Перекрестная ссылка на родственную заявку
По настоящей патентной заявке испрашивается приоритет японской патентной заявки No. 190862/2008, которая зарегистрирована 24 июля 2008 г., японской патентной заявки No. 270294/2008, которая зарегистрирована 20 октября 2008 г., и японской патентной заявки No. 20591/2009, которая зарегистрирована 30 января 2009 г., и полный объем раскрытого содержания всех их включен в настоящее описание в качестве ссылки в виде части раскрытия настоящей заявки.
Известный уровень техники
Область техники, к которой относится изобретение
Настоящее изобретение относится к гену биосинтеза пирипиропена А.
Характеристика известного уровня техники
Как раскрыто в выложенной японской патентной публикации No. 360895/1992 (патентный документ 1) и в Journal of Antibiotics (1993), 46(7), 1168-9 (непатентный документ 1), пирипиропен А обладает ингибиторной активностью в отношении ACAT (ацил-CoA-холестеринацилтрансферазы). Предполагается его применение для лечения заболеваний, вызываемых аккумуляцией холестерина или тому подобного.
Дополнительно в статье в Journal of Synthetic Organic Chemistry, Japan (1998), Vol. 56, No. 6, 478-488 (непатентный документ 2), WO94/09147 (патентный документ 2), выложенной японской патентной публикации No. 184158/1994 (патентный документ 3), выложенной японской патентной публикации No. 239385/1996 (патентный документ 4), выложенной японской патентной публикации No. 259569/1996 (патентный документ 5), выложенной японской патентной публикации No. 269062/1996 (патентный документ 6), выложенной японской патентной публикации No. 269063/1996 (патентный документ 7), выложенной японской патентной публикации No. 269064/1996 (патентный документ 8), выложенной японской патентной публикации No. 269065/1996 (патентный документ 9), выложенной японской патентной публикации No. 269066/1996 (патентный документ 10), выложенной японской патентной публикации No. 291164/1996 (патентный документ 11) и в статье в Journal of Antibiotics (1997), 50(3), 229-36 (непатентный документ 3) раскрыты аналоги и производные пирипиропена, а также их ингибиторная активность в отношении ACAT.
Далее, в статье в Applied and Environmental Microbiology (1995), 61(12), 4429-35 (непатентный документ 4) раскрыто, что пирипиропен А обладает инсектицидной активностью против личинки Helicoverpa armigera. Кроме того, в WO2004/060065 (патентный документ 12) раскрыто, что пирипиропен А обладает инсектицидной активностью против личинки капустной моли и Tenebrio molitor.
Кроме того, в WO2006/129714 (патентный документ 13) и в WO2008/066153 (патентный документ 14) раскрыто, что аналоги пирипиропена обладают инсектицидной активностью против растительной тли.
Более того, в качестве бактерии, продуцирующей пирипиропен А, раскрыты штамм FO-1289 Aspergillus fumigatus в выложенной японской патентной публикации No. 360895/1992 (патентный документ 1); штамм NRRL-3446 Eupenicillium reticulosporum в статье в Applied and Environmental Microbiology (1995), 61(12), 4429-35 (непатентный документ 4); и штамм F1959 Penicillium griseofulvum в патенте WO2004/060065 (патентный документ 12); и штамм PF1169 Penicillium coprobium в Journal of Technical Disclosure 500997/2008 (патентный документ 15).
Также в качестве пути биосинтеза пирипиропена А в статьях в Journal of Organic Chemistry (1996), 61, 882-886 (непатентный документ 5) и в Chemical Review (2005), 105, 4559-4580 (непатентный документ 6) раскрыт предполагаемый путь биосинтеза в штамме FO-1289 Aspergillus fumigatus. В этих документах раскрыто, что в штамме FO-1289 Aspergillus fumigates подструктуры, индивидуально синтезируемые с помощью поликетидсинтазы и пренилтрансферазы, соединяются для синтеза пирипиропена А под действием циклазы.
[Ссылки предшествующего уровня техники]
[Патентные документы]
[Патентный документ 1] выложенная японская патентная публикация No. 360895/1992
[Патентный документ 2] WO94/09147
[Патентный документ 3] выложенная японская патентная публикация No. 184158/1994
[Патентный документ 4] выложенная японская патентная публикация No. 239385/1996
[Патентный документ 5] выложенная японская патентная публикация No. 259569/1996
[Патентный документ 6] выложенная японская патентная публикация No. 269062/1996
[Патентный документ 7] выложенная японская патентная публикация No. 269063/1996
[Патентный документ 8] выложенная японская патентная публикация No. 269064/1996
[Патентный документ 9] выложенная японская патентная публикация No. 269065/1996
[Патентный документ 10] выложенная японская патентная публикация No. 269066/1996
[Патентный документ 11] выложенная японская патентная публикация No. 291164/1996
[Патентный документ 12] WO2004/060065
[Патентный документ 13] WO2006/129714
[Патентный документ 14] WO2008/066153
[Патентный документ 15] Journal of Technical Disclosure 500997/2008
[Непатентные документы]
[Непатентный документ 1] Journal of Antibiotics (1993), 46(7), 1168-9
[Непатентный документ 2] Journal of Synthetic Organic Chemistry, Japan (1998), Vol. 56, No. 6, 478-488
[Непатентный документ 3] Journal of Antibiotics (1997), 50(3), 229-36
[Непатентный документ 4] Applied and Environmental Microbiology (1995), 61(12), 4429-35
[Непатентный документ 5] Journal of Organic Chemistry (1996), 61, 882-886
[Непатентный документ 6] Chemical Review (2005), 105, 4559-4580
[Краткое изложение сущности изобретения]
Изобретатели этого изобретения в настоящий момент выявили нуклеотидную последовательность, кодирующую, по меньшей мере, один полипептид, вовлеченный в биосинтез пирипиропена А. Настоящее изобретение было осуществлено на основе этих данных.
Соответственно, целью настоящего изобретения является обеспечение выделенным новым полинуклеотидом, обладающим нуклеотидной последовательностью, кодирующей, по меньшей мере, один полипептид, вовлеченный в биосинтез пирипиропена А, рекомбинантным вектором, включающим полинуклеотид, и трансформантом, включающим полинуклеотид.
Далее, в соответствии с одним вариантом осуществления настоящего изобретения предлагается выделенный полинуклеотид, который представляет собой
(a) полинуклеотид, обладающий нуклеотидной последовательностью SEQ ID NO:266,
(b) полинуклеотид, обладающий нуклеотидной последовательностью, которая способна гибридизоваться с нуклеотидной последовательностью SEQ ID NO:266 в жестких условиях, или
(c) полинуклеотид, обладающий полинуклеотидной последовательностью, кодирующей, по меньшей мере, одну аминокислотную последовательность, выбранную из SEQ ID NO: 267-274, или по существу эквивалентную им аминокислотную последовательность.
Также в соответствии с другим вариантом осуществления настоящего изобретения предлагается выделенный полинуклеотид, который обладает, по меньшей мере, одной нуклеотидной последовательностью, выбранной из нуклеотидной последовательности в любом из пунктов (1) или (2) ниже:
(1) нуклеотидной последовательностью в любом из пунктов от (a) до (h) ниже:
(a) нуклеотидной последовательностью от 3342 до 5158 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(b) нуклеотидной последовательностью от 5382 до 12777 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(c) нуклеотидной последовательностью от 13266 до 15144 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(d) нуклеотидной последовательностью от 16220 до 18018 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(e) нуклеотидной последовательностью от 18506 до 19296 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(f) нуклеотидной последовательностью от 19779 до 21389 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(g) нуклеотидной последовательностью от 21793 до 22877 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(h) нуклеотидной последовательностью от 23205 до 24773 в нуклеотидной последовательности, представленной SEQ ID NO:266;
(2) нуклеотидной последовательностью, которая способна гибридизоваться с нуклеотидной последовательностью из (1) в жестких условиях.
Далее, в соответствии с другим вариантом осуществления настоящего изобретения предлагается полинуклеотид, кодирующий, по меньшей мере, один полипептид, вовлеченный в биосинтез пирипиропена А.
Кроме того, в соответствии с другим вариантом осуществления настоящего изобретения предлагается полинуклеотид, кодирующий полипептид, обладающий какой-либо одной или более из активностей - активностью поликетидсинтазы, активностью пренилтрансферазы, активностью гидроксилазы, активностью ацетилтрансферазы или активностью аденилатсинтетазы.
Более того, в соответствии с другим вариантом осуществления настоящего изобретения предлагается полинуклеотид, который происходит из штамма PF1169 Penicillium coprobium.
Дополнительно в соответствии с другим вариантом осуществления настоящего изобретения предлагается рекомбинантный вектор, включающий указанный выше полинуклеотид.
Кроме того, в соответствии с другим вариантом осуществления настоящего изобретения предлагается трансформант, включающий указанный выше полинуклеотид.
Дополнительно в соответствии с одним вариантом осуществления настоящего изобретения предлагается способ получения предшественника пирипиропена А, отличающийся культивированием трансформанта, в который полинуклеотид, обладающий нуклеотидной последовательностью, указанной выше в (c) или (d), включен одновременно или отдельно, и выделением предшественника пирипиропена А из пирипиропена E, представленного следующей формулой:
[Химическая формула 1]
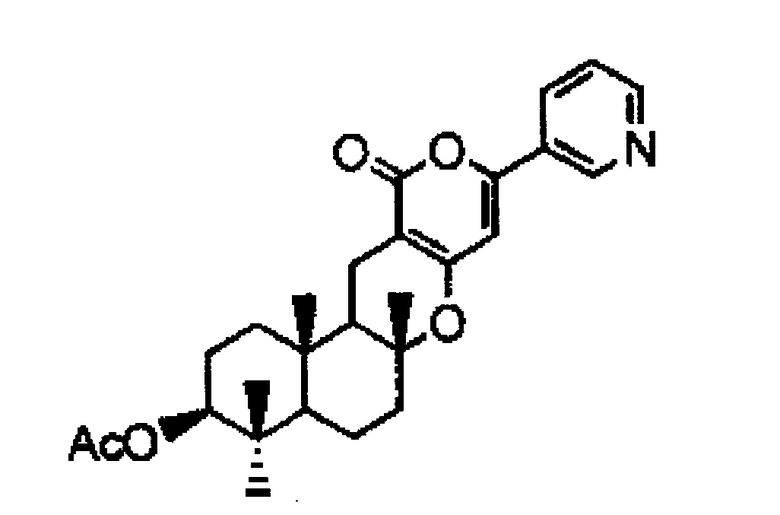
Пирипиропен E
Кроме того, предлагается способ получения, в котором указанный выше предшественник пирипиропена А представляет собой предшественник, представленный следующей формулой (I):
[Химическая формула 2]
Формула (I)
Также предлагается способ получения предшественника пирипиропена А, отличающийся культивированием указанного выше трансформанта, и выделением предшественника пирипиропена А из пирипиропена O, представленного следующей формулой:
[Химическая формула 3]
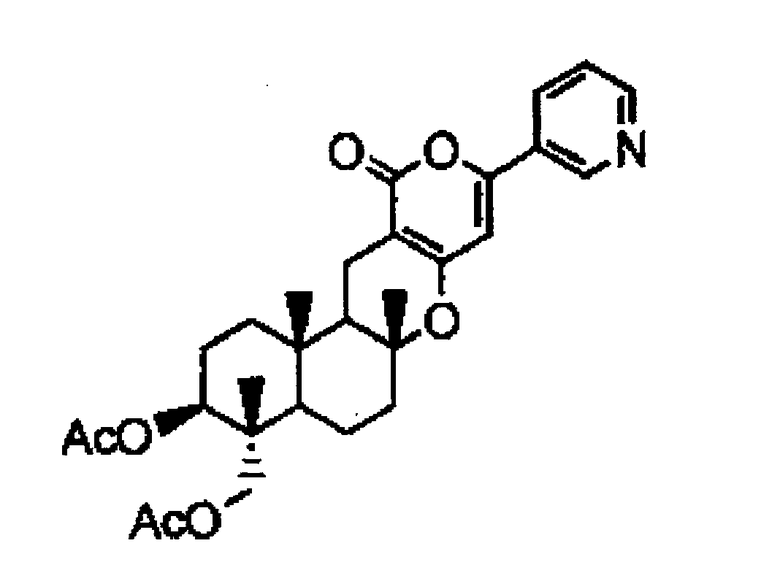
Пирипиропен O
Кроме того, предлагается способ получения, в котором указанный выше предшественник пирипиропена А представляет собой соединение, представленное следующей формулой (II):
[Химическая формула 4]
Формула (II)
В соответствии с одним вариантом осуществления настоящего изобретения предлагается получение нового аналога пирипиропена, улучшение продуктивности бактерии, продуцирующей пирипиропен А, получение нового инсектицидного агента для микроорганизмов, создание новой устойчивости растений к насекомым-вредителям или тому подобное.
[Краткое описание фигур]
[Фигура 1] На фигуре 1 представлен образец электрофореза продуктов ПЦР в агарозном геле. Для электрофореза использовали амплифицированные продукты ПЦР с применением следующих праймеров: M: маркер молекулярной массы (шаг 100 п.о.), дорожка 1: праймеры SEQ ID NO:1 и 2, дорожка 2: праймеры SEQ ID NO:239 и 240, дорожка 3: праймеры SEQ ID NO:237 и 238, дорожка 4: праймеры SEQ ID NO:241 и 242, дорожка 5: праймеры SEQ ID NO:247 и 248, дорожка 6: праймеры SEQ ID NO:251 и 252, дорожка 7: праймеры SEQ ID NO:245 и 246, дорожка 8: праймеры SEQ ID NO:243 и 244, дорожка 9: праймеры SEQ ID NO:249 и 250, дорожка 10: праймеры SEQ ID NO:235 и 236, дорожка 11: праймеры SEQ ID NO:233 и 234, дорожка 12: праймеры SEQ ID NO:227 и 228, дорожка 13: праймеры SEQ ID NO:229 и 230, дорожка 14: праймеры SEQ ID NO:231 и 232.
[Фигура 2] Сходно с фигурой 1 на фигуре 2 представлен образец электрофореза продуктов ПЦР в агарозном геле. Для электрофореза использовали амплифицированные продукты ПЦР с применением следующих праймеров: M: маркер молекулярной массы (шаг 100 п.о.), дорожка 1: праймеры SEQ ID NO:253 и 254, дорожка 2: праймеры SEQ ID NO:257 и 258, дорожка 3: праймеры SEQ ID NO:259 и 260, дорожка 4: праймеры SEQ ID NO:255 и 256, дорожка 5: праймеры SEQ ID NO:261 и 262.
[Фигура 3] Сходно с фигурой 1 на фигуре 3 представлен образец электрофореза продуктов ПЦР в агарозном геле. Для электрофореза использовали амплифицированные продукты ПЦР с применением следующих праймеров: дорожка 1: маркер молекулярной массы (шаг 100 п.о.), дорожка 2: праймеры SEQ ID NO:264 и 265 (амплифицированный фрагмент 400 п.о.).
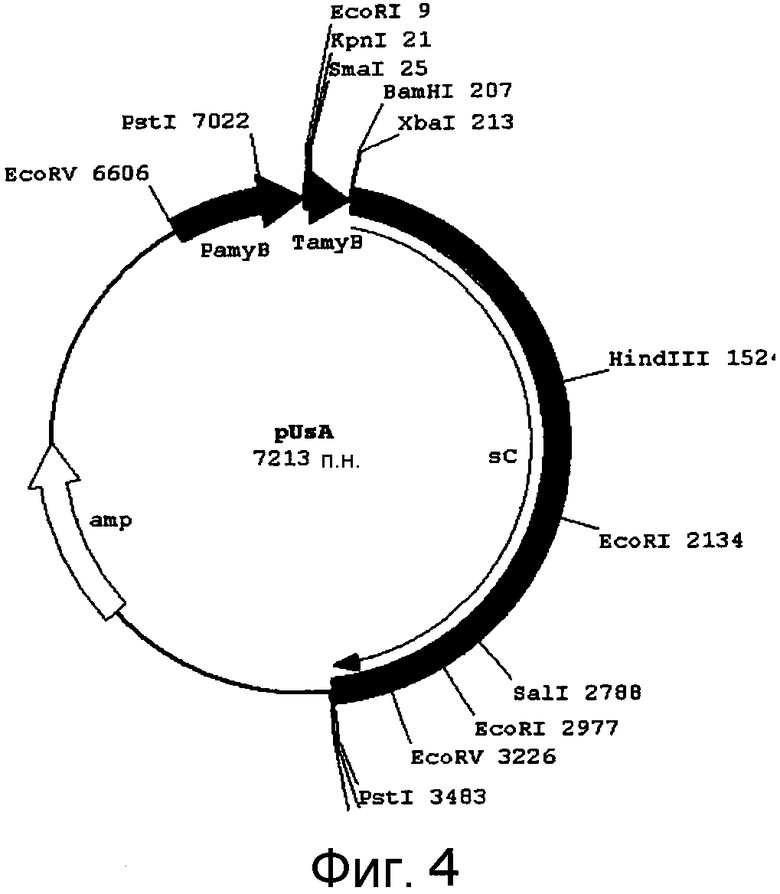
[Фигура 4] На фигуре 4 представлена карта плазмиды pUSA.
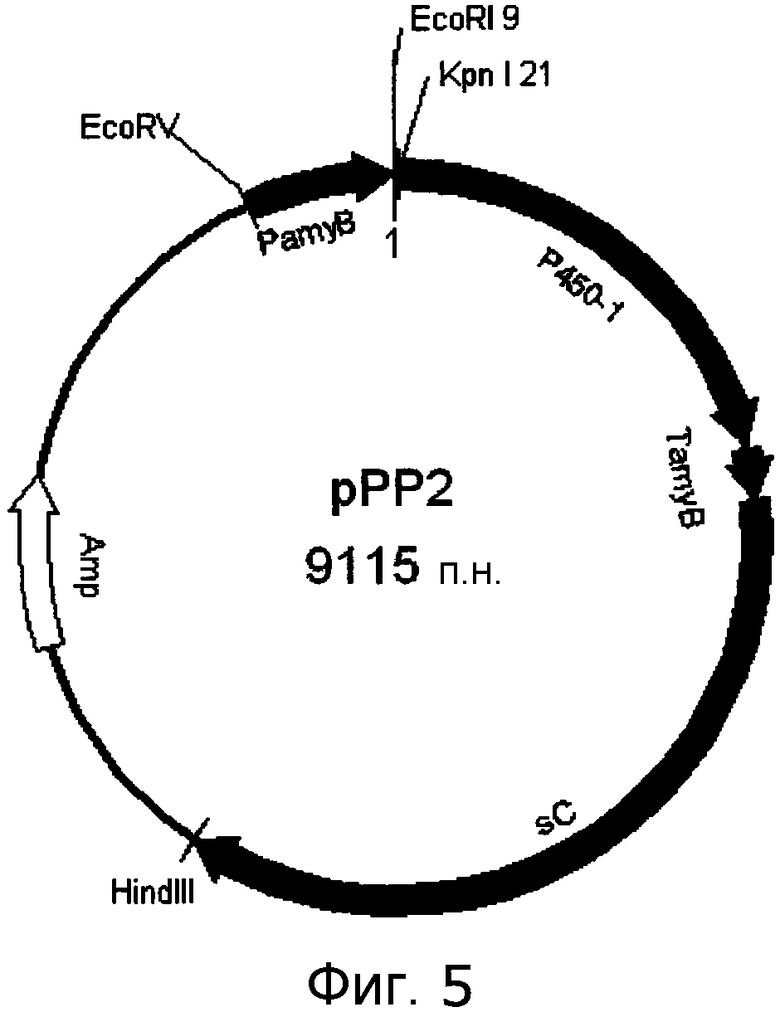
[Фигура 5] На фигуре 5 представлена карта плазмиды pPP2.
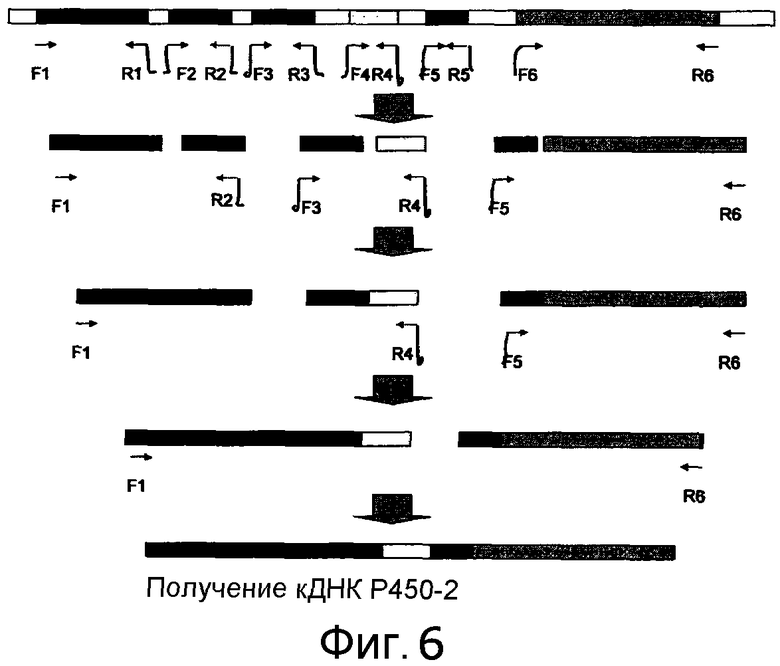
[Фигура 6] На фигуре 6 представлена схема амплификации кДНК P450-2.
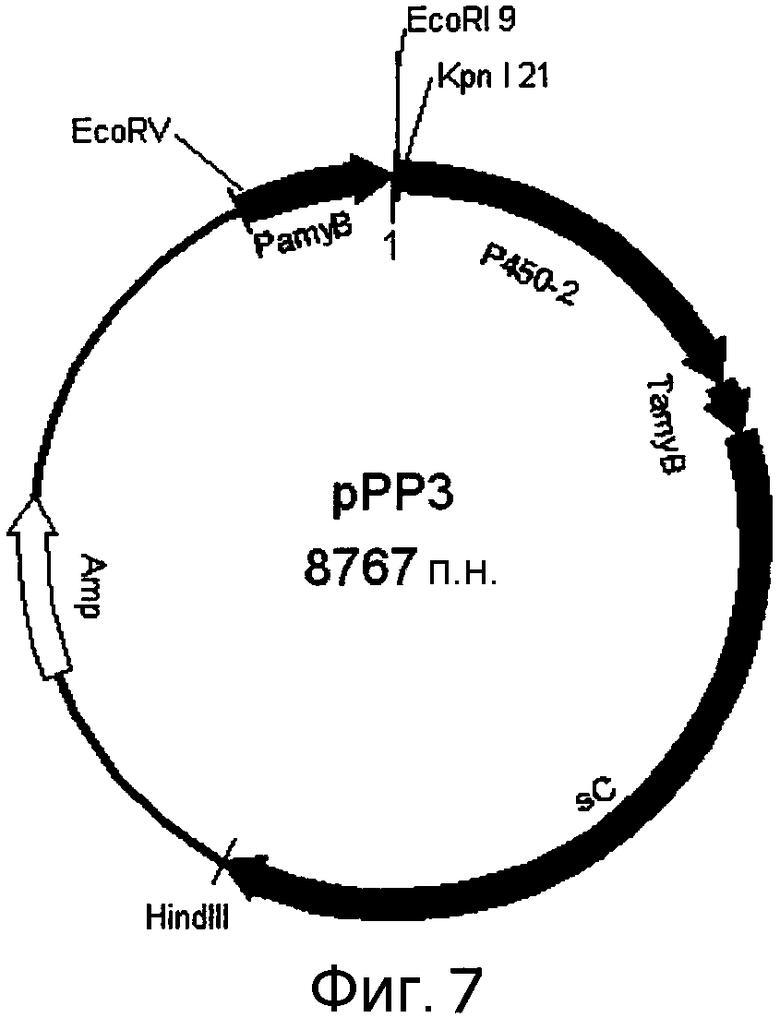
[Фигура 7] На фигуре 7 представлена карта плазмиды pPP3.
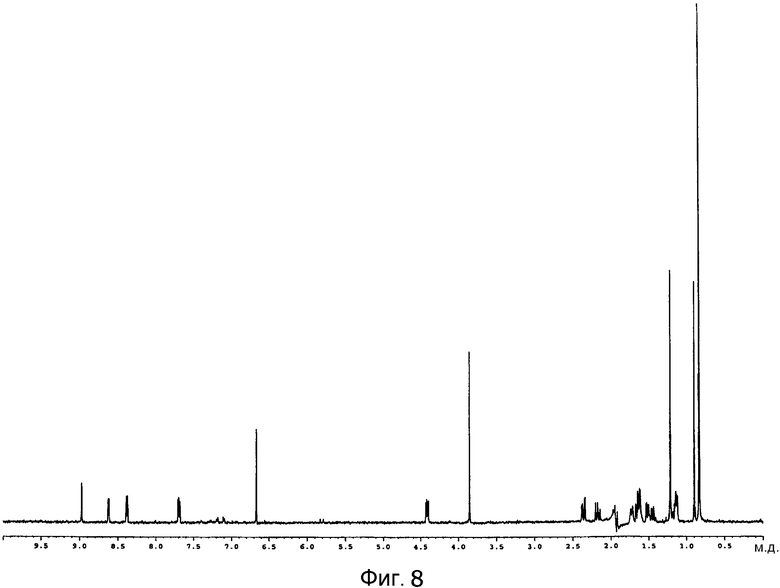
[Фигура 8] На фигуре 8 представлен спектр 1H-ЯМР пирипиропена E в дейтерированном ацетонитриле.
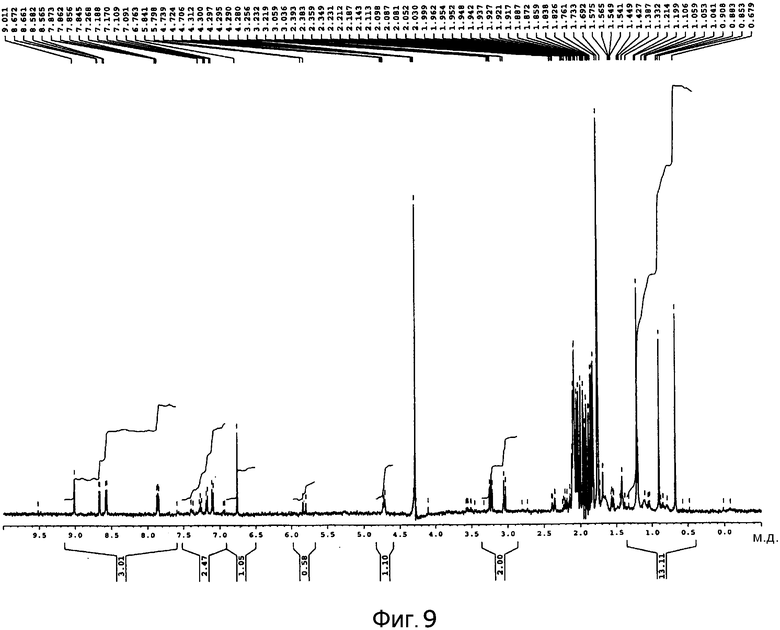
[Фигура 9] На фигуре 9 представлен спектр 1H-ЯМР в дейтерированном ацетонитриле продукта культуры Aspergillus oryzae, трансформированного плазмидой pPP2.
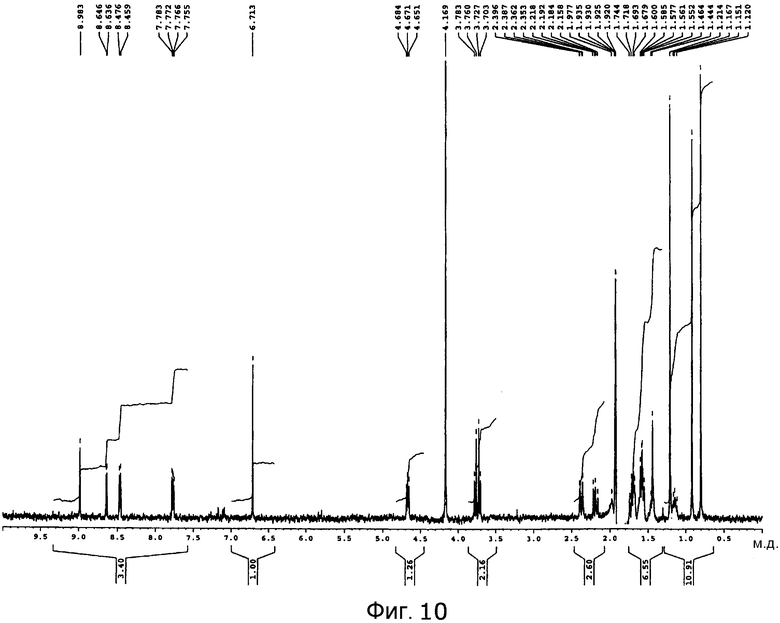
[Фигура 10] На фигуре 10 представлен спектр 1H-ЯМР пирипиропена O в дейтерированном ацетонитриле.
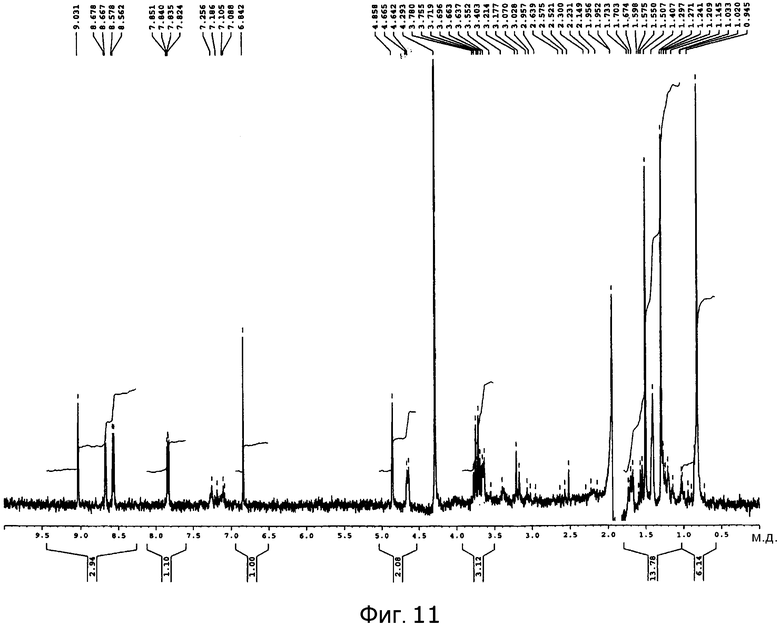
[Фигура 11] На фигуре 11 представлен спектр 1H-ЯМР в дейтерированном ацетонитриле продукта культуры Aspergillus oryzae, трансформированного плазмидой pPP3.
[Подробное описание изобретения]
Депонирование микроорганизмов
Escherichia coli (Escherichia coli EPI300™-T1R), трансформированная плазмидой pCC1-PP1, была депонирована в международном депозитарии запатентованных организмов, государственный институт прогрессивной промышленной науки и технологии (адрес: AIST Tsukuba Central 6, 1-1-1 Higashi, Tsukuba, Ibaraki, Japan, 305-8566), под No. поступления FERM BP-11133 (переведенного из местного депозитария под No. поступления FERM P-21704) (идентификационная ссылка депозиторов: Escherichia coli EPI300™-T1R/pCC1-PP1) 9 октября 2008 г. (исходная дата депонирования).
Aspergillus oryzae, трансформированный плазмидой pPP2, был депонирован в международном депозитарии запатентованных организмов, государственный институт прогрессивной промышленной науки и технологии (адрес: AIST Tsukuba Central 6, 1-1-1 Higashi, Tsukuba, Ibaraki, Japan, 305-8566), под No. поступления FERM BP-11137 (идентификационная ссылка депозиторов: Aspergillus oryzae PP2-1) 23 июня 2009 г.
Aspergillus oryzae, трансформированный плазмидой pPP3, был депонирован в международном депозитарии запатентованных организмов, государственный институт прогрессивной промышленной науки и технологии (адрес: AIST Tsukuba Central 6, 1-1-1 Higashi, Tsukuba, Ibaraki, Japan, 305-8566), под No. поступления FERM BP-11141 (идентификационная ссылка депозиторов: Aspergillus oryzae PP3-2) 3 июля 2009 г.
Выделенный полинуклеотид
В настоящем изобретении представлен выделенный полинуклеотид. Выделенный полинуклеотид по настоящему изобретению представляет собой (a) полинуклеотид, обладающий нуклеотидной последовательностью SEQ ID NO:266; (b) полинуклеотид, обладающий нуклеотидной последовательностью, которая способна гибридизоваться с нуклеотидной последовательностью SEQ ID NO:266 в жестких условиях, или (c) полинуклеотид, обладающий полинуклеотидной последовательностью, кодирующей, по меньшей мере, одну аминокислотную последовательность, выбранную из SEQ ID NO: 267-274, или по существу эквивалентную им аминокислотную последовательность. Указанный выше выделенный полинуклеотид предпочтительно обладает нуклеотидной последовательностью, кодирующей, по меньшей мере, один полипептид, который обладает ферментативной активностью, вовлеченной в биосинтез пирипиропена А.
В настоящем изобретении «по существу эквивалентная аминокислотная последовательность» обозначает аминокислотную последовательность, которая не влияет на активность полипептида, несмотря на тот факт, что одна или более аминокислот изменены в результате замены, делеции, добавки или вставки. Количество измененных аминокислотных остатков составляет предпочтительно от 1 до 40 остатков, более предпочтительно от 1 до нескольких остатков, еще более предпочтительно от 1 до 8 остатков, наиболее предпочтительно от 1 до 4 остатков.
Далее, пример изменения, которое не влияет на активность, включает консервативную замену. Термин «консервативная замена» обозначает замену одного или более аминокислотных остатков другими химически сходными аминокислотными остатками, так что активность полипептида существенно не изменяется. Их примеры включают случаи, когда остаток определенной гидрофобной аминокислоты заменяют остатком другой гидрофобной аминокислоты, и случаи, когда остаток определенной полярной аминокислоты заменяют остатком другой полярной аминокислоты, обладающей теми же зарядами. Функционально сходные аминокислоты, способные на такие замены, известны в данной области техники для каждой аминокислоты. Конкретно, примеры неполярных (гидрофобных) аминокислот включают аланин, валин, изолейцин, лейцин, пролин, триптофан, фенилаланин, метионин и тому подобное. Примеры полярных (нейтральных) аминокислот включают глицин, серин, треонин, тирозин, глутамин, аспарагин, цистеин и тому подобное. Примеры положительно заряженных (основных) аминокислот включают аргинин, гистидин, лизин и тому подобное. Примеры отрицательно заряженных (кислых) аминокислот включают аспарагиновую кислоту, глутаминовую кислоту и тому подобное.
Также выделенный полинуклеотид по настоящему изобретению может представлять собой полинуклеотид, обладающий, по меньшей мере, одной нуклеотидной последовательностью, выбранной из нуклеотидной последовательности в любом из пунктов (1) или (2) ниже:
(1) нуклеотидной последовательностью в любом из пунктов от (a) до (h) ниже:
(a) нуклеотидной последовательностью от 3342 до 5158 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(b) нуклеотидной последовательностью от 5382 до 12777 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(c) нуклеотидной последовательностью от 13266 до 15144 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(d) нуклеотидной последовательностью от 16220 до 18018 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(e) нуклеотидной последовательностью от 18506 до 19296 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(f) нуклеотидной последовательностью от 19779 до 21389 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(g) нуклеотидной последовательностью от 21793 до 22877 в нуклеотидной последовательности, представленной SEQ ID NO:266,
(h) нуклеотидной последовательностью от 23205 до 24773 в нуклеотидной последовательности, представленной SEQ ID NO:266;
(2) нуклеотидной последовательностью, которая способна гибридизоваться с нуклеотидной последовательностью из (1) в жестких условиях.
Полинуклеотид, обладающий, по меньшей мере, одной нуклеотидной последовательностью, выбранной из нуклеотидной последовательности в любом из указанных выше пунктов (1) или (2), предпочтительно кодирует, по меньшей мере, один полипептид, обладающий ферментативной активностью, вовлеченной в биосинтез пирипиропена А.
Термин «жесткие условия» в настоящем изобретении обозначает условия, когда процедура отмывки мембран после гибридизации осуществляется при высоких температурах в растворе с низкими концентрациями соли, например, в условиях отмывки в растворе с концентрацией 2×SSC (1×SSC: 15 мМ цитрата тринатрия, 150 мМ хлорида натрия) и 0,5% SDS при 60°С в течение 20 минут.
Полинуклеотид, обладающий, по меньшей мере, одной нуклеотидной последовательностью, выбранной из нуклеотидной последовательности в любом из указанных выше пунктах (1) или (2) по настоящему изобретению представляет собой полинуклеотид, кодирующий полипептид, обладающий какой-либо одной или более из активностей - активностью поликетидсинтазы, активностью пренилтрансферазы, активностью гидроксилазы, активностью ацетилтрансферазы или активностью аденилатсинтетазы; и, в частности, полинуклеотид, кодирующий полипептид, обладающий активностью гидроксилазы.
Кроме того, в соответствии с одним вариантом осуществления настоящего изобретения указанный выше полинуклеотид представляет собой полинуклеотид, кодирующий полипептид, обладающий активностью в отношении гидроксилирования положения 7 и/или положения 13 указанного выше пирипиропена E или O, или представляет собой полинуклеотид, кодирующий полипептид, обладающий активностью в отношении гидроксилирования положения 11 указанного выше пирипиропена E.
Получение выделенного полинуклеотида
Способ получения выделенного полинуклеотида по настоящему изобретению особенно не ограничен. Например, полинуклеотид может быть выделен из штамма PF1169 Penicillium coprobium или нитчатой бактерии с помощью следующего способа.
Основываясь на гомологии последовательности, полученной методом примера 9 ниже или тому подобного, синтезируют праймеры, способные к специфичной амплификации гена поликетидсинтазы. ПЦР осуществляют для фосмидной геномной библиотеки штамма PF1169 Penicillium coprobium, который получают отдельно с последующей гибридизацией колоний. В результате этого получают рекомбинантный вектор, и определяют последовательность оснований его вставленной ДНК.
Также, основываясь на гомологии последовательности, полученной методом примера 9 ниже или тому подобного, синтезируют праймеры, способные к специфичной амплификации гена пренилтрансферазы. Далее определяют последовательность оснований вставленной ДНК тем же способом, что и описанный выше.
Далее, основываясь на гомологии последовательности, полученной способом примера 9 ниже или тому подобного, синтезируют праймеры, способные к специфичной амплификации любого или обоих гена поликетидсинтазы и гена пренилтрансферазы. Далее определяют последовательность оснований вставленной ДНК тем же способом, что и описанный выше.
Кроме того, основываясь на гомологии последовательности, по меньшей мере, одной нуклеотидной последовательности, выбранной из SEQ ID NO:266, и нуклеотидной последовательности в любом из указанных выше пунктов (1) или (2) по настоящему изобретению, синтезируют праймеры, способные к специфичной амплификации любого одного или более гена поликетидсинтазы, гена пренилтрансферазы, гена гидроксилазы, гена ацетилтрансферазы или гена аденилатсинтетазы, предпочтительно гена гидроксилазы. Далее определяют последовательность оснований вставленной ДНК тем же способом, что и описанный выше.
Кроме того еще, основываясь на аминокислотной последовательности поликетидсинтаз, консервативных среди различных нитчатых бактерий, синтезируют вырожденные праймеры для амплификации и определяют последовательность оснований вставленной ДНК.
Трансформант
В целом примеры способа улучшения продуктивности продукта вторичного метаболизма с помощью рекомбинации генов включают улучшение экспрессии гена, кодирующего белок, катализирующий биосинтетическую реакцию, которая представляет собой реакцию, лимитирующую скорость биосинтеза, улучшение экспрессии гена биосинтеза или разрушение гена, регулирующего экспрессию гена биосинтеза, блокирование необязательной системы вторичного метаболизма и тому подобное. Следовательно, определение гена биосинтеза делает возможным улучшение продуктивности продукта вторичного метаболизма путем лигирования гена в подходящий вектор и введения вектора в бактерию-продуцент.
Между тем, для того чтобы создать новое активное вещество путем рекомбинации генов осуществляют изменение доменов поликетидсинтазы [Ikada and Ohmura, “Protein. Nucleic acid and enzyme” Vol. 43, p. 1265-1277, 1998], [Carreras, C.W. and Santi, D.V., “Current Opinion in Biotechnology”, (UK), 1998, Vol. 9, p. 403-411], [Hutchinson, C.R., “Current Opinion in Microbiology”, (UK), 1998, Vol. 1, p. 319-329], [Katz, L. and McDaniel, R., “Medicinal Research Reviews”, (USA), 1999, Vol. 19, p. 543-558]; разрушение гена биосинтеза; введение модифицированного гена фермента от других организмов [Hutchinson, C.R., “Bio/Technology”, (USA), 1994, Vol. 12, p. 375-380]; и тому подобное. Таким образом, установление гена биосинтеза делает возможным создание нового активного вещества путем лигирования гена с подходящим вектором и введения вектора в бактерию, продуцирующую продукт вторичного метаболизма.
Следовательно, пирипиропен А может быть получен или его продукция может быть улучшена путем лигирования выделенного полинуклеотида по настоящему изобретению с подходящим вектором, введения вектора в хозяина, его экспрессирования, повышения его экспрессии или осуществления разрушения части гена выделенного полинуклеотида с использованием гомологичной рекомбинации и ослабления его функций.
Разрушение гена с использованием гомологичной рекомбинации может быть осуществлено в соответствии с общепринятым способом. Получение вектора, используемого для разрушения гена, и введение вектора в хозяина очевидно для специалистов в данной области техники.
Рекомбинантный вектор по настоящему изобретению предпочтительно включает любой один или более из полинуклеотидов, обладающих нуклеотидной последовательностью SEQ ID NO:266 и указанными выше в (1); полинуклеотид, обладающий нуклеотидной последовательностью, которая способна гибридизоваться с нуклеотидной последовательностью SEQ ID NO:266 и указанными выше в (1) в жестких условиях, или полинуклеотид, обладающий полинуклеотидной последовательностью, кодирующей, по меньшей мере, одну аминокислотную последовательность, выбранную из SEQ ID NO: с 267 по 274 или по существу эквивалентную им аминокислотную последовательность. Более предпочтительно, рекомбинантный вектор по настоящему изобретению представляет собой вектор, в котором указанный выше полинуклеотид включает полинуклеотид, кодирующий полипептид, гидроксилирующий пирипиропен E или O в положении 7 и/или положении 13, и указанный выше полинуклеотид, включающий полинуклеотид, кодирующий полипептид, гидроксилирующий пирипиропен E в положении 11.
Рекомбинантный вектор для введения гена может быть получен путем модификации полинуклеотида, предлагаемого настоящим изобретением, в подходящую форму в зависимости от объекта и лигирования в вектор в соответствии с общепринятым способом, например, способами рекомбинации генов, описанными в руководстве {Sambrook, J. et al., “Molecular cloning: a laboratory manual”, (USA), 2nd Edition, Cold Spring Harbor Laboratory, 1989].
Рекомбинантный вектор, используемый в настоящем изобретении, может быть подходящим образом выбран из вирусных, плазмидных, фосмидных, космидных векторов или тому подобного. Например, когда клеткой-хозяином является E. coli, их примеры включают бактериофаг на основе фага λ и плазмиды на основе pBR и pUC. В случае Bacillus subtilis примеры включают плазмиды на основе pUB. В случае дрожжей примеры включают плазмиды на основе YEp, YRp, YCp и YIp.
Кроме того, предпочтительно, чтобы, по меньшей мере, одна плазмида из используемых плазмид включала маркер селекции для селекции трансформанта. В качестве маркера селекции может быть использован ген, кодирующий устойчивость к лекарственным средствам, и ген, завершающий ауксотрофию. Их конкретные предпочтительные примеры включают при использовании бактерии в качестве хозяина - гены устойчивости к ампициллину, гены устойчивости к канамицину, ген устойчивости к тетрациклину и тому подобное; в случае дрожжей - ген биосинтеза триптофана (TRP1), ген биосинтеза урацила (URA3), ген биосинтеза лейцина (LEU2) и тому подобное; в случае грибов - гены устойчивости к гигромицину, гены устойчивости к биалафосу, гены устойчивости к блеомицину, гены устойчивости к ауреобасидину и тому подобное; и в случае растения - гены устойчивости к канамицину, гены устойчивости к биалафосу и тому подобное.
Кроме того, используемые в настоящем изобретении молекулы ДНК, служащие в качестве экспрессионного вектора, предпочтительно содержат последовательности ДНК, необходимые для экспрессии каждого гена, регуляторные сигналы транскрипции и регуляторные сигналы трансляции, такие как промоторы, сигналы инициации транскрипции, сайты связывания с рибосомой, стоп-сигналы трансляции, терминаторы. Предпочтительные примеры промоторов включают промоторы оперона лактозы, оперона триптофана и тому подобное в Escherichia coli; промоторы гена алкогольдегидрогеназы, гена кислой фосфатазы, гена метаболизма галактозы, гена глицеральдегид-3-фосфатдегидрогеназы или тому подобное в дрожжах; промоторы гена α-амилазы, гена глюкоамилазы, гена целлобиогидролазы, гена глицеральдегид-3-фосфатдегидрогеназы, гена abp1 или тому подобное в грибах; промотор CaMV 35S RNA, CaMV 19S RNA или промотор гена нопалинсинтетазы в растениях.
Хозяин, которому вводят выделенный полинуклеотид по настоящему изобретению, может быть подходящим образом выбран в зависимости от типа используемого вектора из актиномицетов, Escherichia coli, Bacillus subtilis, дрожжей, нитчатых бактерий, клеток растений и тому подобного.
Метод введения рекомбинантного вектора в хозяина может быть выбран в зависимости от тестируемой клетки-хозяина из конъюгационного переноса, трансдукции фагом, а так же методов трансформации, таких как метод с использованием ионов кальция, метод с использованием ионов лития, метод электропорации, метод с использованием ПЭГ, метод Agrobacterium или метод с использованием инжектора частиц.
В случаях, когда в настоящем изобретении в клетки-хозяева вводят множество генов, гены могут находиться в одной молекуле ДНК или раздельно в разных молекулах ДНК. Кроме того, когда клетка-хозяин представляет собой бактерию, каждый ген может быть сконструирован таким образом, чтобы он экспрессировался в виде полицистронной мРНК и вмещался в одну молекулу ДНК.
Трансформант по настоящему изобретению предпочтительно включает любой один или более из полинуклеотидов, имеющих нуклеотидную последовательность SEQ ID NO:266 и упомянутую выше (1); полинуклеотид, имеющий нуклеотидную последовательность, которая способна гибридизоваться с нуклеотидной последовательностью SEQ ID NO:266 и упомянутой выше (1) в жестких условиях, или полинуклеотид, имеющий полинуклеотидную последовательность, кодирующую, по меньшей мере, одну аминокислотную последовательность, выбранную из SEQ ID NO с 267 по 274 или по существу эквивалентную ей аминокислотную последовательность.
Полученный трансформант можно культивировать традиционным способом, и можно исследовать новоприобретенные свойства. В качестве среды можно использовать обычно применяемые компоненты, например, в качестве источников углерода глюкозу, сахарозу, крахмальный сироп, декстрин, крахмал, глицерин, мелассу, животные и растительные масла или тому подобное. Также в качестве источников азота могут быть использованы соевая мука, проростки пшеницы, жидкость от замоченной кукурузы, мука из семян хлопчатника, мясной экстракт, полипептон, мальтоэкстракт, дрожжевой экстракт, сульфат аммония, нитрат натрия, мочевина и тому подобное. Кроме того, при необходимости эффективна добавка натрия, калия, кальция, магния, кобальта, хлора, фосфорной кислоты (гидрофосфат калия или тому подобного), серной кислоты (сульфата магния или тому подобного) или неорганических солей, которые могут образовывать другие ионы. Также при необходимости могут быть добавлены различные витамины, такие как тиамин (гидрохлорид тиамина или тому подобное), аминокислоты, такие как глутаминовая кислота (глутамат натрия или тому подобное) или аспарагин (DL-аспарагин или тому подобное), следовые питательные вещества, такие как нуклеотиды, или агенты селекции, такие как антибиотики. Более того, могут быть подходящим образом добавлены органические вещества или неорганические вещества, которые способствуют росту бактерии и содействуют продукции пирипиропена A.
pH среды составляет, например, приблизительно от pH 5,5 до pH 8. В качестве метода культивирования может быть использовано культивирование на твердой фазе в аэробных условиях, культивирование со встряхиванием, культивирование с барботированием при перемешивании или аэробное культивирование в глубокой части, и особенно аэробное культивирование в глубокой части является наиболее подходящим. Подходящая температура для культивирования составляет от 15°С до 40°С, и во многих случаях рост происходит при приблизительно от 22°С до 30°С. Продукция пирипиропена A варьирует в зависимости от среды и условий культивирования или используемого хозяина. При любом методе культивирования аккумуляция обычно достигает пика от 2 дней до 10 дней. Культивирование прекращают в момент времени, когда аккумуляция пирипиропена A достигает пика, и желаемое вещество выделяют и очищают из культуры.
Для выделения пирипиропена A из культуры его можно экстрагировать и очищать с помощью обычных средств отделения с использованием его свойств, таких как метод экстракции растворителем, метод с использованием ионообменной смолы, метод распределительной колоночной хроматографии, метод гель-фильтрации, диализ, метод осаждения, метод кристаллизации, которые можно использовать раздельно или в соответствующем сочетании.
Метод получения предшественника пирипиропена A
Для выделения пирипиропена A пирипиропен A может быть выделен из предшественника пирипиропена A с помощью известного метода. Пример известного метода включает метод WO2009/022702. Путем культивирования микроорганизма, содержащего вектор, включающий одно или более из указанного выше, предшественник пирипиропена A может быть выделен из пирипиропена E. Предшественник пирипиропена A может представлять собой, например, соединение, представленной упомянутой выше формулой (I).
Предшественник пирипиропена A может также может быть выделен из пирипиропена O путем культивирования микроорганизма, включающего вектор, содержащий одно или более. Примером может служить соединение, представленной упомянутой выше формулой (II).
ПРИМЕРЫ
Настоящее изобретение будет далее подробно иллюстрировано следующими примерами, которые не предназначены для ограничения настоящего изобретения.
Пример 1: Получение геномной ДНК штамма PF1169 Penicillium coprobium
Стерилизованную среду NB (500 мл) помещали в эрленмейеровский сосуд (1 л). Штамм PF1169 Penicillium coprobium (Journal of Technical Discosure No. 500997/2008 (патентный документ 15)), предварительно культивированный в 1/2 агаровой среде CMMY при 28°С в течение 4 дней, добавляли к упомянутой выше среде и культивировали в жидкости при 28°С в течение 4 дней. Производили фильтрование с помощью Miracloth с получением 5 г бактериальных клеток. Из этих бактериальных клеток было получено 30 мкг геномной ДНК в соответствии с руководством, прилагаемым к набору для очистки геномной ДНК Genomic-tip 100/G (производство Qiagen K.K.).
Пример 2: Вырожденные праймеры для амплификации поликетидсинтазы (PKS) и ее амплифицированного фрагмента
Основываясь на аминокислотной последовательности поликетидсинтаз, консервативной для различных нитчатых бактерий, были сконструированы и синтезированы следующие праймеры в качестве вырожденных праймеров для амплификации:
LC1: GAYCCIMGITTYTTYAAYATG (SEQ ID NO:1)
LC2c: GTICCIGTICCRTGCATYTC (SEQ ID NO:2)
(где R=A/G, Y=C/T, M=A/C, I=инозин)
С помощью этих вырожденных праймеров геномной ДНК, полученной в примере 1, и ExTaq полимеразе (производство Takara Bio Inc.) давали провзаимодействовать в соответствии с прилагаемой инструкцией. Был обнаружен амплифицированный фрагмент приблизительно 700 п.н. (см. фигуру 1). Более того, указанный выше амплифицированный фрагмент был проанализирован для установления последовательности расположенных внутри 500 п.н. (SEQ ID NO:3).
Пример 3: Широкомасштабное секвенирование геномной ДНК и поиск гомологии аминокислотной последовательности
Геномную ДНК штамма PF1169 Penicillium coprobium, полученную в примере 1, подвергали широкомасштабному секвенированию и производили поиск гомологичных аминокислотных последовательностей. Конкретно, часть из 50 мкг геномной ДНК предварительно обрабатывали и затем вводили в ДНК-секвенатор Roche 454FLX с получением 103 тысяч фрагментов последовательностей приблизительно 250 п.н. (суммарно 49 м.н. последовательности).
Для этих последовательностей в качестве известных последовательностей для поликетидсинтаз и пренилтрансфераз были отобраны пять следующих последовательностей (последовательности из поликетидсинтаз: Aspergillus(A.) fumigatus PKS 2146 а.к. и Penicillium(P.) griseofluvum синтазы 6-метилсалициловой кислоты 1744 а.к.; а также пренилтрансфераз: Aspergillus(A.) fumigatus пренилтрансферазы, Aspergillus(A.) fumigatus пренилтрансферазы (4-гидроксибензоатоктапренилтрансферазы) и Penicillium(P.) marneffei пренилтрансферазы) и был произведен поиск с помощью программы blastx поиска гомологичных последовательностей с получением в результате 89, 86, 2, 1 и 3 гомологичных последовательностей, соответственно (см. таблицу 1). Далее из гомологичных последовательностей A. fumigatus PKS 2146 а.к. и P. griseofluvum синтазы 6-метилсалициловой кислоты 1744 а.к. было, соответственно, получено 19 и 23 близких последовательностей (непрерывные последовательности A. fumigatus PKS 2146 а.к.: SEQ ID NO: 179-197; непрерывные последовательности P. griseofluvum синтазы 6-метилсалициловой кислоты 1744 а.к.: SEQ ID NO: с 198 по 220) (см. таблицу 1).
гомологичных последователь-ностей
NO.
синтазы
6-метилсалициловой кислоты 1744 а.к.
6-метилсалициловой кислоты 1744 а.к.
Пример 4: Амплификация ПЦР с геномной ДНК
На основании результатов поиска с помощью blastx, полученных в примере 3, были синтезированы 13 типов пар праймеров для поликетидсинтаз, показанных в SEQ ID NO: 227-252. Сходным образом, были синтезированы 5 типов пар праймеров для пренилтрансфераз, показанных в SEQ ID NO: 253-262. При проведении ПЦР для геномной ДНК с использованием этих праймеров наблюдались амплифицированные фрагменты ожидаемого размера для всех пар праймеров (см. фигуру 1 и фигуру 2).
Пример 5: Конструирование фаговой геномной библиотеки
λ-фаговая геномная библиотека штамма PF1169 Penicillium coprobium была сконструирована с помощью набора λBlueSTAR с плечами полусайтов Xho I (производство Takara Bio Inc., Cat. No. 69242-3) в соответствии с прилагаемым руководством. То есть, геномную ДНК частично расщепляли с помощью фермента рестрикции Sau3A1. Фрагмент ДНК приблизительно 20 т.н. (0,5 мкг) лигировали с 0,0 мкг ДНК λBlueSTAR, прилагаемой к набору. Этот раствор лигирования подвергали упаковке in vitro с использованием набора Lambda INN Packaging (производство Nippon Gene Co., Ltd.) на основе руководства, прилагаемого к набору, с получением 1 мл раствора. Этот раствор с упакованными фагами (10 мкл) инфицировали в 100 мкл штамма ER1647 E. coli и культивировали в среде с образованием пятен при 37°С в течение ночи с получением в результате приблизительно 500 клонов пятен. Таким образом, была сконструирована геномная библиотека, состоящая из приблизительно 50000 клонов фагов, в которые было введено от 10 до 20 т.н. геномной ДНК штамма PF1169 Penicillium coprobium путем инфекции.
Пример 6: Скрининг фаговой библиотеки
Для 10000 клонов фаговой библиотеки, полученной в примере 5, производили первичный скрининг путем гибридизации пятен с использованием в качестве зонда продукта ПЦР, амплифицированного с помощью пары праймеров LC1-LC2c, полученных выше. Для промечивания и выявления зонда использовали набор AlkPhos Direct Labelling and Detection System with CDP-Star (производство GE Healthcare, Cat. No. RPN3690). Указанную выше гибридизацию проводили в соответствии с прилагаемыми указаниями.
В результате первичного скрининга осталось 6 клонов в качестве кандидатов. Более того, в результате вторичного скрининга гибридизацией пятен было получено 4 клона. Эти позитивные клоны инфицировали в штамм BM25.8 E. coli, и фаги превращали в плазмиды в соответствии с прилагаемыми указаниями с получением в результате 4 типов плазмид, содержащих желаемую область.
Пример 7: Получение фосмидной геномной библиотеки
Была сконструирована геномная библиотека штамма PF1169 Penicillium coprobium с помощью набора CopyControl Fosmid Library Production (производство EPICENTRE, Cat. No. CCFOS110) в соответствии с прилагаемыми к нему указаниями. То есть, 0,25 мкг фрагмента ДНК приблизительно 40 т.н. геномной ДНК обрабатывали с получением тупых концов и затем включали в фосмидный вектор pCCFOS (производство Epicentre). Этот раствор лигирования подвергали упаковке in vitro с использованием прилагаемого к набору MaxPlax Lambda Packaging Extract в соответствии с прилагаемыми к набору указаниями. Этот раствор с упакованным вирусом (10 мкл) инфицировали в 100 мкл штамма EPI300TM-T1R E. coli и культивировали на среде, содержащей хлорамфеникол, при 37°С в течение ночи и подвергали селекции с получением в результате приблизительно 300 клонов пятен. Таким образом, было получено приблизительно 30000 клонов фосмид, в которые было введено 40 т.н. геномной ДНК штамма PF1169 Penicillium coprobium путем инфекции. Их аликвотировали в 96-луночный планшет с получением приблизительно 50 клонов на лунку. Таким образом была сконструирована геномная библиотека, состоящая из 96 пулов, около 4800 клонов.
Пример 8: Скрининг фосмидной библиотеки
В соответствии с указаниями, прилагаемыми к фосмиде, получали плазмидные ДНК отдельно из 96 пулов библиотеки, полученной в примере 7. С помощью вырожденных праймеров для амплификации поликетидсинтазы, синтезированных в примере 2, проводили ПЦР для 96 пулов этих образцов плазмидной ДНК. В результате из 9 пулов были амплифицированы фрагменты ДНК приблизительно 700 п.н. Затем из позитивных пулов готовили чашку Петри, содержащую колонии из приблизительно 300 клонов или более, и производили повторный скрининг путем гибридизации колоний. В результате из приблизительно 4800 колоний с помощью пары праймеров LC1-LC2c было получено 9 типов фосмид.
Пример 9: Широкомасштабное секвенирование геномной ДНК и поиск гомологии аминокислотной последовательности
Геномную ДНК штамма PF1169 Penicillium coprobium, полученную в примере 1, подвергали широкомасштабному секвенированию и производили поиск гомологичных аминокислотных последовательностей. Конкретно, часть из 50 мкг геномной ДНК подвергали предварительной обработке и затем вводили в ДНК-секвенатор Roche 454FLX с получением 1405 фрагментов последовательностей со средней непрерывной длиной 19,621 т.н. (последовательность в общей сложности длиной 27,568160 м.н.).
Для этих последовательностей в качестве известных последовательностей среди поликетидсинтаз и пренилтрансфераз были отобраны следующие пять последовательностей (последовательности, берущие начало из поликетидсинтаз: Penicillium(P.) griseofluvum синтазы 6-метилсалициловой кислоты 1744 а.к. (P22367) и Aspergillus(A.) fumigatus PKS 2146 а.к. (Q4WZA8); а также пренилтрансфераз: Penicillium(P.) marneffei пренилтрансферазы (Q0MRO8), Aspergillus(A.) fumigatus пренилтрансферазы (Q4WBI5) и Aspergillus(A.) fumigatus пренилтрансферазы (4-гидроксибензоатоктапренилтрансферазы) (Q4WLD0)), и был произведен поиск гомологичных последовательностей с помощью программы blastx с получением в результате 22 (P22367), 21 (Q4WZA8), 2 (Q0MRO8), 3 (Q4WBI5) и 3 (Q4WLD0) гомологичных последовательностей, соответственно.
Пример 10: Скрининг фосмидной библиотеки и анализ последовательности кластеров генов
В соответствии с указаниями, прилагаемыми к фосмидному набору (производство EPICENTRE, набор CopyControl Fosmid Library Production), были отдельно получены плазмидные ДНК из 96 пулов библиотеки, полученной в примере 7. На основании последовательностей, определенных с помощью ДНК-секвенатора Roche 454FLX, был произведен поиск гомологичных аминокислотных последовательностей для выявления областей, примыкающих к поликетидсинтазе и пренилтрансферазе. На основании нуклеотидной последовательности пренилтрансферазы полученной области была синтезирована пара праймеров (No. 27), способная амплифицировать фрагмент ДНК 400 п.н. С помощью этих праймеров была проведена ПЦР для этих 48 пулов образцов плазмидной ДНК. В результате из 11 пулов были амплифицированы ожидаемые фрагменты ДНК приблизительно 400 п.н. (SEQ ID NO:263) (см. фигуру 3). Затем из 6 позитивных пулов получали чашку Петри, содержащую 300 клонов или более, и подвергали повторному скринингу с помощью гибридизации колоний. В результате путем использования пары праймеров 27F+27R (праймер 27F: SEQ ID NO:264, праймер 27R: SEQ ID NO:265) из приблизительно 4800 клонов было получено 4 типа фосмид. Одна из них была названа pCC1-PP1, и была определена вся последовательность вставленного фрагмента (SEQ ID NO:266).
Полученной pCC1-PP1 трансформировали штамм EPI300TM-T1R Escherichia coli (включенный в фосмидный набор) с получением, тем самым, штамма Escherichia coli EPI300TM-T1R/pCC1-PP1.
Когда был произведен поиск гомологии между указанной выше последовательностью SEQ ID NO:266 и каждым из фермента, образующего аденилат; LovB-подобной поликетидсинтазы; цитохром P450 монооксигеназы, интегрального мембранного белка, ФАД-зависимой монооксигеназы, которые являются гидроксилазами; UbiA-подобной пренилтрансферазой; ацетилтрансферазой; белком Tri7 биосинтеза токсина, которые являются ацетилтрансферазами; и транспортирующей катионы АТФазой (все упомянутые выше ферменты берут начало от штамма Af293 Aspergillus fumigatus) при любом поиске наблюдалась высокая гомология 70% или более.
Нуклеотиды 3342-5158 SEQ ID NO:266 кодируют фермент, образующий аденилат, и соответствующий полипептид показан в виде аминокислотной последовательности, представленной SEQ ID NO:267; нуклеотиды 5382-12777 SEQ ID NO:266 кодируют LovB-подобную поликетидсинтазу, и соответствующий полипептид показан в виде аминокислотной последовательности, представленной SEQ ID NO:268; нуклеотиды 13266-15144 SEQ ID NO:266 (в настоящем описании ниже белок, кодируемый этой полинуклеотидной последовательностью (P450-1), обозначается как цитохром P450 монооксигеназа (1)), а нуклеотиды 16220-18018 (в настоящем описании ниже белок, кодируемый этой полинуклеотидной последовательностью (P450-2), обозначается как цитохром P450 монооксигеназа (2)) кодируют цитохром P450 монооксигеназы, и соответствующие полипептиды показаны в виде аминокислотных последовательностей, представленных SEQ ID NO:269 и 270, соответственно; нуклеотиды 18506-19296 SEQ ID NO:266 кодируют интегральный мембранный белок, и соответствующий полипептид показан в виде аминокислотной последовательности, представленной SEQ ID NO:271; нуклеотиды 19779-21389 SEQ ID NO:266 кодируют ФАД-зависимую монооксигеназу, и соответствующий полипептид показан в виде аминокислотной последовательности, представленной SEQ ID NO:272; нуклеотиды 21793-22877 SEQ ID NO:266 кодируют UbiA-подобную пренилтрансферазу, и соответствующий полипептид показан в виде аминокислотной последовательности, представленной SEQ ID NO:273; нуклеотиды 23205-24773 SEQ ID NO:266 кодируют ацетилтрансферазу, и соответствующий полипептид показан в виде аминокислотной последовательности, представленной SEQ ID NO:274; нуклеотиды 25824-27178 SEQ ID NO:266 кодируют белок Tri7 биосинтеза токсина, и соответствующий полипептид показан в виде аминокислотной последовательности, представленной SEQ ID NO:275; и нуклеотиды 27798-31855 SEQ ID NO:266 кодируют транспортирующую катионы АТФазу, и соответствующий полипептид показан в виде аминокислотной последовательности, представленной SEQ ID NO:276.
Пример 11: Гидроксилирование пирипиропена E или пирипиропена O путем трансформации Aspergillus Oryzae
Пирипиропен E, используемый ниже, может быть получен, например, методом культивирования микроорганизма на основе метода, описанного в выложенной японской патентной публикации No. 239385/1996 (патентный документ 4), WO94/09147 или патенте США No. 5597835, или методом общего синтеза, описанного в Tetrahedron Letters, vol. 37, No. 36, 6461-6464, 1996. Также пирипиропен O, используемый ниже, может быть получен, например, методом культивирования микроорганизма на основе метода, описанного в J. Antibiotics 49, 292-298, 1996 или WO94/09147.
(1) Получение экспрессионного вектора для введения в нитчатую бактерию
pUSA (фигура 4) и pHSG399 (Takara Bio Inc.) раздельно гидролизовали с помощью KpnI и лигировали с получением, тем самым, pUSA-HSG. Плазмиду гидролизовали с помощью SmaI и KpnI в указанном порядке и подвергали очистке в геле с получением, тем самым, линейной векторной ДНК, содержащей липкий KpnI конец и тупой SmaI конец.
(2) Получение плазмиды pPP2
При использовании фосмиды pCC1-PP1 в качестве матрицы полинуклеотид указанного выше P450-1 был амплифицирован с помощью пары праймеров P450-1 Kpn F (SEQ ID NO:277)/P450-1 Swa R (SEQ ID NO:278). Очищенный фрагмент ДНК клонировали в pCR-Blunt (Invitrogen, Cat. No. K2700-20). Полученную плазмиду расщепляли с помощью KpnI и SwaI. Указанный выше фрагмент P450-1 лигировали к описанному выше вектору pUSA-HSG с получением, тем самым, плазмиды pPP2, показанной на фигуре 5.
(3) Получение плазмиды pPP3
При использовании фосмиды pCC1-PP1 в качестве матрицы в соответствии со схемой, показанной на фигуре 6, сначала амплифицировали только экзоны с помощью пар праймеров F1 (SEQ ID NO:279)/R1 (SEQ ID NO:280), F2 (SEQ ID NO:281)/R2 (SEQ ID NO:282), F3 (SEQ ID NO:283)/R3 (SEQ ID NO:284), F4 (SEQ ID NO:285)/R4 (SEQ ID NO:286), F5 (SEQ ID NO:287)/R5 (SEQ ID NO:288) и F6 (SEQ ID NO:289)/R6 (SEQ ID NO:290) с получением, тем самым, шести фрагментов. Затем проводили амплификацию с этими фрагментами в качестве матриц с использованием пар праймеров F1/R2, F3/R4 и F5/R6 с получением, тем самым, более длинных фрагментов. Затем путем повторения амплификации с использованием пар праймеров F1/R4 и F1/R6 была получена кДНК, которая не содержит интроны полинуклеотида указанного выше P450-2. Этот фрагмент кДНК был вставлен в pCR-Blunt (Invitrogen, Cat. No. K2700-20), и полученную плазмиду использовали в качестве матрицы для амплификации с помощью пары праймеров infusion F P450-2-cDNA (SEQ ID NO:291)/infusion R P450-2-cDNA (SEQ ID NO:292). На основании указаний к набору с использованием набора In-Fusion Advantage PCR Cloning (Clontech) была получена показанная на фигуре 7 плазмида pPP3.
(4) Трансформация Aspergillus Oryzae (A. oryzae)
A. oryzae (штамм HL-1105) культивировали в агаровой среде CD-Met (содержащей L-метионин 40 мкг/мл) при 30°С в течение одной недели. Из этой чашки Петри были собраны конидии (>108) и высеяны в 100 мл жидкой среды YPD в 500-мл сосуде. Через 20 часов культивирования (30°С, 180 об/мин) были получены бактериальные клетки, имеющие форму шариков мха. Бактериальные клетки собирали с помощью стеклянного фильтра 3G-1, промывали 0,8 М NaCl, и воду тщательно удаляли. Полученное суспендировали с помощью TF раствора I (раствора образования протопласта) и затем встряхивали при 30°С, 60 об/мин в течение 2 часов. С 30-минутным интервалом производили микроскопическое обследование и проверяли наличие протопластов. После этого культуральную среду фильтровали и подвергали центрифугированию (2000 об/мин, 5 минут) для сбора протопластов, которые затем промывали TF раствором II. После промывки добавляли 0,8 объема TF раствора II и 0,2 объема TF раствора III и смешивали с получением, тем самым, суспензии протопластов.
К 200 мкл этой суспензии добавляли 10 мкг плазмидной ДНК (pPP2 или pPP3). Смесь оставляли на льду в течение 30 минут и добавляли TF раствор III (1 мл). Полученную смесь слегка перемешивали и затем оставляли стоять при комнатной температуре в течение 15 минут. После этого в указанные выше протопласты вводили плазмидную ДНК. Для этого добавляли TF раствор II (8 мл) и подвергали центрифугированию (при 2000 об/мин в течение 5 минут). После этого протопласты собирали из 1-2 мл, оставшихся наверху. Выделенный раствор протопластов по каплям наносили на среду восстановления (нижний слой), и приливали среду восстановления (верхний слой). Полученное перемешивали вращением чашки Петри и затем культивировали при 30°С в течение 4-5 дней. Полученные клоны выделяли в среде восстановления (нижний слой), субкультивировали и очищали с получением, тем самым, трансформанта (Aspergillus Oryzae PP2-1 и Aspergillus Oryzae PP3-2).
Готовили указанный выше TF раствор I (раствор образования протопласта) со следующим составом.
После получения указанного выше состава (pH 5,5) производили стерилизацию фильтрованием.
Готовили указанный выше TF раствор II со следующим составом.
После получения указанного выше состава производили стерилизацию автоклавированием.
Готовили указанный выше TF раствор III со следующим составом.
После получения указанного выше состава производили стерилизацию фильтрованием.
Готовили указанную выше среду восстановления со следующим составом.
После получения указанного выше состава (pH 5,5) производили стерилизацию автоклавированием.
Кроме того, готовили использованный выше раствор микроэлементов со следующим составом.
После получения указанного выше состава производили стерилизацию автоклавированием.
(5) Функциональный анализ и дополнительный культуральный тест P450-1
К среде YPD (1% (масс./об.) дрожжевой экстракт, 2% (масс./об.) пептон, 2% (масс./об.) декстроза), содержащей 1% (масс./об.) мальтозу, добавляли 1/100 объема раствора 2 мг/мл пирипиропена E в диметилсульфоксиде с получением среды A. Из флоры Aspergillus oryzae PP2-1, культивируемого в агаровой среде Czapek Dox, собирали его конидии и суспендировали в стерилизованной воде. Эту суспензию конидий доводили до 104 спор/мл. Затем 100 мкл этой доведенной суспензии конидий добавляли к 10 мл среды A и культивировали при встряхивании при 25°С в течение 96 часов. К этому культуральному раствору добавляли 10 мл ацетона, и смесь тщательно перемешивали. После этого ацетон удаляли с помощью центрифужного концентратора. К полученному добавляли 10 мл этилацетата, и полученную смесь тщательно перемешивали, и затем отбирали только слой этилацетата. Высушенный продукт, полученный удалением этилацетата с помощью центрифужного концентратора, растворяли в 1000 мкл метанола. Это использовали в качестве образца и анализировали с помощью ЖХ-МС (Waters, Micromass ZQ, 2996PDA, 2695 Separation module, колонка: Waters XTerra C18 (Ф4,5×50 мм, 5 мкм) и ЖХ-ЯМР (Avance500 производства Burker Daltonik).
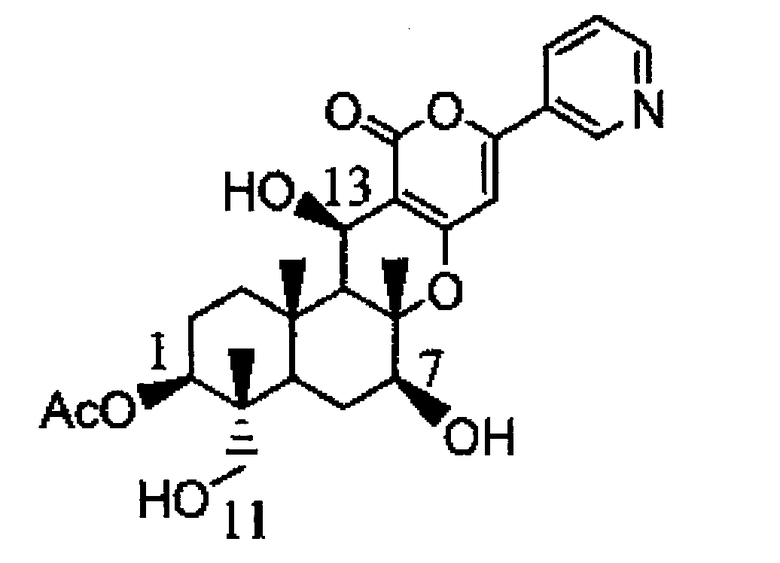
В результате указанного выше ЖХ-МС измерения было подтверждено, что полученное соединение является единственным соединением A, молекулярная масса которого увеличена на 16 по сравнению с пирипиропеном E. Кроме того, по данным измерения ЖХ-ЯМР было подтверждено, что это соединение A представляет собой гидроксид в положении 11 пирипиропена E. Было подтверждено, что указанная выше цитохром P450 монооксигеназа (1) является ферментом, гидроксилирующим пирипиропен E в положении 11 при использовании пирипиропена E в качестве субстрата.
Физико-химические свойства указанного выше соединения A представлены ниже:
1. Масс-спектр: ЭИ-МС 468 M/Z (M+H)+
2. Молекулярная формула: C27H33NO6
3. ВЭЖХ: Колонка: колонка C18 Waters XTerra (5 мкм, 4,5 мм×50 мм), 40°С, подвижная фаза: от 20% водного раствора ацетонитрила до 100% ацетонитрила за 10 минут (линейный градиент), скорость потока: 0,8 мл/мл, детекция: время задержки 6,696 минут при УФ 323 нм.
4. Спектр 1H-ЯМР (CD3CN, 2H: 3,134, 3,157 H-11)
Графики спектра 1H-ЯМР пирипиропена E и спектра 1H-ЯМР по п.4, описанного выше, представлены на фигуре 8 и фигуре 9, соответственно.
(6) Функциональный анализ и дополнительный культуральный тест P450-2
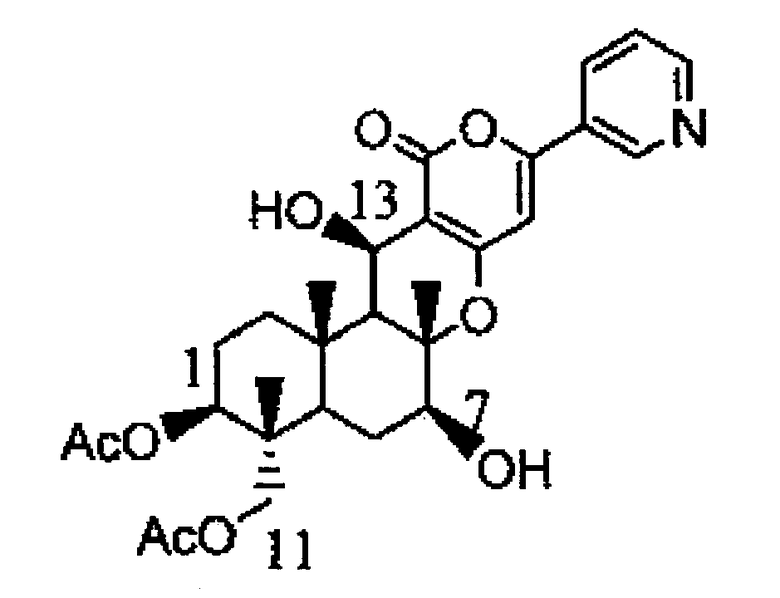
К среде YPD (1% (масс./об.) дрожжевой экстракт, 2% (масс./об.) пептон, 2% (масс./об.) декстроза), содержащей 1% (масс./об.) мальтозу, добавляли 1/100 объема раствора 2 мг/мл пирипиропена E в диметилсульфоксиде с получением среды B и сходным образом добавляли 1/100 объема раствора 2 мг/мл пирипиропена O в диметилсульфоксиде с получением среды C. Из флоры Aspergillus oryzae PP3-2, культивируемого в агаровой среде Czapek Dox, собирали его конидии и суспендировали в стерилизованной воде. Эту суспензию конидий доводили до 104 спор/мл. Затем 500 мкл доведенной суспензии конидий добавляли к 50 мл среды B или среды C и культивировали при встряхивании при 25°С в течение 96 часов. К этому культуральному раствору добавляли 50 мл ацетона, и смесь тщательно перемешивали. После этого ацетон удаляли с помощью центрифужного концентратора. К полученному добавляли 50 мл этилацетата, и полученную смесь тщательно перемешивали, и затем отбирали только слой этилацетата. Высушенный продукт, полученный удалением этилацетата с помощью центрифужного концентратора, растворяли в 1500 мкл метанола. Это использовали в качестве образца и анализировали с помощью ЖХ-МС (производство Waters, Micromass ZQ, 2996PDA, 2695 Separation module, колонка: Waters XTerra C18 (Ф4,5×50 мм, 5 мкм) и ЖХ-ЯМР (производство Burker Daltonik, Avance500). В результате ЖХ-МС измерения в образце, полученном из среды B, было выявлено соединение B, молекулярная масса которого увеличена на 32 по сравнению с пирипиропеном E. Сходным образом в образце, полученном из среды C, было выявлено соединение C, молекулярная масса которого увеличена на 32 по сравнению с пирипиропеном O. Кроме того, по данным измерения ЖХ-ЯМР было подтверждено, что соединение C представляет собой гидроксид в положении 7 и положении 13 пирипиропена O. Было подтверждено, что указанная выше цитохром P450 монооксигеназа (2) является ферментом, гидроксилирующим положение 7 и положение 13 каждого из пирипиропена E или пирипиропена O.
Физико-химические свойства указанного выше соединения B представлены ниже:
1. Масс-спектр: ЭИ-МС 484 M/Z (M+H)+
2. Молекулярная формула: C27H33NO7
3. ВЭЖХ: Колонка: колонка C18 Waters XTerra (5 мкм, 4,5 мм×50 мм), 40°С, подвижная фаза: от 20% водного раствора ацетонитрила до 100% ацетонитрила за 10 минут (линейный градиент), скорость потока: 0,8 мл/мл, детекция: время задержки 5,614 минут при УФ 323 нм.
Физико-химические свойства указанного выше соединения C представлены ниже:
1. Масс-спектр: ЭИ-МС 542 M/Z (M+H)+
2. Молекулярная формула: C29H35NO9
3. ВЭЖХ: Колонка: колонка C18 Waters XTerra (5 мкм, 4,5 мм×50 мм), 40°С, подвижная фаза: от 20% водного раствора ацетонитрила до 100% ацетонитрила за 10 минут (линейный градиент), скорость потока: 0,8 мл/мл, детекция: время задержки 5,165 минут при УФ 323 нм.
4. Спектр 1H-ЯМР (CD3CN, 1H: 4,858 H-13) (CD3CN, 1H: 3,65 H-7)
Графики спектра 1H-ЯМР пирипиропена O и указанного выше соединения C представлены на фигуре 10 и фигуре 11, соответственно.
[Номера регистрации]
FERM BP-11133
FERM BP-11137
FERM BP-11141






















































































































































Представленные изобретения касаются выделенного полинуклеотида, кодирующего полипептид, вовлеченный в биосинтез пирипиропена А, вектора и клетки-хозяина, включающих такой полипептид, и способов получения предшественников пирипиропена А, включающих культивирование клетки-хозяина. Представленный полинуклеотид кодирует полипептид, обладающий какой-либо одной или более из активностей - поликетидсинтазы, пренилтрансферазы, гидроксилазы, ацетилтрансферазы или аденилатсинтазы. Представленные изобретения позволяют синтезировать пирипиропен А, являющийся инсектицидным агентом, и могут быть использованы при формировании устойчивости растений к насекомым-вредителям. 5 н. и 11 з.п. ф-лы, 11 ил., 1 табл., 11 пр.
1. Выделенный полинуклеотид, кодирующий, по меньшей мере, один полипептид, вовлеченный в биосинтез пирипиропена А, который представляет собой
(a) полинуклеотид, обладающий нуклеотидной последовательностью SEQ ID NO: 266,
(b) полинуклеотид, обладающий нуклеотидной последовательностью, которая способна гибридизоваться с нуклеотидной последовательностью SEQ ID NO: 266 в жестких условиях,
(c) полинуклеотид, обладающий полинуклеотидной последовательностью, кодирующей, по меньшей мере, одну аминокислотную последовательность, выбранную из SEQ ID NO: 267-274, или по существу эквивалентную им аминокислотную последовательность, причем в указанной эквивалентной аминокислотной последовательности 1-40 остатков заменены, удалены, добавлены или вставлены по сравнению с SEQ ID NO: 267-274 соответственно или
(d) полинуклеотид, обладающий, по меньшей мере, одной нуклеотидной последовательностью, выбранной из нуклеотидной последовательности в любом из пунктов (1) или (2) ниже:
(1) нуклеотидной последовательностью в любом из пунктов от (а) до (h) ниже:
(a) нуклеотидной последовательностью от 3342 до 5158 в нуклеотидной последовательности, представленной SEQ ID NO: 266,
(b) нуклеотидной последовательностью от 5382 до 12777 в нуклеотидной последовательности, представленной SEQ ID NO: 266,
(c) нуклеотидной последовательностью от 13266 до 15144 в нуклеотидной последовательности, представленной SEQ ID NO: 266,
(d) нуклеотидной последовательностью от 16220 до 18018 в нуклеотидной последовательности, представленной SEQ ID NO: 266,
(e) нуклеотидной последовательностью от 18506 до 19296 в нуклеотидной последовательности, представленной SEQ ID NO: 266,
(f) нуклеотидной последовательностью от 19779 до 21389 в нуклеотидной последовательности, представленной SEQ ID NO: 266,
(g) нуклеотидной последовательностью от 21793 до 22877 в нуклеотидной последовательности, представленной SEQ ID NO: 266,
(h) нуклеотидной последовательностью от 23205 до 24773 в нуклеотидной последовательности, представленной SEQ ID NO: 266;
(2) нуклеотидной последовательностью, которая способна гибридизоваться с нуклеотидной последовательностью из (1) в жестких условиях,
где указанный полинуклеотид кодирует полипептид, обладающий какой-либо одной или более из активностей - активностью поликетидсинтазы, активностью пренилтрансферазы, активностью гидроксилазы, активностью ацетилтрансферазы или активностью аденилатсинтетазы.
2. Полинуклеотид по п.1, который происходит из штамма PF1169 Penicillium coprobium.
3. Полинуклеотид по п.1, кодирующий полипептид, обладающий активностью в отношении гидроксилирования положения 7 и/или положения 13 пирипиропена Е или пирипиропена О.
4. Полинуклеотид по п.1, кодирующий полипептид, обладающий активностью в отношении гидроксилирования положения 11 пирипиропена Е.
5. Экспрессионный вектор, кодирующий полипептид, вовлеченный в биосинтез пирипиропена А, включающий полинуклеотид по п.1.
6. Вектор по п.5, включающий полинуклеотид, кодирующий полипептид, обладающий активностью в отношении гидроксилирования положения 7 и/или положения 13 пирипиропена Е или пирипиропена О.
7. Вектор по п.5, включающий полинуклеотид, кодирующий полипептид, обладающий активностью в отношении гидроксилирования положения 11 пирипиропена Е.
8. Вектор по п.5, где указанный вектор представляет собой плазмиду рСС1-РР1.
9. Вектор по п.5, где указанный вектор представляет собой плазмиду рРР2.
10. Вектор по п.5, где указанный вектор представляет собой плазмиду рРР3.
11. Клетка-хозяин, включающая полинуклеотид по п.1, продуцирующая полипептид, вовлеченный в биосинтез пирипиропена А.
12. Клетка-хозяин по п.11, где указанная клетка-хозяин представляет собой Escherichia coli, включающую плазмиду рСС1-РР1.
13. Клетка-хозяин по п.11, где указанная клетка-хозяин представляет собой Aspergillus oryzae, включающий плазмиду рРР2.
14. Клетка-хозяин по п.12, где указанная клетка-хозяин представляет собой Aspergillus oryzae, включающий плазмиду рРР3.
15. Способ получения предшественника пирипиропена А, который включает культивирование клетки-хозяина по п.13 и выделение предшественника пирипиропена А из пирипиропена Е, указанный предшественник пирипиропена А представляет собой 11-деацетил-пирипиропен О.
16. Способ получения предшественника пирипиропена А, который включает культивирование клетки-хозяина по п.14 и выделение предшественника пирипиропена А из пирипиропена О, где указанный предшественник пирипиропена А представляет собой 7-деацетил-пирипиропен А.
| KR 0100264392, 16.08.2000 | |||
| US 20120046470, 23.02.2012 | |||
| ИНСЕКТИЦИДНЫЕ КОМПОЗИЦИИ, СОДЕРЖАЩИЕ СОЕДИНЕНИЯ, ОБЛАДАЮЩИЕ ИНГИБИРУЮЩЕЙ АКТИВНОСТЬЮ ПРОТИВ АЦИЛ СоА: ХОЛЕСТЕРИНАЦИЛТРАНСФЕРАЗЫ, ИЛИ ИХ СОЛИ В КАЧЕСТВЕ ЭФФЕКТИВНЫХ ИНГРЕДИЕНТОВ | 2003 |
|
RU2305403C2 |

