Известны способы оценки эффективности защиты клеток от апоптоза при активации антиоксидантной активности путем определения роста общего содержания SH-групп белков [Microtubule dynamics and glutathione metabolism in phagocytizing human polymorphonuclear leukocytes / B.R. Burchill, J.M. Oliver, С.B. Pearson et al. // J. of Cell Biology. - 1978. - Vol. 76, №2. - P. 439-447], но данная методика не позволяет оценить уровень восстановленного глутатиона, а следовательно, оценить эффективность защиты клеток от апоптоза. Известен также способ косвенного определения содержания восстановленного глутатиона по активности глутатионпероксидазы [Медицинские лабораторные технологии: В 2-х томах. / Под ред. А.И. Карпищенко - Т. 2 / А.И. Карпищенко. - СПб.: Интермедика. - 1999. - 656 с.], однако, активность этого фермента зависит от конформации активного центра фермента. При изменении конформации активного центра фермента его активность меняется, что не позволяет с высокой точностью оценить уровень восстановленного глутатиона, а, следовательно, оценить эффективность защиты клеток от апоптоза. Известен также способ косвенного определения концентрации восстановленного глутатиона по активности глутатионредуктазы [Медицинские лабораторные технологии: В 2-х томах. / Под ред. А.И. Карпищенко - Т. 2 / А.И. Карпищенко. - СПб.: Интермедика. - 1999. - 656 с], однако активность этого фермента зависит от конформации активного центра фермента. При изменении конформации активного центра фермента его активность меняется, что не позволяет достоверно в любой ситуации оценить с высокой точностью уровень восстановленного глутатиона, а следовательно, оценить эффективность защиты клеток от апоптоза.
Известен также способ определения антиоксидантной активности как защиты клеток от апоптоза по содержанию восстановленного глутатиона, предложенный М.Е. Anderson (1985) в модификации S. Kojima et al. (2004) [Low dose gamma-rays activate immune functions via induction of glutathione and delay tumor growth / S. Kojima, K. Nakayama, H. Ishida // J. Radiat. Res. - 2004. - Vol. 45, №1. - P. 33-39], основаный на взаимодействии восстановленного глутатиона (GSH) с 5,5′-дитио-бис(2-нитробензойной) кислотой (ДТНБ). При этом образуется окисленный глутатион (GSSG), который затем восстанавливается и вновь взаимодействует с ДТНБ. Данный способ является наиболее близким к предлагаемому по технической сущности и достигаемому результату, и выбран в качестве прототипа.
Целью предлагаемого изобретения является повышение эффективности и точности способа.
Указанная цель достигается тем, что дополнительно в инкубационную среду, содержащую лимфоциты, вводится 1,4-дитиоэритритол и аскорбиновая кислота в конечной концентрации 3,0 ммоль и 0,1 ммоль соответственно без которых невозможна последующая защита клеток от апоптоза
Следовательно, только комплексная модернизация способа-прототипа позволяет получить желаемый результат. Только комплексное внесение в инкубационную смесь 1,4-дитиоэритритола и аскорбиновой кислоты в минимальной концентрации 3.0 мМ и 0,1 мМ соответственно позволяет повысить оценку эффективности защиты лимфоцитов от апоптоза, а именно при росте белково-связанного глутатиона на 18-35%, а восстановленного глутатиона на 25-41% считают защиту низкоэффективной, а при росте белково-связанного глутатиона на 36% и более и восстановленного глутатиона на 42% и более считают защиту высокоэффективной.
Антиоксидантная система направлена на эффективную нейтрализацию гидроксирадикалов и снижение токсичной для организма гидроперекиси. Гидроксильный радикал (•OH) участвует в микробицидном и цитотоксическом действии нейтрофилов, моноцитов и Т-лимфоцитов [Окислительный стресс. Прооксиданты и антиоксиданты / Е.Б. Меньщикова, В.З. Панкин, Н.К. Зенков и соавт. - М.: Слово. - 2006. - 556 с.]. Есть два основных механизма синтеза •OH нейтрофилами: первый - образование из пероксида водорода в присутствии металлов переменной валентности в так называемой «реакции Фентона»: Fe2++Н2О2→Fe3++•ОН+ОН- [Лущак В.И. Окислительный стресс и механизмы защиты от него у бактерий / В.И. Лущак // Биохимия. - 2001. - Т. 66, вып. 5. - С. 592-609], второй - в ходе ряда реакций с участием гипогалоидов 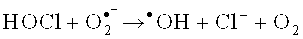
В клетках предусмотрен ряд защитных реакций по блокированию свободных ионов Fe2+ и Cu+. Например, активированные клетки синтезируют лактоферрин, связывающий свободное железо и переводящий его в каталитически неактивную форму, а также продуцируют высокие концентрации таурина, конъюгирующего с гипохлоритом и защищающего клетку от его токсичных эффектов [Taurine chloramines, a product of activated netrophils, inhibits in vitro the genetation of nitric oxide and other macrofage inflammatory mediators / J. Marcinkiewiez, A. Grabowska, J. Bereta et al. // J. Leukocyte Biol. - 1995. - Vol. 58. - P. 667-674].
Образование •ОН показано в ходе микросомального окисления, окисления арахидоновой кислоты, в реакциях с флавиновыми ферментами, убихиноном, пероксинитритом. В ряде исследований клеток in vitro получены свидетельства продукции •ОН данными клетками [Hydroxylation of salicylate by activated neutrophils / W.B. Davis, B.S. Mohammed, D.C. Mays et al. // Biochem Pharmacol. - 1989. - Vol. 38. - P. 4013-4019]. Однако изучение этих реакций зачастую основывалось на использовании ингибиторов и измерении уровня вторичных продуктов. Следовательно, реакции, приписанные •ОН, могли быть вызваны другими оксидантами, в частности 
Генерация •ОН стимулированными клетками в очаге воспаления может существенно лимитироваться отсутствием в среде ионов железа. Исследование реакции Фентона в клетках выявило, что блокирование ионов железа лактоферрином ингибирует непосредственно саму реакцию [Rosen, G.М. Free radicals and phagocytic cells / G.M. Rosen, S. Pou, C.L. Ramos // FASEB J. - 1995. - Vol. 9. - P. 200-211], а утилизация H2O2 миелопероксидазой ограничивает реакцию, даже если железо доступно [Winterbourn, С.С. Myeloperoxidase as an effective inhibitor of hydroxyl radical production: Implications for the oxidative reactions of neutrophils / С.C. Winterbourn // J. Clin. Invest. - 1986. - Vol. 78. - P. 545-557]. Хотя большинство биологических форм железа каталитически неактивно, показана способность клеток к продукции •ОН в присутствии трансферрина, подверженного протеолитической деградации [Phagocyte-derived free radicals stimulated by ingestion of ironrich Staphylococcus aureus: Aspin-trapping study / M.S. Cohen, В.E. Britigan, Y.S. Chai et al. // J. Infect Dis. - 1991. - Vol. 163. - 819-826], или железа в составе Pseudomonas aeruginosa, содержащего сидерофор пиохелин [Possible role of bacterial siderophores in inflammation-Iron bound to the pseudomonas siderophore pyochelin can function as a hydroxyl radical catalyst / T.J. Coffman, C.D. Cox, B.L. Edeker et al. / J. Clin. Invest. - 1990. - Vol. 86. - P. 1030-1038]. Однако M.S. Cohen и соавторы обнаружили, что внутриклеточное железо не всегда доступно: в их экспериментах повышенного образования радикала •ОН не отмечалось, даже если клетки поглощали Staphylococcus aureus, который был преинкубирован с Fe2+ [Phagocyte-derived free radicals stimulated by ingestion of ironrich Staphylococcus aureus: Aspin-trapping study / M.S. Cohen, В.E. Britigan, Y.S. Chai et al. // J. Infect Dis. - 1991. - Vol. 163. - 819-826].
С помощью чувствительных спиновых меток обнаружена наработка гидроксил-радикала клетки in vitro в результате реакции HOCl и
Гидроксильный радикал представляет собой один из наиболее реакционно-способных окислителей и может взаимодействовать почти с любой молекулой клетки. Он модифицирует дезоксирибозу и азотистые основания ДНК, окисляет молекулы белков, углеводов и липидов. Особенно активно •ОН в ходе реакций перекисного окисления липидов атакует фосфолипиды, содержащие в жирнокислотных радикалах ненасыщенные связи, что ведет к образованию гидроперекисей [Дубинина Е.Е. Продукты метаболизма кислорода в функциональной активности клеток (жизнь и смерть, созидание и разрушение). Физиологические и клинико-биохимические аспекты / Е.Е. Дубинина. - СПб.: Медицинская пресса, 2006. - 400 с.; Окислительный стресс. Прооксиданты и антиоксиданты / Е.Б. Меньшикова, В.З. Ланкин, Н.К. Зенков и соавт. - М.: Слово. - 2006. - 556 с.]. Основным компонентом антиокидантной системы является восстановленная форма глутатиона.
Глутатион - трипептид (L-γ-глутамил-L-цистеилглицин) с молекулярной массой 307 Da занимает особое место среди SH-содержащих соединений. Наличие γ-глутамильной связи защищает трипептид от ферментативной деградации. В организме глутатион присутствует в двух формах: окисленной - GSSG и восстановленной - GSH, причем содержание GSH в клетках на несколько порядков выше, чем GSSG [Колесниченко Л.С., 1989; Wu G. et al., 2004; Смирнова Г.В., Октябрьский О.Н., 2005; Марри Р. и соавт., 2009]. По данным P. Pietarinen-Runtti et al. (2000), концентрация GSH в нейтрофилах составляет около 5 нмоль/мг белка. Содержание глутатиона в сыворотке крови здоровых людей незначительно, поэтому клетки основную потребность в GSH обеспечивают путем нематричного синтеза [Wu G. et al., 2004] в ходе двух последовательных реакций, катализируемых γ-глутамилцистеин-синтетазой (КФ 6.3.2.2) и глутатион-синтетазой (КФ 6.3.2.3) [Кулинский В.И., 1990; Смирнова Г.В., Октябрьский О.Н., 2005; Марри Р. и соавт., 2009]. Лимитирующим звеном синтеза является образование γ-глутамилцистеина, зависящее от наличия L-цистеина и его способности окисляться в L-цистин [Зенков Н.К. и соавт., 2001]. В то же время недостаточность глутатион-синтетазы способствует развитию окислительных повреждений в нейтрофилах [Spielberg S.P. et al., 1979].
Глутатион при физиологических значениях рН имеет две анионные карбокси-группы, положительно заряженную аминогруппу и SH-группу цистеинового остатка, которая придает GSH свойства восстановителя и способность быстро обезвреживать свободные радикалы и АФК [Day R.M., 2005; Zhu Y., 2007; Circu C.L. et al., 2009]. Глутатин является типичным тиолом и, участвуя в одноэлектронных восстановительных реакциях, становится GS•, который димеризуется до GSSG, легко реагирующего со свободными SH-группами. Второй тип окислительно-восстановительных превращений с участием GSH - это реакции тиолдисульфидного обмена, которые известны как основной путь образования смешанных дисульфидов глутатиона с белками (белок-SSG) и играют роль в регуляции биологических процессов [Chai Y.C. et al., 1994]. В реакциях третьего типа происходит двухэлектронное окисление глутатиона с образованием интермедиата, который реагирует со второй молекулой GSH (получение GSSG) или иной молекулой (синтез смешанного дисульфида) [Смирнова Г.В., Октябрьский О.Н., 2005].
GSH является стабилизатором мембран [Биленко М.В, 1989; Udupi V., 1992; Trudel S. et al., 2009]. Он защищает клеточные структуры клеток от высокотоксичного OCI - [Carr А.С., Winterbourn С.С., 1997], при этом GSH превращается в глутатион-сульфонамид и дегидроглутатион [Harwood D.T. et al., 2006]. Связывая NO, глутатион образует токсичные для клетки нитрозильные комплексы. Мононитрозоглутатион может активировать апоптоз [Turpaev K.Т. et al., 1997].
Не всегда восстановительного потенциала GSH достаточно для полной нейтрализации прооксидантов. Существует мнение, что взаимодействие GSH с органическими радикалами эффективно только в условиях удаления
Основной антиоксидантный эффект GSH реализует посредством участия в работе ферментов. Глутатион выступает донором водорода при восстановлении Н2О2 и перекисей липидов глутатион-пероксидазами и глутатион-S-трансферазами (ГТ) [Hirayama K., 1989; Sies Н. et al., 1997; Кулинский В.И., 1990; Hayes J.D. et al., 2005; Зенков H.К.. и соавт., 2009; Liu G. et al., 2010]. Высокая активность глутатион-редуктазы и накопление GSH оказывает протекторный эффект в отношении альвеолярных макрофагов, инкубируемых с прооксидантами in vitro [Pietarinen Р.K., 1995].
С изменением окислительно-восстановительного баланса сопряжено большое количество реакций, поэтому поддержание оптимального редокс-состояния цитозоля выступает важным условием нормальной жизнедеятельности клеток. Высокая концентрация глутатиона в цитоплазме, его редокс-активность и возможность поддержания в восстановленном состоянии делают систему GSH/GSSG важнейшим внутриклеточным редокс-буфером [Reed М.С. et al., 2008]. Концентрация GSH в клетке в 500-1000 раз превышает уровень НАДФН и других внутриклеточных редокс-систем, поэтому изменения соотношения GSH/GSSG прямо отражают изменения редокс-статуса клетки [Кулинский В.И., 2007; Aslan М., Canatan D., 2008; Reed М.С, 2008]. Считают, что буферная емкость системы глутатиона защищает репликативную систему клетки, а дефицит GSH приводит к снижению синтеза ДНК и белков [Poot М., 1991; Ланкин В.З., 1997; Day R.M., Suzuki Y.J., 2005; Liu G. et al., 2010], а затем и к апоптозу.
К природным антиоксидантам относят также аскорбиновую кислоту, которая играет важную роль в развитии окислительного стресса в организме.
Аскорбиновая кислота реализует свое антиоксидантное действие в плазме, межклеточной жидкости и на внеклеточном уровне. В организме человека аскорбиновая кислота преимущественно представлена в L-форме. Стрессовые ситуации увеличивают количество метаболитов витамина С в виде дегидроаскорбиновой кислоты.
Аскорбиновая кислота и дегидроаскорбиновая кислота играют активную роль в нескольких процессах, включая защиту от инфекции, повышении иммунности, в процессах заживления ран, а также принимая участие в образовании антистрессовых гормонов. Аскорбат является кофактором дофамин-β-гидроксилазы, которая катализирует синтез норадреналина и других катехоламинов. Аскорбиновая кислота является восстановителем для L-пролингидроксилазы, которая необходима для синтеза коллагена и соединительной ткани в целом. В организме с участием аскорбиновой кислоты происходит регенерация α-токоферола из токофероксильного радикала. Окислительный стресс коррелирует с ухудшением секреции инсулина, а терапия аскорбиновой кислотой прерывает повреждающее действие свободных радикалов, уменьшает степень проявления инсулиновой резистентности [М.И. Балаболкин и соавт., 2003]. Ионы аскорбата являются одним из активных элементов системы антиоксидантной защиты, предохраняя липиды от окисления их пероксидными радикалами. Антиоксидантный эффект аскорбата проявляется при достаточном количестве других антиоксидантов, таких как α-токоферол и глутатион. Глутатион восстанавливает дегидроаскорбиновую кислоту прямым и неферментативным путем до аскорбиновой кислоты. Эта реакция является одним из основных механизмов антиоксидантной системы, часто описываемых как восстановительные циклы - глутатион/глутатиондисульфид и аскорбиновая/дегидроаскорбиновая кислота. При этом клетки периферических тканей поглощают экзогенную дегидроаскорбиновую кислоту и в присутствии глутатиона конвертируют ее в цитоплазме в аскорбиновую кислоту. Восстановление глутатиондисульфида в глутатион катализируется глутатион редуктазой и требует участия NADPH в качестве кофактора. Недостаточность глутатиона снижает содержание аскорбиновой кислоты в тканях и одновременно повышает концентрацию дегидроаскорбиновой кислоты.
При недостатке α-токоферола и глутатиона может превалировать прооксидантный эффект аскорбата и его метаболитов. Прооксидантный эффект аскорбиновой кислоты может наблюдаться не только при недостатке α-токоферола и глутатиона, но и при применении высоких доз аскорбиновой кислоты. Избежать прооксидантного эффекта аскорбиновой кислоты можно в случае создания адекватного внутриклеточного уровня восстановленного глутатиона.
Исходя из вышесказанного, целесообразно использовать в эксперименте комплексное применение аскорбата с протектором SH-групп - а именно 1,4-дитиоэритритолом. Для проникновения внутрь клетки пассивным транспортом происходит превращение аскорбиновой кислоты в дегидроаскорбиновую кислоту, затем последняя подвергается обратимому превращению в аскорбиновую кислоту при участии восстановленного глутатиона. Следовательно, комплексная модернизация способа-прототипа позволяет повысить точность оценки эффективности защиты лимфоцитов от апоптоза.
Каждый вновь введенный в формулу изобретения признак выполняет функцию повышения точности и эффективности способа: дополнительное добавление в инкубационную среду 1,4-дитиоэритритола и аскорбиновой кислоты в низкой концентрации для последующего определения белково-связанного и восстановленного глутатиона. Установлено, что при росте белково-связанного глутатиона на 18-35%, а восстановленного глутатиона на 25-41% считают защиту низкоэффективной, а при росте белково-связанного глутатиона на 36% и более и восстановленного глутатиона на 42% и более считают защиту высокоэффективной.
Роль антиоксидантной системы клетки заключается в снижении токсического эффекта свободных радикалов, в том числе и гидроперекисей липидов.
Антиоксидантную защиту обеспечивает широкий круг веществ, различных по происхождению, физико-химической природе и механизмам действия. Общим их свойством, по определению J.M. Gutteridge (1992), является способность, присутствуя в низких по сравнению с окисляемым субстратом концентрациях, существенно задерживать или ингибировать его окисление. Постоянное образование прооксидантов должно быть уравновешено их инактивацией, поэтому для поддержания гомеостаза необходима адекватная ситуации непрерывная регенерация антиоксидантной способности клеток [Зенков Н.К. и соавт., 2001; Blokhina О. et al., 2003].
Общепринятой номенклатуры антиоксидантов в настоящее время не существует, хотя ряд авторов [Dimascio Р., 1990; Kalra V. et al., 2001; Зайцев В.Г. и соавт., 2003] выделяет два класса: превентивные, снижающие скорость инициации цепной реакции окисления, и гасящие (прерывающие цепь), препятствующие развитию цепной реакции. К превентивным относят каталазу и пероксидазы, разрушающие ROOH, а также агенты, образующие хелатные комплексы с металлами переменной валентности, к прерывающим цепь - фенолы, ароматические амины. В условиях in vivo главными гасящими антиоксидантами являются: витамин Е, нейтрализующий ROO• в липидной фазе мембран [Jore D. et al., 1990; Hong J.H. et al., 2004], фермент СОД, улавливающий
Более известно деление антиоксидантов на ферменты и соединения неферментативной природы. Последние в определенных концентрациях всегда присутствуют в липидной фазе мембран и водных средах организма и расходуются первыми при устранении проявлений окислительного стресса [Droge W., 2002; Blokhina О., 2003]. Ферменты наиболее активно присоединяется к антиоксидантной защите (АОЗ) после включения механизмов индукции [Лущак В.И., 2001]. При возникновении окислительного стресса (ОС) расход антиоксидантов возрастает и меняется экспрессия генов, кодирующих белковые компоненты АОЗ [Дубинина Е.Е., 2006]. Между ферментами и неферментативными элементами АОЗ существует равновесие, причем последние при ряде патологических состояний организма могут выступать в качестве прооксидантов [Зенков Н.К и др., 2001].
Главную роль среди неферментавных антиоксидантных систем защиты отводят глутатиону.
Функционирование клеток связано с уровнем белково-связанного глутатиона. Определение белково-связанного глутатиона основано на способности боргидрата натрия (NaBH4) высвобождать из связи с белками глутатион, который при взаимодействии с ДТНБ образует окрашенное соединение, а именно тио-2-нитробензойную кислоту, водный раствор которой имеет максимум поглощения при длине волны 412 нм [Burchill, B.R. Microtubule dynamics and glutathione metabolism in phagocytizing human polymorphonuclear leukocytes [Text] / B.R. Burchill, J.M. Oliver, C.B. Pearson et al. // J. of Cell Biology. - 1978. - Vol. 76, №2. - P. 439-447].
В настоящее время крайне важно определить уровень белково-связанного глутатиона для защиты клеток от апоптоза.
В настоящее время в лабораторной практике наиболее распространен способ оценки эффективности защиты лимфоцитов от апоптоза и переокисления с помощью определения концентрации белково-связанного и восстановленного глутатиона.
Концентрацию гидроксирадикалов определяют методом, предложенным [Thorn S.R, Elbuken M.E., 1991]. Метод основан на разрушении модельного субстрата 2-дезокси-D-рибозы гидроксильным радикалом, образуемым опсонизированными лимфоцитами.
Содержание восстановленного глутатиона определяют методом, предложенным М.Е. Anderson (1985) в модификации S. Kojima et al. (2004) [Kojima, S. Low dose gamma-rays activate immune functions via induction of glutathione and delay tumor growth / S. Kojima, K. Nakayama, H. Ishida // J. Radiat. Res. - 2004. - Vol. 45, №1. - P. 33-39]. Принцип метода основан на взаимодействии GSH с 5,5′-дитио-бис(2-нитробензойной) кислотой (ДТНБ) с образованием тио-2-нитробензойной кислоты, водный раствор которой имеет максимум поглощения при длине волны 412 нм. При этом образуется GSSG, который восстанавливается глутатионредуктазой до GSH и вновь взаимодействует с ДТНБ. Скорость образования окрашенного продукта пропорциональна содержанию общего глутатиона. Для определения содержания GSSG пробы прединкубируются с блокатором SH-групп 2-винилпиридином («Wako», Япония), который необратимо связывает GSH и, следовательно, скорость образования окрашенного продукта пропорциональна содержанию GSSG.
Лизат клеток готовят на 5% сульфосалициловой кислоте, которая осаждает белки, но не ингибирует активность глутатионредуктазы.
Концентрацию белка в клетках определяют методом [A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding / M.M. Bradford // Analyt. Biochem. - 1976. - Vol. 7, №1, 2. - P. 248-254], основанным на взаимодействии Кумасси голубого G-250 с остатками аргинина и лизина в белках. Свободный краситель красного цвета (максимум поглощения - 495 нм) при образовании комплекса с белком переходит в синюю форму (максимум поглощения - 595 нм).
К 0,1 мл лизата клеток добавляют 1,0 мл раствора Кумасси голубого (100 мг красителя, 50 мл 96° этанола, 100 мл 85% Н3РО4, Н2О до 1,0 л), перемешивают, инкубируют 3 мин при комнатной температуре и измеряют оптическую плотность проб (длина волны 595 нм) против контроля, содержащего 0,1 мл воды и 1,0 мл раствора Кумасси голубого. Содержание белка рассчитывают по калибровочной кривой, построенной по разведениям стандартного раствора альбумина (1,0 мг/мл) и выражают в мг/мл.
В настоящее время крайне важно для защиты клеток от апоптоза и переокисления оценить концентрацию различных форм глутатиона после стимуляции антиоксидантной активности низкими концентрациями 1,4-дитиоэритритола и аскорбиновой кислоты. Для решения этой задачи предложен новый способ защиты клеток от апоптоза после дополнительного добавления в инкубационную среду 1,4-дитиоэритритола и аскорбиновой кислоты в конечной концентрации 3.0 мМ и 0,1 мМ соответственно.
Все сказанное свидетельствует о крайней важности разработки способа защиты клеток от апоптоза, а также для защиты клеток от токсического действия активных форм кислорода.
Популярность указанного выше способа обоснована его высокой чувствительностью, простотой осуществления и достаточной адекватностью получаемых результатов, лежащих в основе определения концентрации гидроксильных радикалов в среде инкубации лимфоцитов.
Существенные признаки, характеризующие изобретение, проявили в заявленной совокупности новые свойства, явным образом не вытекающие из уровня техники в данной области, и не являются очевидными для специалиста.
Идентичной совокупности признаков не обнаружено при изучении патентной и научной медицинской литературы. Данное изобретение может быть использовано в медицинской практике для повышения точности защиты клеток от апоптоза при различных заболеваниях. Таким образом, следует считать предлагаемое изобретение соответствующим условиям патентоспособности: «новизна», «изобретательский уровень», «промышленная применимость».
Метод основан на определении концентрации белок-связанного и восстановленного глутатиона.
Способ осуществляется следующим образом поэтапно:
1. Выделение культуры клеток линии Jurkat.
2. Количественное определение численности жизнеспособных клеток с помощью окраски трипановым синим микроскопическим методом.
Клетки ресуспендируют в 1 мл клеточной взвеси. Отбирают 100 мкл ресуспендированной клеточной суспензии и добавляют 100 мкл 0,1% раствора трипанового синего на физ. растворе, хорошо перемешивают и заполняют камеру Горяева. Предварительно к камере притирают покровное стекло так, чтобы появлялись радужные, ньютоновые кольца (только при этих условиях соблюдался правильный объем камеры). Каплю клеточной взвеси с красителем вносят под притертое покровное стекло. Подсчет клеток производят в 5-ти больших квадратах по диагонали камеры Горяеева. Расчет клеточности лимфоцитов крови производят по формуле
А×106=(число клеток)/4
где А - клеточность лимфоцитов крови.
3. Внесение в инкубационную смесь заявленных добавок.
В культуральную смесь добавляют соединения: 1,4-дитиоэритритол в концентрации 3.0 мМ и аскорбиновую кислоту в конечной концентрации 0,1 мМ.
4. Биохимическое исследование белково-связанного, восстановленного и окисленного глутатиона.
Лизат лимфоцитов готовят на 5% сульфосалициловой кислоте, которая осаждает белки, но не ингибирует активность глутатионредуктазы. Количество общего глутатиона (GSH и GSSG) определяют в пробе содержащей 0,1 М Na-фосфатный буфер (рН=7,5) с 1 мМ ЭДТА, 0,4 мМ НАДФН2, 0,3 мМ ДТНБ и 1 U/мл глутатионредуктазы («Wako», Япония). Окисленный глутатион определяют аналогичным способом в клеточном лизате после предварительной инкубации пробы в течение 30 мин с 10 мМ 2-винилпиридином. Расчет содержания общего и окисленного глутатиона производят с помощью калибровочных графиков, для построения которых используют растворы GSH и GSSG («МР», США) в концентрации от 3 до 100 мкМ, обработанные аналогично опытным пробам. Концентрацию GSH рассчитывают как разницу между концентрацией общего глутатиона и GSSG, выражая результат в нмоль/мг белка.
Определение белково-связанного глутатиона.
После инкубации в экспериментальных условиях (5% и 20% кислорода) в присутствии или отсутствии NEM, DTE, NAC клетки центрифугируют 5 минут при 4°С и 1500 об/мин для их осаждения. Удаляют супернатант. Добавляют 1 мл охлажденного PBS (рН 7,4). Ресуспендируют на вортексе. Центрифугируют 5 минут при 4°С и 1500 об/мин. Удаляют супернатант. Осадок клеток ресуспендируют в 1 мл 5% сульфосалициловой кислоты для получения клеточного лизата. Центрифугируют при 3000 об/мин 10 мин. 1,0 мл осадка белка инкубируют 1 ч при 50°С с 1,0 мл 1% NaBH4». Далее оставшийся белок осаждают добавлением 0,4 мл 30% ТХУ. Пробу инкубируют 15 мин при 50°С. Затем пробу охлаждают 5 мин (0°С). Центрифугируют 10 мин при 3000 об/мин. Супернатант смешивают с 2,5 мл PBS (рН 7,4) и добавляют 2,0 мл ацетона для полного окисления NaBH4. Перемешивают. Центрифугировают 10 мин при 3000 об/мин. Удаляют верхнюю фазу. К нижней фазе добавляют равный объем диэтилового эфира (для удаления ТХУ). Перемешивают. Центрифугировают 10 мин при 3000 об/мин. Затем снова удаляют верхнюю фазу. Далее процедуру отмывки пробы от ТХУ с помощью диэтилового эфира производят 4-х кратно. Затем отбирают 0,1 мл жидкости (из нижней фазы) и смешивают с 0,4 мл 0,01 М фосфатного буфера (рН=7,0). В пробу добавляют 0,1 мл 0,4 мг/мл ДТНБ. Пробу спектрофотометрируют при 412 нм против контроля, содержащего воду вместо раствора осажденного белка. Расчет производят с учетом коэффициента молярной экстинкции 13·103 М-1см-1. Результаты определения концентрации белково-связанного глутатиона выражают в нмоль/мг белка.
Оценка способа оценки эффективности защиты лимфоцитов от апоптоза по способу-прототипу и предлагаемому способу выполнялось 40 раз. Результаты исследования обработаны статистически с использованием пакета программ Stat Soft Statistica 6.0.
При проведении исследования по способу-прототипу уровень восстановленного глутатиона в среде инкубации клеток в норме составил 2.15±0.18 нмоль/мг белка и белково-связанного глутатиона 2.9±0.25 нмоль/мг белка, а при оценке по предлагаемому способу при росте белково-связанного глутатиона на 18-35%, а восстановленного глутатиона на 25-41% считают защиту лимфоцитов от апоптоза низко эффективной, а при росте белково-связанного глутатиона на 36% и более и восстановленного глутатиона на 42% и более считают защиту лимфоцитов от апоптоза высоко эффективной (Табл. 1).
Полученные результаты уровня глутатиона в среде инкубации клеток соответствуют данным литературы [Смирнова Г.В., Октябрьский О.Н., 2005]. Итак, при применении способа-прототипа был получен недостаточно точный результат, не позволивший оценить эффективность защиты лимфоцитов от апоптоза, что связано с отсутствием биохимической стимуляции процесса, а наиболее эффективным и точным был предлагаемый способ. При этом предлагаемый способ прост в исполнении и интерпретации полученных результатов.
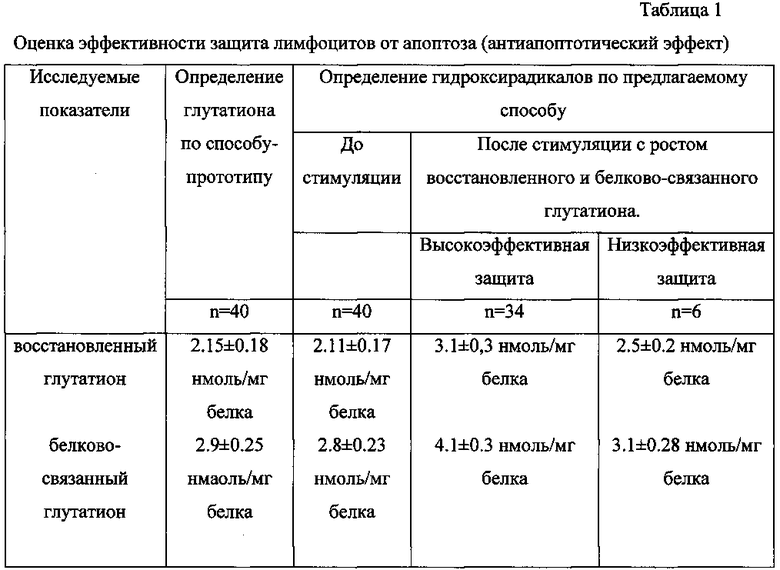
То есть при росте белково-связанного глутатиона на 18-35%, а восстановленного глутатиона на 25-41% считают защиту лимфоцитов от апоптоза низкоэффективной, а при росте белково-связанного глутатиона на 36% и более и восстановленного глутатиона на 42% и более считают защиту лимфоцитов от апоптоза высокоэффективной.
Полученные результаты уровня глутатиона в среде инкубации лимфоцитов соответствуют данным литературы [Смирнова Г.В., Октябрьский О.Н., 2005].
Итак, при применении способа-прототипа был получен недостаточно точный результат, не позволивший оценить эффективность защиты лимфоцитов от апоптоза, что связано с отсутствием биохимической стимуляции процесса, а наиболее эффективным и точным был предлагаемый способ.
При этом предлагаемый способ прост в исполнении и интерпретации полученных результатов.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ЗАЩИТЫ КЛЕТОК ОТ АПОПТОЗА | 2013 |
|
RU2541774C2 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ РАННЕЙ СТАДИИ АПОПТОЗА | 2013 |
|
RU2540500C2 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ ЭФФЕКТИВНОСТИ ЗАЩИТЫ ЛИМФОЦИТОВ ОТ ПЕРЕОКИСЛЕНИЯ | 2013 |
|
RU2525183C1 |
| СПОСОБ ДИАГНОСТИКИ АПОПТОЗА ЛИМФОЦИТОВ | 2013 |
|
RU2545900C2 |
| СПОСОБ ОЦЕНКИ ЭФФЕКТИВНОСТИ СТИМУЛЯЦИИ АНТИОКСИДАНТНОЙ АКТИВНОСТИ | 2013 |
|
RU2516925C1 |
| СПОСОБ СОЗДАНИЯ МОДЕЛИ ПЕРЕКИСНОГО ОКИСЛЕНИЯ ЛИМФОЦИТОВ | 2013 |
|
RU2541771C2 |
| СПОСОБ ГЕНОТИПИРОВАНИЯ ПОЛИМОРФИЗМА rs2551715 ГЕНА ГЛУТАТИОНРЕДУКТАЗЫ У ЧЕЛОВЕКА | 2013 |
|
RU2549688C1 |
| АНТИОКСИДАНТНОЕ, СТРЕСС-И НЕЙРОПРОТЕКТОРНОЕ ФАРМАКОЛОГИЧЕСКОЕ СРЕДСТВО-КОМЕНАТ КАЛИЯ | 2012 |
|
RU2514632C1 |
| АНТИОКСИДАНТНОЕ, СТРЕСС- И НЕЙРОПРОТЕКТОРНОЕ ФАРМАКОЛОГИЧЕСКОЕ СРЕДСТВО - КОМЕНАТ НАТРИЯ | 2012 |
|
RU2506078C1 |
| Способ отбора лиц, работающих в условиях воздействия кремнийсодержащих аэрозолей, для последующего мониторинга состояния бронхолегочной системы | 2023 |
|
RU2813952C1 |
Изобретение касается способа оценки эффективности защиты лимфоцитов от апоптоза, относится к медицине и может быть использовано в биохимии, кардиологии и терапии. Способ включает выделение лимфоцитов, инкубацию клеток 48 часов при температуре 37°С и 5% содержанием СО2, количественное определение жизнеспособности лимфоцитов по включению трипанового синего и биохимическое определение концентрации восстановленного и окисленного глутатиона в лизате лимфоцитов после предварительной инкубации в течение 30 минут с 10 ммоль 2-винилпиридином. Дополнительно в инкубационную среду, содержащую лимфоциты, вводят 1,4-дитиоэритритол и аскорбиновую кислоту в конечной концентрации 3,0 ммоль и 0,1 ммоль соответственно. При росте белково-связанного глутатиона на 18%-35%, а восстановленного глутатиона на 25-41% считают защиту низкоэффективной. При росте белково-связанного глутатиона на 36% и более и восстановленного глутатиона на 42% и более считают защиту высокоэффективной. Использование предлагаемого способа позволяет констатировать защиту клеток от апоптоза. Предлагаемый способ прост в осуществлении и интерпретации полученных результатов. 1 табл.
Способ оценки эффективности защиты лимфоцитов от апоптоза, заключающийся в выделении лимфоцитов, инкубации в течение 48 ч при температуре 37°С и 5% содержании СО2, в количественном определении жизнеспособности лимфоцитов по включению трипанового синего и биохимическом определении концентрации белково-связанного и восстановленного глутатиона в лизате лимфоцитов после предварительной инкубации в течение 30 минут с 10 ммоль 2-винилпиридином, отличающийся тем, что дополнительно в инкубационную среду, содержащую лимфоциты, вводят 1,4-дитиоэритритол и аскорбиновую кислоту в конечной концентрации 3,0 ммоль и 0,1 ммоль соответственно, и при росте окисленного глутатиона на 18-35% и восстановленного глутатиона на 25-41% оценивают защиту лимфоцитов от апоптоза как низкоэффективную, а при росте белково-связанного глутатиона на 36% и более и восстановленного глутатиона на 42% и более оценивают защиту лимфоцитов от апоптоза как высокоэффективную.
| Low dose gamma-rays activate immune functions via induction of | |||
| glutathione and delay tumor growth / S | |||
| Kojima, K | |||
| Nakayama, H | |||
| Ishida // J | |||
| Radiat | |||
| Res | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| - Vol | |||
| Железобетонный фасонный камень для кладки стен | 1920 |
|
SU45A1 |
| - P | |||
| Способ сопряжения брусьев в срубах | 1921 |
|
SU33A1 |
| RU 2013121503 A, 20.11.2014 | |||
| СПОСОБ ПРОГНОЗИРОВАНИЯ УСТОЙЧИВОСТИ ОРГАНИЗМА К СТРЕССОРНОМУ ВОЗДЕЙСТВИЮ | 2006 |
|
RU2322675C1 |
| Chang WK et al | |||
| Glutamine protects activated human T cells from | |||
| apoptosis by up-regulating glutathione | |||

