Настоящее изобретение относится к применению антитела, которое специфично связывается с фосфорилированным фрагментом Tau SEQ ID NO:9 (pS422), для лечения заболеваний головного мозга.
Предшествующий уровень техники
Белок Tau человека (белок, ассоциированный с микротрубочками, Tau (белок нейрофибриллярных клубков, белок парных спиральных филаментов Tau, PHF-Tau) представляет собой нейрональный белок, ассоциированный с микротрубочками, обнаруженный преимущественно в аксонах, функцией которого является стимуляция полимеризации тубулина и стабилизация микротрубочек. Шесть изоформ (изоформа А, В, С, D, Е, F, G, фетальный Tau) обнаружено в головном мозге человека, где самая длинная изоформа содержит 441 аминокислоту (изоформа F, Uniprot P10636-8). Tau и его свойства также описаны Reynolds, C.H. et al., J. Neurochem. 69 (1997) 191-198.
Tau в его гиперфосфорилированной форме является основным компонентом парных спиральных филаментов (PHF), строительного блока нейрофибриллярных повреждений в головном мозге при болезни Альцгеймера (БА). Tau может фосфорилироваться по его остаткам серина или треонина несколькими различными киназами, включающими GSKSbeta, cdk5, MARK и членов семейства MAP киназ.
Таупатии характеризуются аномальным гиперфосфорилированием Tau и представляют собой в соответствии с Iqbal, К. et al. (Biochimica et Biophysica Acta (ВВА) 1739 (2005) 198-210):
- Болезнь Альцгеймера, включая только клубковую форму этого заболевания
- Синдром Дауна, случаи у взрослых
- Комплекс паркинсонизм-деменция (синдром Гуама)
- Деменция боксеров
- Болезнь Пика
- Деменция с аргирофильными зернами
- Лобно-височная деменция
- Кортико-базальная дегенерация
- Паллидо-понто-нигральная дегенерация
- Прогрессирующий надъядерный паралич
- Болезнь Герштмана-Штрауслера-Шейнкера с клубками.
В данное время в Tau из головных мозгов болезни Альцгеймера обнаружено почти 40 сериновых (S)/треониновых (Т) сайтов фосфорилирования (Hanger, D.P. et al., J. Biol. Chem 282 (2007) 23645-23654). Развитие патологии Tau при болезни Альцгеймера обусловлено его состоянием фосфорилирования. Однако большинство из 40 сайтов фосфорилирования не связано с патологией заболевания, поскольку они также обнаружены в Tau, экстрагированном из ткани здорового фетального головного мозга. Только несколько фосфорилирований являются уникальными для болезненного состояния, и предположительно они ответственны за аномальную характеристическую нерастворимость и агрегацию, которая определяет Tau в PHF головного мозга с болезнью Альцгеймера (Morishima-Kawashima, M. et al., J. Biol. Chem 270 (1995) 823-829). Согласно Pel, J.J. et al., Journal of Alzheimer's Disease 14 (2008) 385-392, в существующей литературе представлена ограниченная и неясная информация о том, какие из этих сайтов специфичны для головного мозга при БА. Pei использовал ряд фосфоспецифичных антител к Tau и измерил их уровни в гомогенатах медиальной височной коры от 22 БА и 10 контролей.
Авторы Bussiere, Т. et al. (Acta Neuropathol. 97 (1999) 221-230) описывают, что фосфорилированный серин 422 на белках Tau является патологическим эпитопом, обнаруженным при некоторых заболеваниях с нейрофибриллярной дегенерацией. Augustinack, J.C. et al., (Acta Neuropathol 103 (2002) 26-35) описывают р3422 как коррелирующий с тяжестью нейрональной патологии при болезни Альцгеймера. Guillozet-Bongaarts, A. (J. Neurochem 97 (2006) 1005-1014) описывают фосфорилирование Tau при S422 как составляющее часть процесса созревания PHF. Tau р8422 также обнаружен в сочетании с развивающейся патологией в различных моделях болезни Альцгеймера на трансгенных мышах. Так, Deters, N. et al. упоминают в статье Biochem. Biophys. Res. Commun. 379 (2009) 400-405, что двойные трансгенные мыши Dom5/pR5 проявляют 7-кратно повышенные числа нейронов гиппокампа, которые содержат Tau со специфично фосфорилированным патологическим эпитопом S422. Goetz, J. et al. (Science 293 (2001) 1491-1495) сообщили о появлении Tau, фосфорилированного по S422, в головном мозге трансгенных мышей Tau P301L, инъецированных фибриллами Abeta42.
ЕР 2009104 относится к эпитопам белка Tau, которые встречаются в фосфорилированном состоянии в белке Tau из PHF болезни Альцгеймера, и к применению этих эпитопов для создания антител, специфично обнаруживающих белок Tau болезни Альцгеймера. WO 2002/062851 и US 7446180 относятся к антителам со специфичностью к аномально укороченной форме белка Tau и к диагностическим и терапевтическим аспектам в отношении болезни Альцгеймера и родственных таупатий.
WO 98/22120 относится к способу лечения пациента с болезнью Альцгеймера, включающему стадию введения пациенту антитела против фосфорилированного фрагмента Tau из аминокислот от примерно 207 до примерно 222, аминокислот от примерно 224 до примерно 240 и аминокислот от примерно 390 до примерно 408. Исследования на животных, где фосфорилированный Tau-фрагмент 379-408 [P-Ser396,404] используют для вакцинации трансгенных мышей Tau, упомянуты в статье Asuni, A.A. et al., J. Neuroscience 27 (2007) 9115-9129. US 2008/0050383 относится к способам лечения и предупреждения болезни Альцгеймера или других таупатий у субъекта путем введения фрагмента белка Tau.
Моноклональные антитела против Tau р3422 описаны, например, в ЕР 1876185. Поликпональные антитела против Tau pS422 имеются в продаже (например, ProSci Inc. и Biosource International).
Краткое изложение сущности изобретения
Изобретение включает антитело, связывающееся с Tau, фосфорилированным по серину 422 (Tau pS422), характеризующееся специфичным связыванием с фосфорилированным фрагментом Tau Ser-Ile-Asp-Met-Val-Asp-Ser(PO3H2)-Pro-Gln-Leu-Ala-Thr-Leu-Ala-Asp (SEQ ID NO:9) и с Tau pS422, но не связывающееся с Tau и с фосфорилированным фрагментом MCAK SEQ ID NO:17, для применения при лечении таупатий, и такой способ лечения.
Изобретение включает антитело, связывающееся с Tau, фосфорилированным по серину 422 (Tau pS422), характеризующееся специфичным связыванием с фосфорилированным фрагментом Tau SEQ ID NO:9 и с Tau pS422, но не связывающееся с Tau и с фосфорилированным фрагментом MCAK SEQ ID NO:17, для получения лекарства для лечения таупатий и к такому способу получения.
Изобретение включает антитело, связывающее фосфорилированный Tau, характеризующееся специфичным связыванием с тем же эпитопом, который связывает Mab2.10.3 (антитело анти-Tau pS422).
Изобретение включает применение антитела, связывающего фосфорилированный Tau, характеризующееся специфичным связыванием с с тем же эпитопом, который связывает Mab2.10.3 (антитело анти-Tau pS422), для применения при лечении таупатии или для получения лекарства для лечения таупатии.
Антитело в соответствии с изобретением специфично связывается с Tau р8422 и с агрегированным (фибриллярным), фосфорилированным Tau. Антитело в соответствии с изобретением не связывается с нефосфорилированным Tau, нефосфорилированным фрагментом Таи SEQ ID NO:10 и с фосфорилированным фрагментом MCAK SEQ ID NO:17.
Антитело в соответствии с изобретением предпочтительно принадлежит к подтипу IgG1 человека. В следующей форме осуществления изобретения антитело в соответствии с изобретением предпочтительно принадлежит к подтипу IgG4 человека.
Изобретение включает антитело анти-Tau р8422, характеризующееся тем, что оно включает
a) CDR1H SEQ ID NO:6, CDR2H SEQ ID NO:7 и CDR3H SEQ ID NO:8,
b) CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:24 и CDR3H SEQ ID NO:25,
c) CDR1H SEQ ID NO:31, CDR2H SEQ ID NO:32 и CDR3H SEQ ID NO:33,
d) CDR1H SEQ ID NO:39, CDR2H SEQ ID NO:40 и CDR3H SEQ ID NO:41,
e) CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49,
f) CDR1H SEQ ID NO:55, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49, или
g) CDR1H SEQ ID NO:63, CDR2H SEQ ID NO:64 и CDR3H SEQ ID NO:65.
Предпочтительно антитело характеризуется тем, что оно включает
a) CDR1H SEQ ID NO:6, CDR2H SEQ ID NO:7, CDR3H SEQ ID NO:8 и CDR1L SEQ ID NO:3, CDR2L SEQ ID NO:4, CDR3L SEQ ID NO:5,
b) CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:24 и CDR3H SEQ ID NO:25, и CDR1L SEQ ID NO:27, CDR2L SEQ ID NO:28, CDR3L SEQ ID NO:29,
c) CDR1H SEQ ID NO:31, CDR2H SEQ ID NO:32 и CDR3H SEQ ID NO:33, и CDR1L SEQ ID NO:35, CDR2L SEQ ID NO:36, CDR3L SEQ ID NO:37,
d) CDR1H SEQ ID NO:39, CDR2H SEQ ID NO:40 и CDR3H SEQ ID NO:41, и CDR1L SEQ ID NO:43, CDR2L SEQ ID NO:44, CDR3L SEQ ID NO:45,
e) CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:48, и CDR3H SEQ ID NO:49, и CDR1L SEQ ID NO:51, CDR2L SEQ ID NO:52, CDR3L SEQ ID NO:53,
f) CDR1H SEQ ID NO:55, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49, и CDR1L SEQ ID NO:51, CDR2L SEQ ID NO:60, CDR3L SEQ ID NO:61, или
g) CDR1H SEQ ID NO:63, CDR2H SEQ ID NO:64 и CDR3H SEQ ID NO:65, и CDR1L SEQ ID NO:67, CDR2L SEQ ID NO:68, CDR3L SEQ ID NO:69.
Предпочтительно антитело характеризуется тем, что оно включает a) вариабельную легкую цепь SEQ ID NO:1 и вариабельную тяжелую цепь SEQ ID NO:2,
b) вариабельную легкую цепь SEQ ID NO:26 и вариабельную тяжелую цепь SEQ ID NO:22,
c) вариабельную легкую цепь SEQ ID NO:34 и вариабельную тяжелую цепь SEQ ID NO:30,
d) вариабельную легкую цепь SEQ ID NO:42 и вариабельную тяжелую цепь SEQ ID NO:38,
e) вариабельную легкую цепь SEQ ID NO:50 и вариабельную тяжелую цепь SEQ ID NO:46,
f) вариабельную легкую цепь SEQ ID NO:58 и вариабельную тяжелую цепь SEQ ID NO:54, или
g) вариабельную легкую цепь SEQ ID NO:66 и вариабельную тяжелую цепь SEQ ID NO:62.
Изобретение включает гуманизированный вариант антитела анти-Tau pS422 Mab2.10.3. Изобретение включает химерный вариант антитела анти-Tau pS422 Mab2.10.3. Изобретение включает вариант, истощенный по Т-клеточному эпитопу, антитела анти-Tau pS422 Mab2.10.3. Mab2.10.3 характеризуется его вариабельными цепями SEQ ID NO:1 и SEQ ID NO:2.
Изобретение включает гуманизированный вариант антитела анти-Tau pS422 Mab 005. Изобретение включает химерный вариант антитела анти-Tau pS422 Mab 005. Изобретение включает вариант, истощенный по Т-клеточному эпитопу, антитела анти-Tau pS422 Mab 005. Mab 005 характеризуется его вариабельными цепями SEQ ID NO:26 и SEQ ID NO:22.
Изобретение включает гуманизированный вариант антитела анти-Tau pS422 Mab 019. Изобретение включает химерный вариант антитела анти-Tau pS422 Mab
019. Изобретение включает вариант, истощенный по Т-клеточному эпитопу, антитела анти-Tau pS422 Mab 019. Mab 019 характеризуется его вариабельными цепями SEQ ID NO:34 и SEQ ID NO:30.
Изобретение включает гуманизированный вариант антитела анти-Tau pS422 Mab 020. Изобретение включает химерный вариант антитела анти-Tau pS422 Mab
020. Изобретение включает вариант, истощенный по Т-клеточному эпитопу, антитела анти-Tau pS422 Mab 020. Mab 020 характеризуется его вариабельными цепями SEQ ID NO:42 и SEQ ID NO:38.
Изобретение включает гуманизированный вариант антитела анти-Tau pS422 Mab 085. Изобретение включает химерный вариант антитела анти-Tau pS422 Mab 085. Изобретение включает вариант, истощенный по Т-клеточному эпитопу, антитела анти-Tau pS422 Mab 085. Mab 085 характеризуется его вариабельными цепями SEQ ID NO:50 и SEQ ID NO:46.
Изобретение включает гуманизированный вариант антитела анти-Tau pS422 Mab 086. Изобретение включает химерный вариант антитела анти-Tau pS422 Mab 086. Изобретение включает вариант, истощенный по Т-клеточному эпитопу, антитела анти-Tau pS422 Mab 086. Mab 086 характеризуется его вариабельными цепями SEQ ID NO:58 и SEQ ID NO:54.
Изобретение включает гуманизированный вариант антитела анти-Tau pS422 Mab 097. Изобретение включает химерный вариант антитела анти-Tau pS422 Mab 097. Изобретение включает вариант, истощенный по Т-клеточному эпитопу, антитела анти-Tau pS422 Mab 097. Mab 097 характеризуется его вариабельными цепями SEQ ID NO:66 и SEQ ID NO:62.
Изобретение включает химерный, гуманизированный или истощенный по Т-клеточному эпитопу вариант антитела анти-Tau pS422, включающий CDR1H SEQ ID NO:6, CDR2H SEQ ID NO:7, CDR3H SEQ ID NO:8 и CDR1L SEQ ID NO:3, CDR2L SEQ ID NO:4, CDR3L SEQ ID NO:5 или вариабельную легкую цепь SEQ ID NO:1 и вариабельную тяжелую цепь SEQ ID NO:2.
Изобретение включает химерный, гуманизированный или истощенный по Т-клеточному эпитопу вариант антитела анти-Tau pS422, включающий CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:24 и CDR3H SEQ ID NO:25, и CDR1L SEQ ID NO:27, CDR2L SEQ ID NO:28, CDR3L SEQ ID NO:29, или вариабельную легкую цепь SEQ ID NO:26 и вариабельную тяжелую цепь SEQ ID NO:22.
Изобретение включает химерный, гуманизированный или истощенный по Т-клеточному эпитопу вариант антитела анти-Tau pS422, включающий CDR1H SEQ ID NO:31, CDR2H SEQ ID NO:32 и CDR3H SEQ ID NO:33, и CDR1L SEQ ID NO:35, CDR2L SEQ ID NO:36, CDR3L SEQ ID NO:37 или вариабельную легкую цепь SEQ ID NO:34 и вариабельную тяжелую цепь SEQ ID NO:30.
Изобретение включает химерный, гуманизированный или истощенный по Т-клеточному эпитопу вариант антитела анти-Tau pS422, включающий CDR1H SEQ ID NO:39, CDR2H SEQ ID NO:40 и CDR3H SEQ ID NO:41, и CDR1L SEQ ID NO:43, CDR2L SEQ ID NO:44, CDR3L SEQ ID NO:45 или вариабельную легкую цепь SEQ ID NO:42 и вариабельную тяжелую цепь SEQ ID NO:38.
Изобретение включает химерный, гуманизированный или истощенный по Т-клеточному эпитопу вариант антитела анти-Tau pS422, включающий CDR1H SEQ ID NO:47, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49, и CDR1L SEQ ID NO:51 CDR2L SEQ ID NO:52, CDR3L SEQ ID NO:53 или вариабельную легкую цепь SEQ ID NO:50 и вариабельную тяжелую цепь SEQ ID NO:46.
Изобретение включает химерный, гуманизированный или истощенный по Т-клеточному эпитопу вариант антитела анти-Tau р8422, включающий CDR1H SEQ ID NO:55, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49, и CDR1L SEQ ID NO:51, CDR2L SEQ ID NO:60, CDR3L SEQ ID NO:61 или вариабельную легкую цепь SEQ ID NO:58 и вариабельную тяжелую цепь SEQ ID NO:54.
Изобретение включает химерный, гуманизированный или истощенный по Т-клеточному эпитопу вариант антитела анти-Tau pS422, включающий CDR1H SEQ ID NO:63, CDR2H SEQ ID NO:64 и CDR3H SEQ ID NO:65, и CDR1L SEQ ID NO:67, CDR2L SEQ ID NO:68, CDR3L SEQ ID NO:69 или вариабельную легкую цепь SEQ ID NO:66 и вариабельную тяжелую цепь SEQ ID NO:62.
Изобретение включает способ гуманизации, истощения по Т-клеточному эпитопу или химеризации антитела анти-Tau pS422, характеризующегося тем, что оно включает CDR1H SEQ ID NO:6, CDR2H SEQ ID NO:7, CDR3H SEQ ID NO:8 и CDR1L SEQ ID NO:3, CDR2L SEQ ID NO:4, CDR3L SEQ ID NO:5 или вариабельную легкую цепь SEQ ID NO:1 и вариабельную тяжелую цепь SEQ ID NO:2.
Изобретение включает способ гуманизации, истощения по Т-клеточному эпитопу или химеризации антитела анти-Tau pS422, характеризующегося тем, что оно включает CDR1H SEQ ID NO:6, CDR2H SEQ ID NO:7, CDR3H SEQ ID NO:8 и CDR1L SEQ ID NO:3, CDR2L SEQ ID NO:4, CDR3L SEQ ID NO:5 или вариабельную легкую цепь SEQ ID NO:1 и вариабельную тяжелую цепь SEQ ID NO:2.
Изобретение включает способ гуманизации, истощения по Т-клеточному эпитопу или химеризации антитела анти-Tau pS422, характеризующегося тем, что оно включает CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:24 и CDR3H SEQ ID NO:25, и CDR1L SEQ ID NO:27, CDR2L SEQ ID NO:28, CDR3L SEQ ID NO:29 или вариабельную легкую цепь SEQ ID NO:26 и вариабельную тяжелую цепь SEQ ID NO:22.
Изобретение включает способ гуманизации, истощения по Т-клеточному эпитопу или химеризации антитела анти-Tau pS422, характеризующегося тем, что оно включает CDR1H SEQ ID NO:31, CDR2H SEQ ID NO:32 и CDR3H SEQ ID NO:33, и CDR1L SEQ ID NO:35, CDR2L SEQ ID NO:36, CDR3L SEQ ID NO:37 или вариабельную легкую цепь SEQ ID NO:34 и вариабельную тяжелую цепь SEQ ID NO:30.
Изобретение включает способ гуманизации, истощения по Т-клеточному эпитопу или химеризации антитела анти-Tau pS422, характеризующегося тем, что оно включает CDR1H SEQ ID NO:39, CDR2H SEQ ID NO:40 и CDR3H SEQ ID NO:41, и CDR1L SEQ ID NO:43, CDR2L SEQ ID NO:44, CDR3L SEQ ID NO:45 или вариабельную легкую цепь SEQ ID NO:42 и вариабельную тяжелую цепь SEQ ID NO:38.
Изобретение включает способ гуманизации, истощения по Т-клеточному эпитопу или химеризации антитела анти-Tau pS422, характеризующегося тем, что оно включает CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49, и CDR1L SEQ ID NO:51, CDR2L SEQ ID NO:52, CDR3L SEQ ID NO:53 или вариабельную легкую цепь SEQ ID NO:50 и вариабельную тяжелую цепь SEQ ID NO:46.
Изобретение включает способ гуманизации, истощения по Т-клеточному эпитопу или химеризации антитела анти-Tau pS422, характеризующегося тем, что оно включает CDR1H SEQ ID NO:55, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49, и CDR1L SEQ ID NO:51, CDR2L SEQ ID NO:60, CDR3L SEQ ID NO:61 или вариабельную легкую цепь SEQ ID NO:58 и вариабельную тяжелую цепь SEQ ID NO:54.
Изобретение включает способ гуманизации, истощения по Т-клеточному эпитопу или химеризации антитела анти-Tau pS422, характеризующегося тем, что оно включает CDR1H SEQ ID NO:63, CDR2H SEQ ID NO:64 и CDR3H SEQ ID NO:65, и CDR1L SEQ ID NO:67, CDR2L SEQ ID NO:68, CDR3L SEQ ID NO:69 или вариабельную легкую цепь SEQ ID NO:66 и вариабельную тяжелую цепь SEQ ID NO:62.
Предпочтительно антитело, связывающееся с Tau pS422 и характеризующееся вышеуказанными аминокислотными последовательностями и фрагментами аминокислотных последовательностей, принадлежит к подтипу IgG1 человека. Предпочтительно антитело, связывающееся с Tau pS422 и характеризующееся вышеуказанными аминокислотными последовательностями и фрагментами аминокислотных последовательностей, принадлежит к подтипу IgG4 человека.
Следующей формой осуществления изобретения является фармацевтическая композиция, содержащая антитело в соответствии с изобретением.
Следующей формой осуществления изобретения является применение антитела в соответствии с изобретением для получения фармацевтической композиции.
Следующей формой осуществления изобретения является применение антитела в соответствии с изобретением для лечения таупатии, выбранной из группы, состоящей из болезни Альцгеймера (БА), включая только клубковую форму заболевания, синдрома Дауна (случаев у взрослых), комплекса паркинсонизм-деменция (синдрома Гуама), деменции боксеров, болезни Пика, деменции с аргирофильными зернами, лобно-височной деменции, кортико-базальной дегенерации, паллидо-понто-нигральной дегенерации, прогрессирующего надъядерного паралича и болезни Герштмана-Штрауслера-Шейнкера с клубками.
Следующей формой осуществления изобретения является способ получения фармацевтической композиции, содержащей антитело в соответствии с изобретением.
Следующей формой осуществления изобретения является нуклеиновая кислота, кодирующая вариабельный домен тяжелой цепи и/или вариабельный домен легкой цепи антитела в соответствии с изобретением.
Далее в изобретении предложены экспрессионные векторы, содержащие нуклеиновую кислоту в соответствии с изобретением, способные экспрессировать эту нуклеиновую кислоту в прокариотической или эукариотической клетке-хозяине, и клетки-хозяева, содержащие такие векторы для рекомбинантного продуцирования такого антитела.
Далее изобретение включает прокариотическую или эукариотическую клетку-хозяина, содержащую вектор в соответствии с изобретением.
Далее изобретение включает способ продуцирования рекомбинантного человеческого или гуманизированного антитела в соответствии с изобретением, характеризующийся экспрессией нуклеиновой кислоты в соответствии с изобретением в прокариотической или эукариотической клетке-хозяине и выделением антитела из клетки или из супернатанта клеточной культуры. Далее изобретение включает антитело, которое может быть получено таким рекомбинантным способом.
Далее изобретение включает способ селекции моноклонального антитела в соответствии с изобретением, характеризующийся получением ряда моноклональных антител, связывающихся с Tau pS422, определением специфичного связывания этих антител с фосфорилированным фрагментом Tau SEQ ID NO:9, с Tau и с фосфорилированным фрагментом MCAK SEQ ID NO:17, и селекцией антитела со специфичным связыванием с данным фосфорилированным фрагментом Tau, по меньшей мере 10000-кратным по сравнению с его связыванием с Tau, и со специфичным связыванием с данным фосфорилированным фрагментом Tau, по меньшей мере 100-кратным по сравнению с его связыванием с фосфорилированным фрагментом MCAK Ile-Gln-Lys-Gln-Lys-Arg-Arg-Ser(PO3H2)-Val-Asn-Ser-Lys-Ile-Pro-Ala (SEQ ID NO:17).
Предпочтительно изобретение включает способ селекции моноклонального антитела в соответствии с изобретением, характеризующийся получением ряда моноклональных антител, связывающихся с Tau р8422, определением специфичного связывания этих антител с фосфорилированным фрагментом Tau SEQ ID NO:9, с Tau pS422, с Tau и с фосфорилированным фрагментом MCAK SEQ ID NO:17, и селекцией антитела со специфичным связыванием с данным фосфорилированным фрагментом Tau и с Tau pS422, по меньшей мере 10000-кратным по сравнению с его связыванием с Tau, и со специфичным связыванием с данным фосфорилированным фрагментом Tau, по меньшей мере 100-кратным по сравнению с его связыванием с фосфорилированным фрагментом MCAK Ile-Gln-Lys-Gln-Lys-Arg-Arg-Ser(PC>3H2)-Val-Asn-Ser-Lys-Ile-Pro-Ala (SEQ ID NO:17).
Предпочтительно изобретение включает способ селекции моноклонального антитела в соответствии с изобретением, характеризующийся получением ряда моноклональных антител, связывающихся с Tau pS422, определением специфичного связывания этих антител с фосфорилированным фрагментом Tau SEQ ID NO:9, с Tau pS422, с Tau и с фосфорилированным фрагментом MCAK SEQ ID NO:17, и селекцией антитела со специфичным связыванием с данным фосфорилированным фрагментом Tau и с Tau pS422, по меньшей мере 10000-кратным по сравнению с его связыванием с Tau, и со специфичным связыванием с данным фосфорилированным фрагментом Tau и с Tau pS422, по меньшей мере 100-кратным по сравнению с его связыванием с фосфорилированным фрагментом MCAK Ile-Gln-Lys-Gln-Lys-Arg-Arg-Ser(P03H2)-Val-Asn-Ser-Lys-Ile-Pro-Ala (SEQ ID NO:17).
Антитела в соответствии с изобретением проявляют полезные эффекты для пациентов, нуждающихся в направленной терапии на Tau. Антитела в соответствии с изобретением обладают новыми и изобретательскими свойствами, вызывающими пользу для пациента, страдающего таупатией, в частности БА.
Далее в изобретении предложен способ лечения пациента, страдающего таупатией, в частности БА, включающий введение пациенту, у которого диагностировано такое заболевание (и, следовательно, нуждающемуся в такой терапии), антитела, связывающегося с pS422, в соответствии с изобретением. Антитело предпочтительно вводят в фармацевтической композиции.
Следующей формой осуществления изобретения является способ лечения пациента, страдающего таупатией, в частности БА, характеризующийся введением этому пациенту антитела в соответствии с изобретением.
Далее изобретение включает применение антитела в соответствии с изобретением для лечения пациента, страдающего таупатией, в частности БА, и для получения фармацевтической композиции в соответствии с изобретением. Кроме того, изобретение включает способ получения фармацевтической композиции в соответствии с изобретением.
Далее изобретение включает фармацевтическую композицию, содержащую антитело в соответствии с изобретением, необязательно вместе с буфером и/или адъювантом, полезным для приготовления препаратов антител для фармацевтических целей.
Далее в изобретении предложена фармацевтическая композиция, содержащая антитело в соответствии с изобретением в фармацевтически приемлемом носителе. В одной форме осуществления фармацевтическая композиция может быть включена в готовое изделие или в набор.
Антитело по настоящему изобретению можно применять для диагностики неврологического расстройства, такого как болезнь Альцгеймера, путем обнаружения фосфорилированного полипептида Tau. Антитело по настоящему изобретению можно также применять для специфичного обнаружения Tau р8422 или агрегированного, фосфорилированного Tau.
Подробное описание изобретения
Термин "Tau" в соответствии с изобретением охватывает самую длинную изоформу Tau человека, содержащую 441 аминокислоту (изоформу F, Uniprot Р10636-8).
Термин "фосфорилированный Tau (pTau)" в соответствии с изобретением охватывает фосфорилированную форму самой длинной изоформы Tau человека, содержащей 441 аминокислоту (изоформу F, Uniprot P10636-8), образованную путем фосфорилирования по S422 киназой ERK2.
Термин "агрегированный, фосфорилированный Tau" или "агрегированный (фибриллярный), фосфорилированный Tau" в соответствии с изобретением охватывает агрегированную и фосфорилированную форму самой длинной изоформы Tau человека, содержащей 441 аминокислоту (изоформу F, Uniprot P10636-8), образованную путем фосфорилирования агрегированного Tau киназой ERK2.
Термин "фрагмент Tau" в соответствии с изобретением охватывает фрагмент Tau Ser-Ile-Asp-Met-Val-Asp-Ser-Pro-Gln-Leu-Ala-Thr-Leu-Ala-Asp (SEQ ID NO:10).
Термин "фосфорилированный фрагмент Tau" в соответствии с изобретением охватывает фосфорилированный фрагмент Tau Ser-Ile-Asp-Met-Val-Asp-Ser(PO3H2)-Pro-Gln-Leu-Ala-Thr-Leu-Ala-Asp (SEQ ID NO:9).
Термин "MCAK" в соответствии с изобретением охватывает митотический кинезин человека, ассоциированный с центромерой (кинезиноподобный белок KIF2C, UniProt Q99661)). MCAK_Human (88-102)[95-pSer] представляет собой фосфорилированный фрагмент MCAK, состоящий из аминокислот 88-102, фосфорилированный по серину 95 (SEQ ID NO:17). Этот фосфорилированный фрагмент MCAK не обладает идентичностью или подобием последовательности по сравнению с фосфорилированным фрагментом Tau SEQ ID NO:9. Авторы изобретения обнаружили, что антитела против фосфорилированных фрагментов Tau в соответствии с уровнем техники могут проявлять значительную перекрестную реактивность с неродственными фосфорилированными пептидами и белками человека. Для антител в соответствии с изобретением, которые не связываются с фосфорилированным фрагментом MCAK, такую нежелательную перекрестную реактивность невозможно обнаружить.
Связывание с Tau pS422 и связывание с Tau исследуют с помощью ELISA (твердофазного иммуноферментного анализа) с электролюминесцентным считыванием. Tau или Tau pS422 иммобилизуют при концентрации 2 мкг/мл и добавляют тестируемое антитело (например, человека или мыши). Для обнаружения связанного тестируемого антитела добавляют меченый рутением IgG против человека или против мыши, соответственно, при концентрации 0,5 мкг/мл. Специфичное связывание с Tau pS422 обнаруживают, если отношение сигнала обнаружения Tau pS422 и Tau является по меньшей мере 10000-кратным при максимальном сигнале связывания Tau pS422.
Связывание с фосфорилированным фрагментом Tau SEQ ID NO:9, связывание с нефосфорилированным фрагментом Tau SEQ ID NO:10 и с фосфорилированным фрагментом MCAK SEQ ID NO:17 исследуют с помощью ELISA. Тестируемое антитело инкубируют с иммобилизованным фосфорилированным фрагментом Tau и для сравнения с иммобилизованным нефосфорилированным фрагментом Tau или с фрагментом MCAK. Это антитело является меченым, и метку обнаруживают. Специфичное связывание с фосфорилированным фрагментом Tau обнаруживают, если отношение сигнала обнаружения с использованием фосфорилированного фрагмента Tau и нефосфорилированного фрагмента Tau составляет по меньшей мере 100 при максимальном сигнале связывания фосфорилированного фрагмента Tau, и, если отношение сигнала обнаружения с использованием фосфорилированного фрагмента Tau и фосфорилированного фрагмента MCAK также составляет по меньшей мере 100 при максимальном сигнале связывания фосфорилированного фрагмента Tau.
Термин "эпитоп Mab2.10.3" охватывает эпитоп, локализованный внутри фосфорилированного фрагмента Tau Ser-Ile-Asp-Met-Val-Asp-Ser(PO3H2)-Pro-Gln-Leu-Ala-Thr-Leu-Ala-Asp (SEQ ID NO:9), который специфично распознается Mab2.10.3. Свойство связывания эпитопа антитела к Tau в соответствии с настоящим изобретением определяют с помощью анализа перекрестного блокирования связывания in vitro, такого как анализ перекрестного блокирования связывания Biacore™ in vitro, чтобы определить способность Mab2.10.3 к стерическому затруднению связывания тестируемого антитела с pTau. Для такого анализа Mab2.10.3 иммобилизуют в качестве первого антитела на сенсоре Biacore, после чего осуществляют последовательные впрыскивания pTau и второго антитела, подлежащего тестированию. Если второе антитело не проявляет какого-либо обнаружимого сигнала связывания, это второе антитело связывается с тем же эпитопом, что и Mab2.10.3.
Антитело в соответствии с изобретением связывается с Tau р3422 со сродством от 5×10-8 М-1 до 10-12 M-1, как определено с помощью анализа Biacore™, описанного выше.
Связывание с фибриллярными агрегатами Tau р8422 исследуют с помощью анализа Biacore. Для этого анализа агрегированный Tau р8422 иммобилизуют, и добавляют тестируемое антитело в различных концентрациях, используя коэффициент разведения 2 и максимальную концентрацию 200 нМ. Антитело в соответствии с изобретением связывается с фибриллярным Tau р8422 с Kd от 0,1 до 30, предпочтительно с Kd от 10 до 20 нМ.
Термин "моноклональное антитело или антитело" охватывает различные формы антитела, предпочтительно моноклонального антитела и особенно предпочтительно моноклонального антитела IgG1 или IgG4. Антитело в соответствии с изобретением предпочтительно представляет собой человеческое антитело, гуманизированное антитело, химерное антитело или дополнительно генетически сконструированное антитело, если сохраняются его характеристические свойства в соответствии с изобретением. Антитела, истощенные по Т-клеточному эпитопу, могут быть получены с использованием способов, описанных в WO 98/08097.
"Фрагменты антитела" включают участок полноразмерного антитела, предпочтительно его вариабельный домен или по меньшей мере его антигенсвязывающий сайт. Примеры фрагментов антитела включают диатела, одноцепочечные молекулы антитела и мультиспецифические антитела, образованные из фрагментов антител. Антитела scFv описаны, например, в Huston, J.S., Methods in Enzymol. 203 (1991) 46-88. Кроме того, фрагменты антитела включают одноцепочечные полипептиды, обладающие характеристиками домена VH, а именно способные к сборке вместе с доменом VL, или домена VL, связывающегося с Таи р8422, а именно способные к сборке вместе с доменом VH с образованием функционального антигенсвязывающего сайта. Термин "гуманизированное антитело" относится к антителам, в которых каркасные участки и/или "участки определения комплементарности" (CDR) модифицированы таким образом, что включают CDR иммуноглобулина другого вида по сравнению с видом родительского иммуноглобулина. В предпочтительной форме осуществления
a) CDR1H SEQ ID NO:6, CDR2H SEQ ID NO:7, CDR3H SEQ ID NO:8 и CDR1L SEQ ID NO:3, CDR2L SEQ ID NO:4, CDR3L SEQ ID NO:5,
b) CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:24 и CDR3H SEQ ID NO:25 и CDR1L SEQ ID NO:27, CDR2L SEQ ID NO:28, CDR3L SEQ ID NO:29,
c) CDR1H SEQ ID NO:31, CDR2H SEQ ID NO:32 и CDR3H SEQ ID NO:33 и CDR1L SEQ ID NO:35, CDR2L SEQ ID NO:36, CDR3L SEQ ID NO:37,
d) CDR1H SEQ ID NO:39, CDR2H SEQ ID NO:40 и CDR3H SEQ ID NO:41 и CDR1L SEQ ID NO:43, CDR2L SEQ ID NO:44, CDR3L SEQ ID NO:45,
e) CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49 и CDR1L SEQ ID NO:51, CDR2L SEQ ID NO:52, CDR3L SEQ ID NO:53,
f) CDR1H SEQ ID NO:55, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49 и CDR1L SEQ ID NO:51, CDR2L SEQ ID NO:60, CDR3L SEQ ID NO:61 или
g) CDR1H SEQ ID NO:63, CDR2H SEQ ID NO:64 и CDR3H SEQ ID NO:65 и CDR1L SEQ ID NO:67, CDR2L SEQ ID NO:68, CDR3L SEQ ID NO:69
прививают на каркасный участок человеческого антитела с получением "гуманизированного антитела". См., например, Riechmann, L., et al., Nature 332 (1988) 323-327; и Neuberger, M.S., et al., Nature 314 (1985) 268-270.
"Вариабельный домен" (вариабельный домен легкой цепи (VL), вариабельный домен тяжелой цепи (VH)), как используют в данной заявке, означает каждую из пар доменов легкой и тяжелой цепи, которые вовлечены непосредственно в связывание антитела с антигеном. Вариабельные домены легкой и тяжелой цепи имеют одинаковую общую структуру, и каждый домен включает четыре каркасных (FR) участка, последовательности которых широко консервативны, соединенные тремя "гипервариабельными участками" (или участками определения комплементарности, CDR). Каркасные участки принимают β-складчатую конформацию, а CDR могут образовать петли, соединяющие β-складчатую структуру. Участки CDR в каждой цепи удерживаются в своей трехмерной структуре каркасными участками и вместе с CDR из другой цепи образуют антигенсвязывающий сайт. Участки CDR3 тяжелой и легкой цепи антитела играют особенно важную роль в связывающей специфичности/сродстве антител в соответствии с изобретением и, следовательно, образуют следующий аспект изобретения.
Термин "антигенсвязывающий участок антитела" при использовании в данной заявке относится к аминокислотным остаткам антитела, которые ответственны за связывание антигена. Антигенсвязывающий участок антитела включает аминокислотные остатки из "участков определения комплементарности" или "CDR". "Каркасные" или "FR" участки представляют собой участки вариабельных доменов, иные, чем остатки гипервариабельных участков, как определено в данной заявке. Таким образом, вариабельные домены легкой и тяжелой цепей антитела включают от N- к С-концу домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. В частности, CDR3 тяжелой цепи является участком, который вносит наибольший вклад в связывание антигена и определяет свойства антитела. Участки CDR и FR определяют в соответствии со стандартным определением Кэбота, Kabat et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991) и/или как остатки из "гипервариабельной петли".
Термин "CDR1H" обозначает участок CDR1 вариабельной области тяжелой цепи, вычисленный согласно Кэботу. CDR2L, CDR3H и т.д. означают соответствующие участки из тяжелой (Н) или легкой (L) цепи. Например, антитело, характеризующееся тем, что оно включает CDR1H SEQ ID NO:6, означает, что антитело включает эту аминокислотную последовательность в качестве вариабельного участка тяжелой цепи CDR1 в его вариабельной тяжелой цепи. Например, антитело, характеризующееся тем, что оно включает CDR1H SEQ ID NO:6, CDR2H SEQ ID NO:7, CDR3H SEQ ID NO:8, означает, что антитело включает в его тяжелой цепи в качестве последовательности CDR1 SEQ ID NO:6, в качестве последовательности CDR2 SEQ ID NO:7 и в качестве последовательности CDR3 SEQ ID NO:8.
Термины "нуклеиновая кислота" или "молекула нуклеиновой кислоты", как используют в данной заявке, подразумевают как включающие молекулы ДНК и молекулы РНК. Молекула нуклеиновой кислоты может быть однонитевой или двунитевой, но предпочтительно представляет собой двунитевую ДНК.
Термин "аминокислота", как используют в данной заявке при данном применении, означает группу встречающихся в природе карбокси-α-аминокислот, включающую аланин (трехбуквенный код: ala, однобуквенный код: А), аргинин (arg, R), аспарагин (asn, N), аспарагиновую кислоту (asp, D), цистеин (cys, С), глутамин (gin, Q), глутаминовую кислоту (glu, Е), глицин (gly, G), гистидин (his, Н), изолейцин (ile, I), лейцин (leu, L), лизин (lys, K), метионин (met, M), фенилаланин (phe, F), пролин (pro, Р), серин (ser, S), треонин (thr, Т), триптофан (trp, W), тирозин (tyr, Y) и валин (val, V).
Нуклеиновая кислота "оперативно сцеплена", когда ее помещают в функциональные взаимоотношения с другой аминокислотой. Например, ДНК для препоследовательности или секреторной лидерной последовательности оперативно сцеплена с ДНК для полипептида, если она экспрессируется в виде пребелка, который участвует в секреции полипептида; промотор или энхансер оперативно сцеплен с кодирующей последовательностью, если он влияет на транскрипцию этой последовательности; или сайт связывания рибосомы оперативно сцеплен с кодирующей последовательностью, если он расположен так, что способствует трансляции. В целом "оперативно сцепленный" означает, что последовательности ДНК, которые являются сцепленными, являются коллинеарными, и в случае секреторной лидерной последовательности являются непрерывными и находятся в одной рамке считывания. Однако энхансеры не обязательно должны быть непрерывными. Сцепление осуществляют путем лигирования в удобных сайтах рестрикции. Если такие сайты не существуют, используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с общепринятой практикой.
Как используют в данной заявке, выражения "клетка", "клеточная линия" и "клеточная культура" используют взаимозаменяемо, и все такие обозначения включают потомство. Таким образом, слова "трансформанты" и "трансформированные клетки" включают первичную обсуждаемую клетку и культуры, имеющие от нее происхождение, независимо от числа переносов. Также понятно, что все потомство может быть не в точности идентичным по содержанию ДНК вследствие преднамеренных или случайных мутаций. Включены варианты потомства, обладающие одной и той же функцией и биологической активностью, скрининг на которую осуществляют в исходно трансформированной клетке.
"Участок Fc" антитела не вовлечен непосредственно в связывание антитела с антигеном, но проявляет различные эффекторные функции. "Участок Fc антитела" является термином, хорошо известным специалистам в данной области техники, и определен на основании расщепления антител папаином. В зависимости от аминокислотной последовательности каркасной области их тяжелых цепей антитела или иммуноглобулины делят на классы: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть дополнительно разделены на подклассы (подтипы), например, IgG1, IgG2, IgG3 и IgG4, IgA1 и IgA2. В соответствии с константными областями тяжелой цепи различные классы иммуноглобулинов называют α, δ, ε, γ и µ соответственно. Антитело в соответствии с изобретением предпочтительно включает участок Fc человеческого происхождения, который принадлежит к подтипу IgG1 или IgG4.
Константные легкие и тяжелые цепи человека и константные цепи подтипа IgG1 или IgG4 хорошо известны на уровне техники и описаны, например, Кэботом (см., например, Johnson, G. and Wu, T.T., Nucleic Acids Res. 28 (2000) 214-218). Например, полезная константная область тяжелой цепи человека включает аминокислотную последовательность SEQ ID NO:13 или 14 (IgG1) или SEQ ID NO:15 или 16 (IgG4). Например, полезная константная область легкой цепи человека включает аминокислотную последовательность константную область легкой цепи каппа SEQ ID NO:11. Кроме того, предпочтительно, чтобы антитело имело мышиное происхождение и включало каркас вариабельной последовательности антитела мышиного антитела в соответствии с Кэботом (см., например, Sequences of Proteins of Immunological Interest, Kabat, E.A. et al., 5th edition, DIANE Publishing (1992)).
Изобретение включает способ лечения пациента, нуждающегося в терапии, характеризующийся введением этому пациенту антитела в соответствии с изобретением.
Изобретение включает применение антитела в соответствии с изобретением для терапии.
Изобретение включает применение антитела в соответствии с изобретением для получения лекарства для лечения таупатии, в частности БА.
Изобретение включает применение антитела в соответствии с изобретением для лечения заболеваний головного мозга, предпочтительно для лечения таупатии, в частности БА.
Следующей формой осуществления изобретения является способ продуцирования антитела в соответствии с изобретением, характеризующийся тем, что последовательность нуклеиновой кислоты, кодирующую тяжелую цепь антитела в соответствии с изобретением, и последовательность нуклеиновой кислоты, кодирующую легкую цепь этого антитела, встраивают в один или в два экспрессионных вектора, этот вектор (векторы) встраивают в эукариотическую клетку-хозяина, и кодируемое антитело экспрессируют и выделяют из клетки-хозяина или из супернатанта.
Антитела в соответствии с изобретением предпочтительно продуцируют рекомбинантными способами. Такие способы широко известны на уровне техники и включают экспрессию белка в прокариотических и эукариотических клетках с последующим выделением полипептида антитела и обычно очисткой до фармацевтически приемлемой чистоты. Для экспрессии белка нуклеиновые кислоты, кодирующие легкие и тяжелые цепи или их фрагменты, встраивают в экспрессионные векторы стандартными способами. Экспрессию осуществляют в подходящих прокариотических или эукариотических клетках-хозяевах, таких как клетки СНО, клетки NSO, клетки SP2/0, клетки НЕК293, клетки COS, дрожжи или клетки Е. coli, и антитело выделяют из клеток (из супернатанта или после лизиса клеток).
Рекомбинантное продуцирование антител описано, например, в обзорных статьях Makrides, S.C., Protein Expr. Purif. 17 (1999) 183-202; Geisse, S. et al., Protein Expr. Purif. 8 (1996) 271-282; Kaufman, R.J., Mol. Biotechnol. 16 (2000) 151-160; Werner, R.G., Drug Res. 48 (1998) 870-880.
Антитела могут присутствовать в целых клетках, в клеточном лизате или в частично очищенной или по существу чистой форме. Очистку осуществляют с целью устранения других клеточных компонентов или других контаминантов, например, других клеточных нуклеиновых кислот или белков, стандартными методами (см. Ausubel, F. et al. (eds.), Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York (1987)).
Экспрессия в клетках NSO описана, например, Barnes, L.M. et al., Cytotechnology 32 (2000) 109-123; Barnes, L.M. et al., Biotech. Bioeng. 73 (2001) 261-270. Транзитная экспрессия описана, например, Durocher, Y. et al., Nucl. Acids. Res. 30 (2002) E9. Клонирование вариабельных доменов описано Orlandi, R. et al. Proc. Natl. Acad. Sci. USA 86 (1989) 3833-3837; Carter, P. et al., Proc. Natl. Acad. Sci. USA 89 (1992) 4285-4289; Norderhaug, L. et al., J. Immunol. Methods 204 (1997) 77-87. Предпочтительная система транзитной экспрессии (НЕК 293) описана Schlaeger, E.-J. и Christensen, К. в Cytotechnology 30 (1999) 71-83, и Schlaeger, E.-J. в J. Immunol. Methods 194 (1996) 191-199.
Моноклональные антитела целесообразно выделять из культуральной среды общепринятыми методами очистки иммуноглобулинов, такими как, например, белок А-сефароза, хроматография на гидроксиапатите, гель-электрофорез, диализ или аффинная хроматография. ДНК и РНК, кодирующую моноклональные антитела, легко выделяют и секвенируют, используя общепринятые методы. Клетки гибридомы могут служить в качестве источника такой ДНК и РНК. Когда ДНК выделена, она может быть встроена в экспрессионные векторы, которыми затем трансфицируют клетки-хозяева, такие как клетки НЕК 293, клетки СНО или клетки миеломы, которые в других обстоятельствах не продуцируют белок иммуноглобулин, чтобы получить синтез рекомбинантных моноклональных антител в клетках-хозяевах.
Молекулы нуклеиновых кислот, кодирующие варианты аминокислотной последовательности антитела анти-pS422, получают с помощью ряда способов, известных в данной области техники. Эти способы включают, но не ограничены ими, выделение из природного источника (в случае встречающихся в природе вариантов аминокислотной последовательности) или получение с помощью опосредованного олигонуклеотидами (или сайт-направленного) мутагенеза, ПЦР мутагенеза и кассетного мутагенеза полученного ранее варианта или невариантной версии гуманизированного антитела анти-pS422.
Вариабельные домены тяжелой и легкой цепи в соответствии с изобретением объединяют с последовательностями промотора, инициации трансляции, константной области, 3' нетранслируемой области, полиаденилирования и терминации транскрипции с образованием экспрессионных векторных конструкций. Экспрессионные конструкции тяжелой и легкой цепи могут быть объединены в одном векторе, котрансфицированы, серийно трансфицированы или трансфицированы по отдельности в клетки-хозяева, которые затем сливают с образованием одной клетки-хозяина, экспрессирующей обе цепи.
В другом аспекте в настоящем изобретении предложена композиция, например, фармацевтическая композиция, содержащая моноклональное антитело или их комбинацию, либо его антигенсвязывающий участок по настоящему изобретению, включенные в композицию вместе с фармацевтически приемлемым носителем.
Как используют в данной заявке, "фармацевтически приемлемый носитель" включает любое или все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, изотонические и замедляющие абсорбцию/резорбцию агенты и тому подобное, которые являются физиологически совместимыми. Предпочтительно носитель пригоден для инъекции или инфузии.
Композицию по настоящему изобретению можно вводить с помощью ряда способов, известных в данной области техники. Как понятно специалистам в данной области техники, путь и/или способ введения будет варьировать в зависимости от желаемых результатов.
Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для приготовления стерильных инъекционных растворов или дисперсии. Применение таких сред и агентов для фармацевтически активных веществ известно в данной области техники. Кроме воды, носитель может представлять собой, например, изотонический забуференный физиологический раствор.
Независимо от выбранного пути введения соединения по настоящему изобретению, которые можно применять в подходящей гидратированной форме, и/или фармацевтические композиции по настоящему изобретению готовят в фармацевтически приемлемых лекарственных формах общепринятыми способами, известными специалистам в данной области техники.
Действительные уровни дозировки активных ингредиентов в фармацевтических композициях по настоящему изобретению можно варьировать таким образом, чтобы получить количество активного ингредиента, которое эффективно для достижения желаемого терапевтического ответа для конкретного пациента, композиции и способа введения, но нетоксично для пациента (эффективное количество). Выбранный уровень дозировки будет зависеть от ряда фармакокинетических факторов, включающих активность конкретных применяемых композиций по настоящему изобретению или их эфира, соли или амида, путь введения, время введения, скорость выведения конкретного применяемого соединения, другие лекарства, соединения и/или вещества, используемые в комбинации с конкретными применяемыми композициями, возраст, пол, масса, состояние, общее состояние здоровья и предшествующий анамнез заболевания пациента, подлежащего лечению, и тому подобные факторы, хорошо известные в области медицины.
Изобретение включает применение антител в соответствии с изобретением для лечения пациента, страдающего таупатией, в частности БА.
Изобретение также включает способ лечения пациента, страдающего таким заболеванием, путем введения этому пациенту антитела в соответствии с изобретением.
Далее в изобретении предложен способ получения фармацевтической композиции, содержащей антитело в соответствии с изобретением вместе с фармацевтически приемлемым носителем, и применение антитела в соответствии с изобретением для такого способа.
Далее в изобретении предложено применение антитела в соответствии с изобретением для получения фармацевтического агента, предпочтительно вместе с фармацевтически приемлемым носителем, для лечения пациента, страдающего таупатией, в частности БА.
В изобретении также предложено применение антитела в соответствии с изобретением для получения фармацевтического агента, предпочтительно вместе с фармацевтически приемлемым носителем, для лечения пациента, страдающего раком, в частности, таупатией, в частности, БА.
Приведенные ниже примеры, перечень последовательностей и графические материалы предложены, чтобы способствовать пониманию изобретения, действительный объем которого изложен в прилагаемой формуле изобретения. Понятно, что могут быть произведены модификации в изложенных методах без отклонения от сущности изобретения.
Описание перечня последовательностей
SEQ ID NO:1 Вариабельная легкая цепь Мао 2.10.3
SEQ ID NO:2 Вариабельная тяжелая цепь Mab 2.10.3
SEQ ID NO:3 CDR1L
SEQ ID NO:4 CDR2L
SEQ ID NO:5 CDR3L
SEQ ID NO:6 CDR1H
SEQ ID NO:7 CDR2H
SEQ ID NO:8 CDR3H
SEQ ID NO:9 Фосфорилированный фрагмент Tau
SEQ ID NO:10 Нефосфорилированный SEQ ID NO:9
SEQ ID NO:11 Константная область легкой цепи каппа человека
SEQ ID NO:12 Константная область легкой цепи каппа человека
SEQ ID NO:13 Константная область γ1 человека (аллотип G1m1,17)
SEQ ID NO:14 Константная область γ1 человека (аллотип G1m17)
SEQ ID NO:15 Константная область IgG4 человека
SEQ ID NO:16 Константная область IgG4 человека SPLE-мутант
SEQ ID NO:17 Фосфорилированный фрагмент MCAK fragment (MCAK_Human (88-102)[95-pSer]
SEQ ID NO:18 Праймер
SEQ ID NO:19 Праймер
SEQ ID NO:20 Праймер
SEQ ID NO:21 Праймер
SEQ ID NO:22 Вариабельная тяжелая цепь Mab 005
SEQ ID NO:23 CDRH1
SEQ ID NO:24 CDRH2
SEQ ID NO:25 CDRH3
SEQ ID NO:26 Вариабельная легкая цепь Mab 005
SEQ ID NO:27 CDRL1
SEQ ID NO:28 CDRL2
SEQ ID NO:29 CDRL3
SEQ ID NO:30 Вариабельная тяжелая цепь Mab 019
SEQ ID NO:31 CDRH1
SEQ ID NO:32 CDRH2
SEQ ID NO:33 CDRH3
SEQ ID NO:34 Вариабельная легкая цепь Mab 019
SEQ ID NO:35 CDRL1
SEQ ID NO:36 CDRL2
SEQ ID NO:37 CDRL3
SEQ ID NO:38 Вариабельная тяжелая цепь Mab 020
SEQ ID NO:39 CDRH1
SEQ ID NO:40 CDRH2
SEQ ID NO:41 CDRH3
SEQ ID NO:42 Вариабельная легкая цепь Mab 020
SEQ ID NO:43 CDRL1
SEQ ID NO:44 CDRL2
SEQ ID NO:45 CDRL3
SEQ ID NO:46 Вариабельная тяжелая цепь Mab 085
SEQ ID NO:47 CDRH1
SEQ ID NO:48 CDRH2
SEQ ID NO:49 CDRH3
SEQ ID NO:50 Вариабельная легкая цепь Mab 085
SEQ ID NO:51 CDRL1
SEQ ID NO:52 CDRL2
SEQ ID NO:53 CDRL3
SEQ ID NO:54 Вариабельная тяжелая цепь Mab 086
SEQ ID NO:55 CDRH1
SEQ ID NO:56 CDRH2
SEQ ID NO:57 CDRH3
SEQ ID NO:58 Вариабельная легкая цепь Mab 086
SEQ ID NO:59 CDRL1
SEQ ID NO:60 CDRL2
SEQ ID NO:61 CDRL3
SEQ ID NO:62 Вариабельная тяжелая цепь Mab 097
SEQ ID NO:63 CDRH1
SEQ ID NO:64 CDRH2
SEQ ID NO:65 CDRH3
SEQ ID NO:66 Вариабельная легкая цепь Mab 097
SEQ ID NO:67 CDRL1
SEQ ID NO:68 CDRL2
SEQ ID NO:69 CDRL3
SEQ ID NO:70 Константная область иммуноглобулина кролика (каппа)
SEQ ID NO:71 Константная область иммуноглобулина кролика (гамма)
SEQ ID NO:72 Константная область иммуноглобулина мыши (каппа)
SEQ ID NO:73 Константная область иммуноглобулина мыши (гамма)
Описание графических материалов
Фиг.1: Интрацеребральная локализация pS422 IgG1 после ip введения в мышиной модели TauPS2APP. Конфокальные микрофотографии после множественного флуоресцентного окрашивания первыми антителами против IgG1, конъюгированными с Alexa488, и антителом анти-pTau АТ8, конъюгированным с Alexa555, выявили связанный IgG1 (А) и отложения pTau (В), вместе с DAPI для клеточных ядер (С). Совмещенные изображения показывают
совместно локализованное окрашивание анти-Igd и pTau в некоторых pTau-положительных клетках (D).
Фиг.2: Контроль носителя интрацеребральной локализации IgG1 в мышиной модели TauPS2APP. Конфокальные микрофотографии после множественного флуоресцентного окрашивания среза головного мозга TauPS2APP, показывающие обзор зоны образования СА1 гиппокампа. Множественное окрашивание первыми антителами против IgG1, конъюгированными с Alexa488, отрицательно (А). Антитело анти-pTau АТ8, конъюгированное с Alexa555, выявило отложения pTau (В). Окрашивание DAPI на клеточные ядра (С). Совмещенные изображения показаны в (D). Приемлемой иммунофлуоресценции анти-IgG1 не наблюдали у мышей TauPS2APP после инъекции носителя.
Фиг.3: Внутриклеточная локализация pS422 IgG1 после ip введения в мышиной модели TauPS2APP. Конфокальные микрофотографии после множественного флуоресцентного окрашивания среза головного мозга TauPS2APP, показывающие положительно меченые клетки в пределах префронтальной коры. Множественное окрашивание первыми антителами против IgG1, конъюгированными с Alexa488, и антителом анти-pTau АТ8, конъюгированным с Alexa555, выявило связанный IgG1 (А) и отложения pTau (В). Совмещенные изображения показывают совместно локализованное перинуклеарное и дендритное окрашивание анти-IgG1 и pTau вместе с окрашиванием DAPI на клеточные ядра синим цветом (С).
Фиг 4: Внутриклеточная локализация pS422 IgG1 после ip введения в мышиной модели TauPS2APP. Конфокальные микрофотографии после множественного флуоресцентного окрашивания среза головного мозга TauPS2APP, показывающие положительно меченые клетки в пределах пирамидального слоя зоны СА1 гиппокампа. Множественное окрашивание первыми антителами против IgG1, конъюгированными с Alexa488, и антителом анти-pTau АТ8, конъюгированным с Alexa555, выявило связанный IgG1 (А) и отложения pTau (В). Совмещенные изображения показывают совместно локализованное перинуклеарное и дендритное окрашивание анти-IgG1 и pTau вместе с окрашиванием DAPI на клеточные ядра (С).
Фиг.5: Внутриклеточная локализация pS422 IgG2a после ip введения в мышиной модели TauPS2APP. Конфокальные микрофотографии после множественного флуоресцентного окрашивания среза головного мозга TauPS2APP, показывающие положительно меченые клетки в пределах пирамидального слоя зоны СА1 гиппокампа. Множественное окрашивание первыми антителами против IgG2a, конъюгированными с Alexa488, и антителом анти-pTau AT8, конъюгированным с Alexa555, выявило связанный IgG2a (А) и отложения pTau (В). Совмещенные изображения показывают совместно локализованное перинуклеарное окрашивание анти-IgG2a и pTau вместе с окрашиванием DAPI на клеточные ядра (С).
Фиг.6: Внутриклеточная локализация pS422 IgG2b после ip введения в мышиной модели TauPS2APP. Конфокальные микрофотографии после множественного флуоресцентного окрашивания среза головного мозга TauPS2APP, показывающие положительно меченые клетки в пределах зоны префронтальной коры. Множественное окрашивание первыми антителами против IgG2b, конъюгированными с Alexa488, и антителом анти-pTau AT8, конъюгированным с Alexa555, выявило связанный IgG2b (А) и отложения pTau (В). Совмещенные изображения показывают совместно локализованное перинуклеарное окрашивание анти-IgG2b и pTau, указанное стрелками (С).
Фиг.7: Анализ мыши TauPS2APP. А: Окрашивание серебром по Gallyas сагиттального среза головного мозга 16-месячной мыши TauPS2APP. Типичные клубковые интранейрональные структуры видны в черном цвете и подтверждают положительные нейроны pTau, выявленные иммунофлуоресцентной микроскопией. В: Меченый иммунным окрашиванием золотом ультратонкий срез 16-месячной мыши TauPS2APP. Анти-Tau pS422 mAb специфично связывается с фибриллярными структурами в дендритных отростках в зоне СА1 гиппокампа, как выявлено вторым антителом, конъюгированным с 10 нм коллоидным золотом (стрелка). Размер и плотность меченых фибрилл позволяет предположить, что они сравнимы с парными спиральными филаментами фосфорилированных белков Tau, обнаруженными в дегенерирующих нейронах при болезни Альцгеймера.
Фиг.8: Связывание in vitro антител анти-Tau pS422 со срезами головного мозга БА. Флуоресцентные микрофотографии кортикальных срезов головного мозга человека с БА с моноклональными антителами анти-Tau pS422 (клоны 2.10.3, 2.20.4 и 5.6.11) при указанной концентрации, показывающие положительно меченые отложения pTau. Заметны внутриклеточные отложения pTau, подобные большим нейрофибриллярным клубкам и продольным нитям нейропиля.
Фиг.9: Селективность антител анти-Tau pS422 для Tau pS422 по сравнению с Tau. Измерение с помощью ELISA селективности антител анти-Tau pS422 с использованием планшетов, покрытых Tau (▲) или Tau pS422 (●). Для сравнения Tau-селективное антитело 4/2 показано на верхней левой панели.
Фиг.10: In vitro агрегированный Tau является фибриллярным. Электронная микрофотография отрицательно окрашенного агрегированного Tau.
Фиг.11: Флуоресцентные микрофотографии кортикальных срезов головного мозга человека с БА, окрашенные моноклональными антителами кролика анти-Tau pS422 Mab 005 (первый ряд), Mab 019 (второй ряд), Mab 020 (третий ряд), Mab 085 (четвертый ряд), Mab 086 (пятый ряд) и Mab 097 (шестой ряд), показывающие положительно меченые отложения и филаменты pTau. Серийное разведение IgG показано в каждом ряду, начиная с 2,0 мкг/мл (слева), 0,4 мкг/мл, 0,08 мкг/мл и 0,016 мкг/мл.
Пример 1
Получение и очистка антител
a) Получение антитела
Мышей иммунизировали фрагментом Tau SEQ ID NO:9 (Ser-Ile-Asp-Met-Val-Asp-Ser(PO3H2)-Pro-Gln-Leu-Ala-Thr-Leu-Ala-Asp), который соответствует аминокислотам 416-430 самой длинной изоформы Tau человека. Чтобы дать возможность прямого сочетания посредством тиола с KLH, добавляли цистеин к N-концу фрагмента Tau. Протокол последующей иммунизации, слияния и клонирования и скрининга на специфичные антитела анти-Tau pS422 описан в ЕР 1876185.
b) Очистка клонов 2.10.3, 2.20.4 и 5.6,11
Бесклеточный супернатант культуры гибридомы (250-300 мл) наносили на колонку 25 мл МЕР Hypercell (pall Biosciences), которая была уравновешена 50 мМ Трис-Cl рН 8,0. После промывания буфером для уравновешивания антитело элюировали 30 мМ цитратом натрия, 100 мМ NaCl рН 4,1. Фракции, содержащие антитело, объединяли, а затем подвергали диализу в диализных трубках Spectra-Рог 6-8000 в течение ночи при 4°С против 5 литров 10 мМ Трис-Cl рН 8,0. Диализованный материал наносили на колонкуЮ мл Source 17Q (GE Healthcare), которая была уравновешена в 10 мМ Трис-Cl рН 8,0 (буфер А). После промывания буфером А антитело элюировали градиентом 0-25% буфера В в 10 объемах колонки. Буфер В содержал 10 мМ Трис-Cl, 1 М NaCl рН 8,0. Антитело элюировало примерно при 200 мМ NaCl. Чистоту индивидуальных фракций проверяли с помощью электрофореза в ДСН-ПААГ, и самые чистые фракции объединяли.
За сутки до инъекции мышам каждое антитело подвергали диализу против ФСБ, и концентрацию антитела доводили до 3,2 мг/мл.
Пример 2
Получение Tau, Tau pS422, агрегированного Tau и агрегированного Tau pS422
Tau, содержащий N-концевую слитую метку (His)6-SUMO, экспрессировали в E. coli и очищали ионообменной хроматографией на HiTrapQ (GE Healthcare, Switzerland) с последующей аффинной хроматографией на сефарозе NI-NTA (Qiagen, Switzerland). Затем слитую метку отщепляли расщеплением протеазой SUMO (Invitrogen, Netherlands) с последующей второй стадией хроматографии на сефарозе Ni-NTA для удаления слитой метки.
Tau-pSer422 получали путем инкубации Tau с протеинкиназой ERK2. Молярное отношение ERK2:Tau (примерно 1:10) было выбрано таким образом, чтобы обеспечить максимальный выход фосфорилирования по S422, с последующей инкубацией в течение ночи при 37°С в 10 мМ Трис-Cl рН 8,0, содержащем 1 мМ MgCl2 и 2 мМ АТФ. Затем предположили, что это является стехиометрическим фосфорилированием по сайту 422. Фосфорилирование проверяли с помощью Вестерн-блоттинга, используя собственное моноклональное антитело, специфичное к pS422 в Tau.
Агрегированный (фибриллярный) Tau получали путем инкубации очищенного Tau при конечной концентрации 5 мкМ в 10 мМ Трис-Cl рН 8,0, содержащем 50 мкМ арахидоновую кислоту (Sigma, Switzerland). Инкубацию проводили при 37°С в течение 16 ч. Состояние агрегации проверяли флуоресцентной спектроскопией в присутствии 10 мкМ Thio S (Barghorn and Mandelkow [2002] Biochemistry 41:14885-14896) и электронной микроскопией (фиг.10). Как показано на фиг.10, агрегированный Tau имеет фибриллярный внешний вид. Агрегированный, фосфорилированный Tau получали путем инкубации предварительно агрегированного Tau с киназой ERK2, как описано выше.
Пример 3
Моноклональные антитела анти-Tau pS422 обладают высокой селективностью к Tau, фосфорилированному по S422
а) Пептидный синтез
Пептидные синтезы проводили в автоматическом синтезаторе пептидов, используя химию Fmoc. В повторяющихся циклах пептидные последовательности собирали путем последовательного сочетания соответствующих Fmoc-аминокислот. На каждой стадии сочетания N-концевую Fmoc-группу удаляли обработкой смолы 20% пиперидином в N-метилпирролидоне. Сочетания проводили, используя Fmoc-защищенную аминокислоту (1 ммоль), активированную HBTU/HOBt (1 ммоль каждого) и DIPEA (2 ммоль) в ДМФ. После каждой стадии сочетания не прореагировавшие аминогруппы копировали обработкой смесью уксусной кислоты (0,5 М), DIPEA (0,125 М) и HOBt (0,015 М) в NMP (10 мин вортекс). Между каждыми стадиями смолу промывали N-метилпирролидоном и ДМФ. Включение стерически затрудненных аминокислот осуществляли в автоматических двойных сочетаниях. Для этой цели смолу дважды обрабатывали 1 ммоль активированного строительного блока без стадии копирования между циклами сочетания. В фосфорилированных пептидных последовательностях соответствующее производное серина включали в виде строительного блока Fmoc-Ser(PO(OBzl)OH)-OH. После завершения целевых последовательностей Fmoc-Glu(биотинил-ПЭГ)-ОН (биотин, присоединенный через ПЭГ спейсер) сочетали с пептидом MCAK, используя стандартные условия сочетания аминокислот, тогда как 4× β-аланин (U) и 1× ε-лизин присоединяли к последовательностям фосфо-Tau, используя стандартные условия. Затем DMTr-биотин конъюгировали с пептидами фосфо-Tau с получением биотинилированных целевых последовательностей. После конечного удаления защиты Fmoc (только для MCAK) все пептидные смолы помещали по отдельности в фильтровальные фритты и обрабатывали смесью трифторуксусной кислоты, воды и триизопропилсилана (19 мл:0,5 мл:0,5 мл) в течение 2,5 ч. Растворы для отщепления фильтровали, и пептиды осаждали добавлением холодного (0°С) диизопропилового эфира (300 мл) с получением бесцветных твердых веществ, которые неоднократно промывали диизопропиловым эфиром. Сырые продукты повторно растворяли в смеси уксусная кислота/вода, лиофилизировали, а затем очищали препаративной ВЭЖХ с обращенной фазой, используя градиент ацетонитрил/вода, содержащий 0,1% ТФУ.
HBTU: 2-(1Н-бензотриазол-1-ил)-1,1,3,3-тетраметилурония гексафторфосфат
HOBt: Гидроксибензотриазол
DIPEA: N,N-Диизопропилэтиламин
NMP: N-метил-2-пирролидон
b) Анализ с фосфорилированным пептидом Tau
Пептиды, представляющие последовательность Tau pS422 416-430 (фосфорилированного и нефосфорилированного) и MCAK_Human (88-102)[95-pSer], синтезировали и биотинилировали, чтобы дать возможность покрытия микротитрационного планшета, меченого стрептавидином. Для тестирования на максимальное связывание фосфопептида Tau с аналитическими планшетами различные концентрации фосфопептида Tau в диапазоне от 1 нг/мл до 2000 нг/мл использовали для покрытия. Наконец, 50 нг/мл фосфопептида Tau использовали для покрытия в течение 60 мин при комнатной температуре. Антитела анти-Tau pS422 2.10.3., 2.20.4 и 5.6.11 инкубировали в меченых пептидами микротитрационных планшетах в течение 60 мин при концентрациях вплоть до 1000 нг/мл. После промывания связывание антител обнаруживали, используя антитело IgG Fc против мыши, которое было меченым POD. После инкубации с ABTS® в течение 20 мин при комнатной температуре измеряли поглощение (O.D.) при 405-492 нм. Связывание антитела определяли по EC50. Типичное значение ЕС50 можно было определить в диапазоне от 100 нг/мл для 2.10.3., 8 нг/мл для 2.20.4 и 1 нг/мл для 5.6.11. Нефосфорилированный фрагмент Tau и (MCAK_Human (88-102)[95-pSer]) использовали в качестве контролей. Связывание контрольных пептидов не наблюдалось. Типичные фоновые значения составляли около 30 mE (таблица 1), что составляет примерно 1% максимального значения, измеренного с фосфорилированным фрагментом Tau. Следовательно, Mab2.10.3 связывается с фосфорилированным фрагментом Tau по меньшей мере с 100-кратной селективностью по сравнению с нефосфорилированным пептидом Tau SEQ ID NO:10 и с MCAKJ-fuman (88-102)[95-pSer]. Результаты представлены в таблице 1.
с) Анализ с полноразмерным Tau и Tau pS422
Анализы проводили, используя аналитическую платформу Meso Scale Discovery™ (MSD, Gaithersburg, Maryland, USAMD). 96-луночные микротитрационные планшеты MSD покрывали Tau или Tau pS422 при концентрации 2 мкг/мл в буфере ФСБ в течение 1 ч при комнатной температуре. Затем планшеты блокировали добавлением ФСБ, содержащего 5% БСА и 1% Твин 20, в течение 1 ч. Антитела разводили в буфере Low Cross Buffer (MSD), содержащем 0,1% БСА и 0,1% Твин®20, при концентрации от 30 пг/мл до 20 нг/мл (за исключением клона 2.10.3, где диапазон концентрации составлял от 3 нг/мл до 250 нг/мл) и добавляли в покрытые, блокированные планшеты и инкубировали в течение 3 ч при комнатной температуре. Затем планшеты промывали три раза буфером ФСБ, содержащим 1% Твин. Для обнаружения связанного антитела SULFO-меченый IgG против мыши (MSD) добавляли при концентрации 0,5 мкг/мл, и планшеты инкубировали в течение 1 ч при комнатной температуре. После добавления буфера Read Buffer (MSD) планшеты считывали в считывающем устройстве для планшетов MSD Sector Imager 6000. На фиг.9 показаны данные, полученные с использованием либо Tau, либо Tau pS422 в качестве партнера связывания для моноклональных антител анти-Tau pS422 (клон 5.6.11, 2.20.4 и 2.10.3). Антитела Tau pS422 сравнивали в качестве контроля с антителом анти-Tau (клон 4/2), которое распознает и Tau, и Tau pS422. Путем сравнения связывания с Tau pS422 и Tau было обнаружено, что сигналы связывания с Tau были неотличимы от фоновых сигналов, и, следовательно, связывание с Tau не было обнаружено. Таким образом, было вычислено, что каждое из трех антител анти-Tau pS422 связывается с Tau pS422 по меньшей мере с 10000-кратной селективностью по сравнению с Tau.
Пример 4
Антитело анти-Tau pS422 связывается с фибриллярными агрегатами Tau pS422
Анализ Biacore™ использовали для измерения связывания анти-Tau pS422 (клон 2.10.3) с препаратами агрегированного, фибриллярного Tau pS422. Фосфорилированные фибриллы считают представлением in vitro PHF формы Tau, которая встречается в головном мозге при болезни Альцгеймера. Фибриллярный материал был получен и фосфорилирован, как описано в Примере 2.
Иммобилизация на сенсорных чипах:
Агрегированный Tau pS422 иммобилизовали ковалентно путем аминного сочетания на СМ5-сенсорном чипе. С этой целью агрегированный Tau pS422 разводили в 19 раз в 10 мМ ацетате натрия рН 5,0. Активированную поверхность сенсора приводили в контакт с раствором белка до иммобилизации желаемого уровня белка на чипе. Затем оставшиеся реакционные сложные эфиры N-гидроксисукцинимиды блокировали путем впрыскивания 1 М этаноламина рН 8,0. Было иммобилизовано 450 единиц реакции (RU) белка, чтобы охарактеризовать поведение связывания анти-Tau pS422 (клон 2.10.3). Анализ иммобилизации проводили при КГ, и ФСБ использовали в качестве буфера пробега.
Кинетическое титрование:
Для характеризации антитела буфер пробега заменяли буфером 10 мМ Трис-Cl рН 8,0, 250 мМ NaCl и готовили серию концентраций антитела: 5 различных концентраций, коэффициент разведения 2, максимальная концентрация 200 нМ. Различные растворы антитела впрыскивали последовательно, начиная с самой низкой концентрации. Поверхность сенсора регенерировали 100 мМ H3PO4 после так называемого кинетического титрования. Измерения проводили три раза.
Оценки данных осуществляли с использованием программного обеспечения Biaeval. Вычисление параметров основано на допущении, что связывание антитела агрегированным Tau pS422 является полностью моновалентным. Кинетические и термодинамические параметры, характеризующие связывающее поведение антитела с иммобилизованным белком, представлены в таблице 2.
Таким образом, анти-Tau pS422 (клон 2.10.3) связывается с фибриллярным фосфо-Tau с KD 17 нМ.
Пример 5
Связывание in vitro моноклональных антител анти-Tau pS422 с внутриклеточным pTau в срезах головного мозга пациента с болезнью Альцгеймера
Специфичное и чувствительное иммунологическое окрашивание патологии патологически релевантного pTau в ткани головного мозга при болезни Альцгеймера исследовали с помощью экспериментов по иммуногистохимическому окрашиванию, используя срезы замороженной ткани головного мозга человека от пациентов БА. Оценивание осуществляли, используя замороженную посмертную ткань головного мозга от пациентов БА, у которых нейропатологическим путем была установлена стадия VI по Браак. Срезы замороженной ткани головного мозга получали и обрабатывали для иммуногистохимии без каких-либо химических реактивов, то есть альдегидной фиксацией. Обнаружение отложений pTau осуществляли, используя второе антитело, меченное флуоресцентной меткой, специфичное к IgG мыши, и мониторинг осуществляли с помощью флуоресцентной световой микроскопии.
Кратко, срезы нефиксированной ткани головного мозга, полученные на криостате, из зон коры головного мозга, полученного посмертно от пациентов, у которых была положительно диагностирована болезнь Альцгеймера, метили с помощью непрямой иммунофлуоресценции. В данном описании показаны изображения, полученные с ткани орбитальной фронтальной извилины от пациента БА (пациент 01-05; внутренний банк головного мозга). Последовательную инкубацию в две стадии использовали для обнаружения связанных мышиных антител из различных клонов, которые выявляли с помощью очищенного аффинной хроматографией IgG (H+L) коза против мыши, конъюгированного с Cy3 (Jackson Immuno Research). Получение срезов, окрашивание и флуоресцентную микроскопию осуществляли в соответствии со стандартными методиками.
Специфичное и чувствительное окрашивание патологии pTau очевидно для клонов 2.10.3, 2.20.4 и 5.6.11 (фиг.8). Четко видно насыщенное окрашивание всех типов отложений pTau, а именно нейрофибриллярных клубков, нитей нейропиля и дистрофических нейритов. Минимальная эффективная концентрация 0,05 мкг/мл была определена для всех исследованных клонов, что указывает на высокочувствительное связывание с генуинными отложениями pTau человека.
Обнаружение отложений pTau можно также осуществлять путем использования антитела IgG4 в соответствии с изобретением и меченого флуоресцентной меткой второго антитела, специфичного к IgG4 человека, и осуществлять мониторинг с помощью флуоресцентной световой микроскопии.
Пример 6
Получение тройных трансгенных мышей TauPS2APP
Трансгенных мышей TauPS2APP получали скрещиванием гомозиготных самок PS2APP из линии В6.172Н (Ozmen, L. et al., Neurodegen. Dis. 6 (2009) 29-36) с гомозиготными самцами Tau из линии TauP301L (Goetz et al., Science 293 (2001) 1491-1495). Все полученные в результате трансгенные мыши гетерозиготны по мутации АРР шведский, мутации пресенилина 2 N1411 человека и мутации Tau P301L. Экспрессия трансгенов управляется мышиным промотором Thy-1 в случае АРР человека, а экспрессия трансгенов Tau-прионным промотором в случае трансгена пресенилина 2 человека.
Животных держали при 12-часовом световом цикле, и они получали свободный доступ к корму и воде. Виварий аккредитован Аккредитацией содержания лабораторных животных. Все процедуры проводили, строго придерживаясь швейцарских федеральных законов по защите животных и правил Ассоциации по аттестации и аккредитации содержания лабораторных животных и при личном одобрении местной ветеринарной службы.
Обработка мышей для иммуногистохимического анализа: мышей анестезировали, используя 4% изофлуран, и умерщвляли путем декапитации. Кровь собирали в пробирки, покрытые ЭДТА-NaF (Milian). Головной мозг извлекали, быстро замораживали на сухом льду и хранили при -80°С до дальнейшего использования.
В экспериментах использовали самцов и самок мышей в возрасте 20 месяцев. В этом возрасте усиленное внутриклеточное накопление гиперфосфорилированного Tau и нейрофибриллярная патология четко видны в головном мозге от всех мышей, протестированных к настоящему времени.
Пример 7
Гистология и ультраструктурный анализ отложений pTau в мышиной модели TauPS2APP
У мышей TauPS2APP развивается зависимый от возраста и прогрессирующий фенотип с интранейрональными отложениями pTau со структурами, подобными нейрофибриллярным клубкам, обнаружимыми в нескольких зонах головного мозга. Существенную патологию pTau наблюдают в возрасте 24 месяцев. Типично несколько нейронов, которые являются положительными по внутриклеточному Tau р8422, обнаруживают в пределах префронтальной коры и в слое пирамидальных клеток в зоне СА1 гиппокампа и в зоне, примыкающей к опорной структуре гиппокампа. Было обнаружено, что в этом возрасте pTau распространяется в полиморфный слой и вдоль границы желоба гиппокампа в зону СА3 и фимбрий, а также в краевую полоску.
Окрашивание серебром по Gallyas осуществляли, как описано в статье (Gallyas, F., Acta Morphologica Acad. Sci. Hung. 19 (1971) 1-8), с минорными модификациями, а именно с предварительной инкубацией в 3% периодной кислоте, и после отмывки в 0,5% уксусной кислоте с дополнительной отмывкой в 5% тиосульфате натрия в течение 2 мин. Контрастное окрашивание осуществляли стандартным гематоксилином и эозином. Окрашивание серебром по Gallyas подтвердило появление многочисленных клубковых отложений в гиппокампе и в коре головного мозга мышей TauPS2APP (фиг.7А).
Ультраструктуру отложений pTau исследовали с помощью иммуноэлектронной микроскопии. 16-месячных мышей TauPS2APP перфузировали 2% формальдегидом и 0,5% глутаральдегидом в ФСБ. Срезы головного мозга заключали в Lowicryl HM20 и получали ультратонкие срезы, как описано ранее (Richards, J.G. et al., J. Neurosci. 23 (2003) 8989-9003). Кратко, срезы инкубировали с анти-Tau pS422 при 10 мкг/мл в ФСБ с 2% БСА в течение 1 ч. После 6 отмывок в ФСБ/2% БСА срезы инкубировали со вторым антителом IgG коза против мыши (Amersham, Arlington Heights, IL), конъюгированным с 10 нм золотом, в соотношении 1:20 в ФСБ/2% БСА/0,1% Твин®20 в течение 1 ч и отмывали в ФСБ с 2% БСА. Для контролей авторы изобретения использовали срезы, обработанные нормальной мышиной сывороткой, что приводит в результате к пренебрежимо малому фону менее 10 частиц золота на площади 10 мкм2. Электронные микрофотографии снимали JEOL 1210 при 100 кВ.
Ультраструктурное исследование гиппокампа 16-месячных мышей TauPS2APP с многочисленными pTau-положительными нейронами выявило меченые иммунным окрашиванием золотом фибриллярные отложения, типичные для парных спиральных филаментов (PHF) внутри дендритных отростков (фиг.7В). Ультратонкие срезы, меченые анти-Tau pS422 mAb, выявили специфичное связывание с фибриллярными структурами внутри дендритных отростков в зоне СА1 гиппокампа, как выявлено с помощью второго антитела, конъюгированного с 10 нм коллоидным золотом, видимым в виде черных точек (фиг.7В). Мечение иммунным окрашиванием золотом демонстрирует, что эпитоп pS422 Tau локализован во внутриклеточных фибриллярных структурах в нейронах исследованных мышей TauPS2APP. Размер и плотность меченых фибрилл позволяет предположить, что они сравнимы по структуре с PHF, наблюдаемыми в дегенерирующих нейронах пациентов с БА. Локализация pTau PHF четко демонстрирует внутриклеточное распределение отложений агрегированного pTau у трансгенных мышей TauPS2APP.
Пример 8
Связывание in vivo моноклональных антител анти-Tau р8422 с внутриклеточным pTau в головном мозге мышей pTau мышиной модели болезни Альцгеймера, выявленное с помощью иммуногистохимии и конфокальной лазерной сканирующей микроскопии
Отобранные антитела анти-Tau pS422 (см. таблицу 1) тестировали у 24-месячных тройных трансгенных мышей TauPS2APP и оценивали на связывание с отложениями pTau in vivo. Исследования проводили на мышиной модели TauPS2APP, в которой развивается зависимый от возраста и прогрессирующий фенотип с интранейрональными отложениями pTau со структурами, подобными нейрофибриллярным клубкам, обнаружимыми в нескольких зонах головного мозга, как описано в Примере 4.
Используемые антитела против Tau pS422 вводили i.p. в дозе 20 мг/кг. Иммуногистохимические окрашивания осуществляли через двое суток после ip введения, чтобы обнаружить отложения pTau и связанные антитела подтипа IgG мыши. Группы по три мыши в каждой умерщвляли через двое суток после дозирования. Мышей подвергали глубокой анестезии (~2 минуты в 5% об./об. Forene™ почти до достижения состояния асфиксии), затем вскрывали грудную клетку, и перикардий извлекали и перфузировали ФСБ, и мышей подвергали декапитации, головной мозг делили пополам и быстро замораживали на сухом льду. Парасагиттальные срезы на криостате свежезамороженных головных мозгов резали при толщине 20 мкм криостатом (CM3050S, Leica), закрепляли на предварительно охлажденных стеклах Histobond (Marienfeld, Lauda-Konigshofen, Germany) и хранили при -20°С.
Тройное иммунофлуоресцентное окрашивание наносили, чтобы обнаружить связанные антитела анти-Tau pS422. Срезы гидратировали в ФСБ и обрабатывали 100% ацетоном, предварительно охлажденным до -20°С, в течение 2 мин. Все последующие стадии последовательно проводили в автомате для окрашивания (Autostainer Plus DakoCytomation, High Wycombe, UK) при комнатной температуре. Стекла со срезами головного мозга промывали ФСБ, содержащим 0,01% Твин 20, рН 7,4, в течение 5 минут и блокировали сайты неспецифического связывания последовательной инкубацией в ФСБ с 1% бычьим сывороточным альбумином, 1% овальбумином и 1% нормальной сывороткой козы в течение 20 минут. После отмывки ФСБ и 0,01% Твином 20 стекла инкубировали со специфичными к изотипу IgG идентифицирующими антителами, то есть очищенными аффинной хроматографией IgG1, IgG2a или IgG2b коза против мыши, ковалентно конъюгированными с красителем Alexa Fluor® 488 (А21121, А21131 или А21141, Molecular Probes), при 20 мкг/мл в 1% БСА в ФСБ, рН 7,4 в течение 1 часа. После отмывки ФСБ с 0,01% Твином 20 локализацию pTau оценивали с помощью мечения 5 мкг/мл антитела анти-pTau (АТ-8, Pierce Biotechnology), мышиного моноклонального антитела к фосфорилированному эпитопу S202/205 в белке Tau, которое было ковалентно конъюгировано с красителем Alexa Fluor 555, и наносили его в ФСБ с 1% бычьим сывороточным альбумином, 1% овальбумином и 1% нормальной сывороткой козы в течение 1 часа. После отмывки ФСБ и 0,01% Твином 20 клеточные ядра контрастно окрашивали 4,6'-диамидино-2-фенилидолом (DAPI) при 1 мкг/мл в ФСБ в течение 5 минут. После отмывки ФСБ и 0,01% Твином 20 собственную флуоресценцию липофусцина снижали гашением посредством инкубации в 4 мМ CuSO4 в 50 мМ ацетате аммония, рН 5 в течение 30 минут. После промывания стекол дважды дистиллированной водой и конечной отмывки ФСБ и 0,01% Твином®20 стекла заключали флуоресцентной заливочной средой (S3023 DakoCytomation, High Wycombe, UK). Изображения максимальной проекции снимали последовательно в не перекрывающихся каналах испускания с помощью конфокального лазерного сканирующего микроскопа Leica SP2, AOBS при установке объектива 1 Эри.
Существенная патология pTau отмечена у мышей TauPS2APP в возрасте 24 месяца. Типично несколько нейронов, которые имеют сильный положительный сигнал внутриклеточного Tau р3422, было обнаружено в пределах префронтальной коры и слоя пирамидальных клеток зоны СА1 гиппокампа и зоны, примыкающей к опорной структуре. Было обнаружено, что в этом возрасте pTau распространяется в полиморфный слой и вдоль границы желоба гиппокампа в зону СА3 и фимбрий, а также в краевую полоску.
Иммунологические окрашивания в зонах префронтальной коры и СА1 из гиппокампального образования у мышей TauPS2APP в возрасте 24 месяца показано в изображениях конфокальной микроскопии (фиг.1-6). Антитела к внутриклеточному pTau и введенные антитела на подтип IgG были четко обнаружимы и совместно локализованы в клетках зон головного мозга с существенной патологией pTau. Репрезентативные примеры внутриклеточной локализации IgG1 показаны на фиг.1, 3 и 4 с контролем-носителем (фиг.2). Клетки, положительные по IgG2a (фиг.5) и IgG2b (фиг.6), также были обнаружены в некоторых клетках. Некоторые из pTau-положительных клеток проявляли типичную нейрофибриллярную морфологию, которая напоминает нейрофибриллярные клубки, обнаруженные в дегенерирующих нейронах ткани головного мозга от пациентов с БА. Совместно локализованную иммунологическую реактивность антител подтипа IgG мыши, в частности для IgG1 и pTau, наблюдали в дендритных отростках, типично в префронтальной коре и слое гранулярных клеток в зоне СА1 гиппокампального образования (фиг.3 и 4).
Все протестированные антитела анти-Tau р8422 показали специфичное связывание с внутриклеточными отложениями pTau. Однако степень связывания варьировала между тремя изотипами IgG, как показано в таблице 3.
Пример 9
Хроническая обработка моноклональным антителом анти-Tau pS422 уменьшает патологию Tau в мышиной модели болезни Альцгеймера
Способность мышиного анти-Tau pS422 Mab к уменьшению патологии Tau у мышей в мышиной модели болезни Альцгеймера тестируют путем введения антитела тройным трансгенным мышам Tau.PS2APP в течение периода 4 месяца. Таким образом, группу из 20 тройных трансгенных мышей Tau.PS2APP в возрасте 7 месяцев обрабатывают ip введением один раз в неделю мышиного анти-Tau pS422 Mab (клон 2.10.3). Антитело вводят при концентрации 2 мг/мл в носителе (20 мМ гистидин, 140 мМ NaCl pH 6,0) с получением конечной дозы 20 мг/кг. Вторая группа из 20 тройных трансгенных мышей получает эквивалентный объем носителя. После 4 месяцев всех мышей умерщвляют и извлекают головной мозг, как описано в Примере 5. Затем каждый мозг делят на два полушария, одно из которых готовят для иммуногистохимического анализа, как описано в Примере 5. Второе полушарие используют для количественного определения уровней pTau путем иммунологического анализа, как описано ниже.
1. Экстракция головного мозга мыши для количественных анализов Tau и pTau
Головной мозг взвешивают и гомогенизируют стеклянным гомогенизатором в 10 объемах охлажденного во льду буфера, содержащего 25 мМ Трис-Cl, 800 мМ NaCl, 10% сахарозу, рН 7,5. Затем гомогенаты центрифугируют при 20000 х д. Полученные в результате супернатанты разводят ФСБ/1% БСА/1% Твин®20 1:100 (для анализов Tau) или 1:10 (для анализов pTau).
2. Количественные анализы Tau и pTau
Все анализы проводят, используя аналитическую платформу Meso Scale Discovery (MSD, Gaithersburg, Maryland, USAMD). Иммобилизующие и идентифицирующие антитела метят биотином и SULFO-Tag™, соответственно, в соответствии со стандартными протоколами, предложенными фирмой MSD. В анализах были использованы следующие антитела: (i) 5A6, мышиное антитело, направленное против аминокислот 19-46 в Tau, как описано в Johnson, G.V. et al., J. Neurochem 68 (1997) 430-433; (ii) AT180, мышиное антитело, направленное против рТ231 в Tau [Innogenetics, Belgium]; (iii) мышиное анти-Tau pS422 клон 2.5.2, как описано в ЕР1876185А1; (iv) мышиное анти-Tau клон 4/53, индуцированное против полноразмерного Tau. Антитело 5A6 используют в качестве иммобилизующего антитела, остальные антитела используют в качестве идентифицирующих антител.
96-луночные планшеты MSD, покрытые стрептавидином, блокируют в течение 1 ч при КГ 5% БСА. Планшеты отмывают три раза отмывочным буфером (ФСБ, 0,05% Твин®20). Затем добавляют иммобилизующее антитело, разведенное в аналитическом буфере (ФСБ/1% БСА/1% Твин®20), и планшеты инкубируют при 4°С в течение 30 минут. Затем планшеты отмывают один раз. Затем в каждую лунку добавляют 75 мкл аналитического буфера, после чего добавляют 25 мкл разведенного супернатанта головного мозга и 25 мкл Sulfo-меченого идентифицирующего антитела. Планшеты встряхивают при 4°С в течение 1 ч, а затем отмывают один раз. Наконец, добавляют 170 мкл MSD Read Buffer, и планшеты считывают на MSD Sector Imager 6000.
Чтобы дать возможность количественного определения Tau и Tau pS422 в экстрактах головного мозга мыши, строили стандартные кривые, используя Tau и Tau pS422, полученные, как описано в Примере 2.
О снижении патологии Tau у тройных трансгенных мышей Tau.PS2APP в результате хронической обработки моноклональным антителом анти-Tau pS422 свидетельствовало уменьшение типичной патологии Tau, наблюдаемое у мышей, обработанных антителом, в возрасте 11 месяцев по сравнению с мышами того же возраста, обработанными носителем. Это уменьшение продемонстрировано по сниженному грузу нейрофибриллярных клубков, как обнаружено с помощью иммуногистохимии, и по сниженному уровню различных видов pTau, присутствующих в экстрактах головного мозга, как измерено с помощью количественного иммунологического анализа.
Пример 10
Получение и очистка антител кролика
a) Иммунизация
Для иммунизации использовали кроликов линии NZW из Charles River Laboratories International, Inc. Фосфопептид Tau (416-430)[pS422], подвергнутый сочетанию на KLH, растворяли в буфере K3PO4 рН 7,0 при концентрации 1 мг/мл и смешивали (1:1) с полным адъювантом Фрейнда (ПАФ) до образования стабильной эмульсии. Три кролика получили внутрикожную (i.d.) инъекцию 2 мл эмульсии с последующей второй внутримышечной (i.m.) и третьей подкожной (s.c.) инъекцией, каждая по 1 мл с интервалом в одну неделю. Четвертую i.m. инъекцию 1 мл осуществляли через две недели после двух следующих s.c. инъекций по 1 мл с четырехнедельным интервалом. Образцы по 10 мл периферической цельной крови каждого животного собирали через 4-6 суток после третьей, четвертой, пятой и шестой инъекции и использовали для сортировки отдельных клеток FACS (флуоресцентной сортировки активированных клеток). Дополнительно 0,5 мл сыворотки каждого животного собирали в то же время и использовали для определения специфичного ответа антитела Tau (416-463)[pS422].
b) Ответ антитела
Ответ антитела на иммунизацию определяли путем серийного разведения сывороток, используя ELISA, в котором 30 нг на лунку биотинилированного Tau (416-430)[pS422] инкубировали в 1× ФСБ при 4°С в течение ночи на 96-луночных микротитрационных планшетах (МС1347, Micro Coat Biotechnologie GmbH, Bernried, Germany), предварительно обработанных стрептавидином. Для обнаружения IgG коза против кролика, сшитый с пероксидазой хрена (The Jackson laboratory), использовали при разведении 1:16000. Субстрат ВМ Blue POD, осаждающий тетраметилбензидин (ТМВ), готовый к употреблению раствор от фирмы Roche Diagnostics GmbH, использовали для визуализации. Реакцию останавливали 1 н. HCl и измеряли в Tecan Infinite при 450/690 нм.
с) Клонирование В-клеток
Покрытие планшетов
Стерильные 6-луночные планшеты, покрытые стрептавидином (градации для клеточных культур), инкубировали либо со смесью 3 биотинилированных контрольных пептидов (нефосфорилированный Tau (416-430), MCAK_Human (88-102)[95-pSer] и MAP2_Human (1802-1816)[pSer-1802]), либо с биотинилированным фосфопептидом Tau (416-430)[pS422], каждый в концентрации 0,5-1 мкг/мл в ФСБ при комнатной температуре в течение 1 ч. Планшеты отмывали в стерильном ФСБ три раза перед использованием. 6-луночные планшеты для клеточных культур покрывали 2 мкг/мл KLH (гемоцианином лимфы улитки) в карбонатном буфере (0,1 М бикарбонат натрия, 34 мМ двухзамещенный гидрокарбонат натрия, рН 9,55) в течение ночи при 4°С. Планшеты отмывали в стерильном ФСБ три раза перед использованием.
Выделение мононуклеарных клеток периферической крови кролика (МНПК)
Цельную кровь, содержащую ЭДТА, разводили в два раза 1× ФСБ перед центрифугированием в градиенте плотности на лимфолите для млекопитающих (Cedarlane Laboratories), которое осуществляли для выделения МНПК кролика. МНПК дважды промывали перед окрашиванием антителами.
Среда EL-4 B5
Среда RPMI 1640 (Pan Biootech, Aidenbach, Germany) с добавлением 10% ФСТ (Hyclone, Logan, UT, USA), 2 мМ глутамина, 1% раствора пенициллина/стрептомицина (РАА, Pasching, Austria), 2 мМ пирувата натрия, 10 мМ ГЭПЭС (PAN Biotech, Aidenbach, Germany) и 0,05 мМ бета-меркаптоэтанола (Gibco, Paisley, Scotland)
Истощение макрофагов/моноцитов
Стерильные 6-луночные планшеты (градации для клеточных культур) использовали для истощения макрофагов и моноцитов посредством неспецифической адгезии. Лунки покрывали либо KLH (гемоцианином лимфы улитки), либо стрептавидином и контрольными пептидами. Каждую лунку заполняли максимум 4 мл среды и мононуклеарными клетками периферической крови в количестве вплоть до 6×106 от иммунизированного кролика и давали возможность для связывания в течение 1 ч при 37°С в инкубаторе. 50% клеток в супернатанте использовали для стадии пэннинга; остальные 50% клеток непосредственно подвергали иммунофлуоресцентному окрашиванию и сортировке отдельных клеток.
Пэннинг В-клеток на пептидах
6-луночные планшеты для клеточных культур, покрытые стрептавидином и биотинилированным пептидом Tau (416-430)[р8422], засевали клетками в количестве вплоть до 6×106 на 4 мл среды и давали возможность для связывания в течение 1 ч при 37°С в инкубаторе. Не прикрепленные клетки удаляли осторожной отмывкой лунок 1-2 раза 1× ФСБ. Остальные прикрепленные клетки отделяли трипсином в течение 10 мин при 37°С в инкубаторе, а затем дважды отмывали в среде. Клетки держали на льду до иммунофлуоресцентного окрашивания.
Иммунофлуоресцентное окрашивание и сортировка отдельных клеток
Меченое ФИТЦ (флуоресцеинизотиоцианатом) IgG использовали для сортировки отдельных клеток с помощью AbD Serotec (STAR121F, DQsseldorf, Germany). Для поверхностного окрашивания клетки со стадии истощения и пэннинга инкубировали с меченым ФИТЦ антителом IgG против кролика в ФСБ в течение 30-минутного качания в холодной комнате при 4°С в темноте. После центрифугирования супернатанты удаляли отсасыванием. МНПК подвергали 2 циклам центрифугирования и отмывки охлажденным во льду ФСБ. Наконец, МНПК ресуспендировали в охлажденном во льду ФСБ и сразу подвергали анализам FACS. Перед анализами FACS добавляли пропидиумиодид в концентрации 5 мкг/мл (BD Pharmingen, San Diego, CA, USA) для различения мертвых и живых клеток. FACS проводили, используя прибор Becton Dickinson FACSAria, оборудованный программным обеспечением FACSDiva (BD Biosciences, USA), и отдельные меченые ФИТЦ живые клетки депонировали в 96-луночных планшетах.
В-клеточная культура
В-клеточные культуры получали способом, подобным описанному в статье Zubler, R.H. et al., J Immunol. 134 (1985) 3662-3668. Кратко, отдельные сортированные В-клетки культивировали в 96-луночных планшетах с 210 мкл/лунка среды EL-4 В5 с Pansorbin Cells (1:20000) (Calbiochem (Merck), Darmstadt, Deutschland), 5% супернатантом тимоцитов кролика и облученными гамма-излучением клетками тимомы мыши EL-4-B5 (2×104/лунка) в течение 7 суток при 37°С в атмосфере 5% С02 в инкубаторе. Супернатанты В-клеточной культуры извлекали для скрининга, и клетки сразу собирали для клонирования гена вариабельной области или замораживали при -80°С в 100 мкл буфера RLT (Qiagen, Hilden, Germany).
d) Скрининг В-клеточных клонов
Супернатанты В-клеточной культуры подвергали скринингу на связывание с биотинилированным Tau (416-430)[pS422] с помощью ELISA. Нефосфорилированный Tau (416-430), KLH (гемоцианин лимфы улитки) и необработанный фосфопептид MCAK_Human (88-102)[95-pSer] использовали в качестве контрольных антигенов. Для приготовления планшетов ELISA микротитрационные планшеты, предварительно покрытые стрептавидином, инкубировали с биотинилированным Tau (415-430)[pS422] при 50 нг/мл в течение 1 часа при комнатной температуре. Покрытие KLH или контрольными пептидами осуществляли при 1 мкг/мл. Супернатанты В-клеток разводили от 1:5 до 1:10 и инкубировали в микротитрационных планшетах, покрытых антигеном, в течение 60 мин. После интенсивной отмывки связывание антител кролика обнаруживали, используя идентифицирующее антитело IgG овца против кролика, конъюгированное с дигоксигенином (Chemicon AQ301D). После инкубации с ТМВ при комнатной температуре измеряли поглощение при 370-492 нм. В-клеточные клоны, дающие сигналы выше фоновых с биотинилированным Tau (416-430)[pS422], но не дающие сигналы с KLH и MCAK_Human (88-102)[95-pSer], рассматривали далее и подвергали клонированию гена вариабельной области.
е) ПЦР амплификация V-доменов и секвенирование
Суммарную РНК получали, используя набор для РНК NucleoSpin® 8/96 (Macherey&Nagel; 740709.4, 740698) в соответствии с протоколом фирмы-изготовителя. Все стадии проводили на системе работы с жидкостями epMotion 5075 (Eppendorf). РНК элюировали 60 мкл воды, свободной от РНК-аз. 6 мкл РНК использовали для получения кДНК с помощью обратно-транскриптазной реакции, используя набор для синтеза первой нити Superscript III First-Strand Synthesis SuperMix (Invitrogen 18080-400) и олигоdT-праймер в соответствии с инструкциями фирмы-изготовителя. 4 мкл кДНК использовали для амплификации вариабельных областей тяжелой и легкой цепи иммуноглобулина (VH и VL) со смесью AccuPrime Supermix (Invitrogen 12344-040) в конечном объеме 50 мкл, используя праймеры rbHCfinal.up и rbHCfinal.do для тяжелой цепи и rbLCfinal.up и rbLCfinal.do для легкой цепи (Таблица 4). Условия ПЦР были следующими: "горячий старт" при 94°С в течение 5 мин; 35 циклов 20 с при 94°С, 20 с при 70°С, 45 с при 68°С и конечная элонгация при 68°С в течение 7 мин.
8 мкл из 50 мкл раствора ПЦР наносили на 48 E-Gel 2% (Invitrogen G8008-02). Положительные реакции ПЦР очищали, используя набор NucleoSpin® Extract II (Macherey&Nagel; 740609250) в соответствии с протоколом фирмы-изготовителя, и элюировали в 50 мкл буфера для элюирования. 12 мкл очищенных продуктов ПЦР секвенировали непосредственно в обоих направлениях, используя rbHCfinal.up и rbHCfinal.do для тяжелых цепей и rbLCfinal.up и rbLCfinal.do для легких цепей (Таблица 4).
d) Рекомбинантная экспрессия моноклональных антител кролика и химерные антитела кролик/мышь
Для рекомбинантной экспрессии моноклональных антител кролика продукты ПЦР, кодирующие VH или VL, клонировали в виде кДНК в экспрессионных векторах способом клонирования с выступающими концами (Haun, R.S. et al., Biotechniques 13 (1992) 515-518; Li, M.Z., et al., Nature Methods 4 (2007) 251-256). Линеаризованные экспрессионные плазмиды, кодирующие константную область каппа или гамма кролика, и вставки VL или VH, амплифицировали с помощью ПЦР, используя перекрывающиеся праймеры. Очищенные продукты ПЦР инкубировали с Т4 ДНК полимеразой, которая образует выступающие однонитевые концы. Реакцию останавливали добавлением dCTP. На следующей стадии плазмиду и вставку объединяли и инкубировали с recA, который индуцирует сайт-специфическую рекомбинацию. Рекомбинированные плазмиды трансформировали в E.coli. На следующие сутки выросшие колонии собирали и тестировали на правильно рекомбинированную плазмиду путем получения препарата плазмиды, рестрикционного анализа и секвенирования ДНК. Для экспрессии антитела изолированные плазмиды НС и LC транзитно котрансфицировали в клетки НЕК293, и супернатанты собирали через 1 неделю. Для клонирования и экспрессии химерных антител кролика и мыши области VH и VL амплифицировали с помощью ПЦР и субклонировали в экспрессионных векторах, содержащих константную область каппа мыши или константную область гамма 1 мыши. Химерные плазмиды кролик/мышь НС и LC выделяли, тестировали с помощью рестрикционного анализа и секвенирования ДНК на правильную вставку и транзитно котрансфицировали в клетки НЕК293. Супернатанты собирали через неделю после трансфекции.
е) Очистка антитела
Рекомбинантно экспрессированные моноклональные антитела кролика очищали из супернатантов клеточной культуры на колонках MabSelectSuRe™ (GE Healthcare). Перед нанесением образца колонку уравновешивали 25 ммоль/л Трис-HCl, 25 ммоль/л NaCl, pH 7,4. Элюирование антитела осуществляли 50 ммоль/л ацетата, pH 3,14. Элюированный образец сразу наносили на колонку для обессоливания (Sephadex G25, GE Healthcare) и элюировали в 20 ммоль/л His-HCl, 140 ммоль/л NaCl pH 6,0. Этот буфер также использовали для хранения очищенного антитела. Обычная температура хранения составляла 4°С, комнатную температуру во время процесса очистки и -80°С после деления на аликвоты. Рекомбинантно экспрессированные химерные антитела кролик/мышь из супернатантов клеточных культур очищали на колонках MabSelectSuRe™ (GE Healthcare). Перед нанесением образца колонку уравновешивали 1× ФСБ, pH 7,4. Элюирование антител осуществляли 100 ммоль/л цитрата, pH 3,0. Элюированный образец сразу нейтрализовали 2 моль/л Трис/HCl pH 9,0. Затем антитела наносили на колонку для гель-фильтрации (Superdex 200, GE Healthcare) и элюировали в 20 ммоль/л His-HCl, 140 ммоль/л NaCl pH 6,0. Этот буфер также использовали для хранения очищенных антител. Обычная температура хранения составляла 4°С, комнатную температуру во время процесса очистки и -80°С после деления на аликвоты.
Пример 11
Моноклональные антитела кролика анти-Tau pS422 высокоселективны для Tau, фосфорилированного по pS422, и связываются с фибриллярными агрегатами Tau pS422
a) ELISA
Моноклональные антитела кролика рекомбинантно экспрессировали в клетках НЕК 293. Супернатанты клеточных культур или очищенные антитела кролика тестировали на связывание с биотинилированным Tau (416-430)[pS422], нефосфорилированным Tau (416-430), KLH (гемоцианин лимфы улитки) и необработанным фосфопептидом MCAK_Human (88-102)[95-pSer] с помощью ELISA. Для приготовления планшетов ELISA микротитрационные планшеты, предварительно покрытые стрептавидином, инкубировали с биотинилированным Tau (415-430)[pS422] при 50 нг/мл в течение 1 часа при комнатной температуре. Покрытие KLH или контрольными пептидами осуществляли при 1 мкг/мл. Антитело анти-Tau pS422 кролика (Abeam AB51071) или супернатанты, содержащие антитело кролика, инкубировали в меченых антигеном микротитрационных планшетах в течение 60 мин при различных концентрациях. После интенсивной отмывки связывание антител кролика обнаруживали, используя идентифицирующее антитело IgG овца против кролика, конъюгированное с дигоксигенином. После инкубации с ТМВ при комнатной температуре измеряли поглощение при 370 нм-492 нм. Связывание антитела характеризовали по его значениям ЕС50. Связывание антитела с пептидами, представляющими собой биотинилированный Tau (416-430)[pS422] и нефосфорилированный Tau (416-430), характеризовали по его значениям ЕС50. Перекрестную реактивность с KLH или фосфопептидом MCAK оценивали путем измерения в одной точке при высоких концентрациях, то есть при разведении супернатантов клеточных культур 1:5. Результаты показаны в таблице 5. Было обнаружено, что значения ЕС50 связывания с фосфопептидом Tau более чем в 100 раз ниже, чем значения ЕС50 связывания с пептидом Tau, что указывает на по меньшей мере 100-кратную селективность для фосфорилированного фрагмента Tau по сравнению с нефосфорилированным пептидом Tau. Связывание с KLH и контрольным фосфопептидом MCAK находилось на уровне фона со всеми антителами, что составляет примерно 1<3% измерений максимального значения для фосфопептида Tau.
Тестировали также специфичность к растворимому и агрегированному Tau р8422. Фибриллярные агрегаты Tau р8422 (300 мкг/мл) наносили на микротитрационный планшет на полистироловой основе MaxiSorb (Nunc) в течение ночи при КТ. Таким же способом растворимый полноразмерный Tau и Tau р8422 наносили на микротитрационный планшет MaxiSorb. Добавляли контрольное антитело кролика анти-Tau pS422 (Abeam AB51071) или очищенные антитела кролика и инкубировали в течение 60 мин при концентрациях вплоть до 1000 нг/мл. После интенсивной отмывки связывание антител кролика обнаруживали, используя идентифицирующее антитело IgG овца против кролика, конъюгированное с дигоксигенином (Chemicon AQ301D). После инкубации с ТМВ при комнатной температуре измеряли поглощение при 370 нм-492 нм. Связывание антитела характеризовали по его значениям ЕС50. Результаты показаны в таблице 6.
Моноклональные антитела кролика связывались с белком Tau-pS422 со значениями EC50 ниже 1 нг/мл. Фибриллярный Tau pS422 обнаруживали со значениями EC50 в диапазоне от 0,4 нг/мл до 14 нг/мл. Сигналы связывания с нефосфорилированным полноразмерным белком Tau были неотличимы от фоновых уровней. Таким образом, было установлено, что каждое из антител связывается с Tau pS422 и фибриллярным Tau pS422 по меньшей мере co 100-кратной селективностью по сравнению с Tau.
b) Biacore™
Связывание с агрегатами фибриллярного Tau pS422 дополнительно исследовали и подтверждали с помощью анализа BIAcore™. Измерения проводили, используя прибор BIAcore 3000 при 37°С. Система и буфер для образца представляли собой HBS-EP (10 мМ ГЭПЭС, 150 мМ NaCl, 3,4 мМ ЭДТА, 0,005% полисорбат 20 (об./об.)). Сенсорный чип BIAcore™ CM5 подвергали процедуре предварительного уравновешивания. Затем 0,1% ДСН, 50 мМ NaOH, 10 мМ HCl и 100 мМ H3PO4 впрыскивали за 30 с в проточные ячейки FC1, FC2, FC3 и FC4. Процедуру аминного сочетания проводили в соответствии с инструкциями фирмы-изготовителя, используя эксперт-программу BIAcore 3000™ v. 4.1. После активации EDC/NHS поверхности сенсора нефосфоселективное антитело анти-Tau mAb <TAU>M-4/53-IgG иммобилизовали на сенсорных проточных ячейках FC2, FC3 и FC4. В качестве контроля антитело против СК-ММ (изотип креатинкиназы), распознающее иррелевантный антиген, иммобилизовали на FC1. mAb<TAU>M-4/53-IgG, и антитело против СК-ММ разводили при 30 мкг/мл в 10 мМ NaAc pH 5,0 и впрыскивали со скоростью 10 мкл/мин в течение времени контакта 7 мин для иммобилизации 10000 RU системы иммобилизации антитела. Поверхность инактивировали насыщением 1 М этаноламином. Сенсор уравновешивали 5 циклами с фосфорилированным филаментным белком Tau (исходный раствор 0,3 мг/мл, разведенный 1:100 в HBS-EP) в качестве анализируемого вещества в растворе со скоростью 10 мкл/мин в течение 2 мин. Регенерацию осуществляли 10 мМ глицином pH 2,5 при 30 мкл/мин в течение 3 мин. Принимали, что анализируемое вещество, связывающееся с mAb 4/53, не диссоциирует от филаментов pTau, поскольку диссоциацию филаментов pTau с mAb 4/53 невозможно было наблюдать. Для всех последующих циклов измерения 0,3 мг/мл филаментов pTau разводили 1:100 в буфере HBS-EP и впрыскивали при 10 мкл/мин в течение 1 мин с целью презентирования pTau соответствующим анализируемым антителам в гетерогенном сэндвич-режиме. Анализируемые антитела разводили в буфере HBS-EP до концентрации 100 нМ и впрыскивали в систему при 20 мкл/мин в течение 3 мин. После 3 мин диссоциации поверхность сенсора регенерировали 2 впрыскиваниями 10 мМ глицина pH 2,5 в течение 1 мин при 100 мкл/мин с последующей отмывкой HBS в течение 15 с при 100 мкл/мин. Проводили мониторинг фаз ассоциации и диссоциации взаимодействий. Поскольку анализируемое антитело в растворе является бивалентным, кинетику антитело-pTAU, зависимую от авидитета, характеризовали с помощью двухфазной модели диссоциации, состоящей из стадии быстрой диссоциации, основанной на сродстве, с последующей стадией кинетики, стабилизируемой авидитетом, но ограничивающей скорость, в более поздней диссоциации комплекса. Где возможно, были определены Kd и t/2diss в течение 10 с (ранняя фаза) и 50 с (поздняя фаза) после окончания впрыскивания анализируемого вещества. Измерения кинетики оценивали, используя метод двойной ссылки. Сначала сигнал от контроля FC1 вычитали для коррекции эффекта объема буфера и неспецифического связывания. Затем впрыскивание 0 нМ анализируемого вещества вычитали для коррекции диссоциации первых антител из соответствующей системы иммобилизации. Кинетики скоростей оценивали, используя модель соответствия диссоциации Langmuir 1.1 в соответствии с программным обеспечением оценки Biacore™ v.4.1. Половинное время стабильности комплекса антиген/антитело (мин) вычисляли по формуле ln(2)/60*Kd.
Результаты суммированы в таблице 7.
Пример 12
Связывание моноклональных антител кролика анти-Tau pS422 с внутриклеточным pTau в срезах головного мозга пациентов с болезнью Альцгеймера
Специфичное и чувствительное иммуногистохимическое обнаружение патологии pTau в ткани головного мозга при болезни Альцгеймера моноклональными антителами кролика анти-Tau pS422 исследовали с помощью экспериментов иммунофлуоресцентного окрашивания с использованием срезов замороженной ткани головного мозга пациентов с БА. Метод был, в основном, таким же, как описано в Примере Х (мышиные антитела). IgG кролика обнаруживали с помощью вторых антител коза против кролика, конъюгированных с Alexa Fluor488® (Invitrogen/MolecularProbes АН 034). Специфичное и чувствительное окрашивание отложений и филаментов pTau очевидно для клонов Mab 005, Mab 019, Mab 020, Mab 085, Mab 086 и Mab 097. Заметны внутриклеточные отложения pTau, подобные большим нейрофибриллярным клубкам и продольным нитям нейропиля. Минимальная эффективная концентрация в диапазоне от 0,08 до 0,016 мкг/мл была определена для всех исследованных клонов, что указывает на высокочувствительное связывание с генуинными отложениями pTau человека.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К HER3 И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2560583C2 |
| АНТИТЕЛА ПРОТИВ CXCR4 | 2017 |
|
RU2748233C2 |
| АНТИТЕЛА ПРОТИВ CSF-1R ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2617971C2 |
| АНТИТЕЛА ПРОТИВ CSF-1R ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2617966C2 |
| Антитела к CD43 и их применение для лечения рака | 2016 |
|
RU2694903C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2011 |
|
RU2603078C2 |
| ФОСФОСПЕЦИФИЧНЫЕ АНТИТЕЛА, РАСПОЗНАЮЩИЕ ТАУ | 2012 |
|
RU2639537C2 |
| СОСТАВ, СОДЕРЖАЩИЙ АНТИТЕЛО | 2019 |
|
RU2806628C2 |
| АНТИТЕЛА К ТАУ И СПОСОБЫ ПРИМЕНЕНИЯ | 2014 |
|
RU2661111C2 |
| АНТИИДИОТИПИЧЕСКИЕ АНТИТЕЛА, КОТОРЫЕ ИНДУЦИРУЮТ ИММУННЫЙ ОТВЕТ ПРОТИВ РЕЦЕПТОРА ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА | 1996 |
|
RU2172636C2 |






























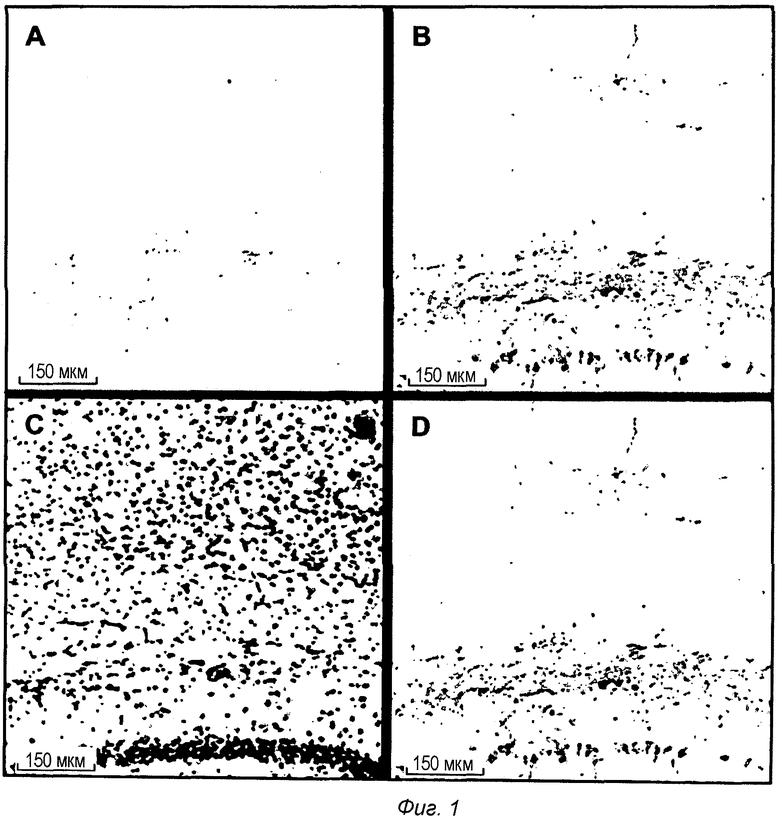
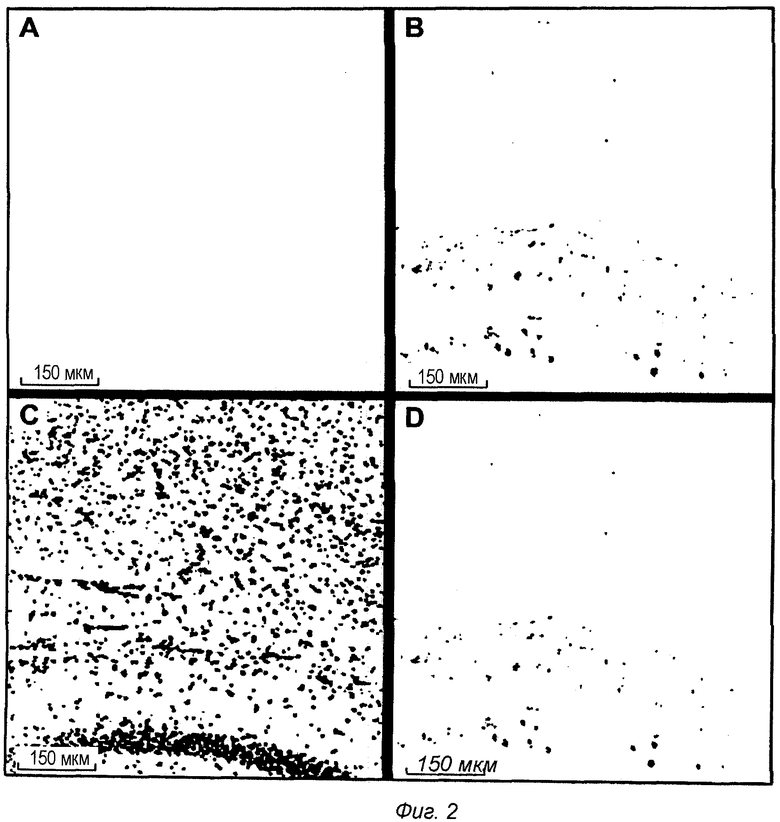
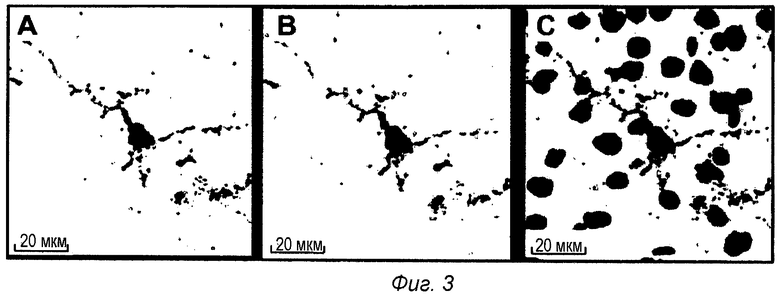
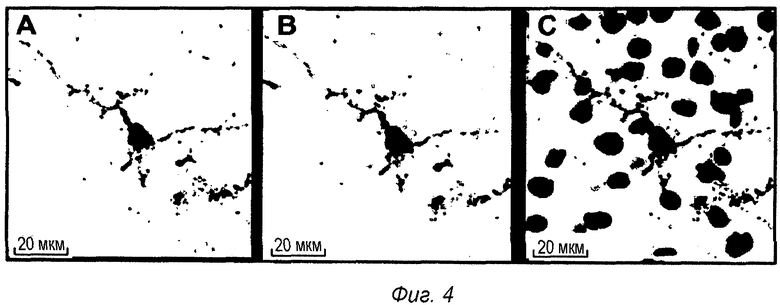
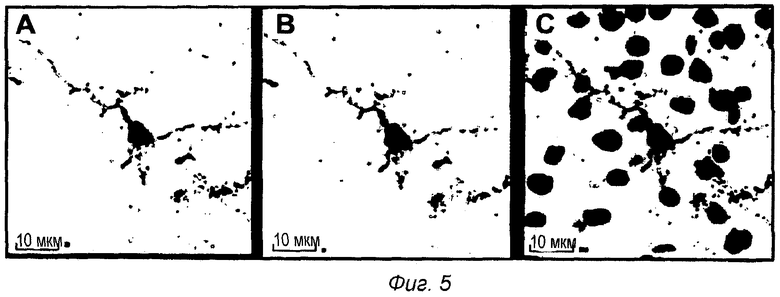
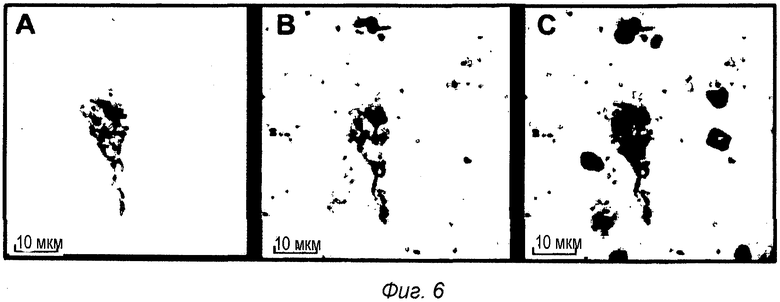
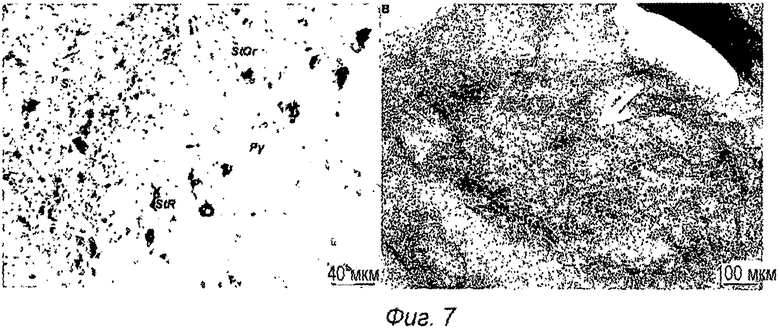
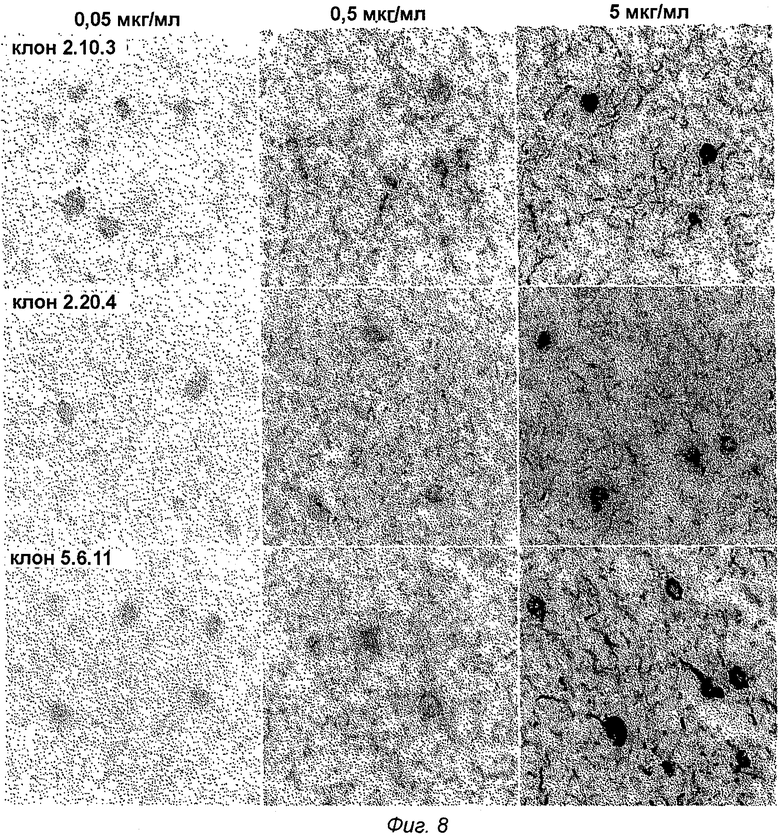
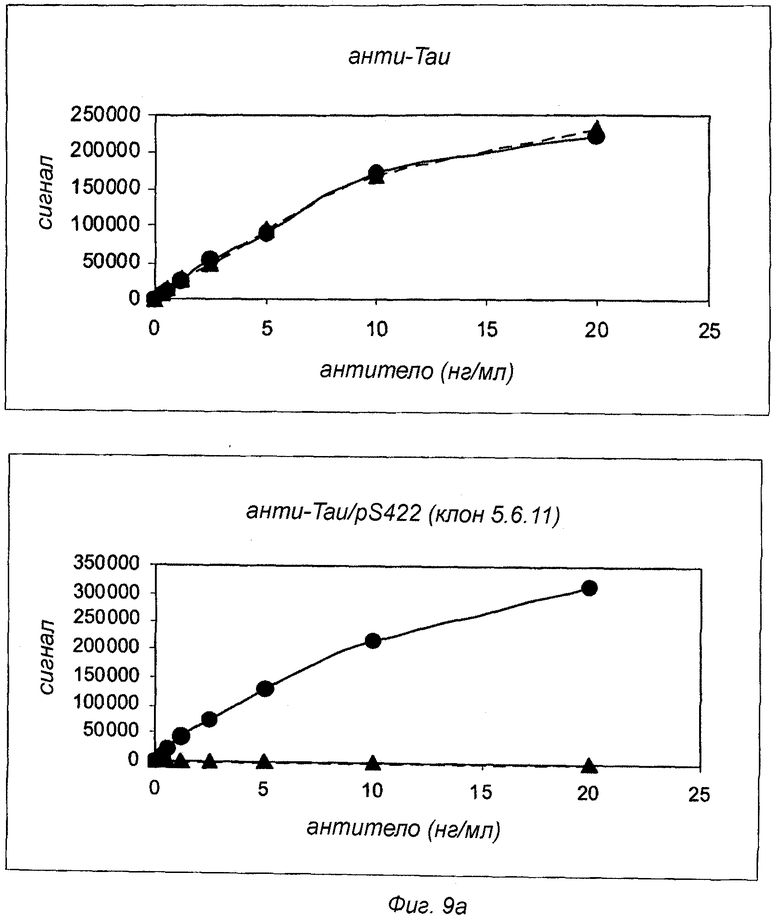
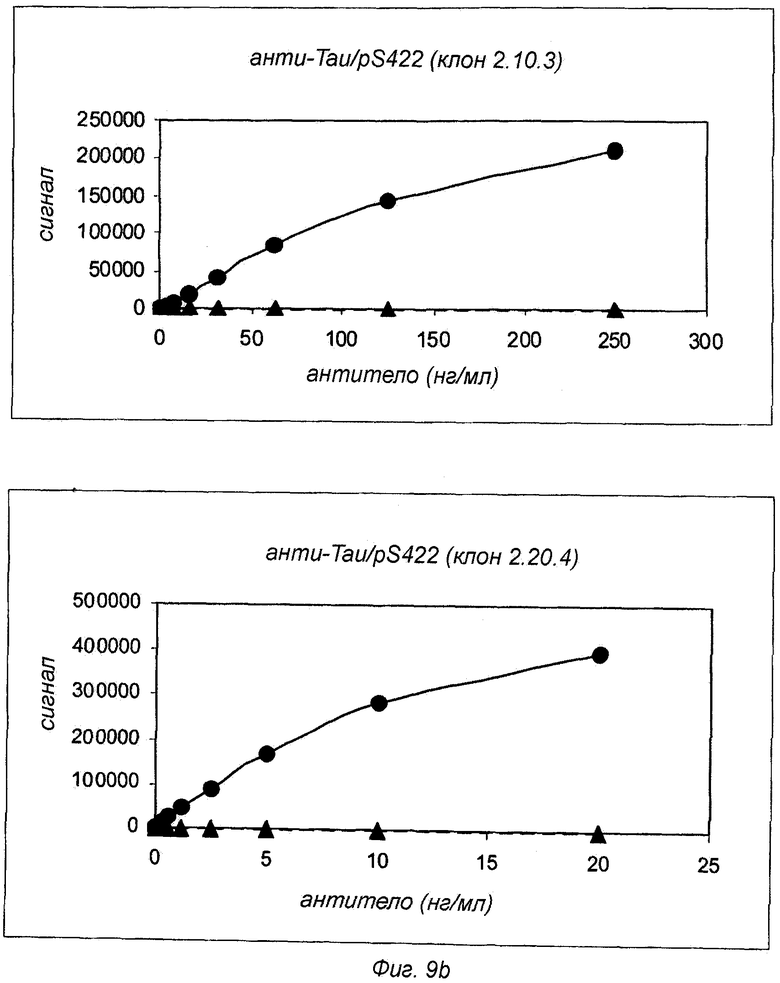


Изобретение относится к биохимии. Описано антитело для лечения таупатии, связывающееся с Tau, фосфорилированным по серину 422 (pS422), характеризующееся тем, что оно специфично связывается с фосфорилированным фрагментом Tau последовательности SEQ ID NO:9 и с Tau pS422, но не связывается с Tau и с фосфорилированным фрагментом МСАК последовательности SEQ ID NO:17. Представлена фармацевтическая композиция для лечения таупатии, а также способ лечения таупатии. Представлен способ получения описанного антитела. Изобретение расширяет арсенал средств для лечения таупатии. 15 н. и 6 з.п. ф-лы, 11 ил., 7 табл., 12 пр.
1. Антитело, специфично связывающееся с фосфорилированным фрагментом Таu последовательности SEQ ID NO:9 (Таu pS422), но не связывающееся с Tau и с фосфорилированным фрагментом МСАК последовательности SEQ ID NO:17, при этом антитело включает:
CDR1H SEQ ID NO:6, CDR1H SEQ ID NO:23, CDR1H SEQ ID NO:31, CDR1H SEQ ID NO:39, CDR1H SEQ ID NO:55 или CDR1H SEQ ID NO:63;
CDR2H SEQ ID NO:7, CDR2H SEQ ID NO:24, CDR2H SEQ ID NO:32, CDR2H SEQ ID NO:40, CDR2H SEQ ID NO:48 или CDR2H SEQ ID NO:64;
CDR3H SEQ ID NO:8, CDR3H SEQ ID NO:25, CDR3H SEQ ID NO:33, CDR3H SEQ ID NO:41, CDR3H SEQ ID NO:49 или CDR3H SEQ ID NO:65;
CDR1L SEQ ID NO:3, CDR1L SEQ ID NO:27, CDR1L SEQ ID NO:35, CDR1L SEQ ID NO:43, CDR1L SEQ ID NO:51, CDR1L SEQ ID NO:59 или CDR1L SEQ ID NO:67;
CDR2L SEQ ID NO:4, CDR2L SEQ ID NO:28, CDR2L SEQ ID NO:36, CDR2L SEQ ID NO:44, CDR2L SEQ ID NO:52, CDR2L SEQ ID NO:60 или CDR2L SEQ ID NO:68;
CDR3L SEQ ID NO:5, CDR3L SEQ ID NO:29, CDR3L SEQ ID NO:37, CDR3L SEQ ID NO:45, CDR3L SEQ ID NO:53, CDR3L SEQ ID NO:61 или CDR3L SEQ ID NO:69.
для применения при лечении таупатии.
2. Антитело для применения по п.1, характеризующееся тем, что это антитело относится к подтипу lgG1 или lgG4 человека.
3. Применение антитела, специфично связывающегося с фосфорилированным фрагментом Tau SEQ ID NO:9 (Таu pS422), но не связывающегося с Таu и с фосфорилированным фрагментом МСАК SEQ ID NO:17, для получения лекарства для лечения таупатии, при этом антитело включает:
CDR1H SEQ ID NO:6, CDR1H SEQ ID NO:23, CDR1H SEQ ID NO:31, CDR1H SEQ ID NO:39, CDR1H SEQ ID NO:55 или CDR1H SEQ ID NO:63;
CDR2H SEQ ID NO:7, CDR2H SEQ ID NO:24, CDR2H SEQ ID NO:32, CDR2H SEQ ID NO:40, CDR2H SEQ ID NO:48 или CDR2H SEQ ID NO:64;
CDR3H SEQ ID NO:8, CDR3H SEQ ID NO:25, CDR3H SEQ ID NO:33, CDR3H SEQ ID NO:41, CDR3H SEQ ID NO:49 или CDR3H SEQ ID NO:65;
CDR1L SEQ ID NO:3, CDR1L SEQ ID NO:27, CDR1L SEQ ID NO:35, CDR1L SEQ ID NO:43, CDR1L SEQ ID NO:51, CDR1L SEQ ID NO:59 или CDR1L SEQ ID NO:67;
CDR2L SEQ ID NO:4, CDR2L SEQ ID NO:28, CDR2L SEQ ID NO:36, CDR2L SEQ ID NO:44, CDR2L SEQ ID NO:52, CDR2L SEQ ID NO:60 или CDR2L SEQ ID NO:68;
CDR3L SEQ ID NO:5, CDR3L SEQ ID NO:29, CDR3L SEQ ID NO:37, CDR3L SEQ ID NO:45, CDR3L SEQ ID NO:53, CDR3L SEQ ID NO:61 или CDR3L SEQ ID NO:69.
4. Применение по п.3, характеризующееся тем, что это антитело относится к подтипу lgG1 или lgG4 человека.
5. Антитело, специфично связывающееся с фосфорилированным фрагментом Tau последовательности SEQ ID NO:9 (Tau pS422), но не связывающееся с Tau и с фосфорилированным фрагментом МСАК последовательности SEQ ID NO:17, характеризующееся тем, что оно включает
a) CDR1H SEQ ID NO:6, CDR2H SEQ ID NO:7 и CDR3H SEQ ID NO:8,
b) CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:24 и CDR3H SEQ ID NO:25,
c) CDR1H SEQ ID NO:31, CDR2H SEQ ID NO:32 и CDR3H SEQ ID NO:33,
d) CDR1H SEQ ID NO:39, CDR2H SEQ ID NO:40 и CDR3H SEQ ID NO:41,
e) CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49,
f) CDR1H SEQ ID NO:55, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49 или
g) CDR1H SEQ ID NO:63, CDR2H SEQ ID NO:64 и CDR3H SEQ ID NO:65 и
a) CDR1L SEQ ID NO:3, CDR2L SEQ ID NO:4, CDR3L SEQ ID NO:5,
b) CDR1L SEQ ID NO:27, CDR2L SEQ ID NO:28, CDR3L SEQ ID NO:29,
c) CDR1L SEQ ID NO:35, CDR2L SEQ ID NO:36, CDR3L SEQ ID NO:37,
d) CDR1L SEQ ID NO:43, CDR2L SEQ ID NO:44, CDR3L SEQ ID NO:45,
e) CDR1L SEQ ID NO:51, CDR2L SEQ ID NO:52, CDR3L SEQ ID NO:53,
f) CDR1L SEQ ID NO:59, CDR2L SEQ ID NO:60, CDR3L SEQ ID NO:61 или
g) CDR1L SEQ ID NO:67, CDR2L SEQ ID NO:68, CDR3L SEQ ID NO:69.
6. Антитело по п.5, характеризующееся тем, что оно включает
a) CDR1H SEQ ID NO:6, CDR2H SEQ ID NO:7, CDR3H SEQ ID NO:8 и CDR1L SEQ ID NO:3, CDR2L SEQ ID NO:4, CDR3L SEQ ID NO:5,
b) CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:24 и CDR3H SEQ ID NO:25 и CDR1L SEQ ID NO:27, CDR2L SEQ ID NO:28, CDR3L SEQ ID NO:29,
c) CDR1H SEQ ID NO:31, CDR2H SEQ ID NO:32 и CDR3H SEQ ID NO:33 и CDR1L SEQ ID NO:35, CDR2L SEQ ID NO:36, CDR3L SEQ ID NO:37,
d) CDR1H SEQ ID NO:39, CDR2H SEQ ID NO:40 и CDR3H SEQ ID NO:41 и CDR1L SEQ ID NO:43, CDR2L SEQ ID NO:44, CDR3L SEQ ID NO:45,
e) CDR1H SEQ ID NO:23, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49 и CDR1L SEQ ID NO:51, CDR2L SEQ ID NO:52, CDR3L SEQ ID NO:53,
f) CDR1H SEQ ID NO:55, CDR2H SEQ ID NO:48 и CDR3H SEQ ID NO:49 и CDR1L SEQ ID NO:59, CDR2L SEQ ID NO:60, CDR3L SEQ ID NO:61 или
g) CDR1H SEQ ID NO:63, CDR2H SEQ ID NO:64 и CDR3H SEQ ID NO:65 и CDR1L SEQ ID NO:67, CDR2L SEQ ID NO:68, CDR3L SEQ ID NO:69.
7. Антитело по п.6, характеризующееся тем, что оно включает
a) вариабельную легкую цепь SEQ ID NO:1 и вариабельную тяжелую цепь SEQ ID NO:2,
b) вариабельную легкую цепь SEQ ID NO:26 и вариабельную тяжелую цепь SEQ ID NO:22,
c) вариабельную легкую цепь SEQ ID NO:34 и вариабельную тяжелую цепь SEQ ID NO:30,
d) вариабельную легкую цепь SEQ ID NO:42 и вариабельную тяжелую цепь SEQ ID NO:38,
e) вариабельную легкую цепь SEQ ID NO:50 и вариабельную тяжелую цепь SEQ ID NO:46,
f) вариабельную легкую цепь SEQ ID NO:58 и вариабельную тяжелую цепь SEQ ID NO:54 или
g) вариабельную легкую цепь SEQ ID NO:66 и вариабельную тяжелую цепь SEQ ID NO:62.
8. Антитело по любому из пп.5-7, характеризующееся тем, что относится к подтипу lgG1 или lgG4 человека.
9. Антитело, специфично связывающееся с фосфорилированным фрагментом Tau последовательности SEQ ID NO:9 (Tau pS422), но не связывающееся с Tau и с фосфорилированным фрагментом МСАК последовательности SEQ ID NO:17, характеризующееся тем, что оно включает CDR1H SEQ ID NO:55, CDR2H SEQ ID NO:48, CDR3H SEQ ID NO:49 и CDR1L SEQ ID NO:59, CDR2L SEQ ID NO:60, CDR3L SEQ ID NO:61.
10. Химерный вариант антитела, связывающийся с Tau, фосфорилированным по серину 422 (Tau pS422), по любому из пп.5-9.
11. Гуманизированный вариант антитела, связывающийся с Tau, фосфорилированным по серину 422 (Tau pS422), по любому из пп.5-9.
12. Истощенный по Т-клеточному эпитопу вариант антитела, связывающийся с Tau, фосфорилированным по серину 422 (Tau pS422), по любому из пп.5-9.
13. Применение антитела по любому из пп.5-12 для получения лекарства для лечения таупатии.
14. Применение по п.13, характеризующееся тем, что антитело относится к подтипу lgG1 или lgG4 человека.
15. Фармацевтическая композиция для лечения таупатии, характеризующаяся тем, что она содержит эффективное количество антитела по любому из пп.5-12.
16. Применение антитела по любому из пп.5-12 для получения фармацевтической композиции для лечения таупатии.
17. Применение антитела по пп.5-12 для лечения таупатии, выбранной из группы, состоящей из болезни Альцгеймера (БА), включая только клубковую форму заболевания, синдрома Дауна (случаев у взрослых), комплекса паркинсонизм-деменция (синдрома Гуама), деменции боксеров, болезни Пика, деменции с аргирофильными зернами, лобно-височной деменции, кортико-базальной дегенерации, паллидо-понто-нигральной дегенерации, прогрессирующего надъядерного паралича и болезни Герштмана-Штрауслера-Шейнкера с клубками.
18. Нуклеиновая кислота, характеризующаяся тем, что она кодирует антитело по любому из пп.5-12.
19. Способ продуцирования рекомбинантного человеческого или гуманизированного антитела по любому из пп.5-12, характеризующийся экспрессией нуклеиновой кислоты по п.18 в прокариотической или эукариотической клетке-хозяине и выделением этого антитела из клетки или супернатанта клеточной культуры.
20. Способ лечения пациента, страдающего таупатией, в частности БА, характеризующийся введением этому пациенту антитела по любому из пп.5-12.
21. Фармацевтическая композиция для лечения таупатии, содержащая терапевтически эффективное количество антитела, специфически связывающегося с фосфорилированным фрагментом Tau последовательности SEQ ID NO:9 (Таu pS422), но не связывающегося с Tau и с фосфорилированным фрагментом МСАК последовательности SEQ ID NO:17, и фармацевтически приемлемый носитель, при этом антитело включает:
CDR1H SEQ ID NO:6, CDR1H SEQ ID NO:23, CDR1H SEQ ID NO:31, CDR1H SEQ ID NO:39, CDR1H SEQ ID NO:55 или CDR1H SEQ ID NO:63;
CDR2H SEQ ID NO:7, CDR2H SEQ ID NO:24, CDR2H SEQ ID NO:32, CDR2H SEQ ID NO:40, CDR2H SEQ ID NO:48 или CDR2H SEQ ID NO:64;
CDR3H SEQ ID NO:8, CDR3H SEQ ID NO:25, CDR3H SEQ ID NO:33, CDR3H SEQ ID NO:41, CDR3H SEQ ID NO:49 или CDR3H SEQ ID NO:65
CDR1L SEQ ID NO:3, CDR1L SEQ ID NO:27, CDR1L SEQ ID NO:35, CDR1L SEQ ID NO:43, CDR1L SEQ ID NO:51, CDR1L SEQ ID NO:59 или CDR1L SEQ ID NO:67;
CDR2L SEQ ID NO:4, CDR2L SEQ ID NO:28, CDR2L SEQ ID NO:36, CDR2L SEQ ID NO:44, CDR2L SEQ ID NO:52, CDR2L SEQ ID NO:60 или CDR2L SEQ ID NO:68;
CDR3L SEQ ID NO:5, CDR3L SEQ ID NO:29, CDR3L SEQ ID NO:37, CDR3L SEQ ID NO:45, CDR3L SEQ ID NO:53, CDR3L SEQ ID NO:61 или CDR3L SEQ ID NO:69.
| ГЛИКОЗАМИНОГЛИКАНЫ, ПРИГОДНЫЕ ДЛЯ ЛЕЧЕНИЯ СТАРЧЕСКОГО СЛАБОУМИЯ | 2000 |
|
RU2248800C2 |
| ЕР 1876185 А1, 01.09.2008 | |||

