Данная заявка испрашивает приоритет предварительной заявки на патент США номер 60/472844, поданной 23 мая 2003 г., и предварительной заявки на патент США номер 60/547975, поданной 26 февраля 2004 г., включенных в данное описание во всей своей полноте в качестве ссылки.
Данное изобретение проведено при правительственной поддержке в рамках NIH Intramural Research Project #Z01-AI-000224. Правительство имеет определенные права на изобретение.
Уровень техники изобретения
Область техники, к которой относится изобретение
Настоящее изобретение относится к новым способам диагностики, прогнозирования, мониторинга прогресса и лечения нарушений, происходящих из-за нарушенной иммунной системы (например, аутоиммунные заболевания, воспалительные заболевания и отторжение трансплантата, и злокачественные опухоли, и инфекционные заболевания), связанных с индуцируемым глюкокортикоидами TNF-рецептором (GITR) и лигандом, ассоциированным с GITR (GITRL), и со связанными с ними модуляторами. Настоящее изобретение далее относится к новым терапевтическим средствам и терапевтическим мишеням и к способу скрининга и оценки тестируемых соединений на предмет вмешательства (лечения) и предотвращения нарушений, происходящих от нарушенной регуляции иммунной системы, что связано с GITR и GITRL.
Уровень техники
В общем, T-лимфоциты ответственны за клеточно-опосредованный иммунитет и играют регуляторную роль путем усиления или подавления ответов других лейкоцитов. Существует представление о том, что Т-лимфоциты играют роль в подавлении иммунных реакций (см., например, Gershon et al. (1970) Immunology 18: 723-35). Однако антигены-мишени для данных супрессорных клеток и механизмы, контролирующие их функцию, пока являются объектами исследования.
Одна из популяций регуляторных T-клеток, которая генерируется в тимусе, отличается от эффекторных Т-клеток экспрессией уникальных мембранных антигенов. Данные регуляторные Т-клетки составляют субпопуляцию CD4+ T-клеток (т.е. T-клеток, которые экспрессируют CD4-антиген), которые совместно экспрессируют CD25-антиген. CD25 также известен как α-цепь рецептора интерлейкина-2 (IL-2R). Совместный перенос или реконституция в Т-клетки CD25+ ассоциированы с предотвращением воспалительных повреждений и аутоиммунитета у различных экспериментальных животных (см. Shevach (2000) Ann. Rev. Immunol. 18: 423-49 и приведенные там ссылки). CD4+CD25+ T-клетки также ассоциированы с ингибированием активации T-клеток in vitro и адаптивного подавления CD4+CD25- T-клеток в совместной культуре (Shevach, выше).
Более двух десятилетий назад было продемонстрировано, что аутореактивные T-клетки избегают механизмов центральной толерантности и существуют на периферии под контролем регуляторных Т-клеток тимусного происхождения. В 1995 г. Sakaguchi и его коллеги продемонстрировали, что малая популяция CD4+ T-клеток, которые в природе экспрессируют α-цепь IL-2R (т.е. CD25), вовлечена в контроль органоспецифических аутореактивных T-клеток (Sakaguchi et al. (1995) J. Immunol. 155: 1151-64). Конкретно они продемонстрировали, что перенос CD4+CD25- T-клеток иммунодефицитным хозяевам вызывает спектр аутоиммунных реакций, которые могут предотвращаться совместным переносом CD4+CD25+ T-клеток (Sakaguchi et al., выше). Последующие исследования связали CD4+CD25+ регуляторные T-клетки с подавлением иммунных реакций на вирусные, бактериальные и протозойные инфекции (Aseffa et al. (2002) J. Immunol. 169:3232-41; Belkaid et al. (2002) Nature 420: 502-07; Hisaeda et al. (2004) Nat. Med. 10:29-30; Kursar et al. (2002) J. Exp. Med. 196:1585-92; Lundgren et al. (2003) Infect. Immun. 71:1755-62; Maloy et al. (2003) J. Exp. Med. 197:111-19). Вместе данные исследования предоставили доказательство того, что удаление CD4+CD25+ T-клеток усилило иммунный ответ. Сделано много попыток для определения активации и супрессии данными CD4+CD25+ T-клетками. Данные клетки представляют собой уникальную родословную клеток тимусного происхождения, которые мощно подавляют эффекторную функцию Т-клеток in vitro и in vivo.
В некоторых исследованиях in vitro выявлено, что CD4+CD25+ клетки подавляют пролиферацию CD4+ T-клеток в ответ и на митогены, и антигены путем выключения транскрипции IL-2 (например, Thornton and Shevach (1998) J. Exp. Med. 188:287-96; Takahashi et al. (1998) Int. Immunol. 10:1969-80). Совместного переноса CD4+CD25+ T-клеток in vivo с аутореактивными CD4+ T-клетками достаточно для подавления индукции и эффекторной фазы органоспецифического аутоиммунитета (Suri-Payer et al. (1999) Eur. J. Immunol. 29:669-77; Suri-Payer et al. (1998) J.Immunol. 160:1212-18). Другие свойства CD4+CD25+ T-клеток включают в себя слабую реакционную способность в ответ на стимуляцию T-клеточного рецептора (TCR) в отсутствие экзогенного IL-2, иммуносупрессию путем межклеточного взаимодействия и необходимость в передаче сигнала с TCR для индукции их супрессорного фенотипа (однако, если они активируются, их супрессорная функция независима от антигенного стимула). Также продемонстрировано, что само приобретение экспрессии CD25, которое может достигаться стимуляцией CD4+CD25- T-клеток, не индуцирует супрессорного фенотипа. Известно, что данные CD4+CD25+ T-клетки существуют у людей (Shevach (2001) J. Exp. Med. 193:F1-F6).
В одном из исследований продемонстрировано, что для генерирования регуляторных CD4+CD25+ T-клеток требуется измененная селекция в тимусе (Jordan et al. (2001) Nat. Immunol. 2:301-06). Кроме того, в исследованиях с нокаут-мышами продемонстрировано, что молекулы, вовлеченные в синтез и реакционную способность IL-2, требуются для генерирования данных клеток; мыши, генетически дефицитные по IL2, или IL2Rβ, или по B7.1 (CD80) и B7.2 (CD86), или по CD28, все характеризуются сильным снижением количества CD4+CD25+ клеток, и в результате этого лимфаденопатией и гиперпролиферацией периферии у некоторых из данных мышей (Papiernik et al. (1998) Int. Immunol. 10:371-78; Salomon et al. (2000) Immunity 12:431-40; Kumanogoh et al. (2001) J. Immunol. 166:353-60).
До недавнего времени в данной области не удавалось определить механизмы, вовлеченные в опосредованное CD4+CD25+ подавление иммунной системы, например антигенная специфичность, молекулы, вовлеченные в приобретение подавления, и молекулы клеточной поверхности или короткодействующие цитокины, вовлеченные в эффекторную фазу подавления; молекулярные мишени CD25+ T-клеток, используемые при модулировании аутоиммунитета, также оставались большей частью неизвестными. В настоящее время было продемонстрировано, путем оценки дифференциальной экспрессии генов с использованием анализа генных чипов на CD4+CD25+ и CD4+CD25- T-клетках, что существуют некоторые дифференциальные зависимые от CD25+ гены (McHugh et al. (2002) Immunity 16:311-23; см. также заявку на патент США 10/194754, включенную в данное описание во всей своей полноте в качестве ссылки). Данные гены, которые, как было определено, преимущественно экспрессируются в CD4+CD25+ T-клетках, могут служить мишенями для терапевтического воздействия и способов скрининга аутоиммунных заболеваний, воспалительных заболеваний и отторжения трансплантата, а также злокачественных опухолей и инфекционных заболеваний.
Значимо то, что один из генов, который, как было определено, дифференциально экспрессировался в CD25+ клетках, является индуцируемым глюкокортикоидами TNF-рецептором (GITR) (McHugh et al., выше). GITR, трансмембранный белковый рецептор клеточной поверхности, является представителем надсемейства рецепторов фактора некроза опухолей (TNFR). Как было продемонстрировано, GITR конститутивно присутствует на неактивированных T-клетках (Gavin et al. (2002) Nat. Immunol. 3:33-41; McHugh et al., выше; Shimizu et al. (2002) Nat. Immunol. 3:135-42). GITR связывается с другим трансмембранным белком, называемым лиганд GITR (GITRL). Антитела-агонисты GITR, как было показано, отменяют супрессорную активность CD4+CD25+ T-клеток, что демонстрирует функциональную роль GITR в регуляции активности данных клеток (McHugh et al., выше). Другое исследование подтвердило, что стимуляция GITR специфичным моноклональным антителом отменяла опосредованную CD4+CD25+ T-клетками супрессию, с индукцией таким образом аутоиммунитета (Shimizu et al., выше). Данные исследования ведут к предположению, что GITR является более надежным маркером CD4+CD25+ T-клеток (Uraushihara et al. (2003) J. Immunol. 171:708-16); однако не только экспрессия GITR характерна для этой субпопуляции, поскольку положительная регуляция GITR также происходит после активации CD4+CD25- T-клеток (McHugh et al., выше; Shimizu et al., выше).
Поскольку GITR, как было показано, значим при регуляции супрессорной активности CD4+CD25+ T-клеток в отношении CD4+CD25- T-клеток, требуется идентифицировать и охарактеризовать новые молекулы, которые взаимодействуют с GITR. Такие новые молекулы, которые взаимодействуют с GITR, описаны в данном описании. Кроме того, предоставлены модуляторы данных молекул.
Сущность изобретения
Настоящее изобретение относится к нуклеотидной и аминокислотной последовательностям нового мышиного гомолога человеческого GITRL. Настоящее изобретение также относится к антителам против мышиного GITRL. Настоящее изобретение также относится к способам как отмены иммуносупрессии путем индукции агонистического связывания GITR-GITRL, так и восстановления или усиления иммуносупрессии путем ингибирования связывания GITR-GITRL, например, путем применения нейтрализующих антител, которые ингибируют активность GITRL (например, которые блокируют взаимодействие между GITR и GITRL). Такая отмена или восстановление/усиление иммуносупрессии благоприятно для лечения различных нарушений, возникающих в результате разрегулированных иммунных реакций, например аутоиммунных нарушений, воспалительных заболеваний и отторжения трансплантата, и злокачественных опухолей и инфекционных заболеваний. Способы по настоящему изобретению направлены на манипулирование GITRL и GITR, включая, но без ограничения, мышиный GITRL и GITR и их гомологи; в состав данных гомологов специфично включен человеческий GITRL и человеческий GITR.
Настоящее изобретение относится к новым выделенным и очищенным полинуклеотидам и полипептидам, относящимся к новому лиганду GITR (GITRL). Изобретение также относится к антителам против GITRL, а также к способам лечения, диагностики, прогнозирования и мониторинга прогресса аутоиммунных нарушений, воспалительных заболеваний и отторжения трансплантата и злокачественных опухолей и инфекционных заболеваний. В одном из вариантов осуществления изобретения описанные способы и молекулы могут применяться для манипуляции исходом иммунного ответа во время лечения заболевания или нарушения, включая аутоиммунные нарушения, воспалительные заболевания и отторжение трансплантата и злокачественные опухоли и инфекционные заболевания. В другом варианте осуществления описанные полинуклеотиды и полипептиды по изобретению, блокирующие или ингибирующие взаимодействие между GITR и GITRL, например, путем отрицательной регуляции экспрессии или активности GITRL или путем связывания с GITRL, но не индуцирующие передачу сигнала с GITR, могут применяться для отмены или усиления супрессии иммунной системы. В другом варианте осуществления взаимодействие между GITR и GITRL может блокироваться или ингибироваться малой молекулой. Специалисту в данной области будет очевидно, что данные типы регуляции (т.е. данные варианты осуществления) являются наиболее благоприятными для лечения аутоиммунных нарушений и некоторых воспалительных заболеваний и сходных или связанных нарушений, а также для лечения отторжения трансплантата. В другом варианте осуществления описанные полинуклеотиды и полипептиды по изобретению, которые индуцируют передачу сигнала с GITR, например, путем положительной регуляции экспрессии или активности GITRL или путем агонистического связывания с GITR, могут использоваться для обращения, блокирования или отмены супрессии иммунной системы. В другом варианте осуществления взаимодействие между GITR и GITRL может усиливаться или имитироваться малой молекулой. Специалисту в данной области будет очевидно, что данные типы регуляции являются наиболее благоприятными для лечения злокачественных опухолей и подобных заболеваний, а также инфекционных заболеваний. Специалисту в данной области также известны вероятные преимущества комбинации данных новых способов лечения с общепринятыми и другими способами лечения.
В другом варианте осуществления изобретение относится к выделенной молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID NO:1 или SEQ ID NO:3. В другом варианте осуществления молекула нуклеиновой кислоты функционально связана, по меньшей мере, с одной последовательностью контроля экспрессии. В другом варианте осуществления предоставляется клетка-хозяин, трансформированная или трансфицированная данной молекулой нуклеиновой кислоты.
В другом варианте осуществления изобретение относится к выделенному аллелю SEQ ID NO:1 или SEQ ID NO:3. В другом варианте осуществления изобретение относится к выделенному гену, включающему в себя нуклеотидную последовательность SEQ ID NO:3.
В другом варианте осуществления изобретение относится к выделенной молекуле нуклеиновой кислоты, которая специфично гибридизуется в условиях высокой жесткости с нуклеотидной последовательностью, приведенной в SEQ ID NO:1 или SEQ ID NO:3, или с комплементарной ей последовательностью.
В другом варианте осуществления изобретение относится к выделенной молекуле нуклеиновой кислоты, которая кодирует белок, содержащий аминокислотную последовательность SEQ ID NO:2 или ее фрагмент, который кодирует активный фрагмент белка. В другом варианте осуществления молекула нуклеиновой кислоты или ее фрагмент функционально связаны, по меньшей мере, с одной последовательностью контроля экспрессии. В другом варианте осуществления предоставляется клетка-хозяин, трансформированная или трансфицированная выделенной молекулой нуклеиновой кислоты или ее фрагмента, функционально связанной, по меньшей мере, с одной последовательностью контроля экспрессии. В другом варианте осуществления изобретение относится к не относящемуся к человеку трансгенному животному, в котором соматические и зародышевые клетки содержат данную выделенную молекулу нуклеиновой кислоты или ее фрагмент. В другом варианте осуществления изобретение относится к не относящемуся к человеку трансгенному животному, в котором соматические и зародышевые клетки содержат ДНК, включающую в себя нуклеотидную последовательность SEQ ID NO:1 или SEQ ID NO:3.
В другом варианте осуществления изобретение относится к выделенному белку, содержащему аминокислотную последовательность, кодируемую выделенной нуклеиновой кислотой, которая специфично гибридизуется в условиях высокой жесткости с нуклеотидной последовательностью, приведенной в SEQ ID NO:1 или SEQ ID NO:3, или с комплементарной ей последовательностью. В другом варианте осуществления изобретение относится к выделенному белку, содержащему аминокислотную последовательность SEQ ID NO:2 или ее активный фрагмент.
В другом варианте осуществления изобретение относится к выделенной молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, комлементарную нуклеотидной последовательности SEQ ID NO:1 или SEQ ID NO:3, или ее фрагмент, где экспрессия данной нуклеиновой кислоты в клетке приводит к сниженной продукции GITRL. В другом варианте осуществления молекула нуклеиновой кислоты или ее фрагмент функционально связаны, по меньшей мере, с одной последовательностью контроля экспрессии. В другом варианте осуществления предоставляется клетка-хозяин, трансформированная или трансфицированная данной выделенной молекулой нуклеиновой кислоты или ее фрагментом, функционально связанной, по меньшей мере, с одной последовательностью контроля экспрессии. В другом варианте осуществления изобретение относится к не относящемуся к человеку трансгенному животному, в котором соматические и зародышевые клетки содержат данную выделенную молекулу нуклеиновой кислоты или ее фрагмент.
В другом варианте осуществления изобретение относится к антисмысловому олигонуклеотиду, комплементарному мРНК, соответствующей молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID NO:1 или SEQ ID NO:3 или ее фрагмент, где олигонуклеотид ингибирует экспрессию GITRL. В другом варианте осуществления изобретение относится к молекуле siРНК (короткой интерферирующей РНК), соответствующей молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID NO:1 или SEQ ID NO:3 или ее фрагмент, где молекула siРНК ингибирует экспрессию GITRL.
В другом варианте осуществления изобретение относится к выделенному антителу, способному к специфичному связыванию выделенного белка, содержащего аминокислотную последовательность, кодируемую выделенной нуклеиновой кислотой, которая специфично гибридизуется в условиях высокой жесткости с нуклеотидной последовательностью, приведенной в SEQ ID NO:1 или SEQ ID NO:3, или с комплементарной ей последовательностью. В другом варианте осуществления антитело нейтрализует активность GITRL. В другом варианте осуществления антитело представляет собой 5F1, имеющее номер ATCC PTA-5336, или 10F12, имеющее номер ATCC PTA-5337. В другом варианте осуществления антитело включает в себя антигенсвязывающие фрагменты 5F1 или 10F12. В другом варианте осуществления изобретение относится к выделенному антителу, способному специфично связывать выделенный белок, содержащий аминокислотную последовательность SEQ ID NO:2 или ее активный фрагмент. В другом варианте осуществления антитело нейтрализует активность GITRL. В другом варианте осуществления антитело представляет собой 5F1, имеющее номер ATCC PTA-5336, или 10F12, имеющее номер ATCC PTA-5337. В другом варианте осуществления антитело содержит антигенсвязывающие фрагменты 5F1 или 10F12.
В другом варианте осуществления изобретение относится к способу скрининга тестируемых соединений, способных ингибировать или блокировать взаимодействие GITRL с GITR, включающему стадии контакта образца, содержащего GITRL и GITR, с тестируемым соединением, и определения того, снижено ли взаимодействие GITRL с GITR в образце по сравнению с взаимодействием GITRL с GITR в образце, не контактировавшем с данным соединением, за счет чего снижение взаимодействия GITRL с GITR в образце, контактировавшем с данным соединением, идентифицирует соединение как то, которое ингибирует или блокирует взаимодействие GITRL с GITR. В другом варианте осуществления идентифицированное соединение применяют в способе лечения субъекта при риске или поставленном диагнозе аутоиммунного заболевания, воспалительного заболевания или отторжения трансплантата, причем данный способ включает стадии выделения из субъекта T-клеток, обработки выделенных T-клеток идентифицированным соединением и переноса Т-клеток обратно субъекту. В другом варианте осуществления идентифицированное соединение применяют в способе лечения субъекта при риске или поставленном диагнозе аутоиммунного заболевания, воспалительного заболевания или отторжения трансплантата, причем данный способ включает введение субъекту идентифицированного соединения. В другом варианте осуществления изобретение относится к способу оценки у субъекта эффективности идентифицированного соединения, включающему стадии выявления первого количества эффекторных Т-клеток у субъекта перед введением субъекту данного соединения, выявления второго числа эффекторных Т-клеток у субъекта после введения субъекту данного соединения и сравнения первого количества и второго количества, причем значимое снижение второго значения количества эффекторных Т-клеток по сравнению с первым значением указывает на то, что данное соединение эффективно при лечении у субъекта аутоиммунного нарушения, воспалительного заболевания или отторжения трансплантата. В другом варианте осуществления эффекторные Т-клетки представляют собой CD4+ T-клетки или CD8+ T-клетки.
В другом варианте осуществления изобретение относится к способу скрининга тестируемых соединений, способных усиливать или имитировать взаимодействие GITRL с GITR, включающему стадии контакта образца, содержащего GITRL и GITR, с тестируемым соединением и определения того, повышается ли взаимодействие GITRL с GITR в образце по сравнению с взаимодействием GITRL с GITR в образце, не контактировавшем с данным соединением, за счет чего такое повышение взаимодействия GITRL с GITR в образце, контактировавшем с данным соединением, идентифицирует данное соединение как то, которое усиливает или имитирует взаимодействие GITRL с GITR. В другом варианте осуществления идентифицированное соединение применяют в способе лечения субъекта при риске или поставленном диагнозе злокачественной опухоли или инфекционного заболевания, причем данный способ включает стадии выделения из субъекта T-клеток, обработки выделенных T-клеток идентифицированным соединением и переноса Т-клеток обратно субъекту. В другом варианте осуществления идентифицированное соединение применяют в способе лечения субъекта при риске или поставленном диагнозе злокачественной опухоли или инфекционного заболевания, причем данный способ включает введение субъекту идентифицированного соединения. В другом варианте осуществления изобретение относится к способу оценки у субъекта эффективности идентифицированного соединения, включающему стадии выявления первого количества эффекторных Т-клеток у субъекта перед введением субъекту данного соединения, выявления второго количества эффекторных Т-клеток у субъекта после введения субъекту данного соединения и сравнения первого количества и второго количества, причем значимое повышение второго значения количества эффекторных Т-клеток по сравнению с первым значением указывает на то, что данное соединение эффективно при лечении у данного субъекта злокачественной опухоли или инфекционного заболевания. В другом варианте осуществления эффекторные Т-клетки представляют собой CD4+ T-клетки или CD8+ T-клетки.
В другом варианте осуществления изобретение относится к способу диагностики у субъекта аутоиммунного нарушения, воспалительного заболевания или отторжения трансплантата, включающему стадии выявления тестируемого количества продукта гена GITRL в образце от субъекта и сравнения тестируемого количества с нормальным количеством продукта гена GITRL в контрольном образце, за счет чего тестируемое количество, значимо превышающее нормальное количество, предоставляет позитивное указание на диагноз аутоиммунного нарушения, воспалительного заболевания или отторжения трансплантата. В другом варианте осуществления изобретение относится к способу диагностики у субъекта злокачественной опухоли или инфекционного заболевания, включающему стадии выявления тестируемого количества продукта гена GITRL в образце от субъекта и сравнения тестируемого количества с нормальным количеством продукта гена GITRL в контрольном образце, причем тестируемое количество, которое значимо ниже нормального количества, предоставляет позитивное указание на диагноз злокачественной опухоли или инфекционного заболевания.
В другом варианте осуществления изобретение относится к способу лечения субъекта при риске или поставленном диагнозе аутоиммунного заболевания, воспалительного заболевания или отторжения трансплантата, включающему введение субъекту антагониста GITR. В другом варианте осуществления способ относится к введению антагониста GITR, так что поддерживается чувствительность эффекторных T-клеток у субъекта к подавлению CD4+CD25+ регуляторными Т-клетками (например, в количестве, эффективном для сохранении такой чувствительности). В другом варианте осуществления антагонист GITR выбран из группы, состоящей из нейтрализующего антитела против GITRL, нейтрализующего антитела против GITR, слитого белка, содержащего GITR, слитого белка, содержащего активный фрагмент GITR, низкомолекулярного антагониста, антисмысловой в отношении GITRL молекулы нуклеиновой кислоты и молекулы нуклеиновой кислоты siРНК против GITRL. В другом варианте осуществления аутоиммунное нарушение или воспалительное заболевание выбрано из группы, состоящей из ревматоидного артрита, энцефаломиелита, остеоартрита, рассеянного склероза, аутоиммунного гастрита, системной красной волчанки, псориаза и других воспалительных дерматозов, диабета I типа, астмы, аллергии и воспалительных заболеваний толстого кишечника, включая болезнь Крона и язвенный колит.
В другом варианте осуществления изобретение относится к способу лечения субъекта при риске или поставленном диагнозе злокачественной опухоли или инфекционного заболевания, включающему введение субъекту агониста GITR. В другом варианте осуществления способ включает введение агониста GITR, так что агонист GITR предоставляет костимуляторный сигнал эффекторным Т-клеткам у субъекта и делает их менее чувствительными к подавлению CD4+CD25+ регуляторными T-клетками субъекта (например, в количестве, эффективном для предоставления такого сигнала). В другом варианте осуществления агонист GITR выбран из группы, состоящей из GITRL, активного фрагмента GITRL, слитого белка, содержащего GITRL, слитого белка, содержащего активный фрагмент GITRL, и агонистического в отношении GITR антитела.
В другом варианте осуществления изобретение относится к способу индукции пролиферации популяции клеток, содержащей эффекторные T-клетки, включающему введение в клеточную популяцию агониста GITR. В другом варианте осуществления агонист GITR выбран из группы, состоящей из GITRL, активного фрагмента GITRL, слитого белка, содержащего GITRL, слитого белка, содержащего активный фрагмент GITRL, и агонистического в отношении GITR антитела. В другом варианте осуществления эффекторные T-клетки представляют собой CD4+ T-клетки или CD8+ T-клетки.
В другом варианте осуществления изобретение относится к способу ингибирования пролиферации популяции клеток, содержащей эффекторные T-клетки, включающему введение в клеточную популяцию антагониста GITR. В другом варианте осуществления антагонист GITR выбран из группы, состоящей из нейтрализующего антитела против GITRL, нейтрализующего антитела против GITR, слитого белка, содержащего GITR, слитого белка, содержащего активный фрагмент GITR, низкомолекулярного антагониста, антисмысловой в отношении GITRL молекулы нуклеиновой кислоты и молекулы нуклеиновой кислоты siРНК против GITRL. В другом варианте осуществления эффекторные T-клетки представляют собой CD4+ T-клетки или CD8+ T-клетки. В другом варианте осуществления антагонист GITR представляет собой 5F1 или 10F12.
В другом варианте осуществления изобретение относится к способу ингибирования или блокирования супрессии клеточной популяции, содержащей эффекторные Т-клетки, в присутствии CD4+CD25+ регуляторных Т-клеток, включающему введение в клеточную популяцию агониста GITR. В другом варианте осуществления способ включает введение агониста GITR, так что агонист GITR предоставляет костимуляторный сигнал эффекторным Т-клеткам у субъекте и делает их менее чувствительными к подавлению CD4+CD25+ регуляторными T-клетками субъекта (например, в количестве, эффективном для предоставления такого сигнала). В другом варианте осуществления агонист GITR выбран из группы, состоящей из GITRL, активного фрагмента GITRL, слитого белка, содержащего GITRL, слитого белка, содержащего активный фрагмент GITRL, и агонистического в отношении GITR антитела. В другом варианте осуществления эффекторные T-клетки представляют собой CD4+ T-клетки или CD8+ T-клетки.
В другом варианте осуществления изобретение относится к способу супрессии клеточной популяции, содержащей эффекторные T-клетки, в присутствии CD4+CD25+ регуляторных T-клеток, включающему введение в клеточную популяцию антагониста GITR. В другом варианте осуществления способ включает введение антагониста GITR, так что поддерживается чувствительность эффекторных T-клеток к подавлению CD4+CD25+ регуляторными T-клетками (например, в количестве, эффективном для поддержания такой чувствительности). В другом варианте осуществления антагонист GITR выбран из группы, состоящей из нейтрализующего антитела против GITRL, нейтрализующего антитела против GITR, слитого белка, содержащего GITR, слитого белка, содержащего активный фрагмент GITR, низкомолекулярного антагониста, антисмысловой в отношении GITRL молекулы нуклеиновой кислоты и молекулы нуклеиновой кислоты siРНК против GITRL. В другом варианте осуществления эффекторные T-клетки представляют собой CD4+ T-клетки или CD8+ T-клетки. В другом варианте осуществления антагонист GITR представляет собой 5F1 или 10F12.
В другом варианте осуществления изобретение относится к способу ингибирования экспрессии GITRL в популяции клеток, включающему обработку данной популяции клеток выделенной молекулой нуклеиновой кислоты, содержащей нуклеотидную последовательность, комплементарную нуклеотидной последовательности SEQ ID NO:1 или SEQ ID NO:3 или ее фрагменту, где экспрессия данной молекулы нуклеиновой кислоты приводит к сниженной продукции GITRL. В другом варианте осуществления изобретение относится к способу ингибирования экспрессии GITRL в популяции клеток, включающему обработку популяции клеток антисмысловым олигонуклеотидом, комплементарным мРНК, соответствующей молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID NO:1 или SEQ ID NO:3 или ее фрагмент, где олигонуклеотид ингибирует экспрессию GITRL.
В другом варианте осуществления изобретение относится к способу ингибирования экспрессии GITRL в популяции клеток, включающему обработку популяции клеток молекулой siРНК, направленной на мРНК, соответствующую молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID NO:1 или SEQ ID NO:3. В другом варианте осуществления изобретение относится к способу ингибирования экспрессии GITRL в популяции клеток, включающему обработку популяции клеток молекулой siРНК, направленной на мРНК, соответствующую выделенной молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, комплементарную нуклеотидной последовательности SEQ ID NO:1 или SEQ ID NO:3.
В другом варианте осуществления изобретение относится к способу ингибирования экспрессии GITRL в популяции клеток, включающему обработку популяции клеток антисмысловым нуклеотидом против молекулы нуклеиновой кислоты, кодирующей GITRL. В другом варианте осуществления изобретение относится к способу ингибирования экспрессии GITRL в популяции клеток, включающему обработку популяции клеток молекулой siРНА, направленной против мРНК, кодирующей GITRL.
В другом варианте осуществления изобретение относится к способу индукции экспрессии GITRL в популяции клеток, включающему обработку популяции клеток путем трансформации или трансфекции популяции клеток выделенной молекулой нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID NO:1 или SEQ ID NO:3, или выделенной молекулой нуклеиновой кислоты, которая кодирует белок, содержащий аминокислотную последовательность SEQ ID NO:2 или ее фрагмент, который кодирует активный фрагмент белка, где данная молекула нуклеиновой кислоты функционально связана, по меньшей мере, с одной последовательностью контроля экспрессии.
В другом варианте осуществления изобретение относится к популяции эффекторных Т-клеток, контактировавших in vitro или ex vivo с агонистом GITR. В другом варианте осуществления агонист GITR выбран из группы, состоящей из GITRL или активного фрагмента GITRL, слитого белка, содержащего GITRL, слитого белка, содержащего активный фрагмент GITRL, низкомолекулярного агониста и агонистического в отношении GITR антитела. В другом варианте осуществления эффекторные Т-клетки представляют собой CD4+ T-клетки или CD8+ T-клетки.
В другом варианте осуществления изобретение относится к способу лечения у субъекта злокачественной опухоли или инфекционного заболевания, причем данный способ включает стадии получения популяции эффекторных T-клеток, обработки данной популяции агонистом GITR и введения обработанной популяции субъекту, пораженному злокачественной опухолью или инфекционным заболеванием. В другом варианте осуществления агонист GITR выбран из группы, состоящей из GITRL, активного фрагмента GITRL, слитого белка, содержащего GITRL, слитого белка, содержащего активный фрагмент GITRL, низкомолекулярного агониста и агонистического в отношении GITR антитела. В другом варианте осуществления субъект подвержен злокачественной опухоли, и обработанную популяцию используют в качестве противоопухолевой вакцины.
В другом варианте осуществления изобретение относится к фармацевтической композиции, содержащей агонист GITR и фармацевтически приемлемый носитель. В другом варианте осуществления агонист GITR выбран из группы, состоящей из GITRL, активного фрагмента GITRL, слитого белка, содержащего GITRL, слитого белка, содержащего активный фрагмент GITRL, низкомолекулярного агониста и агонистического в отношении GITR антитела.
В другом варианте осуществления изобретение относится к фармацевтической композиции, содержащей антагонист GITR и фармацевтически приемлемый носитель. В другом варианте осуществления антагонист GITR выбран из группы, состоящей из нейтрализующего антитела против GITRL, нейтрализующего антитела против GITR, слитого белка, содержащего GITR, слитого белка, содержащего активный фрагмент GITR, низкомолекулярного антагониста, антисмысловой в отношении GITRL молекулы нуклеиновой кислоты и молекулы нуклеиновой кислоты siРНК против GITRL. В другом варианте осуществления антитело содержит антигенсвязывающие фрагменты 5F1 или 10F12.
В другом варианте осуществления изобретение относится к вакцинному адъюванту, содержащему агонист GITR и антиген, выбранный из группы, состоящей из вирусного антигена, бактериального антигена, антигена грибов, паразитарного антигена, ракового антигена, ассоциированного с опухолью антигена или их фрагментов. В другом варианте осуществления агонист GITR выбран из группы, состоящей из GITRL, активного фрагмента GITRL, слитого белка, содержащего GITRL, слитого белка, содержащего активный фрагмент GITRL, низкомолекулярного агониста и агонистического в отношении GITR антитела.
В другом варианте осуществления изобретение относится к вакцинному адъюванту, содержащему антагонист GITR и антиген, выбранный из группы, состоящей из аутоантигена, белка амилоидного пептида, аллоантигена, антигена трансплантата, аллергена и их фрагментов. В другом варианте осуществления антагонист GITR выбран из группы, состоящей из нейтрализующего антитела против GITRL, нейтрализующего антитела против GITR, слитого белка, содержащего GITR, слитого белка, содержащего активный фрагмент GITR, низкомолекулярного антагониста, антисмысловой в отношении GITRL молекулы нуклеиновой кислоты и молекулы нуклеиновой кислоты siРНК против GITRL. В другом варианте осуществления антитело содержит антигенсвязывающие фрагменты 5F1 или 10F12.
В другом варианте осуществления изобретение относится к способу скрининга тестируемых соединений на способность нейтрализовать активность GITRL, включающему стадии контакта с данным соединением образца, содержащего GITRL и нейтрализующее антитело, и определения того, снижается ли взаимодействие GITRL с нейтрализующим антителом в образце относительно взаимодействия GITRL с нейтрализующим антителом в образце, не контактировавшем с данным соединением, причем снижение взаимодействия GITRL с нейтрализующим антителом в образце, контактировавшем с соединением, идентифицирует соединение как то, которое ингибирует или блокирует взаимодействие GITRL с нейтрализующим антителом. В другом варианте осуществления антитело представляет собой 5F1 или 10F12.
В другом варианте осуществления изобретение относится к способу предоставления костимуляторного сигнала популяции клеток, содержащей эффекторные Т-клетки, причем данный способ включает введение агониста GITR. В другом варианте осуществления агонист GITR представляет собой белок, выбранный из группы, состоящей из GITRL или активного фрагмента GITRL, слитого белка, содержащего GITRL, слитого белка, содержащего активный фрагмент GITR, и агонистического антитела против GITR. В другом варианте осуществления эффекторные Т-клетки представляют собой CD4+ T-клетки или CD8+ T-клетки.
Краткое описание чертежей
На фиг.1 показано выравнивание (основанное на матрице аминокислотных замен BLOSUM62; см. Henikoff and Henikoff (1992) Proc. Natl. Acad. Sci. USA 89:10915-19) аминокислотных последовательностей мышиного (m) и человеческого (h) GITRL.
На фиг.2 показаны результаты экспериментов по влиянию связывания GITRL:GITR на пролиферацию CD4+CD25+ T-клеток. Измеряли включение тимидина (CPM) как средство оценки клеточной пролиферации. На фиг.2A показано, что агонистическое антитело против GITR стимулирует пролиферацию CD4+CD25+ T-клеток, но не CD4+CD25- T-клеток. На фиг.2B показано, что клетки YB2/0, экспрессирующие GITRL, стимулируют пролиферацию CD4+CD25+ клеток.
На фиг.3A показано, что GITRL, экспрессированный клетками YB2/0 (~50000), а также агонистическое антитело против GITR (2 мкг/мл) отменяют супрессию (т.е. дают отрицательный процент супрессии), продуцируемую выделенными CD4+CD25+ супрессорными Т-клетками (# супрессоров). На фиг.3B показано, что клетки GITRL-YB2/0 в количествах менее 50000 (т.е. 3000-25000) могли частично отменять супрессию в зависимом от дозы режиме.
На фиг.4 показано, что клетки GITRL-YB2/0 (~50000) не отменяют супрессию, когда вместо свежевыделенных CD4+CD25+ T-клеток используют активированные CD4+CD25+ T-клетки; сходные результаты получены с агонистическим антителом против GITR.
На фиг.5A и 5B показано, что индуцированная GITRL отмена супрессии, продуцированной свежевыделенными CD4+CD25+ T-клетками, может отменяться сама (т.е. происходит восстановление супрессии) в присутствии антитела против GITRL («анти-GITRL»=антитело 5F1). Фиг.5B включает дополнительный эксперимент, где показано, что контрольное антитело («контрольный Ig») не восстанавливает супрессию.
На фиг.6 показано, что антитело против GITRL может только усиливать супрессию в присутствии CD4+CD25+ T-клеток. На фиг.6A показана супрессия пролиферации клеток лимфатических узлов в присутствии антитела против GITRL (5F1). На фиг.6B показано отсутствие супрессорной активности антитела против GITRL, когда популяция клеток лимфоузла лишена CD4+CD25+ T-клеток.
На фиг.7 показано распределение клеток, экспрессирующих GITRL, в лимфоидных тканях. Фиг.7A. Анализ проточной цитометрии проводили на CD11c+ клетках, обогащенных посредством магнитных гранул из селезенки мышей BALB/c, путем окрашивания антителами против CD4, против CD8 и против GITRL. Экспрессию GITRL определяли сравнением интенсивности флуоресценции в разделенных субпопуляциях CD4+, CD8+ и CD4-CD8- (верхняя, средняя и нижняя панели гистограмм соответственно), окрашенных антителом против GITRL (закрашенные гистограммы) с интенсивностью флуоресценции данных клеток, окрашенных антителом контроля изотипа (незакрашенные гистограммы). Фиг.7B. Экспрессию GITRL дендритными клетками селезенки (DC) и B-1 B-клетками определяли путем окрашивания свежевыделенных BALB/c CD11c+ DC селезенки (верхняя панель гистограмм) и CD11clow B220+ плазмацитоидных DC (нижняя панель гистограмм) mAb против GITRL (закрашенные гистограммы) или антителом контроля изотипа (незакрашенные гистограммы) и проведения анализа проточной цитометрии.
Фиг.7C (верхняя панель гистограмм). Интенсивность флуоресценции B220+ B-клеток среди всех спленоцитов (закрашенная гистограмма) и сортированных по CD11b+ B220+ перитонеальных (perC) B-1 B-клеток (незакрашенная гистограмма со сплошной линией), окрашенных антителом против GITRL, по сравнению с интенсивностью флуоресценции клеток, окрашенных контролем изотипа (незакрашенная гистограмма с пунктирной линией). Фиг.7C (нижняя панель гистограмм). Показано сравнение интенсивности флуоресценции окрашенных антителом против GITRL (закрашенная гистограмма) и окрашенных антителом против изотипа (незакрашенная гистограмма) макрофагов perC (CD11b+ B220- клетки). Фиг.7D. Тимоциты окрашивали на предмет экспрессии CD4, CD8 и GITRL или контроля изотипа. Интенсивность флуоресценции клеток CD4+CD8- (верхний левый квадрант), CD4+CD8+ (верхний правый квадрант) и CD4-CD8+ (верхний правый квадрант), окрашенных антителом против GITRL (закрашенная гистограмма), сравнивали с интенсивностью флуоресценции данных клеток, окрашенных антителом контроля изотипа (незакрашенная гистограмма). Фиг.7E. Экспрессию GITRL сортированными CD44+CD25- (R1), CD44+CD25+ (R2), CD44-CD25+ (R3) или CD44-CD25- (R4) тимусными предшественниками определяли сравнением флуоресценции данных клеток, окрашенных антителом против GITRL (закрашенная гистограмма), с флуоресценцией данных клеток, окрашенных антителом контроля изотипа (незакрашенная гистограмма). Фиг.7F. Нестимулированные клетки лимфатического узла окрашивали антителами против CD4, против CD8, против CD25 и/или против GITRL. CD4+CD8- клетки (верхний левый квадрант), а не CD4-CD8+ клетки (нижний правый квадрант) далее разделяли в отношении экспрессии данными клетками CD25. Экспрессию GITRL сортированными CD4+CD8-CD25- (верхняя правая панель гистограмм), CD4+CD8-CD25+ (нижняя правая панель гистограмм) или CD4-CD8+ (нижняя (средняя) панель гистограмм) клетками лимфатического узла определяли сравнением интенсивности флуоресценции данных клеток, окрашенных антителом против GITRL (закрашенные гистограммы), с флуоресценцией данных клеток, окрашенных антителом контроля изотипа (незакрашенные гистограммы). Результаты представляют пять независимых экспериментов.
На фиг.8 показана отрицательная регуляция APC GITRL после стимуляции. Фиг.8A. Экспрессию GITRL очищенными В-клетками селезенки B220+ или общими перитонеальными (PerC) B220+CD11b+ B-1 B-клетками определяли для различных моментов времени после обработки polyI:C (10 мкг/мл), LPS (0,5 мкг/мл), CpG (ODN 1826, 1 мкМ), анти-CD40 и IL-4 (10 мкг/мл и 20 нг/мл соответственно) и анти-IgM (F(ab')2-фрагмент антител козы против IgM, μ-цепь, 1 мкг/мл). Представлена интенсивность флуоресценции окрашенных анти-GITRL стимулированных клеток (закрашенные гистограммы), окрашенных анти-GITRL нестимулированных (среда) клеток (незакрашенные гистограммы со сплошной линией) и клеток, окрашенных антителом контроля изотипа (незакрашенные гистограммы с пунктирной линией). Фиг.8B. Экспрессию GITRL B220+ В-клетками (закрашенная гистограмма), представленными среди всех спленоцитов, обработанных mAb против CD3 (0,5 мкг/мл) после периода культивирования длительностью 48 часов, сравнивали с экспрессией GITRL нестимулированными B220+ B-клетками (незакрашенные гистограммы со сплошной линией) и B220+ B-клетками, окрашенными антителом контроля изотипа (незакрашенная гистограмма с пунктирной линией). Фиг.8C. Экспрессия GITRL (верхние панели гистограмм) и B7.2 (т.е. CD86) (нижние панели гистограмм) очищенными CD11c+ DC после культивирования с LPS (0,5 мкг/мл) и без него в указанные моменты времени. Фиг.8D. Экспрессия GITRL общими спленоцитами, разделенными на экспрессирующие CD4+ или CD8+ клетки после периода культивирования длительностью 48 часов в отсутствие или в присутствии растворимого mAb против CD3 (0,5 мкг/мл). Графики представляют от двух до четырех независимых экспериментов; все эксперименты проводили на тканях, выделенных из мышей BALB/c.
На фиг.9 продемонстрировано влияние блокирования взаимодействия GITR/GITRL на ингибирование пролиферации лимфоцитов. На фиг.9A и 9B отрезки указывают на значения стандартного отклонения. Фиг.9A. Пролиферацию клеток (ось y) лимфатического узла (LN; 1×105) и селезенки (Sp; 0,5×105) с клетками CD25+ или без них (все или Δ25 соответственно) определяли после 72 часов культивирования с различными концентрациями растворимого антитела против CD3 (ось x). Клетки инкубировали в присутствии очищенного mAb против GITRL (10 мкг/мл; закрашенные кружки) или крысиным антителом контроля изотипа IgG2a (10 мкг/мл; незакрашенные кружки). Результаты представлены тремя независимыми экспериментами. Фиг.9B. CD4+CD25- или CD8+ T-клетки культивировали в присутствии 5×104 облученных (3000 R) истощенных по T-клеткам APC и в присутствии 5×104 облученных (8000 R) YB2/0-GITRL (незакрашенные кружки) или контрольных YB2/0 (закрашенные кружки) клеток. Культуры активировали различными концентрациями растворимого mAb против CD3 (ось x) и измеряли пролиферацию (ось y) после 72-часового периода культивирования. Фиг.9C. Среднюю флуоресценцию (ось x) очищенных CD4+CD25- T-клеток, окрашенных антителом против GITR, определяли в различные периоды времени после активации растворимым антителом против CD3 (0,5 мкг/мл) в присутствии облученных (3000R) истощенных по T-клеткам спленоцитов. Результаты представляют, по меньшей мере, два независимых эксперимента.
На фиг.10 продемонстрировано, что экспрессия GITR CD25- T-клетками требуется для отмены супрессии. Фиг.10A. Пролиферацию совместных культур CD4+CD25- T-клеток (5×104) из различных нокаут-мышей и различные количества CD4+CD25+ T-клеток (ось x) из различных нокаут-мышей [(Aa) CD4+CD25-:GITR +/+, CD4+CD25+:GITR +/+; (Ab) CD4+CD25-:GITR +/+, CD4+CD25+:GITR -/-; (Ac) CD4+CD25-:GITR -/-, CD4+CD25+:GITR +/+ и (Ad) CD4+CD25-:GITR -/-, CD4+CD25+:GITR -/-], инкубированные с облученными АРС из мышей дикого типа (5×104) и с растворимым анти-CD3 (0,5 мкг/мл) и с 2 мкг/мл антитела против GITR (закрашенные кружки) или с антителом контроля изотипа (незакрашенные кружки), определяли путем измерения захвата 3Н-тимидина (счет/мин; ось y). Фиг.10В. Пролиферацию совместных культур проводили, как указано выше (фиг.10А), с различными количествами мышиных CD4+CD25+ Т-клеток (ось х) и или (Ва) мышиными CD4+CD25- Т-клетками, или (Вb) крысиными CD4+CD25- Т-клетками в присутствии облученных (3000 R) крысиных АРС. Культуры стимулировали коктейлем антител против крысиного и мышиного анти-CD3 (0,25 мкг/мл каждого) и обрабатывали 2 мкг/мл контроля изотипа (крысиные IgG; незакрашенные кружки) или антителом против GITR (DTA-1; закрашенные кружки). Отрезки указывают на значения стандартного отклонения, рассчитанные из значений пролиферации в трех параллельных культурах. Фиг.10С. Показана флуоресценция (ось х) окрашенных CFSE мышиных CD4+CD25+ (верхние панели) и крысиных CD4+CD25- Т-клеток (нижние панели), совместно культивированных в соотношении супрессор-отвечающая клетка 1:8 с контролем изотипа (крысиный IgG; левые панели) или антителом против GITR (DTA-1; правые панели). Мышиные и крысиные субпопуляции Т-клеток различались путем окрашивания специфичными антителами против CD4. Представлены результаты двух-четырех независимых экспериментов.
На фиг.11 продемонстрировано, что сигналы GITR требуются для преодоления супрессии, опосредованной эндогенными регуляторными Т-клетками. Фиг.11А. Меченные CFSE клетки лимфатического узла (LN) (5×104) из мышей В6 (дикий тип), GITR +/-, CD28 -/- и GITR -/- культивировали в течение 72 часов с различными концентрациями растворимого mAb против CD3 (ось x). Все клетки LN культивировали без экзогенного IL-2 (50 ед/мл) (Аа) и с ним (Ас). Клетки LN, истощенные по CD25+ клеткам (LNΔ25), культивировали без экзогенного IL-2 (50 ед/мл) (Аb) и с ним (Аd). Отрезки, указывающие значения стандартного отклонения, пропущены для ясности. Фиг.11В. Оценку путем проточной цитометрии разведения CFSE сортированными по CD4+ и CD8+ Т-клетками лимфоузла, выделенными из животных CD28 -/-, GITR -/-, GITR +/+ или GITR +/-, проводили после культивирования в течение 72 часов. Данные результаты соответствовали концентрации растворимого анти-CD3, равной 0,62 мкг/мл (как на фиг.11A). Фиг.11C. Анализ путем проточной цитометрии экспрессии CD25 проводили на H-2Db-позитивных CD4+CD25- клетках, которые оставались нестимулированными (незакрашенные гистограммы с пунктирной линией), были получены из мышей GITR -/- (незакрашенные гистограммы со сплошной линией) или были получены из мышей GITR +/+ (закрашенные гистограммы). Экспрессию CD25 клетками CD4+CD25-, полученными из мышей GITR -/- или GITR +/+, определяли через 24 часа культивирования с APC LN из мышей дикого типа, в присутствии анти-CD3 (0,5 мкг/мл) и в отсутствие (левые панели гистограмм) или присутствии (правые панели гистограмм) CD4+CD25+ клеток из мышей BALB/c в соотношении супрессор:отвечающая клетка 1:2. Также определяли экспрессию CD25 в отсутствие (верхние панели гистограмм) или в присутствии (нижние панели гистограмм) 50 ед/мл rhIL-2. Приведенные выше результаты представляют три независимых эксперимента.
На фиг.12 продемонстрировано, что CD28-зависимая совместная стимуляция усиливает экспрессию и реактивность GITR. Фиг.12A. Анализ путем проточной цитометрии экспрессии GITR очищенными CD4+CD25- или CD8+ T-клетками (2,5×104) через 72 часа культивирования со связанным на планшете анти-CD3 и 2 мкг/мл связанного на планшете контролем изотипа хомяка («aCD3») или связанного на планшете анти-CD28 («aCD3+aCD28»). Фиг.12B (левая панель гистограмм). Окрашивание против GITR CD4+CD25- T-клеток, культивируемых в присутствии облученных, истощенных по Т-клеткам спленоцитов и растворимого анти-CD3 (0,5 мкг/мл) в присутствии коктейля антител против CD80/86 (10 мкг/мл каждого) (т.е. антител против B7.1/7.2) или в его отсутствие в течение 72 часов. Фиг.12B (правая панель гистограмм). Окрашивание против GITR CD4+CD25- T-клеток, культивируемых в присутствии облученных, истощенных по Т-клеткам спленоцитов и растворимого анти-CD3 (0,5 мкг/мл) в присутствии коктейля антител против IL-2 и IL-2Ra. Фиг.12C. Пролиферацию оценивали в присутствии или отсутствие mAb против CD80/86 (10 мкг/мл каждого; «aB7») с добавлением mAb против GITR (24 мкг/мл; «DTA-1») или антитела контроля изотипа (2 мкг/мл; «крысиный IgG»). Отрезки указывают на значения стандартного отклонения. Результаты представляют два или три независимых эксперимента.
На фиг.13 продемонстрировано, что связывание GITRL с GITR предоставляет костимуляторный сигнал в отношении эффекторных T-клеток. Фиг.13A. Пролиферацию эффекторных Т-клеток GITR+/TCR+ HT-2 (4×104) отдельно (белые столбцы) или в совместной культуре с 1×104 контрольных клеток YB2/0 (штрихованные столбцы) или экспрессирующих GITRL клеток YB2/0 (закрашенные столбцы), в отсутствие или в присутствии одной или двух гранул анти-CD3 на клетку HT-2, определяли измерением захвата 3H-тимидина (счет/мин; ось y). Фиг.13B. Пролиферацию 4×104 клеток HT-2, совместно культивируемых с двумя гранулами анти-CD3 на клетку, 1×104 экспрессирующих GITRL клеток YB2/0, и возрастающими концентрациями (нг/мл; ось x) антитела против GITRL (5F1.1; закрашенные кружки) или антитела контроля изотипа (rIgG1; незакрашенные кружки) определяли измерением захвата 3H-тимидина (счет/мин; ось y). Фиг.13C. Пролиферацию 4×104 клеток HT-2, совместно культивируемых с двумя гранулами анти-CD3 на клетку, 1×104 экспрессирующих GITRL клеток YB2/0, и возрастающими концентрациями (нг/мл; ось x) четырех различных антител против GITRL: 5F1.1 (закрашенные кружки), MGLT-10 (закрашенные квадраты), MGTL-15 (незакрашенные кружки) или поликлонального антитела (незакрашенные кружки) определяли измерением захвата 3H-тимидина (счет/мин; ось y).
На фиг.14 продемонстрировано, что блокирование связывания GITR-GITRL антителом против GITRL предотвращает адаптивный перенос индуцированного PLP экспериментального аутоиммунного энцефаломиелита (EAE). Оценивали частоту возникновения EAE у мышей. Мышам вводили инъекцией 5×106 спленоцитов, которые выделяли у самок мышей SLJ, иммунизированных 150 мкг пептида PLP, и повторно стимулировали ex vivo в течение 3 суток в трех различных вариантах условий: 10 мкг/мл одного PLP (незакрашенные кружки), 10 мкг/мл PLP и 10 мкг/мл антитела контроля изотипа (CKO1; закрашенные кружки) или 10 мкг/мл PLP и антитела против GITRL (5Fl.1; закрашенные квадраты). Мониторинг возникновения EAE проводили в течение 52 суток (ось x) и считали по шкале от 0 до 5 (ось y).
Подробное описание изобретения
Поскольку антитела против GITR, продуцирующие противодействие подавляющей активности, как оказывается, продуцируют агонистический сигнал, было предсказано, что взаимодействие GITR с GITRL должно также ингибировать подавляющую активность регуляторных Т-клеток. Недостаток подходящих реагентов ранее препятствовал подробному функциональному анализу взаимодействий GITR/GITRL в более физиологических условиях. Идентифицировали мышиный ортолог GITRL и получали антагонистические антитела, которые специфично связываются с мышиным ортологом GITRL, т.е. не характеризуются перекрестным взаимодействием с человеческим GITRL.
С использованием данного реагента оценивали распределение в тканях и регуляцию GITRL. Кроме того, исследовали способность взаимодействий GITR/GITRL регулировать супрессию T-клеток с использованием мышей GITR -/-. Поскольку и CD25-, и CD25+ T-клетки экспрессируют GITR, хотя в различной степени, предыдущие исследования, демонстрирующие ингибирование супрессорной функции после обработки совместных культур агонистическим антителом против GITR, приводили к двусмысленным результатам в отношении клеточной мишени воздействия GITR. С использованием комбинации CD4+CD25+ и CD4+CD25- T-клеток из мышей дикого типа и GITR -/- в экспериментах с совместным культивированием было обнаружено, что связывание GITR на отвечающих CD4+CD25- T-клетках, а не на CD4+CD25+ супрессорных Т-клетках, требовалось для отмены супрессии. В отсутствие CD4+CD25+ T-клеток T-клетки GITR -/- характеризовались пролиферативными реакциями, сходными с таковыми у животных дикого типа, хотя они полностью супрессируются в присутствии физиологических количеств CD4+CD25+ T-клеток. Данные результаты впервые указывают на то, что взаимодействие GITR/GITRL предоставляет ранее не известный сигнал, который делает эффекторные Т-клетки устойчивыми к ингибиторному действию CD4+CD25+ T-клеток. Таким образом, отрицательная регуляция экспрессии GITRL после вторичных воспалительных сигналов может способствовать опосредованной CD4+CD25+ супрессии и предотвращать вредные последствия избыточного ответа эффекторных клеток.
Данное исследование пролило свет на механизмы, лежащие в основе взаимодействия GITR и его лиганда GITRL, особенно касательно действия на CD4+CD25- T-клетки, которые в традиционном понимании являются мишенью супрессорной активности. В итоге с использованием мышей GITR -/- было продемонстрировано, что способность mAb против GITR (агонистического мышиного антитела против GITR) отменять супрессию опосредована его действием на CD4+CD25-, а не на CD4+CD25+ T-клетки (как ранее предполагалось в некоторых исследованиях). APC (антигенпредставляющие клетки) конститутивно экспрессируют GITRL, который отрицательно регулируется после передачи сигнала с Toll-подобного рецептора. Хотя CD4+CD25- Т-клетки мышей GITR -/- были способны развивать пролиферативные реакции, они были неспособны к пролиферации в присутствии физиологических количеств CD4+CD25+ T-клеток. Таким образом, GITRL предоставляет значимый сигнал для CD4+CD25- T-клеток и других эффекторных T-клеток (например, CD8+ T-клеток), делая их устойчивыми к опосредованной CD4+CD25+ регуляции при инициации иммунного ответа. Отрицательная регуляция GITRL воспалительными стимулами может увеличивать чувствительность эффекторных Т-клеток (например, CD4+CD25- T-клеток) к супрессорной активности, например, в ходе злокачественного или инфекционного заболевания.
И в завершение, настоящее изобретение относится к нуклеотидной и аминокислотной последовательностям нового мышиного гомолога человеческого GITRL. Человеческий GITRL идентифицирован (Kwon et al. (1999) J. Biol. Chem. 274:6056-61; Gurney et al. (1999) Curr. Biol. 9:215-18); кроме того, совсем недавно несколько исследовательских групп также сообщили о клонировании мышиного лиганда GITR (Kim et al., 2003; Tone et al., 2003; Yu et al., 2003).
В одном из аспектов настоящее изобретение относится к нуклеотидным последовательностям, аминокислотным последовательностям и их активным фрагментам, и/или слитым белкам, относящимся к новому мышиному гомологу человеческого GITRL. Полинуклеотиды GITRL по изобретению включают в себя полинуклеотиды, которые модулируют экспрессию GITRL, например экспрессирующие векторы, содержащие полинуклеотиды GITRL, которые могут положительно регулировать экспрессию GITRL, и/или антисмысловые и/или относящиеся к РНКi полинуклеотиды GITRL, которые отрицательно регулируют экспрессию GITRL. Также предоставляется применение таких полинуклеотидов для модулирования экспрессии GITRL в клетках и/или животных. В дополнение к полипептидам GITRL, изобретение также относится к другим агонистическим полипептидам, например к активным фрагментам GITRL и/или слитым белкам GITRL, которые способны имитировать GITRL, т.е. индуцировать активность GITR в эффекторных T-клетках. Трансформированные клетки-хозяева и трансгенные животные, содержащие полинуклеотиды GITRL, также входят в объем изобретения.
В другом аспекте предоставляются антитела, которые специфично связываются с новыми мышиными полипептидами GITRL по изобретению (т.е. которые не связываются с человеческим GITRL). В частности, предоставляются нейтрализующие антитела, которые ингибируют активность GITRL (например, антитела, которые препятствуют связыванию GITRL с GITR); о данных антителах можно сказать, что они нейтрализуют активность GITRL (т.е. делают GITRL неэффективным). Нейтрализующие антитела по изобретению включают в себя не относящиеся к человеку и человеческие антитела к GITRL, которые ингибируют активность GITRL, а также химеризованные и/или гуманизированные версии не относящихся к человеку антител по изобретению, которые ингибируют активность GITRL. Также в объем изобретения включены антагонистические антитела, которые могут содержать одну или несколько мутаций, способных функционировать в плане увеличения времени полужизни, стабильности и аффинности антитела или способных функционировать в плане модулирования эффекторных функций антитела.
Другой аспект изобретения относится к анализам по скринингу, в которых полинуклеотиды и полипептиды GITRL, включающие, но без ограничения, их человеческие гомологи, применяют для идентификации соединений, способных модулировать активность GITR в клетке, организме или у субъекта. Изобретение также относится к способам оценки эффективности идентифицированных соединений, причем число T-клеток пациента определяется до и после введения идентифицированного соединения. Кроме того, изобретение относится к способам лечения пациентов или субъектов с использованием идентифицированных соединений.
В дополнение к предоставлению способов скрининга тестируемых соединений, способных модулировать активность GITR, например агонистов GITR или антагонистов GITR, изобретение относится к способам диагностики, прогноза и мониторинга прогресса нарушений, относящихся к нарушенной регуляции иммунной системы, например аутоиммунных заболеваний, воспалительных заболеваний и отторжения трансплантата, и злокачественных опухолей и инфекционных заболеваний.
В данном описании также описаны способы применения GITRL и родственных молекул по изобретению, включая молекулы-агонисты GITR (т.е. полинуклеотиды GITRL, полипептиды GITRL, их активные фрагменты и/или слитые белки, низкомолекулярные агонисты и агонистические в отношении GITR антитела) и молекулы-антагонисты GITR (т.е. ингибиторные полинуклеотиды GITRL, нейтрализующие GITR антитела, нейтрализующие GITRL антитела, низкомолекулярные антагонисты и слитые белки GITR), для теравпетического лечения нарушений, связанных с нарушенной регуляцией иммунной системы. Например, предоставляются способы лечения субъекта при риске или поставленном диагнозе аутоиммунного заболевания, воспалительного заболевания, отторжения трансплантата и/или других воспалительных заболеваний, включающих введение субъекту антагонистов GITR, например нейтрализующего антитела против GITR; также предоставляются способы лечения субъекта при риске или поставленном диагнозе злокачественной опухоли или инфекционных заболеваний, включающие введение субъекту агонистов GITR, например GITRL, или его агонистического слитого белка. Альтернативно, предоставляются способы индукции или ингибирования пролиферации Т-клеток посредством введения агонистов GITR, например GITRL (включая его агонистические слитые белки), или антагонистов GITR, например нейтрализующих антител против GITRL или антагонистических слитых белков GITR соответственно. Сходным образом, также предоставлены способы блокирования или усиления супрессии T-клеток в присутствии CD4+CD25+ T-клеток, включающие введения агонистов GITR, например GITRL (включая их агонистические слитые белки), или антагонистов GITR, например нейтрализующих антител против GITRL соответственно. Популяции T-клеток, обработанных полипептидами GITRL и родственными молекулами (включая их агонистические слитые белки), входят в объем изобретения и могут вводиться субъекту способом лечения злокачественной опухоли или инфекционного заболевания. Предоставлены другие способы лечения, включая способ лечения субъекта при риске или поставленном диагнозе аутоиммунного заболевания, воспалительного заболевания или отторжения трансплантата, антагонистическим соединением, которое снижает активность GITR, и способы лечения субъекта при риске или поставленном диагнозе злокачественной опухоли или инфекционного заболевания агонистическим соединением, которое повышает активность GITR. Фармацевтические композиции, например вакцинные адъюванты, содержащие полинуклеотиды, полипептиды GITRL и родственные молекулы (включая агонистические слитые белки GITRL и антагонистические антитела против GITRL) по изобретению также входят в объем изобретения. Способы по настоящему изобретению направлены на GITRL и GITR, включая, но без ограничения, мышиные GITRL и GITR и их гомологи; в состав данных гомологов специально включены человеческие GITRL и GITR.
Полинуклеотиды и полипептиды GITRL
Настоящее изобретение относится к новым выделенным и очищенным полинуклеотидам и полипептидам, относящимся к новому лиганду GITR (GITRL). Гены, полинуклеотиды, белки и полипептиды по настоящему изобретению включают, но без ограничения, мышиный GITRL и его гомологи.
Например, изобретение относится к очищенным и выделенным полинуклеотидам, кодирующим мышиный GITRL. Предпочтительные последовательности ДНК по изобретению включают в себя геномные, кДНК и химически синтезированные последовательности ДНК.
Нуклеотидная последовательность кДНК, кодирующая данный новый лиганд, обозначенная как кДНК мышиного GITRL, приведена в SEQ ID NO:1. Полинуклеотиды по настоящему изобретению также включают в себя полинуклеотиды, которые гибридизуются в жестких условиях с SEQ ID NO:1 или с комплементарной ей последовательностью, и/или кодируют полипептиды, которые сохраняют существенную биологическую активность (т.е. активные фрагменты) полноразмерного мышиного GITRL. Полинуклеотиды по настоящему изобретению также включают в себя непрерывные части последовательности, приведенной в SEQ ID NO:1, содержащие, по меньшей мере, 21 следующих друг за другом нуклеотидов.
Нуклеотидная последовательность геномной ДНК, кодирующей данный новый лиганд, обозначенная как геномная ДНК мышиного GITRL, приведена в SEQ ID NO:3. Полинуклеотиды по настоящему изобретению также включают в себя полинуклеотиды, которые гибридизуются в жестких условиях с SEQ ID NO:3 или с комплементарной ей последовательностью, и/или кодируют полипептиды, сохраняющие существенную биологическую активность полноразмерного мышиного GITRL. Полинуклеотиды по настоящему изобретению также включают в себя непрерывные части последовательности, приведенные SEQ ID NO:3, содержащие, по меньшей мере, 21 следующих друг за другом нуклеотидов.
Аминокислотная последовательность мышиного GITRL приведена в SEQ ID NO:2. Полипептиды по настоящему изобретению также включают в себя непрерывные части последовательности, приведенной в SEQ ID NO:2, содержащие, по меньшей мере, 7 следующих друг за другом аминокислот. Предпочтительный полипептид по настоящему изобретению включает в себя любую непрерывную часть последовательности, приведенной в SEQ ID NO:2, которая сохраняет существенную биологическую активность полноразмерного мышиного GITRL. Полинуклеотиды по настоящему изобретению также включают в себя, в дополнение к полинуклеотидам мышиного происхождения, описанным выше, полинуклеотиды, которые кодируют аминокислотную последовательность, приведенную в SEQ ID NO:2, или ее непрерывную часть и которые отличаются от описанных выше полинуклеотидов только вследствие хорошо известной вырожденности генетического кода.
Выделенные полинуклеотиды по настоящему изобретению могут использоваться в качестве зондов для гибридизации и праймеров с целью идентификации и выделения нуклеиновых кислот, имеющих последовательности, идентичные в отношении тех, которые кодируют описанные полинуклеотиды, или сходные с ними. Способы гибридизации для идентификации и выделения нуклеиновых кислот включают полимеразную цепную реакцию (ПЦР), способы саузерн-гибридизации, гибридизацию in situ и нозерн-гибридизацию и хорошо известны специалистам в данной области.
Реакции гибридизации могут проводиться в условиях различной жесткости. Жесткость реакции гибридизации заключается в затрудненности, с которой любые две молекулы нуклеиновой кислоты будут гибридизоваться друг с другом. Предпочтительно каждый гибридизующийся полинуклеотид гидридизуется с соответствующим ему полинуклеотидом в условиях сниженной жесткости, более предпочтительно в жестких условиях и наиболее предпочтительно в условиях высокой жесткости. Примеры условий жесткости показаны ниже в таблице 1: условия высокой жесткости соответствуют тем, которые, по меньшей мере, столь же жесткие, как, например, условия A-F; жесткие условия, по меньшей мере, такие же, как, например, условия G-L; условия сниженной жесткости, по меньшей мере, столь же жесткие, как, например, условия M-R.
2 SSPE (1×SSPE представляет собой 0,15M NaCl, 10мМ NaH2PO4 и 1,25мМ EDTA, pH 7,4) в буферах гибридизации и промывки может заменяться на SSC (1×SSC представляет собой 0,15M NaCl и 15мМ цитрат натрия); промывки проводят в течение 15 минут после завершения гибридизации.
TB*-TR*: Температура гибридизации для гибридов, которые, как предсказано, составляют менее 50 пар оснований в длину, должна быть на 5-10°C ниже температуры плавления (Tm) гибрида, где Tm определяется по следующим уравнениям. Для гибридов менее 18 пар оснований в длину Tm (°C)=2 (число оснований A+T)+4 (число оснований G+C). Для гибридов от 18 до 49 пар оснований в длину Tm(°C)=81,5+16,6(log10Nа+)+0,41(% G+C)-(600/N), где N составляет число оснований в гибриде и Na+ представляет собой концентрацию ионов натрия в буфере гибридизации (Na+ для 1Х SSC=0,165M).
Дополнительные примеры условий жесткости для гибридизации полинуклеотидов предоставлены в Sambrook et al., Molecular Cloning: A Laboratory Manual, Chs. 9 & 11, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989), и Ausubel et al., eds., Current Protocols in Molecular Biology, Sects. 2.10 & 6.3-6.4, John Wiley & Sons, Inc. (1995), включенных в данное описание в качестве ссылки.
Выделенные полинуклеотиды по настоящему изобретению могут использоваться в качестве зондов и праймеров для гибридизации с целью идентификации и выделения ДНК, имеющей последовательности, кодирующие аллельные варианты описанных полинуклеотидов. Аллельные варианты представляют собой встречающиеся в природе альтернативные формы описанных полинуклеотидов, которые кодируют полипептиды, идентичные или имеющие значительное сходство с полипептидами, кодирующими описанные полинуклеотиды. Предпочтительно аллельные варианты имеют, по меньшей мере, 90% идентичности по последовательности (более предпочтительно, по меньшей мере, 95% идентичности; наиболее предпочтительно, по меньшей мере, 99% идентичности) с описанными полинуклеотидами.
Выделенные полинуклеотиды по настоящему изобретению могут также использоваться в качестве зондов и праймеров для гибридизации с целью идентификации и выделения молекул ДНК, имеющих последовательности, кодирующие полипептиды, гомологичные описанным полинуклеотидам. Данные гомологи представляют собой полинуклеотиды и полипептиды из видов, отличных от тех, к котором относятся описанные полипептиды и полинуклеотиды, или молекулы из одного вида, но со значительным сходством по последовательности относительно описанных полинуклеотидов и полипептидов. Предпочтительно гомологи полинуклеотидов имеют, по меньшей мере, 50% идентичности по последовательности (более предпочтительно, по меньшей мере, 75% идентичности; наиболее предпочтительно, по меньшей мере, 90% идентичности) с описанными полинуклеотидами, тогда как гомологи полипептидов имеют, по меньшей мере, 30% идентичности по последовательности (более предпочтительно, по меньшей мере, 45% идентичности; наиболее предпочтительно, по меньшей мере, 60% идентичности) с описанными полипептидами. Предпочтительно гомологи описанных полинуклеотидов и полипептидов выделены из видов млекопитающих.
Выделенные полинуклеотиды по настоящему изобретению могут также использоваться в качестве зондов и праймеров для гибридизации с целью идентификации клеток и тканей, которые экспрессируют полипептиды по настоящему изобретению, и условия, в которых они экспрессируются.
Кроме того, выделенные полинуклеотиды по настоящему изобретению могут использоваться для изменения (т.е. усиления, снижения или модификации) экспрессии генов, соответствующих полинуклеотидам по настоящему изобретению, в клетке или организме. Данные соответствующие гены представляют собой геномные последовательности ДНК по настоящему изобретению (например, SEQ ID NO:3), которые транскрибируются с продукцией их РНК, из которых полинуклеотиды кДНК по настоящему изобретению (например, SEQ ID NO:1).
Измененная экспрессия генов по настоящему изобретению, включая, но без ограничения, мышиный GITRL и его гомологи, могут достигаться в клетке или организме путем использования различных ингибиторных полинуклеотидов, например антисмысловых полинуклеотидов (например, антисмысловых молекул нуклеиновой кислоты GITRL) и рибозимов, которые связываются и/или расщепляют мРНК, транскрибированные с генов по изобретению (см., например, Galderisi et al. (1999) J. Cell Physiol. 181:251-57; Sioud (2001) Curr. Mol. Med. 1:575-88). Такие ингибиторные полинуклеотиды могут использоваться при профилактике или лечении аутоиммунных нарушений, воспалительных заболеваний, отторжения трансплантата и сходных или связанных заболеваний.
Антисмысловые полинуклеотиды или рибозимы по изобретению могут быть комплементарными всей кодирующей цепи гена по изобретению или только его части. Альтернативно, антисмысловые полинуклеотиды или рибозимы могут быть комплементарными некодирующей области кодирующей цепи гена по изобретению. Антисмысловые полинуклеотиды или рибозимы могут конструироваться с использованием химического синтеза и реакций ферментативного лигирования с использованием методик, хорошо известных в данной области. Нуклеозидные связи химически синтезированных полинуклеотидов могут модифицироваться для усиления их способности противодействовать опосредованной нуклеазами деградации, а также для усиления их специфичности в отношении последовательности. Такие модификации связей включают, но без ограничения, связи фосфотиоата, метилфосфоната, фосфороамидата, боранофосфата, морфолина и пептидной нуклеиновой кислоты (ПНК) (Galderisi et al., supra; Heasman (2002) Dev. Biol. 243:209-14; Micklefield (2001) Curs. Med. Chem. 8:1157-79). Альтернативно, данные молекулы могут продуцироваться биологически с использованием экспрессирующего вектора, в который субклонирован полинуклеотид по настоящему изобретению в антисмысловой (т.е. обратной) ориентации.
Ингибиторные полинуклеотиды по настоящему изобретению также включают в себя образующие триплекс олигонуклеотиды (TFO), которые связываются с большой бороздкой дуплекса ДНК с высокой специфичностью и активностью (Knauert and Glazer (2001) Hum. Mol. Genet. 10:2243-51). Экспрессия генов по настоящему изобретению может ингибироваться направленным действием TFO, комплементарных регуляторным областям генов (т.е. промоторным и/или энхансерным последовательностям) с образованием структур тройной спирали, которые предотвращают транскрипцию данных генов.
В одном из вариантов осуществления изобретения ингибиторные полинуклеотиды по настоящему изобретению представляют собой молекулы коротких интерферирующих РНК (siРНК) (например, нуклеиновые молекулы siРНК GITRL). Данные молекулы siРНК являются короткими (предпочтительно 19-25 нуклеотидов; наиболее предпочтительно 19 или 21 нуклеотидов), двухцепочечными молекулами РНК, которые вызывают специфическую в плане последовательности деградацию мРНК-мишени. Данная деградация известна как РНК-интерференция (РНКi) (например, Bass (2001) Nature 411:428-29). Впервые выявленная в низших организмах, РНКi эффективно применяется на клетках млекопитающих и, как было недавно показано, предотвращает фульминантный гепатит у мышей, обработанных молекулами siРНК, направленными на мРНК Fas (Song et al. (2003) Nature Med. 9:347-51). Кроме того, недавно было сообщено о том, что интратекально доставленная siРНК блокирует болевые ответы на двух моделях (агонист-индуцированная болевая модель и нейропатическая болевая модель) у крыс (Dorn et al. (2004) Nucleic Acids Res. 32(5):e49).
Молекулы siРНК по настоящему изобретению могут генерироваться путем отжига вместе двух комплементарных одноцепочечных молекул РНК (одна из которых совпадает с частью мРНК-мишени) (Fire et al., патент США № 6506559) или путем применения единственной молекулы РНК-шпильки, которая сворачивается сама на себя с продукцией необходимой двухцепочечной части (Yu et al. (2002) Proc. Natl. Acad. Sci. USA 99:6047-52). Молекулы siРНК могут химически синтезироваться (Elbashir et al. (2001) Nature 411:494-98) или продуцироваться посредством транскрипции in vitro с использованием одноцепочечных ДНК-матриц (Yu et al., supra). Альтернативно, молекулы siРНК могут продуцироваться биологически, кратковременно (Yu et al., выше; Sui et al. (2002) Proc. Natl. Acad. Sci. USA 99:5515-20) или стабильно (Paddison et al. (2002) Proc. Natl. Acad. Sci. USA 99:1443-48), с использованием экспрессирующего(их) вектора(ов), содержащего смысловые и антисмысловые последовательности siРНК. Недавно снижение уровня мРНК-мишени в первичных человеческих клетках эффективным и специфичным в отношении последовательности образом было продемонстрировано с использованием аденовирусных векторов, экспрессирующих РНК-шпильки, которые далее процессируются в siРНК (Arts et al. (2003) Genome Res. 13:2325-32).
Молекулы siРНК, направленные на полинуклеотиды по настоящему изобретению, могут обозначаться на основе критериев, хорошо известных в данной области (например, Elbashir et al. (2001) EMBO J. 20:6877-88). Например, сегмент-мишень мРНК-мишени предпочтительно должен начинаться с AA (наиболее предпочтительно), TA, GA или CA; отношение GC молекулы siРНК предпочтительно должно составлять 45-55%; молекула siРНК предпочтительно не должна содержать три одинаковых нуклеотида в ряд; молекула siРНК предпочтительно не должна содержать семь смешанных G/C в ряд; и сегмент-мишень предпочтительно должен находиться в области ORF мРНК-мишени и предпочтительно должен отстоять, по меньшей мере, на 75 н.п. после инициирующего ATG и, по меньшей мере, на 75 н.п. перед стоп-кодоном. На основе данных критериев или на основе других известных критериев (например, Reynolds et al. (2004) Nature Biotechnol. 22:326-30) молекулы siРНК по настоящему изобретению, направленные на них полинуклеотиды РНК по настоящему изобретению, могут конструироваться рядовыми специалистами в данной области.
Измененная экспрессия генов по настоящему изобретения в организме может также достигаться посредством создания не относящихся к человеку трансгенных животных, в геномы которых введены полинуклеотиды по настоящему изобретению. Такие трансгенные животные включают животных, которые имеют множественные копии гена (т.е. трансгена) по настоящему изобретению. Тканеспецифическая(ие) регуляторная(ые) последовательность(и) может(гут) быть функционально связанной(ыми) с трансгенами для прямой экспрессии полипептида по настоящему изобретению в конкретных клетках или на конкретной стадии развития. Способы получения трансгенных животных посредством манипуляции с эмбрионами и микроинъекции, особенно таких животных, как мышь, стали общепринятыми и хорошо известны в данной области (например, Bockamp et al., Physiol. Genomics, 11:115-32 (2002)).
Измененная экспрессия генов по настоящему изобретению в организме также может достигаться посредством создания животных, эндогенные гены которых, соответствующие полинуклеотидам по настоящему изобретению, разрушены посредством встраивания посторонних нуклеотидных последовательностей (т.е. нокаут-животного). Кодирующая область данного эндогенного гена может быть разрушена с образованием за счет этого нефункционального белка. Альтернативно, вышележащая регуляторная область эндогенного гена может разрушаться или заменяться различными регуляторными элементами, что приводит к измененной экспрессии еще функционального белка. Способы получения нокаут-животных включают гомологичную рекомбинацию и хорошо известны в данной области (например, Wolfer et al., Trends Neurosci., 25:336-40 (2002)).
Выделенные полинуклеотиды по настоящему изобретению могут быть функционально связаны с последовательностью контроля экспрессии и/или лигированы в экспрессирующий вектор для рекомбинантной продукции полипептидов по настоящему изобретению. Основные способы экспрессии рекомбинантных белков хорошо известны в данной области. Такие рекомбинантные белки могут экспрессироваться в растворимой форме для применения нарушений, происходящих из-за нарушенной регуляции иммунной системы; такие нарушения включают, например, злокачественные опухоли и инфекционные заболевания, и аутоиммунные нарушения и воспалительные заболевания, и отторжение трансплантата. Аутоиммунные нарушения и воспалительные заболевания включают, но без ограничения, ревматоидный артрит, энцефаломиелит, остеоартрит, рассеянный склероз, аутоиммунный гастрит, системную красную волчанку и другие воспалительные дерматозы, диабет I типа, астму, аллергию и воспалительные заболевания толстого кишечника, включая болезнь Крона и язвенный колит.
Подразумевается, что используемый экспрессирующий вектор относится к молекуле нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой он связан. Другим типом вектора является плазмида, которая относится к кольцевой двухцепочечной ДНК-петле, в которую могут лигироваться дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, в котором дополнительные сегменты ДНК могут лигироваться в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальную точку начала репликации, и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) могут интегрироваться в геном клетки-хозяева после введения в клетку-хозяин и за счет этого реплицируются вместе с геномом-хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы обозначены в данном описании как рекомбинантные экспрессирующие векторы (или просто экспрессирующие векторы). В общем, экспрессирующие векторы, применяемые в способах рекомбинантной ДНК, часто находятся в виде плазмид. В настоящем описании плазмида и вектор могут употребляться взаимозаменяемо, поскольку плазмида является наиболее часто используемым видом вектора. Однако подразумевается, что изобретение относится к другим видам экспрессирующих векторов, таким как вирусные векторы (например, дефектные по репликации ретровирусы, аденовирусы и аденоассоциированные вирусы), которые выполняют эквивалентные функции.
В другом варианте осуществления полинуклеотиды по настоящему изобретению применяют для создания агонистов GITR, например полипептидов GITRL, включая активные фрагменты и/или их слитые белки, которые также входят в объем изобретения. Например, полипептид GITRL или его активные фрагменты могут сливаться со второй группой, например, иммуноглобулином или его фрагментом (например, его связывающим Fc-фрагментом). В некоторых вариантах осуществления первый полипептид включает в себя полноразмерный полипептид GITRL. Альтернативно, первый полипептид может включать в себя меньше полноразмерного полипептида GITRL. Кроме того, растворимые формы GITRL могут сливаться через «линкерные» последовательности с Fc-частью иммуноглобулина. Другие слитые белки, такие как с глутатион-S-трансфераза (GST), Lex-A, тиоредоксин (TRX) или мальтозосвязывающий белок (MBP), также могут использоваться.
Слитые белки могут дополнительно включать с себя линкерную последовательность, соединяющую фрагмент GITRL или GITRL со второй группой. Применение таких линкерных последовательностей хорошо известно в данной области. Например, слитый белок может включать в себя пептидный линкер, например пептидный линкер длиной примерно 2-20, более предпочтительно менее 10 аминокислот. В одном из вариантов осуществления пептидный линкер может составлять 2 aминокислоты в длину.
В другом варианте осуществления слитый белок включает в себя гетерологичную сигнальную последовательность (т.е. полипептидную последовательность, которая не присутствует в полипептиде, кодируемом нуклеиновой кислотой GITRL) с его N-конца. Например, сигнальная последовательность из другого белка может сливаться с полипептидом GITRL, включая их активные фрагменты и/или слитые белки. В конкретных клетках-хозяевах (например, клетках-хозяевах млекопитающего), экспрессия и/или секреция GITRL может увеличиваться путем использования гетерологичной сигнальной последовательности.
Сигнальный пептид, который может включаться в состав слитого белка, представляет собой MKFLVNVALVFMVVYISYIYA (SEQ ID NO:11). Если требуется, одна или несколько аминокислот может, кроме того, встраиваться между первым полипептидным радикалом, включающим в себя радикал GITRL, и вторым полипептидным радикалом. Второй полипептид предпочтительно растворим. В некоторых вариантах осуществления второй полипептид усиливает период полужизни (например, период полужизни в сыворотке) связанного полипептида. В некоторых вариантах осуществления второй полипептид включает в себя последовательность, которая облегчает ассоциацию слитого полипептида со вторым полипептидом GITRL. В предпочтительном осуществлении второй полипептид включает в себя, по меньшей мере, область иммуноглобулинового пептида. Полипептиды слияния иммуноглобулина известны в данной области и описаны, например, в патентах США № 5516964; 5225538; 5428130; 5514582; 5714147 и 5455165, которые включены в данное описание в качестве ссылки во всей своей полноте.
Химерный белок или слитый белок по изобретению может продуцироваться стандартными способами рекомбинантной ДНК. Например, фрагменты ДНК, кодирующие различные полипептидные последовательности, лигируют вместе в одну рамку считывания общепринятыми способами, например, путем использования тупых концов или липких концов для лигирования, расщепления ферментами рестрикции для обеспечения подходящих концов, подходящего заполнения липких концов, обработки щелочной фосфатазой во избежание нежелательного присоединения и ферментативного лигирования. В другом варианте осуществления ген слияния может быть синтезирован общепринятыми способами, включая автоматизированные ДНК-синтезаторы. Альтернативно, ПЦР-амплификация генных фрагментов может проводиться с использованием якорных праймеров, которые дают начало комплементарным перекрываниям между двумя следующими друг за другом генными фрагментами, которые затем могут отжигаться и повторно амплифицироваться с образованием химерной генной последовательности (см., например, Ausubel et al. (Eds.) CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, John Wiley & Sons, 1992). Более того, многие экспрессирующие векторы, которые кодируют радикал слияния (например, Fc-область иммуноглобулиновой тяжелой цепи), доступны коммерчески. Кодирующая GITRL нуклеиновая кислота может клонироваться в такой экспрессирующий вектор, так что радикал слияния связан в рамку считывания с белком-иммуноглобулином. В некоторых вариантах осуществления слитые полипептиды GITRL существуют в виде олигомеров, таких как димеры или тримеры.
Некоторые клеточные линии могут действовать в виде подходящих клеток-хозяев для рекомбинантной экспрессии полипептидов по настоящему изобретению. Клеточные линии хозяина-млекопитающего включают в себя, например, клетки COS, клетки CHO, клетки 293T, клетки A431, клетки 3T3, клетки CV-1, клетки HeLa, клетки L, клетки BHK21, клетки HL-60, клетки U937, клетки HaK, клетки Jurkat, а также клеточные линии, полученные из культуры in vitro первичной ткани и первичных эксплантатов.
Альтернативно, может быть возможной рекомбинантная продукция полипептидов по настоящему изобретению в низших эукариотах, таких как дрожжи, или в прокариотах. Потенциально подходящие штаммы дрожжей включают в себя Saccharomyces cerevisiae, Schizosaccharomyces pombe, штаммы Kluyveromyces и штаммы Candida. Потенциально подходящие бактериальные штаммы включают в себя Escherichia coli, Bacillus subtilis и Salmonella typhimurium. Если полипептиды по настоящему изобретению получены в дрожжах или бактериях, может быть необходимым модифицировать их, например, путем фосфорилирования или гликозилирования подходящих участков для придания функциональности. Такое ковалентное связывание может осуществляться с использованием хорошо известных химических или ферментативных способов.
Экспрессия в бактериях может приводить к образованию телец включения, содержащих рекомбинантный белок. Таким образом, рефолдинг рекомбинантного белка может потребоваться для продукции активного или более активного материала. Некоторые способы получения правильно свернутых гетерологичных белков из бактериальных телец включения известны в данной области. Данные способы в основном включают солюбилизацию белка из телец включения, с последующей полной денатурацией с использованием хаотропного средства. Когда в первичной аминокислотной последовательности белка присутствуют остатки цистеина, часто является необходимым выполнять рефолдинг в окружении, которое обеспечивает непосредственное образование дисульфидных связей (редокс-система). Основные способы рефолдинга описаны в Kohno (1990) Meth. Enzymol. 185:187-95. В EP 0433225 и в заявке на патент США № 08/163877 описаны другие подходящие способы.
Полипептиды по настоящему изобретению могут также рекомбинантно продуцироваться путем функциональной связи выделенных полинуклеотидов по настоящему изобретению с подходящими последовательностями контроля в одном или нескольких экспрессирующих векторах насекомых, таких как бакуловирусные векторы, и с использованием экспрессирующей системы клеток насекомых. Материалы и способы для экспрессирующих систем на основе бакуловируса/Sf9 доступны коммерчески в виде набора (например, набор MaxBac®, Invitrogen, Carlsbad, Калифорния).
Агонисты GITR, например белок GITRL, его активные фрагменты и/или слитый белок, могут быть получены путем выращивания культуры трансформированных клеток-хозяев в условиях культивирования, необходимых для экспрессии требуемого белка. После рекомбинантной экспрессии подходящих клеток-хозяев полипептиды по настоящему изобретению могут очищаться от культуральной среды или клеточных экстрактов с использованием известных методов очистки, таких как гель-фильтрация и ионообменная хроматография. Растворимые формы агонистов GITR, например белка GITRL, его активных фрагментов и/или слитого белка, могут очищаться из кондиционированной среды. Мембраносвязанные формы, например, белка GITRL по изобретению могут очищаться получением общей мембранной фракции из экспрессирующих клеток и экстракцией из мембран неионным детергентом, таким как Triton X-100. Очистка также может включать аффинную хроматографию со средствами, о которых известно, что они связывают полипептиды по настоящему изобретению. Данные способы очистки также могут быть использованы для очистки полипептидов по настоящему изобретению от других источников, включая природные источники. Как описано выше, агонисты GITR, например белок GITRL, его активные фрагменты и/или слитый белок, могут также экспрессироваться в виде продукта трансгенных животных, например, как компонент молока трансгенных коров, коз, свиней или овец, которые характеризуются соматическими или зародышевыми клетками, содержащими полинуклеотидную последовательность, кодирующую агонисты GITR.
Методы, которые могут быть использованы для очистки агонистов GITR, например белка GITRL, его активных фрагментов и/или слитого белка, известны специалистам в данной области. Например, белок GITRL по изобретению может быть концентрирован с использованием коммерчески доступного фильтра для концентрирования, например, устройства для ульрафильтрации Amicon или Millipore Pellicon. После стадии концентрирования концентрат может быть помещен на очищающий матрикс, такой как среда для гель-фильтрации. Альтернативно, можно использовать анионообменную смолу, например, матрикс или субстрат, имеющие подвешенные диэтиламиноэтиловые (DEAE) или полиэтилениминовые (PEI) группы. Матриксы могут представлять собой акриламид, агарозу, декстран, целлюлозу или другие типы, обычно используемые при очистке белков. Альтернативно, может использоваться катионообменная стадия. Подходящие катионообменники включают различные нерастворимые матриксы, содержащие сульфопропильные или карбоксиметильные группы. Сульфопропильные группы являются предпочтительными (например, колонки с S-Sepharose®). Очистка агонистов GITR, например белка GITRL, его активных фрагментов и/или слитого белка, из культуральной надосадочной жидкости может также включать одну или несколько колоночных стадий на таких аффинных смолах, как конканавалин A-агароза, гепарин-Toyopearl® или Cibacron blue 3GA Sepharose®; или хроматографию гидрофобных взаимодействий с использованием таких смол, как фенилэфирная, бутилэфирная или пропилэфирная; или иммунно-аффинную хроматографию. Наконец, одна или несколько стадий высокоэффективной жидкостной хроматографии с обращенной фазой (ОФ-ВЭЖХ) с применением гидрофобных сред для ОФ-ВЭЖХ, например силикагеля, имеющего навешенные метильные или другие алифатические группы, могут быть использованы для дальнейшей очистки белка GITRL. Аффинные колонки, содержащие антитела против белка GITRL, также могут использоваться в очистке согласно известным способам. Некоторые из предшествующих стадий очистки в различных сочетаниях или с другими известными способами также могут использоваться для предоставления, по существу, очищенного выделенного рекомбинантного белка. Предпочтительно, выделенный белок GITRL очищен так, что он является, по существу, лишенным других белков млекопитающих.
Альтернативно, агонисты GITR, например белок GITRL, его активные фрагменты и/или слитые белки, могут также рекомбинантно экспрессироваться в форме, которая облегчает очистку. Например, данные полипептиды могут экспрессироваться в виде слитых белков с такими белками, как мальтозосвязывающий белок (MBP), глутатион-S-трансфераза (GST) или тиоредоксин (TRX). Наборы для экспрессии и очистки таких слитых белков коммерчески доступны от New England BioLabs (Beverly, Массачусетс), Pharmacia (Piscataway, Нью-Джерси) и Invitrogen соответственно. Агонисты GITR, например белок GITRL, его активные фрагменты и/или слитые белки, могут также быть мечены малым эпитопом и впоследствии идентифицированы с использованием специфического антитела против данного эпитопа. Предпочтительным эпитопом является эпитоп FLAG, который коммерчески доступен от Eastman Kodak (New Haven, Коннектикут).
Агонисты GITR, например белок GITRL, его активные фрагменты и/или слитые белки, могут также продуцироваться путем общепринятого химического синтеза. Способы химического синтеза таких полипептидов хорошо известны специалистам в данной области. Такие химически синтезированные полипептиды могут обладать биологическими свойствами общими с природными, очищенными полипептидами и, таким образом, могут применяться в качестве биологически активных или иммунологических заместителей для природных полипептидов.
Агонисты GITR, например белок GITRL, его активные фрагменты и/или слитые белки, также включают молекулы, структурно отличные от описанных полипептидов (например, которые имеют несколько измененную последовательность), но которые имеют, по существу, те же самые биохимические характеристики, что и описанные полипептиды (например, изменены только в функционально несущественных аминокислотных остатках). Такие молекулы включают в себя встречающиеся в природе аллельные варианты и обдуманно сконструированные варианты, содержащие изменения, замены, перемещения, вставки или делеции. Способы таких изменений, замен, перемещений, вставок или делеций хорошо известны специалистам в данной области. В некоторых вариантах осуществления полипептидный радикал GITRL предоставлен в виде полипептида-варианта GITRL, имеющего мутации во встречающейся в природе последовательности GITRL (дикий тип), что приводит к получению последовательности GITRL, более устойчивой к протеолизу (относительно немутированной последовательности).
Описанные способы получения агонистов GITR, например белка GITRL, его активных фрагментов и/или слитых белков, могут использоваться для получения антагонистов GITR, особенно растворимых белков GITR, их активных фрагментов и/или слитых белков. Специалисту в данной области будет очевидно, что все, что требуется для получения антагонистов GITR, например растворимого GITR, его активных фрагментов и/или слитых белков, будет представлять собой последовательность нуклеиновой кислоты или аминокислотную последовательность GITR, причем обе они известны. С использованием данных последовательностей антагонисты GITR, например растворимый GITR, его активные фрагменты и/или слитые белки, могут генерироваться с использованием способов рекомбинантной ДНК и/или химического синтеза, как описано выше.
Антитела против GITRL
В других вариантах осуществления изобретение относится к антагонистам GITR, таким как антитела или их антигенсвязывающие фрагменты, которые специфично связываются с GITRL, предпочтительно с GITRL млекопитающего (например, мыши) и нейтрализуют активность GITR.
Специалисту в данной области будет очевидно, что используемый здесь термин «антитело» относится к белку, содержащему, по меньшей мере, одну, предпочтительно две вариабельных области тяжелой (H) цепи (обозначаемые как VH), и, по меньшей мере, одну, предпочтительно две вариабельных области легкой (L) цепи (обозначаемые как VL). Области VH и VL могут далее подразделяться на области гипервариабельности, обозначенные «области, определяющие комплементарность» («CDR»), между которыми находятся области, которые являются более консервативными, называемые «каркасные области» («FR»). Содержание FR и CDR точно определено (см., Kabat, E.A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U. S. Department of Health and Human Services, NIH Publication № 91-3242, и Chothia, C. et al. (1987) J. Mol. Biol. 196:901-917, включенные в данное описание в качестве ссылки). Каждый VH и VL состоит из трех CDR и четырех FR, построенных от N-конца до С-конца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4.
Антитело может далее включать в себя константные области легкой и тяжелой цепи с образованием, таким образом, тяжелой и легкой иммуноглобулиновой цепи соответственно. В одном из вариантов осуществления антитело представляет собой тетрамер двух тяжелых иммуноглобулиновых цепей и двух легких иммуноглобулиновых цепей, в котором тяжелые и легкие иммуноглобулиновые цепи связаны между собой, например, дисульфидными связями. Константная область тяжелой цепи состоит из трех доменов, CH1, CH2 и CH3. Константная область легкой цепи состоит из одного домена CL. Вариабельная область легкой и тяжелой цепей состоит из связывающего домена, который взаимодействует с антигеном. Константные области антител обычно опосредуют связывание антитела с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первым компонентом (C1q) классической системы комплемента.
Иммуноглобулин относится к белку, состоящему из одного или нескольких полипептидов, по существу, кодируемых иммуноглобулиновыми генами. Распознаваемые гены человеческого иммуноглобулина включают в себя гены константных областей каппа, лямбда, альфа (IgA1 и IgA2), гамма (IgG1, IgG2, IgG3, IgG4), дельта, эпсилон и мю, а также мириады генов варибельных областей иммуноглобулинов. Полноразмерные «легкие цепи» иммуноглобулинов (примерно 25 кД, или 214 аминокислот) кодируются геном вариабельной области с NH2-конца (примерно 110 аминокислот) и геном константной области каппа или лямбда с COOH-конца. Полноразмерные иммуноглобулиновые «тяжелые цепи» (примерно 50 кД, или 446 аминокислот) сходным образом кодируются геном вариабельной области (примерно 116 аминокислот) и одним из других указанных выше генов константной области, например гамма (кодирует примерно 330 аминокислот). Гены константных областей тяжелых цепей иммуноглобулина кодируют класс антитела, т.е. изотип (например, IgM или IgG1). Антигенсвязывающий фрагмент антитела (или просто «часть антитела» или «фрагмент»), используемый здесь, относится к одному или нескольким фрагментам полноразмерного антитела, которые сохраняют способность специфично связывать антиген (например, CD3). Примеры связывающих фрагментов, охваченные термином «антигенсвязывающий фрагмент» антитела, включают в себя (i) Fab-фрагмент, одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) F(ab')2-фрагмент, двухвалентный фрагмент, состоящий из двух Fab-фрагментов, связанных дисульфидным мостиком в шарнирной области; (iii) Fd-фрагмент, состоящий из доменов VH и CH1; (iv) Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела, (v) dAb-фрагмент (Ward et al., (1989) Nature 341:544-46), который состоит из домена VH; и (vi) выделенную определяющую комплементарность область (CDR). Боле того, хотя два домена Fv-фрагмента, VL и VH, кодируются раздельными генами, их можно соединить с использованием рекомбинантных способов синтетическим линкером, который позволяет им составлять одну белковую цепь, в которой VL и VH области образуют пару с получением одновалентных молекул (известных как одноцепочечные Fv (scFv); см., например, Bird et al. (1988) Science 242:423-26; и Huston et al. (1988) Proc.Natl. Acad. Sci. USA: 85:5879-83). Такие одноцепочечные антитела, как подразумевается, также охватываются термином «антигенсвязывающий фрагмент» антитела. Данные фрагменты антигена получают с использованием общепринятых способов, известных специалистам в данной области, и фрагменты подвергают скринингу при использовании таких же методик, как и для интактных антител.
Специалисту в данной области будет очевидно, что описанные здесь способы получения молекул антитела против полипептидов по настоящему изобретению, например мышиного GITRL, могут также использоваться для получения молекул антитела против других белков, например GITR или человеческого GITRL. Следовательно, способы получения молекул антитела относятся не только к полипептидам по настоящему изобретению, как описано, но также, например, к GITR или человеческому GITRL.
Молекулы антитела против полипептидов по настоящему изобретению, например, нейтрализующие антитела против мышиного GITRL, включающие, но без ограничения, мышиный GITRL и его гомологи, могут использоваться при профилактике или лечении аутоиммунных нарушений, воспалительных заболеваний, отторжения трансплантата и сходных или родственных нарушений. Другие молекулы антитела, например агонистические антитела против GITR, могут использоваться по способам изобретения для лечения злокачественной опухоли, инфекционных заболеваний и сходных или родственных нарушений. Такие молекулы антитела могут продуцироваться способами, хорошо известными специалистам в данной области. Например, моноклональные антитела могут продуцироваться путем образования гибридом хорошо известными способами. Гибридомы, образованные таким способом, затем подвергают скринингу с использованием стандартных способов, таких как сорбционный иммуноферментный анализ (ELISA), для идентификации одной или нескольких гибридом, которые продуцируют антитело, специфично связывающееся с полипептидами по настоящему изобретению. Например, белки GITRL по изобретению могут также использоваться для иммунизации животных для получения поликлональных и моноклональных антител, которые специфично взаимодействуют с белком GITRL и которые могут ингибировать связывание лигандов с рецептором, т.е. GITR. Сходным образом, белки GITR также могут использоваться для получения поликлональных и моноклональных антител, которые специфично взаимодействуют с GITR. Пептидные иммуногены, кроме того, могут содержать остаток цистина с С-конца и могут конъюгироваться с гаптеном, таким как гемоцианин морского блюдечка (KLH). Дополнительные пептидные иммуногены могут генерироваться путем замещения остатков тирозина сульфатированными тирозиновыми остатками. Способы синтеза таких пептидов известны в данной области, например, как в Merrifield (1963) J. Amer. Chem. Soc. 85:2149-54; Krstenansky et al. (1987) FEBS Lett. 211:10. Полноразмерный полипептид по настоящему изобретению может использоваться в качестве иммуногена или, альтернативно, могут использоваться антигенные пептидные фрагменты данных полипептидов. Антигенные пептид из полипептида по настоящему изобретению включает, по меньшей мере, 7 непрерывных аминокислотных остатков и включает эпитоп, так что антитело, полученное против данного пептида, образует специфичный иммунный комплекс с данным полипептидов. Предпочтительно антигенный пептид включает, по меньшей мере, 10 аминокислотных остатков, более предпочтительно, по меньшей мере, 15 аминокислотных остатков, даже более предпочтительно, по меньшей мере, 20 аминокислотных остатков и наиболее предпочтительно по меньшей мере, 30 аминокислотных остатков.
Моноклональные антитела могут быть получены другими способами, известными специалисту в области технологии рекомбинантной ДНК. В качестве альтернативы получения секретирующих моноклональные антитела гибридом моноклональное антитело против полипептида по настоящему изобретению может быть идентифицировано и выделено путем скрининга рекомбинантной комбинаторной библиотеки иммуноглобулина (например, антительного фагового дисплея) полипептидом по настоящему изобретению (например, GITRL) или GITR, с выделением, таким образом, представителей библиотеки иммуноглобулинов, которые связываются с полипептидами по настоящему изобретению или с GITR соответственно. Способы и коммерчески доступные наборы для получения и скрининга библиотек фагового дисплея хорошо известны специалистам в данной области. Кроме того, примеры способов и реагентов, особенно подходящих для применения при получении и скрининге библиотек антительного дисплея, могут быть найдены в литературе. Например, способ «комбинаторного антительного дисплея» был разработан для идентификации и выделения фрагментов антител, имеющих особую специфичность в отношении антигена, и может использоваться для продукции моноклональных антител (для описания комбинаторного антительного дисплея см., например, Sastry et al. (1989) Proc. Natl. Acad. Sci. USA 86:5728; Huse et al. (1989) Science 246:1275; Orlandi et al. 1989 Proc. Natl. Acad. Sci. USA 86:3833). После иммунизации животного иммуногеном, как описано выше, клонировали репертуар антител из полученных в результате объединенных B-клеток. Способы получения последовательности ДНК вариабельных областей разнообразных популяций иммуноглобулиновых молекул с использованием смеси олигомерных праймеров и ПЦР. Например, смешанные олигонуклеотидные праймеры, соответствующие 5'-концевым лидерным (сигнальный пептид) последовательностям и/или последовательностям каркаса 1 (FR1), а также праймеры к консервативной 3'-концевой области могут использоваться для ПЦР-амплицификации вариабельных областей легких и тяжелых цепей из некоторого количества мышиных антител (Larrick et al. (1991) Biotechniques 11:152-56). Аналогичную стратегию можно также использовать для амплификации вариабельных областей легких и тяжелых цепей из человеческих антител (Larrick et al.(1991) Methods: Companion to Methods in Enzymology 2:106-10).
Поликлональные сыворотки и антитела могут продуцироваться иммунизацией подходящего субъекта полипептидом по настоящему изобретению. Титр антитела у иммунизированного субъекта может быть подвергнут мониторингу в течение времени стандартными способами, такими как ELISA с использованием иммобилизованного маркерного белка. Если требуется, молекулы антитела, направленные против полипептида по настоящему изобретению, могут быть выделены из субъекта или культуральной среды и далее очищены хорошо известными методами, такими как хроматография на белке А, с получением фракции IgG.
Фрагменты антител к полипептидам по настоящему изобретению могут быть получены путем расщепления антител по способам, хорошо известным в данной области. Например, иммунологически активные Fab- и F(ab')2-фрагменты могут быть получены обработкой антител ферментом, таким как пепсин.
Кроме того, химерные, гуманизированные и одноцепочечные антитела по настоящему изобретению, включающие в себя относящиеся к человеку и не относящиеся к человеку части, могут продуцироваться с использованием стандартных способов рекомбинантной ДНК и/или рекомбинантной комбинаторной библиотеки иммуноглобулинов. Гуманизированные антитела также могут продуцироваться с использованием трансгенных мышей, не способных экспрессировать эндогенные гены иммуноглобулиновых легких и тяжелых цепей, но способных экспрессировать человеческие гены легких и тяжелых цепей. Например, человеческие моноклональные антитела (mAb), направленные против GITRL, могут генерироваться с использованием трансгенных мышей, несущих иммуноглобулиновые гены человека, а не мышиные иммуноглобулиновые гены. Спленоциты данных трансгенных мышей, иммунизированных интересующим антигеном, могут затем использоваться для продукции гибридом, которые секретируют человеческие mAb со специфичными аффинностями в отношении эпитопов из человеческого белка (см., например, Wood et al., WO 91/00906; Kucherlapati et al., WO 91/10741; Lonberg et al. WO 92/03918; Kay et al., WO 92/03917; Lonberg et al. (1994) Nature 368:856-59; Green et al. (1994) Nat. Genet. 7:13-21; Morrison et al. (1994) Proc. Natl. Acad. Sci. USA 81:6851-55; Bruggeman (1993) Year Immunol 7:33-40; Tuaillon et al. (1993) Proc. Natl. Acad. Sci. USA 90:3720-24; Bruggeman et al. (1991) Eur. J. Immunol. 21:1323-26).
Химерные антитела, включающие химерные иммуноглобулиновые цепи, могут продуцироваться посредством способов рекомбинантной ДНК, известных в данной области. Например, ген, кодирующий константную область Fc молекулы мышиного (или относящегося к другому виду) моноклонального антитела, расщепляют ферментами рестрикции с удалением области, кодирующей мышиный Fc, и замещают эквивалентной частью гена, кодирующего константную область человеческого Fc (см. Robinson et al., публикация Международного патента PCT/US86/02269; Akira, et al., заявка на Европейский патент 184187; Taniguchi, заявка на Европейский патент 171496; Morrison et al., заявка на Европейский патент 173494; Neuberger et al., WO 86/01533; Cabilly et al., патент США № 4816567; Cabilly et al., заявка на Европейский патент 125023; Better et al. (1988) Science 240:1041-43; Liu et al. (1987) Proc. Natl. Acad. Sci. USA 84: 3439-43; Liu et al. (1987) J. Immunol. 139:3521-26; Sun et al. (1987) Proc. Natl. Acad. Sci. USA 84: 214-18; Nishimura et al. (1987) Cancer Res. 47:999-1005; Wood et al. (1985) Nature 314: 446-49; и Shaw et al. (1988) J. Natl. Cancer Inst. 80:1553-59).
Антитело или иммуноглобулиновая цепь могут гуманизироваться способами, известными в данной области. Гуманизированные антитела, включающие в себя гуманизированные иммуноглобулиновые цепи, могут генерироваться путем замены последовательностей вариабельной области Fv, которые непосредственно не вовлечены в связывание антигена, эквивалентными последовательностями из человеческой вариабельной области Fv. Общие способы получения гуманизированных антител предоставлены Morrison (1985) Science 229:1202-07; Oi et al. (1986) BioTechniques 4:214; Queen et al., патенты США № 5585089; 5693761; 5693762, содержание которых включено в данное описание в качестве ссылки. Данные способы включают выделение, манипуляцию и экспрессию последовательностей нуклеиновой кислоты, которые кодируют все или части иммуноглобулиновых вариабельных областей Fv, по меньшей мере, из одной из тяжелой или легкой цепи. Источники таких нуклеиновых кислот хорошо известны специалистам в данной области и, например, могут быть получены из гибридомы, продуцирующей антитело против ранее определенной мишени. Рекомбинантная ДНК, кодирующая гуманизированное антитело, или ее фрагмент, может затем клонироваться в подходящий экспрессирующий вектор.
Гуманизированные молекулы антитела, или иммуноглобулины, или молекулы антитела с пересаженными CDR могут продуцироваться путем пересадки CDR или замены CDR, где может пересаживаться одна, две или все CDR иммуноглобулиновой цепи. См., например, патент США 5225539; Jones et al. (1986) Nature 321:552-25; Verhoeyan et al. (1988) Science 239:1534; Beidler et al. (1988) J. Immunol. 141:4053-60; Winter, патент США № 5225539, содержание которых включено в данное описание в качестве ссылки. Winter описывает способ пересадки CDR, который может применяться для получения гуманизированных антител по настоящему изобретению (заявка на патент Великобритании GB 2188638A; Winter, патент США № 5225539), содержание которых включено в данное описание в качестве ссылки. Все CDR конкретного человеческого антитела могут быть заменены, по меньшей мере, частью не относящегося к человеку CDR, или только некоторые из данных CDR могут быть заменены не относящимися к человеку CDR. Необходимо заменять только такое количество CDR, которое необходимо для связывания гуманизированного антитела к определенному ранее антигену.
Моноклональные, химерные или гуманизированные антитела, модифицированные, например, делецией, добавлением или заменой других частей антитела, например константной области, также входят в объем изобретения. Например, антитело может быть модифицировано следующим образом: (i) делецией константной области; (ii) заменой константной области другой константной областью, например, константной областью, предназначенной для увеличения периода полужизни, стабильности или сродства антитела, или константной областью из другого вида или класса антител; или (iii) модификацией одной или нескольких аминокислот в константной области для изменения, например, числа участков гликозилирования, функции эффекторных клеток, связывания с Fc-рецептором (FcR), фиксации комплемента и т.д.
Способы изменения константной области антитела известны в данной области. Антитела с измененной функцией, например, измененным сродством в отношении эффекторного лиганда, такого как FcR на клетке или C1-компонент комплемента, могут продуцироваться путем замены, по меньшей мере, одного аминокислотного остатка в константной области антитела другим остатком (см., например, EP 388151 A1, патент США 5624821 и патент США 5648260, содержание которых включено в данное описание в качестве ссылки). Сходные типы изменений мышиного (или относящегося к другому виду) иммуноглобулина могут применяться для снижения или элиминации данных функций. Такие изменения известны в данной области.
Например, возможно изменить сродство Fc-области антитела (например, IgG, такого как человеческого IgG) в отношении FcR (например, Fc гамма R1) или сродство в отношении связывания C1q путем замены конкретного(ых) остатка(ов) остатком(ами), имеющим(ими) подходящую функциональную группу на его боковой цепи, или путем введения заряженной функциональной группы, такой как глутамат или аспартат, или ароматического неполярного остатка, такого как фенилаланин, тирозин, триптофан или аланин (см., например, патент США 5624821).
Антитела против GITRL по изобретению могут использоваться для выделения, очистки и/или детекции полипептидов GITRL в надосадочной жидкости, клеточном лизате или на клеточной поверхности. Антитела, описанные в данном изобретении, также могут использоваться в диагностике для мониторинга уровня белка GITRL как часть процедуры клинического тестирования или в клинике для направленного действия на терапевтический модулятор в клетке или ткани, содержащей антиген антитела GITRL. Например, терапевтическое средство, такое как малая молекула, или другое терапевтическое средство по изобретению может связываться с антителом GITRL для направленного действия данного терапевтического средства на клетку или ткань, экспрессирующую GITRL. Связывание нейтрализующих или не нейтрализующих антител (предпочтительно моноклональные антитела) с белком GITRL может также использоваться при лечении состояний, включающих в себя нарушенную регуляцию иммунной системы, например аутоиммунных заболеваний. Данные нейтрализующие моноклональные антитела могут быть способными блокировать связывание GITRL с GITR. Настоящее изобретение далее относится к композициям, содержащим антитело, которое специфично взаимодействует с GITRL. Сходным образом, антитела против GITR могут быть полезны при выделении, очистке и/или детекции GITR, диагностического мониторинга уровня GITR, или клинического направленного действия терапевтического модулятора на клетку или ткань, содержащие GITR. Агонистические антитела против GITR (предпочтительно, моноклональные антитела) могут также использоваться для лечения состояний, включающих нарушенную регуляцию иммунной системы, например, злокачественную опухоль или инфекционные заболевания. Данные агонистические антитела могут индуцировать активность GITR. Таким образом, настоящее изобретение далее относится к композициям, содержащим антитело к GITR.
Анализы по скринингу GITRL
Полинуклеотиды и полипептиды по настоящему изобретению могут использоваться в анализах по скринингу с целью идентификации фармакологических средств или соединений-прототипов средств, которые способны модулировать активность GITRL и, таким образом, GITR в клетке или организме и за счет этого являются регуляторами иммунных реакций. Например, образцы, содержащие GITRL (природный или рекомбинантный), могут контактировать с одним из множества тестируемых соединений (или биологических средств или малых органических молекул), и активность GITRL в каждом из обработанных образцов можно сравнивать с активностью GITRL в необработанных образцах или в образцах, контактировавших с различными тестируемыми соединениями. Такие сравнения определяют, приводит ли какое-либо из тестируемых соединений к: 1) существенно сниженному уровню экспрессии или активности GITRL, что таким образом указывает на ингибитор GITRL (например, соединение, которое сохраняет или усиливает иммунную супрессию), или 2) существенному повышенному уровню экспрессии или активности GITRL, что таким образом указывает на активатор GITRL (например, соединение, которое отменяет иммунную супрессию). В одном из вариантов осуществления идентификацию тестируемых соединений, способных модулировать активность GITRL, проводят с использованием высокопроизводительных анализов скрининга, например анализов BIACORE® (Biacore International AB, Упсала, Швеция), BRET (перенос энергии резонанса биолюминесценции) и FRET (флуоресценции с переносом энергии резонанса), а также ELISA и основанных на клетках анализов.
Малые молекулы
Сниженная активность GITR в организме (или у субъекта), страдающем аутоиммунными нарушениями, воспалительными заболеваниями или отторжением трансплантата (или при их риске), или в клетках из такого организма (или субъекта), вовлеченных в такие нарушения, может также достигаться посредством применения малых молекул (обычно органических малых молекул), которые являются антагонистами, т.е. ингибируют активность GITR. Новые малые молекулы-антагонисты могут идентифицироваться способами скрининга, описанными выше, и могут использоваться в способах лечения по настоящему изобретению, описанных ниже. Наоборот, повышенная активность GITR в организме (или у субъекта), страдающем злокачественной опухолью или инфекционным заболеванием (или при его риске), или в клетке из такого организма (или субъекта), вовлеченных в такие нарушения, может также достигаться посредством применения малых молекул (обычно органических малых молекул), которые являются агонистами, т.е. усиливают активность GITR. Новые малые молекулы-антагонисты могут идентифицироваться способами скрининга, описанными выше, и могут использоваться в способах лечения злокачественной опухоли или инфекционного заболевания, описанных ниже.
Способы диагностики, прогноза и мониторинга прогресса аутоиммунных нарушений и злокачественных опухолей
В данной области хорошо известно, что иммунологические механизмы, исследованные на экспериментальных животных, особенно на мышах, могут переноситься и часто переносятся на иммунную систему человека. По существу, хотя описанные здесь примеры демонстрируют роль GITR в иммунной супрессии CD4+CD25+ регуляторными клетками на мышиной модели, описанные способы диагностики, прогнозирования и мониторинга нарушений, связанных с нарушенной регуляцией иммунной системы, например аутоиммунных нарушений, воспалительных заболеваний и отторжения трансплантата, и злокачественных опухолей и инфекционных заболеваний, могут, в частности, использоваться для диагностики, прогноза и мониторинга таких заболеваний у людей. При воплощении описанных способов специалисту в данной области будет очевидно, что человеческие гомологи GITR и GITRL, а также человеческие агонисты и антагонисты GITR могут использоваться в заявленных способах диагностики, прогнозирования и мониторинга таких нарушений у людей.
Настоящее изобретение относится к способам диагностики, прогноза и мониторинга прогресса аутоиммунных нарушений у субъекта (например, в которых прямо или опосредованно участвует повышение уровня GITRL) путем выявления положительной регуляции активности GITR, например путем выявления положительной регуляции GITRL, включая, но без ограничения, применение таких способов на субъектах-людях. Специалисту в данной области будет очевидно, что данные способы могут применяться в отношении воспалительных заболеваний, а также отторжения трансплантата. Данные способы могут проводиться путем использования предварительно упакованных диагностических наборов, содержащих, по меньшей мере, одно соединение из группы, включающей полинуклеотид GITRL или его фрагменты, полипептид GITRL или его фрагменты (включая слитые белки), или антитела против полипептидов GITRL или их производных, или описанных здесь модуляторов полинуклеотидов и/или полипептидов GITRL, которые могут подходящим образом применяться, например, в клинических условиях. Кроме того, специалисту в данной области будет очевидно, что положительная регуляция GITRL может также выявляться непрямыми способами, такими как подсчет числа иммунных клеток.
«Диагностический» или «диагностирование» означает идентификацию наличия или отсутствия патологического состояния. Диагностические способы включают детекцию положительной регуляции GITRL путем определения тестируемого количества продукта гена GITRL (например, мРНК, кДНК или полипептида, включая их фрагменты) в биологическом образце от субъекта (человека или не относящегося к человеку млекопитающего) и сравнения тестируемого количества с нормальным количеством или интервалом (т.е. количеством или интервалом для индивидуума(ов), о котором(ых) известно, что он(и) не страдает(ют) от аутоиммунных заболеваний) генного продукта GITRL. Хотя конкретный способ диагностики может не предоставлять определительный диагноз аутоиммунных нарушений, бывает достаточно того, что способ предоставляет позитивное указание на то, которое способствует постановке диагноза.
Настоящее изобретение также относится к способам прогноза таких аутоиммунных нарушений путем выявления положительной регуляции активности GITR, например, путем выявления положительной регуляции GITRL. «Прогностический» или «прогнозирование» означает предсказание возможного развития и/или тяжести патологического состояния. Способы прогноза включают определение тестируемого количества продукта гена GITRL в биологическом образце от субъекта и сравнение тестируемого количества с прогностическим количеством или интервалом (т.е. количеством или интервалом для индивидуумов с различной тяжестью аутоиммунных нарушений) продукта гена GITRL. Различные количества продукта гена GITRL в тестируемом образце согласуются с конкретными прогнозами аутоиммунных нарушений. Выявление количества продукта гена GITRL на конкретном прогностическом уровне обеспечивает прогноз для субъекта.
Настоящее изобретение также относится к способам мониторинга прогресса или курса таких аутоиммунных нарушений путем выявления позитивной регуляции активности GITR, например, путем выявления положительной регуляции GITRL. Способы мониторинга включают определение тестируемого продукта гена GITRL в биологических образцах, взятых от субъекта в первый или второй раз, и сравнение данных количеств. Изменение количества продукта гена GITRL от первого раза ко второму указывает на изменение в ходе аутоиммунных заболеваний при снижении количества, указывающем на ремиссию аутоиммунных нарушений, и при повышении количества, указывающем на прогрессию аутоиммунных заболеваний. Такие анализы по мониторингу также могут использоваться для оценки эффективности конкретного терапевтического вмешательства в организм пациентов, которых лечат от аутоиммунных заболеваний.
Повышенная экспрессия GITRL в способах, описанных выше, может выявляться в различных биологических образцах, включая жидкости организма (например, цельную кровь, плазму и мочу), клетки (например, цельные клетки, клеточные фракции и клеточные экстракты) и ткани. Биологические образцы включают срезы ткани, такие как биопсии и замороженные срезы, взятые для гистологических целей. Предпочтительные биологические образцы включают кровь, плазму, лимфу, биопсии ткани, мочу, ЦСЖ (цереброспинальную жидкость), синовиальную жидкость и БАЛ (бронхоальвеолярный лаваж). Следует понимать, что анализ биологических образцов не обязательно требует выделения клеток или ткани от субъекта. Например, подходящие средства для мечения, которые связывают продукты гена GITRL (например, антитела, нуклеиновые кислоты), могут вводиться субъекту и визуализироваться (при связывании с мишенью) с использованием стандартной технологии получения изображения (например, CAT, ЯМР (MRI) и PET).
В диагностических и прогностических анализах по настоящему изобретению продукт гена GITRL выявляют и подвергают количественному анализу с получением тестируемого количества. Затем тестируемое количество сравнивают с нормальным количеством или интервалом. Количество, значительно превышающее нормальное количество или интервал, представляет собой положительный признак в диагностике аутоиммунных нарушений. Конкретные способы выявления и количественного анализа генных продуктов GITRL описано ниже.
Нормальные количества или фоновые уровни продуктов гена GITRL могут определяться для любого конкретного типа образца и популяции. В общем, фоновые (нормальные) уровни белка или мРНК GITRL определяют измерением количества белка или мРНК GITRL в типе биологического образца от нормальных (т.е. здоровых) субъектов. Альтернативно, нормальные уровни генного продукта GITRL могут определяться путем измерения данного количества в здоровых клетках или тканях, взятых от того же субъекта, из которого взяты поврежденные (или, возможно, поврежденные) тестируемые клетки или ткани. Количество генного продукта GITRL (нормальное количество или тестируемое количество) может определять или выражаться в количестве относительно числа клеток, количества общего белка, объема. Для определения количества клеток в образце можно измерить конститутивно экспрессируемый генный продукт или другой генный продукт, экспрессируемый на известном уровне в клетках типа, из которого взят биологический образец.
Следует понимать, что способы анализа по настоящему изобретению необязательно требуют измерения абсолютных значений продукта гена GITRL, поскольку относительного количества достаточно для многих применений данных способов. Также следует понимать, что кроме количества или обилия продуктов гена GITRL варианты продуктов гена GITRL, или его аномальные продукты, или их профили экспрессии (например, мутированные транскрипты, укороченные полипептиды) могут быть идентифицированы путем сравнения нормальных генных продуктов и профилей экспрессии.
Диагностические, прогностические анализы и анализы по мониторингу по настоящему изобретению включают выявление и количественный анализ продуктов гена GITRL в биологических образцах. Продукты гена GITRL включают в себя мРНК GITRL и полипептид GITRL, и оба могут измеряться с использованием способов, хорошо известных специалистам в данной области.
Например, мРНК GITRL может непосредственно выявляться и подвергаться количественному анализу с использованием анализов, основанных на гибридизации, например, на нозерн-гибридизации, гибридизации in situ, дот- и слот-блотов и олигонуклеотидных чипов. Основанные на гибридизации анализы относятся к анализах, в которых зонд из нуклеиновой кислоты гибридизуется с нуклеиновой кислотой-мишенью. В некоторых форматах мишень, зонд или оба иммобилизованы. Иммобилизованная нуклеиновая кислота может представлять собой ДНК, РНК или другой олигонуклеотид или полинуклеотид и может включать в себя встречающиеся в природе или не встречающиеся в природе нуклеотиды, нуклеотидные аналоги или остовы. Способы селекции последовательностей зонда нуклеиновой кислоты для применения по настоящему изобретения основаны на последовательности нуклеиновой кислоты GITRL и хорошо известны в данной области.
Альтернативно, мРНК GITRL может амплифицироваться до выявления и количественного анализа. Такие основанные на амплификации анализы хорошо известны в данной области и включают полимеразную цепную реакцию (ПЦР), ПЦР с обратной транскрипцией (ОТ-ПЦР), ПЦР-сорбционный иммуноферментный анализ (ПЦР-ELISA) и лигазную цепную реакцию (ЛЦР). Праймеры и зонды для продукции и выявления амплифицированных продуктов гена GITRL (например, мРНК или кДНК) могут легко конструироваться и продуцироваться без излишнего экспериментирования специалистами в данной области, основываясь на последовательности нуклеиновой кислоты GITRL. Амплицифированные продукты гена GITRL могут анализироваться непосредственно, например, путем гель-электрофореза; путем гибридизации с зондом нуклеиновой кислоты; путем секвенирования; путем выявления флуоресцентного, фосфоресцентного или радиоактивного сигнала или любым из различных хорошо известных способов. Кроме того, специалистам в данной области известны способы увеличения сигнала продуцируемого амплификацией целевых последовательностей нуклеиновой кислоты. Специалисту в данной области будет очевидно, что какой бы способ амплификации ни применяли, могут использоваться разнообразные количественные способы, известные в данной области (например, количественный ПЦР), если требуется количественный анализ продуктов генов GITRL.
Полипептид GITRL (или его фрагменты) может быть выявлен с использованием различных хорошо известных иммунологических анализов с использованием антител против GITRL, описанных выше. Иммунологические анализы относятся к анализам, в которых используются антитела (например, поликлональные, моноклональные, химерные, гуманизированные, scFv и их фрагменты), которые специфично связываются с полипептидом GITRL (или его фрагментом). Такие хорошо известные иммунологические анализы, подходящие для воплощения настоящего изобретения, включают ELISA, радиоиммунный анализ (RIA), иммунопреципитацию, иммунофлуоресценцию, активированную флуоресценцией сортировку клеток (FACS) и вестерн-блоттинг. Полипептид GITRL также может быть выявлен с использованием меченого GITR.
Специалисту в данной области понятно, что указанные выше способы могут применяться в отношении аутоиммунных заболеваний и других нарушений (например, воспалительных заболеваний), включающих, но без ограничения, ревматоидный артрит, остеоартрит, рассеянный склероз, аутоиммунный гастрит, системную красную волчанку, псориаз и другие воспалительные дерматозы, диабет I типа, астму, аллергию, и воспалительные заболевания кишечника, включая болезнь Крона и язвенный колит.
Специалисту в данной области также будет очевидно, что указанные выше способы или их вариации могут быть также использованы для диагностики, прогнозирования и мониторинга прогресса у субъекта различных злокачественных опухолей и инфекционных заболеваний (например, в которых прямо или опосредованно участвует снижение уровня GITRL) путем выявления отрицательной регуляции активности GITR, например путем выявления отрицательной регуляции GITRL, включая, но без ограничения, применение таких способов на субъектах-людях.
Применение GITRL и родственных молекул для лечения
Авторы полагают, что они являются первыми, кто обнаружил, что связывание GITR на эффекторных T-клетках за счет GITRL или других агонистов GITR предоставляет костимуляторный сигнал в отношении эффекторных T-клеток, в которых данный сигнал делает эффекторные Т-клетки менее чувствительными в отношении супрессии CD4+CD25+ регуляторными Т-клетками и повышает способность эффекторных Т-клеток пролиферировать в ответ на анти-CD3 или другие активирующие сигналы. Хотя для раскрытия данного механизма использовали экспериментальных мышей, в данной области хорошо известно, что иммунологические механизмы, исследованные на экспериментальных мышах, могут переноситься и часто переносятся на иммунную систему человека. Как таковые, описанные способы применения GITRL и родственных молекул, т.е. агонистов GITR или антагонистов GITR, для лечения нарушений, связанных с нарушенной регуляцией иммунной системы, например аутоиммунных нарушений, воспалительных заболеваний и отторжения трансплантата, и злокачественных опухолей и инфекционных заболеваний, могут, в частности, использоваться для диагностики, прогноза и мониторинга таких заболеваний у людей. При воплощении описанных способов специалисту в данной области будет очевидно, что человеческие гомологи GITR и GITRL, а также человеческие агонисты и антагонисты GITR могут использоваться в заявленных способах применения GITRL и родственных GITRL молекул, т.е. агонистов или антагонистов GITR, при лечении аутоиммунных нарушений, воспалительных заболеваний и отторжения трансплантата, и злокачественных опухолей и инфекционных заболеваний у людей.
Описанные здесь родственные GITRL молекулы, т.е. агонисты и антагонисты GITR, включающие модуляторы активности полинуклеотида и/или полипептида GITRL, идентифицированные способами, описанными выше, могут применяться in vitro, ex vivo или включенными в фармацевтические композиции и введенными субъектам in vivo для лечения, например, аутоиммунных заболеваний путем введения антагонистов GITR (например, ингибиторных полинуклеотидов GITRL, низкомолекулярных антагонистов, нейтрализующих антител против GITR и/или нейтрализующих антител против GITRL), или, например, злокачественных опухолей введением агониста GITR (например, полинуклеотидов GITRL, полипептидов GITRL или их слитых белков, низкомолекулярных агонистов и/или агонистических антител против GITR). Такие GITRL и/или связанные с ним молекулы (включая модуляторы) включают, но без ограничения, мышиный GITRL и его гомологи (и антитела к таким молекулам), и такие гомологи включают, но без ограничения, человеческий GITRL. Некоторые подходы фармакогеномики, подлежащие рассмотрению при определении того, следует ли вводить GITRL и/или родственные GITRL молекулы, хорошо известны специалистам в данной области и включают в себя ассоциации в пределах всего генома, подход ген-кандидат и профилирование генной экспрессии. Фармацевтическая композиция по изобретению составляется таким образом, чтобы быть совместимой с предназначенным ей путем введения (например, пероральные композиции, в общем, содержащей инертный разбавитель или съедобный носитель). Другие не ограничивающие примеры путем введения включают парентеральное (например, внутривенное), внутрикожное, подкожное, пероральное (например, ингаляция), чрескожное (местное), введение через слизистые и ректальное введение. Фармацевтические композиции, совместимые с каждым из предназначенных путей введения, хорошо известны в данной области.
Агонисты или антагонисты GITR могут применяться в виде фармацевтических композиций в комбинации с фармацевтически приемлемым носителем. Такие композиции могут содержать, в дополнение к агонистам или антагонистам GITR и носителю, различные разбавители, наполнители, соли, буферы, стабилизаторы, солюбилизаторы и другие вещества, хорошо известные в данной области. Термин «фармацевтически приемлемый» означает нетоксичный материал, который не снижает эффективность биологической активности активного(ых) ингредиента(ов). Характеристики носителя зависят от пути введения.
Фармацевтические композиции по изобретению могут также содержать цитокины, лимфокины или другие кроветворные факторы, такие как M-CSF, GM-CSF, IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL- 14, IL-15, G-CSF, фактор стволовых клеток и эритропоэтин. Фармацевтическая композиция может также включать антитела против цитокинов, как более подробно описано ниже. Фармацевтическая композиция может содержать тромболитические или антитромбозные факторы, такие как активатор плазминогена и фактор VIII. Фармацевтическая композиция может далее содержать противовоспалительные средства, как более подробно описано ниже. Такие дополнительные факторы и/или средства могут быть включены в фармацевтическую композицию с целью продукции синергического эффекта с агонистами или антагонистами GITR или для минимизации побочных эффектов, вызванных агонистами или антагонистами GITR. Наоборот, агонисты или антагонисты GITR могут быть включены в препараты конкретного цитокина, лимфокина, другого фактора гемопоэза, тромболитического или антитромбозного фактора, или противовоспалительного средства для минимизации побочных эффектов цитокина, лимфокина, тромболитического или антитромбозного фактора, или противовоспалительного средства.
Фармацевтическая композиция по изобретению может быть в виде липосомы, в которой сочетаются агонисты или антагонисты GITR, в дополнение к другим фармацевтически приемлемым носителям, с амфипатическими средствами, такими как липиды, которые существуют в агрегированной форме в виде мицелл, нерастворимых монослоев, жидких кристаллов или ламеллярных слоев в водном растворе. Подходящие для липосомного препарата липиды включают, без ограничения, моноглицериды, диглицериды, сульфатиды, лизолецитин, фосфолипиды, сапонин, желчные кислоты и т.д. Получение таких липосомных препаратов находится на уровне техники, существующем в данной области, как описано, например, в патенте США № 4235871; патенте США № 4501728; патенте США № 4837028 и патенте США № 4737323, которые включены в данное описание в качестве ссылки.
Используемый здесь термин «терапевтически эффективное количество» означает общее количество каждого активного компонента фармацевтической композиции или способа, которого достаточно для того, чтобы показать значимое преимущество для пациента, например ослабление симптомов, выздоровление или увеличение скорости выздоровления от таких состояний. При применении в отношении индивидуального активного ингредиента, вводимого отдельно, данный термин относится только к данному ингредиенту. При применении в отношении комбинации, данный термин относится к сочетанным количествам активных ингредиентов, которые приводят к терапевтическому действию, вводятся ли они в сочетании, серийно или одновременно.
При воплощении способа лечения или применения по настоящему изобретению терапевтически эффективное количество агониста GITR (например, полинуклеотида GITRL или полипептида GITRL, экспрессируемого с него) или антагониста GITR (например, нейтрализующего антитела против GITRL или нейтрализующего антитела против GITR) вводят субъекту, например млекопитающему (например, человеку). Агонист или антагонист GITR могут вводиться в соответствии со способом по изобретению отдельно или в комбинации с другими вариантами лечения, такими как варианты лечения с использованием цитокинов, лимфокинов или других факторов гемопоэза или противовоспалительных средств. При совместном введении с одним или несколькими средствами агонисты или антагонисты GITR могут вводиться одновременно со вторым средством или последовательно. При последовательном введении лечащий врач принимает решение по подходящей последовательности введения, например, полипептида GITRL (или его слитого белка) или нейтрализующего антитела против GITRL в комбинации с другими средствами.
При пероральном введении терапевтически эффективного количества агониста или антагониста GITR связующее средство находится в виде таблетки, капсулы, порошка, раствора или эликсира. При введении в виде таблетки фармацевтическая композиция по изобретению может, кроме того, содержать твердый носитель, такой как желатин или адъювант. Таблетка, капсула и порошок содержат примерно от 5 до 95% связующего средства, и предпочтительно примерно от 25 до 90% связующего средства. При введении в жидкой форме может добавляться твердый носитель, такой как вода, нефть, масла животного или растительного происхождения, минеральное масло, соевое масло или кунжутное масло или синтетические масла. Жидкая форма фармацевтической композиции может дополнительно содержать физиологический солевой раствор, раствор декстрозы или другого сахара, или гликоли, такие как этиленгликоль, пропиленгликоль или полиэтиленгликоль. При введении в жидкой форме фармацевтическая композиция содержит примерно от 0,5 до 90% по массе связующего средства и предпочтительно примерно от 1 до 50% по массе связующего средства.
Когда терапевтически эффективное количество агониста или антагониста GITR вводят путем внутривенной, кожной или подкожной инъекции, агонист или антагонист GITR находится в виде свободного от пирогенов, приемлемого для парентерального введения водного раствора. Получение таких приемлемых для парентерального введения белковых растворов, имеющих должные характеристики pH, изотоничности, стабильности и тому подобное, известно специалистам в данной области. Предпочтительная фармацевтическая композиция для внутривенной, кожной или подкожной инъекции должны содержать, в дополнение к агонисту или антагонисту GITR, изотонический носитель, такой как хлорид натрия для инъекций, раствор Рингера для инъекций, декстроза для инъекций, декстроза и хлорид натрия для инъекций, лактатный раствор Рингера для инъекций или другой носитель, известный в данной области. Фармацевтическая композиция по настоящему изобретению может также содержать стабилизаторы, консерванты, буферы, антиоксиданты или другие добавки, известные специалистам в данной области.
Количество агониста или антагониста GITR в фармацевтической композиции по настоящему изобретению зависит от природы и тяжести подлежащего лечению состояния и от природы предварительных способов лечения, которые претерпел пациент. Наконец, лечащий врач может принять решение по количеству агониста или антагониста GITR, которым лечить каждого индивидуального пациента. Сначала лечащий врач будет вводить малые дозы агониста или антагониста GITR и будет наблюдать ответ пациента. Большие дозы агониста или антагониста GITR могут вводиться до получения от пациента оптимального терапевтического эффекта, и в данной точке дозировку в основном далее не повышают. Предполагается, что различные фармацевтические композиции, применяемые для воплощения способов настоящего изобретения, должны содержать примерно от 0,1 мкг до 100 мг, например, полипептида GITRL или нейтрализующего антитела против GITRL на кг массы тела.
Длительность внутривенной (в./в.) терапии с использованием фармацевтической композиции по настоящему изобретению изменяется в зависимости от тяжести подлежащего лечению заболевания и состояния и потенциальной реакции идиосинкразии каждого индивидуального пациента. Предполагается, что длительность каждого применения агониста или антагониста GITR может находиться в интервале от 12 до 24 часов непрерывного в./в. введения. Также предполагается подкожная (п./к.) терапия с использованием фармацевтической композиции по настоящему изобретению. Данные варианты лечения могут проводиться ежесуточно, еженедельно или, более предпочтительно, раз в две недели или ежемесячно. Также предполагается, что агонист или антагонист GITR представляет собой малую молекулу, причем данные способы лечения могут вводиться ежесуточно, два раза в сутки, три разa в сутки и т.д. Наконец, лечащий врач принимает решение по подходящей длительности в./в. или п./к. лечения малой молекулой и временному согласованию лечения с использованием фармацевтической композиции по настоящему изобретению.
Полинуклеотиды и белки по настоящему изобретению, как ожидается, характеризуются одним или несколькими вариантами применения биологической активности (включая ассоциированые с указанными здесь способами анализа), идентифицированными выше. Варианты применения активности, описанные для белков по настоящему изобретению, могут предоставляться путем введения таких белков или путем применения полинуклеотидов, кодирующих такие белки (такие как, например, полинуклеотиды в вариантах генной терапии или векторы, подходящие для введения ДНК).
Применение GITRL и других агонистов GITR для усиления иммунного ответа
В одном из аспектов настоящее изобретение относится к способам повышения пролиферации иммунных клеток, например T-клеток (например, эффекторных T-клеток), путем контакта иммунной клетки или популяции иммунных клеток с агонистом GITR, например полинуклеотидом или полипептидом GITRL по изобретению (например, его слитым белком) и/или агонистическим антителом против GITR, который потенцирует или усиливает активность GITR. Данные способы основаны, по меньшей мере, частично, на том открытии, что агонистическое антитело против GITR отменяет опосредованную CD4+CD25+ T-клетками супрессию пролиферации CD4+CD25- T-клетками (пример 5). Способы также основаны, частично, на том открытии, что связывание с GITR, например, GITRL или агонистического антитела против GITR, индуцирует пролиферацию эффекторных Т-клеток (например, CD4+CD25- и CD8+ T-клеток) (пример 9 и пример 13). Авторы также показали, что связывание с GITR GITRL предоставляет костимуляторный сигнал эффекторным T-клеткам (например, CD4+ и CD8+ T-клеток), с повышением, таким образом, способностей эффекторных T-клеток с преодолением супрессии, опосредованной CD4+CD25+ регуляторными Т-клетками, и пролиферацией в ответ на анти-CD3 (примеры 11 и 13); т.е. связывание GITR, экспрессируемого на эффекторных T-клетках, агонистами GITR (например, полипептидом GITRL, его активными фрагментами и/или агонистическим антителом против GITR) делает эффекторные Т-клетки менее чувствительными к супрессии регуляторными CD4+CD25+ Т-клетками. Соответственно агонисты GITR, стимулирующие активность GITR в эффекторных Т-клетках, могут применяться сами по себе или в комбинации с антигеном, например, в качестве адъюванта (например, вакцинного адъюванта) для положительной регуляции иммунного ответа in vivo, например, для применения при лечении злокачественных опухолей или инфекционных нарушений.
В одном из вариантов осуществления агонисты GITR (например, полинуклеотиды, полипептиды GITRL, их активные фрагменты и/или слитые белки, низкомолекулярные агонисты и/или агонистические антитела против GITR) могут использоваться для лечения различных иммунодефицитов и иммунных нарушений (включая тяжелый комбинированный иммунодефицит (SCID)), например, при положительной регуляции роста и пролиферации Т-клеток. Данные иммунодефициты могут быть генетическими или могут быть вызваны вирусными (например, ВИЧ), а также бактериальными или грибковыми инфекциями, или могут быть результатом аутоиммунных нарушений. Более конкретно, инфекционные заболевания, вызванные вирусной, бактериальной, грибковой или другой инфекцией могут подлежать лечению с использованием белка по настоящему изобретению, и они включают инфекции ВИЧ, вирусами гепатита, герпесвирусами, микобактериями, Leishmania spp., малярии и различные грибковые инфекции, такие как кандидоз. Конечно, в данном отношении белок по настоящему изобретению может также использоваться, когда в основном требуется стимуляция иммунной системы, т.е. при лечении злокачественной опухоли.
Положительная регуляция антигенов антигенпредставляющих клеток (APC) (например, положительная регуляция B7.1, B7.2 и B7.3) как средство положительной регуляции иммунных реакций может также применяться при лечении. Положительная регуляция иммунных реакций может происходить в виде усиления существующего иммунного ответа или возникновения исходного иммунного ответа. Например, усиление иммунного ответа посредством стимуляции антигенпредставляющих функций дендритных клеток может использоваться в случаях вирусной инфекции. Кроме того, системные вирусные заболевания, такие как грипп, обычная простуда и энцефалит, могут ослабляться путем системного введения стимуляторных форм антигенпредставляющих молекул, например антигенов дендритных клеток.
Альтернативно, противовирусные иммунные реакции могут усиливаться у инфицированного пациента путем выделения из пациента Т-клеток, совместного стимулирования Т-клеток ex vivo загруженными вирусным антигеном специализированными APC (например, B-клетками, макрофагами и/или дендритными клетками) и агонистами GITR (например, полинуклеотидами, полипептидами GITRL, их активными фрагментами и/или слитыми белками, низкомолекулярными агонистами и/или агонистическими антителами против GITR). Агонисты GITR (например, полинуклеотиды, полипептиды GITRL, их активные фрагменты, и/или слитые белки, и/или агонистические антитела против GITR) могут подаваться в виде растворимого белка или экспрессироваться APC. Другим способом усиления противовирусных иммунных реакций является выделение инфицированных клеток у пациента, трансфекция их нуклеиновой кислотой, кодирующей белок GITRL по настоящему изобретению, описанный здесь, так что данные клетки экспрессируют на своей поверхности все или часть белка, и повторно вводят трансфицированные клетки пациенту. Теперь инфицированные клетки будут способны доставлять костимуляторный сигнал эффекторным T-клеткам in vivo, т.е. экспрессия белка GITRL или его активного фрагмента инфицированными клетками и связывание такого белка GITRL с GITR на инфицированных Т-клетках может делать эффекторные T-клетки менее чувствительными к супрессии CD4+CD25+ регуляторными клетками T-клетками.
В другом применении положительная регуляция или усиление антигенной функции APC может использоваться в индукции противоопухолевого иммунитета. Опухолевые клетки (например, саркома, меланома, лимфома, лейкоз, нейробластома и карцинома), транфицированные нуклеиновой кислотой, кодирующей, по меньшей мере, один пептид по настоящему изобретению, могут вводиться субъекту с целью преодоления опухолево-специфичной устойчивости у субъекта. Если требуется, опухолевая клетка может трансфицироваться таким образом, чтобы она экспрессировала комбинацию пептидов. Например, опухолевые клетки, полученные от пациента, могут трансфицироваться ex vivo экспрессирующим вектором, направляющим экспрессию агониста GITR (например, полипептидов GITR, их активных фрагментов, и/или слитых белков, и/или агонистических антител против GITR), одним или в комбинации с пептидом, имеющим только B7.2-подобную активность, или в связи с пептидом, имеющим B7.1-подобную активность, и т.д. Трансфицированные опухолевые клетки возвращают пациенту, результатом чего является экспрессия данных пептидов на поверхности трансфицированной клетки. Альтернативно, способы генной терапии могут использоваться для направленного действия на опухолевую клетку с транфекцией in vivo.
Присутствие агониста GITR (например, полипептида GITRL, его активных фрагментов и/или слитых белков, низкомолекулярного агониста и/или агонистического антитела против GITR), в комбинации с пептидом, имеющим активность антигена APC (например, B7.1, B7.2 и т.д.) на поверхности опухолевой клетки, предоставляет необходимый костимуляторный сигнал T-клеткам для индукции опосредованного Т-клетками иммунного ответа против трансфицированных опухолевых клеток. Кроме того, опухолевые клетки, которые лишены молекул MHC класса I или MHC класса II или которые не могут повторно экспрессировать достаточное количество молекул MHC класса I или MHC класса II, могут трансфицироваться нуклеиновыми кислотами, кодирующими весь или часть (например, часть, укороченную по цитоплазматическому домену) белок α-цепь MHC класса I и белок β2-микроглобулин или белок α-цепь MHC класса II и белок β-цепь MHC класса II (или соответствующие нуклеиновые кислоты человеческих HLA) для экспрессии за счет этого белков MHC класса I или MHC класса II (или соответствующих молекул HLA) на клеточной поверхности. Экспрессия подходящего МНС класса I или класса II в связи с агонистом GITR (например, полипептидом GITRL, его активными фрагментами и/или слитыми белками, низкомолекулярным агонистом и/или агонистическим антителом против GITR) и/или пептидом, имеющим активность антигена APC (например, B7.1, B7.2, и т.д.), индуцирует опосредованный Т-клетками иммунный ответ против трансфицированных опухолевых клеток. Необязательно ген, кодирующий антисмысловую конструкцию, которая блокирует экспрессию ассоциированного с MHC класса II белка, например, инвариантной цепи, может также совместно трасфицироваться с ДНК, кодирующей агонист GITR (например, полипептид GITRL, его активные фрагменты и/или слитые белки, низкомолекулярный агонист и/или агонистическое антитело против GITR) и/или пептид, имеющий активность антигена APC, для обеспечения представления ассоциированных с опухолью антигенов и индукции опухолеспецифического иммунитета. Таким образом, индукции опосредованного T-клетками иммунного ответа у субъекта-человека может быть достаточно для преодоления опухолеспецифической толерантности у данного субъекта.
В других вариантах осуществления агонисты GITR (например, полипептид GITRL, его активные фрагменты и/или слитые белки, низкомолекулярные агонисты и/или агонистические антитела против GITR) по изобретению могут применяться в качестве вакцинных адъювантов. Адъюванты представляют собой иммуномодулирующие соединения, которые способны усиливать и/или направлять развитие и профиль иммунных реакций против различных антигенов, которые сами по себе слабоиммуногенны. Цитокины и/или лимфокины могут использоваться в качестве адъювантов. Подходящий выбор адъювантов может индуцировать хорошие гуморальные или клеточные иммунные реакции, которые не будут развиваться в отсутствие адъювантов. В частности, адъюванты имеют значимые эффекты по усилению иммунного ответа на субъединичные и пептидные антигены в вакцинах. Их стимуляторная активность также является преимущественной для развития антигенспецифических иммунных реакций направленных против белковых антигенов. Для различных антигенов, которые требуют сильных реакций в слизистых, высоких титров в сыворотке, индукции CTL (цитотоксических T-лимфоцитов) и интенсивных клеточных реакций, комбинации адъюванта и цитокина/лимфокина предоставляют стимулы, которые не предоставляются большинством антигенных препаратов.
Используемая фраза «вакцинный адъювант» или «вакцинная терапия», как предполагается, означает применение агониста GITR (например, полинуклеотида GITRL, полипептида GITRL, его активного фрагмента, его слитого белка и/или агонистического антитела против GITR) в комбинации с антигеном (например, вирусным, паразитарным и бактериальным полипептидами, белками или пептидами) или другими антигенами (например, полипептидами, белками или пептидами опухолевых и раковых клеток) или полинуклеотиды, кодирующие антиген для усиления, подавления или модулирования иммунного ответа на антиген. Для целей данного определения «комбинация» означает применение в связи, одновременно (комбинированно или некомбинированно) или последовательно с антигеном.
Термин «композиция вакцинного адъюванта» относится к вакцинному адъюванту, который, кроме того, включает иммунологически приемлемые разбавители или носители приемлемым образом для получения инъецируемых жидких растворов или суспензий. Композиция вакцинного адъюванта может дополнительно включать средства, которые дополнительно усиливают иммунный ответ, вызванный агонистом GITR. Например, композиция вакцинного адъюванта может, кроме того, включать 3-O-дезацетилированный монофосфориллипид A (MPL®; Corixa Corporation, Сиэтл, Вашингтон) или монофосфориллипид А и его производные и аналоги. MPL может использоваться в интервале 1-100 мкг/доза.
Антигены, используемые для вакцинной терапии, включают белки, пептиды или полипептиды, полученные из иммуногенных и неиммуногенных белков, а также любое из следующего: сахара, белки, полинуклеотиды, или олигонуклеотиды, или другие макромолекулярные компоненты, или их фрагменты. При использовании в данном разделе «пептид» включает ряд, по меньшей мере, из шести аминокислот и содержит, по меньшей мере, одну антигенную детерминанту, в то время как «полипептид» является более длинной молекулой, чем пептид, но не составляет полноразмерный белок. Используемый здесь термин «фрагмент» включает часть, но меньшую, чем вся молекула, сахара, белка, полинуклеотидов или олигонуклеотидов, или других макромолекулярных компонентов.
Используемый здесь термин «эффективное количество адъюванта» означает дозу комбинации описанных адъювантов, которая подходит для достижения повышенного иммунного ответа у хозяина-позвоночного. Конкретная дозировка зависит от возраста, массы и медицинского состояния хозяина, а также от способа введения антигена.
Композиция вакцинного адъюванта по изобретению может вводиться относящемуся к человеку или не относящемуся к человеку позвоночному различными путями, включая, но без ограничения, интраназальный, пероральный, вагинальный, ректальный, парентеральный, внутрикожный, чрескожный (см., например, Международную публикацию WO 98/20734, которая включена в данное описание в качестве ссылки), внутримышечный, внутрибрюшинный, подкожный, внутривенный и внутриартериальный. Количество антигенного компонента или компонентов антигенной композиции изменяется частично в зависимости от природы антигена, а также от возраста, массы и медицинского состояния субъекта, а также от способа введения. Подходящие дозы легко определят специалисты в данной области. Предпочтительно, но не обязательно, чтобы антиген и комбинация адъювантов вводились в одно и то же время. Число доз и режимов дозирования для антигенной композиции также легко определят специалисты в данной области. В некоторых случаях адъювантные свойства комбинации адъювантов могут снизить число необходимых доз или временной ход режима дозирования.
Комбинации адъювантов по настоящему изобретению подходят для применения в комбинации с разнообразными антигенами из различных патогенных микроорганизмов, включая, но без ограничения, антиген из вирусов, бактерий, грибков или паразитарных микроорганизмов, которые инфицируют людей и не относящихся к человеку позвоночных, или из раковой клетки или опухолевой клетки (например, саркомы, меланомы, лимфомы, лейкоза, нейробоастомы и карциномы). Антиген может включать в себя пептиды или полипептиды, полученные из белков, а также фрагменты любого из следующего: сахара, белки, полинуклеотиды или олигонуклеотиды, раковые или опухолевые клетки или другие макромолекулярные компоненты. В некоторых случаях в антигенную композицию входит более одного антигена.
Требуемые вирусные вакцины, содержащие адъювантные комбинации по данному изобретению, включают вакцины, направленные на профилактику и/или лечение заболевания, вызванного, без ограничений, вирусом иммунодефицита человека, вирусом иммунодефицита обезьян, респираторно-синцитиальным вирусом, вирусом парагриппа типов 1-3, вирусом гриппа, вирусом простого герпеса, цитомегаловирусом человека, вирусом гепатита A, вирусом гепатита В, вирусом гепатита С, человеческим папилломавирусом, вирусом полиомиелита, ротавирусом, калицивирусами, вирусом кори, вирусом свинки, вирусом краснухи, аденовирусом, вирусом бешенства, вирусом собачьей чумы, вирусом чумы крупного рогатого скота, коронавирусом, парвовирусом, вирусом инфекционного ринотрахеита, вирусом кошачьего лейкоза, вирусом кошачьего инфекционного перитонита, вирусом инфекционной болезни фабрициевой сумки птиц, вирусом болезни Ньюкасла, вирусом болезни Марека, вирусом респираторного и репродуктивного синдрома свиней, вирусом артериита лошадей и различными вирусами энцефалита.
Требуемые бактериальные вакцины, содержащие адъювантные комбинации по данному изобретению включают вакцины, направленные на профилактику и/или лечение заболевания, вызванного, без ограничений, Haemophilus influenzae (типируемой и нетипируемой), Haemophilus somnus, Moraxella catarrhalis, Streptococcus pneumoniae, Streptococcus pyogenes, Streptococcus agalactiae, Streptococcus faecalis, Helicobacter pylori, Neisseria naefaifagitidis, Neisseria gonorrhoeae, Chlamydia trachomatis, Chlamydia pneumoniae, Chlamydia psittaci, Bordetella pertussis, Salmonella typhi, Salmonella typhimurium, Salmonella choleraesuis, Escherichia coli, Shigella, Vibrio cholerae, Corynebacterium diphtheriae, Mycobacterium tuberculosis, комплексом Mycobacterium avium-Mycobacterium intracellular, Proteus miabilis, Proteus vulgaris, Staphylococcus aureus, Clostridium tetani, Leptospira interrogans, Borrelia burgdorferi, Pasteurella haemolytica, Pasteurella multocida, Actinobacillus pleuropneumoniae и Mycoplasma gallisepticum.
Требуемые грибковые вакцины, содержащие адъювантные комбинации по данному изобретению, включают вакцины, направленные на профилактику и/или лечение заболевания, вызванного, без ограничений, Aspergillis, Blastomyces, Candida, Coccidiodes, Cryptococcus и Histoplasma.
Требуемые вакцины против паразитов, содержащие адъювантные комбинации по данному изобретению, включают вакцины, направленные на профилактику и/или лечение заболевания, вызванного, без ограничений, Leishmaynia major, Ascaris, Trichuris, Giardia, Schistosoma, Cryptosporidium, Trichomonas, Toxoplasma gondii и Pneumocystis carinii.
Требуемые вакцины для индукции терапевтического или профилактического противоопухолевого эффекта в хозяине-позвоночном, которые содержат комбинации адъювантов по данному изобретению, включают вакцины, использующие раковый антиген или опухолеассоциированный антиген, включая, без ограничений, простатоспецифичный антиген (PSA), простатоспецифичный мембранный антиген (PSMA), раковый эмбриональный антиген (CEA), MUC-1, Her2, CA-125, MAGE-3, EGFR, HELP, GCC, CD66-c, простазин, TMPRSS3, TADG 12 и TADG 15.
В случае HIV и SIV антигенные композиции включают в себя, по меньшей мере, один белок, полипептид, пептид или фрагмент, происходящий из указанного вируса. В некоторых случаях в антигенную композицию включены многие белки HIV или SIV, полипептиды, пептиды и/или их фрагменты.
Препараты комбинации адъювантов по данному изобретению также подходят для включения в качестве адъюванта в полинуклеотидные вакцины (также известные как ДНК-вакцины). Такие вакцины могут далее включать способствующие средства, такие как бупивикаин (см. патент США № 5593972, включенный в данное описание в качестве ссылки).
Способы 1) стимуляции функций антигенпредставляющей клетки, например, функций дендритной клетки; 2) выделения Т-клеток из пациента, совместной стимуляции их ex vivo и введения обратно субъекту; 3) трансфекции опухолевых клеток для индукции противоопухолевого иммунитета и 4) применения вакцинных адъювантов, хорошо известны в данной области (см., например, Cerundolo et al. (2004) Dendritic cells: a journey from laboratory to clinic. Nat. Immunol. 5(1):7-10; Ko et al. (2003) Immunotherapy of malignant diseases. Int. Arch. Allergy Immunol. 132:294-309; Valmori et al. (1999) An antigen-targeted approach to adoptive transfer therapy of cancer. Cancer Res. 59:2167- 73).
Применение антагонистов GITR для снижения активности иммунных клеток
Еще в одном аспекте изобретение относится к способу поддержания чувствительности эффекторных Т-клеток, например CD4+ и CD8+ T-клеток или их популяции, к супрессии CD4+ CD25+ регуляторными Т-клетками. Способ может включать контакт популяции T-клеток с антагонистом GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярный антагонист, нейтрализующее антитело против GITR и/или нейтрализующее антитело против GITRL) в количестве, достаточном для ингибирования активности иммунной клетки или популяции. Антагонисты GITR могут также вводиться субъектам, у которых требуется иммунный ответ. Данные условия включают в себя, например, аутоиммунные нарушения (например, артритные нарушения), воспалительные заболевания или трансплантацию органов.
Данные способы основаны, по меньшей мере, частично на открытии, что снижение активности GITR, например, с использованием нейтрализующего антитела против GITRL восстанавливает опосредованную CD4+CD25+ супрессию (пример 13), т.е. нейтрализующее антитело против GITRL поддерживает чувствительность эффекторных Т-клеток, например CD4+ и CD8+ T-клеток, к супрессии CD4+CD25+ регуляторными Т-клетками. Кроме того, авторы продемонстрировали, что инкубация эффекторных Т-клеток нейтрализующим антителом против GITRL ослабляет заболевание при мышином экспериментальном аутоиммунном энцефалите (EAE) (пример 14). В соответствие с этим антагонисты GITR, т.е. молекулы, которые ингибируют активность GITR (например, антитела против GITRL), могут использоваться для поддержания чувствительности эффекторных T-клеток к супрессии CD4+CD25+ T-клетками in vivo, например, для лечения или профилактики ассоциированных с иммунными клетками патологий, включающих отторжение трансплантата, воспалительные заболевания и аутоиммунные нарушения.
Также могут использоваться способы применения антагонистов GITR, ингибирующих активность (например, пролиферацию, дифференцировку, выживание) эффекторной Т-клетки, и, таким образом, могут использоваться для лечения или профилактики различных иммунных нарушений. Не ограничивающие примеры нарушений, которые могут подлежать лечению или профилактике, включают, но без ограничения, отторжение трансплантата, аутоиммунные заболевания (включая, например, сахарный диабет, артрит (включая ревматоидный артрит, ювенильный ревматоидный артрит, остеоартрит, псориатический артрит), рассеянный склероз, энцефаломиелит, бульбоспинальный паралич, системную красную волчанку, аутоиммунный тироидит, дерматит (включая атопический дерматит и экзематозный дерматит), псориаз, синдром Шегрена, болезнь Крона, афтозную язву, воспаление радужки глаза, конъюнктивит, кератоконъюнктивит, язвенный колит, спондиоартропатию, анкилозирующий спондилит, инфекционно-аллергическую астму, аллергическую астму, кожную красную волчанку, склеродермию, вагинит, проктит, медикаментозный дерматит, лепрозные возвратные реакции, лепрозную узловую эритему, аутоиммунный увеит, аллергический энцефаломиелит, острую некротизирующую геморрагическую энцефалопатию, идиопатическую билатеральную нейросенсорную потерю слуха, апластическую анемию, истинную эритроцитарную аплазию, идиопатическую тромбоцитопению, полихондрит, гранулематоз Вегенера, хронический активный гепатит, синдром Стивенса-Джонсона, идиопатические афты, плоский лишай, болезнь Грейвса, саркоидоз, первичный билиарный цирроз, задний увеит и интерстициальный фиброз легкого), болезнь трансплантат против хозяина и аллергию, такую как атопическая аллергия. Предпочтительные нарушения, которые можно лечить с использованием способов, включающих введение антагонистов GITR, например, нейтрализующего GITRL антитела, включают артритные нарушения (например, ревматоидный артрит, ювенильный ревматоидный артрит, остеоартрит, псориатический артрит и анкилозирующий спондилит (предпочтительно, ревматоидный артрит)), рассеянный склероз, диабет I типа, волчанку (СКВ), IBD, болезнь Крона, астму, васкулит, аллергию, склеродерму и псориаз.
В другом варианте осуществления антагонисты GITR, отдельно или в комбинации с другими описываемыми терапевтическими средствами (например, агонистами TNF), могут использоваться для лечения множественной миеломы и связанных В-лимфоцитарных злокачественных опухолей (Brenne, A. et al. (2002) Blood 99 (10):3756-62).
С использованием антагонистов GITR (например, ингибиторных полинуклеотидов GITRL, низкомолекулярных антагонистов и/или нейтрализующих антител к GITR и/или GITRL) возможно модулировать иммунные реакции некоторыми путями. Отрицательная регуляция может происходить в виде ингибирования или блокирования иммунного ответа, уже прогрессирующего, или может участвовать в профилактике индукции иммунного ответа. Функции активированных Т-клеток могут ингибироваться путем усиления супрессии T-клеточных реакций, или путем индукции специфичной толерантности в T-клетках, или по обоим механизмам. Иммуносупрессия T-клеточных реакций, в основном, представляет собой активный, неспецифичный в отношении антигена процесс, который требует продолжительного воздействия на T-клетки супрессирующего средства. Толерантность, в которой участвует отсутствие реактивности или анергия Т-клеток, отличается от иммуносупрессии тем, что она, в основном, является антигенспецифичной и сохраняется после прекращения воздействия вызывающего толерантность агента. Практически толерантность может демонстрироваться отсутствием Т-клеточного ответа после повторного воздействия специфичного антигена в отсутствие вызывающего толерантность средства.
Отрицательная регуляция или предотвращение одной или нескольких функций антигена антигенпредставляющей клетки (например, B7.1) и, таким образом, предотвращение высокого уровня синтеза лимфокинов активированными Т-клетками может использоваться в ситуациях трансплантации ткани, кожи или органа и при болезни трансплантат против хозяина (GVHD). Например, блокада функции Т-клеток должна приводить к снижению деструкции ткани в тканевых трансплантатах. Обычно в трансплантатах ткани отторжение трансплантата инициируется посредством его распознавания как чужеродного T-клетками с последующей иммунной реакцией, которая разрушает трансплантат. Введение антагониста GITR (например, ингибиторных полинуклеотидов GITRL, низкомолекулярного антагониста, нейтрализующего антитела против GITR и/или нейтрализующего антитела против GITRL) в комбинации с молекулой, которая ингибирует или блокирует взаимодействие лимфоцитарного антигена B7 с его природным(и) лигандом(ами) на иммунных клетках (например, с растворимой мономерной формой пептида, имеющего активность B7.2, отдельно или в связи с мономерной формой пептида, имеющего активность другого В-лимфоцитарного антигена (например, B7.1), или с блокирующим антителом) перед трансплантацией, может приводить к связыванию данной молекулы с природным(и) лигандом(ами) на иммунных клетках без передачи соответствующего костимуляторного сигнала. Блокирование функции лимфоцитарного антигена B7 таким способом предотвращает синтез иммунными клетками, например эффекторными Т-клетками, цитокинов и, таким образом, действует в качестве иммунодепрессанта. Более того, отсутствие костимуляции может также быть достаточным для придания Т-клеткам анергии, с индукцией за счет этого у субъекта толерантности. Индукция долгосрочной толерантности реагентами, блокирующими лимфоцитарный антиген B7, может отменить необходимость повторного введения данных блокирующих реагентов. Для достижения у субъекта достаточной иммуносупрессии или толерантности также может быть необходимым блокировать функцию комбинации В-лимфоцитарных антигенов.
Эффективность конкретных блокирующих реагентов при профилактике отторжения органного трансплантата или GVHD может оцениваться с использованием экспериментальных животных, что предсказывает их эффективность на людях. Примеры подходящих систем, которые могут использоваться, включают аллогенные трансплантаты сердца у крыс и ксеногенные трансплантаты островков поджелудочной железы у мышей, причем оба они применяются для оценки иммуносупрессорных эффектов слитых белков CTLA4Ig in vivo, как описано Lenschow et al. (1992) в Science 257:789-92 и Turka et al. (1992) в Proc. Natl. Acad. Sci. USA 89:11102-05. Кроме того, мышиные модели GVHD (см., например, Paul, ed., Fundamental Immunology, Raven Press, New York, 1989, pp.846-47) могут использоваться для определения эффекта блокирования функции B-лимфоцитарных антигенов in vivo на развитие данного заболевания.
Блокирование функции антигена APC может быть также терапевтически полезным при лечении аутоиммунных заболеваний. Многие аутоиммунные нарушения являются результатом несоответствующей активации Т-клеток, которые способны взаимодействовать с собственной тканью и способствуют продукции цитокинов и аутоантител, вовлеченных в патологию заболеваний. Предотвращение активации аутореактивных Т-клеток может снижать или отменять симптомы заболевания. Введение антагонистов GITR (например, ингибиторных полинуклеотидов GITRL, низкомолекулярных антагонистов и/или нейтрализующих антител против GITR и/или GITRL) в комбинации с реагентами, которые блокируют костимуляцию Т-клеток, разрушая взаимодействия лиганд-рецептор антигенов APC, может использоваться для ингибирования активации и предотвращения продукции аутоантител или полученных из Т-клеток цитокинов, которые могут быть вовлечены в процесс заболевания. Кроме того, антагонисты GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярные антагонисты и/или нейтрализующие антитела против GITR и/или GITRL) в комбинации с блокирующими реагентами могут индуцировать антигенспецифическую толерантность аутореактивных T-клеток, что может приводить к долгосрочной ремиссии заболевания. Эффективность данных средств для предотвращения или ослабления аутоиммунных нарушений может определяться с использованием некоторого количества хорошо охарактеризованных моделей аутоиммунных заболеваний человека на экспериментальных животных. Примеры включают мышиный экспериментальный аутоиммунный энцефалит (EAE), системную красную волчанку у мышей MRL/lpr/lpr или у гибридных мышей NZB, мышиный аутоиммунный коллагеновый артрит, сахарный диабет у мышей NOD и крыс BB и мышиный экспериментальный бульбоспинальный паралич (см., например, Paul, ed., Fundamental Immunology, Raven Press, New York, 1989, pp.840-56).
В одном из вариантов осуществления антагонисты GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярные антагонисты и/или нейтрализующие антитела против GITR и/или GITRL), включая их фармацевтические композиции, вводят в комбинированной терапии, т.е. комбинированными с другими средствами, например терапевтическими средствами, которые могут использоваться для лечения патологических состояний и нарушений, таких как иммунные нарушения и воспалительные заболевания. Термин «в комбинации» в данном контексте означает, что данные средства вводят, по существу, в одно время, вместе или последовательно. При последовательном введении в начале введения второго соединения первое из двух соединений предпочтительно еще выявляется в эффективных концентрациях в участке обработки.
Например, комбинированное лечение может включать в себя один или несколько антагонистов GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярные антагонисты и/или нейтрализующие антитела против GITR и/или GITRL), составленный в один препарат и/или введенные с одним или несколькими дополнительными терапевтическими средствами, например одним или несколькими цитокинами и ингибиторами фактора роста, иммунодепрессантами, противовоспалительными средствами, метаболическими ингибиторами, ингибиторами ферментов и/или цитотоксическими или цитостатическими средствами, как более подробно описано ниже. Более того, один или несколько антагонистов GITR (например, ингибиторных полинуклеотидов против GITRL, низкомолекулярных антагонистов и/или нейтрализующих антител против GITR и/или GITRL), описанных здесь, могут использоваться в комбинации с двумя или более описанными терапевтическими средствами. Такие комбинированные варианты лечения могут предпочтительно использовать более низкие дозировки вводимых терапевтических средств, таким образом, избегая предпочтительных вариантов токсичности или осложнений, ассоциированных с различными видами монотерапии. Более того, описанные здесь терапевтические средства действуют на пути, которые отличаются от пути рецептора GITRL, и, таким образом, усиливают и/или обладают синергией в отношении эффектов антагонистов GITR, т.е. в данной случае эффекторные Т-клетки сохраняют свою чувствительность к супрессии CD4+CD25+ регуляторными Т-клетками.
Предпочтительные терапевтические средства, используемые в комбинации с антагонистом GITRL, представляют собой такие средства, которые препятствуют аутоиммунному и последующему воспалительному ответу на разных стадиях. В одном из вариантов осуществления один или несколько из антагонистов GITR (например, ингибиторных полинуклеотидов GITRL, низкомолекулярных антагонистов и/или нейтрализующих антител против GITR и/или GITRL), описанных здесь, может составляться в общий препарат и/или вводиться с одним или несколькими дополнительными средствами, например с другими антагонистами цитокинов или факторов роста (например, растворимыми рецепторами, пептидными ингибиторами, малыми молекулами, слитыми белками с лигандами); или с антителами или их антигенсвязывающими фрагментами, которые связываются с другими мишенями (например, антителами, которые связываются с другими цитокинами или факторами роста, их рецепторами или другими молекулами клеточной поверхности); и противовоспалительными цитокинами или их агонистами. Не ограничивающие примеры средств, которые могут связываться в комбинации с антагонистами GITR (например, ингибиторными полинуклеотидами против GITRL, низкомолекулярными антагонистами и/или нейтрализующими антителами против GITR и/или GITRL), описанными здесь, включают, но без ограничения, антагонисты одного или нескольких интерлейкинов (IL) или их рецепторов, например антагонисты IL-1, IL-2, IL-6, IL-7, IL-8, IL-12, IL-13, IL-15, IL-16, IL-18 и IL-22; антагонисты цитокинов или факторов роста или их рецепторов, таких как фактор некроза опухоли (TNF), LT, EMAP-II, GM-CSF, FGF и PDGF. Антагонисты GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярные антагонисты и/или нейтрализующие антитела против GITR и/или GITRL) могут также сравниваться, например, с антителами-ингибиторами поверхностных молекул, например, CD2, CD3, CD4, CD8, CD25, CD28, CD30, CD40, CD45, CD69, CD80 (B7.1), CD86 (B7.2), CD90 или их лигандов, включая CD154 (gp39 или CD40L), или LFA-1/ICAM-1 и VLA-4/VCAM-1 (Yusuf-Makagiansar et al. (2002) Med. Res. Rev. 22:146-67). Предпочтительные антагонисты, которые могут использоваться в комбинации с антагонистами GITR (например, ингибиторными полинуклеотидами GITRL, низкомолекулярными антагонистами и/или нейтрализующими антителами против GITR и/или GITRL), описанными здесь, включают в себя антагонисты IL-1, IL-12, TNFα, IL-15, IL-17, IL-18 и IL-22.
Примеры таких средств включают в себя антагонисты IL-12, такие как химерные, гуманизированные, человеческие или генерированные in vitro антитела (или их антигенсвязывающие фрагменты), которые связываются с IL-12 (предпочтительно человеческим IL-12), например антитело, описанное в WO 00/56772; ингибиторы рецептора IL-12, например антитела против человеческого рецептора IL-12; и растворимые фрагменты рецептора IL-12, например человеческого рецептора IL-12. Примеры антагонистов IL-15 включают антитела (или их антигенсвязывающие фрагменты) против IL-15 или его рецептора, например химерные, гуманизированные, человеческие или генерированные in vitro антитела против человеческого IL-15 или его рецептора, растворимые фрагменты рецептора IL-15 и IL-15-связывающие белки. Примеры антагонистов IL-18 включают антитела, например химерные, гуманизированные, человеческие или генерированные in vitro антитела против человеческого IL-18 или его рецептора (или их антигенсвязывающие фрагменты), растворимые фрагменты рецептора IL-18 и IL-18-связывающие белки (IL-18BP, Mallet et al. (2001) Circ. Res. 28). Примеры антагонистов IL-1 включают ингибиторы интерлейкин-1-превращающего фермента (ICE), такие как Vx740, антагонисты IL-1, например, IL-1RA (Anikinra, Amgen), sILlRII (Immunex), и антитела против рецептора IL-1 (или их антигенсвязывающие фрагменты).
Примеры антагонистов TNF включают антитела, например химерные, гуманизированные, человеческие или генерированные in vitro антитела (или их антигенсвязывающие фрагменты) против TNF (например, человеческого TNFα), например, D2E7 (человеческое антитело против TNFα, патент США № 6258562), CDP-571/CDP-870/BAY-10-3356 (гуманизированное антитело против TNFα; Celltech/Pharmacia), cA2 (химерное антитело против TNFα; Remicade™, Centocor); фрагменты антитела против TNF (например, CPD870); растворимые фрагменты рецепторов TNF, например, человеческие рецепторы TNF p55 или p75 или их производные, например, 75 кД TNFR-IgG (слитый белок 75 кД рецептор TNF-IgG, Enbrel™; Immunex; см., например, Arthritis & Rheumatism (1994) 37:S295; J. Invest. Med. (1996) 44:235A), p55 кД TNFR-IgG (слитый белок 75 кД рецептор TNF-IgG (Lenercept)); антагонисты ферментов, например, ингибиторы TNFα-превращающего фермента (TACE) (например, производное альфа-сульфонилгидроксамовой кислоты, WO 01/55112, и N-гидроксиформамидный ингибитор TACE GW 3333, -005 или -022); и TNF-bp/s-TNFR (растворимый TNF-связывающий белок; см., например, Arthritis & Rheumatism (1996) 39 (9) (приложение): S284; Amer. J. Physiol.-Heart and Circulatory Physiology (1995) 268:37-42). Предпочтительными антагонистами TNF являются растворимые фрагменты рецепторов TNF, например человеческие рецепторы TNF p55 или p75, например 75 кД TNFR-IgG, и ингибиторы TNFα-превращающего фермента (TACE).
В других вариантах осуществления описываемые антагонисты GITR могут вводиться в комбинации с одним или несколькими из следующих компонентов: антагонистами IL-13, например растворимые рецепторы IL-13 (sIL-13) и/или антитела против IL-13; антагонистами IL-2, например DAB 486-IL-2 и/или DAB 389-IL-2 (слитые белки IL-2; Seragen; см., например Arthritis & Rheumatism (1993) 36:1223), и/или антитела против IL-2R, например анти-Tac (гуманизированное антитело против IL-2R; Protein Design Labs, Cancer Res. (1990) 50 (5):1495-502). Еще одна комбинация включает антагонисты GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярные антагонисты и/или нейтрализующие антитела против GITR и/или GITRL) в комбинации с неистощающими ингибиторами CD4 (IDEC-CE9.1/SB 210396; неистощающее приматизированное антитело против CD4; IDEC/SmithKline). Другие предпочтительные комбинации включают в себя антагонисты костимуляторного пути CD80 (B7.1) или CD86 (B7.2), включая антитела, растворимые рецепторы или антагонистические лиганды; а также гликопротеиновый лиганд p-селектина (PSGL), противовоспалительные цитокины, например IL-4 (DNAX/Schering); IL-10 (SCH 52000; рекомбинантный IL-10 DNAX/Schering); IL-13 и TGF-β, и их агонисты (например, агонистические антитела).
В других вариантах осуществления один или несколько антагонистов GITR могут составляться в препарат и/или совместно вводиться с одним или несколькими противовоспалительными лекарственными средствами, иммунодепрессантами или метаболическими ингибиторами или ингибиторами ферментов. Не ограничивающие примеры лекарственных средств или ингибиторов, которые могут использоваться в комбинации с антагонистами GITR (например, ингибиторными полинуклеотидами GITRL, низкомолекулярными антагонистами и/или нейтрализующими антителами против GITR и/или GITRL), описанными здесь, включают одно или несколько следующих средств: нестероидные противоспалительные лекарственные средства (NSAID), например ибупрофен, тенидап (см., например, Arthritis & Rheumatism (1996) 39 (9) (supplement): S280)), напроксен (см., например, Neuro. Report (1996) 7:1209-13), мелоксикам, пироксикам, диклофенак и индометацин; сульфазалазин (см., например, Arthritis & Rheumatism (1996) 39 (9) (supplement): S281); кортикостероиды, такие как преднизолон; противовоспалительные средства-супрессоры цитокинов (CSAID); ингибиторы биосинтеза нуклеотидов, например ингибиторы биосинтеза пуринов, антагонисты фолиевой кислоты (например, метотрексат (N-[4-[[(2,4-диамино-6-птеридинил)метил]метиламино]бензоил]-L-глутаминовая кислота); и ингибиторы биосинтеза пиримидина, например ингибиторы дигидрооротатдегидрогеназы DHODH (например, лефлуномид (см., например, Arthritis & Rheumatism (1996) Vol.39, № 9 (supplement), S131; Inflammation Research (1996) 45:103-07). Предпочтительные терапевтические средства для применения в комбинации с антагонистами GITR (например, ингибиторными полинуклеотидами GITRL, низкомолекулярными антагонистами и/или нейтрализующими антителами против GITR и/или GITRL) включают NSAID, CSAID, ингибиторы (DHODH) (например, лефлуномид) и ингибиторы фолиевой кислоты (например, метотрексат).
Примеры дополнительных ингибиторов включают одно или несколько следующих средств: кортикостероиды (пероральные, ингалируемые и для местной инъекции); иммунодепрессанты, например циклоспорин, такролимус (FK-506); и ингибиторы mTOR, например сиролимус (рапамицин) или производные рапамицина, например растворимые производные рапамицина (например, сложноэфирные производные рапамицина, например CCI-779 (Elit (2002) Current Opinion Investig. Drugs 3 (8):1249-53; Huang et al. (2002) Current Opinion Investig. Drugs 3 (2):295-304); средства, препятствующие передаче сигнала провоспалительными цитокинами, такими как TNFα или IL-1 (например, IRAK, NIK, IKK, p38 или ингибиторы MAP-киназы); ингибиторы COX2, например целекоксиб, рофекоксиб и их варианты, см., например, Arthritis & Rheumatism (1996) Vol.39, № 9 (supplement), S81); ингибиторы фосфодиэстеразы, например R973401 (ингибитор фосфодиэстеразы типа IV; см., например, Arthritis & Rheumatism (1996) Vol.39, № 9 (supplement), S282)); ингибиторы фосфолипазы, например ингибиторы цитозольной фосфолипазы 2 (cPLA2) (например, аналоги трифторметилкетона (патент США № 6350892)); ингибиторы фактора роста сосудистого эндотелия или рецептора фактора роста, например ингибитор VEGF и/или ингибитор VEGF-R; и ингибиторы ангиогенеза. Предпочтительными терапевтическими средствами для применения в комбинации с антагонистами GITR (например, ингибиторными полинуклеотидами GITRL, низкомолекулярными антагонистами и/или нейтрализующими антителами против GITR и/или GITRL) являются иммуносупрессоры, например циклоспорин, такролимус (FK-506); ингибиторы mTOR, например сиролимус (рапамицин) или производные рапамицина, например растворимые производные рапамицина (например, сложноэфирные производные рапамицина, например CCI-779); ингибиторы COX2, например целекоксиб и его варианты; и ингибиторы фосфолипазы, например ингибиторы цитозольной фосфолипазы 2 (cPLA2), например аналоги трифторметилкетона.
Дополнительные примеры терапевтических средств, которые могут комбинироваться с антагонистом GITRL, включают в себя одно или несколько из следующих средств: 6-меркаптопурины (6-MP); азатиопринсульфалазин; мезалазин; олсалазин; хлорохин/гидроксихлорохин; пенцилламин; ауротиомалат (внутримышечный и пероральный); азатиоприн; кохицин; агонисты бета-2-адренорецепторов (салбутамол, тербуталин, салметерал); ксантины (теофиллин, аминофиллин); кромогликат; недокромил; кетотифен; ипратропиум и окситропиум; микофенолат мофетил; агонисты аденозина; противотромбозные средства; ингибиторы комплемента; и адренергические средства.
Применение описанных здесь агонистов GITR в комбинации с другими терапевтическими средствами для лечения или профилактики конкретных иммунных нарушений более подробно описано ниже.
Не ограничивающие примеры или средства для лечения или профилактики артритных нарушений (например, ревматоидного артрита, воспалительного артрита, ревматоидного артрита, ювенильного ревматоидного артрита, остеоартрита и псориатического артрита), с которыми могут комбинироваться антагонисты GITR, включают одно или несколько из следующих средств: описанные выше антагонисты IL-12, NSAID; CSAID; антагонисты TNF, например, TNFα, описанные выше; описанные выше неистощающие антитела против CD4; описанные выше антагонисты IL-2; противовоспалительные цитокины, например, IL-4, IL-10, IL-13 и TGFα или их агонисты; описанные выше антагонисты IL-1 или рецептора IL-1; описанные выше ингибиторы фосфодиэстеразы; описанные выше ингибиторы COX-2; илопрост (см., например, Arthritis & Rheumatism (1996) Vol.39, № 9 (supplement), S82); метотрексат; талидомид (см., например, Arthritis & Rheumatism (1996) Vol.39, № 9 (supplement), S282) и родственные талидомиду лекарственные средства (например, Celgen); лефлуномид; ингибитор активации плазминогена, например транексамовая кислота; см., например, Arthritis & Rheumatism (1996) Vol.39, № 9 (supplement), S284); ингибитор цитокинов, например T-614; см., например, Arthritis & Rheumatism (1996) Vol.39, № 9 (supplement), S282); простагландин E1 (см., например, Arthritis & Rheumatism (1996) Vol.39, № 9 (supplement), S282); азатиоприн (см., например, Arthritis & Rheumatism (1996) Vol.39, № 9 (supplement), S281); ингибитор интерлейкин-1-превращающего фермента (ICE); ингибитор zap-70 и/или lck (ингибитор тирозиновой киназы zap-70 или lck); описанный выше ингибитор фактора роста эндотелия сосудов или рецептора фактора роста эндотелия сосудов; кортикостероидные противовоспалительные лекарственные средства (например, SB203580); ингибиторы TNF-конвертазы; IL-11 (см., например, Arthritis & Rheumatism (1996) Vol.39, № 9 (supplement), S296); IL-13 (см., например, Arthritis & Rheumatism (1996) Vol.39, № 9 (supplement), S308); ингибиторы IL-17 (см., например, Arthritis & Rheumatism (1996) Vol.39, № 9 (supplement), S120); золото; пеницилламин; хлорохин; гидроксихлорохин; хлорамбуцил; циклофосфамид; циклоспорин; общее облучение лимфоидной ткани; антитимоцитарный глобулин; CD5-токсины; перорально вводимые пептиды и коллаген; динатрия лобензарит; регулирующие цитокины средства (CRA) HP228 и HP466 (Houghten Pharmaceuticals, Inc.); антисмысловые фосфотиоатные олигодезоксинуклеотиды ICAM-1 (ISIS 2302; Isis Pharmaceuticals, Inc.); растворимый рецептор комплемента 1 (TP10; T Cell Sciences, Inc.); преднизон; орготеин; гликозамингликана полисульфат; миноциклин; антитела против IL2R; морские и растительные липиды (жирные кислоты из рыбы и семян растений; см., например, DeLuca et al. (1995) Rheum. Dis. Clin. North Am. 21:759-77); ауранофин; фенилбутазон; меклофенамовая кислота; флуфенамовая кислота; внутривенный иммуноглобулин; зилейтон; микофеноловая кислота (RS-61443); такролимус (FK-506); сиролимус (рапамицин); амиприлоз (терафектин); кладрибин (2-хлордезоксиаденозин); андазарибин. Предпочтительные комбинации включают один или несколько антагонистов GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярные антагонисты и/или нейтрализующие антитела против GITR и/или GITRL) в комбинации с метотрексатом или лефлуномидом, и, в случаях умеренного или тяжелого ревматоидного артрита, циклоспорин.
Предпочтительные примеры ингибиторов для применения в комбинации с антагонистами GITR для лечения артритных нарушений включают антагонисты TNF (например, химерные, гуманизированные, человеческие или генерированные in vitro антитела или их антигенсвязывающие фрагменты, которые связываются с TNF; растворимые фрагменты рецептора TNF, например человеческого рецептора TNF p55 или p75 или их производные, например 75 кД TNFR-IgG (слитый белок 75 кД рецептор TNF-IgG, Enbrel™), слитый белок p55 кД рецептор TNF-IgG; антагонисты ферментов TNF, например, ингибиторы TNFα-превращающего фермента (TACE)); антагонисты IL-12, IL-15, IL-17, IL-18, IL-22; истощающие T-клетки и В-клетки средства (например, антитела против CD4 или против CD22); низкомолекулярные ингибиторы, например метотрексат и лефлуномид; сиролимус (рапамицин) и их аналоги, например CCI-779; ингибиторы cox-2 и cPLA2; NSAID; ингибиторы p38, ингибиторы TPL-2, Mk-2 и NFkb; RAGE или растворимый RAGE; P-селектин или ингибиторы PSGL-1 (например, низкомолекулярные ингибиторы, антитела против него, например антитела против P-селектина); агонисты рецептора эстрогена бета (ERB) или антагонисты ERB-NFkb. Наиболее предпочтительные дополнительные терапевтические средства, которые могут вводиться совместно и/или входить в один препарат с одним или несколькими антагонистами GITR (например, ингибиторными полинуклеотидами GITRL, низкомолекулярными антагонистами и/или нейтрализующими антителами против GITR и/или GITRL), включают одно или несколько следующих средств: растворимый фрагмент рецептора TNF, например человеческого рецептора TNF p55 или p75 или его производного, например 75 кД TNFR-IgG (слитый белок 75 кД рецептор TNF-IgG, Enbrel™); метотрексат, лефлуномид или сиролимус (рапамицин) и его аналоги, например CCI-779.
Не ограничивающие примеры средств для лечения или профилактики рассеянного склероза, с которыми может комбинироваться антагонисты GITR, включают следующее: интерфероны, например интерферон-альфа-1a (например,Avonex™; Biogen) и интерферон-1b (Betaseron™; Chiron/Berlex); сополимер 1 (Cop-1; копаксон; Teva Pharmaceutical Industries, Inc.); гипербарический кислород; внутривенный иммуноглобулин; кладрибин; антагонисты TNF, описанные выше; кортикостероиды; преднизолон; метилпреднизолон; азатиоприн; циклофосфамид; циклоспорин; метотрексат; 4-аминопиридин и тизанидин. Дополнительные антагонисты, которые могут быть использованы в комбинации с антагонистами GITR (например, ингибиторными полинуклеотидами GITRL, низкомолекулярными антагонистами и/или нейтрализующими антителами против GITR и/или GITRL), включают в себя антитела или антагонисты человеческих цитокинов или факторов роста, например TNF, LT, IL-1, IL-2, IL-6, IL-7, IL-8, IL-12, IL-15, IL-16, IL-18, EMAP-11, GM-CSF, FGF и PDGF. Описанные выше антагонисты GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярные антагонисты и/или нейтрализующие антитела против GITR и/или GITRL) могут комбинироваться с антителами против других молекул клеточной поверхности, например CD2, CD3, CD4, CD8, CD25, CD28, CD30, CD40, CD45, CD69, CD80, CD86, CD90 или их лигандов. Антагонисты GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярные антагонисты и/или нейтрализующие антитела против GITR и/или GITRL) могут также комбинироваться с такими средствами, как метотрексат, циклоспорин, FK506, рапамицин, микофенолат мофетил, лефлуномид, NSAID, например ибупрофен, кортикостероиды, например преднизолон, ингибиторы фосфолиэстеразы, агонисты аденозина, противотромбозные средства, ингибиторы комплемента, адренергические средства, средства, которые препятствуют передаче сигнала провоспалительных цитокинов, ингибиторы IL-1b-превращающего фермента (например, Vx740), анти-P7s, PSGL, ингибиторы TACE, ингибиторы передачи сигнала T-клеток, такие как ингибиторы киназ, ингибиторы металлопротеиназ, сульфалазин, азатиоприн, 6-меркаптопурины, ингибиторы ангиотензин-превращающего фермента, описанные выше растворимые рецепторы цитокинов и их производные и противовоспалительные цитокины (например, IL-4, IL-10, IL-13 и TGF).
Предпочтительные примеры терапевтических средств для лечения рассеянного склероза, с которыми могут комбинироваться антагонисты GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярные антагонисты и/или нейтрализующие антитела против GITR и/или GITRL), включают интерферон-b, например IFNb-1a и IFNb-1b; копаксон, кортикостерлиды, ингибиторы IL-1, ингибиторы TNF, антитела против лиганда CD40 и CD80 и антагонисты IL-12.
Не ограничивающие примеры средств для лечения или профилактики воспалительного заболевания кишечника (например, болезнь Крона, язвенный колит), с которыми может комбинироваться антагонист GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярный антагонист и/или нейтрализующее антитело против GITR и/или GITRL), включают следующее: буденозид; эпидермальный фактор роста; кортикостероиды; циклоспорин, сульфасалазин; аминосалицилаты; 6-меркаптопурин; азатиоприн; метронидазол; ингибиторы липоксигеназы; мезаламин; олсалазин; балсалазид; антиоксиданты; ингибиторы тромбоксана; антагонисты рецептора IL-1; моноклональные антитела против IL-1; моноклональные антитела против IL-6; факторы роста; ингибиторы эластазы; соединения пиридинилимидазола; антагонисты TNF, описанные выше; цитокины IL-4, IL-10, IL-13 и/или TGFβ или их агонисты (например, антитела-агонисты); IL-11; конъюгированные с глюкуронидом или декстраном пролекарства преднизолона, дексаметазона или будезонида; антисмысловые фосфотиоатные олигодезоксинуклеотиды ICAM-1 (ISIS 2302; Isis Pharmaceuticals, Inc.); растворимый рецептор комплемента 1 (TP10; T Cell Sciences, Inc.); мезалазин замедленного высвобождения; метотрексат; антагонисты фактора активации тромбоцитов (PAF); ципрофлоксацин и лигнокаин.
В одном из вариантов осуществления антагонист GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярный антагонист и/или нейтрализующее антитело против GITR, и/или нейтрализующее антитело против GITRL) может использоваться в сочетании с одним или несколькими антителами, направленными на другие мишени, задействованные в регуляции иммунных реакций, например, отторжение трансплантата или болезнь трансплантат против хозяина. Не ограничивающие примеры средств для лечения или профилактики иммунных реакций, с которыми может комбинироваться антагонист GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярный антагонист, нейтрализующее антитело против GITR и/или нейтрализующее антитело против GITRL) по изобретению, включают следующее: антитела против других молекул клеточной поверхности, включая, но без ограничения, CD25 (рецептор интерлейкина-2а), CD11a (LFA-1), CD54 (ICAM-1), CD4, CD45, CD28/CTLA4, CD80 (B7.1) и/или CD86 (B7.2). В еще одном осуществлении антагонист GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярный антагонист, нейтрализующее антитело против GITR и/или нейтрализующее антитело против GITRL) применяют в комбинации с одним или несколькими общими иммунодепрессантами, например циклоспорином A или FK506.
В других вариантах осуществления антагонисты GITR (например, ингибиторные полинуклеотиды GITRL, низкомолекулярный антагонист и/или нейтрализующее антитело против GITRL) применяют в качестве вакцинных адъювантов против аутоиммунных заболеваний, воспалительных заболеваний или отторжения трансплантата. Комбинации адъювантов для лечения данных типов нарушений подходят для применения в сочетании с разнообразными антигенами из аутоантигенов-мишеней, например аутоантигенов, участвующих в аутоиммунитете, например, из основного белка миелина, например из белка амилоидного пептида, или антигенов трансплантата, например аллоантигенов. Антиген может включать в себя пептиды или полипептиды, полученные из белков, а также фрагменты следующих соединений: сахара, белки, полинуклеотиды и олигонуклеотиды, аутоантигены, белок амилоидного пептида, антигены трансплантата, аллергены или другие макромолекулярные компоненты. В некоторых случаях в антигенную композицию включают более одного антигена.
Например, требуемые вакцины для регуляции реакций на аллергены в хозяине-позвоночном, которые содержат адъювантные комбинации по изобретению, включают вакцины, содержащие аллерген или его фрагмент. Примеры таких аллергенов описаны в патенте США № 5830877 и публикации международной патентной заявки № WO99/51259, которые включены в данное описание в качестве ссылки во всей своей полноте, и включают пыльцу, яды насекомых, перхоть животных, споры грибов и лекарственные средства (такие как пенициллин). Вакцины препятствуют продукции антител IgE, известной причине аллергических реакций. В другом примере требуемые вакцины для профилактики или лечения заболевания, характеризующегося отложением амилоида у хозяина-позвоночного, которые содержат адъювантные комбинации по изобретению, включают вакцины, содержащие части белка амилоидного пептида (APP). Данное заболевание называются по-разному: болезнь Альцгеймера, амилоидоз или амилоидогенное заболевание. Таким образом, вакцины по изобретению включают адъювантные комбинации по изобретению плюс пептид Aβ, а также фрагменты пептида Aβ и антитела против пептида Aβ или их фрагменты.
Способы 1) отрицательной регуляции функции антигенпредставляющей клетки и 2) комбинированной терапии для управления иммуносупрессией хорошо известны в данной области (см., например, Xiao et al. (2003) Dendritic cell vaccine design: strategies for eliciting peripheral tolerance therapy of autoimmune diseases. BioDrugs 17:103-11; Kuwana (2002) Induction of anergic and regulatory T cells by plasmacytoid dendritic cells and other dendritic cell subsets. Hum. Immunol. 63:1156-63; Lu et al. (2002) Manipulation of dendritic cells for tolerance induction in transplantation and autoimmune disease. Transplantation 73:S19-S22; Rifle et al. (2002) Dendritic cells and second signal blockade: a step toward allograft tolerance. Transplantation 73:S1-S2; Mancini et al. (2004) The management of immunosuppression: the art and the science. Crit. Care. Nurs.Q. 27:61-64).
Другой аспект настоящего изобретения соответственно относится к наборам для проведения комбинированного введения антагонистов GITR (например, ингибиторных полинуклеотидов GITRL, низкомолекулярных антагонистов и/или нейтрализующих антител против GITR и/или GITRL) с другими терапевтическими соединениями. В одном из вариантов осуществления набор включает одно или несколько связующих средств, составленных в фармацевтический носитель, и, по меньшей мере, одно средство, например терапевтическое средство, составленных, как надлежит, в один или несколько фармацевтических препаратов.
Настоящее изобретение иллюстрируется следующими примерами, относящимися к новой мышиной кДНК, обозначенной как кДНК мышиного GITRL, кодирующей новый полипептидный лиганд, обозначенный как мышиный GITRL, а также новыми антителами против GITRL. Специалисту в данной области будет очевидно, что принципиальные подходы, представленные примерами, могут применяться ко всем гомологам мышиного GITRL.
Примеры
Следующие примеры приведены для облегчения понимания изобретения, но никоим образом не предназначены для ограничения его объема и не должны рассматриваться таким образом. Примеры не включают в себя подробного описания общепринятых способов, таких как проточная цитометрия (например, FACS), ПЦР, нозерн-гибридизация и гибридизация in situ, или способов, используемых при конструировании векторов, встраивании генов, кодирующих полипептиды, в такие векторы и плазмиды, введении таких векторов и плазмид в клетки-хозяева и экспрессии полипептидов с таких векторов и плазмиды в клетках-хозяевах. Такие способы хорошо известны обычным специалистам в данной области.
Пример 1
Идентификация последовательностей ДНК GITRL
Пример 1.1. Идентификация кДНК мышиного GITRL и геномных последовательностей
Два подхода предпринимали для идентификации мышиного гомолога GITRL. В одном подходе аминокислотную последовательность человеческого GITRL (из инвентарного № GenBank AX077015) использовали в поиске Tblastn против данных div1; div2; div3; div4; gbdiv_cu; Celera mouse (cm); и draft_ mouse-dna. Геномную последовательность ga_69772862.cm_4 идентифицировали как одну из возможных соответствующих последовательностей, подлежащих исследованию. Пропущенные аминокислотные последовательности идентифицировали в поиске Tblastn с аминокислотной последовательности человеческого GITRL (инвентарный № GenBank AX077015) против незамаскированного объединенного мышиного генома Celera (cm) с использованием значения ожидания (E)=10 (по умолчанию), 100 и 1000. Геномную последовательность ga_x5j8b7w7wj5_041.cm_aa_2 идентифицировали как геномную последовательность, содержащую пропущенные аминокислотные последовательности.
В другом подходе аминокислотную последовательность человеческого GITRL (из инвентарного номера GenBank NM005092) использовали в поиске Tblastn против незамаскированного объединенного мышиного генома Celera (cm) с использованием значения ожидания (E)=10 (по умолчанию) и 1000. Геномную последовательность ga_x5j8b7w7wj5_041.cm_aa_2 идентифицировали как геномную последовательность, содержащую три области с парой с высоким счетом (HSP).
Предполагаемую мышиную последовательность ДНК конструировали на основе трех областей с HSP, полученных в описанном выше поиске Tblastn. Данную последовательность кДНК редактировали на основе сравнения выравнивания трех человеческих экзонных последовательностей с соответствующей человеческой геномной последовательностью от Celera (ga_x2htbl3vud5_66.ch_r25h_1) и трех производных мышиных предполагаемых экзонных последовательностей с соответствующей мышиной геномной последовательностью от Celera (ga_x5j8b7w7wj5_041.cm_aa_2). Данное редактирование учитывало сплайсинговые соединения человеческой последовательности. Отредактированная последовательность кДНК мышиного GITRL содержала открытую рамку считывания длиной 519 н.п. (кодирующая последовательность 522 н.п.), соответствующую белку длиной 173 аминокислот.
Праймеры конструировали на основе предполагаемых экзонов геномной последовательности мышиного GITRL и использовали для выделения соответствующего физического клона кДНК из библиотеки кДНК мышиного тимуса посредством ПЦР. Последовательности прямого (SEQ ID NO:4) и обратного (SEQ ID NO:5) ПЦР-праймера представляли собой:
5'ATGGAGGAAATGCCTTTGAGAG 3' (прямой праймер) и
5'GAATGGTAGATCAGGCATTAAGATG 3' (обратный праймер).
Полученный в результате фрагмент субклонировали и определяли его ДНК-последовательность с использованием стандартных способов. Полученный в результате фрагмент подтверждал существование кДНК мышиного GITRL, содержащей все три предсказанных экзона (см. ниже). Данный фрагмент продлевали, чтобы включить конечный кодирующий фрагмент (две аминокислоты) кДНК путем ПЦР-амплификации данного полученного в результате клона кДНК.
Последовательности прямого (SEQ ID NO:6) и обратного (SEQ ID NO:7) ПЦР-праймеров для данной стадии представляли собой:
5'TTTAAAGTCGACCCACCATGGAGGAAATGCCTTTGAGAG 3' (прямой праймер), и
5'TTTAAAGAATTCTCATTAAGAGATGAATGGTAGATCAGGCAT 3' (обратный праймер).
Прямой ПЦР-праймер содержал участок SalI, последовательность Kozak для инициации трансляции и ATG, кодирующий инициаторный метионин. Обратный праймер содержал участок EcoRI. Участки SalI и EcoRI использовали для направленного субклонирования и проводили определение последовательности конечного клона кДНК.
Полноразмерная последовательность кДНК мышиного GITRL и ее предсказанная аминокислотная последовательность приведены в SEQ ID NO:1 и SEQ ID NO:2 соответственно.
Выравнивание последовательностей кДНК человеческого GITRL (SEQ ID NO:8) и кДНК мышиного GITRL выявляло 69,6% идентичность. Выравнивание аминокислотных последовательностей человеческого GITRL (SEQ ID NO:9) и мышиного GITRL (фиг.1) выявляло 54,1% идентичности и 60,0% сходства. Данная степень аминокислотной идентичности является сходной с той, которая существует, в общем, между мышиными и человеческими гомологами других лигандов TNFR (Oshima et al. (1998) Int. Immunol. 10:517-26).
Сравнение клонированной последовательности кДНК мышиного GITRL (SEQ ID NO:1) с доступными мышиными базами данных выявило единичный нуклеотидный полиморфизм (SNP) в кодирующей области мышиного GITRL (перестановка A/C в нуклеотидном положении 470 или SEQ ID NO:1 в экзоне 3, которая приводит к замене аспарагина на треонин в аминокислотном положении 157 SEQ ID NO:2).
Сравнение последовательности кДНК мышиного GITRL с последовательностью генома мыши от Celera (ga_x5j8b7w7wj5_041.cm_aa_2), описанной выше, выявило, что локус мышиного GITRL содержит три экзона и два интрона (см. таблицу 2 ниже) с размером и положением экзонов, консервативным в человеческом и мышином GITRL. Геномная ДНК-последовательность мышиного GITRL приведена в SEQ ID NO:3.
Сравнение геномной структуры мышиного GITRL (таблица 2) с геномной структурой человеческого GITRL (см. таблицу 3 ниже) показывает, что размер экзонов и положение интронов консервативны в геномных ДНК-последовательностях человеческого и мышиного GITRL. Геномная ДНК-последовательность человеческого GITRL приведена в SEQ ID NO:10.
Пример 1.2. Профиль гидрофобности мышиного GITRL
Профиль гидрофобности мышиного GITRL определяли посредством TopPred (Claros and von Heijne (1994) Comput. Appl. Biosci. 10:685-6). График коэффициента гидрофобности по аминокислотным остатками мышиного GITRL (SEQ ID NO:2) выявил единичную предполагаемую гидрофобную область, расположенную примерно между аминокислотами 25-50, что сходно с человеческим GITRL. Данный гидрофобный сегмент соответствует предсказанной трансмембранной области для трансмембранных белков типа II.
Пример 2
Экспрессия мышиного GITRL в ткани
Олигонуклеотидные зонды, основанные на последовательности кДНК мышиного GITRL (SEQ ID NO:1) применяли для тестирования нескольких образцов мышиной ткани на предмет экспрессии GITRL путем нозерн-гибридизации и ПЦР в реальном времени (например, Heid et al. (1996) Genome Res. 6:986-94; Mullah et al. (1998) Nucleic Acids Res. 26:1026-31; Giulietti et al. (2001) Methods 25:386-401).
Хотя при нозерн-гибридизации выявлялись едва детектируемые транскрипты в сердце, селезенке, легком, лимфатическом узле, почке и печени, при последующей гибридизации in situ выявляли экспрессию GITRL в сердце, селезенке, лимфатическом узле и тимусе. Экспрессия GITRL в данных тканях, в основном, ограничивалась перикардиальными и эндокардиальными клетками сердца, белой пульпой селезенки, корковой, околокорковой и мозговой зонами лимфатических узлов, и корковой и мозговой зонами тимуса.
Экспрессию GITRL в тимусе, селезенке и лимфатическом узле далее подтверждали путем ПЦР-анализа в реальном времени. GITRL экспрессировался на наиболее высоком уровне в селезенке и в стимулированных липополисахаридом (ЛПС) клетках селезенки, которые являются первичными В-лимфоцитами. Исчезающе низкий уровень экспрессии GITRL выявлялся в желудке, головном мозге и почках. ПЦР-анализ в реальном времени также выявлял транскрипты GITRL в печени, активированных CD25- клетках, активированных CD25+ клетках и активированных конканавалином А клетках лимфатического узла, хотя в меньшей степени, чем в селезенке и ЛПС-бластах. Не выявляли экспрессии GITRL в покоящихся CD25- или CD25+ клетках. ПЦР-анализ в реальном времени зрелых и стимулированных ЛПС дендритных клеток (DC) костно-мозгового происхождения также демонстрировал фоновую экспрессию GITRL незрелыми DC, которая повышалась после стимуляции ЛПС в течение 24 часов, но уменьшалась ниже фона через 48 часов стимуляции ЛПС. Экспрессию GITRL также выявляли в различной степени во всех тестированных линиях эндотелиальных клеток (bEND3, C166, EOMA, MSI и SVEC4-10), и, как было продемонстрировано, она оставалась относительно неизменной, когда клеточные линии стимулировали ЛПС. кДНА GITRL, наоборот, не выявляли путем ПЦР в следующих нестимулированных мышиных клеточных линиях установленного происхождения: линия Т-клеток E10, фетальная тимусная линия T2, плазмацитома T10, тимома EL4, пре-В-клеточные линии BAF3 и PREB, В-клеточная гибридома B9, моноцитарная DA1G, моноцитарная MI, FBMD-1 фетального костного мозга, эмбриональная карцинома P19, MDF из печени и линия эмбриональных стволовых клеток E14.
Пример 3
Функциональная экспрессия рекомбинантного мышиного GITRL
Пример 3.1. Связывание GITRL с GITR клеточной поверхности
Для определения того, кодирует ли выделенная в примере 1 мышиная кДНК функциональный лиганд GITR (GITRL), клетки Cos, экспрессирующие мышиный GITRL, слитый с эпитопом FLAG (GITRL-Flag-Cos), или контрольный мышиный рецептор IL-21, слитый с эпитопом FLAG (IL-21R-Flag-Cos), инкубировали в течение различных отрезков времени с клетками 293T, экспрессирующими мышиный GITR (GITR-293T). Взаимодействие клетка-клетка выявляли путем проточной цитометрии с использованием меченного фикоэритрином антитела против Flag (PE-FLAG) и меченного флуоресцеинизотиоцианатом (FITC) антитела против GITR. Даже через 1 мин после совместного центрифугирования клеток GITR-293T и GITRL-Flag-Cos ~90% клеток GITR-293T (выявляемых посредством FITC-флуоресценции) совместно окрашивались анти-FLAG (что выявлено путем флуоресценции PE), указывая на то, что клетки GITR-293T связывались с клетками GITRL-Flag-Cos, и данное взаимодействие сохранялось в течение всех 60 мин эксперимента. Наоборот, клетки GITR-293T, инкубированные с клетками IL-21R-Flag-Cos, значимо не окрашивались анти-FLAG, даже через 60 мин после совместного центрифугирования. Эти данные демонстрируют, что мышиная кДНК, выделенная в примере 1, кодирует функциональный GITRL, способный связывать GITR клеточной поверхности.
Пример 3.2. Связывание GITRL с растворимым GITR
Способность мышиного GITRL связывать GITR подтверждали путем инкубации клеток Cos, экспрессирующих мышиный GITRL (GITRL-Cos), или клеток Cos с mock-транфекцией рекомбинантным GITR, слитым с Fc-частью человеческого IgG (GITR-Fc) или контрольным человеческим IgG (HIgG). Связывание GITR-Fc с GITRL определяли путем проточной цитометрии с использованием антитела осла против человеческого антитела, конъюгированного с FITC (FITC-Ab). Инкубация клеток GITRL-Cos с GITR-Fc приводила к 3,6-кратному увеличению связывания FITC-Ab (28,8%) по сравнению с инкубацией клеток GITRL-Cos с контрольным HIgG (7,9%). Неокрашенные клетки GITRL-Cos, клетки GITRL-Cos, инкубированные со слитым белком CTLA-4:Fc и FITC-Ab, и клетки GITRL-Cos, инкубированные только с FITC-Ab, не характеризовались флуоресценцией. Ни обработанные клетки Cos, ни клетки Cos с mock-транфекцией не характеризовались какой-либо заметной флуоресценцией. Эти данные демонстрируют, что мышиная кДНК, выделенная в примере 1, кодируют функциональный GITRL, способный связывать растворимый GITR.
Пример 4
Связывание мышиного GITRL с GITR приводит к пролиферации клеток CD4 + CD25 +
Эффект связывания GITRL:GITR на клеточную пролиферацию определяли путем стимуляции ~50000 мышиных T-клеток посредством ~50000 облученных истощенных по Т-клеткам спленоцитов и 100 МЕ/мл IL-2 в течение 65-72 ч в отсутствие или присутствии различных концентраций GITR-связывающего белка. Два GITR-связывающих белки использовали в данных анализах: агонистическое антитело против GITR (см., например, McHugh et al. (2002) Immunity 16:311-23; см. также заявку патент США 10/194754) или мышиный GITRL, экспрессированный на поверхности модифицированных клеток YB2/0 (GITRL-YB2/0). Клеточную пролиферацию анализировали посредством введения 1 мкКи 3H-тимидина в течение последних 6-12 ч культивирования и последующего измерения включения 3H-тимидина с использованием сцинтилляционного счетчика.
Как показано на фиг.2A, CD4+CD25- T-клетки не отвечали на любую концентрацию антитела против GITR. Наоборот, антитело против GITR стимулировало пролиферацию CD4+CD25+ T-клеток во всех тестируемых концентрациях. Например, культивирование CD4+CD25+ T-клеток в присутствии наиболее низких тестируемых титров антитела против GITR (~0,02 мкг/мл) приводило к ~3-кратному повышению включения 3H-тимидина (~15000 имп/мин) по сравнению с клетками, культивированными в отсутствие антитела против GITR. Способность антитела против GITR стимулировать пролиферацию CD4+CD25+ T-клеток достигало плато, равного ~45000 имп/мин при концентрации антитела ~0,3 мкг/мл, соответствующей ~9-кратному повышению включения 3H-тимидина по сравнению с клетками, культивируемыми в отсутствие антитела против GITR.
По аналогии с результатами, полученными с антителом против GITR, клетки GITRL-YB2/0 не стимулировали пролиферацию CD4+CD25- T-клеток (фиг.2B). Наоборот, клетки GITRL-YB2/0 заметно стимулировали пролиферацию CD4+CD25+ T-клеток. Например, культивирование CD4+CD25+ T-клеток в присутствии ~10000 клеток GITRL-YB2/0 приводило к ~4-5-кратному возрастанию включения 3H-тимидина по сравнению с клетками, культивируемыми в присутствии равного количества немодифицированных клеток YB2/0. Увеличение количества клеток YB2/0 до ~50000 приводило к ~15-кратному повышению включения 3H-тимидина по сравнению с клетками, культивируемыми в присутствии равного количества немодифицированных клеток YB2/0 (фиг.2B).
Пример 5
Связывание мышиного GITRL с GITR отменяет опосредованную CD4 + CD25 + T-клетками супрессию CD4 + CD25 - T-клеток
Анализ супрессии T-клеток, применяемый в данных примерах, был описан ранее (см., например, Thornton and Shevach (2000) J. Immunol. 164:183-90; McHugh et al. (2002) Immunity 16:311-23; оба источника включены в данное описание в качестве ссылки). В кратком изложении, ~50000 CD4+CD25- отвечающих T-клеток культивировали в присутствии ~50000 облученных, истощенных по Т-клеткам спленоцитов, 0,5 мг/мл антитела против CD3 и различных количеств свежевыделенных супрессорных CD4+CD25+ T-клеток. Затем анализировали способность ~50000 облученных клеток GITRL-YB2/0 или 2 мкг/мл агонистического антитела против GITR отменять супрессию пролиферации CD4+CD25- путем измерения включения 3H-тимидина на сцинтилляционном счетчике.
Как показано на фиг.3A, CD4+CD25+ клетки снижали пролиферацию CD4+CD25- клеток зависимым от дозы образом. И антитело против GITR, и клетки GITRL-YB2/0 были способны полностью отменять супрессию CD4+CD25- в любом тестируемом интервале количества супрессорных CD4+CD25+ клеток. Таким образом, связывание GITRL с его рецептором GITR, как и связывание агонистического антитела против GITR с GITR, блокировало супрессорную функцию CD4+CD25+ клеток. Способность клеток GITRL-YB2/0 отменять супрессию реализовывалась независимым от дозы образом, так что такое малое количество, как ~3000 клеток GITRL-YB2/0, по меньшей мере, частично отменяло или снижало супрессию в данном анализе (фиг.3B). Ни немодифицированные YB2/0, ни клетки YB2/0, экспрессирующие GITR, не имели заметного эффекта на опосредованную CD4+CD25+ T-клетками супрессию (фиг.3A).
В отличие от результатов, полученных на свежевыделенных CD4+CD25+ T-клетках, связывание GITRL:GITR характеризовалось малым эффектом до его отсутствия в отношении супрессии, опосредованной CD4+CD25+ T-клетками, активированными антителом против CD3, истощенными по Т-клеткам спленоцитами, и IL-2 (фиг.4). Ни добавление антитела против GITR, ни добавление ~50000 клеток GITRL-YB2/0 не могло отменять супрессию, опосредованную ~25000 активированными CD4+CD25+ T-клетками. Когда активированные CD4+CD25+ T-клетки добавляли в анализ (например, ~1500-12500 клеток), однако антитело против GITR и клетки GITRL-YB2/0 могли частично снижать, но не полностью отменять супрессию.
Пример 6
Антитело против GITRL восстанавливает супрессию, опосредованную CD4 + CD25 + T-клетками
Пример 6.1. Выделение антител против мышиного GITRL
Антитела, специфичные в отношении мышиного GITRL, продуцировали путем иммунизации крыс крысиными клетками YB2/0, экспрессирующими мышиную кДНК GITRL (GITRL-YB2/0). С использованием способов, известных в данной области, создавали гибридомы антитела и подвергали скринингу против клеток Phoenix, экспрессирующих мышиный GITRL с использованием проточной цитометрии. Идентифицировали два антитела, 5F1 и 10F12, которые специфично связывались с клетками GITRL-Phoenix и не связывались с контрольными клетками Phoenix с имитацией трансфекции. Данные гибридомы антитела были депонированы в American Type Culture Collection (ATCC) 22 июля 2003 г.; учетный номер ATCC для гибридомы 5F1 - PTA-5336 и PTA-5337 для гибридомы 10F12.
Пример 6
Пример 6.2. Антитела против мышиного GITRL блокируют эффекты GITRL на супрессорную активность CD4+CD25+ T-клеток
Как описано выше в примере 5, клетки YB2/0, экспрессирующие GITRL на их клеточной поверхности, способны отменять супрессию пролиферации CD4+CD25- T-клеток, опосредованную свежевыделенными CD4+CD25+ T-клетками. Для определения того, могут ли антитела против GITRL восстанавливать опосредованную CD4+CD25+ T-клетками супрессию, проводили анализ супрессии T-клеток, как описано в примере 5, в присутствии или в отсутствие антитела против GITRL 5F1 или 10F12 или контрольных антител.
Как показано в примере 5, культивирование отвечающих CD4+CD25- T-клеток и свежевыделенных CD4+CD25+ супрессорных T-клеток в присутствии клеток GITRL-YB2/0 приводило к полной отмене супрессии пролиферации CD4+CD25- (фиг.5A и 5B). Добавление 10% образцов гибридомной культуральной надосадочной жидкости, содержащих антитело против GITRL 5F1, в анализ приводило к частичному (фиг.5B) до почти полного (фиг.5A) восстановления опосредованной CD4+CD25+ супрессии. Добавление антитела против GITRL 10F12 дало сходные результаты. Как и ожидалось, присутствие контрольных антител («контрольного Ig») не имело подходящего эффекта на способность клеток GITRL-YB2/0 отменять супрессию (фиг.5B). Эти данные демонстрируют, что антитела против GITRL блокируют способность GITRL выключать супрессорную активность CD4+CD25+ клетки.
Пример 6.3. Антитела против мышиного GITRL супрессируют T-клеточные реакции только в присутствии CD4+CD25+ T-клеток
Культуры клеток лимфоузлов стимулировали в присутствии различных концентраций агонистического антитела против CD3 перед (фиг.6A) или после истощения (фиг.6B) CD4+CD25+ T-клеток и измеряли пролиферацию путем определения включения 3H-тимидина на сцинтилляционном счетчике. Для определения эффектов антитела против GITRL на пролиферацию 10% образцы гибридомной культуральной надосадочной жидкости, содержащие антитело против GITRL 5F1, добавляли к параллельным культурам.
Как показано на фиг.6A, добавление антитела против GITRL подавляло пролиферацию клеток лимфатического узла, содержащих CD4+CD25+ T-клеток, где клетки стимулировали концентрацией от ~0,075 мкг/мл до ~0,75 мкг/мл антитела против CD3. Супрессию не наблюдали в присутствии антитела против GITRL, когда клетки лимфатического узла, содержащие CD4+CD25+ T-клетки, стимулировали 1,0 мкг/мл антитела против CD3. В отличие от результатов, полученных с культурами клеток лимфоузла, содержащими CD4+CD25+ T-клетки, добавление антитела против GITRL, в общем, не оказывает супрессорного действия на клеточные культуры лимфатического узла, которые истощены по CD4+CD25+ T-клеткам (фиг.6B). Все вместе эти данные указывают на то, что антитело против GITRL блокирует взаимодействие между GITR, экспрессированным на CD4+CD25+ T-клетках, и GITRL, экспрессированном на других клетках, и что блокада данного взаимодействия GITR/GITRL усиливает регуляторную функцию CD4+CD25+ T-клеток по восстановлению иммуносупрессии.
Пример 7
Распределение экспрессирующих GITRL клеток в лимфоидных тканях
Агонистическое антитело против GITRL использовали для оценки экспрессии GITRL в мышиных тканях посредством проточной цитометрии. Свежевыделенные субпопуляции CD11c+ дендритных клеток (DC) селезенки, экспрессирующих только CD4, только CD8 или и CD4, и CD8, конститутивно экспрессировали GITRL на низком уровне (фиг.7A). Однако поверхностная экспрессия GITRL была заметно выше среди CD11clow B220+ плазмацитоподобных дендритных клеток (Nakano et al., 2001). На фиг.7B окрашивание mAb против GITRL или контролем изотипа проводили на указанных субпопуляциях свежевыделенных CD11c+ DC селезенки из мышей BALB/c.
Сходным образом, свежевыделенные B220+ B-клетки селезенки конститутивно экспрессировали GITRL, как и перитонеальные B-1 B-клетки (perC CD11b+B220+), хотя на более высоком уровне (фиг.7C, вверху). Было обнаружено, что покоящиеся перитонеальные макрофаги (perC CD11b+B220-) также экспрессировали данный лиганд (фиг.7C, внизу).
Субпопуляции тимоцитов, прошедшие селекцию, не экспрессировали измеряемого количества GITRL (фиг.7D). В отличие от этого, как показано на фиг.7E, экспрессия GITRL выявлялась на всех субпопуляциях CD4-CD8- тимусных предшественников при экспрессии на наиболее высоком уровне в CD44+CD25+ (R2) и CD44-CD25+ (R3) субпопуляциях.
GITRL не выявляли на нестимулированных клетках лимфоузлов (фиг.7F). GITRL также не выявляли на нестимулированных Т-клетках селезенки (данные не показаны). Эти данные демонстрируют экспрессию GITRL профессиональными антигенпредставляющими клетками (DC, B-клетки и макрофаги; фиг.7A-7C) и тимусными CD4-CD8- клетками-предшественниками (фиг.7E), но не Т-клетками, прошедшими селекцию (фиг.7D), или покоящимися Т-клетками на периферии (нестимулированные клетки лимфатического узла и селезенки; фиг.7F, и данные не показаны). Эти данные соответствовали данным, полученным путем нозерн-гибридизации, гибридизации in situ и ПЦР в реальном времени, как описано в примере 2 (выше).
Пример 8
APC отрицательно регулируют GITRL после стимуляции TLR
Эффекты активации B-клеток на экспрессию GITRL оценивали после обработки лигандами Toll-подобного рецептора (TLR) или анти-CD40 и IL-4, или анти-IgM. Стимуляция В-клеток селезенки (B220+ спленоцитов) или перитонеальных B-1 B-клеток (B220+CD11b+ PerC) приводила к быстрой, но преходящей положительной регуляции GITRL, которая была заметна через 4 часа при большинстве видов обработки (фиг.8A). После 48-60 часов стимуляции экспрессия падала до уровня ниже того, что был до стимуляции, и стабилизировалась. Исключением были обработанные polyI:C B-1 B-клетки из полости брюшины, которые не характеризовались указанной отрицательной регуляцией в течение оцениваемых отрезков времени. Как и ожидалось, в ходе эксперимента во всех группах повышался уровень CD86, и это указывало на то, что наблюдаемая отрицательная регуляция GITRL не являлась вторичной по отношению к клеточной гибели (данные не показаны).
Снижение GITRL, экспрессированного B-клетками, после обработки анти-CD40 и IL-4 указывало на то, что экспрессия может модулироваться после предоставления Т-клеточной помощи. Экспрессию GITRL В-клетками среди всех спленоцитов оценивали после культивирования с антителом против CD3. В данных условиях культивирования экспрессия GITRL на B220+ спленоцитах также отрицательно регулировалась через 48 часов (фиг.8B). Так, физиологический уровень T-клеточной активности также приводил к снижению экспрессии GITRL В-клетками селезенки.
CD11c+ DC селезенки обогащали посредством магнитных гранул и оценивали на предмет экспрессии GITRL через 12 и 36 часа культивирования в присутствии ЛПС. DC, культивированные в среде или ЛПС, экспрессировали GITRL в течение исходных 12 часов, с умеренной положительной регуляцией, индуцируемой ЛПС. Однако через 36 часов экспрессия GITRL не выявлялась на обработанных ЛПС DC, а также на тех, которые культивировали только в среде. На фиг.8C показана экспрессия GITRL (верхние панели гистограммы) и CD86 (B7.2) (нижние панели гистограммы) очищенными CD11c+ DC после культивирования с ЛПС (0,5 мкг/мл) и без него в указанные моменты времени. Как показано, экспрессия CD86 (B7.2) положительно регулировалась, как и ожидалось (Banchereau and Steinman, 1998). Снижение экспрессии GITRL DC селезенки, культивируемыми в среде, указывает на то, что «спонтанного» созревания DC, которое происходит во время культивирования in vitro (Vremec and Shortman, 1997), достаточно для отрицательной регуляции экспрессии GITRL. По этой причине показаны только результаты после стимуляции ЛПС, хотя DC подвергали тем же видам обработки, показанным для В-клеток. Сходно с результатами другого опубликованного отчета (Tone et al., 2003), было обнаружено, что полученные из костного мозга DC экспрессируют GITRL конститутивно и что такая экспрессия лишь минимально снижается после обработки различными лигандами TLR (данные не показаны).
И CD4, и CD8 T-клетки экспрессировали измеряемые уровни GITRL после 48-часового культивирования со спленоцитами в отсутствие («+Med.») или в присутствии («+aCD3») растворимого антитела против CD3 (фиг.8D), что подтверждало полученные ранее результаты ПЦР в реальном времени (см. также пример 2).
Пример 9
Блокада взаимодействия GITR/GITRL ингибирует пролиферацию лимфоцитов
Поскольку GITRL конститутивно экспрессировался APC и поскольку взаимодействия GITR/GITRL, как предполагается, отменяют супрессорные функции CD4+CD25+ T-клеток, тестировали способность антитела против GITRL усиливать супрессию, опосредованную эндогенной популяцией CD4+CD25+ T-клеток, присутствующих во вторичных лимфоидных органах. Проводили сравнение пролиферативных реакций общих клеток лимфатического узла (LN), общих спленоцитов (Sp) и LN или Sp с истощением по CD25+ клеткам (Δ25), причем каждую популяцию культивировали с антителом против GITRL (закрашенные кружки) или с контрольным антителом (IgG крысы; незакрашенные кружки). Добавление антитела против GITRL снижало пролиферативный ответ общих клеток лимфоузла (фиг.9A, вверху слева) и, в меньшей степени, общих спленоцитов (фиг.9A, внизу слева). Однако ингибиторный эффект также был очевидным в культурах, истощенных по CD25+ клетками (фиг.9A, вверху справа (LN); внизу справа (Sp)), и это создает возможность того, что взаимодействия GITR/GITRL предоставляют костимуляторные сигналы для CD25- T-клеток.
Для прямого тестирования данной возможности пролиферативные реакции очищенных CD4+CD25- и CD8+ T-клеток оценивали в присутствии APC и клеток YB2/0, экспрессирующих GITRL. Пролиферация CD4+CD25- и CD8+ T-клеток, по существу, усиливалась в присутствии экспрессирующих GITRL клеток (фиг.9B), что было, в частности, очевидно при низких концентрациях анти-CD3 для CD4+CD25- T-клеток.
То, что антитело против GITR опосредует свои эффекты путем действия на CD4+CD25- T-клетки, может показаться противоречивым, поскольку покоящиеся T-клетки экспрессируют лишь небольшие количества GITR. Однако данное противоречие разрешается демонстрацией того, что экспрессия GITR быстро регулируется на повышение после активации T-клеток (фиг.9C), достигая максимальных уровней между 48 и 72 часами после активации. Данные результаты поддерживают возможность того, что взаимодействия GITR/GITRL могут влиять на активацию CD4+CD25- T-клеток независимо от регуляторных CD4+CD25+ T-клеток.
Пример 10
Отмена супрессии требует экспрессии GITR CD25 - T-клетками
Предшествующие исследования указывали на то, что нефункциональное связывание GITR на CD4+CD25+ T-клетках ингибировало их супрессорные способности (McHugh et al, 2002; Shimizu et al., 2002). Однако поскольку активированные Т-клетки также экспрессируют GITR, авторы попытались определить соответствующую(ые) клеточную(ые) мишень(и) вовлечения GITR, приводящего к отмене супрессии. Пролиферацию измеряли в присутствии или в отсутствие mAb против GITR (DTA-1) в совместных культурах с использованием комбинаций CD4+CD25+ и CD4+CD25- T-клеток из GITR +/+ и GITR -/- мышей (фиг.10). Как сообщалось ранее (Shimizu et al., 2002), когда и CD4+CD25+, и CD4+CD25- T-клетки экспрессировали GITR, добавление mAb против GITR в совместные культуры приводило к повышению пролиферативного ответа по сравнению с совместными культурами, получавшими антитело изотипа (фиг.10A, панель a). Когда CD4+CD25-, но не CD4+CD25+ T-клетки экспрессировали GITR в совместных культурах, добавление антитела против GITR приводило к усилению T-клеточной пролиферации сходно с тем, которое наблюдается, когда CD4+CD25+ T-клетки экспрессируют GITR (фиг.10A, панель b). Однако в совместных культурах CD4+CD25- GITR -/- и CD4+CD25+ GITR +/+ T-клеток добавление антитела против GITR не имело эффекта на пролиферацию (фиг.10A, панель c). Как и ожидалось, добавление антитела против GITR к совместным культурам CD4+CD25- GITR -/- и CD4+CD25+ GITR -/- T-клеток также не имело эффекта на пролиферацию Т-клеток (фиг.10A, панель d). Результаты, сходные с теми, которые описаны выше, были получены с препаратом антитела против mGITR, полученного коммерчески (данные не показаны).
Сильный аргумент в поддержку гипотезы о том, что отмена супрессии являлась последствием нефункционального связывания GITR, экспрессированного CD4+CD25+ T-клетками, был получен в предшествующем исследовании, в котором использовали комбинации крысиных отвечающих клеток и мышиных CD4+CD25+ регуляторных T-клеток (Shimizu et al., 2002). mAb против GITR (DTA-1), использованное в данных исследованиях, получали у крысы, и, следовательно, не связывалось с клетками крысы (id.). Эксперименты, аналогичные тем, что описаны выше, проводили с использованием совместных культур крысиных CD4+CD25- отвечающих клеток, мышиных CD4+CD25+ супрессоров и облученных крысиных APC (фиг.10B, панель b). Совместно культивируемые CD4+CD25-отвечающие клетки, мышиные CD4+CD25+ супрессоры и облученные APC крысы включали в качестве контроля (фиг.10B, панель a). Сходно с данными, полученными от мышей GITR -/-, отмены опосредованной CD4+CD25+ супрессии не происходило (фиг.10B, панель b), кроме ситуации, когда GITR на отвечающей CD25- популяции мог перекрестно связываться (фиг.10B, панель a).
Дальнейший анализ системы крыса/мышь выполняли путем оценки разведения CFSE при совместном культивировании крысиных CD4+CD25- и мышиных CD4+CD25+ T-клеток посредством проточной цитометрии. В присутствии антитела контроля изотипа крысиные CD4+CD25- T-клетки лишь частично супрессировались мышиными CD4+CD25+ T-клетками при совместном культивировании в соотношении супрессоров к отвечающим клеткам 1:8 (фиг.10C, левые панели гистограмм). Однако добавление антитела против GITR приводило к дополнительной экспансии мышиных CD4+CD25+ T-клеток и к последующему повышению супрессии крысиных T-клеток (фиг.10C, правая панель гистограмм). Повышенное разведение CFSE мышиных CD4+CD25+ T-клеток после нефункционального связывания GITR может частично ингибироваться добавлением блокирующих антител против CD25, и это указывало на то, что IL-2, выделяемый вначале отвечающими Т-клетками, также требовался для данной экспансии (данные не показаны). Все вместе данные результаты недвусмысленно демонстрируют, что вовлечение GITR на отвечающих CD4+CD25- T-клетках требуется для преодоления супрессии, опосредованной CD4+CD25+ T-клетками.
Пример 11
Экспрессия сигналов GITR требуется для регуляции и преодоления супрессии, опосредованной эндогенными регуляторными Т-клетками
Поскольку CD28 и GITR, как оказывается, предоставляют костимуляторные сигналы во время активации T-клеток, авторы пытались определить, играют ли они различные роли во время первичного ответа. Авторы сравнили способность GITR и CD28 способствовать T-клеточной пролиферации в присутствии или в отсутствие эндогенных CD4+CD25+ T-клеток лимфоузла и в присутствии или в отсутствие экзогенного IL-2 (фиг.11). Те же образцы, которые использовали для исследования пролиферации, одновременно оценивали на предмет разведения CFSE после 72-часового периода культивирования. В отсутствие экзогенного IL-2, СD4+ и CD8+ T-клетки из GITR -/- и CD28 -/- животных не могли пролиферировать (фиг.11A, панель a). Клетки LN из животных GITR +/- характеризовались промежуточным фенотипом по сравнению с животными дикого типа и GITR -/- (фиг.11A, панель a). Ответ T-клеток из мышей дикого типа значительно усиливался после истощения CD25+ T-клеток, что указывало на то, что супрессия в данных условиях культивирования опосредовалась CD25+ T-клетками, присутствующими в нормальном лимфоузле (фиг.11A, ср. панели a и b}. Наиболее важно то, что в отсутствие CD4+CD25+ T-клеток реакции CD4+ и CD8+ Т-клеток лимфоузла из мышей GITR -/- были сравнимыми с таковыми мышей дикого типа по оценкам включения 3H-тимидина (фиг.11A, панель b) и разведения CSFE (фиг.11B, верхний набор панелей). Однако через 72 часа CD4+ и CD8+ T-клетки из животных CD28 -/- не пролиферировали даже в отсутствие CD4+CD25+ T-клеток (фиг.11A, панель b).
Очень различный профиль ответа наблюдали при добавлении экзогенного IL-2 к культурам интактных клеток лимфоузла. Пролиферация CD4+ и CD8+ T-клеток полностью ингибировалась в отсутствие GITR, что анализировали по захвату 3H-тимидина (фиг.11A, панель c) или по недостаточному разведению CFSE (фиг.11B, средний набор панелей). Измеряемую пролиферацию Т-клеток из CD28 -/- животных, наоборот, выявляли посредством включения 3H-тимидина (фиг.11A, панель c), хотя профиль CFSE демонстрировал, что в основном за измеренную пролиферацию были ответственны CD8+ T-клетки (фиг.11B). В присутствии IL-2 (50 ед/мл) лимфатические узлы, истощенные по CD4+CD25+ T-клеткам от всех животных, характеризовались сходными уровнями пролиферации, что оценено по включению 3H-тимидина (фиг.11A, панель d) и разведению CFSE (фиг.11B, нижний набор панелей). Все вместе данные результаты указывают на то, что дефекты активации T-клеток в GITR -/- и CD28 -/- мышах являются различными.
Неспособность общих Т-клеток, присутствующих в лимфоузлах мышей GITR -/-, пролиферировать в присутствии экзогенного IL-2, указывала на то, что у данных животных может затрагиваться экспрессия высокоаффинного рецептора IL-2. Поскольку экспрессия цепи IL-2Ra первично индуцируется его лигандом (Depper et al., 1985; Malek and Ashwell, 1985), способность активированных анти-CD3 CD4+CD25- T-клеток из мышей GITR -/- экспрессировать данную цепь оценивали в присутствии или в отсутствие CD4+CD25+ T-клеток и в присутствии или в отсутствие IL-2 (фиг.11C). В присутствии регуляторных Т-клеток добавление IL-2 к совместным культурам приводило к усиленной экспрессии CD25 посредством GITR +/+, но не GITR -/- CD4+CD25- T-клеток через 24 часов (фиг.11C, нижняя правая панель гистограммы). Однако способность GITR -/- CD4+CD25- T-клеток претерпевать индуцированную IL-2 экспрессию CD25 может легко восстанавливаться путем удаления CD4+CD25+ T-клеток (фиг.11C, нижняя левая панель гистограммы). Таким образом, ослабление реакционной способности в отношении IL-2 в GITR -/- Т-клетках в присутствии CD4+CD25+ T-клеток, по меньшей мере, частично была следствием их неспособности экспрессировать высокоаффинный рецептор IL-2 в присутствии концентраций экзогенного IL-2, достаточных для индукции экспрессии CD25 на клетках дикого типа.
Пример 12
CD28-зависимая стимуляция усиливает экспрессию и реакционную способность GITR
Хотя данные, представленные выше, указывали на то, что вовлечение GITR или CD28 на CD25- T-клетках предоставляло сигнал, который позволял отвечающим Т-клеткам избегать супрессии, природа сигнала, вовлеченного в данный процесс, оставалась неясной. Частичное сохранение пролиферативных реакций CD28 -/-CD8+ T-клеток под воздействием IL-2 в присутствии CD4+CD25+ T-клеток указывало на то, что передача сигнала CD28/B7 может регулировать чувствительность T-клеток к взаимодействиям GITR/GITRL. Более того, в предшествующих исследованиях было продемонстрировано, что взаимодействия CD28-CD80/CD86 могут усиливать экспрессию некоторых представителей семейства TNFR (Gilfillan et al., 1998; Rogers et al., 2001). Таким образом, авторы оценивали данную способность в отношении GITR. Очищенные CD4+CD25- и CD8+ T-клетки из мышей CD28 -/- или мышей дикого типа оставались нестимулированными или активировались низкими концентрациями связанного с планшетом антитела против CD3 в присутствии или в отсутствие связанного с планшетом антитела против CD28 (фиг.12A). Хотя Т-клетки дикого типа, на которые воздействовало только анти-CD3, лишь слегка положительно регулировали GITR, экспрессия GITR CD4+CD25- и CD8+ T-клетками была сильно усилена включением анти-CD28.
Сходным образом, положительная регуляция экспрессии GITR на CD4+CD25- T-клетках заметно ингибировалась добавлением анти-CD80/CD86 (анти-B7.1/7.2) (фиг.12B, левая панель гистограммы). Ингибирование экспрессии GITR в культурах, содержащих анти-CD80/CD86, не была вторичной по отношению к сниженной продукции IL-2, поскольку положительная регуляция GITR CD4+CD25- T-клетками не предотвращалась в культурах, содержащих mAb анти-IL-2/IL-2R (фиг.12B (правая панель гистограммы)).
Усиленная экспрессия GITR, индуцированная происходящими от CD28 костимуляторными сигналами, была параллельна усиленной реакционной способности на передачу сигнала за счет GITR (фиг.12C). Добавление антитела против GITR, по существу, усиливало пролиферацию CD4+ и CD8+ эффекторных T-клеток из мышей дикого типа (фиг.12C, левые панели). Однако, когда к данным культурам добавляли анти-CD80/CD86, присутствие антитела против GITR лишь слабо повышало пролиферацию GITR +/+ CD4+CD25- T-клеток (фиг.12C, GITR +/+, вверху слева; GITR -/-, вверху справа) и GITR+/+CD8+ T-клеток (фиг.12C, GITR+/+, внизу слева; GITR -/-, внизу справа) в пределах тестируемого интервала концентраций анти-CD3. Обработка анти-GITR не влияла на реакции очищенных CD4+CD25- и CD8+ T-клеток из мышей GITR -/- (фиг.12C, правые панели). Эти данные указывают на то, что опосредованные CD28 сигналы, отличные от костимуляции продукции IL-2, усиливают экспрессию GITR и ослабляют опосредованную GITR передачу сигнала.
Пример 13
Связывание GITRL с GITR предоставляет костимуляторный сигнал эффекторным T-клеткам
Четыреста тысяч эффекторных Т-хелперных клеток HT-2 (GITR+/TCR+) культивировали в отсутствие или в присутствии следующих реагентов: 1) покрытые анти-CD3 гранулы в соотношении 1:1 или 1:2, 2) десять тысяч клеток YB/2, которые не модифицированы для экспрессии GITRL (родительские YB2/0) или модифицированы для экспрессии GITRL (YB2/0 muGITRL), и 3) возрастающие концентрации антитела контроля изотипа или четырех различных антител против GITRL (5F1, MGTL-10, MGTL-15 или поликлональное антитело против GITRL). Пролиферацию измеряли по включению 3H-тимидина в течение последних 5 часов 4-часового периода культивирования. На фиг.13A продемонстрировано, что GITRL усиливает пролиферацию T-клеток, стимулированных анти-CD3. Кроме того, опосредованное GITRL усиление Т-клеточной пролиферации может быть блокировано антителом 5F1, но не антителом контроля изотипа (фиг.13B), и не коммерчески доступными MGTL-10, MGTL-15, и не поликлональными антителами против GITRL (фиг.13C). Эти данные предоставляют дальнейшие доказательства того, что GITRL предоставляет костимуляторный сигнал, а также что 5F1 представляет собой нейтрализующее антитело против GITRL.
Пример 14
Блокирование связывания GITR-GITRL антителом против GITRL предоставляет адаптивный перенос индуцированного PLP экспериментального аутоиммунного энцефаломиелита
Самок мышей SJL в возрасте девяти недель иммунизировали 150 мкг пептида PLP (аминокислоты 139-151) [HSLGKWLGHPDKF (SEQ ID NO:12)] в полном адъюванте Фрейнда. Через десять суток после иммунизации клетки селезенки собирали и повторно стимулировали ex vivo в течение 3 суток 10 мкг/мл PLP (аминокислоты 139-151) в отсутствие (без обработки) или в присутствии 10 мкг/мл антитела (антитело против GITRL или контрольное антитело). После повторной стимуляции 5×106 клеток селезенки адаптивно переносили наивным SJL мышам в возрасте 10 недель и затем мышей подвергали мониторингу в течение 52 суток на предмет экспериментального аутоиммунного энцефаломиелита (EAE), измеряемого по шкале от 0 до 5. EAE развивался у 40% мышей, которые получали клетки селезенки, повторно стимулированные в отсутствие каких-либо антител (без обработки), и у 80% мышей, которые получали клетки селезенки, повторно стимулированные в присутствии контрольного антитела (CK01) (фиг.14). Кроме того, не было существенных различий по шкале заболевания между животными, которые получали клетки селезенки без обработки, и животными, которые получали клетки селезенки, обработанные контрольным антителом. Значимо то, что ни у одной мыши, которые получали клетки селезенки, которые повторно стимулировали в присутствии антитела против GITRL (5F1), не развивался EAE (p=0,0023 по отношению к обработке контрольным антителом) (фиг.14). Эти данные указывают на то, что блокада пути GITR/GITRL ограничивает способность CD25- T-клеток преодолевать супрессию с отрицательным модулированием их способности осуществлять аутоиммунные реакции.
Пример 15
Экспериментальные методики
Пример 15.1. Антитела и реагенты
Все антитела, использованные для проточной цитометрии или функциональных исследований, были от BD-Pharmingen, кроме: меченного тремя цветами aCD4 (клон CT-CD4) и aB220 (клон RA3-6B2), которые приобретали у Caltag (Burlingame, Калифорния), и MGTL-10, MGTL-15 и поликлональные антитела против GITRL, которые приобретали у Alexis Biochemicals (Сан-Диего, Калифорния). Очищенные F(ab')2-фрагменты антител козы против μ-цепи IgM приобретали у Jackson Immunoresearch (West Grove, Пенсильвания). Анти-IL-2 (клон S4B6) использовали в виде асцитной жидкости. Человеческий рекомбинантный IL-2 получали из National Cancer Institute (Frederick, Мэриленд). IL-4, IFN-γ, IL-12 и колонки для обогащения T-клеток приобретали у R&D Systems (Миннеаполис, Миннесота). Poly-I:C и ЛПС приобретали от Sigma. CpG приобретали от InvivoGen (Сан-Диего, Калифорния). Анти-GITRL (клон 5F1; также клон 10F12) и анти-GITR (клон DTA-1) и пептид PLP продуцировали в собственной лаборатории. Магнитные гранулы против B220, CD11c, CD11b, CD8, CD4 и PE приобретали у Miltenyi (Auburn, Калифорния).
Пример 15.2. Мыши
Мышей BALB/c и C57B1/6 (самки в возрасте 6-8 недель) приобретали от вивария NCI Frederick (Frederick, Мэриленд). Мыши CD28 -/- предоставлены д-ром Alfred Singer (NIH/NCI). GITR +/- эмбрионы (Sv129×B6) предоставлены C. Ricarrdi (Perugia University Medical School, Италия) (Ronchetti et al., 2002). Происходящих от них GITR +/- мышей подвергали обратному скрещиванию с мышами C57BL/6 и полученное потомство подвергали скринингу на мутантную аллель посредством ПЦР. Идентифицированное потомство GITR +/- затем скрещивали путем инбридинга с получением GITR -/- мышей. Всех мышей разводили и содержали на фабриках NIH/NIAID в SPF (лишенных специфичных патогенных микроорганизмов) условиях.
Пример 15.3. Клонирование и экспрессия кДНК
Аминокислотную последовательность человеческого GITRL (инвентарный № GenBank NM_005092) использовали для поиска в базе данных Celera на предмет mGITRL. Геномная последовательность ga_x5j8b7w7wj5_041.cm_aa_2 содержала три области пар с высоким счетом (HSP). Основываясь на допущении, что данные области соответствуют экзонам мышиного GITRL, конструировали для ПЦР-амплификации. С прямым праймером (5'-ATGGAGGAAATGCCTTTGAGAG-3') (SEQ ID NO:4) и обратным праймером (5'-GAATGGTAGATCAGGCATTAAGATG-3') (SEQ ID NO:5) амплифицировали клон кДНК из тимусной библиотеки мыши. Полученный в результате фрагмент субклонировали и определяли последовательность ДНК. Последующий полноразмерный клон амплифицировали посредством ПЦР из предыдущей конструкции с использованием праймеров 5'-TTTAAAGTCGACCCACCATGGAGGAAATGCCTTTGAGAG-3' (прямой) (SEQ ID NO:6) и 5'-TTTAAAGAATTCTCATTAAGAGATGAATGGTAGATCAGGCAT-3' (обратный) (SEQ ID NO:7). Полученный в результате фрагмент ПЦР субклонировали в ретровирусный вектор GFP-RV (Ouyang et al., 1998) и проводили определение последовательности конечного клона кДНК. Затем данный вектор трансфицировали в клеточную линию Phoenix. Образцы надосадочной жидкости из трансфицированных клеток Phoenix использовали для трансдукции клеточной линии YB2/0. Экспрессирующие GFP клетки YB2/0 затем сортировали на FACS и сохраняли в культуре. Предсказанная аминокислотная последовательность mGITRL была идентична таковой, полученной в другой группе (Kim et al., 2003), за исключением замены аланина валином в аминокислотном положении 48 их последовательности.
Пример 15.4. Продукция и очистка моноклональных антител
Крыс Lewis иммунизировали один раз s.q. 100×106 клеток YB2/0-GITRL в CFA. Через две недели данных крыс иммунизировали s.q. 100×106 клеток YB2/0-GITRL в IFA. Через две недели крыс стимулировали 50×106 клеток YB2/0-GITRL в PBS. Через четверо суток получали селезенку и проводили слияние клеток, как описано ранее (Coligan et al., 2003). Образцы надосадочной жидкости из полученных в результате гибридом подвергали скринингу путем проточной цитометрии с использованием клеток Phoenix-GITRL и Phoenix. Антитела очищали из образцов клеточной культуральной надосадочной жидкости с использованием загруженных белком G колонок и элюированные антитела диализовали против PBS.
Пример 15.5. Очистка клеток
T-клетки очищали из периферических лимфоузлов мышей. CD25+ T-клетки метили магнитными гранулами и очищали на autoMACS (Miltenyi Biotech, Auburn, Калифорния) по протоколу производителя. Чистота CD25+ клеток обычно составляла от 97 до 99 процентов. Клетки, остававшиеся в негативной фракции, впоследствии метили микрогранулами против CD4 или против CD8 и очищали с использованием метода позитивной селекции на autoMACS. Чистота рутинно составляла 90-95 процентов. Истощенные по T-клеткам суспензии селезенки получали из суспензий с лизированными эритроцитами путем истощения Thy1.2+ клеток с использованием autoMACS. Клетки B220+ очищали из спленоцитов сходным образом с использованием микрогранул против B220 (Miltenyi Biotech, Auburn, Калифорния), и чистота полученных в результате препаратов составляла более 90%. Перитонеальные клетки получали путем промывки полости брюшины (PerC) 10 мл холодного HBSS. Для получения DC селезенки суспензии клеток селезенки подготавливали способом, аналогичным описанному (Vremec et al., 2000). Затем DC селезенки очищали от суспензий с использованием микрогранул против CD11c (Miltenyi Biotech, Auburn, Калифорния). Полученные в результате суспензии DC были обычно 85-90-процентной чистоты. CD4+CD25- клетки крысы получали истощением CD25+ клеток селезенки крысы с использованием PE-антител против крысиного CD25 (OX-39) с последующим применением микрогранул против PE. После истощения CD4+ клетки затем отбирали из истощенной фракции с использованием микрогранул против крысиного CD4.
Пример 15.6. Анализы пролиферации in vitro
Анализы супрессии проводили, как описано (Thornton and Shevach, 1998). В кратком изложении (5×104) клетки совместно культивировали в дополненной FBS RPMI 1640 (Atlanta Biologicals, Atlanta, Джорджия) с облученными (3000 Р), истощенными по T-клеткам клетками селезенки (5×104) в присутствии 0,5 мкг/мл mAb против CD3 (2C11) в 96-луночном плоскодонном планшете. К некоторым культурам добавляли антитела, специфичные в отношении GITR, или крысиные Ig против изотипа добавляли до конечной концентрации 2 мкг/мл. Титрованные количества CD4+CD25+ клеток добавляли в конечном отношении супрессора к отвечающей клетке, равных 0:1, 1:2, 1:4 или 1:8. Культуры подвергали действию 1 мкКи 3H-тимидина в течение конечных 5-8 часов 72-часового культивирования и проводили эксперимент в трех параллелях, кроме указанных иначе случаев. Совместные культуры крысиных и мышиных субпопуляций T-клеток были установлены сходным образом, за исключением того, что облученные (3000 Р), истощенные по CD4 клетки селезенки крысы использовали в качестве APC, и крысиные и мышиные Т-клетки стимулировали с использованием коктейля mAb против крысиного CD3 (0,25 мкг/мл) и против мышиного CD3 (0,25 мкг/мл).
Пример 15.7. Культивирование in vitro клеток лимфоузла и мечение CFSE
Истощенные по CD25+ суспензии клеток лимфоузла получали на autoMACS, как описано выше. Клетки метили CFSE в концентрации 2 мкМ в течение 8 минут на водяной бане с температурой 37°C. Клетки затем промывали в полной RPMI 1640. Клетки (5×104/лунка) затем культивировали в 96-луночных планшетах в присутствии или в отсутствие rhIL-2 (50 ед./мл). Параллельные лунки или обрабатывали 1 мкКи 3H-тимидина в течение конечных 5-8 часов 72-часового культивирования или использовали для анализа разведения CFSE.
Пример 16
Обсуждение
Предположение о роли GITR в функционировании CD4+CD25+ T-клеток возникало из демонстрации того, что поликлональные и моноклональные антитела против GITR отменяли супрессорные эффекты CD4+CD25+ T-клеток при добавлении к совместным культурам CD25+ и CD25- T-клеток (McHugh et al., 2002; Shimizu et al., 2002). CD4+CD25+ Т-клетки оказались наиболее вероятной мишенью реагентов против GITR, поскольку CD25+ T-клетки в свежей культуре экспрессировали GITR на более высоком уровне, чем покоящиеся CD25- T-клетки, и поскольку антитела против GITR вместе с IL-2 препятствовали пролиферации CD25+, но не CD25- T-клеток в отсутствие сигнала TCR. Более того, когда Shimizu et al добавляли крысиное mAb против mGITR, которое не взаимодействовало с клетками крысы, к совместным культурам мышиных CD25+ супрессоров и отвечающих крысиных Т-клеток, наблюдалась отмена супрессии. Данные исследования привели к гипотезе, что вовлечение GITR посредством агонистических антител против GITR и, предположительно, его физиологического лиганда генерировало сигнал, который ингибировал супрессорную активность CD4+CD25+ T-клеток и отменял неспособность к ответу CD25+ T-клеток на экзогенный IL-2.
Планировали эксперименты, в которых пытались расширить и подтвердить данные исследования путем клонирования мышиного GITRL, анализа его распределения в ткани и окончательно определения мишени агонистических антител против GITR с использованием смесей CD25+ и CD25 T-клеток из мышей дикого типа и мышей GITR -/-. Все вместе данные исследования демонстрируют, что антитела против GITR и GITRL препятствуют супрессорным эффектам CD4+CD25+ T-клеток путем предоставления CD25- T-клеткам уникального сигнала, повышающего их барьер для подавления CD4+CD25+ T-клетками. Данные исследования показывают, что GITRL избирательно экспрессируется на клеточной поверхности APC при наиболее высоком уровне экспрессии на B-1 B-лимфоцитах; при промежуточных уровнях на обычных B-2 B-лимфоцитах, макрофагах и B220+ DC; и при низких уровнях на субпопуляциях B220- DC. GITRL уникален среди представителей надсемейства TNF в том, что он экспрессируется на покоящихся APC, и его экспрессия отрицательно регулируется путем включения B-клеточного рецептора CD40 или различных Toll-подобных рецепторов. Другие представители надсемейства TNF (4-1BB-L, OX40-L, LIGHT, CD70, CD30-L) не выявляются на покоящихся APC, и их экспрессия положительно регулируется путем активации APC через стимуляцию Toll-подобных рецепторов (Croft, 2003). В настоящих исследованиях не установлено, сопровождается ли отрицательная регуляция экспрессии GITRL на клеточной поверхности секрецией растворимого GITRL, но Tone et al. (2003) сообщили, что стимуляция ЛПС DC, макрофагов и B-клеток приводит к отрицательной регуляции мРНК GITRL. Экспрессия GITRL на покоящихся APC заметно указывает на то, что он функционирует на ранних этапах процесса активации Т-клеток. Также была показана экспрессия GITRL на других клеточных типах, включая эндотелиальные клетки (Gurney et al., 1999; Kwon et al., 1999), активированные Т-клетки и некоторые субпопуляции тимоцитов DN, и функция данной молекулы на этих более поздних клеточных типах еще подлежит определению.
Способность антител против GITR и экспрессирующих GITRL клеток усиливать активацию CD25- T-клеток отдельно, а также способность анти-GITRL частично ингибировать активацию CD25- T-клеток в отсутствие CD25+ T-клеток потребовала тщательной повторной оценки клеточной(ых) мишени(ей) данных реагентов. Добавление анти-GITR в совместные культуры CD25+ и CD25- T-клеток из мышей дикого типа и мышей GITR -/- продемонстрировало, что мишенью для опосредованной антителом отмены супрессии являлась CD25- T-клетка. Это далее подтверждалось экспериментами с использованием крысиных CD25- отвечающих клеток и мышиных CD25+ супрессорных Т-клеток в совместных культурах, которое решительно демонстрировало, что нефункциональное связывание GITR на субпопуляции CD25+ T-клеток не отменяет их супрессорной функции. Фактически оценка разведения CFSE демонстрирует, что нефункциональное связывание GITR способствует экспансии CD25+ T-клеток в совместных культурах, что окончательно усиливает супрессию крысиных CD25- отвечающих клеток. Поэтому возможно, что усиление пролиферации обработанных анти-GITR крысиных/мышиных совместных культур, о котором сообщалось Shimizu et al. (2002) и которое предполагалось следствием действия крысиных T-клеток, в действительности отражает пролиферацию мышиных CD25+ супрессорных клеток. Прояснить это не могло одно лишь измерение включения 3H-тимидина.
В одном из предшествующих отчетов предполагалось, что дефицитные по GITR T-клетки гиперреактивны в отношении стимуляции TCR (Ronchetti et al., 2002). Однако экстраполяция данных наблюдений на настоящие наблюдения затруднена, поскольку в том отчете не анализировались реакции очищенных CD4+, CD8+ T-лимфоцитов и не оценивалась роль CD4+CD25+ T-клеток. В настоящих исследованиях реакции высокоочищенных CD4+CD25- и CD8+ T-клеток из мышей GITR -/- были сравнимыми с таковыми в контроле. Однако в присутствии физиологических количеств регуляторных Т-клеток CD4+ и CD8+ T-клетки из GITR -/- животных полностью не отвечали на перекрестное связывание с CD3. Данный результат четко демонстрирует, что в отсутствие взаимодействий GITR/GITRL супрессия является доминантной. Действительно, супрессия активации CD4+CD25- T-клеток из GITR -/- мышей была так сильна, что ее не могли преодолеть добавлением высоких концентраций экзогенного IL-2, который обычно отменяет супрессорные эффекты больших количеств CD25+ T-клеток в отношении ответа CD4+CD25- Т-клеток дикого типа (Takahashi et al., 1998; Thornton and Shevach, 1998). Супрессорные эффекты CD4+CD25+ были опосредованы ингибированием продукции IL-2 и экспрессией CD25 популяцией отвечающих клеток CD4+CD25- GITR -/-, что напоминало явление, ранее описанное для эффектов CD4+CD25+ T-клеток на нормальные реакции CD8+ T-клеток (Piccirillo and Shevach, 2001).
Костимуляторные сигналы, доставляемые GITR и CD28, оказались различными, но взаимосвязанными. В отсутствие регуляторных T-клеток реакции CD4+ и CD8+ T-клеток из GITR -/- мышей были идентичными реакциям клеток из мышей дикого типа. Наоборот, при условиях культивирования, используемых в данном исследовании, CD4+ и CD8+ T-клетки из CD28 -/- мышей не реагировали. В отличие от этого, когда к культурам, содержащим регуляторные Т-клетки, добавляли IL-2, реакции CD4+ и CD8+ T-клеток из GITR -/- мышей не восстанавливались, в то время как реакции CD8+ клеток из CD28 -/- мышей частично восстанавливались. С другой стороны, потенциальные кооперативные взаимоотношения и взаимная иерархия в передаче сигнала между CD28 и GITR подтверждались демонстрацией того, что (1) CD4+CD25- и CD8 T-клетки не могли положительно регулировать GITR в отсутствие перекрестного связывания CD28, и (2) антитела против CD80/CD86 заметно ингибировали положительную регуляцию экспрессии GITR и способность реагировать на антитела против GITR. Таким образом, данные результаты указывают на то, что важная дополнительная функция передачи сигнала CD28-CD80/CD86 во время активации T-клеток представляет собой допущение устойчивости T-клеток к опосредованной CD25+ супрессии путем усиления экспрессии и функции GITR.
Сходно с результатами, предложенными в других опубликованных исследованиях (Shimizu et al., 2002; Tone et al., 2003), данные исследования указывают на то, что нефункциональное связывание GITR на CD25- T-клетках в отсутствие регуляторных T-клеток предоставляет некоторую степень совместной стимуляции. Однако нефункциональное связывание GITR не требуется для костимуляции CD25- T-клеток, как для CD28, поскольку CD4+CD25- и CD8 T-клетки из GITR -/- мышей отвечали одинаково с Т-клетками дикого типа. Авторам изобретения удалось наблюдать, что вовлечение GITR на CD4+CD25- и CD8+ эффекторных T-клетках посредством GITRL на ранних этапах иммунного ответа служит главным образом для того, чтобы сделать популяцию эффекторов устойчивой, например менее чувствительной, к супрессорным эффектам CD4+CD25+ T-клеток. В ходе ответа воспалительные сигналы, в конечном счете, приводят к отрицательной регуляции экспрессии GITRL, с повышением таким образом, чувствительности эффекторных клеток к опосредованной CD25+ супрессии. Хотя имеются только ограниченные данные, касающиеся опосредованной CD25+ супрессии иммунных реакций на инфекционные агенты in vivo, некоторые исследования указывают на то, что она действует в фазе ограничения, а не инициации ответа для предотвращения повреждения тканей, вторичного по отношению к избыточному воспалению (Suvas et al., 2003). Следует подчеркнуть, что вовлечение GITR на CD25+ T-клетках в присутствии IL-2 индуцирует их экспансию (McHugh et al., 2002). Остается возможность того, что вовлечение GITR CD25+ T-клетках за счет GITRL на покоящихся APC in vivo приводит к неспецифической экспансии регуляторных Т-клеток в присутствии IL-2, секретируемого эффекторными T-клетками на ранних этапах иммунного ответа. Данная неспецифическая экспансия может быть критической для последующего образования объединенной группы антигенспецифичных супрессорных клеток, которые функционируют в плане ингибирования эффекторной клеточной активности на более поздних этапах ответа.
Авторы изобретения ранее предположили, что манипуляция с взаимодействиями GITR/GITRL может оказаться эффективным путем манипуляции функцией регуляторных T-клеток in vivo (McHugh and Shevach, 2002). Хотя данная концепция была основана на том, что мишенью антитела против GITR является CD4+CD25+ T-клетка, по-прежнему имеется представление, что взаимодействие GITR/GITRL представляет важную терапевтическую мишень. Таким образом, обработка агонистическим антителом против GITR или агонистическим GITRL-Fc должна сделать эффекторные клетки устойчивыми, например, менее чувствительными к супрессорным эффектам CD4+CD25+ T-клеток и должна оказаться эффективной для лечения злокачественных опухолей и инфекционных заболеваний, для усиления иммунных реакций на злокачественные опухоли (в применении отдельно или в комбинации с другими противоопухолевыми способами лечения, например с противоопухолевыми вакцинами) и для усиления иммунных реакций на инфекционные патологические микроорганизмы, включая вирусы, бактерии и т.д. (в применении отдельно или в комбинации с другими способами лечения от инфекционных патологических микроорганизмов, например, с вакцинными адъювантами для ослабления вакцин против инфекционных агентов). Аналогично, ингибирование взаимодействий GITR/GITRL агонистом GITR, например посредством нейтрализующего антитела против GITRL или GITR-Fc, должно снизить порог эффекторных Т-клеток к супрессии и, таким образом, может быть полезно для профилактики и/или лечения аутоиммунных нарушений, воспалительных заболеваний, отторжения трансплантата (или пересаженного органа или ткани) и болезни трансплантат против хозяина.
Ссылки
Следующие ссылки являются полными цитируемыми ссылками с указанием автора и года, встречающихся в описании:
Aseffa, A., Gumy, A., Launois, P., MacDonald, H.R., Louis, J.A., and Tacchini-Cottier, F. (2002). The early IL-4 response to Leishmania major and the resulting Th2 cell maturation steering progressive disease in BALB/c mice are subject to the control of regulatory CD4+CD25+ Т cells. J Immunol 169, 3232-3241.
Banchereau, J., and Steinman, R.M. (1998). Dendritic cells and the control of immunity. Nature 392, 245-252.
Balked, Y., Piccirillo, C.A., Mendez, S., Shevach, E.M., and Sacks, D.L. (2002). CD4+CD25+ regulatory Т cells control Leishmania major persistence and immunity. Nature 420, 502-507.
Coligan, J.E., Frisbee, A.M., Argufies, D.H., Shevach, E.M., and Strobe, W., eds. (2003). Current Protocols in Immunology (John Wiley & Sons, Inc.).
Croft, M. (2003). Co-stimulatory members of the TNFR family: keys to effective T-cell immunity? Nat Rev Immunol 3, 609-620.
Depper, J.M., Leonard, W.J., Dracula, C., Crone, M., Walkman, T.A., and Greene, W.C. (1985). Interlaken 2 (IL-2) augments transcription of the IL-2 receptor gene. Proc Natal Aced Sci USA 82, 4230-4234.
Gavin, M.A., Clarke, S.R., Negro, E., Gallegos, A., and Rude sky, A. (2002). Homeostasis and energy of CD4(+)CD25(+) suppressor T cells in vivo. Nat Immunol 3, 33-41.
Gilfillan, M.C., Noel, P.J., Paddock, E.R., Rainer, S.L., and Thompson, С.В. (1998). Expression of the co stimulatory receptor CD30 is regulated by both CD28 and cytokines. J Immunol 160, 2180-2187.
Godfrey, D.L., Kennedy, J., Soda, Т., and Slotting, A. (1993). A developmental pathway involving four phenotypic ally and functionally distinct subsets of CD3-CD4-CD8-triple-negative adult mouse thymocytes defined by CD44 and CD25 expression. J Immunol 150, 4244-4252.
Gurney, A.L., Marsters, S.A., Huang, R.M., Pitti, R.M., Mark, D.Т., Baldwin, D.Т., Gray, A.M., Dowd, A.D., Brush, A.D., Heldens, A.D, et al. (1999). Identification of a new member of the tumor necrosis factor family and its receptor, a human ortholog of mouse GITR. Curr Biol 9, 215-218.
Hisaeda, H., Maekawa, Y., Iwakawa, D., Okada, H., Himeno, K, Kishihara, K., Tsukumo, S., and Yasutomo, K. (2004). Escape of malaria parasites from host immunity requires CD4(+)CD25(+) regulatory T cells. Nat Med 10, 29-30.
Kim, J.D., Choi, B.K., Bae, J.S., Lee, U.H., Han, I.S., Lee, H.W., Youn, B.S., Vinay, D.S., and Kwon, B.S. (2003). Cloning and characterization of GITR ligand. Genes Immun 4, 564-569.
Kursar, M., Bonhagen, K., Fensterle, J., Kohler, A., Hurwitz, R., Kamradt, Т., Kaufmann, S.H., and Mittucker, H.W. (2002). Regulatory CD4+CD25+ Т cells restrict memory CD8+ Т cell responses. J Exp Med 196, 1585-1592.
Kwon, В., Yu, K.Y., Ni, J., Yu, G.L., Jang, I.K., Kim, Y. J., Xing, L., Liu, D., Wang, S.X., and Kwon, B.S. (1999). Identification of a novel activation-inducible protein of the tumor necrosis factor receptor superfamily and its ligand. J Biol Chem 274, 6056-6061.
Lundgren, A., Sim-Payer, E., Enarsson, K., Svennerholm, A.M., and Lundin, B.S. (2003). Helicobacter pylori-specific CD4+ CD25 high regulatory Т cells suppress memory T-cell responses to H. pylori in infected individuals. Infect Immun 71, 1755-1762.
Malek, Т.R., and Ashwell, J.D. (1985). Interleukin 2 upregulates expression of its receptor on a T cell clone. J Exp Med 161, 1575-1580.
Maloy, K.J., Salaun, L., Cahill, R., Dougan, G., Saunders, N.J., and Powrie, F. (2003). CD4+CD25+ T(R) cells suppress innate immune pathology through cytokine-dependent mechanisms. J Exp Med 197, 111-119.
McHugh, R.S., and Shevach, E.M. (2002). The role of suppressor T cells in regulation of immune responses. J Allergy Clin Immunol 110, 693-702.
McHugh, R.S., Whitters, M.J., Piccirillo, C.A., Young, D.A., Shevach, E.M., Collins, M., and Byrne, M.C. (2002). CD4(+)CD25(+) immunoregulatory T cells: gene expression analysis reveals a functional role for the glucocorticoid-induced TNF receptor. Immunity 16, 311-323.
Nakano, H., Yanagita, M., and Gunn, M.D. (2001). CD1 1c(+)B220(+)Gr-1(+) cells in mouse lymph nodes and spleen display characteristics of plasmacytoid dendritic cells. J Exp Med 194, 1171-1178.
Ouyang, W., Ranganath, S.H., Weindel, K., Bhattacharya, D., Murphy, T.L., Sha, W.C., and Murphy, K.M. (1998). Inhibition of Th1 development mediated by GATA-3 through an IL-4-independent mechanism. Immunity 9, 745-755.
Piccirillo, C.A., and Shevach, E.M. (2001). Cutting edge: control of CD8+ T cell activation by CD4+CD25+ immunoregulatory cells. J Immunol 167, 1137-1140.
Rogers, P.R., Song, J., Gramaglia, I., Killeen, N., and Croft, M. (2001). OХ40 promotes Bcl-xL and Bcl-2 expression and is essential for long-term survival of CD4 Т cells. Immunity 15, 445-455.
Ronchetti, S., Nocentini, G., Riccardi, C., and Pandolfi, P.P. (2002). Role of GITR in activation response of Т lymphocytes. Blood 100, 350-352.
Sakaguchi, S., Sakaguchi, N., Asano, M., Itoh, M., and Toda, M. (1995). Immunologic self-tolerance maintained by activated Т cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J Immunol 155, 1151-1164.
Shimizu, J., Yamazaki, S., Takahashi, Т., Ishida, Y., and Sakaguchi, S. (2002). Stimulation of CD25(+)CD4(+) regulatory Т cells through GITR breaks immunological self-tolerance. Nat Immunol 3, 135-142.
Suvas, S., Kumaraguri, U., Pack, C.D., Lee, S., and Rouse, В.Т. (2003). CD4+CD25+ Т cells regulate virus-specific primary and memory CD8+ Т cell responses. J Exp Med 198, 889-901.
Takahashi, Т., Kuniyasu, Y., Toda, M., Sakaguchi, N., Itoh, M., Iwata, M., Shimizu, J., and Sakaguchi, S. (1998). Immunologic self-tolerance maintained by CD25+CD4+ naturally anergic and suppressive Т cells: induction of autoimmune disease by breaking their anergic/suppressive state, Int Immunol 10, 1969-1980.
Thornton, A.M., and Shevach, E.M. (1998). CD4+CD25+ immunoregulatory Т cells suppress polyclonal Т cell activation in vitro by inhibiting interleukin 2 production. J Exp Med 188, 287-296.
Tone, M., Tone, Y., Adams, E., Yates, S.P., Frewin, M.R., Cobbold, S.P., and Waldmann, H. (2003). Mouse glucocorticoid-induced tumor necrosis factor receptor ligand is costimulatory for Т cells. Proc Natl Acad Sci USA 100, 15059-15064.
Uraushihara, К., Kanai, Т., Ко, К., Totsuka, Т., Makita, S., Iiyama, R., Nakamura, Т., and Watanabe, M. (2003). Regulation of murine inflammatory bowel disease by CD25+ and CD25-CD4+ glucocorticoid-induced TNF receptor family-related gene+ regulatory Т cells. J Immunol 777, 708-716.
Vremec, D., Pooley, J., Hochrein, H., Wu, L., and Shortman, K. (2000). CD4 and CD8 expression by dendritic cell subtypes in mouse thymus and spleen. J Immunol 164, 2978-2986.
Vremec, D., and Shortman, K. (1997). Dendritic cell subtypes in mouse lymphoid organs: cross-correlation of surface markers, changes with incubation, and differences among thymus, spleen, and lymph nodes. J Immunol 159, 565-573.
Yu, К.Y., Kim, H.S., Song, S.Y., Min, S.S., Jeong, J.J., and Youn, B.S. (2003). Identification of a ligand for glucocorticoid-induced tumor necrosis factor receptor constitutively expressed in dendritic cells. Biochem Biophys Res Commun 310, 433-438.

















| название | год | авторы | номер документа |
|---|---|---|---|
| СИСТЕМА И СПОСОБ ПОЛУЧЕНИЯ И ХРАНЕНИЯ АКТИВИРОВАННЫХ ЗРЕЛЫХ ДЕНДРИТНЫХ КЛЕТОК | 2011 |
|
RU2575978C2 |
| РАСТВОРИМЫЙ МЕДИАТОР | 2012 |
|
RU2660580C2 |
| АНТИ-GITR-АНТИТЕЛА | 2010 |
|
RU2595409C2 |
| АНТИ-GITR-АНТИТЕЛА | 2010 |
|
RU2646139C1 |
| АНТИТЕЛА ПРОТИВ ОХ40 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2562874C1 |
| СПОСОБЫ ПОЛУЧЕНИЯ ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР КЛЕТОК | 2015 |
|
RU2751362C2 |
| Моноклональное антитело, которое специфически связывается с GITR | 2019 |
|
RU2734432C1 |
| ИММУНОТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ НА ОСНОВЕ ДРОЖЖЕЙ С BRACHYURY | 2012 |
|
RU2619850C2 |
| ЛЕЧЕНИЕ РАКА С ПОМОЩЬЮ ГУМАНИЗИРОВАННОГО АНТИ-CD19 ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА | 2014 |
|
RU2711975C2 |
| БИОМАРКЕРЫ, ПРОГНОЗИРУЮЩИЕ СПОСОБНОСТЬ К ТЕРАПЕВТИЧЕСКОМУ ОТВЕТУ НА ТЕРАПИЮ ХИМЕРНЫМ РЕЦЕПТОРОМ АНТИГЕНА, И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2743657C2 |

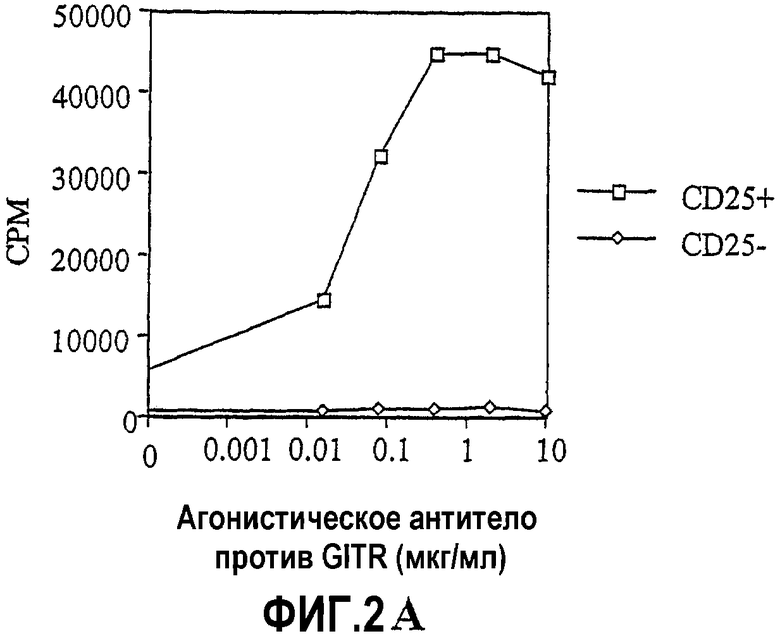
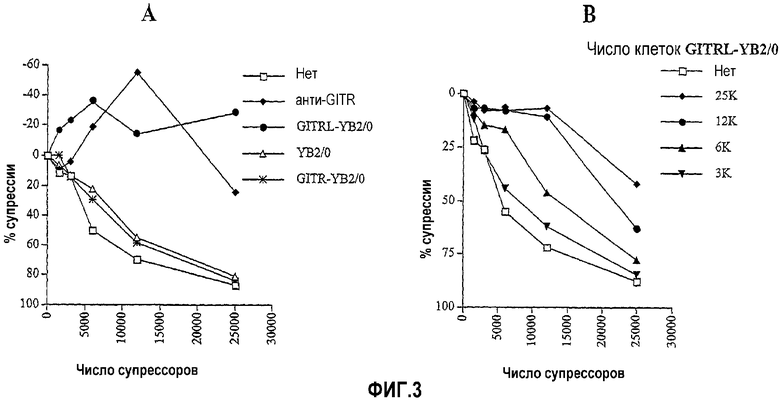
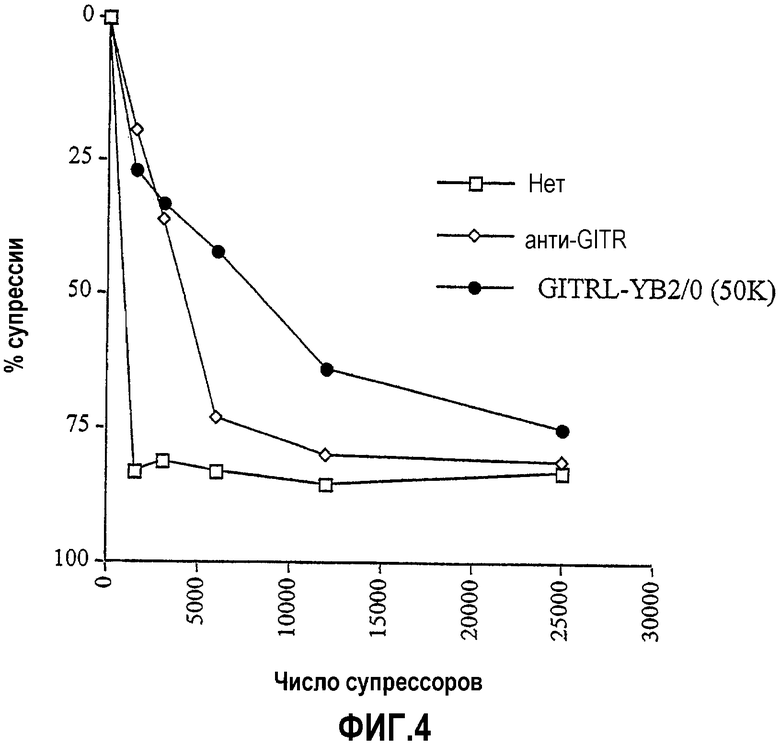
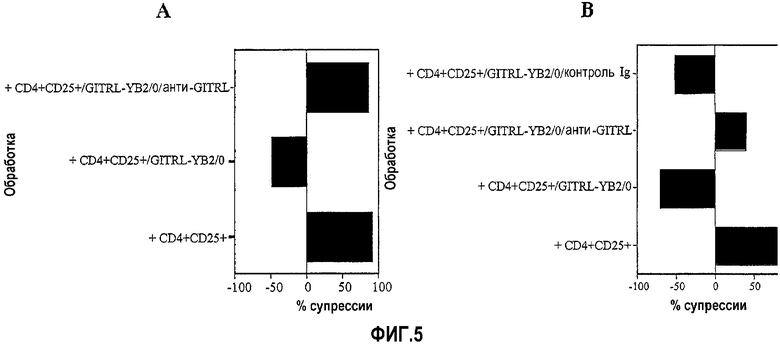
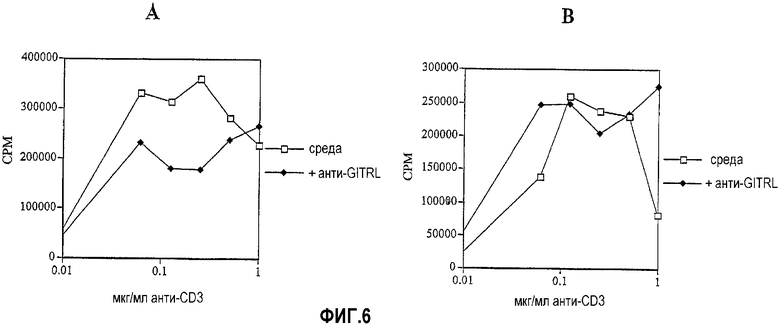
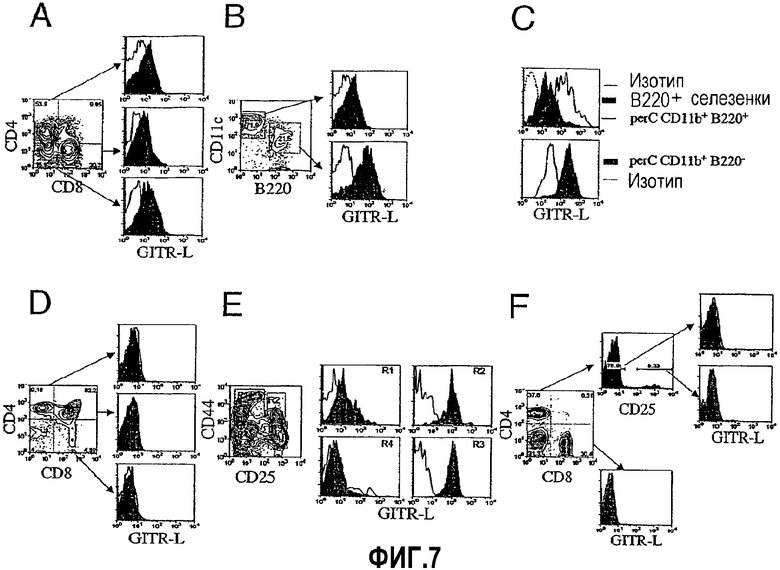
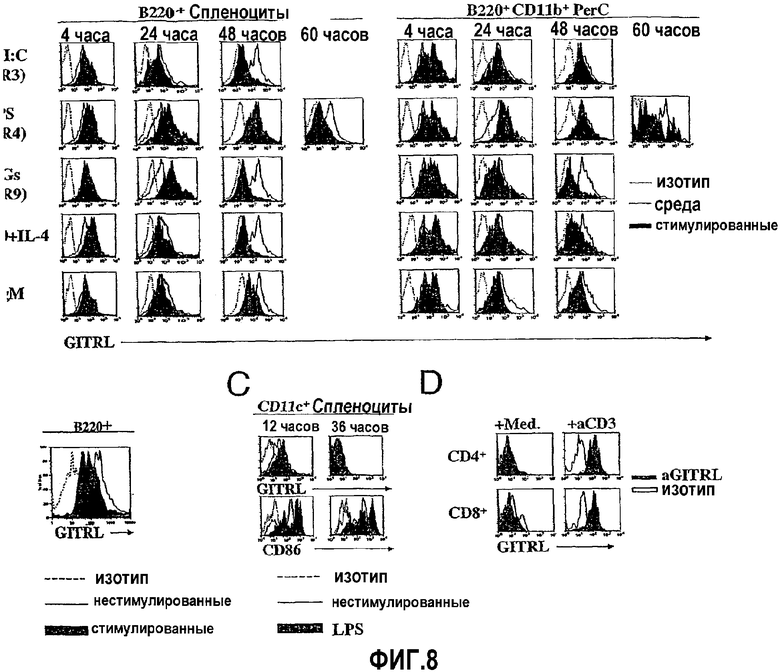
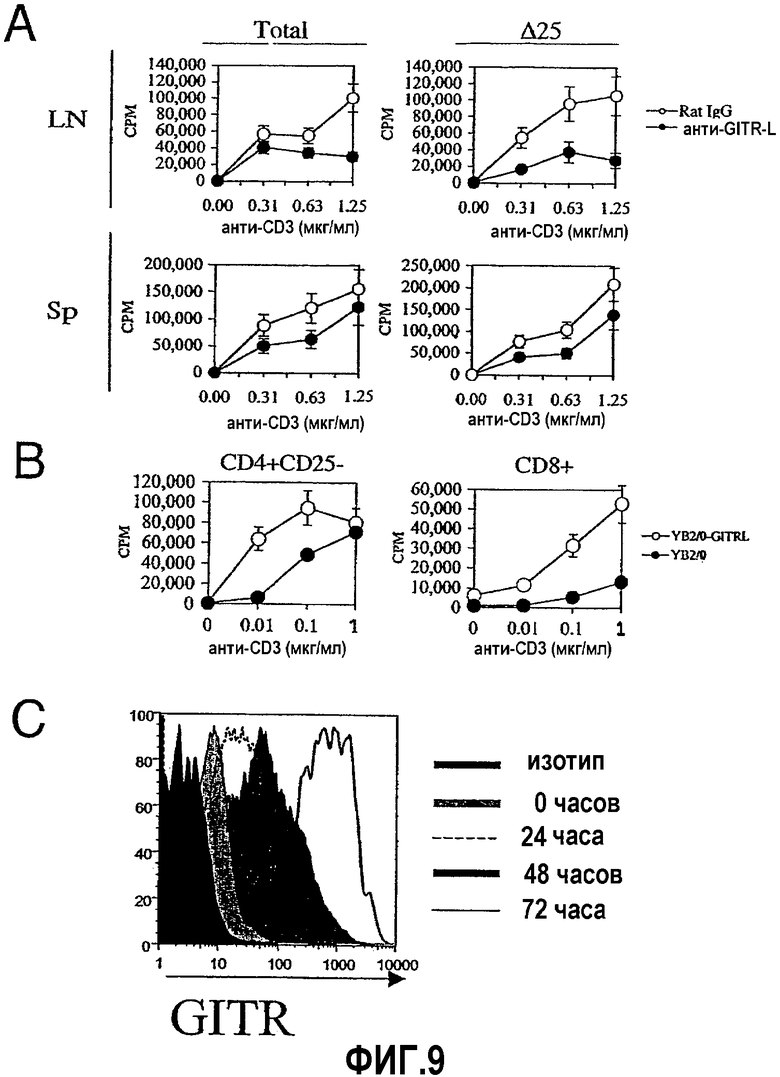
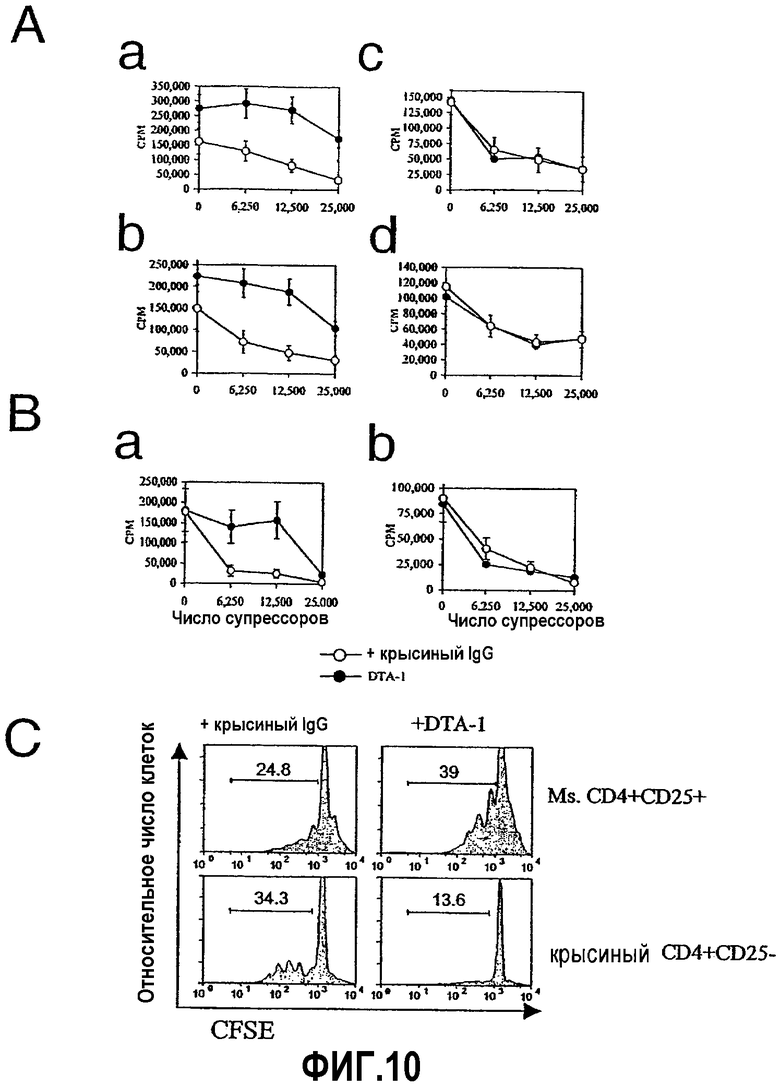
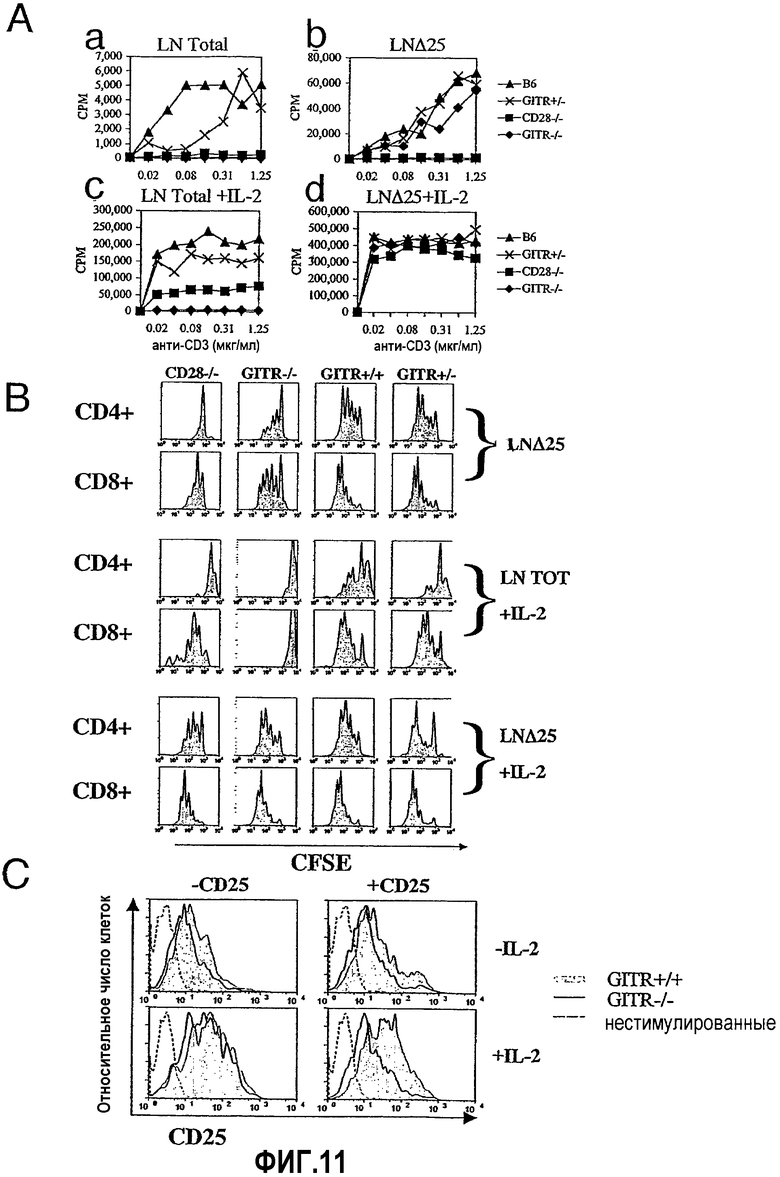
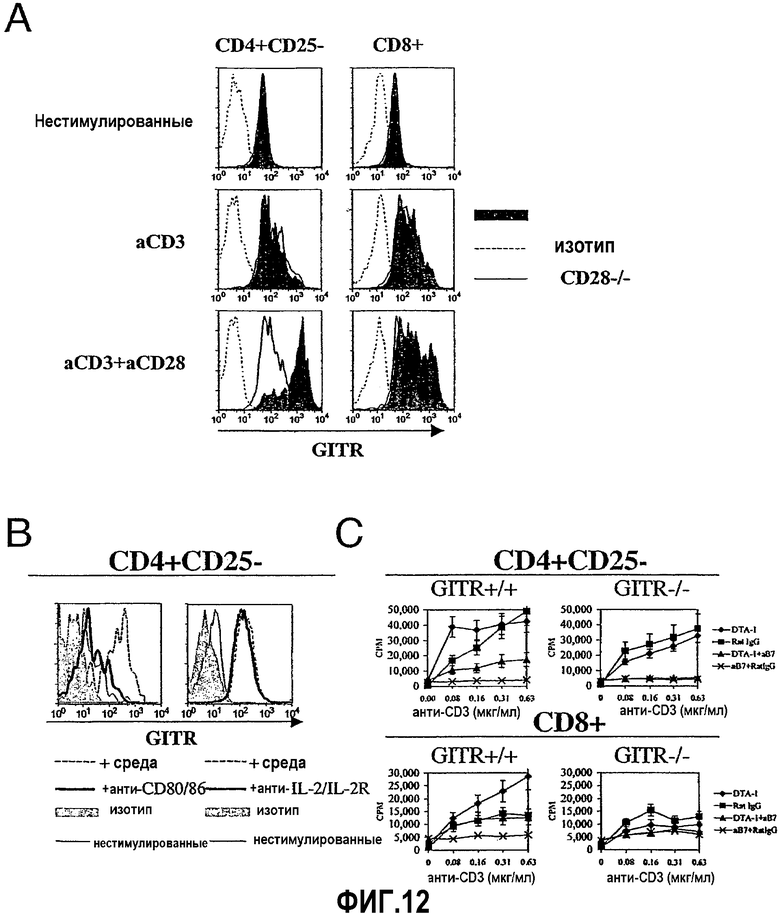
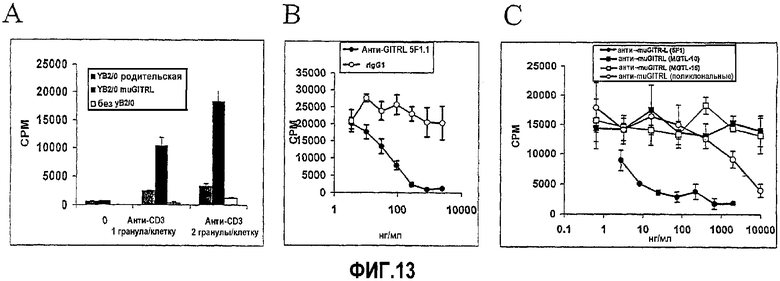
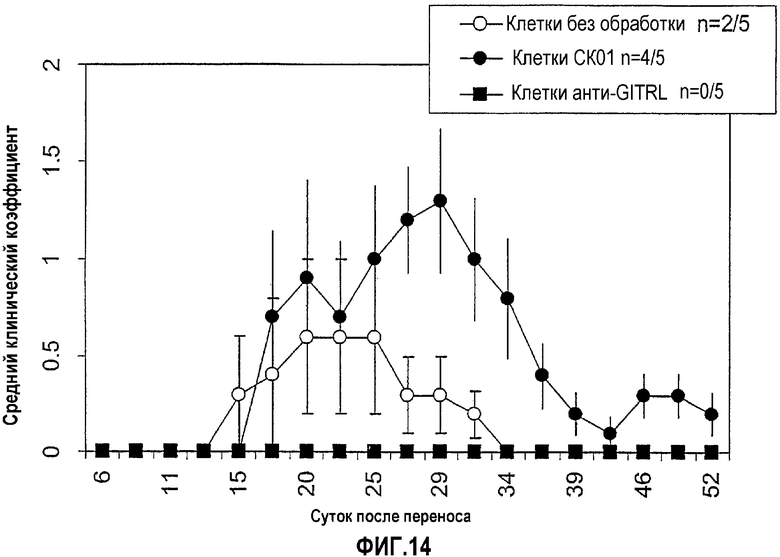
Изобретение относится к биотехнологии. Описан способ лечения субъекта при риске или поставленном диагнозе аутоиммунного заболевания, воспалительного заболевания или отторжения трансплантата, включающий введение субъекту антагониста GITR, в котором антагонист GITR предотвращает связывание GITRL с GITR на эффекторных Т-клетках. Раскрыт способ ингибирования пролиферации популяции клеток, содержащей эффекторные Т-клетки, включающий введение в клеточную популяцию антагониста GITR, в котором антагонист GITR предотвращает связывание GITRL с GITR на эффекторных Т-клетках. Предложен способ супрессии клеточной популяции, содержащей эффекторные Т-клетки, в присутствии CD4+CD25+ регуляторных Т-клеток, включающий введение в клеточную популяцию антагониста GITR, в котором антагонист GITR предотвращает связывание GITRL с GITR на эффекторных Т-клетках. Описана фармацевтическая композиция, содержащая антагонист GITR и фармацевтически приемлемый носитель для лечения аутоиммунных нарушений, воспалительных заболеваний и отторжения трансплантата, в которой антагонист GITR предотвращает связывание GITRL с GITR на эффекторных Т-клетках. Изобретение может быть использовано при лечении нарушений, возникающих в результате разрегулированных иммунных реакций. 8 н. и 8 з.п. ф-лы, 14 ил., 3 табл.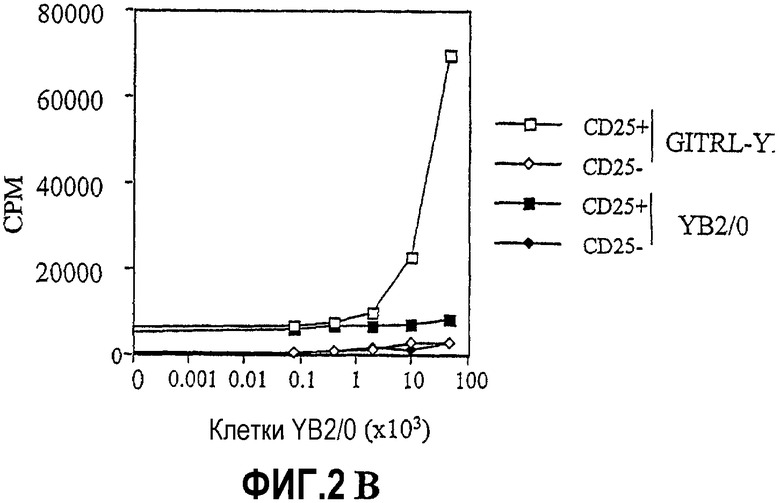
1. Способ лечения субъекта при риске или поставленном диагнозе аутоиммунного заболевания, воспалительного заболевания или отторжения трансплантата, включающий введение субъекту антагониста GITR, в котором антагонист GITR предотвращает связывание GITRL с GITR на эффекторных Т-клетках.
2. Способ по п.1, в котором антагонист GITR выбран из группы, состоящей из нейтрализующего антитела против GITRL, нейтрализующего антитела против GITR, слитого белка, содержащего GITR, слитого белка, содержащего активный фрагмент GITR, и низкомолекулярного антагониста.
3. Способ лечения субъекта при риске или поставленном диагнозе аутоиммунного заболевания, воспалительного заболевания или отторжения трансплантата, включающий введение субъекту антагониста GITR, в котором антагонист GITR выбран из группы, состоящей из антисмысловой в отношении GITRL молекулы нуклеиновой кислоты и молекулы нуклеиновой кислоты siPHK против GITRL.
4. Способ по п.1 или 3, в котором аутоиммунное нарушение или воспалительное заболевание выбрано из группы, состоящей из ревматоидного артрита, энцефаломиелита, остеоартрита, рассеянного склероза, аутоиммунного гастрита, системной красной волчанки, псориаза и других воспалительных дерматозов, диабета I типа, астмы, аллергии, отторжения органного трансплантата, болезни трансплантат против хозяина и воспалительных заболеваний толстого кишечника, включая болезнь Крона и язвенный колит.
5. Способ по п.4, в котором аутоиммунное нарушение представляет собой рассеянный склероз.
6. Способ ингибирования пролиферации популяции клеток, содержащей эффекторные Т-клетки, включающий введение в клеточную популяцию антагониста GITR, в котором антагонист GITR предотвращает связывание GITRL с GITR на эффекторных Т-клетках.
7. Способ по п.6, в котором антагонист GITR выбран из группы, состоящей из нейтрализующего антитела против GITRL, нейтрализующего антитела против GITR, слитого белка, содержащего GITR, слитого белка, содержащего активный фрагмент GITR, и низкомолекулярного антагониста.
8. Способ ингибирования пролиферации популяции клеток, содержащей эффекторные Т-клетки, включающий введение в клеточную популяцию антагониста GITR, в котором антагонист GITR выбран из группы, состоящей из антисмысловой в отношении GITRL молекулы нуклеиновой кислоты и молекулы нуклеиновой кислоты siPHK против GITRL.
9. Способ по п.6 или 8, в котором эффекторные Т-клетки представляют собой CD4+Т-клетки или CD8+Т-клетки.
10. Способ супрессии клеточной популяции, содержащей эффекторные Т-клетки, в присутствии CD4+CD25+ регуляторных Т-клеток, включающий введение в клеточную популяцию антагониста GITR, в котором антагонист GITR предотвращает связывание GITRL с GITR на эффекторных Т-клетках.
11. Способ по п.10, в котором антагонист GITR выбран из группы, состоящей из нейтрализующего антитела против GITRL, нейтрализующего антитела против GITR, слитого белка, содержащего GITR, слитого белка, содержащего активный фрагмент GITR, и низкомолекулярного антагониста.
12. Способ супрессии клеточной популяции, содержащей эффекторные Т-клетки, в присутствии CD4+CD25+ регуляторных Т-клеток, включающий введение в клеточную популяцию антагониста GITR, в котором антагонист GITR выбран из группы, состоящей из антисмысловой в отношении GITRL молекулы нуклеиновой кислоты и молекулы нуклеиновой кислоты siPHK против GITRL.
13. Способ по п.10 или 12, в котором эффекторные Т-клетки представляют собой CD4+Т-клетки или CD8+Т-клетки.
14. Фармацевтическая композиция, содержащая антагонист GITR и фармацевтически приемлемый носитель для лечения аутоиммунных нарушений, воспалительных заболеваний и отторжения трансплантата, в которой антагонист GITR
предотвращает связывание GITRL с GITR на эффекторных Т-клетках.
15. Фармацевтическая композиция по п.14, в которой антагонист GITR выбран из группы, состоящей из нейтрализующего антитела против GITRL, нейтрализующего антитела против GITR, слитого белка, содержащего GITR, слитого белка, содержащего активный фрагмент GITR, и низкомолекулярного антагониста.
16. Фармацевтическая композиция, содержащая антагонист GITR и фармацевтически приемлемый носитель для лечения аутоиммунных нарушений, воспалительных заболеваний и отторжения трансплантата, в которой антагонист GITR
выбран из группы, состоящей из антисмысловой в отношении GITRL молекулы нуклеиновой кислоты и молекулы нуклеиновой кислоты siPHK против GITRL.
| Видоизменение пишущей машины для тюркско-арабского шрифта | 1923 |
|
SU25A1 |
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| ЖЕЛЕЗНОДОРОЖНЫЙ ПУТЬ | 2006 |
|
RU2307885C2 |
| РАСПРЕДЕЛИТЕЛЬНЫЙ ВЕРТИКАЛЬНО-ЗАМКНУТЫЙ | 0 |
|
SU234223A1 |
| «Биология», книга 1 | |||
| Электрическое сопротивление для нагревательных приборов и нагревательный элемент для этих приборов | 1922 |
|
SU1997A1 |
| RU | |||

