Заявка испрашивает приоритет предварительной заявки США №61/177650, поданной 12 мая 2009 г., и предварительной заявки США №61/244638, поданной 22 сентября 2009 г., которые обе включены во всей своей полноте посредством ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам и их фрагментам, которые связываются с Dkk-1 и, в частности, к гуманизированным антителам и их фрагментам, которые связываются с Dkk-1, и, еще более конкретно, к полностью гуманизированным антителам и фрагментам, которые специфически связываются с Dkk-1, в частности, Dkk-1 человека. Предложены нуклеиновые кислоты, кодирующие антитела против Dkk-1 или их фрагменты, а также экспрессирующие векторы и клетки-хозяева, инкорпорирующие эти нуклеиновые кислоты, для осуществления рекомбинантной экспрессии антител против Dkk-1. Также предложены анаболические агенты костной ткани. Также предложены фармацевтические композиции, содержащие антитела или их фрагменты по изобретению. Кроме того, предложены способы лечения заболеваний, состояний и расстройств, таких как поражения костей, которые вызывают уменьшение костной массы. Также предложены способы лечения или предупреждения уменьшения костной массы, способы индуцирования увеличения костной массы и активности Wnt.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Wnt представляют собой секретируемые гликопротеины, которые связываются с рецепторным комплексом, включающим белок, родственный рецептору липопротеинов низкой плотности (LRP5/6), и frizzled белки, и активируют его. Cadigan, K.M. and Y.I. Liu (2005) Journal of Cell Science, 119, 395-402; Nusse, R. (2003) Development, 130(22): 5297-305; и Pinson, K.I. (2000) Nature, 2000, 407(6803): 535-8.
Описано, что Wnt/LRP5 регулирует костную массу, и что активация пути передачи сигнала через Wnt вызывает нарастание костной массы. Boyden, L.M. et al. (2002) N. Engl. J. Med. 346: 1513-1521; Little, R.D. et al. (2002) Am. J. Hum. Genet. 70: 11-19; и Gong, Y. et al. (2001) Cell, 107: 513-523. Передача сигнала через Wnt тесно связана с регуляцией антагонистами, которые включают такие секретируемые молекулы, как Dickkopf 1 (Dkk-1). Tian, E. et al. (2003) N. Engl. J. Med. 349: 2483-2494. Было обнаружено, что фенотип высокой костной массы (НВМ), наблюдаемый у людей, обусловлен наличием единственной точечной мутации в LRP5 (G171V), которая ингибирует способность Dkk-1 связываться с LRP5. Zhang, Y. et al. (2004) Mol. Cell Biol. 24(11): 4677-84.
Решающая роль передачи сигнала через Wnt в процессах образования и роста костей делает Dkk-1 полезной мишенью для лечения заболеваний или состояний, при которых пациент может получить пользу от повышенной остеобластной активности (увеличенной плотности костной массы, усиленного костеобразования без соответствующего повышения резорбции кости), в том числе, например, путем снижения количества переломов, которые происходят, например, в результате нелеченого остеопороза. В WO2006/015373 (US2006/0127393) описаны антитела к Dkk-1 для лечения различных заболеваний, включая нарушения костной ткани. В US2008/0193449 описываются различные антитела, специфичные к Dkk-1, которые ингибируют связывание Dkk-1 с LRP5, композиции, содержащие такие антитела, для стимуляции роста костей и композиции, содержащие такие антитела, для лечения таких нарушений костной ткани, как остеопороз.
Сохраняется необходимость в новых анаболических агентах костной ткани, которые антагонизируют активность Dkk-1, тем самым увеличивая остеобластную активность, для лечения заболеваний, состояний и расстройств, таких как остеопороз, при которых пациент может получить пользу от такого усиленного костеобразования.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам и их фрагментам (например, антигенсвязывающим участкам), которые связываются с Dkk-1, и, в частности, к гуманизированным антителам и их фрагментам, которые связываются с Dkk-1, и, еще более конкретно, к полностью гуманизированным антителам и иммунологически функциональным фрагментам, которые связываются с Dkk-1. Такие антитела и их фрагменты антагонизируют способность Dkk-1 ингибировать активность Wnt. Антитела и их иммунологически функциональные фрагменты антагонизируют способность Dkk-1 ингибировать путь передачи сигнала через Wnt в кости с соответствующим увеличением костной массы. Антитела и их иммунологически функциональные фрагменты включают антитела со структурой, существующей в природе, а также полипептиды, которые имеют антигенсвязывающий домен (например, однодоменное антитело). Антитела и их иммунологически функциональные фрагменты могут быть использованы для лечения разнообразных заболеваний, состояний и расстройств, в том числе связанные с уменьшением костной массы, такие как остеопороз. Антитела и их иммунологически функциональные фрагменты также могут быть использованы для лечения заболеваний, состояний или расстройств, связанных с вторичной потерей костной массы, таких как потеря костной массы, обусловленная, например, длительным лечением кортикостероидами, ингибиторами ароматазы или тиазолидинтионами (TZD) или сопутствующая ему; анорексия; или потеря костной массы, связанная с гипогонадизмом или нефрогенной остеодистрофией.
Некоторые из предложенных антител и их функциональных фрагментов включают:
(а) один или более гипервариабельных участков (CDR) легкой цепи (LC), выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:22;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:24; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:26;
(б) один или более гипервариабельных участков (CDR) тяжелой цепи (НС), выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:30, 49 или 50;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:32 или 51; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:34 или 52; или
(в) один или более CDR из (а) и один или более CDR из (б).
Такие антитела и их иммунологически функциональные фрагменты могут специфически связываться с полипептидом Dkk-1. Некоторые из этих антител и их иммунологически функциональных фрагментов включают один, два, три, четыре, пять или все шесть упомянутых выше CDR. Также предложены консервативные модификации указанных последовательностей.
LC и НС других предложенных антител и их иммунологически функциональных фрагментов имеют по меньшей мере 90% идентичности последовательности по отношению к вышеупомянутым последовательностям.
Кроме того, предложены антитела и их иммунологически функциональные фрагменты, которые имеют LC, где CDR1 имеет аминокислотную последовательность, приведенную в SEQ ID NO:22, CDR2 имеет аминокислотную последовательность, приведенную в SEQ ID NO:24, и/или CDR3 имеет аминокислотную последовательность, приведенную в SEQ ID NO:26.
Кроме того, предложены антитела и их иммунологически функциональные фрагменты, которые имеют НС, где CDR1 имеет аминокислотную последовательность, приведенную в SEQ ID NO:30,49 или 50, CDR2 имеет аминокислотную последовательность, приведенную в SEQ ID NO:32 или 51, и/или CDR3 имеет аминокислотную последовательность, приведенную в SEQ ID NO:34 или 52.
Кроме того, предложены антитела и их иммунологически функциональные фрагменты, которые имеют LC, где CDR1 имеет аминокислотную последовательность, приведенную в SEQ ID NO:22, CDR2 имеет аминокислотную последовательность, приведенную в SEQ ID NO:24, и/или CDR3 имеет аминокислотную последовательность, приведенную в SEQ ID NO:26, и НС, где CDR1 имеет аминокислотную последовательность, приведенную в SEQ ID NO:30, 49 или 50, CDR2 имеет аминокислотную последовательность, приведенную в SEQ ID NO:32 или 51, и/или CDR3 имеет аминокислотную последовательность, приведенную в SEQ ID NO:34 или 52.
Кроме того, предложены антитела и их иммунологически функциональные фрагменты, которые имеют LC, где CDR1 имеет аминокислотную последовательность, приведенную в SEQ ID NO:22, CDR2 имеет аминокислотную последовательность, приведенную в SEQ ID NO:24, и CDR3 имеет аминокислотную последовательность, приведенную в SEQ ID NO:26, и НС, где CDR1 имеет аминокислотную последовательность, приведенную в SEQ ID NO:30, 49 или 50, CDR2 имеет аминокислотную последовательность, приведенную в SEQ ID NO:32 или 51, и CDR3 имеет аминокислотную последовательность, приведенную в SEQ ID NO:34 или 52.
Также предложены антитела и их иммунологически функциональные фрагменты, которые включают: (а) вариабельную область LC (VL), имеющую последовательность, по меньшей мере на 80% идентичную последовательности, приведенной в SEQ ID NO:20; (б) вариабельную область НС (VH), имеющую последовательность, по меньшей мере на 80% идентичную последовательности, приведенной в SEQ ID NO:28; или (в) VL из (а) и VH из (б).
Также предложены антитела и их иммунологически функциональные фрагменты, которые аналогичны по структуре, но VL имеет последовательность, по меньшей мере на 90% идентичную последовательности, приведенной в SEQ ID NO:20, и VH имеет последовательность, по меньшей мере на 90% идентичную последовательности, приведенной в SEQ ID NO:28.
Также предложены антитела и их иммунологически функциональные фрагменты, которые аналогичны по структуре, но VL имеет последовательность, по меньшей мере на 95% идентичную последовательности, приведенной в SEQ ID NO:20, и VH имеет последовательность, по меньшей мере на 95% идентичную последовательности, приведенной в SEQ ID NO:28.
Также предложены антитела и их иммунологически функциональные фрагменты, которые включают VL, которая имеет последовательность, приведенную в SEQ ID NO:20, а VH имеет последовательность, приведенную в SEQ ID NO:28.
Некоторые из предложенных антител и их иммунологически функциональных фрагментов имеют LC, которая содержит или состоит из аминокислотной последовательности, приведенной в SEQ ID NO:38 или SEQ ID NO:42, и/или НС, которая содержит или состоит из аминокислотной последовательности, приведенной в SEQ ID NO:36 или SEQ ID NO:40.
Некоторые из предложенных антител и их иммунологически функциональных фрагментов имеют LC, которая состоит из аминокислотной последовательности, приведенной в SEQ ID NO:38 или SEQ ID NO:42, и НС, которая состоит из аминокислотной последовательности, приведенной в SEQ ID NO:36 или SEQ ID NO:40.
Кроме того, предложены антитела и их иммунологически функциональные фрагменты, которые могут конкурировать с мышиным MabJC18 (Mab относится к моноклональному антителу) за связывание с Dkk-1+-клетками. В одном из воплощений согласно изобретению предложены гуманизированное антитело или его фрагмент, которые специфически связываются с антигеном Dkk-1 человека, где CDR вариабельной области LC (CDR1, CDR2 и CDR3) и CDR вариабельной области НС (CDR1, CDR2 и CDR3) имеют следующие аминокислотные последовательности, и где антитело или фрагмент могут конкурировать с мышиным MabJC18 за связывание с Dkk-1+-клетками: LC: (1) CDR1 (SEQ ID NO:22), (2) CDR2 (SEQ ID NO:24) и (3) CDR3 (SEQ ID NO:26); и НС: (1) CDR1 (SEQ ID NO:30, 49 или 50), (2) CDR2 (SEQ ID NO:32 или 51) и (3) CDR3 (SEQ ID NO:34 или 52).
Согласно изобретению также предложены гуманизированное антитело или его фрагмент, которые специфически связываются с антигеном Dkk-1 человека, где гипервариабельные участки (CDR1, CDR2 и CDR3) вариабельной области LC и гипервариабельные участки (CDR1, CDR2 и CDR3) вариабельной области НС имеют следующие аминокислотные последовательности: LC: (1) CDR1 (SEQ ID NO:22), (2) CDR2 (SEQ ID NO:24) и (3) CDR3 (SEQ ID NO:26); и НС: (1) CDR1 (SEQ ID NO:30, 49 или 50), (2) CDR2 (SEQ ID NO:32 или 51) и (3) CDR3 (SEQ ID NO:34 или 52), в которых каркас вариабельного домена LC содержит каркас зародышевой линии IGKV3-11 человека и область IGKJ4.
Согласно изобретению также предложены гуманизированное антитело или его фрагмент, которые специфически связываются с антигеном Dkk-1 человека, где гипервариабельные участки (CDR1, CDR2 и CDR3) вариабельной области LC и гипервариабельные участки (CDR1, CDR2 и CDR3) вариабельной области НС имеют следующие аминокислотные последовательности: LC: (1) CDR1 (SEQ ID NO:22), (2) CDR2 (SEQ ID NO:24) и (3) CDR3 (SEQ ID NO:26); и НС: (1) CDR1 (SEQ ID NO:30, 49 или 50), (2) CDR2 (SEQ ID NO:32 или 51) и (3) CDR3 (SEQ ID NO:34 или 52), в которых каркас вариабельного домена НС содержит каркас зародышевой линии IGHV3-07 человека (с единственной обратной мутацией: R100T) и область IGHJ6.
Согласно изобретению также предложены гуманизированное антитело или его фрагмент, которые специфически связываются с антигеном Dkk-1 человека, где гипервариабельные участки (CDR1, CDR2 и CDR3) вариабельной области LC и гипервариабельные участки (CDR1, CDR2 и CDR3) вариабельной области НС имеют следующие аминокислотные последовательности: LC: (1) CDR1 (SEQ ID NO:22), (2) CDR2 (SEQ ID NO:24) и (3) CDR3 (SEQ ID NO:26); и НС: (1) CDR1 (SEQ ID NO:30, 49 или 50), (2) CDR2 (SEQ ID NO:32 или 51) и (3) CDR3 (SEQ ID NO:34 или 52), в которых каркас вариабельного домена LC содержит каркас зародышевой линии IGKV3-11 человека и область IGKJ4, а каркас вариабельного домена НС содержит каркас зародышевой линии IGHV3-07 человека (с единственной обратной мутацией: R100T) и область IGHJ6.
Кроме того, предложены слитые белки, содержащие один или более фрагментов или участков антител по изобретению. В одном из воплощений предложен слитый полипептид, который содержит по меньшей мере 10 следующих друг за другом аминокислот вариабельной области LC, приведенной в SEQ ID NO:20, и/или по меньшей мере 10 аминокислот вариабельной области НС, приведенной в SEQ ID NO:28.
В другом воплощении слитый белок содержит вариабельную область LC, приведенную в SEQ ID NO:20, и/или вариабельную область НС, приведенную в SEQ ID NO:28. В другом воплощении слитый полипептид содержит одно или более чем одно из следующего:
(а) гипервариабельных участков (CDR) легкой цепи (LC), выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:22;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:24; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:26; или
(б) одного или более гипервариабельных участков (CDR) тяжелой цепи (НС), выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:30, 49 или 50;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:32 или 51; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:34 или 52.
В другом воплощении слитый белок содержит CDR3 НС (SEQ ID NO:34 или 52) и CDR3 LC (SEQ ID NO:26).
В другом воплощении слитый белок содержит одно или более антител по изобретению и аминокислотную последовательность, с которой оно не соединено в нативной молекуле, такую как, например, гетерологичная последовательность или гомологичная последовательность из другой области. В одном из воплощений гетерологичная последовательность представляет собой метку, выбранную из метки FLAG или метки 6His.
Также предложены антитела и их иммунологически функциональные фрагменты, конъюгированные с агентом, который обеспечивает связывание с твердой подложкой. В одном из воплощений антитела и их фрагменты связаны с агентом, который обеспечивает связывание с твердой подложкой. В одном из воплощений агентом, который обеспечивает связывание с твердой подложкой, является биотин или авидин.
Также предложены антитела и их иммунологически функциональные фрагменты, связанные с агентом мечения. В одном из воплощений агент мечения представляет собой флуоресцентную молекулу или радиоактивную молекулу.
Различные предложенные антитела и их иммунологически функциональные фрагменты могут включать единичную LC или НС либо единичный вариабельный домен легкой цепи и/или единичный вариабельный домен тяжелой цепи. Другие предложенные антитела и фрагменты могут включать две LC и/или две НС, при этом в некоторых воплощениях две LC идентичны друг другу; и/или при этом в некоторых воплощениях две НС идентичны друг другу. Предложенные антитела могут включать, например, моноклональные антитела, человеческое антитело, химерное антитело или гуманизированное антитело. Иммунологически функциональные фрагменты предложенных антител могут включать scFv, Fab, Fab', F(ab')2 или однодоменное антитело, но этим не ограничиваются. В одном из воплощений антитело диссоциирует из комплекса с полипептидом Dkk-1 с Kd примерно 100 пМ или меньше.
Также предложен ряд нуклеиновых кислот, кодирующих антитела и их фрагменты. Некоторые нуклеиновые кислоты, например, кодируют CDR LC с аминокислотной последовательностью, приведенной в SEQ ID NO:22, SEQ ID NO:24 или SEQ ID NO:26, так что кодируемый CDR кодирует антитело или его иммунологически функциональный фрагмент, которые могут специфически связываться с полипептидом Dkk-1. Некоторые нуклеиновые кислоты,. например, кодируют CDR НС с аминокислотной последовательностью, приведенной в SEQ ID NO:30, SEQ ID NO:32 или SEQ ID NO:34, так что кодируемый CDR кодирует антитело или его иммунологически функциональный фрагмент, которые могут специфически связываться с полипептидом Dkk-1. В некоторых воплощениях нуклеиновые кислоты содержат или состоят из последовательности, которая кодирует VL- и/или VH-область антитела или его иммунологически функционального фрагмента, где VL имеет последовательность, идентичную по меньшей мере на 70%, 80%, 90% или 95% последовательности, приведенной в SEQ ID NO:20, a VH имеет последовательность, идентичную по меньшей мере на 70%, 80%, 90% или 95% последовательности, приведенной в SEQ ID NO:28. В некоторых воплощениях нуклеиновые кислоты включают последовательность, которая кодирует VL, содержащую или состоящую из последовательности, приведенной в SEQ ID NO:20, и/или последовательность, которая кодирует VH, содержащую или состоящую из последовательности, приведенной в SEQ ID NO:28. В других воплощениях нуклеиновые кислоты включают последовательности, которые кодируют как VL, так и VH с вышеупомянутыми характеристиками для последовательностей.
Кроме того, предложены полинуклеотиды, кодирующие любое из следующего: (a) huMabJC18 или его варианты, показанные в Таблице 4; (б) фрагмент или участок антитела huMabJC18 или его вариантов, показанных в Таблице 4; (в) легкую цепь антитела huMabJC18 или его вариантов, показанных в Таблице 4; (г) тяжелую цепь антитела huMabJC18 или его вариантов, показанных в Таблице 4; (д) одну или более вариабельных областей из LC и/или НС антитела huMabJC18 или его вариантов, показанных в Таблице 4; (е) один или более CDR (один, два, три, четыре, пять или шесть CDR) антитела huMabJC18 или его вариантов, показанных в Таблице 4; (ж) CDR НЗ из НС антитела huMabJC18; (з) CDR H1 из НС антитела huMabJC18; (и) CDR H2 из НС антитела huMabJC18, где аминокислота G57 представляет собой G или W и/или F58 представляет собой F, L, G, Y, М или V, и/или Q59 представляет собой Q, D, H, G, R или W: (к) CDR Н3 из НС антитела huMabJC18, где аминокислота Т100 представляет собой Т или S, и/или аминокислота L102 представляет собой L или Y, и/или аминокислота Е103 представляет собой Е, R, Q, D или K; (л) CDR L1 из LC антитела huMabJC18, где аминокислота Е27 представляет собой Е, Q, и/или аминокислота D30 представляет собой D или S, и/или аминокислота D31 представляет собой D или S, и/или аминокислота F32 представляет собой F или S, и/или аминокислота G33 представляет собой G или Y, и/или аминокислота I34 представляет собой I или L, и/или аминокислота S35 представляет собой S или А, и/или аминокислота F36 представляет собой F или W, и/или аминокислота I37 представляет собой I или М; (м) CDR L2 из LC антитела huMabJC18, где аминокислота G55 представляет собой G или А, и/или аминокислота S56 представляет собой S или Т; (н) CDR L3 из LC антитела huMabJC18, где аминокислота Q94 представляет собой Q или Н, и/или аминокислота L95 представляет собой L, S, А или G, и/или аминокислота K96 представляет собой K, I, L, W, М или S, и/или аминокислота Е97 представляет собой Е или D, и/или аминокислота V98 представляет собой V или L, и/или аминокислота Р99 представляет собой Р или W, и/или аминокислота Р100 представляет собой Р, S или G, и/или аминокислота Т101 представляет собой Т, Y или L; (о) три CDR из LC антитела huMabJC18 или его вариантов, показанных в Таблице 4; (п) три CDR из НС антитела huMabJC18 или его вариантов, показанных в Таблице 4; (р) три CDR из LC и три CDR из НС антитела huMabJC18 или его вариантов, показанных в Таблице 4; и (с) антитело, содержащее любое из перечисленного от (б) до (с).
Также предложены полинуклеотиды, комплементарные любым таким нуклеиновокислотным последовательностям. Полинуклеотиды могут быть одноцепочечными (кодирующими или антисмысловыми) или двухцепочечными молекулами и могут представлять собой молекулы ДНК (геномной, кДНК или синтетической) или РНК. Молекулы РНК включают, например, молекулы гяРНК (гетерогенной ядерной РНК), которые содержат интроны и соответствуют молекуле ДНК по типу "один-к-одному" ("one-to-one"), и молекулы мРНК, которые не содержат интронов. Дополнительные кодирующие или некодирующие последовательности могут, но это не обязательно, находиться внутри предложенного полинуклеотида, и полинуклеотид может, но это не обязательно, быть связанным с другими молекулами и/или подложками.
Предложены полинуклеотиды, которые могут содержать нативную последовательность (то есть эндогенную последовательность, которая кодирует антитело или его часть) или могут содержать вариант такой последовательности. Полинуклеотидные варианты могут содержать одну или более замен, добавлений, делеций и/или вставок, которые не ослабляют иммунореактивности кодируемого полипептида относительно нативной иммунореактивной молекулы. Варианты предпочтительно демонстрируют по меньшей мере примерно 70% идентичности, более предпочтительно по меньшей мере примерно 80% идентичности, более предпочтительно по меньшей мере примерно 90% и наиболее предпочтительно примерно 95% идентичности с полинуклеотидной последовательностью, которая кодирует нативное антитело или его часть.
Согласно изобретению также предложена молекула ДНК, кодирующая аминокислотную последовательность гуманизированного антитела или его фрагмента, содержащих:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:22;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:24; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:26;
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:30, 49 или 50;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:32 или 51; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:34 или 52; или
(в) один или более CDR из (а) и один или более CDR из (б), в которых каркас вариабельного домена LC содержит каркас зародышевой линии IGKV3-11 человека и область IGKJ4.
Согласно изобретению также предложена молекула ДНК, кодирующая аминокислотную последовательность гуманизированного антитела или его фрагмента, содержащих:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:22;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:24; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:26;
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:30, 49 или 50;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:32 или 51; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:34 или 52; или
(в) один или более CDR из (а) и один или более CDR из (б), в которых каркас вариабельного домена НС содержит каркас зародышевой линии IGHV3-07 человека (с единственной обратной мутацией: R100T) и область IGHJ6.
Согласно изобретению также предложена молекула ДНК, кодирующая аминокислотную последовательность гуманизированного антитела или его фрагмента, содержащих:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:22;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:24; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:26;
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:30, 49 или 50;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:32 или 51; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:34 или 52; или
(в) один или более CDR из (а) и один или более CDR из (б), в которых каркас вариабельного домена LC содержит каркас зародышевой линии IGKV3-11 человека и область IGKJ4, а каркас вариабельного домена НС содержит каркас зародышевой линии IGHV3-07 человека (с единственной обратной мутацией: R100T) и область IGHJ6.
Согласно изобретению также предложена молекула ДНК, кодирующая аминокислотную последовательность гуманизированного антитела или его фрагмента, содержащих:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:22);
(2) CDR2 (SEQ ID NO:24); и
(3) CDR3 (SEQ ID NO:26);
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:30, 49 или 50);
(2) CDR2 (SEQ ID NO:32 или 51); и
(3) CDR3 (SEQ ID NO:34 или 52); или
(в) один или более CDR из (а) и один или более CDR из (б), в которых каркас вариабельного домена LC содержит каркас зародышевой линии IGKV3-11 человека и область IGKJ4, а каркас вариабельного домена НС содержит каркас зародышевой линии IGHV3-07 человека (с единственной обратной мутацией: R100T) и область IGHJ6.
В некоторых воплощениях молекул ДНК нуклеотидные последовательности CDR LC представляют собой последовательности, приведенные ниже:
CDR1: как приведено в SEQ ID NO:21,
CDR2: как приведено в SEQ ID NO:23,
CDR3: как приведено в SEQ ID NO:25.
В некоторых воплощениях молекул ДНК нуклеотидные последовательности CDR НС представляют собой последовательности, приведенные ниже:
CDR1: как приведено в SEQ ID NO:29,
CDR2: как приведено в SEQ ID NO:31,
CDR3: как приведено в SEQ ID NO:33.
В некоторых воплощениях молекул ДНК нуклеотидная последовательность вариабельной области LC приведена в SEQ ID NO:19.
В некоторых воплощениях молекул ДНК нуклеотидная последовательность вариабельной области НС приведена в SEQ ID NO:27.
В некоторых воплощениях молекул ДНК нуклеотидная последовательность LC приведена в SEQ ID NO:37 или SEQ ID NO:41.
В некоторых воплощениях молекул ДНК нуклеотидная последовательность НС приведена в SEQ ID NO:35 или SEQ ID NO:39.
Кроме того, предложены векторы, например, экспрессирующие векторы, содержащие любые из полинуклеотидных последовательностей или молекул ДНК, кодирующих антитела или их фрагменты.
В одном из воплощений предложена молекула ДНК, кодирующая аминокислотную последовательность гуманизированного антитела или его фрагмента, содержащих:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:22;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:24; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:26;
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:30, 49 или 50;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:32 или 51; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:34 или 52; или
(в) один или более CDR из (а) и один или более CDR из (б), в которых каркас вариабельного домена LC содержит каркас зародышевой линии IGKV3-11 человека и область IGKJ4, в форме экспрессирующего вектора.
В одном из воплощений предложена молекула ДНК, кодирующая аминокислотную последовательность гуманизированного антитела или его фрагмента, содержащих:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:22;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:24; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:26;
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:30, 49 или 50;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:32 или 51; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:34 или 52; или
(в) один или более CDR из (а) и один или более CDR из (б), в которых каркас вариабельного домена НС содержит каркас зародышевой линии IGHV3-07 человека (с единственной обратной мутацией: R100T) и область IGHJ6, в форме экспрессирующего вектора.
В одном из воплощений предложена молекула ДНК, кодирующая аминокислотную последовательность гуманизированного антитела или его фрагмента, содержащих:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:22;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:24; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:26;
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:30, 49 или 50;
(2) CDR2 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:32 или 51; и
(3) CDR3 с последовательностью, по меньшей мере на 80% идентичной SEQ ID NO:34 или 52; или
(в) один или более CDR из (а) и один или более CDR из (б), в которых каркас вариабельного домена LC содержит каркас зародышевой линии IGKV3-11 человека и область IGKJ4, а каркас вариабельного домена НС содержит каркас зародышевой линии IGHV3-07 человека (с единственной обратной мутацией: R100T) и область IGHJ6, в форме экспрессирующего вектора.
В одном из воплощений предложена молекула ДНК, кодирующая аминокислотную последовательность гуманизированного антитела или его фрагмента, содержащих:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:22);
(2) CDR2 (SEQ ID NO:24); и
(3) CDR3 (SEQ ID NO:26);
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:30, 49 или 50);
(2) CDR2 (SEQ ID NO:32 или 51); и
(3) CDR3 (SEQ ID NO:34 или 52); или
(в) один или более CDR из (а) и один или более CDR из (б), в которых каркас вариабельного домена LC содержит каркас зародышевой линии IGKV3-11 человека и область IGKJ4, а каркас вариабельного домена НС содержит каркас зародышевой линии IGHV3-07 человека (с единственной обратной мутацией: R100T) и область IGHJ6, в форме экспрессирующего вектора.
Также предложена клетка-хозяин, трансформированная экспрессирующим вектором по изобретению. В одном из воплощений предложена клетка-хозяин, трансформированная экспрессирующим вектором, содержащим молекулу ДНК, кодирующую аминокислотную последовательность гуманизированного антитела или его фрагмента, содержащих:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:22);
(2) CDR2 (SEQ ID NO:24); и
(3) CDR3 (SEQ ID NO:26);
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:30, 49 или 50);
(2) CDR2 (SEQ ID NO:32 или 51); и
(3) CDR3 (SEQ ID NO:34 или 52); или
(в) один или более CDR из (а) и один или более CDR из (б), в которых каркас вариабельного домена LC содержит каркас зародышевой линии IGKV3-11 человека и область IGKJ4, а каркас вариабельного домена НС содержит каркас зародышевой линии IGHV3-07 человека (с единственной обратной мутацией: R100T) и область IGHJ6.
Также предложена клетка-хозяин, содержащая рекомбинантную экспрессирующую систему, кодирующую легкую и тяжелую цепи гуманизированного антитела или его фрагмента, где указанное антитело или указанный фрагмент специфически связываются с антигеном Dkk-1 человека, и где CDR LC и НС имеют следующие аминокислотные последовательности:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:22);
(2) CDR2 (SEQ ID NO:24); и
(3) CDR3 (SEQ ID NO:26);
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:30, 49 или 50);
(2) CDR2 (SEQ ID NO:32 или-51); и
(3) CDR3 (SEQ ID NO:34 или 52); или
(в) один или более CDR из (а) и один или более CDR из (б).
Кроме того, предложен способ получения гуманизированного антитела или его фрагмента, которые специфически связываются с антигеном Dkk-1 человека, и где CDR LC и НС имеют следующие аминокислотные последовательности:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:22);
(2) CDR2 (SEQ ID NO:24); и
(3) CDR3 (SEQ ID NO:26);
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:30, 49 или 50);
(2) CDR2 (SEQ ID NO:32 или 51); и
(3) CDR3 (SEQ ID NO:34 или 52); или
(в) один или более CDR из (а) и один или более CDR из (б), включающий предоставление хозяина, трансформированного либо (1) первым экспрессирующим вектором, который кодирует легкую цепь гуманизированного антитела или его фрагмента, и вторым экспрессирующим вектором, который кодирует тяжелую цепь гуманизированного антитела или его фрагмента; либо (2) единичным экспрессирующим вектором, который кодирует как легкую цепь, так и тяжелую цепь гуманизированного антитела или его фрагмента; и поддержание указанного хозяина в таких условиях, что экспрессируется каждая цепь, и выделение гуманизированного антитела или его фрагмента, образованных в результате сборки экспрессируемых таким образом цепей.
Кроме того, предложен способ получения гуманизированного антитела или его фрагмента, которые специфически связываются с антигеном Dkk-1 человека, и где CDR LC и НС имеют следующие аминокислотные последовательности:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:22);
(2) CDR2 (SEQ ID NO:24); и
(3) CDR3 (SEQ ID NO:26);
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:30, 49 или 50);
(2) CDR2 (SEQ ID NO:32 или 51); и
(3) CDR3 (SEQ ID NO:34 или 52); или
(в) один или более CDR из (а) и один или более CDR из (б), включающий культивирование клетки-хозяина, которая содержит рекомбинантную экспрессирующую систему, кодирующую LC и НС указанного антитела или его фрагмента, и извлечение указанного антитела или его фрагмента.
Согласно изобретению предложено гуманизированное антитело или его фрагмент, полученные культивированием клетки-хозяина, содержащей рекомбинантную экспрессирующую систему, кодирующую легкую и тяжелую цепи гуманизированного антитела или его фрагмента, где указанные антитело или фрагмент специфически связываются с антигеном Dkk-1 человека, и где CDR LC и НС имеют следующие аминокислотные последовательности:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:22);
(2) CDR2 (SEQ ID NO:24); и
(3) CDR3 (SEQ ID NO:26);
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:30, 49 или 50);
(2) CDR2 (SEQ ID NO:32 или 51); и
(3) CDR3 (SEQ ID NO:34 или 52); или
(в) один или более CDR из (а) и один или более CDR из (б), и где указанные гуманизированное антитело или его фрагмент могут конкурировать с muMabJC18 (muMab относится к мышиному Mab) за связывание с антигеном Dkk-1 человека.
Также предложена фармацевтическая композиция, содержащая гуманизированное антитело или его фрагмент, которые специфически связываются с антигеном Dkk-1 человека, и где CDR LC и НС имеют следующие аминокислотные последовательности:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:22);
(2) CDR2 (SEQ ID NO:24); и
(3) CDR3 (SEQ ID NO:26);
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:30, 49 или 50);
(2) CDR2 (SEQ ID NO:32 или 51); и
(3) CDR3 (SEQ ID NO:34 или 52); или
(в) один или более CDR из (а) и один или более CDR из (б), и фармацевтически приемлемый эксципиент, разбавитель или носитель.
Также предложена фармацевтическая композиция, содержащая гуманизированное антитело или его фрагмент, которые специфически связываются с антигеном Dkk-1 человека, и где CDR LC и НС имеют следующие аминокислотные последовательности:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:22);
(2) CDR2 (SEQ ID NO:24); и
(3) CDR3 (SEQ ID NO:26);
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:30, 49 или 50);
(2) CDR2 (SEQ ID NO:32 или 51); и
(3) CDR3 (SEQ ID NO:34 или 52); или
(в) один или более CDR из (а) и один или более CDR из (б), где указанное гуманизированное антитело или его фрагмент могут конкурировать с muMabJC18 за связывание с антигеном Dkk-1 человека, и фармацевтически приемлемый эксципиент, разбавитель или носитель.
Кроме того, предложен способ получения гуманизированного антитела или его фрагмента, включающий культивирование клетки-хозяина, содержащей рекомбинантную экспрессирующую систему, кодирующую LC и НС гуманизированного антитела или его фрагмента, где указанные антитело или фрагмент специфически связываются с антигеном Dkk-1 человека, и где CDR LC и НС имеют следующие аминокислотные последовательности:
(а) один или более CDR LC, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:22);
(2) CDR2 (SEQ ID NO:24); и
(3) CDR3 (SEQ ID NO:26);
(б) один или более CDR НС, выбранных из группы, состоящей из:
(1) CDR1 (SEQ ID NO:30, 49 или 50);
(2) CDR2 (SEQ ID NO:32 или 51); и
(3) CDR3 (SEQ ID NO:34 или 52); или
(в) один или более CDR из (а) и один или более CDR из (б), где указанные гуманизированное антитело или его фрагмент могут конкурировать с muMabJC18 за связывание с антигеном Dkk-1 человека, и извлечение указанного гуманизированного антитела или его фрагмента.
Согласно изобретению также предложен способ лечения или предупреждения уменьшения костной массы, включающий введение пациенту, нуждающемуся в этом, эффективного количества гуманизированных антител и их фрагментов по изобретению. В одном из воплощений пациентом является пациент, страдающий раком, дающим метастазы в кость. В одном из воплощений пациентом является пациент, страдающий множественной миеломой. В одном из воплощений пациент выбран из пациентов, страдающих остеопорозом, остеопенией, болезнью Педжета, периодонтитом, ревматоидным артритом и уменьшением костной массы вследствие иммобилизации. В одном из воплощений пациент страдает остеопорозом. В одном из воплощений пациент страдает дефицитом эстрогенов.
Согласно изобретению также предложен способ индуцирования увеличения костной массы, включающий введение пациенту, нуждающемуся в этом, эффективного количества гуманизированных антител и их фрагментов по изобретению. В одном из воплощений пациентом является пациент, страдающий раком, дающим метастазы в кость. В одном из воплощений пациентом является пациент, страдающий множественной миеломой. В одном из воплощений пациент выбран из пациентов, страдающих остеопорозом, остеопенией, болезнью Педжета, периодонтитом, ревматоидным артритом и уменьшением костной массы вследствие иммобилизации. В одном из воплощений пациент страдает остеопорозом. В одном из воплощений пациентом является реципиент с костным трансплантатом или пациент, страдающий переломом кости. В одном из воплощений пациент имеет вторичную потерю костной массы вследствие: длительного лечения кортикостероидами, ингибиторами ароматазы или TZD; анорексии; или вызванную гипогонадизмом или нефрогенной остеодистрофией.
Согласно изобретению также предложен способ индуцирования активности Wnt, включающий введение пациенту, нуждающемуся в этом, эффективного количества гуманизированного антитела или его фрагмента по изобретению.
Согласно изобретению также предложены нейтрализующие моноклональные антитела против Dkk-1 и их иммунологически функциональные фрагменты. В одном из воплощений антитело или его фрагмент связывается с Dkk-1 человека с Kd меньше 100 пМ. В одном из воплощений антитело или его фрагмент антагонизирует действие Dkk-1 с IC50 меньше 100 нМ в функциональных анализах с использованием клеток. В одном из воплощений антитело или его фрагмент имеет прогнозируемый профиль воздействия на человека с ожидаемым увеличением показателей минеральной плотности кости (BMD) после введения один раз в месяц в дозе не более 250 мг.
Также предложены анаболические агенты костной ткани, которые оказывают антагонизируют действие в отношении способности Dkk-1 ингибировать активность Wnt и, в силу этого, приводят к увеличению костной массы, полезные в лечении заболеваний, состояний и расстройств, таких как остеопороз и заболевания, состояния и расстройства, приводящие к вторичной потере костной массы (например, вследствие: длительного лечения кортикостероидами, ингибиторами ароматазы или TZD; анорексии; или вызванной гипогонадизмом или нефрогенной остеодистрофией), когда такое усиленное костеобразование будет полезно пациенту.
Антитела и их фрагменты по изобретению можно вводить в комбинации с другими фармацевтическими агентами (в частности, агентами, используемыми для лечения или предупреждения первичной и вторичной потери костной массы, уменьшений костной массы и агентами, которые ослабляют прочность кости, которые изложены в данном описании ниже). Комбинированную терапию можно применять любым подходящим способом, как, например: (а) в виде единой фармацевтической композиции, которая содержит гуманизированное антитело или его фрагмент по настоящему изобретению, по меньшей мере один дополнительный фармацевтический агент, изложенный в данном описании, и фармацевтически приемлемый эксципиент, разбавитель или носитель; или (б) в виде двух отдельных фармацевтических композиций, содержащих (1) первую композицию, содержащую гуманизированное антитело или его фрагмент по настоящему изобретению и фармацевтически приемлемый эксципиент, разбавитель или носитель, и (2) вторую композицию, содержащую по меньшей мере один дополнительный фармацевтический агент, изложенный в данном описании, и фармацевтически приемлемый эксципиент, разбавитель или носитель. Фармацевтические композиции могут быть введены одновременно или последовательно и в любом подходящем порядке.
Кроме того, предложены наборы, содержащие антитела и/или их фрагменты по изобретению, или содержащие фармацевтические композиции по изобретению. В одном из воплощений набор дополнительно включает инструкции, описывающие как получить антитело или фрагмент и/или как ввести антитело или фрагмент. В другом воплощении набор используют в качестве диагностического средства для определения того, обладает ли пациент способностью увеличивать костную массу в результате ингибирования пути передачи сигнала через Wnt.
Также предложены два акта депонирования в АТСС (согласно условиям Будапештского договора) от 18 февраля 2009 г. Была депонирована плазмида, несущая DH5α E.coli, которая имеет вариабельную область НС huMabJC18, a именно: PCR2.1 ТОРО из E.coli в хозяине E.coli DH5α, UC 25553, обозначение патентного депонирования в АТСС РТА-9835. Была депонирована плазмида, несущая DH5α Е.coli, которая имеет вариабельную область LC huMabJC18, а именно: PCR2.1 ТОРО из Е.coli в хозяине Е.coli DH5α, UC 25554, обозначение патентного депонирования в АТСС РТА-9836. Все ограничения в отношении доступности плазмид для общественного использования, депонированных с этой целью, будут безоговорочно сняты после выдачи патента на основании описания настоящего изобретения.
Другие признаки и преимущества настоящего изобретения будут очевидны из следующих далее подробного описания и Примеров, которые не следует истолковывать как ограничивающие. Содержания всех ссылок, данных Genbank, патентов и опубликованных патентных заявок, приведенных по всему этому описанию, прямо включены в данное описание во всей своей полноте посредством ссылки.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг.1А и 1В показан эффект моноклонального антитела против Dkk-1 мыши (JC18) в модели интактных мышей. Общая BMD дистальных отделов бедренных костей значительно повышалась в результате лечения с использованием JC18 для всех уровней дозы (Фиг.1А). Применение JC18 также значительно повышало показатели костеобразования для уровня дозы 60 мг/кг у мышей (Фиг.1В).
На Фиг.2 показан эффект мышиной химерной конструкции моноклональное антитело против Dkk-1 (мышиная химерная конструкция представляет собой химерное антитело "человек/мышь") в модели интактных мышей. Мышиная химерная конструкция в дозах от 1 до 30 мг/кг повышала общую BMD по сравнению с разбавителем.
На Фиг.3 показано, что JC18 предотвращает уменьшение костной массы, индуцированное дефицитом эстрогенов в модели подвергнутых OVX (овариэктомии) мышей. Как ожидалось, мыши, обработанные разбавителем, демонстрировали остеопению на 8-й неделе после OVX, что продемонстрировано значительным уменьшением общей BMD. Применение JC18 дозозависимым образом повышало общую BMD, что измерено посредством pQCT (периферической количественной компьютерной томографии) для дистальных отделов бедренных костей, на 7-20% по сравнению с обработкой OVX-мышей разбавителем. У мышей, обработанных JC18 в дозе 15 мг/кг два раза в неделю, общая BMD была не только выше, чем у обработанных разбавителем OVX-мышей, но также сохранялась на уровне, соответствующем ложнооперированным контролем, указывая на то, что применение JC18 в этой дозе и этом режиме введения полностью предотвращало развитие остеопении у OVX-мышей.
На Фиг.4 показана специфичность связывания JC18 с гомологами Dkk-1, Dkk-3 и Dkk-4 человека. Для JC18 продемонстрировано слабое связывание с Dkk-4 человека и отсутствие связывания с Dkk-3 человека вплоть до 5 мкг/мл.
На Фиг.5 показана модель мишень-опосредованного распределения лекарственного средства (TMDD).
На Фиг.6 показан график зависимости концентраций h-DKK-1 (DKK-1 человека) с меткой V5-His в сыворотке крови крысы после однократного внутривенного введения в дозах 1, 5, 10 и 100 мкг/кг. Значения для образцов во временные точки свыше 0,5 ч в дозе 0,001 мг/кг, свыше 1 ч в дозе 0,005 мг/кг и свыше 2 ч в дозах 0,01 и 0,1 мг/кг оказались ниже поддающегося количественному определению предела метода анализа и не были включены в данный график.
На Фиг.7 показан график зависимости измеренных и предсказанных на основании модели концентраций свободного/частично свободного huMabJC18 от времени после еженедельного внутривенного введения huMabJC18 крысам Sprague-Dawley. Символы представляют средние результаты измерений (±SD (стандартное отклонение)), а линии представляют профили, предсказанные на основании этой модели.
На Фиг.8А-Е показаны графики зависимости измеренных и предсказанных на основании модели концентраций свободного Dkk-1 от времени после еженедельного внутривенного введения huMabJC18 крысам Sprague-Dawley. Символы представляют средние результаты измерений (±SE (стандартная ошибка)), а линии представляют профили, предсказанные на основании этой модели. Образцы, в которых были детектированы антитела против huMabJC18, удаляли из анализа.
На Фиг.9А-В показаны графики зависимости измеренных и предсказанных на основании модели концентраций свободного/частично свободного huMabJC18 от времени после однократного внутривенного введения huMabJC18 яванским макакам, (а) Самцы макак, (б) Самки макак. Символы представляют измеренные данные для отдельных макак, а линии представляют профили, предсказанные на основании этой модели.
На Фиг.10А-Е показаны графики зависимости измеренных и предсказанных на основании модели концентраций свободного Dkk-1 от времени после однократного внутривенного введения huMabJC18 яванским макакам. Символы представляют измеренные данные, а линии представляют профили, предсказанные на основании этой модели.
На Фиг.11 показана диаграмма зависимости средних концентраций остеокальцина (±SE) через 1,5 и 2,5 недели после еженедельного внутривенного введения первой дозы huMabJC18 крысам Sprague-Dawley. * указывает, что p<0,05 по сравнению с разбавителем.
На Фиг.12 показан график зависимости предсказанных концентраций свободного Dkk-1 у людей после подкожного введения huMabJC18 1 раз/месяц в течение 6 месяцев. Тестировали три разные дозы: 0,003 мг/кг, которая составляет 1/10 от MABEL (минимального уровня ожидаемого биологического эффекта) или дозу, не дающую никакого эффекта; 0,03 мг/кг, которая равна MABEL, и 3,57 мг/кг, которая представляет собой предсказанную эффективную дозу для остеопороза.
На Фиг.13 показан график зависимости предсказанных концентраций свободного huMabJC18 у людей после подкожного введения huMabJC18 1 раз/месяц в течение 6 месяцев. Тестировали три разные дозы: 0,003 мг/кг, которая представляет собой 1/10 от MABEL или дозу, не дающую никакого эффекта; 0,03 мг/кг, которая равна MABEL, и 3,57 мг/кг, которая представляет собой предсказанную эффективную дозу для остеопороза. Показан ожидаемый предел количественного определения (LOQ) биоаналитического анализа, чтобы указать на нецелесообразность введения дозы ниже предсказанного MABEL.
На Фиг.14 показан график зависимости подсчетов занятости рецепторов для huMabJC18 от предсказанной человеческой дозы, основанных на использовании (1) равновесного метода для стационарного состояния (steady state equilibrium method) (Duff, 2006) и (2) механистической РК/PD (фармакокинетической/фармакодинамической) (TMDD) модели.
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение относится к выделенным антителам и их фрагментам (например, антигенсвязывающим участкам), в частности гуманизированным моноклональным антителам и их фрагментам, которые специфически связываются с Dkk-1. В некоторых воплощениях антитела и их фрагменты по изобретению происходят из конкретных последовательностей тяжелой и легкой цепей зародышевых линий и/или содержат конкретные структурные черты, как например участки CDR, содержащие конкретные аминокислотные последовательности. Согласно данному изобретению предложены, например, выделенные антитела и их фрагменты, способы получения таких антител или их фрагментов и фармацевтические композиции, содержащие данные антитела или их фрагменты. Данное изобретение также относится к способам использования антител и их фрагментов, например для детекции Dkk-1, а также для лечения различных заболеваний, состояний или расстройств, таких как поражения костей, которые вызывают уменьшение костной массы. Также предложены способы лечения или предупреждения уменьшения костной массы, способы индукции увеличения костной массы и способы индукции активности Wnt.
Определения
Если в данном описании не указано иное, научные и технические термины, используемые применительно к настоящему изобретению, будут иметь значения, которые обычно понятны обычным специалистам в данной области. Кроме того, если по контексту не требуется иное, термины, использованные в единственном числе, будут подразумевать включение множественного числа, а термины во множественном числе будут включать в себя единственное число. В большинстве случаев, номенклатуры, используемые применительно к культивированию клеток и тканей, молекулярной биологии, иммунологии, микробиологии, генетике, химии белков и нуклеиновых кислот и гибридизации, и методы, относящиеся ко всему перечисленному выше, изложенные в данном описании, представляют собой методы, хорошо известные и широко используемые в данной области. Способы и методы по настоящему изобретению обычно осуществляют в соответствии с традиционными способами, хорошо известными в данной области и как описано в различных общих и более специальных ссылках, которые приведены и рассмотрены по всему описанию настоящего изобретения, если не указано иное. См., например, Sambrook et al. Molecular Cloning: A Laboratory Manual, 2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989), и Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Associates (1992), и Harlow and Lane Antibodies: A Laboratory Manual Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1990), которые включены в данное описание посредством ссылки. Ферментативные реакции и методы очистки выполняют в соответствии с инструкциями изготовителя, как обычно их осуществляют в данной области или как изложено в данном описании. Терминология, используемая применительно к аналитической химии, химии органического синтеза и медицинской и фармацевтической химии, и лабораторные методики и методы, относящиеся ко всему перечисленному выше, изложенные в данном описании, представляют собой методы, хорошо известные и широко используемые в данной области. Для проведения химических синтезов, химических анализов, получения, изготовления и доставки фармацевтических средств и лечения пациентов можно использовать стандартные способы.
Очевидно, что если не указано иное, следующие термины, используемые в этом описании, будут иметь следующие значения.
Термин "Dkk-1", как он использован в данном описании, включает, например, нативные формы Dkk-1 мыши и человека. Типичные белковые и нуклеотидные последовательности Dkk-1 описаны, например, в US2006/0127393 (WO2006/015373) и US2008/0193449. Термин также включает варианты таких нативных последовательностей, которые дают перекрестную иммунологическую реакцию с этими нативными белками. Эти белки могут ингибировать взаимодействие между LRP5 или LRP6 и Wnt. Термин также может относиться к фрагменту нативной или вариантной формы Dkk-1, содержащему эпитоп, с которым может специфически связываться антитело.
Термин "полинуклеотид" или "нуклеиновая кислота" означает одноцепочечные или двухцепочечные полимеры. Нуклеотиды, входящие в состав полинуклеотида, могут представлять собой рибонуклеотиды или дезоксирибонуклеотиды либо модифицированную форму каждого из этих двух типов нуклеотидов. Указанные модификации включают модификации основания, такие как производные бромуридин и инозин, модификации рибозной составляющей, такие как 2',3'-дидезоксирибоза, и модификации межнуклеотидных связей, такие как фосфоротиоат, фосфородитиоат, фосфороселеноат, фосфородиселеноат, фосфороанилотиоат, фосфоранилидат и фосфороамидат. Термин включает как одно-, так и двухцепочечные формы.
Термин "олигонуклеотид" означает полинуклеотид, содержащий 200 или меньше нуклеотидов. В некоторых воплощениях длина олигонуклеотидов составляет 10-60 оснований. В других воплощениях длина олигонуклеотидов составляет от 12, 13, 14, 15, 16, 17, 18, 19 или 20 до 40 нуклеотидов. Олигонуклеотиды могут быть одноцепочечными или двухцепочечными, например, для использования в конструировании мутантного гена. Олигонуклеотиды по изобретению могут быть смысловыми или антисмысловыми олигонуклеотидами. Олигонуклеотид по изобретению может включать метку, в том числе радиоактивную метку, флуоресцентную метку, гаптен или антигенную метку, для анализов на предмет детекции. Олигонуклеотиды по изобретению можно использовать, например, в виде ПЦР-праймеров, праймеров для клонирования или гибридизационных зондов.
"Выделенная молекула нуклеиновой кислоты" означает молекулу ДНК или РНК геномного, мРНК, кДНК или синтетического происхождения либо некоторую их комбинацию, которая не находится в ассоциации со всем полинуклеотидом или его частью, в котором/которой выделенный полинуклеотид обнаруживается в природе, или которая связана с полинуклеотидом, с которым не связана в природе. Для решения задач этого изобретения следует понимать, что термин "молекула нуклеиновой кислоты, содержащая" конкретную нуклеотидную последовательность, не охватывает интактные хромосомы. Выделенные молекулы нуклеиновой кислоты, "содержащие" специфические нуклеиновокислотные последовательности, могут включать, помимо этих специфических последовательностей, кодирующие последовательности для других белков или их частей в количестве до десяти или даже до двадцати или могут включать функционально связанные регуляторные последовательности, которые регулируют экспрессию кодирующего участка перечисленных нуклеиновокислотных последовательностей, и/или могут включать последовательности векторов.
Если не указано иное, расположенный слева конец любой одноцепочечной полинуклеотидной последовательности, рассмотренной в данном описании, представляет собой 5'-конец; направление в левую сторону двухцепочечных полинуклеотидных последовательностей называется 5'-направление. Направление от 5' к 3' синтеза растущих РНК-транскриптов называется направление транскрипции; участки последовательности на цепи ДНК, имеющие ту же последовательность, что и РНК-транскрипт, которые расположены от 5' до 5'-конца РНК-транскрипта, называются "последовательностями, расположенными в прямом направлении"; участки последовательности на цепи ДНК, имеющие ту же последовательность, что и РНК-транскрипт, которые расположены от 3' до 3'-конца РНК-транскрипта, называются "последовательностями, расположенными в обратном направлении".
Термин "контрольная последовательность" относится к полинуклеотидной последовательности, которая может оказывать воздействие на экспрессию и процессинг кодирующих последовательностей, с которыми она лигирована. Природа таких контрольных последовательностей может зависеть от организма-хозяина. В конкретных воплощениях контрольные последовательности для прокариот могут включать промотор, сайт связывания с рибосомой и последовательность терминации транскрипции. Например, контрольные последовательности для эукариот могут включать промоторы, содержащие один или много сайтов распознавания транскрипционных факторов, последовательности энхансеров транскрипции и последовательность терминации транскрипции. "Контрольные последовательности" по изобретению могут включать лидерные последовательности и/или последовательности партнеров слияния.
Термин "вектор" означает любую молекулу или структуру (например, нуклеиновую кислоту, плазмиду, бактериофаг или вирус), используемую для перенесения информации, кодирующей белки, в клетку-хозяина.
Термин "экспрессирующий вектор" или "экспрессирующая конструкция" относится к вектору, который подходит для трансформации клетки-хозяина и содержит нуклеиновокислотные последовательности, которые направляют и/или регулируют (вместе с клеткой-хозяином) экспрессию одного или более гетерологичных кодирующих участков, функционально связанных с ними. Экспрессирующая конструкция может включать, но этим не ограничивается, последовательности, которые оказывают воздействие или регулируют транскрипцию, трансляцию, и, при наличии интронов, оказывают воздействие на РНК-сплайсинг кодирующего участка, функционально связанного с ними.
Как использовано в данном описании, "функционально связанный" означает, что компоненты, к которым применяется этот термин, находятся в такой взаимосвязи, которая в подходящих условиях позволяет им осуществлять присущие им функции. Например, контрольная последовательность в векторе, который "функционально связан" с кодирующей белок последовательностью, лигирована с ней, так что экспрессия такой кодирующей белок последовательности достигается в условиях, соответствующих транскрипционной активности контрольных последовательностей.
Термин "клетка-хозяин" означает клетку, которая трансформирована или способна к трансформации нуклеиновокислотной последовательностью и в силу этого экспрессирует представляющий интерес ген. Термин включает потомство родительской клетки независимо от того, идентично или нет данное потомство по морфологическим или генетическим характеристикам исходной родительской клетке, при условии, что присутствует представляющий интерес ген.
Термин "трансдукция" означает перенос генов от одной бактерии к другой, обычно посредством бактериофага. "Трансдукция" также относится к приобретению и переносу последовательностей эукариотических клеток ретровирусами.
Термин "трансфекция" означает поглощение клеткой чужеродной или экзогенной ДНК, и клетка считается "трансцифированной", когда экзогенная ДНК введена в пространство, окруженное клеточной мембраной. Ряд методов трансфекции хорошо известен в данной области и изложен в данном описании. См., например, Graham et al., 1973, Virology 52: 456; Sambrook et al., 2001, Molecular Cloning: A Laboratory Manual, Id.; Davis et al., 1986, Basic Methods in Molecular Biology, Elsevier; и Chu et al., 1981, Gene, 13: 197. Такие методы можно использовать для введения одной или более группировок экзогенной ДНК в подходящие клетки-хозяева.
Термин "трансформация" относится к изменению генетических характеристик клетки, и клетка трансформирована, когда она модифицирована таким образом, что содержит новую ДНК или РНК. Например, клетка трансформирована, если она генетически модифицирована из своего нативного состояния в результате введения нового генетического материала посредством трансфекции, трансдукции или других методов. После осуществления трансфекции или трансдукции трансформирующая ДНК может подвергаться рекомбинации с ДНК клетки путем физического интегрирования в хромосому данной клетки, или может сохраняться временно в виде эписомального элемента, не реплицируясь, или может реплицироваться независимо в виде плазмиды. Клетка считается "стабильно трансформированной", когда трансформирующая ДНК реплицируется при делении клетки.
Термины "полипептид" или "белок" означают макромолекулу, имеющую аминокислотную последовательность нативного белка, то есть, белка, продуцируемого природной и не подвергнутой рекомбинации клеткой, или продуцируемого генетически сконструированной или рекомбинантной клеткой, и включают молекулы, имеющие аминокислотную последовательность нативного белка, или молекулы, имеющие делеции, вставки и/или замены одной или более аминокислот нативной последовательности. Термины "полипептид" и "белок" в конкретном случае охватывают антитела против Dkk-1 или последовательности, которые имеют делеции, вставки и/или замены одной или более чем одной аминокислоты антитела против Dkk-1. Термин "полипептидный фрагмент" относится к полипептиду, который имеет делецию на аминоконце, делецию на карбоксильном конце и/или внутреннюю делецию по сравнению с полноразмерным нативным белком. Такие фрагменты также могут содержать модифицированные аминокислоты по сравнению с нативным белком. В некоторых воплощениях длина фрагментов составляет примерно 5-500 аминокислот. Например, длина фрагментов может составлять по меньшей мере 5, 6, 8, 10, 14, 20, 50, 70, 100, 110, 150, 200, 250, 300, 350, 400 или 450 аминокислот. Полезные полипептидные фрагменты для этого изобретения включают иммунологически функциональные фрагменты антител, в том числе связывающие домены. В случае антитела против Dkk-1 полезные фрагменты включают, но этим не ограничиваются, участок CDR, вариабельный домен тяжелой или легкой цепи, часть цепи антитела или только его вариабельную область, включающую два CDR, и тому подобное.
Термин "выделенный белок", приведенный в данном описании, означает, что белок, являющийся объектом изобретения, (1) не содержит по меньшей мере некоторых других белков, с которыми его обычно обнаруживают, (2) по существу не содержит других белков из того же источника, например, из того же вида, (3) экспрессируется клеткой другого вида, (4) отделен по меньшей мере примерно от 50 процентов полинуклеотидов, липидов, углеводов или других веществ, с которыми находится в ассоциации в природе, (5) функционально связан (путем ковалентного или нековалентного взаимодействия) с полипептидом, с которым не находится в ассоциации в природе, или (6) не существует в природе. Геномная ДНК, кДНК, мРНК или другая РНК, имеющая синтетическое происхождение, или любая их комбинация могут кодировать такой выделенный белок. Предпочтительно, чтобы выделенный белок по существу не содержал белков или полипептидов или других примесей, которые встречаются в его природном окружении и которые могли бы служить препятствием его терапевтическому, диагностическому, профилактическому, научному или другому применению.
"Вариант" полипептида (например, антитела) содержит аминокислотную последовательность, где в эту аминокислотную последовательность встроены, из этой аминокислотной последовательности делегированы и/или в этой аминокислотной последовательность заменены один или более аминокислотных остатков по сравнению с другой полипептидной последовательностью. Варианты по изобретению включают слитые белки.
"Производное" полипептида представляет собой полипептид (например, антитело), который модифицирован химически некоторым образом, отличающимся от вариантов со вставкой, делецией или заменой, например, посредством конъюгирования с другой химической группировкой.
"Иммунный ответ", как будет очевидно специалисту в данной области, включает любую детектируемую антигенспецифическую или аллогенную активацию ответа Т-хелперных клеток или цитотоксических Т-клеток, продуцирование антител, опосредованную Т-клетками активацию аллергических реакций и тому подобное, но этим не ограничивается. Термин охватывает действие, например, лимфоцитов, антигенпрезентирующих клеток, фагоцитарных клеток, гранулоцитов и растворимых макромолекул, продуцируемых вышеупомянутыми клетками или печенью (включая антитела, цитокины и комплемент), которое выражается в селективном повреждении, деструкции или элиминировании из организма человека вторгающихся патогенов, клеток или тканей, инфицированных патогенами, раковых клеток, или, в случаях аутоиммунного или патологического воспаления, нормальных клеток или тканей человека.
"Путь передачи сигнала" относится к биохимической взаимосвязи между рядом молекул передачи сигнала, которые играют роль в передаче сигнала от одной части клетки к другой части клетки. Как использовано в данном описании, фраза "рецептор клеточной поверхности" включает, например, молекулы и комплексы молекул, способные к получению сигнала и передаче такого сигнала через плазматическую мембрану клетки.
"Антитело" представляет собой молекулу иммуноглобулина, способную к специфическому связыванию с мишенью, такой как углевод, полинуклеотид, липид, полипептид и т.д., по меньшей мере через один сайт распознавания антигена, локализованный в вариабельной области молекулы иммуноглобулина. Как он использован в данном описании, термин охватывает не только интактные поликлональные или моноклональные антитела, но также их фрагменты (такие как Fab, Fab', F(ab')2, Fv), одноцепочечные (scFv) и однодоменные антитела, включая антитела акулы и верблюжьи антитела) и слитые белки, содержащие участок антитела, и любую другую модифицированную конфигурацию молекулы иммуноглобулина, которая содержит сайт распознавания антигена. Антитело представляет собой антитело любого класса, такое как IgG, IgA или IgM (или их подкласс), и антитело, которое не обязательно относится к какому-либо конкретному классу. В зависимости от аминокислотной последовательности константного домена своих тяжелых цепей в антителах иммуноглобулины могут быть отнесены к разным классам. Существуют пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM и некоторые из них могут быть дополнительно подразделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют разным классам иммуноглобулинов, называются альфа, дельта, эпсилон, гамма и мю, соответственно. Общеизвестны субъединичные структуры и пространственные конфигурации разных классов иммуноглобулинов.
Антитела по изобретению могут происходить только из единственного источника или могут быть "химерными", то есть разные участки антитела могут происходить из двух разных антител. Например, участки CDR могут происходить из крысиного или мышиного источника, тогда как каркасные области V-области происходят из другого животного источника, такого как человек. Антитела или связывающие фрагменты по изобретению могут быть получены в гибридоме, методами рекомбинантной ДНК или путем ферментативного или химического расщепления интактных антител. Если не указано иное, термин "антитело" включает, в дополнение к антителам, содержащим две полноразмерные тяжелые цепи и две полноразмерные легкие цепи, их производные, варианты, фрагменты и мутеины, примеры которых описаны ниже.
Термин "легкая цепь" включает полноразмерную легкую цепь и ее фрагменты, имеющие последовательность вариабельной области, достаточную для обеспечения специфичности связывания. Полноразмерная легкая цепь включает домен вариабельной области, VL, и домен константной области, CL Домен вариабельной области легкой цепи расположен на аминоконце полипептида. Легкие цепи по изобретению включают цепи каппа и цепи лямбда.
Термин "тяжелая цепь" включает полноразмерную тяжелую цепь и ее фрагменты, имеющие последовательность вариабельной области, достаточную для обеспечения специфичности связывания. Полноразмерная тяжелая цепь включает домен вариабельной области, VH, и три домена константной области, СН1, СН2 и СН3. VH-домен расположен на аминоконце полипептида, а СН-домены расположены на карбоксильном конце, причем ближе всех к -СООН-концу расположен СН3. Тяжелые цепи по изобретению могут быть цепями любого изотипа, включая IgG (в том числе подтипы IgG1, IgG2, IgG3 и IgG4), IgA (в том числе подтипы IgA1 и IgA2), IgM. Кроме того, VH- и VL-области могут быть подразделены на участки гипервариабельности, называемые определяющими комплементарность участками (CDR), чередующиеся с более консервативными областями, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от аминоконца к карбоксиконцу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента. Вариабельные и константные области в составе легкой и тяжелой цепей соединены областью "J", состоящей примерно из 12 или более аминокислот, при этом тяжелая цепь также включает область "D", состоящую примерно из 10 или более аминокислот. В целом см. Fundamental Immunology, Ch. 7 (Paul, W., ed., 2nd ed. Raven Press, N.Y. (1989)).
Термин "антигенсвязывающий участок" антитела (или просто "участок антитела"), как он использован в данном описании, относится к одному или более фрагментам антитела, которые сохраняют способность специфически связываться с антигеном (например, α5β1). Показано, что антигенсвязывающая функция антитела может осуществляться фрагментами полноразмерного антитела. Примеры связывающих фрагментов, охватываемых термином "антигенсвязывающий участок" антитела, включают (1) фрагмент Fab, моновалентный фрагмент, состоящий из VL-, VH-, CL- и СН1-доменов; (2) фрагмент F(ab')2, бивалентный фрагмент, содержащий два Fab-фрагмента, соединенных дисульфидной связью в шарнирной области; (3) фрагмент Fd, состоящий из VH- и СН1-доменов; (4) фрагмент Fv, состоящий из VL- и VH-доменов одного плеча антитела; (5) фрагмент dAb (Ward et al. (1989) Nature, 341: 544-546), который состоит из VH-домена; и (6) выделенный определяющий комплементарность участок (CDR). Кроме того, хотя эти два домена Fv-фрагмента, VL и VH, кодируются различными генами, их можно соединить вместе посредством синтетического линкера, используя методы рекомбинации, что дает возможность их получения в виде единой белковой цепи, в которой VL-и VH-области соединяются с образованием моновалентной молекулы (известной как одноцепочечный Fv (scFv); см., например, Bird et al. (1988) Science 242: 423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA, 85: 5879-5883). Подразумевают, что такие одноцепочечные антитела также охватываются термином "антигенсвязывающий участок" антитела. Эти фрагменты антитела можно получить, используя любой подходящий метод, в том числе традиционные методы, известные специалистам в данной области, и скрининг данных фрагментов на предмет их полезности можно провести так же, как и для интактных антител.
"Выделенное антитело", как использовано в данном описании, относится к антителу, по существу не содержащему других антител, обладающих другими антигенными специфичностями (например, выделенное антитело, которое специфически связывается с Dkk-1, по существу не содержит других антител, которые специфически связываются с антигенами, отличными от Dkk-1). Тем не менее, выделенное антитело, которое специфически связывается с Dkk-1, может обладать перекрестной реактивностью в отношении других антигенов, таких как молекулы Dkk-1 из других видов. Помимо этого, выделенное антитело по существу может не содержать другого клеточного материала и/или других химических веществ.
Термин "иммунологически функциональный фрагмент" (или просто "фрагмент") иммуноглобулиновой цепи, как он использован в данном описании, относится к участку легкой цепи или тяжелой цепи антитела, в котором отсутствуют по меньшей мере несколько аминокислот, присутствующих в полноразмерной цепи, но который способен специфически связываться с антигеном. Свойство таких фрагментов быть биологически активными заключается в том, что они связываются специфически с антигеном-мишенью и могут конкурировать с интактными антителами за специфическое связывание с заданным эпитопом. В одном из аспектов изобретения такой фрагмент будет сохранять по меньшей мере один CDR, присутствующий в полноразмерной легкой или тяжелой цепи, и в некоторых воплощениях будет содержать единичную тяжелую цепь и/или легкую цепь или их часть. Эти биологически активные фрагменты могут быть получены методами рекомбинантной ДНК или могут быть получены путем ферментативного или химического расщепления интактных антител. Иммунологически функциональные иммуноглобулиновые фрагменты по изобретению включают Fab, Fab', F(ab')2, Fv, однодоменные антитела и одноцепочечные антитела, но этим не ограничиваются, и могут происходить из любого источника, относящегося к млекопитающим, включая, но этим не ограничиваясь, человека, мышь, крысу, представителя семейства верблюдовых или кролика. Кроме того, предполагается, что функциональный участок антител по изобретению, например, один или более чем один CDR, может быть связан ковалентно со вторым белком или небольшой молекулой с целью создания терапевтического агента, направленного на конкретную мишень в организме, обладающего бифункциональными терапевтическими свойствами или имеющего пролонгированный период полувыведения из сыворотки.
"Fab-фрагмент" состоит из одной легкой цепи и СН1 и вариабельных областей одной тяжелой цепи. Тяжелая цепь молекулы Fab не может образовывать дисульфидную связь с другой молекулой тяжелой цепи.
"Fc"-область содержит два фрагмента тяжелой цепи, содержащие СН1- и СН2-домены антитела. Эти два фрагмента тяжелой цепи удерживаются вместе двумя или более дисульфидными связями и посредством гидрофобных взаимодействий СН3-доменов.
"Fab'-фрагмент" содержит одну легкую цепь и участок одной тяжелой цепи, содержащий VH-домен и СН1-домен, а также область между СН1- и СН2-доменами, так что между двумя тяжелыми цепями двух Fab'-фрагментов может образоваться межцепочечная дисульфидная связь с получением молекулы F(ab')2.
"F(ab')2-фрагмент" содержит две легкие цепи и две тяжелые цепи, содержащие участок константной области между СН1- и СН2-доменами, так что между двумя тяжелыми цепями образуется межцепочечная дисульфидная связь. Таким образом, F(ab')2-фрагмент состоит из двух Fab'-фрагментов, которые удерживаются вместе дисульфидной связью между двумя тяжелыми цепями.
"Fv-область" содержит вариабельные области как от тяжелой, так и легкой цепей, но при отсутствии константных областей.
"Одноцепочечные антитела" представляют собой молекулы Fv, в которых вариабельные области тяжелой и легкой цепей соединены посредством гибкого линкера с образованием единой полипептидной цепи, в результате чего образуется антигенсвязывающий участок. Одноцепочечные антитела подробно рассмотрены, например, в WO 88/01649 и патентах США №№4946778 и 5260203.
"Однодоменное антитело" представляет собой иммунологически функциональный иммуноглобулиновый фрагмент, содержащий только вариабельную область тяжелой цепи или вариабельную область легкой цепи. В некоторых случаях две или более VH-областей соединены ковалентной связью посредством пептидного линкера с целью создания бивалентного однодоменного антитела. Мишенями для таких двух VH-областей бивалентного однодоменного антитела могут служить одинаковые или разные антигены.
"Бивалентное антитело" содержит два антигенсвязывающих сайта. В некоторых случаях эти два антигенсвязывающих сайта имеют одинаковые антигенные специфичности. Тем не менее, бивалентные антитела могут быть биспецифическими (см. ниже).
"Полиспецифическое антитело" представляет собой антитело, мишенью для которого является более чем один антиген или эпитоп.
"Биспецифическое", "с двойной специфичностью" или "бифункциональное" антитело представляет собой гибридное антитело, имеющее два разных антигенсвязывающих сайта. Биспецифические антитела представляют собой вид полиспецифического антитела и могут быть получены рядом способов, включая слияние гибридом или связывание Fab'-фрагментов, но этим не ограничиваясь. См., например, Songsivilai & Lachmann (1990), Clin. Exp. Immunol. 79: 315-321 и Kostelny et al. (1992), J. Immunol. 148: 1547-1553. Эти два связывающих сайта биспецифического антитела будут связываться с двумя разными эпитопами, которые могут располагаться на одинаковых или разных белках-мишенях.
Как использовано в данном описании, "моноклональное антитело" относится к антителу, полученному из популяции по существу гомогенных антител, то есть отдельные антитела, составляющие данную популяцию, идентичны, за исключением возможных природных мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными, направленными против единственного антигенного сайта. Кроме того, в отличие от препаратов поликлональных антител, которые обычно включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против единственной детерминанты на антигене. Определение "моноклональный" указывает на характер антитела, которое получено по существу из гомогенной популяции антител, и его не следует истолковывать в качестве необходимой рекомендации получения антитела каким-либо конкретным способом. Например, моноклональные антитела, которые применяют в соответствии с настоящим изобретением, могут быть получены способом с использованием гибридом, впервые описанным в Kohler and Milstein (1975) Nature 256: 495, или могут быть получены способами с использованием рекомбинантной ДНК, такими как описанные в патенте США №4816567. Моноклональные антитела также могут быть выделены из фаговых библиотек, созданных с использованием методов, описанных в McCafferty et al. (1990) Nature, 348: 552-554, например.
Термин "производные человеческого антитела" относится к любой модифицированной форме человеческого антитела, например, конъюгату антитела и другого агента или антитела.
Подразумевается, что термин "гуманизированное антитело" относится к антителам, в которых последовательности CDR, происходящие из зародышевой линии другого вида млекопитающих, такого как мышь, привиты на последовательности каркасных областей иммуноглобулина человека. Дополнительные модификации каркасной области могут быть выполнены в пределах последовательностей каркасных областей иммуноглобулина человека.
Как использовано в данном описании, "гуманизированное" антитело относится к формам отличных от человеческих (например, мышиных) антител, которые представляют собой химерные иммуноглобулины, иммуноглобулиновые цепи или их фрагменты (такие как Fv, Fab, Fab', F(ab')2 или другие антигенсвязывающие подпоследовательности антител), содержащие минимальную последовательность, происходящую из отличного от человеческого иммуноглобулина. Предпочтительно, гуманизированные антитела представляют собой человеческие иммуноглобулины (реципиентное антитело), в которых остатки от реципиента, определяющего комплементарность участка (CDR), заменены на остатки от CDR вида, не относящегося к человеку (донорное антитело), такого как мышь, крыса или кролик, имеющего желаемую специфичность, аффинность и способность. В некоторых случаях остатки каркасной области (FR) Fv человеческого иммуноглобулина заменены на соответствующие остатки отличного от человеческого иммуноглобулина. Кроме того, гуманизированное антитело может содержать остатки, которые не обнаружены ни в реципиентном антителе, ни в импортируемых последовательностях CDR или каркасных последовательностях, но которые включены для дальнейшего улучшения и оптимизации характеристик антител. В общем случае гуманизированное антитело будет содержать по существу все или по меньшей мере один и обычно два вариабельных домена, в которых все или по существу все участки CDR соответствуют участкам отличного от человеческого иммуноглобулина и все или по существу все области FR представляют собой области консенсусной последовательности человеческого иммуноглобулина. В оптимальном случае гуманизированное антитело также будет содержать по меньшей мере часть константной области или домена (Fc) иммуноглобулина, обычно человеческого иммуноглобулина. Предпочтительны антитела, имеющие Fc-области, модифицированные, как описано в WO 99/58572. Другие формы гуманизированных антител имеют один или более CDR (CDR L1, CDR L2, CDR L3, CDR H1, CDR H2 и/или CDR H3), которые изменены относительно исходного антитела.
Подразумевается, что термины "человеческое антитело" или "полностью человеческое антитело", как они использованы в данном описании, включают антитела, имеющие вариабельные области, в которых как каркасные участки, так и участки CDR происходят из последовательностей иммуноглобулинов зародышевой линии человека. Кроме того, если антитело содержит константную область, то данная константная область также происходит из последовательностей иммуноглобулинов зародышевой линии человека. Человеческие антитела по изобретению или их антигенсвязывающие участки могут включать аминокислотные остатки, которые не кодируются последовательностями иммуноглобулинов зародышевой линии человека (например, мутации, введенные в результате случайного или сайтспецифического мутагенеза in vitro или в результате соматической мутации in vivo). Однако подразумевается, что термин "человеческое антитело", как он использован в данном описании, не включает антитела, в которых последовательности CDR, происходящие из зародышевой линии другого вида млекопитающих, такого как мышь, привиты на последовательности каркасных областей иммуноглобулина человека.
Термины "человеческое моноклональное антитело" или "полностью человеческое моноклональное антитело" относятся к антителам, проявляющим единичную специфичность связывания, которые имеют вариабельные области, в которых как каркасные области, так и участки CDR происходят из последовательностей иммуноглобулинов зародышевой линии человека. В одном из воплощений человеческие моноклональные антитела продуцируются гибридомой, которая включает В-клетку, полученную из трансгенного животного, не являющегося человеком, например, трансгенной мыши, геном которой содержит трансген тяжелой цепи и трансген легкой цепи иммуноглобулина человека, причем В-клетка слита с иммортализованной клеткой.
Термин "рекомбинантное человеческое антитело", как он использован в данном описании, включает все человеческие антитела, которые получают, экспрессируют, создают или выделяют рекомбинантными способами, такие как (а) антитела, выделенные из животного (например, мыши), которое является трансгенным или трансхромосомным для генов человеческого иммуноглобулина или полученной из него гибридомы (описана далее ниже), (б) антитела, выделенные из клетки-хозяина, трансформированной с целью экспрессии человеческого антитела, например, из трансфектомы, (в) антитела, выделенные из рекомбинантной комбинаторной библиотеки человеческих антител, и (г) антитела полученные, экспрессированные, созданные или выделенные любыми другими способами, включающими сплайсинг последовательностей генов человеческих иммуноглобулинов с другими последовательностями ДНК. Такие рекомбинантные человеческие антитела имеют вариабельные области, каркасные области и участки CDR в которых происходят из последовательностей иммуноглобулинов зародышевой линии человека. Однако в некоторых воплощениях такие рекомбинантные человеческие антитела могут быть подвергнуты мутагенезу in vitro (или, когда используется животное, трансгенное в отношении последовательности для Ig человека, соматическому мутагенезу in vivo) и такие аминокислотные последовательности VH- и VL-областей рекомбинантных антител представляют собой последовательности, которые, несмотря на то, что они происходят из VH- и VL-последовательностей зародышевой линии человека и имеют к ним отношение, не могут существовать в природе в репертуаре антител зародышевой линии человека in vivo.
Как использовано в данном описании, "изотип" или "класс" относится к классу антитела (например, IgM или IgG), который кодируется генами константной области тяжелой цепи. Константные домены антител не вовлечены в связывание с антигеном, но проявляют различные эффекторные функции. В зависимости от аминокислотной последовательности константной области тяжелой цепи заданное человеческое антитело или заданный иммуноглобулин могут быть отнесены к одному из пяти основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM. Структуры и пространственные конфигурации разных классов иммуноглобулинов хорошо известны. Известно, что среди различных классов человеческих иммуноглобулинов только IgG1, IgG2, IgG3, IgG4 и IgM человека активируют комплемент. Известно, что IgG1 и IgG3 человека опосредуют ADCC у людей.
При использовании в данном описании, термин "подкласс" относится к дальнейшей спецификации внутри изотипа гена константной области тяжелой цепи, например подклассам IgG1, IgG2, IgG3 или IgG4 в пределах изотипа IgG.
При использовании в данном описании, термин "соединение" или "фармацевтическое соединение" включает антитела, их антигенсвязывающие участки, иммуноконъюгаты и биспецифические молекулы.
Фразы "антитело, разпознающее антиген" и "антитело, специфичное к антигену" используются в данном описании взаимозаменяемо с термином "антитело, которое специфически связывается с антигеном".
Термин "антителозависимая клеточная цитотоксичность" или "ADCC" относится к клеточно-опосредованной реакции, в которой неспецифичные цитотоксические клетки (например, NK клетки, нейтрофилы, макрофаги и т.д.) распознают антитело, связавшееся на поверхности клетки-мишени, и после этого вызывают лизис данной клетки-мишени. Такие цитотоксические клетки, опосредующие ADCC, обычно экспрессируют Fc-рецепторы (FcR). Главные клетки для опосредования ADCC (NK клетки) экспрессируют FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII, FcγRIII и/или FcγRIV. Данные по экспрессии FcR на гематопоэтических клетках суммированы в Ravetch and Kinet, Annu. Rev. Immunol., 9: 457-92 (1991). Чтобы оценить ADCC-активность молекулы, можно провести анализ ADCC in vitro, как, например, описано в патентах США №№5500362 или 5821337. Полезные эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (РВМС) и клетки, являющиеся естественными киллерами (NK). Альтернативно или помимо этого, ADCC-активность представляющих интерес молекул можно оценить in vivo, например, на животной модели, такой как описана в Clynes et al., Proc. Natl. Acad. Sci. (USA), 95: 652-656 (1998).
Термины "Fc-рецептор" или "FcR" используют для описания рецептора, который связывается с Fc-областью антитела, при этом Fc-область содержит шарнирную область и CH2- и CH3-домены тяжелой цепи. Например, FcR может иметь нативную последовательность человеческого FcR. FcR может представлять собой рецептор, который связывается с антителом IgG (гамма-рецептор), и включает рецепторы подкпасов FcγRI, FcγRII, FcγRIII и FcγRIV, в том числе аллельные варианты и формы этих рецепторов, подвергнутые альтернативному сплайсингу. Рецепторы FcγRII включают FcγRIIA ("активирующий рецептор") и FcγRIIB ("ингибирующий рецептор"), которые имеют схожие аминокислотные последовательности, имеющие различия главным образом в их цитоплазматических доменах. Активирующий рецептор FcγRIIA содержит иммунорецепторный тирозинсвязанный активаторный мотив (ITAM) в своем цитоплазматическом домене. Ингибирующий рецептор FcγRIIB содержит иммунорецепторный тирозинсвязанный ингибиторный мотив (ITIM) в своем цитоплазматическом домене (см., Daeron, Annu. Rev. Immunol., 15: 203-234 (1997)). Обзоры по FcR приведены в Ravetch and Kinet, Annu. Rev. Immunol., 9: 457-92 (1991); Capel et al., Immunomethods, 4: 25-34 (1994); и de Haas et al., J. Lab. Clin. Med., 126: 330-41 (1995). Другие FcR, включая те, которые будут идентифицированы в будущем, охватываются приведенным в данном описании термином "FcR". Термин также включает неонатальный рецептор, FcRn, который отвечает за перенос материнских IgG к зародышу (Guyer et al., Immunol., 117: 587 (1976) и Kim et al., J. Immunol., 24: 249 (1994)). Важнейший FcR-связывающий сайт на Ре-фрагментах иммуноглобулина расположен в шарнирной области между CH1- и CH2-доменами. Эта шарнирная область взаимодействует с FcR1-3 на различных лейкоцитах и инициирует атаку мишени этими клетками (Wines et al., J. Immunol., 164: 5313-5318 (2000)). Шарнирная область охватывает последовательности, описанные в патенте США №6165476, но этим не ограничивается.
Термин "способный индуцировать антителозависимую клеточную цитотоксичность" относится к способности агента, такого как антитело, демонстрировать ADCC, что можно измерить с использованием анализа(ов), известного(ых) специалистам в данной области. Такая активность обычно характеризуется связыванием Fc-области с различными FcR. Не ограничиваясь каким-либо конкретным механизмом, специалистам в данной области будет понятно, что способность антитела демонстрировать ADCC может, например, быть обусловлена его подклассом (например, IgG1 или IgG3), мутациями, введенными в Fc-область, или обусловлена модификациями углеводных структур в Fc-области антитела. Такие модификации описаны, например, в публикации патентной заявки США №2007/0092521.
Термин "нейтрализующее антитело" относится к антителу, которое связывается с лигандом, предотвращает связывание данного лиганда с его партнером по связыванию и прерывает биологический ответ, который в противном случае являлся бы результатом связывания лиганда с его партнером по связыванию. При оценке связывания и специфичности антитела или его иммунологически функционального фрагмента, антитело или фрагмент будут по существу ингибировать связывание лиганда с его партнером по связыванию в том случае, когда избыток антитела уменьшает количество партнера по связыванию, связанного с лигандом, по меньшей мере примерно на 20%, 30%, 40%, 50%, 60%, 70%, 80%, 85%, 90%, 95%, 97%, 99% или более (как измерено в анализе конкурентного связывания in vitro). В случае антител к Dkk-1 нейтрализующее антитело будет уменьшать способность Dkk-1 связываться с LRP5 или LRP6, тем самым индуцируя поддающееся измерению увеличение активности Wnt.
Термин "конкурировать", когда он применяется в контексте антител, которые конкурируют за один и тот же эпитоп, означает, что конкуренцию между антителами определяют согласно анализу, в котором тестируемое антитело или иммунологически функциональный фрагмент предотвращает или ингибирует специфическое связывание антитела сравнения с общим антигеном (например, Dkk-1 или его фрагментом). Могут быть использованы многочисленные типы анализов конкурентного связывания, например: твердофазный прямой или непрямой радиоиммуноанализ (RIA), твердофазный прямой или непрямой иммуноферментный анализ (EIA), сэндвич-анализ конкурентного связывания (см., например, Stahli et al. (1983) Methods in Enzymology 9: 242-253); твердофазный прямой EIA с использованием системы биотин-авидин (см., например, Kirkland et al., (1986) J. Immunol. 137: 3614-3619), твердофазный анализ с прямым введением метки, твердофазный сэндвич-анализ с прямым введением метки (см., например, Harlow and Lane (1988) Antibodies, A Laboratory Manual, Cold Spring Harbor Press); твердофазный RIA с прямым введением метки 125I (см., например, Morel et al. (1988) Molec. Immunol. 25: 7-15); твердофазный прямой EIA с использованием системы биотин-авидин (см., например, Cheung, et al. (1990) Virology, 176: 546-552); и RIA с прямым введением метки (Moldenhauer et al. (1990) Scand. J. Immunol., 32: 77-82). Обычно такой анализ включает использование очищенного антигена, связанного с твердой поверхностью, или клеток, несущих что-либо одно из немеченого тестируемого иммуноглобулина и меченого иммуноглобулина сравнения. Конкурентное ингибирование измеряют, определяя количество метки, связавшейся с твердой поверхностью или с клетками в присутствии тестируемого иммуноглобулина. Обычно тестируемый иммуноглобулин присутствует в избытке. Антитела, идентифицированные в конкурентном анализе (конкурирующие антитела), включают антитела, связывающиеся с тем же самым эпитопом, что и антитело сравнения, и антитела, связывающиеся с соседними эпитопами, расположенными достаточно близко к эпитопу, с которым связывается антитело сравнения, создавая при этом пространственные затруднения. Дополнительные подробности, касающиеся методов определения конкурентного связывания, приведены в данном описании в разделе Примеры. Обычно, если конкурирующее антитело присутствует в избытке, оно будет ингибировать специфическое связывание антитела сравнения с общим антигеном по меньшей мере на 40%, 45%, 50%, 55%, 60%, 65%, 70% или 75%. В некоторых случаях связывание ингибируется по меньшей мере на 80%, 85%, 90%, 95% или 97% или более.
Термин "антиген" относится к молекуле или части молекулы, которая может быть связана селективным связывающим агентом, таким как антитело, и помимо этого может быть использована в животном для получения антител, которые могут связываться с этим антигеном. Антиген может содержать один или более эпитопов, которые способны взаимодействовать с разными антителами.
Термин "эпитоп" включает любую детерминанту, способную специфически связываться с иммуноглобулином или с Т-клеточным рецептором. Эпитоп представляет собой участок антигена, который связывается антителом, специфически нацеленным на этот антиген, и, когда антиген представляет собой белок, включает конкретные аминокислоты, которые непосредственно контактируют с антителом. Чаще всего эпитопы располагаются на белках, но в некоторых случаях могут располагаться на молекулах других видов, таких как нуклеиновые кислоты. Эпитопные детерминанты могут включать химически активные поверхностные группировки молекул, такие как аминокислоты, углеводные боковые цепи, фосфорильные или сульфонильные группы, и могут обладать специфическими пространственными структурными характеристиками и/или специфическими характеристиками заряда. В большинстве случаев, антитела, специфичные к конкретному антигену-мишени, предпочтительно будут распознавать эпитоп на данном антигене-мишени в сложной смеси белков и/или макромолекул.
Под фразой "специфически связывается", которая использована в данном описании, понимают соединение, например, белок, нуклеиновую кислоту, антитело и тому подобное, которое распознает конкретную молекулу и связывается с ней, но по существу не распознает другие молекулы в образце или не связывается с ними. Например, антитело или пептидный ингибитор, которые распознают родственный лиганд и связываются с ним (например, антитело против Dkk-1, которое связывается с родственный ему антигеном, Dkk-1) в образце, но по существу не распознают другие молекулы в образце или по существу не связываются с ними. Так, в указанных условиях анализа специфически связывающаяся группировка (например, антитело или его фрагмент (например, его антигенсвязывающий участок)) предпочтительно связывается с конкретной молекулой-мишенью, например Dkk-1, и не связывается в значительной мере с другими компонентами, присутствующими в тестируемом образце. Для отбора антитела, которое специфически связывается с представляющей интерес молекулой, можно использовать различные форматы анализа. Например, твердофазный иммуноанализ ELISA, иммунопреципитация, BIAcore, FACS (метод сортировки флуоресцентно-активированных клеток) и анализ вестерн-блоттинг среди многих других видов анализа являются теми видами анализа, которые могут быть использованы для идентификации антитела, специфически взаимодействующего с Dkk-1. В типичном случае, специфическое или селективное взаимодействие будет по меньшей мере двукратно превосходить фоновый сигнал или шум и в более типичном случае превосходить фон больше чем в 10 раз, в еще более конкретном случае считается, что антитело "специфически связывается" с антигеном, если равновесная константа диссоциации (KD) составляет не более 1 мкМ, например не более 100 нМ и также, например, не более 10 нМ.
Считают, что антитело по изобретению "специфически связывается" со своим антигеном-мишенью, если константа диссоциации (Kd) предпочтительно составляет примерно 100 пМ или меньше, примерно 1×10-9 или меньше или 1×10-8 или меньше.
Подразумевается, что термин "kon", как он использован в данном описании, относится к скорости образования комплекса или скорости ассоциации конкретного взаимодействия антитело-антиген, тогда как подразумевается, что термин "koff", как он использован в данном описании, относится к скорости распада комплекса или скорости диссоциации конкретного взаимодействия антитело-антиген. Подразумевается, что термин "KD", как он использован в данном описании, относится к константе диссоциации, которая получается из отношения koff к kon (т.е. koff/kon) и выражается в виде молярной концентрации (М). Величины KD для антител можно определить, используя общепринятые в данной области методы. Один из методов определения KD для антитела заключается в применении поверхностного плазменного резонанса, обычно с использованием биосенсорной системы, такой как система BIAcore®.
Термин "высокоаффинное" для антитела IgG в большинстве случаев относится к антителу с KD 1×10-7 М или меньше, 5×10-8 М или меньше либо 5×10-9 М или меньше, по отношению к антигену-мишени. Однако, "высокоаффинное" связывание может варьировать для других изотипов антитела. Например, термин "высокоаффинное" связывание для изотипа IgM относится к антителу с KD 10-6 М или меньше, 10-7 М или меньше либо 10-8 М или меньше.
Термин "идентичность" относится к взаимосвязи между последовательностями двух или более молекул полипептида или двух или более молекул нуклеиновой кислоты, которая определяется путем выравнивания и сравнения этих последовательностей. "Процент идентичности" означает процент идентичных остатков между аминокислотами или нуклеотидами в сравниваемых молекулах, и его рассчитывают, исходя из размера самой маленькой из молекул, подвергаемых сравнению. Для этих расчетов с учетом разрывов при выравниваниях (если таковые имеются) необходимо обратиться к конкретной математической модели или компьютерной программе (то есть, "алгоритму"). Методы, которые могут быть использованы для расчета идентичности выравниваемых нуклеиновых кислот или полипептидов, включают методы, описанные в Computational Molecular Biology (Lesk, A. М., ed.) (1988) New York: Oxford University Press; Biocomputing Informatics and Genome Projects (Smith, D. W., ed.) (1993) New York: Academic Press; Computer Analysis of Sequence Data, Part I, (Griffin, A. М. and Griffin, H. G., eds.) (1994) New Jersey: Humana Press; von Heinje, G. (1987) Sequence Analysis in Molecular Biology, New York: Academic Press; Sequence Analysis Primer (Gribskov, M. and Devereux, J., eds.) (1991) New York: M. Stockton Press; и Carillo et al. (1988) SIAM J. Applied Math. 48: 1073.
При расчете процента идентичности последовательности, подлежащие сравнению, выравнивают способом, который дает наибольшее совпадение между последовательностями. Компьютерной программой, используемой для определения процента идентичности, является пакет программ GCG, которая включает GAP (Devereux et al. (1984) Nucl. Acid Res. 12: 387; Genetics Computer Group, University of Wisconsin, Madison, Wisc.). Компьютерный алгоритм GAP используется для выравнивания двух полипептидов или полинуклеотидов, для которых должен быть определен процент идентичности последовательностей. Последовательности выравнивают с целью получения оптимального совпадения их соответствующих аминокислот или нуклеотидов ("сходного участка", который определяется этим алгоритмом). Вместе с данным алгоритмом используют "штраф за разрыв" (который рассчитывают в виде умноженной на 3 средней диагонали, где "средняя диагональ" представляет собой среднюю величину диагонали используемой матрицы сравнения; "диагональ" представляет собой балл или количественный показатель, назначаемый за каждое полное совпадение аминокислот в соответствии с конкретной матрицей сравнения) и "штраф за длину разрыва" (который обычно представляет собой 1/10 долю от штрафа за разрыв), а также матрицу сравнения, такую как РАМ (protein weight matrix) 250 или BLOSUM 62. В некоторых воплощениях стандартная матрица сравнения (см. Dayhoff et al. (1978) Atlas of Protein Sequence and Structure, 5: 345-352 для матрицы сравнения РАМ 250; Henikoff et al. (1992) Proc. Natl. Acad. Sci. USA 89: 10915-10919 для матрицы сравнения BLOSUM 62) также используется данным алгоритмом.
Рекомендуемые параметры для определения процента идентичности среди полипептидов или нуклеотидных последовательностей с использованием программы GAP включают следующие:
алгоритм: Needleman et al. (1970) J. Mol. Biol. 48, 443-453;
матрица сравнения: BLOSUM 62 из Henikoff et al. (1992), выше;
штраф за разрыв: 12 (но с отсутствием штрафа за конечные разрывы);
штраф за длину разрыва: 4;
порог схожести: 0.
В результате применения некоторых схем выравнивания для осуществления выравнивания двух аминокислотных последовательностей можно получить совпадение только на коротком участке этих двух последовательностей, и этот небольшой выровненный участок может обладать высокой идентичностью последовательности даже при отсутствии какой-либо значительной взаимосвязи между этими двумя полноразмерными последовательностями. Соответственно, выбранный метод выравнивания (программа GAP) можно адаптировать, если так будет желательно, для обеспечения выравнивания, которое охватывает по меньшей мере 50 следующих друг за другом аминокислот полипептида-мишени.
При использовании в данном описании, "по существу чистый" означает, что описываемый вид молекулы присутствует в качестве преобладающего вида, то есть с точки зрения молярного содержания является более многочисленным по сравнению с любым другим индивидуальным видом в той же самой смеси. В некоторых воплощениях по существу чистая молекула представляет собой композицию, где целевой вид составляет по меньшей мере 50% (с точки зрения молярного содержания) от всех присутствующих макромолекулярных видов. В других воплощениях по существу чистая композиция будет содержать по меньшей мере 80%, 85%, 90%, 95% или 99% данного вида из расчета на все макромолекулярные виды, присутствующие в этой композиции. В других воплощениях целевой вид очищен по существу до гомогенного состояния, при котором в данной композиции традиционными методами детекции невозможно выявить примесные виды, и такая композиция состоит из единственного детектируемого макромолекулярного вида.
Термин "аминокислота" имеет свое обычное значение в данной области. Двадцать природных аминокислот и их сокращения соответствуют общепринятому использованию. См. работу Immunology--A Synthesis, 2nd Edition, (E. S. Golub and D. R. Gren, eds.), Sinauer Associates: Sunderland, Mass. (1991), включенную в данное описание посредством ссылки для любого назначения. Стереоизомеры (например, D-аминокислоты) двадцати стандартных аминокислот, неприродные аминокислоты, такие как α-,α-дизамещенные аминокислоты, N-алкиламинокислоты и другие нестандартные аминокислоты также могут представлять собой подходящие компоненты для полипептидов по изобретению. Примеры нестандартных аминокислот включают: 4-гидроксипролин, гамма-карбоксиглутамат, эпсилон-N,N,N-триметиллизин, эпсилон-N-ацетиллизин, O-фосфосерин, N-ацетилсерин, N-формилметионин, 3-метилгистидин, 5-гидроксилизин, сигма-N-метиларгинин и другие подобные аминокислоты и иминокислоты (например, 4-гидроксипролин). В системе обозначения полипептидов, используемой в данном описании, направление в левую сторону соответствует направлению в сторону аминоконца, а направление в правую сторону соответствует направлению в сторону карбоксильного конца в соответствии с обычным использованием и соглашением.
Термин "остеопения" относится к пациенту с диагнозом уменьшение костной массы, соответствующее по меньшей мере одному стандартному отклонению в сравнении с обычным пациентом, считающимся имеющим нормальную минеральную плотность кости (BMD). Например, этот показатель можно определить посредством двухэнергетической рентгеновской абсорбциометрии (DEXA), и BMD пациента сравнивают со стандартом соответствующего возраста и пола (Z-критерий). При определении остеопении можно провести измерения BMD для одной или более костей.
Термин "терапевтически эффективное количество" относится к количеству антитела против Dkk-1, которое вызывает терапевтический ответ у млекопитающего. Такие терапевтически эффективные количества легко определит обычный специалист в данной области.
"Гликоформа" относится к сложной олигосахаридной структуре, содержащей сочетания различных углеводных единиц. Такие структуры описаны, например, в Essentials of Glycobiology, Varki et al., eds., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1999), где также приведен обзор стандартной номенклатуры по гликобиологии. Такие гликоформы включают G2, G1, G0, G-1 и G-2, но этим не ограничиваются (см., например, WO 99/22764).
Под "характером гликозилирования" понимают строение углеводных единиц, которые ковалентно присоединены к белку (например, гликоформа), а также сайт(ы), по которому(ым) гликоформа(ы) ковалентно присоединена(ы) к пептидному остову белка, более конкретно к белку иммуноглобулину.
По всей вероятности, антитела, экспрессируемые разными клеточными линиями или в трансгенных животных, при сравнении друг с другом будут содержать разные гликоформы и/или иметь разный характер гликозилирования. Тем не менее, все антитела, кодируемые молекулами нуклеиновых кислот, предложенными в данном изобретении, или содержащие аминокислотные последовательности, предложенные в данном изобретении, составляют часть настоящего изобретения, вне зависимости от гликозилирования таких антител.
При использовании в данном описании, термин "субъект" включает любого человека или животного, не являющегося человеком. Термин "животное, не являющееся человеком" включает всех позвоночных, например, млекопитающих и немлекопитающих, таких как отличные от человека приматы, овцы, собаки, кошки, лошади, коровы, цыплята, земноводные, пресмыкающиеся и т.д.
При использовании в данном описании, термин "лечить" означает снижение частоты, с которой симптомы заболевания проявляются у пациента. Термин включает введение соединений или агентов по настоящему изобретению для предупреждения или задержки начала развития симптомов, осложнений или биохимических показателей заболевания, ослабления симптомов или подавления либо ингибирования дальнейшего развития заболевания, состояния или расстройства. Лечение может представлять собой профилактическое (для предупреждения или задержки начала заболевания или для предупреждения проявления его клинических или субклинических симптомов) или терапевтическое подавление или ослабление симптомов после проявления заболевания.
Различные общие аспекты изобретения описаны подробно далее в следующих ниже подразделах.
Краткий обзор
Согласно настоящему изобретению, предложены новые композиции, содержащие антитела и антигенсвязывающие сайты иммуноглобулинов, специфичные к Dkk-1. Некоторые из этих антител и фрагментов антитела могут давать перекрестную реакцию с Dkk-1 из нескольких источников млекопитающих, в том числе с Dkk-1 крысы, мыши и человека. Некоторые из антител и фрагментов обладают более высокой аффинностью к Dkk-1 из одного вида, нежели из другого. Согласно изобретению, также предложены новые нейтрализующие антитела, включая химерные, гуманизированные и человеческие антитела, а также антитела и их иммунологически функциональные фрагменты. Также описаны нуклеиновые кислоты, кодирующие антитела и фрагменты, а также способы экспрессии антител с использованием этих нуклеиновых кислот. В другом аспекте изобретение относится к молекулам (например, иммунологически функциональным фрагментам и полипептидам), которые способны проявлять иммунологические свойства связывания антигенсвязывающих сайтов антитела.
Антитела и иммунологически функциональные фрагменты, раскрытые в данном изобретении, имеют ряд полезных применений. Некоторые из антител и фрагментов, например, полезны в анализах специфического связывания, при аффинной очистке Dkk-1 или его лигандов и в скрининг-анализах с целью идентификации других антагонистов активности Dkk-1. Некоторые из антител могут применяться для лечения различных заболеваний, которые ассоциированы с активностью Dkk-1. Таким образом, некоторые антитела и фрагменты могут быть использованы в ряде способов лечения, относящихся к кости, таких как повышение BMD, синтез новой кости, лечение системной потери костной массы (например, костных эрозий), репарация костной ткани и лечение различных форм артрита. Тем не менее, некоторые из описанных антител и фрагментов могут быть использованы для лечения ряда разнообразных заболеваний, которые не являются родственными заболеваниям костной системы.
Антитела и фрагменты
Предложены разнообразные селективные связывающие агенты, полезные для регуляции активности Dkk-1. Эти агенты включают, например, антитела и их иммунологически функциональные фрагменты, которые содержат антигенсвязывающий домен (например, одноцепочечные антитела, однодоменные антитела, иммуноадгезины и полипептиды с антигенсвязывающим участком), и специфически связываются с полипептидом Dkk-1 (например, полипептидом Dkk-1 человека, крысы и/или мыши). Некоторые из агентов, например, полезны в ингибировании связывания Dkk-1 с LRP5 и/или LRP6 и поэтому могут быть использованы для стимулирования одной или более активностей, ассоциированных с передачей сигнала через Wnt.
Структура природных антител
Некоторые из предложенных связывающих агентов имеют структуру, обычно ассоциируемую с природными антителами. Структурные единицы этих антител обычно содержат один или более тетрамеров, каждый из которых состоит из двух идентичных двухэлементных структур полипептидных цепей, хотя некоторые виды млекопитающих также продуцируют антитела, имеющие только одну тяжелую цепь. В типичном антителе каждая пара или двухэлементная структура включает одну полноразмерную "легкую" цепь (в некоторых воплощениях примерно 25 кДа) и одну полноразмерную "тяжелую" цепь (в некоторых воплощениях примерно 50-70 кДа). Каждая индивидуальная иммуноглобулиновая цепь состоит из нескольких "иммуноглобулиновых доменов", каждый из которых состоит примерно из 90-110 аминокислот и демонстрирует характерную картину фолдинга. Эти домены представляют собой основные единицы, из которых состоят полипептиды антител. Аминоконцевая часть каждой цепи обычно включает вариабельный домен, который ответственен за антигенное распознавание. Карбоксиконцевая часть эволюционно более консервативна, чем другой конец данной цепи, и называется "константной областью" или "С-областью". Легкие цепи иммуноглобулина человека в большинстве случаев классифицируются как легкие цепи каппа и лямбда, и каждая их них содержит один вариабельный домен и один константный домен. Обычно тяжелые цепи классифицируют как цепи мю, дельта, гамма, альфа или эпсилон, и они определяют изотип антитела как IgM, IgD, IgG, IgA и IgE, соответственно. IgG имеет несколько подтипов, включая IgG1, IgG2, IgG3 и IgG4, но этим не ограничивается. Подтипы IgM включают IgM1 и IgM2. Подтипы IgA включают IgA1 и IgA2. У людей изотипы IgA и IgD содержат четыре тяжелые цепи и четыре легкие цепи; изотипы IgG и IgE содержат две тяжелые цепи и две легкие цепи; а изотип IgM содержит пять тяжелых цепей и пять легких цепей. С-область тяжелой цепи обычно содержит один или более доменов, которые могут отвечать за эффекторную функцию. Количество доменов константной области тяжелой цепи будет зависеть от изотипа. Например, каждая из тяжелых цепей IgG содержит три домена С-области, известные как СН1, СН2 и СН3. Предложенные антитела могут иметь любой из этих изотипов и подтипов.
В полноразмерных легких и тяжелых цепях вариабельные и константные области соединены областью "J", состоящей примерно из 12 или более аминокислот, при этом тяжелая цепь также включает область "D", состоящую примерно из 10 или более аминокислот. См., например, Fundamental Immunology, 2nd ed., Ch. 7 (Paul, W., ed.) (1989) New York: Raven Press (включено сюда посредством ссылки во всей своей полноте для всех назначений). Вариабельные области каждой пары легкая/тяжелая цепь обычно образуют антигенсвязывающий сайт.
Вариабельные области иммуноглобулиновых цепей в большинстве случаев демонстрируют одну и ту же общую структуру, содержащую относительно консервативные каркасные области (FR), соединенные тремя гипервариабельными участками, чаще называемыми как "определяющие комплементарность участки" или CDR. CDR из двух цепей каждой пары тяжелая цепь/легкая цепь, упомянутой выше, обычно ориентированы относительно друг друга благодаря каркасным областям с образованием структуры, которая связывается специфически с конкретным эпитопом на белке-мишени (например, Dkk-1). Расположение этих элементов от N-конца к С-концу природных вариабельных областей обеих легкой и тяжелой цепей обычно соответствует следующему порядку: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Разработана система нумерации для присвоения номера аминокислотам, которые занимают положения в каждом из этих доменов. Эта система нумерации определена в Kabat Sequences of Proteins of Immunological Interest ((1987 and 1991) National Institutes of Health, Bethesda, Md.) или Chothia & Lesk (1987) J. Mol. Biol. 196: 901-917; Chothia et al. (1989) Nature 342: 878-883. Каждую из легких цепей, предложенных настоящим изобретением, можно объединить с любой из тяжелых цепей, предложенных настоящим изобретением, с образованием антитела. В некоторых воплощениях антитела содержат две идентичные легкие цепи и две идентичные тяжелые цепи. Другие предложенные антитела представляют собой варианты антител, образованных путем комбинирования предложенных тяжелых и легких цепей, и содержат легкие и/или тяжелые цепи, каждая из которых идентична по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 97% или 99% аминокислотным последовательностям этих цепей. В некоторых случаях такие антитела включают по меньшей мере одну тяжелую цепь и одну легкую цепь, тогда как в других случаях такие вариантные формы содержат две идентичные легкие цепи и две идентичные тяжелые цепи.
CDR антител
Гипервариабельные участки (CDR) и каркасные области (FR) заданного антитела можно идентифицировать, используя систему, описанную Kabat et al. в Sequences of Proteins of Immunological Interest, 5th Ed., US Dept. of Health and Human Services, PHS, NIH, NIH Publication no. 91-3242 (1991). Некоторые антитела, изложенные в данном описании, содержат одну или более аминокислотных последовательностей, которые идентичны аминокислотным последовательностям или имеют значительную идентичность последовательности с аминокислотными последовательностями одного или более предложенных CDR.
Предложенные антитела и фрагменты могут включать один, два, три, четыре, пять или все шесть CDR, приведенных выше. Некоторые антитела имеют вариантные формы CDR, приведенные в Таблице 4, причем один или более чем один (то есть, 2, 3, 4, 5 или 6) CDR каждый имеет по меньшей мере 80%, 85%, 90% или 95% идентичности последовательности с предложенной конкретной последовательностью CDR.
Конкурирующие антитела и фрагменты
Также предложены антитела и их иммунологически функциональные фрагменты, которые конкурируют с одним из взятых в качестве примера антител или функциональных фрагментов за специфическое связывание с Dkk-1. Такие антитела и фрагменты также могут связываться с тем же эпитопом, что и одно из взятых в качестве примера антител. Ожидается, что антитела и фрагменты, которые конкурируют с тем же самым эпитопом или связываются с ним, как и взятые в качестве примера антитело или фрагмент, будут демонстрировать аналогичные функциональные свойства.
Моноклональные антитела
Предложенные антитела включают моноклинальные антитела, которые связываются с Dkk-1. Моноклональные антитела можно получить, используя любой способ, известный в данной области, например, посредством иммортализации клеток селезенки, собранных у трансгенного животного после завершения схемы иммунизации. Клетки селезенки можно подвергнуть иммортализации, используя любой метод, известный в данной области, например путем слияния их с миеломными клетками с получением гибридомы. Миеломные клетки для использования в процедурах слияния с целью получения гибридомы предпочтительно представляют собой клетки, не продуцирующие антител, характеризуются высокой эффективностью слияния и недостаточностью ферментов, что делает их неспособными расти на некоторых селективных средах, которые поддерживают рост только желаемых слитых клеток (гибридомы). Примеры подходящих клеточных линий для использования в получении слитых мышиных конструкций включают Sp-20, P3-X63/Ag8, P3-X63-Ag8.653, NS1/1.Ag 4 1, Sp210-Ag14, FO, NSO/U, MPC-11, MPC11-X45-GTG 1.7 и S194/5XXO Bul; примеры клеточных линий, используемых в получении слитых крысиных конструкций, включают R210.RCY3, Y3-Ag 1.2.3, IR983F и 4В210. Другими клеточными линиями, полезными для получения клеточных слитых конструкций, являются U-266, GM1500-GRG2, LICR-LON-НМу2 и UC729-6.
В некоторых случаях линию гибридомных клеток получают путем иммунизации животного (например, трансгенного животного, имеющего последовательности человеческого иммуноглобулина) иммуногеном Dkk-1; сбора клеток селезенки от иммунизированного животного; слияния собранных клеток селезенки с линией миеломных клеток, посредством чего образуются гибридомные клетки; создания линии гибридомных клеток из гибридомных клеток и идентификации линии гибридомных клеток, которая продуцирует антитело, связывающееся с полипептидом Dkk-1. Такие линии гибридомных клеток и моноклональные антитела против Dkk-1, продуцируемые ими, охватываются настоящим изобретением.
Моноклональные антитела, секретируемые линией гибридомных клеток, можно очистить, используя любой метод, известный в данной области. Затем можно провести скрининг гибридомы или Mab на предмет идентификации Мао, обладающих конкретными свойствами, такими как способность блокировать Wnt-индуцированную активность. Примеры таких скринингов приведены ниже в разделе Примеры.
Химерные и гуманизированные антитела
Также предложены химерные и гуманизированные антитела, основу которых составляют вышеупомянутые последовательности. Моноклональные антитела для использования в качестве терапевтических агентов перед применением могут быть модифицированы различными способами. Одним из примеров является "химерное" антитело, которое представляет собой антитело, составленное из белковых сегментов от разных антител, соединенных ковалентной связью с получением функциональных легких или тяжелых цепей иммуноглобулина или их иммунологически функциональных частей. В большинстве случаев часть тяжелой цепи и/или легкой цепи идентична или гомологична соответствующей последовательности в антителах, происходящих из конкретного вида или принадлежащих конкретному классу или подклассу антител, тогда как остальная часть цепи(ей) идентична или гомологична соответствующей последовательности в антителах, происходящих из другого вида или принадлежащих другому классу или подклассу антител. О способах, относящихся к химерным антителам, см., например, патент США №4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA, 81: 6851-6855 (1985). CDR-привитие описано, например, в патентах США №№6180370, 5693762, 5693761, 5585089 и 5530101.
Обычно целью получения химерного антитела является создание химерной конструкции, в которой количество аминокислот из видов предполагаемых пациентов является максимальным. Одним из примеров является "CDR-привитое" антитело, в котором антитело содержит один или более гипервариабельных участков (CDR) из конкретного вида или принадлежащих конкретному классу или подклассу антител, тогда как остальная часть цепи(ей) антитела идентична или гомологична соответствующей последовательности в антителах, происходящих из другого вида или принадлежащих другому классу или подклассу антител. С целью применения на людях V-область или отдельные CDR из антитела грызунов часто прививают к человеческому антителу, заменяя природные V-области или CDR человеческого антитела.
Одним из полезных типов химерного антитела является "гуманизированное" антитело. В большинстве случаев гуманизированное антитело получают из моноклонального антитела, первоначально происходящего из животного, не являющегося человеком. Некоторые аминокислотные остатки в этом моноклональном антителе, обычно из частей антитела, не распознающих антиген, модифицированы таким образом, чтобы они были гомологичными соответствующим остаткам в человеческом антителе соответствующего изотипа. Гуманизацию можно осуществить, например, с использованием различных методов, заменяя по меньшей мере часть вариабельной области антитела грызунов на соответствующие области человеческого антитела (см., например, патенты США №№5585089 и 5693762; Jones et al. (1986) Nature, 321: 522-25; Riechmann et al. (1988) Nature, 332: 323-27; Verhoeyen et al. (1988) Science, 239: 1534-36). В некоторых воплощениях для получения гибридных антител константные области видов, отличных от человека, могут быть использованы наряду с вариабельной(ыми) областью(ями) антител человека.
Полностью человеческие антитела
Также предложены полностью человеческие антитела. Существуют способы получения полностью человеческих антител, специфичных к заданному антигену, при которых людей не подвергают действию антигена ("полностью человеческие антитела"). Одним из способов получения полностью человеческих антител является "гуманизация" гуморальной иммунной системы мыши. Введение локусов человеческого иммуноглобулина (Ig) мышам, у которых эндогенные гены Ig инактивированы, является одним из способов получения полностью человеческих моноклональных антител (Mab) в мышах, животных, которые можно иммунизировать любым желаемым антигеном. Используя полностью человеческие антитела, можно минимизировать иммуногенные и аллергические ответы, которые иногда могут вызываться введением людям мышиных Mab или производных мышиных Mab в качестве терапевтических агентов.
Полностью человеческие антитела можно получить путем иммунизации трансгенных животных (обычно мышей), способных продуцировать репертуар человеческих антител при отсутствии продукции эндогенного иммуноглобулина. Используемые для этой цели антигены обычно содержат шесть или более следующих друг за другом аминокислот и возможно конъюгированы с носителем, таким как гаптен. См., например, Jakobovits et al. (1993) Proc. Natl. Acad. Sci. USA, 90: 2551-2555; Jakobovits et al. (1993) Nature, 362: 255-258; и Bruggermann et al. (1993) Year in Immunol., 7: 33. В одном из примеров применения такого способа трансгенных животных получают, делая у них недееспособными локусы эндогенных мышиных иммуноглобулинов, кодирующие тяжелую и легкую цепи иммуноглобулинов мыши, и вводя в мышиный геном большие фрагменты ДНК генома человека, содержащие локусы, которые кодируют тяжелую и легкую цепь белков человека. Частично модифицированных животных, имеющих не полный комплект человеческих иммуноглобулиновых локусов, далее подвергают кроссбридингу с целью получения животного, имеющего все желаемые модификации иммунной системы. После того, как иммуноген введен, эти трансгенные животные вырабатывают антитела, которые являются иммуноспецифичными к данному иммуногену, но содержат аминокислотные последовательности (включая вариабельные области) человека, а не мыши. Для получения более подробной информации о таких способах см., например, WO96/33735 и WO94/02602, которые тем самым включены посредством ссылки. Дополнительные способы, касающиеся использования трансгенных мышей для получения человеческих антител описаны в патентах США №№5545807, 6713610, 6673986, 6162963, 5545807, 6300129, 6255458, 5877397, 5874299 и 5545806; в WO91/10741 и WO90/04036 и в ЕР 546073 В1 и ЕР 546073А1.
Описанные выше трансгенные мыши, обозначенные в данном описании как мыши "HuMab", содержат минилокус гена человеческого иммуноглобулина, который кодирует последовательности неаранжированных тяжелых (µ и γ) и легкой (κ) цепей иммуноглобулина человека вместе с направленными мутациями, которые инактивируют эндогенные локусы µ- и κ-цепей (Lonberg et al. (1994) Nature, 368: 856-859). Соответственно, в ответ на иммунизацию мыши демонстрируют сниженную экспрессию IgM и κ-цепи мыши, и трансгены с введенными тяжелыми и легкими цепями иммуноглобулина человека подвергаются переключению класса (class switching) и соматическим мутациям, генерируя высокоаффинные моноклональные антитела IgG человека с κ-цепью (Lonberg et al., выше; Lonberg and Huszar (1995) Intern. Rev. Immunol., 13: 65-93; Harding and Lonberg (1995) Ann. N. Y Acad. Sci. 764: 536-546). Получение мышей HuMab подробно описано в Taylor et al. (1992) Nucleic Acids Research, 20: 6287-6295; Chen et al. (1993) International Immunology 5: 647-656; Tuaillon et al. (1994) J. Immunol. 152: 2912-2920; Lonberg et al. (1994) Nature 368: 856-859; Lonberg (1994) Handbook of Exp. Pharmacology 113: 49-101; Taylor et al. (1994) International Immunology 6: 579-591; Lonberg and Huszar (1995) Intern. Rev. Immunol. 13: 65-93; Harding and Lonberg, 1995, Ann. N. Y Acad. Sci. 764: 536-546; Fishwild et al. (1996) Nature Biotechnology 14: 845-851; тем самым вышеупомянутые ссылки включены во всей своей полноте для всех назначений посредством ссылки. См. также патенты США №№5545806, 5569825, 5625126, 5633425, 5789650, 5877397, 5661016, 5814318, 5874299 и 5770429; а также патент США №5545807; и WO 93/1227; WO 92/22646; и WO 92/03918. Технологии, применяемые для получения человеческих антител в этих трансгенных мышах, описаны также в WO 98/24893 и Mendez et al. (1997) Nature Genetics 15: 146-156, которые тем самым включены посредством ссылки. Например, для создания человеческих антител против Dkk-1 можно использовать линии трансгенных мышей НСо7 и НСо12.
Используя гибридомную технологию, можно получить и отобрать из трансгенных мышей, которые описаны выше, антигенспецифические человеческие MAb с желаемой специфичностью. Такие антитела можно клонировать и экспрессировать, используя подходящий вектор и подходящую клетку-хозяина, или эти антитела можно собрать из культивируемых гибридомных клеток.
Полностью человеческие антитела также могут происходить из библиотек фагового дисплея (как описано в Hoogenboom et al. (1991) J. Mol. Biol. 227: 381; и Marks et al. (1991) J. Mol. Biol. 222: 581). Методы фагового дисплея имитируют иммунную селекцию посредством проявления репертуаров антител на поверхности нитевидного бактериофага и последующей селекции фага на основании его связывания с выбранным антигеном. Один из таких методов описан в WO99/10494 (включена посредством ссылки), где описано выделение высокоаффинных и функциональных агонистических антител для рецепторов MPL и msk (митоген- и стресс-активируемая протеинкиназа) с использованием такого подхода.
Биспецифические или бифункциональные антитела
Предложенные антитела также включают биспецифические и бифункциональные антитела, которые содержат один или более CDR или одну или более вариабельных областей, как описано выше. Биспецифическое или бифункциональное антитело в некоторых случаях представляет собой искусственное гибридное антитело, имеющее две разные пары тяжелых/легких цепей и два разных сайта связывания. Биспецифические антитела могут быть получены рядом способов, включая, но этим не ограничиваясь, слияние гибридом или связывание Fab'-фрагментов. См., например, Songsivilai & Lachmann (1990) Clin. Exp. Immunol. 79: 315-321; Kostelny et al. (1992) J. Immunol. 148: 1547-1553.
Различные другие формы
Некоторые из предложенных антител или иммунологически функциональных фрагментов представляют собой вариантные формы антител и фрагментов, описанных выше. Природные аминокислоты можно подразделить на классы на основании общих свойств боковой цепи: 1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile; 2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln; 3) кислотные: Asp, Glu; 4) основные: His, Lys, Arg; 5) остатки, влияющие на ориентацию цепи: Gly, Pro; и 6) ароматические: Trp, Tyr, Phe. Консервативные аминокислотные замены могут включать замену члена одного из этих классов на другой член того же класса. Консервативные аминокислотные замены могут охватывать неприродные аминокислотные остатки, которые обычно вводят посредством химического пептидного синтеза, а не синтеза в биологических системах. Сюда же включены пептидомиметики и другие обращенные или инвертированные формы аминокислотных группировок. Неконсервативные замены могут включать замену члена одного из вышеупомянутых классов на другой член из другого класса. Такие вводимые заменой остатки можно ввести в участки антитела, гомологичные человеческим антителам, или в негомологичные участки молекулы.
При осуществлении таких изменений, в соответствии с некоторыми воплощениями, можно учитывать индекс гидрофобности аминокислот. Профиль гидрофобности белка рассчитывают, приписывая каждой аминокислоте численное значение ("индекс гидрофобности") и затем проводя повторное усреднение этих значений по всей пептидной цепи. Каждой аминокислоте приписан индекс гидрофобности на основании характеристик ее гидрофобности и заряда. Они составляют: изолейцин (+4,5); валин (+4,2); лейцин (+3,8); фенилаланин (+2,8); цистеин/цистин (+2,5); метионин (+1,9); аланин (+1,8); глицин (-0,4); треонин (-0,7); серин (-0,8); триптофан (-0,9); тирозин (-1,3); пролин (-1,6); гистидин (-3,2); глутамат (-3,5); глутамин (-3,5); аспартат (-3,5); аспарагин (-3,5); лизин (-3,9); и аргинин (-4,5).
Важность профиля гидрофобности в придании белку согласованной биологической функции в данной области является очевидной (см., например, Kyte et al., 1982, J. Mol. Biol., 157: 105-131). Известно, что некоторые аминокислоты можно заменить на другие аминокислоты, имеющие аналогичный индекс или показатель гидрофобности с сохранением аналогичной биологической активности. При внесении изменений, основанных на индексе гидрофобности, в некоторых воплощениях включена замена аминокислот, индексы гидрофобности которых находятся в пределах ±2. В некоторых аспектах изобретения включены случаи, где индексы гидрофобности находятся в пределах ±1, а в других аспектах изобретения включены случаи, где индексы гидрофобности находятся в пределах ±0,5.
В данной области также очевидно, что замену схожих аминокислот можно эффективно провести на основании гидрофильности, в частности, когда биологически функциональный белок или пептид, созданный таким образом, предполагается использовать в иммунологических воплощениях, как в настоящем случае. В некоторых воплощениях самая высокая локальная средняя гидрофильность белка, которая определяется гидрофильностью расположенных близко к этому месту аминокислот, коррелирует с его иммуногенностью и антигенсвязывающими свойствами или иммуногенностью, то есть с биологическим свойством данного белка.
Данным аминокислотным остаткам приписаны следующие значения гидрофильности: аргинин (+3,0); лизин (+3,0); аспартат (+3,0±1); глутамат (+3,0 1); серин (+0,3); аспарагин (+0,2); глутамин (+0,2); глицин (0); треонин (-0,4); пролин (-0,5±1); аланин (-0,5); гистидин (-0,5); цистеин (-1,0); метионин (-1,3); валин (-1,5); лейцин (-1,8); изолейцин (-1,8); тирозин (-2,3); фенилаланин (-2,5) и триптофан (-3,4). При внесении изменений, основанных на схожих значениях гидрофильности, в некоторых воплощениях включена замена аминокислот, значения гидрофильности которых находятся в пределах ±2, в других воплощениях включены случаи, где значения гидрофильности находятся в пределах ±1, и в следующих воплощениях включены случаи, где значения гидрофильности находятся в пределах ±0,5. В некоторых случаях на основании гидрофильности также можно идентифицировать эпитопы из первичных аминокислотных последовательностей. Эти участки также обозначаются как "коровые области эпитопов (epitopic core regions)".
Специалист в данной области будет способен определить подходящие варианты полипептидов, которые приведены в данном описании, с использованием хорошо известных методов. Специалист в данной области может идентифицировать подходящие области молекулы, которые могут быть изменены без разрушения активности путем мечения участков, не считающихся важными для активности. Специалист в данной области также будет способен идентифицировать остатки и участки молекул, которые считаются консервативными среди схожих полипептидов. В следующих воплощениях консервативным аминокислотным заменам без разрушения биологической активности или без неблагоприятного воздействия на структуру полипептида могут быть подвергнуты даже участки, которые могут быть важны для биологической активности или для структуры.
Помимо этого, специалист в данной области может составить обзор по исследованиям структурно-функциональных свойств, позволяющим идентифицировать остатки в схожих полипептидах, которые являются важными для активности или структуры. С учетом такого сравнения можно предсказать важность аминокислотных остатков в белке, которые соответствуют аминокислотным остаткам, важным для активности или структуры в схожих белках. Специалист в данной области может сделать выбор химически схожих аминокислотных замен для таких предсказанных важных аминокислотных остатков.
Специалист в данной области также может провести анализ пространственной структуры и аминокислотной последовательности по сравнению с подобной структурой в аналогичных полипептидах. С учетом такой информации специалист в данной области может предсказать ориентацию аминокислотных остатков антитела, принимая во внимание его пространственную структуру. Специалист в данной области может решить не вносить радикальных изменений в аминокислотные остатки, которые согласно прогнозу находятся на поверхности белка, поскольку такие остатки могут быть вовлечены в важные взаимодействия с другими молекулами. Вместе с тем, специалист в данной области может создать тест-варианты, содержащие единственную аминокислотную замену в каждом желаемом аминокислотном остатке. Затем можно провести скрининг этих вариантов с использованием анализов Dkk1-нейтрализующей активности (см. примеры ниже) и таким образом получить информацию касательно того, какие аминокислоты могут быть изменены, а какие не должны быть изменены. Другими словами, на основе информации, почерпнутой из таких рутинных экспериментов, специалист в данной области может легко определить положения аминокислот, где следует избегать других замен, или только их одних, или в комбинации с другими мутациями.
Ряд научных публикаций посвящен предсказанию вторичной структуры. См. Moult (1996) Curr. Op. in Biotech. 7: 422-427; Chou et al. (1974) Biochemistry, 13: 222-245; Chou et al. (1974) Biochemistry, 113: 211-222; Chou et al. (1978) Adv. Enzymol. Relat. Areas Mol. Biol. 47: 45-148; Chou et al. (1979) Ann. Rev. Biochem. 47: 251-276; и Chou et al. (1979) Biophys. J., 26: 367-384. Вместе с тем сейчас доступны компьютерные программы, помогающие предсказывать вторичную структуру. Один из способов предсказания вторичной структуры основан на моделировании по гомологии. Например, два полипептида или белка, которые имеют идентичность последовательности более 30% или схожесть более 40%, часто имеют схожие структурные топологии. Происшедший за последнее время рост базы данных белковых структур (PDB) предоставил расширенные возможности для предсказания вторичной структуры, включая предсказания возможного числа вариантов укладки в структуре полипептида или белка. См. Holm et al. (1999) Nucl. Acid. Res., 27: 244-247. Полагают (Brenner et al. (1997) Curr. Op. Struct. Biol., 7: 69-376), что существует ограниченное число вариантов укладки в заданном полипептиде или белке, и что как только критическое число структур будет установлено, структурные предсказания станут значительно более точными.
Дополнительные методы предсказания вторичной структуры включают метод "протягивания (threading)" (Jones (1997) Curr. Opin. Struct. Biol. 7: 377-87; Sippl et al. (1996) Structure, 4: 15-19), "анализ профилей" (Bowie et al. (1991) Science, 253: 164-170; Gribskov et al. (1990) Meth. Enzym., 183: 146-159; Gribskov et al. (1987) Proc. Nat. Acad. Sci., 84: 4355-4358) и метод "эволюционной взаимосвязи" (см. Holm (1999), выше; и Brenner (1997), выше).
В некоторых воплощениях изобретения сделаны аминокислотные замены, которые: (1) снижают чувствительность к протеолизу, (2) снижают чувствительность к окислению, (3) изменяют аффинность связывания в отношении образования белковых комплексов, (4) изменяют аффинности связывания с лигандами или антигенами и/или (5) придают или модифицируют другие физико-химические или функциональные свойства таких полипептидов. Например, в природной последовательности можно сделать единственную или множественные аминокислотные замены (в некоторых воплощениях консервативные аминокислотные замены). Замены могут быть сделаны в той части антитела, которая лежит вне домена(ов), образующего(их) межмолекулярные контакты. В таких воплощениях можно использовать консервативные аминокислотные замены, которые по существу не изменяют структурных характеристик исходной последовательности (например, одну или более замен аминокислот, которые не нарушают вторичную структуру, характерную для исходного или нативного антитела). Примеры принятых в данной области вторичных и третичных структур полипептидов описаны в Proteins, Structures and Molecular Principles (Creighton, Ed.) (1984) W. H. New York: Freeman and Company; Introduction to Protein Structure (Branden and Tooze, eds.) (1991) New York: Garland Publishing; и Thornton et al. (1991) Nature, 354: 105.
Изобретение также охватывает варианты гликозилирования антител по изобретению, где количество и/или тип сайта(ов) гликозилирования изменены по сравнению с аминокислотными последовательностями исходного полипептида. В некоторых воплощениях белковые варианты антитела содержат большее или меньшее количество сайтов N-гликозилирования, чем нативное антитело. Сайт N-гликозилирования характеризуется последовательностью Asn-X-Ser или Asn-X-Thr, где аминокислотный остаток, обозначенный как X, может быть любым аминокислотным остатком за исключением пролина. Замена аминокислотных остатков с целью создания этой последовательности приводит к появлению возможного нового сайта для добавления N-связанной углеводной цепи. Альтернативно, замены, в результате которых эта последовательность удаляется или изменяется, будут препятствовать добавлению N-связанной углеводной цепи, присутствующей в нативном полипептиде. Например, в результате делеции Asn или в результате замены Asn на другую аминокислоту степень гликозилирования может быть снижена. В других воплощениях создан один или более чем один новый сайт N-гликозилирования. Антитела обычно содержат сайт N-гликозилирования в Fc-области.
Дополнительные предпочтительные варианты антител включают цистеиновые варианты, где один или более остатков цистеина в исходной или нативной аминокислотной последовательности делегированы или заменены другой аминокислотой (например, серином). Цистеиновые варианты полезны, среди прочего, когда антитела должны быть подвергнуты рефолдингу с образованием биологически активной конформации. Цистеиновые варианты могут содержать меньше остатков цистеина, чем нативное антитело, и обычно содержат четное количество, чтобы свести к минимуму взаимодействия, обусловленные неспаренными цистеинами.
Описываемые тяжелые и легкие цепи, домены вариабельных областей и CDR могут быть использованы для получения полипептидов, содержащих антигенсвязывающий участок, который может специфически связываться с полипептидом Dkk-1. Например, один или более CDR, приведенных в Таблице 4, может быть встроен в молекулу (например, полипептид) ковалентно или нековалентно с получением иммуноадгезина. Иммуноадгезин может содержать один или более CDR в виде части более длинной полипептидной цепи, может содержать ковалентно связанный один или более CDR с другой полипептидной цепью или может содержать один или более нековалентно связанных CDR. Присутствие одного или более CDR дает возможность иммуноадгезину связываться специфически с конкретным представляющим интерес антигеном (например, полипептидом Dkk-1 или его эпитопом).
Также предложены миметики (например, "пептидные миметики" или "пептидомиметики") на основе доменов вариабельной области и CDR, которые изложены в данном описании. Эти аналоги могут представлять собой пептиды, непептидные структуры или комбинации пептидных и непептидных участков. Fauchere (1986) Adv. Drug Res. 15: 29; Veber and Freidinger (1985) TINS, p.392; и Evans et al. (1987) J. Med. Chem. 30: 1229, которые включены в данное описание посредством ссылки для любого назначения. Пептидные миметики, которые схожи по структуре с терапевтически полезными пептидами, можно использовать для получения аналогичного терапевтического или профилактического эффекта. Такие соединения часто разрабатывают с помощью компьютеризированного молекулярного моделирования. В большинстве случаев пептидомиметиками по изобретению являются белки, которые схожи по структуре с антителом, проявляющим желаемую биологическую активность, такую как упомянутая в данном описании способность специфически связываться с Dkk-1, но у которых одна или более пептидных связей возможно заменена связью, выбранной из: -CH2NH-, -CH2S-, -СН2-СН2-, -СН=СН-(цис и транс), -СОСН2-, -СН(ОН)СН2- и -CH2SO-, с использованием методов, хорошо известных в данной области. Системная замена одной или более аминокислот консенсусной последовательности D-аминокислотой того же типа (например, D-лизином вместо L-лизина) может быть использована в некоторых воплощениях изобретения для создания более стабильных белков. Помимо этого, "изогнутые" пептиды, содержащие консенсусную последовательность или по существу идентичный вариант консенсусной последовательности, могут быть созданы способами, известными в данной области (Rizo and Gierasch (1992) Ann. Rev. Biochem., 61: 387, включено в данное описание посредством ссылки), например, путем добавления внутренних цистеиновых остатков, способных образовывать внутримолекулярные дисульфидные мостики, что приводит к циклизации пептида.
Также предложены производные антител и иммунологически функциональные фрагменты, которые изложены в данном описании. Производное антитела или фрагмент может содержать любую молекулу или любое вещество, которые придают желаемое свойство антителу или фрагменту, такое как увеличенный период полувыведения в условиях конкретного применения. Производное антитела может содержать, например, детектируемую (или метящую) группировку (например, молекулу радиоактивного вещества, вещества, пригодного для колориметрических исследований, молекулу антигена или фермента, детектируемую гранулу (такую как магнитная или обладающая большой электрической плотностью (например, золотая) гранула) или молекулу, которая связывается с другой молекулой (например, биотин или стрептавидин)), терапевтическую или диагностическую группировку (например, радиоактивную, цитотоксическую или фармацевтически активную группировку) или молекулу, которая улучшает пригодность данного антитела для конкретного применения (например, для введения субъекту, такому как человек, или для других применений in vivo или in vitro). Примеры молекул, которые могут быть использованы для получения производных антитела, включают альбумин (например, сывороточный альбумин человека) и полиэтиленгликоль (ПЭГ). Связанные с альбумином и пегилированные производные антител могут быть получены с использованием методов, хорошо известных в данной области. В одном из воплощений антитело конъюгировано или связано иным образом с транстиретином (TTR) или вариантом TTR. TTR или вариант TTR может быть химически модифицирован, например химическим реагентом, выбранным из группы, состоящей из декстрана, поли(н-винилпирролидона), полиэтиленгликолей, гомополимеров пропиленгликоля, сополимеров полипропиленоксида/ этиленоксида, полиоксиэтилированных полиолов и поливиниловых спиртов.
Другие производные включают ковалентные или агрегированные конъюгаты антител против Dkk-1 или их фрагментов с другими белками или полипептидами, как например, полученные в результате экспрессии рекомбинантных слитых белков, содержащих гетерологичные полипептиды, слитые с N-концом или С-концом полипептида антитела против Dkk-1. Например, конъюгированный пептид может представлять собой гетерологичный сигнальный (или лидерный) полипептид, например, лидерный пептид альфа-фактора дрожжей, или такой пептид, как эпитопная метка. Слитые белки, содержащие антитело против Dkk-1, могут содержать пептиды, добавленные для облегчения очистки или идентификации антитела против Dkk-1 (например, поли-His). Полипептид антитела против Dkk-1 также может быть соединен с пептидом FLAG, как описано в Норр et al., Bio/Technology 6: 1204 (1988) и патенте США №5011912. Пептид FLAG обладает высокой антигенностью и обеспечивает обратимое связывание эпитопа специфическим моноклональным антителом (Mab), делая возможным быстрое проведение анализа и облегчая очистку экспрессируемого рекомбинантного белка. Реагенты, полезные для получения слитых белков, в которых пептид FLAG является слитым с заданным полипептидом, имеются в продаже (Sigma, St. Louis, Mo.).
Олигомеры, которые содержат один или более полипептидов антитела против Dkk-1, могут быть использованы в качестве антагонистов Dkk-1. Олигомеры могут быть в форме соединенных ковалентной связью или соединенных нековалентной связью димеров, тримеров или высших олигомеров. Предполагается использовать олигомеры, содержащие два или более полипептидов антитела против Dkk-1, причем одним из примеров является гомодимер. Другие олигомеры включают гетеродимеры, гомотримеры, гетеротримеры, гомотетрамеры, гетеротетрамеры и т.д.
Одно из воплощений относится к олигомерам, содержащим множественные полипептиды антитела против Dkk-1, соединенные посредством ковалентных или нековалентных взаимодействий между пептидными группировками, слитыми с полипептидами антитела против Dkk-1. Такие пептиды могут представлять собой пептидные линкеры (спейсеры) или пептиды, которые обладают свойством стимулировать олигомеризацию. Среди пептидов, которые могут стимулировать олигомеризацию полипептидов антитела против Dkk-1, присоединенных к ним, находятся лейциновые застежки и некоторые полипептиды, происходящие из антител, как описано более подробно ниже.
В конкретных воплощениях олигомеры содержат от двух до четырех полипептидов антитела против Dkk-1. Группировки антитела против Dkk-1 в таком олигомере могут быть в любой из форм, описанных выше, например, в виде вариантов или фрагментов. Предпочтительно, олигомеры содержат полипептиды антитела против Dkk-1, обладающие Dkk1-связывающей активностью.
В одном из воплощений олигомер получают, используя полипептиды, происходящие из иммуноглобулинов. Получение слитых белков, содержащих некоторые гетерологичные полипептиды, слитые с различными участками происходящих из антител полипептидов (включая Fc-домен), описано, например, в Ashkenazi et al. (1991) PNAS USA, 88: 10535; Byrn et al. (1990) Nature, 344: 677; и Hollenbaugh et al. (1992) "Construction of Immunoglobulin Fusion Proteins" в Current Protocols in Immunology, Suppl. 4, pages 10.19.1-10.19.11.
Одно из воплощений настоящего изобретения относится к димеру, содержащему два слитых белка, созданных путем слияния Dkk1-связывающего фрагмента антитела против Dkk-1 с Fc-областью антитела. Такой димер может быть получен, например, посредством встраивания слитого гена, кодирующего слитый белок, в соответствующий экспрессирующий вектор, экспрессии этого слитого гена в клетках-хозяевах, трансформированных рекомбинантным экспрессирующим вектором, и предоставления экспрессированному слитому белку возможности осуществлять сборку подобно молекулам антител, вследствие чего образуются межцепочечные дисульфидные связи между Fc-группировками с получением димера.
Термин "Fc-полипептид", как он использован в данном описании, включает нативные и мутеиновые формы полипептидов, происходящих из Fc-области антитела. Также включены укороченные формы таких полипептидов, содержащих шарнирную область, которая стимулирует димеризацию. Слитые белки, содержащие Fc-группировки (и олигомеры, образованные из них), обеспечивают преимущество, заключающееся в облегчении очистки аффинной хроматографией на колонках с белком А или белком G.
Один из подходящих Fc-полипептидов, описанный в заявке РСТ WO 93/10151 и патентах США №№5426048 и 5262522, представляет собой одноцепочечный полипептид, простирающийся от N-концевой шарнирной области до С-конца нативной Fc-области человеческого антитела IgG1. Другой полезный Fc-полипептид представляет собой Fc-мутеин, описанный в патенте США №5457035 и в Baum et al. (1994) EMBO J. 13: 3992-4001. Аминокислотная последовательность этого мутеина идентична таковой нативной Fc-последовательности, представленной в WO 93/10151, за исключением того, что аминокислота 19 изменена с Leu на Ala, аминокислота 20 изменена с Leu на Glu, а аминокислота 22 изменена с Gly на Ala. Такой мутеин демонстрирует сниженную аффинность в отношении Fc-рецепторов.
В других воплощениях вариабельная часть тяжелых и/или легких цепей антитела против Dkk-1, таких как раскрыто в данном изобретении, может быть заменена на вариабельную часть тяжелой и/или легкой цепи антитела.
Альтернативно, олигомер представляет собой слитый белок, содержащий множественные полипептиды антитела против Dkk-1 вместе с пептидными линкерами (спейсерными пептидами) или без них. Среди подходящих пептидных линкеров имеются линкеры, описанные в патентах США №№4751180 и 4935233.
Другой способ получения олигомерных производных антитела против Dkk-1 включает использование лейциновой застежки. Домены лейциновых застежек представляют собой пептиды, которые стимулируют олигомеризацию белков, в которых они обнаружены. Первоначально лейциновые застежки были идентифицированы в нескольких ДНК-связывающих белках (Landschuiz et al. (1988) Science, 240: 1759) и с тех пор обнаружены в ряде других белков. Среди известных лейциновых застежек существуют природные пептиды и их производные, которые осуществляют димеризацию или тримеризацию. Примеры доменов лейциновых застежек, подходящих для получения растворимых олигомерных белков, описаны в WO 94/10308, а лейциновая застежка, происходящая из сурфактантного белка D из легких (SPD), описана в у Норре et al. (1994) FEBS Letters, 344: 191, включено сюда посредством ссылки. Применение модифицированной лейциновой застежки, которая способствует стабильной тримеризации гетерологичного белка, слитого с ней, описано в Fanslow et al. (1994) Semin. Immunol., 6: 267-78. В одном из подходов рекомбинантные слитые белки, содержащие фрагмент или производное антитела против Dkk-1, слитые с пептидом лейциновой застежки, экспрессируют в подходящих клетках-хозяевах, и из супернатанта культуры извлекают образующиеся растворимые олигомерные фрагменты или производные антитела против Dkk-1.
Нуклеиновые кислоты
Также предложены нуклеиновые кислоты, которые кодируют одну или обе цепи антитела по изобретению или его фрагмент, производное, мутеин или вариант; полинуклеотиды, достаточные для применения в качестве гибридизационных зондов, ПЦР-праймеров или секвенирующих праймеров для идентификации, анализа, подвергания мутациям или амплификации полинуклеотида, кодирующего полипептид; антисмысловые нуклеиновые кислоты для ингибирования экспрессии полинуклеотида и комплементарные последовательности всего вышеупомянутого. Нуклеиновые кислоты могут быть любой длины. Длина их может составлять, например, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 75, 100, 125, 150, 175, 200, 250, 300, 350, 400, 450, 500, 750, 1000, 1500, 3000, 5000 или более нуклеотидов, и/или они могут содержать одну или более чем одну дополнительную последовательность, например, регуляторные последовательности, и/или представлять собой часть более длинной нуклеиновой кислоты, например вектора. Нуклеиновые кислоты могут быть одноцепочечными или двухцепочечными и могут содержать РНК- и/или ДНК-нуклеотиды и их искусственные варианты (например, пептидо-нуклеиновые кислоты). Также предложены нуклеиновые кислоты, кодирующие слитые белки, которые включают эти пептиды.
ДНК, кодирующая полипептиды антител (например, только тяжелую или легкую цепь, вариабельный домен или полноразмерное антитело), может быть выделена из В-клеток мышей, иммунизированных Dkk-1 или его иммуногенным фрагментом. ДНК может быть выделена с использованием традиционных методик, таких как полимеразная цепная реакция (ПЦР). Фаговый дисплей является другим примером известного метода, посредством которого могут быть получены производные антител. В одном из подходов полипептиды, которые являются компонентами представляющего интерес антитела, экспрессируют в любой подходящей рекомбинантной экспрессирующей системе, и экспрессированным полипептидам предоставляют возможность осуществлять сборку с образованием молекул антитела.
Предложены типичные нуклеиновые кислоты, которые кодируют легкую и тяжелую цепи, вариабельные области и CDR антител и иммунологически функциональных фрагментов. Вследствие вырожденности генетического кода каждая из полипептидных последовательностей также кодируется большим числом других нуклеиновокислотных последовательностей помимо приведенных. Согласно настоящему изобретению предложена каждая вырожденная нуклеотидная последовательность, кодирующая каждое антитело по изобретению.
Кроме того, согласно изобретению предложены нуклеиновые кислоты, которые гибридизуются с другими нуклеиновыми кислотами в конкретных условиях гибридизации. Методы гибридизации нуклеиновых кислот хорошо известны в данной области. См., например, Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6. В примерах известного умеренно жесткого условия гибридизации используют раствор для предварительной промывки, содержащий 5-кратный раствор хлорида натрия/цитрата натрия (SSC), 0,5% SDS (додецилсульфата натрия), 1,0 мМ EDTA (рН 8,0), буфер для гибридизации, представляющий собой примерно 50%-ный формамид, 6-кратный SSC, и температуру гибридизации 55°С (или другие аналогичные растворы для гибридизации, как например содержащие примерно 50% формамида, при температуре гибридизации 42°С), и условия промывки при 60°С, в 0,5-кратном SSC, 0,1% SDS. В качестве примеров известных жестких условий гибридизации: гибридизацию проводят в 6-кратном SSC при 45°С, затем следует одна или более промывок в 0,1-кратном SSC в 0,2% SDS при 68°С. Кроме того, специалист в данной области может манипулировать условиями гибридизации и/или промывки с целью повышения или снижения жесткости гибридизации, чтобы нуклеиновые кислоты, содержащие нуклеотидные последовательности, по меньшей мере на 65, 70, 75, 80, 85, 90, 95, 98 или 99% идентичные друг другу, обычно гибридизовались друг с другом.
Основные параметры, влияющие на выбор условий гибридизации, и руководство для разработки подходящих условий приведены, например в Sambrook, Fritsch, and Maniatis (1989) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., главы 9 и 11; и Current Protocols in Molecular Biology (1995) Ausubel et al., eds., John Wiley & Sons, Inc., разделы 2.10 и 6.3-6.4), и могут быть легко определены специалистами обычной квалификации в данной области исходя из, например, длины ДНК и/или нуклеотидного состава ДНК.
Посредством мутаций в нуклеиновую кислоту могут быть внесены изменения, приводящие к изменениям в аминокислотной последовательности полипептида (например, антитела или производного антитела по изобретению), которую она кодирует. Мутации можно вводить, используя любой метод, известный в данной области. В одном из воплощений изменены один или более конкретных аминокислотных остатков с использованием, например, протокола сайт-специфического мутагенеза. В другом воплощении изменены один или более случайно выбранных остатков с использованием, например, протокола случайного мутагенеза. Каким бы образом это ни было сделано, мутантный полипептид может быть экспрессирован и отобран в результате скрининга по желаемому свойству.
Мутации в нуклеиновую кислоту могут быть введены без существенного изменения биологической активности полипептида, который она кодирует. Например, можно выполнить нуклеотидные замены, приводящие к аминокислотным заменам по несущественным аминокислотным остаткам. Альтернативно, в нуклеиновую кислоту могут быть введены одна или более мутаций, которые селективно изменяют биологическую активность полипептида, который она кодирует. Например, мутация может количественно или качественно изменять биологическую активность. Примеры количественных изменений включают повышение, снижение или исчезновение активности. Примеры качественных изменений включают изменение антигенной специфичности антитела.
В другом аспекте согласно настоящему изобретению предложены молекулы нуклеиновой кислоты, которые подходят для применения в качестве праймеров или гибридизационных зондов для детекции нуклеиновокислотных последовательностей по изобретению. Молекула нуклеиновой кислоты по изобретению может содержать только часть нуклеиновокислотной последовательности, кодирующей полноразмерный полипептид по изобретению, например фрагмент, который может быть использован в качестве зонда или праймера, или фрагмент, кодирующий активный участок (например, Dkk1-связывающий участок) полипептида по изобретению.
Зонды, основанные на последовательности нуклеиновой кислоты по изобретению, можно использовать для детекции нуклеиновой кислоты или схожих нуклеиновых кислот, например транскриптов, кодирующих полипептид по изобретению. Зонд может содержать группу мечения, например, радиоактивный изотоп, флуоресцентное соединение, фермент или кофактор фермента. Такие зонды могут быть использованы для идентификации клетки, которая экспрессирует данный полипептид.
В другом аспекте согласно настоящему изобретению предложены векторы, содержащие нуклеиновую кислоту, кодирующую полипептид по изобретению или его часть (например, фрагмент, содержащий один или более CDR или один или более доменов вариабельной области). Примеры векторов включают плазмиды, вирусные векторы, неэписомные векторы млекопитающих и экспрессирующие векторы, например, рекомбинантные экспрессирующие векторы, но этим не ограничиваются. Рекомбинантные экспрессирующие векторы по изобретению могут содержать нуклеиновую кислоту по изобретению в форме, подходящей для экспрессии нуклеиновой кислоты в клетке-хозяине. Рекомбинантные экспрессирующие векторы включают одну или более регуляторных последовательностей, выбранных на основе клеток-хозяев, которые будут использованы для экспрессии, последовательностей, которые функционально связаны с нуклеиновокислотной последовательностью, подлежащей экспрессии. Регуляторные последовательности включают последовательности, которые направляют конститутивную экспрессию нуклеотидной последовательности в клетках-хозяевах многих типов (например, энхансер ранних генов SV40, промотор вируса саркомы Рауса и цитомегаловирусный промотор), последовательности, которые направляют экспрессию нуклеотидной последовательности только в некоторых клетках-хозяевах (например, тканеспецифичные регуляторные последовательности, см. Voss et al. (1986) Trends Biochem. Sci., 11: 287; Maniatis et al. (1987) Science, 236: 1237, включенные в данное описание посредством ссылки во всей своей полноте), и последовательности, которые направляют индуцибельную экспрессию нуклеотидной последовательности в ответ на конкретные обработку или условие (например, металлотиониновый промотор в клетках млекопитающих и чувствительный к tet (тетрациклину) и/или чувствительный к стрептомицину промотор как в прокариотических, так и эукариотических системах (см. там же)). Специалистам в данной области будет очевидно, что конструирование экспрессирующего вектора может зависеть от таких факторов, как выбор клетки-хозяина, подлежащей трансформации, уровень экспрессии желаемого белка и т.д. Экспрессирующие векторы по изобретению могут быть введены в клетки-хозяева, чтобы посредством этого продуцировать белки или пептиды, в том числе слитые белки или пептиды, кодируемые нуклеиновыми кислотами, которые изложены в данном описании.
В другом аспекте согласно настоящему изобретению предложены клетки-хозяева, в которые введен рекомбинантный экспрессирующий вектор по изобретению. Клеткой-хозяином может быть любая прокариотическая клетка (например, Е.coli) или эукариотическая клетка (например, клетки дрожжей, насекомых или млекопитающих (например, СНО-клетки)). Векторная ДНК может быть введена в прокариотические или эукариотические клетки с использованием традиционных методов трансформации или трансфекции. Для осуществления стабильной трансфекции клеток млекопитающих известно, что в зависимости от используемых экспрессирующего вектора и метода трансфекции только небольшая часть клеток может интегрировать чужеродную ДНК в свой геном. Для идентификации и отбора клеток, где произошла такая интеграция, наряду с представляющим интерес геном в большинстве случаев в клетки-хозяева вводят ген, кодирующий селектируемый маркер (например, устойчивости к антибиотикам). Предпочтительные селектируемые маркеры включают маркеры, придающие устойчивость к лекарственным средствам, таким как G418, гигромицин и метотрексат. Клетки, стабильно трансфицированные введенной нуклеиновой кислотой, среди других способов можно идентифицировать, используя селекцию по лекарственному средству (например, клетки, в которые инкорпорирован селектируемый маркерный ген, будут выживать, в то время как другие клетки погибнут).
Получение антител
Предложенные отличные от человеческих антитела могут происходить, например, из любого продуцирующего антитела животного, такого как мышь, крыса, кролик, коза, осел или примат, не являющийся человеком, (такой как обезьяна (например, яванский макак или макак-резус) или человекообразная обезьяна (например, шимпанзе)). Отличные от человеческих антитела можно использовать, например, в клеточной культуре in vitro и для применений, основанных на культивировании клеток, или для любого другого применения, где иммунный ответ на антитело отсутствует или является незначительным, может быть предупрежден, не имеет особого значения или желателен. В некоторых воплощениях изобретения антитела могут быть получены путем иммунизации полноразмерным Dkk-1 или карбоксиконцевой частью молекулы Dkk-1. Антитела могут быть поликлональными, моноклональными или могут быть синтезированы в клетках-хозяевах в результате экспрессии рекомбинантной ДНК.
Полностью человеческие антитела могут быть получены, как описано выше, путем иммунизации трансгенных животных, содержащих локусы человеческих иммуноглобулинов, или в результате селекции библиотеки фагового дисплея, экспрессирующей репертуар человеческих антител.
Моноклональные антитела (Mab) по изобретению могут быть получены рядом методов, включая традиционную методологию получения моноклональных антител, например, стандартный метод гибридизации соматических клеток по Kohler и Milstein ((1975) Nature, 256: 495). Альтернативно, для получения моноклональных антител можно использовать другие методы, например, вирусную или онкогенную трансформацию В-лимфоцитов. Одной из подходящих для получения гибридомы систем с использованием животных является мышиная система, что представляет собой очень известную процедуру. Протоколы иммунизации и методы выделения иммунизированных спленоцитов для слияния известны в данной области. Для осуществления таких методик проводят слияние В-клеток от иммунизированных мышей с подходящим иммортализованным партнером слияния, таким как мышиная линия миеломных клеток. При желании вместо мышей можно иммунизировать также крыс или других млекопитающих и можно проводить слияние В-клеток от таких животных с мышиной линией миеломных клеток, образуя гибридому. Альтернативно, можно использовать линию миеломных клеток из другого источника, не из мыши. Процедуры слияния для получения гибридомы также хорошо известны.
Предложенные одноцепочечные антитела могут образовываться путем соединения фрагментов вариабельного домена тяжелой и легкой цепи (Fv-области) с помощью аминокислотного мостика (короткого пептидного линкера), приводящего к получению единственной полипептидной цепи. Такие одноцепочечные Fv (scFv) могут быть получены путем слияния ДНК, кодирующей пептидный линкер, между молекулами ДНК, кодирующими оба полипептида вариабельных доменов (VL и VH). Полученные полипептиды могут сворачиваться вокруг себя с образованием антигенсвязывающих мономеров или могут образовывать мультимеры (например, димеры, тримеры или тетрамеры) в зависимости от длины гибкого линкера между двумя вариабельными доменами (Kortt et al. (1997) Prot. Eng. 10: 423; Kortt et al. (2001) Biomol. Eng. 18: 95-108). Комбинируя разные VL- и VH-содержащие полипептиды, можно создавать мультимерные scFv, которые связываются с разными эпитопами (Kriangkum et al. (2001) Biomol. Eng., 18: 31-40). Методы, разработанные для получения одноцепочечных антител, включают методы, описанные в патенте США №4946778; Bird (1988) Science, 242: 423; Huston et al. (1988) Proc. Natl. Acad. Sci. USA, 85: 5879; Ward et al. (1989) Nature, 334: 544; de Graafet al. (2002) Methods Mol. Biol. 178: 379-87.
Предложенные в данном описании антитела, которые являются антителами одного подкласса, можно заменить антителами другого подкласса с использованием методов переключения подкласса. Так, IgG-антитела могут происходить из lgM-антитела, например, и наоборот. Такие методы позволяют получать новые антитела, обладающие антигенсвязывающими свойствами заданного антитела (исходного антитела), а также демонстрирующие биологические свойства, ассоциированные с изотипом или подклассом антитела, отличающимся от изотипа или подкласса исходного антитела. Можно использовать методы рекомбинантной ДНК. В таких процедурах можно использовать клонированную ДНК, кодирующую конкретные полипептиды антитела, например ДНК, кодирующую константный домен антитела желаемого изотипа. См., например, Lantto et al. (2002) Methods Mol. Biol., 178: 303-16.
Вместе с тем также известны способы получения антител, обладающих разными свойствами (то есть, различными аффинностями в отношении антигена, с которым они связываются). Один из таких способов, называемый "перестановка цепей", включает проявление репертуаров генов вариабельных доменов иммуноглобулина на поверхности нитчатого бактериофага, часто обозначаемое как фаговый дисплей. Перестановка цепей использована для получения высокоаффинных антител к гаптену 2-фенилоксазол-5-ону, как описано в Marks et al. (1992) BioTechnology, 10: 779.
Для получения антитела против Dkk-1, обладающего функциональными и биохимическими характеристиками, могут быть выполнены консервативные модификации в отношении тяжелых и легких цепей, описанных в Таблице 1 (и соответствующие модификации в отношении кодирующих нуклеиновых кислот). Методы достижения таких модификаций описаны выше.
Антитела и их функциональные фрагменты по изобретению могут быть дополнительно модифицированы различными путями. Например, если они предназначены для применения в терапевтических целях, то они могут быть конъюгированы с полиэтиленгликолем (пегилированы) для пролонгирования периода полувыведения из сыворотки или для усиления доставки белка. Альтернативно, V-область антител, являющихся объектом изобретения, или их фрагментов может быть слита с Fc-областью молекулы другого антитела. Fc-область, используемую для этой цели, можно модифицировать таким образом, чтобы не происходило связывания ее с комплементом, снижая таким образом вероятность индукции лизиса клеток у пациента в тех случаях, когда слитый белок используют в качестве терапевтического агента. Помимо этого антитела, являющиеся объектом изобретения, или их функциональные фрагменты могут быть конъюгированы с сывороточным альбумином человека с целью увеличения периода полувыведения из сыворотки антитела или его фрагмента. Другим полезным партнером слияния для антител по изобретению или их фрагментов является транстиретин (TTR). TTR обладает способностью образовывать тетрамер, поэтому слитый белок антитело-TTR может образовывать поливалентное антитело, что может повышать его авидность связывания.
Альтернативно, важные модификации функциональных и/или биохимических характеристиках антител и фрагментов, изложенных в данном описании, могут быть достигнуты в результате создания замен в аминокислотной последовательности тяжелых и легких цепей, которые значительно отличаются по своему эффекту в отношении сохранения (а) структуры молекулярного остова в области такой замены, например, складчатой или спиральной конформации, (б) заряда или гидрофобности молекулы в сайте-мишени или (в) объемистости боковой цепи. Термин "консервативная аминокислотная замена" может включать в себя замену нативного аминокислотного остатка на остаток, не являющийся нативным, которая оказывает незначительный эффект или не оказывает никакого эффекта на полярность или заряд аминокислотного остатка в этом положении. Кроме того, любой нативный остаток в полипептиде также можно заменить на аланин, как описано ранее для аланин-сканирующего мутагенеза.
Аминокислотные замены (независимо от того, являются ли они консервативными или неконсервативными) антител, являющихся объектом изобретения, могут быть выполнены специалистами в данной области с использованием рутинных методов. Аминокислотные замены можно использовать для идентификации важных остатков антител, предложенных в данном описании, или для увеличения либо уменьшения аффинности этих антител к Dkk-1 человека, или для модификации аффинности связывания других антител против Dkk-1, изложенных в данном описании.
Экспрессия антител против Dkk-1
Антитела против Dkk-1 и иммунологические функциональные фрагменты могут быть получены любым из целого ряда традиционных методов. Например, антитела против Dkk-1 можно получить с применением рекомбинантных экспрессирующих систем, используя любой метод, известный в данной области. См., например, Monoclonal Antibodies, Hybridomas: A New Dimension in Biological Анализы, Kennet et al. (eds.) Plenum Press, New York (1980); и Antibodies: A Laboratory Manual, Harlow and Lane (eds.), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1988).
Антитела по настоящему изобретению могут быть экспрессированы в линиях гибридомных клеток или в клеточных линиях, отличающихся от гибридомы. Для трансформации клетки-хозяина млекопитающих, насекомых или микробов можно использовать экспрессирующие конструкции, кодирующие антитела. Трансформацию можно проводить, используя любой известный метод введения полинуклеотидов в клетку-хозяина, включая, например упаковку полинуклеотида в вирус или бактериофаг и трансдукцию клетки-хозяина данной конструкцией в соответствии с методиками трансфекции, известными в данной области, как подтверждено примерами согласно патентам США №№4399216, 4912040, 4740461 и 4959455. Используемая оптимальная процедура трансформации будет зависеть от того, какой тип клеток-хозяев подлежит трансформации. Способы введения гетерологичных полинуклеотидов в клетки млекопитающих хорошо известны в данной области и включают декстран-опосредованную трансфекцию, осаждение фосфатом кальция, полибрен-опосредованную трансфекцию, слияние протопластов, электропорацию, инкапсулирование полинуклеотида(ов) в липосомы, смешивание нуклеиновой кислоты с положительно-заряженными липидами и прямую микроинъекцию ДНК в ядра, но этим не ограничиваются.
Рекомбинантные экспрессирующие конструкции по изобретению обычно содержат молекулу нуклеиновой кислоты, кодирующую полипептид, содержащий одно или более чем одно из следующего: константную область тяжелой цепи; вариабельную область тяжелой цепи; константную область легкой цепи; вариабельную область легкой цепи; один или более CDR легкой или тяжелой цепи антитела против Dkk-1. Эти нуклеиновокислотные последовательности встроены в соответствующий экспрессирующий вектор с использованием стандартных методов лигирования. Обычно выбирают такой вектор, чтобы он был функциональным в конкретной используемой клетке-хозяине (то есть, вектор совместим с аппаратом клетки-хозяина), что дает возможность осуществления амплификации и/или экспрессии данного гена). В некоторых воплощениях используют векторы, которые задействованы в анализах комплементации фрагментов белка с использованием таких белковых репортеров, как дигидрофолатредуктаза (см., например, патент США №6270964, который тем самым включен посредством ссылки). Подходящие экспрессирующие векторы могут быть закуплены, например, у Invitrogen Life Technologies или BD Biosciences (ранее "Clontech"). Другие полезные векторы для клонирования и экспрессии антител и фрагментов по изобретению включают векторы, описанные в статье Bianchi and McGrew, Biotech. Biotechnol. Bioeng., 84(4): 439-44 (2003), которая тем самым включена посредством ссылки. Дополнительные подходящие экспрессирующие векторы обсуждаются, например, в Methods Enzymol, vol. 185 (D. V. Goeddel, ed.) (1990) New York: Academic Press.
Обычно, экспрессирующие векторы, используемые в любой из клеток-хозяев, содержат последовательности для поддержания их в плазмидах или вирусах и для клонирования и экспрессии экзогенных нуклеотидных последовательностей. Такие последовательности, все вместе обозначаемые как "фланкирующие последовательности", обычно включают что-то одно или более чем одно из следующих функционально связанных нуклеотидных последовательностей: промотор, одну или более энхансерных последовательностей, ориджин репликации, последовательность терминации транскрипции, полную интронную последовательность, содержащую донорный и акцепторный сайт сплайсинга, последовательность, кодирующую лидерную последовательность для секреции полипептида, сайт связывания с рибосомой, последовательность полиаденилирования, полилинкерный участок для встраивания нуклеиновой кислоты, кодирующей полипептид, подлежащий экспрессии, и селектируемый маркерный элемент.
Возможно, что вектор может содержать последовательность, кодирующую "метку", то есть молекулу олигонуклеотида, локализованную на 5'- или 3'-конце кодирующей последовательности, а именно, олигонуклеотидную последовательность, кодирующую поли-His (например, гекса-His) или другую "метку", доступные антитела для которой имеются в продаже, такую как FLAG®, НА (гемаглютинин из вируса гриппа) или myc. Обычно слияние такой метки с белком-антителом происходит в результате экспрессии, и она может служить в качестве средства для аффинной очистки антитела из клетки-хозяина. Аффинную очистку можно проводить, например, посредством колоночной хроматографии, используя антитела против данной метки в качестве аффинной матрицы. Возможно, что впоследствии метка может быть удалена из очищенного полипептида антитела различными способами, например с использованием для ее отщепления некоторых пептидаз.
Фланкирующие последовательности в экспрессирующем векторе могут быть гомологичными (то есть происходить из того же вида и/или штамма, что и клетка-хозяин), гетерологичными (то есть происходить из другого вида, нежели вид и/или штамм клетки-хозяина), гибридными (то есть представлять собой комбинацию фланкирующих последовательностей более чем из одного источника), синтетическими или нативными. В силу этого источником фланкирующей последовательности может быть любой прокариотический или эукариотический организм, любой организм позвоночных или беспозвоночных или любое растение, при условии, что эта фланкирующая последовательность является функциональной в аппарате клетки-хозяина и может активироваться им.
Фланкирующие последовательности, полезные в векторах по данному изобретению, могут быть получены любым из нескольких способов, хорошо известных в данной области. Обычно, фланкирующие последовательности, полезные в данном изобретении, будут идентифицированы заранее посредством картирования и/или посредством переваривания рестрикционными эндонуклеазами и таким образом могут быть выделены из надлежащего тканевого источника с использованием подходящих рестрикционных эндонуклеаз. В некоторых случаях может быть известна полная нуклеотидная последовательность фланкирующей последовательности. В данном случае фланкирующая последовательность может быть синтезирована с использованием изложенных в данном описании методов синтеза нуклеиновых кислот или клонирования.
Если известна вся или только часть фланкирующей последовательности, то ее можно получить с использованием ПЦР и/или путем скрининга геномной библиотеки с подходящим олигонуклеотидом и/или фрагментом фланкирующей последовательности из того же или другого вида. Если фланкирующая последовательность не известна, фрагмент ДНК, кодирующей фланкирующую последовательность, можно выделить из более длинного куска ДНК, которая может содержать, например, кодирующую последовательность или даже другой ген или другие гены. Выделение может быть проведено посредством переваривания рестрикционными эндонуклеазами с получением надлежащего фрагмента ДНК с последующим выделением с использованием очистки на агарозном геле, хроматографии на колонке Qiagen® (Chatsworth, Calif.) или других методов, известных специалисту в данной области. Выбор подходящих ферментов для выполнения этой задачи будет совершенно очевиден специалистам в данной области.
Обычно ориджин репликации составляет часть экспрессирующих векторов прокариот, в частности, тех, которые приобретены на коммерческой основе, и такой ориджин способствует амплификации вектора в клетке-хозяине. Если выбранный вектор не содержит сайт ориджина репликации, его можно синтезировать химически на основании известной последовательности и лигировать в вектор. Например, ориджин репликации из плазмиды pBR322 (New England Biolabs, Beverly, Mass.) подходит для большинства грамотрицательных бактерий, а для клонирующих векторов в клетках млекопитающих полезны разные ориджины (например, SV40, полиомы, аденовируса, вируса везикулярного стоматита (VSV) или папилломавирусов, таких как HPV (папилломавирус человека) или BPV (папилломавирус крупного рогатого скота)). Обычно ориджин репликации млекопитающих не является необходимым для экспрессирующих векторов для млекопитающих (например, часто используют только ориджин SV40, поскольку он содержит ранний промотор).
Экспрессирующие и клонирующие векторы по настоящему изобретению обычно будут содержать промотор, который распознается организмом хозяина и функционально связан с нуклеиновой кислотой, кодирующей антитело против Dkk-1 или его иммунологически функциональный фрагмент. Промоторы представляют собой нетранскрибируемые последовательности, локализованные выше (то есть, 5') стартового кодона структурного гена (в большинстве случаев в пределах примерно 100-1000 п.о.), которые контролируют транскрипцию структурного гена. Традиционно промоторы относят к одному из двух классов: индуцибельным промоторам и конститутивным промоторам. Индуцибельные промоторы инициируют повышенные уровни транскрипции ДНК под своим контролем в ответ на некоторое изменение в условиях культивирования, как например, присутствие или отсутствие питательного вещества или изменение температуры. Конститутивные промоторы, с другой стороны, инициируют непрерывное получение генных продуктов; то есть существует слабый контроль или не существует экспериментального контроля генной экспрессии. Общеизвестно большое количество промоторов, распознаваемых рядом возможных клеток-хозяев. Подходящий промотор функционально связывают с ДНК, кодирующей антитело против Dkk-1, посредством удаления промотора из источника ДНК в результате переваривания рестрикционными ферментами или амплификации промотора с помощью полимеразной цепной реакции и встраивания последовательности желаемого промотора в вектор.
Промоторы, подходящие для использования вместе с хозяевами-дрожжами, также хорошо известны в данной области. Вместе с промоторами дрожжей предпочтительно используют энхансеры дрожжей. Промоторы, подходящие для использования вместе с клетками-хозяевами, представляющими собой клетки млекопитающих, хорошо известны и включают, но этим не ограничиваются, Промоторы, полученные из геномов таких вирусов, как вирус полиомы, вирус птичьей оспы, аденовирус (такой как аденовирус 2), папилломавирус крупного рогатого скота, вирус птичьей саркомы, цитомегаловирус, ретровирусы, вирус гепатита В и наиболее предпочтительно обезьяний вирус 40 (SV40). Другие подходящие Промоторы млекопитающих включают гетерологичные Промоторы млекопитающих, например Промоторы белков теплового шока и актиновый промотор.
Конкретные Промоторы, полезные для практического использования рекомбинантных экспрессирующих векторов по изобретению, включают: участок раннего промотора SV40 (Bemoist and Chambon (1981) Nature, 290: 304-10); промотор CMV (цитомегаловируса); промотор, содержащийся в длинном 3'-концевом повторе вируса саркомы Рауса (Yamamoto, et al. (1980) Cell, 22: 787-97); промотор тимидинкиназы вируса герпеса (Wagner et al. (1981) Proc. Natl. Acad. Sci. USA, 78: 1444-45); регуляторные последовательности гена металлотионина (Brinster et al. (1982) Nature, 296: 39-42); Промоторы прокариотических экспрессирующих векторов, такие как бета-лактамазный промотор (Villa-Kamaroff et al. (1978) Proc. Natl. Acad. Sci. USA, 75: 3727-31); или tac-промотор (DeBoer et al. (1983) Proc. Natl. Acad. Sci. USA, 80: 21-25), но этим не ограничиваются. Также доступны для использования следующие участки транскрипционного контроля у животных, демонстрирующие тканеспецифичность и использованные в трансгенных животных: контрольный участок гена эластазы I, активный в панкреатических ацинарных клетках (Swift et al. (1984) Cell, 38: 63946; Ornitz et al. (1986) Cold Spring Harbor Symp. Quant. Biol., 50: 399409; MacDonald, 1987, Hepatology, 7: 425-515); контрольный участок гена инсулина, активный в панкреатических бета-клетках (Hanahan (1985) Nature 315: 115-22); контрольный участок вируса опухоли молочных желез у мышей, активный в тестикулярных клетках, клетках молочных желез, лимфоидных и тучных клетках (Leder et al. (1986) Cell, 45: 485-95); контрольный участок гена альбумина, активный в печени (Pinkert et al. (1987) Genes and Devel. 1: 268-76); контрольный участок гена альфа-фетопротеина, активный в печени (Krumlauf et al. (1985) Mol. Cell. Biol. 5: 1639-48; Hammer et al. (1987) Science, 235: 53-58); контрольный участок гена альфа 1-антитрипсина, активный в печени (Kelsey et al. (1987) Genes and Devel., 1: 161-71); контрольный участок гена бета-глобина, активный в миелоидных клетках (Mogram et al., 1985, Nature, 315: 338-40; Kollias et al. (1986) Cell, 46: 89-94); контрольный участок гена основного белка миелина, активный в олигодендроцитах в головном мозге (Readhead et al. (1987) Cell, 48: 703-12); контрольный участок гена легкой цепи-2 миозина, активный в скелетной мышце (Sani (1985) Nature, 314: 283-86); контрольный участок гена гонадотропного рилизинг-гормона, активный в гипоталамусе (Mason et al. (1986) Science, 234: 1372-78); и в особенности контрольный участок гена иммуноглобулина, активный в лимфоидных клетках (Grosschedl et al. (1984) Cell, 38: 647-58; Adames et al. (1985) Nature, 318: 533-38; Alexander et al. (1987) Mol. Cell Biol., 7: 1436-44).
Энхансерная последовательность может быть встроена в вектор для усиления транскрипции нуклеиновой кислоты, кодирующей антитело против Dkk-1 или его иммунологически функциональный фрагмент по настоящему изобретению, у высших эукариот. Энхансеры являются цис-действующими элементами ДНК длиной обычно примерно 10-300 п.о., которые действуют на промоторы с целью усиления транскрипции. Энхансеры являются относительно независимыми от ориентации и положения. Они были обнаружены в 5'- и 3'-положении по отношению к транскрипционной единице. Известны несколько энхансерных последовательностей, предоставляемые генами млекопитающих (например, глобина, эластазы, альбумина, альфа-фетопротеина и инсулина). Также можно использовать энхансерную последовательность из вируса. Энхансер SV40, энхансер раннего промотора цитомегаловируса, энхансер полиомы и энхансеры аденовирусов представляют собой типичные элементы, усиливающие активацию эукариотических промоторов. Несмотря на то, что энхансер может быть сплайсирован в вектор в 5'- или 3'-положении по отношению к молекуле нуклеиновой кислоты, его обычно помещают в сайт, находящийся в 5'-положении по отношению к промотору.
В экспрессирующих векторах последовательность терминации транскрипции обычно располагается в 3'-положении по отношению к концу полипептид-кодирующего участка и служит для терминации транскрипции. Последовательность терминации транскрипции, используемая для экспрессии в прокариотических клетках, обычно представляет собой богатый G-C фрагмент, за которым следует поли-Т-последовательность. Хотя данную последовательность легко клонировать из библиотеки или даже приобрести в коммерческих источниках как часть вектора, ее также можно легко синтезировать, используя методы синтеза нуклеиновых кислот, как например, изложенные в данном описании.
Селектируемый маркерный генный элемент кодирует белок, необходимый для выживаемости и роста клетки-хозяина, растущей в селективной культуральной среде. Типичные селектируемые маркерные гены, используемые в экспрессирующих векторах, кодируют белки, которые (а) придают устойчивость к антибиотикам или другим токсинам, например ампициллину, тетрациклину или канамицину, для прокариотических клеток-хозяев; (б) дополняют ауксотрофные дефициты клетки; или (в) снабжают необходимыми питательными веществами, недоступными в составе сред. Примеры селектируемых маркеров включают ген устойчивости к канамицину, ген устойчивости к ампициллину и ген устойчивости к тетрациклину. Бактериальный ген устойчивости к неомицину также может быть использован для селекции как в прокариотических, так и в эукариотических клетках-хозяевах.
Для амплификации гена, который будет экспрессирован, могут быть использованы другие селектируемые гены. Амплификация представляет собой процесс, посредством которого гены, которые не могут экспрессироваться в единственной копии на достаточно высоких уровнях, обеспечивающих выживаемость и рост клеток в некоторых условия селекции, повторяют в тандеме внутри хромосом на последующих стадиях создания рекомбинантных клеток. Примеры амплифицируемых селектируемых маркеров для клеток млекопитающих включают дигидрофолатредуктазу (DHFR) и не содержащую промотора тимидинкиназу. При использовании этих маркеров трансформанты клеток млекопитающих помещают в условия с применением селекционного давления, где только трансформанты уникально адаптированы для выживания благодаря наличию селектируемого гена, представленного в данном векторе. Селекционное давление устанавливают путем культивирования трансформированных клеток в условиях, при которых концентрация селектируемого агента в среде постепенно увеличивается, обеспечивая ввиду этого выживаемость только тех клеток, в которых селектируемый ген амплифицирован. В этих условиях ДНК, ближайшая селектируемому гену, как например ДНК, кодирующая антитело по изобретению, амплифицируется вместе с селектируемым геном. В результате благодаря амплифицированной ДНК синтезируются повышенные количества полипептида против Dkk-1.
Сайт связывания с рибосомой обычно необходим для инициации трансляции мРНК и характеризуется последовательностью Шайна-Дальгарно (прокариоты) или последовательностью Козака (эукариоты). Этот элемент обычно локализован в 3'-положении по отношению к промотору и 5'-положении по отношению к кодирующей последовательности полипептида, подлежащего экспрессии.
В некоторых случаях, например, там, где гликозилирование является желательным в экспрессирующей системе эукариотической клетки-хозяина, для улучшения гликозилирования или выхода могут быть использованы различные пре-последовательности. Например, может быть изменен сайт отщепления пептидазой конкретного сигнального пептида или добавлены про-последовательности, которые также могут оказывать влияние на гликозилирование. Конечный белковый продукт может иметь в положении -1 (по отношению к первой аминокислоте зрелого белка) одну или более дополнительных аминокислот, характерных для экспрессии, которые не могут быть полностью удалены. Например, конечный белковый продукт может иметь один или два аминокислотных остатка в сайте отщепления пептидазой, присоединенных к аминоконцу. Альтернативно, использование нескольких сайтов ферментативного расщепления может приводить к получению слегка укороченной, но все еще активной формы желаемого полипептида, если ферментативное расщепление происходит в этой области внутри зрелого полипептида.
В тех случаях, когда имеющийся в продаже экспрессирующий вектор не содержит некоторых желаемых фланкирующих последовательностей, которые описаны выше, вектор может быть модифицирован посредством индивидуального лигирования этих последовательностей в данный вектор. После того, как вектор выбран и модифицирован, как необходимо, молекулу нуклеиновой кислоты, кодирующую антитело против Dkk-1 или его иммунологически функциональный фрагмент, встраивают в надлежащий сайт данного вектора.
Завершенный вектор, содержащий последовательности, кодирующие антитело по изобретению или его иммунологически функциональный фрагмент, встраивают в подходящую клетку-хозяина для осуществления амплификации и/или экспрессии полипептида. Трансформация экспрессирующим вектором для антитела против Dkk-1 и его иммунологически функциональным фрагментом выбранной клетки-хозяина может быть осуществлена хорошо известными методами, включая такие методы, как трансфекция, инфекция, применение хлорида кальция, электропорация, микроинъекция, липофекция, метод с использованием DEAE-декстрана или другие известные методы. Выбранный метод будет частично зависеть от типа клетки-хозяина, который будет использован. Эти методы и другие подходящие методы хорошо известны специалисту в данной области.
Трансформированная клетка-хозяин при культивировании в соответствующих условиях синтезирует антитело против Dkk-1 или его функциональный фрагмент, которые затем могут быть собраны из культуральной среды (если клетка-хозяин секретирует его в среду) или непосредственно из клетки-хозяина, продуцирующей его (если клетка-хозяин его не секретирует). Выбор соответствующей клетки-хозяина будет зависеть от различных факторов, таких как желаемые уровни экспрессии, модификации полипептида, которые желательны или необходимы для проявления активности (такие как гликозилирование или фосфорилирование), и легкость сворачивания в биологически активную молекулу.
Клеточные линии млекопитающих, доступные в качестве хозяев для осуществления экспрессии, хорошо известны в данной области и включают многие иммортализованные клеточные линии, доступные из Американской коллекции типовых культур (АТСС), такие как клетки яичников китайского хомячка (СНО), клетки HeLa, клетки почки детеныша хомячка (BHK), клетки почки мартышки (COS), клетки гепатоцеллюлярной карциномы человека (например, Hep G2) и ряд других клеточных линий, но ими не ограничиваются. В некоторых воплощениях наилучшую клеточную линию для экспрессии конкретной ДНК-конструкции можно выбрать путем тестирования различных клеточных линий, определяя, которая из них имеет самые высокие уровни экспрессии, и получая антитела, обладающие конститутивными свойствами в отношении связывания с Dkk-1.
Фармацевтические композиции
В другом аспекте согласно настоящему изобретению предложена композиция, например, фармацевтическая композиция, содержащая моноклональное антитело или его антигенсвязывающий участок(и) по настоящему изобретению, или их комбинацию, изготовленные вместе с фармацевтически приемлемым носителем. Такие композиции могут включать одно из или комбинацию (например, два/две или более разных) антител или иммуноконъюгатов или биспецифических молекул по изобретению. Например, фармацевтическая композиция по изобретению может содержать комбинацию антител (или иммуноконъюгатов, или биспецифических молекул), которые связываются с разными эпитопами на антигене-мишени или которые обладают комплементарными активностями.
Фармацевтические композиции по изобретению также можно вводить в виде комбинированной терапии, то есть в комбинации с другими агентами. Например, комбинированная терапия может включать антитело против Dkk-1 по настоящему изобретению в комбинации по меньшей мере с одним другим противовоспалительным или иммуносупрессивным агентом. Примеры терапевтических агентов, которые можно использовать в комбинированной терапии, описаны более подробно ниже в разделе, касающемся применений антител по изобретению.
При использовании в данном описании, "фармацевтически приемлемый носитель" включает любые и все растворители, диспергирующие среды, вещества для нанесения покрытия, антибактериальные и противогрибковые агенты, изотонические и замедляющие всасывание агенты и тому подобное, которые являются физиологически совместимыми. Обычно носитель является подходящим для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии). В зависимости от пути введения активное соединение, то есть антитело, его антигенсвязывающая часть, иммуноконъюгат или биспецифическая молекула, может быть покрыто веществом для его защиты от действия кислот и других природных условий, которые могут инактивировать это соединение.
В некоторых воплощениях антитела по настоящему изобретению могут быть представлены в нейтральной форме (включая цвиттерионные формы) или в виде положительно или отрицательно-заряженных разновидностей. В некоторых случаях, антитела могут находиться в комплексе с противоионом для образования фармацевтически приемлемой соли. Так, фармацевтические соединения по изобретению могут включать одну или более фармацевтически приемлемых солей.
"Фармацевтически приемлемая соль" относится к соли, которая сохраняет желаемую биологическую активность исходного соединения (например, антитела) и не вызывает нежелательных токсикологических эффектов (см., например, Berge, S.M., et al. (1977) J. Pharm. Sci., 66: 1-19). Например, термин "фармацевтически приемлемая соль" включает комплекс, содержащий одно или более антител и один или более противоионов, причем данные противоионы происходят из фармацевтически приемлемых неорганических и органических кислот и оснований.
Примеры таких солей включают соли присоединения кислоты и соли присоединения основания. Соли присоединения кислоты включают соли, происходящие из нетоксичных неорганических кислот, таких как соляная, азотная, фосфорная, серная, бромистоводородная, иодистоводородная, фосфористая и тому подобное, а также их нетоксичных органических кислот, таких как алифатические моно- и дикарбоновые кислоты, фенил-замещенные алкановые кислоты, гидроксиалкановые кислоты, ароматические кислоты, алифатические и ароматические сульфоновые кислоты и тому подобное. Соли присоединения основания включают соли, происходящие из щелочноземельных металлов, таких как натрий, калий, магний, кальций и тому подобное, а также из нетоксичных органических аминов, таких как N,N'-дибензилэтилендиамин, N-метилглюкамин, хлорпрокаин, холин, диэтаноламин, этилендиамин, прокаин и тому подобное.
Фармацевтически приемлемые неорганические основания также включают ионы металлов. Ионы металлов включают, но этим не ограничиваются, ионы солей соответствующих щелочных металлов, солей щелочно-земельных металлов и другие физиологически приемлемые ионы металлов. Соли, происходящие из неорганических оснований, включают соли алюминия, аммония, кальция, кобальта, никеля, молибдена, ванадия, марганца, хрома, селена, олова, меди, трехвалентного железа, двухвалентного железа, лития, магния, трехвалентного марганца, двухвалентного марганца, калия, рубидия, натрия и цинка и в своих обычных валентностях.
Фармацевтически приемлемые полученные присоединением кислоты соли антител по настоящему изобретению могут быть получены из следующих кислот, включая, без ограничения, муравьиную, уксусную, ацетамидобензойную, адипиновую, аскорбиновую, борную, пропионовую, бензойную, камфорную, угольную, цикламовую, дегидрохолевую, малоновую, эдетовую (этилендиаминтетрауксусную), этилсерную, фендизойную (fendizoic), метафосфорную, янтарную, гликолевую, глюконовую, молочную, яблочную, винную, дубильную, лимонную, азотную, глюкуроновую, малеиновую, фолевую, фумаровую, пировиноградную, аспарагиновую, глутаминовую, соляную, бромистоводородную, иодистоводородную, лизин, изолимонную, трифторуксусную, памовую, антраниловую, метансульфоновую (mesylic), оротовую, щавелевую, щавелевоуксусную, олеиновую, стеариновую, салициловую, аминосалициловую, силикат, пара-гидроксибензойную, никотиновую, фенилуксусную, миндальную, эмбоновую, сульфоновую, метансульфоновую, фосфорную, фосфоновую, этансульфоновую, этандисульфоновую, аммоний, бензолсульфоновую, пантотеновую, нафталинсульфоновую, толуолсульфоновую, 2-гидроксиэтансульфоновую, сульфаниловую, серную, азотную, азотистую, монометиловый сложный эфир серной кислоты, циклогексиламиносульфоновую, β-гидроксимасляную, глицин, глицилглицин, какодиловую, диаминокапроновую, камфорсульфоновую, тиоциановую, оксоглутаровую, пиридоксаль-5-фосфат, хлорфеноксиуксусную, ундекановую, N-ацетил-L-аспарагиновую, галактаровую и галактуроновую кислоты.
Фармацевтически приемлемые органические основания включают триметиламин, диэтиламин, N,N'-дибензилэтилендиамин, хлорпрокаин, холин, дибензиламин, диэтаноламин, этилендиамин, меглумин (N-метилглюкамин), прокаин, циклические амины, катионы четвертичного аммония, аргинин, бетаин, кофеин, клемизоль, 2-этиламиноэтанол, 2-диэтиламиноэтанол, 2-диметиламиноэтанол, этандиамин, бутиламин, этаноламин, этилендиамин, N-этилморфолин, N-этилпиперидин, этилглюкамин, глюкамин, глюкозамин, гистидин, гидрабамин, имидазол, изопропиламин, метилглюкамин, морфолин, пиперазин, пиридин, пиридоксин, неодимий, пиперидин, полиаминные смолы, прокаин, пурины, теобромин, триэтиламин, трипропиламин, триэтаноламин, трометамин, метиламин, таурин, холат, 6-амино-2-метил-2-гептанол, 2-амино-2-метил-1,3-пропандиол, 2-амино-2-метил-1-пропанол, алифатические моно- и дикарбоновые кислоты, фенил-замещенные алкановые кислоты, гидрокси-алкановые кислоты, ароматические кислоты, алифатические и ароматические сульфоновые кислоты, стронций, трицин, гидразин, фенилциклогексиламин, 2-(N-морфолино)этансульфоновую кислоту, бис(2-гидроксиэтил)амино-трис(гидроксиметил)метан, N-(2-ацетамидо)-2-аминоэтансульфоновую кислоту, 1,4-пиперазиндиэтансульфоновую кислоту, 3-морфолино-2-гидроксипропан-сульфоновую кислоту, 1,3-бис[трис(гидроксиметил)метиламино]пропан, 4-морфолинпропансульфоновую кислоту, 4-(2-гидроксиэтил)пиперазин-1 -этансульфоновую кислоту, 2-[(2-гидрокси-1,1-бис(гидроксиметил)этил)амино]-этансульфоновую кислоту, N,N-бис(2-гидроксиэтил)-2-аминоэтансульфоновую кислоту, 4-(N-морфолино)бутансульфоновую кислоту, 3-(N,N-бис[2-гидроксиэтил]амино)-2-гидроксипропансульфоновую кислоту, 2-гидрокси-3-[трис(гидроксиметил)метиламино]-1-пропансульфоновую кислоту, 4-(2-гидроксиэтил)пиперазин-1-(2-гидроксипропансульфоновую кислоту), пиперазин-1,4-бис(2-гидроксипропансульфоновой кислоты) дигидрат, 4-(2-гидроксиэтил)-1-пиперазинпропансульфоновую кислоту, N,N-бис(2-гидроксиэтил)глицин, N-(2-гидроксиэтил)пиперазин-N'-(4-бутансульфоновую кислоту), N-[трис(гидроксиметил)метил]-3-аминопропансульфоновую кислоту, N-трис(гидроксиметил)метил-4-аминобутансульфоновую кислоту, N-(1,1-диметил-2-гидроксиэтил)-3-амино-2-гидроксипропансульфоновую кислоту, 2-(циклогексиламино)этансульфоновую кислоту, 3-(циклогексиламино)-2-гидрокси-1-пропансульфоновую кислоту, 3-(циклогексиламино)-1-пропансульфоновую кислоту, N-(2-ацетамидо)иминодиуксусную кислоту, 4-(циклогексиламино)-1-бутансульфоновую кислоту, N-[трис(гидроксиметил)-метил]глицин, 2-амино-2-(гидроксиметил)-1,3-пропандиол и трометамол.
Фармацевтическая композиция по изобретению также может включать фармацевтически приемлемый антиоксидант. Примеры фармацевтически приемлемых антиоксидантов включают: (1) водорастворимые антиоксиданты, такие как аскорбиновая кислота, гидрохлорид цистеина, бисульфат натрия, метабисульфит натрия, сульфит натрия и тому подобное; (2) растворимые в масле антиоксиданты, такие как аскорбилпальмитат, бутилированный гидроксианизол (ВНА), бутилированный гидрокситолуол (ВНТ), лецитин, пропилгалат, альфа-токоферол и тому подобное; и (3) хелатирующие металл агенты, такие как лимонная кислота, этилендиаминтетрауксусная кислота (EDTA), сорбит, винная кислота, фосфорная кислота и тому подобное.
Примеры подходящих водных и неводных носителей, которые могут быть использованы в фармацевтических композициях по изобретению, включают воду, этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и тому подобное) и подходящие их смеси, растительные масла, такие как оливковое масло, и инъекционные органические сложные эфиры, такие как этилолеат. Надлежащую текучесть можно поддерживать, например, путем использования материалов для нанесения покрытия, таких как лецитин, путем поддержания необходимого размера частиц в случае дисперсий и путем использования поверхностно-активных веществ.
Эти композиции также могут содержать такие адъюванты, как консерванты, увлажнители, эмульгирующие агенты и диспергирующие агенты. Профилактика проникновения микроорганизмов может обеспечиваться как процедурами стерилизации, упомянутыми выше, так и посредством введения различных антибактериальных и противогрибковых агентов, например, парабена, хлорбутанола, фенола, сорбиновой кислоты и тому подобного. Также может быть желательно включить в композиции изотонические агенты, такие как сахара, хлорид натрия и тому подобное. Кроме этого, путем введения агентов, которые задерживают всасывание, таких как моностеарат алюминия и желатин, можно добиться пролонгированного всасывания инъекционной фармацевтической формы.
Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для экстемпорального приготовления стерильных инъекционных растворов или дисперсий. Применение таких сред и агентов для фармацевтически активных веществ известно в данной области. За исключением случаев, когда любые традиционные среды или агент несовместимы с активным соединением, их применение в фармацевтических композициях по изобретению предусматривается. В такие композиции также могут быть введены дополнительные активные соединения.
Терапевтические композиции обычно должны быть стерильными и стабильными в условиях производства и хранения. Такие композиция могут быть изготовлены в виде раствора, микроэмульсии, липосомы или другой предписанной структуры, подходящей для высокой концентрации лекарственного средства. Носителем может быть растворитель или дисперсионная среда, содержащие, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и тому подобное) и их подходящие смеси. Надлежащую текучесть можно поддерживать, например, путем использования материалов для нанесения покрытия, таких как лецитин, путем поддержания необходимого размера частиц в случае дисперсий и путем использования поверхностно-активных веществ. Во многих случаях предпочтительно будет включить в композицию изотонические агенты, например, сахара, полиспирты, такие как манит и сорбит, или хлорид натрия. Путем включения в композицию агента, который задерживает всасывание, например, соли моностеарата и желатина, можно добиться пролонгированного всасывания инъекционных композиций.
Стерильные инъекционные растворы можно изготовить путем введения активного соединения в необходимом количестве в соответствующем растворителе вместе с одним из ингредиентов, приведенных выше, или их комбинацией, как необходимо, с последующей стерилизующей микрофильтрацией. В большинстве случаев дисперсии готовят путем введения активного соединения в стерильный разбавитель, который содержит основную дисперсионную среду и необходимые другие ингредиенты из тех, которые приведены выше. В случае стерильных порошков, используемых для приготовления стерильных инъекционных растворов, способы изготовления включают, но этим не ограничиваются, вакуумную сушку и сублимационную сушку (лиофилизацию), в результате которых получают порошок активного ингредиента плюс любого дополнительного желаемого ингредиента из их раствора, предварительно подвергнутого стерильной фильтрации.
Количество активного ингредиента, которое может быть скомбинировано с веществом-носителем для получения разовой лекарственной формы, будет варьировать в зависимости от подвергаемого лечению субъекта и конкретного способа введения. Количество активного ингредиента, которое может быть скомбинировано с веществом-носителем для получения разовой лекарственной формы, в большинстве случаев будет представлять собой такое количество композиции, которое производит терапевтический эффект. В большинстве случаев, из расчета ста процентов, это количество будеть находиться в диапазоне от примерно 0,01 процента до примерно девяноста девяти процентов активного ингредиента, предпочтительно от примерно 0,1 процента до примерно 70 процентов, наиболее предпочтительно от примерно 1 процента до примерно 30 процентов активного ингредиента в комбинации с фармацевтически приемлемым носителем.
Режимы дозирования корректируют с целью получения оптимального желаемого ответа (например, терапевтического ответа). Например, можно вводить разовый болюс, можно вводить несколько разделенных доз в течение периода времени или дозу можно пропорционально снизить или повысить, исходя из необходимости терапевтической ситуации. Особенно предпочтительно изготовление парентеральных композиций в стандартной лекарственной форме для облегчения введения и однородности дозировки. Стандартная лекарственная форма, как использовано в данном описании, относится к физически дискретным единицам, подходящим в качестве однократных дозировок для субъектов, подлежащих лечению; причем каждая единица содержит предварительно отмеренное количество активного соединения, рассчитанное для получения желаемого терапевтического эффекта, вместе с необходимым фармацевтическим носителем. Технические требования для стандартных лекарственных форм по изобретению обуславливаются (а) уникальными характеристиками активного соединения и конкретным терапевтическим эффектом, который предполагается достигнуть, и (б) органичениями, присущими области изготовления такого активного соединения, для лечения чувствительности у индивидуумов, и напрямую зависят от них.
Что касается введения антитела, то дозировка находится в диапазоне от примерно 0,0001 до 100 мг/кг и чаще от 0,01 до 5 мг/кг массы тела хозяина. Например, дозировки могут составлять 0,3 мг/кг массы тела, 1 мг/кг массы тела, 3 мг/кг массы тела, 5 мг/кг массы тела или 10 мг/кг массы тела или находиться в диапазоне от 1 до 10 мг/кг. Типичная схема лечения включает введение один раз в неделю, один раз в две недели, один раз каждые три недели, один раз каждые четыре недели, один раз в месяц, один раз каждые 3 месяца или один раз каждые 3-6 месяцев. Режимы введения для антитела против Dkk-1 или его антигенсвязывающего участка по изобретению включают, например, 1 мг/кг массы тела или 3 мг/кг массы тела посредством подкожного введения, причем для приема данного антитела используют один из следующих режимов дозирования: (1) каждые четыре недели для шести дозировок, затем каждые три месяца; (2) каждые три недели; (3) 3 мг/кг массы тела однократно, затем 1 мг/кг массы тела каждые три недели.
В некоторых воплощениях два или более антител или их фрагментов с разными специфичностями связывания вводят одновременно, в этом случае дозировка каждого вводимого антитела уменьшается в пределах указанных диапазонов. Обычно введение антител осуществляют многократно. Интервалы между разовыми дозировками могут составлять, например, одну неделю, один месяц, каждые три месяца или один год. Интервалы также могут быть неравномерными, как будет предписано согласно измерению уровней антитела относительно антигена-мишени в крови пациента. В некоторых способах дозировку корректируют для достижения концентрации антитела в плазме крови примерно 1-1000 мкг/мл, а в некоторых способах примерно 25-300 мкг/мл.
Альтернативно, антитело можно вводить в виде композиции с непрерывным высвобождением, в этом случае требуется менее частое введение. Дозировку и частоту введения варьируют в зависимости от периода полувыведения антитела у пациента. Как правило, человеческие антитела показывают самый продолжительный период полувыведения, затем следуют гуманизированные антитела, химерные антитела и отличные от человеческих антитела. Дозировка и частота введения могут варьировать в зависимости от того, является ли данное лечение профилактическим или терапевтическим. При профилактических применениях вводят относительно низкую дозировку с относительно редкими интервалами в течение длительного периода времени. Некоторые пациенты продолжают получать лечение весь остаток жизни. При терапевтических применениях иногда бывает необходима относительно высокая дозировка с относительно короткими интервалами введения до тех пор, пока прогрессирование заболевания не снизится или остановится, и предпочтительно до тех пор, пока у пациента не будет достигнуто частичное или полное облегчение симптомов заболевания. В дальнейшем пациент может перейти на профилактический режим.
Действительные уровни дозировок активных ингредиентов в фармацевтических композициях по настоящему изобретению могут быть изменены таким образом, чтобы получить количество активного ингредиента, являющееся эффективным для достижения желаемого терапевтического ответа для конкретного пациента, композиции и способа введения, без оказания токсичного действия на пациента. Выбранный уровень дозировки будет зависеть от ряда фармакокинетических факторов, включая активность конкретных используемых композиций по настоящему изобретению или их сложного эфира, соли или амида, путь введения, продолжительность введения, скорость экскреции конкретного используемого соединения, продолжительность лечения, другие лекарственные средства, соединения и/или вещества, используемые в комбинации с конкретными применяемыми композициями, возраст, пол, массу, состояние, общее состояние здоровья и предшествующую историю болезни пациента, подлежащего лечению, и подобные факторы, хорошо известные в области медицины.
"Терапевтически эффективная дозировка" антитела против Dkk-1 по изобретению предпочтительно приводит к уменьшению тяжести симптомов заболевания, увеличению частоты и продолжительности периодов отсутствия симптомов заболевания или предупреждению ухудшения или нетрудоспособности вследствие заболевания. Обычный специалист в данной области будет способен определить такие количества исходя из таких факторов, как размер субъекта, тяжесть симптомов у субъекта и конкретные выбранные композиция или путь введения.
Дозировку антитела против Dkk-1 можно рассчитать, используя такие методы, как PK/PD модели мишень-опосредованного распределения лекарственного средства (TMDD). Такие модели могут быть использованы для расчета зависимости ответа от концентрации антитела. PK/PD-модель можно использовать для оценки неопосредованного мишенью удаления антител из организма, метаболизма антител и образования и элиминации комплексов. TMDD-модель можно использовать для преобразования данных, определенных с использованием животной модели (например, на крысах или обезьянах), в данные для человека с целью предсказания эффективной дозы. MABEL-модель можно использовать для определения степени экспрессии и скорости метаболизма. Для расчета занятости рецепторов на основании величин KD можно использовать PK/PD-модель. Для определения степени занятости рецепторов учитывают кинетические данные метаболизма мишени и/или комплекса антитело-мишень. PK/PD-модель может быть использована для прогнозирования безопасности и эффективности клинических исследований.
Композицию по настоящему изобретению можно вводить посредством одного или более путей введения, используя один или более чем один из ряда способов, известных в данной области. Как будет очевидно специалисту в данной области, путь и/или способ введения будет варьировать в зависимости от желаемых результатов. Пути введения антител или их антигенсвязывающих участков по изобретению включают внутривенный, внутримышечный, интрадермальный, внутрибрюшинный, подкожный, спинальный или другие парентеральные пути введения, например посредством инъекции или инфузии. Фраза "парентеральное введение", как она использована в данном описании, означает способы введения, отличные от энтерального и местного введения, обычно посредством инъекции, и включает, без ограничения, внутривенную, внутримышечную, интраартериальную, интратекальную, интракапсулярную, внутриглазничную, интракардиальную, интрадермальную, внутрибрюшинную, транстрахеальную, подкожную, субкутикулярную, внутрисуставную, субкапсулярную, субарахноидальную, интраспинальную, эпидуральную и интрастернальную инъекцию и инфузию.
Альтернативно, антитело или его антигенсвязывающую часть по изобретению можно вводить путем, отличным от парентерального, таким как местный, эпидермальный пути или введением через слизистую, например, интраназально, перорально, вагинально, ректально, сублингвально или местно.
Активные соединения могут быть изготовлены вместе с носителями, которые будут защищать соединение от быстрого высвобождения, как например, в композиции с регулируемым высвобождением, включая имплантаты, трансдермальные пластыри и системы доставки с использованием микроинкапсулирования. Могут быть использованы такие биоразлагаемые биосовместимые полимеры, как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, сложные полиортоэфиры и полимолочная кислота. Многие способы изготовления таких композиций запатентованы или в большинстве случаев известны специалистам в данной области. См., например, Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York (1978).
Терапевтические композиции могут быть введены с использованием медицинских устройств, известных в данной области. Например, терапевтическую композицию по изобретению можно вводить, используя безыгольное устройство для подкожных инъекций, как например, устройства, описанные в патентах США №№5399163, 5383851, 5312335, 5064413, 4941880, 4790824 или 4596556. Примеры общеизвестных имплантатов и модулей, полезных в настоящем изобретении, включают: патент США №4487603, в котором описан имплантируемый микро-инфузионный насос для диспергирования лекарственного средства с регулируемой скоростью; патент США №4486194, в котором описано терапевтический устройство для введения лекарственных средств через кожу; патент США №4447233, в котором описан приспособленный для введения лекарств инфузионный насос для доставки лекарственного средства с точно определенной скоростью инфузии; патент США №4447224, в котором описан имплантируемый инфузионный аппарат с изменяющимся потоком для непрерывной доставки лекарственных средств; патент США №4439196, в котором описана осмотическая система доставки лекарственных средств, имеющая многокамерные отделения; и патент США №4475196, в котором описана осмотическая система доставки лекарственных средств. Специалистам в данной области известно много таких других имплантатов, систем доставки и модулей.
В некоторых воплощениях антитела или их фрагменты по изобретению могут быть изготовлены в виде композиции для обеспечения надлежащего распределения in vivo. Например, гематоэнцефалический барьер (ВВВ) не пропускает многие высокогидрофильные соединения. Для обеспечения прохождения ВВВ терапевтическими соединениями по изобретению (если это желательно) они могут быть изготовлены, например, в липосомах. Относительно способов изготовления липосом см., например, патенты США №№4522811, 5374548 и 5399331. Липосомы могут содержать одну или более группировок, которые селективно транспортируются в конкретные клетки или органы, таким образом усиливая направленную доставку лекарственных средств (см., например, V.V. Ranade (1989) J. Clin. Pharmacol., 29: 685). Типичные направляющие группировки включают фолат или биотин (см., например, патент США №5416016, Low et al.); маннозиды (Umezawa et al., (1988) Biochem. Biophys. Res. Commun., 153: 1038); антитела (P.O. Bloeman et al. (1995) FEBS Lett., 357: 140; M. Owais et al. (1995) Antimicrob. Agents Chemother. 39: 180); рецептор сурфактантного белка A (Briscoe et al. (1995) Am. J. Physiol., 1233: 134); p120 (Schreier et al. (1994) J. Biol. Chem., 269: 9090); см. также K. Keinanen; M.L. Laukkanen (1994) FEBS Lett., 346: 123; J.J. Killion; I.J. Fidler (1994) Immunomethods, 4: 273.
Как приведено в данном описании, предложенные фармацевтические композиции можно вводить для профилактического и/или терапевтического лечений. "Эффективное количество" в большинстве случаев относится к количеству, которое является достаточным, но нетоксичным, количеством активного ингредиента (то есть, антитела против Dkk-1 или его иммунологически функционального фрагмента) для достижения желаемого эффекта, который заключается в уменьшении или устранении тяжести и/или частоты симптомов и/или улучшении или исправлении повреждения. "Терапевтически эффективное количество" относится к количеству, которого достаточно для излечения болезненного состояния или симптомов, или которое иным образом предупреждает, замедляет, задерживает или реверсирует прогрессирование заболевания или любого другого нежелательного симптома. "Профилактически эффективное количество" относится к количеству, которое эффективно для предупреждения, замедления или задержки начала болезненного состояния или симптома.
В общем случае токсичность и терапевтическую эффективность антитела или фрагмента можно определить в соответствии со стандартными фармацевтическими методиками в клеточных культурах и/или на экспериментальных животных, включая, например, определение LD50 (дозы, летальной для 50% популяции) и ED50 (дозы, терапевтически эффективной для 50% популяции). Соотношение между дозами, дающими токсический и терапевтический эффекты, представляет собой терапевтический индекс, и его можно выразить в виде соотношения LD50/ED50. Предпочтительными являются композиции, которые демонстрируют высокие терапевтические индексы.
Данные, полученные на основании исследований в клеточной культуре и/или на животных, можно использовать при определении диапазона дозировок для людей. Дозировка активного ингредиента обычно находится в диапазоне концентраций в кровотоке, которые включают ED50 при незначительной токсичности или ее отсутствии. Дозировка может варьировать внутри этого диапазона в зависимости от используемой лекарственной формы и способа введения.
Как упомянуто в данном описании, эффективное количество фармацевтической композиции, содержащей антитела против Dkk-1 или их иммунологически функциональные фрагменты, предназначенной для применения терапевтически или профилактически, будет зависеть, например, от терапевтической ситуации и задач. Специалисту в данной области будет очевидно, что в соответствии с некоторыми воплощениями приемлемые для лечения уровни дозировок, таким образом, будут варьировать в зависимости, частично, от доставляемой молекулы, показания, для которого будет применено антитело против Dkk-1, пути введения и размера (массы тела, поверхности тела или размера органа) и/или состояния (возраста и общего состояния здоровья) пациента. Для получения оптимального терапевтического эффекта врач может определить дозировку и модифицировать способ введения.
Частота введения будет зависеть от фармакокинетических параметров антитела против Dkk-1 или его иммунологически функционального фрагмента в данной композиции. Например, врач будет вводить композицию до тех пор, пока дозировка не достигнет значения, при котором достигают желаемый эффект. Следовательно, такая композиция может быть введена в виде разовой дозы, или в виде двух или более доз (которые могут содержать или не содержать одно и то же количество желаемой молекулы) в течение периода времени, или в виде непрерывной инфузии с помощью имплантируемого устройства или катетера. Лечение может быть непрерывным в течение периода времени или периодическим. Последующее уточнение подходящей дозировки как правило выполняется специалистом обычной квалификации в данной области и находится в пределах задач, обычно выполняемых им. Соответствующие дозировки могут быть установлены благодаря использованию соответствующих данных зависимости доза-ответ.
Для лечения медицинского расстройства, характеризующегося аномальной или избыточной экспрессией Dkk-1, композицию, содержащую являющиеся объектом изобретения антитела против Dkk-1 или их иммунологически функциональные фрагменты, можно вводить пациенту в количестве и в течение времени, которые являются достаточными для индукции непрерывного улучшения по меньшей мере одного индикатора, отражающего тяжесть данного расстройства. Улучшение считается "непрерывным", если пациент демонстрирует улучшение по меньшей мере в двух случаях, разделенных друг от друга по меньшей мере периодом времени от одного до семи суток или в некоторых случаях от одной до шести недель. Соответствующий интервал будет зависеть до некоторой степени от того, какое болезненное состояние подвергается лечению; определение соответствующего интервала для выявления, является ли такое улучшение непрерывным, находится в компетенции квалифицированного врача. Степень улучшения определяют на основании признаков или симптомов, и также можно использовать опросники, которыми снабжают пациента, такие как опросники оценки качества жизни.
Для определения того, являются ли количество и продолжительность лечения достаточными, могут быть оценены различные индикаторы, которые отражают степень заболевания пациента. Начальную величину для выбранного индикатора или индикаторов устанавливают в результате обследования пациента до введения первой дозы антитела. Предпочтительно, начальное обследование выполняют в пределах примерно 60 суток до введения первой дозы. Если антитело вводят для лечения острых симптомов, как например, для лечения сломанной кости, первую дозу вводят так скоро, насколько это возможно практически после того, как произошло повреждение.
Улучшение вызывают путем введения являющихся объектом изобретения антител против Dkk-1 или их иммунологически функциональных фрагментов, пока у пациента не проявится улучшение по сравнению с начальным уровнем для выбранного индикатора или индикаторов. При лечении хронических состояний такую степень улучшения получают путем повторного введения этого лекарственного средства на протяжении периода времени, по меньшей мере одного месяца или более, например, в течение одного, двух или трех месяцев или больше или в течение неопределенного времени. Часто для лечения острых состояний достаточны период времени от одной до шести недель или даже разовая доза. Для повреждений или острых состояний может быть достаточно разовой дозы.
Несмотря на то, что в состоянии заболевания пациента после лечения могут отмечаться улучшения согласно одному или более индикаторам, лечение может быть продолжено в течение неопределенного времени с применением дозы или частоты введения того же самого уровня либо сниженных дозы или частоты. Если лечение сокращено или отменено, позже оно может быть возобновлено на первоначальном уровне, если эти симптомы будут появляться повторно.
Типичные применения антител против Dkk-1
Детекция и скрининг
Антитела против Dkk-1 и их иммунологически функциональные фрагменты, являющиеся объектом изобретения, можно использовать для детекции Dkk-1 в биологических образцах. Такие применения позволяют осуществлять идентификацию клеток или тканей, продуцирующих данный белок, или служат в качестве диагностических средств для детекции патологических состояний, при которых Dkk-1 сверхэкспрессируется или продуцируется в недостаточном количестве.
Предложенные антитела и фрагменты также можно использовать в способах скрининга молекул, которые связываются с Dkk-1. Можно использовать, например, ряд конкурентных способов скрининга. В некоторых методах молекулу Dkk-1 или ее фрагмент, с которыми связывается антитело против Dkk-1, приводят в контакт с антителом или фрагментом, изложенным в данном описании, вместе с другой молекулой (то есть, молекулой-кандидатом). Ослабление связывания между антителом или фрагментом и Dkk-1 является показателем того, что данная молекула связывается с Dkk-1. Связывание антитела или фрагмента можно детектировать, используя различные способы, например, ELISA. Детекцию связывания антител против Dkk-1 или фрагмента с Dkk-1 можно упростить, пометив антитело детектируемой меткой. В некоторых способах проводят дополнительный анализ молекулы, демонстрирующей связывание при первоначальном скрининге, с целью определения того, ингибирует ли она активность Dkk-1 (например, активирует ли данная молекула передачу сигнала через Wnt).
Лечение расстройств, связанных с костной системой
В других аспектах некоторые из предложенных антител и иммунологически функциональных фрагментов могут быть использованы для лечения пациентов с рядом различных заболеваний, включая, например, заболевания, чувствительные к ингибированию активности Dkk-1. Эти антитела и фрагменты также могут быть использованы для лечения заболеваний, чувствительных к индукции передачи сигнала через Wnt. Термин "пациент", как он использован в данном описании, включает в качестве субъектов людей и животных, если не указано иное. Примеры таких заболеваний включают ряд заболеваний, вовлекающих поражение костей, включая состояния с пониженной костной массой, системную потерю костной массы, снижение костеобразования и костные эрозии, но этим не ограничиваются. Некоторые из антител и фрагментов также могут быть использованы в процессах репарации костной ткани.
В некоторых воплощениях антитела или фрагменты имеют терапевтическое применение, выражающееся в стимуляции активности остеобластов и увеличении минеральной плотности кости или костной массы. Таким образом, эти антитела и фрагменты полезны для лечения пациентов, страдающих от различных медицинских расстройств, в которые вовлечено чрезмерное уменьшение костной массы, или пациентов, для которых необходимо образование новой кости, даже в тех случаях, когда активность остеокластов необязательно может быть повышенной. Блокирование активности Dkk-1 приводит к активации остеобластов посредством сигнала, передающегося белками Wnt. Избыточная активность остеокластов ассоциируется с многочисленными остеопеническими нарушениями, которые можно подвергать лечению предложенными антителами и иммунологически функциональными фрагментами, включая остеопению, остеопороз, периодонтит, болезнь Педжета, уменьшение костной массы вследствие иммобилизации, разлагающие кости метастазы и артрит, в том числе ревматоидный артрит, псориатический артрит, анкилозирующий спондилит и другие состояния, в которые вовлечена костная эрозия.
Лечению также могут быть подвергнуты различные другие состояния с пониженной костной массой, включая различные формы остеопороза, в том числе, но этим не ограничиваясь, индуцируемый приемом кортикоидов остеопороз, остеопороз, индуцируемый после трансплантации, остеопороз ассоциированный с химиотерапией (то есть, индуцируемый химиотерапией остеопороз), индуцируемый иммобилизацией остеопороз, остеопороз вследствие отсутствия механической нагрузки и остеопороз, ассоциированный с применением противосудорожных средств. Дополнительные костные болезни, которые могут быть подвергнуты лечению некоторыми из антител или фрагментов, включают костную болезнь, ассоциированную с почечной недостаточностью, и костные болезни, связанные с питанием, ассоциированные с желудочно-кишечным трактом и/или печенью.
Также могут быть подвергнуты лечению разные формы артрита, примеры включают остеоартрит и ревматоидный артрит. Эти антитела и фрагменты также могут быть использованы для лечения системной потери костной массы, ассоциированной с артритом (например, ревматоидным артритом). При лечении артрита пациентам могут быть полезны инъекции антител или их фрагментов, являющихся объектом изобретения, по периферии поврежденного участка или непосредственно в зону поражения. Например, можно сделать инъекцию антитела или его фрагмента вблизи или непосредственно в воспаленный сустав, тем самым стимулируя в этом месте заживление поврежденной кости.
Известно, что при некоторых видах рака возрастает активность остеокластов и индуцируется резорбция кости, таких как рак молочной железы и предстательной железы. Множественная миелома, которая возникает в костном мозге, также ассоциирована с уменьшением костной массы, по всей вероятности вследствие повышенной экспрессии Dkk-1 клетками плазмы, которая затем подавляет костеобразующую активность остеобластов в окружающей области. Снижение активности Dkk-1 в результате введения антител или их иммунологически функциональных фрагментов, являющихся объектом изобретения, может приводить к увеличению активности остеобластов, что служит в качестве противодействия чрезмерной активности остеокластов, тем самым снижая тяжесть вышеупомянутых расстройств, снижая костную эрозию и индуцируя образование новых костей у пациента. Лечение некоторыми специфичными к Dkk-1 антителами или иммунологически функциональными фрагментами может индуцировать значительное увеличение минеральной плотности кости у пациента, страдающего от остеопенического нарушения.
Ингибирование Dkk-1 антителами или иммунологически функциональными фрагментами, изложенными в данном описании, также может быть использовано при различных применениях, касающихся репарации костной ткани. Например, некоторые антитела и фрагменты могут быть полезны в торможении остеолизиса продуктов износа, ассоциированного с искусственными суставами, ускорении репарации переломов кости и способствовании внедрению костных трансплантатов внутрь окружающей его живой кости, в которую они пересажены.
Как изложено в данном описании, антитела против Dkk-1 или их иммунологически функциональные фрагменты могут быть введены по отдельности или в комбинации с другими терапевтическими агентами, например, в комбинации со средствами терапии рака, с агентами, ингибирующими активность остеокластов, или с другими агентами, усиливающими активность остеобластов. Например, антитела по изобретению могут быть введены раковым пациентам, подвергающимся лучевой терапии или химиотерапии. Химиотерапии, применяемые в комбинации с антителами по изобретению, могут включать применение антрациклинов, таксола, тамоксифена, доксорубицина, 5-фторурацила, оксалоплатина, Velcade® ([(1R)-3-метил-1-[[(2S)-1-оксо-3-фенил-2-[(пиразинилкарбонил)амино]пропил]-амино]-бутил]бороновая кислота) и/или других низкомолекулярных лекарственных средств, которые используют в лечении рака. Пациенты с диагнозом рак молочной железы получат пользу от введения ингибитора ароматазы одновременно с комбинированными лечениями, содержащими химиотерапевтический агент и антитело против Dkk-1 или его иммунологически функциональный фрагмент.
Антитела против Dkk-1 и их иммунологически функциональные фрагменты могут быть использованы по отдельности для лечения вышеупомянутых состояний, приводящих к уменьшению костной массы, или в комбинации с терапевтически эффективным количеством агента, стимулирующего рост костей (анаболического агента), или костного антирезорбтивного агента, включая: костные морфогенетические факторы с обозначением от ВМР-1 до BMP-12 (костный морфогенетический белок); трансформирующий ростовой фактор-β и члены семейства TGF-β; факторы роста фибробластов от FGF-1 до FGF-10; ингибиторы интерлейкина-1 (включая IL-1ra, антитела к IL-1 и антитела к рецепторам IL-1); ингибиторы TNFα (включая этанерцепт, адалимумаб и инфликсимаб); ингибиторы лигандов RANK (активатор рецепторов нуклеарного фактора каппа-В) (включая растворимые RANK, остеопротегерин и антагонистические антитела, которые специфически связываются с RANK или лигандом RANK), паратиреоидный гормон, простагландины серии Е, бисфосфонаты и минеральные вещества, способствующие укреплению костей, такие как фторид и кальций, но этим не ограничиваются. Анаболические агенты, которые могут быть использованы в комбинации с антителами по изобретению и их функциональными фрагментами, включают паратиреоидный гормон и инсулиноподобный ростовой фактор (IGF), причем последний агент предпочтительно находится в комплексе с IGF-связывающим белком. Антагонист рецептора IL-1, подходящий для такого комбинированного лечения, описан в WO89/11540, а подходящий растворимый рецептор-1 TNF описан в WO98/01555. Типичные антагонисты лигандов RANK описаны, например, в WO 03/086289, WO 03/002713, патентах США №№6740511 и 6479635. Все вышеупомянутые патенты и патентные заявки тем самым включены сюда посредством ссылки.
Помимо этого, антитела против Dkk-1 могут быть введены пациентам в комбинации с антителами, которые связываются с опухолевыми клетками и индуцируют цитотоксическое и/или цитостатическое воздействие на рост опухоли. Примеры таких антител включают антитела, которые связываются с белками клеточной поверхности Her2, CDC20, CDC33, муциноподобным гликопротеином и рецептором эпидермального ростового фактора (EGFR), присутствующим на опухолевых клетках, и индуцируют цитостатическое и/или цитотоксическое воздействие на опухолевые клетки, проявляющие эти белки. Примеры таких антител включают HERCEPTIN® для лечения рака молочной железы и RITUXAN® для лечения неходжкинской лимфомы и также включают лекарства на основе антител, такие как ERBITUX® и AVASTIN®. Кроме того, комбинированная терапия может включать в качестве противораковой терапии агенты-полипептиды, которые селективно индуцируют апоптоз в опухолевых клетках, такие как TNF-родственный полипептид TRAIL.
Антитела или их иммунологически функциональные фрагменты, являющиеся объектом изобретения, могут быть введены одновременно с другими видами лечения и терапевтическими агентами, вводимыми для того же состояния. "Одновременное введение", как использовано в данном описании, охватывает виды лечения, которые вводят одновременно или последовательно. Антитела против Dkk-1 или их иммунологически функциональные фрагменты могут быть введены с целью профилактики для предупреждения или ослабления начала уменьшения костной массы на ранних стадиях рака (стадиях I или II) или могут быть назначены для облегчения существующего состояния со сниженной костной массой вследствие метастазирования в кость.
Антитела против Dkk-1 по изобретению могут быть использованы для предупреждения и/или лечения роста опухолевых клеток в кости. Рак, который метастазирует в кость, может легко распространяться, поскольку опухолевые клетки стимулируют остеокласты к резорбции внутреннего костного матрикса. Лечение антителом против Dkk-1 или его иммунологически функциональным фрагментом поможет поддержать минеральную плотность кости в месте локализации таких метастазов путем стимуляции повышенной активности остеобластов. Любой рак, который имеет потенциал к метастазированию в кость, можно предупредить или лечить антителом против Dkk-1, вводимым до или после появления метастазов.
Множественная миелома представляет собой пример типа рака, который может быть предотвращен и/или подвергнут лечению антителом против Dkk-1 или его антигенсвязывающим фрагментом. Пораженные болезнью пациенты обычно демонстрируют уменьшение костной массы вследствие повышенной активации остеокластов в локализованных участках кости. Миеломные клетки либо напрямую, либо опосредованно продуцируют лиганд RANK, белок, который активирует остеокласты, вызывая лизис кости, окружающей миеломные клетки, располагающиеся в костно-мозговом пространстве. Нормальные остеокласты, ближайшие к миеломной клетке, в свою очередь продуцируют IL-6, вызывая рост и пролиферацию миеломных клеток. Помимо этого, клетки множественной миеломы продуцируют Dkk-1, тем самым ингибируя активность остеобластов и затем стимулируя активность процессов резорбции кости в этом заболевании. Лечение животного антителом против Dkk-1 или его иммунологически функциональным фрагментом будет инициировать активность остеобластов, тем самым приводя к увеличению костной массы в месте расположения опухолей. Такое лечение может приводить к ослаблению костной боли и может блокировать приводить к ослаблению костной боли и может блокировать дальнейшее метастазирование в кость путем предотвращения резорбтивной активности, в результате проявления которой высвобождаются питательные вещества кости, утилизируемые опухолевыми клетками. При лечении этого заболевания антитело против Dkk-1 или его иммунологически функциональный фрагмент могут быть введены одновременно с антагонистическими антителами, направленными против лиганда RANK, или антителами против IL-6.
Лечение других расстройств
Помимо вышеупомянутых применений, относящихся к поражениям костей, некоторые предложенные антитела и иммунологически функциональные фрагменты могут быть использованы для лечения других заболеваний. Роль Dkk-1 в этих разных заболеваниях частью основывается на его экспрессии в различных других тканях. Антитела и фрагменты, например, могут быть использованы для лечения заболеваний, при которых желательно стимулировать обновление стволовых клеток. Такие заболевания включают диабет, хроническую сердечную недостаточность и различные заболевания мышц (например, дисфункциональную атрофию в результате, например, иммобилизации или постельного режима; старческую дряхлость (саркопению людей пожилого возраста); мышечные дистрофии; кахексию, ассоциированную с раком, СПИДом или воспалением; белково-энергетическую недостаточность при почечной недостаточности/уремии и мышечную атрофию при ожирении), но этим не ограничиваются. Также можно лечить различные воспалительные заболевания, включая, например, болезнь Крона, колит и воспалительное заболевание кишечника. Антитела и фрагменты также могут быть использованы в лечении различных неврологических заболеваний (например, болезни Альцгеймера, болезни Паркинсона и болезни Гентингтона). Глазные болезни (например, макулярная дегенерация и различные ретинопатии) также можно лечить некоторыми из антител и фрагментов. С использованием некоторых антител также можно лечить разные почечные болезни (например, терминальную стадию почечной недостаточности, хроническую болезнь почек, гломерулонефрит, тубулоинтерстициальный нефрит и нефропатию IgA-типа). Помимо этого, также можно лечить различные легочные заболевания (например, хроническую обструктивную болезнь легких, идиопатический легочный фиброз и кистозный фиброз) и различные кожные поражения, в том числе кожные и эпидермальные заболевания. Примеры кожных расстройств, которые могут быть подвергнуты лечению, включают поражение эпителия (тонкого) кишечника (например, индуцированное химиотерапией поражение) и другие заболевания, при которых желательно стимулировать рост и выживаемость эпителия (тонкого) кишечника.
Наборы
Также предложены наборы, которые включают антитело или иммунологически функциональный фрагмент либо фармацевтическую композицию, которые изложены в данном описании. Некоторые наборы включают такие антитело, фрагмент или композицию в контейнере (например, флаконе или ампуле) и также могут включать в себя инструкции по использованию антитела или фрагмента в различных применениях, относящихся к детекции, скринингу и терапии, описанных выше. Антитело, фрагмент или композиция могут быть в различных формах, в том числе, например, в виде части раствора или в виде твердого вещества (например, лиофилизированного порошка). Инструкции могут включать описание того, как приготовить (например, растворить или ресуспендировать) антитело или фрагмент в соответствующей жидкости и/или как ввести антитело или фрагмент для лечения заболеваний, описанных выше (например, поражений костей, таких как сниженная костная масса, системное уменьшение костной массы, ослабление костеобразования и костные эрозии; для обновления стволовых клеток; для лечения воспалительных заболеваний; неврологических заболеваний; глазных болезней; почечных болезней и поражений кожи).
Наборы также могут включать различные другие компоненты, такие как буферы, соли, комплексообразующие ионы металлов и другие агенты, описанные выше в разделе, относящемся к фармацевтическим композициям. Эти компоненты могут быть включены вместе с антителом или фрагментом или могут находиться в раздельных контейнерах. Наборы также могут включать другие терапевтические агенты для введения вместе с антителом или фрагментом. Примеры таких агентов включают, но этим не ограничиваются, агенты для лечения рака, стимулирующие костеобразование агенты и антитела, которые связываются с опухолевыми клетками, и другие агенты, приведенные выше.
Дальнейшее конкретное описание антител против Dkk-1
Модификации Fcγ-области
В рекомбинантных гуманизированных антителах Fcγ-область может быть модифицирована для того, чтобы избежать взаимодействия с Fcγ-рецептором и комплементом иммунной системы. Этот тип модификации был разработан доктором Майком Кларком из Отделения патологии Кембриджского Университета (Dr. Mike Clark from Department of Pathology at Cambridge University), и способы получения таких антител описаны, например, в WO 99/58572, опубликованной 18 ноября 1999 г.Например, константная область может быть сконструирована так, чтобы иметь больше сходства с константными областями иммуноглобулина человека, для избежания иммунного ответа, если антитело используется для лечения людей. См., например, патенты США №№5997867 и 5866692.
Модификации huMabJC18
huMabJC18 представляет собой полностью гуманизированное моноклональное антитело против Dkk-1, Настоящее изобретение охватывает модификации huMabJC18, включая функционально эквивалентные антитела, которые не затрагивают существенно их свойства, и варианты, обладающие повышенной или сниженной активностью и/или аффинностью. Например, как будет очевидно специалистам в данной области, аминокислотная последовательность huMabJC18 может быть подвергнута мутации для получения антитела с желаемой аффинностью связывания с Dkk-1. Модификация полипептидов является обычной практикой в данной области и не нуждается в подробном изложении в данном описании. Примеры модификаций полипептидов представлены в разделе Примеры. Примеры модифицированных полипептиды включают: полипептиды с консервативными заменами аминокислотных остатков, с одной или более делениями или вставками аминокислот, которые не вносят существенных повреждающих изменений в функциональную активность, или использование химических аналогов.
Вставки в аминокислотную последовательность включают: амино- и/или карбоксил-концевые слияния в диапазоне длины от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки внутрь последовательности одного или многих аминокислотных остатков. Примеры терминальных вставок включают антитело с N-концевым остатком метионина или антитело, слитое с эпитопной меткой. Другие варианты молекулы антитела с вставками включают слияние с N- или С-концом антитела фермента или полипептида, что увеличивает период полувыведения антитела из сыворотки крови.
В вариантах с заменами из молекулы антитела удален по меньшей мере один аминокислотный остаток и на его место встроен другой остаток. Сайты, представляющие наибольший интерес для замещающего мутагенеза, включают гипервариабельные участки, но изменения в каркасе (FR) также рассматриваются. Консервативные замены показаны в Таблице 1 под заголовком "консервативные замены". Если такие замены приводят к изменению биологической активности, то могут быть введены более существенные изменения, которые обозначены как "типичные замены" в Таблице 1 или дополнительно описаны ниже в отношении классов аминокислот, и продукты подвергнуты скринингу.
Существенные модификации биологических свойств антител связаны с избирательными заменами, которые значительно различаются по их влиянию на поддержание (а) структурного остова полипептида в области замены, например, такого как конформация "слой" или "спираль", (б) заряда или гидрофобности молекулы в целевом сайте или (в) объема боковой цепи. Природные остатки подразделяют на группы на основании общих свойств боковой цепи:
(1) неполярные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) полярные незаряженные: Cys, Ser, Thr, Asn, Gln;
(3) кислотные (отрицательно заряженные): Asp, Glu;
(4) основные (положительно заряженные): Lys, Arg;
(5) остатки, влияющие на ориентацию цепи: Gly, Pro; и
(6) ароматические: Trp, Tyr, Phe, His.
Неконсервативные замены производят, меняя представителя одного из этих классов на представителя другого класса.
Любой цистеиновый остаток, не вовлеченный в поддержание надлежащей конформации антитела, также может быть заменен, в большинстве случаев на серин, для улучшения стабильности молекулы к окислению и для предупреждения ошибочного перекрестного сшивания. Наоборот, в антитело может быть добавлена связь(и) через остатки цистеина для улучшения его стабильности, в частности когда антитело представляет собой фрагмент антитела, такой как Fv-фрагмент.
Аминокислотные модификации могут варьировать от изменения или модификации одной или более аминокислот, до полной переделки области, такой как вариабельная область. Изменения в вариабельной области могут менять аффинность связывания и/или специфичность. В некоторых воплощениях в пределах домена CDR производят не более чем от одной до пяти консервативных аминокислотных замен. В других воплощениях в пределах домена CDR производят не более чем от одной до трех консервативных аминокислотных замен. В следующих других воплощениях домен CDR представляет собой CDR Н3 и/или CDR L3.
Гликозилирование huMabJC18
Модификации также включают гликозилированные и негликозилированные полипептиды, а также полипептиды с другими посттрансляционными модификациями, такими как, например, Гликозилирование различными сахарами, ацетилирование и фосфорилирование. Антитела гликозилируются в консервативных положениях их константных областей (см., например, Jefferis and Lund (1997) Chem. Immunol., 65: 111-128; Wright and Morrison (1997) TibTECH, 15: 26-32). Олигосахаридные боковые цепи иммуноглобулинов влияют на белковую функцию (см., например, Boyd et al. (1996) Mol. Immunol., 32: 1311-1318; Wittwe and Howard (1990) Biochem., 29: 4175-4180) и на внутримолекулярное взаимодействие между частями гликопротеина, что может оказывать влияние на конформацию и экспонированную в пространство поверхность гликопротеина (Hefferis and Lund, выше; Wyss and Wagner (1996) Current Opin. Biotech., 7: 409-416). Кроме того, олигосахариды могут быть полезны для нацеливания данного гликопротеина на определенные молекулы на основе специфически распознаваемых структур. Также описано, что Гликозилирование антител влияет на антителозависимую клеточную цитотоксичность (ADCC). В частности сообщалось, что СНО-клетки с регулируемой тетрациклином экспрессией β(1,4)-N-ацетилглюкозаминил-трансферазы III (GnTIII), гликозилтрансферазы, катализирующей образование структур с разветвлением при GlcNAc (N-ацетил-D-глюкозамин), обладают повышенной ADCC-активностью (см., например, Umana et al. (1999) Nature Biotech., 17: 176-180).
Гликозилирование антител обычно происходит посредством образования N-связей или O-связей. N-гликозилирование относится к присоединению углеводной группировки к боковой цепи остатка аспарагина. Последовательности трех аминокислот аспарагин-Х-серин, аспарагин-Х-треонин и аспарагин-Х-цистеин, где Х представляет собой любую аминокислоту за исключением пролина, являются последовательностями узнавания для осуществления ферментативного присоединения углеводной группировки к боковой цепи аспарагина. Так, наличие любой из этих последовательностей трех аминокислот в полипептиде создает сайт возможного гликозилирования. O-гликозилирование относится к присоединению одного из сахаров, N-ацетилгалактозамина, галактозы или ксилозы, к гидроксиаминокислоте, чаще всего к серину или треонину, хотя также можно использовать 5-гидроксипролин или 5-гидроксилизин.
Добавление сайтов гликозилирования в антитело удобно выполнять посредством изменения аминокислотной последовательности таким образом, чтобы она содержала одну или более из описанных выше последовательностей трех аминокислот (для сайтов N-гликозилирования). Изменение также может быть осуществлено посредством добавления одного или более остатков серина или треонина к последовательности исходного антитела либо замены одного или более остатков в последовательности исходного антитела на остатки серина или треонина (для сайтов O-гликозилирования).
Характер гликозилирования антител также можно изменить, не изменяя лежащую в их основе нуклеотидную последовательность. Гликозилирование в большой степени зависит от клетки-хозяина, используемой для экспрессии антитела. Так как тип клетки, используемой для экспрессии рекомбинантных гликопротеинов, например, антител как возможных терапевтических средств, редко представляет собой нативную клетку, можно ожидать вариаций в характере гликозилирования антител (см., например, Hse et al. (1997) J. Biol. Chem., 272: 9062-9070).
В дополнение к выбору клетки-хозяина, факторы, которые влияют на гликозилирование в процессе получения рекомбинантных антител, включают режим культивирования, состав среды, плотность культуры, оксигенация, рН, схемы очистки и тому подобное. Для изменения характера гликозилирования, осуществляемого в конкретном организме хозяина, предложены различные способы, включая введение или сверхэкспрессию некоторых ферментов, вовлеченных в образование олигосахаридов (см., например, патенты США №№5047335, 5510261 и 5278299). Углеводные структуры или некоторые типы углеводных структур можно ферментативно удалить из гликопротеина, например, используя эндогликозидазу Н (Endo H). Помимо этого, методами генетической инженерии можно получить рекомбинантную клетку-хозяина, дефектную в отношении процессинга некоторых типов полисахаридов. Эти и подобные им методы хорошо известны в данной области.
Методы сочетания
Как упомянуто выше, другие способы модификации включают использование методов сочетания, известных в данной области, в том числе ферментативных методов, окислительного замещения и хелатообразования, но этим не ограничиваются. Модификации можно использовать, например, для присоединения меток для проведения иммуноанализа. Модифицированные полипептиды huMabJC18 могут быть получены с использованием общепринятых в данной области методик, а их скрининг может быть проведен с использованием стандартных, известных в данной области анализов, некоторые из них описаны ниже в разделе Примеры. Специалистам в данной области будут понятны любые подходящие способы проведения и скрининга таких модификаций.
Модифицированная константная область
В некоторых воплощениях изобретения антитело содержит модифицированную константную область, такую как константная область, которая является иммунологически инертной или частично инертной, например, не инициирует опосредованный комплементом лизис, не стимулирует антителозависимую клеточноопосредованную цитотоксичность (ADCC) или не активирует микроглии; или обладает сниженными активностями (по сравнению с немодифицированным антителом) в отношении одного или более чем одного из следующего: инициирование опосредованного комплементом лизиса, стимулирование антителозависимой клеточноопосредованной цитотоксичности (ADCC) или активация микроглий. Для достижения оптимального уровня и/или оптимальной комбинации эффекторных функций могут быть использованы разные модификации константной области. См., например, Morgan et al., Immunology, 86: 319-324 (1995); Lund et al., J. Immunology, 157: 4963-4969 (1996); Idusogie et al., J. Immunology, 164: 4178-4184 (2000); Tao et al., J. Immunology, 143: 2595-2601 (1989); и Jefferis et al., Immunological Reviews, 163: 59-76 (1998). В одном из воплощений константная область модифицирована так, как описано в Eur. J. Immunol. (1999) 29: 2613-2624; в заявке РСТ № PCT/GB99/01441; и/или патентной заявке UK №9809951.8.
В одном из воплощений антитело содержит константную область тяжелой цепи IgG2a человека, содержащую следующие мутации: А330Р331 на S330S331 (нумерация аминокислот в соответствии с последовательностью IgG2a дикого типа) (Eur. J. Immunol. (1999), 29: 2613-2624). В другом воплощении константная область является агликозилированной. В другом воплощении константная область является агликозилированной в результате мутации гликозилируемого аминокислотного остатка в константной области. Например, сайт N-гликозилирования N297 может быть изменен посредством мутации на A, Q, K или Н. См., например, Tao et al., J. Immunology, 143: 2595-2601 (1989); и Jefferis et al., Immunological Reviews, 163: 59-76 (1998). В одном из воплощениий константная область агликозилирована с участием ферментов (как например, путем удаления углеводов под действием фермента PNGазы (пептид-N-гликаназы)).
Эффекторный домен
Другие модификации антител включают антитела, модифицированные, как описано, например в WO 99/58572, опубликованной 18 ноября 1999 г. Эти антитела содержат, в дополнение к связывающему домену, направленному на молекулу-мишень, эффекторный домен, имеющий аминокислотную последовательность, по существу гомологичную последовательности всего или части константного домена тяжелой цепи человеческого иммуноглобулина. Эти антитела способны к связыванию с молекулой-мишенью без инициирования значительного комплементзависимого лизиса или клеточноопосредованного разрушения мишени. В некоторых воплощениях эффекторный домен способен к специфическому связыванию с FcRn и/или FcγRIIb. В основе этого обычно лежат химерные домены, происходящие из двух или более CH2-доменов тяжелой цепи человеческого иммуноглобулина. Антитела, модифицированные таким образом, в частности подходят для использования при продолжительной терапии антителами, позволяя избежать воспалительных и других неблагоприятных реакций, свойственных традиционной терапии антителами.
Аффинно зрелые формы
В одном из воплощений антитело представляет собой аффинно зрелую форму антитела. Например, аффинно зрелые формы антител могут быть получены с использованием известных в данной области методик (например, Marks et al., 1992, Bio/Technology, 10: 779-783; Barbas et al. (1994) Proc. Nat. Acad. Sci. USA, 91: 3809-3813; Schier et al. (1995) Gene, 169: 147-155; Yelton et al. (1995) J. Immunol., 155: 1994-2004; Jackson et al. (1995) J. Immunol., 154(7): 3310-9; Hawkins et al. (1992) J. Mol. Biol., 226: 889-896).
Библиотека-сканирующий мутагенез
Приведенные далее способы могут быть использованы для регулирования аффинности антител и для определения характеристик CDR. Один из путей определения характеристик CDR антитела и/или изменения (например, улучшения) аффинности связывания полипептида, такого как антитело, носит название "библиотека-сканирующего мутагенеза". Как правило, "библиотека-сканирующий мутагенез" работает следующим образом. В одном или более чем одном из аминокислотных положений в CDR проводят замену, используя две или более (как например, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20) аминокислот, с применением общепризнанных в данной области методов. Это приводит к созданию небольших библиотек клонов (в некоторых воплощениях по одному для каждого анализируемого аминокислотного положения), каждая из которых представляет собой множество из двух или более членов (если в каждом положении проведена замена на две или более аминокислот). Как правило, такая библиотека также включает клон, содержащий нативную (незамененную) аминокислоту. Небольшое количество клонов, например, приблизительно 20-80 клонов (в зависимости от множества составляющих библиотеки), из каждой библиотеки подвергают скринингу на предмет аффинности связывания с полипептидом-мишенью (или другой мишенью для связывания) и идентифицируют кандидатов, характеризующихся улучшенным связыванием, не изменившимся связыванием и ухудшенным связыванием, и кандидатов, у которых связывание отсутствует.
Определение аффинности связывания
Методы определения аффинности связывания хорошо известны в данной области. Аффинность связывания можно определить, используя BIAcore- или ProteOn-анализ, основанный на поверхностном плазменном резонансе, с помощью которого можно идентифицировать примерно 2-кратные или большие различия в аффинности связывания. Эти типы анализов полезны в частности, когда исходное антитело уже характеризуется связыванием с относительно высокой аффинностью, например, с KD примерно 10 нМ или ниже. Скрининг с использованием поверхностного плазменного резонанса BIAcore или ProteOn, изложен в данном описании в разделе Примеры.
Аффинность связывания можно определить, используя биосенсор Kinexa, анализы с применением аналит-связывающих сцинтилляционных гранул, ELISA, иммуноанализ ORIGEN (IGEN), анализ гашения флуоресценции, анализ переноса флуоресценции и/или дисплей с использованием дрожжей. Используя подходящие виды биоанализов, также можно осуществить скрининг по аффинности связывания.
В некоторых воплощениях в каждом аминокислотном положении в CDR проводят замены (в некоторых воплощениях поочередно) на все 20 природных аминокислот, используя общепризнанные в данной области методы мутагенеза (некоторые из них изложены в данном описании). Это приводит к созданию небольших библиотек клонов (в некоторых воплощениях по одному для каждого анализируемого аминокислотного положения), каждая из которых представляет собой множество из 20 членов (если в каждом положении произведена замена на все 20 аминокислот).
В некоторых воплощениях библиотека, подлежащая скринингу, содержит замены в двух или более положениях, которые могут быть в одном и том же CDR или в двух или более CDR. Так, библиотека может содержать замены в двух или более положениях в одном CDR. Библиотека может содержать замену в двух или более положениях в двух или более CDR. Библиотека может содержать замену в 3, 4, 5 или более положениях, причем указанные положения находятся в двух, трех, четырех, пяти или шести CDR. Такую замену можно получить, используя кодоны с низкой вырожденностью. См., например, Таблицу 2 в Balint et al. (1993) Gene, 137(1): 109-18).
CDR может представлять собой CDRH3 и/или CDRL3. CDR может представлять собой один или более чем один из CDRL1, CDRL2, CDRL3, CDRH1, CDRH2 и/или CDRH3. CDR может представлять собой CDR по Kabat, CDR по Chothia или расширенный CDR.
Кандидаты с улучшенным связыванием могут быть отсеквенированы, тем самым идентифицируется мутант с заменой в CDR, которая приводит к улучшенной аффинности (также называемой как "улучшенная" замена). Также могут быть отсеквенированы кандидаты, которые обладают способностью связываться, тем самым идентифицируется замена в CDR, которая сохраняет свойство связываться.
Можно провести множественные раунды скрининга. Например, кандидаты (каждый содержащий аминокислотную замену в одном или более чем одном положении одного или более CDR) с улучшенным связыванием также полезны для конструирования второй библиотеки, содержащей по меньшей мере исходную и замененную аминокислоту в каждом улучшенном положении в CDR (то есть аминокислотном положении в CDR, по которому мутант с заменой показывает улучшенное связывание). Получение и скрининг или селекция этой библиотеки дополнительно обсуждены ниже.
Библиотека-сканирующий мутагенез также обеспечивает средство для определения характеристик CDR, поскольку частота встречаемости клонов с улучшенным связыванием, не изменившимся связыванием, ухудшенным связыванием и с отсутствием связывания также является источником информации, касающейся важности каждого аминокислотного положения для стабильности комплекса антитело-антиген. Например, если положение в CDR сохраняет свойство связываться, когда в нем производится замена на все 20 аминокислот, то такое положение идентифицируют как положение, которое по-видимому не является существенным для связывания с антигеном. И наоборот, если положение в CDR сохраняет свойство связываться только в случае небольшого процента замен, то такое положение идентифицируют как положение, которое является важным для функционирования CDR. Так, методы библиотека-сканирующего мутагенеза дают информацию касательно положений в CDR, в которых могут быть сделаны замены на многие разные аминокислоты (включая все 20 аминокислот), и положений в CDR, в которых не могут быть сделаны замены или в которых могут быть сделаны замены только на несколько аминокислот.
Кандидаты с улучшенной аффинностью можно комбинировать во второй библиотеке, которая включает улучшенную аминокислоту, исходную аминокислоту в этом положении и также может включать дополнительные замены в этом положении в зависимости от множества составляющих библиотеки, которая желательна, либо дает возможность использования желаемого метода скрининга или селекции. Помимо этого, при желании ближайшее аминокислотное положение может быть рандомизировано в отношении по меньшей мере двух или более аминокислот. Рандомизация ближайших аминокислот может обеспечить дополнительную конформационную гибкость мутантного CDR, которая в свою очередь может дать возможность для введения или облегчить введение большого числа улучшающих мутаций. Эта библиотека также может содержать замену по положениям, которые не показывали улучшенной аффинности в первом раунде скрининга.
Скрининг или селекцию второй библиотеки проводят с целью выявления членов библиотеки с улучшенной и/или измененной аффинностью связывания, используя любой метод, известный в данной области, включая скрининг с использованием BIAcore-анализа с применением поверхностного плазменного резонанса и селекцию с использованием любого метода, известного в данной области для селекции, включая фаговый дисплей, дисплей с использованием дрожжей и рибосомный дисплей.
Слитые белки
Как упомянуто выше, изобретение также охватывает слитые белки, содержащие один или более фрагментов или участков из антител (таких как huMabJC18) или полипептидов по данному изобретению. Слитый полипептид huMabJC18 может быть создан способами, известными в данной области, например, путем синтеза или рекомбинантно. Обычно слитые белки huMabJC18 по данному изобретению получают путем получения и экспрессии кодирующего их полинуклеотида, используя рекомбинантные способы, изложенные в данном описании, хотя они также могут быть получены другими способами, известными в данной области, включая, например, химический синтез.
Конъюгирование
Как упомянуто выше, согласно изобретению также предложены композиции, содержащие антитела или полипептиды huMabJC18, конъюгированные (например, соединенные) с агентом (таким как биотин или авидин), который облегчает присоединение к твердой подложке. Для простоты в большинстве случаев ссылка будет делаться на huMabJC18 или антитела с пониманием того, что эти способы применимы к любому из воплощений, касающихся связывания с Dkk-1, изложенных в данном описании. Конъюгирование в большинстве случаев относится к присоединению этих компонентов, которые изложены в данном описании. Такое присоединение (которое в большинстве случаев представляет собой фиксацию этих компонентов в непосредственной близости друг к другу по меньшей мере для введения) может быть достигнуто различными методами. Например, прямое взаимодействие между агентом и антителом возможно, если каждый из них обладает заместителем, способным взаимодействовать друг с другом, Например, нуклеофильная группа, такая как аминогруппа или сульфгидрильная группа, на одном из них может быть способна взаимодействовать с карбонилсодержащей группой, такой как ангидрид или галогенид кислоты, или с алкильной группой, содержащей хорошую уходящую группу (например, галогенид), на другом из них.
Агенты мечения
Как упомянуто выше, антитело или полипептид по данному изобретению может быть соединен с любым подходящим агентом мечения (альтернативно называемым "меткой"), таким как, например, флуоресцентная молекула, радиоактивная молекула или любые другие метки, известные в данной области. В данной области известны метки, которые в большинстве случае дают (или напрямую, или опосредованно) сигнал.
Композиции
Как упомянуто выше, согласно изобретению также предложены композиции (в том числе фармацевтические композиции) и наборы, содержащие, например, huMabJC18, или, как понятно из этого описания, любое(ой) или все из антител и/или полипептидов, изложенных в данном описании.
Полинуклеотиды, векторы (например, экспрессирующие) и клетки-хозяева
Как упомянуто выше в нескольких конкретных приведенных воплощениях, согласно изобретению также предложены выделенные полинуклеотиды, кодирующие антитела и полипептиды по изобретению (включая антитело, содержащее полипептидные последовательности вариабельных областей легкой цепи и тяжелой цепи, показанные на Фиг.1), и векторы и клетки хозяина, содержащие данный полинуклеотид.
В другом аспекте согласно изобретению предложены полинуклеотиды, кодирующие любое из антител (включая фрагменты антитела) и любой из полипептидов, изложенных в данном описании. Как будет очевидно специалистам в данной области, предложенные полинуклеотиды могут быть получены посредством методик, известных в данной области.
В другом аспекте согласно изобретению предложены композиции (включая фармацевтические композиции, описанные более подробно выше и ниже), содержащие любой из полинуклеотидов по изобретению. В одном из воплощений композиция содержит экспрессирующий вектор, содержащий полинуклеотид, кодирующий huMabJC18, который изложен в данном описании. В одном из воплощений композиция содержит экспрессирующий вектор, содержащий полинуклеотид, кодирующий любое(ой) из антител или полипептидов, изложенных в данном описании. В еще одном другом воплощении композиция содержит или один из двух, или оба полинуклеотида, показанные в SEQ ID NO:19 и SEQ ID NO:28. Экспрессирующие векторы и введение полинуклеотидных композиций также изложено в данном описании.
В другом аспекте согласно изобретению предложены способы получения любого из полинуклеотидов, изложенных в данном описании.
Полинуклеотиды, комплементарные к любой из таких последовательностей, также охватываются настоящим изобретением. Полинуклеотиды могут быть одноцепочечными (кодирующими или антисмысловыми) или двухцепочечными и могут представлять собой молекулы ДНК (геномной, кДНК или синтетической) или" РНК. Молекулы РНК включают молекулы гяРНК, которые содержат интроны и молекуле ДНК по типу "один-к-одному", и молекулы мРНК, которые не содержат интронов. Внутри полинуклеотида по настоящему изобретению могут присутствовать, но это не обязательно, дополнительные кодирующие или некодирующие последовательности, и полинуклеотид может быть соединен, но не обязательно, с другими молекулами и/или материалами подложки.
Полинуклеотиды могут содержать нативную последовательность (то есть, эндогенную последовательность, которая кодирует антитело или его часть) или могут содержать вариант такой последовательности. Полинуклеотидные варианты содержат одну или более замен, добавлений, делеций и/или вставок, таких, что иммунореактивность кодируемого полипептида относительно нативной иммунореактивной молекулы не уменьшается. В большинстве случаев можно оценить влияние на иммунореактивность кодируемого полипептида, который изложен в данном описании. Варианты предпочтительно демонстрируют по меньшей мере примерно 70% идентичности, более предпочтительно по меньшей мере примерно 80% идентичности и наиболее предпочтительно по меньшей мере примерно 90% идентичности с полинуклеотидной последовательностью, которая кодирует нативное антитело или его часть.
Отмечается, что две полинуклеотидные или полипептидные последовательности являются "идентичными", если последовательность нуклеотидов или аминокислот в этих двух последовательностях является одной и той же при их выравнивании с целью максимального соответствия, как описано ниже. Сравнения между двумя последовательностями обычно проводят, сравнивая последовательности в пределах окна сравнения, используемого для идентификации, и сравнивая локальные участки схожести последовательностей. "Окно сравнения", как использовано в данном описании, относится к сегменту, состоящему по меньшей мере из примерно 20 следующих друг за другом положений, обычно от 30 до примерно 75 или от 40 до примерно 50, в котором последовательность можно сравнивать с последовательностью сравнения, состоящей из такого же количества следующих друг за другом положений после того, как эти две последовательности оптимально выровнены.
Оптимальное выравнивание последовательностей для проведения сравнения может быть выполнено с применением программы Megalign в наборе комплектов программ Lasergene для биоинформатики (DNASTAR, Inc., Madison, WI) с использованием параметров по умолчанию. Эта программа включает несколько схем выравнивания, описанных в следующих ссылках: Dayhoff, M.O. (1978) A model of evolutionary change in proteins - Matrices for detecting distant relationships. Dayhoff, M.O. (ed.) Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, Washington DC Vol.5, Suppl. 3, pp.345-358; Hein J. (1990) Unified Approach to Alignment and Phylogenes, pp.626-645, Methods in Enzymology vol. 183, Academic Press, Inc., San Diego, CA; Higgins, D.G. and Sharp, P.M. (1989) CABIOS 5: 151-153; Myers, E.W. and Muller W. (1988) CABIOS 4: 11-17; Robinson, E.D., 1971, Comb. Theor., 11: 105; Santou, N., Nes, M. (1987) Mol. Biol. Evol., 4: 406-425; Sneath, P.H.A. and Sokal, R.R. (1973) Numerical Taxonomy the Principles and Practice of Numerical Taxonomy, Freeman Press, San Francisco, CA; Wilbur, W.J. and Lipman, D.J. (1983) Proc. Natl. Acad. Sci. USA 80: 726-730.
Предпочтительно, "процент идентичности последовательностей" определяют путем сравнения двух оптимально выровненных последовательностей в пределах окна сравнения, состоящего по меньшей мере из 20 положений, причем часть полинуклеотидной или полипептидной последовательности в данном окне сравнения может содержать вставки или делеции (то есть разрывы) в количестве 20 процентов или меньше, обычно от 5 до 15 процентов или от 10 до 12 процентов, по сравнению с последовательностями сравнения (которые не содержат вставок или делеции) для оптимального выравнивания этих двух последовательностей. Процент рассчитывают, определяя количество положений в обеих последовательностях, в которых присутствуют идентичные нуклеиновокислотные основания или идентичный аминокислотный остаток, и получая количество совпадающих положений, деля это количество совпадающих положений на общее количество положений в последовательности сравнения (то есть, размер окна) и умножая результат на 100 с получением процента идентичности последовательностей.
Варианты, также или альтернативно, могут быть по существу гомологичными нативному гену или его части или комплементарной последовательности. Такие полинуклеотидные варианты способны гибридизоваться в условиях умеренной жесткости с природной последовательностью ДНК, кодирующей нативное антитело (или комплементарную последовательность).
Подходящие "условия умеренной жесткости" включают предварительную промывку в растворе 5х SSC, 0,5% SDS, 1,0 мМ EDTA (рН 8,0); гибридизацию при 50°С-65°С, 5х SSC, в течение ночи; затем промывку при 65°С в течение 20 минут каждым из 2х, 0,5х и 0,2х SSC, содержащих 0,1% SDS.
При использовании в данном описании, термины "в очень жестких условиях" или "в условиях высокой жесткости" означают, что: (1) используют низкую ионную силу и высокую температуру для промывки, например 0,015 М хлорид натрия/0,0015 М цитрат натрия/0,1% додецилсульфат натрия при 50°С; (2) в процессе гибридизации используют денатурирующий агент, такой как формамид, например, 50%-ый (об./об.) формамид с 0,1% бычьего сывороточного альбумина/0,1% фикола/0,1% поливинилпирролидона/50 мМ натрий-фосфатным буфером при рН 6,5 с 750 мМ хлоридом натрия, 75 мМ цитратом натрия при 42°С; или (3) используют 50%-ный формамид, 5× SSC (0,75 М NaCl, 0,075 М цитрат натрия), 50 мМ фосфат натрия (рН 6,8), 0,1% пирофосфата натрия, 5× раствор Денхардта, обработанную ультразвуком ДНК из молоки лососевых (50 мкг/мл), 0,1% SDS и 10% декстран-сульфата при 42°C с промывками при 42°С в 0,2× SSC (хлорид натрия/цитрат натрия) и 50%-ном формамиде при 55°С, затем промывку высокой жесткости, состоящую из 0,1× SSC, содержащего EDTA, при 55°С. Специалист в данной области будет знать, как отрегулировать температуру, ионную силу и т.д., которые необходимы для согласования желаемых факторов, таких как длина зонда и тому подобное.
Специалистам обычной квалификации в данной области очевидно, что в результате вырожденности генетического кода, существует много нуклеотидных последовательностей, которые кодируют полипептид, изложенный в данном описании. Некоторые из этих полинуклеотидов несут минимальную гомологию с нуклеотидной последовательностью какого-либо нативного гена. Тем не менее, полинуклеотиды, которые варьируют вследствие различий в частоте использования кодонов, специально охватываются настоящим изобретением. Кроме того, в пределах объема настоящего изобретения находятся аллели генов содержащих полинуклеотидные последовательности, приведенные в данном описании. Аллели представляют собой эндогенные гены, которые изменяются в результате одной или более мутаций, таких как делеции, вставки и/или замены нуклеотидов. Полученные мРНК и белок могут, но не обязательно, иметь измененную структуру или функцию. Аллели могут быть идентифицированы с использованием стандартных методов (таких как, например, гибридизация, амплификация и/или сравнение с последовательностями из баз данных).
Полинуклеотиды по изобретению могут быть получены с использованием химического синтеза, рекомбинантных способов или ПЦР. Способы химического полинуклеотидного синтеза хорошо известны в данной области и не нуждаются в подробном изложении в данном описании. Специалист в данной области может использовать последовательности, предложенные в данном описании, и коммерческий синтезатор ДНК для получения желаемой последовательности ДНК.
Для получения полинуклеотидов с использованием рекомбинантных способов можно встроить в подходящий вектор полинуклеотид, содержащий желаемую последовательность, а вектор в свою очередь можно ввести в подходящую клетку-хозяин для репликации и амплификаци, как далее рассмотрено в данном описании. Полинуклеотиды можно вводить в клетки-хозяева, используя любые средства, известные в данной области. Клетки подвергают трансформации, вводя экзогенный полинуклеотид посредством прямого поглощения, эндоцитоза, трансфекции, F-скрещивания или электропорации. Будучи введенным, экзогенный полинуклеотид может поддерживаться внутри клетки в виде неинтегрирующегося вектора (такого как плазмида) или интегрироваться в геном клетки-хозяина. Амплифицированный таким образом полинуклеотид может быть выделен из клетки-хозяина методами, хорошо известными в данной области. См., например, Sambrook et al.(1989).
Альтернативно, ПЦР позволяет осуществлять воспроизводство последовательностей ДНК. Технология ПЦР хорошо известна в данной области и описана в патентах США №№4683195, 4800159, 4754065 и 4683202, а также в работе: PCR: The Polymerase Chain Reaction, Mullis et al. eds., Birkauswer Press, Boston (1994).
РНК может быть получена в результате использования выделенной ДНК в составе соответствующего вектора, введенной в подходящую клетку-хозяина. Когда клетка реплицируется и ДНК транскрибируется в РНК, РНК затем может быть выделена с использованием методов, хорошо известных специалистам в данной области, как это изложено, например, Sambrook et al. (1989).
Подходящие клонирующие векторы могут быть сконструированы в соответствии со стандартными методами или могут быть выбраны из большого числа клонирующих векторов, доступных в данной области техники. Несмотря на то, что выбор клонирующего вектора может варьировать в зависимости от намеченной для использования клетки-хозяина, полезные клонирующие векторы в большинстве случаев будут обладать способностью к саморепликации, могут содержать единственный сайт для конкретной эндонуклеазы рестрикции и/или могут нести гены маркера, который может быть использован для селекции клонов, содержащих данный вектор. Подходящие примеры включают плазмиды и бактериальные вирусы, например, pUC18, pUC19, Bluescript (например, pBS SK+) и его производные, mp18, mp19, pBR322, pMB9, ColE1, pCR1, RP4, фаговые ДНК и челночные векторы, такие как pSA3 и рАТ28. Эти и многие другие клонирующие векторы поставляются фирмами-поставщиками, такими как BioRad, Stratagene и Invitrogen.
Экспрессирующие векторы в большинстве случаев представляют собой способные к репликации полинуклеотидные конструкции, которые содержат полинуклеотид по изобретению. Подразумевается, что экспрессирующий вектор должен быть способен к репликации в клетках-хозяевах либо в виде эписом, либо как составная часть хромосомной ДНК. Подходящие экспрессирующие векторы включают, например плазмиды, вирусные векторы, включая аденовирусы, аденоассоциированные вирусы, ретровирусы, космиды и экспрессирующие векторы, изложенные в WO 87/04462, но этим не ограничиваются. Компоненты вектора в большинстве случаев могут включать одно или более чем одно из следующего: сигнальную последовательность; ориджин репликации; один или более маркерных генов; подходящие элементы транскрипционного контроля (такие как промоторы, энхансеры и терминатор), но этим не ограничиваются. Для экспрессии (то есть, трансляции) также обычно необходимы один или более контролирующих трансляцию элементов, таких как сайты связывания с рибосомой, сайты инициации трансляции и стоп-кодоны.
Векторы, содержащие представляющие интерес полинуклеотиды, могут быть введены в клетку-хозяина с применением любого из ряда соответствующих средств, включая электропорацию, трансфекцию с применением хлорида кальция, хлорида рубидия, фосфата кальция, DEAE-декстрана или других веществ; бомбардировку микрочастицами; липофекцию и инфекцию (например, когда вектор представляет собой инфекционный агент, такой как вирус коровьей оспы). Выбор вводимых векторов или полинуклеотидов часто будет зависеть от особенностей клетки-хозяина.
Согласно изобретению также предложены клетки-хозяева, содержащие любой из полинуклеотидов, изложенных в данном описании. Любые клетки-хозяева, способные к сверхэкспрессии гетерологичных ДНК, могут быть использованы с целью выделения генов, кодирующих представляющие интерес антитело, полипептид или белок. Неограничивающие примеры клеток-хозяев млекопитающих включают клетки COS, HeLa и СНО, но этим не ограничиваются. См. также, например, WO 87/04462. Подходящие клетки-хозяева, отличные от клеток млекопитающих, включают клетки прокариот (таких как E.coli или В.subtillis) и дрожжей (таких как S.cerevisae, S.pombe или K.lactis). Предпочтительно, клетки-хозяева экспрессируют кДНК с уровнем примерно в 5 раз превышающем, более предпочтительно в 10 раз превышающем, еще более предпочтительно в 20 раз превышающем уровень представляющего интерес соответствующего эндогенного антитела или белка, если он присутствует в клетках-хозяевах. Скрининг клеток-хозяев на предмет специфического связывания с Ab1-40 осуществляется, например, с использованием иммуноанализа или FACS. Клетка, сверхэкспрессирующая представляющие интерес антитело или белок, может быть идентифицирована.
Настоящее изобретение далее проиллюстрировано следующими ниже примерами, которые не следует истолковывать как вносящие дополнительные ограничения. Содержания всех Фигур, таблиц и всех ссылок, патентов и опубликованных патентных заявок, приведенных по всему этому описанию, включены прямо в данное описание посредством ссылки во всей своей полноте.
ПРИМЕРЫ
Пример 1. Создание muMabJC18
Получение
А. Иммуноген
Рекомбинантный Dkk-1 человека (rhuDkk-1), полученный от R&D Systems (№ по каталогу 1096-DK/CF, соответствующий аминокислотам (АА) 2-266 Dkk-1 человека, слитого с меткой 10X His на С-конце), конъюгировали с KLH (гемоцианином лимфы улитки), используя набор Imject Immunogen EDC (Pierce) и mcKLH (KLH из марикультуры) (№ по каталогу 77622). Конъюгированный с KLH белок был получен в Genovac GmbH и использован в качестве иммуногена для слитой конструкции, дав начало субклону JC9H3 (JC18).
SEQ ID NO:1. АА последовательность Dkk1-(10His) человека (аминокислоты, выделенные жирным курсивом, соответствуют подлинному N-концу очищенного белка):

SEQ ID NO:2. Nt (нуклеотидная) последовательность Dkk1-(10His) человека (нуклеотидная последовательность, где нуклеотиды, выделенные жирным курсивом, указываются лишь предположительно для данной последовательности, поскольку R&D не дает информации о последовательности):
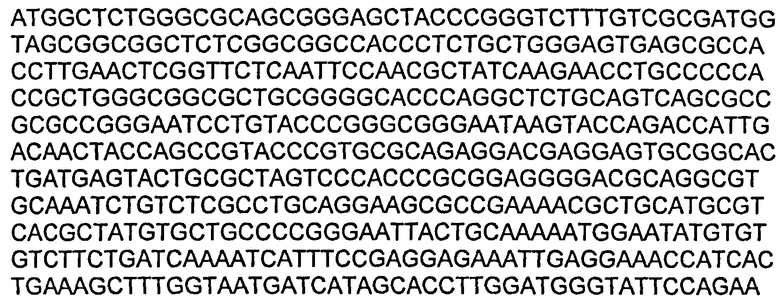

Б. Иммунизация и образование гибридомы
Мышей линии Balb/c иммунизировали внутрибрюшинно (50 мкг/доза/мышь) белком rhuDkk-1, конъюгированным с KLH. Дозу вводили повторно по 3 раза в течение 4 недельного периода. За 2 суток и 1 сутки до проведения слияния мыши получали конечную иммунизирующую дозу 50 мкг белка. Проводили слияние лимфоцитов селезенки с линией несекреторных миеломных клеток sp2/0 и подвергали НАТ-селекции (селекции на среде с гипоксантин-аминоптерин-тимидином), как описано ранее (Galfre and Milstein, Methods Enzymol., 73, 3-46 (1981)). Гибридому, секретирующую Dkk1-специфичные IgG, извлекали, субклонировали и детектировали, используя описанные выше анализы.
Специфичность к Dkk-1 (ELISA-анализы)
А. Детекция
Детекцию Dkk1-специфичных антител осуществляли, используя приведенный далее метод, основанный на ELISA. Белок rhuDkk-1 (R&D Systems, № по каталогу 1096 DK/CF) разбавляли до концентрации 1 мкг/мл в буфере для сенсибилизации поверхностей (Sigma, карбонатный/бикарбонатный буфер для сенсибилизации поверхностей; рН 9,6; № по каталогу С-3041; приготовленный в соответствии с инструкциями производителей). По 100 мкл раствора Dkk-1 добавляли в каждую лунку 96-луночных планшетов Nunc Maxisorp и инкубировали в течение ночи при 4°С. Лунки промывали 4× (250 мкл/лунка) буфером для промывки (0,05% (об./об.)) Твин-20 (Sigma, № по каталогу Р2287) в dPBS (фосфатно-солевой буфер Дульбекко), не содержащем Са/Mg++ (Sigma, № по каталогу D8537)), затем блокировали (200 мкл/лунка) блокирующим буфером (1% PVA (поливиниловый спирт) (Sigma, № по каталогу 363170) (масса/объем) в dPBS (Sigma, № по каталогу D8537)) в течение 2 ч при комнатной температуре (КТ). Блокирующий буфер затем удаляли быстрым декантированием. Антитела разбавляли последовательными 2-кратными серийными разведениями в блокирующем буфере и вносили в каждый планшет в количестве 100 мкл на одну лунку. Планшеты инкубировали в течение 1 часа при КТ и затем промывали, как описано выше.
Детекцию связывания тестируемого антитела с Dkk выполняли, добавляя вторичное антитело, специфичное к видовому антителу. Когда тестировали мышиные антитела, использовали конъюгированные с HRP (пероксидазой хрена) IgG козы против антител мыши (H&L) в количестве 100 мкл/лунка в разведении 1:4000 в блокирующем буфере (Jackson ImmunoResearch, № по каталогу 115-035-146). Когда тестировали гуманизированные антитела, использовали HRP-конъюгированные F(ab')2 IgG козы против человеческих антител в количестве 100 мкл/лунка в разведении 1:4000 в блокирующем буфере (Jackson ImmunoResearch, № по каталогу 109-035-097). Планшеты инкубировали в течение 30 мин при комнатной температуре, затем промывали 2×, как описано выше. В каждую лунку добавляли по 100 мкл свежеприготовленного раствора субстрата (2-компонентный пероксидазный субстрат KPL ABTS, № по каталогу 50-62-00, приготовленный в соответствии с инструкциями производителей) и давали возможность развиваться окраске. После развития окраски измеряли оптическую плотность (OD) в планшетах при 405 и 490 нМ. Величины оптической плотности представляли в виде разности между оптическими плотностями на обеих длинах волн. Для JC18 подемонстрировано связывание с rhuDKK-1 в концентрации до 5 мкг/мл включительно.
Б. Специфичность связывания с другими гомологами Dkk человека
Способность JC18 связываться с другими белками Dkk человека оценивали, используя описанный выше протокол для ELISA за исключением того, что планшеты также сенсибилизировали рекомбинантными Dkk-3 (R&D Systems, № по каталогу 1118-DK/CF) и Dkk-4 человека (R&D Systems, № по каталогу 1269-DK/CF). Для JC18 продемонстрировано слабое связывание с Dkk-4 человека и не продемонстрировано никакого связывания с Dkk-3 человека в концентрации до 5 мкг/мл включительно.
В. Межвидовая перекрестная реактивность
Способность JC18 связываться с белками Dkk других видов оценивали, используя описанный выше протокол для ELJSA за исключением того, что планшеты также сенсибилизировали рекомбинантными Dkk-1 крысы (R&D Systems, № по каталогу 4010-DK/CF), Dkk-1 мыши (R&D Systems, № по каталогу 1765-DK/CF), Dkk-2 мыши (R&D Systems, № по каталогу 2435-DK/CF) и Dkk-4 мыши (R&D Systems, № по каталогу 3105-DK/CF). Для JC18: (1) продемонстрировано связывание с Dkk-1 мыши и крысы, сходное со связыванием с Dkk-1 человека в концентрациях до 5 мкг/мл включительно; (2) не продемонстрировано никакого связывания с Dkk-2 мыши в концентрациях до 5 мкг/мл включительно; и (3) продемонстрировано слабое связывание с Dkk-4 мыши в концентрациях до 5 мкг/мл включительно.
Клонирование и секвенирование
Гомогенизировали один миллион гибридомных клеток, используя спин-колонки QIAshredder, и общую РНК экстрагировали, используя мининабор RNAeasy от QIAGEN. кДНК синтезировали, используя набор Superscript III RT от Invitrogen. Вариабельные области антител против Dkk-1 клонировали, используя наборы праймеров для IgG мыши от Novagen, состоящие из вырожденных праймеров для клонирования генов тяжелой цепи IgG мыши и легких цепей каппа или лямбда мыши. Использовали следующие условия циклов ПЦР: 1 цикл при 94°С в течение 2 мин; пять циклов при 94°С в течение 30 с, 44°С в течение 30 с и 68°С в течение 60 с; затем 25 циклов при 94°С в течение 30 с, 54°С в течение 30 с и 68°С в течение 60 с.Полученные продукты ПЦР клонировали в клонирующий вектор Торо-ТА от Invitrogen и секвенировали. Клонированные последовательности антител подтверждали прямым сравнением с использованием масс-спектрометрического (MS) анализа клонированных последовательностей антител и исходных антител, полученных из асцитов.
Нуклеотидная последовательность и предсказанная аминокислотная последовательность вариабельных областей легкой и тяжелой цепей моноклонального антитела JC18 дикого типа (мышь) приведены ниже.
SEQ ID NO:3. Nt последовательность вариабельной области легкой цепи JC18:

SEQ ID NO:4. АА последовательность вариабельной области легкой цепи JC18:
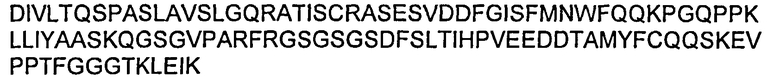
SEQ ID NO:5. Nt последовательность CDR1 вариабельной области легкой цепи JC18:
SEQ ID NO:6. АА последовательность CDR1 вариабельной области легкой цепи JC18:

SEQ ID NO:7. Nt последовательность CDR2 вариабельной области легкой цепи JC18:
SEQ ID NO:8. AA последовательность CDR2 вариабельной области легкой цепи JC18:
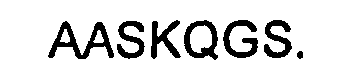
SEQ ID NO:9. Nt последовательность CDR3 вариабельной области легкой цепи JC18:

SEQ ID NO:10. AA последовательность CDR3 вариабельной области легкой цепи JC18:

SEQ ID NO:11. Nt последовательность вариабельной области тяжелой цепи JC18:

SEQ ID NO:12. AA последовательность вариабельной области тяжелой цепи JC18:

SEQ ID NO:13. Nt последовательность CDR1 вариабельной области тяжелой цепи JC18:
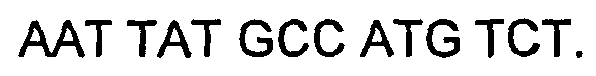
SEQ ID NO:14. AA последовательность CDR1 вариабельной области тяжелой цепи JC18:

SEQ ID NO:15. Nt последовательность CDR2 вариабельной области тяжелой цепи JC18:

SEQ ID NO:16. АА последовательность CDR2 вариабельной области тяжелой цепи JC18:

SEQ ID NO:17. Nt последовательность CDR3 вариабельной области тяжелой цепи JC18:

SEQ ID NO:18. АА последовательность CDR3 вариабельной области тяжелой цепи JC18:

Пример 2. Создание и тестирование гуманизированного моноклонального антитела против Dkk-1 (huMabJC18), полученного в результате мутации muMabJC18
Определение аффинности связывания гуманизированного антитела и его вариантов
В этом примере использованы следующие далее общие способы.
А. Экспрессирующий вектор, использованный для определения характеристик клона
Экспрессия Fab-фрагмента антител протекала под контролем IPTG(изопропилтиогалактозид)-зависимого индуцибельного промотора lacZ, аналогичного описаному в Barbas (2001) Phage display: a laboratory manual, Cold Spring Harbor, NY, Cold Spring Harbor Laboratory Press, pg 2.10. Vector pComb3X, однако, модификации включали добавление и экспрессию следующих дополнительных доменов: константного домена человеческой легкой цепи каппа и константного домена СН1 человеческого иммуноглобулина IgG2a. (С-область цепи Ig гамма-2, номер доступа для белка Р01859; иммуноглобулиновая легкая каппа-цепь (Homo sapiens), номер доступа для белка CAA09181).
Б. Получение Fab в небольшом масштабе
Экспрессию Fab в небольшом масштабе в 96-луночных планшетах осуществляли следующим образом. Начинали с E.coli, трансформированных библиотекой Fab, колонии перекалывали с целью инокуляции как на эталонный планшет (агар LB (бульон Луриа) + ампициллин (50 мкг/мл) + 2% глюкозы), так и рабочий планшет (2 мл/лунка, 96 лунок/планшет, содержащий 1,5 мл LB + ампициллин (50 мкг/мл) + 2% глюкозы). Бактерии в обоих планшетах выращивали при 30°С в течение 8-12 часов. Эталонный планшет хранили при 4°С, а клетки из рабочего планшета собирали в виде осадка после центрифугирования при 5000 об/мин и ресуспендировали в 1 мл LB + ампициллин (50 мкг/мл) + 1 мМ IPTG для индукции экспрессии Fab.
Клетки собирали центрифугированием через 5 ч экспрессии при 30°С, затем ресуспендировали в 500 мкл буфера HBS-EP (100 мМ буфер на основе HEPES (N-2-гидроксиэтил-пиперазин-N-2-этансульфоновая кислота), рН 7,4, 150 мМ NaCl, 0,005% Р20). Лизиса ресуспендированых в HBS-EP клеток достигали в результате одного цикла замораживания (-80°С), затем оттаивания при 37°С. Клеточные лизаты центрифугировали при 5000 об/мин в течение 30 мин для отделения клеточного дебриса от супернатантов, содержащих Fab. Супернатанты фильтровали, используя 96-луночные фильтровальные планшеты Multiscreen HTS (Millipore, № по каталогу MSFBN6B50). Затем супернатанты вводили в устройство BIAcore для плазменного резонанса с целью получения информации об аффинности для каждого Fab. Клоны, экспрессирующие Fab, извлекали из эталонного планшета для секвенирования ДНК и для крупномасштабного получения Fab и подробно характеризовали, как описано ниже.
В. Крупномасштабное получение Fab
Для получения точных кинетических параметров Fab экспрессировали и очищали из культур, полученных в большом количестве. Колбы Эрленмейера, содержащие по 200 мл LB + ампициллин (50 мкг/мл) + 2% глюкозы, инокулировали 5 мл ночной культуры из отобранных Fab-экспрессирующих клонов E.coli. Клоны инкубировали при 30°С до достижения OD550нм 1,0 и затем проводили индукцию, заменяя среду на 200 мл LB + ампициллин (50 мкг/мл) + 1 мМ IPTG. Через 5 ч экспрессии при 30°С клетки собирали в виде осадка после центрифугирования, затем ресуспендировали в 10 мл PBS (забуференного фосфатом физиологического раствора) (рН 8). Лизиса клеток достигали, используя два цикла замораживания/оттаивания (при -80°С и 37°С, соответственно).
Супернатант клеточных лизатов наносили на колонки superflow с Ni-NTA(нитрилацетат)-сефарозой (Qiagen, Valencia. CA), уравновешенные PBS, рН 8, затем промывали PBS, pH 8, в количестве 5 объемов колонки. Индивидуальные Fab элюировали в различных фракциях, используя PBS (pH 8) + 300 мМ имидазол. Фракции, содержащие Fab, объединяли и диализовали против PBS, затем, перед измерением характеристик аффинности, проводили количественное определение, используя ELISA.
Г. Получение целых антител
Для экспрессии целых антител вариабельные области тяжелой и легкой цепи клонировали в экспрессирующие векторы для млекопитающих и, используя липофектамин, проводили трансфекцию клеток HEK (почки эмбриона человека) 293 для осуществления временной экспрессии.
Антитела очищали с использованием белка А, применяя стандартные методы.
BIAcore-анализы
Аффинность huMabJC18 в отношении Dkk1 человека, мыши или крысы определяли, используя систему BIAcore3000™ для поверхностного плазменного резонанса (SPR) (BIAcore Inc., Piscataway, NJ), оборудованную сенсорным чипом СМ5 (BIAcore AB, Uppsala, Sweden). Для анализа взаимодействия huMabJC18 с Dkk1 человека, создавали реакционно-способные поверхности посредством аминного сочетания huMabJC18 стремя проточными ячейками (Fc2, Fc3 и Fc4). СМ5-чипы активировали, используя гидрохлорид N-этил-N'-(3-диметиламинопропил)-карбодиимида (EDC) и N-гидроксисукцинимид (NHS) в соответствии с инструкциями поставщика. IgG huMabJC18 разбавляли в 10 мМ ацетате натрия, pH 4,0, и наносили на активированный чип в концентрации 30 мкг/мл.
Используя различные значения времени протекания сквозь индивидуальные каналы чипа, получали диапазон плотностей для антитела: 200-800 единиц ответного сигнала (RU). Далее поверхности с IgG и веществом сравнения насыщали Fab2 козы против Fc человека (Cappel 55053) для предупреждения неспецифического связывания Dkk-1 с поверхностью чипа. Затем чип блокировали этаноламином. Анализы проводили при 25°С, используя рабочий буфер для BIAcore, состоящий из HBS-EP (0,01 М HEPES, pH 7,4; 0,15 М NaCl; 3 мМ EDTA; 0,005% поверхностно-активного вещества Р20) + BSA (бычий сывороточный альбумин; 2 мг/мл) + СМ(карбоксиметил)- декстрана (2 мг/мл). Серии из пяти трехкратных разведении, начиная с 30,9 нМ концентрации очищенного белка Dkk-1 человека (R&D Systems), вводили в течение 1,5 мин при скорости 100 мкл/мин и диссоциацию регистрировали в течение 8 часов. Кинетические константы скоростей ассоциации (kon) и константы скоростей диссоциации (koff) получали одновременно, аппроксимируя данные к модели связывания Ленгмюра в отношении 1:1 (Karlsson, R. Roos, H. Fagerstam, L. Petersson, B. (1994). Methods Enzymology, 6. 99-110) с использованием программы BIAevaluation. Величины равновесных констант диссоциации (KD) рассчитывали как koff/kon.
Для анализа взаимодействия huMabJC18 с Dkk-1 мыши или крысы, небольшие количества huMabJC18 (обычно 100-400 единиц ответного сигнала) захватывали в индивидуальных проточных ячейках, используя Fab2 козы против Fc человека, которые предварительно иммобилизовали на поверхности СМ5-чипа. Проточная ячейка, насыщенная Fab2 козы против Fc человека, не содержащая гуманизированного антитела, служила в качестве канала сравнения. Dkk-1 (R&D Systems) мыши или крысы в различных концентрациях титровали над поверхностью чипа, используя 70,1 нМ или 30,2 нМ в качестве самой высокой концентрации, соответственно, в сериях с трехкратным разведением. Фазы ассоциации и диссоциации регистрировали при скорости 100 мкл/мин в течение 90 секунд и 30 минут, соответственно. Регенерацию захватывающих поверхностей для всех экспериментов осуществляли двумя 30-секундными импульсными подачами 0,75 мМ H3PO4, за исключением поверхности с huMabJC18, полученной с использованием аминного сочетания, использованной в эксперименте с Dkk-1 человека, которую не регенерировали.
При получении сигналов связывания использовали двойное сравнение и глобальную аппроксимацию к простой модели, применяя программное обеспечение BiaEvaluation v.4.0. Аффинности выводили из соотношения кинетических констант скоростей (KD=koff/kon).
Для скрининг-анализов с целью определения скоростей связывания и диссоциации вариантов huMabJC18 с Dkk-1 человека использовали либо систему BIAcore, либо систему ProteOn XPR36 (BioRad, Inc.). Для BIAcore-анализа биотинилированный Dkk-1 человека захватывали на BIAcore-чипах с SA (стрептавидином) в соответствии с инструкциями поставщика. Биотинилированный белок Dkk-1 человека разбавляли в HBS-EP и впрыскивали над поверхностью чипа в концентрации 5 мкг/мл. Времена протекания сквозь индивидуальные каналы чипа выбирали так, чтобы достичь плотности антигена приблизительно 500-600 единиц ответа (RU). В качестве рабочего буфера использовали буфер HBS-EP. Отфильтрованные клеточные лизаты рекомбинантной Е.coli вводили при высокой температуре (37°С) в течение 1 мин со скоростью 30 мкл/мин, допуская протекание фазы диссоциации продолжительностью 1-5 мин. Поверхности регенерировали, используя 8 мМ NaOH + 8% этанола. В качестве положительного контроля дважды вводили Fab (клон 24) в начале эксперимента и еще раз в конце анализа для оценки воспроизводимости.
В ProteOn-анализе биотинилированный Dkk-1 человека вводили в течение 1 мин в серийных разведениях, подобранных эмпирически для получения различных уровней иммобилизации (150-3000 RU) в пяти каналах сенсорного чипа ProteOn NLC Neutravidin. Шестой канал, остававшийся немодифицированным, служил в качестве поверхности сравнения. Рекомбинантные Fab в отфильтрованных лизатах Е.coli вводили при высокой температуре (37°С) в течение 50 с со скоростью 20 мкл/мин. Рабочий буфер состоял из PBS + BSA (1 мг/мл) + 0,005% Твина-20. Конфигурация системы ProteOn такова, что проба после каждой инъекции протекает через каждый из пяти каналов с Dkk-1, а также через канал сравнения, и можно одновременно вводить шесть образцов Fab. Диссоциацию Fab с Dkk-1-поверхности регистрировали в течение 20 мин. Поверхность чипа регенерировали введением в течение 30 с 8%-го EtOH в 8 мМ NaOH. Клон 24 Fab использовали в качестве контроля. Уровень связывания во время стадии ассоциации и скорость диссоциации для каждого клона находились в хорошем соответствии между различными каналами с лигандом.
Аффинность связывания с Dkk-1 для huMabJC18 и его вариантов
Аминокислотные последовательности huMabJC18 приведены в SEQ ID NO:40 (вариабельная область НС) и SEQ ID NO:42 (вариабельная область LC). Аффинность связывания с Dkk-1 для IgG huMabJC18, определенная с использованием BIAcore, как описано выше, показана в приведенной ниже Таблице 3.
Аминокислотные последовательности CDR вариантов huMabJC18 показаны в приведенной ниже Таблице 4. Все аминокислотные замены, показанные в Таблице 4, описаны относительно последовательности huMabJC18, Данные по аффинности связывания huMabJC18 и его вариантов с h Dkk-1 также представлены в Таблице 4.
CDR представляют собой расширенные CDR, которые включают CDR как в определении по Kabat, так и по Chothia, за исключением CDR H1, который указан в соответствии с определением по Kabat. Аминокислотные остатки нумеруются последовательно (см. SEQ ID NO:28 для нумерации остатков в колонках Н2 и H3 и SEQ ID NO:20 для нумерации остатков в колонках L1, L2 и L3). Все клоны имеют рамку считывания и последовательности CDR H1, идентичные huMabJC18. KD=koff/kon. Все величины koff определяли в режиме скрининга за исключением тех, которые подчеркнуты; их получали в результате глобального анализа серий концентраций hDkk-1, пропускаемых над IgG, захваченными на поверхности BIAcore-чипа или поверхности ProteOn. Ввиду этого подчеркнутые величины KD определяли экспериментально, измеряя kon. Остальные величины KD рассчитаны теоретически на основании оценки для величины kon, равной 1×106 (1/М·с). Поскольку значения констант скоростей ассоциации для всех проанализированных клонов были очень высокими и, следовательно, определялись диффузионными ограничениями (>1×106 и это слишком высокое значение для точного измерения), аффинности вероятнее всего еще выше, чем указано.
Взаимодействие huMabJC18 с гомологами Dkk
HuMabJC18 анализировали на предмет связывания с другими доступными гомологами Dkk, включая Dkk-3 человека, Dkk-4 человека, Dkk-2 мыши и Dkk-4 мыши (R&D Systems). Результаты суммированы в Таблице 5.
Анализы взаимодействия huMabJC18 с гомологами Dkk-1 проводили, как описано выше, используя Fab2 козы против Fc человека в качестве предварительно иммобилизованного захватывающего реагента. Dkk-4 человека, Dkk-2 мыши или Dkk-4 мыши в различных концентрациях титровали над поверхностью чипа, используя 76,3 нМ, 88,8 нМ или 32,3 нМ в качестве самой высокой концентрации, соответственно, в серии с пятикратным разведением. Для Dkk-3 человека не продемонстрировано никакого сигнала специфического связывания.
Dkk1-связывающее антитело JC18
Аффинности связывания мышиного антитела JC18 с Dkk-1 человека, мыши или крысы (R&D Systems), определенные при 25°С в BIAcore-анализе, показаны в приведенной ниже Таблице 6. Для этого анализа проводили аминное сочетание поликлональных анти-мышиных IgG с поверхностью BIAcore-чипа. Захват антитела JC18 против Dkk-1 осуществляли в концентрации 6,53 мкг/мл в течение 1 мин при скорости 5 мкл/мин. Пятикратное серийное разведение Dkk-1 человека, мыши или крысы (R&D Systems) (0,4-250 нМ) вводили в течение 1 мин при скорости 50-100 мкл/мин. Диссоциацию регистрировали в течение 3-5 мин. Регенерацию чипа осуществляли двумя 30-секундными импульсными подачами 100 мМ H3PO4.
Последовательности для huMabJC18
SEQ ID NO:19. Nt последовательность вариабельной области легкой цепи huMabJC18:
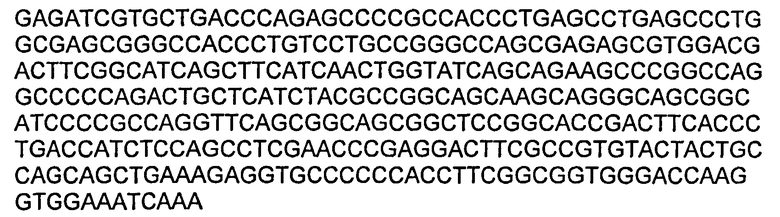
SEQ ID NO:20. АА последовательность вариабельной области легкой цепи huMabJC18:
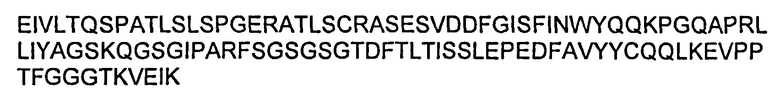
SEQ ID NO:21. Nt последовательность CDR1 вариабельной области легкой цепи huMabJC18:

SEQ ID NO:22. AA последовательность CDR1 вариабельной области легкой цепи huMabJC18:

SEQ ID NO:23. Nt последовательность CDR2 вариабельной области легкой цепи huMabJC18:
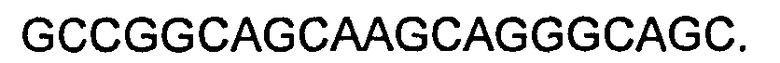
SEQ ID NO:24. AA последовательность CDR2 вариабельной области легкой цепи huMabJC18:
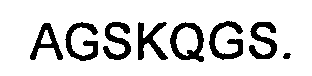
SEQ ID NO:25. Nt последовательность CDR3 вариабельной области легкой цепи huMabJC18:

SEQ ID NO:26. AA последовательность CDR3 вариабельной области легкой цепи huMabJC18:

SEQ ID NO:27. Nt последовательность вариабельной области тяжелой цепи huMabJC18:
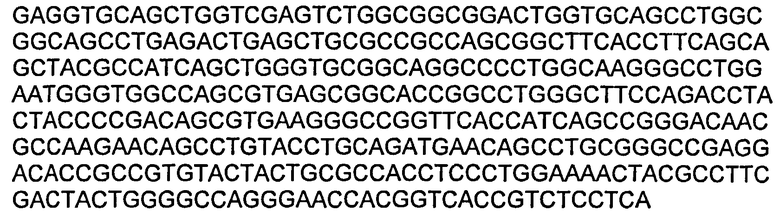
SEQ ID NO:28. AA последовательность вариабельной области тяжелой цепи huMabJC18:

SEQ ID NO:29. Nt последовательность CDR1 вариабельной области тяжелой цепи huMabJC18:
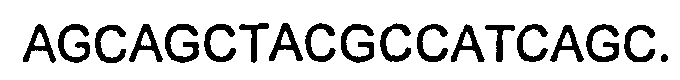
SEQ ID NO:30. AA последовательность CDR1 вариабельной области тяжелой цепи huMabJC18:

SEQ ID NO:49. AA последовательность CDR1 вариабельной области тяжелой цепи huMabJC18 (Kabat).

SEQ ID NO:50. AA последовательность CDR1 вариабельной области тяжелой цепи huMabJC18 (Chothia):

SEQ ID NO:31. Nt последовательность CDR2 вариабельной области тяжелой цепи huMabJC18:
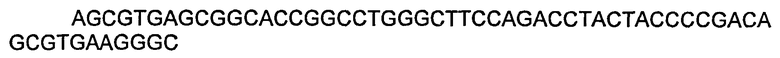
SEQ ID NO:32. AA последовательность CDR2 вариабельной области тяжелой цепи huMabJC18:

SEQ ID NO:51. AA последовательность CDR2 вариабельной области тяжелой цепи huMabJC18 (Chothia):

SEQ ID NO:33. Nt последовательность CDR3 вариабельной области тяжелой цепи huMabJC18:

SEQ ID NO:34. AA последовательность CDR3 вариабельной области тяжелой цепи huMabJC18:

SEQ ID NO:52. AA последовательность (короткая) CDR3 вариабельной области тяжелой цепи huMabJC18:

SEQ ID NO:35. Nt последовательность тяжелой цепи huMabJC18 с лидерной последовательностью:

SEQ ID NO:36. АА последовательность тяжелой цепи huMabJC18 с лидерной последовательностью (мутации *Δа (A329S, P330S) показаны жирным курсивом).

Сайт гликозилирования: НС, N296.
SEQ ID NO:37. Nt последовательность легкой цепи huMabJC18 с лидерной последовательностью:

SEQ ID NO:38. АА последовательность легкой цепи каппа huMabJC18 с лидерной последовательностью:

Изотипы huMabJC18: IgG2(Δа)*, каппа.
Аллотипы huMabJC18: G2(n-), Km3.
SEQ ID NO:39. Nt последовательность тяжелой цепи huMabJC18 без лидерной последовательности:
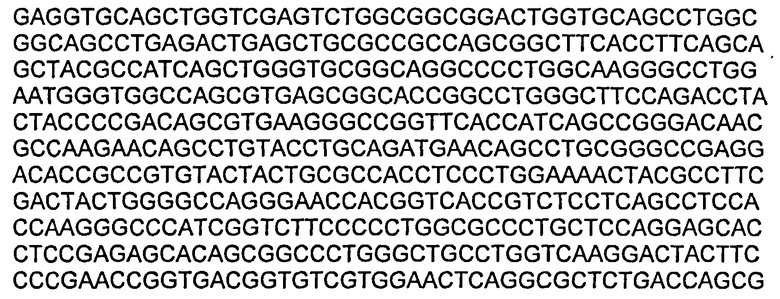

SEQ ID NO:40. АА последовательность тяжелой цепи huMabJC18 без лидерной последовательности (мутации *Δа (A329S, P330S) показаны жирным курсивом):

Сайт гликозилирования: НС, N296.
SEQ ID NO:41. Nt последовательность легкой цепи huMabJC18 без лидерной последовательности:
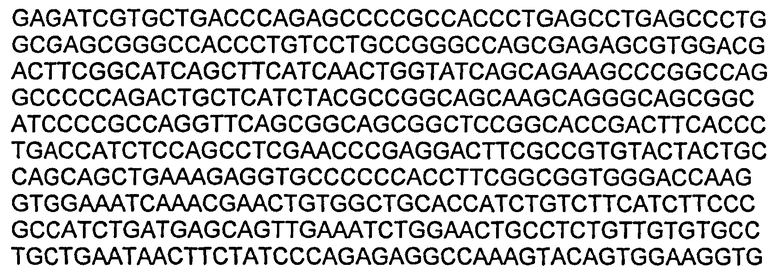

SEQ ID NO:42. АА последовательность легкой цепи каппа huMabJC18 без лидерной последовательности:

Пример 3. Функциональная активность huMabJC18 in vitro: определение влияния JC18 на активность Wnt в функциональном анализе, основанном на использовании клеток
Стабильная клеточная линия U20S (U20S TF), содержащая люциферазный репортерный ген под контролем TK-промотора (тимидинкиназного промотора) с сайтами связывания TCF (фактора Т-клеток), была выбрана для проведения функционального анализа, основанного на использовании клеток. Промотор активируется через Wnt-сигнальный путь, когда клетки обрабатывают Wnt3a-кондиционированными средами. Такая активация антагонизируется Dkk-1. Опосредованное Dkk-1 ингибирование передачи сигнала через Wnt, в свою очередь, может быть реверсировано посредством добавления Dkk1-нейтрализующего mAb.
В частности, JC18 представляет собой моноклональное антитело, которое связывается с Dkk-1 мыши и человека с высокой аффинностью. Его способность реверсировать ингибирование передачи сигнала через Wnt3a, вызываемое Dkk-1, изучали в клетках U2OS TOPFlash. Клетки U2OS получали из АТСС и стабильно трансфицировали плазмидой TOPFlash (Upstate), TCF-люциферазной репортерной конструкцией. Клетки поддерживали при 37°С, 5% CO2 в средах McCoys 5A, дополненных 10% фетальной телячьей сыворотки, 1% Pen-Strep (пенициллина-стрептомицина) и 2 мМ глутамином. Клетки U20S TOPFlash рассевали в обычной ростовой среде с плотностью 31250 клеток/см2 и инкубировали в течение ночи. Клетки обрабатывали белком rhDkk-1 (0,25 мкг/мл) или разбавителем (PBS, Gibco) и mAb против Dkk-1 в различных концентрациях в Optimem (Gibco). После инкубации в течение ночи проводили лизис клеток, используя лизирующий буфер Reporter Lysis Buffer (Promega), и экспрессию люциферазы количественно измеряли, используя люциферазный реагент (Promega).
HuMabJC18, полностью гуманизированные mAb против Dkk-1, демонстрировали удовлетворительную эффективность в функциональном анализе с IC50 1,3 нМ (Таблица 7).
Пример 4. Оценка in vivo эффективности действия на кости muMabJC18 и мышиной химерной конструкции
Самок взрослых мышей C57BL/6 используют для оценки влияния in vivo на скелетную структуру тестируемых антител против Dkk-1. Животных размещают при 24°С в условиях цикла 12 ч свет/12 ч темнота и со свободным доступом к воде и имеющемуся в продаже корму (корму для лабораторных животных (грызунов) Purina 5001, Purina-Mills, St. Louis, МО). Эксперименты проводят в соответствии с утвержденными Pfizer протоколами по уходу за животными, и животных содержат в соответствии с руководством по уходу и использованию лабораторных животных ILAR (Институт исследований на лабораторных животных). Мышей обрабатывают либо разбавителем, либо антителами посредством перорального введения через желудочный зонд один или два раза в неделю в течение нескольких недель. По окончании исследований мышей подвергают эвтаназии и собирают сыворотку крови для измерений уровней свободного антитела и свободного Dkk-1. Помимо этого получают образцы костей для оценки изменений в костной массе и остеогенезе с использованием периферической количественной компьютерной томографии (pQCT), микро-СТ (микрокомпьютерной томографии) и гистоморфометрии.
Правые бедренные кости сканируют с использованием pQCT (Stratec XCT Research M; Norland Medical Sistems, Fort Atkison, WI, USA) с программным обеспечением, версия 5.40. Получают изображения поперечных срезов толщиной 1 мм каждого дистального метафиза бедренной кости на расстоянии 2,5 мм проксимально по отношению к дистальному концу (~1,5 мм до зоны роста, то есть места, богатого губчатой костью) и изображения поперечных срезов толщиной 1 мм каждого диафиза бедренной кости на расстоянии 8 мм проксимально по отношению к дистальному концу (места, богатого кортикальной костью) с размером векселя 0,10 мм. Для кости в целом, трабекулярной и кортикальной кости определяют объемное содержание костной массы, плотность и площадь.
Правые бедренные кости сканируют с использованием установки для микро-СТ (Micro-CT40, Scanco Medical, Auenring 6-8, Bassersdorf, Switzerland) с программным обеспечением, версия 3.1. Для определения объема трабекулярной кости получают изображение поперечного среза дистального метафиза бедренной кости (в общей сложности 50 срезов толщиной по 16 мкм каждый, общая толщина = 0,8 мм) на расстоянии от 2,3 до 3,1 мм проксимально по отношению к дистальному концу (~1,3-2,1 мм от зоны роста).
Левые бедренные кости обрабатывают для гистоморфометрической оценки губчатой кости. Вкратце, левую бедренную кость обезвоживают в возрастающих концентрациях этанола и закрепляют в недекальцифицированном состоянии в метилметакрилате. Используя микротом Reichert-Jung Polycut S (Leica Corp., Heidelberg, Germany), готовят продольные фронтальные срезы толщиной 4 и 10 мкм дистальных отделов бедренной кости. Срезы толщиной 4 мкм окрашивают модифицированным красителем трихромом по Массону, а срезы толщиной 10 мкм оставляют неокрашенными. Все гистоморфометрические измерения проводят на губчатой костной ткани дистального метафиза бедренной кости в области между 0,375 и 0,875 мм проксимально по отношению к месту соединения зоны роста и эпифиза, используя систему анализа изображений (Osteomeasure, Inc., Altanta, GA). Объем губчатой кости в процентном отношении к площади костной ткани и занятую остеокластами поверхность в процентном отношении к общему периметру губчатой кости измеряют в окрашенных срезах толщиной 4 мкм. Рассчитывают число трабекул, плотность и расстояние между ними. Основанные на измерении содержания флуорохромов показатели костеобразования, включая выраженную в процентах площадь поверхности губчатой кости вместе с двойной флуорохромной меткой (минерализующуюся поверхность), уровень аппозиции минералов, скорость костеобразования (соотношение площади поверхности кости и объема костной ткани) получают на неокрашенных срезах толщиной 10 мкм.
Протокол и результаты исследования на модели интактных мышей
А. Влияние мышиного моноклонального антитела (JC18) против Dkk-1 в модели интактных мышей
Самкам взрослых мышей C57BL/6 перорально вводили разбавитель или мышиные IgG1 или JC18 в дозе 10, 30 и 60 мг/кг один раз в неделю в течение 6 недель. При вскрытии от каждой мыши получали как правую, так и левую бедренные кости. Правые бедренные кости анализировали, используя pQCT, a левые бедренные кости анализировали, используя гистоморфометрический метод. Общая BMD дистальных отделов бедренных костей значительно повышалась в результате обработки с использованием JC18 для всех уровней дозы (Фиг.1А). Применение JC18 также значительно повышало показатели костеобразования для уровня дозы 60 мг/кг у мышей (Фиг.1В).
Б. Эффект мышиной химерной конструкции моноклонального антитела против Dkk-1 (мышиная химерная конструкция представляет собой химерное антитело "человек-мышь") в модели интактных мышей.
Интактных самок мышей C57BL/6 в возрасте 4-х месяцев обрабатывали мышиной химерной конструкцией в дозе 0; 0,001; 0,01; 0,1; 1; 3; 10 и 30 мг/кг еженедельно в течение 6 недель. Одну группу мышей обрабатывали мышиной химерной конструкцией в дозе 5 мг/кг дважды в неделю в течение 6 недель. Дистальные отделы правой бедренной кости каждого животного анализировали, используя pQCT. Как показано на Фиг.2, мышиная химерная конструкция в дозах от 1 до 30 мг/кг повышала общую BMD по сравнению с разбавителем.
SEQ ID NO:43. Химерное антитело (hu-mu): AA последовательность химеры huMabJC18L-var (вариабельная область легкой цепи huMabJC18) и каппа-цепи мыши:

SEQ ID NO:44. Химерное антитело (hu-mu): AA последовательность химеры huMabJC18H-var (вариабельная область тяжелой цепи huMabJC18) и константной области IgG1 мыши:

SEQ ID NO:45. Химерное антитело (hu-mu): нуклеотидная последовательность huMabJC18L-var:
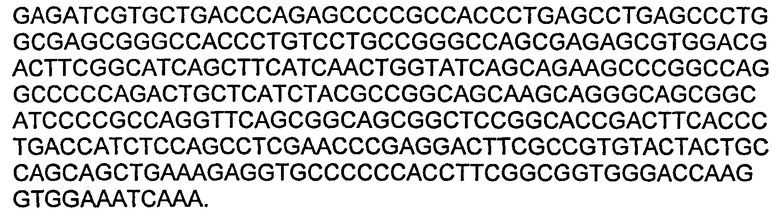
SEQ ID NO:46. Химерное антитело (hu-mu): нуклеотидная последовательность каппа-цепи мыши:

SEQ ID NO:47. Химерное антитело (hu-mu): нуклеотидная последовательность huMabJC18H-var:

SEQ ID NO:48. Химерное антитело (hu-mu): нуклеотидная последовательность СН1-СН2-СН3 мышиного IgG1:

В. Модель овариэктомизированных мышей
Самок взрослых мышей C57BL/6 подвергают ложной хирургической операции или овариэктомии (OVX) для оценки влияния на скелетную структуру тестируемых антител против Dkk-1 в условиях дефицита эстрогенов, что имитирует постменопаузальное уменьшение костной массы. Животных размещают при 24°С в условиях цикла 12 ч свет/12 ч темнота и со свободным доступом к воде и имеющемуся в продаже корму (корму для лабораторных животных (грызунов) Purina 5001, Purina-Mills, St. Louis, МО). Эксперименты проводят в соответствии с утвержденными Pfizer протоколами по уходу за животными, и животных содержат в соответствии с руководством по уходу и использованию лабораторных животных ILAR (Институт исследований на лабораторных животных). Мышей обрабатывают либо разбавителем, либо антителами посредством перорального введения через желудочный зонд один или два раза в неделю в течение нескольких недель. По окончании исследований мышей подвергают эвтаназии и получают образцы костей для оценки изменений в костной массе с использованием периферической количественной компьютерной томографии (pQCT), микро-СТ (микрокомпьютерной томографии) и гистоморфометрии. Методология этих измерений описана в предшествующем разделе.
Протокол и результаты исследования
Мышиное моноклональное антитело (JC18) против Dkk-1 предотвращает уменьшение костной массы, индуцированное дефицитом эстрогенов в модели подвергнутых OVX мышей.
Самок мышей C57BL/6 в возрасте 4-х месяцев подвергали либо ложной хирургической операции, либо овариэктомии (OVX) и обрабатывали либо разбавителем, либо JC18 в дозах 0,3; 1; 3; 10 и 30 мг/кг еженедельно, либо JC18 в дозе 15 мг/кг дважды в неделю в течение 8 недель, начиная на следующие сутки после операции. Как и ожидалось, у мышей, обработанных разбавителем, обнаруживали остеопению через 8 недель после OVX, что продемонстрировано значительным уменьшением общей BMD (Фиг.3). Применение JC18 дозозависимым образом повышало общую BMD, что измерено посредством pQCT для дистальных отделов бедренных костей, на 7-20% по сравнению с обработкой OVX-мышей разбавителем. У мышей, обработанных JC18 в дозе 15 мг/кг два раза в неделю, общая BMD была не только выше, чем у обработанных разбавителем OVX-мышей, но также сохранялась на уровне, соответствующем ложнооперированным контролям, указывая на то, что применение JC18 в этой дозе и этом режиме введения полностью предотвращало развитие остеопении у OVX-мышей (Фиг.3).
Пример 5. Антитело huMabJC18 увеличивало костную массу в модели интактных крыс
Взрослым самкам крыс один раз в неделю внутривенно вводили huMabJC18 в дозе 0; 0,1; 1; 10 или 100 мг/кг в течение 6 недель. Уровень остеокальцина в сыворотке крови, биомаркера костеобразования, возрастал на 30, 26 и 25% у крыс, обработанных huMabJC18 в дозе 1, 10 и 100 мг/кг, соответственно, через 2 недели после обработки и не отличался от контрольной величины через 4 недели после обработки. Уровень СТХ (продуктов деградации коллагена I типа - C-концевых телопептидов) в сыворотке крови, маркера резобции кости, не показал соответствующих изменений после обработки. Такие изменения биомаркеров сыворотки крови являются подтверждением анаболического действия huMabJC18 на кость. BMD дистальных отделов бедренной кости и ВМС (содержание минералов в кости) тела бедренной кости были значительно увеличены на 11 и 16% или 7 и 8%, соответственно, у крыс, обработанных huMabJC18 в дозах 10 и 100 мг/кг, что указывает на увеличение массы губчатой и кортикальной костей у этих животных.
Пример 6. Методы интерпретации системной динамики, касающейся терапии моноклональными антителами, направленными на растворимый антиген Dkk-1. антагонист пути передачи сигнала через Wnt: применение механистической доклинической фармакокинетической/фармакодинамической (PK/PD) модели для выбора начальных доз антитела IqG2. применяемого впервые на человеке (FIH)
Остеопороз представляет собой заболевание кости, характеризующееся низкой минеральной плотностью кости, которое приводит к хрупкости кости и последующим переломам кости. Большинство терапий остеопороза с применением фармакологических средств, включая бисфосфонаты, кальцитонин, HRT (гормонозаместительная терапия) и селективные модуляторы рецепторов эстрогенов (SERM), предупреждают уменьшение костной массы посредством снижения резорбции кости. Восстановление костной массы у пациентов, страдающих остеопорозом, является областью неудовлетворенных медицинских потребностей.
Связывание Dkk-1 с рецептором LRP5/6 и корецептором Kremen-1/2 содействует интернализации рецепторного комплекса, приводящей к демпфированию Wnt-сигнала (Diarra et al. (2007) Nat. Med. 13: 156-163).
Генетическое доказательство центральной роли Wnt-пути в поддержании костной массы проистекает из идентификации как активирующих, так и инактивирующих мутаций Wnt-рецептора LRP5. Инактивация LRP5 приводит к уменьшению костной массы и вызывает у людей аутосомно-рецессивное расстройство: синдром "остеопороз-псевдоглиома" (OPPG) (Gong et al. (2001) Cell, 107: 513-523) и аналогичное фенотипическое проявление у LRP5-нокаутированных мышей (Holmen et al. (2004) J. Bone Miner. Res. 19: 2033-2040).
Индивидуумы с НВМ имеют существенно сниженный риск перелома костей скелета
Ожидается, что Dkk1-нейтрализующее антитело будет увеличивать костную массу вследствие усиленного костеобразования остеобластами и, таким образом, предупреждать переломы при остеопорозе. HuMabJC18 представляет собой гуманизированный прототип моноклонального антитела против Dkk-1 для лечения остеопороза среди других расстройств. Оно связывается с Dkk-1 человека, мыши, крысы и яванского макака in vitro с высокой аффинностью (KD<100 пМ). Оно увеличивает костную массу у интактных мышей и восстанавливает костную массу скелета при остеопении у подвергнутых овариэктомии мышей в модели постменопаузального уменьшения костной массы (Li et al., 2009; рукопись в состоянии подготовки).
Несмотря на то, что в стадии разработки находится большое количество антител, опубликовано лишь небольшое число сообщений, в которых используются доклинические данные для прогнозирования клинической фармакокинетики или эффективной дозы антител (Lobo et al. (2004) J. Pharm. Sci., 93: 2645-2668; Agoram (2009) Br. J. Clin. Pharmacol., 67: 153-160). Аллометрические степенные модели обычно используются для межвидового масштабирования фармакокинетики (РК) антител, когда ожидается линейная PK (Wang et al. (2008) Clin. Pharmacol. Ther., 84: 548-558). Однако в отличие от небольших молекул, взаимодействие антитела с его мишенью часто влияет на РК антитела. Фармакокинетика (PK) и фармакодинамика (PD) тесно связаны, и понимание PK/PD требует знаний об антителе, мишени и взаимодействиях антитело-мишень. Максимальная величина для предсказания экспозиции лекарственного препарата и влияния, оказываемого заданной дозой, вытекает из связи PK с PD-ответом (Agoram et al. (2007) Drug Discov. Today, 12: 1018-1024). Таким образом, механистическое PK/PD моделирование представляет собой рациональное и эффективное средство прогнозирования как PK, так и клинически эффективной дозы антител для человека.
В настоящем исследовании одновременное получение характеристик мишени (Dkk-1) антитела huMabJC18 у крыс и макак сделало возможным глубокое понимание фармакокинетики и фармакодинамики ответа. Все это сочетали со знаниями об уровне мишени у здоровых субъектов по сравнению с больными, скоростях метаболизма мишени и скоростях ассоциации/диссоциации комплекса антитело-мишень для механистических прогнозов PK/PD в клинике.
Цель данного исследования заключалась в сравнении и сопоставлении предсказанных клинических начальных доз антитела IgG2 против Dkk-1 (huMabJC18), полученных в результате следующих вычислений: (1) уровня, не вызывающего видимых неблагоприятных эффектов (NOAEL) в токсикологическом исследовании, (2) минимального уровня ожидаемого биологического эффекта (MABEL) с использованием классического уравнения Даффа и (3) MABEL с использованием модели мишень-опосредованного распределения лекарственного средства (TMDD).
Для определения NOAEL использовали доклинические исследования безопасности на крысах и макаках. Для получения рекомендуемой начальной дозы вводили 100-кратный коэффициент запаса к NOAEL. Классическое уравнение Даффа использовали для расчета степени занятости рецепторов и MABEL определяли как дозу, которая приводила к 10%-ой занятости рецепторов. TMDD-модель использовали для аппроксимации концентраций свободных Dkk-1 и антитела с течением времени в исследованиях на крысах и макаках. Данную модель экстраполировали на людей и MABEL оценивали как дозу, которая давала 10%-ое снижение уровня Dkk-1.
Тестируемые вещества
Моноклональные антитела против Dkk-1 были приготовлены в Genovac (Germany) посредством иммунизации мышей Balb/C полноразмерным рекомбинантным белком Dkk-1 человека (R&D Systems, Minneapolis, MN). Осуществляли гуманизацию и аффинное созревание одиночного мышиного антитела с изотипом IgG1/каппа (JC18), используя стратегию библиотека-сканирующего мутагенеза (Pons et al., 2009; рукопись в состоянии подготовки). По сравнению с исходным мышиным антителом гуманизированное антитело (huMabJC18) демонстрировало увеличение аффинности более чем в 100 раз для Dkk-1 как человека, так и мыши.
HDkk-1-V5-6His (человека) и rDkk-1-TEV-V5-6His (крысы), использованные в кинетических исследованиях Dkk-1, клонировали и экспрессировали в собственной лаборатории и квалифицировали как функциональные, используя TOPFLASH-анализ (Ai et al., 2005).
Эксперимент с использованием BIAcore
Взаимодействие между huMabJC18 и Dkk-1 человека, или мыши, или крысы, или яванского макака анализировали, используя систему BIAcore 3000ТМ, оборудованную сенсорным чипом СМ5 (BIAcore AB, Uppsala, Sweden). В процессе анализа взаимодействия регистрировали фазы ассоциации и диссоциации. При получении сигналов связывания использовали двойное сравнение и глобальную аппроксимацию к простой модели, применяя программное обеспечение BiaEvaluation v.4.0. Аффинности выводили из соотношения кинетических констант скоростей (KD=koff/kon).
Исследования на животных
Все исследования на животных проводили в соответствии с протоколами об уходе и использовании, утвержденными Комитетом по институциональному уходу за животными и их использованию (IACUC).
Кинетические исследования Dkk-1 на крысах
hDkk-1-V5-6His вводили в виде разовой внутривенной болюс-дозы самцам крыс Sprague Dawley (n=3/доза) в количестве 1, 5, 10 и 100 мкг/кг. Отбирали серийные образцы крови перед введением и через 0,083; 0,25; 0,5; 1; 2; 4; 6; 8 и 24 часа после введения. rDkk-1-TEV-V5-6His вводили в виде разовой внутривенной болюс-дозы самцам крыс Sprague Dawley (n=3/доза) в количестве 100 мкг/кг. Отбирали серийные образцы (300 мкл) перед введением и через 0,083; 0,17; 0,33; 0,5; 1; 2; 4 и 6 часов после введения. Сыворотку крови получали центрифугированием и хранили при -20°С до проведения анализа.
Фармакокинетическое/фармакодинамическое исследование на крысах
Эксперименты проводили на самках крыс Sprague Dawley (n=40, массой тела 250-350 г в течение всей продолжительности исследования, Charles River Laboratories, Wilmington, MA). Самкам крыс Sprague-Dawley (n=8/доза) внутривенно в течение шести последовательных недель вводили HuMabJC18 один раз в неделю в дозе 0,1; 1; 10 и 100 мг/кг. Одной группе крыс (n=8) в качестве контроля вводили разбавитель (20 мМ гистидин, рН 6,5, с 140 мМ NaCl).
Серийные образцы крови отбирали перед введением и через 1, 3, 8, 24, 48, 72, 168, 240, 336, 408, 504, 576, 672, 744, 840, 912 и 1008 часов после введения первой дозы у каждой обрабатываемой группы. Сыворотку крови получали центрифугированием и хранили при -20°С до проведения анализа концентраций Dkk-1 и huMabJC18.
Определение минеральной плотности кости у крыс
Для оценки влияния обработки mAb против Dkk-1 на костную массу иссеченные правые бедренные кости сканировали, используя периферическую количественную компьютерную томографию (pQCT, Stratec XCT Research M, Norland Medical Sistems, Fort Atkinson, WI) с программным обеспечением, версия 5.40. Сканировали поперечные срезы толщиной 1 мм каждого дистального метафиза бедренной кости на расстоянии 2,5 мм проксимально по отношению к дистальному концу, сайт, обогащенный губчатой костью, с размером вокселя 0,10 мм. Общую объемную минеральную плотность кости (BMD) определяли, как описано ранее (Ke et al., 2001).
Фармакокинетическое/фармакодинамическое исследование на яванских макаках
Уход за животными и экспериментальные процедуры в данном. исследовании осуществляли в соответствии с Актом США о благополучии животных (US Animal Welfare Act) и состояния регламентировали в соответствии с Руководством по уходу и использованию лабораторных животных (Институт исследований лабораторных животных, 1996).
В этом исследовании использовали пять самцов и пять самок яванских макак (Масаса fascicularis) в возрасте от 2 до 5 лет и массой от 3,2 до 5,2 кг (Charles River Primates, BioResearch Facility, Houston, Texas). Животных (n=1/пол/группа) разделяли на 5 групп. HuMabJC18 вводили посредством медленной внутривенной инъекции в виде разовой дозы 4 группам макак, состоящим из 1 самца и 1 самки, в дозах 0,1; 1; 10 или 100 мг/кг. Оставшейся группе аналогичным образом вводили разбавитель в качестве контроля.
Для каждой обрабатываемой группы посредством бедренной венепункции отбирали образцы цельной крови (~2 мл) до обработки и через 0,1; 0,5; 1; 3; 8; 24; 48; 72; 168; 240; 336; 408; 504; 576 и 672 часа после введения дозы. Сыворотку отделяли от цельной крови центрифугированием, после чего образцы хранили при -80°С до проведения анализа. Позже проводили анализ образцов на предмет концентрации Dkk-1 и huMabJC18.
Анализ свободного Dkk-1
ELISA-система для анализа Dkk-1 человека от Assay Designs (Ann Arbor, Michigan) (№ по каталогу 900-151) была утверждена для измерения общего Dkk-1 в сыворотке крови человека в соответствии с инструкциями производителя с минимальным изменением: рекомбинантный Dkk-1 человека (R&D Systems, Minneapolis, MN) (№ по каталогу 1096-dk-10/cf) использовали в анализе в качестве стандартов.
Чтобы проанализировать содержание свободного Dkk-1 (не связанного с терапевтическими антителами) в сыворотке крови крыс и макак, использовали антитела против Dkk-1 для захвата несвязанного Dkk-1 сыворотки крови, а для остальных стадий использовали реагенты из ELISA-системы для анализа Dkk-1 человека от Assay Designs (№ по каталогу 900-151) в соответствии с инструкцией к набору со следующими изменениями: в качестве стандартов для анализов на крысах и макаках в данном анализе использовали рекомбинантный Dkk-1 крысы (№ по каталогу 4010-dk-10/cf) и Dkk-1 человека (№ по каталогу 1096-dk-10/cf) от R&D Systems (Minneapolis, MN), соответственно.
Образцы сыворотки крови человека от здоровых и больных субъектов закупали у Bioreclamation Inc. (Nassau, NY).
Анализ V5-меченного Dkk-1
Концентрации V5-меченного Dkk-1 в сыворотке крови определяли ELISA-методом. Образцы разбавляли в PBS-буфере (содержащем 3% BSA и 0,05% Твина-20) до конечного минимально необходимого разведения (MRD) в диапазоне от 1/4 до 1/40, используемого для снижения фона и дающего возможность попадания в линейный диапазон анализа концентраций (~0,195-25 нг/мл в планшете для анализа). Калибровочные стандарты и стандарты для качественного контроля hDkk-1-V5-6His или rDkk-1-TEV-V5-6His разбавляли в растворе того же состава, что и образцы. Сенсибилизацию 96-луночных планшетов для твердофазного иммуноферментного анализа (Nalgene Nunc, Rochester, NY) проводили в течение ночи при 4°С, используя 50 мкл раствора антитела против V5 (Invitrogen, Carlsbad, CA) в концентрации 2 мкг/мл, затем промывали PBS-буфером, содержащим 0,05% Твина-20, после чего блокировали PBS-буфером, содержащим 3% BSA и 0,05% Твина-20. Разбавленные образцы и стандарты добавляли в планшеты (50 мкл/лунка) и инкубировали со встряхиванием при комнатной температуре в течение 1 часа. Затем планшеты промывали, используя два цикла промывки, после чего инкубировали вместе с конъюгированным с пероксидазой хрена вторичным антителом против Dkk-1 (из набора от Analysis Designs, Ann Arbor, Ml) в течение 1 часа. После промывки планшеты обрабатывали для развития цветной реакции в течение ~10 мин субстратом 3,3',5,5'-тетраметилбензидином (ТМВ) (KPL, Gaithersburg, MD), затем реакцию останавливали, используя 2 М H2SO4. Величины OD растворов сравнения вычитали как фоновые после прочтения оптической плотности OD при длине волны 450 нМ (с вычитанием при 650 нМ). Калибровочные стандарты V5-меченного Dkk-1 использовали для построения калибровочной кривой, применяя 4-параметрическую аппроксимацию с равномерной взвешенной величиной (uniform weighting), заложенную в SoftMax Pro 4.8. Концентрации в сыворотке крови V5-меченного Dkk-1 в исследуемых образцах определяли интерполяцией с использованием этой калибровочной кривой.
Анализ остеокальцина
Концентрации остеокальцина в сыворотке крови измеряли, используя набор для ELISA-анализа остеокальцина Rat-MIDTM Osteocalcin (Nordic Bioscience Diagnostics A/S, Herlev, Denmark).
Анализ свободных/частично свободных антител huMabJC18 для PK/PD-образцов крыс
Концентрации huMabJC18 в сыворотке крови определяли электрохемилюминисцентным (ECL) методом, используя систему Meso Scale Discovery (MSD) (Gaithersburg, MD). Образцы разбавляли в PBS-буфере, содержащем 1% BSA, до конечного минимально необходимого разведения (MRD) от 1/2 до 1/150, затем дополнительно выполняли 10-4000-кратное разведение, чтобы снизить помехи фона и попасть в линейный диапазон анализов (~5-1300 нг/мл в планшете для анализа). Калибровочные стандарты и стандарты для качественного контроля HuMabJC18 разбавляли в растворе того же состава, что и образцы. Сенсибилизацию 96-луночных MSD-планшетов для сильного связывания (№ по каталогу L11XB-3) проводили в течение ночи при 4°С, используя hDkk-1-V5-6His (полученный в лаборатории) в концентрации 5 мкг/мл. После опрокидывания планшета для удаления раствора для сенсибилизации планшеты блокировали 1%-ным BSA в PBS. В планшеты добавляли разбавленные образцы и стандарты (25 мкл/лунка) и инкубировали со встряхиванием при комнатной температуре в течение 2 часов. Затем планшеты промывали, используя три цикла промывки PBS-буфером, содержащим 0,05% Твина-20, после чего инкубировали с рутенилированным антителом козы против IgG человека от MSD (№ по каталогу R32AJ-1) в течение 1 часа. После промывки в каждую лунку добавляли считывающий буфер Т от MSD (4х) вместе с поверхностно-активным веществом (№ по каталогу R92TC-1) и незамедлительно прочитывали на устройстве для визуализации MSD Sector Imager 6000. Величины для растворов сравнения вычитали как фоновые из величин для образцов. Калибровочные стандарты гуманизированного антитела против Dkk-1 использовали для построения калибровочной кривой, применяя 4-параметрическую аппроксимацию с фактором взвешивания 1/y2, заложенную в программное обеспечение MSD Discovery Workbench, версия 3.0. Концентрации в сыворотке крови гуманизированного антитела против DKK-1 в исследуемых образцах определяли интерполяцией с использованием этой калибровочной кривой.
Анализ свободных/частично свободных антител huMabJC18 для образцов, взятых у макак
Концентрации huMabJC18 в сыворотке крови определяли ELISA-методом. Образцы разбавляли в PBS-буфере (содержащем 3% BSA и 0,05% Твина-20) до конечного минимально необходимого разведения (MRD) в диапазоне от 1/4 до 1/40, затем дополнительно выполняли 10-500-кратное разведение, чтобы снизить помехи фона и попасть в линейный диапазон анализов (~8-100 нг/мл в планшете для анализа). Калибровочные стандарты и стандарты для качественного контроля HuMabJC18 разбавляли в растворе того же состава, что и образцы. Сенсибилизацию 96-луночных планшетов для твердофазного иммуноферментного анализа проводили в течение ночи при 4°С, используя hDkk-1-V5-6His (полученный в лаборатории) в концентрации 1,5 мкг/мл, и затем блокировали PBS (содержащим 3% BSA и 0,05% Твина-20) после промывки PBS-буфером, содержащим 0,05% Твина-20. Разбавленные образцы и стандарты добавляли в планшеты (100 мкл/лунка) и инкубировали со встряхиванием при комнатной температуре в течение 1 часа. Затем планшеты промывали, используя три цикла промывки, после чего инкубировали в течение 1 часа вместе с биотинилированным мышиным антителом против IgG2 человека (Invitrogen, Carlsbad, CA), далее добавляли стрептавидин, конъюгированный с пероксидазой хрена (Jackson ImmunoResearch, West Grove, PA), и планшет затем инкубировали в течение 30 минут. После промывки планшеты обрабатывали для развития цветной реакции в течение ~10 мин субстратом 3,3',5,5'-тетраметилбензидином (ТМВ), затем реакцию останавливали, используя 2 М H2SO4. Величины OD растворов сравнения вычитали как фоновые после прочтения оптической плотности OD при длине волны 450 нМ (с вычитанием при 650 нМ). Калибровочные стандарты гуманизированного антитела против Dkk-1 использовали для построения калибровочной кривой, применяя 4-параметрическую аппроксимацию с равномерной взвешенной величиной, заложенную в SoftMax Pro 4.8. Концентрации в сыворотке крови гуманизированного антитела против DKK-1 в исследуемых образцах определяли интерполяцией с использованием этой калибровочной кривой.
Анализ противолекарственных антител (ADA)
Присутствие противолекарственных антител (ADA) против huMabJC18 у крыс и макак измеряли с применением анализа связывания сшивающего лиганда (LBA), используя платформу от Meso Scale Discovery (MSD) (Gaithersburg, MD). Образцы сыворотки крови (25 мкл), разбавленные 1:10 в используемом для анализа разбавителе (3% BSA, 0,05% Твина 20, PBS), добавляли в 96-луночный MSD-планшет для сильного связывания, который сенсибилизировали, используя huMabJC18 в концентрации 1 мкг/мл в карбонатном буфере, рН 9,6. После инкубации в течение 1 часа при комнатной температуре планшет промывали, в каждую лунку добавляли по 25 мкл меченного рутением huMabJC18 в концентрации 1 мкг/мл и вновь инкубировали в течение 1 часа при комнатной температуре. Планшет промывали и затем добавляли 150 мкл считывающего буфера от MSD (2×), планшет прочитывали на устройстве для визуализации MSD Sector Imager 6000.
Некомпартментальный фармакокинетический анализ
Фармакокинетический анализ проводили, используя вариант программного продукта для отдельных предприятий WinNonLin, версию 5.2 (Pharsight Corp., Cary, NC). Константу скорости заключительной фазы (kel) определяли методом линейной регрессии для профиля зависимости логарифма концентрации в плазме крови от времени. Конечный период полувыведения (t1/2) рассчитывали как 0,693/kel. Величины Cmax, Tmax и Cmin получали непосредственно из зарегистрированных данных. Площадь под кривой зависимости концентрации в сыворотке крови от времени (AUC0-t конечное) рассчитывали, используя метод трапеций с аппроксимацией линейной зависимостью, и экстраполировали к бесконечности (inf) (AUC0-inf), используя kel. Клиренс (CL) рассчитывали, используя соотношение доза/AUC0-inf. Объем распределения (Vc) рассчитывали, используя соотношение CL/kel. Среднюю концентрацию Cave для 1-го интервала дозирования рассчитывали как AUC(1-й интервал дозирования)/tконечное (конечное время).
В кинетическом исследовании Dkk-1 на крысах величина периода полувыведения, указанная для hDkk-1, представляла собой среднее значение для величин периодов полувыведения, определенных при уровнях доз 5, 10 и 100 мкг/кг. Для расчета величины периода полувыведения, соответствующего уровню дозы 1 мкг/кг, не было получено удовлетворительных данных. Период полувыведения для rDkk-1 рассчитывали на основании анализа, проведенного в дозе 100 мкг/кг на группе из 3-х ранее не подверженных экспериментам крыс, которым вводили только одну эту дозу.
Фармакокинетический/фармакодинамический анализ
Был проведен предварительный некомпартментальный анализ (Jusko, 1992) данных зависимости концентрации антитела от времени индивидуально для крыс и макак, выявивший нелинейную фармакокинетику для данного антитела. Для описания PK/PD-профиля huMabJC18 у крыс и макак использовали механистическую модель мишень-опосредованного распределения лекарственного средства (TMDD) (Mager и Jusko, 2001) (Фиг.5). Вкратце, TMDD-модель предполагает, что за наблюдаемый нелинейный характер фармакокинетики отвечает высокоаффинное, с насыщением, связывание антитела (huMabJC18) с мишенью (Dkk-1). Антитело в центральном компартменте (объем V1) связывается (константа скорости, kon) со свободным Dkk-1 с образованием антитело-Dkk1-рецепторного комплекса. Образовавшись, комплекс может диссоциировать (константа скорости, koff), или комплекс антитело-Dkk1 может быть элиминирован (константа скорости, kel, комплекс). Несвязанное антитело также может быть непосредственно элиминировано из центрального компартмента с константой скорости первого порядка (kel). Данная модель была применена для учета распределения антитела по сайтам неспецифического связывания в тканях, которое описывается константами скоростей k12 и k21.
Данная модель описывалась следующим набором дифференциальных уравнений:
(1) 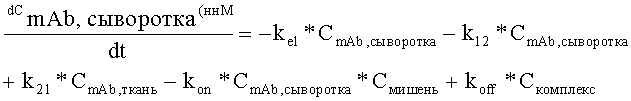
(2) 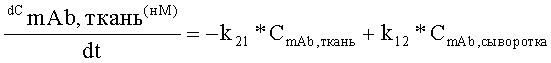
(3) 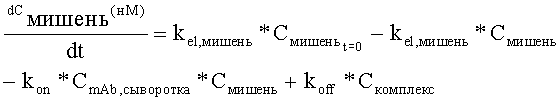
(4) 
где CmAb,сыворотка равна концентрациям свободного huMabJC18 в сыворотке крови, KD=Koff/kon, [mAbсуммарно]=CmAb,сыворотка+Cкомплекс и [мишеньсуммарно]=Cмишень+Cкомплекс.
Эта система дифференциальных уравнений была решена численно с применением программного обеспечения NONMEM, версия V, работающего в оболочке DOS под Windows XP с использованием Compaq Visual Fortran, версия 6.6. Для этой жесткой задачи использовали ADVAN 8 TOL=3. Чтобы оценить концентрацию свободного Dkk-1 при t=0, компартмент свободного Dkk-1 калибровали, используя стандартную дозу при t=0 и устанавливая в положение F3 (свободный Dkk-1). Вариабельность ответа на Dkk-1 у обработанных разбавителем макак характеризовали, используя полиномиальную модель четвертого порядка:
Y=А*ВРЕМЯ^4-B*ВРЕМЯ^3+C*ВРЕМЯ^2-D*ВРЕМЯ+DKK0,
где А, В, С и D представляют собой константы, определенные на основании данных по разбавителю и фиксированные в данной модели, a Dkk0 равняется концентрации Dkk-1 перед введением дозы (t=0 ч). Это вычисление было выполнено в блоке $ERROR в NONMEM с учетом аддитивного эффекта лекарственного средства.
Вариабельность ответа на Dkk-1 у обработанных разбавителем крыс характеризовали, используя простую косинусоидную функцию:
F=Baseline+Amplitude*Cos((Time-Peak Time)*(2*π/24)),
где Baseline = базовая линия; Amplitude = амплитуда; Cos = косинус; Peak Time = время пика (Chakraborty et al. (1999) J. Pharmacokinet. Biopharm., 27: 23-43), с учетом мультипликативного эффекта лекарственного средства.
Критерий согласия оценивали, исходя из точности оценок параметров и матрицы корреляции параметров, полученных с использованием стандартной подпрограммы $COV в NONMEM, визуального контроля случайного разброса взвешенной невязки в зависимости от времени и прогнозируемых концентраций, совместно с визуальным контролем прогноза для отдельного субъекта при сравнении реальных графиков зависимостей концентрации от времени на предмет отсутствия систематических ошибок для любых временных точек).
Моделирование концентраций Dkk-1 и huMabJC18 у пациентов с остеопорозом проводили с применением TMDD-модели, используя программу Berkeley-Madonna (v.8.3.9, University of California, Berkeley, CA). Параметры, использованные с целью моделирования ситуации для человека, показаны в Таблице 14. Параметры, относящиеся к РК, мишени и комплексу, при переходе от крыс и макак к человеку пересчитывали в соответствии с принципами аллометрии, используя простую степенную модель вида: Y=a·BWb, где Y - это представляющий интерес параметр, BW - масса тела, а - аллометрический коэффициент, и b - аллометрическая экспонента. Для вычисления периода полувыведения b принимали равным 0,25, для вычисления констант скоростей элиминирования и всасывания (kel и kel) b принимали равным -0,25, а для вычисления объема распределения b принимали равным 1 (Wang et al., 2008).
Оценки прогнозируемой клинической начальной дозы, полученные с использованием этих трех подходов, различались более чем на 5 порядков по величине. Самая высокая начальная доза была получена на основании вычисления NOAEL (1 мг/кг). Самая низкая начальная доза была получена на основании вычисления MABEL с применением уравнения Даффа (0,00003 мг/кг). Начальная доза, полученная на основании вычисления MABEL с применением TMDD-модели, попадала приблизительно в середину этого диапазона (0,03 мг/кг).
Данные, полученные с использованием BIAcore
Сводка данных по кинетике связывания in vitro huMabJC18 с Dkk-1 человека, мышей, крыс и макак, определенных с применением BIAcore-технологии связывания на поверхности, показана в Таблице 8. HuMabJC18 связывается с Dkk-1 человека, мыши и крысы с высокой аффинностью (Kd<2 пМ, <30 пМ и <100 пМ, соответственно). В большинстве случаев значения констант скоростей ассоциации (kon) были слишком велики, а значения констант скоростей диссоциации (koff) были слишком малы для точного измерения с использованием аппаратуры BIAcore. HuMabJC18 также связывается с Dkk-1 яванского макака, хотя KD невозможно было определить вследствие наличия примесей в данном образце Dkk-1.
Экспрессия Dkk-1 у женщин пременопаузального, постменопаузального периода, у женщин с остеопенией и остеопорозом
Концентрации Dkk-1 в образцах сыворотки крови женщин пременопаузального (n=50), постменопаузального периода (n=50), у женщин с остеопенией (n=50) и остеопорозом (n=50) показаны в Таблице 9. Для этого набора протестированных образцов не было отмечено значительного различия в концентрациях Dkk-1 между женщинами пре- и постменопаузального периода, что является подтверждением того, что возраст не оказывает влияния на концентрации Dkk-1. Наблюдалось значительное различие (p<0,01) между уровнями Dkk-1 у женщин пременопаузального периода (среднее значение 2,2 нг/мл) по сравнению с образцами от женщин с остеопенией (Т-критерий -2,2; среднее значение Dkk-1 9,0 нг/мл) и женщин с остеопорозом (Т-критерий -3,0; среднее значение Dkk-1 10,5 нг/мл). Эти величины были включены в PK/PD-модель для предсказания эффективной дозы huMabJC18 для женщин с остеопорозом.
Кинетика мишени Dkk-1 у крысы
V5-His-меченный (h-) Dkk-1 человека и (r-) Dkk-1 крысы вводили самцам крыс Sprague-Dawley внутривенным болюсным введением в концентрации 1, 5, 10 и 100 мкг/кг (h-Dkk-1) или 100 мкг/кг (r-Dkk-1). Кинетика h-Dkk-1 была линейной во всем диапазоне протестированных доз. Средний период полувыведения составил 22,3±6,4 мин для h-Dkk-1 (Фиг.6) и 34 мин для r-Dkk-1.
Фармакокинетика и фармакодинамика на крысах
Профили зависимости среднего значения концентрации свободного/частично свободного huMabJC18 от времени после еженедельного внутривенного введения самкам крыс Sprague-Dawley показаны на Фиг.7. Параметры некомпартментальной фармакокинетики показаны в Таблице 10. Фармакокинетика huMabJC18 была нелинейной в интервале доз, причем наибольшая нелинейность для AUC наблюдалась для дозы до 10 мг/кг. Это указывает на более высокую скорость клиренса при более низких дозах (0,1 и 1 мг/кг) по сравнению с более высокими дозами (10 и 100 мг/кг). Для более поздних временных точек концентрации huMabJC18 в сыворотке крови у некоторых крыс были меньше, чем ожидалось, или ниже предела количественного определения данного анализа. С использованием качественного анализа противолекарственных антител (ADA) в этих образцах было подтверждено наличие антител крысы против huMabJC18, и данные, полученные на этих крысах, были изъяты из анализа и графиков.
Графики средних значений концентраций свободного Dkk-1 для этого же исследования приведены на Фиг.8А-8Е. Концентрации свободного Dkk-1 быстро уменьшались после введения huMabJC18 крысе. Для всех доз, кроме самой низкой дозы, концентрации свободного Dkk-1 оставались пониженными в продолжение данного исследования.
Фармакокинетика и фармакодинамика на яванских макаках Профили зависимости отдельных концентраций свободного/частично свободного huMabJC18 в сыворотке крови от времени для яванских макак показаны на Фиг.9А и 9В, а параметры некомпартментальной фармакокинетики huMabJC18 для яванского макака показаны в Таблице 11. Фармакокинетика huMabJC18 была нелинейной в диапазоне протестированных доз, при этом самые высокие скорости клиренса и самые низкие величины периодов полувыведения наблюдали при более низких дозах. Период полувыведения huMabJC18 для яванских макак лежал в диапазоне 1-13 суток во всем диапазоне доз.
Утрата экспозиции в группе, получающей дозу 1 мг/кг, примерно на 14-е сутки и для одного макака в группе, получающей дозу 10 мг/кг, примерно на 21-е сутки после введения вероятно обусловлена образованием антител против huMabJC18, и эти данные были изъяты из анализа и графиков. Однако этот факт не мог быть подтвержден с использованием качественного ADA-анализа.
Средние концентрации в сыворотке крови свободного Dkk-1 для этого же исследования на яванских макаках показаны на Фиг.10А-10Е. Концентрация свободного Dkk-1 быстро уменьшалась после введения huMabJC18. Продолжительность супрессии Dkk-1 была дозозависимой, и возвращение к базовой линии наблюдалось при более низких дозах. При самых высоких дозах супрессия Dkk-1 сохранялась во всем интервале дозирования.
PK/PD-моделирование
Механистическую TMDD-модель (Mager и Jusko, 2001) использовали для одновременной аппроксимации концентраций свободного huMabJC18 и свободного Dkk-1 во времени (Фиг.5) как для макак, так и для крыс. Была выбрана эта модель, поскольку она учитывает не связанное с мишенью элиминирование антитела, синтез и метаболизм мишени и образование и элиминирование комплекса. Сравнение наблюдаемых и прогнозируемых PK/PD-профилей huMabJC18 и Dkk-1 для крыс показано на Фиг.7 и 8А-Е, а для макак показано на Фиг.9А-В и 10А-Е. Оценки параметров, полученные с использованием TMDD-модели для крыс и макак, показаны в Таблице 12.
Оценки объема распределения (V1) и периода полувыведения, полученные из PK/PD-модели для крысы и макака, согласуются с фармакокинетикой антитела IgG2 для крысы и макака (Peppard и Orlans, 1980; Hinton et al., 2004). Оценка эффективности in vivo для крысы (Kd=34,8 пМ) согласуется с величиной, полученной in vitro для крысы (Kd меньше 100 пМ, см. приведенные выше данные, полученные с использованием BIAcore). Оценки периода полувыведения Dkk-1 (26 мин для макака и 11 мин для крысы), полученные из kel для Dkk-1, аналогичны величинам, подсчитанным в исследованиях метаболизма V5-His-меченных hDkk-1 и rDkk-1 (22,3 мин и 34 мин, соответственно, см. раздел "Кинетика мишени", приведенный выше). Величина периода полувыведения комплекса (полученная на основании оценки kel для комплекса) лежит в промежутке между оцененным периодом полувыведения антитела huMabJC18 и мишени Dkk-1.
Биомаркеры кости у крыс
Помимо мишени Dkk-1 в исследовании на крысах проводили количественное измерение других биомаркеров, в том числе остеокальцина, который является общепринятым биомаркером костеобразования и минеральной плотности кости (BMD) и утвержденным биомаркером остеопороза (см, например, Primer on the Methabolic Bone Diseases and Disorders of Mineral Methabolism (Rosen CJ ed.) pages 152-163 и 174-179, American Society for Djne and Mineral Research, Washington D.C). График зависимости концентрации остеокальцина от дозы через 1,5 и 2,5 недели после введения первой дозы в исследовании на крысах показан на Фиг.11. Наблюдается статистически значимое (p меньше 0,05) возрастание концентраций остеокальцина в группе с дозой 1 мг/кг/неделя через 2,5 недели после введения первой дозы и в группах с дозой 10 и 100 мг/кг/неделя через 1,5 недели после введения дозы.
Как показано в Таблице 13, общая минеральная плотность кости была значительно повышена у крыс, обработанных huMabJC18 в дозах 10 и 100 мг/кг/неделя, на 11% и 16%, соответственно, по сравнению с разбавителем в качестве контроля.
Самок крыс обрабатывали посредством внутривенной инъекции huMabJC18 в различных дозах еженедельно в течение 6 недель. В конце исследования измеряли BMD.
Моделирование PK/PD для человека
Механизм, основанный на модели PK/PD (TMDD) (Фиг.5), использовали с целью моделирования PK/PD huMabJC18 для людей и с целью предсказания эффективной дозы для лечения остеопороза среди других расстройств. Чтобы получить информацию по моделированию PK/PD для человека, приведенные в литературе величины параметров PK для IgG2 антитела (не опосредованной мишенью) объединяли со знаниями кинетики Dkk-1 как мишени и кинетики комплекса huMabJC18-Dkk-1, полученными на основании PK/PD-моделирования на макаках и крысах (Таблица 14).
Результаты предварительных экспериментов на OVX-мышах в качестве модели заболевания указывали на то, что для статистически значимого повышения минеральной плотности кости (данные не показаны) требовалось 50%-ное снижение уровня Dkk-1. Прогнозируемая эффективная доза huMabJC18 для снижения уровня Dkk-1 на величину более 50% во всем интервале дозирования составляет 3,57 мг/кг при введении один раз в месяц. Данная предполагаемая доза характеризуется ассоциированной с ней неопределенностью, которая обусловлена неопределенностью, связанной с кинетикой эндогенного Dkk-1 и высокой чувствительностью супрессии Dkk-1 (и такой дозы) к этому параметру. Доза, прогнозируемая как дающая минимальный ожидаемый биологический эффект, (MABEL) составила 0,03 мг/кг. Прогнозируется, что эта доза будет временно снижать уровни Dkk-1 на величину меньше 20% с последующим возвращением к базовой линии. Прогнозировалось, что доза 0,003 мг/кг (1/10 от MABEL) не оказывает никакого эффекта. Концентрации Dkk-1 для человека, определенные посредством моделирования, показаны на Фиг.12, и должны представлять собой максимальную величину дозы, устанавливаемой в клинике.
Нелинейная фармакокинетика huMabJC18 прогнозируется в диапазоне доз, охватывающем MABEL (0,03 мг/кг) и прогнозируемую эффективную дозу (3,57 мг/кг). Это обусловлено TMDD и показано на Фиг.13.
Интерпретация свойств мишени
Данные, полученные на мышах, указывают на то, что Dkk-1 является главным регулятором ремоделирования кости (Diarra et al. (2007) Nat. Med., 13: 156-163) и что базовые уровни Dkk-1 у больных (овариэктомизированных, OVX) мышей приблизительно в 5 раз выше по сравнению со здоровыми мышами (собственные данные). Также показано, что уровни Dkk-1 повышены в плазме костного мозга и периферической крови у пациентов с множественной миеломой с повреждением костей (Tian et al., 2003). Анализ чувствительности показал, что прогнозируемая человеческая доза моноклонального антитела против Dkk-1 (huMabJC18) была крайне чувствительна к базовым уровням Dkk-1. Чем выше базовые уровни Dkk-1, тем более высокая концентрация антитела необходима для снижения Dkk-1. Базовые уровни мишени часто известны из предварительных клинических измерений, однако отсутствие такой информации для Dkk-1 вынуждает устанавливать рекомендуемые диапазоны уровней Dkk-1 человека, как для здоровых субъектов, так и группы пациентов (пациентов с остеопенией и остеопорозом), которые могли бы быть использованы для более информативных прогнозирований доз на различных стадиях клинической разработки. Результаты анализа показали, что уровни Dkk-1 были выше у женщин с остеопенией и остеопорозом (Т-критерий <-1) по сравнению со здоровыми женщинами.
Скорость метаболизма лигандов-мишеней может варьировать от нескольких минут до нескольких суток, что может оказывать существенное влияние на эффективную дозу и даже на возможность мишени к изменению с целью получения клинической пользы. Некоторые лиганды характеризуются схожей кинетикой среди разных видов, но для других метаболизм непредсказуем a priori. Meno-Tetang и соавт. (Meno-Tetang and Lowe (2005) Basic Clin. Pharmacol. Toxicol. 96: 182-192) показали, что скорости метаболизма IgE определяются диапазоном от 5-8 ч для мыши до 2,7 суток для человека, что оказывает существенное влияние на прогнозирование влияния на человека антитела против IgE. Анализ чувствительности для Dkk-1 подтвердил, что прогнозируемая доза моноклонального антитела против Dkk-1 (huMabJC18) была чувствительна к периоду полувыведения Dkk-1 при большом молярном избытке антитела, необходимом при более высоких скоростях метаболизма мишени. В литературе не было никакой доступной информации о скоростях метаболизма Dkk-1, и поэтому в собственной лаборатории были проведены эксперименты с использованием VS-His-меченного Dkk-1 человека и крысы, вводимого внутривенно крысам. Эти эксперименты показали, что Dkk-1 имел высокую скорость метаболизма (период полувыведения 22,3 мин для h-Dkk-1 и 34 мин для r-Dkk-1) и что формы Dkk-1 человека и крысы характеризовались схожими скоростями метаболизма у крыс. Высокая скорость метаболизма Dkk-1 позже была подтверждена посредством использования PK/PD-моделирования с целью оценки периода полувыведения Dkk-1 у крыс и макак, которым вводили huMabJC18.
Интерпретация PK/PD для huMabJC18 у крыс и макак
В исследованиях huMabJC18 на крысах и макаках продемонстрирована нелинейная фармакокинетика, характеризующаяся более высокими значениями скоростей клиренса и более короткими периодами полувыведения при низких дозах. Это указывает на мишень-опосредованное распределение лекарственного средства (TMDD), при котором взаимодействие антитела с его фармакологической мишенью влияет на распределение при низких дозах (Tabrizi et al. (2006) Drug Discov. Today, 11: 81-88; Wang et al. (2008) Clin. Pharmacol. Ther., 84: 548-558). Этот путь является путем с насыщением при более высоких дозах ввиду ограниченной экспрессии мишени. В результате насыщения мишень-опосредованного пути типичные для IgG механизмы катаболического клиренса с участием FcRn являются доминирующими, что придает антителу характерный ему продолжительный период полувыведения. Более признано TMDD у моноклональных антител, направленных на белки, экспрессируемые на клеточных мембранах, где опосредованный рецепторами эндоцитоз приводит к элиминированию лекарственного средства. Однако TMDD также можно наблюдать при использовании растворимых мишеней: омализумаб и деносумаб представляют собой антитела для растворимых мишеней (IgE и активатора рецепторов NFkB, соответственно), которые демонстрируют нелинейную кинетику элиминирования (Hayashi et al., 2007; Marathe et al., 2008).
Эмпирические PK/PD-модели, состоящие из PK-модели для описания системных концентраций лекарственного средства, которая используется в качестве задающей функции для описания PD, часто не подходят для характеристики TMDD, поскольку они не учитывают взаимозависимости PK и PD. Одиночная модель, описывающая PK лекарственного средства, динамику мишени и их взаимодействие, была предложена Mager и соавт. (Mager and Jusko (2001) J. Pharmacokinet. Pharmacodyn. 28: 507-532). Эта модель учитывает специфическое и неспецифическое распределение и элиминирование молекул лекарственного средства, а также обеспечивает гибкость при учете динамики мишени. В других случаях, когда PK и PD анализированы одновременно, эта модель использована для обеспечения непосредственной связи между дозой, продолжительностью воздействия и ответом (Meno-Tetang and Lowe (2005) Basic. Clin. Pharmacol. Toxicol., 96: 182-192); Ng et al. (2006) Pharm. Res., 23: 95-103.; Wu et al. (2006) J. Pharm. Sci., 95: 1258-1268). В данном случае TMDD-модель использовали для одновременной аппроксимации концентрации антитела huMabJC18 и мишени Dkk-1 для крыс и макак после в/в (внутривенного) введения huMabJC18 в разных уровнях доз. С использованием этой модели была получена оценка не опосредованной мишенью фармакокинетики huMabJC18, которая должным образом соответствовала типичной фармакокинетике IgG2 для каждого вида (Peppard and Orlans (1980) Immunology, 40: 683-686; Hinton et al. (2004) J. Biol. Chem., 279: 6213-6216). Так, период полувыведения составил 2,5 суток у крысы и 13,6 суток у макака. Объем распределения был приблизительно равен объему плазмы крови у макака (0,052 л/кг), но выше, чем объем плазмы крови у крысы (0,147 л/кг).
Период полувыведения мишени Dkk-1 по оценкам, полученным на основании PK/PD-моделирования, составил 11 мин для крысы и 26 мин для макака. Он был того же порядка по величине, что и период полувыведения V5-His-меченного h-Dkk-1, измеренный у крысы (период полувыведения = 25 мин). Величина периода полувыведения комплекса huMabJC18-Dkk-1 по оценкам лежит между периодом полувыведения huMabJC18 и периодом полувыведения Dkk-1, что вероятно является отражением мишень-опосредованного механизма клиренса. Однако возможно, что связывание мишени с антителом мешает связыванию антитела с FcRn, и исследования с целью проверки этой гипотезы находятся в стадии завершения в собственной лаборатории.
Для крысы наблюдали повышение уровней биомаркеров костеобразования (остеокальцина и минеральной плотности кости) в дополнение к снижению уровня целевого биомаркера Dkk-1. Эти данные явились дополнительной поддержкой возможности использования huMabJC18 для лечения нарушений структуры скелета, таких как остеопороз.
Прогнозирование эффективной дозы для человека и расчет MABEL
Для моноклональных антител становится общепризнанным, что рациональный выбор безопасных доз, впервые применяемых на человеке, на основе PK/PD-моделирования имеет важное значение (см., например, вебсайт правительства UK Publications policy and guidance: Department of Health - Publications). Новый параметр, минимальный уровень ожидаемого биологического эффекта (MABEL) включает в себя экстраполяцию наблюдаемых доклинических PK/PD-данных для клинического прогнозирования на основе подхода PK/PD-моделирования. MABEL предложен для рассмотрения в дополнение к уровню, не вызывающему видимых неблагоприятных эффектов (NOAEL), при планировании уровней впервые применяемых на человеке доз терапевтических средств высокого риска в недавно опубликованных Европейских нормативных рекомендациях (European regulatory guidance) (EMEA (2007) Guideline on strategies to identify and, mitigate risks for first-in-human clinical trials with investigational medicinal products, in (EMEA ed.), London).
Для прогнозирования эффективной дозы huMabJC18 механистическую TMDD-модель, использованную с целью аппроксимации доклинических данных для крысы и макака, адаптировали для моделирований PK/PD человека. Приведенные в литературе величины фармакокинетических параметров для антитела IgG2 (не опосредованных мишенью) объединяли со знаниями кинетики Dkk-1 как мишени и кинетики комплекса huMabJC18-Dkk-1, полученными при моделировании PK/PD для крысы и макака.
Величины, полученные с использованием BIAcore для Dkk-1 человека in vitro, применяли для определения констант скоростей ассоциации и диссоциации комплекса (kon и koff) в данной модели. Анализ чувствительности был проведен в отношении kon и koff, и при использовании максимальных констант скоростей ассоциации (fast on-rates) и минимальных констант скоростей диссоциации (slow off-rates), прогнозируемых для этого антитела у людей, оказалось, что супрессия Dkk-1 и, следовательно, доза не чувствительны к достаточно существенным изменениям Kd. Действительно, при значениях Kd меньше 10 пМ, нет никаких изменений в прогнозируемой дозе. Это происходит потому, что скорость метаболизма комплекса huMabJC18-Dkk-1 больше, чем скорость диссоциации комплекса с рецептором. В таких случаях при выборе кандидата можно обойтись без стадий аффинного созревания.
Для таких моделирований использовали средние значения базовых концентраций Dkk-1, измеренные в образцах от пациентов с остеопорозом (см. раздел "Результаты"). Было сделано предположение, что Dkk-1 характеризуется более низкой скоростью метаболизма у человека по сравнению с макаком или крысой. Поэтому период полувыведения Dkk-1 человека рассчитывали, используя аллометрическое вычисление по данным о величинах периодов полувыведения для макака и крысы. В результате получали оценочный период полувыведения Dkk-1 для людей, равный 49 мин. Следует отметить, что поскольку Dkk-1 представляет собой быстро метаболизируемый антиген, общая супрессия Dkk-1 и, следовательно, доза крайне чувствительны к величине периода полувыведения Dkk-1, используемой при данном моделировании.
С использованием аллометрического вычисления периода полувыведения комплекса, оцениваемого по PK/PD-модели для макака, прогнозируемый период полувыведения комплекса huMabJC18-Dkk-1 у людей составил 2,3 суток (kel=0,3 суток-1). Более короткий период полувыведения комплекса для людей был спрогнозирован с использованием аллометрического вычисления периода полувыведения комплекса, оцениваемого при PK/PD-моделировании на крысах. Однако, по результатам проведенного анализа чувствительности, супрессия Dkk-1 (и следовательно доза) оказалась нечувствительной к изменению периода полувыведения комплекса, прогнозируемого при переходе от крыс к макакам.
Предварительные эксперименты на OVX-мышах как на модели постменопаузального уменьшения костной массы показали, что приблизительно 50%-ное снижение уровней Dkk-1 было связано со статистически значимым повышением минеральной плотности кости (данные не показаны). Поэтому, прогнозы в отношении доз для huMabJC18 были сделаны на основании необходимости супрессии уровней Dkk-1 более чем на 50% по отношению к базовой линии в интервале дозирования (1 месяц). В рамках этой прерогативы и с указанными выше предположениями прогнозируемая эффективная доза huMabJC18 составила 3,57 мг/кг при подкожном введении один раз в месяц. По результатам моделирования прогнозируемая доза, оказывающая минимальный эффект (MABEL), на людей (снижение Dkk-1 <20%) составляет 0,03 мг/кг. Для клиники прогнозируется нелинейная фармакокинетика (Фиг.13), при этом huMabJC18 демонстрирует более сильный клиренс и более короткий период полувыведения при низких дозах вследствие TMDD.
Альтернативный подход для предсказания фармакологии для человека состоит в прогнозировании максимальной занятости рецепторов (RO) с использованием формулы, основанной на теории равновесного взаимодействия лекарственных средств:
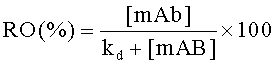
(Фиг.14, (Duff (2006) Expert Scientific Group on Phase One Clinical Trials: Final Report, in Department of Health, UK, London)). Оценка RO с использованием этого способа основывается только на данных о Kd и не учитывает кинетику мишени или комплекса mAb-мишень. Показано, что Dkk-1 характеризуется высокой скоростью метаболизма, и связывание Dkk-1 с huMabJC18 приводит к изменению кинетики мишени (TMDD). В этих условиях RO зачастую невозможно предсказать, исходя из фармакологических равновесных подходов, и показано, что результаты простых расчетов RO на основании Kd существенно превышают прогнозируемую RO для конкретной дозы (Agoram (2009) Br. J. Clin. Pharmacol., 67: 153-160). Что касается Dkk-1, то оценка для RO, рассчитанная с использованием равновесной формулы, составляет ED50 0,00001 мг/кг, в то время как оценка согласно механистической PK/PD-модели составляет ED50 0,01 мг/кг (Фиг.14). Использование расчетов для равновесия могло бы привести к выбору доз, слишком низких для клиники, и замедлить продвижение исследований доз, впервые применяемых на человеке. Другая крайность заключается в том, что, наблюдаемый NOAEL у крысы и макака (100 мг/кг) превышает таковой, необходимый для 100%-го связывания мишени с антителом, что не может быть рекомендовано в качестве оценок начальной клинической дозы. Было сделано заключение, что основанный на PK/PD-модели подход для расчета дозы, соответствующей MABEL, с большей вероятностью является предсказательным для Dkk-1 и обеспечивает эффективность и безопасность клинических исследований.
Наблюдаемый NOAEL (100 мг/кг) превышает уровень, необходимый для 100%-го связывания Dkk-1 с антителом. Другая крайность заключается в том, что MABEL, рассчитанный по уравнению Даффа, вероятно серьезно занижен ввиду отсутствия учета быстрого метаболизма Dkk-1. В связи с этим данные способы не являются в этом случае приемлемыми подходами для установления начальной клинической дозы. Напротив, TMDD-модель обеспечивала отличное описание доклинических данных, которые указывали на роль метаболизма мишени при определении % связывания Dkk-1. Таким образом, оценка MABEL, полученная из TMDD-модели (0,03 мг/кг), будет обеспечивать как безопасность, так и эффективность в исследованиях FIH.
В заключение, huMabJC18 представляет собой гуманизированный прототип антитела против Dkk-1 для лечения остеопороза среди других расстройств. Механистическую TMDD-модель использовали с целью характеристики зависимости ответа от концентрации для huMabJC18 и Dkk-1 у крысы и макака. Эту модель переносили на человека для прогнозирования эффективной дозы и MABEL посредством включения информации об экспрессии мишени и скоростях метаболизма. Было установлено, что эти параметры оказывают большее влияние на зависимость "доза-ответ", чем аффинность для этого механизма.
Информация по депонированию
Авторы заявки провели депонирование вариабельных областей тяжелой и легкой цепи антитела, обозначенного в данном описании как huMAbJC18, вместе с Американской коллекцией типовых культур (АТСС) Manassas, VA, 20110-2209, USA. Как отмечено в изложенном выше описании, вариабельная область НС huMabJC18 была депонирована 19 марта 2009 г. и получила обозначение с № депонирования в АТСС РТА-9835, а вариабельная область LC huMabJC18 была депонирована 19 марта 2009 г. и получила обозначение с № депонирования в АТСС РТА-9836. Эти акты депонирования были проведены в соответствии с положениями Будапештского соглашения о международном признании депонирования микроорганизмов для целей патентной процедуры и нормативного акта, принятого на основании его (Будапештского соглашения). Эти депозиты будут сохранены без ограничения в депозитарии АТСС в течение периода времени 30 лет, или в течение 5 лет после самого последнего запроса, или для эффективной жизни патента, в зависимости от того, какой период длиннее, и будут заменены, если эти депозиты станут нежизнеспособными в течение этого периода. Доступность депонированных материалов не должна быть истолкована как выдача лицензии на практическое применение любых аспектов настоящего изобретения в нарушение прав, предоставленных патентным ведомством какого-либо правительства в соответствии со своими патентными законодательствами. Считается, что специалисту в данной области будет достаточно изложенного выше письменного описания для возможности реализации на практике всех аспектов настоящего изобретения. Объем настоящего изобретения не ограничивается депонированными материалами, поскольку депонированное воплощение предлагается в качестве одной иллюстрации некоторых аспектов данного изобретения, и любые конструкции, которые являются функционально эквивалентными, находятся в пределах объема данного изобретения. Депонирование изложенного в данном описании материала не является признанием того, что изложенного здесь письменного описания недостаточно для практической реализации любого аспекта данного изобретения, в том числе лучшего его варианта, и его не следует истолковывать как ограничение объема формулы изобретения конкретными иллюстрациями, которые оно представляет. Действительно, различные модификации данного изобретения, помимо показанных и изложенных в данном описании, будут очевидны специалистам в данной области из вышеизложенного описания и будут попадать в пределы объема прилагаемой формулы изобретения.
Заголовки разделов, использованные в данном описании, предназначены только для организационных целей, и их никоим образом не следует истолковывать как ограничивающие описываемый объект изобретения. Будет очевидно, что перед температурой, концентрациями, временем и т.д. подразумевается слово "примерно", что рассмотрено в настоящих разъяснениях, так что незначительные и несущественные отклонения находятся в пределах объема разъяснений, приведенных в описании настоящего изобретения. Использование в этой заявке единственного числа включает множественное число, если конкретно не указано иное. Кроме того, применение терминов "содержат", "содержит", "содержащий", "состоят из", "состоит из", "состоящий из", "включают", "включает" и "включающий/включая" не предназначено для ограничения. Следует понимать, что как изложенное выше общее описание, так и следующее после него подробное описание приведены только в иллюстративном и пояснительном смысле и не ограничивают данное изобретение.
Все ссылки, приведенные в данном описании, включая патенты, патентные заявки, статьи, учебники и тому подобное и приведенные там ссылки, при условии, что они не включены ранее, тем самым включены во всей своей полноте посредством ссылки. В том случае, когда один или более чем один из включенных литературных или аналогичных материалов отличается от этой заявки или противоречит ей, в том числе, но не ограничиваясь этим, определенные термины, употребление терминов, описанные методы или тому подобное, этот параметр контролируется данной заявкой.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛЬНЫЕ АГЕНТЫ, СПЕЦИФИЧНЫЕ К CD19 ЧЕЛОВЕКА, И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2773317C2 |
| АНТИТЕЛА ПРОТИВ РЕЦЕПТОРА ФОЛИЕВОЙ КИСЛОТЫ 1, ИХ ИММУНОКОНЪЮГАТЫ И ИСПОЛЬЗОВАНИЕ | 2011 |
|
RU2610663C2 |
| Терапевтические антитела и их применения | 2016 |
|
RU2722381C2 |
| АНТИТЕЛО К CLDN18.2 И ЕГО ПРИМЕНЕНИЯ | 2018 |
|
RU2797709C2 |
| УЛУЧШЕННЫЕ НАНОТЕЛА ПРОТИВ СЫВОРОТОЧНОГО АЛЬБУМИНА ЧЕЛОВЕКА | 2012 |
|
RU2634381C2 |
| АНТИТЕЛА, ОБЛАДАЮЩИЕ СПЕЦИФИЧНОСТЬЮ К BTN2, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2794996C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ ИНТЕРЛЕЙКИНА-31 | 2012 |
|
RU2588462C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ НЕКТИН-4, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2805252C2 |
| Молекулы, связывающиеся с 4-1ВВ | 2011 |
|
RU2710717C2 |
| АНТИТЕЛА ПРОТИВ А2 ТЕНАСЦИНА С И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2584597C2 |



















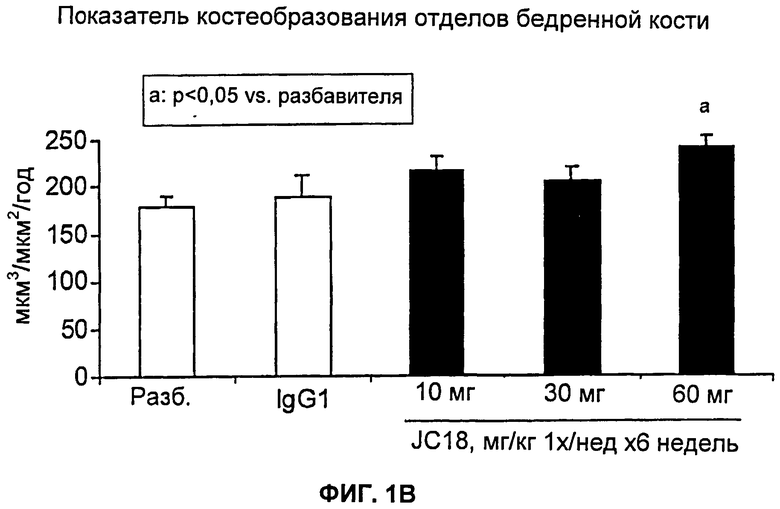

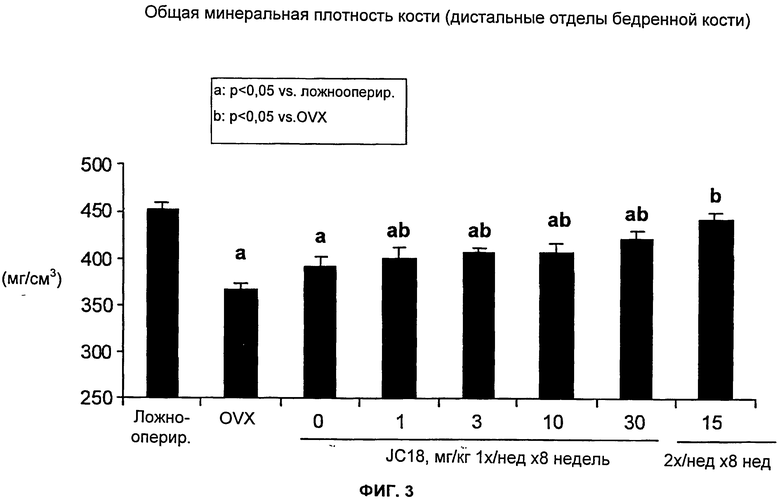
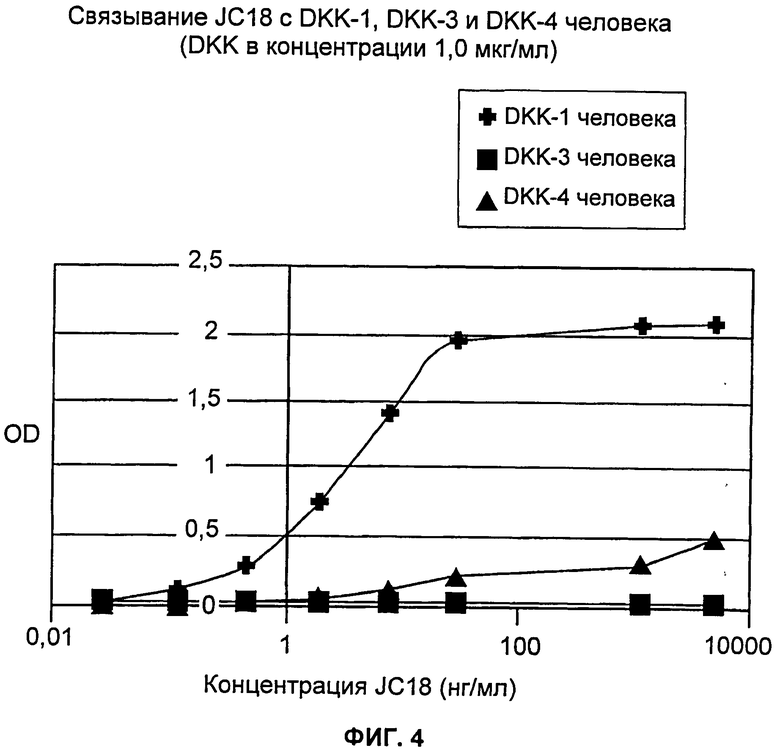
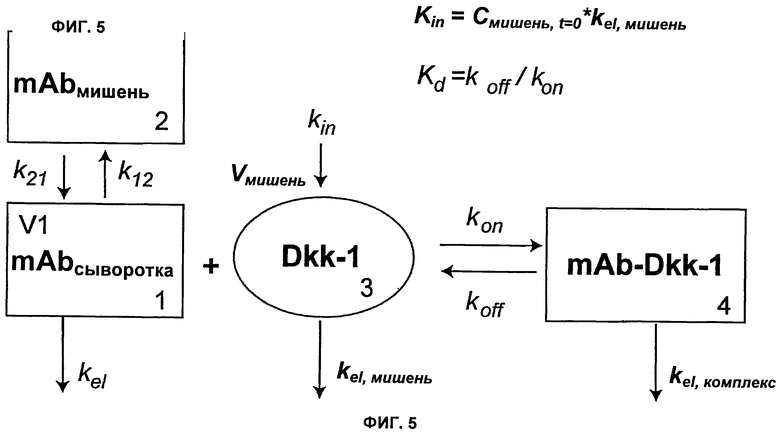
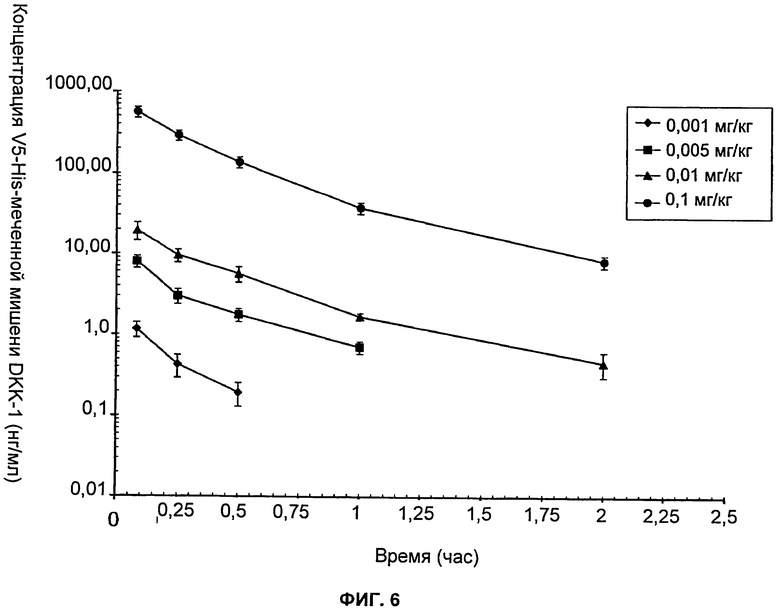
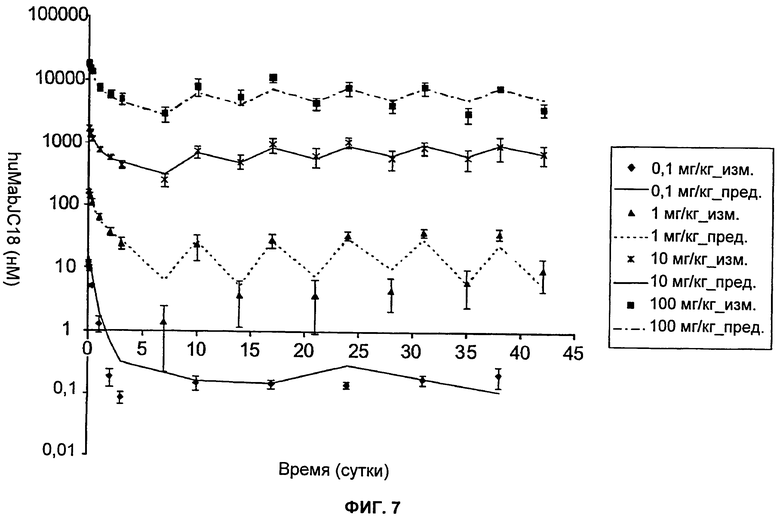
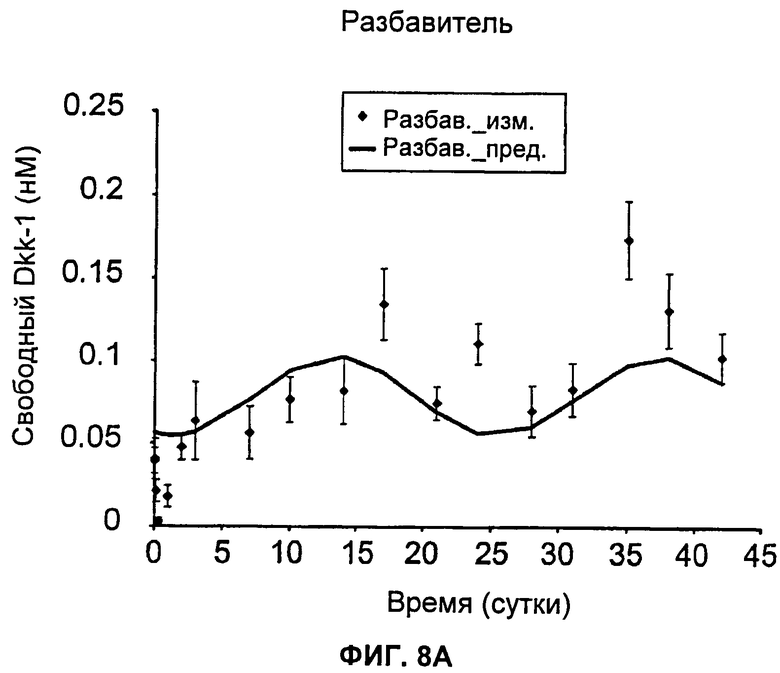
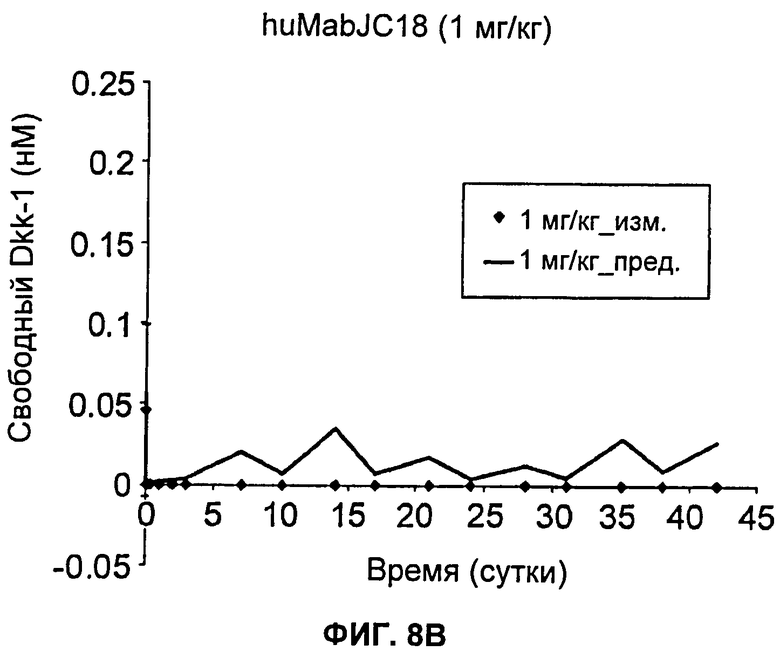
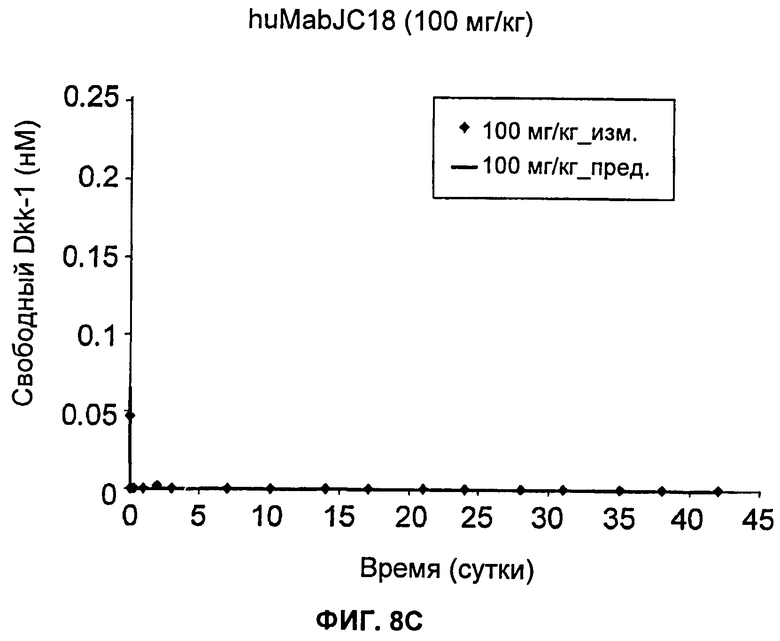
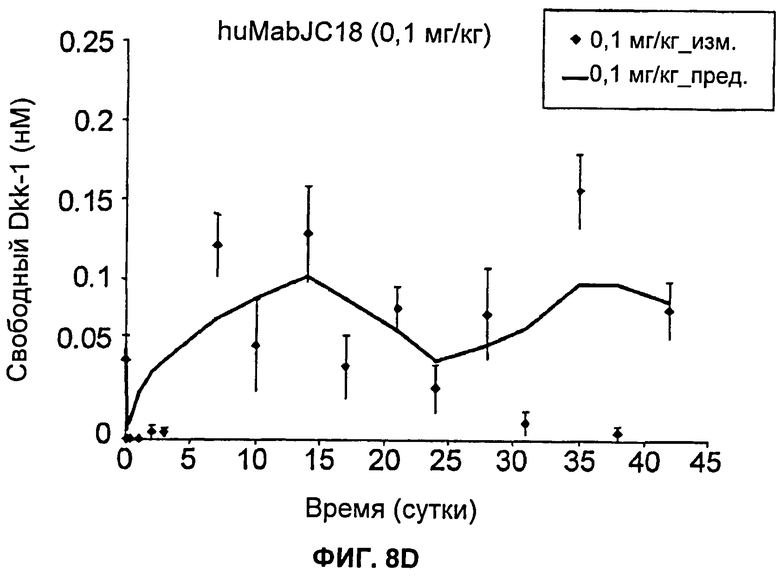
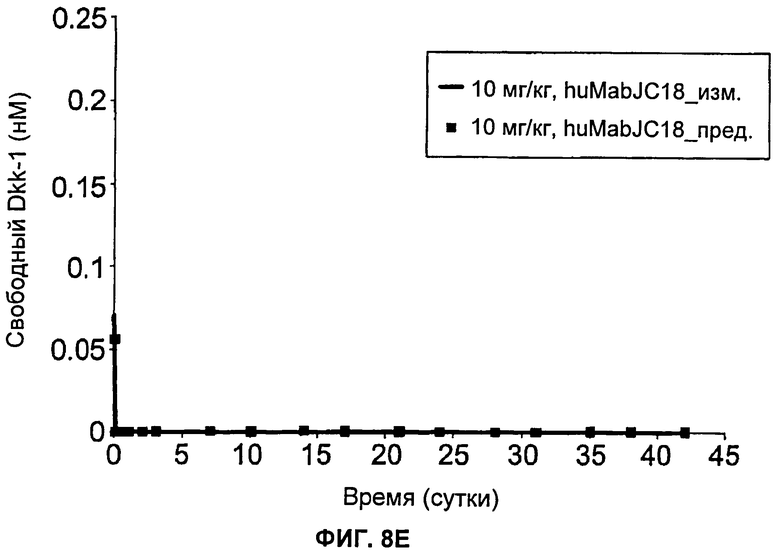
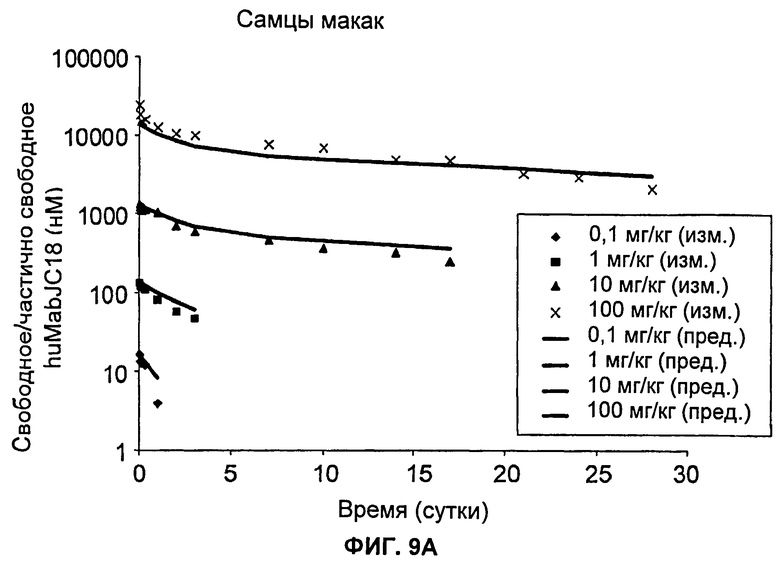
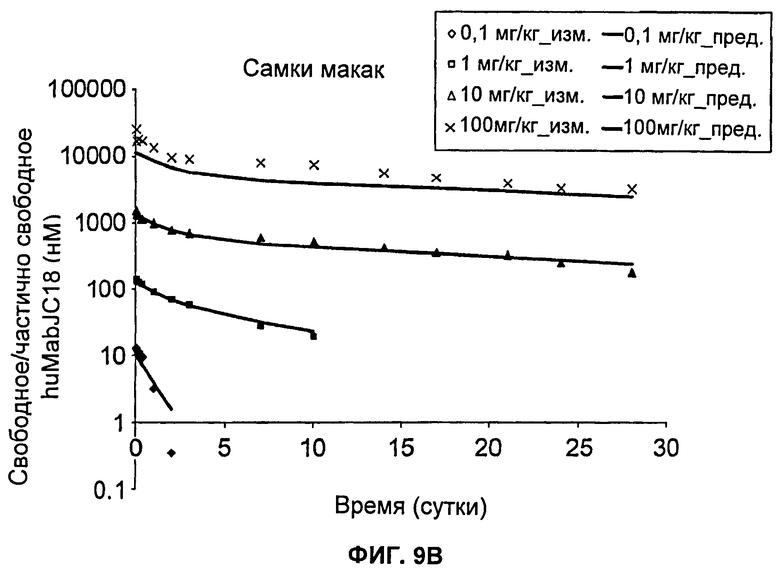

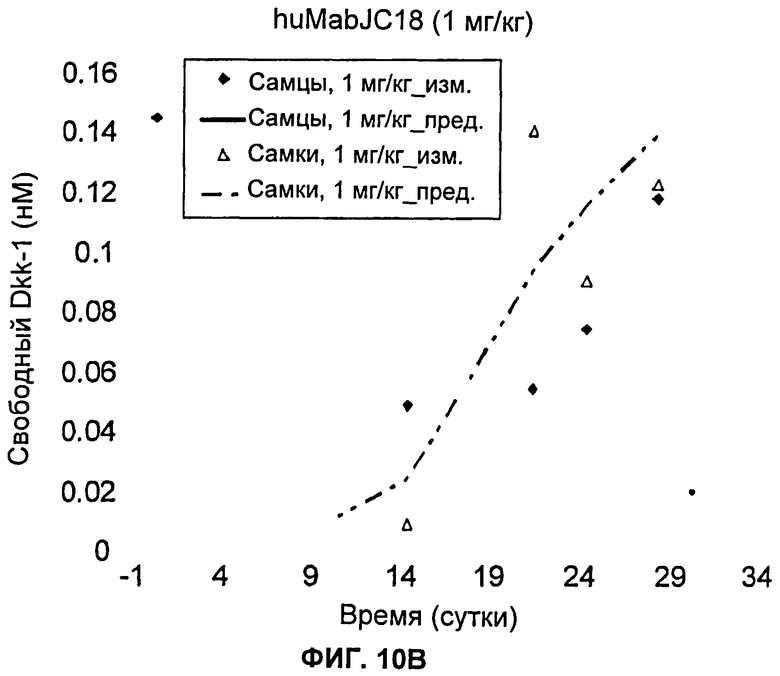
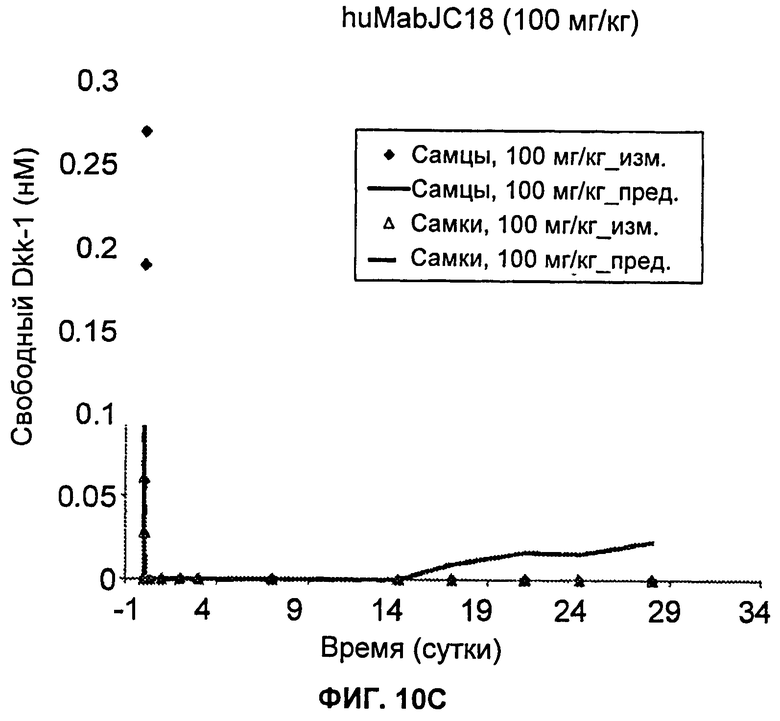
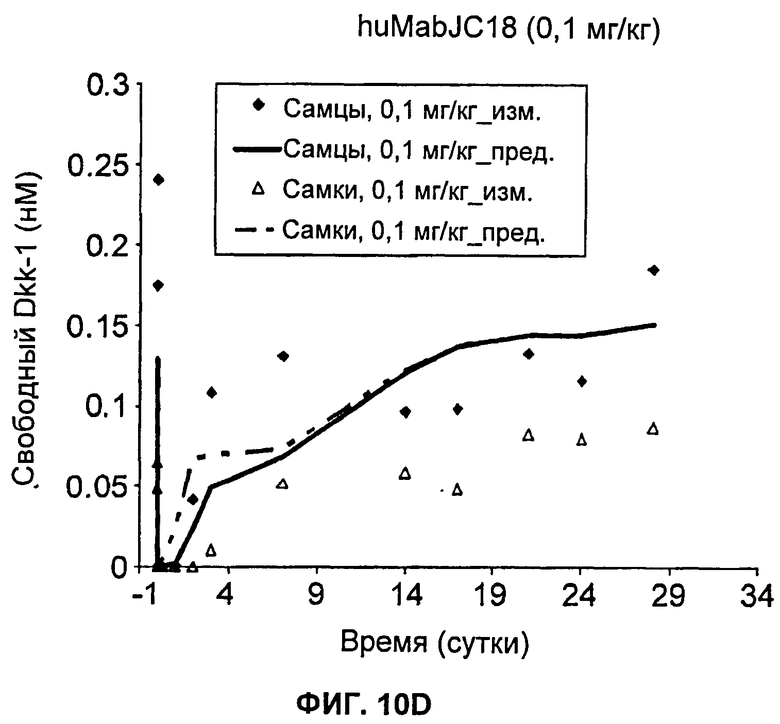
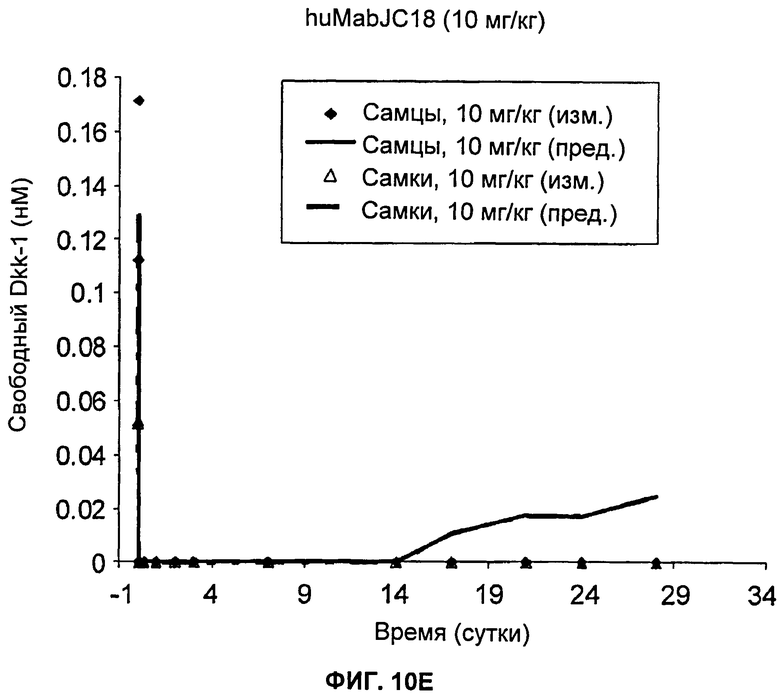

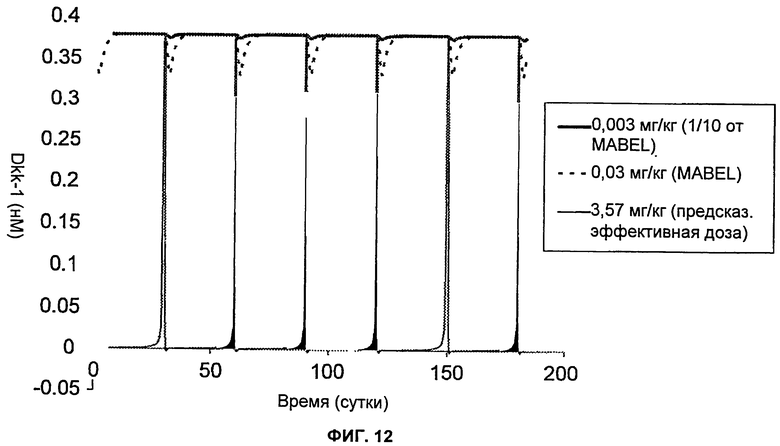
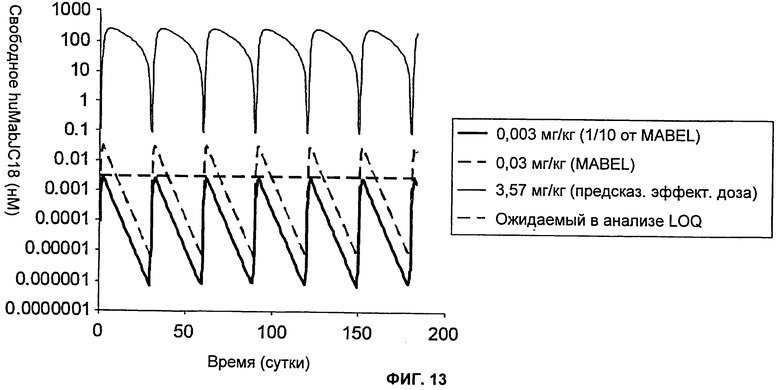
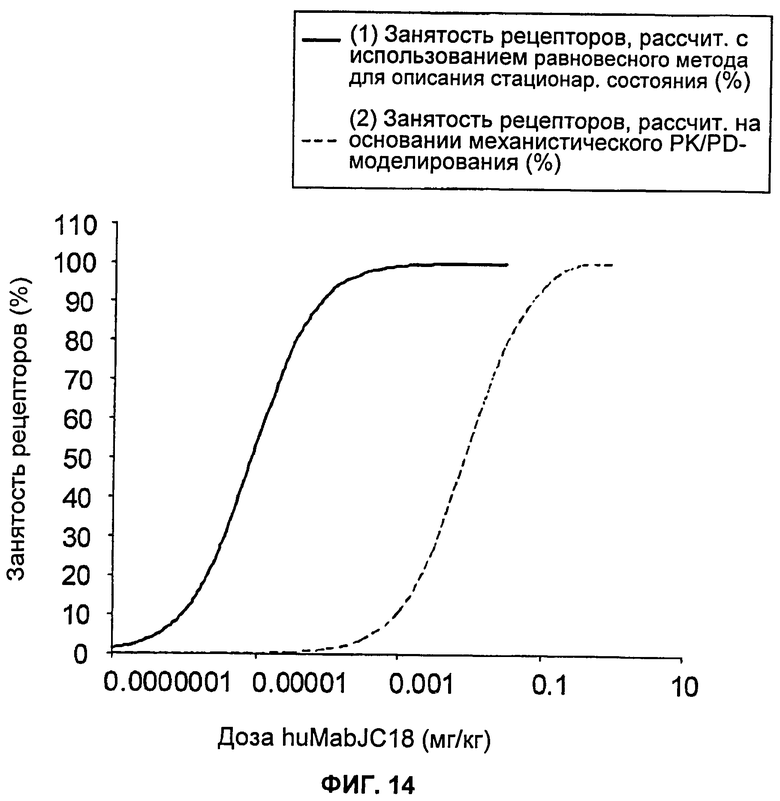
Изобретение относится к области иммунологии. Представлены антитела и их функциональные фрагменты против Dickkopf 1 (Dkk-1), которые выбраны из антител: 1) содержащего CDR1 VH, содержащий аминокислотную последовательность SSYAIS, SYAIS или GFTFSSY; CDR2 VH, содержащий аминокислотную последовательность SVSGTGLGFGTYYPDSVKG или SVSGTGLGFGTY; и CDR3 VH, содержащий аминокислотную последовательность TSLENYAFDY или SLENYAFDY; и CDR1 VL, содержащий аминокислотную последовательность RASESVDDFGISFIN; CDR2 VL, содержащий аминокислотную последовательность AGSKQGS; и CDR3 VL, содержащий аминокислотную последовательность QQLKEVPPT; и 2) антител раскрытых в Таблице 4, содержащейся в материалах заявки. Описаны: нуклеиновые кислоты, кодирующие указанные антитела или их функциональные фрагменты; экспрессирующие векторы, содержащие указанные нуклеиновые кислоты; и клетки для экспрессии указанных антител или функциональных фрагментов, содержащие указанные векторы экспрессии. Предложен способ получения антитела или его функционального фрагмента, включающий стадию культивирования указанной экспрессирующей клетки. Раскрыта композиция, обладающая активностью связывания с Dkk-1, содержащая указанное антитело или его функциональный фрагмент в терапевтически эффективном количестве и фармацевтически приемлемый эксципиент, разбавитель или носитель. Изобретение позволяет расширить арсенал средств для лечения заболеваний, связанных с избыточным взаимодействием Dkk-1 c LRP5/6, которое вызывает активацию Wnt. 7 н. и 7 з.п. ф-лы, 14 ил., 14 табл., 6 пр.
1. Выделенное антитело или его иммунологически функциональный фрагмент, которые специфически связываются с полипептидом Dickkopf 1 (Dkk-1), содержащие
(а) гипервариабельный участок один (CDR1) вариабельной области тяжелой цепи (VH), содержащий аминокислотную последовательность, показанную в SEQ ID NO: 30, 49 или 50; CDR2 VH, содержащий аминокислотную последовательность, показанную в SEQ ID NO: 32 или 51; и CDR3 VH, содержащий аминокислотную последовательность, показанную в SEQ ID NO: 34 или 52; и
(б) CDR1 вариабельной области легкой цепи (VL), содержащий аминокислотную последовательность, показанную в SEQ ID NO: 22; CDR2 VL, содержащий аминокислотную последовательность, показанную в SEQ ID NO: 24; и CDR3 VL, содержащий аминокислотную последовательность, показанную в SEQ ID NO: 26.
2. Антитело или его иммунологически функциональный фрагмент по п. 1, где вариабельная область тяжелой цепи содержит аминокислотную последовательность, показанную в SEQ ID NO: 28, а вариабельная область легкой цепи содержит аминокислотную последовательность, показанную в SEQ ID NO: 20.
3. Антитело или его иммунологически функциональный фрагмент по п. 2, где указанное антитело содержит легкую цепь, содержащую аминокислотную последовательность, показанную в SEQ ID NO: 42 или 38, и тяжелую цепь, содержащую аминокислотную последовательность, показанную в SEQ ID NO: 40 или 36, с С-концевым лизином или без него в SEQ ID NO: 40 или 36.
4. Выделенное антитело или его иммунологически функциональный фрагмент, которые специфически связываются с Dkk-1, содержащие CDR1 VH, CDR2 VH, CDR3 VH, CDR1 VL, CDR2 VL и CDR3 VL, как раскрыто в Таблице 4.
5. Нуклеиновая кислота, кодирующая антитело или иммунологически функциональный фрагмент по любому из пп. 1-3.
6. Экспрессирующий вектор, содержащий нуклеиновую кислоту по п. 5.
7. Выделенная клетка для экспрессии антитела или иммунологически функционального фрагмента по любому из пп. 1-3, содержащая экспрессирующий вектор по п. 6.
8. Способ получения антитела или его иммунологически функционального фрагмента, включающий стадию культивирования клетки по п. 7.
9. Композиция, обладающая активностью связывания с Dkk-1, содержащая антитело или фрагмент по любому из пп. 1-4 в терапевтически эффективном количестве и фармацевтически приемлемый эксципиент, разбавитель или носитель.
10. Антитело или фрагмент по любому из пп. 1-4 для применения в лечении или предупреждении уменьшения костной массы у пациента.
11. Антитело или фрагмент по п. 10, где указанный пациент выбран из пациентов, страдающих раковым заболеванием, дающим метастазы в кости, множественной миеломой, остеопорозом, остеопенией, болезнью Педжета, периодонтитом, ревматоидным артритом и уменьшением костной массы вследствие иммобилизации.
12. Антитело или фрагмент по любому из пп. 1-4 для применения в индуцировании увеличения костной массы у пациента.
13. Антитело или фрагмент по п. 12, где указанный пациент выбран из пациентов, страдающих раковым заболеванием, дающим метастазы в кости, множественной миеломой, остеопорозом, остеопенией, болезнью Педжета, периодонтитом, ревматоидным артритом и уменьшением костной массы вследствие иммобилизации.
14. Антитело или фрагмент по любому из пп. 1-4 для применения в индуцировании активности секретируемых гликопротеинов Wnt у пациента.
| Колосоуборка | 1923 |
|
SU2009A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| O | |||
| AGUILERA, et al., "Epigenetic inactivation of the Wnt antagonist DICKKOPF-1 (DKK-1) gene in human colorectal cancer", Oncogene (2006) 25, pp | |||
| Электролизер с биполярными электродами | 1926 |
|
SU4116A1 |
| Н.М | |||
| ШУБА "Остеопороз - актуальная проблема XXI века: современное представление о патогенезе и терапии", Украхнський ревматологiчний | |||

