Область техники настоящего изобретения
Настоящее изобретение относится к области иммунологии и вирусологии и предполагает использование антител, представляющих собой однодоменные мини-антитела, для профилактики и терапии гриппа, а также к использованию генов этих антител в комбинации с вирусным вектором (вектором на основе рекомбинантного аденовируса) и их экспрессии в организме млекопитающих для профилактики и терапии гриппа.
Предшествующий уровень техники настоящего изобретения
Вирусы гриппа способны вызывать чрезвычайные эпидемические ситуации. Эти вирусы широко распространены в природе и могут поражать людей, многие виды млекопитающих (лошадей, свиней, тюленей и т.д.), а также все виды птиц. Респираторные заболевания, вызываемые вирусами гриппа, в некоторых случаях могут протекать с осложнениями и заканчиваться смертью.
Вирусы гриппа характеризуются большой антигенной вариабельностью. Наиболее подвержены изменениям поверхностные гликопротеины вириона - гемагглютинин и нейраминидаза. Известны два пути их изменения.
Первый - антигенный дрейф. Под давлением популяционного иммунитета мутации, позволяющие уйти из-под контроля иммунной системы, дают вирусу серьезное преимущество и закрепляются. В итоге новые антигенные варианты гемагглютинина и нейраминидазы постоянно сменяют друг друга. В результате возникают эпидемии, т.к. защита от предыдущих штаммов вирусов, в том числе того же самого субтипа, недостаточна для нейтрализации нового штамма. К сожалению, применение современных субъединичных и инактивированных вакцин не решает эту проблему, т.к. обеспечивает хорошую защиту только от штамма, из которого они были получены. Поэтому в настоящее время для защиты населения от новых эпидемических штаммов требуется постоянно создавать новые вакцины.
Второй путь изменения вирусов гриппа - антигенный шифт (сдвиг), т.е. изменение антигенной формулы вируса в результате замены гена(ов) и соответствующего белка(ов). В основе механизма антигенного шифта лежит реассортация или рекомбинация генов, которая может происходить при совместной инфекции двумя и более вирусами [Кочергин-Никитский К.С. // Анализ взаимодействия генов при скрещивании низкопатогенного вируса гриппа птиц подтипа Н5 и высокопродуктивного штамма вируса гриппа человека. М., ГУ НИИ вирусологии им. Д.И. Ивановского РАМН. 2007]. Шифтовые изменения, как правило, затрагивают антигенную структуру гемагглютинина, реже - нейраминидазы. Таким образом, через нерегулярные интервалы времени появляются пандемические варианты вирусов гриппа с новыми антигенными и биологическими свойствами, которые вызывают тяжелые заболевания и гибель людей [Lvov D.K. // Sov. Med. Rev. E. Virol. Rev. 1987. V.2. P.15-37; Webster R.G., Bean W.J., German O.T., et al. // Microbiol. Rev. 1992. V.56. P.152-179]. Например, пандемия «испанки» - вируса гриппа H1N1, в 1918-1920 гг. привела к смерти около 50 млн человек по всему миру. В июне 2009 г. Всемирная организация здравоохранения (ВОЗ) присвоила новому вирусу H1N1, случаи заболевания которым начали выявлять в начале апреля 2009 г., шестую степень угрозы пандемии. Этот вирус представляет собой реассортант, содержащий гены птичьего, человеческого и свиного вирусов [Shinde V., Bridges С.В., Uyeki Т.М. // The New England journal of medicine. 2009. V.360; 25. P.2616-2625]. При возникновении нового пандемического штамма создание вакцины занимает слишком много времени, что не позволяет быстро остановить распространение опасного вируса. Таким образом, несмотря на проводимые широкомасштабные профилактические меры, включающие вакцинацию, во многих странах мира, в том числе в России, ежегодно регистрируются сезонные вспышки гриппа, охватывающие все слои населения от детей до лиц преклонного возраста. Для решения этой проблемы необходимы новые подходы к профилактике и лечению гриппа.
Помимо вакцин для борьбы с гриппом на сегодняшний день существует широкий выбор лекарственных средств. Однако следует отметить, что профилактика подобными средствами не обладает достаточной эффективностью, к тому же многие лекарственные средства имеют широкий перечень противопоказаний и могут вызвать побочные реакции.
В настоящее время для лечения и профилактики гриппа используются препараты нескольких групп, среди которых особое место занимают препараты ингибиторы М2-каналов и ингибиторы нейраминидазы. Из препаратов первой группы в нашей стране в течение многих лет применялся ремантадин, оказывавший выраженный терапевтический и профилактический эффект при гриппе, вызванном вирусом типа А. Однако широкое использование ингибиторов М2-каналов привело к появлению устойчивых штаммов вирусов гриппа, резистентных к ремантадину. Из препаратов второй группы в России зарегистрированы препараты занамивир и осельтамивир. Занамивир не пригоден для широкого использования в клинической практике, т.к. может применяться только в виде ингаляций, что неприемлемо для детей дошкольного возраста и пожилых пациентов. Кроме того, возможен целый ряд нежелательных реакций, включая бронхоспазм и отек гортани. Другой препарат - осельтамивир (Тамифлю) - зарекомендовал себя как высокоэффективный и безопасный препарат, но одним из самых принципиальных недостатков этого препарата является необходимость его раннего применения (наиболее эффективен при приеме в первые 36 часов заболевания).
Проблема своевременного обращения за медицинской помощью и раннего лечения респираторных вирусных инфекций относится к наиболее острым проблемам практического здравоохранения. По данным НИИ гриппа РАМН основная часть пациентов обращаются за медицинской помощью только на 2-3-и сутки, когда становится очевидным, что пациент болен не простой простудой и нельзя ограничиться лечением только жаропонижающими препаратами, однако уже с тяжелыми формами гриппа и вирусными пневмониями пациенты поступают в клиники Института гриппа РАМН только на 5-е сутки. В связи с этим ингибиторы нейраминидазы не могут быть широко использованы в реальных условиях начинающейся эпидемии.
Таким образом, можно заключить, что на сегодняшний день нет эффективных и безопасных препаратов для терапии или профилактики гриппа.
Решением данной проблемы может стать создание препаратов на основе антител, полученных против вируса гриппа типа А. Действие этих препаратов основано на введении в организм антител, полученных к протективному антигену патогена, которые способны защищать организм хозяина от развития инфекции. Такой подход применяется начиная с первой половины 20-го века. До конца 1960-х гг. в СССР сравнительно широко применялась гипериммунная лошадиная сыворотка для профилактики и лечения гриппа. Однако зачастую в качестве побочной реакции данный препарат вызывал аллергию, и в этой связи от него пришлось отказаться. Но сама идея использования содержащихся в крови людей противовирусных антител в настоящее время все еще реализуется в виде препаратов крови. Эти препараты называют иммуноглобулинами. Наиболее эффективен и распространен в России препарат «специфический противогриппозный иммуноглобулин» из крови доноров, специально и многократно вакцинированных против гриппа. Помимо противогриппозных антител, специально стимулируемых вакцинацией, в донорском иммуноглобулине содержатся антитела ко многим широко распространенным инфекционным агентам, в том числе и к респираторным вирусам. В последние годы появились сообщения о формировании антител к введенному человеческому белку. Препарат не лишен и других побочных эффектов: повышения температуры тела, аллергизация организма: от появления сыпи до развития тяжелого состояния и анафилактического шока.
Препараты иммуноглобулинов, полученные генно-инженерными методами, лишены некоторых этих недостатков. Так, на животных показана эффективность моноклональных одноцепочечных антител (scFv), полученных против белка-токсина сибирской язвы [Mabry R., Infect Immun. 2005; 73: 8362-8368.]. Однако при этом используются экстремально высокие дозы белка, что является существенным ограничением. Гуманизированные одноцепочечные однодоменные мини-антитела не отторгаются организмом человека и не вызывают аллергической реакции. Однако данный тип антител имеет ряд недостатков: высокие затраты на их производство и ограниченность генно-инженерных манипуляций.
В настоящее время разработана технология получения с помощью методов генной инженерии еще одного типа антител - рекомбинантных однодоменных мини-антител заданной специфичности. В ее основе лежит получение нуклеотидных последовательностей генов неканонических антител животных семейства верблюдовых (Camelidae). Эти антитела представляют собой димер только одной укороченной (первый константный район СН1 отсутствует) тяжелой цепи иммуноглобулина. Для собственно специфического узнавания и связывания антигена при этом необходим и достаточен лишь один вариабельный домен этого антитела.
Полным эквивалентом термина «однодоменное мини-антитело» для целей настоящего изобретения является вошедшее в широкое употребление обозначение «нанотело», введенное фирмой ABLYNX (NANOBODY™), а также «наноантитело» и «однодоменное наноантитело».
Полученные по этой технологии однодоменные мини-антитела обладают следующими свойствами [Тиллиб С.В. «Верблюжьи мини-антитела» - эффективный инструмент для исследований, диагностики и терапии. Молекулярная биология 2011; 45 (1): 77-85.]:
1) Высокая растворимость и стабильность (в широком диапазоне температур и кислотности среды).
2) Улучшенная проницаемость в клетки за счет малого размера (~2×4 нм, 13-15 кДа).
3) Способность образовывать необычные для классических антител паратипы, позволяющие связываться с углублениями и активными центрами белков.
4) Наличие отработанного способа получения и селекции однодоменных мини-антител.
5) Возможность экономичной наработки в больших количествах. Однодоменные мини-антитела можно нарабатывать в периплазме бактерий Е.coli.
6) Возможность генно-инженерных модификаций для поливалентных однодоменных мини-антител.
7) Низкая иммуногенность.
Еще одним положительным свойством этих антител является возможность их «гуманизации» без заметной потери их специфической активности путем проведения небольшого числа точечных замен аминокислот [Vincke С., Loris R., Saerens D., et al. // J. Biol. Chem. 2009. V.284. №5. P.3273-3284]. Это открывает потенциальную возможность широкого использования однодоменных мини-антител в качестве средств пассивной иммунизации для предотвращения развития различных опасных инфекционных заболеваний [Wesolowski J., Alzogaray V., Reyelt J. et al. Single domain antibodies: promising experimental and therapeutic tools in infection and immunity. Med. Microbiol. Immunol. 2009; 198, 157-174].
Так, было показано (WO 2007/052242; Prendergast Patrick; Composition and method for the treatment of viral infection using camelid heavy chain antibodies), что антитела, полученные против вируса гриппа и отобранные по специфичности к нейраминидазе, способны защищать птиц от развития заболеваний, вызываемых вирусами гриппа различных штаммов. Это техническое решение как наиболее близкое к заявляемому по составу действующего вещества фармацевтической композиции и способу введения его в организм выбрано авторами настоящего за прототип.
Недостатками прототипа являются:
1) Антитела к нейраминидазе вируса гриппа обладают слабой нейтрализующей вирус активностью. Эффективная нейтрализация вируса таким препаратом может обеспечиваться лишь в случае его длительной циркуляции в организме. Это приводит к необходимости введения в организм больших доз препаратов для достижения положительного эффекта, а также к необходимости повторного введения препарата.
2) Для предотвращения возникновения различных иммунных реакций на чужеродный белок (антитело), вводимый в большом количестве вследствие его слабой нейтрализующей вирус активности, используется схема введения, согласно которой препарат вводится дважды, причем второй раз совместно с препаратом, ингибирующим ФНО-α. При этом зачастую повторное введение является затруднительным для пациентов.
Таким образом, в уровне техники существует острая потребность в разработке фармацевтической композиции для профилактики и терапии гриппа на основе специфических (рекомбинантных) антител, характеризующихся высокой нейтрализующей вирус активностью, которые должны продолжительное время поддерживаться/воспроизводиться в организме, что обеспечило бы достаточность однократного введения композиции.
Задачей настоящего изобретения явилось создание:
1) модифицированного однодоменного мини-антитела, которое характеризуется высокой нейтрализующей вирус активностью и за счет этого может эффективно связывать определенный эпитоп гемагглютинина вируса гриппа и тем самым блокировать развитие инфекции вируса гриппа в организме.
2) вектора, обеспечивающего доставку и продолжительную экспрессию в организме гена однодоменного мини-антитела, которое может эффективно связывать определенный эпитоп гемагглютинина вируса гриппа и характеризуется высокой нейтрализующей вирус активностью, что при однократном введении может эффективно блокировать развитие инфекции вируса гриппа в организме.
3) композиции, состоящей из однодоменного мини-антитела и аденовирусного вектора, обеспечивающего доставку и продолжительную экспрессию в организме гена однодоменного мини-антитела, которое может эффективно связывать определенный эпитоп гемагглютинина вируса гриппа и характеризуется высокой нейтрализующей вирус активностью, что при однократном введении может блокировать развитие инфекции вируса гриппа в организме.
Раскрытие настоящего изобретения
Указанная выше задача настоящего изобретения решена тем, что разработка препарата проведена с использованием специально полученных однодоменных мини-антител, связывающих поверхностный белок гемагглютинин вируса гриппа таким образом, что при этом нарушается (блокируется и нейтрализуется) инфекционная активность вируса гриппа А.
Более конкретно, поставленная задача решается настоящим изобретением за счет создания однодоменного мини-антитела против вируса гриппа, специфически связывающегося с определенным эпитопом гемагглютинина вируса гриппа типа A и подавляющего инфекцию вируса гриппа типа А. Также создан вирусный вектор, экспрессирующий однодоменное мини-антитело против вируса гриппа, которое может эффективно связывать определенный эпитоп гемагглютинина вируса гриппа и тем самым блокировать развитие инфекции вируса гриппа в организме. При этом вирусный вектор может быть аденовирусным вектором, экспрессирующим однодоменное мини-антитело против вируса гриппа, которое способно эффективно связывать определенный эпитоп гемагглютинина вируса гриппа и тем самым блокировать развитие инфекции вируса гриппа в организме. Также предлагается композиция, состоящая из эффективного количества однодоменного мини-антитела против вируса гриппа и вирусного вектора, экспрессирующего однодоменное мини-антитело против вируса гриппа, способного эффективно связывать гемагглютинин вируса гриппа и тем самым блокировать развитие инфекции вируса гриппа в организме.
В качестве способов применения для действующего вещества (антитела) были выбраны следующие: использование однодоменных мини-антител в виде суспензии в фармацевтически приемлемом растворителе; использование в качестве носителя для действующего вещества рекомбинантного вирусного вектора на основе аденовируса, экспрессирующего ген однодоменного мини-антитела, способного связывать и/или нейтрализовывать вирус гриппа; использование композиций, содержащих как вирусный вектор с геном однодоменного мини-антитела, так и однодоменное мини-антитело (в виде белка).
Выбор путей реализации с целью получения фармацевтической композиции с заявляемыми свойствами обусловлен следующими факторами:
Настоящее изобретение предусматривает использование в качестве действующего вещества фармацевтической композиции особого однодоменного мини-антитела, способного эффективно связываться с поверхностным белком гемоглютинином вируса гриппа таким образом, что при этом нарушается (блокируется, нейтрализуется) инфекционная активность вируса. Большинство используемых в настоящее время нейтрализующих вирус антител (классических антител, состоящих из 4-х иммуноглобулиновых цепей) имеют сродство именно к белку гемоглютинину вируса гриппа. Антитела против гемагглютинина способны предотвращать проникновение вируса в клетку и таким образом предотвращать развитие инфекции. В отличие от них действие антител против нейраминидазы осуществляется на уровне выхода зрелых частиц вируса из клетки и поэтому способны лишь сократить период заболевания и его интенсивность. Ввиду того что именно антитела к гемагглютинину могут обладать выраженной нейтрализующей вирус активностью и оказывают свое действие против вируса гриппа сразу после попадания его в организм, то есть способны предотвратить развитие инфекции, однодоменные мини-антитела, отобранные против гемагглютинина вируса гриппа и обладающие при этом нейтрализующей вирус активностью, были использованы в качестве действующего вещества для создания фармацевтической композиции, предназначенной для профилактики и терапии гриппа.
Настоящее изобретение относится к способу использования однодоменного мини-антитела, предусматривающему применение вирусных векторов в качестве носителя для действующего вещества. Вирусные векторы представляют собой рекомбинантные вирусы, в геном которых включен целевой ген (ген однодоменного мини-антитела) с набором регуляторных элементов. В качестве вирусных векторов наиболее часто применяют следующие рекомбинантные вирусы: аденовирусы [Шмаров М.М., Тутыхина И.Л., Логунов Д.Ю. и др. Индукция протективного иммунного ответа у мышей, вакцинированных рекомбинантным аденовирусом птиц CELO, экспрессирующим гликопротеин G вируса бешенства. Журнал микробиологии, эпидемиологии и иммунологии 2006; 4: 69-71; Тутыхина И.Л., Шульпин М.И., Чвала И.А. и др. Конструирование рекомбинантных аденовирусов CELO, экспрессирующих ген гемагглютинина вируса гриппа А птиц, и изучение возможности их использования в качестве вакцин для защиты от вируса гриппа А птиц H5N1 и H7N1. Молекулярная генетика, микробиология и вирусология 2011.; Тутыхина И.Л., Шмаров М.М., Логунов Д.Ю. и др. Конструирование и перспективы использования в медицине рекомбинантных аденовирусных наноструктур. Российские нанотехнологии. 2009; 4 (11-12): 82-92; Liu М.А. Immunologic basis of vaccine vectors. Immunity. 2010; 33 (4): 504-15; Lasaro M.O., Ertl H.C. New insights on adenovirus as vaccine vectors. Mol Ther. 2009; 17 (8): 1333-9], ретровирусы [Liu M.A. Immunologic basis of vaccine vectors. Immunity. 2010; 33 (4): 504-15; Pincha M., Sundarasetty B.S., Stripecke R.. Lentiviral vectors for immunization: an inflammatory field. Expert Rev Vaccines. 2010; 9 (3): 309-321; Negri D.R., Michelini Z., Cara A. Toward integrase defective lentiviral vectors for genetic immunization. Curr HIV Res. 2010; 8 (4): 274-281.], аденоассоциированные вирусы [Sun J.Y., Anand-Jawa V., Chatterjee S., Wong K.K. Immune responses to adeno-associated virus and its recombinant vectors. Gene Ther. 2003; 10 (11): 964-76]. Среди существующих систем доставки антигенов вирусные векторы занимают особое место, поскольку обладают следующими свойствами:
- имеют естественный механизм взаимодействия с клеткой и проникновения в клетку;
- транспортируют чужеродный генетический материал в ядро клетки;
- способны обеспечивать длительную экспрессию антигена;
- вирусная оболочка выполняет защитную функцию для генетического материала, кодирующего антиген.
При введении в организм вирусные векторы проникают в различные типы клеток, где осуществляют экспрессию встроенных в ДНК генов. В этой связи вирусные векторы являются широко используемым инструментом для доставки генов в клетки человека и животных [R. Harrop, et al., Advanced Drug Delivery Reviews, V.58, I.8, 2006, P.931-947].
Наряду с вышеперечисленным вирусные векторы отличаются высокой эффективностью экспрессии целевого гена в различных типах клеток, безопасностью для человека и животных.
Таким образом, использование вирусных векторов в качестве носителя для действующего вещества заявляемой композиции позволяет доставлять в организм гены целевых белков, что, в свою очередь, обеспечивает продление периода циркуляции продукта целевого гена в организме.
В качестве носителя используют вирусные векторы, полученные на основе рекомбинантного аденовируса человека 5-го серотипа. Известно, что аденовирусные векторы используются на практике наиболее часто. В настоящее время на основе аденовирусных векторов разрабатываются вакцины против различных бактериальных (туляремия, туберкулез, бруцеллез и др.) и вирусных (вирус иммунодефицита человека, папилломавирус человека, вирус бешенства, вирус Эбола и др.) патогенов человека [Wang J, Thorson L, Stokes RW, Santosuosso M, Huygen K, Zganiacz A, Hitt M, Xing Z. Single mucosal, but not parenteral, immunization with recombinant adenoviral-based vaccine provides potent protection from pulmonary tuberculosis. J Immunol. 2004 Nov 15; 173 (10): 6357-65; Richardson JS, Yao MK, Tran KN, Croyle MA, Strong JE, Feldmann H, Kobinger GP. Enhanced protection against Ebola virus mediated by an improved adenovirus-based vaccine. PLoS One. 2009; 4(4): e5308. Epub 2009 Apr 23; Li WH, Zhang Y, Wang SH, Liu L, Yang F. Recombinant replication-defective adenovirus based rabies vaccine. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2003 Dec; 25 (6): 650-4; Lees CY, Briggs DJ, Wu X, Davis RD, Moore SM, Gordon C, Xiang Z, Ertl HC, Tang DC, Fu ZF. Induction of protective immunity by topic application of a recombinant adenovirus expressing rabies virus glycoprotein. Vet Microbiol. 2002 Apr 2; 85 (4): 295-303]. Так, были получены аденовирусные векторы, содержащие нуклеотидную последовательность гемагглютинина вируса гриппа человека, для дальнейшего использования в качестве вакцины для человека и животных против патогенного штамма вируса гриппа (Van Kampen KR, Shi Z, Gao P, Zhang J, Foster KW, Chen DT, Marks D, Elmets CA, Tang DC. Vaccine. 2005, V 23, p.1029-1036). Ген гемагглютинина вируса гриппа птиц H5N1 включали в ДНК рекомбинантной псевдоаденовирусной наночастицы (РПАН) на основе аденовируса человека 5-го серотипа также с целью использования полученного продукта как вакцины для человека, млекопитающих животных и птиц (Toro Н, Tang DC, Suarez DL, Sylte MJ, Pfeiffer J, Van Kampen KR. Vaccine 2007; Gao W, Soloff AC, Lu X, Montecalvo A, Nguyen DC, Matsuoka Y, Robbins PD, Swayne DE, Donis RO, Katz JM, Barratt-Boyes SM, Gambotto A. J. of Virology, 2006, V.80, №4, p.1959-1964). Также аденовирусные векторы использовали для генной терапии опухолей в случае, когда в их ДНК был клонирован ген опухолевого супрессора р53 (US 7033750). Кроме того, два препарата, представляющих собой аденовирусные векторы на основе аденовируса человека 5-го серотипа, разрешены к применению в онкологии.
Также применение вирусных векторов в качестве носителей наделяет фармацевтическую композицию следующими преимуществами:
- С помощью вирусных векторов может быть решена проблема нестабильности препаратов антител (вводимых в организм в виде белков).
- Вирусные векторы стабильны, могут быть лиофилизованы, могут храниться при 4°C длительное время без потери активности.
- Технологии получения вирусных векторов в промышленных масштабах являются экономически оправданными и позволяют получать препараты с высоким титром. Процесс получения нового вида вирусного вектора занимает несколько недель, что может позволить быстро реагировать на меняющуюся эпидемиологическую обстановку в максимально сжатые сроки [R. Harrop, et al., Advanced Drug Delivery Reviews, V.58, I.8, 2006, Р.931-947].
- Использование вирусных векторов позволяет обеспечивать наработку протективного уровня антител уже через 96 часов после введения. Более того, протективный уровень антител поддерживается в организме не менее 2-х недель [Kasuya K., Mol Ther. 2005 Feb; 11 (2): 237-44; Sofer-Podesta С., Infect Immun. 2009 Apr; 77 (4): 1561-8; Chiuchiolo M.J., J Infect Dis. 2006 Nov 1; 194 (9): 1249-57] в случае аденовирусных векторов.
Использование препаратов, представляющих собой вирусный вектор, несущий в составе ДНК ген однодоменного мини-антитела, способного нейтрализовать вируса гриппа типа актуального штамма, позволяет обеспечить уже через 48-96 часов защиту организма от циркулирующего на данный момент штамма вируса гриппа. Более того, использование фармацевтической композиции, состоящей из вирусного вектора одного типа, несущего ген однодоменного мини-антитела, и белковых препаратов соответствующего однодоменного мини-антитела позволяет обеспечить защиту организма в период непосредственно после введения препарата, длящуюся как минимум 3-4 недели.
Таким образом, целесообразным является использование комплексного препарата, имеющего две составляющие: однодоменных мини-антител против вируса гриппа типа А и рекомбинантного вирусного вектора, экспрессирующего ген однодоменных мини-антител против вируса гриппа типа А.
Такой препарат будет обладать следующими преимущественными характеристиками:
1) Основой препарата являются антитела - наиболее эффективные природные противовирусные вещества.
2) Кратковременный курс приема (1 раз), что снижает риск развития побочных эффектов и непереносимости.
3) Эффективность действия на различных стадиях развития инфекционного процесса.
4) Возможность использования в качестве дополнения к поздней вакцинации лиц из групп риска в первые 2 недели после вакцинации (на период выработки антител).
5) Возможность использования для лиц с иммунодефицитом, которые на вакцинацию могут дать недостаточный иммунный ответ.
6) Возможность использования для пожилых лиц, для которых эффективность вакцинации снижается и достигает 50-70%, как дополнение к вакцинации.
7) Возможность использования для невакцинированных лиц, находящихся в контакте с заболевшими родственниками и соседями.
8) Возможность использования для тех, кто по каким-либо причинам не был вакцинирован вовремя.
9) Стоимость курса применения подобного препарата сравнима со стоимостью одной дозы вакцины.
Настоящее изобретение объединяет преимущества различных упомянутых подходов и относится к однодоменным мини-антителам, связывающимся/нейтрализующим вирус гриппа, а также к способу применения таких антител как путем использования в качестве носителя действующего вещества (антитела) рекомбинантного вирусного вектора, несущего экспрессирующую кассету, содержащую нуклеотидную последовательность, которая кодирует однодоменное мини-антитело, связывающееся с вирусом гриппа, так и путем создания фармацевтической композиции, состоящей из однодоменного мини-антитела против вируса гриппа и рекомбинантного вирусного вектора, экспрессирующего данное однодоменное мини-антитело.
Итак, настоящее изобретение относится к однодоменному мини-антителу, характеризующемуся аминокислотной последовательностью SEQ ID NO: 2, специфически связывающемуся с гемагглютинином вируса гриппа типа A (H5N2) и подавляющему инфекцию вируса гриппа типа A (H5N2), и его функционально активным вариантам. Однодоменное мини-антитело, характеризующееся аминокислотной последовательностью SEQ ID NO: 2, в настоящей заявке иногда именуется своим торговым наименованием «Неогриппаб».
Также настоящее изобретение относится к нуклеотидной последовательности, кодирующей однодоменное мини-антитело, характеризующееся аминокислотной последовательностью SEQ ID NO: 2, и его функционально активные варианты. Частные случаи такой последовательности включают без ограничения SEQ ID NO: 1 и ее генетически вырожденные варианты.
Согласно следующему аспекту настоящее изобретение относится к рекомбинантному вирусному вектору, содержащему такую нуклеотидную последовательность и экспрессирующему однодоменное мини-антитело, характеризующееся аминокислотной последовательностью SEQ ID NO: 2, или его функционально активные варианты. Согласно одному из предпочтительных вариантов осуществления этого аспекта вирусный вектор является аденовирусным вектором. Однако среднему специалисту в данной области техники будет очевидно, что в качестве вирусных векторов могут быть использованы и некоторые другие вирусы, такие как аденоассоциированные вирусы, лентивирусы.
Согласно следующему аспекту настоящее изобретение относится к фармацевтической композиции для подавления развития инфекции вируса гриппа A (H5N2), содержащей эффективное количество раскрытого выше однодоменного мини-антитела и/или вирусного вектора, экспрессирующего такое однодоменное мини-антитело. Среднему специалисту в настоящей области техники будет очевидно, что для разных целей (терапевтических, профилактических, комбинированных) может быть предпочтительно использование фармацевтических композиций, содержащих однодоменное антитело или кодирующий его вирусный вектор как по отдельности, так и в сочетании. Настоящее изобретение охватывает каждый такой вариант.
Таким образом, настоящее изобретение относится к фармацевтической композиции для профилактики или терапии гриппа, представляющей собой суспензию раскрытых выше рекомбинантных вирусных векторов и/или раскрытых выше однодоменных мини-антител в фармацевтически приемлемом растворителе или наполнителе. Такие фармацевтически приемлемые растворители и наполнители хорошо известны из предшествующего уровня техники (Remington′s Pharmaceutical Sciences, 18th edition, A.R. Gennaro, Ed., MackPublishing Company [1990]; Pharmaceutical Formulation Development of Peptides and Proteins, S. Frokjaer and L. Hovgaard, Eds., Taylor & Francis [2000]; и Handbook of Pharmaceutical Excipients, 3rd edition, A. Kibbe, Ed., Pharmaceutical Press [2000]).
Применение такой фармацевтической композиции позволяет добиться транзиторной экспрессии у пациента однодоменных мини-антител, которые за счет своих уникальных свойств, подробно описанных выше, способны обеспечивать достаточно надежную иммунную защиту против вируса гриппа. Среднему специалисту в данной области техники будет понятно, что применение такой фармацевтической композиции эффективно в основном для профилактики, но также и для терапии инфекции.
Кроме того, настоящее изобретение относится к фармацевтической композиции для профилактики или терапии гриппа, представляющей собой суспензию раскрытого выше рекомбинантного вирусного вектора и однодоменных мини-антител, связывающихся с вирусом гриппа того же типа и антигенной формулы, что и однодоменное мини-антитело, кодируемое экспрессирующей кассетой, содержащейся в указанном рекомбинантном вирусном векторе, в фармацевтически приемлемом растворителе или наполнителе.
Применение такой фармацевтической композиции позволяет обеспечить достаточно надежную иммунную защиту против вируса гриппа не только в порядке профилактики, но и после инфицирования пациента вирусом гриппа. Разумеется, чем на более ранней стадии инфекции будет применена фармацевтическая композиция согласно настоящему изобретению, тем более эффективным окажется такое лечение.
Кроме того, настоящее изобретение относится к способу профилактики или терапии гриппа, предусматривающему введение нуждающемуся в этом пациенту профилактически или терапевтически эффективного количества одной из раскрытых выше фармацевтических композиций.
В соответствии с одним из предпочтительных вариантов осуществления настоящего изобретения указанную фармацевтическую композицию вводят интраназально.
В соответствии с более конкретными вариантами осуществления настоящего изобретения интраназальное введение осуществляют в форме капель или в форме спрея (аэрозоля).
Авторы настоящего изобретения исходят из того, что, как известно среднему специалисту в данной области техники, первичные, исходно получаемые последовательности однодоменных мини-антител, могут служить исходными модулями более сложных многомодульных препаратов. Возможно объединение в одном мультивалентном производном двух, трех и более моновалентных первичных однодоменных мини-антител. Эти объединяемые в одну конструкцию однодоменные мини-антитела могут связываться как с одним и тем же эпитопом антигена-мишени, так и с его разными эпитопами, или даже с различными антигенами-мишенями. Возможно также комбинированное объединение в одну конструкцию однодоменных мини-антител и других молекул или лекарств с получением многофункциональных препаратов [Conrath KE, Lauwereys М, Wyns L, Muyldermans S. Camel single-domain antibodies as modular building units in bispecific and bivalent antibody constructs. J Biol Chem. 2001 Mar 9; 276 (10): 7346-50; Zhang J, Tanha J, Hirama T, Khieu NH, To R, Tong-Sevinc H, Stone E, Brisson JR, MacKenzie CR. Pentamerization of single-domain antibodies from phage libraries: a novel strategy for the rapid generation of high-avidity antibody reagents. J Mol Biol. 2004 Jan 2; 335 (1): 49-56; Cortez-Retamozo V, Backmann N, Senter PD, Wernery U, De Baetselier P, Muyldermans S, Revets H. Efficient cancer therapy with a nanobody-based conjugate. Cancer Res. 2004 Apr 15; 64 (8): 2853-7; Baral TN, Magez S, Stijlemans B, Conrath K, Vanhollebeke B, Pays E, Muyldermans S, De Baetselier P. Experimental therapy of African trypanosomiasis with a nanobody-conjugated human trypanolytic factor. Nat Med. 2006 May; 12 (5): 580-4; Coppieters K, Dreier T, Silence K, Haard HD, Lauwereys M, Casteels P, Beirnaert E, Jonckheere H, Wiele CV, Staelens L, Hostens J, Revets H, Remaut E, Elewaut D, Rottiers P. Formatted anti-tumor necrosis factor alpha VHH proteins derived from camelids show superior potency and targeting to inflamed joints in a murine model of collagen-induced arthritis. Arthritis Rheum. 2006 Jun; 54 (6): 1856-66]; мультимеризация с помощью введения дополнительных аминокислотных последовательностей, взаимодействующих белковых доменов, таких как лейциновые зипперы [Harbury Р.В., Zhang T., Kim P.S., et al. A switch between two-, three- and four-stranded coiled coils in GCN4 leucine zipper mutants. Science, 1993, 262: 1401-1407; Shirashi Т., Suzuyama k., Okamoto H. et al. Increased cytotoxicity of soluble Fas ligand by fusing isoleucine zipper motif. Biochem. Biophys. Res. Communic. 2004, 322: 197-202; Chenchik A., Gudkov A., Komarov A., Natarajan V. Reagents and methods for producing bioactive secreted peptides. 2010. US Patent Application 20100305002], последовательностей небольших белков, образующих стабильные комплексы [Deyev SM, Waibel R, Lebedenko EN, Schubiger AP, Plückthun A. Design of multivalent complexes using the barnase*barstar module. Nat Biotechnol. 2003, 21 (12): 1486-92.].
Для модулирования свойств препарата однодоменного мини-антитела, например увеличения времени жизни in vivo (в крови пациента), в состав конечного соединения могут быть введены дополнительные аминокислотные последовательности, последовательности белков (таких, например, как сывороточный альбумин) или другое однодоменное мини-антитело, специфически связывающееся с мажорным и долгоживущим белком в крови человека (например, альбумином или иммуноглобулином) [Gibbs WW. Nanobodies. Sci Am. 2005 Aug; 293 (2): 78-83; Harmsen MM, Van Solt CB, Fijten HP, Van Setten MC. Prolonged in vivo residence times of llama single-domain antibody fragments in pigs by binding to porcine immunoglobulins. Vaccine. 2005 Sep 30; 23 (41): 4926-34; Coppieters K, Dreier T, Silence K, Haard HD, Lauwereys M, Casteels P, Beirnaert E, Jonckheere H, Wiele CV, Staelens L, Hostens J, Revets H, Remaut E, Elewaut D, Rottiers P. Formatted anti-tumor necrosis factor alpha VHH proteins derived from camelids show superior potency and targeting to inflamed joints in a murine model of collagen-induced arthritis. Arthritis Rheum. 2006 Jun; 54 (6): 1856-66].
Среднему специалисту в данной области техники будет очевидно, что такие модификации и прочие варианты антител, лежащих в основе настоящего изобретения, подпадают под объем настоящего изобретения, поскольку являются структурными и функциональными вариантами однодоменных мини-антител. Таким образом, авторы настоящего изобретения понимают под термином "однодоменные мини-антитела" как первичные, исходно получаемые, "минимальные" аминокислотные последовательности однодоменных мини-антител, так и их модификации, полученные в результате упомянутых адаптации или «форматирования» и их варианты.
Термин «вариант антитела» для целей настоящего изобретения означает полипептид, который содержит изменения в аминокислотной последовательности, а именно делеции, вставки, добавления или замены аминокислот, при условии, что при этом сохраняется необходимый уровень активности белка, например, как минимум 10% от активности исходного однодоменного мини-антитела. Ряд изменений в варианте белка зависит от положения или от типа аминокислотного остатка в трехмерной структуре белка. Количество изменений может составлять, например, от 1 до 30, более предпочтительно от 1 до 15, и наиболее предпочтительно от 1 до 5 изменений в последовательности исходного однодоменного мини-антитела. Эти изменения могут иметь место в областях полипептида, которые не являются критичными для его функции. Это становится возможным благодаря тому, что некоторые аминокислоты обладают высокой гомологией друг с другом, и поэтому третичная структура или активность белка не нарушаются при таком изменении. Поэтому в качестве варианта белка может выступать белок, который характеризуется гомологией не менее 70%, предпочтительно не менее 80%, более предпочтительно не менее 90%, и наиболее предпочтительно не менее 95% по отношению к аминокислотной последовательности исходного однодоменного мини-антитела при условии сохранения активности полипептида. Гомология между аминокислотными последовательностями может быть установлена с использованием хорошо известных методов, например с помощью выравнивания последовательностей в компьютерной программе BLAST 2.0, которая вычисляет три параметра: счет, идентичность и сходство.
Замена, делеция, вставка, добавление или замена одного или нескольких аминокислотных остатков будут представлять собой консервативную мутацию или консервативные мутации при условии, что активность белка при этом сохраняется. Примером консервативной мутации(ий) является консервативная замена(ы). "Консервативная аминокислотная замена" представляет собой замену, при которой аминокислотный остаток заменяется аминокислотным остатком, имеющим сходную боковую цепь. В данной области техники определены семейства аминокислот, имеющих сходные боковые цепи. Эти семейства включают в себя аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серии, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, примеры консервативных замен включают замену Ala на Ser или Thr, замену Arg на Gln, His или Lys, замену Asn на Glu, Gln, Lys, His или Asp, замену Asp на Asn, Glu или Gln, замену Cys на Ser или Ala, замену Gln на Asn, Glu, Lys, His, Asp или Arg, замену Glu на Asn, Gln, Lys или Asp, замену Gly на Pro, замену His на Asn, Lys, Gln, Arg или Tyr, замену Ile на Leu, Met, Val или Phe, замену Leu на Ile, Met, Val или Phe, замену Lys на Asn, Glu, Gln, His или Arg, замену Met на Ile, Leu, Val или Phe, замену Phe на Trp, Tyr, Met, Ile или Leu, замену Ser на Thr или Ala, замену Thr на Ser или Ala, замену Trp на Phe или Tyr, замену Tyr на His, Phe или Trp и замену Val на Met, Ile или Leu.
Фрагменты ДНК, которые кодируют по существу тот же функциональный полипептид, могут быть получены, например, путем модификации нуклеотидной последовательности фрагмента ДНК, кодирующего исходное однодоменное мини-антитело, например, посредством метода сайт-направленного мутагенеза, так, что один или несколько аминокислотных остатков в определенном сайте будут делетированы, заменены, вставлены или добавлены. Фрагменты ДНК, модифицированные как описано выше, могут быть получены с помощью традиционных методов обработки с целью получения мутации.
Фрагменты ДНК, которые кодируют по существу тот же функциональный полипептид исходного однодоменного мини-антитела, могут быть получены путем экспрессирования фрагментов ДНК, имеющих мутацию, описанную выше, в соответствующей клетке, и установления активности экспрессируемого продукта.
Организация вариабельных доменов неканонических антител (VHH, наноантител) в значительной степени подобна той, что у вариабельных доменов (VH) классических антител (у человека VH-домены иммуноглобулинов подкласса IgG3 имеют особо выраженную гомологию с VH и VHH верблюдовых). В обоих случаях V-домены состоят из четырех консервативных каркасных участков (FR, «framework regions»), окружающих три гипервариабельных участка (определяющие комплементарность, CDR, от «complementarity determining regions). В обоих случаях домены формируют типичную для V-домена иммуноглобулина пространственную структуру из двух бета-слоев (-листов), один - из четырех аминокислотных цепочек, и второй - из пяти [Padlan Е.А. X-Ray crystallography of antibodies. Adv. Protein Chem. 1996; 49: 57-133. Muyldermans S., Cambillau C., Wyns L. Recognition of antigens by single-domain antibody fragments: the superfluous luxury of paired domains. TIBS 2001; 26: 230-235]. В этой структуре все три гипервариабельных участка кластеризуются с одной стороны V-домена (где они участвуют в узнавании антигена) и располагаются в петлях, соединяющих бета-структуры. Однако имеются и важные отличия, связанные с функционированием VHH в формате одного домена. Так, гипервариабельные участки CDR1 и CDR3 заметно увеличены в случае VHH. Часто в гипервариабельных участках VHH обнаруживаются цистеиновые остатки, причем присутствующие сразу в двух участках (чаще всего в CDR1 и CDR3, реже - в CDR2 и CDR3). При исследовании кристаллических структур VHH было показано, что эти цистеиновые остатки формируют дисульфидные связи, что приводит к дополнительной стабилизации структуры петель данного антигена. Наиболее явным и воспроизводимым отличительным признаком VHH являются четыре замены гидрофобных аминокислотных остатков на гидрофильные во втором каркасном участке (Val37Phe, Gly44Glu, Leu45Arg, Trp47Gly, согласно нумерации Кабат). Этот каркасный участок в случае VH домена является высококонсервативным, обогащен гидрофобными аминокислотными остатками и особо важен для образования связи с вариабельным доменом VL легкой цепи. VHH-домен в этом плане сильно отличается: указанные замены гидрофобных аминокислот на гидрофильные делают невозможной ассоциацию VHH и VL. Эти замены также объясняют высокую растворимость VHH, наноантитела, когда его получают в виде рекомбинантного белка [Тиллиб С.В. «Верблюжьи наноантитела» - эффективный инструмент для исследований, диагностики и терапии. Молекулярная биология 2011; 45 (1): 77-85].
Именно гипервариабельные районы мини-антител определяют их специфическое взаимодействие с антигеном, и именно гомологичные замены аминокислот в этих участках могут приводить к получению несколько различающихся по последовательности мини-антител, которые обладают идентичными или близкими свойствами. Таким образом, среднему специалисту в данной области техники будет очевидно, что под объем настоящего изобретения подпадают не только указанная в приложении последовательность мини-антитела, но и те, которые могут быть получены путем замен аминокислот в гипервариабельных участках (указанных в перечне последовательностей как CDR) на другие, но очень близкие по свойствам, аминокислоты (консервативных замен).
Замена, делеция, вставка или добавление нуклеотидов, описанных выше, также включают мутации, которые имеют место в природе и, например, обусловлены изменчивостью.
Полипептиды однодоменных мини-антител согласно настоящему изобретению могут кодироваться большим множеством молекул нуклеиновых кислот, что является результатом хорошо известного в данной области техники явления вырожденности генетического кода. Суть феномена состоит в том, что любая аминокислота (за исключением триптофана и метионина), входящая в состав природных пептидов, может кодироваться более чем одним триплетным нуклеотидным кодоном (см. таблицу 1). Любая из этих вырожденных кодирующих молекул нуклеиновых кислот может входить в состав экспрессирующих кассет, содержащихся в заявленных в соответствии с настоящим изобретением рекомбинантных псевдоаденовирусных наночастицах, подпадающих под объем настоящего изобретения.
Краткое описание чертежей
На фиг.1 представлены аминокислотная последовательность отобранного однодоменного мини-антитела SEQ ID NO: 2, способного специфически связывать и блокировать вирус гриппа A, и кодирующая ее нуклеотидная последовательность SEQ ID NO: 1. В указанной аминокислотной последовательности подчеркнуты соответственно (слева направо, от N- к С-концу) гипервариабельные участки CDR1, CDR2 и CDR3, которые собственно являются основными определяющими специфичность связывания районами подобных антигенузнающих молекул.
На фиг.2 представлены дополнительные аминокислотные последовательности, которые присоединяли (методом молекулярного клонирования и генно-инженерного встраивания дополнительных последовательностей ДНК, кодирующих соответствующие аминокислотные последовательности) к С-концу исходно отобранного антитела. Указаны четыре дополнительных аминокислотных участка (домена): (1) - линкерный линейный участок (соответствующий верхнему удлиненному шарнирному участку неканонических иммуноглобулинов IgG2a верблюда), (2) - специальный тримеризующий домен (выделен серым цветом) - "ILZ" («isoleucine zipper domain, изолейциновая молния»), а также (3) - НА-таг (пептид из белка гемагглютинина, к которому есть коммерческие антитела) и (4) His-таг (шесть остатков гистидина на самом С-конце белка для эффективной очистки белка с помощью металл-хелатной хроматографии).
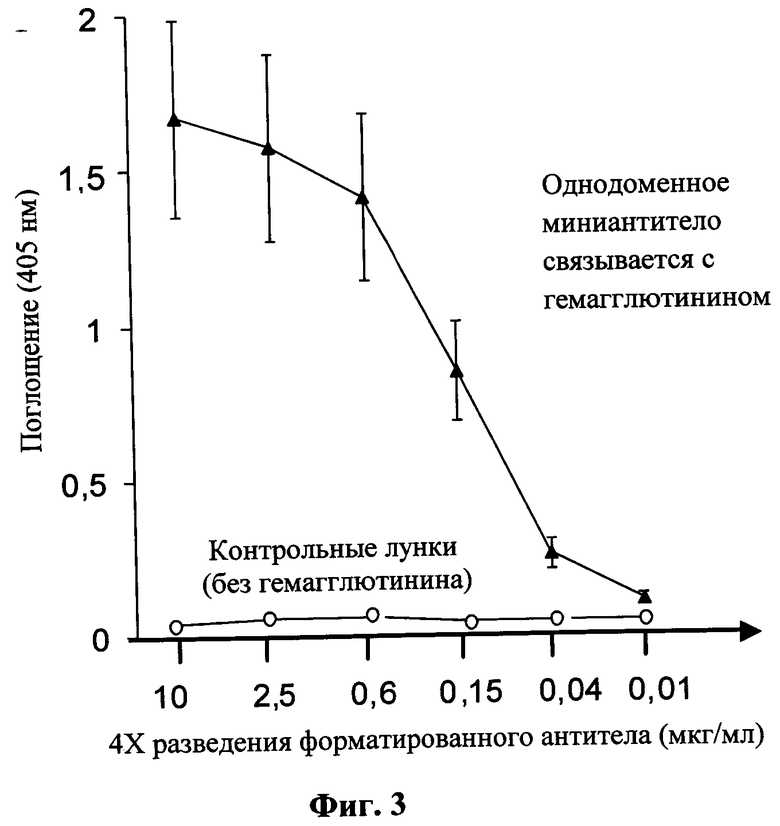
На фиг.3 проиллюстрированы результаты иммуноферментного анализа узнавания отобранным о дно доменным мини-антителом SEQ ID NO: 2 как иммобилизованного в лунках иммунологической плашки препарата белка гемагглютинина (в верхней части рисунка), так и препарата вируса гриппа (в нижней части чертежа). В качестве контрольных использовали ячейки с иммобилизованным белком овальбумином кур.
На фиг.4 проиллюстрированы результаты иммуноферментного анализа узнавания отобранным однодоменным мини-антителом SEQ ID NO: 2 иммобилизованного в лунках иммунологической плашки препарата вируса гриппа. В качестве контрольной использовали ячейку с иммобилизованным белком овальбумином кур.
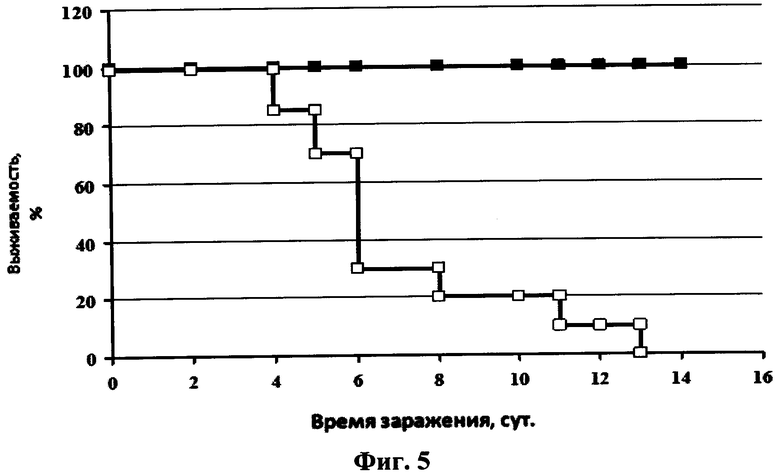
На фиг.5 представлен график зависимости выживаемости мышей после интраназального введения однодоменного мини-антитела против вируса гриппа и последующего заражения их вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2) от времени после заражения. Сплошная линия с маркером в виде квадрата черного цвета - 20 мкл интраназально раствора однодоменного мини-антитела против вируса гриппа в фосфатном буфере в дозе 2,5 мкг/мышь, сплошная линия с маркером в виде квадрата белого цвета - 20 мкл интраназально раствора фосфатного буфера.
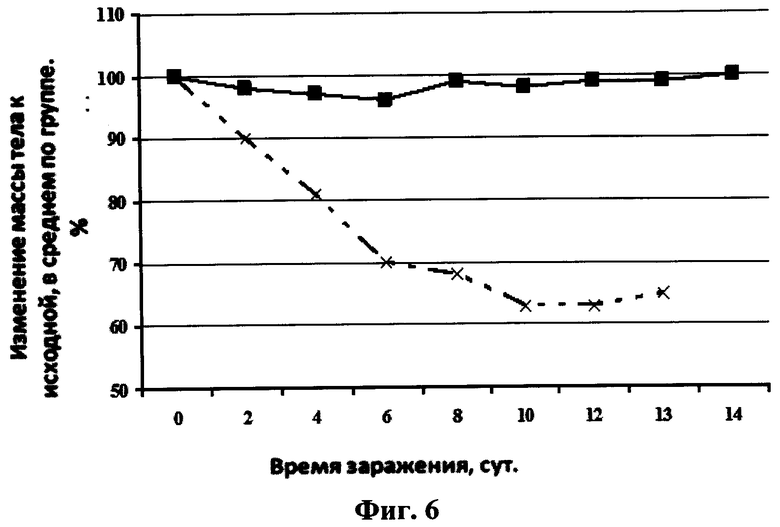
На фиг.6 представлен график зависимости изменения массы тела мышей после интраназального введения однодоменного мини-антитела против вируса гриппа и последующего заражения их вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2) от времени после заражения. Сплошная линия с маркером в виде квадрата черного цвета - 20 мкл интраназально раствора однодоменного мини-антитела против вируса гриппа в фосфатном буфере в дозе 2,5 мкг/мышь, сплошная линия с маркером в виде креста - 20 мкл интраназально раствора фосфатного буфера.
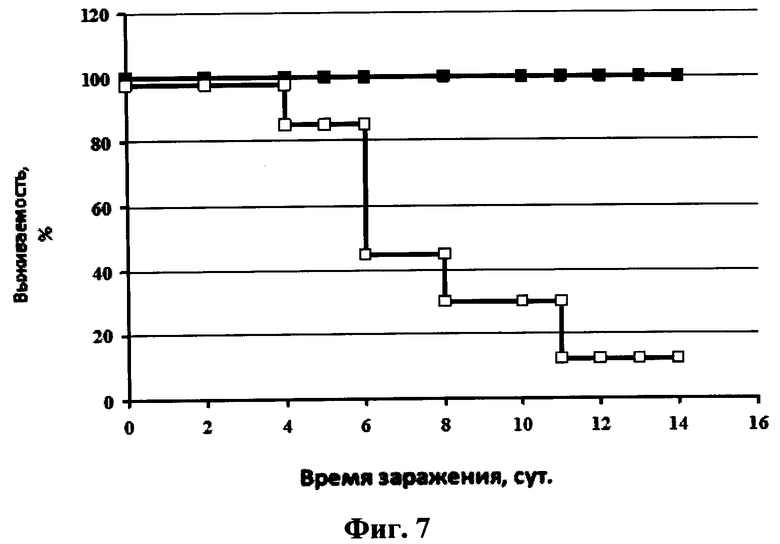
На фиг.7 представлен график зависимости выживаемости мышей после заражения их вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2) и последующего интраназального введения раствора однодоменного мини-антитела против вируса гриппа в фосфатном буфере в дозе 2,5 мкг/мышь. Сплошная линия с маркером в виде квадрата черного цвета - 20 мкл интраназально раствора однодоменного мини-антитела против вируса гриппа в фосфатном буфере в дозе 2,5 мкг/мышь, сплошная линия с маркером в виде квадрата белого цвета - 20 мкл интраназально раствора фосфатного буфера.
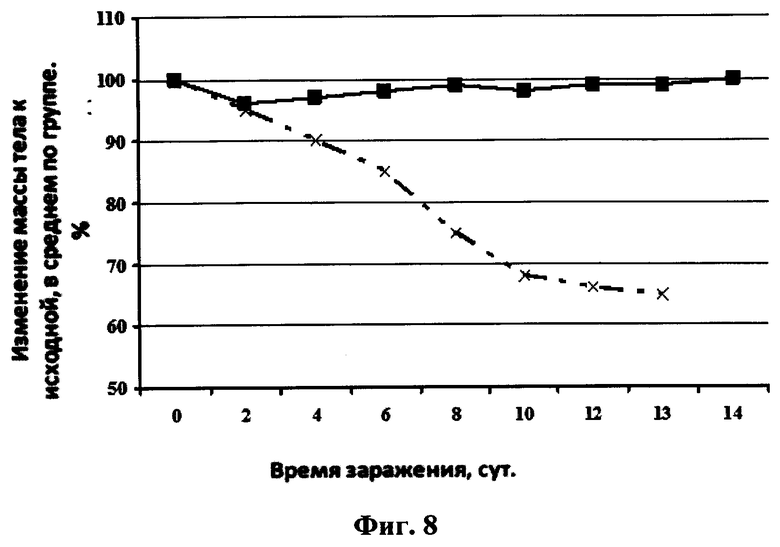
На фиг.8 представлен график зависимости изменения массы тела мышей после заражения их вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2) и последующего интраназального введения раствора однодоменного мини-антитела против вируса гриппа в фосфатном буфере в дозе 2,5 мкг/мышь. Сплошная линия с маркером в виде квадрата черного цвета - 20 мкл интраназально раствора однодоменного мини-антитела против вируса гриппа в фосфатном буфере в дозе 2,5 мкг/мышь, сплошная линия с маркером в виде креста - 20 мкл интраназально раствора фосфатного буфера.
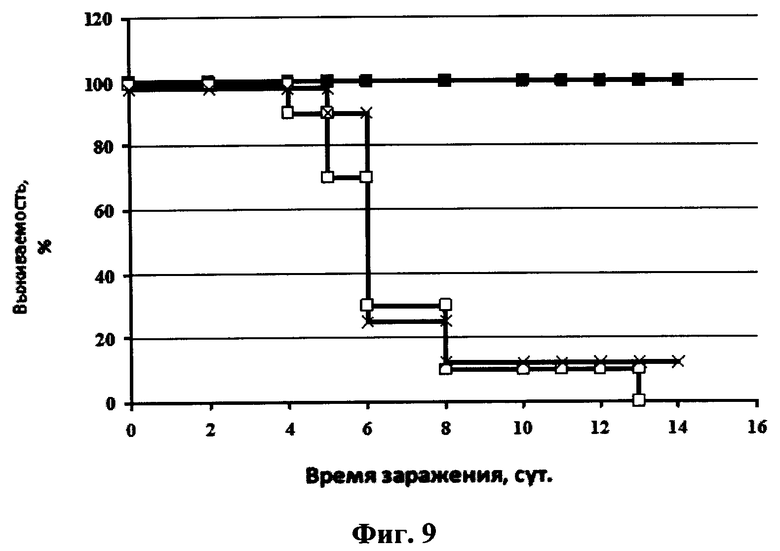
На фиг.9 представлен график зависимости выживаемости мышей после интраназального введения рекомбинантного аденовирусного вектора Ad5-anti-flu и последующего заражения их вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2) от времени после заражения. Сплошная линия с маркером в виде квадрата черного цвета- 20 мкл интраназально препарата рекомбинантных аденовирусных векторов Ad5-anti-flu в дозе 107 БОЕ/мышь, сплошная линия с маркером в виде креста - 20 мкл интраназально препарата рекомбинантных аденовирусных векторов Ad-null, не содержащего вставку целевого гена в экспрессирующей кассете в дозе 107 БОЕ/мышь, сплошная линия с маркером в виде квадрата белого цвета - 20 мкл интраназально 1×PBS.
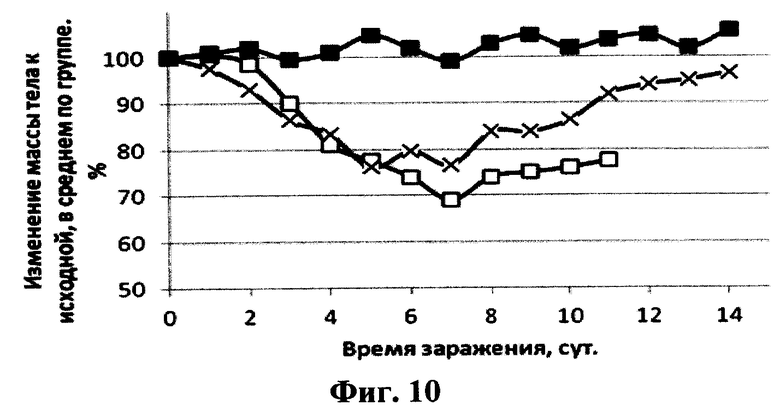
На фиг.10 представлены графики зависимости изменения массы тела мышей после интраназального введения рекомбинантного аденовирусного вектора Ad5-anti-flu и последующего заражения их вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2) от времени после заражения. Сплошная линия с маркером в виде квадрата черного цвета - 20 мкл интраназально препарата рекомбинантных аденовирусных векторов Ad5-anti-flu в дозе 107 БОЕ/мышь, сплошная линия с маркером в виде креста - 20 мкл интраназально препарата рекомбинантных аденовирусных векторов Ad-null, не содержащего вставку целевого гена в экспрессирующей кассете в дозе 107 БОЕ/мышь, сплошная линия с маркером в виде квадрата белого цвета - 20 мкл интраназально 1×PBS.
Примеры осуществления настоящего изобретения
Пример 1
Получение библиотеки вариабельных доменов одноцепочечных антител
Иммунизация
Двугорбого верблюда Camelus bactrianus последовательно иммунизировали 4 раза путем подкожного введения антигенного материала, смешанного с равным объемом неполного адъюванта Фройнда. В качестве антигена использовали препарат инактивированного вируса гриппа птиц A/Mallard duck/PA/10218/84(H5N2) из музея ФГБУ «НИИЭМ им. Н.Ф. Гамалеи» Минздрава России и препарат вируса гриппа птиц H5N2 (штамм А/17/Утка/Потсдам/86/92), полученного от ФГУП НПО «Микроген» Минздрава России, г. Иркутск. Вторую иммунизацию проводили через 3 недели после первой, затем с интервалом в две недели проводили еще две иммунизации. Взятие крови (150 мл) проводили через 6 дней после последней инъекции. Для предотвращения свертывания взятой крови добавляли 50 мл стандартного солевого раствора (PBS), содержащего гепарин (100 ед./мл) и ЭДТА (3 мМ).
Кровь разводили в 2 раза стандартным солевым раствором (PBS), содержащим 1 мМ ЭДТА. 35 мл разбавленного раствора крови наслаивали на ступеньку специальной среды (Histopaque-1077, Sigma) с плотностью 1,077 г/мл объемом 15 мл и проводили центрифугирование в течение 20 мин при 800×g. Мононуклеарные клетки (лимфоциты и моноциты) отбирали из интерфазной зоны плазма/Histopaque, после чего промывали раствором PBS, содержащим 1 мМ ЭДТА.
Суммарную РНК из В-лимфоцитов выделяли с помощью реагента TRIzol (Invitrogen). Затем, на колонке с олиго(d-Т)-целлюлозой из тотальной РНК выделяли поли(А)-содержащую РНК. Концентрацию РНК определяли с помощью биофотометра (Eppendorf) и проверяли качество выделенной РНК с помощью электрофореза в 1,5%-ном агарозном геле с формальдегидом.
Реакцию обратной транскрипции проводили по стандартному протоколу [Sambrook et al., 1989] с использованием обратной транскриптазы H-M-MuLV и праймера олиго(dT)15 в качестве затравки.
Продукты обратной транскрипции использовали в качестве матрицы в двухступенчатой полимеразной цепной реакции и полученные продукты амплификации клонировали по сайтам NcoI(PstI) и NotI в фагмидный вектор, как описано ранее [Hamers-Casterman et al., 1993; Nguyen et al., 2001; Saerens et al., 2004; Rothbauer et al., 2006]. Процедуру селекции проводили также аналогично тем, что описаны в указанных работах. Она базировалась на методе фагового дисплея, в которой в качестве фага-помощника использовали бактериофаг M13KO7 (New England Biolabs, США).
Пример 2
Селекция однодоменных мини-антител, специфически узнающих вирус гриппа A, их форматирование и наработка
Селекцию однодоменных мини-антител проводили методом фагового дисплея с использованием препарата инактивированного вируса гриппа птиц A/Mallard duck/PA/10218/84(H5N2), иммобилизованного на дне лунок 96-луночного ИФА-планшета. Использовали полистироловые иммунологические плашки с высокой сорбцией MICROLON 600 (Greiner Bio-One). Для блокировки использовали 1% БСА (Sigma-Aldrich, США) и/или 1% обезжиренное молоко (Bio-Rad, США) в PBS. Процедуру селекции и последующей амплификации отбираемых фаговых частиц (содержащих ген однодоменного мини-антитела внутри, а экспрессирующееся однодоменное мини-антитело - в составе поверхностного фагового белка pIII) повторяли, как правило, последовательно три раза. Все манипуляции проводили, как описано в публикациях [Тиллиб С.В., Иванова Т.И., Васильев Л.А. 2010. Фингерпринтный анализ селекции «однодоменных мини-антител» методом фагового дисплея с использованием двух вариантов фагов-помощников. Acta Naturae 2010; 2 (3): 100-108; Hamers-Casterman С., Atarhouch Т., Muyldermans S. et al. Nature 1993; 363: 446-448.; Nguyen V.K., Desmyter A., Muyldermans S. Adv. Immunol. 2001; 79: 261-296; Saerens D., Kinne J., Bosmans E., Wernery U., Muyldermans S., Conrath K. J Biol Chem. 2004; 279: 51965-51972; Rothbauer U., Zolghadr K., Tillib S., et al. Nature Methods 2006; 3: 887-889].
Последовательности клонов отобранных однодоменных мини-антител группировали согласно схожести их фингерпринтов, получаемых при электрофоретическом разделении продуктов гидролиза амплифицированных последовательностей однодоменных мини-антител параллельно тремя частощепящими рестрикционными эндонуклеазами (HinfI, MspI, RsaI). Затем, отобранные клоны-представители каждой из различающихся по фингерпринту групп использовали для аналитической наработки однодоменных мини-антител, активность которых проверяли методом иммуноферментного анализа на предмет эффективного связывания с иммобилизованным в ячейке иммунологического планшета препарата инактивированного вируса гриппа птиц A/Mallard duck/PA/10218/84(H5N2).
В результате была отобрана последовательность, кодирующая однодоменное мини-антитело «Неогриппаб» (фиг.1).
Для того чтобы повысить эффективность выделения и детекции однодоменного мини-антитела, а также с целью значительно усилить биологическую активность исходно отобранного моновалентного однодоменного антитела, была использована разработанная нами ранее эффективная процедура форматирования (модифицирования аминокислотной последовательности антитела), подробно описанная в работе Тиллиба и соавт. (Tillib et al., 2013, Antiviral Res. 97: 245-254. Formatted single-domain antibodies can protect mice against infection with influenza virus (H5N2). // Форматированные однодоменные антитела могут защитить мышь от инфицирования вирусом гриппа (H5N2).) Эта процедура включала присоединение к С-концу исходно отобранного антитела четырех дополнительных аминокислотных участков (доменов), представленных на фиг.2: (1) - линкерного линейного участка (соответствующего верхнему удлиненному шарнирному участку особых иммуноглобулинов IgG2a верблюда), (2) - специального тримеризующегося домена - "ILZ" ("isoleucine zipper domain", или, в переводе, «изолейциновой молнии»), (3) - НА-тага (пептида из белка гемагглютинина, к которому есть коммерческие антитела) и (4) His-тага (шести остатков гистидина на самом С-конце белка для эффективной очистки белка с помощью металл-хелатной хроматографии).
Продукция однодоменных мини-антител
Последовательности кодирующей ДНК отобранного однодоменного мини-антитела «Неогриппаб», форматированного как описано выше, клонировали в экспрессионный плазмидный вектор - модифицированный вектор pHEN6 [Conrath KE, Lauwereys М, Galleni М, Matagne A, Frère JM, Kinne J, Wyns L, Muyldermans S. Beta-lactamase inhibitors derived from single-domain antibody fragments elicited in the Camelidae. Antimicrob Agents Chemother. 2001; 45: 2807-12]. Благодаря наличию на N-конце экспрессируемой последовательности сигнального пептида (pelB) нарабатываемый рекомбинантный белок (однодоменное мини-антитело) накапливается в периплазме бактерий, что позволяет эффективно его выделять методом осмотического шока, не разрушая собственно бактериальные клетки. Продукцию однодоменных мини-антител проводили в Е.coli (штамм BL21). Экспрессию индуцировали добавлением 1 мМ индолил-бета-D-галактопиранозида и клетки инкубировали при интенсивном перемешивании в течение 7 часов при 37°C или в течение ночи при 29°C. Однодоменное мини-антитело выделяли из периплазматического экстракта с использованием аффинной хроматографии на Ni-NTA-агарозе с использованием системы для очистки QIAExpressionist (QIAGEN, США).
Полученные в результате элюции белковые фракции концентрировали (до конечной концентрации примерно 1-5 мг/мл) с помощью ультрафильтрационных приспособлений (с размером отсечения 10 кДа) Amicon фирмы Millipore, после чего наслаивали в объеме 0,5-2 мл на колонку для гель-фильтрационной хроматографии (Sephacryl S-100 HR; 1,5×50 см). Хроматографию проводили, используя систему BioLogic LP фирмы Bio-Rad, при постоянной скорости потока 0,3 мл/мин. В качестве буферного раствора использовали стандартный солевой буферный раствор (PBS). Фракции собирали согласно профилю поглощения (при длине волны 280 нм) элюированного белка. Размер тримеризованного форматированного антитела примерно соответствовал размеру (и пику элюции) маркерного белка - бычьего сывороточного альбумина (BSA, 67 кДа). Объединенные отобранные форматированные антитела (из фракций соответствующих пиков элюции) снова концентрировали (как описано выше), дополнительно очищали от эндотоксина на аффинной колонке фирмы Thermo Scientific (Detoxi-Gel Endotoxin Removing Gel), после чего стерилизовали фильтрованием через целлюлоза-ацетатную мембрану (фирмы Nalgene) с порами 0,2 мкм. Очищенные антитела хранили в аликвотах при 4°C или, после добавления 50% глицерина, при -20°C. Для более долгого хранения антитела лиофилизировали.
Демонстрация связывания однодоменного мини-антитела с вирусом гриппа
Способность однодоменного мини-антитела «Неогриппаб» связывать гемагглютинин и целый вирус гриппа проверяли с использованием метода иммуноферментного анализа с иммобилизованным белком гемагглютинином (Н5 из штамма A/American green-winged teal/California/HKWF609/07), Cat. No. 11699-V08H, Sino Biological Inc.) и и инактивированным рекомбинантным вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2), по стандартному протоколу. В качестве контроля использовали лунки с иммобилизованным белком овальбумином кур (неспецифический белок). Детекцию связавшегося с гемагглютинином или с вирусом гриппа форматированного однодоменного мини-антитела проводили с помощью анти-НА антител мыши, вторичных антител к иммуноглобулинам мыши, конъюгированных с пероксидазой хрена, и хромогенного субстрата АБТС (2,2'-азино-бис(3-этилбензтиазолин)-6-сульфоновой кислоты, Sigma).
На фиг.3 и 4 представлены результаты анализа, из которых следует, что однодоменное мини-антитело «НЕОГРИППАБ» специфически связывается как с иммобилизованным белком гемагглютинином (Н5 из штамма А/American green-winged teal/California/HKWF609/07), Cat. No. 11699-V08H, Sino Biological Inc.), так и с препаратом вируса гриппа, но не связывается с овальбумином (или БСА), использованным в качестве блокирующего агента.
Отобранную последовательность однодоменного мини-антитела «Неогриппаб» затем клонировали в геном рекомбинантного аденовирусного вектора.
Пример 3
Создание плазмидной конструкции pShuttle-CMV-Неогриппаб с участками генома аденовируса человека 5-го серотипа для получения рекомбинантного аденовирусного вектора, несущего экспрессирующую кассету, содержащую нуклеотидную последовательность, которая кодирует однодоменное мини-антитело «Неогриппаб», связывающееся с вирусом гриппа
Для получения плазмидной конструкции pShuttle-CMV-Неогриппаб в качестве вектора использовали плазмидную конструкцию pShuttle-CMV с участками генома аденовируса человека 5-го серотипа для получения рекомбинантных аденовирусных векторов, входящую в набор к системе «AdEasy Adenoviral vector system» («Stratagene» Cat. No. 240009). Плазмидную конструкцию pShuttle-CMV гидролизовали по сайту для эндонуклеазы рестрикции EcoRV, а затем осуществляли вставку нуклеотидной последовательности однодоменного мини-антитела с получением, соответственно, конструкции pShuttle-CMV-Неогриппаб. Нуклеотидную последовательность однодоменного мини-антитела «Неогриппаб» для вставки в pShuttle-CMV получали путем гидролиза плазмидной конструкции pAL-Неогриппаб по сайтам для эндонуклеазы рестрикции EcoRV. Наличие гена однодоменного мини-антитела «Неогриппаб» в составе плазмидной конструкции pShuttle-CMV-Неогриппаб подтверждали рестрикционным анализом при использовании эндонуклеаз EcoRI, NotI и EcoRV и методом ПЦР.
Пример 4
Получение рекомбинантного аденовирусного вектора Ad5-Неогриппаб, несущего экспрессирующую кассету, содержащую нуклеотидную последовательность, которая кодирует однодоменное мини-антитело «Неогриппаб», связывающееся с вирусом гриппа
Получение рекомбинантного аденовирусного вектора Ad5-Неогриппаб проводили согласно методике «AdEasy Adenoviral vector system» («Stratagene» Cat. No. 240009). Наличие гена однодоменного мини-антитела Неогриппаб в составе соответствующего рекомбинантного аденовирусного вектора Ad5-Неогриппаб подтверждали методом ПЦР.
Далее определяли титр препарата рекомбинантного аденовирусного вектора Ad5-Неогриппаб методом бляшкообразования на культуре клеток 293 (клетки эмбриональной почки человека).
Пример 5
Нейтрализация вируса гриппа однодоменным мини-антителом «Неогриппаб» в культуре клеток in vitro
Постановку реакции нейтрализации вируса проводили с использованием линии клеток MDCK, культивируемых в 96-луночных культуральных планшетах (Costar, США) до конфлюэнтности клеточного монослоя 80%. Перед проведением реакции в культуральных планшетах с клетками среду заменяли на бессывороточную. Для проведения реакции использовали вирус гриппа птиц A/Mallard duck/PA/10218/84(H5N2) в дозе, равной 100 ТЦД50 на лунку. Разведения вируса и исследуемого антитела «Неогриппаб» производили в бессывороточной среде DMEM. В лунки 96-луночного планшета вносили по 50 мкл суспензии вируса, затем добавляли в каждую лунку равные объемы двукратных разведений сывороток, начиная с 1:10. Планшет инкубировали при 37°C в течение 1 часа. После инкубации смесь вируса и сывороток добавляли в каждую лунку планшета с клетками для адсорбции вируса на клетках. Клетки инкубировали при 37°C в атмосфере 5% CO2 в течение 2 часов. Затем из планшетов отбирали бессывороточную среду и заменяли ее на содержащую эмбриональную сыворотку КРС среду MEM с 0,2% БСА и трипсином в концентрации 1 мкг/мл. Результаты реакции нейтрализации вируса учитывали через 72 ч по наступлению ЦПД в культуре клеток по методу Рида и Менча. Исследуемое антитело «Неогриппаб» обеспечивало нейтрализацию вируса гриппа в интервале концентраций от 0,009 мкг/мл до 2,5 мкг/мл.
Пример 6
Нейтрализация вируса гриппа однодоменным мини-антителом «Неогриппаб», экспрессируемым рекомбинантным аденовирусным вектором Ad5-Неогриппаб в культуре клеток in vitro
Экспрессию однодоменного мини-антитела «Неогриппаб» производили в клетках линии А549. Для этого клетки А549 рассевали на лунки 48-луночного планшета (Costar, США) в количестве 104 клеток на лунку, после чего к клеткам добавляли бессывороточную среду 293AGT («Invitrogen», США) и инкубировали при 37°C в атмосфере 5% CO2 в течение 20-24 ч. Затем клетки трансдуцировали препаратом рекомбинантного аденовирусного вектора Ad5-Неогриппаб в дозе 5 БОЕ/клетку. Клетки инкубировали при 37°C в атмосфере 5% CO2 в течение 2 ч, после чего из лунок отбирали среду и заменяли ее новой порцией среды 293AGT. Трансдуцированные клетки инкубировали в течение 72 ч, затем производили отбор культуральной среды. Таким образом, получали препараты культуральной среды, содержащей экспрессированные рекомбинантным аденовирусным вектором Ad5-Неогриппаб однодоменное мини-антитело «Неогриппаб».
Постановку реакции нейтрализации вируса проводили с использованием линии клеток MDCK, культивируемых в 96-луночных культуральных планшетах (Costar, США) до конфлюэнтности клеточного монослоя 80%. Перед проведением реакции в культуральных планшетах с клетками среду заменяли на бессывороточную. Для проведения реакции использовали вирус гриппа птиц A/Mallard duck/PA/10218/84(H5N2) в дозе, равной 100 ТЦД50 на лунку. Разведения вируса и культуральной среды, содержащей исследуемое антитело «Неогриппаб», производили в бессывороточной среде DMEM. В лунки 96-луночного планшета вносили по 50 мкл суспензии вируса, затем добавляли в каждую лунку равные объемы двукратных разведений культуральных среды, начиная с 1:10. Планшет инкубировали при 37°C в течение 1 ч. После инкубации смесь вируса и культуральной среды добавляли в каждую лунку планшета с клетками для адсорбции вируса на клетках. Клетки инкубировали при 37°C в атмосфере 5% CO2 в течение 2 часов. Затем из планшетов отбирали бессывороточную среду и заменяли ее на содержащую эмбриональную сыворотку КРС среду MEM с 0,2% БСА и трипсином в концентрации 1 мкг/мл. Результаты реакции нейтрализации вируса учитывали через 72 ч по наступлению ЦПД в культуре клеток по методу Рида и Менча. Препараты культуральной среды, содержащие экспрессированное рекомбинантным аденовирусным вектором Ad5-Неогриппаб однодоменное мини-антитело «Неогриппаб», обеспечивали нейтрализацию вируса гриппа в разведении 1:100.
Пример 7
Применение однодоменного мини-антитела «Неогриппаб» для профилактики заболевания гриппом
С целью профилактики заболевания гриппом типа A самок мышей линии Balb/c массой тела 18 г разделяли на 2 группы по 50 мышей в каждой группе:
- первой группе мышей вводили 20 мкл раствора однодоменных мини-антител «Неогриппаб» в фосфатном буфере дозе 2,5 мкг/мл,
- второй группе мышей интраназально вводили 20 мкл фосфатного буфера.
Через 1 час после введения вышеуказанных препаратов мышей заражали 25 ЛД50 адаптированного по своим вирулентным свойствам для модели мышей вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2). Заражение производили интраназально под легким эфирным наркозом. В течение 14 дней после заражения учитывали выживаемость мышей и изменение массы их тела.
Протекция мышей против вируса A/Mallard duck/PA/10218/84(H5N2) составляла 100% в группе, которой вводили препарат однодоменного мини-антитела «Неогриппаб» (см. фиг.5). Также у этих мышей не наблюдалось уменьшение массы тела. Кривые уменьшения массы тела у мышей каждой из двух вышеуказанных групп после инфекции вирусом гриппа представлены на фиг.6.
Таким образом, интраназальное введение однодоменного мини-антитела «Неогриппаб» полностью защищает мышей от заражения вирусом гриппа дозой 25 ЛД50.
Пример 8
Применение однодоменного мини-антитела «Неогриппаб» для терапии гриппа
С целью терапии гриппа типа А 100 самок мышей линии Balb/c массой тела 18 г заражали 25 ЛД50 адаптированного по своим вирулентным свойствам для модели мышей вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2). Заражение производили интраназально под легким эфирным наркозом.
Через 4 часа после введения вышеуказанных препаратов мышей разделяли на 2 группы по 50 мышей в каждой группе:
- первой группе мышей вводили 20 мкл раствора однодоменных мини-антител «Неогриппаб» в фосфатном буфере в дозе 2,5 мкг/мл,
- второй группе мышей вводили 20 мкл интраназально фосфатного буфера.
В течение 14 дней после заражения учитывали выживаемость мышей и изменение массы их тела.
Протекция мышей против вируса A/Mallard duck/PA/10218/84(H5N2) составляла 100% в группе, которой вводили препарат однодоменного мини-антитела «Неогриппаб» (см. фиг.7). Также у этих мышей не наблюдалось уменьшение массы тела. Кривые уменьшения массы тела у мышей каждой из двух вышеуказанных групп представлены на фиг.8.
Таким образом, интраназальное введение однодоменного мини-антитела Неогриппаб полностью защищает мышей от заражения вирусом гриппа дозой 25 ЛД50.
Пример 9
Применение рекомбинантного аденовирусного вектора Ad5-Неогриппаб для профилактики заболевания гриппом
С целью профилактики заболевания гриппом самок мышей линии Balb/c массой тела 18 г разделяли на 3 группы по 50 мышей в каждой группе:
- первой группе мышей интраназально вводили 20 мкл коллоидного раствора рекомбинантного аденовирусного вектора Ad5-Неогриппаб в фосфатном буфере в дозе 107 БОЕ/мышь,
- второй группе мышей интраназально вводили 20 мкл коллоидного раствора рекомбинантного аденовирусного вектора Ad-null, не содержащего вставку целевого гена в экспрессирующей кассете в фосфатном буфере в дозе 107 БОЕ/мышь,
- третьей группе мышей интраназально вводили 20 мкл фосфатного буфера.
Через 72 часа после введения вышеуказанных препаратов мышей заражают 25 ЛД50 адаптированного по своим вирулентным свойствам для модели мышей вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2). Заражение производили интраназально под легким эфирным наркозом. В течение 14 дней после заражения учитывали выживаемость мышей и изменение массы их тела.
Протекция мышей против вируса A/Mallard duck/PA/10218/84(H5N2) составляла 100% в группе, которой вводили препарат рекомбинантного аденовирусного вектора Ad5-Неогриппаб (см. фиг.9). Также у этих мышей не наблюдалось уменьшение массы тела. Кривые уменьшения массы тела у мышей каждой из трех вышеуказанных групп после инфекции вирусом гриппа представлены на фиг.10.
Таким образом, интраназальное введение рекомбинантного аденовирусного вектора Ad5-Неогриппаб полностью защищало мышей от заражения вирусом гриппа в дозе 25 ЛД50.
Пример 10
Применение фармацевтической композиции, состоящей из однодоменного мини-антитела «Неогриппаб» и рекомбинантного аденовирусного вектора Ad5-Неогриппаб для профилактики и терапии гриппа
С целью проведения профилактики и терапии гриппа самок мышей линии Balb/c массой тела 18 г разделяли на 18 групп по 20 мышей в каждой группе:
1, 2 и 3 группа - каждому животному интраназально вводили 20 мкл препарата, содержащего 3 мкг однодоменных мини-антител «Неогриппаб» и 107 БОЕ рекомбинантного аденовирусного вектора Ad5-Неогриппаб,
4, 5 и 6 группа - каждому животному интраназально вводили 20 мкл препарата, содержащего 3 мкг БСА и 107 БОЕ рекомбинантного аденовирусного вектора Ad5-Неогриппаб,
7, 8 и 9 группа - каждому животному интраназально вводили 20 мкл препарата, содержащего 3 мкг однодоменных мини-антител «Неогриппаб» и 107 БОЕ рекомбинантного аденовирусного вектора Ad5-null,
10, 11 и 12 группа - каждому животному интраназально вводили 20 мкл препарата, содержащего 3 мкг БСА и 107 БОЕ рекомбинантного аденовирусного вектора Ad5-null,
13, 14 и 15 группа - каждому животному интраназально вводили 20 мкл фосфатного буфера,
16, 17 и 18 группа - интактные мыши.
В группах 1, 4, 7, 10, 13, 16 через 1 час после введения вышеуказанных препаратов мышей заражали 25 ЛД50 адаптированного по своим вирулентным свойствам для модели мышей вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2). Заражение производили интраназально под легким эфирным наркозом. В течение 14 дней после заражения учитывали выживаемость мышей и изменение их массы тела.
В группах 2, 4, 8, 11, 14, 17 через 72 часа после введения вышеуказанных препаратов мышей заражали 25 ЛД50 адаптированного по своим вирулентным свойствам для модели мышей вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2). Заражение производили интраназально под легким эфирным наркозом. В течение 14 дней после заражения учитывали выживаемость мышей и изменение их массы тела.
В группах 3, 6, 9, 12, 15, 18 через 14 дней после введения вышеуказанных препаратов мышей заражали 25 ЛД50 адаптированного по своим вирулентным свойствам для модели мышей вирусом гриппа птиц A/Mallard duck/PA/10218/84(H5N2). Заражение производили интраназально под легким эфирным наркозом. В течение 14 дней после заражения учитывали выживаемость мышей и изменение их массы тела.
Протекция мышей против вируса A/Mallard duck/PA/10218/84(H5N2) составляла 100% в группах №1-3, 5-8. Также у этих мышей не наблюдалось уменьшение массы тела (см. таблицу 2).
Таким образом, интраназальное введение фармацевтической композиции, состоящей из однодоменных мини-антител «Неогриппаб» и рекомбинантных аденовирусных векторов Ad5-Неогриппаб, полностью защищает от инфекции вируса гриппа типа A при всех выбранных режимах заражения в отличие от использования только антител «Неогриппаб» или только вектора Ad5-Неогриппаб.
| название | год | авторы | номер документа |
|---|---|---|---|
| НАНОАНТИТЕЛО "ANTI-FLU", РЕКОМБИНАНТНЫЕ ВИРУСНЫЕ ВЕКТОРЫ И ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ ДЛЯ ПРОФИЛАКТИКИ И ТЕРАПИИ ГРИППА ТИПА А | 2011 |
|
RU2502745C2 |
| ПРОТИВОГРИППОЗНАЯ ВАКЦИНА ШИРОКОГО СПЕКТРА ДЕЙСТВИЯ ПРОТИВ ПТИЧЬЕГО ГРИППА А НА ОСНОВЕ ЭКТОДОМЕНА БЕЛКА М2 | 2014 |
|
RU2571944C1 |
| Универсальная противогриппозная вакцина | 2015 |
|
RU2618918C2 |
| Штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа Ad5-tetOFF-E3-HA125, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов H1, H2, H5 для создания противогриппозных иммуногенных препаратов, способ получения гена | 2018 |
|
RU2713722C1 |
| РЕКОМБИНАНТНАЯ ПСЕВДОАДЕНОВИРУСНАЯ ЧАСТИЦА НА ОСНОВЕ ГЕНОМА АДЕНОВИРУСА ЧЕЛОВЕКА 5 СЕРОТИПА, ПРОДУЦИРУЮЩАЯ ГЕМАГГЛЮТИНИН ВИРУСА ГРИППА ШТАММА A/Brisbane/59/2007(H1N1) И СПОСОБ ЕЕ ИСПОЛЬЗОВАНИЯ | 2012 |
|
RU2523599C1 |
| Экспрессионный вектор на основе аденовируса человека 5 серотипа, индуцирующий кросс-протективный иммунитет к вирусам гриппа А субтипа Н1, и фармацевтическая композиция на его основе | 2023 |
|
RU2802753C1 |
| Однодоменное антитело для нейтрализации вирусов и его модификации, и способ их применения для экстренной профилактики заболеваний, вызываемых вирусом гриппа А | 2021 |
|
RU2777073C1 |
| Вакцина против гриппа типа А, гриппа типа B и COVID-19 | 2021 |
|
RU2751485C1 |
| СПОСОБ СОЗДАНИЯ РЕКОМБИНАНТНОГО АДЕНОВИРУСА ПТИЦ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИРУСА ГРИППА ПТИЦ Н5N1 | 2006 |
|
RU2326943C1 |
| Экспрессионный вектор на основе аденовируса человека 5 серотипа индуцирующий кросс-протективный иммунитет к вирусам гриппа А субтипа H3 и фармацевтическая композиция на его основе. | 2023 |
|
RU2814189C1 |




Настоящее изобретение относится к вирусологии и предполагает использование однодоменных мини-антител для профилактики и терапии гриппа. Заявлено однодоменное мини-антитело, специфически связывающееся с определенным эпитопом гемагглютинина вируса гриппа типа A (H5N2) и подавляющее инфекцию вируса гриппа типа A (H5N2). Создан аденовирусный вирусный вектор, экспрессирующий однодоменное мини-антитело, которое может эффективно связывать определенный эпитоп гемагглютинина вируса гриппа и тем самым блокировать развитие гриппозной инфекции. Предлагается композиция, состоящая из эффективного количества однодоменного мини-антитела и вирусного вектора, экспрессирующего такое однодоменное мини-антитело. Предлагается способ профилактики или терапии инфекции вируса гриппа A (H5N2), предусматривающий введение нуждающемуся в этом пациенту профилактически или терапевтически эффективного количества фармацевтической композиции интраназально в форме капель или спрея. 5 н. и 4 з.п. ф-лы, 10 ил., 2 табл., 10 пр.
1. Однодоменное мини-антитело, характеризующееся аминокислотной последовательностью SEQ ID NO: 2, специфически связывающееся с гемагглютинином вируса гриппа типа A (H5N2) и подавляющее инфекцию вируса гриппа типа A (H5N2), или его функционально активный вариант.
2. Нуклеотидная последовательность, кодирующая однодоменное мини-антитело по п.1 или его функционально активный вариант.
3. Рекомбинантный вирусный вектор, экспрессирующий однодоменное мини-антитело по п.1 или его функционально активный вариант, содержащий нуклеотидную последовательность по п.2.
4. Вирусный вектор по п.3, отличающийся тем, что он является аденовирусным вектором.
5. Фармацевтическая композиция для профилактики или терапии инфекции вируса гриппа A (H5N2), содержащая профилактически или терапевтически эффективное количество однодоменного мини-антитела по п.1 или его функционально активного варианта и/или вирусного вектора по п.3 и фармацевтически приемлемый носитель.
6. Способ профилактики или терапии инфекции вируса гриппа A (H5N2), предусматривающий введение нуждающемуся в этом пациенту профилактически или терапевтически эффективного количества фармацевтической композиции по п.5.
7. Способ по п.6, в котором указанную фармацевтическую композицию вводят интраназально.
8. Способ по п.7, в котором интраназальное введение осуществляют в форме капель.
9. Способ по п.7, в котором интраназальное введение осуществляют в форме спрея.
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ И СПОСОБЫ | 2008 |
|
RU2468034C2 |
| US 20130022608 A1, 24.01.2013 | |||
| US 2012315323 A1, 13.12.2012 | |||

