Изобретение относится к области иммунологии, биотехнологии и медицины (конкретно - онкологии) и касается структуры синтетического пептидного антигена, используемого в составе конъюгатов с различными белками-носителями для получения антител, специфически распознающих изоформу 2 эукариотического фактора элонгации трансляции 1А (eEF1A2), но не распознающих очень сходную с ней изоформу 1 того же фактора (eEF1A1), и применяемых для селективного выявления изоформы 2, потенциального онкогена и онкомаркера, в различных биологических образцах. Изобретение может быть использовано с целью разработки иммунотест-систем для выявления в биологических образцах злокачественно перерожденных клеток и/или клеток с измененным биохимическим аппаратом контроля запрограммированной гибели (апоптоза) и способных к злокачественному перерождению. Более конкретно, изобретение представляет собой структуру синтетического пептида формулы
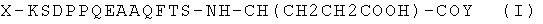 ,
,
где X - ацетильная группа или атом водорода, Y-NH2- или OH-группа, а остальная часть пептидного производного представляет собой фрагмент аминокислотной последовательности eEF1A2 человека, включающий аминокислотные остатки с №330 по №343. Фрагмент идентичной структуры присутствует также в аминокислотных последовательностях eEF1A2 западной гориллы, белощекого хохлатого гиббона, яванского макака и макака-резуса, обыкновенной игрунки, быка, свиньи, лошади, африканского слона, гигантской панды, кролика, китайского хомячка, летучей мыши малой бурой ночницы, грызуна голого землекопа, серого голохвостого опоссума, тасманийского сумчатого дьявола, утконоса. Отличительной характеристикой данной структуры является ее способность после присоединения к белку-носителю (например, бычьему сывороточному альбумину (БСА), яичному альбумину (ЯА), гемоцианину гемолимфы моллюска (ГЦ)) вызывать при введении в организм животного образование антител, специфически распознающих только изоформу 2 eEF1A, но не изоформу 1 того же фактора, а после присоединения к хроматографическому носителю - специфически сорбировать вышеуказанные антитела из раствора и использоваться для их очистки.
Предпосылки создания изобретения.
Злокачественные опухоли являются второй, после сердечно-сосудистых заболеваний, причиной смертности людей в развитых странах. В настоящее время развиваются различные методы терапии злокачественных опухолей, среди которых одним из основных является удаление ткани опухоли хирургическим путем. Однако в ходе разработки стратегии хирургической операции или иной схемы лечения возникают проблемы точной диагностики злокачественности или доброкачественности процесса опухолеобразования, точной локализации границ опухоли и прогноза развития заболевания. Получаемые с целью диагностики образцы тканей опухоли, а также окружающих тканей необходимо исследовать на наличие возможных маркеров и/или факторов злокачественного перерождения клеток, чтобы уточнить диагноз (злокачественная или доброкачественная опухоль), определить необходимость и возможность оперативного вмешательства с целью удаления опухоли, границы иссечения тканей; в ходе операции по иссечению опухоли с помощью выявления таких маркеров необходимо контролировать полноту удаления злокачественно перерожденной ткани. Одним из таких маркеров-факторов злокачественного перерождения клеток является изоформа 2 эукариотического фактора элонгации трансляции 1A.
Эукариотический фактор элонгации трансляции 1 (eEF1A) отвечает за доставку аминоацил-тРНК в А сайт рибосомы и обеспечение корректного кодон-антикодонового взаимодействия, а также может принимать участие и в передаче деацилированной на рибосоме тРНК назад к аминоацил-тРНК синтетазе для повторного аминоацилирования [1]. Кроме того, eEF1A выполняет целый ряд функций, не связанных с белковым синтезом, например взаимодействует с F-актином и тубулином, а также формирует комплекс с кальмодулином, что может иметь значение для процессов фосфорилирования, катализируемых Ca2+/кальмодулин-зависимой киназой [2]. Показано, что eEF1A играет роль, пока еще точно не выясненную, в процессах внутриклеточного протеолиза и апоптоза, а также обладает шапероноподобными свойствами [2].
Существуют две изоформы eEF1A, кодируемые разными генами. Сходство аминокислотных последовательностей этих белков составляет 97%, идентичность - 93%. Экспрессия изоформ eEF1A в организме является тканеспецифичной и происходит по принципу «или-или» [3]. В подавляющем большинстве тканей экспрессируется eEF1A1, и только в миоцитах поперечнополосатых мышц и в нейронах - терминально дифференцированных и долгоживущих клетках - наблюдается экспрессия eEF1A2, хотя в пренатальном периоде в миоцитах и нейронах также наблюдается экспрессия изоформы 1 [4]. Недавно было обнаружено, что изоформа 2 может появляться и в других тканях организма, однако такое ее необычное появление практически всегда связано с онкогенезом. Надэкспрессия eEF1A2 была выявлена в злокачественных опухолях яичников, грудной железы, толстой и прямой кишок, простаты, легких, почек, печени, щитовидной железы, тела матки и некоторых других, а также в культивируемых клеточных линиях, полученных из некоторых из вышеназванных опухолей [5]. В ряде случаев была прямо показана онкогенная роль eEF1A2 - этот белок функционировал как фактор, способствующий иммортализации клеток [6]. В связи с этим особый интерес представляет разработка высокоспецифичных средств детекции изоформы 2 в опухолевых тканях.
Для количественного определения отдельных изоформ одного и того же белка можно использовать моноспецифичные антитела, селективно связывающие только одну изоформу. Известно, что моноспецифичные антитела для выявления отдельных изоформ одного и того же белка можно получить путем иммунизации животных фрагментами этих изоформ, имеющими специфичные для каждой из изоформ аминокислотные последовательности [4]. При этом указанные фрагменты получают обычно путем химического синтеза из производных аминокислот, а затем для повышения иммуногенности конъюгируют с помощью гомо- или гетеробифункциональных реагентов с биополимерами, обычно белками [4, 7]. У иммунизированных животных забирают кровь, получают из нее антисыворотку, а моноспецифичные антитела из полученных антисывороток выделяют аффинной хроматографией на соответствующих пептидных или белковых лигандах, ковалентно связанных с хроматографическими носителями.
Получение моноспецифичных антител против изоформ eEF1A является нетривиальной задачей из-за большого сходства изоформ и консервативности аминокислотных последовательностей eEF1A среди разных видов животных, даже эволюционно удаленных [3]. В литературе упоминалось использование полученных моноспецифичных антипептидных антител против изоформ 1 и 2 eEF1A мышей для выявления данных белков иммуногистохимическим методом и методом иммуноблоттинга [4]. Эти моноспецифичные антитела были получены против фрагментов eEF1A2 217-234 и 439-452, однако подробные характеристики антител в статье не были приведены. Фирма Novus Biologicals (США) производит и продает антитела против фрагмента eEF1A2 360-409 VIDCHTAHIACKFAELKEKIDRRSGKKLEDNPKSLKSGDAAIVEMVPGKP (каталожный № NBP1-55245), позиционируемые как моноспецифичные анти-eEF1A2 антитела (http://ww/v.novusbio.com/EEF1A2-Antibody_NBP1-55245.html?rr=true). Хотя фрагмент 360-409 содержит четыре аминокислотных остатка, различающихся в изоформах 1 и 2 eEF1A (Ile361, Asn390, Ser393 и Glu403 в eEF1A2; Leu361, Gly390, Phe393 и Asp403 в eEF1A1), свыше 90% его последовательности представлено одинаковыми для eEF1A1 и eEF1A2 аминокислотными остатками. Это указывает на высокую вероятность выработки антител, обладающих специфичностью к обеим изоформам eEF1A, в ответ на иммунизацию этим фрагментом. Фирма Novus Biologicals не приводит сравнительного определения специфичности взаимодействия антител против фрагмента 360-409 с изоформами 1 и 2 eEF1A. Другие коммерчески доступные препараты антител, в том числе и моноклональных, не обладают селективностью в отношении той или иной изоформ eEF1A.
Сущность изобретения.
Задачей настоящего изобретения является разработка пептидного производного, которое может быть получено методом химического синтеза, и должно: 1) содержать фрагмент изоформы 2 фактора элонгации трансляции 1A человека, 2) в виде конъюгата с различными белками-носителями вызывать образование антител, специфично взаимодействующих с этой изоформой, но не взаимодействующих с изоформой 1 eEF1A, 3) в виде лиганда, ковалентно связанного с хроматографическим носителем, использоваться для селективной аффинной очистки моноспецифичных анти-eEF1A2 антител. Получаемые в ответ на указанные конъюгаты пептидного производного антитела должны быть пригодны к использованию для селективного иммунохимического выявления изоформы 2 eEF1A в биообразцах.
Указанная задача решается
- описываемым пептидным производным, имеющим формулу (I), содержащим фрагмент 330-343 аминокислотной последовательности белка eEF1A2 человека, а также кролика, быка, лошади и некоторых других видов млекопитающих и имеющим различия в четырех позициях аминокислотной последовательности по сравнению с аналогичным фрагментом изоформы 1 eEF1A, полученным с помощью стандартных приведенных в литературе методов твердофазного пептидного синтеза и синтеза в растворе [8];
- способом выработки в организме животного моноспецифичных антител, взаимодействующих с изоформой 2 фактора элонгации трансляции 1А человека и других млекопитающих (при условии идентичности аминокислотных последовательностей этих белков), но не взаимодействующих с изоформой 1 этого фактора, предусматривающим введение животному эффективной дозы конъюгата пептидного производного (I) с приемлемым белком-носителем, обычно, но не исключительно с бычьим сывороточным альбумином, овальбумином, гемоцианином, в виде композиции с общеизвестными адъювантами;
- специфичным хроматографическим сорбентом для аффинной очистки антител против изоформы 2 eEF1A, представляющим собой вышеуказанное пептидное производное, ковалентно присоединенное к любому подходящему хроматографическому носителю.
Упомянутое пептидное производное, его соли, содержащие приемлемые противоионы (обычно и чаще всего Na+, K+, NH4+, триэтиламмоний, триэтаноламмоний, хлорид, сульфат, фосфат, гидрофосфат или дигидрофосфат, карбонат или бикарбонат, формиат, ацетат, трифторацетат) образуют с белками-носителями с применением гомо- и гетеробифункциональных соединений ковалентно связанные конъюгаты, способные в виде композиций с известными адъювантами вызывать образование антител, специфично взаимодействующих с изоформой 2 eEF1A человека, а также западной гориллы, белощекого хохлатого гиббона, яванского макака и макака-резуса, обыкновенной игрунки, быка, свиньи, лошади, африканского слона, гигантской панды, кролика, китайского хомячка, летучей мыши малой бурой ночницы, грызуна голого землекопа, серого голохвостого опоссума, тасманийского сумчатого дьявола, утконоса, но не взаимодействующих с изоформой 1 этого белка, а в виде ковалентных соединений с подходящими хроматографическими носителями с активированными группами - носители для очистки вышеупомянутых антител методом аффинной хроматографии.
Выбор фрагмента 330-343 eEF1A2 в качестве гаптена для получения антител, специфичных к eEF1A2, но не связывающих eEF1A1, является предпочтительным, поскольку этот довольно короткий фрагмент содержит в своем составе четыре аминокислотных остатка, существенно различающихся в изоформах 1 и 2 eEF1A (Ser331, Gln335, Gln339, Ser342 в eEF1A2 и Asn331, Met335, Gly339, Ala342 в eEF1A1), разделенные двумя-тремя аминокислотными остатками, идентичными в обеих изоформах, то есть не содержит идентичных для обеих изоформ eEF1A четырех-, пяти- и шестичленных пептидных фрагментов, соответствующих по длине линейным B-эпитопам [9], что значительно увеличивает вероятность получения антител, специфичных только к одной изоформе eEF1A. Аминокислотная последовательность фрагмента 330-343 ограничивает выбор способов конъюгации соответствующего данному фрагменту пептидного производного конъюгацией гомобифункциональными реагентами глутаровым альдегидом или диметилимидами дикарбоновых кислот с насыщенной неразветвленной углеводородной цепью различной длины с образованием сшивки между эпсилон-аминогруппой остатка лизина (соответствующего Lys330) и свободными аминогруппами молекулы белка-носителя и конъюгацией с использованием гетеробифункционального реагента N-гидроксисукцинимидного эфира малеимидобензойной кислоты с образованием сшивки между эпсилон-аминогруппой остатка лизина и SH-группами молекулы белка-носителя [7]. Пептидные производные формулы (I), содержащие фрагмент 330-343 eEF1A2 человека, предпочтительнее всего получать известными стандартными методами пептидного синтеза на твердой фазе (в ручном, полуавтоматическом или автоматическом режиме) или в растворе [8] с последующей очисткой осаждением и перекристаллизацией, гель-фильтрацией и/или высокоэффективной жидкостной хроматографией на подходящем хроматографическом сорбенте с обращенной фазой или на ионообменном сорбенте.
Существенным свойством пептидных производных формулы (I) и их приемлемых солей является их способность в виде конъюгата с белком-носителем при введении животным с развитой иммунной системой, например, но не исключительно, млекопитающим, вызывать образование антител, специфично взаимодействующих с изоформой 2 eEF1A человека и ряда других млекопитающих, но не взаимодействующих с изоформой 1 eEF1A этих же видов. В качестве белка-носителя могут использоваться любые растворимые глобулярные белки, о которых известно, что они обладают собственной иммуногенностью для иммунизируемых животных и содержат доступные для воздействия конъюгирующего реагента свободные амино- или SH-группы. Наиболее предпочтительные, но не исчерпывающие все возможные варианты примеры - бычий сывороточный альбумин, яичный альбумин, гемоцианин.
Другим существенным свойством пептидного производного формулы (I) и его приемлемых солей является их способность в виде ковалентно связанных с подходящим хроматографическим носителем, например, но не исключительно, агарозой (предварительно активированной N-гидроксисукцинимидом, бромцианом и т.п. реагентами), выступать в качестве лигандов при аффинной хроматографии моноспецифичных антител против eEF1A2 человека и ряда других млекопитающих.
Данное изобретение иллюстрируется следующими примерами, которые никоим образом не должны рассматриваться как ограничивающие область настоящего изобретения.
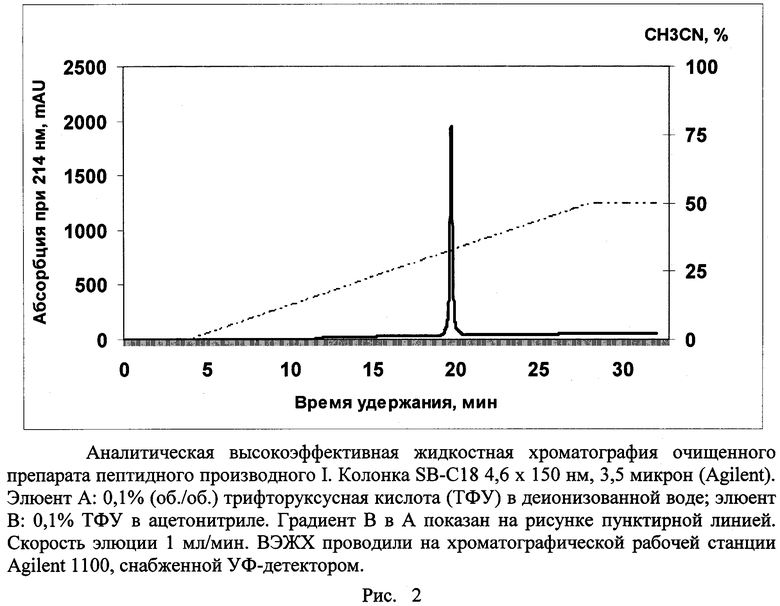
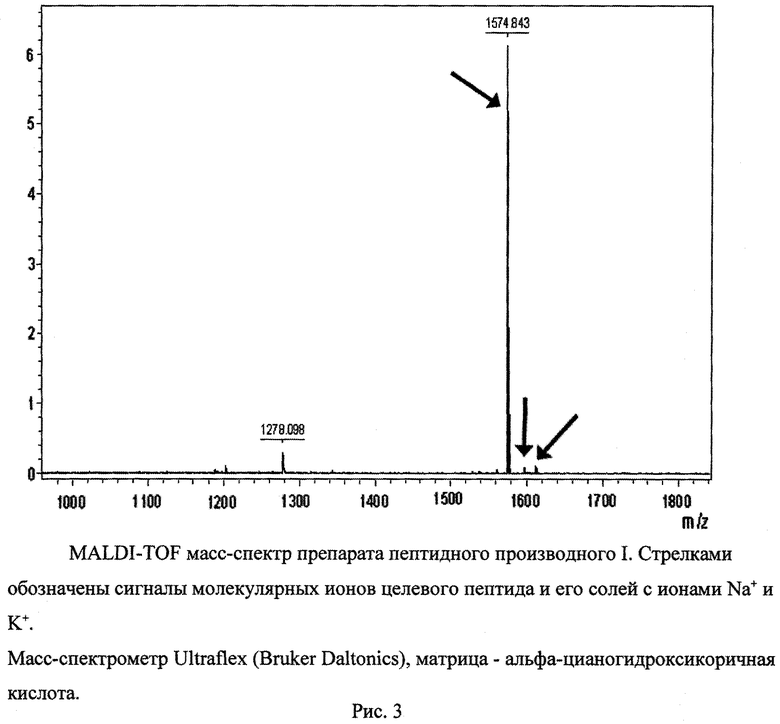
Пример 1. Выбор и синтез пептидного производного для получения анти-eEF1A2 антител. Из базы знаний UniProt извлекали аминокислотные последовательности изоформ 1 и 2 eEF1A человека и выравнивали их с помощью программы CLUSTAL W (рис.1). Выбирали в аминокислотной последовательности eEF1A2 фрагмент длиной не более 14 аминокислотных остатков, содержащий максимальное количество позиций, занимаемых различающимися в eEF1A1 и eEF1A2 аминокислотными остатками, при этом различающиеся аминокислотные остатки не должны были обладать сходными физико-химическими свойствами и не должны были разделяться идентичными для eEF1A1 и eEF1A2 фрагментами длиной более 4 остатков. Данным критериям отвечал фрагмент, соответствующий позициям с 330 по 343. В качестве гаптена использовали пептид альфа-N-Ac-KKSDPPQEAAQFTS-NH-CH(CH2CH2COOH)-CONH2, содержащий фрагмент 330-343 eEF1A2 человека; так как данный пептидный фрагмент извлечен из середины белковой цепи, то блокирование свободных альфа-амино- и альфа-карбоксильных групп предпочтительно. Синтез пептида поводили стандартным методом твердофазного пептидного синтеза в автоматическом режиме, исходя из 9-флуоренилметоксикарбонил-защищенных по альфа-аминогруппе производных аминокислот. Для очистки целевого пептида от примесей реагентов, использовавшихся в ходе синтеза, а также побочных продуктов синтеза применяли стандартные процедуры осаждения и кристаллизации из растворов и высокоэффективной жидкостной хроматографии (ВЭЖХ). После очистки препарат подвергали лиофилизации. Очищенный препарат пептидного производного характеризовали стандартными методами аналитической ВЭЖХ (рис.2) и времяпролетной масс-спектрометрии с матрично-активируемой лазерной десорбцией-ионизацией (MALDI-TOF масс-спектрометрия) (рис.3). Препарат был получен в кристаллической форме в виде натриевой, калиевой и трифторацетатной солей. Препарат растворим в воде и водно-солевых растворах при величинах pH от 6 до 8 до концентрации 1 мг/мл, хорошо растворим в диметилсульфоксиде и ацетонитриле, удовлетворительно - в их смесях с водой и водными солевыми растворами. Лиофилизованный препарат при хранении при -30°C в течение двух лет не менял своих характеристик, приведенных на рис.2 и 3.
Пример 2. Получение конъюгата пептидного производного формулы (I) с бычьим сывороточным альбумином (БСА) проводили по стандартной методике, используя в качестве конъюгирующего реагента глутаровый альдегид [7].
К 3 мл раствора БСА с концентрацией 1 мг/мл в фосфатно-солевом буфере, pH 7,4 (ФСБ) добавляли 5,8 мг пептидного производного (I) в виде раствора в 160 мкл диметилсульфоксида (ДМСО), молярное соотношение белок: пептид 1:36. Смесь охлаждали до 4°C и по каплям при постоянном перемешивании на магнитной мешалке при 4°C добавляли 2%-ный (по объему) раствор глутарового альдегида в воде до конечного объема, вдвое превышающего объем смеси белка с пептидом. По окончании добавления раствора глутарового альдегида смесь перемешивали 1 ч при 4°C, после чего добавляли сухой NaBH4 до конечной концентрации 10 мг/мл (всего 63 мг) и инкубировали при перемешивании 1 ч при комнатной температуре. Далее раствор конъюгата диализовали 36 ч против трех смен по 1 л ФСБ и концентрировали ультрафильтрацией с помощью устройства для центрифужного концентрирования Vivaspin 6 МСО 10,000 ("Sartorius", Швейцария) до конечного объема 2 мл.
Пример 3. Получение конъюгата пептидного производного формулы (I) с гемоцианином с использованием в качестве конъюгирующего реагента глутарового альдегида проводили так же, как в примере 2, но после диализа раствор конъюгата не концентрировали, поскольку гемоцианин и его конъюгаты с пептидами легко выпадают в осадок при концентрации выше 1 мг/мл.
Пример 4. Получение конъюгата пептидного производного формулы (I) с яичным альбумином с использованием в качестве конъюгирующего реагента глутарового альдегида также проводили по стандартной методике [7].
К 7 мл раствора яичного альбумина с концентрацией 1 мг/мл в ФСБ добавляли 10,3 мг пептидного производного (I), растворенного в 290 мкл ДМСО, молярное соотношение белок : пептид 1:40. К этой смеси при 4°C добавляли по каплям при постоянном перемешивании 7 мл 2%-ного раствора глутарового альдегида, через 1 ч перемешивания при 4°C добавляли 150 мг NaBH4, растворенного в 700 мкл ФСБ, и перемешивали 1 ч при комнатной температуре. Затем полученный раствор конъюгата диализовали в течение 36 ч против трех смен по 2 л ФСБ и концентрировали ультрафильтрацией с помощью устройства для центрифужного концентрирования Vivaspin 20 МСО 10,000 ("Sartorius", Швейцария) до конечного объема 3,6 мл.
Пример 5. Получение антител мыши против изоформы 2 eEF1A путем иммунизации конъюгатом пептидного производного (I) с БСА.
5 белым беспородным мышам-самцам в возрасте от 12 до 18 недель четыре раза вводили внутрибрюшинно 50 мкл смеси раствора конъюгата пептидного производного (I) с БСА (1,0 мг/мл) в ФСБ с адъювантом Фрейнда (1:1 по объему). Количество (доза) вводимого конъюгата - 25 мкг на одно животное. Интервалы между инъекциями составляли две недели. Первую и вторую инъекцию проводили смесью раствора конъюгата с полным адъювантом Фрейнда, последующие две - смесью раствора конъюгата с неполным адъювантом Фрейнда. Адъювант Фрейнда добавляли в соотношении 1:1 к растворенным в ФСБ конъюгатам и смешивали до получения однородной эмульсии. Контрольную (неиммунную) сыворотку получали из крови мышей контрольной группы (5 особей), которым вводили ФСБ вместо раствора конъюгата пептидного производного (I) с БСА. Забор крови проводили методом декапитации. После взятия кровь выдерживали 1-3 ч при комнатной температуре, затем оставляли на ночь при 4°C для формирования сгустка. После центрифугирования в течение 15 мин при 2500-3000 об/мин сыворотку крови отбирали в микропробирки и повторно центрифугировали в тех же условиях. Супернатант отбирали и хранили в виде аликвот при -20°C.
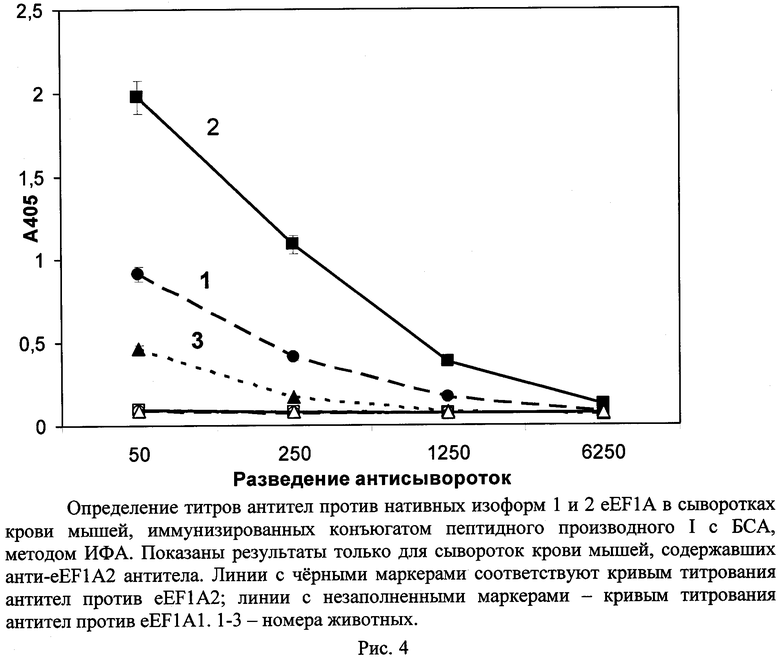
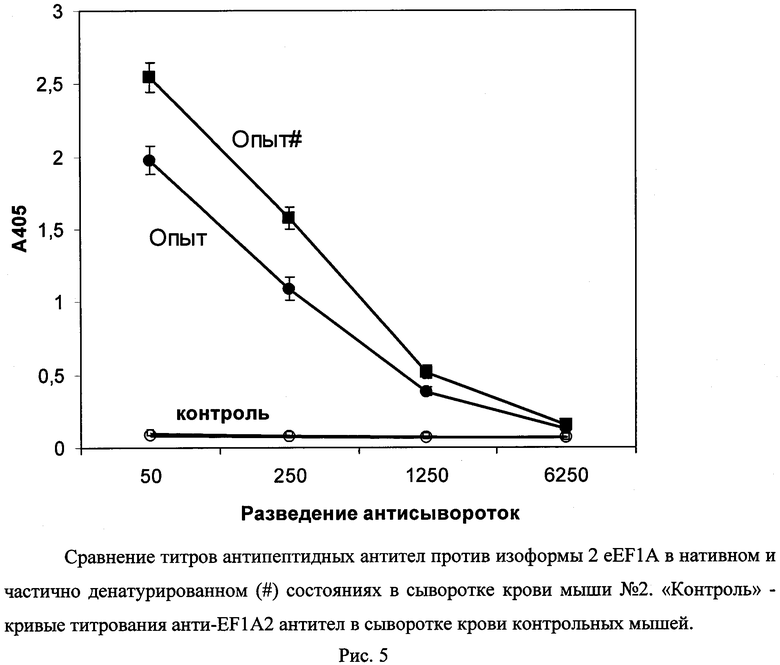
Определение титров антител проводили методом иммуноферментного анализа (ИФА). Для этого белки eEF1A2 или eEF1A1 сорбировали из 0,05М натрий-бикарбонатного буфера, pH 8,5 в лунки предварительно УФ-облученного полистирольного планшета высокой сорбционной емкости ("Costar", США, кат. №9018) в количестве 0,5 мкг белка/лунку. В лунки с сорбированными белками после промывания их 300 мкл ФСБ, содержащим 0,1% Твина 20 (ФСБ-Т), вносили раствор обезжиренного молока (50% молока по объему) в ФСБТ (ФСБТ-молоко) и инкубировали 1 ч при комнатной температуре при перемешивании. После удаления этого раствора в лунки вносили сыворотки крови мышей, разведенные в 50, 250, 1250 и 6250 раз ФСБТ, содержащим 2 мг/мл БСА, и преинкубированные после разведения до внесения в лунки 1 ч при 37°C (для удаления антител против БСА). Инкубировали 2 ч при 37°C. После удаления раствора сыворотки крови лунки планшета промывали, как указано выше, и вносили в них раствор конъюгата пероксидазы хрена с антителами козы против иммуноглобулинов мыши в разведении 1:10000 в ФСБТ-молоке. Инкубировали 1 ч при перемешивании при 37°C, после чего промывали, как указано выше. Далее в лунки планшета вносили по 100 мкл раствора субстрата пероксидазы: ABTS в концентрации 0,5 мг/мл и H2O2 в концентрации 0,01% в 0,08М натрий-цитратном буфере, pH 4,0; раствор готовили непосредственно перед употреблением в склянке темного стекла. Инкубировали 45 мин в темноте при 37°C (при перемешивании), после чего измеряли оптическую плотность растворов в лунках при 405 нм. За величину титра антител принимали такое разведение сыворотки крови иммунизированного животного, для которого соответствующее значение оптической плотности в ИФА превышает двукратное значение оптической плотности контрольного образца сыворотки крови, взятого в таком же разведении. Результаты титрования сывороток крови мышей представлены на рис.4. Видно, что конъюгат пептидного производного (I) вызывал у трех мышей из 5 формирование моноспецифичных анти-eEF1A2 антител; антитела против eEF1A1 в сыворотках крови тех же мышей не выявлялись. Титры анти-eEF1A2 антител в сыворотках крови мышей составляли 1:1250, 1:6250, 1:250 (рис.4). Титры антител против нативного и частично денатурированного eEF1A2 (инкубированного 15 мин при 70°C) в сыворотке крови мыши №2 практически совпадали (рис.5).
Пример 6. Получение антител козы против изоформы 2 eEF1A путем иммунизации конъюгатом пептидного производного (I) с гемоцианином моллюска.
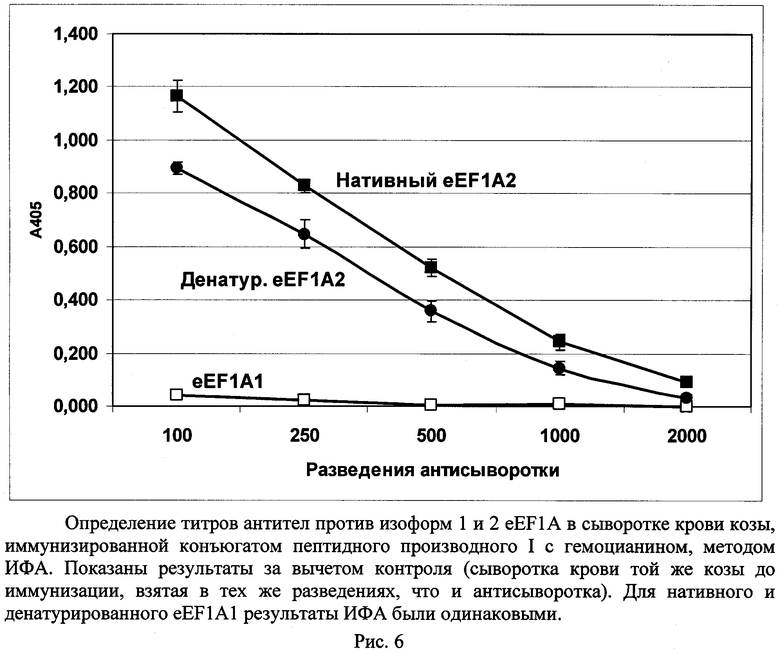
Кастрированному козлу (возраст - 2 года) вводили 3 раза по 2 мл смеси раствора конъюгата пептидного производного (I) с гемоцианином в ФСБ с адъювантом Фрейнда (1:1 по объему) в виде эмульсии, первая инъекция - с полным адъювантом Фрейнда, две последующие - с неполным адъювантом Фрейнда. Доза вводимого конъюгата - 1 мг/инъекцию. Интервал между инъекциями - две недели. Кровь отбирали через одну неделю после последней иммунизации и получали из нее сыворотку, как описано в примере 5. Титр анти-eEF1A2- и анти-eEF1A1-антител определяли, как описано в примере 5, но вместо конъюгата пероксидазы хрена с антителами козы против иммуноглобулинов мыши использовали конъюгат пероксидазы хрена с антителами кролика против иммуноглобулинов козы («Имтек», Россия) в разведении 1:5000. В качестве контрольного препарата использовали сыворотку крови того же козла, полученную из крови, взятой за 1 неделю до начала иммунизации. Из рис.6 видно, что введение козлу конъюгата пептидного производного (I) с гемоцианином моллюска приводит к формированию у животного моноспецифичных анти-eEF1A2 антител; анти-eEF1A1 антитела при этом не обнаруживаются. Титр антител против частично денатурированного белка eEF1A2 (см. пример 5) был немного ниже (1:1000), чем против нативного (1:2000).
Пример 7. Получение антипептидных антител козы против изоформы 2 eEF1A путем иммунизации конъюгатом пептидного производного (I) с яичным альбумином.
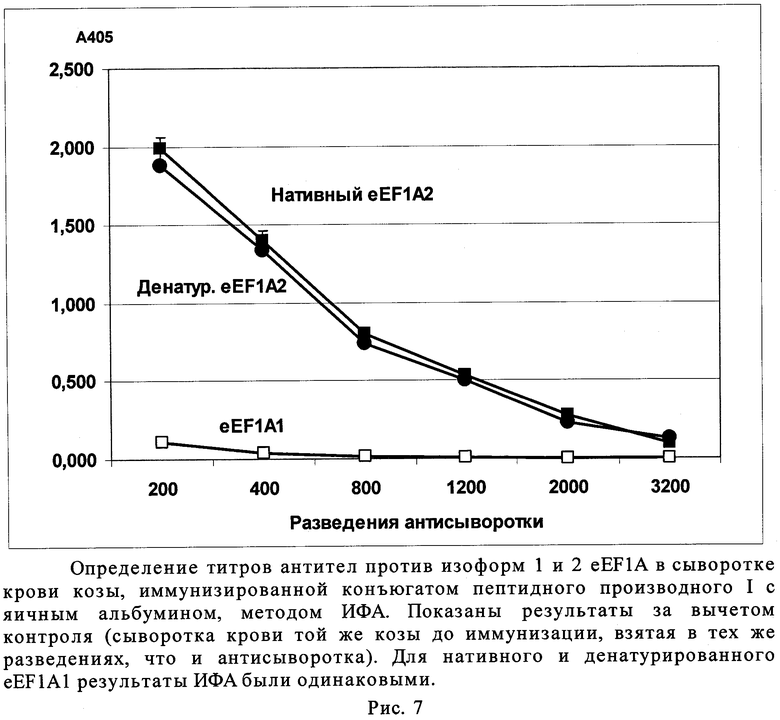
Иммунизацию животного и определение титров анти-eEF1A2- и анти-eEF1A1-антител проводили, как описано в примере 6. Из рис.7 видно, что титры антител против нативного и частично денатурированного eEF1A2, полученные при иммунизации конъюгатом пептидного производного (I) с яичным альбумином, практически совпадают. Антитела против изоформы 1 eEF1A не обнаруживаются.
Пример 8. Получение хроматографического носителя путем конъюгации пептидного производного (I) с гранулами Аффи-геля 15 (Affi-Gel 15, "Bio-Rad", США) и очистка моноспецифичных анти-eEF1A2 антител на этом аффинном носителе.
Аффи-гель 15 ("Bio-Rad", США) представляет собой гранулированную агарозу, модифицированную N-гидроксисукцинимидом. Для защиты от гидролиза N-гидроксисукцинимидных групп гранулы модифицированной агарозы поставляются в виде суспензии в пропаноле-2.
К 1 мл Аффи-геля 15, отфильтрованного от пропанола-2, добавляли 1 мл раствора 7 мг пептидного производного (I) в диметилсульфоксиде и инкубировали 4 ч на качалке при температуре 20°C. По окончании инкубации гель отфильтровывали от раствора пептида. В фильтрате методом аналитической ВЭЖХ на обращенной фазе обнаружены лишь следовые количества пептида, что соответствовало присоединению не менее 4 мкмолей остатков пептидного производного (I) к 1 мл носителя. Далее гель суспендировали в 0,5 мл 2%-ного (по объему) раствора этаноламина в диметилсульфоксиде и инкубировали 1 ч на качалке при той же температуре. По окончании реакции гель отфильтровывали от раствора и уравновешивали 10 объемами 20 мМ трис-HCl буфера, pH 8,0, содержащего 25 мМ NaCl и 0,02% (вес/объем) NaN3 (буфер образца). Упаковывали гель в колонку 0,8×2,1 см (объем 0,8 мл) и промывали 3 объемами буфера образца, контролируя pH на выходе с колонки. На подготовленную колонку наносили порциями по 1 мл препарат иммуноглобулинов G в буфере образца, полученный из сыворотки крови козла, иммунизированного конъюгатом пептидного производного (I) с гемоцианином или с яичным альбумином, путем осаждения сульфатом аммония 50% насыщения и последующей хроматографии на носителях DEAE Affi-gel Blue, Affi-gel Protein А или Affi-gel Protein G, как это рекомендовано в литературе [9]. Нанесение препарата на аффинный носитель прекращали, когда в проскоке с помощью ИФА выявляли антитела, взаимодействующие с eEF1A2, при этом достигалось насыщение аффинного носителя специфичными антителами. После нанесения носитель промывали 5 объемами буфера образца, затем 3 объемами раствора 1М NaCl в буфере образца. Связавшиеся с носителем антитела элюировали 3 объемами 0,1М глицин-HCl буфера, pH 2,5 и тут же добавляли в элюат ½ объема 1М трис-HCl буфера, pH 8,5. Полученный раствор антител диализовали 1 сутки против буфера образца и определяли в нем концентрацию иммуноглобулинов G спектрофотометрически по поглощению света с длиной волны 280 нм (удельная экстинкция раствора иммуноглобулинов G равна 1,43 для 0,1%-ного раствора при длине светового пути 1 см). За один цикл аффинной хроматографии на колонке объемом 1,0 мл очищали до 50 мкг иммуноглобулинов G, связывавшихся с пептидным производным (I). Полученный препарат иммуноглобулинов G тестировали на взаимодействие с изоформами 1 и 2 eEF1A методом ИФА, как описано в примере 5. Чувствительность выявления анти-eEF1A2 антител с помощью данного метода составляла не выше 5 нг в пробе. Анти-eEF1A1 антитела в препарате, полученном после аффинной хроматографии, не выявлялись. Объединенный элюат антител после нескольких экспериментов по аффинной хроматографии концентрировали ультрафильтрацией в 10 раз, смешивали с равным объемом глицерина и хранили при -20°C.
Пример 9. Селективное выявление изоформы 2 eEF1A в лизатах тканей методом иммуноблоттинга с помощью моноспецифичных анти-eEF1A2 антител, полученных против пептидного производного (I).
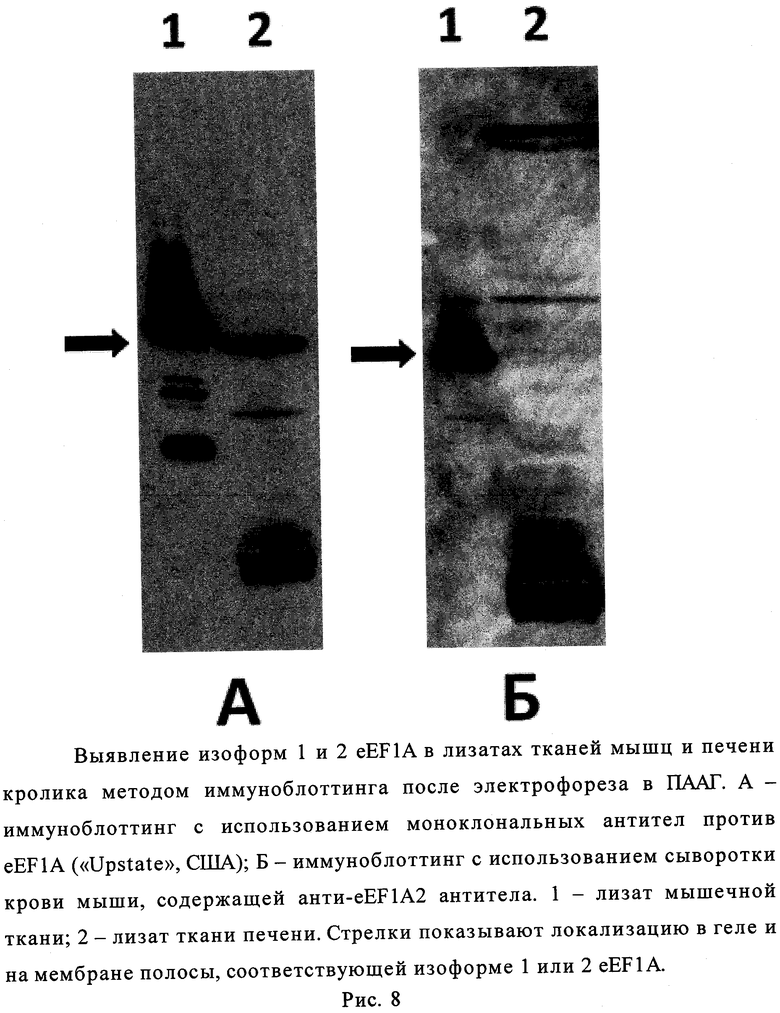
Лизаты тканей печени (источник изоформы 1 eEF1A) и мышц (источник изоформы 2 eEF1A) кролика получали из образцов тканей здоровых животных после эвтаназии. Образцы тканей растирали в жидком азоте в керамической ступке. Затем добавляли лизирующий буфер (10 мМ калий-фосфатный буфер, pH 7,4, содержащий 100 мМ NaCl, 5 мМ MgCl2, 10% (по объему) глицерин, 1% (масса/объем) Nonidet Р-40, 1 мМ дитиотреит, 0,1 мМ фенилметилсульфонилфторид) в количестве 1 мл на 1 мг ткани и инкубировали 30 мин во льду. Затем центрифугировали 30 мин при 14000 g, 4°C. В полученный супернатант добавляли буфер Лэммли для образцов в 4-кратной концентрации (4×) (250 мМ трис-HCl буфер, pH 6,8, 8% (масса/объем) додецилсульфат натрия (ДДС-Na), 40% (по объему) глицерин, 200 мМ дитиотреит, 0,05% (масса/объем) бромфеноловый синий) в количестве 1 объем 4× буфера Лэммли на 3 объема супернатанта. Образцы инкубировали на кипящей водяной бане 2 мин и подвергали диск-электрофорезу в денатурирующих условиях по Лэммли [10]. По окончании электрофореза белки из геля переносили на нитроцеллюлозную мембрану Hybond ECL ("Amersham Bioscience", США) при помощи аппарата для «мокрого» переноса ("Amersham Bioscience") с использованием буфера Towbin при постоянном напряжении 100 В 1 час при 4°C. Детекцию изоформ eEF1A проводили с помощью иммуноферментного анализа. Мембрану блокировали в солевом фосфатном буфере с Tween 20 (PBS-T: 1,47 мМ KH2PO4, 4,29 мМ Na2HPO4·7H2O, 137 мМ NaCl, 2,68 мМ KCl, 0,1% Tween 20, pH 7,3) с добавлением 5% нежирного сухого молока. В качестве препаратов специфичных антител против eEF1A использовали разведенную в 100 раз блокирующим буфером (5%-ный раствор БСА (масса/объем) в 1,47 мМ KH2PO4, 4,29 мМ Na2HPO4, 137 мМ NaCl, 2,68 мМ KCl, 0,1%-ном (масса/объем) Tween 20, 5%-ном обезжиренном сухом молоке, pH 7,3) сыворотку крови иммунной мыши, содержащую антитела против пептидного производного (I), узнающие только eEF1A2, либо препарат моноклональных антител, узнающих обе изоформы eEF1A («Upstate», США) - в качестве контроля. Связавшиеся с изоформами eEF1A антитела проявляли конъюгатом антител кролика против иммуноглобулинов G мыши с пероксидазой хрена («Sigma», США, рабочее разведение 1:5000) с использованием хемилюминесцентного косубстрата пероксидазы SuperSignal West Dura Extended Duration Substrate («Pierce», США, каталожный №34076) в соответствии с рекомендациями фирмы. Антитела, полученные против пептидного производного I, выявляли в лизатах тканей только изоформу 2 eEF1A, но не выявляли изоформу 1 этого фактора (рис.8).
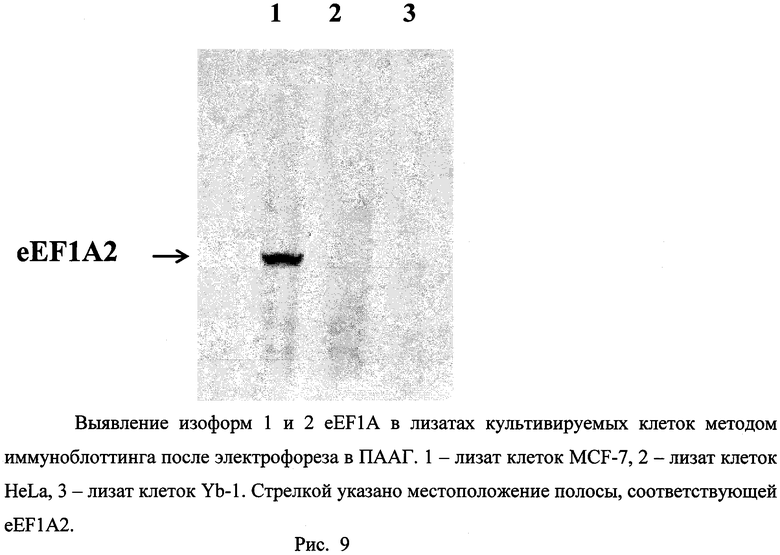
Пример 10. Связывание изоформы 2 eEF1A в лизатах клеточных линий человека моноспецифичными анти-eEF1A2 антителами, полученными против пептидного производного (I), регистрируемое методом иммуноблоттинга .
Клеточные линии MCF7, HeLa выращивали в чашках Петри диаметром 10 см в стандартной среде DMEM с 5% фетальной сыворотки теленка, линию Yb-1 - в среде DMEM/F12 с 5% лошадиной сыворотки, 20 нг/мл эпидермального фактора роста, 0,5 мг/мл гидрокортизона, 1 мг/мл холерного токсина, 10 мкг/мл инсулина в присутствии антибиотиков пенициллина и стрептомицина при 37°C в атмосфере с содержанием CO2 5%. Из чашки Петри, содержащей 5 млн. клеток, удаляли культуральную среду и снимали клетки скребком в 2 мл ФСБ. Полученную суспензию клеток центрифугировали 2 мин при 200 g. Осадок клеток объемом 50 мкл суспендировали в лизирующем буфере (50 мМ трис-HCl буфер, pH 8,0, 150 мМ NaCl, 1% Nonidet Р-40, 0,5% дезоксихолата натрия, 0,1% додецилсульфата натрия) и после прогревания при 70°C в течение 5 мин подвергали электрофорезу в готовых пластинах полиакриламидного геля в денатурирующих условиях. После электрофореза гель отмывали и белки переносили под действием электрического поля на нитроцеллюлозную мембрану (см. пример 9), которую инкубировали в растворе антител козы против eEF1A2, полученных способом, описанным в примере 8, и разведенных в 50 раз блокирующим буфером (см. пример 9). Связавшиеся с изоформами eEF1A антитела проявляли конъюгатом антител кролика против иммуноглобулинов G козы с пероксидазой хрена («Имтек», Россия, рабочее разведение 1:5000) с использованием хемилюминесцентного косубстрата пероксидазы SuperSignal West Dura Extended Duration Substrate («Pierce», США, каталожный №34076) в соответствии с рекомендациями фирмы; время экспозиции блота с раствором субстрата пероксидазы 20 мин. Из рисунка 9 видно, что специфичные анти-eEF1A2 антитела выявляют данную изоформу в лизате клеток MCF-7, где предварительно была выявлена значительная экспрессия мРНК eEF1A2, в лизатах же клеток HeLa (низкий уровень экспрессии мРНК eEF1A2) и Yb-1 (экспрессия мРНК eEF1A2 отсутствует) данной изоформы фактора элонгации трансляции не обнаруживается. При этом имеющаяся в клетках Yb-1 изоформа eEF1A1 с помощью специфичных анти-eEF1A2 антител не выявляется.
Список использованной литературы
1. Negrutskii, В.S., El'skaya, А.V. (1998) Eukaryotic translation elongation factor 1 alpha: structure, expression, functions, and possible role in aminoacyl-tRNA channeling, Prog. Nucleic. Acid Res. Mol. Biol. 60, 47-78.
2. Mateyak, M.K., Kinzy, T.G. (2010) eEF1A: thinking outside the ribosome, J. Biol. Chem. 285, 21209-21213.
3. Kahns, S., Lund, A., Kristensen, P., Knudsen, C.R., Clark, B.F., Cavallius, J., Merrick, W.C. (1998) The elongation factor 1 A-2 isoform from rabbit: cloning of the cDNA and characterization of the protein, Nucleic Acids Res. 26, 1884-1890.
4. Newbery, H.J., Loh, D.H., O'Donoghue, J.E., Tomlinson, V.A.L., Chau, Y-Y., Boyd, J.A., Bergmann, J.H., Braunstein, D., Abbott, C.M. (2007) Translation elongation factor eEF1A2 is essential for postweaning survival in mice. J. Biol. Chem., 282, 28951-28959.
5. Lee, M.-H., Surh, Y.-J. (2009) eEF1A2 as a putative oncogene. Ann. N.Y. Acad. Sci., 1171, 87-93.
6. Li, Z., Qi, C.F., Shin, D.M., Zingone, A., Newbery, H.J., Kovalchuk, A.L., Abbott, С.М., Morse, H.C 3rd. (2010). Eefla2 promotes cell growth, inhibits apoptosis and activates JAK/STAT and АКТ signaling in mouse plasmacytomas. PLoS One, 5(5), e10755.
7. Van Regenmortel, M.H., Muller, S. (1999). Synthetic peptides as antigens. Amsterdam: Elsevier Science, 1999.
8. Lloyd-Williams, P., Albericio, F., Giralt, E. (1997). Chemical approaches to the synthesis of peptides and proteins. New York.: CRC Press LLC.
9. Иммунологические методы (1987). Под ред. Г. Фримеля. М.: «Медицина».
10. Laemmli U.K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227, 680-685.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИЯ НА ОСНОВЕ СИНТЕТИЧЕСКИХ ПЕПТИДОВ И ЛИПИДОВ ДЛЯ ВАКЦИНЫ ПРОТИВ ГЕПАТИТА С | 2015 |
|
RU2675108C2 |
| ТРИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА ПРОТИВ IL-17A, IL-17F И ДРУГОЙ ПРОВОСПАЛИТЕЛЬНОЙ МОЛЕКУЛЫ | 2016 |
|
RU2680011C2 |
| ПЕПТИД АПОВ-РЕЦЕПТОРА, ОТВЕТСТВЕННЫЙ ЗА СВЯЗЫВАНИЕ ЛИПОПРОТЕИНОВ НИЗКОЙ ПЛОТНОСТИ | 1994 |
|
RU2043116C1 |
| АНТИГЕН, АССОЦИИРОВАННЫЙ С РЕВМАТОИДНЫМ АРТРИТОМ | 2013 |
|
RU2649368C2 |
| ИММУНОЦИТОКИНЫ НА ОСНОВЕ IL-15 И IL-R[альфа] ДОМЕНА SUSHI | 2012 |
|
RU2644671C2 |
| ГЛИКОЗИЛИРОВАННЫЕ КОНЪЮГАТЫ МОЛЕКУЛ С ПОВТОРЯЮЩИМСЯ МОТИВОМ | 2010 |
|
RU2574201C2 |
| ПРИМЕНЕНИЕ АНТИТЕЛА ПРОТИВ TAU PS422 ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ ГОЛОВНОГО МОЗГА | 2010 |
|
RU2536247C2 |
| Антигены, ассоциированные с воспалительным заболеванием кишечника | 2012 |
|
RU2612878C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО К PD-L1 | 2017 |
|
RU2665790C1 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ IL-4 И/ИЛИ IL-13, И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2580049C2 |

Изобретение относится к области иммунологии и медицины. Предложено пептидное производное для выработки в организме специфических антител, взаимодействующих с изоформой 2 фактора элонгации трансляции 1A (eEF1A2), но не взаимодействующих с изоформой 1 фактора элонгации трансляции 1A (eEF1A1), содержащее фрагмент аминокислотной последовательности eEF1A2 с 330 по 343 положения. Также рассмотрены основанный на использовании пептидного производного по изобретению способ выработки в организме животного моноспецифичных антител, взаимодействующих с eEF1A2, но не с eEF1A1, и его применение для аффинной очистки таких моноспецифических антител. Изобретение может быть использовано с целью разработки иммунотест-систем для выявления в биологических образцах клеток с измененным биохимическим аппаратом контроля запрограммированной гибели (апоптоза), способных к злокачественному перерождению и образованию злокачественной опухоли. 3 н.п. ф-лы, 9 ил., 10 пр.
1. Пептидное производное для выработки в организме специфических антител, взаимодействующих с изоформой 2 фактора элонгации трансляции 1A человека и других млекопитающих (при условии идентичности аминокислотных последовательностей этих белков), но не взаимодействующих с изоформой 1 фактора элонгации трансляции 1A, общей формулы I
X-KSDPPQEAAQFTS-NH-CH(CH2CH2COOH)-COY
(Х-лизил-серил-глутамил-пролил-пролил-глутаминил-глутамил-аланил-аланил-глутаминил-фенилаланил-треонил-серил-глутамат или -глутаматамид) (I),
или его фармацевтически приемлемая соль, или его сольват,
где X - ацетильная группа или атом водорода, Y-NH2- или OH-группа, а остальная часть пептидного производного представляет собой фрагмент аминокислотной последовательности изоформы 2 фактора элонгации трансляции человека 1A, включающий аминокислотные остатки с №330 по №343.
2. Способ выработки в организме животного моноспецифичных антител, взаимодействующих с изоформой 2 фактора элонгации трансляции 1A человека и других млекопитающих (при условии идентичности аминокислотных последовательностей этих белков), но не взаимодействующих с изоформой 1 этого фактора, предусматривающий введение животному эффективной дозы конъюгата пептидного производного по п.1 с белком-носителем в виде композиции с общеизвестными адъювантами.
3. Применение пептидного производного по п.1 для аффинной очистки моноспецифических антител, взаимодействующих с изоформой 2 фактора элонгации трансляции 1A человека и млекопитающих, но не взаимодействующих с изоформой 1 этого фактора.
| SHARMA S | |||
| et al., "Characterization of a putative ovarian oncogene, elongation factor 1[alpha], isolated by panning a synthetic phage display single-chain variable fragment library with cultured human ovarian cancer cells." Clinical Cancer Research (2007); 13(19): 5889-5896 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| SOARES D.C | |||
| et al., "Structural models of | |||

