Перекрестные ссылки на родственные заявки
Настоящая заявка притязает на преимущество от Предварительной заявки US Serial No.61/118,492, поданной 28 ноября 2008 г.
Область техники, к которой относится изобретение
Настоящее изобретение охватывает вакцины против гриппа, в частности вакцины от птичьего гриппа. Вакцины могут представлять собой рекомбинантные птичьи вакцины.
Уровень техники
Птичий грипп - это грипп, который вызывают вирусы, приспособившиеся к птицам. Вирус птичьего гриппа (AIV) представляет собой РНК-вирус, принадлежащий к семейству Orthomyxoviridae, который классифицируется как вирус гриппа типа А, что относится к его нуклеопротеиду и мембранным белкам. AIV имеет липидную оболочку, в которой есть два особых гликопротеида:гемагглютинин (НА), который облегчает проникновение вируса в клетки хозяина, и нейраминидаза (NA), которая способствует высвобождению его потомства из инфицированных клеток (de Jong et al., J Clin Vrol. 2006, 35 (1):2-13). Подтип H5N1 (вирус, содержащий НА5 и NA1) конкретно связывали с недавними вспышками в Азии, России, на Ближнем Востоке, в Европе и Африке (Olsen et al., Science 2006, 312 (5772):384-388).
Очень патогенный вирус гриппа А подтипа H5N1 и есть новый вирус птичьего гриппа, вызывающий всеобщую озабоченность как потенциальная угроза пандемии. H5N1 уже погубил миллионы домашних птиц во все возрастающем числе стран по всей Азии, Европе и Африке. Эксперты в области здравоохранения озабочены тем, что совместное существование вирусов гриппа человека и вирусов птичьего гриппа (особенно H5N1) создает возможность для обмена генетического материала между видоспецифичными вирусами, возможно, создавая новые вирулентные штаммы гриппа, которые легко передаются и детальны для человека (Food Safety Research Information Office. "A Focus on Avian Influenza". Created May 2006, updated November 2007).
Co времени первой вспышки H5N1 в 1997 г. отмечалось все большее число случаев передачи HPAI H5N1 от птиц к человеку, вызывающих клинически тяжелые и смертельные инфекции. Тем не менее, поскольку существует значительный межвидовой барьер между птицами и человеком, вирус не так легко переходит на людей. Несмотря на то, что миллионы птиц заразились вирусом со времени его открытия, чуть больше 200 человек умерли от птичьего гриппа в Индонезии, Лаосе, Вьетнаме, Румынии, Китае, Турции и России.
Недавно поводились исследования растений в качестве источника получения терапевтических средств, таких как вакцины, антитела и биофармацевтические препараты. Однако получение вакцин, антител, белков и биопрепаратов из растений находится далеко от лечебного процесса, и существует множество препятствий, обычно связанных с таким получением вакцин. Ограничения для успешного производства растительных вакцин включают низкий выход биопродукта или экспрессируемого антигена (Chargelegue et al., Trends in Plant Science 2001, 6, 495-496), нестабильность белков, неустойчивость качества продукта (Schillberg et al., Vaccine 2005, 23, 1764-1769) и недостаточная способность получения вирусоподобных продуктов ожидаемого размера и иммуногенности (Amtzen et al., Vaccine 2005, 23, 1753-1756).
Учитывая подверженность животных, включая человека, к AIV, необходим способ профилактики инфицирования AIV и защиты животных. Соответственно, существует потребность в способах получения эффективных вакцин против гриппа.
Сущность изобретения
Предусмотрены композиции, включающие полипептид гриппа и его фрагменты и варианты. Полипептид или антиген вырабатывается в растениях и является высоко иммуногенным и протективным.
Полипептиды и их фрагменты и варианты могут быть составлены в виде вакцин и/или фармацевтических или иммунологических композиций. Такие вакцины или композиции могут применяться для вакцинации животных и обеспечения защиты против гомологичных и гетерологичных штаммов гриппа.
Способы изобретения включают способы применения, включающие введение животным эффективного количества антигенного полипептида либо его фрагмента или варианта для выработки защитного иммуногенного ответа. Способы также включают способы получения антигенных полипептидов в растениях ряски. После продукции в ряске антигенный полипептид может быть частично или существенно очищен для применения в качестве вакцины или иммунологической композиции.
Также предусмотрены наборы, включающие по меньшей мере один антигенный полипептид либо его фрагмент или вариант и инструкции по применению.
Краткое описание фигур
Следующее подробное описание, представленное для примера, но не предназначенное для ограничения изобретения лишь конкретными описанными воплощениями, лучше всего можно понять в сочетании с прилагаемыми чертежами.
На фиг.1 представлена таблица, в которой приведены SEQ ID NO, соотнесенные с последовательностями полинуклеотидов и белков.
На фиг.2 представлена синтетическая (оптимизированная по кодонам) и мутированная последовательность ДНК, кодирующей гемагглютинин (НА) вируса H5N1 А/курица/Индонезия/7/2003 (SEQ ID NO:1).
На фиг.3 представлены нативные и синтетические/мутированные последовательности белка H5N1 А/курица/Индонезия/7/2003 (НА).
На фиг.4 представлена последовательность кДНК гена НА вируса дикого типа (нативного) А/курица/Индонезия/7/2003 (H5N1) (номер доступа в GenBank EF473080) (SEQ ID NO:3).
На фиг.5 представлены сопоставление белковых последовательностей НА и таблица идентичности последовательностей.
На фиг.6 представлена последовательность вектора MerB01 (SEQ ID NO:6).
На фиг.7 представлена карта вектора MerB01.
На фиг.8 представлены сопоставление последовательностей ДНК и таблица идентичности последовательностей.
На фиг.9 представлены типичная чашка при скрининге положительных трансгенных растений на НА и результаты определения НА.
На фиг.10 представлены результаты определения НА у трансгенных растений, экспрессирующих НА вируса H5N1.
На фиг.11 представлена таблица, в которой приведен ожидаемый выход заданных композиций.
На фиг.12-14 представлены результаты анализа ингибирования гемагглютинации, проведенного с различными антителами.
На фиг.15 представлены SDS-PAGE (окрашивание серебром) и western-блот.
На фиг.16 представлена western-гибридизация с использованием различных сывороток.
На фиг.17 представлен анализ иммунолокализации экспрессированного в Lemna НА с помощью моноклонального антитела против гемагглютинина Н5 вируса гриппа А/Вьетнам/1203/04.
На фиг.18 представлена таблица, в которой приведена схема вакцинации при исследовании иммуногенности.
На фиг.19 представлена сводка данных по защите после провоцирующей пробы с HPAI H5N1.
На фиг.20 представлены титры ингибирования гемагглютинации (log2) от сывороток, взятых на 35-й день у кур, вакцинированных полученным в Lemna НА.
На фиг.21 представлена таблица, в которой приведены сводные серологические данные по образцам, взятым до провоцирующей пробы на 42-й день и после пробы на 56-й день.
Раскрытие сущности изобретения
Предусмотрены композиции, включающие антиген гриппа и его фрагменты и варианты, вызывающие иммуногенный ответ у животных. Антигенные полипептиды либо их фрагменты или варианты могут вырабатываться в растениях ряски. Антигенные полипептиды либо их фрагменты или варианты могут быть составлены в виде вакцин либо фармацевтических или иммунологических композиций и могут применяться для выработки защитного иммунитета у животных. В одном воплощении полипептидным антигеном является полипептид гемагглютинина либо его активный фрагмент или вариант.
Предусматривается, что антигенные полипептиды или антигены изобретения могут представлять собой полные полипептиды либо их активные фрагменты или варианты. Под "активными фрагментами" или "активными вариантами" подразумевается то, что эти фрагменты или варианты сохраняют антигенный характер полипептида. Таким образом, настоящее изобретение охватывает любые полипептиды, антигены, эпитопы или иммуногены гриппа, вызывающие иммуногенный ответ у животных. Полипептид, антиген, эпитоп или иммуноген гриппа может представлять собой любой полипептид, антиген, эпитоп или иммуноген гриппа, как-то белок, пептид либо его фрагмент или вариант, который вызывает, индуцирует или стимулирует иммуногенный ответ у животных.
Антигенным полипептидом, представляющим особый интерес, является гемагглютинин (НА). Гемагглютинин гриппа означает такой тип гемагглютинина, который находится на поверхности вирусов гриппа. Он представляет собой антигенный гликопротеид и отвечает за связывание вируса с инфицируемыми клетками. Существуют различные антигены НА, любой из которых может использоваться при воплощении изобретения. Представляет интерес НА из H5N1, сильно патогенного вируса птичьего гриппа. В частности, НА может быть выделен из H5N1 штамма А/курица/Индонезия/7/2003. Однако при воплощении изобретения можно использовать НА из других вирусов гриппа (т.е. H1-Н16), включая H1, Н3, Н5, Н6, Н7, Н9 и т.п. Также предусматривается, что можно использовать предшественников любых белков НА.
НА представляет собой гомотримерный трансмембранный белок с эктодоменом, состоящим из глобулярной головки и стержня (стволового участка). Оба участка несут N-связанные олигосахариды, которые играют важную роль в биологической функции НА (Schuize I.T., J Infect Dis, 1997. 176 Suppi 1:p.S24-8; Deshpande K.L. et al., PNAS USA, 1987, 84 (1):p.36-40). Среди различных подтипов вируса гриппа А имеется значительная изменчивость в сайтах гликозилирования головки, тогда как олигосахариды стержня более консервативны и необходимы для активности слияния (Ohuchi R. et al., J Virol, 1997, 71 (5):p.3719-25). Гликаны вблизи эпитопов антигенного пептида влияют на распознавание антител (Skehel J.J. et al., PNAS USA, 1984, 81 (6):p.1779-83), а гликаны вблизи от протеолитического сайта модулируют расщепление и влияют на инфекционность вируса гриппа (Deshpande K.L. et al., 1987). Анализ нуклеотидных последовательностей 62 генов Н5 подтверждает гипотезу о том, что дополнительное гликозилирование возле участка связывания с рецептором в глобулярной головке НА является адаптацией вируса вслед за межвидовым переносом от диких птиц, в частности водоплавающих птиц, к домашним птицам (Banks J. et al., Avian Dis, 2003, 47 (3 Suppl):p.942-50).
Для белка НА вируса гриппа идентифицировано более 150 В-клеточных эпитопов, а также 113 CD4+ и 35 CD8+ Т-клеточных эпитопов, однако сообщалось только об ограниченном числе эпитопов для штаммов/подтипов вируса птичьего гриппа (Bui H.H. et al., PNAS USA, 2007, 104 (1):р.246-51). Изучение сайтов аминокислотных замен в естественных и отобранных с помощью моноклональных антител антигенных вариантах показало, что все антигенные сайты находятся на поверхности мембранного дистального домена НА1 и преимущественно окружают сайты связывания с рецептором. Имеются две заметные особенности антигенных сайтов:петлеобразная структура некоторых из них и наличие углеводородных боковых цепей (Skehel J.J. et al., Annu Rev Biochem, 2000, 69:p.531-69). Описана локализация и тонкая структура двух антигенных сайтов Н5 (Kaverin N.V. et al., J Gen Virol, 2002. 83 (Pt 10):p.2497-505). Сайт 1 представляет собой экспонированную петлю, включающую остатки 140-145 НА1, которые соответствуют антигенным сайтам А Н3 и Са2 H1, а сайт 2 состоит из двух подсайтов, причем один (остатки 156 и 157 НА1) соответствует сайту В у подтипа Н3, а другой (остатки 129-133 НА1) соответствует сайту Sa у подтипа H1. Исследование по картированию эпитопов свидетельствует о том, что антигенная структура НА у недавно выделенного H5N1 существенно отличается от малопатогенного штамма Н5 и быстро эволюционирует (Kaverin N.V. et al., J Virol, 2007. 81 (23):p.12911-7). Анализ консервативности эпитопов свидетельствует о том, что для Т-клеточных эпитопов вероятен значительный уровень перекрестной реактивности между штаммами, но он гораздо меньше для Ab-эпитопов. При использовании библиотеки перекрывающихся пептидов впервые был идентифицирован Т-клеточный эпитоп AIV, представляющий собой 15-мерный пептид -Н5246-260 внутри домена НА1, который индуцирует действие Т-клеток у цыплят, иммунизированных против НА Н5 (Haghighi H.R. et al., PLoS ONE, 2009. 4 (11):p.e7772).
Отметим, что в данном раскрытии изобретения и особенно в формуле и/или пунктах такие термины, как "составляют", "составлен", "составляющий" и им подобные могут иметь значения, придаваемые им в Патентном законодательстве США, напр., они могут означать "включают", "включен", "включающий" и им подобные; а такие термины, как "состоящий в основном из" и "состоит в основном из" имеют значения, придаваемые им в Патентном законодательстве США, напр., они разрешают элементы, не указанные прямо, но исключают элементы, встречающиеся на предшествующем уровне техники или затрагивающие основные или новые характеристики изобретения.
Если не указано иначе, все технические и научные термины, используемые в настоящем изобретении, имеют те же самые значения, которые обычно понимаются специалистами в той области, к которой относится данное изобретение. Значения единственного числа включают и значения множественного числа, если из контекста четко не следует иное. Точно так же слово "или" может включать "и", если из контекста четко не следует иное.
Под "животными" подразумеваются млекопитающие, птицы и др. Животные или организмы хозяина включают млекопитающих и человека. Животные могут быть выбраны из группы, состоящей из лошадиных (напр., лошади), псовых (напр., собак, волков, лис, койотов, шакалов), кошачьих (напр., львов, тигров, домашних кошек, диких кошек, других больших кошек и иных кошачьих, включая гепардов и рысей), овечьих (напр., овец), бычьих (напр., крупного рогатого скота), свиных (напр., свиней), птиц (напр., кур, уток, гусей, индеек, перепелок, фазанов, попугаев, зябликов, ястребов, ворон, страусов, эму и казуаров), приматов (напр., просимид, долгопятов, макак, гиббонов, человекообразных обезьян) и рыб. Термин "животные" также включает индивидуальных животных на всех стадиях развития, включая эмбриональные и зародышевые стадии.
Термины "белок", "пептид", "полипептид" и "фрагмент полипептида" в настоящем изобретении применяются взаимозаменяемым образом к полимерам аминокислотных остатков любой длины. Полимеры могут быть линейными или разветвленными, они могут содержать модифицированные аминокислоты или аналоги аминокислот и могут прерываться другими химическими молекулами, чем аминокислоты. Термины также охватывают полимеры аминокислот, подвергнутые модификации естественным образом или вмешательством, к примеру, образованием дисульфидной связи, гликозилированием, липидированием, ацетилированием, фосфорилированием или любой иной манипуляцией или модификацией типа конъюгирования с меченым или биоактивным компонентом.
Антигенные полипептиды по изобретению способны давать защиту от гриппа. То есть они способны стимулировать иммунные ответы у животных. Под "антигеном" или "иммуногеном" подразумевается субстанция, вызывающая специфический иммунный ответ у животного-хозяина. Антиген может составлять целый организм, убитый, ослабленный или живой; субъединица или часть организма; рекомбинантный вектор, содержащий вставку с иммуногенными свойствами; кусок или фрагмент ДНК, способный вызвать иммунный ответ при предоставлении животному-хозяину; полипептид, эпитоп, гаптен или любая их комбинация. С другой стороны, иммуноген или антиген может содержать токсин или антитоксин.
Термин "иммуногенный или антигенный полипептид" в настоящем изобретении охватывает полипептиды, иммунологически активные в том смысле, что при введении хозяину они способны вызывать иммунный ответ гуморального и/или клеточного типа против белка. Предпочтительно фрагмент белка таков, что он обладает практически той же иммунологической активностью, что и целый белок. Таким образом, фрагмент белка по изобретению содержит или состоит в основном или состоит по меньшей мере из одного эпитопа или антигенной детерминанты. Термин "иммуногенный или антигенный" полипептид в настоящем изобретении включает полную последовательности белка, его аналоги или его иммуногенные фрагменты. Под "иммуногенным или антигенным фрагментом" понимается фрагмент белка, который содержит один или несколько эпитопов и тем самым вызывает иммунологический ответ, описанный выше. Такие фрагменты могут быть идентифицированы при помощи любого количества методик картирования эпитопов, хорошо известных в данной области. Напр., см. Epitope Mapping Protocols in Methods in Molecular Biology, Vol.66 (Glenn E.Morris, Ed., 1996). К примеру, линейные эпитопы могут быть определены, напр., путем одновременного синтеза большого количества пептидов на твердых подложках, причем эти пептиды соответствуют частям молекулы белка, и проведения реакции пептидов с антителами, когда пептиды все еще прикреплены к подложкам. Такие методики известны и описаны, к примеру, в U.S. Pat. No.4,708,871; Geysen et al, 1984; Geysen et al., 1986. Точно так же конформационные эпитопы легко идентифицируются путем определения пространственной конформации аминокислот, напр., методом рентгеноструктурного анализа и 2-мерного ядерного магнитного резонанса. Напр., см. Epitope Mapping Protocols, supra. Способы, особенно применимые к белкам Т. parva, полностью описаны в PCT/US2004/022605, включенном путем ссылки во всей полноте.
Как уже обсуждалось, изобретение охватывает активные фрагменты и варианты антигенных полипептидов. Так, термин "иммуногенный или антигенный полипептид" также предусматривает делеции, вставки и замены в последовательности, если только полипептид продолжает вызывать выработку иммунологического ответа, как описано. Термин "консервативная вариация" обозначает замену одного аминокислотного остатка другим биологически сходным остатком либо замену нуклеотида в нуклеотидной последовательности таким образом, чтобы кодируемый аминокислотный остаток не изменялся либо заменялся на другой биологически сходный остаток. В этом смысле особо предпочтительные замены в общем будут консервативными по природе, то есть это замены, которые имеют место внутри семейства аминокислот.К примеру, аминокислоты в общем делятся на четыре семейства:(1) кислые - аспартат и глутамат; (2) основные - лизин, аргинин, гистидин; (3) неполярные - аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан; и (4) незаряженные полярные - глицин, аспарагин, глутамин, цистин, серин, треонин, тирозин. Фенилаланин, триптофан и тирозин иногда классифицируют как ароматические аминокислоты. Примеры консервативных вариаций включают замену одного гидрофобного остатка, такого как изолейцин, валин, лейцин или метионин, на другой гидрофобный остаток, или замену одного полярного остатка на другой полярный остаток, как-то замену аргинина на лизин, глутаминовой кислоты на аспарагиновую кислоту или глутамина на аспарагин и т.д.; или подобную консервативную замену аминокислоты на структурно родственную аминокислоту, которая не будет иметь большого эффекта на биологическую активность. Таким образом, белки с практически такой же аминокислотной последовательностью, что и контрольная молекула, но содержащие замены второстепенных аминокислот, которые практически не влияют на иммуногенность белка, попадают под определение контрольного полипептида. Все полипептиды, образованные такими модификациями, включены в настоящее изобретение. Термин "консервативная вариация" также включает применение замещенных аминокислот вместо незамещенных исходных аминокислот при условии, что антитела против замещенного полипептида также будут обладать иммунореактивностью к незамещенному полипептиду.
Термин "эпитоп" относится к тому сайту на антигене или гаптене, с которым взаимодействуют специфичные В-клетки и/или Т-клетки. Термин также применяется взаимозаменяемым образом с "антигенной детерминантой" или "сайтом антигенной детерминанты". Антитела, распознающие один и тот же эпитоп, могут быть идентифицированы методом простого иммуноанализа, показывающим способность одного антитела блокировать связывание другого антитела с антигеном мишени.
"Иммунологический ответ" на композицию или вакцину - это развитие в организме хозяина клеточного и/или опосредованного антителами иммунного ответа на данную композицию или вакцину. Обычно "иммунологический ответ" включает один или несколько из следующих эффектов:образование антител, В-клеток, хелперных Т-клеток и/или цитотоксических Т-клеток, направленных конкретно на антиген или антигены, включенные в данную композицию или вакцину. Предпочтительно у хозяина должен проявляться терапевтический либо защитный иммунологический ответ таким образом, что будет усиливаться сопротивляемость к новой инфекции и/или будет уменьшаться клиническая тяжесть заболевания. Такая защита будет проявляться уменьшением или отсутствием симптомов, обычно проявляющихся у инфицированного хозяина, более коротким сроком выздоровления и/или снижением титра вируса у инфицированного хозяина.
Синтетические антигены также включены в это определение, например, полиэпитопы, фланкирующие эпитопы и другие рекомбинантные или синтетически полученные антигены. Напр., см. Bergmann et al., 1993; Bergmann et al., 1996; Suhrbier, 1997; Gardner et al., 1998. Иммуногенные фрагменты для целей настоящего изобретения обычно включают по меньшей мере 3 аминокислоты, по меньшей мере 5 аминокислот, по меньшей мере 10-15 аминокислот или 15-25 аминокислот или более аминокислот в молекуле. Не существует критической верхней границы для длины фрагмента, который может составлять почти полную последовательность белка или даже слитый белок, содержащий по меньшей мере один эпитоп белка.
Соответственно, полинуклеотид, экспрессирующий эпитоп, имеет такую минимальную структуру, которая содержит или состоит в основном или состоит из нуклеотидов, кодирующих эпитоп или антигенную детерминанту полипептида гриппа. Полинуклеотид, кодирующий фрагмент полипептида гриппа, может содержать или состоять в основном или состоять из как минимум 15 нуклеотидов, примерно 30-45 нуклеотидов, примерно 45-75 или по меньшей мере 57, 87 или 150 последовательных или смежных нуклеотидов в последовательности, кодирующей полипептид. При воплощении изобретения можно использовать такие методы определения эпитопов, как создание библиотек перекрывающихся пептидов (Hemmer et al., 1998), Pepscan (Geysen et al., 1984; Geysen et al., 1985; Van der Zee R. et al., 1999; Multipin®™. Peptide Synthesis Kits de Chiron) и алгоритмы (De Groot et al., 1999; PCT/US 2004/022605).
Термины "нуклеиновая кислота" и "полинуклеотид" относятся к РНК или ДНК, линейным или разветвленным, однонитчатым или двунитчатым, либо к их гибридам. Термин также охватывает гибриды РНК/ДНК. Неограничивающими примерами полинуклеотидов являются:гены или фрагменты генов, экзоны, интроны, мРНК, тРНК, рРНК, рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, вектора, выделенная ДНК любой последовательности, выделенная РНК любой последовательности, зонды из нуклеиновых кислот и праймеры. Полинуклеотид может содержать модифицированные нуклеотиды, как-то метилированные нуклеотиды и аналоги нуклеотидов, урацил, другие сахара и соединительные группы типа фторорибозы и тиолата и нуклеотидные ветки. Последовательность полинуклеотидов может быть дополнительно модифицирована после полимеризации, как-то посредством конъюгации с меченой компонентой. Другие типы модификаций, охватываемые данным определением, включают кэпы, замены одного или нескольких природных нуклеотидов их аналогами и введение средств для присоединения полинуклеотида к белкам, ионам металлов, меченым компонентам, другим полинуклеотидам или к твердой подложке. Полинуклеотиды могут быть получены химическим синтезом либо происходить из микроорганизма.
Термин "ген" применяется в широком смысле для обозначения любых сегментов полинуклеотидов, связанных с биологической функцией. При этом гены включают интроны и экзоны, как в геномной последовательности, либо только кодирующие последовательности, как в кДНК, и/или регуляторные последовательности, необходимые для их экспрессии. Например, ген также означает фрагмент нуклеиновой кислоты, который экспрессирует мРНК или функциональную РНК либо кодирует определенный белок, а также включает регуляторные последовательности.
Изобретение также включает комплементарные нити полинуклеотидов, кодирующих антиген, эпитоп или иммуноген гриппа. Комплементарная нить может быть полимером любой длины, а также содержать дезоксирибонуклеотиды, рибонуклеотиды и аналоги в любой комбинации.
"Выделенный" биологический компонент (как-то нуклеиновая кислота или белок или органелла) означает такой компонент, который был существенно выделен или очищен от других биологических компонентов в тех клетках организма, в которых этот компонент встречается в природе, например, других хромосомных и внехромосомных ДНК и РНК, белков и органелл. К "выделенным" нуклеиновым кислотам и белкам относятся нуклеиновые кислоты и белки, очищенные стандартными методами очистки. Термин также охватывает нуклеиновые кислоты и белки, полученные по рекомбинантной технологии, а также химическим синтезом.
Термин "очищенный" в настоящем изобретении не требует абсолютной чистоты, скорее он служит относительным термином. Так, например, очищенным препаратом полипептида является такой, в котором полипептид является более обогащенным, чем в своем естественном окружении. То есть этот полипептид отделен от других клеточных компонентов. Под практически очищенным подразумевается то, что было удалено по меньшей мере 60%, 70%, 80%, 90%, 95% или по меньшей мере 98% или больше клеточных компонентов или материалов. Точно так же полипептид может быть частично очищенным. Под частично очищенным подразумевается то, что было удалено менее 60% клеточных компонентов или материалов. То же самое касается полинуклеотидов. Полипептиды, приведенные в настоящем изобретении, могут быть очищены любым известным способом.
Как отмечено выше, антигенными полипептидами либо их фрагментами или вариантами являются антигенные полипептиды гриппа, продуцируемые в ряске. Фрагменты и варианты приведенных полинуклеотидов и кодируемых ими полипептидов также охватываются настоящим изобретением. Под "фрагментом" имеется в виду часть полинуклеотида или часть кодируемой им антигенной последовательности аминокислот.Фрагменты полинуклеотидов могут кодировать фрагменты белков, сохраняющие биологическую активность нативного белка, поэтому они обладают иммуногенной активностью, как отмечено выше. Фрагменты последовательностей полипептидов сохраняют способность вызывать защитный иммунитет у животных.
"Варианты" служат для обозначения существенно сходных последовательностей. У полинуклеотидов варианты включают делеции и/или вставки одного или нескольких нуклеотидов по одному или нескольким сайтам в нативном полинуклеотиде и/или замены одного или нескольких нуклеотидов по одному или нескольким сайтам в нативном полинуклеотиде. В настоящем изобретении "нативный" полинуклеотид или полипептид содержит природную последовательность нуклеотидов или аминокислот, соответственно. Варианты определенного полинуклеотида по изобретению (то есть контрольного полинуклеотида) также можно оценивать путем сравнения степени идентичности последовательности между полипептидом, кодируемым вариантом полинуклеотида, и полипептидом, кодируемым контрольным полинуклеотидом. "Вариант" белка означает белок, полученный из нативного белка путем делеции или вставки одной или нескольких аминокислот по одному или нескольким сайтам в нативном белке и/или замены одной или нескольких аминокислот по одному или нескольким сайтам в нативном белке. Варианты белков, охваченные настоящим изобретением, являются биологически активными, то есть они способны вызывать иммунный ответ.
Гомологи полипептидов гриппа из птиц, свиней, лошадей, кошек, собак, уток, индеек, кур, перепелок и других видов, включая диких животных, входят в рамки настоящего изобретения. В настоящем изобретении термин "гомологи" включает ортологи, аналоги и паралоги. Термин "аналоги" относится к двум полинуклеотидам или полипептидам, имеющим одинаковые или близкие функции, но возникших по отдельности в неродственных организмах. Термин "ортологи" относится к двум полинуклеотидам или полипептидам из разных видов, но возникших из общего предкового гена при расхождении видов. В норме ортологи кодируют полипептиды, имеющие одинаковые или сходные функции. Термин "паралоги" относится к двум полинуклеотидам или полипептидам, возникшим путем дупликации в геноме. Паралоги обычно имеют разные функции, но эти функции могут быть связанными. Аналоги, ортологи и паралоги полипептида гриппа дикого типа могут отличаться от полипептида гриппа дикого типа посттрансляционными модификациями, различиями в аминокислотной последовательности или тем и другим. В частности, гомологи по изобретению обычно проявляют идентичность последовательности по меньшей мере на 80-85%, 85-90%, 90-95% или 95%, 96%, 7%, 98% или 99% со всей или частью последовательности полипептида или полинуклеотида гриппа дикого типа и проявляют сходные функции. Варианты включают аллельные варианты. Термин "аллельные варианты" относится к полинуклеотидам или полипептидам, содержащим полиморфизмы, вызывающие изменения аминокислотной последовательности белка и существующие в природной популяции (напр., вида или разновидности вируса). Такие природные аллельные варианты, как правило, вносят 1-5% вариаций в полинуклеотиде или полипептиде. Аллельные варианты могут быть идентифицированы при секвенировании данной последовательности нуклеиновой кислоты у целого ряда различных видов, что легко выполняется при помощи гибридизационных зондов для идентификации генетических локусов одного и того же гена у этих видов. Предусматривается, что все такие вариации нуклеиновых кислот и возникающие при этом полиморфизмы или вариации аминокислот, которые являются результатом естественной аллельной изменчивости и не изменяют функциональную активность данного гена, входят в рамки изобретения.
В настоящем изобретении термин "производное" или "вариант" относится к полипептидам или нуклеиновым кислотам, кодирующим полипептиды, содержащим одну или несколько консервативных изменений аминокислот или других небольших модификаций, при которых (1) соответствующий полипептид обладает практически эквивалентной функцией в сравнении с полипептидом дикого типа или (2) антитело против этого полипептида проявляет иммунореактивность к полипептиду дикого типа. Эти варианты или производные включают полипептиды, содержащие небольшие модификации первичной аминокислотной последовательности полипептидов гриппа, которые могут давать пептиды, обладающие практически эквивалентной активностью по сравнению с соответствующим немодифициованным полипептидом. Такие модификации могут быть преднамеренными, как при направленном мутагенезе, или спонтанными. Термин "вариант" также предусматривает делеции, вставки и замены в последовательности, если только полипептид продолжает вызывать иммунологический ответ, как описано. Термин "вариант" также включает такие модификации полипептида, при которых природный сигнальный пептид заменяется на гетерологичный сигнальный пептид, что облегчает экспрессирование или секрецию полипептида из организма хозяина. Он также включает такие модификации полипептида, при которых трансмембранный домен и/или цитоплазматический хвост заменяется на сходную гетерологичную последовательность, что облегчает мембранную экспрессию полипептида в организме хозяина.
Термин "консервативная замена" обозначает замену аминокислотного остатка другим биологически сходным остатком или замену нуклеотида в последовательности нуклеиновой кислоты, при которой кодируемый аминокислотный остаток не изменяется или заменяется на другой биологически сходный остаток. В этом отношении особенно предпочтительные замены в общем должны быть консервативными по природе, как описано выше.
Полинуклеотиды изобретения включают последовательности, которые являются результатом вырожденности генетического кода, напр., оптимизированного употребления кодонов у определенного хозяина. В настоящем изобретении "оптимизированные" относятся к полинуклеотидам, подвергнутым генетической инженерии для усиления их экспрессии у данного вида. Для получения оптимизированных полинуклеотидов, кодирующих полипептиды гриппа, последовательность ДНК гена белка вируса гриппа можно модифицировать так, чтобы она:1) содержала кодоны, которые предпочтительны у сильно экспрессируемых генов у данного вида; 2) имела практически такое же содержание А+Т или G+C в составе оснований нуклеотидов, как у данного вида; 3) образовала последовательность инициации данного вида; или 4) устранить последовательности, которые вызывают дестабилизацию, неправильное полиаденилирование, деградацию и экстерминацию РНК либо образуют вторичные структуры типа шпилек или сайты сплайсинга РНК. Усиление экспрессии белка гриппа у данного вида достигается использования частоты распределения употребительности кодонов у эукариот и прокариот или у определенного вида. Термин "частота предпочтительного употребления кодонов" относится к проявляемой клетками определенного хозяина предпочтительности употребления нуклеотидных кодонов для определения данной аминокислоты. Существует 20 природных аминокислот, большинство которых определяются более чем одним кодоном. Поэтому все вырожденные последовательности нуклеотидов включены в изобретение, если только последовательность аминокислот полипептида гриппа, кодируемого последовательностью нуклеотидов, будет функционально неизмененной.
Степень идентичности между двумя последовательностями аминокислот может быть установлена с помощью программы попарного сравнения BLAST и матрикса blosum62 из NCBI (Национального центра информации по биотехнологии), используя стандартные параметры (напр., см. алгоритм BLAST или BLASTX, доступный на сервере NCBI (Bethesda, MD, США), а также в Altschul et al.; при этом в настоящем документе об использовании алгоритма или BLAST или BLASTX и матрикса blosum62 говорится с помощью термина "бласт").
"Идентичность" в отношении последовательностей может относиться к количеству положений с идентичными нуклеотидами или аминокислотами, деленному на количество нуклеотидов или аминокислот в более короткой из двух последовательностей, при этом совмещение двух последовательностей может проводиться в соответствии с алгоритмом Вилбура и Липмана (Wilbur and Lipman), например, при размере окна в 20 нуклеотидов, длине слова в 4 нуклеотида и штрафе за пробел в 4 балла, а анализ с помощью компьютера и интерпретацию данных о последовательности, включая их сопоставление, можно удобно проводить с помощью коммерчески доступных программ (напр., комплекта Intelligenetics™ Suite, Intelligenetics Inc., CA). Когда говорят, что последовательности РНК являются сходными или имеют какую-то степень идентичности или гомологичности последовательности с последовательностями ДНК, то тимидин (Т) в последовательности ДНК считается равным урацилу (U) в последовательности РНК. Таким образом, последовательности РНК входят в рамки изобретения и могут быть выведены из последовательностей ДНК, полагая тимидин (Т) в последовательности ДНК равным урацилу (U) в последовательности РНК.
Степень идентичности или степень схожести между двумя аминокислотными последовательностями либо степень идентичности между двумя нуклеотидными последовательностями можно определить с помощью пакета программ Vector NTI (Invitrogen, 1600 Faraday Ave., Carlsbad, CA).
В следующих документах представлены алгоритмы для сравнения относительной идентичности или гомологичности последовательностей, а кроме того или наряду с вышесказанным положения этих ссылок можно использовать для определения степени гомологичности или идентичности:Needleman SB and Wunsch CD; Smith TF and Waterman MS; Smith TF, Waterman MS and Sadler JR; Feng DF and Dolittle RF; Higgins DG and Sharp PM; Thompson JD, Higgins DG and Gibson TJ; Devereux J, Haeberlie P and Smithies О. И без излишнего экспериментирования специалисты могут воспользоваться многими другими программами или ссылками для определения степени гомологичности.
Реакции гибридизации могут проводиться в условиях различной "жесткости". Условия, повышающие жесткость реакции гибридизации, хорошо известны. Например, см. "Molecular Cloning:A Laboratory Manual", 2nd edition (Sambrook et al., 1989).
Вектор" означает плазмиду из рекомбинантной ДНК или РНК или вирус, содержащие гетерологичный полинуклеотид для доставки в клетки мишени in vitro либо in vivo. Гетерологичный полинуклеотид может содержать представляющие интерес последовательности, предназначенные для профилактики или терапии, и необязательно может иметь вид экспрессионной кассеты. В настоящем изобретении вектор не обязательно способен к репликации в клетках конечной мишени или субъекта. Термин охватывает клонирующие вектора и вирусные вектора.
Термин "рекомбинантный" обозначает полинуклеотиды полусинтетического или синтетического происхождения, которые либо не встречаются в природе, либо соединены с другим полинуклеотидом в такой конфигурации, которая не встречается в природе.
"Гетерологичный" означает происходящий из генетически отличающегося объекта от того объекта, с которым он сравнивается. Например, полинуклеотид может быть вставлен методами генетической инженерии в плазмиду или вектор из другого источника - это и есть гетерологичный полинуклеотид. Промотор, отделенный от своей собственной кодирующей последовательности и оперативно связанный с другой кодирующей последовательностью, является гетерологичным промотором.
Настоящее изобретение касается птичьих вакцин либо фармацевтических или иммунологических композиций, которые могут включать эффективное количество рекомбинантного антигена птичьего гриппа и фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель.
Объект изобретения, раскрытого здесь, частично касается композиций и способов, связанных с неожиданным открытием того, что антиген птичьего гриппа, полученный в растительной системе экспрессии белка, оказался высоко иммуногенным и защищал кур от воздействия гомологичных и гетерологичных штаммов птичьего гриппа.
Композиции
В одном воплощении объектом изобретения, раскрытого здесь, является композиция, содержащая антиген гриппа и фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель.
В одном воплощении объектом изобретения, раскрытого здесь, является композиция, содержащая антиген птичьего гриппа, полученный в системе экспрессии Lemna, и фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель.
В одном воплощении объектом изобретения, раскрытого здесь, является композиция, содержащая антиген птичьего гриппа, полученный в системе экспрессии Lemna, и растительный материал из рода Lemna, и фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель.
В одном воплощении объектом изобретения, раскрытого здесь, является белок, полученный в системе экспрессии Lemna, содержащий антиген птичьего гриппа. Белок может быть гликозилированными.
В одном воплощении объектом изобретения, раскрытого здесь, является белок, полученный в системе экспрессии Lemna, содержащий антиген птичьего гриппа и растительный материал из рода Lemna.
В одном воплощении объектом изобретения, раскрытого здесь, является устойчиво трансформированные растения или растительные культуры, экспрессирующие антиген птичьего гриппа, причем растения или растительные культуры выбираются из рода Lemna.
В одном воплощении, в котором иммунологическая композиция или вакцина от птичьего гриппа является рекомбинантной иммунологической композицией или вакциной, композиция или вакцина содержит рекомбинантный вектор и фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель; рекомбинантный вектор является вектором для растительной экспрессии, который может содержать полинуклеотид, кодирующий полипептид, антиген, эпитоп или иммуноген гриппа. Полипептидом, антигеном, эпитопом или иммуногеном гриппа может быть гемагглютинин, белок матрикса, нейраминидаза, неструктурный белок, нуклеопротеид, полимераза или любой их фрагмент.
В другом воплощении полипептид, антиген, эпитоп или иммуноген гриппа может происходить из птиц, инфицированных штаммом гриппа или птичьего гриппа. В одном воплощении антигеном, эпитопом или иммуногеном птичьего гриппа является белок гемагглютинина (НА) (напр., предшественник НА0, НА1 и/или НА2) H1 или Н2, белок матрикса (напр., белок матрикса M1 или М2), нейраминидаза, неструктурный (NS) белок (напр., NS1 или NS2), нуклеопротеид (NP) и полимераза (напр., полимераза РА, полимераза-1 РВ1 или полимераза-2 РВ2). Вирусы гриппа типа А могут инфицировать людей, птиц, свиней, лошадей, собак, кошек и других животных, но естественным хозяином для этих вирусов являются дикие птицы.
В другом воплощении антигеном птичьего гриппа может быть гемагглютинин (НА) из других подтипов гриппа А (примеры:H1, Н3, Н5, Н6, Н7, Н9). В следующем воплощении антигеном птичьего гриппа может быть НА из изолята H5N1. В другом воплощении антиген H5N1 выделяют из штамма А/курица/Индонезия/7/2003.
Настоящее изобретение касается птичьих вакцин или композиций, которые могут включать эффективное количество рекомбинантного антигена птичьего гриппа и фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель. В одном воплощении антигеном птичьего гриппа может быть гемагглютинин.
В другом воплощении рекомбинантный антиген гриппа экспрессируется в растениях. В следующем воплощении растения представляют собой растения ряски. В следующем воплощении растения представляют собой растения Lemna. В одном воплощении рекомбинантный антиген гриппа может экспрессироваться в запатентованной системе экспрессирования белков в Lemna minor - системе LEX™ фирмы Biolex.
В другом воплощении фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель может представлять собой эмульсию типа вода в масле. В следующем воплощении эмульсия типа вода в масле может представлять собой тройную эмульсию вода/масло/вода (W/O/W). В следующем воплощении фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель может представлять собой эмульсию типа масло в воде.
Изобретение также охватывает полинуклеотиды гриппа, содержащиеся в молекуле вектора или экспрессионного вектора и функционально связанные с элементом промотора и необязательно с энхансером.
В одном аспекте настоящего изобретения предусмотрены полипептиды гриппа, в частности полипептиды птичьего гриппа. В одном аспекте настоящего изобретения предусмотрены полипептиды с последовательностями, приведенными в SEQ ID NO:2, 4, 5, 8, 10, 12 или 14, и их варианты или фрагменты.
В другом аспекте настоящего изобретения предусмотрены полипептиды, у которых последовательности по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96% 97% 98% или 99% идентичны антигенным полипептидам изобретения, в частности полипептидам с последовательностями, приведенными в SEQ ID NO:2, 4, 5, 8, 10, 12 или 14.
В следующем аспекте настоящего изобретения предусмотрены фрагменты и варианты полипептидов гриппа, приведенных выше (SEQ ID NO:2, 4, 5, 8, 10, 12 или 14), которые могут легко быть получены специалистами хорошо известными методами молекулярной биологии.
Варианты представляют собой гомологичные полипептиды, у которых аминокислотные последовательности по меньшей мере на 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичны антигенным полипептидам изобретения, в частности аминокислотным последовательностям, приведенным в SEQ ID NO:2, 4, 5, 8, 10, 12 или 14.
Иммуногенные фрагменты полипептидов гриппа включают по меньшей мере 8, 10, 15 или 20 последовательных аминокислот, по меньшей мере 21 аминокислоту, по меньшей мере 23 аминокислоты, по меньшей мере 25 аминокислот или по меньшей мере 30 аминокислот полипептидов гриппа с последовательностями, приведенными в SEQ ID NO:2, 4, 5, 8, 10, 12 или 14, либо их вариантов. В другом воплощении фрагменты полипептидов гриппа включают специфический антигенный эпитоп, находящийся на полномерном полипептиде гриппа.
В другом аспекте настоящего изобретения предусмотрены полинуклеотиды, кодирующие полипептиды гриппа, как-то полинуклеотиды, кодирующие полипептиды с последовательностями, приведенными в SEQ ID NO:2,4, 5, 8, 10, 12 или 14. В следующем аспекте настоящего изобретения предусмотрены полинуклеотиды, кодирующие полипептиды, у которых последовательности по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичны полипептидам с последовательностями, приведенными в SEQ ID NO:2, 4, 5, 8, 10, 12 или 14, либо их консервативные варианты, аллельные варианты, гомологи или иммуногенные фрагменты, содержащие по меньшей мере 8 или по меньшей мере 10 последовательных аминокислот одного из этих полипептидов, либо комбинации этих полипептидов.
В другом аспекте настоящего изобретения предусмотрены полинуклеотиды с нуклеотидными последовательностями, приведенными в SEQ ID NO:1, 3, 7, 9, 11 или 13, либо их варианты. В следующем аспекте настоящего изобретения предусмотрены полинуклеотиды, у которых последовательности по меньшей мере на 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичны одному из полинуклеотидов с последовательностями, приведенными в SEQ ID NO:1, 3, 7, 9, 11 или 13, либо его вариантам.
Полинуклеотиды изобретения могут содержать дополнительные последовательности, как-то дополнительные кодирующие последовательности в пределах одной и той же единицы транскрипции, контролирующие элементы, как-то промоторы, сайты связывания рибосом, 5'-UTR, 3'-UTR, терминаторы транскрипции, сайты полиаденилирования, дополнительные единицы транскрипции под контролем того же или другого промотора, последовательности, позволяющие клонирование, экспрессирование, гомологическую рекомбинацию и трансформацию клеток хозяина, и любые такие конструкции, которые могут быть нужны для обеспечения воплощений данного изобретения.
В векторах изобретения предпочтительно присутствуют элементы для экспрессии полипептида, антигена, эпитопа или иммуногена гриппа. Как минимум, они включают, состоят в основном или состоят из кодона инициации (ATG), стоп-кодона и промотора, а также необязательно последовательности полиаденилирования для некоторых векторов, как-то плазмид и некоторых вирусных векторов, напр., других вирусных векторов, чем поксвирусы. Если полинуклеотид кодирует фрагмент полипептида, напр., пептид гриппа, то в векторе предпочтительно ATG находится на 5'-конце рамки считывания, а стоп-кодон находится на 3'-конце. Могут присутствовать и другие элементы для контролирования экспрессии, как-то последовательности энхансеров, стабилизирующие последовательности типа интронов и сигнальные последовательности, способствующие секреции белка.
Настоящее изобретение также касается препаратов, содержащих вектора, как-то экспрессирующие вектора, напр., терапевтических композиций. Препараты могут содержать один или несколько векторов, напр., экспрессирующих векторов, как-то экспрессирующих in vivo векторов, включающих и экспрессирующих один или несколько полипептидов, антигенов, эпитопов или иммуногенов гриппа. В одном воплощении вектор содержит или экспрессирует полинуклеотид, который включает, состоит в основном или состоит из полинуклеотида, кодирующего (и предпочтительно экспрессирующего) антиген, эпитоп или иммуноген гриппа, в фармацевтически или ветеринарно приемлемом носителе, эксципиенте или наполнителе. Так, в одном воплощении изобретения другой вектор или другие вектора в препарате включают, состоят в основном или состоят из полинуклеотидов, кодирующих и при соответствующих условиях экспрессирующих один или несколько других белков полипептида, антигена, эпитопа или иммуногена гриппа (напр., гемагглютинина нейраминидазы, нуклеопротеида) или их фрагменты.
В другом воплощении вектор или вектора в препарате включают, состоят в основном или состоят из полинуклеотидов, кодирующих один или несколько белков или фрагментов полипептида, антигена, эпитопа или иммуногена гриппа, причем вектор или вектора экспрессируют эти полинуклеотиды. В другом воплощении препарат содержит один, два или несколько векторов, содержащих полинуклеотиды, кодирующие и экспрессирующие, предпочтительно in vivo, полипептид, антиген, слитый белок или эпитоп гриппа. Изобретение также касается смеси векторов, содержащих полинуклеотиды, кодирующие и экспрессирующие различные полипептиды, антигены, эпитопы или иммуногены гриппа, напр., полипептиды, антигены, эпитопы или иммуногены гриппа из разных видов, как-то человека, лошадей, свиней, собак, кошек в дополнение к видам птиц, включающим кур, уток, индеек, перепелок и гусей.
В следующем воплощении изобретения экспрессирующий вектор представляет собой плазмидный вектор или ДНК-плазмидный вектор, в частности, экспрессирующий in vivo вектор. В одном конкретном неограничивающем примере в качестве вектора для встраивания последовательности полинуклеотида может использоваться плазмида pVR.1020 или 1012 (VICAL Inc.; Luke et al., 1997; Hartikka et al., 1996; напр., см. U.S. Patent Nos.5,846,946 6,451,769). Плазмида pVR1020 происходит из pVR1012 и содержит сигнальную последовательность tPA человека. В одном воплощении сигнал tPA человека включает аминокислоты от М(1) до S(23) из GenBank под номером доступа HUMTPA14. В другом конкретном неограничивающем примере плазмида, используемая в качестве вектора для встраивания последовательности полинуклеотида, может содержать последовательность сигнального пептида IGF1 лошади от аминокислоты М(24) до аминокислоты А(48) из GenBank под номером доступа U28070. Дополнительная информация о ДНК-плазмидах, которые можно принимать во внимание или использовать на практике, приведена, к примеру, в U.S. Patent Nos.6,852,705; 6,818,628; 6,586,412; 6,576,243; 6,558,674; 6,464,984; 6,451,770; 6,376,473 и 6,221,362.
Термин плазмида охватывает любые единицы транскрипции ДНК, содержащие полинуклеотиды по изобретению и элементы, необходимые для их экспрессии in vivo в клетке или клетках нужного хозяина или мишени, причем в этом отношении следует отметить, что в рамки изобретения входят сверхспиральные и не сверхспиральные, кольцевые плазмиды, а также линейные формы.
Каждая плазмида включает или содержит или состоит в основном, наряду с полинуклеотидом, кодирующим антиген, эпитоп или иммуноген гриппа, необязательно слитый с последовательностью гетерологичного пептида, из варианта, аналога или фрагмента, оперативно связанного с промотором или находящегося под контролем промотора или зависимого от промотора. В общем, предпочтительно использовать сильный промотор, функционирующий в клетках эукариот.Сильным промотором может быть самый ранний промотор цитомегаловируса (CMV-IE) человека или мыши либо необязательно имеющий другое происхождение, как-то крысы или морской свинки - суперпромотор (Ni M. et al., Plant J. 7, 661-676, 1995). Промотор CMV-IE может содержать в себе действительный промотор, который может быть связан или не связан с энхансером. Можно обратиться к ЕР-А-260 148, ЕР-А-323 597, U.S. Patent Nos.5,168,062, 5,385,839 и 4,968,615, а также к заявке РСТ No. WO 87/03905. Промотор CMV-IE предпочтительно является промотором CMV-IE человека (Boshart et al., 1985) или CMV-IE мыши.
В более общем смысле промотор имеет вирусное, растительное или клеточное происхождение. Другой сильный вирусный промотор, чем CMV-IE, который может успешно использоваться при воплощении изобретения - это ранний/поздний промотор вируса SV40 или промотор LTR вируса саркомы Рауса. Сильным клеточным промотором, который может успешно использоваться при воплощении изобретения, является промотор какого-нибудь гена цитоскелета, как-то промотор десмина (Kwissa et al., 2000) или промотор актина (Miyazaki et al., 1989).
Можно использовать любые конститутивные, регулируемые или индуцибельные промоторы. Например, к конститутивным промоторам относится промотор маннопинсинтазы из Agrobacterium tumefaciens. С другой стороны, может оказаться выгодным использование промоторов генов теплового шока, промоторов индуцируемых засухой генов, промоторов индуцируемых патогенами генов, промоторов индуцируемых ранами генов и промоторов индуцируемых освещением/темнотой генов. Может оказаться полезным использование промоторов, контролируемых такими регуляторами роста растений, как абсциссовая кислота, ауксины, цитокинины и гиббереллиновая кислота. Также можно выбрать промоторы, обеспечивающие тканеспецифичную экспрессию (напр., промоторы, специфичные для корней, листьев и цветов).
Плазмиды могут содержать и другие элементы контроля экспрессии. Особенно выгодно включение стабилизирующих последовательностей, напр., последовательностей интронов, к примеру, интрона алкогольдегидрогеназы кукурузы (интрона ADHI кукурузы), первого интрона hCMV-IE (заявка РСТ No. WO 1989/01036), интрона II гена р-глобина кролика (van Ooyen et al., 1979). В другом воплощении плазмиды могут содержать 3'-UTR. Таким 3'-UTR может быть 3'-UTR нопалинсинтазы (Nos) Agrobacterium.
Что касается сигнала полиаденилирования (polyA) для плазмид и вирусных векторов помимо поксвирусов, то можно использовать сигнал поли(А) гена бычьего гормона роста (bGH) (см. U.S. 5,122,458), сигнал поли(А) гена β-глобина кролика или сигнал поли(А) вируса SV40.
"Клетки хозяина" означают прокариотические или эукариотические клетки, которые подвергались генетическим изменениям или могут подвергаться генетическим изменениям при применении экзогенных полинуклеотидов типа рекомбинантных плазмид или векторов. При указании генетически измененных клеток термин относится как к первоначально измененным клеткам, так и к их потомству.
В одном воплощении рекомбинантный антиген гриппа экспрессируется в трансгенных растениях ряски. В другом воплощении трансгенные растения представляют собой растения Lemna. В следующем воплощении трансгенные растения представляют собой Lemna minor. В следующем воплощении рекомбинантный антиген гриппа может экспрессироваться в системе экспрессирования белков в Lemna minor - системе LEX™ фирмы Biolex. Подробности о системе экспрессии белков в Lemna minor приведены, к примеру, в U.S. Patent Nos.6,815,184; 7,022,309; 7,160,717; 7,176,024, 6,040,498, 7,161,064 и 7,326,38; содержание которых включено путем ссылки во всей полноте. В воплощениях антигеном гриппа может быть любой полипептид, приведенный в настоящем изобретении, или полипептид, кодируемый любым полинуклеотидом, приведенным в настоящем изобретении.
Способы экспрессирования антигенных полипептидов гриппа в ряске
Итак, в некоторых воплощениях изобретения полипептиды гриппа либо их фрагменты или варианты экспрессируются в ряске. Эти способы включают использование экспрессирующих кассет, которые вводятся в растения ряски любым подходящим методом трансформации, известным в этой области. Полинуклеотиды в этих экспрессирующих кассетах могут подвергаться модификации для усиления экспрессии антигенных полипептидов гриппа либо их фрагментов или вариантов в ряске следующим образом.
Кассеты для экспрессирования в ряске антигенных полипептидов гриппа
Трансгенные растения ряски, экспрессирующие полипептиды гриппа либо их фрагменты или варианты, получают путем трансформации ряски экспрессирующей кассетой, содержащей полинуклеотид, кодирующий полипептид гриппа либо его фрагмент или вариант. При этом полинуклеотид, кодирующий нужный полипептид гриппа либо его фрагмент или вариант, встроен в экспрессирующую кассету и вводится в растения ряски любым подходящим методом трансформации, известным в этой области.
В некоторых воплощениях растения ряски, трансформированные экспрессирующей кассетой, содержащей полинуклеотид, кодирующий нужный полипептид гриппа либо его фрагмент или вариант, также трансформируют экспрессирующей кассетой, обеспечивающей экспрессию другого гетерологичного полипептида, например, другого полипептида гриппа, его фрагмента или варианта. Экспрессирующая кассета, обеспечивающая экспрессию другого гетерологичного полипептида, может быть представлена в том же самом полинуклеотиде (к примеру, в том же самом трансформирующем векторе) для введения в растения ряски или в другом полинуклеотиде (к примеру, в другом трансформирующем векторе) для введения в растения ряски в одно и то же время или в разное время одним и тем же или разными методами введения, например, одним и тем же или разными методами трансформации.
Экспрессирующие кассеты для применения при трансформации ряски включают контролирующие экспрессию элементы, которые как минимум включают участок инициации транскрипции (напр., промотор), функционально связанный с нужным полинуклеотидом, т.е. полинуклеотидом, кодирующим антигенный полипептид гриппа, его фрагмент или вариант.В настоящем изобретении "функционально связанные" в отношении нуклеотидных последовательностей относится к множественным последовательностям нуклеотидов, находящимся в функциональной взаимосвязи друг с другом. Обычно функционально связанные последовательности ДНК являются смежными и, если нужно соединить два кодирующих белки участка, в одной рамке считывания. Такая экспрессирующая кассета снабжена несколькими сайтами рестрикции для встраивания нужного полинуклеотида или полинуклеотидов (напр., одного полинуклеотида, двух полинуклеотидов и т.д.), находящихся под транскрипционным контролем промотора и других контролирующих экспрессию элементов. В определенных воплощениях изобретения подлежащий трансформации полинуклеотид содержит две или несколько экспрессирующих кассет, каждая из которых содержит по меньшей мере один нужный полинуклеотид.
Под "контролирующим экспрессию элементом" имеется в виду регуляторный участок ДНК, обычно содержащий ТАТА-бокс, способный направлять РНК-полимеразу II или в некоторых воплощениях РНК-полимеразу III для инициации синтеза РНК в надлежащем сайте инициации транскрипции для определенной кодирующей последовательности. Контролирующий экспрессию элемент может дополнительно включать другие распознающие последовательности, которые обычно располагаются перед или с 5'-стороны относительно ТАТА-бокса и влияют на (напр., повышают) скорость инициации транскрипции. Кроме того, контролирующий экспрессию элемент может дополнительно содержать последовательности, которые обычно располагаются после или с 3'-стороны относительно ТАТА-бокса и влияют на (напр., повышают) скорость инициации транскрипции.
Участок инициации транскрипции (напр., промотор) может быть нативным или гомологичным либо чужеродным или гетерологичным для растения ряски, а также представлять собой природную последовательность или синтетическую последовательность. Под чужеродным имеется в виду то, что участок инициации транскрипции не встречается в тех организмах ряски дикого типа, в которые он вводится. Под "функциональным промотором" имеется в виду промотор, который, если он функционально связан с последовательностью, кодирующей нужный антигенный полипептид гриппа либо его фрагмент или вариант, способен управлять экспрессией (т.е. транскрипцией и трансляцией) кодируемого полипептида, фрагмента или варианта. Промоторы можно выбирать, исходя из нужного результата. Так, экспрессирующие кассеты изобретения могут содержать конститутивные, индуцибельные, тканеспецифичные или другие промоторы для экспрессии в ряске.
В экспрессирующих кассетах по настоящему изобретению можно использовать любые подходящие промоторы, известные в этой области, включая бактериальные, дрожжевые, грибковые промоторы и промоторы насекомых, млекопитающих и растений. Например, можно использовать промоторы растений, в том числе промоторы ряски. Примеры промоторов включают промотор вируса 35S мозаичности цветной капусты, промоторы опинсинтетазы (напр., nos, mas, ocs и др.), промотор убиквитина, промотор актина, промотор малой субъединицы рибулозобифосфат-(RubP)-карбоксилазы и промотор алкогольдегидрогеназы. Из уровня техники известен промотор малой субъединицы RubP-карбоксилазы ряски (Silverthome et al. (1990) Plant Mol. Biol. 15:49). Подходят и другие промоторы из вирусов, инфицирующих растения, предпочтительно ряску, в том числе промоторы, выделенные из вируса мозаичности колоказии, вируса хлореллы (напр., промотор аденин-метилтрансферазы вируса хлореллы; Mitra et al. (1994) Plant Mol. Biol. 26:85), вируса бронзовости томата, вируса погремковости табака «раттлвирус», вируса некроза табака, вируса кольцевой пятнистости табака, вируса кольцевой пятнистости томата, вируса мозаичности огурца, вируса пеньковидности арахиса, вируса мозаичности люцерны, бацилиформного баднавируса тростника и т.п.
Контролирующие экспрессию элементы, включая промоторы, можно выбрать так, чтобы получить требуемый уровень регуляции. Например, в некоторых случаях может оказаться выгодным использование промотора, придающего конститутивную экспрессию (напр., промотора маннопинсинтазы из Agrobacterium tumefaciens). С другой стороны, в других ситуациях может оказаться выгодным использование таких промоторов, которые активируются в ответ на специфические воздействия окружающей среды (напр., промоторов генов теплового шока, промоторов индуцируемых засухой генов, промоторов индуцируемых патогенами генов, промоторов индуцируемых ранами генов и промоторов индуцируемых освещением/темнотой генов) или регуляторы роста растений (напр., промоторов генов, индуцируемых абсциссовой кислотой, ауксинами, цитокининами и гиббереллиновой кислотой). А также еще можно выбрать промоторы, обеспечивающие тканеспецифичную экспрессию (напр., промоторы, специфичные к корням, листьям и цветкам).
В целом на мощность данного промотора может повлиять комбинация и пространственная организация цис-действующих нуклеотидных последовательностей типа вышележащих активирующих последовательностей. Например, активирующие последовательности нуклеотидов, полученные из гена октопинсинтазы Agrobacterium tumefaciens, могут усиливать транскрипцию из промотора маннопинсинтазы Agrobacterium tumefaciens (см. U.S. Patent 5,955,646 to Gelvin et al.). В настоящем изобретении экспрессирующая кассета может содержать активирующие последовательности нуклеотидов, вставленные выше последовательности промотора для усиления экспрессии нужного антигенного полипептида гриппа либо его фрагмента или варианта. В одном воплощении экспрессирующая кассета включает три вышележащие активирующие последовательности, полученные из гена октопинсинтазы Agrobacterium tumefaciens, функционально связанные с промотором, полученным из гена маннопинсинтазы Agrobacterium tumefaciens (см. U.S. Patent 5,955,646, включенный путем ссылки).
Таким образом, экспрессирующая кассета включает, в направлении транскрипции 5'-3', контролирующий экспрессию элемент, содержащий участок инициации транскрипции и трансляции, полинуклеотид, кодирующий нужный антигенный полипептид гриппа (либо его фрагмент или вариант), и участок терминации транскрипции и трансляции, функционирующий у растений. В соответствии с настоящим изобретением можно использовать любые подходящие последовательности терминации, известные в этой области. Участок терминации может быть нативным к участку инициации транскрипции, нативным к данной кодирующей последовательности или может иметь другое происхождение. Удобные участки терминации доступны из Ti-плазмиды А. tumefaciens, как-то участки терминации октопинсинтетазы и нопалинсинтетазы. Также см. Guerineau et al. (1991) Mol. Gen. Genet. 262:141; Proudfoot (1991) Cell 64:671; Sanfacon et al. (1991) Genes Dev. 5:141; Mogen et al. (1990) Plant Cell 2:1261; Munroe et al. (1990) Gene 91:151; Ballas et al. (1989) Nucleic Acids Res. 17:7891; и Joshi et al. (1987) Nucleic Acids Res. 15:9627. Дополнительные примеры последовательностей терминации последовательность терминации малой субъединицы RubP-карбоксилазы гороха и последовательность терминации вируса 35S мозаичности цветной капусты.
В общем, экспрессирующая кассета должна содержать ген селекционного маркера для селекции трансформированных клеток или тканей ряски. Гены селекционных маркеров включают гены, кодирующие устойчивость к антибиотикам типа тех, что кодируют неомицин-фосфотрансферазу II (NEO) и гигромицин-фосфотрансферазу (НРТ), а также гены, придающие устойчивость к соединениям гербицидов. Гены устойчивости к гербицидам обычно кодируют модифицированный белок мишени, нечувствительный к гербициду, или фермент, который разлагает или детоксифицирует гербицид в растениях до того, как он подействует:см. DeBlock et al. (1987) EMBO J. 6:2513; DeBlock et al. (1989) Plant Physiol. 91:691; Fromm et al. (1990) BioTechnology 8:833; Gordon-Kamm et al. (1990) Plant Cell 2:603. Например, устойчивость к гербицидам типа глифосфата или сульфонилмочевины получали с помощью генов, кодирующих мутантные ферменты мишени 5-енолпирувилшикимат-3-фосфатсинтазу (EPSPS) и ацетолактатсинтазу (ALS). Устойчивость к глюфосинату аммония, бормоксинилу и 2,4-дихлорфеноксиацетату (2,4-D) получали с помощью бактериальных генов, кодирующих фосфинотрицин-ацетилтрансферазу, нитрилазу или 2,4-дихлорфеноксиацетат-монооксигеназу, которые детоксифицируют соответствующие гербициды.
В целях настоящего изобретения гены селекционных маркеров включают гены, кодирующие неомицин-фосфотрансферазу II (Fraley et al. (1986) CRC Critical Reviews in Plant Science 4:1); цианамидгидратазу (Maier-Greiner et al. (1991) Proc. Natl. Acad. Sci. USA 88:4250); аспартаткиназу; дигидропиколинатсинтазу (Peri et al. (1993) BioTechnology 11:715); ген bar (Toki et al. (1992) Plant Physiol. 100:1503; Meagher et al. (1996) Crop Sci. 36:1367); триптофандекарбоксилазу (Goddijn et al. (1993) Plant Mol. Biol. 22:907); неомицин-фосфотрансферазу (NEO; Southern et al. (1982) J. Mol. Appl. Gen. 1:327); гигромицин-фосфотрансферазу (НРТ или HYG; Shimizu et al. (1986) Mol. Cell. Biol. 6:1074); дигидрофолатредуктазу (DHFR; Kwok et al. (1986) Proc. Natl. Acad. Sci. USA 83:4552); фосфинотрицин-ацетилтрансферазу (DeBlock et al. (1987) EMBO J. 6:2513); дегалогеназу 2,2-дихлорпропионовой кислоты (Buchanan-Wollatron et al. (1989) J. Cell. Biochem. 13D:330); синтазу ацетогидроксикислот (U.S. Pat. No.4,761,373 to Anderson et al.; Haughn et al. (1988) Mol. Gen. Genet. 221:266); 5-енолпирувилшикимат-фосфатсинтазу (aroA; Comai et al. (1985) Nature 317:741); галоарилнитрилазу (WO 87/04181 to Stalker et al.); ацетил-КоА-карбоксилазу (Parker et al. (1990) Plant Physiol. 92:1220); дигидроптероатсинтазу (sull; Guerineau et al. (1990) Plant Mol. Biol. 15:127); и 32 кДа полипептид фотосистемы II (psbA; Hirschberg et al. (1983) Science 222:1346 (1983).
Также включены гены, кодирующие устойчивость к гентамицину (напр., aacCl; Wohlleben et al. (1989) Mol. Gen. Genet. 217:202-208); хлорамфениколу (Herrera-Estrella et al. (1983) EMBO J. 2:987); метотрексату (Herrera-Estrella et al. (1983) Nature 303:209; Meijer et al. (1991) Plant Mol. Biol. 16:807); гигромицину (Waldron et al. (1985) Plant Mol. Biol. 5:103; Zhijian et al. (1995) Plant Science 108:219; Meijer et al. (1991) Plant Mol. Biol. 16:807); стрептомицину (Jones et al. (1987) Mol. Gen. Genet. 210:86); спектиномицину (Bretagne-Sagnard et al. (1996) Transgenic Res. 5:131); блеомицину (Hille et al. (1986) Plant Mol. Biol. 7:171); сульфонамидам (Guerineau et al. (1990) Plant Mol. Biol. 15:127); бромоксинилу (Stalker et al. (1988) Science 242:419); 2,4-D (Streber et al. (1989) BioTechnology 7:811); фосфинотрицину (DeBlock et al. (1987) EMBO J. 6:2513); спектиномицину (Bretagne-Sagnard and Chupeau, Transgenic Res. 5:131).
Ген bar придает устойчивость к гербицидам типа глюфосината, как-то фосфинотрицину (РРТ) или биалафосу и др. Как отмечено выше, другие селекционные маркеры, которые могут использоваться в векторных конструкциях, включают ген pat, тоже для устойчивости к биалафосу и фосфинотрицину, ген ALS для устойчивости к имидазолинону, ген НРТ или HYG для устойчивости к гигромицину, ген EPSP-синтазы для устойчивости к глифосату, ген Hm1 для устойчивости к Нс-токсину и другие селективные реагенты, обычно используемые и известные рядовым специалистам. См. Yarranton (1992) Curr. Opin. Biotech. 3:506; Chistopherson et al. (1992) Proc. Natl. Acad. Sci. USA 89:6314; Yao et al. (1992) Cell 71:63; Reznikoif(1992) Mol. Microbiol. 6:2419; Barkley et al. (1980) The Operon 177-220; Hu et al. (1987) Cell 48:555; Brown et al. (1987) Cell 49:603; Figge et al. (1988) Cell 52:713; Deuschle et al. (1989) Proc. Natl. Acad. Sci. USA 86:5400; Fuerst et al. (1989) Proc. Natl. Acad. Sci. USA 86:2549; Deuschle et al. (1990) Science 248:480; Labow et al. (1990) Mol. Cell. Biol. 10:3343; Zambretti et al. (1992) Proc. Natl. Acad. Sci. USA 89:3952; Bairn et al. (1991) Proc. Natl. Acad. Sci. USA 88:5072; Wyborski et al. (1991) Nuc. Acids Res. 19:4647; Hillenand-Wissman (1989) Topics in Mol. and Struct. Biol. 10:143; Degenkolb et al. (1991) Antimicrob. Agents Chemother. 35:1591; Kleinschnidt et al. (1988) Biochemistry 27:1094; Gatz et al. (1992) Plant J. 2:397; Gossen et al. (1992) Proc. Natl. Acad. Sci. USA 89:5547; Oliva et al. (1992) Antimicrob. Agents Chemother. 36:913; Hiavka et al. (1985) Handbook of Experimental Pharmacology 78; и Gill et al. (1988) Nature 334:721. Эти работы включены в настоящее изобретение путем ссылки.
Вышеприведенный список генов селекционных маркеров не является лимитирующим. В настоящем изобретении можно использовать любые гены селекционных маркеров.
Модификация нуклеотидных последовательностей для усиления экспрессии в растениях
Если антигенный полипептид гриппа либо его фрагмент или вариант экспрессируется в ряске, то последовательность экспрессируемого полинуклеотида, кодирующего полипептид гриппа либо его фрагмент или вариант, можно модифицировать для усиления его экспрессии в ряске. Одной из таких модификаций является синтез полинуклеотида с использованием предпочтительных кодонов растений, в частности предпочтительных кодонов ряски. Существуют методы синтеза последовательностей нуклеотидов с предпочтительными кодонами растений. Напр., см. U.S. Patent Nos.5,380,831 and 5,436,391; EP 0359472; ЕР 0385962; WO 91/16432; Perlak et al. (1991) Proc. Natl. Acad. Sci. USA 15:3324; lannacome et al. (1997) Plant Mol. Biol. 34:485; и Murray et al. (1989) Nucleic Acids Res. 17:477, которые включены путем ссылки. Синтез может осуществляться любым методом, известным специалистам. Предпочтительные кодоны можно определить по кодонам, имеющих наибольшую частоту в белках, экспрессирующихся в ряске. Например, частота употребительности кодонов для Lemna minor приведена в следующей таблице.
Lemna minor [gbplnj:4 CDSs (1597 кодонов)
В целях настоящего изобретения "предпочтительными кодонами ряски" являются кодоны, частота употребления которых в ряске превышает 17%. "Предпочтительными кодонами Lemna" в настоящем изобретении являются кодоны, частота употребления которых в роде Lemna превышает 17%. "Предпочтительными кодонами Lemna minor" в настоящем изобретении являются кодоны, частота употребления которых у Lemna minor превышает 17%, причем частота употребления кодонов у Lemna minor взята из Базы данных по употребительности кодонов (GenBank Release 160.0 (June 15, 2007).
Также предусматривается, что оптимизированным или синтетическим может быть весь или любая часть полинуклеотида, кодирующего антигенный полипептид гриппа либо его фрагмент или вариант.Иными словами, можно использовать и полностью оптимизированные, и частично оптимизированные последовательности. Например, предпочтительные кодоны ряски могут составлять 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 87%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%. В одном воплощении предпочтительные кодоны ряски составляют от 90 до 96% кодонов. Кодирующая последовательность полинуклеотида, кодирующего антигенный полипептид гриппа либо его фрагмент или вариант, может содержать кодоны, используемые с частотой как минимум 17% у Lemna gibba или как минимум 17% у Lemna minor. В одном воплощении полипептидом гриппа является полипептид НА, к примеру, полипептид НА, приведенный в SEQ ID NO:2, а экспрессирующая кассета содержит оптимизированную кодирующую последовательность для этого полипептида НА, причем кодирующая последовательность включает предпочтительные кодоны ряски, например, предпочтительные кодоны Lemna minor или Lemna gibba. В одном таком воплощении экспрессирующая кассета включает SEQ ID NO:1, которая содержит предпочтительные кодоны Lemna minor, кодирующий полипептид НА, приведенный в SEQ ID NO:2.
Можно проводить и другие модификации полинуклеотида, кодирующего антигенный полипептид гриппа либо его фрагмент или вариант.Эти модификации включают, но не ограничиваются, элиминацией последовательностей, кодирующих ложные сигналы полиаденилирования, сигналы сайтов сплайсинга экзон-интрон, транспозоноподобные повторы, и других таких хорошо изученных последовательностей, которые могут быть вредными для экспрессии гена. Содержание GC в последовательности можно довести до среднего уровня для ряски, рассчитанного относительно известных генов, экспрессирующихся в этом растении. По возможности полинуклеотид, кодирующий гетерологичный полипептид, можно модифицировать так, чтобы избежать образования вторичных структур мРНК типа шпилек.
Существуют известные различия между оптимальными контекстными последовательностями нуклеотидов инициации трансляции для кодонов инициации трансляции у животных и растений. "Контекстные последовательности нуклеотидов инициации трансляции" в настоящем изобретении относятся к идентичности трех нуклеотидов непосредственно с 5'-стороны от кодона инициации трансляции. "Кодон инициации трансляции" означает кодон, который инициирует трансляцию мРНК, транскрибируемой из данной последовательности нуклеотидов. Состав таких контекстных последовательностей нуклеотидов инициации трансляции может влиять на эффективность инициации трансляции. Например, см. Lukaszewicz et al. (2000) Plant Science 154:89-98; и Joshi et al. (1997) Plant Mol. Biol. 35:993-1001. В настоящем изобретении контекстная нуклеотидная последовательность инициации трансляции для кодона инициации трансляции у полинуклеотида, кодирующего антигенный полипептид гриппа либо его фрагмент или вариант, может быть модифицирована для усиления экспрессии в ряске. В одном воплощении нуклеотидная последовательность модифицируется таким образом, чтобы три нуклеотида непосредственно выше кодона инициации трансляции были "АСС". В другом воплощении этими нуклеотидами являются "АСА".
Экспрессию антигенного полипептида гриппа в ряске можно усилить и при помощи 5'-лидерных последовательностей. Такие лидерные последовательности могут усиливать трансляцию. Лидерные последовательности (лидеры) трансляции известны в этой области и к ним относятся лидеры пикорнавирусов, напр., лидер EMCV (5'-некодирующий участок вируса энцефаломиокардита; EIroy-Stein et al. (1989) Proc. Natl. Acad. Sci USA 86:6126); лидеры потивирусов, напр., лидер TEV (вируса гравировки табака; Allison et al. (1986) Virology 154:9); связывающий белок тяжелой цепи иммуноглобулина человека (BiP; Macajak and Samow (1991) Nature 353:90); нетранслируемый лидер из мРНК белка оболочки вируса мозаичности люцерны (AMV RNA 4; Jobling and Gehrke (1987) Nature 325:622); лидер вируса мозаичности табака (TMV; Gallic (1989) Molecular Biology of RNA, 23:56); лидер вируса гравировки картофеля (Tomashevskaya et al. (1993) J. Gen. Virol. 74:2717-2724); 5'-нетранслируемый участок Fed-1 (Dickey (1992) EMBO J. 11:2311-2317); 5'-нетранслируемый участок RbcS (Silverthome et al. (1990) J. Plant. Mol. Biol. 15:49-58); и лидер вируса хлоротической крапчатости кукурузы (MCMV; Lommel et al. (1991) Virology 81:382). Также см. Della-Cioppa et al. (1987) Plant Physiology 84:965. Лидерные последовательности, содержащие интроны растений, включая последовательность интрона из гена алкогольдегидрогеназы 1 (ADH1) кукурузы, гена каталазы клещевины или гена РАТ1 триптофанового пути Arabidopsis, как было показано, тоже повышают эффективность трансляции у растений (Callis et al. (1987) Genes Dev. 1:1183-1200; Mascarenhas et al. (1990) Plant Mol. Biol. 15:913-920).
В некоторых воплощениях настоящего изобретения перед полинуклеотидом, кодирующим нужный антигенный полипептид гриппа либо его фрагмент или вариант, для повышения эффективности его трансляции вставляется последовательность нуклеотидов, соответствующая нуклеотидам 1222-1775 гена алкогольдегидрогеназы 1 кукурузы (ADH1; номер доступа GenBank X04049). В другом воплощении экспрессирующая кассета содержит лидер из гена малой субъединицы 5 В рибулозобифосфаткарбоксилазы Lemna gibba (лидер RbcS; см. Buzby et al. (1990) Plant Cell 2:805-814).
Предусматривается, что в настоящем изобретении можно использовать любые модификации, усиливающие экспрессию последовательностей нуклеотидов, описанных выше, включая любые одиночные модификации или возможные комбинации модификаций. Выражение "модифицирована для усиления экспрессии" в ряске в настоящем изобретении относится к последовательности полинуклеотида, содержащей любую из этих модификаций либо их комбинаций.
Сигнальные пептиды
Нужный полипептид гриппа в норме или предпочтительно может экспрессироваться в виде секретируемого белка. Секретируемые белки обычно транслируются из предшественников полипептидов, включающих "сигнальный пептид", который взаимодействует с рецепторным белком на мембране эндоплазматического ретикулума (ER), направляя транслокацию растущей цепи полипептида через мембрану в эндоплазматический ретикулум для секреции из клетки. Такой сигнальный пептид обычно отщепляется от предшественника полипептида с образованием "зрелого" полипептида без сигнального пептида. В одном воплощении настоящего изобретения полипептид гриппа либо его фрагмент или вариант экспрессируется в ряске из последовательности полинуклеотида, функционально связанного с последовательностью нуклеотидов, кодирующей сигнальный пептид, направляющий секрецию антигенного полипептида гриппа либо его фрагмента или варианта в культуральную среду. Известны растительные сигнальные пептиды, которые направляют транслокацию белка в эндоплазматический ретикулум (для секреции из клетки). Например, см. U.S. Patent No.6,020,169. В настоящем изобретении для направления экспрессируемого полипептида в ER можно использовать любой растительный сигнальный пептид.
В некоторых воплощениях сигнальным пептидом является сигнальный пептид основной эндохитиназы Arabidopsis thaliana (аминокислоты 14-34 белка с № доступа NCBI ВАА82823), сигнальный пептид экстенсина (Stiefel et al. (1990) Plant Cell 2:785-793), сигнальный пептид α-амилазы риса (аминокислоты 1-31 белка с №доступа NCBI ААА33885; также см. GenBank M24286). В другом воплощении сигнальный пептид соответствует сигнальному пептиду секретируемого белка ряски.
В качестве альтернативы можно использовать сигнальный пептид млекопитающих для направления полученного рекомбинантным способом антигенного полипептида гриппа для секреции из ряски. Было показано, что растительные клетки распознают сигнальные пептиды млекопитающих, направляющие в эндоплазматический ретикулум, причем эти сигнальные пептиды могут направлять секрецию полипептидов не только через плазматическую мембрану, но и через клеточную стенку растений. См. U.S. Patent Nos.5,202,422 and 5,639,947.
В одном воплощении последовательность нуклеотидов, кодирующая сигнальный пептид, подвергается модификации для усиления экспрессии в ряске, используя любые модификации или комбинации модификаций, приведенные выше, для последовательности нужного полинуклеотида.
Секретируемый антигенный полипептид гриппа либо его фрагмент или вариант может быть извлечен из культуальной среды любым стандартным способом, известным в этой области, включая методы хроматографии, электрофореза, диализа, экстракции растворителями и др. При этом можно получить частично или существенно очищенный антигенный полипептид гриппа либо его фрагмент или вариант из культуральной среды.
Трансформированные растения ряски и узелковые культуры ряски
Настоящим изобретением предусмотрены трансформированные растения ряски, экспрессирующие нужный полипептид гриппа либо его фрагмент или вариант.Термин "ряска" относится к представителям семейства Lemnaceae. Это семейство в настоящее время подразделяется на пять родов и 38 видов ряски следующим образом:род Lemna (L. aequinoctialis, L. disperma, L. ecuadoriensis, L. gibba, L. japonica, L. minor, L. miniscula, L. obscura, L. perpusilla, L. tenera, L. trisuica, L. turionifera, L. valdiviana)', род Spirodela (S. intermedia, S. polyrrhiza, S. punctatd); род Wolffia (Wa. angusta, Wa. arrhiza, Wa. australina, Wa. borealis, Wa. brasiliensis, Wa. columbiana, Wa. elongata, Wa. globosa, Wa. microscopica, Wa. neglectd); род Wolfiella (Wl. caudata, Wl. denticulata, Wl. gladiata, Wl. hyalina, Wl. lingulata, Wl. repunda, Wl. rotunda, Wl. neotropica) и род Landoltia (L. punctata). Любые другие рода или виды Lemnaceae, если они существуют, также являются предметом настоящего изобретения. Виды Lemna можно классифицировать по таксономической схеме, описанной в Landolt (1986) Biosystematic Investigation on the Family of Duckweeds: The family of Lemnaceae - A Monograph Study (Geobatanischen Institut ETH, Stiftung Rubel, Zurich).
В настоящем изобретении "растения" включают целые растения, органы растений (напр., вайи (листья), стебли, корни и т.д.), семена, клетки растений и их потомство. Части трансгенных растений входят в рамки изобретения и включают, напр., клетки растений, протопласты растений, культуры тканей растений, из которых можно регенерировать растения, ткани, каллюсы растений, зародыши, а также цветки, семяпочки, стебли, плоды, листья, корни, корешки, узелки и др., возникающие в трансгенных растениях или их потомстве, предварительно трансформированных нужным полинуклеотидом и поэтому состоящих хотя бы частично из трансгенных клеток. В настоящем изобретении термин "растительные клетки" включает клетки семян, зародышей, семяпочек, меристемы, ткани каллюса, листьев, листовидных отростков (пластинок), корней, узелков, побегов, пыльников и пыльцы.
В настоящем изобретении "узелок ряски" означает ткань ряски, содержащую клетки ряски, причем по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% клеток составляют дифференцированные клетки. В настоящем изобретении "дифференцированные клетки" означают клетки, которые отличаются по меньшей мере одним фенотипическим признаком (напр., особой морфологией клеток или экспрессией маркерной нуклеиновой кислоты или белка) от недифференцированных клеток или клеток в других типах тканей. Дифференцированные клетки описанных в настоящем изобретении узелковых культур ряски образуют кафельную гладкую поверхность из сцепленных клеток, слившихся своими клеточными стенками, с узелками, начавшими образовывать зачатки листовидных пластинок, разбросанными по всей ткани. Поверхность ткани узелковых культур содержит эпидермальные клетки, соединенные друг с другом через плазмодесмы.
Характер произрастания ряски идеален для культивирования. Растения быстро размножаются посредством вегетативного почкования новых пластинок, что по внешнему виду аналогично бесполому размножению у дрожжей. Размножение происходит путем вегетативного почкования из клеток меристемы. Область меристемы небольшая и находится на вентральной поверхности пластинки. Клетки меристемы залегают двумя кармашками, по одному с каждой стороны средней жилки. Небольшая область средней жилки также является тем местом, из которого отрастает корешок и отходит стебелек, соединяющий каждую пластинку с материнской пластинкой. Кармашки меристемы защищены тканевыми клапанами. Из этих кармашков поочередно отпочковываются пластинки. Период удвоения зависит от вида и составляет всего лишь 20-24 ч (Landolt (1957) Ber. Schweiz. Bot. Ges. 67:271; Chang et al. (1977) Bull. Inst. Chem. Acad. Sin. 24:19; Datko and Mudd (1970) Plant Physiol. 65:16; Venkataraman et al. (1970) Z Pflanzenphysiol. 62:316). Интенсивное культивирование ряски дает самые высокие скорости прироста биомассы за единицу времени (Landolt and Kandeler (1987) The Family of Lemnaceae-A Monographic Study Vol.2:Phytochemistry, Physiology, Application, Bibliography (Veroffentlichungen des Geobotanischen Institutes ETH, Stiftung Rubel, Zurich), при этом прирост сухой массы составляет 6-15% от сырого веса (Tillberg et al. (1979) Physiol. Plant. 46:5; Landolt (1957) Ber. Schweiz. Bot. Ges. 67:271; Stomp, неопубликованные данные). Содержание белка у ряда видов ряски, выращенных в различных условиях, как сообщалось, составляет 15-45% от сухой массы (Chang et al. (1977) Bull. Inst. Chem. Acad. Sin. 24:19; Chang and Chui (1978) Z. Pflanzenphysiol. 89:91; Porath et al. (1979) Aquatic Botany 7:272; Appenroth et al. (1982) Biochem. Physiol. Pflanz. 177:251). Судя по этим цифрам, уровень продукции белка на литр среды у ряски является величиной того же порядка, что и в дрожжевых системах экспрессии генов.
Трансформированные растения ряски по изобретению можно получить путем введения в нужные растения ряски экспрессирующей конструкции, содержащей полинуклеотид, кодирующий антигенный полипептид гриппа либо его фрагмент или вариант.
Термин "введение" в контексте полинуклеотидов, к примеру, экспрессирующей конструкции, содержащей полинуклеотид, кодирующий антигенный полипептид гриппа либо его фрагмент или вариант, служит для обозначения введения в растения ряски полинуклеотидов таким образом, чтобы полинуклеотид получил доступ внутрь клетки растения ряски. Если вводится более одного полинуклеотида, то эти полинуклеотиды можно собрать в виде одной нуклеотидной конструкции или в виде отдельных нуклеотидных конструкций и расположить в одном и том же или в разных трансформирующих векторах. Соответственно, эти полинуклеотиды могут вводиться в нужные клетки ряски при трансформации за один раз, за несколько раз или, к примеру, в процессе селекции. Композиции и способы изобретения не зависят от конкретного способа введения одного или нескольких полинуклеотидов в растения ряски, лишь бы только полинуклеотиды получили доступ внутрь по меньшей мере одной клетки растения ряски. Методы введения полинуклеотидов в растения известны, в том числе методы кратковременной трансформации, методы устойчивой трансформации и методы, использующие вирусы.
"Кратковременная трансформация" в контексте полинуклеотидов типа полинуклеотида, кодирующего антигенный полипептид гриппа либо его фрагмент или вариант, служит для обозначения того, что полинуклеотид вводится в растения ряски, но не встраивается в геном растений ряски.
Под "устойчивым введением" или "устойчиво введенным" в контексте полинуклеотидов (типа полинуклеотида, кодирующего антигенный полипептид гриппа либо его фрагмент или вариант), вводимых в растения ряски, имеется в виду то, что введенный полинуклеотид устойчиво встраивается в геном ряски, поэтому растения ряски устойчиво трансформируются полинуклеотидом.
"Устойчивая трансформация" или "устойчиво трансформированный" должно означать то, что полинуклеотид, к примеру, полинуклеотид, кодирующий антигенный полипептид гриппа, либо его фрагмент или вариант, введенный в растения ряски, встраивается в геном растения и может наследоваться его потомством, в частности потомством из нескольких последующих поколений. В некоторых воплощениях последующие поколения включают потомство, полученное вегетативным путем (т.е. при бесполом размножении), к примеру, при клональном размножении. В других воплощениях последующие поколения включают потомство, полученное путем полового размножения.
Экспрессирующая конструкция, содержащая полинуклеотид, кодирующий антигенный полипептид гриппа либо его фрагмент или вариант, может вводится в нужные растения ряски по любой методике трансформации, известной специалистам. Подходящие методы введения последовательностей нуклеотидов в растения ряски или растительные клетки или узелки включают микроинъекцию (Crossway et al. (1986) Biotechniques 4:320-334), электропорацию (Riggs et al. (1986) Proc. Natl. Acad. Sci. USA 83:5602-5606), трансформацию с помощью Agrobacterium (U.S. Patent Nos.5,563,055 и 5,981,840, которые оба включены путем ссылки), прямой перенос генов (Paszkowski et al. (1984) EMBO J. 3:2717-2722), баллистическое ускорение частиц (напр., см. U.S. Patent Nos.4,945,050; 5,879,918; 5,886,244; и 5,932,782, которые все включены путем ссылки; и Tomes et al. (1995) "Direct DNA Transfer into Intact Plant Cells via Microprojectile Bombardment," in Plant Cell, Tissue, and Organ Culture:Fundamental Methods, ed. Gamborg and Phillips (Springer-Verlag, Berlin); McCabe et al. (1988) Biotechnology 6:923-926). Из подвергнутых трансформации клеток можно вырастить растения в соответствии со стандартными способами.
Как отмечено выше, устойчиво трансформированную ряску можно получить любым известным методом переноса генов, как-то одним из методов переноса генов, приведенных в U.S. Patent No.6,040,498 или U.S. Patent Application Publication Nos.2003/0115640, 2003/0033630 или 2002/0088027; которые все включены путем ссылки во всей полноте. Растения ряски или узелковые культуры можно эффективно трансформировать экспрессирующей кассетой, содержащей описанную последовательность нуклеиновой кислоты, любым из целого ряда методов, включая перенос генов с помощью Agrobacterium, баллистическую бомбардировку или электропорацию. Можно использовать Agrobacterium tumefaciens или Agrobacterium rhizogenes. Устойчивые трансформанты ряски можно выделить путем трансформирования клеток ряски нужной последовательностью нуклеиновой кислоты вместе с геном придающим устойчивость к селекционному агенту, с последующим культивированием трансфомированных клеток в среде, содержащей селекционного агента. Например, см. U.S. Patent No.6,040,498, содержание которого включено путем ссылки во всей полноте.
Устойчиво трансформированные растения ряски, используемые в этих методах, должны проявлять нормальную морфологию и размножаться половым путем и/или размножаться вегетативным путем (т.е. бесполым размножением), например, клональным размножением. Предпочтительно, трансформированные растения ряски настоящего изобретения содержат одну копию перенесенной нуклеиновой кислоты, содержащей полинуклеотид, кодирующий антигенный полипептид гриппа либо его фрагмент или вариант, а перенесенная нуклеиновая кислота не содержит заметных перестроек. Предусматривается, что трансформированные растения ряски по изобретению могут содержать перенесенную нуклеиновую кислоту в малом количестве копий (т.е. не более 12 копий, не более 8 копий, не более 5 копий или же не более 3 копий, а с другой стороны и менее 3 копий нуклеиновой кислоты на трансформированную клетку).
Трансформированные растения, экспрессирующие антигенный полипептид гриппа либо его фрагмент или вариант, можно культивировать в условиях, подходящих для экспрессирования антигенного полипептида гриппа либо его фрагмента или варианта. Затем антигенный полипептид гриппа либо его фрагмент или вариант можно извлечь из растений ряски, культуральной среды или растений ряски и культуральной среды и, при желании, очистить любым стандартным методом выделения и очистки, известным в этой области, включая хроматографию, электрофорез, диализ, экстракцию растворителями и т.п. Затем антигенный полипептид гриппа либо его фрагмент или вариант можно составить в виде вакцины для терапевтического применения, как описано далее в настоящем изобретении.
Способы получения полипептидов птичьего гриппа
Как описано полностью в настоящем изобретении, в одном воплощении способ получения антигенного полипептида птичьего гриппа включает: (а) культивирование в культуральной среде ряски культуры растений ряски или культуры узелков ряски, причем культура растений ряски или культура узелков ряски устойчиво траснформирована для экспрессии антигенного полипептида, при этом антигенный полипептид экспрессируется из последовательности нуклеотидов, содержащей кодирующую последовательность для данного антигенного полипептида и оперативно связанную кодирующую последовательность для сигнального пептида, направляющего секрецию антигенного полипептида в культуральную среду; и (b) извлечение антигенного полипептида из культуральной среды. Термин извлечение включает выделение из культуральной среды или очистку.
После продукции рекомбинантного полипептида в ряске для очистки белка можно использовать любой доступный метод. Различные стадии включают освобождение белка от небелкового или растительного материала, а затем очистку нужного белка от других белков. Начальные стадии в процессе очистки включают центрифугирование, фильтрование или их комбинации. Белки, секретированные во внеклеточное пространство тканей, можно получить методом вакуумной или центрифужной экстракции. Минимальная обработка может включать и получение неочищенных препаратов. Другие методы включают мацерацию и экстракцию для того, чтобы непосредственно использовать экстракт.
В таких методах очистки нужного белка могут использоваться различия в размерах, физико-химических свойствах и сродстве связывания белков. Такие методы включают хроматографию, в том числе аффинную, эксклюзионную, жидкостную высокого давления, обратнофазовую и анионообменную хроматографию, аффинные метки, фильтрование и т.д. В частности, для очистки экспрессируемого белка можно использовать аффинную хроматографию на иммобилизованных ионах N1, см. Favacho et al. (2006) Protein expression and purification 46:196-203; а также см. Zhou et al. (2007) The Protein J 26:29-37; Wang et al. (2006) Vaccine 15:2176-2185; и WO/2009/076778; которые все включены путем ссылки. В процессе очистки можно использовать протективные реагенты, как-то осмотические, антиоксиданты, ингибиторы фенольного окисления, ингибиторы протеаз и др.
Способы применения
В одном воплощении объектом изобретения, раскрытого здесь, является способ вакцинации животных, включающий введение животному эффективного количества вакцины, которая может содержать эффективное количество рекомбинантного антигена птичьего гриппа и фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель.
Вакцина или композиция содержит рекомбинантный полипептид гриппа. Рекомбинантный полипептид может быть получен в растениях ряски. Рекомбинантный полипептид может быть частично или существенно очищен. Рекомбинантный полипептид может быть гликозилирован.
В одном воплощении объектом изобретения, раскрытого здесь, является способ выработки иммунного ответа, включающий введение птицам вакцины, содержащей экспрессированный антиген птичьего гриппа, при этом вырабатывается иммунный ответ.
В одном воплощении объектом изобретения, раскрытого здесь, является способ выработки иммунного ответа, включающий введение птицам вакцины, содержащей антиген птичьего гриппа, полученный в ряске, и растительный материал из ряски, при этом вырабатывается иммунный ответ.
В одном воплощении объектом изобретения, раскрытого здесь, является способ получения устойчиво трансформированных растений или культуры растений, выбранных из рода Lemna, включающий (а) введение в растения генетической конструкции, содержащей ген антигена птичьего гриппа; и (b) культивирование растений. Способы трансформации ряски доступны и изложены в настоящем изобретении.
В одном воплощении объектом изобретения, раскрытого здесь, является способ получения вакцины или композиции, включающий выделение антигена птичьего гриппа, полученного в системе экспрессии Lemna, и необязательно смешивание с фармацевтически или ветеринарно приемлемым носителем, эксципиентом или наполнителем.
В одном воплощении объектом изобретения, раскрытого здесь, является способ получения вакцины или композиции, включающий смешивание антигена птичьего гриппа, полученного в системе экспрессии Lemna, и растительного материала из рода Lemna, и необязательно добавку фармацевтически и ветеринарно приемлемого носителя, эксципиента или наполнителя.
В следующем воплощении вакцина или композиция может вводиться однодневным или старшим цыплятам.
В одном воплощении изобретения можно использовать режим «прайм-буст», который состоит из по меньшей мере одного первичного введения и по меньшей мере одного повторного введения (бустера), используя по меньшей мере один общий полипептид, антиген, эпитоп или иммуноген. Как правило, иммунологическая композиция или вакцина, используемая при первичном введении, отличается по природе от той, что используется в качестве бустера. Однако следует отметить, что одна и та же композиция может использоваться и при первичном введении, и в качестве бустера. Такая методика введения называется "прайм-буст".
В настоящем изобретении для экспрессии кодирующей последовательности или ее фрагментов, кодирующих антигенный полипептид гриппа либо его фрагмент или вариант, используется рекомбинантный вирусный вектор. В частности, вирусный вектор может экспрессировать последовательность птичьего гриппа, более конкретно ген НА или его фрагмент, кодирующий антигенный полипептид. Предусмотренные настоящим изобретением вирусные вектора включают поксвирусы (напр., вирус коровьей оспы или ослабленный вирус коровьей оспы, птичьи поксвирусы или ослабленные птичьи поксвирусы, напр., поксвирус канарейки, домашней птицы, голубя, дикого голубя, перепелки, ALVAC, TROVAC; напр., см. US 5,505,941, US 5,494,8070), поксвирус енота, поксвирус свиньи и др.), аденовирусы (напр., аденовирус человека, аденовирус собаки), герпесвирусы (напр., герпесвирус собаки, герпесвирус индейки, вирус болезни Марека, вирус инфекционного ларинготрахеита, герпесвирус кошки, герпесвирус быка, герпесвирус свиньи), бакуловирусы, ретровирусы и др. В другом воплощении экспрессирующим вектором из птичьего поксвируса может быть вектор из поксвируса канарейки типа ALVAC. В следующем воплощении экспрессирующим вектором из птичьего поксвируса может быть вектор из поксвируса домашней птицы типа TROVAC. Антигеном, эпитопом или иммуногеном гриппа может быть такой гемагтлютинин, как Н5. Вектором из поксвируса домашней птицы может быть vFP89 или VFP2211. Вектором из поксвируса канарейки может быть vCP2241 (напр., см. US 2008/0107681 и US 2008/0107687). Экспрессируемый антиген птичьего гриппа по изобретению вставляется под контроль специфичного промотора поксвируса, напр., промотора осповакцины 7,5 кДа (Cochran et al., 1985), промотора осповакцины I3L (Riviere et al., 1992), промотора осповакцины НА (Shida, 1986), промотора осповакцины ATI (Funahasi et al., 1988), промотора осповакцины Н6 (Taylor et al., 1988b; Guo et al., 1989; Perkus et al., 1989), среди прочего.
В другом аспекте методики или режима прайм-буст по изобретению вводится композиция, содержащая антиген птичьего гриппа по изобретению, после чего вводится рекомбинантный вирусный вектор, который содержит и экспрессирует антиген птичьего гриппа и/или его варианты или фрагменты in vivo. Аналогичным образом методика прайм-буст может включать введение рекомбинантного вирусного вектора с последующим введением рекомбинантного антигена птичьего гриппа по изобретению. Еще следует отметить, что как первичное, так и вторичное введение может включать рекомбинантный антиген птичьего гриппа по изобретению. Таким образом, рекомбинантный антиген птичьего гриппа по изобретению может вводиться в любом порядке с вирусным вектором или же может применяться сам по себе в качестве и первичной, и вторичной композиции.
В следующем аспекте методики прайм-буст по изобретению вводится композиция, содержащая антиген птичьего гриппа по изобретению, после чего вводится инактивированная вирусная композиция или вакцина, содержащая антиген птичьего гриппа. Аналогичным образом методика прайм-буст может включать введение инактивированной вирусной композиции или вакцины с последующим введением рекомбинантного антигена птичьего гриппа по изобретению. Еще следует отметить, что как первичное, так и вторичное введение может включать рекомбинантный антигенный полипептид по изобретению. Антигенные полипептиды изобретения могут вводиться в любом порядке с инактивированной вирусной композицией или вакциной или же могут применяться сами по себе в качестве и первичной, и вторичной композиции.
Режим прайм-буст включает по меньшей мере одно прайм-введение и по меньшей мере одно буст-введение с использованием по меньшей мере одного общего полипептида и/или его вариантов или фрагментов. Вакцина, используемая при прайм-введении, может отличаться по природе от той, что используется в качестве последующей бустерной вакцины. Прайм-введение может включать одно или несколько введений. Точно так же буст-введение может включать одно или несколько введений.
Объем дозы композиции для заданных видов млекопитающих, напр., объем дозы птичьих композиций на основе вирусных векторов, напр., на основе вирусных векторов не из поксвирусов, обычно составляет от 0,1 до 2,0 мл, от 0,1 до 1,0 мл и от 0,5 мл до 1,0 мл.
Эффективность вакцин можно тестировать через 2-4 недели после последней иммунизации путем введения животным, как-то птицам, пробы вирулентного штамма гриппа, предпочтительно гриппа, принадлежащего к подтипу Н5, как-то штамма H5N1, H5N2, H5N8 или H5N9. Для провоцирующей пробы на эффективность вакцины используются как гомологичные, так и гетерологичные штаммы. Проба может вводиться животным путем распыления, в нос, в глаза, в трахеи и/или перорально. Содержание вируса в пробе может составлять 105-8 EID50 (доза, достаточная для заражения 50% яиц) в объеме, зависящем от способа введения. Например, если вводится распылением, то суспензия вируса в виде аэрозоля дает капельки размером 1-100 мкм, если вводится в нос, в трахеи или перорально, то объем вирусной пробы составляет 0,5 мл, 1-2 мл и 5-10 мл, соответственно. Животных можно наблюдать ежедневно в течение 14 дней после пробы на клинические признаки, к примеру, обезвоживание и липкие отверстия клоаки. Кроме того, группы животных можно подвергнуть эвтаназии и обследовать на патологические признаки легочного и плеврального кровоизлияния, трахеита, бронхита, бронхиолита и бронхопневмонии. Можно взять ротоглоточные мазки у всех животных после пробы для выделения вируса. Наличие или отсутствие вирусных антигенов в дыхательных тканях можно оценивать методом количественной полимеразной цепной реакции с обратной транскриптазой в реальном времени (qRRT-PCR). Можно взять пробы крови до и после провоцирующей пробы и подвергнуть анализу на наличие специфических антител против вируса гриппа H5N1.
Композиции, содержащие рекомбинантные антигенные полипептиды по изобретению, используемые в методиках прайм-буст, содержатся в фармацевтически или ветеринарно приемлемых носителях, разбавителях или эксципиентах. Методики изобретения защищают животных от птичьего гриппа и/или предотвращают прогрессирование заболевания у инфицированных животных.
Различные введения предпочтительно проводятся с интервалом в 1-6 недель. В одном воплощении также предусматривается повторное введение (бустер) раз в год. Возраст животных составляет по меньшей мере один день на время первого введения.
Следует иметь в виду, что настоящее описание приводится для примера и оно не ограничивается этим. Из описания и общих сведений специалист сможет определить количество введений, способ введения и вводимые дозы для каждой методики введения без ненужного экспериментирования.
Настоящим изобретением предусматривается по меньшей мере одно введение животному эффективного количества терапевтической композиции по изобретению. Животными могут быть самцы, самки, беременные самки и новорожденные. Такое введение может проводиться различными способами, в том числе внутримышечно (im), интрадермально (id) или подкожно (sc) либо путем введения в нос или перорально. Терапевтические композиции по изобретению также можно вводить с помощью безигольного прибора (такого, к примеру, как Pigjet, Dermojet, Biojector, Avijet (Merial, GA, США), прибора Vetjet или Vitajet (Bioject, Oregon, США). Другой подход к введению плазмидных композиций заключается в использовании электропорации (напр., см. Tollefsen et al., 2002; Tollefsen et al., 2003; Babiuk et al., 2002; PCT Application No. WO 99/01158). В другом воплощении терапевтическая композиция вводится животным с помощью генной пушки или бомбардировки частицами золота. В предпочтительном воплощении животными являются птицы.
В одном воплощении изобретения предусматривается введение терапевтически эффективного количества композиции для доставки и экспрессии антигена или эпитопа гриппа в клетках мишени. Определение терапевтически эффективного количества является обычным делом для специалистов. В одном воплощении композиция включает экспрессионный вектор, содержащий полинуклеотид, который экспрессирует антиген или эпитоп гриппа, и фармацевтически или ветеринарно приемлемый носитель, наполнитель или эксципиент. В другом воплощении фармацевтически и ветеринарно приемлемый носитель, наполнитель или эксципиент способствует трансфекции или инфекции и/или улучшает сохранность вектора или белка в организме хозяина.
В одном воплощении объектом изобретения, раскрытого здесь, является режим вакцинации и способ детектирования для различения между инфицированными и вакцинированными животными (DIVA). В настоящее время существует два типа вакцин от птичьего гриппа (Al):вакцины из инактивированного целого вируса AI (AIV) и живые рекомбинантные вакцины на основе поксвируса домашней птицы и вируса болезни Newcastle, в которых первичной мишенью для выработки защитного иммунитета оказался гемагглютинин (НА) (Peyre et al., Epidemiol Infect, 2009, 137(1): p.1-21; Bublot et al., Ann N Y Acad Sci, 2006, 1081: p.193-201; Skehel et al., Annu Rev Biochem, 2000, 69: p.531-69). Обычные инактивированные вакцины требуют выращивания AIV в яйцах с зародышами или в клеточной культуре, для чего требуется сильно изолированное помещение с потенциальным риском заражения окружающей среды и персонала. Кроме того, в настоящее время не существует коммерчески доступного теста DIVA, совместимого с применением инактивированных вакцин (Bublot et al., 2006; El Sahly et al., Expert Rev Vaccines, 2008, 7(2): p.241-7; Veits et al., Vaccine, 2008, 26(13): p.1688-96). Была выдвинута стратегия "отличения инфицированных от вакцинированных животных" (DIVA) как возможное решение для окончательного искоренения AIV без массового забоя птиц и соответствующего экономического ущерба, особенно в развивающихся странах (Food and Agriculture Organization of the United Nations (FAO) (2004). FAO, OIE & WHO Technical consultation on the Control of Avian Influenza. Animal Health Special Report). Эта стратегия соединяет выгоды вакцинации (меньше вируса в окружающей среде) со способностью идентифицировать зараженные стаи птиц, что позволяет выполнять и другие меры контроля, включая искоренение. На уровне стаи простой подход заключается в том, чтобы регулярно отслеживать "индикаторных" птиц, оставшихся невакцинированными в каждой вакцинированной стае, но это может вызвать некоторые проблемы управления, особенно при идентификации "индикаторных" в больших стаях. В качестве альтернативы можно проводить тестирование на полевую экспозицию на вакцинированных птицах. Для этого следует использовать системы вакцинации, позволяющие обнаруживать полевую экспозицию на вакцинированных популяциях. В последние годы было разработано несколько систем, включающих применение вакцин, содержащих вирус одного и того же подтипа Н, но отличающихся по N от полевого вируса. Антитела к N полевого вируса действуют как естественные маркеры инфекции, однако будут возникать проблемы, если появится такой полевой вирус, у которого будет другой N-антиген, чем у существующего полевого вируса, или если подтипы с другими N-антигенами уже циркулируют на месте. С другой стороны, применение вакцин, содержащих только НА, позволит использовать классические методы AGID и ELISA основанных на NP или матриксе для выявления инфекции у вакцинированных птиц.
В настоящем изобретении показано, что применение вакцин или композиций настоящего изобретения позволяет выявлять инфицированность гриппом у вакцинированных животных с помощью подходящего диагностического теста, в котором выявляется антительный ответ против других белков гриппа, чем НА, как-то методом иммунодиффузии в агаровом геле или ELISA на NP. В настоящем изобретении показано, что применение вакцин или композиций настоящего изобретения позволяет выявлять инфицированность животных путем различения между инфицированными и вакцинированными животными (DIVA). В настоящем изобретении раскрыт способ диагностики инфицированности гриппом у животных с помощью основанного на NP метода иммуногенной детекции, как, например, ELISA на NP. В одном воплощении объектом изобретения, раскрытого здесь, является способ диагностики инфицированности гриппом у животных, включающий: а) контактирование твердого субстрата, содержащего нуклеопротеид (NP), с образцом, взятым у животного; b) контактирование твердого субстрата с моноклональным антителом (MAb) против NP; и с) детектирование связывания MAb с образцом, захваченным NP на твердом субстрате, при этом степень ингибирования тестируемого образца относительно отрицательного контроля означает то, что субъект инфицирован гриппом, тем самым устанавливается диагноз инфицированности гриппом у субъекта.
Промышленные изделия
В одном воплощении объектом изобретения, раскрытого здесь, является набор для выполнения способа выработки или индуцирования иммунного ответа, который может включать любые из рекомбинантных иммунологических композиций или вакцин от гриппа либо инактивированных иммунологических композиций или вакцин от гриппа и рекомбинантных вирусных композиций или вакцин от гриппа, а также инструкции для выполнения способа.
Другим воплощением изобретения является набор для выполнения способа выработки иммунологического или защитного ответа против гриппа у животных, включающий композицию или вакцину, содержащую антиген птичьего гриппа по изобретению, и рекомбинантную вирусную иммунологическую композицию или вакцину против гриппа, а также инструкции для выполнения способа доставки в эффективном количестве для выработки иммунного ответа у животных.
Другим воплощением изобретения является набор для выполнения способа выработки иммунологического или защитного ответа против гриппа у животных, включающий композицию или вакцину, содержащую антиген птичьего гриппа по изобретению, и инактивированную иммунологическую композицию или вакцину против гриппа, а также инструкции для выполнения способа доставки в эффективном количестве для выработки иммунного ответа у животных.
Следующий аспект настоящего изобретения касается набора для вакцинации типа прайм-буст по настоящему изобретению, как описано выше. Набор может включать по меньшей мере два флакона:первый флакон, содержащий вакцину или композицию для прайм-вакцинации по настоящему изобретению, и второй флакон, содержащий вакцину или композицию для буст-вакцинации по настоящему изобретению. Набор может содержать и дополнительные первые или вторые флаконы для дополнительных прайм-вакцинаций или дополнительных буст-вакцинации.
Изобретением охвачены следующие воплощения. В одном воплощении представлена композиция, содержащая антиген птичьего гриппа либо его фрагмент или вариант и фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель. В другом воплощении представлена вышеописанная композиция, в которой антиген птичьего гриппа либо его фрагмент или вариант включает иммуногенный фрагмент, содержащий по меньшей мере 15 аминокислот антигена птичьего гриппа. В следующем воплощении представлены вышеприведенные композиции, в которых антиген птичьего гриппа либо его фрагмент или вариант вырабатывается в ряске. В одном воплощении представлены вышеприведенные композиции, в которых антиген птичьего гриппа либо его фрагмент или вариант частично очищен. В другом воплощении представлены вышеприведенные композиции, в которых антиген птичьего гриппа либо его фрагмент или вариант существенно очищен. В одном воплощении представлены вышеприведенные композиции, в которых антиген птичьего гриппа либо его фрагмент или вариант представляет собой полипептид птичьего H5N1. В одном воплощении представлены вышеприведенные композиции, в которых полипептид H5N1 представляет собой полипептид гемагглютинина. В одном воплощении представлены вышеприведенные композиции, в которых последовательность антигена птичьего гриппа либо его фрагмента или варианта по меньшей мере на 80% идентична последовательности, приведенной в SEQ ID NO:2, 4, 5, 8, 10, 12 или 14. В одном воплощении представлены вышеприведенные композиции, в которых антиген птичьего гриппа кодируется полинуклеотидом, последовательность которого по меньшей мере на 70% идентична последовательности, приведенной в SEQ ID NO:1, 3, 6, 7, 9, 11 или 13. В одном воплощении представлены вышеприведенные композиции, в которых фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель представляет собой эмульсию типа вода в масле или вода в масле в воде или масло в воде. В другом воплощении представлен способ вакцинации животных, подверженных птичьему гриппу, включающий введение вышеприведенных композиций животным. В одном воплощении представлен способ вакцинации животных, подверженных птичьему гриппу, включающий режим прайм-буст. В одном воплощении представлен существенно очищенный антигенный полипептид, экспрессированный в ряске, причем полипептид включает:аминокислотную последовательность, которая по меньшей мере на 80% идентична полипептиду с последовательностью, приведенной в SEQ ID NO:2, 4, 5, 8, 10, 12 или 14. В любом воплощении животные предпочтительно представляют собой птиц, лошадей, собак, кошек или свиней. В одном воплощении представлен способ диагностики инфицированное™ гриппом у животных. В другом воплощении представлен набор для вакцинации типа прайм-буст, включающий по меньшей мере два флакона, причем первый флакон содержит композицию настоящего изобретения, а второй флакон содержит композицию для буст-вакцинации, включающий композицию, содержащую рекомбинантный конкурирующий вектор, или композицию, содержащую инактивированную вирусную композицию.
Фармацевтически или ветеринарно приемлемые носители или наполнители или эксципенты хорошо известны специалистам. Например, фармацевтически или ветеринарно приемлемым носителем или наполнителем или эксципиентом может быть 0,9% раствор NaCl (напр., физраствор) или фосфатный буфер. Другие фармацевтически или ветеринарно приемлемые носители или наполнители или эксципиенты, которые можно использовать для способов данного изобретения, включают поли-(L-глутамат) или поливинилпирролидон, но не ограничиваются ими. Фармацевтически или ветеринарно приемлемым носителем или наполнителем или эксципиентом может быть любое соединение или комбинация соединений, способствующих введению вектора (или белка, экспрессируемого из вектора по изобретению in vitro); предпочтительно носитель, наполнитель или эксципиент может способствовать трансфекции и/или улучшать сохранность вектора (или белка). Дозы и объемы доз обсуждаются в общем описании, а также могут быть установлены специалистом из данного описания в сочетании с общими познаниями в этой области, без ненужного экспериментирования.
Катионные липиды, содержащие соль четвертичного аммония, которые предпочтительно, но не исключительно подходят для плазмид, - это предпочтительно липиды, имеющие следующую формулу:
где: R1 означает насыщенный или ненасыщенный неразветвленный алифатический радикал, содержащий 12-18 атомов углерода, R2 - другой алифатический радикал, содержащий 2 или 3 атома углерода, а Х - аминогруппа или гидроксил, напр., это DMRIE. В другом воплощении катионный липид может быть связан с нейтральным липидом, напр., DOPE.
Из этих катионных липидов предпочтение отдается DMRIE (N-(2-гидроксиэтил)-N,N-диметил-2,3-бис(тетрадецилокси)-1-пропанаммоний; WO 96/34109), предпочтительно связанному с нейтральным липидом, предпочтительно DOPE (диолеоил-фосфатидилэтаноламин; Behr, 1994), с образованием DMRIE-DOPE.
Предпочтительно смесь плазмиды с адъювантом готовится экстемпорально и предпочтительно одновременно с введением препарата или незадолго до введения препарата; например, незадолго до или перед введением готовится смесь плазмиды с адъювантом, предпочтительно так, чтобы осталось достаточно времени перед введением для того, чтобы смесь образовала комплекс, напр., от 10 до 60 минут перед введением, как-то примерно за 30 минут до введения.
Если присутствует DOPE, то молярное отношение DMRIE:DOPE предпочтительно составляет от 95:5 до 5:95, более предпочтительно примерно 1:1, напр., 1:1.
Соотношение адъювант DMRIE или DMRIE-DOPE:плазмида может составлять по весу 50:1 и 1:10, как-то 10:1 и 1:5, или 1:1 и 1:2, напр., 1:1 и 1:2.
Фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель может представлять собой эмульсию типа вода в масле. Примеры подходящих эмульсий вода в масле включают вакцинные масляные эмульсии типа вода в масле, которые являются стабильными и жидкими при 4°С и содержат:от 6 до 50% об. содержащей антиген водной фазы, предпочтительно от 12 до 25% об. и от 50 до 94% об. масляной фазы, содержащей полностью или частично неметаболизируемое масло (напр., минеральное масло типа парафинового масла) и/или метаболизируемое масло (напр., растительное масло или эфиры жирной кислоты, полиоля или спирта), от 0,2 до 20% об. ПАВ, предпочтительно от 3 до 8% об. ПАВ, которые представляют собой полностью или частично либо в виде смеси полиглицериновых эфиров, которые представляют собой предпочтительно (поли)рицинолеаты глицерина, либо рициновые масла полиоксиэтилена или гидрогенированные рициновые масла полиоксиэтилена. Примеры ПАВ, которые могут использоваться в эмульсиях типа вода в масле, включают этоксилированные эфиры сорбитана (напр., полиоксиэтилен-(20)-сорбитанмоноолеат (Tween 80®) фирмы AppliChem Inc., Cheshire, CT) и эфиры сорбитана (напр., сорбитанмоноолеат (Span 80®) фирмы Sigma Aldrich, St. Louis, МО). Кроме того, касательно эмульсий вода в масле см. также U.S. Patent No.6,919,084, напр., приведенный в нем пример 8, включенный путем ссылки. В некоторых воплощениях содержащая антиген водная фаза включает солевой раствор, содержащий один или несколько буферных реагентов. Примером подходящего буферного раствора является физраствор с фосфатным буфером. В предпочтительном воплощении эмульсия вода в масле может представлять собой тройную эмульсию вода/масло/вода (W/O/W) (U.S. Patent No.6,358,500). Примеры других подходящих эмульсий описаны в U.S. Patent No.7,371,395.
Иммунологические композиции и вакцины по изобретению могут содержать или состоять в основном из одного или нескольких адъювантов. Подходящими адъювантами для применения при воплощения настоящего изобретения являются:(1) полимеры акриловой или метакриловой кислоты, малеинового ангидрида и производных алкенилов, (2) иммуностимулирующие последовательности (ISS), как-то последовательности олигодезоксирибонуклеотидов, содержащие одно или несколько неметилированных звеньев CpG (Klinman et al., 1996; WO 98/16247), (3) эмульсии типа масло в воде, как-то эмульсия SPT, описанная на стр.147 в "Vaccine Design, The Submit and Adjuvant Approach" published by M. Powell, M. Newman, Plenum Press 1995, и эмульсия MF59, описанная на стр.183 той же самой работы, (4) катионные липиды, содержащие соль четвертичного аммония, напр., DDA, (5) цитокины, (6) гидроксид алюминия или фосфат алюминия, (7) сапонин или (8) другие адъюванты, приведенные в любом из процитированных документов и включенные путем ссылки в настоящую заявку, или (9) любые их комбинации или смеси.
Эмульсии типа масло в воде (3), которые особенно пригодны для вирусных векторов, могут основываться на: легком жидком парафиновом масле (типа Европейской Фармакопеи), изопреноидном масле типа сквалана, сквалена, масле, образующемся при олигомеризации алкенов, напр., изобутена или децена, сложных эфирах кислот или спиртов, содержащих неразветвленную алкильную группу, как-то растительных маслах, этилолеате, пропиленгликоле, ди(каприлат/капрате), три(каприлат/капрате) глицерина и диолеате пропиленгликоля, либо эфирах разветвленных жирных спиртов или кислот, особенно изостеариновой кислоты.
Масло используется в комбинации с эмульгаторами для получения эмульсии. Эмульгаторами могут быть неионные детергенты, как-то:эфиры, с одной стороны - сорбитана, маннида (напр., ангидроманнитололеат), глицерина, полиглицерина или пропиленгликоля, а с другой стороны - олеиновой, изостеариновой, рицинолеиновой или гидроксистеариновой кислоты, причем данные эфиры необязательно этоксилированы, либо блочные сополимеры полиоксипропилена, полиоксиэтилена типа Pluronic, напр., L121.
Из адъювантных полимеров типа (1) предпочтение оказывается сшитым полимерам акриловой или метакриловой кислоты, особенно сшитым полиалкиниловыми эфирами Сахаров или полиспиртов. Эти соединения известны под названием карбомеры (Pharmeuropa, vol.8, no.2, June 1996). Специалисты в этой области также могут обратиться к U.S. Patent No.2,909,462, в котором представлены такие акриловые полимеры, сшитые полигидроксильным соединением, содержащим как минимум три гидроксильные группы, предпочтительно не больше восьми таких групп, причем атомы водорода по меньшей мере трех гидроксильных групп замещены ненасыщенными алифатическими радикалами, содержащими как минимум два атома углерода. Предпочтительными радикалами являются те, что содержат 2-4 атома углерода, напр., винилы, аллилы и другие группы с ненасыщенным этиленом. Ненасыщенные радикалы могут содержать и другие заместители типа метила. Особенно подходят препараты, поступающие в продажу под названием Carbopol (BF Goodrich, Ohio, США). Они прошиты аллилсахарозой или аллилпентаэритритолом. Из них следует отметить Carbopol 974Р, 934Р и 971Р.
Что касается сополимеров малеинового ангидрида и производных алкенилов, то предпочтение оказывается EMA (Monsanto), которые являются неразветвленными или сшитыми сополимерами этилена и малеинового ангидрида и прошиты, к примеру, дивиниловым эфиром. Можно также обратиться к J. Fields et al., 1960.
В отношении структуры полимеры акриловой или метакриловой кислоты и EMA предпочтительно образованы основными звеньями, имеющими следующую формулу
где: R1 и R2, которые могут быть одинаковыми или разными, означают Н или СН3;
х = 0 или 1, предпочтительно х=1;
y = 1 или 2, причем х+y=2.
Для EMA х=0 и y=2, а для карбомеров х=y=1.
Эти полимеры растворимы в воде или физиологическом солевом растворе (20 г/л NaCl), а рН можно довести до 7,3-7,4, напр., с помощью гидроксида натрия (NaOH), получая раствор адъюванта, в который можно ввести экспрессионный вектор. Концентрация полимера в конечной иммунологической или вакцинной композиции может составлять от 0,01 до 1,5% масс. (w/v), от 0,05 до 1% масс. (w/v) и от 0,1 до 0,4% масс. (w/v).
Цитокин или цитокины (5) могут находиться в иммунологической композиции или вакцине в виде белка или могут экспрессироваться в организме вместе с их иммуногеном или иммуногенами либо эпитопами. Предпочтение отдается совместной экспрессии цитокина или цитокинов, либо в том же векторе, в котором экспрессируется иммуноген или иммуногены либо эпитопы, либо в отдельном векторе.
Изобретением предусмотрено получение таких комбинированных композиций, например, смешиванием активных компонентов, предпочтительно вместе с адъювантом, носителем, цитокином и/или разбавителем.
Цитокины, которые можно использовать в настоящем изобретении, включают гранулоцитарный колониестимулирующий фактор (G-CSF), гранулоцит/макрофаг колониестимулирующий фактор (GM-CSF), α-интерферон (IFN-α), β-интерферон (IFN-β), γ-интерферон (IFN-y), интерлейкин-1α (IL-1α), интерлейкин-1β (IL-1β), интерлейкин-2 (IL-2), интерлейкин-3 (IL-3), интерлейкин-4 (IL-4), интерлейкин-5 (IL-5), интерлейкин-6 (IL-6), интерлейкин-7 (IL-7), интерлейкин-8 (IL-8), интерлейкин-9 (IL-9), интерлейкин-10 (IL-10), интерлейкин-11 (IL-11), интерлейкин-12 (IL-12), α-фактор некроза опухолей (TNF-α), β-фактор некроза опухолей (TNF-β) и трансформирующий фактор роста β (TGF-β), но не ограничиваются ими. Предусматривается, что цитокины можно вводить совместно и/или последовательно с иммунологической композицией или вакциной настоящего изобретения. Так, например, вакцина настоящего изобретения может содержать и молекулу экзогенной нуклеиновой кислоты, экспрессирущей in vivo подходящий цитокин, напр., цитокин, соответствующий данному вакцинируемому организму или в котором нужно выработать иммунологический ответ (например, птичий цитокин для препаратов, вводимых птицам).
Примеры подходящих эмульсий или адъювантов дополнительно описаны, к примеру, в US 6,235,282; US 6,224,882; US 7,371,395; US 2006/0233831; US 2005/0238660; US 2006/0233831 (все это патенты и патентные заявки фирмы Merial).
Иммунологические композиции или вакцины по изобретению включают или состоят в основном или состоят из эффективного для выработки терапевтической реакции количества одного или нескольких экспрессирующих векторов и/или полипептидов, как изложено в настоящем изобретении, причем эффективное количество можно установить из данного описания, включая документы, включенные в него. и общих познаний в этой области, без ненужного экспериментирования.
В случае иммунологических композиций и/или вакцин на основе плазмидного вектора доза может содержать, состоять в основном или состоять в общем случае из от 1 мкг до 2000 мкг, предпочтительно от 50 мкг до 1000 мкг и более предпочтительно от 100 мкг до 800 мкг плазмиды, экспрессирующей антиген, эпитоп или иммуноген гриппа. При введении иммунологической композиции и/или вакцины на основе плазмидного вектора методом электропорации доза плазмиды обычно составляет от 0,1 мкг до 1 мг, предпочтительно от 1 мкг до 100 мкг, более предпочтительно от 2 мкг до 50 мкг.Объем дозы может составлять от 0,1 до 2 мл, предпочтительно от 0,2 до 1 мл.
Предпочтительно, если антигеном служит гемагглютинин, дозировка измеряется в единицах гемагглютинации (HAU) или в мкг НА. В предпочтительном воплощении дозировка может составлять 655 единиц гемагглютинации (HAU, 0,2 мкг НА) на дозу, 6550 единиц HAU (2,3 мкг НА) на дозу или 65500 единиц HAU на дозу. В некоторых воплощениях дозировка составляет 26200 HAU (9,2 мкг НА) на дозу. Объем может составлять от 0,1 мл до 1,0 мл, предпочтительно от 0,1 до 0,3 мл для однодневных цыплят и от 0,3 до 0,5 мл для старших цыплят.
Иммунологические композиции и/или вакцины содержат в одной дозе от 104 до 1011, предпочтительно от 105 до 1010 и более предпочтительно от 106 до 109 вирусных частиц рекомбинантного аденовируса, экспрессирующего антиген, эпитоп или иммуноген гриппа. В случае иммунологических композиций и/или вакцин на основе поксвируса доза может составлять от 102 БОЕ до 109 БОЕ. Иммунологические композиции и/или вакцины содержат в одной дозе от 105 до 109, предпочтительно от 102 до 108 БОЕ рекомбинантного поксвируса или герпесвиуса, экспрессирующего антиген, эпитоп или иммуноген гриппа.
Объем дозы композиций для заданных видов млекопитающих, напр., объем дозы композиций для птиц на основе вирусных векторов, напр., композиций на основе вирусных векторов не из поксвирусов, обычно составляет от 0,1 до 2,0 мл, от 0,1 до 1,0 мл и от 0,1 мл до 0,5 мл.
Далее изобретение будет дополнительно описано на следующих неограничивающих примерах.
Примеры
Конструирование ДНК-вставок, плазмид и рекомбинантных вирусных или растительных векторов проводили стандартными методами молекулярной биологии, описанными в J. Sambrook et al. (Molecular Cloning:A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1989).
Пример 1. Конструирование плазмид и трансформация растений
В данном исследовании экспрессировали синтетический ген гемагглютинина (НА) из изолята сильно патогенного птичьего гриппа (HPAI) H5N1 А/курица/Индонезия/7/2003 (ck/Indonesia) с помощью запатентованной системы экспрессиования белков в Lemna minor - системы LEX™ фирмы Biolex.
Гемагглютинин (НА) является поверхностным гликопротеидом вируса, который отвечает за прикрепление вируса к терминальным сиаловым кислотам на рецепторах клетки-хозяина и опосредует слияние между частицами вируса и мембранами клетки посредством своего расщепления. Он является ключевым антигеном в реакции хозяина на вирус гриппа и при естественной инфекции, и при вакцинации.
Его предшественник НА0 - белок, содержащий 564 аминокислоты с приблизительной молекулярной массой 77 кДа и обладающий способностью к агглютинации эритроцитов. Имеется 6 предполагаемых сайтов N-гликозилирования в области НА1 и 1 предполагаемый сайт N-гликозилирования в области НА2.
НА высоко экспрессировался в апопластном пространстве растений, имел ожидаемый размер при анализе методом Western-блота и обладал активностью гемагглютинации. Из трансгенной линии Lemna получали неочищенный растительный экстракт для оценки иммуногенности и эффективности на цыплятах SPF. Значительный титр ингибирования гемагглютинации сыворотки при использовании гомологичных и гетерологичных антигенов свидетельствует, что полученный из Lemna НА является сильно иммуногенным. Трехнедельным цыплятам SPF, вакцинированным однократной дозой полученного в Lemna НА, составленного в эмульсии типа вода в масле, вводили провоцирующую пробу с изолятом HPAI H5N1 A/ck/Indonesia/7/2003 или антигенным вариантом A/ck/WestJava/PWT-WU/2006. Вырабатывалась полная или на 80-90% защита от A/ck/Indonesia/7/2003 и A/ck/WestJava/PWT-WU/2006, сответственно. Полная клиническая защита была получена у НА-вакцинированных птиц, обработанных в однодневном возрасте вакциной от птичьего гриппа с вектором из поксвируса домашней птицы (схема прайм-буст). У всех вакцинированных наблюдалось резкое уменьшение ротоглоточного выброса вируса, а метод ELISA на NP в образцах сыворотки четко дифференцировал вакцинированных цыплят от инфицированных. Защита не наблюдалась у цыплят, получавших измельченную НА-экспрессирующую ряску.
В заключение, экспрессированный в Lemna minor НА вызывал сильный иммунный ответ и давал отличную степень защиты против гомологичного H5N1 и варианта H5N1. Трансгенная ряска может стать хорошей альтернативой существующим инактиви-рованным вакцинам с потенциалом для DP/A.
Конструирование плазмид для трансформации растений. Была разработана оптимизированная версия гена гемагглютинина (А) из изолята А/курица/Индонезия/ 7/2003 (H5N1) вируса высоко патогенного птичьего гриппа (HPAI) с предпочтительным использованием кодонов L. minor (содержание GC=63-67%). Синтетический ген НА модифицировали по сайту расщепления между НА1 и НА2 из последовательности высоко патогенного птичьего гриппа (множественные основные аминокислоты: RERRRKKR - SEQ ID NO:17) в последовательность слабо патогенного птичьего гриппа (RETR - SEQ ID NO:18). Нативную сигнальную последовательность НА заменяли на сигнальную последовательность α-амилазы риса (GenBank M24286), слитую с 5'-концом оптимизированной по кодонам кодирующей последовательности H5N1 (SEQ ID NO:1). Добавили сайты рестрикционных эндонуклеаз (5' - EcoRI и 3' - SacI) для клонирования в бинарные векторы Agrobacterium tumefaciens. Оптимизированный к L. minor ген НА клонировали по EcoRI/SacI в модифицированный бинарный вектор pMSP3 A. tumefaciens (Gasdaska J. et al., Bioprocessing J. 3, 50-56, 2003) между химерным промотором октопин- и маннопинсинтазы с 5'-лидером SSU1 RBCS Lemna gibba и терминатором нопалинсинтазы (Nos), получая вектор для трансформации растений MerB01.
Создание и скрининг трансгенной линии. Используя клетки A. tumefaciens C58Z707 (Hepbum A.G. et al., J. Gen. Microbiol. 131, 2961-2969, 1985), трансформированные вектором для трансформации растений MerBOl, были созданы трансгенные растения, представляющие индивидуальные клональные линии, из быстро растущих узелков L. minor, как описано в Yamamoto Y. et al., In vitro Cell. Dev. Biol. 37, 349-353 (2001). Для скрининга трансгенов индивидуальные клональные линии предварительно выдерживали в течение 1 недели с 150-200 мМ m-2s-1 в вентилируемых сосудах для культивирования растений, содержащих среду SH (Schenk R. et al., Can. J. Bot. 50, 199-204, 1972) без сахарозы. Затем 15-20 предварительно обработанных пластинок помещали в вентилируемые сосуды, содержащие свежую среду SH, и давали им расти в течение 2 недель. Образцы тканей из каждой линии замораживали и хранили при -70°С. Впоследствии эти образцы тканей подвергали скринингу на экспрессию НА методом гемагглютинации. Вкратце, замороженные ткани гомогенизировали, центрифугировали и отбирали супернатанты для анализа. Разведения трансгенных образцов инкубировали с 10% раствором эритроцитов индейки (Fitzgerald Industries International) и определяли активность гемагглютинации. Наиболее активные линии, отобранные этим методом при начальном разведении, снова подвергали анализу с использованием больших разведении для определения титра. Образцы сравнивали с рекомбинантным H5N1 в качестве положительного контроля и контролем Lemna дикого типа. Пример скрининга линий представлен на фиг.9.
Пример 2. Создание линии птичьего гриппа H5N1
Для скрининга было создано 150 трансгенных линий птичьего гриппа H5N1. После создания трансгенных линий их подвергали скринингу на экспрессию птичьего гриппа H5N1 в среде и в ткани. Вкратце, растения культивировали в течение 2 недель в небольших исследовательских сосудах и отбирали среды и ткани для анализа. Для анализа тканей замороженные ткани гомогенизировали, центрифугировали и отбирали супернатанты для анализа.
Образцы подвергали скринингу методом гемагглютинации. Вкратце, разведения трансгенных образцов инкубировали с 10% раствором эритроцитов индейки (Fitzgerald Industries International, Concord, MA, США) и определяли активность гемагглютинации. Наиболее активные линии, отобранные этим методом при начальном разведении, снова подвергали анализу с использованием более сильных разведении. Образцы сравнивали с рекомбинантным H5N1 в качестве положительного контроля и растениями Lemna дикого типа в качестве отрицательного контроля. Анализ культуральных сред методом гемагглютинации показал отсутствие активности в одной подгруппе линий, поэтому остальные линии не тестировали этим методом. На фиг.9 представлен репрезентативный планшет при анализе гемагглютинации и результаты метода гемагглютинации при скрининге трансгенных растений (в формате диаграммы и таблицы). Наиболее активные при начальном скрининге линии подращивали так, чтобы получить примерно 1 кг биомассы для дальнейшего изучения.
Пример 3. Продукция гемагглютинина птичьего гриппа H5N1 в Lemna minor
Для характеристики НА H5N1 использовали методы гемагглютинации (ГА), ингибирования гемагглютинации (ИГ), ELISA, SDS-PAGE и western-блота. Рекомбинантный белок также подвергали скринингу против комплекта положительных сывороток цыплят методом ИГ.
Экстрагирование растений. Из линии, содержащей НА H5N1, получали неочищенный тканевой экстракт по методике, описанной ниже. Все операции выполняли при 4°С. Смешивали 100 г замороженной биомассы с 200 мл экстракционного буфера (50 мМ NaPO4, 0,3 М NaCl, 10 мМ ЭДТА, рН 7,4, коктейль из ингибиторов протеаз 1:1000 (Sigma P9599, Sigma, St. Louis, МО, США)), а затем гомогенизировали в измельчителе Waring Blender 4 раза по 20 сек с охлаждением по 10-20 сек между ними. Гомогенат центрифугировали при 14000 g в течение 30 мин при 4°С, очищали пропусканием через марлю для удаления крупных кусков, а затем через фильтр из ацетата целлюлозы (0,22 мкм). Полученный гомогенат хранили при 4°С или на льду для немедленного тестирования. Для дальнейшего анализа гомогенат замораживали порциями при -80°С, чтобы избежать любых циклов замораживания-оттаивания. Общий растворимый белок (TSP) определяли методом Брэдфорда с бычьим сывороточным альбумином в качестве стандарта.
Метод гемагглютинации (ГА). Метод гемагглютинации является допустимым тестом для выявления и количественного определения гемагглютинирующего антигена. В основе метода ГА лежит то, что вирусный гемагглютинин будет связываться с рецепторами на поверхности эритроцитов (RBC), что приведет к их агглютинации. Метод ГА выполняли, используя серийные 2-кратные разведения неочищенных экстрактов на планшетах с U-образным дном Nunc. Инкубировали 50 мкл 10% эритроцитов индейки (Fitzgerald Industries International Inc.) с 50 мкл тестируемых образцов в течение 1 часа при комнатной температуре и определяли титр при наибольшем разведении, при котором еще четко отмечался сгусток. Отрицательные контроли включали ряску дикого типа и PBS, а положительные контроли включали экспрессированный в бакуловирусе рекомбинантный гемагглютинин птичьего гриппа A/Vietnam/1203/2004 (87 мкг/мл).
Отрицательные контроли - PBS и образец ряски дикого типа - не вызывали гемагтлютинации, свидетельствуя, что единственным источником агглютинации был НА H5N1 (фиг.10). Титр НА составлял 64, 12800 и 51200-102400 для инактивированного птичьего гриппа H5N1 ck/Indonesia/03 (мутантного), контрольного рекомбинантного белка НА и неочищенного экстракта, содержащего НА H5N1, соответственно. Результаты свидетельствуют, что даже при разведении в 102400 раз неочищенный экстракт все еще способен агглютинировать RBC и не давать им образовывать плотный осадок. Судя по определению методом ГА, неочищенный экстракт, содержащий НА H5N1, биологически активен и обладает значительно большей активностью, чем инактивированный целый вирус при 108,5 EID50 и контрольный рекомбинантный НА при 87 мкг/мл.
Для начального скрининга использовали коммерческие эритроциты индейки. Для оценки пригодности композиций неочищенный экстракт НА H5N1 анализировали стандартизированным методом ГА. Свежие эритроциты курицы промывали 3 раза с помощью PBS и инкубировали с тестируемыми образцами в течение 30 мин вместо 1 ч. Результаты показали 1-2-кратные отличия в титре НА между стандартным методом ГА и текущим методом ГА. Ожидаемый выход определяли, как показано на фиг.11.
Методы ингибирования гемагглютинации (ИГ) и ELISA. В основе метода ингибирования гемагглютинации лежит то, что взаимодействие специфических антител с гомологичным вирусным гемагглютинином должно ингибировать гемагглютинацию. Распознавание экспрессированного антигена НА специфическими антителами подтверждает антигенность НА.
Агглютинирующая активность неочищенного экстракта НА H5N1 успешно нейтрализировалась всеми положительными на ГА сыворотками, т.е. моноклональным антителом к гемагглютинину Н5 вируса гриппа A/Vietnam/I 203/04 (Rocldand, Gilbertsville, PA), пулом моноклональных антител СР62 и 364/1 против H5N1 (CDC, Atlanta, GA, США), сывороткой цыплят FP2211 и сывороткой цыплят с птичьим гриппом H5N1 ck/Indonesia/03 (мутантным). Отрицательные контроли включали PBS и образец ряски дикого типа, которые не вызывали гемагглютинации (фиг.12-14). Результаты подтверждают то, что в неочищенном экстракте НА H5N1 обладает ожидаемой антигенностью.
Для серологического анализа образцов, собранных при исследовании клинической иммуногенности, проводили тест на ИГ по стандартной методике NVSL. Тестировали на перекрестную реактивность панелью антигенов с сывороткой: H5N1 ветви 2.1 А/курица/Indonesia/7/2003 (Indo/03), H5N1 ветви 2.1 (вариант) A/ck/WestJava/PWT-WIJ72006, H5N1 ветви 2.2 A/WS/Mongolia/244/05, H5N1 ветви 1 A/Vietnam/I 203/2004 (VN/04) и H5N8 A/turkey/Ireland/1378/1983 (Ireland/83). Статистический анализ проводили с помощью SAS V9.1. Блокирующий ферментный иммуносорбентный анализ (bELISA) проводили согласно инструкции производителя (FlockCheck AI MultiS-Screen Antibody Test Kit, IDEXX Laboratories, Westbrook, ME).
SDS-PAGE и Western-блот. Белковые образцы (неочищенные тканевые экстракты) разбавляли буфером для образцов SDS-PAGE, разделяли в 10% бис-трис геле Nu-PAGE (Invitrogen, Carlsbad, CA) и переносили на мембрану PVDF с помощью iBIot фирмы Invitrogen. Мембрану блокировали 1 час при комнатной температуре (или всю ночь при 4°С), инкубировали с моноклональным антителом против гемагглютинина Н5 вируса гриппа A/Vietnam/1203/04 (Rockland) в течение 1 часа при комнатной температуре. После четырех отмывок в PBS с 0,1% Tween-20 мембрану инкубировали 1 час с HRP-конъюгированным вторичным антителом, отмывали 4 раза в PBS с 0,05% Tween-20, а затем проявляли в течение 5 минут с помощью системы субстратов мембранной пероксидазы ТМВ (KPL, Gaitersburg, MD). Анализ изображений проводили с помощью системы анализа инфракрасных изображений Odyssey LICOR 9120 (LICOR, Lincoln, NE).
При окрашивании серебром SDS-PAGE наблюдалась отчетливая полоса при 77 кДа у линии, экспрессирующей НА (фиг.15А). Western-блот с помощью моноклонального антитела против гемагглютинина Н5 вируса гриппа A/Vietnam/1203/04 подтвердил экспрессию НА с ожидаемым молекулярным весом в 77 кДа, тогда как экспрессия Lemna дикого типа оставалась отрицательной (фиг.15 В). На western-блоте в невосстанавливающих условиях и моноклональное антитело против гемагглютинина Н5 вируса гриппа A/Vietnam/1203/04 (Rockland) и пул моноклональных антител СР62 и 364/1 против H5N1 (CDC, Atlanta, GA) распознавали НА H5N1 в виде одной преобладающей полосы с ожидаемым молекулярным весом в 77 кДа, тогда как Lemna дикого типа оставалась отрицательной (фиг.15 с). На фиг.16 также видно распознавание НА сывороткой цыплят FP2211 и сывороткой цыплят с птичьим гриппом H5N1 ck/Indonesia/03 (мутантным) в виде одной ожидаемой полосы в 77 кДа, тогда как Biolex дикого типа оставался отрицательным. Инактивированный целый вирус и рекомбинантный контрольный НА проявляли две полосы в 50 кДа и 28 кДа, указывая на то, что НАО расщеплялся на две субъединицы НА1 и ЯА2. Результаты western-блота согласуются с наблюдениями методом ингибирования гемагглютинации.
Итоги. Результаты анализа гемагглютинации подтвердили биологическую активность НА H5N1 с титром в 51200 единиц гемагглютинина (HAU)/50 мкл, что значительно выше, чем у рекомбинантного НА при 87 мкг/мл и инактивированного птичьего гриппа H5N1 ck/Indonesia/03 (мутантного) при 108,5 EID50. Гемагглютинирующая активность НА H5N1 успешно нейтрализировалась панелью ИГ положительных сывороток, т.е. моноклональным антителом к гемагглютинину Н5 вируса гриппа A/Vietnam/I 203/04 (Rockland), пулом моноклональных антител СР62 и 364/1 против H5N1 (CDC), сывороткой цыплят FP2211 и сывороткой цыплят с птичьим гриппом H5N1 ck/Indonesia/03. Результаты свидетельствуют о том, что каждое антитело распознает антигены в их нативном виде. Экспрессия НА дополнительно проверялась методами SDS-PAGE и western-блота. При окрашивании серебром SDS-PAGE проявлялась полоса в 77 кДа, соответствующая ожидаемому размеру предшественника НА0. На western-блотах НА H5N1 с ожидаемым молекулярным весом в 77 кДа очень хорошо распознавался всеми протестированными МАb и сыворотками цыплят, т.е. моноклональным антителом к гемагглютинину Н5 вируса гриппа A/Vietnam/1203/04 (Rockland), пулом моноклональных антител СР62 и 364/1 против H5N1 (CDC), сывороткой цыплят FP2211 и сывороткой цыплят с птичьим гриппом H5N1 ck/Indonesia/03.
Пример 4. Характеристика экспрессии НА из AIV H5N1 штамма Indonesia, полученного в Lemna (система Biolex), методом иммунолокализации в растениях
Экспрессию НА в тканях Lemna анализировали методом иммунофлуоресценции. Растение фиксировали на предметном стекле в буфере MTSB (50 мМ EGTA, 50 мМ Pipes, 5 мМ MgSO4, pH 7,0) с 4% формальдегидом под вакуумом, затем промывали MTSB + 0,1% Triton X100, а затем водой + 0,1% Triton Х100. Клеточную стенку расщепляли с помощью дриселазы (Driselase) (Sigma-Aldrich, St. Louis, МО) в течение 30 мин при 37°С, снова промывали MTSB+0,1% Triton X100, MTSB+10% DMSO+3% NP40 и MTSB+0,1% Triton X100, а затем блокировали MTSB+3% BSA. После этого обработанное растение инкубировали с моноклональным антителом против гемагглютинина Н5 вируса гриппа A/Vietnam/1203/04 в течение ночи при 4°С и инкубировали 3 часа с конъюгированным с флюоресцеином (FITC) вторичным антителом при комнатной температуре, а предметные стекла исследовали с помощью флуоресцентного микроскопа Nikkon Eclipse 600.
Результаты показали, что у Lemna дикого типа не наблюдалось флуоресцентного фона, тогда как у трансформированных Lemna выявлялся сильный и специфичный сигнал флуоресценции (фиг.17). Они также свидетельствуют, что НА экспрессируется в апопласте растительной ткани, что согласуется с клеточной локализацией мишени для экспрессии НА.
Пример 5. Исследование иммуногенности и провоцирующих проб
Исследования иммуногенности и провоцирующих проб проводились на цыплятах, свободных от специфических патогенов (SPF), вакцинированных в 3-недельном возрасте экспрессированным в Lemna НА с адъювантом. Для каждой вакцинированной группы выделяли по 10 цыплят. В качестве отрицательного контроля для обоих исследований была включена группа А, вакцинированная материалом Lemna дикого типа с адъювантом, а для исследования провоцирующих проб также была включена группа экспериментального рекомбинантного НА, экспрессированного в бакуловирусной системе, с адъювантом. Одна группа (группа 8) получала вакцину против AIV Н5 с вектором из поксвируса домашней птицы (vFP89; см. US 2008/0107681 и US 2008/0107687) в однодневном возрасте за 3 недели до введения экспрессированного в Lemna НА с адъювантом (см. ниже).
Исследование иммуногенности. Цыплят вакцинировали, как описано на фиг.18. Тестировали 6 групп 3-недельных цыплят по двум разным схемам: одно введение (группы 5-7) или два введения (группы 2-4) при трех уровнях дозы (655 HAU, 6550 HAU и 26200 HAU). Схему прайм-буст (группа 8) исследовали на 1-дневных цыплятах, подвергнутых праймингу с помощью TROVAC® (vFP89), экспрессирующего ген НА H5N8 (А/индейка/Ireland(1/1378/83), и бустингу экспрессированным в Lemna НА при 6550 HAU. TROVAC® вводили подкожно в затылочную часть шеи (103 TCID50/0,2 мл на дозу). Эмульсии типа вода в масле из неочищенного экстракта Lemna вводили внутримышечно в лапку (0,3 мл на дозу). На 21-й и 35-й день брали пробы крови для теста на ингибирование гемагглютинации.
Ни один цыпленок не проявлял отрицательной реакции на происходящие из растений вакцины. Иммуногенность определяли по титру ИГ в сыворотках, взятых у вакцинированных цыплят (фиг.20). Цыплята, вакцинированные Lemna дикого типа, давали отрицательный результат при анализе ИГ против всех тестируемых антигенов Н5. Через 21 день после иммунизации вырабатывались специфические антитела в группах НА Lemna, причем средние титры ИГ против гомологичных штаммов Indo/0,3 достигали 4, 6,5 и 8,1 log2 при дозировке в 655 HAU, 6550 HAU и 26200 HAU, соответственно. На 35-й день после вакцинации (p.v.) титры ИГ против Indo/0,3 оставались на уровне 4,7, 6,6 и 7,6 log2 для низких и высоких доз по схеме с одним введением, тогда как по схеме с двумя введениями титры ИГ значительно возрастали до 6,8, 9,4 и 9,5 log2, что свидетельствует о явном буст-эффекте (р<0,005) и эффекте дозы (р<0,005 между низкими и средними/высокими дозами). Этот результат также проявлялся по титру ИГ против гетерологичных штаммов Mong/244/05 и VN/1203/04 при 2,9, 5,4, 6,5 log2 против 5,3, 7,7, 8,5 log2 и 2,6, 3,2, 4,8 против 4,2, 6,0, 6,6 log2 для схемы с одним введением и схемы с двумя введениями при дозировке в 655 HAU, 6550 HAU и 26200 HAU, соответственно. Иммунный ответ был самым сильным против гомологичного штамма H5N1 ветви 2.1 Indo/03, за ним следовала ветвь 2.2 Mong/244/05, а дальше ветвь 1 VN/1203/04 при обеих схемах вакцинации. Также исследовали схему прайм-буст с использованием экспрессирующего НА рекомбинантного поксвируса домашней птицы в качестве прайма при промежуточной дозе НА Lemna в 6550 HAU. На 21-й день после прайма ни для каких антигенов Н5 не наблюдалось титра ИГ, за исключением tk/Ire/83 с титром в 4,0 log2. На 35-й день после буста титр ИГ возрастал до 5,3, 5,6, 5,4 и 9,2 log2 против VN/1203/04, Indo/03, Mong/244/05 и tk/Ire/83, соответственно. Однако по сравнению со схемой с двумя введениями НА Lemna с титрами в 6,0, 9,4, 7,7 и 7,3 log2 антительный ответ оказался слабым, за исключением tk/Ire/83.
Исследование провоцирующих проб. Цыплят вакцинировали согласно фиг.19. Аналогично исследованию иммуногенности, цыплят вакцинировали НА Lemna при трех разных дозах, но при однократной иммунизации (группы 2-3, 5-7), за исключением группы 4 (пероральная вакцинация) и группы 8 (схема прайм-буст).
На 42-й день цыплятам вводили в нос/рот провоцирующую пробу вируса HPAI H5N1 A/ck/Indonesia/07/2003 (группы 1-4) или A/ck/VestJava/PWT-WU/2006 (группы 5-8) при 106,0 EID50 на цыпленка. После введения пробы ежедневно вели наблюдение за заболеваемостью и смертностью цыплят, а больных цыплят считали инфицированными гриппом. Брали ротоглоточные мазки для определения выброса вируса из дыхательных путей на 2-й и 4-й день после пробы в 1,5 мл среды BHI (Becton-Dickinson, Sparks, MD), содержащей антимикробные соединения (100 мкг/мл гентамицина, 100 ед./мл пенициллина и 5 мкг/мл амфотерицина В). У остальных цыплят из всех групп брали кровь для выделения сыворотки в возрасте 42 и 56 дней. Птиц подвергали эвтаназии внутривенным введением пентобарбитала натрия (100 мг/кг массы тела) в возрасте 56 дней.
Ожидалось, что провоцирующая проба с вирусом HPAI H5N1 будет воспроизводимым образом вызывать 100% смертность у интактных цыплят в пределах 2 дней. В обеих группах отрицательного контроля цыплята, вакцинированные Lemna дикого типа и провоцированные штаммом Indo/03, и цыплята, вакцинированные контрольным экспериментальным рекомбинантным НА и провоцированные PWT/06, умерли за этот период (фиг.19). В группах, провоцированных Indo/03, выжили 100% цыплят, однократно вакцинированных внутримышечно полученным в Lemna НА при дозировке в 655 HAU и 6550 HAU. В группах, провоцированных PWT/06, выжили 9 и 8 из 10 цыплят по схеме одного введения при 6550 HAU и 26200 HAU, соответственно. Одну птицу подвергли эвтаназии через 10 дней после пробы (д.п.п.) из-за тяжелой кривошеи в группе 6550 HAU. Схема прайм-буст TROVAC®/Lemna проявляла 100% защиту от этого варианта штамма.
Выброс вируса исследовали методом количественной ОТ-ПЦР в образцах ротоглоточных мазков, взятых их выживших птиц через 2 и 4 дня после пробы. Ротоглоточные мазки тестировали методом количественной полимеразной цепной реакции с обратной транскриптазой в реальном времени (qRRT-PCR) на вирус птичьего гриппа, а пороговые значения циклов qRRT-PCR преобразовывали в эквивалентные инфекционные титры в куриных яйцах с зародышами, исходя из регрессионной прямой, полученной с помощью серии разведении провоцирующего вируса (Lee et al., Journal of Virological Methods 119(2):151-158). Вкратце, из материала ротоглоточных мазков экстрагировали РНК добавлением 250 мкл среды из мазка к 750 мкл Trizol LS (Invitrogen Inc., Carlsbad, CA) с последующим перемешиванием на вибрационной мешалке и инкубацией при комнатной температуре в течение 10 минут, после чего добавляли 200 мкл хлороформа. Образцы опять перемешивали, инкубировали при комнатной температуре в течение 10 минут, а затем центрифугировали 15 минут примерно при 12000 g. Собирали водную фазу и выделяли РНК с помощью набора для выделения вирусной РНК MagMAX AI/ND (Ambion Inc., Austin, TX) в соответствии с инструкцией к набору, используя систему обработки магнитных частиц KingFisher (Thermo Scientific, Waltham, MA). Для получения РНК для количественного стандарта использовали провоцирующие штаммы вируса птичьего гриппа. Запасы вируса в аллантоидной жидкости разбавляли бульоном BHI (Becton-Dickinson) и титровали в куриных яйцах с зародышами на момент разведения стандартными методами (Swayne et al., 2008, Avian influenza. In:Isolation and Identification of Avian Pathogens. 5th ed., pp.128-134). Тотальную вирусную РНК экстрагировали из 10-кратных разведении титрованного вируса, как описано для материала мазков. Проводили qRRT-PCR на ген матрикса гриппа, как описано ранее (Lee et al., 2004). Титры вируса в образцах рассчитывали из стандартных кривых, рассчитанных программой Smart Cycler II (Cepheid Inc., Sunnyvale, CA) или экстраполяцией уравнения стандартной кривой. В группах, провоцированных Indo/03, все цыплята, вакцинированные Lemna дикого типа, оказались положительными с титром вируса в 106,9 EID50o, тогда как выделение вируса в группах НА Lemna резко уменьшалось чуть выше предела обнаружения до 102,9 и 103,1 EID50 при 6550 HAU и 655 HAU, соответственно, на 2-й день, и не обнаруживалось на 4-й день после пробы. В группах, провоцированных штаммом антигенного варианта PWT/06, все цыплята, иммунизированные экспериментальным НА при 5000 HAU, все еще выделяли вирус на уровне 107,1 EID50 на 2-й день после пробы, и только 1, 2 и 1 из 10 птиц выявлялись в группах 6550 HAU, 26200 HAU и TROVAC®/Lemna, соответственно, причем 2 оставались положительными в группах НА Lemna при 6550 и 26200 HAU через 4 дня после пробы, однако вирус оказался возле или ниже предела обнаружения (103,5 EID50) в группе TROVAC /Lemna.
Образцы также исследовали на наличие специфичных к нуклеопротеиду (NP) антител до и после пробы с H5N1 с помощью набора для ELISA (фиг.19). NP-специфичные антитела отсутствовали во всех образцах сыворотки, собранных после иммунизации НА Lemna и до проведения пробы. После пробы с PWT/06 у 9 из 9, 8 из 8 и 8 из 10 образцов проявлялся положительный сигнал в группах 6550 HAU, 26200 HAU и прайм-буст, соответственно.
На фиг.21 представлен серологический анализ образцов, собранных до провоцирующей пробы на 42-й день и после пробы на 56-й день (14 д.п.п.). Ни в группе Lemna дикого типа, ни в пероральной группе не вырабатывался гуморальный иммунитет к штамму Indo/03, a 3 из 10 вакцинированных экспериментальным экспрессированным в бакуловирусе НА проявляли заметный титр антител в 2,4 log2 против штамма VN/04. Все другие группы проявляли положительный иммунный ответ на тестируемые антигены, т.е. Indo/03, VN/04 и Mong/05, что подтверждает данные при исследовании иммуногенности.
После пробы с Indo/03 средний титр ИГ против Indo/03 возрастал с 4,5 до 7,1 log2 и с 6,9 до 8,2 log2 в группах 655 HAU и 6550 HAU, соответственно. Сыворотки также проявляли заметное повышение против PWT/06 из необнаруживаемого до 2,7 log2 и с 2,2 до 3,8 log2. После пробы с PWT/06 средний титр ИГ против гомологичного PWT/06 прыгнул с 2,2 до 6,0 log2, с 2,2 до 6,3 log2 и с 2,7 до 4,9 log2 в группах 6550 HAU, 26200 HAU и прайм-буст, соответственно. Аналогичная тенденция наблюдалась и в титре ИГ против Indo/03, с 6,9 до 8,6 log2, с 6,8 до 9,0 log2 и с 7,1 до 7,7 log2 в тех же группах.
Интересно, что результаты ELISA на NP показали, как и ожидалось, что до провоцирующей пробы не было заметного иммунного ответа на NP. Однако после пробы большинство сывороток защищенных птиц давали положительную реакцию. Этот результат четко показывает, что вакцина с экспрессированным в Lemna НА сама по себе или же режим вакцинации прайм-буст с помощью экспрессирующего НА вектора из поксвируса домашней птицы (так называемая схема прайминг-бустинг) полностью совместима со стратегией DIVA (отличения инфицированных от вакцинированных животных). Применение такой вакцины должно способствовать легкому выявлению инфекций в вакцинированной стае путем проверки на антитела к NP с помощью коммерчески доступного ELISA.
Пример 6. Очистка белка птичьего гриппа из растений ряски
Антигенный полипептид птичьего гриппа либо его фрагмент или вариант очищали путем выделения антигенного полипептида из культуральной среды. Начальная очистка включает центрифугирование для удаления растительного материала и клеточных обломков. После такой частичной очистки неочищенный экстракт можно очистить путем сдвига рН и тепловой обработки с последующим фильтрованием через диатомитовый фильтр. Из этих осветленных экстрактов очищали рекомбинантный НА методом аффинной хроматографии на колонке с фетуином. Очистку можно определять методом денситометрии окрашенных Кумасси голубым гелей SDS-PAGE.
Растительную ткань гомогенизировали в 50 мМ фосфате натрия, 0,3 М хлориде натрия и 10 мМ ЭДТА, рН 7,2, с помощью измельчителя Silverson. Гомогенат подкисляли до рН 4,5 с помощью 1 М лимонной кислоты и центрифугировали при 7500 g в течение 30 минут при 4°С. Значение рН супернатанта доводили до рН 7,2 с помощью 2 М 2-амино-2-(гидроксиметил)-1,3-пропандиола (трис), а затем фильтровали через фильтры на 0,22 мкм. Материал наносили прямо на смолу с белком A mAbSelect SuRe (GE Healthcare), уравновешенную раствором, содержащим 50 мМ фосфата натрия, 0,3 М хлорида натрия и 10 мМ ЭДТА, рН 7,2. После нанесения колонку промывали до уровня фона уравновешивающим буфером с последующей промежуточной промывкой 5 колоночными объемами 0,1 М ацетата натрия, рН 5,0. Связавшиеся антитела элюировали 10 колоночными объемами 0,1 М ацетата натрия, рН 3,0. Элюат с белка А немедленно нейтрализировали с помощью 2 М трис. Для удаления агрегатов элюат с белка А доводили до рН 5,5 и наносили на колонку с керамическим гидроксиапатитом типа I (BioRad, CA, США), уравновешенную 25 мМ фосфата натрия, рН 5,5. После промывки колонки 5 колоночными объемами уравновешивающего буфера белок элюировали за одну стадию с помощью 0,25 М фосфата натрия, рН 5,5. Фракции, содержащие белок по A280, объединяли и хранили при -80°С (Сох К.М. et al., 2006, 24(12): р.1591-7).
Все документы, процитированные или приведенные в процитированных в заявке документах, и все документы, процитированные или приведенные в настоящем изобретении ("приведенные в нем документы"), и все документы, процитированные или приведенные в процитированных в нем документах, вместе с инструкциями производителей, описаниями, спецификациями на продукты и сведениями о продуктах для любых продуктов, указанных в нем или в любом документе, включенном в него путем ссылки, настоящим включаются в него путем ссылки и могут использоваться при воплощении изобретения.
После подробного описания предпочтительных воплощений настоящего изобретения следует иметь в виду, что изобретение, определенное вышеприведенными параграфами, не должно ограничиваться конкретными деталями, изложенными в вышеприведенном описании, так как возможны многие явные его варианты, не отходящие от сути или не выходящие за рамки настоящего изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЕ ВАКЦИНЫ ПРОТИВ ВИРУСА "СИНЕГО ЯЗЫКА", ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2575599C2 |
| ВАКЦИНЫ ПРОТИВ FMDV НА ОСНОВЕ РЕКОМБИНАНТНОГО АДЕНОВИРУСНОГО ВЕКТОРА И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2725495C2 |
| РЕКОМБИНАНТНЫЕ ВЕКТОРЫ, ЭКСПРЕССИРУЮЩИЕ АНТИГЕНЫ ВИРУСА ПТИЧЬЕГО ГРИППА, И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2761869C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ВИРУСНОГО ВЕКТОРА APMV-8 | 2010 |
|
RU2592544C2 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА АФРИКАНСКОЙ КАТАРАЛЬНОЙ ЛИХОРАДКИ | 2007 |
|
RU2446823C2 |
| РЕКОМБИНАНТНЫЕ ВЕКТОРЫ HVT, ЭКСПРЕССИРУЮЩИЕ АНТИГЕНЫ ПАТОГЕНОВ ПТИЦ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2620936C2 |
| НОВЫЕ ЦИТОКИНЫ ПТИЦ И КОДИРУЮЩИЕ ИХ ГЕНЕТИЧЕСКИЕ ПОСЛЕДОВАТЕЛЬНОСТИ | 2008 |
|
RU2562108C2 |
| РЕКОМБИНАНТНЫЕ HVT-ВЕКТОРЫ, ЭКСПРЕССИРУЮЩИЕ МНОЖЕСТВЕННЫЕ АНТИГЕНЫ ПАТОГЕНОВ ПТИЦ, И СОДЕРЖАЩИЕ ИХ ВАКЦИНЫ | 2017 |
|
RU2752836C2 |
| ДНК-ПЛАЗМИДЫ, ОБЛАДАЮЩИЕ ПОВЫШЕННОЙ ЭКСПРЕССИЕЙ И СТАБИЛЬНОСТЬЮ | 2007 |
|
RU2456346C2 |
| КОМПОЗИЦИИ РЕКОМБИНАНТНОГО CDV И ЕЕ ПРИМЕНЕНИЕ | 2011 |
|
RU2567337C2 |















































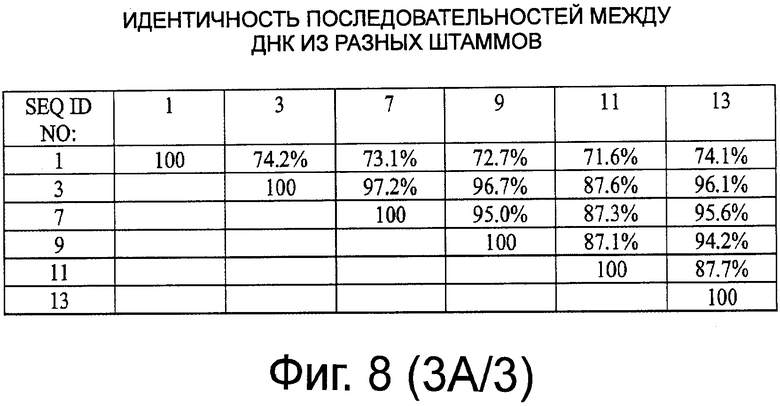





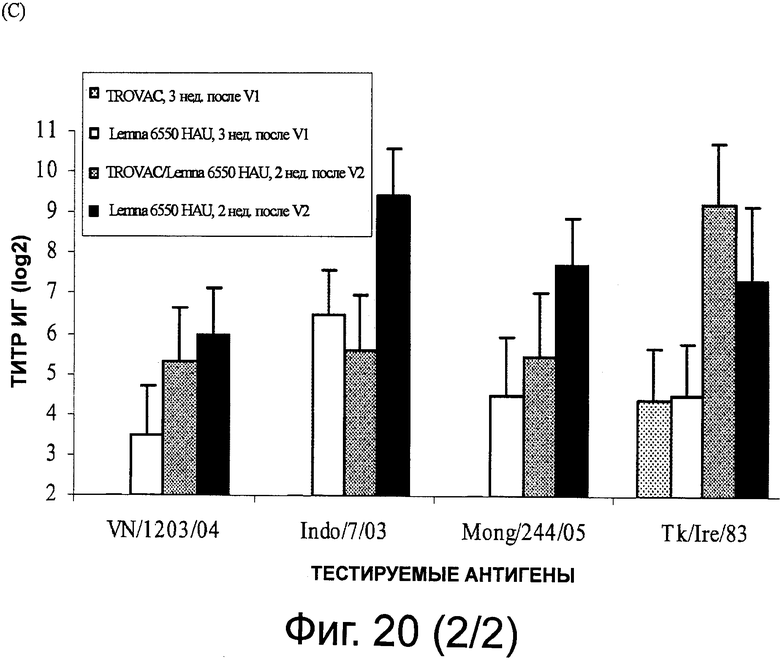
Изобретение относится к области биотехнологии, генной инженерии и вирусологии. Изобретение охватывает вакцины против гриппа, в частности вакцины от птичьего гриппа. Вакцины могут представлять собой субъединичные вакцины на основе гемагглютинина гриппа. Гемагглютинин может экспрессироваться в растениях, в том числе в ряске. Изобретение также охватывает рекомбинантные вектора, кодирующие и экспрессирующие антигены, эпитопы или иммуногены гриппа, которые могут использоваться для защиты животных от гриппа. Оно также охватывает режим вакцинации, совместимый со стратегией DIVA, включая схему прайм-буст с помощью вектора и субъединичных вакцин. Предложенная группа изобретений может быть использована в медицине и ветеринарии. 9 н. и 13 з.п. ф-лы, 21 ил., 6 пр.
1. Композиция, содержащая антиген птичьего гриппа и фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель,
в которой антиген птичьего гриппа содержит полипептид, имеющий последовательность, приведенную в SEQ ID NO: 2, 4, 5, 8, 10, 12 или 14, и где композиция используется для вызова у животного иммунного ответа против гриппа.
2. Композиция по п. 1, в которой антиген птичьего гриппа содержит иммуногенный фрагмент, содержащий по меньшей мере 15 аминокислот полипептида птичьего гриппа.
3. Композиция по п. 1, в которой антиген птичьего гриппа экспрессирован в ряске.
4. Композиция по п. 1, в которой антиген птичьего гриппа частично очищен.
5. Композиция по п. 1, в которой антиген птичьего гриппа существенно очищен.
6. Композиция по п. 1, в которой антигеном птичьего гриппа является полипептид птичьего H5N1.
7. Композиция по п. 6, в которой полипептидом птичьего H5N1 является полипептид гемагглютинина.
8. Композиция по п. 1, в которой фармацевтически или ветеринарно приемлемый носитель, эксципиент или наполнитель представляет собой эмульсию типа вода в масле или эмульсию типа масло в воде.
9. Способ вакцинации животных, подверженных птичьему гриппу, включающий по меньшей мере одно введение эффективного количества композиции по любому из пп. 1-8,
где композиция содержит антиген птичьего гриппа, который содержит полипептид, имеющий последовательность, приведенную в SEQ ID NO: 2, 4, 5, 8, 10, 12 или 14.
10. Способ по п. 9, в котором способ включает введение в режиме прайм-буст.
11. Способ по п. 10, в котором режим прайм-буст включает прайм-введение композиции по любому из пп. 1-8 и буст-введение композиции, включающей, в фармацевтически или ветеринарно приемлемом носителе или эксципиенте, рекомбинантный вирусный вектор, содержащий полинуклеотид для экспрессирования in vivo антигена птичьего гриппа, его варианта или фрагмента для защиты субъекта от гриппа и/или для предотвращения прогрессирования заболевания у инфицированного субъекта.
12. Способ по п. 10, в котором режим прайм-буст включает прайм-введение композиции, включающей, в фармацевтически или ветеринарно приемлемом носителе, разбавителе или эксципиенте, рекомбинантный вирусный вектор, содержащий полинуклеотид для экспрессирования in vivo антигена птичьего гриппа, варианта или фрагмента полипептида птичьего гриппа либо их смеси, и буст-введение композиции по любому из пп. 1-8 для защиты субъекта от гриппа и/или для предотвращения прогрессирования заболевания у инфицированного субъекта.
13. Способ по п. 10, в котором режим прайм-буст включает прайм-введение композиции по любому из пп. 1-8 и буст-введение инактивированной вирусной композиции или вакцины, содержащей антиген птичьего гриппа.
14. Способ по п. 10, в котором режим прайм-буст включает прайм-введение инактивированной вирусной композиции или вакцины, содержащей антиген птичьего гриппа, и буст-введение композиции по любому из пп. 1-8 для защиты субъекта от гриппа и/или для предотвращения прогрессирования заболевания у инфицированного субъекта.
15 Способ по п. 9, в котором животными являются птицы, лошади, псовые, кошачьи или свиньи.
16. Существенно очищенный антиген птичьего гриппа, экспрессированный в ряске, где полипептид является имуногенным и/или протективным, причем полипептид содержит аминокислотную последовательность, имеющую последовательность, приведенную в SEQ ID NO: 2, 4, 5, 8, 10, 12 или 14, где антиген птичьего гриппа гликозилирован.
17. Растение ряски, стабильно трансформированное геном для экспрессии антигена птичьего гриппа,
в котором антиген птичьего гриппа содержит полипептид, имеющий последовательность, приведенную в SEQ ID NO: 2, 4, 5, 8, 10, 12 или 14.
18. Плазмида, содержащая полинуклеотид, кодирующий антиген птичьего гриппа, имеющий последовательность, приведенную в SEQ ID NO: 2, 4, 5, 8, 10, 12 или 14, где плазмида предназначена для трансформации растений.
19. Набор для вакцинации типа прайм-буст, включающий по меньшей мере два флакона, причем первый флакон содержит композицию по любому из пп. 1-8, а второй флакон содержит композицию для буст-вакцинации, включающую композицию, содержащую рекомбинантный конкурирующий вектор, или композицию, содержащую инактивированную вирусную композицию,
где композиция содержит антиген птичьего гриппа, который содержит полипептид, имеющий последовательность, приведенную в SEQ ID NO: 2, 4, 5, 8, 10, 12 или 14.
20. Культура узелков ряски, стабильно трансформированная геном для экспрессии антигена птичьего гриппа,
в которой антиген птичьего гриппа содержит полипептид, имеющий последовательность, приведенную в SEQ ID NO: 2, 4, 5, 8, 10, 12 или 14.
21. Способ получения антигенного полипептида птичьего гриппа, включающий:
(a) трансформацию культуры растений ряски плазмидой, содержащей фрагмент ДНК, кодирующий антиген птичьего гриппа с функционально присоединенной последовательностью, кодирующей сигнальный пептид, который направляет секрецию антигена птичьего гриппа;
(b) культивирование в культуральной среде культуры растений ряски, где культура растений ряски стабильно траснформирована для экспрессии антигена птичьего гриппа; и
(c) извлечение антигена птичьего гриппа, где антиген птичьего гриппа содержит полипептид, имеющий последовательность, приведенную в SEQ ID NO: 2, 4, 5, 8, 10, 12 или 14.
22. Способ получения антигенного полипептида птичьего гриппа, включающий:
(a) трансформацию культуры узелков ряски плазмидой, содержащей фрагмент ДНК, кодирующий антиген птичьего гриппа с функционально присоединенной последовательностью, кодирующей сигнальный пептид, который направляет секрецию антигена птичьего гриппа;
(b) культивирование в культуральной среде культуры узелков ряски, где культура узелков ряски стабильно траснформирована для экспрессии антигена птичьего гриппа; и
(c) извлечение антигена птичьего гриппа, где антиген птичьего гриппа содержит полипептид, имеющий последовательность, приведенную в SEQ ID NO: 2, 4, 5, 8, 10, 12 или 14.
| US 2007286873 A1, 13.12.2007 | |||
| US 2008107681 A1, 08.05.2008 | |||
| ВАКЦИНА ПРОТИВ ГРИППА ПТИЦ ИНАКТИВИРОВАННАЯ ЭМУЛЬСИОННАЯ | 2006 |
|
RU2323741C2 |

