Связанные заявки
По данной заявке испрашивается приоритет по предварительным заявкам США № 60/758426, поданной 12 января 2006 года, 60/759085, поданной 12 января 2006 года, и 60/801991, поданной 18 мая 2006 года, которые включены в настоящее описание в качестве ссылок в полном объеме.
Техническая область
Настоящее изобретение относится к антагонистам OX-2/CD200 (обозначаемым в настоящем документе как CD200) и способам уничтожения или элиминации клеток, сверхэкспрессирующих CD200 у субъекта со злокачественной опухолью или аутоиммунным заболеванием. Способы терапии для лечения злокачественной опухоли представляют собой сочетание двух механизмов. Более конкретно, настоящее изобретение относится к лечению злокачественной опухоли с использованием терапии, которая: (1) препятствует взаимодействию между CD200 и его рецептором, блокируя иммунную супрессию, обеспечивая посредством этого уничтожение злокачественных клеток; и/или (2) непосредственно уничтожает злокачественные клетки посредством либо (a) антителозависимой клеточной цитотоксичности или опосредуемой комплементом цитотоксичности, либо (b) нацеливания на клетки с использованием слитой молекулы, которая включает нацеливающую на CD200 часть. Также настоящее изобретение относится к способу лечения аутоиммунных нарушений посредством терапии, которая повышает антителозависимую клеточную цитотоксичность и/или опосредуемую комплементом цитотоксичность CD200-положительных иммунных клеток.
Предпосылки изобретения
В ответе организма на болезненное состояние, включая злокачественную опухоль, участвуют различные механизмы. Например, CD4+ T-хелперные клетки играют ключевую роль в эффективном иммунном ответе против различных злокачественных опухолей посредством обеспечения эффекторных клеток стимулирующими факторами. Полагают, что цитотоксические T-клетки являются наиболее эффективными клетками для устранения злокачественных клеток, и T-хелперные клетки активируют цитотоксические T-клетки посредством секреции Th1-цитокинов, таких как IL-2 и IFN-γ. Было показано, что при различных злокачественных опухолях T-хелперные клетки обладают измененным фенотипом по сравнению с клетками, встречающимися у здоровых индивидов. Одним из выраженных измененных признаков является сниженная продукция Th1-цитокинов и переключение на продукцию Th2-цитокинов. (См., например, Kiani, et al., Haematologica 88:754-761 (2003); Maggio, et al., Ann Oncol. 13 Suppl 1:52-56 (2002); Ito, et al., Cancer 85:2359-2367 (1999); Podhorecka, et al., Leuk Res 26:657-660 (2002); Tatsumi, et al., J. Exp. Med. 196:619-628 (2002); Agarwal, et al., Immunol. Invest 32:17-30 (2003); Smyth, et al., Ann Surg Oncol. 10:455-462 (2003); Contasta, et al., Cancer Biother Radiopharm 18:549-557 (2003); Lauerova, et al., Neoplasma 49:159-166 (2002)). Было показано, что реверсия этого переключения цитокинов на профиль Th1 усиливает противоопухолевые эффекты T-клеток. (См. Winter, et at., Immunology 108:409-419 (2003); Inagawa, et al., Anticancer Res 18:3957-3964 (1998)).
Механизмы, лежащие в основе способности опухолевых клеток направлять экспрессию цитокинов T-хелперными клетками с Th1 на Th2, включают секрецию цитокинов, таких как IL-10 или TGF-β, а также экспрессию поверхностных молекул, взаимодействующих с клетками иммунной системы. CD200, молекула, экспрессируемая на поверхности дендритных клеток, которая обладает высокой степенью гомологии с молекулами генного семейства иммуноглобулинов, вовлечена в иммунную супрессию (Gorczynski et al., Transplantation 65:1106-1114 (1998)). Например, было показано, что экспрессирующие CD200 клетки могут ингибировать стимуляцию продукции Th1-цитокинов.
Хотя иммунные клетки могут способствовать атаке на злокачественные клетки и их устранению, в определенных случаях, таких как аутоиммунные нарушения, аллергии и отторжение трансплантатов тканей или органов, иммунная система может быть причиной заболевания. В целях ингибирования вредных иммунных реакций в таких случаях пациентам можно вводить иммунодепрессивные средства, такие как кортикостероиды и антагонисты цитокинов. Однако эти общие иммунодепрессанты могут вызывать нежелательные побочные эффекты, включая токсичность и сниженную устойчивость к инфекции. Таким образом, необходимы альтернативные и, возможно, более специфичные способы лечения аутоиммунитета.
Был показан успех нескольких иммуномодулирующих лекарственных средств, включая лекарственные средства на основе антител, в отношении лечения некоторых злокачественных опухолей и аутоиммунных нарушений. Однако существует клиническая потребность в дополнительных лекарственных средствах на основе антител для лечения как злокачественных опухолей, так и аутоиммунных нарушений. Более того, существует связанная необходимость в гуманизированных или иных химерных моноклональных антителах человека/мыши. В широко опубликованных исследованиях у пациентов, которым вводили мышиные моноклональные антитела против TNF (фактора некроза опухоли), развивались ответы против введенных мышиных антител. (Exley A. R., et al., Lancet 335: 1275-1277 (1990)). Этот тип иммунного ответа на схему лечения, обычно называемый ответом человека против мышиных антител (HAMA) (Mirick et al. Q J. Nucl. Med. Mol. Imaging 2004; 48: 251-7), снижает эффективность лечения и даже может сделать лечение полностью неэффективным. Было показано, что гуманизированные или химерные моноклональные антитела человека/мыши значительно снижают ответ HAMA и повышают терапевтическую эффективность лекарственных средств на основе антител. См., например, LoBuglio et al., P.N.A.S. 86:4220-4224 (June 1989). Более того, в клиническом применении могут быть пригодными антитела, в которых конкретные функции являются либо усиленными, либо сниженными.
Сущность изобретения
Настоящее изобретение относится к средствам и способам модулирования функции CD200. Средства, которые модулируют функцию CD200, включают средства, которые модулируют активность и/или экспрессию CD200 и/или его рецептора (CD200R). В некоторых вариантах осуществления средства ингибируют функцию или активность CD200. Таким образом, в некоторых аспектах указанные средства действуют в качестве антагонистов CD200. Некоторые антагонисты могут связываться с CD200 и ингибировать или нарушать взаимодействие CD200 с его рецептором. Другие антагонисты могут связываться с CD200, однако они могут не блокировать взаимодействие CD200:CD200R. Таким образом, антагонисты CD200 включают любое средство, которое способно модулировать эффекты CD200 посредством механизмов, которые могут включать блокирование взаимодействия CD200:CD200R или могут не включать его. Антагонисты CD200 включают, но не ограничиваются ими, полипептиды, низкомолекулярные соединения, металлорганические соединения, олигонуклеотидные конструкции, конструкции RNAi, аптамеры, зеркальные аптамеры (spiegelmers), антисмысловые нуклеиновые кислоты, ингибиторы в виде замкнутой нуклеиновой кислоты (LNA), ингибиторы в виде пептидной нуклеиновой кислоты (PNA), иммуномодулирующие средства, антитела, антигенсвязывающие фрагменты, пролекарства и/или соединения пептидомиметиков.
В определенных вариантах осуществления указанный антагонист представляет собой антитело против CD200. Антитела, как рассматривают в настоящем документе, включают антигенсвязывающие фрагменты, Fab, Fv, scFv, Fab' и F(ab')2, моноклональные и поликлональные антитела, сконструированные антитела (включая химерные, одноцепочечные, полученные пересаживанием CDR, гуманизированные, полностью человеческие антитела и полученные искусственной селекцией антитела) и синтетические или полусинтетические антитела. В некоторых аспектах настоящее изобретение относится к химерным, гуманизированным человеческим и деиммунизированным антителам против CD200 и их антигенсвязывающим фрагментам. В следующих вариантах осуществления описанное в настоящем документе антитело содержит тяжелую цепь, содержащую аминокислотную последовательность, которая является по меньшей мере на 90% идентичной аминокислотной последовательности, выбранной из SEQ ID NO:7, 9, 11 и 20, или ее фрагментам. Также включается антитело, содержащее аминокислотную последовательность, которая является приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной или сходной с аминокислотной последовательностью, представленной в SEQ ID NO:7, 9, 11 и 20, или его фрагменты (включая, но не ограничиваясь ими, фрагменты, соответствующие последовательностям без лидерных последовательностей). Указанное антитело может дополнительно содержать легкую цепь, содержащую аминокислотную последовательность, которая является по меньшей мере на приблизительно 90% идентичной или сходной с аминокислотной последовательностью, выбранной из SEQ ID NO:24, 26, 28 и 32, или ее фрагменты. Аналогично, упомянутая выше аминокислотная последовательность может быть приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной или сходной с аминокислотной последовательностью, представленной в SEQ ID NO:24, 26, 28 и 32, включая ее фрагменты (включая, но не ограничиваясь ими, фрагменты, соответствующие последовательностям без лидерных последовательностей).
В одном варианте осуществления изобретение относится к антителу против CD200, содержащему тяжелую цепь, содержащую аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной аминокислотной последовательности SEQ ID NO:7, а также содержащему легкую цепь, содержащую аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной SEQ ID NO:24. Также антитела включают антитела против CD200, содержащие аминокислотные последовательности, которые являются приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичными или сходными с одной или несколькими аминокислотными последовательностями, представленными в SEQ ID NO:7 и 24, или их фрагментами. Фрагменты включают, но не ограничиваются ими, последовательности, соответствующие последовательностям, указанным в SEQ ID NO:7 и 24 без лидерных последовательностей. Таким образом, изобретение относится к антителу против CD200, содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:6 (включая ее фрагменты и комплементарные ей последовательности), и также содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:23 (включая ее фрагменты и комплементарные ей последовательности). Также включается антитело против CD200, содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:6, включая ее фрагменты и комплементарные ей последовательности, и также содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:23, включая ее фрагменты и комплементарные ей последовательности. Также изобретение относится к антителам против CD200, содержащим аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:6 или 23, включая ее фрагменты и комплементарные ей последовательности.
В другом варианте осуществления изобретение относится к антителу против CD200, содержащему тяжелую цепь, содержащую аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной аминокислотной последовательности SEQ ID NO:9, и также содержащую аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной SEQ ID NO:26. Также включаются антитела против CD200, содержащие аминокислотные последовательности, которые являются приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичными или сходными с одной или несколькими аминокислотными последовательностями, представленными в SEQ ID NO:9 и 26, или их фрагментами. Фрагменты включают, но не ограничиваются ими, последовательности, соответствующие последовательностям, указанным в SEQ ID NO:9 и 26 без лидерных последовательностей. Таким образом, изобретение относится к антителу против CD200, содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:8 (включая ее фрагменты и комплементарные ей последовательности), и также содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:25 (включая ее фрагменты и комплементарные ей последовательности). Также антитела включают антитело против CD200, содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:8, включая ее фрагменты и комплементарные ей последовательности, и также содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:25, включая ее фрагменты и комплементарные ей последовательности. Также изобретение относится к антителам против CD200, содержащим аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:8 или 25, включая ее фрагменты и комплементарные ей последовательности.
В следующем варианте осуществления изобретение относится к антителу против CD200, содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной аминокислотной последовательности SEQ ID NO:11, и также содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной SEQ ID NO:26. Также включаются антитела против CD200, содержащие аминокислотные последовательности, которые являются приблизительно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичными или сходными с одной или несколькими аминокислотными последовательностями, представленными в SEQ ID NO:11 и 26, или их фрагментами. Фрагменты включают, но не ограничиваются ими, последовательности, соответствующие последовательностям, указанным в SEQ ID NO:11 и 26 без лидерных последовательностей. Таким образом, изобретение относится к антителу против CD200, содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:10 (включая ее фрагменты и комплементарные ей последовательности), и также содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:25 (включая ее фрагменты и комплементарные ей последовательности). Также включается антитело против CD200, содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:10, включая ее фрагменты и комплементарные ей последовательности, и также содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:25, включая ее фрагменты и комплементарные ей последовательности. Также изобретение относится к антителам против CD200, содержащим аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%s 95%, 96%, 97%, 98%, 99% или 100% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:10 или 25, включая ее фрагменты и комплементарные ей последовательности.
В дополнительном варианте осуществления изобретение относится к антителу против CD200, содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной аминокислотной последовательности SEQ ID NO:11, и также содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной SEQ ID NO:28. Также включаются антитела против CD200, содержащие аминокислотные последовательности, которые являются приблизительно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичными или сходными с одной или несколькими аминокислотными последовательностями, представленными в SEQ ID NO:11 и 28, или их фрагментами. Фрагменты включают, но не ограничиваются ими, последовательности, соответствующие последовательностям, указанным в SEQ ID NO:11 и 28 без лидерных последовательностей. Таким образом, изобретение относится к антителу против CD200, содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:10 (включая ее фрагменты и комплементарные ей последовательности), и также содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:27 (включая ее фрагменты и комплементарные ей последовательности). Также включается антитело против CD200, содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:10, включая ее фрагменты и комплементарные ей последовательности, и также содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:27, включая ее фрагменты и комплементарные ей последовательности. Также это изобретение относится к антителам против CD200, содержащим аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:10 или 27, включая ее фрагменты и комплементарные ей последовательности.
В другом варианте осуществления изобретение относится к антителу против CD200, содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной аминокислотной последовательности SEQ ID NO:20, и также содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной SEQ ID NO:32. Также включаются антитела против CD200, содержащие аминокислотные последовательности, которые являются приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичными или сходными с одной или несколькими аминокислотными последовательностями, представленными в SEQ ID NO:20 и 32, или их фрагментами. Фрагменты включают, но не ограничиваются ими, последовательности, соответствующие последовательностям, указанным в SEQ ID NO:20 и 32 без лидерных последовательностей. Таким образом, изобретение относится к антителу против CD200, содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:19 (включая ее фрагменты и комплементарные ей последовательности), и также содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:31 (включая ее фрагменты и комплементарные ей последовательности). Также включается антитело против CD200, содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:19, включая ее фрагменты и комплементарные ей последовательности, и также содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:31, включая ее фрагменты и комплементарные ей последовательности. Таким образом, включаются антитела против CD200, содержащие аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% гомологичной или сходной с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:19 или 31, включая ее фрагменты и комплементарные ей последовательности.
Антитела против CD200, представленные в настоящем изобретении, включают антитела и антигенсвязывающие фрагменты с измененной эффекторной функцией(ями) или с ее отсутствием. Антитела включают антитела, которые содержат измененный константный или Fc-участок либо с повышенными, либо со сниженными эффекторными функциями. Также изобретение относится к антителам с измененными эффекторными функциями или их отсутствием, вследствие повышенной или сниженной аффинности связывания, которая может появляться вследствие изменений в вариабельных участках. Измененные эффекторные функции включают, например, повышенную или сниженную способность связывать один или несколько Fc-рецепторов (FcR) или эффекторных клеток, повышенную или сниженную антигензависимую цитотоксичность (ADCC) и/или повышенную или сниженную комплементзависимую цитотоксичность (CDC). Варианты антител включают, но не ограничиваются ими, антитела, в которых константный участок или Fc-участок содержит одну или несколько аминокислотных вставок, делеций и/или замен. В дополнительных вариантах осуществления эти варианты антител содержат константный участок, где источником CH1-участка и шарнирной области является IgG2 человека и источником CH2 и CH3-участков является IgG4 человека. Также включаются антитела, в которых константный или Fc-участок проявляют измененное гликозилирование. Упомянутые выше антитела и антигенсвязывающие фрагменты (включая одноцепочечные антитела) могут быть мышиными, химерными, гуманизированными, полностью человеческими или деиммунизированными и включают антитела, содержащие каркасные области IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgA, IgD или IgE. Более того, указанные антитела, включая их фрагменты и варианты, могут представлять собой блокирующие или неблокирующие антитела или их фрагменты.
В некоторых аспектах изобретение относится к антителам против CD200, которые проявляют сниженную эффекторную функцию или ее отсутствие. Антитела со сниженной эффекторной функцией или ее отсутствием могут содержать вариантный или измененный Fc-участок или константный участок, например, такой как константный участок с заменами, вставками и/или делециями одной или нескольких аминокислот, или константный участок с одним или несколькими изменениями гликозилирования. Вариантный константный участок включает, например, участок, где одна или несколько аминокислот замещены аланином, как в случае мутации Ala-Ala, описанной в настоящем документе, или где изменены, добавлены или удалены одна или несколько углеводных групп. Изменение количества и/или расположения углеводных групп можно проводить с помощью продукции указанного антитела в конкретных типах клеток, в которых посттрансляционные модификации могут быть снижены, могут отсутствовать или могут быть повышены. В одном варианте осуществления эффекторная функция антител против CD200 устраняется посредством замены константного домена IgG1 слитым доменом IgG2/4. Могут быть предусмотрены другие способы устранения эффекторной функции, например, такие как мутация в участках, о которых известно, что они взаимодействуют с FcR, или вставка пептида в шарнирную область, устраняя посредством этого критические участки, требуемые для взаимодействия с FcR.
В некоторых аспектах и способах настоящего изобретения антитела против CD200 с измененными эффекторными функциями или с их отсутствием включают антитела против CD200 с заменами, вставками и/или делециями одной или нескольких аминокислот. В определенных вариантах осуществления такое вариантное антитело против CD200 проявляет сниженную эффекторную функцию или ее отсутствие. В определенных вариантах осуществления вариантный константный участок (указанного отличающегося антитела) обладает по меньшей мере приблизительно 70% гомологией с константным участком или Fc-участком с нативной последовательностью и/или с константным участком или Fc-участком исходного антитела или их фрагментом; в других вариантах осуществления вариантный константный участок или Fc-участок обладает по меньшей мере приблизительно 80% гомологией или сходством с ними; в других вариантах осуществления по меньшей мере приблизительно 90% гомологией или сходством с ними и в дополнительных вариантах осуществления по меньшей мере приблизительно 95% гомологией или сходством с ними. В конкретных вариантах осуществления вариантное антитело содержит конструкцию G2/G4. Таким образом, настоящее изобретение относится к константному участку или Fc-участку антитела против CD200 со сниженной эффекторной функцией или с ее отсутствием, где указанный константный участок содержит тяжелую цепь, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:13, 15, 18, 22, и ее фрагменты. Также настоящее изобретение относится к вариантным константным участкам антитела против CD200, где антитело содержит аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной или сходной с аминокислотной последовательностью выбранной из SEQ ID NO:13, 15, 18, 22, и ее фрагментами. Также изобретение включает антитела, содержащие аминокислотную последовательность, которая является приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичной или сходной с аминокислотной последовательностью, представленной в SEQ ID NO:13, 15, 18, 22, и ее фрагментами. Фрагменты включают, но не ограничиваются ими, последовательности без лидерных последовательностей. Кроме того, в некоторых вариантах осуществления константный участок антитела против CD200 со сниженной эффекторной функцией или с ее отсутствием и содержащий конструкцию G2/G4 кодируется нуклеиновой кислотой, выбранной из группы, состоящей из SEQ ID NO:12, 14, 16, 17 и 21, или ее фрагментами и комплементарными ей последовательностями. В определенных вариантах осуществления антитело против CD200 со сниженной эффекторной функцией или с ее отсутствием кодируется нуклеиновой кислотой, содержащей последовательность нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной или сходной с последовательностью, выбранной из SEQ ID NO:12, 14, 16, 17 и 21, включая ее фрагменты и комплементарные ей последовательности. В других вариантах осуществления вариантное антитело против CD200 кодируется последовательностью нуклеиновой кислоты, содержащей последовательность, которая является приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 91%, 98%, 99% или 100% гомологичной или сходной с последовательностью нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO:12, 14, 16, 17 и 21, включая ее фрагменты и комплементарные ей последовательности. В других вариантах осуществления нуклеиновая кислота, кодирующая вариантное антитело против CD200, содержит последовательность нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO:12, 14, 16, 17 и 21, включая ее фрагменты и комплементарные ей последовательности. Включаются антигенсвязывающие фрагменты и как блокирующие, так и неблокирующие антитела или их фрагменты.
В одном варианте осуществления настоящее изобретение относится к вариантному антителу против CD200, содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной аминокислотной последовательности SEQ ID NO:13, и также содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной SEQ ID NO:28. Также включается антитело против CD200, содержащее одну или несколько аминокислотных последовательностей, которые являются приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичными аминокислотной последовательности, представленной в SEQ ID NO:13 и 28, или ее фрагментам. Фрагменты включают, но не ограничиваются ими, последовательности, соответствующие последовательностям, указанным в SEQ ID NO:13 и 28 без лидерных последовательностей. Таким образом, изобретение относится к вариантному антителу против CD200, содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:12 (включая ее фрагменты и комплементарные ей последовательности), и также содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:27 (включая ее фрагменты и комплементарные ей последовательности). Также включается антитело против CD200, содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:12, включая ее фрагменты и комплементарные ей последовательности, и также содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:27, включая ее фрагменты и комплементарные ей последовательности. Таким образом, включаются антитела против CD200, содержащие аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:12 или 27, включая ее фрагменты и комплементарные ей последовательности.
В другом варианте осуществления изобретение относится к вариантному антителу против CD200, содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной аминокислотной последовательности SEQ ID NO:15, и также содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной SEQ ID NO:24. Также включается антитело против CD200, содержащее аминокислотную последовательность, которая является приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной одной или нескольким аминокислотным последовательностям, представленным в SEQ ID NO:15 и 24, или их фрагментам. Фрагменты включают, но не ограничиваются ими, последовательности, соответствующие последовательностям, указанным в SEQ ID NO:15 и 24, без лидерных последовательностей (например, фрагмент SEQ ID NO:15, начинающийся в аминокислоте 20 или 21). Таким образом, изобретение относится к вариантному антителу против CD200, содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:14 (включая ее фрагменты и комплементарные ей последовательности), и также содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:23 (включая ее фрагменты и комплементарные ей последовательности). Также включается антитело против CD200, содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:14, включая ее фрагменты и комплементарные ей последовательности, и также содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:23, включая ее фрагменты и комплементарные ей последовательности. Таким образом, включаются антитела против CD200, содержащие аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:14 или 23, включая ее фрагменты и комплементарные ей последовательности.
В дополнительном варианте осуществления изобретение относится к вариантному антителу против CD200, содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной аминокислотной последовательности SEQ ID NO:13, и также содержащему аминокислотную последовательность, которая является по меньшей мере на 90% идентичной SEQ ID NO:28. Также включается антитело против CD200, содержащее одну или несколько аминокислотных последовательностей, которые являются приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичными аминокислотной последовательности, представленной в SEQ ID NO:13 и 28, или ее фрагментам. Фрагменты включают, но не ограничиваются ими, последовательности, соответствующие последовательностям, указанным в SEQ ID NO:13 и 28 без лидерных последовательностей. Таким образом, изобретение относится к вариантному антителу против CD200, содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:16 (включая ее фрагменты и комплементарные ей последовательности), и также содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:27 (включая ее фрагменты и комплементарные ей последовательности). Также включается антитело против CD200, содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:16, включая ее фрагменты и комплементарные ей последовательности, и также содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:27, включая ее фрагменты и комплементарные ей последовательности. Таким образом, включаются антитела против CD200, содержащие аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:16 или 27, включая ее фрагменты и комплементарные ей последовательности.
В другом варианте осуществления изобретение относится к вариантному антителу против CD200, содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной аминокислотной последовательности SEQ ID NO:18, и также содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной SEQ ID NO:30. Также включается антитело против CD200, содержащее аминокислотную последовательность, которая является приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной аминокислотной последовательности, представленной в SEQ ID NO:18 и 30, или ее фрагментам. Таким образом, изобретение относится к вариантному антителу против CD200, содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:17 (включая ее фрагменты и комплементарные ей последовательности), и также содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:29 (включая ее фрагменты и комплементарные ей последовательности). Также включается антитело против CD200, содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:17, включая ее фрагменты и комплементарные ей последовательности, и также содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере на 80% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:29, включая ее фрагменты и комплементарные ей последовательности. Таким образом, включаются антитела против CD200, содержащие аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:17 или 29, включая ее фрагменты и комплементарные ей последовательности.
В другом варианте осуществления изобретение относится к вариантному антителу против CD200, содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной аминокислотной последовательности SEQ ID NO:22, и также содержащему аминокислотную последовательность, которая является по меньшей мере приблизительно на 90% идентичной SEQ ID NO:34. Также включается антитело против CD200, содержащее аминокислотную последовательность, которая является приблизительно на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной аминокислотным последовательностям, представленным в SEQ ID NO:22 и 34, или их фрагментам. Таким образом, изобретение относится к вариантному антителу против CD200, содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:21 (включая ее фрагменты и комплементарные ей последовательности), и также содержащему аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая гибридизуется в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:33 (включая ее фрагменты и комплементарные ей последовательности). Также включается антитело против CD200, содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:21, включая ее фрагменты и комплементарные ей последовательности, и также содержащее аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является по меньшей мере приблизительно на 80% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:33, включая ее фрагменты и комплементарные ей последовательности. Таким образом, включаются антитела против CD200, содержащие аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты, которая является приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% гомологичной последовательности нуклеиновой кислоты, представленной в SEQ ID NO:21 и 33, включая ее фрагменты и комплементарные ей последовательности.
Антитела против CD200 с измененной эффекторной функцией также могут проявлять повышенную эффекторную функцию. Повышенные эффекторные функции включают, но не ограничиваются ими, повышенное связывание с одним или несколькими Fc-рецепторами, повышенную способность вызывать ADCC и/или повышенную способность вызывать CDC. Антитела против CD200 с повышенной эффекторной функцией также могут содержать вариантный Fc-участок или константный участок, как описано в настоящем документе. Упомянутые выше антитела против CD200 с измененными эффекторными функциями, более того, могут представлять собой блокирующие или неблокирующие антитела. Например, антитело против CD200 с повышенной эффекторной функцией может связываться с CD200, однако может не блокировать взаимодействие CD200:CD200R. Такое антитело можем быть пригодным при нацеливании эффекторной функции (например, ADCC или CDC) на экспрессирующую CD200 клетку. Как упомянуто ранее, антитела, описанные в настоящем документе, включая упомянутые выше антитела против CD200 с измененной эффекторной функцией(ями), включают мышиные, химерные, гуманизированные, полностью человеческие и деиммунизированные антитела, все из них в их блокирующих и неблокирующих формах, и их фрагменты.
В некоторых аспектах настоящее изобретение относится к способам и композициям для модулирования и уничтожения положительных по CD200 клеток. Положительные по CD200 клетки можно модулировать или уничтожать посредством введения субъекту антагониста CD200. Указанный антагонист может быть нацелен на положительные по CD200 клетки для эффекторной функции, и/или он может нарушать взаимодействие CD200:CD200R. В определенных вариантах осуществления указанный антагонист представляет собой антитело против CD200. Указанное антитело против CD200 может представлять собой антитело, описанное в настоящем описании, включая их любые фрагменты и варианты. Включаются антитела и антигенсвязывающие фрагменты с измененной эффекторной функцией(ями), например, такие как антитела против CD200 со сниженной эффекторной функцией или с ее отсутствием. Также включаются мышиные, химерные, гуманизированные, полностью человеческие и деиммунизированные антитела и антигенсвязывающие фрагменты, включая одноцепочечные антитела. Упомянутые выше антитела могут представлять собой неблокирующие или блокирующие антитела, и они включают антитела, содержащие каркасные области IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgA, IgD или IgE.
Положительные по CD200 клетки вовлечены в определенные типы злокачественных опухолей и определенные аутоиммунные заболевания. Таким образом, положительные по CD200 клетки включают, но не ограничиваются ими, иммунные клетки (например, такие как B-клетки и T-клетки) и злокачественные клетки (например, такие как злокачественные клетки яичника, кожи, легкого, почек, молочной железы, предстательной железы, нейробластомы, лимфомы, миеломы, лейкоза, щитовидной железы и рака плазматических клеток). Также включаются злокачественные клетки из любой ткани или органа, образованных из клеток нервного валика. Таким образом, субъект, нуждающийся в способе модулирования или устранения положительных по CD200 клеток, может представлять собой пациента со злокачественной опухолью или аутоиммунным заболеванием или пациента, которому проведена или который ожидает проведения трансплантации органа.
В одном аспекте настоящее изобретение относится к способам и композициям для лечения аутоиммунного заболевания. Аутоиммунные заболевания, которые можно лечить способами и композициями, представленными в настоящем документе, включают, но не ограничиваются ими, ревматоидный артрит, воспалительное заболевание кишечника (включая язвенный колит и болезнь Крона), системную красную волчанку, рассеянный склероз, тиреоидит Хашимото, пернициозную анемию, болезнь Аддисона, диабет I типа, дерматомиозит, синдром Шегрена, красную волчанку, миастению, синдром Рейтера, болезнь Грэйва, псориаз и аутоиммунные гемолитические заболевания. В некоторых вариантах осуществления пациенту с аутоиммунным заболеванием вводят антагонист CD200, и в определенных вариантах осуществления антагонист представляет собой антитело против CD200. Антитело против CD200 может содержать вариантный константный участок, как описано в настоящем документе. Таким образом, антитело против CD200 может проявлять измененную эффекторную функцию(и), например, такую как повышенная эффекторная функция(и). Указанное антитело может проявлять, например, повышенное связывание с одним или несколькими Fc-рецепторами. Кроме того, указанное антитело может вызывать повышенную ADCC и/или CDC. Более того, указанное антитело может представлять собой либо блокирующее, либо неблокирующее антитело или его фрагмент, и оно может представлять собой либо мышиное, либо химерное, либо гуманизированное, либо полностью человеческое, либо деиммунизированное антитело или его фрагмент.
Злокачественные опухоли, при которых можно использовать описанные способы, включают, но не ограничиваются ими, меланому, рак яичника, рак почек, нейробластому, рак легкого, рак молочной железы, рак предстательной железы, лимфому, миелому, лейкоз и злокачественные опухоли плазматических клеток. Также включаются злокачественные опухоли, образованные из клеток нервного валика, и любые злокачественные опухоли, которые экспрессируют CD200. В определенных вариантах осуществления настоящее изобретение относится к способу лечения гематологических злокачественных опухолей, например, таких как лейкозы, включая хронический лимфоцитарный лейкоз.
В особенно пригодном варианте осуществления терапия злокачественной опухоли в соответствии с этим описанием включает (1) введение антитела против CD200 или антагониста, который препятствует взаимодействию между CD200 и его рецептором для блокирования иммунной супрессии, обеспечивая посредством этого уничтожение злокачественных клеток; и/или (2) введение слитой молекулы, которая включает нацеливающую на CD-200 часть для непосредственного уничтожения злокачественных клеток. Альтернативно антитело непосредственно уничтожает злокачественные клетки через опосредуемую комплементом и/или антителозависимую клеточную цитотоксичность. В различных вариантах осуществления эффекторная функция антитела против CD200 является измененной. В одном конкретном варианте осуществления антитело против CD200 содержит вариантный или измененный константный участок, эффекторная функция которого является сниженной или устранена; такое антитело может быть пригодным для описанных выше способов, например, в (1) и (2).
В определенных вариантах осуществления изобретение относится к слитым молекулам, где антитело против CD200 или антигенсвязывающий фрагмент связаны со второй молекулой. Указанная слитая молекула может содержать, например, низкомолекулярное соединение, полипептид, пептидомиметик, гетероклитический пептид, химиотерапевтическое средство, иммуномодулирующее средство, нацеливающую группу или конструкцию нуклеиновой кислоты (например, антисмысловую, RNAi или нацеливающую на ген конструкцию). Также описание включает антигенсвязывающие фрагменты против CD200, где фрагмент является слитым или иным образом связан с полипептидом, белковым доменом, сывороточным белком, альбумином, PEG (полиэтиленгликолем) или любой другой молекулой, которая будет увеличивать время полужизни указанного фрагмента in vivo. Указанные антигенсвязывающие фрагменты включают, например, Fab, Fv, одноцепочечные фрагменты или scFv, Fab' и F(ab')2.
Также настоящее изобретение относится к способам применения антител против CD200 для определения состояния экспрессии CD200 в образце клеток или ткани, полученном от пациента. Такие способы включают, но не ограничиваются ими, иммуногистохимическое окрашивание образцов тканей и проточный цитометрический анализ окрашенных по CD200 клеток из пациента. Пациент может представлять собой, например, пациента со злокачественной опухолью.
В соответствии со способами и композициями, описанными в настоящем описании, изобретение также относится к способам лечения пациента после трансплантации или аллотрансплантации. Антитело против CD200 или другой антагонист CD200, представленный в настоящем описании, можно вводить пациенту до процесса трансплантации или аллотрансплантации или после этих процессов в целях снижения или устранения положительных по CD200 иммунных клеток, которые могут снизить приживление у пациента трансплантированного органа или ткани. В конкретном варианте осуществления антитело против CD200 с повышенной эффекторной функцией вводят пациенту с трансплантатом.
В следующих вариантах осуществления предусмотрены способы комбинированной терапии, включающие терапию против CD200. Например, пациенту, которому проводят первую терапию, включающую антагонист CD200 (например, антитело против CD200, описанное в настоящем документе), также можно проводить вторую терапию. Антагонист CD200 можно вводить одновременно со второй терапией. Альтернативно антагонист CD200 можно вводить до или после второй терапии. Способы второй терапии включают, но не ограничиваются ими, химиотерапевтические средства, лучевую терапию, вакцины, антибиотики и противовирусные средства и иммуномодулирующие способы лечения.
В другом варианте осуществления, представленном в настоящем описании, предусмотрены способы мониторинга хода терапевтического лечения. Способ включает введение лекарственного средства (например, иммуномодулирующего средства, химиотерапевтического средства и т.д.) и определение уровней CD200 у субъекта по меньшей мере два раза для определения эффективности лечения. Другие способы определения эффективности лечения включают, но не ограничиваются ими, определение наличия злокачественных клеток, общего количества лимфоцитов, размера селезенки, печени и/или лимфатических узлов, количества регуляторных T-клеток, внутриклеточных или сывороточных профилей цитокинов или секреции цитокинов T- или B-клетками, как измеряют посредством ELISPOT - аналитической системы, которая обеспечивает возможность детекции цитокинов или других секретированных молекул в отдельных клетках.
В соответствии с композициями и способами, указанными в настоящих вариантах осуществления, изобретение также относится к любой фармацевтической композиции, содержащей антитело против CD200. Включаются химерные, гуманизированные, человеческие и деиммунизированные антитела против CD200 и антигенсвязывающие фрагменты, включая одноцепочечные антитела. Также включаются мышиные, химерные, гуманизированные, человеческие и деиммунизированные вариантные антитела против CD200 и антигенсвязывающие фрагменты с измененной эффекторной функцией(ями), как описано в настоящем документе. Упомянутые выше антитела и вариантные антитела могут представлять собой либо блокирующие, либо неблокирующие антитела или антигенсвязывающие фрагменты.
В определенных вариантах осуществления у пациентов, для которых является пригодной терапия против CD200, или у пациентов, которые ожидают получения терапии, включающей терапию антагонистом CD200 (включая, например, антитело против CD200), можно проводить скрининг в отношении определенных проводимых у них способов лечения и операций или текущего медицинского состояния. В одном варианте осуществления, например, у пациенток-женщин можно предварительно исследовать наличие беременности и согласие на применение контрацепции, поскольку CD200 играет важную роль в защите против выкидыша. Таким образом, пациенты, получающие указанную терапию, могут быть согласны на применение одного или нескольких способов контрацепции. Указанный пациент может быть согласен на применение одного или нескольких способов контрацепции в течение определенного периода перед началом указанной терапии и/или в ходе указанной терапии. В определенных вариантах осуществления пациентки-женщины получают консультацию, касающуюся рисков в отношении воздействия такой терапии против CD200 на плод. В дополнительных вариантах осуществления можно ожидать, что такие пациенты будут подписывать информированное согласие перед таким лечением. В других аспектах медицинский осмотр в целях консультации пациентов относительно лечения против CD200 может требовать, чтобы такие пациенты применяли механические противозачаточные средства или составы перед проведением лечения против CD200 (см., например, US6908432 и связанные патенты, содержание которых включено в настоящий документ в качестве ссылок). Аналогично, в других вариантах осуществления, пациентов можно обследовать для выявления пациентов, которым ранее проводили хирургическую операцию на головном мозге и/или лучевую терапию на головном мозге; таким пациентам не будут назначать терапию против CD200.
Краткое описание чертежей
На фиг.1 представлена последовательность нуклеиновой кислоты для праймера C7mhHF (SEQ ID NO:1), используемого для получения конструкции G2/G4.
На фиг.2 представлена последовательность нуклеиновой кислоты для праймера Rev Age Pri (SEQ ID NO:2), используемого для получения конструкции G2/G4.
На фиг.3 представлена последовательность нуклеиновой кислоты для праймера C2aB7 rev (SEQ ID NO:3), используемого для получения конструкции G2/G4.
На фиг.4 представлена последовательность нуклеиновой кислоты для lacpri (SEQ ID NO:4), используемого для получения конструкции G2/G4.
На фиг.5 представлена последовательность нуклеиновой кислоты для LeadVHpAX (SEQ ID NO:5).
На фиг.6A-F представлены аминокислотные последовательности и нуклеотидные последовательности для тяжелой и легкой цепей антитела chC2aB7-hG1 (SEQ ID NO:6, 7, 23, 24, 37 и 38). На фиг.6C представлены SEQ ID NO:37 (последовательность нуклеиновой кислоты) и SEQ ID NO:7 (аминокислотная последовательность). SEQ ID NO:7, как представлено на схеме, является непрерывной, однако она изображена с соответствующей нуклеотидной последовательностью, которая включает интроны. На фиг.6F показаны SEQ ID NO:38 (последовательность нуклеиновой кислоты) и SEQ ID NO:24 (аминокислотная последовательность).
На фиг.7A-F представлены аминокислотные последовательности и нуклеотидные последовательности для тяжелой и легкой цепей антитела hB7V4V1-hG1 (SEQ ID NO:8, 9, 25, 26, 39 и 40). На фиг.7C показаны SEQ ID NO:39 (последовательность нуклеиновой кислоты) и SEQ ID NO:9 (аминокислотная последовательность). На фиг.7F показаны SEQ ID NO:40 (последовательность нуклеиновой кислоты) и SEQ ID NO:26 (аминокислотная последовательность).
На фиг.8A-F представлены аминокислотные последовательности и нуклеотидные последовательности для тяжелой и легкой цепей антитела hB7V3V1-hG1 (SEQ ID NO:10, 11, 25, 26, 40 и 41). На фиг.8C показаны SEQ ID NO:41 (последовательность нуклеиновой кислоты) и SEQ ID NO:11 (аминокислотная последовательность). На фиг.8F показаны SEQ ID NO:41 (последовательность нуклеиновой кислоты) и SEQ ID NO:26 (аминокислотная последовательность).
На фиг.9A-F представлены аминокислотные последовательности и нуклеотидные последовательности для тяжелой и легкой цепей антитела hB7V3V2-hG1 (SEQ ID NO:10, 11, 27, 28, 41 и 42). На фиг.9C показаны SEQ ID NO:41 (последовательность нуклеиновой кислоты) и SEQ ID NO:11 (аминокислотная последовательность). На фиг.9F показаны SEQ ID NO:42 (последовательность нуклеиновой кислоты) и SEQ ID NO:28 (аминокислотная последовательность).
На фиг.10A-F представлены аминокислотные последовательности и нуклеотидные последовательности для тяжелой и легкой цепей антитела hB7V3V2-hG2G4 (SEQ ID NO:12, 13, 27, 28, 42 и 43). На фиг.10C показаны SEQ ID NO:43 (последовательность нуклеиновой кислоты) и SEQ ID NO:13 (аминокислотная последовательность). SEQ ID NO:13, как показано на схеме, является непрерывной, однако она изображена с соответствующей нуклеотидной последовательностью, которая включает интроны. На фиг.10F показаны SEQ ID NO:42 (последовательность нуклеиновой кислоты) и SEQ ID NO:28 (аминокислотная последовательность).
На фиг.11A-F представлены аминокислотные последовательности и нуклеотидные последовательности для тяжелой и легкой цепей антитела chC2aB7-hG2G4 (SEQ ID NO:14, 15, 23, 24, 44, 45, 46 и 47). На фиг.11C показаны SEQ ID NO:44 (последовательность нуклеиновой кислоты) и SEQ ID NO:45 (аминокислотная последовательность). SEQ ID NO:45 соответствует аминокислотам 1-337 SEQ ID NO:15. Как показано на схеме, SEQ ID NO:45 является непрерывной, однако изображена с соответствующей нуклеотидной последовательностью, которая включает интроны. На фиг.11F показаны SEQ ID NO:46 (последовательность нуклеиновой кислоты) и SEQ ID NO:47 (аминокислотная последовательность).
На фиг.12A-F представлены аминокислотные последовательности и нуклеотидные последовательности для тяжелой и легкой цепей антитела hB7V3V2-cG2G4 (SEQ ID NO:13, 16, 27, 28, 48 и 49). На фиг.12C показаны SEQ ID NO:48 (последовательность нуклеиновой кислоты) и SEQ ID NO:13 (аминокислотная последовательность). На фиг.12F показаны SEQ ID NO:49 (последовательность нуклеиновой кислоты) и SEQ ID NO:28 (аминокислотная последовательность).
На фиг.13A-D показаны аминокислотные последовательности и нуклеотидные последовательности для тяжелой и легкой цепей антитела chC7-hG2G4 (SEQ ID NO:17, 18, 29 и 30).
На фиг.14A-F показаны аминокислотные последовательности и нуклеотидные последовательности для тяжелой и легкой цепей антитела D1B5-hG1 (SEQ ID NO:19, 20, 31, 32, 50 и 51). На фиг.14C показаны SEQ ID NO:50 (последовательность нуклеиновой кислоты) и SEQ ID NO:20 (аминокислотная последовательность). SEQ ID NO:20, как показано на схеме, является непрерывной, однако она изображена с соответствующей нуклеотидной последовательностью, которая включает интроны. На фиг.14F показаны SEQ ID NO:51 (последовательность нуклеиновой кислоты) и SEQ ID NO:32 (аминокислотная последовательность).
На фиг.15A-D представлены аминокислотные последовательности и нуклеотидные последовательности для тяжелой и легкой цепей антитела G2G4 63L1D (SEQ ID NO:21, 22, 33 и 34).
На фиг.16 представлена последовательность нуклеиновой кислоты прямого праймера для клонирования кДНК CD200 (SEQ ID NO:35).
На фиг.17 представлена последовательность нуклеиновой кислоты обратного праймера для клонирования кДНК CD200 (SEQ ID NO:36).
На фиг.18 показаны эффекты введения гуманизированных антител против CD200 в модели RAJI-CD200/PBL. Гуманизированные антитела против CD200 приводили к ингибированию роста опухоли.
На фиг.19 показаны эффекты введения гуманизированных антител против CD200 с эффекторной функцией и без нее в модели на животных Namalwa_CD200. Антитела без эффекторной функции проявляли эффективность в отношении ингибирования роста опухоли.
На фиг.20 представлена таблица, показывающая уровень экспрессии CD200 в образцах пациента с хроническим лимфоцитарным лейкозом (CLL) по сравнению с нормальными образцами.
На фиг.21 представлены относительные уровни экспрессии CD200, выявленные в линиях злокачественных клеток.
На фиг.22 показан уровень экспрессии антигена CD200 в образцах рака яичника человека относительно уровня экспрессии, выявленного в лимфоцитах периферической крови (PBL) человека.
На фиг.23 показан уровень экспрессии антигена CD200 в образцах пациента с меланомой человека относительно уровня экспрессии, выявленного в PBL.
На фиг.24 показано иммуногистохимическое окрашивание на CD200 в образцах пациента с меланомой.
На фиг.25 показаны эффекты антитела против CD200 на продукцию цитокинов. Уровни продукции IL-2 в анализе смешанной клеточной популяции измеряли в отсутствие и в присутствии антитела против CD200. Используемое антитело представляет собой химерное антитело против CD200 с отсутствием эффекторной функции.
На фиг.26 показаны эффекты введения антител против CD200, с эффекторной функцией или без нее, в модели Namalwa/PBL, в которой опухоли не экспрессируют CD200.
На фиг.27 показан проточный цитометрический анализ экспрессии CD200 на активированных T-клетках. Клетки CD3+ активировали посредством mOKT3, собирали, промывали и подвергали окрашиванию указанными конъюгированными антителами, специфичными к CD25, CD200, CD5, CD4 и CD8 человека. Клетки промывали и анализировали в отношении иммунофлуоресценции на проточном цитометре FacsCaliber с использованием программного обеспечения CellQuest.
На фиг.28 показаны эффекты антител против CD200 на ADCC активированных T-клеток. CD3+ T-клетки человека стимулировали 10 мкг/мл иммобилизованного (нанесенного на планшеты) mOKT3 в течение 72 час. Затем активированные T-клетки подвергали хроматографии для применения в качестве мишеней и инкубировали с очищенными аутологичными CD56+ (NK) клетками в качестве эффекторных клеток. Клетки коинкубировали в течение 4 часов при 37°C с соотношениями эффектор:клетка-мишень 25:1 (A) или 10:1 (B) в присутствии или отсутствие гуманизированного антитела против CD200, способного к опосредованию эффекторной функции (V3V2-G1) или сконструированного для отсутствия эффекторной функции (V3V2-G2G4). Данные представлены в качестве процентного специфичного лизиса. Антитело против CD200 повышало ADCC активированных T-клеток, в то время как антитело против CD200 без эффекторной функции не индуцировало ADCC.
На фиг.29 представлена таблица, показывающая уровень экспрессии CD200 на плазмоцитах.
Подробное описание предпочтительных вариантов осуществления
I. АНТАГОНИСТЫ CD200
CD200 представляет собой высококонсервативный трансмембранный гликопротеин I типа, экспрессируемый на различных типах клеток, включая клетки иммунной системы (например, T-клетки, B-клетки и дендритные клетки (Barclay et al., TRENDS Immunol. 2002: 23)), а также определенные злокачественные клетки, как представлено в настоящем документе. Белок взаимодействует с его рецептором CD200R на миелоидных клетках и субпопуляциях T-клеток (Wright et al. J. Immunol. 2003 (171): 3034-3046 и Wright et al., Immunity 2000 (13): 233-242); взаимодействие CD200:CD200R обеспечивает доставку иммуномодулирующего сигнала в клетки и индуцирует иммуносупрессию, включая ассоциированную с апоптозом иммунную толерантность (Rosenblum et al. 2004 Blood (103): 2691-2698). Таким образом, средства, которые препятствуют функции или активности CD200 и/или его рецептора, могут ингибировать или подвергать реверсии иммунодепрессивные эффекты взаимодействия CD200:CD200R. Кроме того, средства, которые специфично связывают CD200 (но которые могут ингибировать или могут не ингибировать взаимодействие CD200:CD200R), могут запускать последующие процессы, которые подвергают реверсии или устраняют эффекты CD200.
В некоторых аспектах настоящее изобретение относится к антагонистам CD200. Как используют в настоящем документе, термин антагонист включает любое средство, которое способно ингибировать активность, функцию и/или экспрессию CD200 или его рецептора. Примеры включают, но не ограничиваются ими, полипептиды, антитела, низкомолекулярные соединения, аптамеры, зеркальные аптамеры, ингибиторы в виде замкнутых нуклеиновых кислот (LNA), ингибиторы в виде пептидных нуклеиновых кислот (PNA), конструкции нуклеиновых кислот (например, нацеливающие на ген конструкции, антисмысловые конструкции, конструкции для РНК-интерференции (RNAi) и т.д.) и пептидомиметики. В определенных вариантах осуществления антагонист препятствует взаимодействию CD200 и CD200R. В других вариантах осуществления антагонисты CD200 способны снижать иммунодепрессивные эффекты CD200 или способны к нацеливанию на экспрессирующие CD200 клетки для их устранения или элиминации.
В некоторых аспектах антагонисты CD200 представляют собой полипептиды. Полипептиды, используемые в настоящем изобретении, можно конструировать с использованием различных способов, которые известны специалистам в данной области. В одном варианте осуществления полипептиды получают химическим синтезом. В других вариантах осуществления полипептиды представляют собой антитела, сконструированные из фрагмента или нескольких фрагментов одного или нескольких антител. В следующих вариантах осуществления полипептид представляет собой антитело против CD200, как описано в настоящем документе.
Таким образом, в определенных вариантах осуществления антагонисты CD200 представляют собой антитела против CD200. Как используют в настоящем документе, термин "антитела" относится к полным антителам или фрагментам антител, способным связывать CD200 или CD200R. Включаются Fab, Fv, scFv, Fab' и F(ab')2, моноклональные и поликлональные антитела, сконструированные антитела (включая химерные, одноцепочечные, полученные пересадкой CDR, гуманизированные, полностью человеческие антитела и полученные искусственной селекцией антитела) и синтетические или полусинтетические антитела, продуцируемые с использованием фагового дисплея или альтернативных способов. Также включаются антитела, сконструированные или продуцированные таким образом, чтобы они содержали вариантные или измененные константные или Fc-участки либо с повышенной, либо со сниженной способностью связывать одну или несколько эффекторных клеток; такие вариантные антитела включают, но не ограничиваются ими, антитела, в которых константный или Fc-участок содержит измененные паттерны гликозилирования. Небольшие фрагменты, такие как Fv и scFv, обладают преимущественными свойствами для диагностических и терапевтических способов применения вследствие их малого размера и, как следствие, лучшего распределения в тканях. Антитела со сконструированными или вариантными константными или Fc-участками могут быть пригодны для модулирования эффекторных функций, например, таких как ADCC и CDC. Такие антитела со сконструированными или вариантными константными или Fc-участками могут быть пригодны в случаях, например, когда CD200 экспрессируется в нормальной ткани; вариантные антитела против CD200 без эффекторной функции в этих случаях могут вызывать требуемый терапевтический ответ без повреждения нормальной ткани. Более того, антитела, вариантные антитела и их фрагменты могут быть блокирующими (т.е. указанные антитела или фрагменты ингибируют взаимодействие CD200 и CD200R) или неблокирующими (т.е. указанные антитела или фрагменты связываются с CD200, но не блокируют его взаимодействия с CD200R).
Также изобретение относится к антителам против CD200, содержащим тяжелые и легкие цепи, как представлено в настоящем документе, включая тяжелые и легкие цепи, которые являются гомологичными или сходными с тяжелыми и/или легкими цепями, представленными в настоящем документе. "Гомология", или "идентичность", или "сходство" относятся к сходству последовательностей двух пептидов или двух молекул нуклеиновых кислот. Как гомологию, так и идентичность можно определять посредством сравнения положений в каждой последовательности, которые могут быть выравнены для целей сравнения. Когда эквивалентное положение в сравниваемых последовательностях занято тем же основанием или аминокислотой, тогда молекулы являются идентичными в этом положении; когда эквивалентная область занята тем же или сходным аминокислотным остатком (например, сходным по пространственным и/или электронным свойствам), тогда молекулы могут быть названы гомологичными (сходными) в этом положении. Выражение в качестве процентной гомологии/сходства или идентичности относится к функции количества идентичных или сходных аминокислот в положениях, являющихся общими в сравниваемых последовательностях. Термин "гомология" описывает математическое сравнение сходства последовательностей, которое используют для идентификации генов или белков со сходными функциями или мотивами. Как используют в настоящем документе, "идентичность" означает процентное количество идентичных нуклеотидных или аминокислотных остатков в соответствующих положениях в двух или нескольких последовательностях, когда последовательности выравнены для максимизации совпадения последовательностей, т.е. с учетом делеций и вставок. Таким образом, способы определения идентичности разработаны для достижения наибольшего совпадения между тестируемыми последовательностями (см. Computational Molecular Biology, Lesk, A. M., ed., Oxford University Press, New York, 1988; Biocomputing: Informatics and Genome Projects, Smith, D. W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A. M., and Griffin, H. G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; и Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991; и Carillo, H., and Lipman, D., SIAM J. Applied Math., 48: 1073 1988, Devereux, J., et al., Nucleic Acids Research 12(1): 387 (1984), BLASTP, BLASTN, FASTA (Altschul, S. F. et al., J. Molec. Biol. 215: 403-410 (1990) и Altschul et al. Nuc. Acids Res. 25: 3389-3402 (1997)) и BLAST X (BLAST Manual, Altschul, S., et al., NCBI NLM NIH Bethesda, Md. 20894; Altschul, S., et al., J. Mol. Biol. 215: 403-410 (1990))). Последовательность, которая является "неродственной" или "негомологичной", обладает менее чем 40% идентичностью, хотя предпочтительно менее чем 25% идентичностью с последовательностью, представленной в настоящем описании. При сравнении двух последовательностей отсутствие остатков (аминокислот или нуклеиновых кислот) или наличие дополнительных остатков также снижает идентичность и гомологию/сходство.
Таким образом, изобретение относится к антителам, как описано в настоящем документе, без лидерных последовательностей. Таким образом, антитела, представленные в настоящем описании, могут содержать тяжелые и легкие цепи (как описано в настоящем документе), в которых лидерная последовательность либо отсутствует, либо заменена отличающейся лидерной последовательностью. Если используют клетки-хозяева для получения антител, представленных в настоящем описании, то можно выбирать пригодные лидерные последовательности в соответствии с конкретной используемой клеткой-хозяином.
Антитела можно получать способами, хорошо известными в данной области. Например, моноклональные антитела против CD200 можно получать с использованием положительных по CD200 клеток, полипептида CD200 или фрагмента полипептида CD200, в качестве иммуногена, индуцируя, таким образом, иммунный ответ у животных, из которых можно выделять антителопродуцирующие клетки и, в свою очередь, моноклональные антитела. Последовательность таких антител можно определять и можно продуцировать антитела или их варианты рекомбинантными способами. Рекомбинантные способы можно использовать для получения химерных, образованных пересаживанием CDR, гуманизированных и полностью человеческих антител, исходя из последовательности моноклональных антител, а также полипептидов, способных связываться с CD200.
Более того, антитела, полученные из рекомбинантных библиотек ("фаговые антитела"), можно подвергать селекции с использованием положительных по CD200 клеток или полипептидов, полученных из них, в качестве приманки для выделения антител или полипептидов на основе специфичности к мишени. Получение и выделение нечеловеческих и химерных антител против CD200 находится в пределах квалификации специалиста в данной области.
Технологию рекомбинантных ДНК используют для улучшения антител, продуцируемых в нечеловеческих клетках. Таким образом, можно конструировать химерные антитела в целях снижения их иммуногенности при диагностическом и терапевтическом применении. Более того, иммуногенность можно минимизировать гуманизацией антител посредством пересадки CDR и, необязательно, модификации каркасной области. См. патент США № 5225539, содержание которого включено в настоящий документ в качестве ссылок.
Антитела можно получать из сыворотки животного или, в случае моноклональных антител или их фрагментов, продуцировать в клеточной культуре. Технологию рекомбинантных ДНК можно использовать для получения антител в соответствии с общепринятым способом, включая способы в культуре бактериальных клеток или клеток млекопитающих. Выбранная клеточная культуральная система предпочтительно секретирует антительный продукт.
В другом варианте осуществления процесс получения антитела, описанного в настоящем документе, включает культивирование хозяина, например клетки E. coli или млекопитающего, которую трансформировали гибридным вектором. Вектор включает одну или несколько экспрессирующих кассет, содержащих промотор, функционально связанный с первой последовательностью ДНК, кодирующей сигнальный пептид, связанный в правильной рамке считывания со второй последовательностью ДНК, кодирующей белок антитела. Затем белок антитела собирают и выделяют. Необязательно, экспрессирующая кассета может включать промотор, функционально связанный с полицистронными, например бицистронными, последовательностями ДНК, кодирующими белки антитела, каждый из которых по отдельности функционально связан с сигнальным пептидом в правильной рамке считывания.
Размножение гибридомных клеток или клеток-хозяев млекопитающих in vitro проводят в пригодных культуральных средах, которые включают обычные стандартные культуральные среды (например, такие как модифицированная способом Дульбекко среда Игла (DMEM) или среда RPMI 1640), необязательно дополненные сывороткой млекопитающих (например, эмбриональной телячьей сывороткой) или микроэлементами и поддерживающими рост добавками (например, фидерными клетками, такими как нормальные клетки перитонеального экссудата мыши, клетки селезенки, макрофаги костного мозга, 2-аминоэтанол, инсулин, трансферрин, липопротеин низкой плотности, олеиновая кислота или сходные с ними). Размножение клеток-хозяев, которые представляют собой бактериальные клетки или дрожжевые клетки, аналогично проводят в пригодных культуральных средах, известных в данной области. Например, для бактерий пригодные культуральные среды включают среду LE, NZCYM, NZYM, NZM, Terrific Broth, SOB, SOC, 2×YT или минимальную среду M9. Для дрожжей пригодные культуральные среды включают среду YPD, YEPD, минимальную среду или полную минимальную сокращенную среду.
Продукция in vitro приводит к относительно чистым препаратам антител и дает возможность увеличения масштаба продукции для получения больших количеств требуемых антител. Способы культивирования бактериальных клеток, дрожжей, растений или клеток млекопитающих известны в данной области и включают гомогенную суспензионную культуру (например, в пневматическом реакторе или в реакторе с постоянным перемешиванием) и культуру иммобилизованных или захваченных клеток (например, в полых волокнах, микрокапсулах, на агарозных микрогранулах или керамических кассетах).
Большие количества требуемых антител также можно получать посредством размножения клеток млекопитающих in vivo. Для этой цели клетки гибридомы, продуцирующие требуемые антитела, инъецируют гистосовместимым млекопитающим, вызывая рост антителопродуцирующих опухолей. Необязательно, животных перед инъекцией подвергают воздействию углеводорода, особенно минеральных масел, таких как пристан (тетраметилпентадекан). Спустя от одной до трех недель из жидкостей организма этих млекопитающих выделяют антитела. Например, клетки гибридомы, полученные посредством слияния пригодных клеток миеломы с антителопродуцирующими клетками селезенки из мышей Balb/c, или трансфицированные клетки, образованные из гибридомной клеточной линии Sp2/0, которая продуцирует требуемые антитела, инъецируют внутрибрюшинно мышам Balb/c, которым необязательно предварительно вводили пристан. Спустя от одной до двух недель у животных отбирают асцитную жидкость.
Представленные выше и другие способы рассмотрены, например, в Kohler and Milstein, (1975) Nature 256: 495-497; патенте США № 4376110; Harlow and Lane, Antibodies: a Laboratory Manual, (1988) Cold Spring Harbor, описания всех из которых включены в настоящее описание в качестве ссылок. Способы получения рекомбинантных молекул антител описаны в указанных выше ссылках, а также, например, в WO97/08320; патенте США № 5427908; патенте США № 5508717; Smith, 1985, Science, Vol.225, pp.1315-1317; Parmley and Smith, 1988, Gene 73, pp.305-318; De La Cruz et al., 1988, Journal of Biological Chemistry, 263 pp.4318-4322; патенте США № 5403484; патенте США № 5223409; WO88/06630; WO92/15679; патенте США № 5780279; патенте США № 5571698; патенте США № 6040136; Davis et al., 1999, Cancer Metastasis Rev., 18(4): 421-5; Taylor, et al., Nucleic acids Research 20 (1992): 6287-6295; Tomizuka et al., Proc. Natl. Academy of Sciences USA 97(2) (2000): 722-727. Содержание всех этих ссылок включено в настоящий документ в качестве ссылок.
Супернатанты клеточных культур подвергают скринингу в отношении требуемых антител, предпочтительно посредством иммунофлуоресцентного окрашивания положительных по CD200 клеток, посредством иммуноблоттинга, посредством иммуноферментного анализа, например "сэндвич"-анализа или дот-анализа, или радиоиммунного анализа.
Для выделения антител иммуноглобулины в культуральных супернатантах или в асцитной жидкости можно концентрировать, например, посредством осаждения сульфатом аммония, диализом против материала, такого как полиэтиленгликоль, фильтрацией через селективные мембраны или сходные с ними. Если необходимо и/или желательно, антитела очищают общепринятыми способами хроматографии, например гель-фильтрацией, ионообменной хроматографией, хроматографией посредством DEAE-целлюлозы и/или (иммуно-)аффинной хроматографией, например аффинной хроматографией с одним или несколькими поверхностными полипептидами, полученными из положительной по CD200 клеточной линии, или с белком A или G.
Другой вариант осуществления относится к способу получения бактериальной клеточной линии, секретирующей антитела, направленные против CD200, в пригодном млекопитающем. Например, кролика иммунизируют объединенными образцами из положительной по CD200 ткани или клеток или полипептидом CD200 или его фрагментами. Библиотеку фагового дисплея, полученную из иммузированного кролика, конструируют и сортируют посредством пэннинга в отношении требуемых антител в соответствии со способами, хорошо известными в данной области (например, такими как способы, описанные в различных ссылках, включенных в настоящее описание в качестве ссылок).
Также описаны гибридомные клетки, секретирующие моноклональные антитела. Предпочтительные гибридомные клетки являются генетически стабильными, секретируют моноклональные антитела, описанные в настоящем документе, с требуемой специфичностью, и их можно активировать из замороженных культур посредством размораживания и повторного клонирования.
В другом варианте осуществления предусмотрен способ получения гибридомной клеточной линии, секретирующей моноклональные антитела против CD200. В этом способе пригодное млекопитающее, например мышь Balb/c, иммунизируют одним или несколькими полипептидами или антигенными фрагментами CD200 или одним или несколькими полипептидами или антигенными фрагментами, полученными из положительной по CD200 клетки, самими положительными по CD200 клетками или антигенным носителем, содержащим очищенный полипептид, как описано. Антителопродуцирующие клетки иммунизированного млекопитающего выращивают в течение короткого периода времени в культуре или подвергают слиянию с клетками пригодной клеточной линии миеломы. Гибридные клетки, полученные при слиянии, клонируют и отбирают клеточные клоны, секретирующие требуемые антитела. Например, клетки селезенки мышей Balb/c, иммунизированных положительной по CD200 клеточной линией хронического лимфоцитарного лейкоза (CLL), подвергают слиянию с клетками клеточной линии миеломы PAI или клеточной линии миеломы Sp2/0-Ag 14. Затем полученные гибридные клетки подвергают скринингу в отношении секреции требуемых антител и клонируют положительные гибридомные клетки.
Предпочтительным является способ получения гибридомной клеточной линии, отличающийся тем, что мышей Balb/c иммунизируют посредством инъекции подкожно и/или внутрибрюшинно между 106 и 107 клеток положительной по CD200 клеточной линии несколько раз, например от четырех до шести раз, в течение нескольких месяцев, например от двух до четырех месяцев. Клетки селезенки из иммунизированных мышей получают спустя от двух до четырех суток после последней инъекции и подвергают слиянию с клетками клеточной линии миеломы PAI в присутствии обеспечивающего слияние вещества, предпочтительно полиэтиленгликоля. Предпочтительно, клетки миеломы подвергают слиянию с от трех до двадцатикратным избытком клеток селезенки из иммунизированных мышей в растворе, содержащем от приблизительно 30% до приблизительно 50% полиэтиленгликоля с молекулярной массой приблизительно 4000. После слияния клетки размножают в пригодных культуральных средах, как описано в настоящем документе выше, дополненных селективной средой, например средой HAT, через правильные промежутки в целях предотвращения перерастания нормальными клетками миеломы требуемых клеток гибридомы.
Антитела и их фрагменты могут быть "химерными". Химерные антитела и их антигенсвязывающие фрагменты содержат участки из двух или более различных видов (например, мыши и человека). Химерные антитела можно получать с вариабельными участками мыши требуемой специфичности, встроенными в сегменты генов константных доменов человека (например, патент США № 4816567). Таким образом, нечеловеческие антитела можно модифицировать для обеспечения их большей пригодности для клинического применения у человека.
Моноклональные антитела, представленные в настоящем описании, включают "гуманизированные" формы нечеловеческих (т.е. мышиных) антител. Гуманизированные или образованные пересадкой CDR mAb являются особенно пригодными в качестве лекарственных средств для человека, поскольку они не выводятся из кровотока так быстро, как мышиные антитела и, как правило, не вызывают неблагоприятной иммунной реакции. Как правило, гуманизированное антитело обладает одним или несколькими аминокислотными остатками, встроенными в него из нечеловеческого источника. Эти нечеловеческие аминокислотные остатки часто называют "импортными" остатками, которые, как правило, берут из "импортного" вариабельного домена. Способы получения гуманизированных антител, главным образом, хорошо известны в данной области. Например, гуманизацию можно проводить по существу в соответствии со способом Winter и коллег (Jones et al., Nature 321: 522-525 (1986); Reichmann et al., Nature, 332: 323-327 (1988); Verheoeyen et al., Science, 239: 1534-1536 (1988)), посредством замены CDR или последовательностей CDR грызунов соответствующими последовательностями антитела человека. Также см. Staelens et al. 2006 Mol. Immunol. 43: 1243-1257. В конкретных вариантах осуществления гуманизированные формы нечеловеческих (например, мышиных) антител представляют собой антитела человека (реципиентное антитело), в которых остатки гипервариабельной (CDR) области реципиентного антитела заменены остатками гипервариабельного участка из нечеловеческого вида (донорное антитело), такого как мышь, крыса, кролик или не относящийся к человеку примат, имеющие требуемую специфичность, аффинность и способность к связыванию. В некоторых случаях остатки каркасной области иммуноглобулина человека также заменяют соответствующими нечеловеческими остатками (так называемые "обратные мутации"). Кроме того, для изменения аминокислот в выбранных положениях последовательности антитела можно использовать библиотеки фагового дисплея. На свойства гуманизированного антитела также влияют посредством выбора каркасной области человека. Более того, гуманизированные и химерные антитела можно модифицировать, чтобы они содержали остатки, которые не встречаются в реципиентном антителе или в донорном антителе, в целях дальнейшего улучшения свойств антитела, например, таких как аффинность или эффекторная функция.
Также в описании представлены полностью человеческие антитела. Термин "человеческое антитело" включает антитела, имеющие вариабельные и константные участки (при их наличии), образованные из последовательностей иммуноглобулинов эмбрионального типа человека. Антитела человека могут включать аминокислотные остатки, не кодируемые последовательностями иммуноглобулинов эмбрионального типа человека (например, мутации, внесенные случайным или сайт-специфичным мутагенезом in vitro или посредством соматической мутации in vivo). Однако термин "антитело человека" не включает антитела, в которых последовательности CDR, образованные из зародышевой линии других видов млекопитающих, таких как мышь, пересажены в последовательности каркасной области человека (т.е. гуманизированные антитела). Полностью человеческие или человеческие антитела могут быть получены из трансгенной мыши, обладающей генами антитела человека (несущими вариабельные (V), определяющие разнообразие (D), связывающие (J) и константные (C) экзоны), или из клеток человека. Например, в настоящее время является возможным получение трансгенных животных (например, мышей), которые способны при иммунизации продуцировать полный ассортимент антител человека в отсутствие эндогенной продукции иммуноглобулинов (см., например, Jakobovits et al., PNAS, 90: 2551 (1993); Jakobovits et al., Nature, 362: 255-258 (1993); Bruggermann et al., Year in Immunol., 7: 33 (1993); и Duchosal et al. Nature 355: 258 (1992). Штамм трансгенных мышей можно создавать, чтобы они содержали последовательности генов из не подвергнутых реаранжировке генов иммуноглобулинов человека. Последовательности человека могут кодировать как тяжелую, так и легкую цепи антител человека и надлежащим образом функционируют в мышах, подвергаясь реаранжировке для получения широкого ассортимента антител, сходного с ассортиментом у человека. Трансгенных мышей можно иммунизировать белком-мишенью (например, CD200, его фрагментами или клетками, экспрессирующими CD200) для создания разнообразного набора специфичных антител и кодирующей их РНК. Затем нуклеиновые кислоты, кодирующие компоненты цепей антитела в таких антителах, можно клонировать из животного в вектор для воспроизведения. Как правило, клонируют отдельные популяции нуклеиновых кислот, кодирующих последовательности тяжелой и легкой цепей, а затем отдельные популяции подвергают рекомбинации со вставкой в вектор, так что данная копия вектора приобретает случайное сочетание тяжелой и легкой цепей. Вектор конструируют, чтобы он экспрессировал цепи антитела, так чтобы их можно было собирать и воспроизводить на наружной поверхности упаковки дисплея, содержащей вектор. Например, цепи антитела можно экспрессировать в качестве слитых белков с белком оболочки фага из наружной поверхности фага. После этого упаковку фага можно подвергать скринингу в отношении дисплея антител, связывающихся с мишенью.
Кроме того, антитела человека могут быть получены из библиотек фагового дисплея (Hoogenboom et al., J. Mol. Biol., 227: 381 (1991); Marks et al., J. Mol. Biol., 222: 581-597 (1991); Vaughan et al. Nature Biotech 14: 309 (1996)). Можно создавать синтетические фаговые библиотеки, в которых используются случайные сочетания синтетических V-участков антитела человека. Посредством селекции по антигену можно получать полностью человеческие антитела, в которых V-области являются высокосходными по структуре с человеческими. См. патенты США 6794132, 6680209, 4634666 и Ostberg et al. (1983), Hibridoma 2: 361-367, содержание которых включено в качестве ссылки.
Для получения антител человека также см. Mendez et al. Nature Genetics 15: 146-156 (1997), Green и Jakobovits J. Exp. Med. 188: 483-495 (1998), описания которых включены в настоящий документ в качестве ссылок. Кроме того, антитела человека рассмотрены и описаны в патентах США 5939598 и 6673986. Также см. патенты США 6114598, 6075181 и 6162963, которые все поданы 5 июня 1995 года. Также см. патент США 6150584, поданный 2 октября 1996 года, и патенты США 6713610 и 6657103, а также патентные заявки США 10/421011 (US 2003-0229905 A1), 10/455013 (US 2004-0010810 A1), 10/627250 (US 2004-0093622 A1), 10/656623 (US 2006-0040363 A1), 10/658521 (US 2005-0054055 A1), 10/917703 (US 2005-0076395 A1) и 10/978297 (US 2005-0287630 A1). Также см. PCT/US93/06926, поданную 23 июля 1993 года, европейский патент № EP 0463151B1, грант, опубликованный 12 июня 1996 года, международную патентную заявку № WO 94/02602, опубликованную 3 февраля 1994 года, международную патентную заявку № WO 96/34096, опубликованную 31 октября 1996 года, и WO 98/24893, опубликованную 11 июня 1998 года. Описания каждого из цитированных выше патентов, заявок и ссылок включены в настоящий документ в качестве ссылок в полном объеме.
В альтернативном подходе другие компании, включая GenPharm International, Inc., использовали "минилокусный" подход. В минилокусном подходе имитируют экзогенный локус Ig посредством включения фрагментов (отдельных генов) из локуса Ig. Таким образом, образуют конструкцию из одного или нескольких VH-генов, одного или нескольких DH-генов, одного или нескольких JH-генов, константного участка мю и второго константного участка (предпочтительно константного участка гамма) для введения животному. Этот подход описан в патенте США № 5545807, выданном Surani et al., и патентах США №№ 5545806, 5625825, 5625126, 5633425, 5661016, 5770429, 5789650 и 5814318, которые все выданы Lonberg и Kay, патенте США № 5591669, выданном Krimpenfort и Bems, патентах США №№ 5612205, 5721367, 5789215, выданных Berns et al., и патенте США № 5643763, выданном Choi и Dunn, и GenPharm International. Также см. патенты США 5569825, 5877397, 6300129, 5874299, 6255458 и 7041871, описания которых включены в настоящий документ в качестве ссылок. См. также европейский патент № 0546073B1, международные патентные заявки №№ WO 92/03918, WO 92/22645, WO 92/22647, WO 92/22670, WO 93/12227, WO 94/00569, WO 94/25585, WO 96/14436, WO 97/13852 и WO 98/24884, описания которых включены в настоящий документ в качестве ссылок в полном объеме. Кроме того, см. Taylor et al. (1992 Nuc. Acids. Res., 20: 6287), Chen et al. (1993 Int. Immunol. 5: 647), Tuaillon et al. (1993 PNAS U S A. 90: 3720-4), Choi et al., (1993 Nature Genetics 4: 117), Lonberg et al. (1994 Nature 368: 856-859), Taylor et al. (1994 International Immunology 6: 579-591) и Tuaillon et al. (1995 J Immunol. 154: 6453-65), Fishwild et al. (1996 Nature Biotechnology 14: 845) и Tuaillon et al. (2000 Eur J Immunol. 10: 2998-3005), описания которых включены в настоящий документ в качестве ссылок в полном объеме.
В определенных вариантах осуществления предусмотрены деиммунизированные антитела против CD200 или их антигенсвязывающие фрагменты. Деиммунизированные антитела или их антигенсвязывающие фрагменты можно модифицировать таким образом, чтобы обеспечить неиммуногенность или уменьшенную иммуногенность в отношении данных видов антитела или его антигенсвязывающего фрагмента. Деиммунизацию можно проводить посредством модификации антитела или его антигенсвязывающего фрагмента с использованием любого из множества способов, известных специалистам в данной области (см., например, публикации PCT №№ WO 04/108158 и WO 00/34317). Например, антитело или его антигенсвязывающий фрагмент можно деиммунизировать посредством идентификации потенциальных T-клеточных эпитопов и/или B-клеточных эпитопов в аминокислотной последовательности антитела или его антигенсвязывающего фрагмента и удаления одного или нескольких потенциальных T-клеточных эпитопов и/или B-клеточных эпитопов из антитела или его антигенсвязывающего фрагмента, например, с использованием рекомбинантных способов. Затем модифицированное антитело или его антигенсвязывающий фрагмент необязательно можно получать и тестировать для идентификации антител или их антигенсвязывающих фрагментов, которые обладают сохраненным одним или несколькими требуемыми видами биологической активности, например, такими как аффинность связывания, однако обладают сниженной иммуногенностью. Способы идентификации потенциальных T-клеточных эпитопов и/или B-клеточных эпитопов можно проводить с использованием способов, известных в данной области, например, таких как компьютерные способы (см. например, публикацию PCT № WO 02/069232), способы in vitro или in silico, и биологические способы анализа или физические способы (например, такие как определение связывания пептидов с молекулами MHC, определение связывания комплексов пептид:MHC с T-клеточными рецепторами из вида, предназначенного для получения антитела или его антигенсвязывающего фрагмента, тестирование белка или его пептидных частей с использованием трансгенных животных с молекулами MHC из вида, предназначенного для получения антитела или его антигенсвязывающего фрагмента, или тестирование с трансгенными животными преобразованных клетками иммунной системы из вида, предназначенного для получения антитела или его антигенсвязывающего фрагмента, и т.д.). В различных вариантах осуществления деиммунизированные антитела против CD200, описанные в настоящем документе, включают деимунизированные антигенсвязывающие фрагменты, Fab, Fv, scFv, Fab' и F(ab')2, моноклональные антитела, мышиные антитела, сконструированные антитела (например, такие как химерные, одноцепочечные, полученные пересадкой CDR, гуманизированные, полностью человеческие антитела и подвергнутые искусственной селекции антитела), синтетические антитела и полусинтетические антитела.
В следующем варианте осуществления получают рекомбинантную ДНК, содержащую вставку, кодирующую вариабельный домен тяжелой цепи и/или вариабельный домен легкой цепи антител, направленных против CD200, или положительную по CD200 клеточную линию. Термин ДНК включает кодирующие одноцепочечные ДНК, двухцепочечные ДНК, состоящие из указанных кодирующих ДНК и комплементарных им ДНК, или сами эти комплементарные (одноцепочечные) ДНК.
Более того, ДНК, кодирующая вариабельный домен тяжелой цепи и/или вариабельный домен легкой цепи антител, направленных против CD200 или положительной по CD200 клеточной линии, может представлять собой ферментативно или химически синтезированную ДНК, имеющую аутентичную последовательность ДНК, кодирующую вариабельный домен тяжелой цепи и/или вариабельный домен легкой цепи, или ее мутантную форму. Мутантная форма аутентичной ДНК представляет собой ДНК, кодирующую вариабельный домен тяжелой цепи и/или вариабельный домен легкой цепи упомянутых выше антител, в которой одна или несколько аминокислот удалены, вставлены или заменены одной или несколькими другими аминокислотами. Предпочтительно в способах применения с гуманизацией и оптимизацией экспрессии указанная модификация(ии) находится снаружи CDR вариабельного домена тяжелой цепи и/или вариабельного домена легкой цепи антитела. Термин мутантная ДНК также включает молчащие мутантные формы, в которых один или несколько нуклеотидов заменены другими нуклеотидами с новыми кодонами, кодирующими ту же аминокислоту(ы). Термин мутантная последовательность также включает вырожденную последовательность. Вырожденные последовательности являются вырожденными с точки зрения генетического кода в том, что неограниченное количество нуклеотидов заменяют другими нуклеотидами, не вызывая изменения исходно кодируемой аминокислотной последовательности. Такие вырожденные последовательности могут быть пригодными вследствие их отличающихся участков рестрикции и/или частоты конкретных кодонов, которые являются предпочтительными в конкретном хозяине, в частности E. coli, для достижения оптимальной экспрессии вариабельного домена тяжелой цепи мыши и/или вариабельного домена легкой цепи мыши. Подразумевают, что термин мутантная форма включает мутантную форму ДНК, полученную посредством мутагенеза in vitro аутентичной ДНК в соответствии со способами, известными в данной области.
Для сборки полных тетрамерных молекул иммуноглобулинов и экспрессии химерных антител, вставки рекомбинантных ДНК, кодирующие вариабельные домены тяжелой и легкой цепи, подвергают слиянию с соответствующими ДНК, кодирующими константные домены тяжелой и легкой цепей, а затем переносят в пригодные клетки-хозяева, например, после встраивания в гибридные векторы.
Можно использовать рекомбинантные ДНК, включающие вставку, кодирующую вариабельный домен тяжелой цепи мыши антитела, направленного на CD200, или положительную по CD200 клеточную линию, слитые с константным доменом IgG человека, например γ1, γ2, γ3 или γ4; в конкретных вариантах осуществления γ1 или γ4. Также предусмотрены рекомбинантные ДНК, включающие вставку, кодирующую вариабельный домен легкой цепи мыши антитела, направленного на описанную в настоящем документе клеточную линию, слитую с константным доменом человека κ или λ, предпочтительно κ.
Другой вариант осуществления относится к рекомбинантным ДНК, кодирующим рекомбинантный полипептид, где вариабельный домен тяжелой цепи и вариабельный домен легкой цепи связаны посредством спейсера, необязательно содержащего сигнальную последовательность, способствующую процессингу антитела в клетке-хозяине, и/или последовательность ДНК, кодирующую пептид, упрощающий очистку антитела, и/или участок расщепления и/или пептидный спейсер и/или средство. Подразумевают, что ДНК, кодирующая средство, представляет собой ДНК, кодирующую средство, пригодное для диагностического или терапевтического применения. Таким образом, молекулы средства, которые представляют собой токсины или ферменты, особенно ферменты, способные катализировать активацию пролекарств, являются особенно целесообразными. ДНК, кодирующая такое средство, обладает последовательностью ДНК, кодирующей встречающийся в природе фермент или токсин, или ее мутантной формы, и ее можно получать способами, хорошо известными в данной области.
Таким образом, моноклональные антитела или антигенсвязывающие фрагменты, представленные в настоящем изобретении, могут представлять собой голые антитела или антигенсвязывающие фрагменты, которые не конъюгированы с другими средствами, например лекарственным средством или поддающейся детекции меткой. Альтернативно моноклональные антитело или антигенсвязывающий фрагмент могут быть конъюгированы со средством, например, таким как цитотоксическое средство, низкомолекулярное средство, гормон, фермент, фактор роста, цитокин, рибозим, пептидомиметик, химическое вещество, пролекарство, молекула нуклеиновой кислоты, включая кодирующие последовательности (такие как антисмысловые последовательности, RNAi, нацеливающие на ген конструкции и т.д.) или поддающуюся детекции метку (например, контрастное вещество для ЯМР или рентгеновское контрастное вещество, флуоресцентная молекула и т.д.). В определенных вариантах осуществления полипептид против CD200 или антигенсвязывающий фрагмент (например, Fab, Fv, одноцепочечное scFv, Fab' и F(ab')2) является слитым с молекулой, которая увеличивает время полужизни указанного полипептида или антигенсвязывающего фрагмента. Молекулы, которые могут быть связаны с указанным полипептидом против CD200 или антигенсвязывающим фрагментом, включают, но не ограничиваются ими, сывороточные белки, включая альбумин, полипептиды, другие белки или белковые домены, и PEG.
Для экспрессии клонированных генов тяжелой цепи и легкой цепи в клетках млекопитающих доступно несколько возможных векторных систем. Один класс векторов основан на встраивании требуемых последовательностей генов в геном клетки-хозяина. Клетки, которые обладают стабильно встроенной ДНК, можно подвергать селекции посредством одновременного встраивания генов устойчивости к лекарственному средству, таких как gpt E. coli (Mulligan, R. C. and Berg, P., Proc. Natl. Acad. Sci., USA, 78: 2072 (1981)) или Tn5 neo (Southern, P. J. и Berg, P., J. Mol. Appl. Genet., 1: 327 (1982)). Ген селективного маркера может быть либо связанным с последовательностями ДНК гена, подлежащими экспрессии, либо введенным в указанную клетку посредством котрансфекции (Wigler, M. et al., Cell, 16: 77 (1979)). Во втором классе векторов используются элементы ДНК, которые обеспечивают способность к автономной репликации экстрахромосомной плазмиды. Эти векторы могут быть образованы из любых вирусов животных, таких как бычий вирус папилломы (Sarver, N. et al., Proc. Natl. Acad. Sci., USA, 79: 7147 (1982)), вирус полиомы (Deans, R. J. et al., Proc. Natl. Acad. Sci., USA, 81: 1292 (1984)) или вирус SV40 (Lusky, M. and Botchan, M., Nature, 293: 79 (1981)).
Поскольку кДНК иммуноглобулина состоит только из последовательностей, соответствующих зрелой мРНК, кодирующей белок антитела, то для синтеза мРНК иммуноглобулина требуются дополнительные элементы для экспрессии гена, регулирующие транскрипцию гена и процессинг РНК. Эти элементы могут включать сигналы для сплайсинга, промоторы транскрипции, включая энхансеры индуцибельных промоторов и сигналы терминации. Экспрессирующие кДНК векторы, включающие такие элементы, включают векторы, описанные Okayama, H. and Berg, P., Mol. Cell Biol., 3: 280 (1983); Cepko, C. L. et al., Cell, 37: 1053 (1984); и Kaufman, R. J., Proc. Natl. Acad. Sci., USA, 82: 689 (1985).
В определенных вариантах осуществления антитело против CD200 может представлять собой блокирующее или неблокирующее антитело. Как используют в настоящем документе, блокирующее антитело представляет собой антитело, которое блокирует взаимодействие между CD200 и CD200R. Неблокирующее антитело связывается и/или взаимодействует с CD200, но не блокирует взаимодействие с CD200R. Таким образом, в определенных вариантах осуществления антитело против CD200 представляет собой либо блокирующее, либо неблокирующее мышиное, химерное, гуманизированное, человеческое или деиммунизированное антитело.
II. АНТАГОНИСТЫ CD200 C ИЗМЕНЕННЫМИ ЭФФЕКТОРНЫМИ ФУНКЦИЯМИ
Антагонисты CD200 могут быть изменены, чтобы вызывать повышенные или сниженные эффекты относительно исходного или родительского антагониста. Например, антагонист, который связывает CD200, может приводить ко вторичным функциям после связывания с CD200 и, в некоторых случаях, ингибирования взаимодействия CD200:CD200R. Например, антагонист может содержать дополнительные участки связывания для других лигандов, включая рецепторы или внеклеточные белки. Связывание с этими другими лигандами может запускать другие процессы, такие как привлечение или накопление других клеток и активация различных процессов, включая гибель клеток. Таким образом, в некоторых аспектах настоящее изобретение относится к антагонистам CD200, которые приводят к измененным вторичным функциям (или эффекторным функциям, как указано ниже). В определенных вариантах осуществления антагонист CD200 с измененной вторичной или эффекторной функцией(ями) проявляет повышенную, сниженную вторичную или эффекторную функцию(и) или их отсутствие и, кроме того, может блокировать взаимодействие CD200:CD200R или может не блокировать его. В конкретных вариантах осуществления антагонист CD200 с измененной вторичной или эффекторной функцией(ями) представляет собой антитело против CD200.
A) ЭФФЕКТОРНЫЕ ФУНКЦИИ
Взаимодействие антител и комплексов антитело-антиген с клетками иммунной системы влияет на множество видов ответа, обозначаемых в настоящем документе как эффекторные функции. Иллюстративные эффекторные функции включают связывание Fc-рецептора, фагоцитоз, снижение активности рецепторов клеточной поверхности (например B-клеточного рецептора; BCR) и т.д. Другие эффекторные функции включают ADCC, в случае которой антитела связывают Fc-рецепторы на естественных киллерных (NK) клетках или макрофагах, что приводит к клеточной гибели, и CDC, которая представляет собой клеточную гибель, индуцируемую активацией каскада комплемента (рассмотрено в Daeron, Annu. Rev. Immunol. 15: 203-234 (1997); Ward and Ghetie, Therapeutic Immunol. 2: 77-94 (1995); и Ravetch and Kinet, Annu. Rev. Immunol. 9: 457-492 (1991)). Такие эффекторные функции, как правило, требуют комбинирования Fc-участка со связывающим доменом (например, вариабельным доменом антитела) и их можно оценивать с использованием различных способов анализа, например, как описано в настоящем документе.
Несколько эффекторных функций антитела, включая ADCC, опосредуются Fc-рецепторами (FcR), которые связывают Fc-участок антитела. В случае ADCC, NK-клетки или макрофаги связываются с Fc-участком комплекса антитела и обеспечивают лизис клетки-мишени. Перекрестное связывание FcR на NK-клетках запускает опосредуемую перфорином/гранзимом цитотоксичность, в то время как в макрофагах это перекрестное связывание обеспечивает высвобождение медиаторов, таких как оксид азота (NO), TNF-α и реакционно-способные формы кислорода. В случае положительных по CD200 клеток-мишеней, антитело против CD200 связывается с клеткой-мишенью, и Fc-участок направляет эффекторную функцию на клетку-мишень. Аффинность антитела в отношении конкретного FcR и, таким образом, эффекторную активность, опосредуемую антителом, можно модулировать посредством изменения аминокислотной последовательности и/или посттрансляционных модификаций Fc-участка и/или константного участка антитела.
FcR определяют по их специфичности в отношении изотипов иммуноглобулинов; Fc рецепторы для антител IgG обозначают как FcγR, для IgE обозначают как FcεR, для IgA обозначают как FcαR и т.д. Было идентифицировано три подкласса FcγR: FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD16). Вследствие того, что каждый подкласс FcγR кодируется двумя или тремя генами и альтернативный сплайсинг РНК приводит к множеству транскриптов, существует широкое разнообразие изоформ FcγR. Три гена, кодирующих подкласс FcγRI (FcγRIA, FcγRIB и FcγRIC), образуют кластер в области 1q21.1 длинного плеча хромосомы 1; гены, кодирующие изоформы FcγRII (FcγRIIA, FcγRIIB и FcγRIIC), и два гена, кодирующих FcγRIII (FcγRIIIA и FcγRIIIB), образуют кластер в области 1q22. Эти различные подтипы FcR экспрессируются на различных типах клеток (рассмотрено в Ravetch and Kinet, Annu. Rev. Immunol. 9: 457-492 (1991)). Например, у человека FcγRIIIB встречается только на нейтрофилах, в то время как FcγRIIIA встречается на макрофагах, моноцитах, естественных киллерных (NK) клетках и субпопуляции T-клеток. В частности, FcγRIIIA представляет собой единственный FcR, представленный на NK-клетках, одном из типов клеток, вовлеченных в ADCC.
FcγRI, FcγRII и FcγRIII представляют собой рецепторы суперсемейства иммуноглобулинов (IgSF); FcγRI имеет три домена IgSF в его внеклеточном домене, в то время как FcγRII и FcγRIII имеют только два домена IgSF в их внеклеточных доменах. Другим типом Fc-рецепторов является неонатальный Fc-рецептор (FcRn). FcRn является структурно сходным с главным комплексом гистосовместимости (MHC) и состоит из α-цепи, нековалентно связанной с β2-микроглобулином.
Участок связывания антител человека и мыши для FcγR был ранее картирован на так называемой "нижней шарнирной области", состоящей из остатков 233-239 (нумерация посредством индекса EU, как в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)). Woof et al. Molec. Immunol. 23: 319-330 (1986); Duncan et al. Nature 332: 563 (1988); Canfield и Morrison, J. Exp. Med. 173: 1483-1491 (1991); Chappel et al., Proc. Natl. Acad. Sci USA 88: 9036-9040 (1991). Из остатков 233-239 P238 и S239 были типированы, как возможно вовлеченные в связывание.
Другие цитированные ранее участки, возможно вовлеченные в связывание с FcγR, представляют собой: G316-K338 (IgG человека) для FcγRI человека (только посредством сравнения последовательностей; не оценивали мутантные формы с замещением) (Woof et al. Molec Immunol. 23: 319-330 (1986)); K274-R301 (IgG1 человека) для FcγRIII человека (на основе пептидов) (Sarmay et al. Molec. Immunol. 21: 43-51 (1984)); Y407-R416 (IgG человека) для FcγRIII человека (на основе пептидов) (Gergely et al. Biochem. Soc. Trans. 12: 739-743 (1984)); а также N297 и E318 (IgG2b мыши) для FcγRII мыши (Lund et al., Molec. Immunol., 29: 53-59 (1992)).
Эффекторные клетки человека представляют собой лейкоциты, которые экспрессируют один или несколько FcR и выполняют эффекторные функции. В определенных вариантах осуществления клетки экспрессируют по меньшей мере FcγRIII и выполняют эффекторную функцию ADCC. Примеры лейкоцитов человека, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови (PBMC), естественные киллерные (NK) клетки, моноциты, цитотоксические T-клетки и нейтрофилы. Эффекторные клетки можно выделять из их нативного источника, например из крови или PBMC.
При CDC комплекс антитело-антиген связывает комплемент, что приводит к активации каскада комплемента и образованию мембраноатакующего комплекса. Активация классического каскада комплемента начинается посредством связывания первого компонента системы комплемента (C1q) с антителами (соответствующего подкласса), которые связаны с узнаваемым ими антигеном; таким образом, активация каскада комплемента регулируется частично посредством аффинности связывания иммуноглобулина с белком C1q. C1q и две сериновые протеазы, C1r и C1s, образуют комплекс C1, первый компонент каскада CDC. C1q представляет собой шестивалентную молекулу с молекулярной массой приблизительно 460000 и структурой, в которой шесть коллагеновых "стеблей" связаны с шестью участками глобулярных головок. Burton and Woof, Advances in Immunol. 51: 1-84 (1992). Для активации каскада комплемента необходимо связывание C1q по меньшей мере с двумя молекулами IgG1, IgG2 или IgG3, однако только с одной молекулой IgM, связанными с антигенной мишенью (Ward and Ghetie, Therapeutic Immunology 2: 77-94 (1995) p.80). Для оценки активации комплемента можно проводить анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202: 163 (1996).
Было предположено, что в связывание C1q вовлечены различные остатки молекулы IgG, включая остатки Glu318, Lys320 и Lys322 на CH2-домене, аминокислотный остаток 331, расположенный на изгибе вблизи указанной бета-цепи, остатки Lys235 и Gly237, расположенные в нижней шарнирной области, и остатки с 231 по 238, расположенные в N-концевом участке CH2-домена (см., например, Xu et al., J. Immunol. 150: 152A (Abstract) (1993), WO94/29351; Tao et al., J. Exp. Med., 178: 661-667 (1993); Brekke et al., Eur. J. Immunol, 24: 2542-47 (1994); Burton et al.; Nature, 288: 338-344 (1980); Duncan and Winter, Nature 332: 738-40 (1988); патент США № 5648260 и патент США № 5624821). Кроме того, было предположено, что способность IgG связывать C1q и активировать каскад комплемента также зависит от наличия, отсутствия или модификации углеводной группы, расположенной между двумя CH2-доменами (которая обычно присоединена к Asn297) (Ward and Ghetie, Therapeutic Immunology 2: 77-94 (1995). В определенных вариантах осуществления один или несколько из этих остатков могут быть модифицированы, замещены или удалены, или один или несколько аминокислотных остатков могут быть встроены, чтобы повысить или снизить активность антител против CD200, представленных в настоящем документе, в отношении CDC.
B) АНТИТЕЛА ПРОТИВ CD200 С МОДУЛИРОВАННОЙ ЭФФЕКТОРНОЙ ФУНКЦИЕЙ(ЯМИ)
Эффекторные функции, вовлекающие константный участок специфичного к мишени антитела, можно модулировать посредством изменения свойств константного участка или Fc-участка. Измененные эффекторные функции включают, например, модулирование одного или нескольких из следующих видов активности: ADCC, CDC, апоптоз, связывание с одним или несколькими Fc-рецепторами и провоспалительные ответы. Модулирование относится к повышению, снижению или устранению активности по сравнению с активностью второго антитела. В определенных вариантах осуществления второе антитело представляет собой антитело с эффекторной функцией. Второе антитело может представлять собой сконструированное антитело или природное антитело, и оно может быть названо неизмененным, нативным или исходным антителом. В конкретных вариантах осуществления модулирование включает случаи, когда активность устраняется или полностью отсутствует. Кроме того, в некоторых случаях неизмененное антитело может проявлять активность эффекторной функции, сходную или эквивалентную активности антител chC2aB7-hG1 или hB7V3V2-hG1, описанных в настоящем документе. Аналогично, функциональный или неизмененный константный участок или Fc-участок может обладать эффекторной функцией нативного константного домена или Fc-домена; в некоторых случаях константный участок или Fc-участок chC2aB7-hG1 или hB7V3V2-hG1 может соответствовать неизмененным доменам. Для целей, представленных в настоящем документе, chC2aB7-hG1 и hB7V3V2-hG1 представляют собой стандарты, с которыми сравнивают виды активности других антител, при этом hB7V3V2-hG1 является предпочтительным стандартом.
Полипептидный вариант с измененной аффинностью связывания FcR и/или активностью ADCC и/или измененной активностью CDC представляет собой полипептид, который обладает либо повышенной, либо сниженной активностью связывания FcR и/или активностью ADCC и/или активностью CDC по сравнению с нативным или исходным полипептидом или с полипептидом, содержащим Fc или константный участок с нативной последовательностью. Полипептидный вариант, который проявляет повышенное связывание с FcR, связывает по меньшей мере один FcR с большей аффинностью, чем исходный полипептид. Полипептидный вариант, который проявляет сниженное связывание с FcR, связывает по меньшей мере один FcR с меньшей аффинностью, чем исходный полипептид. Такие варианты, которые проявляют сниженное связывание с FcR, могут обладать небольшим или незначительным связыванием с FcR, например, 0-20% связыванием с FcR по сравнению с уровнем связывания с FcR константного участка или Fc-участка иммуноглобулина с нативной последовательностью. Аналогично полипептидный вариант, который проявляет измененную активность ADCC и/или CDC, может проявлять либо повышенную, либо сниженную активность ADCC и/или CDC по сравнению с нативным или исходным полипептидом. Полипептидный вариант, который проявляет сниженную ADCC и/или CDC, может проявлять сниженную активность ADCC и/или CDC или ее отсутствие, как представлено в настоящем документе в качестве примера. В определенных вариантах осуществления исходный или нативный полипептид и его вариант представляют собой антитела или антигенсвязывающие фрагменты. В конкретных вариантах осуществления указанное антитело или антигенсвязывающий фрагмент связывают CD200 и могут блокировать взаимодействие CD200:CD200R или могут не блокировать его.
Fc-участок или константный участок с нативной последовательностью содержат аминокислотную последовательность, идентичную аминокислотной последовательности Fc-участка или константного участка цепи, встречающегося в природе. Вариантный или измененный Fc-участок или константный участок содержат аминокислотную последовательность, которая отличается от последовательности участка тяжелой цепи с нативной последовательностью посредством модификации, вставки или делеции по меньшей мере одной аминокислоты. В определенных вариантах осуществления вариантный или измененный константный участок обладают заменой, вставкой и/или делецией по меньшей мере одной аминокислоты, по сравнению с константным участком с нативной последовательностью или с константным участком исходного полипептида, например, заменами, вставками и/или делециями от приблизительно одной до приблизительно ста аминокислот в константном участке с нативной последовательностью или в константном участке исходного полипептида. В некоторых вариантах осуществления вариантный или измененный константный участок, представленный в настоящем документе, будет обладать по меньшей мере приблизительно 70% гомологией (сходством) или идентичностью с константным участком с нативной последовательностью и/или с константным участком исходного полипептида, и в некоторых случаях по меньшей мере приблизительно 75% и в других случаях по меньшей мере приблизительно 80% гомологией или идентичностью с ним, и в других вариантах осуществления по меньшей мере приблизительно 85%, 90% или 95% гомологией или идентичностью с ним. Вариантный или измененный константный участок также может содержать делеции или вставки одной или нескольких аминокислот. Кроме того, вариантный константный участок может содержать замены, делеции или вставки одной или нескольких аминокислот, которые приводят к измененным посттрансляционным модификациям, включая, например, измененный паттерн гликозилирования.
Вариантные антитела против CD200, как описано в настоящем документе, могут кодироваться последовательностью нуклеиновой кислоты, которая кодирует полипептид со вставками, делециями или заменами одной или нескольких аминокислот относительно нативной или исходной полипептидной последовательности. Более того, вариантные антитела могут кодироваться последовательностями нуклеиновых кислот, которые гибридизуются в строгих условиях с последовательностью нуклеиновой кислоты, кодирующей вариантное антитело против CD200. Множество условий можно использовать для детекции гибридизации, и строгость определяется, главным образом, стадией промывания анализа гибридизации. Как правило, высокие температуры и низкие концентрации солей приводят к высокой строгости, в то время как низкие температуры и высокие концентрации солей приводят к низкой строгости. Гибридизацию низкой строгости проводят посредством промывания, например, приблизительно в 2,0×SSC при 50°С, и гибридизацию высокой строгости проводят приблизительно с 0,2×SSC при 50°С.
Антитела или их антигенсвязывающие фрагменты с измененными эффекторными функциями или с их отсутствием можно получать конструированием или продукцией антител с вариантным константным участком, Fc-участком или участками тяжелой цепи; для получения антител с измененной функцией и/или активностью можно использовать технологии рекомбинантных ДНК и/или клеточной культуры, и условия экспрессии. Например, технологию рекомбинантных ДНК можно использовать для создания замен, делеций или вставок одной или нескольких аминокислот в участках (например, таких как Fc или константные участки), которые влияют на функцию антитела, включая эффекторные функции. Альтернативно изменения в посттрансляционных модификациях, например, таких как паттерны гликозилирования, можно проводить посредством манипулирования с клеточной культурой и условиями экспрессии, в которых продуцируют антитело.
Таким образом, некоторые аспекты и способы, представленные в настоящем описании, относятся к антителам против CD200 с измененными эффекторными функциями, которые содержат замены, вставки и/или делеции одной или нескольких аминокислот. В определенных вариантах осуществления такое вариантное антитело против CD200 проявляет сниженную эффекторную функцию или ее отсутствие. В конкретных вариантах осуществления вариантное антитело содержит конструкцию G2/G4 вместо домена G1 (см., например, фиг.10, 11, 12, 13 и 15).
В дополнение к замене G1-домена конструкцией G2/G4, как представлено в настоящем документе, антитела против CD200 со сниженной эффекторной функцией можно получать посредством внесения других типов замен в аминокислотную последовательность определенных участков антитела. Такие изменения аминокислотной последовательности включают, но не ограничиваются ими, мутацию Ala-Ala, описанную Bluestone et al. (см. WO 94/28027 и WO 98/47531; также см. Xu et al. 2000 Cell Immunol. 200; 16-26). Таким образом, в определенных вариантах осуществления антитела против CD200 с мутациями в константном участке, включая мутацию Ala-Ala, можно использовать для снижения или устранения эффекторной функции. В соответствии с этими вариантами осуществления, константный участок антитела против CD200 содержит мутацию на аланин в положении 234 или мутацию на аланин в положении 235. Кроме того, константный участок может содержать двойную мутацию: мутацию на аланин в положении 234 и вторую мутацию на аланин в положении 235. В одном варианте осуществления антитело против CD200 содержит каркасную область IgG4, где мутация Ala-Ala соответствует мутации(ям) с фенилаланина на аланин в положении 234 и/или мутации с лейцина на аланин в положении 235. В другом варианте осуществления, антитело против CD200 содержит каркасную область IgG1, где мутация Ala-Ala может соответствовать мутации(ям) с лейцина на аланин в положении 234 и/или мутации с лейцина на аланин в положении 235. Альтернативно или дополнительно, антитело против CD200 может обладать другими мутациями, включая точковую мутацию K322A в CH2-домене (Hezareh et al. 2001 J Virol. 75: 12161-8). Антитело с указанной мутацией(ями) в константном участке, более того, может представлять собой блокирующее или неблокирующее антитело.
Изменения шарнирной области также влияют на эффекторные функции. Например, делеция шарнирной области может снизить аффинность к Fc-рецепторам и может снизить активацию комплемента (Klein et al. 1981 PNAS USA 78: 524-528). Таким образом, настоящее изобретение также относится к антителам с изменениями в шарнирной области.
В конкретных вариантах осуществления антитела против CD200 могут быть модифицированы либо для усиления, либо для ингибирования комплементзависимой цитотоксичности (CDC). Модулированная активность CDC может быть достигнута внесением замен, вставок или делеций одной или нескольких аминокислот в Fc-участок антитела (см., например, патент США № 6194551). Альтернативно или дополнительно, в Fc-участок можно вносить остаток(и) цистеина, обеспечивая посредством этого образование межцепочечных дисульфидных связей в этом участке. Полученное таким образом гомодимерное антитело может обладать повышенной или сниженной способностью к интернализации и/или повышенным или сниженным опосредуемым комплементом уничтожением клеток. См. Caron et al., J. Exp. Med. 176: 1191-1195 (1992) и Shopes, B. J. Immunol. 148: 2918-2922 (1992), WO99/51642, Duncan & Winter Nature 322: 738-40 (1988); патент США № 5648260; патент США № 5624821 и WO94/29351. Также можно получать гомодимерные антитела с повышенной противоопухолевой активностью с использованием гетеробифункциональных поперечных линкеров, как описано в Wolff et al. Cancer Research 53: 2560-2565 (1993). Альтернативно можно конструировать антитело, которое обладает двойными Fc-участками и, таким образом, может обладать повышенными способностями в отношении лизиса комплементом и ADCC. См. Stevenson et al. Anti-Cancer Drug Design 3: 219-230 (1989).
Другой потенциальный способ модулирования эффекторной функции антител включает изменение гликозилирования. Эта тема была недавно рассмотрена Raju, который обобщил предполагаемую значимость олигосахаридов, находящихся на IgG человека, с их степенью эффекторной функции (Raju, TS. BioProcess International April 2003, 44-53). В соответствии с Wright and Morrison, микрогетерогенность олигосахаридов IgG человека может влиять на биологические функции, такие как CDC и ADCC, связывание с различными Fc-рецепторами и связывание с белком C1q (Wright A. & Morrison SL. TIBTECH 1997, 15 26-32). Хорошо документировано, что паттерны гликозилирования антител могут отличаться в зависимости от продуцирующей клетки и условий культивирования клеток (Raju, TS. BioProcess International April 2003, 44-53). Такие различия могут приводить к изменениям как эффекторной функции, так и фармакокинетики (Israel et al. Immunology. 1996; 89(4): 573-578; Newkirk et al. P. Clin. Exp. 1996; 106(2): 259-64). Отличия в эффекторной функции могут быть связаны со способностью IgG связываться с Fcγ-рецепторами (FcγR) на эффекторных клетках. Shields et al. показали, что IgG с вариантами в аминокислотной последовательности, который обладает повышенным связыванием с FcγR, может проявлять вплоть до 100% повышенной ADCC при использовании эффекторных клеток человека (Shields et al. J Biol Chem. 2001 276(9): 6591-604). Несмотря на то, что эти варианты включают замены в аминокислотах, не встречающиеся на связывающей поверхности, как природа сахарного компонента, так и его структурный паттерн также могут приводить к выявленным различиям. Кроме того, наличие или отсутствие фукозы в олигосахаридном компоненте IgG может повысить связывание и ADCC (Shields et al. J Biol Chem. 2002; 277(30): 26733-40). IgG, который лишен фукозилированного углевода, связанного с Asn297, проявлял нормальное связывание рецептора с Fcγ-рецептором. Напротив, связывание с рецептором FcγRIIA было улучшено на 50% и сопровождалось повышенной ADCC, особенно при более низких концентрациях антитела.
Работа Shinkawa et al. показала, что антитело к рецептору для IL-5 человека, продуцированное в гибридоме крысы, демонстрировало более чем на 50% более высокую ADCC по сравнению с антителом, продуцированным в клетках яичника китайского хомяка (CHO) (Shinkawa et al. J Biol Chem. 2003 278(5): 3466-73). Анализ моносахаридного состава и олигосахаридного профиля показал, что продуцируемые гибридомой крысы IgG обладают более низким содержанием фукозы, чем продуцируемый CHO белок. Авторы сделали заключение, что отсутствие фукозилирования IgG1 играет критическую роль в усилении активности ADCC.
Другой подход был предпринят Umana et al., которые изменили паттерн гликозилирования chCE7, химерного антитела IgG1 против нейробластомы (Umana et al. Nat Biotechnol. 1999 Feb; 17(2): 176-80). С использованием тетрациклина они регулировали активность фермента гликозилтрансферазы (GnnII), который разрезает олигосахариды, вовлеченные в активность ADCC. Активность ADCC исходного антитела была немного выше фонового уровня. Измерение активности ADCC chCE7, продуцируемого при различных уровнях тетрациклина, показало оптимальный диапазон экспрессии GnTIH для максимальной активности ADCC для chCE7 in vitro. Эта активность коррелировала с уровнем ассоциированного с константным участком разрезанного комплексного олигосахарида. Вновь оптимизированные варианты проявляли значительную активность ADCC. Аналогично Wright и Morrison продуцировали антитела в клеточной линии CHO, дефицитной по гликозилированию (1994 J Exp. Med. 180: 1087-1096), и показали, что антитела, продуцированные в этой клеточной линии, были неспособны к опосредуемому комплементом цитолизу. Таким образом, поскольку известные изменения, которые влияют на эффекторную функцию, включают модификации паттерна гликозилирования или изменение количества гликозилированных остатков, настоящее изобретение относится к антителу против CD200, где гликозилирование изменено либо для повышения, либо для снижения эффекторной функции(ий), включая ADCC и CDC. Измененное гликозилирование включает уменьшение или увеличение количества гликозилированных остатков, а также изменение паттерна или расположения гликозилированных остатков.
Существуют другие подходы для изменения эффекторной функции антител. Например, антителопродуцирующие клетки можно подвергать гипермутагенезу с образованием посредством этого антител с измененными случайным образом нуклеотидными и полипептидным остатками на протяжении всей молекулы антитела (см. WO 2005/011735). Подвергнутые гипермутагенезу клетки-хозяева включают клетки, дефицитные по репарации ошибочно спаренных оснований ДНК. Антитела, полученные таким способом, могут быть менее антигенными и/или обладают благоприятными фармакокинетическими свойствами. Кроме того, такие антитела можно подвергать селекции в отношении свойств, таких как повышенная или сниженная эффекторная функция(ии).
Кроме того, понятно, что эффекторная функция может варьироваться в соответствии с аффинностью связывания антитела. Например, антитела с высокой аффинностью могут быть более эффективными в отношении активации системы комплемента по сравнению с антителами с относительно более низкой аффинностью (Marzocchi-Machado et al. 1999 Immunol. Invest 28: 89-101). Таким образом, антитело можно изменять, чтобы происходило снижение аффинности связывания с его антигеном (например, посредством изменения вариабельных участков антитела способами, такими как замена, вставка или делеция одного или нескольких аминокислотных остатков). Антитело против CD200 со сниженной аффинностью связывания может проявлять сниженные эффекторные функции, включая, например, сниженную ADCC и/или CDC.
III. СПОСОБЫ УСТРАНЕНИЯ ИЛИ ЭЛИМИНАЦИИ КЛЕТОК, СВЕРХЭКСПРЕССИРУЮЩИХ CD200
В соответствии с настоящим изобретением, предусмотрены способы устранения клеток, которые экспрессируют CD200 у субъекта, посредством введения субъекту лекарственного средства, содержащего антагонист CD200. Как упомянуто выше, CD200 экспрессируется на некоторых иммунных клетках; и, как представлено в настоящем описании, также CD200 экспрессируется на некоторых злокачественных клетках. Отличающаяся экспрессия CD200 обеспечивает подход для нацеливания на злокачественные клетки (т.е. положительные по CD200 клетки) в целях терапии. Аналогично можно проводить нацеливание на положительные по CD200 иммунные клетки для их устранения в способах лечения аутоиммунных нарушений.
CD200, через его взаимодействие с CD200R на миелоидных клетках, модулирует иммуносупрессию посредством доставки сигнала, ингибирующего миелоидную активность и/или миграцию. Мыши с нокаутом CD200, например, показывают более активный иммунный ответ после иммуногенных стимулов (Hoek et al. Science 2000), и экспрессирующие CD200 клетки приводят к иммуносупрессии посредством индукции переключения профиля цитокинов в стимулированных иммунных клетках (см. данные, представленные в настоящем документе). Конкретно, положительные по CD200 клетки способны индуцировать переключение продукции цитокинов с Th1 на Th2 в анализах смешанной клеточной популяции. Несмотря на то, что положительные по CD200 клетки способны подавлять иммунный ответ, положительные по CD200 злокачественные клетки, таким образом, могут быть способны избегать атаки иммунными клетками. Однако экспрессию CD200 на мембране злокачественных клеток, а также иммунных клеток, можно использовать для нацеливания на эти клетки при лечении. Например, антагонист CD200 может специфично нацеливаться на положительные по CD200 клетки и препятствовать взаимодействию CD200:CD200R, ингибируя посредством этого иммунную супрессию, а также нацеливать иммунные эффекторные клетки на положительные по CD200 клетки. Варианты осуществления настоящего изобретения, таким образом, относятся к способам нацеливания на положительные по CD200 клетки для их устранения, включающим антагонист, который связывается с CD200 и, в некоторых случаях, препятствует взаимодействию CD200:CD200R.
В определенных вариантах осуществления настоящее изобретение относится к способам усиления иммунного ответа. Такие способы включают введение лекарственного средства, содержащего антагонист CD200, и в конкретных вариантах осуществления антагонист представляет собой антитело против CD200 или антигенсвязывающий фрагмент, как указано в настоящем документе. Без связи с каким-либо конкретным механизмом(ами), блокирующее антитело против CD200, антигенсвязывающий фрагмент, полипептид или другой антагонист могут устранять положительные по CD200 клетки посредством блокирования иммунной супрессии, позволяя посредством этого иммунным клеткам атаковать и элиминировать положительные по CD200 клетки. Альтернативно или в сочетании с упомянутым выше механизмом, антитело против CD200 (как блокирующее, так и неблокирующее) или другой антагонист может привлекать эффекторные клетки или другие лиганды (например, компонент комплемента) к положительной по CD200 клетке, с которой антитело или антагонист связан, и направлять положительную по CD200 клетку на опосредуемую эффектором клеточную гибель.
В одном аспекте настоящее изобретение относится к способам модулирования ADCC и/или CDC положительных по CD200 клеток-мишеней посредством введения мышиного, химерного, гуманизированного или человеческого антитела против CD200 субъекту, нуждающемуся в этом. Изобретение относится к вариантным антителам против CD200, которые вызывают повышенную ADCC и/или CDC, и к вариантным антителам против CD200, которые проявляют сниженную или активность ADCC и/или CDC или ее отсутствие.
В одном варианте осуществления вариантное антитело против CD200 содержит вариантный или измененный Fc-участок или константный участок, где вариантный Fc или константный участок проявляют повышенную эффекторную функцию. Такой указанный вариантный участок может содержать замены, вставки или делеции одной или нескольких аминокислот. Альтернативно или дополнительно, вариантный или измененный Fc-участок или константный участок может содержать посттрансляционные модификации, включая, например, измененный паттерн гликозилирования. Измененный паттерн гликозилирования включает повышение или снижение количества гликозидных связей и/или модификацию положения (т.е. номера аминокислотного остатка) одной или нескольких гликозидных связей.
В другом варианте осуществления изобретение относится к способам устранения или элиминации положительных по CD200 клеток, включающим вариантные антитела против CD200, которые проявляют сниженную активность ADCC и/или CDC или ее отсутствие. В одном варианте осуществления вариантное антитело против CD200 содержит вариантный или измененный Fc-участок или константный участок, где вариантный Fc-участок или константный участок проявляет сниженную эффекторную функцию или ее отсутствие. Такой указанный вариантный или измененный Fc-участок или константный участок может содержать замены, вставки или делеции одной или нескольких аминокислот. Альтернативно или дополнительно, вариантный Fc-участок или константный участок может содержать измененные посттрансляционные модификации, включая, но не ограничиваясь ими, измененный паттерн гликозилирования. Примеры измененных паттернов гликозилирования описаны выше.
В следующем варианте осуществления мышиное, химерное, гуманизированное, человеческое или деиммунизированное антитело против CD200, вводимое пациенту, представляет собой неблокирующее антитело. Неблокирующее антитело против CD200 может представлять собой вариантное антитело, как описано выше, и, следовательно, может проявлять модулированную эффекторную функцию(и). Например, вариантное антитело против CD200 может не блокировать взаимодействие CD200:CD200R и также может содержать вариантный константный участок, который приводит к повышенной эффекторной функции, например, такой как повышенная ADCC.
A) СПОСОБЫ ЛЕЧЕНИЯ ПАЦИЕНТОВ С АУТОИММУННЫМИ НАРУШЕНИЯМИ
В некоторых аспектах изобретение относится к лечению пациентов с аутоиммунными нарушениями лекарственным средством, включающим антагонист CD200. В определенных вариантах осуществления антагонист представляет собой антитело против CD200 или его антигенсвязывающий фрагмент. В других вариантах осуществления антитело против CD200 или его фрагмент представляет собой вариантное антитело против CD200, которое проявляет модулированную эффекторную активность. Например, вариантное антитело может содержать вариантный или измененный константный участок, способный приводить к повышенной или усиленной эффекторной функции, например, такой как ADCC. Кроме того, указанное антитело может представлять собой неблокирующее антитело и может представлять собой мышиное, химерное, гуманизированное, человеческое или деиммунизированное антитело против CD200. Таким образом, способы лечения пациентов с аутоиммунными нарушениями могут включать любой из антагонистов CD200 и антитела, как указано в настоящем описании.
В определенных вариантах осуществления антитела против CD200 или антагонисты CD200 можно использовать для устранения любого типа клетки, которая экспрессирует CD200 на своей поверхности, включая, например, иммунные клетки, такие как T-клетки, B-клетки и дендритные клетки. В одном варианте осуществления антитела против CD200 могут быть пригодными для направленного разрушения иммунных клеток, вовлеченных в нежелательный иммунный ответ, например, такой как иммунные ответы, ассоциированные с аутоиммунным нарушением, трансплантатами, аллергией или воспалительными нарушениями. Иллюстративные аутоиммунные заболевания и нарушения, которые можно лечить антителами против CD200, представленными в настоящем документе, включают, например, воспалительные ответы, такие как воспалительные заболевания кожи, включая псориаз и дерматит (например, атопический дерматит); дерматомиозит; системную склеродермию и склероз; ответы, ассоциированные с воспалительным заболеванием кишечника (такие как болезнь Крона и язвенный колит); респираторный дистресс-синдром (включая взрослый респираторный дистресс-синдром; ARDS); дерматит; менингит; энцефалит; увеит; колит; гломерулонефрит; аллергические состояния, такие как экзема и астма, и другие состояния, вовлекающие инфильтрацию T-клеток и хронические воспалительные ответы; атеросклероз; нарушение адгезии лейкоцитов; ревматоидный артрит; системную красную волчанку (SLE); сахарный диабет (например, сахарный диабет I типа или инсулинзависимый сахарный диабет); рассеянный склероз; синдром Рейно; аутоиммунный тиреоидит; аллергический энцефаломиелит; синдром Шегрена; ювенильный диабет; и иммунные ответы, ассоциированные с острой и замедленной гиперчувствительностью, опосредуемой цитокинами и T-лимфоцитами, как правило, встречающейся при туберкулезе, саркоидоз, полимиозит, гранулематоз и васкулит; пернициозную анемию (болезнь Аддисона); заболевания, вовлекающие диапедез лейкоцитов; воспалительное нарушение центральной нервной системы (ЦНС); синдром множественного повреждения органов; гемолитическую анемию (включая, но не ограничиваясь ими, криоглобулинемию или Кумбс-положительную анемию); миастению gravis; опосредуемые комплексом антиген-антитело заболевания; заболевание, направленное против гломерулярной мембраны; антифосфолипидный синдром; аллергический неврит; болезнь Грэйвса; миестенический синдром Ламберта-Итона; буллезный пемфигоид; пемфигус; аутоиммунные полиэндокринопатии; болезнь Рейтера; синдром ригидности мышц; болезнь Бехчета; гигантоклеточный артрит; связанный с иммунными комплексами нефрит; IgA-нефропатию; IgM-полиневропатии; иммунную тромбоцитопеническую пурпуру (ITP) или аутоиммунную тромбоцитопению и аутоиммунные гемолитические заболевания, тиреоидит Хашимото и т.д.
В соответствии со способами и композициями, описанными в настоящем документе, изобретение также относится к способам лечения пациента, подвергшегося трансплантации или аллотрансплантации. Антитело против CD200 или другой антагонист CD200 по настоящему изобретению можно вводить пациенту до трансплантации или аллотрансплантации или после нее, в целях снижения или устранения положительных по CD200 иммунных клеток, которые могут снизить приживление у пациента трансплантированного органа или ткани. В конкретном варианте осуществления антитело против CD200 с повышенной эффекторной функцией вводят пациенту с трансплантатом. Кроме того, антитело против CD200 представляет собой неблокирующее антитело.
Лекарственные средства, содержащие антагонисты CD200 или антитела, можно вводить пациентам в комбинированных способах лечения. Таким образом, направленное уничтожение некоторых популяций иммунных клеток для лечения или профилактики аутоиммунных нарушений, усиления или продления выживаемости трансплантата, лечения или профилактики аллергии или лечения или профилактики воспалительных нарушений, можно вводить в качестве части комбинированной терапии. Например, пациенту, которому вводят первое лекарственное средство, содержащее антагонист CD200 (например, антитело против CD200, описанное в настоящем документе), также можно вводить второе лекарственное средство. Антагонист CD200 можно вводить одновременно со вторым лекарственным средством. Альтернативно антагонист CD200 можно вводить до или после второго лекарственного средства. Вторые лекарственные средства включают, но не ограничиваются ими, противовоспалительные средства, иммунодепрессивные средства и/или противоинфекционные средства.
Комбинированные способы лечения, представленные в настоящем изобретении, включают, например, антагонист CD200, как описано в настоящем документе, вводимый одновременно или последовательно сериями со стероидами, противомалярийными средствами, аспирином, нестероидными противовоспалительными лекарственными средствами, иммунодепрессантами или цитотоксическими лекарственными средствами. Включаются кортикостероиды (например, преднизон, дексаметазон и преднизолон), метотрексат, метилпреднизолон, макролидные иммунодепрессанты (например, сиролимус и такролимус), ингибиторы митоза (например, азатиоприн, циклофосфамид и метотрексат), продукты метаболизма грибов, которые ингибируют активность T-лимфоцитов (например, циклоспорин), микофенолата мофетил, глатирамера ацетат и цитотоксические и повреждающие ДНК средства (например, хлорамбуцил). В случае аутоиммунных нарушений и пациентов с аллотрансплантатами или трансплантатами, терапию против CD200 можно комбинировать с лекарственными средствами на основе антител, включая даклизумаб, полученное способами генетической инженерии моноклональное антитело IgG1 человека, которое специфично связывается с α-цепью рецептора для интерлейкина-2, а также различные другие антитела, нацеленные на иммунные клетки или другие клетки. Такие комбинированные способы лечения могут быть пригодны для лечения диабета 1 типа, ревматоидного артрита, волчанки и идиопатической тромбоцитопенической пурпуры и других аутоиммунных показаний. Также изобретение относится к лекарственным средствам против аутоиммунных нарушений и для пациентов с трансплантатами, содержащим антагонист CD200 (например, такой как антитела и их варианты, описанные в настоящем изобретении), конъюгированный с одним или несколькими средствами.
B) СПОСОБЫ ЛЕЧЕНИЯ ПАЦИЕНТОВ СО ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛЬЮ
В одном аспекте изобретение относится к способу лечения злокачественной опухоли, в котором субъекту вводят средство, которое прекращает или ингибирует взаимодействие CD200 с его рецептором. Препятствование взаимодействию CD200:CD200R впоследствии вызывает реверсию или ингибирует иммунную супрессию, усиливая, таким образом, иммунный ответ. Возможные средства для препятствования взаимодействию CD200:CD200R включают, например, низкомолекулярные соединения, химические вещества, полипептиды, неорганические молекулы и металлоорганические соединения. Также взаимодействие CD200:CD200R можно ингибировать посредством снижения экспрессии либо мембранного белка, либо его рецептора, посредством антисмысловой, RNAi или генной терапии. Кроме того, полипептид, специфичный к CD200 или CD200R, такой как специфичное к CD200 или CD200R антитело или его фрагменты, может ингибировать иммуносупрессивные эффекты взаимодействия CD200:CD200R.
Злокачественные клетки, которые можно обрабатывать антагонистом CD200, включают любые злокачественные клетки, которые проявляют экспрессию CD200 или активацию CD200. Злокачественные опухоли, в случае которых можно применять терапию против CD200, включают, например, рак яичника, меланому, миелому, нейробластому, рак почек, рак молочной железы, рак предстательной железы, гематологические злокачественные опухоли (например, лимфомы и лейкозы) и злокачественную опухоль плазматических клеток. Также включаются любые злокачественные клетки, образованные из клеток нервного валика. В некоторых вариантах осуществления антагонист CD200 представляет собой антитело против CD200. Такие антитела, используемые в качестве лекарственных средств против злокачественной опухоли, способны препятствовать взаимодействию CD200 и его рецепторов. Это препятствование может блокировать иммуносупрессивный эффект CD200. Посредством улучшения таким образом иммунного ответа, такие антитела могут обеспечивать устранение злокачественных клеток. Антитела против CD200 также могут быть нацелены на злокачественные клетки для опосредуемой эффектором гибели клеток.
В одном варианте осуществления вариантное антитело против CD200, которое проявляет модулированную активность ADCC и/или CDC, можно вводить субъекту с положительными по CD200 злокачественными клетками. Например, вариантное антитело против CD200, используемое для лечения злокачественной опухоли, может проявлять повышенную эффекторную активность по сравнению с исходным или нативным антителом. В другом варианте осуществления вариантное антитело против CD200 проявляет сниженную эффекторную функцию, включая сниженную ADCC, относительно нативного антитела. Указанное антитело может представлять собой мышиное, химерное, гуманизированное, человеческое или деиммунизированное антитело. Злокачественные опухоли, в случае которых для лечения можно использовать вариантное антитело против CD200, включают, но не ограничиваются ими, злокачественные опухоли клеток нервного валика. Также включаются злокачественная опухоль плазматических клеток, рак яичника, рак кожи, рак легкого, рак почек, рак молочной железы, рак предстательной железы, нейробластома, лимфома, миелома и лейкоз.
Антитела по настоящему изобретению можно вводить в качестве лекарственного средства пациентам со злокачественной опухолью, особенно, но не ограничиваясь ими, пациентам с CLL, злокачественной опухолью плазматических клеток, раком яичника, раком кожи, раком легкого, раком почек, раком молочной железы, раком предстательной железы, нейробластомой, лимфомой, миеломой, лейкозом и любой злокачественной опухолью, образованной из клеток нервного валика. В особенно пригодных вариантах осуществления терапия злокачественной опухоли в соответствии с этим описанием включает (1) введение антител против CD200 или антагониста, который препятствует взаимодействию между CD200 и его рецептором для блокирования иммунной супрессии, обеспечивая посредством этого уничтожение злокачественных клеток; и/или (2) введение слитой молекулы, которая включает нацеливающую на CD-200 часть для непосредственного уничтожения злокачественных клеток. Альтернативно антитело непосредственно уничтожает злокачественные клетки через опосредуемую комплементом или антителозависимую клеточную цитотоксичность. Поскольку CD200 также экспрессируется на нормальных клетках, таких как эндотелиальные клетки, хотя и на более низких уровнях, чем на злокачественных клетках, также может быть преимущественным введение антитела против CD200 с константным участком, модифицированным для снижения или устранения ADCC или CDC для ограничения повреждения нормальных клеток. Например, если экспрессия CD200 повышается на некоторых активированных нормальных клетках (например, активированных T-клетках), придавая таким клеткам восприимчивость к уничтожению антителом против CD200 с эффекторной функцией, то, таким образом, также может быть преимущественным применение антитела против CD200, лишенного эффекторной функции, во избежание устранения этих клеток, которые способствуют уничтожению злокачественных клеток.
В конкретном варианте осуществления эффекторную функцию антитела против CD200 устраняют посредством замены константного домена IgG1 слитым доменом IgG2/4. Могут быть предусмотрены другие способы устранения эффекторной функции, например, такие как мутация в областях, о которых известно, что они взаимодействуют с FcR, или встраивание пептида в шарнирную область, устраняя посредством этого критические участки, требуемые для взаимодействия с FcR. Вариантные антитела против CD200 со сниженной эффекторной функцией или ее отсутствием также включают варианты, как описано в настоящем документе ранее.
Упомянутые выше средства для ингибирования или предотвращения взаимодействия CD200:CD200R можно использовать в сочетании с другими способами лечения или с другими средствами. Другие средства включают, но не ограничиваются ими, полипептиды, низкомолекулярные соединения, химические вещества, металлы, металлоорганические соединения, неорганические соединения, молекулы нуклеиновых кислот, олигонуклеотиды, аптамеры, зеркальные аптамеры, антисмысловые нуклеиновые кислоты, ингибиторы на основе замкнутой нуклеиновой кислоты (LNA), ингибиторы на основе пептидной нуклеиновой кислоты (PNA), иммуномодулирующие средства, антигенсвязывающие фрагменты, пролекарства и соединения пептидомиметиков.
В некоторых аспектах настоящее изобретение относится к комбинированным способам лечения, включающим антагонист CD200, включая антитела, описанные в настоящем описании, и иммуномодулирующие соединения, вакцины или химиотерапию. Иллюстративные примеры пригодных иммуномодулирующих средств, которые можно использовать в таких комбинированных способах лечения, включают средства, которые блокируют отрицательную регуляцию T-клеток или антигенпредставляющих клеток (например, антитела против CTLA4, антитела против PD-L1, антитела против PDL-2, антитела против PD-1 и т.п.), или средства, которые усиливают положительную костимуляцию T-клеток (например, антитела против CD40 или антитела против 4-1BB), или средства, которые повышают количество NK-клеток или активность T-клеток (например, антитела против CD200 отдельно или в сочетании с ингибиторами, такими как IMiD, талидомид или аналоги талидомида). Более того, иммуномодулирующая терапия может включать вакцины против злокачественной опухоли, такие как дендритные клетки, нагруженные клетками опухоли, белки, пептиды, РНК или ДНК, полученные из таких клеток, полученные от пациента белки теплового шока (hsp) или общие адъюванты, стимулирующие иммунную систему на различных уровнях, такие как CpG, Luivac, Biostim, Ribominyl, Imudon, Bronchovaxom или любое другое соединение или другой адъювант, активирующий рецепторы врожденной иммунной системы (например, агонист toll-подобного рецептора, антитела против CTLA-4 и т.д.). Также иммуномодулирующая терапия может включать лечение цитокинами, такими как IL-2, GM-CSF и IFN-гамма.
В дополнительных вариантах осуществления перед началом лечения против CD200 устраняют существующие регуляторные T-клетки посредством реагентов, таких как антитело против CD25, флударабин или циклофосфамид. Также посредством терапии против CD200 усиливают терапевтическую эффективность миелоаблятивных способов лечения, с последующей трансплантацией костного мозга или адоптивным переносом T-клеток, реакционноспособных в отношении клеток CLL. В других вариантах осуществления эффективность лечения против CD200 повышают посредством блокирования иммуносупрессивных механизмов с помощью средств, таких как антитела против PDL1 и/или 2, антитела против IL-10, антитела против IL-6 и т.п. Более того, может быть преимущественным устранение плазмацитоидных дендритных клеток, для которых показано, что они являются иммунодепрессивными в окружении злокачественной опухоли. В этих вариантах осуществления, в которых доставка антитела против CD200 предназначена для усиления иммунного ответа посредством блокирования иммунной супрессии, например, также можно использовать вариантное антитело против CD200, лишенное эффекторной функции.
В особенно пригодных вариантах осуществления терапия, которая усиливает иммунный ответ, представляет собой введение полипептида, который связывается с CD200, отдельно или в сочетании с одним из упомянутых ранее иммуномодулирующих способов лечения. Таким образом, антагонист CD200 (включая антитело против CD200, как описано в настоящем документе) можно использовать в сочетании с моноклональным антителом (например, ритуксимабом, трастузумабом, алемтузумабом, цетуксимабом или бевацизумабом), включая конъюгированное моноклональное антитело (например, гемтузумаб озагомицин, ибритутомаб тиуксетан или тозитумомаб).
Более того, комбинирование способы терапии против CD200 с химиотерапевтическими средствами могут быть особенно пригодны для снижения общей опухолевой массы, для ограничения ангиогенеза, для повышения доступности опухоли, для усиления чувствительности к ADCC, для достижения повышенной иммунной функции посредством предоставления большего количества опухолевого антигена или для повышения экспрессии аттрактанта T-клеток LIGHT. Когда лекарственное средство против CD200 вводят субъекту в сочетании с другим общепринятым средством против злокачественной опухоли, либо одновременно, либо последовательно, может быть показано, что терапия против CD200 усиливает терапевтический эффект каждого средства отдельно. Фармацевтические соединения, которые можно использовать для комбинированной противоопухолевой терапии, включают, в качестве иллюстрации: аминоглутетимид, амсакрин, анастрозол, аспарагиназа, bcg, бикалутамид, блеомицин, бусерелин, бусульфан, камптотецин, капецитабин, карбоплатину, кармустин, хлорамбуцил, цисплатину, кладрибин, клодронат, колхицин, циклофосфамид, ципротерон, цитарабин, декарбазин, дактиномицин, даунорубицин, диенэстрол, диэтилстилбестрол, доцетаксел, доксорубицин, эпирубицин, эстрадиол, эстрамустин, этопозид, экземестан, филграстим, флударабин, флудрокортизон, фторурацил, флуоксиместерон, флутамид, гемцитабин, генистеин, гозерелин, гидроксимочевина, идарубицин, ифосфамид, иматиниб, интерферон, иринотекан, летрозол, лейковорин, леупролид, левамизол, ломустин, мехлорэтамин, медроксипрогестерон, магестрол, мелфалан, меркаптопурин, месну, метотрексат, митомицин, митотан, митоксантрон, нилутамид, нокодазол, октреотид, оксалиплатину, паклитаксел, памидронат, пентостатин, пликамицин, порфимер, прокарбазин, ралтитрексед, ритуксимаб, стрептозоцин, сурамин, тамоксифен, темозоломид, тенипозид, тестостерон, тиогуанин, тиотепа, титаноцена дихлорид, топотекан, трастузумаб, третиноин, винбластин, винкристин, виндезин и винорелбин.
Эти химиотерапевтические противоопухолевые соединения можно разделить по их механизму действия на группы, включающие, например, следующие классы средств: антиметаболиты/противоопухолевые средства, такие как аналоги пиримидинов (5-фторурацил, флоксуридин, капецитабин, гемцитабин и цитарабин) и аналоги пуринов, антагонисты фолатов и сходные ингибиторы (меркаптопурин, тиогуанин, пентостатин и 2-хлордезоксиаденозин (кладрибин)); антипролиферативные/антимитотические средства, включая природные продукты, такие как алкалоиды барвинка (винбластин, винкристин и винорелбин), нарушающие образование микротрубочек средства, такие как таксан (паклитаксел, доцетаксел), винкристин, винбластин, нокодазол, эпотилоны и навелбин, эпидиподофиллотоксины (этопозид, тенипозид), повреждающие ДНК средства (актиномицин, амсакрин, антрациклины, блеомицин, бусульфан, камптотецин, карбоплатина, хлорамбуцил, цисплатин, циклофосфамид, цитоксан, дактиномицин, даунорубицин, доксорубицин, эпирубицин, гексаметилмеламиноксалиплатин, ифосфамид, мелфалан, мехлорэтамин, митомицин, митоксантрон, нитрозомочевина, пликамицин, прокарбазин, таксол, таксотер, тенипозид, триэтилентиофосфорамид и этопозид (VP16)); антибиотики, такие как дактиномицин (актиномицин D), даунорубицин, доксорубицин (адриамицин), идарубицин, антрациклины, митоксантрон, блеомицины, пликамицин (митрамицин) и митомицин; ферменты (L-аспарагиназа, которая системно метаболизирует L-аспарагин и устраняет клетки, которые не обладают способностью синтезировать их собственный аспарагин); антитромбоцитарные средства; антипролиферативные/антимитотические алкилирующие средства, такие как азотистые иприты (мехлорэтамин, циклофосфамид и аналоги, мелфалан, хлорамбуцил), этиленимины и метилмеламины (гексаметилмеламин и тиотепа), алкилсульфонаты-бусульфан, нитрозомочевины (кармустин (BCNU) и аналоги, стрептозоцин), тразены-дакарбазинин (DTIC); антипролиферативные/антимитотические антиметаболиты, такие как аналоги фолиевой кислоты (метотрексат); координационые комплексы платины (цисплатин, карбоплатин), прокарбазин, гидроксимочевина, митотан, аминоглутетимид; гормоны, аналоги гормонов (эстроген, тамоксифен, гозерелин, бикалутамид, нилутамид) и ингибиторы ароматазы (летрозол, анастрозол); антикоагулянты (гепарин, синтетические соли гепарина и другие ингибиторы тромбина); фибринолитические средства (такие как тканевой активатор плазминогена, стрептокиназа и урокиназа), аспирин, дипиридамол, тиклопидин, клопидогрел, абциксимаб; средства против миграции; антисекреторные средства (бревелдин); иммунодепрессивные средства (циклоспорин, такролимус (FK-506), сиролимус (рапамицин), азатиоприн, микофенолат (мофетил); иммуномодулирующие средства (талидомид и его аналоги, такие как леналидомид (ревлимид, CC-5013) и CC-4047 (актимид)), циклофосфамид; антиангиогенные соединения (TNP-470, генистеин) и ингибиторы факторов роста (ингибиторы сосудисто-эндотелиального фактора роста (VEGF), ингибиторы фибробластного фактора роста (FGF)); блокатор рецептора для ангиотензина; доноры оксида азота; антисмысловые олигонуклеотиды; антитела (трастузумаб); ингибиторы клеточного цикла и индукторы дифференцировки (третиноин); ингибиторы mTOR, ингибиторы топоизомеразы (доксорубицин (адриамицин), амсакрин, камптотецин, даунорубицин, дактиномицин, энипозид, эпирубицин, этопозид, идарубицин и митоксантрон, топотекан, иринотекан), кортикостероиды (кортизон, дексаметазон, гидрокортизон, метилпреднизолон, преднизон и преднизолон); ингибиторы передающей сигнал фактора роста киназы; индукторы митохондриальной дисфункции и активаторы каспаз; и разрушающие хроматин средства.
В определенных вариантах осуществления фармацевтические соединения, которые можно использовать для комбинированной терапии против ангиогенеза, включают: (1) ингибиторы высвобождения "ангиогенных молекул", такие как bFGF (основной фибробластный фактор роста); (2) нейтрализаторы ангиогенных молекул, такие как антитела против βbFGF; и (3) ингибиторы ответа эндотелиальных клеток на ангиогенные стимулы, включая ингибитор коллагеназы, ингибиторы обновления базальной мембраны, ангиостатические стероиды, полученные из грибов ингибиторы ангиогенеза, тромбоцитарный фактор 4, тромбоспондин, лекарственные средства против артрита, такие как D-пеницилламин и тиомалат золота, аналоги витамина D3, альфа-интерферон и т.п. Для дополнительных предложенных ингибиторов ангиогенеза см. Blood et al., Biochim. Biophys. Acta, 1032: 89-118 (1990), Moses et al., Science, 248: 1408-1410 (1990), Ingber et al., Lab. Invest, 59: 44-51 (1988), и патенты США №№ 5092885, 5112946, 5192744, 5202352 и 6573256. Кроме того, существует широкое множество соединений, которые можно использовать для ингибирования ангиогенеза, например, пептиды или средства, которые блокируют опосредуемый VEGF каскад ангиогенеза, белок эндостатина или его производные, связывающие лизин фрагменты ангиостатина, меланин или меланин-стимулирующие соединения, фрагменты плазминогена (например, Kringles 1-3 плазминогена), субъединицы тропонина, антагонисты витронектина αvβ3, пептиды, образованные из сапозина B, антибиотики или аналоги (например, тетрациклин или неомицин), диеногест-содержащие композиции, соединения, содержащие ингибиторную основу MetAP-2 совместно с пептидом, соединение EM-138, халькон и его аналоги и ингибиторы NAALAD-азы. См., например, патенты США №№ 6395718, 6462075, 6465431, 6475784, 6482802, 6482810, 6500431, 6500924, 6518298, 6521439, 6525019, 6538103, 6544758, 6544947, 6548477, 6559126 и 6569845.
В зависимости от характера комбинированной терапии, введение антитела против CD200 можно продолжать во время введения другого лекарственного средства и/или после этого. Введение антитела можно проводить в качестве однократной дозы или в качестве многократных доз. В некоторых случаях введение антитела против CD200 начинают по меньшей мере за несколько суток до общепринятого лечения, в то время как в других случаях введение начинают либо непосредственно до введения, либо во время введения общепринятого лекарственного средства. В некоторых случаях антитело против CD200 будут вводить после других лекарственных средств, или его можно вводить поочередно с другими лекарственными средствами.
Антитела по настоящему изобретению можно применять для непосредственного уничтожения или устранения злокачественных клеток in vivo. Непосредственное уничтожение вовлекает введение антител (которые являются необязательно слитыми с цитотоксическим лекарственным средством) субъекту, которому требуется такое введение. Поскольку антитела распознают CD200 на злокачественных клетках, то уничтожаются любые такие клетки, с которыми связываются антитела. Когда антитела используют отдельно для уничтожения или устранения злокачественных клеток, такого уничтожения или устранения можно достигать, инициируя эндогенные иммунные функции хозяина, такие как CDC и/или ADCC. Способы анализа для определения наличия уничтожения антителом клеток таким способом находятся в пределах компетенции специалистов в данной области.
Таким образом, в одном варианте осуществления антитела по настоящему описанию можно применять для доставки различных цитотоксических соединений. С антителами по настоящему изобретению может быть слито любое цитотоксическое соединение. Слияние можно проводить химически или генетически (например, посредством экспрессии единой слитой молекулы). Цитотоксическое соединение может представлять собой биологическую молекулу, такую как полипептид, или низкомолекулярное соединение. Как будет понятно специалистам в данной области, в случае низкомолекулярного соединения применяют химическое слияние, в то время как в случае биологических соединений можно применять любо химическое, либо генетическое слияние.
Неограничивающие примеры цитотоксических соединений включают терапевтические лекарственные средства, испускающее излучение соединение, молекулы из растений, грибов или бактерий, биологические белки и их смеси. Цитотоксические лекарственные средства могут представлять собой внутриклеточно действующие цитотоксические лекарственные средства, такие как эмиттеры короткопробежного излучения, включая, например, высокоэнергетические α-эмиттеры короткопробежного излучения. Примерами ферментативно активных токсинов и их фрагментов являются фрагмент дифтерийного токсина A, несвязывающие активные фрагменты дифтерийного токсина, экзотоксин A (из Pseudomonas aeruginosa), цепь рицина A, цепь абрина A, цепь модецина A, альфа-сарцин, некоторые белки Aleurites fordii, некоторые белки Dianthin, белки Phytolacca americana (PAP, PAPII и PAP-S), ингибитор Morodica charantia, курцин, кротин, ингибитор Saponaria officinalis, гелонин, митогиллин, рестриктоцин, феномицин и эномицин, например. Способы получения ферментативно активных полипептидов иммунотоксинов описаны в WO84/03508 и WO85/03508, которые включены в настоящее описание в качестве ссылок. Некоторые цитотоксические группы образованы, например, адриамицином, хлорамбуцилом, дауномицином, метотрексатом, неокарциностатином и платиной.
Способы конъюгации антител с цитотоксическими средствами ранее описаны и находятся в пределах квалификации специалистов данной области.
Альтернативно антитело можно связывать с высокоэнергетическими эмиттерами излучения, например, с радиоизотопом, таким как 131I, γ-эмиттер, который, когда он локализован в области опухоли, приводит к уничтожению диаметром несколько клеток. См., например, S. E. Order, "Analysis, Results, and Future Prospective of the Therapeutic Use of Radiolabeled Antibody in Cancer Therapy", Monoclonal Antibodies for Cancer Detection и Therapy, R. W. Baldwin et al. (eds.), pp.303-316 (Academic Press 1985), которая включена в настоящий документ в качестве ссылки в полном объеме. Другие пригодные радиоизотопы включают α-эмиттеры, такие как 212Bi, 213Bi и 211At, и β-эмиттеры, такие как 186Re и 90Y.
В некоторых вариантах осуществления связывающие CD200 антитела по настоящему изобретению обеспечивают благоприятный эффект, состоящий в блокировании иммунной супрессии при CLL посредством нацеливания на лейкозные клетки непосредственно через CD200. Конкретно, стимуляция иммунной системы может обеспечить устранение клеток CLL из селезенки и лимфатических узлов. Авторам неизвестно о достижении какого-либо успешного устранения клеток CLL из этого микроокружения с помощью средств, которые просто нацелены на B-клетки (таких как алемтузумаб). Напротив, реактивные в отношении CLL T-клетки могут обладать лучшим доступом в эти органы, чем антитела. В других вариантах осуществления прямого уничтожения клеток достигают нацеливанием на клетки CLL посредством антител (Ат) против CD200.
В соответствии с композициями и способами по настоящему изобретению, в особенно пригодных вариантах осуществления комбинирование прямого уничтожения клеток и запуска иммунного ответа в направлении профиля Th1 обеспечивает особенно эффективный подход к лечению злокачественной опухоли. Таким образом, в одном варианте осуществления предусмотрено лечение злокачественной опухоли, где антитело или фрагмент антитела, которые связываются с CD200 и как a) блокируют взаимодействие между CD200 и его рецептором, так и b) прямо уничтожают злокачественные клетки, экспрессирующие CD200, вводят пациенту со злокачественной опухолью. Механизм, посредством которого уничтожают злокачественные клетки, может включать, но не ограничиваться ими, ADCC и/или CDC; слияние с токсином; слияние с радиоактивной меткой; слияние с биологическим веществом, вовлеченным в уничтожение клеток, таким как гранзим B или перфорин; слияние с цитотоксическим вирусом; слияние с цитокином, таким как TNF-α или IFN-α. В альтернативном варианте осуществления лечение злокачественной опухоли вовлекает введение антитела, которое как a) блокирует взаимодействие между CD200 и его рецептором, так и b) усиливает активность цитотоксических T-клеток или NK-клеток против опухоли. Такое усиление активности цитотоксических T-клеток или NK-клеток, например, может быть добавлено посредством слияния антитела с цитокинами, например, такими как IL-2, IL-12, IL-18, IL-13 и IL-5. Кроме того, такое усиление может быть достигнуто посредством введения антитела против CD200 в сочетании с ингибиторами, такими как IMiD, талидомид или аналоги талидомида.
В другом варианте осуществления лечение злокачественной опухоли включает введение антитела, которое как (1) блокирует взаимодействие между CD200 и его рецептором, так и (2) привлекает T-клетки к клеткам опухоли. Привлечение T-клеток может быть достигнуто посредством слияния Ат с хемокинами, такими как MIG, IP-10, I-TAC, CCL21, CCL5 или LIGHT. Также лечение химиотерапевтическими средствами может приводить к требуемой активации LIGHT. Комбинированное действие блокирования иммунной супрессии и уничтожения опухолевых клеток через нацеливание посредством антитела представляет собой уникальный подход, который обеспечивает повышенную эффективность.
Антитела против CD200 в соответствии с настоящим изобретением также можно использовать в качестве диагностического инструмента. Биоптаты или образцы тканей со злокачественными клетками можно тестировать в отношении экспрессии CD200 перед лечением в целях предсказания эффективности терапии против CD200, отдельно или в сочетании с другими средствами или способами (такими как химиотерапевтические средства, лучевая терапия, иммуномодулирующая терапия и т.д.). Например, с использованием крови, полученной от пациентов с гемопоэтическими злокачественными опухолями, можно оценивать экспрессию CD200 на злокачественных клетках посредством анализа FACS с применением антител против CD200 в сочетании с соответствующими маркерами злокачественных клеток, например, такими как CD38 и CD19 на клетках CLL. Для лечения антителами против CD200 можно отбирать пациентов с уровнями CD200, по меньшей мере в 1,4 раза превышающими уровни, встречающиеся на нормальных B-клетках. В качестве другого примера, образцы ткани от пациента можно окрашивать антителом против CD200 для определения экспрессии CD200 в злокачественных и нормальных клетках пациента.
В другом примере применения антител против CD200 по настоящему изобретению, в качестве диагностического или прогностического инструмента, получают биоптаты от пациентов со злокачественной опухолью и определяют экспрессию CD200 посредством анализа FACS с использованием антител против CD200 или посредством иммуногистохимии с использованием антител против CD200. Если опухолевые клетки экспрессируют CD200 на уровнях, которые по меньшей мере в 1,4 раза превышают соответствующие уровни в нормальной ткани, тогда пациентов со злокачественной опухолью отбирают для иммуномодулирующей терапии (включая, но не ограничиваясь ими, терапию, включающую терапию против CD200). В случае злокачественной опухоли, образованной из клеток, которые в норме не экспрессируют CD200, любой поддающийся детекции уровень CD200 в биоптатах злокачественной опухоли указывает на потенциальную полезность терапии против CD200. Иммуномодулирующая терапия может представлять собой терапию против CD200, однако она также может представлять собой любую другую терапию, влияющую на иммунную систему пациента. Примеры пригодных иммуномодулирующих способов терапии включают введение средств, которые блокируют супрессию T-клеток или антигенпредставляющих клеток (например, средств против CTLA4, PD-L1, PDL-2, PD-1), или введение средств, которые усиливают положительную костимуляцию T-клеток (например, средств против CD40 или против 4-1 BB). Более того, иммуномодулирующая терапия может представлять собой вакцины против злокачественных опухолей, такие как гетероклитические пептиды или пептиды опухолевых клеток, которые приводят к цитотоксическим T-клеткам или дендритным клеткам, нагруженным опухолевыми клетками, или введение средств, которые повышают количество NK-клеток или активность T-клеток (например, антитела против CD200 отдельно или в сочетании с ингибиторами, такими как IMiD, талидомид или аналоги талидомида), или введение средств, которые устраняют регуляторные T-клетки (например, антитела против CD200 отдельно или в сочетании с ONTAK) или плазмацитоидные дендритные клетки. Комбинирование со средствами, которые повышают миграцию T-клеток или дендритных клеток, например, такими как любое средство, блокирующее SPARC, также обладает преимуществами. Более того, иммуномодулирующая терапия может представлять собой вакцины против злокачественной опухоли, такие как дендритные клетки, нагруженные клетками опухоли, полученные от пациента экзосомы с опухолевой РНК или опухолевой ДНК, белок опухоли или пептиды опухоли, полученные от пациента белки теплового шока (hsp), hsp, нагруженные опухолевыми антигенами или основные адъюванты, стимулирующие иммунную систему на различных уровнях, такие как CpG, Luivac, Biostim, Ribominyl, Imudon, Bronchovaxom или любое другое соединение, активирующее рецепторы врожденной иммунной системы (например, toll-подобные рецепторы). Также терапия может включать лечение цитокинами, такими как IL-2, GM-CSF и IFN-гамма. Также предусмотрено комбинирование со средствами, восстанавливающими сниженную активность дендритных клеток в опухолевом окружении, например, такими как ингибиторы MAP киназы.
В одном варианте осуществления антитела по настоящему изобретению также можно применять для детекции злокачественных клеток in vivo. Детекцию in vivo проводят посредством мечения антитела, введения меченого антитела субъекту, а затем визуализации субъекта. Примеры меток, пригодных для диагностической визуализации в соответствии с настоящим изобретением, представляют собой радиоактивные метки, такие как 131I, 111In, 123I, 99mTc, 32P, 125I, 3H, 14C и 188Rh, флуоресцентные метки, такие как флуоресцеин и родамин, активные в отношении ядерного магнитного резонанса метки, позитрон-испускающие изотопы, детектируемые сканером позитронной эмиссионной томографии ("PET"), хемилюминесцентные вещества, такие как люциферин, и ферментные маркеры, такие как пероксидаза или фосфатаза. Также можно использовать короткопробежные эмиттеры излучения, такие как изотопы, детектируемые детекторными зондами короткого действия, такими как трансректальный зонд. Антитело можно метить такими реагентами с использованием способов, известных в данной области. Например, см. Wensel and Meares, Radioimmunoimaging and Radioimmunotherapy, Elsevier, N.Y. (1983), которая включена в настоящий документ в качестве ссылки, для указаний, касающихся радиоактивного мечения антител. Также см. D. Colcher et al., "Use of Monoclonal Antibodies as Radiopharmaceuticals for the Localization of Human Carcinoma Xenografts in Athymic Mice", Meth. Enzymol. 121: 802-816 (1986), которая включена в настоящий документ в качестве ссылки.
Меченное радиоактивной меткой антитело в соответствии с настоящим изобретением можно применять для диагностических тестов in vitro. Специфичная активность антитела, его связывающего участка, зонда или лиганда зависит от времени полураспада, изотопной чистоты радиоактивной метки и того, каким образом метка встраивается в биологическое вещество. В иммунологических тестах чем более высокой является специфичная активность, как правило, тем более высокой является чувствительность. Способы мечения антител радиоактивными изотопами, главным образом, известны в данной области.
Меченное радиоактивной меткой антитело можно вводить пациенту, у которого оно локализуется в области злокачественных клеток, несущих антиген, с которым реагирует антитело, и его выявляют или "визуализируют" in vivo с использованием известных способов, таких как радиоизотопное сканирование с использованием, например, гамма-камеры или эмиссионной томографии. См. например, A. R. Bradwell et al., "Developments in Antibody Imaging", Monoclonal Antibodies for Cancer Detection and Therapy, R. W. Baldwin et al., (eds.), pp. 65-85 (Academic Press 1985), которая включена в настоящий документ в качестве ссылки. Альтернативно можно использовать позитронно-эмиссионный поперечный томографический сканер, такой как обозначаемый Pet VI, расположенный в Brookhaven National Laboratory, где радиоактивная метка испускает позитроны (например, 11C, 18F, 15O и 13N).
Меченные флуорофором и хромофором биологические вещества можно получать из стандартных групп, известных в данной области. Поскольку антитела и другие белки поглощают свет, имеющий длину волны вплоть до приблизительно 310 нм, флуоресцентные группы следует выбирать, чтобы они обладали значительным поглощением при длинах волн выше 310 нм и предпочтительно выше 400 нм. Описано множество пригодных флуоресцирующих веществ и хромофоров Stryer, Science, 162: 526 (1968) и Brand, L. et al., Annual Review of Biochemistry, 41: 843-868 (1972), которая включена в настоящий документ в качестве ссылки. Антитела можно метить флуоресцентными хромофорными группами посредством общепринятых способов, таких как способы, описанные в патентах США №№ 3940475, 4289747 и 4376110, которые включены в настоящий документ в качестве ссылок.
В другом варианте осуществления, в соответствии с настоящим изобретением, предусмотрены способы мониторинга течения и/или эффективности терапевтического лечения. Способ включает введение иммуномодулирующего лекарственного средства и определение уровней CD200 у субъекта по меньшей мере два раза для определения эффективности терапии. Например, можно определять уровни CD200 до лечения, и после по меньшей мере одного введения лекарственного средства можно снова определять уровни CD200. Снижение уровней CD200 указывает на эффективное лечение. Измерение уровней CD200 врач может использовать в качестве ориентира для повышения дозируемого количества или частоты лечения. Безусловно, следует понимать, что можно подвергать мониторингу непосредственно уровни CD200 или, альтернативно, можно подвергать мониторингу маркер, который коррелирует с CD200. Другие способы определения эффективности этой терапии включают, но не ограничиваются ими, детекцию злокачественных клеток, общего количества лимфоцитов, размера лимфатических узлов, количества регуляторных T-клеток, профилей цитокинов в сыворотке или внутри клеток, или секрецию цитокинов T- или B-клетками, как измеряют посредством ELISPOT.
C. ДРУГИЕ АНТАГОНИСТЫ CD200
Антагонисты CD200 и полипептиды и/или антитела, используемые в настоящем изобретении, особенно показаны для диагностических и терапевтических способов применения, как описано в настоящем документе. Таким образом, антагонист CD200 и антитела против CD200 и их варианты можно использовать в способах лечения, включая комбинированные способы лечения, в диагностике и прогнозировании заболевания, а также в мониторинге прогрессирования заболевания.
В терапевтических вариантах осуществления настоящего изобретения предусмотрены биспецифичные антитела. Биспецифичные антитела представляют собой моноклональные, предпочтительно человеческие или гуманизированные, антитела, которые обладают специфичностью связывания в отношении по меньшей мере двух различных антигенов. В данном случае одна из специфичностей связывания представляет собой специфичность к антигену CD200 на клетке (например, такой как злокачественная клетка или иммунная клетка), другая специфичность представляет собой специфичность к любому другому антигену и предпочтительно к белку клеточной поверхности или рецептору или субъединице рецептора.
Способы получения биспецифичных антител находятся в пределах квалификации специалистов в данной области. Традиционно рекомбинантная продукция биспецифичных антител основана на коэкспрессии двух пар тяжелая цепь/легкая цепь иммуноглобулина, где две тяжелых цепи обладают различной специфичностью (Milstein and Cuello, Nature, 305: 537-539 (1983)). Вариабельные домены антитела с требуемой специфичностью связывания (участки связывания антитело-антиген) можно подвергать слиянию с последовательностями константных доменов иммуноглобулинов. Слияние предпочтительно проводят с константным доменом тяжелой цепи иммуноглобулина, включая по меньшей мере часть шарнирной области, участки CH2 и CH3. ДНК, кодирующие слитые молекулы тяжелой цепи иммуноглобулина и, если желательно, легкой цепи иммуноглобулина, встраивают в отдельные экспрессирующие векторы и котрансфицируют в пригодный организм-хозяин. Для дополнительных деталей иллюстративных известных в настоящее время способов получения биспецифиных антител см., например, Suresh et al., Methods in Enzymology, 121: 210 (1986); WO 96/27011; Brennan et al., Science 229: 81 (1985); Shalaby et al., J. Exp. Med. 175: 217-225 (1992); Kostelny et al., J. Immunol. 148(5): 1547-1553 (1992); Hollinger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993); и Gruber et al., J. Immunol. 152: 5368 (1994); и Tutt et al., J. Immunol. 147: 60 (1991). Также биспецифичные антитела включают поперечно-сшитые или гетероконъюгированные антитела. Гетероконъюгированные антитела можно получать с использованием любых пригодных способов поперечного сшивания. Пригодные поперечно-сшивающие агенты хорошо известны в данной области и описаны в патенте США № 4676980, совместно с рядом способов поперечного сшивания.
Также описаны различные способы получения и выделения фрагментов биспецифичных антител непосредственно из рекомбинантной культуры клеток. Например, биспецифичные антитела продуцировали с использованием лейциновых молний. Kostelny et al., J. Immunol., 148(5): 1547-1553 (1992). Пептиды лейциновой молнии из белков Fos и Jun можно связывать с Fab'-участками двух различных антител посредством слияния генов. Гомодимеры антитела можно восстанавливать в шарнирной области с образованием мономеров, а затем повторно окислять с образованием гетеродимеров антитела. Также этот способ можно использовать для получения гомодимеров антитела. Технология "димерных антител", описанная Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993), обеспечила альтернативный механизм для получения фрагментов биспецифичных антитела. Фрагменты содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) посредством линкера, который является слишком коротким для обеспечения образования пары между двумя доменами одной цепи. Таким образом, VH- и VL-домены одного фрагмента вынуждены образовывать пары с комплементарными VL- и VH-доменами другого фрагмента, образуя посредством этого два антигенсвязывающих участка. Также описана другая стратегия получения фрагментов биспецифичных антител посредством применения димеров одноцепочечных Fv (scFv). См. Gruber et al., J. Immunol. 152: 5368 (1994). Альтернативно антитела могут представлять собой "линейные антитела", как описано в Zapata et al. Protein Eng. 8(10): 1057-1062 (1995). В кратком изложении, эти антитела содержат пару тандемных сегментов Fd (VH-CH1-VH-CH1), которые образуют пару антигенсвязывающих участков. Линейные антитела могут быть биспецифичными или моноспецифичными.
D. СПОСОБЫ ВВЕДЕНИЯ И СОСТАВЫ
Способ введения антител для антител по настоящему изобретению (любых из чистого антитела, меченого антитела, антитела, слитого с токсином, и т.д.) проводят в соответствии с известными способами, например, инъекцией или инфузией посредством внутривенного, внутрибрюшинного, интрацеребрального, внутримышечного, подкожного, внутриглазного, интраартериального, интратекального, ингаляционного или внутрираневого способов или посредством систем с замедленным высвобождением. Предпочтительно антитело вводят непрерывно инфузией или посредством болюсной инъекции. Антитела можно вводить локальным или системным способом.
Антитела по настоящему изобретению можно получать в смеси с фармацевтически приемлемым носителем. Способы изготовления и введения соединений по настоящей заявке можно найти в "Remington's Pharmaceutical Sciences", Mack Publishing Co., Easton, PA, последнее издание. Эту терапевтическую композицию можно вводить внутривенно или через нос или легкое, предпочтительно в качестве жидкости или аэрозоля порошка (лиофилизированного). Также композицию можно вводить парентерально или подкожно, если желательно. При системном введении терапевтическая композиция должна быть стерильной, по существу не содержащей пирогенов, и находиться в приемлемом для парентерального введения растворе, имеющем точно соответствующие требованиям pH, изотоничность и стабильность. Например, фармацевтический препарат по существу не содержит пирогенных материалов, так чтобы он был пригодным для введения в качестве лекарственного средства для человека. Эти условия известны специалистам в данной области.
Фармацевтические композиции, пригодные для применения, включают композиции, где одно или несколько из антител по настоящему изобретению содержатся в количестве, эффективном для достижения предполагаемой для них цели. Более конкретно, терапевтически эффективное количество означает количество антитела, эффективное для предотвращения, ослабления или смягчения симптомов заболевания или продления выживаемости подлежащего лечению субъекта. Определение терапевтически эффективного количества находится в пределах квалификации специалистов в данной области, особенно с учетом подробного описания, представленного в настоящем документе. Терапевтически эффективные дозировки можно определять с использованием способов in vitro и in vivo.
Несмотря на то, что представленное выше изобретение относится к антителам, в некоторых вариантах осуществления можно использовать полипептиды, образованные из таких антител в соответствии с настоящим описанием.
ПРИМЕРЫ
Модель на мышах и создание CD200+ клеток
Модель Raji/PBL
Мышам NOD.CB17-Prkdc<scid> (Jackson Laboratory) инъецировали 200 мкл RPMI, содержащей 4×106 клеток RAJI (ATCC), подкожно совместно с 0, 1, 5 или 10 миллионами PBL. В группы включали по девять или десять мышей. PBL выделяли из 250 мл цельной крови в градиенте histopaque с последующим лизисом эритроцитов с использованием 0,9% хлорида аммония. Мониторинг роста опухоли проводили три раза в неделю посредством измерения длины и ширины штангенциркулем. Объем опухоли вычисляли, исходя из: длина × ширина × ширина/2.
Различия между группами, в которых инъецировали PBL, по сравнению с группой, в которой вводили только опухолевые клетки, анализировали посредством 2-стороннего непарного t-теста Стьюдента. Значимые отличия были выявлены в группах, в которых вводили 5 или 10 миллионов PBL, но не в группе, в которой вводили 1 миллион PBL, начиная с 32 суток.
Модель PBL Namalwa
Мышам NOD.CB17-Prkdc<scid> (Jackson Laboratory) инъецировали 200 мкл RPMI, содержащей 4×106 клеток Namalwa (ATCC), подкожно совместно с 0, 2 или 10 миллионами PBL. В группы включали по 9 или 10 мышей. PBL выделяли из 250 мл цельной крови в градиенте histopaque с последующим лизисом эритроцитов с использованием 0,9% хлорида аммония. Мониторинг роста опухоли проводили три раза в неделю посредством измерения длины и ширины штангенциркулем. Объем опухоли вычисляли, исходя из: длина × ширина × ширина/2.
Создание стабильных экспрессирующих CD200 клеточных линий
Стабильные экспрессирующие CD200 клеточные линии Raji и Namalwa создавали с использованием лентивирусной экспрессирующей системы Virapower (Invitrogen, Carlsbad, CA). кДНК CD200 выделяли из клеток первичного CLL посредством ОТ-ПЦР с использованием прямого праймера 5'-GACAAGCTTGCAAGGATGGAGAGGCTGGTGA-3' (SEQ ID NO:34) и обратного праймера 5'-GACGGATCCGCCCCTTTTCCTCCTGCTTTTCTC-3' (SEQ ID NO:35). Продукт ПЦР клонировали в вектор Gateway внесением pCR8/GW/TOPO-TA, и отдельные клоны секвенировали. Клоны с правильной последовательностью рекомбинировали как в смысловой, так и в антисмысловой ориентациях в лентивирусные векторы pLenti6/V5/DEST и pLenti6/UbC/V5/DEST с использованием технологии Gateway (Invitrogen, Carlsbad, CA). Главным отличием между этими двумя векторами является промотор, используемый для запуска экспрессии CD200: pLenti6/V5/DEST содержит предранний промотор CMV человека, в то время как pLenti6/UbC/V5/DEST содержит промотор убиквитина C человека.
Псевдотипированные исходные растворы лентивируса VSV-G с высоким титром получали посредством временной котрансфекции клеток 293-FT, по рекомендациям изготовителя. Клетки Raji или Namalwa трансдуцировали ресуспендированием 106 клеток в 1 мл среды для роста, содержащей 12 мкг/мл Polybrene, и добавлением 1 мл исходного раствора лентивируса. После инкубации клеток в течение ночи при 37°C среду, содержащую вирус, удаляли и заменяли 4 мл свежей среды. Через двое суток в инфицированных клетках анализировали экспрессию CD200 посредством проточной цитометрии. Во всех экспериментах ≥70% клеток были CD200+, в то время как CD200 не выявлялся в исходных клеточных линиях и в клетках, трансдуцированных отрицательным контрольным вирусом (антисмысловая последовательность CD200).
Для выделения клональных клеточных линий, которые сверхэкспрессируют CD200, инфицированные клетки подвергали селекции с помощью бластицидина в течение 13 суток. Используемые концентрации бластицидина составляли 6 мкг/мл для клеток Raji или 2 мкг/мл для клеток Namalwa. Затем выделяли стабильные клоны посредством лимитирующего разведения устойчивых к бластицидину клеток в 96-луночных планшетах. Клоны подвергали скринингу в 96-луночном планшете посредством проточной цитометрии с использованием PE-конъюгированных антител мыши против CD200 человека (клон MRC OX104, Serotec) и BD FACSCalibur, оборудованного High Throughput Sampler. После скрининга всего 2000 клонов Raji и 2000 клонов Namalwa, размножали клоны с наиболее высокой экспрессией CD200 для дальнейшей охарактеризации с использованием общепринятых способов.
ПРИМЕР 1
Эффективность гуманизированных вариантов C2aB7 в модели RAJI_CD200/PBL
A) Для оценки того, сохраняют ли гуманизированные варианты C2aB7 их эффективность в моделях опухоли in vivo, химерное C2aB7 (см. публикацию патентной заявки США номер 2005/0129690) и 3 гуманизированных варианта (C2aB7V4V1, C2aB7V3V1 и C2aB7V3V2), а также отрицательное контрольное антитело alxn4100 тестировали в модели RAJI-CD200/PBL. Клетки RAJI, трансдуцированные CD200, инъецировали подкожно мышам NOD.CB17-Prkdc<scid> и оценивали способность PBL снижать рост опухоли в присутствии или отсутствие химерных или гуманизированных антител C2aB7 или контрольного антитела alxn4100 (которое не связывает опухолевые клетки). Антитела в указанных ниже концентрациях вводили первоначально с опухолевыми клетками, а затем два раза в неделю внутривенно. Были подготовлены следующие группы с 10 мышами в каждой:
Группа 1: 4×106 RAJI_CD200 подкожно
Группа 2: 4×106 RAJI_CD200 подкожно + 6×106 PBL
Группа 3: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 5 мг/кг C2aB7
Группа 4: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 20 мг/кг C2aB7V4V1
Группа 5: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 5 мг/кг C2aB7V4V1
Группа 6: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 20 мг/кг C2aB7V3V1
Группа 7: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 5 мг/кг C2aB7V3V1
Группа 8: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 5 мг/кг C2aB7V3V2
Группа 9: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 20 мг/кг alxn4100
Длину и толщину опухоли измеряли 3 раза в неделю, и объем опухоли вычисляли как длина опухоли×ширина×ширина/2. На фиг.18 показано, что, как и ожидалось, экспрессия CD200 на клетках опухоли предотвращала снижение иммунными клетками роста опухоли. Все гуманизированные варианты C2aB7 блокировали рост опухоли вплоть до 97% в дозах 20 мг/кг. Контрольное антитело alxn4100 не влияло на рост опухоли. Эти данные показывают, что все гуманизированные антитела являются высокоэффективными в отношении блокирования роста опухоли.
B) Ускользание от иммунологического надзора посредством CD200
Несмотря на то, что иммунная система человека способна индуцировать иммунный ответ против многих типов злокачественных опухолей, этот ответ является недостаточным для устранения злокачественной опухоли у большинства пациентов, вероятно вследствие ускользания от иммунного надзора через подавление иммунной системы опухолью. Авторы настоящего изобретения идентифицировали, что активация иммуносупрессирующей молекулы CD200 повышается в 1,5-5,4 раз в клетках хронического лимфоцитарного лейкоза (CLL) у всех исследованных пациентов (n=80). Известно, что взаимодействие CD200 с его рецептором изменяет профили цитокинов с Th1 на Th2 в смешанных лимфоцитарных реакциях и приводит к индукции регуляторных T-клеток, которые, как полагают, препятствуют опухолеспецифичному иммунитету эффекторных T-клеток. В этом исследовании авторы настоящего изобретения исследовали, участвует ли экспрессия CD200 на клетках опухоли в ускользании от иммунного надзора, предотвращая посредством этого устранение опухолевых клеток иммунной системы в модели ксенотрансплантации на мышах hu/SCID, и влияет ли введение антагонистического антитела против CD200 на рост опухоли в этой модели.
Клеточные линии неходжкинской лимфомы человека RAJI и Namalwa трансдуцировали посредством CD200 человека и инъецировали подкожно совместно с лимфоцитами периферической крови (PBMC) человека мышам NOD/SCID. Рост опухоли у мышей, которым вводили экспрессирующие CD200 опухолевые клетки, сравнивали с ростом опухоли у мышей, которым вводили клетки опухоли, не экспрессирующие CD200, с течением времени. В последующих экспериментах мышам вводили химерные или гуманизированные антитела против CD200 (диапазон доз от 1 мг/кг до 20 мг/кг) посредством внутривенной инъекции. Лечение начинали либо сразу, либо через 7 суток после инъекции опухолевых клеток.
В отсутствие экспрессии CD200 PBMC снижали рост опухоли RAJI или Namalwa вплоть до 75%. Напротив, рост опухолей RAJI или Namalwa, экспрессирующих CD200 на уровнях, сравнимых с CLL, не снижался посредством PBMC. Введение антител против CD200 в дозе 5 мг/кг приводило практически к полному ингибированию роста опухоли (у 1/10 мышей развивалась небольшая опухоль) в процессе исследования, даже когда введение начинали через 7 суток после инъекции опухолевых клеток.
Наличие CD200 человека на опухолевых клетках ингибирует способность лимфоцитов человека уничтожать опухолевые клетки. Лечение экспрессирующих CD200 опухолей антагонистических антител против CD200 ингибирует рост опухоли, указывая на потенциал в отношении терапии против CD200, как на перспективный подход для CLL.
C) Эффективность C2aB7G1 против конструкций C2aB7G2/G4
Для оценки того, являются ли антитела против CD200 без эффекторной функции (слитые конструкции G2/G4 C2aB7, как описано ниже) равно эффективными или более эффективными, чем конструкции G1, варианты G1 и G2/G4, а также гуманизированный вариант C2aB7 (alxn5200) тестировали в модели Raji_CD200/PBL. Клетки RAJI, трансдуцированные CD200, как описано, подкожно инъецировали мышам NOD.CB17-Prkdc<scid> и оценивали способность PBL снижать рост опухоли в присутствии или отсутствии химерных антител против CD200 c2aB7G1 (c2aB7), c2aB7G2/G4 или гуманизированных вариантов hC2aB7V3V1G1 (V3V1) или hC2aB7V3V2G1 (V3V2), или контрольного антитела alxn4100. Антитела в концентрациях, указанных ниже, вводили первоначально с опухолевыми клетками, а затем два раза в неделю внутривенно. Были подготовлены следующие группы с 10 мышами в каждой:
Группа 1: 4×106 RAJI_CD200 подкожно
Группа 2: 4×106 RAJI_CD200 подкожно + 6×106 PBL
Группа 3: 4×106 RAJI_СD200 подкожно + 6×106 PBL + 20 мг/кг hV3V2-G1
Группа 4: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 5 мг/кг alxn 5200
Группа 5: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 2,5 мг/кг alxn 5200
Группа 6: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 1 мг/кг alxn 5200
Группа 7: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 20 мг/кг chC2aB7G2/G4
Группа 8: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 5 мг/кг chC2aB7G2/G4
Группа 9: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 2,5 мг/кг chC2aB7G2/G4
Группа 10: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 1 мг/кг chC2aB7G2/G4
Группа 11: 4×106 RAJI_CD200 подкожно + 6×106 PBL + 20 мг/кг alxn4100
Длину и толщину измеряли три раза в неделю, и объем опухоли вычисляли как длина×толщина×толщина/2. На фиг.19 показано, что, как и ожидалось, экспрессия CD200 на клетках опухоли предотвращала снижение роста опухоли иммунными клетками. Однако добавление антител против CD200 снижало объем опухоли вплоть до 100%. Несмотря на то, что 20 мг/кг C2aB7G1 приводило к росту небольших опухолей у 6/10 мышей, только у 1 мыши выросли опухоли в группе, в которой вводили 20 мг/кг C2aB7G2/G4, подтверждая, что вариант G2/G4 может приводить к эффективности, лучшей или по меньшей мере равной эффективности варианта G1. Все антитела против CD200, включая гуманизированные варианты, полностью блокировали рост опухоли в дозе 5 мг/кг. Введение контрольного антитела не снижало рост опухоли. Эти данные показывают, что вариант G2/G4 C2aB7 является высокоэффективным в отношении блокирования роста опухоли экспрессирующих CD200 опухолей. Эти данные далее подтверждают, что гуманизированные варианты C2aB7 являются высокоэффективными в отношении блокирования роста опухоли в этой модели.
D) Создание конструкции G2/G4
Плазмиды изменяли в две стадии, сначала заменяя в участке IgG1 участок Age I в CH1-участке человека через стоп-кодон на участок BamH I, расположенный после поли-A-сигнала SV40. C2aB7-6 и cC7 G2G4 (антитело L-SIGN) расщепляли Age I и BamH I и C2aB7-6 обрабатывали CIP. Фрагмент длиной 10315 п.н. из C2AB7-6 и фрагмент длиной 1752 п.н. из cC7 G2G4 очищали электрофорезом и экстракцией из геля. Эти фрагменты лигировали, встраивали посредством электропорации в XL1 Blue E. coli и высевали на планшеты LB/carb/gluc. Колонии выращивали в растворе и выделяли ДНК с использованием колонок Qiagen miniprep. Наличие фрагмента IgG2G4 Age I/BamH I, в противоположность фрагменту IgG1, определяли расщеплением посредством Pvu II, которое приводило к наличию двух полос 267 и 1152 п.н. в противоположность одной полосе 1419 п.н. Для дальнейшего применения выбирали клон 21.
Остальную часть CH1-участка с конца вариабельного участка до участка Age I создавали в форме IgG2/G4 с использованием ПЦР с перекрыванием. Фрагмент ПЦР, содержащий начало участка CH1 до участка Age I, ранее создавали при получении плазмиды cC7 G2G4. В реакции ПЦР использовали праймеры C7mhHF (TCCTCAGCCTCCACCAAGGGCC, SEQ ID NO:1) и Rev Age Pri (GGGCGCCTGAGTTCCACGAC, SEQ ID NO:2) с G2G4 63L1D в качестве матрицы для получения фрагмента 142 п.н. Праймеры C2aB7 rev (GGCCCTTGGTGGAGGCTGAGGAAACTGTGAGAGTGGTGC, SEQ ID NO:3) и lacpri (GCTCCCGGCTCGTATGTTGTGT, SEQ ID NO:4) использовали с Fab C2aB7 в качестве матрицы для получения вариабельного участка тяжелой цепи мыши (и вышележащего материала) во фрагменте приблизительно из 1250 п.н. Эти фрагменты очищали посредством электрофореза и экстракции из геля и использовали в ПЦР с перекрыванием с праймерами Rev Age Pri (GGGCGCCTGAGTTCCACGAC, SEQ ID NO:2) и LeadVHpAX (ATATGAAATATCTGCTGCCGACCG, SEQ ID NO:5) для получения фрагмента 558 п.н., который очищали на колонке для очистки продуктов ПЦР. Этот фрагмент из 558 п.н. и клон 21 расщепляли посредством Xho I и Age I с получением фрагмента из 458 п.н., который очищали электрофорезом и экстракцией из геля. Клон 21 также расщепляли посредством Xho I и Age I, обрабатывали посредством CIP и фрагмент из 11,6 т.п.н. очищали электрофорезом и экстракцией из геля. Эти фрагменты лигировали и вводили посредством электропорации в XL1 Blue E. coli и высевали на планшеты LB/carb/gluc. Было выявлено, что клон C2aB7G2G4.11 обладает ожидаемыми фрагментами рестрикции при расщеплении посредством Pvu II.
Конечную конструкцию C2AB7G2G4.11 секвенировали. Было выявлено, что стоп-кодон TAA легкой цепи подвергся мутации в последовательности TCA, так что к C-концу легкой цепи добавлялось 6 аминокислот. Было выявлено, что она встречалась в клоне варианта IgG1 C2AB7-6, который был исходным для C2AB7G2G4.11. Антитела, содержащие конструкцию G2G4, представлены на фиг.10, 11, 12, 13 и 15.
ПРИМЕР 2
Экспрессия CD200 на злокачественных клетках
A. Определение повышенной активации CD200 у пациентов CLL
Лимфоциты от 15 пациентов с CLL окрашивали конъюгированным с FITC антителом против CD5 (e-bioscience), конъюгированным с APC антителом против CD19 (e-bioscience) и конъюгированным с PE антителом против CD200 (Serotec). Лимфоциты от здоровых доноров окрашивали соответствующим образом. Определяли экспрессию CD200 на клетках CD5+CD19+. Как представлено на фиг.20, несмотря на то, что уровень экспрессии CD200 варьировался среди образцов пациентов с CLL, во всех образцах CLL были показаны повышенные уровни (в диапазоне от 1,6-4 раз) экспрессии CD200 по сравнению с экспрессией CD200 на нормальных B-клетках. Пациентов с CLL, показывающих повышенные уровни экспрессии CD200, отбирали для лечения против CD200 в соответствии с описанными в настоящем документе способами.
B. Анализ FACS на злокачественных клеточных линиях
Экспрессию CD200 оценивали посредством анализа FACS с использованием панели клеточных линий NCI60 от пациентов с меланомой, пациентов с раком предстательной железы, пациентов с глиобластомой, пациентов с астроцитомой, пациентов с нейробластомой, пациентов с раком яичника, пациентов с раком легкого и пациентов с раком почки. C2aB7 метили Zenon-Alexa488 в соответствии с инструкциями изготовителя (Invitrogen). От полумиллиона до 1 миллиона клеток окрашивали 1 мкг меченого антитела в течение 20 мин, с последующим промыванием посредством PBS. Окрашивание клеток оценивали с использованием FACSCalibur (Becton Dickinson). Окрашивание меченных антителом клеток сравнивали с образцами, которые оставались немечеными, и определяли соотношение окрашенные/неокрашенные. На фиг.21 соотношение выше 1, но ниже 2 указано как +/-, соотношение между 2 и 3 представляет собой +, соотношение между 3 и 10 представляет собой ++, соотношение >10 представляет собой +++. Ни одна из тестированных клеточных линий глиобластомы, астроцитомы, рака предстательной железы или рака легкого не экспрессировала CD200, и они не представлены ниже. Четыре из 5 тестированных клеточных линий меланомы, 2/2 клеточных линий рака яичника, 2/3 клеточных линий рака почки, 2/2 клеточных линий нейробластомы и 1/3 клеточных линий рака молочной железы экспрессировали CD200 на поддающихся детекции уровнях на клеточной поверхности, подтверждая, что солидные опухоли также могут использовать CD200 в качестве механизма ускользания от иммунного надзора.
C. RT-QPCR на образцах пациента
Для проверки наличия повышенной активации CD200 не только клеточных линий, но также и на первичных образцах пациента, проводили RT-QPCR и иммуногистохимию (IHC) первичных образцов пациента. Образцы РНК от пациентов с раком яичника и меланомой получали из Cytomix. Получали кДНК и образцы разбавляли 1:100 и 1:1000 в 10 нг/мл РНК дрожжей. Образцы исследовали QPCR с анализом CD200 Hs00245978_m1, как предоставлено ABI. Для нормализации 18S анализ 18S (ABI) проводили с образцами, разбавленными 1:10000. Каждое разведение проводили в двух экземплярах. Образцы рака яичника и меланомы совместно с образцами пациента с CLL нормализовали к 18S, а затем определяли кратность экспрессии относительно нормальных PBL. На фиг.22 показана экспрессия CD200 на образцах рака яичника. Оказалось, что серозные/серозные метастатические/папиллярные серозные образцы обладали наиболее высокой экспрессией CD200, приблизительно от 10 до 20 раз выше нормальных PBL. Экспрессия CD200 была относительно низкой в эндометриоидных образцах, мукоидных образцах и образцах главных клеток, во всех случаях на уровне или ниже уровня нормальной экспрессии в яичнике (в 1-5 раз выше нормальных PBL).
На фиг.23 показаны уровни экспрессии CD200 нескольких образцов метастазов меланомы: в тощую кишку, тонкий кишечник, лимфатический узел, легкое, кожу и головной мозг. Несколько из этих образцов совпадают норма/опухоль, что указано номером (-1 пара или -2 пара). Другие дополнительные образцы без совпадения с нормой также исследовали для сравнения. Образцы тощей кишки показали значительно более высокие уровни экспрессии CD200, чем в нормальном органе, с уровнями в метастатических образцах, в 4-7 раз превышающими уровни в нормальной тощей кишке.
D. Иммуногистохимия первичных образцов пациентов
IHC проводили для 2 замороженных образцов пациента с меланомой (LifeSpan). Для окрашивания использовали Fab-фрагменты D1B5 и C2aB7. Антитело IgG1 использовали в качестве изотипического контроля. Связывание первичных антител исследовали посредством противомышиного вторичного антитела и хромагена DAB.
Как показано на фиг.24, оба тестированных образца меланомы показали выраженное мембранное окрашивание антителами против CD200, однако отсутствие окрашивания изотипическим контролем. Нормальные образцы кожи не показали окрашивания CD200. Эти данные показывают, что CD200 обладает повышенной активацией не только в клеточных линиях меланомы и рака яичника, но также в первичных образцах пациента.
E. Ускользание от иммунного надзора клеток меланомы и опухоли яичника посредством повышенной активации иммуносупрессивной молекулы CD200
Ускользание от иммунного надзора является ключевым признаком прогрессирования злокачественной опухоли. Опухоли могут ускользать от иммунной системы множеством механизмов, каждый из которых является существенным препятствием для иммунотерапии. Внедрение новых и более эффективных форм иммунотерапии будет требовать понимания этих процессов, а также их сходств и отличий в различных злокачественных опухолях. Авторы настоящего изобретения ранее идентифицировали повышенную активацию иммуносупрессивной молекулы CD200 на клетках хронического лимфоцитарного лейкоза. Наличие CD200 подавляет продукцию Th1-цитокинов, требуемую для эффективного цитотоксического T-клеточного ответа. Авторы настоящего изобретения показали в моделях на животных, что экспрессия CD200 клетками опухоли человека предотвращает отторжение лимфоцитами человека опухоли, и введение антагонистического антитела против CD200 ингибирует рост опухоли. В этом исследовании авторы настоящего изобретения оценивали наличие повышенной активации CD200 в других злокачественных опухолях и наличие влияния экспрессии CD200 на этих злокачественных клетках на иммунный ответ.
Относительные уровни транскрипта CD200 количественно определяли посредством RT-QPCR в образцах аденокарциномы яичника (серозные/серозные метастататические/папиллярные серозные, эндометриоидные, мукоидные, главных клеток) и в метастатических образцах пациентов со злокачественной меланомой.
Экспрессию CD200 на клеточной поверхности оценивали посредством IHC в двух замороженных образцах тканей пациентов с меланомой и трех пациентов с карциномой (серозной) по сравнению с нормальной кожей и нормальными яичниками. Экспрессию CD200 на клеточной поверхности злокачественных клеточных линий меланомы SK-MEL-5, SK-MEL-24 и SK-MEL-28 и клеточной линии рака яичника OV-CAR-3 оценивали посредством анализа FACS с использованием PE-меченого антитела против CD200. Эффект экспрессирующих CD200 злокачественных клеточных линий на смешанный профиль цитокинов в лимфоцитарных реакциях оценивали посредством добавления клеток к культуре образованных из моноцитов дендритных клеток с аллогенными T-клетками человека. Продукцию цитокинов (IL-2 и IFN-γ для Th1, IL4 и IL10 для Th2) выявляли в супернатанте посредством ELISA.
Количественный ПЦР показал уровни экспрессии CD200 в образцах серозной аденокарциномы яичника, вплоть до 20 раз превышающие уровни в нормальных PBL, и равные или вплоть до 4 раз превышающие уровни в нормальном яичнике. Экспрессия CD200 была равной или меньшей уровней нормальных яичников в образцах эндометриоидной, мукоидной и образованной из главных клеток аденокарциномы яичника. В метастазах злокачественной меланомы в тощую кишку уровни экспрессии CD200 оказались значительно более высокими, чем в нормальных образцах. В метастазах злокачественной меланомы в легкое 2/6 показали более высокую экспрессию CD200, чем нормальные образцы.
IHC показала выраженное, специфичное, ассоциированное с мембраной окрашивание CD200 на злокачественных клетках обоих пациентов с меланомой. Образец нормальной кожи показал слабое окрашивание эндотелиальных клеток. Среди трех пациентов с раком яичника у одного было показано выраженное окрашивание CD200, у одного было показано умеренно положительное окрашивание и у одного были показаны группы слабо окрашенных опухолевых клеток. Во всех трех случаях в строме было показано выраженное окрашивание.
CD200 высоко экспрессировался на клеточной поверхности злокачественных клеточных линий меланомы SK-MEL-24 и SK-MEL-28, а также на клеточной линии рака яичника OV-CAR-3, и умеренно экспрессировался на клеточной линии меланомы SK-MEL-5. Добавление любой из этих клеточных линий в реакцию смешанных лимфоцитов подавляло продукцию цитокинов Th1, в то время как клеточные линии, не экспрессирующие CD200, не подавляли ее, показывая прямую корреляцию. Добавление антагонистического антитела против CD200 в процессе культивирования отменяло этот эффект.
Клетки меланомы и опухоли яичников могут активировать CD200, подавляя таким образом иммунный ответ. Лечение антагонистическим антителом против CD200 позволяет иммунной системеувеличивать эффективную цитотоксическую реакцию против опухолевых клеток.
F. Эффект экспрессирующих CD200 злокачественных клеточных линий на профили цитокинов в реакциях смешанных лимфоцитов
Способность клеток, сверхэкспрессирующих CD200, переключать ответ цитокинов с TH1-ответа (IL-2, IFN-γ) на Th2-ответ (IL-4, IL-10) оценивали в реакции смешанных лимфоцитов. В качестве источника экспрессирующих CD200 клеток использовали либо трансфицированные CD200 клетки, либо клетки из положительных по CD200 злокачественных клеточных линий.
Реакции смешанных лимфоцитов проводили в 24-луночных планшетах с использованием 250000 дендритных клеток, полученных созреванием из периферических моноцитов человека с использованием IL-4, GM-CSF и IFN-γ и 1×106 отвечающих клеток. Отвечающие клетки представляли собой обогащенные T-клетками лимфоциты, очищенные из периферической крови с использованием Ficoll. Содержание T-клеток повышали инкубацией клеток в течение 1 часа во флаконах для культуры тканей и отбором фракции неприкрепляющихся клеток. 500000 клеток из злокачественных клеточных линий меланомы SK-MEL-1, SK-MEL-24, SK-MEL-28, клеточной линии рака яичника OVCAR3 и клеточной линии неходжкинской лимфомы Namalwa или первичных клеток CLL в качестве положительного контроля добавляли к дендритным клеткам в присутствии или отсутствии 30 мкг/мл антитела против CD200. Супернатанты собирали через 48 и 68 часов и анализировали в них наличие цитокинов.
Цитокины, такие как IL-2, IFN-γ и IL-10, находящиеся в супернатанте культуры тканей, количественно определяли с использованием ELISA. Совпадающие пары иммобилизованного и детектирующего антитела для каждого цитокина получали от R+D Systems (Minneapolis, MN), стандартную кривую для каждого цитокина получали с использованием рекомбинантного цитокина человека. Иммобилизованное антитело против цитокина наносили на планшет в PBS в оптимальной концентрации. После инкубации в течение ночи планшеты промывали и блокировали в течение 1 часа посредством PBS, содержащего 1% BSA и 5% сахарозу. После 3 промываний PBS, содержащим 0,05% Tween, супернатанты добавляли в двукратных или трехкратных разведениях в PBS, содержащем 1% BSA. Захваченные цитокины выявляли посредством соответствующего биотинилированного антитела против цитокина с последующим добавлением конъюгированного с щелочной фосфатазой стреправидина и субстрата SigmaS. Окрашивание оценивали с помощью устройства для считывания планшетов ELISA (Molecular Devices).
Как показано на фиг.25A, наличие клеточных линий с высокой экспрессией CD200 (MEL-24, MEL-28, OVCAR-3) приводило к подавлению Th1-цитокинов, таких как IL-2 и IFN-γ. Напротив, добавление MEL-1 (низкая экспрессия CD200) или Namalwa (отсутствие экспрессии CD200) не влияло на профиль цитокинов. Добавление антитела против CD200 hB7VH3VL2 в количестве 50 мкг/мл полностью восстанавливало Th1-ответ (фиг.25B), указывая на то, что лечение пациентов с меланомой или раком яичника антителом против CD200 может произвести благоприятный терапевтический эффект.
ПРИМЕР 3
Устранение активированных T-клеток посредством C2aB7-G1 и его производных
Для оценки наличия эффекта лечения против CD200 в модели злокачественной опухоли с использованием клеток опухоли, не экспрессирующих CD200, клетки Namalwa и PBL человека инъецировали мышам NOD/SCID и мышам проводили введение, как представлено ниже. В этой модели CD200 представлен только на иммунных клетках, в естественных условиях экспрессирующих CD200, таких как B-клетки и фолликулярные T-хелперные клетки. Схема формирования групп:
10 животных/группа
Группа 1: 4×106 Namalwa подкожно
Группа 2: 4×106 Namalwa подкожно + 8×106 PBL
Группа 3: 4×106 Namalwa подкожно + 8×106 PBL + 20 мг/кг hV3V2-G1
Группа 4: 4×106 Namalwa подкожно + 8×106 PBL + 5 мг/кг hV3V2-G1
Группа 5: 4×106 Namalwa подкожно + 8×106 PBL + 2,5 мг/кг hV3V2-G1
Группа 6: 4×106 Namalwa подкожно + 4×106 PBL
Группа 7: 4×106 Namalwa подкожно + 8×106 PBL + 20 мг/кг chC2aB7-G2G4
Группа 8: 4×106 Namalwa подкожно + 8×105 PBL + 5 мг/кг chC2aB7-G2G4
Группа 9: 4×106 Namalwa подкожно + 8×IO6 PBL + 2,5 мг/кг chC2aB7-G2G4
Группа 10: 4×106 Namalwa подкожно + 4×106 PBL + 20 мг/кг chC2aB7-G2G4
Группа 11: 4×106 Namalwa подкожно + 8×106 PBL + 20 мг/кг alxn4100
В инъекционную смесь была включена 1/10 дозы. Последующее дозирование представляло собой 2x/неделя внутривенно в течение 3 недель.
Длину (L) и ширину (W) опухоли измеряли 3 раза в неделю, и объемы опухолей вычисляли как L×W×W/2. На фиг.26 показано, что, как установлено ранее, одновременная инъекция PBL человека с клетками Namalwa ингибирует рост опухоли. Не было выявлено эффекта chC2aB7-G2G4 на опосредуемое PBL ингибирование роста опухоли. Напротив, введение ALXN5200 (hB7VH3 VL2-G1) блокировало опосредуемое PBL ингибирование роста опухоли. В отсутствие CD200 на клетках опухоли, оказалось, что лечение антителом против CD200 посредством антитела, которое опосредует эффекторную функцию, такого как конструкции G1, устраняет критические эффекторные клетки в популяции PBL. Эти данные подтверждают, что направленная против CD200 терапия против злокачественной опухоли является менее эффективной, когда используют антитело с эффекторной функцией по сравнению с использованием антитела без эффекторной функции. Однако лечение против CD200 с использованием конструкции с эффекторной функцией может производить терапевтически благоприятный эффект в случаях, когда желательным является устранение иммунных клеток, таких как условия трансплантации или аутоиммунные заболевания.
ПРИМЕР 4
Уничтожение T-клеток посредством hB7VH3VL2
Для оценки того, приводит ли инкубация активированных T-клеток с антителами против CD200, содержащими константный участок, опосредующий эффекторную функцию (например G1), к уничтожению T-клеток, T-клетки активировали и проводили анализ уничтожения, как описано ниже.
A. Выделение CD3+ T-клеток
Лимфоциты периферической крови (PBL) человека получали от нормальных здоровых добровольцев посредством центрифугирования в градиенте плотности гепаринизированной цельной крови с использованием системы AccuspinTM System. В каждую пробирку Accuspin (Sigma, St. Louis, MO; car# A2055) добавляли пятнадцать мл Histopaque-1077 (Sigma, St. Louis, MO; каталожный # H8889), которую затем центрифугировали при 1500 об./мин в течение 2 минут, чтобы обеспечить прохождение Histopaque через фритту. Тридцать мл цельной крови наслаивали на фритту и пробирки центрифугировали в течение 15 минут при 2000 об./мин при комнатной температуре без торможения. Границу раздела PBL собирали и мононуклеарные клетки промывали два раза в PBS 2% инактивированной нагреванием эмбриональной телячьей сывороткой (FBS) (Atlas Biologicals, Ft. Collins, CO; каталожный # F-0500-D) при центрифугировании при 1200 об./мин в течение 10 минут. CD3+ T-клетки выделяли пропусканием через колонку HTCC-5 (R&D Systems) в соответствии с инструкциями изготовителя. Элюированные клетки промывали, подсчитывали и ресуспендировали в RPMI 1640, содержащей 5% инактивированную нагреванием сыворотку одного донора, 2 мМ L-глутамин, 10 мМ Hepes и пенициллин/стрептомицин.
B. Активация посредством связанного с планшетом mOKT3
На лунки 12-луночных планшетов (Falcon) наносили 10 мкг/мл mOKT3 (Orthoclone), разведенного в PBS, посредством инкубации в течение ночи при 4°C. Остаточное антитело удаляли, и планшеты осторожно промывали посредством PBS. Очищенные CD3+ T-клетки, выделенные, как описано выше, добавляли в планшеты до конечной концентрации 2×106/лунка в RPMI 1640, содержащей 5% инактивированную нагреванием сыворотку одного донора, 2 мМ L-глутамин, 10 мМ Hepes и пенициллин/стрептомицин. Клетки поддерживали в течение 72 часов при 37°С в инкубаторе с увлажнением, содержащем 5% CO2.
C. Мечение 51 хромом активированных mOKT3 CD3+ клеток-мишеней
В конце периода культивирования активированные mOKT3 клетки CD3+ собирали, промывали и ресуспендировали в количестве 107 клеток/мл в RPMI 1640 без сыворотки. В клетки вводили хром добавлением 125 мКи 51хрома (Perkin Elmer, Billerica, MA)/106 клеток в течение 2 часов при 37°C. Меченые клетки собирали, промывали в RPMI, содержащей 5% инактивированную нагреванием сыворотку одного донора, и ресуспендировали в конечной концентрации 2×105 клеток/мл в той же среде.
D. Получение аутологичных эффекторных NK-клеток
Лимфоциты периферической крови (PBL) человека от того же индивида получали, как описано выше, посредством центрифугирования в градиенте плотности. Границу раздела PBL собирали и мононуклеарные клетки промывали два раза в PBS 2% инактивированной нагреванием эмбриональной телячьей сывороткой (FBS) (Atlas Biologicals, Ft. Collins, CO; каталожный # F-0500-D) с центрифугированием при 1200 об./мин в течение 10 минут. Клетки CD56+ выделяли посредством положительной селекции над конъюгированными с антителами против CD56 магнитными гранулами (Miltenyi Biotec, Auburn, CA, Cat # 120-000-307) в соответствии с инструкциями изготовителя. Элюированные клетки промывали, подсчитывали и ресуспендировали в количестве 1,3×106 клеток/мл в RPMI 1640, содержащей 5% инактивированную нагреванием сыворотку одного донора, 2 мМ L-глутамин, 10 мМ Hepes и пенициллин/стрептомицин. Клетки инкубировали в течение ночи при 37°С в инкубаторе с увлажнением, содержащем 5% CO2, в конечной концентрации 4×106 клеток/лунка в 3 мл той же среды. В конце периода культивирования клетки собирали, промывали, подсчитывали и ресуспендировали в не содержащей сыворотку RPMI, содержащей 2 мМ L-глутамин, 10 мМ Hepes, 2×10-5 M 2-меркаптоэтанол и пенициллин/стрептомицин.
E. Анализ ADCC
Меченные 51Cr активированные mOKT3 CD3+ клетки-мишени, полученные, как описано выше, распределяли в лунки 96-луночного планшета в количестве 104 клеток/лунка в 50 мкл. CD56+ эффекторные клетки собирали, промывали, подсчитывали и ресуспендировали в количестве либо 2,5×106 клеток/мл (для соотношения эффектор:клетка-мишень 25:1), либо 106 клеток/мл (для соотношения эффектор:клетка-мишень 10:1) и распределяли (100 мкл/лунка) в лунки, содержащие клетки-мишени. К эффекторам и мишеням добавляли десятикратные разведения антител против CD200 (V3V2-G1 или V3V2-G2/G4) в конечных концентрациях 10, 1, 0,1 и 0,01 мкг/мл. Контроли анализа включали следующее: 1) эффекторы и мишени в отсутствие антитела (0 Ат); 2) клетки-мишени в отсутствие эффекторов (спонтанный лизис) и 3) эффекторы и мишени, инкубированные с 0,2% Tween-80 (максимальное высвобождение). Культивирование клеток для всех условий проводили по три экземпляра. Клетки инкубировали при 37°C в течение 4 часов в инкубаторе с увлажнением, содержащем 5% CO2. В конце периода культивирования планшеты центрифугировали для осаждения клеток, и 150 мкл клеточного супернатанта переносили в сцинтилляционные флаконы и подсчитывали в гамма-сцинтилляционном счетчике (Wallac). Результаты выражали в качестве процентного лизиса в соответствии со следующей формулой:
(Среднее количество импульсов в минуту в образце (cpm) - средний спонтанный лизис) × 100
(средний максимальный лизис - средний спонтанный лизис)
F. Проточная цитометрия
Сто мкл клеточных суспензий (активированные mOKT3 CD3+ клетки или очищенные CD56+ NK-клетки), полученных, как описано выше, распределяли в 96-луночный планшет с круглым дном (Falcon, Franklin Lakes NJ; каталожный # 353077). Клетки инкубировали в течение 30 минут при 4°C с указанными сочетаниями следующих компонентов: флуоресцеинизотиоцианат (FITC)-, фикоэритрин (PE)-, PerCP-Cy5.5- или аллофикоцианин (APC)-конъюгированные антитела (все от Becton-Dickinson, San Jose, CA); антитело против CD25 человека-FITC (каталожный # 555431); антитело против CD3 человека-APC (каталожный # 555335); антитело против CD200 человека-PE (каталожный # 552475); антитело против CD8 человека-PerCP-Cy5.5 (каталожный # 341051); антитело против CD4 человека-APC (каталожный # 555349); антитело против CD5 человека-APC (каталожный # 555355) и антитело против CD56 человека-APC (каталожный # 341025). Также включали изотипические контроли для каждого меченого антитела. После промывания клеток два раза буфером FACS (центрифугирование при 1800 об./мин в течение 3 минут), клетки ресуспендировали в 300 мкл PBS (Mediatech, Herndon, VA; каталожный # 21-031-CV) и анализировали посредством проточной цитометрии с использованием устройства FacsCaliber и программного обеспечения CellQuest (Becton Dickinson, San Jose, CA).
Как представлено на фиг.27, активированные T-клетки показывают высокую экспрессию CD200 на их поверхности. Активированные T-клетки эффективно уничтожаются в присутствии VH3VL2-G1, но не VH3VL2-G2G4, когда в качестве эффекторных клеток используют NK-клетки (фиг.28). Эти данные показывают, что антитела против CD200 с эффекторной функцией могут элиминировать активированные T-клетки. Такое антитело может обладать терапевтическим применением в условиях трансплантации или для лечения аутоиммунных заболеваний.
В дополнение к регуляторным T-клеткам было показано, что плазмацитоидные дендритные клетки играют отрицательную иммунорегуляторную роль в злокачественной опухоли человека (Wei S, Kryczek I, Zou L, Daniel B, Cheng P, Mottram P, Curiel T, Lange A, Zou W Plasmacytoid dendritic cells induce CD8+ regulatory T cells in human ovarian carcinoma. Cancer Res. 2005 Jun 15; 65(12): 5020-6). Комбинирование терапии, элиминирующей плазмацитоидные дендритные клетки, с терапией против CD200, таким образом, может обладать преимуществом.
ПРИМЕР 5
CD200 на плазматических клетках
Клетки костного мозга от 10 пациентов с множественной миеломой и 3 нормальных доноров получали, первоначально лизируя эритроциты с использованием хлорида аммония. Клетки ресуспендировали в буфере для FACS и метили следующими смесями антител:
*Kappa-FITC/CD38-PE/CD138-PerCP-Cy5.5
*Lambda-FITC/CD38-PE/CD138-PerCP-Cy5.5
*Изотипический контроль-FITC/CD38-PE
*CD200-FITC/CD38-PE
Данные получали с использованием BD FACS Canto и анализировали с использованием программного обеспечения BD DiVA. Анализировали экспрессию CD200 на ярких в отношении CD38 клетках (плазматических клетках). Как представлено на фиг.29, часть плазматических клеток экспрессируют CD200 с высокой интенсивностью у нормальных доноров. У пациентов с множественной миеломой большинство плазматических клеток экспрессируют CD200.
В случае множественной миеломы, аналогично CLL или другим злокачественным опухолям, экспрессирующим CD200, экспрессия CD200 клетками опухоли может препятствовать устранению иммунной системой опухолевых клеток. Следовательно, антагонистическая терапия против CD200 может обеспечить устранение иммунной системой злокачественных клеток. Аблятивная терапия против CD200, нацеленная на плазматические клетки, может оказывать терапевтически благоприятный эффект при аутоиммунных заболеваниях или в условиях трансплантации.
ПРИМЕР 6
CD200 на вирусах
Также CD200 экспрессируется на ряде вирусов, таких как вирус миксомы M141R или вирус герпеса человека 8. Аналогично экспрессии CD200 на клетках опухоли, CD200 на вирусах может препятствовать эффективному выведению вируса иммунной системой. Лечение антагонистическим антителом против CD200 может оказывать терапевтически благоприятный эффект при инфекции экспрессирующим CD200 вирусом, обеспечивая устранение вируса иммунной системой. Альтернативно можно применять аблятивное антитело против CD200.
Будет понятно, что можно проводить различные модификации вариантов осуществления, описанных в настоящем документе. Например, как поймут специалисты в данной области, конкретные последовательности, описанные в настоящем документе, могут быть немного изменены без обязательного неблагоприятного влияния на функциональность полипептида, антитела или фрагмента антитела, применяемых для связывания OX-2/CD200. Например, часто можно проводить замены одной или нескольких аминокислот в последовательности антитела без нарушения функциональности антитела или фрагмента. Таким образом, следует понимать, что полипептиды или антитела, имеющие степень идентичности более 70% с конкретными антителами, описанными в настоящем документе, находятся в пределах объема этого описания. В особенно пригодных вариантах осуществления предусмотрены антитела, имеющие идентичность более приблизительно 80% с конкретными антителами, описанными в настоящем документе. В других пригодных вариантах осуществления предусмотрены антитела, имеющие идентичность более приблизительно 90% с конкретными антителами, описанными в настоящем документе. Таким образом, представленное выше описание не следует истолковывать как ограничивающее, а только как иллюстрацию предпочтительных вариантов осуществления. Специалисты в данной области предусмотрят другие модификации в пределах объема и сущности этого изобретения.
ССЫЛКИ
Представленные ниже ссылки включены в настоящий документ в качестве ссылок для более полного описания уровня техники, к которой относится настоящее изобретение. Любое несоответствие между этими публикациями, представленными ниже, или публикациями, включенными в качестве ссылок выше, и настоящим изобретением следует принимать в пользу настоящего изобретения.
1) Agarwal, et al., (2003). Disregulated expression of the Th2 cytokine gene in patients with intraoral squamous cell carcinoma. Immunol. Invest 32:17-30.
2) Almasri, N.M. et al. (1992). Am J Hemato 140, 259-263.
3) Contasta, et al., (2003). Passage from normal mucosa to adenoma and colon cancer: alteration of normal sCD30 mechanisms regulating TH1/TH2 cell functions. Cancer Biother Radiopharm 18:549-557.
4) Gorczynski, et al., (1998). Increased expression of the novel molecule OX-2 is involved in prolongation of murine renal allograft survival. Transplantation 65:1106-1114.
5) Gorczynski, et al., (2001). Evidence of a role for CD200 in regulation of immune rejection of leukaemic tumour cells in C57BL/6 mice. Clin Exp. Immunol. 126:220-229.
6) Hainsworth, J.D. (2000). Oncologist 2000; 5(5):376-84.
7) Inagawa, et al., (1998). Mechanisms by which chemofherapeutic agents augment the antitumor effects of tumor necrosis factor: involvement of the pattern shift of cytokines from Th2 to Тh1 in tumor lesions. Anticancer Res 18:3957-3964.
8) Ito, et al., (1999). Lung carcinoma: analysis of Т helper type 1 and 2 cells and Т cytotoxic type 1 and 2 cells by intracellular cytokine detection with flow cytometry. Cancer 85:2359-2367.
9) Kiani, et al., (2003). Normal intrinsic Th1/Th2 balance in patients with chronic phase chronic myeloid leukemia not treated with interferon-alpha or imatinib. Haematologica 88:754-761.
10) Lauerova, et al., (2002). Malignant melanoma associates with Th1/Th2 imbalance that coincides with disease progression and immunotherapy response. Neoplasma 49:159-166.
11) Maggio, et al., (2002). Chemokines, cytokines and their receptors in Hodgkin's lymphoma cell lines and tissues. Ann Oncol. 13 Suppl 1:52-56.
12) Nilsson, К. (1992). Burn Cell. 5(1):25-41.
13) Podhorecka, et al., (2002). Т type 1/type 2 subsets balance in B-cell chronic lymphocytic leukemia-the three-color flow cytometry analysis. Leuk Res 26:657-660.
14) Pu, Q.Q. and Bezwoda, W. (2000). Anticancer Res. 20(4):2569-78.
15) Smyth, et al., (2003). Renal cell carcinoma induces prostaglandin E2 and T-helper type 2 cytokine production in peripheral blood mononuclear cells. Ann Surg Oncol. 10:455-462.
16) Tatsumi, et al., (2002). Disease-associated bias in Т helper type 1 (Th1)/Th2 CD4(+) Т cell responses against MAGE-6 in HLA-DRB10401(+) patients with renal cell carcinoma or melanoma. J Exp. Med. 196:619-628.
17) Walls A V et al. (1989). Int. J Cancer 44846-853.
18) Winter, et al., (2003). Tumour-induced polarization of tumour vaccine-draining lymph node Т cells to a type 1 cytokine profile predicts inherent strong immunogenicity of the tumour and correlates with therapeutic efficacy in adoptive transfer studies. Immunology 108:409-419.
19) Cameron, С. M., J. W. Barrett, L. Liu, A. R. Lucas, and G. McFadden. 2005. Myxoma virus M141R expresses a viral CD200 (vOX-2) that is responsible for down-regulation of macrophage and T-cell activation in vivo. J Virol. 79:6052.
20) Foster-Cuevas, M., G. J. Wright, M. J. Puklavec, M. H. Brown, and A. N. Barclay. 2004. Human herpesvirus 8 K14 protein mimics CD200 in down-regulating macrophage activation through CD200 receptor. J Virol. 78:7667.
21) Nicholas, J. 2003. Human herpesvirus-8-encoded signalling ligands and receptors. J Biomed Sci 10:475.
22) Shiratori, I., M. Yamaguchi, M. Suzukawa, K. Yamamoto, L. L. Lanier, T. Saito, and H. Arase. 2005. Down-regulation of basophil function by human CD200 and human herpesvirus-8 CD200. J Immunol. 175:4441.
23) Voigt, S., G. R. Sandford, G. S. Hayward, and W. H. Burn. 2005. The English strain of rat cytomegalovirus (CMV) contains a novel captured CD200 (vOX2) gene and a spliced CC chemokine upstream from the major immediate-early region: further evidence for a separate evolutionary lineage from that of rat CMV Maastricht. J Gen. Virol. 86:263.
24) Zhang, J., J. Wang, C. Wood, D. Xu, and L. Zhang. 2005. Kaposi's sarcoma-associated herpesvirus/human herpesvirus 8 replication and transcription activator regulates viral and cellular genes via interferon-stimulated response elements. J Virol. 79:5640.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ РАЗРУШЕНИЯ КЛЕТОК С ИСПОЛЬЗОВАНИЕМ ЭФФЕКТОРНЫХ ФУНКЦИЙ АНТИ-EphA4 АНТИТЕЛ | 2007 |
|
RU2429246C2 |
| АНТИТЕЛА ПРОТИВ PD-1 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2788616C2 |
| АНТИТЕЛА К БЕЛКУ ПРОГРАММИРУЕМОЙ КЛЕТОЧНОЙ СМЕРТИ 1 | 2018 |
|
RU2812915C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХ С ОСЬЮ PD-1, И ИНГИБИТОРОВ TIGIT | 2014 |
|
RU2702108C2 |
| УЛУЧШЕННЫЕ СПОСОБЫ ЛЕЧЕНИЯ ВАСКУЛЯРИЗИРОВАННЫХ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2015 |
|
RU2692248C2 |
| ЛЕЧЕНИЕ РАКА С ПОМОЩЬЮ АНТИТЕЛА К СЕМАФОРИНУ 4D В КОМБИНАЦИИ С ЭПИГЕНЕТИЧЕСКИМ МОДУЛИРУЮЩИМ АГЕНТОМ | 2018 |
|
RU2783535C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХ С ОСЬЮ PD-1, И ИНГИБИТОРОВ TIGIT | 2014 |
|
RU2825390C2 |
| ЛИГАНД GITR И СВЯЗАННЫЕ С ЛИГАНДОМ GITR МОЛЕКУЛЫ И АНТИТЕЛА И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2369636C2 |
| АНТИТЕЛА К ИНТЕГРИНУ АЛЬФА-2 И ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2545401C2 |
| АНТИТЕЛА, РАСПОЗНАЮЩИЕ СОДЕРЖАЩИЙ УГЛЕВОДЫ ЭПИТОП НА CD-43 И СЕА, ЭКСПРЕССИРУЕМЫХ НА ЗЛОКАЧЕСТВЕННЫХ КЛЕТКАХ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2461572C2 |





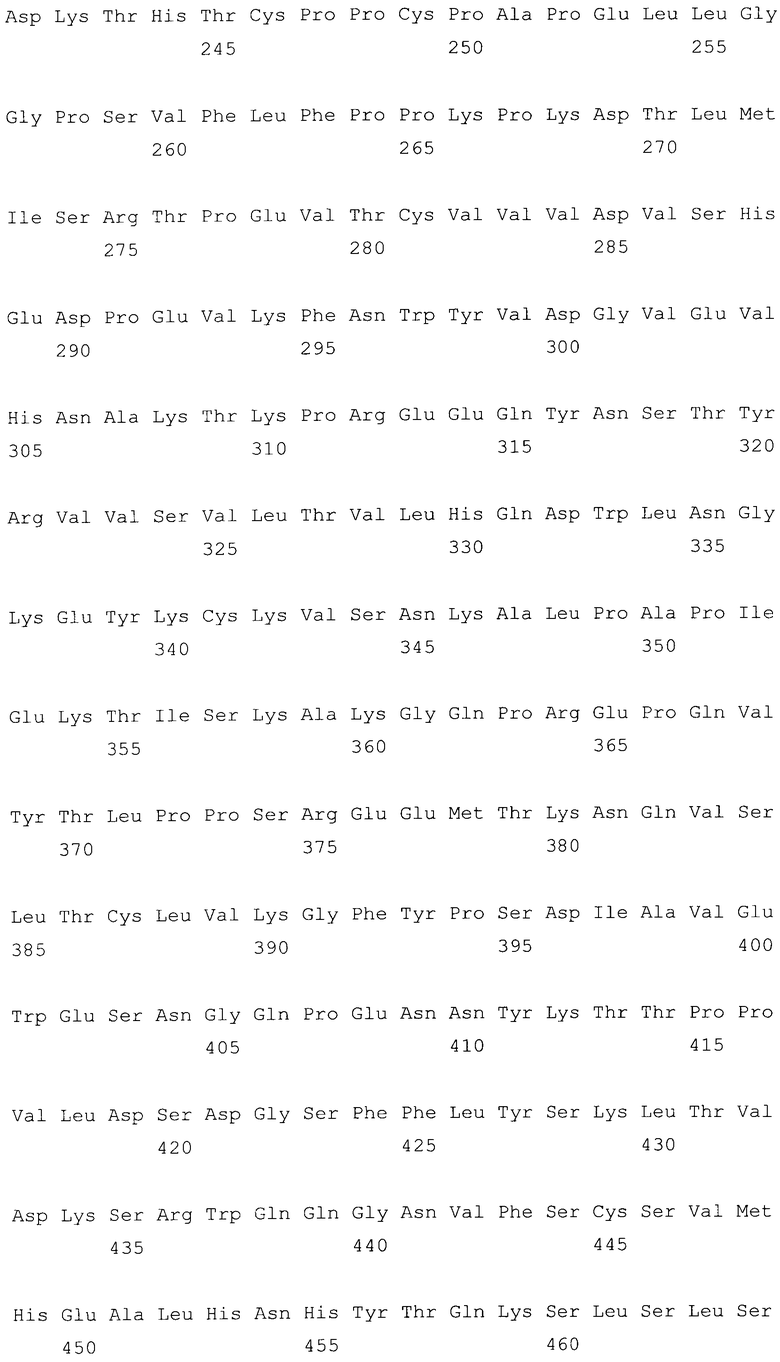






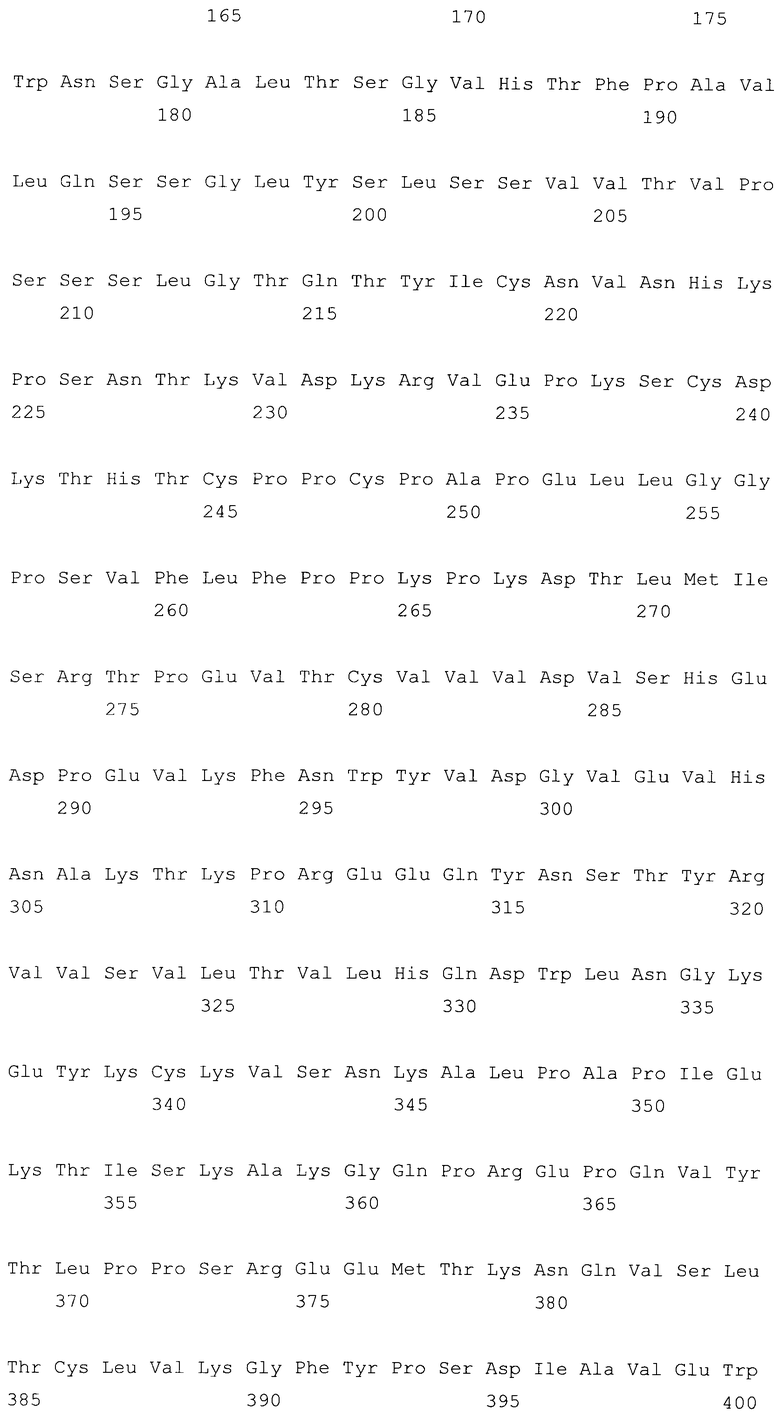













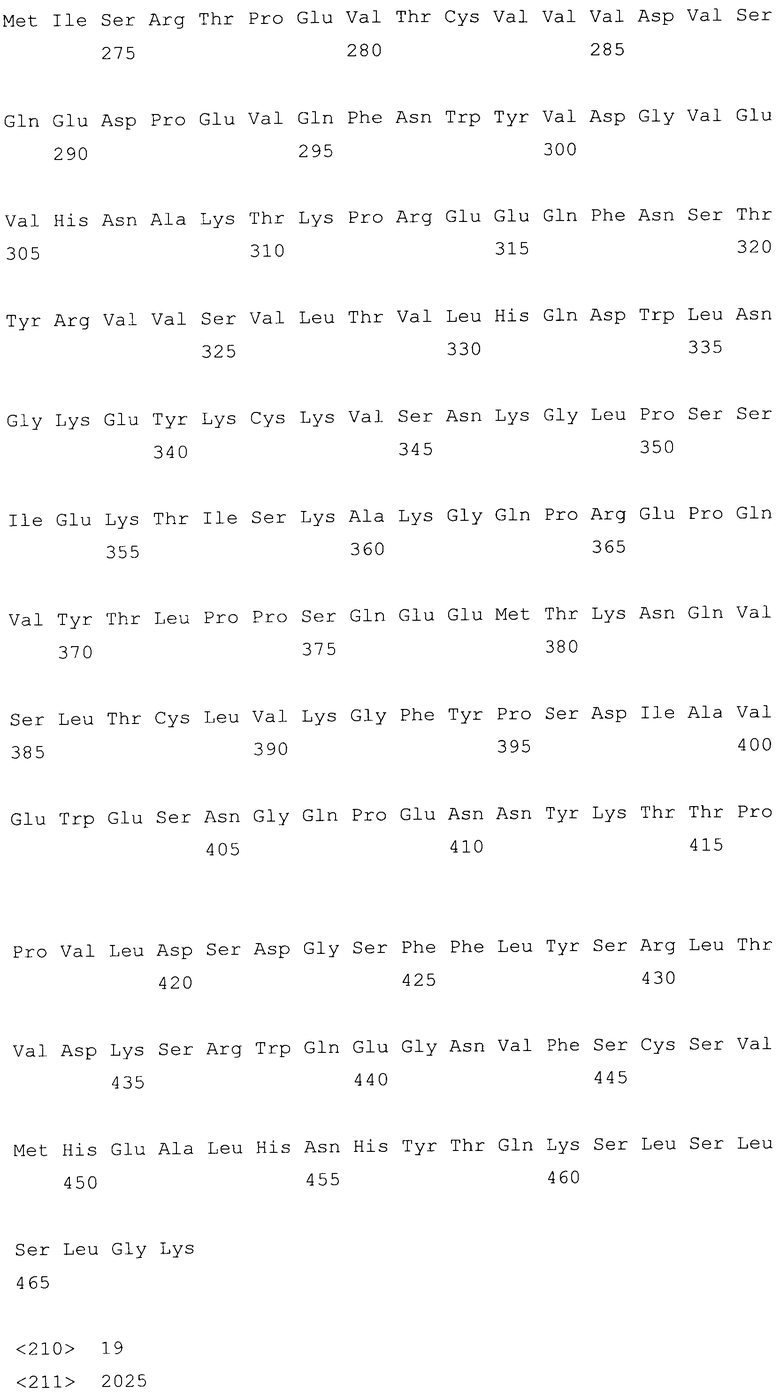


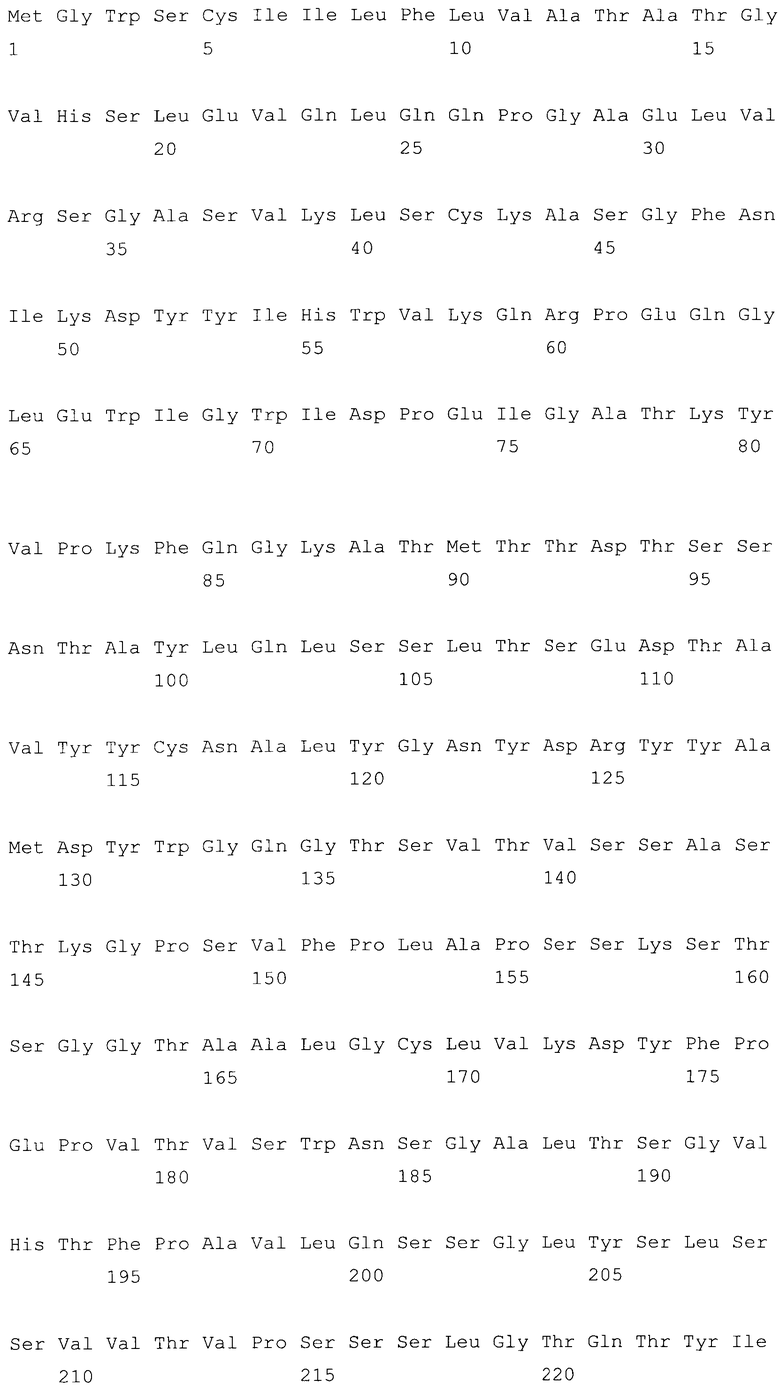
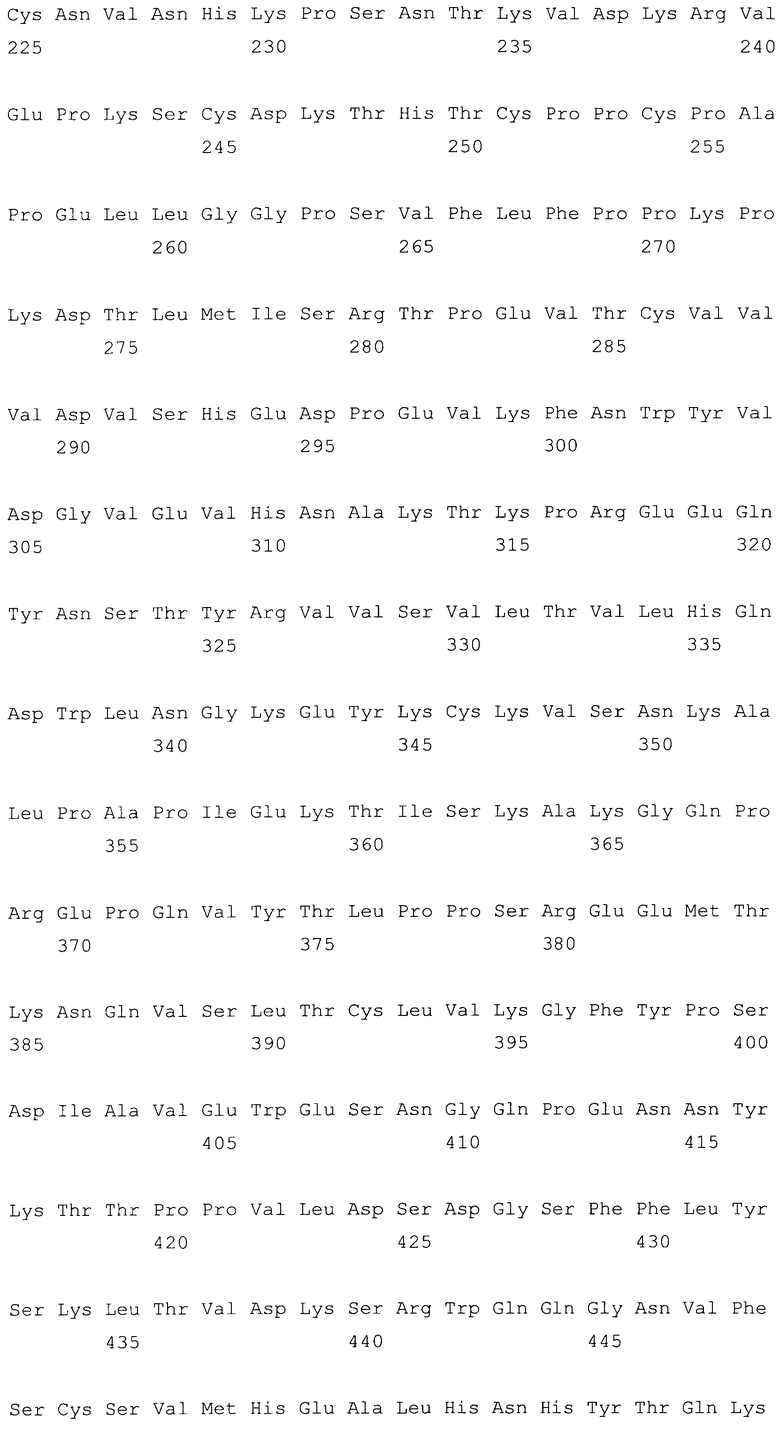




























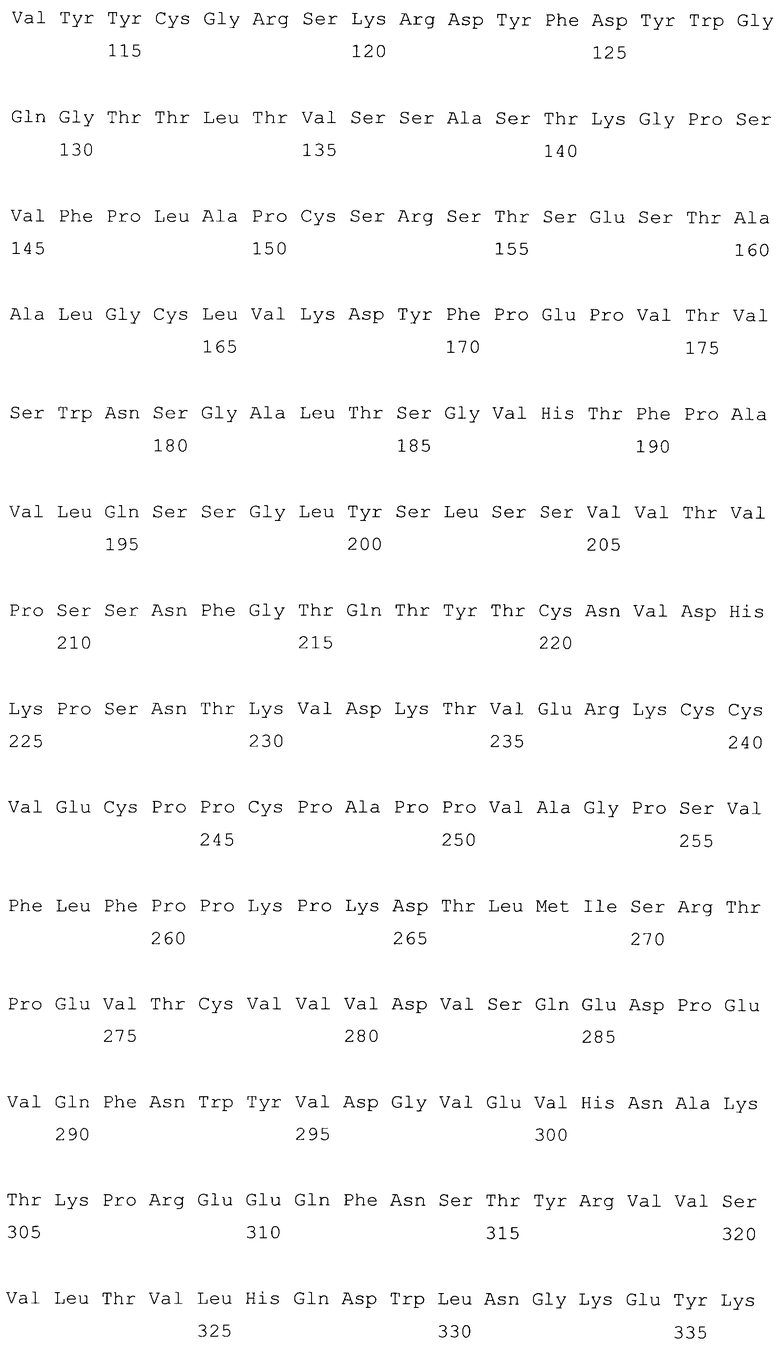








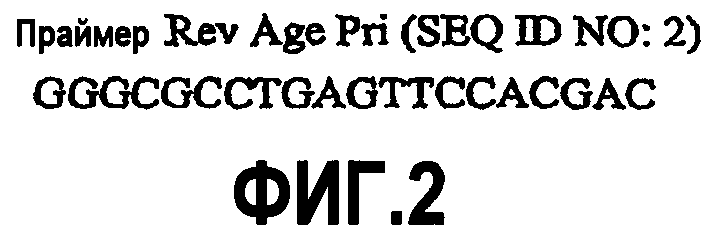
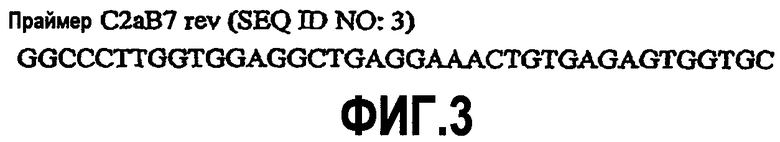




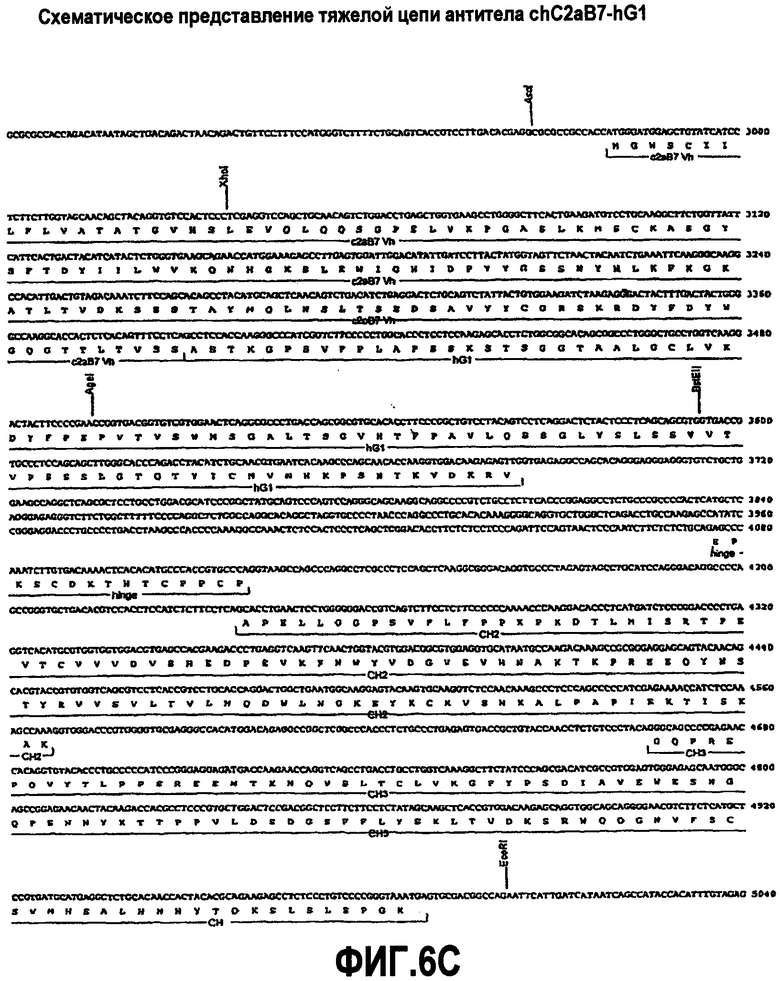


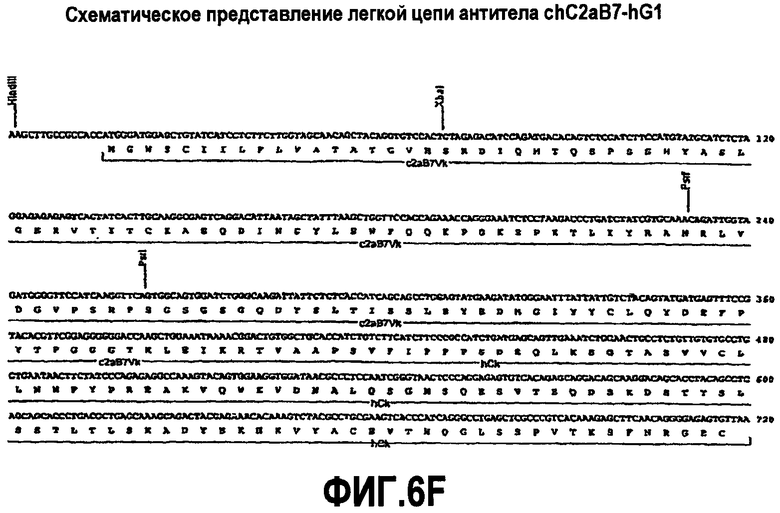


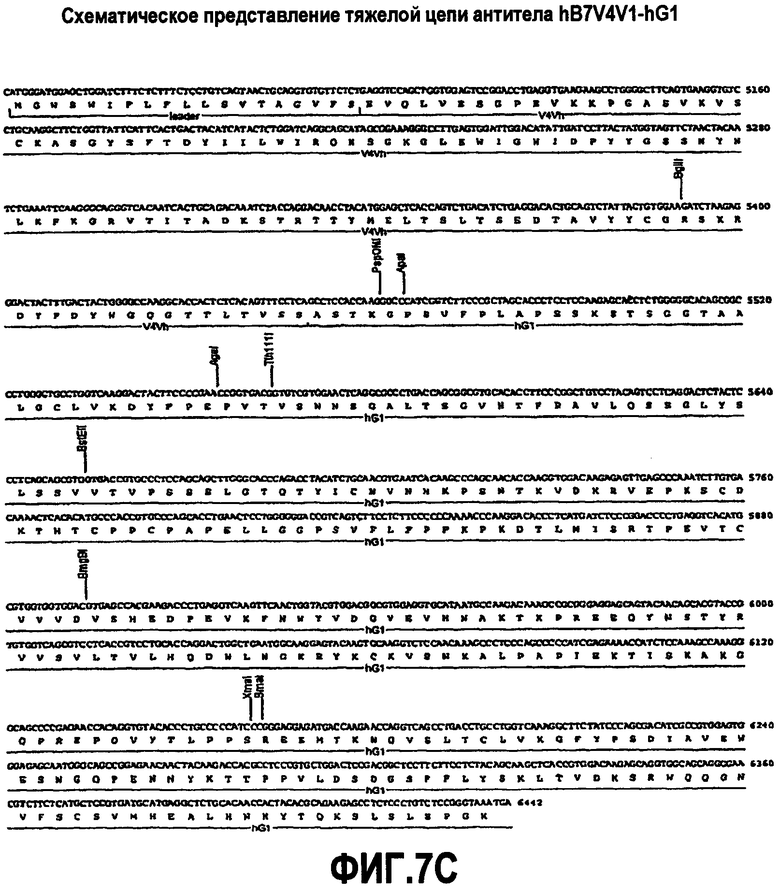

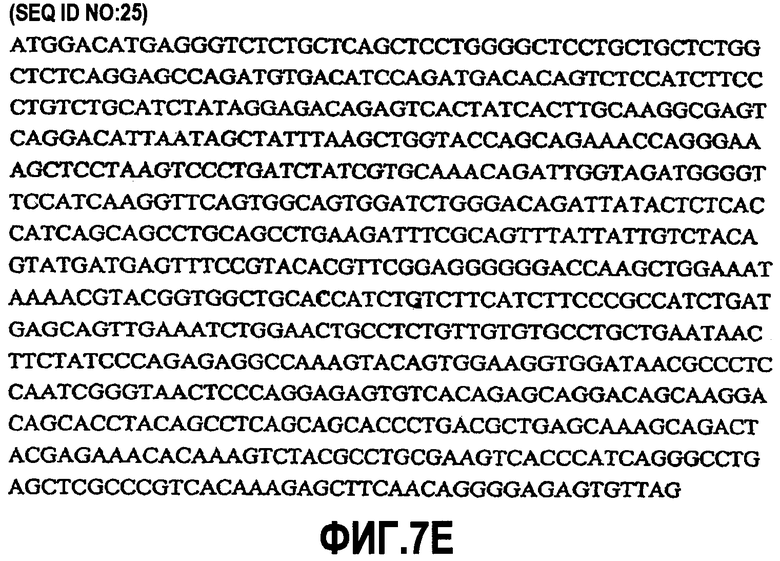
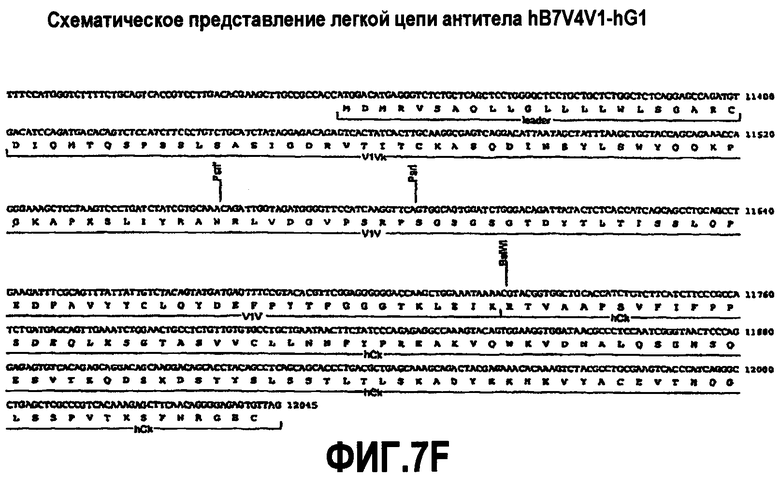



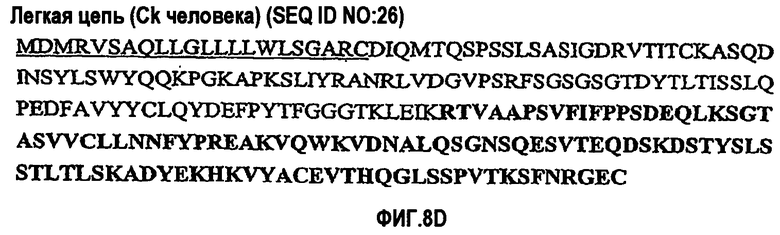
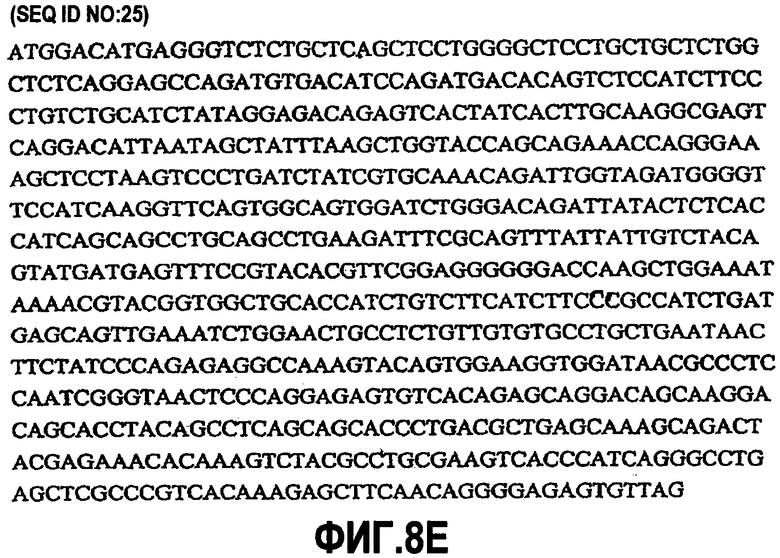
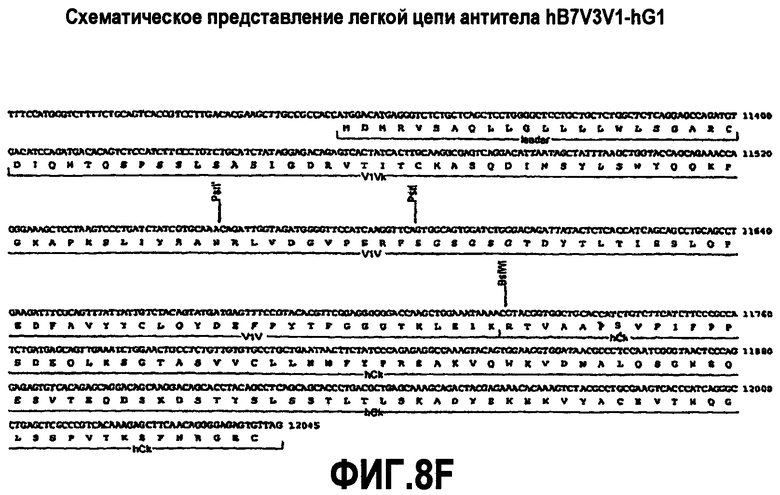





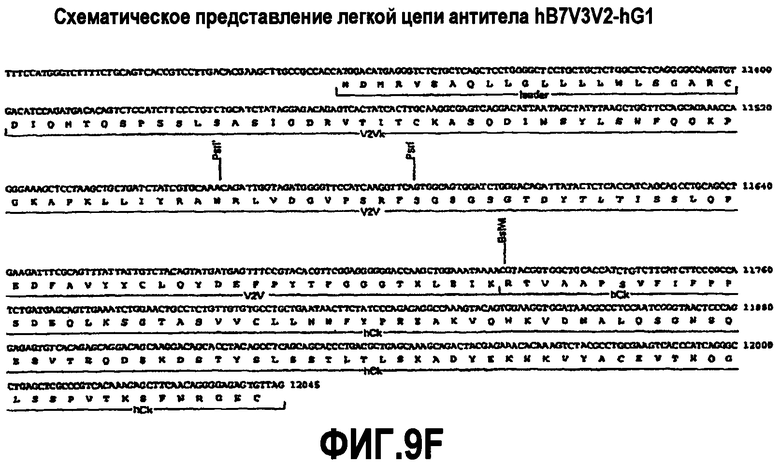


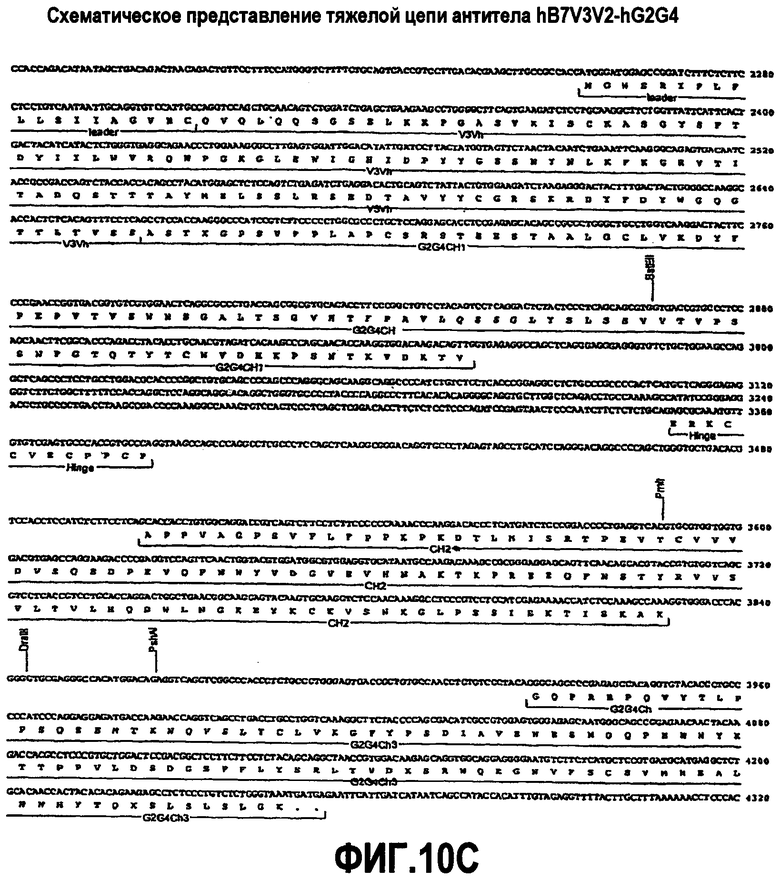

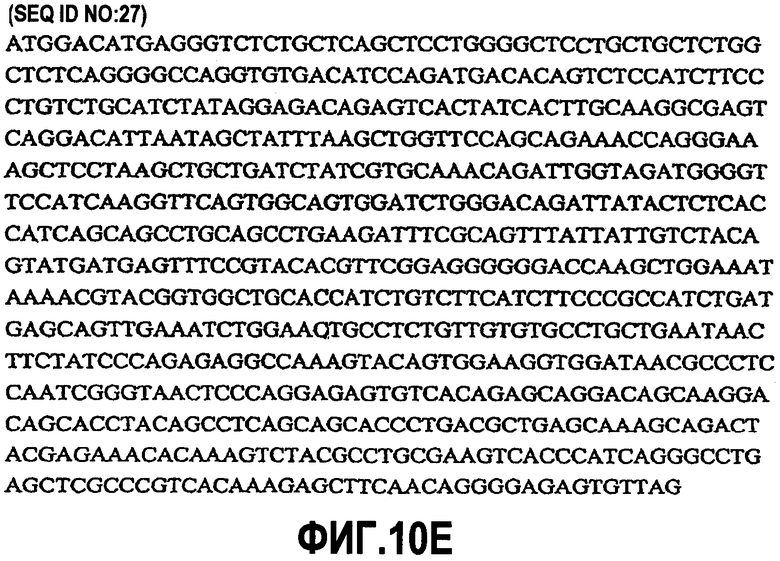
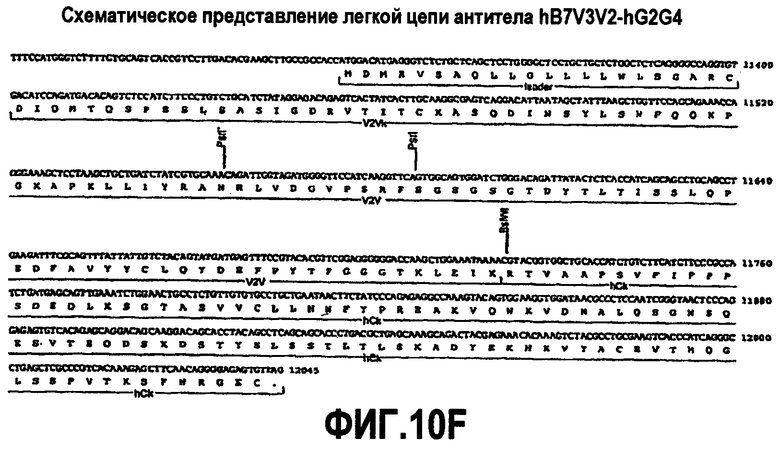





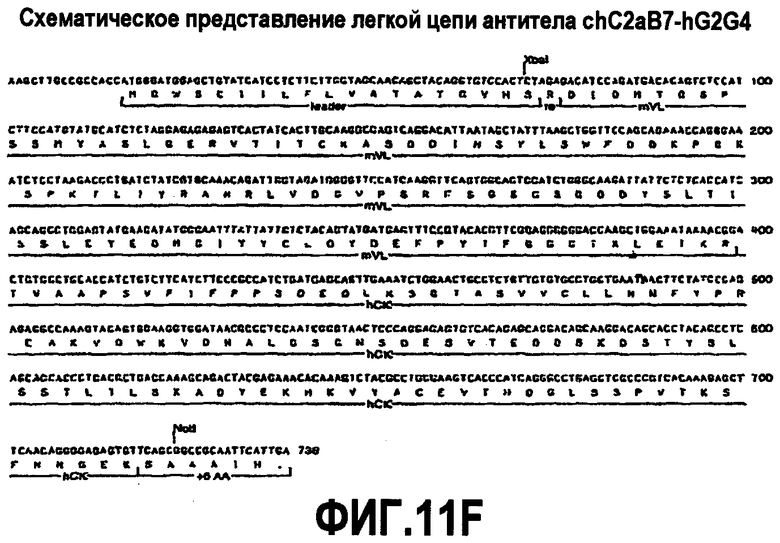





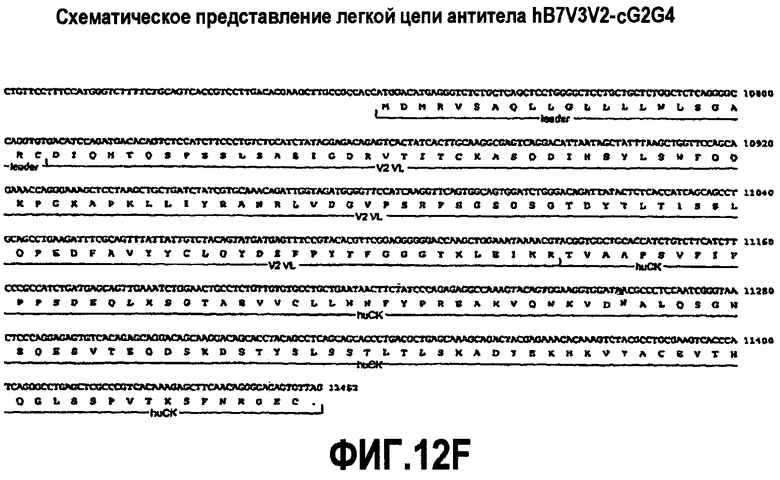



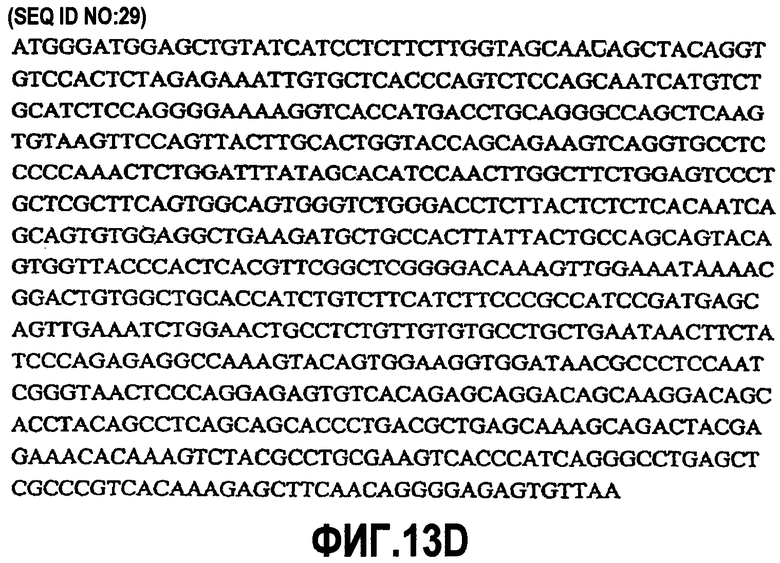


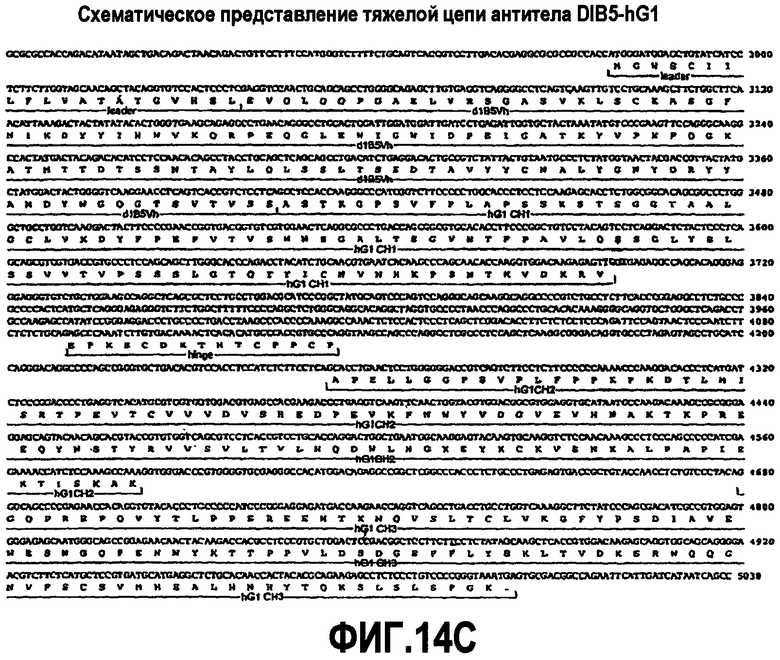

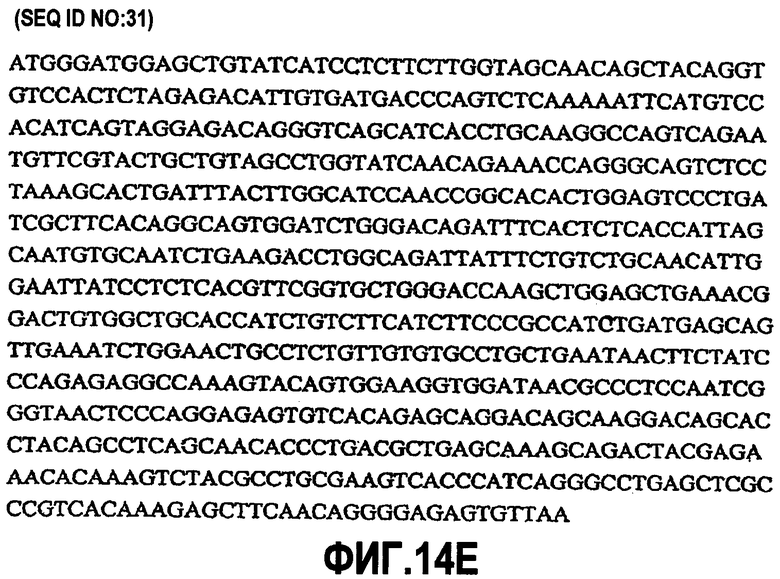
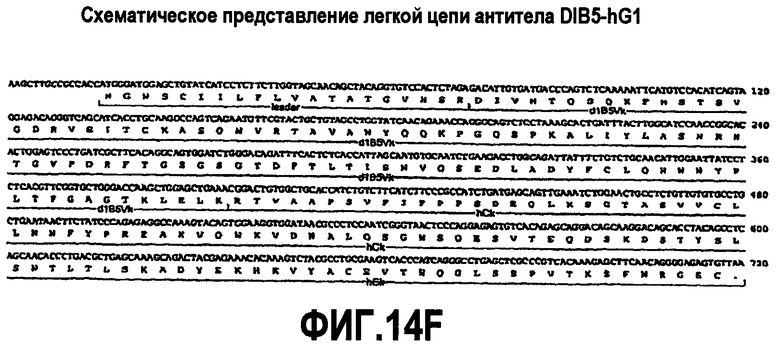




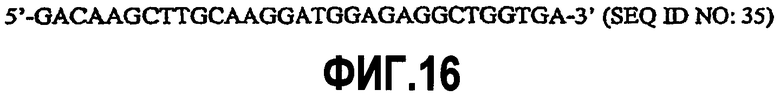

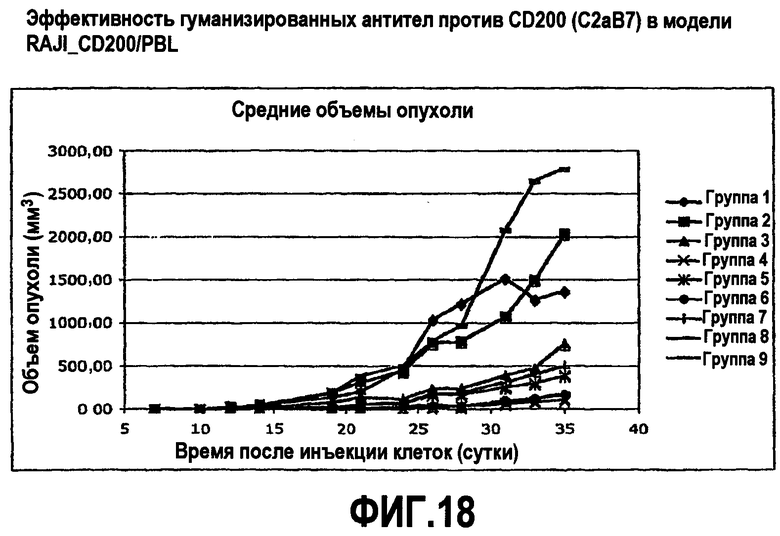
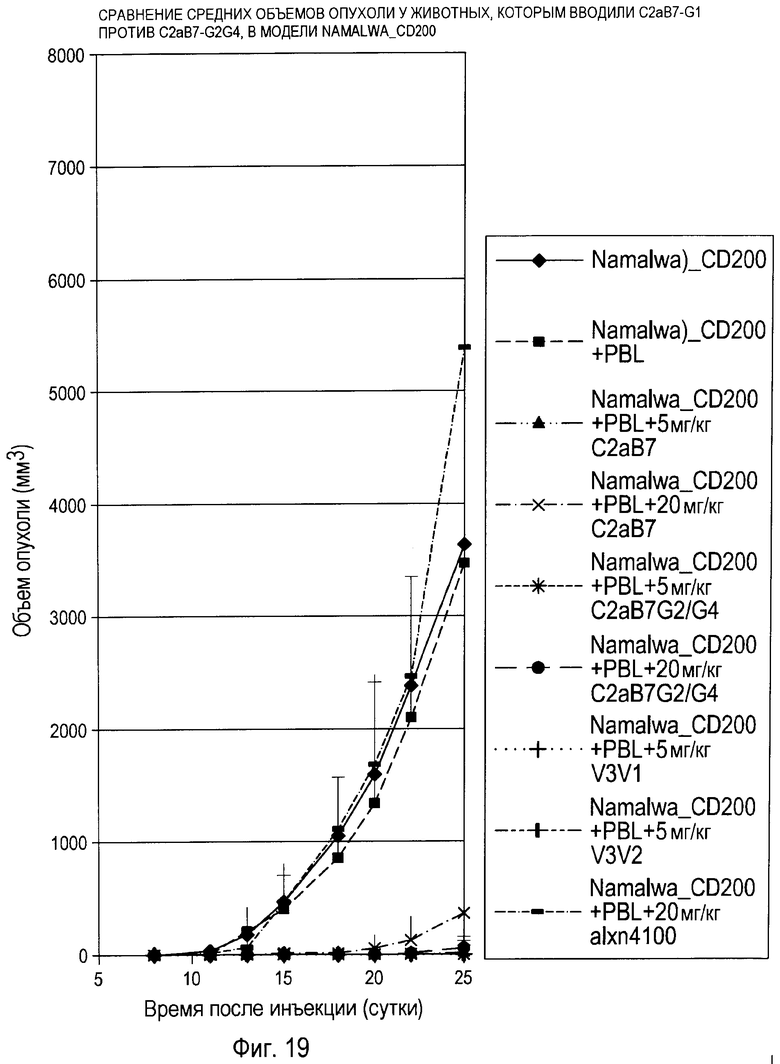
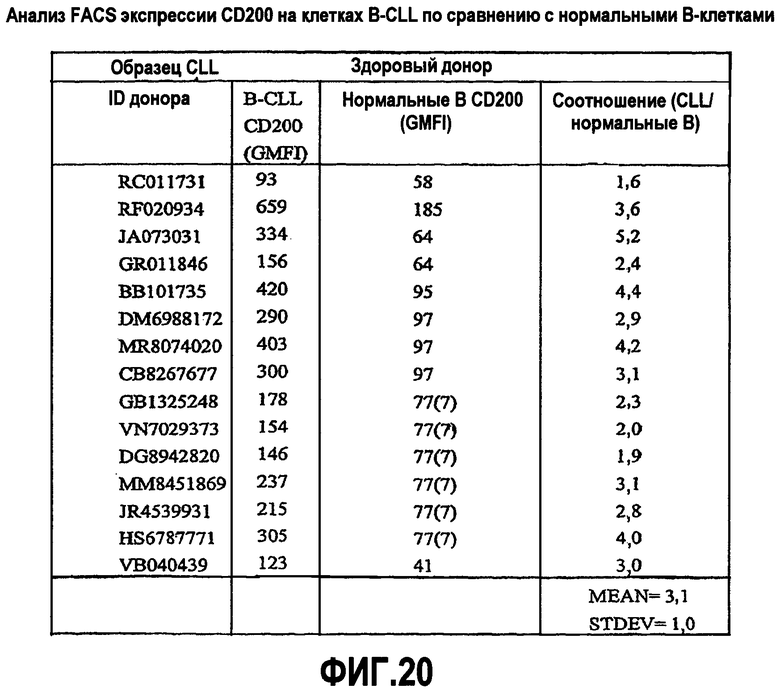

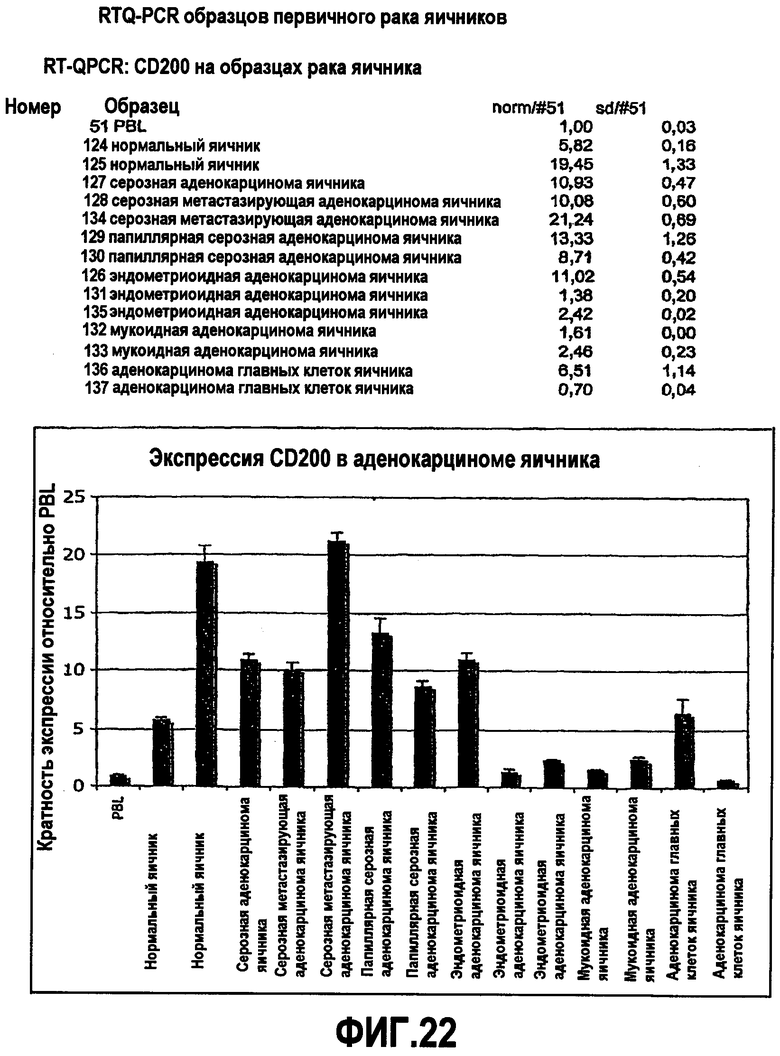
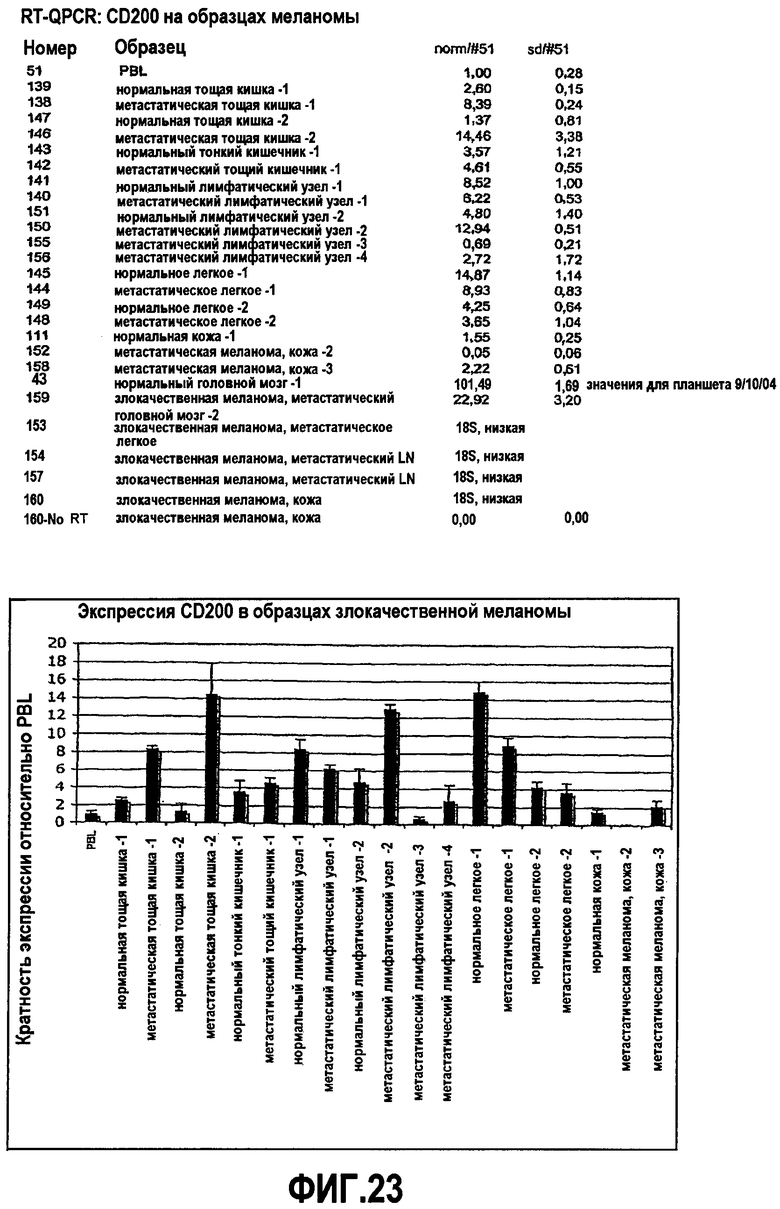
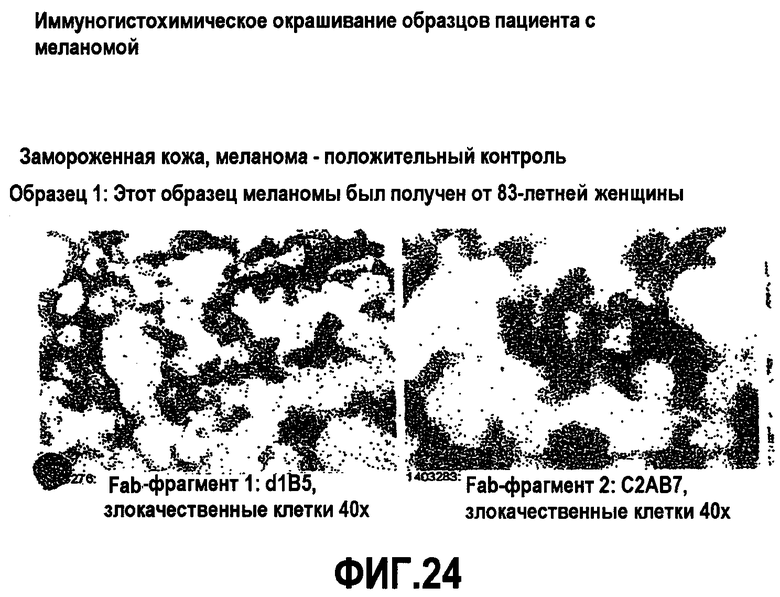
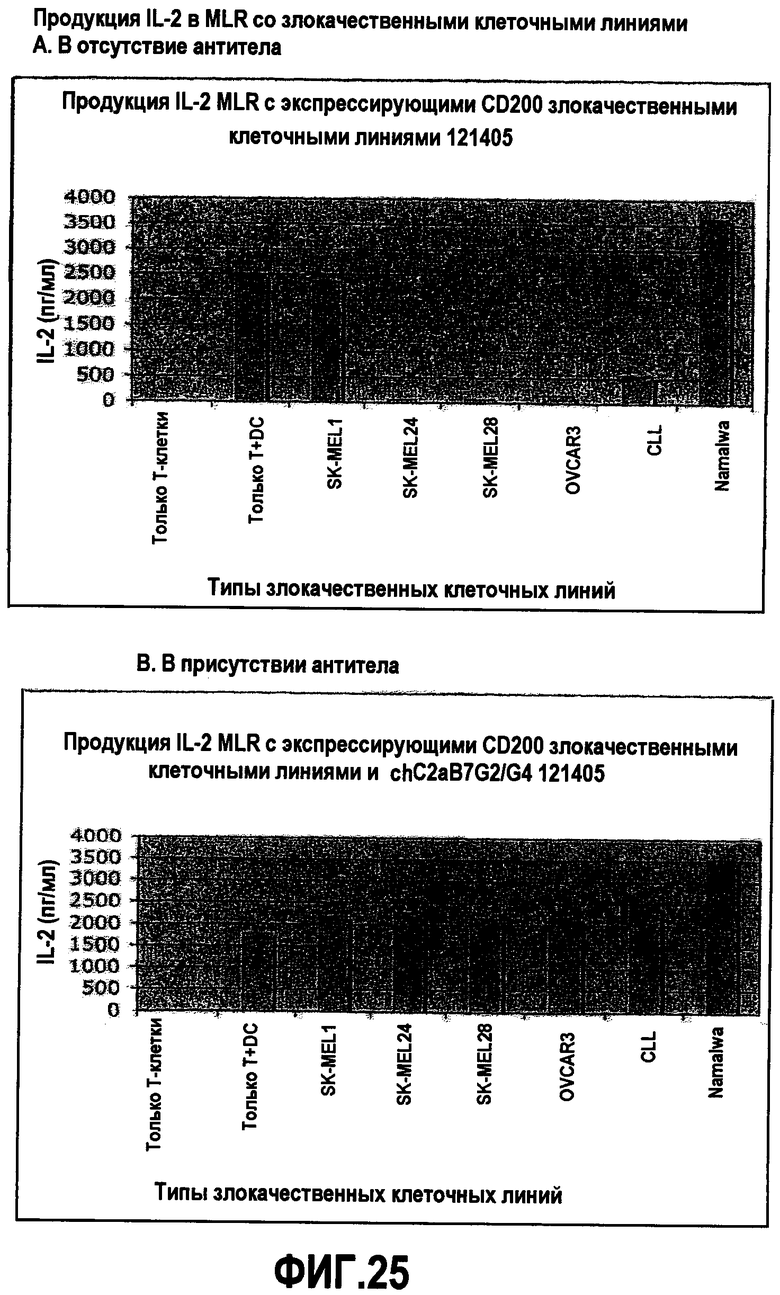
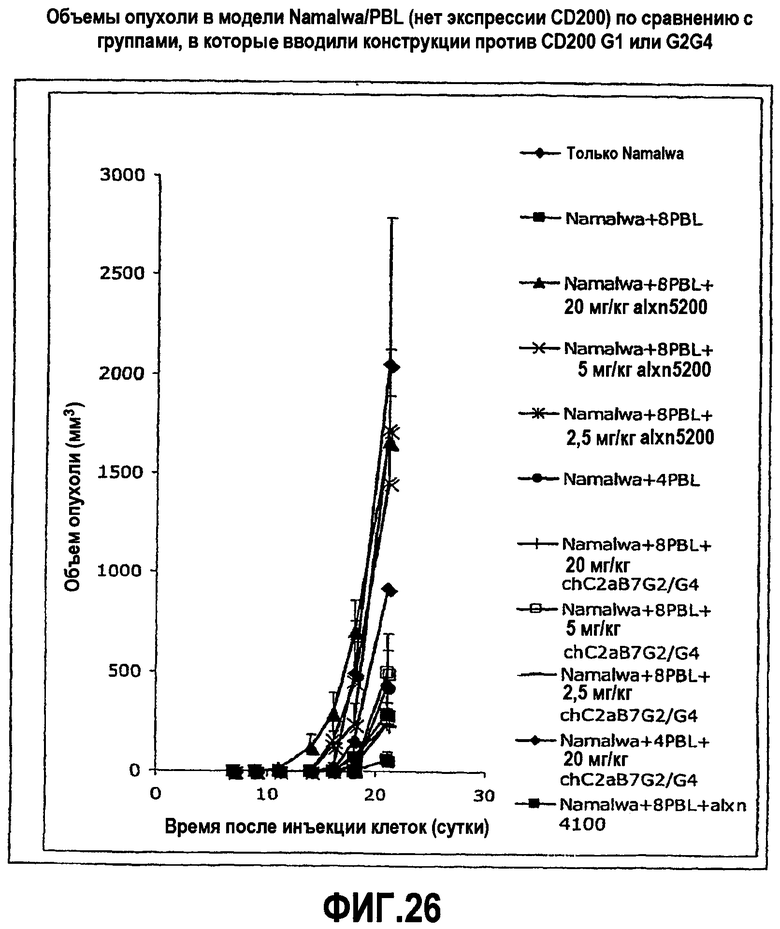


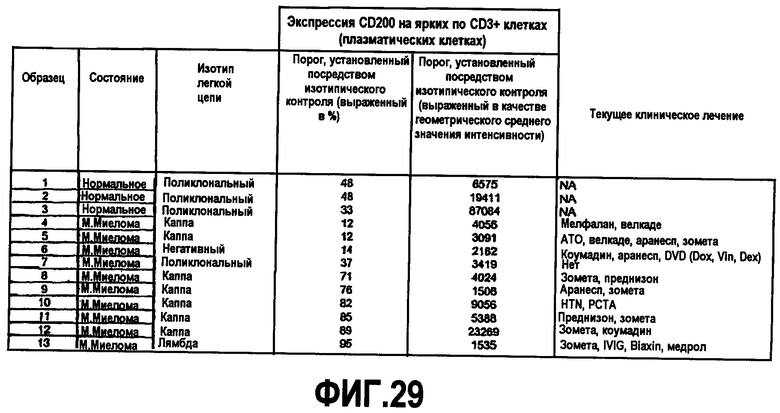
Настоящее изобретение относится к иммунологии и биотехнологии. Предложены: фармацевтическая композиция и способ, основанные на введении пациенту терапевтически эффективного количества антитела против CD200, содержащего константный участок G2/G4, для лечения больного злокачественным новообразованием, содержащим положительные по CD200 злокачественные клетки. При этом указанное антитело против CD200 ингибирует взаимодействие между CD200 и CD200R. Использование изобретения обеспечивает лучшее ингибирование роста опухолей при пониженной эффекторной функции, по сравнению с использованием антитела, содержащего константный участок G1, что может найти применение в медицине. 2 н. и 22 з.п. ф-лы, 29 ил., 6 пр.
1. Фармацевтическая композиция для лечения больного злокачественным новообразованием, содержащим положительные по CD200 злокачественные клетки, причем фармацевтическая композиция содержит фармацевтически приемлемый носитель и эффективное количество антитела против CD200, содержащего константный участок G2/G4, где указанное антитело против CD200 ингибирует взаимодействие между CD200 и CD200R.
2. Фармацевтическая композиция по п.1, где константный участок G2/G4 содержит аминокислоты 137-462 последовательности SEQ ID NO:13.
3. Фармацевтическая композиция по п.1 или 2, где антитело против CD200 представляет собой мышиное антитело, химерное антитело, гуманизированное антитело, деиммунизированное антитело или антитело человека.
4. Фармацевтическая композиция по п.1, где антитело против CD200 содержит:
(i) полипептид легкой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 67-711 последовательности SEQ ID NO:25, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 58-408 последовательности SEQ ID NO:10;
(ii) полипептид легкой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 67-711 последовательности SEQ ID NO:27, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 58-408 последовательности SEQ ID NO:10;
(iii) полипептид легкой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 67-711 последовательности SEQ ID NO:25, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 58-408 последовательности SEQ ID NO:8;
(iv) полипептид легкой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 67-711 последовательности SEQ ID NO:27, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 61-2002 последовательности SEQ ID NO:12;
(v) полипептид легкой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 67-705 последовательности SEQ ID NO:23, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 61-1999 последовательности SEQ ID NO:14;
(vi) полипептид легкой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 67-711 последовательности SEQ ID NO:27, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 58-1391 последовательности SEQ ID NO:16;
(vii) полипептид легкой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 67-705 последовательности SEQ ID NO:31, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 61-427 последовательности SEQ ID NO:19; или
(viii) полипептид легкой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 58-696 последовательности SEQ ID NO:33, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, которая кодируется нуклеотидами 58-2026 последовательности SEQ ID NO:21.
5. Фармацевтическая композиция по п.1, где антитело против CD200 содержит:
(i) полипептид легкой цепи, содержащий аминокислоты 23-236 последовательности SEQ ID NO:26, и полипептид тяжелой цепи, содержащий аминокислоты 20-136 последовательности SEQ ID NO:11;
(ii) полипептид легкой цепи, содержащий аминокислоты 23-236 последовательности SEQ ID NO:28, и полипептид тяжелой цепи, содержащий аминокислоты 20-136 последовательности SEQ ID NO:11;
(iii) полипептид легкой цепи, содержащий аминокислоты 23-236 последовательности SEQ ID NO:26, и полипептид тяжелой цепи, содержащий аминокислоты 20-136 последовательности SEQ ID NO:9; или
(iv) полипептид легкой цепи, содержащий аминокислоты 20-231 последовательности SEQ ID NO:34, и полипептид тяжелой цепи, содержащий аминокислоты 20-472 последовательности SEQ ID NO:22.
6. Фармацевтическая композиция по п.1, где антитело против CD200 содержит:
(a) легкую цепь, содержащую:
(i) CDR1 легкой цепи, содержащий аминокислотную последовательность из остатков 44-51 последовательности SEQ ID NO:24; CDR2 легкой цепи, содержащий аминокислотную последовательность из остатков 70-76 последовательности SEQ ID NO:24; и CDR3 легкой цепи, содержащий аминокислотную последовательность из остатков 109-117 последовательности SEQ ID NO:24;
(ii) аминокислотную последовательность, содержащую остатки 21-127 последовательности SEQ ID NO:24; или
(iii) аминокислотную последовательность, содержащую остатки 21-234 последовательности SEQ ID NO:24; и
(b) тяжелую цепь, содержащую:
(i) CDR1 тяжелой цепи, содержащий аминокислотную последовательность из остатков 46-55 последовательности SEQ ID NO:15; CDR2 тяжелой цепи, содержащий аминокислотную последовательность из остатков 70-86 последовательности SEQ ID NO:15; и CDR3 тяжелой цепи, содержащий аминокислотную последовательность из остатков 119-126 последовательности SEQ ID NO:15;
(ii) аминокислотную последовательность, содержащую остатки 21-137 последовательности SEQ ID NO:15; или
(iii) аминокислотную последовательность, содержащую остатки 21-463 последовательности SEQ ID NO:15.
7. Фармацевтическая композиция по п.1, где антитело против CD200 содержит:
(a) легкую цепь, содержащую:
(i) CDR1 легкой цепи, содержащий аминокислотную последовательность из остатков 44-54 последовательности SEQ ID NO:32; CDR2 легкой цепи, содержащий аминокислотную последовательность из остатков 70-76 последовательности SEQ ID NO:32; и CDR3 легкой цепи, содержащий аминокислотную последовательность из остатков 109-117 последовательности SEQ ID NO:32;
(ii) аминокислотную последовательность, содержащую остатки 21-127 последовательности SEQ ID NO:32; или
(iii) аминокислотную последовательность, содержащую остатки 21-234 последовательности SEQ ID NO:32; и
(b) тяжелую цепь, содержащую:
(i) CDR1 тяжелой цепи, содержащий аминокислотную последовательность из остатков 46-55 последовательности SEQ ID NO:20; CDR2 тяжелой цепи, содержащий аминокислотную последовательность из остатков 70-86 последовательности SEQ ID NO:20; и CDR3 тяжелой цепи, содержащий аминокислотную последовательность из остатков 119-131 последовательности SEQ ID NO:20; или
(ii) аминокислотную последовательность, содержащую остатки 21-142 последовательности SEQ ID NO:20.
8. Фармацевтическая композиция по п.1, где антитело против CD200 содержит:
(a) легкую цепь, содержащую:
(i) аминокислотную последовательность, содержащую остатки 23-129 последовательности SEQ ID NO:28; или
(ii) аминокислотную последовательность, содержащую остатки 23-236 последовательности SEQ ID NO:28; и
(b) тяжелую цепь, содержащую:
(iii) аминокислотную последовательность, содержащую остатки 20-136 последовательности SEQ ID NO:13; или
(iv) аминокислотную последовательность, содержащую остатки 20-462 последовательности SEQ ID NO:13.
9. Фармацевтическая композиция по п.1 или 2, где злокачественное новообразование выбрано из группы, состоящей из рака плазматических клеток, лимфомы, лейкоза, рака яичников, рака кожи, рака легких, рака почек, рака молочной железы, рака предстательной железы, нейробластомы и миеломы.
10. Фармацевтическая композиция по п.1 или 2, где лечение включает введение пациенту второго средства или проведение второй терапии.
11. Фармацевтическая композиция по п.10, где второе средство является химиотерапевтическим средством.
12. Фармацевтическая композиция по п.10, где терапия является радиационной терапией.
13. Способ лечения больного злокачественным новообразованием, содержащим положительные по CD200 злокачественные клетки, где способ включает введение пациенту терапевтически эффективного количества антитела против CD200, содержащего константный участок G2/G4, где указанное антитело против CD200 ингибирует взаимодействие между CD200 и CD200R.
14. Способ по п.13, где константный участок G2/G4 содержит аминокислоты 137-462 последовательности SEQ ID NO:13.
15. Способ по п.13 или 14, где антитело против CD200 представляет собой мышиное антитело, химерное антитело, гуманизированное антитело, деиммунизированное антитело или антитело человека.
16. Способ по п.13, где антитело против CD200 содержит:
(i) полипептид легкой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 67-711 последовательности SEQ ID NO:25, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 58-408 последовательности SEQ ID NO:10;
(ii) полипептид легкой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 67-711 последовательности SEQ ID NO:27, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 58-408 последовательности SEQ ID NO:10;
(iii) полипептид легкой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 67-711 последовательности SEQ ID NO:25, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 58-408 последовательности SEQ ID NO:8;
(iv) полипептид легкой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 67-711 последовательности SEQ ID NO:27, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 61-2002 последовательности SEQ ID NO:12;
(v) полипептид легкой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 61-705 последовательности SEQ ID NO:23, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 61-1999 последовательности SEQ ID NO:14;
(vi) полипептид легкой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 67-711 последовательности SEQ ID NO:27, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 58-1391 последовательности SEQ ID NO:16;
(vii) полипептид легкой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 61-705 последовательности SEQ ID NO:31, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 61-427 последовательности SEQ ID NO:19; или
(viii) полипептид легкой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 58-696 последовательности SEQ ID NO:33, и полипептид тяжелой цепи, содержащий аминокислотную последовательность, кодируемую нуклеотидами 58-2026 последовательности SEQ ID NO:21.
17. Способ по п.13, где антитело против CD200 содержит:
(i) полипептид легкой цепи, содержащий аминокислоты 23-236 последовательности SEQ ID NO:26, и полипептид тяжелой цепи, содержащий аминокислоты 20-136 последовательности SEQ ID NO:11;
(ii) полипептид легкой цепи, содержащий аминокислоты 23-236 последовательности SEQ ID NO:28, и полипептид тяжелой цепи, содержащий аминокислоты 20-136 последовательности SEQ ID NO:11;
(iii) полипептид легкой цепи, содержащий аминокислоты 23-236 последовательности SEQ ID NO:26, и полипептид тяжелой цепи, содержащий аминокислоты 20-136 последовательности SEQ ID NO:9; или
(iv) полипептид легкой цепи, содержащий аминокислоты 20-231 последовательности SEQ ID NO:34, и полипептид тяжелой цепи, содержащий аминокислоты 20-472 последовательности SEQ ID NO:22.
18. Способ по п.13, где антитело против CD200 содержит:
(a) легкую цепь, содержащую:
(i) CDR1 легкой цепи, содержащий аминокислотную последовательность из остатков 44-51 последовательности SEQ ID NO:24; CDR2 легкой цепи, содержащий аминокислотную последовательность из остатков 70-76 последовательности SEQ ID NO:24; и CDR3 легкой цепи, содержащий аминокислотную последовательность из остатков 109-117 последовательности SEQ ID NO: 24;
(ii) аминокислотную последовательность, содержащую остатки 21-127 последовательности SEQ ID NO:24; или
(iii) аминокислотную последовательность, содержащую остатки 21-234 последовательности SEQ ID NO:24; и
(b) тяжелую цепь, содержащую:
(i) CDR1 тяжелой цепи, содержащий аминокислотную последовательность из остатков 46-55 последовательности SEQ ID NO:15; CDR2 тяжелой цепи, содержащий аминокислотную последовательность из остатков 70-86 последовательности SEQ ID NO:15; и CDR3 тяжелой цепи, содержащий аминокислотную последовательность из остатков 119-126 последовательности SEQ ID NO:15;
(ii) аминокислотную последовательность, содержащую остатки 21-137 последовательности SEQ ID NO:15; или
(iii) аминокислотную последовательность, содержащую остатки 21-463 последовательности SEQ ID NO:15.
19. Способ по п.13, где антитело против CD200 содержит:
(a) легкую цепь, содержащую:
(i) CDR1 легкой цепи, содержащий аминокислотную последовательность из остатков 44-54 последовательности SEQ ID NO:32; CDR2 легкой цепи, содержащий аминокислотную последовательность из остатков 70-76 последовательности SEQ ID NO:32; и CDR3 легкой цепи, содержащий аминокислотную последовательность из остатков 109-117 последовательности SEQ ID NO:32;
(ii) аминокислотную последовательность, содержащую остатки 21-127 последовательности SEQ ID NO:32; или
(iii) аминокислотную последовательность, содержащую остатки 21-234 последовательности SEQ ID NO:32; и
(b) тяжелую цепь, содержащую:
(i) CDR1 тяжелой цепи, содержащий аминокислотную последовательность из остатков 46-55 последовательности SEQ ID NO:20; CDR2 тяжелой цепи, содержащий аминокислотную последовательность из остатков 70-86 последовательности SEQ ID NO:20; и CDR3 тяжелой цепи, содержащий аминокислотную последовательность из остатков 119-131 последовательности SEQ ID NO:20; или
(ii) аминокислотную последовательность, содержащую остатки 21-142 последовательности SEQ ID NO:20.
20. Способ по п.13, где антитело против CD200 содержит:
(a) легкую цепь, содержащую:
(i) аминокислотную последовательность, содержащую остатки 23-129 последовательности SEQ ID NO:28; или
(ii) аминокислотную последовательность, содержащую остатки 23-236 последовательности SEQ ID NO:28; и
(b) тяжелую цепь, содержащую:
(iii) аминокислотную последовательность, содержащую остатки 20-136 последовательности SEQ ID NO:13; или
(iv) аминокислотную последовательность, содержащую остатки 20-462 последовательности SEQ ID NO:13.
21. Способ по п.13 или 14, где злокачественное новообразование выбрано из группы, состоящей из рака плазматических клеток, лимфомы, лейкоза, рака яичников, рака кожи, рака легких, рака почек, рака молочной железы, рака предстательной железы, нейробластомы и миеломы.
22. Способ по п.13 или 14, где лечение включает введение пациенту второго средства или проведение второй терапии.
23. Способ по п.22, где второе средство является химиотерапевтическим средством.
24. Способ по п.22, где терапия является радиационной терапией.
| US 20050129690 А1, 16.06.2005 г., | |||
| WO 2005007809 A2, 27.01.2005 | |||
| MUELLER et al., "Humanized porcine vcam-specific monoclonal antibodies with chimeric IgG2/G4 | |||
| сonstant regions block human leukocyte binding to porcine endothelial cells", | |||
| Molecular Immunology, Vol.34, No.6, 1997, стр.441-452 | |||
| WO 1994028027 А1, 08.12.1994 | |||
| Dorai et | |||
| al., |

