УРОВЕНЬ ТЕХНИКИ
Ген aad-1 (первоначально известный из Sphingobium herbicidovorans) кодирует белок арилоксиалканоатдиоксигеназы (AAD-1). Этот признак придает устойчивость к гербицидам 2,4-дихлорфеноксиуксусной кислоте и арилоксифеноксипропионату (обычно называемым «фоп»-гербицидами, таким как квизалофоп) и может быть использован в качестве селектируемого маркера при трансформации растений и в селекционных питомниках. Ген aad-1, как таковой, в связи с устойчивостью растений к гербицидам впервые раскрыт в WO 2005/107437 (также см. US 2009-0093366).
На экспрессию гетерологичных или чужеродных генов у растений влияет то, в каком месте хромосомы встроен чужеродный ген. Это может быть следствием, например, структуры хроматина (например, гетерохроматина) или близости элементов регуляции транскрипции (например, энхансеров) к участку интеграции (Weising с соавторами, Ann. Rev. Genet. 22: 421-477, 1988). Один и тот же ген в одном и том же типе трансгенного растения (или другого организма) может проявлять широкую изменчивость уровня экспрессии при разных событиях. Также могут существовать различия в пространственных или временных картинах экспрессии. Например, различия в относительной экспрессии трансгена в разных растительных тканях могут не соответствовать картинам, ожидаемым, исходя из элементов регуляции транскрипции, присутствующих во введенной генной конструкции.
Таким образом, часто создают и подвергают скринингу большое количество событий, чтобы идентифицировать событие, которое приводит к экспрессии введенного представляющего интерес гена на уровне, достаточном для данной цели. В коммерческих целях обычно осуществляют от нескольких сотен до нескольких тысяч разных событий и осуществляют скрининг таких событий в отношении одного события, которое дает требуемые уровни и картины экспрессии трансгена. Событие, которое дает требуемые уровни и/или картины экспрессии трансгена, применимо для интрогрессии трансгена в другое генетическое окружение в результате полового ауткроссинга с использованием обычных способов селекции. У потомства от таких скрещиваний сохраняются характеристики экспрессии трансгена исходного трансформанта. Такую методику используют для того, чтобы обеспечить надежную экспрессию гена у некоторых сортов, которые хорошо адаптированы к местным условиям роста.
Заявки на выдачу патента США 20020120964 A1 и 20040009504 A1 относятся к генетическому событию PV-GHGT07(1445) в растениях хлопчатника и композициям и способам его выявления. WO 02/100163 относится к событию MONI5985 в растениях хлопчатника и композициям и способам его выявления. WO 2004/011601 относится к событию MON863 в растениях кукурузы и композициям и способам его выявления. WO 2004/072235 относится к событию MON88913 в растениях хлопчатника и композициям и способам его выявления.
WO 2006/098952 относится к событию 3272 у кукурузы. WO 2007/142840 относится к событию MIR162 у кукурузы.
Патент США № 7179965 относится к хлопчатнику, в котором произошло событие cry1F и событие cry1Ac. Кукуруза AAD-1, в которой произошло конкретное событие, раскрытое в настоящем описании, ранее не была описана.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к предпосевному и/или довсходовому внесению гербицида на площади или поле, которое засеяно семенами, в которых имеет место событие AAD-1. В некоторых предпочтительных вариантах в семени имеет место событие DAS-40278-9 кукурузы. В некоторых предпочтительных вариантах гербицид может представлять собой состав, содержащий активный ингредиент 2,4-D. Такие гербициды и составы также можно применять для предпосевного внесения. Дополнительные гербициды, такие как глифосат, можно использовать в комбинации, включая и предпосевное применение.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фигуре 1 показана карта плазмиды pDAS1740.
На фигуре 2 показаны компоненты вставки для DAS-40278-9 (pDAS1740).
На фигуре 3 показана карта рестрикции и компоненты вставки для DAS-40278-9 (pDAS1740).
На фигуре 4 показаны ампликоны, праймеры и методика клонирования в случае ДНК-вставки и границы DAS-40278-9.
Фигура 5 иллюстрирует положения праймеров по отношению к вставке и границам DAS-40278-9.
Фигура 6 иллюстрирует области соединения и инсерцию DAS-40278-9.
Фигура 7 представляет собой диаграмму скрещивания, описанного в примере 7.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Последовательности SEQ ID NO: 1-28 являются праймерами, которые описаны в настоящей публикации.
SEQ ID NO: 29 представляет вставку и фланкирующие последовательности для рассматриваемого события DAS-40278-9.
Последовательности SEQ ID NO: 30-33 являются праймерами для фланкирующих маркеров, которые описаны в примере 4.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение включает предпосевное и/или довсходовое внесение гербицида на площадь или поле, которое затем засевают семенами, в которых имеет место событие AAD-1. В некоторых предпочтительных вариантах в семени имеет место событие DAS-40278-9 кукурузы. В некоторых предпочтительных вариантах гербицид может представлять собой состав, содержащий активный ингредиент 2,4-D. Такие гербициды и составы также можно применять для предпосевного внесения. Дополнительные гербициды, такие как глифосат, можно использовать в комбинации, включая и предпосевное применение. Настоящее изобретение не ограничено кукурузой, а может включать, например, применение хлопчатника и/или сои, содержащих ген aad-1.
Примеры, включенные в настоящее описание, отчасти относятся к предпосевному и/или довсходовому внесению гербицидов. Такие применения не ограничены событием «278». Кроме того, применимость устойчивости, обеспечиваемой генами AAD-1, может быть объяснима с учетом укороченного промежутка времени от применения до нового посева. Это обеспечивает растениеводам намного большую гибкость в составлении графика проводимых ими посевов относительно момента полного уничтожения сорняков («выжигания»). Без использования настоящего изобретения ожидание в течение 7-30 дней или около того после уничтожения сорняков до посева может быть причиной значительной потери урожая. Таким образом, настоящее изобретение обеспечивает преимущества в этом отношении. См., например, пример 13. Любые интервалы между посевами/внесением гербицидов и любые диапазоны концентраций/нормы расхода гербицида(ов), приведенные в качестве примеров или предлагаемые в настоящем описании, можно использовать согласно настоящему изобретению.
Таким образом, настоящее изобретение относится к новым способам применения гербицидов. Такие применения могут включать баковые смеси более чем одного гербицида. Некоторые предпочтительные гербициды для применения согласно настоящему изобретению включают феноксиауксиновый гербицид, такой как 2,4-D; 2,4-DB; MCPA; MCPB. Такие применения можно сочетать с одним или несколькими дополнительными генами устойчивости к гербицидам и соответствующим гербицидом (например, глифосатом и/или глюфосинатом). Один, два, три или больше гербицидов можно использовать в преимущественных комбинациях, которые могут быть очевидны для специалиста в данной области, обладающих эффектом настоящего изобретения. Один или несколько предлагаемых гербицидов можно вносить на поле/площадь перед засеванием семенами согласно настоящему изобретению. Такое внесение можно осуществлять, например, в пределах 14 дней до посева. Один или несколько предлагаемых гербицидов также можно вносить при посеве и/или после посева, но до всходов. Один или несколько предлагаемых гербицидов также можно вносить на почву (для борьбы с сорняками) или поверх сорняков и/или трансгенных растений согласно настоящему изобретению. Три рассматриваемых гербицида можно чередовать или применять в комбинации, например, для борьбы или предотвращения появления сорняков, которые могут быть устойчивы к одному гербициду, но не к другому. Можно использовать разное время внесения трех рассматриваемых типов гербицидов различными путями, которые могут быть известны в данной области.
Таким образом, настоящее изобретение также относится к предпосевному внесению гербицида на площади или поле, которое затем засевают семенами, в которых имеет место событие AAD-1. В некоторых предпочтительных вариантах в семени имеет место событие DAS-40278-9 кукурузы. В некоторых предпочтительных вариантах гербицид может представлять собой состав, содержащий активный ингредиент 2,4-D. Такие гербициды и составы можно применять для предпосевного внесения. Дополнительные гербициды, такие как глифосат, можно использовать в комбинации при предпосевном внесении. В любом из таких вариантов можно использовать, например, кукурузу, хлопчатник и сою.
Ген aad-1 можно комбинировать, например, с признаками, кодирующими резистентность к глифосату (например, резистентными растительными или бактериальными EPSPS, GOX, GAT), резистентность к глюфосинату (например, Pat, bar), резистентность к гербициду, ингибирующему ацетолактатсинтазу (ALS) (например, имидазолинонам [таким как имазетапир], сульфонилмочевине, триазолопиримидинсульфонанилиду, пиримидинилтиобензоатам и другим химическим составам [Csr1, SurA и другим]), резистентность к бромоксинилу (например, Bxn), резистентность к ингибиторам фермента HPPD (4-гидроксифенилпируватдиоксигеназы), резистентность к ингибиторам фитоиндесатуразы (PDS), резистентность к гербицидам, ингибирующим фотосистему II (например, psbA), резистентность к гербицидам, ингибирующим фотосистему I, резистентность к гербицидам, ингибирующим протопорфининогеноксидазу IX (PPO) (например, PPO-1), резистентность к гербицидам на основе фенилмочевины (например, CYP76B1), ферменты, разрушающие дикамбу (см., например, US 20030135879), и другие, которые можно подвергать стэкингу по отдельности или во множестве разных комбинаций для обеспечения возможности эффективно бороться с сорняками или предотвращать смену сорняков и/или резистентность к любому гербициду указанных выше классов.
Что касается дополнительных гербицидов, то некоторые дополнительные предпочтительные ингибиторы ALS (также известной как AHAS) включают триазолопиримидинсульфонанилиды (такие как клорансулам-метил, диклосулам, флорасулам, флуметсулам, метосулам и пенокссулам), пиримидинилтиобензоаты (такие как биспирибак и пиритиобак) и флукарбазон. Некоторые предпочтительные ингибиторы HPPD включают мезотрион, изоксафлутол и сулкотрион. Некоторые предпочтительные ингибиторы PPO включают флумиклорак, флумиоксазин, флуфенпир, пирафлуфен, флутиацет, бутафенацил, карфентразон, сульфентразон и дифениловые эфиры (такие как ацифлуорфен, фомесафен, лактофен и оксифлуорфен).
Гены AAD-1 для применения согласно настоящему изобретению также могут обеспечивать резистентность к соединениям, которые превращаются в феноксиацетатауксиновые гербициды (например, 2,4-DB, MCPB и т.д.). Остаток масляной кислоты, присутствующий в гербициде 2,4-DB, превращается в результате β-окисления в фитотоксичную 2,4-дихлорфеноксиуксусную кислоту. Подобным образом, MCPB превращается в результате β-окисления в фитотоксичный MCPA. Гербициды на основе масляной кислоты сами по себе не являются гербицидными. Они превращаются в соответствующую им кислоту в результате β-окисления в чувствительных растениях, и такая уксуснокислая форма гербицида является фитотоксичной. Растения, неспособные к быстрому β-окислению, не получают вреда от гербицидов на основе масляной кислоты. Однако растения, которые способны к быстрому β-окислению и могут превращать гербицид на основе масляной кислоты в уксуснокислую форму, в дальнейшем оказываются защищены присутствующим AAD-1.
Настоящее изобретение охватывает событие AAD-1 у кукурузы, обозначаемое DAS-40278-9, имеющей семена, депонированные в Американской коллекции типов культур (ATCC) с номером доступа PTA-10244, и полученное из них потомство. Другие аспекты включают растения-потомки, семена и зерно или регенерируемые части растений и семена и потомство кукурузы в случае события DAS-40278-9, а также полученные из них корма и пищевые продукты. Настоящее изобретение также охватывает части растений кукурузы, где имеет место событие DAS-40278-9, которые включают без ограничения пыльцу, семязачаток, цветки, побеги, корни и листья и ядра вегетативных клеток, клеток пыльцы и яйцеклеток. Кроме того, раскрыты растения кукурузы, обладающие устойчивостью к феноксиауксиновым и/или арилоксиалканоатным гербицидам, к новым генетическим составам в случае события DAS-40278-9 у кукурузы и аспектам агрономической эффективности растений кукурузы, имеющих событие DAS-40278-9 кукурузы.
Настоящее изобретение охватывает способы селекции растений и устойчивые к гербицидам растения, включая событие трансформации aad-1 в растениях кукурузы, содержащих полинуклеотидную последовательность, которая описана в настоящей публикации, встроенную в конкретный участок генома клетки кукурузы.
В некоторых вариантах указанное событие/полинуклеотидная последовательность могут быть «подвергнуты стэкингу» с другими признаками, включая, например, другой ген (гены) устойчивости к гербицидам и/или подавляющие насекомых белки. Также в настоящей публикации описаны растения, имеющие единичное событие.
Дополнительные признаки могут быть подвергнуты стэкингу в геноме растения путем скрещивания растений, повторной трансформации трансгенного растения, имеющего событие DAS-40278-9 кукурузы, или добавления новых признаков в результате целенаправленной интеграции посредством гомологичной рекомбинации.
Другие варианты включают эксцизию полинуклеотидных последовательностей, в которых произошло событие DAS-40278-9 кукурузы, включая, например, кассету экспрессию гена pat. После эксцизии полинуклеотидной последовательности модифицированное событие может быть перенаправлено в конкретный участок хромосомы, где дополнительные полинуклеотидные последовательности подвергают стэкингу с событием DAS-40278-9 кукурузы.
В одном варианте изобретение относится к сайту-мишени в хромосоме кукурузы, локализованному в хромосоме 2 в положении примерно 20 сМ между SSR-маркерами UMC 1265 (см. SEQ ID NO: 30 и SEQ ID NO: 31) и MMC0111 (см. SEQ ID NO: 32 и SEQ ID NO: 33), примерно 20 сМ на карте сцепления кукурузы 2008 DAS, при этом сайт-мишень содержит гетерологичную нуклеиновую кислоту. В другом варианте изобретение относится к сайту-мишени в хромосоме кукурузы, содержащему положение, определяемое в последовательности или последовательностью SEQ ID NO: 29, и его остаткам, которые описаны в настоящей публикации, как будет понятно специалисту в данной области.
В одном варианте изобретение относится к способу получения трансгенного растения кукурузы, содержащего вставку гетерологичной нуклеиновой кислоты в хромосоме 2 в положении примерно 20 сМ между SSR-маркерами UMC1265 (см. SEQ ID NO: 30 и SEQ ID NO: 31) и MMC0111 (см. SEQ ID NO: 32 и SEQ ID NO: 33) в положении примерно 20 сМ на карте сцепления кукурузы 2008 DAS. В еще одном варианте встроенная гетерологичная нуклеиновая кислота фланкирована с 5’-стороны всей или частью 5’-фланкирующей последовательности, которая определена в настоящем описании со ссылкой на последовательность SEQ ID NO: 29, и фланкирована с 3’-стороны всей или частью 3’-фланкирующей последовательности, которая определена в настоящем описании со ссылкой на последовательность SEQ ID NO: 29.
Кроме того, в настоящем описании раскрыты анализы для выявления присутствия данного события в образце (например, зерне кукурузы). Анализы могут быть основаны на последовательности ДНК рекомбинантной конструкции, встроенной в геном кукурузы, и на геномных последовательностях, фланкирующих сайт инсерции. Также предлагаются наборы и условия, применимые для проведения анализов.
Также в настоящем описании раскрыто клонирование и анализ последовательностей ДНК полной вставки AAD-1 и ее пограничных областей (в трансгенных линиях кукурузы). Такие последовательности являются уникальными. На основании таких последовательностей вставки и границ создавали специфичные для события праймеры. ПЦР-анализ показал, что такие события можно идентифицировать посредством анализа ПЦР-ампликонов, образуемых с использованием таких специфичных для события наборов праймеров. Таким образом, указанные и другие родственные способы можно применять для того, чтобы однозначно идентифицировать линии кукурузы, в которых имеет место такое событие.
Настоящее изобретение включает применение скрещивания растений и устойчивых к гербицидам растений. Настоящее изобретение включает новые применения событий трансформации растений кукурузы, содержащих рассматриваемые полинуклеотидные последовательности aad-1, которые описаны в настоящей публикации, встроенные в конкретный участок в геноме клетки кукурузы. В некоторых вариантах указанная полинуклеотидная последовательность может быть «подвергнута стэкингу» с другими признаками (такими как другой ген (гены) устойчивости к гербицидам, который кодируют, например, белки, подавляющие насекомых. В некоторых вариантах указанные полинуклеотидные последовательности могут быть вырезаны и затем снова направлены к мишени с дополнительными полинуклеотидными последовательностями. Однако настоящее изобретение включает растения, в которых имеет место единичное событие, которое описано в настоящей публикации.
Кроме того, настоящее изобретение относится к анализам для выявления наличия данного события в образце. Аспекты включают способы конструирования и/или получения любых диагностических молекул нуклеиновой кислоты, приведенных в качестве примера или предлагаемых в настоящем описании, в частности, молекул, полностью или частично основанных на рассматриваемых фланкирующих последовательностях.
Более конкретно, настоящее изобретение отчасти относится к трансгенному событию DAS-40278-9 у кукурузы (также известному как pDAS 1740-278), к применению линий растений, в которых имеют место такие события, и клонированию и анализу последовательностей ДНК такой вставки и/или ее пограничных областей. Линии растений для применения согласно настоящему изобретению можно выявить с использованием последовательностей, раскрытых и предлагаемых в настоящем описании. В некоторых вариантах настоящее изобретение относится к устойчивым к гербицидам линиям кукурузы и к их идентификации. Настоящее изобретение отчасти относится к выявлению наличия данного события, чтобы определить, имеет ли потомство от полового скрещивания представляющее интерес событие. Кроме того, включен способ выявления события, и такой способ применим, например, для того, чтобы обеспечить соблюдение регламента, требующего предпродажного разрешения и нанесения метки на продукты, полученные, например, из рекомбинантных культурных растений. Можно выявлять наличие данного события любым хорошо известным способом выявления нуклеиновой кислоты, таким как полимеразная цепная реакция (ПЦР) или ДНК-гибридизация с использованием зондов нуклеиновых кислот. Специфичный для определенного события ПЦР-анализ обсуждается, например, Windels с соавторами (Med. Fac. Landbouww, Univ. Gent 64/5b: 459462, 1999). Обсуждение относится к идентификации устойчивого к глифосату события 40-3-2 у сои посредством ПЦР с использованием набора праймеров, охватывающих соединение между вставкой и фланкирующей ДНК. Более конкретно, один праймер содержал последовательность из вставки, а второй праймер содержал последовательность из фланкирующей ДНК.
Кукурузу модифицировали с использованием инсерции гена aad-1 из Sphingobium herbicidovorans, который кодирует белок арилоксиалканоатдиоксигеназы (AAD-1). Признак придает устойчивость к гербицидам 2,4-дихлорфеноксиуксусной кислоте и арилоксифеноксипропионату (обычно называемым «фоп»-гербицидами, таким как квизалофоп) и может быть использован в качестве селектируемого маркера при трансформации растений и в селекционных питомниках. Трансформацию кукурузы фрагментом ДНК из плазмиды pDAS1740 переносили, используя скрещивание, получая событие DAS-40278-9.
Образцы геномной ДНК, экстрагированной из двадцати отдельных растений кукурузы, полученных из пяти поколений, по четыре растения на поколение события DAS-40278-9, отбирали для молекулярной характеристики события DAS-40278-9 для AAD-1-кукурузы. Экспрессию белка AAD-1 тестировали, используя набор полосок для быстрого специфичного для AAD-1 теста. Для последующей молекулярной характеристики отбирали только растения, которые тестировали как позитивные в отношении экспрессии белка AAD-1. Саузерн-гибридизация подтвердила, что ген aad-1 присутствует в растениях кукурузы, которые в тесте выявляли как позитивные в отношении экспрессии белка AAD-1, и ген aad-1 был встроен в виде одной интактной копии в такие растения в том случае, когда происходила гибридизация с зондом гена aad-1.
Также в настоящей публикации описана молекулярная характеристика встроенной ДНК в случае события DAS-40278-9 для AAD-1-кукурузы. Событие было получено посредством прокалывания клеток путем встряхивания их в суспензии микроигл (Whiskers) с использованием Fsp I-фрагмента плазмиды pDAS1740. Саузерн-блот-анализ использовали для установления картины интеграции встроенного фрагмента ДНК и определения количества вставок/копий гена aad-1 в событии DAS-40278-9. Были получены данные для демонстрации интеграции и целостности трансгена aad-1, встроенного в геном кукурузы. Осуществляли характеристику интеграции некодирующих областей (предназначенных для регуляции кодирующих областей), таких как промоторы и терминаторы, участки прикрепления к матриксу RB7 Mar v3 и RB7 Mar v4, а также оценивали стабильность вставки трансгена в поколениях. Стабильность встроенной ДНК показана на протяжении пяти отдельных поколений растений. Кроме того, показано отсутствие последовательности остова трансформирующей плазмиды, включая область гена устойчивости к ампициллину (Apr), с использованием зондов, охватывающих почти всю область остова, фланкирующую сайты рестрикции (Fsp I) плазмиды pDAS1740. Подробная физическая карта инсерции получена на основе таких Саузерн-блот-анализов события DAS-40278-9.
Определяли уровни белка AAD-1 в тканях кукурузы. Кроме того, осуществляли анализ состава кукурузного фуража и зерна, чтобы исследовать эквивалентность между изогенной нетрансформированной линией кукурузы и трансгенной линией кукурузы DAS-40278-9 (неопрыскиваемой, опрыскиваемой 2,4-D, опрыскиваемой квизалофопом и опрыскиваемой и 2,4-D и квизалофопом). Агрономические характеристики изогенной нетрансформированной линии кукурузы также сравнивали с кукурузой DAS-40278-9.
Экспрессию в полевых условиях, состав питательных веществ и агрономические испытания нетрансгенного контроля и гибридной линии кукурузы, содержащей арилоксиалканоатдиоксигеназу-1 (AAD-1), проводили в один и тот же год в шести местоположениях в Айове, Иллинойсе (2 местоположения), Индиане, Небраске и Онтарио в Канаде. Уровни экспрессии суммированы в настоящем описании для белка AAD-1 в листьях, пыльце, корне, фураже, целом растении и зерне, представлены результаты агрономических определений и анализа состава образцов фуража и зерна в контроле и растениях кукурузы AAD-1 DAS-40278-9.
Растворимый экстрагируемый белок AAD-1 измеряли, используя способ количественного твердофазного иммуноферментного анализа (ELISA), в листьях, пыльце, корне, фураже, целом растении и зерне кукурузы. Хорошие средние значения экспрессии наблюдали в тканях корня и пыльцы, как более подробно обсуждается в настоящем описании. Значения экспрессии были сходными при всех обработках опрыскиванием, а также в случае делянок, опрыскиваемых и неопрыскиваемых гербицидами 2,4-D и квизалофопом.
Анализ состава, включая пищевые составляющие, минералы, аминокислоты, жирные кислоты, витамины, антипитательные вещества и вторичные метаболиты, проводили для того, чтобы исследовать эквивалентность кукурузы AAD-1 DAS-40278-9 (с обработкой или без обработки гербицидами) контролю. Все результаты анализа состава образцов AAD-1 DAS-40278-9 были такими же хорошими или лучше (биологически и агрономически), чем результаты, полученные на основе контрольных линий и/или обычной кукурузы, как и в случае агрономических данных, собранных с контрольных делянок и делянок с кукурузой AAD-1 DAS-40278-9.
Как упоминается выше в разделе «Уровень техники», введение и интеграция трансгена в геном растений включают некоторые случайные события (поэтому название «событие» используют для данной инсерции, которая экспрессируется). То есть в случае многих методик трансформации, таких как трансформация с помощью Agrobacterium, «генная пушка» и WHISKERS, невозможно предсказать, в каком месте генома будет встроен трансген. Таким образом, идентифицикация фланкирующей геномной ДНК растения с обеих сторон вставки может быть важна для идентификации растения, в котором произошло такое событие инсерции. Например, могут быть сконструированы ПЦР-праймеры, которые создают ПЦР-ампликон, охватывающий область соединения вставки и генома хозяина. Такой ПЦР-ампликон можно использовать для идентификации уникального или отличающегося типа инсерционного события.
Так как «события» исходно являются случайными событиями, то в качестве части настоящего раскрытия, по меньшей мере 2500 семян линии кукурузы, в которой имеет место событие, были депонированы и сделаны доступными для общественности без ограничения (но на которые распространяется патентное право) в Американской коллекции типов культур (ATCC), 10801 University Boulevard, Manassas, VA, 20110. Депозит получил депозитарный номер ATCC PTA-10244 (гибридные семена желтой зубовидной кукурузы (Zea Mays L.): DAS-40278-9; депонированы от имени Dow AgroSciences LLC; Дата получения семян/линии(ий) в ATTC: 10 июля 2009; жизнеспособность подтверждена 17 августа 2009). Указанный депозит был сделан и будет поддерживаться в соответствии и по условиям Будапештского договора в отношении депонирования семян для целей патентной процедуры. Депозит будет поддерживаться без ограничения в депозитарии ATCC, который является общедоступным депозитарием, в течение периода времени, составляющего 30 лет, или пять лет после самого последнего запроса, или в течение срока эффективного действия патента, какой угодно продолжительности, и будет заменен, если он станет нежизнеспособным в течение данного периода.
Депонированные семена являются частью раскрытого изобретения. Ясно, что растения кукурузы могут быть выращены из таких семян, и такие растения являются частью настоящего изобретения. Настоящее изобретение также относится к последовательностям ДНК, находящимся в таких растениях кукурузы, которые применимы для выявления таких растений и их потомства. Способы выявления и наборы согласно настоящему изобретению могут быть направлены на идентификацию любого одного, двух или даже всех трех указанных событий, в зависимости от конечной цели такого теста.
Определения и примеры приведены в настоящем описании, чтобы помочь описать настоящее изобретение и проинструктировать специалистов в данной области в отношении осуществления изобретения на практике. Если не указано иное, термины следует понимать в соответствии с их обычным использованием специалистами в соответствующей области. Использована номенклатура оснований для ДНК, которая указана в 37 CFR § 1.822.
В используемом в настоящем описании смысле термин «потомство» означает потомство любого поколения, полученного от исходного растения, в котором имеет место событие DAS-40278-9 AAD-1 кукурузы.
Трансгенное «событие» получают в результате трансформации растительных клеток гетерологичной ДНК, т.е. конструкцией нуклеиновой кислоты, которая включает представляющий интерес трансген, регенерации популяции растений, полученных в результате инсерции трансгена в геном растения и отбора конкретного растения, характеризуемого инсерцией в конкретное положение в геноме. Термин «событие» относится к исходному трансформанту и потомству трансформанта, которые содержат гетерологичную ДНК. Термин «событие» также относится к потомству, полученному в результате полового ауткроссинга между трансформантом и другим сортом, который содержит геномную/трансгенную ДНК. Даже после повторного возвратного скрещивания с родительской формой встроенная ДНК трансгена и фланкирующая геномная ДНК (геномная/трансгенная ДНК) из трансформированного родителя присутствует в потомстве от скрещивания в том же самом положении в хромосоме. Термин «событие» также относится к ДНК из исходного трансформанта и его потомства, содержащих встроенную ДНК и фланкирующую геномную последовательность непосредственно вблизи со встроенной ДНК, которая, как можно ожидать, может быть передана потомству, которое получает встроенную ДНК, включая представляющий интерес трансген, в результате полового скрещивания одной родительской линии, которая содержит встроенную ДНК (например, исходный трансформант и потомство, полученное в результате самоопыления), и родительской линии, которая не содержит встроенной ДНК.
«Последовательность соединения» охватывает точку, в которой ДНК, встроенная в геном, связана с ДНК из нативного генома кукурузы, фланкирующей точку инсерции, при этом идентификация или выявление одной из таких последовательностей соединения в генетическом материале растения является достаточной для того, чтобы диагностировать событие. Также включены последовательности ДНК, которые охватывают инсерции в описанных в настоящей публикации событиях и сходные длины фланкирующей ДНК. Конкретные примеры таких диагностических последовательностей приведены в настоящем описании; однако другие последовательности, которые перекрывают соединения инсерций или соединения инсерций и геномной последовательности, также являются диагностическими и могут быть использованы согласно настоящему изобретению.
Настоящее раскрытие включает идентификацию таких фланкирующих последовательностей, последовательностей соединения и вставки. Включены соответствующие ПЦР-праймеры и ампликоны. Способы ПЦР-анализа с использованием ампликонов, которые перекрывают встроенную ДНК и ее границы, можно применять для выявления или идентификации запущенных в коммерческое производство трансгенных сортов или линий кукурузы, полученных из данных патентуемых трансгенных линий кукурузы.
Полные последовательности каждой из таких вставок вместе с частями соответствующих фланкирующих последовательностей представлены в настоящем описании в виде SEQ ID NO: 29. Координаты вставки и фланкирующих последовательностей для данного события по отношению к последовательности SEQ ID NO: 29 (всего 8557 пар оснований) помещены ниже. Более подробное обсуждение приведено, например, в примере 3.8. Предпосевные варианты осуществления настоящего изобретения включают применение белка AAD-1, кодируемого остатками 1874-6689 последовательности SEQ ID NO: 29.
Такое событие инсерции и его дополнительные компоненты показаны далее на фигурах 1 и 2. Такие последовательности (в частности, фланкирующие последовательности) являются уникальными. На основании таких последовательностей вставки и границ создавали специфичные для события праймеры. ПЦР-анализ показал, что такие линии кукурузы могут быть идентифицированы в разных генотипах кукурузы с использованием анализа ПЦР-апликонов, создаваемых с использованием таких специфичных для события наборов праймеров. Таким образом, указанные и другие родственные способы можно использовать для однозначной идентификации таких линий кукурузы. Последовательности, указанные в настоящем описании, являются уникальными. Например, поиски с помощью BLAST в базах данных GENBANK не выявили какой-либо значимой гомологии между клонированными последовательностями границ и последовательностями в базе данных.
Способы выявления особенно применимы вместе с селекцией растений для определения того, какие растения в потомстве имеют данное событие после того, как родительское растение, имеющее представляющее интерес событие, скрещивают с растением другой линии, чтобы попытаться придать один или несколько дополнительных признаков потомству. Такие способы ПЦР-анализа полезны для осуществления программ селекции кукурузы, а также для контроля качества, особенно в случае коммерческого производства трансгенного кукурузного зерна. Также могут быть получены и использованы наборы для ПЦР-детекции таких трансгенных линий кукурузы. Способы также могут быть полезны для регистрации продукта и управления производством продукта.
Кроме того, фланкирующие кукурузные/геномные последовательности можно использовать для специфичной идентификации положения каждой вставки в геноме. Такая информация может быть использована для получения молекулярных маркерных систем, специфичных для каждого события. Такие системы можно применять для систем ускоренной селекции и установления данных о сцеплении.
Кроме того, информацию о фланкирующих последовательностях можно использовать для исследования и характеристики процесса интеграции трансгена, характеристики сайта интеграции в геноме, сортировки событий, стабильности трансгенов и фланкирующих его последовательностей и экспрессии гена (особенно в отношении сайленсинга гена, картин метилирования гена, эффектов положения и потенциально связанных с экспрессией элементов, таких как MAR [участки прикрепления к матриксу] и тому подобного).
Настоящее изобретение включает семена, доступные из ATCC с номером депозита PTA-10244. Настоящее изобретение также относится к применению резистентного к гербицидам растения кукурузы, выращенного из семени, депонированного в ATCC под номером доступа PTA-10244. Кроме того, настоящее изобретение относится к применению частей указанного растения, таких как листья, образцы тканей, семена, полученные от указанного растения, пыльца и тому подобное.
Кроме того, настоящее изобретение относится к применению растений, родственных по нисходящей линии, и/или потомков растения, выращенного из депонированного семени, предпочтительно устойчивого к гербицидам растения кукурузы, при этом указанное растение имеет геном, содержащий регистрируемую последовательность соединения геномной ДНК дикого типа/ДНК вставки, которая описана в настоящей публикации. В используемом в настоящем описании смысле термин «кукуруза» означает кукурузу Zea mays и включает все ее сорта, которые можно скрещивать.
Кроме того, настоящее изобретение относится к способам осуществления скрещиваний с использованием растения согласно настоящему изобретению в качестве по меньшей мере одного из родителей. Например, настоящее изобретение относится к применению гибридного растения F1, имеющего в качестве одного или обоих родителей любое из растений, приведенных в настоящем описании в качестве примера. Также в объем настоящего изобретения входит применение семени, полученного от таких гибридов F1 согласно настоящему изобретению. Настоящее изобретение относится к способу получения гибридного семени F1 в результате скрещивания приведенного в качестве примера растения с другим растением (например, инбредным родителем) и сбора полученного в результате гибридных семян. Настоящее изобретение относится к применению приведенного в качестве примера растения, которое является либо родителем женского пола, либо родителем мужского пола. Характеристики полученных в результате растений могут быть улучшены за счет тщательного подбора родительских растений.
Селекцию устойчивого к гербицидам растения кукурузы можно осуществлять, используя первое половое скрещивание первого родительского растения кукурузы, представляющего собой растение кукурузы, выращенное из семени любой из линий, указанных в настоящем описании, и второго родительского растения кукурузы, с получением при этом множества растений-потомков первого поколения; и затем используя отбор растения-потомка первого поколения, которое является резистентным к гербициду (или растения, в котором имеет место по меньшей мере одно из событий согласно настоящему изобретению); и самоопыление растения-потомка первого поколения с получением при этом множества растений-потомков второго поколения; и затем используя отбор из растений-потомков второго поколения растения, которое является резистентным к гербициду (или растения, в котором имеет место по меньшей мере одно из событий согласно настоящему изобретению). Такие стадии могут дополнительно включать возвратное скрещивание растения потомка первого поколения или растения-потомка второго поколения со вторым родительским растением кукурузы или третьим родительским растением кукурузы. Затем можно высевать урожай зерна, содержащий семена кукурузы согласно настоящему изобретению или ее потомства.
Также следует понимать, что два разных трансгенных растения также можно скрещивать с получением потомства, которое содержит два добавленных экзогенных гена, расщепление по которым происходит независимо. Самоопыление соответствующего потомства может давать растения, которые являются гомозиготными по обоим добавленным экзогенным генам. Также предусматривается возвратное скрещивание с родительским растением и ауткроссинг с нетрансгенным растением, как и вегетативное размножение. Другие способы, обычно применяемые для получения разных признаков и культур, известны в данной области. Селекцию на основе возвратного скрещивания использовали для переноса генов просто и в высокой степени наследуемого признака в требуемый гомозиготный сорт культурного растения или инбредную линию, которая является рекуррентным родителем. Источник признака, который необходимо перенести, называют родителем-донором. Ожидается, что получаемое в результате растение обладает свойствами рекуррентного родителя (например, сорта культурного растения) и требуемым признаком, перенесенным от родителя-донора. После начального скрещивания отбирают особи, обладающие фенотипом родителя-донора, и повторно скрещивают (осуществляют возвратное скрещивание) с рекуррентным родителем. Предполагается, что получаемый родитель обладает свойствами рекуррентного родителя (например, сорта культурного растения) и требуемым признаком, перенесенным от родителя-донора.
Молекулы ДНК, раскрытые в настоящем описании, можно применять в качестве молекулярных маркеров в способе скрещивания с использованием маркера (MAB). Молекулы ДНК согласно настоящему изобретению можно применять в способах (такие как AFLP-маркеры, RFLP-маркеры, RAPD-маркеры, SNP и SSR), в которых идентифицируют генетически сцепленные агрономически полезные признаки, которые известны в данной области. Признак резистентности к гербицидам можно проследить в потомстве от скрещивания с растением кукурузы согласно настоящему изобретению (или его потомством и любым другим сортом кукурузы), используя способы MAB. Молекулы ДНК являются маркерами такого признака, и способы MAB, которые хорошо известны в данной области, можно применять для того, чтобы проследить признак (признаки) резистентности к гербицидам в растениях кукурузы, при этом по меньшей мере одна линия кукурузы согласно настоящему изобретению или ее потомство является родителем или предком. Способы согласно настоящему изобретению можно применять для идентификации любого сорта кукурузы, в котором имеет место данное событие.
Способы согласно настоящему изобретению включают способ получения устойчивого к гербицидам растения кукурузы, при этом указанный способ включает скрещивание с растением для применения в настоящем изобретении. Более конкретно, указанные способы могут включать скрещивание двух растений согласно настоящему изобретению или одного растения согласно настоящему изобретению и любого другого растения. Предпочтительные способы дополнительно включают отбор потомства от указанного скрещивания в результате анализа указанного потомства в отношении события, выявляемого согласно настоящему изобретению. Например, настоящее изобретение может включать отслеживание данного события на протяжении циклов скрещивания с растениями, имеющими другие требуемые признаки, такие как агрономические признаки, такие как признаки, оцениваемые в настоящем описании в различных примерах. Растения, в которых имеет место данное событие и которые имеют требуемый признак, могут быть выявлены, идентифицированы, отобраны и оперативно использованы, например, в дальнейших раундах скрещивания. Данное событие/признак также можно сочетать на протяжении скрещиваний и прослеживать согласно настоящему изобретению с признаком (признаками) резистентности к насекомым и/или с дополнительными признаками устойчивости к гербицидам. Одним предпочтительным вариантом таких признаков устойчивости к гербицидам является растение, имеющее данное событие в сочетании с геном, кодирующим резистентность к гербициду дикамба.
Кроме того, AAD-1 отдельно или в сочетании с одним или несколькими дополнительными признаками HTC может быть подвергнута стэкингу с одним или несколькими дополнительными привнесенными признаками (например, резистентностью к насекомым, резистентностью к грибам или устойчивостью к стрессам и т.д.) или признаками, связанными с получаемой продукцией (например, повышенной урожайностью, улучшенным профилем масел, улучшенным качеством волокон и т.д.). Таким образом, настоящее изобретение можно применять для получения полного агрономического комплекса улучшенного качества культуры с возможностью гибкой и экономически эффективной борьбы с любым количеством сельскохозяйственных вредителей.
Способы интеграции полинуклеотидной последовательности в конкретный участок хромосомы растительной клетки в результате гомологичной рекомбинации были описаны в данной области. Например, сайт-специфичная интеграция описана в публикации заявки на выдачу патента США № 2009/0111188 A1, в которой описано применение рекомбиназ или интеграз для того, чтобы опосредовать введение донорной полинуклеотидной последовательности в хромосомную мишень. Кроме того, в международной заявке на выдачу патента № WO 2008/021207 описана опосредованная цинковыми пальцами гомологичная рекомбинация для интеграции одной или нескольких донорных полинуклеотидных последовательностей в конкретные положения генома. Применение рекомбиназ, таких как FLP/FRT, которые описаны в патенте США № 6720475, или CRE/LOX, которые описаны в патенте США № 5658772, можно использовать для интеграции полинуклеотидной последовательности в конкретный участок хромосомы. Наконец, применение мегануклеаз для целенаправленного введения донорных полинуклеотидов в конкретное положение в хромосоме описано Puchta с соавторами, PNAS USA 93 (1996) p. 5055-5060.
Другие различные способы сайт-специфичной интеграции в растительные клетки в общем известны и могут быть применены (Kumar с соавторами, Trands in Plant Sci. 6(4) (2001) pp. 155-159). Кроме того, системы сайт-специфичной рекомбинации, которые были идентифицированы в нескольких прокариотических и низших эукариотических организмах, можно использовать для применения в растениях. Примеры таких систем включают без ограничения: систему рекомбиназ R/RS из плазмиды pSR1 дрожжей Zygosaccharomyces rouxii (Araki с соавторами (1985) J. Mol. Biol. 182: 191-203) и систему Gin/gix фага Mu (Maeser and Kahlmann (1991) Mol. Gen. Genet. 230: 170-176).
В некоторых вариантах осуществления настоящего изобретения может быть желательна интеграция или стэкинг нового трансгена(ов) вблизи существующего трансгенного события. Можно рассматривать трансгенное событие, предпочтительный геномный локус которого был выбран на основе уникальных характеристик, таких как единственный сайт инсерции, нормальное менделевское расщепление и стабильная экспрессия и превосходное сочетание эффективности, включая устойчивость к гербицидам и агрономическую эффективность во многих местоположениях с разными условиями окружающей среды. Вновь интегрированные трансгены должны сохранять характеристики экспрессии трансгена существующих трансформантов. Кроме того, можно разработать анализы для выявления и подтверждения нового события интеграции, так как геномные фланкирующие последовательности и положение в хромосоме нового события интеграции уже идентифицированы. Наконец, интеграция нового трансгена в конкретное положение в хромосоме, которое сцеплено с существующим трансгеном, может упростить интрогрессию трансгенов в другой генетический фон в результате полового ауткроссинга с использованием обычных способов скрещивания.
В некоторых вариантах может быть желательным вырезание полинуклеотидных последовательностей из трансгенной вставки. Например, вырезание трансгена как описано в предварительной заявке на выдачу патента США № 61/297628, где описано применение нуклеаз с цинковыми пальцами для удаления полинуклеотидной последовательности, состоящей из кассеты экспрессии, из интегрированной в хромосому в результате трансгенного события вставки. Полинуклеотидная последовательность, которую удаляют, может представлять собой селектируемый маркер. После вырезания и удаления полинуклеотидной последовательности модифицированное трансгенное событие может быть повторно целенаправленно осуществлено посредством инсерции полинуклеотидной последовательности. Вырезание полинуклеотидной последовательности и последующее повторное целенаправленное осуществление модифицированного трансгенного события дает такие преимущества, как повторное использование селектируемого маркера или возможность преодолеть непредусмотренные изменения в транскриптоме растения в результате экспрессии конкретных генов.
В настоящем описании раскрыт конкретный участок в хромосоме 2 в геноме кукурузы, который является очень хорошим для инсерции гетерологичных нуклеиновых кислот. Также раскрыт молекулярный 5’-маркер, молекулярный 3’-маркер, 5’-фланкирующая последовательность и 3’-фланкирующая последовательность, применимые для идентификации положения участка-мишени в хромосоме 2. Таким образом, настоящее изобретение относится к способам введения представляющих интерес гетерологичных нуклеиновых кислот в такой предварительно установленный участок-мишень или вблизи такого участка-мишени. Настоящее изобретение также охватывает применение семени кукурузы и/или растения кукурузы, содержащего любую гетерологичную нуклеотидную последовательность, встроенную в заявленный участок-мишень или вблизи такого участка. Одним из вариантов осуществления такой целенаправленной интеграции является вырезание и/или замена другой вставкой вместо кассеты экспрессии pat, приведенной в данном описании в качестве примера. В этой связи согласно настоящему изобретению можно использовать, например и без ограничения, целенаправленную гомологичную рекомбинацию.
В используемом в настоящем описании смысле «стэкинг» генов, событий или признаков означает объединение требуемых признаков в одной трансгенной линии. Растениеводы подвергают трансгенные признаки стэкингу за счет осуществления скрещиваний между родителями, каждый из которых имеет требуемый признак, и затем идентифицируя потомство, которое обладает обоими требуемыми признаками. Другой путь стэкинга генов представляет собой перенос во время трансформации двух или более генов в ядро клетки растения одновременно. Другой путь стэкинга генов заключается в повторной трансформации трансгенного растения другим представляющим интерес геном. Например, стэкинг генов можно использовать для объединения двух или более разных признаков, включая, например, два или более разных признаков насекомых, признака(ов) резистентности к насекомым и признака(ов) устойчивости к заболеваниям, двух или более признаков резистентности к гербицидам и/или признака(ов) резистентности к насекомым и признака(ов) резистентности к гербицидам. Применение селектируемого маркера в дополнение к представляющему интерес гену также можно считать стэкингом генов.
«Гомологичная рекомбинация» относится к взаимодействию между любой парой нуклеотидных последовательностей, имеющих соответствующие участки, содержащие сходную нуклеотидную последовательность, посредством которой две нуклеотидные последовательности могут взаимодействовать (рекомбинировать) с образованием новой рекомбинантной последовательности ДНК. Каждый из участков со сходной нуклеотидной последовательностью называют в настоящем описании «последовательностью гомологии». В общем, частота гомологичной рекомбинации возрастает с увеличением длины последовательности гомологии. Таким образом, хотя гомологичная рекомбинация может происходить между двумя нуклеотидными последовательностями, длина которых меньше, чем длина идентичной последовательности, частота рекомбинации (или эффективность) снижается по мере возрастания дивергенции между двумя последовательностями. Рекомбинацию можно осуществлять, используя одну последовательность гомологии в донорной молекуле и молекуле-мишени, создавая при этом продукт рекомбинации, получаемый вследствие «одинарного кроссинговера». Альтернативно, можно помещать две последовательности гомологии в каждой из нуклеотидных последовательностей: донорной последовательности и последовательности-мишени. Рекомбинация между двумя последовательностями гомологии в доноре с двумя последовательностями гомологии в мишени генерирует продукт рекомбинации, получаемый вследствие «двойного кроссинговера». Если последовательности гомологии в донорной молекуле фланкируют последовательность, которую необходимо подвергнуть обработке (например, представляющую интереса последовательность), то рекомбинация вследствие двойного кроссинговера с молекулой-мишенью будет приводить к получению продукта рекомбинации, в котором представляющая интерес последовательность заменяет последовательность ДНК, которая исходно располагалась между последовательностями гомологии в молекуле-мишени. Обмен последовательностями ДНК между мишенью и донором в результате события рекомбинации вследствие двойного кроссинговера называют «заменой последовательности».
Рассматриваемый фермент AAD-1 обеспечивает возможность s трансгенной экспрессии, приводящей к устойчивости к сочетаниям гербицидов, которые обеспечили бы возможность борьбы почти со всеми широколиственными и злаковыми сорняками. AAD-1 может служить в качестве превосходного признака устойчивой к гербицидам культуры (HTC), например, для стэкинга с другими признаками HTC (например, резистентностью к глифосату, резистентностью к глюфосинату, резистентностью к имидазолинонам, резистентностью к бромоксинилу и т.д.) и признаками резистентности к насекомым (Cry1F, Cry1Ab, Cry 34/45 и т.д.). Кроме того, AAD-1 может служить в качестве селектируемого маркера, способствующего селекции первичных трансформантов растений, генетически сконструированных со вторым геном или группой генов.
Признаки HTC согласно настоящему изобретению можно применять в новых сочетаниях с другими признаками HTC (включая без ограничения устойчивость к глифосату). Такие сочетания признаков обеспечивают новые способы борьбы с сорными (и тому подобными) видами благодаря новоприобретенной резистентности или наследственной устойчивости к гербицидам (например, глифосату). Таким образом, кроме признаков HTC новые способы борьбы с сорняками с использованием гербицидов, к которым была получена устойчивость у трансгенных культурных растений благодаря указанному ферменту, входят в объем изобретения.
Кроме того, широко распространены устойчивые к глифосату культуры, выращиваемые по всему миру. Многократное чередование с другими устойчивыми к глифосату культурами, контроль резистентных к глифосату самосевных растений могут быть затруднены в случае чередуемых культур. Таким образом, применение трансгенных признаков согласно настоящему изобретению, подвергаемых по отдельности стэкингу или трансформации в культурные растения, обеспечивает средство борьбы с другими самосевными HTC-культурами.
Предпочтительное растение или семя для применения в настоящем изобретении содержит в своем геноме встроенные последовательности, которые указаны в настоящем описании, вместе с по меньшей мере 20-500 или более следующих друг за другом фланкирующих нуклеотидов с обеих сторон вставки, которые идентифицированы в настоящем описании. Если не указано иное, то ссылка на фланкирующие последовательности относится к последовательностям, определяемым в отношении последовательности SEQ ID NO: 29 (см. таблицу выше). В свою очередь последовательность SEQ ID NO: 29 включает гетерологичную ДНК, встроенную в исходный трансформант, и иллюстративные фланкирующие геномные последовательности непосредственно вблизи встроенной ДНК. Можно ожидать, что все или часть таких фланкирующих последовательностей может быть передана потомству, которое получает встроенную ДНК, включая представляющий интерес трансген, в результате полового скрещивания одной родительской линии, в которой произошло такое событие.
Настоящее изобретение относится к применению культур тканей из регенерированных клеток растения согласно настоящему изобретению. Также изобретение относится к применению растения, регенерированного из такой культуры ткани, особенно, когда указанное растение способно экспрессировать все морфологические и физиологические свойства используемого в качестве примера сорта. Предпочтительные растения для применения в настоящем изобретении могут обладать всеми физиологическими и морфологическими свойствами растения, выращенного из депонированного семени. Настоящее изобретение, кроме того, относится к применению потомства такого семени и к семени, обладающему представляющими интерес качественными свойствами.
Манипуляции (такие как мутация, дополнительная трансфекция и дальнейшее скрещивание) с растениями или семенами или их частями могут приводить к созданию так называемых «производных по существу» сортов. Международный союз по охране новых сортов растений (UPOV) представил следующее руководство по определению того, является ли сорт по существу производным от охраняемого сорта:
[A] Сорт следует считать производным по существу от другого сорта («исходного сорта»), если:
(i) он преимущественно произведен из исходного сорта или из сорта, который сам преимущественно произведен из исходного сорта, с сохранением при этом проявления основных признаков, являющихся результатом реализации генотипа или сочетания генотипов исходного сорта;
(ii) он явно отличается от исходного сорта; и
(iii) за исключением различий, которые являются следствием происхождения, он соответствует исходному сорту по степени проявления основных признаков, являющихся результатом реализации генотипа или комбинации генотипов исходного сорта.
UPOV, Sixth Meeting with International Organizations, Geneva, Oct. 30, 1992; документ подготовлен Бюро Союза.
В используемом в настоящем описании смысле «линия» означает группу растений, в которой наблюдается небольшое генетическое варьирование или нет генетического варьирования среди особей в отношении по меньшей мере одного признака. Некоторые линии могут быть созданы в результате самоопыления в нескольких поколениях и селекции или в результате вегетативного размножения исходного родителя с использованием методики культивирования тканей или клеток.
В используемом в настоящем описании смысле термины «культурный сорт» и «сорт» являются синонимами и относятся к линии, которую применяют для промышленного производства.
«Стабильность» или «стабильный» в отношении данного компонента означает, что компонент сохраняется от поколения к поколению и, предпочтительно, по меньшей мере в течение трех поколений по существу на одном и том же уровне, например, предпочтительно ±15%, более предпочтительно ±10%, наиболее предпочтительно ±5%. На стабильность может влиять температура, местоположение, стресс и время посева. Сравнение последующих поколений в полевых условиях должно показать, что компонент присутствует в аналогичном виде.
«Коммерческую ценность» определяют как наличие хорошей мощности растения и высокой плодовитости, так чтобы культура могла быть использована фермерами в производстве с применением обычного сельскохозяйственного оборудования, и масло, содержащее описанные компоненты, можно было экстрагировать из семени с использованием обычного дробильного и экстрагирующего оборудования. Чтобы представлять ценность с коммерческой точки зрения, урожайность, которую измеряют по массе семян, содержанию масла и общему количеству масла, полученного в расчете на акр (0,4 га), должна быть в пределах 15% от средней урожайности во всем остальном сравнимого коммерческого сорта кукурузы без улучшенных ценных свойств, выращенного в том же регионе.
«Агрономически элитный» означает, что линия обладает требуемыми агрономическими характеристиками, такими как урожайность, зрелость, устойчивость к заболеваниям и тому подобное, в дополнение к устойчивости к насекомым вследствие события(ий) согласно изобретению. Агрономические характеристики, взятые по отдельности или в любой комбинации, которые указаны в примерах ниже, в растении, в котором произошло событие согласно настоящему изобретению, входят в объем настоящего изобретения. Любую или все агрономические характеристики и измеряемые параметры можно использовать для идентификации таких растений, либо в виде точки, либо на любом конце или обоих концах диапазона характеристик, используемых для определения таких растений.
Как будет понятно специалисту в данной области в свете настоящего описания, предпочтительные варианты наборов для выявления, например, могут содержать зонды и/или праймеры, направленные к «последовательностям соединения» и/или содержащие «последовательности соединения» или «переходные последовательности» (где фланкирующая геномная последовательность кукурузы сходится с последовательностью вставки). Например, набор содержит полинуклеотидные зонды, праймеры и/или ампликоны, сконструированные для идентификации одной или обеих последовательностей соединения (где вставка сходится с фланкирующей последовательностью), которые указаны в таблице 1. Один общий замысел состоит в том, чтобы был один праймер, который гибридизуется с фланкирующей областью, и один праймер, который гибридизуется со вставкой. Каждый из таких праймеров часто имеет длину по меньшей мере ~15 остатков. В случае такого расположения праймеры можно использовать для образования/амплификации регистрируемого ампликона, который свидетельствует о наличии события согласно настоящему изобретению. Такие праймеры можно использовать для образования ампликона, который охватывает (и включает) последовательность соединения, которая указана выше.
Праймер(ры), который «садя(и)тся» на фланкирующую последовательность, конструируют так, чтобы он не гибридизовался более чем в пределах примерно 200 оснований или за пределами соединения. Таким образом, могут быть сконструированы типичные фланкирующие праймеры, содержащие по меньшей мере 15 остатков любой нити в пределах 200 оснований во фланкирующих последовательностях от начала вставки. То есть праймеры, содержащие последовательность соответствующего размера в пределах остатков ~1674-1873 и/или ~6690-6890 последовательности SEQ ID NO: 29, входят в объем настоящего изобретения. Подобным образом праймеры для вставки могут быть сконструированы к любому месту на вставке, и для такого конструирования праймеров можно использовать без исключения, например, остатки ~1874-2074 и ~6489-6689.
Специалисту в данной области также будет понятно, что могут быть сконструированы праймеры и зонды для гибридизации в диапазоне стандартных условий гибридизации и/или ПЦР с участком последовательности SEQ ID NO: 29 (или комплемента) и его комплементами, при этом праймер или зонд не является абсолютно комплементарным указанной в качестве примера последовательности. То есть некоторая степень ошибочного спаривания может быть допустима. В случае праймера длиной, например, приблизительно 20 нуклеотидов, обычно примерно один или два нуклеотида не обязательно должны связываться с противоположной нитью, если ошибочно спариваемое основание является внутренним или находится на конце праймера, который противоположен ампликону. Ниже приведены разные подходящие условия гибридизации. Синтетические аналоги нуклеотидов, такие как инозин, также можно использовать в качестве зондов. Также можно использовать зонды на основе пептидо-нуклеиновых кислот (PNA), а также ДНК- и РНК-зонды. Важно, что такие зонды и праймеры являются диагностическими (способными однозначно идентифицировать и отличать) в отношении наличия события согласно настоящему изобретению.
Следует отметить, что могут возникать ошибки в ПЦР-амплификации, которые могут приводить, например, к небольшим ошибкам секвенирования. То есть если не указано иное, последовательности, перечисленные в настоящем описании, определяли, создавая длинные ампликоны с геномной ДНК кукурузы и затем клонируя и секвенируя ампликоны. Нет ничего необычного в том, чтобы найти небольшие различия и минорные несоответствия в последовательностях, образуемых и определяемых таким образом, учитывая множество раундов амплификации, которые необходимы для создания достаточного для секвенирования ампликона с геномных ДНК. Специалисту в данной области следует понимать и иметь в виду, что любые корректировки, необходимые из-за общих ошибок секвенирования или несоответствий таких типов, входят в объем настоящего изобретения.
Также следует отметить, что нет ничего необычного в делетировании некоторых геномных последовательностей, например, когда встраивают последовательность в ходе осуществления определенного события. Таким образом, также могут появляться некоторые различия между рассматриваемыми фланкирующими последовательностями и геномными последовательностями, перечисленными, например, в GENBANK. Некоторые из таких различий обсуждаются ниже в разделе «Примеры». Соответственно, могут быть осуществлены корректировки зондов и праймеров.
Таким образом, применение растения, содержащего полинуклеотид, который имеет определенную степень идентичности с фланкирующими последовательностями и/или последовательностями вставки согласно изобретению, входит в объем настоящего изобретения. Идентичность последовательности согласно настоящему изобретению может означать, что полинуклеотидная последовательность обладает по меньшей мере 65% идентичностью последовательности, предпочтительно по меньшей мере 70% идентичностью последовательности, более предпочтительно по меньшей мере 75% идентичностью последовательности, более предпочтительно по меньшей мере 80% идентичностью и более предпочтительно по меньшей мере 85% 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичностью последовательности с последовательностью, приведенной в качестве примера или описанной в настоящей публикации. Гибридизация и условия гибридизации, которые предлагаются в настоящем изобретении, также можно использовать для определения таких растений и полинуклеотидных последовательностей согласно настоящему изобретению. Последовательность фланкирующих последовательностей плюс последовательности вставки можно подтвердить со ссылкой на депонированное семя.
Компоненты каждой «вставки» показаны на фигурах 1 и 2 и обсуждаются более подробно ниже в примерах. Полинуклеотидные последовательности ДНК таких компонентов или их фрагментов можно использовать в качестве ДНК-праймеров или зондов в способах согласно настоящему изобретению.
В некоторых вариантах предлагаются композиции и способы для выявления наличия трансгена/геномной области инсерции в растениях и семенах и тому подобном растения кукурузы. Предлагаются последовательности ДНК, которые содержат последовательность соединения трансген/геномная область инсерции, предлагаемую в настоящем изобретении (между остатками 1873-1874 и 6689-6690 последовательности SEQ ID NO: 29), их фрагменты и комплементы приведенных в качестве примера последовательностей и их любых фрагментов. Последовательность соединения области инсерции охватывает соединение между гетерологичной ДНК, встроенной в геном, и ДНК из клеток кукурузы, фланкирующей сайт инсерции. Такие последовательности могут быть диагностическими для данного события.
На основании таких последовательности вставки и пограничных последовательностей могут быть созданы специфичные для события праймеры. ПЦР-анализ показал, что линии кукурузы согласно настоящему изобретению могут быть идентифицированы в разных генотипах кукурузы в анализе ПЦР-ампликонов, образованных с использованием таких специфичных для события наборов праймеров. Указанные и другие родственные способы можно использовать для однозначной идентификации таких линий кукурузы. Таким образом, ПЦР-ампликоны, полученные с использованием таких пар праймеров, являются уникальными и могут быть использованы для идентификации таких линий кукурузы.
В некоторых вариантах последовательности, которые содержат непрерывный фрагмент нового трансгена/геномной области инсерции, представляют собой аспект настоящего изобретения. В объем изобретения включены последовательности ДНК, которые содержат полинуклеотиды достаточной длины последовательности вставки трансгена и полинуклеотиды достаточной длины геномной последовательности кукурузы из одного или более из трех указанных выше растений кукурузы и/или последовательности, которые применимы в качестве праймерных последовательностей для получения продукта в виде ампликона, являющегося диагностическим для одного или нескольких из таких растений кукурузы.
Родственные варианты относятся к последовательностям ДНК, которые содержат по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более следующих друг за другом нуклеотидов трансгенной части последовательности ДНК, указанной в настоящем описании (такой как последовательность SEQ ID NO: 29 и ее участки), или ее комплемента и фланкирующую последовательность ДНК кукурузы сходной длины из таких последовательностей или их комплементов. Такие последовательности применимы в качестве ДНК-праймеров в способах амплификации ДНК. Ампликоны, полученные с использованием таких праймеров, являются диагностическими для любых событий, имеющих место в растениях кукурузы, указанных в настоящем описании. Таким образом, изобретение также относится к ампликонам, полученным с использованием таких ДНК-праймеров и гомологичных праймеров.
Настоящее изобретение также может относиться к способам выявления наличия в образце ДНК, которая соответствует событию в растении кукурузы, указанному в настоящем описании. Такие способы могут включать: (a) осуществление контакта образца, содержащего ДНК, с набором праймеров, который при использовании в реакции амплификации нуклеиновой кислоты с ДНК из по меньшей мере одного из таких событий кукурузы дает ампликон, который является диагностическим для указанного события(ий); (b) осуществление реакции амплификации нуклеиновой кислоты с получением таким образом ампликона; и (c) выявление ампликона.
Дополнительные способы выявления могут включать способ выявления присутствия в образце ДНК, соответствующей по меньшей мере одному из указанных событий, при этом указанный способ включает: (a) осуществление контакта образца, содержащего ДНК, с зондом, который гибридизуется в жестких условиях гибридизации с ДНК по меньшей мере из одного из указанных событий у кукурузы и который не гибридизуется в жестких условиях гибридизации с ДНК контрольного растения кукурузы (ДНК, не имеющей отношения к представляющему интерес событию); (b) помещение образца и зонда в жесткие условия гибридизации; и (c) выявление гибридизации зонда с ДНК.
Некоторые варианты включают способы получения растения кукурузы, в котором имеет место событие aad-1 согласно настоящему изобретению, при этом указанный способ включает стадии: (a) полового скрещивания первой родительской линии кукурузы (содержащей кассету экспрессии согласно настоящему изобретению, которая придает указанный признак устойчивости к гербицидам растениям указанной линии) и второй родительской линии кукурузы (в которой отсутствует такой признак устойчивости к гербицидам) с получением при этом множества растений-потомков; и (b) отбор растений-потомков с использованием молекулярных маркеров. Такие способы необязательно могут включать дополнительную стадию возвратного скрещивания растений-потомков со второй родительской линией кукурузы с получением чистосортного растения кукурузы, которое имеет указанный признак устойчивости к насекомым.
Согласно другому аспекту изобретения предлагаются способы определения зиготности потомства от скрещивания с любым одним (или несколькими) из указанных трех событий. Указанные способы могут включать осуществление контакта образца, содержащего ДНК кукурузы, с набором праймеров согласно настоящему изобретению. Указанные праймеры при использовании в реакции амплификации нуклеиновых кислот с геномной ДНК по меньшей мере из одного их указанных событий кукурузы дают первый ампликон, который является диагностическим по меньшей мере для одного из указанных событий кукурузы. Такие способы дополнительно включают осуществление реакции амплификации нуклеиновой кислоты с получением при этом первого ампликона; выявление первого ампликона; и осуществление контакта образца, содержащего ДНК кукурузы, с указанным набором праймеров (указанный набор праймер при использовании в реакции амплификации нуклеиновой кислоты с геномной ДНК из растений кукурузы дает второй ампликон, содержащий нативную геномную ДНК кукурузы, гомологичную области генома кукурузы); и осуществление реакции амплификации нуклеиновой кислоты с получением при этом второго ампликона. Способы дополнительно включают выявление второго ампликона и сравнение первого и второго ампликонов в образце, при этом присутствие обоих ампликонов свидетельствует о том, что образец является гетерозиготным в отношении инсерции трансгена.
Наборы для выявления ДНК могут быть разработаны с использованием композиций, раскрытых в настоящем описании, и способов, хорошо известных в области выявления ДНК. Наборы применимы для идентификации в образце ДНК, связанной с событием у кукурузы согласно изобретению, и могут быть применимы в способах селекции растений кукурузы, содержащих такую ДНК. Наборы содержат последовательности ДНК, гомологичные или комплементарные ампликонам, например, раскрытым в настоящем описании, или последовательности ДНК, гомологичные или комплементарные ДНК, находящейся в генетических элементах трансгена в случае события согласно изобретению. Такие последовательности ДНК могут быть использованы в реакциях амплификации ДНК или в качестве зондов в способе гибридизации ДНК. Наборы также могут содержать реагенты и материалы, необходимые для осуществления способа выявления.
«Зонд» означает изолированную молекулу нуклеиновой кислоты, с которой связана обычная регистрируемая метка или репортерная молекула (такая как радиоактивный изотоп, лиганд, хемилюминесцентный агент или фермент). Такой зонд комплементарен нити нуклеиновой кислоты-мишени, в случае настоящего изобретения нити геномной ДНК одного из указанных событий у кукурузы, либо из растения кукурузы, либо из образца, который содержит ДНК, полученную в результате такого события. Зонды согласно настоящему изобретению включают не только дезоксирибонуклеиновые или рибонуклеиновые кислоты, но также полиамиды и другие вещества для зондов, которые специфично связываются с последовательностью ДНК мишени и могут быть использованы для выявления присутствия такой последовательности ДНК мишени.
«Праймеры» представляют собой изолированные/синтезированные нуклеиновые кислоты, которые отжигаются с комплементарной нитью ДНК-мишени при гибридизации нуклеиновых кислот с образованием гибрида между праймером и нитью ДНК-мишени, затем удлиняются вдоль нити ДНК-мишени полимеразой, например, ДНК-полимеразой. Пары праймеров согласно настоящему изобретению применимы для амплификации последовательности нуклеиновой кислоты мишени, например, посредством полимеразной цепной реакции (ПЦР) или другими обычными способами амплификации нуклеиновых кислот.
Зонды и праймеры обычно имеют 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188, 189, 190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202, 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387, 388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428, 429, 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499 или 500 нуклеотидов или больше. Такие зонды и праймеры специфично гибридизуются с последовательностью-мишенью в условиях высокой жесткости. Предпочтительно, зонды и праймеры согласно настоящему изобретению имеют полное сходство последовательности с последовательностью-мишенью, хотя обычными способами могут быть сконструированы зонды, отличающиеся от последовательности-мишени и сохраняющие способность гибридизоваться с последовательностями-мишенями.
Способы получения и применения зондов и праймеров описаны, например, в Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, ed. Sambrook et al., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989. Пары ПЦР-праймеров могут быть получены из известной последовательности, например, с применением компьютерных программ, предназначенных для этой цели.
Праймеры и зонды, основанные на последовательностях фланкирующей ДНК и вставки, раскрытых в настоящем описании, можно использовать для подтверждения (и в случае необходимости, для корректировки) описанных последовательностей обычными способами, посредством повторного клонирования и секвенирования таких последовательностей.
Зонды и праймеры на основе нуклеиновых кислот согласно настоящему изобретению гибридизуются в жестких условиях с последовательностью ДНК-мишени. Можно использовать любой обычный способ гибридизации или амплификации нуклеиновых кислот для идентификации присутствия в образце ДНК, встроенной в результате трансгенного события. Молекулы нуклеиновых кислот или их фрагменты способны специфично гибридизоваться с другими молекулами нуклеиновой кислоты в определенных условиях. В используемом в настоящем описании смысле говорят, что две молекулы нуклеиновой кислоты способны специфично гибридизоваться друг с другом, если две молекулы способны образовывать структуру нуклеиновой кислоты из двух антипараллельных нитей. Говорят, что молекула нуклеиновой кислоты является «комплементом» другой молекулы нуклеиновой кислоты, если показана их полная комплементарность. В используемом в настоящем описании смысле говорят, что молекулы имеют «полную комплементарность», когда каждый нуклеотид одной из молекул комплементарен нуклеотиду другой. Говорят, что молекулы являются «минимально комплементарными», если они могут гибридизоваться друг с другом со стабильностью, достаточной для обеспечения возможности того, чтобы они оставались отожженными друг с другом по меньшей мере в обычных условиях «низкой жесткости». Подобным образом, говорят, что молекулы являются «комплементарными», если они могут гибридизоваться друг с другом со стабильностью, достаточной для обеспечения возможности того, чтобы они оставались отожженными друг с другом в обычных условиях «высокой жесткости». Обычные условия жесткости описаны Sambrook с соавторами, 1989. Поэтому допустимы отклонения от полной комплементарности, при условии, что такие отклонения не исключают полностью способность молекул образовывать двунитевую структуру. Чтобы молекула нуклеиновой кислоты служила в качестве праймера или зонда, она должна быть только в достаточной степени комплементарна по последовательности, чтобы она могла образовывать стабильную двунитевую структуру в конкретном используемом растворителе и при используемых концентрациях соли.
В используемом в настоящем описании смысле по существу гомологичная последовательность представляет собой последовательность нуклеиновой кислоты, которая будет специфично гибридизоваться с комплементом последовательности нуклеиновой кислоты, с которым ее сравнивают, в условиях высокой жесткости. Термин «жесткие условия» функционально определяют по отношению к гибридизации нуклеиновой кислоты-зонда с нуклеиновой кислотой-мишенью (т.е., с конкретной представляющей интерес последовательностью нуклеиновой кислоты) с использованием конкретного способа гибридизации, обсуждаемого в публикации Sambrook с соавторами, 1989, в разделах 9.52-9.55. См. также публикацию Sambrook с соавторами, 1989, разделы 9.47-9.52 и 9.56-9.58. Соответственно, нуклеотидные последовательности согласно изобретению можно использовать в связи с их способностью избирательно образовывать дуплексные молекулы с комплементарными участками фрагментов ДНК.
В зависимости от предусмотренного применения можно использовать разные условия гибридизации, чтобы добиться разных степеней избирательности зонда по отношению к последовательности-мишени. В случае применений, требующих высокой избирательности, обычно можно использовать относительно жесткие условия для образования гибридов, например, можно выбирать условия относительно низкой концентрации соли и/или высокой температуры, такие как условия, создаваемые при концентрации NaCl от примерно 0,02 М до примерно 0,15 М при температурах от примерно 50°C до примерно 70°C. Жесткие условия, например, могут включать промывку гибридизационного фильтра по меньшей мере два раза буфером для промывки высокой жесткости (0,2X SSC, 0,1% SDS, 65°C). Подходящие условия жесткости, которые стимулируют гибридизацию ДНК, например, 6,0X хлорид натрия/цитрат натрия (SSC) примерно при 45°C с последующей промывкой 2,0×SSC при 50°C известны специалистам в данной области, 6.3.1-6.3.6. Например, концентрация соли на стадии промывки может быть выбрана из условий от низкой жесткости примерно 2,0×SSC при 50°C до высокой жесткости примерно 0,2×SSC при 50°C. Кроме того, температура на стадии промывки может быть увеличена от условий низкой жесткости при комнатной температуре примерно 22°C до условий высокой жесткости примерно при 65°C. И температура и соль могут варьировать, либо температуру, либо концентрацию соли можно поддерживать постоянными, в то время как другую переменную изменяют. Такие селективные условия в малой степени или вообще не допускают несоответствия между зондом и матрицей или нитью-мишенью. Выявление последовательностей ДНК с использованием гибридизации хорошо известно специалистам в данной области, и инструкции, приведенные в патентах США № 4965188 и 5176995, являются примерами способов анализа гибридизации.
В некоторых вариантах нуклеиновая кислота для применения в настоящем изобретении будет специфично гибридизоваться с одним или несколькими праймерами (или ампликонами или другими последовательностями), приведенными в качестве примеров или предлагаемых в настоящем описании, включая их комплементы и фрагменты, в условиях высокой жесткости. В одном аспекте маркерная молекула нуклеиновой кислоты согласно настоящему изобретению имеет последовательность нуклеиновой кислоты, указанную в SEQ ID NO: 3-14, или ее комплементы и/или фрагменты.
В другом аспекте маркерная молекула нуклеиновой кислоты согласно настоящему изобретению имеет от 80% до 100% или от 90% до 100% идентичность последовательности с такими последовательностями нуклеиновых кислот. В следующем аспекте маркерная молекула нуклеиновой кислоты для применения в настоящем изобретении имеет от 95% до 100% идентичность последовательности с такой последовательностью. Такие последовательности можно использовать в качестве маркеров в способах селекции растений, чтобы идентифицировать потомство от генетического скрещивания. Гибридизация зонда с молекулой ДНК мишени может быть выявлена любым из ряда способов, известных специалистам в данной области, при этом такие способы включают без ограничения флуоресцирующие метки, радиоактивные метки, метки на основе антител и хемилюминесцентные метки.
Что касается амплификации последовательности нуклеиновой кислоты мишени (например, в ПЦР) с использованием конкретной пары праймеров для амплификации, то «жесткие условия» представляют собой условия, которые обеспечивают возможность гибридизации пары праймеров только с последовательностью нуклеиновой кислоты мишени, с которой праймер, имеющий соответствующую последовательность дикого типа (или ее комплемент), может связываться и предпочтительно давать уникальный продукт амплификации ампликон.
Термин «специфичный для (последовательности-мишени)» указывает, что зонд или праймер гибридизуется в жестких условиях гибридизации только с последовательностью-мишенью в образце, содержащем последовательность-мишень.
В используемом в настоящем описании смысле «амплифицированная ДНК» или «ампликон» относится к продукту амплификации нуклеиновой кислоты последовательности нуклеиновой кислоты-мишени, которая является частью матрицы нуклеиновой кислоты. Например, чтобы определить, содержит ли растение кукурузы, полученное в результате полового скрещивания, геномную ДНК трансгенного события из растения кукурузы согласно настоящему изобретению, ДНК из образца ткани растения кукурузы может быть подвергнута амплификации нуклеиновой кислоты способом с использованием пары праймеров, которая включает праймер, полученный из фланкирующей последовательности, расположенной в геноме растения вблизи сайта инсерции встроенной гетерологичной ДНК, и второй праймер, полученный из встроенной гетерологичной ДНК, с получением ампликона, который является диагностическим в отношении наличия ДНК, встроенной в результате того события. Ампликон имеет определенную длину и последовательность, которые также являются диагностическими для данного события. Ампликон может иметь длину в диапазоне от суммарной длины пары праймеров плюс одна пара оснований нуклеотидов и/или суммарной длины пары праймеров плюс примерно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188, 189, 190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202, 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387, 388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428, 429, 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499 или 500, 750, 1000, 1250, 1500, 1750, 2000 или более пар оснований нуклеотидов (плюс или минус любое количество из перечисленных выше). Альтернативно, пара праймеров может быть получена из фланкирующей последовательности, расположенной с обеих сторон встроенной ДНК, так, чтобы получить ампликон, который содержит полную нуклеотидную последовательность вставки. Представитель пары праймеров, полученный из геномной последовательности растения, может быть расположен на некотором расстоянии от последовательности встроенной ДНК. Такое расстояние может быть в диапазоне от одной пары оснований нуклеотидов вплоть до примерно двадцати тысяч пар оснований нуклеотидов. Использование термина «ампликон», в частности, исключает димеры праймеров, которые могут быть образованы в термической реакции амплификации ДНК.
Амплификация нуклеиновых кислот может быть осуществлена любым из различных способов амплификации нуклеиновых кислот, известных в данной области, включая полимеразную цепную реакцию (ПЦР). В данной области известно множество способов амлификации, и они описаны наряду с прочими публикациями в патенте США № 4683195 и патенте США № 4683202. Способы ПЦР-амплификации были разработаны для амплификации до 22 т.п.н. геномной ДНК. Такие способы, а также другие способы, известные в области амплификации ДНК, могут быть использованы при практическом осуществлении настоящего изобретения. Последовательность вставки гетерологичной трансгенной ДНК или фланкирующая геномная последовательность в случае данного события кукурузы могут быть подтверждены (и при необходимости скорректированы) посредством амплификации таких последовательностей, связанных с данным событием, с использованием праймеров, полученных из последовательностей, предлагаемых в настоящем изобретении, с последующим стандартным секвенированием ДНК ПЦР-ампликона или клонированной ДНК.
Ампликон, полученный указанными способами, можно выявить, используя множество способов. Электрофорез в агарозном геле и окрашивание бромидом этидия является общеизвестным способом выявления ДНК-ампликонов. Другим таким способом является анализ единиц генетической информации (Genetic Bit Analysis), в случае которого конструируют олигонуклеотиды ДНК, которые перекрывают и близлежащую фланкирующую последовательность геномной ДНК и встроенную последовательность ДНК. Олигонуклеотиды иммобилизуют в лунках планшета для микротитрования. После ПЦР представляющей интерес области (с использованием одного праймера к встроенной последовательности и одного к близлежащей фланкирующей геномной последовательности), однонитевой ПЦР-продукт может быть гибридизован с иммобилизованным олигонуклеотидом и может служить в качестве матрицы для реакции удлинения на одно основание с использованием ДНК-полимеразы и меченых ddNTP, специфичных для ожидаемого следующего основания. Регистрация данных может быть на основе флуоресценции или на основе ELISA. Сигнал свидетельствует о присутствии вставки/фланкирующей последовательности с учетом успешной амплификации, гибридизации и удлинения на одно основание.
Другим способом является способ пиросеквенирования, который описан Winge (Innov. Pharma. Tech. 00: 18-24, 2000). В случае указанного способа конструируют олигонуклеотид, который перекрывает соединение близлежащей геномной ДНК и ДНК вставки. Олигонуклеотид гибридизуют с однонитевым ПЦР-продуктом из представляющей интерес области (один праймер к встроенной последовательности и один к фланкирующей геномной последовательности) и инкубируют в присутствии ДНК-полимеразы, АТФ, сульфурилазы, люциферазы, апиразы, аденозин-5’-фосфосульфата и люциферина. dNTP добавляют по отдельности, и включение приводит к появлению светового сигнала, который измеряют. Световой сигнал свидетельствует о присутствии трансгенной вставки/фланкирующей последовательности с учетом успешной амплификации, гибридизации и удлинения на одно или множество оснований.
Поляризация флуоресценции является другим способом, который можно использовать для выявления ампликона согласно настоящему изобретению. Следуя такому способу, конструируют олигонуклеотид, который охватывает соединение геномной фланкирующей ДНК и встроенной ДНК. Олигонуклеотид гибридизуют с однонитевым ПЦР-продуктом из представляющей интерес области (один праймер ко встроенной ДНК и один к фланкирующей последовательности геномной ДНК) и инкубируют в присутствии ДНК-полимеразы и флуоресцентно-меченного ddNTP. Удлинение на одно основание приводит к включению ddNTP. Включение можно измерить в виде изменения поляризации с использованием флуориметра. Изменение поляризации свидетельствует о присутствии трансгенной вставки/фланкирующей последовательности с учетом успешной амплификации, гибридизации и удлинения на одно основание.
TAQMAN (PE Applied Biosystems, Foster City, Calif.) представляет собой способ выявления и количественной оценки присутствия последовательности ДНК. Коротко, конструируют олигонуклеотидный зонд FRET, который перекрывает соединение геномной фланкирующей ДНК и ДНК вставки. Зонд FRET и ПЦР-праймеры (один праймер к последовательности ДНК вставки и один к фланкирующей геномной последовательности) подвергают циклической реакции в присутствии термостабильной полимеразы и dNTP. В ходе специфичной амплификации ДНК-полимераза Taq удаляет и высвобождает флуоресцирующий остаток от связи с гасящим остатком на зонде FRET. Сигнал флуоресценции свидетельствует о присутствии фланкирующей последовательности/последовательности трансгенной вставки с учетом успешной амплификации и гибридизации.
Способ «молекулярных маяков» описан для применения при выявлении последовательности. Коротко, конструируют олигонуклеотидный зонд FRET, который перекрывает соединение фланкирующей геномной ДНК и ДНК вставки. Результатом уникальной структуры зонда FRET является то, что он имеет вторичную структуру, которая удерживает флуоресцирующий остаток и гасящий остаток в тесной близости. Зонд FRET и ПЦР-праймеры (один праймер к последовательности ДНК вставки и один к фланкирующей геномной последовательности) подвергают циклической реакции в присутствии термостабильной полимеразы и dNTP. После успешной ПЦР-амплификации гибридизация зонда FRET с последовательностью-мишенью приводит к устранению вторичной структуры зонда и пространственному разделению флуоресцирующего и гасящего остатков. Сигнал флуоресценции свидетельствует о присутствии фланкирующей геномной последовательности/последовательности трансгенной вставки с учетом успешной амплификации и гибридизации.
В дополнение к раскрытию положения в геноме кукурузы, которое прекрасно подходит для инсерции, настоящее изобретение также относится к применению семени кукурузы и/или растения кукурузы, содержащих по меньшей мере одну вставку, не имеющую отношения к aad-1, вблизи такого положения в геноме. Один вариант заключается в том, чтобы осуществить замену другой вставкой вместо вставки aad-1, приведенной в настоящем описании в качестве примера. В общем, согласно настоящему изобретению в этой связи можно применять, например, целенаправленную гомологичную рекомбинацию. Методика такого типа является, например, предметом заявки WO 03/080809 A2 и соответствующей опубликованной заявки на выдачу патента США (US 20030232410). Таким образом, настоящее изобретение относится к применению растений и растительных клеток, содержащих гетерологичную вставку (вместо или одновременно с множественными копиями aad-1), фланкированную всей или известной частью фланкирующих последовательностей, указанных в настоящем описании (например, остатки 1-1873 и 6690-8557 последовательности SEQ ID NO: 29).
Ниже приведены некоторые пункты/варианты растений, семян, полинуклеотидов и способов выявления, которые можно применять согласно некоторым вариантам осуществления настоящего изобретения:
1. Трансгенное растение кукурузы, содержащее геном, при этом указанный геном содержит последовательность SEQ ID NO: 29.
2. Семя кукурузы, содержащее геном, в котором имеет место AAD-1-событие DAS-40278-9, которое имеется в семени, депонированном в Американской коллекции типов культур (ATCC) с номером доступа PTA-10244.
3. Семя кукурузы по п.2, в котором указанное семя содержит геном, и указанный геном содержит последовательность SEQ ID NO: 29.
4. Растение кукурузы, полученное выращиванием из семени по п.2, при этом указанное растение содержит последовательность SEQ ID NO: 29.
5. Растение-потомок, полученное от растения кукурузы по п.4, при этом в указанном растении-потомке имеет место AAD-1-событие DAS-40278-9.
6. Устойчивое к гербицидам растение-потомок, полученное от растения кукурузы по п.1, при этом указанное растение-потомок содержит последовательность SEQ ID NO: 29.
7. Трансгенное растение кукурузы, содержащее трансгенную вставку в участке хромосомы кукурузы, являющемся мишенью, расположенном в хромосоме 2 в положении примерно 20 сМ между SSR-маркерами UMC1265, частично амплифицируемым при использовании последовательностей SEQ ID NO: 30 и SEQ ID NO: 31, и MC0111, частично амплифицируемым при использовании последовательностей SEQ ID NO: 32 и SEQ ID NO: 33, при этом участок-мишень содержит гетерологичную нуклеиновую кислоту.
8. Способ получения трансгенного растения кукурузы по п.7, при этом указанный способ включает встраивание гетерологичной нуклеиновой кислоты в положение в хромосоме 2 примерно 20 сМ между SSR-маркерами UMC1265, частично амплифицируемым при использовании последовательностей SEQ ID NO: 30 и SEQ ID NO: 31, и MMC0111, частично амплифицируемым при использовании последовательностей SEQ ID NO: 32 и SEQ ID NO: 33.
9. Часть растения по п.4, при этом указанная часть выбрана из группы, состоящей из пыльцы, семязачатка, цветков, семенных коробочек, волокон, побегов, корней и листьев, при этом указанная часть содержит последовательность SEQ ID NO: 29.
10. Трансгенное растение кукурузы по п.7, в котором указанное растение содержит вставку трансгена в геномной последовательности или фланкированную геномной последовательностью, выбранной из группы, состоящей из остатков 1-1873 последовательности SEQ ID NO: 29 и остатков 6690-8557 последовательности SEQ ID NO: 29.
11. Изолированная молекула полинуклеотида, при этом указанная молекула:
содержит по меньшей мере 15 нуклеотидов и поддерживает гибридизацию в жестких условиях промывки с последовательностью нуклеиновой кислоты, выбранной из группы, состоящей из остатков 1-1873 последовательности SEQ ID NO: 29, остатков 6690-8557 последовательности SEQ ID NO: 29 и их комплементов;
содержит нуклеотидную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 1-33; и/или
гибридизуется в жестких условиях промывки с нуклеотидной последовательностью, выбранной из группы, состоящей из остатков 1863-1875 последовательности SEQ ID NO: 29, остатков 6679-6700 последовательности SEQ ID NO: 29 и их комплементов.
12. Изолированный полинуклеотид по п.11, в котором указанный полинуклеотид содержит нуклеотидную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 1-33.
13. Изолированный полинуклеотид по п.11, в котором указанный полинуклеотид гибридизуется в жестких условиях промывки с нуклеотидной последовательностью, выбранной из группы, состоящей из остатков 1863-1875 последовательности SEQ ID NO: 29, остатков 6679-6700 последовательности SEQ ID NO: 29 и их комплементов.
14. Полинуклеотид по п.13, в котором указанный полинуклеотид представляет собой ампликон, образованный в полимеразной цепной реакции.
15. Способ выявления события в образце кукурузы, содержащем ДНК кукурузы, при этом указанный способ включает осуществление контакта указанного образца по меньшей мере с одним полинуклеотидом, который является диагностическим для события AAD-1 кукурузы DAS-40278-9, которое имеет место в семени, депонированном в Американской коллекции типов культур (ATCC) с номером доступа PTA-10244.
16. Способ по п.15, в котором указанный способ включает осуществление контакта указанного образца с
a) первым праймером, который связывается с фланкирующей последовательностью, выбранной из группы, состоящей из остатков 1-1873 последовательности SEQ ID NO: 29, остатков 6690-8557 последовательности SEQ ID NO: 29 и их комплементов; и
b) вторым праймером, который связывается с последовательностью вставки, содержащей остатки 1874-6689 последовательности SEQ ID NO: 29 или ее комплемента;
подвергание указанного образца полимеразной цепной реакции; и анализ в отношении ампликона, образованного между указанными праймерами.
17. Способ по п.16, в котором указанные праймеры выбраны из группы, состоящей из последовательностей SEQ ID NO: 1-28.
18. Способ по п.15, в котором указанный полинуклеотид содержит по меньшей мере 30 нуклеотидов и гибридизуется в жестких условиях с последовательностью, выбранной из группы, состоящей из остатков 1863-1875 последовательности SEQ ID NO: 29, остатков 6679-6700 последовательности SEQ ID NO: 29 и их комплементов; при этом указанный способ дополнительно включает подвергание указанного образца и указанного полинуклеотида гибридизации в жестких условиях; и анализ указанного образца в отношении гибридизации указанного полинуклеотида с указанной ДНК.
19. Набор для выявления ДНК, содержащий первый праймер и второй праймер по п.17.
20. Набор для выявления ДНК, предназначенный для осуществления способа по п.18.
21. Набор для выявления ДНК, содержащий полинуклеотид по п.13.
22. Полинуклеотид по п.12, при этом указанный полинуклеотид содержит последовательность SEQ ID NO: 29.
23. Способ получения полинуклеотида по п.22.
24. Способ получения трансгенного растения по п.10, в котором указанный способ включает встраивание трансгена в участок ДНК генома кукурузы, при этом указанный участок ДНК имеет 5’-конец, содержащий остатки нуклеотидов 1-1873 последовательности SEQ ID NO: 29, и 3’-конец, содержащий остатки нуклеотидов 6690-8557 последовательности SEQ ID NO: 29.
25. Способ, включающий скрещивание первого растения кукурузы, содержащего последовательность SEQ ID NO: 29, со вторым растением кукурузы с получением третьего растения кукурузы, содержащего геном, и анализ указанного третьего растения кукурузы в отношении присутствия последовательности SEQ ID NO: 29 в указанном геноме.
26. Способ по п.25, в котором указанный способ применяют при селекции растения кукурузы и/или для интрогрессии признака устойчивости к гербицидам в растение кукурузы.
Все патенты, заявки на выдачу патентов, предварительные заявки и публикации, упоминаемые или цитируемые в настоящем описании, включены в виде ссылки в объеме, в котором они не противоречат подробному руководству, приведенному в настоящем описании.
Следующие далее примеры включены с целью иллюстрации способов, применяемых при практическом осуществлении изобретения, и демонстрации некоторых предпочтительных вариантов осуществления изобретения. Такие примеры не следует толковать как ограничивающие. Специалистам в данной области будет понятно, что инструкции, приведенные в следующих примерах, представляют конкретные подходы, применяемые для иллюстрации предпочтительных способов для практического осуществления изобретения. Однако в свете настоящего описания специалистам в данной области будет понятно, что могут быть осуществлены многочисленные изменения указанных конкретных вариантов и все еще получить при этом подобные или сходные результаты, не отходя от сути и не выходя за рамки объема изобретения. Если не указано иное, все проценты указаны по массе и все соотношения в смеси растворителей указаны по объему, если не отмечено иное.
Следующие примеры включают предпосевное и/или довсходовое применение. Такие применения не ограничены событием «278». Кроме того, применимость устойчивости, обеспечиваемой данными генами AAD-1, может быть объяснима с учетом укороченного промежутка времени от применения до нового посева. Это обеспечивает растениеводам намного большую гибкость в составлении графика проводимых ими посевов относительно момента полного уничтожения сорняков («выжигания»). Без применения настоящего изобретения ожидание в течение 7-30 дней или около того после уничтожения сорняков до посева может быть причиной значительной потери урожая. Таким образом, настоящее изобретение обеспечивает преимущества в этом отношении. См., например, пример 13. Любые интервалы между посевами/внесением гербицидов и любые диапазоны концентраций/нормы расхода гербицида(ов), приведенные в качестве примеров или предлагаемые в настоящем описании, можно использовать согласно настоящему изобретению.
Если не указано иное, использованы следующие сокращения.
ПРИМЕРЫ
Пример 1. Трансформация и отбор события AAD1 pDAS 1740-278
Событие AAD1, pDAS 1740-278, получали WHISKER-опосредованной трансформацией линии кукурузы Hi-II. Применяемый способ трансформации описан в заявке на выдачу патента США № 20090093366. FspI-фрагментом плазмиды pDAS1740, также называемой pDAB3812 (фигура 1), трансформировали линию кукурузы. Указанная плазмидная конструкция содержит растительную кассету экспрессии, содержащую RB7 MARv3 :: промотор v2 убиквитина 1 Zea mays // AAD1 v3 // 3’-UTR PER5 Zea mays :: транскрипционная единица растения (PTU) RB 7 MARv4.
Получали многочисленные события. Тем событиям, которые были жизнеспособными и приводили к получению здоровой резистентной к галоксифопу каллусной ткани, присваивали уникальные идентификационные коды, представляющие предполагаемые события трансформации, и сразу же переносили в свежую среду для селекции. Растения регенерировали из ткани, полученной в результате каждого уникального события, и переносили в теплицу.
Брали образцы листьев для молекулярного анализа, чтобы подтвердить присутствие трансгена AAD-1 с использованием Саузерн-блота, подтверждения границ ДНК и подтверждения с помощью геномных маркеров. Позитивные растения T0 переопыляли с инбредными линиями, чтобы получить семя T1. Растения T1 с событием pDAS 1470-278-9 (DAS-40278-9) отбирали, подвергали самоопылению и характеризовали в течение пяти поколений. Одновременно растения T1 подвергали возвратному скрещиванию и осуществляли интрогрессию в элитную зародышевую плазму (XHH13) посредством селекции с помощью маркеров в течение нескольких поколений. Такое событие создавали на основе независимого трансформированного изолята. Событие отбирали на основе его уникальных характеристик, таких как единственный сайт инсерции, нормальное менделевское расщепление и стабильная экспрессия и превосходное сочетание эффективности, включая устойчивость к гербицидам и агрономическую эффективность в широком диапазоне генотипических фонов и во многих местоположениях с разными условиями окружающей среды. Следующие далее примеры содержат данные, которые использовали для характеристики события pDAS-1740-278-9.
Пример 2. Характеристика события pDAS 1740-278-9 с использованием Саузерн-блота
Саузерн-блот-анализ использовали для установления картины интеграции встроенного фрагмента ДНК и определить количество вставок/копий гена aad-1 в случае события pDAS-1740-278-9 (DAS- 40278-9). Получали данные для демонстрации интеграции и целостности трансгена aad-1, встроенного в геном кукурузы.
Данные, полученные с использованием Саузерн-блота, свидетельствуют, что вставка pDAS 1740/Fsp I-фрагмента в случае события DAS-40278-9 происходила в простой интеграции одной интактной копии PTU aad-1 из плазмиды pDAS1740. Подробный Саузерн-блот-анализ проводили, используя зонды, специфичные для гена, промотора, терминатора и других регуляторных элементов, находящихся в области плазмиды, и описанные ферменты рестрикции, которые имеют сайты расщепления, расположенные в пределах плазмиды, и продуцируют гибридизующиеся фрагменты внутри плазмиды или фрагменты, которые охватывают соединение плазмиды с геномной ДНК кукурузы (пограничные фрагменты). Молекулярные массы, показанные на основании данных Саузерн-гибридизации, в случае комбинации данного фермента рестрикции и данного зонда были уникальными для данного события и позволяли установить картины для его идентификации. Такие анализы также показали, что фрагмент плазмиды был встроен в геномную ДНК кукурузы без перестройки PTU aad-1. Идентичные фрагменты гибридизации наблюдали в пяти разных поколениях кукурузы в случае трансгенного события DAS-40278-9, что свидетельствует о стабильности наследования инсерции PTU aad-1 в поколениях. Гибридизация с использованием смеси трех зондов к остову, расположенных вне сайта рестрикции Fsp I в плазмиде pDAS1740, не выявляла никаких специфичных фрагментов ДНК/генов, что свидетельствует об отсутствии гена резистентности к ампициллину и отсутствии других области остова вектора, непосредственно вблизи сайтов рестрикции Fsp I плазмиды pDAS1740 в случае трансгенного события DAS-40278-9 кукурузы. Иллюстративная карта вставки в случае aad-1-события DAS-40278-9 у кукурузы представлена на фигурах 2, 3.
Пример 2.1. Сбор образцом листьев кукурузы и выделение геномной ДНК (гДНК)
гДНК получали из листьев отдельных растений, в которых имело место событие aad-1 кукурузы DAS-40278-9. гДНК экстрагировали из ткани листьев, собранных от отдельных растений, несущих событие aad-1 кукурузы DAS-40278-9. Использовали трансгенные семена кукурузы из пяти разных поколений в случае события DAS-40278-9. Отобрали двадцать отдельных растений кукурузы, по четыре растения на поколение, для события DAS-40278-9. Кроме того, гДНК выделяли из обычного растения кукурузы, XHH13, которое содержит генетический фон, который является типичным для основной линии, не имеющей гена aad-1.
Перед выделением гДНК от каждого растения отбирали вырезанные пробиванием ткани образцы листьев, чтобы тестировать экспрессию белка aad-1 с использованием набора тест-полосок для быстрого анализа (American Bionostica, Swedesboro, NJ) согласно способу, рекомендованному производителем. Каждый вырезанный пробиванием образец листа получал оценку + или - в связи с присутствием или отсутствием aad-1, соответственно. Дальнейшей характеристике подвергали только позитивные растения из пяти поколений в случае события DAS-40278-9.
Образцы листьев кукурузы собирали от отдельных растений, в которых имеет место событие DAS-40278-9, и обычного контроля XHH13. Образцы листьев быстро замораживали в жидком азоте и хранили при температуре примерно -80° вплоть до использования.
Индивидуальную геномную ДНК экстрагировали из замороженной ткани листьев кукурузы, следуя стандартному способу CTAB. При необходимости некоторое количество геномной ДНК дополнительно очищали, используя Qiagen Genomic-Tip (Qiagen, Valencia, CA) в соответствии со способами, рекомендованными производителем. После экстракции количество ДНК определяли спектрофлуориметрически, используя зеленый реагент Pico Green (Invitrogen, Carlsbad, CA). Затем ДНК визуализировали в агарозном геле с подтверждением значений, полученных в анализе с использованием Pico Green, и с определением качества ДНК.
Пример 2.2. Расщепление и разделение ДНК
Для молекулярной характеристики ДНК девять микрограмм (9 мкг) геномной ДНК из образца ДНК в случае события DAS-40278-9 кукурузы и обычного контроля расщепляли, добавляя к каждому образцу ДНК приблизительно от пяти до одиннадцати единиц выбранного фермента рестрикции на микрограмм ДНК и соответствующий буфер для реакции. Каждый образец инкубировали примерно при 37°C в течение ночи. Для расщепления использовали ферменты рестрикции EcoR I, Nco I, Sac I, Fse I и Hind III (New England Biolabs, Ipswich, MA). Позитивный контрольный образец для гибридизации получали, объединяя плазмидную ДНК, pDAS1740 (pDAB3812), с геномной ДНК из обычного контроля в соотношении, примерно эквивалентном 1 копии трансгена на геном кукурузы, и расщепляли, используя такие же способы и фермент рестрикции, что и для тестируемых образцов. ДНК из обычной контрольной кукурузы (XHH13) расщепляли, используя такие же способы и ферменты рестрикции, что и для тестируемых образцов, чтобы использовать в качестве негативного контроля.
Образцы расщепленной ДНК преципитировали, используя Quick-Precip (Edge BioSystems, Gaithersburg, MD) и ресуспендировали в 1x Blue Juice (Invitrogen, Carlsbad, CA), доводя до объема, требуемого для загрузки на гель. Затем образцы ДНК и маркеры размера молекул подвергали электрофорезу в 0,8% агарозном геле, используя 1x TBE-буфер (Fisher Scientific, Pittsburgh, PA), при 55-65 вольтах в течение приблизительно 18-22 часов, чтобы добиться разделения фрагментов. Гели красили бромидом этидия (Invitrogen, Carlsbad, CA) и ДНК визуализировали в ультрафиолетовом (УФ) свете.
Пример 2.3. Саузерн-перенос и обработка мембран
Саузерн-блот анализ осуществляли, по существу, как описано Memelink с соавторами (1994) Southern, Northern, and Western Blot Analysis. Plant Mol. Biol. Manual F1: 1-23. Кратко, после электрофоретического разделения и визуализации фрагментов ДНК гели подвергали депуринизации, используя 0,25 н. HCl (Fisher Scientific, Pittsburgh, PA), в течение примерно 15 минут и затем подвергали действию денатурирующего раствора (AccuGENE, Sigma, St. Louis, MO) в течение приблизительно 30 минут с последующим действием нейтрализующего раствора (AccuGENE, Sigma, St. Louis, MO) в течение по меньшей мере 30 минут. Саузерн-перенос осуществляли в течение ночи на нейлоновые мембраны (Roche Diagnostics, Indianapolis, IN), используя капиллярную систему с 10×SSC (Sigma, St. Louis, MO). После переноса мембраны промывали в растворе 2x SSC и ДНК связывали с мембраной за счет поперечного сшивания под действием УФ. Такой способ приводил к получению Саузерн-блот-мембран, готовых для гибридизации.
Пример 2.4. Мечение и гибридизация ДНК-зондов
Фрагменты ДНК, связанные с нейлоновой мембраной, выявляли, используя меченый зонд. Зонды, используемые для исследования, создавали с помощью основанного на ПЦР включения меченного дигоксигенином (DIG) нуклеотида, [DIG-11]-dUTP, из фрагментов, образованных праймерами, специфичными к элементам гена и другим областям плазмиды pDAS1740. Создание ДНК-зондов посредством ПЦР-синтеза осуществляли, используя набор для синтеза DIG-зондов в ПЦР (Roche Diagnostics, Indianapolis, IN), следуя способам, рекомендованным производителем. Список зондов, используемых для исследования, приведен в таблице 1.
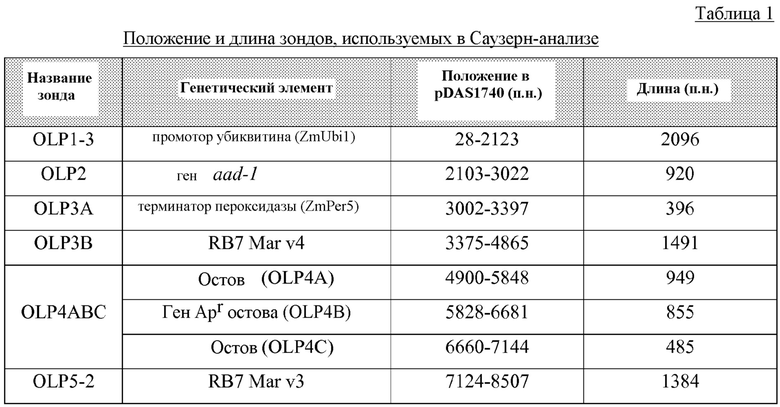
Меченые зонды анализировали, используя электрофорез в агарозном геле, чтобы определить их качество и количество. Затем требуемое количество меченого зонда использовали для гибридизации с ДНК-мишенью на нейлоновых мембранах для выявления специфичных фрагментов способами, описанными для раствора DIG Easy Hyb (Roche Diagnostics, Indianapolis, IN). Кратко, блоты на нейлоновых мембранах с фиксированной на них ДНК кратковременно промывали в 2×SSC и предварительно гибридизовали с 20-25 мл предварительно нагретого раствора DIG Easy Hyb в сосудах для гибридизации примерно при 50°C в течение минимум 30 минут в печи для гибридизации. Затем раствор для предварительной гибридизации сливали и заменяли 20 мл предварительно нагретого раствора DIG Easy Hyb, содержащего требуемое количество специфичных зондов, предварительно денатурированных кипячением в воде в течение 5 минут. Затем осуществляли стадию гибридизации примерно при 40-60°C в течение ночи в печи для гибридизации.
Пример 2.5. Детекция
В конце гибридизации с зондами растворы DIG Easy Hyb, содержащие зонды, сливали в чистые пробирки и хранили при -20°C. Такие зонды могут быть повторно использованы 2-3 раза согласно способу, рекомендованному производителем. Мембранные блоты кратковременно промывали и затем дважды промывали в чистых пластиковых емкостях с использованием буфера для промывки низкой жесткости (2×SSC, 0,1% SDS) в течение примерно 5 минут при комнатной температуре, затем два раза промывали буфером для промывки высокой жесткости (0,1×SSC, 0,1% SDS), каждый раз в течение 15 минут примерно при 65°C. Затем мембранные блоты переносили в другие чистые пластиковые емкости и кратковременно промывали 1x буфером для промывки из набора буферов для промывки и блокирования DIG (Roche Diagnostics, Indianapolis, IN) в течение примерно 2 минут, затем блокировали в 1 x буфере для блокирования в течение минимум 30 минут с последующей инкубацией с антителом против DIG-AP (щелочная фосфатаза) (разведение 1:5000, Roche Diagnostics, Indianapolis, IN) в 1x буфере для блокирования в течение минимум 30 минут. После 2-3 промывок 1x буфером для промывки специфичные ДНК-зонды, которые оставались связанными с мембранными блотами, и DIG-меченые стандарты ДНК визуализировали, используя систему выявления нуклеиновых кислот на основе хемилюминесценции CDP-Star (Roche Diagnostics, Indianapolis, IN), следуя рекомендациям производителя. Блоты экспонировали с пленкой для регистрации хемилюминесценции (Roche Diagnostics, Indianapolis, IN) один или несколько периодов времени (временных точек), чтобы определить гибридизующиеся фрагменты и визуализировать стандарты молекулярного размера. Затем пленки проявляли, используя проявитель All-Pro 100 Plus (Konica SRX-101), и изображения сканировали для сообщения данных. Количество и размеры выявленных полос документировали для каждого зонда. DIG-меченный маркер молекулярной массы II (MWM DIG II), видимый после выявления DIG, как описано, использовали для определения размеров гибридизующихся фрагментов на Саузерн-блотах.
Пример 2.6. Снятие зондов
ДНК-зонды снимали с мембранных блотов после получения данных о Сайзерн-гибридизации, и мембранные блоты можно было повторно использовать для гибридизации с другим ДНК-зондом способами, рекомендованными производителем (DIG Application Manual for Filter Hybridization (2003). Roche Diagnostics). Кратко, после детекции сигнала и экспонирования с пленкой мембранные блоты тщательно промывали водой Milli-Q и затем два раза промывали в буфере для снятия зондов (0,2 н. NaOH, 0,1% SDS) в течение примерно 15 минут при комнатной температуре или при 37°C. Затем мембранные блоты кратковременно промывали в 2×SSC, после чего они были готовы для предварительной гибридизации и гибридизации с другим ДНК-зондом. Мембранные блоты экспонировали с новой пленкой для регистрации хемилюминесценции, чтобы убедиться, что все ДНК-зонды были сняты, перед тем как приступить к следующей гибридизации. Пленки после повторного экспонирования хранили вместе с пакетом данных о предыдущей гибридизации в файле исследования для отчета.
Пример 2.7. Результаты Саузерн-блоттинга
Ожидаемые и наблюдаемые размеры фрагментов в случае конкретного расщепления и зонда, основанных на сайтах для известных ферментов рестрикции pDAS 1740/Fsp I-фрагмента, приведены в таблице 2. В результате указанных расщеплений и гибридизации идентифицировали два типа фрагментов: внутренние фрагменты, в случае которых известные сайты рестрикции фланкировали область зонда и которые полностью находились в пределах pDAS 1740/Fsp I-фрагмента, и пограничные фрагменты, в случае которых сайт для известного фермента локализован на одном конце области зонда, а второй сайт предположительно локализован в геноме кукурузы. Размеры пограничных фрагментов варьировали в зависимости от события, поскольку в большинстве случаев сайты интеграции ДНК-фрагментов являются уникальными для каждого события. Пограничные фрагменты обеспечивают средство для локализации сайта фермента рестрикции относительно интегрированной ДНК и для оценки количества ДНК-инсерций. На основании Саузерн-блот-анализов, проведенных в настоящем исследовании, был сделан вывод о том, что единственная копия интактной PTU aad-1 из плазмиды pDAS 1740/Fsp I встраивалась в геном кукурузы в случае события DAS-40278-9, как подробно указано на карте вставки (фигуры 2, 3).
~6300
~16000
~6300
~16000
# Одинаковые значения в обычном контроле, BC3S1, и в некоторых образцах BC3S2.
1 Ожидаемые размеры фрагментов основаны на карте плазмиды pDAS 1740 (pDAB3812), которая показана на фигуре 1.
2 Наблюдаемые размеры фрагментов оценивали приблизительно на основании таких анализов, и они основаны на указанных размерах фрагментов DIG-меченного маркера молекулярной массы ДНК II. Вследствие включения DIG-молекул для визуализации маркерные фрагменты в эксперименте обычно были на 5-10% больше, чем их фактически указанная молекулярная масса.
Ферменты рестрикции, имеющие уникальный сайт рестрикции в плазмиде pDAS1740, EcoR I, Nco I, Sac I, Fse I/Hind III, были выбраны для характеристики вставки гена aad-1 в случае события DAS-40278-9. Предполагали, что пограничный фрагмент длиной >3382 п.н., >2764 п.н., >4389 п.н. гибридизуется с зондом гена aad-1 после расщепления EcoR I, Nco I и Sac I, соответственно (таблица 2). Наблюдали одну полосу гибридизации aad-1 длиной ~12000 п.н., ~4000 п.н. и ~16000 п.н. при использовании EcoR I, Nco I и Sac I, соответственно, что свидетельствует о наличии единственного сайта инсерции гена aad-1 в геноме кукурузы в случае события DAS-40278-9. Двойное расщепление с использованием Fse I и Hind III было выбрано для высвобождения фрагмента длиной 3361 п.н., который содержит растительную транскрипционную единицу aad-1 (PTU, промотор/ген/терминатор) (таблица 2). Предсказанный фрагмент длиной 3361 п.н. наблюдали при использовании зонда к гену aad-1 после расщепления Fse I/Hind III. Результаты, полученные при расщеплении всеми четырьмя ферментами/сочетанием ферментов образца DAS-40278-9 с последующей гибридизацией с зондом к гену aad-1, показали, что одна копия интактной PTU aad-1 из плазмиды pDAS1740 была встроена в геном кукурузы в случае события DAS-40278-9.
Ферменты рестрикции Nco I, Sac I и Fse I/Hind III отбирали для характеристики области промотора (ZmUbi1) для aad-1 в случае события DAS-40278-9. Расщепления Nco I и Sac I предположительно образуют фрагменты пограничной области длиной >3472 п.н. и >4389 п.н., соответственно, наблюдаемые при гибридизации с ДНК-зондами, специфичными к области промотора ZmUbi1 (таблица 2). Две полосы гибридизации ~6300 п.н. и ~3600 п.н. выявляли с использованием зонда к промотору ZmUbi1 после расщепления Nco I. Однако полоса ~3600 п.н. присутствовала на всех дорожках с образцами, включая обычные контроли, что свидетельствует о том, что полоса длиной ~3600 п.н. является полосой неспецифичного сигнала, возникающего в результате гомологичного связывания зонда к промотору убиквитина (ZmUbi1), полученному из кукурузы, с эндогенным геном ubi кукурузы. Напротив, полосу сигнала ~6300 п.н. выявляли в тестируемых образцах DAS-40278-9, но не выявляли в обычных контролях, что свидетельствует о том, что полоса ~6300 п.н. является специфичной для зонда промотора ZmUbi1 из плазмиды pDAS1740 и, следовательно, является ожидаемой полосой Nco I/ZmUbi1, указанной в таблице 2. Подобным образом выявляли две полосы гибридизации длиной ~3800 п.н. и ~16000 п.н. с использованием зонда промотора ZmUbi1 после расщепления Sac I. Полосу ~3800 п.н. выявляли на всех дорожках с образцами, включая обычные контроли, и поэтому считали, что она является результатом неспецифичной гибридизации зонда к промотору ZmUbi1 с эндогенным геном ubi кукурузы. Полосу гибридизации ~16000 п.н., которая присутствует только в образцах DAS-40278-9, расценивают как ожидаемую полосу Sac I/ZmUbi1. Предполагается, что двойное расщепление с использованием Fse I/Hind III высвобождает фрагмент PTU aad-1 длиной 3361 п.н., который гибридизуется с зондом промотора ZmUbi1 (таблица 2). Такую полосу 3361 п.н. и полосу неспецифичной гибридизации длиной ~6400 п.н. выявляли с использованием зонда к промотору ZmUbi1 после расщепления Fse I/Hind III. Полосу длиной ~6400 п.н. считают результатом неспецифичного связывания зонда к промотору ZmUbi1 с эндогенным геном ubi кукурузы, так как такая полоса присутствует на всех дорожках с образцами, включая обычные контроли. Кроме того, наблюдали другую полосу, очень близкую к полосе ~6400 п.н., в обычном контроле, BC3S1, и в некоторых из образцов BC3S2. Такую дополнительную полосу, очень близкую к ~6400 п.н., также считают неспецифичной, поскольку она присутствует на дорожках с обычными контрольными образцами XHH13 и наиболее вероятно ассоциирована с генетическим фоном XHH13.
Такие же ферменты рестрикции/сочетание ферментов, Nco I, Sac I и Fse I/Hind III, выбирали для характеристики области терминатора (ZmPer5) для aad-1 в случае события DAS-40278-9. Предполагают, что расщепление Nco I образует фрагмент пограничной области длиной >2764 п.н., выявляемый при гибридизации с ДНК-зондами, специфичными по отношению к области терминатора ZmPer5 (таблица 2). Две полосы гибридизации ~4000 п.н. и ~3900 п.н. выявляли с использованием зонда к терминатору ZmPer5 после расщепления Nco I. Полоса ~3900 п.н. присутствовала на всех дорожках с образцами, включая обычные контроли, что свидетельствует о том, что полоса длиной ~3900 п.н. является полосой неспецифичного сигнала, вероятно, вследствие гомологичного связывания зонда к терминатору гена пероксидазы, полученному из кукурузы (ZmPer5), с эндогенным геном per кукурузы. Напротив, сигнальную полосу ~4000 п.н. выявляли в тестируемых образцах DAS-40278-9, но не выявляли в обычных контролях, что свидетельствует о том, что полоса длиной ~4000 п.н. является специфичной для зонда к терминатору ZmPer5 из плазмиды pDAS 1740, и поэтому она является ожидаемой полосой Nco I/ZmPer5, указанной в таблице 2. Предполагали, что пограничный фрагмент длиной >1847 п.н. гибридизуется с зондом к терминатору ZmPer5 после расщепления Sac I. Две полосы гибридизации длиной ~1900 п.н. и ~9000 п.н. выявляли с использованием зонда к терминатору ZmPer5 после расщепления Sac I. Полосу длиной ~9000 п.н. выявляли на всех дорожках с образцами, включая обычные контроли, и поэтому расценивали как неспецифичную гибридизацию зонда к терминатору ZmPer5 с эндогенным геном per кукурузы. Полосу гибридизации ~1900 п.н., которая присутствует только в образцах DAS-40278-9, расценивают как ожидаемую полосу Sac I/ZmPer5. Двойное расщепление с использованием Fse I/Hind III предположительно высвобождает фрагмент PTU aad-1 длиной 3361 п.н., который гибридизуется с зондом к терминатору ZmPer5 (таблица 2). Такую полосу 3361 п.н. и дополнительную полосу неспецифичной гибридизации длиной ~2100 п.н. выявляли с использованием зонда к терминатору ZmPer5 после расщепления Fse I/Hind III. Дополнительная полоса ~2100 п.н. является результатом неспецифичного связывания зонда к терминатору ZmPer5 с эндогенным геном кукурузы, так как такая полоса присутствует на всех дорожках с образцами, включая негативные контроли. Результаты, полученные с использованием таких расщеплений образца DAS-40278-9 с последующей гибридизацией с зондами к промотору ZmUbi1 и терминатору ZmPer5, дополнительно подтвердили, что одна копия интактной PTU aad-1 из плазмиды pDAS1740 встроена в геном кукурузы в случае события DAS-40278-9.
Ферменты рестрикции, Nco I и Sac I, выбирали для характеристики остальных компонентов из pDAS1740/Fsp I-фрагмента в случае события AAD-1 DAS-40278-9 кукурузы (таблица 2). Последовательности ДНК компонентов RB7 Mar v3 и RB7 Mar v4 обладают более 99,7% идентичностью, поэтому ожидали, что ДНК-зонды, специфичные для RB7 Mar v3 или RB7 Mar v4, гибридизуются с ДНК-фрагментами, содержащими любой из вариантов RB7 Mar. Предполагали, что два пограничных фрагмента длиной >2764 п.н. и >3472 п.н. гибридизуются с зондами к RB7 Mar v4 и RB7 Mar v3 после расщепления Nco I (таблица 2). Две полосы гибридизации ~4000 п.н. и ~6300 п.н. наблюдали с использованием зонда либо к RB7 Mar v4, либо к RB7 Mar v3 после расщепления Nco I в образцах DAS-40278-9. Подобным образом предполагали два пограничных фрагмента длиной >1847 п.н. и >4389 п.н. в случае использования зондов RB7 Mar v4 и RB7 Mar v3 после расщепления Sac I (таблица 2). Полосы гибридизации длиной ~1900 п.н. и ~16000 п.н. выявляли в образцах DAS-40278-9 с использованием зонда RB7 Mar v4 или RB7 Mar v3 после расщепления Sac I.
Вместе взятые результаты Саузерн-гибридизации, полученные с использованием таких зондов к элементам, показывают, что ДНК, встроенная в случае события DAS-40278-9 кукурузы, содержит интактную PTU aad-1 вместе с участками прикрепления к матриксу RB7 Mar v3 и RB7 Mar v4 на 5’- и 3’-концах вставки, соответственно.
Пример 2.8. Отсутствие последовательностей остова
Комбинацию трех ДНК-фрагментов в эквимолярном соотношении (таблица 1), охватывающую почти полную Fsp I-область остова (4867-7143 п.н. в плазмиде pDAS1740) плазмиды pDAS1740, использовали в качестве зонда к остову, чтобы охарактеризовать AAD-1-событие у кукурузы DAS-40278-9. Для получения события DAS-40278-9 использовали плазмидный pDAS 1740/Fsp I-фрагмент, поэтому не ожидали специфичной гибридизации с комбинацией зондов к остову (таблица 2) после расщепления любым ферментом рестрикции. Было подтверждено, что сигнал специфичной гибридизации не регистрируется при использовании зонда к остову после расщепления Nco I или Sac I ни в одном из образцов DAS-40278-9. Дорожки с позитивным контролем содержали ожидаемые гибридизующиеся полосы, что свидетельствует о том, что зонды способы гибридизоваться с любами гомологичными ДНК-фрагментами, если они присутствуют в образцах. Данные свидетельствуют о том, что инсерция в случае события DAS-40278-9 кукурузы не содержит никакой последовательности векторного остова вне Fsp I-области из плазмиды pDAS1740.
Образцы листьев из пяти разных поколений для события DAS-40278-9 использовали для проведения Саузерн-блот-анализа с целью молекулярной характеристики. Картину интеграции исследовали, используя переваривание выбранным ферментом рестрикции и комбинации зондов, для характеристики встроенного гена, aad-1, а также некодирующих областей, включая промотор, терминатор экспрессии гена и участки связывания с матриксом.
Саузерн-блот-характеристика ДНК, встроенной в случае события DAS-40278-9, показывает, что единственная интактная копия PTU aad-1 была интегрирована при событии DAS-40278-9. Молекулярные массы, показанные при использовании Саузерн-гибридизации в случае сочетания фермента рестрикции и зонда, были уникальными для события и формировали картины для его идентификации. Картины гибридизации являются идентичными на протяжении всех пяти поколений, что свидетельствует о том, что вставка является стабильной в геноме кукурузы. Гибридизация с зондами, охватывающими область остова за пределами pDAS 1740/Fsp I-фрагмента для трансформации из плазмиды pDAS 1740, подтверждает, что последовательности векторного остова не были включены в событие DAS-40278-9.
Пример 3. Клонирование и характеристика последовательности ДНК вставки и фланкирующих пограничных областей в случае события DAS-40278-9 кукурузы
Чтобы охарактеризовать встроенную ДНК и описать геномный сайт инсерции, определяли последовательности ДНК вставки и пограничных областей в случае события DAS-40278-9. Все было подтверждено 8557 п.н. геномной последовательности в случае события DAS-40278-9, содержащей 1873 п.н. 5’-фланкирующей пограничной последовательности, 1868 п.н. 3’-фланкирующей пограничной последовательности и 4816 п.н. ДНК-вставки. ДНК-вставка длиной 4816 п.н. содержит интактную кассету экспрессии aad-1, неполный MAR v3 длиной 259 п.н. на 5’-конце и неполный MAR v4 длиной 1096 п.н. на 3’-конце. Анализ последовательностей выявил инсерцию 21 п.н. на 5’-участке соединения при интеграции и делецию двух пар нуклеотидов из локуса инсерции генома кукурузы. Инсерция одной пары нуклеотидов обнаружена на 3’-участке соединения при интеграции между геномом кукурузы и вставкой DAS-40278-9. Также обнаружено изменение одного основания (T на C) во вставке в положении 5212 в некодирующей области 3’-UTR. Ни одно из таких изменений не влияло на состав открытой рамки считывания кассеты экспрессии aad-1.
ПЦР-амплификация на основе последовательностей вставки и пограничных последовательностей события DAS-40278-9 подтвердила, что пограничные области происходили из кукурузы, и что области соединения можно использовать для специфичной для события идентификации DAS-40278-9. Анализ последовательности, охватывающей области соединения, показал, что новых открытых рамок считывания (ORF>=200 кодонов) не возникало в результате инсерции ДНК в случае события DAS-40278-9, а также ни одна из геномных открытых рамок считывания не была прервана при интеграции DAS-40278-9 в нативный геном кукурузы. В целом, характеристика последовательностей вставки и пограничных последовательностей AAD-1-события кукурузы DAS-40278-9 показала, что только одна интактная копия кассеты экспрессии aad-1 была интегрирована в нативный геном кукурузы.
Пример 3.1. Экстракция и количественная оценка геномной ДНК
Геномную ДНК экстрагировали из лиофилизированных или свежеприготовленных измельченных тканей листьев, применяя модифицированный способ CTAB. Образцы ДНК растворяли в 1×TE (10 мМ трис, pH 8,0, 1 мМ EDTA) (Fluka, Sigma, St. Louis, MO) и количественно оценивали способом Pico Green согласно инструкциям производителя (Molecular Probes, Eugene, OR). Для ПЦР-анализа образцы ДНК разбавляли водой специальной очистки для молекулярной биологии (5 PRIME, Gaithersburg, MD), чтобы получить в результате концентрацию 10-100 нг/мкл.
Пример 3.2. ПЦР-праймеры
В таблице 3 приведен список последовательностей праймеров, которые использовали для клонирования вставки ДНК и фланкирующих пограничных областей события DAS-40278-9, причем положения и описания представлены на фигуре 4. В таблице 4 приведен список последовательностей праймеров, которые использовали для подтверждения последовательности вставки и пограничных последовательностей. Положения праймера отмечены на фигурах 4 и 5, соответственно. Все праймеры синтезировали в Integrated DNA Technologies, Inc. (Coralville, IA). Праймеры растворяли в воде (5 PRIME, Gaithersburg, MD) до концентрации 100 мкМ для получения маточного раствора и разбавляли водой до концентрации 10 мкМ для получения рабочего раствора.
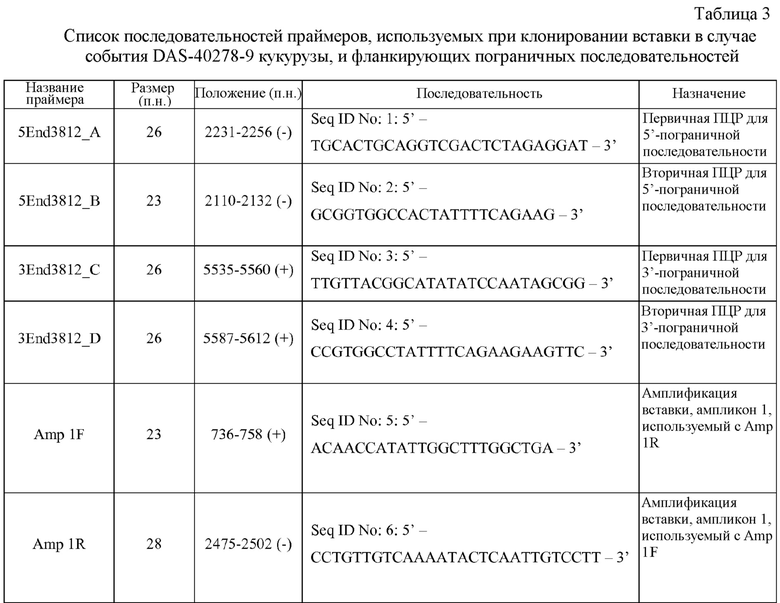




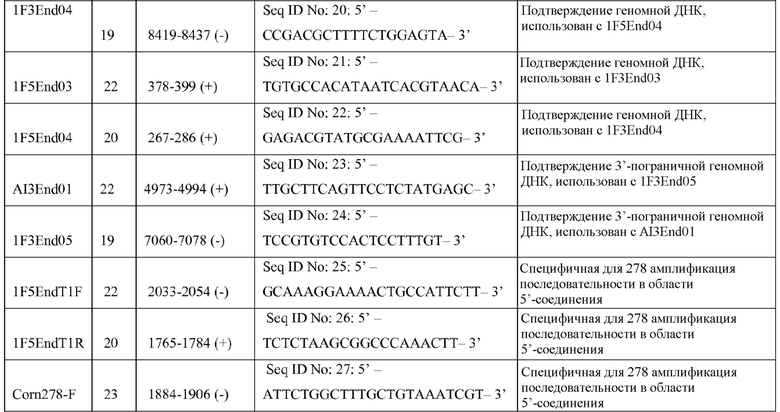

Пример 3.3. Прогулка по геному
Универсальный набор для прогулки по геному (Clontech Laboratories, Inc., Mountain View, CA) использовали для клонирования 5’- и 3’-фланкирующих пограничных последовательностей события DAS-40278-9 кукурузы. Согласно инструкции производителя примерно 2,5 мкг геномной ДНК события AAD-1 кукурузы DAS-40278-9 расщепляли в течение ночи, используя EcoR V, Stu I (оба фермента поставляются в наборе) или Sca I (New England Biolabs, Ipswich, MA). Расщепленную ДНК очищали, используя DNA Clean and ConcentratorTM-25 (ZYMO Research, Orange, CA), с последующим лигированием с адаптерами Genome WalkerTM, чтобы сконструировать библиотеки Genome WalkerTM. Каждую библиотеку Genome WalkerTM использовали в качестве ДНК-матрицы для первичной ПЦР-амплификации с адаптерным праймером AP1, поставляемым в наборе, и каждым специфичным для конструкции праймером 5End3812_A и 3End3812_C. Затем один микролитр после разбавления 1:25 смеси для первичной ПЦР-реакции использовали в качестве матрицы для вторичной ПЦР-амплификации с гнездовым адаптерным праймером AP2 и каждым гнездовым специфичным для конструкции праймером 5End3812_B и 3End3812_D. В ПЦР-амплификации использовали TaKaRa LA TaqTM HS (Takara Bio Inc., Shiga, Japan). В 50 мкл реакционной смеси для ПЦР использовали 1 мкл ДНК-матрицы, 8 мкл 2,5 мМ смеси dNTP, 0,2 мкМ каждого праймера, 2,5 единицы ДНК-полимеразы TaKaRa LA TaqTM HS, 5 мкл 10 x буфера для ПЦР LA II (плюс Mg2+) и 1,5 мкл 25 мМ MgCl2. Конкретные условия ПЦР указаны в таблице 5.
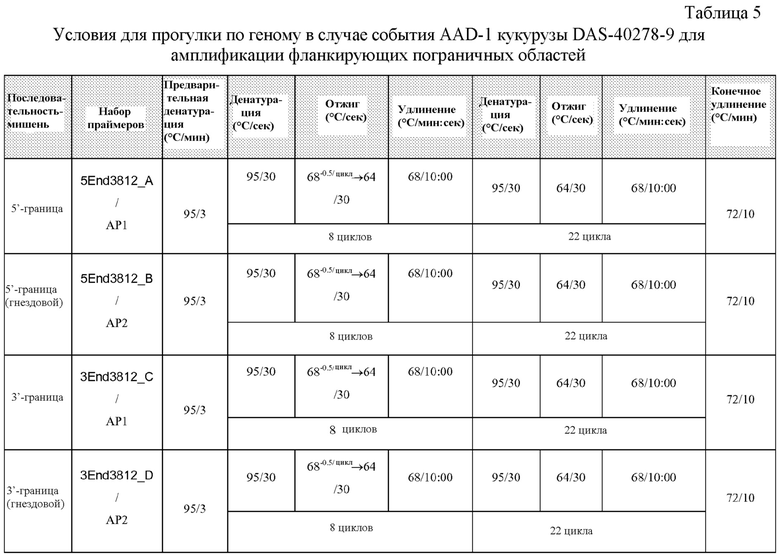
Пример 3.4. Традиционная ПЦР
Стандартную ПЦР использовали для клонирования и подтверждения ДНК-вставки и пограничной последовательности в случае события DAS-40278-9 кукурузы. ДНК-полимеразу TaKaRa LA TaqTM (Takara Bio Inc., Shiga, Japan), HotStarTaq (Qiagen, Valencia, CA), систему для расширенной высокоточной ПЦР (Roche Diagnostics, Inc., Indianapolis, IN) или смесь High-Fidelity PCR Cloning Enzyme and Master Mix Easy-A® (Stratagene, LaJolla, CA) использовали для обычной ПЦР-амплификации согласно способам, рекомендованным производителем. Конкретные условия ПЦР и описания ампликонов приведены в таблице 6А.
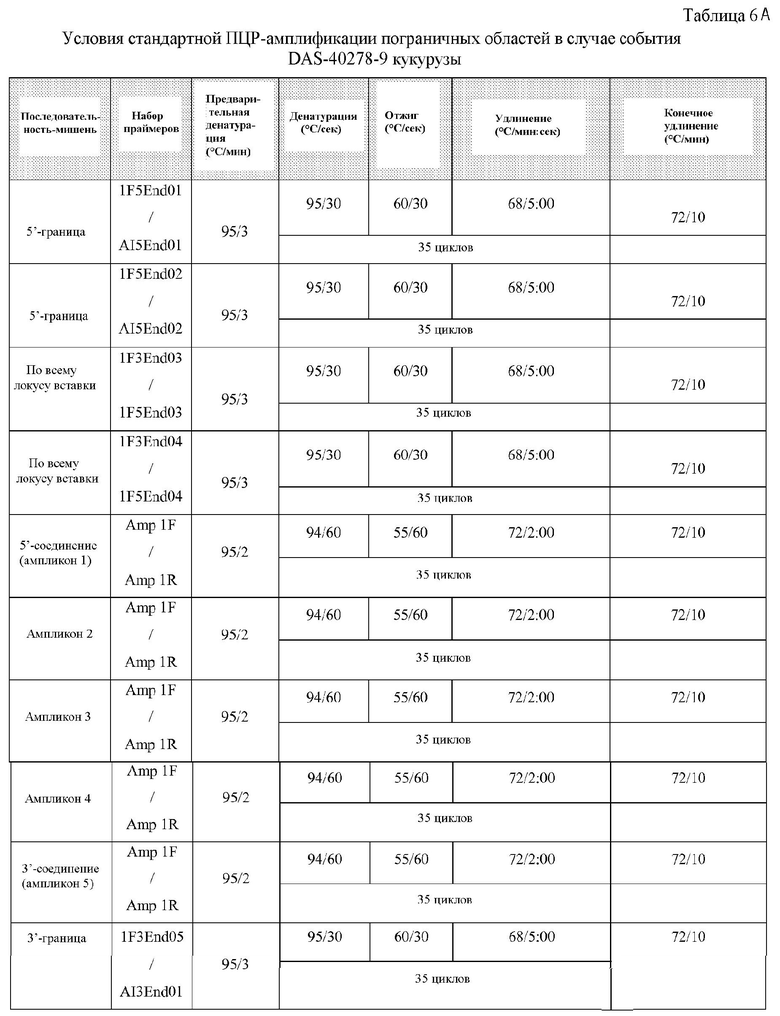
Пример 3.5. Выявление ПЦР-продукта, очистка, субклонирование ПЦР-продуктов и секвенирование
ПЦР-продукты исследовали электрофорезом с использованием 1,2% или 2% E-геля (Invitrogen, Carlsbad, CA) согласно инструкции к продукту. Размер фрагментов оценивали путем сравнения с ДНК-маркерами. При необходимости ПЦР-фрагменты очищали, вырезая фрагменты из 1% агарозного геля в 1x TBE, окрашенного бромидом этидия, используя набор для экстракции из геля QIAquick (Qiagen, Carlsbad, CA).
ПЦР-фрагменты субклонировали в pCR®4-TOPO®, используя набор TOPO TA Cloning® для секвенирования (Invitrogen, Carlsbad, CA) согласно инструкции к продукту. В частности, от двух до пяти микролитров реакционной смеси для клонирования TOPO® использовали для трансформации химически компетентных клеток One Shot TOP 10, следуя инструкции производителя. Клонированные фрагменты подтверждали, получая минипрепараты плазмидной ДНК (набор QIAprep Spin Miniprep, Qiagen, Carlsbad, CA) с последующим рестрикционным расщеплением EcoR I или прямой ПЦР на колониях с использованием праймеров T3 и T7, поставляемых в наборе. Затем плазмидную ДНК или маточные растворы в глицерине выбранных колоний отправляли на секвенирование.
После субклонирования предполагаемые целевые ПЦР-продукты сначала секвенировали, чтобы подтвердить, что были клонированы ожидаемые фрагменты ДНК. Колонии, содержащие соответствующие последовательности ДНК, отбирали для опосредованной праймером прогулки, чтобы определить полные последовательности ДНК. Секвенирование осуществляли в Cogenics (Houston, TX).
Конечную сборку вставки и пограничных последовательностей осуществляли, используя компьютерную программу Sequencher (версия 4.8, Gene Codes Corporation, Ann Arbor, MI). Аннотацию к вставке и пограничным последовательностям события DAS-40278-9 кукурузы готовили, используя Vector NTI (версии 10 и 11, Invitrogen, Carlsbad, CA).
Поиск гомологии осуществляли, используя программу BLAST и базу данных GenBank. Анализ открытых рамок считывания (ORF) с использованием Vector NTI (версия 11, Invitrogen) осуществляли для идентификации ORF (>=200 кодонов) в полных последовательностях вставки и фланкирующих пограничных последовательностях.
Пример 3.6. Пограничная последовательность на 5’-конце
Фрагмент ДНК амплифицировали с каждой библиотеки, полученной при прогулке по геному в случае события DAS-40278-9 у кукурузы, используя набор специфичных гнездовых праймеров для 5’-конца трансгена. ПЦР-продукт длиной примерно 800 п.н. наблюдали в обеих библиотеках, полученных при прогулке по геному в случае события DAS-40278-9 с использованием EcoR V и Stu I. Библиотека, полученная при прогулке по геному с использованием Sca I, давала продукт длиной примерно 2 т.п.н. Фрагменты клонировали в pCR®4-TOPO® и случайным образом отбирали по шесть колоний из каждой библиотеки для окончательного секвенирования, чтобы подтвердить, что вставка содержала ожидаемые последовательности. Полное секвенирование вставок в результате опосредованной праймером прогулки выявило, что фрагменты, амплифицированные с библиотек, полученных при прогулке по геному с использованием Stu I, EcoR V и Sca I в случае события DAS-40278-9 у кукурузы, имели длину 793, 822 и 2132 п.н., соответственно. ДНК-фрагменты, образованные из библиотек, полученных в результате прогулки по геному с использованием Stu I и EcoR V, на 100% совпадали с ДНК-фрагментом, образованным из библиотеки, полученной в результате прогулки по геному с использованием Sca I, что свидетельствует о том, что такие ДНК-фрагменты амплифицированы с 5’-области вставки трансгена. Поиск с использованием BLAST получаемой в результате геномной последовательности кукурузы длиной 1873 п.н. показал высокое сходство с последовательностью клона BAC кукурузы. Кроме того, анализ последовательности соединения инсерции показал, что последовательность длиной 917 п.н. MAR v3 в области 5’-конца укорочена по сравнению с плазмидным pDAS1740/Fsp I-фрагментом, при этом остается 259 п.н. неполного MAR v3 в 5’-области кассеты экспрессии aad-1.
Пример 3.7. Пограничная последовательность на 3’-конце
ДНК-фрагмент размером примерно 3 т.п.н. амплифицировали с библиотеки, полученной в результате прогулки по геному с использованием Stu I в случае события DAS-40278-9 кукурузы, применяя набор гнездовых праймеров, специфичных для 3’-конца трансгена. ДНК-фрагмент клонировали в pCR(R)4-TOPO® и случайным образом отбирали десять колоний для окончательного секвенирования, чтобы подтвердить инсерцию ожидаемых последовательностей. Три клона с ожидаемыми вставками полностью секвенировали, получая ДНК-фрагмент длиной 2997 п.н. Анализ последовательности такого ДНК-фрагмента выявил неполный элемент MAR v4 (пропущено 70 п.н. его 5’-области) и геномную последовательность кукурузы длиной 1867 п.н. Поиск с использованием BLAST показал, что геномная последовательность ДНК длиной 1867 п.н. на 100% совпадала с последовательностью в таком же клоне BAC кукурузы, который идентифицировали с помощью 5’-пограничной последовательности.
Пример 3.8. Последовательности вставки ДНК и соединения
ДНК-вставку и области соединения клонировали для случая события DAS-40278-9 кукурузы, используя основанные на ПЦР способы, которые описаны ранее. Конструировали пять пар праймеров на основе 5’- и 3’-фланкирующих пограничных последовательностей и ожидаемой последовательности трансгена. Всего клонировали и секвенировали пять перекрывающихся ДНК-фрагментов (ампликон 1 длиной 1767 п.н., ампликон 2 длиной 1703 п.н., ампликон 3 длиной 1700 п.н., ампликон 4 длиной 1984 п.н. и ампликон 5 длиной 1484 п.н.) (фигура 4). Полные последовательности вставки и фланкирующих границ собирали на основе перекрывающейся последовательности пяти фрагментов. Конечная последовательность подтверждает присутствие 4816 п.н. вставки ДНК, полученной из pDAS 1740/Fsp I, 1873 п.н. 5’-фланкирующей пограничной последовательности и 1868 п.н. 3’-фланкирующей пограничной последовательности. ДНК-вставка длиной 4816 п.н. содержит интактную кассету экспрессии aad-1, 259 п.н. неполного MAR v3 на 5’-конце и 1096 п.н. неполного MAR v4 на 3’-конце (SEQ ID NO: 29).
По меньшей мере два клона для каждой пары праймеров использовали для опосредованной праймером прогулки, чтобы получить полную информацию о последовательности ДНК-вставки и пограничных с ней последовательностей. Анализ последовательностей показал наличие инсерции 21 п.н. в области соединения с 5’-стороны от места интеграции между геномной ДНК и интегрированным неполным MAR v3 из pDAS1740/Fsp I. Результаты поиска с использованием BLAST и анализа с использованием Vector NTI показали, что ДНК вставки длиной 21 п.н. не проявляет гомологии ни с ДНК какого-либо вида растения, ни с плазмидной ДНК pDAS1740. Инсерция одной пары нуклеотидов была обнаружена в области соединения с 3’-стороны от места интеграции между геномной ДНК кукурузы и неполным MAR v4 из pDAS1740/Fsp I. Интеграция ДНК также приводила к делеции двух нуклеотидов в локусе инсерции генома кукурузы (фигура 6). Кроме того, различие в одном нуклеотиде (T на C) в положении 5212 п.н. наблюдали в нетранслируемой 3’-UTR-области вставки ДНК (SEQ ID NO: 29). Однако ни одно из таких изменений, по-видимому, не является важным для экспрессии aad-1 или создания каких-либо новых ORF (>=200 кодонов) на протяжении областей соединения вставки DAS-40278-9.
Пример 3.9. Подтверждение геномных последовательностей кукурузы
Чтобы подтвердить сайт инсерции трансгена в случае события DAS-40278-9 в геноме кукурузы, осуществляли ПЦР-амплификацию с разными парами праймеров (фигура 4). Геномную ДНК, соответствующую событию DAS-40278-9, и геномную ДНК других трансгенных или нетрансгенных линий кукурузы использовали в качестве матрицы. Два специфичных для aad-1 праймера, AI5End01 и AI5End02, и два праймера, сконструированных в соответствии с 5’-концевой пограничной последовательностью, 1F5End01 и 1F5End02, использовали для амплификации ДНК-фрагмента, охватывающего область гена aad-1 до 5’-концевой пограничной последовательности. Подобным образом, чтобы амплифицировать ДНК-фрагмент, охватывающий область гена aad-1 до 3’-концевой пограничной последовательности, использовали праймер 1F3End05, полученный из 3’-концевой пограничной последовательности, и специфичный для aad-1 праймер AI3End01. ДНК-фрагменты ожидаемых размеров амплифицировали только с геномной ДНК в случае события AAD-1 DAS-40278-9, при этом каждая пара праймеров состояла из одного праймера, локализованного во фланкирующей границе события AAD-1 DAS-40278-9 кукурузы, и одного специфичного для aad-1 праймера. Контрольные образцы ДНК не давали ПЦР-продуктов с такими же парами праймеров, что свидетельствует о том, что клонированные 5’- и 3’-концевые пограничные последовательности действительно представляют собой последовательности, вышележащие и нижележащие от встроенной конструкции гена aad-1, соответственно. Следует отметить, что слабую полосу размером примерно 8 т.п.н. наблюдали во всех образцах кукурузы, включая событие AAD-1 кукурузы DAS-40278-9, событие AAD-1 кукурузы DAS-40474 и нетрансгенную линию кукурузы XHH13, когда для ПЦР-амплификации использовали пару праймеров 1F5End01 и AI5End01. Наблюдаемая слабая полоса (на полученном геле) могла быть результатом неспецифической амплификации в геноме кукурузы при использовании такой пары праймеров.
Чтобы дополнительно подтвердить инсерцию ДНК в геноме кукурузы, использовали два праймера, локализованных в 5’-концевой пограничной последовательности, 1F5End03 и 1F5End04, и два праймера, локализованных в 3’-концевой пограничной последовательности, 1F3End03 и 1F3End04, чтобы амплифицировать ДНК-фрагменты, охватывающие локус инсерции. ПЦР-амплификация либо с парой праймеров 1F5End03/1F3End03, либо с парой праймеров 1F5End04/1F3End04 приводила к получению фрагмента ожидаемого размера примерно 8 т.п.н. из геномной ДНК в случае события AAD-1 кукурузы DAS-40278-9. Напротив, не получали ПЦР-продуктов с геномной ДНК в случае события AAD-1 кукурузы DAS-40474-7 или в случае нетрансгенной линии кукурузы XHH13. С учетом того, что событие AAD-1 кукурузы DAS-40278-9 и событие DAS-40474-7 являются результатом трансформации HiII с последующим возвратным скрещиванием исходных линий с трансгенными событиями с линией кукурузы XHH13, большая часть генома в случае каждого из указанных двух событий теоретически происходит из линии кукурузы XHH13. Весьма вероятно, что только фланкирующие пограничные последовательности вблизи трансгена aad-1 перенесены из исходной геномной ДНК и сохранились в ходе процесса интрогрессии события AAD-1, тогда как другие области геномных последовательностей могли быть заменены геномными последовательностями XHH13. Таким образом, не является неожиданным, что ни одного фрагмента не было амплифицировано с геномной ДНК в случае события AAD-1 кукурузы DAS-40474-7 и XHH13 при использовании либо пары праймеров 1F5End03/1F3End03, либо пары праймеров 1F5End04/1F3End04. Фрагменты длиной примерно 3,1 и 3,3 т.п.н. амплифицировали с использованием пары праймеров 1F5End03/1F3End03 и 1F5End04/1F3End04, соответственно, с геномной ДНК линий кукурузы HiII и B73, но не линии кукурузы A188. Результаты показывают, что пограничные последовательности происходили из генома линии кукурузы B73.
Дополнительное клонирование геномной ДНК кукурузы из B73/HiII осуществляли, чтобы гарантировать достоверность фланкирующих пограничных последовательностей. Амплифицированные в ПЦР фрагменты секвенировали, чтобы подтвердить данные об области вставки ДНК, интегрированной в конкретное положение геномной ДНК B73/HiII. Праймеры конструировали на основе полученной последовательности. Набор праймеров Amp 1F/Amp 5R использовали для амплификации фрагмента длиной 2212 п.н., охватывающего область от 5’- до 3’-соединений из нативного генома B73/HiII без ДНК вставки. Анализ последовательности показал, что существует делеция двух пар нуклеотидов из нативного генома B73 в локусе инсерции трансгена. Анализ последовательностей ДНК из клонированного фрагмента нативного генома B73 выявил одну ORF (>=200 кодонов), расположенную ниже 3’-области соединения при интеграции. Кроме того, нет других ORF на всем протяжении исходного локуса, в который происходила интеграция в случае события AAD-1 кукурузы DAS-40278-9. Поиск с использованием BLAST также подтвердил, что обе 5’-концевая и 3’-концевая пограничные последовательности, имеющиеся в случае события DAS40278-9, локализованы рядом в таком же клоне BAC кукурузы.
Учитывая уникальность области 5’-соединения при интеграции события AAD-1 кукурузы DAS-40278-9, конструировали две пары специфичных ПЦР-праймеров, 1F5EndT1F/1F5EndT1R и Corn278-F/Corn278-R, чтобы амплифицировать такое соединение вставки с геномного растения. Как предполагали, требуемый ДНК-фрагмент был образован только в геномной ДНК в случае события AAD-1 кукурузы DAS-40278-9, но не в других трансгенных или нетрансгенных линиях кукурузы. Таким образом, указанные две пары праймеров можно использовать в качестве идентификаторов, специфичных для события AAD-1 кукурузы DAS-40278-9.
Пример 4. Характеристика генома с помощью фланкирующих SSR-маркеров DAS-40278-9
Чтобы охарактеризовать и описать геномный сайт инсерции, определяли маркерные последовательности, локализованные вблизи вставки. Панель полиморфных SSR-маркеров использовали для идентификации и картирования положения трансгена. Событие pDAS 1740-278 локализовано в хромосоме 2 в положении примерно 20 сМ между SSR-маркерами UMC1265 и MMC0111 примерно в положении 20 сМ на карте сцепления кукурузы 2008 DAS. В таблице 6 суммирована информация о праймерах для указанных двух маркеров, которые обнаружены в непосредственной близости к трансгену pDAS1740-278.

Пример 4.1. Выделение гДНК
гДНК экстрагировали из вырезанных пробиванием ткани образцов листьев, используя набор для тестирования растений DNEasy 96 (Qiagen, Valencia, California). Были осуществлены модификации протокола для автоматизации. Выделенную гДНК количественно оценивали, используя краситель PicoGreen® из Molecular Probes, Inc. (Eugene, OR). Концентрацию гДНК доводили до 5 нг/мкл разбавлением всех образцов с использованием стерильной деионизованной воды.
Пример 4.2. Скрининг гДНК с использованием маркеров
Разбавленную гДНК генотипировали, используя подгруппу маркеров в виде простых повторяющихся последовательностей (SSR). SSR-маркеры синтезировали в Applied Biosystems (Foster City, California) с использованием прямых праймеров, меченных флуоресцирующими метками 6-FAM, HEX/VIC или NED (голубой, зеленый и желтый, соответственно). Маркеры делили на группы или панели на основе их флуоресцирующей метки и размера ампликона, чтобы облегчить мультиплексирование и анализ после ПЦР.
ПЦР осуществляли в 384-луночных планшетах для анализа, при этом каждая реакционная смесь содержала 5 нг геномной ДНК, 1,25X буфер для ПЦР (Qiagen, Valencia, California), 0,20 мкМ каждого прямого и обратного праймера, 1,25 мМ MgCl2, 0,015 мМ каждого из dNTP и 0,3 единицы ДНК-полимеразы HotStart Taq (Qiagen, Valencia, California). Амплификацию осуществляли в системе ПЦР GeneAmp 9700 с двойным 384-луночным модулем (Applied Biosystems, Foster City, California). Использовали следующую программу амплификации: (1) начальная активация Taq при 95°C в течение 12 минут; (2) 30 секунд при 94°C; (3) 30 секунд при 55°C; (4) 30 секунд при 72°C; (5) повторение стадий 2-4 на протяжении 40 циклов; и (6) 30-минутное конечное удлинение при 72°C. Продукты ПЦР для каждой панели SSR-маркеров мультиплексировали вместе, добавляя по 2 мкл каждого продукта ПЦР одного и того же растения к стерильной деионизованной воде до общего объема 60 мкл. Из мультиплексированных продуктов ПЦР 0,5 мкл вносили в 384-луночные планшеты для загрузки, содержащие 5 мкл буфера для загрузки, состоящего из смеси в соотношении 1:100 стандарта размера в парах нуклеотидов GeneScan 500 LIZ и формамида ABI HiDi (Applied Biosystems, Foster City, California). Затем образцы загружали в анализатор ДНК ABI Prism 3730x1 (Applied Biosystems, Foster City, California) для капиллярного электрофореза, используя рекомендации производителя с использованием общего рабочего времени 36 минут. Данные о маркерах собирали, используя компьютерную программу для автоматизированного сбора данных о последовательностях ABI Prism 3730x1, версии 4.0, и извлекали посредством компьютерной программы GeneMapper 4.0 (Applied Biosystems) для характеристики аллелей и определения размеров фрагментов.
Пример 4.3. Результаты анализа SSR-маркеров
Данные о праймерах для фланкирующих маркеров, которые идентифицированы в непосредственной близости с трансгеном, приведены в таблице 6. Два наиболее близко ассоциированных маркера, UMC1265 и MMC0111, локализованы примерно на расстоянии 20 сМ от вставки трансгена в хромосоме 2.
Пример 5. Характеристика белка aad-1 в случае события DAS-40278-9
Характеризовали биохимические свойства рекомбинантного белка aad-1, полученного из трансгенной кукурузы, в которой имеет место событие DAS-40278-9. Использовали электрофорез в полиакриламидном геле с додецилсульфатом натрия (SDS-ПААГ, окрашенный Кумасси синим, и способы выявления гликопротеидов), Вестерн-блот, анализы на иммунодиагностических тест-полосках, времяпролетную масс-спектрометрию с лазерной десорбцией/ионизацией при содействии матрицы (MALDI-TOF MS) и анализ на основе секвенирования белков с использованием тандемной МС для характеристики биохимических свойств белка.
Пример 5.1. Анализ на иммунодиагностических полосках
Присутствие белка aad-1 в ткани листа DAS-40278-9 подтверждали, используя коммерчески полученные иммунодиагностические тест-полоски из American Bionostica. Полоски позволяли различать трансгенные и нетрансгенные растения при тестировании неочищенных экстрактов листьев (данные не показаны). Нетрансгенные экстракты (XHH13) не содержали регистрируемых количеств иммунореактивного белка. Такой результат также был подтвержден в Вестерн-блот-анализе.
Чтобы тестировать экспрессию белка aad-1, осуществляли анализ на иммунодиагностических полосках. Четыре вырезанных пробиванием образца листьев собирали от каждого растения XHH13 (контрольное растение) и растения с событием DAS-40278-9, зажимая ткань крышками в виде защелкивающегося колпачка отдельных помеченных микроцентрифужных пробирок объемом 1,5 мл. После доставки в лабораторию в каждую пробирку добавляли 0,5 мл буфера для экстракции aad-1 (American Bionostica, Swedesboro, NJ), и ткань гомогенизировали, используя одноразовый пестик, с последующим встряхиванием образца в течение ~10 секунд. После гомогенизации в пробирку помещали тест-полоску и обеспечивали возможность проявления в течение ~5 минут. Присутствие или отсутствие белка aad-1 в экстракте растения подтверждали на основе появления (или отсутствия) тестовой линии на иммунодиагностической полоске. После того, как экспрессия белка aad-1 была подтверждена для трансгенного события, собирали ткань стебля кукурузы, лиофилизировали и хранили примерно при -80°C вплоть до использования.
Пример 5.2. Очистка белка aad-1 из кукурузы
Готовили иммунологически очищенный полученный из кукурузы белок aad-1 (молекулярная масса: ~33 кДа) или неочищенные водные экстракты из ткани стебля кукурузы. Все ткани листьев и стеблей собирали и транспортировали в лабораторию следующим образом. Листья срезали с растения ножницами и помещали в тканевые мешки и хранили примерно при -20°C для использования в будущем. Отдельно срезали стебли чуть выше уровня почвы, помещали в тканевые мешки и сразу же замораживали примерно при -80°C в течение ~6 часов. Затем стебли помещали в лиофилизатор на 5 суток, чтобы удалить воду. После полного высушивания тканей их измельчали до получения тонкодисперсного порошка с сухим льдом и хранили примерно при -80°C до того, как они требовались.
Полученный из кукурузы белок aad-1 экстрагировали из лиофилизированной ткани стебля в буфере на основе фосфата (см. компоненты буфера в таблице 7), отвешивая ~30 грамм лиофилизированной ткани в охлажденный стеклянный измельчитель объемом 1000 мл и добавляя 500 мл буфера для экстракции. Ткань измельчали сверху в течение 60 секунд, и растворимые белки собирали центрифугированием образца в течение 20 минут при 30000×g. Осадок повторно экстрагировали, как описано, и надосадки объединяли и фильтровали через 0,45-мкм фильтр. Профильтрованные надосадки загружали примерно при +4°C на иммуноаффинную анти-aad-1-колонку, на которую было конъюгировано моноклональное антитело, полученное в Strategic Biosolution Inc. (мАт 473F1 85.1; очищенное на белке A; партия №: 609.03C-2-4; 6,5 мг/мл (всего ~35,2 мг)) (Windham, ME), конъюгированное с CNBr-активированной сефарозой 4B (GE Healthcare, Piscataway, NJ). Несвязанные белки собирали и колонку тщательно промывали предварительно охлажденным буфером на основе 20 мМ бикарбоната аммония, pH 8,0. Связанные белки элюировали 3,5 М NaSCN (Sigma, St. Louis, MO), 50 мМ трис-буфером (Sigma, St. Louis, MO), pH 8,0. Собирали семь фракций по 5 мл, и фракции с номерами 2→7 диализовали в течение ночи примерно при +4°C против 10 мМ трис-буфера, pH 8,0. Фракции исследовали, используя SDS-ПААГ и Вестерн-блот, и оставшиеся образцы хранили примерно при +4°C вплоть до использования в последующих анализах.
Белок, который связывается с иммуноаффинной колонкой, исследовали в SDS-ПААГ, и результаты показали, что элюированные фракции содержали белок aad-1 с приблизительной молекулярной массой 33 кДа. Кроме того, также осуществляли Вестерн-блоттинг, и результат был позитивным в отношении белка aad-1. Полученный из кукурузы белок aad-1 выделяли из ~30 г лиофилизированного материала стеблей.
Пример 5.3. SDS-ПААГ и Вестерн-блот
Лиофилизированную ткань из стеблей растений с событием DAS-40278-9 и XHH13 (~100 мг) отвешивали в микроцентрифужные пробирки объемом 2 мл и экстрагировали, используя ~1 мл PBST (Sigma, St. Louis, MO), содержащего 10% смесь ингибиторов протеаз растений (Sigma, St. Louis, MO). Экстракцию облегчали, добавляя 4 небольших шарикоподшипника и измельчая образец, используя Geno-Grinder, в течение 1 минуты. После измельчения образцы центрифугировали в течение 5 минут при 20000×g, и надосадки смешивали 4:1 с 5x-буфером для образца по Лэммли (2% SDS, 50 мМ трис, pH 6,8, 0,2 мг/мл бромфенолового синего, 50% (масс./масс.) глицерина, содержащего 10% свежедобавленного 2-меркаптоэтанола) и нагревали в течение 5 минут при ~100°C. После непродолжительного центрифугирования 45 мкл надосадка непосредственно нагружали на SDS-ПААГ-гель BioRad Criterion (Bio-Rad, Hercules, CA), вставленный в камеру для геля Criterion Cell. Позитивный эталонный стандарт полученного из микроорганизмов aad-1 ресуспендировали в концентрации 1 мг/мл в PBST, pH 7,4, и далее разбавляли PBST. Затем образец смешивали с буфером по Лэммли Bio-Rad с 5% 2-меркаптоэтанолом и обрабатывали, как описано ранее. Электрофорез проводили с использованием трис/глицин/SDS-буфера (Bio-Rad, Hercules, CA) при напряжении 150-200 В вплоть до момента, когда фронт красителя появлялся на конце геля. После разделения гель разрезали пополам, и одну половину красили синим красителем для белка Pierce GelCode, а другую половину подвергали электроблоттингу на нитроцеллюлозную мембрану (Bio-Rad, Hercules, CA) с использованием ячейки для электрофоретического переноса Mini trans-blot (Bio-Rad, Hercules, CA) в течение 60 минут при постоянном напряжении 100 вольт. Буфер для переноса содержал 20% метанол и трис/глициновый буфер из Bio-Rad. Для иммунодетекции мембрану исследовали, используя в качестве зонда специфичное для aad-1 поликлональное кроличье антитело (Strategic Biosolution Inc., Newark, DE, очищенное на белке A кроличье поликлональное антитело, партия №: DAS F1 197-15 1, 1,6 мг/мл). Конъюгат антитела козы против IgG (H+L) кролика и щелочной фосфатазы (Pierce Chemical, Rockford, IL) использовали в качестве второго антитела. Субстрат SigmaFast BCIP/NBT использовали для проявления и визуализации полос иммунореактивного белка. Мембрану тщательно промывали водой, чтобы остановить реакцию, и запись результатов осуществляли, используя цифровой сканер (Hewlett Packard, Palo Alto, CA).
В препарате белка aad-1, полученного из P. fluorescens, основная полоса белка, который визуализировали на окрашенных Кумасси SDS-ПААГ-гелях, содержала белок с молекулярной массой приблизительно 33 кДа. Как и ожидали, соответствующий происходящий из кукурузы белок aad-1 (событие DAS-40278-9) был идентичен по размеру экспрессируемым микроорганизмами белкам. Как и следовало ожидать, очищенные из растений фракции в дополнение к белку aad-1 содержали небольшое количество иммунологически неактивных примесей. Совместно очищаемые белки, вероятно, сохранялись на колонке благодаря слабым взаимодействиям с матриксом колонки или утечке моноклонального антитела с колонки в жестких условиях элюирования. Другие исследователи также сообщали о неспецифичной адсорбции пептидов и аминокислот на иммуноадсорбентах на основе активированной бромцианом сефарозы 4B (Kennedy и Barnes, 1983; Holroyde с соавторами, 1976; Podlaski и Stern, 2008).
Полученный из Pseudomonas белок aad-1 давал положительный сигнал ожидаемого размера в Вестерн-блот-анализе с использованием поликлонального антитела. Такой же сигнал также наблюдали в экстракте стеблей трансгенной кукурузы DAS-40278-9. В Вестерн-блот-анализе aad-1 не обнаружили иммунореактивных белков в контрольном экстракте XHH13 и не наблюдали белков с альтернативными размерами (агрегатов или продуктов распада) в трансгенных образцах.
Пример 5.4. Выявление посттрансляционного гликозилирования
Очищенный посредством иммуноаффинной хроматографии полученный из кукурузы белок aad-1 (фракция № 3) смешивали в соотношении 4:1 с 5x буфером по Лэммли. Полученный из микроорганизмов aad-1, соевый ингибитор трипсина, бычий сывороточный альбумин и пероксидазу хрена разбавляли водой Milli-Q до концентрации, примерно соответствующей концентрации полученного из растений aad-1, и смешивали с буфером по Лэммли Bio-Rad. Затем белки нагревали при ~95°C в течение 5 минут и центрифугировали при 20000×g в течение 2 минут, чтобы получить осветленный надосадок. Полученные надосадки непосредственно наносили на гель Bio-RadCriterion и подвергали электрофорезу, используя рабочий буфер XT MES (Bio-Rad, Hercules, CA), по существу, как описано выше, за исключением того, что электрофорез проводили при 170 В в течение ~60 минут. После электрофореза гель разрезали пополам, и одну половину красили синим красителем GelCode на общий белок согласно протоколу производителя. После завершения окрашивания гель сканировали на денситометре Molecular Dynamics, получая непрерывную визуальную регистрацию геля. Другую половину геля красили, используя набор для окрашивания гликопротеидов GelCode (Pierce Chemical, Rockford, IL) согласно протоколу производителя, чтобы визуализировать гликопротеиды. Гликопротеиды (с пределом детекции уже на уровне 0,625 нг на полосу) визуализировали в виде пурпурных полос на светло-розовом фоне. После завершения окрашивания гликопротеидов гели сканировали, используя цифровой сканер Hewlett Packard, получая непрерывную визуальную регистрацию геля. После улавливания изображения окрашивания гликозилирования гели красили, используя GelCode Blue, чтобы подтвердит присутствие негликозилированных белков. Результаты показали, что и полученные из кукурузы и полученные из микроорганизмов белки aad-1 не имели регистрируемых ковалентно связанных углеводов. Полученный результат также был подтвержден по «отпечаткам пальцев» пептидных масс.
Пример 5.5. Отпечатки пальцев пептидных масс, полученных с использованием масс-спектрометрии, и секвенирование aad-1, полученного из кукурузы и Pseudomonas
Проводили масс-спектрометрический анализ aad-1, полученного из Pseudomonas и кукурузы. Белок aad-1, полученный из стебля трансгенной кукурузы (событие DAS-40278-9), подвергали расщеплению в растворе трипсином с последующей MALDI-TOF-МС и ESI-ЖХ/МС.
Массы выявляемых пептидов сравнивали с массами, рассчитанными на основе потенциальных сайтов расщепления протеазами в последовательности полученного из кукурузы белка aad-1. Теоретическое расщепление получали in silico, используя бесплатное программное обеспечение Protein Analysis Worksheet (PAWS) из Proteometrics LLC. Белок aad-1 после денатурации легко расщепляется протеазами и будет давать многочисленные пики пептидов.
В продукте расщепления трипсином полученного из трансгенной кукурузы белка aad-1 (событие DAS-40278-9) выявляемые пептидные фрагменты охватывали почти полную последовательность белка, при этом отсутствовал только один небольшой триптический фрагмент на C-конце белка, от F248 до R253, и один короткий (2 аминокислоты) пептидный фрагмент. Такой анализ подтвердил, что аминокислотная последовательность полученного из кукурузы белка совпадала с аминокислотной последовательностью белка aad-1, полученного из микроорганизмов. Результаты таких анализов показывают, что аминокислотная последовательность полученного из кукурузы белка aad-1 была эквивалентна белку, экспрессируемому P. fluorescens.
Пример 5.5.1. Секвенирование триптических пептидных фрагментов
Кроме отпечатков пальцев пептидных масс секвенировали аминокислотные остатки на N- и C-концах полученного из кукурузы белка aad-1 (иммуноаффинно очищенного из кукурузы, в которой имеет место событие DAS-40278-9) и сравнивали с последовательностью полученного из микроорганизмов белка. Получали последовательности белка, используя тандемную масс-спектрометрию, для первых 11 остатков полученных из микроорганизмов и кукурузы белков (таблица 8). Аминокислотные последовательности обоих белков представляли собой последовательность A’ H A A L S P L S Q R” (SEQ ID NO: 30), что свидетельствует о том, что N-концевой метионин был удален аминопептидазой (таблица 8). N-концевая последовательность белка aad-1, как ожидалось, представляла собой последовательность M’ A H A A L S P L S Q R’2. (SEQ ID NO: 31). Такие результаты свидетельствует, что во время или после трансляции у кукурузы и P. fluorescens N-концевой метионин отщепляется метиониновой аминопептидазой (MAP). MAP быстро отщепляют остатки метионила, когда второй остаток в белке небольшой, такой как Gly, Ala, Ser, Cys, Thr, Pro и Val (Walsh, 2006). Кроме того, что был удален метионин, показано, что небольшая часть N-концевого пептида белка aad-1 была ацетилирована после отщепления N-концевого метионина (таблица 8). С таким результатом часто сталкиваются в случае экспрессируемых эукариотами (растением) белков, так как примерно 80-90% N-концевых остатков модифицированы (Polevoda и Sherman, 2003). Также было показано, что белки с серином и аланином на N-концах чаще всего ацетилированы (Polevoda и Sherman, 2002). Два котрансляционных процесса, отщепление N-концевого остатка метионина и N-концевое ацетилирование, безусловно являются наиболее распространенными модификациями и происходят в случае подавляющего большинства (~85%) эукариотических белков (Polevoda и Sherman, 2002). Однако примеры, демонстрирующие биологическую значимость, ассоциированную с N-концевым ацетилированием, встречаются редко (Polevoda и Sherman, 2000).


Кроме N-ацетилирования также существует небольшое N-концевое укорочение, которое появляется во время очистки получаемого из кукурузы белка aad-1 (таблица 8). Такие «рваные концы» приводили к потере аминокислот A2, H3 и A4 (в различных формах и количествах) из белка, полученного из кукурузы. Полагают, что такое укорочение произошло во время очистки белка aad-1, так как зонд для Вестерн-блота неочищенных экстрактов листьев давал одну выраженную полосу с такой же молекулярной массой, как и полученный из микроорганизмов белок aad-1. Буфер для экстракции для образцов, подвергаемых Вестерн-блоттингу, содержал избыток смеси ингибиторов протеаз, которая включает в себя смесь ингибиторов протеаз с широкой специфичностью для ингибирования сериновых, цистеиновых, аспарагиновых и металлопротеиназ и аминопептидаз.
C-концевую последовательность белков aad-1, полученных из кукурузы и микроорганизмов, определяли, как описано выше, и сравнивали с ожидаемыми аминокислотными последовательностями (таблица 9). Результаты показали, что измеряемые последовательности были идентичны ожидаемым последовательностям, и белки aad-1, полученные из кукурузы и микроорганизмов, были идентичными и не были изменены на C-конце.


Пример 6. Экспрессия в полевых условиях, анализ состава питательных веществ и агрономические характеристики гибридной линии кукурузы, в которой имеет место событие DAS-40278-9
Целью настоящего исследования было определение уровней белка AAD-1, найденного в тканях кукурузы. Кроме того, осуществляли анализ состава кукурузного корма и зерна для исследования эквивалентности изогенной нетрансформированной линии кукурузы и трансгенной линии кукурузы DAS-40278-9 (без опрыскивания, с опрыскиванием 2,4-D, опрыскиванием квизалофопом и опрыскиванием 2,4-D и квизалофопом). Агрономические характеристики изогенной нетрансформированной линии кукурузы также сравнивали с кукурузой DAS-40278-9. Исследования экспрессии в полевых условиях, состава и агрономические испытания проводили в шести исследуемых местоположениях в главных производящих кукурузу регионах в США и Канаде. Такие местоположения представляют регионы с разнообразной агрономической практикой и разными условиями окружающей среды. Такие испытания проводили в Айове, Иллинойсе (2 местоположения), Индиане, Небраске и Онтарио в Канаде.
Средние значения во всех местоположениях для контрольного образца, образцов неопрыснутых AAD-1, образцов AAD-1+квизалофоп, AAD-1+2,4-D и AAD-1+оба вносимых препарата были в известных из литературы пределах для кукурузы. Наблюдали ограниченные значимые различия между неопрыснутой кукурузой AAD-1, кукурузой AAD-1+квизалофоп, AAD-1+2,4-D или AAD-1+оба препарата и контролем, но различия не считали биологически значимыми, поскольку они были небольшими, и результаты были в пределах, обнаруженных для коммерческой кукурузы. Графики результатов определения состава не показывают никаких биологически значимых, связанных с обработкой различий в составе между линиями кукурузы: неопрыснутой AAD-1, AAD-1+квизалофоп, AAD-1+2,4-D или AAD-1+оба препарата, и контрольной линией кукурузы. В заключение, результаты определения состава кукурузы неопрыснутой AAD-1, AAD-1+квизалофоп, AAD-1+2,4-D и AAD-1+оба препарата подтверждают эквивалентность кукурузы AAD-1 (событие DAS 40278-9) с обычными линиями кукурузы.
Пример 6.1. Тестируемые линии кукурузы
Гибридные семена, в которых имеет место событие DAS-40278-9, и контрольные растения в виде обычных гибридных семян с таким же генетическим фоном, как и линия тестируемого материала, но не имеющие события DAS-40278-9, перечислены в таблице 10.
Описанные выше растения кукурузы выращивали в местоположениях в основных регионах возделывания кукурузы в США и Канаде. Шесть полигонов для испытания в полевых условиях, Richland, IA; Carlyle, IL; Wyoming, IL; Rockville, IN; York, NE; и Branchton, Ontario, Canada (обозначенные IA, IL1, IL2, IN, NE и ON), представляют регионы с разнообразной агрономической практикой и разными условиями окружающей среды для кукурузы.
Тестируемые и контрольные семена кукурузы высевали по норме высева примерно 24 семени на ряд с расстоянием между семенами в ряду примерно 10 дюймов (25 см). В каждом месте закладывали по 4 повтора делянок для каждой обработки, при этом каждая делянка имела ряды 2-25 футов (0,6-7,5 м). Делянки распределяли в соответствии с рандомизированным полноблочным (RCB) планом с уникальной рандомизацией в каждом месте. Каждая делянка кукурузы была ограничена 2 рядами нетрансгенного гибрида кукурузы со сходным периодом достижения зрелости. Все место испытания было окружено минимум 12 рядами (или 30 футов (9 м)), нетрансгенного гибрида кукурузы сходной относительной зрелости.
Осуществляли подходящие практические мероприятия по борьбе с насекомыми, сорняками и заболеваниями, чтобы получить соответствующий агрономическим требованиям урожай. Максимальные и минимальные ежемесячные температуры наряду с количеством атмосферных осадков и орошением были средними в каждом местоположении. С такими диапазонами значений обычно сталкиваются при производстве кукурузы.
Пример 6.2. Внесения гербицидов
Обработку гербицидами проводили, используя объем раствора для опрыскивания 20 галлонов на акр (187 л/га). Такое внесение было запланировано для того, чтобы воспроизвести максимальную утвержденную норму в коммерческой практике. В таблице 11 перечислены гербициды, которые были использованы.
b аи = активный ингредиент.
2,4-D (Weedar 64) применяли в виде 3 внесений разбрасыванием сверху для тестирования объектов 4 и 5 (сезонные всего 3 фунта эк/акр). Отдельные внесения проводили до всходов и примерно на стадиях V4 и V8-V8.5. Планируемые нормы для отдельных внесений составляли 1,0 фунт эк/акр в случае Weedar 64 или 1120 г эк/га. Фактические нормы внесения были в диапазоне 1096-1231 г эк/акр.
Квизалофоп (Assure II) применяли в виде одного внесения разбрасыванием сверху для тестирования объектов 3 и 5. Срок внесения был приурочен примерно к стадии роста V6. Планируемая норма внесения составляла 0,0825 фунтов аи/акр для Assure II или 92 г аи/га. Фактические нормы внесения были в диапазоне 90,8-103 г аи/га.
Пример 6.3. Сбор агрономических данные и результаты
Агрономические характеристики регистрировали для всех тестируемых объектов в блоках 2, 3 и 4 в каждом местоположении. В таблице 12 перечислены следующие измеряемые характеристики.
Проводили анализ агрономических данных, собранных для следующих объектов: контрольных, aad-1 неопрыснутых, aad-1+2,4-D, aad-1+квизалофоп и aad-1+оба препарата. В случае анализа в различных регионах не наблюдали статистически значимых различий в значениях для ранних популяций (V1 и V4), мощности, конечных популяций, повреждений культуры, времени выметывания пестичных столбиков, времени пыления, стеблевого полегания, корневого полегания, заболеваемости, повреждения насекомыми, количества дней до созревания, высоты растений и жизнеспособности пыльцы (форме и окраске), судя по суммарным данным анализа в различных регионах (таблица 13). В случае исследования способности сохранять зеленую окраску и высоты до початка наблюдали значимые различия при использовании парных t-критериев между контролем и объектом aad-1+квизалофоп, но они не сопровождались значимыми общими эффектами обработки или p-значениями, скорректированными с учетом средней доли ложных отклонений от гипотезы (FDR) (таблица 13).
(0,303, 0,819)
(0,443, 0,819)
(0,556, 0,819)
(0,393, 0,819)
(0,883, 0,984)
(0,687, 0,863)
(0,423, 0,819)
(0,263, 0,819)
(0,197, 0,819)
(0,161, 0,819)
(0,633, 0,819)
(0,729, 0,889)
(0,747, 0,889)
(0,802, 0,924)
(0,943, 1,00)
(0,521, 0,819)
(1,00, 1,00)
(1,00, 1,00)
(1,00, 1,00)
(0,130, 0,819)
(0,996, 1,00)
(0,781, 0,917)
(0,088, 0,819)
(0,224, 0,819)
(0,564, 0,819)
(0,480, 0,819)
(0,245, 0,819)
(0,245, 0,819)
(0,931, 1,00)
(0,546, 0,819)
(0,439, 0,819)
(0,605, 0,819)
(0,618, 0,819)
(0,409, 0,819)
(0,739, 0,889)
(0,409, 0,819)
(0,809, 0,924)
(0,470, 0,819)
(0,629, 0,819)
(0,629, 0,819)
(1,00, 1,00)
(1,00, 1,00)
(1,00, 1,00)
(0,337, 0,819)
(0,850, 0,960)
(0,306, 0,819)
(0,573, 0,819)
(0,573, 0,819)
(0,612, 0,819)
(0,315, 0,819)
(0,315, 0,819)
(0,612, 0,819)
(0,356, 0,819)
(0,356, 0,819)
(0,356, 0,819)
(0,356, 0,819)
(0,457, 0,819)
(0,457, 0,819)
(0,457, 0,819)
(0,373, 0,819)
(0,250, 0,819)
(0,034m, 0,819)
(0,144, 0,819)
(0,106, 0,819)
(0,383, 0,819)
(0,265, 0,819)
(0,265, 0,819)
(0,265, 0,819)
(0,500, 0,819)
(0,500, 0,819)
(0,736, 0,889)
(0,500, 0,819)
(0,558, 0,819)
(0,302, 0,819)
(0,185, 0,819)
(0,104, 0,819)
(0,206, 0,819)
(0,209, 0,819)
(0,350, 0,819)
(0,286, 0,819)
(0,089, 0,819)
(0,018m, 0,786)
(0,214, 0,819)
(0,016m, 0,786)
b Сравнение обработок с опрыскиванием и без опрыскивания с контролем с использованием t-критерия.
c P-значения, скорректированные с использованием способа оценки средней доли ложных отклонений от гипотезы (FDR).
d Визуальная оценка по шкале 1-9; 9 = высокие растения с крупными крепкими листьями.
e Шкала 0-100%; 0 = нет повреждения и 100 = мертвое растение.
f Количество тепловых единиц, которые были накоплены с момента посева.
g Шкала 0-100%; % пыльцевых зерен с разрушенными стенками.
h Шкала 0-100%; % пыльцевых зерен с интенсивной желтой окраской.
i Визуальная оценка по шкале 1-9, где 1 означает отсутствие видимой зеленой ткани.
j Визуальная оценка по шкале 1-9, где 1 означает слабую устойчивость к заболеваниям.
k Визуальная оценка по шкале 1-9, где 1 означает слабую устойчивость к насекомым.
l NA = статистический анализ не проводили, так как не было вариабельности между повторами или обработками.
m Статистическое различие показано P-значением <0,05.
Пример 6.4. Сбор образцов
Образцы для экспрессии и анализа композиции собирали, как указано в таблице 14.
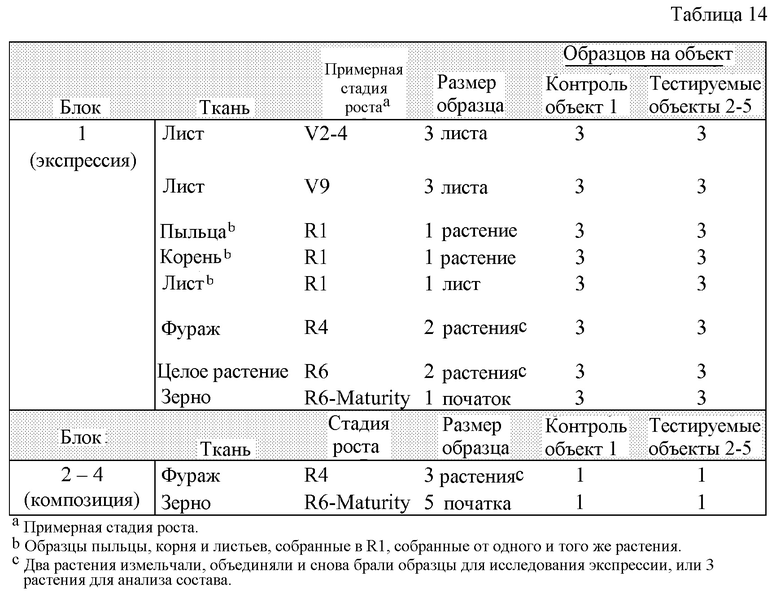
Пример 6.5. Определение белка aad-1 в образцах кукурузы
Образцы кукурузы анализировали в отношении количества белка aad-1. Растворимый экстрагируемый белок aad-1 количественно оценивали, используя набор для твердофазного иммуноферментного анализа (ELISA), приобретенный из Beacon Analytical System, Inc. (Portland, ME).
Образцы тканей кукурузы выделяли из тестируемых растений и готовили для анализа экспрессии, используя грубый размол, лиофилизацию и тонкое измельчение (при необходимости) с помощью измельчителя Geno/Grinder (Certiprep, Metuchen, New Jersey). Для пыльцы не требовалось дополнительной подготовки. Белок aad-1 экстрагировали из тканей кукурузы раствором фосфатно-солевого буфера, содержащим детергент твин-20 (PBST) и содержащим 0,5% бычьего сывороточного альбумина (БСА). В случае пыльцы белок экстрагировали, используя 0,5% буфер PBST/БСА, содержащий 1 мг/мл аскорбата натрия и 2% смесь ингибиторов протеаз. Экстракты из растительных тканей и пыльцы центрифугировали; собирали водный надосадок, при необходимости разбавляли соответствующим буфером и анализировали, используя набор для ELISA aad-1 в формате типа «сэндвича». Набор использовали в ходе следующих стадий. Аликвоту разбавленного образца и биотинилированное моноклональное анти-aad-1-антитело инкубировали в лунках планшета для микротитрования, покрытых иммобилизованным моноклональным анти-aad-1-антителом. Такие антитела связываются с белком aad-1 в лунках и образуют «сэндвич» с белком aad-1, который связан между растворимым и иммобилизованным антителом. Несвязанные образцы и конъюгат затем удаляли из планшета, промывая PBST. В лунки добавляли избыточное количество конъюгата стрептавидин-фермент (щелочная фосфатаза) для инкубации. В конце периода инкубации несвязанные реагенты удаляли из планшета промывкой. Последующее добавление субстрата фермента приводило к образованию окрашенного продукта. Так как aad-1 был связан в сэндвиче из антител, уровень развития окраски был связан с концентрацией aad-1 в образце (т.е., более низкие концентрации остатка приводили к меньшему развитию окраски). Поглощение при 405 нм измеряли, используя устройство для считывания планшетов Molecular Devices V-max или Spectra Max 190. Получали калибровочную кривую и вычисляли концентрацию aad-1 в неизвестных образцах из уравнения полиномиальной регрессии, используя компьютерную программу Soft-MAX ProTM, которая была совместима с устройством для считывания планшетов. Образцы анализировали, используя лунки в двух повторах, при этом сообщали среднюю концентрацию в двух повторах лунок.
Суммарные данные о концентрации белка aad-1 (усредненной для разных регионов) в различных формах кукурузы показаны в таблице 15. Средняя концентрация белка aad-1 была в диапазоне от 2,87 нг/мг сухой массы в корне на стадии R1 до 127 нг/мг в пыльце. Результаты определения экспрессии для неопрыскиваемых и опрыскиваемых делянок были сходными. Белок aad-1 не выявляли ни в одном из контрольных образцов, за исключением одного контрольного образца корня из Индианы.
отклонение
b Значения в скобках в пределах между LOD способа и пределом количественного определения (LOQ).
Пример 6.6. Анализ состава
Образцы фуража и зерна кукурузы анализировали в отношении содержания питательных веществ, используя различные тесты. Анализы, осуществляемые в случае фуража, включали анализ зольности, общего жира, влажности, белков, углеводов, грубых волокон, кислотного детергентного волокна, нейтрального детергентного волокна, кальция и фосфора. Анализы, осуществляемые в случае зерна, включали анализы основных компонентов пищи (зольных веществ, общего жира, влажности, белков, углеводов, грубых волокон, кислотного детергентного волокна), нейтрального детергентного волокна (NDF), минералов, аминокислот, жирных кислот, витаминов, вторичных метаболитов и антипитательных веществ. Результаты анализа питательных веществ в фураже и зерне кукурузы сравнивали со значениями, сообщенными в литературе (см.; Watson, 1982 (4); Watson, 1984 (5); ILSI Crop Composition Database, 2006 (6); OECD Consensus Document on Compositional Considerations for maize, 2002 (7); и Codex Alimentarius Commission 2001 (8)).
Пример 6.6.1. Анализ питательных компонентов, волокон и минералов в фураже
Проводили анализ белков, жира, зольности, влажности, углеводов, ADF, NDF, кальция и фосфора в образцах кукурузного фуража, полученного из следующих растений кукурузы: контрольных, неопрыскиваемых aad-1, aad-1+квизалофоп, aad-1 +2,4-D и aad-1+оба препарата. Суммарные результаты по всем регионам показаны в таблице 16. В случае анализа по всем регионам и анализа в отдельных регионах все средние значения для питательных компонентов, волокон и минералов были в пределах, известных из литературы. Не наблюдали статистических различий в анализе по различным регионам между контрольными и трансгенными объектами в отношении содержания влаги, ADF, NDF, кальция и фосфата. В отношении белка и зольности наблюдали значимые различия при использовании парных t-критериев для неопрыскиваемых AAD-1 (белок), aad-1+квизалофоп (белок) и aad-1+оба препарата (зольность), но при этом не наблюдали значимых общих эффектов обработки или p-значений, скорректированных в соответствии с FDR. В отношении исследования жира наблюдали и значимые различия при использовании парного t-критерия и скорректированное p-значение в случае aad-1+квизалофоп по сравнению с контролем, но не наблюдали значимого общего эффекта обработки. В отношении углеводов наблюдали статистически значимый общий эффект обработки, парный t-критерий и FDR-скорректированное p-значение при сравнении aad-1+квизалофоп и контроля. Также в отношении углеводов наблюдали значимое различие при использовании парного t-критерия в случае неопрыскиваемых растений aad-1, но без значимого FDR-скорректированного p-значения. Такие различия не являются биологически значимыми, так как результаты, полученные в разных регионах, для таких аналитов были в диапазонах, о которых сообщается в литературе для кукурузы, и отличия от контроля были небольшими (<23%).
(Pr>F)b
(0,016e, 0,066)
(0,010e,
0,051)
(0,285, 0,450)
(0,245, 0,402)
(0,202, 0,357)
(0,005e,
0,028e)
(0,233, 0,391)
(0,093, 0,213)
(0,742, 0,859)
(0,525,
0,708)
(0,673, 0,799)
(0,019e, 0,069)
(0,651, 0,782)
(0,988,
0,988)
(0,699, 0,820)
(0,501, 0,687)
(0,015e, 0,061)
(0,006e,
0,034e)
(0,262, 0,424)
(0,538, 0,708)
(0,925, 0,970)
(0,833,
0,925)
(0,677, 0,800)
(0,851, 0,937)
(0,169, 0,322)
(0,242,
0,402)
(0,809, 0,911)
(0,978, 0,985)
(0,532, 0,708)
(0,930,
0,970)
(0,815, 0,911)
(0,150, 0,296)
(0,198, 0,354)
(0,427,
0,615)
(0,288, 0,450)
(0,608, 0,762)
b Общий эффект обработки, оцениваемый с использованием F-критерия.
c Сравнение трансгенных обработок с контролем с использованием t-критерия.
d P-значения, скорректированные с использованием способа оценки средней доли ложных отклонений от гипотезы (FDR).
e Статистическое различие показано P-значением <0,05.
Пример 6.6.2. Анализ питательных компонентов и волокон в зерне
Суммарные результаты анализа питательных компонентов (белка, жира, золы, влажности, холестерина и углеводов) и волокон (ADF, NDF и общих пищевых волокон) в зерне кукурузы во всех местоположениях показаны в таблице 17. Все результаты в отношении питательных компонентов и волокон были в известных из литературы диапазонах, и не наблюдали значимых различий в анализе в разных местоположениях между контролем и трансгенными объектами в отношении жира, золы, NDF и общих пищевых волокон. В отношении содержания влаги наблюдали значимый общий эффект обработки, но он не сопровождался значимыми парными t-критериями или FDR-скорректированными p-значениями. В отношении ADF наблюдали значимый парный t-критерий в случае aad-1+оба препарата, но не наблюдали значимого общего эффекта обработки или FDR-скорректированного p-значения. В отношении белка и углеводов получали значимые различия с использованием парных t-критериев, скорректированные p-значения и общие эффекты обработки в случае неопрыснутых aad-1, aad-1+квизалофоп и aad-1+оба препарата. Так как такие различия были небольшими (<12%) и все значения были в известных из литературы диапазонах, такие различия не считали биологически важными.
(Pr>F)b
(0,002e, 0,016e)
(0,0004e, 0,013e)
(0,061, 0,161)
(0,002e, 0,015e)
(0,238, 0,397)
(0,095, 0,215)
(0,955, 0,977)
(0,427, 0,615)
(0,178, 0,330)
(0,364, 0,557)
(0,982, 0,985)
(0,397, 0,587)
(0,406, 0,594)
(0,056, 0,152)
(0,117, 0,254)
(0,114, 0,250)
(0,002e, 0,015e)
(0,001e,
0,013e)
(0,074, 0,185)
(0,003e, 0,019e)
(0,130, 0,269)
(0,197,
0,354)
(0,078, 0,193)
(0,035e, 0,106)
(0,455, 0,638)
(0,120, 0,254)
(0,121, 0,254)
(0,552, 0,708)
(0,164, 0,313)
(0,195, 0,353)
(0,487, 0,679)
(0,215, 0,370)
b Общий эффект обработки, оцениваемый с использованием F-критерия.
c Сравнение трансгенных обработок с контролем с использованием t-критерия.
d P-значения, скорректированные с использованием способа оценки средней доли ложных отклонений от гипотезы (FDR).
e Статистическое различие показано P-значением <0,05.
f NR = не сообщается.
g NA = статистический анализ не проводили, так как большинство данных было <LOQ.
Пример 6.6.3. Анализ минералов в зерне
Осуществляли анализ образцов зерна кукурузы в отношении минералов кальция, хрома, меди, йода, железа, магния, марганца, молибдена, фосфора, калия, селена, натрия и цинка. Суммарные результаты, полученные во всех местоположениях, показаны в таблице 18. Все результаты были в известных из литературы диапазонах. В случае анализа в разных местоположениях не наблюдали значимых различий в отношении кальция, меди, железа и калия. Средние результаты для хрома, йода, селена и натрия были ниже предела количественного определения таким способом. В отношении магния и фосфора наблюдали значимые парные t-критерии для неопрыснутых растений aad-1 и растений aad-1+квизалофоп, но они не сопровождались значимыми общими эффектами обработки или FDR-скорректированными p-значениями. В отношении марганца и молибдена наблюдали значимый парный t-критерий для неопрыснутых растений aad-1, но не обнаружили FDR-скорректированного p-значения и общего эффекта обработки. В случае aad-1+оба препарата наблюдали значимый парный t-критерий в отношении цинка, но значимое FDR-скорректированное p-значение или общий эффект обработки отсутствовали. Кроме того, такие отличия от контроля были небольшими (<13%), и все значения были в известных из литературы диапазонах в том случае, когда таковые были доступны.
(Pr>F)b
(0,146, 0,289)
(0,505, 0,687)
(0,944, 0,977)
(0,898, 0,957)
(0,655, 0,782)
(0,890,
0,957)
(0,817, 0,911)
(0,749, 0,863)
(0,086, 0,206)
(0,310,
0,482)
(0,801, 0,911)
(0,145, 0,289)
(0,010f, 0,051)
(0,017f,
0,066)
(0,145, 0,289)
(0,070, 0,177)
(0,025f, 0,082)
(0,884, 0,957)
(0,942, 0,977)
(0,505, 0,687)
(0,020f, 0,072)
(0,067, 0,173)
(0,206, 0,362)
(0,046, 0,132)
(0,012f, 0,057)
(0,035f,
0,106)
(0,055, 0,150)
(0,085, 0,206)
(0,330, 0,510)
(0,655,
0,782)
(0,722, 0,839)
(0,454, 0,638)
(0,183, 0,336)
(0,108,
0,238)
(0,627, 0,768)
(0,027f, 0,085)
b Общий эффект обработки, оцениваемый с использованием F-критерия.
c Сравнение трансгенных обработок с контролем с использованием t-критерия.
d P-значения, скорректированные с использованием способа оценки средней доли ложных отклонений от гипотезы (FDR).
e NA = статистический анализ не проводили, так как большинство данных было <LOQ.
f Статистическое различие показано P-значением <0,05.
Пример 6.6.4. Анализ аминокислот в зерне
Образцы кукурузы анализировали в отношении содержания аминокислот в контроле, в неопрыснутых растениях кукурузы aad-1, в растениях aad-1+квизалофоп, aad-1+2,4-D и aad-1+оба препарата, и суммарные результаты по всем регионам и на полях в отдельных местоположениях показаны в таблице 19. Уровни всех аминокислот были в диапазонах, которые сообщаются в литературе, и не наблюдали значимых различий в анализе в разных местоположениях в отношении аргинина, лизина и тирозина. Значимые различия наблюдали в отношении нескольких аминокислот в анализе в разных местоположениях. В таких случаях содержание аминокислот в контроле было ниже, чем в aad-1-трансгенных линиях, что может быть связано с более низким содержанием общего белка в контрольном зерне по сравнению с линиями aad-1. В случае неопрыснутого исследуемого объекта aad-1 обнаружили значимые общие эффекты обработки наряду со значимыми парными t-критериями и получили FDR-скорректированные p-значения для всех аминокислот, за исключением аргинина, глицина, лизина, триптофана и тирозина. В случае aad-1+квизалофоп обнаружили значимые общие эффекты обработки наряду со значимыми парными t-критериями и получили FDR-скорректированные p-значениями в случае всех аминокислот за исключением аргинина, цистеина, глицина, лизина, триптофана и тирозина. В случае объекта aad-1+2,4-D обнаружили значимые общие эффекты обработки наряду со значимыми парными t-критериями (со значимыми FDR-скорректированными p-значениями) для всех аминокислот, за исключением аргинина, аспарагиновой кислоты, глицина, гистидина, лизина, тирозина и валина. В случае объекта aad-1+оба препарата обнаружили значимые общие эффекты обработки наряду со значимыми парными t-критериями и FDR-скорректированными p-значениями для всех аминокислот, за исключением аргинина, глицина, лизина, серина, триптофана и тирозина. Хотя было много различий, наблюдаемых для аминокислот, различия были небольшими (<15%), но не наблюдались между разными местоположениями, и все средние значения были в диапазонах, сообщаемых в литературе.
(Pr>F)b
(0,0005e, 0,013e)
(0,0005e, 0,013e)
(0,021e, 0,074)
(0,001e, 0,013e)
(0,286, 0,450)
(0,139,
0,283)
(0,929, 0,970)
(0,897, 0,957)
(0,002e, 0,015e)
(0,003e,
0,021e)
(0,060, 0,160)
(0,004e, 0,027e)
(0,009e, 0,050e)
(0,020e,
0,072)
(0,018e, 0,067)
(0,005e, 0,028e)
(0,0003e, 0,013e)
(0,0004e, 0,013e)
(0,017e, 0,067)
(0,001e, 0,013e)
(0,018e, 0,067)
(0,013e,
0,059)
(0,217, 0,371)
(0,016e, 0,066)
(0,001e, 0,013e)
(0,002e,
0,014e)
(0,036, 0,109)
(0,002e, 0,014e)
(0,001e, 0,014e)
(0,001e,
0,014e)
(0,044e, 0,127)
(0,001e, 0,013e)
(0,0003e, 0,013e)
(0,0003e, 0,013e)
(0,013e, 0,059)
(0,001e, 0,013e)
(0,210, 0,367)
(0,128,
0,265)
(0,879, 0,956)
(0,102, 0,226)
(0,001e, 0,013e)
(0,001e,
0,013e)
(0,014e, 0,061)
(0,001e, 0,014e)
(0,001e, 0,013e)
(0,001e,
0,013e)
(0,023e, 0,077)
(0,001e, 0,013e)
(0,0004e, 0,013e)
(0,001e,
0,013e)
(0,012e, 0,059)
(0,001e, 0,014e)
(0,002e, 0,014e)
(0,001e,
0,014e)
(0,042e, 0,122)
(0,015e, 0,061)
(0,001e, 0,014e)
(0,001e,
0,013e)
(0,023e, 0,077)
(0,003e, 0,020e)
(0,067, 0,173)
(0,025e,
0,082)
(0,014e, 0,060)
(0,029e, 0,092)
(0,535, 0,708)
(0,214,
0,370)
(0,907, 0,964)
(0,500, 0,687)
(0,002e, 0,014e)
(0,002e,
0,014e)
(0,054, 0,148)
(0,001e, 0,014e)
b Общий эффект обработки, оцениваемый с использованием F-критерия.
c Сравнение трансгенных обработок с контролем с использованием t-критериев.
d P-значения, скорректированные с использованием способа оценки средней доли ложных отклонений от гипотезы (FDR).
e Статистическое различие показано P-значением <0,05.
Пример 6.6.5. Анализ жирных кислот в зерне
Осуществляли анализ образцов зерна кукурузы в отношении жирных кислот. Суммарные результаты, полученные в разных местоположениях, показаны в таблице 20. Все результаты для образцов контрольной кукурузы, неопрыснутых aad-1-растений кукурузы, aad-1+квизалофоп, aad-1+2,4-D и aad-1+оба препарата, анализируемых в отношении жирных кислот, были в диапазонах, опубликованных в литературе. Результаты для каприловой (8:0), каприновой (10:0), лауриновой (12:0), миристиновой (14:0), миристолевой (14:1), пентадекановой (15:0), пентадеценовой (15:1), гептадекановой (17:0), гептадеценовой (17:1), гамма-линоленовой (18:3), эйкозадиеновой (20:2), эйкозатриеновой (20:3) и арахидоновой (20:4) кислот были ниже предела количественного определения (LOQ) способа. В анализе в разных местоположениях не выявлены различия для 16:0 пальмитиновой, 16:1 пальмитолевой, 18:0 стеариновой, 18:2 линолевой, 18:3 линоленовой и 20:0 арахидоновой кислот. В отношении 18:1 олеиновой и 20:1 эйкозеновой кислот наблюдали значимые парные t-критерии в случае неопрыснутой кукурузы aad-1 (18:1) и кукурузы aad-1+2,4-D (18:1 и 20:1), но они не сопровождались значимыми общими эффектами обработки или FDR-скорректированными p-значениями. В отношении 22:0 бегеновой кислоты обнаружили значимый общий эффект обработки и значимые парные t-критерии в случае aad-1+2,4-D и aad-1+оба препарата, но значимые FDR-скорректированные p-значения отсутствовали.
(Pr>F)c
(0,618, 0,763)
(0,280,
0,445)
(0,617, 0,763)
(0,544, 0,708)
(0,804, 0,911)
(0,551,
0,708)
(0,555, 0,708)
(0,392, 0,582)
(0,119, 0,254)
(0,437,
0,626)
(0,259, 0,421)
(0,598, 0,756)
(0,013g, 0,059)
(0,178,
0,329)
(0,015g, 0,061)
(0,092, 0,213)
(0,189, 0,345)
(0,144,
0,289)
(0,453, 0,638)
(0,119, 0,254)
(0,537, 0,708)
(0,202,
0,357)
(0,842, 0,932)
(0,266, 0,428)
(0,061, 0,161)
(0,341,
0,525)
(0,153, 0,297)
(0,175, 0,328)
(0,089, 0,210)
(0,497,
0,687)
(0,013g, 0,059)
(0,121, 0,254)
(0,093, 0,213)
(0,887,
0,957)
(0,011g, 0,054)
(0,023g, 0,077)
b Объединенный диапазон.
c Общий эффект обработки, оцениваемый с использованием F-критерия.
d Сравнение трансгенных обработок с контролем с использованием t-критериев.
e P-значения, скорректированные с использованием способа оценки средней доли ложных отклонений от гипотезы (FDR).
f NA = статистический анализ не проводили, так как большинство данных было <LOQ.
g Статистическое различие показано P-значением <0,05.
Пример 6.6.6. Анализ витаминов в зерне
Определяли уровни витаминов A, B1, B2, B5, B6, B12, C, D, E, ниацина и фолиевой кислоты в образцах зерна контрольной кукурузы, неопрыснутой кукурузы aad-1, aad-1+квизалофоп, aad-1+2,4-D и aad-1+оба препарата. Суммарные результаты, полученные для всех местоположений, показаны в таблице 21. Витамины B12, D и E нельзя было количественно измерить используемыми способами анализа. Все средние результаты, полученные в отношении витаминов, были сходны со значениями, о которых сообщается в литературе. Результаты, полученные в отношении витаминов, диапазоны для которых не указаны в литературе (витамины B5 и C), были сходны с полученными контрольными значениями (отличие от контроля <22%). При анализе в разных местоположениях не наблюдали значимых различий, за исключением витаминов B1, C и ниацина. Используя парные t-критерии, наблюдали значимые различия в отношении витамина B1 между контролем и неопрыснутыми растениями aad-1, aad-1+квизалофоп и aad-1+оба препарата, но они не сопровождались значимыми общими эффектами обработки или FDR-скорректированными p-значениями. В отношении витамина C наблюдали значимый общий эффект обработки наряду со значимыми парными t-критериями и FDR-скорректированными p-значениями в случае aad-1+квизалофоп и aad-1+2,4-D. Подобным образом в отношении ниацина наблюдали значимый общий эффект обработки наряду со значимыми парными t-критериями и FDR-скорректированными p-значениями в случае aad-1+квизалофоп и aad-1+оба препарата. Значимый парный t-критерий получен в случае aad-1+2,4-D в отношении ниацина, но он не сопровождался значимым общим эффектом обработки или FDR-скорректированным p-значением. Так как не наблюдали различий между разными местоположениями, и значения были в известных из литературы диапазонах (в том случае, когда таковые имелись), различия не считали биологически значимыми.
(Pr>F)b
(0,372, 0,566)
(0,967,
0,983)
(0,770, 0,883)
(0,221, 0,376)
(0,041e, 0,121)
(0,013e,
0,059)
(0,375, 0,567)
(0,032e, 0,100)
(0,443, 0,631)
(0,600,
0,756)
(0,227, 0,383)
(0,543, 0,708)
(0,623, 0,766)
(0,391,
0,582)
(0,968, 0,983)
(0,424, 0,615)
(0,859, 0,938)
(0,652,
0,782)
(0,652, 0,782)
(0,088, 0,210)
(0,268, 0,429)
(0,005e,
0,028e)
(0,004e, 0,026e)
(0,068, 0,173)
(0,050, 0,140)
(0,002e,
0,017e)
(0,018e, 0,067)
(0,002e, 0,016e)
(0,779, 0,890)
(0,403,
0,592)
(0,931, 0,970)
(0,916, 0,970)
b Общий эффект обработки, оцениваемый с использованием F-критерия.
c Сравнение трансгенных обработок с контролем с использованием t-критериев.
d P-значения, скорректированные с использованием способа оценки средней доли ложных отклонений от гипотезы (FDR).
e Статистическое различие показано P-значением <0,05.
f NR = не сообщается.
g NA = статистический анализ не проводили, так как большинство данных было <LOQ.
Пример 6.6.7. Анализ антипитательных веществ и вторичных метаболитов в зерне
Определяли уровни вторичных метаболитов (кумаровой кислоты, феруловой кислоты, фурфурола и инозитола) и антипитательных веществ (фитиновой кислоты, раффинозы и ингибитора трипсина) в образцах зерна кукурузы в контроле, в неопрыснутых растениях aad-1, aad-1+квизалофоп, aad-1+2,4-D и aad-1+оба препарата. Суммарные результаты, полученные для всех местоположений, показаны в таблицах 22 и 23. В случае анализа в разных местоположениях все значения были в известных из литературы диапазонах. Не наблюдали значимых различий между результатами, полученными для объектов aad-1 и контрольных объектов в анализе в разных местоположениях в отношении инозитола и ингибитора трипсина. Результаты для фурфурола и раффинозы были ниже предела количественного определения данным способом. Значимые парные t-критерии наблюдали в отношении кумаровой кислоты (неопрыснутые aad-1, aad-1+2,4-D и aad-1+оба препарата) и феруловой кислоты (aad-1+квизалофоп и aad-1+оба препарата). Такие различия не сопровождались значимыми общими эффектами обработки или FDR-скорректированными p-значениями, и результаты были сходны с контролем (различие <10%). Значимый общий эффект обработки, парный t-критерий и FDR-скорректированное p-значение получены для фитиновой кислоты (неопрыснутые aad-1). Так как все результаты были в известных из литературы диапазонах и сходны с контролем (различие <11%), такие различия не считали биологически важными.
(Pr>F)b
(0,038e, 0,113)
(0,090,
0,211)
(0,022e, 0,074)
(0,029e, 0,091)
(0,051, 0,141)
(0,010e,
0,051)
(0,080, 0,196)
(0,019e, 0,069)
(0,548, 0,708)
(0,973,
0,984)
(0,612, 0,763)
(0,526, 0,708)
b Общий эффект обработки, оцениваемый с использованием F-критерия.
c Сравнение трансгенных обработок с контролем с использованием t-критериев.
d P-значения, скорректированные с использованием способа оценки средней доли ложных отклонений от гипотезы (FDR).
e Статистическое различие показано P-значением <0,05.
f NA = статистический анализ не проводили, так как большинство данных было <LOQ.
(Pr>F)b
(0,003e, 0,020e)
(0,099,
0,224)
(0,245, 0,402)
(0,158, 0,304)
(0,954, 0,977)
(0,631,
0,770)
(0,387, 0,582)
(0,813, 0,911)
b Общий эффект обработки, оцениваемый с использованием F-критерия.
c Сравнение трансгенных обработок с контролем с использованием t-критериев.
d P-значения, скорректированные с использованием способа оценки средней доли ложных отклонений от гипотезы (FDR).
e Статистическое различие показано P-значением <0,05.
f NA = статистический анализ не проводили, так как большинство данных было <LOQ.
Пример 7. Дополнительные агрономические испытания
Оценивали агрономические характеристики линии кукурузы 40278 по сравнению с почти изолинейной кукурузой в разных условиях среды. В обработку включали 4 генетически разных гибрида и соответствующие им почти изолинейные контрольные гибриды, которые тестировали всего в 21 местоположении.
Четыре тестируемых гибрида представляли собой гибриды, имеющие зрелость от средней до поздней в диапазоне от 99 до 113 дней относительной зрелости. В эксперименте A тестировали событие DAS-40278-9 в генетическом фоне в случае конверсии инбредной C x BC3S1. Такой гибрид имел относительную зрелость 109 дней и его тестировали в 16 местоположениях (таблица 24). В эксперименте B тестировали гибридный фон в случае конверсии инбредной E x BC3S1, гибрид с относительной зрелостью 113 дней. Такой гибрид тестировали в 14 местоположениях, используя группу местоположений, немного отличающуюся от местоположений в эксперименте A (таблица 24). В экспериментах C и D тестировали гибридные фоны в случае конверсии BC2S1 x инбредной D и конверсии BC2S1 x инбредной F, соответственно. Оба указанных гибрида имеют относительную зрелость 99 дней, и их тестировали в одних и тех же 10 местоположениях.
{TC «Таблица 11. Местоположения, используемые в агрономических испытаниях в эксперименте 2» \f D \1" 1"}
Для каждого испытания использовали рандомизированный полноблочный план с двумя повторами для каждого местоположения и двумя делянками рядов. Длина рядов составляла 20 футов (6 м) и каждый ряд засевали по 34 семени на ряд. Использовали стандартную для регионов агрономическую практику при проведении испытаний.
Данные собирали и анализировали, используя восемь агрономических характеристик: высоту растений, высоту до початка, стеблевое полегание, корневое полегание, конечную популяцию, влажность зерна, натуральную массу и урожайность. Параметры, высота растений и высота до початка, дают информацию об общем виде гибридов. Агрономические характеристики стеблевое полегание и корневое полегание в процентах определяют возможности сбора урожая гибрида. Оценка конечной популяции позволяет измерять качество семян и сезонные условия выращивания, которые влияют на урожай. Влажность зерна в процентах при сборе определяет зрелость гибрида, и урожайность (бушелей/акр с корректировкой на влажность) и натуральная масса (масса в фунтах бушеля кукурузы с корректировкой на 15,5% влажности) описывают воспроизводительную способность гибрида.
Дисперсионный анализ проводили для разных местоположений полей, используя линейную модель. Объект и местоположение включали в модель в качестве фиксированных эффектов. Исследовали смешанные модели, включающие местоположение и местоположение по объектам в качестве случайных эффектов, но местоположение по объектам объясняло только небольшую часть дисперсии, и компонента дисперсии часто значимо не отличалась от нуля. В случае стеблевого и корневого полегания использовали логарифмическое превращение, чтобы стабилизировать дисперсию, однако средние и диапазоны сообщали в единицах исходной шкалы. Различия признавали значимыми при 95% доверительном уровне. Значимость общего эффекта обработки оценивали, используя t-критерий.
Результаты таких испытаний агрономических характеристик можно найти в таблице 2. Не обнаружено статистически значимых различий ни для одного из четырех гибридов 40278 по сравнению с изолинейными контролями (p<0,05) в отношении параметров: высоты до початка, стеблевого полегания, корневого полегания, влажности зерна, натуральной массы и урожайности. Оценка конечной популяции и высота растений были статистически разными в экспериментах A и B, соответственно, но не наблюдали сходных различий при сравнении с другими тестированными гибридами 40278. Некоторая наблюдаемая изменчивость может быть следствием низких уровней генетической вариабельности, сохраняющейся в результате возвратного скрещивания растений с событием DAS-40278-9 с элитными инбредными линиями. Общий диапазон значений измеряемых параметров во всех случаях был в пределах значений, полученных для традиционных гибридов кукурузы, и нельзя было прийти к заключению о повышенной засоренности сорняками. В заключение, данные анализа агрономических характеристик показывают, что кукуруза 40278 биологически эквивалентна обычной кукурузе.
Пример 8. Применение сайта инсерции события DAS-40278-9 кукурузы для целенаправленной интеграции
Соответствующая агрономическая эффективность трансгена в случае события DAS-40278-9 кукурузы на протяжении нескольких поколений в полевых условиях свидетельствует, что идентифицированные области вокруг сайта инсерции в случае события DAS-40278-9 кукурузы обеспечивают хорошие положения в геноме для целенаправленной интеграции других представляющих интерес трансгенных генов. Такая целенаправленная интеграция позволяет преодолевать проблемы, связанные с так называемым «эффектом положения» и риском возникновения мутации в гене при интеграции трансгена в хозяине. Дополнительные преимущества такой целенаправленной интеграции включают без ограничения уменьшение большого количества событий трансформации, которые необходимо подвергать скринингу и тестировать до получения трансгенного растения, которое имеет требуемый уровень экспрессии трансгена, к тому же без проявления аномалий в результате случайной инсерции трансгена в важный локус в геноме хозяина. Кроме того, такая целенаправленная интеграция позволяет осуществлять стэкинг трансгенов, который делает выведение элитных линий растений с обоими генами более эффективным.
Используя представленные инструкции, специалист сможет направить представляющие интерес полинуклеиновые кислоты в тот же сайт инсерции в хромосоме 2, что и в случае события DAS-40278-9 кукурузы, или в сайт, расположенный непосредственно вблизи сайта инсерции в случае события DAS-40278-9 кукурузы. Один из таких способов раскрыт в международной заявке на выдачу патента № WO2008/021207, которая включена в настоящее описание в виде ссылки в полном объеме.
Кратко, до 20 т.п.н. геномной последовательности, фланкирующей с 5’-стороны сайта инсерции, и до 20 т.п.н. геномной последовательности, фланкирующей с 3’-стороны сайта инсерции (части, которые указаны со ссылкой на последовательность SEQ ID NO: 29), используют для фланкирования представляющего интерес гена или генов, которые предполагается встроить в геномное положение в хромосоме 2 посредством гомологичной рекомбинации. Представляющий интерес ген или гены могут быть помещены в точно такой же сайт инсерции, как в случае события DAS-40278-9 кукурузы или могут быть помещены в ином месте в пределах областей длиной 20 т.п.н. вокруг сайтов инсерции события DAS-40278-9 кукурузы, чтобы обеспечить подходящий уровень экспрессии трансгена без неблагоприятного влияния на растение. ДНК-векторы, содержащие представляющий интерес ген или гены и фланкирующие последовательности, могут быть доставлены в растительные клетки одним из нескольких способов, известных специалистам в данной области, включая без ограничения трансформацию, опосредованную Agrobacterium. Инсерция векторов донорной ДНК в участок-мишень события DAS-40278-9 кукурузы может быть дополнительно усилена одним из нескольких способов, включая без ограничения коэкспрессию или повышающую регуляцию усиливающих рекомбинацию генов или понижающую регуляцию генов, подавляющих эндогенную рекомбинацию. Кроме того, в данной области известно, что двунитевое расщепление специфичных последовательностей в геноме можно использовать для увеличения частоты гомологичной рекомбинации, таким образом инсерция в сайт инсерции события DAS-40278-9 кукурузы и его фланкирующие области может быть усилена за счет экспрессии природных или сконструированных специфичных для последовательности эндонуклеаз для расщепления таких последовательностей. Таким образом, используя инструкции, приведенные в настоящем описании, можно встроить любую гетерологичную нуклеиновую кислоту в хромосому 2 кукурузы в участок-мишень, расположенный между 5’-молекулярным маркером, обсуждаемым в примере 4, и 3’-молекулярным маркером, обсуждаемым в примере 4, предпочтительно в пределах последовательности SEQ ID NO: 29 и/или в ее областях, которые обсуждаются в настоящем описании.
Пример 9. Эксцизия кассеты экспрессии гена pat из события DAS-40278-9 кукурузы
Удаление кассеты экспрессии селектируемого маркерного гена является предпочтительным для целенаправленной инсерции в геномный локус события DAS-40278-9 кукурузы. Удаление селектируемого маркера pat из события DAS-40278-9 кукурузы позволяет повторно использовать селектируемый маркер pat в целенаправленной интеграции полинуклеиновых кислот в хромосому 2 в последующих поколениях кукурузы.
Применяя описанные инструкции, специалист способен вырезать представляющие интерес полинуклеиновые кислоты из события DAS-40278-9 кукурузы. Один из таких способов раскрыт в предварительной заявке на выдачу патента США № 61/297628, которая включена в настоящее описание в виде ссылки в полном объеме.
Кратко, конструируют специфичные для последовательности эндонуклеазы, такие как нуклеазы с цинковыми пальцами, которые узнают, связывают и расщепляют специфичные последовательности ДНК, которые фланкируют кассету экспрессии гена. Нуклеазы с цинковыми пальцами доставляют в растительную клетку посредством скрещивания родительского растения, которое содержит кассеты экспрессии трансгенных нуклеаз с цинковыми пальцами, со вторым родительским растением, в котором имеет место событие DAS-40278-9 кукурузы. Полученное потомство выращивают до состояния зрелости и анализируют в отношении утраты кассеты экспрессии pat посредством нанесения на листья гербицида, который содержит глюфосинат. Растения-потомки, которые не являются резистентными к гербициду, подтверждают на молекулярном уровне и выращивают для самоопыления. Эксцизия и удаление кассеты экспрессии pat молекулярно подтверждают у потомства, полученного в результате такого самоопыления. Используя инструкции, представленные в настоящем описании, любую гетерологичную нуклеиновую кислоту можно вырезать из хромосомы 2 кукурузы в участке-мишени, расположенном между 5’-молекулярным маркером и 3’-молекулярным маркером, которые обсуждаются в примере 4, предпочтительно в пределах последовательности SEQ ID NO: 29 или в ее указанных областях.
Пример 10. Устойчивость к резким внезапным переломам стебля
Резкий внезапный перелом стебля относится к повреждению стеблей кукурузы сильными ветрами после применения гербицидов-регуляторов роста, обычно в период быстрого роста. Тесты с использованием механического «удара», в которых используют прут, чтобы физически толкнуть кукурузу, имитируя повреждение сильными ветрами, проводили на гибридной кукурузе, в которой имеет место событие DAS-40278-9, и контрольных растениях, в которых не происходило событие DAS-40278-9. Обработки осуществляли в четырех разных географических местоположениях и повторяли четыре раза (имело место исключение в случае одного испытания, которое повторяли только три раза). Делянки состояли из восьми рядов: по четыре ряда для каждого из двух гибридов, при этом два ряда для кукурузы, в которой имеет место событие DAS-40278-9, и два ряда для кукурузы без события. Каждый ряд имел длину двадцать футов (6 м). Растения кукурузы выращивали до стадии развития V4 и вносили коммерческий гербицид, содержащий 2,4-D (Weedar 64, Nufarm Inc., Burr Ridge, IL) при норме 1120 г эк/га, 2240 г эк/га и 4480 г эк/га. Через семь дней после внесения гербицида осуществляли тест с механический ударом. Тест с механическим ударом для исследования резких внезапных переломов заключался в сбивании 4-футовым (1,2 м) прутом двух рядов кукурузы для имитации повреждения ветром. Высота прута и скорость перемещения были подобраны так, чтобы обеспечить низкий уровень поломки стеблей (10% или меньше) в случае необработанных растений, чтобы гарантировать тест достаточно жесткий, чтобы показать различие между обработками. Проведение обработки для получения резких переломов осуществляли в направлении, противоположном наклону кукурузы.
В двух местоположениях, где проводили испытания, дули сильные ветры и грозы через 2-3 дня после внесения гербицида 2,4-D. Два дня подряд гроза была в Huxley IA. Сообщали о скорости ветра от 2 до 17 м сек-1 с порывами 33 м сек-1 в месте полевой делянки. Направление ветра было переменным. В один из дней сообщалось о грозе в Lanesboro MN. О ветрах высокой скорости сообщалось в месте указанной полевой делянки. Кроме того, при обеих грозах шел дождь. Сочетанием дождя и ветра объясняли резкие внезапные переломы, о которых сообщается.
Оценки резких переломов в тесте с механическими ударами (и суровой погодой) осуществляли благодаря визуальной оценке процента повреждения. Перед механической обработкой прутом, вызывающим резкие переломы, осуществляли оценку густоты стояния растений гибридной кукурузы, в которой имеет место событие DAS-40278-9, и контролей. Через несколько дней после обработки прутом для резких переломов повторно оценивали густоту стояния растений на делянке. Определяли процент полегания и процент пониженной густоты стояния растения на делянке (таблица 26). Данные, полученные в испытаниях, показали, что гибридная кукуруза, в которой имело место событие DAS-40278-9, менее предрасположена к резким внезапным переломам по сравнению с нулевыми растениями после применения 2,4-D.
Средний % полегания через 7-8 дней после внесения1
4XP811XTR)
4XP811XTR)
BE9515XT)
BE9515XT)
1120 г эк/га
2240 г эк/га
4480 г эк/га
танные
Средний3 % полегания через 11-14 дней после внесения
4XP811XTR)
BE9515XT)
1120 г эк/га
2240 г эк/га
4480 г эк/га
танные
Средний3 % снижения густоты стояния через 11-14 дней после внесения
4XP811XTR)
BE9515XT)
1120 г эк/га
2240 г эк/га
4480 г эк/га
танные
2 Обработки, повторяемые четыре раза по рандомизированному полноблочному плану (только одно испытание осуществляли в трех повторах).
3 Средние значения, скорректированные с учетом необработанных растений (средние для необработанных принимали за нулевые значения).
Пример 11. Анализ белка в зерне
Зерно с повышенным содержанием общего белка получали от гибридной кукурузы, в которой имело место событие DAS-40278-9, по сравнению с контрольными растениями, не имеющими такого события. Проводили два последовательных многоцентровых полевых испытания, которые включали неопрыскиваемые растения и обработанные гербицидами растения, при этом использовали сочетания трех разных гербицидов. В 7 из 8 статистических сравнений в случае события DAS-40278-9 получали зерно со значимо более высоким содержанием общего белка (таблица 27). Полученные данные были подтверждены в анализах отдельных аминокислот.
2,4-D
2,4-D
Пример 12. Дополнительные агрономические испытания
Агрономические характеристики гибридной кукурузы, в которой имело место событие DAS-40278-9, сравниваемой с почти изолинейной кукурузой, собирали в многоцентровых полевых испытаниях, в разных географических условиях в течение сезона выращивания. Данные собирали и анализировали в отношении агрономических характеристик, как описано в примере 7. Результаты для гибридных линий кукурузы, в которых имело место событие DAS-40278-9, по сравнению с нулевыми растениями перечислены в таблице 28. Кроме того, агрономические характеристики для гибридных линий кукурузы, в которых имело место событие DAS-40278-9, и нулевых растений, опрыснутых гербицидом квизалофопом (280 г эк/га) на стадии развития V3 и опрыснутых 2,4-D (2240 г эк/га) на стадии развития V6, описаны в таблице 29.
акр, указанных в тысячах)
Пример 13. Предпосевное и/или довсходовое внесение
Предпосевные внесения гербицидов для «выжигания» сорняков предназначены для уничтожения сорняков, которые появились после перезимовки или ранней весной перед посевом данной культуры. Обычно такие внесения применяют в системах, где не применяют подготовку почвы вспахиванием или проводят ограниченную вспашку, в которых не происходит физическое удаление сорняков перед посевом. Поэтому гербицидная программа должна осуществлять борьбу с очень широким спектром широколиственных и злаковых сорняков, присутствующих во время посева. Глифосат, грамоксон и глюфосинат являются примерами неизбирательных, непоследействующих гербицидов, широко используемых для предпосевного внесения «выжигающих» гербицидов.
Однако с некоторыми сорняками трудно бороться в этот период сезона по одной или нескольким из следующих причин: наследственной нечувствительности вида или биотипа сорняка к гербициду, относительно крупного размера озимых однолетних сорняков и холодной погоды, ограничивающей поглощение и активность гербицидов. Несколько вариантов гербицидов доступны для приготовления баковых смесей с такими гербицидами с целью расширения спектра сорняков и повышения активности по отношению к сорнякам, когда неизбирательные гербициды являются слабыми. Примером может быть применение баковой смеси 2,4-D с глифосатом для борьбы с Conyza canadensis (мелколепестник канадский). Глифосат можно применять в дозе от 420 до 1680 г эк/га, чаще от 560 до 840 г эк/га для предпосевной борьбы по типу выжигания с большинством имеющихся сорняков; однако 280-1120 г эк/га 2,4-D можно применять для борьбы с многими широколиственными видами сорняков (например, с мелколепестником канадским).
2,4-D является предпочтительным гербицидом, поскольку он эффективен против очень широкого круга широколиственных сорняков, эффективен даже при низких температурах, и является очень недорогим. Однако если последующей культурой является чувствительная двудольная культура, остатки 2,4-D в почве (хотя и недолго живущие) могут негативно влиять на культуру. Культуры, которые содержат ген aad-1, устойчивы к 2,4-D и не подвержены негативному влиянию остатков 2,4-D в почве. Повышенная гибкость и пониженная стоимость вносимых компонентов в баковые смеси (или коммерческие готовые смеси) будут улучшать параметры борьбы с вредителями и повышать надежность применений по типу выжигания в случае ситуаций, когда не применяют подготовку почвы вспахиванием или проводят ограниченную вспашку.
Кукуруза aad-1
Трансгенную гибридную кукурузу (pDAS 1740-278), содержащую ген aad-1, который кодирует белок арилоксиалканоатдиоксигеназы (AAD-1), оценивали в отношении устойчивости к довсходовому внесению 2,4-D на поле. Испытания проводили в нескольких местоположениях в Миссисипи, Индиане и Миннесоте, используя рандомизированный полноблочный план с тремя повторами двурядных делянок длиной примерно 6 м в каждом местоположении. Обрабатываемые гербицидами делянки закладывали в паре с необрабатываемыми делянками, чтобы обеспечить точную оценку всхожести и раннего сезонного роста. Гербицидные обработки по 1120, 2240 и 4480 г эк/га 2,4-D-амина проводили сразу после посева, но до всходов культуры (через 0-2 дня после посева). Информация о почве и атмосферных осадках при таких испытаниях приведена в таблице 30.
Примерно через 16-21 дней после посева и внесения 2,4-D повреждение составляло в среднем от 12 до 31% в случае обычного контрольного гибрида при нормах, повышенных от 1120 до 4480 г эк/га 2,4-D-амина. Повреждение гибридной кукурузы, содержащей pDAS 1740-278, было в диапазоне от 3 до 9% в том же диапазоне норм внесения. Предлагаемые в настоящее время нормы целевого внесения 2,4-D для трансгенной гибридной кукурузы (pDAS 1740-278), содержащей ген aad-1, составляют 1120 г эк/га или ниже в случае 2,4-D. Результаты полевых испытаний показывают, что гибридная кукуруза, содержащая pDAS 1740-278, имеет высокую устойчивость к обработкам гербицидом 2,4-D при нормах, более чем в два-четыре раза превышающих предлагаемые нормы для целевого применения, с минимальными повреждениями (таблица 31).
b вносили через 0-2 дня после посева до всходов;
c Оценки через 16-21 день после внесения.
Довсходовое внесение 2,4-D-амина применяют при нормах 1120, 2240, 4480 г эк/га за 7 дней, 15 дней или 30 дней до посева гибридной кукурузы, содержащей ген aad-1, и обычных контрольных гибридов. Довсходовое внесение применяют, используя известные в данной области способы, на полевых делянках, которые расположены в географически разных областях. Обрабатываемые гербицидами делянки закладывают в паре с необрабатываемыми делянками, чтобы обеспечить точную оценку всхожести и раннего сезонного роста. Примерно через 16-21 день после посева и внесений 2,4-D за 7, 15 или 30 дней до посева измеряют повреждение обычных контрольных гибридов и гибридной кукурузы, содержащей aad-1. Результаты полевых испытаний показывают, что гибридная кукуруза, содержащая aad-1, обладает высокой устойчивостью к довсходовым обработкам гербицидом 2,4-D за 7, 15 или 30 дней до посева.
Хлопчатник aad-1
Трансгенный хлопчатник, содержащий ген aad-1, который кодирует белок арилоксиалканоатдиоксигеназы (AAD-1), оценивали в отношении устойчивости к довсходовому внесению 2,4-D на поле. Испытания состояли из трех повторов, и их проводили в нескольких местоположениях в Миссисиппи, Джорджии, Теннесси и Арканзасе. Использовали рандомизированный полноблочный план, по которому отдельный ряд отделен защитным рядом, длиной примерно 10 футов (3 м) (20 футов (6 м) в испытании в Миссисиппи). Семена высевали, используя по 8 семян на фут (0,3 м), затем растения вручную прореживали до 3,5 растений/фут в ряду. Обрабатываемые гербицидами делянки закладывали в паре с необрабатываемыми делянками, чтобы обеспечить точную оценку всхожести и раннего сезонного роста. Все делянки получали по меньшей мере ½ дюйма (12,5 мм) дождевой или ирригационной воды в пределах 24 часов после внесения. Гербицидные обработки по 560, 1120, 2240 и 4480 г эк/га 2,4-D-амина (WEEDAR 64, Nufarm, Burr Ridge, II) проводили сразу после посева, но перед всходами культуры. Информация о почве и атмосферных осадках при таких испытаниях приведена в таблице 32.
Довсходовое внесение 2,4-D по норме 560, 1120 и 2240 г эк/га значимо не влияло на густоту стояния трансгенных растений хлопчатника, содержащих ген aad-1, по сравнению с необработанной делянкой.
Примерно через 7-8 дней после посева и довсходового применения 2,4-D в концентрациях 560, 1120, 2240 и 4480 г эк/га сообщали о снижении густоты стояния на 3%, отсутствии снижения, снижении на 19% и 34%, соответственно, в случае хлопчатника, содержащего aad-1, по сравнению со снижением густоты стояния в необработанном контроле. Через 11-14 дней после посева и внесения 2,4-D в концентрациях 560, 1120, 2240 и 4480 г эк/га сообщали о снижении густоты стояния на 5%, 2%, 14% и 25%, соответственно, в случае хлопчатника, содержащего aad-1, по сравнению со снижением густоты стояния в необработанном контроле.
Довсходовое внесение 2,4-D в концентрации 560, 1120, 2240 и 4480 г эк/га не вызывало эпинастии. Хлороз наблюдали на уровне <5% при визуальной оценке через 7-8 дней после внесения в случае обработок 2240 и 4480 г эк/га. Хлороз не выявляли в случае обработок 560 и 1120 г эк/га на 7-8 день после внесения. Кроме того, хлороз не выявляли в хлопке, содержащем aad-1, ни при одной из концентраций 560, 1120, 2240 и 4480 г эк/га в оставшийся период сезона.
Довсходовое внесение 2,4-D в концентрациях 560 и 1120 г эк/га приводило к минимальному ингибированию роста менее чем на 2% (которое статистически не значимо по сравнению с необработанной контрольной делянкой). Полученные результаты были постоянны на протяжении испытания: 7-8, 11-14, 27-30 и 52-57 дней после внесения. Довсходовое внесение 2,4-D в концентрации 2240 г эк/га приводило к ингибированию роста менее чем на 10%. Довсходовое внесение 2,4-D в концентрации 4480 г эк/га приводило к ингибированию роста на 27%, 28%, 17% и 8,3% при оценке на 7-8, 11-14, 27-30 и 52-57 дни после внесения.
Повреждение, вызванное довсходовым внесением 2,4-D в концентрациях 560 и 1120 г эк/га, значимо не отличалось от необработанной контрольной делянки. Оценки повреждений в случае растений хлопчатника aad-1, обработанных 2,4-D в концентрации 2240 г эк/га, были в диапазоне от 3% до 15% в течение всего испытания. Оценки повреждений в случае растений хлопчатника aad-1, обработанных 2,4-D в концентрации 4480 г эк/га, были в диапазоне от 8% до 34% в ходе всего испытания.
Отличия в высоте растений, характере плодоношения и урожайности не были выявлены у хлопчатника aad-1, который обрабатывали, используя довсходовое внесение 2,4-D в концентрациях 560, 1120, 2240 и 4480 г эк/га.
Довсходовое внесение 2,4-D-амина осуществляли, используя нормы внесения 560, 1120, 2240, 4480 г эк/га, за 7 дней, 15 дней или 30 дней до посева хлопчатника, содержащего ген aad-1, и контрольного хлопчатника. Довсходовое внесение осуществляли, используя известные в данной области способы, на полевых делянках, которые расположены в географически разных местоположениях. Обрабатываемые гербицидами делянки закладывают в паре с необрабатываемыми делянками, чтобы обеспечить точную оценку всхожести и раннего сезонного роста. После посева и внесения 2,4-D за 7, 15 или 30 дней до посева измеряли повреждение хлопчатника, содержащего ген aad-1, и контрольного хлопчатника. Результаты полевого тестирования показывают, что хлопчатник, содержащий ген aad-1, обладает устойчивостью к довсходовым обработкам гербицидом 2,4-D за 7, 15 или 30 дней до посева.
В настоящем примере обсуждаются некоторые из многочисленных доступных вариантов. Специалисты в области борьбы с сорняками в качестве примеров отметят множество других внесений, включая без ограничения грамоксон+2,4-D или глюфосинат+2,4-D с использованием продуктов, описанных в федеральном идентификаторе гербицидов (CPR, 2003), и применений, описанных в руководстве по защите растений Agriliance (2003). Специалистам в данной области также будет известно, что описанный выше пример может быть применен по отношению к любой 2,4-D-чувствительной (или чувствительной к другому феноксиауксиновому гербициду) культуре, которая может быть защищена геном AAD-1 (v3) при стабильной трансформации.
| название | год | авторы | номер документа |
|---|---|---|---|
| СОБЫТИЕ DAS-40278-9 AAD-1, РОДСТВЕННЫЕ ЛИНИИ ТРАНСГЕННОЙ КУКУРУЗЫ И ИХ СОБЫТИЕ-СПЕЦИФИЧЕСКАЯ ИДЕНТИФИКАЦИЯ | 2010 |
|
RU2614120C2 |
| ДЕТЕКЦИЯ AAD-1 ОБЪЕКТА DAS-40278-9 | 2010 |
|
RU2577143C2 |
| СОБЫТИЕ 416 AAD-12, РОДСТВЕННЫЕ ЛИНИИ ТРАНСГЕННОЙ СОИ И ИХ СОБЫТИЕСПЕЦИФИЧНАЯ ИДЕНТИФИКАЦИЯ | 2010 |
|
RU2603252C2 |
| ЛИНИИ ТРАНСГЕННОЙ СОИ, ГЕНЕТИЧЕСКОЕ СОБЫТИЕ 8264.42.32.1, УСТОЙЧИВОЕ К ГЕРБИЦИДАМ С ПАКЕТИРОВАННЫМИ ГЕНАМИ НА ЕГО ОСНОВЕ, И ИХ ДЕТЕКТИРОВАНИЕ | 2012 |
|
RU2636021C2 |
| СПОСОБЫ ПОВЫШЕНИЯ УРОЖАЙНОСТИ РЕЗИСТЕНТНЫХ К 2,4-D СЕЛЬСКОХОЗЯЙСТВЕННЫХ КУЛЬТУР | 2013 |
|
RU2628504C2 |
| ПАКЕТИРОВАННОЕ СОБЫТИЕ 8264.44.06.1 С УСТОЙЧИВОСТЬЮ К ГЕРБИЦИДАМ, СВЯЗАННЫЕ ТРАНСГЕННЫЕ ЛИНИИ СОИ И ИХ ДЕТЕКТИРОВАНИЕ | 2011 |
|
RU2608650C2 |
| ДЕТЕКТИРОВАНИЕ AAD-12-СОБЫТИЯ 416 У СОИ | 2010 |
|
RU2573898C2 |
| УСТОЙЧИВЫЙ К НАСЕКОМЫМ-ВРЕДИТЕЛЯМ И ТОЛЕРАНТНЫЙ К ГЕРБИЦИДАМ СЕЛЕКЦИОННЫЙ ГИБРИД ТРАНСФОРМАНТА СОИ pDAB9582.814.19.1 и pDAB4468.04.16.1 | 2012 |
|
RU2627161C2 |
| СПОСОБЫ ОБНАРУЖЕНИЯ ДНК ДЛЯ САЙТ-СПЕЦИФИЧЕСКОЙ НУКЛЕАЗНОЙ АКТИВНОСТИ | 2013 |
|
RU2678001C2 |
| ТРАНСГЕННЫЙ ОБЪЕКТ СОИ MON 87708 И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2010 |
|
RU2624025C2 |












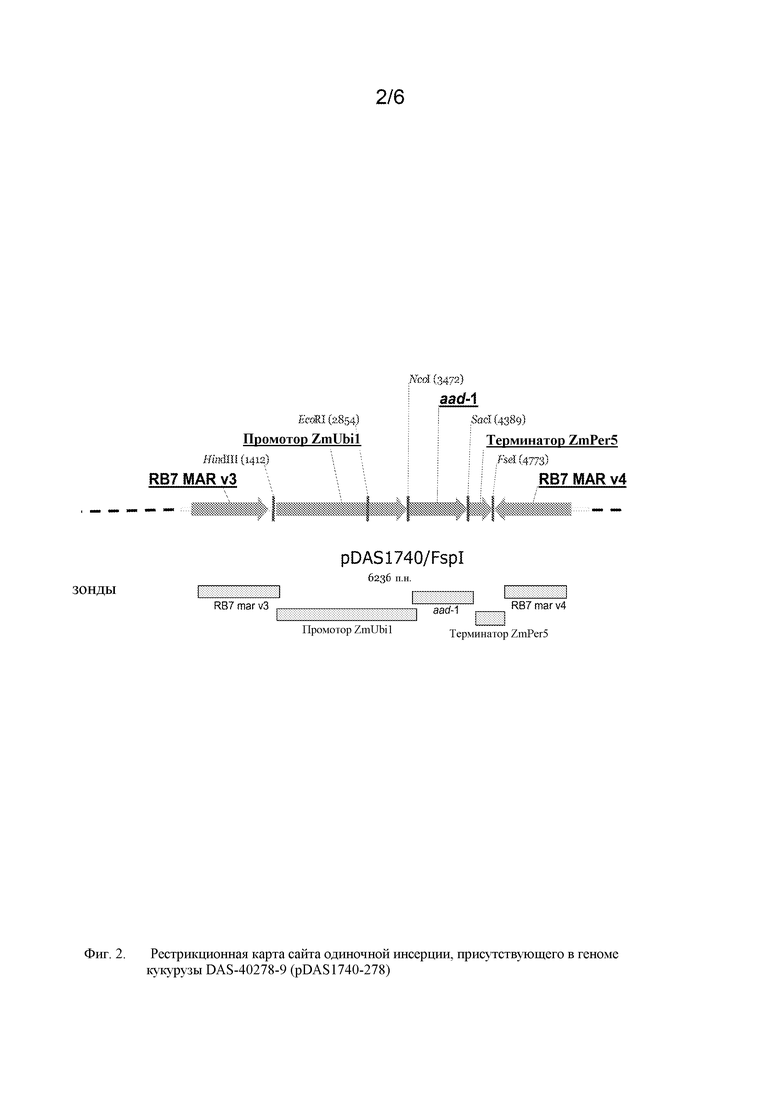
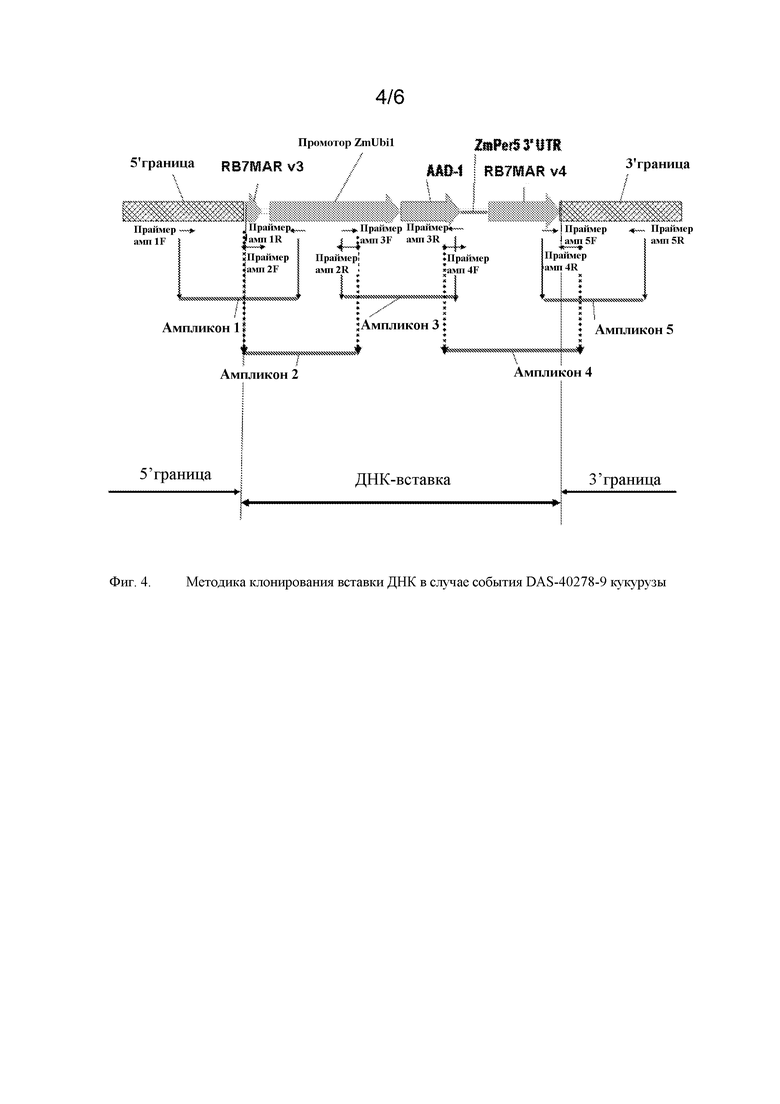


Изобретение относится к области биохимии, в частности к способу борьбы с сорняками, включающему посев семян на определенной площади и внесение арилоксиалканоатного гербицида на указанной площади за 30 дней до посева семян на указанной площади, причем указанные семена содержат белок арилоксиалканоатдиоксигеназу AAD-1, кодируемый последовательностью SEQ ID NO: 29. Изобретение позволяет эффективно бороться с сорняками на определенной площади, содержащей культурные растения. 11 з.п. ф-лы, 7 ил., 33 табл., 13 пр.
1. Способ борьбы с сорняками, включающий посев семян на определенной площади и внесение арилоксиалканоатного гербицида на указанной площади за 30 дней до посева семян на указанной площади, причем указанные семена содержат белок арилоксиалканоатдиоксигеназу AAD-1, кодируемый последовательностью SEQ ID NO: 29.
2. Способ по п. 1, в котором указанный гербицид выбран из группы, состоящей из 2,4-D, 2,4-DB, МСРА, МСРВ и арилоксифеноксипропионата.
3. Способ по п. 1, в котором указанная площадь представляет собой поле и указанный гербицид вносят в течение 7 дней, 11 дней, 14 дней или 15 дней до посева указанных семян.
4. Способ по п. 1, используемый для борьбы с резистентными к глифосату сорняками на указанной площади, при этом из указанных семян выращивают растения, дополнительно обладающие признаком устойчивости к глифосату, и указанный способ включает внесение арилоксиалканоатного гербицида по меньшей мере на части указанной площади.
5. Способ по п. 1, в котором указанным гербицидом является феноксиауксин.
6. Способ по п. 4, в котором по меньшей мере один из указанных сорняков является резистентным к глифосату самосевом другого вида, отличного от указанного растения.
7. Способ по п. 1, в котором указанные сорняки находятся на площади, подвергаемой культивации, причем на указанной площади имеется множество растений, выращиваемых из указанных семян, причем указанный способ дополнительно включает внесение арилоксиалканоатного гербицида поверх растений.
8. Способ по п. 7, включающий довсходовое, послевсходовое или довсходовое и послевсходовое нанесение гербицида на растения, части растений и/или культивируемую площадь.
9. Способ по п. 1, в котором указанным арилоксиалканоатным гербицидом является 2,4-D-амин и указанный 2,4-D-амин вносят с нормой расхода в диапазоне, выбранном из группы, состоящей из 560, 1120, 2240 и 4480 г эк/га.
10. Способ по п. 1, в котором указанным арилоксиалканоатным гербицидом является 2,4-D-амин и указанный 2,4-D-амин вносят с нормой расхода 280-1120 г эк/га.
11. Способ по п. 1, в котором указанное семя получено из растения кукурузы.
12. Способ по п. 1, в котором указанное семя содержит SEQ ID NO: 29, как имеет место в семени, депонированном в Американской коллекции типов культур (АТСС) с номером доступа РТА-10244.
| HERMAN R.A | |||
| et al., Compositional safety of event DAS-40278-9 (AAD-1) herbicide-tolerant maize, GM Crops, 2010, 1-5, 294-311 | |||
| WO 03024227 A2, 27.03.2003 | |||
| US 2009093366 A1, 09.04.2009 | |||
| ТРАНСГЕННЫЕ РАСТЕНИЯ, ЭКСПРЕССИРУЮЩИЕ ГЕН АЦЕТИЛ-СОА-КАРБОКСИЛАЗЫ | 1996 |
|
RU2187555C2 |

