ПРИОРИТЕТ
По этой заявке испрашивается приоритет на приоритет по дате подачи временной заявки на патент США с регистрационным № 61/582138, поданной 30 декабря 2011 года. По этой заявке также испрашивается приоритет по дате подачи временной заявки на патент США с регистрационным № 61/617252, поданной 29 марта 2012 года.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение, в общем, относится к области молекулярной биологии растений, и более конкретно, к области стабильной экспрессии множественных генов в трансгенных растениях.
УРОВЕНЬ ТЕХНИКИ
Многие виды растений могут быть трансформированы трансгенами из других видов, чтобы ввести агрономически требуемые свойства или характеристики, например, повышающие такое качество, как пищевая ценность, увеличивающие урожайность, придающие устойчивость к насекомым-вредителям или заболеваниям, повышающие засухоустойчивость и стрессоустойчивость, улучшающие качества садово-огородной продукции (такие как пигментация и рост), придающие устойчивость к гербицидам, делающих возможным получение промышленно применимых соединений и/или материалов из растений и/или делающих возможным получение фармацевтических средств. Введение трансгены в растительные клетки и последующая регенерация фертильных трансгенных растений, которые содержат стабильно интегрированную копию трансгена, можно применять для получения трансгенных растений, которые обладают требуемыми свойствами.
Контроль и регуляция генной экспрессии может осуществляться разными механизмами. Инициация транскрипции гена является основным механизмом регуляции генной экспрессии. Инициация транскрипции обычно контролируется полинуклеотидными последовательностями, локализованными в 5'-фланкирующей области или области, лежащей выше транскрибируемого гена. Такие последовательности вместе называют промоторами. Промоторы обычно содержат сигналы для РНК-полимеразы к началу транскрипции, так что может быть получена матричная РНК (мРНК). Зрелая мРНК транслируется рибосомой, при этом происходит синтез белков. ДНК-связывающие белки специфично взаимодействуют с промоторными последовательностями ДНК, стимулируя образование транскрипционного комплекса, и инициируют процесс генной экспрессии. Существует множество эукариотических промоторов, выделенных из растений и охарактеризованных, которые являются функциональными, запуская экспрессию трансгена в растениях. Промоторы, которые влияют на генную экспрессию в ответ на стимулы окружающей среды, доступность питательных веществ или неблагоприятные условия, включая тепловой шок, анаэробиоз или присутствие тяжелых металлов, были выделены и охарактеризованы. Также существуют промоторы, которые контролируют генную экспрессию во время развития или специфичным для ткани или органа образом. Кроме того, были выделены и охарактеризованы прокариотические промоторы из бактерий и вирусов, которые являются функциональными, запуская экспрессию трансгена в растениях.
Типичный эукариотический промотор состоит из минимального промотора и других цис-элементов. Минимальный промотор АО существу представляет собой область TATA-бокса, где ДНК-полимераза II (polII), TATA-связывающий белок (TBP) и TBP-ассоциированные факторы (TAF) могут связываться, инициируя транскрипцию. Однако в большинстве случаев другие элементы последовательности, отличные от TATA-мотива, необходимы для точной транскрипции. Было обнаружено, что такие элементы последовательности (например, энхансеры) повышают общий уровень экспрессии близлежащих генов, часто независимым от положения и/или ориентации образом. Другие последовательности вблизи участка старта транскрипции (например, последовательности INR) некоторых генов polII могут обеспечивать альтернативный участок связывания для факторов, которые также вносят вклад в активацию транскрипции, даже альтернативно обеспечивать участки связывания корового промотора для транскрипции в случае промоторов, в которых отсутствуют функциональные TATA-элементы. См., например, Zenzie-Gregory с соавторами (1992) J. Biol. Chem. 267: 2823-30.
Другие элементы регуляции генов включают последовательности, которые взаимодействуют со специфичными ДНК-связывающими факторами. Такие мотивы последовательностей иногда называют цис-элементами, и они обычно зависят от положения и ориентации, хотя они могут быть обнаружены с 5ʹ- или 3ʹ-стороны от колирующей последовательности гена или в интроне. Такие цис-элементы с которыми связываются специфичные для ткани или специфичные для стадии развития факторы транскрипции, отдельно или в сочетании, могут определять пространственно-временную картину экспрессии с промотора на уровне транскрипции. Расположение вышележащих цис-элементов с последующим минимальным промотором обычно обеспечивает полярность конкретного промотора. Промоторы растений, которые были клонированы и широко использованы как для фундаментальных исследований, так и для биотехнологических применений, обычно являются однонаправленными, управляющими только одним геном, который был слит с их 3'-концом (т.е., ниже). См., например, публикации Xie с соавторами (2001) Nat. Biotechnol. 19(7): 677-9; патент США № 6388170.
Множество цис-элементов (или «вышележащих регуляторных последовательностей») было идентифицировано в растительных промоторах. Такие цис-элементы широко варьируют по типу регуляции, которую они оказывают по отношению к оперативно связанным генам. Некоторые элементы действуют, увеличивая транскрипцию оперативно связанных генов в ответ на сигналы окружающей среды (например, температуру, влажность и ранение). Другие цис-элементы могут отвечать на сигналы в ходе развития (например, прорастание, созревание семян и цветение) или на пространственную информацию (например, тканевую специфичность). См., например, Langridge с соавторами (1989) Proc. Natl. Acad. Sci. USA 86: 3219-23. Тип регуляции конкретными промоторными элементами обычно является качеством, присущим промотору; т.е., гетерологичный ген под контролем такого промотора вероятно должен экспрессироваться в соответствие с регуляцией нативного гена, из которого был выделен промоторный элемент. Такие элементы обычно также можно заменить другими элементами и сохранить при этом характерный присущий им тип регуляции генной экспрессии.
Часто необходимо введение множества генов в растения для метаболической инженерии и объединения признаков, и такие гены часто регулируются идентичными или гомологичными промоторами. Однако вероятно возникает основанный на гомологии сайленсинг генов (HBGS) в случае, когда множество введенных трансгенов имеют гомологичные промоторы, которые ими управляют. См., например, Mol с соавторами: (1989) Plant Mol. Biol. 13: 287-94. Сообщалось, что HBGS часто возникает в трансгенных растениях. См., например, Vaucheret and Fagard (2001) Trends Genet. 17: 29-35. Было предложено несколько механизмов для объяснения явления HBGS, и характерным признаком всех таких механизмов является тот факт, что гомология последовательностей в промоторе запускает механизмы клеточного узнавания, которые приводят к сайленсингу повторяющихся генов. См., например, Matzke and Matzke (1995) Plant Physiol. 107:679-85; Meyer and Saedler (1996) Ann. Rev. Plant Physiol. Plant Mol. Biol. 47:23-48; Fire (1999) Trends Genet. 15:358-63; Hamilton and Baulcombe (1999) Science 286:950-2; и Steimer et al. (2000) Plant Cell 12:1165-78.
Методики, позволяющие избегать HBGS в трансгенных растениях, часто заключаются в разработке синтетических промоторов, которые функционально эквивалентны, но имеют минимальную гомологию последовательностей. В случае применения таких синтетических промоторов для экспрессии трансгенов в культурных растениях, они помогают избегать или уменьшать HBGS. См., например, публикации Mourrain с соавторами, (2007) Planta 225(2): 365-79; Bhullar с соавторами (2003) Plant Physiol. 132: 988-98. Такие промоторы могут быть созданы встраиванием известных цис-элементов в новый или синтетический участок ДНК или альтернативно «перестановкой доменов», при этом домены одного промотора заменяют функционально эквивалентными доменами других гетерологичных промоторов.
Таким образом, остается необходимость в конструкциях и способах для эффективной стабильной экспрессии множественных трансгенов с минимальным риском рекомбинации или утраты трансгенов в ходе селекции или множества поколений трансгенных растений.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В настоящем описании описаны способы превращения полярного промотора Ubi1 в синтетические двунаправленные промоторы, так чтобы один синтетический промотор мог управлять экспрессией двух генов, фланкирующих промотор. В некоторых вариантах способ превращения полярного промотора Ubi1 в синтетический двунаправленный промотор может включать, например и без ограничения, идентификацию нуклеотидной последовательности минимального промоторного элемента промотора Ubi1; и/или получение нуклеиновой кислоты, содержащей две нуклеотидные последовательности минимального промоторного элемента Ubi1, ориентированные в противоположных направлениях. В конкретных вариантах нуклеиновая кислота может содержать две нуклеотидные последовательности минимального промоторного элемента Ubi1, ориентированные в противоположных направлениях, так что конец каждого минимального промоторного элемента, который расположен ближе всего к соответствующему нативному гену Ubi1, находится дальше от другого минимального промоторного элемента, чем конец нуклеиновой кислоты, который соседствует с кодирующей последовательностью, оперативно связанной с промоторным элементом. В некоторых примерах минимальный промоторный элемент Ubi1 выделяют из кукурузы. Дополнительные элементы нативного промотора Ubi1, которые могут быть сконструированы для включения в синтетический двунаправленный промотор, включают интроны Ubi1, экзоны Ubi1 и/или всю или часть области, лежащей выше промотора Ubi1. В некоторых примерах синтетический двунаправленный промотор может содержать больше одной из описанных выше областей.
Также в настоящем описании описаны минимальные промоторы Ubi1, которые могут быть применимы при конструировании синтетических промоторов (например, синтетических двунаправленных промоторов), и особенно синтетических промоторов, получаемых описанными выше способами. В некоторых вариантах синтетический двунаправленный промотор представляет собой промотор, который способен контролировать транскрипцию оперативно связанной нуклеотидной последовательности в растительной клетке. Например, в конкретных вариантах синтетический двунаправленный промотор может обладать способностью контролировать транскрипцию в растительной клетке двух оперативно связанных нуклеотидных последовательностей, которые фланкируют промотор.
Конкретные варианты осуществления изобретения относятся к клеткам (например, растительным клеткам), содержащим минимальный промотор Ubi1 или его функциональный эквивалент. Например, конкретные варианты включают клетку, содержащую синтетический промотор (например, синтетический двунаправленный промотор), который содержит минимальный промотор Ubi1 или его функциональный эквивалент. Растительные клетки согласно конкретным вариантам могут быть представлены в виде культуры клеток, ткани, части растения и/или целого растения. Таким образом, растение (например, однодольное или двудольное), содержащее клетку, которая содержит минимальный промотор Ubi1 или его функциональный эквивалент, включено в некоторые варианты осуществления изобретения.
Некоторые варианты осуществления изобретения относятся к средствам для инициации транскрипции независимым от направления образом. Средства для инициации транскрипции независимым от направления образом включают минимальный промотор Ubi1 SEQ ID NO: 1. Некоторые варианты осуществления изобретения относятся к средствам для инициации транскрипции двух представляющих интерес оперативно связанных нуклеотидных последовательностей. Средства для инициации транскрипция двух представляющих интерес оперативно связанных нуклеотидных последовательностей включают синтетический двунаправленный промотор Ubi1 SEQ ID NO: 5.
Вышеуказанные и другие отличительные признаки будут более понятными из следующего подробного описания нескольких вариантов осуществления изобретения, которое следует далее со ссылкой на прилагаемые фигуры.
Также изобретение относится к конструкциям и способам экспрессии множественных генов в растительных клетках и/или тканях растений. Конструкции по изобретению содержат по меньшей мере один двунаправленный промотор, связанный с несколькими кассетами экспрессии генов. В некоторых вариантах в конструкциях и способах по изобретению используют двунаправленный промотор, основанный на минимальном коровом промоторном элементе гена убиквитина-1 Zea mays или его функциональном эквиваленте. В некоторых вариантах конструкции и способы по изобретению обеспечивают возможность экспрессии от трех до двадцати генов.
В одном аспекте изобретение относится к синтетическому полинуклеотиду, содержащему минимальный коровый промоторный элемент из гена убиквитина-1 Zea mays или Zea luxurians. В одном варианте минимальный коровый промоторный элемент содержит полинуклеотидную последовательность, которая по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентична последовательности SEQ ID NO: 1 или ее комплементу. В следующем или альтернативном варианте минимальный коровый промоторный элемент содержит полинуклеотидную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 1 и 15-39. В следующем варианте минимальный коровый промоторный элемент содержит последовательность SEQ ID NO: 1 или ее комплемент. В следующем варианте минимальный коровый промоторный элемент по существу состоит из последовательности SEQ ID NO: 1 или ее комплемента. В другом варианте синтетический полинуклеотид дополнительно содержит экзон из гена убиквитина-1 и интрон из гена убиквитина-1. В следующем варианте экзон или интрон получены из гена убиквитина-1 Zea mays или Zea luxurians.
В другом варианте синтетический полинуклеотид дополнительно содержит вышележащую регуляторную последовательность из гена убиквитина-1. В следующем варианте вышележащая регуляторная последовательность содержит полинуклеотидную последовательность, которая по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентична последовательности SEQ ID NO: 4 или ее комплементу. В следующем варианте вышележащая регуляторная последовательность содержит последовательность SEQ ID NO: 4 или ее комплемент. В следующем варианте вышележащая регуляторная последовательность по существу состоит из последовательности SEQ ID NO: 4 или ее комплемента.
В другом варианте синтетический полинуклеотид дополнительно содержит по меньшей мере один элемент, выбранный из списка, содержащего вышележащую регуляторную последовательность (URS), энхансерный элемент, экзон, интрон, участок старта транскрипции, TATA-бокс и консенсусный элемент теплового шока. В другом варианте синтетический полинуклеотид дополнительно содержит представляющую интерес нуклеотидную последовательность, оперативно связанную с минимальным коровым промоторным элементом. В другом варианте синтетический полинуклеотид дополнительно содержит элемент, выбранный из группы, состоящей из вышележащей регуляторной последовательности (URS), энхансерного элемент, экзона, интрона, участка старта транскрипции, TATA-бокса, консенсусного элемента теплового шока и их сочетания. В другом варианте синтетический полинуклеотид дополнительно содержит представляющую интерес нуклеотидную последовательность, оперативно связанную с минимальным коровым промоторным элементом.
В другом варианте синтетический полинуклеотид дополнительно содержит второй минимальный коровый промоторный элемент из Zea mays или Zea luxurians, при этом два минимальных коровых промоторных элемента находятся в полинуклеотиде в обратной комплементарной ориентации по отношению друг к другу. В следующем или альтернативном варианте синтетический полинуклеотид дополнительно содержит экзон из гена убиквитина-1 и интрон из гена убиквитина-1. В следующем варианте синтетический полинуклеотид содержит полинуклеотидную последовательность, которая по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентична последовательности SEQ ID NO: 3 или ее комплементу. В следующем варианте синтетический полинуклеотид содержит последовательность SEQ ID NO: 3 или ее комплемент. В следующем варианте синтетический полинуклеотид по существу состоит из последовательности SEQ ID NO: 3 или ее комплемента.
В следующем или альтернативном варианте синтетический полинуклеотид дополнительно содержит вышележащую регуляторную последовательность из гена убиквитина-1. В следующем варианте вышележащая регуляторная последовательность содержит полинуклеотидную последовательность, которая по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентична последовательности SEQ ID NO: 4 или ее комплементу. В следующем варианте вышележащая регуляторная последовательность содержит последовательность SEQ ID NO: 4 или ее комплемент. В следующем варианте вышележащая регуляторная последовательность по существу состоит из последовательности SEQ ID NO: 4 или ее комплемента.
В другом варианте синтетический полинуклеотид, содержащий два минимальных коровых промоторных элемента, дополнительно содержит по меньшей мере один элемент выбранный из списка, содержащего вышележащую регуляторную последовательность (URS), экзон, интрон, участок старта транскрипции, TATA-бокс, консенсусный элемент теплового шока и нуклеотидную последовательность старта и/или остановки трансляции. В следующем или альтернативном варианте синтетический полинуклеотид, содержащий два минимальных коровых промоторных элемента, дополнительно содержит элемент, выбранный из группы, состоящей из вышележащей регуляторной последовательности (URS), экзона, интрона, участка старта транскрипции, TATA-бокса, консенсусного элемента теплового шока, нуклеотидной последовательности старта и/или остановки трансляции и их сочетания. В следующем варианте синтетический полинуклеотид содержит последовательность SEQ ID NO: 5 или ее комплемент. В следующем варианте синтетический полинуклеотид по существу состоит из последовательности SEQ ID NO: 5 или ее комплемента.
В другом варианте синтетический полинуклеотид, содержащий два минимальных коровых промоторных элемента, содержит первую представляющую интерес нуклеотидную последовательность, оперативно связанную с одним из минимальных коровых промоторных элементов. В следующем варианте синтетический полинуклеотид, содержит вторую представляющую интерес нуклеотидную последовательность, оперативно связанную с минимальным коровым промоторным элементом, который оперативно не связан с первой представляющей интерес нуклеотидной последовательностью.
В одном варианте синтетического полинуклеотида по изобретению экзон получен из гена убиквитина-1 видов Zea. В одном варианте предлагаемого синтетического полинуклеотида экзон получен из гена убиквитина-1 Zea mays или Zea luxurians. В другом варианте интрон получен из гена убиквитина-1 видов Zea. В другом варианте интрон получен из гена убиквитина-1 Zea mays или Zea luxurians. В следующем или альтернативном варианте видом Zea является Zea mays. В другом варианте видом Zea является Zea luxurians.
В другом аспекте изобретение относится к способу получения трансгенной клетки. Способы включают трансформацию клетки синтетическим полинуклеотидом, описанным в настоящем описании. В одном варианте клетка является растительной клеткой. В другом аспекте изобретение относится к растительной клетке, содержащей синтетический полинуклеотид, описанный в настоящем описании. В другом аспекте изобретение относится к растению, содержащему растительную клетку, содержащую синтетический полинуклеотид, описанный в настоящем описании.
В другом аспекте изобретение относится к способу экспрессии представляющей интерес нуклеотидной последовательности в растительной клетке. Способ включает введение в растительную клетку представляющей интерес нуклеотидной последовательности, оперативно связанной со средствами для инициации транскрипции независимым от направления образом. В другом аспекте изобретение относится к способу экспрессии представляющей интерес нуклеотидной последовательности в растительной клетке. Способ включает введение в растительную клетку представляющей интерес нуклеотидной последовательности, оперативно связанной со средствами для инициации транскрипции двух оперативно связанных представляющих интерес нуклеотидных последовательностей. В следующем варианте способ включает введение в растительную клетку нуклеиновой кислоты, содержащей: (a) представляющую интерес нуклеотидную последовательность, оперативно связанную со средствами для инициации транскрипции двух оперативно связанных представляющих интерес нуклеотидных последовательностей; и (b) вторую представляющую интерес нуклеотидную последовательность, оперативно связанную со средствами для инициации транскрипции двух оперативно связанных представляющих интерес нуклеотидных последовательностей.
В следующем или альтернативном варианте средства для инициации транскрипции двух оперативно связанных представляющих интерес нуклеотидных последовательностей включают последовательность SEQ ID NO: 5 или ее комплемент. В следующем или альтернативном варианте средства для инициации транскрипции двух оперативно связанных представляющих интерес нуклеотидных последовательностей включают последовательность SEQ ID NO: 5. В другом варианте средства для инициации транскрипции двух оперативно связанных представляющих интерес нуклеотидных последовательностей включают комплемент последовательности SEQ ID NO: 5. В другом варианте нуклеиновую кислоту вводят в растительную клетку так, чтобы целенаправленно встроить представляющую интерес нуклеотидную последовательность, оперативно связанную со средствами для инициации транскрипции двух представляющих интерес оперативно связанных нуклеотидных последовательностей в предварительно определяемый участок в ДНК растительной клетки. В следующем или альтернативном варианте представляющую интерес нуклеотидную последовательность, оперативно связанную со средствами для инициации транскрипции двух представляющих интерес оперативно связанных нуклеотидных последовательностей целенаправленно встраивают в предварительно определяемый участок, используя рекомбинацию, опосредованную нуклеазами с цинковыми пальцами.
В другом аспекте изобретение относится к конструкции нуклеиновой кислоты для экспрессии множественных генов в растительных клетках и/или тканях. Конструкция нуклеиновой кислоты содержит (a) двунаправленный промотор; и (b) две кассеты экспрессии генов на противоположных концах двунаправленного промотора; при этом по меньшей мере одна из кассет экспрессии генов содержит два или больше генов, связанных через переключатель трансляции.
В одном варианте конструкция нуклеиновой кислоты не содержит вирусной последовательности. В другом варианте двунаправленный промотор не содержит вирусной последовательности. В другом варианте двунаправленный промотор содержит по меньшей мере один энхансер. В другом варианте двунаправленный промотор не содержит энхансера. В другом варианте конструкция нуклеиновой кислоты содержит бинарный вектор для опосредованной Agrobacterium трансформации.
В одном варианте двунаправленный промотор содержит элемент, выбранный из группы, состоящей из цис-элемента или вышележащей регуляторной последовательности (URS), энхансерного элемента, экзона, интрона, участка старта транскрипции, TATA-бокса, консенсусного элемента теплового шока и их сочетания. В следующем или альтернативном варианте двунаправленный промотор содержит вышележащую регуляторную последовательность (URS) из гена убиквитина. В следующем варианте двунаправленный промотор содержит (i) промотор, отличный от промотора гена убиквитина, и (ii) вышележащую регуляторную последовательность (URS) из гена убиквитина.
В другом варианте двунаправленный промотор содержит минимальный коровый промоторный элемент из гена убиквитина-1 Zea mays или Zea luxurians. В другом варианте двунаправленный промотор дополнительно содержит второй минимальный коровый промотор из Zea mays или Zea luxurians, при этом два минимальных коровых промоторных элемента находятся в обратной комплементарной ориентации по отношению друг к другу. В следующем варианте минимальный коровый промоторный элемент содержит полинуклеотидную последовательность по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% идентичную последовательности SEQ ID NO: 1 или ее комплементу. В следующем или альтернативном варианте минимальный коровый промоторный элемент содержит полинуклеотидную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 1 и 15-39. В следующем варианте минимальный коровый промоторный элемент содержит полинуклеотидную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 1 и 15-34. В следующем варианте минимальный коровый промоторный элемент содержит полинуклеотидную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 1 и 15-29. В следующем варианте минимальный коровый промоторный элемент содержит полинуклеотидную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 1 и 15-24. В следующем варианте минимальный коровый промоторный элемент содержит полинуклеотидную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 1 и 15-19. В следующем варианте минимальный коровый промоторный элемент содержит полинуклеотидную последовательность SEQ ID NO: 1.
В следующем или альтернативном варианте двунаправленный промотор содержит экзон из гена убиквитина-1 и/или интрон из гена убиквитина. В следующем варианте двунаправленный промотор содержит полинуклеотид по меньшей мере на 75%, 80%, 85%, 90%, 95% или 100% идентичный последовательности SEQ ID NO: 3 или ее комплементу. В следующем варианте двунаправленный промотор содержит полинуклеотид SEQ ID NO: 3 или его комплемент. В другом варианте двунаправленный промотор содержит интрон из гена алкогольдегидрогеназы. В одном варианте конструкцией нуклеиновой кислоты стабильно трансформированы трансгенные растения. В одном варианте растения являются однодольными растениями. В другом варианте растения являются двудольными растениями. В другом варианте растения не являются однодольными растениями. В другом варианте растения не являются двудольными растениями.
В следующем или альтернативном варианте двунаправленный промотор содержит вышележащую регуляторную последовательность из гена убиквитина. В следующем варианте вышележащая регуляторная последовательность из гена убиквитина содержит полинуклеотид с последовательностью по меньшей мере на 75%, 80%, 85%, 90%, 95% или 100% идентичной последовательности SEQ ID NO: 4 или ее комплементу. В следующем варианте вышележащая регуляторная последовательность из гена убиквитина содержит полинуклеотид SEQ ID NO: 4 или его комплемент. В другом варианте двунаправленный промотор содержит полинуклеотид по меньшей мере на 75%, 80%, 85%, 90%, 95% или 100% идентичный последовательности SEQ ID NO: 5 или ее комплементу. В другом варианте двунаправленный промотор содержит полинуклеотид SEQ ID NO: 5 или его комплемент.
В одном варианте обе кассеты экспрессии генов содержат два или более генов, связанных через переключатель трансляции. В следующем или альтернативном варианте переключатель трансляции выбран из группы, состоящей из внутреннего участка связывания рибосомы (IRES), альтернативного сайта сплайсинга, сайта расщепления рибозимом, полинуклеотидной последовательности, кодирующей пептид 2A, полинуклеотидной последовательности, кодирующей 2A-подобный пептид, полинуклеотидной последовательности, кодирующей интеин, полинуклеотидной последовательности, кодирующей сайт расщепления протеазой и их сочетания. В следующем или альтернативном варианте переключатель трансляции содержит цис-действующий элемент гидролазы (CHYSEL). В следующем варианте CHYSEL представляет собой последовательность пептида 2A или 2A-подобного пептида. В другом варианте ген, расположенный выше переключателя трансляции, не содержит стоп-кодона трансляции. В другом варианте конструкция нуклеиновой кислоты позволяет или делает возможной экспрессию по меньшей мере четырех генов. В следующем варианте все четыре гена являются трансгенами. В другом варианте конструкция нуклеиновой кислоты обеспечивает возможность экспрессии от трех до двадцати генов. В другом варианте конструкция нуклеиновой кислоты обеспечивает возможность экспрессии от четырех до восьми генов. В следующем или альтернативном варианте гены являются трансгенами. В другом варианте по меньшей мере одна кассета экспрессии содержит полинуклеотидную последовательность, кодирующую слитый белок. В следующем варианте слитый белок содержит от трех до пяти генов.
В некоторых вариантах экспрессия генов с двунаправленного промотора по меньшей мере в четыре раза выше, чем с однонаправленного промотора. В некоторых вариантах экспрессия генов с двунаправленного промотора от трех до десяти раз выше, чем с однонаправленного промотора. В некоторых вариантах экспрессия генов с двунаправленного промотора в четыре-восемь раз выше, чем с однонаправленного промотора. В некоторых вариантах маркерный ген для селекции помещают на дальнем конце от промотора (т.е., на 3ʹ-конце кассеты экспрессии генов ниже другого гена).
В другом аспекте изобретение относится к способу получения трансгенного растения, включающему трансформацию растительной клетки конструкцией нуклеиновой кислоты, предлагаемой в настоящем описании. В другом аспекте изобретение относится к способу получения трансгенной клетки, включающему трансформацию клетки конструкцией нуклеиновой кислоты, предлагаемой в настоящем описании. В другом аспекте изобретение относится к растительной клетке, содержащей конструкцию нуклеиновой кислоты согласно настоящему изобретению. В следующем или альтернативном варианте конструкцией нуклеиновой кислоты стабильно трансформирована растительная клетка. В другом аспекте изобретение относится к трансгенному растению, содержащему конструкцию нуклеиновой кислоты согласно настоящему изобретению. В следующем или альтернативном варианте конструкцией нуклеиновой кислоты стабильно трансформированы клетки трансгенного растения. В другом аспекте изобретение относится к способу экспрессии множественных генов в растительных клетках и/или тканях, включающему введение в растительные клетки и/или ткани конструкции нуклеиновой кислоты согласно настоящему изобретению. В следующем или альтернативном варианте растительные клетки и/или ткани стабильно трансформированы конструкцией нуклеиновой кислоты, предлагаемой в настоящем описании. В другом аспекте изобретение относится к бинарному вектору для опосредованной Agrobacterium трансформации. В одном варианте бинарный вектор содержит конструкцию нуклеиновой кислоты согласно настоящему изобретению. В другом варианте бинарный вектор содержит синтетический полинуклеотид согласно настоящему изобретению. В другом аспекте изобретение относится к применению двунаправленного промотора согласно настоящему изобретению для экспрессии множественных трансгенов в растениях.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ И ПОСЛЕДОВАТЕЛЬНОСТЕЙ
На фиг. 1 показан примерный (не в масштабе) промотор кукурузы Ubi1 (ZmUbi1), который содержит приблизительно 900 п.н. вышерасположенного элемента, локализованного с 5ʹ-стороны от участка старта транскрипции (TSS). Вышерасположенный элемент содержит TATA-бокс (локализованный приблизительно в положении -30 п.н. от TSS) и два перекрывающихся консенсусных элемента теплового шока (локализованных приблизительно -200 п.н. от TSS). Указанный промотор также содержит примерно 1100 п.н. с 3ʹ-сторонв от области TSS. Такая 3ʹ-область содержит близлежащую лидерную последовательность (экзона ZmUbi1) и интрон.
На фиг. 2 показан примерный вариант предлагаемого синтетического двунаправленного промотора Ubi1, который содержит минимальный коровый элемент minUbi1P, клонированный выше (в обратной комплементарной ориентации) промотора ZmUbi1.
На фиг. 3 показано примерное схематичное изображение кассет экспрессии генов yfp и GUS, каждый из которых оперативно связан с синтетическим двунаправленным промотором Ubi1.
На фиг. 4 показана типичная карта плазмиды pDAB105801.
На фиг. 5 показана типичная карта плазмиды pDAB 108706.
На фиг. 6 показана типичная карта плазмиды pDAB101556.
На фиг. 7A показана последовательность SEQ ID NO: 1, которая содержит область длиной 215 п.н. минимального корового промотора Ubi1 Zea mays (minUbi1P). На фиг. 7B показана последовательность SEQ ID NO: 2, которая содержит интрон Ubi1 Z. mays.
На фиг. 8A показана последовательность SEQ ID NO: 3, которая содержит обратный комплемент полинуклеотида, содержащего минимальный коровый промотор minUbi1P Z. mays (подчеркнут); лидер Ubi1 Z. mays (экзон ZmUbi1; жирным шрифтом); и интрон Ubi1 Z. mays (строчными буквами). На фиг. 8B показана последовательность SEQ ID NO: 4, которая содержит участок вышележащего элемента Ubi1 Z. mays, и такой элемент (и/или его обратный комплемент) может быть локализован в синтетическом промоторе Ubi1, при этом элемент minUbi1P расположен рядом с его 5ʹ- или 3’-концом.
На фиг. 9 показана последовательность SEQ ID NO: 5, которая содержит примерный синтетический двунаправленный промотор Ubi1, при этом обратный комплемент первого minUbi1P и второго minUbi1P подчеркнуты.
На фиг. 10 показана последовательность SEQ ID NO: 6, которая содержит примерную нуклеиновую кислоту, содержащую кассеты экспрессии генов yfp и GUS, управляемые синтетическим двунаправленным промотором Ubi1.
Последовательность SEQ ID NO: 7 содержит прямой праймер: 5ʹ-GATGCCTCAG TGGGAAAGG-3ʹ. Последовательность SEQ ID NO: 8 содержит обратный праймер YFP: 5ʹ-CCATAGGTGA GAGTGGTGAC AA-3ʹ. Последовательность SEQ ID NO: 9 содержит прямой праймер инвертазы: 5ʹ-TGGCGGACGA CGACTTGT-3ʹ. Последовательность SEQ ID NO: 10 содержит обратный праймер инвертазы: 5ʹ-AAAGTTTGGA GGCTGCCGT-3ʹ. SEQ ID NO: 11 содержит зонд инвертазы: 5ʹ-CGAGCAGACC GCCGTGTACT TCTACC-3ʹ. Последовательность SEQ ID NO: 12 содержит прямой праймер AAD1: 5ʹ-TGTTCGGTTC CCTCTACCAA-3ʹ. Последовательность SEQ ID NO: 13 содержит обратный праймер AAD1: 5ʹ-CAACATCCAT CACCTTGACT GA-3ʹ. Последовательность SEQ ID NO: 14 содержит зонд AAD1: 5ʹ-CACAGAACCG TCGCTTCAGC AACA-3ʹ (также см. таблицу 7).
На фиг. 11 показан типичный Вестерн-блот-анализ, подтверждающий стабильную экспрессию YFP и GUS, управляемую двунаправленной промоторной конструкцией убиквитина-1 Z. mays (pDAB 108706), в растениях кукурузы T0. В типичных растениях наблюдали стабильную экспрессию YFP в листе, управляемую минимальным коровым промоторным элементом Min-UbiP1. Количество белка, которое было продуцировано, показано в частях на миллион (ч/млн).
На фиг. 12 показан типичный Вестерн-блот-анализ, показывающий стабильную экспрессию YFP и GUS с контрольной конструкции, содержащей промотор ZmUbi1, который управляет только экспрессией YFP (pDAB101556); кодирующая последовательность GUS не входит в состав такой конструкции. Количество белка, которое было продуцировано указано в частях на миллион (ч/млн).
На фиг. 13A показаны примерные конструкции блоков кассет четырех генов pDAB105843 [видны две кассеты AAD1-2A-YFP (или Phiyfp) плюс Cry34-2A-Cry35] и pDAB105846 [видны две кассеты YFP (или Phiyfp)-2A-AAD1 плюс Cry34-2A-Cry35]. Заштрихованные стрелки указывают направление транскрипции с двунаправленного промотора. Ubi1-minP содержит 200-нуклеотидную последовательность выше участка старта транскрипции промотора Ubi1 кукурузы. Ubi1-URS содержит вышерасположенную регуляторную область промотора Ubi1 кукурузы, состоящую из последовательности выше участка старта транскрипции, за исключением 200-нуклеотидного минимального промотора (показан в виде стрелки). Ubi1-Int содержит интрон промотора Ubi1 кукурузы. На фиг. 13B показаны дополнительные примерные бинарные конструкции блоков четырехгенных кассет из pDAB108717 и pDAB108718.
На фиг. 14 показаны примерные схематичные представления конструкций с множественными генами, предлагаемых в настоящем описании. Переключение трансляции показано с использованием специального символа.
На фиг. 15 показаны типичные карты плазмид pDAB105818 и pDAB 105748.
На фиг. 16 показаны типичные карты плазмид pDAB105803 и pDAB105840.
На фиг. 17 показаны типичные карты плазмид pDAB105841 и pDAB105842.
На фиг. 18 показаны типичные карты плазмид pDAB105843 и pDAB101917.
На фиг. 19 показана типичная карта плазмиды pDAB108717.
На фиг. 20 показаны типичные карты плазмид pDAB105844 и pDAB105845.
На фиг. 21 показаны типичные карты плазмид pDAB105846 и pDAB108718.
На фиг. 22 показаны типичные данные экспрессии белка для Cry35 pDAB108717 (фиг. 22A) и pDAB108718 (фиг. 22B).
На фиг. 23 показана последовательность нуклеиновой кислоты для кассет экспрессии генов pDAB108717, при этом указан каждый ген и элемент.
На фиг. 24 A-E показаны дополнительные минимальные коровые промоторы (min-Ubi1P или Ubi1-minP) с последовательностями SEQ ID NO: 15-39.
На фиг. 25 показаны примерные последовательности для желтых флуоресцирующих белков Phialidium sp. SL-2003 (Phiyfp, SEQ ID NO: 50; и Phiyfpv3, SEQ ID NO: 51).
На фиг. 26 показаны примерные варианты синтетического двунаправленного промотора Ubi1 и предлагаемые конструкции, включая pDAB108706 (ZMUbi двунаправленный (-200)) и pDAB108707 (ZMUbi двунаправленный (-90)). pDAB101556 (контроль ZmUbi1-YFP) и pDAB108716 (ZMUbi1 без минимального промотора) служат в качестве контрольных конструкций с однонаправленными промоторами.
На фиг. 27A показаны примерные результаты экспрессии (V6) с четырех конструкций, показанных на фиг. 26 для белка YFP (ЖХМС) в нг/см2. На фиг. 27B показаны примерные результаты относительной экспрессии (V6) с четырех конструкций, показанных на фиг. 26, для РНК YFP.
На фиг. 28A показаны примерные результаты экспрессии (V6) с четырех конструкций, показанных на фиг. 26 для белка GUS (ЖХМС) в нг/см2. На фиг. 28B показаны примерные результаты относительной экспрессии (V6) с четырех конструкций, показанных на фиг. 26, для РНК GUS.
На фиг. 29A показаны примерные результаты экспрессии (V6) с четырех конструкций, показанных на фиг. 26, для белка AAD1 (ЖХМС) в нг/см2. На фиг. 29B показаны примерные результаты относительной экспрессии (V6) с четырех конструкций, показанных на фиг. 26, для РНК AAD1.
На фиг. 30A показан статистический анализ результатов экспрессии (V6) с четырех конструкций, показанных на фиг. 26, для белка YFP (ЖХМС) в нг/см2. Средние значения для pDAB108707, pDAB108706, pDAB101556 и pDAB108716 составляют 57,63, 52,66, 49,75 и 0, соответственно. На фиг. 30B показан статистический анализ результатов относительной экспрессии (V6) с четырех конструкций, показанных на фиг. 26, для РНК YFP. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 составляют 9,96, 8,07, 6,95 и 1,01, соответственно.
На фиг. 31A показан статистический анализ результатов экспрессии (V6) с четырех конструкций, показанных на фиг. 26, для белка GUS (ЖХМС) в нг/см2. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 составляют 151,27, 143,22, 0 и 213,17, соответственно. На фиг. 31B показан статистический анализ результатов относительной экспрессии (V6) с четырех конструкций, показанных на фиг. 26, для РНК GUS. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 составляют 0,65, 0,78, 0 и 3,03 соответственно.
На фиг. 32A показан статистический анализ результатов экспрессии (V6) с четырех конструкций, показанных на фиг. 26, для белка AAD1 (ЖХМС) в нг/см2. Средние значения для pDAB 108706, pDAB108707, pDAB101556 и pDAB108716 составляют 710,88, 1417,01, 856,58 и 1795,43, соответственно. На фиг. 32B показан статистический анализ результатов относительной экспрессии (V6) с четырех конструкций, показанных на фиг. 26, для РНК AAD1. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 составляют 1,33, 1,37, 1,93 и 2,93, соответственно.
На фиг. 33A, 33B и 33C показаны примерные результаты экспрессии (V10) с четырех конструкций, показанных на фиг. 26, для белка YFP, AAD1 и GUS (ЖХМС) в нг/см2, соответственно.
На фиг. 34A, 34B и 34C показан статистический анализ результатов экспрессии (V10) с четырех конструкций, показанных на фиг. 26, для белка YFP, GUS и AAD1 (ЖХМС) в нг/см2, соответственно. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 в случае YFP (фиг. 34A) составляют 71,77, 81,81, 49,58 и 23,01, соответственно. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 в случае GUS (фиг. 34B) составляют 109,63, 98,25, 0 и 138,02, соответственно. Средние значения для pDAB 108706, pDAB 108707, pDAB101556 и pDAB108716 в случае AAD1 (фиг. 34C) составляют 666,11, 597,80, 715,12 и 1002,84, соответственно.
На фиг. 35A, 35B и 35C показаны примерные результаты экспрессии (R3) с четырех конструкций, показанных на фиг. 26, для белка YFP, GUS и AAD1 (ЖХМС) в нг/см2, соответственно.
На фиг. 36A, 36B и 36C показан статистический анализ результатов экспрессии (R3) с четырех конструкций, показанных на фиг. 26, для белка YFP, GUS и AAD1 (ЖХМС) в нг/см2, соответственно. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 в случае YFP (фиг. 36A) составляют 91,38, 49,49, 21,67 и 0,40, соответственно. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 в случае GUS (фиг. 36B) составляют 5,52, 16,81, 1,07 и 46,60, соответственно. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 в случае AAD1 (фиг. 36C) составляют 156,71, 153,44, 165,40 и 197,80, соответственно.
На фиг. 37A показаны примерные результаты относительной экспрессии (V6) РНК Cry34 с четырех конструкций pDAB105748 (ZMUbi1-YFP), pDAB105818 (ZMUbi1-Cry34/ZMUbi1-Cry35/ZMUbi1-AAD1), pDAB108717 (YFP/AAD-1-ZMUbi1 двунаправленный-Cry34-Cry35) и pDAB108718 (AAD1/YFP-ZMUbi1 двунаправленный-Cry34-Cry35). На фиг. 37B показаны примерные результаты относительной экспрессии (V6) белка Cry34 (ЖХМС) с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 38A показаны примерные результаты относительной экспрессии (V6) РНК AAD1 с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718. На фиг. 38B показаны примерные результаты относительной экспрессии (V6) белка AAD1 (ЖХМС) с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 39A показаны примерные результаты относительной экспрессии (V6) РНК YFP с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718. На фиг. 39B показаны примерные результаты относительной экспрессии (V6) белка YFP (ЖХМС) с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 40A показаны примерные результаты относительной экспрессии (V6) РНК Cry35 с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718. На фиг. 40B показаны примерные результаты относительной экспрессии (V6) белка Cry35 (ELISA) с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 41 показаны примерные результаты относительной экспрессии (V6) РНК PAT с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 42A показан статистический анализ результатов экспрессии (V6) РНК Cry34 с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB 108718 со средними значениями 0, 2,42, 2,67 и 2,25, соответственно. На фиг. 42B показан статистический анализ результатов экспрессии (V6) белка Cry34 с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718 со средними значениями 0, 596,94, 2044,73 и 719,18, соответственно.
На фиг. 43A показан статистический анализ результатов экспрессии (V6) РНК AAD1 с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB 108718 со средними значениями 0, 1,98, 2,68 и 2,03, соответственно. На фиг. 43B показан статистический анализ результатов экспрессии (V6) белка AAD1 с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718 со средними значениями 0, 2237,54, 5763,88 и 2379,15, соответственно.
На фиг. 44A показан статистический анализ результатов экспрессии (V6) РНК YFP с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718 со средними значениями 3,59, 0, 2,78 и 1,95, соответственно. На фиг. 44B показан статистический анализ результатов экспрессии (V6) белка YFP с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718 со средними значениями 1420,69, 251,68, 1154,04 и 706,04, соответственно.
На фиг. 45A показан статистический анализ результатов экспрессии (V6) РНК Cry35 с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718 со средними значениями 0, 1,12, 3,74 и 3,20, соответственно. На фиг. 45B показан статистический анализ результатов экспрессии (V6) белка Cry35 с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB 108718 со средними значениями 0, 283,54, 635,83 и 90,97, соответственно.
На фиг. 46 показан статистический анализ результатов экспрессии (V6) РНК PAT с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB 108718 со средними значениями 1,56, 0,07, 1,46 и 1,01, соответственно.
На фиг. 47A, 47B, 47C и 47D показаны примерные результаты экспрессии белка (V10) YFP, AAD1, Cry34 и Cry35, соответственно, с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 48A, 48B, 48C и 48D показан статистический анализ результатов экспрессии белка (V10) YFP, AAD1, Cry34 и Cry35, соответственно, с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718. Средние значения для YFP (фиг. 48A) составляют 1033,47, 27,51, 136,18 и 119,06, соответственно. Средние значения для AAD1 (фиг. 48B) составляют 80,89, 1323,80, 1544,69 и 802,50, соответственно. Средние значения для Cry34 (фиг. 48C) составляют 0, 246,05, 1089,18 и 769,81, соответственно. Средние значения для Cry35 (фиг. 48D) составляют 0, 90,75, 106,09 и 88,80, соответственно.
На фиг. 49A, 49B, 49C, и 49D показаны примерные результаты экспрессии белка (R3) YFP, AAD1, Cry34 и Cry35, соответственно, с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 50A, 50B, 50C, и 50D показан статистический анализ результатов экспрессии белка (R3) YFP, AAD1, Cry34 и Cry35, соответственно, с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и DAB108718. Средние значения для YFP (фиг. 50A) составляют 2589,63, 43,62, 1305,27 и 1727,96, соответственно. Средние значения для AAD1 (фиг. 50B) составляют 244,41, 1803,99, 1642,44 и 1279,17, соответственно. Средние значения для Cry34 (фиг. 50C) составляют 422,45, 7258,15, 9285,74 и 7544,75, соответственно. Средние значения для Cry35 (фиг. 50D) составляют 0, 373,35, 441,11 и 348,45, соответственно.
На фиг. 51 показаны примерные результаты, полученные с использованием Вестерн-блота для экспрессии белков Cry34, Cry35 и AAD1 с pDAB108718 и pDAB108717.
СПОСОБ (СПОСОБЫ) ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Разработка трансгенных продуктов становится все более сложной, требуя расположения множества трансгенов в виде пирамиды в одном локусе. Традиционно каждый трансген обычно требует наличия уникального промотора для экспрессии, поэтому требуется множество промоторов для экспрессии разных трансгенов в одном блоке генов. Кроме увеличения размера генного блока, это часто приводит к многократному использованию одного и того же промотора, чтобы получить сходные уровни картин экспрессии разных трансгенов, которые контролируют один и тот же признак. Известно, что конструкции, содержащие множество генов, управляемых одним и тем же промотором, вызывают сайленсинг генов, тем самым делая получение трансгенных продуктов менее эффективным в полевых условиях. Избыток сайтов связывания факторов транскрипции (TF) вследствие повторения промоторов может вызывать истощение по эндогенным TF, приводя к инактивации транскрипции.
Предлагаются конструкции и способы объединения двунаправленной промоторной системы с бицистронной организацией генов либо на одном, либо на обоих концах промотора, например, с применением последовательности 2A вируса Thosea asigna. Белок 2A, который имеет длину только 16-20 аминокислот, расщепляет полибелок на его карбоксильном конце. Такое свойство «саморасщепления» или «рибосомного перепрыгивания» пептида 2A или 2A-подобного пептида может быть использовано для процессинга искусственных полибелков, продуцируемых в трансгенных растениях. В одном варианте гены Cry34 и Cry35 сливают в одной кассете экспрессии генов, при этом гены YFP (или Phiyfp) и AAD1 сливают в другой кассете экспрессии генов (с одной открытой рамкой считывания (ORF), при этом копию гена белка 2A помещают между двумя генами в каждом сочетании). Например, каждая из таких кассет экспрессии гена (или пары генов) может быть помещена на любом конце двунаправленного промотора, чтобы управлять 4 трансгенами с использованием одного промотора. Таким образом, конструкции и способы, предлагаемые в настоящем описании, применимы для избегания повторного использования одного и того же промотора и значимо снижают размер коммерческих конструкций. Кроме того, управление четырьмя или большим количеством генов с одного промотора также делает возможной коэкспрессию генов, контролирующих один признак.
Растительные промоторы, используемые для фундаментальных исследований или биотехнологических применений, обычно являются однонаправленными, управляющими только одним геном, который был слит с их 3ʹ-концом (ниже). Часто необходимо введение множества генов в растения для метаболической инженерии и объединения признаков и поэтому обычно требуется множество промоторов в будущих трансгенных культурных растениях для управления экспрессией множества генов. Требуется разработка методик, которые помогут сэкономить на количестве используемых промоторов и позволят одновременно осуществлять корегуляцию экспрессии пирамиды генов. В некоторых вариантах предлагаемые двунаправленные промоторы могут управлять транскрипцией множества транскрипционных единиц, включая интерферирующие РНК, искусственные микро-РНК или последовательности РНК-шпильки.
Одним из способов уменьшения количества используемых промоторов является применение переключателей, активирующих важную транскрипцию, которые могут управлять транскрипцией в обоих направлениях. Такие промоторы называют двунаправленными промоторами. Могут быть сконструированы синтетические промоторы, чтобы ограничить уровень гомологии среди множества промоторов, которые необходимо использовать для генетической инженерии культурных растений, что поможет избежать основанного на гомологии сайленсинга генов. Искусственно сконструированные двунаправленные промоторы могут быть ценными средствами для разработки трансгенных растений. В некоторых случаях сообщалось о двунаправленном функционировании промоторов в растениях, включая промотор 35S CaMV 35S и промотор маннопинсинтазы (mas). Однако применимость таких промоторов для предсказуемой, стабильной и одновременной экспрессии генов в двух направлениях не была исследована.
Другой способ для согласованной экспрессии множества генов заключается в кодировании одной открытой рамкой считывания полибелкового предшественника, содержащего короткий промежуточный мотив, обладающий свойствами самопроцессинга, между двумя кодирующими последовательностями. Автокаталитический процессинг полибелкового предшественника приводит к высвобождению множества независимых белков, результатом которого является их синхронизированная согласованная экспрессия. Синтетическая последовательность подвергающегося самогидролизу пептида 2A была использована как в растительных, так и в животных системах для экспрессии двух трансгенов. Последовательность пептида 2A используется несколькими известными вирусами и состоит из 16-20 аминокислот. Такая последовательность пептида 2A подвергается саморасщеплению (или рибосомному перепрыгиванию) котрансляционно за счет модификации активности рибосомы, что обеспечивает возможность гидролиза 2A между двумя белками, приводя к высвобождению двух белковых продуктов.
Предлагаются конструкции и способы сочетания подхода на основе двунаправленного промотора с процессингом полибелка с использованием промежуточных синтетических мотивов, при этом легко может быть достигнута экспрессия по меньшей мере 4 трансгенов с использованием одного промотора. Гены Cry34 и Cry35 и гены YFP (или Phiyfp) и AAD1 были слиты в виде кассет экспрессии генов или генных пар в одной открытой рамке считывания (ORF) с копией гена белка 2A, помещенной между генами. Пары генов могут быть помещены на любом конце двунаправленного промотора для управления четырьмя трансгенами с использованием одного промотора. Конструкции и/или способы, предлагаемые в настоящем описании, применимы для избегания повторного использования одного и того же промотора, при том избегая потенциальных проблем, связанных с сайленсингом генов. Кроме того, такой способ конструирования трансгенов может значимо уменьшить размер блоков трансгенов, содержащих множество трансгенов. Управление четырьмя или большим количеством генов с одного промотора также позволяет осуществлять коэкспрессию генов, контролирующих одни признак, обеспечивающую долговременную эффективность трансгенных продуктов.
Разработка трансгенных растений становится все более сложной и обычно требует создания блоков из множества трансгенов в одном локусе. См. публикацию Xie с соавторами, (2001) Nat. Biotechnol. 19(7): 677-9. Так как каждый трансген обычно требует наличия уникального промотора для экспрессии, требуется множество промоторов для экспрессии разных трансгенов в одном блоке генов. Кроме увеличения размера генного блока, это часто приводит к многократному использованию одного и того же промотора для получения сходных уровней картин экспрессии разных трансгенов. Такой способ часто проблематичен, так как экспрессия множества генов, управляемых одним и тем же промотором, может приводить к сайленсингу генов или HBGS. Избыток сайтов связывания факторов транскрипции (TF) в повторяющихся промоторах может вызывать истощение по эндогенным TF и приводить к пониженной регуляции транскрипции. Сайленсинг трансгенов вероятно будет нежелательным образом влиять на эффективность трансгенного растения, получаемого для экспрессии трансгенов. Повторяющиеся последовательности в трансгене могут приводить к гомологичной рекомбинации генов внутри локуса и в результате к перестройкам полинуклеотида.
Растительные промоторы, используемые для фундаментальных исследований или биотехнологических применений, обычно являются однонаправленными и регулируют только один ген, который был слит с их 3ʹ-концом (ниже). Для получения трансгенных растений с различными требуемыми признаками или характеристиками, может быть полезным уменьшение количества промоторов, которые используют для управления экспрессией трансгенов, которые кодируют требуемые признаки и характеристики. Часто требуется введение множества трансгенов в растения для метаболической инженерии и объединения признаков и поэтому обычно требуется множество промоторов для управления экспрессией множества генов. Благодаря разработке одного синтетического двунаправленного промотора, который может управлять экспрессией двух трансгенов, которые фланкируют промотор, общее количество промоторов, необходимых для разработки трансгенных культурных растений, может быть уменьшено с уменьшением при этом повторного использования одного и того же промотора, уменьшением размера трансгенных конструкций и/или снижением вероятности HBGS.
В вариантах осуществления настоящего изобретения применяют способ, в котором однонаправленный промотор из гена убиквитина-1 кукурузы (например, ZmUbi1) используют для конструирования синтетического двунаправленного промотора, так что один промотор может управлять экспрессией двух генов, по одному на каждом конце промотора. Способы, применяемые в настоящем описании, могут включать идентификацию минимального корового промоторного элемента Ubi1 (minUbi1P) из гена ZmUbi1 и разработку такого элемента в новых контекстах для конструирования определенных синтетических двунаправленных промоторов. Синтетические двунаправленные промоторы, такие как промоторы, которые могут быть созданы способом согласно некоторым вариантам осуществления изобретения, могут обеспечить возможность специалистам в данной области создавать блоки трансгенов в растительных клетках и растениях с уменьшением при этом многократного использования одного и того же промотора и уменьшением размера трансгенных конструкций. Кроме того, регуляция экспрессии двух генов с одного синтетического двунаправленного промотора также может обеспечить коэкспрессию двух генов в одних и тех же условиях, таких, которые могут быть применимы, например, в том случае, когда каждый из двух генов вносит вклад в один признак хозяина. В некоторых случаях сообщалось о применении двунаправленных промоторов в растениях, включая промотор CaMV 35 (Barfield and Pua (1991), Plant Cell Rep. 10(6-7): 308-14; Xie с соавторами (2001), и промотор маннопинсинтазы (mas) (Velten с соавторами, (1984) EMBO J. 3(12): 2723-30; Langridge с соавторами (1989) Proc. Natl. Acad. Sci. USA 86: 3219-23).
Инициация транскрипции и модулирование экспрессии растительных генов управляется различными элементами последовательности ДНК, которые вместе в определенном порядке расположены в промоторе. Эукариотические промоторы состоят из минимального корового промоторного элемента (minP) и дополнительных вышележащих регуляторных последовательностей (URS). Коровый промоторный элемент представляет собой минимальный участок непрерывной последовательности ДНК, который достаточен для управления точной инициацией транскрипции. Коровые промоторы в растениях также содержат канонические области, ассоциированные с инициацией транскрипции, такие как CAAT- и TATA-боксы. Элемент TATA-бокса обычно расположен примерно на 20-35 нуклеотидов выше сайта инициации транскрипции.
Активация minP зависит от URS, с которой связываются различные белки и затем взаимодействуют с комплексом инициации транскрипции. URS содержат последовательности ДНК, которые определяют пространственно-временную картину экспрессии с промотора, содержащего URS. Полярность промотора часто определяется ориентацией minP, тогда как URS является биполярной (т.е., она функционирует независимо от ориентации). Например, синтетический однонаправленный полярный промотор 35S CaMV может быть превращен в двунаправленный промотор слиянием minP с 5'-концом промотора в противоположной ориентации. См., например, публикацию Xie с соавторами, (2001) Nat. Biotechnol. 19(7): 677-9.
Некоторые сокращения в описании перечислены в таблице 1.
Сокращения, используемые в описании
В конкретных примерах некоторых вариантов модифицированные элементы промотора Ubi1 кукурузы (ZmUbi1), полученного из инбредной линии Z. mays B73, используют для конструирования синтетических двунаправленных промоторов, которые могут функционировать в растениях, обеспечивая характеристики регуляции экспрессии, которые являются уникальными по сравнению с ранее имеющимися двунаправленными промоторами. Такой промотор ZmUbi1, исходно полученный из B73, содержит последовательности, локализованные в геноме кукурузы в пределах 899 оснований с 5ʹ-стороны от участка старта транскрипции, и далее в пределах примерно 1093 оснований с 3ʹ-стороны от участка старта транскрипции. Christensen с соавторами (1992), Plant Mol. Biol. 18(4): 675-89 (описан ген ZmUbi1 B73). Модифицированный промотор ZmUbi1, полученный из B73, который использован в некоторых примерах, представляет собой промотор длиной примерно 2 т.п.н., который содержит TATA-бокс; два перекрывающихся консенсусных элемента теплового шока; лидерную последовательность длиной 82 или 83 нуклеотида (в зависимости от эталонной нити) непосредственно вблизи с участком старта транскрипции, которую называют в настоящем описании экзоном ZmUbi1; и интрон длиной 1015-1016 нуклеотидов (см., например, фиг. 1). Другие варианты промотора убиквитина кукурузы, полученные из видов Zea и генотипов Zea mays, могут иметь высокую консервативность последовательности вблизи элемента minP, состоящего из TATA-элемента и вышележащих консенсусных элементов теплового шока. Таким образом, варианты осуществления изобретения приведены в качестве примеров с использованием такой короткой (~200 нуклеотидов) высоко консервативной области (например, SEQ ID NO: 1) промотора ZmUbi1 в качестве минимального корового промоторного элемента для конструирования синтетических двунаправленных растительных промоторов.
В используемом в настоящем описании смысле единственное число включает указание множественного числа, если контекст ясно и однозначно не диктует иное.
В используемом в настоящем описании смысле фраза «обратное скрещивание» относится к способу, в котором селекционер скрещивает гибридное потомство снова с одним из родителей, например, гибрид первого поколения F1 с одним из родительских генотипов гибрида F1.
В используемом в настоящем описании смысле фраза «интрон» относится к любой последовательности нуклеиновой кислоты, входящей в состав гена (или экспрессируемой представляющей интерес нуклеотидной последовательности), которая транскрибируется, но не транслируется. Интрон отличается от 5'-концевой нетранслируемой лидерной последовательности, которая не считается частью гена. Интроны включают нетранслируемую последовательность нуклеиновой кислоты в экспрессируемой последовательности ДНК, а также соответствующую последовательность в транскрибированных с нее молекулах РНК.
В используемом в настоящем описании смысле фраза «изолированный» относится к биологическому компоненту (включая нуклеиновую кислоту или белок), который был по существу отделен, получен отдельно или очищен от других биологических компонентов в клетке организма, в которой компонент присутствует в природе (т.е., от других хромосомных и внехромосомных ДНК и РНК и белков), при осуществлении химического или функционального изменения компонента (например, нуклеиновая кислота может быть выделена из хромосомы в результате разрушения химических связей, связывающих нуклеиновую кислоту с остальной ДНК в хромосоме). Молекулы нуклеиновой кислоты и белки, которые были «выделены», включают молекулы нуклеиновых кислот и белки, очищенные стандартными способами очистки. Фраза «выделенные» также охватывает нуклеиновые кислоты и белки, полученные в результате основанной на рекомбинации экспрессии в клетке-хозяине, а также химически синтезированные молекулы нуклеиновых кислот, белки и пептиды.
В используемом в настоящем описании смысле фраза «экспрессия генов» относится к процессу, при котором информация, закодированная в единице транскрипции нуклеиновой кислоты (включая, например, геномную ДНК) превращается в функционирующую, не функционирующую или структурную часть клетки, часто включая синтез белка. На экспрессию генов могут влиять внешние сигналы; например, воздействие на клетку, ткань или организм агента, который повышает или понижает экспрессию генов. Экспрессия гена также может регулироваться в любом месте на пути от ДНК к РНК и к белку. Регуляция экспрессии генов происходит, например, посредством контролей, действующих на транскрипцию, трансляцию, транспорт и процессинг РНК, распад интермедиаторных молекул, таких как мРНК, или посредством активации, инактивации, компартментализации или распада специфичных белковых молекул после того, как они были созданы, или сочетанием указанного. Экспрессия генов может быть измерена на уровне РНК или на уровне белка любым способом, известным в данной области, включая без ограничения Нозерн-блот, ОТ-ПЦР, Вестерн-блот или анализ (анализы) активности белка in vitro, in situ или in vivo.
В используемом в настоящем описании смысле фраза «основанный на гомологии сайленсинг генов» (HBGS) относится к родовому термину, который охватывает как транскрипционный сайленсинг генов, так и посттранскрипционный сайленсинг генов. Сайленсинг локуса-мишени несцепленным локусом сайленсинга может быть результатом ингибирования транскрипции (транскрипционный сайленсинг генов; TGS) или распада мРНК (посттранскрипционный сайленсинг генов; PTGS) вследствие образования двунитевой РНК (днРНК), соответствующей промотору или транскрибируемым последовательностями, соответственно. Вовлечение разных клеточных компонентов в каждый процесс свидетельствует о том, что индуцированные днРНК TGS и PTGS вероятно являются следствием расхождения общего древнего механизма. Однако строгое сравнение TGS и PTGS было трудно осуществить, так как обычно оно основано на анализе отдельных локусов сайленсинга. Отдельный трансгенный локус может быть описан с точки зрения запуска как TGS, так и PTGS, вследствие образования днРНК, соответствующей промоторным и транскрибируемым последовательностям разных генов-мишеней. См., например, Mourrain с соавторами. (2007) Planta 225: 365-79. Вероятно, что миРНК являются фактическими молекулами, которые запускают TGS и PTGS она гомологичных последовательностях: миРНК может запускать в такой модели сайленсинг и метилирование гомологичных последовательностей в цис- и в трансположении за счет распространения метилирования трансгенных последовательностей на эндогенный промотор.
В используемом в настоящем описании смысле фраза «молекула нуклеиновой кислоты (или «нуклеиновая кислота» или «полинуклеотид») относится к полимерной форме нуклеотидов, которая может включать как смысловую, так и антисмысловую нити РНК, кДНК, геномной ДНК, и синтетическим формам и смешанным полимерам из описанных выше. Нуклеотид может относиться к рибонуклеотиду, дезоксирибонуклеотиду или модифицированной форме любого типа нуклеотида. Термин «молекула нуклеиновой кислоты» в используемом в настоящем описании смысле является синонимом терминов «нуклеиновая кислота» и «полинуклеотид». Молекула нуклеиновой кислоты обычно имеет длину по меньшей мере 10 оснований, если не указано иное. Термин может относиться к молекуле РНК или ДНК неопределенной длины. Термин включает однонитевые и двунитевые формы ДНК. Молекула нуклеиновой кислоты может включать как встречающиеся в природе, так и модифицированные нуклеотиды, связанные вместе встречающимися в природе и/или не встречающимися в природе нуклеотидными связями.
Молекулы нуклеиновой кислоты могут быть модифицированы химически или биохимически или могут содержать неприродные или дериватизованные основания нуклеотидов, что будет легко понятно специалистам в данной области. Такие модификации включают, например, метки, метилирование, замену одного или нескольких встречающихся в природе нуклеотидов аналогом, межнуклеотидные модификации (например, незаряженные связи: например, метилфосфонаты, фосфотриэфиры, фосфорамидаты, карбаматы и т.д.; заряженные связи: например, фосфоротиоаты, фосфородитиоаты и т.д.; боковые остатки: например, пептиды; интеркаляторы: например, акридин, псорален и т.д.; хелаторы; алкиляторы и модифицированные связи: например, альфа-аномерные нуклеиновый кислоты и т.д.). Термин «молекула нуклеиновой кислоты» также охватывает любую топологическую конформацию, включая однонитевые, двунитевые, частично дуплексные, триплексные, имеющие форму шпильки, кольцевые и блокированные конформации.
Транскрипция происходит в направлении от 5ʹ- к 3ʹ-концу вдоль нити ДНК. Это означает, что РНК образуется в результате последовательного добавления рибонуклеотид-5ʹ-трифосфатов к 3ʹ-концу растущей цепи (с необходимым удалением пирофосфата). Как в линейной, так и в кольцевой молекуле нуклеиновой кислоты дискретные элементы (например, конкретные нуклеотидные последовательности) могут быть названы как «вышележащие» относительно следующего элемента, если они связаны или могут быть связаны с той же нуклеиновой кислотой в 5ʹ-направлении от данного элемента. Подобным образом, дискретные элементы могут быть «нижележащими» относительно другого элемента, если они связаны или могут быть связаны с той же нуклеиновой кислотой в 3ʹ-направлении от данного элемента.
В используемом в настоящем описании смысле фраза «положение основания» относится к расположению данного основания или нуклеотидного остатка в указанной нуклеиновой кислоте. Указанная нуклеиновая кислота может быть определена по выравниванию (см. ниже) с эталонной нуклеиновой кислотой.
В используемом в настоящем описании смысле фраза «гибридизация» относится к процессу, при котором олигонуклеотиды и их аналоги гибридизуются с использованием водородных связей, которые включают водородные связи согласно правилам спаривания Уотсона-Крика, Хугстена или обратного спаривания Хугстена между комплементарными основаниями. Обычно молекулы нуклеиновых кислот состоят из азотистых оснований, которые являются либо пиримидинами (цитозин (C), урацил (U) и тимин (T)) или пуринами (аденин (A) и гуанин (G)). Такие азотистые основания образуют водородные связи между пиримидином и пурином, и связывание пиримидина с пурином называют «спариванием оснований». Более конкретно, A будет образовывать водородную связь с T или U, а G будет связываться с C. «Комплементарное» относится к спариванию оснований, которое происходит между двумя отдельными последовательностями нуклеиновых кислот или двумя отдельными областями одной и той же последовательности нуклеиновой кислоты.
В используемом в настоящем описании смысле фразы «специфично гибридизуемые» и «специфично комплементарные» относятся к такой комплементарности, при которой происходит стабильное и специфичное связывание между олигонуклеотидом и ДНК- или РНК-мишенью. Олигонуклеотид не должен быть обязательно комплементарным на 100% с последовательностью своей мишени, чтобы быть специфично гибридизуемым. Олигонуклеотид способен специфично гибридизоваться, когда связывание олигонуклеотида с молекулой-мишенью ДНК или РНК мешает нормальному функционированию ДНК- или РНК-мишени, и существует достаточная степень комплементарности, чтобы избежать неспецифичного связывания олигонуклеотида с последовательностями, которые не являются мишенями в тех условиях, в которых требуется специфичное связывание, например, в физиологических условиях в случае анализов или систем in vivo. Такое связывание называют специфичной гибридизацией.
Условия гибридизации, результатом которых является конкретная степень жесткости, могут варьировать в зависимости от природы выбранного способа гибридизации и состава и длины гибридизующихся последовательностей нуклеиновых кислот. В общем, температура гибридизации и ионная сила (особенно концентрация Na+ и/или Mg2+) буфера для гибридизации будут вносить вклад в жесткость гибридизации, хотя количество промывок также влияет жесткость. Расчеты, имеющие отношение к условиям гибридизации, необходимым для достижения конкретной степени жесткости, обсуждаются в публикации Sambrook с соавторами, Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989, в главах 9 и 11.
В используемом в настоящем описании смысле фраза «жесткие условия» охватывает условия, в которых гибридизация будет происходить только в том случае, если существует менее 50% ошибочных спариваний между гибридизуемой молекулой и ДНК-мишенью. «Жесткие условия» включают дополнительные конкретные уровни жесткости. Таким образом, в используемом в настоящем описании смысле условия «умеренной жесткости» представляют собой условия, в которых молекулы с более чем 50% несовпадений последовательностей не будут гибридизоваться; условия «высокой жесткости» представляют собой условия, в которых последовательности с более чем 20% несовпадений не буду гибридизоваться; и условия «очень высокой жесткости» представляют собой условия, в которых последовательности с более чем 10% несовпадений не будут гибридизоваться.
В конкретных вариантах жесткие условия могут включать гибридизацию при 65ºC с последующими промывками при 65ºC с использованием 0,1х SSC/0,1% SDS в течение 40 минут.
Ниже перечислены типичные не ограничивающие условия гибридизации:
Очень высокая жесткость: гибридизация в 5x SSC-буфере при 65ºC в течение 16 часов; промывка два раза в 2x SSC-буфере при комнатной температуре по 15 минут каждая; и промывка два раза в 0,5x SSC-буфере при 65ºC по 20 минут каждая.
Высокая жесткость: Гибридизация в 5x-6x SSC-буфере при 65-70ºC в течение 16-20 часов; промывка два раза в 2x SSC-буфере при комнатной температуре в течение 5-20 минут каждая; и промывка два раза в 1x SSC-буфере при 55-70ºC в течение 30 минут каждая.
Умеренная жесткость: Гибридизация в 6x SSC-буфере при температуре от комнатной температуре до 55ºC в течение 16-20 часов; промывка по меньшей мере два раза в 2x-3x SSC-буфере при температуре от комнатной температуры до 55ºC в течение 20-30 минут каждая.
В конкретных вариантах специфично гибридизуемые молекулы нуклеиновых кислот могут оставаться связанными в условиях очень высокой жесткости. В указанных и дополнительных вариантах специфично гибридизуемые молекулы нуклеиновых кислот могут оставаться связанными в условиях очень высокой жесткости. В указанных и дополнительных вариантах специфично гибридизуемые молекулы нуклеиновых кислот могут оставаться связанными в условиях гибридизации умеренной жесткости.
В используемом в настоящем описании смысле фраза «олигонуклеотид» относится к короткому полимеру нуклеиновой кислоты. Олигонуклеотиды могут быть образованы расщеплением более длинных фрагментов нуклеиновой кислоты или полимеризацией отдельных предшественников нуклеотидов. Автоматизированные синтезаторы позволяют осуществлять синтез олигонуклеотидов длиной до нескольких сотен пар оснований. Так как олигонуклеотиды могут связываться с комплементарной нуклеотидной последовательностью, их можно использовать в качестве зондов для выявления ДНК или РНК. Олигонуклеотиды, состоящие из ДНК (олигодезоксирибонуклеотиды), можно использовать в ПЦР, способе амплификации небольших последовательностей ДНК. В ПЦР олигонуклеотид обычно называют «праймером», который позволяет ДНК-полимеразу удлинять олигонуклеотид и реплицировать комплементарную нить.
В используемом в настоящем описании смысле фраза «идентичность последовательностей» или «идентичность» относится к контексту, описывающему две последовательности нуклеиновой кислоты или полипептидные последовательности, и может относиться к остаткам в двух последовательностях, которые являются одинаковыми при выравнивании для максимального соответствия на протяжении конкретного окна сравнения.
В используемом в настоящем описании смысле фраза «идентичность последовательностей в процентах» относится к значению, определяемому при сравнении двух оптимально выровненных последовательностей (например, последовательностей нуклеиновых кислот и аминокислотных последовательностей) в окне сравнения, при этом часть последовательности в окне сравнения может содержать добавления или делеции (т.е., пробелы) по сравнению с эталонной последовательностью (которая не содержит добавлений или делеций) для оптимального выравнивания двух последовательностей. Процент вычисляют посредством определения количества положений, в которых идентичный нуклеотид или аминокислотный остаток встречается в обеих последовательностях, получая количество совпадающих положений, деления количества совпадающих положений на общее количество положений в окне сравнения, и умножения результата на 100 с получением процента идентичности последовательностей.
Способы выравнивания последовательностей для сравнения хорошо известны в данной области. Описаны различные программы и алгоритмы выравнивания, например, в публикациях: Smith and Waterman (1981) Adv. Appl. Math. 2:482; Needleman and Wunsch (1970) J. Mol. Biol. 48:443; Pearson and Lipman (1988) Proc. Natl. Acad. Sci. U.S.A. 85:2444; Higgins and Sharp (1988) Gene 73:237-44; Higgins and Sharp (1989) CABIOS 5:151-3; Corpet et al. (1988) Nucleic Acids Res. 16:10881-90; Huang et al. (1992) Comp. Appl. Biosci. 8:155-65; Pearson et al. (1994) Methods Mol. Biol. 24:307-31; Tatiana et al. (1999) FEMS Microbiol. Lett. 174:247-50. Подробное описание способов выравнивания последовательностей и вычисления гомологии можно найти, например, в публикации Altschul с соавторами (1990), J. Mol. Biol. 215:403-10.
Основное средство поиска, основанное на локальном выравнивании (BLAST™) Национального центра биотехнологической информации (NCBI) (Altschul с соавторами (1990)) доступно из нескольких источников, включая Национальный центр биотехнологической информации (National Center for Biotechnology Information (Bethesda, MD)), и из интернета для применения вместе с несколькими программами анализа последовательностей. Описание того, как определить идентичность последовательностей с использованием такой программы, доступно в интернете в разделе «помощь» для BLAST™. В случае сравнений последовательностей нуклеиновых кислот можно использовать функцию «Blast 2 sequences» программы BLAST™ (Blastn), применяя параметры по умолчанию. В случае последовательностей нуклеиновых кислот с еще большим сходством с эталонными последовательностями будет наблюдаться увеличение идентичности в процентах при оценивании таким способом.
В используемом в настоящем описании смысле фраза «оперативно связанные» относится к контексту, в котором первая последовательность нуклеиновой кислоты оперативно связана со второй последовательностью нуклеиновой кислоты, при этом первая последовательность нуклеиновой кислоты находится в функциональной взаимосвязи со второй последовательностью нуклеиновой кислоты. Например, промотор оперативно связан с кодирующей последовательностью, когда промотор влияет на транскрипцию или экспрессию кодирующей последовательности. В случае получения путем рекомбинации оперативно связанные последовательности нуклеиновых кислот обычно являются смежными и, при необходимости связать две кодирующие белки области, находятся в одной и той же рамке считывания. Однако элементы не должны быть обязательно смежными, чтобы являться оперативно связанными.
В используемом в настоящем описании смысле фраза «промотор» относится к области ДНК, которая обычно локализована выше (по отношению к 5ʹ-области гена), которая необходима для транскрипции. Промоторы могут обеспечивать возможность правильной активации или репрессии гена, который они контролируют. Промотор может содержать специфичные последовательности, которые распознаются факторами транскрипции. Такие факторы могут связываться с последовательностями промоторной ДНК и приводят к привлечению РНК-полимеразы, фермента, который синтезирует РНК с кодирующей области гена.
В используемом в настоящем описании смысле фраза «трансформирует» или «трансдуцирует» относится к процессу, при котором вирус или вектор трансформирует клетку молекулами нуклеиновой кислоты. Клетка «трансформирована» молекулой нуклеиновой кислоты, «трансдуцированной» в клетку, когда молекула нуклеиновой кислоты становится стабильно реплицируемой клеткой, либо при включении молекулы нуклеиновой кислоты в клеточный геном, либо в результате эписомной репликации. В используемом в настоящем описании смысле термин «трансформация» охватывает все способы, благодаря которым молекула нуклеиновой кислоты может быть введена в такую клетку. Примеры включают без ограничения: трансформацию вирусными векторами; трансформацию плазмидными векторами; электропорацию (Fromm с соавторами (1986), Nature 319: 791-3); липофекцию (Feigner с соавторами (1987), Proc. Natl. Acad. Sci. USA 84: 7413-7); микроинъекцию (Mueller с соавторами, (1978) Cell 15: 579-85); опосредованный Agrobacterium перенос (Fraley с соавторами, (1983) Proc. Natl. Acad. Sci. USA 80: 4803-7); прямой захват ДНК; опосредованную прокалыванием микроиглами (whisker) трансформацию; и бомбардировку микрочастицами (Klein с соавторами (1987), Nature 327: 70).
В используемом в настоящем описании смысле фраза «трансген» относится к экзогенной последовательности нуклеиновой кислоты. В одном примере трансген представляет собой последовательность гена (например, гена устойчивости к гербицидам), гена, кодирующего промышленно или фармацевтически применимое соединение, или ген, кодирующий требуемый сельскохозяйственный признак. В еще одном примере трансген представляет собой антисмысловую последовательность нуклеиновой кислоты, при этом экспрессия антисмысловой последовательности нуклеиновой кислоты ингибирует экспрессию последовательности нуклеиновой кислоты-мишени. Трансген может содержать регуляторные последовательности, оперативно связанные с трансгеном (например, промотор). В некоторых вариантах представляющая интерес последовательность нуклеиновой кислоты является трансгеном. Однако в других вариантах представляющая интерес последовательность нуклеиновой кислоты является экзогенной последовательностью нуклеиновой кислоты, при этом требуются дополнительные геномные копии эндогенной последовательности нуклеиновой кислоты, или последовательность нуклеиновой кислоты, которая находится в антисмысловой ориентации по отношению к последовательности молекулы нуклеиновой кислоты-мишени в организме хозяина.
В используемом в настоящем описании смысле фраза «вектор» относится к молекуле нуклеиновой кислоты, которую вводят в клетку, получая при этом трансформированную клетку. Вектор может содержать последовательности нуклеиновой кислоты, которые обеспечивают его репликацию в клетке-хозяине, такие как начало репликации. Примеры включают без ограничения плазмиду, космиду, бактериофаг или вирус, который несет экзогенную ДНК в клетку. Вектор также может включать один или несколько генов, антисмысловых молекул и/или селектируемых маркерных генов и других генетических элементов, известных в данной области. Вектор может трансдуцировать, трансформировать или инфицировать клетку, вызывая при этом в клетке экспрессию молекул нуклеиновой кислоты и/или белков, кодируемых вектором. Вектор необязательно может включать вещества, помогающие достигать входа молекулы нуклеиновой кислоты в клетку (например, липосому).
В используемом в настоящем описании смысле фраза «растение» охватывает растения и части растений, включая без ограничения растительные клетки и ткани растений, такие как листья, стебли, корни, цветки, пыльцу и семена. Классы растений, которые можно использовать в настоящем описании, обычно находятся в широком диапазоне, включая высшие и низшие растения, поддающиеся мутагенезу, включая покрытосеменные (однодольные и двудольные растения), голосеменные, папоротникообразные и многоклеточные водоросли. Таким образом, «растение» включает двудольные растения и однодольные растения. Примеры двудольных растений включают табак, Arabidopsis, сою, томат, папайю, канолу, подсолнечник, хлопчатник, люцерну, картофель, виноград, голубиный горох, горох, Brassica, турецкий горох, сахарную свеклу, рапс, арбуз, дыню, перец, арахис, кабачок, редис, шпинат, тыкву крупноплодную, брокколи, капусту, морковь, цветную капусту, сельдерей, китайскую капусту, огурец, баклажан и латук. Примеры однодольных растений включают кукурузу, рис, пшеницу, сахарный тростник, ячмень, рожь, сорго, орхидеи, бамбук, банан, рогоз, лилии, овес, лук, просо и тритикале.
В используемом в настоящем описании смысле фраза «материал растений» относится к листьям, стеблям, корням, цветкам или частям цветков, фруктам, пыльце, яйцеклеткам, зиготам, семенам, черенкам, культурам клеток и тканей или любой другой части или продукту растения. В некоторых вариантах материал растений включает семядолю и лист.
В используемом в настоящем описании смысле фраза «переключение трансляции» относится к механизму, находящемуся на конце гена, обеспечивающему трансляцию гена, расположенного непосредственно ниже. Механизм переключения трансляции может функционировать на уровне нуклеиновой кислоты (например, вирусный или эукриотический внутренний участок связывания рибосомы (IRES), альтернативный сайт сплайсинга или участок расщепления рибозимом) или на уровне пептида/белка (например, пептид 2A, 2A-подобный пептид, пептид интеина или сайт расщепления протеазой).
Такие механизмы переключения трансляции на уровне нуклеиновой кислоты или на уровне пептида/белка хорошо известны в данной области. См., например, публикации li, Z., H. M. Schumacher, et al. (2010) J Biotechnol 145(1): 9-16; Chen, Y., K. Perumal, et al. (2000) Gene Expr 9(3): 133-143; Dinkova, T. D., H. Zepeda, et al. (2005) Plant J 41(5): 722-731; Dorokhov, Y. L., M. V. Skulachev, et al. (2002) Proc Natl Acad Sci U S A 99(8): 5301-5306; Fernandez-Miragall, O. and C. Hernandez (2011) PLoS One 6(7): e22617; Groppelli, E., G. J. Belsham, et al. (2007) J Gen Virol 88(Pt 5): 1583-1588; Ha, S. H., Y. S. Liang, et al. (2010) Plant Biotechnol J 8(8): 928-938; Karetnikov, A. and K. Lehto (2007) J Gen Virol 88(Pt 1): 286-297; Karetnikov, A. and K. Lehto (2008) Virology 371(2): 292-308; Khan, M. A., H. Yumak, et al. (2009) J Biol Chem 284(51): 35461-35470; и Koh, D. C., S. M. Wong, et al. (2003) J Biol Chem 278(23): 20565-20573, содержание которых включено в настоящее описание в виде ссылки в полном объеме. Конструкции экспрессии множества генов, содержащие модифицированные интеины, раскрыты в патентах США № 7026526 и 7741530, а также в заявке на выдачу патента США 2008/0115243.
В используемом в настоящем описании смысле фраза «селектируемый маркер» или «селектируемый маркерный ген» относится к гену, который необязательно использован при трансформации растения, например, чтобы защитить растительные клетки от селективного агента или обеспечить резистентность/устойчивость к селективному агенту. Только те клетки или растения, которые получили функциональный селектируемый маркер, способны делиться или расти в условиях присутствия селективного агента. Примеры селективных агентов могут включать, например, антибиотики, включая спектиномицин, неомицин, канамицин, паромомицин, гентамицин и гигромицин. Такие селектируемые маркеры включают ген неомицинфосфотрансферазы (npt II), который экспрессирует фермент, придающий резистентность к антибиотику канамицину, и гены резистентности к родственным антибиотикам неомицину, паромицину, гентамицину и G418, или ген гигромицинфосфотрансферазы (hpt), который экспрессирует фермент, придающий резистентность к гигромицину. Другие гены селектируемых маркеров могут включать гены, кодирующие резистентность к гербицидам, включая Bar (резистентность к BASTA® (аммонию глюфосината) или фосфинотрицину (PPT)), ацетолактатсинтазу (ALS, резистентность к таким ингибиторам, как сульфонилмочевины (SU), имидазолиноны (IMI), триазолопиримидины (TP), пиримидинилоксибензоаты (POB) и сульфониламинокарбонилтриазолиноны, которые предотвращают первую стадию синтеза аминокислот с разветвленной цепью), глифосату, 2,4-D и резистентность или чувствительность к металлам. Фраза «позитивные по маркеру» относится к растениям, которые были трансформированы так, что содержат селектируемый маркерный ген.
Различные селектируемые или регистрируемые маркеры могут быть включены в выбранный экспрессирующий вектор, что позволяет осуществлять идентификацию и селекцию трансформированных растений или трансформантов. Имеется множество способов для подтверждения экспрессии селектируемых маркеров в трансформированных растениях, включая, например, секвенирование ДНК и ПЦР (полимеразную цепную реакцию), Саузерн-блоттинг, РНК-блоттинг, иммунологические способы выявления белка, экспрессированного с вектора, например, преципитируемого белка, который опосредует резистентность к фосфинотрицину, или других белков, таких как белки репортерных генов β-глюкуронидазы (GUS), люциферазы, зеленого флуоресцирующего белка (GFP), DsRed, β-галактозидазы, хлорамфениколацетилтрансферазы (ХАТ), щелочной фосфатазы и тому подобных (см. Sambrook с соавторами, Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Press, N.Y., 2001).
Селектируемые маркерные гены используют для селекции трансформированных клеток или тканей. Селектируемые маркерные гены включают гены, кодирующие резистентность к антибиотикам, такие как гены, кодирующие неомицинфосфотрансферазу II (NEO) и гигромицинфосфотрансферазу (HPT), а также гены, придающие резистентность к гербицидным соединениям. Гены резистентности к гербицидам обычно кодируют модифицированный белок-мишень, нечувствительный к гербициду, или фермент, который разрушает или осуществляет детоксикацию гербицида в растении до того, как он может начать действовать. Например, резистентность к глифосату может быть получена с использованием генов, кодирующих мутантные ферменты-мишени, 5-енолпирувилшикимат-3-фосфатсинтазу (EPSPS). Гены и мутанты EPSPS были раскрыты в патентах США № 4940835, 5188642, 5310667, 5633435, 5633448 и 6566587. Резистентность к аммонию глюфосината, бромоксинилу и 2,4-дихлорфеноксиацетату (2,4-D) получали, используя бактериальные гены, кодирующие фосфинотрицинацетилтрансферазу, нитрилазу или 2,4-дихлорфеноксиацетатмонооксигеназу, которые детоксифицируют соответствующие гербициды. Ферменты/гены резистентности/устойчивости к глюфосинату были раскрыты в патентах США № 5273894, 5276268, 5550318 и 5561236. Ферменты/гены резистентности к 2,4-D были раскрыты ранее в патентах США № 6100446 и 6153401, а также в заявках на выдачу патентов US 2009/0093366 и WO 2007/053482. Ферменты/гены нитрилазы были ранее раскрыты в патенте США № 4810648.
Другие гербициды могут ингибировать точку роста или меристему, включая имидазолинон или сульфонилмочевину, и были описаны гены резистентности/устойчивости к таким гербицидам: гены синтазы ацетогидроксикислот (AHAS) и ацетолактатсинтазы (ALS). Гены и мутанты AHAS были раскрыты в патентах США № 4761373, 5304732, 5331107, 5853973 и 5928937. Гены и мутанты ALS были раскрыты в патентах США № 5013659 и 5141870.
Гены резистентности к глифосату включают мутантные гены 5-енолпирувилшикимат-3-фосфатсинтазы (EPSP) (в результате введения рекомбинантных нуклеиновых кислот и/или различных форм мутагенеза in vivo нативных генов EPSP), гены aroA и гены глифосатацетилтрансферазы (GAT), соответственно). Гены резистентности к другим фосфоносоединениям включают гены резистентности к глюфосинату (гены фосфотрицинацетилтрансферазы (PAT) из видов Streptomyces, включая Streptomyces hygroscopicus и Streptomyces viridichromogenes), и пиридинокси- или феноксипропионовые кислоты и циклогексоны (гены, кодирующие ингибитор ACC-азы). Гены ацетил-кофермент A-карбоксилазы (ACC-азы), определяющие резистентность/устойчивость к гербицидам, были описаны в патентах США 5162602 и 5498544.
Молекула ДНК, кодирующая мутант гена aroA, может быть получена из ATCC с номером доступа 39256, и нуклеотидная последовательность мутантного гена раскрыта в патенте США № 4769061 Comai, в Европейской заявке на выдачу патента № 0 333 033, Kumada с соавторами, и в патенте США № 4975374 Goodman с соавторами, где раскрыты нуклеотидные последовательности генов глутаминсинтетазы, которые придают резистентность к таким гербицидам, как L-фосфинотрицин. Нуклеотидная последовательность гена PAT представлена в европейской заявке № 0 242 246 Leemans с соавторами. Также в публикации DeGreef с соавторами, Bio/Technology 7: 61 (1989), описано получение трансгенных растений, которые экспрессируют химерные гены bar, кодирующие активность PAT. Примерами генов, придающих резистентность к феноксипропионовым кислотам и циклогексонам, включая сетоксидим и галоксифоп, являются гены Acc1-S1, Acc1-S2 и Acc1-S3, описанные Marshall с соавторами, Theon. Appl. Genet. 83: 435 (1992). Гены GAT, способные придавать резистентность к глифосату, описаны в WO 2005012515, Castle с соавторами. Гены, придающие резистентность к гербицидам 2,4-D, fop и пиридилоксиауксину, описаны в WO 2005107437 и заявке на выдачу патента США с регистрационным номером 11/587893.
Другие гербициды могут ингибировать фотосинтез, включая триазин (гены psbA и 1s+) или бензонитрил (ген нитрилазы). В публикации Przibila с соавторами, Plant Cell 3: 169 (1991) описана трансформация Chlamydomonas плазмидами, кодирующими мутантные гены psbA. Нуклеотидные последовательности генов нитрилазы раскрыты в патенте США № 4810648 Stalker, и молекулы ДНК, содержащие такие гены, доступны из ATCC номера доступа 53435, 67441 и 67442. Клонирование и экспрессия ДНК, кодирующей глутатион-S-трансферазу, описаны Hayes с соавторами, Biochem. J. 285: 173 (1992).
В целях настоящего изобретения селектируемые маркерные гены включают без ограничения гены, кодирующие: неомицинфосфотрансферазу II (Fraley с соавторами, (1986) CRC Critical Reviews в Plant Science, 4: 1-25); цианамидгидратазу (Maier-Greiner с соавторами, (1991) Proc. Natl. Acad. Sci. USA, 88: 4250-4264); аспартаткиназу; дигидродипиколинатсинтазу (Perl с соавторами, (1993) Bio/Technology, 11: 715-718); триптофандекарбоксилазу (Goddijn с соавторами, (1993) Plant Mol. Bio., 22: 907-912); дигидропиколинатсинтазу и десенсибилизированную аспартаткиназу (Perl et al. (1993) Bio/Technology, 11: 715-718); ген bar (Toki с соавторами, (1992) Plant Physiol., 100: 1503-1507 и Meagher с соавторами (1996), Crop Sci., 36: 1367); ген триптофандекарбоксилазы (Goddijn с соавторами (1993) Plant Mol. Biol., 22:907-912); неомицинфосфотрансферазы (NEO) (Southern с соавторами (1982) J. Mol. Appl. Gen., 1: 327); гигромицинфосфотрансферазы (HPT или HYG) (Shimizu с соавторами (1986) Mol. Cell Biol., 6: 1074); дигидрофолатредуктазы (DHFR) (Kwok с соавторами, (1986) PNAS USA 4552); фосфинотрицинацетилтрансферазы (DeBlock с соавторами (1987) EMBO J., 6: 2513); дегалогеназы 2,2-дихлорпропионовой кислоты (Buchanan-Wollatron с соавторами, (1989) J. Cell. Biochem. 13D: 330); синтазы ацетогидроксикислоты (Anderson с соавторами, патент США № 4761373; Haughn с соавторами, (1988) Mol. Gen. Genet. 221: 266); 5-енолпирувилшикиматфосфатсинтазы (aroA) (Comai с соавторами, (1985) Nature 317: 741); галогенарилнитрилазы (Stalker с соавторами, опубликованная заявка PCT WO87/04181); ацетил-кофермент A-карбоксилазы (Parker с соавторами, (1990) Plant Physiol. 92: 1220); дигидроптероатсинтазы (sul I) (Guerineau с соавторами, (1990) Plant Mol. Biol. 15: 127); и полипептида фотосистемы II 32 кД (psbA) (Hirschberg с соавторами, (1983) Science, 222: 1346).
Также включены гены, кодирующие резистентность к: хлорамфениколу (Herrera-Estrella с соавторами, (1983) EMBO J., 2: 987-992); метотрексату (Herrera-Estrella с соавторами, (1983) Nature, 303: 209-213; Meijer с соавторами, (1991) Plant Mol Bio., 16: 807-820 (1991); гигромицину (Waldron с соавторами, (1985) Plant Mol. Biol., 5: 103-108; Zhijian с соавторами, (1995) Plant Science, 108: 219-227 и Meijer с соавторами, (1991) Plant Mol. Bio. 16: 807-820); стрептомицину (Jones с соавторами, (1987) Mol. Gen. Genet., 210: 86-91); спектиномицину (Bretagne-Sagnard с соавторами, (1996) Transgenic Res., 5: 131-137); блеомицину (Hille с соавторами, (1986) Plant Mol. Biol., 7: 171-176); сульфонамиду (Guerineau с соавторами, (1990) Plant Mol. Bio., 15: 127-136); бромоксинилу (Stalker с соавторами, (1988) Science, 242: 419-423); 2,4-D (Streber с соавторами, (1989) Bio/Technology, 7: 811-816); глифосату (Shaw с соавторами, (1986) Science, 233: 478-481); и фосфинотрицину (DeBlock с соавторами, (1987) EMBO J., 6: 2513-2518).
Авторы не считают приведенный выше список генов селектируемых маркеров и репортерных генов ограничивающим. Любой репортерный ген или ген селектируемого маркера включен в настоящее изобретение. При необходимости такие гены могут быть секвенированы способами, известными в данной области.
Синтезируют гены репортеров и селектируемых маркеров для оптимальной экспрессии в растении. То есть, кодирующая последовательность гена модифицирована для усиления экспрессии в растениях. Синтетический маркерный ген конструируют так, чтобы он экспрессировался в растениях на более высоком уровне, получая в результате более высокую эффективность трансформации. Способы оптимизации синтеза генов доступны в данной области. В действительности, несколько генов были оптимизированы для повышения экспрессии генного продукта в растениях.
Последовательность маркерного гена может быть оптимизирована для экспрессии в конкретном виде растений или альтернативно может быть оптимизирована для оптимальной экспрессии в семействах растений. Предпочтительные для растений кодоны могут быть определены на основании кодонов, встречающихся с наиболее высокой частотой в белках, экспрессируемых в наибольшем количестве в конкретном представляющем интерес виде растения. См., например, EPA 0359472; EPA 0385962; WO 91/16432; Perlak et al. (1991) Proc. Natl. Acad. Sci. USA, 88:3324-3328; и Murray et al. (1989) Nucleic Acids Research, 17: 477-498; патент U.S. № 5380831; и патент U.S. № 5436391. Таким образом нуклеотидные последовательности могут быть оптимизированы для экспрессии в любом растении. Известно, что вся или любая часть генной последовательности может быть оптимизированной или синтетической. То есть, также можно использовать полностью оптимизированные или частично оптимизированные последовательности.
Гены, которые придают резистентность к гербициду:
A. Резистентность/устойчивость синтазы ацетогидроксикислоты (AHAS) и ацетолактатсинтазы (ALS) к гербицидам имидазолинону или сульфонилмочевине. Гены и мутанты AHAS раскрыты в патентах США № 4761373, 5304732, 5331107, 5853973 и 5928937. Гены и мутанты ALS раскрыты в патентах США № 5013659 и 5141870.
B. Гены ацетил-кофермент A-карбоксилазы (ACC-азы) резистентности/устойчивости к гербицидам циклогександионам и/или арилоксифеноксипропановой кислоте (включая галоксифоп, диклофоп, феноксипроп, флуазифоп, хизалофоп) описаны в патентах США 5162602 и 5498544.
C. Гены резистентности/устойчивости к глифосату. Ген 5-енолпирувил-3-фосфошикиматсинтазы (ES3P-синтазы) описан в патенте США № 4769601. Гены 5-енолпирувмлшикимат-3-фосфатсинтазы (EPSPS) и мутанты описаны в патентах США № 4940835, 5188642, 5310667, 5633435, 5633448 и 6566587.
D. Гены резистентности/устойчивости к глюфосинату (биалафосу, фосфинотрицину (PPT)). Ген фосфинотрицинацетилтрансферазы (Pat) был описан в патентах США № 5273894, 5276268 и 5550318; и ген резистентности к биалафосу (Bar) был описан в патентах США № 5561236 и 5646024, 5648477 и 7112665. Ген глутаминсинтетазы (GS) был описан в патенте США № 4975372 и Европейской заявке на выдачу патента EP 0333033 A1.
E. Гены гидроксифенилпируватдиоксигеназы (HPPD), определяющие резистентность/устойчивость к гербицидам изоксазолу, дикетонитрилам и/или трикетонам, включая сулькотрион и мезотрион, описаны в патентах США № 6268549 и 6069115.
F. Гены резистентности/устойчивости к 2,4-D. Ген 2,4-D-монооксигеназы описан в патентах США № 6100446 и 6153401. Дополнительные гены резистентности/устойчивости к 2,4-D раскрыты в US 2009/0093366 и WO 2007/053482.
G. Ген имидазолглицеринфосфатдегидратазы (IGPD) против гербицидов имидазола и/или триазола описан в патенте США № 5541310. Гены ферментов, разрушающих дикамбу (оксигеназы, ферредоксина и редуктазы), против гербицида дикамба раскрыты в патентах США № 7022896 и 7105724.
H. Гены против гербицидов, которые ингибируют фотосинтез, включая триазин (гены psbA и 1s+) или бензонитрил (ген нитилазы). См., например, публикацию Przibila с соавторами, Plant Cell 3: 169 (1991), в которой раскрыта трансформация Chlamydomonas плазмидами, кодирующими мутантные гены psbA. Нуклеотидные последовательности генов нитрилазы раскрыты в патенте США № 4810648, и молекулы ДНК, содержащие такие гены, доступны из ATCC с номерами доступа 53435, 67441 и 67442. Клонирование и экспрессия ДНК, кодирующей глутатион-S-трансферазу, описаны Hayes с соавторами, Biochem. J. 285: 173 (1992).
Если специально не указано иное, все технические и научные термины, используемые в настоящем описании, имеют такое же значение, которое обычно подразумевают специалисты в области, к которой относится изобретение. Определения обычных терминов молекулярной биологии можно найти, например, в публикациях: Lewin, Genes V, Oxford University Press, 1994 (ISBN 0-19-854287-9); Kendrew с соавторами, (ed.), The Encyclopedia of Molecular Biology, Blackwell Science Ltd., 1994 (ISBN 0-632-02182-9); и Meyers (ed.), Molecular Biology and Biotechnology: A Comprehensive Desk Reference, VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8).
Предлагаются молекулы нуклеиновой кислоты, содержащие синтетическую нуклеотидную последовательность, которая может функционировать в качестве двунаправленного промотора. В некоторых вариантах синтетический двунаправленный промотор может быть оперативно связан с одной или двумя представляющими интерес нуклеотидными последовательностями. Например, синтетический двунаправленный промотор может быть оперативно связан с одной или двумя представляющими интерес нуклеотидными последовательностями (например, двумя генами, по одному на каждом конце промотора), так, чтобы регулировать транскрипцию по меньшей мере одной представляющей интерес (например, одной или двух) нуклеотидной последовательности(ей). Благодаря включению URS из промотора в синтетический двунаправленный промотор могут быть достигнуты конкретные картины экспрессии и регуляции (например, такие, которые наблюдают в случае генов под контролем нативного промотора) представляющей интерес нуклеотидной последовательности, которая оперативно связана с синтетическим двунаправленным промотором.
Некоторые варианты осуществления изобретения проиллюстрированы в настоящем описании на примере включения минимального корового промоторного элемента из однонаправленного промотора гена убиквитина-1 кукурузы (ZmUbi1) в молекулярном контексте, отличном от контекста нативного промотора, с целью конструирования синтетического двунаправленного промотора. Такой минимальный коровый промоторный элемент называют в настоящем описании «minUbi1P», и он имеет блину приблизительно 200 нуклеотидов. Секвенирование и анализ элементов minUbi1P из множества видов Zea и генотипов Z. mays показал, что функциональные элементы minUbi1P являются высоко консервативными, так что элемент minUbi1P может сохранять свою функцию как инициатора транскрипция, если он имеет, например, по меньшей мере примерно 75%; по меньшей мере примерно 80%; по меньшей мере примерно 85%; по меньшей мере примерно 90%; по меньшей мере примерно 91%; по меньшей мере примерно 92%; по меньшей мере примерно 93%; по меньшей мере примерно 94%; по меньшей мере примерно 95%; по меньшей мере примерно 96%; по меньшей мере примерно 97%; по меньшей мере примерно 98%; по меньшей мере примерно 99%; и/или по меньшей мере примерно 100% идентичность последовательности с элементом minUbi1P, имеющим последовательность SEQ ID NO: 1. Характеристики элементов minUbi1P, которые могут быть полезны в некоторых вариантах осуществления изобретения, могут включать, например и без ограничения, указанную выше высокую консервативность нуклеотидной последовательности; присутствие по меньшей мере одного TATA-бокса; и/или присутствие по меньшей мере одного (например, двух) консенсусного элемента(ов) теплового шока. В конкретных элементах minUbi1P более одного консенсусного элемента теплового шока могут перекрываться в последовательности minUbi1P.
Способ введения элемента minUbi1P в молекулярном контексте, отличном от контекста нативного промотора, с целью конструирования синтетического двунаправленного промотора, может включать изменение ориентации элемента minUbi1P в нуклеиновой кислоте на противоположное по отношению к остальной последовательности промотора, включая его нативный минимальный коровый промотор. Таким образом, синтетический двунаправленный промотор может содержать первый элемент minUbi1P, включенный с 5ʹ-стороны от второго минимального корового промоторного элемента (например, второго элемента minUbi1P) в промоторе в обратной ориентации, так что он может быть оперативно связан с представляющей интерес нуклеотидной последовательностью, локализованной с 5’-стороны от первого элемента minUbi1P. Например, первый элемент minUbi1P может быть введен на 5’-конце промотора ZmUbi1 в обратной ориентации.
Синтетический двунаправленный промотор Ubi1 также может содержать один или несколько дополнительных элементов последовательности, кроме по меньшей мере одного элемента minUbi1P. В некоторых вариантах синтетический двунаправленный промотор Ubi1 может содержать URS промотора; экзон (например, лидер или сигнальный пептид); интрон; спейсерную последовательность; и/или сочетания одного или нескольких из вышеуказанных элементов. Например, и без ограничения, синтетический двунаправленный промотор Ubi1 может содержать последовательность URS из промотора Ubi1 (например, промотора Ubi1 кукурузы); экзон, кодирующий лидерный пептид из гена Ubi1; интрон из гена Ubi1 gene; и сочетания указанных элементов.
В некоторых примерах, содержащих синтетический двунаправленный промотор Ubi1, который включает URS промотора, URS может быть выбрана так, чтобы она придавала конкретные регуляторные свойства синтетическому промотору. Известные промоторы широко варьируют по типу регуляции, которую они осуществляют по отношению к оперативно связанным генам (например, ответы на условия окружающей среды, сигналы при развитии и пространственную информацию), и URS, включенная в гетерологичный промотор, обычно сохраняет тип регуляции, который URS осуществляет по отношению к нативному промотору и оперативно связанному гену(ам). Langridge с соавторами, (1989), выше. Примеры эукариотических промоторов, которые были охарактеризованы и могут содержать URS, входящую в состав синтетического двунаправленного промотора Ubi1, согласно некоторым вариантам, включают, например и без ограничения: промоторы, описанные в патентах США № 6437217 (промотор RS81 кукурузы); 5641876 (промотор актина риса); 6426446 (промотор RS324 кукурузы); 6429362 (промотор PR-1 кукурузы); 6232526 (промотор A3 кукурузы); 6177611 (конститутивные промоторы кукурузы); 6433252 (промотор олеозина L3 кукурузы); 6429357 (промотор актина 2 риса и интрон актина 2 риса); 5837848 (специфичный для корня промотор); 6294714 (индуцируемые светом промоторы); 6140078 (индуцируемые солями промоторы); 6252138 (индуцируемые патогенами промоторы); 6175060 (индуцируемые недостаточностью фосфора промоторы); 6388170 (двунаправленные промоторы); 6635806 (промотор гамма-коиксина); и промотора описанные в заявке на выдачу патента США с регистрационным номером 09/757089 (промотор альдолазы хлоропластов кукурузы).
Дополнительные примеры прокариотических промоторов включают промотор нопалинсинтазы (NOS) (Ebert с соавторами, (1987) Proc. Natl. Acad. Sci. USA 84(16): 5745-9); промотор октопинсинтазы (OCS) (который несут индуцирующие опухоли плазмиды Agrobacterium tumefaciens); промоторы колимовирусов, такие как промотор 19S вируса мозаики цветной капусты (CaMV) (Lawton с соавторами, (1987) Plant Mol. Biol. 9: 315-24); промотор 35S CaMV (Odell с соавторами, (1985) Nature 313: 810-2); 35S-промотор вируса мозаики норичника (Walker с соавторами, (1987) Proc. Natl. Acad. Sci. USA 84(19): 6624-8); промотор синтазы сахарозы (Yang and Russell (1990) Proc. Natl. Acad. Sci. USA 87: 4144-8); промотор R-генного комплекса (Chandler с соавторами, (1989) Plant Cell 1: 1175-83); CaMV35S (патенты США № 5322938, 5352605, 5359142 и 5530196); FMV35S (патенты США № 6051753 и 5378619); промотор PC1SV (патент США № 5850019); промотор SCP1 (патент США № 6677503); и промоторы AGRtu.nos (номер доступа в GenBank V00087; Depicker с соавторами, (1982) J. Mol. Appl. Genet. 1: 561-73; Bevan с соавторами, (1983) Nature 304: 184-7), и тому подобные.
В некоторых вариантах синтетический двунаправленный промотор Ubi1 может дополнительно содержать экзон в дополнение к элементу(ам) minUbi1P. Например, в конкретных вариантах может требоваться направление к цели или перенос полипептида, кодируемого представляющей интерес нуклеотидной последовательностью, оперативно связанной с промотором, в конкретное субклеточное положение и/или компартмент. В указанных и других вариантах кодирующая последовательность (экзон) может быть включена в молекулу нуклеиновой кислоты между элементом minUbi1P и нуклеотидной последовательностью, кодирующей полипептид. Такие элементы могут быть расположены согласно решению специалиста так, чтобы синтетический двунаправленный промотор Ubi1 стимулировал экспрессию полипептида (или одной или обеих кодирующих полипептиды последовательностей, которые оперативно связаны с промотором), содержащего пептид, кодируемый введенной кодирующей последовательностью, в функциональной взаимосвязи с остальной частью полипептида. В конкретных примерах может быть введен экзон, кодирующий лидерный, транзитный или сигнальный пептид (например, лидерный пептид Ubi1).
Пептиды, которые могут кодироваться экзонов, введенным в синтетический двунаправленный промотор Ubi1, включают, например, и без ограничения: экзон лидера убиквитина (например, Ubi1); и транзитный пептид хлоропластов (CTP) (например, CTP EPSPS A. thaliana (Klee с соавторами, (1987) Mol. Gen. Genet. 210: 437-42), и CTP EPSPS Petunia hybrida (della-Cioppa с соавторами, (1986) Proc. Natl. Acad. Sci. USA 83: 6873-7)), как проиллюстрировано на примере целенаправленной доставки в хлоропласты монооксигеназы дикамбы (DMO) в международной публикации PCT № WO 2008/105890.
Интроны также могут быть введены в синтетический двунаправленный промотор Ubi1 в некоторых вариантах осуществления изобретения, например, между элементом minUbi1P и представляющей интерес нуклеотидной последовательностью, которая оперативно связанна с промотором. В некоторых примерах интрон, введенный в синтетический двунаправленный промотор Ubi1, может представлять собой без ограничения 5ʹ-UTR, которая функционирует в качестве лидерной последовательности трансляции, которая присутствует в полностью процессированной мРНК выше стартовой последовательности трансляции (такая лидерная последовательность трансляции может влиять на процессинг первичного транскрипта в мРНК, на стабильность мРНК и/или на эффективность трансляции). Примеры лидерных последовательностей трансляции включают лидеры белков теплового шока кукурузы и петунии (патент США № 5362865), лидеры белков оболочки вирусов растений, лидеры рибулозо-1,5-бифосфаткарбоксилазы/оксигеназы растений и другие. См., например, Turner and Foster (1995) Molecular Biotech. 3(3): 225-36. Не ограничивающие примеры 5ʹ-UTR включают GmHsp (патент США № 5659122); PhDnaK (патент США № 5362865); AtAnt1; TEV (Carrington and Freed (1990) J. Virol. 64: 1590-7); и AGRtu.nos (номер доступа в GenBank V00087; и Bevan с соавторами, (1983) Nature 304: 184-7).
Дополнительные последовательности, которые необязательно могут быть введены в синтетический двунаправленный промотор Ubi1, включает, например и без ограничения: 3ʹ-нетранслируемые последовательности; 3ʹ-области терминации транскрипции; и области полиаденилирования. Они представляют собой генетические элементы, располагаемые ниже представляющей интерес нуклеотидной последовательности (например, представляющей интерес последовательности, которая оперативно связана с синтетическим двунаправленным промотором Ubi1), и включают полинуклеотиды, которые обеспечивают сигнал полиаденилирования, и/или другие регуляторные сигналы, способные влиять на транскрипцию, процессинг мРНК или экспрессию генов. Сигнал полиаденилирования может функционировать в растениях, вызывая добавление полиадениловых нуклеотидов к 3ʹ-концу предшественника мРНК. Последовательность полиаденилирования может быть получена из природного гена, из различных генов растений или из генов T-ДНК. Не ограничивающим примером 3ʹ-области терминации транскрипции является 3ʹ-область нопалинсинтазы (nos 3ʹ; Fraley с соавторами, (1983) Proc. Natl. Acad. Sci. USA 80: 4803-7). Пример применения разных 3ʹ-нетранслируемых областей приведен в публикации Ingelbrecht с соавторами, (1989), Plant Cell 1: 671-80. Не ограничивающие примеры сигналов полиаденилирования включают сигнал полиаденилирования гена RbcS2 Pisum sativum (Ps.RbcS2-E9; Coruzzi с соавторами, (1984) EMBO J. 3: 1671-9) и AGRt nos (номер доступа в GenBank E01312).
В некоторых вариантах синтетический двунаправленный промотор Ubi1 содержит одну или несколько нуклеотидных последовательностей, которые облегчают направление нуклеиновой кислоты, содержащей промотор, в конкретный локус-мишень в геноме целевого организма. Например, могут быть введены одна или несколько последовательностей, которые являются гомологичными участкам геномной последовательности ДНК хозяина (например, редким или уникальным последовательностям геномной ДНК). В некоторых примерах такие гомологичные последовательности могут направлять рекомбинацию и интеграцию нуклеиновой кислоты, содержащей синтетический двунаправленный промотор Ubi1, в участок гомологичной ДНК в геноме-хозяина. В конкретных примерах синтетический двунаправленный промотор Ubi1 содержит одну или несколько нуклеотидных последовательностей, которые облегчают направление нуклеиновой кислоты, содержащей промотор, в редкое или уникальное положение в геноме хозяина с использованием сконструированных нуклеазных ферментов, которые узнают последовательность в редком или уникальном положении, и способствуют интеграции в такое редкое или уникальное положение. Такая система целенаправленной интеграции с использованием эндонуклеаз типа цинковых пальцев в качестве нуклеазного фермента описана в заявке на выдачу патента США № 13/011735.
Нуклеиновые кислоты, содержащие синтетический двунаправленный промотор Ubi1, могут быть получены с использованием любого способа, известного в данной области, включая, например и без ограничения: RCA; ПЦР-амплификацию; ОТ0ПЦР-амплификацию; OLA и SNuPE. Указанные и другие эквивалентные способы хорошо известны специалистам в данной области и дополнительно подробно описаны, например и без ограничения: Sambrook с соавторами, Molecular Cloning: A Laboratory Manual, 3rd Ed., Cold Spring Harbor Laboratory, 2001; и Ausubel с соавторами, Current Protocols in Molecular Biology, John Wiley and Sons, 1998. Все публикации, цитированные выше, включают указанные выше руководства вместе с приведенными в них любыми чертежами, фигурами и/или таблицами.
Доставка и/или трансформация: Настоящее изобретение также относится к способам трансформации клетки молекулой нуклеиновой кислоты, содержащей синтетический двунаправленный промотор Ubi1. Согласно некоторым вариантам можно использовать любой из большого количества способов, хорошо известных в данной области для введения молекул нуклеиновых кислоты в растения, для трансформации растения молекулой нуклеиновой кислоты, содержащей синтетический двунаправленный промотор Ubi1, например, для введения одного или нескольких синтетических двунаправленных промоторов Ubi1 в геном растения-хозяина и/или для дополнительного введения одной или нескольких представляющих интерес молекул нуклеиновых кислот, оперативно связанных с промотором.
Подходящие способы трансформации растений включают любой способ, которым ДНК может быть введена в клетку, например и без ограничения: электропорацию (см., например, патент США 5384253); бомбардировку микрочастицами (см., например, патенты США 5015580, 5550318, 5538880, 6160208, 6399861 и 6403865); опосредованную Agrobacterium трансформацию (см., например, патенты США 5635055, 5824877, 5591616, 5981840 и 6384301); и трансформацию протопластов (см., например, патент США 5508184). Благодаря применению способов, таких как описанные выше способы, клетки фактически любого вида растения могут быть стабильно трансформированы, и такие клетки можно развивать в трансгенные растения способами, известными специалистам в данной области. Например, способы, которые могут быть особенно применимы в контексте трансформации хлопчатника, описаны в патентах США 5846797, 5159135, 5004863 и 6624344; способы трансформации растений Brassica, в частности, описаны, например, в патенте США 5750871; способы трансформации сои описаны, например, в патенте США 6384301; и способы трансформации кукурузы описаны, например, в патентах США 7060876 и 5591616 и международной публикации PCT WO 95/06722.
После осуществления доставки экзогенной нуклеиновой кислоты в реципиентную клетку обычно идентифицируют трансформированную клетку для дальнейшего культивирования и регенерации растения. Чтобы повысить возможность идентификации трансформантов, может требоваться использование селектируемого или подвергаемого скринингу маркерного гена в векторе для трансформации, используемом для создания трансформанта. В данном случае популяцию потенциально трансформированных клеток можно анализировать, экспонируя клетки с агентом или агентами для селекции, или клетки можно подвергать скринингу в отношении требуемого признака маркерного гена.
Клетки, которые выживают при воздействии на них агента для селекции, или клетки, которые были оценены как позитивные в скрининговом анализе, можно культивировать в среде, которая поддерживает регенерацию растений. В некоторых вариантах любая подходящая среда для культуры ткани растений (например, среды MS и N6) может быть модифицирована введением дополнительных веществ, таких как регуляторы роста. Ткань можно поддерживать в основной среде в регуляторами роста до тех пор, пока не будет получено достаточно ткани, чтобы попытаться начать регенерацию растения, или осуществляют многократные раунды селекции вручную до тех пор, пока морфология ткани не станет подходящей для регенерации (например, по меньшей мере 2 недели), затем ее переносят в среду, способствующую образованию побегов. Культуры периодически переносят вплоть до момента, когда происходит образование достаточного количества побегов. После образования побегов их переносят в среду, способствующую образованию корней. После образования достаточного количества корней растения могут быть перенесены в почву для дальнейшего роста и созревания.
Чтобы подтвердить присутствие требуемой молекулы нуклеиновой кислоты, содержащей синтетический двунаправленный промотор Ubi1 в регенерирующих растениях, можно осуществить разнообразные анализы. Такие анализы включают, например: молекулярные биологические анализы, такие как Саузерн-блоттинг и Нозерн-блоттинг и ПЦР; биохимические анализы, такие как выявление присутствия белкового продукта, например, иммунологическими способами (ELISA и/или Вестерн-блоты) или по осуществлению ферментативной функции; анализы частей растений, такие как анализы листьев и корней; и анализы фенотипа целого регенерированного растения.
События целенаправленной интеграции могут быть подвергнуты скринингу, например, при ПЦР-амплификации с использованием, например, олигонуклеотидных праймеров, специфичных для представляющих интерес молекул нуклеиновых кислот. Подразумевается, что ПЦР-генотипирование включает без ограничения амплификацию в полимеразной цепной реакции (ПЦР) геномной ДНК, полученной из выделенной ткани каллуса растения-хозяина, предположительно содержащей представляющую интерес молекулу нуклеиновой кислоты, интегрированную в геном, с последующим стандартным клонированием и анализом последовательности продуктов ПЦР-амплификации. Способы ПЦР-генотипирования хорошо описаны (см., например, Rios с соавторами, (2002) Plant J. 32: 243-53), и могут быть применимы по отношению к геномной ДНК, полученной из любого вида растения или типа ткани, включая культуры клеток. Сочетания олигонуклеотидных праймеров, которые связываются как с последовательностью-мишенью, так и с вводимыми последовательностями, можно использовать последовательно или совместно в реакциях ПЦР-амплификации. Могут быть получены олигонуклеотидные праймеры, сконструированные для отжига с сайтом-мишенью, введенными последовательностями нуклеиновой кислоты и/или сочетаниями таких последовательностей. Таким образом, методика ПЦР-генотипирования может включать, например, и без ограничения: амплификацию специфичных последовательностей генома растения; амплификацию множества специфичных последовательностей генома растения; амплификацию неспецифичных последовательностей генома растения; и сочетания любых указанных выше способов. Специалист в данной области может разработать дополнительные сочетания праймеров и реакций амплификации, чтобы подробно исследовать геном. Например, может быть сконструирован набор прямых и обратных олигонуклеотидных праймеров для отжига с последовательностью(ями) нуклеиновой кислоты, специфичной для мишени вне границ вводимой последовательности нуклеиновой кислоты.
Могут быть сконструированы прямые и обратные олигонуклеотидные праймеры для специфичного отжига с введенной молекулой нуклеиновой кислоты, например, с последовательностью, соответствующей кодирующей области в представляющей интерес нуклеотидной последовательности, входящей в ее состав, или другими частями молекулы нуклеиновой кислоты. Такие праймеры могут быть использованы вместе с праймерами, описанными выше. Олигонуклеотидные праймеры могут быть синтезированы в соответствии с требуемой последовательностью и являются коммерчески доступными (например, из Integrated DNA Technologies, Inc., Coralville, IA). После амплификации может быть осуществлено клонирование и секвенирование или прямой анализ последовательности продуктов амплификации. Специалист в данной области может предусмотреть альтернативные способы для анализа продуктов амплификации, созданных при ПЦР-генотипировании. В одном варианте в ПЦР-амплификациях используют олигонуклеотидные праймеры, специфичные для генной мишени.
Некоторые варианты осуществления настоящего изобретения также относятся к клеткам, содержащим синтетический двунаправленный промотор Ubi1, например, который может присутствовать в конструкции нуклеиновой кислота. В конкретных примерах синтетический двунаправленный промотор Ubi1 согласно некоторым вариантам осуществления изобретения может быть использован в качестве регуляторной последовательности для регуляции экспрессии трансгенов в растительных клетках и растениях. В некоторых таких примерах применение синтетического двунаправленного промотора Ubi1, оперативно связанного с представляющей интерес нуклеотидной последовательностью (например, трансгеном) может способствовать уменьшению количеств гомологичных промоторов, необходимых для регуляции экспрессии заданного количества представляющих интерес нуклеотидных последовательностей и/или уменьшению размера конструкции нуклеиновой кислоты (кислот), необходимой для введения заданного количества представляющих интерес нуклеотидных последовательностей. Кроме того, применение синтетического двунаправленного промотора Ubi1 может обеспечить возможность совместной экспрессии двух оперативно связанных представляющих интерес нуклеотидных последовательностей в одних и тех же условиях (т.е., условиях, в которых промотор является активным). Такие примеры могут быть особенно применимы, например, когда каждая из двух представляющих интерес оперативно связанных нуклеотидных последовательностей вносит вклад в один признак трансгенного хозяина, содержащего представляющие интерес нуклеотидные последовательности, и коэкспрессия представляющих интерес нуклеотидных последовательностей преимущественно влияет на экспрессию признака трансгенного хозяина.
В некоторых вариантах трансгенное растение, содержащее один или несколько синтетических двунаправленных промоторов Ubi1 и/или одну или несколько представляющих интерес нуклеотидных последовательностей, может иметь один или несколько требуемых признаков, придаваемых (например, вводимых, усиливаемых или формируемых с участием) экспрессией представляющей интерес нуклеотидной последовательности(тей) в растении. Такие признаки могут включать, например, и без ограничения: резистентность к насекомым, другим вредителям и вызывающим заболевания агентам; устойчивость к гербицидам; повышенную стабильность, урожайность или сроки хранения; устойчивость к условиям окружающей среды; получение фармацевтических средств; получение промышленных продуктов; и повышение содержания питательных веществ. В некоторых примерах требуемый признак может быть результатом трансформации растения молекулой нуклеиновой кислоты, содержащей синтетический двунаправленный промотор Ubi1, оперативно связанный с представляющей интерес нуклеотидной последовательностью. В некоторых примерах требуемый признак можно придавать растению, получаемому в виде растения-потомка, в результате селекции, и такой признак можно придавать с использованием одной или нескольких представляющих интерес нуклеотидных последовательностей, оперативно связанных с синтетическим двунаправленным промотором Ubi1, которые передаются в растение от родительского растения, содержащего представляющую интерес нуклеотидную последовательность, оперативно связанную с синтетическим двунаправленным промотором Ubi1.
Трансгенным растением согласно некоторым вариантам осуществления изобретения может быть любое растение, которое можно трансформировать молекулой нуклеиновой кислоты согласно изобретению, или которое получают скрещиванием с растением, трансформированным молекулой нуклеиновой кислоты согласно изобретению. Соответственно растение может быть двудольным или однодольным. Не ограничивающие примеры двудольных растений для применения в некоторых примерах включают: люцерну; бобы; брокколи; канолу; капусту; морковь; цветную капусту; сельдерей; китайскую капусту; хлопчатник; огурец; баклажан; латук; дыню; горох; перец; арахис; картофель; тыкву; редис; рапс; шпинат; сою; кабачок; сахарную свеклу; подсолнечник; табак; томат и арбуз. Не ограничивающие примеры однодольных растений для применения в некоторых примерах включают: кукурузу; лук; рис; сорго; пшеницу; рожь; просо; сахарный тростник; овес; тритикале; просо и газонные травы.
В некоторых вариантах трансгенное растение можно применять или культивировать любым способом, при этом требуется присутствие синтетического двунаправленного промотора Ubi1 и/или оперативно связанной представляющей интерес нуклеотидной последовательности. Соответственно, такие трансгенные растения могут быть сконструированы так, чтобы они, наряду с прочим, имели один или несколько требуемых признаков благодаря трансформации молекулами нуклеиновых кислот согласно изобретению, и их можно возделывать и/или культивировать любым способом, известным специалистам в данной области.
Хотя изобретение описано со ссылкой на конкретные способы и варианты осуществления, будет понятно, что могут быть осуществлены различные модификации и изменения, не выходя за рамки объема изобретения.
Следующие примеры приведены для иллюстрации некоторых конкретных признаков и/или вариантов. Примеры не следует рассматривать как ограничивающие изобретение конкретными иллюстративными признаками или вариантами.
ПРИМЕРЫ
ПРИМЕР 1: Трансформация и экспрессия
Трансформация Agrobacterium tumefaciens: Бинарным вектором pDAB108706 трансформировали штамм Agrobacterium tumefaciens DAt13192 тройной системы (предварительная заявка на выдачу патента США № 61/368965). Бактериальные колонии выделяют и затем выделяют ДНК бинарной плазмиды и подтверждают, используя расщепление ферментами рестрикции.
Трансформация кукурузы: стерилизация початков и выделение зародышей. Чтобы получить незрелые зародыши кукурузы, растения Zea mays (сорт B104) выращивают в теплице и подвергают самоопылению или опылению между сибсами, чтобы получить початки. Початки собирают примерно через 9-12 дней после опыления. В день эксперимента поверхность початков стерилизуют, погружая в 20% раствор хозяйственного отбеливателя, который содержит 5% гипохлорита натрия, и встряхивая в течение 20-30 минут, с последующими тремя промывками в стерильной воде. После стерилизации незрелые зиготические зародыши (1,5-2,2 мм) асептически вырезают из каждого початка и случайным образом распределяют в микроцентрифужные пробирки, содержащие жидкую среду для инфекции (основная среда LS, 4,43 г/л; раствор витамина N6 [1000X], 1,00 мл/л; L-пролин, 700,0 мг/л; сахароза, 68,5 г/л; глюкоза, 36,0 г/л; 2,4-D, 1,50 мг/л. В случае указанного набора экспериментов для каждой обработки используют объединенные зародыши из 2-3 початков.
Инициация культуры Agrobacterium: Исходные культуры в глицерине Agrobacterium, содержащие бинарные векторы, описанные выше, высевают штрихом на чашках с минимальной средой AB, содержащих соответствующие антибиотики, и выращивают при 20ºC в течение 3-4 суток. Отбирают отдельную колонию и высевают штрихом на чашки YEP, содержащие такие же антибиотики, и инкубируют при 28ºC в течение 1-2 суток.
Культура и совместная культура Agrobacterium: В день эксперимента колонии Agrobacterium собирают из YEP-чашки, суспендируют в 10 мл среды для инфекции в одноразовой пробирке объемом 50 мл, и плотность клеток доводят до OD600 = 0,2-0,4 нм, используя спектрофотометр. Культуры Agrobacterium помещают в ротационный встряхиватель при 100 об./мин, при комнатной температуре, в это время осуществляют вырезание зародышей. Незрелые зиготические зародыши размером 1,5-2,2 мм выделяют из стерилизованных зерен кукурузы и помещают в 1 мл среды для инфекции и промывают один раз такой же средой. Суспензию Agrobacterium (2 мл) добавляют в каждую пробирку и пробирки переворачивают примерно 20 раз, затем встряхивают в течение 10-15 минут. Зародыши переносят в среду для совместного культивирования (соли MS, 4,33 г/л; L-пролин, 700,0 мг/л; миоинозит, 100,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба - 3,30 мг/л; сахароза, 30,0 г/л; Gelzan™, 3,00 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л, AgNO3, 15,0 мг/л; ацетосирингон, 100 мкМ), ориентируют щитком вверх и инкубируют в течение 3-4 суток в условиях освещения при 25ºC.
Временная экспрессия GUS и YFP/Phiyfp: Временную экспрессию YFP/Phiyfp и GUS можно исследовать в трансформированных зародышах и через 3 дня после совместного культивирования с Agrobacterium. Зародыши исследуют с помощью стереомикроскопа (Leica Microsystems, Buffalo Grove, IL), используя YFP-фильтр и источник света 500 нм. Зародыши, в которых наблюдают экспрессию YFP/Phiyfp, отбирают для гистохимического анализа GUS. Раствор для окрашивания GUS готовят, как описано Maniatis с соавторами (1989), и зародыши инкубируют в 1 мл раствора в течение 24 часов при 37ºC. Зародыши исследуют в отношении временной экспрессии GUS под микроскопом.
Отбор каллусов и регенерация растений с предполагаемыми событиями: После периода совместного культивирования зародыши переносят в среду для стадии покоя (MS-соли, 4,33 г/л; L-пролин, 700,0 мг/л; миоинозит, 100,0 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота] 500,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба, 3,30 мг/л; сахароза, 30,0 г/л; гельзан 2,30 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л; AgNO3, 15,0 мг/л; карбенициллин, 250,0 мг/л) без селективного агента и инкубируют при 24-часовом освещении с интенсивностью света 50 мкмоль⋅м-2⋅сек-1 в течение 7 суток при 28ºC. Зародыши переносят в среду для селекции 1 (MS-соли, 4,33 г/л; L-пролин, 700,0 мг/л; миоинозит, 100,0 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота] 500,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба, 3,30 мг/л; сахароза, 30,0 г/л; Gelzan™ 2,30 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л; AgNO3, 15,0 мг/л; карбенициллин, 250,0 мг/л), содержащую 100 нМ галоксифопа и инкубируют при 24-часовом освещении с интенсивностью света 50 мкмоль⋅м-2⋅сек-1 в течение 7 суток при 28ºC.
Зародыши с пролиферирующими эмбриогенными каллусами переносят в среду для селекции 2 (MS-соли, 4,33 г/л; миоинозит, 100,0 мг/л; L-пролин, 700,0 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота] 500,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба, 3,30 мг/л; сахароза, 30,0 г/л; Gelzan™ 2,30 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л; AgNO3, 15,0 мг/л; карбенициллин, 250,0 мг/л), содержащую 500 нМ галоксифопа, и инкубируют при 24-часовом освещении с интенсивностью света 50 мкмоль⋅м-2⋅сек-1 еще в течение 14 суток при 28ºC. Описанная стадия селекции обеспечивает возможность трансгенному каллусу дальше пролиферировать и дифференцироваться. Период селекции каллусов продолжается три недели. Пролиферирующие эмбриогенные каллусы переносят в среду для регенерации 1 (MS-соли, 4,33 г/л; миоинозит, 100,0 мг/л; L-пролин, 350,0 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота] 250,0 мг/л; ферментативный гидролизат казеина 50,0 мг/л; NAA 0,500 мг/л; ABA 2,50 мг/л; BA 1,00 мг/л; сахароза, 45,0 г/л; Gelzan™ 2,50 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л; AgNO3, 1,00 мг/л; карбенициллин, 250,0 мг/л), содержащую 500 нМ галоксифопа, и культивируют при 24-часовом освещении с интенсивностью света 50 мкмоль⋅м-2⋅сек-1 в течение 7 суток при 28ºC. Эмбриогенные каллусы с побегами/почками переносят в среду для регенерации 2 (MS-соли, 4,33 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л; миоинозит, 100,0 мг/л; сахароза, 60,0 г/л; геллановая камедь G434TM 3,00 г/л; карбенициллин, 250,0 мг/л), содержащую 500 нМ галоксифопа. Культуры инкубируют при 24-часовом освещении с интенсивностью света 50 мкмоль⋅м-2⋅сек-1 в течение 7-10 суток при 28ºC. Небольшие побеги с первичными корнями переносят в среду для удлинения побегов и образования корней (MS-соли, 4,33 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л; миоинозит, 100,0 мг/л; сахароза, 60,0 г/л; геллановая камедь G434TM 3,00 г/л; карбенициллин, 250,0 мг/л) в боксах MAGENTATM (Sigma-Aldrich, St. Louis, MO) и инкубируют в условиях 16/8 часов освещение/темнота в течение 7 суток при 28ºC. Предполагаемые трансгенные растеньица анализируют в отношении количества копий трансгенов и переносят в теплице.
ПРИМЕР 2: Конструирование синтетического двунаправленного промотора Ubi1 и вектора pDAB108706
Иллюстративное схематичное изображение промотора убиквитина-1 кукурузы (Ubi1) показано на фиг. 1. Промотор Ubi1 клонировали из кукурузы. Плазмиду, содержащую промотор Ubi1, амплифицировали в ПЦР, используя систему ПЦР-амплификации высокой точности. Область длиной приблизительно 200 нуклеотидов промотора Ubi1 кукурузы идентифицировали как минимальный коровый промотор Ubi1 Zea mays Ubi1 (minUbi1P) (SEQ ID NO: 1). Затем minUbi1P с последовательностью SEQ ID NO: 1 добавляли к полинуклеотиду (SEQ ID NO: 2), содержащему экзон убиквитина-1 Zea mays (экзон ZmUbi1 экзон) и интрон убиквитина-1 Zea mays (интрон ZmUbi1), используя способы клонирования, общеизвестные в данной области, для получения полинуклеотида SEQ ID NO: 3. Затем полученный полинуклеотид клонировали в обратной ориентации выше нуклеиновой кислоты, содержащей промотор Ubi1 кукурузы (включая вышележащую регуляторную последовательность Ubi1 (URS)); SEQ ID NO: 4), получая синтетический двунаправленный промотор Ubi1 SEQ ID NO: 5 (см., например, фиг. 5).
Последовательности, кодирующие репортерные гены, клонировали ниже каждого конца синтетического двунаправленного промотора Ubi1. Последовательность, кодирующую желтый флуоресцирующий белок (yfp), встраивали ниже фрагмента полинуклеотида, который содержит промоторные элементы minUbi1P, экзон ZmUbi1 и интрон ZmUbi1. Кроме того, лежащую ниже лидерную последовательность, содержащую полинуклеотидную последовательность стоп-кодона рамки 3 и консенсусную полинуклеотидную последовательность кукурузы (Козак), добавляли к фрагменту, содержащему промоторные элементы minUbi1P, экзон ZmUbi1 и интрон ZmUbi1. Последовательность, кодирующую uidA (GUS), встраивали ниже синтетического двунаправленного промотора Ubi1 в обратной ориентации по отношению к последовательности yfp (SEQ ID NO: 6; см., например, фиг. 3). Полученный полинуклеотид, содержащий синтетический двунаправленный промотор Ubi1, оперативно связанный с генами yfp и GUS, клонировали в плазмиде pDAB105801.
Бинарный вектор, который содержал кассеты экспрессии генов GUS и yfp из плазмиды pDAB105801, получали, используя реакцию GATEWAYTM L-R CLONASETM (Invitrogen, Carlsbad, CA). Полученный вектор, pDAB108706, содержит кассеты экспрессии генов GUS, yfp и aad-1 в области T-нити (см., например, фиг. 5).
ПРИМЕР 3: Временная экспрессия генов, оперативно связанных с синтетическим двунаправленным промотором убиквитина 1
Получали изображения типичных примеров временной экспрессии YFP и GUS в зародышах Zea mays, трансформированных pDAB108706. Обе стороны двунаправленного промотора ZmUbi1 могут вызывать высокий уровень экспрессии оперативно связанных последовательностей, кодирующих yfp и GUS. Уровни экспрессии YFP сравнимы с уровнями экспрессии GUS. Полученные наблюдения подтверждают, что обе стороны двунаправленного промотора ZmUbi1 являются биологически функциональными. Кроме того, элемент minUbi1P синтетического двунаправленного промотора Ubi1 может экспрессировать YFP на уровнях экспрессии, сходных с уровнями экспрессии в каллусе Zea mays, трансформированном бинарной плазмидой (pDAB101556), которая содержала однонаправленный промотор ZmUbi1, управляющий кодирующей последовательностью yfp. Экспрессия YFP или GUS не выявлена в незрелых зародыш, используемых в качестве негативного контроля, которые не были трансформированы бинарной конструкцией и не содержали последовательностей, кодирующих yfp или GUS.
ПРИМЕР 4: Стабильная экспрессия генов, оперативно связанных с синтетическим двунаправленным промотором убиквитина 1
Можно исследовать картины клеток каллуса Zea mays, которые стабильно трансформированы бинарным вектором pDAB108706, который содержит последовательность, кодирующую yfp. Такие клетки получали из зародышей Z. mays, которые были размножены на среде для селекции 2. Двунаправленный промотор ZmUbi1 может вызвать высокий уровень экспрессии последовательности, кодирующей yfp. Полученные результаты подтверждают, что минимальный промоторный элемент Min-UbiP1 двунаправленного промотора ZmUbi1 способен экспрессировать репортерный ген в стабильно трансформированных клетках каллуса Z. mays. Уровни экспрессии белка YFP сходны с уровнями экспрессии YFP в каллусе Z. mays, трансформированном контрольным бинарным вектором, который содержал однонаправленный промотор ZmUbi1, управляющий последовательностью, кодирующей yfp (pDAB101556). Экспрессию YFP или GUS не выявляли в каллусе, используемом в качестве негативного контроля, который не был трансформирован бинарной конструкцией и не содержал последовательности, кодирующей yfp или GUS.
ПРИМЕР 5: Оценка количества копий трансгенов с использованием ПЦР в реальном времени TaqMan®
Зародыши Zea mays трансформировали бинарным вектором, содержащим двунаправленный промотор ZmUbi1, pDAB108706, а другие растения трансформировали контрольным бинарным вектором, pDAB101556. Присутствие трансгенов yfp в геноме обеих групп растений Z. mays подтверждали в анализе с использованием гидролизных зондов. Получали стабильно трансформированные трансгенные молодые растения Z. mays, которые развивались из каллуса, и анализировали, чтобы идентифицировать события наличия небольшого количества копий (1-2 копии) полноразмерных вставок T-нити из бинарного вектора pDAB108706 и контрольного бинарного вектора pDAB101556. Идентифицированные молодые растения переносили в теплицу и выращивали.
Систему Roche Light Cycler480™ использовали для определения количества копий трансгенов в случае событий трансформации бинарным вектором pDAB108706 и в случае контрольных событий трансформации бинарным вектором pDAB101556. В данном способе используют биплексную реакцию TaqMan®, в которой применяют олигонуклеотиды, специфичные к гену yfp и к эндогенному эталонному гену инвертазы Z. mays (номер доступа в Genbank: U16123.1), в одном анализе. Количество копий и зиготность определяли, измеряя интенсивность yfp-специфичной флуоресценции относительно специфичной для инвертазы флуоресценции, которые сравнивали с известными стандартами количества копий.
В Z. mays, трансформированных бинарным вектором pDAB108706, специфичный для гена yfp фрагмент ДНК амплифицировали с использованием одного набора праймеров/зондов TaqMan®, содержащего зонд, меченый флуоресцирующим красителем FAM, и инвертазу амплифицировали с использованием второго набора праймеров/зондов TaqMan®, содержащего зонд, флуоресцентно меченый HEX (таблица 2). Смесь для ПЦР-реакции готовили, как указано в таблице 3, и специфичные для генов фрагменты ДНК амплифицировали согласно условиям, указанным в таблице 4. Количество копий и зиготность образцов определяли, измеряя относительную интенсивность флуоресценции, специфичной для репортерного гена, yfp, по отношению к флуоресценции, специфичной для эталонного гена инвертазы, по сравнению с известными стандартами количества копий.
Прямые и обратные нуклеотидные праймеры и флуоресцирующие зонды (синтезированные в Integrated DNA Technologies, Coralville, IA)
Стандарты создавали разбавлением вектора pDAB108706 в геномной ДНК Z. mays B104 (гДНК), чтобы получить стандарты с известным соотношением pDAB108706:гДНК. Например, готовили образцы, имеющие одну, две и четыре копии векторной ДНК на одну копию гДНК Z. mays B104. Разведения с одной и двумя копиями pDAB108706, смешанными со стандартом гДНК Z. mays B104, подтверждали по сравнению с контрольным событием для Z. mays, в случае которого известно, что растение является гемизиготным, и контрольным событием для Z. mays, в случае которого известно, что растение является гомозиготным (событие Z. mays 278; см. международную публикацию патента PCT № WO 2011/022469 A2). Биплексный анализ TaqMan®, в котором используют олигонуклеотиды, специфичные к гену AAD1, и олигонуклеотиды, специфичные к эндогенному эталонному гену Z. mays, гену инвертазы, осуществляли посредством амплификации и выявления фрагмента ДНК, специфичного для гена AAD1 с использованием одного набора праймеров/зондов TaqMan®, содержащего зонд, меченый флуоресцирующим красителем FAM, и посредством амплификации и выявления фрагмента ДНК, специфичного для гена инвертазы, с использованием второго набора праймеров/зондов TaqMan®, содержащего зонд, флуоресцентно меченый HEX (таблица 2). Реакционную смесь TaqMan® для AAD1 готовили, как указано в таблице 3, и специфичные фрагменты амплифицировали согласно условиям, указанным в таблице 4.
Смесь для ПЦР-реакции Taqman
Уровень флуоресценции, которая была генерирована в случае каждой реакции, анализировали, используя термоциклер Roche LightCycler 480™ согласно инструкциям производителя. Флуоресцирующий остаток FAM возбуждали при оптической плотности 465/510 нм, и флуоресцирующий остаток HEX возбуждали при оптической плотности 533/580 нм. Количество копий определяли сравнением значений мишень/эталон для неизвестных образцов (результат, полученный с использованием LightCycler 480™) со значениями мишень/эталон четырех стандартов с известным количеством копий (ноль, 1 копия (геми-), 2 копии (гомо-) и 4 копии).
Условия для ПЦР-амплификации в термоциклере
Результаты анализа количества копий трансгенов в трансгенных растениях, полученных в результате трансформация конструкцией с двунаправленным промотором ZmUbi1 (pDAB108706), и в трансгенных растениях, полученных в результате трансформации контрольной конструкцией YFP с однонаправленным промотором ZmUbi1 (pDAB101556), показаны в таблице 5. Только растения с 1-2 копиями трансгена yfp переносили в теплицу для дальнейших анализов экспрессии.
Оценка количества копий трансгенов в трансгенных растениях, полученных в случае конструкции с двунаправленным промотором и контрольной конструкции
ПРИМЕР 6: Стабильная экспрессия в целых растениях генов, оперативно связанных с синтетическим двунаправленным промотором убиквитина 1
Целые растения, которые содержали небольшое количество копий T-ДНК бинарной плазмиды pDAB 108706, и растения, которые содержали небольшое количество копий контрольной бинарной плазмиды pDAB101556, выращивали в теплице. Анализировали типичные примеры стабильной экспрессии YFP в тканях листьев и корней трансгенных растений кукурузу T0, полученных из зародышей Z. mays, трансформированных pDAB108706. Двунаправленный промотор ZmUbi1 может вызывать высокий уровень экспрессии последовательностей, кодирующих yfp, как в тканях листьев, так и в тканях корней. Микроскопический анализ также подтверждает, что минимальный коровый элемент Min-UbiP1 в двунаправленном промоторе ZmUbi1 может управлять контрольной бинарной плазмидой (pDAB101556), которая содержит однонаправленный промотор ZmUbi1, управляющий экспрессией последовательности, кодирующей yfp. В таких контрольных растениях также наблюдали стабильную экспрессию YFP в тканях листьев и в тканях корней.
ПРИМЕР 7: Вестерн-блот-анализ генов, оперативно связанных с синтетическим двунаправленным промотором убиквитина 1
Общий растворимый белок: Брали образцы от трансформированных растений кукурузы T0 на стадии развития V6. Всего отбирали по четыре вырезанных пробойником образца листа из самого молодого распустившегося листа в пробирку Matrix и помещали в бокс Matrix. В качестве негативного контроля четыре вырезанных пробойником образца листа от двух нетрансформированных растений кукурузы B104 на стадии развития V6 отбирали в пробирку Matrix. Стальные шарики помещали в пробирки Matrix с образцами и затем добавляли 400 мкл PBST в каждую пробирку. Пробирки закрывали крышками и белок экстрагировали, разбивая шариками при 1500 об./мин в течение 5 минут в измельчителе тканей Kleco™. Обломки осаждали центрифугированием.
5 мкл образца из каждой пробирки разбавляли до 25 мкл в PBST в 96-луночном планшете для микротитрования. Такие образцы анализировали в отношении общего растворимого белка, используя набор для анализа белка BCA (Thermo Scientific Pierce, Rockford, IL), согласно инструкциям производителя. Стандарты бычьего сывороточного альбумина (БСА), прилагаемые в наборе, анализировали в двух повторах, и средние значения использовали для создания стандартной кривой, которую затем использовали для вычисления общего растворимого белка в каждом образце. Общий растворимый белок для каждого образца затем нормализовали в мг/мкл.
Протокол Вестерн-блоттинга
Вестерн-блот-анализ YFP/Phiyfp: В 96-луночном планшете для микротитрования каждый образец экстрагированного белка объемом 5 мкл разбавляли, используя 5 мкл 2x буфера Лэммли + 2-β-меркаптоэтанол. Контрольные образцы очищенного YFP/Phiyfp в буфере HEPES (50 мМ HEPES, 200 мМ KCl, 10% глицерин) приобретали из Axxora (San Diego, CA). Образцы готовили в том же планшете, разбавлением 1:1 буфером Лэммли, получая стандартную кривую со следующими концентрациями: 0,5 нг/мкл, 0,25 нг/мкл и 0,125 нг/мкл. Образцы нагревали в термоциклере при 95ºC в течение 30 минут и затем охлаждали при 4ºC. Затем готовили гели Bio-Rad Criterion gel™, используя буфер MES/SDS. Образцам давали возможность нагреться до комнатной температуры и по 10 мкл образца загружали в каждую лунку двух гелей. Кроме того, образцы очищенного YFP/Phiyfp, используемые для стандартной кривой, и маркер в виде лэддера белков загружали в лунки геля. Гели подвергали электрофорезу при 150 В и 150 мА в течение 90 минут. После разгона покрытия с гелей снимали и белки переносили на нитроцеллюлозную мембрану, используя систему iBlot™ (Invitrogen). Белок переносили из геля на мембрану при 20 В в течение 10 минут. Нитроцеллюлозную мембрану удаляли и помещали в блокирующий буфер StartingBlock T20™ на ночь при 4ºC. Затем блокирующий буфер удаляли и мембрану обрабатывали, используя протокол, указанный в таблице 6.
Связывание антител выявляли, используя систему детекции хемилюминесценции Amersham ECL™ plus, следуя инструкциям производителя. Пленку экспонировали 10 минут и 30 минут. Экспонированную в течение 10 минут пленку использовали для количественной оценки белка, и пленку, сверэкспонированную в течение 30 минут, использовали для подтверждения отсутствия белка в B104 и других контрольных образцах. Мембрану приклеивали с задней стороны экспонируемой пленки и белок оценивали количественно, используя анализ плотностей пикселей. Плотность пикселей очищенных белковых стандартов сначала использовали для получения стандартной кривой, которую применяли для количественной оценки белка в образцах. Хотя на мембране видны полосы мономера и димера PhiYFP даже в очищенном стандарте, только мономер PhiYFP использовали для количественной оценки экспрессии белка. Затем значения, полученные для белка, нормализовали в нг/мкл. Отношение нормализованного общего растворимого белка (TSP) к PhiYFP вычисляли в единицах нг YFP/мг TSP, или альтернативно в частях на миллион (ч/млн).
Вестерн-блот-анализ GUS: Экспрессию белка GUS количественно оценивали способом, подобным способу оценки PhiYFP со следующими исключениями: образец экстракта объемом 10 мкл разбавляли 1:1 2x буфером Лэммли + 2-β-меркаптоэтанол, денатурировали при 95ºC в течение 30 минут и затем по 15 мкл загружали на гель. На обработанные мембраны с пленкой (экспозиция 1 минута) сверху наслаивали мембрану для оценки плотности пикселей.
Результаты Вестерн-блот-анализа 12 трансгенных растений кукурузы T0, полученных из зародышей Z. mays, трансформированных бинарным вектором pDAB 108706, показаны на фиг. 11. Двунаправленный промотор ZmUbi1 демонстрировал высокую экспрессию последовательностей, кодирующих yfp и GUS, из ткани листа. Такие наблюдения подтверждают, что минимальный промоторный элемент Min-UbiP1 двунаправленного промотора ZmUbi1, приводил к экспрессии YFP на уровнях экспрессии, сходных с уровнями экспрессии в каллусе Z. mays, трансформированном бинарной плазмидой, содержащей однонаправленный промотор ZmUbi1, управляющий последовательностью, кодирующей yfp (pDAB101556; см. фиг. 12).
ПРИМЕР 8: Конструирование блоков кассет четырех генов
Конструкцию плазмиды pDAB105803 использовали в качестве исходной плазмиды для создания блока кассет четырех генов (aad1-2a-Phiyfp и cry34-2a-cry35), управляемого одним двунаправленным промотором убиквитина-1 Zea mays. Типичная карта плазмиды pDAB105803 показана на фиг. 16, такая плазмида содержит Двунаправленный промотор убиквитина-1 Zea mays.
Фрагмент aad1-2a-Phiyfp, полученный из плазмиды pDAB105841, клонировали в разрезанном BamHI и SacI векторном остове плазмиды pDAB105803, используя способы клонирования, общеизвестные в данной области. При этом получали промежуточную плазмиду pDAB105842 (фиг. 17). NotI/XbaI-расщепленный фрагмент cry34(8V6)-2a-cry35, полученный из плазмиды pDAB105840, клонировали между сайтами NotI/SpeI плазмиды pDAB105842, чтобы сконструировать плазмиду pDAB105843. Плазмида pDAB105843 содержит кассеты генов cry34(8V6)-2a-cry35 и aad1-2a-Phiyfp с каждой стороны от двунаправленного промотора ZmUbi1 (фиг. 18).
Бинарный вектор, содержащий двунаправленный промотор ZmUbi1 и кассеты экспрессии генов cry34(8V6)-2a-cry35 и Phiyfp-2a-aad1 из плазмиды pDAB105842, создавали, применяя реакцию GATEWAY L-R CLONASE (Invitrogen, Carlsbad, CA) с использованием плазмиды назначения pDAB101917. Полученный вектор, pDAB108717, содержит кассеты экспрессии генов cry34(8V6)-2a-cry35, aad1-2a-Phiyfp и PAT в границах T-ДНК (фиг. 19).
ПРИМЕР 9: Конструирование второго блока кассет четырех генов
Конструкцию плазмиды pDAB105803 использовали для создания второго блока кассет четырех генов (Phiyfp-2a-aad1 и cry34-2a-cry35), управляемого одним двунаправленным промотором убиквитина-1 Zea mays. Фрагмент Phiyfp-2a-aad1, полученный из плазмиды pDAB105844, клонировали в разрезанном BamHI и SacI векторном остове плазмиды pDAB105803, используя способы клонирования, общеизвестные в данной области. При этом получали промежуточную плазмиду pDAB105845 (фиг. 20). Расщепленный NotI/XbaI фрагмент cry34(8V6)-2a-cry35, полученный из плазмиды pDAB105840, клонировали между сайтами NotI/SpeI плазмиды pDAB105845, конструируя плазмиду pDAB105846 (фиг. 21). Плазмида pDAB105846 содержит кассеты генов cry34(8V6)-2a-cry35 и Phiyfp-2a-aad1 с каждой стороны от двунаправленного промотора ZmUbi1.
Бинарный вектор, содержащий двунаправленный промотор ZmUbi1 и кассеты генов cry34(8V6)-2a-cry35 и Phiyfp-2a-aad1 из плазмиды pDAB105846, создавали, используя реакцию GATEWAY L-R CLONASE (Invitrogen, Carlsbad, CA) с использованием плазмиды назначения pDAB101917. Полученный вектор, pDAB108718, содержит кассеты экспрессии генов cry34(8V6)-2a-cry35, Phiyfp-2a-aad1 и PAT в границах T-ДНК (фиг. 21).
ПРИМЕР 10: Трансформация штамма Agrobacterium tumefaciens DAt13192
Бинарными векторами pDAB108717 и pDAB108718 трансформировали Agrobacterium tumefaciens, тройной штамм DAt13192 (см. предварительную заявку на выдачу патента США № 61/368965). Выделяли бактериальные колонии, и ДНК бинарной плазмиды экстрагировали и подтверждали, используя расщепления ферментами рестрикции.
ПРИМЕР 11: Трансформация кукурузы
Стерилизация початков и выделение зародышей: Чтобы получить незрелые зародыши кукурузы, растения Zea mays (сорт B104) выращивали в теплице и подвергали самоопылению или опылению между сибсами, чтобы получить початки. Початки собирали примерно через 9-12 дней после опыления. В день эксперимента поверхность початков стерилизовали, погружая в 20% раствор хозяйственного отбеливателя, который содержит 5% гипохлорита натрия, и встряхивая в течение 20-30 минут, с последующими тремя промывками в стерильной воде. После стерилизации незрелые зиготические зародыши (1,5-2,2 мм) асептически вырезали из каждого початка и случайным образом распределяли в микроцентрифужные пробирки, содержащие жидкую среду для инфекции (основная среда LS, 4,43 г/л; раствор витаминов N6 [1000X], 1,00 мл/л; L-пролин, 700,0 мг/л; сахароза, 68,5 г/л; глюкоза, 36,0 г/л; 2,4-D, 1,50 мг/л. В случае указанного набора экспериментов для каждой обработки использовали объединенные зародыши из 2-3 початков.
Инициация культуры Agrobacterium: Исходные культуры в глицерине штаммов Agrobacterium, содержащих бинарные векторы, описанные выше, высевали штрихом на чашках с минимальной средой AB, содержащих соответствующие антибиотики, и выращивали при 20ºC в течение 3-4 суток. Отбирали отдельную колонию и высевали штрихом на чашки YEP, содержащие такие же антибиотики, и инкубировали при 28ºC в течение 1-2 суток.
Культура и совместная культура Agrobacterium: В день эксперимента колонии Agrobacterium собирали из YEP-чашки, суспендировали в 10 мл среды для инфекции в одноразовой пробирке объемом 50 мл, и плотность клеток доводили до OD600 = 0,2-0,4 нм, используя спектрофотометр. Культуры Agrobacterium помещали в ротационный встряхиватель при 115 об./мин, при комнатной температуре, в это время осуществляли вырезание зародышей. Незрелые зиготические зародыши размером 1,5-2,2 мм выделяли из стерилизованных зерен кукурузы и помещали в 1 мл среды для инфекции и промывали один раз такой же средой. Суспензию Agrobacterium (2 мл) добавляли в каждую пробирку и пробирки переворачивали примерно 20 раз, затем встряхивали в течение 10-15 минут. Зародыши переносили в среду для совместного культивирования (соли MS, 4,33 г/л; L-пролин, 700,0 мг/л; миоинозит, 100,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба - 3,30 мг/л; сахароза, 30,0 г/л; Gelzan™, 3,00 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л, AgNO3, 15,0 мг/л; ацетосирингон, 100 мкМ), ориентировали щитком вверх и инкубируют в течение 3-4 суток в условиях освещения при 25ºC.
Временная экспрессия YFP/Phiyfp: Временную экспрессию YFP/Phiyfp можно было наблюдать в трансформированных зародышах через 3 дня после совместного культивирования с Agrobacterium. Зародыши исследовали с помощью стереомикроскопа (Leica Microsystems, Buffalo Grove, IL), используя YFP-фильтр и источник света 500 нм.
Отбор каллусов и регенерация растений с предполагаемыми событиями: После периода совместного культивирования зародыши переносили в среду для стадии покоя (MS-соли, 4,33 г/л; L-пролин, 700,0 мг/л; миоинозит, 100,0 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота] 500,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба, 3,30 мг/л; сахароза, 30,0 г/л; Gelzan™ 2,30 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л; AgNO3, 15,0 мг/л; карбенициллин, 250,0 мг/л), содержащую 3 мг/л биалафоса, и инкубировали при 24-часовом освещении с интенсивностью света 50 мкмоль⋅м-2⋅сек-1 в течение 7 суток при 28ºC. Зародыши переносили в среду для селекции 1 (MS-соли, 4,33 г/л; L-пролин, 700,0 мг/л; миоинозит, 100,0 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота] 500,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба, 3,30 мг/л; сахароза, 30,0 г/л; Gelzan™ 2,30 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л; AgNO3, 15,0 мг/л; карбенициллин, 250,0 мг/л), содержащую 3 мг/л биалафоса, и инкубировали при 24-часовом освещении с интенсивностью света 50 мкмоль⋅м-2⋅сек-1 в течение 7 суток при 28ºC.
Зародыши с пролиферирующими эмбриогенными каллусами переносили в среду для селекции 2 (MS-соли, 4,33 г/л; миоинозит, 100,0 мг/л; L-пролин, 700,0 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота] 500,0 мг/л; ферментативный гидролизат казеина 100,0 мг/л; дикамба, 3,30 мг/л; сахароза, 30,0 г/л; Gelzan™ 2,30 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л; AgNO3, 15,0 мг/л; карбенициллин, 250,0 мг/л), содержащую 5 мг/л биалафоса, и инкубировали при 24-часовом освещении с интенсивностью света 50 мкмоль⋅м-2⋅сек-1 еще в течение 14 суток при 28ºC. Описанная стадия селекции обеспечивает возможность трансгенному каллусу дальше пролиферировать и дифференцироваться. Период селекции каллусов продолжался три недели. Пролиферирующие эмбриогенные каллусы переносили в среду для регенерации 1 (MS-соли, 4,33 г/л; миоинозит, 100,0 мг/л; L-пролин, 350,0 мг/л; MES [(2-(н-морфолино)этансульфоновая кислота), свободная кислота] 250,0 мг/л; ферментативный гидролизат казеина 50,0 мг/л; NAA 0,500 мг/л; ABA 2,50 мг/л; BA 1,00 мг/л; сахароза, 45,0 г/л; Gelzan™ 2,50 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л; AgNO3, 1,00 мг/л; карбенициллин, 250,0 мг/л), содержащую 3 мг/л биалафоса, и культивировали при 24-часовом освещении с интенсивностью света 50 мкмоль⋅м-2⋅сек-1 в течение 7 суток при 28ºC.
Эмбриогенные каллусы с побегами/почками переносили в среду для регенерации 2 (MS-соли, 4,33 г/л; модифицированные MS-витамины [1000X], 1,00 мл/л; миоинозит, 100,0 мг/л; сахароза, 60,0 г/л; геллановая камедь G434™ 3,00 г/л; карбенициллин, 250,0 мг/л), содержащую 3 мг/л биалафоса. Культуры инкубировали при 24-часовом освещении с интенсивностью света 50 мкмоль⋅м-2⋅сек-1 в течение 7-10 суток при 28ºC. Небольшие побеги с первичными корнями переносили в среду для удлинения побегов и образования корней (MS-соли, 4,33 г/л; раствор витаминов N6 [1000X], 1,00 мл/л; миоинозит, 100,0 мг/л; сахароза, 30,0 г/л; агар 5,50 г/л; в камере Phytatray и инкубировали в условиях 16/8 часов освещенности/темноты при 90 мкмоль⋅м-2⋅сек-1 в течение 7 суток при 28ºC. Затем отбирали здоровые предполагаемые трансгенные растеньица и инкубировали в условиях 16/8 часов освещенности/темноты при 200 мкмоль⋅м-2⋅сек-1 еще в течение 2-5 суток при 25ºC и анализировали в отношении количества копий трансгенов и переносили в теплицу.
ПРИМЕР 12: Временная экспрессия Phiyfp
Осуществляли временную экспрессию Phiyfp в зародышах Zea mays, трансформированных pDAB108717. Двунаправленный промотор ZmUbi1 может вызывать экспрессию Phiyfp с кассеты экспрессии генов aad1-2a-Phiyfp, при этом в случае нетрансформированного зародыша не наблюдали никакой флуоресценции Phiyfp. Сходные уровни экспрессии Phiyfp можно наблюдать в случае зародышей Zea mays, трансформированных бинарной плазмидой pDAB105748 (фиг. 15), содержащей однонаправленный промотор Ubi1 Zea mays (Zm), управляющий одной последовательностью, кодирующей Phiyfp, проявляя при этом ожидаемый уровень экспрессии YFP/Phiyfp. Временную экспрессию Phiyfp можно наблюдать в зародышах Zea mays, трансформированных pDAB108718, при этом двунаправленный промотор ZmUbi1 может экспрессировать Phiyfp с кассеты экспрессии генов Phiyfp-2a-aad1.
ПРИМЕР 13: Экспрессия Phiyfp а стабильно трансформированной кукурузе
Экспрессия Phiyfp в стабильно трансформированном каллусе Zea mays, управляемая двунаправленным промотором ZmUbi1: В зародышах Zea mays, трансформированных бинарным вектором pDAB108717, содержащим кассету экспрессии генов aad1-2a-Phiyfp, наблюдали хорошую экспрессию Phiyfp. Двунаправленный промотор ZmUbi1 может вызывать высокий уровень экспрессии Phiyfp. Полученные результаты подтверждают, что минимальный промоторный элемент Min-UbiP1 двунаправленного промотора ZmUbi1 способен экспрессировать репортерный ген, например, Phiyfp или YFP. Уровни экспрессии белка Phiyfp сходны с уровнями экспрессии в каллусе Zea mays, трансформированном контрольным бинарным вектором, который содержал однонаправленный промотор ZmUbi1, управляющий последовательностью, кодирующей Phiyfp (pDAB105748). Экспрессию Phiyfp не выявляли в каллусе, используемом в качестве негативного контроля, который не был трансформирован бинарной конструкцией и не содержал последовательностей, кодирующих Phiyfp.
В зародышах Zea mays, трансформированных бинарным вектором pDAB108718, который содержит кассету экспрессии генов Phiyfp-2a-aad1, наблюдали хорошую экспрессию Phiyfp. Двунаправленный промотор ZmUbi1 может вызывать высокий уровень экспрессии Phiyfp. Полученные результаты подтверждают, что минимальный промоторный элемент Min-UbiP1 двунаправленного промотора ZmUbi1 способен экспрессировать репортерный ген, например Phiyfp или YFP.
ПРИМЕР 14: Оценка количества копий трансгенов
Оценка количества копий трансгенов с использованием ПЦР в реальном времени TaqMan™: Растения Zea mays трансформировали бинарными векторами, содержащими двунаправленный промотор ZmUbi1, pDAB108717 и pDAB108718, а другие растения трансформировали контрольным бинарным вектором, pDAB105748. Присутствие кодирующей последовательности (Phiyfp, aad1, cry34, cry35, Pat) в геноме растений Z. mays, трансгенных по pDAB108717 и pDAB108718, подтверждали в анализе с использованием гидролизных зондов TaqMan Растения, трансгенные по контрольному вектору pDAB105748, анализировали в отношении присутствия последовательности Phiyfp. Получали стабильно трансформированные трансгенные молодые растения Z. mays, которые развивались из каллуса, и анализировали, чтобы идентифицировать события, при которых растения содержали небольшое количество копий (1-2 корпии) полноразмерных вставок T-нити из бинарных векторов pDAB108717 и pDAB108718 и контрольного бинарного вектора pDAB105748. Идентифицированные молодые растения переносили в теплицу и выращивали.
Систему Roche Light Cycler480™ использовали для определения количества копий трансгенов в случае событий трансформации бинарным вектором pDAB108717 и pDAB108718. В данном способе используют биплексную реакцию TaqMan®, в которой применяют олигонуклеотиды, специфичные к кодирующей последовательности и к эндогенному эталонному гену инвертазы Z. mays (номер доступа в Genbank: U16123.1), в одном анализе. Количество копий и зиготность определяли, измеряя интенсивность флуоресценции, специфичной для кодирующей последовательности, относительно специфичной для инвертазы флуоресценции, которые сравнивали с известными стандартами количества копий.
Прямой и обратный нуклеотидные праймеры и флуоресцирующие зонды (синтезированные в Integrated DNA Technologies, Coralville, IA)
В случае образцов Z. mays, трансформированных бинарными векторами pDAB108717 и pDAB108718, фрагмент ДНК, специфичный для кодирующей последовательности, амплифицируют с использованием одного набора праймеров/зондов TaqMan®, содержащего зонд, меченый флуоресцирующим красителем FAM, и инвертазу амплифицировали с использованием второго набора праймеров/зондов TaqMan®, содержащего зонд, флуоресцентно меченый HEX (таблица 7). Смесь для ПЦР-реакции готовили, как указано в таблице 8, и специфичные для генов фрагменты ДНК амплифицировали согласно условиям, указанным в таблице 9. Количество копий и зиготность образцов определяли, измеряя относительную интенсивность флуоресценции, специфичной для кодирующей последовательности, по отношению к флуоресценции, специфичной для эталонного гена инвертазы, по сравнению с известными стандартами количества копий.
Стандарты создавали разбавлением вектора (pDAB108717 и pDAB108717) в геномной ДНК Z. mays B104 (гДНК), чтобы получить стандартны с известным соотношением вектор:гДНК. Например, готовили образцы, имеющие одну, две и четыре копии векторной ДНК на одну копию гДНК Z. mays B104. Разведения с одной и двумя копиями вектора, смешанными со стандартом гДНК Z. mays B104, подтверждали по сравнению с контрольным событием для Z. mays, в случае которого известно, что растение является гемизиготным, и контрольным событием для Z. mays, в случае которого известно, что растение является гомозиготным (событие Z. mays 278; см. международную публикацию патента PCT № WO 2011/022469 A2). Биплексный анализ TaqMan®, в котором используют олигонуклеотиды, специфичные для кодирующей последовательности гена, и олигонуклеотиды, специфичные к эндогенному эталонному гену Z. mays, гену инвертазы, осуществляли посредством амплификации и выявления фрагмента ДНК, специфичного для кодирующей последовательности гена, с использованием одного набора праймеров/зондов TaqMan®, содержащего зонд, меченый флуоресцирующим красителем FAM, и посредством амплификации и выявления фрагмента ДНК, специфичного для гена инвертазы, с использованием второго набора праймеров/зондов TaqMan®, содержащего зонд, флуоресцентно меченый HEX). Согласно таблице 7, реакционную смесь TaqMan® для кодирующей последовательности готовили, как указано в таблице 8, и специфичные фрагменты амплифицировали согласно условиям, указанным в таблице 9.
Смесь для ПЦР-реакции Taqman®
Уровень флуоресценции, генерируемой в каждой реакции, анализировали, используя термоциклер Roche LightCycler 480™ согласно инструкциям производителя. Флуоресцирующий остаток FAM возбуждали при оптической плотности 465/510 нм, и флуоресцирующий остаток HEX возбуждали при оптической плотности 533/580 нм. Количество копий определяли сравнением значений мишень/эталон для неизвестных образцов (результат, полученный с использованием LightCycler 480™) со значениями мишень/эталон четырех стандартов с известным количеством копий (например, ноль, 1 копия (геми-), 2 копии (гомо-) и 4 копии).
Условия для ПЦР-амплификации в термоциклере
Результаты анализа количества копий трансгенов в трансгенных растениях, полученных в результате трансформация конструкцией с двунаправленным промотором ZmUbi1 (pDAB108717 и pDAB 108718), и в трансгенных растениях, полученных в результате трансформации контрольной конструкцией Phiyfp с однонаправленным промотором ZmUbi1 (pDAB105748), суммированы в таблице 10. Только растения с 1-2 копиями трансгенов переносили в теплицу для дальнейших анализов экспрессии.
Оценка количества копий трансгенов в трансгенных растениях, полученных с использованием конструкций с двунаправленным промотором и контрольных конструкций
ПРИМЕР 15: Стабильная экспрессия Phiyfp в растениях кукурузы T0
Стабильная экспрессия Phiyfp в растениях Zea mays T0, управляемая двунаправленным промотором ZmUbi1: Можно исследовать зародыши Zea mays, трансформированные бинарным вектором pDAB108717, содержащим кассету экспрессии генов aad1-2a-Phiyfp. Двунаправленный промотор ZmUbi1 может вызывать высокий уровень экспрессии Phiyfp как в тканях побегов, так и в тканях корней. Результаты подтверждают, что минимальный промоторный элемент Min-UbiP1 двунаправленного промотора ZmUbi1 способен экспрессировать репортерный ген, например, Phiyfp или YFP, который слит в виде двойного цистрона с aad1 с использованием последовательности 2A. Уровни экспрессии белка Phiyfp сходны с уровнями экспрессии в зародышах Z. mays, трансформированных контрольным бинарным вектором, который содержит однонаправленный промотор ZmUbi1, управляющий последовательностью, кодирующей Phiyfp (pDAB105748). Экспрессию Phiyfp не выявляли в растениях, используемых в качестве негативного контроля, которые не были трансформированы бинарной конструкцией и не содержали последовательностей, кодирующих Phiyfp.
Можно наблюдать экспрессию Phiyfp в тканях листьев и корней растений Zea mays T0, трансгенных по бинарному вектору pDAB108718, который содержит кассету экспрессии генов Phiyfp-2a-aad1. Двунаправленный промотор ZmUbi1 может вызывать высокий уровень экспрессии Phiyfp. Результаты подтверждают, что минимальный промоторный элемент Min-UbiP1 двунаправленного промотора ZmUbi1 способен экспрессировать репортерный ген, например Phiyfp или YFP, слитый с aad-1 с использованием последовательности 2A.
ПРИМЕР 16: Анализ белков Cry34, Cry35 и AAD1
Образцы растений собирали в колонки 1-10 бокса Matrix в конических пробирках объемом 1,5 мл, в которые клали 1 стальной шарик и затем добавляли PBST + 0,5% БСА (0,6 мл). Затем осуществляли разбивание шариком в боксе для измельчения образца в измельчителе Geno в течение 5 минут при 1500 об./мин, затем центрифугировали при 3700 об./мин в течение 7 минут при 4ºC.
Анализ ELISA Cry34/35: В отдельном 96-луночном планшете с глубокими лунками образец экстракта разбавляли 1:200 в PBST + 1% реагент Blotto. Затем по два объема 25 мкл разбавленного образца переносили в отдельные 96-луночные планшеты, в которых были разнесены анти-Cry34 и анти-Cry35 (Meso Scale Discovery). В 11 и 12 колонки каждого планшета добавляли стандартные концентрации Cry34 и Cry35 в PBST + 1% реагент Blotto (25 мкл). Затем планшеты инкубировали, встряхивая при комнатной температуре в течение одного часа. Затем планшеты промывали PBST (3×300 мкл). Затем в каждую лунку добавляли 25 мкл раствора конъюгированного с SulfoTAG анти-Cry34 и анти-Cry35 и инкубировали, встряхивая при комнатной температуре в течение одного часа. Затем планшеты промывали PBST (3×300 мкл). Затем добавляли буфер для регистрации Read Buffer T (Meso Scale Discovery) в объеме 150 мкл и планшет сразу же подвергали регистрации, используя считывающее устройство SECTOR® 6000. Концентрации белков в образце можно вычислять, используя стандартную кривую для соответствующего белка, получаемую с использованием данных с того же самого планшета.
Анализ ELISA AAD-1: В отдельном 96-луночном планшете с глубокими лунками образец экстракта разбавляли 1:20 в PBST + 0,5% БСА. Затем по два объема 200 мкл разбавленного образца переносили в отдельные 96-луночные планшеты, которые были покрыты анти-AAD1 (предоставленными Acadia Bioscience LLC). В 11 и 12 колонки каждого планшета добавляли стандартные концентрации AAD1 в PBST + 0,5% БСА (200 мкл). Затем в каждую лунку добавляли биотинилированного анти-AAD1 в объеме 50 мкл и планшеты инкубировали, встряхивая при комнатной температуре в течение одного часа. Затем планшеты промывали PBST (5×300 мкл). Затем в каждую лунку добавляли 100 мкл раствора конъюгата стрептавидина-щелочной фосфатазы и инкубировали, встряхивая при комнатной температуре в течение 30 минут. Затем планшеты промывали PBST (5×300 мкл). Затем добавляли субстрат (п-нитрофенилфосфат, PNPP) в объеме 100 мкл и инкубировали, встряхивая при комнатной температуре в течение 45 минут. Затем планшеты регистрировали при A405 на устройстве для считывания планшетов SpectraMax M5 (Molecular Devices). Концентрации белков в образце можно вычислить, используя стандартную кривую, получаемую с использованием данных с того же самого планшета.
ПРИМЕР 17: Анализ белков растений кукурузы T0
Анализ белков растений кукурузы T0, экспрессия которых управляется конструкцией с двунаправленным промотором убиквитина-1 Zea mays (pDAB108717): Типичные анализы ELISA 11 трансгенных растений кукурузы T0, получаемых из зародышей Zea mays, трансформированных pDAB108717, которая содержит cry34-2a-cry35 и aad1-2a-Phiyfp, суммированы в таблице 11. Двунаправленный промотор ZmUbi1 вызывает высокий уровень экспрессии кодирующих последовательностей как Cry34, так и Cry35, в листьях. Неожиданно данные, полученные для белка, показывают наличие более высокой, до 4-кратной, экспрессии Cry34 с двунаправленной конструкции pDAB108717, по сравнению с конструкцией, управляемой однонаправленным промотором ZmUbi1. Также неожиданно наблюдали сходную в 8-10 раз более высокую экспрессию белков Cry35 и AAD1 с двунаправленной конструкции pDAB108717 по сравнению с конструкцией, управляемой однонаправленным промотором ZmUbi1. Полученные данные показывают, что один двунаправленный промотор убиквитина 1 Zm в конструкции pDAB108717 может экспрессировать множество генов (например, Cry34, Cry35 и AAD1) на неожиданно более высоких уровнях по сравнению с растениями Zea mays, трансформированными бинарной плазмидой, которая содержит однонаправленный промотор ZmUbi1, управляющий такими же генами, в которых каждая кодирующая последовательность управляется независимым промотором ZmUbi1.
Корреляция экспрессии Cry34 и Cry35 в растениях кукурузы T0, управляемой конструкцией с двунаправленным промотором убиквитина-1 Zea mays (pDAB108717): Анализ корреляции между белками Cry34 и Cry35 в 11 трансгенных растениях кукурузы T0, полученных из зародышей Zea mays, трансформированных pDAB108717, которая содержит cry34-2a-cry35, показан ан фиг. 23A. Очень высокая корреляция (R-квадрат = 0,98) демонстрирует строгую корегуляцию экспрессии Cry34 и Cry 35 с кассеты экспрессии генов cry34-2a-cry35, управляемой двунаправленным промотором ZmUbi1.
Экспрессия Cry34/Cry35/AAD1 в трансгенных по pDAB108717 растениях кукурузы T0
Анализ белков растений кукурузы T0, экспрессия которых управляется конструкцией с двунаправленным промотором убиквитина-2 Zea mays (pDAB 108718): Типичный анализ ELISA 11 трансгенных растений кукурузы T0, полученных из зародышей Zea mays, трансформированных pDAB108718, которая содержит cry34-2a-cry35, суммирован в таблице 12. Двунаправленный промотор ZmUbi1 вызывал высокую экспрессию последовательностей, кодирующих как Cry34, так и Cry35, в листьях. Данные, полученные в отношении белков, демонстрируют в несколько раз более высокую экспрессию белков Cry34, Cry35 и AAD1 с двунаправленной конструкции pDAB108718 по сравнению с конструкцией, управляемой однонаправленным промотором ZmUbi1. Полученные данные подтверждают, что двунаправленный промотор убиквитина-1 Zea mays в конструкции pDAB108718 вызывает экспрессию нескольких генов (например, Cry34, Cry35 и AAD1) на неожиданно более высоких уровнях по сравнению с растениями Zea mays, трансформированными бинарной плазмидой, которая содержит однонаправленный промотор ZmUbi1, управляющий такими же генами, при этом каждая кодирующая последовательность управляется независимым промотором ZmUbi1.
Корреляция экспрессии Cry34 и Cry35 в растениях кукурузы T0, управляемой конструкцией с двунаправленным промотором убиквитина-1 Zea mays (pDAB108718): Анализ корреляции между белками Cry34 и Cry35 в 11 трансгенных растениях кукурузы T0, полученных из зародышей Zea mays, трансформированных pDAB108718, которая содержит cry34-2a-cry35, показан на фиг. 23B. Очень высокая корреляция (R-квадрат = 0,98) демонстрирует строгую корегуляцию экспрессии Cry34 и Cry 35 с кассеты экспрессии генов cry34-2a-cry35, управляемой двунаправленным промотором ZmUbi1.
Экспрессия Cry34/Cry35/AAD1 в трансгенных pDAB108718-растениях кукурузы T0
ПРИМЕР 18: Стекинг трансгенов: синтетические двунаправленные промоторы (данные T1)
Экспрессия генов в растениях T1, управляемая конструкциями с двунаправленным промотором убиквитина-1 Zea mays: от десяти до двенадцати событий введения одной копии на конструкцию отбирали для анализа, за исключением того, что для контрольной конструкции pDAB108716 было только одно событие. Тестировали пять растений/событий для стадии V6 и три растения/события для стадий V10-12 и R3. Анализы белков осуществляли, используя ЖХМС или ELISA.
Конструкции, используемые в данном примере, показаны на фиг. 26. pDAB108706 (ZMUbi, двунаправленный (-200)) и pDAB108707 (ZMUbi, двунаправленный (-90)) представляют собой конструкции с типичным двунаправленным промотором согласно настоящему изобретению; pDAB101556 (ZmUbi1-YFP, контроль) и pDAB108716 (ZMUbi1, без минимального промотора) служат в качестве контрольных конструкций с однонаправленными промоторами.
Примерные результаты экспрессии (V6) с четырех конструкций для белка YFP (ЖХМС) в нг/см2 показаны на фиг. 27A, и примерные результаты относительной экспрессии (V6) с четырех конструкций для РНК YFP показаны на фиг. 27B.
Примерные результаты экспрессии (V6) с четырех конструкций для белка GUS (ЖХМС) в нг/см2 показаны на фиг. 28A, и примерные результаты относительной экспрессии (V6) с четырех конструкций для РНК GUS показаны на фиг. 28B.
Примерные результаты экспрессии (V6) с четырех конструкций для белка AAD1 (ЖХМС) в нг/см2 показаны на фиг. 29A, и примерные результаты относительной экспрессии (V6) с четырех конструкций для РНК AAD1 показаны на фиг. 29B.
Статистический анализ результатов экспрессии (V6) с четырех конструкций для белка YFP protein (ЖХМС) в нг/см2 показан на фиг. 30A, и средние значения для pDAB108707, pDAB108706, pDAB101556, и pDAB108716 составляют 57,63, 52,66, 49,75 и 0, соответственно. Статистический анализ результатов относительной экспрессии (V6) с четырех конструкций для РНК YFP показан на фиг. 30B, и средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 составляют 9,96, 8,07, 6,95 и 1,01, соответственно.
Статистический анализ результатов экспрессии (V6) с четырех конструкций для белка GUS (ЖХМС) в нг/см2 показан на фиг. 31A, и средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 составляют 151,27, 143,22, 0 и 213,17, соответственно. Статистический анализ результатов относительной экспрессии (V6) с четырех конструкций для РНК GUS показан на фиг. 31B, и средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 составляют 0,65, 0,78, 0 и 3,03, соответственно.
Статистический анализ результатов экспрессии (V6) с четырех конструкций для белка AAD1 (ЖХМС) в нг/см2 показан на фиг. 32A, и средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 составляют 710,88, 1417,01, 856,58 и 1795,43, соответственно. Статистический анализ результатов относительной экспрессии (V6) с четырех конструкций для РНК AAD1 показан на фиг. 32B, и средние значения для pDAB 108706, pDAB108707, pDAB101556 и pDAB108716 составляют 1,33, 1,37, 1,93 и 2,93, соответственно.
На фиг. 33A, 33B и 33C показаны примерные результаты экспрессии (V10) с четырех конструкций для белка YFP, AAD1 и GUS (ЖХМС) в нг/см2, соответственно.
На фиг. 34A, 34B и 34C показан статистический анализ результатов экспрессии (V10) с четырех конструкций для белка YFP, GUS и AAD1 (ЖХМС) в нг/см2, соответственно. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 в случае YFP (фиг. 34A) составляют 71,77, 81,81, 49,58 и 23,01, соответственно. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 в случае GUS (фиг. 34B) составляют 109,63, 98,25, 0 и 138,02, соответственно. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 в случае AAD1 (фиг. 34C) составляют 666,11, 597,80, 715,12 и 1002,84, соответственно.
На фиг. 35A, 35B и 35C показаны примерные результаты экспрессии (R3) с четырех конструкций для белка YFP, GUS и AAD1 (ЖХМС) в нг/см2, соответственно.
На фиг. 36A, 36B и 36C показан статистический анализ результатов экспрессии (R3) с четырех конструкций для белка YFP, GUS и AAD1 (ЖХМС) в нг/см2, соответственно. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 в случае YFP (фиг. 36A) составляют 91,38, 49,49, 21,67 и 0,40, соответственно. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 в случае GUS (фиг. 36B) составляют 5,52, 16,81, 1,07 и 46,60, соответственно. Средние значения для pDAB108706, pDAB108707, pDAB101556 и pDAB108716 в случае AAD1 (фиг. 36C) составляют 156,71, 153,44, 165,40 и 197,80, соответственно.
Результаты показывают, что двунаправленные промоторы Ubi1 кукурузы согласно настоящему изобретению могут вызывать высокий уровень экспрессии GUS и YFP, при этом экспрессия YFP с двунаправленного промотора Ubi1 кукурузы сходна с экспрессией YFP, управляемой однонаправленным Ubi1 кукурузы. Результаты также свидетельствуют о том, что двунаправленная транскрипция не оказывает значимого влияния на экспрессию GUS (экспрессию GUS по сравнению с конструкциями, в которых отсутствует минимальный промотор, без экспрессии YFP).
ПРИМЕР 19: Сочетание двунаправленного промотора и бицистронной последовательности 2A для управления четырьмя трансгенами с одного промотора (данные T1)
Экспрессия генов в растениях T1, управляемая конструкциями с двунаправленным промотором убиквитина-1 Zea mays: от десяти до двенадцати событий введения одной копии на конструкцию отбирали для анализа, за исключением того, что для контрольных конструкций было четыре или пять событий на конструкцию. Тестировали пять растений/событий для стадии V6 и три растения/события для стадий V10-12 и R3. Анализы белков осуществляли, используя ЖХМС или ELISA.
На фиг. 37A показаны примерные результаты относительной экспрессии (V6) РНК Cry34 с четырех конструкций pDAB105748 (ZMUbi1-YFP), pDAB105818 (ZMUbi1-Cry34/ZMUbi1-Cry35/ZMUbi1-AAD1), pDAB108717 (YFP/AAD-1-ZMUbi1 двунаправленный-Cry34-Cry35) и pDAB108718 (AAD1/YFP-ZMUbi1 двунаправленный-Cry34-Cry35). На фиг. 37B показаны примерные результаты относительной экспрессии (V6) белка Cry34 (ЖХМС) с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 38A показаны примерные результаты относительной экспрессии (V6) РНК AAD1 с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718. На фиг. 38B показаны примерные результаты относительной экспрессии (V6) белка AAD1 (ЖХМС) с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 39A показаны примерные результаты относительной экспрессии (V6) РНК YFP с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718. На фиг. 39B показаны примерные результаты относительной экспрессии (V6) белка YFP (ЖХМС) с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 40A показаны примерные результаты относительной экспрессии (V6) РНК Cry35 с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718. На фиг. 40B показаны примерные результаты относительной экспрессии (V6) белка Cry35 (ELISA) с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 41 показаны примерные результаты относительной экспрессии (V6) РНК PAT с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 42A показан статистический анализ результатов экспрессии (V6) РНК Cry34 с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB 108718 со средними значениями 0, 2,42, 2,67 и 2,25, соответственно. На фиг. 42B показан статистический анализ результатов экспрессии (V6) белка Cry34 с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718 со средними значениями 0, 596,94, 2044,73 и 719,18, соответственно.
На фиг. 43A показан статистический анализ результатов экспрессии (V6) РНК AAD1 с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB 108718 со средними значениями 0, 1,98, 2,68 и 2,03, соответственно. На фиг. 43B показан статистический анализ результатов экспрессии (V6) белка AAD1 с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718 со средними значениями 0, 2237,54, 5763,88 и 2379,15, соответственно.
На фиг. 44A показан статистический анализ результатов экспрессии (V6) РНК YFP с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718 со средними значениями 3,59, 0, 2,78 и 1,95, соответственно. На фиг. 44B показан статистический анализ результатов экспрессии (V6) белка YFP с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718 со средними значениями 1420,69, 251,68, 1154,04 и 706,04, соответственно.
На фиг. 45A показан статистический анализ результатов экспрессии (V6) РНК Cry35 с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718 со средними значениями 0, 1,12, 3,74 и 3,20, соответственно. На фиг. 45B показан статистический анализ результатов экспрессии (V6) белка Cry35 с тех же самых четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB 108718 со средними значениями 0, 283,54, 635,83 и 90,97, соответственно.
На фиг. 46 показан статистический анализ результатов экспрессии (V6) РНК PAT с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB 108718 со средними значениями 1,56, 0,07, 1,46 и 1,01, соответственно.
На фиг. 47A, 47B, 47C и 47D показаны примерные результаты экспрессии белка (V10) YFP, AAD1, Cry34 и Cry35, соответственно, с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 48A, 48B, 48C и 48D показан статистический анализ результатов экспрессии белка (V10) YFP, AAD1, Cry34 и Cry35, соответственно, с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 49A, 49B, 49C, и 49D показаны примерные результаты экспрессии белка (R3) YFP, AAD1, Cry34 и Cry35, соответственно, с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и pDAB108718.
На фиг. 50A, 50B, 50C, и 50D показан статистический анализ результатов экспрессии белка (R3) YFP, AAD1, Cry34 и Cry35, соответственно, с четырех конструкций pDAB105748, pDAB105818, pDAB108717 и DAB108718.
На фиг. 51 показаны примерные результаты, полученные с использованием Вестерн-блота для экспрессии белков Cry34, Cry35 и AAD1 с pDAB108718 и pDAB108717.
Результаты показывают, что все четыре трансгена в конструкциях, управляемых одним промотором, являются функциональными с хорошими уровнями экспрессии. Три гена (Cry34/Cry35/AAD1) в блоке с двунаправленным Ubi1 имеют высокие уровни экспрессии, которые сходны с уровнями экспрессии, обеспечиваемыми в блоке генов, отдельно управляемых Ubi1 (DExT).
Хотя выше обсуждается несколько иллюстративных аспектов и вариантов осуществления, специалистам в данной области будут понятны некоторые их модификации, преобразования, добавления и дополнительные сочетания. Поэтому подразумевается, что следующая прилагаемая формула изобретения и введенные далее пункты формулы изобретения следует интерпретировать как включающие все такие модификации, преобразования, добавления и дополнительные сочетания, которые составляют истинную суть и входят в объем изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ И КОНСТРУКТ ДЛЯ СИНТЕТИЧЕСКОГО ДВУНАПРАВЛЕННОГО РАСТИТЕЛЬНОГО ПРОМОТОРА SCBV | 2012 |
|
RU2627595C2 |
| СИНТЕТИЧЕСКИЕ ПРОИСХОДЯЩИЕ ИЗ BRASSICA ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТОВ | 2013 |
|
RU2626535C2 |
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТА ИЗ BRASSICA | 2013 |
|
RU2636035C2 |
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТОВ | 2013 |
|
RU2632648C2 |
| ТРАНЗИТНЫЙ ПЕПТИД ХЛОРОПЛАСТОВ | 2013 |
|
RU2636036C2 |
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТОВ | 2013 |
|
RU2720973C2 |
| МОЛЕКУЛЫ НУКЛЕИНОВЫХ КИСЛОТ, КОТОРЫЕ ПРИДАЮТ УСТОЙЧИВОСТЬ К НАСЕКОМЫМ-ВРЕДИТЕЛЯМ ОТРЯДА ЖЕСТКОКРЫЛЫХ | 2011 |
|
RU2639549C2 |
| МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ, КОТОРЫЕ ВОЗДЕЙСТВУЮТ НА СУБЪЕДИНИЦУ С ВАКУОЛЯРНОЙ АТФАЗЫ И ПРИДАЮТ УСТОЙЧИВОСТЬ К ЖЕСТКОКРЫЛЫМ НАСЕКОМЫМ-ВРЕДИТЕЛЯМ | 2011 |
|
RU2644669C2 |
| УСТОЙЧИВЫЕ К ГЛИФОСАТУ РАСТЕНИЯ И СПОСОБЫ, СВЯЗАННЫЕ С НИМИ | 2013 |
|
RU2636037C2 |
| НОВЫЙ КЛАСС ГЕНОВ УСТОЙЧИВОСТИ К ГЛИФОСАТУ | 2013 |
|
RU2721122C2 |


































































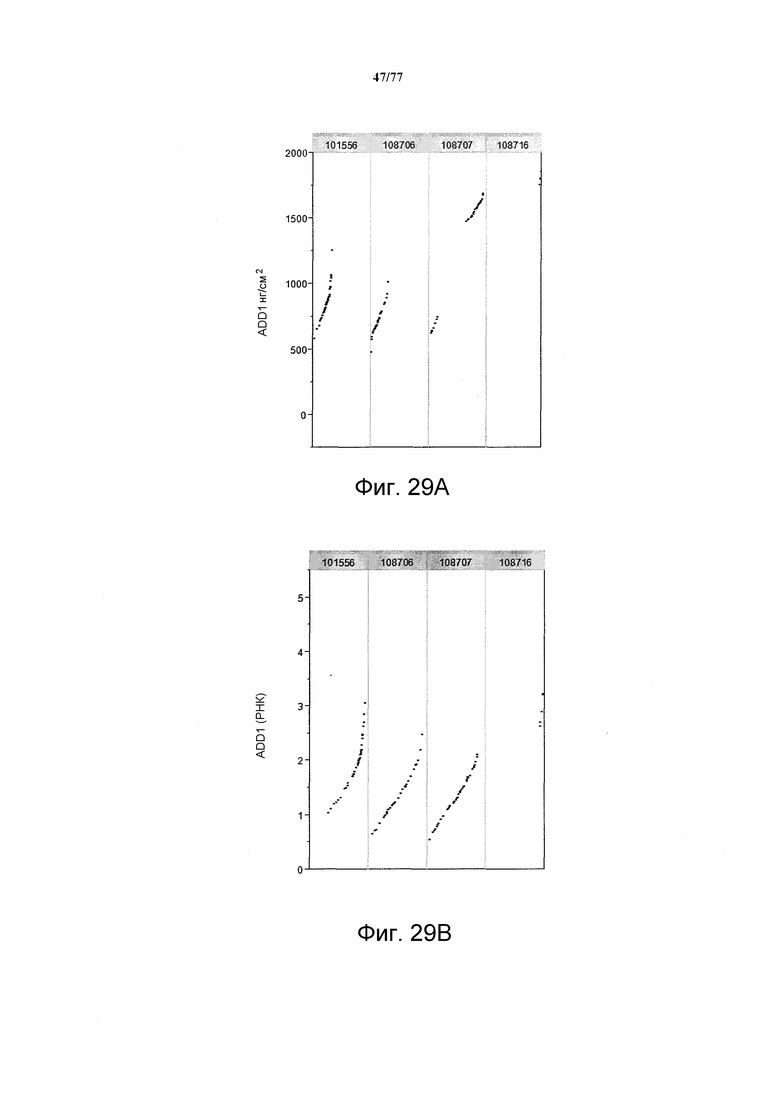







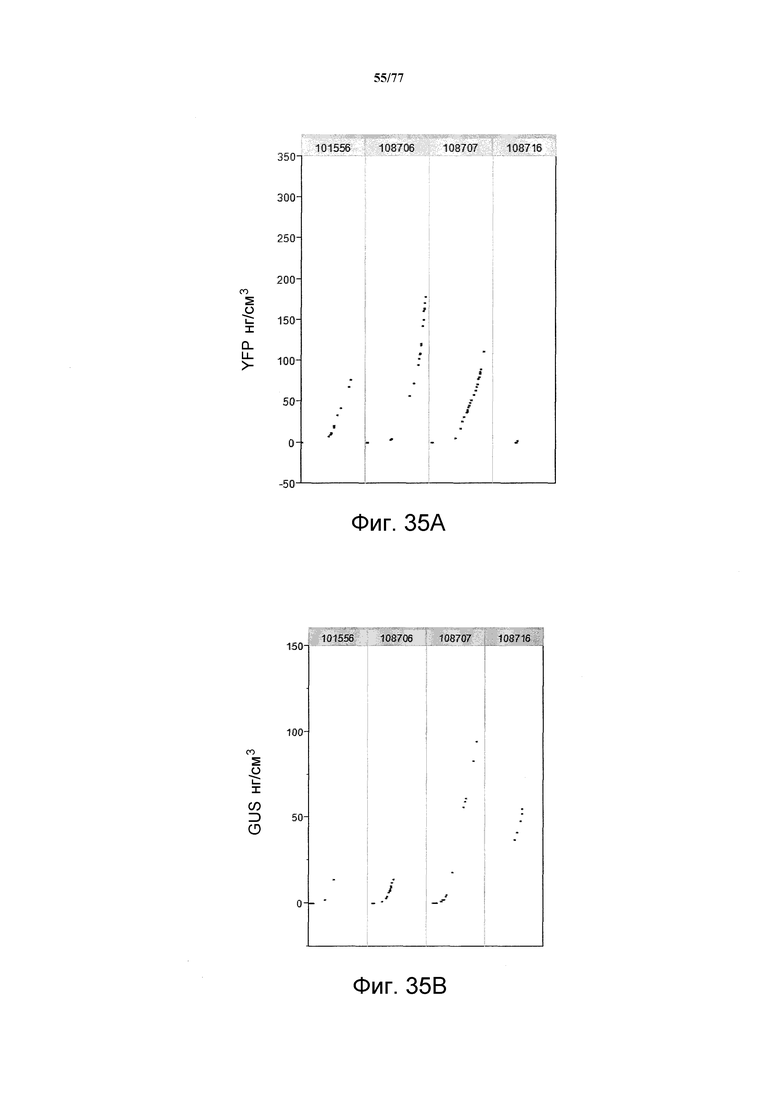









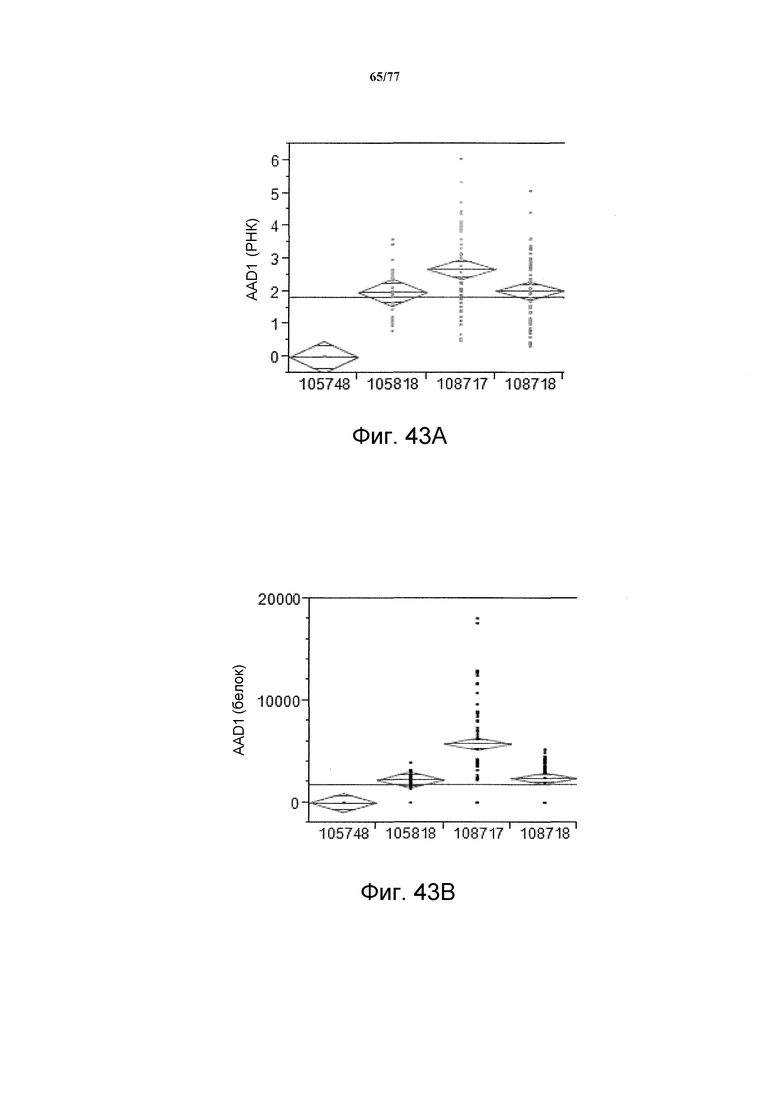






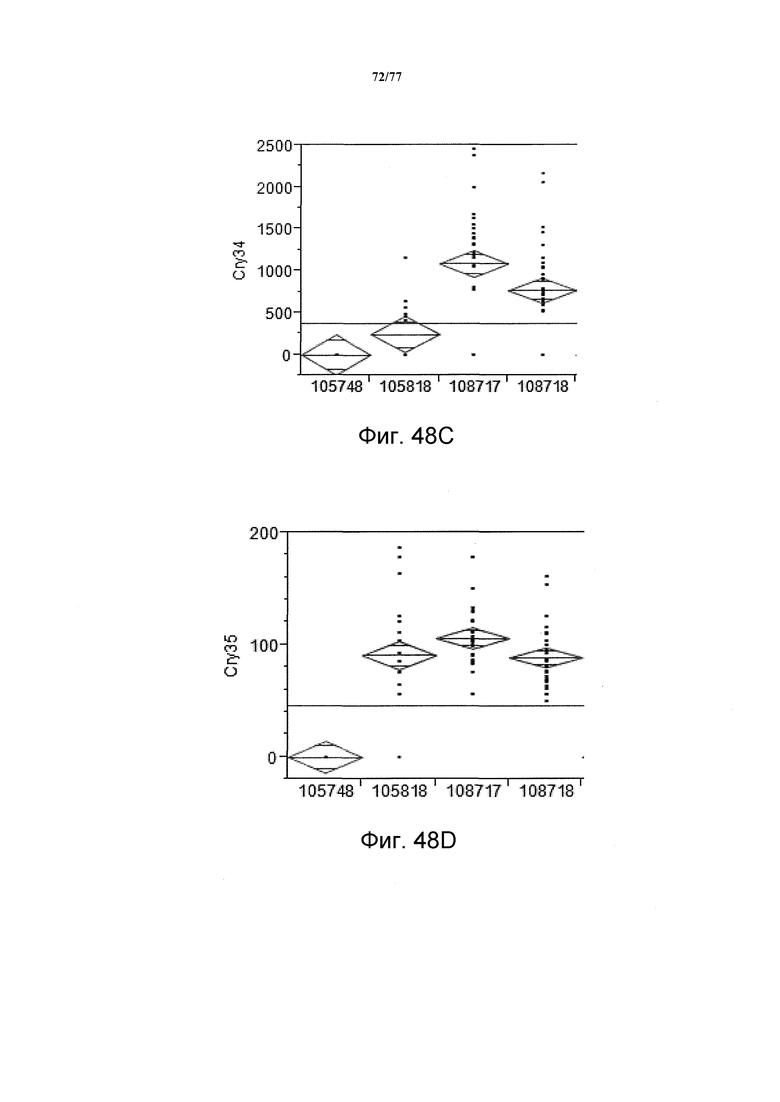





Изобретение относится к области биохимии, в частности к синтетическому двунаправленному промоторному полинуклеотиду для повышения экспрессии гетерологичных белков в растительной клетке, а также к синтетическому полинуклеотиду для экспрессии двух гетерологичных белков в растительной клетке. Также раскрыт способ получения трансгенной растительной клетки, экспрессирующей два гетерологичных белка с использованием вышеуказанного синтетического полинуклеотида. Изобретение также относится к растительной клетке для экспрессии гетерологичного белка, содержащей вышеуказанный полинуклеотид, а также к растению, ее содержащему. Изобретение позволяет эффективно осуществлять экспрессию гетерологичных белков в растительной клетке. 5 н. и 2 з.п. ф-лы, 51 ил., 12 табл., 19 пр. 
1. Синтетический двунаправленный промоторный полинуклеотид для повышения экспрессии гетерологичных белков в растительной клетке, характеризующийся двумя минимальными коровыми промоторными элементами, имеющими нуклеотидную последовательность SEQ ID NO: 1, причем два минимальных коровых промоторных элемента находятся в синтетическом полинуклеотиде в обратной комплементарной ориентации по отношению друг к другу.
2. Синтетический полинуклеотид для экспрессии двух гетерологичных белков в растительной клетке, причем полинуклеотид характеризуется:
первой нуклеотидной последовательностью, кодирующей гетерологичный белок;
второй нуклеотидной последовательностью, кодирующей гетерологичный белок; и
двумя минимальными коровыми промоторными элементами, имеющими нуклеотидную последовательность SEQ ID NO: 1, в двунаправленном промоторе, причем минимальные коровые промоторы находятся в двунаправленном промоторе в обратной комплементарной ориентации по отношению друг к другу,
причем первая нуклеотидная последовательность оперативно связана с двунаправленным промотором в направлении 5' синтетического полинуклеотида по отношению к двунаправленному промотору, и
причем вторая нуклеотидная последовательность оперативно связана с двунаправленным промотором в направлении 3' синтетического полинуклеотида по отношению к двунаправленному промотору.
3. Способ получения трансгенной растительной клетки, экспрессирующей два гетерологичных белка, включающий трансформацию клетки полинуклеотидом по п. 2.
4. Способ по п. 3, дополнительно включающий регенерацию растения из трансгенной растительной клетки.
5. Растительная клетка для экспрессии гетерологичного белка, содержащая полинуклеотид по п. 2.
6. Растение для экспрессии гетерологичного белка, содержащее растительную клетку по п. 5.
7. Способ по п. 3, в котором полинуклеотид направляют в предварительно определяемый участок в геноме растительной клетки, используя рекомбинацию, опосредованную нуклеазами типа цинковых пальцев.
| US 2009038025 A1, 05.02.2009 | |||
| WO 2006013072 A2, 09.02.2006 | |||
| СПОСОБ ПРОИЗВОДСТВА ИНСТАНТ-ПОРОШКА ДЛЯ ПОЛУЧЕНИЯ АРОМАТИЗИРОВАННОГО ТОПИСОЛНЕЧНОГО НАПИТКА | 2008 |
|
RU2385129C1 |
| DONALD RG et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| ЭКСПРЕССИЯ ПРОМОТОРА СИНТАЗЫ АЦЕТООКСИКИСЛОТ В ИНТРОДУЦИРОВАННЫХ ГЕНАХ РАСТЕНИЙ | 1995 |
|
RU2197527C2 |

