Изобретение относится к области биотехнологии и касается способа быстрого выявления серологического родства микробов, имеющих общие детерминанты в составе липополисахаридов, по изменениям в подвижности бактерий рода Azospirillum в присутствии поликлональных антител к О-антигенам.
Известны способы исследования серологических свойств липополисахаридов (ЛПС, О-антигенов) азоспирилл (Azospirillum), которые могут быть использованы для определения серологической специфичности этих бактерий (Коннова О.Н., Бойко А.С., Бурыгин Г.Л., Федоненко Ю.П., Матора Л.Ю., Коннова С.А., Игнатов В.В. Химические и серологические исследования липополисахаридов бактерий рода Azospirillum // Микробиология. - 2008. - Т.77. №3. - С.350-357).
Известны способы определения титра антител, вызывающих агглютинацию клеток (Бойко А.С., Смолькина О.Н., Федоненко Ю.П., Здоровенко Э.Л., Качала В.В., Коннова С.А., Игнатов В.В. Особенности структуры О-полисахаридов азоспирилл серогруппы I // Микробиология. - 2010. - Т.79. №2. - С.219-227).
Известны способы определения наличия антигенных перекрестов экстрактов ЛПС в реакциях иммунопреципитации в геле или твердофазного иммуноферментного анализа (ИФА) с использованием специфических антител (Федоненко Ю.П., Бойко А.С., Здоровенко Э.Л., Коннова С.А., Шашков А.С., Игнатов В.В., Книрель Ю.А. Структурные особенности О-специфических полисахаридов бактерий Azospirillum серогруппы III // Биохимия. - 2011. - Т.76. Вып.7. - С.976-982).
Использование данных методов позволило провести исследование серологических свойств и структур (хемотипов) липополисахаридов (ЛПС, О-антигенов) и выделить на сегодняшний день три серогруппы азоспирилл.
Однако для постановки реакций иммунопреципитации в геле и проведения твердофазного иммуноферментного анализа (ИФА) необходимо выделение бактериальных липополисахаридов. Кроме того, для ИФА или для определения титра антител, вызывающих агглютинацию клеток, необходимы планшеты соответствующего типа, а в случае иммунопреципитации требуются пластины агарозного геля. Время регистрации результатов с начала постановки реакции без учета времени подготовки образцов составляет от 3-х до 24-х часов в случае всех трех методов.
Ближайшим аналогом заявляемого решения является способ регистрации изменений в подвижности бактерий рода Azospirillum в присутствии поликлональных антител на поверхностные структуры бактерий для выявления общих детерминант в составе клеточной мембраны и чехла, покрывающего полярный жгутик, обеспечивающий подвижность бактерий, а также при исследовании углеводных антигенных детерминант, представленных в составе клеточной поверхности близкородственных штаммов (Sp107 и Sp245) (Бурыгин Г.Л., Широков А.А., Шелудько А.В., Кацы Е.И., Щеголев С.Ю., Матора Л.Ю. Выявление чехла на поверхности полярного жгутика Azospirillum brasilense // Микробиология. 2007. Т.76. №6. С.822-829; Шелудько А.В. Генетико-физиологические аспекты социального поведения ассоциативных бактерий Azospirillum brasilense // Автореф. дис. … д-ра биол. наук. - Саратов: ИБФРМ РАН, 2010. - 48 с.).
Однако известный способ предполагает для определения процента подвижных клеток и их скорости движения у тестируемых штаммов проведения записи видеофайлов подвижности бактерий при добавлении и без добавления антител с последующим компьютерным анализом видеоизображения, поскольку используются антитела в концентрациях, значительно снижающих показатели подвижности, но не останавливающих движение всех клеток в бактериальной суспензии. Последнее обстоятельство значительно увеличивает время регистрации результатов, ограничивая число штаммов, которые можно проанализировать в одном эксперименте. Данный способ предполагает дополнительную операцию по отмыванию клеток от культуральной жидкости и суспендирования их в фосфатном буфере при подготовке бактериальных суспензий.
Задачей, на решение которой направлено заявляемое техническое решение, является оптимизация подходов выявления серологического родства микробов и условий регистрации изменений в подвижности бактерий рода Azospirillum в присутствии поликлональных антител к О-антигенам.
Технический результат, достигаемый при использовании заявляемого изобретения, заключается в сокращении времени на выявление серологически родственных штаммов бактерий рода Azospirillum непосредственно в жидких бактериальных культурах, независимо от использованных для культивирования сред и условий подготовки клеточных суспензий.
Поставленная задача решается тем, что в СПОСОБЕ ВЫЯВЛЕНИЯ БАКТЕРИЙ РОДА AZOSPIRILLUM, ИМЕЮЩИХ ОБЩИЕ АНТИГЕННЫЕ ДЕТЕРМИНАНТЫ В СОСТАВЕ ЛИПОПОЛИСАХАРИДОВ, включающем: - получение специфических антител на липополисахариды; - определение “рабочей" концентрации антител на липополисахариды "модельного штамма", серологическое родство с которым необходимо установить; - выращивание культур исследуемых штаммов бактерий рода Azospirillum на жидких средах при температуре 30°С и подготовку бактериальных суспензий на основе выращенных бактерий рода Azospirillum; - добавление антител в "рабочей" концентрации к бактериальным суспензиям с последующим приготовлением препаратов для световой микроскопии при комнатной температуре; - оценку подвижности бактерий рода Azospirillum при микроскопии препаратов; - выявление бактерий, имеющих общие детерминанты в составе полисахаридов, по снижению подвижности бактерий рода Azospirillum в присутствии антител на липополисахариды "модельного штамма", согласно заявляемому решению в качестве "рабочей" концентрации антител используют удвоенную концентрацию антител, в присутствии которой неподвижны все клетки гомологичного модельного штамма бактерий рода Azospirillum в поле зрения микроскопа; "рабочую" концентрацию антител, в присутствии которой неподвижны все клетки гомологичного штамма бактерий рода Azospirillum, сохраняют неизменной, независимо от партии полученных антител; для определения "рабочей" концентрации антител проводят оценку процента подвижных клеток гомологичного штамма бактерий рода Azospirillum в поле зрения микроскопа; в качестве исследуемых штаммов бактерий рода Azospirillum используют 18-часовые культуры, разведенные 50 мМ фосфатно-солевым буфером (ФСБ, рН 7) до значений A600=0.3-0.5 (1=1 см), без отмывания клеток от культуральной жидкости; о наличии микробов, имеющих общие детерминанты в составе полисахаридов, судят но остановке движения бактерий рода Azospirillum в присутствии антител на липополисахариды "модельного штамма".
Сущность заявляемого решения заключается в том, что способ включает определение "рабочей" концентрации антител, предварительно полученных одним из известных способов, на липополисахариды "модельного штамма" серогруппы, принадлежность к которой требуется установить, выращивание культур тестируемых микроорганизмов и подготовку бактериальных суспензий, добавление антител к бактериальным суспензиям с последующим приготовлением препаратов для световой микроскопии, оценку подвижности микроорганизмов при микроскопии препаратов.
О наличии серологического родства (принадлежности к серогруппе) судят по остановке движения клеток в присутствии антител на липополисахариды "модельного штамма". Штаммы, клетки которых неподвижны в присутствии антител на липополисахариды "модельного штамма", можно считать серологически близкими.
Преимущество изобретения заключается в быстром и простом способе выявления микробов, имеющих общие антигенные детерминанты (то есть обладающих серологическим родством) в составе О-антигена липополисахаридов, преобладающих компонентов внешней мембраны грамотрицательных бактерий, как правило, определяющих серологическую специфичность бактерий, что является существенным в иммутотаксономических исследованиях бактерий рода Azospirillum.
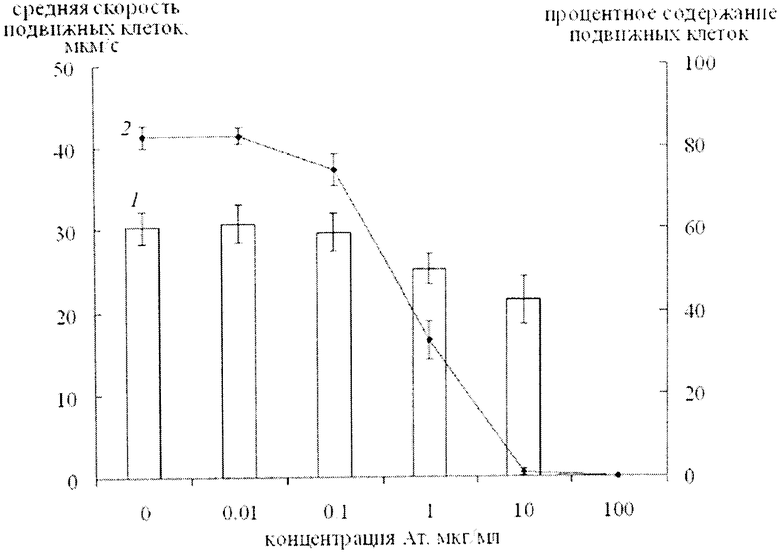
Изобретение поясняется чертежом, на котором представлен график изменения средней скорости движения (1) и количества (2) подвижных клеток A. brasilense Sp245 с увеличением концентрации антител на липополисахарид этого штамма.
Описание сущности технического решения.
Бактерии рода Azospirillum из семейства Rhodospirillaceae в естественных условиях обитают в основном на корнях высших растений, произрастающих в разнообразных климатических условиях. Интерес к изучению этих бактерий объясняется их способностью стимулировать рост широкого круга растений, в том числе и агрономически значимых. Прогресс в их исследовании в качестве потенциальных стимуляторов роста растений в значительной степени зависит от развития быстрых и надежных способов качественного определения данных микроорганизмов. Выявляющие системы на основе антител являются весьма удобным средством мониторинга разнообразных микроорганизмов. На клетках азоспирилл, поддерживаемых в жидких средах, имеется один полярно расположенный жгутик, обуславливающий подвижность и хемотаксис - важные факторы для эффективной колонизации растений данными бактериями. Полярный жгутик бактерий A. brasilense покрыт чехлом, в формировании которого показано участие молекул липополисахаридов (ЛПС), что может позволить быстро выявлять антигенные перекресты между штаммами при таксономических исследованиях на основании изменения характеристик подвижности микроорганизмов в присутствии антител на ЛПС.
Заявляемый способ включает получение одним из известных способов специфических антител на липополисахариды, определение "рабочей" концентрации антител на липополисахариды "модельного штамма" серогруппы, принадлежность к которой требуется установить, выращивание тестируемых культур микроорганизмов и подготовку бактериальных суспензий, добавление антител к бактериальным суспензиям с последующим приготовлением препаратов для световой микроскопии, оценку подвижности микроорганизмов при микроскопии препаратов. О наличии серологического родства (принадлежности к серогруппе) свидетельствует остановка движения клеток в присутствии антител на липополисахариды "модельного штамма".
Штаммы, клетки которых неподвижны в присутствии антител на липополисахариды "модельного штамма", можно считать серологически близкими.
Антитела получают одним из известных способов, например для получения специфических антител на ЛПС готовят суспензию грамотрицательных бактерий в 2%-ном растворе глутарового альдегида, после чего проводят иммунизацию кроликов обработанной суспензией с последующим забором крови и выделением из нее антител в виде фракции иммуноглобулинов класса G.
Для определения "рабочей концентрации" антител проводят оценку подвижных клеток "модельного штамма" в присутствии этих антител в поле зрения микроскопа. Клетки 18-часовых жидких культур "модельного штамма", выращенные при 30°С, осаждают центрифугированием в течение 15 мин при 5000 g, удаляют надосадочную жидкость, затем суспендируют в 50 мМ фосфатно-солевом буфере (ФСБ) с рН 7) и повторно центрифугируют в течение 15 мин при 5000 g, осадок ресуспендируют в ФСБ до значений А600=0.5 (1=1 см). Полученную суспензию клеток используют для определения характеристик подвижности клеток (процент подвижных клеток и их средняя скорость движения) в присутствии различных концентраций штаммоспецифических антител к ЛПС (диапазон конечных концентраций антител 0.01-1000 мкг/мл с шагом, кратным 10). Для этого определяют концентрацию полученных и очищенных антител (мкг/мл), затем антитела в концентрациях 0.02, 0.2, 2, 20, 200 и 2000 мкг/мл последовательно в соотношении 1:1 быстро смешивают на покровном стекле с суспензией бактерий (в итоге концентрация антител станет 0.01, 0.1, 1, 10, 100 и 1000 мкг/мл), покровное стекло каплей вниз помещают на предметное стекло с лункой (препарат "висячая капля"). При микроскопии препарата "висячая капля" проводят запись видеофайлов подвижности бактерий. Определяют процент подвижных клеток и их среднюю скорость движения, используя доступную программу анализа видеоизображения. Результаты трех независимых экспериментов используют для построения диаграммы зависимости количества подвижных клеток (в процентах) и скорости их движения (мкм/с) от концентрации штаммоспецифических антител на ЛПС. На диаграмме выбирают концентрацию антител, значительно снижающую процент подвижных клеток и их скорость движения. Результаты подвижности бактерий в присутствии этого количества антител можно использовать при исследовании углеводных антигенных детерминант, представленных в составе поверхности интактных клеток. Удвоенную концентрацию антител, в присутствии которой неподвижны все клетки в поле зрения микроскопа, используют в качестве "рабочей" для быстрого определения серологического родства.
Процедуру определения "рабочей концентрации" антител для определения серологического родства можно упростить, исключив этап получения видеозаписи с последующим определением скорости движения клеток, ограничившись оценкой процента подвижных клеток в поле зрения при микроскопии, поскольку данный показатель является наиболее выраженным.
Определение "рабочей концентрации" для каждой партии антител достаточно провести один раз. Необходимо отметить, что независимо от партии антител (антител, полученных после иммунизации разных животных, или антител, выделенных в разное время из сыворотки одного животного) концентрация антител, в присутствии которой неподвижны все клетки гомологичного штамма (то есть "рабочая" концентрация), остается неизменной, а концентрация антител, значительно снижающая процент подвижных клеток и их скорость движения, может варьировать. Как показала практика, "рабочая" концентрация (то есть концентрация антител, останавливающая движение всех клеток) для всех антител, полученных после иммунизации разных животных, или антител, выделенных в разное время из сыворотки одного животного, была одной и той же.
Следующим этапом является оценка подвижности клеток тестируемых штаммов в присутствии рабочей концентрации антител.
- 18 ч культуры исследуемых штаммов, выращенных при 30°С, разводят 50 мМ фосфатно-солевым буфером (ФСБ, рН 7) до значений A600=0.3-0.5 (1=1 см). Количество жизнеспособных клеток при данных значениях A600 составляет 108 в мл.
- Далее проводят микроскопию разведенных культур и оценивают подвижность бактерий без добавления антител. Как правило, в случае бактерий рода Azospirillum, выращенных в жидких средах, 60-90% клеток в популяции подвижны.
- Затем разведенную бактериальную культуру в соотношении 1:1 (оптимальное соотношение 2 к 2 мкл) быстро смешивают на покровном стекле с антителами в рабочей концентрации, покровное стекло каплей вниз помещают на предметное стекло с лункой (препарат "висячая капля"). Препарат "висячая капля" микроскопируют с использованием объектива с широким охватом ноля зрения.
- При микроскопии препарата "висячая капля" оценивают процент подвижных клеток в поле зрения микроскопа. По результатам трех независимых наблюдений делают заключение о влиянии антител на подвижность бактерий.
Клеточные агрегаты, возникающие в результате взаимодействия антител с корпускулярными антигенами, не принимаются во внимание. Необходимо отметить, что некоторая доля бактерий (менее 10%) в жидких средах образует агрегаты независимо от присутствия в среде антител. Агрегация характерна для азоспирилл и может усиливаться вследствие воздействия на бактериальную популяцию каких-либо внешних факторов. Штаммы, клетки которых неподвижны в присутствии антител на липополисахариды "модельного штамма", можно считать серологически близкими. Процент подвижных клеток в присутствии рабочей концентрации антител на ЛПС в случае таких штаммов составляет не более 10%, что может являться результатом снижения количества свободных антител при их взаимодействии с полисахаридами, экскретируемыми бактериями в культуральную жидкость.
Таким образом, наблюдаемая при микроскопии остановка 90% двигающихся (подвижных) клеток в присутствии антител на липополисахариды "модельного штамма" свидетельствует о наличии серологического родства (принадлежности к серогруппе).
- В результате общее время на выявление бактерий - от смешивания антител с бактериальной культурой до регистрации результатов без учета времени подготовки образцов - составляет от 1-3 мин.
В табл.1 представлены характеристики подвижности клеток A. brasilense в присутствии антител на ЛПС штаммов Sp7 и Sp245 в фосфатно-солевом буфере.
мкм/с
мкм/с
В табл.2 представлены результаты взаимодействия антител на ЛПС штаммов, относящихся к серогруппам I и II, с клетками и экстрактами липополисахаридов.
На чертеже представлен график изменения средней скорости движения (1) и количества (2) подвижных клеток A. brasilense Sp245 с увеличением концентрации антител на липополисахарид этого штамма.
Заявляемый способ осуществляют следующим образом.
В модельных экспериментах проведен анализ подвижности 23-х штаммов, относящихся к Azospirillum brasilense, Azospirillum lipoferum, Azospirillum irakense и Azospirillum sp., в присутствии антител на ЛПС штаммов Sp7 или Sp245, относящихся к разным серогруппам.
Определили "рабочую" концентрацию антител на ЛПС, предварительно полученных одним из известных способов. Бактерии "модельного штамма" выращивали на жидкой синтетической малатной среде, содержащей в 1 л: малат Na - 5 г, KH2PO4 - 0.4 г, K2HPO4 - 0.4 г, NaCl - 0.1 г, MgSO4 - 0.2 г, FeSO4·7H2O - 0.02 г, Na2MoO4·2H2O - 0.002 г, NH4Cl - 1 г (рН 6.8-7.0). Далее клетки 18-ч жидких культур, выращенных при 30°С, осаждали центрифугированием в течение 15 мин при 5000 g, удаляли надосадочную жидкость, затем суспендировали в 50 мМ фосфатно-солевом буфере (ФСБ, рН 7) и повторно центрифугировали в течение 15 мин при 5000 g, осадок ресуспендировали в ФСБ до значений А600=0.5 (1=1 см). Полученную суспензию клеток использовали для определения характеристик подвижности клеток (процент подвижных клеток и их среднюю скорость движения) в присутствии различных концентраций штаммоспецифических антител к ЛПС (диапазон конечных концентраций антител 0.01-1000 мкг/мл с шагом, кратным 10). Для этого при комнатной температуре антитела в концентрациях 0.02, 0.2, 2, 20, 200 и 2000 мкг/мл последовательно в соотношении 1:1 (2 и 2-м мкл) быстро смешивали на покровном стекле с суспензией бактерий, покровное стекло каплей вниз помещали на предметное стекло с лункой (препарат "висячая капля"). Микроскопию препарата проводили при комнатной температуре, используя объектив с широким охватом поля зрения. При микроскопии препарата "висячая капля" записывали видеофайлы подвижности бактерий и определяли процент подвижных клеток и их среднюю скорость движения, используя доступную программу анализа видеоизображения. По результатам трех независимых экспериментов строили диаграммы зависимости количества подвижных клеток (в процентах) и скорости их движения (мкм/с) от концентрации штаммоспецифических антител на ЛПС. На фиг.1 представлен график изменения средней скорости движения (1) и количества подвижных (2) клеток А. brasilense Sp245 с увеличением концентрации антител на липополисахарид этого штамма. На диаграмме выбирали концентрацию антител, значительно снижающую процент подвижных клеток и их скорость движения. Результаты подвижности бактерий в присутствии этого количества антител использовали при исследовании углеводных антигенных детерминант, представленных в составе клеточной поверхности. Концентрация антител на ЛПС штамма Sp245, значительно снижающая процент подвижных клеток и их скорость движения, составляет 10 мкг/мл. При концентрации этих антител 100 мкг/мл все клетки Sp245 неподвижны (фиг.1). Удвоенную концентрацию антител, в присутствии которой неподвижны все клетки в поле зрения микроскопа, использовали в качестве "рабочей" для быстрого определения серологического родства. Таким образом, для быстрого определения серологического родства со штаммом Sp245 "рабочая концентрация" антител на ЛПС этого штамма составляет 200 мкг/мл. В случае антител на ЛПС штамма Sp245 концентрация антител, в присутствии которой неподвижны все клетки гомологичного штамма оставалась неизменной (100 мкг/мл при этом "рабочая" концентрация составляла 200 мкг/мл), независимо от партии антител, а концентрация антител, значительно снижающая процент подвижных клеток и их скорость движения варьировала от 10 до 30 мкг/мл. Определение "рабочей концентрации" антител для определения серологического родства проводили также по упрощенной схеме, ограничившись оценкой процента подвижных клеток в поле зрения микроскопа, поскольку данный показатель является наиболее выраженным. Для определения "рабочей концентрации" антител на ЛПС штамма Sp7 использовали данный подход. Так, процент подвижных клеток Sp7 в присутствии антител на ЛПС этого штамма составляет 80-90%, 20-30, 5-10 и 0% в присутствии соответственно 1, 10, 100 и 1000 мкг/мл белка. Таким образом, для быстрого определения серологического родства со штаммом Sp7 "рабочая концентрация" антител на ЛПС этого штамма составляет 2000 мкг/мл, а концентрация антител, значительно снижающая процент подвижных клеток и их скорость движения, составляет 100-300 мкг/мл. В случае антител на ЛПС штамма Sp7 концентрация антител, в присутствии которой неподвижны все клетки гомологичного штамма, также оставалась неизменной. Таким образом, определение рабочей концентрации для каждой партии антител достаточно провести один раз.
В случае ряда штаммов бактерии, выращенные на жидкой синтетической малатной среде, осаждали центрифугированием в течение 15 мин при 5000 g, удаляли надосадочную жидкость, затем суспендировали в 50 мМ фосфатно-солевом буфере (ФСБ, рН 7) и повторно центрифугировали в течение 15 мин при 5000 g, осадок ресуспендировали в ФСБ до значений А600=0.5 (1=1 см). Суспензию бактерий в ФСБ в соотношении 1:1 (2 к 2 мкл) быстро смешивали на покровном стекле последовательно с антителами к ЛПС Sp7 или Sp245. Концентрация антител в итоге составляла величину, значительно снижающую процент подвижных клеток и их скорость движения у штаммов Sp7 или Sp245 (100 и 10 мкг/мл соответственно). Покровное стекло каплей вниз помещали на предметное стекло с лункой (препарат "висячая капля"). При микроскопии препарата "висячая капля" записывали видеофайлы подвижности бактерий и определяли процент подвижных клеток и их среднюю скорость движения, используя программу анализа видеоизображения. В серии экспериментов по исследованию подвижности клеток азоспирилл 9-ти штаммов определены штаммы (результаты представлены в табл. 1), подвижность которых снижалась в присутствии антител к ЛПС Sp7 и Sp245 в концентрации 100 и 10 мкг/мл, соответственно (концентрации Ат, значительно снижающие показатели подвижности гомологичных штаммов). Однако подвижность клеток штамма Sp107, относящегося к серогругше I в присутствии 10 мкг/мл антител к ЛПС Sp245, изменялась незначительно (табл. 1). ЛПС двух близкородственных штаммов A. brasilense Sp245 и Sp107 в тесте иммунодиффузии демонстрировали лишь минорные антигенные различия, выражающиеся в образовании небольшой шпоры при слиянии полос преципитации. Наличие данной шпоры свидетельствовало о том, что в составе ОПС1 штамма Sp107 отсутствуют некие антигенные детерминанты, присутствующие в составе ОПС1 штамма Sp245. Вполне вероятно, это может объяснять лишь незначительное изменение подвижности клеток Sp107 в присутствии использованной концентрации Ат (10 мкг/мл) на ЛПС Sp245 (табл. 1). Ингибирующего эффекта удалось достичь увеличением концентрации Ат на ЛПС Sp245 до 100 мкг/мл (концентрация антител ингибирующая подвижность всех клеток штамма Sp245), поскольку это приводило к увеличению количества антител на общие для Sp245 и Sp107 детерминанты (табл. 2). Таким образом, антитела в концентрациях, останавливающих подвижность всех клеток гомологичного штамма, добавленные к исследуемым бактериям, ингибируют подвижность клеток независимо от топких особенностей организации клеточной поверхности азоспирилл.
Для оценки подвижности клеток тестируемых штаммов в присутствии рабочей концентрации антител (антитела в конечной концентрации, останавливающие подвижность всех клеток гомологичного штамма) бактерии выращивали на жидкой синтетической малатной среде (см. выше) или богатой среде LB, содержащей в 1 л: триптон - 10 г, дрожжевой экстракт - 5 г, NaCl - 5 г (рН 7.5). 18 ч культуры исследуемых штаммов, выращенных при 30°С, разводили 50 мМ фосфатно-солевым буфером (ФСБ, рН 7) до значений А600=0.3-0.5 (1=1 см). Количество жизнеспособных клеток при данных значениях А600 составляло 108 в мл. Этап отмывания клеток не проводили. Далее при комнатной температуре проводили микроскопию разведенных культур и оценивали подвижность бактерий без добавления антител. Подвижные клетки в популяции исследуемых штаммов составляли не менее 60%. При комнатной температуре разведенную бактериальную суспензию в соотношении 1:1 (2 и 2 мкм) быстро смешивали на покровном стекле последовательно с антителами к ЛПС Sp7 или Sp245 в концентрации 2000 и 200 мкг/мл соответственно (в итоге концентрация антител составляла 1000 и 100 мкг/мл). Покровное стекло каплей вниз помещали на предметное стекло с лункой (препарат "висячая капля"). Микроскопию препарата проводили при комнатной температуре, используя объектив с широким охватом поля зрения, оценивая процент подвижных клеток в присутствии антител. По результатам трех независимых наблюдений делали заключение о влиянии антител на подвижность бактерий. Результаты представлены в табл. 2. Этапы записи видеофайлов и их обработки при помощи программы компьютерного анализа видеоизображения не проводили. Антитела на ЛПС Sp7 (серотип II) или Sp245 (серотип I) останавливали движение бактерий только тех штаммов, клетки которых агглютинировали с антителами (табл. 2). Также выявлена корреляция между наличием антигенных перекрестов в реакции иммунодиффузии и снижением подвижности клеток в присутствии антител на ЛПС Sp7 или Sp245 у штаммов Sp7, CD, SR14, SR55, SR80 или Sp245, Sp107, S27, SR15, SR75, SR81 и SpRG20a (табл. 2). Из семи штаммов, взаимодействующих с Ат к ЛПС Sp245, для A. brasilense Sp245, Sp107, S27, SR75 и A. lipoferum RG20a, установлена идентичность структуры повторяющегося звена О-специфических полисахаридов (ОПС), представленного линейным пента-D-рамнаном. ОПС штамма SR15 является D-рампаном, но с тетрасахаридным повторяющимся звеном. Полученные в работе данные позволяют дополнить серогруппу I штаммом Azospirillum sp.SR81. Штаммы Azospirillum серогруппы II характеризуются наличием гетерополисахаридных ОПС, дающих перекрестную реакцию с антителами к ЛПС типового штамма A. brasilense Sp7. Из пяти штаммов, взаимодействующих с Ат к ЛПС Sp7, для трех описана структура повторяющегося звена ОПС. Результаты данной работы позволяют дополнить серотип II штаммом A. brasilense SR14. Необходимо отметить, что результаты влияния антител на подвижность бактерий заметны уже через 1 мин после приготовления препаратов для микроскопии (препарат "висячая капля"). Клетки штаммов, подвижных в присутствии антител, продолжали двигаться даже после 30-60 мин инкубации с антителами. Влияние антител к ЛПС на подвижность клеток не зависит от среды культивирования бактерий и способа подготовки бактериальной суспензии. Так, влияние антител на подвижность клеток в бактериальных культурах (не отмытых от среды культивирования) или отмытых и суспендированных в ФБ азоспирилл, не различалось. Влияние антител на бактерии, выращенные как на жидкой синтетической малатной среде, так и на богатой среде LB, было одинаковым. Клеточные агрегаты, возникающие в результате взаимодействия антител с корпускулярными антигенами, не принимались во внимание. Необходимо отметить, что некоторая доля бактерий (менее 10%) в жидких средах образует агрегаты независимо от присутствия в среде антител. Агрегация характерна для азоспирилл и может усиливаться вследствие воздействия на бактериальную популяцию каких-либо внешних факторов. Таким образом, заявляемый способ, заключающийся в ингибировании подвижности бактерий при добавлении к клеточной суспензии антител на ЛПС, может быть использован при определении штаммов, демонстрирующих антигенные перекресты с относящимися к разным серологическим типам штаммами Sp7 (серотип II) и Sp245 (серотип I). Результаты влияния антител на подвижность согласуются с данными таких тестов, как агглютинация клеток антителами и выявления наличия антигенных перекрестов в реакции иммунодиффузии.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКОВ КЛЕТОЧНОЙ ПОВЕРХНОСТИ | 2017 |
|
RU2670001C1 |
| СОСТАВ ДЛЯ ДОСТАВКИ АКТИВНЫХ ВЕЩЕСТВ К КОРНЯМ ПШЕНИЦЫ | 2007 |
|
RU2347366C2 |
| ШТАММ КУЛЬТИВИРУЕМЫХ ГИБРИДНЫХ КЛЕТОК ЖИВОТНЫХ Mus. musculus L - ПРОДУЦЕНТ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ, СПЕЦИФИЧНЫХ К О-АНТИГЕНУ ХОЛЕРНЫХ ВИБРИОНОВ О1 СЕРОГРУППЫ | 2010 |
|
RU2425874C1 |
| СПОСОБ ПОВЫШЕНИЯ ВСХОЖЕСТИ СЕМЯН ПШЕНИЦЫ, ПОРАЖЕННЫХ КЛОПОМ ВРЕДНОЙ ЧЕРЕПАШКИ | 2006 |
|
RU2314692C1 |
| Способ биодеградации малахитового зеленого (варианты) | 2017 |
|
RU2676153C2 |
| Штамм гибридных культивируемых клеток животных MUS мUSсULUS L. - продуцент моноклональных антител к бактериям рода BRUceLLa | 1990 |
|
SU1752764A1 |
| ШТАММ КУЛЬТИВИРУЕМЫХ ГИБРИДНЫХ КЛЕТОК ЖИВОТНЫХ Mus. Musculus L - ПРОДУЦЕНТ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ, СПЕЦИФИЧНЫХ К ЛПС ХОЛЕРНЫХ ВИБРИОНОВ О139 СЕРОГРУППЫ | 2010 |
|
RU2425875C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЛИПОПОЛИСАХАРИДОВ | 2002 |
|
RU2237719C2 |
| Штамм культивируемых гибридных клеток животных Mus. musculus - продуцент моноклональных антител к мембранному белку, общему для типичных и атипичных холерных вибрионов 01 серогруппы | 2022 |
|
RU2785463C1 |
| БЕЛОК NMB0928 И ЕГО ПРИМЕНЕНИЕ В ФАРМАЦЕВТИЧЕСКИХ КОМПОЗИЦИЯХ | 2004 |
|
RU2335505C2 |
Изобретение касается способа выявления серологического родства микробов, имеющих общие детерминанты в составе липополисахаридов. Способ включает получение специфических антител на липополисахариды. Определение "рабочей" концентрации антител на липополисахариды "модельного штамма", серологическое родство с которым необходимо установить. Выращивание культур исследуемых штаммов бактерий рода Azospirillum и добавление антител в "рабочей" концентрации к бактериальным суспензиям с последующим приготовлением препаратов для световой микроскопии. Оценку подвижности бактерий рода Azospirillum. Выявление бактерий, имеющих общие детерминанты в составе полисахаридов, по снижению подвижности бактерий рода Azospirillum в присутствии антител на липополисахариды "модельного штамма". В качестве "рабочей" концентрации антител используют удвоенную концентрацию антител, в присутствии которой неподвижны все клетки гомологичного модельного штамма бактерий рода Azospirillum. Представленное изобретение позволяет сократить время на выявление серологически родственных штаммов бактерий рода Azospirillum непосредственно в жидких бактериальных культурах. 1 ил., 2 табл.
Способ выявления бактерий рода Azospirillum, имеющих общие антигенные детерминанты в составе липополисахаридов, включающий: - получение специфических антител на липополисахариды; - определение "рабочей" концентрации антител на липополисахариды "модельного штамма", серологическое родство с которым необходимо установить; - выращивание культур исследуемых штаммов бактерий рода Azospirillum на жидких средах при температуре 30°C и подготовку бактериальных суспензий на основе выращенных бактерий рода Azospirillum; - добавление антител в "рабочей" концентрации к бактериальным суспензиям с последующим приготовлением препаратов для световой микроскопии при комнатной температуре; - оценку подвижности бактерий рода Azospirillum при микроскопии препаратов; - выявление бактерий, имеющих общие детерминанты в составе полисахаридов, по снижению подвижности бактерий рода Azospirillum в присутствии антител на липополисахариды "модельного штамма", отличающийся тем, что в качестве "рабочей" концентрации антител используют удвоенную концентрацию антител, в присутствии которой неподвижны все клетки гомологичного модельного штамма бактерий рода Azospirillum в поле зрения микроскопа; "рабочую" концентрацию антител, в присутствии которой неподвижны все клетки гомологичного штамма бактерий рода Azospirillum, сохраняют неизменной независимо от партии полученных антител; для определения «рабочей» концентрации антител проводят оценку процента подвижных клеток гомологичного штамма бактерий рода Azospirillum в поле зрения микроскопа; в качестве исследуемых штаммов бактерий рода Azospirillum используют 18-часовые культуры, разведенные 50 мМ фосфатно-солевым буфером (ФСБ, рН 7) до значений А600=0.3-0.5 (1=1 см), без отмывания клеток от культуральной жидкости; о наличии микробов, имеющих общие детерминанты в составе полисахаридов, судят по остановке движения бактерий рода Azospirillum в присутствии антител на липополисахариды "модельного штамма".
| Г.Л.БУРЫГИН и др., Выявление чехла на поверхности полярного жгутика Azospirillum Brasilense, Микробиология, 2007, том 76,N6, стр.822-829 | |||
| ШЕЛУДЬКО А.В., Генетико-физиологические аспекты социального поведения ассоциативных бактерий Azospirillum Brasilense, Автореферат дисс., Саратов, 2010, стр.20-21 | |||
| MATORA LIU et al., Antigenic identity of the |

