Область техники, к которой относится изобретение
Настоящее изобретение относится к области иммунологии и вирусологии и предполагает использование антител, представляющих собой тримеризованные однодоменные антитела, для профилактики и терапии бешенства.
Предшествующий уровень техники
Бешенство - особо опасная болезнь животных всех видов и человека, характеризующаяся тяжелым поражением центральной нервной системы и летальным исходом. Ежегодно в мире от бешенства умирают 45-60 тысяч человек, и приблизительно 10 миллионов людей, получивших повреждения от животных, обращаются за антирабической помощью. Особая опасность бешенства состоит в том, что до настоящего времени не найдено эффективных средств лечения уже развившегося заболевания.
В России показатель обращаемости за антирабической помощью в среднем по стране составляет 300,0 на 100 тысяч населения. Ежегодно более 250 тысяч человек подвергаются риску заражения вирусом бешенства и нуждаются в проведении им специфической профилактики.
Тенденция осложнения эпизоотической обстановки в России пока сохраняется и даже усиливается. Характерно одновременное возрастание числа случаев бешенства как диких, так и домашних плотоядных. Подъем эпизоотии природного типа несомненен. Нарастает эпизоотологическая и эпидемиологическая значимость бешенства волков, лис, енотовидных собак. В эпизоотические цепи все чаще включаются дикие куньи, зарегистрированы единичные случаи бешенства лосей, бобров, ондатр, хомяков, зайцев, ежей, серых крыс и мышей.
Основным способом борьбы с бешенством животных является специфическая профилактика с применением вакцинных препаратов и антирабического иммуноглобулина. Согласно рекомендуемому ВОЗ лечению бешенства в условии III категории контакта (единственный или множественные трансдермальные укусы или царапины, облизывание поврежденной кожи; загрязнение слизистых оболочек слюной при облизывании, контакты с летучими мышами) наряду с применением вакцинного препарата назначается немедленное введение антирабического иммуноглобулина. Однако существующие на сегодняшний день препараты, содержащие антирабический иммуноглобулин (гетерологичный (лошадиный) и гомологичный (человеческий)), не отвечают современным требованиям безопасности и эффективности. Поэтому получение новых инновационных средств борьбы с вирусом бешенства является актуальной задачей для повышения уровня защиты населения от вируса бешенства и снижения числа летальных исходов инфицирования.
Одним из наиболее эффективных на сегодняшний день подходов для создания безопасных и эффективных препаратов для борьбы с вирусными инфекциями является использование вируснейтрализующих антител. В настоящее время в России зарегистрирован и закупается для государственных нужд только один препарат иммуноглобулина антирабического из сыворотки крови человека производства Фармацевтической компании «Сычуаньская Юанда Шуян» (Китай). Аналогичных препаратов отечественного производства нет.
По сравнению с иммуноглобулинами, получаемыми из сыворотки человека, которому вводили вакцинный препарат инактивированного вируса бешенства, рекомбинантные моноклональные антитела или фрагменты антител, которые можно получать в культурах клеток в строго контролируемых и воспроизводимых условиях, являются более безопасными, лучше охарактеризованными и стандартизированными и больше подходят для производства и применения в качестве терапевтических средств.
В качестве ближайшего аналога технического решения, составляющего основу настоящего изобретения, можно привести следующий пример. Патентная заявка US 2013/0039926 A1 («Human antibodies against rabies and uses thereof» - «Антитела человека против бешенства и их использование», опубликованная 14 февраля 2013 года) описывает получение рекомбинантных моноклональных человеческих антител и их антиген-связывающих фрагментов (Fab), способных специфически взаимодействовать с основным поверхностным вирусным антигеном, гликопротеином G, вируса бешенства и в результате такого взаимодействия противодействовать способности вируса инфицировать клетки.
Данное техническое решение, как наиболее близкое к заявляемому по составу действующего вещества и способу его использования, выбрано авторами настоящего изобретения за прототип.
Недостатками прототипа являются:
- Использование в качестве продуцента антител клеток млекопитающих, в частности миеломы мышей, которые сложны в культивировании и хранении, требовательны к качеству используемых реагентов, могут быть инфицированы патогенами, опасными для человека (вирусы, микоплазмы и др.) и должны постоянно контролироваться на их отсутствие, что в конечном итоге отражается на высокой стоимости моноклональных антител;
- Использование традиционной гибридомной технологии на мышах вызывает необходимость в проведении дополнительных генно-инженерных манипуляций для гуманизации по генам иммуноглобулинов с целью приведения аминокислотных последовательностей заявляемых антител в соответствие с человеческими, однако, это не отменяет посттрансляционных модификаций, характерных именно для иммуноглобулинов мыши, а не человека. Полученные таким образом антитела могут вызывать нежелательную иммунную реакцию при их введении в организм человека.
- Моноклональные антитела имеют большой размер получаемых антител, что влечет за собой пониженную проницаемость в ткани;
- Структурные особенности накладывают ограничение на узнавание некоторых «скрытых» эпитопов, находящихся, например, в углублениях, щелях малого размера в структуре белков, что сказывается на снижении биологической активности моноклональных антител.
Таким образом, в уровне техники существует потребность в разработке новых антител - антиген-узнающих молекул, лишенных указанных выше недостатков и специфически узнающих вирус бешенства, а также способных ингибировать его инфицирующую активность.
Раскрытие изобретения
Задачей настоящего изобретения явилось создание нового однодоменного антитела, которое будет эффективно связывать и нейтрализовать вирус бешенства.
Техническая задача решается за счет того, что получено тримеризованное однодоменное антитело, специфически связывающее основной поверхностный (капсидный) белок, гликопротеин G, вируса бешенства и способное нейтрализовать вирус бешенства.
С целью получения более стабильного, компактного, экономичного в производстве, удобного для всевозможных последующих его модификаций и разнообразных путей комбинированного использования антитела против вируса бешенства была использована технология генерирования библиотеки однодоменных антител и последующего отбора последовательностей, кодирующих однодоменное антитело с заданной специфичностью с помощью метода фагового дисплея.
В основе использованной технологии лежит получение нуклеотидных последовательностей генов неканонических антител животных семейства верблюдовых (Camelidae). Эти антитела представляют собой димер только одной укороченной (первый константный район CH1 отсутствует) тяжелой цепи иммуноглобулина. Для собственно специфического узнавания и связывания антигена при этом необходим и достаточен лишь один вариабельный домен этого антитела.
Полным эквивалентом термина «однодоменное антитело» для целей настоящего изобретения является вошедшее в широкое употребление обозначение «нанотело», «NANOBODY», а также «наноантитело» и «однодоменное наноантитело».
Полученные по этой технологии однодоменные антитела обладают следующими улучшенными свойствами [Тиллиб С.В. «Верблюжьи мини-антитела» - эффективный инструмент для исследований, диагностики и терапии. Молекулярная биология. 2011; 45(1): 77-85]:
1) Наличие отработанного заявителем способа получения и селекции однодоменных антител, в основе которого лежит получение нуклеотидных последовательностей генов неканонических антител животных семейства верблюдовых (Camelidae);
2) Возможность экономичной наработки в больших количествах, так как продуцентом является бактерия Е.Coli, что говорит о промышленной применимости изобретения;
3) Биологическая безопасность антител, так как культура бактерии Е.Coli не может контаминироваться патогенами эукариотов;
4) Низкая иммуногенность, так как однодоменное антитело состоит только из вариабельного фрагмента тяжелой цепи, ответственного за связывание с антигеном и не содержит других доменов, на которые образуются нежелательные иммунные реакции.
5) Однодоменные антитела имеют улучшенную проницаемость в клетки за счет малого размера (~2×4 нм, 13-15 кДа), что очень важно при инфицировании вирусом бешенства.
6) Высокая растворимость и стабильность (в широком диапазоне температур и кислотности среды) позволяет готовить на основе однодоменных антител препараты для медицинского применения.
Хотя получаемые (в бактериальной системе экспрессии) однодоменные антитела не содержат посттрансляционных модификаций, еще одним положительным свойством этих антител является возможность относительно экономичной их «гуманизации» без заметной потери их специфической активности путем проведения небольшого числа точечных замен аминокислот [Vincke С., Loris R., Saerens D., et al. // J. Biol. Chem. 2009. V.284. №5. P.3273-3284]. Это открывает потенциальную возможность широкого использования однодоменных антител в качестве средств пассивной иммунизации для предотвращения развития различных опасных инфекционных заболеваний [Wesolowski J., Alzogaray V., Reyelt J. et al. Single domain antibodies: promising experimental and therapeutic tools in infection and immunity. Med. Microbiol. Immunol. 2009; 198, 157-174].
Поставленная задача решается настоящим изобретением за счет создания тримеризованного однодоменного антитела, специфически связывающегося с поверхностным (капсидным) белком - гликопротеином G, вируса бешенства и нейтрализующего инфекционную активность вируса бешенства.
Способ получения тримеризованного однодоменного антитела, специфически связывающегося с гликопротеином G вируса бешенства и нейтрализующим инфекционную активность вируса бешенства, включает следующие принципиальные этапы:
1) индукция образования специфических антител в результате 5-этапной иммунизации представителя семейства Верблюдовых (двугорбого верблюда) субстанцией, приготовленной на основе смешанного с адьювантом (Gerbu LQ) препарата инактивированного вируса бешенства;
2) клонирование последовательностей антиген-узнающих доменов - особых одноцепочечных антител, синтезирующихся в лимфоцитах иммунизированного животного, в фагмидном векторе, использующемся в последующей процедуре фагового дисплея, получение специфически обогащенной библиотеки последовательностей, кодирующих однодоменные антитела;
3) селекция методом модифицированного фагового дисплея (функционального отбора фаговых частиц, несущих на поверхности экспрессируемое антитело, а внутри - ДНК, кодирующую это антитело) клонов антител, связывающихся с заданным антигеном, как с целым вирусом бешенства, так и с гликопротеином G вируса бешенства, из полученной библиотеки однодоменных антител;
4) проведение переклонирования и генно-инженерных модификаций кодирующих последовательностей отобранных однодоменных антител с целью их эффективной наработки в бактериальной системе экспрессии, последующей эффективной очистки (в качестве продуцента действующего вещества используются бактерии E.coli), а также для увеличения их биологической активности (благодаря гомотримеризации);
5) демонстрация функциональной активности отобранных и форматированных тримеризованных антител, а именно способности специфически узнавать гликопротеин G вируса бешенства и вирус бешенства, а также нейтрализовать in vitro инфекционную активность вируса бешенства.
Принципиально важным этапом получения из исходно отобранных методом фагового дисплея мономерных однодоменных антител модифицированных производных, обладающих существенно более высокой биологической активностью (нейтрализация вируса бешенства), является присоединение к концу первичного антитела дополнительных последовательностей (доменов), которые включают: а) линейный участок (линкерный район, соответствующий длинному шарнирному участку верблюжьего антитела («hinge», h), который важен для разделения антиген-узнающей функции от функций других доменов, б) принципиально важный для усиления нейтрализующей вирус функции тримеризующийся домен (вариант лейциновой молнии, ILZ), а также (на самом конце) участки, кодирующие два олигопептидных тага: в) HA-таг и г) (His)6-таг. Последние два участка существенно облегчают, соответственно, детекцию и очистку данного тримеризованного одноцепочечного антитела. Подробное описание использования тримеризующего домена с целью повышения биологической активности одноцепочечного антитела было описано в нашей недавней работе (Tillib S., Ivanova T.I., Vasilev L.A., Rutovskaya M.V., Saakyan S.A., Gribova I.Y., Tutykhina I.L., Sedova E.S., Lysenko A.A., Shmarov M.M., Logunov D.Y., Naroditsky B.S., Gintsburg A.L. Formatted single-domain antibodies can protect mice against infection with influenza virus (H5N2). Antiviral Research 2013, 97: 245-254. // Форматированные однодоменные антитела могут защитить мышь от летальной инфекции, вызванной вирусом гриппа).
Авторы настоящего изобретения исходят из того, что, как известно среднему специалисту в данной области техники, первичные, исходно получаемые последовательности однодоменных антител могут быть затем адаптированы или «форматированы» различным образом для последующего практического использования.
Так, однодоменные антитела могут быть исходными модулями (блоками) более сложных многомодульных препаратов. Возможно объединение в одном мультивалентном производном двух, трех и более моновалентных первичных однодоменных антител. Эти объединяемые в одну конструкцию однодоменные антитела могут связываться как с одним и тем же эпитопом антигена-мишени, так и с его разными эпитопами или даже с различными антигенами-мишенями. Возможно также комбинированное объединение в одну конструкцию однодоменных антител и других молекул или лекарств с получением многофункциональных препаратов; мультимеризация с помощью введения дополнительных аминокислотных последовательностей, взаимодействующих белковых доменов, таких как лейциновые зипперы, или последовательностей небольших белков, образующих стабильные комплексы. Для модулирования свойств препарата однодоменного антитела, например увеличения времени жизни или совершенствования способа очистки, в состав конечного соединения могут быть введены дополнительные аминокислотные последовательности. Среднему специалисту в данной области техники будет очевидно, что такие модификации и прочие варианты антител, лежащих в основе настоящего изобретения, подпадают под объем настоящего изобретения, поскольку являются структурными и функциональными вариантами однодоменных антител. Таким образом, автор настоящего изобретения понимает под термином "однодоменные антитела" как первичные, исходно получаемые, "минимальные" аминокислотные последовательности однодоменных антител, так и их модификации, полученные в результате упомянутых, адаптации или «форматирования» и их варианты. Термин «вариант антитела» для целей настоящего изобретения означает полипептид, который содержит изменения в аминокислотной последовательности, а именно делеции, вставки, добавления или замены аминокислот, при условии, что при этом сохраняется необходимый уровень активности белка, например, как минимум 10% от активности исходного однодоменного антитела. Ряд изменений в варианте белка зависит от положения или от типа аминокислотного остатка в трехмерной структуре белка. Для вариабельного домена антитела (или однодоменного антитела) такими принципиальными положениями являются три гипервариабельных участка, называемые CDR1, CDR2 и CDR3, фланкированные более консервативными каркасными участками. Особенно важную роль для специфического узнавания в данном случае играет CDR3, тогда как все другие участки существенно менее характеристичные и воспроизводящиеся с высокой гомологией в других генах вариабельных доменов иммуноглобулинов.
Количество изменений может составлять, например, от 1 до 30, более предпочтительно от 1 до 15 и наиболее предпочтительно от 1 до 5 изменений в последовательности исходного однодоменного антитела. Эти изменения могут иметь место в областях полипептида, которые не являются критичными для его функции. Это становится возможным благодаря тому, что некоторые аминокислоты обладают высокой гомологией друг с другом и поэтому третичная структура или активность белка не нарушаются при таком изменении. Поэтому в качестве варианта белка может выступать белок, который характеризуется гомологией не менее 70%, предпочтительно не менее 80%, более предпочтительно не менее 90% и наиболее предпочтительно, не менее 95% по отношению к аминокислотной последовательности исходного однодоменного антитела при условии сохранения активности полипептида.
Гомология между аминокислотными последовательностями может быть установлена с использованием хорошо известных методов, например с помощью выравнивания последовательностей в компьютерной программе BLAST 2.0, которая вычисляет три параметра: счет, идентичность и сходство.
Замена, делеция, вставка, добавление или замена одного или нескольких аминокислотных остатков будут представлять собой консервативную мутацию или консервативные мутации при условии, что активность белка при этом сохраняется.
Примером консервативной мутации является консервативная замена. "Консервативная аминокислотная замена" представляет собой замену, при которой аминокислотный остаток заменяется аминокислотным остатком, имеющим сходную боковую цепь. В данной области техники определены семейства аминокислот, имеющих сходные боковые цепи. Эти семейства включают в себя аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серии, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Поскольку гипервариабельные районы однодоменных антител определяют их специфическое взаимодействие с антигеном, то именно гомологичные замены аминокислот в этих участках могут приводить к получению несколько различающихся по последовательности однодоменных антител, которые обладают идентичными или близкими свойствами. Таким образом, среднему специалисту в данной области техники будет очевидно, что под объем настоящего изобретения подпадают не только указанные в приложении последовательности однодоменных антител, но и те, которые могут быть получены путем замен аминокислот в гипервариабельных участках (указанных в перечне последовательностей как CDR) на другие, но очень близкие по свойствам, аминокислоты (консервативных замен).
Фрагменты ДНК, которые кодируют по существу тот же функциональный полипептид, могут быть получены, например, путем модификации нуклеотидной последовательности фрагмента ДНК, кодирующего исходное однодоменное антитело, например, посредством метода сайт-направленного мутагенеза, так, что один или несколько аминокислотных остатков в определенном сайте будут делетированы, заменены, вставлены или добавлены. Фрагменты ДНК, модифицированные, как описано выше, могут быть получены с помощью традиционных методов обработки с целью получения мутации.
Фрагменты ДНК, которые кодируют по существу тот же функциональный полипептид исходного однодоменного антитела, получают путем экспрессирования фрагментов ДНК, имеющих мутацию, описанную выше, и установления активности экспрессируемого продукта.
Замена, делеция, вставка или добавление нуклеотидов, описанных выше, также включают мутации, которые имеют место в природе и, например, обусловлены изменчивостью.
Полипептиды однодоменных антител согласно настоящему изобретению могут кодироваться большим множеством молекул нуклеиновых кислот, что является результатом хорошо известного в данной области техники явления вырожденности генетического кода. Суть феномена состоит в том, что любая аминокислота (за исключением триптофана и метионина), входящая в состав природных пептидов, может кодироваться более чем одним триплетным нуклеотидным кодоном. Любая из этих вырожденных кодирующих молекул нуклеиновых кислот может входить в состав кассет, экспрессирующих однодоменные антитела, заявленные в соответствии с настоящим изобретением и подпадающие под объем настоящего изобретения.
Описание фигур
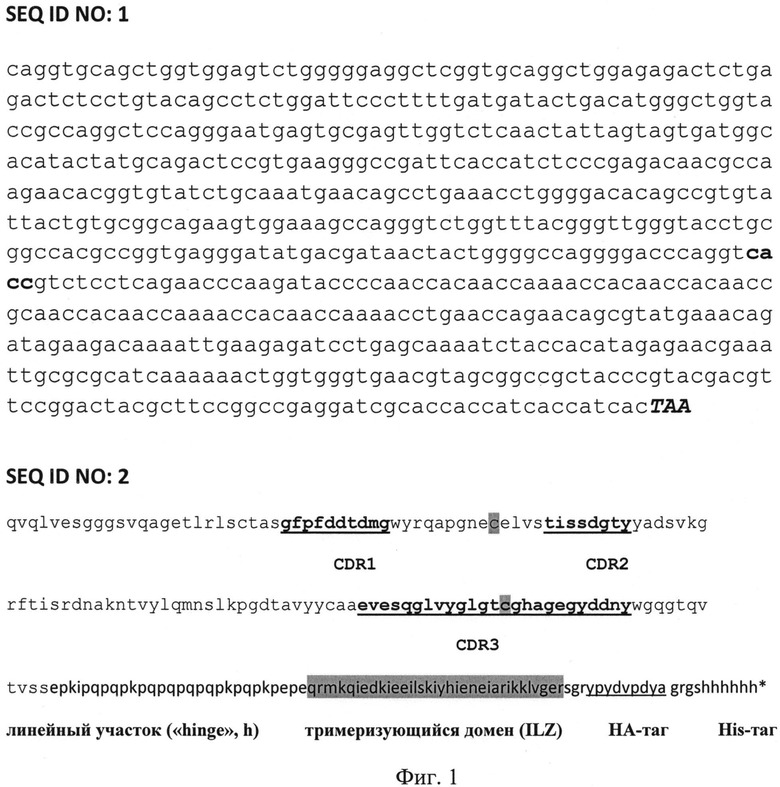
На фиг.1 представлены:
SEQ ID NO: 2 - аминокислотная последовательность отобранного тримеризованного однодоменного антитела, способного специфически связывать и нейтрализовать вирус бешенства,
SEQ ID NO: 1 - нуклеотидная последовательность, кодирующая аминокислотную последовательность отобранного тримеризованного однодоменного антитела, способного специфически связывать и нейтрализовать вирус бешенства.
В указанной последовательности SEQ ID NO: 2 выделены жирным и подчеркнуты соответственно (слева направо, от N- к C-концу) гипервариабельные участки CDR1, CDR2 и CDR3, которые являются определяющими (особенно важен CDR3) для специфического узнавания данным одноцепочечным антителом поверхностного белка вируса бешенства. В аминокислотной последовательности выделены также два остатка цитозина, которые, исходя из многих предшествующих структурных исследований подобных молекул, могут образовывать дополнительную неканоническую Cys-Cys-связь и таким образом существенно стабилизировать структуру антитела.
Добавленный при тримеризации исходного однодоменного антитела фрагмент содержит линейный участок (линкерный район, соответствующий длинному шарнирному участку верблюжьего антитела («hinge», h), принципиально важный для усиления нейтрализующей вирус функции тримеризующийся домен (вариант лейциновой молнии, ILZ, выделен серым цветом), а также (на самом конце) участки, кодирующие два олигопептидных тага: HA-таг и (His)6-таг. На конце находится терминирующий кодон TAA (в аминокислотной последовательности ему соответствует звездочка).
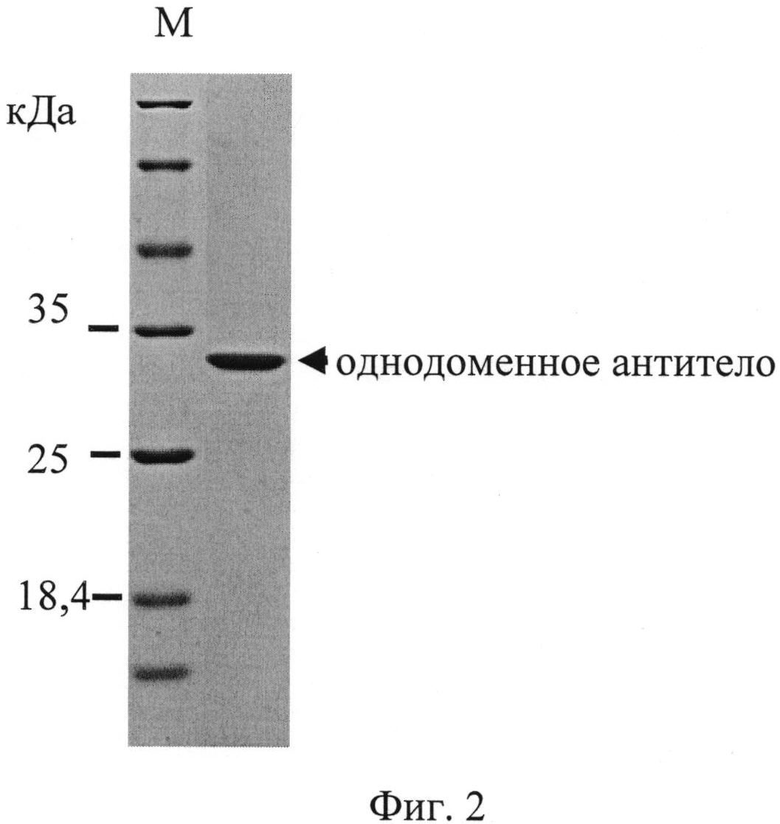
На фиг.2 представлены данные анализа очищенного тримеризованного однодоменного антитела, специфически связывающегося с вирусом бешенства, с помощью электрофореза по Лэммли в разделяющем 14% полиакриламидном геле в денатурирующих и восстановительных условиях (с додецил сульфатом натрия, SDS, и с дитиотреитолом, ДТТ). В левой дорожке нанесен маркер молекулярного веса.
Тримеризованное однодоменное антитело было наработано в периплазме бактерий и затем почищено (в том числе с помощью аффинной металл-хелатной хроматографии и последующей гель-фильтрационной хроматографии) до практически индивидуально белкового продукта, мигрирующего в геле в зоне белков размером примерно 30 кДа (реальный размер - примерно 22 кДа; антитело движется в геле замедленно вследствие особенностей структуры, в частности из-за наличия концевых положительно заряженных шести остатков гистидина (His-таг)).
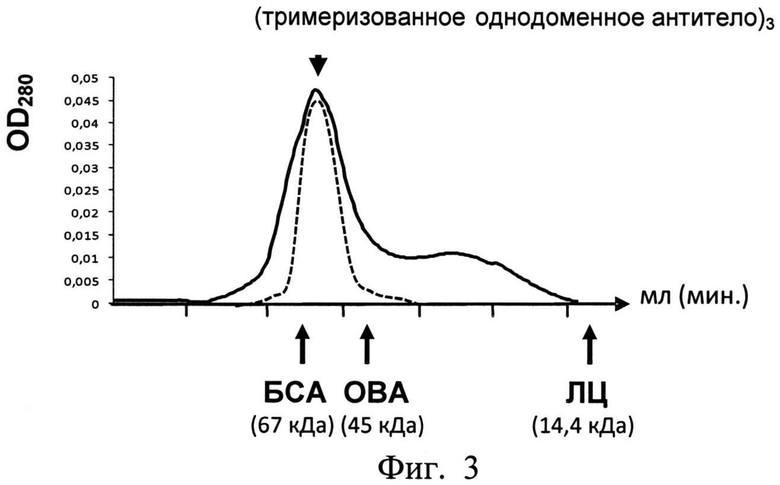
На фиг.3 показан профиль гель-фильтрационной хроматографии очищенного тримеризованного однодоменного антитела, пропущенного через колонку Sephacryl S-100 HR 1.5×50 в стандартном буферно-солевом растворе PBS (в нормальных физиологических условиях без денатурации). Белки с известным молекулярным весом были использованы как маркеры: Бычий сывороточный альбумин, БСА - 67 кДа; Овальбумин, ОВА - 45 кДа; Лизоцим, ЛЦ - 14,4 кДа. Пунктирной линией обозначен профиль очищенного препарата антитела, полученный в результате повторного раунда гель-фильтрации. Препарат антитела элюируется во фракциях, соответствующих тримеризованному антителу.
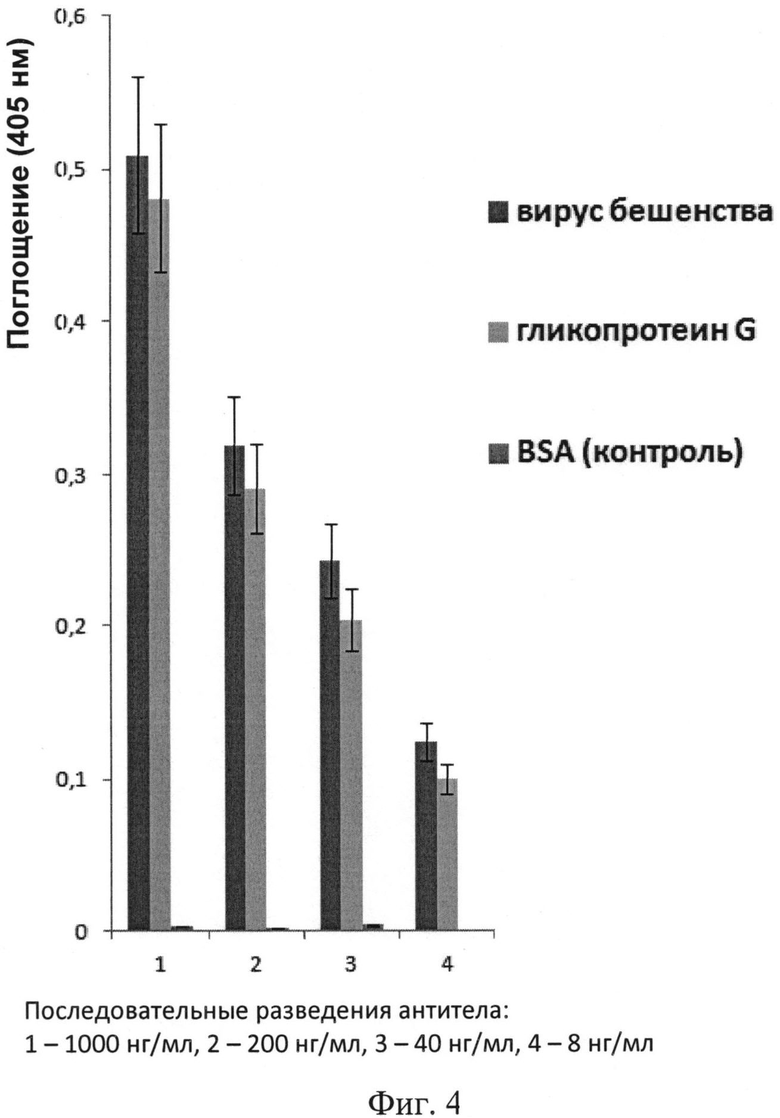
На фиг.4 проиллюстрированы результаты иммуноферментного анализа связывания тримеризованных однодоменных антител с иммобилизованными в лунке планшета препаратом инактивированного вируса бешенства и белком гликопротеином G вируса бешенства. В качестве контроля использовали лунки с иммобилизованным белком бычьим сывороточным альбумином (BSA). Тримеризованное однодоменное антитело брали в последовательных разведениях с фактором 5, начиная с концентрации 1 мкг/мл. Представлены полученные данные поглощения при длине волны 405 нм (ось ординат), отражающие эффективность связывания тримеризованных одноцепочечных антител с иммобилизованными антигенами. Представлены усредненные полученные данные. Отрезками в верхней части столбцов показаны диапазоны отклонений результатов в индивидуальных ячейках.
Примеры осуществления настоящего изобретения
Пример 1.
Получение библиотеки вариабельных доменов неканонических одноцепочечных антител.
Двугорбого верблюда Camelus bactrianus последовательно иммунизировали 5 раз путем подкожного введения антигенного материала. Вторую иммунизацию проводили через 3 недели после первой, затем с интервалом в 10-14 дней проводили еще три иммунизации.
В качестве антигена использовали вакцину антирабическую из штамма "Щелково-51" инактивированную жидкую культуральную (Рабиков) (ФГУП "Щелковский биокомбинат", Россия).
Взятие крови (150 мл) проводили через 5 дней после последней инъекции. Для предотвращения свертывания взятой крови добавляли 50 мл стандартного солевого раствора (PBS), содержащего гепарин (100 ед./мл) и ЭДТА (3 мМ).
Кровь разводили в 2 раза стандартным солевым раствором (PBS), содержащим 1 мМ ЭДТА. 35 мл разбавленного раствора крови наслаивали на ступеньку специальной среды (Histopaque-1077, Sigma) с плотностью 1,077 г/мл объемом 15 мл и проводили центрифугирование в течение 20 мин при 800×g. Мононуклеарные клетки (лимфоциты и моноциты) отбирали из интерфазной зоны плазма/Histopaque, после чего промывали раствором PBS, содержащим 1 мМ ЭДТА.
Суммарную РНК из В-лимфоцитов выделяли с помощью реагента TRIzol (Invitrogen). Затем, на колонке с олиго(dT)-целлюлозой из тотальной РНК выделяли поли(A)-содержащую РНК. Концентрацию РНК определяли с помощью биофотометра (Eppendorf) и проверяли качество выделенной РНК с помощью электрофореза в 1,5%-ном агарозном геле с формальдегидом.
Реакцию обратной транскрипции проводили с использованием обратной транскриптазы H-M-MuLV и праймера олиго(dt)15 в качестве затравки по стандартному протоколу фирмы-производителя фермента (Fermentas/Thermo Fisher Scientific).
Продукты обратной транскрипции использовали в качестве матрицы в двухступенчатой полимеразной цепной реакции, и полученные продукты амплификации клонировали по сайтам Ncol(Pstl) и Notl в фагмидный вектор (pHEN4), как описано ранее [Hamers-Casterman C., Atarhouch Т., Muyldermans S. et al. Nature 1993; 363: 446-448.; Nguyen V.K., Desmyter A., Muyldermans S. Adv. Immunol. 2001; 79: 261-296; Saerens D., Kinne J., Bosmans E., Wernery U., Muyldermans S., Conrath K. J Biol Chem. 2004; 279: 51965-51972; Rothbauer U., Zolghadr K., Tillib S., et al. Nature Methods 2006; 3: 887-889]. Процедуру селекции проводили также аналогично тем, что описаны в указанных работах. Она базировалась на методе фагового дисплея, в которой в качестве фага-помощника использовали бактериофаг M13KO7 (New England Biolabs, США).
В результате проведенной работы была получена библиотека однодоменных антител.
Пример 2.
Селекция однодоменных антител, специфически узнающих вирус бешенства (гликопротеин G), их тримеризация (модификация, форматирование).
Селекцию однодоменных антител проводили методом фагового дисплея с использованием препарата инактивированного вируса бешенства, а на финальной стадии также и гликопротеина G вируса бешенства (из коммерческого набора PLATELLATM RABIES II KIT, Bio-Rad, Франция), иммобилизованного на дне лунок 96-луночного ИФА-планшета. Использовали полистироловые иммунологические плашки с высокой сорбцией MICROLON 600 (Greiner Bio-One). Для блокировки использовали 1% БСА (Sigma-Aldrich, США) и/или 1% обезжиренное молоко (Bio-Rad, США) в PBS. Процедуру селекции и последующей амплификации отбираемых фаговых частиц (содержащих ген однодоменного антитела внутри, а экспрессирующееся однодоменное антитело - в составе поверхностного фагового белка pIII) повторяли, как правило, последовательно три раза. Все манипуляции проводили, как описано в публикациях Тиллиб С.В., Иванова Т.И., Васильев Л.А. 2010. Фингерпринтный анализ селекции «однодоменных мини-антител» методом фагового дисплея с использованием двух вариантов фагов-помощников. Acta Naturae 2010; 2 (3): 100-108; Hamers-Casterman С, Atarhouch Т., Muyldermans S. et al. Nature 1993; 363: 446-448.; Nguyen V.K., Desmyter A., Muyldermans S. Adv. Immunol. 2001; 79: 261-296; Saerens D., Kinne J., Bosmans E., Wernery U., Muyldermans S., Conrath K. J Biol Chem. 2004; 279: 51965-51972; Rothbauer U., Zolghadr K., Tillib S., et al. Nature Methods 2006; 3: 887-889].
Последовательности клонов отобранных однодоменных антител группировали согласно схожести их фингерпринтов, получаемых при электрофоретическом разделении продуктов гидролиза амплифицированных последовательностей однодоменных антител параллельно тремя частощепящими рестрикционными эндонуклеазами (Hinfl, Mspl, Rsal). Затем, отобранные клоны-представители каждой из различающихся по фингерпринту групп использовали для аналитической наработки однодоменных антител, активность которых проверяли методом иммуноферментного анализа на предмет эффективного связывания с иммобилизованным в ячейке иммунологического планшета препарата инактивированного вируса гриппа птиц A/Mallard duck/PA/10218/84(H5N2).
В результате была отобрана последовательность, кодирующая однодоменное антитело (фиг.1, SEQ ID NO: 2).
Для того чтобы повысить эффективность выделения и детекции однодоменного антитела, а также с целью значительно усилить биологическую активность исходно отобранного моновалентного однодоменного антитела, была использована разработанная нами ранее эффективная процедура тримеризации (форматирования, модифицирования аминокислотной последовательности антитела), подробно описанная в работе Тиллиба и соавт. (Tillib et al., 2013, Antiviral Res. 97: 245-254. Formatted single-domain antibodies can protect mice against infection with influenza virus (H5N2) // Форматированные однодоменные антитела могут защитить мышь от инфицирования вирусом гриппа (H5N2)). Эта процедура включала присоединение к С-концу исходно отобранного антитела четырех дополнительных аминокислотных участков (доменов), представленных на фиг.1 - линкерного линейного участка (соответствующего верхнему удлиненному шарнирному участку особых иммуноглобулинов lgG2a верблюда), - специального тримеризующегося домена - "ILZ" ("isoleucine zipper domain", или, в переводе, «изолейциновой молнии»), - НА-тага (пептида из белка гемагглютинина, к которому есть коммерческие антитела) и His-тага (шести остатков гистидина на самом С-конце белка для эффективной очистки белка с помощью металл-хелатной хроматографии).
Пример 3.
Наработка тримеризованных однодоменных антител, специфически узнающих вирус бешенства (гликопротеин G).
Последовательность кодирующей ДНК отобранного и тримеризованного, как описано выше, одноцепочечного антитела, указанного на фиг.1, клонировали в экспрессионный плазмидный вектор - модифицированный вектор pHEN6 [Conrath КЕ, Lauwereys М, Galleni М, Matagne A, Frere JM, Kinne J, Wyns L, Muyldermans S. Beta-lactamase inhibitors derived from single-domain antibody fragments elicited in the Camelidae. Antimicrob Agents Chemother. 2001; 45: 2807-12]. Благодаря наличию на N-конце экспрессируемой последовательности сигнального пептида (peIB) нарабатываемый рекомбинантный белок (однодоменное антитело) накапливается в периплазме бактерий, что позволяет эффективно его выделять методом осмотического шока, не разрушая собственно бактериальные клетки. Наработку однодоменных антител проводили в Е.coli (штамм BL21). Экспрессию индуцировали добавлением 1 мМ индолил-бета-D-галактопиранозида и клетки инкубировали при интенсивном перемешивании в течение 7 часов при 37°C или в течение ночи при 28°С. Однодоменное антитело выделяли из периплазматического экстракта с использованием аффинной хроматографии на Ni-NTA-агарозе с использованием системы для очистки QIAExpressionist (QIAGEN, США).
Полученные в результате элюции белковые фракции концентрировали (до конечной концентрации примерно 1-5 мг/мл) с помощью ультрафильтрационных приспособлений (с размером отсечения 10 кДа) Amicon фирмы Millipore, после чего наслаивали в объеме 0,5-2 мл на колонку для гель-фильтрационной хроматографии (Sephacryl S-100 HR; 1,5×50 см). Хроматографию проводили, используя систему BioLogic LP фирмы Bio-Rad, при постоянной скорости потока 0,3 мл/мин. В качестве буферного раствора использовали стандартный солевой буферный раствор (PBS). Фракции собирали согласно профилю поглощения элюированного белка при длине волны 280 нм (фиг.3). Размер тримеризованного одноцепочечного антитела примерно соответствовал размеру (и пику элюции) маркерного белка - бычьего сывороточного альбумина (BSA, 67 кДа). Объединенные отобранные тримеризованные антитела (из фракций соответствующих пиков элюции) снова концентрировали (как описано выше), дополнительно очищали от эндотоксина на аффинной колонке фирмы Thermo Scientific (Detoxi-Gel Endotoxin Removing Gel), после чего стерилизовали фильтрованием через целлюлоза-ацетатную мембрану (фирмы Nalgene) с порами 0,2 мкм. Получаемый препарат тримеризованного одноцепочечного антитела на различных стадиях анализировали с помощью электрофореза в 14% SDS-полиакриламидном геле (фиг.2). Очищенные антитела хранили в аликвотах при 4°C или, после добавления 50% глицерина, при -20°C. Для более долгого хранения антитела лиофилизировали.
Пример 4.
Демонстрация специфического связывания полученного по изобретению тримеризованного одноцепочечного антитела с вирусом бешенства и с гликопротеином G вируса бешенства.
Способность полученного тримеризованного однодоменного антитела связывать как целый (инактивированный, но сохранивший целостность капсида) вирус бешенства (вакцинный штамм Щелково-51), так и конкретный его белок, гликопротеин G (из коммерческого набора PLATELLATM RABIES II KIT, Bio-Rad, Франция), иммобилизованные (из раствора с концентрацией примерно 4 мкг белка в 1 мл) в лунках иммунологического планшета, проверяли с использованием метода иммуноферментного анализа с использованием стандартного протокола. В качестве контроля использовали лунки с иммобилизованным белком бычьим сывороточным альбумином (неспецифический белок). Тримеризованное однодоменное антитело брали в последовательных разведениях с фактором 5, начиная с концентрации 1 мкг/мл. Детекцию связавшегося с гликопротеином G или с вирусом бешенства форматированного однодоменного антитела проводили с помощью анти-НА антител мыши, вторичных антител к иммуноглобулинам мыши, конъюгированных с пероксидазой хрена, и хромогенного субстрата АБТС (2,2′-азино-бис(3-этилбензтиазолин)-6-сульфоновой кислоты, Sigma). На фиг.4 представлены результаты анализа, из которых следует, что тримеризованное однодоменное антитело специфически связывается как с иммобилизованным вирусом бешенства, так и иммобилизованным белком гликопротеином G вируса бешенства, как в случае изолированного белка, так и, по-видимому, в случае, когда этот белок находится в составе вирусного капсида, но не связывается с бычьим сывороточным альбумином (BSA), использованным в качестве блокирующего агента.
Пример 5.
Демонстрация способности тримеризованного однодоменного антитела нейтрализовать вирус бешенства in vitro (т.е. вируснейтрализующей активности на культуре клеток).
Определение вируснейтрализующей активности тримеризованного однодоменного антитела проводили по общеизвестной утвержденной методике «Методические указания по определению вируснейтрализующих антител к вирусу бешенства в сыворотках крови животных в реакции нейтрализации в культуре клеток ВНК-21 (FAVN)», 2011 год.
Для сопоставления вируснейтрализующей активности тримеризованного однодоменного антитела по изобретению использовали широко известный в России китайский коммерческий антирабический иммуноглобулин («Сычуаньская Юанда Шуян Фармацевтическая компания», Китай). Проводить сравнение с прототипом не представлялось возможным из-за неполного раскрытия технологии получения антител авторами патента, а также отсутствия подобных антител в коммерческом доступе.
Предварительно была установлена концентрация белка в препарате китайского иммуноглобулина спектрофотометрически и проведено титрование на культуре клеток по вышеуказанной стандартной методике. По результатам титрования было установлено, что при разведении иммуноглобулина до 300 мкг/мл и выше уровень вируснейтрализующих антител в препарате является защитным (0,5 МЕ/мл). Соответственно, данную концентрацию взяли исходной для обоих исследуемых препаратов.
Далее проводили титрование в реакции нейтрализации тримеризованного однодоменного антитела и китайского иммуноглобулина одновременно. Результаты титрования приведены в таблице.
Таким образом, уровень вируснейтрализующих антител при концентрации белка 300 мкг/мл у китайского иммуноглобулина составил 0,5 МЕ/мл, а у тримеризованного однодоменного антитела при той же концентрации составил 9720 МЕ/мл, что в 19440 раз выше, чем у китайского иммуноглобулина.
В результате эксперимента было установлено, что тримеризованное однодоменное антитело способно нейтрализовать вирус бешенства in vitro и защищать культуру клеток от инфицирования вирусом бешенства при концентрации 300 мкг/мл и выше. Причем вируснейтрализующая активность тримеризованного однодоменного антитела по изобретению выше, чем у коммерческого иммуноглобулина в 19440 раз.
Промышленная применимость
Все представленные примеры подтверждают промышленную применимость заявленного тримеризованного однодоменного антитела для профилактики и терапии бешенства и подтверждают выполнение поставленной технической задачи.
| название | год | авторы | номер документа |
|---|---|---|---|
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПАССИВНОЙ ИММУНИЗАЦИИ ПРОТИВ БЕШЕНСТВА, ФАРМАЦЕВТИЧЕСКИЙ НАБОР, СПОСОБ ПРИМЕНЕНИЯ ФАРМАЦЕВТИЧЕСКОГО НАБОРА | 2016 |
|
RU2661028C2 |
| Рекомбинантный слитый белок, специфически связывающий и нейтрализующий вирус бешенства, способ его получения и применение | 2023 |
|
RU2829507C1 |
| Генетическая конструкция на основе оптимизированного гена консенсусного гликопротеина вируса бешенства для профилактики бешенства | 2018 |
|
RU2707544C1 |
| Нуклеотидная последовательность, оптимизированная для экспрессии в бактериях консенсусного гликопротеина вируса бешенства | 2018 |
|
RU2717255C1 |
| НЕЙТРАЛИЗУЮЩЕЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СВЯЗЫВАЮЩЕЕСЯ С ГЛИКОПРОТЕИНОМ G ВИРУСА БЕШЕНСТВА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО, И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ | 2017 |
|
RU2718835C2 |
| ПРЕПАРАТ НА ОСНОВЕ НЕЙТРАЛИЗУЮЩИХ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ, СВЯЗЫВАЮЩИХСЯ С ГЛИКОПРОТЕИНОМ ВИРУСА БЕШЕНСТВА | 2018 |
|
RU2711553C1 |
| ПРОТИВОВИРУСНОЕ ОДНОДОМЕННОЕ МИНИ-АНТИТЕЛО, НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ, ЭКСПРЕССИРУЮЩИЙ РЕКОМБИНАНТНЫЙ ВИРУСНЫЙ ВЕКТОР, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И СПОСОБ ПРОФИЛАКТИКИ ИЛИ ТЕРАПИИ ГРИППА ТИПА А | 2013 |
|
RU2536956C1 |
| НАНОАНТИТЕЛО, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩЕЕ БЕЛОК СЕА, СПОСОБ ЕГО ИСПОЛЬЗОВАНИЯ ДЛЯ ДЕТЕКЦИИ ЭТОГО БЕЛКА | 2012 |
|
RU2493166C1 |
| ОДНОДОМЕННОЕ МИНИ-АНТИТЕЛО aHER2/aSKBR3-1, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩЕЕ РЕЦЕПТОР ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА HER2/ERBB2/neu И СПОСОБНОЕ ЧЕРЕЗ ЭТО ВЗАИМОДЕЙСТВИЕ ПРОНИКАТЬ ВНУТРЬ КЛЕТКИ-МИШЕНИ (ИНТЕРНАЛИЗОВАТЬСЯ), СПОСОБ ПОЛУЧЕНИЯ ДАННОГО АНТИТЕЛА И СПОСОБ ДЕТЕКЦИИ БЕЛКА HER2/ERBB2/neu И КЛЕТОК, ЭКСПРЕССИРУЮЩИХ ЭТОТ БЕЛОК В ПОВЫШЕННОМ КОЛИЧЕСТВЕ, С ПОМОЩЬЮ МИНИ-АНТИТЕЛА aHER2/aSKBR3-1 | 2012 |
|
RU2522929C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ ФРАГМЕНТЫ (Fab) ПРОТИВ ВИРУСА БЕШЕНСТВА, ИЗОЛИРОВАННЫЙ ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ Fab ПРОТИВ ВИРУСА БЕШЕНСТВА, КЛЕТКА ДРОЖЖЕЙ, ТРАНСФОРМИРОВАННАЯ ФРАГМЕНТОМ ДНК, И СПОСОБ ПОЛУЧЕНИЯ Fab ПРОТИВ ВИРУСА БЕШЕНСТВА С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ | 2010 |
|
RU2440412C2 |
Настоящее изобретение относится к области иммунологии и вирусологии и предполагает использование антител, представляющих собой тримеризованные однодоменные антитела, для профилактики и терапии бешенства. Создано тримеризованное однодоменное антитело, специфически связывающееся с поверхностным (капсидным) белком - гликопротеином G, вируса бешенства, имеющее аминокислотную последовательность SEQ ID NO: 2, аминокислотная последовательность которого кодируется нуклеотидной последовательностью SEQ ID NO: 1, и при этом нейтрализующее инфекционную активность вируса бешенства. 2 з.п. ф-лы, 4 ил., 1 табл.
1. Тримеризованное однодоменное антитело, специфически связывающееся с гликопротеином G вируса бешенства, имеющее аминокислотную последовательность SEQ ID NO: 2.
2. Тримеризованное однодоменное антитело по п.1, аминокислотная последовательность которого кодируется нуклеотидной последовательностью SEQ ID NO: 1.
3. Тримеризованное однодоменное антитело по п.1, отличающееся тем, что оно нейтрализует вирус бешенства.
| US 2013039926 A1, 14.02.2013 | |||
| US 2009041777 A1, 12.02.2009 | |||
| СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ ВИРУСА БЕШЕНСТВА НЕЙТРАЛИЗУЮЩИЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА И НУКЛЕИНОВЫЕ КИСЛОТЫ И СВЯЗАННЫЕ С НИМИ СПОСОБЫ | 2001 |
|
RU2272809C2 |

