Изобретение относится к способу тестирования предполагаемого или известного иммуномодулирующего лекарственного средства для активации T-клеток, который включает стадию приведения в контакт культуры мононуклеарных клеток периферической крови (PBMC) с предварительно определяемым количеством предполагаемого или известного иммуномодулирующего лекарственного средства in vitro и наблюдения активации T-клеток в PBMC культуре с использованием системы считывания при контакте с предполагаемым или известным иммуномодулирующим лекарственным средством, где плотность клеток в предварительной культуре PBMC корректируют так, чтобы сделать возможным межклеточный контакт PBMC, и где предварительную культуру PBMC культивируют в течение по меньшей мере 12 ч. Изобретение дополнительно относится к способу тестирования ослабляющих цитокиновый шторм лекарственных средств in vitro.
В данном описании процитировано множество документов, включая патентные заявки и руководства изготовителя. Раскрытие этих документов, хотя и не считается важным для патентоспособности данного изобретения, при этом включено посредством ссылки в полном объеме. Более конкретно, все цитированные документы включены посредством ссылки в такой же степени, как если бы было указано, что каждый отдельный документ специально и отдельно включен посредством ссылки.
Иммунотерапевтические лекарственные средства, которые модулируют активность лимфоцитов, доклинически оценивают в двух системах: животные модели, обычно грызуны и, если доступны, приматы и культуры мононуклеарных клеток периферической крови (PBMC) человека.
PBMC обычно используют, поскольку, во-первых, они содержат все субпопуляции лимфоцитов, а также моноциты, и, во-вторых, они легко доступны из венозной крови, которую берут у здоровых доноров или у пациентов. Стимуляцию этих PBMC in vitro рассматривают в качестве эффективного индикатора активностей иммуномодулирующего лекарственного средства, ожидаемых in vivo.
Определенные активирующие T-клетки средства, в частности моноклональные антитела (МАТ), затрагивающие T-клеточный антигенный рецептор (TCR), такие как OKT3, первое МАТ, использованное в клинике для иммуносупрессии, вызывают системное высвобождение провоспалительных цитокинов (Abramowicz et al., 1989). Среди них наиболее опасными являются TNF, интерферон-γ (IFN-γ) и IL-2. У пациентов, получающих терапию МАТ, контроль такого синдрома высвобождения цитокинов или «цитокинового шторма» обычно достигается лечением кортикостероидами в высоких дозах.
TGN1412 представляет собой гуманизированное моноклональное антитело (МАТ) подкласса IgG4 со специфичностью к костимулирующей молекуле CD28, экспрессируемой T-клетками человека. Его называют «суперагонистом CD28» (CD28SA), поскольку в отличие от классических МАТ со специфичностью к CD28 оно может активировать T-лимфоциты без одновременного вовлечения T-клеточного антигенного рецептора (TCR) (Hunig, 2007). TGN1412 разработано ныне не существующей TeGenero AG, Wurzburg.
Во время первого исследования у человека, которое проводило независимое Parexel Clinical Trial Unit в Northwick Park Hospital, London, 13 марта 2006 года, внутривенное применение TGN1412 в дозе 100 мкг/кг массы тела у здоровых людей-добровольцев вело к угрожающему для жизни синдрому высвобождения цитокинов, который поддавался контролю только после перевода добровольцев в отделение интенсивной терапии больницы (Suntharalingam et al., 2006).
В доклинической работе, представленной спонсором TeGenero AG, не представлены свидетельства о таком «цитокиновом шторме» в аналогичной модели на крысах с использованием суперагониста со специфичностью к CD28 крысы, и на яванских макаках, получавших сам TGN1412 в дозах, превышавших вплоть до 50 раз дозы для людей-добровольцев (Duff, 2006). Кроме того, добавление TGN1412 в культуры PBMC человека также не приводило к высвобождению цитокинов. Все ключевые эксперименты на макаках и культурах PBMC повторяли в British National Institute for Biological Standards и Control (NIBSC), действующего от лица правительственной Expert Scientific Group on Phase One Clinical Trials, и подтверждали безопасное поведение TGN1412 в этих системах (Duff, 2006). Через три года после провала испытаний TNG1412 до сих пор не выяснено, почему этот анализ in vitro не предупредил о цитокиновом шторме, который перенесли люди-добровольцы.
Неспособность грызунов и яванских макаков высвобождать токсичные системные цитокины после инъекции CD28SA очевидно обусловлен межвидовыми различиями в реактивности интактной иммунной системы на такие средства, и в отношении таких различий сделаны конкретные предположения (Gogishvili et al., 2009; Nguyen et al., 2006; Schraven и Kalinke, 2008).
Эти находки указывают на то, что вследствие видоспецифических паттернов реакции даже животные модели на приматах не всегда являются надежными средствами прогноза реактивности человека на новые лекарственные средства.
Человек имеет приблизительно 1012 T-лимфоцитов, и менее чем 1% от них циркулирует в крови в тот или иной момент. Следовательно, неспособность культивируемых PBMC отвечать на TGN1412 обусловлена или функциональным дефектом в этих клетках по сравнению с теми, что находятся в лимфоидных тканях (которые, очевидно, отвечали высвобождением цитокинов у добровольцев), или потребностью в типе клеток, которые присутствуют в лимфоидных органах, но не в крови, для опосредованной TGN1412 активации T-лимфоцитов.
Следовательно, необходимо понять механизм воспроизведения цитокинового шторма, наблюдавшегося у людей-добровольцев в лондонском исследовании TGN1412, в клеточной культуре и тестировать ее чувствительность к фармакологической супрессии.
В более широком смысле, неспособность известных культур PBMC человека отвечать на растворимый TGN1412 высвобождением цитокинов указывает на то, что эта система не отвечает на все активирующие лимфоциты средства таким же образом, как отвечает интактная иммунная система человека внутри организма. Коррекция этого дефекта может не только сделать возможным подробный анализ эффектов суперагонистов CD28 человека (SA), таких как TGN1412, но также может обнаружить реактивность других, по-видимому, безвредных лекарственных средств во время доклинической разработки.
Соответственно, техническая проблема изобретения состоит в том, чтобы предоставить улучшенное средство для тестирования иммуномодулирующих лекарственных средств in vitro, в частности CD28SA, в отношении возможного цитокинового шторма. Дополнительная цель изобретения состоит в том, чтобы предоставить средство для тестирования лекарственных средств, подходящих для ослабления цитокинового шторма.
Для решения первой указанной технической проблемы в изобретении предложен способ тестирования предполагаемого или известного иммуномодулирующего лекарственного средства для активации T-клеток, который содержит стадию приведения в контакт культуры мононуклеарных клеток периферической крови (PBMC) и предварительно определяемого количества предполагаемого или известного иммуномодулирующего лекарственного средства in vitro и наблюдения активации T-клеток в культуре PBMC с использованием системы считывания при контакте с предполагаемым или известным иммуномодулирующим лекарственным средством, где плотность клеток в предварительной культуре PBMC корректируют так, чтобы сделать возможным межклеточный контакт PBMC, и где предварительную культуру PBMC культивируют в течение по меньшей мере 12 ч.
В предпочтительном варианте осуществления изобретения система считывания наблюдает высвобождение по меньшей мере одного цитокина из PBMC в культуре PBMC, наблюдает пролиферацию клеток или представляет собой другую подходящую систему считывания, такую как изменения в экспрессии гена, экспрессии белка и/или посттрансляционных модификациях.
Термин «иммуномодулирующее лекарственное средство» предпочтительно обозначает какое-либо терапевтическое средство, которое способно активировать или подавлять иммунную систему, например, посредством активации или ингибирования функций лимфоцитов, в частности функций T-клеток, такого как ингибирование или активация T-клеток. Конкретные примеры иммуномодулирующих лекарственных средств предоставлены в настоящем документе ниже.
Термин «активация T-клеток» предпочтительно определяет механизмы активации T-клеток, которые могут незначительно варьировать между различными типами T-клеток. Однако «двухсигнальная модель» в CD4+ T-клетках применима к T-клеткам большинства типов. Более подробно, активация CD4+ T-клеток типично происходит через вовлечение как T-клеточного рецептора, так и CD28 на поверхности T-клетки посредством кодируемой главным комплексом гистосовместимости антигенпредставляющей молекулы и связанного с ним антигенного пептида и членов семейства B7 на поверхности антигенпредставляющей клетки (APC), соответственно. Оба межклеточных контакта, как правило, необходимы для образования эффективного иммунного ответа. Например, в отсутствие костимуляции CD28, передача только сигналов T-клеточного рецептора может привести к анэргии T-клеток. Дополнительные нисходящие пути передачи сигнала как от CD28, так и от T-клеточного рецептора, включают многие дополнительные белки, известные специалистам. Активацию T-клеток можно определить посредством высвобождения цитокинов и/или клеточной пролиферации, в частности, пролиферации T-клеток, как описано в настоящем документе ниже.
Термин «мононуклеарные клетки периферической крови (PBMC)» предпочтительно определяет любые клетки крови, которые имеют круглое ядро, например лимфоциты и моноциты. Эти клетки крови рециркулируют между тканями и кровью и являются определяющим компонентов в борьбе иммунной системы с инфекцией и адаптации к возбудителям. Популяция лимфоцитов в PBMC типично состоит из T-клеток (CD4 и CD8 положительные ~75%), B клеток и NK клеток (вместе ~25%). PBMC можно получить из образцов цельной крови способами, хорошо известными в данной области. Например, PBMC можно получить из цельной крови, используя градиент фиколла. Фиколл представляет собой гидрофильный полисахарид, который разделяет слои крови по плотности, где моноциты и лимфоциты формируют слой между плазмой и раствором фиколла. Помимо цельной гепаринизированной крови в качестве начального материала можно использовать лейкоцитарную пленку (слой белых клеток поверх эритроцитов, полученных центрифугированием цельной крови) и содержимое лейкоредукционной камеры, получаемое в качестве побочного продукта тромбоцитарной массы в банках крови.
Термин «предварительно культивированный» обозначает, что культуру PBMC культивируют в отсутствие иммуномодулирующих лекарственных средств и до контакта с такими лекарственными средствами, подлежащими тестированию. Способы культивирования PBMC хорошо известны в данной области, а также описаны в настоящем документе ниже.
Термин «наблюдать» включает качественное, полуколичественное и количественное измерение концентрации по меньшей мере одного цитокина или пролиферации (клеток), предпочтительно пролиферации T-клеток, или других клеточных ответов, таких как изменения экспрессии гена, экспрессии белка и/или посттрансляционных модификаций, используя известные в данной области способы.
Изобретение основано на том удивительном открытии, что культура PBMC, которую получают стандартными способами, и, кроме того, предварительно культивируют в течение предварительно определяемого периода времени перед контактом с иммуномодулирующим лекарственным средством, внезапно проявляет чувствительность в отношении высвобождения цитокинов, запускаемого посредством контакта с иммуномодулирующими лекарственными средствами, которые не запускают высвобождение цитокинов в отсутствие процесса предварительного культивирования. Кроме того, изобретение основано на том открытии, что этому эффекту предварительного культивирования содействуют межклеточные контакты PBMC в течение предварительно определяемого периода времени. Другими словами, когда добавляют иммуномодулирующее лекарственное средство, культура PBMC не должна быть свежеполученной.
Следовательно, изобретение можно использовать для предсказания реактивности индивидуумов к иммуномодулирующим лекарственным средствам, таким как TGN1412, и, как объяснено более подробно далее, способности иммуносупрессорных лекарственных средств, таких как кортикостероиды, контролировать нежелательные реакции. Также его можно использовать для более глубокого понимания механизма действия CD28SA (т.е. суперагонистического специфичного к CD28 моноклонального антитела). Как указано выше, CD28 экспрессирован на поверхности T-клеток, а костимуляция через CD28 и TCR обычно необходима для активации T-клеток в физиологических условиях, принимая во внимание, что также возможна непосредственная активация T-клеток посредством суперагонистических антител к CD28. Кроме того, изобретение можно использовать в скрининге потенциала активации T-клеток у предполагаемых иммуномодулирующих лекарственных средств, включая высвобождение цитокинов. Поскольку PBMC, собранные после предварительного культивирования при высокой плотности, наиболее вероятно отражают состояние реактивности T-клеток, которые находят в лимфоидных органах, их также следует использовать в сочетании с активирующим средством для тестирования иммуносупрессорных лекарственных средств, поскольку циркулирующие T-клетки можно более легко супрессировать вследствие их «неактивного» состояния, что ведет к ложным результатам относительно эффективности таких лекарственных средств.
В предпочтительном варианте осуществления изобретения стадию предварительного культивирования осуществляют посредством хранения культуры PBMC в течение по меньшей мере 24 ч, предпочтительно по меньшей мере 36 ч, более предпочтительно по меньшей мере 45 ч, при температуре от 35°C до 40°C, предпочтительно при температуре от 36°C до 38°C, в отсутствие иммуномодулирующих лекарственных средств, и до контакта с иммуномодулирующим лекарственным средством, подлежащим тестированию.
В другом предпочтительном варианте осуществления изобретения плотность клеток культуры PBMC во время стадии предварительного культивирования составляет по меньшей мере 2×106/мл, предпочтительно по меньшей мере 5×106/мл, более предпочтительно по меньшей мере 107/мл. С точки зрения плотности клеток на поверхности сосуда с культурой ткани она должна составлять по меньшей мере 4×105/см2, предпочтительно по меньшей мере 106/см2, наиболее предпочтительно по меньшей мере 2×106/см2. Предоставленные значения применяют непосредственно к сосуду, состоящему из плоских лунок. В круглых лунках или конических лунках общая плотность будет отличаться тем, что плотность клеток выше на дне лунки и ниже в верхних частях лунки. Соответственно, приведенные выше плотности клеток, измеренные в объеме, также относятся к частичным объемам в сосудах любого типа, т.е. приведенные плотности следует обеспечивать в частичном объеме, например, по меньшей мере порядка 10 мкл от присутствующего общего объема предварительной культуры. Также изобретение охватывает любой другой способ достижения плотностей клеток для жизнеспособных клеток, как заявлено. Минимальное число клеток, находящихся в межклеточном контакте, предпочтительно составляет по меньшей мере 50000.
В дополнительном предпочтительном варианте осуществления иммуномодулирующее лекарственное средство представляет собой иммуностимулирующее лекарственное средство, такое как антитело, предпочтительно моноклональное антитело. В частности, моноклональное антитело может представлять собой суперагонистическое специфичное к CD28 человека моноклональное антитело. Тестируемое иммуномодулирующее лекарственное средство может быть выбрано из группы, но без ограничения, состоящей из лектинов, таких как конканавалин A (ConA) или фитогемагглютинин (PHA), природных экстрактов, содержащих лектины, таких как экстракт эхинацеи (например, Echinacin, Madaus, Germany) или экстракт омелы (например, Lektinof, Madaus, Germany), суперантигенов, таких как стафилококковый энтеротоксин B (SEB), стафилококковый энтеротоксин A (SEA), токсин синдрома токсического шока-1 (TSST-1), стафилококковых пирогенов, таких как стафилококковый пирогенный энтеротоксин B (SPEB), суперантигенов, продуцируемых микоплазмами, такими как Mycoplasma arthritidis, или бактериями черной чумы, такими как Yersinia pseudotuberculosis, суперантигенов, продуцируемых определенными патогенными вирусами, такими как ЭБВ или ВИЧ-1.
В другом предпочтительном варианте осуществления изобретения наблюдаемый цитокин выбирают из группы, состоящей из TNF, IFN-γ, IL-1β, IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL12p70, IL-13, IL-14, IL-15, IL-16, IL-17, IL-21, IL-35 и LT и их сочетаний.
Альтернативно можно наблюдать пролиферацию клеток, где пролиферация имеет место, если число клеток увеличивается за единицу времени на предварительно определяемое количество. Это количество может выбрать специалист исходя из обычных соображений. Клетки, которые пролиферируют в этом анализе, предпочтительно представляют собой PBMC и более предпочтительно T-клетки.
В дополнительном предпочтительном варианте осуществления способа тестирования предполагаемого или известного иммуномодулирующего лекарственного средства для активации T-клеток культура PBMC содержит T-клетки памяти.
В данной области хорошо известно, что T-клетки памяти являются субпопуляцией антигенспецифических T-клеток, которые ранее встречались со своим когнатным антигеном и отвечали на него. Таким образом, термин «обученная антигеном T-клетка» альтернативно применяют в данной области. Кроме того, в этом отношении предпочтительно, чтобы T-клетки памяти представляли собой CD45RO+ T-клетки. Авторы настоящего изобретения обнаружили, что цитокины, высвобождаемые PBMC в способе по изобретению, преимущественно высвобождают T-клетки памяти, которые содержатся в PBMC.
В дополнительном предпочтительном варианте осуществления, обладающем особой значимостью в отношении второй указанной цели, предварительно культивированную культуру PBMC дополнительно приводят в контакт с предполагаемым лекарственным средством для ослабления высвобождения по меньшей мере одного цитокина (иммуносупрессорное лекарственное средство), одновременно с контактом культуры PBMC с известным иммуномодулирующим, в частности стимулирующим, лекарственным средством, или впоследствии после предварительно определяемого периода времени, или прежде за предварительно определяемый период времени, где дополнительно наблюдают высвобождение цитокинов. Предварительно определяемый период времени может находиться в диапазоне от 10 с до 12 ч, предпочтительно в диапазоне от 10 с до 1 ч. В том случае, когда культуру PBMC дополнительно приводят в контакт с иммуносупрессорным лекарственным средством, следует понимать, что иммуносупрессорный эффект можно определить по сравнению с состоянием, в котором культуру PBMC приводят в контакт с только предполагаемым или известным иммуномодулирующим лекарственным средством. Иммуносупрессорное лекарственное средство также можно приводить в контакт с предварительно культивированной культурой PBMC только для того, чтобы определить, например, его супрессорный эффект, оказываемый на естественную, латентную активацию T-клеток или на неестественную, повышенную активацию T-клеток, например, обусловленную иммунным нарушением или факторами окружающей среды.
Предполагаемое лекарственное средство для ослабления высвобождения цитокинов предпочтительно выбирают из группы, состоящей из кортикостероидов, включая в качестве неограничивающих примеров дексаметазон или метилпреднизолон.
Предполагаемое лекарственное средство для ослабления высвобождения цитокинов, кроме того, может представлять собой другое иммуносупрессорное лекарственное средство, выбранное из группы, состоящей из рапамицина или ингибиторов кальциневрина, таких как циклоспорин A, воклоспорин или такролимус.
В этом предпочтительном варианте осуществления возможно проводить тестирование применимости конкретного лекарственного средства, предназначенного для ослабления цитокинового шторма (иммуносупрессорное лекарственное средство), для этой цели in vitro, в частности, в тех случаях, когда цитокиновый шторм возникает in vivo, несмотря на введение иммуномодулирующего лекарственного средства в концентрации, которую прежде всего сочтут предотвращающей цитокиновый шторм. В частности, этот вариант изобретения предоставляет возможность создания совместимой пары иммуномодулирующего лекарственного средства и ослабляющего лекарственного средства и предоставляет надежное средство для управления неожиданным цитокиновым штормом в экспериментах in vivo, в частности, в клинических исследованиях с участием человека.
В этом отношении термин «цитокиновый шторм», который в данной области также известен как «гиперцитокинемия», предпочтительно определяет системную воспалительную реакцию у пациента, которая среди прочего отличается гипотензией, пирексией и/или дрожью, и потенциально ведет к смерти. Цитокиновый шторм предположительно обусловлен неконтролируемым механизмом положительной обратной связи между цитокинами и иммунными клетками, который ведет к чрезвычайно высоким уровням различных цитокинов.
В дополнительном варианте осуществления изобретение относится к кортикостероиду, рапамицину и/или ингибитору кальциневрина, для использования в лечении, ослаблении или профилактике цитокинового шторма при введении специфического к CD28 человека суперагонистического моноклонального антитела.
В предпочтительном варианте осуществления антитело представляет собой TGN1412.
Аминокислотная последовательность антитела TGN1412 описана в WO-A2 2006/050949.
В дополнительном предпочтительном варианте осуществления кортикостероидом является дексаметазон и/или метилпреднизолон.
В дополнительном предпочтительном варианте осуществления ингибитором кальциневрина является циклоспорин A, воклоспорин и/или такролимус.
Далее приведено подробное объяснение изобретения посредством примеров и фигур. На чертежах:
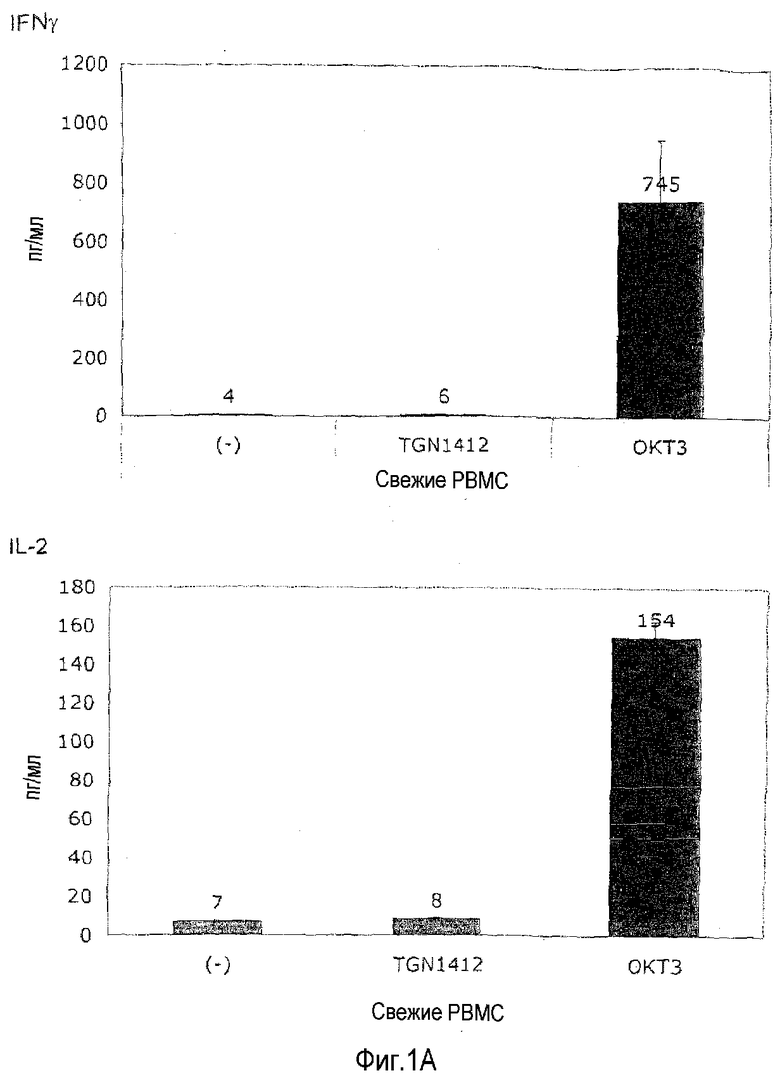
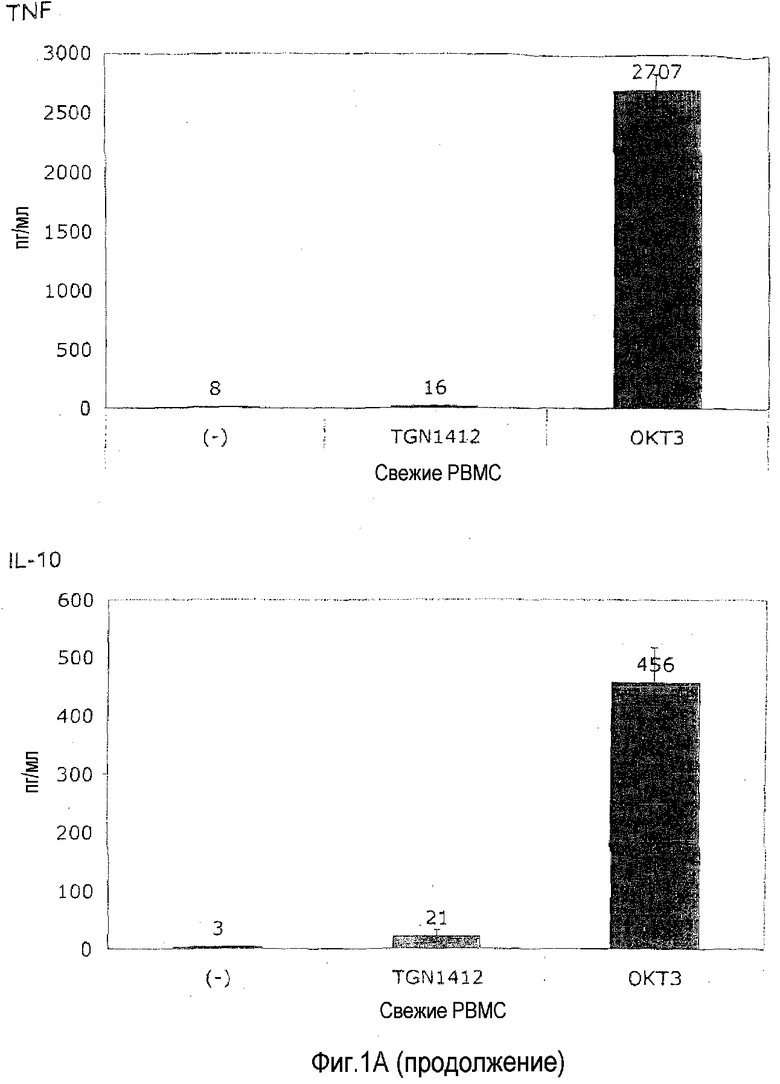
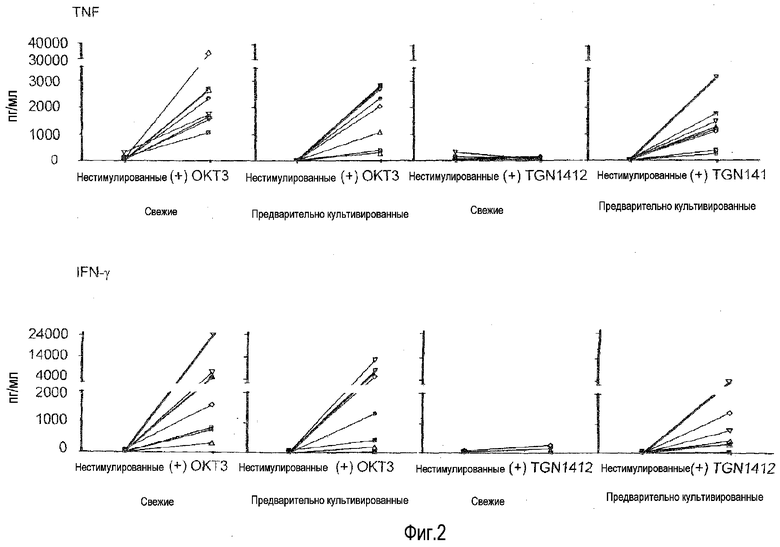
фиг.1: индукция высвобождения цитокинов посредством CD3-специфического МАТ OKT3 и суперагониста CD28 TGN1412 из PBMC человека.
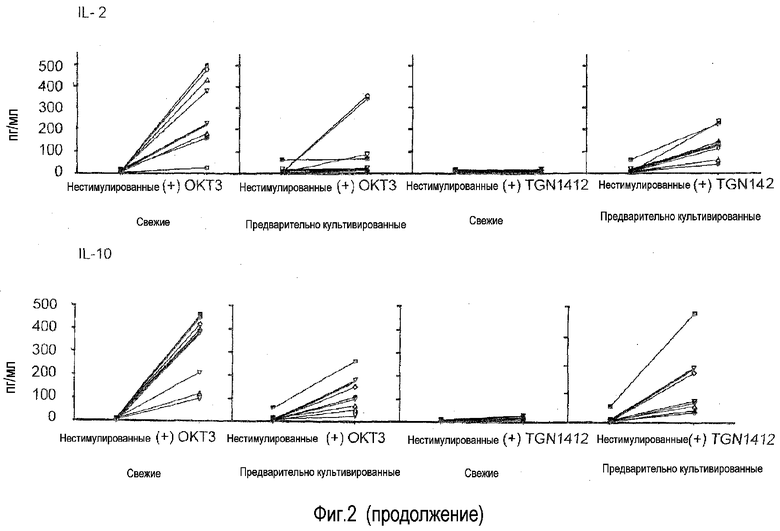
фиг.2: последовательный переход из TGN1412-нереактивного в TGN1412-реактивное состояние посредством предварительного культивирования в течение 2 суток.
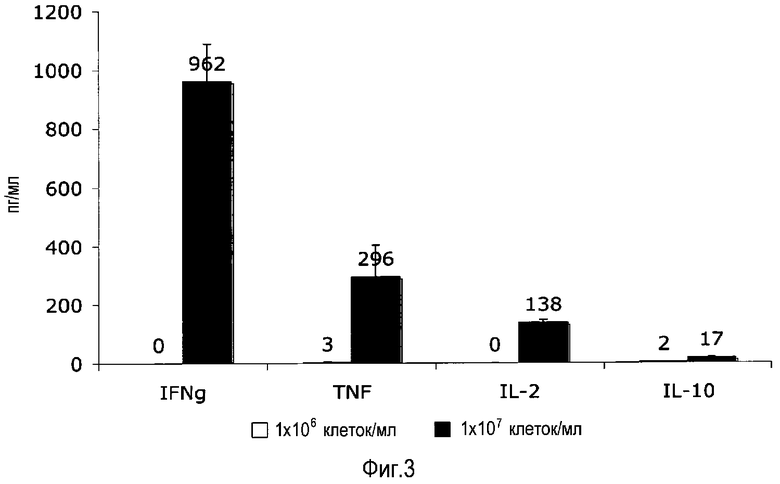
фиг.3: для приобретения TGN1412-реактивности необходима высокая плотность клеток во время предварительной культуры.
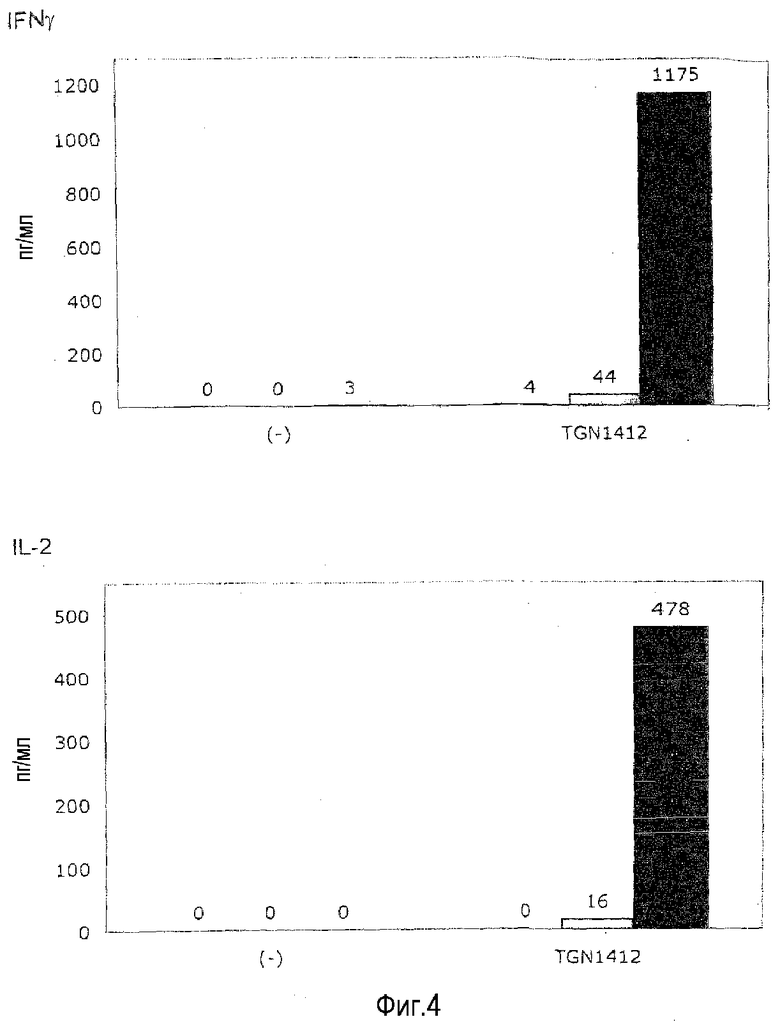
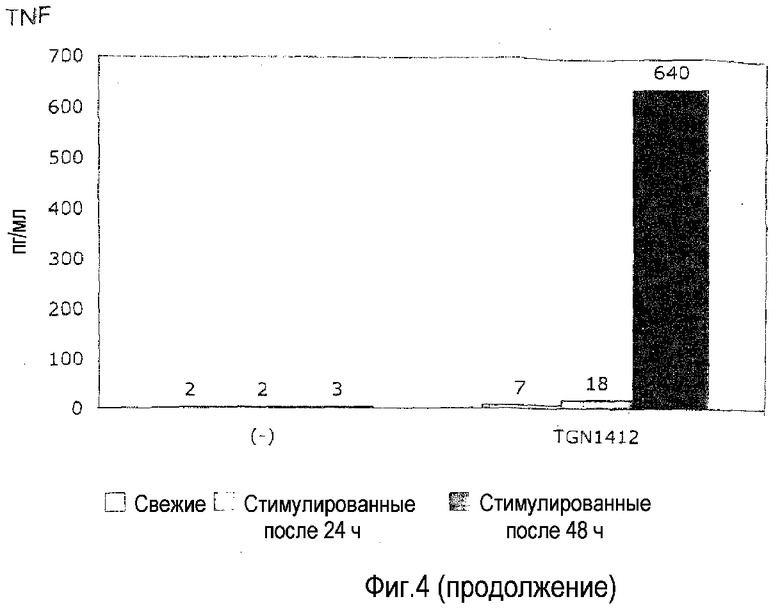
фиг.4: для приобретения TGN1412-реактивности в предварительных культурах с высокой плотностью необходимо инкубирование в течение 2 суток.
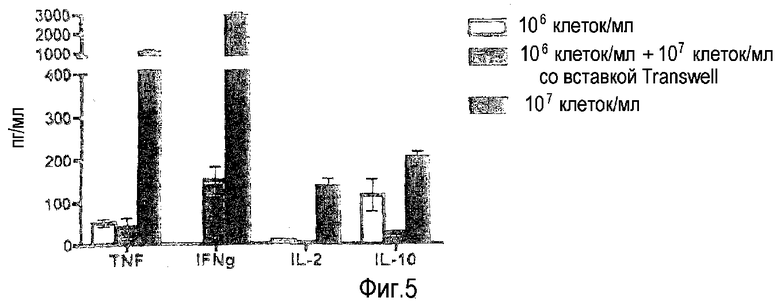
фиг.5: для приобретения TGN1412-реактивности в культурах с высокой плотностью необходим межклеточный контакт.
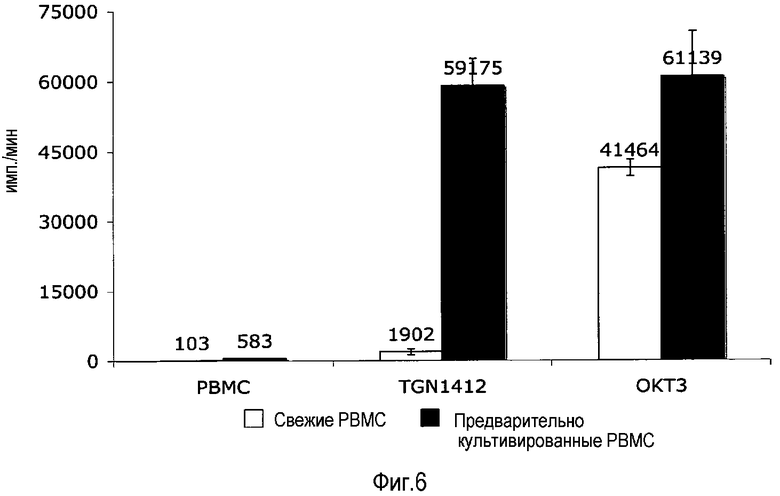
фиг.6: предварительно культивированные, а не свежие PBMC отвечают на TGN1412 пролиферацией.
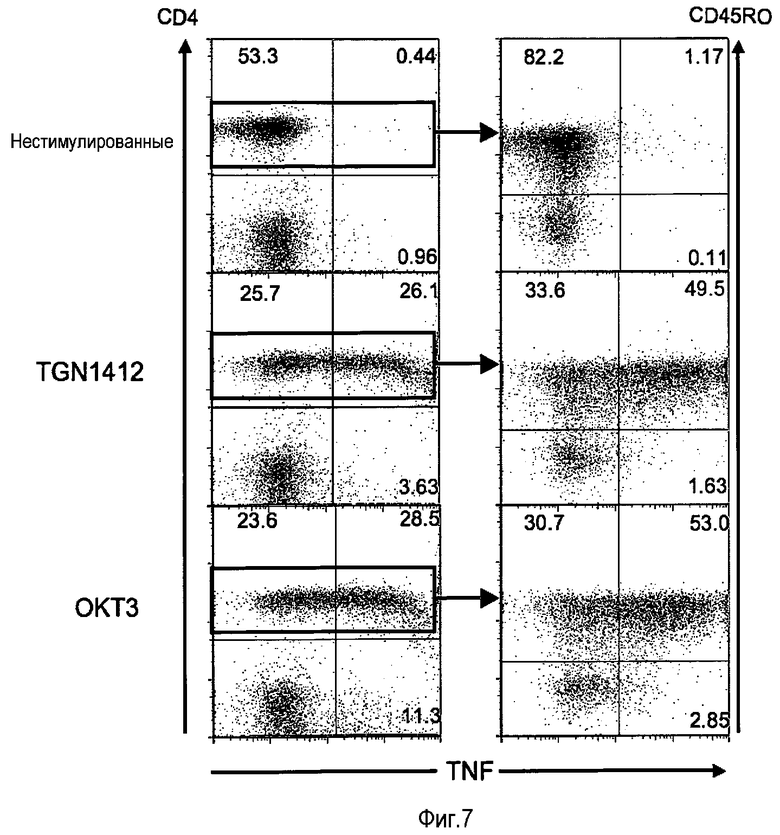
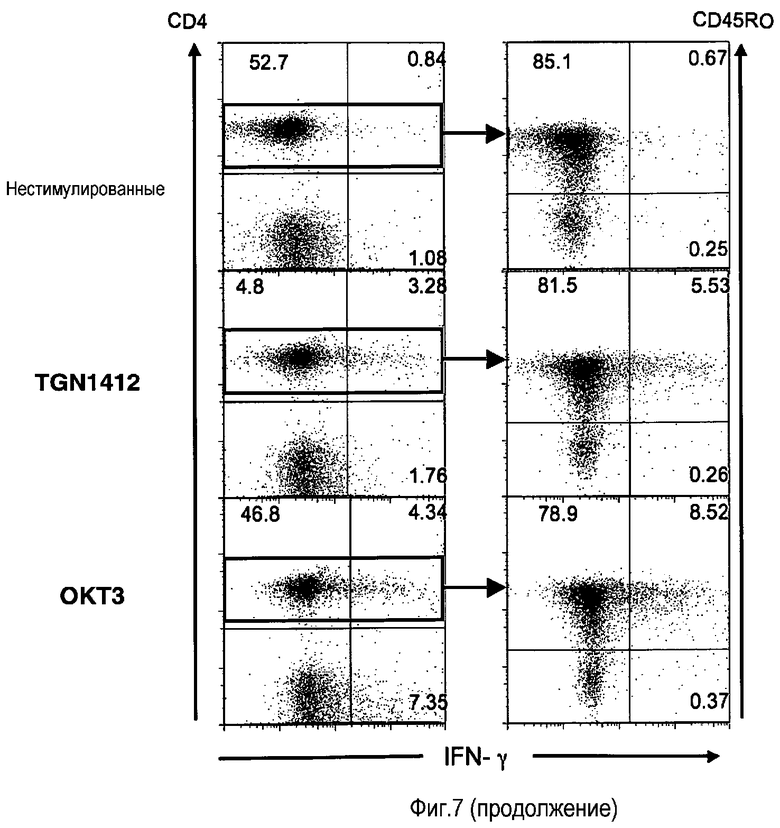
фиг.7: CD45RO CD4 T-клетки (памяти) являются основным источником провоспалительных цитокинов, высвобождаемых посредством как OKT3, так и TGN1412.
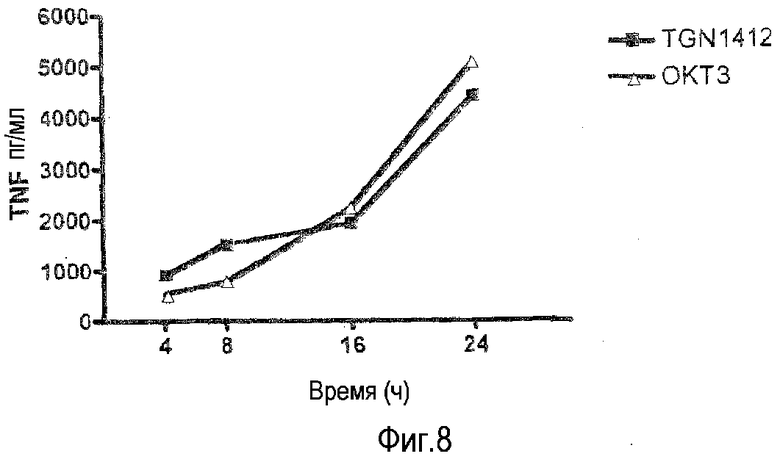
фиг.8: сравнимая кинетика высвобождения TNF из предварительно культивированных PBMC при высокой плотности, индуцированная OKT3 и TGN1412.
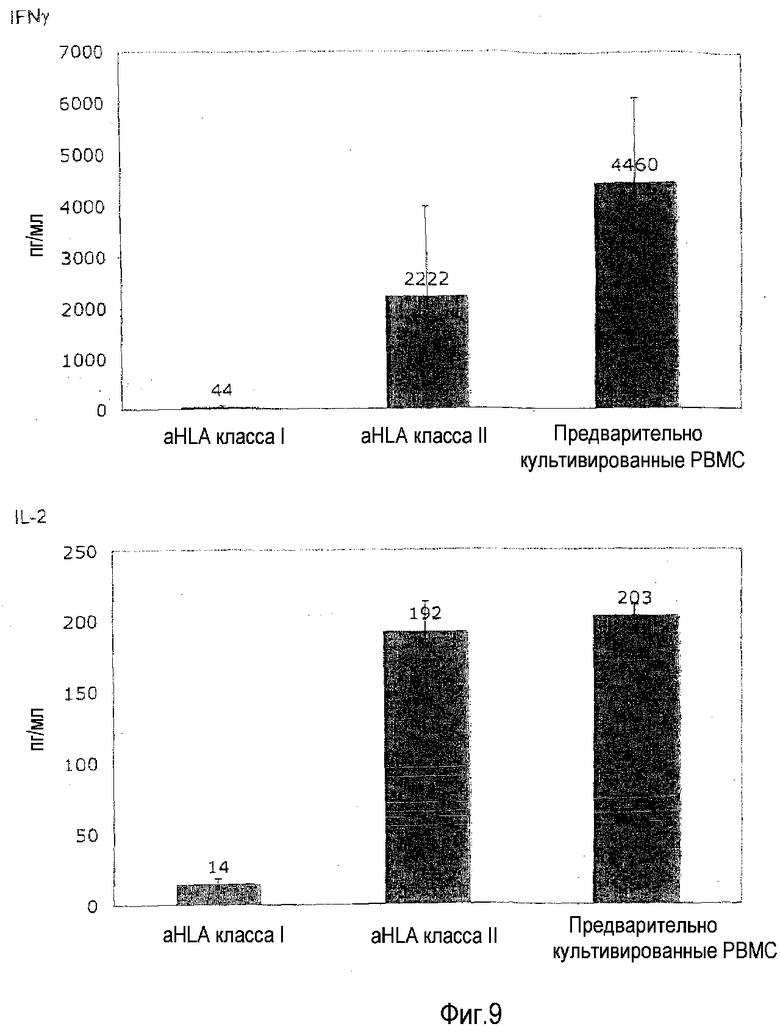
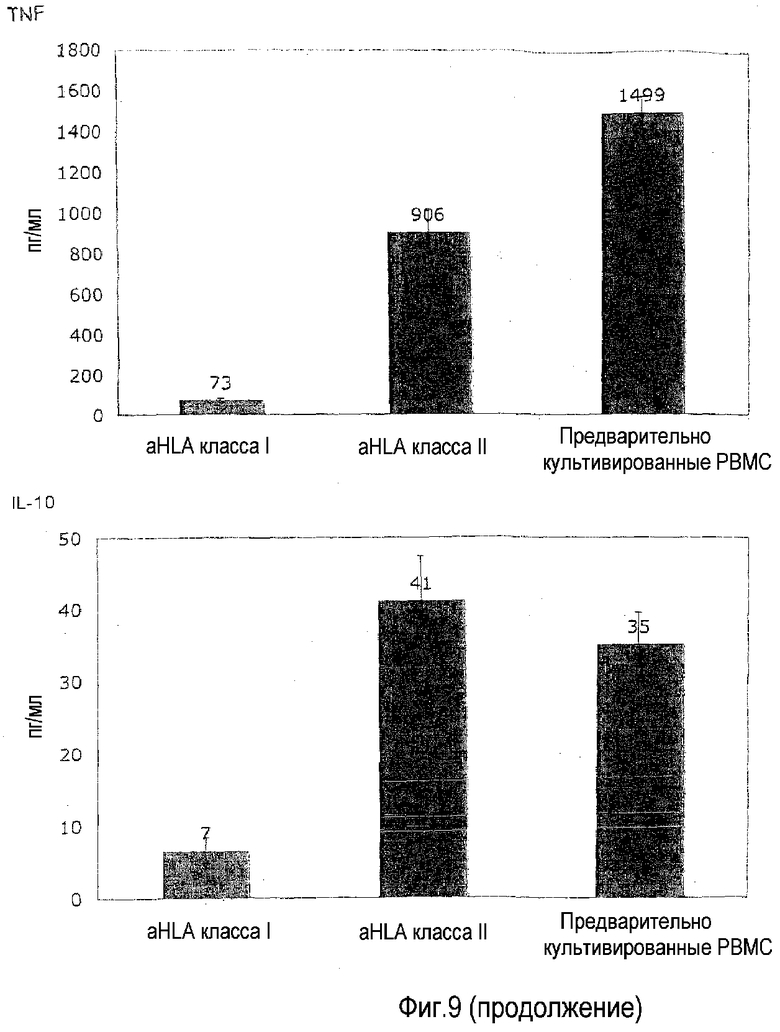
фиг.9: приобретение TGN1412-реактивности блокируется посредством МАТ к HLA-антигенам.
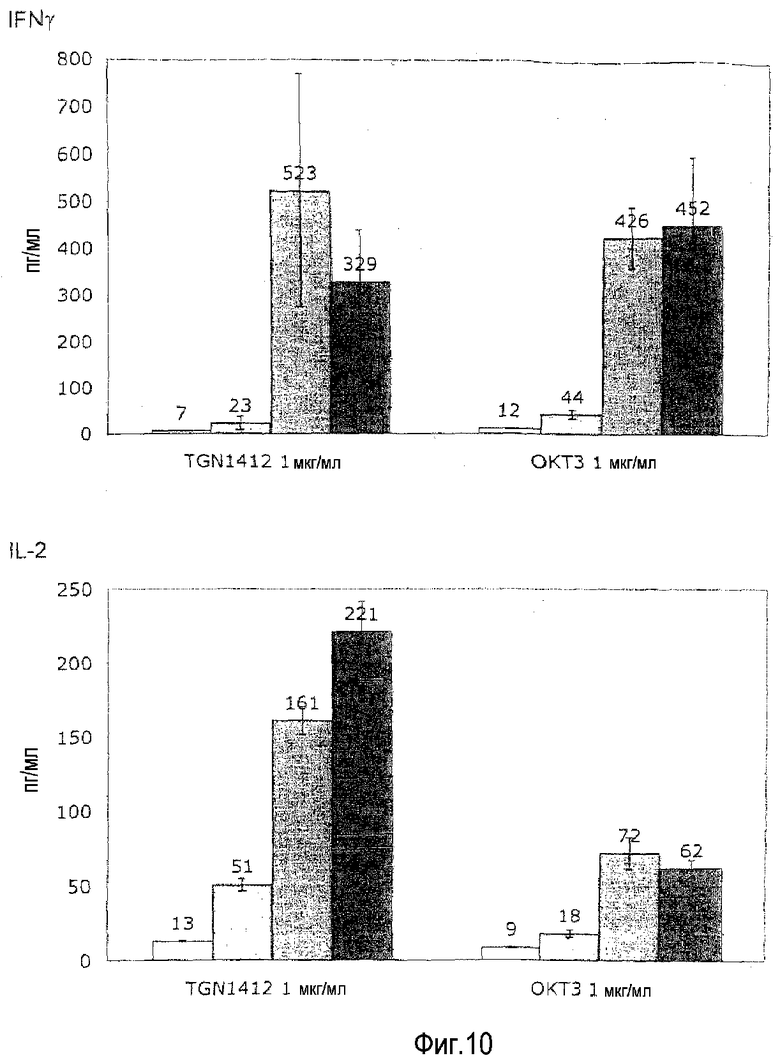
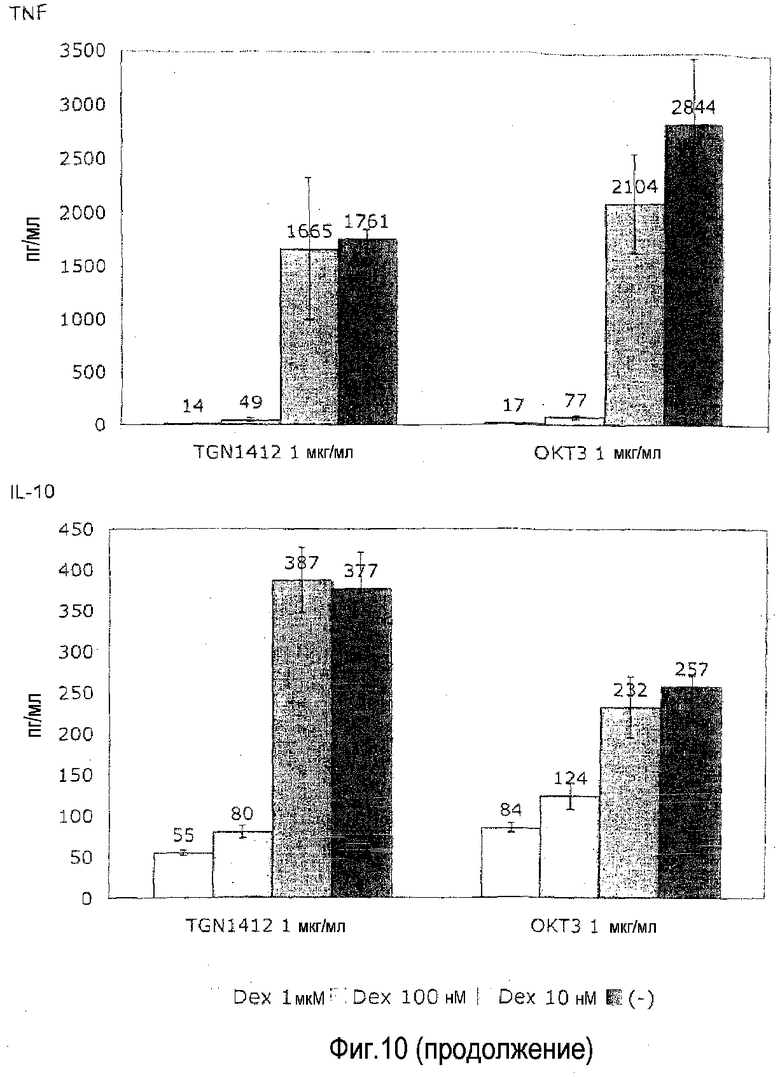
фиг.10: сравнимая чувствительность высвобождения цитокинов, индуцированная посредством OKT3 и TGN1412, к опосредованной кортикостероидами супрессии.
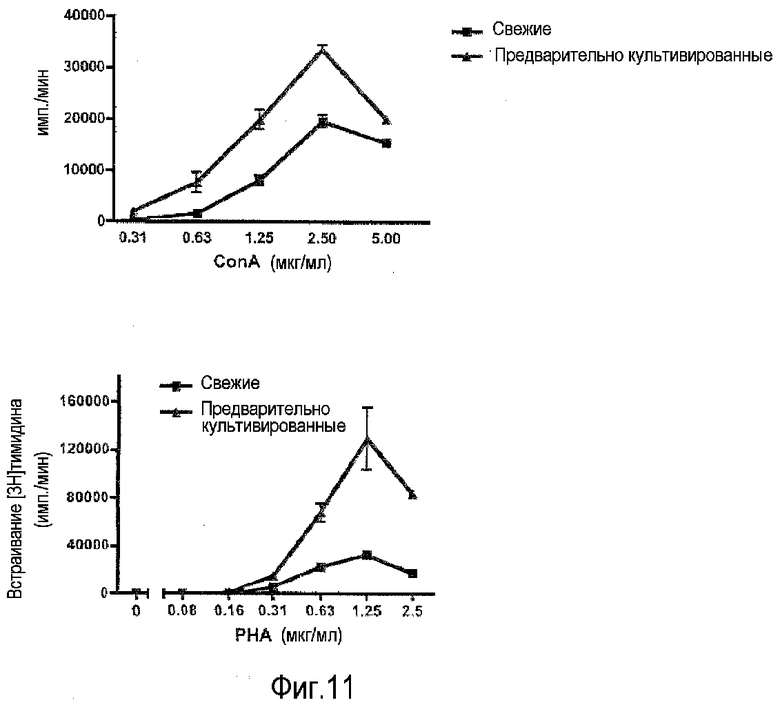
фиг.11: усиление пролиферативного ответа T-клеток на ConA и PHA в предварительно культивированных PBMC.
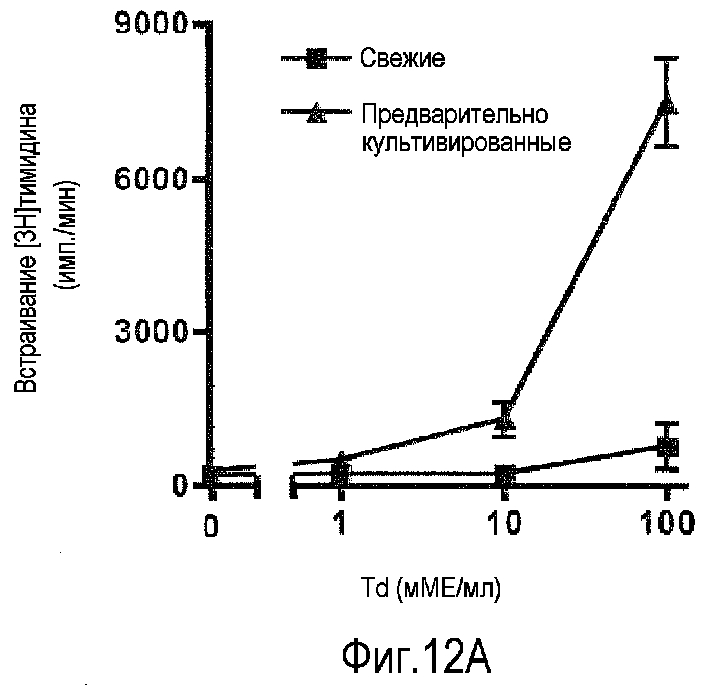
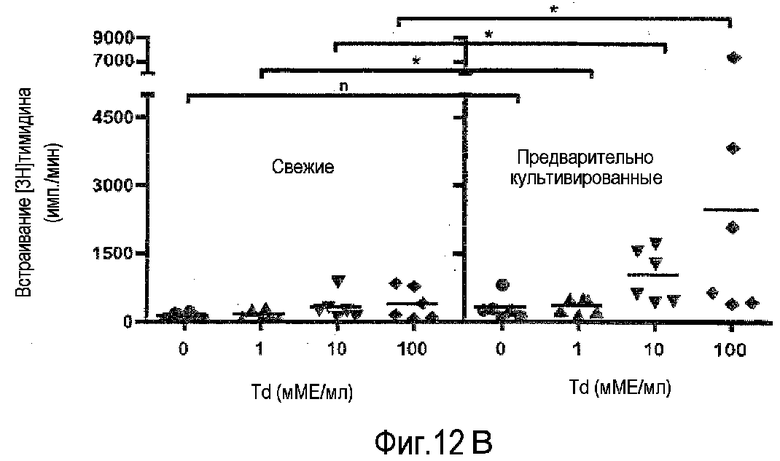
фиг.12: усиление повторного ответа на столбнячный/дифтерийный анатоксины в предварительно культивированных PBMC (A), компиляция данных от шести индивидуумов-доноров (B).
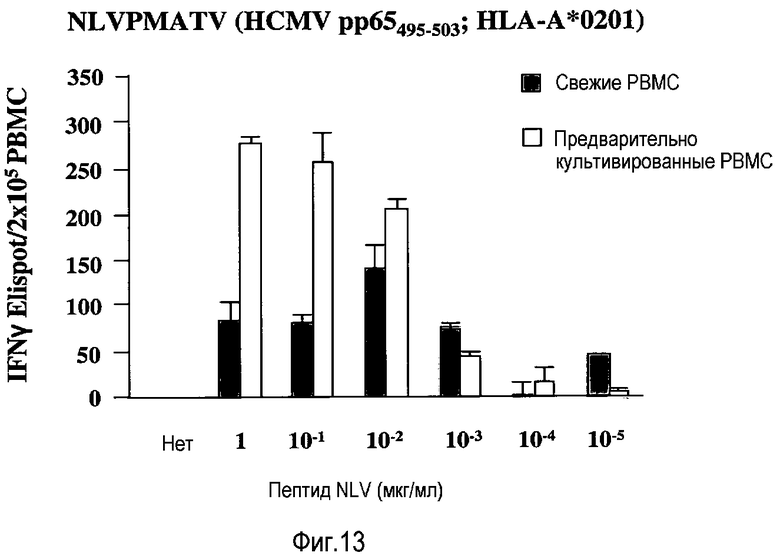
фиг.13: усиление пролиферативного ответа T-клеток на вирусный пептид в предварительно культивированных PBMC.
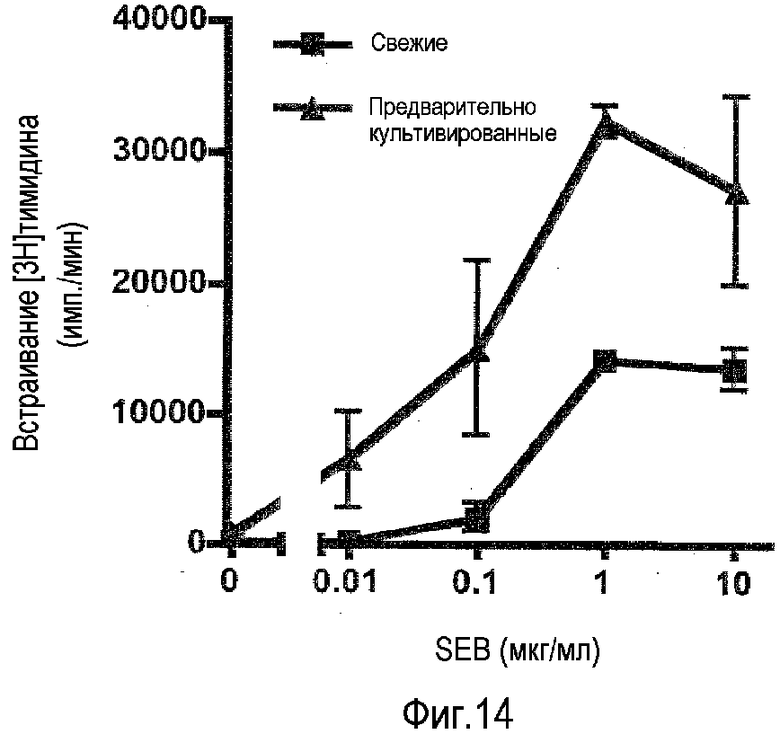
фиг.14: усиление пролиферативного ответа T-клеток на стафилококковый эндотоксин B в предварительно культивированных PBMC.
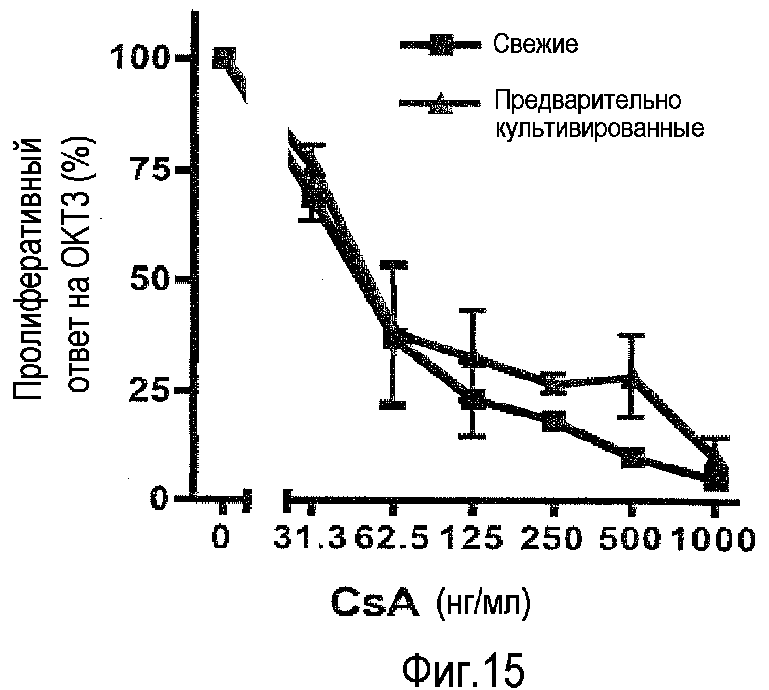
фиг.15: сниженная супрессия пролиферативного ответа T-клеток на высокие дозы CsA в предварительно культивированных PBMC.
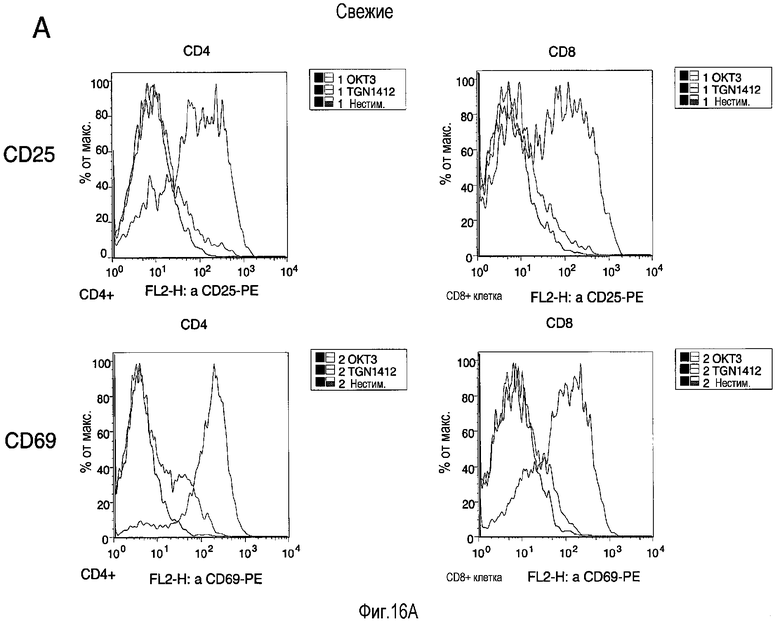
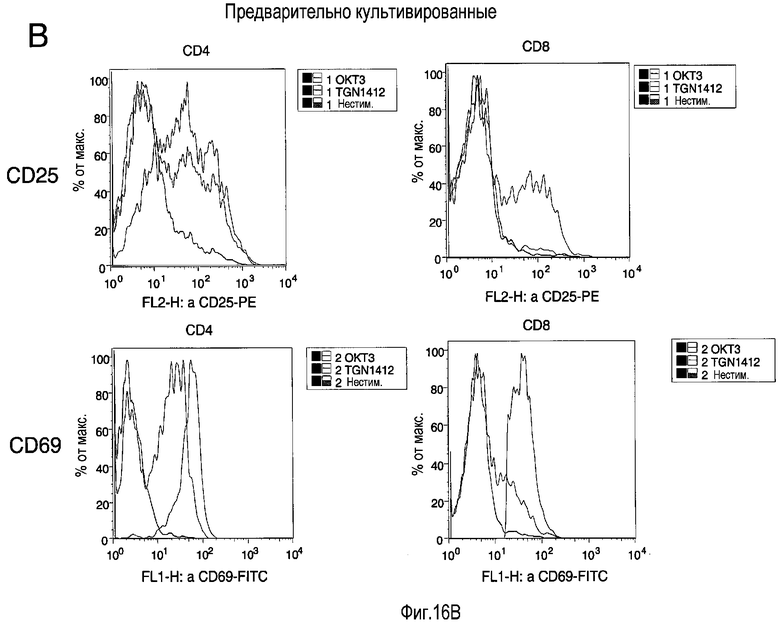
фиг.16: индукция маркеров активации на свежих (A) и предварительно культивированных (B) T-клетках посредством TGN1412 и OKT3 стимуляции. Использовали свежие (A) или предварительно культивированные в условиях с высокой плотностью в течение 2 суток (B) PBMC, собранные и повторно стимулированные в стандартных условиях (с низкой плотностью) с использованием или 1 мкг/мл OKT3 (черная линия), 1 мкг/мл TGN1412 (серая линия), или без стимулирующего антитела (закрашенная гистограмма). После 18 ч клетки собирали и окрашивали с использованием МАТ, специфичных к CD4, CD8, CD25 и CD69. Приведены гистограммы для CD25 (верхняя) и CD69 (нижняя), отобранных по CD4 (слева) или CD8 T-клетки (справа).
Изобретение проиллюстрировано примером.
Пример 1: Сравнительный пример
Для индукции высвобождения цитокинов в настоящем изобретении, а также в этом сравнительном примере использовали стандартную систему стимуляции PBMC, как ее используют исследователи по всему миру для изучения ответа PBMC человека на иммуномодулирующие средства. В этой системе использовали свежеполученные PBMC, выделенные из гепаринизированной венозной крови посредством центрифугирования в градиенте плотности (Lymphocyte Separation Medium LSM 1077, PAA Laboratories, Pasching, Germany) по инструкциям производителя. Альтернативно, свежую лейкоцитарную массу, полученную из камер системы редукции лейкоцитов (Caridian, Gambro BCT, Lakewood, CO, USA) в качестве побочного продукта при получении тромбоцитарной массы (Dietz et al., 2006), используют в качестве начального материала для очистки фиколлом с по существу такими же результатами. PBMC культивировали в 96-луночных планшетах для культуры ткани (Greiner bio-one, Frickenhausen, Germany), в которых 2×105 клеток стимулировали в 0,2 мл обогащенной среды для культивирования RPMI 1640 (GIBCO/Invitrogen, Long Island, NY, USA) с добавлением 10% аутологичной сыворотки или коммерчески доступной смешанной AB сыворотки (PAA Laboratories) с по существу такими же результатами. При использовании коммерчески доступной смешанной AB сыворотки ее следует предварительно тестировать в сравнении с аутологичной сывороткой для того, чтобы гарантировать, что получают сравнимые результаты.
В этой системе тканевой культуры свежевыделенные PBMC стимулировали растворимым TGN1412, поставляемым компанией TheraMab GmbH, Wurzburg. Оно представляло собой партию с тем же качеством GMP, что использовали во время лондонского исследования (Suntharalingam et al., 2006). В качестве положительного контроля для индукции высвобождения цитокинов использовали OKT3 клинической чистоты («Muromonomab», Janssen-Cilag, Neuss, Germany), о котором хорошо известно, что оно запускает высвобождение цитокинов как in vitro, так и у пациентов (Abramowicz et al., 1989). Через 24 часа панель цитокинов, включая основные про- и противовоспалительные факторы, обнаруженные в плазме добровольцев из исследования TGN1412 (Suntharalingam et al., 2006), анализировали посредством технологии анализа цитокинов на бусах (CBA) (Becton Dickinson, Mountain View, CA, USA) по инструкциям производителя.
Концентрация использованного как OKT3, так и TGN1412 составляла 1 мкг/мл, что находится в диапазоне оцениваемой концентрации, достигаемой в циркуляции у добровольцев во время лондонского исследования TGN1412 (Duff, 2006). Обширное титрование обоих антител, не представленное здесь, показало, что эта концентрация находится в оптимальном диапазоне для биологических ответов.
На фиг.1A показано, что растворимый TGN1412 не способен индуцировать высвобождение цитокинов в свежих PBMC. В отличие от этого OKT3 высоко эффективен в отношении индукции TNF, IFN-γ и IL-2, о которых известно, что все они вносят вклад в патологические проявления синдрома высвобождения цитокинов, а также противовоспалительного цитокина IL-10. PBMC от здорового донора выделяли посредством центрифугирования в плотности фиколла и культивирования в 0,2 мл среды RPMI1640 с добавлением 10% AB сыворотки в течение 24 ч при плотности 106/мл в 96-луночных плоскодонных планшетах для культуры ткани. Цитокины в супернатантах анализировали после 24 часов инкубирования посредством анализа цитокинов на бусах. Моноклональные антитела использовали в конечной концентрации 1 мкг/мл. Приведены среднее для трех значений и стандартное отклонение.
Неспособность TGN1412 и способность OKT3 индуцировать высвобождение этих и других цитокинов (не показано) в такой стандартной культуре PBMC воспроизвели минимум у 10 индивидуумов-доноров, и это согласуется с данными, поданными компанией TeGeneroAG и воспроизведенными в отчете Scientific Expert Group о безопасности I фазы клинических исследований (Duff, 2006).
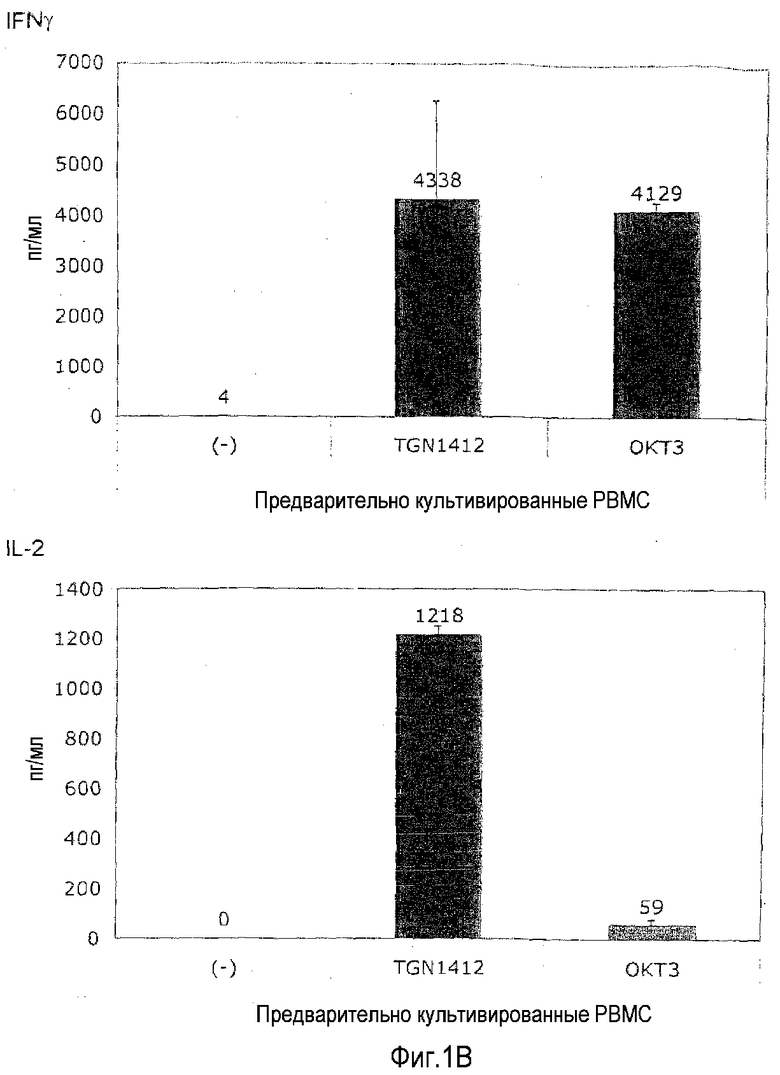
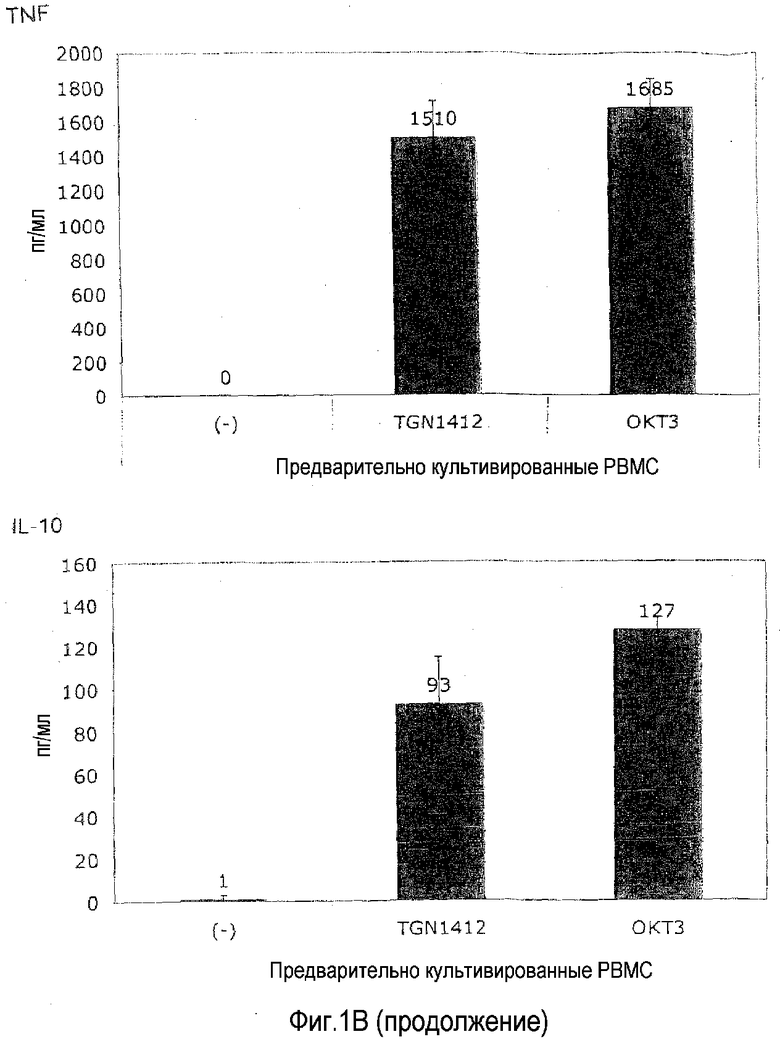
Пример 2: Ответ на TGN1412 после предварительного культивирования
Для эксперимента на фиг.1B PBMC от здорового донора культивировали в 1,5 мл среды в течение 2 суток при плотности 107/мл в 24-луночных плоскодонных планшетах для культуры ткани перед отмыванием и доведением до 106/мл. Используя эти клетки, выполняли тот же эксперимент, что описан в примере 1.
На фиг.1B показано, что отвечаемость на TGN1412 восстанавливалась, к удивлению, просто посредством предварительного культивирования PBMC в течение 2 суток без явной стимуляции. Когда 4 декабря 2008 года получали клетки, число полученных PBMC превышало необходимое для текущего эксперимента, и лишние клетки хранили в культуральной среде в течение 2 суток при 37°C. Когда эти клетки использовали для абсолютно такого же эксперимента, как выполняли ранее с использованием свежих клеток (фиг.1A), происходило что-то совершенно неожиданное: теперь TGN1412 индуцировал высвобождение цитокинов в степени, сравнимой с OKT3. На фиг.1B приведен пример такого эксперимента.
В связи с этим дополнительно исследовали воспроизводимость и основополагающего механизма.
Пример 3: Воспроизводимость ключевого наблюдения
На фиг.2 резюмирован эффект предварительного культивирования, оказываемый на реактивность на TGN1412 у 7 здоровых индивидуумов-доноров. Приведены данные для 7 здоровых индивидуумов-доноров, каждый обозначен символом. Условия для стимуляции антителом и для предварительного культивирования аналогичны фиг.1. Несмотря на то, что существуют специфичные для доноров вариации в ответах как на OKT3, так и на TGN1412, ясно, что во всех случаях свежие клетки доноров не могли отвечать на стимуляцию TGN1412 высвобождением цитокинов и что это рефракторное состояние утрачивали после 2 суток предварительного культивирования. Специфические для доноров вариации были ожидаемы, как проиллюстрировано посредством значительных различий в величине цитокинового шторма, перенесенного добровольцами в лондонском исследовании TGN1412 (Suntharalingam et al., 2006).
Пример 4: Оптимизация нового способа
На фиг.3-5 описаны параметры, определяющие приобретение чувствительности к TGN1412 в T-клетках периферической крови.
Для эксперимента с фиг.3 культивировали PBMC при плотности 106/мл (левые столбцы) или 107/мл (правые столбы) в течение 2 суток перед стимуляцией посредством TGN1412 при плотности 106/мл, как описано на фиг.1.
Для эксперимента с фиг.4 свежие PBMC (левые столбцы из тройки) и PBMC, предварительно культивированные при плотности 107/мл в течение 24 (средние столбцы из тройки) или 48 ч (правые столбцы из тройки), стимулировали OKT3 или TGN1412 в течение 24 ч в условиях, приведенных на фиг.1.
Для эксперимента с фиг.5 PBMC культивировали в течение 2 суток при высокой плотности (107/мл, правые столбцы из тройки) или при низкой плотности (106/мл, левые и средние столбцы из тройки) в 1,5 мл культурах. В группе, представленной в виде средних столбцов из троек, эти культуры с низкой плотностью дополнительно содержали вставку с полупроницаемой мембраной, на которой дополнительные PBMC культивировали при высокой плотности (106/мл ниже мембраны, 107/мл выше мембраны, средние столбцы из троек). Клетки повторно стимулировали и анализировали, как приведено на фиг.1.
Как показано выше, PBMC приобретают чувствительность к TGN1412 посредством предварительного культивирования в среде с 10% аутологичной или коммерчески доступной AB сывороткой. Авторы настоящего изобретения тестировали роль следующих параметров в приобретении чувствительности к TGN1412.
Плотность клеток. В отличие от стандартных анализов стимуляции PBMC, где клетки культивировали при плотности 106/мл или 2×105/см2 культуральной лунки, «стоянку» в течение 2 суток осуществляли при в 10 раз более высокой плотности. На фиг.3 показано, что предварительное культивирование PBMC при высокой (107/мл или 2×106/см2), но не при низкой (106/мл или 2×105/см2) плотности, индуцирует реактивность на TGN1412 во вторичной культуре. В связи с требованиями к плотности клеток и культуре в течение 2 суток, половину среды для культивирования осторожно заменяли свежей предварительно нагретой средой после 24 ч для того, чтобы обеспечить клетки достаточными питательными веществами.
Время. На фиг.4 показано, что полную реактивность на TGN1412 (сравнимую с реактивностью на OKT3) достигали после 2 суток предварительного культивирования. Предварительное культивирование в течение 1 суток вело только к незначительному повышению реактивности.
Необходимость межклеточного контакта. Необходимость высокой плотности клеток во время предварительного культивирования PBMC для приобретения реактивности на TGN1412 может быть обусловлена потребностью в межклеточном контакте и/или действием растворимых факторов, которые должны достичь определенной концентрации, чтобы содействовать созреванию до реактивного состояния. Используя систему Transwell (Corning incorporated, Lowell, MA, USA), в которой клетки, культивировавшиеся при высокой плотности, отделяли от тех клеток, которые культивировали при низкой плотности, посредством мембраны с порами 8 мкм, допускающей диффузию растворимых факторов, на фиг.5 показано, что необходим межклеточный контакт.
Пример 5: дополнительные характеристики нового способа
На фиг.6 показано, что предварительно культивированные, но не свежевыделенные PBMC, пролиферируют в ответ на TGN1412. Свежие и предварительно культивированные PBMC получали и культивировали, как описано для фиг.1A и B соответственно. На 3 сутки добавляли 1 мкКи 3H-тимидина, культуры собирали через 16 ч и обрабатывали для подсчета импульсов в жидкости. Помимо высвобождения цитокинов поликлональная активация T-клеток приводит к пролиферации, которую можно измерить в виде инкорпорированной радиоактивности, инкорпорированной здесь в виде меченного тритием тимидина. Как видно на фиг.6, OKT3 стимулировало пролиферацию как свежих, так и предварительно культивированных PBMC, тогда как TGN1412 был только способен индуцировать пролиферацию в предварительно культивированных PBMC. Таким образом, пролиферацию также можно использовать в качестве считываемых данных.
На фиг.7 показано, что в предварительно культивированных PBMC TGN1412 высвобождает провоспалительные цитокины из CD4 клеток памяти. PBMC, предварительно культивированные в течение 2 суток при высокой плотности, стимулировали в течение 18 часов, используя 1 мкг/мл OKT3 или TGN1412. Вместо 18 ч, как правило, возможна стимуляция в течение периода времени от 15 до 20 ч, предпочтительно от 16 до 18 ч. Во время последних 4 часов культуры, 5 мкг/мл брефелдина A добавляли для блокирования клеточного экспорта цитокинов. После окрашивания поверхностей клеток конъюгированным с флуорохромом МАТ к CD4 и CD45RO (маркер клеток памяти), клетки фиксировали, пермеабилизировали и окрашивали МАТ к TNF - левые изображения - или IFN-γ - правые изображения - (все из BD Pharmingen, Mountain View, CA, USA).
На проточном цитометре BD Calibur получали 15000 событий отбора живых клеток, а данные анализировали с использованием программного обеспечения FlowJo (Three Star Inc., Ashland, OR, USA). Цитокиновый шторм, вызываемый посредством OKT3 у пациентов, является хорошо известным феноменом, и сопроводительная информация, предоставляемая с этим лекарственным средством (Muromonomab, Janssen-CILAG), в прямой форме предупреждает об этом синдроме. Чтобы сравнить клеточный источник двух ключевых провоспалительных цитокинов TNF и IFN-γ в ответ на OKT3 и TGN1412, их задерживали в клетках посредством блокады их транспорта через аппарат Гольджи, используя лекарственное средство брефелдин A (Sigma Aldrich, Steinheim, Germany), и выявляли посредством внутриклеточного окрашивания фиксированных пермеабилизированных клеток флуоресцентными МАТ со специфичностью к TNF и IFN-γ соответственно (Becton Dickinson, Mountain View, CA, USA). Параллельно определяли фенотип клеточной поверхности стимулированных PBMC. Таким образом, идентифицировали основную продуцирующую цитокины субпопуляцию T-клеток, CD4 T-клетки, и затем делили на субпопуляции нативных клеток (CD45RO-) и клеток памяти (CD45RO+). На фиг.7 показано, что как в свежих, так к в предварительно культивированных PBMC OKT3 и TGN1412 запускали образование цитокинов преимущественно в субпопуляции CD4 клеток памяти.
На фиг.8 показано, что высвобождение TNF из предварительно культивированных PBMC придерживается такой же кинетики, при индукции посредством или OKT3, или TGN1412. PBMC, предварительно культивированные при высокой плотности, получали, как на фиг.1B, и стимулировали с использованием 1 мкг/мл OKT3 или TGN1412. Супернатанты собирали в отмеченные моменты времени и содержание TNF анализировали, как на фиг.1. Высвобождение TNF, наиболее важного провоспалительного цитокина в «цитокиновом шторме», in vivo придерживается той же кинетики, когда индуцируют посредством или OKT3, или TGN1412 (Abramowicz et al., 1989; Suntharalingam et al., 2006). Таким образом, для двух МАТ сравнивали кинетику высвобождения TNF в предварительно культивированных PBMC и обнаружили, что они по существу одинаковы.
Пример 6: основополагающий механизм для индукции реактивности на TGN142 во время предварительного культивирования при высокой плотности: распознавание собственного MHC
В этом примере, не ограничиваясь теорией, предоставлена функциональная информация об изобретении. Исходя из работы, выполненной на мышах, предполагали, что распознавание молекул главного комплекса гистосовместимости (MHC; у человека HLA) антигенными рецепторами (TCR) T-клеток в лимфоидных органах (лимфатические узлы, селезенка и т.д.) подготавливает TCR к усиленной передаче сигнала во время последующей встречи с антигеном (Stefanova et al., 2002). Этот процесс обозначают как «MHC-сканирование» и он описывает постоянное взаимодействие TCR тканевых T-клеток с молекулами HLA на их соседних клетках независимо от того, какие пептиды они содержат (Stefanova et al., 2002). Это взаимодействие ведет к подготовке TCR и сборке сигнальной платформы, которая облегчает полную активацию T-клеток, когда TCR встречает подходящий антигенный пептид. Этот процесс в данной области также называют «тонизирующей» передачей сигнала TCR. Поскольку в лаборатории авторов настоящего изобретения ранее показано, что передача сигналов суперагониста CD28 происходит через амплификацию слабых TCR сигналов (Dennehy et al., 2007), была выдвинута гипотеза, что потеря этого подготавливающего сигнала делает циркулирующие T-клетки (которые не имеют межклеточного контакта, в отличие от ситуации в лимфоидных органах) невосприимчивыми к стимуляции TGN1412. Ее тестировали, включая блокирующие МАТ, которые реагируют со всеми молекулами HLA класса I и класса II человека во время двухсуточного предварительного культивирования (МАТ 646-2.6 и Tű39, Becton-Dickenson) в концентрации 10 мкг/мл.
На фиг.9 показано, что специфические к HLA класса I, и в меньшей степени к HLA класса II, МАТ способны блокировать приобретение TGN1412 реактивности, что иллюстрирует необходимость распознавания HLA посредством TCR клеток, приобретающих эту реактивность. Предварительное культивирование PBMC с высокой плотностью выполняли, как задано на фиг.1B. В некоторые предварительные культуры с высокой плотностью добавляли МАТ к HLA класса I или HLA класса II в концентрации 10 мкг/мл.
Эта находка весьма убедительно подсказывает, что взаимодействие TCR с молекулами HLA в плотно упакованном лимфоидном органе является предпосылкой для сильной реактивности на TGN1412, которую перенесли добровольцы в лондонском исследовании, и что эту ситуацию имитирует культура с высокой плотностью in vitro, таким образом, восстанавливающая реактивность циркулирующих T-клеток, которые утратили межклеточный контакт, до реактивности T-клеток в лимфоидных органах. Это также объясняет, почему ответ на OKT3 наблюдали не только в предварительно культивированных, но также в свежих PBMC. Поскольку в отличие от TGN1412 OKT3 обращается к самому TCR, нет необходимости в «подготовке» этого рецептора посредством взаимодействии с HLA-молекулами. Дифференциальная способность МАТ, специфичных к HLA класса I и класса II, блокировать приобретение реактивности на TGN1412 объясняется тем фактом, что более 90% доступных HLA молекул в культуре PBMC относятся к классу I.
Пример 7: тестирование способности кортикостероидов контролировать опосредованное TGN1412 высвобождение цитокинов
Индуцированный OKT3 цитокиновый шторм обычно контролируют посредством профилактики или вмешательства с использованием кортикостероидов в высоких дозах (Goldman et al., 1989). До сегодняшнего дня не было возможности тестировать чувствительность индуцированного TGN1412 высвобождения цитокинов к кортикостероидам, поскольку не существовало тестовой системы. Таким образом, авторы настоящего изобретения использовали новый способ для сравнения чувствительности высвобождения цитокинов в свежих и предварительно культивированных PBMC к дексаметазону («Dex», Sigma-Aldrich).
На фиг.10 показано, что индуцированное TGN1412 высвобождение цитокинов чувствительно к кортикостероидам. Предварительно культивированные в высокой плотности клетки получали и стимулировали посредством МАТ, как описано в фиг.1B. Где указано, дексаметазон включали в заданных конечных концентрациях, а цитокины измеряли после 24 ч культивирования.
Высвобождение цитокинов, индуцированное посредством как OKT3, так и TGN1412 полностью подавляли посредством наивысшей используемой дозы (1 мкМ), и, кроме того, почти полностью подавляли при вдесятеро меньшей дозе (100 нМ). Это убедительно подсказывает, что индуцированное TGN1412 высвобождение цитокинов можно контролировать подходящей лекарственной терапией кортикостероидами, которую в клинике используют для пациентов, которых лечат OKT3.
Пример 8: реактивность на митогенные лектины
Митогенные лектины конканавалин A (ConA) и фитогемагглютинин (PHA) обычно используют для тестирования реактивности T-клеток в препаратах PBMC. Эти лектины действуют через один или несколько гликопротеинов клеточной поверхности, где они связываются с остатками сахаров. На фиг.11 показано, что для обоих лектинов пролиферативный ответ T-клеток значительно усиливается, если PBMC предварительно культивировали в течение 2 суток, следуя приведенному выше протоколу.
Пример 9: антигенспецифический вторичный ответ
В результате преднамеренной вакцинации или естественной инфекции частота T-лимфоцитов, специфичных к данному микробному антигену, повышается до уровня, при котором повторная стимуляция in vitro делает возможным обнаружение пролиферативных ответов. Классическими примерами таких антигенов являются дифтерийный и столбнячный анатоксины. Такие препараты обычно используют для оценки статуса вакцинации индивидуумов. Для этой цели Sanofi Pasteur предлагает два анатоксина в виде одного препарата. На фиг.12A проиллюстрировано, что действительно вторичный ответ на столбнячный/дифтерийный анатоксины значительно усиливается посредством предварительного культивирования PBMC при высокой плотности в течение 2 суток. На фиг.12B компилированы данные от шести индивидуумов-доноров. * обозначает статистическую значимость p<0,05.
Пример 10: ответ CD8 T-клеток на пептид, полученный из цитомегаловируса
Вторичные ответы на столбнячный и дифтерийный анатоксины обращены к CD4 T-клеткам. Чтобы протестировать, влияет ли предварительное культивирование при высокой плотности клеток также на вторичный ответ CD8 T-клеток, PBMC положительного по HLA A2 индивидуума, ранее подвергавшегося воздействию цитомегаловируса, стимулировали пептидом, представленным посредством HLA A2 CD8 T-лимфоцитам индивидуума. В качестве считываемых данных выбирали число секретирующих IFN-γ CDS T-клеток, поскольку известно, что такие CD8 T-клетки памяти быстро секретируют IFN-γ при стимуляции через антигенный рецептор. Количественное определение осуществляли с использованием так называемого анализа ELISPOT, где PBMC культивировали на фильтровальной мембране, покрытой реагирующим с IFN-γ моноклональным антителом. Таким образом, IFN-γ, захваченный во время 24 ч инкубационного периода, затем визуализировали с использованием второго меченого ферментом МАТ и превращения бесцветного в цветной субстрат. Как показано на фиг.13, предварительное культивирование PBMC в течение 2 суток при высокой плотности клеток значительно увеличивало частоту продуцирующих IFN-γ клеток в ответ на вирусный пептид.
Пример 11: ответ на «суперантигенный» стафилококковый эндотоксин B
Некоторые бактерии секретируют так называемые суперантигены, которые способны стимулировать значительно большую часть T-лимфоцитов (5-20%) через их антигенные рецепторы, чем в случае стандартных антигенов (1/100,000 у неиммунизированного индивидуума). Соответственно пролиферативный ответ на такие суперантигены можно измерить in vitro, используя PBMC. На фиг.14 показано, что предварительное культивирование PBMC при высокой плотности в течение 2 суток также значительно усиливало чувствительность этого анализа.
Пример 12: тестирование иммуносупрессорных лекарственных средств
Активность иммуномодулирующих лекарственных средств можно неверно оценить при использовании свежих вместо предварительно культивированных PBMC. Циклоспорин A (CsA) представляет широко используемое иммуносупрессорное лекарственное средство, которое действует на пути передачи сигнала антигенного рецептора T-клетки. Соответственно сниженную способность передачи сигналов циркулирующих T-клеток можно перевести в повышенную чувствительность к этому лекарственному средству. На фиг.15 показано, что это действительно верно: несмотря на то, что и свежие, и предварительно культивированные PBMC (из того же исходного препарата) отвечали на стимуляцию TCR с использованием OKT3, предварительно культивированные PBMC не так просто супрессировать посредством высоких доз CsA, как свежие PBMC (p<0,03).
Пример 13: индукция маркеров активации T-клеток на клеточной поверхности
На фиг.16 показана индукция маркеров активации на свежих (A) и предварительно культивированных (B) T-клетках посредством стимуляции TGN1412 и OKT3. PBMC использовали свежими (A) или предварительно культивировали в условиях высокой плотности в течение 2 суток (B), собирали и повторно стимулировали в стандартных условиях (при низкой плотности) с использованием одного из 1 мкг/мл OKT3 (черная линия), 1 мкг/мл TGN1412 (серая линия) или без стимулирующего антитела (заполненная гистограмма). После 18 ч клетки собирали и окрашивали с использованием МАТ, специфичных к CD4, CD8, CD25 и CD69. Гистограммы приведены для CD25 (верх) и CD69 (низ), отобранных по CD4 (слева) или CD8 T-клеткам (справа).
Источники информации
1. Abramowicz, D. et al., Transplantation 47, 606-608 (1989).
2. Dennehy, K.M. et al., J Immunol 178, 1363-1371 (2007).
3. Dietz, A.B. et al., Transfusion 46, 2083-2089 (2006).
4. Duff, G.W. C Expert Scientific Group on Phase One Clinical Trials Final Report (Norwich, UK, Stationary Office 2006).
5. Gogishvili, T. et al., PLoS ONE 4, e4643 (2009).
6. Goldman, M. et al., Lancet 2, 802-803 (989).
7. Hunig, T., Adv Immunol 95, 111-148 (2007).
8. Nguyen, D.H. et al., Proc Natl Acad Sci USA 103, 7765-7770 (2006).
9. Schraven, B. et al., Immunity 28, 591-595 (2008).
10. Stefanova, I. et al., Nature 420, 429-434 (2002).
11. Suntharalingam, G. et al., N Engl J Med 355, 1018-1028 (2006).
| название | год | авторы | номер документа |
|---|---|---|---|
| ФАРМАЦЕВТИЧЕСКИЙ ПРЕПАРАТ | 2009 |
|
RU2542512C2 |
| ПРОИЗВОДНЫЕ 5-(4-МЕТАНСУЛЬФОНИЛФЕНИЛ)ТИАЗОЛА ДЛЯ ЛЕЧЕНИЯ ОСТРЫХ И ХРОНИЧЕСКИХ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ | 2009 |
|
RU2495031C2 |
| УЛУЧШЕННЫЕ СПОСОБЫ ПРОИЗВОДСТВА СРЕДСТВ АДОПТИВНОЙ КЛЕТОЧНОЙ ТЕРАПИИ | 2015 |
|
RU2741899C2 |
| АНТИТЕЛА ПРОТИВ αβTCR | 2012 |
|
RU2630656C2 |
| СПОСОБ УСИЛЕНИЯ ИММУННОГО ОТВЕТА НА АНТИГЕН У МЛЕКОПИТАЮЩИХ | 2004 |
|
RU2341289C2 |
| КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ АРТРИТА | 2009 |
|
RU2563360C2 |
| СПОСОБ ЛЕЧЕНИЯ АУТОИММУННОГО ЗАБОЛЕВАНИЯ (ВАРИАНТЫ) | 2009 |
|
RU2539110C2 |
| СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ | 2009 |
|
RU2540013C2 |
| ПРОБИОТИЧЕСКИЙ ШТАММ Bifidobacterium longum, КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ТАКОЙ ШТАММ, И ЕГО ПРИМЕНЕНИЕ | 2008 |
|
RU2473681C2 |
| ИММУНОМОДУЛИРУЮЩИЕ ПРОБИОТИЧЕСКИЕ МОЛОЧНО-КИСЛЫЕ БАКТЕРИИ | 2007 |
|
RU2535974C2 |
Изобретение относится к медицине и описывает способ тестирования предполагаемого или известного иммуномодулирующего лекарственного средства для активации Т-клеток, который включает стадию приведения в контакт культуры мононуклеарных клеток периферической крови (РВМС) с предварительно определяемым количеством предполагаемого или известного иммуномодулирующего лекарственного средства in vitro и наблюдение активации Т-клеток в культуре РВМС, используя систему считывания, при контакте с предполагаемым или известным иммуномодулирующим лекарственным средством, где плотность клеток культуры РВМС во время стадии предварительного культивирования составляет по меньшей мере 2×106/мл, предпочтительно по меньшей мере 5×106/мл, более предпочтительно по меньшей мере 107/мл, или по меньшей мере 4×105/cм2, предпочтительно по меньшей мере 106/см2, наиболее предпочтительно по меньшей мере 2×106/cм2, при этом предварительную культуру РВМС культивируют в течение по меньшей мере 12 ч. Изобретение обеспечивает улучшенное средство для тестирования иммуномодулирующих лекарственных средств in vitro. 9 з.п. ф-лы, 16 ил., 13 пр.
1. Способ тестирования предполагаемого или известного иммуномодулирующего лекарственного средства для активации Т-клеток, который включает стадию приведения в контакт культуры мононуклеарных клеток периферической крови (РВМС) с предварительно определяемым количеством предполагаемого или известного иммуномодулирующего лекарственного средства in vitro и наблюдение активации Т-клеток в культуре РВМС, используя систему считывания, при контакте с предполагаемым или известным иммуномодулирующим лекарственным средством, где плотность клеток культуры РВМС во время стадии предварительного культивирования составляет по меньшей мере 2×106/мл, предпочтительно по меньшей мере 5×106/мл, более предпочтительно по меньшей мере 107/мл, или по меньшей мере 4×105/cм2, предпочтительно по меньшей мере 106/см2, наиболее предпочтительно по меньшей мере 2×106/cм2, при этом предварительную культуру РВМС культивируют в течение по меньшей мере 12 ч.
2. Способ по п.1, где система считывания наблюдает в культуре РВМС высвобождение по меньшей мере одного цитокина из РВМС, наблюдает пролиферацию клеток или представляет собой другую подходящую систему считывания, такую как система считывания изменений экспрессии гена, экспрессии белка и/или посттрансляционных модификаций.
3. Способ по п.1, где стадию предварительного культивирования осуществляют посредством хранения культуры РВМС в течение по меньшей мере 24 ч, предпочтительно по меньшей мере 36 ч, более предпочтительно по меньшей мере 45 ч, при температуре от 35°С до 40°С, предпочтительно от 36°С до 38°С, в отсутствие иммуномодулирующих лекарственных средств и до контакта с иммуномодулирующим лекарственным средством, подлежащим тестированию.
4. Способ по п.1, где иммуномодулирующее лекарственное средство представляет собой иммуностимулирующее лекарственное средство, такое как антитело, предпочтительно моноклональное антитело, в частности специфичное к CD28 человека суперагонистическое моноклональное антитело, или выбранное из группы, состоящей из пектинов, таких как конканавалин А (СоnА) или фитогемагглютинин (РНА), природных экстрактов, содержащих пектины, таких как экстракт эхинацеи или экстракт омелы, суперантигенов, таких как стафилококковый энтеротоксин В (SЕВ), стафилококковый энтеротоксин A (SEA), токсин синдрома токсического шока-1 (TSST-1), стафилококковых пирогенов, таких как стафилококковый пирогенный энтеротоксин В (SРЕВ), суперантигенов, продуцируемых микоплазмами, такими как Mycoplasma arthritidis, или бактериями черной чумы, такими как Yersinia pseudotuberculosis, суперантигенов, продуцируемых определенными патогенными вирусами, такими как ЭБВ или ВИЧ-1.
5. Способ по п.1, где наблюдаемый цитокин выбирают из группы, состоящей из TNF, IF-γ, IL-1β, IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL12p70, IL-13, IL-14, IL-15, IL-16, IL-17, IL-21, IL-35 и LT и их сочетаний.
6. Способ по п.1, где предварительно культивированную культуру РВМС дополнительно приводят в контакт с предполагаемым лекарственным средством для ослабления высвобождения по меньшей мере одного цитокина, одновременно с приведением в контакт культуры РВМС с известным иммуномодулирующим, в частности стимулирующим, лекарственным средством или впоследствии после предварительно определяемого периода времени или прежде за предварительно определяемый период времени, где дополнительно наблюдают высвобождение цитокинов.
7. Способ по п.6, где предварительно определяемый период времени находится в диапазоне от 10 с до 12 ч, предпочтительно в диапазоне от 10 с до 1 ч.
8. Способ по п.6, где предполагаемым лекарственным средством для ослабления высвобождения цитокинов является кортикостероид.
9. Способ по п.6, где предполагаемым лекарственным средством для ослабления высвобождения цитокинов является иммуносупрессорное лекарственное средство, выбранное из группы, состоящей из рапамицина или ингибиторов кальциневрина, таких как циклоспорин А, воклоспорин или такролимус.
10. Способ по п.1, где РВМС получены из образца цельной крови индивидуума, при этом способ полезен для предсказания реактивности индивидуума к иммуномодулирующему лекарственному средству.
| Ganesh Suntharalingam et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Приспособление для увеличения сцепной силы тяги паровозов и других повозок | 1919 |
|
SU355A1 |
| Ледовских, Мария Владимировна | |||
| Сравнительное изучение капсулированных препаратов циклоспорина А (контроль качества и терапевтический лекарственный мониторинг) | |||
| Автореферат диссертации, 2006. | |||

