Данная заявка заявляет приоритет согласно предварительной заявке на патент США номер 60/943, 994, поданной 14 июня, 2007, полное содержание которой включено здесь в частности путем ссылки.
Область изобретения
Настоящее изобретение относится в целом к конечным продуктам гликирования ("AGE") и в частности к определенным слитым белкам, которые содержат рецептор конечных продуктов гликирования ("RAGE"). Слитые белки по изобретению связываются с AGE и другими лигандами RAGE (например, S100 и HMGB1), и композиции, содержащие слитые белки по изобретению, могут использоваться для лечения заболеваний.
Предшествующий уровень техники
Конечные продукты гликирования (AGE) являются результатом неферментативного гликирования и окисления белков. Они появляются в вызванных стрессом условиях, приводящих к аутоиммунным заболеваниям соединительной ткани, и могут формироваться в воспаленной ткани вследствие окисления или миелопероксидазного пути. AGE включены в ряд обусловленных диабетом осложнений. Например, характерные структурные изменения при диабетической нефропатии, утолщенная клубочковая базальная мембрана и мезангиальное расширение сопровождаются накоплением AGE, приводя к гломерулосклерозу и интерстициальному фиброзу. Пролонгированная инфузия AGE в недиабетических крысах приводит к развитию подобных морфологических изменений и значительной протеинурии. Было показано, что ингибиторы AGE, такие как аминогуанидин, предотвращают диабетическую нефропатию у диабетических животных моделей, и, как недавно было продемонстрировано, делают то же самое в одном клиническом исследовании на пациентах с диабетом. Также AGE являются вполне утвержденной терапевтической мишенью при диабетической ретинопатии. Обширные исследования на диабетических мышах и крысах демонстрируют пользу ингибирования образования AGE при лечении этого заболевания.
Атеросклероз значительно ускорен у пациентов с диабетом и связан с повышенным риском смертности от сердечно-сосудистых и цереброваскулярных заболеваний. Исследования на животных и человеке наводят на мысль, что AGE играют значительную роль в формировании и развитии атеросклерозных поражений. Увеличенное накопление AGE в диабетических сосудистых тканях связано с изменениями в функционировании эндотелиальных клеток, макрофагов и гладкомышечных клеток.
AGE взаимодействуют с рецепторами на поверхности клеток на моноцитах, макрофагах, эндотелиальных клетках микроциркуляторного русла, гладкомышечных клетках, мезангиальных клетках и нейронах. Рецептор для конечных продуктов гликирования (RAGE) является членом иммуноглобулинового суперсемейства рецепторов клеточной поверхности. RAGE состоит из трех внеклеточных иммуноглобулин-подобных доменов, трансмембранного домена и цитоплазматического домена, который вовлечен в сигнальную систему. RAGE связывает множество лигандов помимо AGE, включая S100/кальгранулины, амфотерин/HMGB1 и амилоидные фибриллы. RAGE действует через сигнальный каскад, включающий NF-κВ. Экспрессия RAGE активируется в присутствии лигандов RAGE и увеличивается в суставах субъектов с ревматоидным артритом (RA).
RAGE имеет секретируемую изоформу, не имеющую трансмембранный домен, названную растворимым RAGE (sRAGE). Как было показано, введение of sRAGE восстанавливает заживление ран (Goova, et al. (2001) Am. J. Pathol. 159, 513-525) и подавляет диабетический атеросклероз (Park, et al. (1998) Nat Med. 4(9): 1025-31). Слитые белки, состоящие из лиганд-связывающего элемента RAGE, и иммуноглобулинового элемента, обсуждаются в WO 2004/016229 А2 (Wyeth, Madison, NJ) и публикации патентной заявки США 2006/0057679 A1 (O'Keefe, Т. et al.).
Существует потребность в новых способах лечения AGE-опосредованных заболеваний, таких как заболевания, которые ассоциированы с увеличенным количеством AGE. Эта потребность и другие удовлетворяются настоящим изобретением.
Краткое изложение сущности изобретения
Настоящее изобретение обеспечивает материалы и способы лечения заболеваний, ассоциированных с увеличенным количеством AGE. В одном варианте осуществления настоящее изобретение предоставляет слитый белок, содержащий, по меньшей мере, один полипептид, содержащий: (а) первую аминокислотную последовательность, по меньшей мере, на 95% идентичную лиганд-связывающему домену рецептора конечного продукта гликирования (RAGE) млекопитающего, причем первая аминокислотная последовательность способна связывать лиганд RAGE; и (b) вторую аминокислотную последовательность, по меньшей мере, на 95% идентичную константному домену тяжелой цепи иммуноглобулина IgG4 человека или его фрагменту; где первая аминокислотная последовательность содержит, по меньшей мере, одну мутацию по сравнению с лиганд-связывающим доменом RAGE дикого типа. В одном варианте осуществления изобретения слитый белок по изобретению может также содержать линкерную последовательность между первой аминокислотной последовательностью и второй аминокислотной последовательностью. В некоторых вариантах осуществления лиганд-связывающий домен RAGE может быть из RAGE млекопитающего, например, RAGE человека. Подходящий лиганд-связывающий домен RAGE млекопитающего может содержать аминокислоты 1-344 из SEQ ID NO:6 или аминокислоты 24-344 из SEQ ID NO:6. В одном варианте осуществления изобретения слитый белок по изобретению может содержать аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 и SEQ ID NO:8. В одном варианте осуществления изобретения выделенный слитый белок по изобретению содержит SEQ ID NO:6 или SEQ ID NO:8. В другом варианте осуществления изобретения выделенный слитый белок по изобретению состоит из SEQ ID NO:6 или SEQ ID NO:8. В некоторых вариантах осуществления изобретения слитые белки по изобретению могут также содержать линкер между аминокислотной последовательностью RAGE и аминокислотной последовательностью IgG4. Настоящее изобретение также рассматривает молекулы нуклеиновых кислот (например, молекулы ДНК или РНК), кодирующие слитые белки изобретения, а также клетки-хозяева, экспрессирующие молекулы нуклеиновых кислот, кодирующие слитые белки по изобретению.
Настоящее изобретение также предоставляет фармацевтическую композицию, содержащую слитый белок по изобретению и фармацевтически приемлемый наполнитель или разбавитель.
Настоящее изобретение обеспечивает способы лечения заболеваний, опосредованных AGE. Такие заболевания включают любое заболевание, характеризующееся увеличенным количеством AGE у субъекта, например, млекопитающего, такого как человек. Способы лечения AGE-опосредованного заболевания содержат введение субъекту, имеющему AGE-опосредованное заболевание, терапевтически эффективного количества фармацевтической композиции, содержащей слитый белок по изобретению. Примеры заболеваний, которые могут излечиваться способами по изобретению, включают, но не ограничиваются этим, диабетическую нефропатию, ревматоидный артрит и аутоиммунные заболевания, такие как дерматит, гломерулонефрит, рассеянный склероз, симпатическая офтальмия, аутоиммунное легочное воспаление, инсули-нозависимый сахарный диабет, аутоиммунные глазные воспаления, системная красная волчанка, резистентность к инсулину, ревматоидный артрит, диабетическая ретинопатия и склеродермия. Любой слитый белок по изобретению может быть использован при осуществлении на практике способов по изобретению. В одном варианте осуществления способы по изобретению могут осуществляться, используя слитый белок, содержащий SEQ ID NO:6 или SEQ ID NO:8. В другом варианте осуществления способы по изобретению могут осуществляться, используя слитый белок, который состоит из SEQ ID NO:6 или SEQ ID NO:8.
В другом варианте осуществления изобретения настоящее изобретение предоставляет способы снижения уровней лиганда, связывающегося посредством RAGE, у млекопитающего (например, человека), нуждающегося в этом. Такие способы могут предусматривать введение млекопитающему RAGE лиганд-снижающего количества слитого белка по изобретению.
В других вариантах осуществления изобретение предоставляет рекомбинантный вектор экспрессии, содержащий последовательности ДНК по изобретению; клетку-хозяина, трансформированную, трансдуцированную или трансфицированную этим вектором; и способ продуцирования слитого белка, который предусматривает культивирование клетки-хозяина, трансформированной, трансдуцированной или трансфицированной нуклеиновой кислотой, кодирующей слитый белок по изобретению в условиях, пригодных для осуществления экспрессии слитого белка.
Изобретение также предоставляет композиции, содержащие настоящий слитый белок или его фрагменты. В некоторых вариантах осуществления изобретение включает композиции, содержащие настоящий слитый белок или его фрагменты, к которым прямо или опосредованно присоединены радиоизотоп, хелатор, токсин, флуорохром, биотин, пептидные эпитопы, такие как his-тэги, myc-тэги, или сахара. Другие варианты осуществления изобретения включают настоящий слитый белок, слитый с другим белком с целью изменения периода полувыведения из организма или биологической функции, и гликозилированные варианты слитого белка.
Эти и другие аспекты настоящего изобретения станут ясны при обращении к следующему подробному описанию.
Краткое описание чертежей
Фиг.1 представляет собой столбчатую диаграмму, демонстрирующую влияние иллюстративного слитого белка RAGE-Ig на лейкостаз на мышиной модели со стрептозотоцин-индуцированным диабетом.
Фиг.2A-2D представляют собой столбчатые диаграммы, демонстрирующие влияние иллюстративного слитого белка RAGE-Ig на проницаемость сосудов сетчатки в различных слоях сетчатки на мышиной модели со стрептозотоцин-индуцированным диабетом.
Фиг.3 представляет собой столбчатую диаграмму, демонстрирующую влияние иллюстративного слитого белка RAGE-Ig на нитрование белков сетчатки на мышиной модели со стрептозотоцин-индуцированным диабетом.
Фиг.4 представляет собой столбчатую диаграмму, демонстрирующую влияние иллюстративного слитого белка RAGE-Ig на экспрессию ICAM в сетчатке на мышиной модели со стрептозотоцин-индуцированным диабетом.
Фиг.5А представляет собой столбчатую диаграмму, демонстрирующую влияние иллюстративного слитого белка RAGE-Ig на число бесклеточных капилляров, измеренных на квадратный мм ткани сетчатки у диабетических мышей после 10 месяцев диабета. Фиг.5В представляет собой столбчатую диаграмму, демонстрирующую влияние иллюстративного слитого белка RAGE-Ig на число «теней» перицитов на 1000 капиллярных клеток, измеренных у диабетических мышей после 10 месяцев диабета.
Фиг.6 представляет собой столбчатую диаграмму, демонстрирующую влияние иллюстративного слитого белка RAGE-Ig на 50% порог реакции на прикосновение у диабетических мышей после 10 месяцев диабета.
Фиг.7 предоставляет блок-схему, демонстрирующую экспериментальный протокол Примера 3.
Фиг.8 является линейным графиком, демонстрирующим влияние иллюстративного слитого белка RAGE-Ig на массу тестируемых животных для мышиной модели с артритом, индуцированным коллагеном II типа.
Фиг.9 является столбчатой диаграммой, демонстрирующей влияние иллюстративного слитого белка RAGE-Ig на число случаев артрита для мышиной модели с артритом, индуцированным коллагеном II типа.
Фиг.10 является столбчатой диаграммой, демонстрирующей влияние иллюстративного слитого белка RAGE-Ig на начало артрита для мышиной модели с артритом, индуцированным коллагеном II типа.
Фиг.11 является линейным графиком, демонстрирующим влияние иллюстративного слитого белка RAGE-Ig на число случаев артрита в виде временной функции для мышиной модели с артритом, индуцированным коллагеном II типа.
Фиг.12 является линейным графиком, демонстрирующим влияние иллюстративного слитого белка RAGE-Ig на тяжесть артрита в виде временной функции для мышиной модели с артритом, индуцированным коллагеном II типа.
Фиг.13 является линейным графиком, демонстрирующим влияние иллюстративного слитого белка RAGE-Ig на число лап с артритом, измеренное в виде временной функции для мышиной модели с артритом, индуцированным коллагеном II типа.
Фиг.14A-14D являются микрофотографиями, демонстрирующими влияние иллюстративного слитого белка RAGE-Ig на морфологию суставов в виде функции увеличивающегося количества слитого белка для мышиной модели с артритом, индуцированным коллагеном II типа.
Фиг.15 является столбчатой диаграммой, демонстрирующей влияние иллюстративного слитого белка RAGE-Ig на синовит (черные столбцы) и паннус (серые столбцы) для мышиной модели с артритом, индуцированным коллагеном II типа.
Фиг.16 является столбчатой диаграммой, демонстрирующей влияние иллюстративного слитого белка RAGE-Ig на краевую эрозию (черные столбцы) и изменения архитектуры (серые столбцы) для мышиной модели с артритом, индуцированным коллагеном II типа.
Фиг.17 является столбчатой диаграммой, демонстрирующей влияние иллюстративного слитого белка RAGE-Ig на полную гистологическую оценку артрита для мышиной модели с артритом, индуцированным коллагеном II типа.
Фиг.18 является столбчатой диаграммой, демонстрирующей влияние иллюстративного слитого белка RAGE-Ig на потерю белка матрикса сустава для мышиной модели с артритом, индуцированным коллагеном II типа.
Фиг.19A-19D являются микрофотографиями срезов, окрашенных толуидиновым синим, демонстрирующими влияние иллюстративного слитого белка RAGE-Ig на потерю белка матрикса сустава для мышиной модели с артритом, индуцированным коллагеном II типа.
Подробное описание изобретения
Определения
Используемые здесь термины "рецептор конечного продукта гликирования" или RAGE относятся к белкам, имеющим аминокислотные последовательности, которые по существу похожи на нативные аминокислотные последовательности RAGE млекопитающих, и функцию связывания одного или более лигандов RAGE специфическим способом лиганд-рецептор. Термины "конечный продукт гликирования" и "AGE" относятся к гетерогенной группе молекул, образованных неферментативной реакцией восстанавливающих сахаров со свободными аминогруппами белков, липидов и нуклеиновых кислот, как описано выше.
При использовании здесь "лиганд-связывающий домен RAGE" или "RAGE-LBD" относится к любому белку RAGE млекопитающего или любой части белка RAGE млекопитающего, которая сохраняет способность связывать лиганд RAGE специфическим способом лиганд-рецептор. В частности, без ограничения, лиганд-связывающий домен RAGE включает полипептид, имеющий один или более внеклеточных доменов трансмембранного белка RAGE. Согласно Таблице 6, подходящий RAGE-LBD может содержать, по меньшей мере, аминокислоты 1-99, или аминокислоты 24-99, или аминокислоты 1-208, или аминокислоты 24-208, или аминокислоты 1-301, или аминокислоты 24-301, или аминокислоты 1-344, или аминокислоты 24-344 из SEQ ID NO:6.
Термин «выделенный» при использовании в контексте данного описания для определения чистоты слитого белка означает, что белок по существу не содержит другие белки, с которыми он был соединен во время продуцирования, включая, без ограничения, по существу отсутствие других белков, присутствующих во время экспрессии слитого белка в клеточной культуральной среде. Например, выделенный белок по изобретению может содержать 1- 25%, 20-25%, 15-20%, 10-15%, 5-10%, 1-5% или менее чем около 2 масс.% примесного белкового остатка процессов продуцирования. Композиции, содержащие выделенные белки по изобретению, однако, могут содержать другие белки, добавленные в качестве стабилизаторов, носителей, наполнителей или дополнительных лекарственных средств.
При использовании здесь "белок" и "полипептид" являются взаимозаменяемыми.
При использовании здесь "лечение" заболевания или нарушения относится к улучшению, по меньшей мере, одного признака или симптома заболевания или нарушения у субъекта.
Термин "нуклеиновая кислота" относится к полинуклеотидам, таким как дезоксирибонуклеиновая кислота (ДНК), и, где подходит, рибонуклеиновая кислота (РНК). Следует также понимать, что этот термин включает, как эквиваленты, аналоги или РНК, или ДНК, полученные из нуклеотидных аналогов, и, когда применимо к описанному варианту осуществления изобретения, одно- (смысловые или антисмысловые) и двухцепочечные полинуклеотиды.
Термин "или" при использовании здесь означает и используется взаимозаменяемо с термином "и/или", если контекст не указывает ясно иного.
Термин "процент идентичности " относится к идентичности последовательности между двумя аминокислотными последовательностями или между двумя нуклеотидными последовательностями. Процент идентичности может определяться сравнением положения в каждой последовательности, которая может быть подвергнута выравниванию с целью сравнения. Выражение в виде процента идентичности относится к функции числа идентичных аминокислот или нуклеиновых кислот в положениях, общих для сравниваемых последовательностей. Могут использоваться различные алгоритмы выравнивания и/или программы, включая FASTA, BLAST или ENTREZ. FASTA и BLAST доступны как часть пакета программ GCG для анализа последовательностей (University of Wisconsin, Madison, Wis.) и могут использоваться, например, с параметрами по умолчанию. ENTREZ обеспечивается the National Center for Biotechnology Information, National Library of Medicine, National Institutes of Health, Bethesda, Md. В одном из вариантов осуществления процент идентичности двух последовательностей может быть определен программой GCG с весом гэпа 1, например, каждый аминокислотный гэп оценивается, если было одно аминокислотное или нуклеотидное несовпадение между двумя последовательностями.
Идентичность последовательности можно определить, сравнивая референсную последовательность или подпоследовательность референсной последовательности с изучаемой последовательностью (например, нуклеотидной последовательностью, аминокислотной последовательностью и т.д.). Референсная последовательность и изучаемая последовательность оптимально выравниваются относительно произвольного числа остатков, названного окном сравнения. Для того, чтобы получить оптимальное выравнивание, добавления или делеции, такие как гэпы, могут вводиться в изучаемую последовательность. Процент идентичности последовательности определяется установлением числа положений, в которых один и тот же остаток присутствует в обеих последовательностях, и делением числа совпадающих положений на общую длину последовательностей в окне сравнения и умножением на 100 с получением процентов. В дополнение к числу совпадающих положений число и размер гэпов также учитывается при расчете процента идентичности последовательности.
Идентичность последовательностей обычно определяют, используя компьютерные программы. Типичной программой является программа BLAST (Basic Local Alignment Search Tool), доступная в National Center for Biotechnology Information (NCBI, http://www.ncbi.nlm.nih.gov/). Эта программа сравнивает сегменты в изучаемой последовательности с последовательностями в базе данных для определения статистической значимости совпадений, затем устанавливает и сообщает только те совпадения, которые являются более значащими, чем пороговый уровень. Подходящей версией программы BLAST является та, которая учитывает гэпы, например, версия 2.Х (Altschul, et al.. Nucleic Acids Res 25(17):3389-402, 1997). Дополнительные подходящие программы идентификации белков с идентичностью последовательностей по отношению к белкам по изобретению включают, но не ограничиваются этим, PHI-BLAST (Pattern Hit Initiated BLAST, Zhang, et al., Nucleic Acids Res 26(17):3986-90, 1998) и PSI-BLAST (Position-Specific Iterated BLAST, Altschul, et al., Nucleic Acids Res 25(17):3389-402, 1997). Эти программы доступны открыто на веб-сайте NCBI, указанном выше, и могут использоваться со стандартными настройками для определение идентичности последовательностей согласно изобретению.
Слитые белки
Настоящее изобретение обеспечивает выделенный слитый белок, содержащий, по меньшей мере, один полипептид, содержащий: (а) первую аминокислотную последовательность, по меньшей мере, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную лиганд-связывающему домену рецептора конечного продукта гликирования (RAGE) млекопитающего, причем первая аминокислотная последовательность способна связывать лиганд RAGE; и (b) вторую аминокислотную последовательность, по меньшей мере, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную константному домену тяжелой цепи иммуноглобулина IgG4 человека или его фрагменту, где первая аминокислотная последовательность содержит, по меньшей мере, одну мутацию, или по меньшей мере две мутации, или по меньшей мере три мутации, или 1-4 мутации, или 1-10 мутаций по сравнению с лиганд-связывающим доменом RAGE дикого типа. Примерами мутаций, которые могут быть сделаны в первой аминокислотной последовательности, являются те, которые увеличивают стабильность слитого белка, например, получая лиганд-связывающий домен RAGE, более устойчивый к протеолитической деградации, как, например, те, которые делают слитый белок более устойчивым к фурин-подобным протеазам. Подходящие фрагменты второй аминокислотной последовательности включают фрагменты, которые сохраняют способность увеличивать период полувыведения из сыворотки слитых белков, частью которых они являются, по сравнению с периодом полувыведения из сыворотки той же первой аминокислотной последовательности в отдельности. Предпочтительно первую аминокислотную последовательность и вторую аминокислотную последовательность получают из лиганд-связывающего домена RAGE человека и IgG4 человека.
Слитые белки по изобретению могут содержать одну или более аминокислотных последовательностей в дополнение к лиганд-связывающему домену RAGE и константному домену IgG4 или его фрагменту. Например, слитый белок по изобретению может содержать линкерную последовательность, которая может вставляться между лиганд-связывающим доменом RAGE и последовательностью IgG. Слитые белки по изобретению могут содержать одну или более тэг-последовательностей, например, тэг-последовательности для очистки, такие как 6-гистидины. Слитые белки по изобретению могут содержать один или более эпитопов, распознаваемых коммерчески доступными антителами, например, c-myc (EQKLISEEDL, SEQ ID NO:9) и гемагглютинин (YPYDVPDYA, SEQ ID NO:10), полученный из эпитопного тэга белка гемагглютинина гриппа.
Любой белок RAGE млекопитающего, известный специалисту в данной области техники, может использоваться при осуществлении настоящего изобретения. Предпочтительно внеклеточный домен белка RAGE может использоваться для определения лиганд-связывающего домена, который может быть мутирован и использоваться как первая аминокислотная последовательность слитого белка. Подходящий пример белков RAGE млекопитающего включает, но не ограничивается этим, домены RAGE примата, человека (например, GenBank регистрационный номер NP_001127 и NP_751947), мыши (например, GenBank регистрационный номер NP_031451), собаки (например, GenBank регистрационный номер AAQ81297), крысы (например, GenBank регистрационный номер NP_445788), кошки, крупного рогатого скота (например, GenBank регистрационный номер AAI20128), овцы, лошади и свиньи (например, GenBank регистрационный номер AAQ73283).
Аминокислотные последовательности RAGE, содержащие одно или более изменений или модификаций по сравнению с последовательностью дикого типа, могут использоваться в настоящем изобретении. Такие изменения или модификации включают, но не ограничиваются этим, точечные мутации, делеции с N-конца, делеции с С-конца, внутренние делеции и их сочетания. Любое изменение или модификация могут вводиться в последовательность RAGE для применения в настоящем изобретении, при условии, что получающийся белок сохраняет биологическую активность, например, способность связывать один или более лигандов RAGE. Слитые белки по изобретению также включают белки с или без эндогенных паттернов гликозилирования, включая, без ограничения, слитые белки, в которых первая аминокислотная последовательность получена из лиганд-связывающего домена RAGE млекопитающего с или без сопутствующего гликозилирования с нативным паттерном связывающего домена.
Любой подходящий Fc-участок IgG может использоваться при осуществлении данного изобретения, предпочтительно, из молекулы IgG4, например, аминокислотные остатки 149-473 из GenBank рег. номер ААН25985. Участок IgG для применения в настоящем изобретении может быть Fc-участком IgG4 Fc и может содержать один или более СН2 и СН3 участков молекулы IgG4.
Примеры подходящих слитых белков представлены в следующих таблицах.
Таблица 1 предоставляет нуклеотидную последовательность генной последовательности слитого белка человеческого RAGE-IgG4 Fc.
Таблица 1: Слитая генная последовательность человеческого RAGE-IgG4 Fc (SEQ ID NO:1).

Текст с жирным шрифтом означает кодирующую последовательность для сигнальной последовательности RAGE, текст с обычным шрифтом означает кодирующую последовательность для человеческого RAGE, и подчеркнутый текст означает кодирующую последовательность для Fc-участка IgG4.
Таблица 2: Аминокислотная последовательность слитого белка человеческого RAGE-IgG4Fc (SEQ ID NO:2).
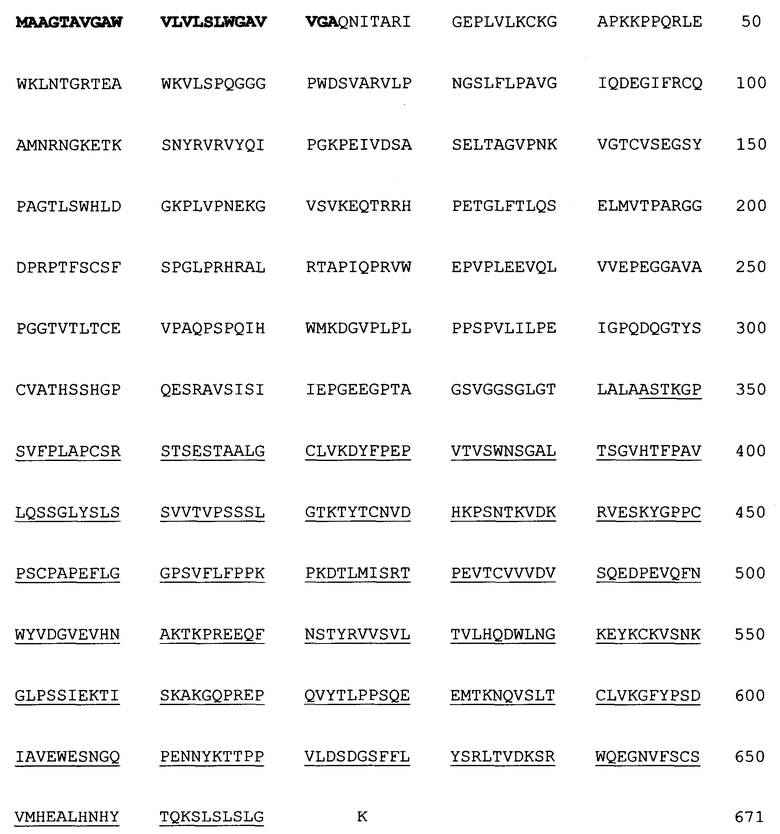
Текст с жирным шрифтом означает аминокислотную последовательность сигнальной последовательности RAGE, текст с обычным шрифтом означает аминокислотную последовательность человеческого RAGE, и подчеркнутый текст означает аминокислотную последовательность Fc-участка IgG4.
Таблица 3: Слитая генная последовательность человеческий RAGE-линкер-IgG4 Fc (SEQ ID NO:3).


Текст с жирным шрифтом означает кодирующую последовательность для сигнальной последовательности RAGE, текст с обычным шрифтом означает кодирующую последовательность для человеческого RAGE, подчеркнутый двойной линией текст означает кодирующую последовательность для пептидного линкера, и подчеркнутый одной линией текст означает кодирующую последовательность для Fc-участка IgG4.
Таблица 4: Аминокислотная последовательность слитого белка человеческий RAGE-линкер-IgG4Fc (SEQ ID NO:4).

Текст с жирным шрифтом означает аминокислотную последовательность сигнальной последовательности RAGE, текст с обычным шрифтом означает аминокислотную последовательность человеческого RAGE, подчеркнутый двойной линией текст означает аминокислотную последовательность пептидного линкера, и подчеркнутый одной линией текст означает аминокислотную последовательность для Fc-участка IgG4.
Таблица 5: Слитая генная последовательность человеческого RAGE варианта-IgG4 Fc (SEQ ID NO:5).

Текст с жирным шрифтом означает кодирующую последовательность для сигнальной последовательности RAGE, текст с обычным шрифтом означает кодирующую последовательность для варианта RAGE человека, выделенные жирным шрифтом буквы, подчеркнутые волнистой линией, означают участки точечных мутаций, введенных в последовательность варианта hRAGE, и подчеркнутый текст означает кодирующую последовательность для Fc-участка IgG4.
Таблица 6: Аминокислотная последовательность слитого белка человеческого RAGE варианта-IgG4Fc (SEQ ID NO:6).

Текст с жирным шрифтом означает аминокислотную последовательность сигнальной последовательности RAGE, текст с обычным шрифтом означает аминокислотную последовательность варианта RAGE человека, выделенные жирным шрифтом буквы, подчеркнутые волнистой линией, означают участки точечных мутаций, введенных в вариант hRAGE, и подчеркнутый текст означает аминокислотную последовательность для Fc-участка IgG4.
Таблица 7: Слитая генная последовательность человеческий RAGE вариант-линкер-IgG4 Fc (SEQ ID NO:7).

Текст с жирным шрифтом означает кодирующую последовательность для сигнальной последовательности RAGE, текст с обычным шрифтом означает кодирующую последовательность для варианта RAGE человека, выделенные жирным шрифтом буквы, подчеркнутые волнистой линией, означают участки точечных мутаций, введенных в последовательность варианта hRAGE, подчеркнутый двойной линией текст означает последовательность, кодирующую пептидный линкер, и подчеркнутый текст означает кодирующую последовательность для Fc-участка IgG4.
Таблица 8: Аминокислотная последовательность человеческий RAGE вариант-линкер-IgG4 Fc (SEQ ID NO:8).

Текст с жирным шрифтом означает аминокислотную последовательность сигнальной последовательности RAGE, текст с обычным шрифтом означает аминокислотную последовательность варианта RAGE человека, выделенные жирным шрифтом буквы, подчеркнутые волнистой линией, означают участки точечных мутаций, введенных в вариант hRAGE, подчеркнутый двойной линией текст означает аминокислотную последовательность пептидного линкера, и подчеркнутый текст означает аминокислотную последовательность для Fc-участка IgG4.
ЭКСПРЕССИЯ СЛИТЫХ БЕЛКОВ RAGE
Слитые белки по изобретению могут быть продуцированы в любой системе экспрессии белка, известной специалисту в данной области техники, например, эукариотических системах экспрессии, бактериальных системах экспрессии и вирусных системах экспрессии. Ряд систем хозяин-вектор экспрессии могут использоваться для экспрессии слитого белка по изобретению. Такие системы хозяев представляют собой носители, в которых слитые белки по изобретению могут продуцироваться, и из которых они впоследствии могут быть очищены. Такие системы включают, но не ограничиваются этим, микроорганизмы, такие как бактерии, дрожжи, клетки насекомых или растительные клетки. RAGE, экспрессированные в системах экспрессии дрожжей или млекопитающих, например, клетки COS-7, могут быть схожими или слегка отличаться по молекулярной массе и паттерну гликозилирования от нативных молекул, в зависимости от системы экспрессии. Экспрессия ДНК RAGE в бактериях, таких как Е. coli, дает негликозилированные молекулы. Различные паттерны гликозилирования могут быть получены с использованием бакуловирусных систем экспрессии в клетках насекомых. Функциональные мутантные аналоги RAGE млекопитающих, имеющие инактивированные участки N-гликозилирования, могут быть получены олигонуклеотидным синтезом и методиками лигирования или сайт-специфического мутагенеза. Эти аналогичные белки могут продуцироваться в гомогенной форме с пониженным содержанием углеводов с хорошим выходом при использовании дрожжевых систем экспрессии.
Молекулы нуклеиновых кислот, кодирующие слитые белки по изобретению, могут быть получены, а нуклеотидная последовательность этих полинуклеотидов может быть определена любым способом, известным в данной области техники. Принимая во внимание изложенные здесь и известные полипептидные последовательности RAGE и их идентифицированные или идентифицируемые лиганд-связывающие элементы, и известные последовательности константных доменов тяжелых цепей IgG, нуклеотидные последовательности, кодирующие эти полипептиды, могут быть определены с использованием способов, хорошо известных в данной области техники, т.е. нуклеотидные кодоны, известные как кодирующие определенные аминокислоты, могут собираться таким образом, чтобы получить нуклеиновую кислоту, которая кодирует слитый белок по изобретению. Нуклеотидные кодоны могут выбираться, исходя из используемой системы экспрессии, например, выбирая кодоны, которые соответствуют более многочисленным молекулам tRNA, присутствующим в системе экспрессии, при этом может экспрессироваться более высокий уровень слитого белка. Такой полинуклеотид, кодирующий слитый белок, может собираться из химически синтезированных олигонуклеотидов (например, как описано в Kutmeier et. Al., Biotechniques 17:242(1994), что, вкратце, включает синтез перекрывающихся олигонуклеотидов, содержащих части последовательности, кодирующей слитый белок, отжиг и лигирование этих олигонуклеотидов, и затем амплификацию дотированных олигонуклеотидов с использованием полимеразной(ых) цепной(ых) реакции (реакций) (ПЦР).
Рекомбинантная экспрессия слитого белка по изобретению (включая другие молекулы, содержащие или альтернативно состоящие из фрагментов слитого белка или их вариантов) может требовать конструирования вектора (векторов) экспрессии, содержащего полинуклеотид, который кодирует слитый белок. После того как получен полинуклеотид, кодирующий слитый белок по изобретению, вектор (векторы) для продуцирования слитого белка может быть получен технологией рекомбинантных ДНК, используя методики, хорошо известные в данной области техники. Такие векторы экспрессии, содержащие кодирующие последовательности RAGE-Fc, могут также содержать соответствующие транскрипционные и трансляционные контрольные сигналы /последовательности, например, сайты связывания рибосомы (например, последовательности Козака), участки внутренней посадки рибосомы (IRES) и участки полиаденилирования и т.д.
Молекулы нуклеиновых кислот, кодирующие слитые белки по изобретению, могут переноситься в клетки млекопитающих с использованием ретровирусных векторов, дефектных по репликации (например, векторы, полученные из вируса мышиного лейкоза Молони (MLV) или ВИЧ), и псевдотипироваться белком G вируса везикулярного стоматита (VSV-G) для стабильной вставки единичных копий молекул нуклеиновых кислот, кодирующих слитый белок по изобретению, в делящиеся клетки. Ретровирусные векторы доставляют гены, закодированные как РНК, которые, после введения в клетку, обратно транскрибируются в ДНК и стабильно интегрируются в геном клетки-хозяина. Множественные генные вставки в одну клетку могут увеличивать экспрессию и секрецию слитого белка. Множественные раунды заражения могут также увеличивать число интегрированных генных копий и следовательно количество экспрессированного слитого белка. Интегрированный(е) ген(ы), кодирующий(е) слитый белок сохраняется (сохраняются) в клетках во время клеточного деления благодаря их присутствию в геноме.
В некоторых вариантах осуществления изобретения настоящее изобретение предоставляет стабильную клеточную линию, которая экспрессирует слитые белки по изобретению. Один подходящий способ для быстрого получения стабильных, экспрессирующих белок на высоком уровне клеточных линий млекопитающих заключается в использовании системы экспрессии GPEx™ (Gala Biotech, дочерняя компания Catalent Pharma Solutions, Middleton, WL, Bleck, Gregory Т., Bioprocessingjournal.com September/October 2005 p1-7). Такой способ может содержать получение псевдотипированного ретровирусного вектора, дефектного по репликации, на основе MMLV, и преобразование клеток млекопитающих (например, клеток СНО) этим вектором. Вектор может интегрироваться в геном клеток, давая таким образом стабильную клеточную линию.
ОЧИСТКА ВЫДЕЛЕННОГО СЛИТОГО БЕЛКА
Выделенные слитые белки по изобретению могут быть получены культивированием подходящих систем хозяин /вектор для экспрессии рекомбинантных продуктов трансляции настоящих последовательностей ДНК, которые затем очищаются из культуральных сред или клеточных экстрактов с использованием методик, хорошо известных в данной области техники.
Например, супернатанты из систем, которые секретируют рекомбинантный белок в культуральные среды, могут сначала концентрироваться, используя коммерчески доступный концентрирующий фильтр для белка, например, ультрафильтровальную установку Amicon или Millipore Pellicon. После стадии концентрирования концентрат может быть нанесен на подходящую матрицу для очистки. Например, подходящая аффинная матрица может содержать, например, AGE, или лектин, или Белок А, или Белок G, или молекулу антитела, связанную с подходящей подложкой. Альтернативно, может использоваться анионообменная смола, например, матрица или субстрат, имеющие «подвешенные» диэтиламиноэтильные (DEAE) группы. Матрицы могут быть акриламидными, агарозными, декстрановыми, целлюлозными или других типов, обычно применяемых в очистке белка. Альтернативно, может использоваться катионообменная стадия. Подходящие катионообменники включают различные нерастворимые матрицы, содержащие сульфопропильные или карбоксиметильные группы. Сульфопропильные группы являются предпочтительными.
Рекомбинантный белок, полученный в бактериальной культуре, обычно выделяют первоначальной экстракцией из клеточных осадков, за которой следуют стадии одного или более концентрирований, высаливания, водной ионообменной хроматографии или гель-фильтрации. В заключение может применяться высокоэффективная жидкостная хроматография (ВЭЖХ) для конечных стадий очистки. Микробные клетки, использованные для экспрессии рекомбинантного RAGE млекопитающего, могут разрушаться любым подходящим способом, включая цикл замораживания и оттаивания, разрушение ультразвуком, механическое разрушение или применение агентов, лизирующих клетки.
Ферментация дрожжей, которые экспрессируют слитый белок по изобретению в виде секретируемого белка, значительно упрощает очистку. Секретируемый рекомбинантный белок, получающийся в результате крупномасштабной ферментации, может быть очищен способами, аналогичными способам, раскрытым Urdal et al. (J. Chromatog. 296:171, 1984). Эта ссылка описывает две последовательные стадии обращенно-фазовой ВЭЖХ для очистки рекомбинантного GMCSF человека на колонке для препаративной ВЭЖХ.
ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ
Слитые белки по изобретению могут быть составлены в виде смеси, пригодной для введения субъекту, нуждающемуся в этом, например, могут быть составлены в виде фармацевтических композиций. Композиции по изобретению могут содержать один или более фармацевтически приемлемых носителей, наполнителей или разбавителей. Используемый здесь термин "фармацевтически приемлемый носитель" включает любой и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, изотонические и замедляющие всасывание агенты и т.п., которые являются физиологически совместимыми. В одном из вариантов осуществления изобретения носитель является пригодным для парентерального введения. Носитель может быть пригодным для введения в центральную нервную систему (например, интраспинально или внутрицеребрально). Альтернативно, носитель может быть пригодным для внутривенного, подкожного, внутрибрюшинного или внутримышечного введения. В другом варианте осуществления изобретения носитель является пригодным для перорального введения. Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для экстемпорального приготовления стерильных инъецируемых растворов или дисперсии. Применение таких сред и агентов для фармацевтически активных веществ хорошо известно в данной области техники. За исключением случаев, когда любые стандартные среды или агент являются несовместимыми со слитыми белками по изобретению, применение их в фармацевтических композициях по изобретению является предполагаемым. Дополнительные активные соединения могут также быть включены в композиции.
Пригодные носители обычно являются нетоксичными по отношению к реципиентам в применяемых дозах и концентрациях. Как правило, приготовление фармацевтических композиций по изобретению включает в себя комбинирование слитого белка по изобретению с одним или более буферами, антиоксидантами, такими как аскорбиновая кислота, низкомолекулярные полипептиды (менее чем около 10 остатков), белки, аминокислоты, углеводы, включая глюкозу, трегалозу, сахарозу или декстрины, хелатирующие агенты, такие как ЭДТА, глутатион, и другими стабилизаторами и наполнителями. Нейтральный забуференный физиологический раствор или физиологический раствор, смешанный с неспецифическим сывороточным альбумином, являются типичными подходящими разбавителями.
ТЕРАПЕВТИЧЕСКОЕ ВВЕДЕНИЕ СЛИТЫХ БЕЛКОВ ПО ИЗОБРЕТЕНИЮ
Настоящее изобретение предполагает введение слитых белков по изобретению в форме фармацевтической композиции, содержащей слитый белок по изобретению и фармацевтически приемлемый разбавитель или носитель, субъекту (например, млекопитающему, в частности, человеку), нуждающемуся в этом. Настоящее изобретение также предоставляет способ лечения заболевания человека такими композициями.
Как правило, способы по изобретению могут предусматривать введение фармацевтической композиции, содержащей фармацевтически эффективное количество слитого белка по изобретению. Применяемое фармацевтически эффективное количество может изменяться в соответствии с такими факторами как стадия заболевания, возраст, пол и масса индивидуума.
Фармацевтически эффективное количество слитого белка по изобретению может составлять от около 1 мкг слитого белка /1 кг массы тела субъекта до около 500 мг слитого белка /1 кг массы тела субъекта, или от около 10 мкг слитого белка /1 кг массы тела субъекта до около 500 мг слитого белка /1 кг массы тела субъекта, или от около 100 мкг слитого белка /1 кг массы тела субъекта до около 500 мг слитого белка /1 кг массы тела субъекта, или от около 1 мг слитого белка /1 кг массы тела субъекта до около 500 мг слитого белка /1 кг массы тела субъекта, или от около 10 мг слитого белка /1 кг массы тела субъекта до около 500 мг слитого белка /1 кг массы тела субъекта, или от около 100 мг слитого белка /1 кг массы тела субъекта до около 500 мг слитого белка /1 кг массы тела субъекта, или от около 100 мкг слитого белка /1 кг массы тела субъекта до около 25 мг слитого белка /1 кг массы тела субъекта, или от около 1 мг слитого белка /1 кг массы тела субъекта до около 25 мг слитого белка /1 кг массы тела субъекта, или от около 5 мг слитого белка /1 кг массы тела субъекта до около 25 мг слитого белка /1 кг массы тела субъекта, или от около 10 мг слитого белка /1 кг массы тела субъекта до около 25 мг слитого белка /1 кг массы тела субъекта, или от около 15 мг слитого белка /1 кг массы тела субъекта до около 25 мг слитого белка /1 кг массы тела субъекта, или от около 100 мкг слитого белка /1 кг массы тела субъекта до около 10 мг слитого белка /1 кг массы тела субъекта, или от около 1 мг слитого белка /1 кг массы тела субъекта до около 10 мг слитого белка /1 кг массы тела субъекта, или от около 2.5 мг слитого белка /1 кг массы тела субъекта до около 10 мг слитого белка /1 кг массы тела субъекта, или от около 5 мг слитого белка /1 кг массы тела субъекта до около 10 мг слитого белка /1 кг массы тела субъекта, или от около 7.5 мг слитого белка /1 кг массы тела субъекта до около 10 мг слитого белка /1 кг массы тела субъекта.
В некоторых вариантах осуществления изобретения фармацевтически эффективное количество слитого белка по изобретению может составлять 0.5 мг слитого белка /1 кг массы тела субъекта, 1 мг слитого белка /1 кг массы тела субъекта, 2 мг слитого белка /1 кг массы тела субъекта, 3 мг слитого белка /1 кг массы тела субъекта, 4 мг слитого белка /1 кг массы тела субъекта, 5 мг слитого белка /1 кг массы тела субъекта, 6 мг слитого белка /1 кг массы тела субъекта, 7 мг слитого белка /1 кг массы тела субъекта, 8 мг слитого белка /1 кг массы тела субъекта, 9 мг слитого белка /1 кг массы тела субъекта или 10 мг слитого белка /1 кг массы тела субъекта.
Единичная лекарственная форма относится к физически дискретным единицам, подходящим в качестве единичных доз млекопитающим, подвергаемым лечению; при этом каждая единица содержит заданное количество слитого белка по изобретению, рассчитанное для получения желаемого терапевтического эффекта, совместно с требуемым фармацевтическим носителем. Единичная лекарственная форма слитого белка по изобретению может составлять от около 1 мг до около 1000 мг, от около 25 мг до около 1000 мг, от около 50 мг до около 1000 мг, от около 100 мг до около 1000 мг, от около 250 мг до около 1000 мг, от около 500 мг до около 1000 мг, от около 100 мг до около 500 мг, от около 200 мг до около 500 мг, от около 300 мг до около 500 мг или от около 400 мг до около 500 мг. Единичная доза слитого белка по изобретению может составлять около 100 мг, 200 мг, 300 мг, 400 мг, 500 мг, 600 мг или 700 мг.
Композиции по изобретению могут содержать слитые белки по изобретению на уровне от около 0.1 масс.% до около 20 масс.%, от около 0.1 масс.% до около 18 масс.%, от около 0.1 масс.% до около 16 масс.%, от около 0.1 масс.% до около 14 масс.%, от около 0.1 масс.% до около 12 масс.%, от около 0.1 масс.% до около 10 масс.%, от около 0.1 масс.% до около 8 масс.%, от около 0.1 масс.% до около 6 масс.%, от около 0.1 масс.% до около 4 масс.%, от около 0.1 масс.% до около 2 масс.%, от около 0.1 масс.% до около 1 масс.%, от около 0.1 масс.% до около 0.9 масс.%, от около 0.1 масс.% до около 0.8 масс.%, от около 0.1 масс.% до около 0.7 масс.%, от около 0.1 масс.% до около 0.6 масс.%, от около 0.1 масс.% до около 0.5 масс.%, от около 0.1 масс.% до около 0.4 масс.%, от около 0.1 масс.% до около 0.3 масс.% или от около 0.1 масс.% до около 0.2 масс.% от общей массы композиции.
Фармацевтические композиции по изобретению могут содержать один или более слитых белков по изобретению на уровне от около 1 масс.% до около 20 масс.%, от около 1 масс.% до около 18 масс.%, от около 1 масс.% до около 16 масс.%, от около 1 масс.% до около 14 масс.%, от около 1 масс.% до около 12 масс.%, от около 1 масс.% до около 10 масс.%, от около 1 масс.% до около 9 масс.%, от около 1 масс.% до около 8 масс.%, от около 1 масс.% до около 7 масс.%, от около 1 масс.% до около 6 масс.%, от около 1 масс.% до около 5 масс.%, от около 1 масс.% до около 4 масс.%, от около 1 масс.% до около 3 масс.% или от около 1 масс.% до около 2 масс.% от общей массы композиции. Фармацевтические композиции по изобретению могут содержать один или более слитых белков по изобретению на уровне около 0.1 масс.%, около 0.2 масс.%, около 0.3 масс.%, около 0.4 масс.%, около 0.5 масс.%, около 0.6 масс.%, около 0.7 масс.%, около 0.8 масс.%, около 0.9 масс.%, около 1 масс.%, около 2 масс.%, около 3 масс.%, около 4 масс.%, около 5 масс.%, около 6 масс.%, около 7 масс.%, около 8 масс.% или около 9 масс.% в расчете на общую массу композиции.
Режимы дозирования можно регулировать, чтобы обеспечить оптимальный терапевтический ответ. Например, можно ввести один болюс, можно ввести несколько разделенных доз в течение времени, или доза может быть пропорциональна уменьшена или увеличена, как предписано потребностями в соответствии с терапевтической ситуацией. В частности удобно составить парентеральные композиции в единичной лекарственной форме для облегчения введения и однородности дозы. Композиции по изобретению могут быть составлены и введены внутривенной, внутримышечной или подкожной инъекцией. В некоторых вариантах осуществления композиции по изобретению могут вводиться подкожно или внутримышечно.
В некоторых вариантах осуществления изобретения режим дозирования может включать введение повторных доз, например, введение еженедельной дозы. Режимы лечения могут включать еженедельную дозу в течение одного периода времени (например, в течение четырех недель), за которым следует менее частый "поддерживающий" режим дозирования (например, один раз в месяц или один раз в два месяца).
Режимы дозирования могут регулироваться для достижения желательных терапевтических исходов.
Способы по изобретению включают способы подавления AGE-зависимых воспалительных реакций у людей, предусматривающие введение эффективного количества фармацевтической композиции, содержащей один или более слитых белков по изобретению.
Способы по изобретению включают способы ингибирования AGE-опосредованной биологической активности, предусматривающие введение фармацевтической композиции, содержащей один или более слитых белков по изобретению. Как обсуждалось выше, AGE вовлечены в ряд заболеваний или состояний, таких как аутоиммунные заболевания. Аутоиммунные нарушения, заболевания или состояния, которые могут лечиться, облегчаться, обнаруживаться, диагностироваться, прогнозироваться или наблюдаться при использовании слитого белка по изобретению, включают, но не ограничиваются этим, дерматит, гломерулонефрит, рассеянный склероз, симпатическую офтальмию, аутоиммунное легочное воспаление, инсулинозависимый сахарный диабет, аутоиммунные глазные воспаления, системную красную волчанку, резистентность к инсулину, ревматоидный артрит, диабетическую ретинопатию и склеродермию.
Другие нарушения, которые могут лечиться или предупреждаться способами по изобретению, могут характеризоваться, как правило, как включающие любое нарушение, при котором пораженная клетка показывает увеличенную экспрессию RAGE или одного или более лигандов RAGE, или любое нарушение, которое поддается лечению (т.е. один или более симптомов могут быть устранены или облегчены) путем снижения функции RAGE. Например, функция RAGE может снижаться введением агента, который мешает взаимодействию между RAGE и лигандом RAGE.
Увеличенная экспрессия RAGE связана с несколькими патологическими состояниями, такими как диабетическая васкулопатия, нефропатия, ретинопатия, нейропатия и другие нарушения, включая болезнь Альцгеймера и иммунные/воспалительные реакции стенок кровяных сосудов. Лиганды RAGE продуцируются в ткани, пораженной многочисленными воспалительными нарушениями, включая артрит (такой как ревматоидный артрит). Отложения амилоида в тканях вызывает ряд токсических эффектов в клетках и являются признаком заболеваний, называемых амилоидозами. RAGE связывается с бета-складчатым фибриллярным веществом, таким, который обнаружен в бета-амилоидном пептиде, А-бета, амилине, сывороточном амилоиде А и прионных пептидах (prion-derived peptides). RAGE также экспрессируется на повышенных уровнях в тканях, имеющих амилоидные структуры. Соответственно, RAGE вовлечен в амилоидные нарушения. RAGE-амилоидное взаимодействие, как полагают, приводит к окислительному стрессу, ведущему к нейрональной дегенерации.
Множество лигандов RAGE, и в частности лиганды из семейств S100/кальгранулина и амфотерина (HMGB1), продуцируются в воспаленных тканях. Это наблюдение справедливо для острого воспаления, такого как наблюдаемое в ответ на введение липополисахаридов (как, например, при сепсисе), и для хронического воспаления, такого как наблюдаемое в различных формах артрита, язвенного колита, воспалительных заболеваниях кишечника и т.д. Сердечно-сосудистые заболевания и в частности те, которые являются результатом атеросклеротических бляшек, также, как полагают, имеют значительный воспалительный компонент. Такие заболевания включают окклюзионные, тромботические и эмболические заболевания, такие как стенокардия, нарушения, связанные с нестабильными бляшками (fragile plaque disorder), и эмболический инсульт соответственно. Опухолевые клетки также показывают увеличенную экспрессию лиганда RAGE, в частности амфотерина, указывающую на то, что рак является также RAGE-связанным нарушением. Кроме того, окислительные эффекты и другие аспекты хронического воспаления могут способствовать возникновению определенных опухолей.
AGE являются терапевтической мишенью для ревматоидного артрита и других воспалительных заболеваний.
Соответственно RAGE-связанные нарушения, которые можно лечить композициями по изобретению, включают в дополнение к аутоиммунным нарушениям, обсуждаемым выше: амилоидозы (такие как болезнь Альцгеймера), болезнь Крона, острые воспалительные заболевания (такие как сепсис), шок (например, септический шок, геморрагический шок), сердечно-сосудистые заболевания (например, атеросклероз, инсульт, нарушения, связанные с нестабильными бляшками, стенокардия и рестеноз), диабет (и в частности сердечно-сосудистые заболевания у страдающих диабетом), осложнения при диабете, нарушения, связанные с прионами, рак, васкулит и другие васкулитные синдромы, такие как некротизирующие васкулиты, нефропатии, ретинопатии и нейропатии.
Следующие примеры предоставляются только в иллюстративных целях и никоим образом не предназначены для ограничения объема настоящего изобретения.
Примеры
В следующих примерах эксперименты на мышах проводили со слитым белком, содержащим внеклеточные домены мышиного RAGE (аминокислотные остатки 1-342), слитые с шарниром, СН2 и СН3 доменами Fc-фрагмента тяжелой цепи мышиного IgG2a. Конструкция экспрессировалась в клетках СНО при использовании системы экспрессии GPEx™. Последовательность использованного мышиного RAGE представлена в следующей таблице.
Таблица 9 (SEQ ID NO:11) Последовательность мышиного RAGE

где сигнальный пептид RAGE = одинарное подчеркивание, внеклеточный домен RAGE = не подчеркнут, шарнирная область мышиного IgG2a = двойное подчеркивание, СН2-участок мышиного IgG2a = пунктирное прдчеркивание и СН3 участок мышиного IgG2a = волнистое подчеркивание.
ПРИМЕР 1
Действие слитых белков RAGE по изобретению на стрептозотоцин-индуцированный диабет у мышей.
Стрептозотоцин-индуцированный диабет у мышей является в данной области техники общепризнанной моделью ретинальных изменений, вызванных диабетом (см. Obrosova IG, Drel VR, Kumagai AK, Szabo C, Pacher P, Stevens MJ. Early diabetes-induced biochemical changes in the retina: comparison of rat and mouse models. Diabetologia. 2006 Oct: 49(10):2525-33.)
Настоящий эксперимент включал 5 лечебных групп, содержащих 15 C57BL/6 мышей на группу: 1) контроль без диабета; 2) диабетический контроль, содержащий мышей, обработанных стрептозотоцином при 45 мг/кг в течение 5 последующих дней перед началом исследования для вызывания диабета; 3) мыши, обработанные стрептозотоцином, которые также получали 10 мкг/день mRAGE-IgG2aFc, инъецированного интраперитонеально, 3 инъекции/неделю; 4) мыши, обработанные стрептозотоцином, которые также получали 100 мкг/день mRAGE-IgG2aFc, инъецированного интраперитонеально, 3 инъекции/неделю; и 5) мыши, обработанные стрептозотоцином, которые также получали 300 мкг/день mRAGE-IgG2aFc, инъецированного интраперитонеально, 3 инъекции/неделю.
Во время исследования у мышей оценивали массу тела, уровень глюкозы в крови, гликогемоглобин (GHb), альбуминурию и тактильную чувствительность как показатель функции сенсорных нервов. Мышей умерщвляли в конце исследования и оценивали проницаемость сосудов сетчатки, используя флуоресцентный зонд, адгезию лейкоцитов к капиллярам сетчатки и NF-κВ-регулируемую экспрессию белка (СОХ-2, ICAM, iNOS).
Результаты двухмесячного исследования
Изучали воздействия слитого белка RAGE-Ig на развитие изменений, вызванных диабетом, в физиологии сетчатки и метаболизме у мышей C57B1/6J. Слитый белок вводили интраперитонеально в 3 различных концентрациях (10 мкг, 100 мкг и 300 мкг) три раза в неделю. Не наблюдалось никаких неблагоприятных воздействий при любой дозе лекарственного средства на прибавление в весе или общее самочувствие у диабетических мышей. Уровни глюкозы в крови у неголодных мышей составляли 155±24 мг/дл (среднее±станд. отклонение), 358±38,417±36, 376±36 и 370±55 у контроля без диабета, диабетического контроля, диабетической +10 мкг слитого белка RAGE-Ig, диабетической +100 мкг слитого белка RAGE-Ig и диабетической+300 мкг слитого белка RAGE-Ig групп соответственно.
Параметры, связанные с ретинопатией, измеренные в краткосрочных исследованиях, представляли собой (1) лейкостаз, (2) проницаемость для эндогенного альбумина у сосудов сетчатки, (3) нитрование белков сетчатки и (4) экспрессию ретинальных ICAM и СОХ-2.
1. Лейкостаз.
Методы: При двухмесячном диабете кровь удаляли из сосудистой системы анестезированных животных путем полной перфузии с фосфатно-солевым буфером (PBS) через сердечный катетер. Животных затем перфузировали лектином конканавалином А, связанным с флуоресцеином (20 мкг/мл в PBS; Vector Laboratories, Burlingame, CA), как описано ранее (см. Joussen et al., FASEB J. 2004 Sep; 18(12):1450-2). Изображение плоско закрепленных сетчаток получали посредством флуоресцентной микроскопии, и подсчитывали число лейкоцитов, прилипших к стенке сосуда.
Результат: Значительное увеличение лейкостаза было продемонстрировано у мышей, у которых был диабет в течение 2 месяцев, по сравнению с мышами без диабета (Р<0.05). Лейкостаз не ингибировался в любой из групп, обработанных слитым белком RAGE-Ig (см. Фиг.1).
2. Проницаемость сосудов
Методы: При двухмесячном диабете получали путем замораживания срезы глаз (10 мкм), фиксировали в метаноле в течение 10 минут и промывали четырехкратно в PBS. Каждый срез инкубировали с антителами овцы против сывороточного альбумина мыши (sheep anti-mouse serum albumin) (Abeam, Cambridge MA; AB8940; 1:2000 разведение) в течение 2 ч. После промывки срезы инкубировали с FTIC-меченным вторичным антителом (АВ 6743; 1:1000 разведение) в течение 90 мин. На основании флуоресцентной микроскопии среднее количество флуоресценции определяли в 3 различных участках для каждого из 4 слоев сетчатки (внутренний плексиформный слой, внутренний ядерный слой, наружный плексиформный слой, наружный ядерный слой). Количество флуоресценции в каждом участке являлось средним из 10 рандомизированных измерений, и количество флуоресценции в каждом слое сетчатки представляет собой среднюю величину флуоресценции в каждом из 3 различных участков внутри этого слоя.
Результаты:
Диабет приводил к значительному увеличению флуоресценции в бессосудистой сетчатке (т.е. вследствие просачивания альбумина из сосудов) в каждом из 4 исследуемых слоев сетчатки. Результаты представлены на Фиг.2 (2А - внутренний плексиформный слой, 2В - внутренний ядерный слой, 2С - наружный плексиформный слой, 2D - наружный ядерный слой). Чтобы определить альбумин во внутреннем и внешнем ядерных слоях, мы преднамеренно измеряли в тонком пространстве между ядрами, таким образом эти цифры могут не быть столь же строгими, как цифры из плексиформных слоев, в которых не было ядер, искажающих измерения.
3. Нитрование белков сетчатки
Методы: При двухмесячном диабете выделяли и гомогенизировали сетчатки. Получали дот-блоты посредством блоттинга 50 мкг гомогената белков из каждого животного на нитроцеллюлозную мембрану. Мембраны блокировали молоком (5%), промывали и окрашивали с использованием иммунной метки, используя антитела к нитротиразину (Upstate Biotechnology, Inc. #05-233; 1:500 разведение), в течение 2 часов и затем окрашивали вторичным антителом (конъюгат козьих антител к IgG мыши с HRP (пероксидазой хрена), Bio-Rad; 1:1000 разведение) в течение 1 часа. После усиленной промывки окрашивание с использованием иммунной метки, обнаруженное посредством антитела, визуализировали усиленной хемилюминесценцией (ECL, Santa Cruz Biotechnology, Santa Cruz, CA). Зависимую от окрашивания с использованием иммунной метки хемилюминесценцию регистрировали на пленке, и проводили количественный анализ плотности окрашенных с использованием иммунной метки пятен. Результаты выражают как процент от значений, обнаруживаемых в контролях без диабета.
Результаты:
Результаты представлены на Фиг.3. Ретинальные гомогенаты из диабетических мышей показали ожидаемое увеличение нитрования белков. Терапия ингибировала эту посттрансляционную модификацию дозозависимым способом. Считается, что нитрование белков является параметром и окислительного, и нитратного стрессов.
4. Экспрессия ретинальных ICAM и СОХ-2
Способы: Сетчатки изолировали и разрушали ультразвуком, и супернатант использовали как полный экстракт сетчатки. Образцы (50 мкг) фракционировали при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE), переносили с помощью электроблоттинга на нитроцеллюлозную мембрану и мембраны блокировали Tris-забуференным физиологическим раствором, содержащим 0.02% Tween 20 и 5% обезжиренного молока. Применяли антитела для ICAM-1 (1:200 разведение; Santa Cruz Biotechnology) и СОХ-2 с последующим применением вторичного антитела в течение 1 часа. После промывки результаты визуализировали усиленной хемилюминесценцией.
Результаты:
Результаты показаны на Фиг.4. Поскольку экспрессия ICAM-1 на эндотелиальных клетках играет критическую роль в адгезии лейкоцитов к стенкам сосудов (лейкостаз), мы измеряли влияние диабета и терапии на экспрессию ICAM-1 в сетчатке. Двухмесячный диабет приводил к значительному увеличению экспрессии ретинальных ICAM-1. Введение слитого белка RAGE-Ig приводило к дозозависимому уменьшению экспрессии ICAM, и самая высокая доза значительно ингибировала эту экспрессию.
Проявление окрашенной с использованием иммунной метки полосы, соответствующей молекулярной массе СОХ-2, не увеличилось при диабете и не изменилось у животных, получающих терапию (не показано).
Конечные точки, использованные в этом краткосрочном исследовании воздействий слитого белка RAGE-Ig, были выбраны в силу того, что они все, как было обнаружено, связаны с развитием ранних (дегенеративных) стадий диабетической ретинопатии, т.е. различные терапии, которые, как было обнаружено, ингибируют вызванную диабетом дегенерацию капилляров сетчатки, также ингибируют эти дефекты.
Ингибирование RAGE действительно ингибировало патологии, связанные с сосудистой проницаемостью и нитратным стрессом в сетчатке. Нитратный стресс также рассматривается как маркер окислительного стресса. Ингибитор RAGE, однако, не ингибировал патологий, связанных с лейкостазом.
ПРИМЕР 2
Воздействие слитых белков RAGE по изобретению на долгосрочный стрептозотоцин-индуцированный диабет у мышей.
Стрептозотоцин-индуцированный диабет у мышей является в данной области техники общепризнанной моделью ретинальных изменений, вызванных диабетом (см. Obrosova IG, Drel VR, Kumagai AK, Szabo C, Pacher P, Stevens MJ. Early diabetes-induced biochemical changes in the retina: comparison of rat and mouse models. Diabetologia. 2006 Oct: 49(10):2525-33).
Долгосрочные исследования включали 5 групп обработки, содержащих 25 мышей C57BL/6 на группу: 1) недиабетический контроль; 2) диабетический контроль, содержащий мышей, обработанных стрептозотоцином при 45 мг/кг в течение 5 последующих дней перед началом исследования для вызывания диабета; 3) мыши, обработанные стрептозотоцином, которые также получали 10 мкг/день mRAGE-IgG2aFc, инъецированного интраперитонеально, 3 инъекции/неделю; 4) мыши, обработанные стрептозотоцином, которые также получали 100 мкг/день mRAGE-IgG2aFc, инъецированного интраперитонеально, 3 инъекции/неделю; и 5) мыши, обработанные стрептозотоцином, которые также получали 300 мкг/день mRAGE-IgG2aFc, инъецированного интраперитонеально, 3 инъекции/неделю.
Во время исследования у мышей оценивали массу тела, уровень глюкозы в крови, гликогемоглобин (GHb), альбуминурию и тактильную чувствительность как показатель функции сенсорных нервов. Мышей умерщвляли в конце исследования и оценивали количественную гистопатологию и нейродегенерацию сетчатки.
Параметры, связанные с ретинопатией, измеренные в долгосрочном исследовании, представляли собой (1) бесклеточные капилляры, (2) «тени» перицитов и (3) ганглиозные клетки. В качестве маркера периферической нейропатии в долгосрочном исследовании измеряли также чувствительность лапы к легкому прикосновению.
Гистопатология сетчатки, вызванная диабетом
После 10 месяцев диабета глаза фиксировали в формалине и одну сетчатку выделяли из каждого животного, промывали в проточной воде в течение ночи и расщепляли в течение 2 ч в растворе неочищенного трипсина, как мы сообщали ранее. Ретинальную сосудистую систему выделяли мягким отделением нервных клеток посредством "щетки", полученной из одного волоса. После полной чистки от нервных клеток выделенную сосудистую систему раскладывали на предметном стекле микроскопа, высушивали в течение ночи, окрашивали гематоксилином и Шифф-йодной кислотой, обезвоживали и накрывали покровным стеклом. Дегенеративные (бесклеточные) капилляры количественно анализировали в 6-7 участках, соответствующих средней периферии сетчатки (mid-retina) (200X увеличение), замаскированным образом. Бесклеточные капилляры идентифицировали как сосудистые трубочки с размерами капилляров, не имеющие ядер где-либо вдоль своей длины, и описывались на квадратный миллиметр площади сетчатки. Тени перицитов оценивали из распространения выступающих «бугорков» на капиллярных базальных мембранах, из которых исчезли перициты. Исследовали, по меньшей мере, 1000 капиллярных клеток (эндотелиальные клетки и перициты) в 5 участках средней периферии сетчатки (400Х увеличение) замаскированным образом. Тени на любом уже бесклеточном сосуде исключали.
Для изучения влияния диабета на нейродегенерацию сетчатки подсчитывали клетки в слое ганглиозных клеток. Фиксированные формалином глаза заключали в парафин, делали саггитальные срезы сетчатки, проходя через оптический нерв, и окрашивали гематоксилином-эозином. Число клеток в ганглиозном слое подсчитывали на двух участках (средней периферии сетчатки и области сетчатки в заднем полюсе глаза около диска зрительного нерва) с обеих сторон зрительного нерва. Аналогичные участки с двух сторон зрительного нерва усреднялись и выражались на единицу длины.
Результаты. Как предполагалось на основании предшествующей работы, долгосрочный диабет приводил к значительному увеличению числа дегенеративных, бесклеточных капилляров в сетчатке (Фиг.5А). Все дозы слитого белка RAGE-Ig значительно ингибировали эту дегенерацию капилляров, без наличия какого-либо влияния на тяжесть гипергликемии. Диабет также имел тенденцию увеличивать дегенерацию перицитов («тени» перицитов), но результаты не достигали статистической значимости (Фиг.5В). Мы ранее обнаружили, что потерю перицитов гораздо более трудно обнаружить у диабетических мышей С57В1/6 по сравнению с диабетическими крысами или более крупными видами, и сейчас мы рассматриваем ее как ненадежный параметр сосудистого заболевания для этой модели. Возможно, в результате неспособности обнаружить значительную потерю перицитов в контроле с диабетом мы не обнаружили какого-либо влияния слитого белка RAGE-Ig на потерю перицитов у этих мышей.
Диабет не вызывал уменьшение числа клеток в ретинальном слое ганглиозных клеток (т.е. нейродегенерацию) у этих мышей С57В1/6. Этот вывод согласовывался с предшествующим исследованием этой мышиной модели. В отсутствие влияния диабета на нейродегенерацию сетчатки, мы не способны оценить, воздействовал бы или нет ингибитор на нейродегенерацию.
Чувствительность на легкое прикосновение (признак периферической нейропатии).
Пациенты с диабетической нейропатией могут показывать ряд аберрантных ощущений, включающих спонтанную боль, боль, вызываемую легким прикосновением, и гипералгезию. Существуют накапливающиеся данные, что грызуны с диабетом воспроизводят эту гипералгезию и проявляют тактильную аллодинию. У грызунов это измеряется как порог тактильной чувствительности лапы.
Способы: Мышей (8-месячный диабет) переносили в испытательную клетку с дном из проволочной сетки и оставляли акклиматизироваться в течение 10-15 минут. Использовали волоски фон Фрея для определения 50% порога механического отдергивания для отдергивания стопы. Комплект волосков с логарифмически увеличивающейся жесткостью, начиная с волоска, который имел массу изгиба 0.6 г, применялся последовательно к подошвенной поверхности правой задней лапы с давлением, которое вызывало изгибание волоска. Подъем лапы регистрировали, как положительный ответ, и выбирали более легкий волосок для следующего измерения. Если ответ отсутствовал через 5 секунд, затем использовали следующий более тяжелый волосок. Этот процесс продолжали до тех пор, пока не были сделаны четыре измерения после первоначального изменения в поведении, или до тех пор, пока не наблюдались пять последовательных отрицательных (6 г) или четыре последовательных положительных (0.4 г) ответа. Получающуюся последовательность положительных и отрицательных оценок использовали для расчета 50% порога реакции отдергивания.
Результаты: Диабет значительно увеличивал чувствительность лапы к легкому прикосновению, подразумевая, что требуется более низкое давление для диабетических животных, чтобы отдернуть их лапы, чем в случае животных без диабета (Фиг.6). Этот вызванный диабетом дефект значительно ингибировался при каждой дозе слитого белка sRAGE-Ig.
Ретинопатия: Исследования, проведенные с использованием слитого белка RAGE-Ig, осуществляли для 2 сроков диабета: (1) долгосрочные (10 месяцев) исследования для оценки влияния терапии на долгосрочную гистопатологию диабетической ретинопатии, которая развивается у мышей, и (2) 2-3 месячные исследования для оценки физиологических и молекулярных воздействий терапии, которые предположительно лежат в основе воздействий на долгосрочную гистопатологию. Физиологические и молекулярные конечные точки, исследуемые по отношению к воздействиям слитого белка RAGE-Ig, выбирались ввиду того, что все, как было обнаружено в других исследованиях, соотносятся с (и вероятно причинно связаны с) развитием ранних (дегенеративных) стадий диабетической ретинопатии. Все три дозы терапии ясно и существенно ингибировали вызванную диабетом дегенерацию сосудистой системы сетчатки. Аналогично, все три дозы лекарственного средства, по-видимому, ингибировали также вызванное диабетом увеличение проницаемости сетчатки у этих мышей. Эти выводы представляют собой большое клиническое значение, поскольку ранние (непролиферативные) стадии диабетической ретинопатии все еще определяют на основе сосудистой патологии (сосудистой неперфузии и дегенерации и увеличенной проницаемости).
Влияние терапии на измеренные молекулярные и физиологические конечные точки в сетчатках от диабетических мышей было смешанным. Ингибирование RAGE действительно ингибировало нарушения, связанные с нитратным стрессом, маркером окислительного стресса в сетчатке. Ингибитор RAGE, однако, не ингибировал нарушения, связанные с лейкостазом. Отсутствие воздействия терапии на лейкостаз является неожиданным, поскольку другая группа недавно сообщала, что их sRAGE действительно ингибировал увеличение лейкостаза при диабете. Доказательство, которое мы получили со времени начала наших исследований с использованием слитого белка RAGE-Ig (Diabetes 57:1387-93, 2008), однако, показывает, что воздействия лекарственной терапии на ретинальный лейкостаз при диабете не предсказывают воздействие терапии на дегенерацию капилляров сетчатки при диабете. Следовательно, отсутствие воздействия терапии на ретинальный лейкостаз никоим образом не уменьшает значение обнаруженных воздействий лекарственного средства.
Неожиданно оказалось, что существует показатель «доза-эффект» лекарственного средства в отношении экспрессии ICAM-1 и нитрования белков в сетчатках диабетических животных, тогда как этот показатель «доза-эффект» не являлся выраженным для проницаемости капилляров сетчатки и дегенерации. По-видимому, можно было бы предположить, что ни ICAM, ни нитрование не вовлечены в ретинальные сосудистые нарушения при диабете, хотя мы имеем данные при использовании ICAM-1-нокаутных животных, которые опровергают это заключение.
Очевидно, что это лекарственное средство действительно попадало в сетчатку, действительно проявляло биологические эффекты и действительно демонстрировало значительную способность данного лекарственного средства ингибировать, по меньшей мере, ранние сосудистые поражения при диабетической ретинопатии.
Сенсорная нейропатия: Другими авторами постулировано, что конечные продукты гликирования (AGE) и взаимодействие этих AGE с RAGE вызывают окислительный стресс, активируют NF-kB и различные NF-kB-опосредованные провоспалительные гены в нервах, и увеличивают неврологическую дисфункцию, включая измененное болевое ощущение. Настоящие данные находятся в соответствии с фактами, что RAGE-опосредованная сигнальная система вносит вклад в развитие, по меньшей мере, нескольких аспектов диабетической нейропатии, и обеспечивают доказательства того, что слитый белок sRAGE-Ig ингибирует этот процесс в долгосрочном исследовании.
ПРИМЕР 3
Оценка слитого белка RAGE-Ig при использовании мышиной модели с артритом, индуцированным коллагеном II типа.
Иммунизация чувствительных линий мышей коллагеном II типа, основного компонента хряща сустава, вызывает прогрессирующий воспалительный артрит (Wooley et al. Journal of Experimental Medicine 1981; 154:688-700). Индуцированный коллагеном артрит (CIA) клинически характеризуется эритемой и эдемой, причем ширина пораженной лапы обычно увеличивается на 100%. Клинический оценочный индекс разработан для оценки прогрессирования болезни по отношению к деформации сустава и спондилиту (Wooley, Methods In Enzymology 1988; 162:361-373). Гистопатология пораженных суставов показывает синовит, формирование паннуса и эрозию суставов и костей, которые могут также быть представлены индексом. Иммунологические лабораторные наблюдения включают высокие уровни антител к коллагену II типа и гипер-гаммаглобулинемию. Эта модель в настоящее время хорошо адаптирована для исследования иммунотерапевтических подходов к заболеванию суставов (Staines et al. British Journal of Rheumatology 1994; 33(9):798-807) и успешно используется для изучения как биологических, так и фармакологических агентов для лечения ревматоидного артрита (RA) (Wooley et al. Arthritis Rheum 1993; 36:1305-1314, and Wooley et al. Journal of Immunology 1993; 151:6602-6607).
Антагонизм рецептора RAGE признается как потенциальная терапевтическая мишень ревматоидного артрита. Блокада RAGE у мышей с вызванным коллагеном артритом приводила к подавлению клинических и гистологических признаков артрита, и уменьшение интенсивности заболевания было связано с уменьшением уровней TNFα, IL-6 и матричных металлопротеиназ ММР-3, ММР-9 и ММР-13 в ткани лап с артритом (Hofmann et al. Genes Immun 2002; 3(3):123-135). Это показывает, что вызванный коллагеном артрит чувствителен к RAGE-направленной терапии.
Этот эксперимент может оценить влияние слитого белка RAGE-Ig на CIA при трех дозах, введенных со времени иммунизации коллагеном типа II. План исследования показан на Фиг.7.
Сорок мышей DBA/1 LacJ в возрасте 8-10 недель получали из Jackson Labs и акклиматизировали в виварии в течение минимум 10 дней перед проведением эксперимента. Все животные весили >16 г на время начала эксперимента. Мышей разделяли на четыре лечебных группы: 1) 100 мкл стерильного PBS интраперитонеальной инъекцией ежедневно; 2) 100 мкл стерильного PBS, содержащего 10 мкг слитого белка RAGE-Ig, интраперитонеальной инъекцией ежедневно; 3) 100 мкл стерильного PBS, содержащего 100 мкг слитого белка RAGE-Ig, интраперитонеальной инъекцией ежедневно; и 4) 100 мкл стерильного PBS, содержащего 300 мкг слитого белка RAGE-Ig, интраперитонеальной инъекцией ежедневно.
Через три дня после первоначального введения дозы всех мышей инъецировали 100 мкг бычьего коллагена II типа в полном адъюванте Фрейнда (FCA) внутрикожно в основание хвоста. Мышей контролировали ежедневным осмотром на предмет начала болезни, которое регистрировали. Мышей еженедельно взвешивали и отмечали состояние общего самочувствия. Животных, пораженных артритом, клинически оценивали пять раз в неделю до 10 недель после иммунизации, и измерения лап проводили три раза в неделю. Мышей без признаков артрита через десять недель после иммунизации рассматривали как отрицательный результат заболевания.
РЕЗУЛЬТАТЫ
Общее самочувствие и токсичность. Никаких острых токсических случаев не встречалось во время исследования, и все животные выжили в ходе эксперимента. Лечение хорошо переносилось, и никаких признаков побочных явлений, таких как потускнение шерсти или зуд, не наблюдалось. Массы мышей (Фиг.8) демонстрируют небольшие изменения по массе в ходе исследования, что является типичным вследствие преходящей потери массы у отдельных животных, соответствующей началу болезни. Никакие из этих изменений между группами не достигали статистической значимости.
Число случаев и начало артрита. Конечное число случаев коллагенового артрита в исследовании показано на Фиг.9. Контрольные мыши достигали 100% начала, что не является необычным в классической модели коллагенового артрита, где обычный диапазон числа случаев 80%-100%. Мыши, обработанные 10 мкг RAGE в день, достигали числа случаев 80%, что не являлось значительным уменьшением числа случаев. Мыши, обработанные 100 мкг RAGE в день, проявляли 60% число случаев, что значительно ниже, чем у контрольной группы (p<0.05). Неожиданно число случаев артрита у мышей, обработанных 300 мкг RAGE, составило 100%, и таким образом было подобно числу случаев контроля.
Среднее значение (и стандартная ошибка среднего (SEM)) периода начала заболевания представлено на Фиг.10. Начало заболевания было типичным в контрольной группе, со средним периодом первого проявления артрита 38.6. Начало заболевания у мышей, обработанных или 10 мкг, или 100 мкг RAGE, номинально задерживалось до 42.5, что не достигало статистической значимости. Однако начало заболевания значительно откладывалось (p<0.05) у мышей, обработанных 300 мкг RAGE. Следовательно, хотя мыши при высокой дозе не проявляли уменьшения числа случаев, время до развития клинически явного артрита заметно возрастало.
Модуляцию начала заболевания обработкой RAGE можно легко оценить при помощи графика зависимости числа случаев заболевания от времени (Фиг.11). Типичное свойство быстрого начала заболевания для CIA обнаружено в контрольной группе, тогда как мыши, обработанные RAGE в дозе или 10 мкг, или 100 мкг, приводили к задержке начала заболевания и к более низкому конечному числу случаев артрита. В течение приблизительно восьми недель у мышей, обработанных 300 мкг RAGE, заболевание развивалось по похожей схеме, но группа животных с поздним артритом приводила к высокому числу случаев заболевания, но с отсроченным началом заболевания.
Тяжесть заболевания и прогрессирование. Анализ совокупной оценки для сустава у обработанных и контрольных животных выявил значительное воздействие терапии с использованием RAGE на тяжесть вызванного коллагеном артрита (Фиг.12). У контрольных мышей обнаруживалось типичное хроническое прогрессирующее заболевание с явным увеличением кумулятивного индекса артрита. В отличие от этого, мыши, обработанные RAGE в любой дозе, проявляли явное уменьшение совокупной оценки артрита. Разница между контрольной и обработанной группами достигала высокого уровня статистической значимости (p<0.001) с 43 дня после иммунизации, и эта разница поддерживалась на всем протяжении исследования. Хотя терапия с использованием RAGE при 100 мкг/день достигала самой низкой совокупной оценки артрита, не существовало значительной разницы между RAGE группами по отношению к оценке артрита, предполагая, что достигался «пороговый» эффект, а не классический дозозависимый эффект.
Анализ влияния терапии с использованием RAGE на число лап с артритом (Фиг.13) действительно демонстрирует значительное влияние на прогрессирование заболевания. Кроме того, обнаруживалось значительное влияние на число пораженных лап с 43 дня. Уровень значимости варьировал от p<0.001 до p<0.025, что может показывать, что влияние RAGE было более выраженным в отношении тяжести заболевания, чем прогрессирования артрита; однако, максимальное число пораженных лап (40) является более ограниченным, чем максимальная совокупная оценка заболевания (120). Кроме того, не было никаких значительных изменений между группами, обработанными RAGE, хотя группа со 100 мкг RAGE действительно проявляла более высокий уровень замедления артрита.
Эти результаты предполагают, что введение белка RAGE оказывает заметное влияние на вызванный коллагеном артрит при введении с использованием профилактической схемы. Явные токсические эффекты от инъекции RAGE отсутствовали при любой дозе, и, по-видимому, лечение являлось очень хорошо переносимым. Общее число случаев заболевания значительно снижалось у мышей, получающих 100 мкг ежедневно, и задержка начала заболевания наблюдалась у мышей, обработанных 300 мкг/день. Однако наиболее очевидный признак клинической активности обнаруживался в уменьшении оценки заболевания и числа лап с артритом, где большое расхождение между мышами, обработанными RAGE, и контрольными животными обнаруживалось с 43 дня после иммунизации. На данном этапе контрольные животные испытывали типичное прогрессирование тяжелого артрита, тогда как обработка при помощи RAGE во всех дозах замедляло прогрессирование болезни.
Гистопатологическая оценка: Конечности всех мышей удаляли при окончании клинического оценочного исследования и хранили в нейтральном забуференном растворе формалина. Суставы декальцинировали в течение 18 дней в 10% муравьиной кислоте, дегидратировали и заключали в парафиновые блоки. Срезы нарезали вдоль продольной оси, закрепляли и окрашивали или гематоксилином и эозином, или толуидиновым синим. Препараты разрезали приблизительно по средней линии и затем сагиттальные центральные образцы закрепляли для оценки. Это обеспечивало состоятельную географическую оценку. Пять-десять образцов закрепляли (обычно 4-6 образцов на предметное стекло). После окрашивания предметные стекла необратимо соединяли с покровными стеклами. Минимум 3 отдельных среза на препарат оценивали слепым методом, при этом оценщик не осведомлен о назначении в группе. На передних конечностях все локтевые, лучезапястные и метакарпальные суставы оценивали, в то время как все коленные, голеностопные и метатарзальные суставы оценивались на задних лапах. Цифры не оценивали, так как процедура получения срезов уничтожает большинство проксимальных межфаланговых суставов. Предметные стекла оценивали на присутствие синовита, формирование паннуса, краевых эрозий, изменений архитектуры (главным образом, подвывиха) и деструкции. Общая оценка на основе этих коллективных точек, затем, была присвоена каждому срезу. Оценочная система основывалась на следующем:
Синовит рассматривали по толщине синовиальной мембраны и оценивали по бальной шкале следующим образом: 0 - для толщины менее чем 3 клетки; 1 - для толщины 3-5 клеток; 2 - для толщины 6-10 клеток; 3 - для толщины 10-20 клеток; и 4 - для толщины 20-30 клеток.
Формирование паннуса оценивали следующим образом: 0 - при отсутствии формирования паннуса; 1 - при наличии микроворсинок; 2 - при явном прикреплении паннуса; 3 - при заметном прикреплении паннуса; и 4 - при суставной щели, заполненной паннусом.
Краевые эрозии оценивали следующим образом: 0 - при отсутствии видимых эрозий; 1 - при незначительном дефекте (indentation) в области прикрепления капсулы; 2 - при явных эрозиях хряща; 3 - при эрозиях, распространяющихся в субхондральную кость; и 4 - при обширной эрозии кости и хряща.
Изменения архитектуры оценивают следующим образом: 0 - при нормальной архитектуре сустава; 1 - при отечных изменениях; 2 - при незначительном подвывихе суставных поверхностей; 3 - при крупном подвывихе суставных поверхностей; 4 - при полном фиброзе и образовании коллагеновых сшивок.
Общая оценка отображает: 0 - при классическом нормальном внешнем виде сустава; 1 - при незначительных изменениях; в соответствии с ремиссией; может быть клинически нормальным; 2 - при определенном воспалительном артрите; 3 - обширное воспалительное эрозивное заболевание; и 4 - при деструктивном эрозивном артрите.
Деградация хрящевого и костного матрикса. Серийные срезы окрашивали на компоненты хрящевого матрикса, используя гистохимический краситель толуидиновый синий. Срезы с толуидиновым синим оценивали на потерю протеогликанов. Окрашивание суставной поверхности сравнивали с окрашиванием пластинки роста и оценивали следующим образом: 0 - при отсутствии потери протеогликанов; нормальное окрашивание толуидиновым синим; 1 - при незначительной потере протеогликанов; некоторая потеря окрашивания у поверхностной зоны хряща; 2 - при средней потере протеогликанов; слабое окрашивание поверхностной зоны хряща; 3 - при значительной потере протеогликанов; отсутствие окрашивания толуидиновым синим поверхностной зоны хряща; и 4 - при крупной потере протеогликанов; отсутствие окрашивания толуидиновым синим глубокой зоны хряща.
РЕЗУЛЬТАТЫ
Гистологические выводы по артриту, вызванному коллагеном. Срезы оценивали на воспалительные и эрозивные параметры заболевания. Внешний вид артрита (Фиг.14) показывает типичную патологию воспалительного эрозивного заболевания на этот момент времени в контрольной группе (обработана PBS) с типичными признаками артрита - синовиальной гипертрофией и гиперплазией, заметном прикреплении паннуса и краевыми эрозиями.
Обработка слитым белком RAGE-Ig при 10 мкг/мл (Фиг.14В) приводила к умеренным изменениям в воспалительных и эрозивных параметрах с общим улучшением в проявлениях эрозий и нарушенных поверхностях хряща. Обработка слитым белком RAGE-Ig при 100 мкг/мл (Фиг.14С) приводила к уменьшению формирования паннуса и эрозий по сравнению с контролем, и общее отличие было вполне заметно. Однако введение слитого белка RAGE-Ig при 300 мкг/мл (Фиг.14D) приводило к артриту, который проявлялся менее серьезно, чем патология, наблюдаемая в контроле с физиологическим раствором, но тем не менее был довольно серьезен, с синовиальной гипертрофией и гиперплазией, с заметным прикреплением паннуса и краевыми эрозиями.
Анализ оценок воспаления (Фиг.15) выявил снижение воспаления у мышей, обработанных слитым белком RAGE-Ig во всех дозах, при сравнении с контрольными (обработанными физиологическим раствором) животными. Однако синовит значительно уменьшился (p<0.05) только в группе 100 мкг/мл, и формирование паннуса демонстрировало подобные снижения в оценке (p<0.03). Снижения воспалительных параметров заболевания, обнаруживаемые при использовании слитого белка RAGE-Ig или при 10 мкг/мл, или 300 мкг/мл, были недостаточными, чтобы достичь статистической значимости.
Оценка изменений в эрозивных признаках (эрозии и изменения в архитектуре суставов), вызванного коллагеном артрита, показала похожую схему эффектов. Значительное снижение (p<0.01) эрозий суставов наблюдалось в группе, обработанной слитым белком RAGE-Ig при 100 мкг/мл, при сравнении с контрольными (обработанными физиологическим раствором) животными (Фиг.16), тогда как снижения, наблюдаемые у мышей, обработанных слитым белком RAGE-Ig при 10 мкг/мл и 300 мкг/мл, не достигали значимости.
Комбинация гистопатологических параметров в общей гистологической оценке артрита (Фиг.17) отражала выводы об индивидуальных патологических параметрах. Наблюдались значительные отличия между контрольными (физиологический раствор) обработанными животными и мышами, обработанными слитым белком RAGE-Ig при 100 мкг/мл (p<0.02), а общая оценка для мышей, обработанных 10 мкг/мл, как раз достигала значимости (p=0.05), между тем никаких значительных снижений в общей оценке заболевания не наблюдалось при применении слитого белка RAGE-Ig при 300 мкг/мл.
Срезы, окрашенные толуидиновым синим, исследовали, чтобы определить, влиял ли слитый белок RAGE-Ig на потерю белков матрикса из сустава с артритом. Данные (показанные на Фиг.18 и 19) наводят на мысль, что слитый белок RAGE-Ig действительно защищал от потери протеогликанов, но этот эффект является статистически значимым (p<0.05) только при дозе 100 мкг/мл дозы. Контрольная группа с PBS показывает значительную потерю матрикса хряща (протеогликаны и коллагены), и заметная потеря окрашивания проксимальной поверхности хряща наблюдается у мышей, обработанных слитым белком RAGE-Ig при 300 мкг/мл. В отличие от этого, существует хорошая сохранность белка матрикса при введении слитого белка RAGE-Ig при 10 мкг/мл или 100 мкг/л.
Гистологические выводы подтверждают клинические данные, которые показывают, что обработка вызванного коллагеном артрита слитым белком RAGE-Ig приводила к воздействию на число случаев и тяжесть заболевания. Гистологические параметры достигали максимума уровней статистической значимости у мышей, обработанных 100 мкг/мл, и достигали статистической значимости в общей патологии у мышей, обработанных 10 мкг/мл. Слитый белок RAGE-Ig при 100 мкг/мл обеспечивал хорошую сохранность структуры сустава, и значительное уменьшение всех признаков артрита при оценивании. Общая оценка заключалась в том, что слитый белок RAGE-Ig блокировал эрозивную фазу артрита, поскольку на степень воспалительных изменений оказывалось меньше влияния, чем на вторичные параметры заболевания. Мыши, обработанные слитым белком RAGE-Ig при 300 мкг/мл, не защищались в той же степени, как при более низких дозах, в свою очередь вызывая возможность супрессивного ответа на этот уровень введения белка. В целом, эти выводы находятся в соответствии с клиническими результатами наблюдений, полученными в этом исследовании, и демонстрируют, что слитый белок RAGE-Ig может проявлять противоартритный эффект.
Хотя изобретение описано подробно и со ссылкой на специфические варианты осуществления изобретения, специалисту в данной области техники будет очевидно, что различные изменения и модификации могут быть сделаны в нем без отклонения от его объема и сущности, и такие изменения и модификации могут осуществляться в пределах объема приложенной формулы изобретения. Все патенты и публикации включены здесь путем ссылки в том же объеме, как если бы каждая отдельная публикация специфически и индивидуально указывалась как включенная путем ссылки во всей своей полноте.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИПЕПТИДЫ ДЛЯ СВЯЗЫВАНИЯ С "РЕЦЕПТОРОМ КОНЕЧНЫХ ПРОДУКТОВ ГЛИКИРОВАНИЯ", А ТАКЖЕ ИХ КОМПОЗИЦИИ И СПОСОБЫ, В КОТОРЫХ ОНИ ПРИНИМАЮТ УЧАСТИЕ | 2010 |
|
RU2558301C2 |
| СПОСОБ И ДИАГНОСТИЧЕСКИЙ НАБОР ДЛЯ ИЗМЕРЕНИЯ КОЛИЧЕСТВА БЕЛКА БЕТТА ig-h3 | 2002 |
|
RU2281512C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ КОНЕЧНЫХ ПРОДУКТОВ ГЛУБОКОГО ГЛИКИРОВАНИЯ (RAGE) И ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2518351C2 |
| ПОЛИПЕПТИДЫ И ПОЛИНУКЛЕОТИДЫ И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ ИММУНОЛОГИЧЕСКИХ НАРУШЕНИЙ И РАКА | 2012 |
|
RU2623161C2 |
| ПЕПТИДЫ НАПРАВЛЕННОГО ДЕЙСТВИЯ НА VEGFR-1/NRP-1 | 2008 |
|
RU2488592C2 |
| СПОСОБЫ ЛЕЧЕНИЯ СОСТОЯНИЙ АНТИТЕЛАМИ, КОТОРЫЕ СВЯЗЫВАЮТ РЕЦЕПТОР КОЛОНИЕСТИМУЛИРУЮЩЕГО ФАКТОРА 1 (CSF1R) | 2012 |
|
RU2670743C9 |
| ИММУНОСУПРЕССОРНЫЕ ПОЛИПЕПТИДЫ И НУКЛЕИНОВЫЕ КИСЛОТЫ | 2008 |
|
RU2506275C2 |
| Новые гибридные ActRIIB белки-ловушки лигандов для лечения заболеваний, связанных с мышечной атрофией | 2016 |
|
RU2733492C2 |
| Белок, связанный с гликопротеином человека, композиция, фармацевтическая композиция, вектор экспрессии | 2016 |
|
RU2773819C2 |
| НОВЫЕ IL-17-СВЯЗЫВАЮЩИЕ СОЕДИНЕНИЯ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2010 |
|
RU2550272C2 |


























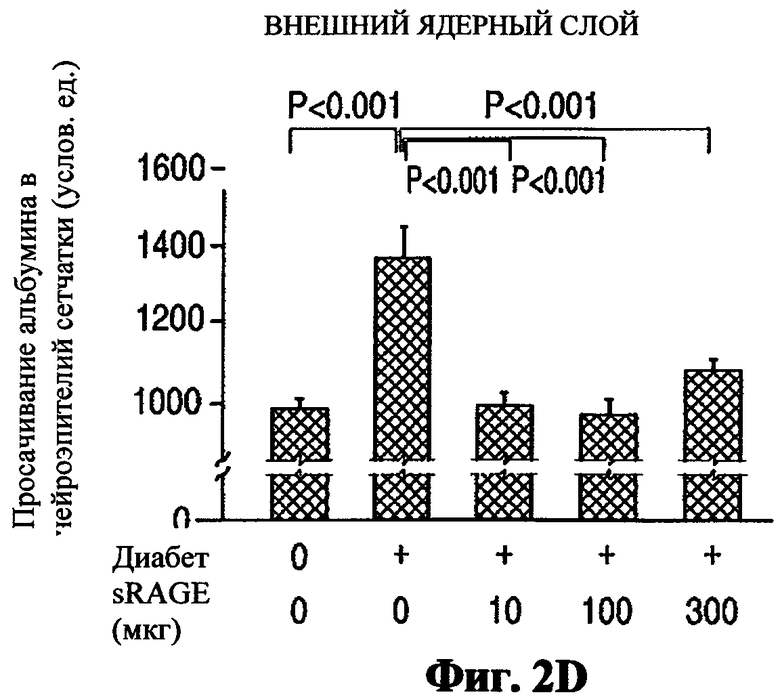










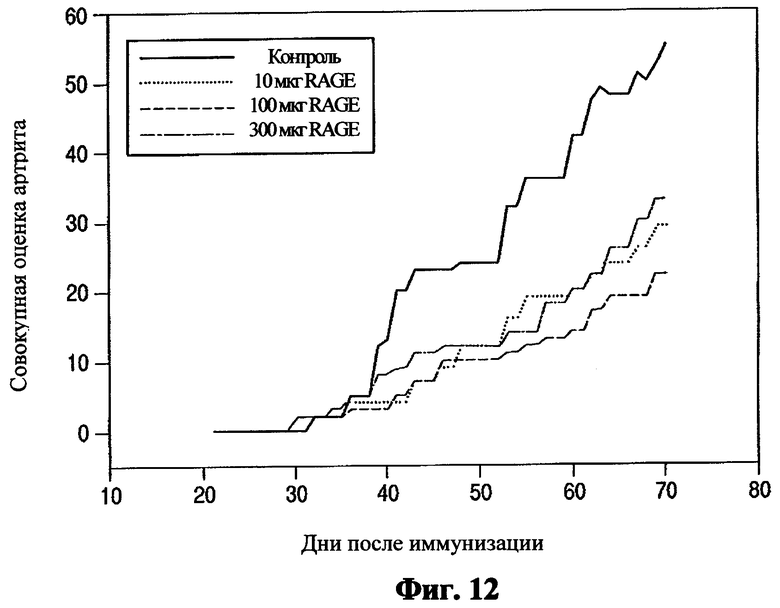







Настоящее изобретение относится к области биохимии. Предложен слитый белок для лечения заболеваний, опосредованных конечными продуктами гликирования (AGE), состоящий из фрагмента варианта рецептора конечных продуктов гликирования (RAGE) человека, имеющего две точечные мутации H217R и R221H, и фрагмента константного домена иммуноглобулина человека IgG4, при необходимости соединенных линкером. Кроме того, рассмотрены нуклеиновая кислота и рекомбинантная клетка-хозяин для получения слитого белка, а также фармацевтическая композиция для лечения AGE-опосредованных заболеваний, содержащая слитый белок. Данное изобретение обеспечивает пониженную агрегацию слитого белка. 4 н. и 9 з.п. ф-лы, 19 ил., 3 пр., 9 табл.
1. Слитый белок для лечения AGE-опосредованных заболеваний, состоящий из:
(a) первой аминокислотной последовательности, выбранной из группы, состоящей из аминокислот 1-301, аминокислот 24-301, аминокислот 1-344 или аминокислот 24-344 из SEQ ID NO: 6, причем первая аминокислотная последовательность способна связывать лиганд RAGE; и
(b) второй аминокислотной последовательности, состоящей из (1) аминокислотной последовательности константного домена тяжелой цепи иммуноглобулина человека IgG4 или (2) аминокислотной последовательности фрагмента тяжелой цепи иммуноглобулина человека IgG4, содержащего шарнирный, CH2 и CH3 участки,
где первая аминокислотная последовательность и вторая аминокислотная последовательность при необходимости соединены линкером.
2. Слитый белок по п.1, где первая аминокислотная последовательность состоит из аминокислот 1-344 из SEQ ID NO: 6.
3. Слитый белок по п.1 или 2, где вторая аминокислотная последовательность состоит из аминокислотной последовательности константного домена тяжелой цепи иммуноглобулина человека IgG4.
4. Слитый белок по п.1, где слитый белок состоит из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 6 и SEQ ID NO: 8.
5. Слитый белок по п.4, где слитый белок состоит из аминокислотной последовательности SEQ ID NO: 6.
6. Слитый белок по п.1, дополнительно содержащий линкер между первой аминокислотной последовательностью и второй аминокислотной последовательностью.
7. Слитый белок по любому из пп.1-6 для применения в способе снижения уровней лиганда, связывающегося посредством RAGE, у млекопитающего.
8. Нуклеиновая кислота для получения слитого белка по любому из пп.1-6, где указанная нуклеиновая кислота кодирует слитый белок по любому из пп.1-6.
9. Рекомбинантная клетка-хозяин для получения слитого белка по любому из пп.1-6, содержащая нуклеиновую кислоту по п.8.
10. Фармацевтическая композиция для лечения AGE-опосредованных заболеваний, содержащая от 0,1% до 20 мас.% слитого белка по любому из пп.1-6 от общей массы фармацевтической композиции и фармацевтически приемлемый носитель.
11. Фармацевтическая композиция по п.10, содержащая слитый белок по п.5.
12. Фармацевтическая композиция по любому из пп.10 или 11 для применения в лечении диабетической нефропатии или ревматоидного артрита, или аутоиммунного заболевания.
13. Фармацевтическая композиция по любому из пп.10-12 для применения в лечении дерматита, гломерулонефрита, рассеянного склероза, симпатической офтальмии, аутоиммунного легочного воспаления, инсулинозависимого сахарного диабета, аутоиммунных глазных воспалений, системной красной волчанки, резистентности к инсулину, ревматоидного артрита, диабетической ретинопатии и склеродермии.
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| СЕВЕРИНА Т | |||
| И | |||
| и др., "Результаты применения тиоктацида в лечении диабетической нейропатии у больных сахарным диабетом 1 типа", Сахарный диабет (2000), 4: 33-35 | |||

