Настоящее изобретение относится к способу оценки биологической активности яиц гельминтов, прежде всего яиц гельминтов р. Trichuris, предпочтительно яиц Trichuris suis, на различных стадиях их жизненного цикла. Способ позволяет осуществлять контролируемое получение препаратов яиц гельминтов в качестве эффективных компонентов терапевтических лекарственных средств и гарантировать их безопасное и терапевтически эффективное применение для человека.
Известно, что паразитарные инфекции оказывают воздействие на активацию иммунной системы животного-хозяина (см. обзор D.М. McKay, Parasitology, 132, 2006, cc.1-12). Такая активация влияет также на встречаемость и протекание аутоиммунных заболеваний. Результаты эпидемиологических исследований показали, что в регионах с высокими коэффициентами заражения червями аутоиммунные заболевания являются более редкими, чем в регионах, в которых коэффициенты такого заражения является более низкими вследствие лучших гигиенических условий. Изучение профилей цитокинов в организме пациентов, страдающих болезнью Крона, представляющей собой хроническое воспалительное заболевание кишечника, показало, что иммунные Th2-клетки могут подвергаться стимуляции в результате заражения гельминтами. Болезнь Крона, т.е. Th1-доминантное аутоиммунное заболевание, можно предупреждать или оказывать воздействие на это заболевание путем заражения гельминтами (Summers и др., Am J Gastroenterol, 98, 2003, cc.2034-2041).
Уже в 1971 г. Beer (Br. Med. J., 3, 1971, cc.41) опубликовал данные о том, что Trichuris suis может служить в качестве нематоды, пригодной для осуществления целенаправленного умеренного заражения человека, не индуцирующего патогенные действия, которые сопровождают, например, заражение патогенным для человека возбудителем Trichuris trichiura Результаты более современных исследований продемонстрировали, что заражение человека Trichuris suis, по-видимому, является лишь кратковременным, и очевидно, что черви изгоняются до того, как может произойти их размножение. Однако положительные результаты клинических исследований применения Trichuris suis при хронических воспалительных заболеваниях кишечника свидетельствуют о том, что указанное кратковременное заражение человека может запускать обладающую терапевтической эффективностью модуляцию иммунной системы (Summers и др., Am. J. Gastroenterol., 98, 2003, cc.2034-2041).
Жизненный цикл Trichuris suis начинается с откладывания неэмбрионированных яиц (стадия L0), которые выделяются из инфицированного животного вместе с фекалиями. В почве в течение периода времени, составляющего 3-6 месяцев, происходит эмбрионирование до образования личинки первой стадии L1. В течение первых нескольких дней после достижения стадии L1 в яйце можно наблюдать движения личинки, после чего яйца переходят в состояние покоя, в котором они могут оставаться годами, не теряя при этом своей инфективности. После поглощения оральным путем пригодным хозяином личинки, находящиеся на стадии L1, вылупляются из яйца, попадая в полость кишечника, и в течение нескольких часов проникают в слизистую оболочку слепой кишки и ободочной кишки. Находясь в слизистой оболочке кишечника, личинки проходят в течение нескольких последующих недель дальнейшие стадии развития (L2-L4), после чего они в виде взрослой личинки, находящейся на стадии L5, снова выходят в полость кишечника (Beer, Parasitol., 67, 1973, cc.253-262).
Для получения яиц Trichuris suis (Trichuris suis ova = TSO) для фармацевтических целей неэмбрионированные TSO (находящиеся на стадии L0), которые были отложены in vitro или in situ, выделяют и очищают с помощью пригодных методов (WO 1999/33479, WO 2007/076868, WO 2007/134761). Затем в контролируемых лабораторных условиях происходит эмбрионирование с образованием биологически активных TSO (находящихся на стадии L1), которые представляют собой обладающий фармацевтической активностью компонент фармацевтического продукта.
Яйца гельминтов, которые предназначены для терапевтических применений, можно классифицировать как биологические фармацевтические продукты. Основным параметром, который необходимо тестировать при производстве и применении биологических фармацевтических продуктов, является биологическая активность, которая в конечном итоге представляет собой меру терапевтической эффективности фармацевтического продукта. Без анализа биологической активности нельзя ни рационально разрабатывать и контролировать технологический процесс, ни определять дозу фармацевтического продукта, необходимую для достижения терапевтического действия и безопасную для пациента в случае его планируемого применения на человеке. Поэтому в качестве фармацевтических продуктов можно безопасно и эффективно применять только те препараты яиц гельминтов, биологическая активность которых охарактеризована в достаточной степени.
До настоящего времени были известны три аналитических метода, которые можно было применять для Trichuris suis, однако они не позволяли в достаточной степени охарактеризовать биологическую активность.
1. Оценка коэффициента эмбрионирования (Kringel и др., Vet. Parasitol., 139, 2006, cc.132-139, параграф 2.2 (Trichuris suis); а также Johnson и др., Intl. J. Parasitol., 28, 1998, cc.627-633 (Ascaris suum)): Исследование проводят с помощью микроскопа. На основе морфологических критериев оценивают, содержат ли яйца интактные полностью сформированные личинки, находящиеся на стадии L1. Однако в отличие от молекулярно-биологического способа, предлагаемого в настоящем изобретении, этот метод, основанный только на морфологической оценке, не позволяет сделать уверенное заключение о том, что личинки действительно полностью эмбрионированы. Еще одно ограничение заключается в том, что сам по себе коэффициент эмбрионирования не несет информацию о жизнеспособности и биологической активности эмбрионированных TSO, находящихся на стадии L1. Кроме того, для проведения морфологической оценки требуется наличие определенного опыта у исследователя и субъективная классификация пограничных случаев при проведении морфологической оценки ограничивает правильность и точность этого метода.
2. Определение коэффициента заражения: В случае Trichuris suis анализ проводят на свиньях. В определенный момент времени определяют количество личинок, присутствующих в слизистой оболочке кишечника, и сопоставляют с введенной дозой TSO (Kringel и др., Vet. Parasitol., 139, 2006, cc.132-139, а также Summers и др., Gastroenterology, 128, 2005, cc.825-832 или Johnson и др., Intl. J. Parasitol., 28, 1998, cc.627-633 (Ascaris suum)). Проблема определения коэффициента заражения заключается в том, что он зависит не только от функциональной способности яиц гельминтов, но в равной степени также и от индивидуальных факторов организма хозяина, таких, например, как иммунная система, кишечная флора и кишечная функция лабораторных животных. Таким образом, определение коэффициента заражения на лабораторных животных неизбежно сопряжено с высокой вариабельностью, и его нельзя стандартизовать вследствие того, что система тестирования основана на использовании природных объектов, что существенно ограничивает ее применимость для тестирования биологической активности. В случае Trichuris suis проведение теста по оценке инфективности на свиньях связано с длительным инкубационным периодом, составляющим по меньшей мере три недели, и вследствие высокой вариабельности оно требует большого количества животных, что как с этической, так и с экономической точки зрения делает проблематичным применение этого теста как стандартного.
Кроме того, Kringel с соавторами описали выделение яиц из фекалий животных для доказательства биологической активности яиц гельминтов, которыми были заражены животные. Для этого необходимо выжидать в течение 7-8 недель после заражения животных, чтобы вылупившиеся личинки созрели до достижения стадии L5, спарились и сами отложили яйца. При этом отсутствует прямая количественная связь, позволяющая судить о коэффициенте заражения или биологической активности взрослых личинок по количеству яиц в фекалиях. Метод, разработанный Kringel с соавторами, существенно отличается от предлагаемого в настоящем изобретении способа определения коэффициента вылупляемости in vivo, поскольку согласно методу Kringel с соавторами анализируют яйца следующего поколения.
3. В WO 2007/134761 описан метод определения жизнеспособности яиц Trichuris suis, в котором моделируют in vitro прохождение яиц через желудочно-кишечный тракт свиньи и при этом создают условия для созревания яиц.
Однако описанные методы не являются достаточными для оценки биологической активности препаратов яиц гельминтов с точностью, необходимой для продуктов медицинского назначения.
Так, одна из основных проблем, связанных с биологическим фармацевтическим продуктом, биологическую активность которого тестируют на животных, заключается в стандартизации препарата. Моделирование на животных, такое как указанное выше заражение свиней, является чрезвычайно дорогостоящим и требует значительного количества времени. Поэтому при создании настоящего изобретения был разработан способ, в котором осуществляют различные типы анализов, которые касаются различных аспектов развития яиц гельминтов и их биологической активности. Для того чтобы сделать надежное заключение, необходимо провести по меньшей мере один из тестов, предлагаемых в изобретении, для подлежащей анализу партии, однако предпочтительно провести по меньшей мере три, более предпочтительно по меньшей мере четыре или даже пять из предлагаемых в настоящем изобретении тестов, в которых определяют в каждом случае соответствующие параметры. Результаты индивидуальных тестов рассматриваются в совокупности, причем пригодный препарат гельминтов в каждом тесте должен удовлетворять заранее определенным предельным значениям, для того, чтобы можно было сделать вывод о том, что препарат яиц гельминтов пригоден для фармацевтического применения.
Таким образом, существует выраженная необходимость в разработке способа, пригодного для промышленного применения, который позволяет осуществлять полный и надежный анализ биологической активности TSO и яиц других гельминтов на различных стадиях развития и тем самым характеризовать различные биологические функции яиц гельминтов. Только такой способ позволяет осуществлять контролируемое производство пригодного для продажи медицинского продукта.
Задача настоящего изобретения заключается в осуществлении полного анализа биологической активности яиц гельминтов с помощью способа, более подробно представленного ниже в настоящем описании и позволяющего, с одной стороны, обеспечивать строгий контроль процесса производства и, с другой стороны, обеспечивать возможность безопасного и терапевтически эффективного применения на пациенте.
Принимая во внимание сложность проблемы и недостатки известных в настоящее время методов, становится очевидным, что надежное определение биологической активности нельзя осуществлять с помощью метода, заключающегося в осуществлении только одного вида анализа.
Таким образом, объектом изобретения является способ определения биологической активности яиц Trichuris, которые содержат полностью эмбрионированные личинки, заключающийся в том, что осуществляют по меньшей мере одну из следующих оценок:
оценку индуцируемой температурой активности яиц Trichuris с помощью биохимических и/или молекулярно-биологических методов, прежде всего измерение содержания АТФ, определение индуцибельности генной экспрессии в эмбрионированных яйцах Tricuris,
оценку с помощью микроскопа подвижности личинки Tricuris в яйце в течение длительного периода наблюдения после активации посредством предварительной инкубации при повышенных температурах и/или
оценку коэффициента вылупляемости личинок Trichuris в организме лабораторного животного, при этом интактные эмбрионированные яйца, выделенные из содержимого кишечника, оценивают количественно путем сравнения с внутренним стандартом.
Так, была разработана система, включающая пять видов анализов, согласно которой в каждом случае анализируют различные биологические функции гельминтов на определенной фазе их жизненного цикла. Хотя может оказаться достаточным использовать лишь один метод анализа в процессе осуществления отдельных стадий производства фармацевтического препарата, как правило, для определения биологической активности конечного продукта следует осуществлять по меньшей мере 3 из перечисленных ниже в таблице 1 видов анализа.
Для надежной характеризации биологической активности пригодных для фармацевтического применения яиц гельминтов предпочтительно осуществляют весь способ полностью, включая по меньшей мере три, предпочтительно 4 и более предпочтительно все 5 его стадий. В частности, может оказаться достаточным осуществлять лишь отдельные стадии способа для характеризации конкретных аспектов биологической активности или для мониторинга определенных подстадий процесса производства.
Пять отдельных стадий способа основаны на принципах тестирования, которые уже были описаны в другой связи. Адаптация способа с целью его применения на эмбрионированных яйцах гельминтов, прежде всего на эмбрионированных яйцах Trichuris suis, является новой, и она требует разработки ряда новых подстадий, не описанных для настоящего времени. Способ в целом, основанный на исследовании пяти различных аспектов биологической активности и тем самым позволяющий осуществлять исчерпывающую характеризацию, также является новым, и до настоящего времени не было описано его применение для яиц гельминтов, прежде всего для яиц Trichuris suis. Предпочтительной общей отличительной чертой трех из пяти описанных стадий способа является активация находящихся в состоянии покоя личинок Trichuris путем предварительной инкубации при повышенной температуре. Другая отличительная особенность рассматриваемого способа заключается в том, что его можно осуществлять на интактных жизнеспособных яйцах гельминтов, и в том, что измеряемые параметры зависят только от активности яиц гельминтов, но не от связанных с хозяином факторов или от индивидуальных способностей специалиста, осуществляющего тест, и вследствие этого рассматриваемый способ является более объективным, чем методы, применявшиеся для настоящего времени. Благодаря этому оказывается возможным осуществлять стандартизацию, которая необходима для фармацевтического применения.
С помощью предлагаемого способа оказывается возможным надежно определять биологическую активность препаратов яиц гельминтов с точностью, требуемой для фармацевтического продукта. Точное определение биологической активности яиц гельминтов согласно способу, предлагаемому в настоящем изобретении, представляет собой необходимую стадию при производстве пригодного для применения фармацевтического продукта.
Ниже более подробно описаны 5 видов анализа, представляющие собой составную часть способа.
а) Оценка количества копий геномной ДНК
При каждом делении клетки в процессе эмбрионального развития набор хромосом удваивается и разделяется между двумя дочерними клетками. Следовательно, количество копий геномной ДНК в яйце можно рассматривать в качестве коррелята количества соматических клеток в развивающейся личинке. Таким образом, оценку количества копий геномной ДНК можно использовать для определения статуса эмбрионального развития развивающейся личинки на любой стадии. Это является очевидным преимуществом по сравнению с описанным ранее методом (оценка коэффициента эмбрионирования с помощью микроскопа, см. выше), с помощью которого можно анализировать только конечный результат эмбрионирования, продолжающегося в течение примерно 13 недель, и который не дает никакой информации о том, закончено ли полностью в действительности эмбриональное развитие (т.е. соответствует ли количество клеток полностью сформированной личинке на стадии L1). Эмбрионирование представляет собой составную часть фармацевтического процесса получения яиц гельминтов. Оценка количества копий геномной ДНК впервые позволяет осуществлять анализ в режиме сопровождения процесса производства. С одной стороны, это дает возможность рациональным путем проводить усовершенствование процесса производства и анализировать эффект изменений процесса, а, с другой стороны, осуществлять стандартным путем мониторинг в течение фазы эмбрионирования.
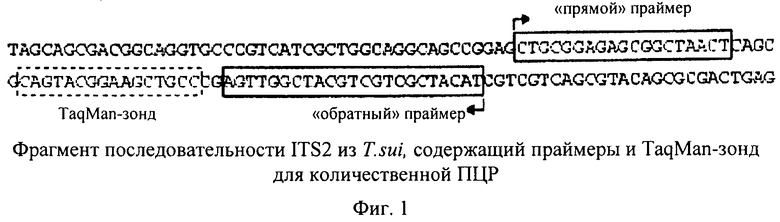
С помощью метода количественной ПЦР можно оценивать количество копий определенной нуклеотидной последовательности в образце. Чем большее количество копий определенной нуклеотидной последовательности присутствует в образце, тем быстрее процесс амплификации выходит на плато. Количество копий можно оценивать сравнительно точно с использованием калибровочных кривых. Одним из методов, который, как правило, применяют для этой цели, является ПЦР в реальном времени или так называемая TaqMan-ПЦР (ПЦР по методу TaqMan). Специалисту в данной области известны альтернативные методы амплификации нуклеиновых кислот, и их также можно применять для определения количества копий геномной ДНК. Точное количество копий, как правило, определяют с использованием соответствующих калибровочных кривых.
Предпочтительно в качестве предпочтительного маркера для геномной ДНК гельминтов можно применять область ITS1-5.8S-ITS2, кодирующую часть рибосомной РНК. Последовательность ITS1-5.8S-ITS2 уже была описана для Trichuris suis (Cutillas и др., Parasitol Res, 100, 2001, cc.383-389), а также для других видов Trichuri и других гельминтов. Описан также тест по оценке жизнеспособности, согласно которому осуществляют мониторинг эмбрионального развития Ascaris suum с помощью количественной ПЦР области IST-1 (Pecson и др., Appl. Envir. Microbiol., 72, 2006, cc.7864-7872).
Однако указанный тест предназначен для другой цели, а именно для тестирования эффективности методов инактивации гельминтов.
Количественная ПЦР (кПЦР), с помощью которой амплифицируют частичную последовательность из области ITS1-5.8S-ITS2, является наиболее пригодной для оценки количества копий маркерной области и, следовательно, количества копий геномной ДНК. Поскольку элементы ITS1 и ITS2 являются видоспецифическими, то этот метод можно применять помимо анализа стадии эмбрионирования также и для качественной поверки идентичности организма. Такой тест не был описан в указанных выше прототипах прежде всего потому, что ранее не было известно, что T.suis на стадии L1 содержит примерно 1000 клеток согласно оценке на основе количества копий генома (см. пример 1).
На фиг.1 представлена соответствующая последовательность гена Trichuris suis, которую можно использовать, например, для оценки количества копий геномной ДНК. Соответствующая последовательность гена представлена в SEQ ID NO:1. Последовательность «прямого» праймера представлена в SEQ ID NO:2, последовательность «обратного» праймера представлена в SEQ ID NO:3, а последовательность Taqman-зонда представлена в SEQ ID NO:4. Специалисту в данной области должно быть очевидно, что для оценки количества копий можно использовать также и другие участки из генома Trichuris suis. Предпосылкой для возможности использования последовательности является то, что она должна представлять собой нуклеотидную последовательность, которая является специфической для Trichuris suis, и что сходные последовательности не встречаются в других организмах, которые могут присутствовать в качестве примесей в образце, предназначенном для анализа.
Оценку количества копий геномной ДНК можно использовать также и для другой цели. Таким путем можно подтверждать, что представляющий интерес организм присутствует в препарате и с использованием соответствующих других последовательностей можно определять, присутствуют ли примеси в виде других организмов.
Важным аспектом, который нельзя упускать из виду при оценке количества копий геномной ДНК, является то, что яйца гельминтов необходимо разрушать перед проведением измерений. Как проиллюстрировано в примерах, для данной цели наиболее пригодным оказался «гомогенизатор Поттера», причем в предпочтительном варианте осуществления изобретения используют объем 2 мл и устанавливают ширину зазора от 0,01 до 0,03 мм. Гомогенизацию предпочтительно осуществляют в течение периода времени, составляющего от 5 до 15 мин, предпочтительно примерно 10 мин. Разрушение клеток всегда осуществляют тогда, когда клеточные компоненты должны находиться в форме, пригодной для соответствующей стадии анализа.
б) Оценка метаболической активности
Многие методы оценки жизнеспособности клеток основаны на анализе метаболической активности. Аденозинтрифосфат играет центральную роль в клеточном метаболизме в качестве переносчика энергии и аккумулятора энергии, и его можно использовать в качестве маркера при оценке внутриклеточной метаболической активности. В настоящем изобретении продемонстрировано, что обнаружение этого нуклеотида можно использовать также для оценки жизнеспособности различных организмов, таких как личинки, находящиеся на стадии L1 Trichuris suis.
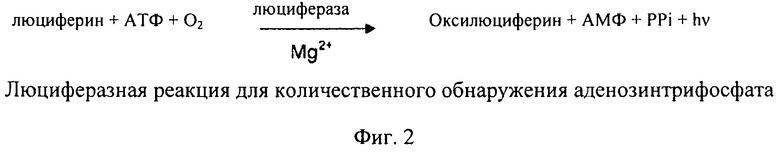
Количественную оценку аденозинтрифосфата в биологических системах можно осуществлять на основе измерения люминесценции с помощью люциферазной реакции. Люминесценцию определяют как испускание «холодного» света. Люминесцентные системы основаны на химической, биохимической или электрохимической активации субстратов, которые при возвращении в свое исходное состояние испускают часть энергии возбуждения в форме света. При обнаружении аденозинтрифосфата применяют люциферазы, выделенные из светляков (Photinus pyralis; светляк). Эукариотический фермент катализирует окисление люциферина в оксилюциферин в присутствии аденозинтрифосфата, кислорода и ионов магния с испусканием света. Уравнение реакции представлено на фиг.2.
Предпосылками для успешного осуществления обнаружения аденозинтрифосфата в эмбрионированных жизнеспособных яйцах Trichuris suis являются стимуляция синтеза аденозинтрифосфата в личинках, находящихся на стадии L1, на уровне, достаточном для измерения люминесценции, полное разрушение яиц с помощью пригодного метода гомогенизации для высвобождения сформированного внутри клетки нуклеотида, а также количественное и при этом беспрепятственное улавливание аденозинтрифосфата из сложной матрицы. При создании изобретения неожиданно был установлен неописанный до настоящего времени факт, что содержание АТФ в личинках, находящихся на стадии L1, до завершения эмбрионального развития является низким и только после соответствующей активации путем инкубации в течение продолжительного периода времени при определенной температуре возрастает до постоянного высокого уровня. Кроме того, для того, чтобы различать живые и мертвые личинки, находящиеся на стадии L1, необходим эффективный метод инактивации, такой чтобы можно было воспроизводимым образом определять исходный уровень аденозинтрифосфата в убитых организмах.
Согласно изобретению для анализа метаболической активности популяций яиц гельминтов предпочтительно оценивают содержание АТФ.
Для анализа метаболической активности индивидуальных яиц гельминтов можно применять также методы окрашивания с использованием производных тетразолия, которые первоначально были разработаны для свободно доступных клеток животных в клеточных культурах или для гистологических срезов. Производные тетразолия восстанавливаются в обладающих метаболической активностью клетках в результате воздействия митохондриальных ферментов с образованием окрашенных формазанов, которые откладываются в клетках. Решающее значение для возможности применения методов окрашивания, разработанных для свободно доступных клеток животных, для многоклеточных личинок гельминтов, находящихся на стадии L1, в яйцах, имеет предварительная обработка яиц, которая дает возможность субстрату проникать в них, не влияя на жизнеспособность находящейся в яйце личинки. Личинки Trichuris suis, находящиеся на стадии L1, окружены жесткой оболочкой, которая значительно ограничивает обмен веществом с окружающей средой и препятствует проникновению субстанций, необходимых для осуществления анализа.
Было установлено, что для щадящего разрушения оболочки яйца предпочтительно следует применять метод обработки яиц хлорноватистой кислотой, который уже был описан ранее (Beer, Parasitol., 67, 1973, cc.263-278), и эффективность указанной обработки при необходимости можно повышать путем последующего ферментативного расщепления с помощью хитиназы и протеазы.
Для того чтобы иметь возможность оценить метаболическую активность, необходимо определять «нулевое значение». В данном случае речь идет о величине, путем сравнения с которой оценивают соответствующую метаболическую активность. В предпочтительном варианте осуществления изобретения «нулевое значение» определяют на неактивных яйцах гельминтов.
Установлено, что криоинактивация (быстрое замораживание и хранение при -80°C в течение 24 ч) является методом, пригодным для инактивации яиц. Было установлено, что аденозинтрифосфатный сигнал от образцов, инактивированных таким путем, можно снижать до уровня фонового шума. Получение криоинактивированных образцов яиц осуществляют в забуференном фосфатом физиологическом растворе хлорида натрия. Очевидно, что криоинактивацию можно применять также в сочетании с другими методами обнаружения.
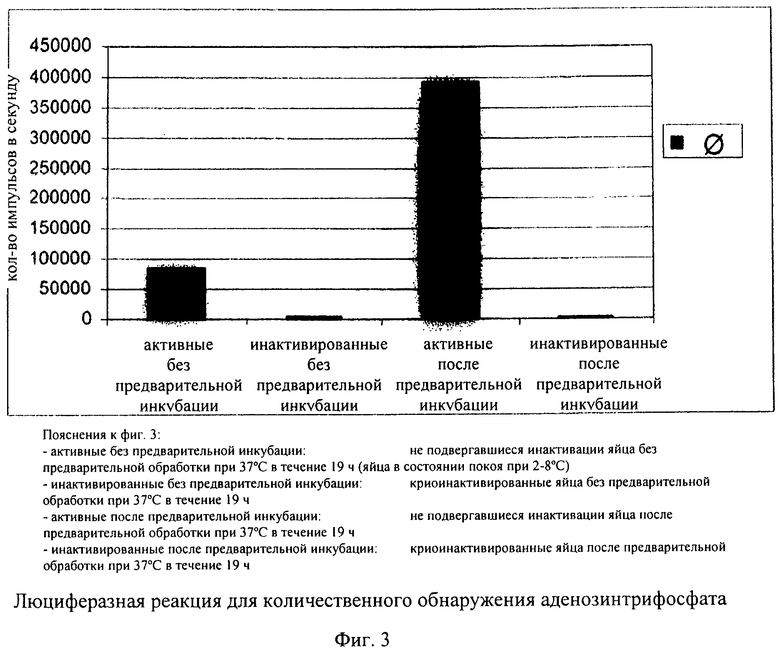
На фиг.3 продемонстрировано в качестве примера воздействие криоинактивации при количественном обнаружении аденозинтрифосфата с помощью люциферазной реакции. С использованием криоинактивации можно оценивать конкретно воздействие предварительной инкубации. Установлено, что в случае инактивированных (убитых) яиц гельминтов даже после предварительной инкубации было невозможно выявить увеличение уровня аденозинтрифосфата. В отличие от этого, биологически активные яйца гельминтов обладают активностью, которую выявляли по увеличению уровня аденозинтрифосфата после предварительной обработки при 37°С в течение периода времени, составляющего 19 ч.
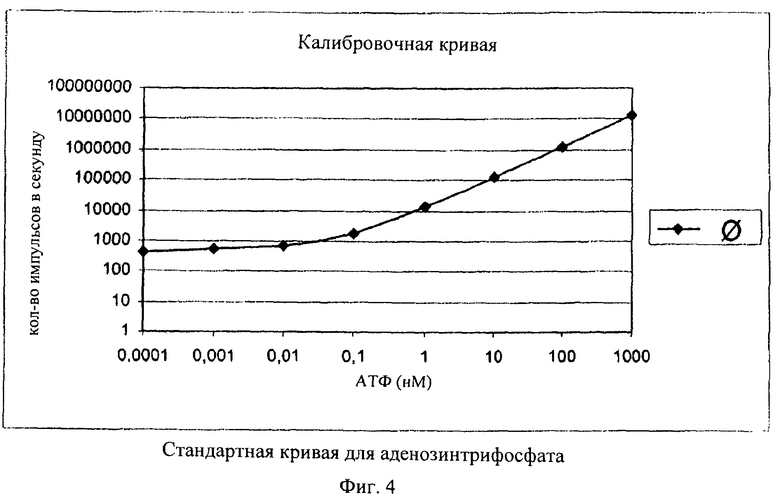
В одном из предпочтительных вариантов осуществления изобретения для каждой партии использовали по 5500 яиц в 550 мкл раствора. После предварительной инкубации яйца отделяли центрифугированием (5 мин при 500 об/мин) и ресуспендировали в 225 мкл буфера для лизиса. Для высвобождения содержимого клеток, включающего аденозинтрифосфат, образовавшийся при инкубации, яйца гомогенизировали с использованием гомогенизатора Поттера (объем 2 мл, ширина зазора 0,01-0,03 мм) в течение 10 мин. Только данный метод позволял осуществлять полное разрушение яиц и обеспечивать количественное высвобождение нуклеотида. Для инактивации расщепляющих аденозинтрифосфат гидролаз разрушение яиц осуществляли с использованием буфера для лизиса (компонент: фосфорная кислота, рН 2). Из каждой индивидуальной партии отбирали с помощью пипетки по 50 мкл и вносили в 96-луночные микропланшеты и каждую партию разводили с помощью еще 100 мкл фосфатного буфера. Люциферазную реакцию начинали путем добавления 100 мкл поступающего в продажу набора для анализа аденозинтрифосфата. После инкубации в течение 2 мин осуществляли считывание лунок микропланшета с помощью люминометра и определяли люминесценцию (фиг.3). Количественную оценку осуществляли с использованием стандартной кривой для аденозинтрифосфата в фосфатном буфере (1,0 мкМ, 0,1 мкМ, 10 нМ, 1 нМ, 0,1 нМ, 10 пМ, 1,0 пМ, 0,1 пМ). Типичная калибровочная кривая для АТФ проиллюстрирована на фиг.4.
Путем целенаправленного добавления аденозинтрифосфата к гомогенизированным криоинактивированным образцам эмбрионированных яиц можно достигать выхода, составляющего 84%, в линейном диапазоне стандартной кривой (от 1 до 1000 нМ). При этом не было выявлено соответствующего воздействия сложного матрикса гомогената яиц на люциферазную реакцию и люминесценцию.
Данный пример четко демонстрирует, что измерение люминесценции можно использовать для того, чтобы отличать жизнеспособные эмбрионированные яйца Trichuris suis от инактивированных и следовательно нежизнеспособных яиц. Однако тесты можно успешно осуществлять только при использовании комбинации оптимизированных параметров способа, включающей активацию образца, гомогенизацию образца и получение пригодных контролей с помощью оптимизированного метода инактивации. Пригодное содержание АТФ в биологически активных яйцах гельминтов составляет по меньшей мере примерно 0,01 пмоля АТФ на яйцо.
в) Анализ индуцибельной генной экспрессии
Принцип, положенный в основу этого метода, заключается в осуществлении индукции генной экспрессии, которая может иметь место только в живых клетках и которая позволяет различать живые и мертвые клетки. Кроме того, индуцибельность генной экспрессии, по-видимому, является предпосылкой для того, чтобы покоящаяся личинка, находящаяся на стадии L1, могла проходить через различные состояния активации, необходимые для индукции последующих стадий жизненного цикла (вылупление, внедрение в слизистую оболочку и т.д.).
По-видимому, наиболее предпочтительным является анализ экспрессии белков теплового шока, поскольку ее можно легко индуцировать путем повышения температуры и образующаяся матричная РНК (мРНК), как правило, обладает стабильностью в отношении быстрого расщепления. При создании настоящего изобретения впервые удалось обнаружить в Trichuris suis последовательность гена, обладающую высокой степенью гомологии с последовательностью белка теплового шока hsp70 из гельминта Caenorabhditis elegans.
Согласно одному из предпочтительных вариантов осуществления настоящего изобретения анализируют генную экспрессию белка теплового шока Т.suis. На фиг.5 приведено сравнение белковой последовательности Trichuris suis (SEQ ID NO:5) с аминокислотной последовательностью гомологичного белка из Caenorabhditis elegans (SEQ ID NO:6).
Индуцибельную генную экспрессию можно выявлять путем оценки содержания матричной РНК, кодирующей определенный индуцибельный белок. Из аминокислотной последовательности можно выводить нуклеотидную последовательность матричной РНК. Для специалиста в данной области не представляет трудностей выбор пригодных «прямого» и «обратного» праймеров и размещение между ними последовательности Taqman-зонда. После этого с помощью ПЦР в реальном времени можно обнаруживать индукцию гена. Для информативного обнаружения необходимой предпосылкой является анализ также и контрольного образца, содержащего убитые яйца червей. Кроме того, данный анализ следует применять для белка, который легко и надежно поддается индукции. Одним из примеров является белок теплового шока, однако для обнаружения можно с равным успехом использовать также и другие индуцируемые определенными стимулами гены.
Обнаружение экспрессируемой матричной РНК на уровне индивидуального яйца можно осуществлять также с помощью метода флуоресцентной in situ гибридизации (FISH) с использованием пригодного генного зонда. Для быстрого получения надежных результатов наиболее предпочтительно можно затем анализировать популяцию яиц с помощью проточной цитометрии.
Последовательность зонда, пригодного для гибридизации in situ, представлена на фиг.6 (SEQ ID NO:7).
г) Оценка подвижности личинок в интактных яйцах гельминтов
Принцип, положенный в основу этого анализа, заключается в обнаружении движений тела в качестве параметров функциональности личинки, находящейся на стадии L1, в яйце. Подвижность личинки является предпосылкой для ее вылупления в соответствующих условиях окружающей среды. В случае свободно живущих личинок и червей, таких как C.elegans, анализ подвижности можно использовать для надежного доказательства жизнеспособности и функциональной способности.
При создании настоящего изобретения неожиданно и впервые на примере Trichuris suis было установлено, что путем точного регулирования температуры окружающей среды можно индуцировать слабые движения также и у личинок гельминтов, которые находятся в яйце, а не находятся в свободном состоянии. Согласно известным к настоящему времени данным считалось, что после завершения эмбрионирования личинки переходят в состояние покоя и активируются и вылупливаются только после их поглощения пригодным хозяином. Движения личинок в яйце являются очень медленными и их можно обнаружить только тогда, когда согласно настоящему изобретению их анализируют с помощью цейтрафера (аппарат для замедленной киносъемки) под микроскопом в течение продолжительного периода времени. Тест на обнаружение подвижности с помощью микроскопа можно автоматизировать с использованием программного обеспечения для анализа изображений. Коэффициент подвижности, представляющий собой параметр биологической активности яиц, рассчитывают по следующей формуле:
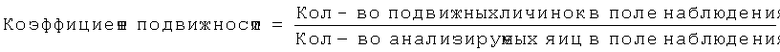
При оценке подвижности личинок для успешного применения способа решающее значение имеет точная настройка температуры. Прежде всего, яйца подвергают предварительной инкубации в течение периода времени, составляющего от 2 до 30 ч, предпочтительно от 4 до 20 ч при точно установленной температуре, находящейся в пределах от 36 до 42°С, предпочтительно от 37 до 41°С и наиболее предпочтительно при 39,5°С. После осуществления указанной предварительной инкубации личинки помещают под пригодный микроскоп, поддерживая при этом температуру по меньшей мере приблизительно на установленном уровне. После этого осуществляют наблюдение с использованием замедленной киносъемки в течение периода времени, составляющего от 2 мин до 4 ч, предпочтительно от 30 мин до 2 ч при температуре от 36 до 42°С, предпочтительно от 38 до 40°С.
На фиг.7 представлены результаты оценки коэффициента подвижности согласно изобретению, свидетельствующие о том, что в случае инактивированных яиц подвижность полностью отсутствует, а в отличие от этого в случае активных яиц червей коэффициент подвижности увеличивается с течением времени.
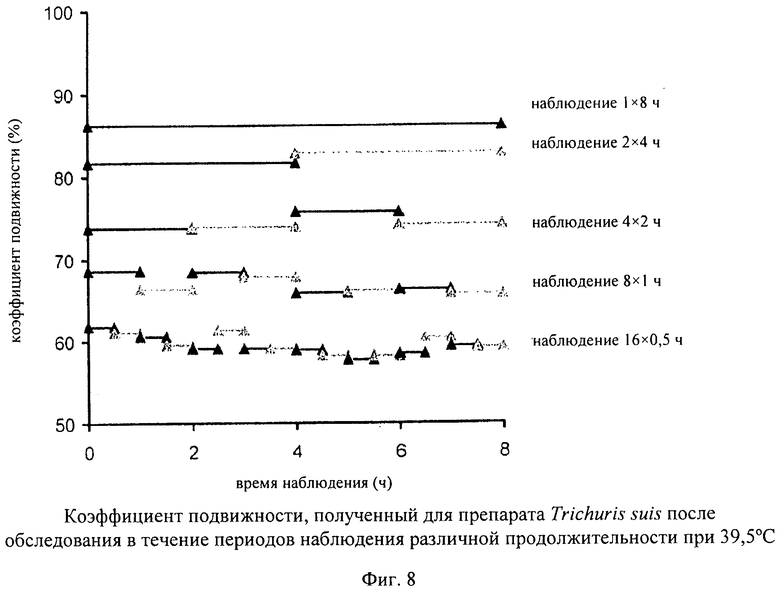
На фиг.8 представлены данные о коэффициенте подвижности для различных периодов наблюдения.
д) Оценка коэффициента вылупляемости в кишечнике
Принцип, положенный в основу этой стадии способа, заключается в оценке коэффициента вылупляемости личинок гельминтов в кишечнике лабораторных животных или в альтернативном варианте в содержимом кишечника, которое ранее было изъято из организма лабораторных животных.
Согласно одному из вариантов осуществления изобретения коэффициент вылупляемости можно оценивать в содержимом кишечника, которое изъято из организма лабораторных животных, как правило, предпочтительно из верхнего отдела ободочной кишки или из нижнего отдела двенадцатиперстной кишки. В этом случае можно оценивать коэффициент вылупляемости в содержимом кишечника вместо того, чтобы определять коэффициент вылупляемости непосредственно в организме лабораторного животного. Кроме того, способ можно осуществлять также путем анализа яиц, которые прошли через кишечник и выделены из фекалий. Это дает то преимущество, что не требуется убивать лабораторное животное и его можно использовать несколько раз для определения коэффициента вылупляемости.
На разработанной при создании изобретения стадии способа содержимое кишечника анализируют по истечении относительно короткого промежутка времени после инокуляции и подсчитывают количество интактных яиц, а также оболочек яиц, оставшихся после вылупления. Важное преимущество по сравнению с известным из существующего уровня техники методом оценки инфективности заключается в том, что в данном случае по вылуплению анализируют только первую стадию жизненного цикла, на которую факторы, зависящие от хозяина, оказывают лишь слабое влияние.
При создании изобретения неожиданно впервые было установлено для Trichuris suis, что поддающееся количественной оценке вылупление личинок происходит не только в организме свиньи, представляющем собой пригодного хозяина, но также и в организме кролика. Следовательно, например, для Trichuris suis можно использовать лабораторное животное с меньшими размерами кишечника, что значительно облегчает обнаружение микроскопически малых яиц и оболочек яиц.
При создании изобретения неожиданно было установлено, что можно осуществлять стойкое окрашивание оболочек яиц с помощью поступающих в продажу флуоресцентных красителей, разработанных для маркировки белков. Связывание флуоресцентных красителей с оболочкой яйца существенно облегчает обнаружение яиц, поскольку содержимое кишечника можно анализировать с помощью флуоресцентного микроскопа.
Еще одной технической инновацией является применение неэмбрионированных или инактивированных яиц гельминтов в качестве определенного внутреннего стандарта. Для таких яиц характерно такое же время прохождения через кишечник или время пребывания в кишечнике, но в отличие от эмбрионированных и биологически активных яиц они остаются интактными при прохождении через кишечник. Путем маркировки яиц, предназначенных для применения в качестве внутреннего стандарта, с помощью второго (отличного от первого) флуоресцентного красителя можно легко различать внутренний стандарт и яйца, подлежащие анализу.
Для анализа определяют количество интактных яиц внутреннего стандарта, маркированных флуоресцентным красителем 1, и пустых оболочек яиц в предназначенном для анализа образце и количество интактных яиц, маркированных флуоресцентным красителем 2, в содержимом кишечника в определенный момент времени после инокуляции. Коэффициент вылупляемости в кишечнике в пересчете на количество интактных (из которых не вылупились личинки) яиц можно рассчитывать согласно приведенной ниже формуле.
Формула для расчета коэффициента вылупляемости:

где:
- [IE] = количество интактных яиц в образце, подлежащем анализу (маркированных красителем 1),
- [IS] = количество интактных яиц внутреннего стандарта (маркированных красителем 2),
- i = образец содержимого кишечника,
- 0 = образец инокулята.
В приведенной ниже таблице 2 обобщены данные о ПЦР-анализе, а также предпочтительные виды оценки и предпочтительные получаемые при этом значения, которые представляют собой предельные значения, позволяющие судить, обладает ли анализируемый препарат требуемой биологической активностью.
В предпочтительном варианте осуществления изобретения новый способ тестирования заключается в том, что оценивают индуцируемую температурой способность к активации эмбрионированных яиц гельминтов in vitro. Измеряют два дополняющих друг друга параметра (коэффициент подвижности и среднее содержание АТФ), которые в совокупности дают балл активности in vitro. Балл активности in vitro хорошо коррелирует с биологической активностью in vivo. Однако он может служить только прогностическим фактором для биологической активности, если используемые для его определения личинки являются полностью эмбрионированными. В этом случае применяют метод количественной ПЦР, с помощью которого измеряют количество копий генома T.suis в яйце. С помощью осуществляемого параллельно нового теста по оценке вылупления in vivo удостоверяются, что анализируемые яйца действительно обладают активностью в физиологических условиях. Ниже представлена схема осуществления предпочтительного варианта способа:
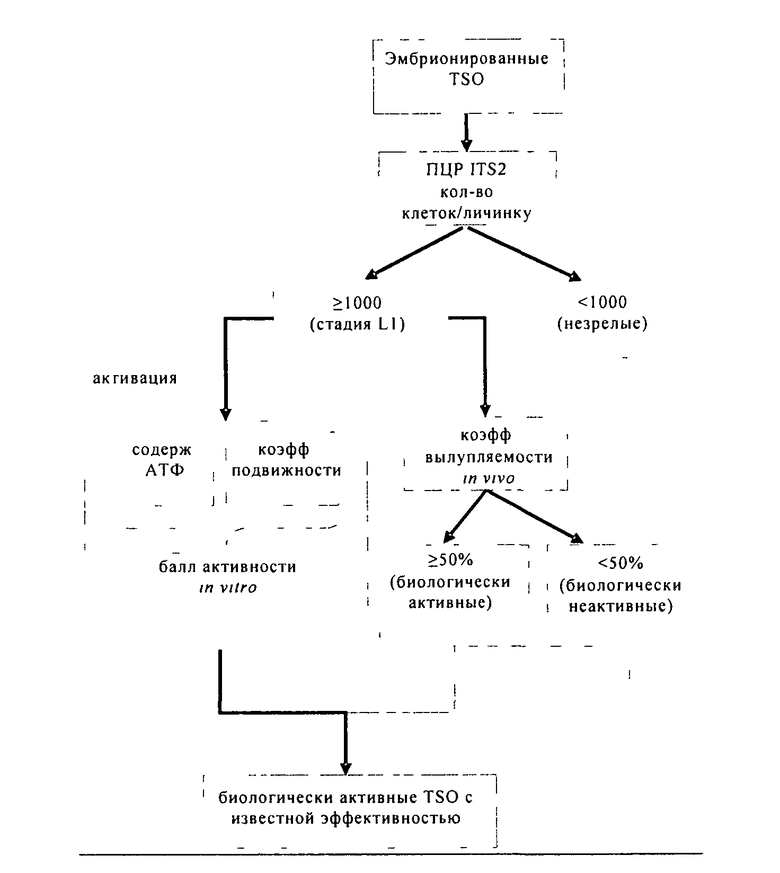
Важным аспектом настоящего изобретения является оценка индуцируемой температурой способности к активации in vitro.
TSO, находящиеся на стадии L1 (L1-TSO), после завершения эмбрионирования переходят в состояние покоя, в котором они могут находиться в течение нескольких лет, не теряя жизнеспособности, до тех пор пока после поглощения хозяином они не перейдут в активированное состояние и не начнут вылупляться. При создании изобретения неожиданно был установлен неописанный до настоящего времени факт, что яйца можно активировать также in vitro всего лишь путем инкубации в течение определенного периода времени в определенном диапазоне («окне») температур без использования других факторов. Активированное состояние яиц количественно оценивают путем определения двух дополнительных параметров: подвижности личинок, находящихся внутри интактных яиц, и содержания АТФ в лизатах активированных перед этим яиц. Неожиданно был установлен факт, который находится в противоречии с известными из научной литературы данными, заключающийся в том, что указанная в настоящем описании предварительная инкубация индуцирует слабые движения личинки в яйце. Движения личинок продолжаются в течение нескольких часов, при этом не наблюдается гибель личинок или вылупление из яйца. Поэтому в предпочтительном варианте способа, предлагаемого в изобретении, применяют соответствующую предварительную инкубацию яиц, приводящую к активации. Путем наблюдения с помощью микроскопа с использованием замедленной киносъемки за достаточно большим количеством яиц оказывается возможным надежно оценивать долю яиц в популяции, которые содержат активные подвижные личинки (т.е. определять коэффициент подвижности). Однако, поскольку данный способ не позволяет выявлять различия между состояниями с различной активностью для индивидуальной личинки, т.е. различные уровни подвижности/активности, то его объединяют с определением содержания АТФ, которое представляет собой количественную меру общей активности анализируемой популяции яиц. Оба анализируемых параметра, а именно коэффициент подвижности, представляющий собой прямую, но дискретную переменную, и содержание АТФ, представляющее собой косвенную, но непрерывную переменную, взаимно дополняют друг друга, позволяя надежно оценивать активность in vitro, которая хорошо коррелирует с активностью in vivo. На основе обоих параметров можно рассчитывать балл активности in vitro, который хорошо коррелирует с активностью яиц in vivo.
Предлагаемый в изобретении способ in vitro существенно отличается от известных в настоящее время методов. Определение коэффициента эмбрионирования, когда яйца визуально анализируют только с целью определения присутствия полностью сформированных личинок (Kringel с соавторами), в отличие от способа, предлагаемого в настоящем изобретении, не позволяет сделать никакого заключения о жизнеспособности или активности личинок. Оценка коэффициента вылупляемости in vitro путем моделирования прохождения через желудочно-кишечный тракт представляется существенно более сложным и подверженным воздействию случайных факторов методом по сравнению с предлагаемым в настоящем изобретении способом. Кроме того, для теста по оценке вылупляемости in vitro не опубликованы данные о корреляции с активностью in vivo.
В предпочтительном варианте осуществления изобретения оценивают состояние эмбрионирования. Балл активности in vitro дает значимые результаты, которые можно использовать для предсказания ситуации in vivo, только в том случае, когда личинки полностью прошли через стадию эмбрионального развития. Не полностью развитые личинки в определенных условиях дают позитивную реакцию в тесте на индуцируемую температурой способность к активации, однако они не являются функциональными и не обладают способностью вылупляться в хозяине и развиваться согласно нормальному жизненному циклу личинки. Согласно известным к настоящему времени методам полное эмбриональное развитие оценивают с помощью микроскопа на основе морфологических критериев. Такая оценка требует наличия большого опыта, она является ненадежной и по своей природе ее можно осуществлять только на индивидуальных яйцах. В отличие от этого способ, предлагаемый в настоящем изобретении, позволяет получать объективную оценку количества копий генома в большой популяции яиц, на основе чего можно рассчитывать среднее значение соматических клеток на одну личинку.
Результаты теста на индуцируемую температурой способность к активации предпочтительно подтверждают с помощью нового теста in vivo, который позволяет надежно определять коэффициент вылупляемости в физиологических условиях с помощью простой и быстрой процедуры. При создании изобретения неожиданно было установлено, что в кишечном тракте имеет место поддающееся количественной оценке вылупление личинок из биологически активных яиц, в то время как предварительно инактивированные яйца проходят через кишечник в неизмененном виде. Согласно новому разработанному способу долю активных TSO в препарате оценивают косвенным путем, для чего подсчитывают только неактивные яйца, прошедшие без изменений через кишечный тракт лабораторного животного. Однако количественную оценку можно успешно осуществлять только в том случае, когда используют новый разработанный внутренний стандарт, представляющий собой флуоресцентномеченные неактивные яйца, которые также проходят без изменений через кишечный тракт. Кроме того, был установлен удивительный и неожиданный факт, что поддающееся количественной оценке вылупление биологически активных TSO может иметь место также в кишечнике кролика. Таким образом, существует животное, которое можно использовать в качестве животной модели вместо свиньи, содержание которого требует меньших затрат и является более простым и которое имеет существенно меньший объем кишечника, что делает более легким выделение микроскопически малых яиц.
Предлагаемый в изобретении тест in vivo отличается от известного теста по оценке инфективности, проводимого на свиньях. Оценка коэффициента вылупляемости занимает всего лишь 1-3 дня, в то время как продолжительность теста по оценке инфективности составляет несколько недель. Кроме того, при оценке коэффициента вылупляемости анализируют только первую стадию жизненного цикла T.suis в хозяине, которая слабо зависит от индивидуальных факторов организма хозяина. В противоположность этому коэффициент заражения на свиньях предпочтительно оценивают через 3-4 недели после заражения, и он подвержен естественной биологической вариабельности, которая объясняется, прежде всего, тем, что каждый хозяин имеет отличную от других индивидуальную сформированную иммунную систему.
Еще одним преимуществом представленного в настоящем описании анализа является то, что яйца можно выделять из фекалий животных и поэтому для осуществления теста не требуется убивать животных.
Примеры
Пример 1:
На основе последовательности ITS2 Trichuris suis был разработан метод количественной ПЦР с использованием TaqMan-системы. Последовательность-мишень, праймер и Taqman-зонд проиллюстрированы ниже. Последовательность-мишень выбирали таким образом, чтобы она позволяла отличать Trichuris suis от других видов Trichuris, последовательность которых также известна. Тем самым тест помимо того, что он дает информацию о количестве копий ITS2, обеспечивает также качественное доказательство того, что анализируемый организм действительно представляет собой Trichuris suis.
Для приготовления образца яйца (1000 яиц в 500 мкл раствора) гомогенизировали в гомогенизаторе Поттера (объем 2 мл; ширина зазора 0,01-0,03 мм) в течение 10 мин. Путем исследования под микроскопом неожиданно было установлено, что с помощью этого метода оказывается возможным полностью разрушать оболочки яиц, в то время как использование других методов, обычно применяемых при выделении генов, для разрушения яиц Trichuris suis оказывалось безуспешным. После добавления 1,54 мкг ДНК спермы рыбы гомогенат яиц подвергали взаимодействию с поступающим в продажу набором для выделения генов (набор DNeasy для выделения из крови и ткани; фирма Qiagen) согласно инструкциям производителя. С использованием изолята ДНК (общий объем 50 мкл) проводили ПЦР согласно протоколу, представленному ниже в таблице.
Эффективность амплификации, обеспечиваемая данным методом анализа, составляет от 92 до 98%, а чувствительность достаточно велика для того, чтобы было можно определить количество копий ITS2-гена в одном яйце.
С помощью метода ПЦР анализировали количество ITS2-копий в неэмбрионированных и полностью эмбрионированных яйцах Trichuris suis. Результаты представлены в приведенной ниже таблице.
ПЦР-анализ показал, что количество копий ITS2 в эмбрионированных TSO примерно в 1000 выше, чем количество копий ITS2 в неэмбрионированных TSO. Поскольку неэмбрионированное яйцо (стадия L0) можно рассматривать как индивидуальную клетку, то это означает, что личинка T.suis, находящаяся на стадии L1, содержит примерно 1000 соматических клеток. До настоящего времени для T.suis количество соматических клеток в личинке, находящейся на стадии L1, не было известно. Для родственного организма C.elegans было установлено, что количество соматических клеток составляет 959, что хорошо согласуется с установленным при создании настоящего изобретения количеством клеток, составляющим примерно 1000.
Для проверки того, можно ли с помощью рассматриваемого способа тестирования анализировать другие стадии развития личинки, проводили эмбрионирование TSO в течение 4 недель и при этом регулярно осуществляли ПЦР-анализ ITS2 (таблица 5).
Данный пример убедительно демонстрирует, что способ можно применять для характеризации различных стадий развития личинки от L0 до L1 на основе количества соматических клеток. Следовательно, данный способ пригоден прежде всего для осуществления анализа процесса эмбрионирования в режиме сопровождения. При производстве препаратов яиц гельминтов, по-видимому, наиболее предпочтительным является процесс, при котором в течение первых 28 дней эмбрионирования относительное количество копий достигает 50-1200.
Пример 2:
Для обнаружения аденозинтрифосфата можно применять поступающие в продажу наборы, которые помимо прочего содержат необходимые для осуществления реакции люциферазы, а также применяемый в качестве субстрата люциферин и соли магния. Необходимо осуществлять стимуляцию образования аденозинтрифосфата путем предварительной инкубации эмбрионированных яиц. Содержание аденозинтрифосфата в жизнеспособных эмбрионированных яйцах, находящихся в состоянии покоя, не позволяет достаточно надежно отличать их от мертвых яиц (фиг.3). Установлено, что способ позволяет это осуществлять в том случае, когда яйца инкубируют в течение периода времени, составляющего 19 ч, при 37°С. Существенное преимущество разработанного способа анализа заключается в том, что для такой инкубации не требуется приготовление образца. Можно непосредственно использовать предназначенные для анализа суспензии яиц вне зависимости от качественного и количественного состава среды. Можно применять даже сильнокислые среды, имеющие значения рН <2, без приготовления образца, и они не оказывают отрицательного воздействия на стимуляцию аденозинтрифосфата в яйцах. Благодаря применению инкубации оказывается возможным однозначно различать живые и мертвые личинки, находящиеся на стадии F1 (фиг.3).
Пример 3:
С использованием праймеров, выведенных из последовательности белка теплового шока hsp70, которая была обнаружена в Caenorhabditis elegans, и гомологичных последовательностей из Trichuris muris и Trichuris vulpis, оказалось возможным амплифицировать последовательность гена в РНК-изоляте из Trichuris suis. Новая последовательность из Trichuris suis после трансляции в белковую последовательность на 65% совпадала с белковой последовательностью белка hsp70 из С.elegans (фиг.5).
Для предполагаемого белка теплового шока из Т.suis был разработан метод количественной ОТ-ПЦР. Количественную оценку осуществляли путем сравнения с конститутивно экспрессируемой 188-рРНК. Активные эмбрионированные яйца Т.suis подвергали тепловому шоку (20 мин при 45°С) и экспрессию белка теплового шока сравнивали с экспрессией в необработанных активных яйцах Т.suis.
Анализ относительных уровней экспрессии четко продемонстрировал, что с помощью теплового шока можно индуцировать экспрессию мРНК, сходной с мРНК hsp70 в T.suis (таблица 6). Следовательно, в тесте по оценке активности можно применять указанный ген, экспрессию которого можно эффективно индуцировать в соответствующих экспериментальных условиях.
Обнаружение активной генной экспрессии в индивидуальном яйце осуществляют с помощью метода флуоресцентной in situ гибридизации (FISH). Для обнаружения предполагаемого белка теплового шока создавали несущий флуоресцентную метку ДНК-зонд, пример которого представлен на фиг.6 (SEQ ID NO:7).
После введения флуоресцентной метки в активные яйца популяцию яиц можно анализировать также с помощью проточной цитометрии. Преимущество этого подхода по сравнению с исследованием с помощью микроскопа заключается в том, что данный анализ требует для своего осуществления меньше времени и дает более объективные результаты.
Данный пример убедительно демонстрирует, что представленный в настоящем описании способ позволяет анализировать индукцию генной экспрессии в яйцах (индивидуальных) гельминтов. Таким образом, впервые создан способ, с помощью которого можно анализировать способность к активации личинок гельминтов, находящихся в состоянии покоя на стадии L1.
Если для проверки биологической активности препаратов яиц гельминтов используют индукцию генной экспрессии, то в качестве критерия приемлемого качества препарата можно считать наличие 25-100% позитивных яиц, предпочтительно 50-100% позитивных яиц гельминтов.
Пример 4:
4.1 Зависимость коэффициента подвижности от условий анализа
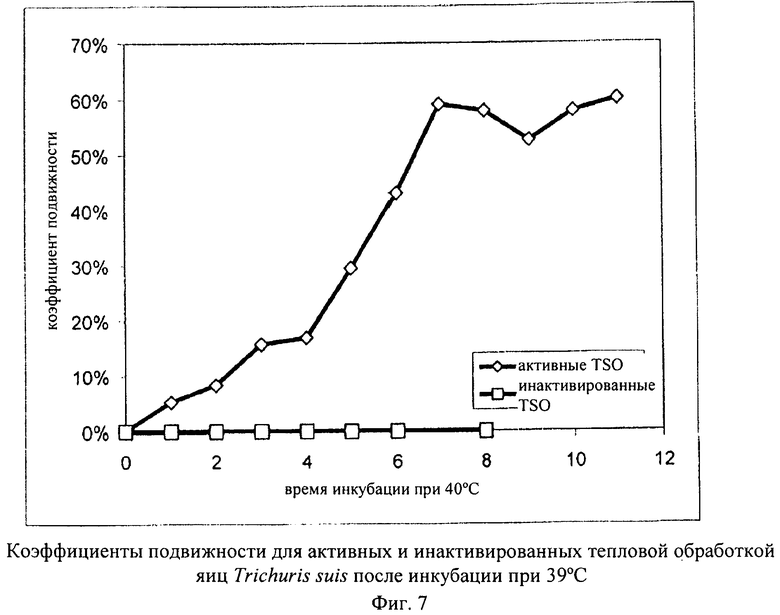
Активные яйца Trichuris suis и яйца Trichuris suis, инактивированные путем инкубации при 48°С в течение 72 ч, инкубировали в течение 11 ч при 39°С. Для каждого варианта через каждый час проводили в течение 5 мин обследование яиц и определяли коэффициент подвижности. На фиг.7 представлены результаты, характеризующие изменение коэффициента подвижности с течением времени.
Данный пример убедительно демонстрирует, что для активации личинок из состояния покоя и возможности измерения постоянного высокого коэффициента подвижности необходима относительно продолжительная предварительная инкубация при повышенных температурах (в данном случае: 7 ч при 39°С). Этот результат оказался неожиданным, поскольку до настоящего времени полностью эмбрионированные личинки на стадии L1, находящиеся в яйце, классифицировали как неподвижные (Parasitology, 67, 1973, cc.253-262). По-видимому, то, что подвижность личинки в яйце не была обнаружена, объясняется тем, что для этого требуется продолжительная предварительная инкубация и то, что медленные движения можно выявить только с помощью замедленной киносъемки.
Сравнение активных и неактивированных яиц однозначно свидетельствует о том, что анализ подвижности представляет собой простой путь, с помощью которого можно различать биологически активные и неактивные яйца Trichuris suis.
Наряду с фазой активации на измеряемый коэффициент подвижности оказывает влияние продолжительность периода наблюдения (длина «окна наблюдения»), поскольку личинки двигаются несинхронно и, следовательно, удлинение «окна наблюдения» повышает вероятность обнаружения спорадических движений в живом яйце. Об этом свидетельствуют результаты анализа (фиг.7), в котором осуществляли наблюдение за препаратом Trichuris suis после фазы активации (8 ч при 37°С) в течение промежутков времени различной продолжительности при температуре от 39,5 до 40°С. Результаты анализа продемонстрировали также, что подвижность остается стабильной в течение всего периода наблюдения, составлявшего 8 ч.
Таким образом, фаза активации при строго контролируемой температуре и длинное «окно наблюдения» представляют собой характерные и новые отличительные признаки способа, предлагаемого в настоящем изобретении. В примере 4 продемонстрировано, что у личинок, находящихся в покое на стадии L1, которые еще находятся в яйце, можно индуцировать и оценивать движения в соответствующих температурных условиях и условиях «окна наблюдения». С помощью рассматриваемого способа можно простым путем обнаруживать подвижность, являющуюся предпосылкой для вылупления, и, следовательно, представляющую собой параметр биологической активности яиц, и однозначно различать активные и неактивные личинки, находящиеся на стадии L1.
4.2 Предпочтительные условия тестирования
Установлено, что предпочтительными являются следующие условия:
образец, содержащий 15000 яиц гельминтов в 300 мкл фосфатного буфера, рН 7,4 переносят в 96-луночный планшет (площадь дна: 0,31 см). При такой плотности посева в поле зрения микроскопа при 200-кратном увеличении попадают примерно 80-150 яиц. После инкубации в течение 8 ч при 37°С (фаза активации) температуру повышают до 39,5°С и последовательно выбирают 4 поля наблюдения, каждое из которых обследуют в течение 2 ч. При этом на пленку регистрируют по 3 изображения в минуту. После этого определяют количество яиц и количество двигающихся личинок и рассчитывают коэффициент подвижности. Коэффициент подвижности для образца рассчитывают в виде среднего значения по 4 индивидуальным измерениям.
4.3 Воспроизводимость и точность - сравнение с тестом по оценке инфективности
Для оценки воспроизводимости теста по оценке подвижности в описанных выше условиях анализировали 4 аналогичных образца из партии Trichuris suis в 4 различных дня. Результаты измерений представлены в приведенных ниже таблицах. Для сравнения представлены результаты 4 серий измерений с помощью теста по оценке инфективности, которые были проведены с той же партией Trichuris suis.
Коэффициенты подвижности, полученные в 4 независимых сериях измерений, отличались друг от друга максимум на 4,2%, в отличие от этого максимальная разница между коэффициентами инфективности, полученными в 4 независимых сериях измерений, составляла 35,5%. Точность индивидуальных измерений в тесте по оценке подвижности составляла от 1,9 до 6,3%, в отличие от этого в тесте по оценке инфективности она составляла от 9,6 до 58,4%. Таким образом, тест по оценке подвижности превосходит тест по оценке инфективности как с точки зрения воспроизводимости, так и с точки зрения точности.
4.4. Корреляция между подвижностью и биологической активностью (достоверность) - сравнение с тестом по оценке инфективности
Для проверки корреляции между подвижностью и биологической активностью приготавливали смеси, содержащие активные TSO и инактивированные тепловой обработкой TSO, и анализировали с помощью теста по оценке подвижности в условиях, описанных в разделе 4.2. Тепловую инактивацию осуществляли путем выдерживания яиц при температуре 48°С в течение 72 ч. Для получения объективных результатов образцы перед проведением измерений зашифровывали.
Относительную биологическую активность рассчитывали в виде частного, полученного делением измеренного коэффициента подвижности для образца на определенный перед этим коэффициент подвижности для активных TSO. Достоверность измерения определяли путем сравнения относительной биологической активности и имеющей место в действительности доли активных яиц в образце.
Аналогичную серию измерений с использованием TSO из той же самой партии проводили с помощью теста по оценке инфективности. Лабораторных животных заражали различными количествами активных TSO с известным коэффициентом инфективности. Инактивированные яйца в смесь не добавляли. Результаты проиллюстрированы в приведенной ниже таблице.
С помощью проиллюстрированного в настоящем описании теста по оценке подвижности можно корректно определять относительную биологическую активность образца TSO с погрешностью <10% от ожидаемой величины во всем ее диапазоне от 0 до 100%. Точность измерения (разброс измеренных величин) составляла менее 5% в диапазоне от 50 до 100% относительной биологической активности. При определении более низких величин относительной биологической активности точность составляла менее 15%.
Результаты теста по оценке инфективности находились в пределах примерно 25-32% от ожидаемой величины в диапазоне от 0 до 100% относительной биологической активности. Разброс измеренных величин составлял от 28 до 80%.
Данный пример убедительно демонстрирует, что коэффициент подвижности линейно коррелирует с уровнем относительной биологической активности. Тест по оценке подвижности существенно превосходит тест по оценке инфективности с точки зрения достоверности и точности.
Тест по оценке подвижности можно применять для определения биологической активности препаратов яиц гельминтов, когда она находится в диапазоне от 30 до 100%, предпочтительно от 60 до 100%.
Пример 5:
В качестве примера кроликам вводили оральным путем различные смеси эмбрионированных, неэмбрионированных и инактивированных эмбрионированных яиц. Через 8 ч содержимое кишечника лабораторных животных анализировали с помощью микроскопа и подсчитывали количества интактных яиц и пустых оболочек яиц. Поскольку в данном опыте яйца не были флуоресцентномеченными, то коэффициент вылупляемости рассчитывали на основе соотношения количеств интактных эмбрионированных и неэмбрионированных яиц (с использованием приведенной выше формулы, в которой [IE] = количество эмбрионированных яиц и [IS] = количество неэмбрионированных яиц). В приведенной ниже таблице представлены результаты опыта, а также рассчитанные на их основе коэффициенты вылупляемости:
Сравнение данных, полученных для групп 1-3, продемонстрировало, что в организме кролика может происходить вылупление личинок только из активных эмбрионированных яиц (о чем свидетельствует выраженное уменьшение количества интактных эмбрионированных яиц и присутствие большого количества пустых оболочек яиц). В отличие от этого инактивированные эмбрионированные яйца сохранялись в кишечнике в неизмененном состоянии (группа 2). Это является убедительным доказательством того, что предлагаемый в изобретении способ позволяет отличать биологически активные и инактивированные яйца. Кроме того, установлено, что неэмбрионированные яйца также остаются интактными после прохождения через кишечник (группа 3). Это дает возможность применять неэмбрионированные яйца, которые проходят через кишечник лабораторных животных без расщепления, в качестве внутреннего стандарта. Такой внутренний стандарт необходим, поскольку из содержимого кишечника выделяют только часть яиц, и, следовательно, абсолютное количество выделенных яиц не является информативным. Для расчета коэффициента вылупляемости используют приведенную выше формулу с учетом внутреннего стандарта. Расчет по этой формуле показал, что коэффициент вылупляемости для проанализированных в данном опыте яиц Trichuris suis составлял 67,9%, в то время как коэффициент вылупляемости для инактивированных яиц Trichuris suis был равен 0% (строго математически: -12,6%).
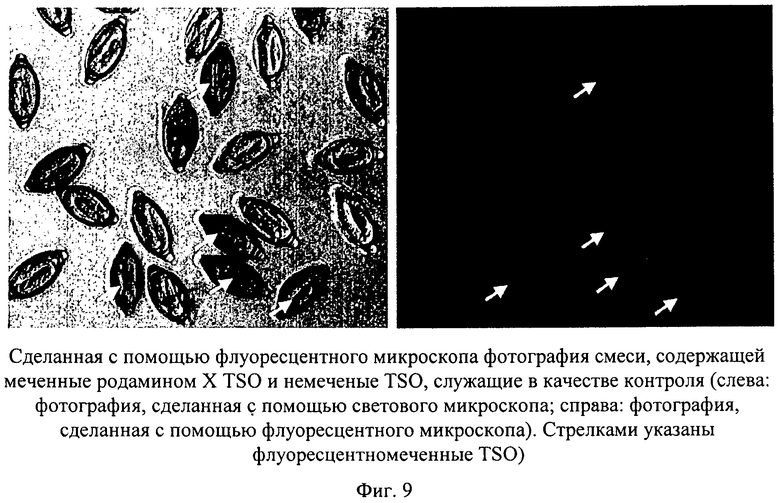
Для обеспечения более эффективного выявления и осуществления более четкой дифференциации яиц в образце и яиц, служащих в качестве внутреннего стандарта, осуществляли ковалентное мечение яиц Trichuris suis флуоресцентным зондом. Для примера в качестве флуоресцентного зонда использовали сукцинимидиловый эфир родамина X, который согласно спецификации производителя может взаимодействовать с первичными аминогруппами белков. 20 мкл 1М раствора бикарбоната натрия добавляли к суспензии, содержащей 1000 яиц Trichuris suis в 200 мкл фосфатного буфера, рН 7,4, и затем осуществляли взаимодействие с 5 мкл 0,5%-ного раствора сукцинимидилового эфира родамина Х в ДМСО. Раствор инкубировали в течение 1 ч при комнатной температуре при встряхивании. Затем яйца очищали, осуществляя в общей сложности 8 раз центрифугирование (10 мин при 500 об/мин) и замену супернатантного буфера. С помощью флуоресцентного микроскопа было четко выявлено окрашивание в красный цвет оболочек яиц (фиг.9).
Флуоресцентные метки можно с успехом применять для оценки с помощью микроскопа коэффициента вылупляемости. Это позволяет также осуществлять объективный и быстрый анализ методом проточной цитометрии с возбуждением флуоресценции.
В целом данный пример убедительно демонстрирует, что с помощью способа, предлагаемого в настоящем изобретении, можно количественно анализировать фазу вылупления и применять его для оценки биологической активности. Оценку биологической активности препаратов яиц гельминтов можно осуществлять на основе коэффициентов вылупляемости, когда они находятся в диапазоне примерно 25-100%.
Пример 6:
Корреляция индуцируемой температурой способности к активации in vitro с биологической активностью in vivo
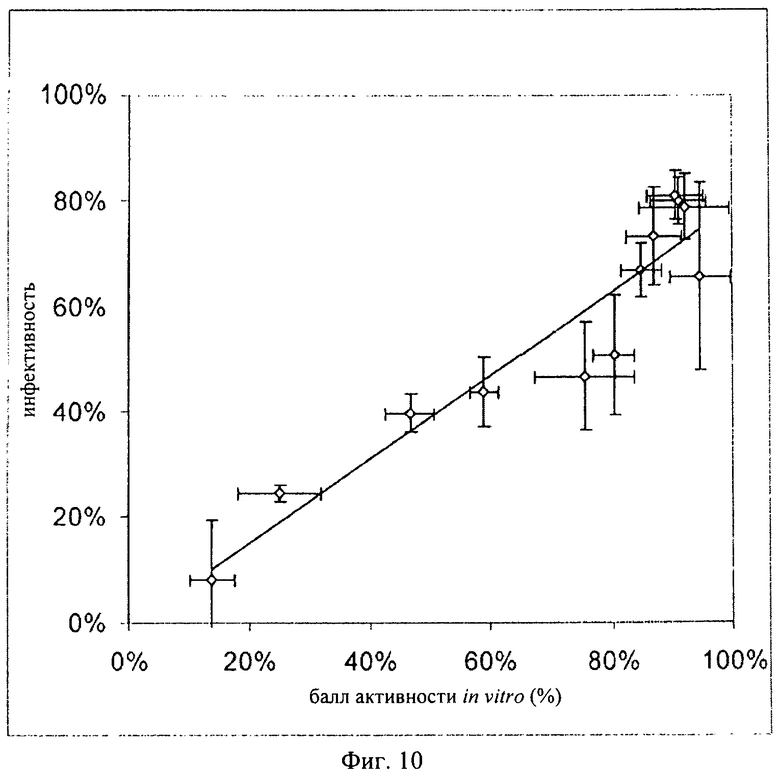
Для определения корреляции индуцируемой температурой способности к активации с биологической активностью in vivo анализировали 12 образцов TSO различного качества. Прежде всего, образцы анализировали в отношении полноты эмбрионального развития с помощью метода ПЦР. Затем после осуществления индуцируемой температурой активации определяли коэффициент подвижности и содержание АТФ. Параллельно для каждого из 12 образцов осуществляли тест по оценке инфективности на свиньях. Тест по оценке инфективности осуществляли каждый раз на 5 свиньях согласно методу, описанному Kringel с соавторами. Два определенных in vitro параметра, а именно коэффициент подвижности и содержание АТФ, объединяли в один балл активности in vitro:
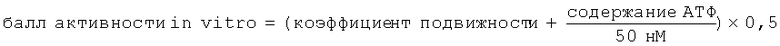
Корреляция между баллом активности in vitro и инфективностью продемонстрирована на фиг.10.
Данный пример демонстрирует хорошую корреляцию между баллом активности in vitro и биологической активностью in vivo. Небольшие отклонения для индивидуальных образцов, по-видимому, обусловлены скорее недостатками теста in vivo, которому присущи погрешности, обусловленные биологической вариабельностью (факторы, связанные с хозяином).
Анализ образца с помощью теста по оценке инфективности занимает 6 недель, требует использования 5 свиней и является очень трудоемким. Представленный в настоящем описании тест in vitro, заключающийся в осуществлении трех из пяти предлагаемых в настоящем изобретении анализов, занимает 1-2 дня, является сравнительно нетрудоемким и требует сравнительно небольших материальных затрат.





Изобретение касается способа определения биологической активности эмбрионированных яиц Trichuris. Охарактеризованный способ включает осуществление по меньшей мере 3-х анализов, выбранных из:
- оценки и/или подтверждения стадии эмбрионального развития яиц с помощью метода количественной ПЦР с использованием пригодных маркерных последовательностей для определения количества копий геномной ДНК,
- оценки метаболической активности эмбрионированных яиц с помощью биохимических и/или молекулярно-биологических методов,
- оценки индуцибельности генной экспрессии в эмбрионированных яйцах,
- оценки подвижности личинок Trichurs с помощью микроскопа в течение продолжительных периодов наблюдения после предварительной инкубации при повышенных температурах и/или
- оценки коэффициента вылупляемости личинок Trichuris в организме лабораторного животного. Представленное изобретение может быть использовано при получении безопасных фармацевтических продуктов, терапевтически эффективных при применении у человека. 14 з.п. ф-лы, 10 ил., 11 табл., 6 пр.
1. Способ определения биологической активности эмбрионированных яиц Trichuris для обеспечения контролируемого получения терапевтически эффективных фармацевтических продуктов и таким образом гарантии их безопасности и терапевтически эффективного применения для человека, в котором осуществляют по меньшей мере 3 из следующих анализов:
а) оценку и/или подтверждение стадии эмбрионального развития яиц Trichuris с помощью метода количественной ПЦР с использованием пригодных маркерных последовательностей для определения количества копий геномной ДНК,
б) оценку метаболической активности эмбрионированных яиц Trichuris с помощью биохимических и/или молекулярно-биологических методов,
в) оценку индуцибельности генной экспрессии в эмбрионированных яйцах Trichuris,
г) оценку с помощью микроскопа подвижности личинок Trichuris, включая личинки находящихся на стадии покоя в яйце, в течение продолжительных периодов наблюдения после предварительной инкубации при повышенных температурах и/или
д) оценку коэффициента вылупляемости личинок Trichuris в организме лабораторного животного, при этом осуществляют количественное сравнение интактных эмбрионированных яиц, выделенных из фекалий, с внутренним стандартом.
2. Способ по п.1а), в котором с помощью метода количественной ПЦР с использованием пригодных специфических для Trichuris suis последовательностей оценивают количество копий геномной ДНК.
3. Способ по п.1б), отличающийся тем, что для оценки метаболической активности эмбрионированных яиц Trichuris измеряют содержание АТФ.
4. Способ по п.3, отличающийся тем, что перед осуществлением измерений люминесценции осуществляют предварительную инкубацию яиц Trichuris в следующих условиях:
аа) при температуре от 36 до 42°С,
бб) в течение промежутка времени, составляющего от 2 до 30 ч,
вв) в суспензионной среде, имеющей значение pH от 0,1 до 3.
5. Способ по п.1б), отличающийся тем, что яйца Trichuris сначала обрабатывают средством для предварительной обработки, выбранным из хлорноватистой кислоты, хитиназы и/или протеазы, и затем окрашивают солями тетразолия.
6. Способ по п.1в), отличающийся тем, что оценивают индуцибельность белка теплового шока.
7. Способ по п.1в), отличающийся тем, что обнаружение экспрессии осуществляют на основе гибридизации с флуоресцентномеченным нуклеотидным зондом.
8. Способ по п.1в), отличающийся тем, что гибридизацию обнаруживают с помощью проточной цитометрии.
9. Способ по п.1г), отличающийся тем, что подвижность личинок гельминтов, находящихся в яйце, оценивают с помощью микроскопа в течение периодов времени, составляющих от 2 мин до 8 ч, с использованием замедленной киносъемки.
10. Способ по п.1д), отличающийся тем, что подлежащие анализу яйца гельминтов маркируют флуоресцентными зондами и внутренние стандарты маркируют имеющими другую окраску флуоресцентными зондами.
11. Способ по п.1д), отличающийся тем, что в качестве системы для анализа применяют содержимое кишечников кроликов и/или свиней.
12. Способ по п.1д), отличающийся тем, что в качестве внутреннего стандарта для определения коэффициента вылупляемости применяют неэмбрионированные или инактивированные яйца Trichuris.
13. Способ по любому из пп.1-12, отличающийся тем, что осуществляют по меньшей мере четыре анализа, выбранные из анализов, указанных в 1а), 1б), 1в), 1г) и/или 1д).
14. Способ по любому из пп.1-13, отличающийся тем, что перед осуществлением оценки яйца Trichuris подвергают предварительной инкубации в течение периода времени, составляющего от по меньшей мере 30 мин до 24 ч, в строго стандартизованных условиях, причем предварительная инкубация при необходимости включает также регулирование имеющих существенное значение параметров, прежде всего температуры.
15. Способ по любому из пп.1-14, отличающийся тем, что по меньшей мере 50% яиц Trichuris suis являются биологически активными.
| WO 2007134761 A1, 29.11.2007 | |||
| WO 2007076868 A2, 12.07.2007 | |||
| RU 2008122346 A, 10.02.2010 |

