Область техники, к которой относится изобретение
Данное изобретение относится к новому терапевтическому агенту для мезотелиомы.
Уровень техники
Мезотелиома представляет собой опухоль, которая встречается в мезотелии, покрывающем поверхность плевры, брюшины и перикарда, которые, соответственно, обволакивают органы грудной полости, такие как легкие и сердце, и брюшные органы, такие как пищеварительный тракт и печень. В случае диффузной плевральной мезотелиомы боль в груди вызывается инвазией межреберных нервов на плевральной стороне грудной стенки, и, вследствие роста опухоли, могут возникать респираторные и циркуляторные нарушения и накопление плевральной жидкости в плевре на этой же стороне органа [Takagi, Journal of Clinical and Experimental Medicine, (March Supplement), «Respiratory Diseases», pp.469-472, 1999]. В конечном счете, происходит пролиферация опухоли в соседние медиастинальные органы, прогрессирующая до прямой инвазии сердца или развития опухоли в брюшную полость через диафрагму, или может быть ее развитие вне грудной полости в результате дополнительного лимфатического или циркуляторного (через кровь) метастазирования (Takagi, там же).
Сообщалось, что в США диффузная плевральная мезотелиома встречается у 3000 субъектов ежегодно, число случаев начало заметно увеличиваться в 1980-ых годах, и часто ее наблюдают у людей в возрасте шестидесяти лет, причем встречаемость у мужчин является приблизительно в пять раз более высокой, чем встречаемость у женщин [Takagi, Journal of Clinical and Experimental Medicine, (March Supplement), «Respiratory Diseases», pp.469-472, 1999]. Согласно недавним сообщениям в США и Европе, встречаемость мезотелиомы демонстрирует тенденцию быстрого увеличения и, на основании эпидемиологической статистики из Великобритании в 1995 году, предсказывается, что количество смертей от мезотелиомы будет продолжать увеличиваться на протяжении следующих 25 лет, а также указывалось, что, в самом плохом из возможных сценариев, существует риск, что она будет ответственной за 1% всех смертей среди людей, родившихся в 1940-ых годах (Nakano, Respiration, Vol.18, №9, pp.916-925, 1999).
Многочисленные различные классификации клинических стадий заболевания использовали для мезотелиомы, и поскольку используемые способы классификации стадии заболевания были различными, предыдущие терапевтические сообщения о мезотелиоме встречались с трудностями для сравнения результатов лечения (Nakano, Respiration, Vol.18, №9, pp.916-925, 1999). Международная классификация TNM (Система клинико-патологического анализа опухолей) для злокачественной плевральной мезотелиомы была разработана в 1995 году International Mesothelioma Interest Group (IMIG) (Nakano, Respiration, Vol.18, №9, pp.916-925,1999).
Кроме того, злокачественная мезотелиома имеет причинную связь с подверганием воздействию асбеста, и это было также продемонстрировано в экспериментах на животных [Tada, Journal of Clinical and Experimental Medicine (March Supplement), «Respiratory Diseases», pp.406-408, 1999]. Асбест, который ингалировали в дыхательные пути, достигает местоположения непосредственно под плеврой, где со временем развивается опухоль вследствие продолжительного раздражения в течение, по меньшей мере, приблизительно 20 лет, и эта опухоль распространяется тонким слоем по всей поверхности плевры [Takagi, Journal of Clinical and Experimental Medicine, (March Supplement), «Respiratory Diseases», pp.469-472, 1999]. Таким образом, хотя злокачественная мезотелиома классифицируется как связанное с асбестом заболевание, не все злокачественные мезотелиомы обусловлены асбестом, и хорошо документированное подвергание воздействию асбеста наблюдается только у приблизительно половины из всех пациентов [Tada, Journal of Clinical and Experimental Medicine (March Supplement), «Respiratory Diseases», pp.406-408,1999].
Злокачественная плевральная мезотелиома является устойчивой к лечению, ассоциирована с крайне плохим прогнозом и требует немедленного принятия контрмер (Nakano, Respiration, Vol.18, №9, pp.916-925, 1999). Например, хотя антагонист фолиевой кислоты, метотрексат (МТХ), имеет удовлетворительную степень эффективности, равную 37%, при лечении единственной большой дозой в комбинации с лейковином, его применение не распространилось вследствие технической трудности, связанной с нанесением на мезотелиому, которое вызывает сохранение большого количества плевральной жидкости [Nakano, Journal of Clinical and Experimental Medicine (March Supplement), «Respiratory Diseases», pp.570-673, 2003]. Кроме того, хотя плевролегочное иссечение и плевроэктомию выполняли для диффузной плевральной мезотелиомы, существует повышенная вероятность возникновения рецидива после такого лечения, и частота послехирургического локального рецидива, в частности, является высокой (35-43%) [Takagi, Journal of Clinical and Experimental Medicine, (March Supplement), «Respiratory Diseases», pp.469-472,1999].
Известно, что многочисленные клеточные линии мезотелиомы человека и несколько клеточных линий мезотелиомы мыши экспрессируют IL-6 т vitro, и сообщалось, что у мышей с трансплантированной им линией клеток АВ22 мышиной мезотелиомы, которая экспрессирует высокий уровень IL-6, IL-6 детектировали в сыворотке перед началом роста раковых клеток, перед клиническими симптомами и изменениями в лимфоцитарной ткани периферической крови (Bielefeldt-Ohmann, Cancer Immunol. Immunother. 40, 241-250, 1995). Кроме того, уровни IL-6 в сыворотке у пациентов со злокачественной плевральной мезотелиомой являются более высокими в сравнении с пациентами с легочной аденомой, осложненной плевральным выпотом, это же относится к тромбоцитозу, который является одним из клинических симптомов злокачественной плевральной мезотелиомы - известно, что существует явная корреляция между уровнями IL-6 в сыворотке и количеством тромбоцитов (Nakano, British Journal of Cancer, 77, №6,: 907-912, 1998). Кроме того, опухолевые клетки пациентов с плевральной мезотелиомой экспрессируют высокие уровни IL-6, и сообщалось, что уровни IL-6 в сыворотке увеличиваются перед смертью (Higashihara, Cancer, Vol.70, №. 8, pp.2105-2108,1992).
Bielefeldt-Ohmann, et al. сообщали, что в результате введения дважды в неделю крысиных антител против мышиного IL-6 (6 В4) мышам с трансплантированной АВ22, наблюдались эффекты, которые значительно уменьшали возникновение и прогрессирование клинических симптомов (Bielefeldt-Ohmann, Cancer Immunol. Immunother. 40, 241-250, 1995). Однако, согласно сообщению Bielefeldt, анти-IL-6 антитело не оказывает прямое ингибирующее рост воздействие на АВ22 in vitro, не наблюдались различия в посмертных аутопсиях (результатах вскрытия) мышей, подвергнутых лечению анти-IL-6 антителом, и мышей, не обработанных указанным антителом, и наблюдались большое число опухолей значительного размера даже у леченых мышей (Bielefeldt-Ohmann, Cancer Immunol. Immunother. 40, 241-250, 1995). To есть, ингибирование роста мезотелиомы анти-IL-6 антителом не было известно ни in vitro, ни in vivo.
Takagi, Journal of Clinical and Experimental Medicine, (March Supplement), «Respiratory Diseases», pp.469-472, 1999
Nakano, Respiration, Vol.18, №9, pp.916-925, 1999
Tada, Journal of Clinical and Experimental Medicine (March Supplement), «Respiratory Diseases», pp.406-408, 1999
Nakano, Journal of Clinical and Experimental Medicine (March Supplement), «Respiratory Diseases», pp.570-673, 2003
Bielefeldt-Ohmann, Cancer Immunol. Immunother. 40,241-250,1995
Higashihara, Cancer, Vol.70, №8, pp.2105-2108,1992.
Раскрытие изобретения
Известно, что антагонисты IL-6 действуют непосредственно на мезотелиому и демонстрируют ингибирующие рост воздействия. Целью данного изобретения является обеспечение нового терапевтического агента для мезотелиомы, который содержит антагонист IL-6 в качестве его активного ингредиента.
В результате проведения интенсивных исследований по разработке нового терапевтического агента для мезотелиомы, который ингибирует рост клеток мезотелиомы, автор данного изобретения сделал новое открытие, что рост клеток мезотелиомы может быть ингибирован с помощью ингибирования или прерывания передачи сигнала, связанного с IL-6, что и привело к реализации данного изобретения.
Таким образом, данное изобретение обеспечивает терапевтический агент для мезотелиомы, который содержит антагонист интерлейкина-6 (IL-6) в качестве его активного ингредиента.
Вышеупомянутой мезотелиомой является, например, плевральная мезотелиома и, более конкретно, злокачественная плевральная мезотелиома. В злокачественную плевральную мезотелиому включена диффузная плевральная мезотелиома.
Вышеупомянутым антагонистом IL-6 являетсяантитело к IL-6-рецептору и, предпочтительно моноклональное антитело к IL-6-рецептору. Вышеупомянутое антитело к IL-6-рецептору является особенно предпочтительно моноклональным антителом к IL-6-рецептору человека, таким как антитело РМ-1, или моноклональным антителом к мышиному IL-6-рецептору, таким как антитело MR16-1. Вышеупомянутым антителом к IL-6-рецептору является предпочтительно рекомбинантное антитело.
Вышеупомянутое антитело к IL-6-рецептору может быть химерным антителом, гуманизированным антителом или антителом человека. В данном изобретении, особенно предпочтительным антителом является гуманизированное антитело РМ-1.
Данное изобретение может также быть в указанных ниже формах:
(1) Применение антитела к рецептору интерлейкина-6 (IL-6) для получения терапевтического агента для мезотелиомы.
(2) Применение по (1) выше, в котором мезотелиома является плевральной мезотелиомой.
(3) Применение по (2) выше, где плевральная мезотелиома является злокачественной плевральной мезотелиомой.
(4) Применение по (1) выше, где антителом к IL-6-рецептору является моноклональное антитело к IL-6-рецептору.
(5) Применение по (1) выше, где антителом к IL-6-рецептору является моноклональное антитело к IL-6-рецептору человека.
(6) Применение по (1) выше, где антителом к IL-6-рецептору является моноклональное антитело к IL-6-рецептору мыши.
(7) Применение по любому из (1)-(5) выше, где антителом к IL-6-рецептору является рекомбинантное антитело.
(8) Применение по (5) выше, где моноклональным антителом к IL-6-рецептору человека является антитело РМ-1.
(9) Применение по (6) выше, где моноклональным антителом к IL-6-рецептору мыши является антитело MR 16-1.
(10) Применение по любому из (4)-(9) выше, где антителом к IL-6-рецептору является химерное антитело, гуманизированное антитело или человеческое антитело к IL-6-рецептору.
(11) Применение по (10) выше, где гуманизированным антителом к IL-6-рецептору является гуманизированное антитело РМ-1.
Данное изобретение может также принимать указанные ниже формы.
(1) Способ лечения мезотелиомы, предусматривающий введение антитела к рецептору интерлейкина-6 (IL-6) субъекту, нуждающемуся в таком лечении.
(2) Способ по (1) выше, где мезотелиома является плевральной мезотелиомой.
(3) Способ по (2) выше, где плевральная мезотелиома является злокачественной плевральной мезотелиомой.
(4) Способ по (1) выше, где антителом к IL-6-рецептору является моноклональное антитело к IL-6-рецептору.
(5) Способ по (1) выше, где антителом к IL-6-рецептору является моноклональное антитело к IL-6-рецептору человека.
(6) Способ по (1) выше, где антителом к IL-6-рецептору является моноклональное антитело к IL-6-рецептору мыши.
(7) Способ по любому из (1)-(5) выше, где антителом к IL-6-рецептору является рекомбинантное антитело.
(8) Способ по (5) выше, где моноклональным антителом к IL-6-рецептору человека является антитело РМ-1.
(9) Способ по (6) выше, где моноклональным антителом к IL-6-рецептору мыши является антитело MR16-1.
(10) Способ по любому из (4)-(9) выше, где антителом к IL-6-рецептору является химерное антитело, гуманизированное антитело или человеческое антитело к IL-6-рецептору.
(11) Способ по (10) выше, где гуманизированным антителом к IL-6-рецептору является гуманизированное антитело РМ-1.
Краткое описание фигур
Фиг.1 является диаграммой, которая отображает результаты примера 1, показывающей продуцирование IL-6 различными клеточными линиями злокачественной мезотелиомы.
Фиг.2 является диаграммой, которая отображает результаты примера 2, показывающей продуцирование (или его отсутствие) IL-6-рецептора (IL-6R) различными клеточными линиями злокачественной мезотелиомы. Кроме того, GAPDH показывает количество мРНК GAPDH (глицеральдегид-3-фосфатдегидрогеназы), используемой в качестве внешнего контроля.
Фиг.3 является диаграммой, которая показывает результаты примера 3, где продуцирование фактора роста эндотелия сосудов (VEGF) клеточными линиями Н2050 (А) и Н2452 (В) злокачественной мезотелиомы индуцируется IL-6 и IL-6R.
Фиг.4 является диаграммой, которая показывает результаты примера 4, в котором эксперимент, сходный с примером 3, проводили для клеточной линии Н28 злокачественной мезотелиомы, показывающей, что эта клеточная линия продуцирует фактор роста эндотелия сосудов (VEGF), не требуя индукции IL-6/IL-6R.
Фиг.5 является диаграммой, которая показывает результаты примера 5, где фосфорилирование STAT3, связанное со стимуляцией IL-6, усиливается в клеточной линии Н2025, в которой продуцирование VEGF индуцируется IL-6, и наоборот, фосфорилирование STAT3, вследствие стимуляции IL-6, не наблюдается в клеточной линии Н28, в которой продуцирование VEGF не индуцируется IL-6.
Фиг.6 является диаграммой, которая показывает результаты примера 6, где в противоположность экспрессии SOCS3 вследствие стимуляции IL-6 и IL-6R, индуцируемой в клеточной линии Н2052, в которой продуцирование VEGF индуцируется IL-6, SOCS3 неиндуктивно экспрессируется в клеточной линии Н28, в которой продуцирование VEGF не индуцируется IL-6.
Фиг.7 является схемой, показывающей структуру промотора плазмид pGL3-VEGF и pGL3-VEGFmut вместе с его окружением, используемого в примере 7.
Фиг.8 является диаграммой, которая показывает результаты примера 7, где в системе, в которой промотор VEGF связан с репортерным геном люциферазы, в случае изменения сайта связывания pSTAT3 в промоторе VEGF, не происходит активация промотора VEGF IL-6.
Фиг.9 является схемой, показывающей механизм индукции промотора VEGF (продуцирования VEGF) вследствие стимуляции IL-6 в клеточной линии Н2052, как предсказано из результатов примеров 5-7.
Фиг.10 является диаграммой, которая показывает результаты примера 8, где рост клеточных линий Н2052 увеличивается в зависимости от концентрации IL-6 в присутствии IL-6R, где график (А) показывает результат в отсутствие растворимого рецептора IL-6, а график (В) показывает результат, полученный в присутствии растворимого рецептора IL-6.
Фиг.11 является диаграммой, которая показывает результаты примера 9, где стимуляция роста клеточной линии Н2052 IL-6 и IL-6R ингибируется MRA (гуманизированным антителом РМ-1).
Фиг.12 является диаграммой, которая показывает результаты примера 10, где рост клеточной линии Н226 увеличивается в зависимости от концентрации IL-6 в присутствии IL-6R и MRA демонстрирует ингибирующее воздействие на этот рост.
Фиг.13 является диаграммой, которая отражает результаты примера 8, показывающие, что рост клеточной линии Н226 увеличивается в зависимости от концентрации IL-6 в присутствии IL-6R.
Фиг.14 является диаграммой, которая показывает результаты примера 11, где стимуляция IL-6 и IL-6R роста клеточных линий злокачественной мезотелиомы Н2052 и Н226 ингибируется гуманизированным антителом РМ-1.
Фиг.15 является диаграммой, которая показывает результаты примера 12, где индукция продуцирования VEGF стимуляцией IL-6 в клеточных линиях злокачественной мезотелиомы MSTO, Н226, Н2052 и Н2452 ингибируется гуманизированным антителом РМ-1.
Фиг.16 является диаграммой, которая показывает результаты примера 13, где фосфорилирование STAT3, индуцируемое стимуляцией IL-6/IL-6R в клеточных линиях Н2052 и Н2452, ингибируется гуманизированным антителом РМ-1.
Фиг.17 является диаграммой, которая показывает результаты примера 14, где активация роста клеточных линий Н2052 и Н226 стимуляцией IL-6 и растворимым IL-6R не ингибируется анти-VEGF антителом.
Осуществление изобретения
IL-6 является цитокином, также называемым стимулирующим В-клетки фактором 2 (BSF2) или интерфероном (32. Первоначально было обнаружено, что IL-6 является фактором дифференцировки, участвующим в активации В-лимфоцитов (Hirano Т. et al.„ Nature, 324, 73-76, 1986), после чего было определено, что он является мультифункциональным цитокином, оказывающим воздействие на функции различных клеток (Akira S. et al.. Adv. In Immunology, 54, 1-78, 1993). Сообщалось также, что IL-6 индуцирует созревание Т-клеток (Lotz M. et al., J. Exp.Med., 167, 1253-1258,1988).
IL-6 реализует свою биологическую активность с использованием различных белков на клетках. Одним из них является лиганд-связывающий белок IL-6-рецепторов, имеющий молекулярную массу приблизительно 80 кД (Taga Т. et al., J. Exp.Med., 166, 967-981, 1987; Yamasaki К. et al.. Science, 241, 825-828, 1987). Кроме существования в мембрано-связанной форме, которая экспрессируется на клеточной мембране проникновением через эту мембрану, IL-6-рецепторы существуют также в виде растворимых IL-6-рецепторов, состоящих в основном из внеклеточного региона.
Другой из этих мембрано-связанных белков gpl30, имеющий молекулярную массу приблизительно 130 кД, участвует в передаче сигнала, не опосредованной связыванием лиганда. IL-6 и IL-6-рецепторы образуют комплекс IL-6/IL-6-penenTop и в результате последующего связывания с gpl30 биологическая активность IL-6 передается в клетки (Taga Т. etal.. Cell, 58, 573-581, 1989).
Антагонистами IL-6 являются вещества, которые ингибируют передачу биологической активности IL-6. Известные примеры этих антагонистов IL-6 включают в себя антитело к IL-6 (анти-IL-6 антитело), антитело к IL-6-рецепторам (анти-IL-6-рецептор антитело), антитело к gpl30 (анти-gp130 антитело), варианты IL-6 и неполные пептиды IL-6 или IL-6-рецептора.
Несколько сообщений появлялись в отношении анти-IL-6-рецептор антитела (Novick D. et al„ Hybridoma, 10, 137-146, 1991; Huang Y.W. et al., Hybridoma, 12, 621-630, 1993; публикация не прошедшего экспертизу Международного патента №WO 95-09873; публикация не прошедшего экспертизу патента Франции №FR 2694767 и патент США №US 521628). Известно, что гуманизированное антитело РМ-1 было получено трансплантацией гипервариабельного участка (CDR, область, определяющая комплементарность) одной из форм мышиного анти-РМ-1 антитела (Hirata Y. et al., J. Immunol., 143, 2900-2906, 1989) в человеческое антитело (публикация не прошедшего экспертизу Международного патента №WO 92-19759).
Вышеупомянутым антагонистом IL-6 является предпочтительно антитело к IL-6-рецептору и более предпочтительно моноклональное антитело к IL-6-рецептору человека или моноклональное антитело к IL-6-рецептору мыши. Примером вышеуказанного моноклонального антитела к IL-6-рецептору человека является антитело РМ-1, тогда как примером моноклонального антитела к IL-6-рецептору мыши является антитело MR 16-1. Вышеупомянутым антителом является предпочтительно химерное антитело, гуманизированное антитело или человеческое антитело, примером которых является гуманизированное антитело РМ-1.
Имеются определенные ограничения в отношении происхождения, типа или формы антагониста IL-6, используемого в данном изобретении, при условии, что он ингибирует рост клеток мезотелиомы и применим в качестве активного ингредиента терапевтического агента для лечения мезотелиомы.
Антагонистами IL-6 являются вещества, которые ингибируют биологическую активность IL-6 прерыванием передачи сигнала IL-6. Антагонистами IL-6 являются предпочтительно вещества, которые оказывают ингибирующее воздействие на связывание IL-6, IL-6-рецептора и gp130. Примеры антагонистов IL-6 включают в себя aHTH-IL-6 антитело, aнти-IL-6-рецептор антитело, анти-gpl30 антитело, варианты IL-6, варианты растворимого IL-6-рецептора, неполные пептиды IL-6-рецептора и низкомолекулярные вещества, демонстрирующие сходную активность.
AHTH-IL-6-рецептор антитело, используемое в данном изобретении, может быть получено в форме поликлонального антитела или моноклонального антитела с использованием известных способов. Моноклональное антитело, происходящее из млекопитающего, является особенно предпочтительным в качестве анти-IL-6-рецептор антитела, используемого в данном изобретении. Примеры моноклонального антитела, происходящего из клеток млекопитающего, включают в себя антитела, продуцируемые гибридомами, и антитела, продуцируемые в хозяине, трансформированном экспрессирующим вектором, который содержит ген антитела, с использованием методов генной инженерии. В результате связывания этого антитела с IL-6-рецепторами, связывание IL-6 с IL-6-рецепторами ингибируется, с прерыванием посредством этого передачи биологической активности IL-6 клеткам.
Примерами таких антител являются антитело MR16-1 (Tamura Т. et al., Proc. Natl. Acad. Sci. USA, 90, 11924-11928, 1993), антитело РМ-1 (Hirata Y. et al., J. Immunol., 143, 2900-2906, 1989), антитело AUK12-20, антитело AUK64-7 и антитело AUK146-15 (публикация Международной патентной заявки №WO 92-19759). Среди них, в качестве такого антитела особенно предпочтительным является антитело РМ-1.
Кроме того, продуцирующая антитело РМ-1 гибридомная клеточная линия была депонирована согласно международному Будапештскому Договору под именем FERM ВР-2998 12 июля 1989 года в Международном Депозитарии запатентованных организмов Национального Института передовой промышленной науки и технологии (Tsukuba Central 6, 1-1-1 Higashi, Tsukuba, Ibaraki, Japan) в виде РМ-1. Кроме того, продуцирующая антитело MR 16-1 гибридомная клеточная линия была депонирована согласно международному Будапештскому Договору под именем FERM ВР-5875 13 марта 1997 года в Международном Депозитарии запатентованных организмов Национального Института передовой промышленной науки и технологии (Tsukuba Central 6, 1-1-1 Higashi, Tsukuba, Ibaraki, Japan) в виде гибридомы крыса-мышь MR16-1.
Продуцирующая моноклональное анти-IL-6R. антитело гибридома может быть в основном получена способом, описанным ниже с использованием известной технологии. А именно, эта гибридома может быть получена с использованием IL-6-рецептора в качестве сенсибилизирующего антигена, иммунизацией им в соответствии с обычными способами иммунизации, слиянием полученных иммуноцитов с известными клетками-хозяевами в соответствии с обычными способами слияния клеток и затем скринингом клеток, которые продуцируют моноклональное антитело, в соответствии с обычными способами скрининга.
Более конкретно, получение анти-1b-6-рецептор антитела может проводиться способом, описанным ниже. Например, IL-6-рецептор человека, используемый в качестве сенсибилизирующего антигена для получения антитела, получают с использованием генной или аминокислотной последовательностей IL-6-рецептора, описанных в публикации Европейской патентной заявки №ЕР 325474, тогда как IL-6-рецептор мыши получают с использованием генной или аминокислотной последовательностей IL-6-рецептора, описанных в публикации патентной заявки Японии, №3-155795.
Имеются два типа белка IL-6-рецептора, состоящие из белка, который экспрессируется на клеточной мембране, и белка, который высвобождается из клеточной мембраны (растворимый IL-6-рецептор) (Yasukawa К. et al., J. Biochem., 108, 673-676, 1990). Растворимый IL-6-рецептора по существу состоит из внеклеточного региона IL-6-рецептора, который связывается с клеточной мембраной, и отличается от мембрано-связанного IL-6-рецептора тем, что он не имеет проникающего через клеточную мембрану региона или проникающего через клеточную мембрану региона и внутриклеточного региона. В качестве белка IL-6-рецептора может быть использован любой IL-6-рецептор при условии, что он может быть использован в качестве сенсибилизирующего антигена для получения анти-IL-6-рецептор антитела, используемого в данном изобретении.
После вставки последовательности гена IL-6-рецептора в известную систему экспрессирующего вектора для трансформации подходящих клеток-хозяев, желаемый IL-6-рецептор очищают известными способами из клеток-хозяев или культурального супернатанта с последующим использованием этого очищенного IL-6-рецептора в качестве сенсибилизирующего антигена. Кроме того, слитый белок, состоящий из белка IL-6-рецептора и другого белка, может быть также использован в качестве сенсибилизирующего антигена.
Escherichia coli (Е. со/г), содержащая плазмиду pIBIBSF2R, которая содержит кДНК, кодирующую IL-6-рецептор человека, была депонирована согласно международному Будапештскому Договору под депозитным номером FERM ВР-2232 9 января 1989 года в Международном Депозитарии запатентованных организмов Национального Института передовой промышленной науки и технологии (Tsukuba Central 6, 1-1-1 Higashi, Tsukuba, Ibaraki, Japan) в виде НВ 101-pIBIBSF2R.
Хотя и нет особых ограничений в отношении млекопитающего, иммунизируемого сенсибилизирующим антигеном, его предпочтительно выбирают с учетом совместимости с клетками-хозяевами, использованными для слияния клеток, обычные примеры млекопитающих включают в себя грызунов, таких как мыши, крысы и хомячки.
Иммунизацию животного сенсибилизирующим антигеном проводят в соответствии с известными способами. В качестве типичного примера такого способа, иммунизацию предпочтительно проводят инъекцией сенсибилизирующего антигена в брюшную полость или под кожу млекопитающего. Более конкретно, суспензию сенсибилизирующего антигена, разведенную подходящим количеством забуференного фосфатом солевого раствора (ЗФР) или физиологического солевого раствора, смешивают с подходящим количеством обычного адъюванта, такого как полный адъювант Фрейнда, необязательно с последующим эмульгированием, и введением несколько раз млекопитающему каждые 4 дня - 21 день. Кроме того, может быть использован подходящий носитель при иммунизации сенсибилизирующим антигеном.
После подтверждения, что иммунизация была проведена таким образом, и уровень антител в сыворотке увеличился до желаемого уровня, иммуноциты извлекают из этого млекопитающего и используют для слияния клеток. Предпочтительным примером иммуноцитов, используемых для слияния клеток, являются клетки селезенки.
Различные известные клеточные линии, такие как P3X6Ag8.653 (Keamey J.F. et al., J. Immunol., 123, 1548-1550, 1979), P3X63Ag8U.l (Current Topics in Microbiology and Imminiology, 81, 1-7, 1978), NS-1 (Kohler G. and Milstein C., Eur. J. Immunol., 6, 511-519, 1976), MPC-11 (Nargulies D.H. et al.. Cell, 8, 405-415, 1976), SP2/0 (Shulman M. et al., Nature, 276,269-270, 1978), F0 (de St. Groth S.F. et al\ J. Immunol. Methods, 35,1-21,1980), S194 (Trowbridge I.S., J. Exp.Med., 148, 313-323, 1978) или R210 (Galfre G. et aL, Nature, Til, 131-133, 1979), уже использовали подходящим образом в качестве клеток миеломы млекопитающего, служащих в качестве других клеток-хозяев, слитых с вышеупомянутыми иммуноцитами.
Слияние вышеупомянутых иммуноцитов и миеломных клеток может в основном проводиться в соответствии с известными способами, такими как способ Milstein et al, (Kohler G. and Milstein С., Methods Enzymol., 73, 3-46, 1981).
Более конкретно, вышеупомянутое слияние клеток проводят, например, в обычной питательной культуральной жидкости в присутствии стимулятора слияния клеток. Примеры стимуляторов слияния, которые используют, включают в себя полиэтиленгликоль (ПЭГ) и вирус Сендай (HVJ) и, необязательно, может быть также добавлено вспомогательное вещество, такое как диметилсульфоксид, для увеличения эффективности слияния.
Соотношение используемых иммуноцитов и миеломных клеток равно предпочтительно 1:10 (миеломные клетки: иммуноциты). Примеры культуральных жидкостей, которые могут быть использованы для вышеупомянутого слияния клеток, включают в себя культуральную жидкость RPMI 1640 для выращивания вышеупомянутых миеломных клеток, культуральную жидкость MEM и другие обычные культуральные жидкости, используемые для этого типа культивирования клеток. Кроме того, сывороточная добавка, такая как фетальная телячья сыворотка (ФТС), может быть также использована в комбинации с вышеупомянутой культуральной жидкостью.
Слияние клеток проводят тщательным смешиванием заранее определенных количеств вышеупомянутых иммуноцитов и миеломных клеток в вышеупомянутой культуральной жидкости, добавлением раствора ПЭГ, такого как раствор ПЭГ, имеющего среднюю молекулярную массу приблизительно 1000-6000, предварительно нагретого до 37°С, обычно при концентрации 30-60% (масса/объем), и затем перемешиванием для образования слитых клеток (гибридом). Далее, агенты слияния клеток и т.д., которые могут быть вредными для развития гибридомы, могут быть удалены повторением процедуры, состоящей из последовательного добавления подходящей культуральной жидкости и затем центрифугирования для удаления супернатанта.
Гибридому отбирают культивированием в обычной селективной культуральной жидкости, такой как культуральная жидкость HAT (культуральная жидкость, содержащая гипоксантин, аминоптерин и тимидин). Культивирование в указанной культуральной жидкости HAT продолжают в течение периода времени, который составляет обычно несколько дней - несколько недель, который является достаточным для разрушения клеток, не являющихся гибридомой-мишенью (неслитых клеток). Затем гибридому, которая продуцирует антитело-мишень, подвергают скринингу и клонируют проведением обычных способов конечного разведения.
Наряду с получением вышеупомянутой гибридомы с помощью иммунизации антигеном животных, отличающихся от человека, желаемое человеческое антитело, имеющее связывающую активность в отношении желаемого антигена или антиген-экспрессирующих клеток, может быть получено сенсибилизацией лимфоцитов человека желаемым белком-антигеном или антиген-экспрессирующими клетками in vitro, и затем слиянием сенсибилизированных В-лимфоцитов с миеломными клетками, такими как U266 (см. публикацию патентной заявки Японии №1-59878). Кроме того, желаемое человеческое антитело может быть также получено введением антигена или антиген-экспрессирующих клеток в трансгенное животное, имеющее набор генов антител человека (см. международные публикации патентных заявок с номерами WO 93/12227, WO 92/03918, WO 94/02602, WO 94/25585, WO 96/34096 или WO 96/33735).
Гибридома, которая продуцирует моноклональное антитело, полученная таким образом, может субкультивироваться в обычной культуральной жидкости и храниться в жидком азоте в течение продолжительного периода времени.
Способ, в котором гибридому культивируют в соответствии с обычными способами и получают в форме культурального супернатанта, или способ, в котором гибридому выращивают введением млекопитающему, который совместим с ней, с последующим получением формы асцитов, может быть использован для получения моноклонального антитела из этой гибридомы. Первый способ является более подходящим для получения антитела с высокой чистотой, тогда как последний способ является подходящим для получения антитела в большом объеме.
Например, получение продуцирующей анти-IL-6-рецептор антитело гибридомы может проводиться в соответствии со способом, описанным в публикации патентной заявки Японии №3-139293. Оно может проводиться с использованием способа, в котором РМ-1-продуцирующую гибридому, депонированную согласно международному Будапештскому Договору под депозитным номером FERM ВР-2998 12 июля 1989 года в Международном Депозитарии запатентованных организмов Национального Института передовой промышленной науки и технологии (Tsukuba Central 6, 1-1-1 Higashi, Tsukuba, Ibaraki, Japan), инъецируют в брюшную полость мышей Balb/c для получения асцитов с последующей очисткой антитела РМ-1 из этих асцитов, или способа, в котором указанную гибридому культивируют в среде, такой как 10% фетальная телячья сыворотка, среда RPMI 1640, содержащая 5% BM-Condimed HI (Boehringer Mannheim), гибридомная среда SFM (Gibco-BRL) или среда PFHM-11 (Gibco-BRL), с последующей очисткой антитела РМ-1 из культурального супернатанта.
Рекомбинантное антитело, полученное с использованием технологии рекомбинации генов - клонированном гена антитела из гибридомы, включением его в подходящий вектор и введения в хозяина - может быть использовано в качестве моноклонального антитела в данном изобретении (см., например, Borrebaeck C.A.K. and Larrick J.W., Therapeutic Monoclonal Antibodies, publishing in the United Kingdom by MacMillan Publishers Ltd., 1990).
Более конкретно, мРНК, которая кодирует вариабельную (V) область антитела, выделяют из клеток, таких как гибридомные клетки, которые продуцируют желаемое антитело. Выделение мРНК проводят с помощью получения вначале тотальной РНК в соответствии с известными способами, такими как гуанидин-центрифугирование (Chirgwin J.M. et al., Biochemistry, 18, 5294-5299, 1979) или AGPC (Chomczynski P. et al., Anal. Biochem., 162, 156-159, 1987), из которой затем получают фракцию мРНК с использованием набора для очистки мРНК (Pharmacia) и т.д. Кроме того, мРНК может быть получена непосредственно с использованием набора для очистки мРНК QuickPrep (Pharmacia).
кДНК вариабельной (V) области антитела синтезируют из полученной мРНК с использованием обратной транскриптазы. Синтез кДНК может проводиться с использованием, например, набора AMV Reverse Transcriptase First-Strand cDNA Synthesis Kit. Кроме того, набор 5'-Ampli Finder Race Kit (Clontech) и 5'-RACE-cnoco6 с использованием ПЦР (Frohman M.A. et al., Proc. Natl. Acad. Sci. USA, 85, 8998-9002,1988) могут быть использованы для синтеза и амплификации кДНК. Желаемый ДНК-фрагмент очищают из полученного ПЦР-продукта и связывают с ДНК вектора. Далее, из этого связанного продукта получают рекомбинантный вектор и вводят его в Е. coli и т.д. с последующим отбором колонии (клона) для получения желаемого рекомбинантного вектора. Последовательность оснований этой желаемой ДНК подтверждают затем известным способом секвенирования, таким как дезокси-способ.
После получения ДНК, которая кодирует V-область желаемого антитела, ее связывают с ДНК, которая кодирует константную область желаемого антитела (С-область), с последующим включением в экспрессирующий вектор. Альтернативно, ДНК, кодирующая V-область этого антитела, может быть включена в экспрессирующий вектор, который содержит ДНК С-области этого антитела.
Для получения антитела, используемого в данном изобретении, ген антитела, который будет описан позднее, включают в экспрессирующий вектор под контроль, например, энхансера или промотора. Затем, клетки-хозяина трансформируют этим экспрессирующим вектором для создания возможности экспрессии этого антитела.
В данном изобретении, может быть использовано полученное из рекомбинантного гена антитело, которое было искусственно изменено с целью снижения межвидовой антигенности в отношении человека, примерами которого являются химерное антитело, гуманизированное антитело и человеческое антитело. Эти измененные антитела могут быть получены с использованием известных способов.
Химерные антитела получают связыванием ДНК, кодирующей V-область антитела, полученной, как описано выше, с ДНК, кодирующей С-область антитела человека, с последующим включением этого связанного продукта в экспрессирующий вектор и продуцированием посредством встраивания в хозяина (см. публикацию Европейской патентной заявки №ЕР 125023 или публикацию Международной патентной заявки WO 92-19759). Химерное антитело, которое применимо в данном изобретении, может быть получено с использованием этого известного способа.
Например, плазмиды, содержащие ДНК, которые кодируют V-области L-цепи и Н-цепи химерного антитела РМ-1, были названы pPM-k3 и pPM-hl, соответственно, и Escherichia coli, сохраняющая эти плазмиды, была депонированы на основании международного Будапештского Договора под названиями NCIMB40366 и NCIMB40362, соответственно, 12 февраля 1991 года в Национальных Коллекциях Industrial and Marine Bacteria Limited (23 St Machar Drive, Aberdeen AB2 1RY, Scotland, Commonwealth of Great Britain and Northern Ireland).
Гуманизированные антитела, которые также называют реконструированными антителами, получают трансплантацией гипервариабельного участка (CDR) антитела млекопитающих, не являющихся человеком, таких как мыши, в соответствующую область антитела человека, такие способы обычной рекомбинации генов являются известными (см. публикацию Европейской патентной заявки №ЕР 125023 или публикацию Международной патентной заявки WO 92-19759).
Более конкретно, ДНК-последовательность, сконструированную таким образом, что она связывает CDR антитела мыши с каркасным участком (FR, framework region) вариабельной области антитела человека, синтезируют при помощи ПЦР с множества олигонуклеотидов, продуцируемых таким образом, чтобы они имели перекрывающиеся части на своих концах. Затем полученную ДНК связывают с ДНК, кодирующей С-область антитела человека, с последующим включением в экспрессирующий вектор и встраиванием в хозяина для получения этой ДНК в данном хозяине (см. публикацию Европейской патентной заявки №EU 239400 или публикацию Международной патентной заявки №WO 92-19759.
FR, с которым гипервариабельная область образует удовлетворительный сайт связывания антигена, выбирается из FR антитела человека, связанного посредством CDR. Аминокислоты каркасного района вариабельной области антитела могут быть заменены таким образом, что гипервариабельная область этого реконструированного антитела человека образует подходящий сайт связывания антигена (Sato К.. et al., Cancer Res, 53, 851-856, 1993).
С-область антитела человека используют для химерного антитела и гуманизированного антитела. Примером С-области человека является Сγ, и, например, могут быть использованы Сγ1, Сγ2, Сγ3 или Сγ4. Кроме того, С-область антитела человека может быть модифицирована для улучшения стабильности антитела или его продуцирования.
Химерные антитела состоят из вариабельной области антитела, происходящей из млекопитающего, отличающегося от человека, и константной (С) области, происходящей из антитела человека, тогда как гуманизированные антитела состоят из гипервариабельной области антитела, происходящего из млекопитающего, отличающегося от человека, и каркасного участка и С-области, происходящих из антитела человека, и, поскольку их антигенность в человеке уменьшается, они применимы в качестве антител, используемых в данном изобретении.
Предпочтительным конкретным примером гуманизированного антитела, используемого в данном изобретении, является гуманизированное антитело РМ-1 (см. публикацию Международной патентной заявки №WO 92-19759).
Наряду с описанным ранее способом, другой способ получения антитела человека состоит в получении антитела человека при помощи пэннинга (panning technique -метод сортировки антител на колонке) с использованием библиотеки антител человека. Например, фаг, который связывается с антигеном, может быть также выбран с использованием вариабельной области антитела человека в виде одноцепочечного антитела (scFv) и экспрессией на поверхности фага с использованием метода фагового дисплея (phage display). Затем ДНК-последовательность, которая кодирует вариабельную область антитела человека, которая связывается с этим антигеном, может быть определена анализом генов отобранного фага. После идентификации ДНК-последовательности этого scFv, которая связывается с антигеном, может быть получен экспрессирующий вектор, который эквивалентен этой последовательности, для получения антитела человека. Эти способы являются уже общеизвестными, и может быть сделана ссылка на WO 92/01047, WO 92/20791, WO 93/06213, WO 93/11236, WO 93/19172, WO 95/01438 и WO 95/15388.
Ген антитела, сконструированный описанным выше способом, может быть экспрессирован и получен известными способами. В случае клеток млекопитающих, ген антитела может быть экспрессирован ДНК или вектором, который содержит указанную ДНК, в котором функционально связаны обычно используемый применимый промотор, ген антитела, который должен быть получен, и сигнал полиаденилирования на 3'-конце. Примером промотора/энхансера является ранний промотор/энхансер цитомегаловируса человека.
Кроме того, примеры других промоторов/энхансеров, которые могли бы использоваться для экспрессии антитела, используемого в данном изобретении, включают в себя промоторы/энхансеры вирусов, такие как промоторы/энхансеры ретровируса, полиомавируса, аденовируса или вируса 40 обезьяны (SV40), а также промоторы/энхансеры, происходящие из клеток млекопитающих, такие как фактор элонгации 1а человека (HEFIa).
Например, в случае использования промотора/энхансера SV40, антитело может быть легко экспрессировано в соответствии со способом Mulligan et al., (Mulligan R.C. et al.. Nature, 277, 108-114, 1979), или в случае промотора/энхансера HEFIa, антитело может быть легко экспрессировано в соответствии со способом Mizushima et al., (Mizushima S. and Nagata S., Nucleic Acids Res., 18, 5322,1990).
В случае Escherichia coli, антитело может быть экспрессировано функциональным связыванием обычно используемого применимого промотора с сигнальной последовательностью для секреции антитела и геном антитела, который должен быть экспрессирован. Примеры промоторов включают в себя промотор lacZ и промотор агаВ. В случае применения промотора lacZ, антитело должно экспрессироваться в соответствии со способом Ward et al., (Ward E.S. et al.. Nature, 341, 544-566, 1989; Ward E.S. et al., FASEB J., 6, 2422-2447, 1992), или, в случае использования промотора агаВ, антитело должно экспрессироваться в соответствии со способом Better et al., (Better M. et al.. Science, 240, 1041-1043, 1988).
Сигнальная последовательность pelB (Lei S.P. et al., J, Bacterial., 169, 4379-4383, 1987) должна быть использована как сигнальная последовательность для секреции антитела в случае его продуцирования в периплазме Е. coli. После отделения антитела, продуцированного в периплазме, это антитело используют для подходящего рефолдинга (повторного сворачивания) структуры антитела (см., например, WO 96/30394).
Примеры источников репликации, которые могут быть использованы, включают в себя источники репликации, происходящие из SV40, полиомавируса, аденовируса или папилломавируса коров (BPV), а для увеличения числа копий гена в системе клетки-хозяина этот экспрессирующий вектор может содержать селективный маркер, такой как ген аминогликозидфосфотрансферазы (АРН), ген тимидинкиназы (ТК), ген ксантин-гуанинфосфорибозилкиназы (Ecogpt) или ген дигидрофолатредуктазы (dhfr).
Любая произвольная система продуцирования может быть использована для получения антитела в данном изобретении. Эта система продуцирования для получения антитела может быть системой in vitro или системой in vivo. Примеры систем продуцирования in vitro включают в себя системы продуцирования, которые используют эукариотические клетки, и системы продуцирования, которые используют прокариотические клетки.
В случае использования эукариотических клеток, эта система продуцирования может использовать клетки животных, клетки растений или клетки грибов. Известные примеры клеток животных включают в себя: (1) клетки млекопитающих, такие как СНО, COS, миеломные клетки, ВНК (клетки почки детеныша хомячка) клетки HeLa и Vero, (2) клетки земноводных, такие как фолликулярные клетки африканской древесницы, и (3) клетки насекомых, такие как клетки sf9, sf21 и Тп5. Известные примеры клеток растений включают в себя клетки, происходящие из Nicotiana tabacum, и они должны культивироваться в каллусах. Известные примеры клеток грибов включают в себя клетки дрожжей, такие как виды Saccharomyces, в том числе Saccharomyces cerevisiae, и плесени, такие как виды Aspergillus, в том числе Aspergillus niger.
В случае использования прокариотических клеток, применяют систему продуцирования, которая использует бактериальные клетки. Известные примеры бактериальных клеток включают в себя Е. coli и Bacillus subtilis.
Антитело получают встраиванием гена желаемого антитела в эти клетки трансформацией, и затем трансформированные клетки культивируют in vitro. Культивирование проводят в соответствии с известными способами. Например, для культуральной жидкости могут быть использованы DMEM, MEM, RPMI 1640 или IMDM, в сывороточная добавка, такая как фетальная телячья сыворотка (ФТС), может быть также использована в комбинации. Кроме того, антитело может быть получено in vivo перенесением клеток, в которые был вставлен ген антитела, в брюшную полость и т.д. животного.
С другой стороны, примеры продуцирования in vivo включают в себя системы продуцирования, которые используют животных, и системы продуцирования, которые используют растения. В случае применения животного, примеры систем продуцирования включают в себя системы продуцирования с использованием млекопитающих или насекомых.
Примеры млекопитающих, которые могут быть использованы, включают в себя коз, свиней, овец, мышей и кроликов (Vicki Glaser, SPECTRUM Biotechnology Applications, 1993). Тутовый шелкопряд может быть использован в случае насекомых. В случае применения растений, могут быть, например, использованы растения табака.
Ген антитела вводят в этих животных или в эти растения с последующим продуцированием и извлечением антитела, продуцированного в телах животных и в растениях. Например, ген антитела вставляют в промежуточном местоположении в ген, который кодирует белок, уникально продуцируемый в молоке, такой как [3-казеин козы, для получения в форме слитого белка. ДНК-фрагмент, который кодирует этот слитый белок, в который был вставлен ген антитела, инъецируют затем в эмбрион козы, и затем этот эмбрион козы вводят в самку козы. Затем желаемое антитело получают из молока, продуцируемого трансгенной козой или ее потомством, которое родилось от козы, получившей этот эмбрион. Может быть использован подходящий гормон в трансгенной козе для увеличения количества молока, содержащего желаемое антитело, продуцируемое трансгенной козой (Ebert K.M. et al., Bio/Technology, 12, 699-702, 1994).
Кроме того, в случае применения тутового шелкопряда, его инфицируют бакуловирусом, в который был вставлен ген желаемого антитела, с последующим получением желаемого антитела из жидкости тела тутового шелкопряда (Maeda S. et al., Nature, 315, 592-594, 1985). Кроме того, в случае применения растения табака, ген желаемого антитела вставляют в экспрессирующий вектор растений, такой как pMON 530, после чего этот вектор вводят в бактерии, такие как Agrobacterium tumefaciens. Затем эту бактерию используют для инфицирования растения табака, такого как Nicotiana tabacum, для получения желаемого антитела из листьев табака (Jilian K. -C. Ма, et al., Eur. J. Immunol., 24, 131-138, 1994).
Как было описано выше, в случае продуцирования антитела системами продуцирования in vitro или in vivo, хозяин может быть одновременно трансфецирован раздельным включением ДНК, кодирующей тяжелую цепь (Н-цепь) или легкую цепь (L-цепь), в отдельных экспрессирующих векторах, или трансформацией хозяина включением ДНК, кодирующей Н-цепь и L-цепь, в едином экспрессирующем векторе (см. публикацию Международной патентной заявки №WO 94-11523).
Антителом, используемым в данном изобретении, может быть фрагмент антитела или его модифицированный продукт, при условии, что он может быть предпочтительно использован в данном изобретении. Примеры фрагментов антител включают в себя Fab, F(ab')2, Fv- или Н-цепь и одноцепочечный Fv (scFv), в котором Fv или Fv- или Н-цепь и L-цепь связаны подходящим линкером.
Более конкретно, после образования фрагмента антитела обработкой антитела ферментом, таким как папаин или пепсин, или конструированием гена, который кодирует эти фрагменты антител, и вставки его в экспрессирующий вектор, он экспрессируется в подходящих клетках-хозяевах (см., например, Со М. S. et al., J. Immunol., 152, 2968-2976, 1994; Better М. & Horwitz A.H., Methods in Enzymology, 178, 476-496, 1989; Plueckthun A. & Skerra A., Methods in Enzymology, 178, 476-496, 1989; Lamoyi E., Methods in Enzymology, 121, 652-663, 1989; Rousseaux J. et al., Methods in Enzymology, 121, 663-666, 1989; BirdR.E. etal., TIBTECH, 9, 132-137, 1991).
Вышеупомянутый scFv получают связыванием V-области Н-цепи и V-области L-цепи линкером и предпочтительно пептидным линкером (Huston J.S. et al., Proc. Natl. Acad. Sci. USA, 85, 5879-5883, 1988). V-область Н-цепи и V-область L-цепи в scFv могут иметь любые происхождения, описанные для вышеупомянутых антител. Примером пептидного линкера, используемого для связывания V-областей, является произвольный одноцепочечный пептид, состоящий из 12-19 аминокислотных остатков.
ДНК, которая кодирует scFv, получают с использованием ДНК, кодирующей вышеупомянутые Н-цепь или V-область Н-цепи антитела, и ДНК, кодирующей L-цепь или V-область L-цепи, в качестве матриц, амплификации ДНК-части, кодирующей желаемую аминокислотную последовательность среди этих последовательностей, при помощи ПЦР с использованием пары праймеров, которая определяет оба конца, и затем амплификации в комбинации с парой праймеров, которая определяет ДНК, кодирующую пептидную линкерную часть, а также оба ее конца, таким образом, что каждая Н-цепь и L-цепь является связанной.
Кроме того, после получения ДНК, которые кодируют scFv, могут быть получены экспрессирующие векторы, которые содержат их, а также хозяева, которые были трансформированы указанными экспрессирующими векторами, в соответствии с обычными способами. Кроме того, scFv могут быть получены в соответствии с обычными способами с использованием этих хозяев.
Эти фрагменты антител могут быть получены из хозяина получением и экспрессией их генов, как это описано ранее. Термин «антитело», используемый в данном изобретении, включает в себя и эти фрагменты антител.
Антитела, связанные с различными типами молекул, такими как полиэтиленгликоль (ПЭГ), могут быть использованы для получения модифицированных продуктов антител. Термин «антитело», используемый в данном изобретении, включает в себя и эти модифицированные продукты антител. Эти модифицированные продукты антител могут быть получены проведением химической модификации полученных антител. Эти способы уже были разработаны в данной области.
Как было описано ранее, экспрессируемое антитело может быть отделено от внутренней части и от наружной части клеток или выделено из хозяина и очищено до гомогенности. Отделение и очистка антитела, используемая в данном изобретении, может проводиться аффинной хроматографией. Примеры колонок, используемых для аффинной хроматографии, включают в себя белок А-колонку и белок G-колонку. Примеры носителей, используемых для белок А-колонки, включают в себя Hyper D, POROS и Sepharose F.F. Другие способы отделения и очистки, используемые с обычными белками, могут быть также использованы, и в отношении этих способов нет ограничений.
Антитело, используемое в данном изобретении, может быть выделено и очищено подходящим выбором и объединением, например, хроматографии, другой, чем вышеупомянутая аффинная хроматография, фильтрования, ультрафильтрации, высаливания или диализа. Примеры других типов хроматографии включают в себя ионообменную хроматографию, гидрофобную хроматографию и гель-фильтрацию. Эти типы хроматографии могут быть применены к высокоэффективной жидкостной хроматографии (ВЖХ). Кроме того, может быть также использована обращенно-фазовая ВЖХ.
Концентрация антитела, полученного, как описано выше, может быть измерена с помощью измерения оптической плотности, ELISA и т.д. А именно, в случае измерения оптической плотности, после достаточного разведения в ЗФР(-), оптическую плотность измеряют при 280 нм с последующим расчетом концентрации на основе того, что OD 1,35 соответствует 1 мг/мл. Кроме того, измерение концентрации при помощи ELISA может проводиться способом, описанным ниже. А именно, 100 мкл козьего антитела против IgG человека (TAG), разведенного до 1 мкг/мл 0,1 М бикарбонатным буфером (рН 9,6), добавляют в 96-луночный планшет (Nimc) с последующим инкубированием в течение ночи при 4°С для иммобилизации этого антитела на планшете. После блокирования добавляют 100 мкл подходящим образом разведенного антитела, используемого в данном изобретении, пробу, содержащую антитело, или стандарт IgG человека (Cappel), с последующим инкубированием в течение 1 часа при комнатной температуре.
После промывания этого планшета, добавляют 100 мкл разбавленного в 5000 раз меченого щелочной фосфатазой антитела против IgG человека- (BIOSOURCE) с последующим инкубированием в течение 1 часа при комнатной температуре. После промывания планшета, добавляют раствор субстрата с последующим инкубированием и затем измерением оптической плотности при 405 нм с использованием Microplate Reader модели 3550 (BioRad) для расчета концентрации желаемого антитела.
Вариант IL-6, используемый в данном изобретении, является веществом, которое имеет связывающую активность с IL-6-рецептором, но не передает биологическую активность IL-6. То есть, хотя вариант IL-6 конкурирует с IL-6 за связывание с IL-6-рецептором, передача сигнала IL-6 прерывается, так как вариант IL-6 не передает биологическую активность IL-6.
Вариант IL-6 получают введением мутации, приводящей к замене аминокислотного остатка в аминокислотной последовательности IL-6. Хотя нет ограничений в отношении происхождения IL-6, служащего в качестве основы варианта IL-6, IL-6 человека является предпочтительным в отношении антигенности и т.д.
Более конкретно, это выполняют предсказанием вторичной структуры аминокислотной последовательности IL-6 с использованием известной программы молекулярного моделирования, такой как WHATIF (Vriend et al., J. Mol. Graphics, 8, 52-56, 1990) и затем оцениванием эффектов для всех аминокислотных остатков, которые должны быть заменены. После определения подходящего замененного аминокислотного остатка, с использованием затем вектора, содержащего последовательность оснований, которая кодирует ген IL-6, в качестве матрицы, и введением мутации таким образом, что эта аминокислота заменяется, при помощи ПЦР, проводимой обычным образом, получают ген, который кодирует вариант IL-6. Он может быть затем включен в подходящий экспрессирующий вектор, необязательно, с получением варианта IL-6 в соответствии с вышеупомянутыми способами экспрессии рекомбинантного антитела, получения и очистки.
Конкретные примеры вариантов IL-6 описаны в Brakenhoff et al., J. Biol. Chem., 269, 86-93,1994; Savino et al., EMBOJ., 13,1357-1367, 1994; WO 96-18648 и WO-96-17869.
Неполный пептид IL-6 или неполный пептид IL-6-рецептора, используемый в данном изобретении, является веществом, которое имеет связывающую активность в отношении IL-6-рецептора или IL-6, соответственно, и не передает биологическую активность IL-6. То есть, неполный пептид IL-6 или неполный пептид IL-6-рецептора специфически ингибирует связывание IL-6 с IL-6-рецептором захватом IL-6-рецептора или IL-6 связыванием с ними. В результате, передача сигнала IL-6 прерывается, так как биологическая активность IL-6 не передается.
Неполный пептид IL-6 или неполный пептид IL-6-рецептора является пептидом, который состоит из аминокислотной последовательности части или целого района, участвующего в связывании между IL-6 и IL-6-рецептором, в аминокислотной последовательности IL-6 или IL-6-рецептора. Такой пептид состоит обычно из 10-80, предпочтительно 20-50 и более предпочтительно 20-40 аминокислотных остатков,
Неполный пептид IL-6 или неполный пептид IL-6-рецептора может быть получен способом, в котором район, участвующий в связывании между IL-6 и IL-6-рецептором, идентифицируют в аминокислотной последовательности IL-6 или IL-6-рецептора, и часть этой последовательности или вся последовательность обычно является известной, причем примеры этого способа включают в себя способы генной инженерии и способы синтеза пептидов.
Неполный пептид IL-6 или неполный пептид IL-6-рецептора может быть получен с использованием способов генной инженерии включением ДНК, которая кодирует желаемый пептид, в экспрессирующий вектор, и получением желаемого пептида в соответствии с вышеупомянутыми способами экспрессии рекомбинантного антитела, получения и очистки.
Неполный пептид IL-6 или неполный пептид IL-6-рецептора может быть получен с использованием способов синтеза пептидов при помощи способа, обычно используемого в синтезе пептидов, такого как способ твердофазного синтеза или способ жидкофазного синтеза.
Более конкретно, это должно проводиться в соответствии со способом, описанным в Zokuiyakuhin-no-Kaihatsu, Vol.14, Peptide Synthesis, N. Yajima ed., Hirokawa Shoten Publishing (1991). В случае твердофазного синтеза, используют способ, в котором, например, пептидную цепь удлиняют альтернативным повторением реакции, в которой аминокислоту, соответствующую С-концу синтезируемого пептида, связывают с подложкой, которая является нерастворимой в органическом растворителе, а аминокислоты, в которых α-аминогруппы и функциональные группы боковых цепей защищены подходящими защитными группами, последовательно связывают в последовательности от С-конца к N-концу, и реакции, в которой указанные защитные группы α-аминогрупп или пептид, связанный со смолой, удаляют. Способы твердофазного синтеза пептидов в общих чертах подразделяют на Вос-способ и Fmoc-способ в зависимости от типа используемых защитных групп.
После синтеза желаемого пептида таким образом, проводят реакцию удаления защитных групп и реакцию отсоединения пептидной цепи от подложки. В реакции отсоединения пептидной цепи в Вое-способе обычно используют фторид водорода или трифторметансульфоновую кислоту, тогда как в Fmoc-способе обычно используют ТФУ.
В Вос-способе, например, вышеупомянутую смолу с защищенным пептидом обрабатывают во фториде водорода в присутствии анизола. Затем защитные группы удаляют и пептид отсоединяют от подложки с последующим извлечением пептида. Затем неочищенный пептид получают лиофилизацией этого продукта. С другой стороны, в Fmoc-способе, например, реакцию удаления защитных групп и реакцию отсоединения пептидной цепи от подложки проводят с использованием той же самой процедуры, которая описана выше для ТФУ.
Полученный неочищенный пептид может быть выделен и очищен применением ВЖХ. Элюция должна проводиться при оптимальных условиях с использованием растворителя на основе смеси вода-ацетонитрил, обычно используемого для очистки белка. Затем, фракцию, соответствующую пику полученного профиля хроматографии, отделяют и лиофилизируют. Затем пептидную фракцию, очищенную таким образом, идентифицируют анализом молекулярной массы с использованием масс-спектрометрии, анализом аминокислотного состава или анализом последовательности аминокислот.
Конкретные примеры неполных пептидов IL-6 и неполных пептидов IL-6-рецептора описаны в публикации патентной заявки Японии №2-188600, публикации патентной заявки Японии №7-324097, публикации патентной заявки Японии №8-311098 и патенте США №US 5210075.
Ингибирующая передачу сигнала IL-6 активность антагониста IL-6, используемого в данном изобретении, может оцениваться обычно применяемьми способами. Более конкретно, IL-6-зависимую миеломную линию человека (S6B45, КРММ2), линию клеток КТЗ Т-лимфомы Леннерта или IL-6-зависимую клеточную линию МН60 или BSF2 культивируют с последующим добавлением IL-6 при одновременном присутствии антагониста IL-6 и измерением поглощения 3H-тимидина IL-6-зависимыми клетками.
Кроме того, 125-меченый IL-6, связанный с экспрессирующими IL-6-рецептор клетками, измеряют культивированием экспрессирующих IL-6-рецептор клеток в форме клеток U226 с последующим добавлением 125-меченого IL-6 и одновременным добавлением антагониста IL-6. В вышеупомянутой системе анализа группа отрицательного контроля, которая не содержит антагониста IL-6, обеспечена, наряду с группой, в которой присутствует антагонист IL-6, и сравнение результатов, полученных из этих двух групп, позволяет оценить ингибирующую IL-6 активность антагониста IL-6.
Как будет показано в примерах, которые будут описаны позднее, поскольку ингибирующие рост воздействия на клетки мезотелиомы наблюдали для анти-IL-6-рецептор антитела, было предположено, что анти-IL-6-рецептор антитело и другие антагонисты IL-6 применимы в качестве терапевтических агентов для мезотелиомы.
Субъектом лечения в данном изобретении является млекопитающее. Млекопитающим-мишенью этого лечения предпочтительно является человек.
Терапевтический агент для мезотелиомы или ингибитор роста клеток мезотелиомы данного изобретения может вводиться системно или локально, перорально или парентерально. Например, могут быть выбраны внутривенная инфузия или другая форма внутривенного введения, внутримышечная инъекция, интраторакальная инъекция, интраперитонеальная инъекция, подкожная инъекция, суппозитории, клизма или пероральные покрытые энтеросолюбильным покрытием пилюли и т.д., и способ введения может быть подходящим образом выбран в зависимости от возраста и симптомов пациента. Эффективную дозу выбирают в диапазоне 0,01 мг - 100 мг/кг массы тела на одно введение. Альтернативно, доза 1-1000 мг и предпочтительно 5-50 мг может быть выбрана на одного пациента.
Предпочтительные доза и способ введения в случае анти-IL-6-рецептор антитела, являются, например, такими, что количество свободного антитела, присутствующее в крови, равно эффективной дозе, конкретным примером чего является способ, посредством которого его вводят с использованием такого способа, как внутривенное капельное вливание или другая форма внутривенного введения или подкожная инъекция и т.д. в соответствии с такой схемой введения, как дважды в неделю, один раз в неделю, один раз каждые две недели или один раз каждые четыре недели в виде единственного введения или в дозах, разделенных на несколько введений при дозе 0,5 мг - 40 мг и предпочтительно 1 мг - 20 мг в месяц (4 недели) на 1 кг массы тела. Схема введения может быть скорректирована, например, удлинением интервала введения с двух раз в неделю или одного раза в неделю до одного раза каждые две недели, одного раза каждые три недели или одного раз каждые четыре недели, при наблюдении состояния пациента и тенденций величин анализа крови.
Терапевтический агент для мезотелиомы или ингибитор роста клеток мезотелиомы может содержать фармацевтически приемлемый носитель или вспомогательный агент в зависимости от способа введения. Примеры таких носителей и вспомогательных агентов (добавок) включают в себя воду, фармацевтически приемлемые органические растворители, коллаген, поливиниловый спирт, поливинилпирролидон, карбоксивиниловый полимер, натрий-карбоксиметилцеллюлозу, полиакрилат натрия, аргинат натрия, водорастворимый декстран, натрий-карбоксиметилкрахмал, пектин, метилцеллюлозу, этилцеллюлозу, ксантановую камедь, аравийскую камедь, казеин, желатин, агар, диглицерин, пропиленгликоль, полиэтиленгликоль, вазелин, парафин, стеариловый спирт, стеариновую кислоту, человеческий сывороточный альбумин (ЧСА), маннит, сорбит, лактозу и поверхностно-активные вещества, приемлемые для использования в качестве фармацевтических добавок. Хотя используемые добавки выбирают подходящим образом или комбинируют из вышеупомянутых добавок в соответствии с лекарственной формой, они не ограничиваются указанными выше добавками.
Примеры
Хотя следующее описание обеспечивает подробное объяснение данного изобретения при помощи примеров и ссылочных примеров, данное изобретение не ограничивается этими примерами.
Пример 1 - Продуцирование IL-6 клеточными линиями злокачественной мезотелиомы
Культивирование клеточных линий MSTO, Н2052, Н28, Н226 и Н2452 злокачественной мезотелиомы начинали при концентрации клеток 5 х 104 на лунку в 24-луночном планшете, содержащем культуральную жидкость (RPMI, содержащую 10% фетальную телячью сыворотку, ФТС), эту культуральную жидкость заменяли на следующий день, после чего эти клетки культивировали еще в течение 3 дней с последующим измерением концентраций IL-6 в культуральном супернатанте с использованием полностью автоматизированной системы хемилюминесцентного иммуноферментного анализа (Fujirebio, Lumipulse) и коррекции на количество клеточного белка. Эти результаты показаны на фиг.1. Этот эксперимент повторяли три раза. Хотя клеточная линия Н28 не продуцировала IL-6, четыре другие линии продуцировали IL-6. Клеточные линии Н2054 и Н226 продуцировали особенно высокие уровни IL-6.
Пример 2 - Экспрессия IL-6R в клеточных линиях злокачественной мезотелиомы Уровни экспрессии IL-6-рецептора измеряли для пяти клеточных линий злокачественной мезотелиомы на уровне мРНК. Клетки КТ-3 использовали для положительного контроля, а синовиоциты использовали для отрицательного контроля. Эти клетки культивировали в течение 48 часов в RPMI, содержащей 10% ФТС, и мРНК, кодирующую IL-6-рецептор (IL-6R), в этих клетках измеряли ПЦР с обратной транскрипцией (ОТ-ПЦР) с использованием системы GeneAmp PCR (Applied Biosystems) в качестве детектирующего устройства. Эти результаты показаны на фиг.2. Фигура 2 (нижняя часть) показывает количество мРНК GAPDH (глицеральдегид-3-фосфатдегидрогеназы), используемой в качестве внутреннего контроля.
Согласно этим результатам, авторы считают, что клетки злокачественной мезотелиомы едва ли экспрессируют какое-либо количество IL-6-рецептора. Поскольку плевральная жидкость в случаях злокачественной плевральной мезотелиомы состоит из кровянистой плевральной жидкости, авторы предполагают, что растворимый IL-6-рецептор присутствует в больших количествах, и это предполагает, что он участвует в передаче возбуждающей реакции IL-6.
Пример 3 - Индукция продуцирования VEGF стимуляцией IL-6 (1) Опухолевые клетки клеточных линий Н2052 и Н2452 злокачественной мезотелиомы культивировали в трех сериях в 24-луночных планшетах при начальной концентрации клеток 5 × 104 на лунку в среде RPMI 1640. На следующий день жидкость культуры клеток заменяли с последующим началом стимуляции (1) рекомбинантным IL-6 (10 нг/мл), (2) смесью рекомбинантный IL-6 (10 нг/мл)+рекомбинантный растворимый IL-6R (100 нг/мл) и (3) ингибированием анти-IL-6-рецептор антителом, гуманизированным антителом РМ-1 (см. WO 92/19759) (25 мкг/мл) (RPMI использовали для контроля в виде среды). После культивирования в течение дополнительных 3 дней измеряли концентрации фактора роста эндотелия сосудов (VEGF) в культуральных супернатантах с использованием набора Quantikine Human VEGF Immunoassay Kit (R & D System) и корректировали на количество клеточного белка.
Эти результаты показаны на фиг.3. Как видно из этих диаграмм, продуцирование VEGF индуцировалось стимуляцией IL-6 в обеих клеточных линиях Н2052 и Н2452.
Пример 4 - Индукция продуцирования VEGF стимуляцией IL-6 (2) Эксперимент примера 3 повторяли с использованием клеточной линии злокачественной мезотелиомы Н28. Эти результаты показаны на фиг.4. Как видно из этих результатов, хотя клеточная линия Н28 продуцирует высокие уровни VEGF, она не отвечала на стимуляцию IL-6.
Пример 5 - Фосфорилирование STAT3 стимуляцией IL-6
Клеточную линию Н2025, в которой продуцирование VEGF индуцируется IL-6, и клеточную линию Н28, в которой продуцирование VEGF не индуцируется IL-6, инкубировали в присутствии рекомбинантного IL-6 (10 нг/мл) и растворимого рекомбинантного IL-6-рецептора (100 нг/мл) вместе с сигнальным трансдуктором и активатором транскрипции 3 (STAT3) с последующим анализом фосфорилирования STAT3 в p-STAT3 при 0 часов, при 0,5 ч и 1 ч инкубирования при помощи Вестерн-блоттинга.
Эти результаты показаны на фиг.5.
Как видно из фиг.5, значительное фосфорилирование STAT3 наблюдали для клеточной линии Н2025 через 30 минут после IL-6-стимуляции. В отличие от этого, только слабое фосфорилирование STAT3 наблюдали в ответ на IL-6-стимуляцию для клеточной линии Н28. Отметим, что p-STAT3 на фиг.5 показывает фосфорилированный STAT3, тогда как STAT3 показывает сумму фосфорилированного и нефосфорилированного STAT3.
Пример 6 - Индукция SOCS3 стимуляцией IL-6
Клеточную линию Н2025, в которой продуцирование VEGF индуцируется IL-6, и клеточную линию Н28, в которой продуцирование VEGF не индуцируется IL-6, инкубировали в присутствии рекомбинантного IL-6 (10 нг/мл) и растворимого рекомбинантного IL-6-рецептора (100 нг/мл) с последующим измерением уровней экспрессии супрессоры индукции цитокиновой сигнализации 3 (SOCS3) и глицеральдегид-3-фосфатдегидрогеназы (GAPDH) измерением уровней мРНК, которые кодируют их, с последующей ОТ-ПЦР-амплификацией при 0 ч, 2 ч и 4 ч инкубации. Эти результаты показаны на фиг.6.
Как видно из фиг.6, индукцию мРНК SOCS3 наблюдали спустя 2 часа после стимуляции IL-6 в клеточной линии Н2052. Непрерывно высокую экспрессию SOCS3 наблюдали в клеточной линии Н28.
Пример 7 - Анализ промотора VEGF
Получали плазмиду pGL3-VEGF, которая контролирует экспрессию гена люциферазы промотором VEGF, и плазмиду pGL3-VEGFmut, которая контролирует экспрессию гена люциферазы мутированным промотором VEGF (фиг.7).
50000 клеток Н2052, каждую, трансфецировали 1 мкг pGL3-VEGF или pGL3-VEGFmut и трансфецированные клетки стимулировали рекомбинантным IL-6 (10 нг/мл) и рекомбинантным растворимым IL-6-рецептором (100 нг/мл). После стимуляции в течение 2 дней исследовали люциферазную активность, индуцированную от промотора VEGF и мутированного промотора VEGF. Эти результаты показаны на фиг.8.
Как видно из фиг.8, в случае удаления сайта связывания фосфорилированного STAT3 из промотора VEGF активацию промотора VEGF вследствие стимуляции IL-6 уже не наблюдали. На основании результатов вышеупомянутых примеров 5, 6 и 7, авторы считают, что увеличенное продуцирование VEGF вследствие стимуляции IL-6 в клеточной линии Н2052 было опосредовано системой JAK-STAT. Эта предсказанная схема показана на фиг.9.
Пример 8 - Стимуляция роста Н2052 и Н226 добавлением IL-6 и растворимого IL-6-рецептора
Клетки клеточной линии Н2052 распределяли в 96-луночном планшете, содержащем среду RPMI, содержащую 10% ФТС, при 500 клетках на лунку с последующим культивированием в пяти сериях в течение 6-7 дней в присутствии или в отсутствие рекомбинантного растворимого IL-6-рецептора при 100 нг/мл и в присутствии различных концентраций (0, 1, 10 или 100 нг/мл) IL-6. Эти результаты показаны на фиг.10 и фиг.13. Как видно из этих диаграмм, клеточная линия Н2052 и клеточная линия Н226 растут зависимым от концентрации IL-6 образом в присутствии рекомбинантного растворимого IL-6-рецептора (100 нг/мл).
Пример 9 - Ингибирование стимуляции роста клеток Н2052 вследствие добавления IL-6 и IL-6R антителом к IL-6R (анти-IL-6R антителом)
Для определения, ингибируется ли стимуляция роста клеток Н2052 IL-6 и IL-6-рецептором антителом к рецептору IL-6, клетки Н2052 распределяли в 96-луночном планшете, содержащем среду RPMI, содержащую 10% ФТС, при 500 клетках на лунку с последующим культивированием в трех сериях в течение 7 дней в присутствии рекомбинантного растворимого IL-6-рецептора при 100 нг/ил и в присутствии различных концентраций (0, 1 или 25 мкг/мл) гуманизированного антитела РМ-1. После культивирования величину роста клеток Н2052 (OD 450) измеряли с использованием MTS-анализа. Эти результаты показаны на фиг.11. В результате, было определено, что антитело к рецептору IL-6 ингибировало рост зависимым от концентрации образом. С другой стороны, ингибитрующую активность не наблюдали в случае добавления IgGI человека вместо MRA в такой же концентрации, какая была в контроле.
Пример 10- Стимуляция роста клеток Н226 добавлением IL-6 и растворимого IL-6-рецептора и ингибирование анти-Л-бР антителом
Клетки клеточной линии Н226 распределяли в 96-луночном планшете, содержащем среду RPMI, содержащую 10% ФТС, при 500 клетках на лунку с последующим культивированием в трех сериях в течение 7 дней в присутствии рекомбинантного растворимого IL-6-рецептора при 100 нг/ил и в присутствии различных концентраций (0, 1, 10 или 100 нг/мл) IL-6. Эти результаты показаны на фиг.12. Как видно из этой диаграммы, клетки Н226, которые продуцируют высокие уровни IL-6, росли зависимым от концентрации IL-6 образом в присутствии рекомбинантного растворимого IL-6-рецептора (100 нг/мл) таким же образом, что и клетки Н2052, продуцирующие высокие уровни IL-6.
Пример 11 - Ингибирование стимуляции роста клеточной линии Н2052 и клеточной линии Н226, индуцированной добавлением IL-6 и растворимого IL-6-рецептора, анти-IL-6-рецептор антителом
Клетки клеточной линии Н2052 и клеточной линии Н226 распределяли в 96-луночном планшете, содержащем среду RPMI, содержащую 10% ФТС, при 500 клетках на лунку с последующим культивированием в пяти сериях в течение 6 дней в присутствии IL-6 (10 нг/мл) и растворимого IL-6-рецептора (100 нг/мл) в присутствии различных концентраций (0, 1 мкг/мл, 5 мкг/мл) гуманизированного антитела РМ-1. После культивирования скорости роста клеток клеточной линии Н2052 и клеточной линии Н226 измеряли MTS-анализом. Скорости роста клеток показали, во сколько раз рост клеток увеличивался в сравнении с отсутствием добавления IL-6/sIL-6R. Эти результаты показаны на фиг.14. В результате, анти-IL-6-рецептор антитело полностью ингибировало стимулирующее рост воздействие, индуцированное добавлением IL-6/sIL-6R в клеточной линии Н2052 и клеточной линии Н226. С другой стороны, ингибирующее рост воздействие не наблюдали в случае добавления тех же самых концентраций IgGI человека (Sigma) в качестве контроля вместо анти-IL-6-рецептор антитела.
Пример 12 - Ингибирование индукции продуцирования VEGF, индуцированного стимуляцией IL-6, анти-IL-6-рецептор антителом
Проводили исследование ингибирования индукции продуцирования VEGF, индуцированного стимуляцией IL-6 в клеточных линиях MSTO, Н226, Н2052 и Н2452 мезотелиомы, анти-IL-6-рецептор антителом, в условиях, описанных в примере 3, за исключением концентраций гуманизированного антитела РМ-1 (0,1 мкг/мл, 5 мкг/мл). Эти результаты показаны на фиг.15. В результате, анти-IL-6-рецептор антитело полностью ингибировало индукцию продуцирования VEGF, индуцированного стимуляцией IL-6/sIL-6R. С другой стороны, ингибирующее воздействие на индукцию продуцирования VEGF, не наблюдали в случае добавления тех же самых концентраций IgGI человека (Sigma) в качестве контроля вместо анти-IL-6-рецептор антитела.
Пример 13 - Ингибирование фосфорилирования STAT3 anmu-IL-6-рецептор антителом
Проводили исследования для определения, ингибируется ли фосфорилирование STAT3, индуцированное стимуляцией IL-6 (10 нг/мл) и растворимым IL-6-рецептором (100 нг/мл), 5 мкг/мл анти-IL-6-рецептор антителом в клеточной линии Н2052 и клеточной линии Н2452, которые продуцируют VEGF. Эти результаты показаны на фиг.16. В результате, анти-IL-6-рецептор антитело значительно ингибировало образование фосфорилированного STAT3 (p-STAT3), которое индуцируется стимуляцией IL-6/sIL-6R в клетках злокачественной мезотелиомы. С другой стороны, значительное ингибирование фосфорилирования STAT3 не наблюдали в случае добавления той же самой концентрации IgGI человека (Sigma) в качестве контроля вместо анти-1b-6-рецептор антитела.
Пример 14 - Влияние анти-VEGF антитела на рост клеток злокачественной мезотелиомы
Клетки клеточной линии Н2052 и клеточной линии Н226 распределяли в 96-луночном планшете при 500 клетках на лунку в среде RPMI, содержащей 10% ФТС, с последующим культивированием в пяти сериях в течение 6-7 дней в присутствии IL-6 (10 нг/мл) и рекомбинантного растворимого IL-6-рецептора (100 нг/мл) в присутствии 1 мкг/мл анти-VEGF антитела. После культивирования величины роста исследовали MTS-анализом. Ту же самую концентрацию IgGI человека (Sigma) использовали для контроля вместо анти-VEGF антитела. Эти результаты показаны на фиг.17. В результате, анти-VEGF антитело не ингибировало рост клеток, индуцированный стимуляцией IL-6/sIL-6R. На основании этого результата, было явно показано, что воздействие на рост клеток злокачественной мезотелиомы, индуцированное стимуляцией IL-6/sIL-6R, не было опосредовано VEGF.
Ссылочный пример 1 - Получение растворимого IL-6-рецептора человека Растворимый IL-6-рецептор получали при помощи ПЦР с использованием плазмиды pBSF2R.236, которая содержит ДНК, кодирующую IL-6-рецептор, полученной в соответствии со способом Yamasaki, et al. (Yamasaki, K. et aL, Science. 241, 825-828, 1988). Затем плазмиду pBSF2R.236 расщепляли рестриктазой SphI с получением кДНК IL-6-рецептора, который затем вставляли в mpl8 (Amersham). В кДНК IL-6-рецептора вводили мутацию при помощи ПЦР с использованием системы мутагенеза in vitro (Amersham) с применением синтетического олиго-праймера, сконструированного таким образом, чтобы вставить стоп-кодон в кДНК IL-6-рецептора. В результате этой процедуры стоп-кодон вставлялся в местоположение аминокислоты 345, и получали кДНК, которая кодирует растворимый IL-6-рецептор.
Для экспрессии кДНК, кодирующей растворимый IL-6-рецептор, в клетках СНО ее связывали с плазмидой pSV (Pharmacia) с получением плазмиды pSVL344. Затем кДНК растворимого IL-6-рецептора, разрезанную HindIII-SalI, встраивали в плазмиду pECEdhfr, содержащую кДНК dhfr, для получения экспрессирующей плазмиды pEXEdhfi-344 клеток СНО.
Затем 10 мкг плазмиды pEXEdhfr344 использовали для трансфекции линии dhfr-СНО клеток DXB-11 (Uriaub, G. et al., Proc. Natl. Acad. Sci. USA. 77, 4216-4220, 1980) в соответствии со способом осаждения фосфатом кальция (Chen, С.et al., Mol. Cell. Biol. 7, 2745-2751, 1987). Затем трансфецированные клетки СНО культивировали в течение 3 недель в не содержащей нуклеозидов селективной культуральной среде аМЕМ, содержащей 1 мМ глутамин, 10% диализованную ФТС, 100 Е/мл пенициллина и 100 мкг/мл стрептомицина.
Затем отобранные клетки СНО подвергали скринингу способом конечных разведении с получением единственного клона клеток СНО. Этот клон клеток СНО размножали с метотрексатом при концентрации от 20 нМ до 200 нМ с получением линии клеток СНО 5Е27, продуцирующей растворимый IL-6-рецептор человека. Линию клеток СНО 5Е27 культивировали в среде Дульбекко в модификации Искова (IMDM, Gibco), содержащей 5% ФТС.После получения культурального супернатанта концентрацию растворимого IL-6-рецептора в этом культуральном супернатанте измеряли при помощи ELISA. В результате было подтверждено, что растворимый IL-6-рецептор присутствует в этом культуральном супернатанте.
Ссылочный пример 2 - Получение анти-IL-6-человека антитела Мышей Balb/c иммунизировали 10 мкг рекомбинантного IL-6 (Hirano, Т. et al., Immunol. Lett. 17, 41, 1988) вместе с полным адъювантом Фрейнда и иммунизацию продолжали один раз в неделю, пока анти-1Е-6 антитело не детектировали в сыворотке. Иммуноциты иссекали из локальных лимфатических узлов и сливали с миеломной клеточной линией P3U1 с использованием полиэтиленгликоля 1500. Гибридому отбирали по способу Oi, et al. с использованием культуральной жидкости HAT (Selective Methods in Cellular Immunology, W.H. Freeman and Co., San Francisco, 351, 1980) для получения гибридомы, которая продуцировала анти-IL-6 человека антитело.
Анализ связывания IL-6 выполняли способом, описанным ниже, на гибридоме, которая продуцировала aнти-IL-6 человека-антитело. А именно, гибкий поливиниловый 96-луночный планшет (Dynatech Laboratories, Alexandria, VA) покрывали в течение ночи при 4°С 100 мкл козьего антитела против мышиного Ig (10 мкг/мл, Cooper Biomedical, Malvem, PA) в 0,1 M карбонат-гидрокарбонатном буфере (рН 9,6). Затем этот планшет обрабатывали в течение 2 часов при комнатной температуре 100 мкл ЗФР, содержащим 1% бычий сывороточный альбумин (БСА).
После промывания планшета ЗФР 100 мкл культурального супернатанта гибридомы добавляли в каждую лунку с последующим инкубированием в течение ночи при 4°С.Планшет промывали и в каждую лунку добавляли 125I-меченый рекомбинантный IL-6 при 2000 имп/мин на 0,5 нг на лунку с последующим измерением радиоактивности каждой лунки после промывания при помощи гамма-счетчика (Beckman Gamma 9000, Beckman Instruments, Fullerton, CA). 32 из 216 гибридомных клонов были положительными согласно анализу связывания IL-6. В конце концов, из этих клонов получали стабильный клон MH166.BSF2. Анти-IL-6 антитело МН166, продуцируемое указанной гибридомой, имело подтип IgGlK.
Затем, нейтрализующую активность антитела МН166 в отношении роста гибридомы исследовали с использованием IL-6-зависимого мышиного гибридомного клона MH60.BSF2. Клетки MH60.BSF2 распределяли в лунки микропланшета при 1 × 104200 мкл на лунку с последующим добавлением пробы, содержащей антитело МН166, и культивированием в течение 48 часов и затем дополнительным культивированием в течение 6 часов после добавления 3H-тимидина (New England Nuclear, Boston, MA) при 0,5 мкКи на лунку. Затем эти клетки помещали на фильтровальную бумагу из стекловолокна и обрабатывали автоматическим харвестером клеток (Labo Mash Science, Tokyo, Japan). В качестве контроля использовали кроличье анти-IL-6 антитело.
В результате антитело МН166 зависимьм от объема образом ингибировало поглощение 3H-тимидина клетками MH60.BSF2, индуцированными IL-6. Таким образом, было ясно показано, что антитело MH60.BSF2 нейтрализует активность IL-6.
Ссылочный пример 3 - Получение антитела против IL-6-рецептора человека Анти-IL-6-рецептор антитело МТ18, полученное по способу Hirata, et al, (Hirata, Y. et al., J. Immunol. 143, 2900-2906, 1989), связывали с активированной Сефарозой 4 В (Pharmacia Fine Chemicals, Piscataway, NJ) в соответствии с прилагаемым протоколом для очистки IL-6-рецептора (Yamasaki, К. et al.. Science 241, 825-828, 1988). Клеточную линию миеломы человека U266 солюбилизировали гидрохлоридом п-парааминофенилметансульфонилфторида (Wako Chemicals), содержащим 1% дигитонин (Wako Chemicals), 10 мМ триэтаноламин (рН 7,8) и 0,15 M Nad (дигитониновый буфер), и затем смешивали с антителом МТ18, связанным с гранулами Сефарозы 4 В. Затем эти гранулы промывали шесть раз дигитониновьм буфером с получением частично очищенного IL-6-рецептора для иммунизации.
Мышь Balb/c иммунизировали четыре раза каждые 10 дней вышеупомянутым частично очищенным IL-6-рецептором, полученным из 3 х 109 клеток U266, с последующим получением гибридомы в соответствии с обычными способами. Связывающую активность культурального супернатанта гибридомы из лунок с реакцией положительного роста на IL-6-рецептор исследовали согласно способу, описанному ниже. 5 × 107 клеток U266 метили 35S-метионином (2,5 мКи) и солюбилизировали вышеупомянутым дигитониновым буфером. Затем эти солюбилизированные клетки U266 смешивали с антителом МТ18, связанным с гранулами Сефарозы 4 В, имеющими объем 0,04 мл, с последующими промыванием шесть раз дигитониновым буфером, элюцией 35S-метионин-меченого IL-6-рецептора 0,25 мл дигитонинового буфера (рН 3,4) и нейтрализацией 0,025 мл 1 М Триса (рН 7,4).
0,05 мл гибридомного культурального супернатанта смешивали с 0,01 мл Белок G-Сефарозы (Pharmacia). После промывания эту Сефарозу инкубировали с 0,005 мл раствора 35S-метионин-меченого IL-6-рецептора, полученного, как описано выше. Затем иммунопреципитат анализировали электрофорезом в ДСН-ПААГ и гибридомный культуральный супернатант, который реагирует с IL-6-рецептором, исследовали. В результате был получен гибридомный клон РМ-1 (PERM BP-2998) с положительной реакцией. Антитело, продуцируемое из гибридомы РМ-1, имеет подтип IgGlK.
Ингибирующую связывание IL-6 активность антитела, продуцируемого гибрид омой РМ-1, в отношении IL-6-рецептора человека исследовали с использованием клеточной линии U266 миеломы человека. Рекомбинантный IL-6-человека получали из Е. coli (Hirano, Т. et al., Immunol. Lett. 17, 41-45, 1988) и метили 125I с использованием реагента Болтона-Хантера (New England Nuclear, Boston, MA) (Taga. T. et al., J. Exp.Med. 166,967-981,1987).
4 × 105 клеток U266 культивировали с 70% (об./об.) культуральным супернатантом гибридомы РМ-1 и 14000 имп/мин 125I-меченого IL-6 в течение часа. 70 мкл пробы наносили на 300 мкл ФТС в микроцентрифужной полиэтиленовой пробирке на 400 мкл с последующим центрифугированием и измерением радиоактивности этих клеток.
В результате, было ясно определено, что антитело, продуцируемое гибридомой РМ-1, ингибирует связывание IL-6 с IL-6-рецептором.
Ссылочный пример 4 - Получение антитела против IL-6-рецептора мыши
Моноклональное антитело к IL-6-рецептору мыши получали в соответствии со способом, описанным в Saito, Т. et al.,J. Immunol. 147,168-173, 1991.
Клетки СНО, продуцирующие растворимый IL-6-рецептор мыши, культивировали в культуральной жидкости IMDM, содержащей 10% ФТС, с последующей очисткой растворимого IL-6-рецептора мыши из культурального супернатанта с использованием аффинной колонки, в которой антитело RS12 (см. Saito, Т. et а1., там же) против мышиного IL-6-рецептора было иммобилизовано в геле Affigel 10 (BioRad).
50 мкг полученного растворимого IL-6-рецептора мыши смешивали с полным адъювантом Фрейнда и инъецировали в брюшную полость крысы Wistar. Эту крысу дополнительно иммунизировали с неполным адъювантом Фрейнда, начиная с момента времени спустя две недели. Затем клетки селезенки крысы собирали в день 45 и, после слияния 2 × 108 клеток с 1 × 107 клеток клеточной линии P3U1 мышиной миеломы с использованием 50% ПЭГ1500 (Boehringer Mannheim) в соответствии с обычными способами, эти слитые клетки подвергали скринингу на гибридому со средой HAT.
После добавления гибридомного культурального супернатанта на планшет, покрытый кроличьим антителом против IgG крысы (Cappel), растворимому IL-6-рецептору мыши давали реагировать. Затем гибридому, которая продуцировала антитело к растворимому IL-6-рецептору мыши, подвергали скринингу при помощи ELISA с использованием кроличьего антитела против IL-6-рецептора мыши и меченого фосфатазой овечьего антитела против кроличьего IgG. Гибридомные клоны, в которых было подтверждено продуцирование антитела, подвергали последующему скринингу два раза для получения единственного гибридомного клона. Этот клон был назван MR16-1.
Нейтрализующую активность во время передачи сигнала мышиным IL-6 этого продуцируемого гибридомой антитела исследовали по поглощению 3H-тимидина с использованием клеток MH60.BSF2 (Matsuda, Т. et al., J. Immunol. 18, 951-956, 1988). 1 × 104 клеток MH60.BSF2/200 мкл на лунку получали в 96-луночном планшете. К этому планшету добавляли 10 пг/мл IL-6 мыши и 12,3-1000 нг/мл антитела MR16-1 или антитела RS12 и культивировали в течение 44 часов при 37°С и 5% СO2 с последующим добавлением 1 мкКи на лунку 3H-тимидина. Затем поглощение 3H-тимидина измеряли спустя 4 часа. В результате, антитело MR16-1 ингибировало поглощение 3H-тимидина клетками MH60.BSF2.
Таким образом, было определено, что антитело, продуцируемое гибридомой MR16-1 (FERM ВР-5875), ингибирует связывание IL-6 с IL-6-рецептором.
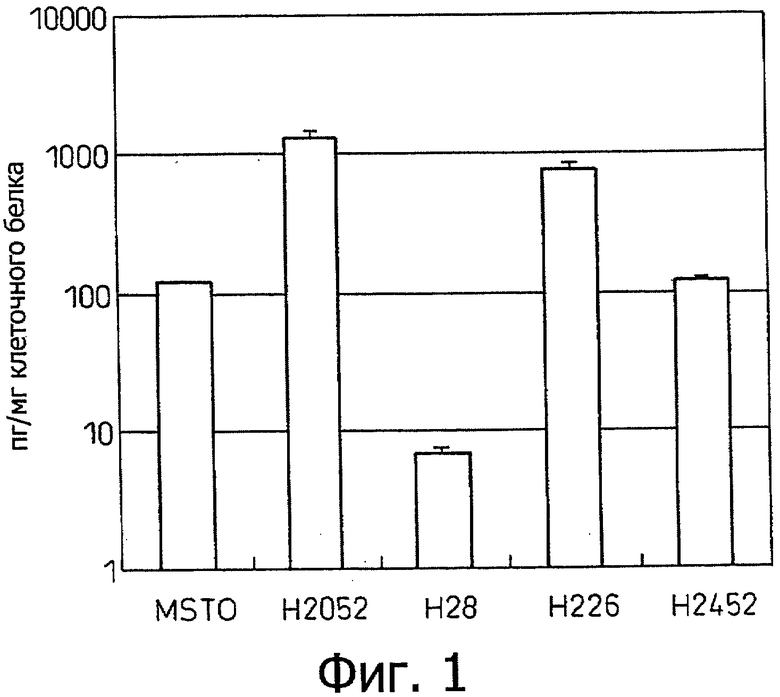

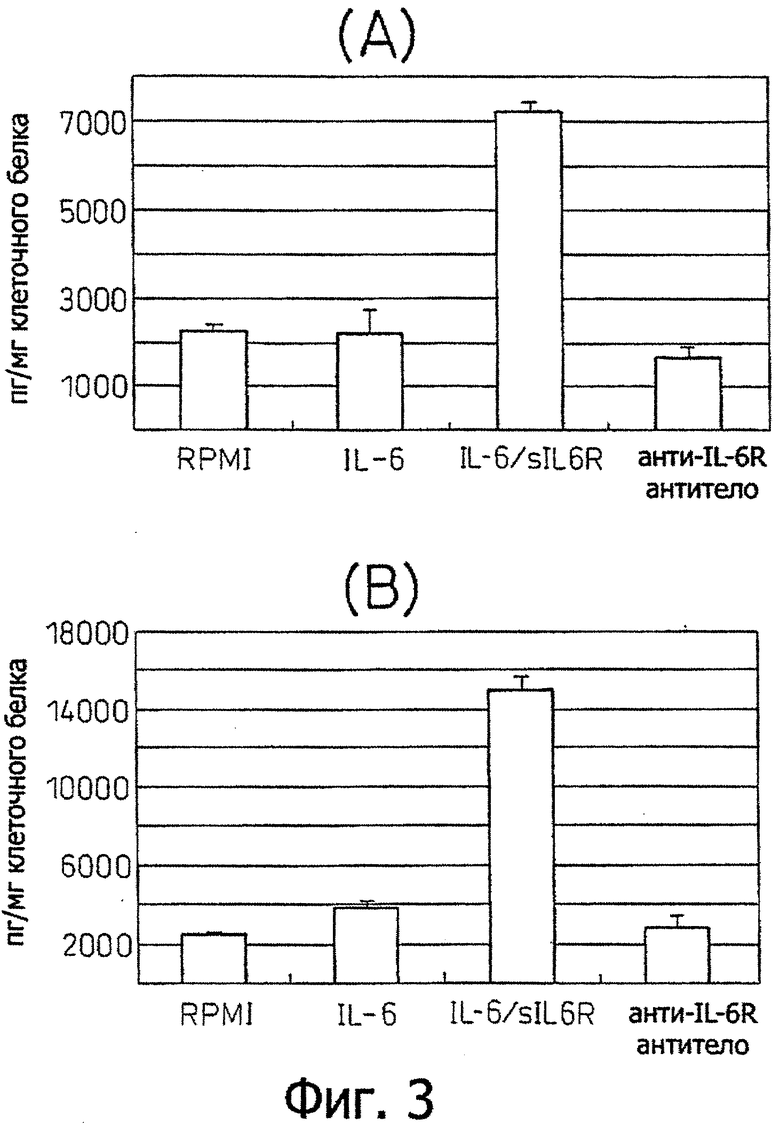
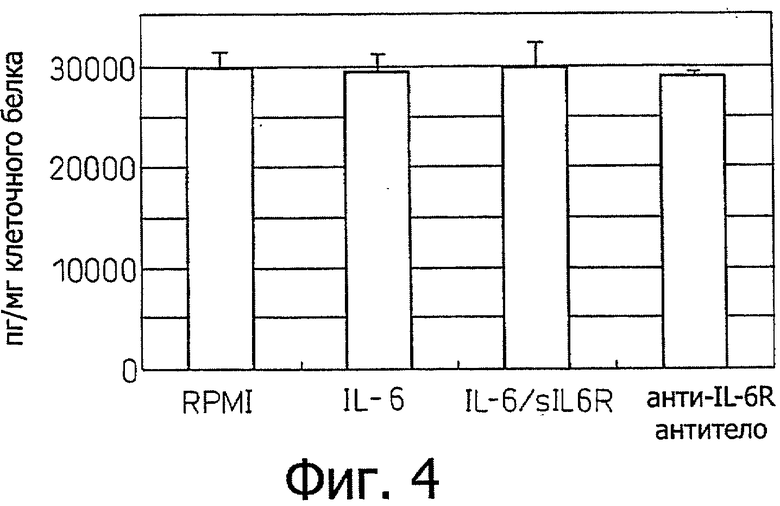


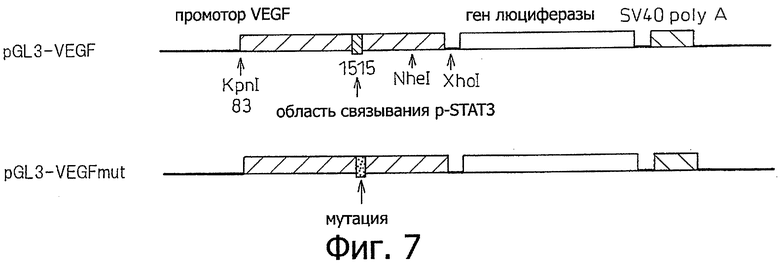




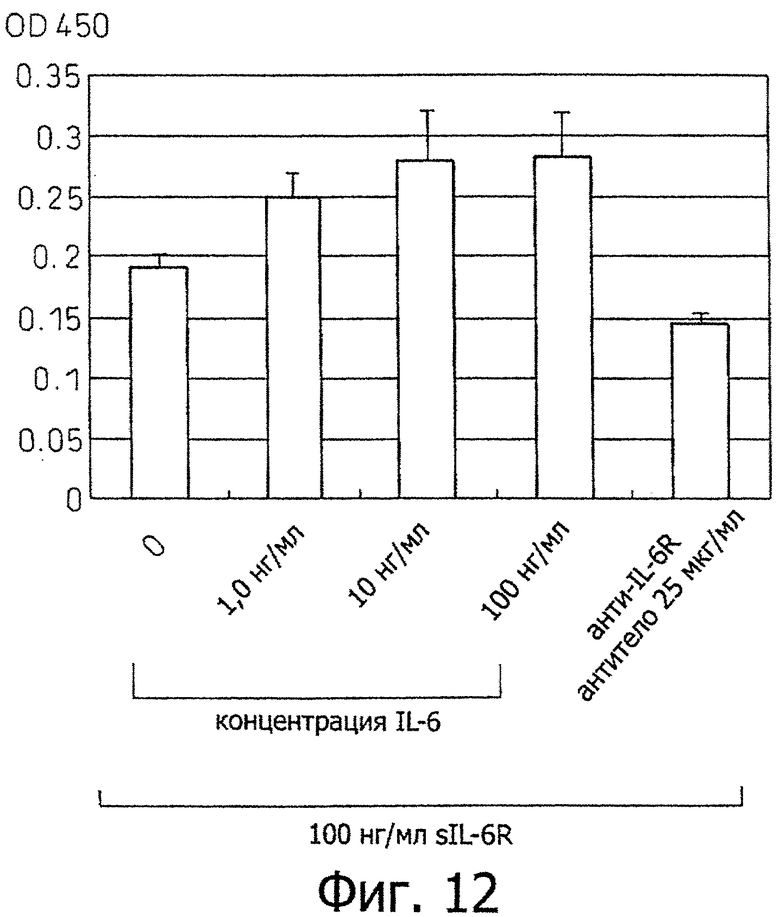

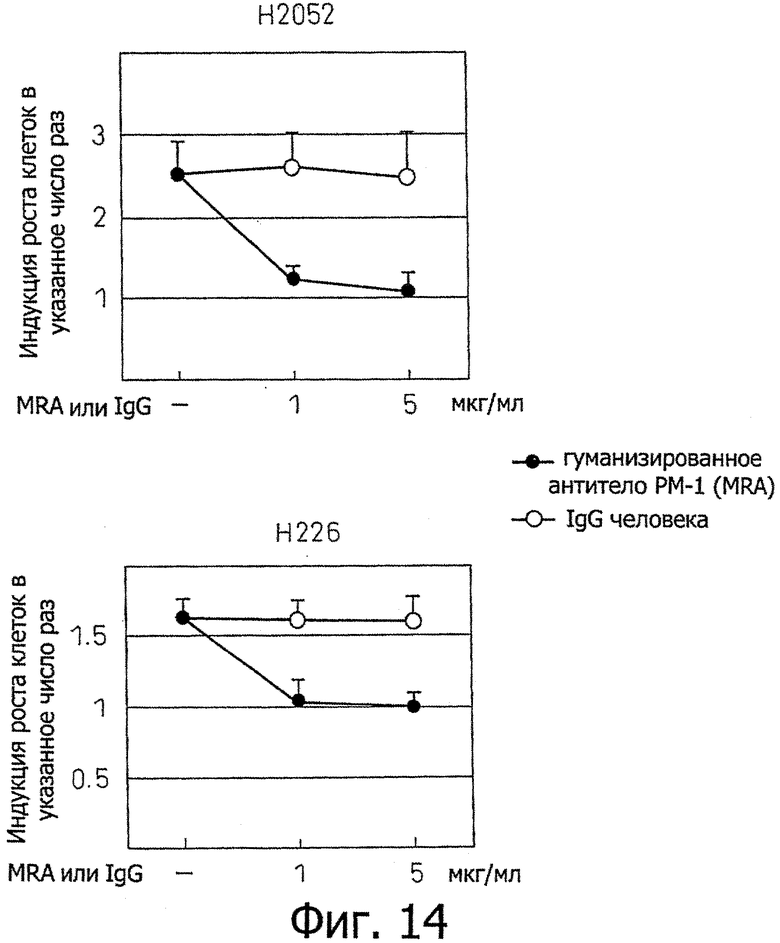
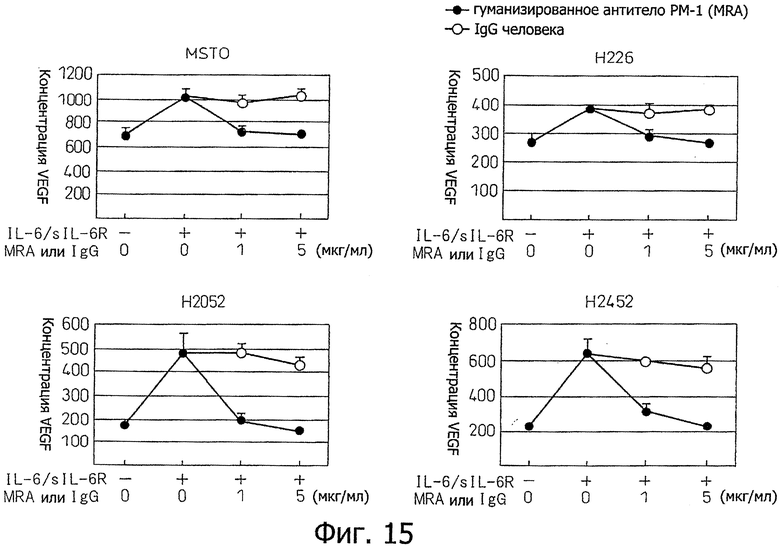
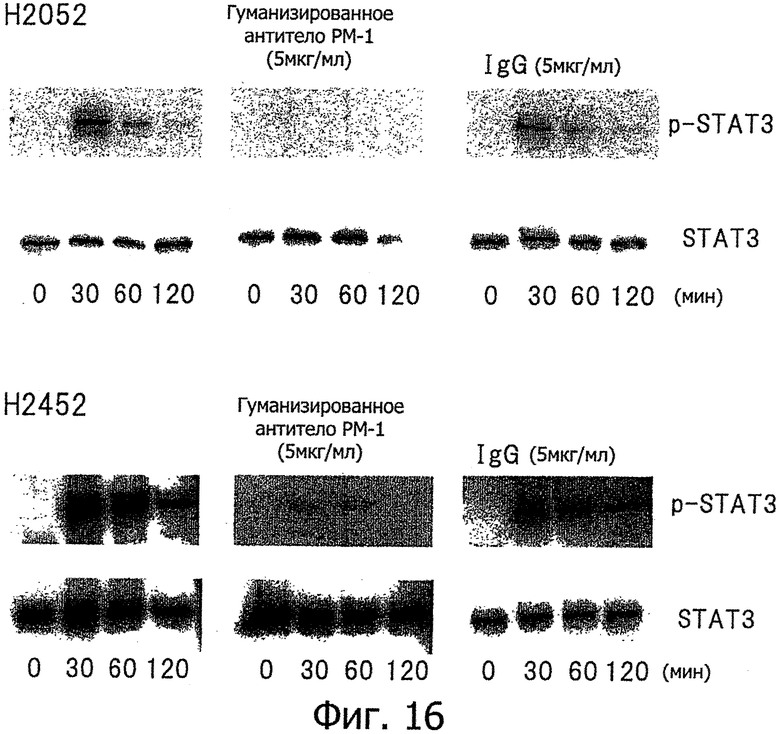
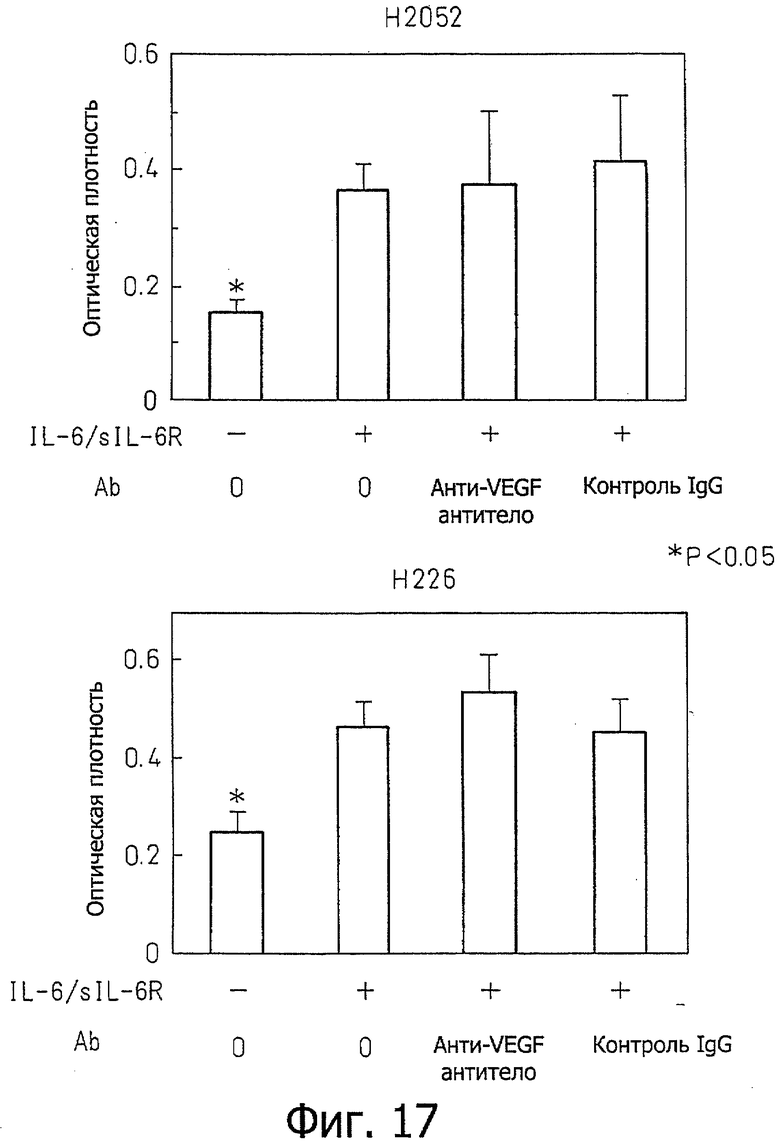
Изобретение относится к лечению мезотелиомы. Терапевтический агент для мезотелиомы, который содержит антитело к рецептору интерлейкина-6 (IL-6) в качестве его активного ингредиента, где мезотелиома является плевральной мезотелиомой, где плевральная мезотелиома является злокачественной плевральной мезотелиомой, где антителом к IL-6-рецептору является моноклональное антитело к IL-6-рецептору, где антителом к IL-6-рецептору является моноклональное антитело к IL-6-рецептору человека, где антителом к IL-6-рецептору является моноклональное антитело к IL-6-рецептору мыши, где антителом к IL-6-рецептору является рекомбинантное антитело, где моноклональным антителом к IL-6-рецептору человека является антитело РМ-1, где моноклональным антителом к IL-6-рецептору мыши является антитело MR 16-1, где антителом к IL-6-рецептору является химерное антитело, гуманизированное антитело или человеческое антитело, где антителом к IL-6-рецептору является химерное антитело, гуманизированное антитело или человеческое антитело, где гуманизированным антителом к IL-6-рецептору является гуманизированное антитело РМ-1, где гуманизированным антителом к IL-6-рецептору является гуманизированное антитело РМ-1. Изобретение обеспечивает расширение арсенала средств лечения мезотелиомы. 12 з.п. ф-лы, 17 ил., 14 пр.
1. Терапевтический агент для лечения мезотелиомы, который содержит антитело к рецептору интерлейкина-6 в качестве его активного ингредиента.
2. Терапевтический агент для лечения мезотелиомы по п.1, где мезотелиома является плевральной мезотелиомой.
3. Терапевтический агент для лечения мезотелиомы по п.2, где плевральная мезотелиома является злокачественной плевральной мезотелиомой.
4. Терапевтический агент для лечения мезотелиомы по п.1, где антителом к IL-6-рецептору является моноклональное антитело к IL-6-рецептору.
5. Терапевтический агент для лечения мезотелиомы по п.1, где антителом к IL-6-рецептору является моноклональное антитело к IL-6-рецептору человека.
6. Терапевтический агент для лечения мезотелиомы по п.1, где антителом к IL-6-рецептору является моноклональное антитело к IL-6-рецептору мыши.
7. Терапевтический агент для лечения мезотелиомы по любому из пп.1-5, где антителом к IL-6-рецептору является рекомбинантное антитело.
8. Терапевтический агент для лечения мезотелиомы по п.5, где моноклональным антителом к IL-6-рецептору человека является антитело РМ-1.
9. Терапевтический агент для лечения мезотелиомы по п.6, где моноклональным антителом к IL-6-рецептору мыши является антитело MR 16-1.
10. Терапевтический агент для лечения мезотелиомы по любому из пп.4-6 и 8-9, где антителом к IL-6-рецептору является химерное антитело, гуманизированное антитело или человеческое антитело.
11. Терапевтический агент для лечения мезотелиомы по п.7, где антителом к IL-6-рецептору является химерное антитело, гуманизированное антитело или человеческое антитело.
12. Терапевтический агент для лечения мезотелиомы по п.10, где гуманизированным антителом к IL-6-рецептору является гуманизированное антитело РМ-1.
13. Терапевтический агент для лечения мезотелиомы по п.11, где гуманизированным антителом к IL-6-рецептору является гуманизированное антитело РМ-1.
| BIELEFELDT-OHMANN H | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ ИЛИ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, ВЫЗЫВАЕМЫХ ОБРАЗОВАНИЕМ IL-6 | 1995 |
|
RU2147442C1 |
| EP 0409607 А2, 23.01.1991 | |||

