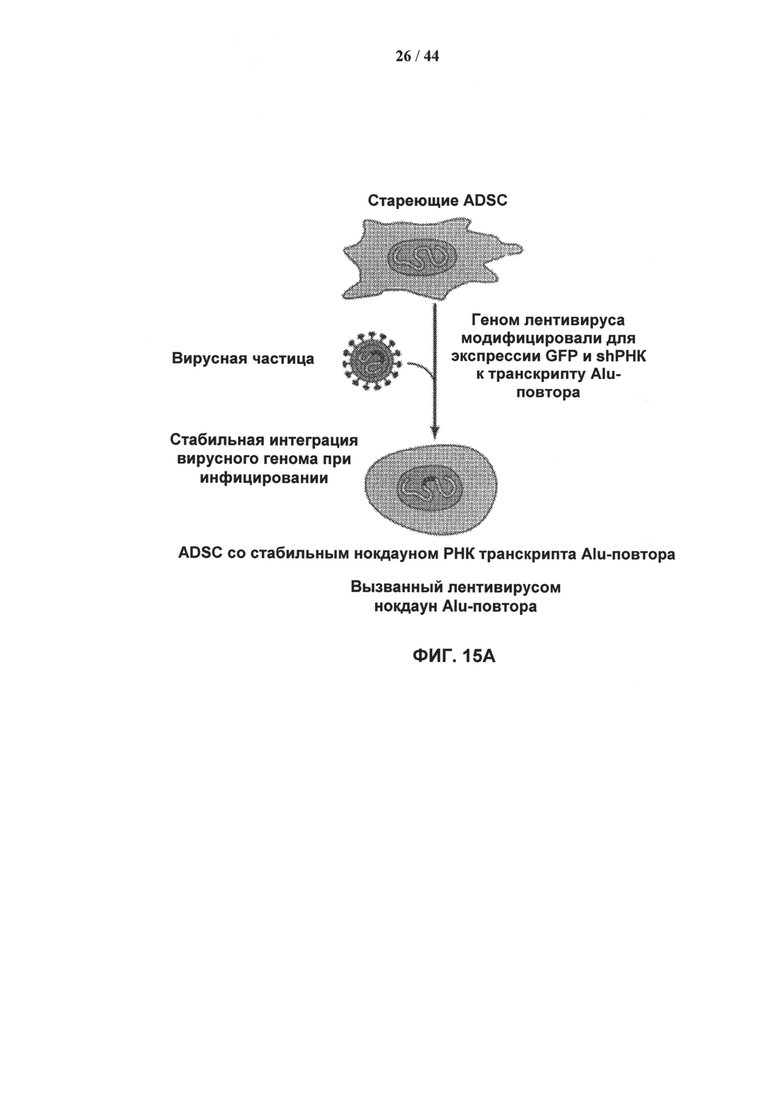

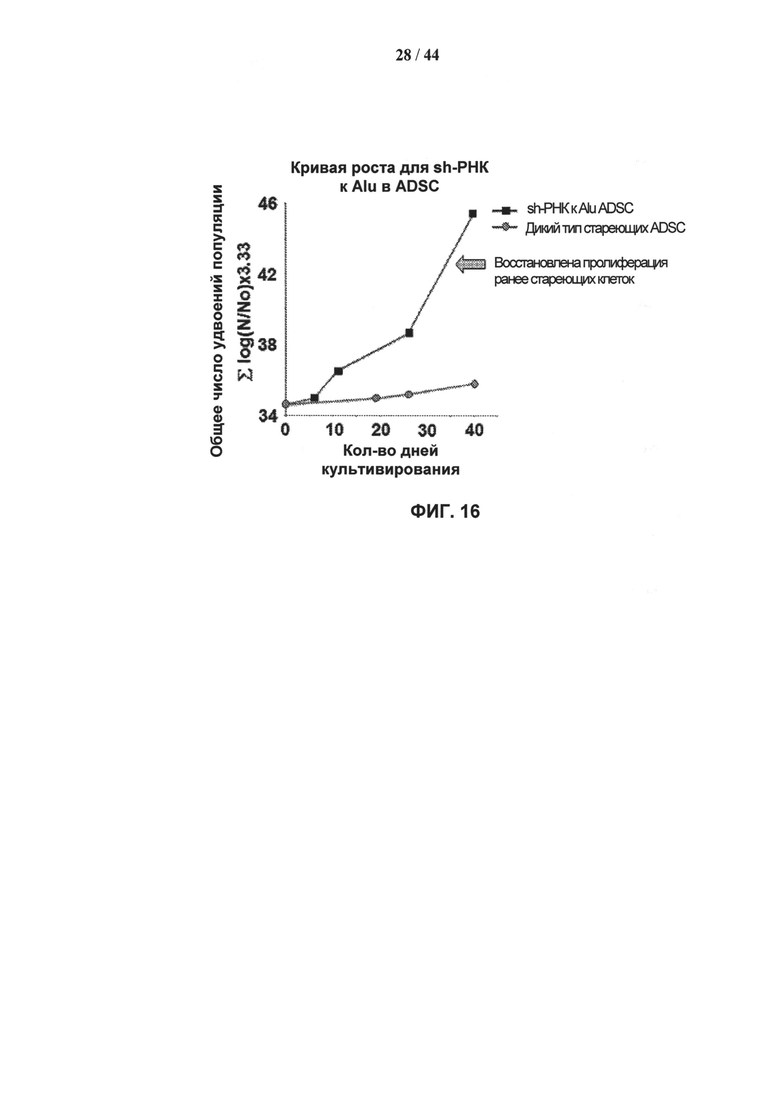


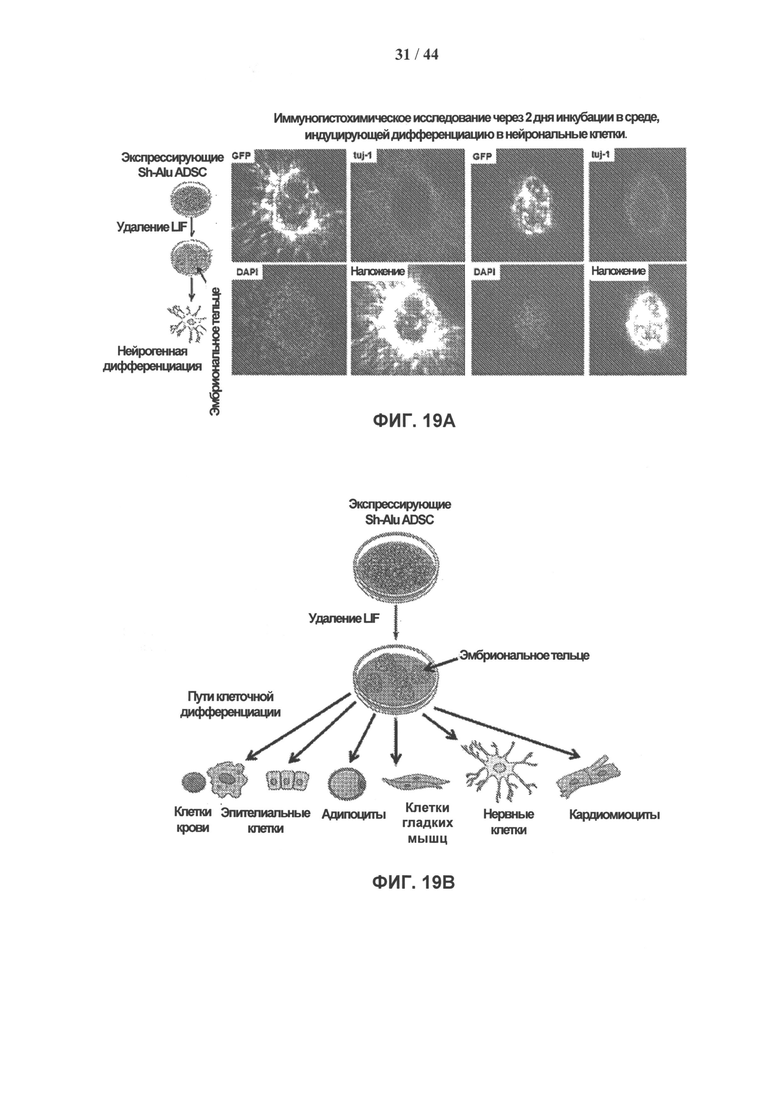
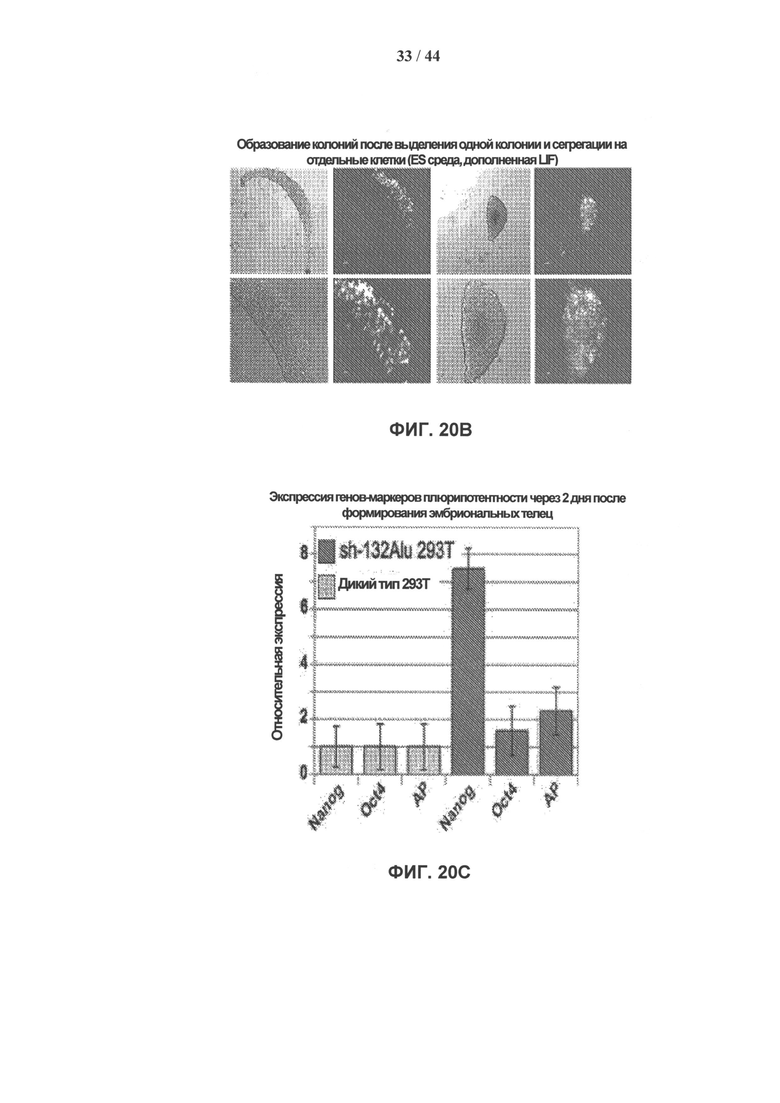
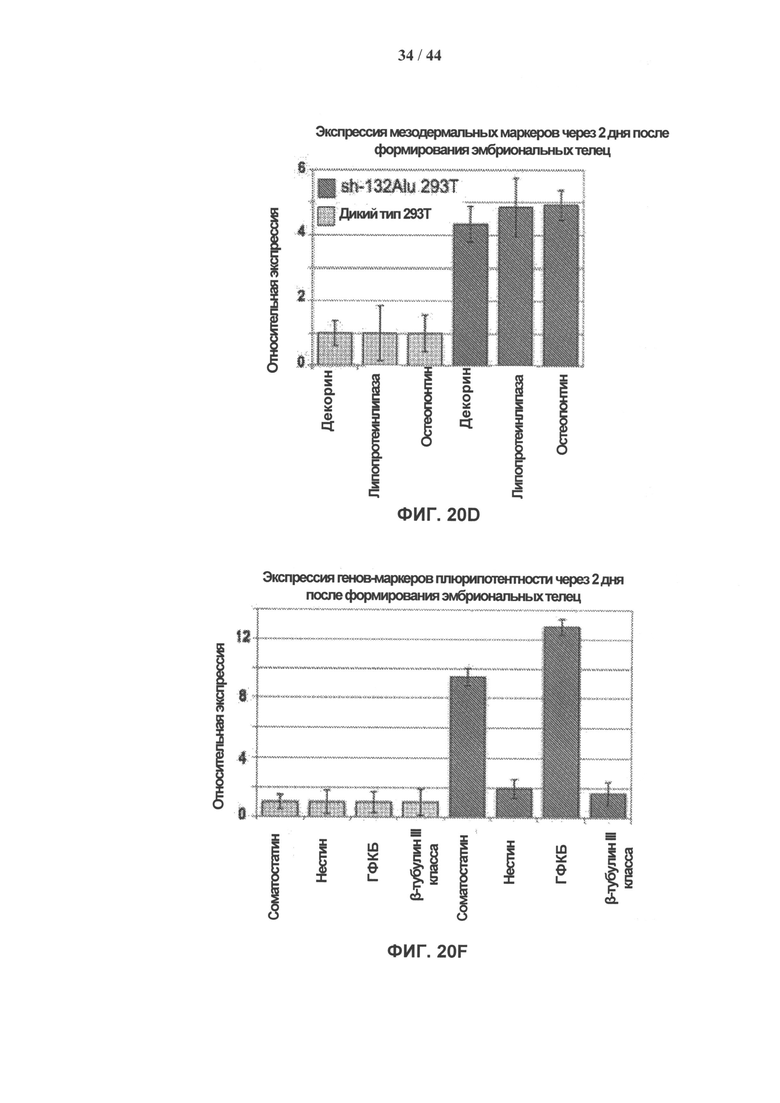
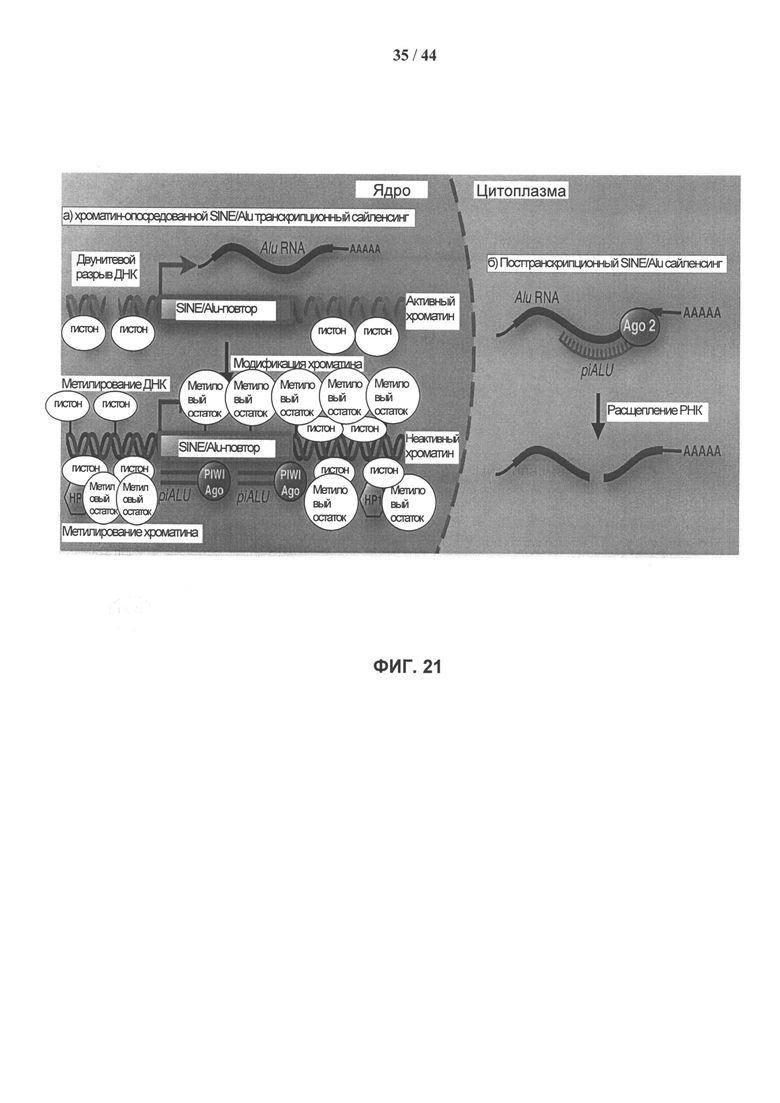
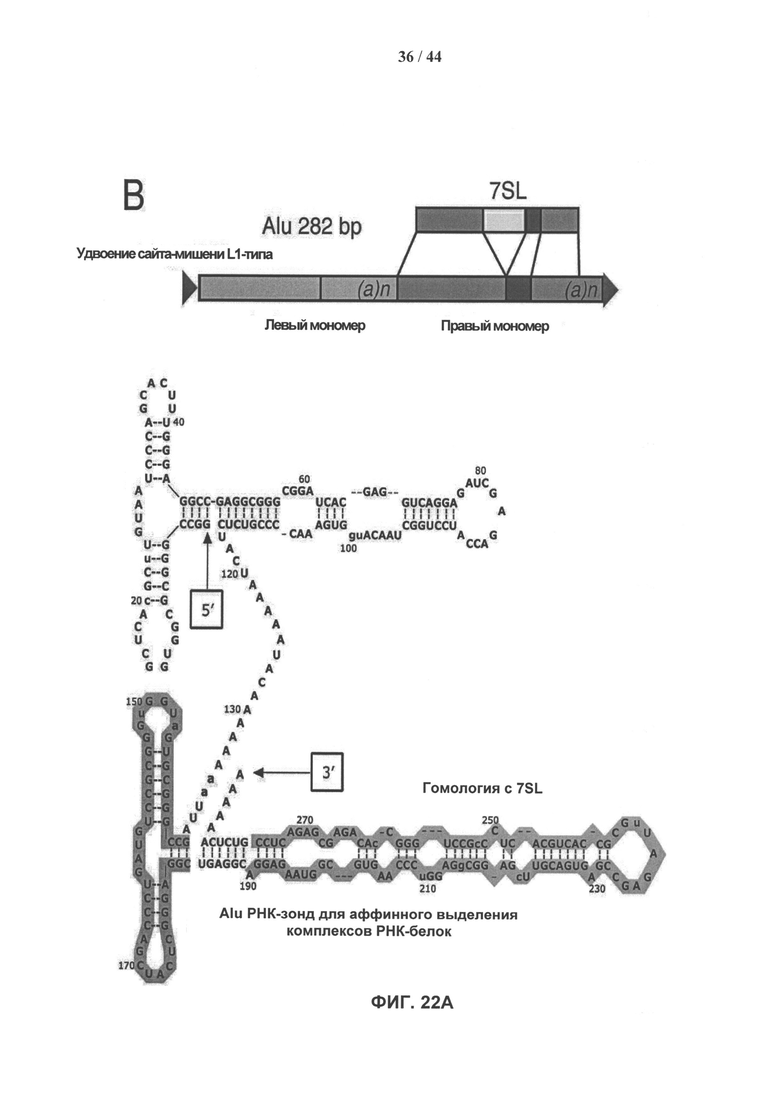
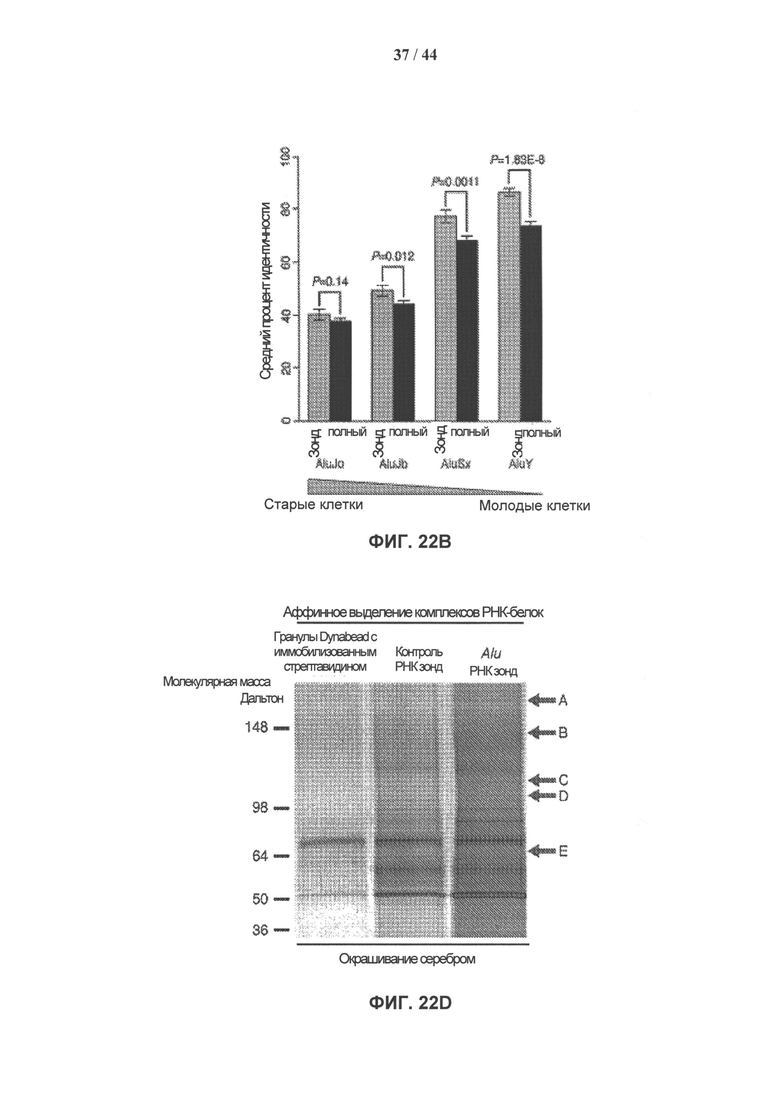
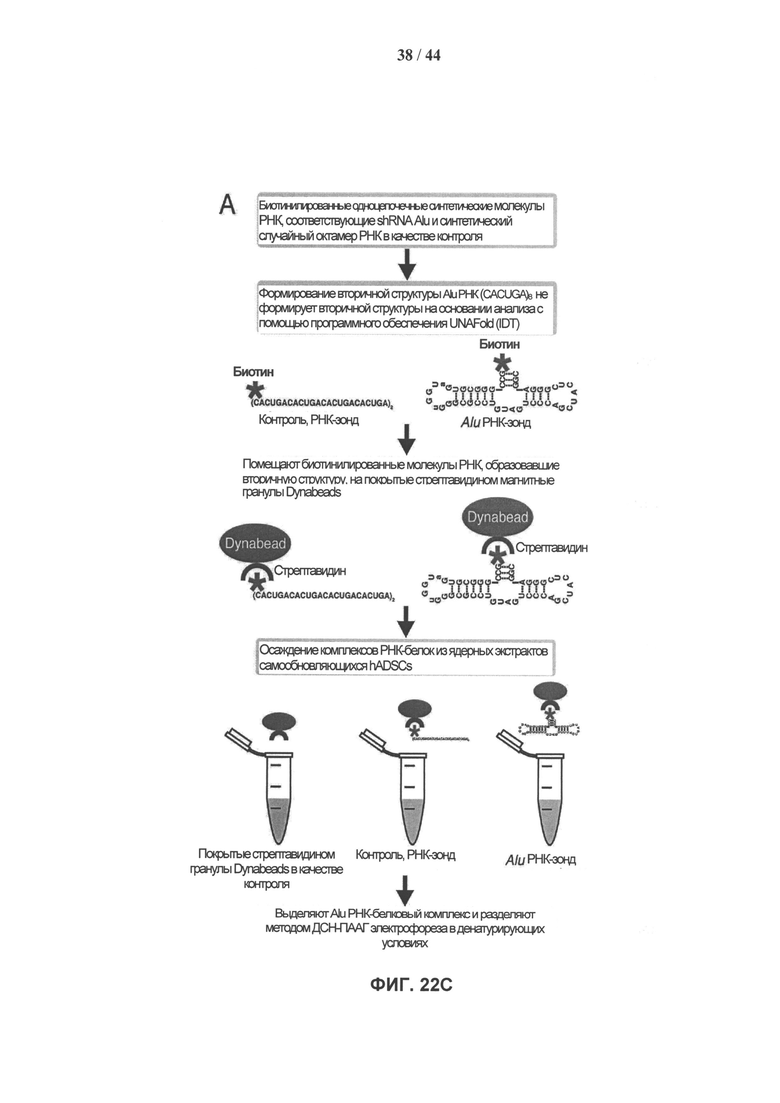

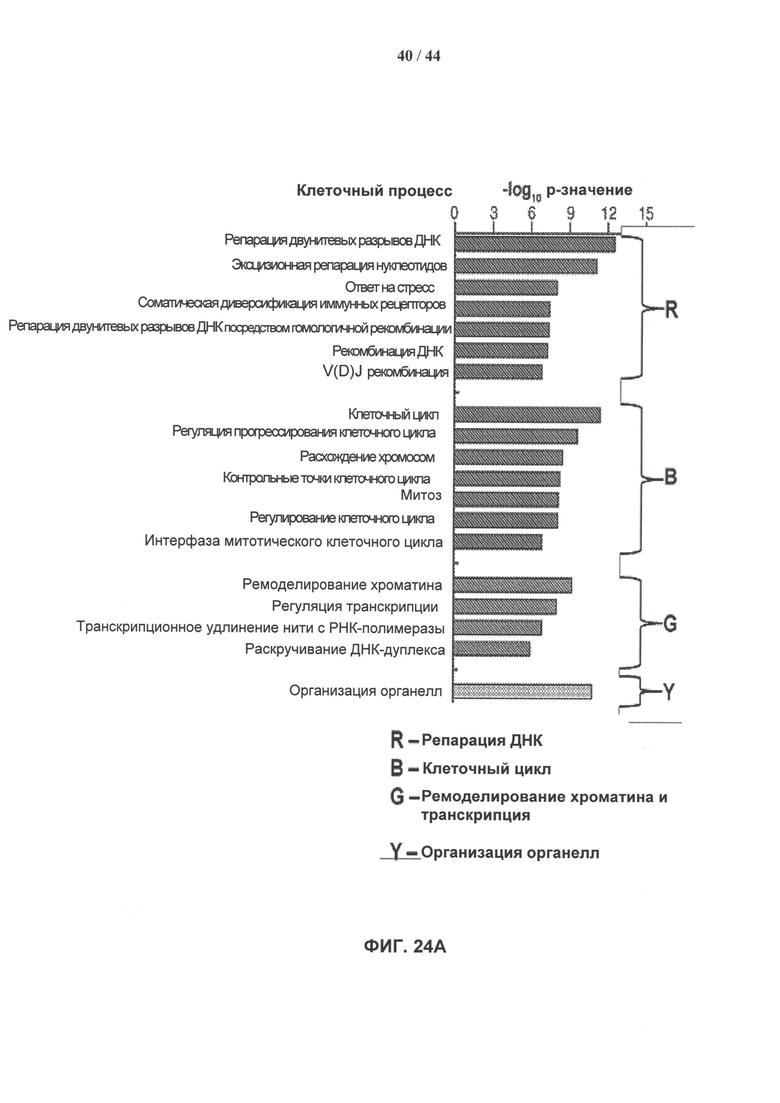
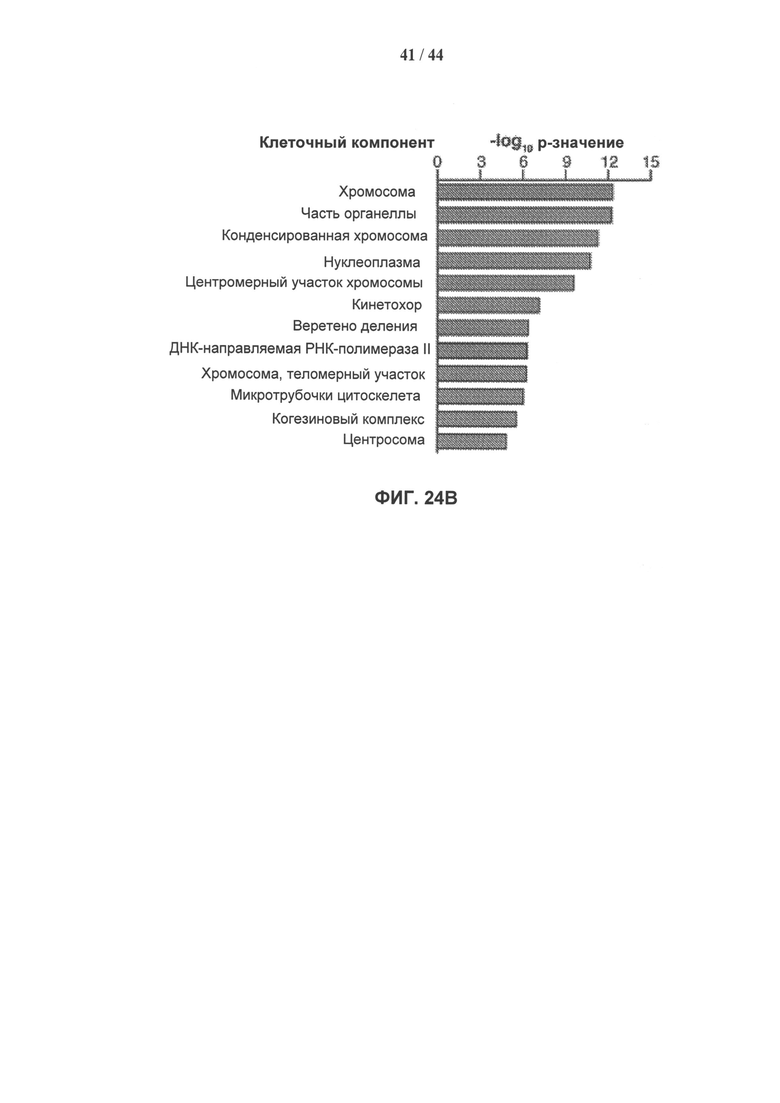

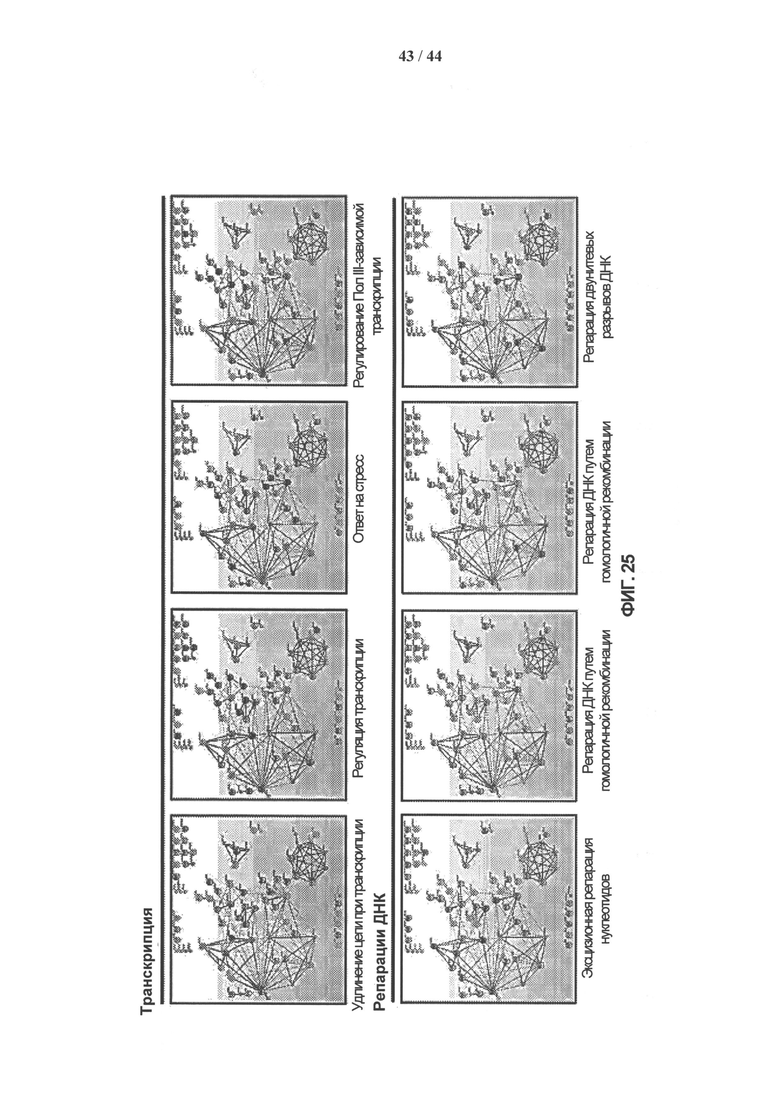

Изобретение относится к области биотехнологии. Описан способ индукции, и/или восстановления, и/или поддержания фенотипа без признаков старения или его аспектов, в частности способности к пролиферации и/или плюрипотентности в клетке млекопитающего. Способ включает снижение уровня или активности транскриптов ретротранспозонов SINE/Alu в клетке в количестве, достаточном для индукции или восстановления способности к пролиферации и/или плюрипотентности указанной клетки млекопитающего. 3 н. и 22 з.п. ф-лы, 58 ил., 9 табл., 3 пр.
1. Способ индукции, восстановления или поддержания способности к пролиферации и/или плюрипотентности клетки млекопитающего, за исключением эмбриональной стволовой клетки человека, причем указанный способ включает: снижение уровня или активности транскриптов ретротранспозонов SINE/Alu за счет введения в указанную клетку конструкции, которая содержит или кодирует молекулу малой интерферирующей РНК, представляющую собой миРНК или мшРНК, нацеленную на транскрипты ретротранспазонов SINE/Alu, в количестве, достаточном для индукции или восстановления способности к пролиферации и/или плюрипотентности указанной клетки млекопитающего и для разрушения мРНК-мишени.
2. Способ по п. 1, отличающийся тем, что указанный способ включает одно или более из следующего:
восстановление способности к пролиферации и/или плюрипотентности клетки млекопитающего, характеризующейся сниженной или утраченной способностью к пролиферации и/или сниженной или утраченной плюрипотентностью;
индукцию способности к пролиферации и/или плюрипотентности клетки млекопитающего, которая не обладает такой способностью; поддержание способности к пролиферации и/или плюрипотентности клетки млекопитающего.
3. Способ по п. 1, отличающийся тем, что указанная клетка млекопитающего включает клетку, выбранную из группы, состоящей из стволовой клетки, необновляющейся клетки-предшественника, окончательно дифференцированной клетки и стареющей клетки.
4. Способ по п. 3, отличающийся тем, что указанная стволовая клетка млекопитающего включает клетку, выбранную из группы, состоящей из стволовой клетки пуповинной крови, взрослой стволовой клетки и индуцированной плюрипотентной стволовой клетки (ИПСК), за исключением эмбриональной стволовой клетки человека.
5. Способ по п. 3, отличающийся тем, что указанная стволовая клетка млекопитающего представляет собой стволовую клетку, полученную из ткани, выбранной из группы, состоящей из жировой ткани человека, костного мозга человека, нервной ткани человека, гладкомышечной ткани человека, кардиомиоцитов человека, эндотелиальной ткани человека и эпителиальной ткани человека.
6. Способ по п. 3, отличающийся тем, что указанная клетка проявляет один или более признаков старения.
7. Способ по п. 3, отличающийся тем, что указанная клетка детерминирована к какому-либо пути дифференцировки клеток.
8. Способ по п. 7, отличающийся тем, что указанная клетка детерминирована к дифференцировке в клеточный тип, выбранный из группы, состоящей из эктодермы, мезодермы и энтодермы.
9. Способ по п. 7, отличающийся тем, что указанная клетка детерминирована к дифференцировке в клеточный тип, выбранный из группы, состоящей из жировых клеток человека, клеток крови человека, нервных клеток человека, гладкомышечных клеток человека, адипоцитов человека, хондроцитов человека, кардиомиоцитов человека, эндотелиальных клеток человека и эпителиальных клеток человека.
10. Способ по п. 9, отличающийся тем, что указанная малая интерферирующая РНК включает молекулу, выбранную из группы, состоящей из одноцепочечной РНК, спаренной двухцепочечной РНК (дцРНК), малой шпилечной РНК (мшРНК, shPHK).
11. Способ по п. 9, отличающийся тем, что указанная конструкция содержит sh-132Alu.
12. Способ по п. 9, отличающийся тем, что указанная конструкция обеспечивает стабильную отрицательную регуляцию транскриптов ретротранспозонов SINE/Alu.
13. Способ по п. 9, отличающийся тем, что указанная конструкция включает вектор.
14. Способ по любому из пп. 1-13, отличающийся тем, что указанный способ не включает нацеленную повышающую регуляцию Oct3/4 и/или Sox2, и/или Klf4, и/или с- тус, и обеспечение одной или более гетерологичных конструкций в указанной клетке, кодирующих Oct3/4, и/или Sox2, и/или Klf4, и/или с-тус.
15. Клетка млекопитающего, полученная при осуществлении способа индукции, восстановления и поддержания способности к пролиферации и/или плюрипотентности по любому из пп. 1-14 и предназначенная для экспрессии, содержит конструкцию, которая снижает уровень или активность транскриптов SINE/Alu в указанной клетке, в результате чего указанная клетка восстановила или поддерживает способность к пролиферации и/или плюрипотентности, и/или демонстрирует уменьшение признаков старения по сравнению со стволовой клеткой того же типа, не содержащей указанной конструкции, при этом указанная конструкция содержит или кодирует молекулу малой интерферирующей РНК, представляющую собой миРНК или мшРНК, нацеленную на транскрипты ретротранспазонов SINE/Alu.
16. Клетка по п. 15, отличающаяся тем, что малая интерферирующая РНК включает молекулу, выбранную из группы, состоящей из одноцепочечной РНК, спаренной двухцепочечной РНК (дцРНК), малой шпилечной РНК (мшРНК, shPHK).
17. Клетка по п. 15, отличающаяся тем, что указанная конструкция обеспечивает стабильную отрицательную регуляцию транскриптов SINE/Alu.
18. Клетка по п. 15, отличающаяся тем, что указанная клетка включает клетку, выбранную из группы, состоящей из эмбриональной стволовой клетки, за исключением эмбриональной стволовой клетки человека, стволовой клетки пуповинной крови, взрослой стволовой клетки и ИПСК.
19. Клетка по п. 18, отличающаяся тем, что указанная клетка представляет собой стволовую клетку, полученную из ткани, выбранной из группы, состоящей из жировой ткани человека, костного мозга человека, нервной ткани человека, гладкомышечной ткани человека, остеобластов и остеокластов человека, ткани спинного мозга человека, кардиомиоцитов человека, эндотелиальной ткани человека и эпителиальной ткани человека.
20. Клетка по любому из пп. 15-19, отличающаяся тем, что указанная клетка выбрана из группы, состоящей из:
клетки, которая ранее представляла собой или получена из клетки, которая имеет сниженную или утраченную способность к пролиферации и/или сниженную или утраченную плюрипотентность;
клетки, которая ранее представляла собой или получена из стволовой клетки, проявляющей один или более признаков старения;
клетки, которая ранее представляла собой или получена из стволовой клетки, детерминированной к какому-либо пути дифференцировки клеток; и клетки, которая ранее представляла собой или получена из необновляющейся клетки-предшественника.
21. Клетка по любому из пп. 15-20, отличающаяся тем, что указанная клетка детерминирована к дифференцировке в клеточный тип, выбранный из группы, состоящей из жировой клетки человека, клеток крови человека, нервных клеток человека, гладкомышечных клеток человека, остеобластов и остеокластов человека, клеток поджелудочной железы человека, адипоцитов человека, хондроцитов человека, кардиомиоцитов человека, эндотелиальных клеток человека и эпителиальных клеток человека.
22. Клетка по любому из пп. 15-20, отличающаяся тем, что указанная клетка не содержит конструкцию, нацеленную на повышающую регуляцию или кодирующую Oct3/4, и/или Sox2, и/или Klf4, и/или с-тус.
23. Способ получения дифференцированной клетки, причем указанный способ включает:
получение клетки по любому из пп. 15-22; и
культивирование указанной клетки в условиях, которые индуцируют или допускают дифференцировку в эмбриоидное тельце и/или окончательно дифференцированную клетку, а именно в среде, не содержащей фактор ингибирования лейкемии.
24. Способ трансдифференцировки клетки млекопитающего из клетки первого типа или линии в клетку второго типа или линии, причем указанный способ включает:
трансформацию дифференцированной клетки в плюрипотентную клетку способом по любому из пп. 1-14;
культивирование указанной клетки в условиях, которые индуцируют или допускают дифференцировку указанной клетки;
отбор клеток, которые дифференцировались в клетки второго типа или линии; и
культивирование клеток второго типа или линии, при этом клетки первого типа или линии представляют собой клетку мезодермального типа, или адипоцит, или клетку костного мозга, или выбранную из группы, состоящей из клетки крови, эмбриональной клетки, за исключением эмбриональной клетки человека, эпителиальной клетки, адипоцита, гладкомышечной клетки, нервной клетки, панкреатической бета-клетки и кардиомиоцита, а клетки второго типа или линии представляют собой клетку нейроэктодермального типа.
25. Способ по п. 24, отличающийся тем, что культивирование указанной клетки в условиях, которые индуцируют или допускают дифференцировку указанной клетки, включает культивирование указанной клетки в среде, не содержащей или с пониженным содержанием фактора ингибирования лейкемии (LIF).
| US 2006177825 A1, 10.08.2006 | |||
| US 2010221827 A1, 02.09.2010 | |||
| US 2009099060 A1, 16.04.2009 | |||
| СПОСОБ ПОЛУЧЕНИЯ ПЛЮРИПОТЕНТНЫХ КЛЕТОК | 2009 |
|
RU2399667C1 |

