Область техники
Настоящее изобретение относится к области биотехнологии, иммунологии, медицины. Оно касается рекомбинантного белка на основе однодоменных антител, специфически связывающего и нейтрализующего вирус бешенства; штамма-продуцента рекомбинантного белка (CHO-AntiRabD-ABD) на основе линии эукариотических клеток яичника китайского хомячка CHO; применение рекомбинантного белка для нейтрализации вируса бешенства у млекопитающих в схеме экстренной лечебно-профилактической иммунизации. Изобретение может быть использовано в фармацевтической промышленности для получения лекарственных препаратов для пассивной иммунизации против бешенства.
Предшествующий уровень развития
Бешенство - вакциноконтролируемое зоонозное вирусное заболевание, поражающее центральную нервную систему. Заболевание вызывает вирус бешенства Rabies virus, относящийся к роду Lyssavirus семейства Rhabdoviridae. На этапе появления клинических симптомов его летальность составляет 100%. По данным ВОЗ передача вируса бешенства людям почти в 99% случаев происходит от домашних собак. Инфекция передается через слюну, обычно при укусах, царапинах или прямом контакте со слизистыми оболочками (например, глаза, ротовая полость или открытые раны). Бешенство присутствует на всех континентах за исключением Антарктиды, причем 95% случаев смерти людей происходят в странах Азии и Африки. (ВОЗ, Информационный бюллетень Бешенство, 19 января 2023 г.).
В России, по данным Роспотребнадзора, ежегодно за антирабической помощью обращается около 400 тысяч человек, из них около 250 тысяч человек нуждаются в проведении специфического лечения. Социально/экономическое значение проблемы бешенства неуклонно растет вследствие формирования новых природных очагов инфекции. В 2021 году зарегистрировано 6 случаев гидрофобии, в 2020 году - 7 случаев гидрофобии среди населения Российской Федерации. По поводу укусов, ослюнения и оцарапывания животными в 2021 году в медицинские организации обратилось более 333 тыс. человек (227,7 на 100 тыс. населения), в 2020 - более 338 тыс. (230,5 на 100 тыс. населения). Из них укусы диких животных составили 2021 году - 10 332 (7,05 на 100 тыс. населения), в 2020 году 8 227 случаев (5,61 на 100 тыс. населения) (https://www.rospotrebnadzor.ru/activities/recommendations/details.php?ELEMENT_ID=20827). Экономический ущерб от укусов, ослюнений, оцарапываний животными, потенциальными источниками возбудителя, в 2019 году по некоторым подсчетам составил более 4 млрд рублей (Михеева М.А., Михеева И.В. Динамика рейтинга экономического ущерба от инфекционных болезней как критерий эффективности эпидемиологического контроля. Журнал микробиологии, эпидемиологии и иммунобиологии. 2020; 97(2): 174-181).
До настоящего времени эффективных средств и методов лечения уже развившегося заболевания бешенством не разработано. Однако даже после контакта с подозрительным животным можно предотвратить заболевание бешенством с помощью методики, называемой «пост экспозиционная профилактика» (также экстренная ПЭП, экстренная лечебно-профилактическая иммунизация). Возможность ее осуществления основана на длительном инкубационном периоде бешенства, за время которого после введения антирабических препаратов формируется иммунитет и заболевания не происходит. Инкубационный период обычно длится 2-3 месяца, но может варьировать от недели до года в зависимости от таких факторов, как место проникновения вируса в организм и вирусная нагрузка (ВОЗ, Информационный бюллетень Бешенство, 19 января 2023 г.). При этом меры постэкспозиционной профилактики зависят от степени контакта с животным, которое предположительно может быть заражено бешенством. ВОЗ требует проведение мер ПЭП при контактах категории II (промывание раны и срочная вакцинация) или III (промывание раны, срочная вакцинация и введение антирабического иммуноглобулина/моноклональных антител).
Таким образом, при разработке мер ПЭП учтено, что индукция активного иммунитета требует некоторого времени и в случае контактов III категории, вызывающих более серьезные опасения, в схему добавляется пассивная иммунизация антирабическим иммуноглобулином (АИГ, RIG) или моноклональными антителами (mAb), действие которых наступает практически с момента введения. Иммуноглобулин против бешенства обеспечивает пассивную защиту путем нейтрализации вируса в ране в период до появления антител, индуцированных вакциной через 7-10 дней после начала первичного постконтактного курса.
В настоящее время в мире чаще используются иммуноглобулин антирабический человеческий (HRIG), полученный из плазмы крови иммунизированных людей-доноров, и иммуноглобулин антирабический лошадиный (ERIG) из сыворотки крови иммунизированных лошадей. Данные препараты содержат высокий титр специфичных к вирусу бешенства иммуноглобулинов.
Известен человеческий антирабический иммуноглобулин высокой степени очистки (патент CN 101450967A «High-purity rabies human immunoglobulin and raw material production method thereof» - Антирабический человеческий иммуноглобулин высокой степени очистки и сырье для его получения, методы использования). Полученный по такому способу препарат характеризуется тем, что содержание иммуноглобулина составляет более 98% от общего содержания белка; титр антирабических антител - равен или больше, чем 150 МЕ/мл, содержание белка - меньше, чем 120 г/л. При этом содержание мономера и димера иммуноглобулина Ig G составляет более чем 99%. Процесс получения такого иммуноглобулина достаточно трудоемок и включает несколько этапов: 1) подготовка сырья на основе крови с титром антирабических антител 10 МЕ/мл крови; 2) фракционирование холодным этанолом, очистка; 3) ионообменная хроматография; 4) инактивация вирусов инкубацией при низком pH; 5) мембранная фильтрация для удаления вирусов; 6) ультрафильтрация, дозирование, стерилизация.
Известен препарат гетерологичного антирабического иммуноглобулина для внутривенного и внутримышечного введения и способ его получения (Патент РФ 2339401). В патенте предложен вирусбезопасный способ получения гетерологичного антирабического иммуноглобулина из высокоспецифичной гетерологичной антирабической сыворотки каприлатно-спиртовым методом с пепсиновой обработкой, который позволяет получить высокоочищенный гетерологичный антирабический иммуноглобулин для внутривенного и внутримышечного введения. При этом в качестве исходного сырья используют высокоспецифичную антирабическую сыворотку, полученную от иммунизации животных-продуцентов очищенным вирусом бешенства, выращенным на культуре клеток Vero или ПСХ.
Однако поликлональные антирабические иммуноглобулины (АИГ) крови (лошадиный и человеческий) имеют существенные недостатки:
- ограниченная производственная мощность, малый масштаб производства (особенно для АИГ из крови человека), отсутствие волонтеров-доноров или проблемы с животными-донорами;
- проблемы достижения качества - различия между партиями, влияющие на эффективность и безопасность, дополнительно содержат ненейтрализующие антитела;
- низкая безопасность - риск передачи инфекций, в т.ч. зоонозов через кровь доноров;
- возможная реактогенность, введение гетерологичного для человека лошадиного иммуноглобулина может сопровождаться развитием аллергических реакций, в том числе анафилактическим шоком и сывороточной болезнью;
- проблема интерференции с антирабическими вакцинами, пассивные антитела могут нарушать продукцию вакцинных антител;
- человеческий АИГ может мешать иммунному ответу на живые вакцины, такие как корь, эпидемический паротит, полиомиелит или краснуху, их нельзя вводить в течение 3 месяцев после АИГ;
- ограниченное проникновение в головной мозг из-за наличия гематоэнцефалического барьера (ГЭБ) и крупного размера антител (≈150 КДа). (Kumar N.N., Pizzo M.E., Nehra G. et al. Passive immunotherapies for central nervous system disorders: current delivery challenges and new approaches // Bioconjug Chem. 2018 Dec 19;29(12):3937-3966).
Моноклональные антитела из-за наличия недостатков у классических иммуноглобулинов рассматриваются в качестве предпочтительного варианта для разработок и внедрения в практику на современном этапе (Fan L., Zhang L., Li J., Zhu F. Advances in the progress of monoclonal antibodies for rabies. Hum Vaccin Immunother. 2022 Dec 31;18(1):2026713). В мире препараты на основе антирабических моноклональных антител находятся на различных стадиях разработки. Индия является первой страной, где лицензированы два препарата на основе рекомбинантных антирабических mAb - Rabishield (SII RMab, одно гомологичное гуманизированное Mab) и Twinrab™ (RabiMabs, гетерологичный коктейль из 2-х мышиных Mabs).
Известно нейтрализующее моноклональное антитело, связывающееся с гликопротеином G вируса бешенства, фрагменты ДНК, кодирующие указанное антитело, и антигенсвязывающий фрагмент (Патент РФ 2718835). Предложенное авторами нейтрализующее моноклональное антитело или его антигенсвязывающий фрагмент, селективно связывающее гликопротеин вируса бешенства, включает вариабельные участки тяжелой (VH) и легкой (VL) цепей. При этом вариабельный домен тяжелой цепи иммуноглобулина (VH) включает последовательность гипервариабельных регионов CDRH1, CDRH2 и CDRH3, а вариабельный домен легкой цепи иммуноглобулина (VL) включает последовательность гипервариабельных регионов CDRL1, CDRL2 и CDRL3. Изобретение позволяет получить антитело с высокой аффинностью.
Известно биспецифическое антитело против вируса бешенства и его применение (Патент РФ 2764740). Биспецифическое антитело содержит два антиген-связывающих фрагмента, которые связываются с эпитопом I и эпитопом III G-белка вируса бешенства, и обладает нейтрализующей вирус бешенства активностью. Также изобретение относится к фармацевтической композиции, включающей указанное антитело, и способу предупреждения или лечения бешенства.
Однако препараты антирабических моноклональных антител (MAт) также имеют существенные недостатки:
- негуманизированные МАт можно использовать только один раз в жизни человека из-за формирования иммунного ответа к ним, а гуманизация является сложной технологической задачей.
- достаточно высокие показатели интерференции с иммунным ответом на вакцину, сравнимые с АИГ (Investigation of SYN023 interference with vaccine-induced immunity in these animal studies found reductions in RVNA titers at doses at and above 0.1 mg/kg that were similar to reductions observed with RIG. A phase I study conducted in China found the ARV MAbs to be safe, with no serious adverse events reported (Ding et al., 2020)). Выбор подходящей дозы для коктейля mAb является сложной задачей, так как слишком высокая доза может повлиять на иммунный ответ на вакцину и, таким образом, увеличить риск развития бешенства.
Решением проблемы может стать разработка препаратов на основе однодоменных антител (VHH). Средняя молекулярная масса однодоменных антител составляет 15 кДа, что в десять раз меньше, чем обычное антитело, это позволяет легче преодолевать гематоэнцефалический барьер (Горшкова Е.Н., Василенко Е.А., Тиллиб С.В. и др. Однодоменные антитела и биоинженерные препараты на их основе: новые возможности для диагностики и терапии // Медицинская иммунология. 2016. Т. 18. № 6. С. 505-520., Elodie Pothin, Dominique Lesuisse and Pierre Lafaye. Brain delivery of single-domain antibodies: a focus on VHH and VNAR). Антигенсвязывающий участок формируется только одним вариабельным доменом VHH, подобные белковые структуры практически не вызывают иммунного ответа (не являются иммуногенами для организма пациента), VHH не вызывают Fc - зависимой интерференции с антирабической вакциной. В мире разработки на основе антирабических однодоменных антителах находятся на различных стадиях, но о зарегистрированных к применению препаратах не известно.
Однако однодоменные антитела (VHH) имеют существенный недостаток для применения в качестве пассивной антирабической ПЭП.
Из-за небольшого размера VHH имеют короткий период полувыведения из плазмы крови, равный 1,16 часа (Terryn S., Francart A., Lamoral S. et al. Protective effect of different anti-rabies virus VHH constructs against rabies disease in mice. PLoS One. 2014 Oct 27;9(10):e109367) по сравнению с несколькими неделями полувыведения для обычных антител (Paul, P.S.; Mengeling, W.L.; Pirtle, E.C. Duration and biological half-life of passively acquired colostral antibodies to porcine parvovirus. Am. J. Vet. Res. 1982, 43, 1376-1379). Таким образом, возникает большой иммунный разрыв между однократным введением антирабических VHH и образованием вакцинных антител на 7 день после проведения ПЭП, что может вызвать заболевание бешенством у покусанного человека.
Из патента РФ № 2661028 известен один из подходов к пролонгированию фармакокинетики антирабического однодоменного антитела. Авторы изобрели фармацевтическую композицию для экстренной профилактики бешенства, включающую два препарата. Препарат 1 содержит вирус-нейтрализующие антирабические однодоменные антитела, специфичные к гликопротеину G вируса бешенства, действие которых начинается с 0,5 часов после введения и продолжается вплоть до 12 часов. Пролонгирует действие однодоменных антител Препарат 2, содержащий рекомбинантные псевдоаденовирусные частицы, несущие ген вирус-нейтрализующих антирабических однодоменных антител, действие которых начинается с 6-ти часов после введения в организм и продолжается вплоть до 7 дней с поддержанием защитного уровня антител рекомендованного ВОЗ (0,5 МЕ/мл в сыворотке крови). Таким образом, недостаток однодоменных антител связанный с их быстрым выведением из организма компенсируется экспрессией данных антител с помощью аденовирусных векторов. Однако основным недостатком предложенного решения является высокая стоимость фармацевтической композиции из-за необходимости производить второй препарат с использованием дорогостоящей технологии наработки вирусных векторов. При этом в случае наличия предсуществующих антител к аденовирусному вектору возможно снижение эффективности экстренной профилактики бешенства.
В Российской Федерации для экстренной ПЭП бешенства применяются два типа препаратов - антирабический иммуноглобулин лошадиный (гетерологичный) отечественного производства, а также антирабический иммуноглобулин человеческий (гомологичный) импортного производства. Из импортных препаратов зарегистрированы два препарата: Ребинолин («Камада», Лтд., Израиль) и Иммуноглобулин антирабический из сыворотки крови человека («Сычуаньская Юанда Шуян Фармацевтическая компания», ООО, Китай). Препараты моноклональных и однодоменных антирабических антител в РФ не зарегистрированы к применению.
Таким образом, в уровне техники существует потребность в разработке недорого средства для экстренной профилактики бешенства на основе однодоменных антител (VHH), обладающего улучшенными фармакокинетическими параметрами и не оказывающего влияния на эффективность антирабической вакцины.
Раскрытие сущности изобретения
Технической задачей изобретения является расширение арсенала средств для пассивной иммунизации млекопитающих в схеме экстренной лечебно-профилактической иммунизации против бешенства.
Технический результат заключается в получении рекомбинантного слитого белка, на основе антирабического однодоменного антитела, с пролонгированным периодом полувыведения, который можно применять для пассивной иммунизации в схеме экстренной лечебно-профилактической иммунизации против бешенства, без снижения эффективности антирабической вакцины.
Указанный технический результат достигается тем, что разработан рекомбинантный слитый белок, на основе однодоменного антитела, специфически связывающий и нейтрализующий вирус бешенства, где указанный белок представлен аминокислотной последовательностью SEQ ID NO:1.
Существует вариант изобретения, в котором разработанный рекомбинантный слитый белок кодируется нуклеотидной последовательностью SEQ ID NO:2.
Кроме того, технический результат достигается тем, что разработан штамм-продуцент рекомбинантного слитого белка CHO-AntiRabD-ABD, полученный на основе линии клеток яичника китайского хомячка, в геном которых интегрирована экзогенная последовательность, которая представлена экспрессионной кассетой, включающей SV40-промотор, ген устойчивости к пуромицину и сигнал полиаденилирования, экспрессионной кассетой, включающей EF1-промотор, ген рекомбинантного разработанного слитого белка, сигнал полиаденилирования вируса SV40, причем место интеграции представлено нуклеотидной последовательностью SEQ ID NO: 3 единая направляющая РНК к которому закодирована последовательностью SEQ ID NO: 4.
Также технический результат достигается тем, что разработан способ получения рекомбинантного слитого белка, включающий следующие стадии:
- культивирование штамма - продуцента в подходящих условиях для продуцирования и накопления разработанного рекомбинантного слитого белка;
- очистка рекомбинантного слитого белка из культуральной жидкости.
Кроме того, технический результат достигается тем, что разработано применение рекомбинантного слитого белка, заключающееся в его введении млекопитающим в количестве, достаточном для эффективной нейтрализации вируса бешенства.
Разработан вариант применения рекомбинантного слитого белка, который предусматривает однократное введение млекопитающему, что обеспечивает нейтрализацию вируса бешенства на протяжении не менее 7 суток.
Другой вариант применения рекомбинантного слитого белка отличается тем, что, введение млекопитающим проводят одновременно с антирабической вакциной.
Краткое описание фигур
На фиг. 1 схематически представлена кодирующая последовательность SEQ ID NO:2 рекомбинантного слитого белка AntiRabD-ABD, которая в направлении от 5́'- конца к 3 - концу включает последовательности лидерного пептида IgGκ мыши (IgGκSP), антирабического домена, гибкого глицин-сериновый линкера (G2S)3 и альбумин - связывающего домена.
На фиг. 2 изображена схема рекомбинантного слитого белка, специфически связывающего и нейтрализующего вирус бешенства (SEQ ID NO:1). Светло - серым цветом выделена последовательность антирабического домена (AntiRabD), темно-серым цветом выделена последовательность альбумин-связывающего домена (ABD), между ними находится последовательность гибкого пептидного глицин-серинового линкера (G2S)3.
На фиг. 3 изображены составные части кодирующей последовательности единой направляющей РНК (нРНК) SEQ ID NO: 3 к месту интеграции в геноме клеток СНО (SEQ ID NO:4). Спейсерная часть (криспрРНК) отмечена серым цветом (20 п.н.), последовательность транс-активирующей РНК (тракрРНК), взаимодействующей с нуклеазой Cas9 из Streptococcus pyogenes (SpCas9) не имеет цветовой метки.
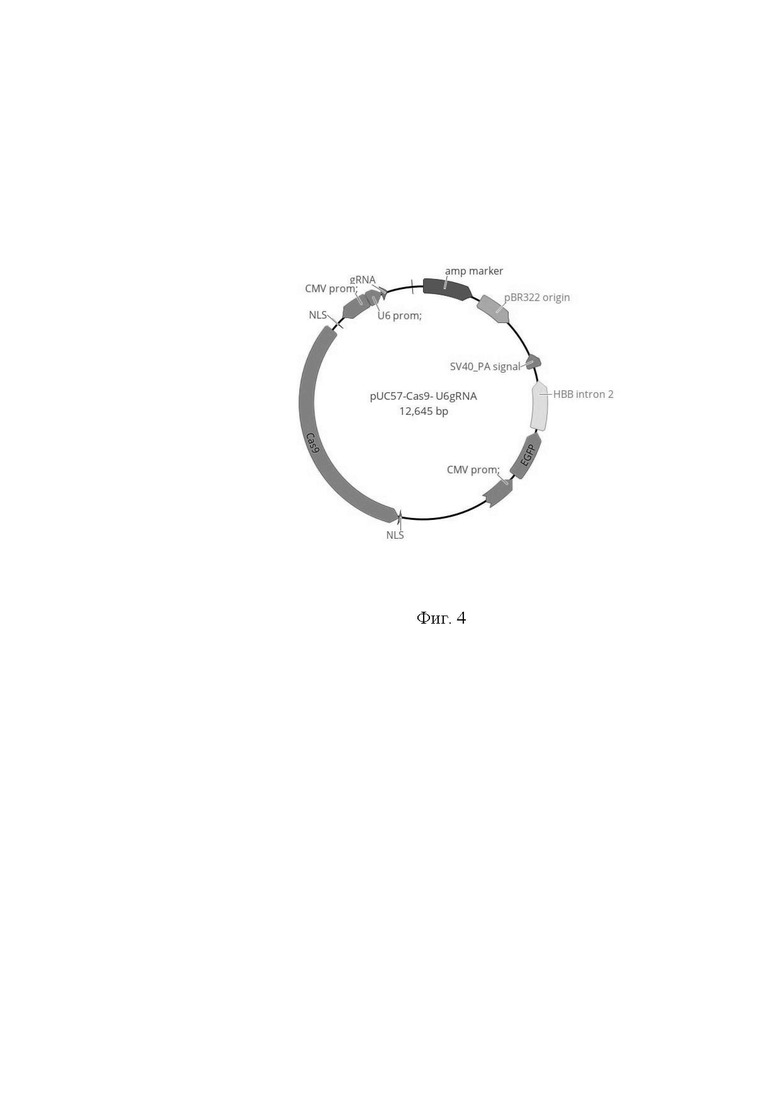
На фиг. 4 изображена схема созданной генетической конструкции pUC57-Cas9-U6gRNA, содержащей ген нуклеазы Cas9 из Streptococcus pyogenes (SpCas9) и единой направляющей РНК (нРНК, gRNA) к гену ЛДГА клеточной линии CHO. Последовательность нРНК находится под контролем промотора U6.
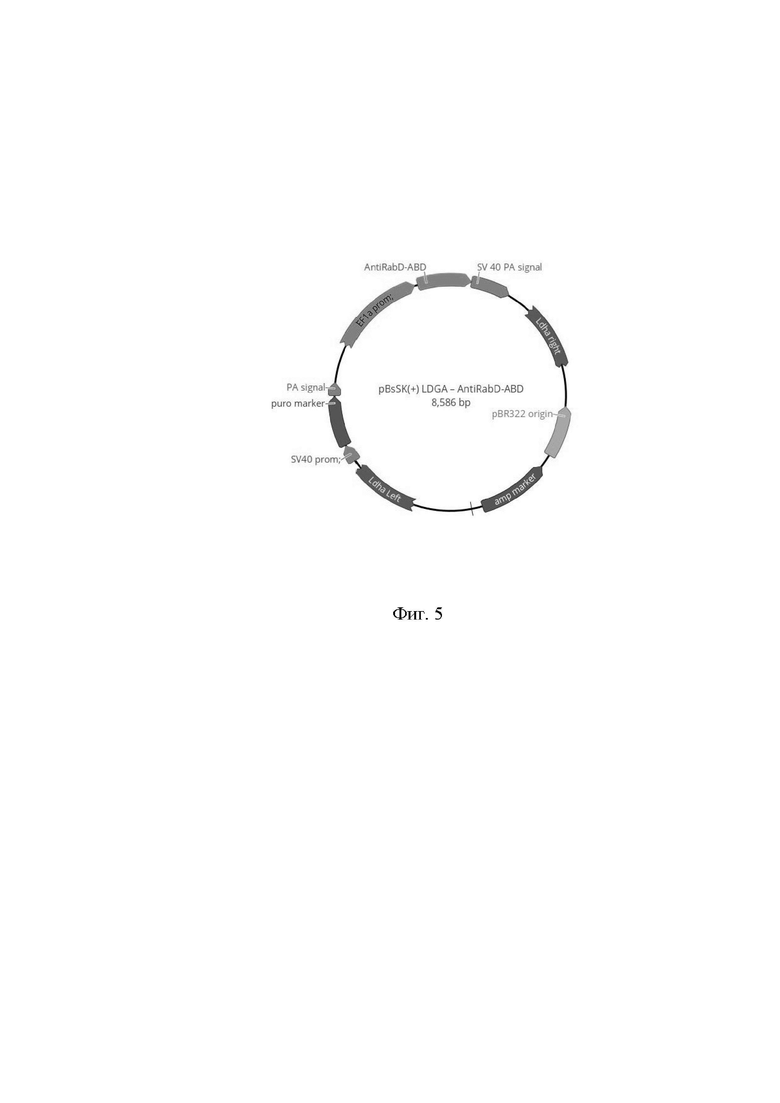
На фиг. 5 изображена схема созданной генетической конструкции pBsSK(+) LDGA - AntiRabD-ABD, содержащей экспрессионную кассету состоящую из EF1-промотора, гена рекомбинантного слитого белка SEQ ID NO: 2 и SV40 поли(А) терминатора; экспрессионную кассету состоящую из SV40-промотора, гена устойчивости к пуромицину и сигнал полиаденилирования; правое и левое плечо гомологии первого экзона гена ЛДГА клеточной линии CHO, обрамляющие две экспрессионные кассеты.
На фиг. 6 представлена схема расположения элементов экзогенной последовательности, находящейся в месте интеграции SEQ ID NO: 4 первого экзона гена ЛДГА в геноме клетки CHO-AntiRabD-ABD. Экзогенная последовательность содержит экспрессионную кассету, включающей SV40-промотор, ген устойчивости к пуромицину и синтетический сигнал полиаденилирования, экспрессионную кассету, включающую EF1-промотор, ген рекомбинантного слитого белка SEQ ID NO: 2, сигнал полиаденилирования вируса SV40.
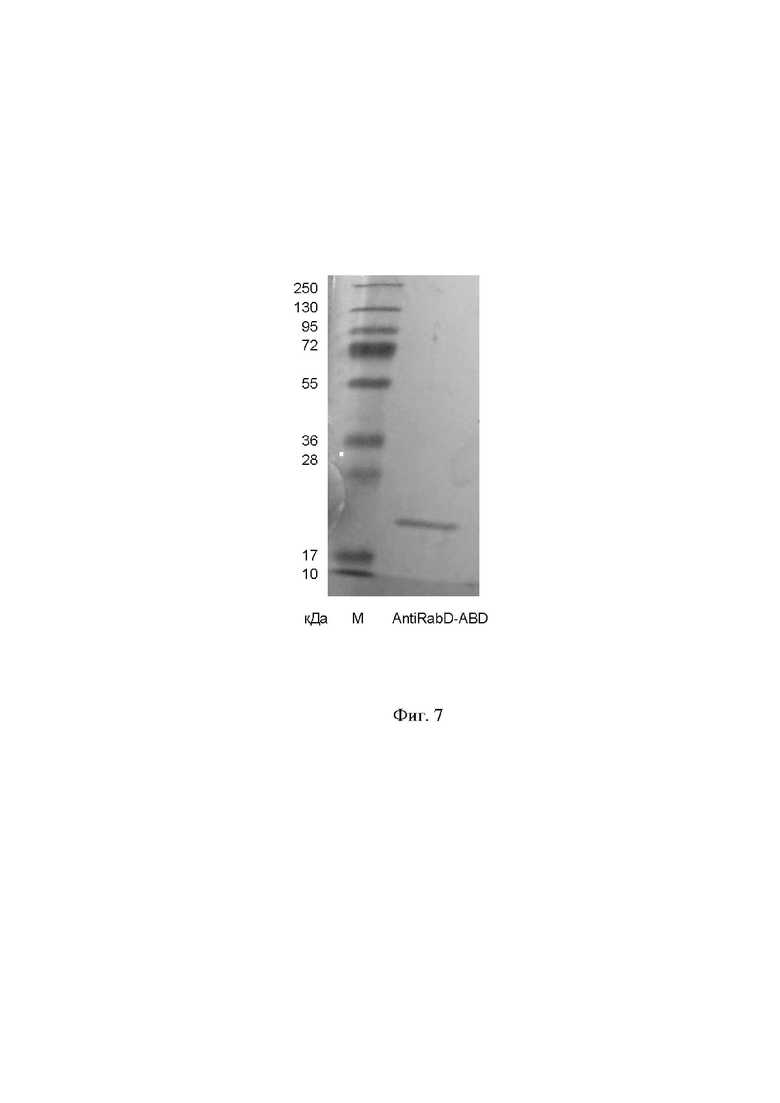
На фиг. 7 представлен электрофорез очищенного рекомбинантного слитого белка, специфически связывающего и нейтрализующего вирус бешенства, по Леммли в 10% ПААГ. На дорожку наносили 1,0 мкг белка. М - маркер PageRuler Plus Prestained Protein Ladder #26619 ThermoScientific. Окраска проводилась с использованием EZBlue Gel Staining Reagent. На дорожке, подписанной AntiRabD-ABD, детектирована полоса по молекулярной массе соответствующая рекомбинантному слитому белку (21,3 кДа).
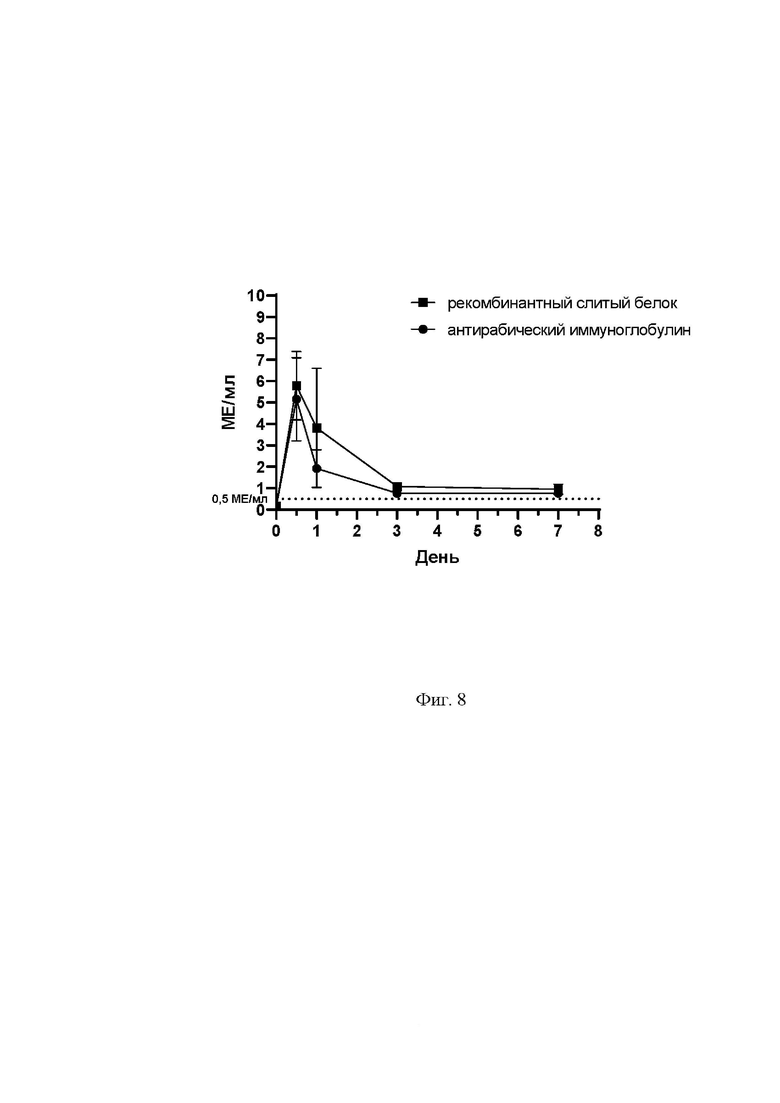
На фиг. 8 представлен график, отражающий динамику нейтрализующей активности против вируса бешенства (МЕ/мл) у рекомбинантного слитого антитела, измеренную на протяжении 7 дней в сыворотке крови крыс методом FAVN. Группе опыт вводили препарат рекомбинантного слитого антитела (квадратный маркер обозначает среднее значение по группе, вертикальные линии - разброс значений), группе сравнения вводили антирабический иммуноглобулин (круглый маркер обозначает среднее значение по группе, вертикальные линии - разброс значений). На графике точечной линией отмечен защитный уровень антирабических антител в сыворотке крови, рекомендуемый ВОЗ (0,5 МЕ/мл).
Реализация изобретения
Авторы настоящего изобретения создали методами генной инженерии и биотехнологии уникальный рекомбинантный слитый белок, на основе однодоменного антитела, специфически связывающий и нейтрализующий вирус бешенства. Изобретение имеет преимущества перед существующими аналогами.
Рекомбинантный слитый белок по изобретению имеет пролонгированный период полувыведения по сравнению с однодоменным антителом (VHH). Это достигается за счет создания конструкции, в которой антирабический домен (однодоменное антитело) рекомбинантного белка слит с альбумин-связывающимся доменом протеина G грамположительной бактерии рода Streptococcus (ABD), который продляет период полувыведения рекомбинантного слитного белка из сыворотки крови за счет связывания с альбумином, как минимум до 7 дней, то есть до момента образования антител в ответ на антирабическую вакцину.
Основными технологическими преимуществами эукариотического штамма - продуцента CHO в сравнении с бактериальной системой экспрессии (н-р, E.coli) является отсутствие необходимости очистки синтезированного белка от бактериальных эндотоксинов, а также наличие эффективного фолдинга и полноценных посттрансляционных модификаций.
Созданный авторами изобретения штамм-продуцент CHO-AntiRabD-ABD получен с помощью системы направленного редактирования генома CRISPR/Cas9, что позволило интегрировать кодирующую последовательность рекомбинантного слитого белка, специфически связывающего и нейтрализующего вирус бешенства в геном клеток для стабильной продукции белка. Стабильные штаммы-продуценты предпочтительны для крупномасштабного производства рекомбинантных белков.
Находящаяся с С-конца рекомбинантного слитого белка последовательность альбумин-связывающего домена дает возможность получать высокую степень очистки белка методом аффинной хроматографии с использованием иммобилизованного сывороточного альбумина человека.
Заявленное авторами изобретение позволяет упростить и удешевить способ получения антирабического средства на основе однодоменных антител, описанное в патенте РФ № 2661028, за счет создания пролонгирующей фармакокинетику модификации однодоменного антитела с получением уникального рекомбинантного слитого белка. В сравнении с известным решением уже после однократного введения рекомбинантного слитого белка млекопитающему антирабическая защита присутствует на период не менее 7 суток, что не требует дополнительного введения препарата рекомбинантных псевдоаденовирусных частиц, экспрессирующих данное антитело. В результате существенно снижается стоимость производства антирабического средства из-за отсутствия необходимости в выпуске второго препарата на основе аденовирусных векторов. Помимо этого, устраняется вероятность снижения эффективности экстренной профилактики бешенства, в случае наличия высокого уровня предсуществующих антител к аденовирусному вектору в организме.
Таким образом, предлагаемый по изобретению рекомбинантный слитый белок, на основе однодоменного антитела, специфически связывающий и нейтрализующий вирус бешенства является инновационным и способен расширить линейку существующих антирабических средств пассивной профилактики для применения в стандартной схеме лечебно-профилактической иммунизации против бешенства.
Осуществление изобретения подтверждается следующими примерами, которые при этом не ограничивают его объем.
Пример 1
Получение последовательности антирабического домена, нейтрализующего вирус бешенства
В примере рассмотрен многоэтапный процесс получения и отбора уникальной кодирующей последовательности антирабического домена, как первого функционального фрагмента созданного авторами рекомбинантного слитого белка. За счет отсутствия Fc-фрагмента у получаемого антирабического домена, будет исключена Fc-зависимая интерференция с иммунным ответом на одновременно введенную антирабическую вакцину. (Stockinger, B., & Lemmel, E.-M. (1978). Fc receptor dependency of antibody mediated feedback regulation: On the mechanism of inhibition. Cellular Immunology, 40(2), 395-403. doi:10.1016/0008-8749(78)90347-7).
Задача по получению антирабического домена решалась путем получения библиотеки вариабельных доменов (син. однодоменное антитело, VHH, наноантитело) неканонических антител верблюда с дальнейшим отбором методом фагового дисплея домена с требуемой специфичностью. Методы решения задачи известны специалистам в данной области техники и не раскрываются здесь более подробно.
Первым этапом работ было получение неканонических антител к вирусу бешенства, для этого проводили 5-кратную иммунизацию двугорбого верблюда путем подкожных инъекций препаратом инактивированного вируса бешенства, смешанного с адъювантом (Gerbu LQ). Введение осуществляли последовательно, вторую иммунизацию проводили через 21 день после первой, далее интервал составлял 10-14 дней. Через 5 дней после последней иммунизации у животного проводили взятие крови и выделяли мононуклеарные клетки периферической крови из которых получали РНК, на основе которой синтезировали кДНК. На матрице полученной кДНК ставили гнездовую ПЦР, позволяющую амплифицировать последовательности антигенсвязывающих доменов неканонических антител верблюда. В результате была получена библиотека ампликонов нуклеотидных последовательностей антигенсвязывающих доменов.
Вторым этапом было проведено клонирование последовательностей антигенсвязывающих доменов в фагмидный вектор, использующийся далее в процедуре фагового дисплея, и получена специфически обогащенная библиотека последовательностей, кодирующих антигенсвязывающие домены.
На следующем этапе из полученной фаговой библиотеки была проведена селекция методом фагового дисплея (функционального отбора фаговых частиц, несущих на поверхности экспрессируемые антигенсвязывающие домены, а внутри - ДНК, кодирующую эти антигенсвязывающие домены) клонов фагов, связывающихся с инактивированным вирусом бешенства. Отобранные ДНК клоны были секвенированы и таким образом получена кодирующая последовательность антигенсвязывающего домена, специфически связывающегося с вирусом бешенства (далее антирабический домен, AntiRabD).
Далее кодирующую последовательность антирабического домена химически синтезировали (ЗАО «Евроген», РФ), клонировали в экспрессионную плазмиду и трансдуцировали E.coli для наработки белка. Рекомбинантный белок очищали хроматографическими методами и использовали в эксперименте по определению вируснейтрализующей активности антирабического домена.
Нейтрализующую активность детектировали в реакции вирусной нейтрализации с флюоресцентной детекцией (FAVN, Fluorescent antibody virus neutralization) по известной среднему специалисту методике (OIE Manual of diagnostic tests and vaccines for terrestrial animals (mammals, birds and bees), Chapter 3.1.17. Rabies (infection with rabies virus) and other lyssaviruses. -2018.- p. 591-594).
Результаты реакции вирусной нейтрализации с флюоресцентной детекцией показали наличие вируснейтрализующей активности в отношении вируса бешенства у полученного уникального антирабического домена.
Таким образом, в ходе работ авторами была создана уникальная последовательность первого функционального фрагмента рекомбинантного слитого белка, т.е. антирабический домен, который способен специфически связывать и нейтрализовать вирус бешенства, не вступая в Fc-зависимую интерференцию с одновременно введенной антирабической вакциной.
Пример 2
Создание кодирующей последовательности рекомбинантного слитого белка, связывающего и нейтрализующего вирус бешенства
В ходе данной работы кодирующая последовательность антирабического домена, полученного в примере 1, объединялась с последовательностью второго функционального фрагмента рекомбинантного слитого белка с целью удлинения периода полувыведения из организма субъекта.
Задача по созданию кодирующей последовательности рекомбинантного слитого белка по изобретению решалась методами in cilico путем объединения нуклеотидной последовательности антирабического домена из примера 1 с нуклеотидной последовательностью альбумин-связывающего домена протеина G грамположительной бактерии рода Streptococcus (ABD), между ними была встроена нуклеотидная последовательность гибкого пептидного глицин-серинового линкера (G2S)3. Данная модификация влияет на фармакокинетику и позволяет пролонгировать период полувыведения антирабического домена из крови субъекта за счет связывания с альбумином, что позволяет поддерживать нейтрализующую активность (по стандарту ВОЗ не менее 0,5 МЕ/мл крови) минимум до 7 суток, то есть до момента образования антител в ответ на введенную антирабическую вакцину. Гибкий пептидный глицин-сериновый линкер (G2S)3 в составе рекомбинантного слитого белка позволяет сохранить специфическую активность антирабического домена и способность второго фрагмента связывать альбумин сыворотки крови. Дополнительно с 5́'- конца нуклеотидной последовательности рекомбинантного слитого белка была встроена нуклеотидная последовательность лидерного пептида IgGκ мыши (IgGκSP) для обеспечения секреции рекомбинантного белка клетками CHO. С целью эффективной экспрессии гена рекомбинантного слитого белка в эукариотической клеточной системе, проводили соответствующую оптимизацию кодонов его нуклеотидной последовательности, широко известную специалистам. Конструктивные части созданной кодирующей последовательности рекомбинантного слитого белка, специфически связывающего и нейтрализующего вирус бешенства изображены на фиг. 1.
Созданная кодирующая рекомбинантный слитый белок нуклеотидная последовательность представлена в перечне под номером SEQ ID NO: 2, что однако не ограничивает объем изобретения и возможности представления в других вариантах на основе факта вырожденности генетического кода. Аминокислотная последовательность рекомбинантного слитого белка, специфически связывающего и нейтрализующего вирус бешенства, представлена в перечне последовательностей под номером SEQ ID NO: 1.
Таким образом, созданная авторами изобретения уникальная последовательность SEQ ID NO: 2 кодирует аминокислотную последовательность SEQ ID NO: 1 рекомбинантного слитого белка, специфически связывающего и нейтрализующего вирус бешенства, с пролонгированным периодом полувыведения из организма субъекта и без интерференции с антирабической вакциной.
Пример 3
Получение штамма-продуцента рекомбинантного слитого белка, специфически связывающего и нейтрализующего вирус бешенства
В примере представлен ход работ по конструированию штамма на основе эукариотической линии клеток CHO (Chinese hamster ovary, клетки яичника китайского хомячка), стабильно продуцирующего рекомбинантный слитый белок из примера 1.
Поставленная задача решалась путем введения в геном эукариотической линии клеток CHO экзогенной последовательности SEQ ID NO: 2, кодирующей рекомбинантный слитый белок, с использованием известной специалистам системы направленного редактирования генома CRISPR/Cas9 (Barrangou, R., Doudna, J. Applications of CRISPR technologies in research and beyond. Nat Biotechnol 34, 933-941 (2016). https://doi.org/10.1038/nbt.3659).
Схема получения штамма-продуцента включала следующие этапы:
1) Выбор локуса в геноме клеток CHO для встраивания экзогенной последовательности;
2) Подбор места интеграции и конструирование последовательности сайт-специфической единой направляющей РНК (нРНК);
3) Создание генетической конструкции, несущей гены нРНК и Cas9;
4) Создание генетической конструкции, несущей ген рекомбинантного слитого белка SEQ ID NO: 2;
5) Проведение направленной интеграции экзогенной последовательности в геном CHO с помощью системы CRISPR/Cas9, путем трансфекции двумя полученными на предыдущих этапах генетическими конструкциями;
6) Селекция и отбор клонов клеток CHO, продуцирующих рекомбинантный слитый белок SEQ ID NO1.
На первом этапе работ был проведен выбор локуса в геноме клеток CHO для встраивания экзогенной последовательности, кодирующей рекомбинантный слитый белок по изобретению. Для работы с геномом CHO использовался вариант так называемого референсного генома Cricetulus griseus (GenBank: GCF_000419365.1). В качестве мишени был выбран ген ЛДГА кодирующий субъединицу А лактат дегидрогеназы. Ранее было показано, что в этот ген возможно встраивание генов интереса, без каких либо нежелательных последствий для жизнедеятельности клетки (Lee J.S., Kallehauge T.B., Pedersen L.E., Kildegaard H.F. Site-specific integration in CHO cells mediated by CRISPR/Cas9 and homology-directed DNA repair pathway. Sci. Rep. 2015 Feb 25;5:8572).
На втором этапе методом биоинформационного анализа был проведен подбор места интеграции экзогенной последовательности в выбранный локус клеток линии CHO с использованием общедоступных компьютерных программ и ресурсов Geneious 9 (Biomatters, Inc.), NCBI BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) и CRISPR RGEN Tools (http://www.rgenome.net/cas-offinder/). Была выбрана последовательность места интеграции 5'-GAAACGTAAAAGTAATACCGAGG-3' (РАМ - сайт подчеркнут), находящаяся в первом экзоне гена ЛДГА, которая представлена в перечне последовательностей под номером SEQ ID NO: 4 (без РАМ-сайта).
Для направленного внедрения экзогенной последовательности в геном клеток CHO на следующем этапе был проведен дизайн единой направляющей РНК (нРНК, gRNA) к месту интеграции. Была создана кодирующая последовательность единой нРНК, которая состояла из 5'-концевой спейсерной части (криспрРНК) и последовательности транс-активирующей РНК (тракрРНК), взаимодействующей с нуклеазой Cas9 из Streptococcus pyogenes (SpCas9). Кодирующая последовательность единой нРНК представлена в перечне под номером SEQ ID NO: 3, а также на фигуре 3.
Созданная последовательность, кодирующая единую нРНК (96 п.о.) была химически синтезирована (ЗАО «Евроген», РФ). Для проведения клонирования использовали вектор pUC57-Cas9, который содержал экспрессионную кассету, состоящую из CMV-промотора и гена Cas9 из Streptococcus pyogenes (SpCas9) с сигналами ядерной локализации, а также векторную часть pUC57-Cas9. Последовательность, кодирующая единую нРНК была клонирована в вектор pUC57-Cas9 по сайтам XbaI/Bsu15I под контроль промотора U6 (специальный промотор для клеточной РНК полимеразы III). В результате работ была создана генетическая конструкция pUC57-Cas9- U6gRNA (фиг. 4).
Далее была создана вторая генетическая конструкция, с целевым геном. Последовательность, кодирующая рекомбинантный слитый белок SEQ ID NO: 2 была химически синтезирована (ЗАО «Евроген», РФ). Для проведения клонирования использовали вектор pBsSK(+) LDGA, который содержал экспрессионную кассету состоящую из EF1-промотора и SV40 поли(А); экспрессионную кассету состоящую из SV40-промотора, гена устойчивости к пуромицину и синтетического сигнала полиаденилирования; правое и левое плечо гомологии первого экзона гена ЛДГА, обрамляющие две экспрессионные кассеты; векторную часть плазмиды pBluescript II SK(+). Последовательности плечей гомологии первого экзона гена ЛДГА длиной ~800 нуклеотидов были получены амплификацией с помощью специфических праймеров из генома клеточной линии CHO и клонированы в плазмиду pBluescript II SK(+). Последовательность, кодирующая рекомбинантный слитый белок SEQ ID NO: 2 была клонирована в вектор pBsSK(+) LDGA по сайтам XbaI/Bsu15I под контроль промотора EF1. В результате работ была создана генетическая конструкция pBsSK(+) LDGA - AntiRabD-ABD (фиг. 5).
На пятом этапе работы была проведена трансфекция клеток линии CHO двумя созданными генетическими конструкциями pUC57-Cas9- U6gRNA и pBsSK(+) LDGA - AntiRabD-ABD. Клетки CHO культивировали в среде DMEM c добавлением до 10% эмбриональной телячьей сыворотки. За сутки до трансфекции клетки были посеяны в лунки 6-ти луночного планшета в количестве 1×106 клеток на лунку. Для трансфекции использовался реагент Lipofectamin 2000 (Invitrogen, США). При трансфекции использовали 4 мкг ДНК двух вышеуказанных генетических конструкций в равном соотношении и 12 мкл реагента Lipofectamine 2000. Через 24 часа оценивали эффективность трансфекции клеток при помощи флуоресцентной микроскопии по свечению зеленого флуоресцентного белка GFP.
На шестом этапе работ были проведены селекция и отбор клонов клеток CHO, продуцирующих рекомбинантный слитый белок SEQ ID NO 1.
Через 48 часов после трансфекции проводили селекцию клонов в которые произошло встраивание экзогенной ДНК. Поскольку при использовании системы редактирования CRISPR/Cas9 в геном клетки CHO-AntiRabD-ABD встраиваются экспрессионная кассета с целевым геном и экспрессионная кассета с геном устойчивости к антибиотику (фигура 6), селекцию проводили с использованием антибиотика пуромицин (7 мкг/мл культуральной среды).
После воздействия пуромицином, клетки подвергнутые трансфекции обрабатывали трипсином, смешивали со свежей средой DMEM (10% сыворотки, 300 мкг/л L-глутамина, 7 мкг/мл пуромицина) и рассевали на 96-луночный культуральный планшет. Через 4-5 суток клетки снимали трипсином из тех лунок, в которых образовались колонии и переносили на культуральные чашки с диаметром 3 см. После того как клетки в культуральных чашках образовывали монослой, часть клеток отбирали для проведения ПЦР анализа для детекции целевого гена. В результате были отобраны 8 клонов, в которых присутствовала нуклеотидная последовательность рекомбинантного слитого белка SEQ ID NO: 2. У отобранных клонов из культуральных чашек брали пробы культуральной жидкости и определяли уровень продукции рекомбинантного слитого белка методами электрофореза и иммуноблоттинга. Один клон с самой высокой экспрессией был адаптирован для роста в суспензионной культуре в среде SFM4CHO (HyClone) для крупномасштабной продукции белка.
Штамм линии клеток яичника китайского хомячка CHO-AntiRabD-ABD был депонирован в коллекции ФГБУ «НИЦЭМ им. Н.Ф. Гамалеи» Минздрава России.
Таким образом, в результате работ был получен штамм линии клеток яичника китайского хомячка CHO-AntiRabD-ABD - стабильный продуцент рекомбинантного слитого белка, специфически связывающего и нейтрализующего вирус бешенства.
Пример 4
Способ получения рекомбинантного слитого белка, специфически связывающего и нейтрализующего вирус бешенства
В данном примере рассмотрен способ получения рекомбинантного слитого белка по изобретению, описывающий процесс культивирования штамма-продуцента CHO-AntiRabD-ABD, накопления и очистки рекомбинантного белка.
В ходе работ были подобраны оптимальные технологические условия культивирования штамма-продуцента CHO-AntiRabD-ABD. Выращивание штамма - продуцента CHO-AntiRabD-ABD и синтез рекомбинантного слитого белка осуществляли в суспензионной культуре на бессывороточной среде пригодной для выращивания культуры клеток CHO, например SFM4CHO (HyClone), в атмосфере с 8% концентрацией CO2 в культивирования с перемешиванием 120 об/мин, в течение 3 суток.
Далее для проведения очистки полученную суспензионную культуру штамма-продуцента CHO-AntiRabD-ABD выдерживали без перемешивания 30 минут для осаждения клеток и отбирали культуральную надосадочную жидкость.
Выделение и очистку рекомбинантного слитого белка осуществляли многостадийно. Полученную культуральную жидкость центрифугировали при 5000 тыс/об в течение 15 минут и концентрировали с помощью тангенциальной фильтрации с использованием мембраны с номинальным отсечением по молекулярной массе 10 кДа. Далее проводили отделение рекомбинантного слитого белка методом афинной хроматографии с использованием в качестве сорбента сефарозы (NHS activeted Sephar 4 fast flow) с иммобилизированным человеческим сывороточным альбумином, для чего полученный концентрат с предыдущей стадии разбавляли стартовым буфером, наносили на хроматографическую колонку и элюировали. Полученный элюат стерилизовали через мембранный фильтр с отверстиями размером 0,22 мкм с получением концентрированного раствора очищенного рекомбинантного слитого белка AntiRabD-ABD.
Экспрессия рекомбинантного слитого белка SEQ ID NO: 1 штаммом-продуцентом CHO-AntiRabD-ABD была оценена методом электрофореза в 10% полиакриламидном геле по Леммли. Показанные на фиг. 7 результаты электрофореза подтверждают синтез штаммом-продуцентом CHO-AntiRabD-ABD рекомбинантного слитого белка, специфически связывающего и нейтрализующий вирус бешенства.
Аминокислотная последовательность полученного рекомбинантного слитого белка была подтверждена методом масс-спектрометрии и соответствовала SEQ ID NO: 1.
Таким образом, предложенный способ получения с использованием штамма - продуцента CHO-AntiRabD-ABD позволяет проводить выращивание, накопление и очистку рекомбинантного слитого белка по изобретению.
Пример 5
Изучение нейтрализующей активности рекомбинантного слитого белка in vitro против вируса бешенства
В ходе работ было проведено изучение вирус-нейтрализующей активности полученного в предыдущем примере рекомбинантного слитого белка в отношении вируса бешенства в реакции вирусной нейтрализации с флюоресцентной детекцией (FAVN).
Сущность метода FAVN заключается в нейтрализации in vitro постоянного количества вируса бешенства (штамм CVS) специфическим антителом, в данном случае рекомбинантным слитым белков, с внесением в чувствительную культуру клеток (культура клеток почка сирийского хомячка ВНК-21).
Постановку реакции вирусной нейтрализации с флюоресцентной детекцией осуществляли по общеизвестной методике (OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. vol. 1. - 6nd ed. - Paris, 2008. - Chap. 2.1.13. - P. 304-323).
В работе использовали следующие образцы:
1. Рекомбинантный слитый белок AntiRabD-ABD (1 мг/мл).
2. Стандартный образец иммуноглобулина человека против бешенства ВОЗ (WHO, International Laboratory For Biological Standards, Copenhagen, Denmark: Second International Standard for Rabies Immunoglobulin) с активностью 30 МЕ/мл.
3. Стандартный образец активности антирабического иммуноглобулина человека Европейской фармакопеи (European Pharmacopoeia Reference Standard Human rabies immunoglobulin BPR, Strasburg. Batch: 1.1, Id: 004X14) с активностью 91 ME/ml.
Реакцию проводили в культуре клеток BHK-21/клон 13 в 96-луночных плоскодонных планшетах. Клетки добавляли к предварительно проинкубированной вместе смеси исследуемых образцов и вируса бешенства CVS-11. Через три дня окрашивали зафиксированную клеточную культуру с помощью флуоресцентного антирабического конъюгата (FITC Anti-Rabies Monoclonal Globulin, Fujirebio Diagnostics Inc., США). Инфицированные клетки учитывали, осматривая всю поверхность каждой лунки. Результат оценивали качественным образом: отсутствие флуоресцирующих клеток - отрицательный результат, присутствие одной и более флуоресцирующих клеток - положительный результат. Титры исследуемых образцов выражали в МЕ/мл, сравнивая полученный для рекомбинантного слитого белка результат со стандартными образцами. Для этого стандартный образец ВОЗ предварительно был разведен до значения активности 0,50 МЕ/мл, а Международный стандарт Европейской фармакопеи - 2,0 МЕ/мл.
Нейтрализующая активность исследуемого образца в сравнении со стандартами рассчитывалась по формуле Spearman-Kärber (ВОЗ, 1996):

Полученные результаты реакции вирусной нейтрализации с флюоресцентной детекцией представлены в таблице 1.
Таблица 1 - Результаты определения методом FAVN нейтрализующей активности рекомбинантного слитого белка AntiRabD-ABD против вируса бешенства CVS-11 в сравнении со стандартными образцами
AntiRabD-ABD
ЕФ
Таким образом, при определении антирабической нейтрализующей активности рекомбинантного слитого белка AntiRabD-ABD в реакции вирусной нейтрализации с флюоресцентной детекцией (FAVN) было установлено, что образец AntiRabD-ABD с концентрацией белка 1 мг/мл обладает вируснейтрализующей активностью 1047,6 МЕ/мл. Полученные данные свидетельствует о наличии у рекомбинантного слитого белка высокого уровня нейтрализующей активности против вируса бешенства.
Результаты проведенного эксперимента свидетельствуют о наличии нейтрализующей активности против вируса бешенства у рекомбинантного слитого белка (SEQ ID NO: 1).
Пример 6
Нейтрализующая активность рекомбинантного слитого белка против вируса бешенства на лабораторных животных
В данном исследовании оценивали in vivo способность рекомбинантного слитого белка нейтрализовать вирус бешенства по показателю выживаемость. Определение значения нейтрализующей активности проводили в реакции нейтрализации (РН) вируса бешенства на мышах с использованием штамма фиксированного вируса бешенства CVS-11 по известному специалистам методу P. Atanasiu (Методы лабораторных исследований по бешенству. Женева: ВОЗ, 1975).
Суть метода состоит в нейтрализации постоянной дозы предварительно протитрованного вируса бешенства последовательными разведениями испытуемого образца на мышах.
Перед проведением реакции нейтрализации определяли LD50 вируса бешенства, для чего готовили 10-кратные разведения 10% мозговой суспензии вируса и вводили мышам интрацеребрально по 0,03 мл, каждое разведение вводили 6 животным. Регистрировали гибель мышей со специфическими для бешенства признаками с 6 по 14 день после заражения. Расчет LD50 вируса проводили по методу Reed и Muench (Методы лабораторных исследований по бешенству. Женева: ВОЗ, 1975). Для реакции нейтрализации использовали суспензию мозга, содержащую 300 LD50/0,03 мл.
В качестве препарата сравнения использовали коммерческий антирабический иммуноглобулин (положительный контроль) с активностью 150 МЕ/мл, представляющий собой поликлональный антирабический иммуноглобулин человека.
Для проведения реакции нейтрализации рекомбинантный слитый белок AntiRabD-ABD и антирабический иммуноглобулин выровняли путем разведения стерильным фосфатным буфером до концентрации 1 МЕ/мл каждый, сделали серию разведений с шагом 1:2, смешали равные объемы растворов исследуемых веществ с вирусом бешенства штамма CVC-11 (из расчета 150 LD50/мышь), инкубировали 1 час при температуре +37°С, далее вводили интрацеребрально по 30 мкл полученной смеси мышам Balb/c весом 8-10 г. Каждое разведение вводили группе из 6 мышей. Гибель мышей оценивали в период с 6 по 14 день после заражения.
Результаты эксперимента по оценке вируснейтрализующей активности на мышах отражены в таблице 2.
Таблица 2 - Результаты оценки реакции нейтрализации вируса бешенства рекомбинантным слитым белом AntiRabD-ABD по выживаемости мышей (%) после заражения штаммом CVS-11 в инфицирующей дозе 150 LD50
МЕ/мл
Как следует из таблицы 2, в реакции нейтрализации на мышах было установлено, что рекомбинантный слитый белок обладает нейтрализующими (протективными) свойствами против тест-штамма вируса бешенства штамма CVS-11 с активностью 150 ЛД50/мышь. В разведениях от 1:2 до 1:64 из 10 мышей в опыте выжили все 10, т.е. выживаемость составила 100%, что сравнимо с положительным контролем. При этом защитными титрами специфических антител к вирусу бешенства можно считать диапазон от 1 МЕ/мл вплоть до 0,0155 МЕ/мл как для рекомбинантного слитого белка AntiRabD-ABD, так и для коммерческого антирабического иммуноглобулина.
Таким образом, установлено, что при оценке на лабораторных животных рекомбинантный слитый белок AntiRabD-ABD обладает нейтрализующей активностью против вируса бешенства.
Пример 7
Динамика нейтрализующей активности против вируса бешенства рекомбинантного слитого белка
В ходе работы была исследована динамика изменения нейтрализующей активности против вируса бешенства рекомбинантного слитого белка по изобретению в различные сроки после внутримышечного введения на модели крыс.
Нейтрализующая активность против бешенства измерялась в реакции вирусной нейтрализации с флюоресцентной детекцией (FAVN) (OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Vol. 1. - 6nd ed. - Paris, 2008. - Chap. 2.1.13. - P. 304-323). Критерием оценки являлись значения нейтрализующей активности в сыворотке крови крыс, выраженные в международных единицах (МЕ). При оценке учитывался установленный ВОЗ защитный уровень антирабических антител в сыворотке крови 0,5 МЕ/мл.
В исследовании использовались беспородные крысы, самки, весом около 300 грамм. Группы состояли из 5 животных. Забор крови у крыс проводили из хвостовой вены в следующие временные точки: до введения препарата, 12 часов, 1 день, 3 день, 7 день.
Лабораторным животным вводили в качестве испытуемого препарата - рекомбинантный слитый белок, препаратом сравнения служил зарегистрированный в качестве лекарственного средства антирабический иммуноглобулин. Исходный препарат рекомбинантного слитого белка содержал 1048 МЕ/мл, антирабический иммуноглобулин содержал 150 МЕ/мл. Для получения исследуемых доз исходные препараты разводили в необходимом объеме стерильным фосфатным буфером, расчет величины дозы проводился с учетом эквивалентного переноса дозы для человека (МЕ/кг) на крысу.
Выбор уровней доз в эксперименте обосновывался предварительной оценкой нейтрализующей активности препаратов на крысах с последующим определением относительной активности рекомбинантного слитого белка (испытуемый препарат) к антирабическому глобулину (препарат сравнения). Было установлено, что через 12 часов после введения уровень нейтрализующей активности рекомбинантного слитого белка в сыворотке крови после внутримышечного введения 100 МЕ на крысу соответствует уровню нейтрализующей активности антирабического глобулина в сыворотке крови после внутримышечного введения 25 МЕ на крысу.
Таким образом, крысам группы опыт вводили препарат рекомбинантного слитого белка однократно внутримышечно в дозе 100 МЕ. Группе сравнения вводили антирабический иммуноглобули в дозе 25 МЕ на крысу. Группе отрицательный контроль вводили фосфатный буфер (PBS) в равном объеме (200 мкл) на крысу.
Полученные результаты эксперимента представлены в таблице 3 и на фиг. 8.
Таблица 3 - Динамика изменения во времени нейтрализующей активности против вируса бешенства рекомбинантного слитого белка в сравнительном опыте на крысах, оцененная методом FAVN
(до введения)
Как следует из таблицы 3 и фиг. 8, период сохранения нейтрализующей активности рекомбинантного слитого белка в сыворотке крови крыс в защитном титре более 0,5 МЕ/мл составляет не менее 7 дней после введения. Защитный титр антирабических антител (препарата сравнения) после введения также сохранялся не менее 7 дней после введения. Таким образом, динамика нейтрализующей активности рекомбинантного слитого белка была сравнима с зарегистрированным антирабическим иммуноглобулином, что позволяет применять рекомбинантный слитый белок по изобретению в качестве средства для пассивной иммунизации против бешенства.
В итоге, было показано, что рекомбинантный слитый белок может применяться млекопитающим для эффективной нейтрализации вируса бешенства в качестве средства для пассивной иммунизации против бешенства.
Промышленная применимость
Изобретение может применяться в фармацевтической промышленности для производства препарата для пассивной иммунизации против бешенства и использоваться медицинскими работниками в рамках оказания стандартной лечебно-профилактической иммунизации против бешенства в лечебно-профилактических учреждениях страны.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="Слитый белок.xml"
softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2023-12-19">
<ApplicationIdentification>
<IPOfficeCode></IPOfficeCode>
<ApplicationNumberText></ApplicationNumberText>
<FilingDate>2023-12-14</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>20231214</ApplicantFileReference>
<ApplicantName languageCode="ru">федеральное государственное
бюджетное учреждение "Национальный исследовательский центр
эпидемиологии и микробиологии имени почетного академика
Н.Ф.Гамалеи" Министерства здравоохранения Российской
Федерации</ApplicantName>
<ApplicantNameLatin>The National Research Center for Epidemiology
and Microbiology named after Honorary Academician N.F. Gamaleya of
the Ministry of Health of the Russian Federation</ApplicantNameLatin>
<InventionTitle languageCode="ru">Рекомбинантный слитый белок,
специфически связывающий и нейтрализующий вирус бешенства, способ его
получения и применение</InventionTitle>
<SequenceTotalQuantity>4</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>179</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..179</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>QVQLVESGGDSVQAGGSLRLSCAASGLTVRNNCMGWFRQSPGKEREAVA
AEGFDRTYYADSVKGRFTFSQDNAKNTVYLQMNSLKPEDTAMYYCAARSCGYGLLSRPEYYDYWGQGTQV
TVSSGGSGGSGGSGLAEAKVLANRELDKYGVSDFYKRLINKAKTVEGVEALKLHILAALP</INSDSeq_
sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>615</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..615</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atggagaccgacacactgctcttgtgggtcctgcttctctgggtgccag
gcagcaccggagatcaggtgcaactggttgagagtggaggcgacagcgtgcaggctggcggatccctgag
gttgagctgtgcagcctctggactgacagtgaggaacaattgcatgggatggtttcggcagagccctggg
aaggagagagaagcagtggctgcagagggcttcgacagaacctactatgccgattccgtcaaagggaggt
ttaccttcagccaagacaatgccaagaacactgtgtatttgcagatgaacagtctcaaacccgaggacac
agccatgtattactgcgctgcacggagctgtggctatggactcttgtccagacctgagtactacgactat
tggggacaaggcacccaggtgacggtctctagcggaggcagcggagggagcggcggttccggactggccg
aagcaaaagtgctggctaaccgcgagctggataagtatggcgtatcagatttttacaaacgcctcattaa
caaagcgaagactgttgaaggggtcgaggcattgaagctccatattctggctgcgctgcccggtggcggg
tccggt</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>96</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..96</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gaaacgtaaaagtaataccggttttagagctagaaatagcaagttaaaa
taaggctagtccgttatcaacttgaaaaagtggcaccgagtcggtgc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>20</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..20</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>genomic DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q8">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Cricetulus barabensis</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gaaacgtaaaagtaataccg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПАССИВНОЙ ИММУНИЗАЦИИ ПРОТИВ БЕШЕНСТВА, ФАРМАЦЕВТИЧЕСКИЙ НАБОР, СПОСОБ ПРИМЕНЕНИЯ ФАРМАЦЕВТИЧЕСКОГО НАБОРА | 2016 |
|
RU2661028C2 |
| ТРИМЕРИЗОВАННОЕ ОДНОДОМЕННОЕ АНТИТЕЛО, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩЕЕСЯ С ГЛИКОПРОТЕИНОМ G ВИРУСА БЕШЕНСТВА, НЕЙТРАЛИЗУЮЩЕЕ ВИРУС БЕШЕНСТВА | 2013 |
|
RU2533802C1 |
| Генетическая конструкция на основе оптимизированного гена консенсусного гликопротеина вируса бешенства для профилактики бешенства | 2018 |
|
RU2707544C1 |
| РЕКОМБИНАНТНЫЕ АНТИТЕЛА И КОМПОЗИЦИИ, А ТАКЖЕ СПОСОБЫ ИХ СОЗДАНИЯ И ИСПОЛЬЗОВАНИЯ | 2002 |
|
RU2292353C2 |
| Генетическая (рекомбинантная) ДНК-конструкция, содержащая кодон-оптимизированный ген гликопротеина (белка G) вируса бешенства с консенсусной аминокислотной последовательностью, которая составлена с учетом аминокислотных последовательностей белка G, выделяемого из штаммов вируса бешенства, циркулирующих на территории Российской Федерации | 2015 |
|
RU2626605C2 |
| Однодоменные наноантитела против шиповидного белка вируса SARS-CoV-2 | 2021 |
|
RU2794141C2 |
| ОДНОДОМЕННЫЕ АНТИТЕЛА К БЕЛКУ GP ВИРУСА ЭБОЛА ДЛЯ ИММУНОТЕРАПИИ ЛИХОРАДКИ ЭБОЛА | 2015 |
|
RU2644202C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ ФРАГМЕНТЫ (Fab) ПРОТИВ ВИРУСА БЕШЕНСТВА, ИЗОЛИРОВАННЫЙ ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ Fab ПРОТИВ ВИРУСА БЕШЕНСТВА, КЛЕТКА ДРОЖЖЕЙ, ТРАНСФОРМИРОВАННАЯ ФРАГМЕНТОМ ДНК, И СПОСОБ ПОЛУЧЕНИЯ Fab ПРОТИВ ВИРУСА БЕШЕНСТВА С ИСПОЛЬЗОВАНИЕМ ДРОЖЖЕЙ | 2010 |
|
RU2440412C2 |
| Тяжелоцепочечные моноклональные антитела, специфически связывающиеся с S белком вируса SARS-CoV-2, и способ их применения для терапии заболеваний, вызываемых различными вариантами вируса SARS-CoV-2 | 2024 |
|
RU2836313C1 |
| Нуклеотидная последовательность, оптимизированная для экспрессии в бактериях консенсусного гликопротеина вируса бешенства | 2018 |
|
RU2717255C1 |




Изобретение относится к области биотехнологии, иммунологии, медицины. Предложены рекомбинантный слитый белок, на основе однодоменного антитела, специфически связывающий и нейтрализующий вирус бешенства, штамм линии клеток яичника китайского хомячка, продуцирующий указанный белок, способ получения указанного белка и применение указанного белка в качестве средства для пассивной иммунизации против бешенства. Изобретение позволяет получить рекомбинантный слитый белок с пролонгированным периодом полувыведения, который можно применять для пассивной иммунизации в схеме экстренной лечебно-профилактической иммунизации против бешенства, без снижения эффективности антирабической вакцины. 4 н. и 3 з.п. ф-лы, 8 ил., 3 табл., 7 пр.
1. Рекомбинантный слитый белок, на основе однодоменного антитела, специфически связывающий и нейтрализующий вирус бешенства, где указанный белок представлен аминокислотной последовательностью SEQ ID NO: 1.
2. Рекомбинантный слитый белок по п.1, отличающийся тем, что аминокислотная последовательность SEQ ID NO: 1 кодируется нуклеотидной последовательностью SEQ ID NO: 2.
3. Штамм линии клеток яичника китайского хомячка CHO-AntiRabD-ABD - продуцент рекомбинантного слитого белка по п.1, в геном которых интегрирована экзогенная последовательность, которая содержит экспрессионную кассету, включающую SV40-промотор, ген устойчивости к пуромицину и сигнал полиаденилирования, экспрессионную кассету, включающую EF1-промотор, ген с SEQ ID NO: 2, кодирующий рекомбинантный слитый белок с SEQ ID NO: 1, сигнал полиаденилирования вируса SV40, где экзогенная последовательность была направлена в геном клеток CHO в место интеграции с SEQ ID NO: 4 путем трансфекции клеток линии CHO двумя генетическими конструкциями pBsSK(+) LDGA – AntiRabD-ABD и pUC57-Cas9-U6gRNA, где генетическая конструкция pBsSK(+) LDGA - AntiRabD-ABD содержит экспрессионную кассету, состоящую из SV40-промотора, гена устойчивости к пуромицину и сигнала полиаденилирования, и экспрессионную кассету, состоящую из EF1-промотора, гена с SEQ ID NO: 2, кодирующего рекомбинантный слитый белок SEQ ID NO: 1, сигнала полиаденилирования вируса SV40, а генетическая конструкция pUC57-Cas9-U6gRNA содержит экспрессионную кассету, состоящую из CMV-промотора, гена Cas9 из Streptococcus pyogenes (SpCas9) с сигналами ядерной локализации, гена с последовательностью SEQ ID NO: 3 под контролем промотора U6, кодирующего единую направляющую РНК к месту интеграции с SEQ ID NO: 4.
4. Способ получения рекомбинантного слитого белка по п.1, включающий следующие стадии:
- культивирование штамма-продуцента по п.3 в подходящих условиях для продуцирования и накопления рекомбинантного слитого белка по п.1;
- очистка рекомбинантного слитого белка по п.1 из культуральной жидкости.
5. Применение рекомбинантного слитого белка по п.1 в качестве средства для пассивной иммунизации против бешенства, заключающееся в его введении млекопитающим в количестве, достаточном для эффективной нейтрализации вируса бешенства.
6. Применение рекомбинантного слитого белка по п.5, отличающееся тем, что однократное введение млекопитающему обеспечивает нейтрализацию вируса бешенства на протяжении не менее 7 суток.
7. Применение рекомбинантного слитого белка по любому из пп.5-6, отличающееся тем, что введение млекопитающему проводят одновременно с антирабической вакциной.
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПАССИВНОЙ ИММУНИЗАЦИИ ПРОТИВ БЕШЕНСТВА, ФАРМАЦЕВТИЧЕСКИЙ НАБОР, СПОСОБ ПРИМЕНЕНИЯ ФАРМАЦЕВТИЧЕСКОГО НАБОРА | 2016 |
|
RU2661028C2 |
| ТРИМЕРИЗОВАННОЕ ОДНОДОМЕННОЕ АНТИТЕЛО, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩЕЕСЯ С ГЛИКОПРОТЕИНОМ G ВИРУСА БЕШЕНСТВА, НЕЙТРАЛИЗУЮЩЕЕ ВИРУС БЕШЕНСТВА | 2013 |
|
RU2533802C1 |
| CN 101450967 A, 10.06.2009 | |||
| WO 2020029860 A1, 13.02.2020 | |||
| НЕЙТРАЛИЗУЮЩЕЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СВЯЗЫВАЮЩЕЕСЯ С ГЛИКОПРОТЕИНОМ G ВИРУСА БЕШЕНСТВА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО, И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ | 2017 |
|
RU2718835C2 |
| ГОРШКОВА Е.Н | |||
| и др | |||
| Однодоменные антитела и биоинженерные препараты на их основе: новые возможности для диагностики и терапии | |||
| Медицинская иммунология | |||
| Токарный резец | 1924 |
|
SU2016A1 |

