Перекрестные ссылки на связанные заявки
Эта заявка PCT заявляет о приоритетном преимуществе предварительной заявки США № 62/787711, поданной 2 января 2019 г., и 62/838190, поданной 24 апреля 2019 г., каждая из которых полностью включена в настоящий документ ссылкой.
Ссылка на список последовательностей приведена в электронном виде
Содержимое представленного в электронном виде списка последовательностей в текстовом файле ASCII (Имя: 3763_016PC02_SeqListing_ST25.txt; Размер: 90 231 байт; и Дата создания: 30 декабря 2019 г.), поданного вместе с заявкой, полностью включено в настоящий документ ссылкой.
Заявление о государственной поддержке
Эта работа была поддержана Программой инновационных промышленных технологий (10081300, Разработка нового терапевтического средства на основе моноклонального антитела через механизм ингибирования образования глиальных рубцов при ишемическом инсульте), финансируемой Министерством торговли, промышленности и энергетики (MOTIE, Корея).
Область техники, к которой относится изобретение
В настоящем раскрытии представлены антитела (например, деиммунизированные или с созревшей аффинностью), которые специфически связываются с представителем A5 семейства 19 со сходством последовательностей (FAM19A5), композициями, содержащими такие антитела, и способом применения таких антител для предотвращения или лечения нарушений или заболеваний, например, вызванных повреждением центральной нервной системы у объекта.
Предшествующий уровень техники
FAM19A5 является представителем подсемейства белков TAFA, которое состоит из пяти высокогомологичных небольших белков. Tang T. Y. et al., Genomics 83(4):727-34 (2004). Эти белки содержат консервативные остатки цистеина в фиксированных положениях и отдаленно родственны воспалительному белку макрофагов 1-альфа (MIP-1-альфа), представителю семейства CC-хемокинов. Белки TAFA преимущественно экспрессируются в определенных областях головного и спинного мозга. Считается, что эти белки вырабатываются и секретируются взрослыми нервными стволовыми клетками в процессах нейрогенеза.
FAM19A5 преимущественно экспрессируется в головном мозге позвоночных, и считается, что FAM19A5 важен для развития, дифференциации, формирования всей центральной нервной системы и может использоваться для профилактики или лечения повреждений и/или заболеваний центральной нервной системы. Патентная публикация США № 2015/0118230.
Хотя ингибирование FAM19A5 может играть важную роль в лечении центральной нервной системы, все еще существует потребность в разработке антител, которые специфически связываются с FAM19A5 и которые способны модулировать активность FAM19A5, особенно тех антител, которые можно использовать у людей без побочных эффектов.
Краткое изложение раскрытия
В настоящем документе предложено выделенное антитело или его антигенсвязывающая часть, которая специфически связывает человеческий белок представителя A5 семейства 19 со сходством последовательностей (FAM19A5) («анти-FAM19A5 антитело»), включающее CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где CDR1, CDR2 и CDR3 тяжелой цепи содержат аминокислотные последовательности, указанные в SEQ ID NO: 5, 6 и 7, соответственно, каждая из которых необязательно содержит одну, две, три, четыре или пять мутаций, при этом CDR1, CDR2 и CDR3 легкой цепи содержат аминокислотные последовательности, указанные в SEQ ID NO: 8, 9 и 10, соответственно, где, по меньшей мере, одну из CDR1, CDR2 и CDR3 легкой цепи, содержащей одну, две, три, четыре или пять мутаций, и при этом антитело имеет пониженную иммуногенность у человека по сравнению с референсным антителом, содержащим VH, указанную в SEQ ID NO: 11, и VL, указанную в SEQ ID NO: 12.
В некоторых воплощениях CDR3 тяжелой цепи анти-FAM19A5 антитела, раскрытого в данном документе, включает аминокислотную последовательность, указанную в SEQ ID NO: 7.
В некоторых воплощениях CDR1 тяжелой цепи анти-FAM19A5 антитела включает аминокислотную последовательность, указанную в SEQ ID NO: 5, с одной или двумя мутациями. В некоторых воплощениях мутации включают замену треонина в аминокислоте 3 из SEQ ID NO: 5 на кислую аминокислоту. В дополнительных воплощениях мутации включают замену серина в аминокислоте 5 из SEQ ID NO: 5 на кислую аминокислоту. В некоторых воплощениях кислая аминокислота включает аспарагиновую кислоту или глутаминовую кислоту.
В некоторых воплощениях CDR2 тяжелой цепи анти-FAM19A5 антитела включает аминокислотную последовательность, указанную в SEQ ID NO: 6, с одной, двумя, тремя, четырьмя или пятью мутациями. В некоторых воплощениях мутации включают замену аргинина в аминокислоте 16 из SEQ ID NO: 6 на основную аминокислоту. В некоторых воплощениях основная аминокислота включает лизин. В дополнительных воплощениях мутации включают одну или несколько из следующих: (а) замену глицина в аминокислоте 6 из SEQ ID NO: 6 на кислую аминокислоту; (b) замену серина в аминокислоте 7 в SEQ ID NO: 6 на кислую аминокислоту; (c) замену серина в аминокислоте 8 из SEQ ID NO: 6 на кислую аминокислоту; (d) замену треонина в аминокислоте 9 SEQ ID NO: 6 на кислую аминокислоту; и (e) замену аргинина в аминокислоте 16 из SEQ ID NO: 6 на основную аминокислоту. В некоторых воплощениях кислая аминокислота включает аспарагиновую кислоту или глутаминовую кислоту. В некоторых воплощениях основная аминокислота включает лизин.
В некоторых воплощениях CDR3 легкой цепи анти-FAM19A5 антитела, раскрытого в данном документе, включает аминокислотную последовательность, указанную в SEQ ID NO: 10, с одной, двумя, тремя, четырьмя или пятью мутациями. В некоторых воплощениях мутации включают одну или несколько из следующих: (а) замену серина в аминокислоте 6 из SEQ ID NO: 10 на кислую аминокислоту или алифатическую аминокислоту; (b) замену аспарагина в аминокислоте 7 из SEQ ID NO: 10 на кислую аминокислоту или на гидроксил или сера/селенсодержащую аминокислоту; (c) замену глицина в аминокислоте 8 из SEQ ID NO: 10 на кислую аминокислоту или на гидроксил или сера/селенсодержащую аминокислоту; (d) замену глицина в аминокислоте 9 из SEQ ID NO: 10 на кислую аминокислоту или на гидроксил или сера/селенсодержащую аминокислоту; и (e) замену изолейцина в аминокислоте 10 из SEQ ID NO: 10 на основную аминокислоту. В некоторых воплощениях кислая аминокислота включает аспарагиновую кислоту или глутаминовую кислоту. В некоторых воплощениях гидроксильная или сера/селенсодержащая аминокислота включает серин. В дополнительных воплощениях основная аминокислота включает гистидин.
В некоторых воплощениях CDR1 легкой цепи анти-FAM19A5 антитела по настоящему раскрытию включает аминокислотную последовательность, указанную в SEQ ID NO: 8, с одной, двумя, тремя или четырьмя мутациями. В некоторых воплощениях мутации включают одну или несколько из следующих: (а) замену тирозина в аминокислоте 6 из SEQ ID NO: 8 на кислую аминокислоту; (b) замену аргинина в аминокислоте 7 из SEQ ID NO: 8 на кислую аминокислоту; (c) замену глицина в аминокислоте 8 из SEQ ID NO: 8 на кислую аминокислоту; и (d) замену серина в аминокислоте 9 из SEQ ID NO: 8 на кислую аминокислоту. В некоторых воплощениях кислая аминокислота включает глутаминовую кислоту или глутамин.
В некоторых воплощениях CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 9, с одной, двумя, тремя или четырьмя мутациями. В некоторых воплощениях мутации включают одну или несколько из следующих: (а) замену глутаминовой кислоты в аминокислоте 1 из SEQ ID NO: 9 на кислую аминокислоту; (b) замену серина в аминокислоте 2 из SEQ ID NO: 9 на кислую аминокислоту; (c) замену аспарагина в аминокислоте 3 из SEQ ID NO: 9 на кислую аминокислоту, основную аминокислоту или алифатическую аминокислоту; и (d) замену лизина в аминокислоте 4 из SEQ ID NO: 9 на кислую аминокислоту или алифатическую аминокислоту. В некоторых воплощениях кислая аминокислота включает глутамин, аспарагин, аспарагиновую кислоту или глутаминовую кислоту. В некоторых воплощениях основная аминокислота включает гистидин. В дополнительных воплощениях алифатическая аминокислота включает лейцин. В некоторых воплощениях мутации включают замену серина у аминокислоты 2 из SEQ ID NO: 9 на кислую аминокислоту. В некоторых воплощениях кислая аминокислота включает аспартат, глутамат, аспарагин, глутамин или их комбинации. В некоторых воплощениях кислая аминокислота представляет собой аспарагин.
Также в настоящем документе предлагается выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, содержащая CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 5; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 13; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 20; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 10.
Также в настоящем документе предлагается выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, содержащая CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 21; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 22; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 23.
Также в настоящем документе предлагается выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, содержащая CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 21; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 24; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 23.
Также в настоящем документе предлагается выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, содержащая CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 25; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 23.
Также в настоящем документе предлагается выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, содержащая CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 24; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 23.
Также в настоящем документе предлагается выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, содержащая CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 26; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 27.
Также в настоящем документе предлагается выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, содержащая CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 28; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 29.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, которая идентична, по меньшей мере, на около 80%, по меньшей мере, около 85%, по меньшей мере, около 90%, по меньшей мере, около 95%, по меньшей мере, около 96%, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% или около 100% указанной аминокислотной последовательности в SEQ ID NO: 11 и/или где VL включает аминокислотную последовательность, которая идентична, по меньшей мере, на около 80%, по меньшей мере, около 85%, по меньшей мере, около 90%, по меньшей мере, около 95%, по меньшей мере, около 96%, при, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% или около 100% аминокислотной последовательности, указанной в SEQ ID NO: 12.
В некоторых воплощениях анти-FAM19A5 антитело перекрестно конкурирует с референсным антителом, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где: (a) VH содержит аминокислотную последовательность, указанную в SEQ ID NO: 33, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 38; (b) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 39; (c) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 41; (d) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 40; (e) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 42; (f) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 43; или (g) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 44.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, связывается с тем же эпитопом FAM19A5 человека, что и референсное антитело, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где: (a) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 33, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 38; (b) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 39; (c) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 41; (d) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 40; (e) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 42; (f) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 43; или (g) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 44.
В некоторых воплощениях эпитоп FAM19A5 человека включает аминокислотную последовательность, указанную в SEQ ID NO: 90, 91 или 92.
Настоящее раскрытие дополнительно обеспечивает выделенное антитело или его антигенсвязывающую часть, которая специфически связывает человеческий белок представителя A5 семейства 19 со сходством последовательностей (FAM19A5) («анти-FAM19A5 антитело»), содержащий CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где CDR1, CDR2 и CDR3 тяжелой цепи содержат аминокислотные последовательности, указанные в SEQ ID NO: 16, 17 и 18, соответственно, каждая из которых необязательно содержит одну, две или три мутации, при этом CDR1, CDR2 и CDR3 легкой цепи содержат аминокислотные последовательности, указанные в SEQ ID NO: 30, 31 и 32, соответственно, где, по меньшей мере, одну из CDR1, CDR2 и CDR3 содержит одну, две или три мутации, и при этом антитело имеет пониженную иммуногенность у человека и/или более высокую аффинность связывания с человеческим белком FAM19A5 по сравнению с референсным антителом, содержащим VH, указанный в SEQ ID NO: 35, и VL, указанный в SEQ ID NO: 45.
В некоторых воплощениях CDR3 тяжелой цепи анти-FAM19A5 антитела включает аминокислотную последовательность, указанную в SEQ ID NO: 18, 128 или 129.
В некоторых воплощениях CDR1 тяжелой цепи анти-FAM19A5 антитела включает аминокислотную последовательность, указанную в SEQ ID NO: 16. В других воплощениях CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 19.
В некоторых воплощениях CDR2 тяжелой цепи анти-FAM19A5 антитела включает аминокислотную последовательность, указанную в SEQ ID NO: 18.
В некоторых воплощениях CDR3 легкой цепи анти-FAM19A5 антитела включает аминокислотную последовательность, указанную в SEQ ID NO: 32.
В некоторых воплощениях CDR2 легкой цепи анти-FAM19A5 антитела включает аминокислотную последовательность, указанную в SEQ ID NO: 31.
В некоторых воплощениях CDR1 легкой цепи анти-FAM19A5 антитела, раскрытого в данном документе, включает аминокислотную последовательность, указанную в SEQ ID NO: 30, с одной мутацией. В некоторых воплощениях мутация включает замену серина в аминокислоте 4 из SEQ ID NO: 30 на алифатическую аминокислоту. В некоторых воплощениях алифатическая аминокислота включает валин.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, которая идентична, по меньшей мере, на около 80%, по меньшей мере, около 85%, по меньшей мере, около 90%, по меньшей мере, около 95%, по меньшей мере, около 96%, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% или около 100% указанной аминокислотной последовательности в SEQ ID NO: 35 и/или где VL включает аминокислотную последовательность, которая идентична, по меньшей мере, на около 80%, по меньшей мере, около 85%, по меньшей мере, около 90%, по меньшей мере, около 95%, по меньшей мере, около 96%, при, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% или около 100% аминокислотной последовательности, указанной в SEQ ID NO: 45.
В некоторых воплощениях анти-FAM19A5 антитело перекрестно конкурирует с референсным антителом, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 36, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46. В других воплощениях анти-FAM19A5 антитело перекрестно конкурирует с референсным антителом, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 37, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46. В дополнительных воплощениях анти-FAM19A5 антитело перекрестно конкурирует с референсным антителом, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 130, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46. В некоторых воплощениях анти-FAM19A5 антитело перекрестно конкурирует с референсным антителом, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 131, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
В некоторых воплощениях анти-FAM19A5 антитело выбрано из группы, состоящей из IgG1, IgG2, IgG3, IgG4, их варианта и любой их комбинации. В некоторых воплощениях анти-FAM19A5 антитело представляет собой химерное антитело, человеческое антитело или гуманизированное антитело. В некоторых воплощениях анти-FAM19A5 антитело содержит Fab, a Fab', F(ab')2, Fv или одноцепочечный Fv (scFv).
В некоторых воплощениях раскрытое в данном документе анти-FAM19A5 антитело представляет собой scFv. В некоторых воплощениях scFv содержит VH и VL, где: (a) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 33, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 38; (b) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 39; (c) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 41; (d) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 40; (e) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 42; (f) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 43; (g) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 44; (h) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 36, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46; (i) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 37, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46; (j) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 130, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46; или (k) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 131, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
В некоторых воплощениях анти-FAM19A5 антитело проявляет одно или несколько из следующих свойств: (а) связывается с растворимым FAM19A5 человека с KD 10 нМ или менее, что измерено с помощью иммуноферментного анализа (ELISA); (b) связывается с мембраносвязанным FAM19A5 человека с KD 10 нМ или менее, что измерено с помощью ELISA; (c) уменьшает, обращает вспять, задерживает и/или предотвращает начало реактивного глиоза; (d) подавляет чрезмерную пролиферацию реактивных астроцитов; (e) снижает экспрессию протеогликанов хондроитинсульфата, включая нейрокан и нейрон-глиальный антиген 2 (NG2); (f) увеличивает экспрессию c-fos и pERK в ядрах нейронов; (g) способствует выживанию нейронов; (h) увеличивает экспрессию GAP43 в нейронах; (i) способствует отрастанию аксона; (j) вызывает нормализацию кровеносных сосудов, например, в опухоли; (k) подавляет рост опухоли; (l) усиливает проникновение иммунных клеток в опухоль; (m) усиливает инфильтрацию нейрональных клеток в опухоль; (n) усиливает фагоцитарную активность макрофага или микроглии; (o) увеличивает потенциал митохондриальной мембраны макрофага или микроглии; (p) снижает рекрутирование миелоидных супрессорных клеток (MDSC) в опухоль; (q) уменьшает некроз и отек опухоли; (r) снижает проницаемость ткани опухоли; и (s) увеличивает скорость кровотока в опухоли.
В настоящем документе также представлены нуклеиновые кислоты, кодирующие анти-FAM19A5 антитела, раскрытые в настоящем документе, векторы, содержащие нуклеиновую кислоту, клетки, содержащие вектор, и иммуноконъюгаты, содержащие анти-FAM19A5 антитела по настоящему раскрытию. Композиции, содержащие анти-FAM19A5 антитела, нуклеиновые кислоты, векторы, клетки или иммуноконъюгаты согласно настоящему раскрытию, и носитель также раскрыты в данном документе. В настоящем раскрытии также представлены наборы, содержащие анти-FAM19A5 антитела, нуклеиновые кислоты, векторы, клетки или иммуноконъюгаты в соответствии с настоящим раскрытием, а также инструкцию по применению.
В настоящем документе также предлагается способ получения антитела, которое специфически связывается с человеческим белком FAM19A5, включающий культивирование клеток, описанных в настоящем документе, в подходящих условиях и выделение антитела.
Настоящее раскрытие дополнительно обеспечивает способ лечения заболевания или состояния у объекта, нуждающегося в этом, включающий введение анти-FAM19A5 антитела, нуклеиновой кислоты, вектора, клетки или иммуноконъюгата, описанных в данном документе. В некоторых воплощениях заболевание или состояние включает опухоль, фиброз, глаукому, расстройство настроения, ретинопатию, возрастную дегенерацию желтого пятна или невропатическую боль. В некоторых воплощениях заболевание или состояние представляет собой опухоль.
В некоторых воплощениях изобретения опухоль включает меланому, рак поджелудочной железы, глиому, рак молочной железы, лимфому, рак легкого, рак почки, рак предстательной железы, фибросаркому, аденокарциному толстой кишки, рак печени или рак яичников. В некоторых воплощениях глиома представляет собой мультиформную глиобластому (GBM).
В некоторых воплощениях анти-FAM19A5 антитело, нуклеиновая кислота, вектор, клетка или иммуноконъюгат по настоящему раскрытию вызывает нормализацию кровеносных сосудов. В некоторых воплощениях нормализация кровеносных сосудов сопровождается изменениями свойств кровеносных сосудов, включая повышенную коннективность, увеличенную толщину стенки, уменьшенный диаметр сосуда, более правильное направление и характер распределения сосудов, увеличенное количество сосудов, уменьшение утечки и проницаемости, увеличенное покрытие перицитов и близость к кровеносным сосудам, повышенную оксигенацию или их комбинации.
В некоторых воплощениях анти-FAM19A5 антитело, нуклеиновая кислота, вектор, клетка или иммуноконъюгат по настоящему раскрытию подавляет рост опухоли.
В некоторых воплощениях анти-FAM19A5 антитело, нуклеиновая кислота, вектор, клетка или иммуноконъюгат, раскрытые в настоящем документе, усиливают инфильтрацию иммунных клеток в опухоль. В некоторых воплощениях иммунные клетки содержат макрофаги, дендритные клетки, Т-лимфоциты, В-лимфоциты, клетки-естественные киллеры (NK) или их комбинации. В некоторых воплощениях иммунные клетки дополнительно проявляют гипертрофию. В некоторых воплощениях усиленная инфильтрация иммунных клеток в опухоль сопровождается усиленной инфильтрацией нейрональных клеток в опухоль. В некоторых воплощениях нейронные клетки включают астроциты, глиальные клетки или их комбинации.
В некоторых воплощениях анти-FAM19A5 антитело, нуклеиновая кислота, вектор, клетка или иммуноконъюгат усиливают фагоцитарную активность макрофага или микроглии. В некоторых воплощениях анти-FAM19A5 антитело, нуклеиновая кислота, вектор, клетка или иммуноконъюгат увеличивает потенциал митохондриальной мембраны макрофага или микроглии.
В некоторых воплощениях анти-FAM19A5 антитело, нуклеиновая кислота, вектор, клетка или иммуноконъюгат, раскрытые в настоящем документе, снижают привлечение к опухоли клеток-супрессоров миелоидного происхождения (MDSC). В некоторых воплощениях анти-FAM19A5 антитело, нуклеиновая кислота, вектор, клетка или иммуноконъюгат уменьшают некроз и отек в опухоли. В некоторых воплощениях анти-FAM19A5 антитело, нуклеиновая кислота, вектор, клетка или иммуноконъюгат снижают тканевую проницаемость опухоли. В некоторых воплощениях анти-FAM19A5 антитело, нуклеиновая кислота, вектор, клетка или иммуноконъюгат увеличивают скорость кровотока в опухоли.
В некоторых воплощениях способ лечения заболевания или нарушения дополнительно включает введение дополнительного терапевтического агента. В некоторых воплощениях дополнительный терапевтический агент включает химиотерапию, иммунотерапию, лучевую терапию или их комбинации.
Краткое описание чертежей
На фиг. 1A, 1B и 1C представлен анализ связывания отдельных клонов scFv с белком FAM19A5. Поглощение измеряли при 405 нм. Номера клонов указаны на оси X. фиг. 1A, 1B и 1C показан анализ 96 клонов из 4-го, 5-го или 5-го цикла биопэннинга, полученных от первого цыпленка, второго цыпленка и третьего цыпленка, соответственно. Для каждого из клонов, показанных на фиг. 1A, 1B и 1C вертикальные полосы соответствуют белку FAM19A5, белку отрицательного контроля, белку гемагглютинина и BSA (слева направо). На каждой из фиг. 1A, 1B и 1C, заключенные в рамку клоны представляют собой те 8 клонов, которые были отобраны для дальнейшего анализа (см. Пример 4).
На фиг. 2A и 2B представлены приблизительный размер и связывающая способность различных scFv против FAM19A5, соответственно. Размер антител показан с помощью SDS-Page, а способность связывания измерена с помощью ELISA. Показанные антитела включают (слева направо): 1-28, 1-85, 2-13, 2-14, 2-20, 2-29, 3-2 и 3-26. На фиг. 2B, для каждого из показанных антител полоса слева представляет связывание антител с белком FAM19A5. Полоса справа представляет собой отрицательный контроль (измеренное связывание в присутствии только блокирующего буфера, т.е. без рекомбинантного белка FAM19A5). Столбец, обозначенный как «только 2-й», представляет другой отрицательный контроль, чтобы показать фоновый уровень анализа (измеренное связывание в отсутствие первичных анти-FAM19A5 антител).
На фиг. 3A и 3B представлено сравнение способности различных анти-FAM19A5 антител нейтрализовать экспрессию FAM19A5 в глиальных клетках мыши и человека, соответственно. Нейтрализация показана как процентное снижение экспрессии FAM19A5, которое показано как средняя интенсивность флуоресценции (MFI). Процент снижения можно рассчитать по следующей формуле: 100% - [((MFI FAM19A5 + анти-FAM19A5 антитело)/(MFI FAM19A5 + контрольное антитело)) x 100]. Процент снижения для каждого из антител показан в скобках.
На фиг. 4 представлены аминокислотные последовательности эпитопов F1-F6 (конъюгированных с BSA) и их расположение на полипептиде FAM19A5 человека. Показанная верхняя аминокислотная последовательность представляет собой изоформу 2 FAM19A5 дикого типа (без сигнального пептида). Вторая показанная аминокислотная последовательность представляет собой ту же последовательность, но остатки цистеина были мутированы на серин для снижения неспецифической активности во время синтеза пептида.
На фиг. 5 представлены результаты ELISA для связывания антитела 3-2 с фрагментами эпитопа с F1 по F6. Самый левый столбец («FAM19A5») представляет собой положительный контроль и показывает связывание антитела 3-2 против FAM19A5 с целым белком FAM19A5. Группы «нерелевантный белок» и «только блокирующий» (т.е. только блокирующий буфер, т.е. отсутствие белка FAM19A5) представляют собой отрицательные контроли. Для каждой из групп полоса слева соответствует контролю изотипа, а полоса справа соответствует антителу 3-2.
На фиг. 6A, 6B, 6C, 6D, 6E, 6F, 6G, 6H, 6I и 6J представлены результаты анализа аланинового сканирования, показывающие специфические аминокислотные остатки во фрагменте эпитопа F2, которые важны для связывания различных вариантов антител 3-2 к белку FAM19A5. На фиг. 6A, 6B, 6C и 6D показаны результаты для следующих антител, полученных в клетках HEK293F: (A) антитело 3-2 дикого типа, (B) антитело 1-30, (C) антитело 1-32 и (D) антитело 6-10, соответственно. Фиг. 6E, 6F, 6G, 6H, 6I и 6J показывают результаты для следующих антител, полученных в клетках CHO: (E) антитело 1-17, (F) антитело 1-30, (G) антитело 1-32, (H) антитело 4-11, (I) антитело 6-10 и (J) антитело с низким PI, соответственно. Как обсуждалось в примере 7, были получены мутантные пептиды, где каждый из мутантных пептидов имел замену аланином в одном аминокислотном остатке внутри эпитопного фрагмента F2. Связывание антител с различными мутантными пептидами измеряли с помощью ELISA.
Фиг. 7 идентифицирует потенциальные сайты иммуногенности в вариабельной области легкой цепи (VL) (верхние три ряда) и вариабельной области тяжелой цепи (VH) (нижние три ряда) антитела 3-2. VL соответствует SEQ ID NO: 12, а VH соответствует SEQ ID NO: 11. Также показаны последовательности человеческих зародышевых линий, используемых для каркасных областей антитела 3-2: VL = лямбда иммуноглобулина, вариабельная 1-51 (IGLV1-51*02) и лямбда иммуноглобулина, соединяющая 2 (IGLJ2*01); VH = тяжелая цепь иммуноглобулина вариабельная 3-64 (IGHV3-64*04) и тяжелая цепь иммуноглобулина, соединяющая 1 (IGHJ1*01). Используемые зародышевые линии человека представляли собой человеческие зародышевые линии (IgBLAST, NCBI), наиболее гомологичные клону 3-2. Показаны неразборчивые пептиды, связывающие MHC класса II с высоким и умеренным потенциалом иммуногенности, определенным анализом ITOPE™. В частности, пептиды, связывающие MHCII с высоким потенциалом иммуногенности, представляют собой следующие: (i) пептид № 3: остатки 85-93 SEQ ID NO: 12 (IYYCGSWDS); (ii) Пептид № 9: остатки 64-72 SEQ ID NO: 11 (VRGRATISR); и (iii) пептид № 10: остатки 79-87 SEQ ID NO: 11 (VRLQLNNPG). Пептиды, связывающие MHCII с умеренным потенциалом иммуногенности, представляют собой следующие: (i) остатки 16-24 SEQ ID NO: 12 (VKITCSGGG); (ii) Пептид № 2: остатки 48-56 SEQ ID NO: 12 (IYESNKRPS); (iii) Пептид № 4: остатки 18-26 SEQ ID NO: 11 (LSLVCKASG); (iv) Пептид № 5: остатки 19-27 SEQ ID NO: 11 (SLVCKASGF); (v) Пептид № 6: остатки 32-40 SEQ ID NO: 11 (FNMFWVRQA); (vi) Пептид № 7: остатки 45-53 SEQ ID NO: 11 (LEYVAQISS); (vii) Пептид № 8: остатки 48-56 SEQ ID NO: 11 (VAQISSSGS); и (viii) пептид № 11: остатки 81-89 SEQ ID NO: 11 (LQLNNPGAE). Гомологичные пептиды из базы данных Т-клеточных эпитопов следующие: Пептид № 5, Пептид № 6 и Пептид № 9. Всего было идентифицировано 11 связывающих пептидов, которые обозначены как пептиды №1- №11. «P1» для каждого из связывающих пептидов указывает первое якорное положение.
Фиг. 8 идентифицирует потенциальные сайты иммуногенности в вариабельной области легкой цепи (VL) (верхние три ряда) и вариабельной области тяжелой цепи (VH) (нижние три ряда) антитела 2-13. VL соответствует SEQ ID NO: 45, а VH соответствует SEQ ID NO: 35. Также показаны последовательности человеческих зародышевых линий, используемых для каркасных областей антитела 2-13: VL = лямбда иммуноглобулина, вариабельная 3-27 (IGLV3-27*01) и лямбда иммуноглобулина, соединяющая 2 (IGLJ2*01); VH = тяжелая цепь иммуноглобулина вариабельная 3-64 (IGHV3-64*04) и тяжелая цепь иммуноглобулина, соединяющая 1 (IGHJ1*01). Используемые зародышевые линии человека были наиболее гомологичными клону 2-13 (IgBLAST, NCBI). Показаны неразборчивые пептиды, связывающие MHC класса II с высоким и умеренным потенциалом иммуногенности, определенным анализом ITOPE™. В частности, пептиды, связывающие MHCII с высоким потенциалом иммуногенности, представляют собой следующие: (i) Пептид № 1: остатки 16-24 SEQ ID NO: 45 (VKITCSGGS); (ii) Пептид № 2: остатки 80-88 SEQ ID NO: 45 (VYFCGTEDI); и (iii) пептид № 8: остатки 79-87 SEQ ID NO: 35 (VRLQLNNLR). Пептиды, связывающие MHCII с умеренным потенциалом иммуногенности, представляют собой следующие: (i) Пептид № 3: остатки 18-26 SEQ ID NO: 35 (LSLVCKASG); (ii) Пептид № 4: остатки 20-28 SEQ ID NO: 35 (LVCKASGFT); (iii) пептид № 5: остатки 36-44 SEQ ID NO: 35 (WVRQTPGKG); (iv) Пептид № 6: остатки 47-55 SEQ ID NO: 35 (YVAEITNDG); (v) Пептид № 7: остатки 64-72 SEQ ID NO: 35 (VKGRATISR); (vi) Пептид № 9: остатки 81-89 SEQ ID NO: 35 (LQLNNLRAE); и (vii) пептид № 10: остатки 86-94 SEQ ID NO: 35 (LRAEDTGTY). Гомологичные пептиды из базы данных Т-клеточных эпитопов следующие: Пептид №4 и Пептид №5. Было идентифицировано всего 10 связывающих пептидов, которые обозначены как пептиды №1- №10. «P1» для каждого из связывающих пептидов указывает первое якорное положение.
На фиг. 9A, 9B и 9C представлен анализ связывания деиммунизированных антител 3-2. На фиг. 9А схематически показано, где были сделаны различные мутации аминокислот для деиммунизации антитела 3-2. На фиг. 9B представлено сравнение последовательностей вариабельной области легкой цепи (VL) и вариабельной области тяжелой цепи (VH) среди следующих: (i) антитело 3-2 дикого типа («клон 3-2»), (ii) полностью деиммунизированное антитело 3-2 («полностью деиммунизированный клон 3-2») и (iii) деиммунизированное за исключением одной аминокислоты в CDR2 тяжелой цепи) антитело («деиммунизированный клон 3-2»). Аминокислотные остатки с высоким и умеренным потенциалом иммуногенности, определенные анализом ITOPE™, заключены в рамки и обозначены цифрами «1» и «2», соответственно. Фиг. 9C показывает сравнение связывания (i) антитела 3-2 дикого типа, (ii) полностью деиммунизированного антитела 3-2 и (iii) деиммунизированного, за исключением одной аминокислоты, с CDR2 тяжелой цепи («деиммунизированный клон-3-2») антитело к белку FAM19A5, что измерено с помощью ELISA. Каждый фаг, презентирующий одноцепочечный вариабельный фрагмент (scFv), добавляли в лунки микротитрационного планшета, покрытого FAM19A5 (■) или анти-НА антителом (□). Фоновый сигнал измеряли в контрольных лунках, покрытых BSA. Лунки исследовали HRP-конъюгированным анти-M13 антителом. Измеряли оптическую плотность при 405 нм. Результаты представлены в виде среднего значения ± стандартное отклонение, полученных на основании четырех повторов экспериментов.
На фиг.10A и 10B представлен анализ двух различных деиммунизированных антител 2-13: (i) полностью деиммунизированного и (ii) деиммунизированного, за исключением одной аминокислоты в CDR2 тяжелой цепи) антитела («деиммунизированного клон 2-13»). На фиг. 10A представлено сравнение вариабельной области легкой цепи (VL) (верхние три строки) и вариабельной области тяжелой цепи (VH) (нижние три строки) для двух деиммунизированных антител 2-13 к антителам 2-13 дикого типа («клон 2-13»). Аминокислотные остатки с высоким и умеренным потенциалом иммуногенности, определенные анализом ITOPE™, заключены в рамки и обозначены цифрами «1» и «2», соответственно. Фиг. 10B показывает сравнение связывания (i) антитела 2-13 дикого типа, (ii) полностью деиммунизированного антитела 2-13 и (iii) деиммунизированного, за исключением одной аминокислоты, с CDR2 тяжелой цепи ((деиммунизированный клон 2-13) антитела к белку FAM19A5, что измерено с помощью ELISA. Каждый фаг, презентирующий одноцепочечный вариабельный фрагмент (scFv), добавляли в лунки микротитрационного планшета, покрытого FAM19A5 (■) или анти-НА антителом (□). Фоновый сигнал измеряли в контрольных лунках, покрытых BSA. Лунки исследовали HRP-конъюгированным анти-M13 антителом. Измеряли оптическую плотность при 405 нм. Результаты представлены в виде среднего значения ± стандартное отклонение, полученных на основании четырех повторов экспериментов.
На фиг. 11 представлено сравнение связывающей способности различных вариантов деиммунизированного клона 2-13. Идентичность различных вариантов антител представлена по оси абсцисс. Каждый аминокислотный остаток CDRL1, CDRL2, CDRL3, CDRH1 и CDRH2 был заменен глутаминовой кислотой и аспарагиновой кислотой. Реактивность 70 вариантов антител анализировали фаговым иммуноферментным анализом. Каждый отображаемый scFv фаг добавляли в лунки микротитрационного планшета, покрытого FAM19A5 (■) или анти-НА антителом (□). Фоновый сигнал измеряли в контрольных лунках, покрытых BSA. Лунки исследовали HRP-конъюгированным анти-M13 антителом. Измеряли оптическую плотность при 405 нм. Результаты показаны как среднее значение ± стандартное отклонение, полученное на основании трех повторов экспериментов.
Фиг. 12 представлена схема способов, используемых для создания деиммунизированных анти-FAM19A5 антител с улучшенными физико-химическими свойствами.
Фиг. 13А и 13В представлено сравнение физико-химических свойств нескольких деиммунизированных вариантов 3-2 с антителом 3-2 дикого типа (т.е. недеиммунизированным) («исходным»). Показанные варианты антител включают: (i) антитело с низкой изоэлектрической точкой («Low_PI»), (ii) 1-17, (iii) 1-30, (iv) 1-32, (v) 4-11 и (vi) 6-10. На фиг. 13А показан анализ связывания антител с белком FAM19A5. Результаты представлены как среднее значение ± стандартное отклонение. На фиг. 13B показаны данные растворимости (балл CamSol) и гидрофобности (балл GRAVY).
Фиг. 14A и 14B представлены выравнивание последовательностей вариабельных областей тяжелой цепи (фиг. 14A) и вариабельных областей легкой цепи (фиг. 14B) для различных вариантов деиммунизированных антител 3-2. Показанные деиммунизированные вариантные антитела связаны с антителом 3-2 в том смысле, что эти антитела эквивалентны антителу 3-2, за исключением того, что они были деиммунизированы для снижения иммуногенности при введении людям. Аминокислотные остатки с высоким и умеренным потенциалом иммуногенности, определенные анализом ITOPE™, заключены в рамки и обозначены цифрами «1» и «2», соответственно. Также отмечены другие представляющие интерес аминокислотные остатки: «3» = различия в ближайшей последовательности зародышевой линии человека; «4» = потенциальное метилирование аргинина или лизина; «5» = потенциальное окисление триптофана или метионина; «6» = потенциальное дезамидирование аспарагина; «7» = потенциальная изомеризация аспартата; «8» = вставка редкой аминокислоты; и «9» = свободный цистеин или неканоническая пара цистеина. Такие аминокислотные остатки могут приводить к вариантам продуктов во время естественного клеточного процессирования и деградации.
На фиг. 15 представлено выравнивание последовательностей вариабельных областей легкой цепи (верхние три ряда) и вариабельных областей тяжелой цепи (нижние три ряда) для различных деиммунизированных и/или созревших аффинных вариантов антител 2-13. Эти вариантные антитела родственны антителу 2-13 в том смысле, что они эквивалентны антителу 2-13, за исключением того, что они были деиммунизированы и/или подверглись созреванию аффинности. Аминокислотные остатки с высоким и умеренным потенциалом иммуногенности, определенные анализом ITOPE™, заключены в рамки и обозначены цифрами «1» и «2», соответственно. Также отмечены другие представляющие интерес аминокислотные остатки: «3» = потенциальное метилирование аргинина или лизина; «4» = потенциальное окисление триптофана или метионина; «5» = потенциальное дезамидирование аспарагина; «6» = потенциальная изомеризация аспартата; «7» = вставка редкой аминокислоты; и «8» = свободный цистеин или неканоническая пара цистеина. Такие аминокислотные остатки могут приводить к вариантам продуктов во время естественного клеточного процессирования и деградации.
На фиг. 16A, 16B и 16C представлен уровень экспрессии двух вариантов (2-13D-37-1.5W-41 и 2-13D-37-3W-16) деиммунизированного антитела 2-13D-37, измеренный с помощью SDS-PAGE и вестерн-блоттинга. Фиг. 16A и 16B показывают результаты с использованием SDS-PAGE для антител из культуральной среды и очищенного белка. Фиг. 16C показывает результаты, полученные с использованием вестерн-блоттинга. На каждой из фиг. 16A, 16B и 16C, дорожки «1» и «2» соответствуют антителам 2-13D-37-1.5W-41 и 2-13D-37-3W-16, соответственно. На дорожках «1» и «2» «А» и «В» соответствуют состояниям до центрифугирования и после центрифугирования, соответственно. На фиг. 16B, левая панель показывает уменьшающуюся SDS-PAGE, а правая панель показывает невосстанавливающую SDS-PAGE.
На фиг. 17А и 17В представлен анализ двух вариантов (2-13D-37-1.5W-41 и 2-13D-37-3W-16) деиммунизированного антитела 2-13D-37, которые были получены в результате созревания аффинности. На фиг. 17A показано выравнивание аминокислотной последовательности вариабельной области легкой цепи (верхние три прямоугольника) и вариабельной области тяжелой цепи (нижние три прямоугольника) для следующих антител: деиммунизированный клон 2-13D-37, деиммунизированный клон 2-13D-37-1.5W-41, и деиммунизированный клон 2-13D-37-3W-16. Аминокислотные остатки с высоким и умеренным потенциалом иммуногенности, определенные анализом ITOPE™, заключены в рамки и обозначены цифрами «1» и «2», соответственно. Также отмечены другие представляющие интерес аминокислотные остатки: «3» = потенциальное окисление триптофана или метионина; «4» = потенциальное дезамидирование аспарагина; «5» = потенциальная изомеризация аспартата; и «6» = неканоническая пара цистеина. Такие аминокислотные остатки могут приводить к вариантам продуктов во время естественного клеточного процессирования и деградации. Фиг. 17B показывает анализ связывания деиммунизированного клона 2-13D-37, деиммунизированного клона 2-13D-37-1.5W-41 и деиммунизированного клона 2-13D-37-3W-16 с белком FAM19A5. Каждый фаг, презентирующий одноцепочечный вариабельный фрагмент (scFv), добавляли в лунки микротитрационного планшета, покрытого FAM19A5 (■) или анти-НА антителом (□). Фоновый сигнал измеряли в контрольных лунках, покрытых BSA. Лунки исследовали HRP-конъюгированным анти-M13 антителом. Измеряли оптическую плотность при 405 нм.
На фиг. 18 представлена схема общего рабочего процесса анализа HDX-MS, использованного в Примере 11.
На фиг. 19 представлена таблица, суммирующая процент покрытия, достигнутый в различных экспериментальных условиях, протестированных для анализа HDX-MS, описанного в Примере 11. Был отрегулирован один или несколько из следующих параметров: (i) концентрация образца, (ii) условия гашения, (iii) концентрация пепсина и/или продолжительность гидролиза, и (v) время выдерживания гашения (минуты). В зависимости от анализа использовали колонки с иммобилизованным пепсином C18 или C8 с расщеплением в режиме реального времени (включено) или в автономном режиме (выключено), как показано на фиг. 19.
На фиг. 20A и 20B представлены результаты гидролиза белка FAM19A5 пепсином с использованием оптимизированных условий, описанных в примере 11. Фиг. 20A показывает процент покрытия и избыточности 44 пептидов, которые были идентифицированы относительно зрелого белка FAM19A5 (SEQ ID NO: 101, т.е. SEQ ID NO: 2 минус сигнальный пептид, который соответствует первым 25 аминокислотам). Каждая из горизонтальных полос представляет собой отдельный пептид. На фиг. 20B представлены аминокислотные последовательности 44 пептидов, включая их начальный и конечный сайты относительно SEQ ID NO: 101, массу одного иона (MHP) и время удерживания (RT).
Фиг. 21 показывает процент покрытия и избыточность 22 пептидов, которые были идентифицированы при расщеплении белка FAM19A5 пепсином после мечения дейтерием, как описано в Примере 11. Каждая из горизонтальных полос представляет собой отдельный пептид.
На фиг. 22A, 22B, 22C, 22D и 22E представлено сравнение поглощения дейтерия между единичным (только антиген, «1») и комплексом антиген-антитело (2-13) («2») в зависимости от времени. По оси ординат показано максимальное поглощение дейтерия (если пептид содержал пролин, максимальное значение составляло [(количество аминокислот - 1) - (количество пролина)]). По оси абсцисс отложена продолжительность мечения дейтерием. Фиг. 22A предоставляет данные для следующих пептидов: (i) FLKEGQL (SEQ ID NO: 102) (верхний левый график), (ii) FLKEGQLAAGTCE (SEQ ID NO: 103) (верхний правый график), (iii) LKEGQLAAG (SEQ ID NO: 104) (средний левый график), (iv) LKEGQLAAGTCEI (SEQ ID NO: 105) (средний правый график), (v) LKEGQLAAGTCEIVTL (SEQ ID NO: 106) (нижний левый график) и (vi) AAGTCEI (SEQ ID NO: 107) (нижний правый график). Фиг. 22B предоставляет данные для следующих пептидов: (i) RDSSQPPRTIARQTARCAC (SEQ ID NO: 108) (верхний левый график), (ii) QPPRTIARQTA (SEQ ID NO: 109) (верхний правый график), (iii) ACRKGQIAGTTRARPAC (SEQ ID NO: 110) (средний левый график), (iv) ACRKGQIAGTTRARPACVD (SEQ ID NO: 111) (средний правый график), (v) ACRKGQIAGTTRARPACVDA (SEQ ID NO: 112) (нижний левый график) и (vi) ARIIKTKQWC (SEQ ID NO: 113) (нижний правый график). Фиг. 22C предоставляет данные для следующих пептидов: (i) ARIIKTKQWCDM (SEQ ID NO: 114) (верхний левый график), (ii) ARIIKTKQWCDML (SEQ ID NO: 115) (верхний правый график), (iii) ARIIKTKQWCDMLPCL (SEQ ID NO: 116) (средний левый график), (iv) RIIKTKQWCDM (SEQ ID NO: 117) (средний правый график), (v) RIIKTKQWCDML (SEQ ID NO: 118) (нижний левый график) и (vi) WCDMLPCL (SEQ ID NO: 119) (нижний правый график). Фиг. 22D предоставляет данные для следующих пептидов: (i) LPCLEGEG (SEQ ID NO: 120) (верхний левый график), (ii) PCLEGEG (SEQ ID NO: 121) (верхний правый график), (iii) PCLEGEGCD (SEQ ID NO: 122) (средний левый график), (iv) PCLEGEGCDL (SEQ ID NO: 123) (средний правый график), (v) EGEGCDL (SEQ ID NO: 124) (нижний левый график) и (vi) EGEGCDLL (SEQ ID NO: 125) (нижний правый график). Фиг. 22E предоставляет данные для следующих пептидов: (i) LLINRSGWTCTQPGGRIKTTT (SEQ ID NO: 126) (левый график) и (ii) LINRSGWTCTQPGGRIKTTT (SEQ ID NO: 127) (правый график). На каждом из графиков дополнительная информация о пептидах (т.е. начальный и конечный сайты (SEQ ID NO: 101, т.е. SEQ ID NO: 2 минус сигнальный пептид, который соответствует первым 25 аминокислотам) и размер) представлены в верхнем углу каждого графика.
На фиг. 23A, 23B и 23C показаны ключевые аминокислотные остатки вдоль белка FAM19A5 со значительными различиями в поглощении дейтерия (т.е. более ± 05 Да) между отдельными и сложными образцами. На фиг. 23A представлен анализ бабочкообразной диаграммы поглощения дейтерия отдельным антигеном (только антиген, верхний график) и комплексом антиген-антитело (2-13) (нижний график). Фиг. 23B представляет собой график разницы в поглощении дейтерия между отдельными и комплексными образцами. Фиг. 23C показывает данные как сумму разницы в поглощении дейтерия менее 15 Да. На фиг. 23A, 23B и 23C каждая из линий представляет различную продолжительность мечения дейтерием: (i) «1» (033 минуты или 20 секунд), (ii) «2» (10 минут), (iii) «3». (60 минут) и (iv) «4» (240 минут). Также каждая точка соответствует отдельному пептиду. На фиг. 23B и 23C показаны начальный и конечный сайты (SEQ ID NO: 101) пептидов с различиями в поглощении дейтерия более чем ± 05 Да между отдельным антигеном (только антиген) и комплексом антиген-пептид. Пунктирная рамка на фиг. 23B и 23C показывает различия в поглощении дейтерия менее ± 05 Да.
Фиг. 24 представлен анализ тепловой карты, показывающий области белка FAM19A5 со значительными различиями в поглощении дейтерия между одиночными (только антиген) и комплексными образцами антиген-антитело (2-13). Остатки, обведенные пунктирной красной линией, представляют ключевые аминокислотные остатки: (i) CRKGQIAGTTRAR (аминокислотные остатки 38-50 SEQ ID NO: 101 или аминокислотные остатки 63-75 SEQ ID NO: 2) и (ii) PACVDARIIKTKQW (аминокислотные остатки 51-64 SEQ ID NO: 101 или аминокислотные остатки 76-85 SEQ ID NO: 2).
На фиг. 25 представлена трехмерная структура белка FAM19A5 и расположение ключевого связывающего эпитопа для антитела 2-13.
Подробное описание раскрытия
В настоящем документе раскрыто выделенное моноклональное антитело или его антигенсвязывающая часть, которое специфически связывается с белком представителя A5 семейства 19 со сходством последовательностей (FAM19A5) («анти-FAM19A5 антитело») и проявляет одно или несколько свойств, раскрытых в данном документе. В частности, анти-FAM19A5 антитело было деиммунизировано для снижения иммуногенности у человека.
Чтобы облегчить понимание представленного в данном документе раскрытия, определен ряд терминов и фраз. Дополнительные определения приведены в подробном описании.
I. Определения
В этом раскрытии термины в единственном числе охватывают и множественное число; например, «антитело» означает одно или несколько антител. По существу, термины, «один или несколько» и «по меньшей мере один» могут использоваться в данном документе взаимозаменяемо.
Кроме того, «и/или» в данном контексте следует понимать как конкретное раскрытие каждой из двух указанных особенностей или компонентов с другим или без другого. Таким образом, термин «и/или», используемый в данном документе во фразе, такой как «A и/или B», предназначен для включения «A и B», «A или B», «A» (отдельно) и «B» (отдельно). Аналогичным образом, термин «и/или», используемый во фразе, такой как «A, B и/или C», предназначен для охвата каждого из следующих аспектов: A, B и C; А, В или С; А или С; А или В; B или C; А и С; А и В; B и C; А (отдельно); B (отдельно); и C (отдельно).
Понятно, что везде, где аспекты описаны в данном документе на языке «включающий», также предусмотрены аналогичные аспекты, описанные в терминах «состоящий из» и/или «состоящий по существу из».
Если не указано иное, все технические и научные термины, используемые в данном документе, имеют то же значение, которое обычно понимается специалистом в данной области техники, к которой относится это раскрытие. Например, the Concise Dictionary of Biomedicine and Molecular Biology, Juo, Pei-Show, 2nd ed., 2002, CRC Press; The Dictionary of Cell and Molecular Biology, 3rd ed., 1999, Academic Press; и the Oxford Dictionary Of Biochemistry And Molecular Biology, Revised, 2000, Oxford University Press, предоставляют специалистам общий словарь многих терминов, используемых в этом раскрытии.
Единицы, префиксы и символы обозначаются в их принятой форме Système International de Unites (SI). Числовые диапазоны включают числа, определяющие диапазон. Если не указано иное, аминокислотные последовательности написаны слева направо в ориентации от аминоконца до карбоксиконца. Заголовки, представленные в данном документе, не являются ограничениями различных аспектов раскрытия, которое может быть получено путем ссылки на описание в целом. Соответственно, термины, определенные непосредственно ниже, более полно определены со ссылкой на описание в целом.
Термин «около» используется в данном документе для обозначения примерно, приблизительно, приближенно или в области. Когда термин «около» используется в сочетании с числовым диапазоном, он изменяет этот диапазон, расширяя границы выше и ниже указанных числовых значений. В общем, термин «около» может изменять числовое значение выше и ниже указанного значения с отклонением, например, на 10 процентов, в большую или меньшую сторону (выше или ниже).
Термин «представитель A5 семейства 19 со сходством последовательностей» или «FAM19A5» относится к белку, который принадлежит к семейству TAFA (также известному как семейство FAM19) из пяти высокогомологичных белков и преимущественно экспрессируется в головном и спинном мозге. FAM19A5 также известен как TAFA5 или хемокиноподобный белок TAFA-5.
У человека ген, кодирующий FAM19A5, расположен на хромосоме 22. Существует несколько изоформ FAM19A5 человека (UniProt: Q7Z5A7), которые, как считается, получаются путем альтернативного сплайсинга: изоформа 1 (UniProt: Q7Z5A7-1), состоящая из 132 аминокислот, изоформа 2 (UniProt: Q7Z5A7-2), которая состоит из 125 аминокислот и изоформы 3 (UniProt: Q7Z5A7-3), которая состоит из 53 аминокислот. Считается, что белок FAM19A5 человека существует как в мембраносвязанной, так и в растворимой (секретируемой) форме. Считается, что изоформа 1 представляет собой мембранный белок с одной трансмембранной областью. Изоформа 2, о которой сообщалось в Tang T. Y. et al., Genomics 83 (4): 727-34 (2004) как секретируемый белок (растворимый), содержит сигнальный пептид в положениях аминокислот 1-25. Изоформа 1 считается мембранным белком и предсказана на основе данных EST. Ниже приведены аминокислотные последовательности трех известных изоформ FAM19A5 человека.
(I) Изоформа 1 (UniProt: Q7Z5A7-1, трансмембранный белок): эта изоформа была выбрана в качестве канонической последовательности.
MAPSPRTGSR QDATALPSMS STFWAFMILA SLLIAYCSQL AAGTCEIVTL DRDSSQPRRT IARQTARCAC RKGQIAGTTR ARPACVDARI IKTKQWCDML PCLEGEGCDL LINRSGWTCT QPGGRIKTTT VS (SEQ ID NO: 1)
(II) Изоформа 2 (UniProt: Q7Z5A7-2, растворимый белок):
MQLLKALWAL AGAALCCFLV LVIHAQFLKE GQLAAGTCEI VTLDRDSSQP RRTIARQTAR CACRKGQIAG TTRARPACVD ARIIKTKQWC DMLPCLEGEG CDLLINRSGW TCTQPGGRIK TTTVS (SEQ ID NO: 2)
(III) Изоформа 3 (UniProt: Q7Z5A7-3):
MYHHREWPAR IIKTKQWCDM LPCLEGEGCD LLINRSGWTC TQPGGRIKTT TVS (SEQ ID NO: 3)
Термин «FAM19A5» включает любые варианты или изоформы FAM19A5, которые естественным образом экспрессируются клетками. Соответственно, описанные в данном документе антитела могут перекрестно реагировать с различными изоформами одного и того же вида (например, разными изоформами FAM19A5 человека) или перекрестно реагировать с FAM19A5 других видов, кроме человека (например, FAM19A5 мыши). Альтернативно, антитела могут быть специфичными для FAM19A5 человека и не могут проявлять перекрестную реактивность с другими видами. FAM19A5 или его любые варианты и изоформы можно либо выделить из клеток или тканей, которые их экспрессируют в природе, либо получить рекомбинантно. Полинуклеотид, кодирующий человеческий FAM19A5, имеет учетный номер GenBank BC039396 и следующую последовательность:
Таблица 1A. Полинуклеотидная последовательность FAM19A5 человека
(Учетный номер GenBank BC039396)
Термины «антитело» и «антитела» являются терминами из уровня техники и могут использоваться в данном документе взаимозаменяемо и относятся к молекуле с сайтом связывания антигена, которая специфически связывает антиген. Используемые в данном документе термины включают целые антитела и любые антигенсвязывающие фрагменты (т.е. «антигенсвязывающие части») или их отдельные цепи. «Антитело» относится в одном варианте к гликопротеину, содержащему, по меньшей мере, две тяжелые (H) цепи и две легкие (L) цепи, соединенные между собой дисульфидными связями, или его антигенсвязывающую часть. В другом воплощении «антитело» относится к одноцепочечному антителу, содержащему единственный вариабельный домен, например, домен VHH. Каждая тяжелая цепь состоит из вариабельного участка тяжелой цепи (обозначенного в настоящем документе как VH) и константного участка тяжелой цепи. В некоторых встречающихся в природе антителах константная область тяжелой цепи состоит из трех доменов: СН1, СН2 и СН3. В некоторых встречающихся в природе антителах каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно VL) и константной области легкой цепи. Константная область легкой цепи содержит один домен, CL.
Области VH и VL могут быть далее разделены на области гипервариабельности, называемыми областями, отвечающими за комплементарность связывания (CDR), которые перемежаются областями, которые являются более консервативными, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от аминоконца до карбокси-конца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Вариабельные области тяжелой и легкой цепей содержат домен связывания который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (C1q) классической системы комплемента.
Термин «нумерация по Kabat» и подобные термины известны в данной области и относятся к системе нумерации аминокислотных остатков в вариабельных областях тяжелой и легкой цепей антитела или его антигенсвязывающей части. В некоторых аспектах CDR антитела можно определить в соответствии с системой нумерации Kabat (см., например, Kabat EA & Wu TT (1971) Ann NY Acad Sci 190: 382-391 and Kabat EA et al., (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242). Используя систему нумерации Kabat, CDR в молекуле тяжелой цепи антитела обычно присутствуют в положениях аминокислот с 31 по 35, которые необязательно могут включать одну или две дополнительные аминокислоты, следующие за 35 (обозначенные в схеме нумерации Kabat как 35A и 35B) (CDR1), в положениях аминокислот с 50 по 65 (CDR2) и в положениях аминокислот с 95 по 102 (CDR3). Используя систему нумерации Kabat, CDR в молекуле легкой цепи антитела обычно присутствуют в положениях аминокислот с 24 по 34 (CDR1), положениях аминокислот с 50 по 56 (CDR2) и положениях аминокислот с 89 по 97 (CDR3). В конкретном воплощении CDR описанных в данном документе антител были определены в соответствии со схемой нумерации Kabat.
Фразы «нумерация положений аминокислот, как в Kabat», «положение Kabat» и их грамматические варианты относятся к системе нумерации, используемой для вариабельных доменов тяжелой цепи или вариабельных доменов легкой цепи при компиляции антител в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991). При использовании этой системы нумерации фактическая линейная аминокислотная последовательность может содержать меньшее количество или дополнительные аминокислоты, соответствующие укорочению или вставке в FW или CDR вариабельного домена. Например, вариабельный домен тяжелой цепи может включать вставку одной аминокислоты (остаток 52a по Kabat) после остатка 52 из H2 и вставленные остатки (например, остатки 82a, 82b и 82c и т.д. по Kabat) после остатка 82 тяжелой цепи FW. См. таблицу 1B.
Таблица 1B
Нумерацию остатков по Kabat можно определить для данного антитела путем выравнивания областей гомологии последовательности антитела со «стандартной» пронумерованной последовательностью по Kabat. Chothia вместо этого ссылается на расположение структурных петель (Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987)). Конец петли CDR-H1 по Chothia при нумерации с использованием соглашения о нумерации Kabat варьирует между H32 и H34 в зависимости от длины петли (это связано с тем, что схема нумерации по Kabat размещает вставки в H35A и H35B; если 35A или 35B отсутствуют, петля заканчивается в 32, если присутствует только 35А, петля заканчивается в 33; если присутствуют оба 35А и 35В, петля заканчивается в 34). Гипервариабельные участки AbM представляют собой компромис между CDR по Kabat и структурными петлями Chothia и используюся программным обеспечением Oxford Molecular's AbM antibody modeling.
IMGT (ImMunoGeneTics) также обеспечивает систему нумерации для вариабельных областей иммуноглобулина, включая CDR. См., например, Lefranc, M.P. et al., Dev. Comp. Immunol. 27: 55-77 (2003), который включен в данный документ ссылкой. Система нумерации IMGT была основана на выравнивании более 5000 последовательностей, структурных данных и характеристик гипервариабельных петель и позволяет легко сравнивать вариабельные и CDR-области для всех видов. Согласно схеме нумерации IMGT, VH-CDR1 находится в положениях с 26 по 35, VH-CDR2 находится в положениях с 51 по 57, VH-CDR3 находится в положениях с 93 по 102, VL-CDR1 находится в положениях с 27 по 32, VL-CDR2 находится в положениях от 50 до 52, а VL-CDR3 находится в положениях от 89 до 97.
Для всех положений аминокислот константной области тяжелой цепи, обсуждаемых в настоящем описании, нумерация соответствует индексу EU, впервые описанному в Edelman et al., 1969, Proc. Natl. Acad. Sci. USA 63 (1): 78-85, описывающая аминокислотную последовательность миеломного белка EU, который является первым секвенированным lgG1 человека. Индекс EU из Edelman et al. также изложен в Kabat et al., 1991, Sequences of Proteins of Immunological Interest, 5th Ed., United States Public Health Service, National Institutes of Health, Bethesda. Таким образом, фразы «индекс EU в соответствии с Kabat» или «индекс EU из Kabat» и «положение… в соответствии с индексом EU, изложенным в Kabat» и их грамматические варианты относятся к системе нумерации остатков, основанной на человеческом lgG1 EU-антителе из Edelman et al. как изложено в Kabat 1991.
Система нумерации, используемая для вариабельных доменов (как тяжелой цепи, так и легкой цепи) и аминокислотной последовательности константной области легкой цепи, изложена в Kabat 1991.
Антитела могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA или IgY), любого класса (например, IgD, IgG2, IgG3, IgG4, IgA1 или IgA2) или любого подкласса (например, IgG1, IgG2, IgG3 и IgG4 у людей и IgG1, IgG2a, IgG2b и IgG3 у мышей) молекулы иммуноглобулина. Иммуноглобулины, например IgG1, существуют в виде нескольких аллотипов, которые отличаются друг от друга не более чем несколькими аминокислотами. Описанное в данном документе антитело может принадлежать к любому из общеизвестных изотипов, классов, подклассов или аллотипов. В некоторых воплощениях описанные в данном документе антитела относятся к подклассу IgG1, IgG2, IgG3 или IgG4 или к любому их гибриду. В некоторых воплощениях антитела относятся к подклассу IgG2, IgG4 или IgG2/IgG4.
«Антитело» включает, например, как встречающиеся в природе, так и не встречающиеся в природе антитела; моноклональные и поликлональные антитела; химерные и гуманизированные антитела; человеческие и нечеловеческие антитела; полностью синтетические антитела; одноцепочечные антитела; моноспецифические антитела; мультиспецифические антитела (включая биспецифические антитела); тетрамерные антитела, содержащие две молекулы тяжелой цепи и две молекулы легкой цепи; мономер легкой цепи антитела; мономер тяжелой цепи антитела; димер легкой цепи антитела, димер тяжелой цепи антитела; пару легкой цепи антитела-тяжелой цепи антитела; интратела; гетероконъюгированные антитела; моновалентные антитела; одноцепочечные антитела; верблюжьи антитела; аффи-боди; антиидиотипические (анти-Id) антитела (включая, например, анти-анти-Id антитела) и однодоменные антитела (sdAbs), которые включают связывающие молекулы, состоящие из одного мономерного вариабельного домена антитела, которые полностью способны к связыванию антигена (например, домен VH или домен VL). Harmen M. M. and Haard H. J. Appl Microbiol Biotechnol. 77(1): 13-22 (2007)).
Термин «антигенсвязывающая часть» антитела в контексте настоящего описания относится к одному или нескольким фрагментам антитела, которые сохраняют способность специфически связываться с антигеном (например, FAM19A5 человека). Такие «фрагменты» составляют, например, от около 8 до около 1500 аминокислот в длину, соответственно от около 8 до около 745 аминокислот в длину, соответственно от около 8 до около 300, например от около 8 до около 200 аминокислот, или около от 10 до около 50 или 100 аминокислот в длину. Было показано, что антигенсвязывающая функция антитела может быть осуществлена с помощью фрагментов полноразмерного антитела. Примеры связывающих фрагментов, охватываемых термином «антигенсвязывающая часть» антитела, например, описанного в данном документе анти-FAM19A5 антитела, включают (i) фрагмент Fab, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) фрагмент F(ab')2, двухвалентный фрагмент, содержащий два фрагмента Fab, связанных дисульфидным мостиком в шарнирной области; (iii) фрагмент Fd, состоящий из доменов VH и CH1; (iv) фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела, и дисульфидно-связанных Fv (sdFv) (v) фрагмент dAb (Ward et al., (1989) Nature 341: 544-546), который состоит из домена VH; и (vi) выделенную область, определяющую комплементарность (CDR) или (vii) комбинацию двух или более выделенных CDR, которые необязательно могут быть соединены синтетическим линкером. Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, они могут быть соединены с использованием рекомбинантных методов с помощью синтетического линкера, который позволяет им быть в виде единой белковой цепи, в которой VL и пары областей VH образуют одновалентные молекулы (известные как одноцепочечный Fv (scFv); см., например, Bird et al., (1988) Science 242: 423-426; и Huston et al., (1988) Proc. Natl. Acad. Sci. USA 85: 5879-5883). Такие одноцепочечные антитела также предназначены для охватывания термином "антигенсвязывающая часть" антитела. Эти фрагменты антител получаются с помощью традиционных методов известных специалистам в данной области, и фрагменты проверяют на пригодность также как и интактные антитела. Антигенсвязывающие части могут быть получены методами рекомбинантной ДНК или ферментативным или химическим расщеплением интактных иммуноглобулинов.
Используемые в данном документе термины «вариабельная область» и «вариабельный домен» используются взаимозаменяемо и являются общими в данной области техники. Вариабельная область обычно относится к части антитела, как правило, к части легкой или тяжелой цепи, обычно около амино-концевых аминокислот с 110 по 120 в зрелой тяжелой цепи и с около 90 по 115 аминокислот в зрелой легкой цепи, которые сильно различаются по последовательности среди антител и используются для связывания и специфичности конкретного антитела в отношении его конкретного антигена. Вариабельность последовательности сосредоточена в тех областях, которые называются областями, определяющими комплементарность (CDR), в то время как более высококонсервативные области в вариабельном домене называются каркасными областями (FR).
Безотносительно какого-либо конкретного механизма или теории, считается, что CDR легкой и тяжелой цепей в первую очередь ответственны за взаимодействие и специфичность антитела с антигеном. В некоторых воплощениях вариабельная область представляет собой вариабельную область человека. В некоторых воплощениях вариабельная область включает CDR грызунов или мышей и каркасные области (FR) человека. В конкретных воплощениях вариабельная область представляет собой вариабельную область приматов (например, приматов, отличных от человека). В некоторых воплощениях вариабельная область содержит CDR грызунов или мышей и каркасные области (FR) приматов (например, приматов, отличных от человека).
Используемый в данном документе термин «тяжелая цепь» (HC), когда он используется в отношении антитела, может относиться к любому отдельному типу, например, альфа (α), дельта (δ), эпсилон (ε), гамма (γ) и мю (μ) на основе аминокислотной последовательности константного домена, которая дает классы антител IgA, IgD, IgE, IgG и IgM, соответственно, включая подклассы IgG, например, IgG1, IgG2, IgG3 и IgG4.
Используемый в данном документе термин «легкая цепь» (LC) при использовании в отношении антитела может относиться к любому отдельному типу, например, каппа (κ) или лямбда (λ) на основе аминокислотной последовательности константных доменов. Аминокислотные последовательности легкой цепи хорошо известны в данной области. В конкретных воплощениях легкая цепь представляет собой легкую цепь человека.
Термины «VL» и «домен VL» используются взаимозаменяемо для обозначения вариабельной области легкой цепи антитела.
Термины «VH» и «домен VH» используются взаимозаменяемо для обозначения вариабельной области тяжелой цепи антитела.
Используемые в данном документе термины «константная область» и «константный домен» являются взаимозаменяемыми и имеют общепринятое значение в данной области техники. Константная область представляет собой часть антитела, например, карбоксильную концевую часть легкой и/или тяжелой цепи, которая не участвует напрямую в связывании антитела с антигеном, но может проявлять различные эффекторные функции, такие как взаимодействие с рецептором Fc. Константная область молекулы иммуноглобулина обычно имеет более консервативную аминокислотную последовательность по сравнению с вариабельным доменом иммуноглобулина.
«Область Fc» (область кристаллизующегося фрагмента), или «домен Fc», или «Fc» относится к С-концевой области тяжелой цепи антитела, которая опосредует связывание иммуноглобулина с тканями или факторами хозяина, включая связывание с Fc-рецепторами, расположенными на различных клетках иммунной системы (например, эффекторных клетках) или с первым компонентом (C1q) классической системы комплемента. Таким образом, Fc-область включает константную область антитела, за исключением первого иммуноглобулинового домена константной области (например, CH1 или CL). В изотипах антител IgG, IgA и IgD Fc-область включает два идентичных фрагмента белка, происходящих из второго (CH2) и третьего (CH3) константных доменов двух тяжелых цепей антитела; Fc-области IgM и IgE содержат три константных домена тяжелой цепи (домены CH 2-4) в каждой полипептидной цепи. Для IgG Fc-область включает домены иммуноглобулина Cγ2 и Cγ3 и шарнир между Cγ1 и Cγ2. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут варьировать, Fc-область тяжелой цепи человеческого IgG обычно определяется как простирающаяся от аминокислотного остатка в положении C226 или P230 (или аминокислоты между этими двумя аминокислотами) до карбоксильного конца тяжелой цепи, где нумерация соответствует индексу EU, как в Kabat. Домен CH2 Fc-области человеческого IgG простирается от около аминокислоты 231 до около аминокислоты 340, тогда как домен CH3 расположен на C-концевой стороне домена Cm в Fc-области, то есть он простирается от около аминокислоты 341 до около аминокислоты 447 в IgG. В данном контексте Fc-область может представлять собой Fc с нативной последовательностью, включая любой аллотипический вариант, или вариант Fc (например, не встречающийся в природе Fc). Fc также может относиться к этой области отдельно или в контексте полипептида, содержащего Fc, такого как «связывающий белок, содержащий Fc-область», также называемый «гибридный белок Fc» (например, антитело или иммуноадгезия).
«Область Fc с нативной последовательностью» или «Fc с нативной последовательностью» включает аминокислотную последовательность, которая идентична аминокислотной последовательности Fc-области, встречающейся в природе. Fc-области человека с нативной последовательностью включают Fc-область человеческого IgG1 с нативной последовательностью; Fc-область человеческого IgG2 с нативной последовательностью; Fc-область человеческого IgG3 с нативной последовательностью; и Fc-область человеческого IgG4 с нативной последовательностью, а также их природные варианты. Нативная последовательность Fc включает различные аллотипы Fc (см., например, Jefferis et al., (2009) mAbs 1: 1; Vidarsson G. et al., Front Immunol. 5: 520 (опубликовано онлайн 20 окт. 2014 г.)).
«Рецептор Fc» или «FcR» представляет собой рецептор, который связывается с Fc-областью иммуноглобулина. FcR, которые связываются с антителом IgG, включают рецепторы семейства FcγR, с учетом аллельных вариантов и альтернативно сплайсированных форм этих рецепторов. Семейство FcγR состоит из трех активирующих (FcγRI, FcγRIII и FcγRIV у мышей; FcγRIA, FcγRIIA и FcγRIIIA у людей) и одного ингибиторного (FcγRIIB) рецептора. Человеческий IgG1 связывается с большинством человеческих рецепторов Fc и вызывает наиболее сильные эффекторные функции Fc. Он считается эквивалентным мышиному IgG2a в отношении типов активирующих Fc-рецепторов, с которыми связывается. Напротив, человеческий IgG4 вызывает наименьшие эффекторные функции Fc. Vidarsson G. et al., Front Immunol. 5: 520 (опубликовано онлайн 20 окт. 2014 г.).
Константной областью можно манипулировать, например, с помощью рекомбинантной технологии, чтобы устранить одну или несколько эффекторных функций. «Эффекторная функция» относится к взаимодействию Fc-области антитела с Fc-рецептором или лигандом или к биохимическому событию, которое возникает в результате этого. Примеры «эффекторных функций» включают связывание C1q, комплемент-зависимую цитотоксичность (CDC), связывание с рецептором Fc, эффекторные функции, опосредованные FcγR, такие как ADCC и зависимый от антител клеточно-опосредованный фагоцитоз (ADCP), а также отрицательную регуляцию рецептора клеточной поверхности (например, В-клеточного рецептора; BCR). Такие эффекторные функции обычно требуют, чтобы Fc-область была объединена со связывающим доменом (например, вариабельным доменом антитела). Соответственно, термин «константная область без функции Fc» включает константные области с пониженной или отсутствующей одной или несколькими эффекторными функциями, опосредованными Fc-областью.
Эффекторные функции антитела можно уменьшить или избежать с помощью различных подходов. Эффекторные функции антитела можно уменьшить или избежать, используя фрагменты антитела, лишенные Fc-области (например, такие как Fab, F(ab')2, одноцепочечный Fv (scFv) или sdAb, состоящий из мономерного домена VH или VL). В качестве альтернативы, так называемые агликозилированные антитела могут быть получены путем удаления сахаров, которые связаны с определенными остатками в Fc-области, для снижения эффекторных функций антитела при сохранении других ценных атрибутов Fc-области (например, пролонгированного периода полувыведения и гетеродимеризации). Агликозилированные антитела могут быть получены, например, путем удаления или изменения остатка, к которому присоединен сахар, ферментативного удаления сахаров, продуцирования антитела в клетках, культивируемых в присутствии ингибитора гликозилирования, или путем экспрессии антитела в клетках, неспособных к гликозилированию белков (например, бактериальных клетках-хозяевах). См., например, публ. США № 20120100140. Другой подход заключается в использовании Fc-областей из подкласса IgG, которые обладают пониженной эффекторной функцией, например, антитела IgG2 и IgG4 характеризуются более низкими уровнями эффекторных функций Fc, чем IgG1 и IgG3. Остатки, наиболее проксимальные к шарнирной области в домене CH2 части Fc, отвечают за эффекторные функции антител, поскольку он содержит в значительной степени перекрывающийся сайт связывания для рецепторов C1q (комплемента) и IgG-Fc (FcγR) на эффекторных клетках врожденной иммунной системы. Vidarsson G. et al., Front Immunol. 5: 520 (опубликовано онлайн 20 окт. 2014 г.). Соответственно, антитела со сниженными эффекторными функциями Fc или без них могут быть получены путем создания, например, химерной Fc-области, которая включает домен CH2 из антитела IgG изотипа IgG4 и домен CH3 из антитела IgG изотипа IgG1 или химерной Fc-области, которая включает шарнирную область из IgG2 и область CH2 из IgG4 (см., например, Lau C. et al., J. Immunol. 191: 4769-4777 (2013)), или Fc-области с мутациями, которые приводят к измененным эффекторным функциям Fc, например, уменьшению или отсутствию функций Fc. Такие Fc-области с мутациями известны в данной области. См., например, публ. США № 20120100140 и заявки РСТ и США, процитированные в ней и в An et al., mAbs 1: 6, 572-579 (2009); раскрытие которых включено ссылкой во всей полноте.
«Шарнир», «шарнирный домен» или «шарнирная область» или «шарнирная область антитела» относится к домену константной области тяжелой цепи, которая соединяет домен CH1 с доменом CH2 и включает верхнюю, среднюю и нижнюю части шарнира (Roux et al., J. Immunol. 1998 161: 4083). Шарнир обеспечивает различные уровни гибкости между связывающими и эффекторными областями антитела, а также обеспечивает сайты для межмолекулярных дисульфидных связей между двумя константными областями тяжелой цепи. В данном контексте шарнир начинается на Glu216 и заканчивается на Gly237 для всех изотипов IgG (Roux et al., 1998 J Immunol 161: 4083). Последовательности шарниров IgG1, IgG2, IgG3 и IgG4 дикого типа известны в данной области. См., например, Kabat EA et al., (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242; Vidarsson G. et al., Front Immunol. 5: 520 (опубликовано онлайн 20 окт. 2014 г.).
Термин «домен CH1» относится к константной области тяжелой цепи, связывающей вариабельный домен с шарниром в константном домене тяжелой цепи. При использовании в данном документе, домен CH1 начинается в A118 и заканчивается в V215. Термин «домен CH1» включает домены CH1 дикого типа, а также их естественные варианты (например, аллотипы). Последовательности домена CH1 из IgG1, IgG2, IgG3 и IgG4 (включая дикий тип и аллотипы) известны в данной области. См., например, Kabat EA et al., (1991) выше и Vidarsson G. et al., Front Immunol. 5: 520 (опубликовано онлайн 20 окт. 2014 г.). Примеры доменов CH1 включают домены CH1 с мутациями, которые модифицируют биологическую активность антитела, например, период полувыведения, например, описанные в публ. США № 20120100140 и патентах и публикациях США, а также публикациях РСТ, процитированных в ней.
Термин «домен CH2» относится к константной области тяжелой цепи, связывающей шарнир с доменом CH3 в константном домене тяжелой цепи. В данном контексте домен CH2 начинается с P238 и заканчивается на K340. Термин «домен CH2» включает домены CH2 дикого типа, а также их естественные варианты (например, аллотипы). Последовательности домена CH2 IgG1, IgG2, IgG3 и IgG4 (включая дикий тип и аллотипы) известны в данной области. См., например, Kabat EA et al., (1991) выше и Vidarsson G. et al., Front Immunol. 5: 520 (опубликовано онлайн 20 окт. 2014 г.). Типичные домены CH2 включают домены CH2 с мутациями, которые модифицируют биологическую активность антитела, например, период полувыведения и/или сниженную эффекторную функцию Fc, например, описанные в публ. США № 20120100140 и патентах и публикациях США, а также публикациях РСТ, процитированных в ней.
Термин «домен CH3» относится к константной области тяжелой цепи, которая является С-концом по отношению к домену CH2 в константном домене тяжелой цепи. В данном контексте домен CH3 начинается с G341 и заканчивается на K447. Термин «домен CH3» включает домены CH3 дикого типа, а также их естественные варианты (например, аллотипы). Последовательности домена CH3 IgG1, IgG2, IgG3 и IgG4 (включая дикий тип и аллотипы) известны в данной области. См., например, Kabat EA et al., (1991) выше и Vidarsson G. et al., Front Immunol. 5: 520 (опубликовано онлайн 20 окт. 2014 г.). Примеры доменов CH3 включают домены CH3 с мутациями, которые модифицируют биологическую активность антитела, например, период полувыведения, например, описанные в публ. США № 20120100140 и патентах и публикациях США, а также публикациях РСТ, процитированных в ней.
Используемый в данном документе термин «изотип» относится к классу антител (например, IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgD и антитела IgE), которые кодируются генами константной области тяжелой цепи.
«Аллотип» относится к встречающимся в природе вариантам в пределах конкретной группы изотипа, причем эти варианты отличаются несколькими аминокислотами (см., например, Jefferis et al., (2009) mAbs 1: 1). Описанные в данном документе антитела могут быть любого аллотипа. Аллотипы IgG1, IgG2, IgG3 и IgG4 известны в данной области. См., например, Kabat EA et al., (1991) выше; Vidarsson G. et al., Front Immunol. 5: 520 (опубликовано онлайн 20 окт. 2014 г.); и Lefranc MP, mAbs 1: 4, 1-7 (2009).
Фразы «антитело, распознающее антиген» и «антитело, специфичное к антигену» используются в данном документе взаимозаменяемо с термином «антитело, которое специфически связывается с антигеном».
Термин «выделенное антитело» в контексте настоящего описания предназначен для обозначения антитела, которое по существу не включает других антител, обладающих другой антигенной специфичностью (например, выделенное антитело, которое специфически связывается с FAM19A5, по существу не включает антител, которые специфически связывают антигены, кроме FAM19A5). Однако выделенное антитело, которое специфически связывается с эпитопом FAM19A5, может обладать перекрестной реактивностью с другими белками FAM19A5 из разных видов.
«Аффинность связывания» обычно относится к силе общей суммы нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). Если не указано иное, в контексте настоящего описания термин «аффинность связывания» относится к внутренней аффинности связывания, которая отражает взаимодействие 1: 1 между членами пары связывания (например, антителом и антигеном). Аффинность молекулы X к ее партнеру Y обычно можно представить константой диссоциации (KD). Аффинность можно измерить и/или выразить рядом способов, известных в данной области, включая, без ограничения указанным, константу равновесной диссоциации (KD) и константу равновесной ассоциации (KA). KD рассчитывается как частное koff/kon и выражается как молярная концентрация (M), тогда как KA рассчитывается как частное kon/koff. kon относится к константе скорости ассоциации, например, антитела с антигеном, а koff относится к диссоциации, например, антитела с антигеном. Kon и koff могут быть определены методами, известными среднему специалисту в данной области, такими как иммуноанализы (например, твердофазный иммуноферментный анализ (ELISA)), BIAcore® или анализ кинетического исключения (KinExA).
Используемые в данном документе термины «специфически связывает», «специфически распознает», «специфическое связывание», «избирательное связывание» и «селективно связывает» являются аналогичными терминами в контексте антител и относятся к молекулам (например, антителам), которые связываться с антигеном (например, эпитопом или иммунным комплексом), поскольку такое связывание понятно специалисту в данной области. Например, молекула, которая специфически связывается с антигеном, может связываться с другими пептидами или полипептидами, как правило, с более низкой аффинностью, как определено, например, с помощью иммуноанализа, BIAcore®, прибора KinExA 3000 (Sapidyne Instruments, Boise, ID) или других известных в данной области анализов. В конкретном воплощении молекулы, которые специфически связываются с антигеном, связываются с антигеном с KA, которая составляет, по меньшей мере, 2 log, 25 log, 3 log, 4 log или больше, чем KA, когда молекулы связываются с другим антигеном.
Антитела обычно специфически связываются со своим когнатным антигеном с высокой аффинностью, что отражается константой диссоциации (KD) от 10-5 до 10-11 M или меньше. Обычно считается, что любая KD, превышающая примерно 10-4 М, указывает на неспецифическое связывание. В данном контексте антитело, которое «специфически связывается» с антигеном, относится к антителу, которое связывается с антигеном и практически идентичными антигенами с высокой аффинностью, что означает, что KD составляет 10-7 М или меньше, предпочтительно 10-8 М или меньше, даже более предпочтительно 10-9 М или меньше, и наиболее предпочтительно от 10-8 М до 10-10 М или меньше, при определении, например, с помощью иммуноанализа (например, ELISA) или технологии поверхностного плазмонного резонанса (SPR) в приборе BIACORE 2000 с использованием заранее определенного антигена, но не связывается с высокой аффинностью с неродственными антигенами.
Используемый в данном документе термин «антиген» относится к любому природному или синтетическому иммуногенному веществу, такому как белок, пептид или гаптен. Антигеном может быть FAM19A5 или его фрагмент.
В контексте настоящего описания термин «эпитоп» относится к локализованной области антигена, с которой антитело может специфически связываться. Эпитоп может представлять собой, например, смежные аминокислоты полипептида (линейный или непрерывный эпитоп), или эпитоп может, например, происходить вместе из двух или более несмежных областей полипептида или полипептидов (конформационных, нелинейных, прерывистый или несмежный эпитоп). Эпитопы, образованные из смежных аминокислот, обычно, но не всегда, сохраняются при воздействии денатурирующих растворителей, тогда как эпитопы, образованные в результате третичной укладки, обычно теряются при обработке денатурирующими растворителями. Эпитоп обычно включает, по меньшей мере, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или 20 аминокислот в уникальной пространственной конформации. Способы определения того, какие эпитопы связаны с данным антителом (например, картирование эпитопов), хорошо известны в данной области и включают, например, иммуноблоттинг и анализы иммунопреципитации, в которых перекрывающиеся или смежные пептиды из (например, из FAM19A5) проверяются на реактивность с данным антителом (например, анти-FAM19A5 антителом). Способы определения пространственной конформации эпитопов включают методы в данной области техники и методы, описанные в данном документе, например, рентгеновскую кристаллографию, 2-мерный ядерный магнитный резонанс и HDX-MS (см., например, Epitope Mapping Protocols in Methods in Molecular Biology, Vol. 66, G. E. Morris, Ed. (1996)).
В некоторых воплощениях эпитоп, с которым связывается антитело, может быть определен, например, с помощью ЯМР-спектроскопии, исследований кристаллографии дифракции рентгеновских лучей, анализов ELISA, обмена водорода/дейтерия в сочетании с масс-спектрометрией (например, жидкостной хроматографией с электрораспылительной масс-спектрометрией), массивом сканирующие анализы на основе олигопептидов и/или картирование мутагенеза (например, картирование с помощью сайт-направленного мутагенеза). Для рентгеновской кристаллографии кристаллизация может быть выполнена с использованием любого из известных в данной области способов (например, Giege R et al., (1994) Acta Crystallogr D Biol Crystallogr 50 (Pt 4): 339-350; McPherson A (1990)) Eur J Biochem 189: 1-23; Chayen NE (1997) Structure 5: 1269-1274; McPherson A (1976) J Biol Chem 251: 6300-6303). Кристаллы Антитело:антиген могут быть изучены с использованием хорошо известных методов дифракции рентгеновских лучей и могут быть уточнены с использованием компьютерного программного обеспечения, такого как X-PLOR (Yale University, 1992, распространяется Molecular Simulations, Inc.; см., например, Meth Enzymol (1985) volumes 114 & 115, eds Wyckoff HW et al.; U.S. 2004/0014194) и BUSTER (Bricogne G (1993) Acta Crystallogr D Biol Crystallogr 49 (Pt 1): 37-60; Bricogne G (1997) Meth Enzymol 276A: 361-423, ed Carter CW; Roversi P et al., (2000) Acta Crystallogr D Biol Crystallogr 56 (Pt 10): 1316-1323). Исследования картирования мутагенеза могут быть выполнены с использованием любого способа, известного специалисту в данной области. См., например, Champe M. et al., (1995) J Biol Chem 270: 1388-1394 и Cunningham BC & Wells JA (1989) Science 244: 1081-1085 для описания методов мутагенеза, включая методы мутагенеза аланинового сканирования.
Термин «картирование эпитопа» относится к процессу идентификации молекулярных детерминант распознавания антитело-антиген.
Термин «связывается с одним и тем же эпитопом» применительно к двум или более антителам означает, что антитела связываются с одним и тем же сегментом аминокислотных остатков, как определено данным способом. Методы определения того, связываются ли антитела с «одним и тем же эпитопом на FAM19A5» с описанными в данном документе антителами, включают, например, методы картирования эпитопа, такие как рентгеновский анализ кристаллов комплексов антиген: антитело, который обеспечивает атомное разрешение эпитопа и масс-спектрометрия водородно-дейтериевого обмена (HDX-MS). Другие способы контролируют связывание антитела с фрагментами антигена или мутированными вариантами антигена, где потеря связывания из-за модификации аминокислотного остатка в последовательности антигена часто считается признаком эпитопного компонента. Кроме того, также могут использоваться вычислительные комбинаторные способы для картирования эпитопов. Эти способы основаны на способности представляющего интерес антитела к аффинному выделению специфических коротких пептидов из библиотек комбинаторных пептидов фагового дисплея. Ожидается, что антитела, имеющие одинаковые последовательности VH и VL или одинаковые последовательности CDR1, 2 и 3, будут связываться с одним и тем же эпитопом.
Антитела, которые «конкурируют с другим антителом за связывание с мишенью», относятся к антителам, которые ингибируют (частично или полностью) связывание другого антитела с мишенью. Конкурируют ли два антитела друг с другом за связывание с мишенью, т.е. ингибирует ли и в какой степени одно антитело связывание другого антитела с мишенью, можно определить с помощью известных экспериментов по конкуренции. В некоторых воплощениях антитело конкурирует с другим антителом и ингибирует его связывание с мишенью, по меньшей мере, на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100%. Уровень ингибирования или конкуренции может быть различным в зависимости от того, какое антитело является «блокирующим антителом» (т.е. холодным антителом, которое сначала инкубируют с мишенью). Конкурентные анализы можно проводить, как описано, например, в Ed Harlow and David Lane, Cold Spring Harb Protoc; 2006; doi: 10.1101/pdb.prot4277 или в 11 главе "Using Antibodies" by Ed Harlow and David Lane, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, USA 1999. Конкурирующие антитела связываются с одним и тем же эпитопом, перекрывающимся эпитопом или с соседними эпитопами (что, например, подтверждается стерическими препятствиями).
Другие анализы конкурентного связывания включают: твердофазный прямой или непрямой радиоиммуноанализ (RIA), твердофазный прямой или непрямой иммуноферментный анализ (EIA), сэндвич-конкурентный анализ (см. Stahli et al., Methods in Enzymology 9: 242 (1983)); твердофазный прямой биотин-авидиновый ИФА (см. Kirkland et al., J. Immunol. 137: 3614 (1986)); твердофазный анализ с прямой меткой, твердофазный сэндвич-анализ с прямым мечением (см. Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Press (1988)); твердофазный РИА с прямой меткой с использованием метки I-125 (см. Morel et al., Mol. Immunol. 25 (1): 7 (1988)); твердофазный прямой биотин-авидиновый EIA (Cheung et al., Virology 176: 546 (1990)); и RIA с прямым мечением (Moldenhauer et al., Scand. J. Immunol. 32:77 (1990)).
«Биспецифическое» или «бифункциональное антитело» представляет собой искусственное гибридное антитело, имеющее две разные пары тяжелой/легкой цепи и два разных сайта связывания. Биспецифичные антитела могут быть получены с помощью различных способов, включающих слияние гибридом или соединение Fab'-фрагментов. См., например, Songsivilai & Lachmann, Clin. Exp., Immunol. 79:315-321 (1990); Kostelny et al., J. Immunol. 148, 1547-1553 (1992).
Используемый в данном документе термин «моноклональное антитело» относится к антителу, которое проявляет единственную специфичность связывания и аффинность к конкретному эпитопу, или к композиции антител, в которой все антитела проявляют единственную специфичность связывания и аффинность к конкретному эпитопу. Соответственно, термин «человеческое моноклональное антитело» относится к антителу или композиции антител, которые проявляют единственную специфичность связывания и которые имеют вариабельные и необязательные константные области, полученные из последовательностей иммуноглобулина зародышевой линии человека. В одном воплощении человеческие моноклональные антитела продуцируются гибридомой, которая включает В-клетку, полученную от трансгенного животного, не являющегося человеком, например, трансгенной мыши, имеющего геном, содержащий трансген тяжелой цепи человека и трансген легкой цепи, слитую с иммортализованной клеткой.
Термин «рекомбинантное человеческое антитело» в контексте настоящего описания включает все человеческие антитела, которые получены, экспрессированы, созданы или выделены рекомбинантными способами, такие как (а) антитела, выделенные из животного (например, мыши), которое является трансгенным или трансхромосомным для генов иммуноглобулинов человека или гибридомы, полученной из них, (b) антитела, выделенные из клетки-хозяина, трансформированной для экспрессии антитела, например, из трансфектомы, (c) антитела, выделенные из рекомбинантной, комбинаторной библиотеки человеческих антител, и (d) антитела получены, экспрессированы, созданы или выделены любыми другими способами, которые включают сплайсинг последовательностей гена иммуноглобулина человека с другими последовательностями ДНК. Такие рекомбинантные человеческие антитела содержат вариабельные и константные области, которые используют определенные последовательности иммуноглобулинов зародышевой линии человека, кодируемые генами зародышевой линии, но включают последующие перестройки и мутации, которые происходят, например, во время созревания антител. Как известно в данной области (см., например, Lonberg (2005) Nature Biotech. 23 (9): 1117-1125), вариабельная область содержит антигенсвязывающий домен, который кодируется различными генами, которые перестраиваются с образованием антитела, специфичного к чужеродному антигену. Помимо реаранжировки, вариабельная область может быть дополнительно модифицирована путем множественных замен одной аминокислоты (называемых соматической мутацией или гипермутацией) для увеличения аффинности антитела к чужеродному антигену. Константная область изменится при дальнейшем ответе на антиген (т.е. при переключении изотипа). Следовательно, перестроенные и соматически мутированные молекулы нуклеиновых кислот, которые кодируют полипептиды иммуноглобулинов легкой цепи и тяжелой цепи в ответ на антиген, не могут быть идентичными по последовательности с исходными молекулами нуклеиновой кислоты, а вместо этого будут по существу идентичными или похожими (т.е. иметь, по меньшей мере, 80% идентичности).
«Человеческое» антитело (HuMAb) относится к антителу, имеющему вариабельные области, в которых как каркасная область, так и области CDR происходят из последовательностей иммуноглобулина зародышевой линии человека. Кроме того, если антитело содержит константную область, эта константная область также происходит из последовательностей иммуноглобулина зародышевой линии человека. Описанные в данном документе антитела могут включать аминокислотные остатки, не кодируемые последовательностями иммуноглобулина зародышевой линии человека (например, мутации, введенные случайным или сайт-специфическим мутагенезом in vitro или соматической мутацией in vivo). Однако, при использовании в настоящем документе, подразумевается, что термин «человеческое антитело» не включает антитела, в которых последовательности CDR, полученные из зародышевой линии других видов млекопитающих, таких как мыши, были пересажены в человеческие каркасные последовательности. Термины «человеческие» антитела и «полностью человеческие» антитела используются как синонимы.
«Гуманизированное» антитело относится к антителу, в котором некоторые, большая часть или все аминокислоты вне доменов CDR нечеловеческого антитела заменены соответствующими аминокислотами, полученными из человеческих иммуноглобулинов. В одном воплощении гуманизированной формы антитела некоторые, большая часть или все аминокислоты вне доменов CDR были заменены аминокислотами из человеческих иммуноглобулинов, тогда как некоторые, большая часть или все аминокислоты в одной или нескольких областях CDR не изменены. Небольшие добавления, делеции, вставки, замены или модификации аминокислот допустимы до тех пор, пока они не отменяют способность антитела связываться с конкретным антигеном. «Гуманизированное» антитело сохраняет антигенную специфичность, аналогичную исходному антителу.
Используемый в данном документе термин «деиммунизированный» или «деиммунизация» относится к процессу, в котором антитело или его антигенсвязывающая часть модифицируют для снижения его иммуногенности, например, у человека. Например, последовательности вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL) исходного антитела могут быть проанализированы, и из каждой V-области может быть создана «карта» Т-клеточных эпитопов человека, показывающая расположение эпитопов в отношение к определяющим комплементарность участкам (CDR) и другим ключевым остаткам в последовательности. Индивидуальные Т-клеточные эпитопы из карты Т-клеточных эпитопов анализируют, чтобы идентифицировать альтернативные аминокислотные замены с низким риском изменения активности конечного антитела. Разработан ряд альтернативных последовательностей VH и VL, включающих комбинации аминокислотных замен, и эти последовательности впоследствии включаются в ряд FAM19A5-специфических антител или их антигенсвязывающей части для применения в раскрытых в данном документе способах диагностики и лечения, которые затем тестируют на функциональность. Полные гены тяжелой и легкой цепей, содержащие модифицированные V- и человеческие С-области, затем клонируют в экспрессирующие векторы, а последующие плазмиды вводят в клеточные линии для получения целого антитела. Затем антитела сравнивают в соответствующих биохимических и биологических анализах и определяют оптимальный вариант. Антитело можно деиммунизировать с использованием способов, описанных в данном документе, или любыми другими способами, известными в данной области. См., например, WO 98/52976 или WO 00/34317.
«Химерное антитело» относится к антителу, в котором вариабельные области происходят из одного вида, а константные области происходят из другого вида, например из антитела, в котором вариабельные области происходят из антитела мыши, а константные области происходят из антитела человека.
Термин «перекрестная реакция», используемый в данном документе, относится к способности описанного в данном документе антитела связываться с FAM19A5 другого вида. Например, описанное в данном документе антитело, которое связывает человеческий FAM19A5, также может связывать другой вид FAM19A5 (например, мышиный FAM19A5). В данном контексте перекрестная реактивность может быть измерена путем определения специфической реактивности с очищенным антигеном в анализах связывания (например, SPR, ELISA) или связывания или иного функционального взаимодействия с клетками, физиологически экспрессирующими FAM19A5. Способы определения перекрестной реактивности включают стандартные анализы связывания, как описано в данном документе, например, анализ поверхностного плазмонного резонанса (SPR) Biacore® с использованием прибора Biacore® 2000 SPR (Biacore AB, Упсала, Швеция) или методы проточной цитометрии.
Термин «встречающийся в природе», используемый в данном документе по отношению к объекту, относится к тому факту, что объект может быть найден в природе. Например, полипептидная или полинуклеотидная последовательность, присутствующая в организме (включая вирусы), которая может быть выделена из природного источника и которая не была намеренно модифицирована человеком в лаборатории, является встречающейся в природе.
«Полипептид» относится к цепи, содержащей, по меньшей мере, два последовательно связанных аминокислотных остатка без верхнего предела длины цепи. Один или несколько аминокислотных остатков в белке могут содержать модификацию, такую как, помимо прочего, гликозилирование, фосфорилирование или образование дисульфидной связи. «Белок» может включать один или несколько полипептидов.
Используемый в данном документе термин «молекула нуклеиновой кислоты» включает молекулы ДНК и молекулы РНК. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной и может быть кДНК.
Используемый в данном документе термин «вектор» предназначен для обозначения молекулы нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой она связана. Одним из типов вектора является «плазмида», которая относится к кольцевой двухцепочечной петле ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Некоторые векторы проявляют способность к автономной репликации в клетке-хозяине в которую они введены (например, бактериальные вектора имеющие бактериальную точку начала репликации и эписомальные вектора млекопитающих). Другие векторы (например, неэписомальные векторы млекопитающих) интегрируются в геном клетки-хозяина при введении в клетку-хозяин, и таким образом, реплицируются вместе с хозяйским геномом. Более того, некоторые векторы способны управлять экспрессией генов, с которыми они функционально связаны. Такие векторы называются в данном документе «рекомбинантными экспрессирующими векторами» (или просто «экспрессирующими векторами»). В общем, экспрессирующие векторы, применимые в методах рекомбинантных ДНК, часто имеют форму плазмид. В настоящем описании термины «плазмида» и «вектор» могут использоваться взаимозаменяемо, поскольку плазмида является наиболее часто используемой формой вектора. Однако также включены другие формы экспрессирующих векторов, такие как вирусные векторы (например, дефектные по репликации ретровирусы, аденовирусы и аденоассоциированные вирусы), которые выполняют эквивалентные функции.
Термин «рекомбинантная клетка-хозяин» (или просто «клетка-хозяин») в контексте настоящего описания предназначен для обозначения клетки, которая содержит нуклеиновую кислоту, которая в природе не присутствует в клетке, и может быть клеткой, в которую введен рекомбинантный экспрессирующий вектор. Следует понимать, что такие термины предназначены для обозначения не только определенного объекта, но также и потомства такой клетки. Поскольку определенные модификации могут происходить в последующих поколениях из-за мутаций или влияний окружающей среды, такое потомство не может фактически быть идентичным родительской клетке, но по-прежнему входит в объем термина «клетка-хозяин», который используется в данном документе.
Используемый в данном документе термин «связанный» относится к ассоциации двух или более молекул. Связь может быть ковалентной или нековалентной. Связь также может быть генетической (т.е. рекомбинантно слитой). Такие связи могут быть достигнуты с использованием широкого ряда признанных в данной области методов, таких как химическая конъюгация и получение рекомбинантных белков.
Используемый в данном документе термин «введение» относится к физическому введению терапевтического агента или композиции, содержащей терапевтический агент, объекту с использованием любого из различных способов и систем доставки, известных специалистам в данной области. Предпочтительные пути введения описанных в данном документе антител включают внутривенный, внутрибрюшинный, внутримышечный, подкожный, спинномозговой или другие парентеральные пути введения, например, путем инъекции или инфузии. Фраза «парентеральное введение» в контексте настоящего описания означает способы введения, отличные от энтерального и местного, обычно путем инъекции, и включает, без ограничения указанным, внутривенные, внутрибрюшинные, внутримышечные, внутриартериальные, интратекальные, внутрилимфатическые, внутриочаговые, внутрикапсулярные, интраорбитальные, внутрисердечные, интрадермальные, транстрахеальные, подкожные, субкутикулярные, внутрисуставные, субкапсулярные, субарахноидальные, внутриспинальные, эпидуральные и внутригрудинные инъекции и инфузии, а также электропорацию in vivo. Альтернативно, описанное в данном документе антитело можно вводить непарентеральным путем, например местным, эпидермальным или слизистым путем, например интраназально, перорально, вагинально, ректально, сублингвально или местно. Введение также может выполняться, например, один раз, множество раз и/или в течение одного или нескольких продолжительных периодов.
Термины «лечить», «лечиться» и «лечение», при использовании в данном документе, относятся к любому типу вмешательства или выполняемого процесса, или введению активного агента объекту с целью обращения, облегчения, улучшения, подавления или замедления или предотвращения прогрессирования, развития, тяжести или рецидива симптома, осложнения, состояния или биохимических показателей, связанных с заболеванием. Лечение может относиться к объекту, страдающему заболеванием, или объекту, у которого нет заболевания (например, для профилактики).
Используемый в данном документе термин «объект» включает любого человека или животное, не являющееся человеком. Термин «животное, не являющееся человеком» включает всех позвоночных, например, млекопитающих и не млекопитающих, таких как не являющиеся человеком приматы, овцы, собаки, коровы, куры, земноводные, рептилии и т.д.
Используемый в данном документе термин «начало глиоза» или «начало реактивного глиоза» включает начало или инициацию глиоза. Глиоз - это неспецифическое реактивное изменение глиальных клеток в центральной нервной системе (ЦНС, например, головной и/или спинной мозг) в ответ на травму или повреждение, например, травму, цереброспинальное повреждение, опухоль головного мозга, инфекцию, ишемию, инсульт, аутоиммунные ответы и/или нейродегенеративные заболевания и включают пролиферацию или гипертрофию нескольких различных типов глиальных клеток, включая астроциты, микроглию и олигодендроциты. Начало глиоза может привести к образованию рубца, который препятствует регенерации аксонов в травмированной или поврежденной части ЦНС. Вредные эффекты начала глиоза включают необратимое или постоянное повреждение нейронов и/или предотвращение восстановления окружающих нейронов. Соответственно, термины «отсрочить начало глиоза» и «отсрочить начало реактивного глиоза» включают ингибирование, замедление, подавление или предотвращение начала или возникновения глиоза и связанных с ним пагубных эффектов на ЦНС.
Используемый в данном документе термин «чрезмерная пролиферация реактивных астроцитов» включает аномальное увеличение количества астроцитов из-за разрушения близлежащих нейронов, например, в результате повреждения ЦНС, травмы, травмы, цереброспинального повреждения, опухоли головного мозга, инфекции, ишемии, инсульта, аутоиммунных реакций и/или нейродегенеративного заболевания. Чрезмерное разрастание реактивных астроцитов может привести к пагубным эффектам в ЦНС, включая образование рубцов, которые препятствуют регенерации аксонов в части ЦНС, которая была травмирована или повреждена, обострение воспаления, выработку и высвобождение нейротоксичных уровней активных форм кислорода, высвобождение потенциального эксайтотоксичного глутамата, потенциального вклада в генез припадков, нарушения функции гематоэнцефалического барьера, цитотоксического отека во время травмы и инсульта, возможности хронической цитокиновой активации астроцитов, способствующей хронической боли, и вторичной дегенерации после повреждения ЦНС. Sofroniew, Michael V. (2009) Trends in Neurosciences, 32(12):638-47; McGraw, J. et al. (2001) Journal of Neuroscience Research 63(2):109-15; и Sofroniew, M. V. (2005) The Neuroscientist 11(5): 400-7. Соответственно, термины «подавление чрезмерной пролиферации реактивных астроцитов» включают ингибирование, замедление, подавление, сдерживание или предотвращение чрезмерной или аномальной пролиферации реактивных астроцитов и связанных с этим пагубных эффектов на ЦНС.
Используемый в данном документе термин «протеогликаны хондроитинсульфата» включает протеогликаны, состоящие из белкового ядра и хондроитинсульфата. Хондроитинсульфатные протеогликаны, также известные как CSPG, представляют собой молекулы внеклеточного матрикса, широко экспрессирующиеся в развивающейся и взрослой ЦНС. CSGP играют ключевую роль в развитии нервной системы и формировании глиальных рубцов, и они ингибируют регенерацию аксонов после повреждения в ЦНС. Известные CSPG включают аггрекан (CSPG1), версикан (CSPG2), нейрокан (CSPG3), CSPG4 (или нейронглиальный антиген 2 (NG2)), CSPG5, SMC3 (CSPG6, структурное поддержание хромосом 3), бревикан (CSPG7) и CD44 (CSPG8, кластер дифференцировки 44), фосфаканневрокан (CSPG3). Rhodes, K. E. and Fawcett, J. W. (2004) Journal of Anatom. 204(1):33-48. Таким образом, термин «снижение экспрессии протеогликанов хондроитинсульфата» включает снижение, ингибирование, снижение уровня одного или нескольких CSGP или снижение активности или превращение в неактивный один или несколько CSGP. В некоторых воплощениях термин включает уменьшение, ингибирование, снижение уровня нейрокана, NG2 или обоих, или снижение активности, или превращение в неактивный нейрокан, NG2 или обоих.
Используемый в данном документе термин «нейрон» включает электрически возбудимые клетки, которые обрабатывают и передают информацию посредством электрических и химических сигналов. Нейроны являются основными компонентами головного и спинного мозга ЦНС и ганглиев периферической нервной системы (PNS) и могут соединяться друг с другом, образуя нейронные сети. Типичный нейрон состоит из тела клетки (сомы), дендритов и аксона. Сома (тело клетки) нейрона содержит ядро. Дендриты нейрона - это клеточные продолжения с множеством ветвей, где происходит большая часть входных данных в нейрон. Аксон представляет собой более тонкий, похожий на кабель выступ, идущий от сомы и несущий нервные сигналы от сомы и определенные типы информации обратно к соме. Термин «способствовать возобновлению роста нейронов» включает стимуляцию, промотирование, увеличение или активацию роста нейронов, предпочтительно после травмы или повреждения.
Используемый в данном документе термин «c-fos» включает протоонкоген c-fos, который быстро индуцируется стимуляцией нейромедиатора. c-fos существует у многих видов, включая мышь и человека. Ген и белок c-fos известны и охарактеризованы. См. Curran, T, The c-fos proto-oncogene, pp 307-327 (The Oncogene Handbook, Reddy EP et al., (eds.) Elsevier)(1988). Экспрессию c-fos можно определить способами, известными в данной области, например нозерн-блоттингом, количественной ПЦР или иммуногистохимией. Термин «увеличивает экспрессию c-fos» включает повышение уровня мРНК c-fos, белка c-fos или активности белка c-fos.
Используемый в данном документе термин «pERK» включает фосфорилированную киназу, регулируемую внеклеточными сигналами. Киназа, регулируемая внеклеточными сигналами, или ERK, включает ERK1 и ERK2, является представителем семейства митоген-активируемых протеинкиназ (MAPK). ERK активируется посредством фосфорилирования своей вышестоящей киназой с образованием pERK, которая затем активирует нижележащие мишени. ERK участвует в нейронной и синаптической пластичности, лежащей в основе обучения, а также в повышенной чувствительности к боли и памяти. Ji R.R. et al., Nat Neurosci (1999) 2:1114-1119. Ген, белок, фосфорилирование и активация ERK известны и охарактеризованы, а экспрессия ERK и pERK может быть определена способами, известными в данной области (например, нозерн-блоттинг, количественная ПЦР или иммуногистохимия). См. Gao Y. J. and Ji R. R., Open Pain J. (2009) 2:11-17. Термин «увеличивает экспрессию pERK» включает повышение уровня мРНК ERK, белка ERK или активности pERK.
Используемый в данном документе термин «GAP43», также известный как «белок 43, ассоциированный с ростом», представляет собой белок, специфичный для нервной ткани, который способствует образованию, регенерации и пластичности нейритов. Benowitz L. I. and Routtenberg A. (1997) Trends in Neurosciences 20 (2): 84-91; Aarts L. H. et al., (1998) Advances in Experimental Medicine and Biology 446: 85-106. GAP43 человека кодируется геном GAP43. Последовательность полипептида GAP43 человека (UniProt: KB - P17677) и последовательность кДНК, кодирующая полипептид, известны в данной области. Kosik K. S. et al., (1988) Neuron 1(2):127-32; Ng S. C. et al., (1988) Neuron 1(2):133-9. Экспрессию GAP43 можно определить методами, известными в данной области (например, нозерн-блоттинг, количественная ПЦР или иммуногистохимия). Термин «увеличение GAP43 в нейронах» включает усиление или увеличение уровня мРНК GAP43, белка GAP43 или увеличение активности белка GAP43.
Используемый в данном документе термин «терапевтически эффективное количество» относится к количеству лекарственного средства, отдельно или в комбинации с другим терапевтическим агентом, эффективному для «лечения» заболевания или нарушения у объекта или снижения риска, потенциала, возможности или возникновения заболевания или расстройства (например, поражение центральной нервной системы). «Терапевтически эффективное количество» включает количество лекарственного средства или терапевтического агента, которое обеспечивает некоторое улучшение или пользу для объекта, имеющего или подверженного риску заболевания или расстройства (например, повреждение центральной нервной системы, такое как черепно-мозговая травма или другое заболевание, описанное в данном документе). Таким образом, «терапевтически эффективное» количество - это количество, которое снижает риск, потенциал, возможность или возникновение заболевания или расстройства или обеспечивает некоторое облегчение, смягчение и/или уменьшение, по меньшей мере, одного показателя (например, начало реактивного глиоза) и/или уменьшение, по меньшей мере, одного клинического симптома заболевания или расстройства.
II. Анти-FAM19A5 антитела
В настоящем документе раскрыты антитела, например моноклональные антитела, которые характеризуются конкретными функциональными особенностями или свойствами. Например, антитела, которые специфически связывают человеческий FAM19A5, мутированы (например, путем замены или удаления) путем измлечения и/или модификации областей или остатков, которые являются высокоиммуногенными для человека (т.е. деиммунизированы). Соответственно, антитела, раскрытые в данном документе, т.е. анти-FAM19A5 антитело, обладают пониженной иммуногенностью при введении объекту-человеку по сравнению с референсным антителом (например, соответствующим антителом, кторое не было деиммунизировано, например, антителом 3-2 или 2 -13).
Кроме того, описанные в данном документе антитела обладают одним или несколькими из следующих функциональных свойств:
(а) связываются с растворимым FAM19A5 человека с KD 10 нМ или менее;
(b) связываются с мембраносвязанным FAM19A5 человека с KD 10 нМ или менее;
(c) уменьшают, обращают вспять, задерживают и/или предотвращают начало реактивного глиоза;
(d) подавляют чрезмерную пролиферацию реактивных астроцитов
(e) снижают экспрессию протеогликанов хондроитинсульфата, включая нейрокан и нейрон-глиальный антиген 2 (NG2);
(е) увеличивают экспрессию c-fos и pERK в ядрах нейронов;
(g) обеспечивают выживания нейронов.
(h) увеличивают экспрессию GAP43 в нейронах; и
(i) обеспечивают отрастание аксона.
В некоторых воплощениях анти-FAM19A5 антитело было деиммунизировано таким образом, что антитело является менее иммуногенным при введении объекту-человеку по сравнению с референсным антителом (например, соответствующим антителом, которое не было деиммунизировано, например, антителом 3-2 или 2-13). В некоторых воплощениях иммуногенность антитела снижена, по меньшей мере, на около 5%, по меньшей мере, на около 10%, по меньшей мере, на около 15%, по меньшей мере, на около 20%, по меньшей мере, на около 25%, по меньшей мере, на около 30%, по меньшей мере, около 35%, по меньшей мере, около 40%, по меньшей мере, около 45%, по меньшей мере, около 50%, по меньшей мере, около 60%, по меньшей мере, около 70%, по меньшей мере, около 80%, по меньшей мере, около 90% или, по меньшей мере, около 100% по сравнению с референсным антителом (например, соответствующим антителом, которое не было деиммунизировано, например, антителом 3-2 или 2-13). В некоторых воплощениях процесс деиммунизации не изменяет аффинность связывания антитела.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, претерпело созревание аффинности, так что антитело связывается с белком FAM19A5 с большей аффинностью по сравнению с референсным антителом (например, соответствующим антителом, которое не претерпело созревания аффинности, например, антителом 2 -13). В некоторых воплощениях аффинность связывания антитела увеличена, по меньшей мере, на около 5%, по меньшей мере, около 10%, по меньшей мере, около 15%, по меньшей мере, около 20%, по меньшей мере, около 25%, по меньшей мере, на около 30%, по меньшей мере, около 35%, по меньшей мере, около 40%, по меньшей мере, около 45%, по меньшей мере, около 50%, по меньшей мере, около 60%, по меньшей мере, около 70%, по меньшей мере, около 80%, по меньшей мере, около 90%, или по меньшей мере, около 100% или более по сравнению с референсным антителом (например, соответствующим антителом, которое не претерпело созревания аффинности, например, антителом 2-13). В некоторых воплощениях процесс созревания аффинности не изменяет иммуногенность антитела.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, было деиммунизировано и претерпело созревание аффинности, так что по сравнению с референсным антителом (например, соответствующим антителом, которое не было деиммунизировано или не претерпело созревания аффинности, например, антителом 3-2 или 2-13), антитело не только менее иммуногенно при введении человеку, но также связывается с белком FAM19A5 с большей аффинностью. В некоторых воплощениях иммуногенность антитела снижена, по меньшей мере, на около 5%, по меньшей мере, около 10%, по меньшей мере, около 15%, по меньшей мере, около 20%, по меньшей мере, около 25%, по меньшей мере, около 30%, по меньшей мере, около 35%, по меньшей мере, около 40%, по меньшей мере, около 45%, по меньшей мере, около 50%, по меньшей мере, около 60%, по меньшей мере, около 70%, по меньшей мере, около 80%, по меньшей мере, около 90% или, по меньшей мере, около 100% по сравнению с референсным антителом. В некоторых воплощениях аффинность связывания антитела увеличена, по меньшей мере, на около 5%, по меньшей мере, около 10%, по меньшей мере, около 15%, по меньшей мере, около 20%, по меньшей мере, около 25%, по меньшей мере, около 30%, по меньшей мере, около 35%, по меньшей мере, около 40%, по меньшей мере, около 45%, по меньшей мере, около 50%, по меньшей мере, около 60%, по меньшей мере, около 70%, по меньшей мере, около 80%, по меньшей мере, около 90%, или по меньшей мере, на около 100% или более по сравнению с референсным антителом.
В некоторых воплощениях анти-FAM19A5 антитело специфически связывается с растворимым FAM19A5 человека или мембраносвязанным человеческим с высокой аффинностью, например, с KD 10-7 М или меньше, 10-8 М (10 нМ) или меньше, 10-9 M (1 нМ) или меньше, 10-10 M (01 нМ) или меньше, 10-11 M или меньше, или 10-12 M или меньше, например, 10-12 M - 10-7 M, 10-11 M - 10-7 M, 10-10 M - 10-7 M, или 10-9 M - 10-7 M, e.g., 10-12 M, 5 X 10-12 M, 10-11 M, 5 X 10-11 M, 10-10 M, 5 X 10-10 M, 10-9 M, 5 X 10-9 M, 10-8 M, 5 X 10-8 M, 10-7 M, или 5 X 10-7 M. Стандартные анализы для оценки способности антитела к связыванию с FAM19A5 человека различных видов известны в данной области, включая, например, ELISA, вестерн-блоттинг и RIA. Подходящие анализы подробно описаны в Примерах. Кинетику связывания (например, аффинность вязывания) антител также можно оценить стандартными анализами, известными в данной области, такими как ELISA, анализ Biacore® или KinExA. Анализы для оценки эффектов антител на функциональные свойства FAM19A5 (например, связывание лиганда) более подробно описаны ниже и в примерах.
В некоторых воплощениях анти-FAM19A5 антитело специфически связывается с растворимым FAM19A5 человека с KD, например, как определено с помощью ELISA, 10-7 M или меньше, 10-8 M (10 нМ) или меньше, 10-9 M (1 нМ) или меньше, 10-10 М или меньше, от 10-12 М до 10-7 М, от 10-11 М до 10-7 М, от 10-10 М до 10-7 М, от 10-9 М до 10-7 M или от 10-8 M до 10-7 M. В некоторых воплощениях анти-FAM19A5 антитело специфически связывается с растворимым FAM19A5 с KD 10 нМ или менее, например, от 01 до 10 нМ, от 01 до 5 нМ. от 01 до 1 нМ, от 05 до 10 нМ, от 05 до 5 нМ, от 05 до 1 нМ, от 1 до 10 нМ, от 1 до 5 нМ или от 5 до 10 нМ. В некоторых воплощениях анти-FAM19A5 антитело специфически связывается с растворимым FAM19A5 человека с KD около 1 пМ, около 2 пМ, около 3 пМ, около 4 пМ, около 5 пМ, около 6 пМ, около 7 пМ, около 8 пМ, около 9 пМ, около 10 пМ, около 20 пМ, около 30 пМ, около 40 пМ, около 50 пМ, около 60 пМ, около 70 пМ, около 80 пМ, около 90 пМ, около 100 пМ, около 200 пМ, около 300 пМ, около 400 пМ, около 500 пМ, около 600 пМ, около 700 пМ, около 800 пМ, или около 900 пМ, или около 1 нМ, около 2 нМ, около 3 нМ, около 4 нМ, около 5 нМ, около 6 нМ, около 7 нМ, около 8 нМ, или около 9 нМ, или около 10 нМ, около 20 нМ, около 30 нМ, около 40 нМ, около 50 нМ, около 60 нМ, около 70 нМ, около 80 нМ или около 90 нМ, как определено с помощью ELISA.
В некоторых воплощениях анти-FAM19A5 антитело специфически связывается с мембраносвязанным человеком с KD, например, как определено с помощью ELISA, 10-7 M или меньше, 10-8 M (10 нМ) или меньше, 10-9 M (1 нМ) или меньше, 10-10 M или меньше, от 10-12 M до 10-7 M, от 10-11 M до 10-7 M, от 10-10 M до 10-7 M, от 10-9 M до 10 -7 M или от 10-8 M до 10-7 M. В некоторых воплощениях анти-FAM19A5 антитело специфически связывается с мембраносвязанным FAM19A5 человека с KD 10 нМ или менее, как определено с помощью ELISA, например, между 01 и 10 нМ, от 01 до 5 нМ, от 01 до 1 нМ, от 05 до 10 нМ, от 05 до 5 нМ, от 05 до 1 нМ, от 1 до 10 нМ, от 1 до 5 нМ или от 5 до 10 нМ. В некоторых воплощениях анти-FAM19A5 антитело специфически связывается с мембраносвязанным FAM19A5 человека с KD примерно 1 пМ, примерно 2 пМ, примерно 3 пМ, примерно 4 пМ, примерно 5 пМ, примерно 6 пМ, примерно 7 пМ, примерно 8 пМ, около 9 пМ, около 10 пМ, около 20 пМ, около 30 пМ, около 40 пМ, около 50 пМ, около 60 пМ, около 70 пМ, около 80 пМ, около 90 пМ, около 100 пМ, около 200 пМ, примерно 300 пМ, примерно 400 пМ, примерно 500 пМ, примерно 600 пМ, примерно 700 пМ, примерно 800 пМ, или примерно 900 пМ, или примерно 1 нМ, примерно 2 нМ, примерно 3 нМ, примерно 4 нМ, примерно 5 нМ, около 6 нМ, около 7 нМ, около 8 нМ, или около 9 нМ, или около 10 нМ, около 20 нМ, около 30 нМ, около 40 нМ, около 50 нМ, около 60 нМ, около 70 нМ, около 80 нМ или около 90 нМ, как определено с помощью ELISA.
Анти-FAM19A5 антитело по настоящему раскрытию может задерживать или ингибировать начало глиоза, например, задерживать, замедлять или подавлять начало или начало неспецифического реактивного изменения глиальных клеток в центральной нервной системе (ЦНС, например, мозга и/или спинного мозга) в ответ на травму или повреждение, например, травму, цереброспинальное повреждение, опухоль головного мозга, инфекцию, ишемию, инсульт, аутоиммунные реакции и/или нейродегенеративное заболевание.
Анти-AM19A5 антитело по настоящему изобретению может задерживать, ингибировать, замедлять, супрессировать, сдерживать или предотвращать чрезмерную или аномальную пролиферацию реактивных астроцитов и связанные с ней пагубные эффекты на ЦНС. Например, анти-FAM19A5 антитело по настоящему изобретению может ингибировать или предотвращать аномальное увеличение количества астроцитов из-за разрушения нейронов, например, в результате повреждения ЦНС, травмы, травмы, цереброспинального повреждения, опухоли головного мозга, инфекции, ишемии, инсульта, аутоиммунных реакций и/или нейродегенеративного заболевания, ингибирует или предотвращают образование рубцов в ЦНС, ингибирует или снижает высвобождение нейротоксических уровней активных форм кислорода или высвобождение потенциально эксайтотоксического глутамата, уменьшает или ингибирует судороги, боль и/или вторичную дегенерацию после травмы ЦНС. Анти-AM19A5 антитело по настоящему изобретению может содействовать, стимулировать, увеличивать или активировать повторный рост нейронов и/или аксонов, предпочтительно после поражения или повреждения ЦНС.
Анти-AM19A5 антитело по настоящему изобретению может ингибировать экспрессию протеогликанов хондроитинсульфата, включая протеогликаны, состоящие из ядра белка и хондроитинсульфата (CSGP), таких как аггрекан (CSPG1), версикан (CSPG2), нейрокан (CSPG3), CSPG4 (или нейрон-глиальный антиген 2 (NG2)), CSPG5, SMC3 (CSPG6, структурное поддержание хромосом 3), бревикан (CSPG7), CD44 (CSPG8, кластер дифференцировки 44) и фосфаканневрокан (CSPG3). В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию ингибирует, снижает или уменьшает уровень нейрокана и/или NG2, или активность нейрокана и/или NG2.
Анти-AM19A5 антитело по настоящему изобретению может увеличивать экспрессию c-fos и pERK в ядре нейронов, например, увеличивать мРНК, белок и/или активность белка c-fos и pERK. Анти-AM19A5 антитело по настоящему изобретению также может увеличивать или усиливать уровень экспрессии мРНК GAP43, белка GAP43 или увеличивать или усиливать активность белка GAP43.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где CDR1, CDR2 и CDR3 тяжелой цепи содержат набор аминокислотных последовательностей далее в SEQ ID NO: 5, 6 и 7, соответственно, каждая из которых необязательно содержит одну, две, три, четыре или пять мутаций, при этом CDR1, CDR2 и CDR3 легкой цепи содержат аминокислотные последовательности, указанные в SEQ ID NO: 8, 9 и 10, соответственно, где, по меньшей мере, одну из CDR1, CDR2 и CDR3 легкой цепи, содержащий одну, две, три, четыре или пять мутаций, и где антитело имеет пониженную иммуногенность у человека по сравнению с референсным антителом, содержащим VH, указанную в SEQ ID NO: 11, и VL, указанную в SEQ ID NO: 12.
В некоторых воплощениях мутации, включенные в антитело, представляют собой замену, делецию и/или вставку. В одном воплощении мутация представляет собой замену, например консервативную замену. «Консервативная замена» (также называемая консервативной заменой) в контексте настоящего описания означает аминокислотную замену, которая изменяет данную аминокислоту на другую аминокислоту с аналогичными биохимическими свойствами (например, заряд, гидрофобность и размер). Хотя существует множество способов классификации аминокислот, их часто разделяют на шесть основных групп на основе их структуры и общих химических характеристик их R-групп.
Таблица 2. Классы аминокислот
И, наоборот, радикальное замещение или радикальная замена - это замена аминокислоты, при которой происходит замена исходной аминокислоты конечной аминокислотой с различными физико-химическими свойствами. В некоторых воплощениях аминокислотная мутация в антителе FAM19A5 представляет собой радикальную замену. В других воплощениях аминокислотная мутация в антителе FAM19A5 представляет собой комбинацию консервативной замены и радикальной замены.
В некоторых воплощениях CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7. В некоторых воплощениях CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7, с одной, двумя или тремя мутациями.
В некоторых воплощениях CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 5, необязательно с одной или двумя мутациями. В некоторых воплощениях мутации включают замену треонина в аминокислоте 3 из SEQ ID NO: 5 на кислую аминокислоту. В дополнительных воплощениях мутации включают замену серина в аминокислоте 5 из SEQ ID NO: 5 на кислую аминокислоту. В некоторых воплощениях кислая аминокислота включает аспарагиновую кислоту или глутаминовую кислоту.
В некоторых воплощениях CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 6, необязательно с одной, двумя, тремя, четырьмя или пятью мутациями. В некоторых воплощениях мутации включают замену аргинина в аминокислоте 16 из SEQ ID NO: 6 на основную аминокислоту. В некоторых воплощениях основная аминокислота включает лизин. В некоторых воплощениях мутации включают одну или несколько из следующих:
(а) замену глицина в аминокислоте 6 SEQ ID NO: 6 на кислую аминокислоту;
(b) замену серина в аминокислоте 7 в SEQ ID NO: 6 на кислую аминокислоту;
(c) замену серина в аминокислоте 8 в SEQ ID NO: 6 на кислую аминокислоту;
(d) замену треонина в аминокислоте 9 SEQ ID NO: 6 на кислую аминокислоту; и
(e) замену аргинина в аминокислоте 16 SEQ ID NO: 6 на основную аминокислоту.
В некоторых воплощениях кислая аминокислота включает аспарагиновую кислоту или глутаминовую кислоту. В некоторых воплощениях основная аминокислота включает лизин.
В некоторых воплощениях CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 10, необязательно с одной, двумя, тремя, четырьмя или пятью мутациями. В некоторых воплощениях мутации включают одну или несколько из следующих:
(а) замену серина в аминокислоте 6 из SEQ ID NO: 10 на кислую аминокислоту или алифатическую аминокислоту;
(b) замену аспарагина в аминокислоте 7 из SEQ ID NO: 10 на кислую аминокислоту или на гидроксил или сера/селенсодержащую аминокислоту;
(c) замену глицина в аминокислоте 8 из SEQ ID NO: 10 на кислую аминокислоту или на гидроксил или сера/селенсодержащую аминокислоту;
(d) замену глицина в аминокислоте 9 из SEQ ID NO: 10 на кислую аминокислоту или на гидроксил или сера/селенсодержащую аминокислоту; и
(e) замену изолейцина в аминокислоте 10 из SEQ ID NO: 10 на основную аминокислоту.
В некоторых воплощениях кислая аминокислота включает аспарагиновую кислоту или глутаминовую кислоту. В некоторых воплощениях гидроксильная или сера/селенсодержащая аминокислота включает серин. В дополнительных воплощениях основная аминокислота включает гистидин.
В некоторых воплощениях CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8, с одной, двумя, тремя или четырьмя мутациями. В некоторых воплощениях мутации включают одну или несколько из следующих:
(а) замену тирозина в аминокислоте 6 SEQ ID NO: 8 на кислую аминокислоту;
(b) замену аргинина в аминокислоте 7 в SEQ ID NO: 8 на кислую аминокислоту;
(c) замену глицина в аминокислоте 8 SEQ ID NO: 8 на кислую аминокислоту; и
(d) замену серина в аминокислоте 9 в SEQ ID NO: 8 на кислую аминокислоту.
В некоторых воплощениях кислая аминокислота включает глутаминовую кислоту или глутамин.
В некоторых воплощениях CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 9, с одной, двумя, тремя или четырьмя мутациями. В некоторых воплощениях мутации включают одну или несколько из следующих:
(а) замену глутаминовой кислоты в аминокислоте 1 из SEQ ID NO: 9 на кислую аминокислоту;
(b) замену серина в аминокислоте 2 SEQ ID NO: 9 на кислую аминокислоту;
(c) замену аспарагина в аминокислоте 3 SEQ ID NO: 9 на кислую аминокислоту, основную аминокислоту или алифатическую аминокислоту; и
(d) замену лизина в аминокислоте 4 из SEQ ID NO: 9 на кислую аминокислоту или алифатическую аминокислоту.
В некоторых воплощениях кислая аминокислота включает глутамин, аспарагин, аспарагиновую кислоту или глутаминовую кислоту. В некоторых воплощениях основная аминокислота включает гистидин. В дополнительных воплощениях алифатическая аминокислота включает лейцин.
В некоторых воплощениях мутации включают замену серина в аминокислоте 2 из SEQ ID NO: 9 на кислую аминокислоту. В некоторых воплощениях кислая аминокислота включает аспартат, глутамат, аспарагин, глутамин или их комбинации. В дополнительных воплощениях кислая аминокислота представляет собой аспарагин.
В некоторых воплощениях изобретения анти-FAM19A5 антитело, раскрытое в настоящем документе, содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 5; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 13; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 20; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 10.
В некоторых воплощениях анти-FAM19A5 антитело содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 21; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 22; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 23.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи содержит указанную аминокислотную последовательность в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 21; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 24; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 23.
В некоторых воплощениях анти-FAM19A5 антитело содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 25; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 23.
В некоторых воплощениях изобретения анти-FAM19A5 антитело, раскрытое в настоящем документе, содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 24; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 23.
В некоторых воплощениях анти-FAM19A5 антитело содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 26; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 27.
В некоторых воплощениях анти-FAM19A5 антитело содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 28; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 29.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию является гуманизированным. В других воплощениях гуманизированное анти-FAM19A5 включает каркасную область человеческого антитела. В некоторых воплощениях анти-FAM19A5 антитело содержит одну или несколько (например, одну, две, три, четыре, пять, шесть, семь или более) мутаций в каркасной области (например, FR1, FR2, FR3 и FR4 из VH и/или FR1, FR2, FR3 и FR4 из VL) антитела.
В некоторых воплощениях анти-FAM19A5 антитело включает мутацию в FR1 из VH. В некоторых воплощениях мутация включает аминокислотную замену в остатке 19 (например, серин на основную аминокислоту, например, аргинин) и/или в остатке 21 (например, валин на гидроксил или сера/селенсодержащую аминокислоту, например, серин) из SEQ ID NO: 11.
В некоторых воплощениях раскрытое в данном документе анти-FAM19A5 антитело включает мутацию в FR2 из VH. В некоторых воплощениях мутация включает аминокислотную замену в остатке 49 (например, аланина на гидроксил или сера/селенсодержащую аминокислоту, например серин) из SEQ ID NO: 11.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит мутацию в FR3 из VH. В некоторых воплощениях мутация включает аминокислотную замену в остатке 79 (например, валин на алифатическую аминокислоту, например, лейцин), остаток 80 (например, аргинин на ароматическую аминокислоту, например, тирозин), остаток 83 (например, лейцин на гидроксил или сера/селенсодержащую аминокислоту, например, метионин), остаток 85 (например, аспарагин на гидроксил или сера/селенсодержащую аминокислоту, например, серин), остаток 86 (например, пролин на алифатическую аминокислоту, например, лейцин) и/или остаток 87 (например, глицин на основную аминокислоту, например, аргинин) из SEQ ID NO: 11.
В некоторых воплощениях анти-FAM19A5 антитело включает мутацию в FR2 из VL. В некоторых воплощениях мутация включает делецию аминокислотного остатка 39 из SEQ ID NO: 12.
В некоторых воплощениях раскрытое в данном документе анти-FAM19A5 антитело включает мутацию в FR3 из VL. В некоторых воплощениях мутация включает аминокислотную замену в остатке 81 (например, аспарагиновой кислоты на алифатическую аминокислоту, например, глицин) и/или в остатке 85 (например, изолейцин на кислую аминокислоту, например, аспарагиновую кислоту) из SEQ ID NO: 12.
В некоторых воплощениях анти-FAM19A5 антитело согласно настоящему раскрытию включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, которая идентична, по меньшей мере, на около 80%, по меньшей мере, около 85%, по меньшей мере, около 90%, по меньшей мере, около 95%, по меньшей мере, около 96%, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% или около 100% аминокислотной последовательности, указанной как SEQ ID NO: 11, и/или где VL включает аминокислотную последовательность, которая идентична, по меньшей мере, около 80%, по меньшей мере, около 85%, по меньшей мере, около 90%, по меньшей мере, около 95%, по меньшей мере, около 96%, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% или около 100% аминокислотной последовательности, указанной как SEQ ID NO: 12, где антитело имеет пониженную иммуногенность по сравнению с референсным антителом, содержащим VH, указанной как SEQ ID NO: 11, и VL, указанной как SEQ ID NO: 12.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, перекрестно конкурирует с референсным антителом, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где:
(а) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 33, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 38;
(b) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 39;
(c) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 41;
(d) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 40;
(e) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 42;
(f) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 43; или
(g) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 44.
В некоторых воплощениях анти-FAM19A5 антитело связывается с тем же эпитопом FAM19A5 человека, что и референсное антитело, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где:
(а) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 33, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 38;
(b) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 39;
(c) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 41;
(d) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 40;
(e) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 42;
(f) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 43; или
(g) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 44.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где CDR1, CDR2 и CDR3 тяжелой цепи содержат аминокислотные последовательности, указанные в SEQ ID NO: 16, 17 и 18, соответственно, каждая из которых необязательно содержит одну, две или три мутации, при этом CDR1, CDR2 и CDR3 легкой цепи содержат аминокислотные последовательности, указанные в SEQ ID NO: 30, 31 и 32, соответственно, где, по меньшей мере, одна из CDR1, CDR2 и CDR3 легкой цепи содержит одну, две или три мутации, и где антитело имеет пониженную иммуногенность у человека и/или более высокую аффинность связывания с человеческим белком FAM19A5, по сравнению с референсным антителом, содержащим VH, указанную в SEQ ID NO: 35, и VL, указанную в SEQ ID NO: 45.
В некоторых воплощениях CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 18. В некоторых воплощениях CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 18, необязательно с одной или двумя мутациями. В некоторых воплощениях мутация включает одно или несколько из следующего: (a) замену треонина в аминокислоте 2 из SEQ ID NO: 18 на гидроксил или сера/селенсодержащую аминокислоту; и (b) замену глутаминовой кислоты в аминокислоте 4 из SEQ ID NO: 18 на алифатическую аминокислоту. В некоторых воплощениях гидроксильная или сера/селенсодержащая аминокислота включает серин. В некоторых воплощениях алифатическая аминокислота включает валин. В некоторых воплощениях мутация включает одно или несколько из следующего: (а) замену треонина в аминокислоте 2 из SEQ ID NO: 18 на кислую кислоту; и (b) замену глутаминовой кислоты в аминокислоте 4 из SEQ ID NO: 18 на алифатическую аминокислоту. В некоторых воплощениях кислая кислота включает аспарагин. В некоторых воплощениях алифатическая аминокислота включает аланин.
В некоторых воплощениях CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 16. В некоторых воплощениях CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 16, необязательно с одной мутацией. В некоторых воплощениях мутация включает замену треонина в аминокислоте 3 из SEQ ID NO: 16 на кислую аминокислоту. В некоторых воплощениях кислая аминокислота включает аспарагиновую кислоту.
В некоторых воплощениях CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 17.
В некоторых воплощениях CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 32. В некоторых воплощениях CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 31. В некоторых воплощениях CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 31, с одной, двумя или тремя мутациями.
В дополнительных воплощениях CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 30, необязательно с одной мутацией. В некоторых воплощениях мутация включает замену серина в аминокислоте 4 из SEQ ID NO: 30 на алифатическую аминокислоту. В некоторых воплощениях алифатическая аминокислота включает валин.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 16; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 17; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 18; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 80; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 31; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 32.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 19; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 17; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 18; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 80; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 31; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 32.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 19; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 17; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 128; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 80; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 31; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 32.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 19; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 17; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 129; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 80; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 31; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 32.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию является гуманизированным. В других воплощениях гуманизированное анти-FAM19A5 включает каркасную область человеческого антитела. В некоторых воплощениях анти-FAM19A5 антитело содержит одну или несколько (например, одну, две, три, четыре, пять, шесть, семь или более) мутаций в каркасной области (например, FR1, FR2, FR3 и FR4 из VH и/или FR1, FR2, FR3 и FR4 из VL) антитела.
В некоторых воплощениях раскрытое в данном документе анти-FAM19A5 антитело включает мутацию в FR1 из VH. В некоторых воплощениях мутация включает аминокислотную замену в остатке 19 (например, серин на основную аминокислоту, например, аргинин), остатке 21 (например, валин на гидроксил или сера/селенсодержащую аминокислоту, например серин) и/или остатке 23 (например, от лизина до гидроксила или серу/селенсодержащей аминокислоты, например серин) из SEQ ID NO: 35.
В некоторых воплощениях анти-FAM19A5 антитело включает мутацию в FR2 из VH. В некоторых воплощениях мутация включает аминокислотную замену в остатке 40 (например, тирозин на алифатическую аминокислоту, например, аланин) и/или остатке 49 (например, аланин на гидроксил или сера/селенсодержащую аминокислоту, например, серин) из SEQ ID NO: 35.
В некоторых воплощениях анти-FAM19A5 антитело включает мутацию в FR3 из VH. В некоторых воплощениях мутация включает аминокислотную замену в остатке 79 (например, валин на алифатическую аминокислоту, например, лейцин), остатке 80 (например, аргинин на ароматическую аминокислоту, например, тирозин), остатке 83 (например, лейцин в гидроксил или сера/селенсодержащую аминокислоту, например, метионин), и/или остатке 85 (например, аспарагин в гидроксил или сера/селенсодержащую аминокислоту, например серин) из SEQ ID NO: 35.
В некоторых воплощениях анти-FAM19A5 антитело включает мутацию в FR1 из VL. В некоторых воплощениях мутация включает аминокислотную замену в остатке 16 (например, валин на алифатическую аминокислоту, например, аланин) из SEQ ID NO: 45.
В некоторых воплощениях анти-FAM19A5 антитело включает мутацию в FR2 из VL. В некоторых воплощениях мутация включает делецию аминокислотного остатка 34 из SEQ ID NO: 45.
В некоторых воплощениях раскрытое в данном документе анти-FAM19A5 антитело включает мутацию в FR3 из VL. В некоторых воплощениях мутация включает аминокислотную замену в остатке 76 (например, аспарагиновой кислоты на кислую аминокислоту, например, глутаминовую кислоту), остатке 80 (например, валина на кислую аминокислоту, например, аспарагиновую кислоту) и/или остатке 82 (например, от фенилаланина до ароматической аминокислоты, например, тирозина) SEQ ID NO: 45.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, которая идентична, по меньшей мере, на около 80%, по меньшей мере, около 85%, по меньшей мере, около 90%, по меньшей мере, около 95%, по меньшей мере, около 96%, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% или около 100% указанной аминокислотной последовательности в SEQ ID NO: 35 и/или где VL включает аминокислотную последовательность, которая идентична, по меньшей мере, на около 80%, по меньшей мере, около 85%, по меньшей мере, около 90%, по меньшей мере, около 95%, по меньшей мере, около 96%, при, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% или около 100% аминокислотной последовательности, указанной в SEQ ID NO: 45.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию перекрестно конкурирует с референсным антителом, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где (i) VH содержит аминокислотная последовательность, указанную в SEQ ID NO: 36, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46; (ii) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 37, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46; (iii) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 130, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46; или (iv) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 131, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, связывается с тем же эпитопом FAM19A5 человека, что и референсное антитело, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где (i) VH содержит аминокислотная последовательность, указанная в SEQ ID NO: 36, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46; (ii) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 37, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46; (iii) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 130, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46; или (iv) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 131, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
В некоторых воплощениях анти-FAM19A5 антитело согласно настоящему раскрытию конкурирует за связывание (или ингибирует связывание) эпитопа FAM19A5 человека с референсным антителом (например, антителом 3-2 или 2-13).
В некоторых воплощениях анти-FAM19A5 антитело ингибирует связывание такого референсного антитела (например, антитела 3-2 или 2-13) с FAM19A5 человека, по меньшей мере, на около 10%, по меньшей мере, около 20%, по меньшей мере, около 30%, по меньшей мере, около 40%, по меньшей мере, около 50%, по меньшей мере, около 60%, по меньшей мере, около 70%, по меньшей мере, около 80%, по меньшей мере, около 90% или, по меньшей мере, около 100%. Конкурирующие антитела связываются с одним и тем же эпитопом, перекрывающимся эпитопом или с соседними эпитопами (что, например, подтверждается стерическими препятствиями). Конкурируют ли два антитела друг с другом за связывание с мишенью, можно определить с помощью экспериментов по конкуренции, известных в данной области, таких как RIA и EIA.
Методы определения того, связываются ли два антитела с одним и тем же эпитопом, включают, например, способы картирования эпитопа, такие как рентгеновский анализ кристаллов комплексов антиген: антитело, который обеспечивает атомное разрешение эпитопа и масс-спектрометрия обмена водорода/дейтерия (HDX-MS), способы мониторинга связывания антитела с фрагментами антигена или мутированными вариациями антигена, где потеря связывания из-за модификации аминокислотного остатка в последовательности антигена часто считается критерием эпитопного компонента, вычислительные комбинаторные способы для картирования эпитопа.
Анти-AM19A5 антитело, которое может быть использовано в раскрытых в данном документе способах, может связываться, по меньшей мере, с одним эпитопом зрелого FAM19A5 человека, как определено, например, путем связывания антител с фрагментами FAM19A5 человека. В некоторых воплощениях анти-FAM19A5 антитело связывается, по меньшей мере, с одним эпитопом, который имеет аминокислотную последовательность TLDRDSSQPRRTIARQTARC (SEQ ID NO: 90 или аминокислотные остатки с 42 по 61 из SEQ ID NO: 2), или связывается с фрагментом расположенный в аминокислотной последовательности SEQ ID NO: 90, например, эпитоп, имеющий, по меньшей мере, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот SEQ ID NO: 90. В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, связывается с одной или несколькими аминокислотами, соответствующими аминокислотным остаткам с 44 по 52 (т.е. DRDSSQPRR) из SEQ ID NO: 2, например, аминокислотным остаткам 45, 46, 50, 51, и 52 (RD---PRR), например, аминокислотные остатки 45, 50, 51 и 52 (т.е. R----PRR), например, аминокислотные остатки 43, 50 и 51 (т.е. R----PR).
В некоторых воплощениях анти-FAM19A5 антитело связывается, по меньшей мере, с одним эпитопом, который имеет аминокислотную последовательность TARCACRKGQIAGTTRARPA (SEQ ID NO: 91 или аминокислотные остатки 58-77 из SEQ ID NO: 2), или связывается с фрагментом расположенный в аминокислотной последовательности SEQ ID NO: 91, например, эпитоп, имеющий, по меньшей мере, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот SEQ ID NO: 91. В некоторых воплощениях анти-FAM19A5 антитело связывается с одной или несколькими аминокислотами, соответствующими аминокислотным остаткам с 63 по 75 (т.е. CRKGQIAGTTRAR) из SEQ ID NO: 2. В некоторых воплощениях анти-FAM19A5 антитело связывается, по меньшей мере, с одним эпитопом, который имеет аминокислотную последовательность ARPACVDARIIKTKQWCDML (SEQ ID NO: 92 или аминокислотные остатки с 74 по 93 из SEQ ID NO: 2), или связывается с фрагментом расположенным в аминокислотной последовательности SEQ ID NO: 92, например, эпитоп, имеющий, по меньшей мере, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот SEQ ID NO: 92. В некоторых воплощениях анти-FAM19A5 антитело связывается с одной или несколькими аминокислотами, соответствующими аминокислотным остаткам 76-89 (т.е. PACVDARIIKTKQW) из SEQ ID NO: 2.
В некоторых воплощениях, по меньшей мере, один эпитоп имеет аминокислотную последовательность, которая идентична, по меньшей мере, 90%, по меньшей мере, около 95%, по меньшей мере, около 96%, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% или около 100% с SEQ ID NO: 90, 91 или 92.
В некоторых воплощениях анти-FAM19A5 антитело или его антигенсвязывающая часть связывается только с эпитопом FAM19A5 человека, который представляет собой SEQ ID NO: 89, 90, 91, 92, 93 или 94, или фрагментом, расположенным в пределах аминокислотной последовательности из SEQ ID NO: 89, 90, 91, 92, 93 или 94, например, эпитопом, имеющим 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот из SEQ ID NO: 89, 90, 91, 92, 93 или 94.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию связывается с SEQ ID NO: 90 или его фрагментом в своей нативной конформации (т.е. неденатурированной). В некоторых воплощениях анти-FAM19A5 антитело или его антигенсвязывающая часть связывается как с гликозилированным, так и с негликозилированным FAM19A5 человека.
В некоторых воплощениях анти-FAM19A5 антитело связывается с одним или несколькими дополнительными эпитопами FAM19A5. В некоторых воплощениях один или несколько дополнительных эпитопов FAM19A5 выбраны из QLAAGTCEIVTLDR (SEQ ID NO: 89, эпитоп F1), TLDRDSSQPRRTIARQTARC (SEQ ID NO: 90, эпитоп F2), TARCACRKGQIAGTTRARPA (SEQ ID NO: 91), эпитопа ARPACVDARIIKTKQW CDML (SEQ ID NO: 92, эпитоп F4), CDMLPCLEGEGCDLLINRSG (SEQ ID NO: 93, эпитоп F5) или NRSGWTCTQPGGRIKTTTVS (SEQ ID NO: 94, эпитоп F6) или фрагмента, расположенного в пределах аминокислотной последовательности SEQ NO: 89, SEQ ID NO: 90, SEQ ID NO: 91, SEQ ID NO: 92, SEQ ID NO: 93 или SEQ ID NO: 94, или любой их комбинации. Фрагмент, расположенный в пределах аминокислотной последовательности SEQ ID NO: 89, SEQ ID NO: 90, SEQ ID NO: 91, SEQ ID NO: 92, SEQ ID NO: 93 или SEQ ID NO: 94, включает фрагмент, имеющий 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот любой из SEQ ID NO: 89, SEQ ID NO: 90, SEQ ID NO: 91, SEQ ID NO: 92, SEQ ID NO: 93 или SEQ ID NO: 94. В некоторых воплощениях один или несколько дополнительных эпитопов FAM19A5 выбраны из SEQ ID NO: 89, 90, 91, 92, 93 или 94 или фрагмента, расположенного в пределах аминокислотной последовательности SEQ ID NO: 89, 90, 91, 92, 93 или 94, например, фрагмента, имеющего 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот SEQ ID NO: 89, 90, 91, 92, 93 или 94, или любой их комбинации. В некоторых воплощениях анти-FAM19A5 антитело или его антигенсвязывающая часть согласно настоящему описанию связывается с любым из одного или нескольких дополнительных эпитопов в их нативной конформации (т.е. неденатурированной). В некоторых воплощениях анти-FAM19A5 антитело или его антигенсвязывающая часть связывается как с гликозилированным, так и с негликозилированным одним или несколькими дополнительными эпитопами FAM19A5.
В некоторых воплощениях в настоящем документе предлагается антитело или его антигенсвязывающий фрагмент, который связывается с FAM19A5 (например, FAM19A5 человека) с 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более высокой аффинностью, чем аффинность к другому белку семейства FAM19A, что измерено, например, с помощью иммуноанализа (например, ELISA), поверхностного плазмонного резонанса или анализа кинетического исключения. В некоторых воплощениях анти-FAM19A5 антитело или его антигенсвязывающий фрагмент связывается с FAM19A5 (например, FAM19A5 человека) без перекрестной реактивности с другим белком семейства FAM19A, что измерено, например, с помощью иммуноанализа.
В некоторых воплощениях анти-FAM19A5 антитела по настоящему раскрытию не являются нативными антителами или не являются встречающимися в природе антителами. Например, в некоторых воплощениях анти-FAM19A5 антитела имеют посттрансляционные модификации, которые отличаются от модификаций антител, которые встречаются в природе, такие как наличие большей, меньшей посттрансляционной модификации или посттрансляционной модификации другого типа.
Аминокислотные последовательности CDR из VH и VL типичных антител по настоящему изобретению представлены в таблицах 3 и 4, соответственно. Аминокислотные последовательности VH и VL представлены в таблицах 5 и 6, соответственно.
Таблица 3. Аминокислотные последовательности CDR вариабельной области тяжелой цепи (идентифицированные с использованием IMGT)
(«3-2»)
(SEQ ID NO: 7)
(«2-13»)
Таблица 4. Аминокислотные последовательности CDR вариабельной области легкой цепи (идентифицированные с использованием IMGT)
(«3-2»)
(SEQ ID NO: 10)
(«2-13»)
Таблица 5. Аминокислотная последовательность вариабельной области тяжелой цепи
(«3-2»)
(«2-13»)
Таблица 6. Аминокислотная последовательность вариабельной области легкой цепи
(«3-2»)
(«2-13»)
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит вариабельные области тяжелой и легкой цепи, где вариабельная область тяжелой цепи (VH) включает CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 33 и/или вариабельная область легкой цепи (VL) включает CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 38. В других воплощениях анти-FAM19A5 антитело содержит VH и VL, где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 33, и/или VL включает аминокислотную последовательность, указанную в SEQ ID NO: 38.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит вариабельные области тяжелой и легкой цепи, где вариабельная область тяжелой цепи (VH) включает CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 34 и/или вариабельная область легкой цепи (VL) включает CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 39. В других воплощениях анти-FAM19A5 антитело содержит VH и VL, где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и/или VL включает аминокислотную последовательность, указанную в SEQ ID NO: 39.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит вариабельные области тяжелой и легкой цепи, где вариабельная область тяжелой цепи (VH) включает CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 34 и/или вариабельная область легкой цепи (VL) включает CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 41. В других воплощениях анти-FAM19A5 антитело содержит VH и VL, где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и/или VL включает аминокислотную последовательность, указанную в SEQ ID NO: 41.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит вариабельные области тяжелой и легкой цепи, где вариабельная область тяжелой цепи (VH) включает CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 34 и/или вариабельная область легкой цепи (VL) включает CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 40. В других воплощениях анти-FAM19A5 антитело содержит VH и VL, где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и/или VL включает аминокислотную последовательность, указанную в SEQ ID NO: 40.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит вариабельные области тяжелой и легкой цепи, где вариабельная область тяжелой цепи (VH) включает CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 34 и/или вариабельная область легкой цепи (VL) включает CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 42. В других воплощениях анти-FAM19A5 антитело содержит VH и VL, где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и/или VL включает аминокислотную последовательность, указанную в SEQ ID NO: 42.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит вариабельные области тяжелой и легкой цепи, где вариабельная область тяжелой цепи (VH) включает CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 34 и/или вариабельная область легкой цепи (VL) включает CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 43. В других воплощениях анти-FAM19A5 антитело содержит VH и VL, где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и/или VL включает аминокислотную последовательность, указанную в SEQ ID NO: 43.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит вариабельные области тяжелой и легкой цепи, где вариабельная область тяжелой цепи (VH) включает CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 34 и/или вариабельная область легкой цепи (VL) включает CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 44. В других воплощениях анти-FAM19A5 антитело содержит VH и VL, где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и/или VL включает аминокислотную последовательность, указанную в SEQ ID NO: 44.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию включает вариабельные области тяжелой и легкой цепи, где вариабельная область тяжелой цепи (VH) включает CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 36 и/или вариабельная область легкой цепи (VL) включает CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 46. В других воплощениях анти-FAM19A5 антитело содержит VH и VL, где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 36, и/или VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит вариабельные области тяжелой и легкой цепи, где вариабельная область тяжелой цепи (VH) включает CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 37 и/или вариабельная область легкой цепи (VL) включает CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 46. В других воплощениях анти-FAM19A5 антитело содержит VH и VL, где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 37, и/или VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему изобретению содержит вариабельные области тяжелой и легкой цепи, где вариабельная область тяжелой цепи (VH) включает CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 130 и/или вариабельная область легкой цепи (VL) включает CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 46. В других воплощениях анти-FAM19A5 антитело содержит VH и VL, где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 37, и/или VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит вариабельные области тяжелой и легкой цепи, где вариабельная область тяжелой цепи (VH) включает CDR1, CDR2 и CDR3 тяжелой цепи из SEQ ID NO: 131 и/или легкую вариабельная область цепи (VL) включает CDR1, CDR2 и CDR3 легкой цепи из SEQ ID NO: 46. В других воплощениях анти-FAM19A5 антитело содержит VH и VL, где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 37, и/или VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит VH и VL, где VH включает аминокислотную последовательность, которая идентична, по меньшей мере, на около 80%, по меньшей мере, около 85%, по меньшей мере, около 90%, по меньшей мере, около 95%, по меньшей мере, около 96%, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% или около 100% аминокислотной последовательности, указанной в SEQ ID NO: 33, 34, 36, 37, 130 или 131.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит VH и VL, где VL включает аминокислотную последовательность, которая идентична, по меньшей мере, на около 80%, по меньшей мере, около 85%, по меньшей мере, около 90%, по меньшей мере, около 95%, по меньшей мере, около 96%, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% или около 100% аминокислотной последовательности, указанной в SEQ ID NO: 38, 39, 40, 41, 42, 43, 44 или 46.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию содержит VH и VL, где:
(а) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 33, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 38;
(b) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 39;
(c) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 41;
(d) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 40;
(e) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 42;
(f) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 43;
(g) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 44;
(h) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 36, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46;
(i) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 37, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46;
(j) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 130, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46; или
(k) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 131, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
Домен VH или одна или несколько его CDR, описанных в данном документе, могут быть связаны с константным доменом для образования тяжелой цепи, например, полноразмерной тяжелой цепи. Точно так же домен VL или один или несколько его CDR, описанных в данном документе, могут быть связаны с константным доменом для образования легкой цепи, например, полноразмерной легкой цепи. Полноразмерная тяжелая цепь и полноразмерная легкая цепь объединяются с образованием полноразмерного антитела.
Соответственно, в конкретных воплощениях в настоящем документе предлагается антитело, содержащее легкую цепь и тяжелую цепь антитела, например, отдельные легкую цепь и тяжелую цепь. Что касается легкой цепи, в конкретном воплощении легкая цепь антитела, описанного в данном документе, представляет собой легкую каппа-цепь. В другом конкретном воплощении легкая цепь антитела, описанного в данном документе, представляет собой легкую лямбда-цепь. В еще одном конкретном воплощении легкая цепь антитела, описанного в настоящем документе, представляет собой легкую каппа-цепь человека или легкую лямбда-цепь человека. В конкретном воплощении описанное в данном документе антитело, которое специфически связывается с полипептидом FAM19A5 (например, FAM19A5 человека), содержит легкую цепь, которая содержит любые описанные в данном документе аминокислотные последовательности VL или CDR из VL, и где константная область легкой цепи включает аминокислотная последовательность константной области легкой цепи каппа человека. В конкретном воплощении описанное в данном документе антитело, которое специфически связывается с полипептидом FAM19A5 (например, FAM19A5 человека), содержит легкую цепь, которая содержит описанные в данном документе аминокислотные последовательности VL или CDR из VL, и где константная область легкой цепи включает аминокислотная последовательность константной области легкой цепи лямбда человека. Неограничивающие примеры последовательностей константной области человека были описаны в данной области, например, см. Патент № 5693780 и Kabat EA et al. (1991) выше.
Что касается тяжелой цепи, в некоторых воплощениях тяжелая цепь антитела, описанного в настоящем документе, может быть тяжелой цепью альфа (α), дельта (δ), эпсилон (ε), гамма (γ) или мю (μ). В другом конкретном воплощении тяжелая цепь описанного антитела может включать человеческую альфа (α), дельта (δ), эпсилон (ε), гамма (γ) или мю (μ) тяжелую цепь. В одном воплощении описанное в данном документе антитело, которое специфически связывается с FAM19A5 (например, с FAM19A5 человека), включает тяжелую цепь, которая содержит описанную в данном документе аминокислотную последовательность VH или CDR из VH, и где константная область тяжелой цепи содержит аминокислотную последовательность константной области гамма (γ) тяжелой цепи человека. В другом воплощении описанное в данном документе антитело, которое специфически связывается с FAM19A5 (например, с FAM19A5 человека), включает тяжелую цепь, которая включает аминокислотную последовательность VH или CDR из VH, описанную в данном документе, и где константная область тяжелой цепи включает аминокислотную последовательность кислота тяжелой цепи человека, описанная в данном документе или известная в данной области. Неограничивающие примеры последовательностей константной области человека были описаны в данной области, например, см. Патент № 5693780 и Kabat EA et al. (1991) выше.
В некоторых воплощениях описанное в данном документе антитело, которое специфически связывается с FAM19A5 (например, FAM19A5 человека), содержит домен VL и домен VH, содержащий VH или CDR из VH и VL и CDR из VL, описанные в данном документе, и при этом константные области содержат аминокислотные последовательности константных областей молекулы иммуноглобулина IgG, IgE, IgM, IgD, IgA или IgY или молекулы иммуноглобулина IgG, IgE, IgM, IgD, IgA или IgY человека. В другом конкретном воплощении описанное в данном документе антитело, которое специфически связывается с FAM19A5 (например, FAM19A5 человека), включает домен VL и домен VH, содержащие любые аминокислотные последовательности, описанные в данном документе, и где константные области содержат аминокислотные последовательности константной области молекулы иммуноглобулина IgG, IgE, IgM, IgD, IgA или IgY, любого подкласса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) молекулы иммуноглобулина. В некоторых воплощениях константные области содержат аминокислотные последовательности константных областей человеческого IgG, которые встречаются в природе, включая подклассы (например, IgG1, IgG2, IgG3 или IgG4) и аллотипы (например, G1m, G2m, G3m и nG4m) и их варианты. См., например, Vidarsson G. et al. Front Immunol. 5: 520 (опубликовано онлайн 20 окт. 2014 г.) и Jefferis R. and Lefranc MP, mAbs 1:4, 1-7(2009). В некоторых воплощениях константные области содержат аминокислотные последовательности константных областей человеческих IgG1, IgG2, IgG3 или IgG4 или их вариантов.
В некоторых воплощениях анти-FAM19A5 антитело, раскрытое в настоящем документе, не имеет эффекторных функций Fc, например, комплемент-зависимой цитотоксичности (CDC) и/или антителозависимого клеточного фагоцитоза (ADCP). Эффекторные функции опосредуются Fc-областью, а остатки, наиболее проксимальные к шарнирной области в домене CH2 Fc-области, отвечают за эффекторные функции антител, поскольку этот домен содержит в значительной степени перекрывающийся сайт связывания для C1q (комплемент) и рецепторов IgG-Fc (FcγR) на эффекторных клетках врожденной иммунной системы. Кроме того, антитела IgG2 и IgG4 имеют более низкие уровни эффекторных функций Fc, чем антитела IgG1 и IgG3. Эффекторные функции антитела можно уменьшить или избежать с помощью различных подходов, известных в данной области, включая (1) использование фрагментов антитела, лишенных Fc-области (например, таких как Fab, F(ab')2, одноцепочечный Fv (scFv). или sdAb, состоящий из мономерного домена VH или VL); (2) создание агликозилированных антител, которые могут быть получены, например, путем удаления или изменения остатка, к которому присоединен сахар, ферментативного удаления сахаров, продуцирования антитела в клетках, культивируемых в присутствии ингибитора гликозилирования, или путем экспрессии антитела в клетках, неспособных гликозилировать белки (например, бактериальные клетки-хозяева, см., например, публ. США № 20120100140); (3) использование Fc-областей из подкласса IgG, которые обладают пониженной эффекторной функцией (например, Fc-области из антител IgG2 или IgG4 или химерной Fc-области, содержащей домен CH2 из антител IgG2 или IgG4, см., например, публ. No. 20120100140 и Lau C. et al., J. Immunol. 191: 4769-4777 (2013)); и (4) создание Fc-области с мутациями, которые приводят к снижению функций Fc или их отсутствию. См., например, публ. США № 20120100140 и заявки США и РСТ, процитированные в ней, и An et al., mAbs 1: 6, 572-579 (2009).
Таким образом, в некоторых воплощениях изобретения анти-FAM19A5 антитело, раскрытое в настоящем документе, представляет собой Fab, Fab', F(ab')2, Fv, одноцепочечный Fv (scFv) или sdAb, состоящий из мономерного домена VH или VL. Такие фрагменты антител хорошо известны в данной области и описаны выше.
В некоторых воплощениях анти-FAM19A5 антитело представляет собой одноцепочечный Fv. Аминокислотные последовательности типичного scFv против FAM19A5 представлены в Таблице 7 ниже.
Таблица 7. Аминокислотные последовательности анти-FAM19A5 scFv
(«3-2»)
(«2-13»)
В некоторых воплощениях изобретения анти-FAM19A5 антитело, раскрытое в настоящем документе, содержит Fc-область с пониженной эффекторной функцией Fc или без нее. В некоторых воплощениях константные области содержат аминокислотные последовательности Fc-области человеческого IgG2 или IgG4, в некоторых воплощениях анти-FAM19A5 антитело имеет изотип IgG2/IgG4. В некоторых воплощениях анти-FAM19A5 антитело содержит химерную Fc-область, которая включает домен CH2 из антитела IgG изотипа IgG4 и домен CH3 из антитела IgG изотипа IgG1 или химерную Fc-область, которая включает шарнирную область из IgG2 и области CH2 из IgG4 или Fc-области с мутациями, которые приводят к снижению или отсутствию функций Fc. Области Fc с пониженной эффекторной функцией Fc или без нее включают области, известные в данной области. См., например, Lau C. et al., J. Immunol. К191: 4769-4777 (2013); An et al., MAbs 1: 6, 572-579 (2009); и публ. США № 20120100140 и патенты и публикации США, а также публикации РСТ, процитированные в них. Также Fc-области с пониженной эффекторной функцией Fc или без нее могут быть легко созданы специалистом в данной области.
III. Молекулы Нуклеиновых Кислот
Другой аспект, описанный в данном документе, относится к одной или нескольким молекулам нуклеиновой кислоты, которые кодируют любое из описанных в данном документе антител. Нуклеиновые кислоты могут присутствовать в целых клетках, в клеточном лизате или в частично очищенной или практически чистой форме. Нуклеиновая кислота является «выделенной» или «по существу чистой» при очистке от других клеточных компонентов или других примесей, например, других клеточных нуклеиновых кислот (например, другой хромосомной ДНК, например, хромосомной ДНК, которая связана с выделенной ДНК в природе) или белков стандартными методами, включая обработку щелочью/SDS, разделение в CsCl, колоночную хроматографию, рестрикционные ферменты, электрофорез в агарозном геле и другие, хорошо известные в данной области техники. См. F. Ausubel, et al., Ed. (1987) Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York. Описанная в данном документе нуклеиновая кислота может быть, например, ДНК или РНК и может содержать или не может содержать интронные последовательности. В некоторых воплощениях нуклеиновая кислота представляет собой молекулу кДНК.
Описанные в данном документе нуклеиновые кислоты могут быть получены с использованием стандартных методов молекулярной биологии. Для антител, экспрессируемых гибридомами (например, гибридомами, полученными из трансгенных мышей, несущих гены иммуноглобулинов человека, как описано ниже), кДНК, кодирующие легкую и тяжелую цепи антитела, продуцируемого гибридомой, могут быть получены стандартными методами ПЦР-амплификации или клонирования кДНК. Для антител, полученных из библиотеки генов иммуноглобулинов (например, с использованием методов фагового дисплея), нуклеиновая кислота, кодирующая антитело, может быть извлечена из библиотеки.
Определенные молекулы нуклеиновых кислот, описанные в настоящем документе, кодируют последовательности VH и VL различных анти-FAM19A5 антител по настоящему изобретению. Типичные последовательности ДНК, кодирующие последовательности VH и VL таких антител, представлены в таблицах 8 и 9, соответственно.
Таблица 8. Полинуклеотидная последовательность вариабельной области тяжелой цепи
(«3-2»)
(«2-13»)
Таблица 9. Полинуклеотидная последовательность вариабельной легкой цепи
(«3-2»)
(«2-13»)
Раскрытый в данном документе способ получения анти-FAM19A5 антитела может включать экспрессию тяжелой цепи и легких цепей в линии клеток, содержащей нуклеотидные последовательности, кодирующие тяжелую и легкую цепи, с помощью сигнального пептида. Клетки-хозяева, содержащие эти нуклеотидные последовательности, включены в настоящий документ.
После получения фрагментов ДНК, кодирующих сегменты VH и VL, с этими фрагментами ДНК можно дополнительно провести дополнительные манипуляции стандартными методами рекомбинантных ДНК, например, для преобразования генов вариабельной области в гены полноразмерных цепей антитела, в гены фрагментов Fab или в ген scFv. В этих манипуляциях фрагмент ДНК, кодирующий VL или VH, функционально связан с другим фрагментом ДНК, кодирующим другой белок, таким как константная область антитела или гибкий линкер. Термин «функционально связанный», используемый в данном контексте, предназначен для обозначения того, что два фрагмента ДНК соединены таким образом, что аминокислотные последовательности, кодируемые двумя фрагментами ДНК, остаются в рамке считывания.
Выделенная ДНК, кодирующая область VH, может быть преобразована в ген полноразмерной тяжелой цепи путем функционального связывания ДНК, кодирующей VH, с другой молекулой ДНК, кодирующей константные области тяжелой цепи (шарнир, CH1, CH2 и/или CH3). Последовательности генов константной области тяжелой цепи человека известны в данной области (см., например, Kabat, E.A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242) и фрагменты ДНК, охватывающие эти области, можно получить с помощью стандартной ПЦР-амплификации. Константная область тяжелой цепи может быть константной областью IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD, например, константной областью IgG2 и/или IgG 4. Для гена тяжелой цепи Fab-фрагмента ДНК, кодирующая VH, может быть функционально связана с другой молекулой ДНК, кодирующей только константную область СН1 тяжелой цепи.
Выделенная ДНК, кодирующая область VL, может быть преобразована в ген полноразмерной легкой цепи (а также ген легкой цепи Fab) путем функционального связывания ДНК, кодирующей VL, с другой молекулой ДНК, кодирующей константную область легкой цепи, CL. Последовательности генов константной области легкой цепи человека известны в данной области (см., например, Kabat, E.A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242) и фрагменты ДНК, охватывающие эти области, можно получить с помощью стандартной ПЦР-амплификации. Константная область легкой цепи может быть константной областью каппа или лямбда.
Для создания гена scFv фрагменты ДНК, кодирующие VH и VL, функционально связаны с другим фрагментом, кодирующим гибкий линкер, например, кодирующим аминокислотную последовательность (Gly4 -Ser) 3, так что последовательности VH и VL могут быть экспрессированы. как непрерывный одноцепочечный белок, с областями VL и VH, соединенными гибким линкером (см., например, Bird et al. (1988) Science 242: 423-426; Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85: 5879-5883; McCafferty et al. (1990) Nature 348:552-554). Аминокислотные последовательности типичных scFv против FAM19A5 представлены в таблицах 7.
В некоторых воплощениях настоящего изобретения предлагается вектор, содержащий выделенную молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, кодирующую антитело. В других воплощениях векторы можно использовать для генной терапии.
Подходящие векторы для раскрытия включают экспрессирующие векторы, вирусные векторы и плазмидные векторы. В одном воплощении вектор представляет собой вирусный вектор.
Используемый в данном документе экспрессирующий вектор относится к любой конструкции нуклеиновой кислоты, которая содержит необходимые элементы для транскрипции и трансляции вставленной кодирующей последовательности или, в случае вирусного вектора РНК, необходимые элементы для репликации и трансляции при введении в подходящую клетку-хозяина. Экспрессирующие векторы могут включать плазмиды, фагмиды, вирусы и их производные.
Экспрессирующие векторы по настоящему описанию могут включать полинуклеотиды, кодирующие антитело или описанные в данном документе. В одном воплощении кодирующие последовательности антитела функционально связаны с последовательностью контроля экспрессии. В данном контексте две последовательности нуклеиновой кислоты функционально связаны, когда они ковалентно связаны таким образом, чтобы позволить каждой компонентной последовательности нуклеиновой кислоты сохранять свою функциональность. Считается, что кодирующая последовательность и последовательность контроля экспрессии гена функционально связаны, если они ковалентно связаны таким образом, чтобы обеспечить экспрессию или транскрипцию и/или трансляцию кодирующей последовательности под влиянием или контролем последовательности контроля экспрессии гена. Две последовательности ДНК считаются функционально связанными, если индукция промотора в 5'-генной экспрессионной последовательности приводит к транскрипции кодирующей последовательности и если природа связи между двумя последовательностями ДНК (1) не приводит к введению мутации со сдвигом рамки считывания, (2) не мешает способности промоторной области управлять транскрипцией кодирующей последовательности, или (3) не мешает способности соответствующего транскрипта РНК транслироваться в белок. Таким образом, последовательность экспрессии гена была бы функционально связана с кодирующей последовательностью нуклеиновой кислоты, если бы последовательность экспрессии гена была способна влиять на транскрипцию этой кодирующей последовательности нуклеиновой кислоты, так что полученный транскрипт транслировался в искомое антитело.
Вирусные векторы включают, без ограничения указанным, последовательности нуклеиновых кислот из следующих вирусов: ретровируса, такого как вирус мышиного лейкоза Молони, вирус мышиной саркомы Харви, вирус опухоли молочной железы мышей и вирус саркомы Рауса; лентивируса; аденовируса; аденоассоциированного вируса; вирусов типа SV40; полиомавирусов; вирусов Эпштейна-Барр; вирусов папилломы; вируса герпеса; вируса осповакцины; вируса полиомиелита; и РНК-вируса, такой как ретровирус. Можно без проблем использовать другие векторы, хорошо известные в данной области. Некоторые вирусные векторы основаны на нецитопатических эукариотических вирусах, в которых несущественные гены заменены представляющим интерес геном. Нецитопатические вирусы включают ретровирусы, жизненный цикл которых включает обратную транскрипцию геномной вирусной РНК в ДНК с последующей провирусной интеграцией в клеточную ДНК хозяина. Ретровирусы были одобрены для клинических исследований генной терапии человека. Наиболее полезными являются те ретровирусы, которые являются репликативно-дефектными (т.е. способные управлять синтезом требуемых белков, но неспособные создавать инфекционную частицу). Такие генетически измененные ретровирусные экспрессирующие векторы имеют общее применение для высокоэффективной трансдукции генов in vivo. Стандартные протоколы для получения ретровирусов с дефицитом репликации (включая стадии включения экзогенного генетического материала в плазмиду, трансфекцию линии упаковывающих клеток плазмидой, получение рекомбинантных ретровирусов упаковочной линией клеток, сбор вирусных частиц из сред тканевых культур, и инфицирование клеток-мишеней вирусными частицами) представлены в Kriegler, M., Gene Transfer and Expression, A Laboratory Manual, WH Freeman Co., New York (1990) and Murry, E. J., Methods in Molecular Biology, Vol. 7, Humana Press, Inc., Cliffton, N.J. (1991).
В одном воплощении вирус представляет собой аденоассоциированный вирус, вирус с двухцепочечной ДНК. Аденоассоциированный вирус может быть сконструирован репликативно-дефектным, и он способен инфицировать широкий спектр типов и видов клеток. Кроме того, этот вирус имеет такие преимущества, как устойчивость к нагреванию и липидным растворителям; высокие частоты трансдукции в клетках различных линий, включая гемопоэтические клетки; и отсутствие подавления суперинфекции, что позволяет проводить множественные серии трансдукций. По имеющимся сведениям, аденоассоциированный вирус может быть интегрирован в человеческую клеточную ДНК сайт-специфическим образом; таким образом минимизирование вероятности вставочного мутагенеза и вариабельности экспрессии вставленных генов характерной для ретровирусной инфекции. Кроме того, инфекции аденоассоциированным вирусом дикого типа отслеживали в тканевой культуре в течение больше чем 100 пассажей в отсутствии селективного давления, что подтверждает, что геномная интеграция аденоассоциированного вируса является относительно стабильным событием. Аденоассоциированные вирус может также действовать экстрахромасомально.
В других воплощениях вектор получен из лентивируса. В некоторых воплощениях вектор представляет собой вектор рекомбинантного лентивируса, способного инфицировать неделящиеся клетки.
Геном лентивируса и провирусная ДНК обычно содержат три гена, обнаруженные в ретровирусах: gag, pol и env, которые фланкированы двумя последовательностями длинных концевых повторов (LTR). Ген gag кодирует внутренние структурные (матрикс, капсид и нуклеокапсид) белки; ген pol кодирует РНК-направленную ДНК-полимеразу (обратную транскриптазу), протеазу и интегразу; а ген env кодирует гликопротеины вирусной оболочки. 5 'и 3' LTR служат для стимуляции транскрипции и полиаденилирования РНК вириона. LTR содержит все другие цис-действующие последовательности, необходимые для репликации вируса. Лентивирусы имеют дополнительные гены, включая vif, vpr, tat, rev, vpu, nef и vpx (в HIV-1, HIV-2 и/или SIV).
Рядом с 5' LTR находятся последовательности, необходимые для обратной транскрипции генома (сайт связывания праймера тРНК) и для эффективной инкапсидации вирусной РНК в частицы (сайт Psi). Если последовательности, необходимые для инкапсидации (или упаковки ретровирусной РНК в инфекционные вирионы), отсутствуют в вирусном геноме, цис-дефект предотвращает инкапсидацию геномной РНК.
Однако полученный мутант остается способным управлять синтезом всех белков вириона. В описании предложен способ получения рекомбинантного лентивируса, способного инфицировать неделящуюся клетку, включающий трансфекцию подходящей клетки-хозяина двумя или более векторами, несущими функции упаковки, а именно gag, pol и env, а также rev и tat. Как будет раскрыто в данном документе ниже, векторы, лишенные функционального гена tat, желательны для определенных применений. Таким образом, например, первый вектор может обеспечивать нуклеиновую кислоту, кодирующую вирусный gag и вирусный pol, а другой вектор может предоставлять нуклеиновую кислоту, кодирующую вирусную env, для получения упаковывающей клетки. Введение вектора, обеспечивающего гетерологичный ген, обозначенный в данном документе как вектор-переносчик, в эту упаковывающую клетку дает клетку-продуцент, которая высвобождает инфекционные вирусные частицы, несущие интересующий чужеродный ген.
В соответствии с указанной выше конфигурацией векторов и чужеродных генов второй вектор может обеспечивать нуклеиновую кислоту, кодирующую ген вирусной оболочки (env). Ген env может происходить практически из любого подходящего вируса, включая ретровирусы. В некоторых воплощениях белок env представляет собой амфотропный белок оболочки, который позволяет трансдукцию клеток человека и других видов.
Примеры генов env, полученных из ретровирусов, включают, без ограничения указанным: вирус лейкоза мышей Молони (MoMuLV или MMLV), вирус саркомы мышей Харви (HaMuSV или HSV), вирус опухоли молочной железы мышей (MuMTV или MMTV), вирус лейкоза гиббонов (GaLV или GALV), вирус иммунодефицита человека (HIV) и вирус саркомы Рауса (RSV). Также можно использовать другие гены env, такие как белок G вируса везикулярного стоматита (VSV) (VSV G), гены вирусов гепатита и гриппа.
Вектор, обеспечивающий последовательность вирусной нуклеиновой кислоты env, функционально связан с регуляторными последовательностями, описанными в другом месте в настоящем документе.
В некоторых воплощениях вектор включает лентивирусный вектор, в котором гены вирулентности HIV env, vif, vpr, vpu и nef были удалены без ущерба для способности вектора трансдуктировать неделящиеся клетки.
В некоторых воплощениях вектор включает лентивирусный вектор, который содержит делецию области U3 3'-LTR. Удаление области U3 может быть полным или частичным.
В некоторых воплощениях лентивирусный вектор по раскрытию, содержащий описанную в данном документе нуклеотидную последовательность FVIII, может быть трансфицирован в клетку с (а) первой нуклеотидной последовательностью, содержащей гены gag, pol или gag и pol, и (b) второй нуклеотидной последовательностью, содержащей гетерологичный ген env; где в лентивирусном векторе отсутствует функциональный ген tat. В других воплощениях клетка дополнительно трансфицируется четвертой нуклеотидной последовательностью, содержащей ген rev. В некоторых воплощениях в лентивирусном векторе отсутствуют функциональные гены, выбранные из vif, vpr, vpu, vpx и nef или их комбинации.
В некоторых воплощениях лентивирусный вектор содержит одну или несколько нуклеотидных последовательностей, кодирующих белок gag, респонсивный элемент Rev, центральный полипуриновый тракт (cPPT) или любую их комбинацию.
Примеры лентивирусных векторов раскрыты в WO9931251, W09712622, W09817815, W09817816 и WO9818934, которые полностью включены в настоящий документ ссылкой.
Другие векторы включают плазмидные векторы. Плазмидные векторы подробно описаны в данной области и хорошо известны специалистам в данной области. См., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press, 1989. В последние несколько лет было обнаружено, что плазмидные векторы особенно полезны для доставки генов в клетки in vivo из-за их неспособности реплицироваться в геноме хозяина и интегрироваться в него. Однако эти плазмиды, имеющие промотор, совместимый с клеткой-хозяином, могут экспрессировать пептид из гена, функционально кодируемого в плазмиде. Некоторые широко используемые плазмиды, доступные от коммерческих поставщиков, включают pBR322, pUC18, pUC19, различные плазмиды pcDNA, pRC/CMV, различные плазмиды pCMV, pSV40 и pBlueScript. Дополнительные примеры конкретных плазмид включают pcDNA3.1, каталожный номер V79020; pcDNA3.1/hygro, каталожный номер V87020; pcDNA4/myc-His, каталожный номер V86320; и pBudCE4.1, каталожный номер V53220, все от Invitrogen (Карлсбад, Калифорния). Другие плазмиды хорошо известны обычным специалистам в данной области. Кроме того, плазмиды могут быть специально разработаны с использованием стандартных методов молекулярной биологии для удаления и/или добавления определенных фрагментов ДНК.
IV. Получение антител
Антитела или их фрагменты, которые иммуноспецифически связываются с FAM19A5 (например, человеческий FAM19A5), могут быть получены любым способом, известным в данной области техники для синтеза антител, например, химическим синтезом или методами рекомбинантной экспрессии. В описанных в данном документе способах используются, если не указано иное, обычные методы молекулярной биологии, микробиологии, генетического анализа, рекомбинантной ДНК, органической химии, биохимии, ПЦР, синтеза и модификации олигонуклеотидов, гибридизации нуклеиновых кислот и в смежных областях в пределах квалификации в данной области. Эти методы описаны, например, в цитируемых в данном документе ссылках и полностью объяснены в литературе. См., например, Maniatis T et al., (1982) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press; Sambrook J et al., (1989), Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press; Sambrook J et al., (2001) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY; Ausubel FM et al., Current Protocols in Molecular Biology, John Wiley & Sons (1987 and annual updates); Current Protocols in Immunology, John Wiley & Sons (1987 and annual updates) Gait (ed.) (1984) Oligonucleotide Synthesis: A Practical Approach, IRL Press; Eckstein (ed.) (1991) Oligonucleotides and Analogues: A Practical Approach, IRL Press; Birren B et al., (eds.) (1999) Genome Analysis: A Laboratory Manual, Cold Spring Harbor Laboratory Press.
В конкретном воплощении описанное в данном документе антитело представляет собой антитело (например, рекомбинантное антитело), полученное, экспрессируемое, созданное или выделенное любыми способами, которые включают создание, например, посредством синтеза, генной инженерии последовательностей ДНК. В некоторых воплощениях такое антитело содержит последовательности (например, последовательности ДНК или аминокислотные последовательности), которые в природе не существуют в репертуаре зародышевой линии антитела животного или млекопитающего (например, человека) in vivo. В некоторых воплощениях изобретения анти-FAM19A5 антитела, раскрытые в настоящем документе, были деиммунизированы.
Как описано в примерах (например, в примере 2), анти-FAM19A5 антитела первоначально были получены путем иммунизации цыплят синтетическим пептидом FAM19A5. Следовательно, чтобы минимизировать риск иммуногенности при введении людям, анти-FAM19A5 антитела (например, 3-2 и 2-13) были модифицированы, чтобы они больше напоминали иммуногенные последовательности человеческих антител. В некоторых воплощениях деиммунизированные анти-FAM19A5 антитела, раскрытые в настоящем документе, обладают сходной аффинностью связывания с FAM19A5 человека по сравнению с их соответствующими антителами-аналогами, которые не были деиммунизированы. В некоторых воплощениях анти-FAM19A5 антитела, раскрытые в настоящем документе, также претерпели созревание аффинности. Способы деиммунизации антитела раскрыты в данном документе, а также известны в данной области.
В определенном аспекте в настоящем документе предоставляется способ получения антитела или его антигенсвязывающего фрагмента, которое иммуноспецифически связывается с FAM19A5 (например, FAM19A5 человека), включающий культивирование клетки или клетки-хозяина, описанных в настоящем документе. В определенном аспекте в настоящем документе предусмотрен способ получения антитела или его антигенсвязывающего фрагмента, которое иммуноспецифически связывается с FAM19A5 (например, FAM19A5 человека), включающий экспрессию (например, рекомбинантную экспрессию) антитела или его антигенсвязывающего фрагмента с использованием клетки или клетки-хозяина, описанных в данном документе (например, клетки или клетки-хозяина, содержащих полинуклеотиды, кодирующие антитело, описанное в данном документе). В конкретном воплощении клетка представляет собой выделенную клетку. В конкретном воплощении экзогенные полинуклеотиды были введены в клетку. В конкретном воплощении способ дополнительно включает стадию очистки антитела или его антигенсвязывающего фрагмента, полученного из клетки или клетки-хозяина.
Способы получения поликлональных антител известны в данной области (см., например, главу 11 в: Short Protocols in Molecular Biology, (2002) 5th Ed., Ausubel FM et al., Eds., John Wiley and Sons, New York).
Моноклональные антитела могут быть получены с использованием широкого ряда методов, известных в данной области, включая использование технологий гибридом, рекомбинантных и фаговых дисплеев или их комбинации. Например, моноклональные антитела могут быть получены с использованием методов гибридом, включая методы, известные в данной области и описанные, например, в Harlow E & Lane D, Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling GJ et al., in: Monoclonal Antibodies and T-Cell Hybridomas 563 681 (Elsevier, N.Y., 1981). Используемый в данном документе термин «моноклональное антитело» не ограничивается антителами, полученными с помощью гибридомной технологии. Например, моноклональные антитела могут быть получены рекомбинантно из клеток-хозяев, экзогенно экспрессирующих описанное в данном документе антитело или его фрагмент, например легкую цепь и/или тяжелую цепь такого антитела.
В конкретных воплощениях, «моноклональное антитело» в контексте настоящего описания представляет собой антитело, продуцируемое одной клеткой (например, гибридомой или клеткой-хозяином, продуцирующей рекомбинантное антитело), где антитело иммуноспецифически связывается с FAM19A5 (например, FAM19A5 человека), как определено., например, с помощью ELISA или другого анализа связывания антигена или конкурентного связывания, известного в данной области техники или в Примерах, представленных в настоящем документе. В конкретных воплощениях моноклональное антитело может представлять собой химерное антитело или гуманизированное антитело. В некоторых воплощениях моноклональное антитело представляет собой моновалентное антитело или поливалентное (например, бивалентное) антитело. В конкретных воплощениях моноклональное антитело представляет собой моноспецифическое или мультиспецифическое антитело (например, биспецифическое антитело). Описанные в данном документе моноклональные антитела могут быть, например, получены гибридомным методом, как описано в Kohler G & Milstein C (1975) Nature 256: 495, или могут быть выделены, например, из фаговых библиотек, используя, например, способы, описанные в данном документе. Другие способы получения клональных клеточных линий и экспрессируемых ими моноклональных антител хорошо известны в данной области (см., например, главу 11 в: Short Protocols in Molecular Biology, (2002) 5th Ed., Ausubel FM et al, выше).
Способы получения и скрининга специфических антител с использованием гибридомной технологии являются обычными и хорошо известны в данной области. Например, в гибридомном способе мышь или другое подходящее животное-хозяин, такое как овца, коза, кролик, крыса, хомяк или обезьяна макака, иммунизируют для выработки лимфоцитов, которые продуцируют или способны продуцировать антитела, которые будут специфически связываться с белком (например, человеческим FAM19A5), используемым для иммунизации. Альтернативно лимфоциты можно иммунизировать in vitro. Затем лимфоциты сливают с клетками миеломы с использованием подходящего агента слияния, такого как полиэтиленгликоль, с образованием гибридомной клетки (Goding JW (Ed), Monoclonal Antibodies: Principles and Practice, pp. 59-103 (Academic Press, 1986)). Кроме того, для иммунизации животного может быть использован метод RIMMS (многократная повторная иммунизация) (Kilpatrick KE et al., (1997) Hybridoma 16: 381-9, полностью включено ссылкой).
В некоторых воплощениях мышей (или других животных, таких как куры, крысы, обезьяны, ослы, свиньи, овцы, хомяки или собаки) можно иммунизировать антигеном (например, FAM19A5, таким как FAM19A5 человека), и как только детектируется иммунный ответ, например, антитела, специфичные к антигену, детектируются в сыворотке мыши, селезенку мыши собирают и выделяют спленоциты. Затем спленоциты сливают с помощью хорошо известных методов с любыми подходящими клетками миеломы, например клетками из линии клеток SP20, доступной в Американской коллекции типовых культур (ATCC®) (Манассас, Вирджиния), с образованием гибридом. Гибридомы отбирают и клонируют путем ограниченного разведения. В некоторых воплощениях лимфатические узлы иммунизированных мышей собирают и сливают с клетками миеломы NSO.
Полученные таким образом клетки гибридомы засевают и выращивают в подходящей культуральной среде, которая предпочтительно содержит одно или несколько веществ, которые ингибируют рост или выживание неслитых исходных клеток миеломы. Например, если исходные миеломные клетки лишены фермента гипоксантин-гуанинфосфорибозилтрансферазы (HGPRT или HPRT), то культуральная среда для гибридом, как правило, будет включать гипоксантин, аминоптерин и тимидин («HAT-среда»), вещества, которые предотвращают рост дефицитных по HGPRT клеток.
В конкретных воплощениях используются клетки миеломы, которые эффективно сливаются, поддерживают стабильную продукцию антител на высоком уровне выбранными продуцирующими антитела клетками и чувствительны к среде, такой как среда HAT. Среди этих линий миеломных клеток есть линии миеломных клеток мыши, такие как линия клеток NSO или линии, полученные из опухолей мышей MOPC-21 и MPC-11, доступные в Центре распределения клеток Института Солка, Сан-Диего, Калифорния, США, и клетки SP-2 или X63-Ag8.653 доступны из Американской коллекции типовых культур, Роквилл, Мэриленд, США. Клеточные линии миеломы человека и гетеромиеломы мыши и человека также были описаны для получения человеческих моноклональных антител (Kozbor D (1984) J Immunol 133: 3001-5; Brodeur et al., Monoclonal Antibody Production Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York, 1987)).
Культуральную среду, в которой растут клетки гибридомы, исследуют на выработку моноклональных антител, направленных против FAM19A5 (например, FAM19A5 человека). Специфичность связывания моноклональных антител, продуцируемых гибридомными клетками, определяют способами, известными в данной области, например, иммунопреципитацией или анализом связывания in vitro, таким как радиоиммуноанализ (RIA) или иммуноферментный анализ (ELISA).
После идентификации гибридомных клеток, продуцирующих антитела искомой специфичности, аффинности и/или активности, клоны можно субклонировать с помощью процедур предельных разведений и выращивать стандартными способами (Goding JW (Ed), Monoclonal Antibodies: Principles and Practice, выше). Подходящие культуральные среды для этой цели включают, например, среду D-MEM или RPMI 1640. Кроме того, гибридомные клетки можно выращивать in vivo как асцитные опухоли у животных.
Моноклональные антитела, секретируемые субклонами, подходящим образом отделяют от культуральной среды, асцитной жидкости или сыворотки с помощью обычных процедур очистки иммуноглобулинов, таких как, например, протеин A-сефароза, гидроксилапатитная хроматография, гель-электрофорез, диализ или аффинная хроматография.
Описанные в данном документе антитела включают фрагменты антител, которые распознают специфический FAM19A5 (например, человеческий FAM19A5) и могут быть получены любым методом, известным специалистам в данной области. Например, описанные в данном документе фрагменты Fab и F(ab')2 могут быть получены протеолитическим расщеплением молекул иммуноглобулина с использованием ферментов, таких как папаин (для получения фрагментов Fab) или пепсин (для получения фрагментов F(ab')2). Фрагмент Fab соответствует одному из двух идентичных плеч молекулы антитела и содержит полную легкую цепь, спаренную с доменами VH и CH1 тяжелой цепи. Фрагмент F(ab')2 содержит два антигенсвязывающих плеча молекулы антитела, связанных дисульфидными связями в шарнирной области.
Кроме того, описанные в данном документе антитела или их антигенсвязывающие фрагменты также могут быть получены с использованием различных способов фагового дисплея, известных в данной области. В способах фагового дисплея функциональные домены антител отображаются на поверхности фаговых частиц, которые несут кодирующие их полинуклеотидные последовательности. В частности, последовательности ДНК, кодирующие домены VH и VL, амплифицируются из библиотек кДНК животных (например, библиотек человеческой или нечеловеческой кДНК, таких как библиотеки кДНК пораженных тканей мыши или курицы). ДНК, кодирующие домены VH и VL, рекомбинируют вместе с линкером scFv с помощью ПЦР и клонируют в фагмидный вектор. Вектор электропорируют в E. coli, и E. coli инфицируют фагом-помощником. Фаг, используемый в этих способах, обычно представляет собой нитчатый фаг, включая fd и M13, а домены VH и VL обычно рекомбинантно слиты либо с геном фага III, либо с геном VIII. Фаг, экспрессирующий антигенсвязывающий домен, который связывается с конкретным антигеном, может быть выбран или идентифицирован с помощью антигена, например, с использованием меченого антигена или антигена, связанного или захваченного на твердой поверхности или грануле. Примеры способов фагового дисплея, которые можно использовать для получения описанных в данном документе антител, включают способы, раскрытые в Brinkman U et al., (1995) J Immunol Methods 182: 41-50; Ames RS et al., (1995) J Immunol Methods 184: 177- 186; Kettleborough CA et al., (1994) Eur J Immunol 24: 952-958; Persic L et al., (1997) Gene 187: 9-18; Burton DR & Barbas CF (1994) Advan Immunol 57: 191-280; заявка PCT № PCT/GB91/001134; Международные публикации № WO 90/02809, WO 91/10737, WO 92/01047, WO 92/18619, WO 93/1 1236, WO 95/15982, WO 95/20401 и WO 97/13844; и Пат. США № 5698426, 5223409, 5403484, 5580717, 5427908, 5750753, 5821047, 5571698, 5427908, 5516637, 5780225, 5658727, 5733743 и 5969108.
Как описано в приведенных выше источниках, после отбора фага кодирующие области антитела из фага могут быть выделены и использованы для создания целых антител, включая человеческие антитела, или любого другого искомого антигенсвязывающего фрагмента, и экспрессированы в любом желаемом хозяине, включая клетки млекопитающих, клетки насекомых, клетки растений, дрожжи и бактерии, например, как описано ниже. Методы рекомбинантного получения фрагментов антител, таких как фрагменты Fab, Fab 'и F(ab')2, также могут быть использованы с использованием способов, известных в данной области, таких как раскрытые в публикации РСТ № WO 92/22324; Mullinax RL et al., (1992) BioTechniques 12(6): 864-9; Sawai H et al., (1995) Am J Reprod Immunol 34: 26-34; и Better M et al., (1988) Science 240: 1041- 1043.
В одном аспекте для создания полных антител праймеры ПЦР, включая нуклеотидные последовательности VH или VL, сайт рестрикции и фланкирующую последовательность для защиты сайта рестрикции, могут использоваться для амплификации последовательностей VH или VL из матрицы, например, клонов scFv. Используя методы клонирования, известные специалистам в данной области, амплифицированные ПЦР домены VH можно клонировать в векторы, экспрессирующие константную область VH, а амплифицированные ПЦР домены VL можно клонировать в векторы, экспрессирующие константную область VL, например, константные области каппа или лямбда человека. Домены VH и VL также можно клонировать в один вектор, экспрессирующий необходимые константные области. Векторы конверсии тяжелой цепи и векторы конверсии легкой цепи затем котрансфицируют в клеточные линии для получения стабильных или транзиторных клеточных линий, которые экспрессируют полноразмерные антитела, например, IgG, с использованием методов, известных специалистам в данной области.
Химерное антитело - это молекула, в которой разные части антитела происходят из разных молекул иммуноглобулина. Например, химерное антитело может содержать вариабельную область моноклонального антитела животного, не являющегося человеком (например, мыши, крысы или курицы), слитого с константной областью человеческого антитела. Способы получения химерных антител известны в данной области. См., например, Morrison SL (1985) Science 229: 1202-7; Oi VT & Morrison SL (1986) BioTechniques 4: 214- 221; Gillies SD et al., (1989) J Immunol Methods 125: 191-202; и Пат. США № 5807715, 4816567, 4816397 и 6331415.
Гуманизированное антитело способно связываться с заранее определенным антигеном и включает каркасную область, имеющую по существу аминокислотную последовательность человеческого иммуноглобулина и CDR, имеющие по существу аминокислотную последовательность нечеловеческого иммуноглобулина (например, иммуноглобулина мыши или курицы). В конкретных воплощениях гуманизированное антитело также включает, по меньшей мере, часть константной области иммуноглобулина (Fc), обычно иммуноглобулина человека. Антитело также может включать области СН1, шарнира, СН2, СН3 и СН4 тяжелой цепи. Гуманизированное антитело можно выбрать из любого класса иммуноглобулинов, включая IgM, IgG, IgD, IgA и IgE, и любого изотипа, включая IgG1, IgG2, IgG3 и IgG4. Гуманизированные антитела могут быть получены с использованием различных методов, известных в данной области, включая, без ограничения указанным, трансплантацию CDR (Европейский патент № EP 239400; Международная публикация No. WO 91/09967; и Пат. США № 5225539, 5530101 и 5585089), венирование или перекладка (европейские патенты № EP 592106 и EP 519596; Padlan EA (1991) Mol Immunol 28 (4/5): 489-498; Studnicka GM et al. (1994) Prot Engineering 7 (6): 805-814; и Roguska MA et al., (1994) PNAS 91: 969-973), шаффлинг цепей (Пат. США № 5565332) и методы, раскрытые, например, в Пат. США № 6407213, США. Пат. США № 5766886, международная публикация № WO 93/17105; Tan P et al., (2002) J Immunol 169: 1119-25; Caldas C et al., (2000) Protein Eng. 13(5): 353-60; Morea V et al., (2000) Methods 20(3): 267- 79; Baca M et al., (1997) J Biol Chem 272(16): 10678-84; Roguska MA et al., (1996) Protein Eng 9(10): 895 904; Couto JR et al., (1995) Cancer Res. 55 (23 Supp): 5973s-5977s; Couto JR et al., (1995) Cancer Res 55(8): 1717-22; Sandhu JS (1994) Gene 150(2): 409-10 и Pedersen JT et al., (1994) J Mol Biol 235(3): 959-73. См. также Публ.заявки США № US 2005/0042664 A1 (Фев. 24, 2005), которая полностью включена в настоящий документ ссылкой.
Были описаны способы создания мультиспецифических (например, биспецифических антител), см., например, Пат. США № 7951917; 7183076; 8227577; 5837242; 5989830; 5869620; 6132992 и 8586713.
Однодоменные антитела, например антитела, лишенные легких цепей, можно получить способами, хорошо известными в данной области. См. Riechmann L & Muyldermans S (1999) J Immunol 231: 25-38; Nuttall SD et al., (2000) Curr Pharm Biotechnol 1 (3): 253-263; Muyldermans S, (2001) J Biotechnol 74 (4): 277-302; Патент № 6005079; и международные публикации № WO 94/04678, WO 94/25591 и WO 01/44301.
Кроме того, антитела, которые иммуноспецифически связываются с антигеном FAM19A5, могут, в свою очередь, использоваться для создания антиидиотипических антител, которые «имитируют» антиген, с использованием методов, хорошо известных специалистам в данной области. (см., например, Greenspan NS & Bona CA (1989) FASEB J 7(5): 437-444; и Nissinoff A (1991) J Immunol 147(8): 2429-2438).
В конкретных воплощениях описанное в данном документе антитело, которое связывается с тем же эпитопом FAM19A5 (например, FAM19A5 человека), что и описанное в данном документе анти-FAM19A5 антитело, представляет собой человеческое антитело или его антигенсвязывающий фрагмент. В конкретных воплощениях описанное в данном документе антитело, которое конкурентно блокирует (например, дозозависимым образом) антитела, описанные в настоящем документе, от связывания с FAM19A5 (например, с FAM19A5 человека), представляет собой человеческое антитело или его антигенсвязывающий фрагмент.
Человеческие антитела можно получить с использованием любого способа, известного в данной области. Например, можно использовать трансгенных мышей, которые не способны экспрессировать функциональные эндогенные иммуноглобулины, но могут экспрессировать гены иммуноглобулинов человека. В частности, генные комплексы тяжелых и легких цепей иммуноглобулинов человека можно вводить случайным образом или путем гомологичной рекомбинации в эмбриональные стволовые клетки мыши. Альтернативно, вариабельная область человека, константная область и область разнообразия могут быть введены в эмбриональные стволовые клетки мыши в дополнение к генам тяжелой и легкой цепей человека. Гены мышиного иммуноглобулина тяжелой и легкой цепей можно сделать нефункциональными по отдельности или одновременно с введением локусов иммуноглобулина человека путем гомологичной рекомбинации. В частности, гомозиготная делеция JH-области предотвращает эндогенную выработку антител. Модифицированные эмбриональные стволовые клетки наращивают и микроинъецируют в бластоцисты для получения химерных мышей. Затем химерных мышей разводят для получения гомозиготного потомства, которое экспрессирует человеческие антитела. Трансгенных мышей иммунизируют обычным способом выбранным антигеном, например, полным антигеном или частью антигена (например, FAM19A5). Моноклональные антитела, направленные против антигена, можно получить от иммунизированных трансгенных мышей с использованием общепринятой гибридомной технологии. Трансгены человеческого иммуноглобулина, содержащиеся в трансгенных мышах, перестраиваются во время дифференцировки В-клеток, а затем претерпевают переключение классов и соматические мутации. Таким образом, применение такого метода, возможно для продуцирования терапевтически полезных антител IgG, IgA, IgM и IgE. Для обзора этой технологии получения человеческих антител см. Lonberg N & Huszar D (1995) Int Rev Immunol 13: 65-93. Подробное обсуждение этой технологии получения человеческих антител и человеческих моноклональных антител и протоколов получения таких антител см., например, в международных публикациях № WO 98/24893, WO 96/34096 и WO 96/33735; и Пат. США № 5413923, 5625126, 5633425, 5569825, 5661016, 5545806, 5814318 и 5939598. Примеры мышей, способных продуцировать человеческие антитела, включают Xenomouse™ (Abgenix, Inc.; Пат. США № 6 075 181 и 6 150 184), HuAb-Mouse™ (Mederex, Inc./Gen Pharm; Пат. США № 5545806 и 5569825), Trans Chromo Mouse™ (Kirin) и KM Mouse™ (Medarex/Kirin).
Человеческие антитела, которые специфически связываются с FAM19A5 (например, человеческим FAM19A5), могут быть получены различными способами, известными в данной области, включая способы фагового дисплея, описанные выше, с использованием библиотек антител, полученных из последовательностей иммуноглобулинов человека. См. также Пат. США № 4 444 887, 4 716 111 и 5 885 793; и международные публикации № WO 98/46645, WO 98/50433, WO 98/24893, WO 98/16654, WO 96/34096, WO 96/33735 и WO 91/10741.
В некоторых воплощениях человеческие антитела можно получить с использованием гибридом мыши и человека. Например, лимфоциты периферической крови человека, трансформированные вирусом Эпштейна-Барр (EBV), могут быть слиты с клетками миеломы мыши для получения гибридом мышь-человек, секретирующих человеческие моноклональные антитела, и эти гибридомы мышь-человек могут быть подвергнуты скринингу для определения тех, которые секретируют человеческие моноклональные антитела, которые иммуноспецифически связываются с антигеном-мишенью (например, FAM19A5, таким как FAM19A5 человека)). Такие способы известны и описаны в данной области, см., например, Shinmoto H et al., (2004) Cytotechnology 46: 19-23; Naganawa Y et al., (2005) Human Antibodies 14: 27-31.
V. Способы конструирования антител
Как обсуждалось выше, анти-FAM19A5 антитело, имеющее последовательности VH и VL, описанные в данном документе, можно использовать для создания нового анти-FAM19A5 антитела путем модификации последовательностей VH и/или VL или прикрепленных к ним константных областей. Таким образом, в другом аспекте, описанном в данном документе, структурные особенности анти-FAM19A5 антитела, описанного в данном документе, используются для создания структурно родственных анти-FAM19A5 антител, которые сохраняют, по меньшей мере, одно функциональное свойство антител, описанных в данном документе, такое как связывание с FAM19A5 человека. Например, исходным материалом для способа конструирования являются последовательности VH и/или VL, представленные в данном документе, или одна или несколько их областей CDR. Для создания сконструированного антитела нет необходимости фактически готовить (т.е. экспрессировать в виде белка) антитело, имеющее одну или несколько последовательностей VH и/или VL, представленных в данном документе, или одну или несколько их участков CDR. Скорее, информация, содержащаяся в последовательности(ях), используется в качестве исходного материала для создания последовательности(ей) «второго поколения», полученной из исходной последовательности(ей), а затем последовательность(и) «второго поколения» подготавливается и экспрессируется в виде белка.
Соответственно, в настоящем документе представлены способы получения анти-FAM19A5 антитела, включающие:
(a) обеспечение: (i) последовательности вариабельной области тяжелой цепи, содержащей последовательность CDR1, CDR2 и/или CDR3, как указано в таблице 3, или CDR1, CDR2 и/или CDR3 вариабельной области тяжелой цепи, как указано в таблице 5; и (ii) последовательность вариабельной области легкой цепи, содержащую последовательность CDR1, CDR2 и/или CDR3, как указано в таблице 4, или CDR1, CDR2 и/или CDR3 вариабельной области тяжелой цепи, как указано в таблице 6;
(b) изменение, по меньшей мере, одного аминокислотного остатка в последовательности вариабельной области тяжелой цепи и/или последовательности вариабельной области легкой цепи для создания, по меньшей мере, одной измененной последовательности антитела; и
(c) экспрессию измененной последовательности антитела в виде белка.
Для получения и экспрессии измененной последовательности антитела можно использовать стандартные методы молекулярной биологии.
В некоторых воплощениях антитело, кодируемое измененной последовательностью(ями) антитела, представляет собой антитело, которое сохраняет одно, некоторые или все функциональные свойства анти-FAM19A5 антител, описанных в настоящем документе, которые включают:
(1) снижение иммуногенности у людей;
(2) связывание с растворимым FAM19A5 человека, например, с KD 10 нМ или менее (например, от 001 до 10 нМ), например, при измерении с помощью Biacore;
(3) связывание с мембраносвязанным FAM19A5 человека, например, с KD 10 нМ или менее (например, от 001 до 1 нМ), например, при измерении с помощью ELISA;
(4) связывание с мембраносвязанным FAM19A5 человека, например, с EC50 1 нМ или менее (например, от 001 до 1 нМ), например, при измерении с помощью ELISA;
(5) уменьшение, обращение вспять, задержку и/или предотвращение начала реактивного глиоза;
(6) подавление чрезмерной пролиферации реактивных астроцитов;
(7) снижение экспрессии протеогликанов хондроитинсульфата, включая нейрокан и нейрон-глиальный антиген 2 (NG2);
(8) увеличение экспрессии c-fos и pERK в ядрах нейронов;
(9) стимуляция выживания нейронов;
(10) увеличение экспрессии GAP43 в нейронах;
(11) стимуляция отрастания аксона; и
(12) конкуренция в любом направлении или в обоих направлениях за связывание с FAM19A5 человека с анти-FAM19A5 антителом, раскрытым в данном документе.
Измененное антитело может проявлять одно или несколько, два или более, три или более, четыре или более, пять или более, шесть или более, семь или более, восемь или более, девять или более, десять или более, одиннадцать или все из функциональных свойств, указанных выше в (1) - (12). Функциональные свойства измененных антител можно оценить с помощью стандартных анализов, доступных в данной области техники и/или описанных в данном документе, таких как те, что изложены в примерах (например, ELISA, FACS).
В некоторых воплощениях способов конструирования антител, описанных в данном документе, мутации могут быть введены случайным образом или выборочно вдоль всей или части кодирующей последовательности анти-FAM19A5 антитела, и полученные модифицированные анти-FAM19A5 антитела могут быть проверены на связывающую активность и/или другие функциональные свойства, как описано в данном документе. Мутационные способы описаны в данной области техники. Например, в публикации РСТ WO 02/092780 от Short описаны способы создания и скрининга мутаций антител с использованием насыщающего мутагенеза, сборки синтетического лигирования или их комбинации. В качестве альтернативы, публикация РСТ WO 03/074679 Lazar et al. описывает способы использования компьютерных методов скрининга для оптимизации физико-химических свойств антител.
VI. Клетки и векторы
В определенных аспектах в настоящем документе представлены клетки (например, клетки-хозяева), экспрессирующие (например, рекомбинантно) антитела, описанные в настоящем документе (или их антигенсвязывающий фрагмент), которые специфически связываются с FAM19A5 (например, FAM19A5 человека) и родственными полинуклеотидами и экспрессирующими векторами. В настоящем документе предусмотрены векторы (например, экспрессирующие векторы), содержащие полинуклеотиды, содержащие нуклеотидные последовательности, кодирующие анти-FAM19A5 антитела или фрагмент для рекомбинантной экспрессии в клетках-хозяевах, например, в клетках млекопитающих. В настоящем документе также представлены клетки-хозяева, содержащие такие векторы для рекомбинантной экспрессии анти-FAM19A5 антител, описанных в настоящем документе (например, человеческого или гуманизированного антитела). В конкретном аспекте в настоящем документе представлены способы получения описанного в данном документе антитела, включающие экспрессию такого антитела из клетки-хозяина.
Рекомбинантная экспрессия антитела, описанного в настоящем документе (например, полноразмерного антитела, тяжелой и/или легкой цепи антитела, или одноцепочечного антитела, описанного в настоящем документе), которое специфически связывается с FAM19A5 (например, FAM19A5 человека), включает создание экспрессирующего вектора, включающего полинуклеотид, кодирующий антитело. После получения полинуклеотида, кодирующего молекулу антитела, тяжелую и/или легкую цепь антитела или его фрагмент (например, вариабельные домены тяжелой и/или легкой цепи), описанные в данном документе, вектор для продуцирования молекулы антитела может могут быть получен с помощью технологии рекомбинантных ДНК с использованием методов, хорошо известных в данной области. Таким образом, в данном документе описаны способы получения белка путем экспрессии полинуклеотида, содержащего нуклеотидную последовательность, кодирующую антитело или фрагмент антитела (например, легкую цепь или тяжелую цепь). Способы, хорошо известные специалистам в данной области, можно использовать для конструирования экспрессирующих векторов, содержащих кодирующие последовательности антитела или фрагмента антитела (например, легкой цепи или тяжелой цепи) и соответствующие сигналы контроля транскрипции и трансляции. Эти способы включают, например, in vitro методы рекомбинантных ДНК, синтетические методы и in vivo генетическую рекомбинацию. Также предлагаются реплицируемые векторы, содержащие нуклеотидную последовательность, кодирующую молекулу антитела, описанную в данном документе, тяжелую или легкую цепь антитела, вариабельный домен тяжелой или легкой цепи антитела или его фрагмента, или CDR тяжелой или легкой цепи, функционально связанные с промотором. Такие векторы могут, например, включать нуклеотидную последовательность, кодирующую константную область молекулы антитела (см., например, международные публикации № WO 86/05807 и WO 89/01036; и пат. США № 5122464) и вариабельные домены антитела могут быть клонированы в такой вектор для экспрессии всей тяжелой, всей легкой цепи или обеих целых тяжелой и легкой цепей.
Экспрессирующий вектор может быть перенесен в клетку (например, клетку-хозяин) обычными методами, и полученные клетки затем можно культивировать обычными методами для получения антитела, описанного в данном документе (например, антитела, содержащего VH и/или VL, или одно или более VH и/или VL CDR анти-FAM19A5 антитела согласно настоящему раскрытию) или его фрагмента. Таким образом, в настоящем документе представлены клетки-хозяева, содержащие полинуклеотид, кодирующий антитело, описанное в данном документе, или его фрагменты, или его тяжелую или легкую цепь, или его фрагмент, или одноцепочечное антитело, описанное в данном документе, функционально связанный с промотором для экспрессии таких последовательностей в клетке-хозяине. В некоторых воплощениях для экспрессии двухцепочечных антител векторы, кодирующие как тяжелую, так и легкую цепи по отдельности, могут совместно экспрессироваться в клетке-хозяине для экспрессии всей молекулы иммуноглобулина, как подробно описано ниже. В некоторых воплощениях клетка-хозяин содержит вектор, содержащий полинуклеотид, кодирующий как тяжелую цепь, так и легкую цепь антитела, описанного в данном документе, или его фрагмент. В конкретных воплощениях клетка-хозяин содержит два разных вектора, первый вектор, содержащий полинуклеотид, кодирующий тяжелую цепь или вариабельную область тяжелой цепи описанного в данном документе антитела, или его фрагмент, и второй вектор, содержащий полинуклеотид, кодирующий легкую цепь или вариабельную область легкой цепи антитела, описанного в данном документе, или его фрагмент. В других воплощениях первая клетка-хозяин содержит первый вектор, содержащий полинуклеотид, кодирующий тяжелую цепь или вариабельную область тяжелой цепи антитела, описанного в данном документе, или его фрагмент, а вторая клетка-хозяин содержит второй вектор, содержащий полинуклеотид, кодирующий легкую цепь или вариабельную область легкой цепи антитела, описанного в данном документе. В конкретных воплощениях вариабельная область тяжелой цепи/тяжелой цепи, экспрессируемая первой клеткой, ассоциирована с вариабельной областью легкой цепи/легкой цепи второй клетки с образованием описанного в данном документе анти-FAM19A5 антитела или его антигенсвязывающего фрагмента. В определенных воплощениях в настоящем документе представлена популяция клеток-хозяев, включающая такую первую клетку-хозяин и такую вторую клетку-хозяин.
В конкретном воплощении в настоящем документе представлена популяция векторов, включающая первый вектор, содержащий полинуклеотид, кодирующий вариабельную область легкой цепи/легкой цепи анти-FAM19A5 антитела, описанного в настоящем документе, и второй вектор, содержащий полинуклеотид, кодирующий тяжелую цепь/вариабельная область тяжелой цепи анти-FAM19A5 антитела, описанного в данном документе.
Для экспрессии описанных в данном документе молекул антител можно использовать различные системы хозяин-экспрессирующий вектор. Такие системы хозяин-экспрессия представляют собой носители, с помощью которых могут быть получены и впоследствии очищены представляющие интерес кодирующие последовательности, но также представляют собой клетки, которые при трансформации или трансфекции соответствующими нуклеотидными кодирующими последовательностями могут экспрессировать описанную в данном документе молекулу антитела in situ. Они включают, без ограничения указанным, микроорганизмы, такие как бактерии (например, E. coli и B. subtilis), трансформированные рекомбинантной ДНК бактериофага, плазмидной ДНК или экспрессирующими векторами космидной ДНК, содержащими кодирующие последовательности антитела; дрожжи (например, Saccharomyces Pichia), трансформированные рекомбинантными экспрессирующими векторами дрожжей, содержащими кодирующие последовательности антитела; системы клеток насекомых, инфицированные экспрессирующими векторами рекомбинантного вируса (например, бакуловируса), содержащими кодирующие последовательности антител; системы растительных клеток (например, зеленые водоросли, такие как Chlamydomonas reinhardtii), инфицированные экспрессирующими векторами на основе рекомбинантного вируса (например, вирусом мозаики цветной капусты, CaMV; вирус табачной мозаики, TMV) или трансформированные экспрессирующими векторами на основе рекомбинантных плазмид (например, плазмида Ti), содержащими последовательностями, кодирующими антитела; или клеточные системы млекопитающих (например, COS (например, COS1 или COS), CHO, BHK, MDCK, HEK 293, NSO, PER.C6, VERO, CRL7030, HsS78Bst, HeLa и NIH 3T3, HEK-293T, HepG2, SP210, R1.1, BW, LM, BSC1, BSC40, YB/20 и клетки BMT10), несущие рекомбинантные экспрессионные конструкции, содержащие промоторы, полученные из генома клеток млекопитающих (например, промотор металлотионеина) или из вирусов млекопитающих (например, поздний промотор аденовируса; 7.5K промотор вируса осповакцины). В конкретном воплощении клетки для экспрессии описанных в данном документе антител или их антигенсвязывающего фрагмента представляют собой клетки СНО, например клетки СНО из системы CHO GS System™ (Lonza). В конкретном воплощении клетки для экспрессии описанных в данном документе антител представляют собой клетки человека, например, линии клеток человека. В конкретном воплощении экспрессирующий вектор млекопитающих представляет собой pOptiVEC™ или pcDNA3.3. В конкретном воплощении бактериальные клетки, такие как Escherichia coli, или эукариотические клетки (например, клетки млекопитающих), особенно для экспрессии всей молекулы рекомбинантного антитела, используются для экспрессии молекулы рекомбинантного антитела. Например, клетки млекопитающих, такие как клетки яичника китайского хомячка (СНО), в сочетании с вектором, таким как главный элемент промотора немедленно-раннего гена из цитомегаловируса человека, представляют собой эффективную систему экспрессии антител (Foecking MK & Hofstetter H (1986) Gene 45: 101-5; and Cockett MI et al., (1990) Biotechnology 8(7): 662-7). В некоторых воплощениях описанные в данном документе антитела продуцируются клетками СНО или клетками NSO. В конкретном воплощении экспрессия нуклеотидных последовательностей, кодирующих описанные в данном документе антитела, которые иммуноспецифически связываются с FAM19A5 (например, человеческим FAM19A5), регулируется конститутивным промотором, индуцибельным промотором или тканеспецифическим промотором.
В бактериальных системах можно преимущественно выбрать ряд экспрессирующих векторов в зависимости от применения, предназначенного для экспрессируемой молекулы антитела. Например, когда должно быть получено большое количество такого антитела, для создания фармацевтических композиций молекулы антитела могут быть желательны векторы, которые управляют экспрессией продуктов слияния на высоком уровне, которые легко очищаются. Такие векторы включают, без ограничения указанным, экспрессирующий вектор E. coli pUR278 (Ruether U & Mueller-Hill B (1983) EMBO J 2: 1791-1794), в котором кодирующая последовательность антитела может быть лигирована индивидуально в вектор в рамке с кодирующей областью lac Z, так что образуется гибридный белок; векторы pIN (Inouye S & Inouye M (1985) Nuc Acids Res 13: 3101-3109; Van Heeke G & Schuster SM (1989) J Biol Chem 24: 5503-5509); и т.п. Например, векторы pGEX также можно использовать для экспрессии чужеродных полипептидов в виде гибридных белков с глутатион-5-трансферазой (GST). Как правило, такие гибридные белки растворимы и могут быть легко очищены от лизированных клеток путем адсорбции и связывания с матриксными гранулами глутатион-агарозы с последующей элюцией в присутствии свободного глутатиона. Векторы pGEX предназначены для включения сайтов расщепления протеазой тромбином или фактором Ха, так что клонированный продукт целевого гена может высвобождаться от группы GST.
В системе насекомых вирус ядерного полиэдроза Autographa californica (AcNPV), например, можно использовать в качестве вектора для экспрессии чужеродных генов. Вирус растет в клетках Spodoptera frugiperda. Кодирующую последовательность антитела можно клонировать индивидуально в несущественные области (например, ген полиэдрина) вируса и поместить под контроль промотора AcNPV (например, промотора полиэдрина).
В клетках-хозяевах млекопитающих можно использовать ряд вирусных систем экспрессии. В случаях, когда аденовирус используется в качестве экспрессирующего вектора, представляющая интерес кодирующая последовательность антитела может быть лигирована с комплексом контроля транскрипции/трансляции аденовируса, например, с поздним промотором и трехкомпонентной лидерной последовательностью. Затем этот химерный ген может быть вставлен в геном аденовируса путем рекомбинации in vitro или in vivo. Вставка в несущественную область вирусного генома (например, область E1 или E3) приведет к получению рекомбинантного вируса, который является жизнеспособным и способным экспрессировать молекулу антитела в инфицированных хозяевах (см., например, Logan J & Shenk T (1984).) PNAS 81 (12): 3655-9). Для эффективной трансляции вставленных кодирующих последовательностей антител также могут потребоваться специфические сигналы инициации. Эти сигналы включают кодон инициации ATG и соседние последовательности. Кроме того, кодон инициации должен находиться в фазе с рамкой считывания искомой кодирующей последовательности, чтобы гарантировать трансляцию всей вставки. Эти экзогенные контрольные сигналы трансляции и кодоны инициации могут иметь различное происхождение, как природное, так и синтетическое. Эффективность экспрессии можно повысить путем включения соответствующих элементов энхансера транскрипции, терминаторов транскрипции и т.д. (см., например, Bitter G et al., (1987) Methods Enzymol. 153: 516-544).
Кроме того, может быть выбран штамм клетки-хозяина, который модулирует экспрессию встроенных последовательностей или модифицирует и обрабатывает продукт гена желаемым специфическим образом. Такие модификации (например, гликозилирование) и процессинг (например, расщепление) белковых продуктов могут быть важны для функции белка. Различные клетки-хозяева обладают характерными и специфическими механизмами посттрансляционной обработки и модификации белков и генных продуктов. Соответствующие клеточные линии или хозяйские системы могут быть выбраны для гарантии корректной модификации или процессирования экспрессируемого инородного белка. С этой целью можно использовать эукариотические клетки-хозяева, которые обладают клеточным механизмом для правильного процессинга первичного транскрипта, гликозилирования и фосфорилирования продукта гена. Такие клетки-хозяева млекопитающих включают, без ограничения указанным, клетки CHO, VERO, BHK, Hela, MDCK, HEK 293, NIH 3T3, W138, BT483, Hs578T, HTB2, BT20 и T47D, NSO (линия клеток миеломы мыши, которая не продуцирует эндогенно любые цепи иммуноглобулина), CRL7030, COS (например, COS 1 или COS), PER.C6, VERO, HsS78Bst, HEK-293T, HepG2, SP210, R1.1, BW, LM, BSC 1, BSC40, YB/20, BMT10 и HsS78Bst. В некоторых воплощениях описанные в данном документе анти-FAM19A5 антитела продуцируются в клетках млекопитающих, таких как клетки СНО.
В конкретном воплощении описанные в данном документе антитела или их антигенсвязывающие части имеют пониженное содержание фукозы или не содержат фукозы. Такие антитела можно получить с использованием методов, известных специалистам в данной области. Например, антитела могут экспрессироваться в клетках, дефицитных или лишенных способности к фукозилированию. В конкретном примере клеточные линии с нокаутом обоих аллелей 16-фукозилтрансферазы можно использовать для получения антител или их антигенсвязывающих частей с пониженным содержанием фукозы. Система Potelligent® (Lonza) является примером такой системы, которую можно использовать для получения антител или их антигенсвязывающих частей с пониженным содержанием фукозы.
Для длительного продуцирования рекомбинантных белков с высоким выходом могут быть созданы стабильные экспрессирующие клетки. Например, могут быть сконструированы клеточные линии, которые стабильно экспрессируют описанное в данном документе анти-FAM19A5 антитело и его антигенсвязывающую часть. В конкретных воплощениях клетка, представленная в настоящем документе, стабильно экспрессирует вариабельный домен легкой цепи/легкой цепи и вариабельный домен тяжелой цепи/тяжелой цепи, которые связываются с образованием антитела, описанного в данном документе, или его антигенсвязывающей части.
В определенных аспектах, вместо использования экспрессирующих векторов, которые содержат вирусные источники репликации, клетки-хозяева можно трансформировать ДНК, контролируемой соответствующими элементами контроля экспрессии (например, промотором, энхансером, последовательностями, терминаторами транскрипции, сайтами полиаденилирования и т.п.) и селективным маркером. После введения чужеродной ДНК/полинуклеотида сконструированным клеткам можно дать возможность расти в течение 1-2 дней в обогащенной среде, а затем перевести на селективную среду. Селективный маркер в рекомбинантной плазмиде придает устойчивость к отбору и позволяет клеткам стабильно интегрировать плазмиду в свои хромосомы и расти с образованием фокусов, которые, в свою очередь, можно клонировать и размножать в клеточные линии. Этот способ можно успешно использовать для конструирования клеточных линий, которые экспрессируют анти-FAM19A5 антитело, описанное в данном документе, или его связывающую часть антитела. Такие сконструированные клеточные линии могут быть особенно полезны при скрининге и оценке композиций, которые прямо или косвенно взаимодействуют с молекулой антитела.
Можно использовать ряд систем отбора, включая, помимо прочего, гены тимидинкиназы вируса простого герпеса (Wigler M. et al., (1977) Cell 11 (1): 223-32), гипоксантингуанинфосфорибозилтрансферазы (Szybalska EH & Szybalski W. (1962) PNAS 48 (12): 2026-2034) и аденинфосфорибозилтрансферазы (Lowy I et al., (1980) Cell 22 (3): 817-23) можно использовать в tk-, hgprt- или aprt-клетках, соответственно. Кроме того, устойчивость к антиметаболитам может быть использована в качестве основы отбора для следующих генов: dhfr, который придает устойчивость к метотрексату (Wigler M et al., (1980) PNAS 77 (6): 3567-70; O'Hare K et al., (1981) PNAS 78: 1527-31); gpt, который придает устойчивость к микофеноловой кислоте (Mulligan RC & Berg P (1981) PNAS 78 (4): 2072-6); neo, который придает устойчивость к аминогликозиду G-418 (Wu GY & Wu CH (1991) Biotherapy 3: 87-95; Tolstoshev P (1993) Ann Rev Pharmacol Toxicol 32: 573-596; Mulligan RC (1993) Science 260: 926-932; и Morgan RA & Anderson WF (1993) Ann Rev Biochem 62: 191-217; Nabel GJ & Feigner PL (1993) Trends Biotechnol 11 (5): 211-5); и hygro, который придает устойчивость к гигромицину (Santerre RF et al., (1984) Gene 30 (1-3): 147-56). Способы, широко известные в области технологии рекомбинантных ДНК, могут применяться рутинно для выбора искомого рекомбинантного клона, и такие способы описаны, например, в Ausubel FM et al., (eds.), Current Protocols in Molecular Biology, John Wiley & Sons, NY (1993); Kriegler M, Gene Transfer and Expression, A Laboratory Manual, Stockton Press, NY (1990); и в главах 12 и 13 Dracopoli NC et al. (eds.), Current Protocols in Human Genetics, John Wiley & Sons, NY (1994); Colbere-Garapin F et al., (1981) J Mol Biol 150: 1-14, которые полностью включены в настоящий документ ссылкой.
Уровни экспрессии молекулы антитела можно повысить путем амплификации вектора (для обзора см. Bebbington CR & Hentschel CCG, The use of vectors based on gene amplification for the expression of cloned genes in mammalian cells in DNA cloning, Vol. 3 (Academic Press, New York, 1987)). Когда маркер в векторной системе, экспрессирующей антитело, поддается амплификации, увеличение уровня ингибитора, присутствующего в культуре клетки-хозяина, увеличит количество копий маркерного гена. Поскольку амплифицированная область связана с геном антитела, продуцирование антитела также будет увеличиваться (Crouse GF et al., (1983) Mol Cell Biol 3: 257-66).
Клетка-хозяин может быть котрансфицирована двумя или более экспрессирующими векторами, описанными в данном документе, первый вектор кодирует полипептид, полученный из тяжелой цепи, а второй вектор, кодирующий полипептид, полученный из легкой цепи. Два вектора могут содержать идентичные селектируемые маркеры, которые обеспечивают равную экспрессию полипептидов тяжелой и легкой цепей. Клетки-хозяева можно котрансфицировать разными количествами двух или более экспрессирующих векторов. Например, клетки-хозяева можно трансфицировать любым из следующих соотношений первого экспрессирующего вектора и второго экспрессирующего вектора: 1:1, 1:2, 1:3, 1:4, 1:5, 1:6, 1:7, 1:8, 1:9, 1:10, 1:12, 1 :15, 1:20, 1:25, 1:30, 1:35, 1:40, 1:45 или 1:50.
В качестве альтернативы можно использовать один вектор, который кодирует и способен экспрессировать полипептиды как тяжелой, так и легкой цепи. В таких ситуациях легкая цепь должна располагаться перед тяжелой цепью, чтобы избежать избытка токсичной свободной тяжелой цепи (Proudfoot NJ (1986) Nature 322: 562-565; и Kohler G (1980) PNAS 77: 2197-2199). Кодирующие последовательности тяжелой и легкой цепей могут включать кДНК или геномную ДНК. Экспрессирующий вектор может быть моноцистронным или мультицистронным. Конструкция мультицистронной нуклеиновой кислоты может кодировать 2, 3, 4, 5, 6, 7, 8, 9, 10 или более или в диапазоне 2-5, 5-10 или 10-20 генов/нуклеотидных последовательностей. Например, конструкция бицистроновой нуклеиновой кислоты может содержать в следующем порядке промотор, первый ген (например, тяжелую цепь антитела, описанного в данном документе) и второй ген (например, легкую цепь антитела, описанного в данном документе). В таком экспрессирующем векторе транскрипция обоих генов может управляться промотором, тогда как трансляция мРНК из первого гена может осуществляться с помощью кэп-зависимого механизма сканирования, а трансляция мРНК из второго гена может осуществляться посредством кэп-независимого механизма сканирования, например, с помощью IRES.
После того, как описанная в данном документе молекула антитела была получена путем рекомбинантной экспрессии, ее можно очистить любым способом, известным в данной области техники для очистки молекулы иммуноглобулина, например, с помощью хроматографии (например, с помощью ионного обмена, аффинностнной хроматографии, особенно аффинностнной хроматографии к специфическому антигену после протеина A и эксклюзионной хроматографии), центрифугирования, дифференциальной растворимости или любого другого стандартного метода очистки белков. Кроме того, описанные в данном документе антитела могут быть слиты с гетерологичными полипептидными последовательностями, описанными в данном документе или иным образом известными в данной области, для облегчения очистки.
В конкретных воплощениях антитело или его антигенсвязывающая часть, описанные в настоящем документе, выделяют или очищают. Как правило, выделенное антитело - это антитело, которое по существу не содержит других антител с антигенной специфичностью, отличной от специфичности выделенного антитела. Например, в конкретном воплощении препарат антитела, описанный в настоящем документе, по существу не содержит клеточного материала и/или химических предшественников. Выражение «по существу не содержит клеточного материала» включает препараты антитела, в которых антитело отделено от клеточных компонентов клеток, из которых оно выделено или получено рекомбинантно. Таким образом, антитело, которое практически не содержит клеточного материала, включает препараты антитела, содержащие менее примерно 30%, 20%, 10%, 5%, 2%, 1%, 05% или 01% (по сухой массе) гетерологичного белка (также называемого в данном документе «контаминирующим белком») и/или варианты антитела, например, различные посттрансляционные модифицированные формы антитела или другие различные версии антитела (или связывающие антитело части). Когда антитело получают рекомбинантным способом, оно также обычно практически не содержит культуральной среды, т.е. культуральная среда составляет менее чем около 20%, 10%, 2%, 1%, 05% или 01% от объема белкового препарата. Когда антитело получают путем химического синтеза, оно, как правило, практически не содержит химических предшественников или других химикатов, т.е. оно отделено от химических предшественников или других химикатов, которые участвуют в синтезе белка. Соответственно, такие препараты антитела содержат менее примерно 30%, 20%, 10% или 5% (по сухой массе) химических предшественников или соединений, отличных от представляющего интерес антитела. В конкретном воплощении описанные в данном документе антитела выделяют или очищают.
VII. Количественные анализы
Описанные в данном документе антитела можно тестировать на связывание с FAM19A5, например, с помощью стандартного ELISA. Вкратце, микротитрационные планшеты покрывают очищенным FAM19A5 в концентрации 1-2 мкг/мл в PBS, а затем блокируют 5% бычьим сывороточным альбумином в PBS. Разведения антител (например, разведения плазмы от мышей, иммунизированных FAM19A5) добавляют в каждую лунку и инкубируют в течение 1-2 часов при 37°C. Планшеты промывали PBS/Tween, а затем инкубировали вторичным реагентом (например, для человеческих антител, поликлональным реагентом козьих антител против Fc человеческого IgG), конъюгированным с пероксидазой хрена (HRP) в течение 1 часа при 37°C. После промывки планшеты проявляли с помощью субстрата ABTS (Moss Inc, продукт: ABTS-1000) и анализировали на спектрофотометре при OD 415-495. Затем сыворотки от иммунизированных мышей подвергали дополнительному скринингу с помощью проточной цитометрии на предмет связывания с линией клеток, экспрессирующей человеческий FAM19A5, но не с контрольной линией клеток, которая не экспрессирует FAM19A5. Вкратце, связывание анти-FAM19A5 антител оценивали путем инкубации клеток СНО, экспрессирующих FAM19A5, с анти-FAM19A5 антителом в разведении 1:20. Клетки промывали и детектировали связывание с PE-меченными антителами против IgG человека. Анализы проточной цитометрии выполняли с использованием проточной цитометрии FACS (Becton Dickinson, Сан-Хосе, Калифорния). Предпочтительно для слияния использовали мышей с наивысшими титрами.
Анализ ELISA, как описано выше, можно использовать для скрининга антител и, таким образом, гибридом, которые продуцируют антитела, которые проявляют положительную реактивность с иммуногеном FAM19A5. Гибридомы, которые продуцируют антитела, которые связываются, предпочтительно с высокой аффинностью, с FAM19A5, затем могут быть субклонированы и дополнительно охарактеризованы. Затем можно выбрать один клон от каждой гибридомы, который сохраняет реактивность родительских клеток (с помощью ELISA), для создания банка клеток и для очистки антител.
Для очистки анти-FAM19A5 антител отобранные гибридомы можно выращивать в двухлитровых вращающихся колбах для очистки моноклональных антител. Надосадочные жидкости можно фильтровать и концентрировать перед аффинной хроматографией с протеином A-сефарозой (Pharmacia, Пискатауэй, Нью-Джерси). Элюированный IgG можно проверить с помощью гель-электрофореза и высокоэффективной жидкостной хроматографии для обеспечения чистоты. Буферный раствор можно заменить на PBS, и концентрацию можно определить по OD 280 с использованием коэффициента экстинкции 143. Моноклональные антитела можно разделить на аликвоты и хранить при -80°C.
Чтобы определить, связываются ли выбранные моноклональные анти-FAM19A5 антитела с уникальными эпитопами, каждое антитело можно биотинилировать с использованием коммерчески доступных реагентов (Pierce, Рокфорд, Иллинойс). Связывание биотинилированных MAb можно обнаружить с помощью зонда, меченного стрептавидином. Конкурентные исследования с использованием немеченых моноклональных антител и биотинилированных моноклональных антител могут быть выполнены с использованием планшетов для ELISA, покрытых FAM19A5, как описано выше.
Чтобы определить изотип очищенных антител, изотипический ELISA может быть выполнен с использованием реагентов, специфичных для антител определенного изотипа. Например, для определения изотипа человеческого моноклонального антитела лунки микротитровальных планшетов могут быть покрыты 1 мкг/мл иммуноглобулина человека в течение ночи при 4°C. После блокирования 1% BSA в планшетах начинали реакцию с 1 мкг/мл или меньше тестируемых моноклональных антител или очищенных контролей изотипа при температуре окружающей среды в течение одного-двух часов. Затем в лунках можно начать реакцию взаимодействия с зондами из щелочной фосфатазы конъюгированной либо с человеческим IgG1, либо с человеческим IgM. Планшеты проявляли и анализировали, как описано выше.
Чтобы проверить связывание моноклональных антител с живыми клетками, экспрессирующими FAM19A5, можно использовать проточную цитометрию, как описано в примерах. Вкратце, клеточные линии, экспрессирующие мембраносвязанный FAM19A5 (выращенные в стандартных условиях роста), смешивали с различными концентрациями моноклональных антител в PBS, содержащем 01% BSA, при 4°C в течение 1 часа. После промывания клетки реагировали с меченным флуоресцеином антителом против IgG в тех же условиях, что и при окрашивании первичным антителом. Образцы могут быть проанализированы с помощью прибора FACScan с использованием свойств рассеяния света и бокового рассеяния для анализа отдельных клеток и определения связывания меченых антител. Альтернативный анализ с использованием флуоресцентной микроскопии может быть использован (в дополнение или вместо) анализа проточной цитометрии. Клетки можно окрашивать точно так, как описано выше, и исследовать с помощью флуоресцентной микроскопии. Этот способ позволяет визуализировать отдельные клетки, но может иметь пониженную чувствительность в зависимости от плотности антигена.
Анти-FAM19A5 антитела можно дополнительно протестировать на реактивность с антигеном FAM19A5 с помощью вестерн-блоттинга. Вкратце, клеточные экстракты из клеток, экспрессирующих FAM19A5, могут быть получены и подвергнуты электрофорезу в полиакриламидном геле с додецилсульфатом натрия. После электрофореза отделенные антигены переносятся на нитроцеллюлозные мембраны, блокируются 20% сывороткой мыши и исследуются с помощью тестируемых моноклональных антител. Связывание IgG можно определить с помощью анти-IgG щелочной фосфатазы и проявить с помощью таблеток субстрата BCIP/NBT (Sigma Chem. Co., Сент-Луис, Миссури).
Способы анализа аффинности связывания, перекрестной реактивности и кинетики связывания различных анти-FAM19A5 антител включают стандартные анализы, известные в данной области, например, анализ поверхностного плазмонного резонанса (SPR) Biacore™ с использованием прибора Biacore™ 2000 SPR (Biacore AB, Упсала, Швеция).
В одном воплощении антитело специфически связывается с растворимой формой FAM19A5 человека. В одном воплощении антитело специфически связывается с мембраносвязанной формой FAM19A5 человека. Антитело может специфически связываться с конкретным эпитопом FAM19A5 (например, с SEQ ID NO: 90 или фрагментом в SEQ ID NO: 90). В некоторых воплощениях антитело специфически связывает человеческий FAM19A5, предпочтительно с высокой аффинностью, и не реагирует перекрестно с другими членами подсемейства белков FAM19.
VIII. Биспецифические молекулы
Описанные в данном документе антитела можно использовать для образования биспецифических молекул. Анти-AM19A5 антитело или его антигенсвязывающие части могут быть дериватизированы или связаны с другой функциональной молекулой, например, с другим пептидом или белком (например, с другим антителом или лигандом для рецептора) для создания биспецифической молекулы, которая связывается, по меньшей мере, с два разных сайта связывания или молекулы-мишени. Цитокины, такие как IL-6, CNTF, LIF, EGF и TGFα, были задействованы как триггеры начала глиоза и/или реактивного астроглиоза (Balasingam et al., J. Neurosci. 14 (2): 846-56 (1994); Winter et al., Proc. Natl. Acad. Sci. U. S. A. 20; 92(13):5865-9 (1995)) путем активации белка передатчика сигнала и активатор транскрипции 3 (STAT3), который затем регулирует многие аспекты реактивного астроглиоза после повреждения ЦНС. Herrmann J. E. et al., J. Neurosci. 28(28): 7231-7243 (2008). Например, отсутствие или снижение STAT3 приводит к ослабленной активации глиального фибриллярного кислого белка (GFAP), отсутствию гипертрофии астроцитов и увеличению распространения воспаления, увеличению объема поражения и частичному ослаблению восстановления моторики после повреждения ЦНС. Herrmann J. E. et al., J. Neurosci. 28(28): 7231-7243 (2008). Таким образом, например, анти-FAM19A5 антитело может быть связано с антителом или scFv, которое специфически связывается с любым белком, который участвует в ингибировании начала глиоза и/или чрезмерной пролиферации реактивного астроглиоза для комбинированного лечения, например, антителом к IL- 6, CNTF, LIF, EGF или TGFα.
Кроме того, анти-FAM19A5 антитело может быть связано с антителом или scFv, которое лечит заболевание или нарушение, включая повреждение центральной нервной системы (например, черепно-мозговое повреждение, цереброспинальное повреждение, инсульт или опухоль головного мозга), цереброспинальное системное повреждение, дегенеративное заболевание головного мозга (например, болезнь Хантингтона, болезнь Паркинсона, болезнь Альцгеймера, рассеянный склероз, ALS), дегенеративное цереброспинальное или нервное расстройство или невропатическую боль у объекта (см. заболевания или нарушения в Разделе XII ниже). Например, анти-FAM19A5 антитело может быть связано с антителом или scFv, например, натализумабом (TYSABRI®), алемтузумабом (LEMTRADA®), которыми лечат рассеянный склероз.
Описанное в данном документе антитело фактически может быть дериватизировано или связано с более чем одной другой функциональной молекулой для создания мультиспецифических молекул, которые связываются более чем с двумя различными сайтами связывания и/или молекулами-мишенями; такие мультиспецифические молекулы также охватываются термином «биспецифическая молекула» в том смысле, в котором он в данном документе используется. Для создания биспецифической молекулы, описанной в данном документе, описанное в данном документе антитело может быть функционально связано (например, путем химического связывания, генетического слияния, нековалентной ассоциации или иным образом) с одной или несколькими другими связывающими молекулами, такими как другое антитело, его связывающая часть антитела, пептид или миметик связывания, так что получается биспецифическая молекула. В одном воплощении биспецифическая молекула связывается с FAM19A5 и VEGF. В другом воплощении биспецифическая молекула связывается с FAM19A5 и EGF.
Соответственно, в настоящем документе представлены биспецифические молекулы, содержащие, по меньшей мере, одну первую специфичность связывания для FAM19A5 и вторую специфичность связывания для второго целевого эпитопа. В воплощении, описанном в данном документе, в котором биспецифическая молекула является мультиспецифической, молекула может дополнительно включать третью специфичность связывания.
В одном воплощении биспецифические молекулы, описанные в настоящем документе, содержат в качестве специфичности связывания, по меньшей мере, одно антитело или его связывающую часть антитела, включая, например, Fab, Fab', F(ab')2, Fv или одноцепочечный Fv (scFv). Антитело также может быть димером легкой цепи или тяжелой цепи или любым его минимальным фрагментом, таким как Fv или одноцепочечная конструкция, как описано в Ladner et al., пат. США № 4946778, содержание которого специально включено ссылкой.
Хотя человеческие моноклональные антитела являются предпочтительными, другие антитела, которые можно использовать в биспецифических молекулах, описанных в данном документе, представляют собой мышиные, химерные и гуманизированные моноклональные антитела.
Описанные в данном документе биспецифические молекулы могут быть получены конъюгированием составляющих специфичностей связывания с использованием способов, известных в данной области. Например, каждая специфичность связывания биспецифической молекулы может быть получена отдельно, а затем конъюгирована друг с другом. Когда специфичностями связывания являются белки или пептиды, для ковалентной конъюгации можно использовать различные связывающие или перекрестно сшивающие агенты. Примеры сшивающих агентов включают протеин A, карбодиимид, N-сукцинимидил-S-ацетилтиоацетат (SATA), 55'-дитиобис(2-нитробензойную кислоту) (DTNB), о-фенилендималеимид (oPDM), N- сукцинимидил-3- (2-пиридилдитио) пропионат (SPDP) и сульфосукцинимидил-4-(N-малеимидометил) циклогаксан-1-карбоксилат (сульфо-SMCC) (см., например, Karpovsky et al., (1984) J. Exp. Med., 160: 1686; Liu, MA et al., (1985) Proc. Natl. Acad. Sci. USA 82:8648). Другие способы включают способы, описанные в Paulus (1985) Behring Ins. Mitt. No. 78, 118-132; Brennan et al., (1985) Science 229:81-83), и Glennie et al., (1987) J. Immunol. 139: 2367-2375). Предпочтительными конъюгирующими агентами являются SATA и сульфо-SMCC, оба доступны у Pierce Chemical Co. (Рокфорд, Иллинойс).
Когда специфичностями связывания являются антитела, они могут быть конъюгированы посредством сульфгидрильного связывания С-концевых шарнирных областей двух тяжелых цепей. В особенно предпочтительном воплощении шарнирная область модифицируется, чтобы содержать нечетное количество сульфгидрильных остатков, предпочтительно один, перед конъюгацией.
Альтернативно, обе специфичности связывания можно кодировать в одном и том же векторе, экспрессировать и собирать в одной и той же клетке-хозяине. Этот способ особенно полезен, когда биспецифическая молекула представляет собой mAb x mAb, mAb x Fab, mAb x (scFv) 2, Fab x F(ab')2 или лиганд x гибридный белок Fab. Биспецифическое антитело может включать антитело, содержащее scFv на С-конце каждой тяжелой цепи. Описанная в данном документе биспецифическая молекула может быть одноцепочечной молекулой, содержащей одно одноцепочечное антитело и детерминанту связывания, или одноцепочечной биспецифической молекулой, содержащей две детерминанты связывания. Биспецифические молекулы могут включать, по меньшей мере, две одноцепочечные молекулы. Способы получения биспецифических молекул описаны, например, в пат. США № 5260203; пат. США № 5455030; пат. США № 4881175; пат. США № 5 132 405; пат. США № 5091513; пат. США № 5476786; пат. США № 5 013 653; пат. США № 5 258 498; и пат. США № 5482858.
Связывание биспецифических молекул с их конкретными мишенями может быть подтверждено с использованием признанных в данной области способов, таких как твердофазный иммуноферментный анализ (ELISA), радиоиммуноанализ (RIA), анализ FACS, биоанализ (например, ингибирование роста) или анализ вестерн-блоттинга. Каждый из этих анализов обычно определяет присутствие комплексов белок-антитело, представляющих особый интерес, с использованием меченого реагента (например, антитела), специфичного для интересующего комплекса.
IX. Диагностика
В одном воплощении фрагмент, присоединенный к антителу против FAM19A5, выбран из группы, состоящей из связывающего фрагмента, метящего фрагмента и биологически активного фрагмента.
Описанные в данном документе антитела можно использовать для диагностических целей, включая анализ образцов и визуализацию in vivo, и для этой цели антитело (или его связывающая часть) можно конъюгировать с подходящим детектируемым агентом с образованием иммуноконъюгата. Для диагностических целей подходящими агентами являются детектируемые метки, которые включают радиоизотопы, для визуализации всего тела и радиоизотопы, ферменты, флуоресцентные метки и другие подходящие метки антител для тестирования образцов.
Детектируемые метки могут быть любого из различных типов, используемых в настоящее время в области диагностики in vitro, включая метки в виде частиц, включая золи металлов, такие как коллоидное золото, изотопы, такие как I125 или Tc99, представленные, например, с пептидным хелатирующим агентом N2S2, N3S. или типа N4, хромофоры, включая флуоресцентные маркеры, люминесцентные маркеры, фосфоресцентные маркеры и т.п., а также ферментные метки, которые превращают данный субстрат в детектируемый маркер, и полинуклеотидные метки, которые выявляются после амплификации, такой как полимеразная цепная реакция. Подходящие ферментные метки включают пероксидазу хрена, щелочную фосфатазу и т.п. Например, метка может быть ферментом щелочной фосфатазой, обнаруживаемой путем измерения наличия или образования хемилюминесценции после преобразования 12-диоксетановых субстратов, таких как адамантилметоксифосфорилоксифенилдиоксетан (AMPPD), динатрий 3-(4-(метоксиспиро{l, 2-диоксетан-32'-(5'-хлор)трицикло {3.3.1.1 37}декан}-4-ил)фенилфосфат (CSPD), а также CDP и CDP-star® или другие люминесцентные субстраты, хорошо известные специалистам в данной области, например хелаты подходящих лантаноидов, таких как тербий (III) и европий (III). Средство детекции определяется выбранной меткой. Внешний вид этикетки или продуктов ее реакции может быть достигнут невооруженным глазом, если этикетка представляет собой твердые частицы и накапливается на соответствующих уровнях, или с помощью таких инструментов, как спектрофотометр, люминометр, флуориметр и т.п. в соответствии со стандартной практикой.
Описанные в данном документе антитела также могут быть конъюгированы с терапевтическим агентом с образованием иммуноконъюгата, такого как конъюгат антитело-лекарственное средство (ADC). Подходящие терапевтические агенты включают агенты, модулирующие начало глиоза и/или реактивного астроглиоза, и/или лечат дегенеративные заболевания головного мозга, повреждение центральной нервной системы или невропатическую боль. Терапевтические агенты для лечения дегенеративных заболеваний головного мозга включают лекарства для лечения болезни Хантингтона, болезни Паркинсона, болезни Альцгеймера, рассеянного склероза и бокового амиотрофического склероза (ALS). Сюда входят лекарственные средства, обычно используемые для лечения таких дегенеративных заболеваний головного мозга, например, лекарственные средства, описанные ниже в Разделе XII.
Иммуноконъюгаты можно получить способами, известными в данной области. Предпочтительно, способы конъюгации приводят к связям, которые являются по существу (или почти) неиммуногенными, например пептидными (т.е. амидными), сульфидными (стерически затрудненными), дисульфидными, гидразоновыми и эфирными связями. Эти связи почти неиммуногенны и демонстрируют разумную стабильность в сыворотке (см., например, Senter, P.D., Curr. Opin. Chem. Biol. 13 (2009) 235-244; WO 2009/059278; WO 95/17886).
В зависимости от биохимической природы фрагмента и антитела можно использовать разные стратегии конъюгации. В случае, если фрагмент является природным или рекомбинантным и содержит от 50 до 500 аминокислот, в учебниках есть стандартные процедуры, описывающие химию синтеза белковых конъюгатов, которым может легко следовать квалифицированный специалист (см., например, Hackenberger, CPR, и Schwarzer, D., Angew. Chem. Int. Ed. Engl. 47 (2008) 10030-10074). В одном воплощении используется реакция малеинимидо-фрагмента с остатком цистеина в антителе или фрагменте. Это особенно подходящая химия связывания используется в случае, например, Fab или Fab'-фрагмента антитела. Альтернативно, в одном воплощении проводят связывание с С-концевым концом антитела или фрагмента. С-концевая модификация белка, например, Fab-фрагмента, например, может быть выполнена, как описано (Sunbul, M. and Yin, J., Org. Biomol. Chem. 7 (2009) 3361-3371).
В общем, сайт-специфическая реакция и ковалентное связывание основаны на превращении природной аминокислоты в аминокислоту с реакционной способностью, которая ортогональна реакционной способности других присутствующих функциональных групп. Например, конкретный цистеин в контексте редкой последовательности может ферментативно превращаться в альдегид (см. Frese, M.A., and Dierks, T., ChemBioChem. 10 (2009) 425-427). Также возможно получить желаемую модификацию аминокислоты, используя специфическую ферментативную реактивность определенных ферментов с природной аминокислотой в данном контексте последовательности (см., например, Taki, M. et al., Prot. Eng. Des. Sel. 17 (2004) 119-126; Gautier, A. et al., Chem. Biol. 15 (2008) 128-136; а катализируемое протеазой образование связей C - N используется Bordusa, F., Highlights in Bioorganic Chemistry (2004) 389-403).
Сайт-специфическая реакция и ковалентное связывание также могут быть достигнуты путем селективной реакции концевых аминокислот с соответствующими модифицирующими реагентами. Реакционную способность N-концевого цистеина с бензонитрилами (см. Ren, H. et al., Angew. Chem. Int. Ed. Engl. 48 (2009) 9658-9662) можно использовать для достижения сайт-специфичного ковалентного связывания. Нативное химическое лигирование также может зависеть от C-концевых остатков цистеина (Taylor, E. Vogel; Imperiali, B, Nucleic Acids and Molecular Biology (2009), 22 (Protein Engineering), 65-96).
ЕР 1074 563 описывает способ конъюгации, который основан на более быстрой реакции цистеина в отрезке отрицательно заряженных аминокислот с цистеином, расположенным на отрезке положительно заряженных аминокислот.
Фрагмент также может быть синтетическим пептидом или имитатором пептида. В случае химического синтеза полипептида во время такого синтеза могут быть включены аминокислоты с ортогональной химической реакционной способностью (см., например, de Graaf, A.J. et al., Bioconjug. Chem. 20 (2009) 1281-1295). Поскольку на кону большое разнообразие ортогональных функциональных групп, которые могут быть введены в синтетический пептид, конъюгация такого пептида с линкером является стандартной химией.
Чтобы получить моно-меченый полипептид, конъюгат со стехиометрией 1:1 можно отделить хроматографией от других побочных продуктов конъюгации. Эту процедуру можно облегчить, используя меченый красителем член пары связывания и заряженный линкер. Используя этот вид меченого и сильно отрицательно заряженного члена пары связывания, моноконъюгированные полипептиды легко отделяются от немеченых полипептидов и полипептидов, которые несут более одного линкера, поскольку для разделения можно использовать разницу в заряде и молекулярной массе. Флуоресцентный краситель может быть использован для очистки комплекса от несвязанных компонентов, таких как меченое одновалентное связующее.
X. Фармацевтические композиции
В настоящем документе представлены композиции, содержащие описанное в данном документе антитело или его антигенсвязывающую часть, имеющую искомую степень чистоты в физиологически приемлемом носителе, эксцепиенте или стабилизаторе (Remington's Pharmaceutical Sciences (1990) Mack Publishing Co., Истон, Пенсильвания). Приемлемые носители, эксцепиенты или стабилизаторы нетоксичны для реципиентов в используемых дозировках и концентрациях и включают буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол и м-крезол); полипептиды с низкой молекулярной массой (менее примерно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбитол; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как TWEEN®, PLURONICS® или полиэтиленгликоль (PEG).
В конкретном воплощении фармацевтические композиции содержат антитело или его антигенсвязывающую часть, биспецифическую молекулу или иммуноконъюгат, описанные в настоящем документе, и, возможно, одно или несколько дополнительных профилактических или терапевтических средств в фармацевтически приемлемом носителе. В конкретном воплощении фармацевтические композиции содержат эффективное количество антитела или его антигенсвязывающей части, описанного в данном документе, и, возможно, одно или несколько дополнительных профилактических терапевтических агентов в фармацевтически приемлемом носителе. В некоторых воплощениях антитело является единственным активным ингредиентом, включенным в фармацевтическую композицию. Описанные в данном документе фармацевтические композиции могут быть полезны для усиления, индукции или активации активности FAM19A5 и лечения состояния, такого как повреждение центральной нервной системы, дегенеративное заболевание головного мозга или невропатическая боль.
Фармацевтически приемлемые носители, используемые в парентеральных препаратах, включают водные носители, неводные носители, антимикробные агенты, изотонические агенты, буферы, антиоксиданты, местные анестетики, суспендирующие и диспергирующие агенты, эмульгирующие агенты, связывающие или хелатирующие агенты и другие фармацевтически приемлемые вещества. Примеры водных носителей включают инъекцию хлорида натрия, инъекцию Рингера, инъекцию изотонической декстрозы, инъекцию стерильной воды, инъекцию декстрозы и инъекцию лактата Рингера. Неводные парентеральные носители включают жирные масла растительного происхождения, хлопковое масло, кукурузное масло, кунжутное масло и арахисовое масло. Противомикробные агенты в бактериостатических или фунгистатических концентрациях могут быть добавлены к парентеральным препаратам, упакованным в многодозовые контейнеры, которые включают фенолы или крезолы, ртуть, бензиловый спирт, хлорбутанол, сложные эфиры метил- и пропил-пара-гидроксибензойной кислоты, тимеросал, хлорид бензалкония и хлорид бензетония. Изотонические агенты включают хлорид натрия и декстрозу. Буферы включают фосфат и цитрат. Антиоксиданты включают бисульфат натрия. Местные анестетики включают гидрохлорид прокаина. Суспендирующие и диспергирующие агенты включают карбоксиметилцеллюлозу натрия, гидроксипропилметилцеллюлозу и поливинилпирролидон. Эмульгирующие агенты включают полисорбат 80 (TWEEN® 80). Агенты, связывающие ионы металлов, или хелатирующие агенты, включают EDTA. Фармацевтические носители также включают этиловый спирт, полиэтиленгликоль и пропиленгликоль для смешивающихся с водой носителей; и гидроксид натрия, соляную кислоту, лимонную кислоту или молочную кислоту для регулирования pH.
Фармацевтическая композиция может быть составлена для любого пути введения объекту. Конкретные примеры способов введения включают интраназальный, пероральный, парентеральный, интратекальный, интрацеребровентрикулярный, легочный, подкожный или внутрижелудочковый. В данном документе также рассматривается парентеральное введение, характеризующееся подкожной, внутримышечной или внутривенной инъекцией. Инъекционные препараты могут быть приготовлены в обычных формах, либо в виде жидких растворов или суспензий, твердых форм, подходящих для растворения или суспендирования в жидкости перед инъекцией, либо в виде эмульсий. Инъекционные составы, растворы и эмульсии также содержат один или несколько эксцепиентов. Подходящими эксцепиентами являются, например, вода, физиологический раствор, декстроза, глицерин или этанол. Кроме того, при желании вводимые фармацевтические композиции могут также содержать небольшие количества нетоксичных вспомогательных веществ, таких как смачивающие или эмульгирующие агенты, буферные агенты pH, стабилизаторы, усилители растворимости и другие подобные агенты, такие как, например, натрий ацетат, монолаурат сорбитана, олеат триэтаноламина и циклодекстрины.
Препараты для парентерального введения антитела включают стерильные растворы, готовые к инъекции, стерильные сухие растворимые продукты, такие как лиофилизированные порошки, готовые к смешиванию с растворителем непосредственно перед использованием, включая таблетки для подкожных инъекций, стерильные суспензии, готовые к инъекции, стерильные сухие нерастворимые продукты, готовые к смешиванию с носителем непосредственно перед использованием и стерильными эмульсиями. Растворы могут быть водными или неводными.
При внутривенном введении подходящие носители включают физиологический раствор или фосфатно-солевой буферный раствор (PBS), а также растворы, содержащие загущающие и солюбилизирующие агенты, такие как глюкоза, полиэтиленгликоль и полипропиленгликоль и их смеси.
Смеси для местного применения, содержащие антитело, готовят, как описано для местного и системного введения. Полученная смесь может представлять собой раствор, суспензию, эмульсии и т.п. и может быть приготовлена в виде кремов, гелей, мазей, эмульсий, растворов, эликсиров, лосьонов, суспензий, настоек, паст, пен, аэрозолей, орошений, спреев, суппозиториев, повязок, кожных пластырей или любых других составов, подходящих для местного применения.
Антитело или его антигенсвязывающая часть, описанные в данном документе, могут быть составлены в виде аэрозоля для местного применения, такого как ингаляция (см., например, Пат. США № 4044126, 4414209 и 4364923, которые описывают аэрозоли для доставки стероида, полезного для лечения воспалительных заболеваний, в частности астмы). Эти составы для введения в дыхательные пути могут быть в форме аэрозоля или раствора для распылителя или в виде мелкодисперсного порошка для инсуффляции, отдельно или в комбинации с инертным носителем, таким как лактоза. В таком случае частицы состава в одном воплощении будут иметь диаметр менее 50 микрон, в одном воплощении менее 10 микрон.
Антитело или его антигенсвязывающая часть, описанные в данном документе, могут быть составлены для местного или местного применения, например, для местного нанесения на кожу и слизистые оболочки, например, в глаза, в форме гелей, кремов и лосьонов, а также для применения в глазе или для интрацистернального или интраспинального применения. Предусматривается местное введение для трансдермальной доставки, а также для введения в глаза или слизистую оболочку или для ингаляционной терапии. Также можно вводить назальные растворы антитела отдельно или в комбинации с другими фармацевтически приемлемыми эксцепиентами.
Трансдермальные пластыри, включая устройства для ионтофоретического и электрофоретического действия, хорошо известны специалистам в данной области и могут использоваться для введения антитела. Например, такие пластыри раскрыты в Пат. США № 6267983, 6261595, 6256533, 6167301, 6024975, 6010715, 5985317, 5983134, 5948433 и 5860957.
В некоторых воплощениях фармацевтическая композиция, содержащая антитело или его антигенсвязывающую часть, описанная в настоящем документе, представляет собой лиофилизированный порошок, который можно восстановить для введения в виде растворов, эмульсий и других смесей. Его также можно восстановить и приготовить в виде твердых веществ или гелей. Лиофилизированный порошок получают растворением антитела или его антигенсвязывающей части, описанных в данном документе, или его фармацевтически приемлемого производного в подходящем растворителе. В некоторых воплощениях лиофилизированный порошок является стерильным. Растворитель может содержать эксцепиент, улучшающий стабильность, или другой фармакологический компонент порошка или восстановленного раствора, приготовленного из порошка. Эксцепиенты, которые можно использовать, включают, без ограничения указанным, декстрозу, сорбит, фруктозу, кукурузный сироп, ксилит, глицерин, глюкозу, сахарозу или другой подходящий агент. Растворитель также может содержать буфер, такой как цитрат, фосфат натрия или калия, или другой такой буфер, известный специалистам в данной области техники, в одном воплощении при примерно нейтральном pH. Последующая стерильная фильтрация раствора с последующей лиофилизацией в стандартных условиях, известных специалистам в данной области, обеспечивает желаемый состав. В одном воплощении полученный раствор будет распределен по флаконам для лиофилизации. Каждый флакон будет содержать одну или несколько доз соединения. Лиофилизированный порошок можно хранить в соответствующих условиях, например при температуре от около 4°C до комнатной.
Восстановление этого лиофилизированного порошка водой для инъекций обеспечивает состав для парентерального введения. Для восстановления лиофилизированный порошок добавляют в стерильную воду или другой подходящий носитель. Точное количество зависит от выбранного соединения. Такое количество можно определить опытным путем.
Антитела или их антигенсвязывающие части, биспецифическая молекула или иммуноконъюгат, описанные в настоящем документе, и другие композиции, представленные в настоящем документе, также могут быть составлены для нацеливания на конкретную ткань, рецептор или другую область тела объекта, подлежащего лечению. Многие такие способы нацеливания хорошо известны специалистам в данной области. Все такие способы нацеливания рассматриваются в настоящем документе для использования в настоящих композициях. Неограничивающие примеры способов нацеливания см., например, в Пат. США № 6316652, 6274552, 6271359, 6253872, 6139865, 6131570, 6120751, 6071495, 6060082, 6048736, 6039975, 6004534, 5985307, 5972366, 5900252, 5840674, 5759542 и 5709874. В конкретном воплощении антитело или его антигенсвязывающая часть, описанные в данном документе, нацелены на лечение повреждения центральной нервной системы, дегенеративного заболевания головного мозга или невропатической боли.
Композиции, используемые для введения in vivo, могут быть стерильными. Это легко достигается путем фильтрации, например, через стерильные фильтрующие мембраны.
XI. Наборы
В настоящем документе предлагаются наборы, содержащие одно или несколько описанных в данном документе антител или их антигенсвязывающие части, биспецифические молекулы или их иммуноконъюгаты. В конкретном воплощении в настоящем документе предоставляется фармацевтическая упаковка или набор, включающий один или несколько контейнеров, заполненных одним или несколькими ингредиентами описанных в данном документе фармацевтических композиций, такими как одно или несколько антител, представленных в настоящем документе, или их антигенсвязывающая часть, необязательно инструкция по использованию. В некоторых воплощениях наборы содержат фармацевтическую композицию, описанную в данном документе, и любое профилактическое или терапевтическое средство, такое как те, которые описаны в данном документе.
XII. Терапевтические применения и способы
В настоящем документе также представлены способы смягчения поражения или повреждения ЦНС у нуждающегося в этом объекта (например, человека), включающие введение объекту анти-FAM19A5 антитела, биспецифической молекулы или иммуноконъюгата, описанных в настоящем документе, или их композиции.
В других аспектах, представленных в настоящем документе, представлены способы ингибирования, замедления, подавления, сдерживания, уменьшения, обращения или предотвращения начала или инициации глиоза и связанных с ним пагубных эффектов на ЦНС у объекта, включающие введение объекту анти- FAM19A5 антитела, описанного в данном документе. В некоторых воплощениях в настоящем документе представлены способы ингибирования, замедления, супрессии, сдерживания, уменьшения, обращения или предотвращения чрезмерной или аномальной пролиферации реактивных астроцитов и связанных с ней пагубных эффектов на ЦНС у объекта, включающие введение объекту анти-FAM19A5-антитела по настоящему раскрытию. В некоторых воплощениях в настоящем документе представлены способы снижения, ингибирования или снижения экспрессии протеогликанов хондроитинсульфата (включая уровень нейрокана, NG2 или обоих), или снижения активности, или превращения в неактивный нейрокан, NG2 или обоих в объект, включающий введение объекту анти-FAM19A5 антитела, описанного в данном документе. В некоторых воплощениях в настоящем документе представлены способы стимуляции, промотирования, увеличения или активации роста нейронов, предпочтительно после травмы или повреждения у объекта, включающие введение объекту анти-FAM19A5 антитела, как описано в данном документе. В других воплощениях в настоящем документе представлены способы повышения уровня активности мРНК c-fos, белка c-fos или белка c-fos, а также повышения уровня активности мРНК ERK, белка ERK или pERK, предпочтительно в ядре нейронов у объекта, нуждающегося в этом, включающий введение объекту анти-FAM19A5 антитела по настоящему раскрытию. В определенных воплощениях в настоящем документе представлены способы усиления или повышения уровня мРНК GAP43, белка GAP43 или повышения активности белка GAP43, предпочтительно в нейронах, у объекта, нуждающегося в этом, включающие введение объекту анти-FAM19A5 антитела по настоящему раскрытию. В определенных воплощениях в настоящем документе представлены способы увеличения или стимулирования выживания нейронов и/или стимулирования возобновления роста аксона у объекта, нуждающегося в этом, включающие введение объекту анти-FAM19A5 антитела, раскрытого в настоящем документе. В некоторых воплощениях объект представляет собой человека, предпочтительно человека, имеющего поражение или повреждение нейрона, например, в результате повреждения ЦНС, травмы, ранения, цереброспинального повреждения, опухоли головного мозга, инфекции, ишемии, инсульта, аутоиммунных реакций и/или нейродегенеративной болезни.
В некоторых аспектах в настоящем документе также представлены способы лечения заболевания, расстройства или состояния у объекта, нуждающегося в этом, включающие введение объекту анти-FAM19A5 антитела согласно настоящему раскрытию. В некоторых воплощениях заболевание, нарушение или состояние включает повреждение центральной нервной системы, повреждение спинномозговой системы, дегенеративное заболевание головного мозга, дегенеративное цереброспинальное или нервное расстройство или невропатическую боль. В некоторых воплощениях повреждение центральной нервной системы представляет собой черепно-мозговую травму, цереброспинальное повреждение, инсульт, опухоль головного мозга или их комбинацию. В некоторых воплощениях дегенеративное заболевание головного мозга представляет собой болезнь Хантингтона, болезнь Паркинсона, болезнь Альцгеймера, рассеянный склероз, боковой амиотрофический склероз (ALS) или их комбинацию. Таким образом, в определенных воплощениях в настоящем документе раскрыт способ лечения черепно-мозговой травмы, цереброспинального повреждения, инсульта, опухоли головного мозга или их комбинации у объекта, нуждающегося в этом, включающий введение объекту раскрытого анти-FAM19A5 антитела, раскрытого в данном документе, или их композицию. В некоторых воплощениях в настоящем документе раскрывается способ лечения болезни Гентингтона, болезни Паркинсона, болезни Альцгеймера, рассеянного склероза, ALS у объекта, который в этом нуждается, включающий введение объекту анти-FAM19A5 антитела, раскрытого в настоящем документе, или его композиции. В некоторых воплощениях объектом является человек.
В некоторых воплощениях анти-FAM19A5 антитело можно вводить в комбинации с одним или несколькими дополнительными агентами для лечения повреждения центральной нервной системы (например, черепно-мозговой травмы, цереброспинального повреждения, инсульта или опухоли головного мозга), повреждения цереброспинальной системы, дегенеративного заболевания головного мозга (например, болезни Хантингтона, болезни Паркинсона, болезни Альцгеймера, рассеянного склероза, ALS), дегенеративного цереброспинального или нервного расстройства или невропатической боли.
В некоторых воплощениях заболевание, нарушение или состояние включает опухоль, фиброз, глаукому, ретинопатию, возрастную дегенерацию желтого пятна или расстройство настроения. В некоторых воплощениях заболевание, нарушение или состояние включает опухоль. В некоторых воплощениях изобретения опухоль включает меланому, рак поджелудочной железы, глиому (например, мультиформную глиобластому (GBM)), рак молочной железы, лимфому, рак легкого, рак почки, рак предстательной железы, фибросаркому, аденокарциному толстой кишки, рак печени или рак яичников.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию вызывает нормализацию кровеносных сосудов, например, внутри опухоли. В некоторых воплощениях нормализация кровеносных сосудов сопровождается изменениями свойств кровеносных сосудов, включая повышенную коннективность, увеличенную толщину стенки, уменьшенный диаметр сосудов, более правильное направление и характер распределения сосудов, увеличенное количество сосудов, уменьшение утечки и проницаемости, увеличенное покрытие перицитов и близость к кровеносным сосудам, повышенную оксигенацию или их комбинации.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию подавляет рост опухоли. В некоторых воплощениях рост опухоли подавляется, по меньшей мере, на 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% по сравнению со стандартом (например, рост опухоли у объекта, который не получал анти-FAM19A5 антитело).
В некоторых воплощениях анти-FAM19A5 антитело усиливает проникновение иммунных клеток в опухоль. В некоторых воплощениях инфильтрация иммунных клеток в опухоль усиливается/увеличивается, по меньшей мере, на 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% по сравнению со стандартом (например, объект с злокачественной опухолью, который не получал анти-FAM19A5 антитело). В некоторых воплощениях иммунные клетки содержат макрофаги, дендритные клетки, Т-лимфоциты, В-лимфоциты, клетки-естественные киллеры (NK) или их комбинации. В некоторых воплощениях иммунные клетки проявляют гипертрофию. В некоторых воплощениях инфильтрация иммунных клеток в опухоль сопровождается усиленной инфильтрацией нейрональных клеток в опухоль. В некоторых воплощениях нейронные клетки включают астроциты, глиальные клетки или их комбинации.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию усиливает фагоцитарную активность макрофага или микроглии. В некоторых воплощениях анти-FAM19A5 антитело увеличивает потенциал митохондриальной мембраны макрофага или микроглии. В некоторых воплощениях фагоцитарная активность или потенциал митохондриальной мембраны повышается или увеличивается, по меньшей мере, на 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% по сравнению со стандартом (например, объект со злокачественной опухолью, который не получал анти-FAM19A5 антитело).
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию снижает некроз и отек в опухоли. В других воплощениях анти-FAM19A5 антитело снижает тканевую проницаемость опухоли. В некоторых воплощениях некроз и отек или проницаемость ткани опухоли снижается, по меньшей мере, на 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, или 100% по сравнению со стандартом (например, объект со злокачественной опухолью, который не получал анти-FAM19A5 антитело).
В некоторых воплощениях анти-FAM19A5 антитело увеличивает скорость кровотока в опухоли. В некоторых воплощениях скорость кровотока увеличивается, по меньшей мере, на 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% по сравнению со стандартом (например, объект со злокачественной опухолью, который не получал анти-FAM19A5 антитело).
В некоторых воплощениях способ лечения опухоли включает введение дополнительного терапевтического агента. В некоторых воплощениях дополнительный терапевтический агент включает химиотерапию, иммунотерапию, лучевую терапию или их комбинации. В некоторых воплощениях иммунотерапия включает терапию моноклональным антителом, химерным антигенным рецептором (CAR), терапию Т-клетками, терапию NK-клетками, терапию дендритными клетками (DC), адоптивный перенос клеток (ACT), модулятор иммунных контрольных точек, цитокином, противоопухолевой вакциной, адъювантом, онколитическим вирусом или их комбинацией. В некоторых воплощениях химиотерапия включает темозоломид, гемцитабин, паклитаксел, карбоплатин, цисплатин, элотуксумаб, леналидомид, дексаметазон, оксалиплатин или их комбинации.
В некоторых воплощениях вводят терапевтически эффективное количество анти-FAM19A5 антитела по настоящему раскрытию или его композиции. При обработке объекта (например, человека) терапевтически эффективное количество описанного в данном документе анти-FAM19A5 антитела зависит от таких факторов, как возраст, пол, тяжесть заболевания.
В некоторых воплощениях анти-FAM19A5 антитело по настоящему раскрытию или его композицию вводят внутривенно, перорально, парентерально, транстекально, интратекально, внутрицеребровентрикулярно, легочно, подкожно, внутрикожно, внутримышечно или внутрижелудочково.
Следующие ниже примеры предлагаются в качестве иллюстрации, а не в качестве ограничения.
ПРИМЕРЫ
Пример 1. Экспрессия и очистка белка FAM19A5 человека
Рекомбинантный человеческий белок FAM19A5 получали и очищали, как описано ниже, и очищенный белок использовали в скрининговом анализе антител, основанном на анализе аффинности связывания. Сначала плазмиду LPS-hT, экспрессирующую ген FAM19A5, трансформировали в бактерии и индуцировали сверхэкспрессию белка. После получения белок FAM19A5 очищали с помощью аффинной хроматографии Ni-NTA (Qiagen, Валенсия, Калифорния, США). Используя постепенно повышающуюся концентрацию имидазола, His-меченный белок FAM19A5 удаляли из Ni-колонки. Экспрессию белка в растворе измеряли с помощью красителя кумасси бриллиантовый синий R-250. Захвативший только FAM19A5 имидазол-содержащий раствор использовали для концентрирования белка FAM19A5 с помощью PBS. Когда концентрирование было завершено, как чистоту, так и концентрацию белка FAM19A5 измеряли с помощью вестерн-блоттинга. Концентрированный белок впоследствии использовали для скрининга антител, специфичных к FAM19A5.
Пример 2. Получение библиотек анти-FAM19a5 антител
1. Иммунизация
Белок FAM19A5 использовали в качестве антигена для иммунизации цыплят белого леггорна. 50 мкг конъюгата синтетического пептида KLH смешивали с 750 мкл фосфатно-солевого буфера (PBS) и инкубировали при 37°C в течение 30 минут. После этого удаляли токсин в 2% сквален-эндотоксине MPL (виды монофосфорилированного липида A) и микобактерии (микобактерии) компонентов клеточной стенки TDW и CWS, содержащих адъювант эмульсии вода в масле (RIBI + MPL + TDM + CWS. адъювант, Sigma, Сент-Луис, Миссури, США) в эмульгированном виде, который затем вводили цыплятам подкожно. Цыплят иммунизировали всего четыре раза с интервалом примерно 2-3 недели между иммунизациями. Титр антител, полученных от иммунизированных животных, измеряли с помощью иммуноблоттинга с использованием лизатов клеток HEK293T, которые сверхэкспрессировали белок FAM19A5.
2. Получение библиотеки одноцепочечных вариабельных фрагментов (scFv) из иммунизированной курицы
Используя реагент TRI (Invitrogen, Карлсбад, Калифорния, США), РНК экстрагировали из селезенки, костного мозга и фабрициевой сумки иммунизированных цыплят, описанных выше. Праймеры Олиго-dT и систему синтеза первой цепи Superscript™ III (Invitrogen) использовали для синтеза кДНК первой цепи. Для кДНК, полученной из иммунной системы животных, использовали Expand High Fidelity PCR System (Roche Molecular Systems, Индиана, США) для получения библиотеки одноцепочечных вариабельных областей. В каждой реакции 1 мкл кДНК, 60 пмоль каждого праймера, 10 мкл 10-кратного реакционного буферного раствора, 8 мкл 25 мМ dNTP (Promega, Мадисон, Висконсин, США) и 05 мкл ДНК-полимеразы Taq смешивали с водой. Конечный объем составлял 100 мкл. Реакцию ПЦР проводили с использованием следующих условий: 30 циклов (i) 15 секунд при 94°C, (ii) 30 секунд при 56°C и (iii) 90 секунд при 72°C, с последующим заключительным удлинением в течение 10 минут при 72°C. Продукты ПЦР, содержащие фрагмент длиной около 350 п.н., загружали на 15% агарозный гель и после электрофореза использовали набор для экстракции QIAGEN Gel II (QIAGEN, Валенсия, Калифорния, США) для очистки нуклеотидного фрагмента. Очищенный продукт ПЦР определяли количественно, считывая при OD 260 нм. (1 единица OD = 50 мкг/мл).
VH и VL первого продукта из второй ПЦР случайным образом соединяли с помощью ПЦР с перекрывающимися праймерами (ПЦР с перекрывающимися праймерами). Каждую реакцию ПЦР смешивали со 100 нг очищенного продукта VL и VH, 60 пмоль каждого праймера, 10 мкл 10-кратного реакционного буфера, 8 мкл 25 мМ dNTP, 05 мкл ДНК-полимеразы Taq и воды в конечном объеме 100 мкл. ПЦР выполняли в следующих условиях: 25 циклов (i) 15 секунд при 94°C, (ii) 30 секунд при 56°C и (iii) 2 минуты при 72°C с последующим окончательным удлинением в течение 10 минут при 72°C. Продукты ПЦР, содержащие одноцепочечный фрагмент вариабельной области длиной около 700 п.н., загружали в 15% агарозный гель и после электрофореза использовали набор для экстракции геля QIAGEN II (QIAGEN) для очистки нуклеотидного фрагмента. Очищенный продукт ПЦР определяли количественно, считывая при OD 260 нм. (1 единица OD = 50/мл).
3. Библиотека, лигирование и трансформация
Фрагмент scFv продукта ПЦР и вектор pComb3X - SS (Исследовательский институт Скриппса, Калифорния, США) расщепляли рестрикционным ферментом Sfi. 10 мкг очищенного перекрывающегося продукта PCT смешивали с 360 единицами Sif I (мкг ДНК на 16 единиц, Roche Molecular Systems, Плезантон, Калифорния, США), 20 мкл 10-кратного реакционного буфера и водой до конечного объема 200 мкл. 20 мкг вектора pComb3X-SS смешивали со 120 единицами Sfi I (мкг ДНК на 6 единиц), 20 мкл 10-кратного реакционного буферного раствора и водой до конечного объема до 200 мкл. Смесь гидролизовали при 50°C в течение 8 часов. После этого расщепленный продукт, содержащий фрагмент scFv (примерно 700 п.н.) и вектор (примерно 3400 п.н.), загружали в 1% агарозный гель и очищали с использованием набора для экстракции геля II QIAGEN (QIAGEN, Валенсия, Калифорния, США). 1400 нг Sfi I-рестрицированного вектора pComb3X и 700 нг расщепленных фрагментов scFv смешивали с 5x лигазным буфером, 10 мкл ДНК-лигазы Т4 (Invitrogen, Карлсбад, Калифорния, США) и водой до конечного объема 200 мкл. Смесь инкубировали при 16°C в течение 16 часов для лигирования.
После осаждения этанолом осадок ДНК растворяли в 15 мкл воды. Для создания библиотеки образец лигирования трансформировали в штамм E. coli ER2738 (New England Biolabs Inc., Хитчин, Норт-Хартфордшир, SG4 OTY, Англия, Великобритания) посредством электропорации с использованием vibrator gene (Gene pulser: Bio-Rad Laboratories, Геркулес, Калифорния, США). Клетки смешивали в 5 мл среды Super Broth (SB) и инкубировали при перемешивании со скоростью 250 об/мин в течение одного часа при 37°C. Затем 3 мкл 100 мг/мл канамицина добавляли к 10 мл среды SB. Для определения размера библиотеки 01 мкл, 1 мкл и 10 мкл образца культуры наносили на чашки с агаром Luria Broth (LB), содержащие 50 мкг/мл канамицина. После перемешивания в течение 1 часа к культуре LB добавляли 45 мкл 100 мг/мл канамицина и дополнительно перемешивали еще 1 час. Затем к среде LB добавляли 2 мл фага-помощника VCM13 в воде (> 1011 КОЕ/мл) вместе с предварительно нагретым LB (183 мл), содержащим 925 мкл 100 мг/мл канамицина. Эту смесь перемешивали при 250 об/мин при 37°С в течение дополнительных 2 часов. Затем к культуре добавляли 280 мкл (50 мг/мл) канамицина и перемешивали в течение ночи при 37°С. На следующий день осадок бактерий центрифугировали с использованием высокоскоростной центрифуги (Beckman, JA-10 ротор) при 3000 g, 4°C. После этого бактериальный осадок использовали для экстракции фагмидной ДНК, а надосадочную жидкость переносили в стерильные центрифужные флаконы. Следующие 8 граммов полиэтиленгликоля-8000 (PEG-8000, Sigma) и 6 граммов хлорида натрия добавляли (NaCl, Merck) к надосадочной жидкости и затем выдерживали в течение 30 минут на льду. После этого надосадочную жидкость центрифугировали 15 минут при 15000 g, 4°C. Затем надосадочную жидкость отбрасывали, а осадок фага суспендировали в трис-буферном растворе (TBS), содержащим 1% BSA - восстановление.
Пример 3. Пэннинг библиотеки (биопэннинг) на иммобилизованном антигене
Биопэннинг выполняли с использованием магнитных гранул (Dynabeads M-270 Epoxy, Invitrogen). При комнатной температуре приблизительно 1 × 107 гранул покрывали 5 мкг рекомбинантного белка FAM19A5 путем перемешивания при вращении гранул и белка вместе в течение 20 часов при комнатной температуре. После нанесения покрытия гранулы промывали 4 раза фосфатно-солевым буферным раствором (PBS) и блокировали на один час в PBS, содержащем 3% BSA, при комнатной температуре. Затем покрытые гранулы культивировали в течение двух часов при комнатной температуре с scFv, презентируемым на фаге, описанным выше. Для удаления любого фага, который не связался с покрытыми антигеном гранулами, гранулы промывали 0,05% Tween20/PBS. Затем связанные фаги элюировали 50 мкл 01 М глицина/хлористого водорода (01 М глицин-HCl, pH 22) и нейтрализовали 3 мкл 2 М Tris с хлористым водородом (трис-HCl, pH 91). Эти содержащие фаг надосадочные жидкости использовали для заражения клеток E.coli ER2738, а фаг-помощник VCSM13 использовали для амплификации и спасения в течение ночи. Также ввод (ввод) и продуцирование (вывод) титрами фага из инфицированных фагом культур определяли путем блоттинга инфицированных фагом культур на чашках с агаром LB, содержащим 50 мкг/мл канамицина. На следующий день для осаждения фагов использовали PEG-8000 и NaCl, которые впоследствии использовали для биопэннинга. Биопэннинг выполняли в общей сложности до пяти раз, повторяя описанный выше процесс. При каждой амплификации фаги подвергали скринингу и отбирали на предмет высокой аффинности к белку FAM19A5.
Пример 4. Отбор клона с помощью фагового ELISA.
Для анализа клонов, выбранных биопэннингом, отдельные клоны случайным образом выбирали из фаг-презентируемых scFv и подтверждали с помощью ELISA, что клоны связываются с рекомбинантным белком FAM19A5. Рекомбинантный белок FAM19A5 разводили в 01 М буфере NaHCO3, и 100 нг/лунку белка использовали для покрытия 96-луночных микротитровальных планшетов при 4°C в течение 16 часов. На следующий день планшеты блокировали 3% BSA/PBS при 37°C в течение 1 часа. Затем фаговую надосадочную жидкость смешивали с 6% BSA/PBS и культивировали в течение 2 часов при 37°C. Затем планшеты, содержащие надосадочную жидкость, промывали 0,05% Tween-20/PBS. Конъюгированное с HRP антитело M13 (a-M13-HRP, Pierce Chemical Co, Rockford, IL, USA) разбавляли до 1/5000. 50 мкл разведенного антитела добавляли в планшеты и инкубировали в течение 1 часа при 37°C. После инкубации и промывки в планшеты добавляли 005 М цитратный буферный раствор, 1 мкг/мл 22'-азино-бис (3-этилбензотиазолин-6-сульфоновой кислоты) (ABTS, Amresco, Солон, Огайо, США.) и 0,1% H2O2 для проявления цвета. Оптическую плотность для каждой лунки измеряли при 405 нм.
Из 96 клонов, которые были первоначально идентифицированы, для дальнейшего анализа отобрали 8 клонов scFv, имеющих уникальные последовательности CDR3 тяжелой цепи (HCDR3) и сильное связывание с белком FAM19A5. ФИГ. 1А-1С.
ПРИМЕР 5. ПРОДУЦИРОВАНИЕ АНТИ-FAM19A5-IGG2/4 АНТИТЕЛА
Анти-FAM19A5 ScFv субклонировали в экспрессирующий вектор для млекопитающих. В последовательности гена scFv FAM19A5 ген Cκ человека соединяли с вариабельным доменом легкой цепи, а гены CH1, CH2 и CH3 изотипического иммуноглобулина человека IgG2/4 соединяли с вариабельной областью тяжелой цепи. Антитело, имеющее каждую легкую цепь и каждую тяжелую цепь, синтезировали путем добавления сайтов рестрикции (Genscript, США). Синтезированный ген вставляли в экспрессирующий вектор для клеток млекопитающих, имеющий модифицированный сайт рестрикции для облегчения клонирования. Сначала в вектор вставляли ген легкой цепи с помощью рестрикционных ферментов Hind III и Xba I (New England Biolabs, Великобритания), а затем добавляли в вектор ген тяжелой цепи с помощью рестрикционных ферментов NheΙ и BamHΙ (New England Biolabs, Великобритания).
Для экспрессии и очистки анти-FAM19A5 антитела-IgG2/4 использовали систему инъекции для трансфекции клеток млекопитающих и сверхэкспрессии. Приблизительно 2 мкг/мл экспрессирующего вектора для млекопитающих смешивали с 4 мкг полиэтиленимина (PEI, Polysciences, Уоррингтон, Пенсильвания, США) в 150 мМ хлориде натрия (NaCl, Merck), что соответствует 1/10 объема клеточной культуры. Смесь оставляли на 15 минут при комнатной температуре. Смесь добавляли к клеткам HEK293F (2 × 106 клеток/мл, Invitrogen), которые затем инкубировали в культуральной среде для экспрессии Freestyle™ 293, содержащей 100 Ед/мл пенициллина и стрептомицина (Invitrogen), при 7% CO2 и 37°C и перемешивали при 135 об/мин в течение шести дней. Для очистки экспрессированных анти-FAM19A5 антител IgG2/4 из надосадочной жидкости клеточной культуры использовали аффинную гель-хроматографию на гранулах с протеином A (RepliGen, Уолтем, Массачусетс, США). Хроматографию с протеином А проводили с помощью гель-электрофореза в градиенте 4 ~ 12% Bis-Tris. Размер и выход белка подтверждали окрашиванием кумасси бриллиантовым синим. Связывающую способность антител измеряли с помощью анализа ELISA.
Как показано на фиг. 2А, различные протестированные антитела (т.е. 1-28, 1-85, 2-13, 2-14, 2-20, 2-29, 3-2 и 3-26) были сопоставимы по размеру. За исключением антитела 1-85, протестированные антитела были способны связываться с белком FAM19A5 в различной степени. Фиг. 2B.
ПРИМЕР 6. АНАЛИЗ НЕЙТРАЛИЗУЮЩЕЙ СПОСОБНОСТИ АНТИ-FAM19A5 АНТИТЕЛ
Для дальнейшей оценки функциональных свойств антител использовали следующие способы:
1. Получение рекомбинантного гибридного белка FAM19A5 с Fc кролика
Для конструирования экспрессирующих векторов с FAM19A5 химически синтезировали гены, кодирующие человеческий FAM19A5 (Genscript, Пискатауэй, Нью-Джерси, США). Ген был субклонирован в модифицированный экспрессирующий вектор для млекопитающих, кодирующий шарнирную область человеческого IgG1 и домены CH2-CH3 кроличьего IgG в 3'-области, как сообщалось ранее. См. Han, J., et al., Exp Mol Med. 48(11):e271 (2016).
Экспрессирующий вектор, кодирующий гибрид Fc кролика с FAM19A5, трансфицировали в клетки HEK293F (Invitrogen, Карлсбад, Калифорния, США), используя линейный полиэтиленимин 25 кДа (Polyscience, Уоррингтон, Пенсильвания, США), как сообщалось ранее. См. Boussif, O., et al., Proc Natl Acad Sci U S. A. 92 (16): 7297-301 (1995). Гибридный белок кролика с FAM19A5 очищали из культуральных надосадочных жидкостей транзиторно трансфицированных клеток HEK293F с использованием колонки с протеином А-сефарозой (Repligen, Уолтем, Массачусетс, США) в соответствии с инструкциями производителя.
2. Получение рекомбинантного гибридного белка анти-FAM19A5 scFv с Cκ человека
Гены выбранных клонов субклонировали в модифицированный вектор pCEP4, кодирующий домен Cκ (константный домен κ легкой цепи иммуноглобулина человека) в 5'-области, как сообщалось ранее. См. Lee, Y., et al., Exp Mol Med. 46:e114 (2014). Экспрессирующие векторы, кодирующие гибрид анти-FAM19A5 scFv человека, трансфицировали в клетки HEK293F (Invitrogen), как описано выше. Гибридный белок scFv-hCκ очищали из культуральных надосадочных жидкостей транзиторно трансфицированных клеток HEK293F с использованием колонки с протеином А-сефарозой (Repligen, Уолтем, Массачусетс, США) в соответствии с инструкциями производителя.
3. Нейтрализующая эффективность анти-FAM19A5 антител на глиальных клетках
Для оценки нейтрализующей эффективности антитела подтверждали анализом проточной цитометрии, как описано ранее. См. Kim, M., et al., PLoS One. 7 (4): e35100 (2012). Глиальные клетки мыши и человека высевали в 96-луночный планшет с v-образным дном (Corning Inc., Корнинг, Нью-Йорк, США) с конечной плотностью 3 × 105 клеток на лунку. Клетки обрабатывали 1 мкМ рекомбинантного FAM19A5 с Fc кролика и 5 мкМ гибридного белка Сκ человека с анти-FAM19A5 scFv в буфере для проточной цитометрии [1% (масс./об.) БСА в PBS, содержащем 0,05% (масс./об.) азида натрия] при 37°C в течение 1 ч. После промывания буфером для проточной цитометрии клетки затем инкубировали с конъюгированным с Alexa Fluor 488 антителом против кроличьего IgG (Fc-специфическим) (Jackson Immuno Research Inc., Пенсильвания, США) в течение 1 ч при 37°C в темноте. После дополнительной промывки тем же буфером клетки ресуспендировали в 300 мкл PBS и анализировали проточной цитометрией с использованием прибора FACSCANTO™ II (BD Bioscience, Сан-Хосе, Калифорния, США), оснащенного 488-нм лазером. Данные анализировали с помощью программного обеспечения FlowJo (TreeStar, Ашленд, Орегон, США).
Как показано на фиг. 3A, все протестированные антитела (т.е. 1-28, 1-85, 2-13, 2-14, 2-20, 2-29, 3-2 и 3-26) были способны ингибировать взаимодействие FAM19A5 с первичными глиальными клетками мыши в различной степени. В клеточной линии глиобластомы человека антитело 2-20 не могло нейтрализовать взаимодействие FAM19A5 с клеточной линией. ФИГ. 3B. Другие антитела были способны нейтрализовать активность FAM19A5, хотя и в разной степени. Антитела 1-28, 2-13 и 3-2 показали наибольшую нейтрализующую эффективность в клеточной линии глиобластомы человека.
ПРИМЕР 7. АНАЛИЗ КАРТИРОВАНИЯ ЭПИТОПА С ИСПОЛЬЗОВАНИЕМ ФРАГМЕНТОВ ЭПИТОПА FAM19A5 F1-F6
Из-за его способности нейтрализовать экспрессию FAM19A5 как на мышиных, так и на человеческих глиальных клетках, антитело 3-2 было выбрано для анализа картирования эпитопа. Перекрывающиеся пептидные фрагменты (F1-F6, см. Фиг. 4) человеческого белка FAM19A5 были синтезированы и конъюгированы с BSA. Связывание различных анти-FAM19A5 антител с BSA-конъюгированными пептидными фрагментами F1-F6 определяли с помощью анализа ELISA. Вкратце, FAM19A5 фрагмент F1-F6 (разбавленный до 1 мкг/мл в 50 мМ карбонатном буфере (Biosesang) или до 20 мкг/мл для анализа высокой концентрации) использовали для покрытия лунок 96-луночных иммунопланшетов (Thermo Scientific) (100 мкл/лунку) в течение ночи при 4°C, а затем дважды промывали 1X PBS. Затем планшеты блокировали блокирующим буфером (100 мкл/лунку) в течение 1 часа при комнатной температуре. Во время 1-часовой инкубации соответствующие анти-FAM19A5 антитела разбавляли до 1 мкг/мл (или 20 мкг/мл для анализа с высокой концентрацией) в буфере для разбавления. После промывки планшетов (2 раза с использованием 1x PBS) разбавленные анти-FAM19A5 антитела добавляли в соответствующие лунки и планшеты инкубировали при комнатной температуре в течение 1 часа. Затем планшеты промывали в общей сложности пять раз промывочным буфером. Затем субстрат ODP (приготовленный растворением одной таблетки ODP (O-фенилендиамин дигидрохлорид, Thermo) в 9 мл стерилизованной деионизированной воды и 1 мл 10-кратного стабильного пероксидно-стабильного буфера (Thermo)) был добавлен в каждую лунку, и реакции изменения цвета давали возможность протекать в течение 10 минут. Данную реакцию останавливали добавлением в лунки 100 мкл 2 н. H2SO4 (Daejung). Величину поглощения каждой лунки определяли при 492 нм, используя 96-луночный ридер для микропланшетов (Molecular Device).
Как показано на фиг. 5, антитело 3-2 прочно связывается с фрагментом эпитопа F2 с минимальным связыванием с другими фрагментами.
Затем, чтобы идентифицировать конкретные аминокислотные остатки внутри эпитопного фрагмента F2, с которыми связывается антитело 3-2, был проведен сканирующий аланиновый анализ. Как показано на фиг. 6А, когда аминокислотные остатки R4, D5, P9, R10 или R11 были изменены на аланин, способность антитела 3-2 связываться с фрагментом эпитопа F2 значительно снижалась. Аналогичный анализ был проведен для нескольких деиммунизированных вариантов антитела 3-2 (подробности относительно процесса деиммунизации см. в Примере 8). Как показано на фиг. 6B, 6C, 6D, 6F, 6G, 6I и 6J аминокислотные остатки R4, P9, R10 и R11 важны для связывания антител 1-30, 1-32 и 6-10 с FAM19A5. Для антител 1-17 и 4-11 важны аминокислотные остатки R4, P9 и R10 (см. Фиг. 6E и 6H).
ПРИМЕР 8. ДЕИММУНИЗАЦИЯ АНТИ-FAM19A5 АНТИТЕЛА
1. Оценка иммуногенности in silico
Чтобы снизить риск иммуногенности при введении человеку, был проведен анализ in silico для выявления конкретных областей с высокой иммуногенностью внутри анти-FAM19A5 антител 3-2 и 2-13.
Чтобы идентифицировать неразборчивые пептиды, связывающие MHC класса II, был проведен анализ iTopeTM (Abzena plc., Великобритания) на перекрывающиеся 9-мерные пептиды, охватывающие всю последовательность. Возможные Т-клеточные эпитопы были спрогнозировны путем анализа взаимодействия с 34 различными аллелями MHC класса II. Как показано на фиг. 7 и 8, соответственно, клон 3-2 содержал в общей сложности 11 связывающих пептидов, а клон 2-13 содержал всего 10 неразборчивых пептидов, не связанных с зародышевой линией, связывающих MHC класса II.
Кроме того, неразборчивые связывающие пептиды MHC класса II анализировали с помощью TCEDTM (Abzena plc., Великобритания), которая представляет собой базу данных CD4+ Т-клеточных эпитопов, созданную с помощью анализа стимуляции Т-клеток против более чем 10000 пептидов. Клон 2-13 показал, что два неразборчивых пептида, связывающих MHC класса II, обладают высокой гомологией с известными Т-клеточными эпитопами. Фиг. 8 (выделено зеленым). Клон 3-2 имел три неразборчивых пептида, связывающих МНС класса II, с высокой гомологией с известными Т-клеточными эпитопами. Фиг. 7 (выделено зеленым).
2. Создание композитной библиотеки антител
Композитная библиотека антител была разработана так, чтобы избежать эпитопов CD4+ Т-клеток, связанных с иммуногенностью. Чтобы выбрать наиболее гомологичную зародышевую линию человека для конструирования библиотеки, каркас клона 2-13 и клона 3-2 анализировали в базе данных IgBLAST (NCBI). Для клона 2-13 были выбраны гены человеческой зародышевой линии IGLV1-51*02, IGLJ2*01, IGHV3-64*04 и IGHJ1*01 (Фиг. 9A и 9B). Для клона 2-13 были выбраны гены человеческой зародышевой линии IGLV3-27*01, IGLJ2*01, IGHV3-64*04 и IGHJ1*01 (Фиг. 10).
Библиотека была сконструирована для замены определенных неидентичных аминокислотных остатков в областях связывания MHC класса II клона 2-13 и клона 3-2 на соответствующую аминокислоту в наиболее гомологичной последовательности зародышевой линии человека. Для создания библиотеки сайт-направленного мутагенеза, как описано ранее, была проведена последовательная полимеразная цепная реакция (ПЦР) удлинения путем перекрывания олигонуклеотидов, кодирующих вырожденные кодоны, покрывающие аминокислоты как человека, так и курицы. Baek DS, Kim YS. Biochem Biophys Res Commun. 463(3):414-20 (2015). Продукт ПЦР субклонировали в фагмидный вектор и трансформировали в штамм Escherichia coli ER2738 (New England BioLabs, Ипсвич, Массачусетс, США) для получения фага, как описано ранее. Han, J., et al., Exp Mol Med. 48(11):e271 (2016).
3. Биопэннинг и отбор клонов
Для выделения положительных клонов проводили биопэннинг и иммуноферментный анализ на фаг, как описано ранее (Barbas CF., Phage display: a laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2001). Отбирали клоны фага с реактивностью против FAM19A5, и их нуклеотидные последовательности определяли секвенированием по Сэнгеру. Наконец, были отобраны клоны scFv с наименьшим количеством неразборчивых связывающих эпитопов MHC класса II с сохраненной аффинностью к FAM19A5.
Как показано на фиг. 9B, 7 неразборчивых эпитопов связывания MHC класса II в клоне 3-2 были удалены в деиммунизированном клоне 3-2. Удаленные эпитопы соответствуют связывающим пептидам № 2, 3, 4, 5, 9, 10 и 11, как показано на фиг. 7. Неразборчивые эпитопы, связывающие MHC класса II в CDRH1, не могли быть удалены (т.е. связывающий пептид № 6 на фиг. 7). Кроме того, неразборчивый эпитоп связывания MHC класса II в HFR2 нельзя было удалить, потому что остаток p3, расположенный в CDRH2, играет критическую роль в активности связывания (т.е. связывающий пептид № 8 на фиг. 7). Как показано на фиг. 9C, полная деиммунизация антитела 3-2 путем мутации всех вышеуказанных остатков приводила к тому, что антитело 3-2 не могло связываться с белком FAM19A5. Однако, когда аминокислотный остаток Q50 VH не был мутирован, деиммунизированное антитело 3-2 было способно связываться с белком FAM19A5, как и антитело 3-2 дикого типа.
Как показано на фиг. 10A, 7 неразборчивых эпитопов связывания MHC класса II в клоне 2-13 были удалены в деиммунизированном клоне 2-13. Удаленные эпитопы соответствуют связывающим пептидам №1, 2, 3, 4, 5, 8 и 9, как показано на фиг. 8. Неразборчивый связывающий эпитоп MHC класса II в HFR2 нельзя было удалить, потому что остаток p4, расположенный в CDRH2, играет критическую роль в связывающей активности (т.е. связывающий пептид № 6 на фиг. 8). Как и в случае с антителом 3-2, полная деиммунизация антитела 2-13 также отрицательно влияет на связывание с белком FAM19A5. Фиг. 10B.
Приведенные выше результаты позволяют предположить, что можно снизить иммуногенность анти-FAM19A5 антител без отрицательного воздействия на их способность связываться с белком FAM19A5.
ПРИМЕР 9. СОЗРЕВАНИЕ АФФИННОСТИ ДЕИММУНИЗИРОВАННОГО АНТИТЕЛА 2-13
Электростатическое взаимодействие важно в ассоциации антитела и антигена. Lee J.Y., et al., Nat Commun. 7:13354 (2016). Каждую аминокислоту в 5 CDR, за исключением CDRH3 деиммунизированного клона 2-13, заменяли на глутаминовую кислоту и аспарагиновую кислоту для введения искусственно отрицательно заряженных R-групп. В общей сложности 70 генов, кодирующих точечные мутированные scFv, были химически синтезированы (Integrated DNA Technologies, Коралвилл, Айова, США) и субклонированы в фагмидный вектор для конструирования мутантных scFv, отображающих фаг, как описано ранее. Yoon, A., et al., PLoS One. 11(1):e0146907 (2016).
Мутанты scFv с вариантом заряда, презентированные фагом, были спасены и подвергнуты фаговому иммуноферментному анализу, как описано ранее (Barbas CF. Phage display: a laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2001). Как показано на фиг. 11, все клоны проявляли активность связывания FAM19A5 в различной степени. Два клона (36.H1-3TE и 37.H1-3TD) проявляли более высокую аффинность связывания, чем деиммунизированный клон 2-13. Сравнение последовательностей VH и VL (i) антитела 2-13 дикого типа, (ii) деиммунизированного клона 2-13 антитела и (iii) деиммунизированного антитела 2-13 с созревшей аффинностью (2-13D- 37) представлены на фиг. 15.
ПРИМЕР 10. СОЗРЕВАНИЕ АФФИННОСТИ ДЕИММУНИЗИРОВАННОГО АНТИТЕЛА 2-13D-37
Для создания библиотек сайт-направленного мутагенеза CDR-H3 ген, кодирующий деиммунизированный клон 2-13D-37 scFv, использовали в качестве матрицы для ПЦР. Три последовательных остатка в CDR-H3 были рандомизированы с олигонуклеотидами, кодирующими вырожденные кодоны NNK (N = A, T, G или C, K = G или T), как описано ранее (Lee Y, Kim H, Chung J. Exp Mol Med. 2014;46:e114). Рандомизированные кодоны вводили в CDR-H3 с помощью ПЦР. Амплифицированные фрагменты scFv с помощью ПЦР с перекрывающимися праймерами субклонировали в фагмидный вектор и трансформировали в ER2738 (New England BioLabs) для получения фага, как описано выше. Всего было создано восемь рандомизированных библиотек CDR-H3.
Биопэннинг и фаговый иммуноферментный анализ проводили для выделения положительных клонов, как описано ранее (Barbas CF. Phage display: a laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2001; см. также Пример 8). Отбирали клоны фага с более высокой реактивностью против FAM19A5, и их нуклеотидные последовательности определяли секвенированием по Сэнгеру. Размер и уровень экспрессии клонов-кандидатов (т.е. 2-13D-37-1.5W-41 и 2-13D-37-3W-16), измеренные с помощью SDS-PAGE и вестерн-блоттинга, представлены на фиг. 16B-16C. Сравнение последовательностей клонов-кандидатов (т.е. 2-13D-37-1.5W-41 и 2-13D-37-3W-16) с деиммунизированным клоном 2-13D-37 представлено на фиг. 17А. По сравнению с антителом 2-13D-37 эти деиммунизированные антитела с созревшей аффинностью проявляли большее связывание с белком FAM19A5, что измерено с помощью ELISA (Фиг. 17B).
ПРИМЕР 11. УЛУЧШЕНИЕ ФИЗИКО-ХИМИЧЕСКИХ СВОЙСТВ ДЕИММУНИЗИРОВАННОГО АНТИТЕЛА 3-2
Для улучшения физико-химических свойств деиммунизированных анти-FAM19A5 антител была создана библиотека фагового дисплея с использованием рандомизированных последовательностей из CDR1, CDR2 и CDR3 легкой цепи и из CDR1 и CDR2 тяжелой цепи антитела 3-2. Для создания библиотек CDR сайт-направленного мутагенеза ген, кодирующий деиммунизированный клон 3-2 scFv, использовали в качестве матрицы для ПЦР. Четыре или пять последовательных остатков в CDR были рандомизированы с олигонуклеотидами, кодирующими вырожденные кодоны NNK (N = A, T, G или C, K = G или T), как описано ранее. Lee, Y., et al., J. Exp Mol Med. 46:e114 (2014). Рандомизированные кодоны были введены в пять CDR, кроме CDRH3, с помощью ПЦР. Амплифицированные фрагменты scFv с помощью ПЦР с перекрывающимися праймерами субклонировали в фагмидный вектор и трансформировали в ER2738 (New England BioLabs) для получения фага, как описано выше. Фиг. 12.
Биопэннинг и фаговый иммуноферментный анализ проводили для выделения положительных клонов, как описано ранее (Barbas CF. Phage display: a laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2001). Отбирали клоны фага с реактивностью против FAM19A5, и их нуклеотидные последовательности определяли секвенированием по Сэнгеру. Балл собственной растворимости и балл GRAVY отдельных клонов рассчитывали, как описано ранее. Sormanni, P., et al., Sci Rep. 7(1):8200 (2017). Были отобраны клоны с улучшенной растворимостью, демонстрирующие аналогичную реактивность против FAM19A5: (i) антитело с низкой изоэлектрической точкой («Low_PI»), (ii) 1-17, (iii) 1-30, (iv) 1-32, (v) 4-11 и (vi) 6-10. Результаты показаны на фиг. 13A и 13B. Сравнение последовательностей VH и VL различных деиммунизированных вариантов антител 3-2 к антителам 3-2 дикого типа представлено на фиг. 14A и 14B, соответственно.
ПРИМЕР 12. HDX-MX КАРТИРОВАНИЕ ЭПИТОПА АНТИТЕЛА 2-13
Масс-спектрометрию с обменом водород/дейтерий (HDX-MS) использовали, как описано ниже, для зондирования связывающего эпитопа FAM19A5 человека на клон 2-13 анти-FAM19A5-антитела. См. Фиг. 18.
1. Оптимизация условий максимального охвата FAM19A5.
Перед экспериментами по картированию эпитопов были проведены эксперименты без дейтерирования для создания списка общих пептидов для белка FAM19A5. Автономное и неавтономное расщепление пепсином использовали для гидролиза белка FAM19A5. Houde, D., et al., Methods Mol Biol 988: 269-89 (2013). При неавтономном расщеплении использовали колонку с иммобилизованным пепсином (~ 20°C), и эффективность реакции регулировали путем регулирования скорости потока. В автономном режиме белок FAM19A5 расщепляли вручную пепсином при 4°C в течение 5 минут, а затем смесь загружали в колонку для жидкостной хроматографии для анализа. Для определения условия максимального покрытия белком один или несколько из следующих параметров были скорректированы: фосфат калия, фосфат натрия, время выдержки гашения, восстанавливающий реагент (TCEP), концентрация мочевины и концентрация пепсина. Программное обеспечение Waters PLGS использовалось для анализа исходных данных масс-спектрометрии (МС), полученных для определения того, было ли получено максимальное покрытие белком. Чтобы подтвердить воспроизводимость экспериментов, каждое из экспериментальных условий было повторено, по меньшей мере, дважды, и были подтверждены пептиды, которые были получены. На фиг. 19 показаны различные использованные экспериментальные условия и достигнутый процент покрытия.
Максимальное покрытие наблюдали при следующих условиях: 1M TCEP, 2M мочевина (pH 266) и пепсин в концентрации 1: 2. Как показано на фиг. 20A и 20B, используя эти условия, было идентифицировано 44 пептидных фрагмента, которые покрывали приблизительно 97% белка FAM19A5.
2. Картирование эпитопа FAM19A5
Для получения максимального (100%) связывания (KD = 1 нМ) даже при 15-кратном разведении комплекса антитело-белок в буфере для мечения образцы комплекса антиген-антитело инкубировали в течение не менее 3 часов до мечения водородом/дейтерием. После мечения образцы доводили до того же объема, используя равновесный буфер.
Чтобы начать реакцию мечения, 25 мкл подготовленных образцов (только антиген, только антитело или комплекс антиген-антитело), разведенных в буфере для мечения 1:15, смешивали с раствором для мечения D2O (18 мкМ). Реакции проводили в течение разных периодов времени: 0 мин (т.е. недейтерированная), 20 секунд, 10 минут, 60 минут и 240 минут. Для недейтерированной реакции приготовленный образец вместо этого смешивали с уравновешивающим буфером. В конце каждого периода мечения реакцию останавливали гасящим буфером. Затем образцы встряхивали, немедленно замораживали в жидком азоте и хранили при -80°C до анализа.
Перед анализом с помощью масс-спектрометра хранимым замороженным образцам давали возможность расплавиться, а затем расщепляли пепсином в течение приблизительно 5 минут на льду (т.е. автономный способ, описанный выше). Полученные относительные уровни дейтерия были построены в зависимости от времени обмена с использованием программного обеспечения DynamX 3.0™ (Waters). Окончательные данные о покрытии и избыточности последовательности показаны на фиг. 21.
Сравнение данных по поглощению дейтерия (одиночный антиген/антитело против комплекса антиген-антитело) для каждого из идентифицированных пептидов (см. Фиг. 21) показано на ФИГ. 22A-22E и 23A-23C. Как показано, значительное поглощение дейтерия наблюдалось для следующих пептидных последовательностей: (i) ACRKGQIAGTTRARPAC (остатки 37-53 SEQ ID NO: 101), (ii) ACRKGQIAGTTRARPACVD (остатки 37-55 SEQ ID NO: 101), (iii) ACRKGQIAGTTRARPACVDA (остатки 37-56 SEQ ID NO: 101), (iv) ARIIKTKQWC (остатки 56-65 SEQ ID NO: 101), (v) ARIIKTKQWCDM (остатки 56-67 SEQ ID NO: 101), (vi) ARIIKTKQWCDML (остатки 56-68 SEQ ID NO: 101), (vii) ARIIKTKQWCDMLPCL (остатки 56-71 SEQ ID NO: 101), (viii) RIIKTKQWCDM (остатки 57-67 SEQ ID NO: 101), и (ix) RIIKTKQWCDML (остатки 57-68 SEQ ID NO: 101).
Используя приведенные выше результаты, была построена тепловая карта для идентификации областей в белке FAM19A5 с наиболее значимой разницей в поглощении дейтерия между образцом одного антиген/антитело и комплексом антиген-антитело. Как показано на фиг. 24 аминокислотные остатки 38-50 (CRKGQIAGTTRAR) и 51-64 (PACVDARIIKTKQW) SEQ ID NO: 101 были идентифицированы как важные связывающие остатки для антитела 2-13. На фиг. 25 показано расположение этих остатков в трехмерной структуре белка FAM19A5.
Следует понимать, что раздел «Подробное описание», а не разделы «Сущность изобретения» и «Реферат», предназначен для использования для интерпретации формулы изобретения. Разделы «Сущность изобретения» и «Реферат» могут излагать один или несколько, но не все примерные воплощения настоящего раскрытия, предполагаемые автором(ами), и, таким образом, никоим образом не предназначены для ограничения настоящего раскрытия и прилагаемой формулы изобретения.
Настоящее раскрытие было описано выше с помощью функциональных структурных элементов, иллюстрирующих реализацию определенных функций и их взаимосвязи. Границы этих функциональных структурных элементов были определены в данном документе произвольно для удобства описания. Альтернативные границы могут быть определены при условии, что указанные функции и их взаимосвязи будут выполнены надлежащим образом.
Вышеприведенное описание конкретных воплощений настолько полно покажет общий характер раскрытия, что другие могут, применяя знания в пределах квалификации в данной области, легко модифицировать и/или адаптировать для различных приложений такие конкретные воплощения, без проведения излишних экспериментов, без отклонения из общей концепции настоящего раскрытия. Следовательно, предполагается, что такие адаптации и модификации находятся в пределах значения и диапазона эквивалентов раскрытых воплощений, основанных на замысле и руководстве, представленных в данном документе. Следует понимать, что фразеология или терминология в данном документе предназначена для описания, а не ограничения, так что терминология или фразеология настоящего описания должна интерпретироваться квалифицированным специалистом в свете замысла и руководства.
Широта и объем настоящего раскрытия не должны ограничиваться каким-либо из описанных выше примерных воплощений, а должны определяться только в соответствии со следующей формулой изобретения и ее эквивалентами.
Все публикации, патенты, патентные заявки, интернет-сайты и учетные номера/последовательности баз данных (включая как полинуклеотидные, так и полипептидные последовательности), процитированные в настоящем документе, включены в настоящий документ ссылкой во всей своей полноте для всех целей в той же степени, как если бы каждая отдельная публикация, патент, заявка на патент, интернет-сайт или учетный номер/последовательность из базы данных были специально и индивидуально указаны для включения ссылкой.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> НЬЮРЭКЛ САЙЕНС КО., ЛТД.
СЕУЛ НЭШНЛ ЮНИВЕРСИТИ АрэндДиБи ФАУНДЕЙШН
<120> АНТИТЕЛА ПРОТИВ ПРЕДСТАВИТЕЛЯ A5 СЕМЕЙСТВА 19 СО СХОДСТВОМ
ПОСЛЕДОВАТЕЛЬНОСТЕЙ И СПОСОБ ИХ ПРИМЕНЕНИЯ
<130> 3763.016PC02/C-K/DKC
<150> US 62/787,711
<151> 2019-01-02
<150> US 62/838,190
<151> 2019-04-24
<160> 137
<170> PatentIn version 3.5
<210> 1
<211> 132
<212> PRT
<213> Homo sapiens
<400> 1
Met Ala Pro Ser Pro Arg Thr Gly Ser Arg Gln Asp Ala Thr Ala Leu
1 5 10 15
Pro Ser Met Ser Ser Thr Phe Trp Ala Phe Met Ile Leu Ala Ser Leu
20 25 30
Leu Ile Ala Tyr Cys Ser Gln Leu Ala Ala Gly Thr Cys Glu Ile Val
35 40 45
Thr Leu Asp Arg Asp Ser Ser Gln Pro Arg Arg Thr Ile Ala Arg Gln
50 55 60
Thr Ala Arg Cys Ala Cys Arg Lys Gly Gln Ile Ala Gly Thr Thr Arg
65 70 75 80
Ala Arg Pro Ala Cys Val Asp Ala Arg Ile Ile Lys Thr Lys Gln Trp
85 90 95
Cys Asp Met Leu Pro Cys Leu Glu Gly Glu Gly Cys Asp Leu Leu Ile
100 105 110
Asn Arg Ser Gly Trp Thr Cys Thr Gln Pro Gly Gly Arg Ile Lys Thr
115 120 125
Thr Thr Val Ser
130
<210> 2
<211> 125
<212> PRT
<213> Homo sapiens
<400> 2
Met Gln Leu Leu Lys Ala Leu Trp Ala Leu Ala Gly Ala Ala Leu Cys
1 5 10 15
Cys Phe Leu Val Leu Val Ile His Ala Gln Phe Leu Lys Glu Gly Gln
20 25 30
Leu Ala Ala Gly Thr Cys Glu Ile Val Thr Leu Asp Arg Asp Ser Ser
35 40 45
Gln Pro Arg Arg Thr Ile Ala Arg Gln Thr Ala Arg Cys Ala Cys Arg
50 55 60
Lys Gly Gln Ile Ala Gly Thr Thr Arg Ala Arg Pro Ala Cys Val Asp
65 70 75 80
Ala Arg Ile Ile Lys Thr Lys Gln Trp Cys Asp Met Leu Pro Cys Leu
85 90 95
Glu Gly Glu Gly Cys Asp Leu Leu Ile Asn Arg Ser Gly Trp Thr Cys
100 105 110
Thr Gln Pro Gly Gly Arg Ile Lys Thr Thr Thr Val Ser
115 120 125
<210> 3
<211> 53
<212> PRT
<213> Homo sapiens
<400> 3
Met Tyr His His Arg Glu Trp Pro Ala Arg Ile Ile Lys Thr Lys Gln
1 5 10 15
Trp Cys Asp Met Leu Pro Cys Leu Glu Gly Glu Gly Cys Asp Leu Leu
20 25 30
Ile Asn Arg Ser Gly Trp Thr Cys Thr Gln Pro Gly Gly Arg Ile Lys
35 40 45
Thr Thr Thr Val Ser
50
<210> 4
<211> 1501
<212> DNA
<213> Homo sapiens
<400> 4
ggcggcggag gatggcgcgc gcggggcccg cacgtggagg ccggcgcggg ggcgcgggca 60
gggccggctg ctgagacgcg ctgctgcccc ccgcgcgggc gccgcggctt caatggcgcc 120
atcgcccagg accggcagcc ggcaagatgc gaccgccctg cccagcatgt cctcaacttt 180
ctgggcgttc atgatcctgg ccagcctgct catcgcctac tgcagtcagc tggccgccgg 240
cacctgtgag attgtgacct tggaccggga cagcagccag cctcggagga cgatcgcccg 300
gcagaccgcc cgctgtgcgt gtagaaaggg gcagatcgcc ggcaccacga gagcccggcc 360
cgcctgtgtg gacgcaagaa tcatcaagac caagcagtgg tgtgacatgc ttccgtgtct 420
ggagggggaa ggctgcgact tgttaatcaa ccggtcaggc tggacgtgca cgcagcccgg 480
cgggaggata aagaccacca cggtctcctg acaaacacag cccctgaggg ggccccggga 540
gtggccttgg ctccctggag agcccacgtc tcagccacag ttctccactc gcctcggact 600
tcacccgttc tctgccgccc gcccactccg tttccctgtg gtccgtgaag gacggcctca 660
ggccttggca tcctgagctt cggtctgtcc agccgacccg aggaggccgg actcagacac 720
ataggcgggg ggcggcacct ggcatcagca atacgcagtc tgtgggagcc cggccgcgcc 780
cagcccccgc cgaccgtggc gttggccctg ctgtcctcag aggaggagga ggaggaggca 840
gctccggcag ccacagaagg ctgcagccca gcccgcctga gacacgacgc ctgccccagg 900
ggactgtcag gcacagaagc ggcctcctcc cgtgccccag actgtccgaa ttgcttttat 960
tttcttatac tttcagtata ctccatagac caaagagcaa aatctatctg aacctggacg 1020
caccctcact gtcagggtcc ctggggtcgc ttgtgcgggc gggagggcaa tggtggcaga 1080
gacatgctgg tggccccggc ggagcggaga gggcggccgt ggtggaggcc tccaccccag 1140
gagcaccccg cacaccctcg gaggacgggc ttcggctgcg cggaggccgt ggcacacctg 1200
cgggaggcag cgacggcccc cacgcagacg ccgggaacgc aggccgcttt attcctctgt 1260
acttagatca acttgaccgt actaaaatcc ctttctgttt taaccagtta aacatgcctc 1320
ttctacagct ccatttttga tagttggata atccagtatc tgccaagagc atgttgggtc 1380
tcccgtgact gctgcctcat cgatacccca tttagctcca gaaagcaaag aaaactcgag 1440
taacacttgt ttgaaagaga tcattaaatg tattttgcaa agcccaaaaa aaaaaaaaaa 1500
a 1501
<210> 5
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy Chain CDR1 - 3-2; 1-7A-IT Abs
<400> 5
Gly Phe Thr Phe Ser Ser Phe Asn Met Phe
1 5 10
<210> 6
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy Chain CDR2 - 3-2 Ab
<400> 6
Gln Ile Ser Ser Ser Gly Ser Ser Thr Asn Tyr Ala Pro Ala Val Arg
1 5 10 15
Gly
<210> 7
<211> 22
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy Chain CDR3 - 3-2; 1-7A-IT; Low-PI; 1-30; 1-17; 1-32; 4-11;
6-10 Abs
<400> 7
Ser Ser Tyr Asp Cys Pro Tyr Gly His Cys Ser Ser Gly Val Asp Ser
1 5 10 15
Ala Gly Glu Ile Asp Ala
20
<210> 8
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR1 - 3-2; 1-7A-IT; 1-17; 1-32; 4-11; 6-10 Abs
<400> 8
Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr Gly
1 5 10
<210> 9
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR2 - 3-2 Ab
<400> 9
Glu Ser Asn Lys Arg Pro Ser
1 5
<210> 10
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR3 - 3-2; 1-7A-IT Abs
<400> 10
Gly Ser Trp Asp Ser Ser Asn Gly Gly Ile
1 5 10
<210> 11
<211> 131
<212> PRT
<213> Artificial Sequence
<220>
<223> VH - 3-2 Ab
<400> 11
Ala Val Thr Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly
1 5 10 15
Ala Leu Ser Leu Val Cys Lys Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Asn Met Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val
35 40 45
Ala Gln Ile Ser Ser Ser Gly Ser Ser Thr Asn Tyr Ala Pro Ala Val
50 55 60
Arg Gly Arg Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Val Arg
65 70 75 80
Leu Gln Leu Asn Asn Pro Gly Ala Glu Asp Thr Gly Thr Tyr Tyr Cys
85 90 95
Ala Lys Ser Ser Tyr Asp Cys Pro Tyr Gly His Cys Ser Ser Gly Val
100 105 110
Asp Ser Ala Gly Glu Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile
115 120 125
Val Ser Ser
130
<210> 12
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> VL - 3-2 Ab
<400> 12
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Ala Pro Gly Ser Ala Pro Val Thr Leu Ile
35 40 45
Tyr Glu Ser Asn Lys Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala
65 70 75 80
Asp Asp Glu Ala Ile Tyr Tyr Cys Gly Ser Trp Asp Ser Ser Asn Gly
85 90 95
Gly Ile Phe Gly Ala Gly Thr Thr Leu Thr Val Leu
100 105
<210> 13
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy Chain CDR2 - 1-7A-IT Ab
<400> 13
Gln Ile Ser Ser Ser Gly Ser Ser Thr Asn Tyr Ala Pro Ala Val Lys
1 5 10 15
Gly
<210> 14
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy Chain CDR1 - Low-PI; 1-30; 1-17; 1-32; 4-11; 6-10 Abs
<400> 14
Gly Phe Asp Phe Glu Ser Phe Asn Met Phe
1 5 10
<210> 15
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy Chain CDR2 - Low-PI; 1-30; 1-17; 1-32; 4-11; 6-10 Abs
<400> 15
Gln Ile Ser Ser Ser Glu Glu Asp Glu Asn Tyr Ala Pro Ala Val Lys
1 5 10 15
Gly
<210> 16
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy Chain CDR1 - 2-13; 2-13D Abs
<400> 16
Gly Phe Thr Phe Ser Ser His Gly Met Phe
1 5 10
<210> 17
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy Chain CDR2 - 2-13; 2-13D; 2-13D-37 Abs
<400> 17
Glu Ile Thr Asn Asp Gly Ser Gly Thr Asn Tyr Gly Ser Ala Val Lys
1 5 10 15
Gly
<210> 18
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy Chain CDR3 - 2-13; 2-13D; 2-13D-37 Abs
<400> 18
Ser Thr Tyr Glu Cys Pro Gly Gly Phe Ser Cys Trp Gly Asp Thr Gly
1 5 10 15
Gln Ile Asp Ala
20
<210> 19
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Heavy Chain CDR1 - 2-13D-37 Ab
<400> 19
Gly Phe Asp Phe Ser Ser His Gly Met Phe
1 5 10
<210> 20
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR2 - 1-7A-IT
<400> 20
Glu Asn Asn Lys Arg Pro Ser
1 5
<210> 21
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR1 - Low-PI; 1-30 Abs
<400> 21
Ser Gly Gly Gly Ser Glu Glu Glu Gln Tyr Tyr Tyr Gly
1 5 10
<210> 22
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR2 - Low-PI Ab
<400> 22
Glu Asp Glu Glu Arg Pro Ser
1 5
<210> 23
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR3 - Low-PI; 1-30; 1-17; 1-32 Abs
<400> 23
Gly Ser Trp Asp Ser Glu Asp Glu Asp His
1 5 10
<210> 24
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR2 - 1-30; 1-32 Abs
<400> 24
Gln Asp Glu Glu Arg Pro Ser
1 5
<210> 25
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR2 - 1-17 Ab
<400> 25
Glu Asp Glu Gln Arg Pro Ser
1 5
<210> 26
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR2 - 4-11 Ab
<400> 26
Glu Asp His Glu Arg Pro Ser
1 5
<210> 27
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR3 - 4-11 Ab
<400> 27
Gly Ser Trp Asp Ser Ser Asp Glu Asp His
1 5 10
<210> 28
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR2 - 6-10 Ab
<400> 28
Gln Asp Leu Leu Arg Pro Ser
1 5
<210> 29
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR3 - 6-10 Ab
<400> 29
Gly Ser Trp Asp Ser Leu Ser Ser Ser His
1 5 10
<210> 30
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR1 - 2-13 Ab
<400> 30
Ser Gly Gly Ser Tyr Ser Tyr Gly
1 5
<210> 31
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR2 - 2-13; 2-13D; 2-13D-37 Abs
<400> 31
Trp Asp Asp Glu Arg Pro Ser
1 5
<210> 32
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR3 - 2-13; 2-13D; 2-13D-37 Abs
<400> 32
Gly Thr Glu Asp Ile Ser Gly Thr Ala Gly Val
1 5 10
<210> 33
<211> 131
<212> PRT
<213> Artificial Sequence
<220>
<223> VH - 1-7A-IT Ab
<400> 33
Ala Val Thr Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly
1 5 10 15
Ala Leu Arg Leu Ser Cys Lys Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Asn Met Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val
35 40 45
Ser Gln Ile Ser Ser Ser Gly Ser Ser Thr Asn Tyr Ala Pro Ala Val
50 55 60
Lys Gly Arg Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Tyr Cys
85 90 95
Ala Lys Ser Ser Tyr Asp Cys Pro Tyr Gly His Cys Ser Ser Gly Val
100 105 110
Asp Ser Ala Gly Glu Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile
115 120 125
Val Ser Ser
130
<210> 34
<211> 131
<212> PRT
<213> Artificial Sequence
<220>
<223> VH - Low-PI; 1-30; 1-17; 1-32; 4-11; 6-10 Abs
<400> 34
Ala Val Thr Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly
1 5 10 15
Ala Leu Arg Leu Ser Cys Lys Ala Ser Gly Phe Asp Phe Glu Ser Phe
20 25 30
Asn Met Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val
35 40 45
Ser Gln Ile Ser Ser Ser Glu Glu Asp Glu Asn Tyr Ala Pro Ala Val
50 55 60
Lys Gly Arg Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Tyr Cys
85 90 95
Ala Lys Ser Ser Tyr Asp Cys Pro Tyr Gly His Cys Ser Ser Gly Val
100 105 110
Asp Ser Ala Gly Glu Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile
115 120 125
Val Ser Ser
130
<210> 35
<211> 129
<212> PRT
<213> Artificial Sequence
<220>
<223> VH - 2-13 Ab
<400> 35
Ala Val Thr Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly
1 5 10 15
Ala Leu Ser Leu Val Cys Lys Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Gly Met Phe Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Tyr Val
35 40 45
Ala Glu Ile Thr Asn Asp Gly Ser Gly Thr Asn Tyr Gly Ser Ala Val
50 55 60
Lys Gly Arg Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Val Arg
65 70 75 80
Leu Gln Leu Asn Asn Leu Arg Ala Glu Asp Thr Gly Thr Tyr Phe Cys
85 90 95
Ala Arg Ser Thr Tyr Glu Cys Pro Gly Gly Phe Ser Cys Trp Gly Asp
100 105 110
Thr Gly Gln Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser
115 120 125
Ser
<210> 36
<211> 129
<212> PRT
<213> Artificial Sequence
<220>
<223> VH - 2-13D Ab
<400> 36
Ala Val Thr Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly
1 5 10 15
Ala Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Gly Met Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val
35 40 45
Ser Glu Ile Thr Asn Asp Gly Ser Gly Thr Asn Tyr Gly Ser Ala Val
50 55 60
Lys Gly Arg Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Phe Cys
85 90 95
Ala Arg Ser Thr Tyr Glu Cys Pro Gly Gly Phe Ser Cys Trp Gly Asp
100 105 110
Thr Gly Gln Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser
115 120 125
Ser
<210> 37
<211> 129
<212> PRT
<213> Artificial Sequence
<220>
<223> VH - 2-13D-37 Ab
<400> 37
Ala Val Thr Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly
1 5 10 15
Ala Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Asp Phe Ser Ser His
20 25 30
Gly Met Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val
35 40 45
Ser Glu Ile Thr Asn Asp Gly Ser Gly Thr Asn Tyr Gly Ser Ala Val
50 55 60
Lys Gly Arg Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Phe Cys
85 90 95
Ala Arg Ser Thr Tyr Glu Cys Pro Gly Gly Phe Ser Cys Trp Gly Asp
100 105 110
Thr Gly Gln Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser
115 120 125
Ser
<210> 38
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> VL - 1-7A-IT Ab
<400> 38
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Glu Asn Asn Lys Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Ser Asn Gly Gly
85 90 95
Ile Phe Gly Ala Gly Thr Thr Leu Thr Val Leu
100 105
<210> 39
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> VL - Low-PI Ab
<400> 39
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Glu Glu Glu Gln Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Glu Asp Glu Glu Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Glu Asp Glu Asp
85 90 95
His Phe Gly Ala Gly Thr Thr Leu Thr Val Leu
100 105
<210> 40
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> VL - 1-30 Ab
<400> 40
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Glu Glu Glu Gln Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Gln Asp Glu Glu Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Glu Asp Glu Asp
85 90 95
His Phe Gly Ala Gly Thr Thr Leu Thr Val Leu
100 105
<210> 41
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> VL - 1-17 Ab
<400> 41
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Glu Asp Glu Gln Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Glu Asp Glu Asp
85 90 95
His Phe Gly Ala Gly Thr Thr Leu Thr Val Leu
100 105
<210> 42
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> VL - 1-32 Ab
<400> 42
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Gln Asp Glu Glu Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Glu Asp Glu Asp
85 90 95
His Phe Gly Ala Gly Thr Thr Leu Thr Val Leu
100 105
<210> 43
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> VL - 4-11 Ab
<400> 43
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Glu Asp His Glu Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Ser Asp Glu Asp
85 90 95
His Phe Gly Ala Gly Thr Thr Leu Thr Val Leu
100 105
<210> 44
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> VL - 6-10 Ab
<400> 44
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Gln Asp Leu Leu Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Leu Ser Ser Ser
85 90 95
His Phe Gly Ala Gly Thr Thr Leu Thr Val Leu
100 105
<210> 45
<211> 104
<212> PRT
<213> Artificial Sequence
<220>
<223> VL - 2-13 Ab
<400> 45
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Ser Tyr Ser Tyr Gly Trp Phe Gln Gln
20 25 30
Lys Ser Pro Gly Ser Ala Leu Val Thr Val Ile Tyr Trp Asp Asp Glu
35 40 45
Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ala Leu Ser Gly Ser
50 55 60
Thr Asn Thr Leu Thr Ile Thr Gly Val Gln Ala Asp Asp Glu Ala Val
65 70 75 80
Tyr Phe Cys Gly Thr Glu Asp Ile Ser Gly Thr Ala Gly Val Phe Gly
85 90 95
Ala Gly Thr Thr Leu Thr Val Leu
100
<210> 46
<211> 103
<212> PRT
<213> Artificial Sequence
<220>
<223> VL - 2-13D; 2-13D-37 Abs
<400> 46
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Ala
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Val Tyr Ser Tyr Gly Trp Phe Gln Gln
20 25 30
Lys Pro Gly Ser Ala Leu Val Thr Val Ile Tyr Trp Asp Asp Glu Arg
35 40 45
Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ala Leu Ser Gly Ser Thr
50 55 60
Asn Thr Leu Thr Ile Thr Gly Val Gln Ala Glu Asp Glu Ala Asp Tyr
65 70 75 80
Tyr Cys Gly Thr Glu Asp Ile Ser Gly Thr Ala Gly Val Phe Gly Ala
85 90 95
Gly Thr Thr Leu Thr Val Leu
100
<210> 47
<211> 257
<212> PRT
<213> Artificial Sequence
<220>
<223> scFv - 3-2 Ab
<400> 47
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Ala Pro Gly Ser Ala Pro Val Thr Leu Ile
35 40 45
Tyr Glu Ser Asn Lys Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala
65 70 75 80
Asp Asp Glu Ala Ile Tyr Tyr Cys Gly Ser Trp Asp Ser Ser Asn Gly
85 90 95
Gly Ile Phe Gly Ala Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser
100 105 110
Arg Ser Ser Gly Gly Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val
115 120 125
Thr Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu
130 135 140
Ser Leu Val Cys Lys Ala Ser Gly Phe Thr Phe Ser Ser Phe Asn Met
145 150 155 160
Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val Ala Gln
165 170 175
Ile Ser Ser Ser Gly Ser Ser Thr Asn Tyr Ala Pro Ala Val Arg Gly
180 185 190
Arg Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Val Arg Leu Gln
195 200 205
Leu Asn Asn Pro Gly Ala Glu Asp Thr Gly Thr Tyr Tyr Cys Ala Lys
210 215 220
Ser Ser Tyr Asp Cys Pro Tyr Gly His Cys Ser Ser Gly Val Asp Ser
225 230 235 240
Ala Gly Glu Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser
245 250 255
Ser
<210> 48
<211> 256
<212> PRT
<213> Artificial Sequence
<220>
<223> scFV - 1-7A-IT Ab
<400> 48
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Glu Asn Asn Lys Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Ser Asn Gly Gly
85 90 95
Ile Phe Gly Ala Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser Arg
100 105 110
Ser Ser Gly Gly Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val Thr
115 120 125
Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu Arg
130 135 140
Leu Ser Cys Lys Ala Ser Gly Phe Thr Phe Ser Ser Phe Asn Met Phe
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val Ser Gln Ile
165 170 175
Ser Ser Ser Gly Ser Ser Thr Asn Tyr Ala Pro Ala Val Lys Gly Arg
180 185 190
Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Tyr Cys Ala Lys Ser
210 215 220
Ser Tyr Asp Cys Pro Tyr Gly His Cys Ser Ser Gly Val Asp Ser Ala
225 230 235 240
Gly Glu Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser Ser
245 250 255
<210> 49
<211> 256
<212> PRT
<213> Artificial Sequence
<220>
<223> scFv - Low-PI Ab
<400> 49
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Glu Glu Glu Gln Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Glu Asp Glu Glu Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Glu Asp Glu Asp
85 90 95
His Phe Gly Ala Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser Arg
100 105 110
Ser Ser Gly Gly Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val Thr
115 120 125
Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu Arg
130 135 140
Leu Ser Cys Lys Ala Ser Gly Phe Asp Phe Glu Ser Phe Asn Met Phe
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val Ser Gln Ile
165 170 175
Ser Ser Ser Glu Glu Asp Glu Asn Tyr Ala Pro Ala Val Lys Gly Arg
180 185 190
Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Tyr Cys Ala Lys Ser
210 215 220
Ser Tyr Asp Cys Pro Tyr Gly His Cys Ser Ser Gly Val Asp Ser Ala
225 230 235 240
Gly Glu Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser Ser
245 250 255
<210> 50
<211> 256
<212> PRT
<213> Artificial Sequence
<220>
<223> scFv - 1-30 Ab
<400> 50
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Glu Glu Glu Gln Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Gln Asp Glu Glu Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Glu Asp Glu Asp
85 90 95
His Phe Gly Ala Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser Arg
100 105 110
Ser Ser Gly Gly Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val Thr
115 120 125
Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu Arg
130 135 140
Leu Ser Cys Lys Ala Ser Gly Phe Asp Phe Glu Ser Phe Asn Met Phe
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val Ser Gln Ile
165 170 175
Ser Ser Ser Glu Glu Asp Glu Asn Tyr Ala Pro Ala Val Lys Gly Arg
180 185 190
Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Tyr Cys Ala Lys Ser
210 215 220
Ser Tyr Asp Cys Pro Tyr Gly His Cys Ser Ser Gly Val Asp Ser Ala
225 230 235 240
Gly Glu Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser Ser
245 250 255
<210> 51
<211> 256
<212> PRT
<213> Artificial Sequence
<220>
<223> scFv - 1-17 Ab
<400> 51
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Glu Asp Glu Gln Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Glu Asp Glu Asp
85 90 95
His Phe Gly Ala Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser Arg
100 105 110
Ser Ser Gly Gly Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val Thr
115 120 125
Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu Arg
130 135 140
Leu Ser Cys Lys Ala Ser Gly Phe Asp Phe Glu Ser Phe Asn Met Phe
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val Ser Gln Ile
165 170 175
Ser Ser Ser Glu Glu Asp Glu Asn Tyr Ala Pro Ala Val Lys Gly Arg
180 185 190
Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Tyr Cys Ala Lys Ser
210 215 220
Ser Tyr Asp Cys Pro Tyr Gly His Cys Ser Ser Gly Val Asp Ser Ala
225 230 235 240
Gly Glu Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser Ser
245 250 255
<210> 52
<211> 256
<212> PRT
<213> Artificial Sequence
<220>
<223> scFv - 1-32 Ab
<400> 52
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Gln Asp Glu Glu Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Glu Asp Glu Asp
85 90 95
His Phe Gly Ala Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser Arg
100 105 110
Ser Ser Gly Gly Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val Thr
115 120 125
Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu Arg
130 135 140
Leu Ser Cys Lys Ala Ser Gly Phe Asp Phe Glu Ser Phe Asn Met Phe
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val Ser Gln Ile
165 170 175
Ser Ser Ser Glu Glu Asp Glu Asn Tyr Ala Pro Ala Val Lys Gly Arg
180 185 190
Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Tyr Cys Ala Lys Ser
210 215 220
Ser Tyr Asp Cys Pro Tyr Gly His Cys Ser Ser Gly Val Asp Ser Ala
225 230 235 240
Gly Glu Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser Ser
245 250 255
<210> 53
<211> 256
<212> PRT
<213> Artificial Sequence
<220>
<223> scFv - 4-11 Ab
<400> 53
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Glu Asp His Glu Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Ser Asp Glu Asp
85 90 95
His Phe Gly Ala Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser Arg
100 105 110
Ser Ser Gly Gly Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val Thr
115 120 125
Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu Arg
130 135 140
Leu Ser Cys Lys Ala Ser Gly Phe Asp Phe Glu Ser Phe Asn Met Phe
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val Ser Gln Ile
165 170 175
Ser Ser Ser Glu Glu Asp Glu Asn Tyr Ala Pro Ala Val Lys Gly Arg
180 185 190
Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Tyr Cys Ala Lys Ser
210 215 220
Ser Tyr Asp Cys Pro Tyr Gly His Cys Ser Ser Gly Val Asp Ser Ala
225 230 235 240
Gly Glu Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser Ser
245 250 255
<210> 54
<211> 256
<212> PRT
<213> Artificial Sequence
<220>
<223> scFv - 6-10 Ab
<400> 54
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Gly Ser Tyr Ala Gly Ser Tyr Tyr Tyr
20 25 30
Gly Trp Tyr Gln Gln Lys Pro Gly Ser Ala Pro Val Thr Leu Ile Tyr
35 40 45
Gln Asp Leu Leu Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ser
50 55 60
Thr Ser Gly Ser Thr Ala Thr Leu Thr Ile Thr Gly Val Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Ser Leu Ser Ser Ser
85 90 95
His Phe Gly Ala Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser Arg
100 105 110
Ser Ser Gly Gly Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val Thr
115 120 125
Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu Arg
130 135 140
Leu Ser Cys Lys Ala Ser Gly Phe Asp Phe Glu Ser Phe Asn Met Phe
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val Ser Gln Ile
165 170 175
Ser Ser Ser Glu Glu Asp Glu Asn Tyr Ala Pro Ala Val Lys Gly Arg
180 185 190
Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Tyr Cys Ala Lys Ser
210 215 220
Ser Tyr Asp Cys Pro Tyr Gly His Cys Ser Ser Gly Val Asp Ser Ala
225 230 235 240
Gly Glu Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser Ser
245 250 255
<210> 55
<211> 251
<212> PRT
<213> Artificial Sequence
<220>
<223> scFv - 2-13 Ab
<400> 55
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Val
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Ser Tyr Ser Tyr Gly Trp Phe Gln Gln
20 25 30
Lys Ser Pro Gly Ser Ala Leu Val Thr Val Ile Tyr Trp Asp Asp Glu
35 40 45
Arg Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ala Leu Ser Gly Ser
50 55 60
Thr Asn Thr Leu Thr Ile Thr Gly Val Gln Ala Asp Asp Glu Ala Val
65 70 75 80
Tyr Phe Cys Gly Thr Glu Asp Ile Ser Gly Thr Ala Gly Val Phe Gly
85 90 95
Ala Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser Arg Ser Ser Gly
100 105 110
Gly Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val Thr Leu Asp Glu
115 120 125
Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu Ser Leu Val Cys
130 135 140
Lys Ala Ser Gly Phe Thr Phe Ser Ser His Gly Met Phe Trp Val Arg
145 150 155 160
Gln Thr Pro Gly Lys Gly Leu Glu Tyr Val Ala Glu Ile Thr Asn Asp
165 170 175
Gly Ser Gly Thr Asn Tyr Gly Ser Ala Val Lys Gly Arg Ala Thr Ile
180 185 190
Ser Arg Asp Asn Gly Gln Ser Thr Val Arg Leu Gln Leu Asn Asn Leu
195 200 205
Arg Ala Glu Asp Thr Gly Thr Tyr Phe Cys Ala Arg Ser Thr Tyr Glu
210 215 220
Cys Pro Gly Gly Phe Ser Cys Trp Gly Asp Thr Gly Gln Ile Asp Ala
225 230 235 240
Trp Gly His Gly Thr Glu Val Ile Val Ser Ser
245 250
<210> 56
<211> 250
<212> PRT
<213> Artificial Sequence
<220>
<223> scFv - 2-13D Ab
<400> 56
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Ala
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Val Tyr Ser Tyr Gly Trp Phe Gln Gln
20 25 30
Lys Pro Gly Ser Ala Leu Val Thr Val Ile Tyr Trp Asp Asp Glu Arg
35 40 45
Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ala Leu Ser Gly Ser Thr
50 55 60
Asn Thr Leu Thr Ile Thr Gly Val Gln Ala Glu Asp Glu Ala Asp Tyr
65 70 75 80
Tyr Cys Gly Thr Glu Asp Ile Ser Gly Thr Ala Gly Val Phe Gly Ala
85 90 95
Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser Arg Ser Ser Gly Gly
100 105 110
Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val Thr Leu Asp Glu Ser
115 120 125
Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu Arg Leu Ser Cys Ser
130 135 140
Ala Ser Gly Phe Thr Phe Ser Ser His Gly Met Phe Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gly Leu Glu Tyr Val Ser Glu Ile Thr Asn Asp Gly
165 170 175
Ser Gly Thr Asn Tyr Gly Ser Ala Val Lys Gly Arg Ala Thr Ile Ser
180 185 190
Arg Asp Asn Gly Gln Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Gly Thr Tyr Phe Cys Ala Arg Ser Thr Tyr Glu Cys
210 215 220
Pro Gly Gly Phe Ser Cys Trp Gly Asp Thr Gly Gln Ile Asp Ala Trp
225 230 235 240
Gly His Gly Thr Glu Val Ile Val Ser Ser
245 250
<210> 57
<211> 250
<212> PRT
<213> Artificial Sequence
<220>
<223> scFv - 2-13D-37 Ab
<400> 57
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Ala
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Val Tyr Ser Tyr Gly Trp Phe Gln Gln
20 25 30
Lys Pro Gly Ser Ala Leu Val Thr Val Ile Tyr Trp Asp Asp Glu Arg
35 40 45
Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ala Leu Ser Gly Ser Thr
50 55 60
Asn Thr Leu Thr Ile Thr Gly Val Gln Ala Glu Asp Glu Ala Asp Tyr
65 70 75 80
Tyr Cys Gly Thr Glu Asp Ile Ser Gly Thr Ala Gly Val Phe Gly Ala
85 90 95
Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser Arg Ser Ser Gly Gly
100 105 110
Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val Thr Leu Asp Glu Ser
115 120 125
Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu Arg Leu Ser Cys Ser
130 135 140
Ala Ser Gly Phe Asp Phe Ser Ser His Gly Met Phe Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gly Leu Glu Tyr Val Ser Glu Ile Thr Asn Asp Gly
165 170 175
Ser Gly Thr Asn Tyr Gly Ser Ala Val Lys Gly Arg Ala Thr Ile Ser
180 185 190
Arg Asp Asn Gly Gln Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Gly Thr Tyr Phe Cys Ala Arg Ser Thr Tyr Glu Cys
210 215 220
Pro Gly Gly Phe Ser Cys Trp Gly Asp Thr Gly Gln Ile Asp Ala Trp
225 230 235 240
Gly His Gly Thr Glu Val Ile Val Ser Ser
245 250
<210> 58
<211> 393
<212> DNA
<213> Artificial Sequence
<220>
<223> VH -3-2 Ab
<400> 58
gccgtgacgt tggacgagtc cgggggcggc ctccagacgc ccggaggagc gctcagcctc 60
gtctgcaagg cctccgggtt caccttcagc agcttcaaca tgttctgggt gcgacaggcg 120
cccggcaagg ggctggaata cgtcgctcaa attagcagca gtggtagtag cacaaactac 180
gcacccgcgg tgaggggccg tgccaccatc tcgagggaca acgggcagag cacagtgagg 240
ctgcagctga acaaccccgg ggctgaagac accggcacct actactgcgc caaaagtagt 300
tatgactgtc cttacggtca ttgtagtagt ggtgttgata gtgctggtga gatcgacgca 360
tggggccacg ggaccgaagt catcgtctcc tcc 393
<210> 59
<211> 393
<212> DNA
<213> Artificial Sequence
<220>
<223> VH - 1-7A-IT Ab
<400> 59
gccgtgacgt tggatgaatc cgggggcggc ctccagacgc ccggaggagc gctccgcctc 60
agctgcaagg cctctgggtt caccttcagc agcttcaaca tgttctgggt gcgacaggcg 120
cccggcaagg ggctggaata cgtctcgcag attagcagca gtggtagtag cacaaactac 180
gcacccgcgg tgaaaggccg tgccaccatc tcgagggaca acgggcagag cacactgtat 240
ctgcagatga acagcctgcg cgctgaagac accggcacct actactgcgc caaaagtagt 300
tatgactgtc cttacggtca ttgtagtagt ggtgttgata gtgctggtga gatcgacgca 360
tggggccacg ggaccgaagt catcgtctcc tcc 393
<210> 60
<211> 393
<212> DNA
<213> Artificial Sequence
<220>
<223> VH - Low-PI Ab
<400> 60
gccgtgacgt tggatgaatc cgggggcggc ctccagacgc ccggaggagc gctccgcctc 60
agctgcaagg cctctgggtt caccttcagc agcttcaaca tgttctgggt gcgacaggcg 120
cccggcaagg ggctggaata cgtctcgcag attagcagca gtggtagtag cacaaactac 180
gcacccgcgg tgaaaggccg tgccaccatc tcgagggaca acgggcagag cacactgtat 240
ctgcagatga acagcctgcg cgctgaagac accggcacct actactgcgc caaaagtagt 300
tatgactgtc cttacggtca ttgtagtagt ggtgttgata gtgctggtga gatcgacgca 360
tggggccacg ggaccgaagt catcgtctcc tcc 393
<210> 61
<211> 393
<212> DNA
<213> Artificial Sequence
<220>
<223> VH - 1-30 Ab
<400> 61
gccgtgacgt tggatgaatc cgggggcggc ctccagacgc ccggaggagc gctccgcctc 60
agctgcaagg cctctggctt tgattttgaa agcttcaaca tgttctgggt gcgacaggcg 120
cccggcaagg ggctggaata cgtctcgcag attagcagca gtgaagaaga tgaaaactac 180
gcacccgcgg tgaaaggccg tgccaccatc tcgagggaca acgggcagag cacactgtat 240
ctgcagatga acagcctgcg cgctgaagac accggcacct actactgcgc caaaagtagt 300
tatgactgtc cttacggtca ttgtagtagt ggtgttgata gtgctggtga gatcgacgca 360
tggggccacg ggaccgaagt catcgtctcc tcc 393
<210> 62
<211> 393
<212> DNA
<213> Artificial Sequence
<220>
<223> VH - 1-17 Ab
<400> 62
gccgtgacgt tggatgaatc cgggggcggc ctccagacgc ccggaggagc gctccgcctc 60
agctgcaagg cctctggctt tgattttgaa agcttcaaca tgttctgggt gcgacaggcg 120
cccggcaagg ggctggaata cgtctcgcag attagcagca gtgaagaaga tgaaaactac 180
gcacccgcgg tgaaaggccg tgccaccatc tcgagggaca acgggcagag cacactgtat 240
ctgcagatga acagcctgcg cgctgaagac accggcacct actactgcgc caaaagtagt 300
tatgactgtc cttacggtca ttgtagtagt ggtgttgata gtgctggtga gatcgacgca 360
tggggccacg ggaccgaagt catcgtctcc tcc 393
<210> 63
<211> 393
<212> DNA
<213> Artificial Sequence
<220>
<223> VH - 1-32 Ab
<400> 63
gccgtgacgt tggatgaatc cgggggcggc ctccagacgc ccggaggagc gctccgcctc 60
agctgcaagg cctctggctt tgattttgaa agcttcaaca tgttctgggt gcgacaggcg 120
cccggcaagg ggctggaata cgtctcgcag attagcagca gtgaagaaga tgaaaactac 180
gcacccgcgg tgaaaggccg tgccaccatc tcgagggaca acgggcagag cacactgtat 240
ctgcagatga acagcctgcg cgctgaagac accggcacct actactgcgc caaaagtagt 300
tatgactgtc cttacggtca ttgtagtagt ggtgttgata gtgctggtga gatcgacgca 360
tggggccacg ggaccgaagt catcgtctcc tcc 393
<210> 64
<211> 393
<212> DNA
<213> Artificial Sequence
<220>
<223> VH - 4-11 Ab
<400> 64
gccgtgacgt tggatgaatc cgggggcggc ctccagacgc ccggaggagc gctccgcctc 60
agctgcaagg cctctggctt tgattttgaa agcttcaaca tgttctgggt gcgacaggcg 120
cccggcaagg ggctggaata cgtctcgcag attagcagca gtgaagaaga tgaaaactac 180
gcacccgcgg tgaaaggccg tgccaccatc tcgagggaca acgggcagag cacactgtat 240
ctgcagatga acagcctgcg cgctgaagac accggcacct actactgcgc caaaagtagt 300
tatgactgtc cttacggtca ttgtagtagt ggtgttgata gtgctggtga gatcgacgca 360
tggggccacg ggaccgaagt catcgtctcc tcc 393
<210> 65
<211> 393
<212> DNA
<213> Artificial Sequence
<220>
<223> VH - 6-10 Ab
<400> 65
gccgtgacgt tggatgaatc cgggggcggc ctccagacgc ccggaggagc gctccgcctc 60
agctgcaagg cctctggctt tgattttgaa agcttcaaca tgttctgggt gcgacaggcg 120
cccggcaagg ggctggaata cgtctcgcag attagcagca gtgaagaaga tgaaaactac 180
gcacccgcgg tgaaaggccg tgccaccatc tcgagggaca acgggcagag cacactgtat 240
ctgcagatga acagcctgcg cgctgaagac accggcacct actactgcgc caaaagtagt 300
tatgactgtc cttacggtca ttgtagtagt ggtgttgata gtgctggtga gatcgacgca 360
tggggccacg ggaccgaagt catcgtctcc tcc 393
<210> 66
<211> 387
<212> DNA
<213> Artificial Sequence
<220>
<223> VH - 2-13 Ab
<400> 66
gccgtgacgt tggacgagtc cgggggcggc ctccagacgc ccggaggagc gctcagcctc 60
gtctgcaagg cctccgggtt caccttcagc agccatggca tgttctgggt gcgacagacg 120
cccggcaagg ggttggaata tgtcgctgaa attaccaatg atggtagtgg cacaaactac 180
gggtcggcgg tgaagggccg tgccaccatc tcgagggaca acgggcagag cacagtgagg 240
ctgcagctga acaacctcag ggctgaggac accggcacct acttctgcgc cagatctact 300
tatgaatgtc ctggtggttt tagttgttgg ggtgatactg gtcaaataga cgcatggggc 360
cacgggaccg aagtcatcgt ctcctcc 387
<210> 67
<211> 387
<212> DNA
<213> Artificial Sequence
<220>
<223> VH - 2-13D Ab
<400> 67
gccgtgacgt tggacgagtc cgggggcggc ctccagacgc ccggaggagc gctccgcctt 60
agctgcagcg cctccgggtt caccttcagc agccatggca tgttctgggt gcgacaggcg 120
cccggcaagg ggttggaata tgtctcggag attaccaatg atggtagtgg cacaaactac 180
gggtcggcgg tgaagggccg tgccaccatc tcgagggaca acgggcagag cacactgtat 240
ctgcagatga acagcctcag ggctgaggac accggcacct acttctgcgc cagatctact 300
tatgaatgtc ctggtggttt tagttgttgg ggtgatactg gtcaaataga cgcatggggc 360
cacgggaccg aagtcatcgt ctcctcc 387
<210> 68
<211> 387
<212> DNA
<213> Artificial Sequence
<220>
<223> VH - 2-13D-37 Ab
<400> 68
gccgtgacgt tggacgagtc cgggggcggc ctccagacgc ccggaggagc gctccgcctt 60
agctgcagcg cctccgggtt cgatttcagc agccatggca tgttctgggt gcgacaggcg 120
cccggcaagg ggttggaata tgtctcggag attaccaatg atggtagtgg cacaaactac 180
gggtcggcgg tgaagggccg tgccaccatc tcgagggaca acgggcagag cacactgtat 240
ctgcagatga acagcctcag ggctgaggac accggcacct acttctgcgc cagatctact 300
tatgaatgtc ctggtggttt tagttgttgg ggtgatactg gtcaaataga cgcatggggc 360
cacgggaccg aagtcatcgt ctcctcc 387
<210> 69
<211> 324
<212> DNA
<213> Artificial Sequence
<220>
<223> VL - 3-2 Ab
<400> 69
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgtcaa gatcacctgc 60
tccgggggtg gcagctatgc tggaagttac tattatggct ggtaccagca gaaggcacct 120
ggcagtgccc ctgtcactct gatctatgaa agcaacaaga gaccctcgga catcccttca 180
cgattctccg gttccacatc tggctccaca gccacactaa ccatcactgg ggtccaagcc 240
gatgacgagg ctatctatta ctgtgggagc tgggacagta gcaatggtgg tatatttggg 300
gccgggacaa ccctgaccgt ccta 324
<210> 70
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> VL - 1-7A-IT Ab
<400> 70
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgtcaa gatcacctgc 60
tccgggggtg gcagctatgc tggaagttac tattatggct ggtatcagca gaagcctggc 120
agtgcccctg tcactctgat ctatgaaaac aacaagagac cctcggacat cccttcacga 180
ttctccggtt ccacatctgg ctccacagcc acactaacca tcactggggt ccaagccggc 240
gacgaggctg attattactg tgggagctgg gacagtagca atggtggtat atttggggcc 300
gggacaaccc tgaccgtcct a 321
<210> 71
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> VL - Low-PI Ab
<400> 71
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgtcaa gatcacctgc 60
tccgggggtg gcagctatgc tggaagttac tattatggct ggtatcagca gaagcctggc 120
agtgcccctg tcactctgat ctatgaaaac aacaagagac cctcggacat cccttcacga 180
ttctccggtt ccacatctgg ctccacagcc acactaacca tcactggggt ccaagccggc 240
gacgaggctg attattactg tgggagctgg gacagtagca atggtggtat atttggggcc 300
gggacaaccc tgaccgtcct a 321
<210> 72
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> VL - 1-30 Ab
<400> 72
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgtcaa gatcacctgc 60
tccgggggtg gcagcgaaga agaacagtac tattatggct ggtatcagca gaagcctggc 120
agtgcccctg tcactctgat ctatcaggat gaagaaagac cctcggacat cccttcacga 180
ttctccggtt ccacatctgg ctccacagcc acactaacca tcactggggt ccaagccggc 240
gacgaggctg attattactg tgggagctgg gacagtgaag atgaagatca ttttggggcc 300
gggacaaccc tgaccgtcct a 321
<210> 73
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> VL 1-17 Ab
<400> 73
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgtcaa gatcacctgc 60
tccgggggtg gcagctatgc tggaagttac tattatggct ggtatcagca gaagcctggc 120
agtgcccctg tcactctgat ctatgaagat gaacagagac cctcggacat cccttcacga 180
ttctccggtt ccacatctgg ctccacagcc acactaacca tcactggggt ccaagccggc 240
gacgaggctg attattactg tgggagctgg gacagtgaag atgaagatca ttttggggcc 300
gggacaaccc tgaccgtcct a 321
<210> 74
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> VL 1-32 Ab
<400> 74
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgtcaa gatcacctgc 60
tccgggggtg gcagctatgc tggaagttac tattatggct ggtatcagca gaagcctggc 120
agtgcccctg tcactctgat ctatcaggat gaagaaagac cctcggacat cccttcacga 180
ttctccggtt ccacatctgg ctccacagcc acactaacca tcactggggt ccaagccggc 240
gacgaggctg attattactg tgggagctgg gacagtgaag atgaagatca ttttggggcc 300
gggacaaccc tgaccgtcct a 321
<210> 75
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> VL 4-11 Ab
<400> 75
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgtcaa gatcacctgc 60
tccgggggtg gcagctatgc tggaagttac tattatggct ggtatcagca gaagcctggc 120
agtgcccctg tcactctgat ctatgaagac cacgagagac cctcggacat cccttcacga 180
ttctccggtt ccacatctgg ctccacagcc acactaacca tcactggggt ccaagccggc 240
gacgaggctg attattactg tgggagctgg gacagtagcg atgaagatca ttttggggcc 300
gggacaaccc tgaccgtcct a 321
<210> 76
<211> 321
<212> DNA
<213> Artificial Sequence
<220>
<223> VL 6-10 Ab
<400> 76
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgtcaa gatcacctgc 60
tccgggggtg gcagctatgc tggaagttac tattatggct ggtatcagca gaagcctggc 120
agtgcccctg tcactctgat ctatcaggat ctgctgagac cctcggacat cccttcacga 180
ttctccggtt ccacatctgg ctccacagcc acactaacca tcactggggt ccaagccggc 240
gacgaggctg attattactg tgggagctgg gacagtctga gcagcagcca ttttggggcc 300
gggacaaccc tgaccgtcct a 321
<210> 77
<211> 312
<212> DNA
<213> Artificial Sequence
<220>
<223> VL 2-13 Ab
<400> 77
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgtcaa gataacctgc 60
tccgggggta gctatagcta tggctggttc cagcagaagt ctcctggcag tgcccttgtc 120
actgtgatct actgggatga tgagagaccc tcggacatcc cttcacgatt ctccggtgcc 180
ctatccggct ccacaaacac attaaccatc actggggtcc aagccgacga cgaggctgtc 240
tatttctgtg ggactgaaga catcagcggc actgctggtg tatttggggc cgggacaacc 300
ctgaccgtcc tg 312
<210> 78
<211> 309
<212> DNA
<213> Artificial Sequence
<220>
<223> VL 2-13D Ab
<400> 78
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgcgaa gataacctgc 60
tccgggggtg tgtatagcta tggctggttc cagcagaagc ctggcagtgc ccttgtcact 120
gtgatctact gggatgatga gagaccctcg gacatccctt cacgattctc cggtgcccta 180
tccggctcca caaacacatt aaccatcact ggggtccaag ccgaagacga ggctgattat 240
tattgtggga ctgaagacat cagcggcact gctggtgtat ttggggccgg gacaaccctg 300
accgtcctg 309
<210> 79
<211> 309
<212> DNA
<213> Artificial Sequence
<220>
<223> VL 2-13D-37 Ab
<400> 79
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgcgaa gataacctgc 60
tccgggggtg tgtatagcta tggctggttc cagcagaagc ctggcagtgc ccttgtcact 120
gtgatctact gggatgatga gagaccctcg gacatccctt cacgattctc cggtgcccta 180
tccggctcca caaacacatt aaccatcact ggggtccaag ccgaagacga ggctgattat 240
tattgtggga ctgaagacat cagcggcact gctggtgtat ttggggccgg gacaaccctg 300
accgtcctg 309
<210> 80
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Light Chain CDR1 - 2-13D; 2-13D-37 Abs
<400> 80
Ser Gly Gly Val Tyr Ser Tyr Gly
1 5
<210> 81
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> EP1 Epitope
<400> 81
Ile Val Thr Leu Asp
1 5
<210> 82
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> EP2 Epitope
<400> 82
Asp Ser Ser Gln Pro
1 5
<210> 83
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> EP3 Epitope
<400> 83
Arg Thr Ile Ala Arg
1 5
<210> 84
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> EP4 Epitope
<400> 84
Ala Arg Cys Ala Cys Arg Lys
1 5
<210> 85
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<223> EP5 Epitope
<400> 85
Ala Arg Pro Ala
1
<210> 86
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> EP6 Epitope
<400> 86
Lys Thr Lys Gln Trp Cys Asp Met Leu
1 5
<210> 87
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> EP7 Epitope
<400> 87
Gly Cys Asp Leu Leu Ile Asn Arg
1 5
<210> 88
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> EP8 Epitope
<400> 88
Thr Cys Thr Gln Pro Gly Gly Arg
1 5
<210> 89
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope F1
<400> 89
Gln Leu Ala Ala Gly Thr Cys Glu Ile Val Thr Leu Asp Arg
1 5 10
<210> 90
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope F2
<400> 90
Thr Leu Asp Arg Asp Ser Ser Gln Pro Arg Arg Thr Ile Ala Arg Gln
1 5 10 15
Thr Ala Arg Cys
20
<210> 91
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope F3
<400> 91
Thr Ala Arg Cys Ala Cys Arg Lys Gly Gln Ile Ala Gly Thr Thr Arg
1 5 10 15
Ala Arg Pro Ala
20
<210> 92
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope F4
<400> 92
Ala Arg Pro Ala Cys Val Asp Ala Arg Ile Ile Lys Thr Lys Gln Trp
1 5 10 15
Cys Asp Met Leu
20
<210> 93
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope F5
<400> 93
Cys Asp Met Leu Pro Cys Leu Glu Gly Glu Gly Cys Asp Leu Leu Ile
1 5 10 15
Asn Arg Ser Gly
20
<210> 94
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Epitope F6
<400> 94
Asn Arg Ser Gly Trp Thr Cys Thr Gln Pro Gly Gly Arg Ile Lys Thr
1 5 10 15
Thr Thr Val Ser
20
<210> 95
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 95
Ser Gly Gly Gly Gly Ser Ser Gly Gly Gly Gly Ser
1 5 10
<210> 96
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> 2-13 binding epitope 1
<400> 96
Cys Arg Lys Gly Gln Ile Ala Gly Thr Thr Arg Ala Arg
1 5 10
<210> 97
<211> 14
<212> PRT
<213> Artificial Sequence
<220>
<223> 2-13 binding epitope 2
<400> 97
Pro Ala Cys Val Asp Ala Arg Ile Ile Lys Thr Lys Gln Trp
1 5 10
<210> 98
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> binding epitope
<220>
<221> BINDING
<222> (1)..(2)
<223> Antibody specifically binds to respective residue
<220>
<221> BINDING
<222> (6)..(8)
<223> Antibody specifically binds to respective residue
<400> 98
Arg Asp Ser Ser Gln Pro Arg Arg
1 5
<210> 99
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> binding epitope
<220>
<221> BINDING
<222> (1)..(1)
<223> Antibody Specifically binds to respective residue
<220>
<221> BINDING
<222> (6)..(8)
<223> Antibody Specifically binds to respective residue
<400> 99
Arg Asp Ser Ser Gln Pro Arg Arg
1 5
<210> 100
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> binding epitope
<220>
<221> BINDING
<222> (1)..(1)
<223> Antibody Specifically binds to respective residue
<220>
<221> BINDING
<222> (7)..(8)
<223> Antibody Specifically binds to respective residue
<400> 100
Arg Asp Ser Ser Gln Pro Arg Arg
1 5
<210> 101
<211> 100
<212> PRT
<213> Artificial Sequence
<220>
<223> Mature Human FAM19A5 Isoform 2
<400> 101
Gln Phe Leu Lys Glu Gly Gln Leu Ala Ala Gly Thr Cys Glu Ile Val
1 5 10 15
Thr Leu Asp Arg Asp Ser Ser Gln Pro Arg Arg Thr Ile Ala Arg Gln
20 25 30
Thr Ala Arg Cys Ala Cys Arg Lys Gly Gln Ile Ala Gly Thr Thr Arg
35 40 45
Ala Arg Pro Ala Cys Val Asp Ala Arg Ile Ile Lys Thr Lys Gln Trp
50 55 60
Cys Asp Met Leu Pro Cys Leu Glu Gly Glu Gly Cys Asp Leu Leu Ile
65 70 75 80
Asn Arg Ser Gly Trp Thr Cys Thr Gln Pro Gly Gly Arg Ile Lys Thr
85 90 95
Thr Thr Val Ser
100
<210> 102
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #1 (residues 2-8 of SEQ ID NO: 101)
<400> 102
Phe Leu Lys Glu Gly Gln Leu
1 5
<210> 103
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #2 (residues 2-14 of SEQ ID NO: 101)
<400> 103
Phe Leu Lys Glu Gly Gln Leu Ala Ala Gly Thr Cys Glu
1 5 10
<210> 104
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #3 (residues 3-11 of SEQ ID NO: 101)
<400> 104
Leu Lys Glu Gly Gln Leu Ala Ala Gly
1 5
<210> 105
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #4 (residues 3-15 of SEQ ID NO: 101)
<400> 105
Leu Lys Glu Gly Gln Leu Ala Ala Gly Thr Cys Glu Ile
1 5 10
<210> 106
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #5 (residues 3-18 of SEQ ID NO: 101)
<400> 106
Leu Lys Glu Gly Gln Leu Ala Ala Gly Thr Cys Glu Ile Val Thr Leu
1 5 10 15
<210> 107
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #6 (residues 9-15 of SEQ ID NO: 101)
<400> 107
Ala Ala Gly Thr Cys Glu Ile
1 5
<210> 108
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #7 (residues 20-38 of SEQ ID NO: 101)
<400> 108
Arg Asp Ser Ser Gln Pro Pro Arg Thr Ile Ala Arg Gln Thr Ala Arg
1 5 10 15
Cys Ala Cys
<210> 109
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #8 (residues 24-34 of SEQ ID NO: 101)
<400> 109
Gln Pro Pro Arg Thr Ile Ala Arg Gln Thr Ala
1 5 10
<210> 110
<211> 17
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #9 (residues 37-53 of SEQ ID NO: 101)
<400> 110
Ala Cys Arg Lys Gly Gln Ile Ala Gly Thr Thr Arg Ala Arg Pro Ala
1 5 10 15
Cys
<210> 111
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #10 (residues 37-55 of SEQ ID NO: 101)
<400> 111
Ala Cys Arg Lys Gly Gln Ile Ala Gly Thr Thr Arg Ala Arg Pro Ala
1 5 10 15
Cys Val Asp
<210> 112
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #12 (residues 56-65 of SEQ ID NO: 101)
<400> 112
Ala Cys Arg Lys Gly Gln Ile Ala Gly Thr Thr Arg Ala Arg Pro Ala
1 5 10 15
Cys Val Asp Ala
20
<210> 113
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #12 (residues 56-65 of SEQ ID NO: 101)
<400> 113
Ala Arg Ile Ile Lys Thr Lys Gln Trp Cys
1 5 10
<210> 114
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #13 (residues 56-67 of SEQ ID NO: 101)
<400> 114
Ala Arg Ile Ile Lys Thr Lys Gln Trp Cys Asp Met
1 5 10
<210> 115
<211> 13
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #14 (residues 56-68 of SEQ ID NO: 101)
<400> 115
Ala Arg Ile Ile Lys Thr Lys Gln Trp Cys Asp Met Leu
1 5 10
<210> 116
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #15 (residues 56-71 of SEQ ID NO: 101)
<400> 116
Ala Arg Ile Ile Lys Thr Lys Gln Trp Cys Asp Met Leu Pro Cys Leu
1 5 10 15
<210> 117
<211> 11
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #16 (residues 57-67 of SEQ ID NO: 101)
<400> 117
Arg Ile Ile Lys Thr Lys Gln Trp Cys Asp Met
1 5 10
<210> 118
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #17 (residues 57-68 of SEQ ID NO: 101)
<400> 118
Arg Ile Ile Lys Thr Lys Gln Trp Cys Asp Met Leu
1 5 10
<210> 119
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #18 (residues 64-71 of SEQ ID NO: 101)
<400> 119
Trp Cys Asp Met Leu Pro Cys Leu
1 5
<210> 120
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #19 (residues 68-75 of SEQ ID NO: 101)
<400> 120
Leu Pro Cys Leu Glu Gly Glu Gly
1 5
<210> 121
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #20 (residues 69-75 of SEQ ID NO: 101)
<400> 121
Pro Cys Leu Glu Gly Glu Gly
1 5
<210> 122
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #21 (residues 69-77 of SEQ ID NO: 101)
<400> 122
Pro Cys Leu Glu Gly Glu Gly Cys Asp
1 5
<210> 123
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #22 (residues 69-78 of SEQ ID NO: 101)
<400> 123
Pro Cys Leu Glu Gly Glu Gly Cys Asp Leu
1 5 10
<210> 124
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #23 (residues 72-78 of SEQ ID NO: 101)
<400> 124
Glu Gly Glu Gly Cys Asp Leu
1 5
<210> 125
<211> 8
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #24 (residues 72-79 of SEQ ID NO: 101)
<400> 125
Glu Gly Glu Gly Cys Asp Leu Leu
1 5
<210> 126
<211> 21
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #25 (residues 78-98 of SEQ ID NO: 101)
<400> 126
Leu Leu Ile Asn Arg Ser Gly Trp Thr Cys Thr Gln Pro Gly Gly Arg
1 5 10 15
Ile Lys Thr Thr Thr
20
<210> 127
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Peptide Sequence #26 (residues 79-98 of SEQ ID NO: 101)
<400> 127
Leu Ile Asn Arg Ser Gly Trp Thr Cys Thr Gln Pro Gly Gly Arg Ile
1 5 10 15
Lys Thr Thr Thr
20
<210> 128
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> 2-13D-37-1.5W-41 Ab VH-CDR3
<400> 128
Ser Ser Tyr Val Cys Pro Gly Gly Phe Ser Cys Trp Gly Asp Thr Gly
1 5 10 15
Gln Ile Asp Ala
20
<210> 129
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> 2-13D-37-3W-16 Ab VH-CDR3
<400> 129
Ser Asn Tyr Ala Cys Pro Gly Gly Phe Ser Cys Trp Gly Asp Thr Gly
1 5 10 15
Gln Ile Asp Ala
20
<210> 130
<211> 129
<212> PRT
<213> Artificial Sequence
<220>
<223> 2-13D-37-1.5W-41 Ab VH
<400> 130
Ala Val Thr Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly
1 5 10 15
Ala Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Asp Phe Ser Ser His
20 25 30
Gly Met Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val
35 40 45
Ser Glu Ile Thr Asn Asp Gly Ser Gly Thr Asn Tyr Gly Ser Ala Val
50 55 60
Lys Gly Arg Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Phe Cys
85 90 95
Ala Arg Ser Ser Tyr Val Cys Pro Gly Gly Phe Ser Cys Trp Gly Asp
100 105 110
Thr Gly Gln Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser
115 120 125
Ser
<210> 131
<211> 129
<212> PRT
<213> Artificial Sequence
<220>
<223> 2-13D-37-3W-16 Ab VH
<400> 131
Ala Val Thr Leu Asp Glu Ser Gly Gly Gly Leu Gln Thr Pro Gly Gly
1 5 10 15
Ala Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Asp Phe Ser Ser His
20 25 30
Gly Met Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Val
35 40 45
Ser Glu Ile Thr Asn Asp Gly Ser Gly Thr Asn Tyr Gly Ser Ala Val
50 55 60
Lys Gly Arg Ala Thr Ile Ser Arg Asp Asn Gly Gln Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Gly Thr Tyr Phe Cys
85 90 95
Ala Arg Ser Asn Tyr Ala Cys Pro Gly Gly Phe Ser Cys Trp Gly Asp
100 105 110
Thr Gly Gln Ile Asp Ala Trp Gly His Gly Thr Glu Val Ile Val Ser
115 120 125
Ser
<210> 132
<211> 250
<212> PRT
<213> Artificial Sequence
<220>
<223> scFv - 2-13D-37-1.5W-41 Ab
<400> 132
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Ala
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Val Tyr Ser Tyr Gly Trp Phe Gln Gln
20 25 30
Lys Pro Gly Ser Ala Leu Val Thr Val Ile Tyr Trp Asp Asp Glu Arg
35 40 45
Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ala Leu Ser Gly Ser Thr
50 55 60
Asn Thr Leu Thr Ile Thr Gly Val Gln Ala Glu Asp Glu Ala Asp Tyr
65 70 75 80
Tyr Cys Gly Thr Glu Asp Ile Ser Gly Thr Ala Gly Val Phe Gly Ala
85 90 95
Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser Arg Ser Ser Gly Gly
100 105 110
Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val Thr Leu Asp Glu Ser
115 120 125
Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu Arg Leu Ser Cys Ser
130 135 140
Ala Ser Gly Phe Asp Phe Ser Ser His Gly Met Phe Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gly Leu Glu Tyr Val Ser Glu Ile Thr Asn Asp Gly
165 170 175
Ser Gly Thr Asn Tyr Gly Ser Ala Val Lys Gly Arg Ala Thr Ile Ser
180 185 190
Arg Asp Asn Gly Gln Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Gly Thr Tyr Phe Cys Ala Arg Ser Ser Tyr Val Cys
210 215 220
Pro Gly Gly Phe Ser Cys Trp Gly Asp Thr Gly Gln Ile Asp Ala Trp
225 230 235 240
Gly His Gly Thr Glu Val Ile Val Ser Ser
245 250
<210> 133
<211> 250
<212> PRT
<213> Artificial Sequence
<220>
<223> scFv - 2-13D-37-3W-16 Ab
<400> 133
Ala Leu Thr Gln Pro Ser Ser Val Ser Ala Asn Pro Gly Glu Thr Ala
1 5 10 15
Lys Ile Thr Cys Ser Gly Gly Val Tyr Ser Tyr Gly Trp Phe Gln Gln
20 25 30
Lys Pro Gly Ser Ala Leu Val Thr Val Ile Tyr Trp Asp Asp Glu Arg
35 40 45
Pro Ser Asp Ile Pro Ser Arg Phe Ser Gly Ala Leu Ser Gly Ser Thr
50 55 60
Asn Thr Leu Thr Ile Thr Gly Val Gln Ala Glu Asp Glu Ala Asp Tyr
65 70 75 80
Tyr Cys Gly Thr Glu Asp Ile Ser Gly Thr Ala Gly Val Phe Gly Ala
85 90 95
Gly Thr Thr Leu Thr Val Leu Gly Gln Ser Ser Arg Ser Ser Gly Gly
100 105 110
Gly Gly Ser Ser Gly Gly Gly Gly Ser Ala Val Thr Leu Asp Glu Ser
115 120 125
Gly Gly Gly Leu Gln Thr Pro Gly Gly Ala Leu Arg Leu Ser Cys Ser
130 135 140
Ala Ser Gly Phe Asp Phe Ser Ser His Gly Met Phe Trp Val Arg Gln
145 150 155 160
Ala Pro Gly Lys Gly Leu Glu Tyr Val Ser Glu Ile Thr Asn Asp Gly
165 170 175
Ser Gly Thr Asn Tyr Gly Ser Ala Val Lys Gly Arg Ala Thr Ile Ser
180 185 190
Arg Asp Asn Gly Gln Ser Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg
195 200 205
Ala Glu Asp Thr Gly Thr Tyr Phe Cys Ala Arg Ser Asn Tyr Ala Cys
210 215 220
Pro Gly Gly Phe Ser Cys Trp Gly Asp Thr Gly Gln Ile Asp Ala Trp
225 230 235 240
Gly His Gly Thr Glu Val Ile Val Ser Ser
245 250
<210> 134
<211> 387
<212> DNA
<213> Artificial Sequence
<220>
<223> 2-13D-37-1.5W-41 Ab VH
<400> 134
gccgtgacgt tggacgagtc cgggggcggc ctccagacgc ccggaggagc gctccgcctt 60
agctgcagcg cctccgggtt cgatttcagc agccatggca tgttctgggt gcgacaggcg 120
cccggcaagg ggttggaata tgtctcggag attaccaatg atggtagtgg cacaaactac 180
gggtcggcgg tgaagggccg tgccaccatc tcgagggaca acgggcagag cacactgtat 240
ctgcagatga acagcctcag ggctgaggac accggcacct acttctgcgc cagatcttct 300
tatgtttgtc ctggtggttt tagttgttgg ggtgatactg gtcaaataga cgcatggggc 360
cacgggaccg aagtcatcgt ctcctcc 387
<210> 135
<211> 387
<212> DNA
<213> Artificial Sequence
<220>
<223> 2-13D-37-3W-16 Ab VH
<400> 135
gccgtgacgt tggacgagtc cgggggcggc ctccagacgc ccggaggagc gctccgcctt 60
agctgcagcg cctccgggtt cgatttcagc agccatggca tgttctgggt gcgacaggcg 120
cccggcaagg ggttggaata tgtctcggag attaccaatg atggtagtgg cacaaactac 180
gggtcggcgg tgaagggccg tgccaccatc tcgagggaca acgggcagag cacactgtat 240
ctgcagatga acagcctcag ggctgaggac accggcacct acttctgcgc cagatctaat 300
tatgcttgtc ctggtggttt tagttgttgg ggtgatactg gtcaaataga cgcatggggc 360
cacgggaccg aagtcatcgt ctcctcc 387
<210> 136
<211> 309
<212> DNA
<213> Artificial Sequence
<220>
<223> 2-13D-37-1.5W-41 Ab VL
<400> 136
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgcgaa gataacctgc 60
tccgggggtg tgtatagcta tggctggttc cagcagaagc ctggcagtgc ccttgtcact 120
gtgatctact gggatgatga gagaccctcg gacatccctt cacgattctc cggtgcccta 180
tccggctcca caaacacatt aaccatcact ggggtccaag ccgaagacga ggctgattat 240
tattgtggga ctgaagacat cagcggcact gctggtgtat ttggggccgg gacaaccctg 300
accgtcctg 309
<210> 137
<211> 309
<212> DNA
<213> Artificial Sequence
<220>
<223> 2-13D-37-3W-16 Ab VL
<400> 137
gccctgactc agccgtcctc ggtgtcagca aacccaggag aaaccgcgaa gataacctgc 60
tccgggggtg tgtatagcta tggctggttc cagcagaagc ctggcagtgc ccttgtcact 120
gtgatctact gggatgatga gagaccctcg gacatccctt cacgattctc cggtgcccta 180
tccggctcca caaacacatt aaccatcact ggggtccaag ccgaagacga ggctgattat 240
tattgtggga ctgaagacat cagcggcact gctggtgtat ttggggccgg gacaaccctg 300
accgtcctg 309
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К C10ORF54 И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714232C2 |
| АНТИ-DR5-АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2735956C1 |
| АНТИТЕЛА К C10ORF54 И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2819627C1 |
| АНТИТЕЛО К РЕЦЕПТОРУ ТРОМБОЦИТАРНОГО ФАКТОРА РОСТА (PDGF) И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2778023C1 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
| НОВЫЕ АНТИ-HLA-А2 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2805674C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО АНТИ-EGFR/АНТИ-4-1ВВ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2831836C2 |
| АНТИТЕЛА ПРОТИВ СИГНАЛ-РЕГУЛЯТОРНОГО БЕЛКА АЛЬФА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2771964C2 |
| МОЛЕКУЛЫ АНТИ-SIGLEC-9 АНТИТЕЛА | 2020 |
|
RU2833330C1 |
| АНТИТЕЛО ДЛЯ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ С N-КОНЦЕВЫМ ДОМЕНОМ ЛИЗИЛ-ТРНК-СИНТЕТАЗЫ, ЭКСПОНИРОВАННЫМ НА ВНЕКЛЕТОЧНОЙ МЕМБРАНЕ | 2019 |
|
RU2781304C1 |



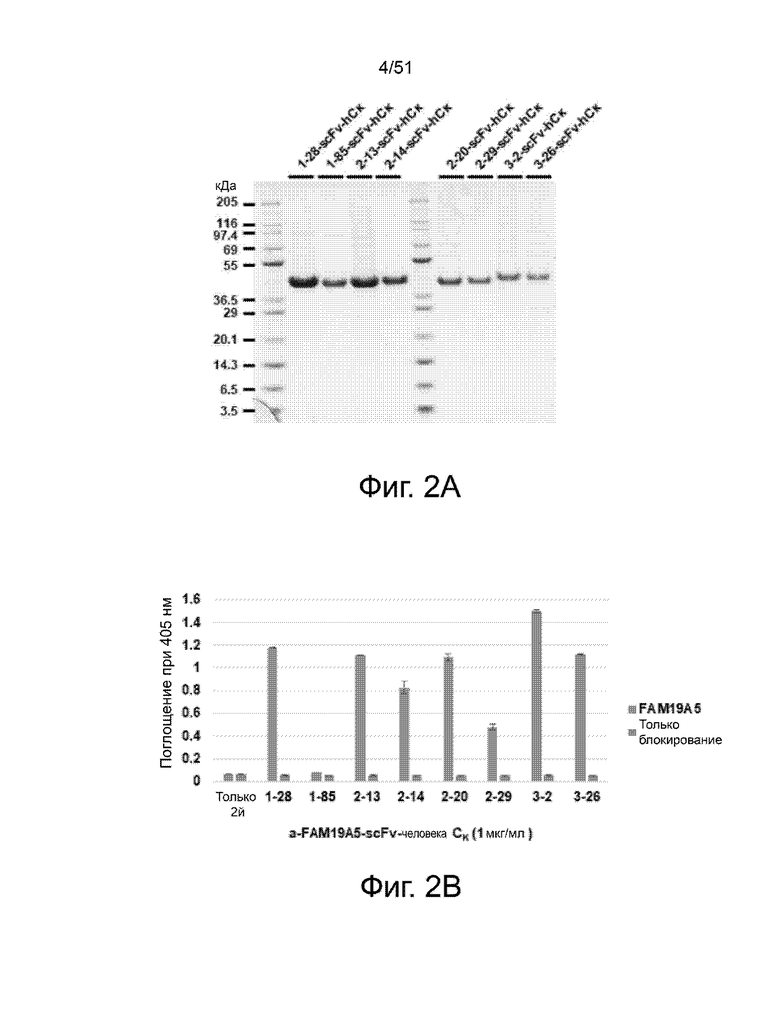
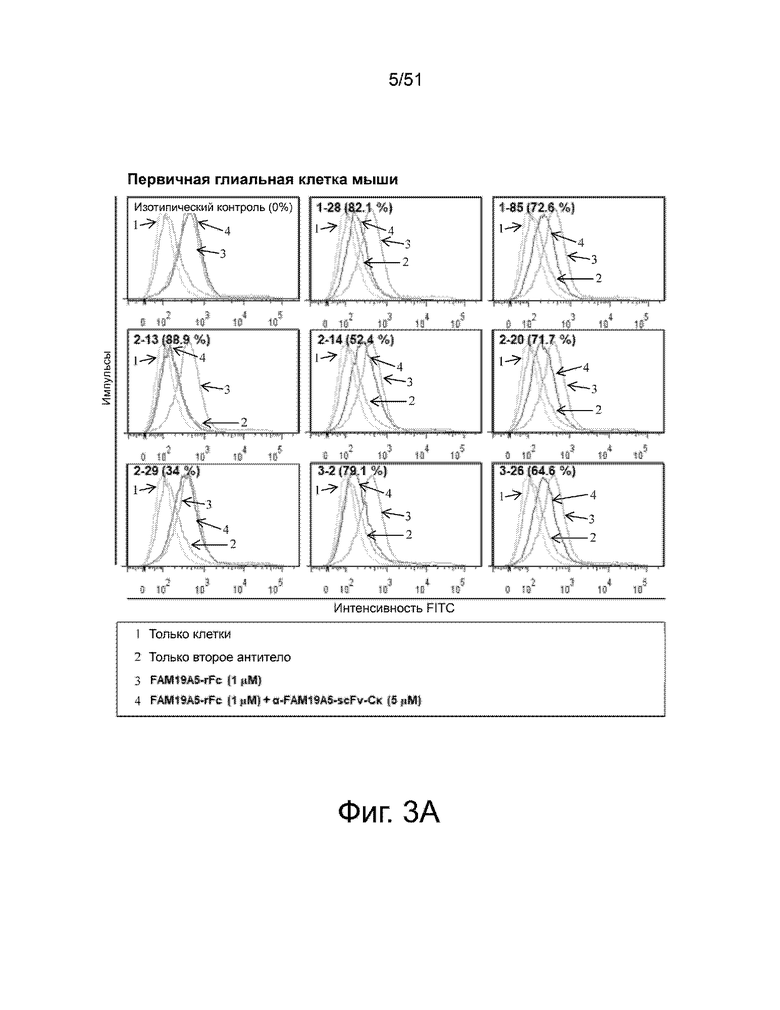
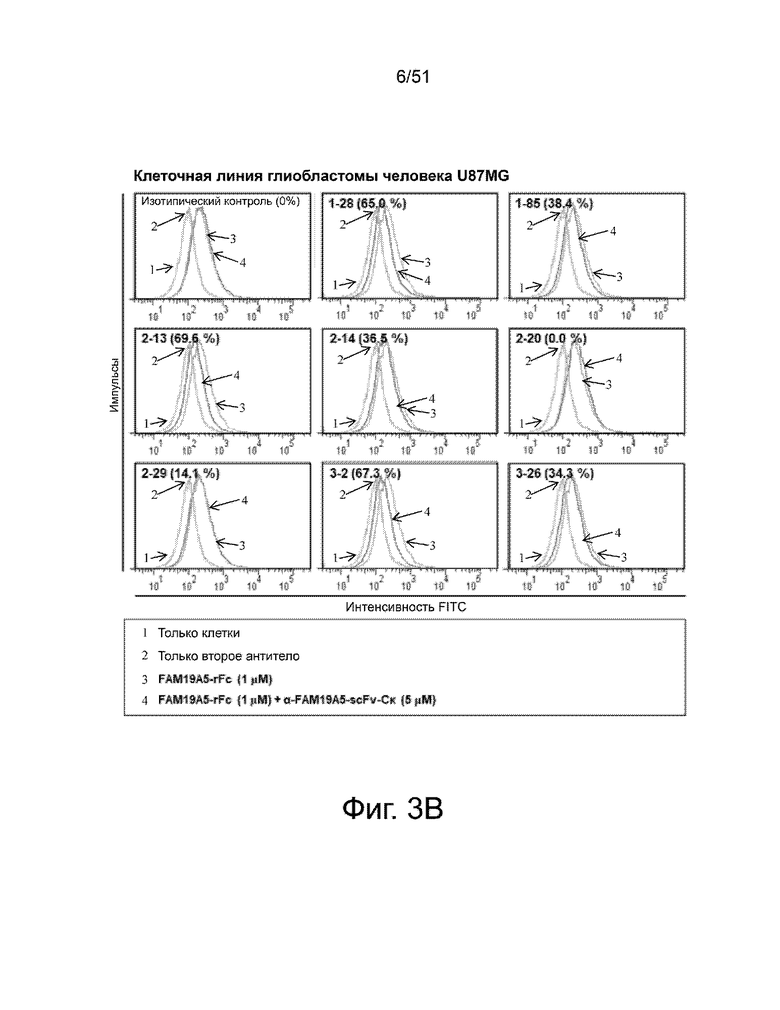








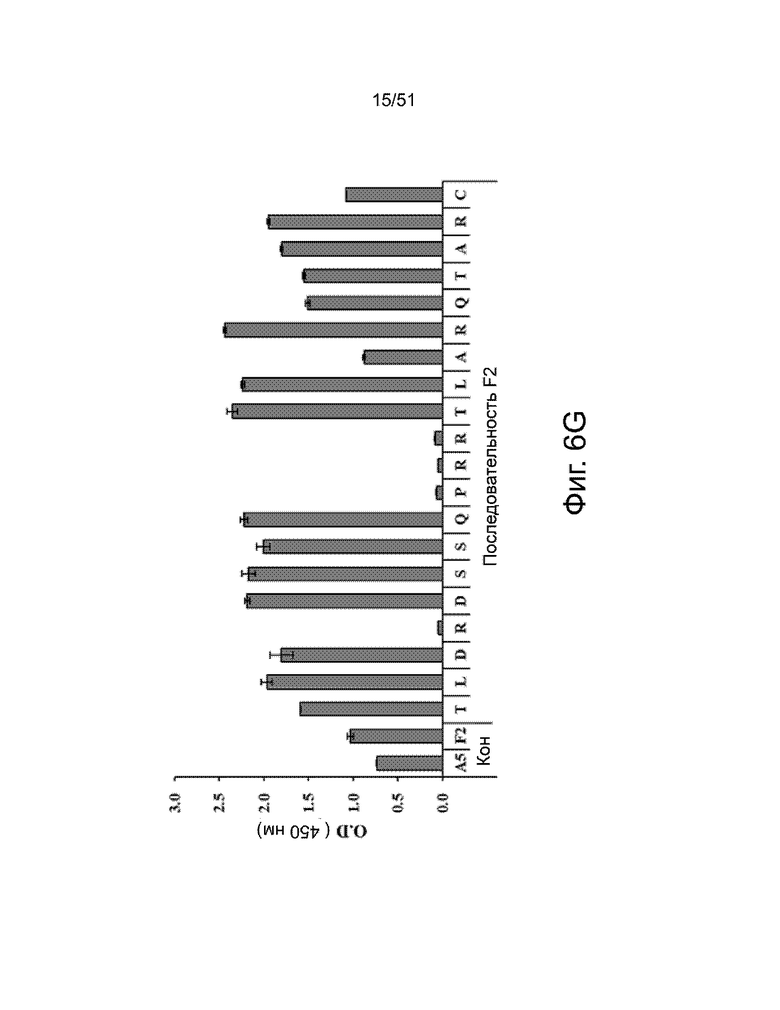





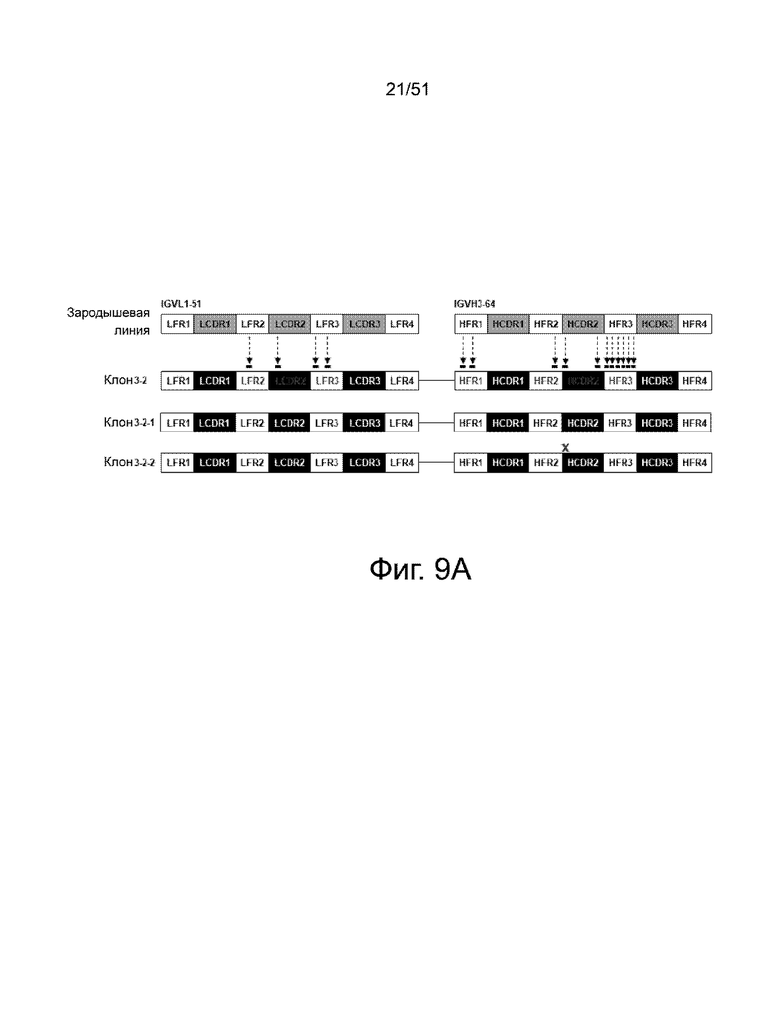

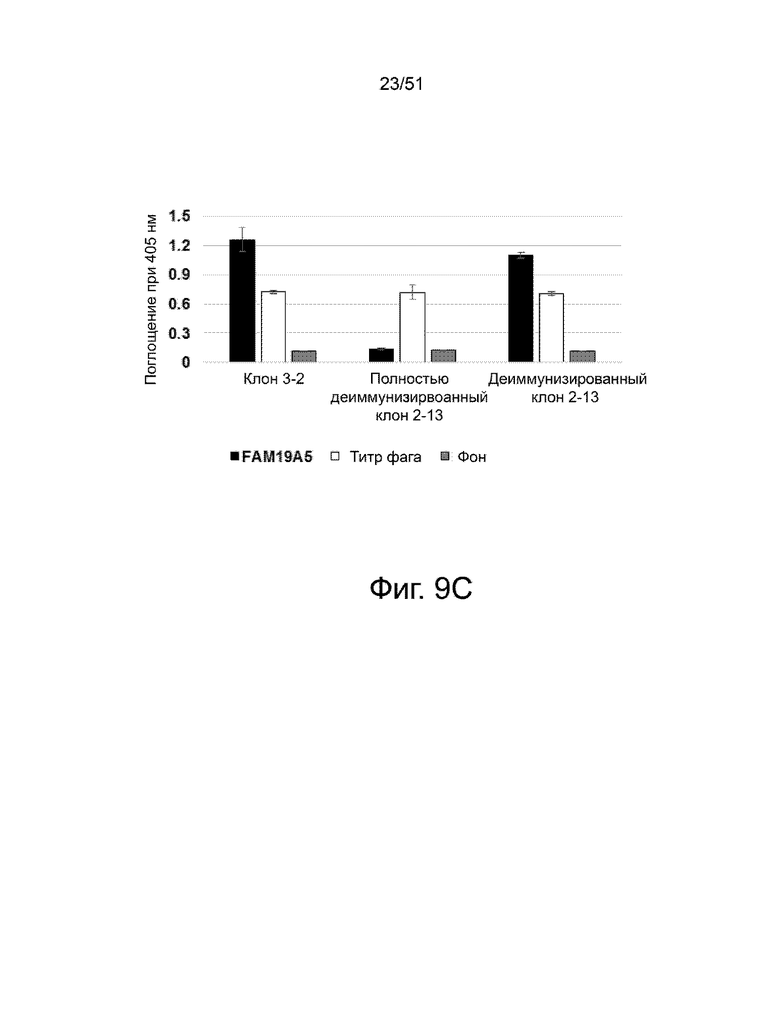

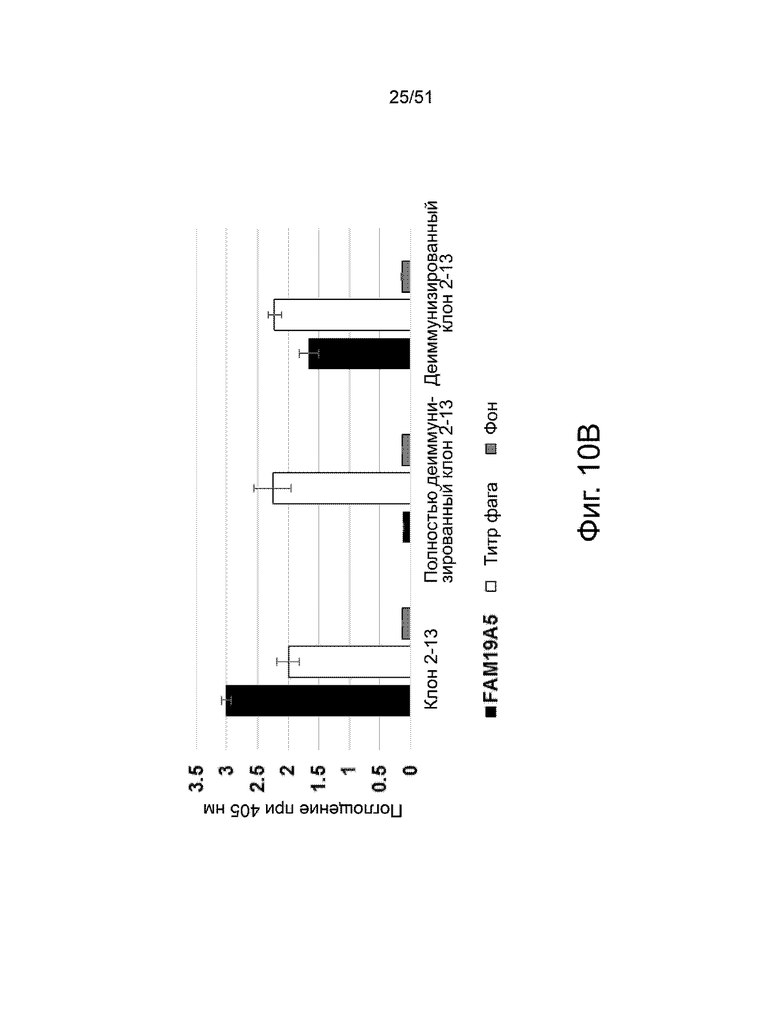
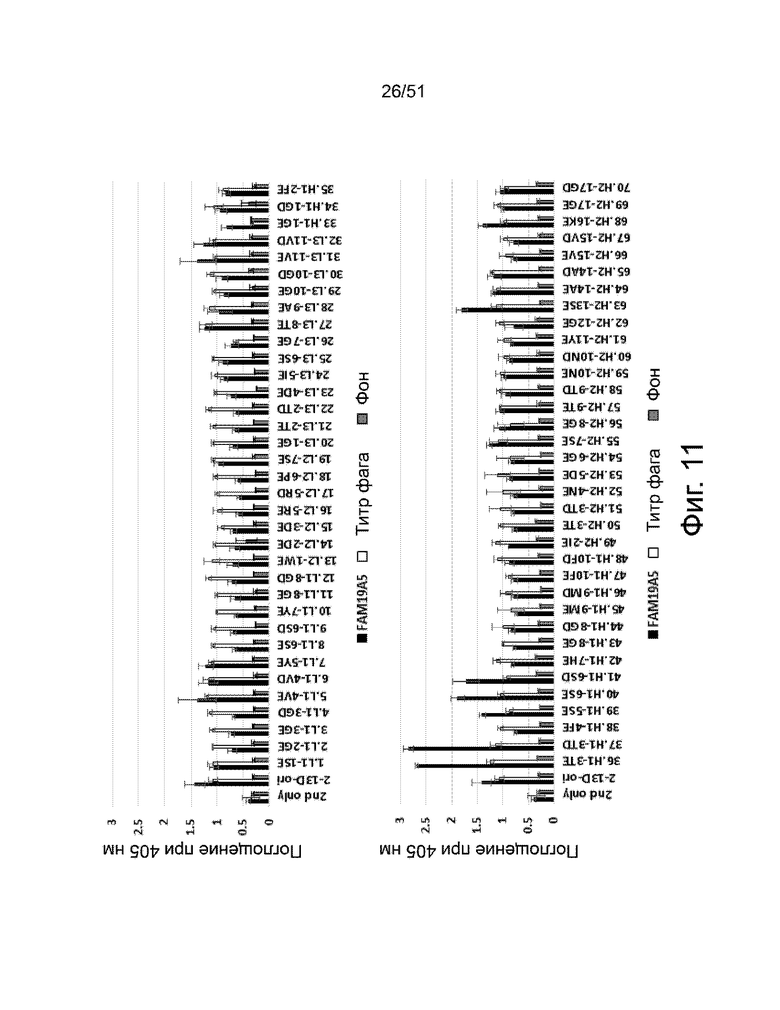

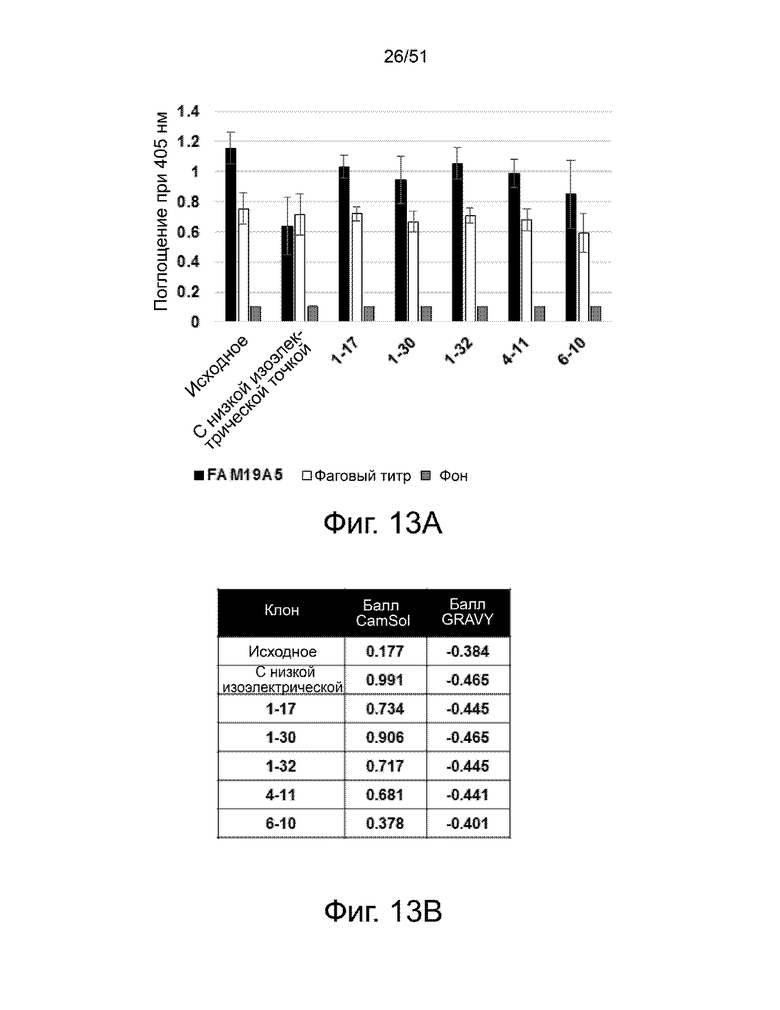


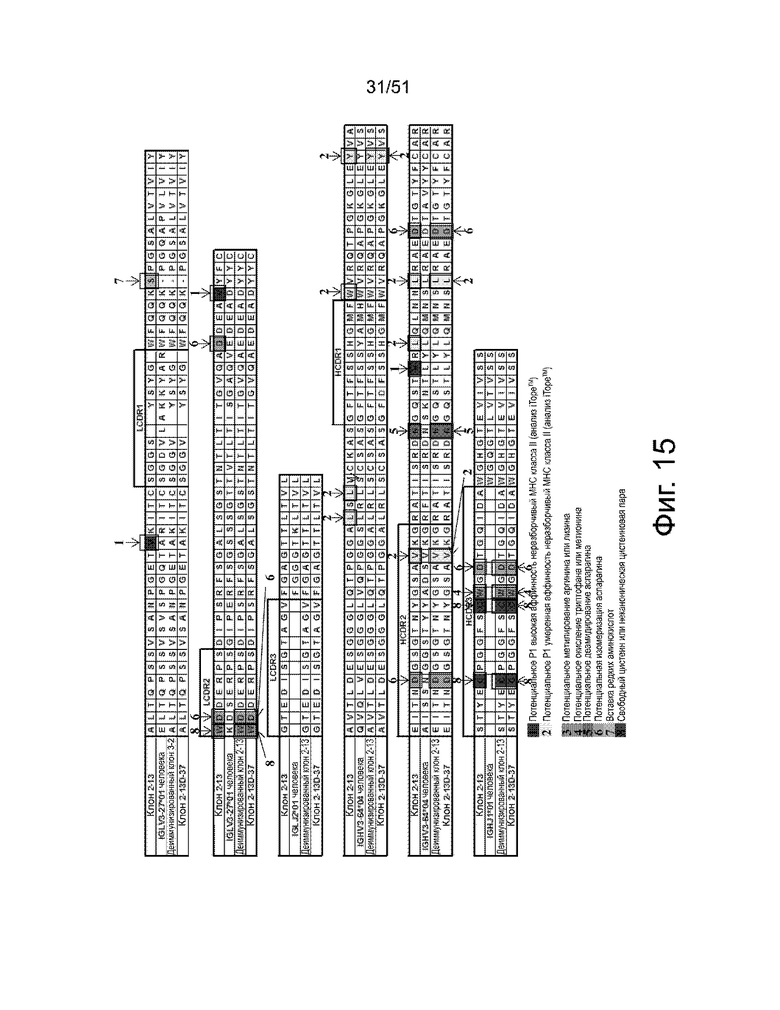
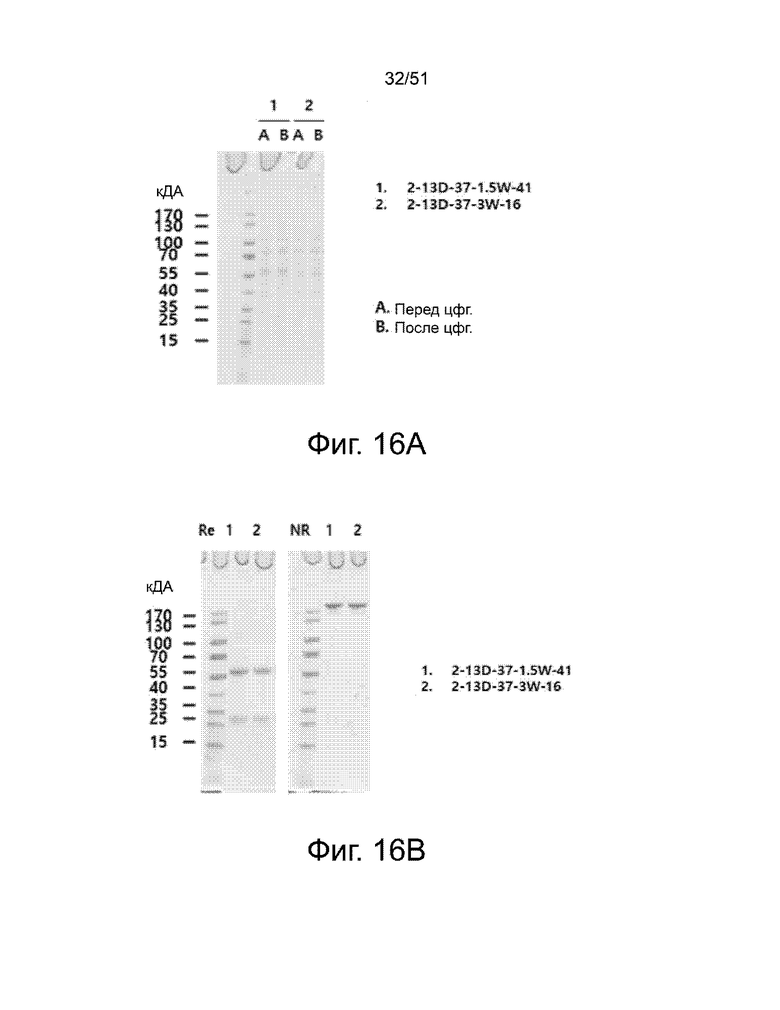











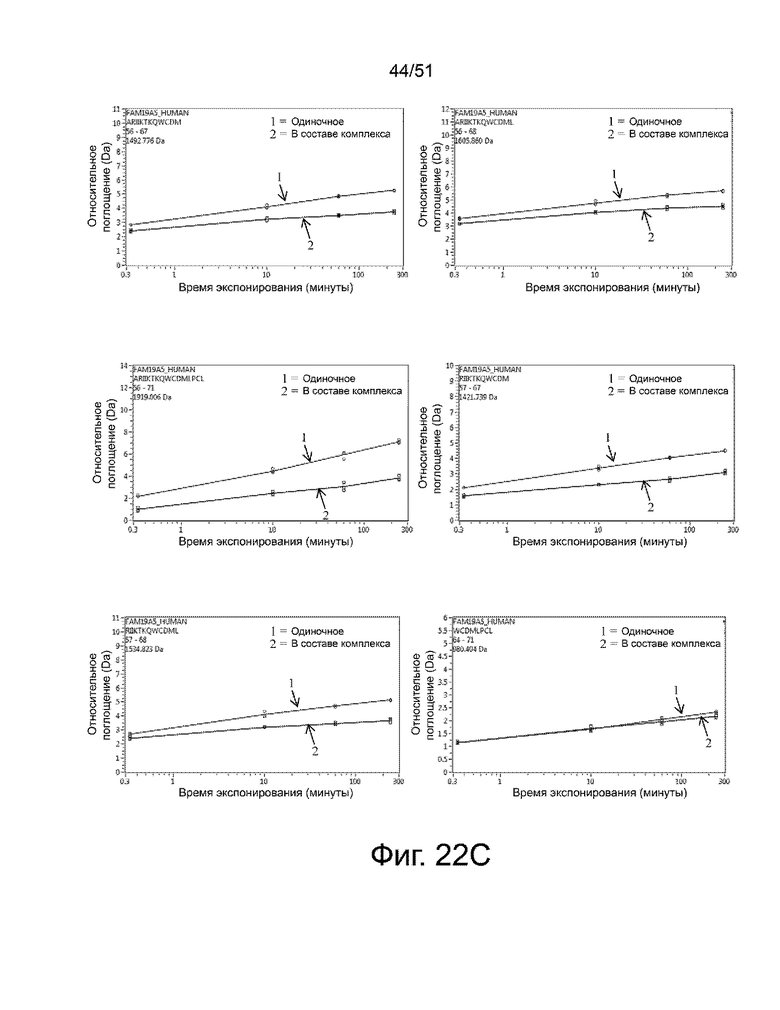




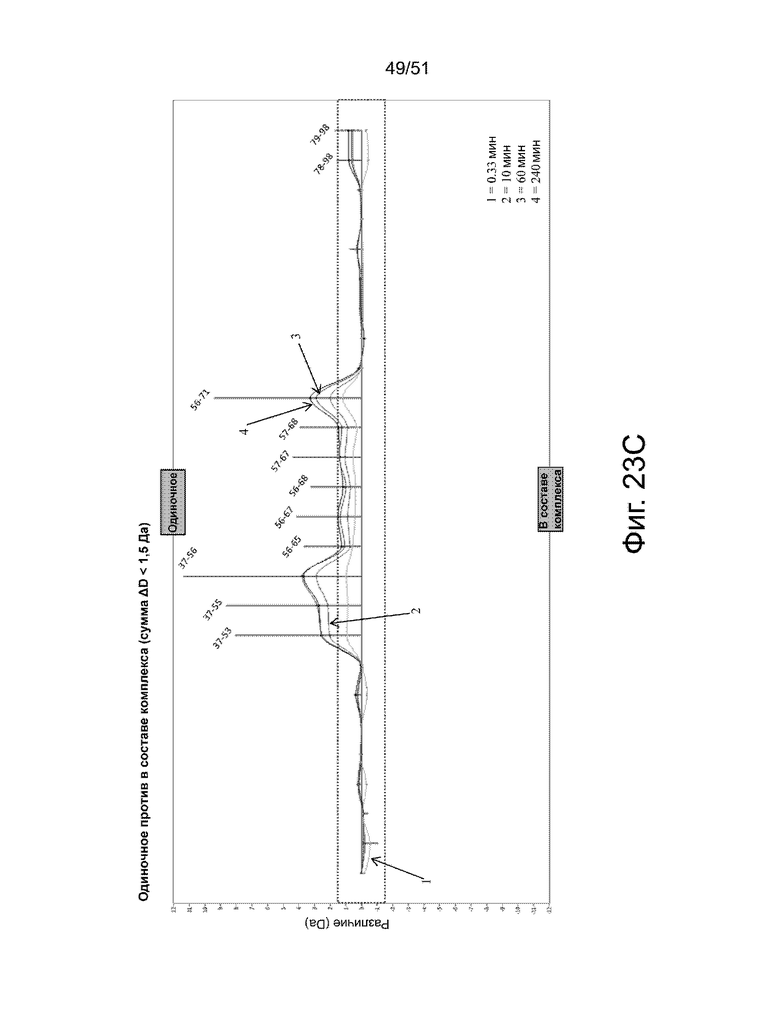
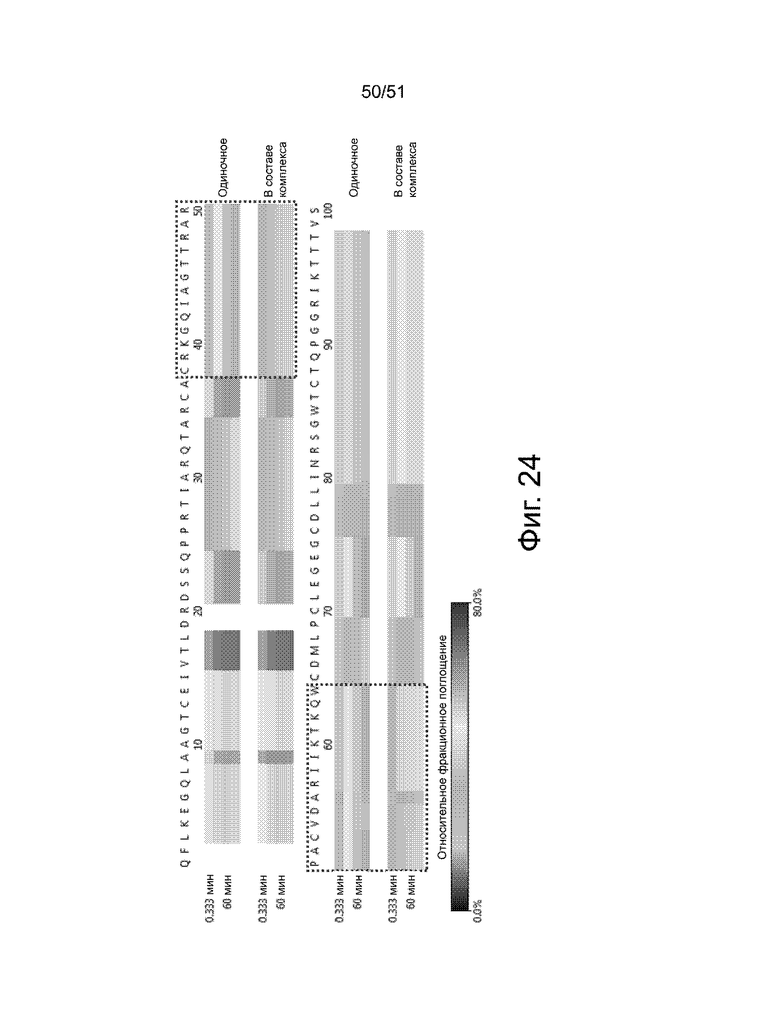

В настоящем изобретении представлены антитела, которые специфически связываются с FAM19A5 человека, а также нуклеиновая кислота, кодирующая антитело, экспрессирующий вектор, клетка для экспрессии антитела, композиция для ингибирования активности FAM19A5, набор для ингибирования активности FAM19A5, способ получения антитела, которое специфически связывается с человеческим белком FAM19A5, и способ лечения расстройств, таких как повреждение центральной нервной системы, дегенеративное заболевание головного мозга или невропатическая боль, путем введения антитела, которое специфически связывается с FAM19A5 человека. 16 н. и 21 з.п. ф-лы, 25 ил., 9 табл., 12 пр.
1. Выделенное антитело или его антигенсвязывающая часть, которое специфически связывает белок члена A5 семейства со сходством последовательностей 19 (FAM19A5) человека («анти-FAM19A5 антитело»), включающее CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где
VH-CDR1 выбрана из SEQ ID NO: 5, 14, 16 или 19;
VH-CDR2 выбрана из SEQ ID NO: 13, 15 или 17;
VH-CDR3 выбрана из SEQ ID NO: 7, 18, 128 или 129; и
VL-CDR1 выбрана из SEQ ID NO: 8, 21 или 80;
VL-CDR2 выбрана из SEQ ID NO: 20, 22, 24, 25, 26, 28 или 31;
VL-CDR3 выбрана из SEQ ID NO: 10, 23, 27, 29 или 32, и
где антитело имеет пониженную иммуногенность у человека по сравнению с референсным антителом, содержащим VH, указанную в SEQ ID NO: 11, и VL, указанную в SEQ ID NO: 12.
2. Выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, включающее CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 5; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 13; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 20; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 10.
3. Выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, включающее CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 21; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 22; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 23.
4. Выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, включающее CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 21; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 24; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 23.
5. Выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, включающее CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 25; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 23.
6. Выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, включающее CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 24; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 23.
7. Выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, включающее CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 26; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 27.
8. Выделенное антитело или его антигенсвязывающая часть, которая специфически связывается с человеческим белком FAM19A5, включающее CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи, где: (i) CDR1 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 14; (ii) CDR2 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 15; (iii) CDR3 тяжелой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 7; (iv) CDR1 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 8; (v) CDR2 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 28; и (vi) CDR3 легкой цепи включает аминокислотную последовательность, указанную в SEQ ID NO: 29.
9. Антитело по любому из пп. 1-8, содержащее вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, которая идентична по меньшей мере на около 80%, по меньшей мере около 85%, по меньшей мере около 90%, по меньшей мере около 95%, по меньшей мере около 96%, по меньшей мере около 97%, по меньшей мере около 98%, по меньшей мере около 99% или около 100% указанной аминокислотной последовательности в SEQ ID NO: 11, и/или где VL включает аминокислотную последовательность, которая идентична по меньшей мере на около 80%, по меньшей мере около 85%, по меньшей мере около 90%, по меньшей мере около 95%, по меньшей мере около 96%, по меньшей мере около 97%, по меньшей мере около 98%, по меньшей мере около 99% или около 100% аминокислотной последовательности, указанной в SEQ ID NO: 12.
10. Антитело по любому из пп. 1-9, отличающееся тем, что указанное антитело перекрестно конкурирует с референсным антителом, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), причем:
(а) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 33, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 38;
(b) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 39;
(c) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 41;
(d) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 40;
(e) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 42;
(f) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 43; или
(g) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 44.
11. Антитело по любому из пп. 1-10, отличающееся тем, что указанное антитело связывается с тем же эпитопом FAM19A5 человека, что и контрольное антитело, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), причем:
(а) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 33, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 38;
(b) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 39;
(c) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 41;
(d) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 40;
(e) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 42;
(f) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 43; или
(g) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 44.
12. Антитело по п. 11, отличающееся тем, что эпитоп FAM19A5 человека включает аминокислотную последовательность, указанную в SEQ ID NO: 90, 91 или 92.
13. Выделенное антитело или его антигенсвязывающая часть, которое специфически связывает белок члена A5 семейства со сходством последовательностей 19 (FAM19A5) человека («анти-FAM19A5 антитело»), включающее CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи,
где CDR1 тяжелой цепи содержит аминокислотную последовательность, выбранную из SEQ ID NO: 16 или 19, CDR2 тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 17 и CDR3 тяжелой цепи содержит аминокислотную последовательность, выбранную из SEQ ID NO: 18, 128 или 129,
где CDR1 легкой цепи содержит аминокислотную последовательность, выбранную из SEQ ID NO: 30 и SEQ ID NO: 30, в которой аминокислота серин в 4 положении SEQ ID NO: 30 заменена на валин, CDR2 легкой цепи содержит аминокислотную последовательность SEQ ID NO: 31 и CDR3 легкой цепи содержит аминокислотную последовательность SEQ ID NO: 32, и
где антитело имеет пониженную иммуногенность в отношении человека и/или более высокую аффинность связывания с человеческим белком FAM19A5 по сравнению с референсным антителом, содержащим VH, указанным в SEQ ID NO: 35, и VL, указанным в SEQ ID NO: 45.
14. Антитело по п. 13, содержащее вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, которая идентична по меньшей мере на около 80%, по меньшей мере около 85%, по меньшей мере около 90%, по меньшей мере около 95%, по меньшей мере около 96%, по меньшей мере около 97%, по меньшей мере около 98%, по меньшей мере около 99% или около 100% указанной аминокислотной последовательности в SEQ ID NO: 35, и/или где VL включает аминокислотную последовательность, которая идентична по меньшей мере на около 80%, по меньшей мере около 85%, по меньшей мере около 90%, по меньшей мере около 95%, по меньшей мере около 96%, по меньшей мере около 97%, по меньшей мере около 98%, по меньшей мере около 99% или около 100% аминокислотной последовательности, указанной в SEQ ID NO: 45.
15. Антитело по любому из пп. 13, 14, отличающееся тем, что указанное антитело перекрестно конкурирует с референсным антителом, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 36, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
16. Антитело по любому из пп. 13, 14, отличающееся тем, что указанное антитело перекрестно конкурирует с референсным антителом, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 37, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
17. Антитело по любому из пп. 13, 14, отличающееся тем, что указанное антитело перекрестно конкурирует с референсным антителом, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 130, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
18. Антитело по любому из пп. 13, 14, отличающееся тем, что указанное антитело перекрестно конкурирует с референсным антителом, которое включает вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где VH включает аминокислотную последовательность, указанную в SEQ ID NO: 131, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
19. Антитело по любому из пп. 1-18, отличающееся тем, что антитело выбрано из группы, состоящей из IgG1, IgG2, IgG3, IgG4, его варианта и любой их комбинации.
20. Антитело по любому из пп. 1-19, которое представляет собой химерное антитело, человеческое антитело или гуманизированное антитело.
21. Антитело по любому из пп. 1-20, содержащее Fab, Fab', F(ab')2, Fv или одноцепочечный Fv (scFv).
22. Антитело по п. 21, которое представляет собой scFv.
23. Антитело по п. 22, отличающееся тем, что scFv включает VH и VL, причем:
(а) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 33, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 38;
(b) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 39;
(c) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 41;
(d) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 40;
(e) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 42;
(f) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 43;
(g) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 34, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 44;
(h) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 36, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46;
(i) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 37, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46;
(j) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 130, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46; или
(k) VH включает аминокислотную последовательность, указанную в SEQ ID NO: 131, и VL включает аминокислотную последовательность, указанную в SEQ ID NO: 46.
24. Антитело по любому из пп. 1-23, которое демонстрирует одно или несколько из следующих свойств:
(а) связывание с растворимым FAM19A5 человека с KD 10 нМ или менее, что измерено с помощью иммуноферментного анализа (ELISA);
(b) связывание с мембраносвязанным FAM19A5 человека с KD 10 нМ или менее, что измерено с помощью ELISA;
(c) уменьшение, обращение вспять, задержка и/или предотвращение начала реактивного глиоза;
(d) подавление чрезмерной пролиферации реактивных астроцитов;
(e) снижение экспрессии протеогликанов хондроитинсульфата, включая нейрокан и нейрон-глиальный антиген 2 (NG2);
(е) увеличение экспрессии c-fos и pERK в ядрах нейронов;
(g) обеспечение выживания нейронов;
(h) увеличение экспрессии GAP43 в нейронах;
(i) обеспечение отрастания аксона;
(j) индукция нормализации кровеносных сосудов, например, в опухоли;
(k) подавление роста опухоли;
(l) усиление инфильтрации иммунных клеток в опухоль;
(m) усиление инфильтрации нейрональных клеток в опухоль;
(n) усиление фагоцитарной активности макрофага или микроглии;
(o) увеличение потенциала митохондриальной мембраны макрофага или микроглии;
(p) уменьшение рекрутирования миелоидных супрессорных клеток (MDSC) в опухоль;
(q) уменьшение некроза и отека опухоли;
(r) уменьшение проницаемости ткани опухоли; и
(s) увеличение скорости кровотока в опухоли.
25. Нуклеиновая кислота, кодирующая антитело по любому из пп. 1-24.
26. Экспрессирующий вектор, содержащий нуклеиновую кислоту по п. 25.
27. Клетка для экспрессии антитела по любому из пп.1-24, содержащая вектор по п. 26.
28. Композиция для ингибирования активности FAM19A5, включающая антитело по любому из пп. 1-24, нуклеиновую кислоту по п. 25, вектор по п. 26 или клетку по п. 27 и фармацевтически приемлемый носитель.
29. Набор для ингибирования активности FAM19A5, включающий антитело по любому из пп. 1-24, нуклеиновую кислоту по п. 25, вектор по п. 26, клетку по п. 27, а также инструкцию по применению.
30. Способ получения антитела, которое специфически связывается с человеческим белком FAM19A5, включающий культивирование клетки по п. 27 в подходящих условиях и выделение антитела.
31. Способ лечения заболевания или состояния у объекта, нуждающегося в этом, включающий введение объекту антитела по любому из пп. 1-24, нуклеиновой кислоты по п. 25, вектора по п. 26, клетки по п. 27, где заболевание или состояние выбрано из повреждения центральной нервной системы, дегенеративного заболевания головного мозга и невропатической боли.
32. Способ по п. 31, отличающийся тем, что антитело, нуклеиновая кислота, вектор, клетка или иммуноконъюгат индуцируют нормализацию кровеносных сосудов.
33. Способ по п. 32, отличающийся тем, что нормализация кровеносных сосудов сопровождается изменениями свойств кровеносных сосудов, включая повышенную связность, увеличенную толщину стенки, уменьшенный диаметр сосуда, более регулярное направление и характер распределения сосудов, увеличенное количество сосудов, уменьшение утечки и проницаемости, увеличенное покрытие перицитов и близость к кровеносным сосудам, повышенную оксигенацию или их комбинации.
34. Способ по любому из пп. 31-33, отличающийся тем, что антитело, нуклеиновая кислота, вектор, клетка или иммуноконъюгат усиливают фагоцитарную активность макрофага или микроглии.
35. Способ по любому из пп. 31-34, отличающийся тем, что антитело, нуклеиновая кислота, вектор, клетка или иммуноконъюгат увеличивают потенциал митохондриальной мембраны макрофага или микроглии.
36. Способ по любому из пп. 31-35, включающий введение дополнительного терапевтического агента.
37. Способ по п. 36, отличающийся тем, что дополнительный терапевтический агент включает химиотерапию, иммунотерапию, лучевую терапию или их комбинации.
| US 9579398 B2, 28.02.2017 | |||
| WO 2018083538 A1, 11.05.2018 | |||
| WANG Y | |||
| et al., Novel Adipokine, FAM19A5, Inhibits Neointima Formation After Injury Through Sphingosine-1-Phosphate Receptor 2, Circulation, 2018, Vol.138, N1, pp.48-63 | |||
| ПРОКОФЬЕВА Л.В | |||
| и др., Курс лекций по общей фармакологии: учебно-методическое пособие, Ульяновск: УлГУ, 2017, 155 с., |

