ОБЛАСТЬ ТЕХНИКИ
В этом документе представлен способ уменьшения агрегации иммуноглобулинов в концентрированных растворах путем внесения обратной мутации (обратных мутаций) в четвертую каркасную область, т.е. в J-сегмент тяжелой цепи иммуноглобулина, с заменой на гидрофильный аминокислотный остаток или аминокислотный остаток, который присутствует в природном иммуноглобулине эмбрионального типа.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Из-за химических и физических свойств иммуноглобулинов, таких как молекулярная масса и доменная структура, включая вторичные модификации, технология их производства и выделения сильно осложнены. Концентрированные растворы иммуноглобулинов необходимы не только для лекарственных препаратов, приготовленных в соответствии с определенной рецептурой, но и для применения в качестве промежуточных продуктов в технологии производства и выделения целевого продукта для достижения малых объемов с целью обеспечения большей экономичности транспортировки и хранения.
Также сильно концентрированный раствор необходим для получения готовой лекарственной формы, содержащей иммуноглобулин. Например, для применения иммуноглобулина подкожно необходимы концентрации более 100 мг/мл, т.е. примерно 150 мг/мл. Но по меньшей мере некоторые иммуноглобулины имеют склонность к агрегации при нефизиологических высоких концентрациях 100 мг/мл или более.
В WO 2009/000098 сообщается о создании генно-инженерными методами и оптимизации одноцепочечных антител на основе их последовательности.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В этом документе представлен способ уменьшения агрегации иммуноглобулинов в концентрированном растворе. Было обнаружено, что две или даже одна обратная мутация в четвертой каркасной области тяжелой цепи иммуноглобулина может снизить образование агрегатов, создавая условия для получения сильно концентрированного раствора вариантного иммуноглобулина с обратными мутациями, имеющего пониженное содержание агрегатов.
Следовательно, в этом документе представлены отдельные особенности способа снижения агрегации иммуноглобулина в растворе, способа модификации иммуноглобулина, способа получения иммуноглобулина и способа гуманизирования иммуноглобулина.
Все описанные в этом документе способы содержат следующие этапы:
a) выравнивание аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина по отношению к референсной аминокислотной последовательности с целью достижения максимального уровня идентичности аминокислотных последовательностей;
b) определение в выровненных аминокислотных последовательностях положений с отличающимися аминокислотными остатками в аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина и в референсной аминокислотной последовательности;
c) модификацию аминокислотной последовательности иммуноглобулина путем замены одного или по меньшей мере одного аминокислотного остатка в аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина в положении, определенном на этапе b), на соответствующий остаток треонина или серина, который присутствует в референсной аминокислотной последовательности, при этом в замещенном положении аминокислотный остаток является треонином или серином в референсной аминокислотной последовательности и при этом аминокислотный остаток не является треонином или серином в аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина;
d) возможно, предложение оптимальной последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность модифицированного иммуноглобулина.
В одном из вариантов воплощения определяется только различие по остаткам треонина, и по ним вносятся изменения. В другом варианте воплощения референсная аминокислотная последовательность для четвертой каркасной области иммуноглобулина является аминокислотной последовательностью xxxxTTxTxSS (SEQ ID NO:01), где x обозначает любой аминокислотный остаток, кроме треонина и серина. В дополнительном варианте воплощения референсная аминокислотная последовательность является аминокислотной последовательностью xxxxTxxTxxx (SEQ ID NO:11), где x в положениях 1, 2, 3, 4, 7 и 9 обозначает любой аминокислотный остаток, кроме треонина и серина, а x в положении 6 обозначает треонин и x в положениях 10 и 11 обозначает серин, при этом различие по остаткам треонина в положении 5 и/или 8 в референсной аминокислотной последовательности относительно аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина определяется на этапе b) и модифицируется на этапе с). В одном из вариантов воплощения референсная аминокислотная последовательность является аминокислотной последовательностью эмбрионального типа человека. В одном из вариантов воплощения всех описанных в этом документе особенностей способы включают следующие этапы:
a) выравнивание аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина по отношению к аминокислотной последовательности WGQGTLVTVSS (SEQ ID NO:02), или аминокислотной последовательности WGRGTLVTVSS (SEQ ID NO:03), или аминокислотной последовательности WGQGTMVTVSS (SEQ ID NO:04), или аминокислотной последовательности WGQGTTVTVSS (SEQ ID NO:05), или аминокислотной последовательности WGKGTTVTVSS (SEQ ID NO:06) с целью достижения максимального уровня идентичности аминокислотных последовательностей,
b) определение в выровненных аминокислотных последовательностях положений с отличающимися аминокислотными остатками в аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина и в референсной аминокислотной последовательности;
c) модификацию иммуноглобулина путем замены аминокислотного остатка в аминокислотной последовательности четвертой каркасной области тяжелой цепи в положении, определенном на этапе b), на соответствующий остаток треонина или серина, который присутствует в референсной аминокислотной последовательности, при этом в данном положении аминокислотный остаток является треонином или серином в референсной аминокислотной последовательности и при этом аминокислотный остаток в данном положении не является треонином или серином в аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина;
а) возможно, предложение оптимальной последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность модифицированного иммуноглобулина.
В одном из вариантов воплощения описанных в этом документе особенностей способы включают в качестве первого этапа этап создания или определения аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина. В одном варианте воплощения референсная последовательность является аминокислотной последовательностью WGQGTLVTVSS (SEQ ID NO:02). В другом варианте воплощения референсная аминокислотная последовательность является последовательностью ххххТххТххх (SEQ ID NO:11). В одном из вариантов воплощения определяют различие в расположении остатков треонина в положении 5 и/или 8 референсной аминокислотной последовательности. В одном из вариантов воплощения определяется только различие по остаткам треонина, и по ним вносятся изменения.
Одной из особенностей, описанных в этом документе, является то, что способ представляет собой способ получения иммуноглобулинов, включающий следующие этапы:
a) выравнивание аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина по отношению к референсной аминокислотной последовательности xxxxTTxTxSS (SEQ ID NO:01) или ххххТххТххх (SEQ ID NO:11) с целью достижения максимального уровня идентичности аминокислотных последовательностей,
b) определение в выровненных аминокислотных последовательностях положений с отличающимися аминокислотными остатками в положении 5 и/или 6 и/или 8 и/или 10 и/или 11 в аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина и в референсной аминокислотной последовательности;
c) модификацию аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина путем замены аминокислотного остатка в аминокислотной последовательности четвертой каркасной области тяжелой цепи в положении, определенном на этапе b), на соответствующий остаток треонина или серина, который присутствует в референсной аминокислотной последовательности, при этом в данном положении аминокислотный остаток является треонином или серином в референсной аминокислотной последовательности и при этом аминокислотный остаток в данном положении не является треонином или серином в аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина;
d) культивирование клетки млекопитающего, содержащей нуклеиновую кислоту, кодирующую аминокислотную последовательность модифицированной тяжелой цепи иммуноглобулина, и нуклеиновую кислоту, кодирующую аминокислотную последовательность соответствующей легкой цепи, для экспрессии тяжелой и легкой цепей иммуноглобулина;
е) выделение иммуноглобулина из этой клетки млекопитающего или культуральной среды и, таким образом, получение иммуноглобулина.
В одном варианте воплощения референсная аминокислотная последовательность является аминокислотной последовательностью WGQGTLVTVSS (SEQ ID NO:02), или аминокислотной последовательностью WGRGTLVTVSS (SEQ ID NO:03), или аминокислотной последовательностью WGQGTMVTVSS (SEQ ID NO:04), или аминокислотной последовательностью WGQGTTVTVSS (SEQ ID NO:05), или аминокислотной последовательностью WGKGTTVTVSS (SEQ ID NO:06). В одном из вариантов воплощения этап модификации аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина - это замена аминокислотного остатка в аминокислотной последовательности четвертой каркасной области тяжелой цепи в положении, определенном на этапе b), который является треонином в референсной аминокислотной последовательности и который не является треонином в аминокислотной последовательности четвертой каркасной области тяжелой цепи, на соответствующий остаток треонина, который присутствует в референсной аминокислотной последовательности. В другом варианте воплощения модификацию производят по меньшей мере по одному выявленному положению. В одном варианте воплощения референсная аминокислотная последовательность является аминокислотной последовательностью WGQGTLVTVSS (SEQ ID NO:02). В одном из вариантов воплощения иммуноглобулин является иммуноглобулином человека или гуманизированным иммуноглобулином. В другом варианте воплощения клетка млекопитающего является клеткой СНО или клеткой НЕК. В другом варианте воплощения определяют различие в расположении остатков треонина в положении 5 и/или 8 референсной аминокислотной последовательности.
Еще одной из особенностей, описанных в этом документе, является способ гуманизирования иммуноглобулина, включающий следующие этапы:
a) выравнивание аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина по отношению к референсной аминокислотной последовательности xxxxTTxTxSS (SEQ ID NO:01) или ххххТххТххх (SEQ ID NO:11) с целью достижения максимального уровня идентичности аминокислотных последовательностей,
b) определение в выровненных аминокислотных последовательностях положений с отличающимися аминокислотными остатками в аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина и в референсной аминокислотной последовательности;
c) гуманизирование иммуноглобулина путем замены аминокислотного остатка в аминокислотной последовательности четвертой каркасной области тяжелой цепи в положении, определенном на этапе b), на соответствующий остаток треонина или серина, который присутствует в референсной последовательности, при этом в данном положении аминокислотный остаток является треонином или серином в референсной аминокислотной последовательности и при этом аминокислотный остаток в данном положении не является треонином или серином в аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина;
d) возможно, предложение оптимальной последовательности нуклеиновой кислоты, кодирующей гуманизированный иммуноглобулин.
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Было обнаружено, что замена двух или даже одного аминокислотного остатка треонина и/или серина на маленький гидрофобный или неполярный аминокислотный остаток, такой как изолейцин или аланин, в аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина увеличивает склонность иммуноглобулина к образованию агрегатов в растворе, особенно в растворах с высокой концентрацией соли и/или высокой концентрацией иммуноглобулина. При обратной замене замещенных аминокислотных остатков на находящиеся в этом положении в природном белке остатки (остаток) треонина и/или серина склонность к образованию агрегатов в растворе, особенно в концентрированных растворах, заметно снижается.
Одной из особенностей изобретения является то, что описанный в этом документе способ является способом уменьшения агрегации иммуноглобулина в растворе, включающим следующие этапы:
- определение того, был ли заменен один или несколько остатков треонина и/или серина в аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина, путем сравнения аминокислотной последовательности с референсной аминокислотной последовательностью для аминокислотной последовательности четвертой каркасной области;
- обратное изменение аминокислотной последовательности иммуноглобулина путем модификации по меньшей мере одного из замещенных остатков треонина и/или серина обратно на присутствующий в референсной последовательности остаток треонина и/или серина и, тем самым, уменьшение агрегации иммуноглобулина в растворе.
Термин "выравнивание" означает процесс расположение рядом друг с другом двух или более аминокислотных последовательностей для достижения максимального уровня идентичности и консервативности аминокислотных последовательностей. Он включает определение позиционной гомологии молекулярных последовательностей, в том числе размещение рядом друг с другом аминокислот или нуклеотидов в гомологичных последовательностях. В результате сравниваемые последовательности представлены в виде, демонстрирующем области с наибольшим статистическим сходством.
"Максимальный уровень идентичности аминокислотной последовательности" по отношению к референсной аминокислотной последовательности определяется как процент аминокислотных остатков в кандидатной аминокислотной последовательности, которые идентичны аминокислотным остаткам референсной аминокислотной последовательности, после выравнивания последовательностей и внесения пропусков, в случае необходимости, для достижения максимального процента идентичности последовательностей, и без учета каких-либо консервативных замен как части идентичности последовательностей. Выравнивание с целью определения идентичности аминокислотных последовательностей может быть достигнуто различными способами, например, с применением общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры для выравнивания аминокислотных последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых аминокислотных последовательностей. Однако с целью использования в этом документе значения "% идентичности аминокислотных последовательностей" получают с помощью компьютерной программы для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 был разработана компанией Genentech, Inc., и исходный код вместе с документацией для пользователя был зарегистрирован в Бюро регистрации авторских прав США (Вашингтон, округ Колумбия, 20559, США), где он зарегистрирован под номером TXU510087 регистрации авторского права в США. Программа ALIGN-2 свободно доступна в компании Genentech, Inc. (Южный Сан-Франциско, Калифорния, США) или может быть скомпилирована из исходного кода. Программу ALIGN-2 необходимо компилировать для применения в операционной системе UNIX, в том числе - Digital UNIX V4.0D. Все параметры сравнения последовательностей устанавливаются программой ALIGN-2 и не изменяются.
В случае применения ALIGN-2 для сравнения аминокислотных последовательностей % идентичности аминокислотных последовательностей для данной аминокислотной последовательности А с, относительно или в сравнении с данной аминокислотной последовательностью В (что иначе можно выразить фразой: данная аминокислотная последовательность А, которая обладает определенным % идентичности аминокислотных последовательностей с, относительно или в сравнении с данной аминокислотной последовательностью В) вычисляется следующим образом:
100×Х/Y,
где Х - это количество аминокислотных остатков, оцененных как идентичные совпадения, согласно программе для выравнивания последовательностей ALIGN-2, в произведенном этой программой выравнивании А и В, и где Y - это суммарное количество аминокислотных остатков в В. Понятно, что если длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, то % идентичности аминокислотной последовательности А относительно В не будет равен % идентичности аминокислотной последовательности В относительно А.
Дополнительной особенностью изобретения является то, что описанный в этом документе способ является способом модификации иммуноглобулина, включающим следующие этапы:
- определение того, был ли заменен один или несколько остатков треонина и/или серина в аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина, путем сравнения аминокислотной последовательности с референсной аминокислотной последовательностью для аминокислотной последовательности четвертой каркасной области;
- обратное изменение аминокислотной последовательности иммуноглобулина путем модификации по меньшей мере одного из замещенных остатков треонина и/или серина обратно на присутствующий в референсной последовательности остаток треонина и/или серина и, тем самым, модификацию иммуноглобулина.
Одной из особенностей изобретения является то, что в этом документе описан способ гуманизирования иммуноглобулина, включающий следующие этапы:
- определение посредством сравнения аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина, иммуноглобулина с пересаженной областью CDR, иммуноглобулина с уменьшенным или отсутствующим эпитопом для T-клеток, или их варианта, с референсной аминокислотной последовательностью для четвертой каркасной области того, был ли заменен один или несколько остатков треонина и/или серина на другой аминокислотный остаток;
- обратное изменение аминокислотной последовательности иммуноглобулина путем модификации по меньшей мере одного из замещенных остатков треонина и/или серина обратно на присутствующий в референсной последовательности остаток треонина и/или серина и, тем самым, гуманизирование иммуноглобулина.
Термин "аминокислотная последовательность четвертой каркасной области" означает аминокислотную последовательность четвертой каркасной области тяжелой цепи иммуноглобулина. Эта аминокислотная последовательность начинается с аминокислотного остатка, расположенного сразу же после С-конца области 3, определяющей комплементарность (CDR3) тяжелой цепи иммуноглобулина, и заканчивается последним аминокислотным остатком вариабельного домена тяжелой цепи. В одном из вариантов воплощения аминокислотные остатки в составе CDR3 тяжелой цепи иммуноглобулина определяют согласно системе Кабата (Kabat).
Термин "химерный иммуноглобулин" означает иммуноглобулин, содержащий аминокислотные остатки, происходящие из иммуноглобулина первого биологического вида, и аминокислотные остатки, происходящие из иммуноглобулина второго биологического вида, не идентичного первому биологическому виду. Если акцепторным биологическим видом является человек, то химерный иммуноглобулин представляет собой "гуманизированный иммуноглобулин". В большинстве случаев гуманизированный иммуноглобулин происходит от иммуноглобулина человека (реципиентного или акцепторного иммуноглобулина), в котором аминокислотные последовательности одной или нескольких гипервариабельных областей, определенных согласно системе Кабата и/или Хотиа и/или другим системам нумерации, заменены на аминокислотную последовательность гипервариабельной области из иммуноглобулина биологического вида, отличного от человека (донорного иммуноглобулина). Гуманизированный иммуноглобулин, в котором целые гипервариабельные области, согласно системе Кабата и/или Хотиа и/или другим системам нумерации, в акцепторном иммуноглобулине человека заменены соответствующими аминокислотными остатками донорного иммуноглобулина биологического вида, отличного от человека, обозначают как "иммуноглобулин с пересаженной областью CDR". Примерами донорных иммуноглобулинов, не принадлежащих человеку, являются иммуноглобулины мыши, крысы, кролика, собаки, хомяка, овцы или приматов, кроме человека, обладающие требуемой специфичностью и аффинностью в отношении представляющего интерес антигена (см., например, Morrison, S.L., et al., Proc. Natl. Acad. Sci. USA 81 (1984) 6851-6855; US 5202238; US 5204244). В некоторых случаях остатки каркасной области (FR) иммуноглобулина человека заменяют соответствующими остатками иммуноглобулина из вида, отличного от человека. Дополнительно, гуманизированные иммуноглобулины могут содержать дополнительные модификации, например аминокислотные остатки, которые не обнаружены в акцепторном иммуноглобулине или в донорном иммуноглобулине. Такие модификации приводят к получению вариантов таких реципиентных или донорных иммуноглобулинов, которые гомологичны, но не идентичны, соответствующей исходной последовательности. Гуманизированный иммуноглобулин, возможно, также будет содержать по меньшей мере часть консервативной области иммуноглобулина, в типичном случае, таковой области иммуноглобулина человека.
В одном из вариантов воплощения иммуноглобулин является химерным иммуноглобулином, или иммуноглобулином с пересаженной областью CDR, или иммуноглобулином с уменьшенным или отсутствующим эпитопом для Т-клеток, или их вариантом. В одном из вариантов воплощения иммуноглобулин содержит консервативную область тяжелой цепи иммуноглобулина человека или ее вариант. В дополнительном варианте воплощения консервативная область иммуноглобулина человека принадлежит к подклассу lgG1, или подклассу lgG4, или подклассу lgG2, или подклассу lgG3, или имеет последовательность SEQ ID NO:07, или SEQ ID NO:08, или является их вариантом.
В одном из вариантов воплощения консервативная область модифицирована таким образом, что не может быть обнаружено связывания с рецептором Fey (например, FcyRIIIa) и/или связывания с Clq, как указано ниже. В одном из вариантов воплощения консервативная область является консервативной областью иммуноглобулина человека и либо принадлежит к подклассу lgG4 человека, либо является мутантной Fc-частью из подкласса lgG1 человека. В другом варианте воплощения консервативная область принадлежит к подклассу lgG1 человека, содержащему мутации L234A и L235A (положения определены относительно полной длины аминокислотной последовательности тяжелой цепи lgG1 человека, т.е., включая вариабельный домен). Хотя lgG4 демонстрирует сниженное связывание с рецептором Fey (FcyRIIIa), иммуноглобулины из других подклассов lgG демонстрируют сильное связывание. Однако Pro238, Asp265, Asp270, Asn297 (потеря углевода в Fc), Pro329, Leu234, Leu235, Gly236, Gly237, lle253, Ser254, Lys288, Thr307, Gln311, Asn434 и/или His435 - это остатки, которые, при их изменении, также обеспечивают пониженное связывание с рецептором Fey (Shields, R.L, et al, J. Biol. Chem. 276 (2001) 6591-6604; Lund, J., et al., FASEB J. 9 (1995) 115-119; Morgan, A, et al, Immunology 86 (1995) 319-324; EP 0 307 434). В одном из вариантов воплощения консервативная область по отношению к связыванию с рецептором Fey принадлежит к подклассу lgG4 или к подклассу lgG1 или lgG2, с мутацией по L234, L235 и/или D265, и/или содержит мутацию PVA236. В одном из вариантов воплощения мутация является S228P, L234A, L235A, L235E и/или PVA236 (PVA236 означает, что аминокислотная последовательность ELLG (указанная с помощью однобуквенного кода обозначения аминокислот) в положении аминокислот с 233 по 236 в составе lgG1 или EFLG в составе lgG4 заменена на PVA). В дополнительном варианте воплощения мутация является S228P в lgG4 и L234A и L235A в lgG1. Консервативная область иммуноглобулина непосредственно участвует в АЗКЦ (антителозависимой клеточно-опосредованной цитотоксичности) и КЗЦ (комплемент-зависимой цитотоксичности). Иммуноглобулин, который не связывается с рецептором Fey и/или фактором комплемента C1q, не вызывает антителозависимую клеточно-опосредованную цитотоксичность (АЗКЦ) и/или комплемент-зависимую цитотоксичность (КЗЦ).
Термин "иммуноглобулин с отсутствующим эпитопом для Т-клеток" означает иммуноглобулин, который был модифицирован с целью прекращения или снижения иммуногенности путем удаления эпитопов для Т-клеток человека (пептидной последовательности в составе белка, способной связываться с молекулами МНС класса II). При этом способе идентифицируют взаимодействия между боковыми цепочками аминокислот пептида и специфическими карманами связывания в желобке связывания на МНС класса II. Обнаруженные иммуногенные области изменяют для устранения иммуногенности. Такие способы в целом описаны, например, в WO 98/52976 или WO 98/08097.
Термин "их вариант" означает иммуноглобулин, содержащий консервативные модификации последовательности или имеющий измененный характер гликозилирования. Термин "консервативные модификации последовательности" означает аминокислотные замены, в том числе таковые, при которых аминокислотный остаток заменяется аминокислотным остатком, имеющим схожую боковую цепь. Семейства аминокислотных остатков, имеющих схожую боковую цепь, определены в соответствующей области знаний. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), разветвленными у бета-атома углерода боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Еще один тип варианта иммуноглобулина имеет измененный характер гликозилирования иммуноглобулина. Под изменением подразумевается делеция одного или нескольких сайтов гликозилирования, обнаруженных в иммуноглобулине, и/или внедрение одного или нескольких сайтов гликозилирования, которые отсутствуют в иммуноглобулине. Гликозилирование иммуноглобулинов в типичном случае является N-гликозилированием. N-гликозилированием называют присоединение углеводного фрагмента к боковой цепочке аспарагинового остатка. Трипептидные последовательности аспарагин-X-серин и аспарагин-X-треонин и аспарагин-Х-цистеин, где Х может быть любой аминокислотой, служат последовательностями распознавания для ферментативного присоединения углеводного фрагмента к боковой цепочке аспарагина.
В одном из вариантов воплощения референсная последовательность является последовательностью SEQ ID NO:02-06, или последовательностью SEQ ID NO:01, или последовательностью SEQ ID NO:11. В другом варианте воплощения определение и обратная мутация относятся к замещенным остаткам треонина.
Далее изобретение иллюстрируется примерами антитела против IL13Rα1 и антитела против OX40L. Соответствующие аминокислотные последовательности и способы получения описаны в WO 2006/072564 и WO 2006/029879 (которые включены сюда в качестве ссылок). Эти антитела используются исключительно для целей иллюстрации особенностей, описанных в этом документе, и не представляют собой какого-либо ограничения. Объем изобретения изложен в прилагаемой формуле изобретения.
Антитело против IL13Rα1 далее обозначается как исходное антитело и имеет аминокислотную последовательность четвертой каркасной области тяжелой цепи WGQGTLVIVSS (SEQ ID NO:09). Сравнение с аминокислотной последовательностью SEQ ID NO:01 и SEQ ID NO:11 приведено ниже.

Можно видеть, что различие между аминокислотными последовательностями заключается в остатке изолейцина в положении 8 в SEQ ID NO:09, присутствующем вместо остатка треонина, имеющегося в SEQ ID NO:01 или SEQ ID NO:11. Следовательно, было получено вариантное антитело, у которого остаток изолейцина заменен в результате обратной мутации на остаток треонина.
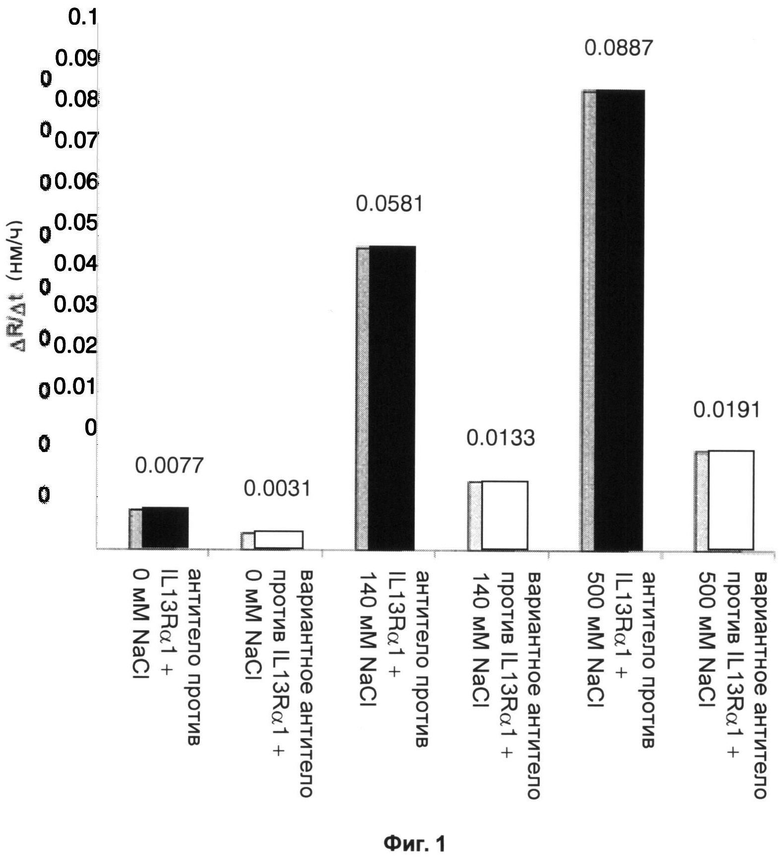
Тогда как исходное антитело демонстрирует высокие уровни образования агрегатов, вариантное антитело с обратной заменой имеет заметно сниженную склонность к образованию агрегатов. На фиг.1 показаны скорости увеличения размеров (нм/ч) агрегатов исходного антитела против IL13Rα1 при концентрации антитела 30 мг/мл при 50°С и различных значениях ионной силы. Также на фиг.1 представлены скорости, полученные для вариантного антитела против IL13Ra1, в котором остаток изолейцина в положении аминокислоты 115 (соответствующем положению 8 в SEQ ID NO:01, 11 и 09) в аминокислотной последовательности четвертой каркасной области тяжелой цепи был заменен обратной мутацией на треонин, и это является единственным его отличием от исходного антитела против IL13Rα1. Единичная обратная мутация снижает скорость увеличения размера агрегатов при всех испытанных значениях ионной силы.
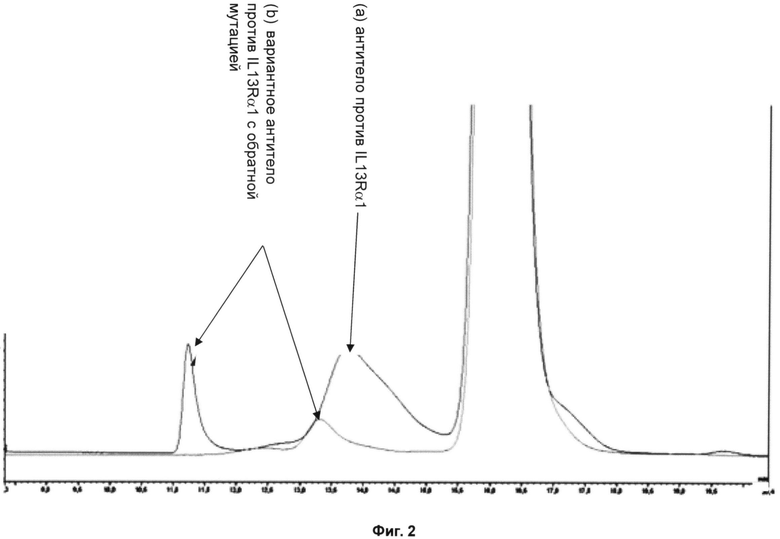
На фиг.2 приведены аналитические хроматограммы, полученные с помощью гель-фильтрации, для двух проб, хранившихся в течение 9 нед. при 40°С и имеющих одинаковую технологию приготовления. Соответствующие данные перечислены в таблице 1 далее.
Можно видеть, что склонность к образованию агрегатов у вариантного антитела с обратной мутацией снижена более чем на 50% по сравнению с исходным антителом.
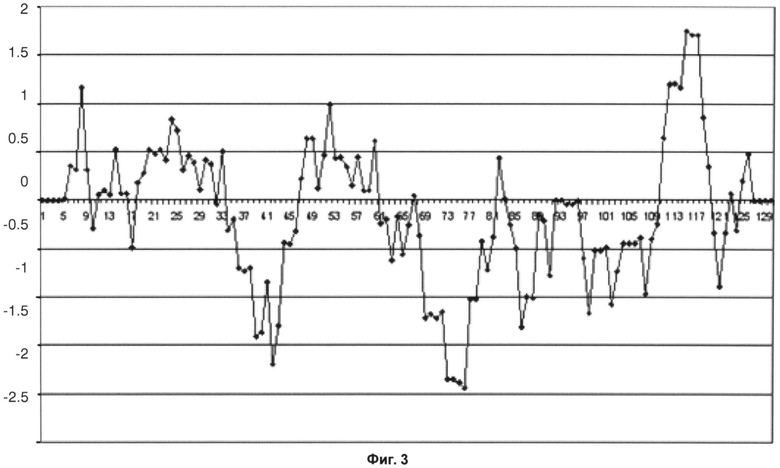
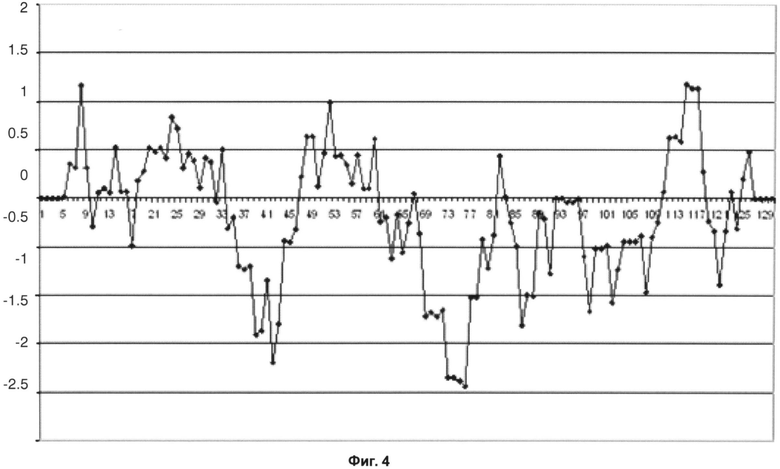
На фиг.3 показан график Кайта-Дулиттла для остатков тяжелой цепи с 1 по 130 с окном анализа или усреднения размером в 9 остатков для исходного антитела против IL13Ra1. В четвертой каркасной области график демонстрирует самые высокие значения гидрофобности для всей тяжелой цепи. На фиг.4 приведен график Кайта-Дулиттла для тяжелой цепи вариантного антитела против IL13Ra1. Можно видеть, что в вариантном антителе с обратной мутацией значения гидрофобности четвертой каркасной области заметно снижены.
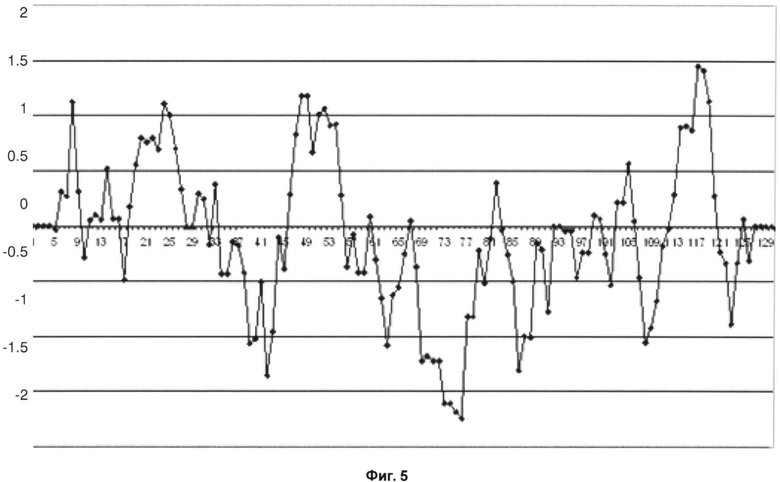
На фиг.5 показан график Кайта-Дулиттла для остатков тяжелой цепи с 1 по 130 с окном анализа/усреднения размером в 9 остатков для антитела против OX40L.
Видно, что и в данном случае самое высокое значение гидрофобности также наблюдается в четвертой каркасной области тяжелой цепи. Проанализировав аминокислотную последовательность четвертой каркасной области тяжелой цепи, можно определить аминокислотную замену, как показано ниже.

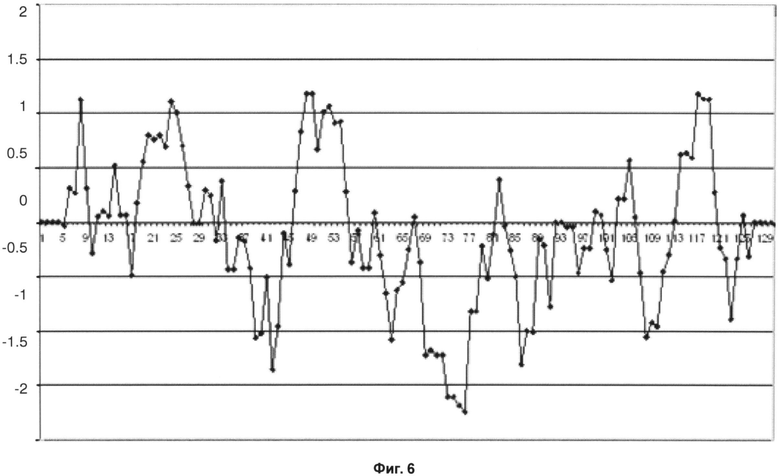
Обратная мутация этой единственной аминокислоты с заменой на встречающийся в природе в антителе эмбрионального типа остаток треонина снизила гидрофобность антитела, как показано на графике Кайта-Дулиттла на фиг.6.
В следующих примерах листинг последовательностей и фигуры приведены исключительно для понимания настоящего изобретения, полный объем которого изложен в прилагаемой формуле изобретения. При этом понимается, что в пределах сущности изобретения в изложенных методиках могут быть произведены модификации.
ОПИСАНИЕ ЛИСТИНГА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO:01 xxxxTTxTxSS - референсная аминокислотная последовательность
SEQ ID NO:02 WGQGTLVTVSS - частичная аминокислотная последовательность FR4/J-элемента эмбрионального типа.
SEQ ID NO:03 WGRGTLVTVSS - частичная аминокислотная последовательность FR4/J-элемента эмбрионального типа.
SEQ ID NO:04 WGQGTMVTVSS - частичная аминокислотная последовательность FR4/J-элемента эмбрионального типа.
SEQ ID NO:05 WGQGTTVTVSS - частичная аминокислотная последовательность FR4/J-элемента эмбрионального типа.
SEQ ID NO:06 WGKGTTVTVSS - частичная аминокислотная последовательность FR4/J-элемента эмбрионального типа.
SEQ ID NO:07 аминокислотная последовательность консервативной области тяжелой цепи IgG1 человека.
SEQ ID NO:08 аминокислотная последовательность консервативной области тяжелой цепи IgG4 человека.
SEQ ID NO:09 WGQGTLVIVSS - аминокислотная последовательность антитела против IL13Rα1.
SEQ ID NO:10 WGQGALVTVSS - аминокислотная последовательность антитела против OX40L.
SEQ ID NO:11 ххххТххТххх - референсная аминокислотная последовательность.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1 - Скорость увеличения размера частиц (ΔR) в единицу времени (Δt) в растворе 30 мг/мл антитела против IL13Rα1 (а) и вариантного антитела с обратной заменой (b), в обоих случаях - в 20 мМ гистидиновом буферном растворе, рН6, с 0, 140 и 500 мМ NaCl, при 50°С, определенная по динамическому рассеянию света.
Фиг.2 - Аналитические хроматограммы, полученные при гель-фильтрации двух растворов 30 мг/мл антитела против IL13Rα1 и формы с обратной заменой, в обоих случаях - в присутствии 500 мМ NaCl в течение 15 ч при 10°С.
Фиг.3 - График Кайта-Дулиттла для остатков тяжелой цепи с 1 по 130 с окном анализа размером в 9 остатков для исходного антитела против IL13Rα1.
Фиг.4 - График Кайта-Дулиттла для остатков тяжелой цепи с 1 по 130 с окном анализа размером в 9 остатков для вариантного антитела против IL13Rα1.
Фиг.5 - График Кайта-Дулиттла для остатков тяжелой цепи с 1 по 130 с окном анализа размером в 9 остатков для исходного антитела против OX40L.
Фиг.6 - График Кайта-Дулиттла для остатков тяжелой цепи с 1 по 130 с окном анализа размером в 9 остатков для вариантного антитела против OX40L.
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Материалы и методу
Аналитическая гель-фильтрация
Содержание высокомолекулярных форм, мономеров антител и низкомолекулярных форм определяли с помощью аналитической гель-фильтрации на колонке TSK 3000S WXL 7,8×300 мм (Tosoh, Штутгарт, Германия). В качестве элюента использовали буферный раствор, содержащий 200 мМ KH2PO4 и 250 мМ KCl, с рН7,0, при скорости потока 0,5 мл/мин. В каждом прогоне наносили белок в количестве 150 мкг. Детекцию производили по поглощению УФ при длине волны 280 нм.
Динамическое рассеяние света (DTS)
Динамическое рассеяние света - это неинвазивная методика измерения размера частиц, в типичном случае в диапазоне размеров менее одного микрона. В настоящем изобретении применяли прибор Zetasizer Nano S (Malvern Instruments, Вустершир, Великобритания) с температуроконтролируемой кварцевой кюветой (25°С) для мониторинга диапазона размеров от 1 нм до 6 мкм. Интенсивность рассеянного света лазера определяли под углом 173°. Интенсивность колеблется со скоростью, зависящей от скорости диффузии частиц, которая, в свою очередь, зависит от размера частиц. Следовательно, данные о размере частиц можно получить с помощью анализа колебания интенсивности рассеянного света (Dahneke, B.E. (ed), Measurement of Suspended Particles by Quasielectric Light Scattering, Wiley Inc. (1983); Pecora, R., Dynamic Light Scattering: Application of Photon Correlation Spectroscopy, Plenum Press (1985)). Распределение размеров вычисляли по интенсивности, используя режим множественного сужения в программном обеспечении DTS (Malvern).
Пример 1
Получение вариантного антитела против IL13Rα1
Гены, кодирующие легкую и тяжелую цепи антитела против IL13Rα1, раздельно внедрили в экспрессионные векторы для клеток млекопитающих. Таким образом были соединены сегменты генов, кодирующие вариабельную область легкой цепи антитела против IL13Rα1 (VL) и консервативную область легкой цепи κ человека (CL), а также сегменты генов для вариабельной области тяжелой цепи антитела против IL13Rα1 (VH) и консервативной области тяжелой цепи y1 человека (СН1-шарнир-СН2-СН3). Общая информация относительно нуклеотидных последовательностей легкой и тяжелой цепей человека, кодоны которых применялись, дана в публикации: Kabat, E.A., Wu, Т.Т., Perry, H.M., Gottesman, К.S., and Foeller, С.Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No. 91-3242 (1991). Транскрипционная единица легкой цепи κ антитела против IL13Rα1 состоит из следующих элементов:
- предранние энхансер и промотор цитомегаловируса человека (HCMV),
- синтетическая 5'-нетранслируемая область, включающая последовательность Козака,
- сигнальная последовательность тяжелой цепи иммуноглобулина мыши, включающая интрон сигнальной последовательности,
- клонированная кДНК вариабельной области легкой цепи антитела против IL13Rα1, с добавлением уникального сайта рестрикции BsmI на 5'-конце и донорного сплайс-сайта и уникального сайта рестрикции NotI на 3'-конце,
- геномная ДНК человека с геном консервативной области цепи κ, включающая энхансер гена lg-κ мыши во 2 интроне (Picard, D., and Schaffner, W., Nature 307 (1984) 80-82), и
- сигнальная последовательность полиаденилирования ("полиА") цепи κ иммуноглобулина человека.
Транскрипционная единица тяжелой цепи y1 антитела против IL13Rα1 состоит из следующих элементов:
- предранние энхансер и промотор цитомегаловируса человека (HCMV),
- синтетическая 5'-нетранслируемая область, включающая последовательность Козака,
- модифицированная сигнальная последовательность тяжелой цепи иммуноглобулина мыши, включающая интрон сигнальной последовательности,
- клонированная кДНК вариабельной области тяжелой цепи антитела против IL13Rα1, с добавлением уникального сайта рестрикции BsmI на 5'-конце и донорного сплайс-сайта и уникального сайта рестрикции NotI на 3'-конце,
- геномная ДНК человека с геном консервативной области тяжелой цепи y1, включающая энхансер гена Ig-µ мыши (Neuberger, M.S., EMBO J. 2 (1983) 1373-1378),
- сигнальная последовательность полиаденилирования ("полиА") цепи y1 иммуноглобулина человека.
Помимо экспрессионной кассеты для легкой цепи κ или тяжелой цепи y1 антитела против IL13Rα1, эти плазмиды содержат:
- ген устойчивости к гигромицину,
- ориджин репликации, oriP, вируса Эпштейна-Барр (EBV),
- ориджин репликации из вектора pUC18, который позволяет данной плазмиде реплицироваться в Е.coli, и
- ген а β-лактамазы, который придает Е.coli устойчивость к ампициллину.
Экспрессионная плазмида, кодирующая тяжелую цепь y1 вариантного антитела против IL13Rα1, была создана посредством сайт-направленного мутагенеза экспрессионных плазмид исходного антитела с применением набора QuickChange™ Site-Directed mutagenesis (Stratagene). Нумерация аминокислот соответствует системе EU (Edelman, G.M., et al., Proc. Natl. Acad. Sci. USA 63 (1969) 78-85; Kabat, E.A., Wu, T.T, Perry, H.M., Gottesman, K.S, and Foeller, C, (1991) Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No. 91-3242).
Стабильно трансфецированный клон клеток СНО продуцирует вариантное антитело против IL13Ra1c выходом 130 мг/л. Выделение целевого продукта производили с применением трех последовательных хроматографических этапов: хроматография на колонке с белком А, катионо-обменная хроматография и анионо-обменная хроматография.
Пример 2
Определение скорости увеличения размера агрегатов с применением динамического рассеяния света
Для слежения за процессом агрегации с течением времени производили измерения динамического рассеяния света (DLS) с равными временными интервалами. Высокие концентрации солей стабилизируют гидрофобные взаимодействия; следовательно, ожидается, что связанная с гидрофобностью агрегация будет более явно выражена при высоких концентрациях солей. Изменение среднего размера частиц (среднего радиуса Z) измеряли как показатель агрегации белков (фиг.1). Пробы диализовали против буферного раствора, содержащего различные количества NaCl (20 мМ His/His-HCl с рН6,0+0/140/500 мМ NaCl) при концентрации белка 30 мг/мл. Измерения динамического рассеяния света проводили на спектрофотометре для прочтения многолуночных планшетов Wyatt DynaPro в 394-луночных планшетах для микротитрования при температуре 50°С.
Пример 3
Испытание стабильности вариантного антитела против IL13Rα1
Индукцию образования высокомолекулярных соединений проводили путем диализа проб против буферного раствора 20 мМ His/His-HCl с рН6,0, содержащего 0 или 500 мМ NaCl, с последующей инкубацией при 10°С в течение 15 ч. Образование высокомолекулярных соединений, в сравнении с необработанными пробами, контролировали с помощью эксклюзионной ВЭЖХ (фиг.2).







Настоящее изобретение относится к области биохимии. Предложен способ получения последовательности нуклеиновой кислоты, кодирующей иммуноглобулин, включающий выравнивание аминокислотной последовательности четвертой каркасной области (FR4) тяжелой цепи иммуноглобулина с референсной последовательностью ххххТххТххх и, в случае выявления в положении 5 и/или 8 указанной последовательности маленьких гидрофобных или неполярных аминокислотных остатков, отличных от треонина, их замену на треонин. Также рассмотрен способ получения иммуноглобулина, основанный на использовании способа получения последовательности нуклеиновой кислоты, кодирующей иммуноглобулин. Данное изобретение обеспечивает снижение склонности иммуноглобулинов к образованию агрегатов в растворе, благодаря чему может найти применение в производстве лекарственных средств. 2 н. и 5 з.п. ф-лы, 6 ил., 1 табл., 3 пр.
1. Способ получения последовательности нуклеиновой кислоты, кодирующей иммуноглобулин, включающий следующие этапы:
a) выравнивание аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина по отношению к референсной последовательности ххххТххТххх (SEQ ID NO: 11) с целью достижения максимального уровня идентичности аминокислотных последовательностей,
b) определение присутствия маленьких гидрофобных или неполярных аминокислотных остатков в положении 5 и/или 8 выровненных аминокислотных последовательностей четвертой каркасной области тяжелой цепи;
c) модификация иммуноглобулина путем замены аминокислотного остатка, определенного на этапе b), в четвертой каркасной области тяжелой цепи, который является треонином в референсной последовательности и при этом не является треонином в четвертой каркасной области тяжелой цепи, на соответствующий остаток треонина, присутствующий в референсной последовательности;
d) предложение последовательности нуклеиновой кислоты, кодирующей иммуноглобулин.
2. Способ получения иммуноглобулина, включающий следующие этапы:
a) выравнивание аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина по отношению к референсной последовательности ххххТххТххх (SEQ ID NO: 11) с целью достижения максимального уровня идентичности аминокислотных последовательностей,
b) определение присутствия маленьких гидрофобных или неполярных аминокислотных остатков в положении 5 и/или 8 выровненных аминокислотных последовательностей четвертой каркасной области тяжелой цепи;
c) модификация иммуноглобулина путем замены аминокислотного остатка, определенного на этапе b), в четвертой каркасной области тяжелой цепи, который является треонином в референсной последовательности и при этом не является треонином в четвертой каркасной области тяжелой цепи, на соответствующий остаток треонина, как в референсной последовательности;
d) предложение последовательности нуклеиновой кислоты, кодирующей аминокислотную последовательность модифицированного иммуноглобулина;
e) культивирование клетки млекопитающего, содержащей нуклеиновую кислоту, кодирующую аминокислотную последовательность модифицированной тяжелой цепи иммуноглобулина, и нуклеиновую кислоту, кодирующую аминокислотную последовательность соответствующей легкой цепи, для экспрессии тяжелой и легкой цепей иммуноглобулина;
f) выделение иммуноглобулина из клетки млекопитающего или культуральной среды и, таким образом, получение иммуноглобулина.
3. Способ по любому из пп.1, 2, отличающийся тем, что референсная последовательность представляет собой последовательность WGQGTLVTVSS (SEQ ID NO: 02), или последовательность WGRGTLVTVSS (SEQ ID NO: 03), или последовательность WGQGTMVTVSS (SEQ ID NO: 04), или последовательность WGQGTTVTVSS (SEQ ID NO: 05), или последовательность WGKGTTVTVSS (SEQ ID NO: 06).
4. Способ по любому из пп.1, 2, отличающийся тем, что способ включает в качестве первого этапа предложение или определение аминокислотной последовательности четвертой каркасной области тяжелой цепи иммуноглобулина.
5. Способ по любому из пп.1, 2, отличающийся тем, что иммуноглобулин является иммуноглобулином человека или гуманизированным иммуноглобулином.
6. Способ по любому из пп.1, 2, отличающийся тем, что иммуноглобулин является химерным иммуноглобулином, или иммуноглобулином с пересаженной областью CDR, или иммуноглобулином с отсутствующим эпитопом для Т-клеток, или их вариантом.
7. Способ по п. 2, отличающийся тем, что клетка млекопитающего является клеткой CHO или клеткой HEK.
| Колосоуборка | 1923 |
|
SU2009A1 |
| "Engineering scFvs for improved stability", Methods Mol Biol, 2003; 207:239-247US 2009/118127 A1, 07.05.2009WO 2004/020579 A2, 11.03.2004NIEBA L | |||
| et al., "Disrupting the hydrophobic patches at the antibody variable/constant domain interface: improved in vivo folding and physical | |||

