ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способу и диагностическому набору для измерения количества белка β ig-h3. В частности, оно относится к способу измерения количества белка β ig-h3 в жидкостях организма с помощью реакции специфического связывания белка β ig-h3 или рекомбинантных белков, содержащих домен fas-1 белка β ig-h3 (включая их фрагменты или производные), с лигандами, а также оно относится к набору для диагностики заболеваний почек, печени, ревматоидного артрита или сердечно-сосудистых заболеваний, содержащему белок β ig-h3 или рекомбинантные белки, соответствующие домену fas-1 белка β ig-h3 (включая их фрагменты или производные), и их лиганды.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
β ig-h3 представляет собой белок внеклеточного матрикса, который индуцируется под действием TGF-β в клетках разных видов, в том числе в клетках меланомы человека, эпителиальных клетках молочной железы, кератиноцитах и легочных фибробластах. TGF-β (трансформирующий фактор роста-β) участвует в процессах роста и дифференцировки во многих видах клеток, причем у млекопитающих обнаружено три вида TGF-β (TGF-β 1, TGF-β 2 и TGF-β 3). Известно, что TGF-β выполняет много сложных функций, таких как регуляция роста, регуляция иммунного ответа, стимуляция костеобразования, в том числе специфических макромолекул хрящевой ткани, стимуляция заживления ран и др. (Bennett, N. T. et al., Am. J. Surg. 1993,165, 728). TGF-β экспрессируется в эпителиальных клетках во время заживления ран, возможно, для стимуляции экспрессии интегрина в кератиноцитах в процессе регенерации эпителиальных клеток. Последние исследования по экспрессии TGF-β показали, что мРНК TGF-β 3 экспрессируется как в эпителии нормальной кожи, так и в эпителии, восстанавливающемся после острых или хронических повреждений, тогда как мРНК TGF-β 1 экспрессируется только в регенерированном эпителии после острых повреждений, а мРНК TGF-β 2 не экспрессируется совсем (Schmid, P. et al., J. Pathol., 1993, 171, 191). Хотя конкретная теория для механизма осуществления вышеописанных процессов еще не установлена, полагают, что TGF-β играет ключевую роль в регенерации эпителия.
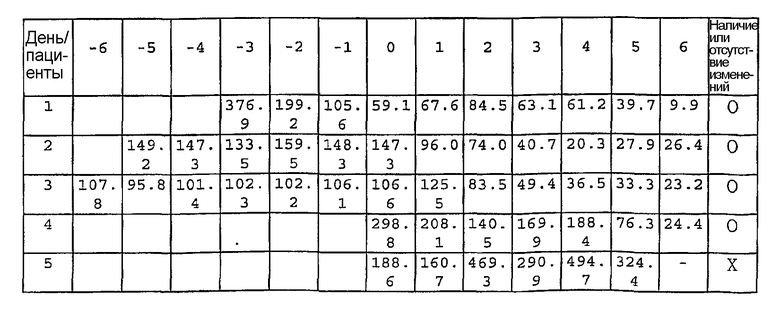
β ig-h3, индуцируемый TGF-β ген h3, впервые обнаружен Stonier et al. А именно, β ig-h3 обнаружен в результате анализа данных, полученных при дифференциальном скрининге библиотеки кДНК клеточной линии A549, клеточной линии аденокарциномы легких человека, обработанной TGF-β 1; показано, что через 2 дня после обработки TGF-β 1 уровень β ig-h3 увеличивается в 20 раз (Stonier, J. et al., DNA cell Biol., 1992, 11, 511). Секвенирование ДНК также показало, что β ig-h3 состоит из 683 аминокислот, представленных в SEQ ID NO: 1, имеет аминоконцевую секреторную последовательность и карбоксиконцевой фрагмент Arg-Gly-Asp (RGD), обеспечивающий лигандное распознавание лиганда некоторых интегринов.
β ig-h3 содержит 4 гомогенных внутренних повторяющихся домена наряду с мотивом RGD, который присутствует в мембранных или секреторных белках млекопитающих, насекомых, морских ежей, растений, дрожжей, бактерий и др. как высоко консервативная последовательность. Такие белки, как периостин, фасциклин I, HLC-2 морского ежа, водорослей CAM и MPB70 микобактерии, также содержат вышеуказанную консервативную последовательность (Kawamoto, T. et al.,Biochem. Biophys. Acta., 1998, 1395,288). Гомогенный домен (далее называемый "домен fas-1"), высоко консервативный в данных белках, состоит из 110-140 аминокислот и включает в себя две очень консервативные ветви (H1 и H2), каждая из которых состоит из 10 аминокислот. β ig-h3, периостин и фасциклин I имеют 4 домена fas-1, HCL-2 имеет 2, а MPB70 - только 1 домен fas-1. Известно, что некоторые из указанных белков, в качестве молекул клеточной адгезии, опосредуют присоединение и отсоединение клеток, хотя биологические функции данных белков до конца еще не выяснены. Например, β ig-h3, периостин и фасциклин I опосредуют присоединение фибробластов, остеобластов и нервных клеток, соответственно, а водорослей CAM, как установлено, является молекулой клеточной адгезии, постоянно присутствующей в зародышах вольвокса (LeBaron, R. G. et al., J. Invest. Dermatol., 104, 844, 1995; Horiuchi, K. et al., J. Bone Miner. Res., 1999, 14, 1239; Huber, O. et al., EMBO J., 1994, 13, 4212).
Очищенный белок β ig-h3 стимулирует адгезию и рост фибробластов кожи, но препятствует адгезии клеток A549, HeLa и WI-38 в среде, не содержащей сыворотки. В особенности β ig-h3 препятствует росту, колониеобразованию и появлению опухолевых клеток. Действительно, рост опухолевой клетки у мышей nude, полученной трансфицированием яйцеклеток китайского хомячка экспрессирующим вектором β ig-h3, значительно уменьшается, что четко установлено в патентах США №5714588 и №5599788. Кроме того, в данных патентах также приводится способ стимуляции роста и адгезии фибробластов в районе поврежденной области путем приведения раны в контакт с требующимся количеством β ig-h3. Следовательно, как молекула клеточной адгезии, в значительной степени индуцируемая TGF-β в клетках разных типов, β ig-h3 играет важную роль в процессах клеточного роста, клеточной дифференцировки, заживления ран, морфогенеза и клеточной адгезии.
Хотя β ig-h3 является эффективным полезным веществом, оно обеспечивается не полностью, так как в организме человека образуется только минимальное количество β ig-h3. Для решения данной проблемы был разработан способ получения β ig-h3 путем экспрессии его в системе эукариотической клетки с использованием методов генной инженерии. В данном случае, однако, клетки, продуцирующие β ig-h3, растут гораздо медленнее, чем другие клетки, что затрудняет получение достаточного количества клеток, продуцирующих β ig-h3. Следовательно, авторы настоящего изобретения разработали способ очистки, который позволяет осуществлять высокопродуктивную экспрессию рекомбинантных белков, содержащих целый белок β ig-h3 или какой-либо из его доменов, с использованием E. coli в качестве хозяина, подтвердили, что данные рекомбинантные белки поддерживают клеточную адгезию и клеточный рост, и подали заявку на патент (патентная заявка Кореи №2000-25664).
Активность β ig-h3 как молекулы клеточной адгезии впервые была описана для фибробластов кожи человека и затем раскрыта для хондроцитов, перитонеальных фибробластов, а также для человеческих фибробластов MRC5. Было предположено, что клеточно-адгезивная активность β ig-h3 опосредуется мотивом RGD, находящимся на карбоксильном конце β ig-h3 в первые дни. Однако позднее было опубликовано, что для стимуляции роста хондроцитов мотив RGD не требуется, а зрелый β ig-h3, в котором мотив RGD утрачивается в результате процессинга карбоксильного конца, может препятствовать клеточной адгезии. В результате было подтверждено, что мотив RGD не является необходимым медиатором для обеспечения клеточно-адгезивной активности β ig-h3. Последние исследования дополнительно подтвердили, что β ig-h3 стимулирует клеточную адгезию и рост, особенно рост фибробластов, независимо от интегрина α1β1, хотя присутствие мотива RGD в β ig-h3 не является обязательным для клеточного роста, опосредованного β ig-h3 (Ohno, S., et al., Biochim. Biophys. Acta, 1999, 1451, 196). Кроме того, было доказано, что пептиды H1 и H2, присутствующие в β ig-h3, не влияют на клеточную адгезию, опосредованную β ig-h3, это позволило предположить, что некоторые аминокислоты, необходимые для клеточной адгезии, расположены не в H1 и H2, а в других участках β ig-h3. Для подтверждения вышесказанного с помощью компьютера анализировали гомологию повторяющегося домена fas-1 белка β ig-h3 и доменов fas-1 других белков, в результате было показано, что в β ig-h3 существует много других консервативных аминокислот, кроме H1 и H2, которые участвуют в клеточной адгезии.
Поэтому авторы настоящего изобретения предприняли попытки обнаружить консервативный мотив, участвующий в активности, касающейся клеточной адгезии и отлипания клеток, и получить содержащий его пептид. В результате авторы настоящего изобретения получили пептиды NKDIL, EPDIM, а также их производные, которые, работая вместе с интегрином 3β 1, опосредуют клеточную адгезию и отлипание клеток, для получения пептидов использовали второй и четвертый домены β ig-h3, известного как молекула клеточной адгезии, а также они раскрыли, что две очень консервативные аминокислоты, аспарагиновая кислота (Asp) и изолейцин (Ile), расположенные вблизи участка H2 во втором и четвертом доменах β ig-h3, необходимы для проявления активности, касающейся клеточной адгезии и отлипания клеток, что позволило им подать заявку на патент (патентная заявка Кореи №2000-25665).
На сегодняшний день не существует публикаций, свидетельствующих о том, что β ig-h3 непосредственно связан с заболеваниями, однако, по-видимому, β ig-h3 связан с некоторыми видами рака человека. Взаимосвязь экспрессии β ig-h3 и развития заболеваний почек, печени, ревматоидного артрита и сердечно-сосудистых заболеваний еще не выяснена, возможность использовать белок β ig-h3 для диагноза заболеваний путем измерения количества белка β ig-h3 в жидкостях организма тоже еще не обсуждалась.
Таким образом, авторы настоящего изобретения разработали способ измерения количества β ig-h3 с использованием рекомбинантного белка, полученного путем соединения нескольких β ig-h3 или четвертого домена fas-1 белка β ig-h3, в качестве стандартного белка, а также содержащий его диагностический набор. Авторы настоящего изобретения завершили работу подтверждением того, что способ и набор по настоящему изобретению можно эффективно использовать как чувствительный способ диагностики степени поражения или развития заболеваний почек, печени, ревматоидного артрита и сердечно-сосудистых заболеваний.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Целью настоящего изобретения является предложение способа измерения количества белка β ig-h3 с использованием белка β ig-h3 или рекомбинантных белков, включающих в себя домены fas-1 белка β ig-h3, а также диагностического набора, содержащего упомянутые белки.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 приведена диаграмма, изображающая структуру рекомбинантного белка β ig- h3,
I, II, III и IV: каждый домен,
 и
и  консервативный участок основной последовательности
консервативный участок основной последовательности
A: β ig-h3; B: человеческий β ig-h3; C: мышиный β ig-h3
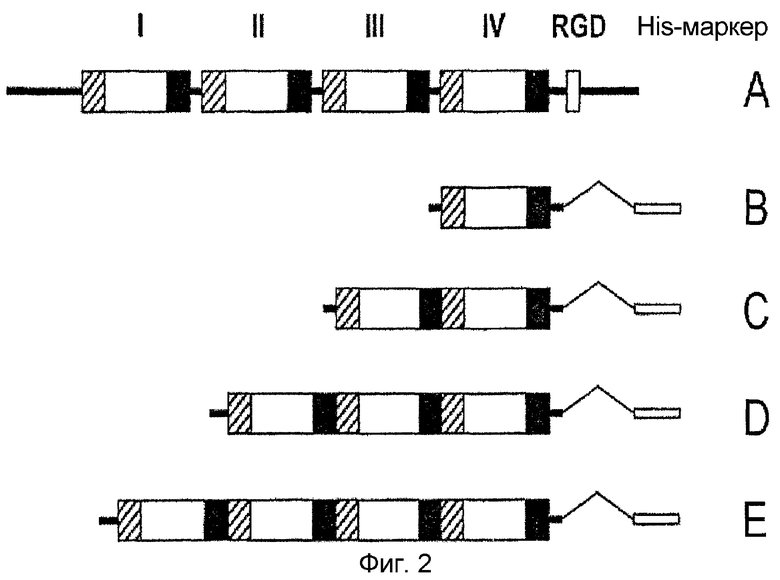
На фиг. 2 приведена диаграмма, изображающая геометрическую структуру рекомбинантных белков β ig-h3 D-IV, полученных путем повторения доменов IV белка β ig-h3,
A: β ig-h3;B:
β ig-h3 D-IV(1x);
C: β ig-h3 D-IV (2x);
D: β ig-h3 D-IV(3x);
E: β ig-h3 D-IV (4x)
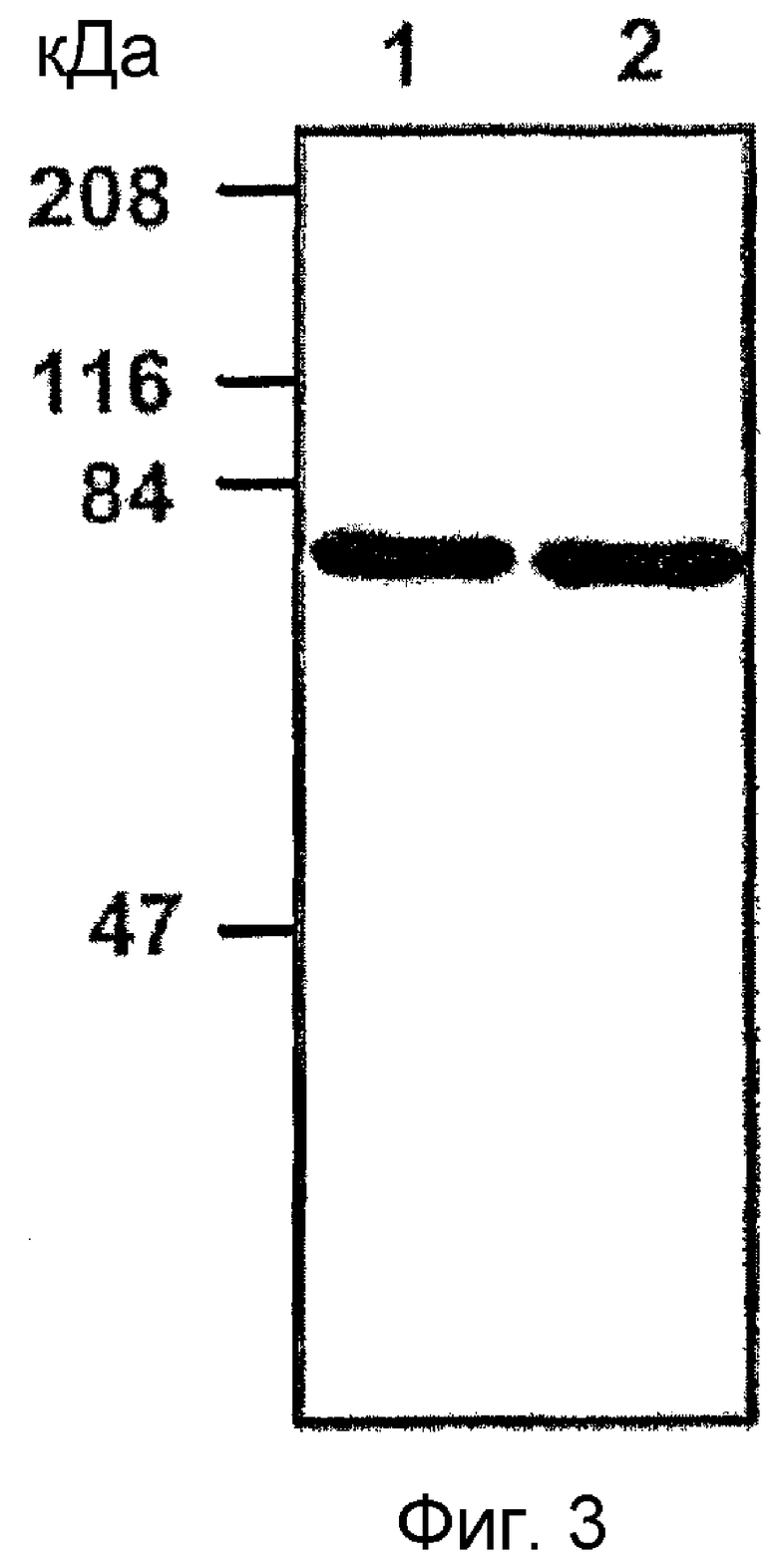
На фиг. 3 приведена фотография электрофореза выделенного рекомбинантного белка β ig-h3,
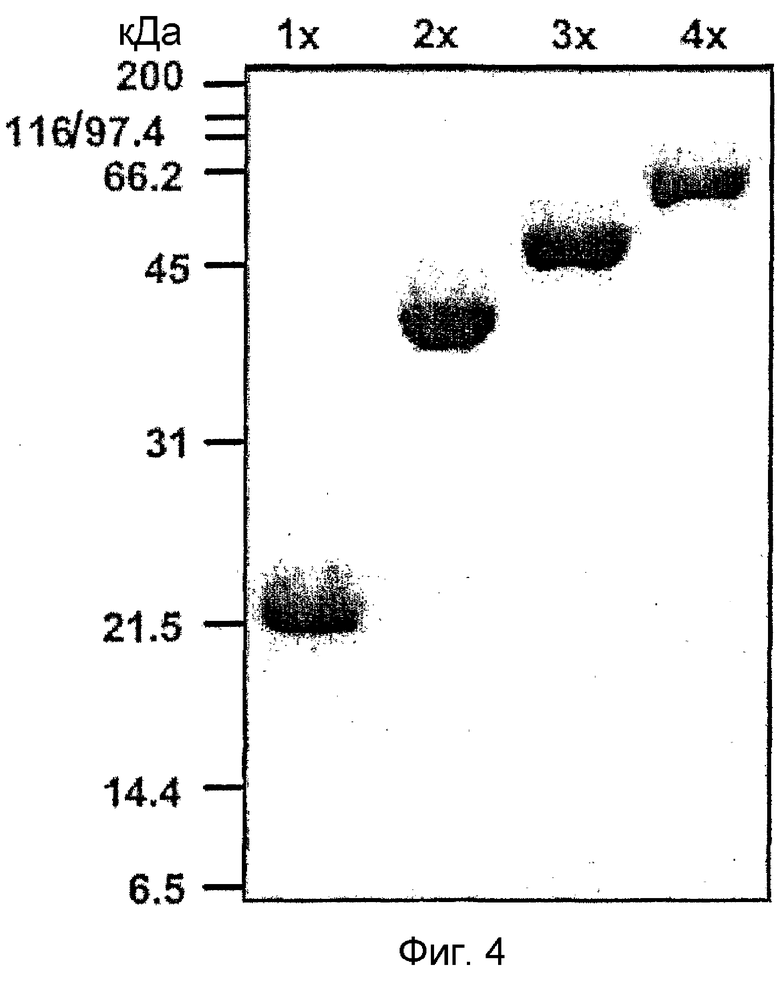
На фиг. 4 приведена фотография электрофореза белков β ig-h3 D-IV (1x, 2x, 3x, 4x),
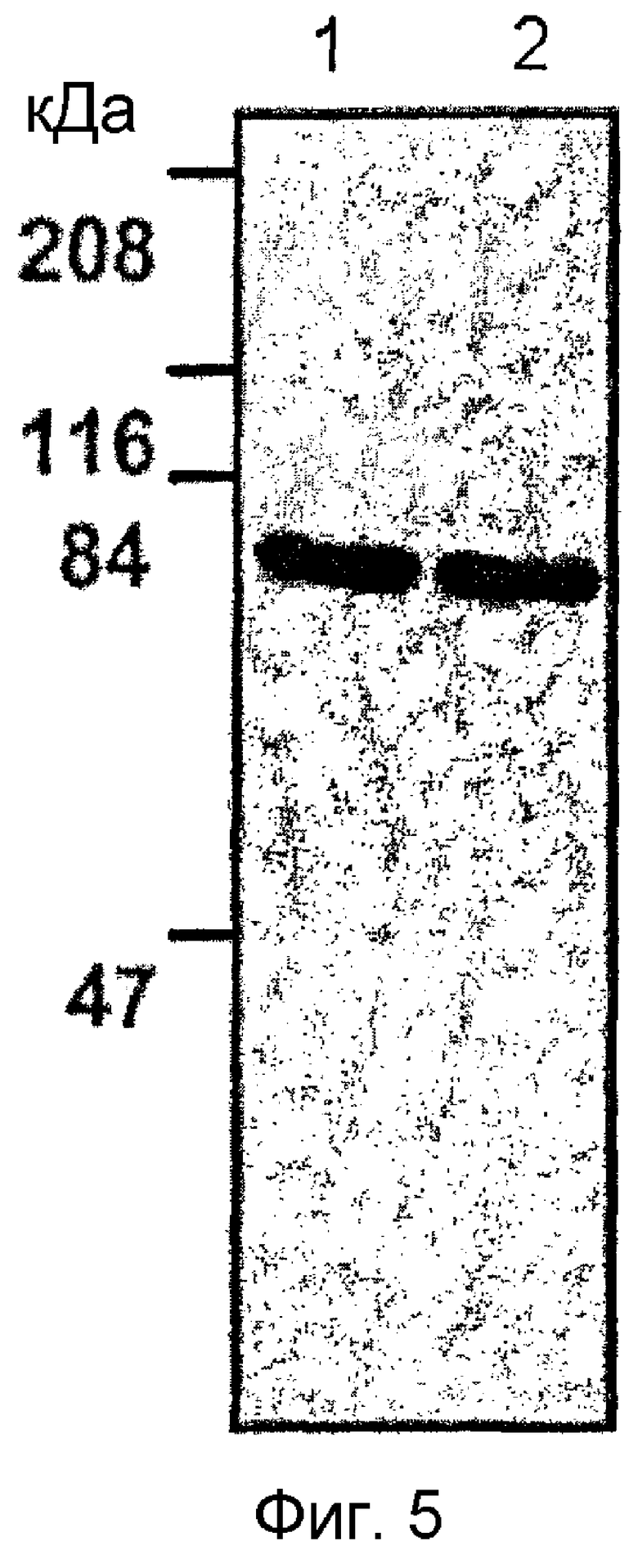
На фиг. 5 приведена фотография, показывающая результат вестерн-блоттинга с использованием первичных антител, с помощью которого идентифицируют человеческий β ig-h3 и мышиный β ig-h3,
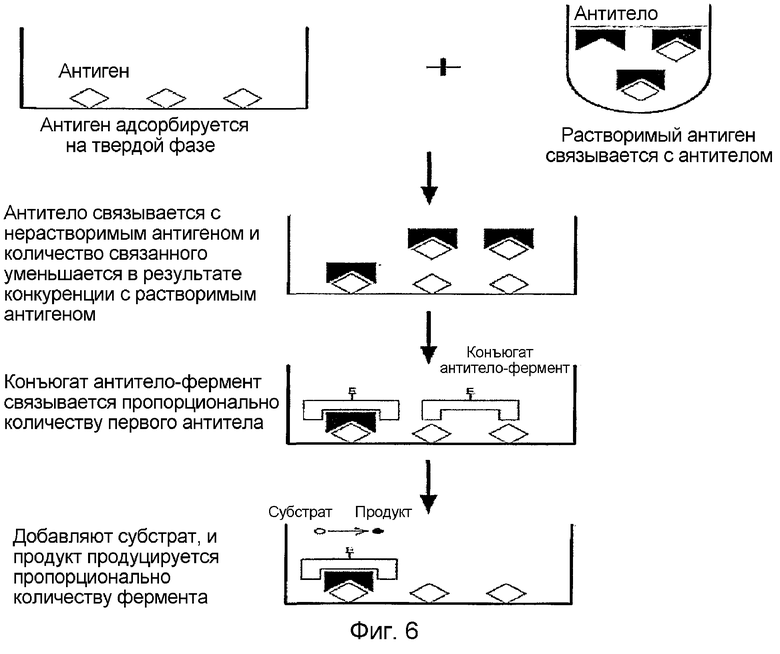
На фиг. 6 приведена диаграмма, демонстрирующая принцип твердофазного иммуноферментного анализа (ELISA)
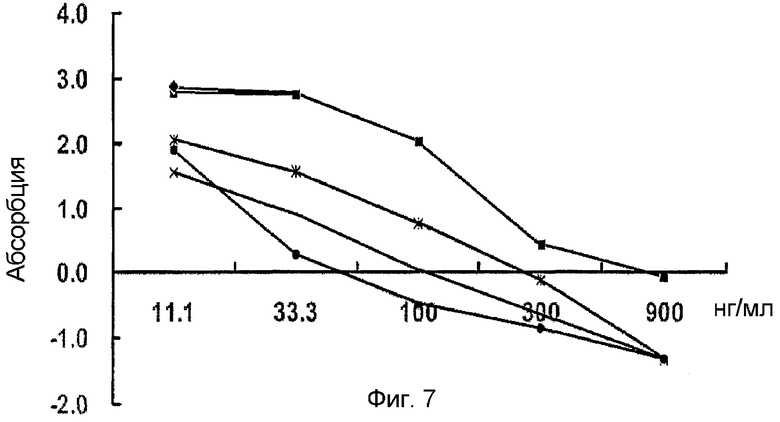
На фиг. 7 приведен график, демонстрирующий количественные соотношения первичных антител,
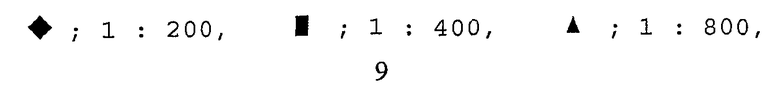
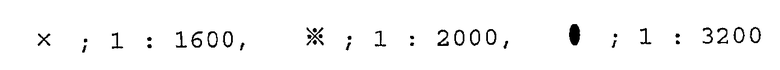
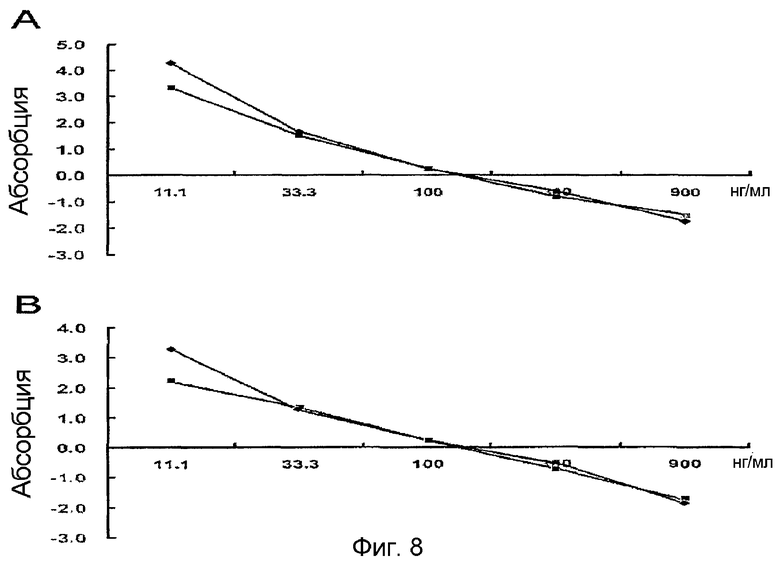
На фиг. 8 приведен график, демонстрирующий количественные соотношения вторичных антител,
A: фиксированные первичные антитела, 1:1600;
В: фиксированные первичные антитела, 1:2000;
 : разбавленные вторичные антитела, 1:1000;
: разбавленные вторичные антитела, 1:1000;
 : разбавленные вторичные антитела, 1:2000;
: разбавленные вторичные антитела, 1:2000;
 : разбавленные вторичные антитела, 1:3000
: разбавленные вторичные антитела, 1:3000
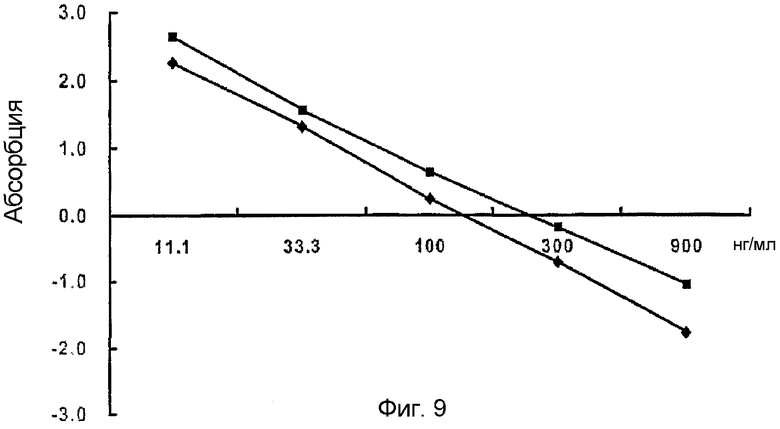
На фиг. 9 приведен график, демонстрирующий покрывающую концентрацию человеческого белка β ig-h3,
: 0,5 мкг/мл,: 1,0 мкг/мл
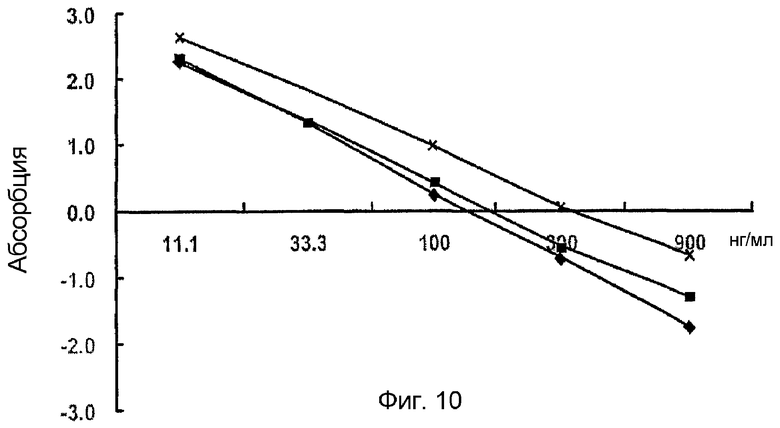
На фиг. 10 приведен график, демонстрирующий, что в качестве стандартного белка можно использовать и человеческий, и мышиный белок β ig-h3, что подтверждается перекрестным анализом,
: покрывающая концентрация человеческого белка β ig-h3 0,5 мкг/мл, первичные антитела против человеческого β ig-h3 1:2000, вторичные антитела 1:2000;
: покрывающая концентрация человеческого белка β ig-h3 0,5 мкг/мл, первичные антитела против мышиного β ig-h3 1:2000, вторичные антитела 1:2000;
 : покрывающая концентрация мышиного белка β ig-h3 0,5 мкг/мл, первичные антитела против человеческого β ig-h3 1:2000, вторичные антитела 1:2000;
: покрывающая концентрация мышиного белка β ig-h3 0,5 мкг/мл, первичные антитела против человеческого β ig-h3 1:2000, вторичные антитела 1:2000;
 : покрывающая концентрация мышиного белка β ig-h3 0,5 мкг/мл, первичные антитела против мышиного β ig-h3 1:2000, вторичные антитела 1:2000
: покрывающая концентрация мышиного белка β ig-h3 0,5 мкг/мл, первичные антитела против мышиного β ig-h3 1:2000, вторичные антитела 1:2000
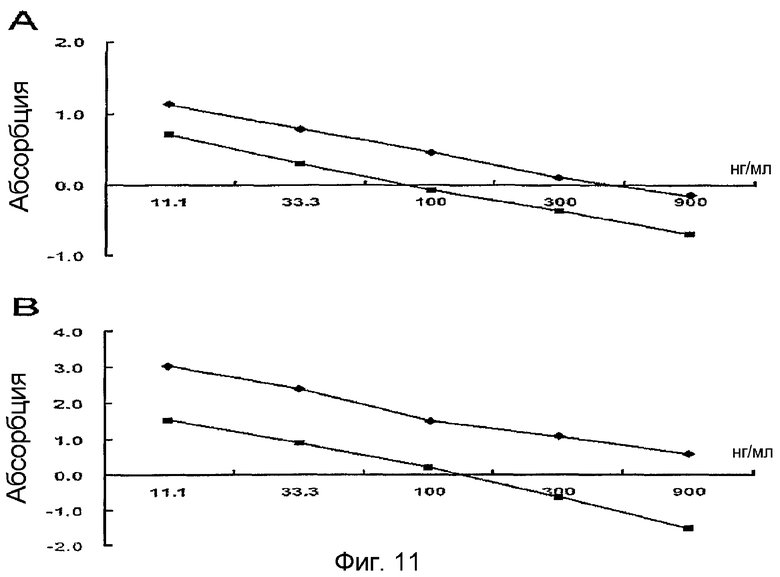
На фиг. 11 приведен график, демонстрирующий, что в качестве стандартного белка можно использовать рекомбинантный белок β ig-h3 D-IV(1x) и рекомбинантный белок β ig-h3 D-IV(4x), что подтверждается перекрестным анализом,
(А): покрывающая концентрация β ig-h3 D-IV(1x) 0,5 мкг/мл, первичные антитела против человеческого β ig-h3 1:2000, вторичные антитела 1:2000;
(А): покрывающая концентрация β ig-h3 D-IV(4x) 0,5 мкг/мл, первичные антитела против человеческого β ig-h3 1:2000, вторичные антитела 1:2000;
(В): покрывающая концентрация β ig-h3 D-IV(1x) 0,5 мкг/мл, первичные антитела против мышиного β ig-h3 1:2000, вторичные антитела 1:2000;
(В): покрывающая концентрация β ig-h3 D-IV(4x) 0,5 мкг/мл, первичные антитела против мышиного β ig-h3 1:2000, вторичные антитела 1:2000
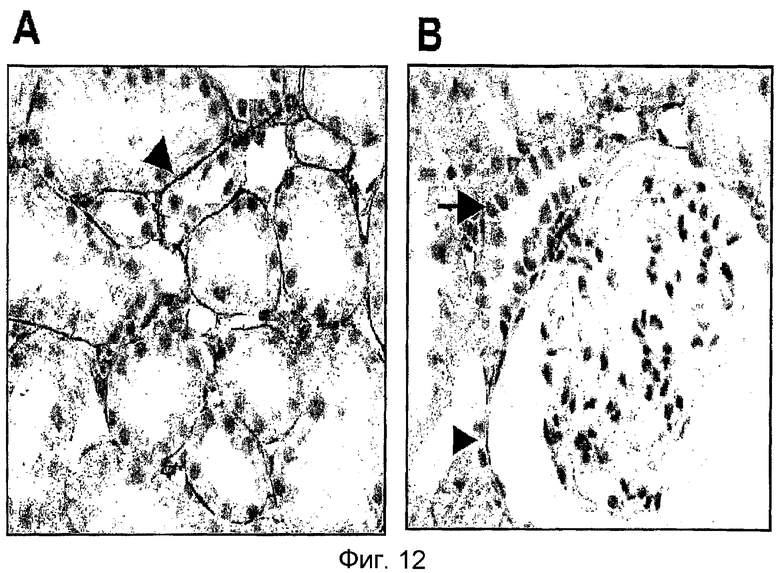
На фиг. 12 приведена фотография иммуногистохимического окрашивания, демонстрирующая характер экспрессии β ig-h3 в почечной ткани,
 (А): характер экспрессии на базальной мембране проксимальной канальцевой клетки S3;
(А): характер экспрессии на базальной мембране проксимальной канальцевой клетки S3;
(B); характер экспрессии на базальной мембране боуменовой капсулы клубочка;
→ (B); характер экспрессии на базальной мембране корковой толстой восходящей сегментной клетки
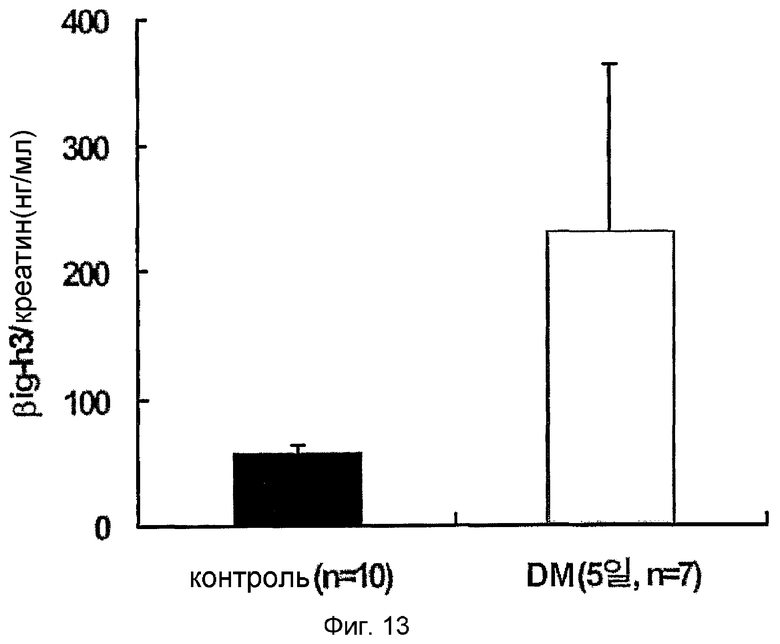
На фиг. 13 приведен график, демонстрирующий уровни β ig-h3 в моче крыс с индуцированным диабетом,
: контрольная группа;
 : крысы с диабетом, индуцированным стрептозотоцином
: крысы с диабетом, индуцированным стрептозотоцином
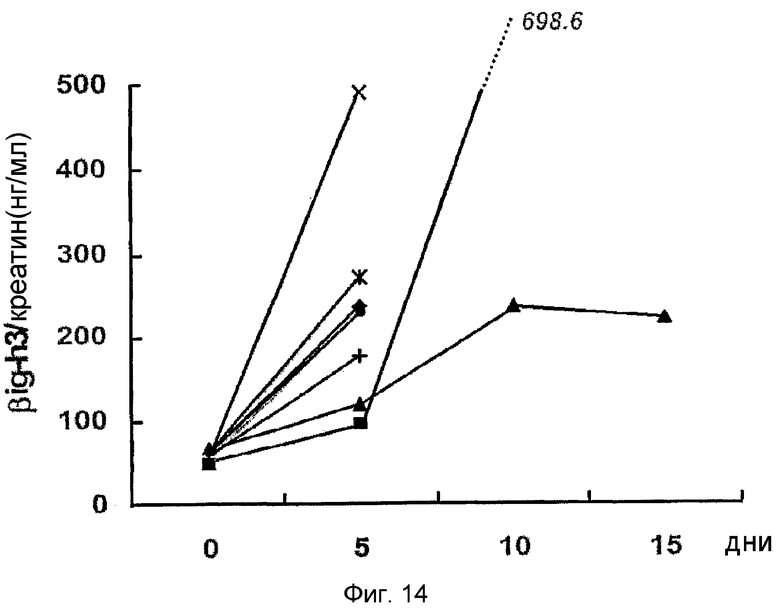
На фиг. 14 приведен график, демонстрирующий индивидуальный уровень β ig-h3 в моче крыс с индуцированным диабетом фиг. 13
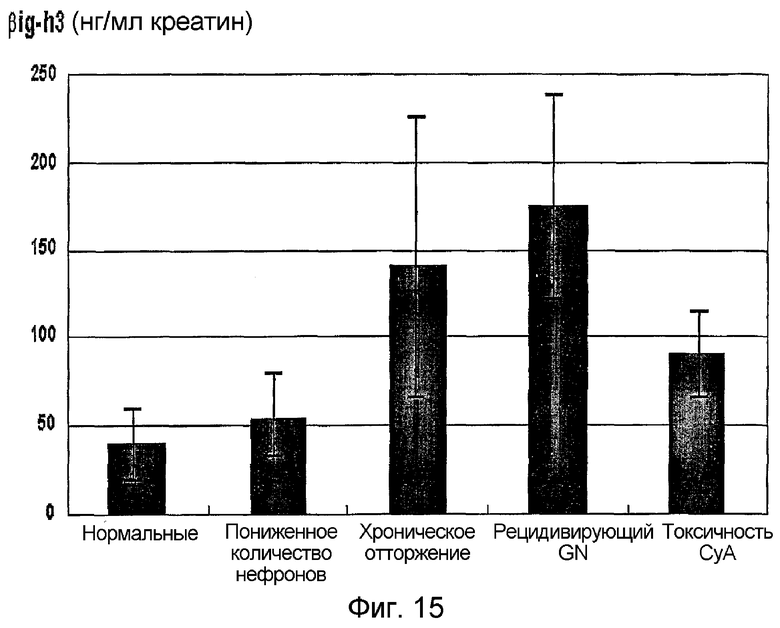
На фиг. 15 приведен график, демонстрирующий уровень β ig-h3 в моче, полученной от каждой нормальной крысы, крысы с пониженным количеством нефронов, крысы с хроническим отторжением, крысы с рецидивирующим GN и крысы с токсичностью CyA
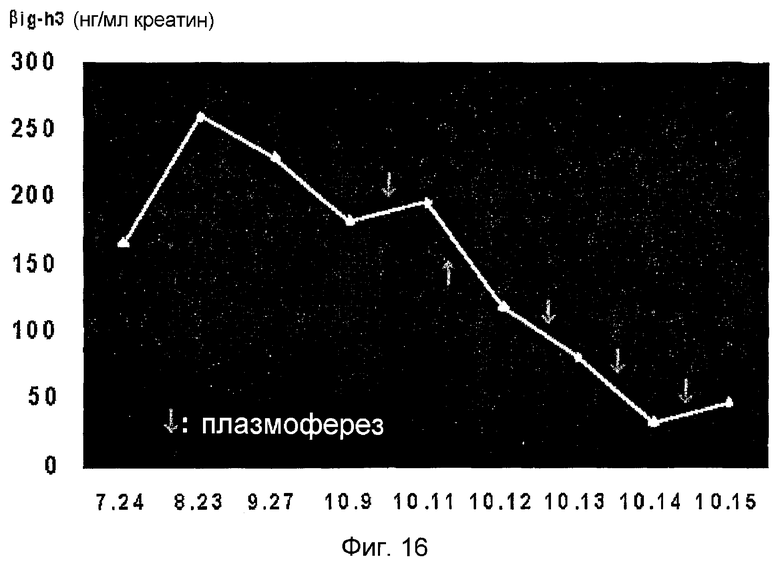
На фиг. 16 приведен график, демонстрирующий различные концентрации белка β ig-h3 посуточно, которые измеряют, используя образцы мочи пациентов, которые подвергаются плазмоферезу в то время как очаговый сегментарный гломерулосклероз (FSGS) повторно развивается после трансплантации почки,
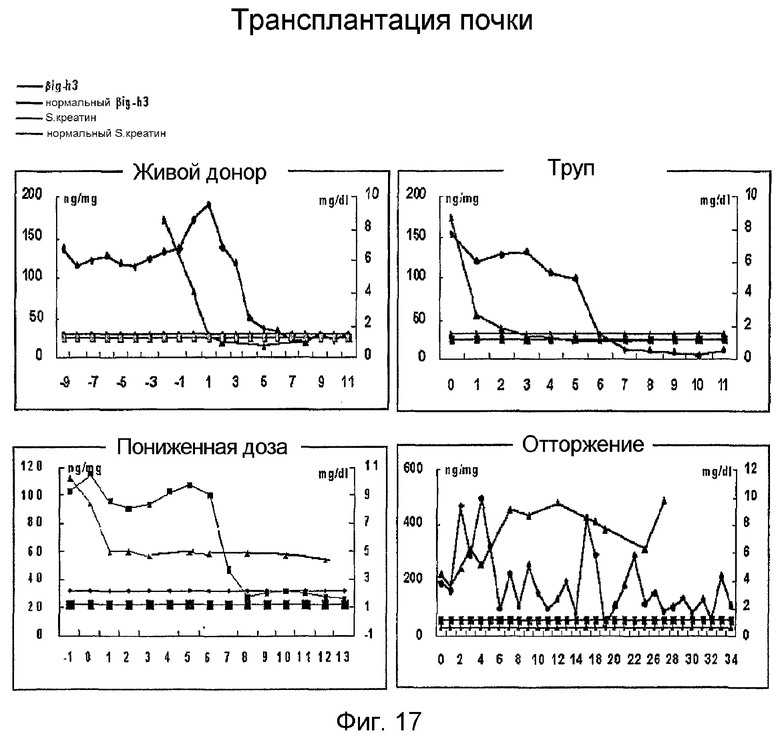
На фиг. 17 приведен график, демонстрирующий концентрации белка β ig-h3 в моче, взятой от живого донора, от трупа, от пациента с недостаточностью и отторжением, которые измеряли до и после трансплантации почки
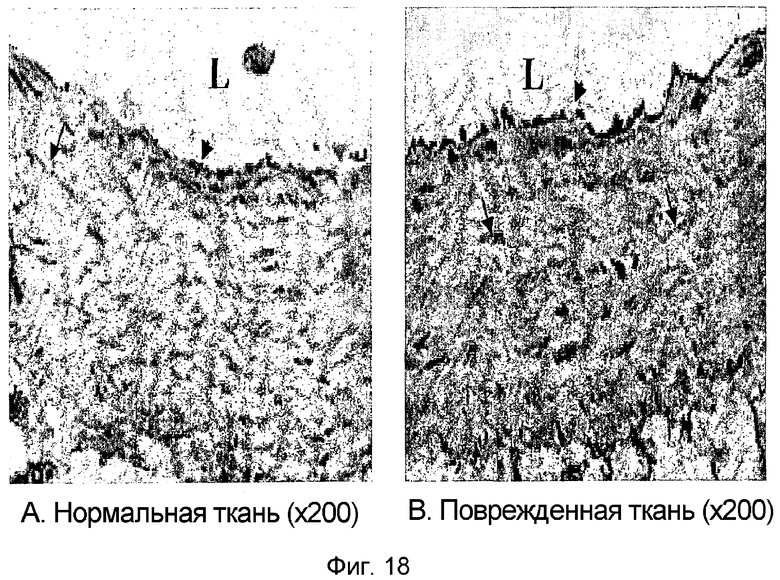
На фиг. 18 приведена фотография иммуногистохимического окрашивания, демонстрирующая характер экспрессии белка β ig-h3 в поврежденных кровеносных сосудах мыши с индуцированным диабетом,
A: нормальные кровеносные сосуды;
B: поврежденные кровеносные сосуды,
L: просвет
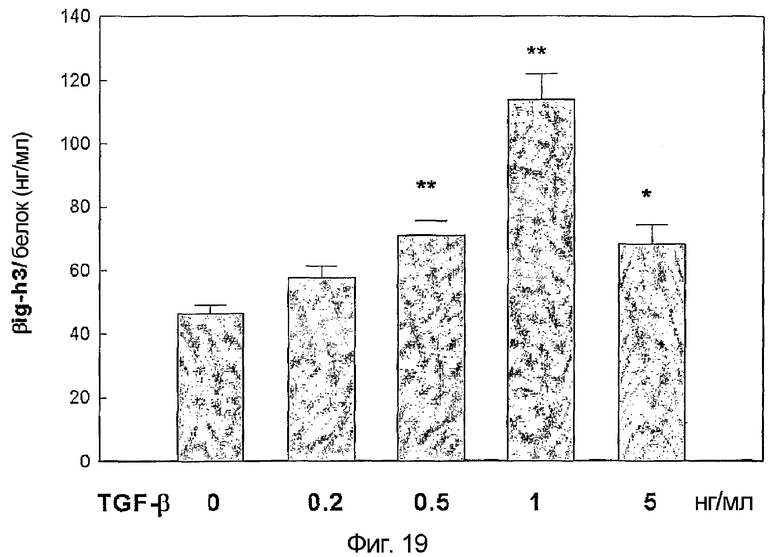
На фиг. 19 приведен график, демонстрирующий характер экспрессии белка β ig-h3 в культуре клеток гладкой мускулатуры сосудов,
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Для достижения вышеуказанной цели настоящее изобретение относится к способу измерения количества белка β ig-h3.
Настоящее изобретение также относится к набору для диагностики заболеваний почек, печени, ревматоидного артрита или сердечно-сосудистых заболеваний, содержащему упомянутый белок.
Другие признаки настоящего изобретения будут описаны ниже.
Способ измерения количества β ig-h3 согласно настоящему изобретению включает в себя следующие стадии:
1) Получение белка β ig-h3 или рекомбинантных белков, содержащих домен fas-1 белка β ig-h3, их фрагментов или производных;
2) Получение специфических лигандов для вышеуказанных рекомбинантных белков, их фрагментов или производных, полученных на описанной выше стадии 1; и
3) Измерение количества белка β ig-h3 в образцах по способу, в котором используется реакция связывания лигандов, полученных на описанной выше стадии 2, с рекомбинантными белками, их фрагментами или производными, полученными на описанной выше стадии 1.
На стадии 1 белком β ig-h3 является либо человеческий β ig-h3, имеющий аминокислотную последовательность, представленную SEQ ID NO: 3, либо мышиный β ig-h3, имеющий аминокислотную последовательность, представленную SEQ ID NO: 5. Структурные элементы человеческого и мышиного белков β ig-h3 приведены на фиг. 1. Заштрихованный участок и заштрихованный накрест участок на фиг. 1 обозначают высоко консервативные последовательности повторяющегося домена fas-1 I, II, III и IV, а пустой участок обозначает мотив RGD.
Белок β ig-h3 имеет 4 домена fas-1. Для домена fas-1 β ig-h3 вышеприведенной стадии 1, предпочтительно выбрать один или более двух из четырех доменов fas-1 белка β ig-h3 и более предпочтительно использовать 4ый домен fas-1. 4ый Домен fas-1 можно использовать либо отдельно, либо в виде рекомбинантного белка, в котором повторяются несколько доменов fas-1, соединенных между собой. Для получения рекомбинантного белка нужно соединить 1-10 доменов fas-1, причем более предпочтительно использовать от 1 до 4 доменов fas-1. В предпочтительных воплощениях настоящего изобретения авторы предлагают примеры использования только 4ого домена fas-1, и, соответственно, рекомбинантные белки, полученные соединением 2, 3 и 4 четвертых доменов fas-1 белка β ig-h3.
Авторы настоящего изобретения получили белки, представленные последовательностями SEQ ID NO:7, NO:8, NO:9 и NO:10, содержащие 4ый домен fas-1, который состоит из 502ой-632ой аминокислот белка β ig-h3 и повторяется соответственно один, два, три или четыре раза, и назвали их "β ig-h3 D-IV(1x)", "β ig-h3 D-IV(2x)", "β ig-h3 D-IV(3x)" и "β ig-h3 D-IV(4x)" (см. фиг. 4).
В качестве фрагментов рекомбинантного белка можно использовать эпитоп белка β ig-h3, в котором происходит реакция специфического связывания с лигандом, а также любую другую часть белка, содержащую пептиды, гидролизующиеся под действием протеазы. Производные рекомбинантного белка могут быть получены в результате образования ковалентной связи, например, путем фосфорилирования и гликозилирования, а также нековалентной связи, в том числе ионной связи, координационной связи, водородной связи или связи ван дер Ваальса. Если фрагменты производных вышеуказанных рекомбинантных белков могут специфически связываться с лигандами, их можно отнести к категории белков по настоящему изобретению.
Для получения стандартного белка по настоящему изобретению конструирование экспрессирующего вектора и трансформацию можно проводить традиционным методом.
На стадии 2 специфичность связывания лигандов с β ig-h3, доменом fas-1 белка β ig-h3, их фрагментами или производными можно подтвердить путем наблюдения реакции связывания лигандов с белком или рекомбинантным белком стадии 1. Существует много видов лигандов, например, антитело, РНК, ДНК, органические соединения, включающие в себя липид, белок или органические соли, или неорганические соединения, включающие в себя ионы металлов или неорганические соли, а предпочтительный лиганд представляет собой первичное антитело против β ig-h3 или домена fas-1 белка β ig-h3 стадии 2, полученное с использованием в качестве антигена белка или рекомбинантного белка (в том числе фрагментов или производных) стадии 1. Первичное антитело может быть получено традиционным методом, причем можно использовать моноклональное антитело или поликлональное антитело.
На стадии 3 количество белка β ig-h3, входящего в состав образца, измеряют с помощью реакции специфического связывания лиганда с белком β ig-h3, его фрагментами или производными. Если происходит реакция связывания с лигандом, можно использовать даже кусочки данных фрагментов или производных. Предпочтительно используют количественный анализ посредством реакции связывания антиген-антитело, в которой в качестве антигена используют белок β ig-h3. Более предпочтительно, выбирают один метод из группы, состоящей из иммуноблотинга (Current Protocols in Molecular Biology, vol 2, chapter 10.8; David et al., Cells (a Laboratory manual), vol 1, chapter 73), иммунопреципитации (Current Protocols in Molecular Biology, vol 2, chapter 10.16; Cells (a Laboratory manual), vol 1, chapter 72), ELISA (Current Protocols in Molecular Biology, vol 2, chapter 11.2; ELISA Theory and Practice, John R. Crowther; The ELISA Guidebook, John R. Crowther), RIA (радиоиммунного анализа) (Nuklearmedizin 1986 Aug; 25 (4): 125-127, Tumor markers as target substances in theradioimmunologic detection of malignancies. von Kleist S; Mariani G. Ann Oncol 1999; 10 Suppl 4: 37-40), метода белковых чипов (Daniel Figeys et. al,Electrophoresis 2001, 22, 208-216; Albala JS. Expert Rev Mol Diagn 2001 Jul; 1 (2): 145-152), экспресс-анализа (Kasahara Y and Ashihara Y, Clinica Chimica Acta 267 (1997), 87-102; патентная заявка Кореи №2000-46639) или микрочипов (Vivian G. cheung et al, Nature genetics 1999, 21, 15-19; Robert J. Lipshutz et al, Nature genetics 1999,21, 20-24; Christine Debouck and Peter N. Goodfellow, Nature genetics 1999, 21, 48-50; DNA Microarrays, M. Schena), причем наиболее предпочтительным методом является ELISA. Массовый анализ образцов также можно проводить, используя биологический микрочип и автоматическую систему микрочипов, наряду с ELISA, на основе которых может быть разработан простой метод самодиагностики с использованием мочи.
В соответствии с предпочтительными воплощениями настоящего изобретения, способ измерения количества белка β ig-h3 путем конкурентного анализа с использованием ELISA включает в себя следующие стадии:
1) Нанесение белка β ig-h3 или рекомбинантного белка, содержащего домен fas-1 β ig-h3, его фрагментов или производных, на основу;
2) Взаимодействие антитела против белка вышеописанной стадии 1, его фрагментов или производных с образцом;
3) Добавление реагента вышеописанной стадии 2 к покрывающему белку стадии 1, проведение реакции и затем промывание; и
4) Добавление вторичного антитела к реагенту вышеописанной стадии 3, проведение следующей реакции и измерение ОП.
В качестве основы на вышеописанной стадии 1 можно использовать все виды традиционно используемых основ, в особенности, нитроцеллюлозную мембрану, поливиниловый планшет (например, 96-луночный планшет), планшет из полистирола и стеклянную пластинку.
Вторичное антитело вышеописанной стадии 4 метят окрашивающими ферментами, флуоресцентными веществами, светящимися веществами, радиоизотопами или хелатами металлов. Все традиционно используемые для мечения вещества пригодны для данного изобретения, а предпочтительными окрашивающими ферментами являются пероксидаза, щелочная фосфатаза, β-D-глактозидаза, малатдегидрогеназа, нуклеаза стафилококка, пероксидаза хрена, каталаза и ацетилхолинэстераза. В качестве флуоресцентных веществ предпочтительно используют флуоресцеинизотиоцианат, фикобилипротеин, родамин, фикоэритрин, фикоцианин, ортофталевый альдегид и др.
Кроме окрашивающих ферментов или флуоресцентных веществ для мечения вторичных антител предпочтительно используют светящиеся вещества, такие как изолюминол, люцигенин, люминол, сложный эфир акридиновой кислоты, имидазол, соль акридиновой кислоты, люциферин, люцифераза и экворин, или радиоизотопы, такие как 125I, 127I, 131I, 14C, 3H, 32P и 35S. Кроме того, антитело может конъюгировать с микромолекулярными гаптенами, такими как биотин, динитрофенил, пиридоксил или флуорескамин.
В случае применения на стадии 4 окрашивающих ферментов, для измерения активности фермента следует использовать окрашивающие субстраты, в качестве которых можно использовать любое вещество, способствующее развитию окраски фермента, связанного со вторичным антителом. В качестве окрашивающих субстратов предпочтительно используют 4-хлор-1-нафтол (4CN), диаминобензидин (DAB), аминоэтилкарбазол (AEC), 2,2'-азино-бис(3-этилбензотиазолин-6-сульфоновую кислоту) (ABTS), o-фнилендиамин (OPD) и тетраметилбензидин (TMB).
В качестве образцов вышеописанной стадии 2 можно использовать все виды жидкостей организма пациентов, страдающих от заболеваний, связанных с β ig-h3. В особенности, предпочтительными являются моча, кровь или синовиальная жидкость пациентов, страдающих от заболеваний почек, печени, ревматоидного артрита или сердечно-сосудистых заболеваний.
Для того чтобы подтвердить, является ли способ измерения количества белка β ig-h3 по настоящему изобретению корректным, авторы в качестве стандартного белка использовали рекомбинантный белок, содержащий мышиный β ig-h3 или 4ый домен fas-1 белка β ig-h3, и сравнивали результат с полученным при использовании человеческого β ig-h3 в качестве стандартного белка.
Для способа измерения β ig-h3 по настоящему изобретению определяли оптимальную покрывающую концентрацию человеческого белка β ig-h3 и количественное соотношение антител. Наилучшим количественным соотношением первичных антител против человеческого β ig-h3 являлось 1:1600 и 1:2000 (см. фиг. 7), а наилучшим количественным соотношением вторичных антител являлось 1:2000 (см. фиг. 8). Соответствующая концентрация белка β ig-h3 составляла 1,0 мкг/мл и 0,5 мкг/мл, но более предпочтительной покрывающей концентрацией являлась 0,5 мкг/мл (см. фиг. 9).
Следовательно, авторы настоящего изобретения за оптимальную покрывающую концентрацию человеческого β ig-h3 приняли 0,5 мкг/мл, наилучшей степенью разбавления для первичных антител против человеческого β ig-h3 и для вторичных антител считали 1:2000, соответственно.
Авторы настоящего изобретения также определили концентрацию белка и количественное соотношение первичных и вторичных антител, используя мышиный β ig-h3, рекомбинантный β ig-h3 D-IV(1x), β ig-h3 D-IV(2x), β ig-h3 DIV(3x) и β ig-h3 D-IV(4x). А именно, получили покрывающую концентрацию каждого белка 0,5 мкг/мл, разбавили первичные антитела против человеческого β ig-h3 и вторичные антитела до соотношения 1:2000 соответственно и провели количественный анализ. Первичные антитела против мышиного β ig-h3 и вторичные антитела разбавили до соотношения 1:2000 и также провели количественный анализ.
В результате во всех случаях были получены графики с прямолинейной зависимостью, это говорит о том, что соотношения являются наилучшими, а интервал измерения составлял 11 нг/мл - 900 нг/мл для всех случаев, это означает, что не существует большой разницы в интервалах измерения (см. фиг. 11 и фиг. 12).
Приведенные выше результаты подтверждают, что стандартным белком может быть любой из человеческого β ig-h3, мышиного β ig-h3, рекомбинантного β ig-h3 D-IV(1x), ig-h3 D-IV(2x), ig-h3 D-IV(3x) и ig-h3 D-IV(4x), а в качестве первичных антител можно использовать либо антитела против человеческого β ig-h3, либо антитела против мышиного β ig-h3.
В данном изобретении предпочтительная покрывающая концентрация стандартного белка составляет 0,1-2,0 мкг/мл, более предпочтительно 0,5-1,0 мкг/мл. Предпочтительный индекс разведения первичных и вторичных антител находится в интервале 1:400-1:3200, более предпочтительно он составляет 1:2000.
Настоящее изобретение относится к набору для диагностики заболеваний почек, печени, ревматоидного артрита или сердечно-сосудистых заболеваний, который позволяет диагностировать заболевания путем измерения количества белка β ig- h3 в жидкостях организма пациентов.
Диагностический набор по настоящему изобретению включает в себя белок β ig-h3 или рекомбинантные белки, содержащие домен fas-1 белка β ig-h3 (включая их фрагменты или их производные), и их лиганды. В данное время в качестве предпочтительных специфических лигандов используют антитела против белка β ig-h3 или доменов fas-1 белка β ig-h3. Данный набор может дополнительно включать буферный раствор, вторичное антитело, раствор для промывания или окрашивающий субстрат.
Диагностический набор по настоящему изобретению подходит для диагноза различных заболеваний, таких как заболевания почек, печени, ревматоидный артрит или сердечно-сосудистые заболевания, путем измерения количества белка β ig-h3 в жидкостях организма.
Заболевания почек можно диагностировать путем измерения количества белка β ig-h3 на основе того факта, что экспрессия β ig-h3 индуцируется под действием TGF-β, который играет важную роль в развитии почечных заболеваний. Для подтверждения вышесказанного измеряли количество β ig-h3 в моче больных диабетом. Полученное в результате количество β ig-h3 в моче пациентов с диабетическими почечными заболеваниями, включая микроальбуминурию, было в пять раз выше, чем у нормальных субъектов. У некоторых больных диабетом, не имеющих почечных заболеваний, количество β ig-h3 также выше, чем у нормальных субъектов. Учитывая вышеприведенные результаты, можно сделать вывод, что уровень β ig-h3 в моче отражает степень поражения почек, а высокий уровень β ig-h3 у некоторых больных диабетом, не имеющих заболеваний почек, позволяет предположить, что их почки уже поражены в некоторой степени, хотя клинические симптомы еще отсутствуют. Следовательно, измерение количества β ig-h3 в моче пациентов является высокочувствительным и важным диагностическим методом, который может отражать поражение почек на ранней стадии.
Для того чтобы подтвердить, действительно ли концентрация β ig-h3 в моче больного диабетом может отражать поражение почки на ранней стадии, измеряли концентрацию β ig-h3 у животного, страдающего диабетом. В результате концентрация β ig-h3 увеличилась в 4 раза через 5 дней после индуцирования диабета (см. фиг. 13). Изменение концентрации β ig-h3 наблюдали у каждого субъекта после индуцирования диабета, приводящему к сильному увеличению концентрации β ig-h3 в моче (см. фиг. 14). На 5 день после индуцирования диабета анализы крови, мочи и креатина были нормальными, и ткани почек казались нормальными. Таким образом, сильное увеличение количества β ig-h3 в моче позволяет предположить, что уже существует минимальное поражение почек, которое не определяется традиционными методами.
Кроме того, авторы настоящего изобретения подтвердили взаимосвязь поражения почек и концентрации β ig-h3 путем измерения количества β ig-h3 в моче у пациентов, подвергавшихся трансплантации почки, до и после операции. Высокая концентрация β ig-h3, наблюдающаяся у пациента до операции, постепенно уменьшалась после успешной операции. Но у пациента № 5, у которого функция почки не восстанавливалась даже после операции, концентрация β ig-h3 оставалась высокой (см. фиг. 2). Все вышеприведенные результаты подтверждают, что концентрация β ig-h3 чувствительно отражает степень поражения почек.
Авторы настоящего изобретения также измерили концентрацию β ig-h3 в моче пациентов с почечной недостаточностью. Концентрация β ig-h3 в моче у всех пациентов с почечной недостаточностью была высокой. Таким образом, опять было подтверждено, что количество β ig-h3 в моче чувствительно отражает поражение почки даже на ранней стадии, следовательно, измерение количества β ig-h3 является очень важным методом диагностики различных заболеваний почек (см. таблицу 3).
Очень важно определить, развивается ли у пациента хронический гепатит в цирроз печени, однако пока не существует метода, который позволяет это сделать. Самым критическим фактором для развития цирроза печени является TGF-β. Следовательно, при развитии цирроза печени уровень β ig-h3, экспрессия которого индуцируется TGF-β, в крови, возможно, будет увеличиваться. В таком случае, количество β ig-h3 также может отражать степень поражения циррозом печени. Фактически доказано, что экспрессия β ig-h3 возрастает при увеличении тяжести цирроза печени, которую устанавливают с помощью иммуногистологического анализа тканей печени пациентов с гепатитом. Авторы настоящего изобретения подразделили состояние пациентов на несколько степеней и стадий на основе результатов биопсии, полученных для пациентов с хроническим гепатитом, и определили концентрацию β ig-h3 в крови для каждой стадии и степени. У пациентов с хроническим гепатитом концентрация β ig-h3 в крови выше, чем у нормальных людей. Доказано, что концентрация β ig-h3 на ранней стадии и низшей степени выше, чем на более поздней стадии и высшей степени (см. таблицу 5). Степень 3 и стадия 3 характеризуются тяжелым состоянием цирроза печени, перешедшего пик в своем развитии. Между тем, степени 1 и 2 и стадии 1 и 2 соответствуют состоянию пациента, при котором очень интенсивно развивается воспалительный процесс. Следовательно, концентрация β ig-h3 отражает интенсивность цирроза печени, так что его развитие можно отслеживать, регулярно измеряя концентрацию β ig-h3 в крови.
Также измеряли концентрацию β ig-h3 в синовиальной жидкости пациентов с ревматоидным артритом и остеоартритом. Концентрация β ig-h3 в синовиальной жидкости пациентов с ревматоидным артритом была в два раза выше, следовательно, измерение концентрации β ig-h3 в синовиальной жидкости может служить эффективным способом диагностики остеоартрита и ревматоидного артрита (см. таблицу 6).
Кроме того, для подтверждения взаимосвязи экспрессии β ig-h3 и сосудистых заболеваний исследовали характер экспрессии β ig-h3 в нормальных и поврежденных кровеносных сосудах диабетических мышей с помощью иммуногистохимических методов. Результаты показали, что белок β ig-h3 в поврежденных кровеносных сосудах диабетических мышей экспрессируется гораздо интенсивнее, чем в нормальных кровеносных сосудах (см. фиг. 18). Поскольку TGF-β, который играет важную роль в развитии сосудистых заболеваний, индуцирует экспрессию β ig-h3, исследовали индуцирование экспрессии β ig-h3 под действием TGF-β 1 в клетках гладкой мускулатуры сосудов, образующих кровеносные сосуды. Полученные результаты подтвердили, что экспрессия β ig-h3 увеличивается по мере увеличения количества TGF-β 1 (см. фиг. 19).
Экспрессия β ig-h3 в крови и тканях отражает их поражение. Следовательно, было доказано, что способ измерения количества белка β ig-h3 настоящего изобретения можно эффективно использовать для диагностики различных сосудистых заболеваний.
Таким образом, диагностический набор для измерения количества белка β ig-h3 по настоящему изобретению может применяться с высокой эффективностью, так как он позволяет установить степень поражения и развития заболеваний почек, печени, ревматоидного артрита или сердечно-сосудистых заболеваний.
ПРИМЕРЫ
Практические и в настоящий момент предпочтительные воплощения данного изобретения являются иллюстративными, как показано в нижеследующих примерах.
Однако следует понимать, что специалисты в данной области, исходя из приведенного описания, могут осуществлять модификации и улучшения в пределах сущности и объема настоящего изобретения.
Пример 1: Получение стандартных белков и первичных антител
<1-1> Выделение человеческого β ig-h3 и мышиного β ig-h3
Авторы настоящего изобретения получили человеческий и мышиный белки β ig-h3. Структурные элементы человеческого и мышиного белков β ig-h3 приведены на фиг. 1. Заштрихованный участок и заштрихованный накрест участок на фиг. 1 обозначают высоко консервативные последовательности повторяющегося домена fas-1 I, II, III и IV, а пустой участок обозначает мотив RGD.
кДНК β ig-h3 (pBS β ig-h3; полученную путем клонирования кДНК клеток папилломы кожи человека), имеющую базовую последовательность, представленную SEQ ID NO: 2, клонированную в векторе pBluescript SK (-), расщепляют с помощью Nde I и Bgl II, получая фрагменты ДНК с тупыми концами. Вышеуказанные фрагменты ДНК субклонируют в сайтах EcoR V и EcoR I вектора pET-29 (полученного от Novagen). Выделяют белок, имеющий аминокислотную последовательность из 69-653 аминокислот β ig-h3, представленную SEQ ID NO: 3, и называют его человеческий β ig-h3.
Затем кДНК β ig-h3 расщепляют с помощью BamH I и Xho I, получая фрагменты ДНК, имеющие базовую последовательность, представленную SEQ ID NO: 4. Вышеуказанные фрагменты ДНК субклонируют в сайтах BamH I и Xho I вектора pET-29. Выделяют белок, имеющий аминокислотную последовательность из 23-641 аминокислот β ig-h3, представленную SEQ ID NO: 5, и называют его мышиный β ig-h3.
Клетки E. coli BL21 (DE3) трансформируют для осуществления экспрессии человеческого и мышиного белков β ig-h3. Трансформированные клетки культивируют в среде LB, содержащей канамицин (50 мкг/мл), при 37°C до тех пор, пока OD595 не достигнет значений 0,5-0,6. В процессе культивирования экспрессию белка β ig-h3 индуцируют обработкой 1 мМ изопропил-β-D-(-)тиогалактопиранозида (IPTG) при 37°C в течение 3 часов.
Клетки E. coli ресуспендируют в буфере для клеточного лизиса (50 мМ Tris-HCl, pH 8,0, 100 мМ NaCl, 1 мМ EDTA, 1% тритон X-100, 1 мМ фенилметансульфонидфторид (называемый далее "PMSF") и 0,5 мМ DTT) и затем разрушают ультразвуком. Процедуру повторяют 5 раз.
Вышеуказанный раствор центрифугируют и нерастворимые внутриклеточные включения, содержащие β ig-h3 растворяют в буферном растворе 20 мМ трис-HCl, содержащем 0,5 M NaCl, 5 мМ имидазол и 8 М мочевину. Белки очищают, используя смолу Ni-NTA (Qiagen). Белки очищают, подвергая их по очереди диализу в буферном растворе 20 мМ трис-HCl, содержащем 0,5 M NaCl и мочевину, начиная с высокой концентрации до низкой концентрации; результаты подтверждают с помощью SDS-PAGE.
Результаты подтверждают, что человеческий и мышиный белки β ig-h3 по настоящему изобретению являются чистыми (фиг. 2).
<1-2> Конструирование и выделение β ig-h3 D-IV(1x) и β ig-h3 D-IV(4x)
Фрагмент ДНК, представленный SEQ ID NO: 6, кодирующей 4ый домен, который соответствует 498-637 аминокислотам человеческого β ig-h3, представленного SEQ. ID. No 1, амплифицируют методом ПЦР. Продукт ПЦР клонируют в векторе pET-29 для конструирования экспрессирующего вектора 4ого домена. Авторы настоящего изобретения называют экспрессирующий вектор 4ого домена "β ig-h3 D-IV".
Базовую последовательность, которая соответствует 4ому домену, синтезируют методом ПЦР, и 3'-конец продукта ПЦР затупляют, используя фрагмент Кленова. Данный продукт ПЦР вставляют в сайт EcoR V вышеуказанного экспрессиирующего вектора pβ ig-h3 D-IV, и называют pβ ig-h3 D-IV(2x). Вставленный фрагмент pβ ig-h3 D-IV(2x) расщепляют с помощью EcoR V и Xho I, и 3'-конец продукта ПЦР затупляют, используя фрагмент Кленова. Данный фрагмент вставляют в сайт EcoR V β ig-h3 D-IV, и называют pβ ig-h3 D-IV(3x). Фрагмент, имеющий тупой 3'-конец также вставляют в сайт EcoR V pβ ig-h3 D-IV(2x), и называют pβ ig-h3 D-IV (4x) (фиг. 3). Для очистки белков на смоле Ni-NTA (Qiagen) получают His-маркер путем присоединения 6 гистидиновых остатков к карбоксильному концу фрагмента ДНК.
Клетки E. coli BS21 (DE3) трансформируют экспрессирующими векторами. Трансформированные клетки культивируют в среде LB, содержащей канамицин (50 мкг/мл). Клетки E. coli ресуспендируют в буфере для клеточного лизиса (50 мМ Tris-HCl, pH 8,0, 100 мМ NaCl, 1 мМ EDTA, 1% тритон X-100, 1 мМ фенилметансульфонидфторид (называемый далее "PMSF") и 0,5 мМ DTT) и затем разрушают ультразвуком. Процедуру повторяют 5 раз. Вышеуказанный раствор центрифугируют, получая супернатанты. Белки выделяют из супернатантов, используя смолу Ni-NTA (Qiagen), и идентифицируют их с помощью SDS-PAGE.
Полученные результаты подтверждают, что экспрессируются следующие белки: β ig-h3 D-IV(1x), имеющий аминокислотную последовательность, представленную SEQ ID NO: 7, β ig-h3 D-IV (2x), имеющий аминокислотную последовательность, представленную SEQ ID NO: 8, β ig-h3 D-IV(3x), имеющий аминокислотную последовательность, представленную SEQ ID NO: 9, и β ig-h3 D-IV (4x), имеющий аминокислотную последовательность, представленную SEQ ID NO: 10. Все вышеуказанные белки содержат 4ый домен человеческого β ig-h3 (фиг. 4).
<1-3> Получение и выделение первичных антител
Первичные антитела получают, используя в качестве антигена человеческий белок β ig-h3 и мышиный белок β ig-h3, выделенные в примере <1-1>. Белки вводят кроликам подкожно в область спины. Для первой инъекции используют смесь 200 мкг белков с полным адъювантом Фрейнда. Для инъекций 2-5 используют смесь 100 мкг белков с неполным адъювантом Фрейнда; инъекции проводят с 3-недельными интервалами. Венозную кровь собирают и оставляют на 2 часа при комнатной температуре. После центрифугирования (10000xg, 10 минут) получают супернатанты, содержащие первичные антитела. Для последующего использования супернатанты хранят при -20°C (фиг. 5).
Пример 2: Определение покрывающей концентрации человеческого белка β ig-h3 и количественного соотношения антител
<2-1> Определение количественного соотношения первичных антител
Для определения количественного соотношения первичных антител и человеческого белка β ig-h3, человеческий β ig-h3 разбавляют (0,5 мкг/мл) 20 мМ карбонатным-бикарбонатным раствором (pH 9,6, содержащим 0,02% азида натрия). Раствор, содержащий β ig-h3, добавляют в каждую лунку 96-луночного планшета (200 мкл/лунку) и выдерживают для образования покрытия в течение ночи при 4°С. Первичные антитела против человеческого β ig-h3 серийно разбавляют разбавляющим раствором (физиологический раствор с фосфатным буфером/Tween 80) в соотношениях 1:200, 1:400, 1:800, 1:1600, 1:2000 и 1:3200 и добавляют к покрытому 96-луночному планшету. К ним также добавляют вторичные антитела (1:5000) и оставляют их взаимодействовать при комнатной температуре в течение полутора часов. Затем добавляют раствор субстрата (полученный путем растворения о-фенилендиамина в метаноле (10 мг/мл), разбавления дистиллированной водой в соотношении 1:100 и смешивания с 10 мкл 30% раствора пероксида водорода) и оставляют взаимодействовать при комнатной температуре в течение 1 часа. Реакцию останавливают добавлением 50 мкл 8 N раствора серной кислоты и проводят ELISA (O D 492 нм).
Полученные результаты показывают, что наилучшее количественное соотношение первичных антител против человеческого β ig-h3 составляет 1:1600 и 1:2000 (фиг. 7).
<2-2> Определение количественного соотношения вторичных антител
Для того чтобы определить количественное соотношение вторичных антител, планшет покрывают человеческим белком β ig-h3 (0,5 мкг/мл). Добавляют первичные антитела против человеческого β ig-h3 (1:1600 и 1:2000). Добавляют вторичные антитела (1:1000, 1:2000 и 1:3000 соответственно) и оставляют взаимодействовать. ELISA проводят по способу вышеописанного примера <2-1>.
Полученные результаты показывают, что наилучшее количественное соотношение первичных антител составляет 1:2000 (фиг. 8).
<2-3> Определение покрывающей концентрации человеческого белка β ig-h3
Для того чтобы определить покрывающую концентрацию человеческого белка β ig-h3, первичные антитела против человеческого β ig-h3 разбавляют в соотношении 1:2000, вторичные антитела разбавляют в соотношении 1:2000, планшет покрывают человеческим белком β ig-h3 с концентрацией 0,5 мкг/мл и 1,0 мкг/мл соответственно и затем проводят ELISA.
Полученные результаты показывают, что подходящей концентрацией человеческого белка β ig-h3 являются и 1,0 мкг/мл, и 0,5 мкл/мл, но 0,5 мкг/мл является более предпочтительной покрывающей концентрацией, так как в данном случае значение R2 приближается к 1 наилучшим образом (фиг. 9).
Исходя из вышеприведенных результатов авторы настоящего изобретения установили, что оптимальной покрывающей концентрацией стандартного белка человеческого β ig-h3 является 0,5 мкг/мл, а наилучшей степенью разбавления первичных антител против человеческого β ig-h3 и вторичных антител является 1:2000, соответственно.
Значения, полученные из вышеописанных результатов, подвергают логарифмическому преобразованию по формуле Робарда (Robard, 1971), приведенной ниже как <Математическая формула 1>. В результате получают кривую в интервале от 11 нг/мл до 900 нг/мл, который является вероятным интервалом измерений. Также показано, что, используя вышеописанные условия реакции, измерение можно проводить, даже начиная с 10 нг/мл (фиг. 10).
<Математическая формула 1>
log b= log×eb/(100-b)
В вышеприведенной формуле b обозначает процент OD в лунке, которая не содержит антигена ни при какой концентрации.
Пример 3: Измерение количественного интервала мышиного β ig-h3, рекомбинантных β ig-h3 D-IV (1x) и β ig-h3 D-IV (4x) путем перекрестного анализа
Авторы настоящего изобретения также определяют концентрацию белка и количественное соотношение первичных и вторичных антител, используя мышиный β ig-h3, рекомбинантные β ig-h3 D-IV(1x) и β ig-h3 D-IV(4x). А именно, для проведения экспериментов получают покрывающую концентрацию каждого белка 0,5 мкг/мл и количественное соотношение первичных антител против человеческого β ig-h3 и вторичных антител 1:2000. Количественное соотношение первичных антител против мышиного β ig-h3 и вторичных антител также устанавливают равным 1:2000.
Полученные в результате графики, имеющие во всех случаях прямолинейную зависимость, подтверждают, что соотношение является наилучшим, а интервал измерения от 11 нг/мл до 900 нг/мл означает, что не существует большой разницы в интервалах измерений (фиг. 11 и фиг. 12).
Полученные результаты подтверждают, что стандартным белком может быть любой из человеческого β ig-h3, мышиного β ig-h3, рекомбинантных β ig-h3 D-IV(1x) и β ig-h3 D-IV(4x), а в качестве первичных антител можно использовать либо антитела против человеческого β ig-h3, либо антитела против мышиного β ig-h3.
Пример 4: Взаимосвязь между заболеваниями почек и экспрессией β ig-h3
<4-1> Измерение β ig-h3 у диабетиков
Авторы настоящего изобретения доказывают взаимосвязь между заболеваниями почек и экспрессией β ig-h3 исходя из того, что TGF-β, который играет важную роль в развитии заболеваний почек, индуцирует экспрессию β ig-h3. Чтобы подтвердить эту взаимосвязь, измеряют количество β ig-h3 в моче диабетиков. А именно, смешивают 110 мкл мочи диабетика и 110 мкл раствора первичных антител (1:1000) в круглодонном планшете, после чего культивируют при 37°С в течение 1 часа. 200 мкл полученной ранее смеси вносят в планшет, покрытый β ig-h3, и оставляют взаимодействовать при комнатной температуре в течение 30 минут. Реакцию останавливают добавлением стоп-раствора, содержащего вторичное антитело и субстрат, и проводят ELISA (OD 492 нм).
Концентрация β ig-h3 в моче диабетиков
Полученное в результате количество β ig-h3 в моче пациентов с диабетическими почечными заболеваниями, включая микроальбуминурию, было в пять раз выше, чем у нормальных субъектов. У некоторых больных диабетом, не имеющих почечных заболеваний, количество β ig-h3 также выше, чем у нормальных субъектов. Учитывая вышеприведенные результаты, можно сделать вывод, что уровень β ig-h3 в моче отражает степень поражения почек, а высокий уровень β ig-h3 у некоторых больных диабетом, не имеющих заболеваний почек, позволяет предположить, что их почки уже поражены в некоторой степени, хотя клинические симптомы еще отсутствуют. Следовательно, измерение количества β ig-h3 в моче пациентов является высокочувствительным и важным диагностическим методом, который может отражать поражение почек на ранней стадии.
<4-2> Измерение β ig-h3 у диабетической животной модели
Для того чтобы подтвердить, действительно ли концентрация β ig-h3 в моче диабетика может отражать поражение почки на ранней стадии, авторы настоящего изобретения измеряют количество β ig-h3 у животного, страдающего диабетом.
Диабет индуцируют у крыс Sprague-Dawley (SD) путем введения стрептозотоцина (60 мг/кг), лекарственного средства, индуцирующего диабет, в брюшную полость крыс. Индуцирование диабета подтверждают измерением глюкозы в крови крыс. На пятый день после индуцирования диабета у крыс берут мочу и измеряют количество β ig-h3 по способу примера <4-1>
Результаты показывают, что через 5 дней после индуцирования диабета количество β ig-h3 увеличивается в 4 раза (56,9±6,4 нг/мг креатина: 230,4±131,8 нг/мг креатина, фиг. 13). Изменение концентрации β ig-h3 наблюдают у каждого субъекта после индуцирования диабета, приводящему к сильному увеличению концентрации β ig-h3 в моче (см. фиг. 14). На 5 день после индуцирования диабета анализы крови, мочи и креатина были нормальными, и ткани почек казались нормальными. Таким образом, сильное увеличение количества β ig-h3 в моче на пятый день после индуцирования диабета позволяет предположить, что уже существует минимальное поражение почек, которое не определяется традиционными методами.
<4-3> Измерение β ig-h3 у пациентов, оперированных по поводу трансплантации почки
Авторы настоящего изобретения подтверждают корреляцию поражения почек и количества β ig-h3 путем измерения количества β ig-h3 в моче у пациентов до и после трансплантации почки. Результаты приведены в таблице 2.
Таблица 2
Изменения концентрации β ig-h3 у пациентов до и после трансплантации почки
Результаты показывают, что высокое количество β ig-h3, наблюдающееся у пациента до операции, постепенно уменьшается после успешной операции. Но у пациента № 5, у которого функция почки не восстановилась даже после операции, количество β ig-h3 остается высоким. Все вышеприведенные результаты подтверждают, что количество β ig-h3 чувствительно отражает степень поражения почек.
<4-4> Измерение β ig-h3 у пациентов с почечной недостаточностью
Авторы настоящего изобретения измеряют количество β ig-h3 в моче пациентов с почечной недостаточностью. Результаты показывают, что в моче у всех указанных пациентов присутствует высокое количество β ig-h3 (таблица 3).
Концентрации β ig-h3 в моче пациентов с почечной недостаточностью
<4-5> Измерение β ig-h3 у пациентов с заболеваниями, связанными с почками
Для того, чтобы определить, действительно ли β ig-h3 по-разному экспрессируется у пациентов с почечными заболеваниями, авторы настоящего изобретения измеряют концентрацию β ig-h3 в моче у пациентов с нормальными симптомами после трансплантации почки, у пациентов, у которых трансплантированная почка имеет меньший размер, у пациентов с хроническим отторжением, пациентов с рецидивирующим пиелитом и у пациентов с циклосфориновой токсичностью, используя способ примера <4-1>.
Результаты показывают, что в среднем концентрация β ig-h3 у пациентов с нормальными симптомами после трансплантации почки составляет 39,4 нг/мг креатина, тогда как у пациентов с хроническим отторжением, рецидивирующим пиелитом и циклосфориновой токсичностью концентрация β ig-h3 сильно увеличена (140,8, 175,4 и 90,9 нг/мг креатина, соответственно) (фиг. 15, таблица 4).
после трансплантации почки (n=47)
18,2
23,0
81,1
65,8
22,4
мальная
мальная
Авторы настоящего изобретения также определяют, уменьшается ли вновь повышенная концентрация β ig-h3 у пациентов с рецидивирующими почечными заболеваниями, если проводится лечение. Результаты показывают, что концентрация β ig-h3 в моче у пациентов, подвергающихся лечению рецидивирующего пиелита после трансплантации почки, постепенно уменьшается, позволяя предположить, что концентрация β ig-h3 в моче уменьшается в процессе лечения. Таким образом, концентрация β ig-h3 может свидетельствовать о реакции на лечение (фиг. 16).
<4-6> Анализ влияния трансплантации почки на концентрацию β ig-h3
Для того, чтобы исследовать изменения концентрации β ig-h3 в моче после трансплантации почки, авторы настоящего изобретения каждый день измеряют концентрацию β ig-h3 в моче у пациентов после трансплантации почки.
Результаты показывают, что концентрация β ig-h3 в моче у пациентов после успешной трансплантации почки, независимо от того, взята ли почки у живого субъекта или у субъекта, погибшего в результате смерти мозга, постепенно уменьшается. А именно, в случае получения почки от живого субъекта, концентрация β ig-h3 в моче возвращается к нормальному уровню в течение 4-5 дней после трансплантации, а в случае получения почки от субъекта, погибшего в результате смерти мозга, концентрация β ig-h3 возвращается к нормальному уровню в течение 6-7 дней (фиг. 17).
Кроме того, концентрация β ig-h3 в моче пациентов, получивших маленькую почку, после трансплантации возвращается к нормальному уровню, хотя уровень креатина у них в крови еще остается высоким, что позволяет предположить, что трансплантированная почка работает нормально, хотя она не может фильтровать продукты жизнедеятельности достаточно хорошо из-за маленького размера. В любом случае, концентрация β ig-h3 отражающая поражение почки, возвращается к нормальному уровню (фиг. 17). Между тем, как концентрация β ig-h3 в моче у пациентов с неудачной трансплантацией почки сильно колеблется.
На основании полученных результатов концентрацию β ig-h3 в моче можно считать эффективным маркером для диагноза почечных заболеваний на ранних стадиях, для установления развития почечных заболеваний и для определения эффекта лечения, так как концентрация β ig-h3 хорошо отражает поражение почек.
В результате авторы настоящего изобретения подтверждают, что концентрация β ig-h3 чувствительно отражает поражение почек на ранних стадиях и имеет большое значение для диагноза различных почечных заболеваний.
Пример 5: Взаимосвязь заболеваний печени и экспрессии β ig-h3
Очень важно определить, развивается ли у пациента хронический гепатит в цирроз печени, однако пока не существует метода, который позволяет это сделать. Самым критическим фактором для развития цирроза печени является TGF-β. Следовательно, при развитии цирроза печени уровень β ig-h3, экспрессия которого индуцируется TGF-β, в крови, возможно, будет увеличиваться. В таком случае, количество β ig-h3 также может отражать степень поражения циррозом печени. Фактически доказано, что экспрессия β ig-h3 возрастает при увеличении тяжести цирроза печени, которую устанавливают с помощью иммуногистологического анализа тканей печени пациентов с гепатитом. Авторы настоящего изобретения подразделяют состояние пациентов на несколько степеней и стадий на основе результатов биопсии, полученных для пациентов с хроническим гепатитом, и определяют концентрацию β ig-h3 в крови для каждой стадии и степени. А именно, авторы настоящего изобретения собирают кровь у пациентов с хроническим гепатитом и измеряют количество β ig-h3 по способу примера <4-1>. Результаты приведены в таблице 5.
Концентрации β ig-h3 в крови у пациентов с хроническим гепатитом
(Норма)
(n=172, ± 28,5)
(Норма)
(n=172,± 28,5)
(n=16,± 30,6)
(n=20,± 30,2)
(n=43,± 72,8)
(n=36,± 79,1)
(n=7,± 21,9)
(n=10,± 21,9)
Результаты показывают, что у пациентов с хроническим гепатитом концентрация β ig-h3 в крови выше, чем у нормальных людей, и что концентрация β ig-h3 на ранней стадии и степени (1 и 2) выше, чем на более поздней стадии и степени (3). Степень 3 и стадия 3 характеризуются тяжелым состоянием цирроза печени, перешедшего пик в своем развитии. Между тем, степени 1 и 2 и стадии 1 и 2 соответствуют состоянию пациента, при котором очень интенсивно развивается воспалительный процесс. Следовательно, концентрация β ig-h3 отражает интенсивность цирроза печени, так, что его развитие можно отслеживать, регулярно измеряя концентрацию β ig-h3 в крови.
Пример 6: Взаимосвязь ревматоидного артрита и экспрессии β ig-h3
Авторы настоящего изобретения подтверждают корреляцию ревматоидного артрита и экспрессии β ig-h3, измеряя количество β ig-h3 в синовиальной жидкости пациентов с остеоартритом и ревматоидным артритом по способу примера <4-1> (таблица 6).
Концентрации β ig-h3 в синовиальной жидкости
Результаты показывают, что концентрация β ig-h3 в синовиальной жидкости пациентов с ревматоидным артритом была в два раза выше, следовательно, измерение концентрации β ig-h3 в синовиальной жидкости может служить эффективным способом диагностики остеоартрита и ревматоидного артрита.
Пример 7: Взаимосвязь сердечно-сосудистых заболеваний и экспрессии β ig-h3
< 7-1 > Измерение β ig-h3 в поврежденных кровеносных сосудах у мышей с индуцированным диабетом
Для подтверждения взаимосвязи экспрессии β ig-h3 и сердечно-сосудистых заболеваний авторы настоящего изобретения исследуют характер экспрессии β ig-h3 в нормальных и поврежденных кровеносных сосудах диабетических мышей с помощью иммуногистохимических методов.
Результаты показали, что белок β ig-h3 в поврежденных кровеносных сосудах диабетических мышей экспрессируется гораздо интенсивнее, чем в нормальных кровеносных сосудах (фиг. 18).
<7-2> Измерение экспрессии β ig-h3, индуцированной TGF-β в клетках гладкой мускулатуры сосудов
На основании того, что экспрессия β ig-h3 индуцируется TGF-β, который играет важную роль в развитии сосудистых заболеваний, авторы настоящего изобретения попытались подтвердить корреляцию экспрессии β ig-h3 и сердечно-сосудистых заболеваний. А именно, авторы настоящего изобретения определяют характер экспрессии β ig-h3, индуцированной TGF-β в клетках гладкой мускулатуры сосудов, образующих кровеносные сосуды, по способу примера <4-1>.
Результаты подтверждают, что экспрессия β ig-h3 возрастает по мере увеличения количества TGF-β (фиг. 19).
Полученные результаты подтверждают, что экспрессия β ig-h3 в крови и тканях отражает их поражение. Следовательно, способ измерения количества белка β ig-h3 по настоящему изобретению можно эффективно использовать для диагностики различных сердечно-сосудистых заболеваний.
ПРОМЫШЛЕННОЕ ПРИМЕНЕНИЕ
Как описано выше, способ измерения количества белка β ig-h3 по настоящему изобретению, в котором в качестве стандартного белка используются человеческий β ig-h3, мышиный β ig-h3, β ig- h3 D-IV(1x) или β ig-h3 D-IV (4x), является недорогим и очень точным при измерении концентрации β ig-h3 в различных жидкостях организма. Количество β ig-h3 позволяет с высокой чувствительностью диагностировать связанные с TGF-β заболевания, такие как заболевания почек, печени, ревматоидный артрит и сердечно-сосудистые заболевания на ранних стадиях, следовательно, способ по настоящему изобретению можно эффективно использовать для определения поражения и развития данных заболеваний, а также для их диагностики.


















Изобретение относится к области медицины и касается способа диагностики заболеваний, связанных со сверхэкспрессией белка β ig-h3 и диагностического набора для его осуществления. Сущность изобретения заключается в измерении уровня белка β ig-h3 в жидкости организма пациента с использованием антитела, специфического в отношении белка β ig-h3, и диагностике заболеваний почек, печени, ревматоидного артрита или сердечно-сосудистых заболеваний, а также набора для диагностики поражений этих органов. Преимущество изобретения заключается в разработке экспресс-метода для предварительной диагностики заболеваний почек, печени, ревматоидного артрита или сердечно-сосудистых заболеваний. 2 н. и 10 з.п. ф-лы, 5 табл., 19 ил.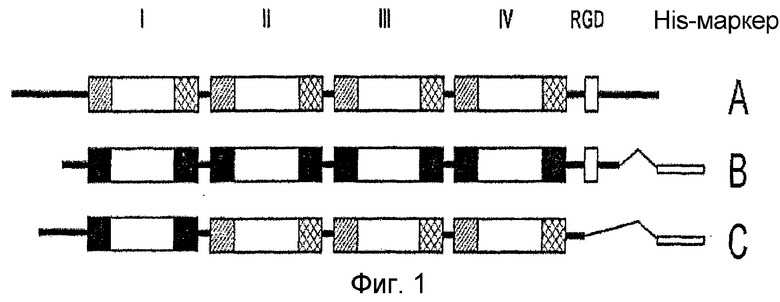
1) Нанесение рекомбинантных белков β ig-h3, содержащих 4-е домены fas-1, их фрагментов или производных, на основу;
2) Взаимодействие антитела против белка β ig-h3, его фрагмента или производного с жидкостью организма пациента;
3) Добавление реагента вышеописанной стадии 2 к покрывающему белку стадии 1, проведение реакции и затем промывание покрытия; и
4) Добавление вторичного антитела к реагенту вышеописанной стадии 3 для осуществления дополнительной реакции и затем измерение ОП.
| US 5714588 А, 03.02.1998 | |||
| US 5444164 A, 22.08.1995 | |||
| US 5599,788 А, 04.02.1997. |

