Настоящее изобретение относится к детекции и лечению ревматоидного артрита (РА). Изобретение включает применение связывающего элемента, который связывает изоформу ED-A фибронектина, в частности, связывающего элемента, который связывает домен ED-A фибронектина.
Ревматоидный артрит (РА) является хроническим воспалительным и деструктивным заболеванием суставов, которое поражает 0,5-1% населения в индустриальном мире и обычно приводит к существенной потере трудоспособности и последующему ухудшению качества жизни.
Считается, что ангиогенез в синовиальной мембране пациентов с РА является важной ранней стадией в патогенезе и в сохранении заболевания (Taylor, 2002). Как и в неопластическом заболевании, ангиогенез питает распространение синовиальной мембраны (Walsh et al., 1998). Вероятно, рост кровеносных сосудов способствует пролиферации воспалительного синовиального паннуса, а также вхождению воспалительных лейкоцитов в синовиальную ткань. Синовиальная мембрана пациентов с РА содержала увеличенные количества фактора роста фибробластов (FGF-2) и фактора роста эндотелия сосудов (VEGF) (Koch, 2003). Сывороточные концентрации VEGF коррелируют с активностью заболевания и уменьшаются, когда синовит успешно подавляется терапией (Taylor, 2002).
Фибронектин (FN) является гликопротеином и широко экспрессируется в различных нормальных тканях и жидкостях тела. Он является компонентом внеклеточного матрикса (ECM) и играет роль во многих биологических процессах, включающих клеточную адгезию, клеточную миграцию, гемостаз, тромбоз, заживление ран, дифференцировку ткани и злокачественную трансформацию.
Различные изоформы FN генерируются альтернативным сплайсингом трех областей (ED-A, ED-B, IIICS) первичного транскрипта пре-мРНК FN, процессом, который модулируется цитокинами и внеклеточным рН (Balza 1988; Carnemolla 1989; Borsi 1990; Borsi 1995). Фибронектин содержит два глобулярных экстрадомена типа-III, которые подвергаются альтернативному сплайсингу: ED-A и ED-B (ffrench-Constant 1995, Hynes 1990, Kaspar et al. 2006). ED-A фибронектина мыши и фибронектина человека являются на 96,7% идентичными (только 3 аминокислоты отличаются между этими двумя состоящими из 90 аминокислот последовательностями, смотрите фиг.2).
Имеются сообщения об экспрессии ED-А фибронектина в опухолевых клетках и в солидных опухолях на уровне мРНК в раке молочной железы (Jacobs et al. 2002, Matsumoto et al. 1999) и раке печени (Oyama et al. 1989, Tavian et al. 1994) и на уровне выделенного белка в фибросаркоме, рабдосаркоме и меланоме (Borsi et al. 1987).
На иммуногистохимическом уровне, присутствие ED-A детектировали во внеклеточном матриксе (ECM) одонтогенных опухолей (Heikinheimo et al. 1991) и гепатоцеллюлярной карциномы (Koukoulis et al. 1995). В отличие от этого, ED-A детектировали в строме злокачественных неоплазм молочной железы (Koukoulis et al. 1993) и в кровеносных сосудах и базальных мембранах хорошо дифференцированного рака почки (Lohi et al. 1995). Однако, в менее дифференцированном раке почки (Lohi et al. 1995) и папиллокарциноме щитовидной железы (Scarpino et al. 1999) ED-A детектировали в кровеносных сосудах, базальных мембранах и строме опухоли. Сообщалось также о присутствии ED-A в сосудистой сети глиом (Borsi et al. 1998). Таким образом, распространение экспрессии ED-A, о котором сообщалось для различных типов опухолей, является высоко вариабельным.
Направленная доставка на основе антител к местам ангиогенеза является привлекательной терапевтической стратегией для лечения рака, но она является в значительной степени неисследованной в отношении хронических воспалительных заболеваний. Авторы настоящего изобретения демонстрировали ранее, что домен ED-B фибронектина, маркер ангиогенеза, экспрессируется в псориатических повреждениях у пациентов и в мышиной модели псориаза, а также в артритных лапках в коллаген-индуцированной модели ревматоидного артрита мыши. С использованием как радиоактивных, так и флуоресцентных способов, было обнаружено, что моноклональное антитело L19, специфическое в отношении EDB, селективно локализуется в местах воспаления in vivo после внутривенного введения. Эти результаты позволяют предположить терапевтический потенциал в отношении селективной доставки на основе L19 биологически активных соединений в местам воспаления (Trachsel, 2007; PCT/EP2007/004044).
Перед гибридизацией in situ было показано, что в образцах артрита человека может присутствовать домен фибронектина, отличный от ED-B, а также ED-A (Berndt et al., 1998; Kriegsmann et al., 2004).
В настоящем документе авторы настоящего изобретения показывают, что анти-EDA-антитело, такое как описанное здесь антитело F8, способно давать более сильное распределение окрашивания на артритных пробах человека в сравнении с анти-EDB-антителом L19 и анти-тенасцин-C-антителами F16 и G11.
Кроме того, было обнаружено, что при использовании как радиоактивных, так и флуоресцентных способов моноклональное антитело человека F8, специфическое в отношении ED-A, селективно локализуется в местах воспаления in vivo после внутривенного введения.
Таким образом, ED-A фибронектина может быть использован в качестве сосудистого маркера ревматоидного артрита.
Связывающие молекулы, такие как молекулы антител, которые связывают A-FN и/или ED-A фибронектина, представляют новые агенты, которые могут быть использованы для приготовления лекарственного средства для лечения ревматоидного артрита (РА).
Настоящее изобретение обеспечивает применение связывающего элемента, например, молекулы антитела, которая связывает изоформу Экстра-Домена-А (ED-A) фибронектина (A-FN), для приготовления лекарственного средства для лечения ревматоидного артрита. Настоящее изобретение обеспечивает также применение связывающего элемента, например, молекулы антитела, которая связывает ED-A фибронектина, для приготовления лекарственного средства для лечения ревматоидного артрита.
Настоящее изобретение дополнительно обеспечивает применение связывающего элемента, например, молекулы антитела, которая связывает изоформу ED-A фибронектина, для доставки к местам ревматоидного артрита молекулы, конъюгированной с этим связывающим элементом. Настоящее изобретение обеспечивает также применение связывающего элемента, например, молекулы антитела, которая связывает ED-A фибронектина, для доставки к местам ревматоидного артрита молекулы, конъюгированной с этим связывающим элементом. Этот связывающий элемент может быть использован для приготовления лекарственного средства для доставки такой молекулы.
Настоящее изобретение обеспечивает применение связывающего элемента, например, молекулы антитела, которая связывает изоформу ED-A фибронектина, для производства диагностического продукта для применения в диагностике ревматоидного артрита. Настоящее изобретение обеспечивает также применение связывающего элемента, например, молекулы антитела, которая связывает ED-A фибронектина, для производства диагностического продукта для применения в диагностике ревматоидного артрита.
Настоящее изобретение обеспечивает дополнительно способ детекции или диагностики ревматоидного артрита у человека или животного, предусматривающий:
(a) введение этому человеку или животному связывающего элемента, например, молекулы антитела, которая связывает ED-A фибронектина, и
(b) определение присутствия или отсутствия этого связывающего элемента в местах ревматоидного артрита тела человека или животного;
где локализация связывающего элемента в местах ревматоидного артрита указывает на наличие ревматоидного артрита.
Настоящее изобретение обеспечивает способ лечения ревматоидного артрита у индивидуума, предусматривающий введение индивидууму терапевтически эффективного количества лекарственного средства, содержащего связывающий элемент, например, молекулу антитела, которая связывает изоформу ED-A фибронектина. Настоящее изобретение обеспечивает также способ лечения ревматоидного артрита у индивидуума, предусматривающий введение индивидууму терапевтически эффективного количества лекарственного средства, содержащего связывающий элемент, например, молекулу антитела, которая связывает ED-A фибронектина.
Настоящее изобретение обеспечивает композицию, содержащую связывающий элемент, например, молекулу антитела, которая связывает изоформу ED-A фибронектина, для применения в способе лечения ревматоидного артрита у индивидуума, предусматривающем введение индивидууму терапевтически эффективного количества лекарственного средства, содержащего связывающий элемент, например, молекулу антитела, которая связывает изоформу ED-A фибронектина. Настоящее изобретение обеспечивает также композицию, содержащую связывающий элемент, например, молекулу антитела, которая связывает ED-A фибронектина, для применения в способе лечения ревматоидного артрита у индивидуума, предусматривающем введение индивидууму терапевтически эффективного количества лекарственного средства, содержащего связывающий элемент, например, молекулу антитела, которая связывает ED-A фибронектина.
Настоящее изобретение обеспечивает способ доставки молекулы к новообразованной сосудистой сети участков ревматоидного артрита у человека или животного, предусматривающий введение человеку или животному связывающего элемента, например, молекулы антитела, которая связывает изоформу ED-A фибронектина, где связывающий элемент конъюгирован с этой молекулой. Настоящее изобретение обеспечивает также способ доставки молекулы к новообразованной сосудистой сети участков ревматоидного артрита у человека или животного, предусматривающий введение человеку или животному связывающего элемента, например, молекулы антитела, которая связывает ED-A фибронектина, где связывающий элемент конъюгирован с этой молекулой.
Связывающим элементом для применения в настоящем изобретении может быть антитело, которое связывает изоформу ED-A фибронектина и/или ED-A фибронектина, содержащее один или несколько определяющих комплементарность областей (CDR) антитела H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 или G9 или их вариантов. Предпочтительно, связывающим элементом для применения в настоящем изобретении является антитело, которое связывает изоформу ED-A фибронектина и/или ED-A фибронектина, содержащее один или несколько определяющих комплементарность областей (CDR) антитела B2, C5, D5, C8, F8, B7 или G9 или их вариантов. Наиболее предпочтительно, связывающим элементом для применения в настоящем изобретении является антитело, которое связывает изоформу ED-A фибронектина и/или ED-A фибронектина, содержащее один или несколько определяющих комплементарность областей (CDR) антитела F8 или их вариантов.
Связывающий элемент для применения в настоящем изобретении может содержать набор CDR H и/или L антитела H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 или G9 или набор CDR H и/или L антитела H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 или G9 с десятью или менее, например, одной, двумя, тремя, четырьмя или пятью аминокислотными заменами в указанном наборе CDR H и/или L. Предпочтительно, связывающий элемент для применения в настоящем изобретении содержит набор CDR H и/или L антитела B2, C5, D5, C8, F8, B7 или G9 с десятью или менее, например, одной, двумя, тремя, четырьмя или пятью аминокислотными заменами в указанном наборе CDR H и/или L. Предпочтительно, связывающий элемент для применения в настоящем изобретении содержит набор CDR H и/или L антитела F8 с десятью или менее, например, одной, двумя, тремя, четырьмя или пятью аминокислотными заменами в указанном наборе CDR H и/или L.
Замены могут быть потенциально произведены в любом остатке в этих наборах CDR и могут находиться в CDR1, CDR2 и/или CDR3.
Например, связывающий элемент для применения в настоящем изобретении может содержать один или несколько CDR, описанных здесь, например, CDR3, и необязательно также CDR1 и CDR2, для образования набора CDR.
Связывающий элемент для применения в настоящем изобретении может также содержать молекулу антитела, например, молекулу антитела человека. Связывающий элемент обычно содержит домен VH и/или VL антитела. Домены VH связывающих элементов также обеспечены для применения в настоящем изобретении. В каждом из этих доменов VH и VL находятся определяющие комплементарность области ("CDR") и каркасные области ("FR"). Домен VH содержит набор HCDR, а домен VL содержит набор LCDR. Молекула антитела может содержать домен VH антитела, содержащий CDR1, CDR2 и CDR3 и каркас VH. Она может альтернативно или также содержать домен VL антитела, содержащий CDR1, CDR2 и CDR3 и каркас VL. Здесь описаны домены VH и VL и CDR антител H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 и G9. Все последовательности VH и VL, последовательности CDR, наборы CDR и наборы HCDR и наборы LCDR, описанные здесь, представляют варианты связывающего элемента для применения в настоящем изобретении. В настоящем документе, "набор CDR" содержит CDR1, CDR2 и CDR3. Так, набор HCDR относится к HCDR1, HCDR2 и HCDR3, а набор LCDR относится к LCDR1, LCDR2 и LCDR3. Если нет другого указания, "набор CDR" включает HCDR и LCDR.
Связывающий элемент для применения в настоящем изобретении может содержать домен VH антитела, содержащий определяющие комплементарность области HCDR1, HCDR2 и HCDR3 и каркас, где HCDR1 является SEQ ID NO:3, 23, 33, 43, 53, 63, 73, 83, 93, 103 или 113, и где необязательно HCDR2 является SEQ ID NO:4 и/или HCDR3 является SEQ ID NO:5. Предпочтительно, HCDR1 является SEQ ID NO:23, 33, 43, 53, 73, 83 или 103. Наиболее предпочтительно, HCDR1 является SEQ ID NO:83.
Обычно, домен VH спарен с доменом VL для обеспечения антигенсвязывающего сайта антитела, хотя, как обсуждается дополнительно ниже, домен VH или VL, один, может быть использован для связывания антигена. Таким образом, связывающий элемент для применения в настоящем изобретении может дополнительно содержать домен VL антитела, содержащий определяющие комплементарность области LCDR1, LCDR2 и LCDR3 и каркас, где LCDR1 является SEQ ID NO:6, 26, 36, 46, 56, 66, 76, 86, 96, 106 или 116 и где, необязательно, LCDR2 является SEQ ID NO:7 и/или LCDR3 является SEQ ID NO:8. Предпочтительно, LCDR1 является SEQ ID NO:26, 36, 46, 56, 76, 86 или 106. Наиболее предпочтительно, LCDR1 является SEQ ID NO:86.
Связывающий элемент для применения в настоящем изобретении может быть выделенной молекулой антитела для ED-A фибронектина, содержащей домен VH и домен VL, где домен VH содержит каркас и набор определяющих комплементарность областей HCDR1, HCDR2 и HCDR3 и где домен VL содержит определяющие комплементарность области LCDR1, LCDR2 и LCDR3 и каркас, и где
HCDR1 имеет аминокислотную последовательность SEQ ID NO:3, 23, 33, 43, 53, 63, 73, 83, 93, 103 или 113,
HCDR2 имеет аминокислотную последовательность SEQ ID NO:4,
HCDR3 имеет аминокислотную последовательность SEQ ID NO:5,
LCDR1 имеет аминокислотную последовательность SEQ ID NO:6, 26, 36, 46, 56, 66, 76, 86, 96, 106 или 116;
LCDR2 имеет аминокислотную последовательность SEQ ID NO:7; и
LCDR3 имеет аминокислотную последовательность SEQ ID NO:8.
Один или несколько CDR или набор CDR антитела могут быть трансплантированы в каркас (например, каркас человека) для обеспечения молекулы антитела для применения в настоящем изобретении. Каркасные области могут содержать последовательности сегментов генов зародышевой линии человека. Таким образом, этот каркас может быть включен в клетки зародышевой линии, посредством чего один или несколько остатков в этом каркасе изменяются для соответствия остаткам в эквивалентном положении в наиболее сходном каркасе зародышевой линии человека. Связывающий элемент для применения в настоящем изобретении может быть выделенной молекулой антитела, имеющей домен VH, содержащий набор HCDR в каркасе зародышевой линии человека, например, DP47. Обычно, связывающий элемент имеет также домен VL, содержащий набор LCDR, например, в каркасе зародышевой линии человека. Этим каркасом зародышевой линии человека домена VL может быть DPK22.
Домен VH для применения в настоящем изобретении может иметь аминокислотную последовательность SEQ ID NO:1, 21, 31, 41, 51, 61, 71, 81, 91, 101 или 111. Предпочтительно, домен VH для применения в настоящем изобретении имеет аминокислотную последовательность SEQ ID NO:21, 31, 41, 51, 71, 81 или 101. Наиболее предпочтительно, домен VH для применения в настоящем изобретении имеет аминокислотную последовательность SEQ ID NO:81. Домен VL для применения в настоящем изобретении может иметь аминокислотную последовательность SEQ ID NO:2, 22, 32, 42, 52, 62, 72, 82, 92, 102 или 112. Предпочтительно, домен VL для применения в настоящем изобретении имеет аминокислотную последовательность SEQ ID NO:22, 32, 42, 52, 72, 82 или 102. Наиболее предпочтительно, домен VL для применения в настоящем изобретении имеет аминокислотную последовательность SEQ ID NO:82.
Связывающий элемент для применения в настоящем изобретении может быть одноцепочечным Fv или содержать одноцепочечный Fv (scFv), содержащий домен VH и домен VL, соединенные пептидным линкером. Квалифицированный в данной области специалист может выбрать подходящие длину и последовательность линкера, например, длину по меньшей мере 5 или 10 аминокислот и до длины приблизительно 15, 20 или 25 аминокислот. Этот линкер может иметь аминокислотную последовательность GSSGG (SEQ ID NO:28). scFv может состоять из аминокислотной последовательности SEQ ID NO:9 или содержать аминокислотную последовательность SEQ ID NO:9.
Одноцепочечный Fv (scFv) может содержаться в мини-иммуноглобулине или малом иммунопротеине (SIP), например, как описано в (Li et al., 1997). SIP может содержать молекулу scFv, слитую с доменом CH4 секреторной изоформы IgE-S2 антитела IgE человека (εS2-CH4; Batista et al., 1996) с образованием молекулы антитела в виде гомодимерного мини-иммуноглобулина.
Альтернативно, связывающий элемент для применения в настоящем изобретении может содержать антигенсвязывающий сайт в молекуле не-антитела, обычно обеспечиваемый одним или несколькими CDR, например, набором CDR, в белковом каркасе, не являющемся антителом. Связывающие элементы, включающие молекулы не-антитела и молекулы антитела, описаны более подробно в настоящем документе.
Связывающий элемент для применения в настоящем изобретении, может быть конъюгирован с молекулой, которая имеет биоцидную, цитотоксическую, иммуносупрессивную или противовоспалительную активность. Интерлейкин-10 является предпочтительной молекулой для конъюгации со связывающим элементом в соответствии с настоящим изобретением. Кроме того, связывающий элемент для применения в настоящем изобретении может быть конъюгирован с радиоактивным изотопом, детектируемой меткой или фотосенсибилизатором.
Эти и другие аспекты настоящего изобретения описаны более подробно ниже.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фиг.1 показывает результаты иммуногистохимии на артритных пробах человека с использованием антител, направленных на маркеры ангиогенеза. Более темное окрашивание указывает сильную экспрессию этого антигена, визуализированную белыми стрелками. F8 является молекулой антитела, которое связывает ED-A, описанный здесь, L19 является молекулой антитела, которое связывает ED-B (например, Pini et al. 1998), F16 и G11 являются молекулами, которые связывают домены А1 и С Тенасцина-С, соответственно (WO2006/050834).
Фиг.2 показывает результаты анализа иммунофлуоресценции на артритных пробах человека с использованием молекулы антитела F8, направленного против домена ED-A фибронектина. Белое окрашивание указывает сильную экспрессию этого антигена.
Фиг.3 показывает выравнивание между А: ED-A человека (верхняя последовательность) и B: ED-A мыши (нижняя последовательность). Звездочки указывают положения аминокислот, в которых аминокислоты ED-A человека и ED-A мыши являются идентичными.
Фиг.4А показывает нуклеотидную последовательность тяжелой цепи (VH) анти-ED-A-антитела Н1 (SEQ ID NO:12). Нуклеотидная последовательность CDR1 тяжелой цепи анти-ED-A-антитела Н1 подчеркнута. Нуклеотидная последовательность CDR2 тяжелой цепи анти-ED-A-антитела H1 показана курсивом и подчеркнута. Нуклеотидная последовательность CDR3 тяжелой цепи анти-ED-A-антитела H1 показана жирным шрифтом и подчеркнута.
Фиг.4В показывает нуклеотидную последовательность линкерной последовательности анти-ED-A-антитела H1 (SEQ ID NO:14).
Фиг.4С показывает нуклеотидную последовательность легкой цепи (VL) анти-ED-A-антитела H1 (SEQ ID NO:13). Нуклеотидная последовательность CDR1 легкой цепи анти-ED-A-антитела Н1 подчеркнута. Нуклеотидная последовательность CDR2 легкой цепи анти-ED-A-антитела H1 показана курсивом и подчеркнута. Нуклеотидная последовательность CDR3 легкой цепи анти-ED-A-антитела H1 показана жирным шрифтом и подчеркнута.
Фиг.5А показывает аминокислотную последовательность тяжелой цепи (VH) анти-ED-A-антитела Н1 (SEQ ID NO:1). Аминокислотная последовательность CDR1 тяжелой цепи (SEQ ID NO:3) анти-ED-A-антитела Н1 подчеркнута. Аминокислотная последовательность CDR2 тяжелой цепи (SEQ ID NO:4) анти-ED-A-антитела H1 показана курсивом и подчеркнута. Аминокислотная последовательность CDR3 тяжелой цепи анти-ED-A-антитела H1 показана жирным шрифтом и подчеркнута.
Фиг.5В показывает аминокислотную последовательность линкерной последовательности анти-ED-A-антитела H1 (SEQ ID NO:11).
Фиг.5С показывает аминокислотную последовательность легкой цепи (VL) анти-ED-A-антитела H1 (SEQ ID NO:2). Аминокислотная последовательность CDR1 легкой цепи (SEQ ID NO:6) анти-ED-A-антитела Н1 подчеркнута. Аминокислотная последовательность CDR2 легкой цепи (SEQ ID NO:7) анти-ED-A-антитела H1 показана курсивом и подчеркнута. Нуклеотидная последовательность CDR3 легкой цепи (SEQ ID NO:8) анти-ED-A-антитела H1 показана жирным шрифтом и подчеркнута.
Фиг.6 показывает последовательность конструкции нуклеиновой кислоты, включающей кодирующую последовательность для F8-IL10. Эта структура является HINDIII-секреторная последовательность-F8-(14aa-линкер)-линкер (SSSSG)3-IL10-Стоп-NotI, показанной следующим образом: сайт рестрикции HINDIII является подчеркнутым, последовательность, кодирующая сигнал секреции, показана курсивом, VH-кодирующая последовательность F8 показана жирным шрифтом после последовательности сигнала секреции, последовательность, кодирующая линкер из 14 аминокислот, показана строчными буквами, VL-кодирующая последовательность F8 показана жирным шрифтом после линкерной последовательности из 14 аминокислот, линкерная последовательность (SSSSG)3, которая следует после кодирующей последовательности F8, подчеркнута и показана курсивом, последовательность, кодирующая IL-10, подчеркнута двойной линией; затем следует стоп-кодон, показанный строчными буквами, с последующим сайтом рестрикции NotI, который является подчеркнутым.
Фиг.7 показывает аминокислотную последовательность конъюгата антитело scFv (F8)-IL-10, включающего линкеры, имеющего структуру: VH-линкер-VL-линкер-IL-10. Домены VH и VL показаны жирным шрифтом, линкер scFv показан строчными буквами, линкер между scFv и IL-10 показан строчными буквами и курсивом, последовательность IL-10 подчеркнута.
Фиг.8 иллюстрирует клонирование, экспрессию и очистку F8-IL10 и HyHel10-IL10:
Фиг.8а показывает схематическое представление вектора pcDNA3.1, содержащего элементы слитых белков F8-IL10. Часть IL10 человека слита с С-концом фрагмента антитела scFv линкером из 15 аминокислот (SSSSG)3. Для секреции рекомбинантных белков необходима последовательность секреции на N-конце.
Фиг.8b показывает результаты анализа электрофорезом в ДСН-ПААГ очищенных слитых белков: Дорожка 1, маркер молекулярной массы; дорожки 2 и 3, F8-IL10 при невосстанавливающих и восстанавливающих условиях. Ожидается, что мономерный белок имеет молекулярную массу 46 кДа.
Фиг.8c показывает профиль вытеснительной хроматографии очищенного F8-IL10 (Супердекс 200). Пик элюции при объеме удерживания 13 мл соответствует нековалентной гомодимерной форме F8-IL10, меньший пик при объеме удерживания 14 мл соответствует мономерной фракции.
Фиг.8d показывает результаты анализа активности F8-IL10. Активность F8-IL10 сравнивали с активностью рекомбинантного IL10 человека на клетках МС/9.
ТЕРМИНОЛОГИЯ
Фибронектин
Фибронектин является антигеном, подверженным альтернативному сплайсингу, и известен ряд альтернативных изоформ фибронектина, как описано в настоящем документе. Экстра Домен-А (EDA или ED-A) известен также как ED, повтор А экстра типа III (EIIIA) или EDI. Последовательность ED-A человека была опубликована Kornblihtt et al. (1984), Nucleic Acids Res. 12, 5853-5868 и Paolella et al. (1988), Nucleic Acids Res. 16, 3545-3557. Последовательность ED-A человека доступна также в базе данных SwissProt в виде аминокислот 1631-1720 (фибронектин типа-III 12; экстра домен 2) аминокислотной последовательности, депонированной под номером доступа P02751. Последовательность ED-A мыши доступна из базы данных SwissProt в виде аминокислот 1721-1810 (фибронектин типа-III 13; экстра домен 2) аминокислотной последовательности, депонированной под номером доступа P11276.
Изоформа ED-A фибронектина (A-FN) содержит Экстра Домен-A (ED-A). Последовательность A-FN человека может быть расшифрована из соответствующей последовательности-предшественника фибронектина человека, которая доступна из базы данных SwissProt под номером доступа P02751. Последовательность A-FN мыши может быть расшифрована из соответствующей последовательности-предшественника фибронектина человека, которая доступна из базы данных SwissProt под номером доступа P11276. A-FN может быть изоформой ED-A фибронектина человека. ED-A может быть Экстра Доменом-А фибронектина человека.
ED-A является содержащей 90 аминокислот аминокислотной последовательностью, которая встраивается в фибронектин (FN) альтернативным сплайсингом и расположена между доменом 11 и 12 FN (Borsi et al., 1987, J. Cell Biol., 104, 595-600). ED-A в основном отсутствует в плазматической форме FN, но имеется в изобилии во время эмбриогенеза, при ремоделировании ткани, фиброзе, трансплантации сердца и росте солидных опухолей.
Альтернативный сплайсинг
Альтернативным сплайсингом называют наличие отличающегося характера сплайсинга первичного РНК-транскрипта ДНК с получением отличающихся мРНК. После вырезания интронов, отбор может определить, какие экзоны сплайсируются вместе с образованием этой мРНК. Альтернативный сплайсинг приводит к получению различных изоформ, содержащих различные экзоны и/или различные количества экзонов. Например, одна изоформа может содержать дополнительную аминокислотную последовательность, соответствующую одному или нескольким экзонам, которые могут содержать один или несколько доменов.
Связывающий элемент
Этот термин обозначает один элемент пары молекул, которые связываются друг с другом. Элементы пары связывания могут быть природно полученными или полностью или частично синтетически полученными. Один элемент пары молекул имеет площадь на его поверхности, или впадину, которая связывается с конкретной пространственной и полярной организацией или комплементарна конкретной пространственной и полярной организации другого элемента пары молекул. Примерами типов пар связывания являются пары антиген-антитело, биотин-авидин, гормон-рецептор гормона, рецептор-лиганд, фермент-субстрат. Настоящее изобретение относится к реакциям типа антиген-антитело.
Связывающий элемент обычно содержит молекулу, имеющую антигенсвязывающий сайт. Например, связывающий элемент может быть молекулой антитела или белком, не являющимся антителом, который содержит антигенсвязывающий сайт.
Антигенсвязывающий сайт может быть обеспечен посредством расположения определяющих комплементарность областей (CDR) на каркасах белка, не являющегося антителом, такого как фибронектин или цитохром В и т.д. (Haan & Maggos, 2004; Koide 1998; Nygren 1997), или рандомизацией или мутированием аминокислотных остатков петли в белковом каркасе для придания связывающей специфичности в отношении желаемой мишени. Каркасы для конструирования новых связывающих сайтов в белках обсуждались подробно Nygren et al. (1997). Белковые каркасы для миметиков антител описаны в WO/0034784, который включен в настоящее описание в качестве ссылки в полном объеме, в котором авторы изобретения описывают белки (миметики антител), которые включают домен фибронектина типа III, имеющий по меньшей мере одну рандомизированную петлю. Подходящий каркас, в который можно трансплантировать один или несколько CDR, например, набор HCDR, может быть обеспечен любым элементом-доменом суперсемейства генов иммуноглобулинов. Этот каркас может быть белком человека или белком не человека. Преимуществом каркаса белка не антитела заключается в том, что он может обеспечивать антигенсвязывающий сайт в молекуле каркаса, который меньше и/или может быть легче получен, чем по меньшей мере некоторые молекулы антител. Малый размер связывающего элемента может придавать полезные физиологические свойства, такие как способность входить в клетки, проникать глубоко в ткани или достигать мишеней внутри других структур или связываться во впадинах белка антигена-мишени. Применение антигенсвязывающих сайтов в каркасах белков не антител обсуждается в Wess, 2004. Типичными являются белки, имеющие стабильный скелет молекулы и одну или несколько вариабельных петель, в которых аминокислотная последовательность этой петли или этих петель является специфически или случайным образом мутированной для создания антигенсвязывающего сайта, который связывает антиген-мишень. Такие белки включают IgG-связывающие домены белка А из S. aureus, трансферрин, тетранектин, фибронектин (например, 10-ый домен фибронектина типа III) и липокалины. Другие подходы включают синтетические "Микротела" (Selecore GmbH), которые основаны на циклотидах - малых белках, имеющих внутримолекулярные дисульфидные связи.
Кроме последовательностей антител и/или антигенсвязывающего сайта, связывающий элемент для применения в настоящем изобретении может содержать другие аминокислоты, например, образующие пептид или полипептид, такие как уложенный домен, или придающие этой молекуле другую функциональную характеристику, наряду со способностью связывать антиген. Связывающие элементы для применения в настоящем изобретении могут нести детектируемую метку или могут быть конъюгированы с токсином или нацеливающей частью или ферментом (например, через пептидильную связь или линкер). Например, связывающий элемент может содержать каталитический сайт (например, в домене фермента), а также антигенсвязывающий сайт, где этот антигенсвязывающий сайт связывается с антигеном и, следовательно, нацеливает этот каталитический сайт на этот антиген. Этот каталитический сайт может ингибировать биологическую функцию этого антигена, например, посредством расщепления.
Хотя, как отмечалось, CDR могут находиться на каркасах не антител, структурой, несущей CDR или набор CDR, будет обычно последовательность тяжелой или легкой цепи антитела или ее существенная часть, в которой расположены CDR или набор CDR в местоположении, соответствующем CDR или набору CDR природных вариабельных доменов VH и VL антитела, кодируемых реаранжированными генами иммуноглобулина. Эти структуры и местоположения вариабельных доменов иммуноглобулина могут быть определены со ссылкой на Kabat 1987 и более поздние переиздания, доступные в настоящее время в Интернете (в immuno.bme.nwu.edu или могут быть найдены на "Kabat" с использованием любой поисковой системы).
Под терминами область CDR или CDR имеются в виду гипервариабельные области тяжелой и легкой цепей иммуноглобулина, как определено Kabat et al. (1987), (Kabat 1991a и более поздние издания). Антитело обычно содержит 3 CDR тяжелой цепи и 3 CDR легкой цепи. Термины CDR или CDR во множественном числе используются в настоящем описании для указания, согласно этому случаю, одной из этих областей или нескольких или даже всех из этих областей, которые содержат большинство аминокислотных остатков, ответственных за связывание посредством аффинности антитела в отношении антигена или эпитопа, который оно узнает.
Среди шести коротких CDR-последовательностей, третий CDR тяжелой цепи (HCDR3) имеет вариабельность большего размера (большее разнообразие, в основном обусловленное механизмом расположения генов, которые его вызывают). Он может быть таким коротким, как 2 аминокислоты, хотя самым длинным размером является 26. Функционально, HCDR3 играет роль отчасти в определении специфичности этого антитела (Segal 1974; Amit 1986; Chothia 1987; Chothia 1989; Caton 1990; Sharon 1990a; Sharon 1990b; Kabat et al., 1991b).
Молекула антитела
Этот термин описывает иммуноглобулин, природный или частично или полностью синтезированный. Этот термин относится также к любому полипептиду или белку, содержащему антигенсвязывающий сайт антитела. Должно быть понятно здесь, что настоящее изобретение не относится к антителам в природной форме, то есть они не находятся в их природном окружении, но они могут быть выделены или получены очисткой из природных источников или получены генетической рекомбинацией или химическим синтезом, и что они могут тогда содержать неприродные аминокислоты, как будет описано далее. Фрагменты антител, которые содержат антигенсвязывающий сайт антитела, включают, но не ограничиваются ими, такие молекулы антител, как Fab, Fab’, Fab’-SH, scFv, Fv, dAb, Fd и диатела.
Можно использовать моноклональные и другие антитела и использовать способы технологии рекомбинантных ДНК для получения других антител или химерных молекул, которые связывают антиген-мишень. Такие способы могут включать введение ДНК, кодирующей вариабельную область иммуноглобулина, или CDR, антитела в константные области или константные области плюс каркасные области другого иммуноглобулина. Смотрите, например, EP-A-184187, GB 2188638A или EP-A-239400 и большой объем последующей литературы. Гибридома или другая клетка, продуцирующая антитело, может быть подвергнута генетической мутации или другим изменениям, которые могут изменять или могут не изменять связывающую специфичность полученных антител.
Поскольку антитела могут быть модифицированы различными путями, термин "молекула антитела" должен пониматься как термин, включающий любой связывающий элемент или любое связывающее вещество, имеющие антигенсвязывающий сайт антитела с требуемыми специфичностью и/или связыванием с антигеном. Таким образом, этот термин включает фрагменты и производные антител, в том числе любой полипептид, содержащий антигенсвязывающий сайт антитела, независимо от того, является ли он природным или полностью или частично синтетическим. Таким образом, в этот термин включены химерные молекулы, содержащие антигенсвязывающий сайт антитела, или, эквивалентно, слитые с другим полипептидом (например, произведенным из другого вида или принадлежащим к другому классу или подклассу антител). Клонирование и экспрессия химерных антител описаны в EP-A-0120694 и EP-A-0125023 и большом объеме последующей литературы.
Дополнительные способы, доступные в области конструирования антител, сделали возможным выделение антител человека и гуманизированных антител. Например, гибридомы человека могут быть получены, как описано Kontermann & Dubel (2001). Фаговый дисплей, другой установленный способ генерирования связывающих элементов, был описан подробно во многих публикациях, таких как WO92/01047 (обсуждаемый дополнительно ниже), и патентах США US5969108, US5565332, US5733743, US5858657, US5871907, US5872215, US5885793, US5962255, US6140471, US6172197, US6225447, US6291650, US6492160, US6521404 и Kontermann & Dubel (2001). Могут быть использованы трансгенные мыши, в которых гены антител мыши инактивированы и функционально заменены генами антител человека с оставлением интактными других компонентов иммунной системы мыши (Mendez 1997).
Синтетические молекулы антител могут быть созданы экспрессией из генов, генерированных с использованием олигонуклеотидов, синтезированных и собранных в подходящих экспрессирующих векторах, например, как описано Knappik et al. (2000) или Krebs et al. (2001).
Было показано, что фрагменты полного антитела могут выполнять функцию связывания антигенов. Примерами связывающих фрагментов являются (i) Fab-фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) Fd-фрагмент, состоящий из доменов VH и CH1; (iii) Fv-фрагмент, состоящий из доменов VL и VH одного антитела; (iv) dAb-фрагмент (Ward 1989; McCafferty 1990; Holt 2003), который состоит из домена VH или VL; (v) выделенные области CDR; (vi) F(ab’)2-фрагмент, бивалентный фрагмент, содержащий два связанных Fab-фрагмента, (vii) одноцепочечные молекулы Fv (scFv), в которых домен VH и домен VL связаны пептидным линкером, который позволяет этим двум доменам ассоциироваться с образованием антигенсвязывающего сайта (Bird 1988; Huston 1988); (viii) биспецифические одноцепочечные димеры Fv (PCT/US92/09965) и (ix) "диатела", мультивалентные или мультиспецифические фрагменты, сконструированные слиянием генов (WO94/13804; Holliger 1993a). Молекулы Fv, scFv или диател могут быть стабилизированы включением дисульфидных мостиков, связывающих домены VH и VL (Reiter 1996). Могут быть также генерированы минитела, содержащие scFv, присоединенный к домену CH3 (Hu 1996). Другими примерами связывающих фрагментов являются Fab’, которые отличаются от Fab-фрагментов добавлением нескольких остатков на карбоксильном конце домена СН1 тяжелой цепи, в том числе одного или нескольких цистеинов из шарнирной области антитела, и Fab’-SH, который является Fab’-фрагментом, в котором остаток (остатки) цистеина константных доменов несут свободную тиоловую группу.
Фрагменты антител для применения в настоящем изобретении могут быть получены из любой из описанных здесь молекул антител, например, молекул антител, содержащих домены VH и/или VL или CDR любых из описанных здесь антител, такими способами, как расщепление ферментами, такими как пепсин или папаин, или расщепление дисульфидных мостиков химическим восстановлением. Другим путем, фрагменты антител настоящего изобретения могут быть получены способами генетической рекомбинации, также хорошо известными лицу с квалификацией в данной области, или еще пептидным синтезом с использованием, например, автоматического синтезатора пептидов, такого как синтезаторы, поставляемые компанией Applied Biosystems, и т.д., или синтезом и экспрессией нуклеиновых кислот.
Функциональные фрагменты антител согласно настоящему изобретению включают любой функциональный фрагмент, полупериод существования которого увеличен химической модификацией, особенно ПЭГилированием, или включением в липосому.
dAb (состоящее из одного домена антитело) является малым мономерным антигенсвязывающим фрагментом антитела, а именно, вариабельной областью тяжелой или легкой цепи антитела (Holt 2003). VH-dAb встречается природно в семействе верблюдовых (например, верблюде, ламе) и может быть получено иммунизацией животного из семейства верблюдовых антигеном-мишенью, выделением антиген-специфических В-клеток и прямого клонирования генов dAb из индивидуальных В-клеток. dAb могут быть также продуцированы в культуре клеток. Их малый размер, хорошая растворимость и термостабильность делает их особенно физиологически применимыми и подходящими для отбора и созревания аффинности. Связывающий элемент настоящего изобретения может быть dAb, содержащим домен VH или VL, по существу установленным здесь, или домен VH или VL, содержащий набор CDR, по существу установленный здесь.
В данном контексте, фраза "по существу установленные" относится к характеристике (характеристикам) релевантных CDR домена VH или VL связывающих элементов, описанных здесь, которые будут либо идентичными, либо в высокой степени сходными с указанными областями, последовательность которых установлена здесь. Как описано здесь, фраза "в высокой степени сходные" в отношении указанной области одного или нескольких вариабельных доменов обозначает, что от 1 до приблизительно 5, например, от 1 до 4, в том числе 1-3 или 1 или 2, или 3 или 4 аминокислотных замен могут быть произведены в этом CDR и/или домене VH или VL.
Биспецифические или бифункциональные антитела образуют вторую генерацию моноклональных антител, в которых две различные вариабельных области объединены в одной и той же молекуле (Holliger 1999). Их применение демонстрировалось как в диагностической области, так и в области терапии на основе их способности рекрутировать новые эффекторные функции или поражать несколько молекул на поверхности опухолевых клеток. Когда необходимо использовать биспецифические антитела, они могут быть общепринятыми биспецифическими антителами, которые могут быть получены различными путями (Holliger 1993b), например, получены химически или из гибридных гибридом, или могут быть любым из вышеупомянутых биспецифических фрагментов антител. Эти антитела могут быть получены химическими способами (Glennie 1987; Repp 1995) или соматическими способами (Staerz 1986; Suresh 1986), но также способами генетической инженерии, которые позволяют усиливать гетеродимеризацию и, следовательно, облегчать процесс очистки желаемого антитела. (Merchand 1998). Примеры биспецифических антител включают антитела, полученные способом BiTE™, в котором могут быть использованы связывающие домены двух антител с разной специфичностью, причем они могут быть непосредственно связаны через короткие гибкие пептиды. Этот способ объединяет два антитела на короткой единой полипептидной цепи. Могут быть сконструированы диатела и scFv без области Fc, с использованием только вариабельных доменов, что потенциально уменьшает эффекты антиидиотипической реакции.
Биспецифические антитела могут быть сконструированы в виде полного IgG, в виде биспецифического F(ab’)2, в виде Fab’PEG, в виде диател или также в виде биспецифического scFv. Кроме того, два биспецифических антитела могут быть связаны с использованием рутинных способов, известных в данной области, с образованием тетравалентных антител.
Биспецифические диатела, в отличие от биспецифических полных антител, могут быть также применимы, так как они могут быть легко сконструированы и экспрессированы в E. coli. Диатела (и многие другие полипептиды, такие как фрагменты антител) подходящих специфичностей связывания могут быть легко отобраны с использованием фагового дисплея (WO94/13804) из библиотек. Если одно плечо диатела должно сохраняться константным, например, со специфичностью, направленной против антигена-мишени, то может быть изготовлена библиотека, в которой другое плечо варьируется, и может быть отобрано антитело подходящей специфичности. Биспецифические полные антитела могут быть получены альтернативными способами генной инженерии, описанными в Ridgeway 1996.
В данной области доступны различные способы получения антител против антигена-мишени. Эти антитела могут быть моноклональными антителами, особенно антителами человека, мышиными, химерными или гуманизированными антителами, которые могут быть получены в соответствии со стандартными способами, хорошо известными лицу с квалификацией в данной области.
Обычно, для получения моноклональных антител или их функциональных фрагментов, в частности, мышиного происхождения, можно сослаться на способы, которые описаны, в частности, в справочнике "Антитела" (Harlow and Lane 1988), или на способ получения из гибридом, описанный Kohler and Milstein, 1975.
Моноклональные антитела могут быть получены, например, из клетки животного, иммунизированного против A-FN или одного из его фрагментов, содержащих эпитоп, узнаваемый указанными моноклональными антителами, например, фрагмента, содержащего ED-A или состоящего из ED-A, или пептидного фрагмента ED-A. Эти A-FN или один из его фрагментов могут быть, в частности, получены в соответствии с обычными рабочими способами, посредством генетической рекомбинации с использованием исходной последовательности нуклеиновой кислоты, содержащейся в кДНК-последовательности, кодирующей A-FN или его фрагмент.
Моноклональные антитела могут быть, например, очищены на аффинной колонке, на которой был предварительно иммобилизован A-FN или один из его фрагментов, содержащий эпитоп, узнаваемый указанными моноклональными антителами, например, фрагмент, содержащий ED-A или пептидный фрагмент ED-A или состоящий из ED-A или пептидного фрагмента ED-A. Моноклональные антитела могут быть очищены хроматографией на белке А и/или G, с последующей очисткой ионообменной хроматографией или без последующей очистки ионообменной хроматографией, нацеленной на элиминацию остаточных белковых примесей, а также ДНК и LPS, самих по себе, с последующей вытеснительной хроматографией на Сефарозном геле или без вытеснительной хроматографии на Сефарозном геле для элиминации потенциальных агрегатов вследствие присутствия димеров или других мультимеров. Все из этих способов могут использоваться одновременно или последовательно.
Антигенсвязывающий сайт
Этот термин описывает часть молекулы, которая связывается со всем антигеном-мишенью или с частью антигена-мишени или комплементарна всему антигену-мишени или части антигена-мишени. В молекуле антитела этот термин относится к антигенсвязывающему сайтом, и он содержит часть антитела, которая связывается со всем антигеном-мишенью или с частью антигена-мишени или комплементарна всему антигену-мишени или части антигена-мишени. Когда антиген является большим, антитело может связываться только с конкретной частью этого антигена, которую называют эпитопом. Антигенсвязывающий сайт антитела может быть обеспечен одним или несколькими вариабельными доменами антитела. Антигенсвязывающий сайт может содержать вариабельную область легкой цепи антитела (VL) и вариабельную область тяжелой цепи антитела (VH).
Выделенные
Этот термин относится к состоянию, в котором связывающие элементы для применения в настоящем изобретении или нуклеиновая кислота, кодирующая такие связывающие элементы, будут обычно находиться в соответствии с настоящим изобретением. Таким образом, связывающие элементы, домены VH и/или VL настоящего изобретения, могут быть обеспечены в выделенном и/или очищенном виде, например, из их природного окружения, по существу в чистой или гомогенной форме, или, в случае нуклеиновой кислоты, в виде свободной или по существу свободной нуклеиновой кислоты или генов, имеющих происхождение, другое, чем последовательность, кодирующая полипептид с требуемой функцией. Выделенные элементы и выделенная нуклеиновая кислота будут свободны или по существу свободны от материала, с которым они природно ассоциированы, такого как другие полипептиды или нуклеиновые кислоты, с которыми их обнаруживают в их природном окружении, или окружении, в котором они получены (например, в культуре клеток), когда такое получение выполняют технологией рекомбинантных ДНК, практикуемой in vitro или in vivo. Элементы и нуклеиновые кислоты могут быть приготовлены с разбавителями или адъювантами и все еще быть выделенными для практических целей - например, эти элементы будут нормально смешиваться с желатином или другими носителями при использовании для покрытия микротитрационных планшетов для применения в иммуноанализах, или будут смешиваться с фармацевтически приемлемыми носителями или разбавителями при использовании в диагностике или терапии. Связывающие элементы могут быть гликозилированы, либо природно, либо при помощи систем гетерологичных эукариотических клеток (например, клеток CHO или NS0 (ECACC 85110503) или они могут быть (например, при получении экспрессией в прокариотических клетках) негликозилированными.
Гетерогенные препараты, содержащие молекулы антител, могут быть также использованы в настоящем изобретении. Например, такие препараты могут быть смесями антител с полноразмерными тяжелыми цепями и тяжелыми цепями, лишенными С-концевого лизина, с различными степенями гликозилирования и/или с дериватизованными аминокислотами, например, циклизацией N-концевой глутаминовой кислоты с образованием остатка пироглутаминовой кислоты.
Один или несколько связывающих элементов в отношении антигена, например, A-FN или ED-A фибронектина, могут быть получены приведением в контакт библиотеки связывающих элементов по настоящему изобретению и антигена или его фрагмента, например, фрагмента, содержащего ED-A или пептидный фрагмент ED-A или состоящего из ED-A или пептидного фрагмента ED-A, и отбором одного или нескольких связывающих элементов этой библиотеки, способных связывать этот антиген.
Библиотека антител может быть подвергнута скринингу с использованием Итеративного скрининга колоний на фильтре (ICFS). В ICFS, бактерии, содержащие ДНК, кодирующую несколько связывающих специфичностей, выращивают в жидкой среде и после достижения стадии экспоненциального роста несколько миллиардов их распределяют на другой носитель, состоящий из предварительно обработанного подходящим образом мембранного фильтра, который инкубируют до появления полностью конфлюэнтных колоний бактерий. Второй улавливающий субстрат состоит из другого мембранного фильтра, предварительно увлажненного и покрытого желаемым антигеном.
Затем этот улавливающий мембранный фильтр помещают на чашку, содержащую подходящую культуральную среду, и покрывают фильтром для выращивания с поверхностью, покрытой бактериальными колониями, ориентированными вверх. Полученный таким образом сэндвич инкубируют при комнатной температуре в течение приблизительно 16 часов. Таким образом можно получить экспрессию генов, кодирующих фрагменты антител scFv, имеющие распределяющее действие, так что эти фрагменты, связывающиеся специфически с антигеном, который присутствует на улавливающей мембране, улавливаются. Затем эту улавливающую мембрану обрабатывают для указания связанных фрагментов антител scFv при помощи колориметрических способов, обычно используемых для этой цели.
Положения окрашенных пятен на улавливающем фильтре позволяет вернуться обратно к соответствующим бактериальным колониям, которые присутствуют на мембране для выращивания и продуцируют улавливаемые фрагменты антител. Такие колонии собирают и выращивают, и бактерии (несколько миллионов) распределяют на новый культуральный мембранный фильтр, повторяя вышеописанные процедуры. Затем проводят аналогичные циклы, пока положительные сигналы на улавливающей мембране соответствуют отдельным положительным колониям, каждая из которых представляет потенциальный источник фрагментов моноклонального антитела, направленных против антигена, используемого в этом отборе. ICFS описан, например, в WO0246455, который включен здесь в качестве ссылки. Библиотека может быть также дисплеем на частицах или молекулярных комплексах, например, реплицируемых генетических упаковках таких частиц бактериофагов (например, T7), или другими системами дисплея in vitro, причем каждая частица или каждый молекулярный комплекс содержит нуклеиновую кислоту, кодирующую вариабельный домен VH антитела, визуально воспроизведенный на них, и необязательно также визуально воспроизведенный домен VL, если он присутствует. Фаговый диплей описан в WO92/01047 и, например, в патентах США US5969108, US5565332, US5733743, US5858657, US5871907, US5872215, US5885793, US5962255, US6140471, US6172197, US6225447, US6291650, US6492160 и US6521404, каждый из которых включен здесь в качестве ссылки в его полном виде.
После отбора связывающих элементов, способных связывать этот антиген и визуально воспроизводить на бактериофагах или других частицах или молекулярных комплексах библиотеки, нуклеиновая кислота может быть извлечена из бактериофагов или других частиц или молекулярных комплексов, визуально воспроизводящих указанный отобранный связывающий элемент. Такая нуклеиновая кислота может быть использована в последующем продуцировании связывающего элемента или вариабельного домена VH или VL антитела экспрессией из нуклеиновой кислоты с последовательностью нуклеиновой кислоты, полученной из бактериофага или другой частицы или молекулярного комплекса, визуально воспроизводящих указанный отобранный связывающий элемент.
Вариабельный домен VH антитела с аминокислотной последовательностью вариабельного домена VH указанного отобранного связывающего элемента может быть обеспечен в выделенной форме, как и связывающий элемент, содержащий такой домен VH.
Дополнительно может быть тестирована способность связывания A-FN или ED-A фибронектина или другого антигена-мишени или изоформы, например, способность конкуренции, например, с любым из анти-ED-A-антител H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 или G9 за связывание с A-FN или фрагментом A-FN, например, ED-A фибронектина.
Связывающий элемент для применения в настоящем изобретении может связывать специфически A-FN и/или ED-A фибронектина. Связывающий элемент настоящего изобретения может связывать A-FN и/или ED-A фибронектина с той же самой аффинностью, что и анти-ED-A-антитело H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 или G9, например, в формате scFv, или с лучшей аффинностью. Связывающий элемент для применения в настоящем изобретении может связывать A-FN и/или ED-A фибронектина с KD 3×10-8 M или с лучшей аффинностью. Предпочтительно, Связывающий элемент для применения в настоящем изобретении может связывать A-FN и/или ED-A фибронектина с KD 2×10-8 M или с лучшей аффинностью. Более предпочтительно, Связывающий элемент для применения в настоящем изобретении может связывать A-FN и/или ED-A фибронектина с KD 1,7×10-8 M или с лучшей аффинностью. Еще более предпочтительно, Связывающий элемент для применения в настоящем изобретении может связывать A-FN и/или ED-A фибронектина с KD 1,4×10-8 M или с лучшей аффинностью. Наиболее предпочтительно, Связывающий элемент для применения в настоящем изобретении может связывать A-FN и/или ED-A фибронектина с KD 3×10-9 M или с лучшей аффинностью.
Связывающий элемент настоящего изобретения может связываться с тем же самым эпитопом на A-FN и/или ED-A фибронектина, что и анти-ED-A-антитело H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 или G9.
Связывающий элемент для применения в настоящем изобретении может не обнаруживать какого-либо значимого связывания с молекулами, другими, чем A-FN и/или ED-A фибронектина. В частности, связывающий элемент может не связывать другие изоформы фибронектина, например, изоформу ED-B или изоформу IIICS фибронектина.
Варианты молекул антител, описанных здесь, могут быть получены и использованы в настоящем изобретении. Способы, необходимые для получения замен в аминокислотных последовательностях CDR, доменах VH или VL антитела и связывающих элементах обычно являются доступными в данной области. Могут быть получены вариантные последовательности с заменами, которые, как может быть предсказано или может не быть предсказано, оказывают минимальное или благоприятное действие на активность, и тестированы на способность связывания A-FN и/или ED-A фибронектина и/или любое другое желаемое свойство.
Варианты аминокислотной последовательности вариабельных доменов любого из доменов VH и VL, последовательности которых специально описаны здесь, могут быть использованы в соответствии с настоящим изобретением, как обсуждалось. Конкретные варианты могут включать одно или несколько изменений последовательности (добавление, делецию, замену и/или инсерцию аминокислотного остатка), могут быть меньшими, чем приблизительно 20 изменений, меньшими, чем приблизительно 15 изменений, меньшими, чем приблизительно 10 изменений или меньшими, чем приблизительно 5 изменений, могут быть 5, 4, 3, 2 изменениями или 1 изменением. Изменения могут быть произведены в одном или нескольких каркасных областях и/или одной или нескольких CDR. Эти изменения обычно не приводят к потере функции, так что связывающий элемент может сохранять способность связывания A-FN и/или ED-A фибронектина. Например, он может сохранять то же самое количественное связывание, что и связывающий элемент, в котором не произведено изменение, например, как измерено в анализе, описанном здесь. Связывающий элемент, содержащий измененную таким образом аминокислотную последовательность, может иметь улучшенную способность связывать A-FN и/или ED-A фибронектина.
Новые области VH или VL, несущие произведенные из CDR последовательности, для применения в настоящем изобретении могут быть генерированы с использованием случайного мутагенеза одного или нескольких генов выбранных генов VH и/или VL для генерирования мутаций в полном вариабельном домене. В некоторых вариантах одна или две аминокислотные замены производят в полном вариабельном домене или наборе CDR. Другим способом, который может быть использован, является прямой мутагенез в отношении областей CDR генов VH или VL.
Как отмечалось выше, аминокислотная последовательность CDR, по существу установленная здесь, может находиться в виде CDR в вариабельном домене антитела человека или его существенной части. Последовательности HCDR3, по существу установленные здесь, представляют варианты настоящего изобретения, и, например, каждая из них может находиться в виде HCDR3 в вариабельном домене тяжелой цепи антитела человека или его существенной части.
Вариабельные домены, используемые в настоящем изобретении, могут быть получены или произведены из любой зародышевой линии или реаранжированного вариабельного домена человека, или могут быть синтетическим вариабельным доменом на основе консенсусных или фактических последовательностей известных вариабельных доменов человека. Вариабельный домен может быть произведен из антитела не человека. Последовательность CDR для применения в настоящем изобретении (например, CDR3) может быть введена в спектр вариабельных доменов, лишенных CDR (например, CDR3), с использованием технологии рекомбинантных ДНК. Например, Marks et al. (1992) описывают способы получения спектров (репертуаров) вариабельных доменов антител, в которых консенсусные праймеры, направленные на 5’-конец или смежные с 5’-концом зоны вариабельного домена, используют вместе с консенсусными праймерами относительно третьей каркасной области генов VH человека, для обеспечения спектра вариабельных доменов VH, лишенных CDR3. Marks et al. дополнительно описывают, как этот спектр может быть объединен с CDR3 конкретного антитела. С использованием аналогичных способов, произведенные из CDR3 последовательности настоящего изобретения могут быть перетасованы со спектрами доменов VH или VL, лишенных CDR3, и эти перетасованные полные домены VH или VL объединяются с родственными доменами VL или VH с получением связывающих элементов для применения в настоящем изобретении. Затем этот спектр может быть визуально воспроизведен в подходящей системе-хозяине, такой как система фагового дисплея WO92/01047, который включен здесь в качестве ссылки в его полном виде, или любой системе, описанной в последующем объеме литературы, в том числе Kay, Winter & McCafferty (1996) так что могут быть отобраны подходящие связывающие элементы. Спектр может состоять из любого количества от 104 индивидуальных элементов и выше, например, до по меньшей мере 105, по меньшей мере 106, по меньшей мере 107, по меньшей мере 108, по меньшей мере 109 или по меньшей мере 1010 элементов.
Подобным образом, один или более или все три CDR могут быть трансплантированы в спектр доменов VH или VL, которые затем подвергают скринингу на связывающий элемент или связывающие элементы для A-FN и/или ED-A фибронектина.
Могут быть использованы один или более из HCDR1, HCDR2 и HCDR3 антитела H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 или G9, или набор HCDR и/или один или более из X LCDR1, LCDR2 и LCDR3 антитела H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 или G9 или набор LCDR антитела H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 или G9.
Подобным образом, могут быть использованы другие домены VH и VL, наборы CDR и наборы HCDR и/или наборы LCDR, описанные здесь.
A-FN и/или ED-A фибронектина могут быть использованы в скрининге на связывающие элементы, например, полипептиды антител, для применения в приготовлении лекарственного средства для лечения ревматоидного артрита. Этот скрининг может быть скринингом спектра доменов, как описано в другом месте здесь.
Основная часть вариабельного домена иммуноглобулина может содержать по меньшей мере три области CDR, вместе с их промежуточными каркасными областями. Эта часть может также включать по меньшей мере приблизительно 50% как первых, так и четвертых каркасных областей, причем эти 50% являются С-концевыми 50% первой каркасной области и N-концевыми 50% четвертой каркасной области. Дополнительными остатками на N-концевой или С-концевой стороне основной части вариабельного домена могут быть остатки, в норме не ассоциированные с природно-встречающимися областями вариабельных доменов. Например, конструирование связывающих элементов настоящего изобретения, выполняемое способами рекомбинантных ДНК, может приводить к введению N- или C-концевых остатков, кодируемых линкерами, введенными для облегчения клонирования или других стадий манипулирования. Другие стадии манипулирования включают введение линкеров для присоединения вариабельных доменов, описанных в другом месте здесь, к дополнительным белковым последовательностям, включающим константные области антитела, другие вариабельные домены (например, для получения диател) или детектируемые/функциональные метки, как обсуждается более подробно в другом месте здесь.
Хотя связывающие элементы могут содержать пару доменов VH и VL, отдельные связывающие домены на основе последовательностей доменов VH или VL могут быть также использованы в настоящем изобретении. Известно, что отдельные домены иммуноглобулина, в частности, домены VH, способны связывать антигены-мишени специфически образом. Например, смотрите обсуждение dAb выше.
В случае любого из отдельных связывающих доменов, эти домены могут быть использованы для скрининга на комплементарные домены, способные образовывать двухдоменный связывающий элемент, способные связывать A-FN и/или ED-A фибронектина. Это может быть достигнуто способами скрининга фагового дисплея с использованием так называемого иерархического двойного комбинаторного подхода, описанного в WO92/01047, включенного здесь в качестве ссылки в его полном виде, в котором отдельную колонию, содержащую клон либо H-, либо L-цепи, используют для инфицирования полной библиотеки клонов, кодирующих другую цепь (L или H), и полученный двухцепочечный связывающий элемент отбирают в соответствии со способами фагового дисплея, такими как способы, описанные в этой ссылке. Этот способ описан также в Marks 1992.
Связывающие элементы для применения в настоящем изобретении, могут дополнительно содержать константные области или их части, например, константные области или их части антитела человека. Например, домен VL может быть присоединен на его С-конце к константным доменам легкой цепи, включающим цепи Сκ или Cλ, например, Cλ. Подобным образом, связывающий элемент на основе домена VH может быть присоединен на его С-конце ко всей или части (например, домену СН1) тяжелой цепи иммуноглобулина, произведенной из любого изотипа антитела, например, IgG, IgA, IgE и IgM и любого из подклассов изотипа, в частности, IgG1 и IgG4. Любой синтетический или другой вариант константной области, которая имеет эти свойства и стабилизирует вариабельные области, также применим в вариантах настоящего изобретения.
Связывающие элементы для применения в настоящем изобретении могут быть помечены детектируемой или функциональной меткой. Меткой может быть любая молекула, которая продуцирует сигнал или может быть индуцирована для продуцирования сигнала, в том числе, но не только, флуоресцирующие агенты, радиоактивные метки, ферменты, хемилюминесцирующие агенты или фотосенсибилизаторы. Таким образом, связывание может быть детектировано и/или измерено детектированием флуоресценции или люминесценции, радиоактивности, активности фермента или поглощения света. Детектируемые метки могут быть присоединены к антителам для применения в настоящем изобретении с использованием общепринятых химических способов, известных в данной области.
Имеются многочисленные способы, при помощи которых метка может производить сигнал, детектируемый наружными средствами, например, визуальным обследованием, с использованием электромагнитной радиации, нагревания и химических реагентов. Эта метка может быть также связана с другим связывающим элементом, который связывает антитело для применения в этом исследовании, или с носителем.
Меченые связывающие элементы, например, scFv, меченые детектируемой меткой, могут быть использованы диагностически in vivo, ex vivo или in vitro, и/или терапевтически.
Например, радиоактивно меченые связывающие элементы (например, связывающие элементы, конъюгированные с радиоизотопом) могут быть использованы в радиодиагностике и радиотерапии. Радиоизотопы для применения в настоящем изобретении включают такие изотопы, как 94mTc, 99mTc, 186Re, 188Re, 203Pb, 67Ga, 68Ga, 47Sc, 111In, 97Ru, 62Cu, 64Cu, 86Y, 88Y, 90Y, 121Sn, 161Tb, 153Sm, 166Ho, 105Rh, 177Lu, 123I, 124I, 125I и 131I.
Например, Связывающий элемент для применения в настоящем изобретении, меченый детектируемой меткой, может быть использован для детектирования, диагностики или мониторинга ревматоидного артрита у человека или животного.
Связывающий элемент настоящего изобретения может быть использован для приготовления диагностического продукта для применения в диагностике ревматоидного артрита.
Настоящее изобретение обеспечивает способ детекции или диагностики ревматоидного артрита у человека или животного, предусматривающий:
(a) введение человеку или животному связывающего элемента по изобретению, например, меченого детектируемой меткой, который связывает изоформу ED-A фибронектина, и
(b) определение присутствия или отсутствия связывающего элемента в новообразованной сосудистой сети тела человека или животного;
где локализация этого связывающего элемента в новообразованной сосудистой сети у человека или животного является указанием на наличие ревматоидного артрита.
При мечении связывающего элемента детектируемой меткой присутствие или отсутствие детектируемой метки может быть определено детектированием этой метки.
Конъюгат или слияние между связывающим элементом для применения в настоящем изобретении и молекулой, которая проявляет биоцидное, цитотоксическое, иммуносупрессивное или противовоспалительное действие на клетках-мишенях в повреждениях, и антителом, направленным против компонента внеклеточного матрикса, который присутствует в таких повреждениях, могут быть использованы в настоящем изобретении. Например, этой конъюгированной молекулой может быть inter alia интерлейкин-10, противовоспалительное или другое лекарственное средство, фотосенсибилизатор или радионуклид. Такие конъюгаты могут быть использованы терапевтически, например, для лечения ревматоидного артрита, как описано здесь.
Получение и применение слияний или конъюгатов связывающих элементов с биоцидными или цитотоксическими молекулами описано, например, в WO01/62298, который включен здесь в качестве ссылки.
Настоящее изобретение обеспечивает способ лечения ревматоидного артрита, предусматривающий введение индивидууму терапевтически эффективного количества лекарственного средства, содержащего Связывающий элемент для применения в настоящем изобретении.
Этот связывающий элемент может быть конъюгатом (i) молекулы, которая проявляет противовоспалительное действие на клетках-мишенях посредством клеточного взаимодействия, противовоспалительной молекулы, IL-10, TGF-бета или другим лекарственным средством, и (ii) связывающего элемента для изоформы ED-A фибронектина и/или ED-A фибронектина.
Этот связывающий элемент может быть конъюгатом (i) молекулы, которая проявляет иммуносупрессивное или противовоспалительное действие, и (ii) связывающего элемента для изоформы ED-A фибронектина и/или ED-A фибронектина.
Этот связывающий элемент может быть конъюгатом (i) интерлейкина-10 (IL10) или TGF-β и (ii) связывающего элемента изоформы ED-A фибронектина и/или ED-A фибронектина. Такой связывающий элемент применим в аспектах настоящего изобретения, описанных здесь, касающихся лечения ревматоидного артрита.
Настоящее изобретение обеспечивает использование связывающего элемента для применения в настоящем изобретении для приготовления лекарственного средства для лечения ревматоидного артрита.
Связывающий элемент может быть конъюгирован или слит с молекулой, которая проявляет биоцидное, цитотоксическое, иммуносупрессивное или противовоспалительное действие, как описано здесь. Связывающий элемент может быть конъюгатом (i) молекулы, которая проявляет биоцидное или цитотоксическое действие на клетках-мишенях посредством клеточного взаимодействия или имеет иммуносупрессивное или противовоспалительное действие, и (ii) связывающего элемента для фибронектина человека в соответствии с настоящим изобретением.
Здесь описан также конъюгат (i) молекулы, которая проявляет биоцидное или цитотоксическое действие на клетках-мишенях посредством клеточного взаимодействия или имеет иммуносупрессивое или противовоспалительное действие, и (ii) связывающего элемента для фибронектина человека в соответствии с применением в настоящем изобретении. Такой конъюгат предпочтительно содержит слитый белок, содержащий биоцидную, цитотоксическую, иммуносупрессивную или противовоспалительную молекулу и указанный связывающий элемент, или, когда этот связывающий элемент является двухцепочечным или мультицепочечным, слитый белок, содержащий биоцидную, цитотоксическую, иммуносупрессивную или противовоспалительную молекулу и компонент полипептидной цепи указанного связывающего элемента. Предпочтительно, этот связывающий элемент является одноцепочечным полипептидом, например, молекулой одноцепочечного антитела, такой как scFv.
В настоящем изобретении может быть использован слитый белок, содержащий иммуносупрессивную или противовоспалительную молекулу и молекулу одноцепочечного Fv-антитела.
Иммуносупрессивная или противовоспалительная молекула, которая проявляет свое действие на клетках-мишенях посредством клеточного взаимодействия, может взаимодействовать непосредственно с клетками-мишенями, может взаимодействовать с мембраносвязанным рецептором на клетке-мишени или нарушать электрохимический потенциал клеточной мембраны. В примере предпочтительного варианта этой молекулой является IL-10.
Как обсуждается дополнительно ниже, этот специфический связывающий элемент является предпочтительно антителом или содержит антигенсвязывающий сайт антитела. Предпочтительно, этот специфический связывающий элемент может быть одноцепочечным полипептидом, таким как одноцепочечное антитело. Это делает возможным удобное получение слитого белка, содержащего одноцепочечное антитело и иммуносупрессивную или противовоспалительную молекулу (например, интерлейкин-10 или TGF-бета). Антигенсвязывающий сайт антитела может быть обеспечен посредством ассоциации домена VH антитела и домена VL антитела в отдельных полипептидах, например, в полном антителе или во фрагменте антитела, таком как Fab или диатело. Когда специфический связывающий элемент является двухцепочечной или мультицепочечной молекулой (например, Fab или полным антителом, соответственно), иммуносупрессивная или противовоспалительная молекула может быть конъюгирована в виде слитого полипептида с одной или несколькими полипептидными цепями в этом специфическом связывающем элементе.
Связывающий элемент может быть конъюгирован с иммуносупрессивной или противовоспалительной молекулой посредством пептидной связи, т.е. в слитом полипептиде, содержащем указанную молекулу и специфический связывающий элемент, или компонент его полипептидной цепи (смотрите, например, Trachsel et al.). Другие способы конъюгации включают химическую конъюгацию, в частности, сшивание с использованием бифункционального реагента (например, с использованием DOUBLE-REAGENTS™ Cross-linking Reagents Selection Guide, Pierce).
Здесь описана также выделенная нуклеиновая кислота, кодирующая Связывающий элемент для применения в настоящем изобретении. Нуклеиновая кислота может включать ДНК и/или РНК. Нуклеиновая кислота может кодировать CDR или набор CDR или домен VH или домен VL или антигенсвязывающий сайт молекулы антитела, например, scFv или IgG, например, IgG1, определенные здесь. Эти нуклеиновые кислоты могут кодировать описанные здесь домены VH и/или VL.
Кроме того, здесь описаны конструкции в форме плазмид, векторов, транскрипционных или экспрессионных кассет, которые содержат по меньшей мере один из полинуклеотидов, описанных выше.
Описана также рекомбинантная клетка-хозяин, которая содержит одну или несколько конструкций, описанных выше. Описана нуклеиновая кислота, кодирующая любой CDR или набор CDR или домен VH или домен VL или антигенсвязывающий сайт антитела или молекулу антитела, например, scFv или IgG1 или IgG4, а также способ получения кодируемого продукта, предусматривающий экспрессию из кодирующей нуклеиновой кислоты. Экспрессия может удобным образом достигаться культивированием при подходящих условиях рекомбинантных клеток-хозяев, содержащих эту нуклеиновую кислоту. После получения посредством экспрессии, домен VH или VL или связывающий элемент могут быть выделены и/или очищены с использованием любого подходящего способа и затем использованы соответствующим образом.
Нуклеиновая кислота может содержать ДНК или РНК и может быть полностью или частично синтетической. Ссылка на нуклеотидную последовательность, установленную здесь, включает молекулу ДНК с указанной последовательностью и включает молекулу РНК с указанной последовательностью, в которой U заменяет T, если нет других указаний.
Описан также способ получения вариабельного домена VH, предусматривающий вызывание экспрессии из кодирующей нуклеиновой кислоты. Такой способ может предусматривать культивирование клеток-хозяев в условиях, подходящих для продуцирования указанного вариабельного домена VH антитела.
Способ получения может включать стадию выделения и/или очистки этого продукта. Способ получения может включать приготовление этого продукта в виде композиции, включающей по меньшей мере один дополнительный компонент, такой как фармацевтически приемлемый эксципиент.
Системы для клонирования и экспрессии полипептида во множестве различных клеток-хозяев хорошо известны. Подходящие клетки-хозяева включают бактерии, клетки млекопитающих, клетки растений, мицелиальные грибы, дрожжи и бакуловирусные системы и клетки трансгенных растений и животных. Экспрессия антител и фрагментов антител в прокариотических клетках хорошо установлена в данной области. В отношении обзора смотрите, например, Plϋckthun 1991. Обычным бактериальным хозяином является E. coli.
Экспрессия в эукариотических клетках в культуре также доступна специалистам с квалификацией в данной области в качестве одной возможности получения связывающего элемента, например, Chadd & Chamow (2001), Andersen & Krummen (2002), Larrick & Thomas (2001). Линии клеток млекопитающих, доступные в данной области для экспрессии гетерологичного полипептида, включают клетки яичника китайского хомячка (СНО), клетки HeLa, клетки почки детеныша хомячка, клетки мышиной меланомы NS0, клетки миеломы крысы YB2/0, клетки эмбриональной почки человека, клетки эмбриональной сетчатки человека и многие другие.
Могут быть выбраны или сконструированы подходящие векторы, содержащие подходящие регуляторные последовательности, включающие последовательности промотора, последовательности терминатора, последовательности полиаденилирования, последовательности энхансера, маркерные гены и другие последовательности, по мере необходимости. Векторами могут быть плазмиды, например, фагмида, или вирусы, например, ‘фаги, по мере необходимости. В отношении дополнительных подробностей смотрите, например, Sambrook & Russell (2001). Многие известные способы и протоколы для манипулирования нуклеиновой кислотой, например, в получении конструкций нуклеиновых кислот, мутагенезе, секвенировании, введении ДНК в клетки и экспрессии генов, и для анализа белков описаны подробно в Ausubel 1999.
Клетка-хозяин может содержать нуклеиновую кислоту, описанную здесь. Такая клетка-хозяин может находиться in vitro или может находиться в культуре. Такая клетка-хозяин может находиться in vivo. Присутствие in vivo клетки-хозяина может сделать возможной внутриклеточную экспрессию связывающего элемента для применения в настоящем изобретении в виде "интрател" или внутриклеточных антител. Интратела могут быть использованы для генотерапии.
Описан также способ, предусматривающий введение описанной здесь нуклеиновой кислоты в клетку-хозяина. Это введение может использовать любой доступный способ. Для эукариотических клеток, подходящие способы могут включать кальций-фосфатную трансфекцию, DEAE-декстрановый способ, электропорацию, опосредованную липосомами трансфекцию и трансдукцию с использованием ретровируса или другого вируса, например, вируса коровьей оспы, или, для клеток насекомых, бакуловируса. Введение нуклеиновой кислоты в клетку-хозяина, в частности, в эукариотическую клетку, может использовать вирусную или основанную на плазмиде систему. Плазмидная система может поддерживаться эписомно или может быть включена в клетку-хозяина или в искусственную хромосому. Включение может быть либо случайной, либо нацеленной интеграцией одной или нескольких копий в единственный локус или во множественные локусы. Для бактериальных клеток, подходящие способы могут включать кальций-хлоридную трансформацию, электропорацию и трансфекцию с использованием бактериофага.
За этим включением может следовать индуцирование или позволение экспрессии из этой нуклеиновой кислоты, например, культивированием клеток-хозяев в условиях, подходящих для экспрессии этого гена. Очистка экспрессируемого продукта может достигаться способами, известными квалифицированному в данной области специалисту.
Эта нуклеиновая кислота может быть интегрирована в геном (например, в хромосому) клетки-хозяина. Интеграция может быть усилена включением последовательностей, которые стимулируют рекомбинацию с геномом, в соответствии со стандартными способами.
Описан также способ, которые предусматривает использование конструкции, указанной выше, для экспрессии связывающего элемента или полипептида, описанного выше.
Связывающие элементы для применения в настоящем изобретении предназначены для применения в способах диагностики или лечения у человека или животного в качестве субъекта, например, у человека. Связывающие элементы для применения в настоящем изобретении могут быть использованы в диагностике или лечении ревматоидного артрита.
Таким образом, настоящее изобретение обеспечивает способы лечения, предусматривающие введение обеспеченного связывающего элемента, фармацевтические композиции, содержащие такой связывающий элемент, и применение такого связывающего элемента в приготовлении лекарственного средства для введения, например, в способе приготовления лекарственного средства или фармацевтической композиции, предусматривающем приготовление связывающего элемента с фармацевтически приемлемым эксципиентом. Фармацевтически приемлемые носители хорошо известны и будут адаптироваться лицом с квалификацией в данной области в зависимости от характера и способа введения выбранного активного соединения (выбранных активных соединений).
Связывающие элементы для применения в настоящем изобретении будут обычно вводиться в форме фармацевтической композиции, которая может содержать по меньшей мере один компонент, наряду со связывающим элементом. Таким образом, фармацевтическая композиция по настоящему изобретению и для применения в соответствии с настоящим изобретением может содержать, наряду с активным ингредиентом, фармацевтически приемлемый эксципиент, носитель, буфер, стабилизатор или другие материалы, хорошо известные квалифицированным в данной области специалистам. Такие материалы должны быть нетоксичными и не должны препятствовать эффективности активного ингредиента. Точная природа носителя или другого материала будет зависеть от способа введения, который может быть пероральным способом, использующим ингаляцию способом или инъекцией, например, внутривенной.
Фармацевтические композиции для перорального введения, такие как, например, наночастицы и т.д., также предполагаются в настоящем изобретении. Такие пероральные готовые формы могут быть в форме таблетки, капсулы, порошка, жидкости или быть в полутвердой форме. Таблетка может содержать твердый носитель, такой как желатин или адъювант. Жидкие фармацевтические композиции обычно содержат жидкий носитель, такой как вода, вазелиновое масло, масла животных или растений, минеральное масло или синтетическое масло. Могут быть включены физиологический солевой раствор, декстроза или другой раствор сахарида или гликоли, такие как этиленгликоль, пропиленгликоль или полиэтиленгликоль.
Для внутривенной инъекции или инъекции в месте поражения активный ингредиент будет в форме парентерально приемлемого водного раствора, который является апирогенным и имеет подходящие рН, изотоничность и стабильность. Специалисты с соответствующей квалификацией в данной области вполне смогут приготовить подходящие растворы с использованием, например, изотонических носителей, таких как инъекционный раствор хлорида натрия, инъекционный раствор Рингера, лактатсодержащий раствор Рингера. Если требуется, могут быть использованы консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки. Многие способы приготовления фармацевтических готовых форм известны квалифицированным в данной области специалистам. Смотрите, например, Robinson, 1978.
Композиция может вводиться отдельно или а комбинации с другими способами лечения, одновременно или последовательно, или в виде объединенного препарата с другим терапевтическим агентом или другими терапевтическими агентами, в зависимости от подлежащего лечению состояния.
Связывающий элемент для применения в настоящем изобретении может быть использован в виде части комбинированной терапии вместе с дополнительным лекарственным компонентом. Комбинированные способы лечения могут быть использованы для обеспечения существенных синергических эффектов, в частности, комбинация связывающего элемента для применения в настоящем изобретении с одним или несколькими другими лекарственными средствами. Связывающий элемент для применения в настоящем изобретении может вводиться одновременно или последовательно или в виде объединенного препарата с другим терапевтическим агентом или другими терапевтическими агентами, для лечения одного или нескольких состояний, перечисленных здесь.
Например, Связывающий элемент для применения в настоящем изобретении может быть использован в комбинации с существующим терапевтическим агентом для лечения ревматоидного артрита.
Существующие терапевтические агенты для лечения ревматоидного артрита включают IL-10, TGF-β, фотосенсибилизаторы и цитотоксические лекарственные средства.
Связывающий элемент для применения в настоящем изобретении и один или несколько вышеуказанных дополнительных лекарственных компонентов могут быть использованы в приготовлении лекарственного средства. Это лекарственное средство может быть средством для раздельного или комбинированного введения индивидууму и, соответственно, может содержать связывающий элемент и дополнительный компонент в виде объединенного препарата или в виде раздельных препаратов. Раздельные препараты могут быть использованы для облегчения раздельного и последовательного или одновременного введения и для возможности введения этих компонентов различными способами, например, пероральным и парентеральным введением.
В соответствии с настоящим изобретением, обеспеченные композиции могут вводиться млекопитающим. Введение может быть введением в "терапевтически эффективном количестве", обнаруживающим пользу для пациента. Такой пользой может быть по меньшей мере уменьшение интенсивности по меньшей мере одного симптома. Таким образом, "лечением ревматоидного артрита" называют уменьшение интенсивности по меньшей мере одного симптома. Фактически вводимое количество и скорость и временной ход введения будут зависеть от природы и тяжести состояния, подлежащего лечению, конкретного животного, получающего лечение, клинического состояния этого индивидуального пациента, причины нарушения, участка доставки композиции, типа связывающего элемента, способа введения, схемы введения и других факторов, известных врачам-практикам. Прописывание лечения, например, решения в отношении дозы и т.д., находится в рамках ответственности общих врачей-практиков и других медицинских докторов и может зависеть от тяжести симптомов и/или прогрессирования подлежащего лечению заболевания. Подходящие дозы антитела хорошо известны в данной области (Ledermann 1991 и Bagshawe 1991. Могут быть использованы конкретные указанные здесь дозы или дозы, описанные в Physician's Desk Reference (2003), подходящие для типа вводимого лекарственного средства. Терапевтически эффективное количество или подходящая доза связывающего элемента для применения в настоящем изобретении может быть определена сравнением его активности in vitro и активности in vivo в модели животного. Способы экстраполяции эффективных доз в мышах и других тест-животных относительно человека известны. Точная доза будет зависеть от ряда факторов, в том числе от того, используется это антитело для диагностики, предупреждения или для лечения, от размера и местоположения обрабатываемой зоны, точного характера антитела (например, полное антитело, фрагмент или диатело) и природы любой детектируемой метки или другой молекулы, присоединенной к этому антителу. Обычная доза антитела будет находиться в диапазоне 100 мкг - 1 г для системных применений и 1 мкг - 1 мг для местных применений. Может вводиться начальная более высокая доза с последующим введением одной или нескольких более низких доз. Антитело может быть полным антителом, например, изотипа IgG1 или IgG4. Эта доза является дозой для одной обработки взрослого пациента, которая может пропорционально корректироваться для детей и младенцев, а также корректироваться для других форматов антител пропорционально молекулярной массе. Обработки могут повторяться с интервалами один раз в день, два раза в неделю, с недельными или месячными интервалами, по решению врача. Обработки могут проводиться каждые две - каждые четыре недели для подкожного введения и каждые четыре - каждые восемь недель для внутривенного введения. В некоторых вариантах осуществления настоящего изобретения, лечение является периодическим, и период между введениями составляет приблизительно две недели или более, например, приблизительно три недели или более, приблизительно четыре недели или более или приблизительно один раз в месяц. В других вариантах осуществления настоящего изобретения, лечение может предоставляться до и/или после операции и лекарство может вводиться или наноситься непосредственно в анатомическом участке хирургического вмешательства.
Дополнительные аспекты и варианты осуществления настоящего изобретения будут очевидными квалифицированным в данной области специалистам при условии прочтения данного описания, включающего следующие экспериментальные примеры.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
РЕЗУЛЬТАТЫ
Гистохимический анализ артритных проб человека
Экспрессию доменов EDA и EDB фибронектина, а также доменов А1 и С тенасцина-С исследовали иммуногистохимией на артритных пробах человека с использованием антител F8, L19, F16 и G11, соответственно.
На фиг.1 более темное окрашивание указывает экспрессию соответствующих антигенов (показанную белыми стрелками). Анти-EDA-антитело F8 приводило к более сильному окрашиванию, следовательно, все дальнейшие эксперименты выполняли с этим антителом.
Выполняли эксперименты с использованием иммунофлуоресценции с антителом F8, которые показали хорошее периваскулярное окрашивание (видимое в виде белых структур на фиг.2).
Моноклональное антитело F8 человека накапливается в участках артрита у мышей
Авторы изобретения исследовали нацеливающую эффективность in vivo F8 в формате миниантител (SIP) (Borsi et al. 2002) в мышиной модели CIA (Courtney et al. 1980) с использованием как флуоресценции, так и радиоактивности для детектирования антител. SIP-формат состоит из фрагмента антитела scFv, связанного с доменом CH4 IgE человека, с получением гомодимерного белка с размером 80 кДа.
Артритным мышам инъецировали SIP(F8), поглощающий в ближней инфракрасной области красителем Alexa 750. Спустя двадцать четыре часа после внутривенной инъекции, животных визуализировали с использованием блока для формирования изображения для инфракрасной флуоресценции (Birchler et al., 1999), выявляющего сильное и селективное накапливание антитела в повреждениях, присутствующих в артритной конечности, видимое в виде белых светящихся лапок с отеком 2-ой степени в передних лапках этой мыши.
Спустя двадцать четыре часа после внутривенной инъекции SIP(F8), радиоактивно меченого 125I, мышей умерщвляли и лапки визуализировали авторадиографией (фосфор-визуализацией) Преимущественное накопление радиоактивности наблюдали в воспаленных конечностях мышей, инъецированных SIP(F8), видимых в виде черного окрашивания при авторадиографии. Одна лапка обнаруживала артритный оценку 2 (отек всей лапки). Другую лапку классифицировали как артрит 1-ой степени (отек отдельных пальцев).
Активность слияния анти-ED-A-антитело-интерлейкин-10
Молекулу антитела F8 в scFv-формате конъюгировали со слитым белком с интерлейкином-10 (IL-10). Биологическую активность этого слитого белка сравнивали с биологической активностью IL-10 человека в анализе, определяющем способность индуцировать IL-4-зависимую пролиферацию клеток MC/9 (Thompson et al., 1991). Эти результаты показаны на фиг.8(d).
МАТЕРИАЛЫ И СПОСОБЫ
Иммуногистохимический анализ артритных проб человека
Замороженные срезы артритных проб человека фиксировали охлажденным на льду ацетоном в течение 10 минут, блокировали фетальной телячьей сывороткой в течение 30 минут и окрашивали на маркеры новообразованной сосудистой сети (фибронектин ED-A и ED-B, домен А1 и С тенасцина-С). Антитела F8, L19, F16 и G11 использовали в качестве myc-меченых scFv в концентрации 10 мкг/мл и инкубировали в течение 1 часа. Первичные антитела ко-инкубировали с анти-myc-антителом 9E10 в концентрации 7 мкг/мл. В качестве антитела третьего детектирования использовали кроличье антитело против мышиного IgG (Dako, Denmark) и APAAP Mouse Monoclonal (Dako, Denmark) а концентрациях 5 и 50 мкг/мл, соответственно, в течение 1 часа в каждом случае. Таблетки Fast Red Tablets (Sigma, Switzerland) использовали для проявления окрашивания с инкубированием в течение 15 минут. Предметные стекла контрастировали гематоксилином в течение 2 минут, промывали водой, заделывали средой для заделки препаратов Glycergel (Dako, Denmark) и анализировали при помощи микроскопа Axiovert S100 TV (Zeiss, Switzerland).
Анализ иммунофлуоресценции на артритных пробах человека
Замороженные срезы артритных проб человека фиксировали охлажденным на льду ацетоном в течение 10 минут, блокировали фетальной телячьей сывороткой в течение 30 минут и окрашивали на домен EDA фибронектина. Антитело F8 использовали в качестве myc-меченого scFv в концентрации 10 мкг/мл и инкубировали в течение 1 часа. Первичное антитело ко-инкубировали с анти-myc-антителом 9E10 в концентрации 7 мкг/мл. В качестве антитела третьего детектирования использовали флуоресцентное антитело против Alexa 596 мыши (Molecular Probes, Denmark) в концентрации 10 мкг/мл в течение 1 часа в каждом случае. Предметные стекла контрастировали Hoechst 33342, заделывали средой для заделки препаратов Glycergel (Dako, Denmark) и анализировали при помощи микроскопа AxioScop 2MOT+ (Zeiss, Switzerland).
Модель животного
Самцов мышей DBA/1 (8-12-недельных) иммунизировали внутрикожной инъекцией при основании хвоста 200 мкг бычьего коллагена типа II (MD Biosciences), эмульгированного с равными объемами полного адъюванта Фрейнда (MD Biosciences). Спустя 2 недели после первой иммунизации эту процедуру повторяли, но использовали неполный адъювант Фрейнда (MD Biosciences) для эмульгирования коллагена. Мышей обследовали один раз в день и каждую мышь, которая обнаруживала эритему и/или отек лапок в 1 или нескольких конечностях, передавали для исследования изображения или лечения.
Артрит подвергали мониторингу с использованием 2 показателей заболевания (клиническая оценка и отек лапок). Для клинической оценки каждую конечность классифицировали ежедневно не слепым образом. (0 = нормальные, 1 = отек 1 или нескольких пальцев на одной и той же конечности, 2 = отек всей лапки), получая максимально возможную оценку 8 на одно животное. Отек лапки оценивали в каждый второй день с использованием калипера для измерения толщины каждой конечности под анестезией с использованием изофлурана. Среднюю величину всех 4 лапок брали в качестве величины лапок для каждого животного.
Ближнее-инфракрасное-изображение артритных лапок
Селективное накопление SIP(F8) у артритных мышей тестировали анализом ближнего-инфракрасного-изображения, как описано Birchler et al. (1999). Вкратце, очищенный SIP(F8) метили Alexa750 (Molecular Probes, Leiden, The Netherlands) в соответствии с рекомендациями изготовителя и 100 мкг меченого белка инъецировали в хвостовую вену артритных мышей. Мышей анестезировали Кетамином 80 мг/кг и Медетомидином 0,2 мг/кг и визуализировали в блоке формирования изображения в ближней-инфракрасной области спектра спустя 24 часа после инъекции (Trachsel et al. 2007; Birchler et al. 1999).
Эксперименты по биораспределению
Эффективность нацеливания in vivo SIP(F8) у артритных мышей оценивали анализом биораспределения, описанным ранее (Borsi et al. 2002; Tarli et al., 1999). Вкратце, очищенный SIP(F8) радиоиодинировали и 10 мкг белка, соответствующие мкКи 125I, инъецировали в хвостовую вену артритных мышей. Мышей умерщвляли спустя 24 часа после инъекции и лапки экспонировали в течение 1 часа и считывали в приборе формирования изображения (phosphorimager) (Fujifilm BAS-5000), как описано ранее (Trachsel et al. 2007).
Антитела
Выделение фрагмента scFv(L19) анти-ED-B-антитела описано ранее (Pini et al. 1998). Исходное анти-ED-A-антитело выделяли из библиотеки ETH-2 с использованием опубликованных процедур (Giovannoni, Nucleic. Acid Research, 2001, 29(5): E27). Созревание аффинности исходного анти-ED-A-антитела, дающее высокую аффинность анти-ED-A-антител, описано в следующем разделе.
Созревание аффинности исходного анти-ED-A-антитела
Исходное анти-ED-A-антитело (ролизведенное из ETH-2 антитело) использовали в качестве матрицы для конструирования библиотеки созревания аффинности. Вариабельность последовательности в VH CDR1 (зародышевой линии DP47) и VL CDR1 (зародышевой линии DPK22) этой библиотеки вводили при помощи ПЦР с использованием частично вырожденных праймеров 5’- CTGGAGCCTGGCGGACCCAGCTCATMNNMNNMNNGCTAAAGGTGAAT CCAGA-3’ (SEQ ID NO:17) для VH и 5’-CCAGGTTTCTGCTGGTACCAGGCTAA MNNMNNMNNGCTAACACTCTGACTGGCCCTGC-3’ (SEQ ID NO:18) для VL (все олигонуклеотиды приобретали из Operon Biotechnologies, Cologne, Germany), в процессе, который генерирует случайные мутации в положениях 31, 32 и 33 VH CDR1 и в положениях 31, 31a и 32 VL CDR1. Комбинации VHVL собирали в формате scFv ПЦР-сборкой с использованием праймеров LMB3long (5’-CAGGAAACAGCTATGACCATGATTAC-3’) (SEQ ID NO:19) и fdseqlong (5’-GACGTTAGTAAATGAATTTTCTGTATGAGG-3’) (SEQ ID NO:20), с использованием очищенных из геля сегментов VH и VL в качестве матриц. Собранные фрагменты VH-VL дважды расщепляли NcoI/NotI и клонировали в расщепленный NcoI/NotI фагмидный вектор pHEN1 (Hoogenboom et al., 1991). Полученный продукт лигирования электропорировали в электрокомпетентные клетки TG-1 E. coli в соответствии с (Viti et al., 2000) с получением библиотеки, содержащей 1,5×107 отдельных клонов антител, которую подвергали скринингу на антитела, которые связывают ED-A с улучшенной аффинностью.
Отбор анти-ED-A-антител
Описанную выше библиотеку антител подвергали скринингу на антитела, которые связывали ED-A с большей аффинностью, чем исходное анти-ED-A-антитело, с использованием анализа BIAcore. Антиген (11A12), используемый в анализе BIAcore, содержал ED-A-домен фибронектина человека и имеет следующую аминокислотную последовательность (SEQ ID NO:120):
MRSYRTEIDKPSQMQVTDVQDNSISVKWLPSSSPVTGYRVTTTPKNGPGPTKTKTAGPDQ
TEMTIEGLQPTVEYWSVYAQNPSGESQPLVQTAVTNIDRPKGLAFTDVDVDSIKIAWES
PQGQVSRYRVTYSSPEDGIHELFPAPDGEEDTAELQGLRPGSEYTVSWALHDDMESQPL
IGTQSTAIPAPTDLKFTQVTPTSLSAQWTPPNVQLTGYRVRVTPKEKTGPMKEINLAPDS
SSVWSGLMVATKYEVSVYALKDTLTSRPAQGWTTLENVRSHHHHHH
Нуклеотидная последовательность антигена (11A12) (SEQ ID NO:121) является следующей:
atgagatcctaccgaacagaaattgacaaaccatcccagatgcaagtgaccgatgttcaggacaacagcattagtgtcaagtggctgccttcaagttcccctgttactggttacagagtaaccaccactcccaaaaatggaccaggaccaacaaaaactaaaactgcaggtccagatcaaacagaaatgactattgaaggcttgcagcccacagtggagtatgtggttagtgtctatgctcagaatccaagcggagagagtcagcctctggttcagactgcagtaaccaacattgatcgccctaaaggactggcattcactgatgtggatgtcgattccatcaaaattgcttgggaaagcccacaggggcaagtttccaggtacagggtgacctactcgagccctgaggatggaatccatgagctattccctgcacctgatggtgaagaagacactgcagagctgcaaggcctcagaccgggttctgagtacacagtcagtgtggttgccttgcacgatgatatggagagccagcccctgattggaacccagtccacagctattcctgcaccaactgacctgaagttcactcaggtcacacccacaagcctgagcgcccagtggacaccacccaatgttcagctcactggatatcgagtgcgggtgacccccaaggagaagaccggaccaatgaaagaaatcaaccttgctcctgacagctcatccgtggttgtatcaggacttatggtggccaccaaatatgaagtgagtgtctatgctcttaaggacactttgacaagcagaccagctcagggagttgtcaccactctggagaatgtcagatctcatcaccatcaccatcactaa
Нуклеотидную последовательность этого антигена амплифицировали при помощи ПЦР с использованием праймеров, содержащих сайты рестрикции BamHI и BglII при 5’ и 3’, соответственно. Полученный ПЦР-продукт и вектор pQE12 (QIAGEN) расщепляли рестрикционными эндонуклеазами BamHI и BglII и затем лигировали в реакции, содержащей отношение инсерта к вектору 3:1. Полученный вектор секвенировали для подтверждения правильности этой последовательности.
Получение антигена
Предварительную культуру клеток TG1 в 10 мл 2TY, Amp, 1% глюкозы электропорировали в присутствии 1 мкл ДНК-минипрепарата 11A12. Затем эту предварительную культуру разводили 1:100 (8 мл в 800 мл 2TY, Amp, 0,1% глюкозы) и выращивали до OD600 0,4-0,6 и затем индуцировали IPTG в течение ночи. На следующий день эти клетки откручивали и супернатант фильтровали (Millipore 0,22 мкм). После центрифугирования и осветления культурального бульона, 11A12 очищали с использованием колонки Hitrap на FPLC (жидкостной хроматографии быстрого разрешения). Ni/ колонку регенерировали следующим образом: колонку промывали 5 колоночными объемами (КО) Н2О с последующим нанесением 3 КО 0,5 M ЭДТА/0,2 M Трис pH 8 для вымывания старого никеля из этой колонки. Затем эту колонку промывали 5 КО Н2О. Затем колонку повторно загружали 2 КО 100 мМ NiSO4 с последующим промыванием колонки несколькими КО Н2О. Затем эту колонку уравновешивали 5 КО лизисного буфера (20 мМ имидазол/250 мМ NaCl/ЗФР рН 7,4). Лизат клеток фильтровали (Millipore 0,45 мкм) и наносили на эту колонку (вручную). Затем эту колонку опять помещали на FPLC и лизисному буферу давали протекать, пока УФ-сигнал не становился стабильным (постоянным), приблизительно 3 КО. Затем начинали программу элюции: градиент 0-100% буфера для элюции (400 мМ имидазол/250 мМ NaCl/ЗФР рН 7,4) в 5 КО. Фракции, содержащие элюированный антиген, объединяли и диализовали против ЗФР в течение ночи.
Экспрессия и очистка анти-ED-A-антител
Анти-ED-A-антитела экспрессировали и очищали следующим образом: Предварительную культуру клеток TG1 в 10 мл 2TY, Amp, 1% глюкозы электропорировали в присутствии 1 мкл ДНК-минипрепарата одного из анти-ED-A-антител. Затем эту предварительную культуру разводили 1:100 (8 мл в 800 мл 2TY, Amp, 0,1% глюкозы) и выращивали до OD600 0,4-0,6 и затем индуцировали IPTG в течение ночи. На следующий день эти клетки откручивали и супернатант фильтровали (Millipore 0,22 мкм). Эти scFv очищали на Белок А-Сефарозной колонке и использовали триэтиленмеламин для элюции scFv из этой колонки. Фракции, содержащие элюированные scFv, диализовали против ЗФР в течение ночи при 4оС. Затем фракции scFv помещали на колонку Супердекс 75 с ЗФР, протекающим при скорости 0,5 мл/мин и собирали фракции 0,25 мл. Эти мономерные фракции использовали для анализа BIAcore.
Анализ BIAcore™ 1
Чип (микропроцессор) BIAcore™ промывали в течение ночи при скорости тока 5 мкл/мин буфером HBS-EP BIACORE™, 0,01 M Hepes pH 7,4, 0,15 M NaCl, 3 мМ ЭДТА, 0,005% поверхностно-активное вещество P20 (тот же самый буфер использовали для этого анализа). Антиген (11A12) разводили до концентрации 50 мкг/мл в ацетатном буфере (рН 4,0) и СООН-группы на этом чипе активировали инъекцией 50 мкл смеси N-гидроксисукцинимида (NHS) и этил-N-(диметиламинопропил)карбодиимида (EDC). 40 мкл антигена 11A12 инъецировали на этот чип и оставшиеся свободные СООН-группы блокировали 30 мкл этаноламина. После фильтрования через фильтр 0,22 мкм, 20 мкл каждого индивидуального бактериального супернатанта инъецировали на этот чип и взаимодействие с этим антигеном подвергали мониторингу в реальном времени.
Анализ BIAcore™ 2
Величины kon, koff и KD исходного анти-ED-A-антитела и анти-ED-A-антител B2, C5, D5, C8, F8, B7 и G9 оценивали с использованием резонанса поверхностных плазмонов. Этот чип уравновешивали в течение ночи буфером, использованным во время анализа, при скорости тока буфера 5 мкл/мин. Всю процедуру нанесения покрытия выполняли при этой скорости тока. Антиген 11A12 разводили 1:25 ацетатным буфером рН 4,00 (обеспеченным BIACORE™) до конечной концентрации 20 мкг/мл. Затем смешивали NHS и EDC и 50 мкл инъецировали для активации СООН-групп на чипе СМ5. Затем выполняли инъекцию 40 мкл этого антигена (длящуюся приблизительно 40 секунд). Затем 30 мкл этаноламина инъецировали для блокирования реактивности возможных свободных СООН.
Каждую пробу анализировали при скорости тока 20 мкл/мин. Инъецировали 20 мкл неразведенного мономерного белка (полученного из гель-фильтрации). Время диссоциации было приблизительно 200 секунд. Затем инъецировали 10 мкл HCl 10 мМ для регенерации чипа. Это инъецирование мономерного белка повторяли при различных разведениях, т.е. разведении 1:2 (в ЗФР) с последующей регенерацией с использованием HCl. Затем выполняли третью инъекцию этого белка, при разведении 1:4, опять с регенерацией HCl. Величины kon, koff и KD для каждого анти-ED-A-антитела оценивали с использованием программного обеспечения BIAevaluation.
Отбор анти-ED-A-антител
Анализ BIAcore™ 1
Этот анализ BIAcore™ давал график для каждого анти-ED-A-антитела, которое анализировали для выведения аффинности антитела в отношении этого антигена следующим образом. Ось х каждого графика соответствует времени и ось y соответствует единицам резонанса (мере, которая указывает аффинность связывания тестируемого антитела в отношении антигена, нанесенного в виде покрытия на чип BIAcore™). Каждый график показывал 3 пика и 1 падение, которые соответствуют изменениям буфера и, следовательно, не влияют на интерпретацию этих результатов.
Восходящая часть каждого графика представляет фазу ассоциации. Чем круче является кривая в этой части графика, тем быстрее является ассоциация антитела с антигеном. Нисходящая часть каждого графика представляет фазу диссоциации антитела от этого антигена. Чем более плоской является кривая в этой части графика, тем медленнее является диссоциация антитела от антигена.
Анти-ED-A-антитела H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 и G9, все, показали более плоскую кривую диссоциации, чем исходное анти-ED-A-антитело, из которого они произведены, что свидетельствует о том, что они связывают ED-A и, следовательно, также A-FN, с более высокой аффинностью, чем исходное анти-ED-A-антитело. Графики для антител E5, F1, F8 и H1 показали наиболее плоские кривые диссоциации из всех тестируемых анти-ED-A-антител. Кривые ассоциации антител H1, C5, D5, E5, C8, F8 и F1 были более плоскими, чем кривые ассоциации, наблюдаемые для исходного анти-ED-A-антитела, в то время как кривая ассоциации, наблюдаемая для антител B2, B7, E8 и G9 была такой же крутой, что и кривая ассоциации, наблюдаемая для исходного анти-ED-A-антитела. Однако, при использовании бактериальных супернатантов IPTG-индуцированных клеток TG-1 E. coli для анализа BIAcore™ антител H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 и G9, концентрация проб тестируемого антитела была неизвестной, но, наиболее вероятно, более низкой, чем концентрация пробы исходного анти-ED-A-антитела, используемого для сравнения. Таким образом, кривая ассоциации антител H1, B2, C5, D5, E5, C8, F8, Fl, B7, E8 и G9 может быть искусственно низкой вследствие низкой концентрации антитела в пробах, использованных для анализа BIAcore™. Однако, поскольку концентрация не влияет существенным образом на диссоциацию антитела от его антигена-мишени в анализе BIAcore™, эти плоские кривые, наблюдаемые для антител H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 и G9, показывают, что эти антитела связывают ED-A по меньшей мере с равной и, возможно, более высокой аффинностью, чем исходное анти-ED-A-антитело.
Анализ BIAcore 2
Величины kon, koff и KD для каждого анти-ED-A-антитела оценивали с использованием программного обеспечения BIAevaluation. Величины kon, koff и KD исходного анти-ED-A-антитела и анти-ED-A-антител B2, C5, D5, C8, F8, B7 и G9 для антигена 11A12 подробно показаны в таблице 2. Анти-ED-A-антитела B2, C5, D5, C8, F8, B7 и G9, все, имеют лучшие величины KD в отношении антигена 11A12, чем исходное анти-ED-A-антитело, из которого они произведены, что свидетельствует о том, что они связывают ED-A и, следовательно, также A-FN, с более высокой аффинностью, чем исходное анти-ED-A-антитело.
Последовательности
Анти-ED-A-антитела H1, B2, C5, D5, E5, C8, F8, F1, B7, E8 и G9, все, являются scFv-антителами, и они были секвенированы с использованием общепринятых способов. Нуклеотидная последовательность анти-ED-A-антитела H1 показана на фиг.3. Аминокислотная последовательность анти-ED-A-антитела H1 показана на фиг.4.
Предпочтительные нуклеотидные последовательности, кодирующие VH и/или VL анти-ED-A-антител B2, C5, D5, E5, C8, F8, F1, B7, E8 и G9, являются идентичными нуклеотидным последовательностям, кодирующим VH и/или VL анти-ED-A-антитела Н1, за исключением того, что нуклеотидные последовательности, кодирующие CDR1 легкой (VL) и тяжелой (VH) цепи Н1, заменены нуклеотидными последовательностями, кодирующими CDR1 легкой (VL) и тяжелой (VH) цепи, приведенными в списке таблицы 1 для соответствующего антитела.
Предпочтительные нуклеотидные последовательности, кодирующие VH и/или VL диатела анти-ED-A-scFv F8, являются идентичными нуклеотидным последовательностям, кодирующим VH и/или VL анти-ED-A-антитела Н1, за исключением того, что нуклеотидные последовательности, кодирующие CDR1 легкой (VL) и тяжелой (VH) цепи Н1, заменены нуклеотидными последовательностями, кодирующими CDR1 легкой (VL) и тяжелой (VH) цепи, приведенными в списке таблицы 1 для анти-ED-A-антитела F8. Предпочтительной нуклеотидной последовательностью, кодирующей линкер, связывающий VH и VL диатела анти-ED-A-scFv F8, является gggtccagtggcggt (SEQ ID NO:29).
Анти-ED-A-антитела B2, C5, D5, E5, C8, F8, F1, B7, E8 и G9 имеют идентичные аминокислотные последовательности относительно анти-ED-A-антитела Н1, за исключением того, что аминокислотные последовательности CDR1 легкой (VL) и тяжелой (VH) цепи Н1 заменены аминокислотными последовательностями CDR1 легкой (VL) и тяжелой (VH) цепи, приведенными в списке таблицы 1 для соответствующего антитела. Аминокислотная последовательность диатела анти-ED-A-scFv F8 является идентичной аминокислотным последовательностям анти-ED-A-антитела Н1, за исключением того, что аминокислотные последовательности CDR1 легкой (VL) и тяжелой (VH) цепи Н1 заменены аминокислотными последовательностями CDR1 легкой (VL) и тяжелой (VH) цепи, приведенными в списке таблицы 1 для анти-ED-A-антитела F8, и аминокислотная последовательность линкера в Н1 заменена аминокислотной последовательностью линкера GSSGG (SEQ ID NO:28).
Аминокислотная последовательность домена VH анти-ED-A-антитела В2 (SEQ ID NO:21) идентична аминокислотной последовательности домена VH анти-ED-A-антитела Н1, за исключением того, что VH CDR1 H1 заменен SEQ ID NO:23.
Аминокислотная последовательность домена VH анти-ED-A-антитела С5 (SEQ ID NO:41) идентична аминокислотной последовательности домена VH анти-ED-A-антитела Н1, за исключением того, что VH CDR1 H1 заменен SEQ ID NO:43.
Аминокислотная последовательность домена VH анти-ED-A-антитела D5 (SEQ ID NO:51) идентична аминокислотной последовательности домена VH анти-ED-A-антитела Н1, за исключением того, что VH CDR1 H1 заменен SEQ ID NO:53.
Аминокислотная последовательность домена VH анти-ED-A-антитела Е5 (SEQ ID NO:61) идентична аминокислотной последовательности домена VH анти-ED-A-антитела Н1, за исключением того, что VH CDR1 H1 заменен SEQ ID NO:63.
Аминокислотная последовательность домена VH анти-ED-A-антитела С8 (SEQ ID NO:71) идентична аминокислотной последовательности домена VH анти-ED-A-антитела Н1, за исключением того, что VH CDR1 H1 заменен SEQ ID NO:73.
Аминокислотная последовательность домена VH анти-ED-A-антитела F8 (SEQ ID NO:81) идентична аминокислотной последовательности домена VH анти-ED-A-антитела Н1, за исключением того, что VH CDR1 H1 заменен SEQ ID NO:83. Домен VH анти-ED-A-диатела F8 имеет ту же самую аминокислотную последовательность, что и домен VH анти-ED-A-антитела F8 (т.е. SEQ ID NO:81).
Аминокислотная последовательность домена VH анти-ED-A-антитела F1 (SEQ ID NO:91) идентична аминокислотной последовательности домена VH анти-ED-A-антитела Н1, за исключением того, что VH CDR1 H1 заменен SEQ ID NO:93.
Аминокислотная последовательность домена VH анти-ED-A-антитела В7 (SEQ ID NO:101) идентична аминокислотной последовательности домена VH анти-ED-A-антитела Н1, за исключением того, что VH CDR1 H1 заменен SEQ ID NO:103.
Аминокислотная последовательность домена VH анти-ED-A-антитела Е8 (SEQ ID NO:111) идентична аминокислотной последовательности домена VH анти-ED-A-антитела Н1, за исключением того, что VH CDR1 H1 заменен SEQ ID NO:113.
Аминокислотная последовательность домена VH анти-ED-A-антитела G9 (SEQ ID NO:31) идентична аминокислотной последовательности домена VH анти-ED-A-антитела Н1, за исключением того, что VH CDR1 H1 заменен SEQ ID NO:33.
Аминокислотная последовательность домена VL анти-ED-A-антитела В2 (SEQ ID NO:22) идентична аминокислотной последовательности домена VL анти-ED-A-антитела Н1, за исключением того, что VL CDR1 H1 заменен SEQ ID NO:26.
Аминокислотная последовательность домена VL анти-ED-A-антитела С5 (SEQ ID NO:42) идентична аминокислотной последовательности домена VL анти-ED-A-антитела Н1, за исключением того, что VL CDR1 H1 заменен SEQ ID NO:46.
Аминокислотная последовательность домена VL анти-ED-A-антитела D5 (SEQ ID NO:52) идентична аминокислотной последовательности домена VL анти-ED-A-антитела Н1, за исключением того, что VL CDR1 H1 заменен SEQ ID NO:56.
Аминокислотная последовательность домена VL анти-ED-A-антитела Е5 (SEQ ID NO:62) идентична аминокислотной последовательности домена VL анти-ED-A-антитела Н1, за исключением того, что VL CDR1 H1 заменен SEQ ID NO:66.
Аминокислотная последовательность домена VL анти-ED-A-антитела С8 (SEQ ID NO:72) идентична аминокислотной последовательности домена VL анти-ED-A-антитела Н1, за исключением того, что VL CDR1 H1 заменен SEQ ID NO:76.
Аминокислотная последовательность домена VL анти-ED-A-антитела F8 (SEQ ID NO:82) идентична аминокислотной последовательности домена VL анти-ED-A-антитела Н1, за исключением того, что VL CDR1 H1 заменен SEQ ID NO:86. Домен VL анти-ED-A-диатела F8 имеет ту же самую аминокислотную последовательность, что и домен VL анти-ED-A-антитела F8 (т.е. SEQ ID NO:82).
Аминокислотная последовательность домена VL анти-ED-A-антитела F1 (SEQ ID NO:92) идентична аминокислотной последовательности домена VL анти-ED-A-антитела Н1, за исключением того, что VL CDR1 H1 заменен SEQ ID NO:96.
Аминокислотная последовательность домена VL анти-ED-A-антитела В7 (SEQ ID NO:102) идентична аминокислотной последовательности домена VL анти-ED-A-антитела Н1, за исключением того, что VL CDR1 H1 заменен SEQ ID NO:106.
Аминокислотная последовательность домена VL анти-ED-A-антитела Е8 (SEQ ID NO:112) идентична аминокислотной последовательности домена VL анти-ED-A-антитела Н1, за исключением того, что VL CDR1 H1 заменен SEQ ID NO:116.
Аминокислотная последовательность домена VL анти-ED-A-антитела G9 (SEQ ID NO:32) идентична аминокислотной последовательности домена VL анти-ED-A-антитела Н1, за исключением того, что VL CDR1 H1 заменен SEQ ID NO:36.
Необязательно, аминокислота в положении 5 домена VH анти-ED-A-антител H1, B2, C5, D5, E5, C8, F8, F1, B7, E8, G9 и диатела scFv F8 может быть остатком лейцина (L), а не остатком валина (V), как показано на фиг.4А. Кроме того, или альтернативно, аминокислота в положении 18 домена VL анти-ED-A-антител H1, B2, C5, D5, E5, C8, F8, F1, B7, E8, G9 и диатела scFv F8 может быть остатком аргинина (R), а не остатком лизина (K), как показано на фиг.4С.
Клонирование, получение и характеристика F8-IL10
Ген IL-10 человека амплифицировали при помощи ПЦР с использованием следующих праймерных последовательностей:
обратного антисмыслового праймера,
5’-TCGGGTAGTAGCTCTTCCGGCTCATCGTCCAGCGGCAGCCCAGGCCAGGGCACC-3’;
и прямого смыслового праймера,
5’-TTTTCCTTTTGCGGCCGCtcattaGTTTCGTATCTTCATTGTCATGTA-3’,
который имел часть содержащего 15 аминокислот линкера (SSSSG)3 на его N-конце и стоп-кодон и сайт рестрикции NotI на его С-конце.
ДНК, кодирующую одноцепочечный вариабельный фрагмент (F8), амплифицировали с сигнальным пептидом с использованием следующих пар праймеров:
обратный антисмысловой праймер,
5’-CCCAAGCTTGTCGACCATGGGCTGGAGCC-3’
и прямой смысловой праймер,
5’-GAGCCGGAAGAGCTACTACCCGATGAGGAAGAGAATTCTTTGATTTCCACCTTGGTCCCTTG- 3’.
С использованием этой стратегии, сайт рестрикции HindIII встраивали при N-конце, а комплементарную часть линкерной последовательности встраивали при С-конце.
Затем одноцепочечный Fv и фрагменты IL-10 собирали с использованием ПЦР и клонировали в сайты рестрикции HindIII и NotI экспрессирующего вектора млекопитающих pcDNA3.1(+).
Клетки CHO-S стабильно трансфицировали этими описанными ранее плазмидами и отбор проводили в присутствии G418 (0,5 г/л).
Клоны G418-резистентных клеток подвергали скринингу на экспрессию слитого белка при помощи ELISA с использованием рекомбинантного EDA фибронектина человека в качестве антигенов и Белка А для детектирования.
Эти слитые белки очищали из среды культуры клеток аффинной хроматографией с использованием колонок Белка А.
Размер этих слитых белков анализировали в восстанавливающих и невосстанавливающих условиях при помощи электрофореза в ДСН-ПААГ и в нативных условиях FPLC-гель-фильтрацией на вытеснительной колонке Superdex S-200 (Amersham Pharmacia Biotech, Dübendorf, Switzerland).
Анализ активности
Биологическую активность hIL10 определяли по его способности индуцировать IL-4-зависимую пролиферацию клеток MC/9 (Thompson et al., 1991) с использованием колориметрического анализа восстановления красителя MTT.
10000 клеток MC/9 (ATCC, Manassas, USA) на лунку в 200 мкл среды, содержащей 5 пг (0,05 Единиц) мышиного IL4/мл (eBiosciences) в 96-луночном микротитрационном планшете обрабатывали в течение 48 часов варьирующимися количествами IL10 человека. Стандарт hIL10 и слитый белок F8-IL10 использовали при максимуме 100 нг/мл IL10-эквивалентов и серийно разводили. Добавляли 10 мкл 5 мг/мл МТТ (Sigma) и инкубировали в течение 3-5 часов. Затем эти клетки центрифугировали, лизировали ДМСО и считывали показания в виде оптической плотности при 570 нм.
ССЫЛКИ
Все ссылки, цитируемые в любом месте в этом описании, в том числе ссылки, цитируемые в любом месте выше, включены тем самым в качестве ссылки в их полном виде и для всех целей.
Amit et al. (1986), Science, 233: 747-753.
Andersen et al. (2002) Current Opinion in Biotechnology 13: 117.
Ausubel et al. (1999) 4th eds., Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, John Wiley & Sons.
Bagshawe K. D. et al. (1991) Antibody, Immunoconjugates and Radiopharmaceuticals 4: 915-922.
Balza et al. (1988), FEBS Lett., 228: 42-44.
Batista et al. J. Exp. Med., 184: 2197-205, 1996.
Berndt et al. Histochem Cell Biol 1998, 109 (3): 249-255.
Birchler et al. (1999), J. Immunol. Methods, 231, 239-248.
Bird et al. (1988) Science, 242, 423-426.
Borsi et al. (1987), J. Cell. Biol., 104, 595-600.
Borsi et al. (1990), FEBS Lett., 261: 175-178.
Borsi et al. (1995), J. Biol.Chem., 270: 6243-6245.
Borsi et al. (1998), Exp. Cell Res., 240, 244-251.
Borsi et al. (2000) Int J Cancer, 102 (1): 75-85.
Brack et al. (2006), Clin. Cancer Res., 12, 3200-3208.
Carnemolla et al. (1989), J. Cell. Biol., 108: 1139-1148.
Caton et al. (1990), J. Immunol., 144: 1965-1968.
Chadd et al. (2001), Current Opinion in Biotechnology 12: 188-194.
Chothia et al. (1987), J. Mol. Biol., 196: 901-917.
Chothia et al. (1989), Nature, 342: 877-883.
Courtney et al. (1980) Nature, 14;283 (5748): 666-8.
Devos et al. (1983), Nucl. Acids Res. 11: 4307-4323.
ffrench-Constant (1995), Exp. Cell Res., 221, 261-271.
Giovannoni, Nucleic. Acid Research, 2001, 29(5): E27.
Glennie M J et al., 1987 J. Immunol. 139, 2367-2375.
Haan et al. (2004), BioCentury, 12(5): A1-A6.
Hanahan et al. (2000), Cell 100, 57-70.
Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor N. Y., pp. 726, 1988
Heikinheimo et al. (1991), Virchows Arch. B Cell Pathol. Incl. Mol. Pathol., 61, 101-109.
Holliger and Bohlen 1999 Cancer and metastasis rev. 18: 411-419.
Holliger et al. (1993a), Proc. Natl. Acad. Sci. USA 90 6444-6448.
Holliger et al. (1993b), Current Opinion Biotechnol 4, 446-449.
Holt et al. (2003) Trends in Biotechnology 21, 484-490.
Hoogenboom et al. (1991), Nucleic Acids Res., 19 (15) 4133-7.
Hu et al. (1996), Cancer Res., 56, 3055-3061.
Huston et al. (1988) PNAS USA, 85, 5879-5883.
Hynes, R. O. (1990). Fibronectins (New York: Springer-Verlag).
Jacobs et al. (2002), Hum. Pathol., 33, 29-38.
Kabat et al. (1987) Sequences of Proteins of Immunological Interest. 4th Edition. US Department of Health and Human Services.
Kabat et al. (1991a), Sequences of Proteins of Immunological Interest, 5th Edition. US Department of Health and Human Services, Public Service, NIH, Washington. (a)
Kabat et al. (1991b), J. Immunol., 147: 1709-1719.
Kaspar et al. (2006), Int. J. Cancer, 118, 1331-1339.
Knappik et al., (2000) J. Mol. Biol. 296, 57-86.
Koch et al. Ann Rheum Dis 2003, 62 Suppl 2: 1160-67.
Kohler and Milstein, Nature, 256: 495-497, 1975.
Koide et al. (1998), Journal of Molecular Biology, 284: 1141-1151.
Kontermann et al. (2001), S, Antibody Engineering, Springer-Verlag New York, LLC; ISBN: 3540413545.
Kornblihtt et al. (1984), Nucleic Acids Res. 12, 5853-5868.
Koukoulis et al. (1993), J. Submicrosc. Cytol. Pathol., 25, 285-295.
Koukoulis et al. (1995), Ultrastruct. Pathol., 19, 37-43.
Krebs et al. (2001), Journal of Immunological Methods, 254 67-84.
Kriegsmann et al. (2004) Rheumatol Int, 24(1): 25-33.
Larrick JW and Thomas DW (2001) Current Opinion in Biotechnology 12: 411-418.
Ledermann J. A. et al. (1991) Int. J. Cancer 47: 659-664.
Li et al. Protein Engineering, 10: 731-736, 1997.
Lohi et al. (1995), Int. J. Cancer, 63, 442-449.
Maeda et al. (1983) Biochem. Biophys. Res. Comm. 115: 1040-1047.
Matsumoto et al. (1999), Jpn. J. Cancer Res., 90, 320-325.
McCafferty et al., (1990) Nature, 348, 552-554.
Mendez, M. et al., (1997) Nature Genet, 15(2): 146-156.
Merchand et al., 1998 Nature Biotech. 16: 677-681
Neri, D., and Bicknell, R. (2005), Nat Rev Cancer 5, 436-446.
Nygren et al. (1997), Current Opinion in Structural Biology, 7: 463-469.
Oyama et al. (1989), J. Biol. Chem., 264, 10331-10334.
Paolella et al. (1988), Nucleic Acids Res. 16, 3545-3557.
Pini et al. (1998), J. Biol. Chem., 273, 21769-21776.
Plückthun (1991), Bio/Technology 9: 545-551.
Reiter et al. (1996), Nature Biotech, 14, 1239-1245.
Repp et al., 1995 J. Hemat. 377-382.
Ridgeway et al. (1996), Protein Eng., 9, 616-621.
Robinson ed., Sustained and Controlled Release Drug Delivery Systems, Marcel Dekker, Inc., New York, 1978.
Roesli et al. (2006), Nature Protocols, 1, 192-199.
Ruf et al. (1991) J. Biol. Chem. 226: 15719-15725.
Rybak et al. (2005), Nat. Methods, 2, 291-298.
Rybak et al. (2006), ChemMedChem., 2, 22-40.
Sambrook and Russell, Molecular Cloning: a Laboratory Manual: 3rd edition, 2001, Cold Spring Harbor Laboratory Press.
Scarpati et al. (1987) Biochemistry 26: 5234-5238.
Scarpino et al. (1999) J. Pathol. 188, 163-167.
Scheurer et al. (2005), Proteomics 5, 3035-3039.
Segal et al. (1974), PNAS, 71: 4298-4302.
Sharon et al. (1990a), PNAS, 87: 4814-4817.
Sharon et al. (1990b), J. Immunol., 144: 4863-4869.
Silacci et al. (2003), Proteomics, 5, 2340-2350.
Staerz U. D. and Bevan M. J. 1986 PNAS 83
Suresh et al. (1986) Method Enzymol. 121: 210-228
Taniguchi et al. (1983) Nature 302, 305-310;
Tarli et al. Blood 1999, 94 (1): 192-198.
Tavian et al. (1994), Int. J. Cancer, 56, 820-825.
Taylor, Arthritis Res 2002, 4 Suppl 3: S99-107.
Terrana et al. (1987), Cancer Res. 47, 3791-3797.
Thompson et al. (1991), J Exp Med, 173: 507-510.
Thorpe (2004), Clin. Cancer Res., 10, 415-427.
Trachsel et al. (2006), Adv. Drug Deliv. Rev., 58, 735-754.
Trachsel et al. Arthritis Res Ther 2007, 9(1): R9.
Viti et al. (2000), Methods Enzymol., 326, 480-505.
Walsh et al. (1998) Am J Pathol, 152 (3): 691-702.
Ward et al. (1989), Nature 341, 544-546.
Wess In: BioCentury, The Bernstein Report on BioBusiness, 12(42), A1-A7, 2004.
Нуклеотидные и аминокислотные последовательности CDR1 тяжелой цепи (VH) и легкой цепи (VL) анти-ED-A-антител со зрелой аффинностью
Данные оценивания BIAcore
| название | год | авторы | номер документа |
|---|---|---|---|
| Антигены, ассоциированные с воспалительным заболеванием кишечника | 2012 |
|
RU2612878C2 |
| ИММУНОКОНЪЮГАТЫ IL2 И МУТАНТНОГО TNF | 2017 |
|
RU2758139C2 |
| ANTI-CXCR4 АНТИТЕЛА И КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВО | 2014 |
|
RU2675516C2 |
| АНТИТЕЛА ПРОТИВ KIT И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2681730C2 |
| ТЕРАПЕВТИЧЕСКИЕ DLL4-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2010 |
|
RU2570639C2 |
| Терапевтические антитела и их применения | 2016 |
|
RU2722381C2 |
| СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ АГЕНТЫ ПРОТИВ В7-Н1 | 2010 |
|
RU2571204C2 |
| СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ АГЕНТЫ ПРОТИВ В7-Н1 | 2010 |
|
RU2706200C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СПЕЦИФИЧНЫЕ К ИЛ-21, И ОБЛАСТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2708336C2 |
| КОМПОЗИЦИИ IL-12, НАЦЕЛЕННЫЕ НА EDB | 2019 |
|
RU2758143C1 |

















































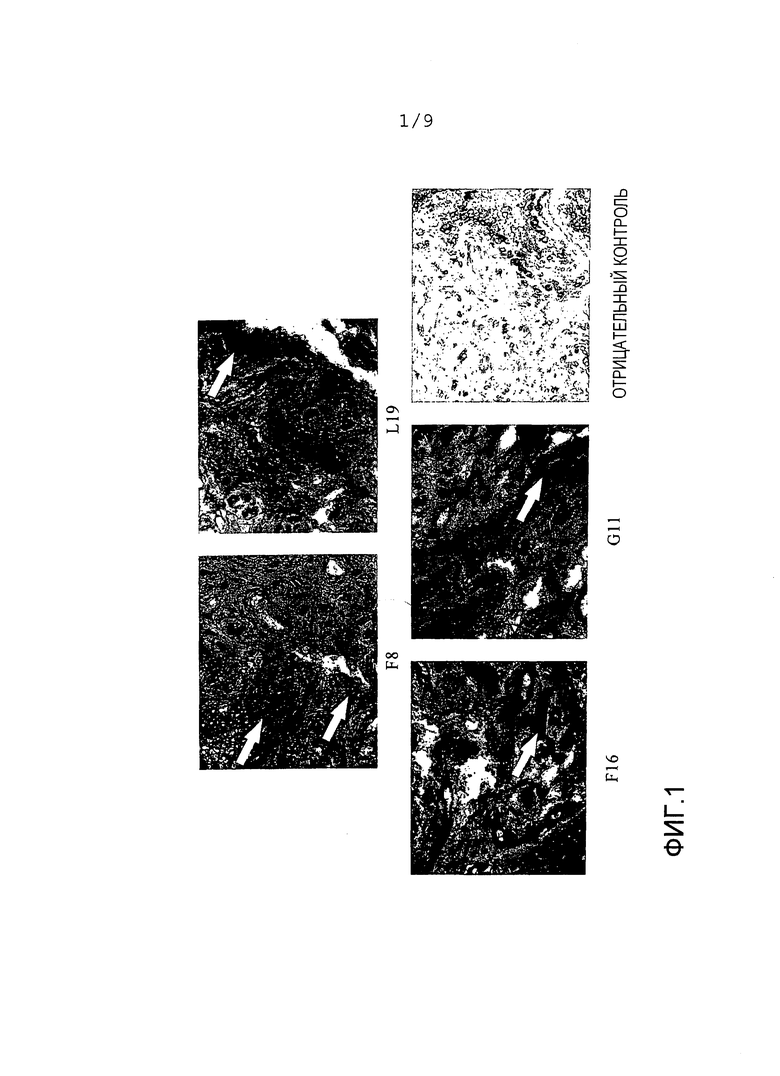







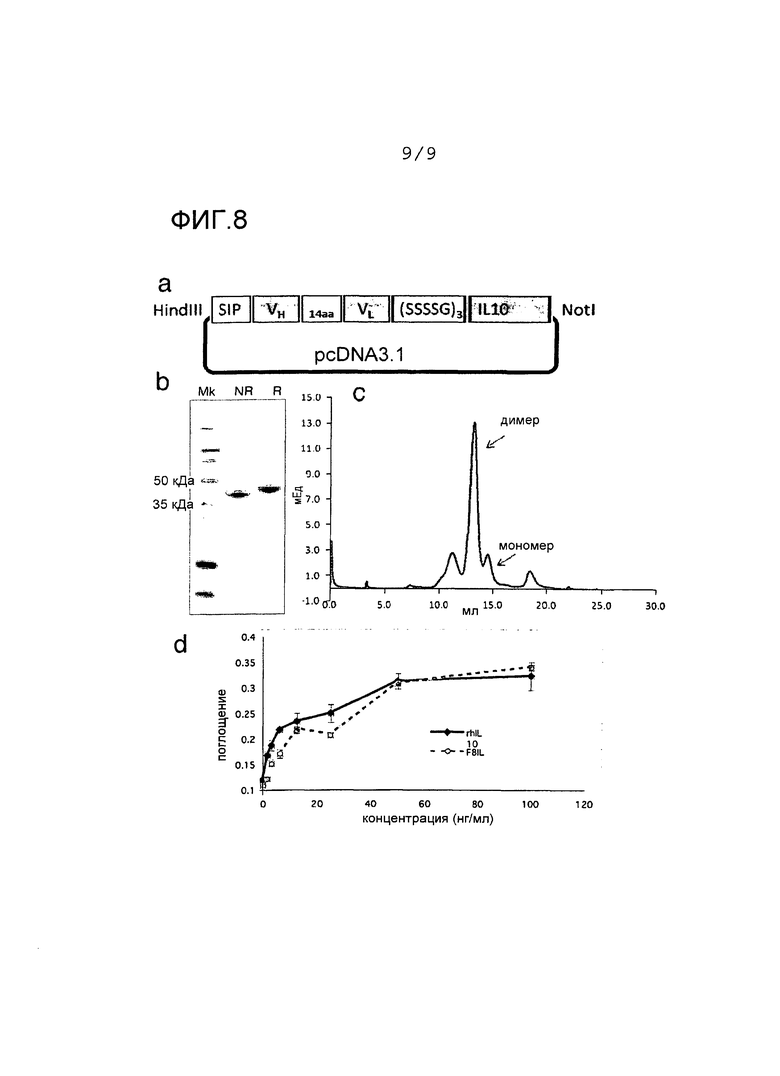
Настоящее изобретение относится к области биотехнологии и фармацевтики, в частности к конъюгатам антитела для лечения ревматоидного артрита у пациента. Антитело, входящее в состав указанных конъюгатов, специфически связывает Экстра-Домен-A (ED-A) фибронектина и конъюгировано с интерлейкином-10. Настоящее изобретение также раскрывает нуклеиновую кислоту, кодирующую указанный конъюгат, клетку-хозяина для его продуцирования, фармацевтическую композицию для лечения ревматоидного артрита у пациента, а также способ получения указанного конъюгата и способ лечения ревматоидного артрита, способ доставки интерлейкина-10 к участкам ревматоидного артрита и способ диагностирования ревматоидного артрита у пациента с его использованием. Настоящее изобретение обеспечивает успешное лечение ревматоидного артрита за счёт селективной доставки интерлейкина-10 к местам локализации заболевания. 15 н. и 20 з.п. ф-лы, 8 ил., 2 табл., 1 пр.
1. Конъюгат антитела для лечения ревматоидного артрита у пациента, содержащий антитело или его антигенсвязывающий фрагмент, которые специфически связывают Экстра-Домен-A (ED-A) фибронектина, конъюгированные с интерлейкином-10, где указанное антитело содержит домен VH и домен VL,
где указанный домен VH содержит набор определяющих комплементарность областей HCDR1, HCDR2 и HCDR3,
где указанная HCDR1 содержит аминокислотную последовательность SEQ ID NO: 83,
указанная HCDR2 содержит аминокислотную последовательность SEQ ID NO: 4, и
указанная HCDR3 содержит аминокислотную последовательность SEQ ID NO: 5; и
где указанный домен VL содержит набор определяющих комплементарность областей LCDR1, LCDR2 и LCDR3,
где указанная LCDR1 содержит аминокислотную последовательность SEQ ID NO: 86,
указанная HCDR2 содержит аминокислотную последовательность SEQ ID NO: 7, и
указанная HCDR3 содержит аминокислотную последовательность SEQ ID NO: 8.
2. Конъюгат антитела по п. 1, где указанный домен VH и/или указанный домен VL содержит каркас зародышевой линии человека.
3. Конъюгат антитела по п. 1, где указанное антитело содержит:
(i) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 81; или
(ii) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 81, где
аминокислота в положении 5 SEQ ID NO:81 представляет собой остаток лейцина (L), а не остаток валина (V).
4. Конъюгат антитела по п. 1, где указанное антитело содержит:
(i) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 82;
(ii) домен VL, содержащий аминокислотную последовательность SEQ ID NO: 82, где аминокислота в положении 18 SEQ ID NO:82 представляет собой остаток аргинина (R), а не остаток лизина (K);
(iii) домен VL, содержащий аминокислоты 1-108 SEQ ID NO:82; или
(iv) домен VL, содержащий аминокислоты 1-108 SEQ ID NO:82, где аминокислота в положении 18 SEQ ID NO:82 представляет собой остаток аргинина (R), а не остаток лизина (K).
5. Конъюгат антитела по п. 1, где указанное антитело содержит:
(i) домен VH, содержащий аминокислотную последовательность SEQ ID NO: 81, где
аминокислота в положении 5 SEQ ID NO:81 представляет собой остаток лейцина (L), а не остаток валина (V); и
(ii) домен VL, содержащий аминокислоты 1-108 SEQ ID NO:82, где аминокислота в положении 18 SEQ ID NO:82 представляет собой остаток аргинина (R), а не остаток лизина (K).
6. Конъюгат антитела по любому из пп. 1-5, где указанный фрагмент антитела содержит одноцепочечный Fv (scFv) или представляет собой диатело.
7. Конъюгат антитела по любому из пп. 1-5, где указанное антитело содержит малый иммунопротеин (SIP).
8. Конъюгат антитела по любому из пп. 1-5, где указанное антитело дополнительно конъюгировано с детектируемой меткой.
9. Конъюгат антитела по любому из пп. 1-5, где указанное антитело дополнительно содержит последовательность myc-tag.
10. Конъюгат антитела по любому из пп. 1-5, где указанное антитело дополнительно конъюгировано с радиоизотопом.
11. Конъюгат антитела по любому из пп. 1-5, где указанный интерлейкин-10 представляет собой интерлейкин-10 человека.
12. Конъюгат антитела по п. 6, где указанный интерлейкин-10 представляет собой интерлейкин-10 человека.
13. Конъюгат антитела по п. 7, где указанный интерлейкин-10 представляет собой интерлейкин-10 человека.
14. Конъюгат антитела по любому из пп. 1-5, где указанное антитело конъюгировано с указанным интерлейкином-10 через пептидный линкер.
15. Конъюгат антитела по п. 14, где указанный пептидный линкер содержит 15 аминокислот.
16. Конъюгат антитела по п. 15, где указанный пептидный линкер содержит аминокислотную последовательность (SSSSG)3.
17. Конъюгат антитела по любому из пп. 1-5, где антитело содержит scFv или диатело, и где указанный домен VH конъюгирован с указанным доменом VL через пептидный линкер.
18. Конъюгат антитела по п. 17, где указанный пептидный линкер содержит 5-25 аминокислот.
19. Конъюгат антитела по п. 18, где указанный пептидный линкер содержит 5 аминокислот.
20. Конъюгат антитела для лечения ревматоидного артрита у пациента, содержащий:
(i) антитело или его антигенсвязывающий фрагмент, которые специфически связывают ED-A фибронектина, содержащие домен VH или домен VL,
где указанный домен VH содержит набор определяющих комплементарность областей HCDR1, HCDR2 и HCDR3,
где указанная HCDR1 содержит аминокислотную последовательность SEQ ID NO: 83,
указанная HCDR2 содержит аминокислотную последовательность SEQ ID NO: 4, и
указанная HCDR3 содержит аминокислотную последовательность SEQ ID NO: 5; и
где указанный домен VL содержит набор определяющих комплементарность областей LCDR1, LCDR2 и LCDR3,
где указанная LCDR1 содержит аминокислотную последовательность SEQ ID NO: 86,
указанная HCDR2 содержит аминокислотную последовательность SEQ ID NO: 7, и
указанная HCDR3 содержит аминокислотную последовательность SEQ ID NO: 8,
где антитело содержит scFv или представляет собой диатело;
где указанный домен VH конъюгирован с указанным доменом VL через пептидный линкер из 5 аминокислот; и
(ii) интерлейкин-10 человека,
где указанный домен VL указанного антитела конъюгирован с интерлейкином-10 человека через пептидный линкер, содержащий аминокислотную последовательность (SSSSG)3.
21. Конъюгат антитела для лечения ревматоидного артрита у пациента, содержащий:
(i) антитело или его антигенсвязывающий фрагмент, которые специфически связывают ED-A фибронектина, содержащие домен VH или домен VL,
(a) где указанный домен VH содержит аминокислотную последовательность SEQ ID NO: 81, где аминокислота в положении 5 SEQ ID NO:81 представляет собой остаток лейцина (L), а не остаток валина (V); и
(b) где указанный домен VL содержит аминокислоты 1-108 SEQ ID NO:82, где аминокислота в положении 18 SEQ ID NO:82 представляет собой остаток аргинина (R), а не остаток лизина (K),
где антитело содержит scFv или представляет собой диатело;
где указанный домен VH конъюгирован с указанным доменом VL через пептидный линкер из 5 аминокислот; и
(ii) интерлейкин-10 человека,
где указанный домен VL указанного антитела конъюгирован с интерлейкином-10 человека через пептидный линкер, содержащий аминокислотную последовательность (SSSSG)3.
22. Конъюгат антитела для лечения ревматоидного артрита у пациента, состоящий из:
(i) антитела или его антигенсвязывающего фрагмента, которые специфически связывают ED-A фибронектина, содержащие домен VH или домен VL,
где указанный домен VH содержит набор определяющих комплементарность областей HCDR1, HCDR2 и HCDR3,
где указанная HCDR1 содержит аминокислотную последовательность SEQ ID NO: 83,
указанная HCDR2 содержит аминокислотную последовательность SEQ ID NO: 4, и
указанная HCDR3 содержит аминокислотную последовательность SEQ ID NO: 5; и
где указанный домен VL содержит набор определяющих комплементарность областей LCDR1, LCDR2 и LCDR3,
где указанная LCDR1 содержит аминокислотную последовательность SEQ ID NO: 86,
указанная HCDR2 содержит аминокислотную последовательность SEQ ID NO: 7, и
указанная HCDR3 содержит аминокислотную последовательность SEQ ID NO: 8,
где антитело представляет собой диатело;
где указанный домен VH конъюгирован с указанным доменом VL через пептидный линкер из 5 аминокислот; и
(ii) интерлейкин-10 человека,
где указанный домен VL указанного антитела конъюгирован с интерлейкином-10 человека через пептидный линкер, содержащий аминокислотную последовательность (SSSSG)3.
23. Конъюгат антитела для лечения ревматоидного артрита у пациента, состоящий из:
(i) антитела или его антигенсвязывающего фрагмента, которые специфически связывают ED-A фибронектина, состоящие из домена VH или домена VL,
(а) где указанный домен VH состоит из аминокислотной последовательности SEQ ID NO: 81, где аминокислота в положении 5 SEQ ID NO:81 представляет собой остаток лейцина (L), а не остаток валина (V); и
(b) где указанный домен VL состоит из аминокислот 1-108 SEQ ID NO:82, где аминокислота в положении 18 SEQ ID NO:82 представляет собой остаток аргинина (R), а не остаток лизина (K),
где антитело представляет собой диатело;
где указанный домен VH конъюгирован с указанным доменом VL через пептидный линкер из 5 аминокислот; и
(ii) интерлейкин-10 человека,
где указанный домен VL указанного антитела конъюгирован с интерлейкином-10 человека через пептидный линкер, содержащий аминокислотную последовательность (SSSSG)3.
24. Конъюгат антитела по п. 11, где указанный интерлейкин-10 человека содержит аминокислотную последовательность из аминокислот 258-417 согласно Фиг. 7.
25. Выделенная нуклеиновая кислота, кодирующая конъюгат антитела по любому из пп. 1-5 или 20-23.
26. Клетка-хозяин для продуцирования конъюгата антитела по любому из пп. 1-5 или 20-23, содержащая нуклеиновую кислоту по п. 25.
27. Способ получения конъюгата антитела по любому из пп. 1-5 или 20-23, включающий культивирование клетки-хозяина, содержащей нуклеиновую кислоту, кодирующую конъюгат антитела, по любому из пп. 1-5 или 20-23 в условиях для продуцирования белка.
28. Способ по п. 27, также содержащий выделение указанного конъюгата антитела.
29. Фармацевтическая композиция для лечения ревматоидного артрита у пациента, содержащая конъюгат антитела по любому из пп. 1-5 или 20-23 и фармацевтически приемлемый носитель.
30. Способ лечения ревматоидного артрита у пациента, нуждающегося в этом, включающий введение указанному пациенту конъюгата антитела по любому из пп. 1-5 или 20-23.
31. Применение конъюгата антитела по любому из пп. 1-5 или 20-23 для приготовления лекарственного средства для лечения ревматоидного артрита.
32. Способ доставки интерлейкина-10 к участкам ревматоидного артрита у пациента, включающий введение указанному пациенту конъюгата антитела по любому из пп. 1-5 или 20-23.
33. Применение конъюгата антитела по любому из пп. 1-5 или 20-23 для получения лекарственного средства для доставки интерлейкина-10 к участкам ревматоидного артрита у пациента.
34. Способ диагностирования ревматоидного артрита у пациента, включающий введение указанному пациенту конъюгата антитела по п. 8 и детектирование детектируемой метки, где локализация этой детектируемой метки связывающего элемента в новообразованной сосудистой сети у человека или животного является указанием на наличие ревматоидного артрита.
35. Применение конъюгата антитела по п. 8
для производства диагностического продукта для диагностики ревматоидного артрита у пациента.
| US 2007202044 A1, 30.08.2007 | |||
| ANDRES F | |||
| MURO, et al | |||
| Regulated splicing of the fibronectin EDA exon is essential for proper skin wound healing and normal lifespan, The Journal of Cell Biology, 2003, Vol.162, No.1, pp.149-160 | |||
| DIETMAR BERNDORF, et al | |||
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| ВОДОПОДЪЕМНОЕ УСТРОЙСТВО ДЛЯ НАПОЛНЕНИЯ ЖЕЛЕЗНОДОРОЖНЫХ БАКОВ И ПОЛЬЗОВАНИЕМ ПАРА ОТ ПАРОВОЗОВ | 1927 |
|
SU7396A1 |
| ОПОСРЕДОВАННАЯ РЕЦЕПТОРАМИ ДОСТАВКА ГЕНОВ С ИСПОЛЬЗОВАНИЕМ ВЕКТОРОВ НА ОСНОВЕ БАКТЕРИОФАГОВ | 1998 |
|
RU2209088C2 |

