В настоящем документе описываются антитела, специфично связывающиеся с рецептором TfR (TfR). Такие антитела полезны, в частности, при лечении пролиферативных и воспалительных расстройств, например лимфом или лейкозов. Конкретно говоря, данное описание относится к определенным гуманизированным антителам против TfR, не уступающим по своим свойствам соответствующим исходным мышиным антителам A24, или их химерному варианту, включающему константную область человеческого иммуноглобулина G1 (IgG1).
Уровень техники
Рецептор TfR (обозначаемый CD71, или Т-9, а в настоящем документе - TfR) - это трансмембранный гликопротеин, представляющий собой гомодимер из двух связанных дисульфидной связью полипептидных цепей, состоящих из 760 аминокислотных остатков, с молекулярной массой приблизительно 90 кДа. TfR играет ключевую роль в регуляции поглощения железа и в клеточном росте (Gill et al., N. Engl. J. Med., 332,1744-1748, 1995 - Hermine et al., N. Engl. J. Med. , 332, 1749-1751, 1995). Когда трансферрин, несущий два иона железа (Fe3+), связывается на клеточной поверхности со своим рецептором, происходит его интернализация через ямки, окаймлённые клатрином, в кислые везикулы, где комплекс TfR с железом диссоциирует. После высвобождения рецептор и апотрансферрин возвращаются на клеточную поверхность.
TfR постоянной экспрессируется на клеточной плазматической мембране ткане, которые постоянно обновляются, например предшественников клеток крови в костном мозге, гепатоцитов в печени, кератиноцитов в эпидермисе и энтероцитов в криптах эпителия тонкого кишечника.
В ряде исследований показано, что экспрессия TfR более интенсивна в злокачественных тканях, чем в соответствующих нормальных (Gatter et al., J Clin Pathol., 36,539-545, 1983 - Faulk et al, Lancet., 2,390-392, 1980 - Shindelman et al., Int J Cancer, 27,329-334, 1981). В нескольких работах сообщалось о терапевтических подходах, основанных на использовании антител против рецептора TfR или самого TfR, конъюгированных с лекарственными препаратами, для ликвидации злокачественных клеток.
Также предлагалось использовать антитела против рецептора TfR, чтобы блокировать взаимодействие между трансферрином и его рецептором, вследствие чего предотвращается поглощение железа клетками, а это ведет к снижению уровня железа и отрицательной регуляции клеточного роста. Но хотя во многих публикациях описывалось получение антител против рецептора TfR, сообщения о моноклональных антителах (mAb) против рецептора TfR, обладающих антипролиферативной активностью весьма немногочисленны.
В работе Траубриджа и Лопеза (Trowbridge I.S., Lopez F. Proc. Natl. Acad. Sci. USA, 79, 1175-1179, 1982) сообщается о свойствах моноклональных антител, обозначенных 42/6 и относящихся к IgA (k), которые блокируют связывание TfR с его рецептором и способны подавлять клеточный рост в линиях Т-клеточного лейкоза человека in vitro путем задержки клеток в фазе S клеточного цикла. Антитела 42/6 и продуцирующие их гибридомы (ATCC HB-8094) описываются в патенте США № 4,434,156.
Лесли с сотрудниками (Lesley J.F. et al. Mol Cell Biol. 5, 1814-21, 1985) изучали влияние моноклональных антител против мышиного рецептора TfR, относящихся к классу IgG или IgM, на связывание TfR и на пролиферацию мышиных лимфомных клеток in vitro. По наблюдениям этих авторов, IgM подавляли клеточный рост, а IgG - нет, хотя последние были способны вызывать снижение уровня экспрессии и деградацию TfR. Однако IgG, перекрестно связанные антителами против иммуноглобулинов, могли подавлять клеточный рост. Позже та же команда (Lesley J.F. et al., Exp Cell Res., 182,215-33, 1989) изучали влияние моноклональных антител против рецептора TfR классов IgG и IgM и их одно- и двухвалентных фрагментов на рост мышиных лимфомных клеток и экспрессию TfR. Они сообщают о том, что это влияние зависит от степени перекрестного связывания рецептора TfR антителами, определяемого валентностью антител. Одновалентные фрагменты молекулы антитела не имеют существенного эффекта; двухвалентные антительные фрагменты вызывали снижение уровня экспрессии рецептора на клеточной поверхности, не влияя на его интернализацию и рециклизацию и не изменяя клеточный рост; поливалентные IgM вызывали накопление комплексов рецептора с антителами на клеточной поверхности, блокируя интернализацию и в результате значительно подавляя клеточный рост.
Из приведенных выше сведений, по-видимому, следует, что антипролиферативные свойства антител против TfR весьма различны у разных антител и не могут быть предсказаны на основании способности данного антитела блокировать связывание TfR с его рецептором.
В ранее опубликованной работе (Moura I.C. et al., J Exp Med, 194, 417-425, 2001), авторы данного изобретения сообщали о моноклональных мышиных IgG (IgG2κ), обозначенных A24, которые связывались с человеческим рецептором TfR.
В публикации WO2005/111082 описываются мышиные антитела A24, способные блокировать пролиферацию Т-клеток; они, по-видимому, эффективнее, чем ранее описанные моноклональные антитела 42/6, подавляют пролиферацию Т-клеток. A24 препятствуют связыванию Tf с TfR в конкуретном формате. A24 также снижают уровень экспрессии рецептора TfR и нарушают его рециклизацию. Кроме того, ex vivo A24 способны блокировать пролиферацию злокачественных Т-клеток, вызывающих как острую, так и хроническую форму Т-клеточного лейкоза взрослых (ATL); см. Moura I.C. et al., Blood, 103,5, 1838-45,1 March 2004, Callens C. et al., 2010; J. Exp. Med., Vol 207 No 4, pp731-750. Сообщалось также, что эти антитела способны предотвращать развитие мантийноклеточной лимфомы как in vitro, так и in vivo (Lepelletier Y. et al. Cancer Res 2007; 67:1145-1154; Callens С. et al. 2008, Leukemia, 22, 42-48).
В настоящее время считается обязательным, чтобы антитела мышиного происхождения для введения людям были гуманизированы - во избежание иммуногенных процессов. Однако когда авторы данного изобретения проводили первую гуманизацию антител A24, у всех гуманизированных вариантов обнаружилась утрата свойств, относящихся к связыванию и апоптозу. Авторам данного изобретения пришлось путем нескольких процедур гуманизации сконструировать специфические варианты A24, в которых соединялись сохраненные функциональные свойства исходных антител A24 с предсказанным снижением иммуногенности для человека.
Раскрытие изобретения
Таким образом, настоящее раскрытие касается выделенного антитела против TfR или к белку, содержащему связывающую антиген часть антитела против TfR, и содержащему что-либо из
(a) вариабельный полипептид тяжелой цепи, который включает HCDR1 последовательности SEQ ID NO:1, HCDR2 последовательности SEQ ID NO:2, HCDR3 последовательности SEQ ID NO:3, и вариабельную легкую цепь полипептида, который включает LCDR1 последовательности SEQ ID NO:4, LCDR2 последовательности SEQ ID NO:5, и LCDR3 последовательности SEQ ID NO:6;
(b) вариабельный полипептид тяжелой цепи, который включает HCDR последовательности SEQ ID NO:1, HCDR2 последовательности SEQ ID NO:2, HCDR3, последовательности SEQ ID NO:3, и вариабельный полипептид легкой цепи, который включает LCDR1 последовательности SEQ ID NO:4, LCDR2 последовательности SEQ ID NO:8, и LCDR3 последовательности SEQ ID NO:6;
(c) вариабельный полипептид тяжелой цепи, который содержит VH последовательности SEQ ID NO:11, и вариабельный полипептид легкой цепи, который содержит VL последовательности SEQ ID NO:13;
(d) вариабельный полипептид тяжелой цепи, который содержит VH последовательности SEQ ID NO:11, и вариабельный полипептид легкой цепи, который содержит VL последовательности SEQ ID NO:14;
(e) вариабельный полипептид тяжелой цепи, который содержит VH последовательности SEQ ID NO:11, и вариабельный полипептид легкой цепи, который содержит VL последовательности SEQ ID NO:15;
(f) вариабельный полипептид тяжелой цепи, который содержит VH последовательности SEQ ID NO:12, и вариабельный полипептид легкой цепи, который содержит VL последовательности SEQ ID NO:13;
(g) вариабельный полипептид тяжелой, который содержит VH последовательности SEQ ID NO:12, и вариабельный полипептид легкой цепи антитела, который составляет область VL, имеющую последовательность SEQ ID NO:14;
(h) вариабельный полипептид тяжелой цепи, который содержит VH последовательности SEQ ID NO:12, и вариабельный полипептид легкой цепи, который содержит VL последовательности SEQ ID NO:15;
причем указанное антитело против TfR или указанный белок против TfR специфично связываются с TfR последовательности SEQ ID NO:16.
В одном из конкретных воплощений данного изобретения связывание указанного антитела или белка с рецептором TfR характеризуется KD, составляющей 10 нM или меньше, предпочтительно KD составляет 1 нM или меньше.
В другом конкретном воплощении данного изобретения указанное антитело или белок вызывает в клетках линии HL-60 апоптоз, уровень которого такой же или больше уровня апоптоза, индуцируемого соответствующими химерными антителами, содержащими вариабельные области исходного мышиного антитела, в которых VH последовательности SEQ ID NO:9 и VL последовательности SEQ ID NO:10.
Предпочтительно такие антитела против TfR по данному изобретению является гуманизированным антителом против TfR.
В другом конкретном воплощении, которое можно объединить с предыдущими воплощениями, указанное антитело против TfR включает константную область человеческого иммуноглобулина изотипа IgG4 или мутантную либо химически модифицированную константную область, с которой указанное антитело не обеспечивает антителозависимую опосредованную клетками цитотоксическую активность (ADCC) или эта активность понижена по сравнению с соответствующим антителом, в котором имеется константная область дикого типа изотипа IgG1. В качестве альтернативы указанное антитело против TfR или белок содержат константную область человеческого иммуноглобулина изотипа IgG1 или мутантную либо химически модифицированную константную область, где указанная мутантная либо химически модифицированная константная область обеспечивает повышенную ADCC указанного антитела по сравнению с соответствующим антителом с константной областью дикого типа изотипа IgG1.
Примеры антител по данному изобретению включают моноклональные гуманизированные антитела mAb1 - mAb16 против TfR, описанные ниже (см. таблицу 1).
Также в настоящем документе описываются выделенные антитела против TfR или белки, как определены выше, для применения в качестве средств медикаментозного лечения или для диагностики, например для применения при лечении опухоли. Предпочтительно указанная опухоль является гематологической опухолью, например лимфомой или лейкозом.
Или же указанные изолированные антитела против TfR или белки можно использовать при лечении заболеваний, связанных с инфекцией Т-лимфотропным вирусом человека типа 1 (HTLV-1); для снижения вирусной нагрузки при воспалительных расстройствах, связанных с инфекцией HTLV-1, в том числе тропического спастического пареза/миелопатии, ассоциированного с HTLV-1 (HAM/TSP), полимиозита и артрита.
Настоящее раскрытие также касается фармацевтической композиции, содержащей антитела против TfR или белки, как определено выше, в сочетании с одним или более фармацевтически приемлемыми эксципиентами, разбавителями или носителями. Фармацевтическая композиция по данному изобретению может также включать дополнительно другие активные ингредиенты.
В одном из конкретных воплощений данного изобретения указанная композиция представляет собой лиофилизованный препарат или преднаполненный шприц либо преднапоненную ампулу, содержащие терапевтически приемлемое количество антител против TfR или белков, как определено выше.
Настоящее раскрытие также касается изолированного полинуклеотида, кодирующего по меньшей мере вариабельную область (области) тяжелой и/или легкой цепи антитела или белка, как определено выше; вектору для клонирования или экспрессии, содержащему одну или более таких нуклеиновых кислот, или к вектору для клонирования или экспрессии для продуцирования антител против TfR, как определено выше, в клетке-хозяине.
В одном воплощении вектор для клонирования или экспрессии содержит по меньшей мере одну из следующих нуклеиновых кислот, кодирующих полипептиды тяжелой и легкой цепей какого-либо из антител mAb1 - mAb16, как описано в примерах ниже.
Настоящее раскрытие также касается клетки-хозяина, содержащей один или более описанных выше векторов для клонирования или экспрессии.
Раскрытие также касается способа получения антител против TfR или белка, как определено выше, включающему: (i) культивирование клетки-хозяина согласно раскрытию для экспрессии указанного антитела или белка указанной клеткой-хозяином, и при необязательно (ii) очистку указанных антитела или белка и (iii) выделение указанных антитела или белка.
Подписи под фигурами
Фиг. 1 - график, демонстрирующие отсутствие индукции апоптоза в клетках первыми шестью гуманизированными вариантами антител INA01 (INA01 варианты 1-6) по сравнению с химерными антителами INA01 с вариабельными областями исходного антитела A24, детектированное с помощью набора для индукции апоптоза в HL-60.
Фиг. 2 - график, демонстрирующие улучшение эффекторной функции у гуманизированных вариантов второго «поколения» (INA01 варианты 7-11); однако все еще ниже, чем химерные антитела INA01 с вариабельными областями исходного антитела A24, детектированное с помощью набора для индукции апоптоза в HL-60.
Фиг. 3 - график, демонстрирующие эффективную индукцию апоптоза в клетках HL-60 антителами третьего «поколения» (INA01 варианты 12-17), которые в этом отношении превзошли химерные антителами INA01 с вариабельными областями исходного антитела A24.
Фиг. 4 - выход при продуцировании антител mAb1- mAb16: количество (в миллиграммах) каждого из продуцируемых антител mAb1- mAb16 показано для двух разных партий.
Фиг. 5 изображает выравнивание последовательностей VL по данному изобретению с VL исходных A24 (фиг. 5a) и выравнивание последовательностей VH по данному изобретению с VH исходных A24 (фиг. 5b).
Осуществление изобретения
Определения
Чтобы облегчить понимание настоящего раскрытия, вначале определены некоторые термины. Дополнительные определения приведены по всему разделу «Осуществление изобретения».
Термин «иммунный ответ/иммунологическая реакция» относится к действию, например, лимфоцитов, клеток, представляющих антиген, фагоцитирующих клеток, гранулоцитов и растворимых макромолекул, продуцируемых указанными клетками или печенью (включая антитела, цитокины и комплемент), в результате которого избирательно повреждаются или разрушаются или удаляются из человеческого организма проникшие в него патогенные агенты, клетки или ткани, зараженные патогенными агентами, раковые клетки или - в случае аутоиммунных реакций или патологического воспалительного процесса - нормальные человеческие клетки или ткани.
Термин «путь передачи сигнала/сигнальный путь» или «сигнальная активность» относится к причинно-следственно связанным биохимическим взаимодействиям, которые, как правило, инициируются белок-белковым взаимодействием, например связыванием фактора роста с соответствующим рецептором, приводящим к передаче сигнала от одной части клетки к другой. Как правило, передача сигнала включает специфичное фосфорилирование одного или более остатков тирозина, серина или треонина в одном или более белках в ряду реакций, обеспечивающих передачу сигнала. Предконечные процессы передачи сигнала обычно включают определенные события в клеточном ядре, приводящие к изменениям в экспрессии генов.
Термин «рецептор TfR/CD71/ТfR относятся к человеческому белку, аминокислотная последовательность которого представлена в настоящем документе как SEQ ID NO: 16 (если не указано иного).
Термин «антитело» в настоящем документе включает цельные антительные молекулы и любые их фрагменты, связывающие антиген (то есть содержащие антигенсвязывающий участок) или одиночные полипептидные цепи молекул антител.
Природные антитела (иммуноглобулины) представляют собой гликопротеины, включающие по меньшей мере две тяжелых (H) и две легких (L) полипептидных цепи, соединенных дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области (сокращенно обозначаемой VH) и константной области, состоящей из трех доменов - CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области (сокращенно обозначаемой VL) легкой цепи и константной области легкой цепи, состоящей из одного домена - CL. Вариабельные области тяжелых и легких цепей подразделяются на гипервариабельные участки, называемые также участками, определяющими комплементарность (CDR), которые перемежаются более консервативными участками, называемыми каркасными (FR). В каждой цепи (как VH, так и VL) имеются три участка CDR и четыре участка FR, располагающиеся в следующем порядке от N-конца к С-концу: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области в тяжелых и легких цепях содержат связывающий домен, который взаимодействует с антигеном. Константные области антительных молекул, возможно, опосредуют связывание иммуноглобулина с тканями или с факторами своего организма, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента.
Термин «антигенсвязывающий участок» антитела (или проще «антигенный участок») в настоящем документе относится к одному или более фрагментам молекулы антитела, сохраняющим способность специфично связываться с антигеном (то есть с определенным участком рецептора TfR). Показано, что функция связывания антигена может выполняться фрагментами полноразмерной молекулы антитела. Примеры фрагментов антительной молекулы, связывающих антиген, охватываются термином «антигенсвязывающий участок» и включают фрагмент Fab (одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH); фрагмент F(ab)2 (двухвалентный фрагмент, содержащий два фрагмента Fab, соединенных дисульфидным мостиком в шарнирной области); фрагмент Fd (состоящий из доменов VH и CH1); фрагмент Fv (состоящий из доменов VL и VH одной «ветви» молекулы антитела); фрагмент Unibody, состоящий из одной ветви с модифицированной тяжелой цепью IgG, например IgG4, в шарнирной области, фрагментом однодоменного (Ward et al., 1989 Nature 341:544-546), или фрагмент наноантитела, состоящий из домена VH; выделенные участки, определяющие комплементарность (CDR), или любые слитые белки, содержащие антигенсвязывающий участок.
Далее, хотя два домена фрагмента Fv - VL и VH - кодируются отдельными генами, эти фрагменты могут быть соединены генно-инженерным путем с помощью синтетического линкера, что позволяет получить их как одноцепочечный белок, в котором пара вариабельных областей VL и VH образуют одновалентную молекулу (известную как одноцепочечный Fv, или scFv); см., например, Bird et al., 1988 Science 242:423-426; Huston et al., 1988 Proc. Natl. Acad. Sci. 85:5879-5883). Такие одноцепочечные антитела также охватываются термином «антигенсвязывающий участок» антитела. Указанные антительные фрагменты получают, используя обычно применяемые для подобных целей методы, известные специалистам в данной области техники; возможности использования и эффективность полученных фрагментов проверяют так же, как интактных антител.
Термин «выделенное антитело» в настоящем документе относится к антителам, которые практически не содержат других антител, обладающих иной антигенной специфичностью (например, выделенное антитело, специфично связывающееся с рецептором TfR, практически свободно от антител, специфично связывающихся с другими антигенами, отличными от рецептора TfR). Выделенное антитело, специфично связывающееся с рецептором TfR, может, однако, проявлять перекрестную реактивность с другими антигенами, например с молекулами TfR других видов живых организмов. Кроме того, выделенное антитело практически не содержит иного клеточного материала и/или содержащихся в клетках химических веществ.
Термин «моноклональные антитела» или «композиция моноклональных антител» в настоящем документе относится к препарату антительных молекул одного молекулярного состава. В композиции по данному изобретению моноклональные антитела обладают одинаковыми специфичностью связывания и сродством к определенному эпитопу.
Термин «изотип» в настоящем документе относится к классам антител (например, IgM, IgE, IgG, например IgG1 или IgG4), определяемым генами константной области тяжелой цепи.
В настоящем документе выражения «антитело, распознающее антиген» и «антитело, специфичное в отношении антигена» употребляются как взаимозаменяемые с термином «антитело, специфично связывающееся с антигеном».
В настоящем документе антитело или белок, которые «специфично связываются с антигеном», например «специфично связываются с TfR», относится к антителу или белку, связывание которых с указанным антигеном (например, с человеческим рецептором TfR последовательности SEQ ID NO:16), характеризуется KD 100 нМ или меньше, 10 нМ или меньше, 1 нМ или меньше.
Термин «KD» в настоящем документе относится к константе диссоциации, которая является отношением Kd к Ka (то есть Kd/Ka) и выражается как молярная концентрация (М). Значения KD для антител можно определить хорошо известными в данной области техники методами. Одним из них является метод поверхностного плазмонного резонанса с использованием биосенсоров, например системы Biacore®. Определение KD для антител против TfR с помощью системы Biacore® описано в разделе «Примеры» ниже.
Термин « Kassoc» или «Ka» в настоящем документе относится к скорости ассоциации для конкретного взаимодействия антитела с антигеном, в то время как термин «Kdis» или «Kd» в настоящем документе относится к скорости диссоциации конкретного комплекса антиген-антитело.
Термин «аффинность» в настоящем документе относится к силе взаимодействия между антителом и антигеном в отдельно взятом участке связывания антигена. В пределах каждого участка связывания антигена между вариабельной областью «ветви» молекулы антитела и множеством участков молекулы антигена имеют место слабые нековалентные взаимодействия; чем больше этих взаимодействий, тем выше аффинность, то есть прочнее связывание в данном антигенсвязывающем участке антитела
Термин «авидность» в настоящем документе относится к количественной характеристике общей стабильности или силе взаимодействия комплекса антиген-антитело. Она зависит от трех главных факторов: аффинности антитела к эпитопам; валентности антигена и антитела; структурной организации взаимодействующих участков. В итоге этими факторами определяется специфичность антитела, то есть вероятность того, что данное антитело свяжется с определенным эпитопом антигена. В одном из конкретных воплощений данного изобретения указанное антитело против TfR является двухвалентным антителом.
Термин «линия клеток HL-60» в настоящем документе относится к линии промиелоцитов, выделенной Collins et al. (PNAS 1978, 75:2458-1462) и описанной также в Gallagher R. et al. (Blood, 1979, 54:713-733), например, доступной в коллекции ATCC® под номером CCL-240™ каталога.
Термин «антитело A24» в настоящем документ относится к антителу, описанному в WO2005/111082.
Термин «антителозависимая опосредованная клетками цитотоксичность» или «ADCC» в настоящем документе относится к активности, приводящей к уничтожению клеток. Активность ADCC может быть измерена при помощи имеющихся в продаже аналитических наборов, например, ADCC Reporter Bioassay производства Promega под номером Ref# G7015, и как кратко описано в разделе «Примеры».
Термин «субъект» в настоящем документе включает человека и животное, не являющееся человеком. Термин «животное» включает всех позвоночных, например, млекопитающих и представителей других классов, например, приматов, не являющихся человеком, овец, собак, кошек, крупный рогатый скот, кур, земноводных, пресмыкающихся и др.
Термин «оптимизированный» в настоящем документе относится к полинуклеотидам, нуклеотидная последовательность которых изменена таким образом, чтобы она кодировала аминокислотную последовательность кодонами, предпочтительными для продукции в клетке или организме, как правило, в эукариотической клетке, например, клетке яичника китайского хомячка (CHO) или в человеческой клетке. Оптимизированная нуклеотидная последовательность составляется так, чтобы полностью или максимально возможно сохранить аминокислотную последовательность, кодировавшуюся исходной нуклеотидной последовательностью. Аминокислотные последовательности, кодируемые оптимизированными нуклеотидными последовательностями, тоже называются оптимизированными.
Степень идентичности в процентах двух последовательностей в настоящем документе является функцией числа идентичных положений в этих двух последовательностях (т.е. % идентичности = (число идентичных положений : общее число положений) х 100) с учетом числа пропусков (гэпов) и длины каждого пропуска, которые необходимо вводить для наилучшего выравнивания двух последовательностей. Сравнение последовательностей и определение степени идентичности двух последовательностей осуществляют с использованием математических алгоритмов, как описано ниже.
Степень идентичности в процентах двух аминокислотных последовательностей можно определять с помощью алгоритма, описанного в работе E. Meyers, W. Miller (Comput. Appl. Biosci., 4:11-17, 1988), который включен в программу ALIGN (версия 2.0), используя матрицу весов замен аминокислотных остатков PAM120, приняв штраф за длину пропуска 12 и штраф за пропуск 4. Или же степень идентичности в процентах двух аминокислотных последовательностей можно определять с помощью алгоритма Needleman и Wunsch (J. Mol, Biol. 48:444-453, 1970), который включен в программу GAP в пакете программ GCG (см. http://www.gcg.com), используя матрицу Blossom 62 или PAM250 и приняв штраф за открытие пропуска 16, 14, 12, 10, 8, 6 или 4 и штраф за продолжение пропуска 1, 2, 3, 4, 5 или 6.
Степень идентичности в процентах двух нуклеотидных последовательностей можно также определить с помощью, например, таких алгоритмов, как программа BLASTN для нуклеотидных последовательностей, используя длину слова по умолчанию (W) 11, математическое ожидание (E) 10, M=5, N=4, и сравнивая обе цепи.
Рекомбинантные антитела
Антитела по настоящему описанию включают гуманизированные рекомбинантные антитела mAb1-mAb16, выделенные и структурно охарактеризованные по аминокислотным последовательностям вариабельных областей тяжелых и легких цепей и изотипу человеческой константной области, как представлено в таблице 1 ниже.
Таблица 1. Аминокислотные последовательности вариабельных областей тяжелых (VH) и легких (VL) цепей mAb1-mAb16
(VH4)
(VH4)
(VH5)
(VH5)
(VH4)
(VH4)
(VH5)
(VH5)
(VH4)
(VH4)
(VH5)
(VH5)
(VH4)
(VH4)
(VH5)
(VH5)
Соответствующие аминокислотные и кодирующие нуклеотидные последовательности константной области изотипных областей IgG4, IgG1 и их мутантные варианты IgG1 AA и IgG1 N297A, которые использовались для получения mAb1- mAb16, хорошо известны в данной области техники.
Аминокислотные последовательности полноразмерных легких и тяжелых цепей и соответствующие кодирующие их нуклеотидные последовательности для mAb1 представлены в таблице 2 ниже.
Таблица 2. Аминокислотные последовательности полноразмерных легких и тяжелых цепей и кодирующие их нуклеотидные последовательности ДНК
Легкая цепь - SEQ ID NO:17
Легкая цепь - SEQ ID NO:19
Примеры аминокислотных последовательностей участков VH CDR1 (или HCDR1), VH CDR2 (или HCDR2), VH CDR3 (или HCDR3), VL CDR1 (или LCDR1); VL CDR2 (или LCDR2); VL CDR3s (или HCDR3) некоторых антител по настоящему описанию представлены в таблице 3.
В таблице 3 участки CDR некоторых антител по настоящему описанию определены по системе нумерации Чотиа (Chothia C., Lesk A.M. 1987, J Mol Biol 196, 901-917).
Чтобы облегчить чтение настоящего документа, участки CDR далее называются HCDR1, HCDR2, HCDR3, LCDR1, LCDR2, LCDR3, соответственно.
Таблица 3. Участки CDR антител mAb1 - mAb16 и референсное антитело A24 (нумерация по системе Чотиа)
mAb5
mAb9
mAb13
mAb6 mAb10 mAb14
mAb7 mAb11 mAb15
mAb8 mAb12 mAb16
В одном из воплощений данного изобретения в выделенных рекомбинантных антителах имеются: вариабельная область тяжелой цепи, содержащая HCDR1 SEQ ID NO:1, HCDR2 SEQ ID NO:2 HCDR3 SEQ ID NO:3; вариабельная область легкой цепи, содержащая LCDR1 SEQ ID NO:4, LCDR2 SEQ ID NO: 5 или 8 и LCDR3 SEQ ID NO: 6, причем указанные антитела специфично связываются с рецептором трансферрина последовательности SEQ ID NO:16.
В конкретных воплощениях данного изобретения выделенные рекомбинантные антитела по настоящему описанию содержат либо:
(a) вариабельный полипептид тяжелой цепи, содержащий область HCDR1 SEQ ID NO:1, HCDR2 SEQ ID NO:2, HCDR3 SEQ ID NO:3, и вариабельный полипептид легкой цепи, содержащий область LCDR1 SEQ ID NO:4, LCDR2 SEQ ID NO:5 и LCDR3 SEQ ID NO:6,
(b) вариабельный полипептид тяжелой цепи, содержащий область HCDR1 SEQ ID NO:1, HCDR2 SEQ ID NO:2, HCDR3 SEQ ID NO:3, и вариабельный полипептид легкой цепи, содержащий LCDR1 SEQ ID NO:4, LCDR2 SEQ ID NO:8 и LCDR3 SEQ ID NO:6, либо
(c) вариабельный полипептид тяжелой цепи, содержащий VH SEQ ID NO:11 и вариабельный полипептид легкой цепи VL SEQ ID NO:13,
(d) вариабельный полипептид тяжелой цепи, содержащий VH SEQ ID NO:11 и вариабельный полипептид легкой цепи VL SEQ ID NO:14,
(e) вариабельный полипептид тяжелой цепи, содержащий VH SEQ ID NO:11 и вариабельный полипептид легкой цепи VL SEQ ID NO:15,
(f) вариабельный полипептид тяжелой цепи, содержащий VH SEQ ID NO:12 и вариабельный полипептид легкой цепи VL SEQ ID NO:13,
(g) вариабельный полипептид тяжелой цепи, содержащий VH SEQ ID NO:12 и вариабельный полипептид легкой цепи VL SEQ ID NO:14,
(h) вариабельный полипептид тяжелой цепи, содержащий VH SEQ ID NO:12 и вариабельный полипептид легкой цепи VL SEQ ID NO:15,
причем указанные антитела специфично связываются с рецептором TfR, имеющего аминокислотную последовательность SEQ ID NO:16.
В одном из конкретных воплощений данного изобретения указанные рекомбинантные антитела против TfR, определенные выше, обладают одним или более из следующих свойств:
(i) они связываются с рецептором трансферрина с KD = 10 нM или меньше, предпочтительно с KD = 1 нM или меньше, как измерено методом поверхностного плазмонного резонанса (SPR), например, как описано в разделе «Примеры» ниже;
(ii) связывание с рецептором трансферрина с EC50 = 0,1 мкг/мл или меньше, предпочтительно EC50 = 0,05 мкг/мл или меньше, измеренный методом твердофазного иммуноферментного анализа (ELISA), как описано в разделе «Примеры» ниже;
(iii) они вызывают апоптоз в клетках линии HL-60, уровень которого такой же или выше уровня апоптоза, измеренного в отношении соответствующих референсных химерных антител, имеющих вариабельные области исходного мышиного антитела VH SEQ ID NO:9 и VL SEQ ID NO:10, например, как измерено путем определения индукции апоптоза с использованием HL-60. Как правило, для такого определения берут рекомбинантные антитела по данному изобретению в концентрации 10 мкг/мл и сравнивают с индукцией апоптоза в клетках HL-60 таким же количеством референсных химерных антител с вариабельными областями исходного мышиного антитела A24, содержащими VH SEQ ID NO:9 и VL SEQ ID NO:10. Индукция апоптоза в HL-60 испытываемого антитела эквивалентна таковой референсного антитела, если доля в процентах (+)-клеток, померенная в случае испытываемого антитела, незначительно меньше, чем доля (+)-клеток, померенная в случае референсного антитела.
В настоящем документе формулировка «соответствующее» референсное химерное антитело относится к референсному антителу, в котором константная область определенного изотипа на 100% идентична константной области того же изотипа антитела, которое должно быть протестировано по определенному свойству, например, на индукцию апоптоза.
В некоторых воплощениях данного изобретения, которые можно объединить с описанными выше воплощениями, предлагаемое в настоящем документе антитело является фрагментом антитела, определенных выше антител.
Фрагменты антител включают, не ограничиваясь перечисленным здесь, фрагменты
Fab, Fab', Fab'-SH, F(ab')2, Fv, UniВody и scFv, диантитела, отдельные домены или наноантитела и другие фрагменты.
Термин «диантитела» относится к небольшим антительным фрагментам, в которых имеются два участка связывания антигена и которые содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одну полипептидную цепь (VH-VL). Благодаря использованию линкера, который слишком короткий, чтобы могло произойти образование пары из двух доменов одной и той же цепи, домены заставляют образовывать пару с комплементарными доменами другой цепи и создавать два антигенсвязывающие сайта.
Однодоменные антитела - это антительные фрагменты, содержащие полный либо частичный вариабельный домен тяжелой цепи или полный либо частичный вариабельный домен легкой цепи молекулы антитела. В некоторых воплощениях данного изобретения однодоменные антитела являются человеческими однодоменными антителами (Domantis, Inc., Waltham, MA; см., например, патент США № 6,248,516 B1).
Антительные фрагменты можно получать различными методами, в том числе, но не ограничиваясь перечисленным, путем протеолитического расщепления интактных молекул антител, а также путем продуцирования рекомбинантными клетками-хозяевами, как описано в настоящем документе.
В некоторых воплощениях данного изобретения предлагаемые антитела являются гуманизированными антителами. Как правило, антитела не человеческого происхождения гуманизируют, чтобы снизить их иммуногенность для людей; при этом гуманизированные антитела обладают такой же (или большей) аффинностью, какая была присуща исходным нечеловеческим антителам. В предпочтительных воплощениях данного изобретения описанные в настоящем документе антитела являются гуманизированными антителами, исходным для которых послужило антитело A24.
Как правило, гуманизированнное антитело содержит один или более вариабельных доменов, в которых CDRs (или их части) происходят из нечеловеческого антитела, например, мышиного антитела A24, а FRs (или их части) происходят из аминокислотных последовательностей человеческих антител. При необходимости гуманизированное антитело также содержит по меньшей мере часть константной области человеческого антитела. В некоторых воплощениях данного изобретения в FRs некоторые аминокислотные остатки заменены на соответственные аминокислотные остатки из нечеловеческих антител (например, из антител A24, из которых происходят аминокислотные последовательности участков CDR); это нужно, например, для повышения специфичности или аффинности. В некоторых конкретных воплощениях данного изобретения некоторые остатки CDR в гуманизированных антителах тоже содержат замены, например, для восстановления или повышения специфичности или аффинности.
Обзор гуманизированных антител и методов их получения можно найти, например, в Almagro и Fransson, Front. Biosci. 13: 1619-1633 (2008); они описаны также, например, в Riechmann et al., Nature 332:323-329 (1988) и Queen et al., Proc. Natl Acad. Sci. USA 86: 10029-10033 (1989); в патентах США №№ 5,821,337, 7,527,791, 6,982,321 и 7,087,409; Kashmiri S.V.S. et al, Methods 36:25-34 (2005) (описывает прививание определяющих специфичность участком (SDR); Padlan, Mol. Immunol. 28:489-498 (1991) (описывающих «изменение поверхности»); Dall'Acqua et al., Methods 36:43-60 (2005) и Osbourn et al, Methods 36:61-68 (2005) и Klimka et al., Br. J. Cancer, 83 :252-260 (2000) (описывающие метод «направленного отбора» к перегруппировке FR).
Рекомбинантное антитело по настоящему описанию предпочтительно является гуманизированным «молчащим» антителом, предпочтительно антителом изотипа IgG1 или IgG4.
В настоящем документе термин «молчащее» антитело относится к антителу, которое не проявляет активности или проявляет низкую активность в отношении активности ADCC, как измерено методом на отделение ADCC.
В одном из воплощений данного изобретения термин «нулевая или низкая активность ADCC» означает, что молчащее антитело показывает активность ADCC, которая составляет по меньшей мере менее 10%, например, менее 50% от активности ADCC, наблюдаемой в отношении соответствующего антитела человеческого изотипа IgG1 дикого типа.
Молчание эффекторных функций может быть достигнуто мутацией в константной части Fc антител, и описано в уровне техники: Strohl 2009 (AA & N297A); Baudino 2008, D265A (Baudino et al., J. Immunol. 181 (2008): 6664-69, Strohl CO Biotechnology 20 (2009): 685-91). Примеры молчащих антител IgG1 включают так называемые мутанты АА, содержащие мутации L234A и L235A в аминокислотной последовательности Fc IgG1. Другое молчащее антитело IgG1 содержит мутацию N297A, из-за которой антитела дегликозилированы или не гликозилированы.
Антитела с мутантными аминокислотными последовательностями можно получить путем мутагенеза (например, путем сайт-направленного мутагенеза или ПЦР-опосредованного мутагенеза) кодирующих нуклеотидных последовательностей молекул с последующей проверкой кодируемых измененных антител на сохранение функции (например, функций, указанных выше), применяя функциональные аналитические методы, описанные в настоящем документе.
Антитела с консервативными модификациями
В некоторых воплощениях данного изобретения антитело (или связывающем белке, содержащем его антигенсвязывающий участок) имеет вариабельную область тяжелой цепи, включающую HCDR1, HCDR2 и HCDR3, и вариабельную область легкой цепи, включающую LCDR1, LCDR2 и LCDR3, причем одна или более из этих последовательностей CDR имеет определенную последовательность на основе антител mAb1 - mAb16, описанных в настоящем документе, или их консервативных модификаций; при этом указанные антитело или белок сохраняют нужные функциональные свойства антител против TfR по данному изобретению.
Нужные функциональные свойства антител против TfR включают, не ограничиваясь:
(i) связывание с рецептором трансферрина с KD = 10 нM или меньше, предпочтительно с KD = 1 нM или меньше, например, как померено SPR, например, с использованием Biacore(R);
(ii) связывание с рецептором трансферрина с EC50 = 0,1 мкг/мл или меньше, предпочтительно EC50 = 0,05 мкг/мл или меньше, например, как померено ELISA, как описано в разделе «Примеры» ниже;
(iii) индукция апоптоза в клетках линии HL-60, уровень которого такой же или выше уровня апоптоза, индуцируемого соответствующим референсным химерным антителом, имеющим вариабельные области исходного мышиного антитела VH SEQ ID NO:9 и VL SEQ ID NO:10, например, как померено путем определения индукции апоптоза с использованием HL-60. Как правило, для такого определения берут рекомбинантные антитела по данному изобретению в концентрации 10 мкг/мл и сравнивают с индукцией апоптоза в клетках линии HL-60 таким же количеством референсного химерного антитела с вариабельными областями исходного мышиного A24, содержащими VH SEQ ID NO:9 и VL SEQ ID NO:10. Индукция апоптоза в клетках линии HL-60 испытываемым антителом эквивалентна таковой референсного антитела, если доля в процентах (+)-клеток, померенная в случае испытываемого антитела, незначительно меньше, чем доля (+)-клеток, померенная в случае референсного антитела.
В настоящем документе термин «консервативные модификации последовательности» относится к таким аминокислотным заменам, когда аминокислотный остаток заменяется на другой, имеющий сходную боковую цепь. Группы аминокислот со сходными боковыми цепями известны в данной области техники. Эти группы включают: аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин); аминокислоты с кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота); аминокислоты с полярными боковыми группами, не несущими электрического заряда (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан); аминокислоты с неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), аминокислоты с разветвлением при β-углеродном атоме (например, треонин, валин, изолейцин); аминокислоты с ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, один или более аминокислотных остатков в участках CDR антител, описанных в настоящем документе, могут быть заменены на остатки других аминокислот из той же группы аминокислот со сходными боковыми цепями, и измененное антитело может быть проверено на сохранение функции, применяя функциональные аналитические методы, описанные в настоящем документе.
Введение модификаций в антитела по данному изобретению осуществляется стандартными методами, известными в данной области техники, например путем сайт-направленного мутагенеза и ПЦР-опосредованного мутагенеза.
Конструирование каркасных участков или фрагментов Fc
Сконструированные антитела по данному изобретению включают антитела, в которых модификации затрагивают аминокислотные остатки каркасных участков в VH и/или VL, например для улучшения свойств антитела. Как правило, модификации в каркасных участках делаются для того, чтобы снизить иммуногенность антитела. Например, один из подходов здесь состоит в «обратном мутировании» одного или более аминокислотных остатков в каркасных участках на соответствующие последовательности зародышевой линии. Говоря конкретнее, антитело, претерпевшее соматическую мутацию, может содержать в каркасных участках аминокислотные остатки, отличные от тех, что имелись в последовательности зародышевой линии, из которой происходит данное антитело. Такие остатки можно идентифицировать путем сравнения аминокислотных последовательностей каркасных участков антитела с таковыми зародышевой линии, от которой происходит данное антитело. Для того чтобы вернуть аминокислотную последовательность к конфигурации зародышевой линии, произошедшие соматические мутации «обратно мутируют» в зародышевую линию, например, путем сайт-направленного мутагенеза или ПЦР-опосредованного мутагенеза. Такие антитела с «обратными мутациями» также входят в объем данного изобретения.
Другой тип модификаций каркасных участков подразумевает мутирование одного или более аминокислотных остатков в каркасных участках или даже в одном или более участков CDRs, для ликвидации Т-клеточных эпитопов с целью уменьшения таким образом иммуногенности антитела.
В частности, в компании Antitope (Кембридж, Великобритания) разработан спектр методов для оценки и ликвидации иммуногенности, базирующихся на идентификации локализации Т-клеточных эпитопов в антителах, предназначенных для терапевтического применения. Эти методы кратко описаны ниже.
<%=X=%> iTope™ - технология in silico для предсказания связывания пептидов с аллельными вариантами человеческих MHC класса II (Perry et al., 2008, Drugs in R&D, 9(6):385-396).
<%=X=%> TCED™ - база данных известных Т-клеточных эпитопов, идентифицированных с использованием аналитического метода картирования Т-клеточных эпитопов EpiScreen™ специально для вариабельных областей антител (Bryson et al., 2010, Biodrugs 24(1):1-8). В этой базе данных с помощью программы BLAST выявляют общие мотивы последовательностей (Altschul S.F. et al., 1997, Nucleic Acids Res. (1997) 25:3389-3402).
В дополнение к описанным выше модификациям в каркасных участках или в участках CDR, антитела по данному изобретению могут быть сконструированы таким образом, чтобы включать модификации в области Fc, как правило, с целью изменения одного или более функциональных свойств антитела, например, времени полужизни в сыворотке крови, связывания комплемента, связывания рецептора Fc и/или антителозависимой опосредованной клетками цитотоксичности.
Также антитело по данному изобретению может быть модифицировано химически (например, к молекуле антитела присоединяют одну или более химических группировок) или модифицировано для изменения его гликозилирования, опять-таки, с целью изменения одного или более функциональных свойств антитела. Все эти воплощения данного изобретения подробнее описаны ниже.
В настоящем документе термины «константная область данного изотипа» и «область Fc» употребляется взаимозаменяемо для определения С-концевой части тяжелой цепи иммуноглобулина, включая нативную область Fc и варианты области Fc. Константная область тяжелой цепи человеческого IgG, как правило, определяется как содержащая аминокислотные остатки от положения C226 или от положения P230 до карбоксильного конца полипептидной цепи в молекуле IgG. Для нумерации аминокислотных остатков области Fc используется индекс EU по Кабату. С-концевой лизин (остаток K447) области Fc может быть удален, например, в процессе получения антител или их очистки. Соответственно, в антительную композицию по данному изобретению могут входить группы молекул антител, в которых нет ни одного остатка K447, группы молекул антител, в которых ни один остаток К447 не удален, и смешанные группы, в которых одни молекулы антител содержат остаток K447, а другие нет.
В одном из конкретных воплощений данного изобретения шарнирный участок области CH1 модифицирован таким образом, что в нем изменилось количество остатков цистеина, например стало меньше или больше. Эта модификация описана в патенте США № 5,677,425 (Bodmer M.W. et al.). Количество остатков цистеина в шарнирном участке области CH1 изменяют с целью, например, способствовать сборке легких и тяжелых цепей или повысить либо понизить стабильность антитела.
В другом воплощении данного изобретения в шарнирном участке области Fc осуществлена мутация для уменьшения биологического времени полужизни антитела. Говоря конкретнее, в поверхностной зоне доменов CH2-CH3 шарнирного участка области Fc создают одну или более мутации аминокислот, обусловливающих нарушение связывания антитела с белком А Staphylococcus (SpA) по сравнению со связыванием SpA нативным шарнирным доменом Fc. Этот подход описан в патенте США № 6,165,745 (Ward E.S. et al.).
В другом воплощении данного изобретения антитела модифицируют для удлинения биологического времени полужизни. При этом возможны различные подходы. Например, в молекуле антитела осуществляют одну или более из следующих мутаций: T252L, T254S, T256F, как описано в патенте США № 6,277,375 (Ward E.S.). Или же для увеличения биологического времени полужизни антитела изменяют область CH1 или CL, чтобы антитело содержало эпитоп связывания рецептора реутилизации, в формировании которого участвуют две петли домена CH2 области Fc IgG, как описано в патентах США №№ 5,869,046 и 6,121,022 (Presta L.G. et al.).
В других воплощениях данного изобретения область Fc изменяют путем замены по меньшей мере одного аминокислотного остатка на иной, чтобы изменить эффекторные функции антитела. Например, одна или более аминокислот могут быть заменены другим аминокислотным остатком так, что изменится аффинность антитела в отношении эффекторного лиганда, но способность к связыванию с антигеном останется, как у исходного антитела. Необходимый для эффекторной функции лиганд, аффинность к которому изменяют, может быть, например, рецептором Fc или компонентом C1 системы комплемента. Этот подход описан подробно в патентах США №№ 5,624,821 и 5,648,260 (оба за авторством Winter et al.).
В еще одном воплощении данного изобретения один или более аминокислотных остатков области Fc заменяют на остаток другой аминокислоты так, чтобы изменилось связывание антитела с белком C1q системы комплемента и/или снизилась либо исчезла цитотоксичность, зависимая от комплемента (CDC). Этот подход описан подробно в патенте США № 6,194,551 (Idusogie Е.Е. et al.).
В другом воплощении данного изобретения один или более аминокислотных остатков области Fc заменяют на остатки других аминокислот так, чтобы изменилась способность антитела связываться с комплементом. Этот подход описан подробно в публикации РСТ WO 94/29351 Bodmer M.W. et al.
В еще одном воплощении данного изобретения модифицируют область Fc, чтобы повысить способность антитела вызывать антителозависимую опосредованную клетками цитотоксичность (ADCC) и/или увеличить аффинность антитела в отношении рецептора Fcγ, путем модификации одной и более аминокислот. Этот подход описан подробнее в публикации РСТ WO 00/42072 (Presta L.G.). Также картированы участки связывания человеческого IgG1 с рецепторами FcγRl, FcγRII, FcγRIII и FcRn и описаны варианты с улучшенным связыванием (см. Shields, R.L. et al., 2001, J. Biol. Chem. 276:6591-6604).
В других воплощениях данного изобретения модифицируют один или более аминокислотных остатков области Fc для снижения способности антитела вызывать антителозависимую опосредованную клетками цитотоксичность (ADCC) и/или уменьшить аффинность антитела в отношении рецептора Fcγ. Такие антитела с ослабленными эффекторными функциями, в частности, со сниженной ADCC, включают молчащие антитела.
В некоторых воплощениях данного изобретения используется домен Fc изотипа IgG1. В некоторых конкретных воплощениях данного изобретения используется мутантный вариант фрагмента Fc IgG1, например молчащий Fc IgG1, который снижает или уничтожает способность слитых полипептидов вызывать антителозависимую опосредованную клетками цитотоксичность (ADCC) и/или связываться с рецептором Fcγ. Например, молчащий мутантный вариант изотипа IgG1 - это IgG1, в котором в положениях 234 и 235 остаток лейцина заменен на остаток аланина, как описано в работе Hezareh М. et al., J. Virol., 2001, Dec;75(24):12161-8.
В некоторых воплощениях данного изобретения домен Fc является мутантным вариантом Fc с ослабленной эффекторной функцией, в котором не происходит гликозилирования в положении 297 домена Fc. Например, в домене Fc имеет место замена остатка аспарагина в положении 297. Например, остаток N297 заменяется на остаток глицина или аланина.
В другом воплощении данного изобретения модификация антитела затрагивает его гликозилирование. Например, может быть получено негликозилированное антитело (то есть к молекуле иммуноглобулина не присоединены остатки сахаров.). Измененное гликозилирование нужно, например, для повышения аффинности антитела в отношении данного антигена. Такие модификации гликозилирования можно осуществить, например, путем изменения одного или нескольких сайтов гликозилирования в аминокислотной последовательности антитела. Например, можно сделать одну или более аминокислотных замен, из-за которых исчезнет один или более сайтов гликозилирования в каркасных участках вариабельных областей, так что гликозилирования в этом сайте происходить не будет. В результате дегликозилирования может возрасти аффинность антитела к данному антигену. Этот подход описан подробнее в патентах США №№ 5,714,350 и 6,350,861 (Co M.S. et al.).
В дополнение или в качестве варианта антитело можно изменить так, что оно будет иметь измененный тип гликозилирования, например, как в гипофукозилированном антителе, в котором количество остатков сахара фукозы понижено, или в антителе, в котором увеличено разделенных пополам структур GlcNac. Показано, что такой измененный профиль гликозилирования приводит к увеличению способности антител вызывать ADCC. Такие модификации углеводов могут быть осуществлены путем, например, экспрессии антител в клетках-хозяевах с измененным механизмом гликозилирования. Клетки с измененным механизмом гликозилирования описаны в данной области техники и их можно использовать в качестве клеток-хозяев для экспрессии рекомбинантных антител по данному изобретению и получать таким образом антитела с измененным гликозилированнием. Например, в ЕР 1 176 195 Hang et al. описывается линия клеток, в которых функционально дефектен ген FUT8, кодирующий фукозилтрансферазу, в результате чего антитела, экспрессирующиеся в клетках этой линии, оказываются гипофукозилированными. Соответственно, в одном из воплощений данного изобретения получают рекомбинантные антитела путем экспрессии в клетках линии, отличающейся пониженным уровнем фукозилирования иммуноглобулинов, например в клетках млекопитающих с недостаточной экспрессией гена FUT8, кодирующего фукозилтрансферазу. В публикации PCT WO 03/035835 Presta L.G. описывается вариант Lecl3 клеточной линии CHO, отличающийся пониженной способностью к присоединению остатка фукозы к углеводам, связанным с остатком аспарагина в положении 297, что также приводит к недостаточному фукозилированию антител, экспрессирующихся в этих клетках-хозяевах (см. также Shields R.L. et al., 2002 J. Biol. Chem. 277:26733-26740). В публикации PCT WO 99/54342 Umana Р. et al. описываются линии клеток, в которых генно-инженерным путем достигнута экспрессия гликозилтрансфераз, модифицирующих гликопротеины (например, β(1,4)-N- ацетилглюкозаминилтрансфераза III (GnTIII)), в результате чего в антителах, экспрессирующихся в этих клетках, повышено содержание разделенных пополам структур GlcNac, что приводит к повышенной активности ADCC антител (см. также Umana Р. et al., 1999, Nat. Biotech. 17:176-180).
Другая модификация антител, входящая в объем данного изобретения, состоит в пэгилировании, или хэзилировании, или сходных подходах. Пэгилирование, например, может увеличить биологическое время полужизни антитела (например, в сыворотке крови). Для пэгилирования антитела, антитело или ее фрагмент связывают с полиэтиленгликолем (PEG) обычно посредством реакции, например, реакционноспособного эфирного или альдегидного производного PEG в условиях, в которых одна или более групп PEG связываются с антителом или ее фрагментом. Присоединение PEG возможно путем реакции ацилирования или алкилирования с использованием реакционноспособных молекул PEG (или аналогичных реакционноспособных водорастворимых полимеров). В настоящем документе термин «полиэтиленгликоль» относится к любой форме PEG, которую использовали для присоединения к другим белкам, например к моно(C1-C10)алкокси- или арилоксиполиэтиленгликолю или полиэтиленгликоль-малеимиду. В некоторых воплощениях данного изобретения присоединению PEG подлежит не гликозилированное антитело. Способы пэгилирования белков известны в данной области техники и применимы к антителам по данному изобретению; см., например ЕР 0 154 316 (Nishimura et al.) и ЕР 0 401 384 (Ishikawa et al.).
Другая модификация антител, входящая в объем данного изобретения, состоит в конъюгировании или слиянии по меньшей мере антигенсвязывающего участка антитела по данному изобретению с сывороточным белком, например человеческим сывороточным альбумином или его фрагментом, для увеличения времени полужизни получающейся молекулы. Такой подход описан, например, в ЕР № 0 322 094 Ballance et al.
Еще одна возможность - это слияние по меньшей мере антигенсвязывающего участка антитела по данному изобретению с другим белком, способным связываться с каким-либо сывороточным белком, например с человеческим сывороточным альбумином или его фрагментом, для увеличения времени полужизни получающейся молекулы. Такой подход описан, например, в ЕР 0 486 525 Nygren et al.
В одном из конкретных воплощений данного изобретения эффекторная функция или функция активации комплемента антитела по данному изобретению снижалась или исчезала по сравнению с антителами дикого типа того же изотипа. В одном из аспектов данного изобретения эффекторную функцию антител по данному изобретению снижали или ликвидировали, применяя метод, выбираемый из: снижения уровня гликозилирования антитела; изменения изотипа антитела на другой, от природы отличающийся низким или нулевым уровнем эффекторной функции; модификации области Fc. В относящихся к этому конкретных воплощениях данного изобретения указанный изотип, отличающийся низким или нулевым уровнем эффекторной функции, является изотипом IgG4.
Молукулы нуклеиновых кислот, кодирующие антитела по данному изобретению
В настоящем документе также описываются молекулы нуклеиновых кислот, кодирующие антитела против TfR или относящиеся к ним белки по данному изобретению. Примеры нуклеотидных последовательностей для вариабельных областей легкой цепи - это те, что кодируют аминокислотные последовательности вариабельных областей легкой цепи любого из антител mAb1 - mAb16; эти последовательности нетрудно получить из таблиц 1 и 2 и используя генетический код, при необходимости с учетом предпочтения кодонов в зависимости от вида клетки-хозяина.
Настоящее описание также относится к молекулам нуклеиновых кислот, происходящим от упомянутых выше нуклеотидных последовательностей, оптимизированных для экспрессии белков в клетках млекопитающих, например в клетках линии СНО.
Указанные нуклеиновые кислоты могут присутствовать в целой клетке, в лизате клеток, либо они могут быть в частично очищенном или в практически чистой форме. Нуклеиновая кислота является «выделенной» или «практически чистой», если она очищена от других клеточных компонентов или иных контаминантов, например, других клеточных нуклеиновых кислот или белков, стандартными методами, в том числе путем обработки щелочным раствором с додецилсульфатом натрия (SDS), центрифугирования в градиенте плотности CsCl, колоночной хроматографии, электрофореза в агарозном геле и других хорошо известных в данной области техники методов (см. Ausubel F. еt al., ed. 1987. Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York). Нуклеиновая кислота по данному изобретению представлена, например, ДНК либо РНК и содержит либо не содержит интронных последовательностей. В одном из воплощений данного изобретения нуклеиновые кислоты входят в состав вектора, например вектора для фагового дисплея или рекомбинантного плазмидного вектора.
Нуклеиновые кислоты по данному изобретению можно получать, используя стандартные молекулярно-биологические методы. Когда получены фрагменты ДНК, кодирующие, например, VH и VL, к ним применяют стандартные приемы технологии рекомбинантной ДНК, например чтобы превратить гены вариабельной области в гены полноразмерной цепи антитела, в гены фрагментов Fab или scFv. Для этого фрагмент ДНК, кодирующий VL и VH (например, VL и VH по таблице 1) функционально объединяют с другой молекулой ДНК или с фрагментом, кодирующим другой белок, например, константную область антитела или гибкий линкер. Термин «функционально связан» в контексте данного изобретения означает, что данные два фрагмента ДНК соединены так, что кодируемые ими аминокислотные последовательности остаются в рамке считывания, или же так, что экспрессия данного белка оказывается регулируемой желаемым промотором.
Выделенная ДНК, кодирующая вариабельную область тяжелой цепи, может быть превращена в ген полноразмерной тяжелой цепи путем функционального связывания с другой молекулой ДНК, кодирующей константную область тяжелой цепи (CH1, CH2 и CH3). Последовательности генов константных областей тяжелых цепей человеческих антител известны в данной области техники (см., например, Kabat, E. A., et al., 1991, Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242), и фрагменты ДНК, включающие эти нуклеотидные последовательности, можно получить стандартным методом амплификации ПЦР. Константная область тяжелой цепи антитела по данному изобретению может быть константной областью IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD. В некоторых воплощениях данного изобретения константную область тяжелой цепи выбирают из изотипов IgG1, например человеческого IgG1. В других воплощениях данного изобретения константную область тяжелой цепи выбирают из изотипов IgG4, например человеческого IgG4. Для гена тяжелой цепи фрагмента Fab ДНК, кодирующая VH, может быть функционально связана с другой молекулой ДНК, кодирующей только участок CH1 константной области тяжелой цепи.
Выделенная ДНК, кодирующая область VL, может быть превращена в ген полноразмерной легкой цепи (равно как и в ген легкой цепи фрагмента Fab) путем функционального связывания ДНК, кодирующей область VL, с другой молекулой ДНК, кодирующей константную область легкой цепи, CL. Нуклеотидные последовательности генов константной области легкой цепи человека известны в данной области техники (см., например, Kabat, E. A., et al., 1991, Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242), и фрагменты ДНК, включающие эти области, можно получить стандартным методом амплификации ПЦР. Константная область легкой цепи может быть «каппа» либо «лямбда» константной областью.
Чтобы получить ген scFv, фрагменты ДНК, кодирующие VH и VL, функционально связывают с другим фрагментом, кодирующим гибкий линкер, например, кодирующим аминокислотную последовательность (Gly4-Ser)3, так что последовательности VH и VL могут экспрессироваться в виде непрерывного одноцепочечного белка, в котором последовательности VL и VH соединены гибким линкером (см., например, Bird R.E. et al., 1988, Science, 242:423-426; Huston J.S. et al., 1988, Proc. Natl. Acad. Sci. USA, 85:5879-5883; McCafferty J. et al., 1990, Nature, 348:552-554).
Получение трансфертом, продуцирующих моноклональные антитела
Антитела по данному изобретению можно получать клетке-хозяине, являющейся трансфектомой, например сочетая приемы технологии рекомбинантной ДНК и методы трансфекции генов, как это известно в данной области техники (см., например, Morrison, S. (1985) Science 229:1202).
Например, для экспрессии антител или их фрагментов ДНК, кодирующие частичные или полноразмерные легкие и тяжелые цепи, можно получить стандартными молекулярно-биологическими или биохимическими методами (например, путем химического синтеза ДНК, амплификации ПЦР или клонирования с использованием гибридом, в которых экспрессируются нужные антитела), и эти ДНК можно включить в состав экспрессионных векторов, так что введенные гены окажутся функционально связанными с нуклеотидными последовательностями, регулирующими транскрипцию и трансляцию. В контексте данного изобретения выражение «функционально связанный» означает, что ген антитела лигирован в вектор таким образом, что имеющиеся в этом векторе нуклеотидные последовательности, регулирующие транскрипцию и трансляцию, регулируют также транскрипцию и трансляцию гена антитела. Экспрессионные вектор и регуляторные последовательности подбирают так, чтобы они были совместимы с используемой экспрессионной клеткой-хозяином. Ген легкой и ген тяжелой цепей антитела могут быть встроены в разные векторы или, чаще, оба гена включают в состав одного и того же экспрессионного вектора. Гены антител встраивают в экспрессионный вектор стандартными методами (например, лигируя комплементарные сайты рестрикции во фрагменте гена антитела, или в отсутствие сайтов рестрикции лигируя тупые концы ДНК). Вариабельные области легких и тяжелых цепей антител, описанных в настоящем документе, можно использовать для формирования генов полноразмерных антитела любого изотипа путем встраивания их в экспрессионные векторы, в которых уже имеются нуклеотидные последовательности, кодирующие константные области легких и тяжелых цепей нужного изотипа; в результате сегмент VH оказывается функционально связанным с сегментом CH в векторе, и сегмент VL оказывается функционально связанным с сегментом CL. В дополнение к этому или вместо этого рекомбинантный экспрессионный вектор может кодировать сигнальный пептид, который способствует секреции полипептидных цепей антитела из клетки-хозяина. Ген цепи антитела может быть встроен в вектор таким образом, что сигнальный пептид оказывается в одной рамке считывания с геном цепи антитела, в результате сигнальный пептид будет присоединен к N-концу цепи антитела. Указанным сигнальным пептидом может быть сигнальный пептид иммуноглобулина или гетерологичный сигнальный пептид (то есть сигнальный пептид белка, не являющегося иммуноглобулином).
Помимо генов цепи антитела описанные в настоящем документе рекомбинантные экспрессионные векторы несут нуклеотидные последовательности, регулирующие экспрессию генов цепи антитела в клетке-хозяине. Термин «последовательность, регулирующая экспрессию» включает промоторы, энхансеры и другие управляющие экспрессией элементы (например, сигналы полиаденилирования), которые регулируют транскрипцию или трансляцию генов цепи антитела. Такие регуляторные последовательности описаны, например, в Goeddel D.V. (Gene Expression Technology. Methods in Enzymology, 185, Academic Press, Сан-Диего, шт. Калифорния, 1990). Специалистам в данной области техники должно быть ясно, что конструкция экспрессионного вектора, в том числе, выбор регуляторных последовательностей зависит от таких факторов, как выбор подлежащей трансформации клетки-хозяина, уровня экспрессии желаемого белка и др. Регуляторные последовательности для экспрессии в клетках млекопитающих включают вирусные элементы, обеспечивающие высокий уровень экспрессии в клетках млекопитающих, например, промоторы и/или энхансеры цитомегаловируса (CMV), полиомавируса обезьян 40 (SV40) и других полиомавирусов, аденовируса (например, аденовирусный большой поздний промотор - AdMLP) и вирус полиомы. Или же можно использовать невирусные регуляторные последовательности, например, промотор гена убиквитина или промотор гена Р-глобина. Также используются регуляторные элементы, включающие нуклеотидные последовательности из разных источников, например, промотор SRα системы, которая содержит последовательности из промотора ранних генов SV40, и длинный концевой повтор вируса Т-клеточного лейкоза человека типа 1 (Takebe Y. et al., 1988 Mol. Cell. Biol. 8:466-472).
Помимо генов цепи антитела и регуляторных последовательностей рекомбинантный экспрессионный вектор по данному изобретению может содержать дополнительные последовательности, например те, которые регулируют репликацию вектора в клетке-хозяине (например, сайт инициации репликации) и гены селектируемых маркеров. Гены отбираемых маркеров помогают отбирать клетки-хозяева с введенным вектором (см., например, патенты США №№ 4,399,216, 4,634,665 и 5,179,017, все - Axel et al.). Например, часто селектируемый маркерный ген придает клетке-хозяину, в который ген был введен, устойчивость к лекарствам, например, G418, гигромицину или метотрексату. Селектируемые маркерные гены включают ген дигидрофолатредуктазы (DHFR) (для использования в dhfr-клетках-хозяевах с амплификацией/отбором по метотрексату), и ген neo (для отбору по G418).
Для экспрессии легких и тяжелых цепей антитела экспрессионный вектор (вектора), кодирующий легкие и тяжелые цепи антитела, вводят путем трансфекции в клетку-хозяина, применяя стандартные методы. Различные формы термина «трансфекция» охватывает весьма разнообразные методы, обычно используемые для введения экзогенной ДНК в прокариотическую или эукариотическую клетку-хозяина, например, электропорацию, осаждение фосфатом кальция, трансфекцию с использованием диэтиламиноэтил-декстрана и проч. Теоретически возможно, чтобы антитела по данному изобретению экспрессировались как в прокариотических, так и в эукариотических клетках-хозяевах. Экспрессия антител в эукариотических клетках, например, в клетках млекопитающих, дрожжей или мицелиальных грибов, ценна потому, что таким эукариотическим клеткам, в особенности клеткам млекопитающих, более, нежели прокариотическим клеткам, свойственна сборка и секреция должным образом сформированных и иммунологически активных антител.
В одном из конкретных воплощений данного изобретения вектор для клонирования или экспрессии по данному изобретению содержит одну из нуклеотидных последовательностей, кодирующих тяжелые и легкие цепи антител mAb1 - mAb16, функционально связанные с подходящими промоторными последовательностями.
Клетки млекопитающих для экспрессии рекомбинантных антител по данному изобретению включают клетки яичника китайского хомячка (СНО клетки) (в том числе dhfr- клетки СНО, описанные Urlaub и Chasin, 1980, Proc. Natl. Acad. Sci. USA, 77:4216-4220, которые используют с селектируемым маркером DHFR, как описано, например, в Kaufman R.J. и Sharp P.A., 1982 Mol. Biol. 159:601-621), клеточную линию CHOK1 dhfr+, мышиные миеломные клетки NSO, клетки COS и SP2, например, клеточную линию GS CHO вместе с системой экспрессии гена GS Xceed™ (Lonza).
Когда рекомбинантные экспрессионные векторы, кодирующие гены антител, введены в клетки млекопитающих, антитела получают путем культивирования этих клеток-хозяев в течение промежутка времени, достаточного для экспрессии антител и при необходимости секреции антител в культуральную среду, где выращиваются клетки-хозяева. После секреции образовавшихся антител их выделяют из культуральной среды и очищают, применяя стандартные методы очистки белков (см., например, Abhinav et al., 2007, Journal of Chromatography, 848: 28-37).
В одном из конкретных воплощений данного изобретения клетка-хозяин - это клетка-хозяин, трансфицированная экспрессионным вектором, имеющим кодирующие последовательности, пригодные для экспрессии антител mAb1-mAb16 соответственно и функционально связанные с подходящими промоторными последовательностями.
Указанные клетки-хозяева можно затем культивировать в подходящих условиях для экспрессии и продуцирования антитела по данному изобретению, выбираемого из группы, состоящей из mAb1-mAb16.
Иммуноконъюгаты
В другом аспекте данного изобретения предлагаются описанные в настоящем документе антитела против TfR или их фрагменты, конъюгированные с терапевтическим веществом, таким как, цитотоксин, лекарственное вещество (например, иммунодепрессант) или радиотоксин. Такие иммуноконъюгаты называются в настоящем документе иммуноконъюгатами. Иммуноконъюгаты, в состав которых входит один или более цитотоксинов, называются «иммунотоксинами». Цитотоксины, или цитотоксические агенты, включают любые агенты, губительные для живых клеток (то есть убивающие) их. Примеры цитотоксинов включают цитохалазин В, грамицидин D, этидийбромид, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол, пуромицин и их аналоги и гомологичные соединения. Терапевтические агенты также включают, например, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил, декарбазин), вещества, используемые при абляции (например, мехлорэтамин, тиотепа, хлорамбуцил, мелфалан, кармустин (BSNU), ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин С, цис-дихлордиаминплатину (II) (DDP, цисплатин) антрациклины (например, даунорубицин, ранее известный под названием дауномицин, и доксорубицин), антибиотики (например, дактиномицин, ранее известный под названием актиномицин), блеомицин, митрамицин, антрамицин (AMC)), антимитотические агенты (например, винкристин и винбластин).
Цитотоксины можно конъюгировать с антителами по данному изобретению, используя методы связывания с помощью линкеров, известные в данной области техники. Примеры линкеров, используемых для получения конъюгатов цитотоксина с антителом, включают, не ограничиваясь, гидразоны, тиоэфиры, сложные эфиры, дисульфиды и содержащие пептиды линкера. Выбирают, например, такой линкер, который подвержен расщеплению при низких рН, присущих содержимому лизосом, или расщеплению протеазами, например протеазами, которые экспрессируются преимущественно в опухолевых тканях, такими как катепсины (например, катепсины В, C и D).
Для дальнейшего ознакомления с различными цитотоксинами, линкерами и методами конъюгирования терапевтических агентов с антителами см. работу Panowksi S. et al. 2014, Jan 1; 6(1): 34-4, где можно найти также обзор конъюгатов антител с лекарственными веществами.
Антитела по данному изобретению могут образовывать конъюгаты также с цитотоксическими радиофармацевтическими препаратами, также именуемыми как радиоиммуноконъюгаты. Примеры радиоактивных изотопов, пригодных для образования конъюгатов с антителами для диагностических или терапевтических целей, включают, не ограничиваясь, иод-131, индий-111, иттрий-90 и лютеций-177. Методы получения радиоиммуноконъюгатов хорошо известны в данной области техники.
Биспецифичные или полиспецифичные молекулы
Другой аспект данного изобретения, раскрытый в настоящем документе, касается биспецифичных или полиспецифичных молекул, включая антитела против TfR по данному изобретению. Возможно получить производное антитела или присоединить антительную молекулу к другой функциональной молекуле, например, к другому пептиду или белку (например, к другому антителу или лиганду рецептора) для того, чтобы получилась биспецифичная молекула, связывающаяся с по меньшей мере двумя различными связывающими сайтами или молекулами-мишенями. Собственно говоря, можно получить производное антитела или присоединить антитело к более чем одной другой функциональной молекуле, чтобы получить полиспецифичные молекулы, связывающиеся с более чем двумя различными связывающими сайтами и/или молекулами-мишенями; в настоящем документе такие полиспецифичные молекулы также охватываются термином «биспецифичная молекула». Для создания биспецифичной молекулы антитело по данному изобретению может быть функционально связано (например, путем химического связывания, слияния на генетическом уровне, нековалентной ассоциации или как-либо иначе) с одной или более другой связывающей молекулой, например с другим антителом, фрагментом антитела, пептидом или миметику связывания, в результате чего образуется биспецифичная молекула.
Соответственно, настоящее описание охватывает биспецифичные молекулы, содержащие по меньшей мере один первый участок, специфично связывающийся с TfR, например, один антигенсвязывающий участок какого-либо из антител mAb1-mAb16, и второй связывающий участок, специфично связывающийся со вторым эпитопом-мишенью. Например, второй эпитоп-мишень является еще одним эпитопом TfR, отличным от первого эпитопа-мишени.
Кроме того, в тех воплощениях данного изобретения, в которых биспецифичные молекулы являются полиспецифичными, они могут помимо участков, специфично связывающихся с первым и вторым эпитопами-мишенями, также включать третью связывающую специфичность.
В одном из воплощений данного изобретения описанные в настоящем документе биспецифичные молекулы содержат в качестве способного к специфичному связыванию компонента по меньшей мере одно антитело или фрагмент антитела, в том числе, например, фрагмент Fab, Fab', F(ab')2, Fv, UniВody или одноцепочечные Fv. Входящее в состав биспецифичной молекулы антитело может быть представлено также димером легкой и тяжелой цепей или каким-либо минимальным его фрагментом, например, Fv или одноцепочечной конструкцией, описанной в патенте США № 4,946,778 Ladner et al.
В биспецифичных антителах, описанных в настоящем документе, могут использоваться также другие антитела, а именно мышиные, химерные и гуманизированные моноклональные.
Биспецифичные молекулы по настоящему описанию получают путем соединения компонентов, способных к специфичному связыванию, применяя известные в данной области техники методы. Например, каждая специфичность связывания биспецифичной молекулы может быть получена отдельно, а затем объединена с другими. Когда специфичности связывания являются белками или пептидами, для их ковалентного соединения могут использоваться различные связывающие и поперечно-сшивающие агенты. Примеры поперчено-сшивающих агентов включают протеин А, карбодиимид, N-сукцинимидил-S-ацетилтиоацетат (SATA), 5,5'-дитиобис(2-нитро)бензойную кислоту (DTNB), орто-фенилендималеимид (oPDM), N- сукцинимидил -3-(2-пиридилдитио)пропионат (SPDP) и сульфосукцинимидил-4-(N-малеимидометил) циклогексан-l-карбоксилат (сульфо-SMCC) (см., например, Karpovsky et al., 1984 J. Exp. Med. 160:1686; Liu M.A. et al., 1985 Proc. Natl. Acad. Sci. USA 82:8648). Другие методы описаны в Paulus, 1985 Behring Ins. Mitt. No. 78,118-132; Brennan M. et al., 1985 Science, 229:81-83) и Glennie et al., 1987, J. Immunol.,139: 2367-2375).
Или же обе связывающие специфичности могут кодироваться одним и тем же вектором и экспрессироваться и собираться в одной и той же клетке-хозяине. Этот подход особенно полезен, когда биспецифичная молекула представляет собой слитые белки mAb x mAb, mAb x Fab, Fab x F(ab')2 или лиганд x Fab. Биспецифичная молекула по данному изобретению может быть одноцепочечной, содержащей одну полипептидную цепь антитела и связывающую детерминанту, или одноцепочечной биспецифичной молекулой, содержащей две связывающие детерминанты.
Связывание биспецифичных молекул с их специфическими мишенями можно подтвердить, например, методами твердофазного иммуноферментного анализа (ELISA), радиоиммунологического анализа (REA), проточной цитометрии FACS, биологического анализа (например, по подавлению пролиферации клеток и апоптозу) или вестерн-блоттинга. Все эти методы позволяют, как правило, выявить присутствие искомых комплексов белок-антитело с помощью меченых реагентов (например, антител), специфичных в отношении искомого комплекса.
Антитела по данному изобретению можно также использовать для получения искусственных рецепторов на Т-клетках (так называемых химерных Т-клеточных рецепторов, или химерных рецепторов антигена, сокращенно CAR). Например, вариабельные области антител можно использовать для формирования scFv, которые присоединяют посредством спейсера к трансмембранному домену и сигнальному внутриклеточному домену Т-клеточного рецептора (например, к CD3 зета), и они могут продуцироваться на поверхности Т-клеток. Такие CARs можно использовать при терапии методом создани\ адаптивного иммунитета путём переноса иммунокомпетентных клеток от донора к реципиенту, например, для лечения пролиферативных расстройств.
Фармацевтические композиции
В другом аспекте данного изобретения предлагается композиция, например, фармацевтическая композиция, содержащая одно из описанных в настоящем документе антител или их комбинацию, например, одно антитело, выбираемое из группы, состоящей из антител mAb1-mAb16, и содержащая также фармацевтически приемлемый носитель. Такие композиции могут включать одно антитело или их комбинацию (например, два или более разных антител), или иммуноконъюгаты. или биспецифичные молекулы, описанные выше.
Фармацевтические композиции, описанные в настоящем документе, можно вводить как часть комбинированной терапии, то есть сочетать с другими агентами. Например, комбинированная терапия может включать антитело против TfR по данному изобретению, например, одно антитело, выбираемое из группы, состоящей из антител mAb1-mAb16, в сочетании с по меньшей мере одним противовирусным, противовоспалительным или иным антипролиферативным агентом. Примеры терапевтических агентов, которые можно использовать в комбинированной терапии, подробнее описаны ниже, где речь идет о применении антител по данному изобретению.
В настоящем документе термин «фармацевтически приемлемый носитель» включает любые растворители, дисперсионную среду, покрытия, антибактериальный или противогрибковый агент, изотонические агенты, агенты, задерживающие абсорбцию, и прочие подобные физиологически совместимые ингредиенты. Указанный носитель должен быть пригоден для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии). В одном из воплощений данного изобретения носитель пригоден для подкожного введения. В зависимости от пути и способа введения композиции в организм активное вещество (то есть антитела, иммуноконъюгаты или биспецифичные молекулы) может быть заключено в какой-либо материал, защищающий это вещество от воздействия кислот и других естественных факторов, которые могут инактивировать данное вещество.
Одним из примеров фармацевтически приемлемого носителя является стерильный физиологический раствор, забуференный фосфатом (PBS). В данной области техники хорошо известны другие пригодные носители (см., например, Gennaro (ed.), Remington's Pharmaceutical Sciences (Mack Publishing Company, 19th ed. 1995)). В состав композиции могут также входить один или более наполнителей, консервантов, солюбилизаторов, забуферивающих агентов, альбумин, чтобы предотвратить потери белка на поверхности емкостей, и др.
Форма фармацевтической композиции по данному изобретению, путь и способ ее введения, дозировка и режим введения обычно зависят от состояния, подлежащего лечению, степени тяжести заболевания, возраста, массы тела и пола индивида и проч.
Фармацевтические композиции по данному изобретению могут быть представлены препаратами для местного, перорального, парентерального, интраназального, внутривенного, внутримышечного, подкожного или внутриглазничного и иного введения.
Фармацевтические композиции по данному изобретению предпочтительно содержат носители, которые фармацевтически приемлемы для препаратов, предназначенных для введения путем инъекций. В частности, это могут быть изотонические, стерильные, солевые растворы (однозамещенного или двухзамещенного фосфата натрия, хлоридов натрия, калия, кальция или магния, другие подобные компоненты или смеси таких солей) или же сухие препараты, в частности, лиофилизованные композиции, которые после добавления стерильной воды или физиологического раствора (в зависимости от конкретного случая) становятся растворами, пригодными для введения путем инъекций.
Дозы для введения подбираются в зависимости от различных параметров и, в частности, зависит от пути и способа введения, характера патологии или же от желаемой продолжительности лечения.
Для получения композиций по данному изобретению эффективное количество антител можно растворить или диспергировать в фармацевтически приемлемом носителе или водной среде.
Формы фармацевтических композиций по данному изобретению, пригодные для введения путем инъекций, включают стерильные водные растворы или дисперсии; препараты, включающие кунжутное масло, арахисовое масло или водный раствор пропиленгликоля; стерильные порошки или лиофилизаты для приготовления стерильных растворов или дисперсий для введения путем инъекций непосредственно перед их использованием. В любом случае форма композиции по данному изобретению должна быть стерильной и жидкой в такой степени, чтобы ее нетрудно было бы вводить с помощью шприца. Они должны быть стабильными в условиях изготовления и хранения, и защищены от загрязнения микроорганизмами, например бактериями и грибками.
Растворы активных веществ, представленных свободными основаниями или фармацевтически приемлемыми солями, готовят на воде, при необходимости смешанной с поверхностно-активным агентом, например гидроксипропилцеллюлозой. Дисперсии готовят на глицерине, жидком полиэтиленгликоле и их смесях, а также на маслах. При обычных условиях хранения и применения эти препараты должны содержать консервант для предотвращения размножения микроорганизмов.
Антитело по изобретению включают в состав описанной выше композиции в нейтральной или солевой форме. Фармацевтически приемлемые в данном случае соли включают соли присоединения кислоты (образованные с участием свободных аминогрупп белка) и соли с неорганическими кислотами, например соляной или фосфорной, либо соли с органическими кислотами, например уксусной, щавелевой, виннокаменной, миндальной и проч. Можно использовать соли, образованные с участием свободных карбоксильных групп белка с неорганическими основаниями, например с гидроокисями натрия, калия, аммония, кальция или железа, а также с органическими основаниями, например с изопропиламином, триметиламином, гистидином, прокаином и др.
Носитель в композиции по данному изобретению может быть растворителем или дисперсионной средой, содержащей, например воду, этиловый спирт, многоатомный спирт (например, глицерин, пропиленгликоль, жидкий полиэтиленгликоль и др.) и их пригодные смеси, а также растительные масла. Необходимая текучесть носителя достигается, например, путем использования покрытий, например лецитина, путем поддержания должного размера частиц в случае дисперсий и путем использования поверхностно-активных агентов. Предотвратить активность микроорганизмов можно с помощью различных антибактериальных и противогрибковых агентов, например парабенов, хлорбутилового спирта, фенола, сорбиновой кислоты, тимеросала и др. Во многих случаях предпочтительно включать в состав композиции по данному изобретению изотонические агенты, например, сахара или хлорид натрия. Для того, чтобы при инъекции вводимая композиция дольше всасывалась в ткани в месте введения, используют агенты, замедляющие всасывание, например моностеарат алюминия и желатин.
Для получения стерильных растворов для введения путем инъекций активные вещества в требуемом количестве растворяют в подходящем растворителе с различными другими ингредиентами, перечисленными выше, при необходимости, после чего стерилизуют путем фильтрации. Для получения дисперсий, как правило, объединяют стерилизованные активные ингредиенты со стерильным носителем, содержащим основную дисперсионную среду и другие необходимые ингредиенты из числа упомянутых выше. Для получения стерильных порошков для приготовления стерильных инъекционных растворов предпочтительны такие методы, как высушивание в вакууме и лиофилизация, которые позволяют получить порошок активного ингредиента вместе с любым дополнительно нужным дополнительным ингредиентом из их раствора, стерилизованного фильтрацией.
Также рассматриваются более концентрированные или высококонцентрированные растворы для инъекций, в которых используется в качестве растворителя диметилсульфоксид (DMSO), который способствует очень быстрому проникновению активных агентов в нужное место и созданию высокой их концентрации на небольшой площади опухоли.
После приготовления раствор для инъекций вводят способом, совместимым с дированной формой, и в таком количестве, которое является терапевтически эффективным. Композиции по данному изобретению нетрудно вводить в организм в различных дозированных формах, например, в форме описанных выше растворов для инъекций, а также можно использовать капсулы, обеспечивающие модифицированное высвобождение активных ингредиентов, и другие формы.
Для парентерального введения водного раствора, например, раствор должен быть подходящим образом забуфферен, если необходимо, и должна быть обеспечена изотоничность жидкого разбавителя достаточным количеством соли или глюкозы. Такие водные растворы особенно подходят для внутривенного, внутримышечного, подкожного и внутрибрюшинного введения. В этой связи стерильные водные среды, которые могут быть использованы, известны в данной области техники в свете настоящего раскрытия. Например, одна доза композиции по данному изобретению может быть растворена в 1 мл раствора хлорида натрия и либо добавлена в жидкость для подкожного введения до 1000 мл, либо введена в нужное место (см., например, "Remington's Pharmaceutical Sciences" 15th Edition, pages 1035-1038, 1570-1580). Дозировка может варьировать по необходимости в зависимости от состояния субъекта, подлежащего лечению. Персонал, ответственный за введение, должен в любом случае определять подходящую дозу конкретно для данного субъекта.
Антитела по данному изобретению входят в состав терапевтической смеси в количестве от около 0,0001 мг до 1,0 мг или от около 0,001 мг до 0,1 мг, или от 0,1 мг до 1,0 мг или даже от 1,0 мг до 10 мг. Возможно также многократное введение.
Помимо лекарственных форм для парентерального введения, например, внутривенного или внутримышечного, другие фармацевтически приемлемые формы включают, например, таблетки или иные твердые формы для перорального применения; капсулы длительного высвобождения, и любая другая используемая сейчас форма.
В некоторых воплощениях данного изобретения для введения антител в клетки используются липосомы и/или наночастицы. Получение и применение липосом и/или наночастиц известно специалистам в данной области техники.
Заключение веществ в нанокапсулы происходит стабильным и воспроизводимым образом. Во избежание побочных эффектов, возможных из-за перегрузки внутреннего пространства клетки полимерами, такие в высшей степени мелкие частицы (размером около 0,1 мкм) получают, как правило, используя полимеры, способные к деградации in vivo. Для применения по данному изобретению пригодны, в частности, биоразлагаемые наночастицы из полиалкил-цианоакрилата, которые легко изготавливать.
Липосомы образуются из фосфолипидов, диспергированных в водной среде и спонтанно образующих многослойные двухслойные концентрические везикулы (также называемые многослойные везикулы (MLVs)). MLVs, как правило, имеют от 25 нм до 4 мкм в диаметре. При их обработке ультразвуком образуются небольшие однослойные везикулы (SUV) диаметром от 200 до 500 Å, в середине которых находится водный раствор. Физические свойства липосом зависят от рН, ионной силы и присутствия двухвалентных катионов.
Применения и способы использования антител или белков по данному изобретению
Антитела или белки по данному изобретению можно использовать в диагностических и терапевтически целях in vitro и in vivo. Например, их можно вводить в культивируемые клетки, например in vitro или in vivo, или субъекту, например in vivo, для лечения, предотвращения или диагностики различных расстройств.
Антитела по данному изобретению способны не только подавлять пролиферацию клеток, но и вызывать апоптоз интенсивно пролиферирующих клеток, например Т-лимфоцитов.
В настоящем документе рассматривается применение антител против TfR или белков по данному изобретению в качестве лекарственного средства, в частности, для использования при лечении, профилактике или диагностировании расстройств, связанных с пролиферацией клеток, например опухолей с высоким уровнем экспрессии рецептора TfR, говоря конкретнее - гематологических опухолей, например лимфом, Т-клеточного лейкоза взрослых (ATL), мантийноклеточной лимфомы (MCL), болезни Ходжкина, В-крупноклеточной лимфомы, периферической Т-клеточной лимфомы, острого лейкоза (миелоидного и лимфоидного), а также солидных опухолей, например почечной карциномы, рака легких (мелкоклеточного) и др.
Также рассматривается применения антител по данному изобретению при лечении, профилактике или диагностировании расстройств, связанных с HTLV-1, в частности, для снижения вирусной нагрузки при воспалительных расстройствах, связанных с инфекцией HTLV-1, в том числе, тропического спастического пареза/миелопатии (HAM/TSP), полимиозита и артрита.
Настоящее описание также относится к способам уменьшения или подавления поглощения трансферрина клетками крови у человека путем введения композиции, содержащей терапевтически эффективную дозу антител по данному изобретению.
Антитела или белки по данному изобретению для описанных выше применений вводят в организм как отдельно взятый активный ингредиент, или в сочетании с, например, адъювантом, или в комбинации с другими лекарственными средствами, например, с противовирусными, противовоспалительными или цитотоксическими, антипролиферативными, химиотерапевтическими или противоопухолевыми агентами и другими для лечения или предотвращения заболеваний, упомянутых выше.
Например, антитела по данному изобретению для описанных выше применений можно использовать в комбинации с азидотимидином, α-интерфероном, моноклональными антителами против В-лимфоцитарного антигена CD20, моноклональными антителами против рецептора к интерлейкину-2 CD25, химиотерапевтическими агентами.
Подходящие антинеопластические агенты включают, не ограничиваясь, алкилирующие агенты (например, циклофосфамид, мехлорэтамин, хлорамбуцил, мелфалан, нитрозомочевину, темозоломид), антрациклины (например, даунорубицин, доксорубицин, эпирубицин, идарубицин, митоксантрон, валрубицин), таксаны (например, паклитаксел, доцетаксел), эпотилоны, ингибиторы топоизомеразы I (например, иринотекан или топотекан), ингибиторы топоизомеразы II (например, этопозид, тенипозид или тафлупозид), аналоги нуклеотидов и их предшественников (например, азацитидин, азатиоприн, капецитабин, цитарабин, фторурацил, гемцитабин, гидроксимочевину, меркаптопурин, метотрексат или тиогуанин), пептидные антибиотики (например, карбоплатин, цисплатин и оксалиплатин), ретиноиды (например, третиноин, алитретиноин, бексаротен), винкаалкалолиды и их производные (например, винбластин, винкристин, виндезин, винорелбин), агенты, напрвленные на определенные мишени, например ингибиторы киназ (например, ибрутиниб, иделализиб, эрлотиниб, гефитиниб, иматиниб, вемурафениб, висмодегиб), ингибиторы протеасом (например, бортезомиб, карфилзомиб), ингибиторы гистондеацетилазы (например, вориностат или ромидепсин).
Соответственно сказанному выше в настоящем описании предлагается еще один аспект данного изобретения:
а именно, способ, как описано выше, включающий совместное введение, например, одновременное или последовательное, терапевтически эффективного количества антител против TfR или белка по данному изобретению и по меньшей мере одного иного лекарственного вещества, причем указанное второе лекарственное вещество является противовирусным или антипролиферативным агентом, например, из числа упомянутых выше.
В одном из воплощений данного изобретения описанные в настоящем документе антитела или белки используются для определения уровня TfR или количества клеток, содержащих рецептор TfR. Это достигается, например, в результате контактирования образца (например, образца in vitro) и контрольного образца с антителами против TfR в условиях, в которых возможно образование комплекса между антителом и TfR. Любые комплексы, образовавшиеся между антителом и TfR, детектируют, и сравнивают полученные данные для изучаемого и контрольного образцов. Например, с использованием композиций по данному изобретению можно осуществлять анализ хорошо известными в данной области техники стандартными методами, например, методом твердофазного иммуноферментного анализа (ELISA) или проточной цитометрии.
Соответственно в одном из аспектов настоящим описанием предлагаются также способы выявления присутствия TfR (например, антигена человеческого TfR) в образце или измерения количества TfR, включающие контактирование образца и контрольного образца с антителом или белком по данному изобретению или с их антигенсвязывающим участком, специфично связывающимися с TfR, в условиях, в которых возможно образование комплекса антитела или его части с TfR. Затем выявляется образовавшийся комплекс, при этом разница в образовании комплекса между образцом и контрольным образцом показательна для присутствия TfR в образце.
В объем настоящего описания также входят наборы, состоящие из композиций (например, антитела, белков, гуманизированных антител, конъюгированных антител и полиспецифичных молекул), раскрытых в настоящем документе, и инструкции по их использованию. Такой набор содержит по меньшей мере один дополнительный реагент или одно или более дополнительных антител или белков (например, антител, обладающих дополняющей активностью, посредством которой связывается с эпитопом антигена-мишени, отличного от эпитопа, с которым связывается первое антитело). Наборы по данному изобретению, как правило, включают инструкцию по применению данного препарата, в которой указано, как пользоваться содержимым набора. Термин «инструкция» в настоящем документе включает любую напечатанную или записанную информацию, приложенную к набору или вложенную в него, или как-то иначе его сопровождающую. Набор по данному изобретению может также содержать средства для определения того, относится ли данный пациент к группе индивидов, чувствительных к лечению антителами к TfR, описанными выше.
Далее данное изобретение, будучи полностью описано, иллюстрируется приведенными ниже примерами, которые не имеют ограничительного характера, а служат только для иллюстрации.
Примеры
Методы
1. Определение аффинности путем твердофазного иммуноферментного анализа (ELISA)
Для определения специфичности антител по данному изобретению метод ELISA применяется согласно следующему протоколу. Лунки 96-луночного планшета покрывают рекомбинантным человеческим TfR (предоставлен R&D Systems, номер по каталогу 2474-TR) и оставляют на ночь, после чего дважды промывают и затем инкубируют с различными десятикратными разведениями (начиная с концентрации 50 мкг/мл до 0,001 мкг/мл включительно) антитела (мышиного и гуманизированного моноклонального) и инкубируют в течение 1 часа при комнатной температуре. Затем промывают два раза и инкубируют со вторыми антителами (козьими против человеческого IgG) в течение 1 часа при комнатной температуре в темноте, после чего промывают два раза и инкубируют с раствором 3,3',5,5'-тетраметилбензидина (ТМВ) в течение 10 минут, затем измеряют поглощение на длине волны 450 нм.
2. Определение аффинности методом поверхностного плазмонного резонанса с использованием системы Biacore
Чтоб определить значения KD, применяли метод поверхностного плазмонного резонанса, используя систему Biacore™, как описано ниже.
Человеческий TfR с гистидиновой меткой (предоставлен R&D Systems, номер по каталогу 2474-TR) связывали на сенсорном чипе на основе карбоксиметилированного декстрана (CM5) с ковалентно иммобилизованными Penta-His mAb. Проводили инжекцию антител в десяти различных концентрациях (от 1 нм до 500 нM) по поверхности anti-His/TfR-His в течение 4 минут, и диссоциация комплекса происходила в течение 5 минут. Получаемые профили по ассоциации и диссоциации анализировали, используя нелинейный метод наименьших квадратов и однократное экспоненциальное сглаживание временных рядов пакета программ BIAevalution.
3. Определение ADCC
Антителозависимая опосредованная клетками цитотоксичность является предпочтительным механизмом для уничтожения раковых клеток-мишеней с помощью лекарственных агентов на основе антител. Антитело связывается с антигеном-мишенью на клеточной поверхности. Когда эффекторная часть Fc связанного с мишенью антитела связывается также с рецептором FcγRIIIa на поверхности эффекторной клетки (преимущественно естественных киллерных клеток), происходит множественное перекрестное связывание клеток двух типов, что ведет к активизации ADCC. Конечным этапом этого пути активации является гибель клетки-мишени, на чем и базируются биологические методы определения ADCC, в которых в качестве эффекторных клеток используются донорские мононуклеары периферической крови (PBMCs) или субпопуляции естественных киллеров (NK) клеток. Набор ADCC Reporter Bioassay Complete Kit (Promega, G7015) включает все эти компоненты и реагенты, необходимые для ADCC Reporter Bioassay, основанного на антителах против CD20. ADCC Reporter Bioassay использует также альтернативный вариант получения данных на раннем этапе пути активации ADCC: активации генной транскрипции в эффекторной клетке с участием NFAT (ядерного фактора активированных Т-клеток). Кроме того, ADCC Reporter Bioassay использует в качестве эффекторных клеток рекомбинантные клетки Т-лимфобластного лейкоза человека Jurkat, в которых стабильно экспрессируется рецептор FcγRIIIa и элемент отклика NFAT, инициирующий экспрессию люциферазы светлячка. Биологическая активность антител в ADCC подсчитывается по продукции люциферазы, образующейся в результате активации пути NFAT; люциферазную активность в эффекторных клетках считают, измеряя люминесценцию. В качестве положительного контроля для сравнения с гуманизированными моноклональными антителами в указанном наборе предлагаются антитела против CD20.
4. Определение активированной Т-лимфоцитами индукции апоптоза в HL-60 и PHA
Для оценки апоптоза клетки инкубировали в 24-луночном планшете в концентрации 100.103 клеток на лунку в объеме 400 мкл. Затем клетки инкубировали 3 и 4 суток в присутствии нескольких разведений мышиного либо гуманизированного антитела (концентрация от 200 мкг/мл до 3 мкг/мл). По истечении указанного времени клетки центрифугировали и метили в присутствии аннексина V и TRO-PO 3 в течение 3-15 минут, после чего проводили проточную цитометрию.
5.Анализ продуктивности (400ml)
Для определения продуктивности синтезировали и клонировали гены VH в экспрессионный вектор pXCIgG (ΔK) для тяжелой цепи. Легкую цепь предварительно субклонировали в векторе pXCKappa. Осуществляли временную экспрессию 16 антител в клетках CHO K1SV GS-KO в объеме 400 мл (в колбе для культивирования со встряхиванием). Полученные антитела очищали путем аффинной хроматографии с использованием протеина А. Концентрации антител определяли, используя предсказанное значение коэффициента экстинкции (1,49) для всех вариантов антител. Целостность вариантов антител оценивали путем электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) в восстанавливающих и в не восстанавливающих условиях.
Примеры mAb1 - mAb16
Примеры mAb1 - mAb16, представленных в таблице 1, могут быть получены с помощью стандартных процедур, осуществляемых для получения и очистки рекомбинантных антител.
Например, нужные кодирующие нуклеотидные последовательности можно клонировать в продуцируюший вектор для рекомбинантной экспрессионный в продуцирующих клетках млекопитающих подходящей линии.
В нижеследующих таблицах 4 и 5 представлены аминокислотные и нуклеотидные последовательности, используемые при осуществлении данного изобретения, в частности, для получения нуклеиновых кислот, экспрессионных векторов и антител, описанных в настоящем документе.
Таблица 4. Краткое описание аминокислотных и нуклеотидных последовательностей, используемых при осуществлении данного изобретения
Таблица 5. Краткое описание аминокислотных и нуклеотидных последовательностей, используемых при осуществлении данного изобретения
Результаты
Конструирование функциональных гуманизированных антител A24
Гуманизация
Процедуру гуманизации осуществляли следующим образом:
1. Идентифицировали домены и области исходного антитела.
2. Последовательности исходного антитела выравнивали с рядом референсных последовательностей.
3. Строили трехмерную структурную модель исходного белка.
4. На основании собранных данных делали первоначальную оценку возможности замены в каждом положении и выделяли группы положений.
5. Строили структурные модели вариантов гуманизированных антител при необходимости и сравнивали с моделью исходного белка для структурного анализа потенциальных замен.
6. Путем анализа положений, различающихся в исходной последовательности и в акцепторной последовательности, проводили прививание CDR. Важные положения, как правило, сохраняли и замену делали только тогда, когда она представлялась сравнительно благоприятной исходя из потенциального влияния на аффинность и совместимость с каркасными участками акцепторной последовательности.
7. Составляли окончательный набор комбинаций вариантов гуманизированных легких и тяжелых цепей.
Описание последовательностей
Для определения содержания доменов каждой аминокислотной цепи и приблизительных границ каждого домена использовали Базу данных консервативных доменов (CDD) (Marchler-Bauer et al., 2011). Границы вариабельного домена и границы участков, определяющих комплементарность, (CDRs) устанавливали в точности согласно ряду обычно используемых определений (Kabat E.A., Wu T.T., 1991, Chothia C., Lesk A.M.,1987; обновление см. Al-Lazikani B. et al., 1997, Honegger А., Plücktuhn А., 2001). Использовали доработанное определение CDR по Чотиа (Al-Lazikani В.. et al. 1997) и нумерацию положений по Чотиа (Chothia С., 1987).
Выравнивание последовательностей
Проводили множественное выравнивание исходной аминокислотной последовательности с мышиными и человеческими последовательностями зародышевой линии с помощью программы MAFFT (Katoh К., et al., 2002); входы в каждое выравнивание осуществляли соответственно степени идентичности данной последовательности и исходной последовательности (SeqID). Сравниваемые множества сократили до уникального набора последовательностей путем кластеризации по 100% SeqID и исключения избыточных вводов.
Идентификация аминокислотных остатков в критически важных положениях
Fv антител имеют ряд критически важных положений, которые образуют межцепочечное взаимодействие VH/VL или входят в дискретный набор канонических структур для пяти CDR (Chothia С., Lesk А.М., 1987, Martin А.С., Thornton J.M., 1996, Al Laziniki В. et al. 1997); прежде, чем предлагать замены в этих положениях, их следует рассмотреть подробно. На основе выравнивания исходной аминокислотной последовательности с человеческими последовательностями зародышевой линии были идентифицированы наиболее совпадающие вводы. Идентификация оптимальной последовательности зародышевой линии человека как акцепторной основывалась на критериях, приведенных ниже.
1. Идентичность последовательностей в каркасных участках
2. Идентичные или сравнимые аминокислотные остатки, обеспечивающие межцепочечное взаимодействие
3. Канонические конформации петель CDRs исходной последовательности
4. В экспрессирующихся антителах имеет место комбинация легких и тяжелых цепей зародышевой линии
Построение трехмерной модели
Моделирование структуры области Fv для исходного мышиного антитела и его вариантов проводили с использованием программной платформы Lonza. Фрагменты структурных матриц-кандидатов для каркасных участков (FR) и участков, определяющих комплементарность, (CDRs), а также полноразмерных Fv сравнивали, ранжировали и отбирали из собственной базы данных по антителам на основании идентичности их последовательностей искомой последовательности и на основании кристаллографических количественных показателей матричных структур, например разрешения (в ангстремах(Å)).
Для структурного выравнивания матриц CDRs и FR в матрицу CDR включали по пять аминокислотных остатков с каждой стороны матрицы CDR. Выравнивание фрагментов достигалось на основании перекрывающихся участков и полученного структурного выравнивания последовательностей. Фрагменты матриц вместе с результатами выравнивания обрабатывали с помощью программы MODELLER (Sali А. et al., 1993).
Этот протокол создал конформационные ограничения, проистекающие из множества выровненных структурных матриц. Сборку структур, удовлетворяющих этим ограничениям, осуществляли, объединяя метод градиентной оптимизации и алгоритм имитации связывания. Из результатов сборки выбирали одну или более модельных структур по энергетической функции, исходя из оценки структуры белковой молекулы и соответствия конформационным ограничениям. Модели анализировали и боковые цепи аминокислотных остатков в положениях, которые в искомой последовательности и в матрице оказались различными, оптимизировали, используя алгоритм оптимизации расположения боковых цепей и минимизации энергии молекулярной системы.
Для достижения конформационной вариабельности участков CDR, а также упаковки ядра белковой молекулы, локальной упаковки доменов и областей и анализа поверхности белковой молекулы использовали комплекс визуализационных и вычислительных средств, чтобы отобрать одну или более предпочтительных моделей.
Сравнение смоделированных структур
Строили по отдельности структурные модели областей Fv исходного и гуманизированного антител, как описано ранее, чтобы убедиться в том, что модели вариантов получились без неустранимого смещения в сторону модельной структуры исходного антитела. Однако высокая степень идентичности последовательностей гуманизированных вариантов и исходного антитела нередко приводит к тому, что для многих моделей выбираются одинаковые структурные матрицы.
Для оценки влияния различных аминокислотных замен на аффинность и стабильность антител применяли ряд структурных критериев. Ключевыми критериями здесь являются доступность для растворителя, локальная упаковка атомов и расположение замены относительно предсказанного участка связывания антигена или контактирования одноцепочечных Fv в димере. В случаях неблагоприятной сольватации, плохих межатомных контактов или плохого расположения неподходящего аминокислотного остатка в критически важном положении последовательности потенциальная замена отвергалась. Чтобы найти подходящие замены, использовались также другие критерии, например электростатические эффекты, паттерн имеющихся или потенциальных водородных связей. Некоторые положения аминокислотной последовательности больше других подходят для замен, поскольку совокупность критически важных положений играет роль в поддержании канонических CDR, укладке ядер отдельных доменов или областей междоменного взаимодействия.
Оценка потенциальных аминокислотных замен
Проводили оценку всех различавшихся положений аминокислотных последовательностей каркасных участков или близлежащих предсказанных эпитопов исходного антитела и антитела-акцептора на основании потенциального влияния этих положений на сродство связывания и стабильность.
В этой оценке имеют значение многие факторы, что проистекает из необходимости учитывать как аффинность, так и стабильность антитела. Эти факторы следующие:
● Положения, ответственные за связывание с антигеном
● Критически важные положения
○ Консервативные остатки в области межцепочечного взаимодействия VH/VL
● Положения, определяющие канонический класс CDR
● Удаленность от CDRs
● Консервативность или вариабельность в данном положении при выравнивании с референсной последовательностью
● Доступность для растворителя
● Локальная упаковка атомов
● Локальная вторичная структура
● Электростатические эффекты
● Паттерн водородных связей
● Паттерн потенциальных водородных связей
● Посттрансляционные модификации
○ N-гликозилирование
○ Деамидирование
Критически важные положения первоначально определяют как положения в CDRs по Чотиа, как находящиеся в критически важных местах области межцепочечного взаимодействия VH/VL; в положениях, способствующих определенной конформации CDR или в положениях, высоко консервативных при выравнивании с референсной последовательностью. Многие положения являются высоко консервативными и в них могут находиться аминокислотные остатки только одного типа или лишь немногих типов.
Отбор оптимальных каркасных участков акцептора
Проводили выравнивание последовательностей, сравнивая вариабельные домены исходного антитела и соответственные последовательности зародышевой линии человека. С учетом идентичности полной последовательности, соответствия положений аминокислотной последовательности, образующих поверхностные участки молекулы антитела, и сходной принадлежности участков CDR к каноническим классам в легкой цепи все равно имелись несколько каркасных участков, в равной мере подходящих в качестве акцепторных; нередко один из них был более подходящим для одной области, но менее подходящим для другой. В итоге были выбраны два каркасных участка легкой цепи - VK6-A26 и VK3-L6.
Тяжелая цепь наилучшим образом соответствовала зародышевой линии человека VH7-7-4.1. В семействе последовательностей зародышевой линии VH7 имеется только один представитель, который можно включить в семейство VH1 (Knappik А. et al., 2000). Поскольку зародышевая линия VH7 встречается, по всей вероятности, реже, чем зародышевая линия VH1, мы сочли предпочтительным использовать последнюю как акцепторный каркасный участок. Однако когда были проанализированы положения с потенциальными обратными мутациями, стало ясно, что в последовательности зародышевой линии VH7 уже имеются подходящие аминокислотные остатки. Поэтому для акцепторных каркасных участков использовали две зародышевых линии последовательностей вариабельной области тяжелой цепи иммуноглобулинов (IGHV) - VH7-7-4.1 и VH1-1-03.
Гены сегмента J сравнивали с последовательностями исходного A24 по FR4 и J сегментам. Наилучшее совпадение обнаружилось для JK4 и JH4 легкой и тяжелой цепей соответственно, и их поэтому выбрали в качестве акцепторных каркасных участков сегмента J.
Гуманизация
Был составлен перечень всех положений, в которых имелись различия между каркасным участком исходного и акцепторного антител. Все эти положения проанализировали как по отдельности, так и в свете других потенциальных аминокислотных замен. Каждое положение ранжировали, и сделали предположения о том, какие аминокислотные остатки подлежат замене в гуманизированных вариантах антител. Для легкой цепи были предложены три гуманизированных цепи: одна с использованием акцептора VK3-L6 и две с VK6-A2, из которых в одной имелись дополнительные существенные положения, сохранившиеся из исходной последовательности (обратные мутации). Для тяжелой цепи были предложены две гуманизированных цепи - по одной для каждого акцептора. Отметим, что прививка в VH7-7-4.1 является более консервативной; см. таблицу 6.
Таблица 6. Гуманизированные цепи
Первое «поколение» 6 гуманизированных антител не сохранило функциональные свойства A24
Основываясь на методах, описанных в предыдущем разделе настоящего документа, мы вначале сконструировали три варианта гуманизированной легкой цепи A24 (далее обозначаются VL1, VL2 и VL3) и два варианта гуманизированной тяжелой цепи A24 (далее обозначаются VH1 и VH2). Соответствующие VH и VL A24 далее обозначаются VH0 и VL0.
Мы получили следующие 6 гуманизированных антител с приведенными ниже комбинациями гуманизированных VL и VH и изотипом IgG1 человека:
INA01 Гуманизированное антитело, вариант 1 - VH1/VL1
INA01 Гуманизированное антитело, вариант 2 - VH2/VL1
INA01 Гуманизированное антитело, вариант 3 - VH1/VL2
INA01 Гуманизированное антитело, вариант 4 - VH2/VL2
INA01 Гуманизированное антитело, вариант 5 - VH1/VL3
INA01 Гуманизированное антитело, вариант 6 - VH2/VL3
С этими 6 гуманизированными антителами было проведено определение индукции апоптоза в клетках HL-60, как описано выше, и полученные данные сравнили с данными по индукции апоптоза для химерной формы референсного мышиного антитела A24 (VH0/VL0) того же изотипа IgG1 человека. Эти результаты представлены на фиг 1.
Как видно из фиг. 1, оказалось, что все испытанные антитела утратили, как ни странно, свойства связывания. Это тем более удивительно, если учесть тот факт, что в некоторых из этих вариантов все 6 участков CDR были такими же, как в исходном мышином антителе A24.
Получение 5 новых гуманизированных антител
Мы сконструировали новую тяжелую цепь (VH3) и получили следующие гуманизированные антитела:
INA01 Гуманизированное антитело, вариант 7 - VH0/VL1
INA01 Гуманизированное антитело, вариант 8 - VH3/VL1
INA01 Гуманизированное антитело, вариант 9 - VH3/VL2
INA01 Гуманизированное антитело, вариант 10 - VH3/VL3
INA01 Гуманизированное антитело, вариант 11 - VH3/VL0
И с этими пятью гуманизированными антителами было проведено определение индукции апоптоза в HL-60, как описано выше, и полученные данные сравнили с данными по индукции апоптоза для химерной формы INA01, соответствующей антителу с вариабельными областями исходного мышиного A24 изотипа IgG1 человека. Эти результаты представлены на фиг. 2.
Как видно из фиг. 2, эти гуманизированные антитела утратили, по меньшей мере частично, способность вызывать апоптоз.
Получение 6 новых гуманизированных антител
Исходя из результатов, полученных для трех новых указанных выше антител, мы сконструировали три новых легких цепи и две новых тяжелых цепи.
В частности, было проведено анализ in silico, чтобы предсказать изменения аминокислотных остатков (в каркасных участках и в участках CDR), которые приводили бы к снижению иммуногенности, но чтобы при этом сохранялись нужные свойства исходного антитела A24.
В этом анализе использовали iTope™ для анализа происходящих из А24 последовательностей на смешанное высокоаффинное связывание с белками МНС класса II человека. Считается, что смешанное выскоаффинное связывание с белками МНС класса II коррелирует с присутствием Т-клеточных эпитопов (Hill J.A. et al., 2003, Arthritis Res. Ther. (2003) 1:R40-R48), хотя Т-клеточный ответ может быть инициирован также средой и низкоаффинным связыванием. Таким образом, технология iTope™ послужила для первоначального - но весьма полезного - отбора «с низким разрешением» с целью выяснить расположение Т-клеточных эпитопов. Кроме того, последовательности анализировали путем поиска с помощью программ BLAST по базе данных Т-клеточных эпитопов TCED™, чтобы выявить эпитопы Т-клеток, ранее идентифицированные при анализе с помощью EpiScreen™ последовательностей других белков.
Как видно из таблицы 7, in silico анализ iTope™ показал, что при аминокислотных заменах L53R и/или S55T в LCDR2 иммуногенность снижается.
Таблица 7
iTope™
МНС II
Мы поэтому решили проверить 3 новых легких цепи - одну (VL4) с двумя аминокислотными заменами в LCDR2 по сравнению с LCDR2 антитела A24, одну (VL5) с только одной аминокислотной заменой в LCDR2 и одну (VL6) с такой же аминокислотной последовательностью LCDR2, что и в A24.
Были получены следующие шесть новых гуманизированных антител с константной областью изотипа IgG1:
INA01 вариант 12 VH4/VL4
INA01 вариант 13 VH5/VL4
INA01 вариант 14 VH4/VL5
INA01 вариант 15 VH5/VL5
INA01 вариант 16 VH4/VL6
INA01 вариант 17 VH5/VL6
На фиг. 5a изображено выравнивание VL4, VL5, VL6 с VL антитела A24.
На фиг. 5b изображено выравнивание VH4 и VH5 с VH антитела A24.
Мы проверили способность этих гуманизированных антител вызывать апоптоз на основании 72-ух часового анализа с HL-60, как описано выше анализ, и по сравнению с химерными INA01, соответствующими антителу с вариабельными областями исходного мышиного антитела A24 и изотипом IgG1 человека. Полученные результаты представлены на фиг. 3.
Как видно на фиг. 3, теперь все 6 новых гуманизированных антител проявляли свойства, сходные с таковыми химерных INA01, соответствующих антителу с вариабельными областями исходного мышиного антитела A24 и изотипом IgG1 человека, или даже превосходящими их.
В частности, антитела варианта 12 и варианта 13 с двумя аминокислотными заменами в LCDR2 обладали большей способностью вызывать апоптоз, нежели исходное химерное антитело INA01.
Превращение IgG
Для экспрессии полноразмерного IgG фрагменты вариабельных доменов тяжелых (VH) и легких цепей (VL) четырех лидирующих вариантов 12, 14, 15 и 17 субклонировали из векторов для экспрессии Fab в экспрессионные векторы, подходящие для человеческого IgG4, человеческого IgG1 дикого типа, человеческого IgG1L234AL235A и человеческого IgG1N297A, указанный человеческий IgG1 L234AL235A обозначили «AA», в результате были получены экспрессионные векторы для продуцирования 16 антител по данному изобретению, mAb1-mAb16, представленных в приведенной ниже таблице 8.
Таблица 8. Вариабельные области и область Fc IgG антител mAb1-mAb16
Временная экспрессия и очистка человеческого IgG
Клетки трансфицировали экспрессионным вектором, содержавшим ДНК, кодирующую тяжелые и легкие цепи IgG mAb1- mAb16.
Результаты продукции антител представлены на фиг. 4. Результаты подтверждают, что антитела изотипа IgG4 продуцируются с большим выходом, чем антитела изотипа IgG1, мутантные IgG1 и мутантные молчащие IgG1.
Данные о свойствах антител mAb1
Таблица 9. Свойства mAb1
(KD нM по данным SPR)
в клетках HL-60
Примечательно, что антитела по данному изобретению имеют KD и EC50 менее 10 нM и даже меньше 1 нM, а способность вызывать апоптоз, измеренный с помощью HL-60, более выражена, нежели у A24, поэтому особенно подходят для использования в качестве лекарства.
Кроме того, антитела по данному изобретению обладают предпочтительными доступными для улучшения качествами, в частности, в отношении продуцирования эукариотическими клетками и введения в организм человека в связи с ожидаемой пониженной иммуногенностью.
Кодирующие последовательности, кодирующие вариабельные области, нетрудно переносить в подходящие экспрессионные вектора и клеточные линии для получения молчащих антител IgG1, например, содержащих вариант Fc IgG1 с мутацией L234A L235A или вариант Fc IgG1 с мутацией N297A.
Или же кодирующие последовательности, кодирующие вариабельные области, нетрудно переносить в подходящие экспрессионные вектора и клеточные линии для получения антител с высокой активностью ADCC, например, содержащие Fc IgG1 дикого типа и/или с разделенным пополам GlcNac, или с низким фукозилированием обладающие гликана в положении аминокислоты N297.
--->
Перечень последовательностей
<110> INATHERYS
<120> Anti-TfR antibodies and their use in treating proliferative and
inflammatory disorders
<130> INA001
<160> 23
<170> PatentIn version 3.5
<210> 1
<211> 7
<212> PRT
<213> Homo sapiens
<400> 1
Gly Tyr Thr Phe Thr Asn Gln
1 5
<210> 2
<211> 6
<212> PRT
<213> Homo sapiens
<400> 2
Asn Thr Tyr Thr Gly Glu
1 5
<210> 3
<211> 8
<212> PRT
<213> Homo sapiens
<400> 3
Glu Gly Trp Asp Ser Met Asp Tyr
1 5
<210> 4
<211> 10
<212> PRT
<213> Homo sapiens
<400> 4
Ser Ala Ser Ser Ser Val Asn Tyr Met His
1 5 10
<210> 5
<211> 7
<212> PRT
<213> Homo sapiens
<400> 5
Ser Thr Ser Asn Arg Ala Thr
1 5
<210> 6
<211> 9
<212> PRT
<213> Homo sapiens
<400> 6
Gln Gln Arg Ser Ser Tyr Pro Leu Thr
1 5
<210> 7
<211> 7
<212> PRT
<213> Homo sapiens
<400> 7
Ser Thr Ser Asn Leu Ala Ser
1 5
<210> 8
<211> 7
<212> PRT
<213> Homo sapiens
<400> 8
Ser Thr Ser Asn Arg Ala Ser
1 5
<210> 9
<211> 117
<212> PRT
<213> Homo sapiens
<400> 9
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Gln
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Ile Asn Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Ile Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Met Ala Thr Tyr Phe Cys
85 90 95
Val Arg Glu Gly Trp Asp Ser Met Asp Tyr Trp Gly Gln Gly Thr Ser
100 105 110
Val Thr Val Ser Ser
115
<210> 10
<211> 107
<212> PRT
<213> Homo sapiens
<400> 10
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Trp Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg
100 105
<210> 11
<211> 117
<212> PRT
<213> Homo sapiens
<400> 11
Gln Val Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Gln
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Ile Asn Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Val Ile Ser Leu Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Ser Ser Leu Lys Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Val Arg Glu Gly Trp Asp Ser Met Asp Tyr Trp Gly Gln Gly Thr Ser
100 105 110
Val Thr Val Ser Ser
115
<210> 12
<211> 136
<212> PRT
<213> Homo sapiens
<400> 12
Met Glu Trp Ser Trp Val Phe Leu Phe Phe Leu Ser Val Thr Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Gln Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Ile Asn Ala
65 70 75 80
Asp Asp Phe Lys Gly Arg Phe Val Ile Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Ser Asn Leu Lys Asn Glu Asp Thr Ala Val
100 105 110
Tyr Phe Cys Val Arg Glu Gly Trp Asp Ser Met Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Ser Val Thr Val Ser Ser
130 135
<210> 13
<211> 107
<212> PRT
<213> Homo sapiens
<400> 13
Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Asn Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Ser Thr Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 14
<211> 107
<212> PRT
<213> Homo sapiens
<400> 14
Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Asn Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Ser Thr Ser Asn Arg Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 15
<211> 107
<212> PRT
<213> Homo sapiens
<400> 15
Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Asn Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
65 70 75 80
Asp Ala Ala Val Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg
100 105
<210> 16
<211> 760
<212> PRT
<213> Homo sapiens
<400> 16
Met Met Asp Gln Ala Arg Ser Ala Phe Ser Asn Leu Phe Gly Gly Glu
1 5 10 15
Pro Leu Ser Tyr Thr Arg Phe Ser Leu Ala Arg Gln Val Asp Gly Asp
20 25 30
Asn Ser His Val Glu Met Lys Leu Ala Val Asp Glu Glu Glu Asn Ala
35 40 45
Asp Asn Asn Thr Lys Ala Asn Val Thr Lys Pro Lys Arg Cys Ser Gly
50 55 60
Ser Ile Cys Tyr Gly Thr Ile Ala Val Ile Val Phe Phe Leu Ile Gly
65 70 75 80
Phe Met Ile Gly Tyr Leu Gly Tyr Cys Lys Gly Val Glu Pro Lys Thr
85 90 95
Glu Cys Glu Arg Leu Ala Gly Thr Glu Ser Pro Val Arg Glu Glu Pro
100 105 110
Gly Glu Asp Phe Pro Ala Ala Arg Arg Leu Tyr Trp Asp Asp Leu Lys
115 120 125
Arg Lys Leu Ser Glu Lys Leu Asp Ser Thr Asp Phe Thr Gly Thr Ile
130 135 140
Lys Leu Leu Asn Glu Asn Ser Tyr Val Pro Arg Glu Ala Gly Ser Gln
145 150 155 160
Lys Asp Glu Asn Leu Ala Leu Tyr Val Glu Asn Gln Phe Arg Glu Phe
165 170 175
Lys Leu Ser Lys Val Trp Arg Asp Gln His Phe Val Lys Ile Gln Val
180 185 190
Lys Asp Ser Ala Gln Asn Ser Val Ile Ile Val Asp Lys Asn Gly Arg
195 200 205
Leu Val Tyr Leu Val Glu Asn Pro Gly Gly Tyr Val Ala Tyr Ser Lys
210 215 220
Ala Ala Thr Val Thr Gly Lys Leu Val His Ala Asn Phe Gly Thr Lys
225 230 235 240
Lys Asp Phe Glu Asp Leu Tyr Thr Pro Val Asn Gly Ser Ile Val Ile
245 250 255
Val Arg Ala Gly Lys Ile Thr Phe Ala Glu Lys Val Ala Asn Ala Glu
260 265 270
Ser Leu Asn Ala Ile Gly Val Leu Ile Tyr Met Asp Gln Thr Lys Phe
275 280 285
Pro Ile Val Asn Ala Glu Leu Ser Phe Phe Gly His Ala His Leu Gly
290 295 300
Thr Gly Asp Pro Tyr Thr Pro Gly Phe Pro Ser Phe Asn His Thr Gln
305 310 315 320
Phe Pro Pro Ser Arg Ser Ser Gly Leu Pro Asn Ile Pro Val Gln Thr
325 330 335
Ile Ser Arg Ala Ala Ala Glu Lys Leu Phe Gly Asn Met Glu Gly Asp
340 345 350
Cys Pro Ser Asp Trp Lys Thr Asp Ser Thr Cys Arg Met Val Thr Ser
355 360 365
Glu Ser Lys Asn Val Lys Leu Thr Val Ser Asn Val Leu Lys Glu Ile
370 375 380
Lys Ile Leu Asn Ile Phe Gly Val Ile Lys Gly Phe Val Glu Pro Asp
385 390 395 400
His Tyr Val Val Val Gly Ala Gln Arg Asp Ala Trp Gly Pro Gly Ala
405 410 415
Ala Lys Ser Gly Val Gly Thr Ala Leu Leu Leu Lys Leu Ala Gln Met
420 425 430
Phe Ser Asp Met Val Leu Lys Asp Gly Phe Gln Pro Ser Arg Ser Ile
435 440 445
Ile Phe Ala Ser Trp Ser Ala Gly Asp Phe Gly Ser Val Gly Ala Thr
450 455 460
Glu Trp Leu Glu Gly Tyr Leu Ser Ser Leu His Leu Lys Ala Phe Thr
465 470 475 480
Tyr Ile Asn Leu Asp Lys Ala Val Leu Gly Thr Ser Asn Phe Lys Val
485 490 495
Ser Ala Ser Pro Leu Leu Tyr Thr Leu Ile Glu Lys Thr Met Gln Asn
500 505 510
Val Lys His Pro Val Thr Gly Gln Phe Leu Tyr Gln Asp Ser Asn Trp
515 520 525
Ala Ser Lys Val Glu Lys Leu Thr Leu Asp Asn Ala Ala Phe Pro Phe
530 535 540
Leu Ala Tyr Ser Gly Ile Pro Ala Val Ser Phe Cys Phe Cys Glu Asp
545 550 555 560
Thr Asp Tyr Pro Tyr Leu Gly Thr Thr Met Asp Thr Tyr Lys Glu Leu
565 570 575
Ile Glu Arg Ile Pro Glu Leu Asn Lys Val Ala Arg Ala Ala Ala Glu
580 585 590
Val Ala Gly Gln Phe Val Ile Lys Leu Thr His Asp Val Glu Leu Asn
595 600 605
Leu Asp Tyr Glu Arg Tyr Asn Ser Gln Leu Leu Ser Phe Val Arg Asp
610 615 620
Leu Asn Gln Tyr Arg Ala Asp Ile Lys Glu Met Gly Leu Ser Leu Gln
625 630 635 640
Trp Leu Tyr Ser Ala Arg Gly Asp Phe Phe Arg Ala Thr Ser Arg Leu
645 650 655
Thr Thr Asp Phe Gly Asn Ala Glu Lys Thr Asp Arg Phe Val Met Lys
660 665 670
Lys Leu Asn Asp Arg Val Met Arg Val Glu Tyr His Phe Leu Ser Pro
675 680 685
Tyr Val Ser Pro Lys Glu Ser Pro Phe Arg His Val Phe Trp Gly Ser
690 695 700
Gly Ser His Thr Leu Pro Ala Leu Leu Glu Asn Leu Lys Leu Arg Lys
705 710 715 720
Gln Asn Asn Gly Ala Phe Asn Glu Thr Leu Phe Arg Asn Gln Leu Ala
725 730 735
Leu Ala Thr Trp Thr Ile Gln Gly Ala Ala Asn Ala Leu Ser Gly Asp
740 745 750
Val Trp Asp Ile Asp Asn Glu Phe
755 760
<210> 17
<211> 233
<212> PRT
<213> Homo sapiens
<400> 17
Met Ser Val Pro Thr Gln Val Leu Gly Leu Leu Leu Leu Trp Leu Thr
1 5 10 15
Asp Ala Arg Cys Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser
20 25 30
Val Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser
35 40 45
Val Asn Tyr Met His Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Arg
50 55 60
Leu Leu Ile Tyr Ser Thr Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Thr Leu Thr Ile Ser Ser
85 90 95
Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Ser
100 105 110
Tyr Pro Leu Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr
115 120 125
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
130 135 140
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
145 150 155 160
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
165 170 175
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
180 185 190
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
195 200 205
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
210 215 220
Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 18
<211> 462
<212> PRT
<213> Homo sapiens
<400> 18
Met Glu Trp Ser Trp Val Phe Leu Phe Phe Leu Ser Val Thr Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Gln Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Ile Asn Ala
65 70 75 80
Asp Asp Phe Lys Gly Arg Phe Val Ile Ser Leu Asp Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Ser Ser Leu Lys Ala Glu Asp Thr Ala Val
100 105 110
Tyr Phe Cys Val Arg Glu Gly Trp Asp Ser Met Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
225 230 235 240
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
275 280 285
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
355 360 365
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
450 455 460
<210> 19
<211> 724
<212> DNA
<213> Homo sapiens
<400> 19
aagcttgccg ccaccatgtc cgtgcctacc caggtgctgg gactgctgct gctgtggctg 60
accgatgcca ggtgccagat cgtgctgacc cagtctcctg ccaccctgtc tgtgtctccc 120
ggcgagagag ctaccctgtc ctgctccgcc tcctcctccg tgaactacat gcactggttc 180
cagcagaagc ccggccagtc ccccagactg ctgatctact ccacctccaa ccgggccacc 240
ggcatccctg ccagattttc cggctctggc tccggcacct cctataccct gaccatctcc 300
agcctggaac ccgaggactt cgccgtgtac tactgccagc agcggtcctc ctaccccctg 360
acctttggcc agggcaccaa gctggaaatc aagcgtacgg tggccgctcc cagcgtgttc 420
atcttccccc caagcgacga gcagctgaag agcggcaccg ccagcgtggt gtgtctgctg 480
aacaacttct accccaggga ggccaaggtg cagtggaagg tggacaacgc cctgcagagc 540
ggcaacagcc aggagagcgt caccgagcag gacagcaagg actccaccta cagcctgagc 600
agcaccctga ccctgagcaa ggccgactac gagaagcaca aggtgtacgc ctgtgaggtg 660
acccaccagg gcctgtccag ccccgtgacc aagagcttca acaggggcga gtgctgatga 720
attc 724
<210> 20
<211> 1411
<212> DNA
<213> Homo sapiens
<400> 20
aagcttgccg ccaccatgga atggtcctgg gtgttcctgt tcttcctgtc cgtgaccacc 60
ggcgtgcact cccaggtgca gctggtgcag tctggccccg agctgaagaa acctggcgcc 120
tccgtgaagg tgtcctgcaa ggcttccggc tacaccttta caaaccaggg catgaactgg 180
gtcaagcagg cccctggcaa gggcctgaag tggatgggct ggatcaacac ctacaccggc 240
gagcccatca acgccgacga cttcaagggc agattcgtga tctccctgga cacctccgcc 300
tccaccgcct acctgcagat cagctctctg aaggccgagg ataccgccgt gtacttctgc 360
gtgcgggaag gctgggactc catggactat tggggccagg gcacctccgt gaccgtgtct 420
agcgcttcta caaagggccc aagcgtgttc cccctggccc cctgctccag aagcaccagc 480
gagagcacag ccgccctggg ctgcctggtg aaggactact tccccgagcc cgtgaccgtg 540
tcctggaaca gcggagccct gaccagcggc gtgcacacct tccccgccgt gctgcagagc 600
agcggcctgt acagcctgag cagcgtggtg accgtgccca gcagcagcct gggcaccaag 660
acctacacct gtaacgtgga ccacaagccc agcaacacca aggtggacaa gagggtggag 720
agcaagtacg gcccaccctg ccccccctgc ccagcccccg agttcctggg cggacccagc 780
gtgttcctgt tcccccccaa gcccaaggac accctgatga tcagcagaac ccccgaggtg 840
acctgtgtgg tggtggacgt gtcccaggag gaccccgagg tccagttcaa ctggtacgtg 900
gacggcgtgg aggtgcacaa cgccaagacc aagcccagag aggagcagtt taacagcacc 960
taccgggtgg tgtccgtgct gaccgtgctg caccaggact ggctgaacgg caaagagtac 1020
aagtgtaagg tctccaacaa gggcctgcca agcagcatcg aaaagaccat cagcaaggcc 1080
aagggccagc ctagagagcc ccaggtctac accctgccac ccagccaaga ggagatgacc 1140
aagaaccagg tgtccctgac ctgtctggtg aagggcttct acccaagcga catcgccgtg 1200
gagtgggaga gcaacggcca gcccgagaac aactacaaga ccaccccccc agtgctggac 1260
agcgacggca gcttcttcct gtacagcagg ctgaccgtgg acaagtccag atggcaggag 1320
ggcaacgtct ttagctgctc cgtgatgcac gaggccctgc acaaccacta cacccagaag 1380
agcctgagcc tgtccctggg ctgatgaatt c 1411
<210> 21
<211> 9
<212> PRT
<213> Homo sapiens
<400> 21
Ile Tyr Ser Thr Ser Asn Arg Ala Thr
1 5
<210> 22
<211> 9
<212> PRT
<213> Homo sapiens
<400> 22
Ile Tyr Ser Thr Ser Asn Arg Ala Ser
1 5
<210> 23
<211> 9
<212> PRT
<213> Mus musculus
<400> 23
Ile Tyr Ser Thr Ser Asn Leu Ala Ser
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОНЪЮГАТЫ АНТИТЕЛ С ЛЕКАРСТВЕННЫМИ ВЕЩЕСТВАМИ И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ РАКОВЫХ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2799547C2 |
| СРЕДСТВО ДЛЯ ИНГИБИРОВАНИЯ ПОГЛОЩЕНИЯ ЖЕЛЕЗА КЛЕТКАМИ | 2019 |
|
RU2817146C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С TFR | 2019 |
|
RU2810756C2 |
| АНТИТЕЛА ПРОТИВ ТРАНСФЕРРИНОВОГО РЕЦЕПТОРА, ОБЛАДАЮЩИЕ ИНДИВИДУАЛИЗИРОВАННОЙ АФФИННОСТЬЮ | 2016 |
|
RU2729416C2 |
| АНТИТЕЛА ПРОТИВ ТРАНСФЕРРИНОВОГО РЕЦЕПТОРА, ОБЛАДАЮЩИЕ ИНДИВИДУАЛИЗИРОВАННОЙ АФФИННОСТЬЮ | 2016 |
|
RU2831724C2 |
| АНТИТЕЛО, СПОСОБНОЕ СВЯЗЫВАТЬСЯ С ТИМИЧЕСКИМ СТРОМАЛЬНЫМ ЛИМФОПОЭТИНОМ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2825304C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДУПРЕЖДЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2017 |
|
RU2766586C2 |
| КОНЪЮГАТ АНТИТЕЛА ПРОТИВ CEA И АНАЛОГА ЭКСАТЕКАНА И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2020 |
|
RU2833323C1 |
| АНТИТЕЛО ПРОТИВ СТОЛБНЯЧНОГО ТОКСИНА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2815280C1 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |

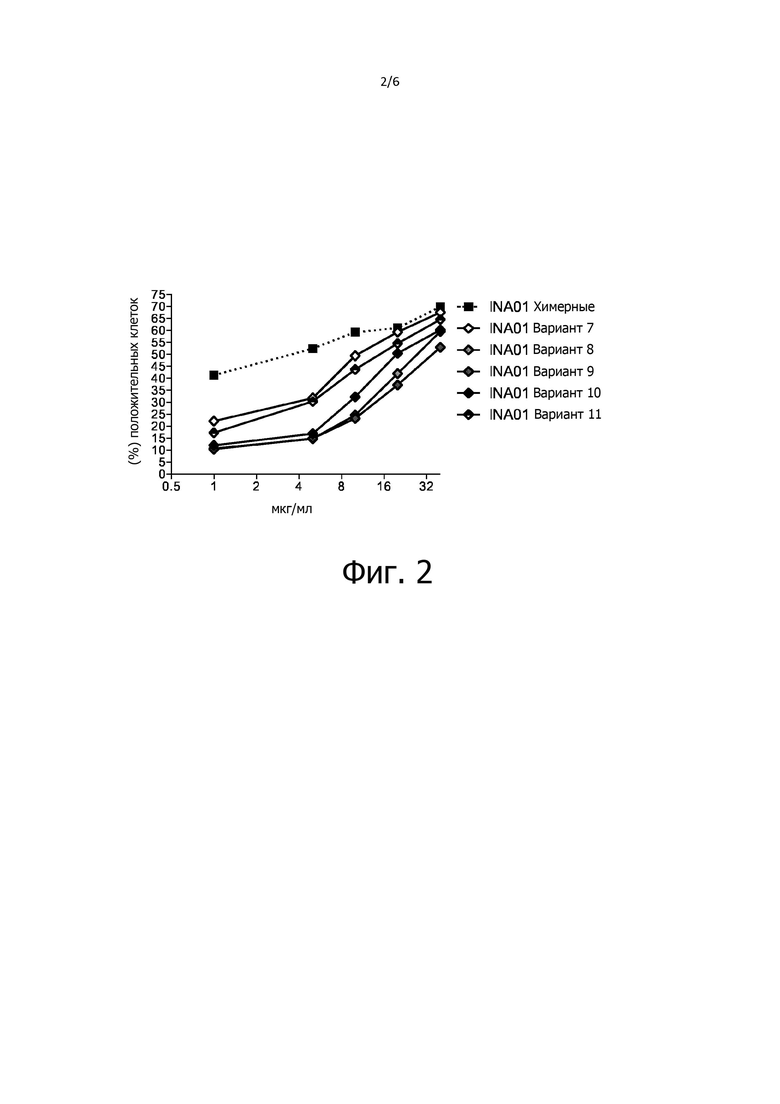
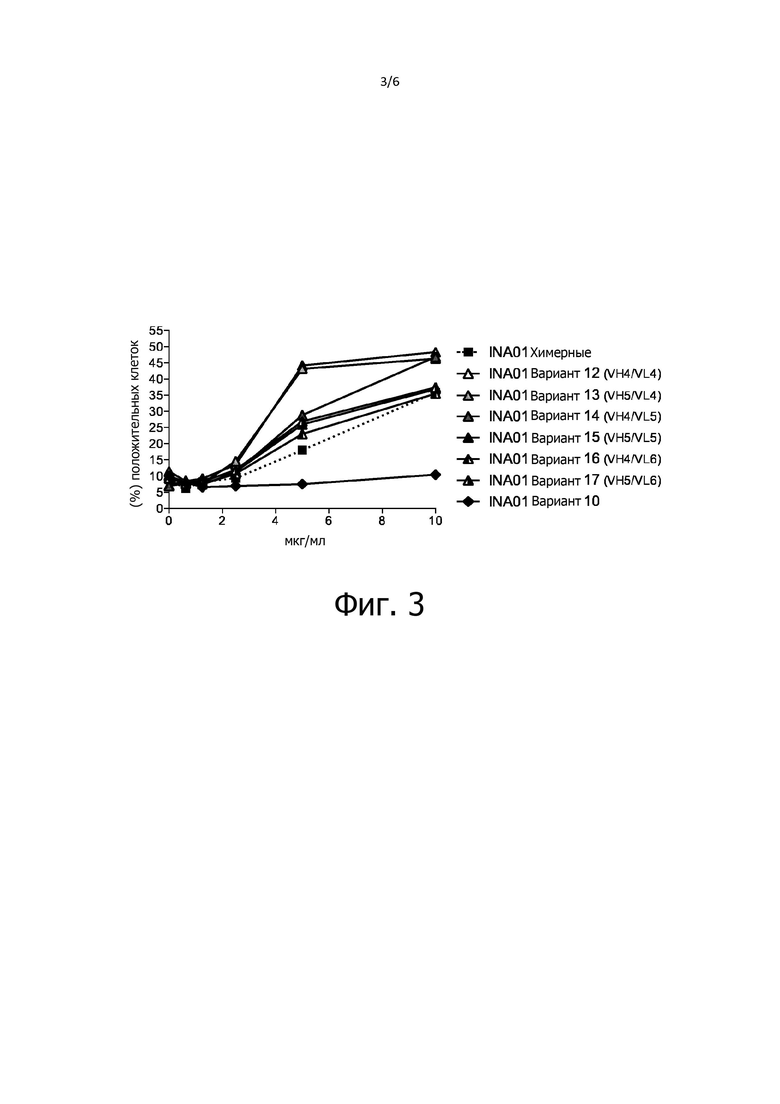
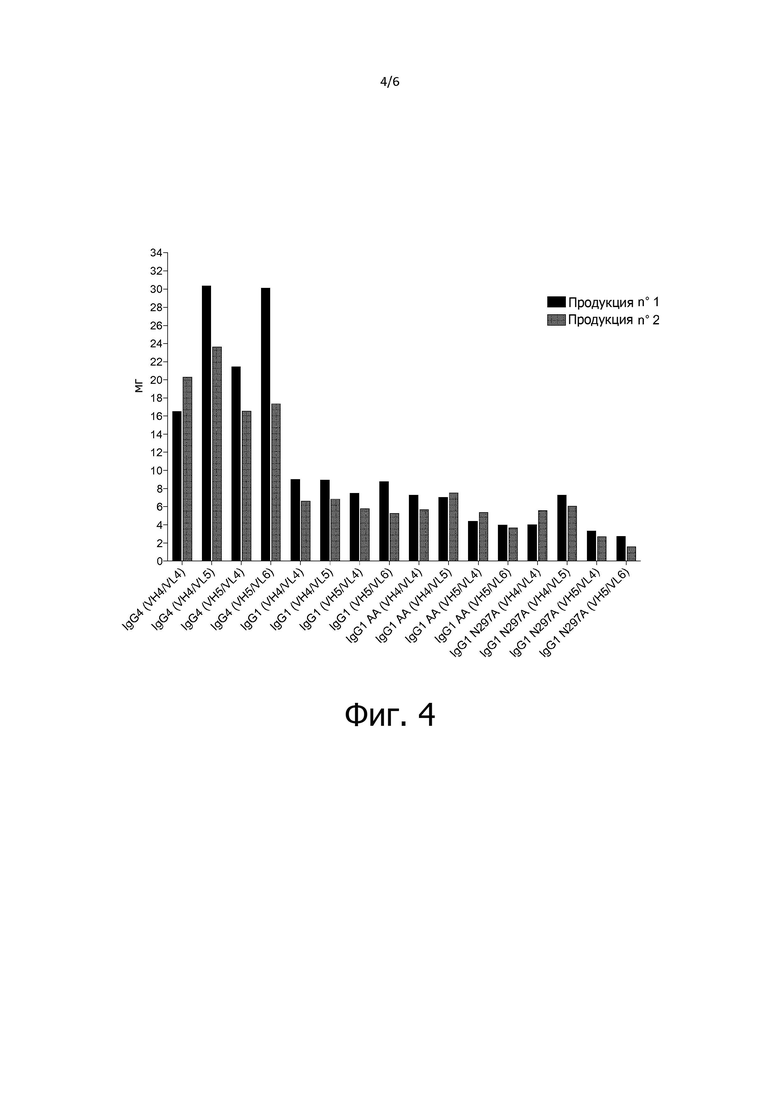


Изобретение относится к области биохимии, в частности к выделенному гуманизированному антителу против рецептора трансферрина (TfR) или антигенсвязывающему фрагменту гуманизированного антитела против TfR, способу его получения, а также к иммуноконъюгату, фармацевтической композиции и лиофилизованному препарату, содержащим вышеуказанное антитело. Также раскрыт вектор, содержащий, по меньшей мере, одну нуклеиновую кислоту, кодирующую вышеуказанное антитело против TfR, а также раскрыта клетка-хозяин, содержащая вышеуказанный вектор. Изобретение также относится к способу обнаружения рецептора трансферрина в образце, включающему контактирование образца с вышеуказанным антителом. Изобретение позволяет эффективно осуществлять лечение опухоли у субъекта. 8 н. и 9 з.п. ф-лы, 9 табл., 5 ил.
1. Выделенное гуманизированное антитело против рецептора трансферрина (TfR) или антигенсвязывающий фрагмент гуманизированного антитела против TfR, включающие что-либо из
(a) вариабельного полипептида тяжелой цепи, содержащего HCDR1 с SEQ ID NO:1, HCDR2 с SEQ ID NO:2, HCDR3 с SEQ ID NO:3, и вариабельного полипептида легкой цепи, содержащего LCDR1 с SEQ ID NO:4, LCDR2 с SEQ ID NO:5 и LCDR3 с SEQ ID NO:6;
(b) вариабельного полипептида тяжелой цепи, содержащего HCDR1 с SEQ ID NO:1, HCDR2 с SEQ ID NO:2, HCDR3 с SEQ ID NO:3, и вариабельного полипептида легкой цепи, содержащего LCDR1 с SEQ ID NO:4, LCDR2 с SEQ ID NO:8 и LCDR3 с SEQ ID NO:6;
(c) вариабельного полипептида тяжелой цепи, содержащего VH с SEQ ID NO:11, и вариабельного полипептида легкой цепи, содержащего VL с SEQ ID NO:13;
(d) вариабельного полипептида тяжелой цепи, содержащего VH, с SEQ ID NO:11, и вариабельного полипептида легкой цепи, содержащего область VL с SEQ ID NO:14;
(e) вариабельного полипептида тяжелой цепи, содержащего VH с SEQ ID NO:12, и вариабельного полипептида легкой цепи, содержащего VL с SEQ ID NO:13; или
(f) вариабельного полипептида тяжелой цепи, содержащего VH с SEQ ID NO:12, и вариабельного полипептида легкой цепи, содержащего VL с SEQ ID NO:14;
причем указанное антитело против TfR специфично связывается с рецептором трансферрина (TfR), имеющим последовательность SEQ ID NO:16.
2. Выделенное гуманизированное антитело против TfR по п. 1, причем указанное антитело связывается с рецептором трансферрина с KD меньше чем 10 нM .
3. Выделенное гуманизированное антитело против TfR по п. 1 или 2, которое вызывает апоптоз в клетках линии HL-60 на том же уровне, что и соответствующее референсное химерное антитело, имеющее VH с SEQ ID NO:9 и VL с SEQ ID NO:10, или на большем уровне.
4. Выделенное гуманизированное антитело против TfR по пп. 1, 2 или 3, содержащее константную область человеческого иммуноглобулина изотипа IgG4 или мутантную, или химически модифицированную константную область, причем указанная мутантная или химически модифицированная константная область обусловливает пониженную или нулевую активность ADCC указанного антитела по сравнению с соответствующим антителом с константной областью дикого типа изотипа IgG1.
5. Выделенное гуманизированное антитело против TfR по пп. 1, 2 или 3, содержащее константную область человеческого иммуноглобулина изотипа IgG1 или мутантную, или химически модифицированную константную область, причем указанная мутантная или химически модифицированная константная область обусловливает повышенную активность ADCC указанного антитела по сравнению с соответствующим антителом с константной областью дикого типа изотипа IgG1.
6. Выделенное гуманизированное антитело против TfR по любому из пп 1-5, которое содержит тяжелую цепь с SEQ ID NO:18 и легкую цепь с SEQ ID NO:17.
7. Иммуноконъюгат для применения при лечении опухоли, содержащий выделенное гуманизированное антитело против TfR по любому из пп. 1-6, конъюгированное с цитотоксином или радиотоксином.
8. Выделенное гуманизированное антитело против TfR по любому из пп. 1-6 или иммуноконъюгат по п. 7 для применения при лечении опухоли, например гематологической опухоли, а конкретнее лимфомы или лейкоза.
9. Выделенное гуманизированное антитело против TfR по любому из пп. 1-6 или иммуноконъюгат по п. 7, отличающиеся тем, что содержатся в преднаполненном шприце.
10. Выделенное гуманизированное антитело против TfR по любому из пп. 1-6 или иммуноконъюгат по п. 7, отличающиеся тем, что содержатся в преднаполненной ампуле.
11. Фармацевтическая композиция для применения при лечении опухоли у субъекта, нуждающегося в этом, содержащая эффективное количество гуманизированного антитела против TfR по любому из пп. 1-6, 8-10 или иммуноконъюгат по любому из пп. 7-10 в комбинации с одним или более фармацевтически приемлемым наполнителем, разбавителем или носителем, необязательно содержащая другие активные ингредиенты.
12. Лиофилизованный препарат для применения при лечении опухоли у субъекта, нуждающегося в этом, содержащий антитело против TfR по любому из пп. 1-6 или иммуноконъюгат по п. 7.
13. Вектор для экспрессии для рекомбинантного продуцирования гуманизированного антитела против TfR по любому из пп. 1-6 в клетке-хозяине, который содержит по меньшей мере одну нуклеиновую кислоту, кодирующую указанное антитело против TfR.
14. Вектор для экспрессии по п. 13, содержащий по меньшей мере одну нуклеиновую кислоту, которая кодирует антитело против TfR, определенное в п. 6.
15. Клетка-хозяин для продуцирования антитела против TfR по любому из пп. 1-6, содержащая вектор для экспрессии по п. 13 или 14.
16. Способ получения антитела против TfR по любому из пп. 1-6, включающий: (i) культивирование клетки-хозяина по п. 15 для экспрессии указанного антитела этой клеткой-хозяином; необязательно (ii) очистку указанного антитела и (iii) выделение указанного антитела.
17. Способ обнаружения рецептора трансферрина в образце, причем указанный способ включает (i) контактирование образца с антителом по любому из пп. 1-6 в условиях, обеспечивающих образование комплекса между этим антителом и рецептором трансферрина, и (ii) обнаружение указанного комплекса.
| WO2005111082 A1, 24.11.2005 | |||
| WO2014020140 A1, 06.02.2014 | |||
| MOURA I.C | |||
| et al., Identification of the transferrin receptor as a novel immunoglobulin (Ig)A1 receptor and its enhanced expression on mesangial cells in IgA nephropathy, THE JOURNAL OF EXPERIMENTAL MEDICINE, 2001, vol.194, no.4, pp.417-425 | |||
| HOOGENBOOM H.R | |||
| et al., Cloning and expression |

