Эта заявка испрашивает приоритет, основанный на предварительной патентной заявке 61/183777, которая была зарегистрирована в Бюро США по патентам и торговым знакам 3 июня 2009 г., и полное раскрытие которой включено в этот документ путем ссылки.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение, в целом, относится к высокопроизводительному анализу признаков растений. В некоторых вариантах осуществления изобретение относится к способам высокопроизводительного анализа и количественной оценки растительных белков, определенных посредством масс-спектрометрии с разовым впрыскиванием образцов смешанного белка, извлеченного из растений и растительной ткани. Далее, некоторые варианты осуществления изобретения относятся к способам анализа генотипов трансгенных растений и поддержки экспрессии в этих растениях желательных растительных признаков.
УРОВЕНЬ ТЕХНИКИ
Расширяющееся применение технологии рекомбинантных ДНК для получения трансгенных растений в коммерческих и индустриальных целях требует развития высокопроизводительных способов анализа линий трансгенных растений. Такие способы нужны для сохранения сортов трансгенных растений в последовательной череде поколений, для предупреждения утечки трансгенов в окружающую среду, а также для помощи в быстром получении трансгенных растений с желательными или оптимальными фенотипами. Кроме того, современные руководства по безопасной оценке ГМ (генетически модифицированных) растений, предназначенных для употребления человеком, требуют их характеристики на уровне ДНК и белка при сравнении между родительской и модифицированной культурой. См. Sesikeran and Vasanthi (2008) Asia Pac. J. Clin. Nutr. 17 Suppl. 1:241-44. Новые экспериментальные сорта ГМ-растений, состоят из все более сложных генетических модификаций, включая, среди прочего, стопочные гены и признаки.
Современные способы анализа трансгенных растений, предпочитаемые в данной области техники, включают: методики, основанные на ДНК (например, ПЦР), ОТ-ПЦР, применение генов-репортеров, саузерн-блоттинг и иммунохимию. Всем перечисленным методологиям присущи различные недостатки, вследствие чего весьма желательно найти более совершенный способ, который определенно способен быстро и недорого идентифицировать, а также количественно оценить множественные продукты трансгенов высокопроизводительным образом посредством анализа ограниченного образца из трансгенного растения.
Методики анализа трансгенных растений, основанные на ДНК, страдают несколькими заметными недостатками. Несмотря на тот факт, что трансформация, опосредованная Agrobacterium, считается наиболее предпочтительным способом генетической трансформации растений, генотипы растений, трансформированных Agrobacterium, трудно анализировать, применяя методики, основанные на ПЦР. См. Nain et al. (2005) Plant Mol. Biol. Rep. 23:59-65. Присутствие в трансформированных тканях даже следовых количеств Agrobacterium приводит к обманчивым результатам ПЦР. Форматы Id. амплификации ДНК также требуют эмпирического тестирования ген-специфических праймеров и условий термоциклирования. Наиболее важно, что подходы к скринингу трансгенных растений, основанные на ДНК, фактически не определяют экспрессию белкового продукта гена. Сходным образом, ОТ-ПЦР или нозерн-блоттинг можно использовать для подтверждения присутствия трансгенных транскриптов в трансгенном растительном материале. См. Alwine et al. (1977) Proc. Nat. Acad. Sci. 74:5350-54; Toplak et al. (2004) Plant Mol. Biol. Rep. 22:237-50. Ни один из этих способов не подтверждает наличия фактической экспрессии белка в исходном растительном материале. Эти методики также требуют применения радиоактивных материалов и/или большего количества тканей и большого времени обработки.
Гены-репортеры, такие как гены, кодирующие флуоресцентные белки, также могут быть котрансформированы в геномы трансгенных растений, то есть, обеспечить инструмент для идентификации трансформантов. Однако гены-репортеры являются лишь косвенными индикаторами генетической рекомбинации. Экспрессия конструкции гена-репортера не подтверждает экспрессию сопутствующего трансгена. Далее, либо ген-репортер, либо трансген могут быть утрачены в последующих поколениях растения-хозяина, тем самым разрывая связь между наличием репортера и представляющего интерес трансгена. Сходным образом, трансгены могут уходить из растения-хозяина в соседние растения, например, в результате перекрестного опыления, без одновременного ухода гена-репортера. Когда в трансгенном растении сложены стопкой множественные гены, необходимо ввести в его геном равное количество генов-репортеров для анализа трансгенного протеома, поскольку функция гена-репортера является только косвенным индикатором функции трансгена, то есть, изменения экспрессии одного трансгена в ответ на присутствие добавочного трансгена могут оказаться не выявленными.
В отличие от способов, описанных выше, для идентификации продуктов экспрессии трансгенов в трансгенном растении можно использовать иммунохимию. Хотя иммунохимия полезна в этих целях, данный способ требует высокой степени очистки белковых образцов для выработки антител. Полученные в результате антитела должны быть протестированы на специфичность, и для этого необходимо создать условия анализа, специфичные в отношении реагентов. Ограничениями полезности этого способа являются высокий уровень экспрессии и очистки, необходимый для проведения иммунохимического анализа, а также связанные с этим проблемы удаления загрязнений из растительной ткани.
Для анализа протеома трансгенного растения также можно использовать масс-спектрометрию. Однако признанные в данной области техники спектрометрические методики требуют, чтобы сложные смеси растительных белков, прежде всего, были разделены двумерным гель-электрофорезом. Rajagopal and Ahern (2001) Science 294(5551):2571-73. См. также Domon and Aebersold (2006) Science 312(5771):212-17, 214. Единичные полосы из разделенного в геле образца белка впоследствии можно расщеплять протеазой и подвергать масс-спектрометрии для идентификации уникального белка, первоначально представленного в нерасщепленной полосе. См., например, Chang et al. (2000) Plant Physiol. 122(2):295-317. Стадия разделения в геле при использовании этого способа является длительным процессом, который затрудняет применение масс-спектрометрии в высокопроизводительных приложениях.
В данной области техники существует потребность в высоко-производительном способе выявления и количественного определения продуктов экспрессии трансгенов в растениях, который не требовал бы очищенного или высокоэкспрессируемого белка, а также использования специфических по отношению к методике реактивов. Такой способ будет полезным подспорьем для растениеводов, занимающихся трансгенными растениями, в сохранении сорта целевого трансгенного растения в ряду последовательных поколений при половом или бесполом размножении. Такой способ также будет полезен для быстрого анализа растений, полученных в результате процедуры трансформации, с целью идентификации таких полученных растений, которые являются трансгенными растениями, экспрессирующими встроенный белок в нужных тканях. Далее, такой способ можно применять для быстрого скрининга растений, подверженных риску контаминации трансгенами из трансгенного растения, для достижения биологической изоляции трансгенного растения.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Специфический вариант осуществления изобретения включает высокопроизводительный способ выявления и количественной оценки присутствия двух или более представляющих интерес белков с известными аминокислотными последовательностями в образце растительного происхождения. Способ включает обеспечение первого впрыскивания смешанного образца растительного происхождения, содержащего белки, и расщепление белков из смешанного образца растительного происхождения на пептиды. В альтернативном варианте способ включает предварительную стадию расщепления белков из смешанного образца растительного происхождения на пептиды с последующим первым впрыскиванием пептидов. Затем пептиды разделяют и ионизируют. Для пептидов получают синхронные масс-спектральные данные и определяют присутствие или отсутствие двух или более представляющих интерес белков.
Еще один вариант осуществления изобретения включает высокопроизводительный способ выявления присутствия двух или более представляющих интерес белков с известными аминокислотными последовательностями в образце растительного происхождения. Способ включает получение масс-спектральных данных для двух или более представляющих интерес белков и осуществление первого впрыскивания смешанного образца растительного происхождения, содержащего белки.
Белки смешанного образца растительного происхождения расщепляют на пептиды, а затем эти пептиды разделяют и ионизируют. Для пептидов получают синхронные масс-спектральные данные. Синхронные масс-спектральные данные сравнивают с масс-спектральными данными, полученными для двух или более представляющих интерес белков, тем самым, определяя наличие или отсутствие двух или более представляющих интерес белков.
Еще один вариант осуществления изобретения включает способ сохранения генотипа ассортимента трансгенных растений. Способ включает: (i) получение масс-спектральных данных для одного или более ожидаемых продуктов трансгенной экспрессии в ряду трансгенных растений, (ii) проведение первого впрыскивания смешанного образца, содержащего белки, из первого поколения ассортимента трансгенных растений, (iii) расщепление белков из смешанного образца растительного происхождения на пептиды, (iv) разделение пептидов, (v) ионизацию пептидов, (vi) получение синхронных масс-спектральных данных для пептидов и сравнение синхронных масс-спектральных данных с масс-спектральными данными, предоставленными для ожидаемых продуктов трансгенной экспрессии, тем самым, определяя наличие или отсутствие ожидаемых продуктов трансгенной экспрессии в первом поколении ассортимента трансгенных растений, (vii) проведение первого впрыскивания смешанного образца, содержащего белки, из второго поколения ассортимента трансгенных растений, (viii) повторение стадий (iii)-(vi) со смешанным образцом, содержащим белки, из второго поколения ассортимента трансгенных растений, и (ix) отказ от размножения второго поколения ассортимента трансгенных растений в том случае, если наличие ожидаемого продукта (продуктов) трансгенной экспрессии не было подтверждено по масс-спектральным данным для пептидов из смешанного белкового образца второго поколения ассортимента трансгенных растений, тем самым, сохраняя генотип ассортимента трансгенных растений.
КРАТКОЕ ОПИСАНИЕ ФИГУР
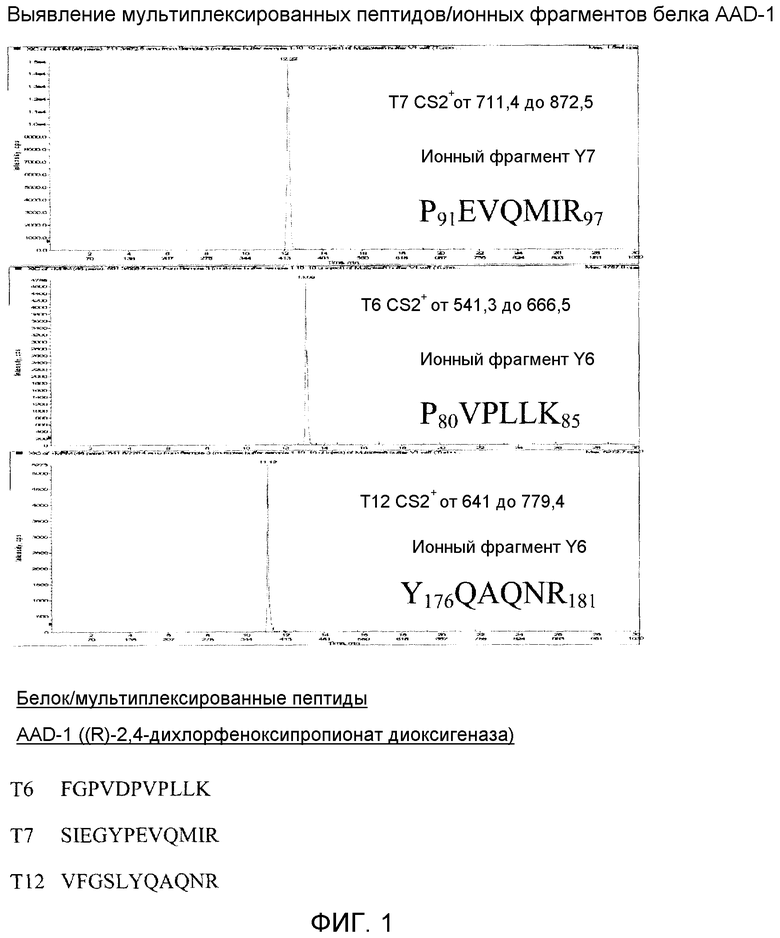
Фиг. 1 показывает ЖХ\МС\МС мультиплексное выявление AAD-1.
Фиг. 2 показывает ЖХ\МС\МС мультиплексное выявление AAD-12.
Фиг. 3 показывает ЖХ\МС\МС мультиплексное выявление Cry1F.
Фиг. 4 показывает ЖХ\МС\МС мультиплексное выявление Cry34.
Фиг. 5 показывает ЖХ\МС\МС мультиплексное выявление Cry35.
Фиг. 6 показывает ЖХ\МС\МС мультиплексное выявление ФАТ.
Фиг. 7 показывает ЖХ\МС\МС мультиплексное выявление Cry1F, экспрессированного в ткани инбредной кукурузы.
Фиг. 8 показывает ЖХ\МС\МС мультиплексное выявление Cry34, экспрессированного в ткани инбредной кукурузы.
Фиг. 9 показывает ЖХ\МС\МС мультиплексное выявление Cry35, экспрессированного в ткани инбредной кукурузы.
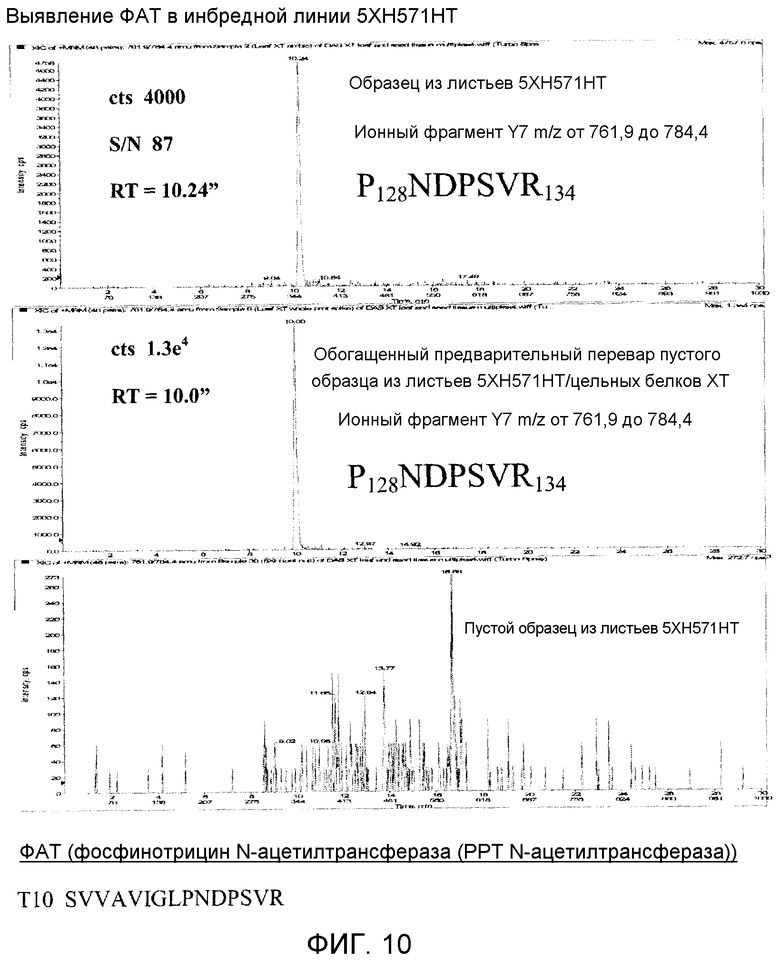
Фиг. 10 показывает ЖХ\МС\МС мультиплексное выявление ФАТ, экспрессированного в ткани инбредной кукурузы.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Специфичность, присущая масс-спектрометрическому подходу при идентификации единичного белка в смешанном образце, уникальна в том, что для идентификации представляющего интерес белка нужна только последовательность представляющего интерес белка. По сравнению с другими форматами мультиплексирования масс-спектрометрия уникальна, будучи способной использовать всю длину первичной аминокислотной последовательности белка, чтобы нацелиться на уникальные идентифицирующие участки первичной аминокислотной последовательности и, практически, исключить неспецифическое выявление. В некоторых вариантах осуществления настоящего изобретения протеолитический фрагмент или набор протеолитических фрагментов, который уникальным образом идентифицирует представляющий интерес белок (белки), применяют для выявления представляющего интерес белка (белков) в смешанном белковом образце.
В общих чертах, специфические способы, предложенные изобретением, позволяют осуществить прямой мониторинг и количественное определение любого белка (белков) из представляющей интерес ткани, для чего требуется только предварительное знание аминокислотной последовательности белка (белков). Способ может быть реализован высокопроизводительным образом без необходимости разрабатывать/тестировать реактив, специфический по отношению к реакции. Наличие очищенного белка в качестве реактива для разработки способа неизменно приносит успех, хотя для разработки способа этого анализа вполне достаточно иметь белок, степень очистки которого составляет, например, 60%. Способы, описанные в представленном раскрытии изобретения, могут исключить для компетентного специалиста необходимость разрабатывать требующие напряжения сил способы приготовления высокоочищенных образцов белка с целью их применения для выработки антител. Таким образом, способы, предложенные настоящим изобретением, помогут сэкономить время и ресурсы. Кроме того, эти способы обеспечивают большее разнообразие белковых реактивов, пригодных для мультиплексного анализа, поскольку усилия для удаления, например, последних 30-40% загрязнений могут стать ненужными. Раскрытые в этом документе способы также представляют необходимое улучшение по сравнению с мультиплексными подходами, основанными на амплификации ДНК, которые требуют эмпирического тестирования геноспецифических праймеров, соблюдения условий термоциклирования и, кроме того, создают ситуации, в которых любая утрата целевой специфичности может значительно снизить точность способа вследствие экспоненциальной амплификации, необходимой для выявления ДНК.
В отдельных вариантах осуществления способы, предлагаемые настоящим изобретением, позволяют осуществить обратную инженерию белка, чтобы определить, почему специфический белок обладает уникальными свойствами, причем реальный белок для начала исследования не требуется. Например, модификации последовательности и/или посттрансляционные модификации целевых белков, которые ассоциированы с желательными или нежелательными призраками растения, могут быть идентифицированы по масс-спектральным данным смешанного белкового образца из растения, экспрессирующего желательный или нежелательный признак.
В некоторых вариантах осуществления изобретения раскрытые в этом документе способы позволяют провести количественное определение или оценку соотношения различных белков в смешанном белковом образце посредством одноразового масс-спектрометрического анализа в противоположность индивидуальному многоразовому измерению каждого представляющего интерес белка и компиляции индивидуальных результатов в общий результат для одного образца.
В других вариантах осуществления настоящее изобретение также предлагает способы, полезные для развития и использования технологии трансгенных растений. Более конкретно, раскрытые в этом документе способы могут быть использованы для сохранения генотипов трансгенных растений в ряду поколений. Кроме того, некоторые варианты осуществления раскрытых в этом документе способов могут быть использованы для обеспечения высокопроизводительного анализа нетрансгенных растений, которые подвержены риску контаминации трансгенами от соседних растений, например, посредством перекрестного опыления. Благодаря этим вариантам осуществления изобретения может быть облегчено и/или достигнуто биологическое предотвращение распространения трансгенов. В других вариантах осуществления раскрытые в этом документе способы могут быть использованы для скрининга результатов процедуры трансформации растений высоко-производительным образом с целью идентификации трансформантов, которые проявляют желательные свойства экспрессии.
I. Сокращения
Настоящее изобретение будет описано с употреблением следующих сокращений:
AAD-1: (R)-2,4-дихлорфеноксипропионат диоксигеназа
AAD-12: (S)-2,4-дихлорфеноксипропионат/альфа-кетоглутарат диоксигеназа
ИСД: Индуцированная столкновениями диссоциация
Cry1F: Пестицидный кристаллический белок Cry1Fa (инсектицидный дельта-эндотоксин Cry1F(a)
Cry34: Кристаллический белок ET79 (Bacillus thuringiensis)
Cry35: Cry35Ab-подобный (Bacillus thuringiensis)
КЭ: Капиллярный электрофорез
ДНК: Дезоксирибонуклеиновая кислота
ELISA: Ферментный иммуносорбентный анализ
УИП: Усиленный ион-продукт
ГМ: Генетически модифицированный
ЗЗИ: Захват, зависящий от информации
ЖХ\МС\МС: Жидкостная хроматография - тандемная масс-спектрометрия
ММР: Мониторинг множественных реакций
МС: Масс-спектрометрия
МСМС: Тандемная масс-спектрометрия
ФАТ: Фосфинотрицин N-ацетилтрансфераза (PPT N-ацетилтрансфераза)
ПЦР: Полимеразная цепная реакция
ОТ-ПЦР: Полимеразная цепная реакция с обратной транскриптазой
II. Термины
Для того чтобы облегчить просмотр различных вариантов осуществления изобретения, описанных в этом документе, дается следующее разъяснение специфических терминов:
Биологическое сдерживание: При использовании в этом документе термин "биологическое сдерживание" относится к ограничению перемещения генетически модифицированных растений или их генетического материала в нежелательные ареалы. Этот термин включает физическое, физико-химическое, биологическое сдерживание, а также другие формы сдерживания, которые предотвращают выживание, распространение или репродукцию генетически модифицированных растений в природную окружающую среду или в искусственные условия выращивания.
Смешанный белковый образец: При использовании в этом документе термин "смешанный белковый образец" употребляется для дифференциации с очищенным белковым образцом. Сложный белковый образец содержит множественные белки и дополнительно может содержать другие загрязнения.
Масс-спектрометрия: При использовании в этом документе общий термин "масс-спектрометрия" относится к любому подходящему способу, устройству или конфигурации масс-спектрометрии, включая, например, ионизацию электрораспылением (ИЭР), МС с ионизацией способом лазерной десорбции в присутствии матрицы (ИЛДПМ), времяпролетную МС с ИЛДПМ, МС с ИЛДПМ при атмосферном давлении, МС с ИЛДПМ в вакууме, тандемную МС или любую комбинацию этих способов. Устройства для масс-спектрометрии измеряют молекулярную массу молекулы (как функцию соотношения между массой и зарядом молекулы), определяя траекторию полета молекулы через множество магнитных и электрических полей. Соотношение между массой и зарядом является физической величиной, широко используемой в электродинамике заряженных частиц. Соотношение между массой и зарядом определенного пептида априорно может рассчитать компетентный специалист в данной области. Две частицы с разным соотношением между массой и зарядом, подвергаясь воздействию одинаковых электрических и магнитных полей, не будут двигаться в вакууме по одной и той же траектории. Настоящее изобретение включает, среди всего прочего, применение жидкостной хроматографии высокого разрешения (ЖХВР) с последующим анализом пептидов способом тандемной МС.
Инструменты для масс-спектрометрии состоят из трех модулей: Источника ионов, который разбивает молекулы образца на ионы, масс-анализатора, который сортирует ионы по их массе, прикладывая электромагнитные поля, и детектора, который измеряет величину индикаторного количества и, тем самым, поставляет данные для расчета относительного содержания каждого представленного иона. Такая методика имеет как качественные, так и количественные варианты применения. К их числу относятся идентификация неизвестных соединений, определение изотопного состава элементов в молекуле, определение структуры соединения посредством наблюдения его фрагментации и количественное определение соединения в образце.
Подробный обзорный анализ методологий и устройств масс-спектрометрии можно найти в следующих источниках, которые включены в тот документ путем ссылки: Carr and Annan (1997) Overview of peptide and protein analysis by mass spectrometry. В: Current Protocols in Molecular Biology, под редакцией Ausubel, et al. New York: Wiley, p. 10.21.1-10.21.27; Paterson and Aebersold (1995) Electrophoresis 16: 1791-1814; Patterson (1998) Protein identification and characterization by mass spectrometry. В: Current Protocols in Molecular Biology, под редакцией Ausubel, et al. New York: Wiley, p. 10.22.1-10.22.24; and Domon and Aebersold (2006) Science 312(5771):212-17.
Мультиплексный: Применительно к термину, употребляемому в этом документе, белки и/или пептиды "мультиплексированы", когда в одном и том же образце представлены два или более представляющих интерес белка и/или пептида.
Признак растения: При использовании в этом документе термин "признак растения" может относиться к любому единичному качественному или количественному признаку растения.
Пептид: Пептиды представляют собой короткие полимеры, образующиеся в результате соединения друг с другом α-аминокислот в определенном порядке. Пептиды также можно получить в результате расщепления полипептидов, например, белков, протеазой.
Белок: Белки представляют собой органические соединения аминокислот, выстроенных линейной цепочкой и соединенных друг с другом пептидными связями между карбоксильными группами и аминогруппами остатков смежных аминокислот. Последовательность аминокислот в белке определяется последовательностью гена, которая закодирована в генетическом коде. В общем, генетический код точно определяет 20 стандартных аминокислот, однако в некоторых организмах генетический код может включать селеноцистеин, а в некоторых археях пирролизин. Аминокислотные остатки в белке часто бывают химически модифицированы посредством посттрансляционной модификации, которая может происходить или до использования белка в клетке, или в качестве составной части механизмов управления. Аминокислотные остатки в белке также можно модифицировать преднамеренно, применяя методики, известные компетентному специалисту. При использовании в этом документе термин "белок" охватывает линейные цепочки, содержащие природные аминокислоты, синтетические аминокислоты, модифицированные аминокислоты или комбинации любых перечисленных типов аминокислот.
Одноразовое впрыскивание: При использовании в этом документе термин "одноразовое впрыскивание" относится к начальной стадии операции в устройстве для МС или ЖХ-МС. Когда белковый образец вводят в устройство одним впрыскиванием, весь образец вводится в один прием.
Стопочный/стэкинг: При использовании в этом документе термин "стопочный" относится к наличию множественных гетерологичных полинуклеотидов, встроенных в геном растения.
Тандемная масс-спектрометрия: При тандемной масс-спектрометрии родительский ион, полученный из представляющей интерес молекулы, может быть отфильтрован в инструменте для МС, а затем фрагментирован с выходом одного или более дочерних ионов, которые анализируют (выявляют и/или количественно определяют) во второй процедуре МС.
Трансгенное растение: При использовании в этом документе термин "трансгенное растение" включает ссылку на растение, которое содержит в своем геноме гетерологичный полинуклеотид. Обычно гетерологичный полинуклеотид стабильно интегрирован в геном, то есть, этот полинуклеотид передается в последующие поколения потомства. Гетерологичный полинуклеотид может быть интегрирован в геном самостоятельно или как часть рекомбинантной экспрессирующей кассеты. Термин "трансгенный" при употреблении в этом документе включает любую клетку, клеточную линию, каллюс, ткань, часть растения или целое растение, генотип которых был изменен присутствием гетерологичной нуклеиновой кислоты, включая как растения с первоначально измененным генотипом, так и растения, полученные при перекрестном опылении или бесполом размножении из исходного трансгенного растения.
III. Выбор трансгенного белка для мультиплексного анализа
Способами, предлагаемыми настоящим изобретением, можно проанализировать любой белок, введенный в растение посредством технологии трансгенной экспрессии. Белки, пригодные для мультиплексного анализа, в соответствии с изобретением могут придавать трансгенному растению такой фенотипический признак, который делает его более совершенным по сравнению с нетрансгенным аналогом. Неограничивающими примерами желательных признаков, которые могут быть приданы трансгенным растениям, являются устойчивость к гербицидам, устойчивость к стрессу, связанному с воздействием окружающей среды, повышенная урожайность, улучшенная пищевая ценность, улучшенный срок хранения, измененное содержание масла, измененный состав масла, измененное содержание сахара, измененное содержание крахмала, выработка растительного фармацевтического средства, выработка продуктов промышленного назначения (например, полигидроксил-алканоатов: макромолекулярных полиэфиров, считающихся идеальными для замещения пластиков, полученных из нефти) и потенциал для биоремедиации. Кроме того, с применением способов, описанных в этом раскрытии изобретения, может быть проанализирована экспрессия одного или более трансгенных белков в одном растительном виде. Добавление двух и более генов или желательных признаков в один представляющий интерес биологический вид или их модуляция известны как стэкинг генов (эффект образования стопки). Кроме того, экспрессия одного или более трансгенных белков может быть одновременно проанализирована на основе раскрытого в этом документе мультиплексного анализа с одним или более эндогенных белков растения.
Предпочтение конкретным белкам, выбранным для анализа, определяется по усмотрению специалиста. Такими белками могут быть, но, не ограничиваясь ими, белки растений, животных, дрожжей и т.п., кроме того, это могут быть белки, не обнаруживаемые в нетрансформированной клетке или обнаруживаемые в трансформированной клетке. Особенно удобными белками, экспрессирующимися в трансгенных растениях, являются такие белки, которые придают этим растениям устойчивость к гербицидам, насекомым или вирусам, а также гены, которые обеспечивают повышенную пищевую ценность, более высокую урожайность, засухоустойчивость, утилизацию азота, выработку полезных в промышленном отношении соединений, лучшие характеристики переработки растения или потенциал для биоремедиации. Примеры полезных белков включают белки, кодируемые инсектицидным геном Bacillus thuringiensis, которые придают устойчивость к насекомым, а также ген 5'-энолпирувил-3'-фосфошикимат-синтазы (EPSPS) и его любые варианты, придающие устойчивость к глифосатным гербицидам. Как будет легко понятно компетентному специалисту, любой белок, придающий растению желательный признак, может экспрессироваться в растительной клетке с применением технологии рекомбинантных ДНК.
IV. Отбор пептидных фрагментов
В соответствии с представленным раскрытием изобретения целевые белки можно идентифицировать, определяя наличие протеолитических пептидных фрагментов целевых белков в спектрах МС из сложной смеси растительных белков. Протеазные ферменты расщепляют белки в специфических аминокислотных последовательностях, которые могут быть легко определены в пределах всей последовательности целевого белка. Следовательно, набор пептидных фрагментов, полученных при расщеплении одной или более протеаз, можно логически вывести априорно. Затем, исходя из первичной аминокислотной последовательности пептида, можно определить логически выведенное уникальное соотношение между его массой и зарядом.
Пептидные фрагменты также можно определить эмпирически по спектрам МС расщепленных целевых белков. Специфический вариант осуществления изобретения может быть связан с отбором пептида, последовательность которого является внутренней по отношению к предсказанным сайтам протеолитического расщепления. Для эмпирического отбора пептидных фрагментов с целью идентификации целевого белка образцы расщепленного белка можно подвергнуть масс-спектрометрии, чтобы определить пептидные фрагменты, которые обеспечивают необходимую чувствительность и специфичность выявления для мультиплексной идентификации белка. Обычно предпочтительными для отбора являются самые обильные ионизированные пептиды. Обильное содержание специфического ионизированного пептида является функцией массы пептида и эффективности ионизации пептида.
Представляющие интерес белки в стопочном продукте будут экспрессироваться на разном уровне по отношению друг к другу. Выявленные пептиды будут получены из индивидуальных белков, экспрессированных в разных и иногда неизвестных концентрациях. Выявление пептидов и белков будет варьироваться в диапазоне от аттомолярных до микромолярных концентраций. Чистота тестируемых образцов будет варьироваться от очищенных стандартных белков в буферных растворах до неочищенного матрикса растения, экстрагированного из различных представляющих интерес тканей. Если целевое выявление потребует уменьшения сложности образца матрикса перед выявлением, то пептидные фрагменты будут идентифицированы в неочищенном матриксе растения так же, как и частично очищенный образец матрикса ткани.
V. Целенаправленный анализ ММР
При идентификации одного или более пептидных фрагментов целевого белка, подлежащих определению в мультиплексном анализе, разрабатывают целенаправленный анализ ММР для целевого белка. Предпочтительно, чтобы целенаправленный анализ ММР для данного целевого белка мог идентифицировать те предварительно выбранные пептиды, которые представляют собой наиболее обильные ионные фрагменты. Таким образом, каждый целевой пептид, подлежащий определению в мультиплексном анализе, может иметь специфический анализ ММР, который разработан для идентификации уникальных ионных фрагментов этого целевого пептида. Мультиплексный анализ осуществляется через одновременный анализ ММР, который включает уникальные фрагменты пептида по типу идентификаторов, специфичные для двух или более целевых белков в образце растительной ткани. Предпочтительно, чтобы мультиплексный анализ мог одновременно идентифицировать те специфические пептиды целевого белка, которые демонстрируют эффективную ионизацию и фрагментацию при анализе, как в присутствии, так и в отсутствие сложного матрикса. Целевой белок, включенный в мультиплексный анализ, может быть идентифицирован с одной или более специфических родительских/дочерних пар ионного перехода.
Затем можно провести одноразовый мультиплексный анализ МС/МС для множественных целевых белков в сложной белковой смеси. Благодаря чувствительности и специфичности масс-спектрометрии, сложный белковый образец, подлежащий мультиплексному анализу МС/МС, не должен быть столь же чистым или обильным, как образец, анализируемый с применением традиционных методик, таких как иммунохимия или ПЦР. Однако сложный белковый образец, подлежащий мультиплексному анализу МС/МС, можно приготовить в соответствии с условиями экстракции, оптимизированными для робастного аналитического выполнения мультиплексного способа. Белковые образцы можно приготовить в соответствии с такими методиками, как, среди всего прочего, солевая экстракция (например, бикарбонатом аммония), солевая экстракция в присутствии мочевины и экстракция детергентом (например, CHAPS) или другие методики по типу обогащения.
Растительный материал, из которого можно приготовить мультиплексированный белковый образец, выбирается по усмотрению компетентного специалиста. Подходящий материал может включать, например, ткань или клетки трансгенного растения, ткань растения или клетки, полученные в результате процедуры генетической трансформации, ткань или клетки нетрансгенных растений, анализируемые на присутствие контаминирующего трансгена, или растительные материалы, подозрительные на трансгенное происхождение растения.
Мультиплексный анализ МС проводят на сложном белковом образце. Сложный белковый образец впрыскивают в ионизационную камеру для МС, в которой вырабатывается первый (родительский) ион. Родительский ион может быть выявлен прямым образом в первой МС или он может быть выделен посредством первой МС, фрагментирован на характерные дочерние ионы, а один или более дочерних ионов могут быть выявлены во второй МС (МС/МС).
Ионы можно выявлять с применением нескольких режимов выявления. Например, отобранные ионы можно выявить, применяя режим селективного мониторинга ионов (СМИ), который включает мониторинг множественных реакций (ММР) или мониторинг избранной реакции (МИР). В альтернативном варианте ионы можно выявлять, применяя режим сканирования.
Соотношение между массой и зарядом можно определить, применяя квадрупольный масс-анализатор. Например, в таком инструменте как "квадруполь" или "квадрупольная ионная ловушка" ионы в осциллирующем радиочастотном поле испытывают силовое воздействие, пропорциональное потенциалу DC (постоянного тока) между электродами, амплитуде RF (радиочастотного) сигнала и m/z (отношения массы к заряду). Напряжение и амплитуду можно подобрать таким образом, чтобы только ионы, имеющие особое соотношение m/z, проходили всю длину квадруполя, а все другие ионы отклонялись от прямого направления. Таким образом, инструменты квадруполя могут действовать как "масс-фильтр" и "масс-детектор" для ионов, впрыснутых в инструмент.
Индуцированную столкновениями диссоциацию ("ИСД") часто применяют, чтобы генерировать дочерние ионы для дальнейшего выявления. При ИСД родительские ионы получают энергию через столкновения с инертным газом, таким как аргон, и впоследствии фрагментируются в ходе процесса, известного как "мономолекулярный распад". В родительском ионе должна быть накоплена достаточная энергия для того, чтобы определенные связи внутри иона могли разорваться вследствие накопленной вибрационной энергии.
При выполнении МС/МС родительские ионы отбирают в первом анализе МС. Затем эти отобранные родительские ионы переправляют в камеру для столкновений, чтобы генерировать специфические дочерние ионы пептида для идентификации и количественного определения. При определенных условиях ионизации/фрагментации родительские и дочерние ионы вырабатываются воспроизводимым образом, что придает методике МС/МС исключительно мощные аналитические возможности.
В типичном случае масс-спектрометр обеспечивает пользователя сканированием ионов, то есть, относительной распространенностью каждого соотношения m/z в данном диапазоне (например, от 10 до 1200 атомных единиц массы). Результаты исследования анализируемого вещества, то есть, масс-спектр, могут быть связаны с количеством анализируемого вещества в исходном образце многими способами, известными в данной области техники. Например, при том условии, что параметры отбора образцов и анализа тщательно контролируются, относительную распространенность данного иона можно сравнить с таблицей, которая преобразует эту относительную распространенность в абсолютное количество исходной молекулы. В альтернативном варианте может быть осуществлен прогон с образцами молекулярных стандартов (например, внутренних стандартов и внешних стандартов), и в результате получена стандартная кривая, основанная на ионах, генерированных из этих стандартов. При использовании такой стандартной кривой относительную распространенность данного иона можно преобразовать в абсолютное количество исходной молекулы. Среднему специалисту в данной области техники должны быть хорошо известны многие другие способы, позволяющие соотнести определенное количество иона с определенным количеством исходной молекулы.
Выбор способа ионизации можно определить на основе подлежащего измерению анализируемого вещества, типа образца, типа детектора, выбора положительного или отрицательного режима и т.д. Ионы можно вырабатывать с применением множества способов, включая, но, не ограничиваясь ими, электронную ионизацию, химическую ионизацию, быструю бомбардировку атомов, десорбцию электрическим полем, ионизацию способом лазерной десорбции в присутствии матрицы (ИЛДПМ), ионизацию способом лазерной десорбции с усилением поверхностью (ИЛДУП), ионизацию способом электрораспыления с десорбцией (ИЭРД), фотонную ионизацию, ионизацию способом электрораспыления и индуктивно связанную плазму. Ионизация способом электрораспыления относится к способам, в которых раствор пропускают через короткую капиллярную трубку с приложенным к ее концу высоким положительным или отрицательным электрическим потенциалом. Раствор, достигающий конца трубки, распыляется (небулизируется), превращаясь в струю или аэрозоль из очень мелких капелек раствора в парах растворителя. Эта аэрозольная дымка капелек протекает через испарительную камеру, которую нагревают для предупреждения конденсации и для полного испарения растворителя. По мере того как капельки становятся меньше, плотность поверхностного электрического заряда увеличивается вплоть до того момента, когда в результате естественного отталкивания между одинаковыми зарядами начнут высвобождаться ионы, а также нейтральные молекулы.
Вытекающий поток ЖХ можно напрямую и автоматически (т.е., "линейно") впрыснуть в электрораспылительное устройство. В некоторых вариантах осуществления изобретения белки, содержащиеся в вытекающем потоке ЖХ, прежде всего, ионизируются электрораспылением, превращаясь в родительский ион. Первый квадруполь МСМС настроен как масс-фильтр для родительских ионов мультиплексированных целевых белков.
Родительские ионы, проходящие через первый квадруполь, затем ионизируются и/или фрагментируются перед попаданием во второй квадруполь. В некоторых вариантах осуществления изобретения ионы сталкиваются с молекулами инертного газа в процессе индуцированной столкновениями диссоциации (ИСД). Подходящие инертные газы включают, например, аргон, гелий, азот и т.д. Желательно, чтобы родительские ионы мультиплексированных целевых белков были фрагментированы на дочерние ионы, которые и подлежат последующему выявлению.
VI. Сохранение сорта трансгенного растения
Способы, представленные в настоящем раскрытии изобретения, можно использовать для сохранения сортового генотипа трансгенного растения. Сложные белковые образцы, приготовленные из следующего поколения потомков трансгенного растения можно подвергнуть мультиплексному анализу МСМС, чтобы определить присутствие или отсутствие представляющего интерес белка. Сложные белковые образцы могут содержать трансгенные белки, экспрессированные в трансгенном растении. Размножая те растения, а которых было подтверждено наличие представляющего интерес трансгенного белка, можно гарантированно обеспечить экспрессию представляющего интерес трансгенного белка в последующих поколениях. Подобным образом, можно отказаться от размножения в следующем поколении тех растений, в которых не было подтверждено наличие представляющего интерес трансгенного белка.
VII. Результаты скрининга процедуры трансформации растения
Способы, представленные в настоящем раскрытии изобретения, также можно использовать для быстрого скрининга результатов процедуры трансформации растения, осуществляемого высокопроизводительным образом. Вследствие вариабельности генотипа и характеристик экспрессии ГМ растений и растительных клеток, полученных в результате рекомбинации ДНК, растения и растительные клетки, полученные в процедуре трансформации растения, не обязательно будут иметь аналогичный или сходный профиль экспрессии введенных трансгенных белков, например, гетерологичных белков. Кроме того, эндогенные белки могут проявлять в различной степени измененный профиль экспрессии. В некоторых вариантах осуществления настоящего изобретения сложные белковые образцы, могут быть приготовлены из растений, растительных тканей или растительных клеток, полученных в процедуре трансформации растения. Затем приготовленные сложные белковые образцы можно подвергнуть мультиплексному анализу МС/МС. Затем спектры МС различных образцов анализируют для идентификации тех образцов, которые демонстрируют желательные характеристики экспрессии. Затем исходное растение, растительную ткань или растительные клетки идентифицированных образцов можно размножить для отбора желательных характеристик экспрессии.
VIII. Достижение биологического сдерживания трансгенов
Трансгены могут выходить из трансгенного растения в окружающую среду и интегрироваться в геном окружающих растений, например, посредством перекрестного опыления. В большинстве случаев это нежелательно. В некоторых вариантах осуществления способы, представленные в этом раскрытии изобретения, применяются для того, чтобы достичь биологического удержания трансгенов в трансгенном растении. В этих и дальнейших вариантах осуществления изобретения сложные белковые образцы могут быть приготовлены из растения (растений), растительной ткани (тканей) или растительных клеток, которые подвержены риску контаминации генетическим материалом из трансгенного растения. Затем приготовленные сложные белковые образцы можно подвергнуть мультиплексному анализу МС/МС. После этого спектры МС различных образцов анализируют, чтобы определить растения, содержащие целевой трансгенный белок, например, трансгенный белок, экспрессируемый в трансгенном растении. Присутствие целевого белка в образце связывают с проникновением трансгена в анализируемое растение (растения), растительную ткань (ткани) или растительные клетки. Биологического сдерживания трансгенов можно достичь, ликвидируя, сдерживая или другим образом ограничивая рост и размножение контаминированного растения (растений), растительной ткани (тканей) или растительных клеток.
В дополнение к описанным в этом документе специфическим примерам возможные варианты осуществления изобретения поддаются различным модификациям и допускают альтернативные формы. Таким образом, варианты осуществления изобретения не ограничиваются раскрытыми конкретными формами. Скорее можно говорить о том, что сфера изобретения охватывает все модификации, эквиваленты и альтернативы, изложенные в прилагаемой формуле изобретения.
ПРИМЕРЫ
ПРИМЕР I
Для разработки мультиплексного анализа ЖХ\МС\МС были отобраны шесть отдельных трансгенных белков (Cry1F, Cry34, Cry35, AAD-1, AAD-12 и ФАТ). Индивидуальные белки были выявлены и идентифицированы посредством масс-спектроскопии в одноразовом впрыскивании сложной белковой смеси.
Первый вариант мультиплексного анализа был проведен посредством протеолитического расщепления шести белков по отдельности с последующим увеличением концентрации полученных белковых пептидов в протеолитически расщепленных экстрактах растительной ткани и применением ЖХ\МС\МС для выявления специфических предшественников/ионных фрагментов для каждого из шести белков в одноразовом впрыскивании. Методика, разработанная для этого первого варианта мультиплексного анализа, впоследствии была применена для мультиплексного выявления четырех белков, экспрессированных в современном селекционном материале инбредной кукурузы.
ПРИМЕР II
Мультиплексное выявление ЖХ\МС\МС у растения
В таблице 1 перечислены полученные концентрации каждого отдельного исходного белка и результирующие разбавления каждого белка при расщеплении трипсином. Перед расщеплением протеазой (трипсином) все белки были забуферены 25 mM бикарбоната аммония, pH 7,9 (SIGMA) для обеспечения эффективных условий расщепления. Аликвотную пробу каждого исходного белка (см. таблицу 1) переносили в стерильную пробирку Эппендорфа емкостью 1,5 мл и доводили до объема 100 мкл с применением 25 mM бикарбоната аммония, pH 7,9. Для замены буфера каждого белка были использованы опресняющие спин-колонки Zeba (Pierce # 89882) в соответствии с рекомендациями производителя. Были проведены три спиновых промывания колонки Zeba с применением в каждом случае 300 мкл 25 mM бикарбоната аммония, pH 7,9 и раскручиванием на скорости 1500 g в течение 1 минуты. Затем аликвотную пробу 100 мкл каждого белка из образца наносили на поверхность смолы колонки Zeba и раскручивали на скорости 1500 g в течение 2 минут. Затем этот забуференный материал прямым образом использовали для расщепления трипсином и получения пептидных фрагментов белка. Начальное расщепление шести белков трипсином не включало предусмотренного протоколом стадии алкилирования аминокислотных остатков цистеина. Эта стадия алкилирования может быть включен позже, но с целью высокопроизводительного анализа, причем пропуск стадии алкилирования может сберечь время и силы для заключительного анализа. Каждый забуференный образец белка 100 мкл был внесен в 5 mM DTT с добавлением 1 мкл 0,5 M DTT, затем денатурирован нагреванием при температуре 95°C в течение 20 минут и охлажден до комнатной температуры (25°C). Модифицированный трипсин степени чистоты для секвенирования был ресуспендирован в 25 mM бикарбоната аммония, pH 7,9, до концентрации 0,4 мкг/мкл. Трипсиновый фермент добавляли к каждому образцу белка, доводя конечное соотношение фермента к субстрату до диапазона от 1:20 до 1:50. Расщепление трипсином проводили в термоциклере, используя температурный профиль 37°C в течение 16 часов, а затем охлаждали материал до 4°C. После расщепления трипсином к каждому перевару белка добавляли 3 мкл 10% муравьиной кислоты.
Концентрации и разбавления белка для каждого мультиплексного ЖХ\МС\МС анализа
трипсинового перевара 6 белков
(мл)
Первоначально каждый из шести белковых образцов, расщепленных трипсином, анализировали способом ИЭР-ЖХ\МС\МС, чтобы определить триптические пептидные фрагменты, способные обеспечить достаточную чувствительность и специфичность для одновременной мультиплексной идентификации всех шести белков в одном анализе. Для разработки методики был применен масс-спектрометр Applied Biosystems MDS Sciex 4000 Q Trap hybrid triple quad, (Foster City, модель CA # 1004229-V), использующий источник турбо V ИЭР, смонтированный с датчиком TSI. Образцы вводили в масс-спектрометр через систему ЖХВР Agilent 1100. Таблица 2 включает информацию о специфическом номере модели и версии программного обеспечения для разных инструментальных компонентов.
Информация о модели/версии программного обеспечения для инструментальных компонентов
Хроматографию с обращенной фазой проводили, применяя систему ЖХВР Agilent 1100, оснащенную 4 мкм колонкой Phenomenex Jupiter Proteo 50×2,0 мм с условием загрузки 95% A (H2O/0,1% муравьиной кислоты)/5% B (ацетонитрила/0,1% муравьиной кислоты) в течение 1 минуты и с градиентом до 90% B в течение 20 минут. Колонку регенерировали с 2-минутной поддержкой на уровне 90% B, а затем повторно уравновешивали до 5% B в течение 5 минут. С целью начального скрининга отдельных белковых пептидов в колонку для анализа загружали приблизительно 10-50 фмоль каждого белка.
Для каждого из шести мультиплексированных белков был проведен анализ способом ЗЗИ с включением двух сканов УИП по двум наиболее обильным ионам, выявленным из списка преобразований ММР, специфических для каждого белка. Эти данные по фрагментированию были информативными для выбора ионных фрагментов, демонстрирующих наиболее высокое содержание в каждом выбранном пептиде. Как правило, для дальнейшей разработки методики отбирали три высших ионных фрагмента из выявленного пептида предшественника. Для осуществления этого ЗЗИ система Sciex 4000 Q TRAP задействовала следующие условия: вольтаж IS 5500, DP 75, EP 10, CXP 12, CUR 10, CAD HIGH, TEM 450, GS1 35, GS2 35, блок RES Ql и блок RES Q3. Значения КЭ для каждого пептида тестировали эмпирически с использованием оптимальной величины для каждого пептида. Используя данные пептидной фрагментации, накопленные в результате анализа индивидуальных ЗЗИ для каждого из шести белков, затем мы провели анализ ММР для каждого отдельного белка, чтобы идентифицировать в каждом белке ионы предшественники с хорошей эффективностью ионизации. Для каждого мультиплексного белка был использован список ММР всех триптических пептидов, чтобы создать индивидуальный способ анализа ММР. Используя данные индивидуального анализа белков способом ММР в качестве критерия эффективности ионизации, мы выбрали для каждого белка пептиды, служащие ионами предшественниками в мультиплексном формате. Мультиплексные пептиды перечислены в таблице 3.
Пептиды, отобранные для начального 6-мультиплексного способа ЖХ\МС\МС
ПРИМЕР III
Кроме того, был разработан одиночный целевой мультиплексный анализ ММР с методикой ЖХ\МС\МС для всех шести белков DAS с использованием данных о предшественнике/ионном фрагменте, полученных в результате анализа индивидуальных белков. Эти белки сначала были мультиплексно выявлены способом ЖХ\МС\МС при подкреплении буфером бикарбоната аммония (25 mM, pH 7,9) с использованием материала трипсинового перевара для каждого из шести белков.
Приблизительно 5-20 фмоль каждого белка впрыскивали в колонку для начального 6-мультиплексоного анализа усиленного буфера.
ФИГУРЫ 1-6 показывают экстрагированные ионные хроматограммы для каждого из шести белков, выявленных в одноразовом впрыскивании. Более конкретно, Фиг. 1 показывает мультиплексное выявление AAD-1 способом ЖХ\МС\МС. Данные представляют собой экстрагированную ЖХ\МС\МС ионную хроматограмму для AAD-1 ((R)-2,4-дихлорфеноксипропионат диоксигеназы), выявленной в одноразовом впрыскивании вместе с AAD-12, Cry1F, Cry34, Cry35 и ФАТ в экстракте из семян кукурузы.
Фиг. 2 показывает мультиплексное выявление AAD-12 способом ЖХ\МС\МС. Данные представляют собой экстрагированную ЖХ\МС\МС ионную хроматограмму для AAD-12 ((S)-2,4-дихлорфеноксипропионат/альфа кетоглуратат диоксигеназы), выявленной в одноразовом впрыскивании вместе с AAD-1, Cry1F, Cry34, Cry35 и ФАТ в экстракте из семян кукурузы.
Фиг. 3 показывает мультиплексное выявление Cry1F способом ЖХ\МС\МС. Данные представляют собой экстрагированную ЖХ\МС\МС ионную хроматограмму для Cry1F (пестицидного кристаллического белка Cry1Fa (инсектицидного дельта-эндотоксина Cry1F(a))), выявленного в одноразовом впрыскивании вместе с AAD-1, AAD-12, Cry34, Cry35 и ФАТ в экстракте из семян кукурузы.
Фиг. 4 показывает мультиплексное выявление Cry34 способом ЖХ\МС\МС. Данные представляют собой экстрагированную ЖХ\МС\МС ионную хроматограмму для Cry34 (кристаллического белка ET79 [Bacillus thuringiensis]), выявленного в одноразовом впрыскивании вместе с AAD-1, AAD-12, Cry1F, Cry35 и ФАТ в экстракте из семян кукурузы.
Фиг. 5 показывает мультиплексное выявление Cry35 способом ЖХ\МС\МС. Данные представляют собой экстрагированную ЖХ\МС\МС ионную хроматограмму для Cry35 (Cry35Ab-подобного [Bacillus thuringiensis]), выявленного в одноразовом впрыскивании вместе с AAD-1, AAD-12, Cry1F, Cry34 и ФАТ в экстракте из семян кукурузы.
Фиг. 6 показывает мультиплексное выявление ФАТ способом ЖХ\МС\МС. Данные представляют собой экстрагированную ЖХ\МС\МС ионную хроматограмму для Cry35 (фосфинотрицин N-ацетилтрансферазы (PPT N-ацетилтрансферазы)), выявленной в одноразовом впрыскивании вместе с AAD-1, AAD-12, Cry1F, Cry34 и Cry35 в экстракте из семян кукурузы.
После подтверждения мультиплексного выявления всех шести белков способом ЖХ\МС\МС смесь пептидов с бикарбонатом аммония была введена в расщепленный протеазой экстракт из ткани семян кукурузы для выявления в межклеточном матриксе семян. Использованная ткань семян происходила из современного инбредного материала. Усиленный образец был разбавлен в соотношении 1:10 бикарбонатом аммония, а затем в соотношении 1:2 в экстракте из семян кукурузы, при этом приблизительно от 0,2 до 1 фмоль каждого белка было впрыснуто в колонку и выявлено с применением мультиплексного способа ЖХ\МС\МС.
ПРИМЕР IV
Четыре отдельных трансгенных белка (Cry1, Cry34, Cry35 и ФАТ) были выявлены и идентифицированы посредством масс-спектроскопии в одноразовом впрыскивании сложной белковой смеси из инбредных растительных материалов (5XH751XT). Белок был выявлен как в ткани листьев, так и в ткани семян.
Описанная выше 6-мультиплексная методика была использована для выявления присутствия четырех отдельных трансгенных белков (Cry1F, Cry34, Cry35 и ФАТ) в одноразовом впрыскивании сложного белкового образца из растительного материала инбредной линии (5XH751XT). В этой попытке был разработан 4-мультиплексный способ с одноразовым впрыскиванием для определения присутствия четырех белков как в ткани семян кукурузы, так и в ткани листьев кукурузы. Экспериментальный контроль включал сравнение с пустым экстрактом 5XH751, обогащенным цельными трансгенными белками перед ферментативным расщеплением и сравнение с ферментативно расщепленным пустым экстрактом 5XH751.
Для этого эксперимента также была продумана методика экстракции, необходимая для робастного аналитического осуществления мультиплексного способа. Поскольку мультиплексное выявление одновременно измеряет множественные белки в каждом индивидуальном образце, применяемый способ экстракции белков должен быть эффективным для всех белков в целях их точного измерения. Как показано на ФИГУРЕ 11, условия экстракции были протестированы на ткани листьев и семян кукурузы. В первой попытке понять, как эти четыре разных белка проявят себя в отношении единого способа экстракции с учетом таких факторов, как стабильность, растворимость и гидрофобность белка, были протестированы три самостоятельных условия экстракции.
Привлекательность простой экстракции белка бикарбонатом аммония заключается в возможности непосредственно перейти от стадии экстракции к расщеплению протеазой, не беспокоясь о супрессии сигнала в связи с буферным компонентом при анализе способом МС. Это может снизить стоимость, но, что представляется более важным, существенно уменьшить затраты времени на подготовку и возможную изменчивость, связанную с заменой буфера. Для того чтобы определить, нужен ли для выявления всех четырех белков в растительных тканях способ агрессивной солюбилизации, был протестирован буфер 8M с мочевиной. Был использован буфер PBS-T, поскольку он традиционно применяется для методик выявления ELISA.
Материал 5XH751XT из ткани листьев: Ткань листьев собирали из выращенного в теплице растения V6-V7 и измельчали под жидким азотом. Испытательный вес каждого образца листьев составлял 1,5 г. Каждый экстрагирующий буфер (Фиг. 11) был протестирован при соотношении между буфером и образцом 2:1 (3 мл). Образцы перемешивали вихревым движением в течение 2 минут, затем центрифугировали 2 минуты и собирали супернатанты.
Материал 5XH751XT из ткани семян: Получали ткань зрелых семян и измельчали ее в шаровой мельнице. Испытательный вес каждого образца семян составлял 1,5 г. Каждый экстрагирующий буфер (Фиг. 11) был протестирован при соотношении между буфером и образцом 2:1 (3 мл). Образцы перемешивали вихревым движением в течение 2 минут, затем центрифугировали 2 минуты и собирали супернатанты.
Образцы, экстрагированные в 8M мочевине или PBS-T, были забуферены в 25 mM бикарбонате аммония с применением центробежных фильтров Pierce Zeba. Для всех образцов 50 мкл тканевого экстракта расщепляли 10 мкг трипсина (удаление) в 110 мкл общего объема бикарбоната аммония. Расщепление белка проводили при 37°C в течение 16 часов с последующим охлаждением до 4°C. В качестве положительного контроля для каждого из четырех белков, подлежащих мультиплексному выявлению, была использована приготовленная из очищенных белковых стандартов смесь типа коктейля, содержащая приблизительно 50 мкг/мл каждого индивидуального цельного белка. Конечное разведение коктейля 1:10 было использовано как с пустым образцом экстракта листьев, так и с пустым образцом экстракта семян для положительного контроля матрикса в приблизительной концентрации 5 мкг/мл. Эти усиленные пустые образцы служили положительным контролем, позволяя точно оценить время удержания пептидов при ЖХ и данные фрагментирования последовательности по методике МС/МС при сравнении с пустым тканевым контролем (отрицательный контроль) и экстрактами 5XH751XT из ткани кукурузы. Поскольку все четыре белка, подлежащих мультиплексному выявлению в материале 5XH751XT, были белками, также использованными для разработки вышеописанного 6-мультиплексного способа ЖХ/МС/МС, не требовалась никакая дополнительная разработка для определения того, какие специфические пептиды/ионные фрагменты белка надо выявлять и при каких инструментальных условиях.
Вышеописанный 6-мультиплексный способ был использован для анализа тканевых экстрактов листьев и семян 5XH751XT. ФИГУРЫ 7-10 представляют подвыборку данных, полученных при 4-мультиплексном анализе одноразового впрыскивания способом LC-МС/МС для выявления белков Cry1F, Cry34, Cry35 и ФАТ, экспрессируемых в ткани листьев и семян инбредной кукурузы.
Более конкретно, Фиг. 7 показывает мультиплексное выявление способом ЖХ-МС/МС белка Cry1F, экспрессированного в ткани инбредной кукурузы. Данные представляют собой спектры МС/МС, специфичные для белка Cry1F, выявленного мультиплексным анализом в ткани листьев кукурузы 5XH751, экстрагированной 25 mM бикарбонатом аммония. Были выявлены три T22-специфических для Cry1F ионных фрагмента, но здесь показан только один фрагмент. Также показаны положительный и отрицательный контроль для транзиции МС/МС.
Фиг. 8 показывает мультиплексное выявление способом ЖХ-МС/МС белка Cry34, экспрессированного в ткани инбредной кукурузы. Данные представляют собой спектры МС/МС, специфичные для белка Cry34, выявленного мультиплексным анализом в ткани листьев кукурузы 5XH751, экстрагированной 25 mM бикарбонатом аммония. Также показаны положительный и отрицательный контроль для транзиции МС/МС. Были выявлены пять T7-специфических для Cry34 ионных фрагментов, но здесь показан только один из них. Легкий сдвиг по времени удержания между образцом 5XH751XT и усиленным положительным контролем Cry34 не является неожиданным для белка, элюированного в ранней, более гидрофобной области обращенно-фазового градиента. Пик, показанный в контрольной панели пустого образца ткани листьев не является специфическим пиком, о чем свидетельствует большой (~7 минут) сдвиг по времени удержания в колонке. Ни один из остальных четырех ионных фрагментов Cry34 не имел специфических пиков.
Фиг. 9 показывает мультиплексное выявление способом ЖХ-МС/МС белка Cry35, экспрессированного в ткани инбредной кукурузы. Данные представляют собой спектры МС/МС, специфичные для белка Cry35, выявленного мультиплексным анализом в ткани листьев кукурузы. Также показаны положительный и отрицательный контроль для транзиции МС/МС. Показаны данные для ткани листьев кукурузы 5XH751, экстрагированной 25 mM бикарбонатом аммония. Были выявлены три T9-специфических для Cry35 ионных фрагмента, но здесь показан только один из них.
Фиг. 10 показывает мультиплексное выявление способом ЖХ/МС/МС белка ФАТ, экспрессированного в ткани инбредной кукурузы. Данные представляют собой спектры МС/МС, специфичные для белка ФАТ, выявленного мультиплексным анализом в ткани листьев кукурузы. Также показаны положительный и отрицательный контроль для транзиции МС/МС.
Для каждого белка был выявлен и фрагментирован специфический пептид предшественник с тремя-пятью соответствующими ионными фрагментами, выявленными для надежного подтверждения белковой последовательности. Каждая из фигур 7, 8, 9 и 10 демонстрирует спектры МС/МС, специфичные для одного из четырех белков, выявленных мультиплексным анализом в ткани кукурузы. Каждая фигура также демонстрирует специфический положительный и отрицательный контроль для конкретной транзиции МС/МС. Два из четырех белков были выявлены в ткани семян. Неспособность выявить два из четырех белков, вероятно, была связана с низкой экспрессией, поскольку исторически данные демонстрируют сниженную экспрессию этих четырех белков в ткани семян по сравнению с тканью листьев.
ПРИМЕР V
Сорт трансгенного растения поддерживают, подтверждая присутствие двух представляющих интерес белков в следующем поколении этого сорта растения. Отбор трансгенного растения основан на присутствии двух представляющих интерес трансгенных белков (A и B). Приготовляют образцы белков A и B, которые подвергают анализу способом ЖХ/МС/МС. По выявленным спектрам МС проводят отбор пептидных ионных фрагментов родительских белков для целевого анализа МС/МС.
Из первого поколения трансгенного растения готовят сложный белковый образец для мультиплексного анализа способом ЖХ/МС/МС, в котором идентифицируют отобранные пептидные ионные фрагменты белков A и B, определяя присутствие отобранных пептидных ионных фрагментов в спектрах МС. Затем готовят сложные белковые образцы из следующего поколения трансгенного растения. Сложные белковые образцы, полученные из этих растений второго поколения, подвергают мультиплексному анализу способом ЖХ/МС/МС. Для поддержания сорта трансгенного растения размножают те растения следующего поколения, из которых были получены сложные белковые образцы, демонстрирующие такие спектры МС, в которых можно идентифицировать оба белка (A и B) по присутствию отобранных пептидных ионных фрагментов. Для поддержания сорта трансгенного растения не размножают те растения следующего поколения, из которых были получены сложные белковые образцы, демонстрирующие такие спектры МС, в которых не удалось идентифицировать оба белка (A и B).
ПРИМЕР VI
Трансформанты, полученные в процедуре трансформации растения, скринируют, чтобы определить присутствие двух представляющих интерес целевых белков. Два целевых белка (A и B) подвергают анализу способом ЖХ/МС/МС. По выявленным спектрам МС проводят отбор пептидных ионных фрагментов целевых родительских белков для целенаправленного анализа МС/МС.
В процедуре трансформации растения получают предполагаемые трансформанты. Из каждого предполагаемого трансформанта получают сложные белковые образцы. Сложные белковые образцы, полученные из этих предполагаемых трансформантов, подвергают мультиплексному анализу способом ЖХ/МС/МС, в котором определяют экспрессию целевых белков по присутствию отобранных пептидных ионных фрагментов. Те трансформанты, из которых были получены сложные белковые образцы, демонстрирующие спектры МС с желательными характеристиками экспрессии белков A и B, определяемыми по присутствию отобранных пептидных ионных фрагментов, размножают.
ПРИМЕР VII
Биологическое сдерживание трансгенов в трансгенном растении. Два целевых белка (A и B), экспрессируемых трансгенным растением, подвергают анализу способом ЖХ/МС/МС. По выявленным спектрам МС проводят отбор пептидных ионных фрагментов целевых родительских белков для целенаправленного анализа МС/МС.
Собирают растительный материал из растений, выросших в окружающей среде по соседству с трансгенным растением. Из этого растительного материала готовят сложные белковые образцы. Сложные белковые образцы, полученные из этого растительного материала, подвергают мультиплексному анализу способом ЖХ/МС/МС, в котором определяют экспрессию контаминирующего трансгена, выявляя присутствие отобранных пептидных ионных фрагментов экспрессируемого трансгенного белка. Те растения, из которых был получен материал, демонстрирующий по результатам мультиплексного анализа способом ЖХ/МС/МС трансгенным белком, уничтожают.
В этот документ путем ссылки включены во всей полноте следующие источники:
Alwine et al. (1977) Proc. Nat. Acad. Sci. 74:5350-54; Baldwin (2004) Mol. Cell. Proteomics 3(1): Carr and Annan (1997) Overview of peptide and protein analysis by mass spectrometry. В: Current Protocols in Molecular Biology, под редакцией Ausubel, et al. New York: Wiley, p. 10.21.1-10.21.27; Chang et al. (2000) Plant Physiol. 122(2):295-317; Domon and Aebersold (2006) Science 312(5771):212-17; Nain et al. (2005) Plant Mol. Biol. Rep. 23:59-65; Patterson (1998) Protein identification and characterization by mass spectrometry. В: Current Protocols in Molecular Biology, под редакцией Ausubel, et al. New York: Wiley, p. 10.22.1-10.22.24; Paterson and Aebersold (1995) Electrophoresis 16: 1791-1814; Rajagopal and Ahern (2001) Science 294(5551):2571-73; Sesikeran and Vasanthi (2008) Asia Pac. J. Clin. Nutr. 17 Suppl. 1:241-44; and Toplak et al. (2004) Plant Mol. Biol. Rep. 22:237-50.
















Изобретение относится к биотехнологии, а именно к способу выявления присутствия двух или более представляющих интерес белков с известными аминокислотными последовательностями в образце растительного происхождения (варианты) и способу сохранения генотипа сорта трансгенного растения. Способы включают расщепление всех белков из образца растительного происхождения специфичной по последовательности протеазой с получением пептидов. Разделяют пептиды в один прием в приборе для жидкостной хроматографии - масс-спектрометрии (ЖХ-МС). Ионизируют пептиды в приборе для ЖХ-МС. Проводят одновременное получение масс-спектральных данных для пептидов в приборе для ЖХ-МС. Сравнивают указанные одновременные масс-спектральные данные для пептидов с ожидаемыми масс-спектральными данными от представляющих интерес белков с известными аминокислотными последовательностями. Осуществляют определение присутствия или отсутствия представляющих интерес белков. Предложенное изобретение позволяет с высокой эффективностью выявления присутствия двух или более представляющих интерес белков растительного происхождения. 3 н. и 14 з.п. ф-лы, 10 ил., 3 табл., 7 пр.
1. Высокопроизводительный способ выявления присутствия двух или более представляющих интерес белков с известными аминокислотными последовательностями в образце растительного происхождения, где указанный способ включает:
(i) расщепление всех белков из образца растительного происхождения специфичной по последовательности протеазой с получением пептидов;
(ii) разделение пептидов в один прием в приборе для жидкостной хроматографии - масс-спектрометрии (ЖХ-МС);
(iii) ионизацию пептидов в приборе для ЖХ-МС; и
(iv) одновременное получение масс-спектральных данных для пептидов в приборе для ЖХ-МС; и
(v) сравнение указанных одновременных масс-спектральных данных для пептидов с ожидаемыми масс-спектральными данными от представляющих интерес белков с известными аминокислотными последовательностями, чем осуществляется определение присутствия или отсутствия представляющих интерес белков.
2. Способ по п. 1, в котором пептиды, разделяемые на стадии (ii), разделяют посредством колоночной хроматографии.
3. Способ по п. 1, в котором колоночная хроматография представляет собой жидкостную колоночную хроматографию.
4. Способ по п. 1, в котором белки, расщепляемые на стадии (i), расщепляют в один прием перед впрыскиванием.
5. Способ по п. 1, в котором пептиды, ионизируемые на стадии (iii), ионизируют в один прием.
6. Способ по п. 1, в котором масс-спектральные данные по соотношению между массой и зарядом пептидов, соответствующих двум или более представляющим интерес белкам, получают в один прием.
7. Способ по п. 1, в котором два или более представляющих интерес белков представляют собой два представляющих интерес белка.
8. Способ по п. 1, в котором два или более представляющих интерес белков представляют собой четыре представляющих интерес белка.
9. Способ по п. 1, в котором образец растительного происхождения представляет собой образец из трансгенного растения.
10. Способ по п. 1, в котором образец растительного происхождения представляет собой образец из трансгенного растения, а два или более представляющих интерес белков являются ожидаемыми продуктами экспрессии трансгена в трансгенном растении.
11. Высокопроизводительный способ выявления присутствия двух или более представляющих интерес белков с известными аминокислотными последовательностями в образце растительного происхождения, где указанный способ включает:
(i) получение масс-спектральных данных для двух или более представляющих интерес белков;
(ii) получение сложного образца растительного происхождения, содержащего неразделенные белки и расщепление всех белков на пептиды специфичной по последовательности протеазой;
(iii) разделение пептидов в один прием в приборе для ЖХ-МС;
(iv) ионизацию пептидов в приборе для ЖХ-МС;
(v) одновременное получение масс-спектральных данных для пептидов в приборе для ЖХ-МС; и
(vi) сравнение одновременных масс-спектральных данных с масс-спектральными данными, полученными для двух или более представляющих интерес белков, чем определяется присутствие или отсутствие двух или более представляющих интерес белков.
12. Способ по п. 11, в котором пептиды, расщепляемые на стадии (iii), расщепляют в один прием.
13. Способ по п. 11, в котором пептиды, ионизируемые на стадии (iv), ионизируют в один прием.
14. Способ по п. 11, в котором образец растительного происхождения представляет собой образец из трансгенного растения.
15. Способ сохранения генотипа сорта трансгенного растения, где указанный способ включает:
(i) получение масс-спектральных данных для одного или более ожидаемых продуктов экспрессии трансгена в сорте трансгенного растения;
(ii) получение сложного образца, содержащего неразделенные белки, из первого поколения сорта трансгенного растения и (ii') расщепление всех из неразделенных белков на пептиды специфичной по последовательности протеазой;
(iii) разделение пептидов в один прием в приборе для ЖХ-МС;
(iv) ионизацию пептидов в приборе для ЖХ-МС;
(v) одновременное получение масс-спектральных данных для пептидов в приборе для ЖХ-МС и сравнение одновременных масс-спектральных данных с масс-спектральными данными, полученными для ожидаемых продуктов экспрессии трансгена, чем определяется присутствие или отсутствие ожидаемых продуктов экспрессии трансгена в первом поколении сорта трансгенного растения;
(vi) проведение первого впрыскивания сложного образца, содержащего белки, из второго поколения сорта трансгенного растения;
(vii) повторение стадий (ii′)-(v) со сложным образцом, содержащим белки, из второго поколения сорта трансгенного растения; и
(viii) отказ от размножения второго поколения сорта трансгенного растения, если присутствие ожидаемого продукта (продуктов) экспрессии трансгена не может быть подтверждено масс-спектральными данными для пептидов из сложного белкового образца из второго поколения сорта трансгенного растения, чем поддерживается генотип сорта трансгенного растения.
16. Способ по п. 15, в котором все из неразделенных белков расщепляют в один прием.
17. Способ по п. 15, в котором пептиды, ионизируемые на стадии (iv), ионизируют в один прием.
| WO2007132164 A2, 22.11.2007 | |||
| US2005153380 A1, 14.07.2005 | |||
| BHUSHAN S | |||
| ET AL., "Catalysis, Subcellular Localization, Expression and Evolution of the Targeting Peptides Degrading Protease, AtPreP2", PLANT AND CELL PHYSIOLOGY, 2005, v | |||
| Способ изготовления звездочек для французской бороны-катка | 1922 |
|
SU46A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Аппарат для сжигания нефти | 1920 |
|
SU985A1 |
| WIENKOOP S | |||
| ET AL., "Cell-specific protein profiling in Arabidopsis thaliana trichomes: | |||

