ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
В настоящей заявке заявлен приоритет по предварительной заявке Соединенных Штатов с серийным No. 61/252074, поданной 15 октября 2009, которая в полном объеме включена в настоящее описание в виде ссылки.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение в целом связано с новыми полипептидами, определяемыми здесь как химерные полипептиды фактора роста фибробластов (FGF), с новой ДНК, кодирующей химерные полипептиды FGF, и с рекомбинантной продукцией химерных полипептидов FGF, а также со способами, композициями и количественными анализами с использованием химерных полипептидов FGF для терапевтического лечения связанных с метаболизмом расстройств и других состояний, и кроме того, для изготовления фармацевтически активных композиций, содержащих химерные полипептиды FGF, композиций, имеющих терапевтические и фармакологические свойства, включая свойства, ассоциированные с лечением связанных с метаболизмом расстройств и других состояний.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Семейство факторов роста фибробластов (FGF) включает в себя подсемейства FGF19, которые включают в себя человеческие FGF21, FGF23 и FGF19, а также мышиный FGF15. В отличие от других членов семейства FGF, которые обычно действуют на свою исходную ткань паракринным образом, члены подсемейства FGF19 действуют на специфические отдаленные ткани эндокринным способом. Эффекты, оказываемые членами семейства FGF, являются результатом их гепарин-зависимого связывания с одним или несколькими членами семейства тирозинкиназных рецепторов FGF (FGFR). Указанное семейство рецепторов включает в себя четыре члена, каждый из которых имеет тирозинкиназный домен, FGFR1, FGFR2, FGFR3 и FGFR4, а также два варианта сплайсинга каждого из FGFR1, FGFR2 и FGFR3. Указанные варианты сплайсинга, которые возникают в экзоне 3 рецепторов FGFR1, FGFR2 и FGFR3, обозначают как варианты "b" и "c" (то есть FGFR1b, FGFR2b, FGFR3c, FGFR1c, FGFR2c и FGFR3c, которые известны также как FGFR1(III)b, FGFR2(III)b, FGFR3(III)c, FGFR1(III)c, FGFR2(III)c и FGFR3(III)c, соответственно).
Члены подсемейства FGF19 вовлечены в регуляцию различных тканеспецифических процессов метаболизма у млекопитающих. Особый интерес представляет FGF19, который, как было показано, нацелен и воздействует как на адипоциты, так и на гепатоциты. Например, мыши, обработанные рекомбинантным человеческим FGF19 (rhFGF19), несмотря на то, что находятся на диете с высоким уровнем потребления жира, проявляют повышенную скорость метаболизма, повышенный уровень окисления липидов, пониженный дыхательный коэффициент и потерю веса. Более того, такие мыши обнаруживают более низкие сывороточные уровни лептина, инсулина, холестерина и триглицеридов, а также нормальные уровни глюкозы в крови, несмотря на то, что находятся на диете с высоким уровнем потребления жира, и на отсутствие потери аппетита. Мыши с ожирением, у которых отсутствовал лептин, но был встроен трансген FGF19, отличались потерей веса, более низкими уровнями холестерина и триглицеридов, и у них не развивался диабет. При инъецировании rhFGF19 мышам с ожирением и диабетом, у которых отсутствовал лептин, у них проявлялась обратимость характеристических свойств метаболизма в виде потери веса и понижения глюкозы в крови (Fu, L. et al., Endocrinology 145(6), 2594-2603 (2004); Tomlinson, E. et al., Endocrinology 143(5), 1741-1747 (2002)).
Другой член подсемейства FGF19, полипептид FGF21, экспрессируется главным образом печенью и вызывает эффекты метаболизма, сходные с таковыми FGF19, такие как повышение метаболизма через его воздействие на жировую ткань, потеря веса, понижение уровней глюкозы в крови и резистентность к ожирению и диабету (Kharitonenkov, A. et al., J. Clin. Invest. 115(6), 1627-1635 (2005)). FGF21-трансгенные мыши также были резистентны к обусловленному диетой ожирению. Более того, у грызунов-моделей диабета введение FGF21 понижало уровни глюкозы и триглицеридов в крови.
Было также показано, что FGF21 участвует в регуляции пути обмена гормона роста (GH). Анаболические эффекты GH опосредованы инсулиноподобным фактором роста 1 (IGF-1), который главным образом продуцируется печенью. GH вызывает транскрипцию IGF-1, таким образом повышая его уровни в циркуляции через активацию Janus-киназы 2 (JAK2) посредством рецептора GH. Активированная JAK2 фосфорилирует члены семейства сигнальных трансдукторов и активаторов транскрипции (STAT), которые при фосфорилировании подвергаются транслокации в ядро и связываются с регуляторными элементами генов-мишеней, включая гены IGF-1. В частности, было показано, что STAT5 в его фосфорилированной форме играет заметную роль в таком ответе.
Влияние GH на уровни IGF-1, вероятнее всего, противостоит эффектам голода или ограничения пищи - состояниям, которые приводят к понижению уровней транскрипции IGF-1 и циркуляции IGF-1 (Thissen, J.P. et al., Endocr. Rev. 15, 80-101 (1994)). Указанные эффекты в отношении IGF-1 могут иметь место за счет понижения уровней фосфорилированного STAT5. В частности, у голодающих крыс, которым инъецируют GH, уровни печеночного фосфорилированного STAT5 ниже, чем у не голодающих крыс (Beauloye, V. et al., Endocrinology 143, 792-800 (2002)). FGF21, который индуцируется в печени в условиях голодания или ограничения пищи, может быть посредником указанного эффекта. Было показано, что FGF21-трансгенные мыши имеют более низкие уровни IGF-1 и фосфорилированного STAT5 (Inagaki, T. et al., Cell Metabolism 8, 77-83 (2008)).
Метаболические эффекты FGF19 и FGF21 осуществляются через их связывание с FGFR1c-, FGFR2c- и FGFR3c-рецепторами, из которых связывание с FGFR1c-, FGFR2c-рецепторами является наиболее значимым. Более того, связывание FGF19 и FGF21 с указанными рецепторами требует ко-рецептора Klotho-бета. Несмотря на превалирование указанных FGFR-рецепторов, метаболические эффекты FGF19 и FGF21 являются адипоцит-специфическими, в силу необходимости участия ко-рецептора Klotho-бета, который имеет тканеспецифическую локализацию.
Было показано также, что FGF19 вызывает эффекты, которые отличаются от эффектов, вызываемых FGF21. Например, было показано, что FGF19 регулирует продукцию желчи печенью через его специфические в отношении печеночной ткани эффекты. В ответ на возникающую после приема пищи продукцию желчи FGF19 осуществляет отрицательную регуляцию продукции желчи путем торможения транскрипции гена холестерин-7-альфа-гидролазы (CYP7A1), фермента, ограничивающего скорость синтеза желчных кислот, и путем стимуляции наполнения желчного пузыря. Кроме того, FGF19 обнаруживает митогенные эффекты, которые не наблюдаются в отношении FGF21. Например, у FGF19-трансгенных мышей развивается аденокарцинома печени в результате повышенной пролиферации и дисплазии гепатоцитов, и у rhFGF19-обработанных мышей проявляется пролиферация гепатоцитов (Nicholes, K. et al., Am J Pathol 160, 2295-2307 (2000).)
Обнаружено, что указанные дополнительные активности FGF19 опосредованы его связыванием с FGFR4. FGF19 может связываться с FGFR4 как Klotho-бета-зависимым, так и Klotho-бета-независимым путем. Несмотря на то, что было показано, что FGF21 может также связываться с FGFR4 Klotho-бета-зависимым путем, никаких четких результатов по передаче сигнала от связывания FGF21 с FGFR4 получено не было.
Необходимо развивать новые терапевтические возможности для лечения связанных с метаболизмом расстройств, таких как диабет, ожирение, высокий уровень сахара в крови и других сходных нарушений. Необходимо также развивать новые терапевтические возможности для таких связанных с метаболизмом расстройств, при которых будут исключены или ослаблены эффекты нежелательного роста или пролиферативного потенциала (например, вызывающего опухоли потенциала) такой терапии. Необходимо также развивать новые терапевтические возможности для таких связанных с метаболизмом расстройств, при которых будет исключен или ослаблен потенциал в отношении резистентности к гормону роста.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В первом аспекте настоящее изобретение относится к химерному полипептиду фактора роста фибробластов человека 19 (hFGF19). В некоторых воплощениях настоящего изобретения последовательность химерного полипептида включает C-концевую часть, которая содержит C-концевую часть последовательности полипептида hFGF19, и N-концевую часть, которая содержит N-концевую часть последовательности полипептида hFGF21. В определенных воплощениях C-концевая часть последовательности полипептида hFGF19 имеет длину от приблизительно 45 до 185 остатков, N-концевая часть последовательности полипептида hFGF21 имеет длину от приблизительно 7 до приблизительно 140 остатков.
В другом воплощении настоящего изобретения предложен химерный полипептид hFGF19, при этом последовательность полипептида включает C-концевую часть, которая содержит C-концевую часть последовательности полипептида hFGF21, и N-концевую часть, которая содержит N-концевую часть последовательности полипептида hFGF19. В некоторых воплощениях C-концевая часть последовательности полипептида hFGF21 имеет длину от приблизительно 8 до приблизительно 145 остатков, а N-концевая часть последовательности полипептида hFGF19 имеет длину от приблизительно 45 до приблизительно 175 остатков.
В другом воплощении настоящего изобретения предложен химерный полипептид hFGF19, где последовательность указанного химерного полипептида включает первую полипептидную последовательность, имеющую хоть какую-нибудь последовательность, идентичную последовательности полипептида hFGF19, и где часть первой полипептидной последовательности замещена частью второй полипептидной последовательности, при этом указанная вторая полипептидная последовательность имеет хоть какую-нибудь последовательность, идентичную последовательности полипептида hFGF21, так, чтобы замещенная часть первой полипептидной последовательности имела длину от приблизительно 3 до приблизительно 185 остатков.
В некоторых воплощениях настоящего изобретения предложен химерный полипептид hFGF19, где последовательность указанного химерного полипептида включает первую полипептидную последовательность, имеющую хоть какую-нибудь последовательность, идентичную последовательности полипептида hFGF19, и где часть первой полипептидной последовательности замещена более чем одной частью второй полипептидной последовательности, при этом указанная вторая полипептидная последовательность имеет хоть какую-нибудь последовательность, идентичную последовательности полипептида hFGF21. В некоторых воплощениях химерный полипептид hFGF19 включает замену петли β1-β2 первого полипептида, замену сегмента β10-β12 первого полипептида и/или замену пяти остатков WGDPI первого полипептида петлей β1-β2 второго полипептида, сегментом β10-β12 второго полипептида и/или соответствующей последовательностью GQV второго полипептида.
В определенных воплощениях настоящего изобретения химерный полипептид hFGF19 включает последовательность
HPIPDSSPLLQFGGQVRQRYLYTSGPHGLSSCFLRIRADGVVDCARGQSAHSLLEIKAVALRTVAIKGVHSVRYLCMGADGKMQGLLQYSEEDCAFEEEIRPDGYNVYRSEKHRLPVSLSSAKQRQLYKNRGFLPLSHFLPMLPMVPEEPEDLRGHLESDMFSSPLETDSMDPFGLVTGLEAVRSPSFEK (SEQ ID NO: 5).
В определенных воплощениях химерный полипептид hFGF19 согласно настоящему изобретению слит со вторым полипептидом, при этом второй полипептид является участком Fc иммуноглобулина, аналогом участка Fc иммуноглобулина и одним или несколькими фрагментами участка Fc иммуноглобулина. В определенных воплощениях иммуноглобулин выбран из группы, состоящей из IgG-1, IgG-2, IgG-3, IgG-4, IgA-1, IgA-2, IgE, IgD и IgM. В некоторых воплощениях участок Fc является человеческим или гуманизированным. В некоторых воплощениях C-конец химерного полипептида hFGF19 слит с N-концом второго полипептида. В некоторых воплощениях C-конец химерного полипептида hFGF19 слит с N-концом второго полипептида посредством линкера, при этом указанный линкер выбран из группы, состоящей из линкера [Gly]n, линкера [Gly3Ser]m и линкера [Gly4Ser]m, где n является целым числом от 1 до 30, а m является целым числом от 1 до 6.
Настоящее изобретение включает в себя химерные полипептиды hFGF19, которые имеют физиологическое время полужизни, которое является, по меньшей мере, или приблизительно таким же, что и у нативного hFGF19. Настоящее изобретение включает в себя химерные полипептиды hFGF19, которые имеют физиологическое время полужизни, которое является, по меньшей мере, или приблизительно таким же, что и у нативного hFGF21.
В определенных воплощениях химерный полипептид hFGF19 не активирует существенным образом FGFR4 ни Klotho-бета-независимым, ни Klotho-бета-зависимым образом. В определенных воплощениях химерный полипептид hFGF19 активирует FGFR1c Klotho-бета-зависимым образом.
В определенных воплощениях химерный полипептид hFGF19, при введении его индивиду, не уменьшает безжировую массу индивида. В определенных воплощениях химерный полипептид hFGF19, при введении его индивиду, не вызывает существенного уменьшения безжировой массы индивида. В определенных воплощениях химерный полипептид hFGF19, при введении его индивиду, не уменьшает плотность костей индивида. В определенных воплощениях химерный полипептид hFGF19, при введении его индивиду, не вызывает существенного уменьшения плотности костей индивида. В определенных воплощениях химерный полипептид hFGF19, при введении его индивиду, не понижает функциональную активность сердца индивида. В определенных воплощениях химерный полипептид hFGF19, при введении его индивиду, по существу, не понижает функциональную активность сердца индивида.
В определенных воплощениях химерный полипептид hFGF19 не понижает или не вызывает существенного понижения количества фосфорилированного полипептида STAT5 in vivo. В определенных воплощениях химерный полипептид hFGF19, при введении его индивиду, не понижает или не вызывает существенного понижения количества фосфорилированного полипептида STAT5 у индивида. В определенных воплощениях, при введении индивиду химерного полипептида hFGF19, количество фосфорилированного полипептида STAT5 у индивида понижается, но при этом указанное количество фосфорилированного полипептида STAT5 превышает количество фосфорилированного полипептида STAT5 при введении нативного hFGF21 указанному индивиду. В определенных воплощениях, при введении химерного полипептида hFGF19 индивиду, количество фосфорилированного полипептида STAT5 составляет от 100% до 5%, от 100% до 10%, от 100% до 20%, от 100% до 30%, от 100% до 40%, от 100% до 50%, от 100% до 60%, от 100% до 70%, от 100% до 80%, от 100% до 90% или от 100% до 95% от количества фосфорилированного полипептида STAT5 у индивида без такого введения. В определенных воплощениях, при введении химерного полипептида hFGF19 индивиду, понижение количества фосфорилированного полипептида STAT5 является меньшим, чем понижение количества фосфорилированного полипептида STAT5 при введении нативного hFGF21. Например, понижение фосфорилированного полипептида STAT5, при введении химерного полипептида hFGF19 индивиду, представляет собой любую из степеней понижения: от 0% до 5%, от 0% до 5%, от 0% до 10%, от 0% до 20%, от 0% до 30%, от 0% до 40%, от 0% до 50%, от 0% до 60%, от 0% до 70%, от 0% до 80%, от 0% до 90% или от 0% до 95% от количества фосфорилированного полипептида STAT5 при введении нативного hFGF21.
В определенных воплощениях химерный полипептид hFGF19 не понижает или не вызывает существенного понижения количества циркулирующего инсулиноподобного фактора роста 1 (IGF-1) in vivo. В определенных воплощениях химерный полипептид hFGF19, при введении его индивиду, не понижает или не вызывает существенного понижения количества циркулирующего IGF-1 у индивида. В определенных воплощениях, при введении индивиду химерного полипептида hFGF19, количество циркулирующего IGF-1 понижается, но при этом указанное количество циркулирующего IGF-1 превышает количество фосфорилированного полипептида STAT5 при введении нативного hFGF21 указанному индивиду. В определенных воплощениях, при введении индивиду химерного полипептида hFGF19, количество циркулирующего IGF-1 составляет от 100% до 5%, от 100% до 10%, от 100% до 20%, от 100% до 30%, от 100% до 40%, от 100% до 50%, от 100% до 60%, от 100% до 70%, от 100% до 80%, от 100% до 90% или от 100% до 95%, от количества циркулирующего IGF-1 у индивида без такого введения. В определенных воплощениях, при введении индивиду химерного полипептида hFGF19, понижение количества циркулирующего IGF-1 является меньшим, чем понижение количества циркулирующего IGF-1 при введении нативного hFGF21. Например, понижение циркулирующего IGF-1, при введении индивиду химерного полипептида hFGF19, составляет от 0% до 5%, от 0% до 10%, от 0% до 20%, от 0% до 30%, от 0% до 40%, от 0% до 50%, от 0% до 60%, от 0% до 70%, от 0% до 80%, от 0% до 90% или от 0% до 95%, от понижения количества циркулирующего IGF-1 при введении нативного hFGF21.
В другом аспекте настоящее изобретение относится к фармацевтической композиции, содержащей терапевтически эффективное количество химерного полипептида hFGF19 согласно настоящему изобретению; и приемлемый фармацевтический носитель.
В другом аспекте настоящее изобретение относится к способам лечения индивида, у которого проявляется одна или более из следующих патологий: ожирение, диабет 1 типа, диабет 2 типа, высокое содержание глюкозы в крови, метаболический синдром, атеросклероз, гиперхолестеринемия, инсульт, остеопороз, остеоартрит, дегенеративное заболевание суставов, мышечная атрофия, саркопения, понижение безжировой массы тела, облысение, морщинистость, повышенная усталость, пониженная выносливость, пониженная функция сердца, дисфункция иммунной системы, злокачественная опухоль, болезнь Паркинсона, старческое слабоумие, болезнь Альцгеймера и пониженная когнитивная функция, где указанный способ включает введение указанному индивиду терапевтически эффективное количество фармацевтической композиции согласно настоящему изобретению.
В другом аспекте настоящее изобретение относится к способу понижения уровней глюкозы в крови индивида, нуждающегося в таком лечении, при этом указанный способ включает введение указанному индивиду терапевтически эффективного количества фармацевтической композиции согласно настоящему изобретению.
В другом аспекте настоящее изобретение относится к изолированной молекуле нуклеиновой кислоты, содержащей молекулу ДНК, имеющую, по меньшей мере, 80%, по меньшей мере, 90%, по меньшей мере, 95% или, по меньшей мере, 99% степень идентичности последовательности молекулы ДНК, кодирующей полипептид, имеющий аминокислотные остатки от приблизительно 1 до приблизительно 190 последовательности SEQ ID NO: 5 или комплементарной ей последовательности.
В другом аспекте настоящее изобретение относится к изолированной молекуле нуклеиновой кислоты, содержащей следующую последовательность:
CACCCCATCCCTGACTCCAGTCCTCTCCTGCAATTCGGGGGCCAAGTCCGGCAGCGGTACCTCTACACCTCCGGCCCCCACGGGCTCTCCAGCTGCTTCCTGCGCATCCGTGCCGACGGCGTCGTGGACTGCGCGCGGGGCCAGAGCGCGCACAGTTTGCTGGAGATCAAGGCAGTCGCTCTGCGGACCGTGGCCATCAAGGGCGTGCACAGCGTGCGGTACCTCTGCATGGGCGCCGACGGCAAGATGCAGGGGCTGCTTCAGTACTCGGAGGAAGACTGTGCTTTCGAGGAGGAGATCCGCCCAGATGGCTACAATGTGTACCGATCCGAGAAGCACCGCCTCCCGGTCTCCCTGAGCAGTGCCAAACAGCGGCAGCTGTACAAGAACAGAGGCTTTCTTCCACTCTCTCATTTCCTGCCCATGCTGCCCATGGTCCCAGAGGAGCCTGAGGACCTCAGGGGCCACTTGGAATCTGACATGTTCTCTTCGCCCCTGGAGACCGACAGCATGGACCCATTTGGGCTTGTCACCGGACTGGAGGCCGTGAGGAGTCCCAGCTTTGAGAAG (SEQ ID NO:7) или ее часть.
В определенных воплощениях изолированная нуклеиновая кислота согласно настоящему изобретению дополнительно содержит последовательность, кодирующую аминокислотные остатки, соответствующие участку Fc иммуноглобулина.
В другом аспекте настоящее изобретение относится к системе экспрессии, содержащей молекулу нуклеиновой кислоты согласно настоящему изобретению. В другом аспекте настоящее изобретение относится к клетке-хозяину, содержащей систему экспрессии или нуклеиновую кислоту согласно настоящему изобретению.
В другом аспекте настоящее изобретение относится к изолированному полипептиду, кодируемому молекулой нуклеиновой кислоты согласно настоящему изобретению.
В другом аспекте настоящее изобретение относится к способу продуцирования изолированного полипептида, включающему культивирование клетки-хозяина согласно изобретению в условиях, подходящих для экспрессии кодируемого полипептида, и извлечение кодируемого полипептида из клеточной культуры. В другом аспекте настоящее изобретение относится к изолированному полипептиду, продуцируемому способом согласно изобретению.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКОГО МАТЕРИАЛА
На ФИГ. 1 приведена аминокислотная последовательность человеческого полипептида FGF19 (SEQ ID NO: 1) и человеческого пре-FGF19-полипептида (SEQ ID NO:3);
На ФИГ. 2 приведена аминокислотная последовательность человеческого полипептида FGF21 (SEQ ID NO:2) и человеческого пре-FGF21-полипептида (SEQ ID NO:4);
На ФИГ. 3 представлены иллюстративные результаты анализа связывания рецептора с использованием химерных полипептидов FGF19 согласно настоящему изобретению;
На ФИГ. 4 представлены иллюстративные результаты анализа связывания рецептора с использованием химерных полипептидов FGF19 согласно настоящему изобретению;
На ФИГ. 5 представлены иллюстративные результаты анализа специфичной для печени экспрессии генов с помощью химерных полипептидов FGF19 согласно настоящему изобретению;
На ФИГ. 6 представлены иллюстративные результаты анализа адипоцит-специфической экспрессии генов с помощью химерных полипептидов FGF19 согласно настоящему изобретению;
На ФИГ. 7 представлены иллюстративные результаты анализа понижения уровней глюкозы в крови с помощью химерных полипептидов FGF19 согласно настоящему изобретению;
На ФИГ. 8 представлены иллюстративные результаты анализа теста толерантности к глюкозе с помощью химерных полипептидов FGF19 согласно настоящему изобретению;
На ФИГ. 9 представлены иллюстративные результаты анализа активности с помощью иммуноглобулиновых Fc слияний химерных полипептидов FGF19 согласно настоящему изобретению; и
На ФИГ. 10-14 представлены иллюстративные результаты анализа рецепторной специфичности с использованием химерных полипептидов FGF19 согласно настоящему изобретению;
На ФИГ. 15 представлены иллюстративные результаты применения химерного полипептида FGF19 согласно настоящему изобретению в отношении уровней фосфорилированного белка Stat5.
На ФИГ. 16 представлены иллюстративные результаты применения химерного полипептида FGF19 согласно настоящему изобретению в отношении суммарной метаболической активности клеток.
На фигуре 17 показано, что FGFR4 необходим для регуляции желчных кислот ("BA", bile acid), но не для повышения толерантности к глюкозе под действием FGF19. На фигуре 17A приведен уровень глюкозы в тесте на толерантность к глюкозе у мышей дикого типа ("WT"), обработанных FGF19 или PBS, и у FGFR4- нокаутных ("KO") мышей. *p<0,05. **p<0,01. Значение p для площади под кривой (AUC) составляло p<0,02 (WT) и p<0,005 (KO). N=6~8. На фигуре 17B представлены различные параметры метаболизма (вес тела (г), отношение веса печени к весу тела (%), инсулин сыворотки (нг/мл), бета-гидроксибутират ("BHB") в сыворотке (мг/л), сывороточный лактат (мг/дл) и сывороточный триглицерид (мг/дл)) у мышей дикого типа, обработанных FGF19 или PBS, и у FGFR4- нокаутных ("KO") мышей, при их умерщвлении на день 7. После 3-часового голодания мышей умерщвляли, и получали сыворотку. N=6~8. На фигуре 17C представлен анализ сывороточного состава BA у мышей WT, обработанных FGF19 или PBS, и у FGFR4- нокаутных мышей. Показаны только основные виды BA. CA: холевая кислота, DCA: дезоксихолевая кислота, MCA: мурихолевая кислота, T-: конъюгированные с таурином. На фигуре 17D показана относительная экспрессия различных печеночных генов (Egr-1, c-Fos, альфа-фетопротеина, Cyp7a1, Cyp8b, Cyp27a1, Cyp7b и GK) у мышей WT, обработанных FGF19 или PBS, и у мышей FGFR4-KO, определяемая методом количественной ПЦР (qPCR) в режиме реального времени. N=6~8. Значениями p для фигур 17B-17D являются: <0,05, **<0,005 (сравнение PBS и FGF19), #<0,05, ##<0,005 (сравнение WT и FGFR4-KO).
На фигуре 18 представлена идентификация вариантов FGF19 с пониженной активностью FGFR4. На фигуре 18A представлена относительная активность люциферазы светлячка, нормированная к люциферазной активности renilla (показана как относительная единица люциферазной активности ("RLU")), из GAL-Elk1-анализа люциферазы, с помощью крысиных клеток L6, трансфицированных KLB [полипептид Klotho-бета] и FGFR1c или FGFR4 и инкубируемых со средой, содержащей повышенные концентрации FGF19(O) или FGF21(▲). На фигуре 18B приведено графическое представление (в масштабе) FGF19 (вверху), FGF21 (внизу) и различных химерных белков с аминокислотным составом слева. На основе результатов GAL-Elk-1-анализов, приведенных на Фиг. 18C, каждую химеру классифицировали в класс (I), (II) или (III), как показано справа. Химеры, которые при использовании кондиционированной среды не проявляли активность FGFR1c, эквивалентную FGF21 или FGF19, здесь не показаны. На фигуре 18C показана активация FGFR1c или FGFR4 в GAL-Elk-1-анализе, при использовании крысиных клеток L6, ко-трансфицированных KLB и/или FGFR (FGFR1c или FGFR4) и инкубируемых в присутствии кондиционированной среды из-под клеток 293, транзитно трансфицированных различными конструкциями FGF (см. фигуру 18B с приведенными на ней аминокислотными составами используемых конструкций FGF). Результаты представлены в виде кратности индукции по отношению к контрольной среде, кондиционированной ложно-трансфицированными клетками. На фигуре 18D представлена кратность индукции для случая активации FGFR в GAL-Elk-1-анализе люциферазы с использованием крысиных клеток L6, трансфицированных FGFR1c, FGFR4 + KLB, или FGFR4 и инкубируемых со средой, содержащей повышенные концентрации очищенного FGF19(O) или FGF19v(▼) (конструкция #4 на фигурах 18B и 18C). На фигуре 18E представлены результаты твердофазного анализа связывания FGF19 и FGF19v с FGFR4, слитым с фрагментом Fc. Схематическая диаграмма указанных экспериментов приведена справа. На фигуре 18F показано, что анти-FGF19-антитело, используемое на Фиг. 18E, распознавало FGF19 и FGF19v с неразличимой аффинностью (контрольный эксперимент ELISA). Схематическая диаграмма указанных экспериментов приведена справа.
На фигуре 19 представлены RLU в GAL-Elk-1-анализе люциферазы в крысиных клетках L6, трансфицированных KLB и FGFR2c или FGFR3c и инкубируемых в присутствии кондиционированной среды, содержащей повышенные концентрации FGF19(O) или FGF21(▲). Клетки L6 были ко-трансфицированы экспрессирующими векторами для KLB и указанного FGFR вместе с GAL-Elk-1, SV40-люциферазой Renilla и Gal-чувствительным люциферазным репортером. Трансфицированные клетки инкубировали со средой, содержащей повышенные концентрации FGF19 или FGF21, в течение 6 часов перед анализом люциферазной активности. Активацию транскрипции устанавливали по относительной люциферазной активности, нормированной к люциферазной активности Renilla, и выражали в относительных единицах люциферазной активности (RLU). На данной фигуре показано, что FGF21 и FGF19 активируют FGFR2c и FGFR3c в присутствии KLB.
На фигуре 20 представлена активация FGFR1c или FGFR4 в GAL-Elk-1-анализе с помощью клеток L6, ко-трансфицированных KLB и/или FGFR (FGFR1c или FGFR4) и инкубируемых в присутствии кондиционированной среды из-под клеток 293, транзитно трансфицированных различными конструкциями FGF, показанными на оси X (см. фигуру 18B с приведенными на ней аминокислотными составами используемых конструкций FGF). Результаты представлены в виде кратности индукции по отношению к контрольной среде, кондиционированной ложно-трансфицированными клетками. Числовые значения, указанные на оси X, соответствуют нумерации конструкций, представленных на Фиг. 18B.
На фигуре 21 показаны эффекты FGF19v у кормленых ужином мышей. На фигуре 21A показана относительная экспрессия разных генов (c-Fos, Egr-1, GK, БТШ и Cyp7a1) у мышей FVB, которым (через хвостовую вену) инъецировали FGF21, FGF19 или FGF19v в количестве 1 мг/кг, или PBS в контроле; значения p: *<0,05, **<0,01, ***<0,001 (по сравнению с PBS). Через 4 часа после инъекции из каждой мыши выделяли печеночную мРНК и подвергали анализу qPCR в режиме реального времени в отношении указанных генов. На фигуре 21B показана относительная экспрессия разных генов (c-Fos, Egr-1, GK, SHP (БТШ) и Cyp7a1) у мышей WT или FGFR4 KO (KO) (N=5-7), которым в/б инъецировали FGF21 или FGF19 в количестве 1 мг/кг, или PBS в контроле. Голодавшим течение ночи мышам в/б инъецировали белок FGF, или PBS в контроле. Через 4 часа после инъекции из каждой мыши выделяли печеночную мРНК и подвергали анализу qPCR в режиме реального времени в отношении указанных генов; значения p: *<0,05, **<0,01, ***<0,001. На фигуре 21C представлена пролиферация клеток HepG2, обработанных FGF21, FGF19 или FGF19v при различных концентрациях (10 нг/мл, 60 нг/мл и 200 нг/мл), измеряемых по интенсивности флуоресценции (× 105) в анализе «безъякорного» клеточного роста, то есть не зависящего от прикрепления клеток. Пролиферацию клеток HepG2 в мягком агаре оценивали на основе превращения резазурина (Alamer Blue), нефлуоресцентного индикаторного красителя, в резоруфин. На фигуре 21D показана кратность изменения BrdU+ гепатоцитов у мышей FVB, которым инфузировали FGF19 или FGF19v. N=6, *p<0,01, ***p<5E-5 (по сравнению с PBS), ##p<0,0002 (по сравнению с FGF19). Мышам FVB имплантировали осмотический насос для реализации непрерывной инфузии указанного белка FGF в количестве 1 нг/час (~0,8 мг/кг/день) (день 0). Мыши получали также ежедневную (q.d.) инъекцию белка FGF 1 мг/кг/день и 30 мг/кг/день BrdU (два раза в день, b.i.d.), начиная со дня 1. На день 7 печени вырезали и осуществляли их анти-BrdU-окрашивание. Результаты представлены в виде кратности индукции против ложно-обработанных животных по числу BrdU-положительных гепатоцитов на анализируемой площади. На фигуре 21E приведены репрезентативные изображения при исследованиях, представленных на фигуре 21C. На фигуре 21F показана относительная экспрессия разных генов ((c-Fos, Egr-1, альфа-фетопротеина, GK, Cyp7a1 и Cyp8b) у мышей, используемых на фигурах 21D и 21E. N=6. *p<0,05, **p<0,005, ***p<0,001 (по сравнению с PBS), #p<0,05, ##p<0,005 (по сравнению с FGF19).
На фигуре 22 показано, что FGF19v и FGF21 оказывают сходные метаболические эффекты и облегчают гипергликемию у мышей ob/ob. 11-недельным мышам ob/ob подкожно имплантировали осмотический насос для реализации непрерывной инфузии 1 нг/час указанного белка FGF (0,4 мг/кг/день) или PBS в контроле (N=7). На фигуре 22A приведены вес тела (г) и глюкоза крови (мг/дл) у мышей ob/ob, которым через осмотический насос инфузировали 1 нг/час FGF21 или FGF19v (0,4 мг/кг/день) или PBS в контроле (N=7). Осмотический насос имплантировали на день 0. На фигуре 22B приведены уровни глюкозы в крови (мг/дл) у мышей ob/ob, которым инфузировали FGF21, FGF19v или PBS в контроле, в случае беспорядочного питания и после того как в течение ночи мыши голодали. На фигуре 22C приведены уровни неэстерифицированных жирных кислот ("NEFA") в сыворотке у мышей ob/ob, которым инфузировали FGF21, FGF19v или PBS в контроле, на день 8. На фигуре 22D представлены результаты теста толерантности к глюкозе, проводимого у мышей ob/ob, которым инфузировали FGF21, FGF19v или PBS в контроле, на день 6. Мыши в течение ночи голодали, и им вводили в/б инъекцию глюкозы в количестве 1 г/кг во время t=0. На фигуре 22E представлено отношение веса органа к весу тела (%) у мышей ob/ob, которым инфузировали FGF21, FGF19v или PBS в контроле, на день 8. На фигуре 22F представлена относительная экспрессия различных генов (альфа-фетопротеина, IGFBP2, SCD-1, Cyp7A, Cyp8B, UCP-1, MCAD и SREBP-1c) из разных тканей (печень, бурая жировая ткань ("BAT") и белая жировая ткань ("WAT")) у мышей ob/ob, которым инфузировали FGF21, FGF19v или PBS в контроле, определяемая методом qPCR. Значения p: *<0,05, **<0,005, ***<0,0005 (по сравнению с PBS в контроле).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новым химерным полипептидам FGF19. В некоторых воплощениях настоящего изобретения полипептидная последовательность химерного FGF19 включает часть последовательности полипептида FGF19 и часть последовательности полипептида FGF21. В определенных предпочтительных воплощениях полипептид FGF19 является видоизмененным человеческим полипептидом FGF19 (hFGF19), последовательность которого определяется последовательностью SEQ ID NO: 1. В определенных предпочтительных воплощениях полипептид FGF21 является видоизмененным человеческим полипептидом FGF21 (hFGF21), последовательность которого определяется последовательностью SEQ ID NO: 2. В другом аспекте настоящее изобретение относится к новым химерным полипептидам FGF19, которые дополнительно слиты с иммуноглобулиновым доменом, таким как домен Fc.
В другом аспекте настоящее изобретение относится к новым химерным полипептидам FGF19, которые имеют измененную рецепторную специфичность. В определенных предпочтительных воплощениях химерный полипептид FGF19 согласно настоящему изобретению не активирует существенным образом FGFR4 ни Klotho-бета-зависимым, ни Klotho-бета-независимым образом. В определенных воплощениях химерный полипептид hFGF19 активирует FGFR1c Klotho-бета-зависимым образом. В определенных воплощениях химерный полипептид FGF19 согласно настоящему изобретению активирует, по меньшей мере, один из рецепторов FGFR1c, FGFR2c или FGFR3c Klotho-бета-зависимым образом.
В некоторых воплощениях химерный полипептид FGF19 согласно настоящему изобретению может обладать одним или несколькими из следующих предпочтительных признаков: указанный полипептид, при его введении, по существу не индуцирует пролиферацию гепатоцитов у индивида, указанный полипептид, при его введении, по существу не индуцирует резистентность к гормону роста у индивида, указанный полипептид не содержит остатка, который является полиморфным в популяции, указанный полипептид имеет физиологическое время полужизни in vivo, которое является, по меньшей мере, или приблизительно таким же, что и у нативного полипептида FGF19 (такого как нативный полипептид hFGF19), указанный полипептид имеет физиологическое время полужизни in vivo, которое является, по меньшей мере, или приблизительно таким же, что и у нативного полипептида FGF21 (такого как нативный полипептид hFGF21), указанный полипептид, при его введении, по существу не способствует понижению безжировой массы у индивида, указанный полипептид, при его введении, по существу не способствует уменьшению плотности костей у индивида, и указанный полипептид, при его введении, по существу не понижает функциональную активность сердца у индивида.
Дополнительные предпочтительные признаки химерного полипептида FGF19 согласно настоящему изобретению могут включать в себя одно или более из следующих свойств: указанный полипептид, в случае необходимости такого лечения, понижает уровень глюкозы в крови индивида; указанный полипептид активирует, по меньшей мере, один из рецепторов, FGFR1c, FGFR2c или FGFR3c, Klotho-бета-зависимым образом; указанный полипептид не активирует существенным образом FGFR4 Klotho-бета-зависимым образом; указанный полипептид, при введении его индивиду, не активирует существенным образом FGFR4 Klotho-бета-независимым образом; указанный полипептид не понижает или по существу не понижает количество фосфорилированного полипептида STAT5; при введении указанного полипептида индивиду, количество фосфорилированного полипептида STAT5 у индивида понижается, но при этом указанное количество фосфорилированного полипептида STAT5 превышает количество фосфорилированного полипептида STAT5 при введении нативного hFGF21 указанному индивиду; при введении указанного полипептида вводят индивиду, указанное понижение количества фосфорилированного полипептида STAT5 у индивида является меньшим, чем понижение количества фосфорилированного полипептида STAT5 при введении нативного hFGF21 указанному индивиду; указанный полипептид, при введении его индивиду, не понижает или по существу не понижает количество циркулирующего полипептида IGF-1; при введении указанного полипептида индивиду, количество IGF-1 у индивида понижается, но при этом количество циркулирующего полипептида IGF-1 превышает количество циркулирующего IGF-1 при введении нативного hFGF21 указанному индивиду; при введении указанного полипептида индивиду, указанное понижение количества циркулирующего IGF-1 у индивида является меньшим, чем понижение количества циркулирующего IGF-1 при введении нативного hFGF21 указанному индивиду; и указанный полипептид, при введении его индивиду, не понижает или по существу не понижает количество циркулирующего полипептида IGF-1 у индивида, имеющего нормальное количество или выше нормального количества GH.
В определенных воплощениях химерный полипептид FGF19 согласно изобретению обладает преимущественными свойствами активации, по меньшей мере, одного из рецепторов FGFR1c, FGFR2c или FGFR3c Klotho-бета-зависимым образом, и не активирует существенным образом FGFR4 ни Klotho-бета-зависимым, ни Klotho-бета-независимым образом. В определенных воплощениях химерный полипептид FGF19 согласно изобретению обладает преимущественными свойствами в отношении активации, по меньшей мере, одного из рецепторов FGFR1c, FGFR2c или FGFR3c Klotho-бета-зависимым образом, не активирует существенным образом FGFR4 ни Klotho-бета-зависимым, ни Klotho-бета-независимым образом, и понижает количество фосфорилированного полипептида STAT5 у индивида, но при этом указанное количество фосфорилированного полипептида STAT5 превышает количество фосфорилированного полипептида STAT5 при введении указанному индивиду нативного hFGF21. В определенных воплощениях химерный полипептид FGF19 согласно изобретению обладает преимущественными свойствами в отношении активации, по меньшей мере, одного из рецепторов FGFR1c, FGFR2c или FGFR3c Klotho-бета-зависимым образом и не активирует существенным образом FGFR4 ни Klotho-бета-зависимым, ни Klotho-бета-независимым образом, а также не содержит остаток, который является полиморфным в данной популяции. В определенных воплощениях химерный полипептид FGF19 согласно изобретению обладает преимущественными свойствами в отношении активации, по меньшей мере, одного из рецепторов FGFR1c, FGFR2c или FGFR3c Klotho-бета-зависимым образом, не активирует существенным образом FGFR4 ни Klotho-бета-зависимым, ни Klotho-бета-независимым образом, понижает у индивида количество фосфорилированного полипептида STAT5, но при этом указанное количество фосфорилированного полипептида STAT5 превышает количество фосфорилированного полипептида STAT5 при введении указанному индивиду нативного hFGF21, а также не содержит остаток, который является полиморфным в данной популяции.
В другом аспекте настоящее изобретение относится к новому химерному полипептиду FGF19, который имеет измененную рецепторную специфичность. В определенных предпочтительных воплощениях химерный полипептид FGF19 согласно изобретению не активирует существенным образом FGFR4 ни Klotho-бета-зависимым, ни Klotho-бета-независимым образом. В определенных воплощениях химерный полипептид hFGF19 согласно изобретению активирует, по меньшей мере, один из рецепторов FGFR1c, FGFR2c или FGFR3c Klotho-бета-зависимым образом.
В другом аспекте химерный полипептид FGF19 не влияет на резистентность к гормону роста у индивида по сравнению с резистентностью к гормону роста, вызываемой нативным FGF21. В другом аспекте химерный полипептид FGF19 по существу не влияет на резистентность к гормону роста у индивида по сравнению с резистентностью к гормону роста, вызываемой нативным FGF21. В определенных воплощениях химерный полипептид FGF19 не понижает или по существу не понижает количество фосфорилированного полипептида STAT5 у индивида. В определенных воплощениях, при введении химерного полипептида hFGF19 индивиду, количество фосфорилированного полипептида STAT5 у индивида понижается, но при этом указанное количество фосфорилированного полипептида STAT5 превышает количество фосфорилированного полипептида STAT5 при введении нативного hFGF21 указанному индивиду. В определенных воплощениях, при введении химерного полипептида hFGF19 индивиду, понижение количества фосфорилированного полипептида STAT5 является меньшим, чем понижение количества фосфорилированного полипептида STAT5 при введении указанному индивиду нативного hFGF21. В определенных воплощениях химерный полипептид FGF19 не понижает или по существу не понижает количество циркулирующего инсулиноподобного фактора роста 1 (IGF-1). В определенных воплощениях, при введении химерного полипептида hFGF19 индивиду, количество циркулирующего IGF-1 понижается, но при этом количество циркулирующего IGF-1 превышает количество циркулирующего IGF-1 при введении указанному индивиду нативного hFGF21. В определенных воплощениях, при введении химерного полипептида hFGF19 индивиду, понижение количества циркулирующего IGF-1 является меньшим, чем понижение количества циркулирующего IGF-1 при введении нативного hFGF21.
В другом аспекте настоящее изобретение относится к новым изолированным молекулам нуклеиновой кислоты, имеющим последовательность, которая кодирует химерный полипептид FGF19 согласно настоящему изобретению, к новым системам экспрессии, которые включают в себя молекулу нуклеиновой кислоты согласно настоящему изобретению, и к клеткам-хозяевам, которые включают в себя молекулу нуклеиновой кислоты согласно изобретению или систему экспрессии согласно изобретению.
В другом аспекте настоящее изобретение включает в себя антитела, которые могут специфически связываться с химерным полипептидом FGF19 согласно настоящему изобретению.
В другом аспекте настоящее изобретение относится к фармацевтическим композициям, которые включают в себя химерный полипептид FGF19 согласно настоящему изобретению и фармацевтически приемлемый носитель.
В другом аспекте настоящее изобретение относится к способам лечения связанного с метаболизмом расстройства у индивида путем введения химерного полипептида FGF19 согласно настоящему изобретению или содержащей его подходящей фармацевтической композиции. В другом аспекте настоящее изобретение относится к способам осуществления, по меньшей мере, одного или более из следующих эффектов у индивида: понижения уровня глюкозы в крови, понижения степени ожирения, увеличения скорости метаболизма, усиления окисления липидов, снижения веса, понижения уровней глюкозы, лептина, инсулина, холестерина и/или триглицеридов в сыворотке крови, излечения диабета и других эффектов в отношении метаболизма, при этом такие эффекты являются результатом введения указанному индивиду терапевтического количества химерного полипептида FGF19 согласно изобретению или содержащей его фармацевтической композиции.
I. Химерные полипептиды FGF19 с N-концевыми последовательностями полипептида FGF21
В одном из аспектов настоящего изобретения полипептидная последовательность химерного FGF19 включает C-концевую часть и N-концевую часть. При этом N-концевая часть полипептидной последовательности химерного FGF19 включает N-концевую часть последовательности полипептида FGF21, а C-концевая часть полипептидной последовательности химерного FGF19 включает C-концевую часть последовательности полипептида FGF19. В некоторых воплощениях вышеизложенного C-концевая часть последовательности полипептида FGF19 и N-концевая часть полипептидной последовательности химерного FGF21 соединены встык, образуя смежные последовательности. В некоторых воплощениях вышеизложенного C-концевая часть последовательности полипептида FGF19 и N-концевая часть полипептидной последовательности химерного FGF21 соединены, образуя смежные последовательности, путем перекрывания в совокупности 1, 2, 3 или более остатков между указанными двумя частями. В некоторых альтернативных воплощениях C-концевая часть последовательности полипептида FGF19 и N-концевая часть полипептидной последовательности химерного FGF21 имеют встроенный между ними спейсер из 1, 2, 3, 4, 5 или более аминокислотных остатков.
В определенных предпочтительных воплощениях полипептид FGF19 является человеческим полипептидом FGF19 (hFGF19), последовательность которого определяется последовательностью SEQ ID NO: 1 (Фиг. 1). В некоторых воплощениях C-концевая часть последовательности полипептида hFGF19 имеет длину от приблизительно 45 до приблизительно 185 остатков, однозначно включая участки длины последовательностей С-концевых частей hFGF19, представленных в таблице 1. В некоторых воплощениях C-концевая часть полипептидной последовательности химерного FGF19 включает C-концевую часть полипептидной последовательности, которая, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97% или, по меньшей мере, на 99% идентична полипептидной последовательности hFGF19. В некоторых воплощениях полипептид FGF19 является прошедшим стадию пре-процессинга человеческим полипептидом FGF19 (hFGF19), который включает в себя свою сигнальную последовательность и последовательность которого определяется последовательностью SEQ ID NO: 3 (Фиг. 1).
В определенных предпочтительных воплощениях полипептид FGF21 является человеческим полипептидом FGF21 (hFGF21), последовательность которого определяется последовательностью SEQ ID NO:2 (Фиг. 2). В некоторых воплощениях N-концевая часть последовательности полипептида hFGF21 имеет длину от приблизительно 7 до приблизительно 140 остатков, однозначно включая участки последовательности N-концевых частей hFGF19, представленных в таблице 2. В некоторых воплощениях N-концевая часть полипептидной последовательности химерного FGF19 включает N-концевую часть полипептидной последовательности, аминокислотная последовательность которой, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97% или, по меньшей мере, на 99% идентична полипептидной последовательности hFGF21. В некоторых воплощениях полипептид FGF21 является прошедшим стадию пре-процессинга человеческим полипептидом FGF21 (hFGF21), который включает свою сигнальную последовательность, и последовательность которого определяется последовательностью SEQ ID NO:4 (Фиг. 2).
Здесь C-концевая часть, N-концевая часть, замещенная часть или замещенная часть полипептидной последовательности, такой как часть полипептидный последовательности hFGF19 или hFGF21, имеет первое положение и последнее положение. Указанные положения соответствуют положениям в полипептидной последовательности, с которых начинается отсчет части, о которой идет речь. Таким образом, последовательность определенной части является непрерывной последовательностью аминокислот, которая начинается с положения или примерно с такого положения в полипептидной последовательности, которое соответствует первому положению, а кончается в положении или примерно в таком положении в полипептидной последовательности, которое соответствует последнему положению. В некоторых воплощениях последнее положение C-концевой части полипептида соответствует последнему или примерно последнему остатку полипептида. В некоторых воплощениях первое положение N-концевой части полипептида соответствует первому или примерно первому остатку полипептида.
Примеры C-концевых частей последовательности полипептида hFGF19, о которых идет речь в настоящем изобретении, включают в себя, не ограничиваясь ими, такие части, которые имеют первое положение, которое соответствует положению или является приблизительно одним из положений 10, 11, 25, 26, 27, 28, 30, 33, 35, 37, 40, 41, 42, 43, 44, 45, 52, 53, 54, 56, 57, 58, 59, 72, 73, 74, 79, 80, 81, 143, 144, 145 или 146 последовательности SEQ ID NO: 1. Каждая типичная C-концевая часть также имеет последнее положение, которое соответствует положению или находится около положения 194 последовательности SEQ ID NO: 1. В таблице 1 показаны полипептидные последовательности иллюстративных C-концевых частей полипептида hFGF19. Здесь рассматриваются также и другие аналогичные части полипептида hFGF19.
Иллюстративные C-концевые части последовательности полипептида hFGF19
Примеры N-концевых частей полипептидной последовательности hFGF21, о которых идет речь в настоящем изобретении, включают, без ограничения, такие части, которые имеют последнее положение, соответствующее или приблизительно соответствующее любому из положений 7, 8, 20, 21, 22, 23, 25, 27, 29, 31, 34, 35, 36, 37, 38, 39, 46, 47, 48, 50, 51, 52, 53, 66, 67, 68, 73, 74, 75, 135, 136, 137 и 138 SEQ ID NO: 2. Каждая иллюстративная N-концевая часть имеет также первое положение, соответствующее или приблизительно соответствующее положению 1 последовательности SEQ ID NO: 2.
В таблице 2 приведен список иллюстративных N-концевых положений полипептида hFGF21. Здесь рассматриваются также и другие аналогичные части полипептида hFGF21.
Иллюстративные N-концевые части последовательности полипептида hFGF21
Подразумевается, что C-концевая часть, N-концевая часть или любая другая часть определяемой здесь полипептидной последовательности может независимо и необязательно включать в себя от одного до пяти или более дополнительных остатков в определенном первом или последнем положении. Например, C-концевая часть полипептида, имеющего на месте первого положения или приблизительно первого положения остаток 100, может, независимо, (i) необязательно включать в себя 1, 2, 3, 4, 5 или более дополнительных остатков, N-концевых по отношению к остатку в положении 100, (ii) необязательно включать в себя 1, 2, 3, 4, 5 или более дополнительных остатков, C-концевых по отношению к последнему остатку, (iii) необязательно начинаться в положении 1, 2, 3, 4, 5 или более остатков, C-концевых по отношению к остатку в положении 100, или (iv) необязательно кончаться в положении 1, 2, 3, 4, 5 или более остатков, N-концевых по отношению к последнему остатку. Один или более дополнительных остатков, если они имеются, могут быть, а могут и не быть теми же самыми, что и остатки в соответствующем положении в данном полипептиде.
В некоторых воплощениях химерного полипептида FGF19 согласно настоящему изобретению N-концевая часть его последовательности включает в себя последовательность, которая выбрана из N-концевых частей последовательности полипептида hFGF21, перечисленных в таблице 2, а C-концевая часть его последовательности включает последовательность, которая выбрана из C-концевых частей последовательности полипептида hFGF19, перечисленных в таблице 1. В некоторых воплощениях выбранная N-концевая часть hFGF21 и выбранная C-концевая часть hFGF19 выбраны независимо по отношению друг к другу. В некоторых воплощениях часть N-концевой последовательности hFGF21 и часть С-концевой последовательности hFGF19 выбраны таким образом, чтобы С-конец части N-концевой последовательности и N-конец части C-концевой последовательности имели в совокупности, по меньшей мере, 1, по меньшей мере, 2 или, по меньшей мере, 3 или более остатков. В некоторых воплощениях последовательность химерного полипептида FGF19 содержит последовательность, в которой C-концевая часть последовательности полипептида FGF19 и N-концевая часть последовательности химерного полипептида FGF21 соединены встык, образуя смежные последовательности, без встроенных между ними аминокислот. В некоторых воплощениях вышеизложенного последовательность химерного полипептида FGF19 содержит последовательность, в которой C-концевая часть полипептидной последовательности FGF19 и N-концевая часть химерной полипептидной последовательности FGF21 соединены последовательно, с перекрыванием 1, 2, 3 или более остатков в совокупности между двумя указанными частями. В некоторых альтернативных воплощениях последовательность химерного полипептида FGF19 содержит последовательность, которая включает C-концевую часть последовательности полипептида FGF19 и N-концевую часть последовательности химерного полипептида FGF21, и дополнительно включает встроенный между ними спейсер из 1, 2, 3, 4, 5 или более аминокислотных остатков.
Иллюстративные последовательности химерных полипептидов FGF19 согласно настоящему изобретению представлены в таблице 3, где их N-концевая часть включает N-концевую часть последовательности полипептида hFGF21, а их C-концевая часть включает C-концевую часть последовательности полипептида hFGF19.
Иллюстративные полипептидные последовательности химерных FGF19
GVHSVRYLCMGADGKMQGLLQYSEEDCAFEEEIRPDGYNVYRSEKHRLPVSLSSAKQRQLYKNRGFLPLSHFLPMLPMVPEEPEDLRGHLESDMFSSPL ETDSMDPFGLVTGLEAVRSPSFEK
II. Химерные полипептиды FGF19 с C-концевыми последовательностями полипептида FGF21
Во втором аспекте настоящего изобретения полипептидная последовательность химерного FGF19 включает C-концевую часть и N-концевую часть. N-концевая часть полипептидной последовательности химерного FGF19 включает N-концевую часть полипептидной последовательности FGF19, а C-концевая часть полипептидной последовательности химерного FGF19 включает C-концевую часть полипептидной последовательности FGF21. В некоторых воплощениях вышеизложенного C-концевая часть полипептидной последовательности FGF21 и N-концевая часть полипептидной последовательности химерного FGF19 соединены последовательно, встык. В некоторых воплощениях вышеизложенного C-концевая часть полипептидной последовательности FGF21 и N-концевая часть полипептидной последовательности химерного FGF19 соединены последовательно, с перекрыванием 1, 2, 3 или более остатков в совокупности между указанными двумя частями. В некоторых альтернативных воплощениях C-концевая часть полипептидной последовательности FGF21 и N-концевая часть полипептидной последовательности химерного FGF19 имеют встроенный между ними спейсер из 1, 2, 3, 4, 5 или более аминокислотных остатков.
В определенных предпочтительных воплощениях полипептид FGF19 является человеческим полипептидом FGF19 (hFGF19), последовательность которого определяется последовательностью SEQ ID NO: 1. В некоторых воплощениях N-концевая часть последовательности полипептида hFGF19 имеет длину от приблизительно 45 до приблизительно 175 остатков, однозначно включая участки последовательности N-концевых частей hFGF19, представленных в таблице 4. В некоторых воплощениях N-концевая часть полипептидной последовательности химерного FGF19 включает N-концевую часть полипептидной последовательности, аминокислотная последовательность которого, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97% или по меньшей мере, на 99% идентична последовательности полипептида hFGF19.
В определенных предпочтительных воплощениях полипептид FGF21 является человеческим полипептидом FGF21 (hFGF21), последовательность которого определяется последовательностью SEQ ID NO: 2. В некоторых воплощениях С-концевая часть последовательности полипептида hFGF21 имеет длину от приблизительно 8 до приблизительно 145 остатков, однозначно включая участки последовательности С-концевых частей hFGF19, представленных в таблице 5. В некоторых воплощениях С-концевая часть полипептидной последовательности химерного FGF21 включает С-концевую часть полипептидной последовательности, которая, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97% или, по меньшей мере, на 99% идентична полипептидной последовательности полипептида hFGF21.
Примеры N-концевых частей последовательности полипептида hFGF19, о которых идет речь в настоящем изобретении, включают, но не ограничены перечисленным, такие которые имеют последнее положение, соответствующее или приблизительно соответствующее любому из положений 9, 10, 24, 25, 26, 27, 39, 40, 41, 42, 43, 44, 51, 52, 53, 55, 56, 57, 58, 71, 72, 73, 78, 79, 80, 142, 143, 144 и 145 в SEQ ID NO: 1. Каждая иллюстративная N-концевая часть имеет также последнее положение, соответствующее или приблизительно соответствующее положению 1 в SEQ ID NO: 1. В таблице 4 представлены полипептидные последовательности иллюстративных N-концевых частей полипептида hFGF19. Здесь рассматриваются также и другие аналогичные части полипептида hFGF19.
Иллюстративные N-концевые части последовательности полипептида hFGF19
Примеры C-концевых частей последовательности полипептида hFGF21, о которых идет речь в настоящем изобретении, включают в себя, без ограничения, такие части, которые имеют последнее положение, которое соответствует или приблизительно соответствует любому из положений 8, 9, 21, 22, 23, 24, 35, 36, 37, 38, 39, 40, 47, 48, 49, 51, 52, 53, 54, 67, 68, 69, 146, 147, 148 и 149 последовательности SEQ ID NO: 2. Каждая иллюстративная C-концевая часть имеет также последнее положение, которое соответствует или приблизительно соответствует положению 181 последовательности SEQ ID NO: 2. В таблице 5 представлен перечень иллюстративных С-концевых частей полипептида hFGF21. Здесь рассматриваются также и другие аналогичные части полипептида hFGF21.
Иллюстративные С-концевые части последовательности полипептида hFGF21
В некоторых воплощениях химерного полипептида FGF19 согласно настоящему изобретению N-концевая часть его последовательности включает последовательность, которая выбрана из частей последовательности полипептида hFGF19, перечисленных в таблице 4, а C-концевая часть его последовательности включает последовательность, которая выбрана из частей последовательности полипептида hFGF21, перечисленных в таблице 5. В некоторых воплощениях выбранная N-концевая часть hFGF19 и выбранная C-концевая часть hFGF21 выбраны независимо по отношению друг к другу. В некоторых воплощениях часть N-концевой последовательности hFGF19 и часть C-концевой последовательности hFGF21 выбраны таким образом, чтобы C-конец части N-концевой последовательности и N-конец части C-концевой последовательности имели в совокупности, по меньшей мере, 1, по меньшей мере, 2, или, по меньшей мере, 3 или более остатков. В некоторых воплощениях последовательность химерного полипептида FGF19 содержит последовательность, в которой N-концевая часть последовательности полипептида FGF19 и C-концевая часть последовательности химерного полипептида FGF21 соединены, образуя смежные последовательности, без встроенных между ними аминокислот. В некоторых воплощениях последовательность химерного полипептида FGF19 содержит последовательность, в которой N-концевая часть последовательности полипептида FGF19 и C-концевая часть последовательности химерного полипептида FGF21 соединены, в совокупности образуя смежные последовательности с перекрыванием 1, 2, 3 или более остатков между двумя указанными последовательностями. В некоторых альтернативных воплощениях последовательность химерного полипептида FGF19 содержит последовательность, которая включает N-концевую часть последовательности полипептида FGF19 и C-концевую часть последовательности химерного полипептида FGF21, а также дополнительно включает встроенный между ними спейсер из 1, 2, 3, 4, 5 или более аминокислотных остатков.
Иллюстративные последовательности химерных полипептидов FGF19 согласно настоящему изобретению представлены в таблице 6, где их N-концевая часть включает N-концевую часть последовательности полипептида hFGF19, а их C-концевая часть включает C-концевую часть последовательности полипептида hFGF21.
Иллюстративные последовательности химерных полипептидов FGF19
PLLQFGGQVRQRYLYTDDAQQTEAHLEIREDGTVGGAADQSPESLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
LYTDDAQQTEAHLEIREDGTVGGAADQSPESLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
IREDGTVGGAADQSPESLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
DGTVGGAADQSPESLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
QSPESLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
SLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
GVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
LCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
FLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
(Конструкция 8, представленная на фигуре 18B)
LPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
(Конструкция 9, представленная на фигуре 18B)
KTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSE
AHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
(Конструкция 10, представленная на фигуре 18B)
QLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
(Конструкция 11, представленная на фигуре 18B)
PESLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
(Конструкция 12, представленная на фигуре 18B)
TVGGAADQSPESLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPL
SMVGPSQGRSPSYAS
(Конструкция 13, представленная на фигуре 18B)
EDGTVGGAADQSPESLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
(Конструкция 14, представленная на фигуре 18B)
DDAQQTEAHLEIREDGTVGGAADQSPESLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVY
QSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
(Конструкция 15, представленная на фигуре 18B)
LLQFGGQVRQRYLYTDDAQQTEAHLEIREDGTVGGAADQSPESLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYAS
III. Химерные полипептиды FGF19 с замещающими последовательностями полипептида FGF21
В третьем аспекте настоящего изобретения полипептидная последовательность химерного FGF19 включает первую полипептидную последовательность, где часть первой полипептидной последовательности замещена частью второй полипептидной последовательности. В предпочтительных воплощениях первый полипептид является человеческим полипептидом FGF19 (hFGF19), последовательность которого определяется последовательностью SEQ ID NO: 1. В некоторых воплощениях первой полипептидной последовательностью является последовательность полипептида, аминокислотная последовательность которого, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97% или, по меньшей мере, на 99% идентична полипептидной последовательности полипептида hFGF19.
В определенных предпочтительных воплощениях второй полипептид является человеческим полипептидом FGF21 (hFGF21), последовательность которого определяется последовательностью SEQ ID NO: 2. В некоторых воплощениях последовательность полипептида FGF21 представляет собой полипептидную последовательность, аминокислотная последовательность которой, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97% или, по меньшей мере, на 99% идентична последовательности полипептида hFGF21.
В некоторых воплощениях последовательность химерного полипептида FGF19 включает последовательность полипептида hFGF19, в которой часть, которая подлежит замещению, выбрана из группы, которая включает, без ограничения, части, которые имеют (i) первое положение, которое соответствует или приблизительно соответствует любому из положений 1, 10, 11, 17, 18, 21, 22, 25, 26, 27, 28, 40, 41, 42, 43, 44, 45, 52, 53, 54, 56, 57, 58, 59, 63, 72, 73, 74, 79, 80, 81, 143, 144, 145 и 146 в SEQ ID NO: 1, и (ii) последнее положение, которое соответствует или приблизительно соответствует любому из положений 9, 10, 24, 25, 26, 27, 29, 31, 32, 34, 36, 39, 40, 41, 42, 43, 44, 51, 52, 53, 55, 56, 57, 58, 66, 71, 72, 73, 78, 79, 80, 142, 143, 144, 145 и 194 в SEQ ID NO: 1, так, чтобы последнее положение было C-концевым по отношению к первому положению, и чтобы указанные положения были выбраны независимо по отношению друг к другу. В таблице 7 представлен перечень иллюстративных положений последовательности полипептида hFGF19, которые подлежат замене частью последовательности полипептида hFGF21.
Иллюстративные замещенные части последовательности полипептида hFGF19
Замещенная часть последовательности полипептида hFGF19 замещена частью последовательности полипептида hFGF21. Иллюстративные замещающие части последовательности полипептида hFGF21 включают в себя, но не ограничены перечисленным, части, которые имеют (i) первое положение, которое соответствует или приблизительно соответствует любому из положений 1, 8, 9, 13, 14, 17, 18, 21, 22, 23, 24, 35, 36, 37, 38, 39, 40, 47, 48, 49, 51, 52, 53, 54, 58, 67, 68, 69, 74, 75, 76, 136, 137, 138 и 139 в SEQ ID NO: 2, и (ii) последнее положение, которое соответствует или приблизительно соответствует любому из положений 7, 8, 20, 21, 22, 23, 24, 25, 27, 29, 31, 34, 35, 36, 37, 38, 39, 46, 47, 48, 50, 51, 52, 53, 61, 66, 67, 68, 73, 74, 75, 135, 136, 137, 138 и 181 в SEQ ID NO: 2, так, чтобы последнее положение было C-концевым по отношению к первому положению, и чтобы указанные положения были выбраны независимо одно от другого. В таблице 8 представлен список иллюстративных частей последовательности полипептида hFGF21, которыми должна быть заменена часть последовательности полипептида hFGF19.
Иллюстративные заменяющие части из последовательности полипептида hFGF21
В некоторых воплощениях химерный полипептид FGF19 включает полипептидную последовательность hFGF19 с частью его последовательности, такой как последовательность, выбранная из частей последовательности полипептида hFGF19, перечисленных в таблице 7, замещенных частей последовательности полипептида hFGF21, таких как части последовательности, перечисленные в таблице 8. В некоторых воплощениях выбранная N-концевая часть hFGF19 и выбранная C-концевая часть hFGF21 выбраны независимо друг от друга. В некоторых воплощениях часть hFGF19, подлежащая замене, включает N-концевой остаток полипептида hFGF19. В некоторых воплощениях часть hFGF19, подлежащая замене, включает C-концевой остаток полипептида hFGF19. В некоторых воплощениях замещающая часть hFGF21 включает N-концевой остаток полипептида hFGF21. В некоторых воплощениях замещающая часть hFGF21 включает C-концевой остаток полипептида hFGF21. В некоторых воплощениях часть hFGF19, подлежащая замене, включает N-концевой остаток полипептида hFGF19, и замещающая часть hFGF21 также включает N-концевой остаток полипептида hFGF21. В некоторых воплощениях часть hFGF19, подлежащая замене, включает C-концевой остаток полипептида hFGF19, и замещающая часть hFGF21 также включает C-концевой остаток полипептида hFGF21.
В некоторых воплощениях часть последовательности hFGF19 и часть последовательности hFGF21 выбраны таким образом, чтобы, по меньшей мере, один из их соответственных концов (например, N-конец части hFGF19 и N-конец части hFGF21, C-конец части hFGF19 и C-конец части hFGF21, или же и то, и другое) имели в совокупности, по меньшей мере, 1, по меньшей мере, 2 или, по меньшей мере, 3 или более остатков на указанных соответствующих концах. В некоторых воплощениях последовательность химерного полипептида FGF19 содержит последовательность, в которой соответствующая часть последовательности полипептида hFGF21 образует смежную последовательность с оставшейся последовательностью полипептида hFGF19 с перекрыванием 1, 2, 3 или более остатков в совокупности между двумя указанными последовательностями.
В некоторых воплощениях последовательность химерного полипептида FGF19 включает последовательность, в которой часть последовательности полипептида hFGF21 замещена в последовательности полипептида hFGF19, так, чтобы полипептидные последовательности hFGF19 и hFGF21 прилегали друг к другу, и между ними не содержалось добавочных, встроенных аминокислот. В некоторых альтернативных воплощениях последовательность химерного полипептида FGF19 содержит последовательность, в которой часть последовательности полипептида hFGF21 замещена в последовательности полипептида hFGF19, так, чтобы полипептидная последовательность химерного FGF19 дополнительно включала в себя встроенный спейсер из 1, 2, 3, 4, 5 или более аминокислотных остатков между последовательностями hFGF19 и hFGF21.
Иллюстративные последовательности химерных полипептидов FGF19 согласно настоящему изобретению, в которых часть hFGF19 заменена частью hFGF21, представлены в таблице 9.
Иллюстративные полипептидные последовательности химерного FGF19
В некоторых воплощениях любого из химерных полипептидов hFGF19, химерный полипептид hFGF19 включает первую полипептидную последовательность, имеющую, по меньшей мере, определенную степень идентичности последовательности таковой полипептида hFGF19, и где часть первой полипептидной последовательности замещена более чем одной частью второй полипептидной последовательности, при этом вторая полипептидная последовательность имеет, по меньшей мере, определенную степень идентичности с последовательностью полипептида hFGF21. В некоторых воплощениях химерный полипептид hFGF19 дополнительно содержит замену петли β1-β2 первого полипептида, замену сегмента β10-β12 первого полипептида и/или замену пяти остатков WGDPI (SEQ ID NO: 287) первого полипептида петлей β1-β2 второго полипептида, сегментом β10-β12 второго полипептида и/или соответствующей последовательностью GQV второго полипептида. В некоторых воплощениях химерный полипептид hFGF19 дополнительно содержит замену петли β1-β2 (аминокислотные остатки 50-57 полипептида FGF19 (SGPHGLSS (SEQ ID NO: 288)) полипептида FGF19 петлей β1-β2 (аминокислотные остатки 51-57 полипептида FGF21 (DDAQQTE (SEQ ID NO: 289)) полипептида FGF21. В некоторых воплощениях химерный полипептид hFGF19 дополнительно содержит замену сегмента β10-β12 (аминокислотные остатки 146-162 полипептида FGF19 (SSAKQRQLYKNRGFLPL (SEQ ID NO: 290)) полипептида FGF19 сегментом β10-β12 (аминокислотные остатки 147-161 полипептида FGF21 (PGNKSPHRDPAPRGP (SEQ ID NO: 291)) полипептида FGF21. В некоторых воплощениях химерный полипептид hFGF19 дополнительно содержит замену аминокислотных остатков 38-42 (WGDPI (SEQ ID NO: 287)) полипептида FGF19 аминокислотными остатками 41-43 (GQV) полипептида FGF21.
Химерные полипептиды FGF19 согласно настоящему изобретению, и в частности, содержащие их фармацевтически активные композиции и способы применения указанных химерных полипептидов FGF19 в терапевтическом лечении одного или более из заболеваний, состояний и т.д., перечисленных или описанных в настоящей заявке или известных в данной области, имеют определенные преимущества над применением либо нативного FGF19 (например, hFGF19), либо нативного FGF21 {например, hFGF21).
В некоторых воплощениях химерные полипептиды FGF19 могут быть менее иммуногенными, чем один или оба из их нативных родительских FGF. Нативный FGF19 и/или FGF21 (такой как hFGF19 или hFGF21) могут присутствовать в популяции более чем в одной аллельной вариации, где есть, по меньшей мере, один аминокислотный остаток, который отличается между указанными аллельными формами. Например, известно, что hFGF21 в зрелой форме имеет полиморфизм в положении 146, где указанным остатком в разных аллелях может быть лейцин (как на Фиг. 2 и в SEQ ID NO: 2) или пролин. Такой полиморфизм может ограничивать применимость нативного hFGF21 в качестве терапевтической композиции. Например, введение полипептида FGF19 индивиду, где эндогенный FGF19 индивида имеет последовательность, отличную от введенного FGF19, может привести к иммунной реакции индивида на введенный hFGF21. Таким образом, в некоторых воплощениях химерный полипептид FGF19 согласно изобретению может включать в себя часть последовательности полипептида hFGF21 и часть последовательности полипептида hFGF19, где обе части включают в себя только части соответствующих полипептидных последовательностей, которые не полиморфны. Это может быть осуществлено, например, путем замены части полиморфной последовательности FGF аналогичной, неполиморфной частью другого полипептида FGF. Например, химерный полипептид FGF19 согласно изобретению, такой как cFGF21/19-2 (см. таблицу 3), который содержит часть полипептида hFGF21, но не включает в себя положение 146, не имеет полиморфизма в указанном положении. Таким образом, химерные полипептиды FGF19 согласно изобретению могут быть преимущественно менее иммуногенными и, таким образом, могут быть преимущественно более подходящими для введения широкому кругу индивидов.
В некоторых воплощениях химерные полипептиды FGF19 могут быть менее опухолегенными, чем один или оба из соответствующих им нативных FGF. В частности, химерные полипептиды FGF19 могут быть менее опухолегенными, чем нативный hFGF19. Нативный hFGF19, как обсуждалось здесь выше, проявляет свою потенциальную опухолегенную активность через его связывание с FGFR4. Такая опухолегенная активность представляется независимой от метаболических эффектов hFGF19, которые, также как и в случае hFGF21, осуществляются через Klotho-бета-зависимое связывание с FGFR1c, FGFR2c и/или FGFR3c. В некоторых воплощениях химерные полипептиды FGF19 согласно изобретению включают в себя часть последовательности полипептида hFGF19, который не содержит FGFR4-эффекторный мотив, и взамен этого замещены соответствующей последовательностью из hFGF21. В некоторых воплощениях химерные полипептиды FGF19 согласно изобретению уже больше по существу не связываются с FGFR4 и/или по существу не активируют FGFR4. В некоторых воплощениях химерные полипептиды FGF19 согласно настоящему изобретению уже больше по существу не связываются с рецептором и/или по существу не активируют рецептор, такой как FGFR4, Klotho-бета-независимым образом. Таким образом, химерные полипептиды FGF19 согласно настоящему изобретению могут быть преимущественно менее иммуногенными и, таким образом, могут быть преимущественно более подходящими для введения широкому кругу индивидов.
В некоторых воплощениях химерные полипептиды FGF19 могут не вызывать резистентность к гормону роста (GH) или проявлять существенно меньшую активность в связи с резистентностью в отношении GH, чем один или оба из соответствующих им нативных FGF. В некоторых воплощениях химерные полипептиды FGF19 могут иметь меньшую активность в связи с резистентностью к GH, чем нативный FGF21, такой как нативный hFGF21. В норме GH оказывает ростовые и метаболические эффекты, которые опосредованы инсулиноподобным фактором роста 1 (IGF-1). Связывание GH с его рецептором приводит к активации Janus-киназы 2 (JAK2), которая затем фосфорилирует белок STAT5. Фосфорилированный полипептид STAT5 переносится в ядро и связывается с чувствительными регуляторными элементами гена, которые вызывают экспрессию IGF-1.
Эффекты GH могут быть ослаблены у индивидов в результате повышения уровней FGF21 или вследствие голода или ограничения питания (лечебной диеты), что тоже повышает уровни FGF21. Эффекты резистентности к GH у индивидов включают в себя экономию энергии, выраженную вялость, пониженную температуру тела, пониженную физическую активность, ингибирование роста, потерю безжировой массы и индукцию синтеза кетоновых тел. Нативный hFGF21 вызывает резистентность к GH путем, например, понижения уровня IGF-1, который в норме индуцируется гормоном роста. Не желая связывать себя какой бы то ни было теорией, авторы изобретения тем не менее полагают, что указанная активность hFGF21 в связи с резистентностью к GH может быть опосредована его способностью понижать количество фосфорилированного полипептида STAT5 и, как результат, уменьшать транслокацию фосфорилированного полипептида STAT5 в ядро и, таким образом, уменьшать экспрессию IGF-1, тем самым вызывая сопротивление эффектам гормона роста.
В некоторых воплощениях химерные полипептиды FGF19 согласно настоящему изобретению не вызывают понижения или по существу не вызывают понижения количества фосфорилированного полипептида STAT5. Таким образом, химерные полипептиды FGF19 согласно настоящему изобретению могут проявлять меньшую активность в связи с резистентностью к GH или по существу отсутствие такой активности и, таким образом, могут преимущественно быть более подходящими для введения широкому кругу индивидов.
В некоторых воплощениях химерные полипептиды FGF19 по существу не вызывают «безъякорный» рост клеток. В некоторых воплощениях химерные полипептиды FGF19 могут по существу не вызывать повышения метаболической активности и/или пролиферации клеток в окружении, требующем «безъякорного» роста клеток. В некоторых воплощениях химерные полипептиды FGF19 могут вызывать «безъякорный» рост клеток в степени, которая меньше, чем соответствующая стимуляция «безъякорного» роста клеток, вызываемая нативным FGF19. В некоторых воплощениях химерные полипептиды FGF19 могут вызывать повышенную метаболическую активность и/или пролиферацию клеток в окружении, требующем «безъякорного» роста клеток, в меньшей степени, чем соответствующая стимуляция «безъякорного» роста клеток, вызываемая нативным FGF19. Поскольку «безъякорный» рост клеток является одной из определяющих характеристик трансформированных клеток, такие химерные полипептиды FGF19 согласно настоящему изобретению, которые не вызывают стимуляции «безъякорного» роста клеток или по существу не вызывают повышенной метаболической активности и/или пролиферации клеток в окружении, требующем «безъякорного» роста клеток, могут в меньшей степени быть способны вызвать стимуляцию или усиление избирательного роста и/или метаболической активности трансформированных клеток и, таким образом, могут преимущественно быть более подходящими для введения широкому кругу индивидов.
IV. Определения
Используемые здесь термины "полипептид FGF19", "белок FGF19" и "FGF19" относятся к полипептиду, имеющему аминокислотную последовательность, которая является той же самой, что и нативная последовательность члена семейства 19 факторов роста фибробластов. Члены такого семейства включают в себя 194-членную аминокислотную последовательность человеческого FGF19 (hFGF19), представленную последовательностью SEQ ID NO: 1 и приведенную на ФИГ. 1. Полипептид FGF19 может быть выделен из природных объектов или его можно получить рекомбинантным способом и/или путем синтеза. Полипептид FGF19 относится, в частности, к возникающим природным образом усеченным или секретируемым формам, возникающим природным образом вариантным формам (например, формы, образованные в результате альтернативного сплайсинга), а также возникающим природным образом аллельным вариантам FGF19. Полипептид FGF19 также относится, в частности, как к невидоизмененным, так и к видоизмененным формам FGF19, таким, например, как 216-членная аминокислотная последовательность человеческого пре-полипептида FGF19, представленная последовательностью SEQ ID NO: 3 и приведенная на ФИГ. 1.
Используемые здесь термины "полипептид FGF21", "белок FGF21" и "FGF21" относятся к полипептиду, имеющему аминокислотную последовательность, которая является той же самой, что и нативная последовательность члена семейства 21 факторов роста фибробластов. Члены такого семейства включают в себя 181-членную аминокислотную последовательность человеческого FGF21 (hFGF21), представленную последовательностью SEQ ID NO: 2 и приведенную на ФИГ. 2. Полипептид FGF21 может быть выделен из природных объектов или его можно получить рекомбинантным способом и/или путем синтеза. Полипептид FGF21 относится, в частности, к возникающим природным образом усеченным или секретируемым формам, возникающим природным образом вариантным формам (например, формы, образованные в результате альтернативного сплайсинга), а также возникающим природным образом аллельным вариантам FGF21. Полипептид FGF21 также относится, в частности, как к невидоизмененным, так и к видоизмененным формам FGF21, таким, например, как 209-членная аминокислотная последовательность человеческого пре-полипептида FGF21, представленная последовательностью SEQ ID NO: 4 и приведенная на ФИГ. 2.
Используемые здесь термины "полипептид FGF" и "белок FGF" относятся к полипептиду, имеющему нативную последовательность члена семейства FGF, такому как человеческий FGF1, FGF2, FGF3, FGF4, FGF5, FGF6, FGF7, FGF8, FGF9, FGF10, FGF11, FGF 12, FGF13, FGF 14, FGF 16, FGF17, FGF18, FGF19, FGF20, FGF21, FGF22, FGF23, а также их гомологи, встречающиеся у млекопитающих. Нативная последовательность данного полипептида FGF может быть выделена из природного источника или может быть получена рекомбинантным и/или синтетическим путем. Нативная последовательность данного полипептида FGF относится, в частности, к возникающим природным образом усеченным или секретируемым формам, возникающим природным образом вариантным формам (например, формы, образованные в результате альтернативного сплайсинга), а также к возникающим природным образом их аллельным вариантам, а кроме того, как к невидоизмененным, так и к видоизмененным формам FGF.
Используемые здесь термины "химерный полипептид" и "химерный белок" относятся к полипептиду, имеющему последовательность, которая включает, по меньшей мере, часть полноразмерной последовательности первой полипептидной последовательности и, по меньшей мере, часть полноразмерной последовательности второй полипептидной последовательности, где первый и второй полипептиды являются разными полипептидами. Химерный полипептид относится также к полипептидам, которые включают в себя две или более не-непрерывные части, полученные из одного и того же полипептида. Химерный полипептид относится также к полипептидам, имеющим, по меньшей мере, одну замену, при этом указанный химерный полипептид включает первую полипептидную последовательность, в которой часть первой полипептидной последовательности замещена частью второй полипептидной последовательности.
Термин "часть", используемый здесь в отношении данной полипептидной последовательности, относится к непрерывному отрезку последовательности данного полипептида, которая короче, чем полноразмерная последовательность данного полипептида. Часть данного полипептида может быть определена по его первому положению и его последнему положению, при этом каждое из указанных первого и последнего положений соответствует положению в последовательности данного полипептида, где положение в последовательности, соответствующее первому положению, находится на N-конце по отношению к положению в последовательности, соответствующему последнему положению, и где последовательность указанной части является непрерывной последовательностью аминокислотных остатков в данном полипептиде, которая начинается в последовательности с положения, соответствующего первому положению, и кончается в последовательности положением, соответствующим последнему положению. Часть может быть определена также, исходя из указания положения в последовательности данного полипептида и протяженности остатков относительно данного указанного положения, при этом последовательность указанной части является непрерывной последовательностью аминокислотных остатков в данном полипептиде, который имеет определенную длину и который расположен в данном полипептиде относительно определенного положения.
Используемый здесь термин "N-концевая часть" данной полипептидной последовательности представляет собой непрерывный отрезок данной полипептидной последовательности, который начинается с N-концевого остатка данной полипептидной последовательности или около него. N-концевая часть данного полипептида может быть определена по длине. Сходным образом, используемый здесь термин "C-концевая часть" данной полипептидной последовательности представляет собой непрерывный отрезок данной полипептидной последовательности, который кончается C-концевым остатком данной полипептидной последовательности или около него. C-концевая часть данного полипептида может быть определена по длине.
Используемые здесь термины "химерный полипептид FGF" и "химерный белок FGF" относятся к полипептиду, имеющему последовательность, которая включает, по меньшей мере, часть первой последовательности полипептида FGF и часть второй последовательности полипептида FGF, где первый и второй полипептиды FGF отличаются друг от друга. Химерный полипептид FGF относится также к полипептидам, которые включают в себя две или более не-непрерывных части, полученных из одного и того же полипептида FGF. Химерный полипептид FGF относится также к полипептидам, имеющим, по меньшей мере, одну замену, при этом указанный химерный полипептид FGF включает первую последовательность полипептида FGF, в которой часть первой последовательности полипептида FGF замещена частью второй последовательности полипептида FGF.
Используемые здесь термины "химерный полипептид FGF19" и "химерный белок FGF19" относятся к химерному полипептиду FGF, имеющему последовательность, которая включает, по меньшей мере, часть последовательности полипептида FGF19 и часть второй полипептидной последовательности. Например, химерный полипептид FGF19 включает полипептиды, в которых указанная вторая полипептидная последовательность является последовательностью полипептида FGF21.
Химерный полипептид FGF19 включает также полипептиды, которые содержат две или более не-непрерывных части, полученных из последовательности полипептида FGF19. Химерный полипептид FGF19 включает также полипептиды, имеющие, по меньшей мере, одну замену, где последовательность химерного полипептида FGF19 включает последовательность полипептида FGF19, в которой часть последовательности полипептида FGF19 замещена частью второй полипептидной последовательности. В таких случаях химерный полипептид FGF19 однозначно охватывает полипептиды, в которых замещающая часть представляет собой часть последовательности полипептида FGF21.
Химерный полипептид FGF19 включает в себя также полипептид, последовательность которого состоит только из частей, полученных либо из последовательности полипептида FGF19, либо из второй последовательности, такой как последовательность полипептида FGF21. Если специально не указано иначе, химерный полипептид FGF19 не ограничен и, если специально не указано иначе, не подразумевается, что он ограничен соответствующим порядком или локализацией последовательности полипептида FGF19 по отношению к любым другим частями последовательности внутри последовательности химерного полипептида FGF19.
"Процент (%) идентичности аминокислотных последовательностей" в отношении идентифицируемых здесь полипептидных последовательностей определяется как процент аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в ссылочной последовательности, после выравнивания последовательностей и, если необходимо, введения гэпов, для достижения максимального процента идентичности последовательностей, и без учета каких бы то ни было консервативных замен как части идентичности указанной последовательности. Выравнивание с целью определения процента идентичности аминокислотных последовательностей может быть достигнуто различными путями, которые известны специалистам в данной области, например, с помощью общедоступного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN, ALIGN-2 или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры для оценки выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания в пределах полноразмерных последовательностей, подлежащих сравнению.
В данном контексте % идентичности аминокислотных последовательностей данной аминокислотной последовательности A другой последовательности, с другой последовательностью или в сравнении с данной аминокислотной последовательностью B (которая альтернативно может быть названа данной аминокислотной последовательностью A, которая имеет или содержит % идентичности аминокислотной последовательности, с другой последовательностью или в сравнении с данной аминокислотной последовательностью B) вычисляется следующим образом: 100, умноженное на частное от деления X/Y, где X является числом аминокислотных остатков, засчитанных как идентичные совпадения программой по выравниванию данной последовательности с таковыми согласно программам по выравниванию последовательностей A и B, и где Y является суммарным числом аминокислотных остатков в B. Следует учесть, что когда длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, % идентичности аминокислотной последовательности A к B не будет равен % идентичности аминокислотной последовательности B к A.
"Процент (%) идентичности последовательности нуклеиновых кислот" в отношении идентифицируемых здесь последовательностей нуклеиновых кислот, кодирующих полипептиды, определяется как процент нуклеотидов в последовательности-кандидате, которые идентичны нуклеотидам в кодирующей полипептид последовательности нуклеиновой кислоты, после выравнивания указанных последовательностей и, если необходимо, введения гэпов, для достижения максимального процента идентичности последовательностей. Выравнивание в целях определения процента идентичности последовательностей нуклеиновых кислот может быть достигнуто различными путями, которые известны специалистам в данной области, например, с помощью общедоступного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN, ALIGN-2 или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры для оценки выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания в пределах полноразмерных последовательностей, подлежащих сравнению.
В данном контексте % идентичности данной последовательности нуклеиновой кислоты С другой последовательности, с другой последовательностью или в сравнении с данной последовательностью нуклеиновой кислоты D (которая альтернативно может быть названа данной последовательностью нуклеиновой кислоты С, которая имеет или содержит % идентичности последовательности нуклеиновой кислоты, с другой последовательностью или в сравнении с данной последовательностью нуклеиновой кислоты D) вычисляется следующим образом: 100, умноженное на частное от деления W/Z, где W является числом нуклеотидов, засчитанных как идентичные совпадения программой по выравниванию данной последовательности с таковыми согласно программам по выравниванию последовательностей C и D, и где Z является суммарным числом нуклеотидов в D. Следует учесть, что когда длина последовательности нуклеиновой кислоты С не равна длине последовательности нуклеиновой кислоты D, % идентичности последовательности нуклеиновой кислоты С к D не будет равен % идентичности последовательности нуклеиновой кислоты D к С.
Термин "изолированный", или "выделенный", используемый для обозначения различных используемых здесь полипептидов, означает полипептид, который идентифицирован и выделен и/или восстановлен из компонента его природного окружения. Предпочтительно, чтобы изолированный полипептид был свободен от ассоциации со всеми компонентами, с которыми он ассоциирован в природе. Примесными компонентами его природного окружения являются материалы, которые обычно мешают при диагностических или терапевтических применениях указанного полипептида, и они могут включать в себя ферменты, гормоны и другие растворенные вещества белковой или небелковой природы. В предпочтительных воплощениях указанный полипептид должен быть очищен (1) до степени, достаточной для получения, по меньшей мере, 15 остатков N-концевой или внутренней аминокислотной последовательности, путем применения секвенатора с вращающимся стаканом, или (2) до гомогенности, определяемой методом ДСН-ПААГ в невосстанавливающих или восстанавливающих условиях с использованием Кумасси или, предпочтительно, серебряного красителя. Изолированный полипептид включает в себя полипептид in situ внутри рекомбинантных клеток, поскольку, по меньшей мере, один из компонентов его природного окружения будет отсутствовать. Однако обычно изолированный полипептид получают в результате, по меньшей мере, одной стадии очистки.
Термин "изолированная" молекула нуклеиновой кислоты, кодирующей полипептид, является молекулой нуклеиновой кислоты, которая идентифицирована и отделена, по меньшей мере, от одной примесной молекулы нуклеиновой кислоты, с которой она обычно ассоциирована в природном источнике кодирующей полипептид нуклеиновой кислоты. Предпочтительно, чтобы изолированная нуклеиновая кислота была свободна от ассоциации со всеми компонентами, с которыми она ассоциирована в природе. Изолированная молекула нуклеиновой кислоты, кодирующей полипептид, является молекулой нуклеиновой кислоты, которая отлична от таковой, которая встречается в природе. Следовательно, изолированные молекулы нуклеиновых кислот отличаются от молекулы нуклеиновой кислоты, кодирующей полипептид, поскольку она существует в природных клетках. Однако изолированная молекула нуклеиновой кислоты, кодирующей полипептид, включает в себя кодирующие полипептид молекулы нуклеиновых кислот, содержащиеся в клетках, которые обычно экспрессируют полипептид, где указанная молекула нуклеиновой кислоты, например, имеет хромосомную локализацию, отличную от таковой в природных клетках.
Используемый здесь термин "контрольные последовательности" относится к последовательностям ДНК, необходимым для экспрессии оперативно связанной кодирующей последовательности в организме конкретного хозяина. Контрольные последовательности, которые являются подходящими для прокариотов, включают в себя, например, промотор, необязательно, последовательность оператора и часть связывания рибосомы. Известно, что в эукариотических клетках используются промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота является "оперативно связанной", когда она находится в функциональном соотношении с другой последовательностью нуклеиновой кислоты. Например, ДНК для предпоследовательности или секреторной лидерной последовательности оперативно связана с ДНК для полипептида, если он экспрессируется в виде пре-белка, который участвует в секреции полипептида; промотор или энхансер оперативно связан с кодирующей последовательностью, если он воздействует на транскрипцию последовательности; или часть связывания рибосомы оперативно связан с кодирующей последовательностью, если он расположен таким образом, чтобы облегчить трансляцию. Обычно "оперативно связанный" означает, что последовательности ДНК связаны, образуя непрерывную последовательность, а в случае секреторной лидерной последовательности, являются непрерывными и находятся в фазе считывания. Однако энхансеры необязательно должны быть непрерывными. Связывание сопровождается лигированием в подходящих участках рестрикции. Если таких участков не существует, в соответствии с общепринятой практикой, используются искусственные олигонуклеотидные адапторы или линкеры.
Термин "антитело" используеся здесь в самом широком смысле и в частности, охватывает, например, одноцепочечные моноклональные антитела (включая агонисты, антагонисты и нейтрализующие антитела), композиции антител с полиэпитопной специфичностью, одноцепочечные антитела и фрагменты антител (см. ниже). Используемый здесь термин "моноклональное антитело" относится к антителу, полученному из популяции по существу гомогенных антител, то есть индивидуальные антитела, содержащиеся в такой популяции, являются идентичными, за исключением возможных возникших природным образом мутаций, которые могут присутствовать в минорных количествах.
В отношении связывания полипептида, антитела, олигопептида или другой органической молекулы с молекулой-мишенью или распознающим рецептором, используемый здесь термин "специфическое связывание" или "специфически связывается с", или является "специфичным в отношении" конкретного полипептида или эпитопа на конкретном полипептиде-мишени или распознающем рецепторе, означает связывание, которое измеримо отличается от неспецифического связывания. Специфическое связывание может быть измерено, например, путем определения связывания молекулы по сравнению со связыванием контрольной молекулы, которая обычно является молекулой со сходной структурой, но которая не обладает связывающей активностью. Например, специфическое связывание может быть определено конкуренцией с контрольной молекулой, которая подобна молекуле-мишени, как, например, в случае избытка немеченой молекулы-мишени. В таком случае специфическое связывание показано, если связывание меченой мишени с зондом специфически ингибируется избытком немеченой мишенью. Используемый здесь термин "специфическое связывание" или "специфически связывается с", или является "специфичным в отношении" конкретного полипептида или эпитопа на конкретном полипептиде-мишени или распознающем рецепторе, означает связывание, которое может быть заингибировано, например, молекулой, имеющей Kd с указанной мишенью, составляющую, по меньшей мере, приблизительно 10-4 M, альтернативно, по меньшей мере, приблизительно 10-5 M, альтернативно, по меньшей мере, приблизительно 10-6 M, альтернативно, по меньшей мере, приблизительно 10-7 M, альтернативно, по меньшей мере, приблизительно 10-8 M, альтернативно, по меньшей мере, приблизительно 10-9 M, альтернативно, по меньшей мере, приблизительно 10-10 M, альтернативно, по меньшей мере, приблизительно 10-11 M, альтернативно, по меньшей мере, приблизительно 10-12 M или более. В одном из воплощений используемый здесь термин "специфическое связывание" относится к связыванию, при котором молекула связывается с конкретным полипептидом или эпитопом на конкретном полипептиде или распознающем рецепторе, без существенного связывания с любым другим полипептидом или эпитопом или рецептором полипептида.
"Жесткость" реакций гибридизации легко определяется рядовыми специалистами в данной области, и обычно она определяется эмпирически, в зависимости от длины зонда, температуры промывания и концентрации соли. Обычно более длинные зонды требуют более высоких температур для надлежащего аннилирования, тогда как более короткие зонды требуют более низких температур. Обычно гибридизация зависит от способности денатурированной ДНК к вторичному отжигу, когда комплементарные цепи находятся в окружении, имеющем температуру ниже их температуры плавления. Чем выше степень требуемой гомологии между зондом и гибридизующейся последовательностью, тем выше относительная температура, которая может быть использована. В результате отсюда следует, что более высокие температуры будут способствовать более жестким условиям реакции, тогда как более низкие температуры будут уменьшать степень жесткости. Для ознакомления с дополнительными подробностями и объяснением понятия жесткости реакций гибридизации см. Ausubel et al., Current Protocols in Molecular Biology, Wiley Interscience Publishers (1995).
Здесь термины "жесткие условия", или "условия высокой жесткости", могут быть идентифицированы как условия, при которых: (1) для промывания используется низкая ионная сила и высокая температура, например 0,015 M хлорид натрия/0,0015 M цитрат натрия/0,1% додецилсульфат натрия при 50°C; (2) в процессе гибридизации используется денатурирующее средство, такое как формамид, например, 50% (об./об.) формамид с 0,1% бычьим сывороточным альбумином/0,1% фиколл/0,1% поливинилпирролидон/50 мМ натрий-фосфатный буфер при pH 6,5 с 750 мМ хлоридом натрия, 75 мМ цитратом натрия при 42°C; или (3) используется 50% формамид, 5×SSC (0,75 M NaCl, 0,075 M цитрат натрия), 50 мМ фосфат натрия (pH 6,8), 0,1% пирофосфат натрия, 5x раствор Денхардта, обработанная ультразвуком ДНК спрмы лосося (50 мкг/мл), 0,1% ДСН и 10% декстрансульфат при 42°C, с промываниями при 42°C в 0,2×SSC (хлорид натрия/цитрат натрия) и 50% формамид при 55°C, с последующим высокожестким промыванием, состоящим из 0,1×SSC, содержащего ЭДТА при 55°C.
"Умеренно жесткие условия" могут быть идентифицированы так, как описано у Sambrook et al., Molecular Cloning: A Laboratory Manual, New York: Cold Spring Harbor Press, 1989, и включают в себя использование промывочного раствора и менее жестких условий гибридизации (например, температуры, ионной силы и % ДСН), чем те, которые описаны выше. Примером умеренно жестких условия является инкубация в течение ночи при 37°C в растворе, содержащем 20% формамид, 5×SSC (150 мМ NaCl, 15 мМ тринатриевый цитрат), 50 mM мМ фосфат натрия (pH 7,6), 5x раствор Денхардта, 10% декстрансульфат и 20 мг/мл денатурированной расщепленной ДНК спермы лосося, с последующей отмывкой фильтров 1×SSC приблизительно при 37-50°C. Специалистам в данной области должно быть понятно, как доводить температуру, ионную силу и т.д. до необходимых значений, чтобы привести их в соответствие с такими факторами как длина зонда и т.д.
Используемый здесь термин "меченый эпитоп" относится к полипептиду, слитому с "маркерным полипептидом". Маркерный полипептид имеет достаточное количество остатков, чтобы обеспечить эпитоп, против которого может быть получено антитело, но при этом является достаточно коротким, чтобы не препятствовать активности полипептида, с которым он слит. Предпочтительно, чтобы маркерный полипептид также был достаточно уникальным, так, чтобы антитело по существу перекрестно не реагировало с другими эпитопами. Подходящие маркерные полипептиды обычно имеют, по меньшей мере, шесть аминокислотных остатков и, как правило, от приблизительно 8 до 50 аминокислотных остатков (предпочтительно, от приблизительно 10 до 20 аминокислотных остатков).
Используемый здесь термин "иммуноадгезин" означает антителоподобные молекулы, которые сочетают активность связывания гетерологичного белка ("адгезин") с эффекторными функциями константных доменов иммуноглобулина. Структурно иммуноадгезины включает слияние аминокислотной последовательности с требуемой специфичностью связывания, которая отличается от специфичности распознавания антигена и связывающей части антитела (то есть являются "гетерологичными"), и последовательности константного домена иммуноглобулина. Адгезиновая часть иммуноадгезиновой молекулы обычно является непрерывной аминокислотной последовательностью, содержащей, по меньшей мере, часть связывания рецептора или лиганда. Последовательность константного домена иммуноглобулина в иммуноадгезине может быть получена из любого иммуноглобулина, такого как подтипы IgG-1, IgG-2, IgG-3 или IgG-4, а также IgA (включая IgA-1 и IgA-2), IgE, IgD или IgM.
Термины "активный" или "активность" здесь относятся к химерному полипептиду FGF19, который сохраняет, по меньшей мере, одну биологическую и/или иммунологическую активность нативного или возникшего природным путем полипептида FGF19 и/или полипептида FGF21, в частности, нативного или возникшего природным путем полипептида hFGF19 и/или полипептида hFGF21. "Биологическая" активность относится к биологической функции (либо ингибиторной, либо стимуляторной, либо кооперативной), вызванной нативным или возникшим природным путем полипептидом FGF19 и/или полипептидом FGF21, отличной от способности вызывать продуцирование антитела против антигенного эпитопа, свойственного нативному или возникшему природным путем полипептиду FGF19 и/или полипептиду FGF21. "Биологическая" активность может также относиться к функции нативного или возникшего природным путем полипептида FGF19 и/или полипептида FGF21, такой как способность связываться с одним или несколькими из соответствующих распознающих его рецепторов. "Иммунологическая" активность относится к способности вызывать продуцирование антитела против антигенного эпитопа, свойственного нативному или возникшему природным путем полипептиду FGF19 и/или полипептиду FGF21. Предпочтительная биологическая активность включает в себя любую одну или более из следующих иллюстративных активностей: усиление метаболизма (или скорости метаболизма) у индивида, уменьшение веса тела индивида, уменьшение ожирения у индивида, уменьшение поглощения глюкозы адипоцитами, усиление высвобождения лептина из адипоцитов, уменьшение триглицеридов у индивида, уменьшение свободных жирных кислот у индивида, Klotho-бета-зависимое связывание с распознающим FGF рецептором и Klotho-бета-независимое связывание с распознающим FGF рецептором. Очевидно, что некоторые из активностей полипептидов FGF19 и/или FGF21 индуцированы непосредственно самим полипептидом, а некоторые индуцированы опосредованно, однако каждая из них является результатом присутствия полипептида FGF19 и/или полипептида FGF21, и в отсутствие указанного полипептида такого результата бы не было.
Термин "антагонист" используется здесь в самом широком смысле и включает любую молекулу, которая частично или полностью блокирует, ингибирует или нейтрализует биологическую активность описанного здесь нативного или химерного полипептида. Сходным образом, термин "агонист" используется здесь в самом широком смысле и включает любую молекулу, которая имитирует биологическую активность описанного здесь нативного или химерного полипептида. Подходящие агонистические или антагонистические молекулы включают в себя, в частности, агонистические или антагонистические антитела или фрагменты антител, фрагменты или варианты аминокислотных последовательностей нативных полипептидов, пептидов, малых органических молекул и т.д. Способы идентификации агонистов или антагонистов полипептида могут включать в себя приведение полипептида в контакт с агонистической или антагонистической молекулой-кандидатом и измерение детектируемого изменения в одной или нескольких биологических активностях, в норме ассоциированных с указанным полипептидом.
Здесь "лечение" или "обработка" относятся к способу получения благотворных или требуемых результатов, включая клинические результаты. В контексте настоящего изобретения благотворные или требуемые клинические результаты включают в себя, но не ограничены перечисленным, один или более из следующих результатов: ослабление одного или более симптомов, возникающих в результате заболевания, уменьшение степени выраженности заболевания, стабилизацию заболевания (например, предотвращение или задержку ухудшения заболевания), задержку или замедление процесса прогрессирования заболевания, улучшение состояния болезни, уменьшение дозы одного или более других лекарственных средств, необходимых для лечения заболевания и/или повышение качества жизни.
Здесь "задержка" прогрессирования означает отсрочку, помеху, замедление, торможение, стабилизацию и/или отсрочку развития заболевания. Такая задержка может иметь различную длительность, в зависимости от истории болезни и/или индивида, подвергаемого лечению.
В некоторых воплощениях описанные здесь способы лечения улучшают (например, уменьшают количество случаев, сокращают длительность течения, ослабляют или облегчают степень тяжести) одного или более симптомов заболевания.
"Симптомом" является любое патологическое явление или отклонение от нормы в структуре, функции или ощущении, испытываемом индивидом.
Как описано в настоящей заявке, "эффективное количество" полипептида, антитела, их агониста или антагониста является количеством, достаточным для осуществления специально заявленного предназначения. "Эффективное количество" может быть определено эмпирически и общеизвестным способом, в зависимости от заявленного предназначения.
Используемый здесь термин "терапевтически эффективное количество" относится к количеству антитела, полипептида или других лекарственных средств, эффективных для "лечения" заболевания или расстройства у индивида или млекопитающего. См. здесь идентификацию термина "лечение".
"Хроническое" введение относится к введению средства (средств) в непрерывном режиме, в отличие от острого режима, с тем, чтобы поддерживать первоначальный терапевтический эффект (активность) в течение длительного периода. "Периодическое" введение относится к лечению, которое проводят не безостановочно, а которое носит циклический характер.
Здесь "млекопитающее" в контексте лечения относится к любому животному, классифицируемому как млекопитающее, включая человека, домашних и сельскохозяйственных животных, а также зверей, спортивных животных или домашних любимцев, таких как собаки, кошки, крупный рогатый скот, лошади, овцы, свиньи, козы, кролики и т.д. Предпочтительно, чтобы млекопитающее являлось человеком.
"Индивидом" является любое млекопитающее, предпочтительно, человек.
"Ожирение" относится к состоянию, при котором млекопитающее имеет индекс массы тела (BMI, Body Mass Index), который рассчитывается как вес (кг), деленный на рост2 (в метрах), составляющий, по меньшей мере, 25,9. Обычно индивиды с нормальным весом имеют BMI от 19,9 до менее чем 25,9. Здесь ожирение может иметь любую причину, будь то генетическая или внешняя причина. Примеры заболеваний, которые могут приводить к ожирению или могут быть причиной ожирения, включают в себя переедание и булимию, поликистозное заболевание яичника, краниофарингиому, синдром Прадера-Вилли, синдром Фрелиха, диабет II типа, дефицит гормона роста у индивидов, нормальный вариант низкорослости, синдром Тернера и другие патологические состояния, при которых происходит понижение метаболической активности или уменьшение расхода энергии в состоянии покоя как процент от массы безжировой ткани, например, у детей с острой лимфобластной лейкемией.
"Состояния, связанные с ожирением" относятся к состояниям, которые являются результатом ожирения или которые усиливаются ожирением, например, но не ограничиваясь дерматологическими расстройствами, такие как инфекции, варикозное расширение вен, акантокератодермия и экзема, непереносимость физической нагрузки, сахарный диабет, резистентность к инсулину, гипертония, гиперхолестеринемия, холелитиаз, остеоартрит, повреждение опорно-двигательного аппарата, тромбоэмболическая болезнь, злокачественная опухоль и ишемическая болезнь сердца (или сердечно-сосудистое заболевание), в частности, такие сердечно-сосудистые заболевания, которые ассоциированы с высоким уровнем триглицеридов и свободных жирных кислот у индивида.
Введение "в сочетании с" или "совместно с" одним или несколькими дополнительными терапевтическими средствами включает в себя одновременное, совместное, последовательное и поочередное введение в любом порядке.
Здесь термин "носители" включает в себя приемлемые фармацевтические носители, эксципиенты или стабилизаторы, которые являются нетоксичными в отношении клетки или млекопитающего, подвергаемого их действию в используемых дозах и концентрациях. Часто физиологически приемлемый носитель забуференным водным раствором. Примеры приемлемых физиологических носителей включают в себя буферы, такие как фосфатный, цитратный и буферы с добавлением других органических кислот; антиоксиданты, включая аскорбиновую кислоту; полипептиды с низким молекулярным весом (менее чем приблизительно из 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатообразующие соединения, такие как ЭДТА; сахарные спирты, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионные поверхностно-активные вещества, такие как TWEENTM, полиэтиленгликоль (ПЭГ) и PLURONICTM.
Обработка антител папаином продуцирует два идентичных антиген-связывающих фрагмента, называемых "Fab"-фрагментами, и оставшийся фрагмент "Fc", обозначение, отражающее способность к быстрой кристаллизации. Fab"-фрагмент состоит из полной L-цепи, вместе с областью вариабельного домена H-цепи (VH), а также первого константного домена тяжелой цепи (CH1). Каждый Fab-фрагмент является моновалентным в отношении связывания антигена, то есть он имеет один антиген-связывающий участок. Обработка антител пепсином дает один крупный F(ab')2-фрагмент, который в общих чертах соответствует двум связанным дисульфидными мостиками Fab-фрагментам, имеющим двухвалентную антиген-связывающую активность, и все еще способен к перекрестному связыванию антигена. Fab'-фрагменты отличаются от Fab-фрагментов тем, что они имеют несколько дополнительных остатков на карбокси-конце CH1-домена, включая один или более цистеинов из шарнирной области антитела. Здесь Fab'-SH служит обозначением для Fab', в котором цистеиновый остаток(остатки) константных доменов несет свободную тиоловую группу. F(ab')2-фрагменты антитела первоначально продуцировались как пары Fab'-фрагментов с шарнирными цистеинами между ними. Известны также и другие виды химического сцепления фрагмента антитела.
"Fv" является минимальным фрагментом антитела, который содержит полный антиген-распознающий и антиген-связывающий участок. Такой участок состоит из димера вариабельных доменов одной тяжелой и одной легкой цепи, находящихся в тесной нековалентной ассоциации друг с другом. Именно в такой конфигурации все три области CDR каждого вариабельного домена взаимодействуют, образуя антиген-связывающий участок на поверхности VH-VL-димера. Совместно указанные шесть областей CDR придают антителу специфичность антигенного связывания. Однако даже один вариабельный домен (или половина Fv-фрагмента, содержащая только три области CDR, специфичные в отношении антигена) обладает способностью распознавать и связывать антиген, хоть и с меньшей аффинностью, чем полный связывающий участок.
"Fc"-фрагмент содержит карбокси-концевые области обеих H-цепей, удерживаемых дисульфидными связями. Эффекторные функции антител определяются последовательностями в области Fc, которая также является частью, распознаваемой рецепторами Fc (FcR), обнаруживаемыми на поверхности определенных типов клеток.
Fab-фрагмент содержит также константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Fab-фрагменты отличаются от Fab'-фрагментов добавлением нескольких остатков на карбокси-конце тяжелой цепи CH1-домена, включая один или более цистеинов из шарнирной области антитела.
"Легкие цепи" антител (иммуноглобулинов) из любого вида позвоночных могут быть отнесены к одному из двух совершенно разных типов, называемых каппа- и лямбда-цепями, на основании аминокислот последовательностей их константных доменов.
В зависимости от константного домена их тяжелых цепей, иммуноглобулины могут быть отнесены к разным классам. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и несколько из них могут быть дополнительно разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA и IgA2.
"Одноцепочечные Fv", или "sFv"-фрагменты антитела содержат VH- и VL-домены антитела, при этом указанные домены представлены единственной полипептидной цепью. Предпочтительно, чтобы полипептид Fv дополнительно содержал полипептидный линкер между VH- и VL-доменами, который дает возможность sFv-фрагменту образовывать структуру, требуемую для связывания антигена. В качестве обзора по sFv-фрагментам см. Pluckthun, The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994).
Используемый здесь термин "диатела" относится к малым фрагментам антитела с двумя антиген-связывающими участками, при этом указанные фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в составе одной и той же полипептидной цепи (VH-VL). С помощью линкера, который является слишком коротким, чтобы допустить спаривание между указанными двумя доменами на одной и той же цепи, указанные домены вынуждены спариваться с комплементарными доменами другой цепи и образовывать два антиген-связывающих участка. Диатела более подробно описаны, например, в Европейской патентной публикации ЕР 0404097; в Международной патентной публикации PCT WO 93/11161; а также у Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993).
"Изолированное" антитело представляет собой антитело, которое идентифицировано и выделено и/или восстановлено из компонента его природного окружения. Примесными компонентами его природного окружения являются вещества, которые могут служить помехой при диагностических или терапевтических применениях такого антитела и могут включать в себя ферменты, гормоны и другие растворимые вещества белковой и небелковой природы. В предпочтительных воплощениях антитело будет очищено (1) более чем на 95% по массе антитела, что определяется методом Лоури, и наиболее предпочтительно, более чем на 99% по массе, (2) до степени, достаточной для получения, по меньшей мере, 15 остатков N-концевой или внутренней аминокислотной последовательности, с применением секвенатора с вращающимся стаканом, или (3) до гомогенности, определяемой методом ДСН-ПААГ в невосстанавливающих или восстанавливающих условиях с использованием Кумасси или, предпочтительно, серебряного красителя. Изолированное антитело включает в себя антитело in situ внутри рекомбинантных клеток, поскольку, по меньшей мере, один из компонентов его природного окружения будет отсутствовать. Однако обычно изолированное антитело получают в результате, по меньшей мере, одной стадии очистки.
Используемое здесь слово "метка" относится к детектируемому соединению или композиции, которая напрямую или опосредованно конъюгирована с антителом, с тем, чтобы получилось "меченое" антитело. Метка может сама по себе быть детектируемой (например, радиоактивные метки или флуоресцентные метки) или, в случае ферментативных меток, может катализировать химическое изменение субстратного соединения или композиции, которое и является детектируемым.
Под "твердой фазой" подразумевается не-водный матрикс, к которому может прикрепиться антитело согласно настоящему изобретению. Примеры твердых фаз включают в себя таковые, полученные частично или полностью из стекла (например, стекло с контролируемым размером пор), полисахаридов (например, агароза), полиакриламидов, полистирола, поливинилового спирта и силиконов. В определенных воплощениях, в зависимости от контекста, твердая фаза может включать в себя лунку аналитического планшета; в других случаях это очистительная колонка (например, колонка аффинной хроматографии). Указанный термин включает в себя также и прерывистую твердую фазу из дискретных частиц, такую как те, что описаны в патенте США No. 4275149.
"Липосома" представляет собой малую везикулу, составленную из разных типов липидов, фосфолипидов и/или поверхностно-активного вещества, которые используются для доставки лекарственного средства (такого как химерный полипептид FGF19 или антитело к нему) в организме млекопитающего. Компоненты липосомы обычно располагаются в определенном порядке, образуя двухслойную систему, сходную с таковой в биологических мембранах.
Здесь термин "приблизительно" в отношении величины или параметра включает в себя (и описывает) вариации, которые связаны указанными величиной или параметром как таковыми. Например, обозначение "приблизительно X" включает в себя обозначение "X".
В настоящем описании и в прилагаемой формуле изобретения формы единственного числа включают в себя также и множественное число, если конкретный контекст определенно не диктует иное. Очевидно, что аспекты и вариации описанного здесь изобретения включают в себя аспекты и варианты, связанные с терминами "состоящий" и/или "по существу состоящий".
V. Химерные варианты FGF19
Помимо описанных здесь химерных полипептидов FGF19, предполагается, что могут быть получены химерные варианты полипептидов FGF19 (или "химерные варианты FGF19"). Химерные варианты FGF19 могут быть получены путем введения соответствующих изменений нуклеотидов в ДНК, кодирующую химерный или нативный полипептид FGF19 или FGF21, и/или путем синтеза требуемого химерного варианта FGF19. Специалистам в данной области должно быть понятно, что аминокислотные изменения могут изменить посттрансляционный процессинг химерного варианта FGF19, например, может измениться число или положение гликозилированных участков или может измениться характер мембранного заякоривания.
Могут быть осуществлены изменения в химерных полипептидах FGF19 согласно настоящему изобретению или в разных их доменах, например, с использованием любой из технологий и методических указаний в отношении консервативных и неконсервативных мутаций, изложенных, например, в патенте США No. 5364934. Вариации могут представлять собой замену, делецию или вставку одного или более кодонов, кодирующих химерный полипептид FGF19, что приводит к изменению аминокислотной последовательности химерного полипептида FGF19. Вариации могут касаться одного или более кодонов, кодирующих химерный полипептид FGF19, который получен из нативного полипептида FGF19 или FGF21.
Вариация необязательно достигается путем замены, по меньшей мере, одной аминокислоты другой аминокислотой в одном или в нескольких из доменов химерного полипептида FGF19. Методическим руководством в определении того, вставка, замена или делеция какого именно аминокислотного остатка может быть осуществлена без неблагоприятного воздействия в отношении требуемой активности, может служить сравнение последовательности химерного полипептида FGF19 с последовательностью известных гомологичных белковых молекул и минимизация числа изменений аминокислотной последовательности, осуществляемых в высокогомологичных областях. Аминокислотные замены могут быть результатом замещения одной аминокислоты другой аминокислотой, имеющей подобные структурные и/или химические свойства, например, замещения лейцина серином, то есть результатом консервативных аминокислотных замен. Вставки или делеции могут необязательно быть заключены в интервале приблизительно от 1 до 5 аминокислот. Допустимые вариации могут определяться систематическим осуществлением вставок, делеций или замен аминокислот в последовательности и тестированием полученных в результате вариантов в отношении активности, проявляемой полноразмерной или зрелой нативной последовательностью.
Здесь представлены фрагменты химерного полипептида FGF19 (или "химерные фрагменты FGF19"). Такие фрагменты могут быть усеченными на N-конце или C-конце или могут быть лишены внутренних остатков, например, по сравнению с полноразмерным нативным белком. Определенные фрагменты лишены аминокислотных остатков, которые не являются необходимыми для требуемой биологической активности химерного полипептида FGF19.
Химерные фрагменты FGF19 могут быть получены с помощью любой из многочисленных общеупотребительных технологий. Требуемые полипептидные фрагменты могут быть синтезированы химическим путем. Альтернативный подход включает получение химерных фрагментов FGF19 путем ферментативного переваривания, например, путем обработки белка ферментом, известным своей способностью расщеплять белки на участках, определяемых конкретными аминокислотными остатками, или путем переваривания ДНК соответствующими рестрикционными ферментами и выделения нужного фрагмента. Другая технология включает выделение и амплификацию ДНК-фрагмента, кодирующего искомый полипептидный фрагмент, путем полимеразной цепной реакции (ПЦР). Олигонуклеотиды, которыми определяются требуемые концы ДНК-фрагмента, используются в качестве 5'- и 3'-праймеров при ПЦР. Предпочтительно, чтобы фрагменты химерного полипептида FGF19 обладали, по меньшей мере, одной биологической и/или иммунологической активностью нативного полипептида FGF19, такого как полипептид hFGF19, представленный на фиг. 1 (SEQ ID NO: 1), или нативного полипептида FGF21, такого как полипептид hFGF21, представленный на фиг. 2 (SEQ ID NO: 2).
В конкретных воплощениях представляющие интерес консервативные замены приведены в таблице 10 под рубрикой предпочтительных замен. Если такие замены приводят к изменению биологической активности, тогда производят более существенные изменения, которые представлены в таблице 10 под рубрикой иллюстративных замен, или же которые производят с учетом классов аминокислот, дополнительно описанных ниже, и осуществляют скрининг полученных продуктов.
Предпочтительные замены аминокислотных остатков
Существенные модификации в отношении функции или иммунологической идентичности химерного полипептида FGF19 выполнены путем селективных замен, которые значительно отличаются их эффектом в отношении поддержания (a) структуры полипептидного остова в области произведенной замены, например, складчатой структуры или спиральной конформации, (b) заряда или гидрофобности молекулы на участке, где произведена замена, или (c) размера боковой цепи. Природные остатки разделены на группы на основе свойств их боковых цепей:
(1) гидрофобные: норлейцин, met, ala, val, leu, ile;
(2) нейтральные гидрофильные: cys, ser, thr;
(3) кислые: asp, glu;
(4) основные: asn, gln, his, lys, arg;
(5) остатки, которые влияют на ориентацию цепи: gly, pro; и
(6) ароматические: trp, tyr, phe.
Неконсервативные замены повлекут за собой замену члена одного из указанных классов членом другого класса. Такие замещенные остатки также могут быть включены в замещенные консервативные участки или, более предпочтительно, в оставшиеся (неконсервативные) участки.
Вариации могут быть осуществлены с помощью методов, известных в данной области, таких как олигонуклеотид-опосредованный (сайт-направленный) мутагенез, аланиновое сканирование и РЦП-мутагенез. Сайт-направленный мутагенез [Carter et al., Nucl. Acids Res., 13: 4331 (1986); Zoller et al., Nucl. Acids Res., 10: 6487 (1987)], кассетный мутагенез [Wells et al., Gene, 34: 315 (1985)], рестрикционный селективный мутагенез [Wells et al., Philos. Trans. R. Soc. London SerA, 317: 415 (1986)] или другие известные технологии могут быть предприняты в отношении клонированной ДНК для получения ДНК, кодирующей химерный вариантный полипептид FGF19.
Сканирующий аминокислотный анализ также может быть предпринят для идентификации одной или более аминокислот на всем протяжении непрерывной последовательности. Среди предпочтительных сканирующих аминокислот можно отметить относительно малые, нейтральные аминокислоты. Такие аминокислоты включают в себя аланин, глицин, серин и цистеин. В указанной группе аланин обычно является предпочтительной сканирующей аминокислотой, поскольку он элиминирует боковую группу за пределами бета-углерода и с меньшей вероятностью может повлиять на конформацию основной цепи варианта [Cunningham and Wells, Science, 244: 1081-1085 (1989)]. Аланин предпочтителен еще и потому, что он является наиболее часто встречающейся аминокислотой. Кроме того, он часто обнаруживается как в скрытых, так и в находящихся на поверхности положениях [Creighton, The Proteins, (W.H. Freeman & Co., N.Y.); Chothia, J. Mol. Biol., 150: 1 (1976)]. Если замена аланина не приводит к образованию адекватных количеств варианта, может быть использована изостерическая аминокислота.
VI. Модификации химерного FGF19
Ковалентные модификации химерного полипептида FGF19 являются включенными в объем, охватываемый настоящим изобретением. Один тип ковалентной модификации включает реактивные нацеленные аминокислотные остатки-мишени химерного полипептида FGF19 с органическим дериватизирующим агентом, который способен реагировать с некоторыми боковыми цепями или с N- или C-концевыми остатками химерного полипептида FGF19. Дериватизация бифункциональными агентами полезна, например, для перекрестного связывания химерного полипептида FGF19 с водонерастворимым матриксом подложки или поверхности для применения в способах очистки антител, и наоборот. Широко используемые перекрестно-связывающие агенты включают в себя, например, 1,1-бис(диазоацетил)-2-фенилэтан, глутаральдегид, сложные эфиры N-гидроксисукцинимида, например, сложные эфиры 4-азидосалициловой кислоты, сложные гомобифункциональные имидоэфиры, включая сложные дисукцинимидиловые эфиры, такие как 3,3'-дитиобис(сукцинимидилпропионат), бифункциональные малеимиды, такие как бис-N-малеимидо-1,8-октан и такие агенты как метил-3-[(пара-азидофенил)дитио]пропиоимидат.
Другие модификации включают в себя дезаминирование глутаминиловых и аспарагиниловых остатков, соответственно, до глутамильного и аспартильного остатков, гидроксилирование пролина и лизина, фосфорилирование гидроксильных групп серильных или треонильных остатков, метилирование α-аминогрупп боковых цепей лизина, аргинина и гистидина [T. E. Creighton, Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco, pp. 79-86 (1983)], ацетилирование N-концевого амина и амидирование любой C-концевой карбоксильной группы.
Другой тип ковалентной модификации химерного полипептида FGF19, включенный в объем, охватываемый настоящим изобретением, включает изменение характера гликозилирования полипептида. Здесь под "изменением характера гликозилирования нативного полипептида" подразумевается делеция одного или более углеводных фрагментов, обнаруживаемых в соответствующем нативном полипептиде FGF19 и/или в полипептидной последовательности FGF21 (либо путем удаления участка, лежащего в основе гликозилирования, или либо путем удаления гликозилирования химическим и/или ферментативным путями), и/или добавление одного или более участков гликозилирования, которые отсутствуют в нативном полипептиде FGF19 и/или в полипептидной последовательности FGF21. Кроме того, указанная фраза подразумевает количественные изменения в гликозилировании нативных белков, включающие в себя изменение в природе и соотношениях различных имеющихся в наличии углеводных фрагментов.
Добавление гликозилированных участков к химерному полипептиду FGF19 может быть осуществлено путем изменения аминокислотной последовательности. Изменение может быть осуществлено, например, путем добавления или замены одного или более сериновых или треониновых остатков к химерному полипептиду FGF19 (для O-связанных участков гликозилирования). Аминокислотная последовательность химерного полипептида FGF19 необязательно может быть изменена через изменения на уровне ДНК, в частности, путем видоизменения ДНК, кодирующей химерный полипептид FGF19 на преселективной основе, так, чтобы образовывались кодоны, которые будут транслироваться в требуемые аминокислоты.
Другим способом увеличения числа углеводных фрагментов на химерном полипептиде FGF19 химическое или ферментативное присоединение гликозидов к полипептиду. Такие способы описаны в данной области, например, в международной заявке WO 87/05330, опубликованной 11 сентября 1987, а также в публикации Aplin and Wriston, CRC Crit. Rev. Biochem., pp. 259-306 (1981).
Удаление углеводных фрагментов, присутствующих на химерном полипептиде FGF19, может быть осуществлено химическим или ферментативным способом или же путем мутационной замены кодонов, кодирующих аминокислотные остатки, которые служат мишенями для гликозилирования. Технологии химического дегликозилирования известны в данной области и описаны, например, Hakimuddin, et al., Arch. Biochem. Biophys., 259: 52 (1987), а также Edge et al., Anal. Biochem., 118131 (1981). Ферментативное расщепление углуглеводных фрагментов на полипептидах может быть достигнуто с помощью различных эндо- и экзогликозидаз, как описано Thotakura et al., Meth. Enzymol., 138: 350 (1987).
Другой тип ковалентной модификации химерного полипептида FGF19 включает связывание химерного полипептида FGF19 с одной из разновидностей небелковых полимеров, например, с полиэтиленгликолем (ПЭГ), полипропиленгликолем или полиоксиалкиленами, способом, описанным в патенте США No. 4640835; 4496689; 4301144; 4670417; 4791192 или 4179337.
Химерный полипептид FGF19 согласно настоящему изобретению может быть модифицирован также и путем слияния химерного полипептида FGF19 с другим, гетерологичным полипептидом или аминокислотной последовательностью.
В одном из воплощений такая химерная молекула содержит слияние химерного полипептида FGF19 с меченым полипептидом, который обеспечивает эпитоп, с которым может избирательно связываться антитело против метки. Такой эпитоп-метку обычно помещают на амино- или карбокси-концах химерного полипептида FGF19. Наличие таких форм меченого эпитопа химерного полипептида FGF19 можно детектировать с помощью антитела против меченого полипептида. Кроме того, метка-эпитоп [или маркерный эпитоп] дает возможность легко очистить химерный полипептид FGF19, используя аффинную очистку с помощью антитела против метки или другого типа аффинность к матриксу, который связывается с меткой эпитопа. Различные меченые полипептиды и соответствующие им антитела хорошо известны в данной области. Примеры включают в себя полигистидиновую (poly-his) или полигистидин-глициновую (poly-his-gly) метки; flu-HA-меченый полипептид и его антитело 12CA5 [Field et al., Mol. Cell. Biol., 8: 2159-2165 (1988)]; c-myc-метка и ее антитела 8F9, 3C7, 6E10, G4, B7 и 9E10 [Evan et al., Molecular and Cellular Biology, 5: 3610-3616 (1985)]; и метка гликопротеина D (gD) простого вируса герпеса и его антитело [Paborsky et al., Protein Engineering, 3(6): 547-553 (1990)]. Другие меченые полипептиды включают в себя меченый пептид [Hopp et al., BioTechnology, 6: 1204-1210 (1988)]; пептид с KT3-эпитопом [Martin et al., Science, 255: 192-194 (1992)]; пептид с α-тубулиновым эпитопом [Skinner et al., J. Biol. Chem., 266: 15163-15166 (1991)]; пептидная метка белка 10 гена T7 [Lutz-Freyermuth et al., Proc. Natl. Acad. Sci. USA, 87: 6393-6397 (1990)].
В альтернативном воплощении полипептид согласно настоящему изобретению может содержать слияние химерного полипептида FGF19 с иммуноглобулином или с конкретной областью иммуноглобулина. Для двухвалентной формы химерной молекулы (называемой также "иммуноадгезином") такое слияние может быть осуществлено с участком Fc иммуноглобулина, аналогом участка Fc иммуноглобулина и одним или несколькими фрагментами участка Fc иммуноглобулина. В некоторых воплощениях указанный иммуноглобулин выбран из группы, состоящей из IgG-1, IgG-2, IgG-3, IgG-4, IgA-1, IgA-2, IgE, IgD и IgM. В некоторых воплощениях указанный участок Fc является человеческим или гуманизированным.
В некоторых воплощениях слиты C-конец химерного полипептида FGF19 и N-конец участка Fc. В некоторых воплощениях слиты N-конец химерного полипептида FGF19 и C-конец участка Fc. В некоторых воплощениях слияние иммуноглобулина включает в себя шарнирную область, области CH2 и CH3, или шарнирную область, области CH1, CH2 и CH3 молекулы IgG1. Для получения иммуноглобулиновых слияний см. также патент США No. 5428130, опубликованный 27 июня 1995. В некоторых воплощениях C-конец химерного полипептида FGF19 слит с N-концом участка Fc через линкер, при этом линкер выбран из группы, состоящей из линкера [Gly]n, линкера [Gly3Ser]m и линкера [Gly4Ser]m, где n представляет собой целое число от 1 до 30, и m представляет собой целое число от 1 до 6.
VII. Применение и способы с применением химерных полипептидов FGF19
Химерные полипептиды FGF19 и их описанные здесь модуляторы могут быть использованы также в качестве терапевтических средств. Химерные полипептиды FGF19 и их модуляторы согласно настоящему изобретению могут быть составлены в соответствии с известными способами получения композиций для фармацевтического применения, согласно которым описанные здесь химерные полипептиды FGF19 комбинируют в смеси с наполнителем, приемлемым фармацевтическим носителем. Терапевтические композиции получают для хранения путем смешивания активного ингредиента, имеющего требуемую степень чистоты, необязательно с физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)), в виде лиофилизированных композиций или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы в используемых дозах и концентрациях являются нетоксичными для реципиентов и включают в себя буферы, такие как фосфатный, цитратный буферы, а также буферы на основе других органических кислот; антиоксиданты, включая аскорбиновую кислоту; низкомолекулярные (состоящие менее чем приблизительно из 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желати или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон, аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатообразующие соединения, такие как ЭДТА; сахарные спирты, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионные поверхностно-активные вещества, такие как TWEENТМ, PLURONICSТМ или ПЭГ.
Композиции, предназначенные для использования in vivo, должны быть стерильными. Этого легко достичь путем фильтрации через стерильные фильтрующие мембраны до или после лиофилизации, с последующим восстановлением влагосодержания.
Указанные здесь терапевтические композиции обычно заключены в контейнер, имеющий стерильное отверстие для доступа, например, мешок с раствором для внутривенного введения или пузырек, имеющий пробку, поддающуюся прокалыванию иглой для подкожных впрыскиваний.
Путь введения соответствует известным способам, например, инъекции или инфузии путем внутривенного, внутрибрюшинного, внутрицеребрального, внутримышечного, внутриглазного, внутриартериального введения или введения внутрь пораженных тканей, местного введения или введения с использованием систем с замедленным высвобождением.
Дозировки и необходимые концентрации лекарственного средства в фармацевтических композициях согласно настоящему изобретению могут варьировать, в зависимости от конкретно предусматриваемого применения. Определение соответствующей дозировки или пути введения хорошо известны рядовым практикующим врачам. Эксперименты на животных являются надежным способом определения эффективных доз для лечения человека. Межвидовая отработка эффективных доз может быть предпринята в соответствии с принципами, сформулированными Mordenti, J. and Chappell, W. "The use of interspecies scaling in toxicokinetics" In Toxicokinetics and New Drug Development, Yacobi et al., Eds., Pergamon Press, New York 1989, pp. 42-96.
При введении in vivo химерного полипептида FGF19 или его агониста или антагониста, нормальные количественные дозировки могут варьировать от приблизительно 10 нг/кг вплоть до 100 мг/кг веса тела млекопитающего или более в день, предпочтительно, от приблизительно 1 мкг/кг/день до 10 мг/кг/день, в зависимости от пути введения. Руководства в отношении конкретных дозировок и способов введения представлены в литературе; см., например, патенты США No. 4657760; 5206344; или 5225212. Считается, что в случае разных лекарственных соединений и разных расстройств будут эффективны разные композиции, что если при введении, направленном на один орган или ткань, используется один путь, то для другого органа или ткани может быть необходим другой путь введения.
Когда требуется введение химерного полипептида FGF19 или его модулятора с замедленным высвобождением в композиции с характеристиками высвобождения, подходящими для лечения какого-нибудь заболевания или расстройства, требующего введения химерного полипептида FGF19 или модулятора, предусмотрена микроинкапсуляция. Микроинкапсуляция рекомбинантных белков для замедленного высвобождения успешно осуществлена в случае с человеческим гормоном роста (rhGH), интерфероном- (rhlFN-), интерлейкином-2 и MN rgpl20, см. Johnson et al., Nat. Med., 2:795-799 (1996); Yasuda, Biomed. Ther., 27: 1221-1223 (1993); Hora et al., Bio/Technology 8: 755-758 (1990); Cleland, "Design and Production of Single Immunization Vaccines Using Polylactide Polyglycolide Microsphere Systems," in Vaccine Design: The Subunit and Adjuvant Approach, Powell and Newman, eds, (Plenum Press: New York, 1995), pp. 439-462; WO 97/03692, WO 96/40072, WO 96/07399; а также патент США No. 5654010.
Композиции с замедленным высвобождением указанных белков разрабатывали с помощью сополимера молочной и гликолевой кислоты (PLGA), в силу его биодоступности и его подверженности биодеградации в широких пределах. Продукты деградации PLGA, молочная и гликолевая кислоты, могут быстро выводиться из человеческого организма. Более того, подверженность указанного полимера биодеградации можно регулировать, от месяцев до лет, в зависимости от его молекулярной массы и состава. Lewis. "Controlled release of bioactive agents from lactide/glycolide polymer," в: M. Chasin and R. Langer (Eds.), Biodegradable Polymers as Drug Delivery Systems (Marcel Dekker: New York, 1990), pp. 1-41.
Терапевтические средства и композиции, содержащие предложенный здесь химерный полипептид FGF19, могут иметь целый ряд различных применений. Указанные применения включают в себя лечение индивида с ожирением или с состоянием, ассоциированным с ожирением. В одном из аспектов химерный полипептид FGF19 вводят индивиду, в случае необходимости, в количестве, эффективном для лечения указанного состояния. Предпочтительно, чтобы состояние было таким, при котором требуется, по меньшей мере, одно из следующих воздействий: понижение уровня глюкозы в крови, усиление метаболизма, снижение массы тела, уменьшение количества телесного жира, понижение уровня триглицеридов, уменьшение количества свободных жирных кислот, повышение уровня высвобождения глюкозы из адипоцитов и/или повышение уровня высвобождения лептина из адипоцитов. Каждый из указанных параметров может быть измерен стандартными способами, например, путем измерения поглощения кислорода с целью определения скорости метаболизма, с использованием шкал для определения веса, а также путем определения измеряемого размера для определения телесного жира. Кроме того, наличие и количество триглицеридов, свободных жирных кислот, глюкозы и лептина может быть определено стандартными способами. Указанные применения включают в себя лечение индивида, страдающего от одного или более из следующих расстройств: диабета 1 типа, диабета 2 типа, высокого содержания глюкозы в крови, метаболического синдрома, атеросклероза, гиперхолестеринемии, инсульта, остеопороза, остеоартрита, дегенеративного заболевания суставов, мышечной атрофии, саркопении, снижения безжировой массы тела, облысения, морщинистости, повышенной усталости, пониженной выносливости, пониженной функции сердца, дисфункции иммунной системы, злокачественной опухоли, болезни Паркинсона, старческого слабоумия, болезни Альцгеймера и пониженной когнитивной функции.
Химерный полипептид FGF19 и композиции, содержащие химерный полипептид FGF19, предпочтительно применяются in vivo. Однако, как описано ниже, их можно вводить и in vitro, например, в способах, описанных ниже в отношении скрининга модуляторов химерного полипептида FGF19. Хотя и очевидно, что химерный полипептид FGF19 может также быть идентифицирован с использованием животных моделей и образцов, полученных от индивидов.
Настоящее изобретение включает также аспекты, при которых химерный полипептид FGF19 согласно настоящему изобретению или содержащую его фармацевтическую композицию вводят индивиду в сочетании со вторым средством, при этом указанным вторым средством предпочтительно является фармакологическое средство. В некоторых воплощениях химерный полипептид FGF19 согласно настоящему изобретению или содержащую его фармацевтическую композицию вводят в терапевтически эффективном количестве в сочетании с терапевтически эффективным количеством второго средства. В некоторых воплощениях химерный полипептид FGF19 или содержащую его фармацевтическую композицию вводят в сочетании со вторым средством, то есть соответствующие периоды введения являются частью единого режима введения. В некоторых воплощениях химерный полипептид FGF19 или содержащую его фармацевтическую композицию вводят одновременно, то есть соответствующие периоды введения друг с другом перекрываются. В некоторых воплощениях химерный полипептид FGF19 или содержащую его фармацевтическую композицию и указанное второе средство вводят неодновременно, то есть соответствующие периоды введения друг с другом не перекрываются. В некоторых воплощениях химерный полипептид FGF19 или содержащую его фармацевтическую композицию и указанное второе средство вводят последовательно, то есть химерный полипептид FGF19 или содержащую его фармацевтическую композицию вводят до и/или после введения второго средства. В некоторых воплощениях химерный полипептид FGF19 или содержащую его фармацевтическую композицию и указанное второе средство вводят одновременно в виде отдельных композиций. В некоторых воплощениях химерный полипептид FGF19 или содержащую его фармацевтическую композицию и указанное второе средство вводят одновременно в виде части одной и той же композиции.
В некоторых воплощениях указанное второе средство является другим химерным полипептидом FGF19 согласно настоящему изобретению. В некоторых воплощениях указанное второе средство является противовоспалительным средством, противодиабетическим средством и/или средством, понижающим уровень холестерина, относящимся к классу "статинов". В некоторых воплощениях указанным вторым средством является инсулин. В некоторых воплощениях указанный инсулин является быстродействующим инсулином, инсулином кратковременного действия, инсулином регулярного действия, инсулином промежуточного действия или инсулином длительного действия. В некоторых воплощениях указанный инсулин представляет собой и/или включает препараты хумалог, лиспро, новолог, апидра, хумалин, аспарт, инсулин регулярного действия, NPH, лента, ультралента, лантус, гларгин, левемир или детемир. В некоторых воплощениях указанным вторым активным средством является статин. В некоторых воплощениях указанный статин представляет собой и/или включает аторвастатин (например, липитор или торваст), церивастатин (например, липобай или бейкол), флувастатин (например, лескол), ловастатин (например, мевакор, алтокор или алтопрев) мевастатин, питавастатин (например, ливало или питава), правастатин (например, правахол, селектин или липостат) розувастатин (например, крестор), симвастатин (например, зокор или липекс), виторин, адвикор, безилат, кадуэт или симкор.
В другом аспекте настоящего изобретения химерный полипептид FGF19 согласно настоящему изобретению или содержащую его фармацевтическую композицию вводят индивиду в сочетании со вторым способом лечения, предпринимаемым в отношении указанного индивида, при этом указанный второй способ лечения включает хирургическое вмешательство. В некоторых воплощениях химерный полипептид FGF19 согласно настоящему изобретению или содержащую его фармацевтическую композицию вводят индивиду в сочетании со вторым способом, или приемом, терапии. В некоторых воплощениях химерный полипептид FGF19 или содержащую его фармацевтическую композицию вводят в сочетании со вторым способом терапии, то есть указанное введение и указанная терапия являются частью единого режима введения. В некоторых воплощениях химерный полипептид FGF19 или содержащую его фармацевтическую композицию вводят одновременно со вторым способом терапии, то есть соответствующие периоды введения и терапии друг с другом перекрываются. В некоторых воплощениях химерный полипептид FGF19 или содержащую его фармацевтическую композицию вводят неодновременно со вторым способом терапии, то есть соответствующие периоды введения и терапии друг с другом не перекрываются. В некоторых воплощениях химерный полипептид FGF19 или содержащую его фармацевтическую композицию вводят последовательно со вторым способом терапии, то есть химерный полипептид FGF19 или содержащую его фармацевтическую композицию вводят до и/или после указанной второй терапии. В некоторых воплощениях химерный полипептид FGF19 или содержащую его фармацевтическую композицию вводят одновременно с указанной второй терапией.
Описанные здесь химерные полипептиды FGF19 могут также быть использованы в качестве маркеров молекулярных весов в способах электрофореза белков.
Химерные полипептиды FGF19 и молекулы нуклеиновых кислот согласно настоящему изобретению могут также быть использованы для типирования тканей, при этом химерные полипептиды FGF19 согласно настоящему изобретению могут быть по-разному экспрессированы в одной ткани по сравнению с другой тканью. Молекулы нуклеиновых кислот химерного полипептида FGF19 найдут применение в области получения зондов для ПЦР, назерн-блоттинга, саузерн-блоттинга и вестерн-блоттинга.
Химерные полипептиды FGF19 согласно настоящему изобретению, которые связываются с другим белком (например, одним из FGFR), могут быть использованы в анализах по идентификации других белков или молекул, участвующих в связывающем взаимодействии. Используя такие методы, можно идентифицировать ингибиторы связывающего взаимодействия рецепторов/лигандов. Белки, участвующие в таких связывающих взаимодействиях, могут быть использованы также для скрининга пептидов или низкомолекулярных ингибиторов или агонистов связывающего взаимодействия. Кроме того, химерный полипептид FGF19 может быть использован для выделения соответствующего лиганда(лигандов). Анализы скрининга могут быть построены таким образом, чтобы можно было обнаружить лидирующие соединения, которые имитируют биологическую активность нативного полипептида FGF19, нативного полипептида FGF21, химерного полипептида FGF19 или рецептора FGF19 и/или FGF21. Такие анализы скрининга будут включать в себя анализы, позволяющие осуществлять высокопроизводительный скрининг химических библиотек, делая их практически подходящими для идентификации низкомолекулярных лекарственных средств-кандидатов. Предполагаемые низкомолекулярные лекарственные средства включают в себя искусственные органические или неорганические соединения. Указанные анализы могут быть предприняты в различных форматах, включая анализы белок-белкового взаимодействия, анализы биохимического скрининга, иммуноанализы и анализы на основе клеток, которые в данной области хорошо разработаны.
В качестве альтернативного подхода для идентификации рецепторов, меченый химерный полипептид FGF19 может быть фотоаффинным методом связан с клеточной мембраной или препаратами экстрактов, которые экспрессируют рецепторную молекулу. Перекрестно-связанный материал разделяли на составляющие компоненты путем ПААГ-ЭФ и снимали на рентгеновскую пленку. Меченый комплекс, содержащий рецептор, можно было вычленить, разделить на пептидные фрагменты и подвергнуть белковому микросеквенированию. Аминокислотную последовательность, полученную в результате микросеквенирования, можно было использовать для создания набора вырожденных олигонуклеотидных зондов для скрининга библиотеки кДНК с целью идентификации геноа, кодирующего предполагаемый рецептор.
В одном из описанных здесь воплощений, когда предпринимаются анализы конкурентного связывания, в качестве конкурента применяются FGFR1c, FGFR2c, FGFR3c и/или FGFR4c, или антитело против химерного полипептида FGF19.
VIII. Антитела против химерного полипептида FGF19
1. Поликлональные антитела
Антитела против химерного полипептида FGF19 могут включать в себя поликлональные антитела. Способы получения поликлональных антител известны специалистам в данной области. Поликлональные антитела могут быть получены у млекопитающего, например, в результате одной или нескольких инъекций иммунизирующего средства и, если это необходимо, также и адъюванта. Обычно иммунизирующее средство и/или адъювант вводят млекопитающему в виде многочисленных подкожных или внутрибрюшинных инъекций. Иммунизирующее средство может включать в себя химерный полипептид FGF19 или слитый с ним белок. Это может оказаться полезным для конъюгации иммунизирующего средства с белком, известным своей иммуногенностью в отношении иммунизируемого млекопитающего. Примеры таких иммуногенных белков включают в себя, но не ограничены перечисленным, гемоцианин лимфы улитки, сывороточный альбумин, бычий тиреоглобулин и ингибитор трипсина из соевых бобов. Примеры адъювантов, которые могут быть использованы, включают в себя полный адъювант Фрейнда и адъювант MPL-TDM (монофосфориллипид A, синтетический дикориномиколят трегалозы). Протокол иммунизации может быть выбран специалистом в данной области без излишнего экспериментирования.
2. Моноклональные антитела
Альтернативно, антитела против химерного полипептида FGF19 могут быть моноклональными антителами. Моноклональные антитела могут быть получены гибридомными способами, такими как способы, описанные Kohler and Milstein, Nature, 256: 495 (1975). В гибридомном способе, мышь, хомяка или другое подходящее животное-хозяин обычно иммунизируют иммунизирующим средством, чтобы получить лимфоциты, которые продуцируют или способны продуцировать антитела, которые будут специфически связываться с иммунизирующим средством. Альтернативно, лимфоциты могут быть иммунизированы in vitro. В предпочтительных воплощениях антитело против химерного полипептида FGF19 специфически связывается с полипептидом согласно настоящему изобретению. В более предпочтительных воплощениях специфически-связывающееся антитело не связывается с нативным полипептидом FGF19 или нативным полипептидом FGF21.
Иммунизирующее средство обычно будет включать в себя химерный полипептид FGF19 или слитый с ним белок. Обычно используются либо лимфоциты периферической крови ("PBL"), если требуются клетки человеческого происхождения, либо клетки селезенки или клетки лимфатических узлов, если требуются клетки млекопитающего, не являющегося человеком. Затем лимфоциты сливают с иммортализованной клеточной линией с помощью подходящего средства для слияния, такого как полиэтиленгликоль, с образованием гибридомной клетки [Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, (1986) pp. 59-103]. Линии иммортализованных клеток обычно представляют собой трансформированные клетки млекопитающих, в частности, миеломные клетки грызунов, бычьи клетки и клетки человеческой природы. Обычно используются миеломные линии клеток крысы или мыши. Гибридомные клетки могут культивироваться в подходящей культуральной среде, которая предпочтительно содержит одно или более веществ, которые ингибируют рост или выживаемость не слитых иммортализованных клеток. Например, если родительские клетки лишены фермента гипоксантингуанинфосфорибозилтрансферазы (HGPRT или HPRT), в таких случаях обычно культуральная среда будет включать в себя гипоксантин, аминоптерин и тимидин ("среда HAT"), при этом указанные вещества препятствуют росту HGPRT-дефицитных клеток.
Предпочтительными иммортализованными линиями клеток обычно являются такие линии клеток, которые эффективно образуют слияния, поддерживают стабильно высокий уровень экспрессии антитела селективными антитело-продуцирующими клетками и являются чувствительными к такой среде как среда HAT. Более предпочтительными иммортализованными линиями клеток являются мышиные миеломные линии, которые могут быть получены, например, из Центра распределения клеток Института Солка, Сан-Диего, Калифорния, и из Американской Коллекции Типовых Культур, Manassas, Ванкувер. Описано также использование линий клеток миеломы человека и гетеромиеломы человек-мышь для получения человеческих моноклональных антител [Kozbor, J. Immunol., 133: 3001 (1984); Brodeur et al., Monoclonal Antibody Production Технологии and Applications, Marcel Dekker, Inc., New York, (1987) pp. 51-63].
Затем культуральную среду, в которой культивируют гибридомные клетки, можно анализировать на предмет определения наличия моноклональных антител против химерного полипептида FGF19. Предпочтительно, чтобы специфичность связывания моноклональных антител, продуцируемых гибридомными клетками, определялась путем иммунопреципитации или путем анализа связывания in vitro, такого как радиоиммуноанализ (RIA) или твердофазный иммуноферментный анализ (ELISA). Такие технологии и методы исследования известны в данной области. Аффинность связывания моноклонального антитела может быть определена, например, анализом Скэтчарда, см. Munson and Pollard, Anal. Biochem., 107: 220 (1980).
После того как идентифицированы требуемые гибридомные клетки, клоны могут быть подвергнуты субклонированию с помощью методов последовательного разведения и стандартных методов выращивания клеток [Goding, выше]. Подходящие культуральные среды для этой цели включают в себя, например, модифицированную Дульбекко среду Игла и среду RPMI-1640. Альтернативно, гибридомные клетки можно выращивать in vivo у млекопитающего в виде асцитных клеток.
Моноклональные антитела, секретируемые азанными субклонами, могут быть изолированы или очищены от культуральной среды или асцитной жидкости с помощью общеизвестных процедур очистки иммуноглобулинов, таких, например, как протеин A-сефароза, хроматография на гидроксилапатите, гель-электрофорез, диализ или аффинная хроматография.
Моноклональные антитела могут быть также получены с помощью технологий рекомбинантных ДНК, например, описанных в патенте США No. 4816567. ДНК, кодирующую моноклональные антитела согласно изобретению, могут быть легко изолированы и секвенированы с помощью общепринятых методов (например, с использованием олигонуклеотидные зонды, которые способны специфически связываться с генами, кодирующими тяжелую и легкую цепи мышиных антител). Гибридомные клетки согласно настоящему изобретению служат в качестве предпочтительного источника такой ДНК. Сразу же после выделения указанную ДНК можно встроить в экспрессирующие векторы, которые затем трансфицируют в клетке-хозяине, такие как обезьяньи клетки COS, клетки яичника китайского хомячка (CHO) или миеломные клетки, которые иначе не продуцируют иммуноглобулины, чтобы добиться синтеза моноклональных антител в рекомбинантных клетках-хозяевах. Указанную ДНК также можно модифицировать, например, путем замены кодирующей последовательности человеческими константными доменами тяжелой и легкой цепей вместо гомологичных мышиных последовательностей [Патент США No. 4816567; Morrison et al., выше] или путем ковалентного связывания с иммуноглобулин-кодирующей последовательностью полной или частичной кодирующей последовательности не-иммуноглобулинового полипептида. Такой не-иммуноглобулиновый полипептид может быть использован вместо константных доменов антитела согласно изобретению или может быть использован вместо вариабельных доменов одного антиген-связывающего участка антитела согласно изобретению для создания двухвалентного химерного антитела.
Антитела могут быть моновалентными антителами. Способы получения моновалентных антител хорошо известных в данной области. Например, один из способов включает рекомбинантную экспрессию легкой цепи и модифицированной тяжелой цепи иммуноглобулина. Тяжелую цепь обычно укорачивают в любой точке области Fc, с тем, чтобы предотвратить перекрестное связывание с тяжелой цепью. Альтернативно, релевантные цистеиновые остатки заменяют другим аминокислотным остатком или удаляют, с тем, чтобы предотвратить перекрестное связывание с тяжелой цепью.
Методы in vitro также являются подходящими для получения моновалентных антител. Расщепление антител с целью получения их фрагментов, в частности, Fab-фрагментов, может быть осуществлено с помощью общеизвестных в данной области технологий.
3. Человеческие и гуманизированные антитела
Антитела против химерного полипептида FGF19 согласно изобретению могут дополнительно включать в себя гуманизированные антитела или человеческие антитела. Гуманизированные формы не-человеческих (например, мышиных) антител являются иммуноглобулинами, цепями иммуноглобулинов или их фрагментами (такими как Fv, Fab, Fab', F(ab')2 или другими антиген-связывающими последовательностями антител), которые содержат минимальную последовательность, полученную из не-человеческого иммуноглобулина. Гуманизированные антитела включают в себя человеческие иммуноглобулины (реципиентское антитело), в которых остатки из определяющей комплементарность области (CDR) реципиента заменены остатками из области CDR вида, не относящегося к человеческому (донорское антитело), например, мыши, крысы или кролика, обладающими требуемой специфичностью, аффинностью и емкостью. В некоторых случаях остатки рамки считывания Fv человеческого иммуноглобулина заменены соответствующими остатками, обнаруживаемыми у вида, не относящегося к человеческому. Гуманизированные антитела могут также включать в себя остатки, которые не обнаруживаются ни в последовательностях реципиентского антитела, ни в заимствованных последовательностях CDR или рамки считывания. Как правило, гуманизированное антитело будет содержать по существу все из, по меньшей мере, одного, а обычно двух, вариабельных доменов, в которых все или по существу все из CDR-областей соответствуют таковым не-человеческого иммуноглобулина, и все или по существу все из FR-областей являются областями консенсусной последовательности человеческого иммуноглобулина. Оптимально гуманизированное антитело будет также содержать, по меньшей мере, участок константной области (Fc) иммуноглобулина, обычно человеческого иммуноглобулина [Jones et al., Nature, 321: 522-525 (1986); Riechmann et al., Nature, 332: 323-329 (1988); а также Presta, Curr. Op. Struct. Biol., 2: 593-596 (1992)].
Способы гуманизации не-человеческих антител хорошо известны в данной области. Обычно гуманизированное антитело имеет один или более аминокислотных остатков, встроенных в него из организма, не являющегося человеческим организмом. Указанные не-человеческие аминокислотные остатки часто называют "импортными" остатками, которые обычно взяты из "импортного" вариабельного домена. В основном гуманизация может быть предпринята в соответствии со способом, описанным Winter и соавторами [Jones et al., Nature, 321: 522-525 (1986); Riechmann et al., Nature, 332: 323-327 (1988); Verhoeyen et al., Science, 239: 1534-1536 (1988)], путем замены участков CDR или последовательностей CDR соответствующими последовательностями человеческого антитела. Соответственно, такие "гуманизированные" антитела являются химерными антителами (патент США No. 4816567), в которых по существу менее чем интактный человеческий вариабельный домен заменен соответствующей последовательностью, полученной из вида, не являющегося человеком. На практике гуманизированные антитела обычно являются человеческими антителами, в которых некоторые остатки CDR и, возможно, некоторые остатки FR замещены остатками из аналогичных участков антител грызунов.
Человеческие антитела могут также быть получены с помощью различных технологий, известных в данной области, включая библиотеки фагового дисплея [Hoogenboom and Winter, J. Mol. Biol., 227:381 (1991); Marks et al., J. Mol. Biol. 222: 581 (1991)]. Технологии, описанные Cole с соавт. и Boerner с соавт., также доступны для получения человеческих моноклональных антител (Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77 (1985), а также Boerner et al., J. Immunol., 147(l): 86-95 (1991)]. Сходным образом, человеческие антитела могут быть получены путем встраивания локусов человеческого иммуноглобулина трансгенным животным, например, мышам, у которых гены эндогенного иммуноглобулина частично или полностью инактивированы. В результате наблюдается продукция человеческого антитела, которое похоже на человеческие антитела во всех отношениях, включая реаранжировку генов, их сборку и репертуар антител. Указанный подход описан, например, в патенте США No. 5545807; 5545806; 5569825; 5625126; 5633425; 5661016, а также в следующих научных публикациях: Marks et al., Bio/Technology 10, 779-783 (1992); Lonberg et al., Nature 368 856-859 (1994); Morrison, Nature 368, 812-13 (1994); Fishwild et al., Nature Biotechnology 14, 845-51 (1996); Neuberger, Nature Biotechnology 14, 826 (1996); Lonberg and Huszar, Intern. Rev. Immunol. 13 65-93 (1995).
4. Биспецифические антитела
Биспецифические антитела являются моноклональными, предпочтительно, человеческими или гуманизированными, антителами, которые обладают специфичностями связывания, по меньшей мере, в отношении двух различных антигенов. В данном случае одна из указанных специфичностей связывания направлена на FGF19, а другая направлена на любой другой антиген, и предпочтительно, на белок или рецептор, или же субъединицу рецептора клеточной поверхности.
Способы получения биспецифических антител известны в данной области. Рекомбинантная продукция биспецифических антител основана на совместной экспрессии двух пар иммуноглобулиновых тяжелых цепей/легких цепей, где указанные две тяжелые цепи имеют различные специфичности [Milstein and Cuello, Nature, 305: 537-539 (1983)]. В силу случайного расхождения генов иммуноглобулиновых тяжелых цепей и легких цепей, указанные гибридомы (квадромы) продуцируют потенциальную смесь из десяти различных молекул антител, из которых только одна имеет правильную биспецифическую структуру. Очистка правильной молекулы обычно осуществляется постадийной аффинной хроматографией. Сходные приемы описаны в международной заявке WO 93/08829, опубликованной 13 мая 1993, а также в публикации Traunecker et al., EMBO J., 10: 3655-3659 (1991).
Вариабельные домены антитела с требуемыми специфичностями связывания (комбинированные участки связывания антиген-антитело) могут быть слиты с последовательностями константных доменов иммуноглобулина. Такое слияние предпочтительно с константным доменом тяжелой цепи иммуноглобулина, включающей в себя, по меньшей мере, часть шарнирной части, областей CH2 и CH3. Предпочтительно иметь первую константную область тяжелой цепи, содержащую участок, необходимый для связывания легкой цепи, присутствующей, по меньшей мере, в одном из слияний. ДНК, кодирующие слияния тяжелой цепи иммуноглобулина, и, в случае необходимости, легкую цепь иммуноглобулина, встраивают в отдельные экспрессирующие векторы и совместно трансфицируют в соответствующий организм-хозяин. В отношении дальнейших подробностей выработки биспецифических антител см., например, Suresh et al., Methods in Enzymology, 121: 210 (1986).
В соответствии с другим подходом, описанным в международной заявке WO 96/27011, поверхность контакта между парой молекул антител может быть сконструирована с тем, чтобы максимизировать процент гетеродимеров, которые получены из культуры рекомбинантных клеток. Предпочтительная поверхность контакта включает в себя, по меньшей мере, часть области CH3 константного домена антитела. В указанном способе одна или более боковых цепей из малых аминокислот из области поверхности контакта первой молекулы антитела заменены более крупными боковыми цепями (например, тирозином или триптофаном). Компенсаторно на поверхности контакта второй молекулы антитела создают "полости" размером, идентичным или сходным с размером крупной боковой цепи (крупным боковым цепям), путем замены боковых цепей из крупных аминокислот более мелкими (например, аланином или треонином). Это обеспечивает механизм повышения выхода гетеродимеров над такими нежелательными конечными продуктами как гомодимеры.
Биспецифические антитела могут быть получены в виде полноразмерных антител или фрагментов антител (например, F(ab')2- биспецифических антител). Технологии получения биспецифических антител из фрагментов антител описаны в литературе. Например, биспецифические антитела могут быть получены с помощью химической связи. Brennan et al., Science 229: 81 (1985) описывают методологию, согласно которой интактные антитела подвергают протеолитическому расщеплению, с получением F(ab')2- фрагментов. Указанные фрагменты восстанавливают в присутствии дитиолового комплексообразующего агента арсенита натрия для стабилизации вицинальных дитиолов и предотвращения образования межмолекулярных бисульфидных связей. Затем полученные Fab'-фрагменты преобразуют в производные тионитробензоата (TNB). Затем одно из производных Fab'-TNB превращают в Fab'-тиол путем восстановления меркаптометиламином и смешивают с эквимолярным количеством другого производного Fab'-TNB, с образованием биспецифического антитела. Полученные биспецифические антитела могут быть использованы в качестве средств для селективной иммобилизации ферментов.
Fab'-фрагменты могут быть непосредственно получены из E. coli и химически спарены, с образованием биспецифических антител. Shalaby et al., J. Exp. Med. 175: 217-225 (1992) описывают продукцию молекулы F(ab')2 полностью гуманизированного биспецифического антитела. Каждый Fab'-фрагмент был отдельно секретирован из E. coli и подвергнут прямому химическому спариванию in vitro с образованием биспецифического антитела. Образованное таким образом биспецифическое антитело было способно связываться с клетками, экспрессирующими в повышенном количестве рецептор ErbB2, и с нормальными человеческими T-клетками, а также запускать литическую активность человеческих цитотоксических лимфоцитов против человеческих злокачественных опухолей молочные железы в качестве мишени.
Описаны также различные технологии получения и выделения фрагментов биспецифических антител непосредственно из культуры рекомбинантных клеток. Например, биспецифические антитела были получены с помощью лейциновых «молний». Kostelny et al., J. Immunol. 148(5): 1547-1553 (1992). Пептиды лейциновых «молний» из белков Fos и Jun соединяли с Fab'-фрагментами двух различных антител путем слияния генов. Гомодимеры антитела восстанавливали в шарнирной области, с образованием мономеров, а затем реоксидировали, с получением гетеродимеров антитела. Указанный способ может быть использован также для продуцирования гомодимеров антитела. Технология получения "диатела", описанная Hollinger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993), обеспечила альтернативный механизм получения фрагментов биспецифического антитела. Указанные фрагменты включают в себя вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) посредством линкера, который является слишком коротким, чтобы допустить спаривание между указанными двумя доменами на одной и той же цепи. Соответственно, VH- и VL-домены одного фрагмента вынуждены спариваться с VL- и VH-доменами другого фрагмента, образуя таким образом два антиген-связывающих участка. Описана также и другая стратегия получения фрагментов биспецифических антител с помощью одноцепочечных Fv (sFv)-димеров. См. Gruber et al., J. Immunol. 152: 5368 (1994). Предусмотрены антитела более чем с двумя валентностями. Например, могут быть получены триспецифические антитела. Tutt et al., J. Immunol. 147: 60 (1991).
Здесь иллюстративные биспецифические антитела могут связываться с разными эпитопами на данном полипептиде FGF19. Альтернативно, плечо полипептида против химерного полипептида FGF19 может быть соединено с плечом, которое связывается с триггерной молекулой на лейкоците, такой как молекула рецептора T-клеток (например, CD2, CD3, CD28 или B7), или рецепторов Fc IgG (FcγR), таких как FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD 16), с тем, чтобы сфокусировать клеточные защитные механизмы на клетках, экспрессирующих конкретный полипептид FGF19. Биспецифические антитела могут быть использованы также для локализации цитотоксических средств на клетках, которые экспрессируют конкретный полипептид FGF19. Указанные антитела имеют FGF19-связывающее плечо, а также плечо, которое связывается с цитотоксическим средством или радионуклидным хелатообразующим соединением, таким как EOTUBE, DPTA, DOTA или TETA. Другое представляющее интерес биспецифическое антитело связывается с полипептидом FGF19 и дополнительно связывается с тканевым фактором (TF).
5. Антитела-гетероконъюгаты
Антитела-гетероконъюгаты также входят в объем, охватываемый настоящим изобретением. Антитела-гетероконъюгаты состоят из двух ковалентно связанных антител. Была, например, вынесена на рассмотрение возможность того, чтобы такие антитела нацеливали клетки иммунной системы на нежелательные клетки [патент США No. 4676980], а также были использованы для лечения ВИЧ-инфекции [WO 91/00360; WO 92/200373; EP 03089]. Рассматривается возможность того, что такие антитела могут быть получены in vitro с помощью известных методов химического синтеза белков, включая методы, в которых используются поперечно-сшивающие агенты. Например, иммунотоксины могут быть сконструированы с помощью реакции дисульфидного обмена или путем образования тиоэфирной связи. Примеры соответствующих реагентов для такой цели включают в себя иминотиолат и метил-4- меркаптобутиримидат, а также и другие, описанные, например, в патенте США No. 4676980.
6. Конструирование эффекторной функции
Может оказаться желательным модифицировать антитело согласно изобретению в отношении эффекторной функции, таким образом, чтобы усилить, например, эффективность антитела. Например, цистеиновый остаток(остатки) может быть встроен в область Fc, создавая таким образом возможность образования в указанной области межцепьевой дисульфидной связи. Полученное таким образом гомодимерное антитело может иметь улучшенную способность к интернализации и/или повышенную комплемент-опосредованную киллерную функцию в отношении клеток и антитело-зависимую клеточную цитотоксичность (ADCC). См. Caron et al., J. Exp Med., 176: 1191-1195 (1992) и Shopes, J. Immunol., 148: 2918-2922 (1992). В одном из примеров гомодимерные антитела с повышенной противоопухолевой активностью могут быть получены также с помощью гетеробифункциональных поперечных линкеров, как описано у Wolff et al. Cancer Research, 53: 2560-2565 (1993). Альтернативно, антитело может быть сконструировано таким образом, чтобы оно имело двойственные области Fc и могло, таким образом, иметь повышенные активности в отношении комплемент-зависимого лизиса и в отношении ADCC. См. Stevenson et al., Anti-Cancer Drug Design, 3: 219-230 (1989).
7. Иммуноконъюгаты
Настоящее изобретение относится также к иммуноконъюгатам, содержащим антитело, конъюгированное с цитотоксическим средством, таким как химиотерапевтическое средство, токсин (например, ферментативно активный токсин бактериального, грибкового, растительного или животного происхождения, или его фрагменты), или радиоактивный изотоп (то есть радиоактивный конъюгат).
Химиотерапевтические средства, применимые для получения таких иммуноконъюгатов, описаны выше. Ферментативно активные токсины и их фрагменты, которые могут быть использованы, включают в себя дифтерийную A-цепь, не связывающиеся активные фрагменты дифтерийного токсина, A-цепь эндотоксина (из Pseudomonas aeruginosa), A-цепь рицина, A-цепь абрина, A-цепь модецина, альфа-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор momordica charantia, курцин, кротин, ингибитор sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. Для продуцирования радиоконъюгированных антител доступны различные ралионуклиды. Примеры включают в себя 212Bi, 131I, 131In, 90Y и 186Re.
Конъюгаты антитела и цитотоксического средства образуются с помощью разнообразных бифункциональных связующих с белком веществ, таких как N-сукцинимидил-3-(2-пиридилдитиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат HCL), активные сложные эфиры (такие как дисукцинимидил суберат), альдегиды (такие как глутаральдегид), бис-азидо-соединения (такие как бис(пара-азидобензоил) гександиамин), производные бис-диазония (такие как бис-(пара-диазонийбензоил)-этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат), и бис-активные фтористые соединения (такие как l,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин может быть получен, как описано у Vitetta et al., Science, 238: 1098 (1987). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилен-триаминпентауксусная кислота (MX-DTPA) является иллюстративным хелатообразующим соединением для конъюгации радионуклида с антителом. См. WO94/11026.
В другом воплощении антитело может быть конъюгировано с "рецептором" (таким как стрептавидин) для использования в пре-таргетировании опухолей, где указанный конъюгат антитело-рецептор вводят индивиду, с последующим удалением несвязанного конъюгата из кровотока с помощью осветляющего вещества, а затем вводят "лиганд" (например, авидин), который конъюгирован с цитотоксическим средством (например, радионуклидом).
8. Иммунолипосомы
Описанные здесь антитела могут быть составлены также в виде иммунолипосом. Липосомы, содержащие антитело, получают, используя методы, известные в данной области, такие как методы, описанные у Epstein et al., Proc. Natl. Acad. Sci. USA, 82: 3688 (1985); Hwang et al., Proc. Natl. Acad. Sci. USA, 77: 4030 (1980); и Патенты США No. 4485045 и 4544545. Липосомы с повышенным временем циркуляции описаны в патенте США No. 5013556.
Особенно полезные липосомы могут быть получены способом выпаривания с обращенной фазой, с липидной композицией, содержащей фосфатидилхолин, холестерин и ПЭГ-дериватизированный фосфатидилэтаноламин (ПЭГ-ФЭА). Липосомы экструдировали через фильтры с определенным размером пор, с получением липосом определенного диаметра. Fab'-фрагменты антитела согласно настоящему изобретению могут быть конъюгированы с липосомами, как описано у Martin et al., J. Biol. Chem., 257: 286-288 (1982), через реакцию обмена дисульфидами.
9. Фармацевтические композиции антител
Антитела, специфически связывающиеся с идентифицируемым здесь полипептидом FGF19, так же как и с другими молекулами, идентифицируемыми посредством описываемых выше анализов скрининга, могут быть назначены для лечения различных расстройств в виде фармацевтических композиций.
Если полипептид FGF19 является внутриклеточным, и в качестве ингибиторов используются полноразмерные антитела, интернализирующиеся антитела являются предпочтительными. Однако для доставки антитела или фрагмента антитела в клетки могут быть использованы также липофекция или липосомы. Когда используются фрагменты, предпочтительным является наименьший ингибиторный фрагмент, который специфически связывается со связывающим доменом белка-мишени. Например, на основе последовательностей вариабельной области антитела могут быть сконструированы пептидные молекулы, которые сохраняют способность связываться с последовательностью белка-мишени. Такие пептиды могут быть синтезированы химическим путем и/или получены на основе технологии рекомбинантной ДНК. См., например, Marasco et al., Proc. Natl. Acad. Sci. USA, 90: 7889-7893 (1993). Здесь композиция может содержать также более одного активного соединения, если это необходимо для лечения в случае конкретных показаний, предпочтительно, соединений с комплементарными активностями, которые не оказывают отрицательного влияния друг на друга. Альтернативно, или дополнительно, композиция может включать в себя средство, которое усиливает ее функцию, такое, например, как цитотоксический агент, цитокин, химиотерапевтическое средство или рост-ингибирующее средство. Такие молекулы присутствуют, соответственно, в сочетаниях и в количествах, которые являются эффективными для преследуемых здесь целей.
Активные ингредиенты могут также быть заключены в микрокапсулы, полученные, например, с помощью технологий коацервации или путем полимеризации на границе раздела фаз, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли-(метилметакрилатные) микрокапсулы, соответственно, в коллоидных системах доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсиях. Такие технологии описаны в Руководстве Remington's Pharmaceutical Sciences, выше.
Композиции, используемые для введения in vivo, должны быть стерильными. Этого легко достичь путем фильтрации через стерильные фильтрующие мембраны.
Могут быть получены препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают в себя полупроницаемые матрицы твердых гидрофобных полимеров, содержащие антитело, которые представлены в виде профилированных изделий, например пленок или микрокапсул.
Примеры матриц с замедленным высвобождением включают в себя полимеры сложного эфира, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды (патент США No. 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, недеградируемый этиленвинилацетат, деградируемые сополимеры молочной кислоты и гликолевой кислоты, такие как LUPRON DEPOTTM (инъецируемые микросферы, состоящие из сополимера молочной кислоты - гликолевой кислоты и лейпролидацетата), и поли-D-(-)-3-гидроксимасляную кислоту. В то время как такие полимеры как этиленвинилацетат и молочная кислота - гликолевая кислота способствуют высвобождению молекул в течение периода более 100 дней, определенные гидрогели высвобождают белки в течение более коротких периодов. Когда инкапсулированные антитела остаются в организме в течение долгого периода, они могут подвергнуться денатурации или агрегации в результате воздействия влажности при 37°C, с последующей потерей биологической активности и возможными изменениями иммуногенности. Для стабилизации могут быть разработаны рациональные стратегии, в зависимости от рассматриваемого механизма. Например, если обнаружено, что механизм агрегации связан с образованием межмолекулярных S-S-связей через тио-дисульфидный обмен, стабилизации можно достичь путем модификации сульфгидрильных остатков, лиофилизации из кислотных растворов, путем контролирования содержания влаги, использования соответствующих добавок и разработки специфических композиций полимерной матрицы.
G. Применение антител
Антитела против химерного полипептида FGF19 согласно изобретению имеют различные применения. Например, анти-FGF19-антитела могут быть использованы в диагностических анализах, связанных с химерным полипептидом FGF19, например, для детектирования его экспрессии в специфических клетках, тканях или сыворотке. Могут быть использованы различные диагностические технологии анализов, известных в данной области, таких как анализы конкурентного связывания, прямой или непрямой сэндвич-анализы и анализы иммунопреципитации, проводимые либо в гетерогенной, либо в гомогенной фазах [Zola, Monoclonal Antibodies: A Manual of Технологии, CRC Press, Inc. (1987) pp. 147-158]. Антитела, используемые в диагностических анализах, могут быть маркированы детектируемым фрагментом. Детектируемый фрагмент должен быть способен к продуцированию, либо напрямую, либо опосредованно, детектируемого сигнала. Например, указанный детектируемый фрагмент может быть радиоизотопом, таким как 3H, 14C, 32P, 35S или 125I, флуоресцентным или хемилюминесцентным соединением, таким как флуоресцеин изотиоцианат, родамин или люциферин, или ферментом, таким как щелочная фосфатаза, бета-галактозидаза или пероксидаза хрена. Может быть использован любой известный в данной области способ конъюгации антитела с детектируемым фрагментом, включая способы, описанные Hunter et al., Nature, 144: 945 (1962); David et al., Biochemistry, 13: 1014 (1974); Pain et al., J. Immunol. Meth., 40: 219 (1981); а также Nygren, J. Histochem. and Cytochem., 30: 407 (1982).
Антитела против химерного полипептида FGF19 полезны также для аффинной очистки химерного полипептида FGF19 от рекомбинантных клеточных культур или природных источников. При таком процессе антитела против химерного полипептида FGF19 иммобилизуют на соответствующей подложке, такой как смола сефадекс или фильтровальная бумага, с помощью методов, хорошо известных в данной области. Затем иммобилизованное антитело приводят в контакт с образцом, содержащим подлежащий очистке химерный полипептид FGF19, и после этого подложку промывают соответствующим растворителем, который удалит практически весь материал в образце, за исключением химерного полипептида FGF19, который связан с иммобилизованным антителом. В конце подложку промывают другим подходящим растворителем, который высвободит химерный полипептид FGF19 из комплекса с антителом.
IX. Получение химерного полипептида FGF19
Приведенное ниже описание связано главным образом с продуцированием химерного полипептида FGF19 путем культивирования клеток, трансформированных или трансфицированных вектором, содержащим нуклеиновую кислоту, кодирующую химерный полипептид FGF19. Конечно же, подразумевается, что альтернативные способы, которые хорошо известны в данной области, также могут быть использованы для получения химерного полипептида FGF19. Например, химерный полипептид FGF19 или его части могут быть получены путем прямого пептидного синтеза с использованием твердофазных технологий [см., например, Stewart et al., Solid-Phase Peptide Synthesis, W.H. Freeman Co., San Francisco, Calif. (1969); Merrifield, J. Am. Chem. Soc, 85: 2149-2154 (1963)]. Синтез белков in vitro может быть осуществлен ручными методами или же с использованием автоматики; автоматический синтез может быть осуществлен, например, с помощью пептидного синтезатора компании Applied Biosystems (Foster City, Калифорния), руководствуясь инструкциями производителя. Различные участки химерного полипептида FGF19 могут быть отдельно синтезированы химическим путем и соединены с использованием химических или ферментативных методов, с получением полноразмерного химерного полипептида FGF19.
1. Выделение ДНК, кодирующей химерный полипептид FGF19
Фрагменты кДНК, кодирующие химерные полипептиды FGF согласно изобретению, могут быть получены с помощью методологии ПЦР с использованием кДНК, кодирующей, по меньшей мере, часть нативного полипептида FGF19 и, по меньшей мере, часть нативного полипептида FGF21 в качестве матриц. Например, в одном случае фрагмент кДНК, кодирующий N-концевую часть полипептида FGF21, и фрагмент кДНК, кодирующий C-концевую часть полипептида FGF19, амплифицируют отдельно и очищают, используя стандартные способы, такие как ПЦР с последующим электрофорезом в агарозном геле. Последовательности праймеров создаются таким образом, чтобы имело место перекрывание по 18 нуклеотидам на 3'-конце кДНК-фрагмента FGF21 и на 5'-конце кДНК-фрагмента FGF19. Вторую амплификацию с помощью ПЦР проводят с использованием смеси из двух фрагментов кДНК в качестве матриц, получая в результате кДНК, которая кодирует химерный полипептид фрагмента FGF21 и фрагмента FGF19. Полученный в результате фрагмент кДНК расщепляют под действием соответствующих рестрикционных ферментов, очищают путем электрофореза в агарозном геле и клонируют в плазмидный вектор pRK5.sm (плазмидный вектор на основе pUC, содержащий CMV-промотор для экспрессии в клетках млекопитающих), используя для этого стандартные методики. Последовательность полученной в результате плазмиды подтверждают сэндвич-методом секвенирования ДНК.
ДНК, кодирующая химерный полипептид FGF19, может быть получена из библиотеки кДНК, полученной из ткани, о которой известно, что в ней имеет место экспрессия мРНК FGF19 и/или FGF21, и что она экспрессируется на детектируемом уровне. Соответственно, человеческая ДНК FGF19 и/или FGF21 может быть получена обычным способом из библиотеки кДНК, полученной из человеческой ткани, так, как описано в примерах. Ген, кодирующий FGF19 и/или FGF21, также может быть получен из геномной библиотеки или с помощью известных процедур синтеза (например, автоматического синтеза нуклеиновых кислот).
Библиотеки могут быть подвергнуты скринингу с помощью зондов (таких как антитела против FGF19 и/или FGF21 или олигонуклеотидов, содержащих, по меньшей мере, приблизительно 20-80 оснований), сконструированных для идентификации представляющего интерес гена или кодируемого им белка. Скрининг кДНК или геномной библиотеки с помощью выбранного зонда может проводиться с использованием стандартных процедур, таких, которые описаны Sambrook et al., Molecular Cloning: A Laboratory Manual (New York: Cold Spring Harbor Laboratory Press, 1989). Альтернативные способы связаны с выделением гена, кодирующего FGF19, и с применением методологии ПЦР [Sambrook et al., выше; Dieffenbach et al., PCR Primer: A Laboratory Manual (Cold Spring Harbor Laboratory Press, 1995)].
В приведенных ниже примерах описаны технологии скрининга библиотеки кДНК. Олигонуклеотидные последовательности, выбранные в качестве зондов, должны быть достаточно длинными и достаточно однозначно идентифицируемыми, чтобы ложно-позитивные результаты были сведены к минимуму. Предпочтительно, чтобы олигонуклеотиды были мечены таким образом, чтобы их можно было детектировать при гибридизации с ДНК в подвергаемой скринингу библиотеке. Способы мечения хорошо известны в данной области и предусматривают применение радиоактивных меток, таких как 32P-меченная АТФ, биотинилирование или ферментативное мечение. Условия гибридизации, включая условия умеренной жесткости и высокой жесткости, освещены в Руководстве Sambrook et al., см. выше.
Последовательности, идентифицируемые в таких способах скрининга библиотек, можно сравнивать и выравнивать с другими известными последовательностями, депонированными и доступными в общественных базах данных, таких как GenBank, или других приватных базах данных по последовательностям. Идентичность последовательностей (либо на аминокислотном, либо на нуклеотидном уровне) в рамках определенных областей молекулы или на всем протяжении полноразмерной последовательности может быть определена с помощью методов, известных в данной области и описанных в настоящей заявке.
Нуклеиновая кислота, имеющая последовательность, кодирующую белок, может быть получена путем скрининга селективных библиотек кДНК или геномных библиотек с помощью дедуктивной аминокислотной последовательности, описанной здесь впервые, и, если это необходимо, с помощью общепринятых процедур удлинения праймера, как описано у Sambrook et al., выше, для детекции предшественников и процессинга промежуточных копий мРНК, которые могли не подвергнуться обратной транскрипции в кДНК.
2. Отбор и трансформация клеток-хозяев
Клетки-хозяева трансфицируют или трансформируют описанными здесь экспрессирующим или клонирующим векторами для продуцирования химерного полипептида FGF19 и культивируют в соответствующей питательной среде, модифицированной в соответствии с индуцирующими промоторами, селективными трансформантами или амплифицирующимися генами, кодирующими требуемые последовательности. Условия культивирования, такие как среда, температура, pH и пр., могут, без излишнего экспериментирования, быть подобраны специалистами в данной области. В общем, принципы, протоколы и практические рекомендации для максимализации продуктивности клеточных культур можно найти у Mammalian Cell Biotechnology: a Practical Approach, M. Butler, ed. (IRL Press, 1991) and Sambrook et al., см. выше.
Способы трансфекции эукариотических клеток и трансформации прокариотических клеток известны рядовым специалистам в данной области, например, CaCl2, CaPO4, посредством липосом и электропорации. В зависимости от используемой клетки-хозяина, трансформацию осуществляют, пользуясь стандартными технологиями, подходящими для конкретно выбранных клеток. Кальциевую обработку с использованием хлорида кальция, как описано у Sambrook et al., выше, или электропорацию обычно осуществляют в случае прокариотов. Инфицирование с помощью Agrobacterium tumefaciens осуществляют при трансформации определенных клеток растений, как описано у Shaw et al., Gene, 23: 315 (1983), а также в международной заявке WO 89/05859, опубликованной 29 июня 1989. В случае клеток млекопитающих, лишенных таких клеточных стенок, может быть использован метод Graham and van der Eb, Virology, 52: 456-457 (1978) кальциево-фосфатного осаждения. Общие аспекты трансфекции системы клеток-хозяев млекопитающих описаны в патенте США No. 4399216. Трансформацию дрожжевых клеток обычно осуществляют согласно методу Van Solingen et al., J. Bact., 130: 946 (1977) и Hsiao et al., Proc. Natl. Acad. Sci. (USA), 76: 3829 (1979). Однако могут быть использованы также и другие способы включения ДНК в клетки, такие как способы с использованием микроинъецирования в ядра, электропорации, слияния бактериального протопласта с интактными клетками, или же с использованием поликатионов, например, полибрена, полиорнитина. В отношении различных технологий трансформации клеток млекопитающих см. Keown et al., Methods in Enzymology, 185: 527-537 (1990) и Mansour et al., Nature, 336: 348-352 (1988).
Здесь подходящие для клонирования или экспрессии ДНК в векторах клетки-хозяева включают в себя прокариотические клетки, клетки дрожжей или клетки высших эукариот. Подходящие прокариоты включают в себя, но не ограничены перечисленным, эубактерии, такие как грамотрицательные или грамположительные организмы, например, энтеробактерии, такие как E. coli. Различные штаммы E. coli являются общедоступными, к ним относятся штамм K12 MM294 E. coli (ATCC 31,446); E. coli X11776 (ATCC 31,537); штамм W3110 E. coli (ATCC 27,325) и K5772 (ATCC 53,635). Другие подходящие прокариотические клетки-хозяева включают в себя энтеробактерии, такие как Escherichia, например, E. coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например, Salmonella typhimurium, Serratia, например, Serratia marcescans, и Shigella, а также палочковидные бактерии, такие как B. subtilis и B. licheniformis (например, B. licheniformis 41P, описанные в документе DD 266710, опубликованном 12 апреля 1989), псевдомонады, такие как P. Aeruginosa и Streptomyces. Приведенные примеры являются иллюстративными, но не ограничивающими объем притязаний. Штамм W3110 является особенно предпочтительным хозяином или родительским штаммом, поскольку он является общим штаммом-хозяином для продуктов ферментации рекомбинантной ДНК. Предпочтительно, чтобы клетка-хозяин секретировала минимальные количества протеолитических ферментов. Например, штамм W3110 может быть модифицирован таким образом, чтобы вызвать генетическую мутацию в генах, кодирующих белки, эндогенные для клетки-хозяина, и примеры таких клеток-хозяев включают в себя штамм W3110 1A2 E. coli, который имеет полный генотип tonA; штамм W3110 9E4 E. coli, который имеет полный генотип tonA ptr3; штамм W3110 27C7 E. coli (ATCC 55,244), который имеет полный генотип tonA ptr3 phoA E15 (argF-lac)169 degP ompT kanr; штамм W3110 37D6 E. coli, который имеет полный генотип tonA ptr3 phoA E15 (argF-lac)169 degP ompT rbs7 ilvG kanr; штамм W3110 40B4 E. coli, который является штаммом 37D6 с делеционной мутацией degP неустойчивости к канамицину; и штамм E. Coli, имеющий мутантную периплазматическую протеиназу, описанную в патенте США No. 4946783, опубликованном 7 августа 1990. Альтернативно, подходящими являются способы клонирования in vitro, например, ПЦР или другие полимеразные реакции нуклеиновых кислот.
Помимо прокариот, эукариотические микроорганизмы, такие как мицелиальные грибы или дрожжи, также являются подходящими хозяевами для клонирования или экспрессии FGF19-кодирующих векторов. Saccharomyces cerevisiae является широко используемым низшим эукариотическим микроорганизмом-хозяином. Другие включают в себя Schizosaccharomyces pombe (Beach and Nurse, Nature, 290: 140 [1981]; EP 139383, опубликованный 2 мая 1985); клетки-хозяева Kluyveromyces (патент США No. 4943529; Fleer et al., Bio/Technology, 9: 968-975 (1991)), такие, например, как K. lactis (MW98-8C, CBS683, CBS4574; Louvencourt et al., J. Bacterid., 154(2): 737-742 [1983]), K. fragilis (ATCC 12,424), K. bulgaricus (ATCC 16,045), K. wickeramii (ATCC 24, 178), K. waltii (ATCC 56,500), K. drosophilarum (ATCC 36,906; Van den Berg et al., Bio/Technology, 8: 135 (1990)), K. Thermotolerans и K. marxianus; yarrowia (EP 402226); Pichia pastoris (EP 183070; Sreekrishna et al., J. Basic Microbiol., 28: 265-278 [1988]); Candida; Trichoderma reesia (EP 244234); Neurospora crassa (Case et al., Proc. Natl. Acad. Sci. USA, 76:5259-5263 [1979]); шванниомицеты, такие как Schwanniomyces occidentalis (EP 394538 опубликованный 31 октября 1990); и мицелиальные грибы, такие, например, как Neurospora, Penicillium, Tolypocladium (WO 91/00357 опубликованная 10 января 1991), и клетки-хозяева Aspergillus, такие как A. nidulans (Ballance et al., Biochem. Biophys. Res. Commun., 112: 284-289 [1983]; Tilburn et al., Gene, 26: 205-221 [1983]; Yelton et al., Proc. Natl. Acad. Sci. USA, 81: 1470-1474 [1984]) и A. niger (Kelly and Hynes, EMBO J., 4: 475-479 [1985]). Метилотрофные дрожжи в данном случае являются подходящими и включают в себя, но не ограничены перечисленным, дрожжи, способные расти на метаноле, выбранные из видов, состоящих из Hansenula, Candida, Kloeckera, Pichia, Saccharomyces, Torulopsis и Rhodotorula. Список специфических видов, которые являются иллюстративными для указанного класса дрожжей, можно найти у C. Anthony, The Biochemistry of Methylotrophs, 269 (1982).
Подходящие для экспрессии гликозилированного химерного полипептида FGF19 клетки-хозяева получают из многоклеточных организмов. Примеры клеток беспозвоночных включают клетки насекомых, таких как Drosophila S2 и Spodoptera Sf9, а также клетки растений. Примеры используемых линий клеток-хозяев млекопитающих включают клетки яичника китайского хомячка (CHO) и клетки COS. Более специфические примеры включают линию клеток CV1 почки обезьяны, трансформированных SV40 (COS-7, ATCC CRL 1651); линию эмбриональных клеток человека (клетки 293 или 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol., 36: 59 (1977)); клетки яичника китайского хомячка/-DHFR (CHO, Urlaub and Chasin, Proc. Natl. Acad. Sci. USA, 77: 4216 (1980)); мышиные клетки Сертоли (TM4, Mather, Biol. Reprod., 23: 243-251 (1980)); клетки легких человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); и мышиные клетки опухоли молочной железы (MMT 060562, ATCC CCL51). Подразумевается, что выбор подходящих клеток-хозяев находится в рамках компетенции специалистов в данной области.
3. Отбор и применение репликативного вектора
Нуклеиновая кислота (например, кДНК или геномная ДНК), кодирующая химерный полипептид FGF19, может быть встроена в репликативный вектор для клонирования (амплификация ДНК) или для экспрессии. Различные векторы являются общедоступными. Например, вектор может быть представлен в виде плазмиды, космиды, вирусной частицы или фага. Подходящие последовательности нуклеиновых кислот могут быть встроены в указанный вектор путем применения различных методик. Обычно ДНК встраивают в подходящий участок (участки) рестрикционной эндонуклеазы с помощью известных в данной области технологий. Компоненты вектора обычно включают в себя, но не ограничены перечисленным, одну или более из сигнальных последовательностей, ориджин репликации, один или более маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции. При конструировании подходящих векторов, содержащих один или более из указанных компонентов, используются стандартные технологии лигирования, которые известны специалистам в данной области.
Химерный полипептид FGF19 может быть получен рекомбинантным путем не только непосредственно, но также и в результате слияния полипептида с гетерологичным полипептидом, который представлять собой сигнальную последовательность или другой полипептид, имеющий специфический участок расщепления на N-конце зрелого белка или полипептида. Обычно сигнальная последовательность может быть компонентом вектора, или же она может быть частью ДНК, кодирующей химерный полипептид FGF19, которую встраивают в вектор. Сигнальная последовательность может быть исходной сигнальной последовательностью FGF19 или FGF21, таких как hFGF19 или hFGF21. Таким образом, в указанных воплощениях химерный полипептид FGF19 согласно настоящему изобретению может включать в себя, по меньшей мере, N-концевую часть пре-hFGF19, например, по меньшей мере, остатки 1-22 последовательности SEQ ID NO: 3. В таких воплощениях химерный полипептид FGF19 согласно настоящему изобретению может включать в себя, по меньшей мере, N-концевую часть пре-hFGF1, например, по меньшей мере, остатки 1-28 последовательности SEQ ID NO: 4.
Сигнальная последовательность может быть прокариотической сигнальной последовательностью, выбранной, например, из группы щелочной фосфатазы, пенициллиназы, lpp или теплостабильных лидерных последовательностей энтеротоксина II. В случае секреции в дрожжевых клетках сигнальной последовательностью может быть, например, лидерная последовательность инвертазы дрожжей, лидерная последовательность альфа-фактора (включая лидерные последовательности альфа-фактора Saccharomyces и Kluyveromyces, последняя из которых описана в патенте США No. 5010182), или лидерная последовательность кислой фосфатазы, лидерная последовательность глюкоамилазы C. albicans (EP 362179, опубликованный 4 апреля 1990), или сигнальная последовательность, описанная в документе WO 90/13646, опубликованном 15 ноября 1990. При экспрессии в клетках млекопитающих могут быть использованы сигнальные последовательности млекопитающих для прямой секреции белка, такие как сигнальные последовательности из секретируемых полипептидов близкородственного или же одного и того же вида, а также вирусные секреторные лидерные последовательности.
Как экспрессирующий, так и клонирующий векторы содержат последовательность нуклеиновой кислоты, которая позволяет вектору реплицироваться в одной или нескольких избранных клетках-хозяевах. Такие последовательности хорошо известны для бактерий, дрожжей и вирусов. Ориджин репликации из плазмиды pBR322 является подходящим для грамотрицательных бактерий, 2µ-плазмидный ориджин является подходящим для дрожжей, а различные вирусные ориджины (вируса SV40, полиомы, аденовируса, VSV или BPV) являются подходящими для клонирующих векторов в клетках млекопитающих.
Обычно экспрессирующий и клонирующий векторы содержат ген селекции, называемый также селективным маркером. Типичные селективные гены кодируют белки, которые (a) придают резистентность к антибиотикам или другим токсинам, например, к ампициллину, неомицину, метотрексату или тетрациклину, (b) дополнительный ауксотрофный дефицит или (c) доставляют критические питательные вещества, не доступные из комплексной среды, например, ген, кодирующий D-аланинрацемазу для палочковидных бактерий.
Примерами подходящих селективных маркеров для клеток млекопитающих являются такие маркеры, которые дают возможность идентифицировать клетки, компетентные в отношении поглощения FGF19-кодирующей нуклеиновой кислоты, например, DHFR или тимидинкиназа. Подходящей клеткой-хозяином при использовании DHFR дикого типа является линия клеток CHO, дефицитных в отношении активности DHFR, получаемых и размножаемых так, как описано Urlaub et al., Proc. Natl. Acad. Sci. USA, 77: 4216 (1980). Подходящим селективным геном для применения в дрожжах является ген trp1, присутствующий в дрожжевой плазмиде YRp7 [Stinchcomb et al., Nature, 282: 39 (1979); Kingsman et al., Gene, 7: 141 (1979); Tschemper et al., Gene, 10: 157 (1980)]. Ген trp1 наделяет селективным маркером мутантный штамм дрожжей, лишенных способности расти в среде с триптофаном, например, ATCC No. 44076 или PEP4-1 [Jones, Genetics, 85: 12 (1977)].
Обычно экспрессирующий и клонирующий векторы содержат промотор, оперативно связанный с кодирующей FGF19 последовательностью нуклеиновой кислоты, для прямого синтеза мРНК. Промоторы, распознаваемые разными потенциальными клетками-хозяевами, хорошо известны. Промоторы, подходящие для применения в прокариотических клетках-хозяевах, включают в себя системы промоторов β-лактамазы и лактозы [Chang et al., Nature, 275: 615 (1978); Goeddel et al., Nature, 281: 544 (1979)], щелочной фосфатазы, систему промоторов триптофана (trp) [Goeddel, Nucleic Acids Res., 8: 4057 (1980); EP 36,776], и гибридные промоторы, такие как промотор tac [deBoer et al., Proc. Natl. Acad. Sci. USA, 80: 21-25 (1983)]. Промоторы для применения в бактериальных системах будут содержать также последовательность Шайна-Дальгарно (S.D.), оперативно связанную с ДНК, кодирующей FGF19.
Примеры подходящих промоторных последовательностей для применения в дрожжевых клетках-хозяевах включают в себя промоторы для 3-фосфоглицераткиназы [Hitzeman et al., J. Biol. Chem., 255: 2073 (1980)] или других гликолитических ферментов [Hess et al., J. Adv. Enzyme Reg., 7: 149 (1968); Holland, Biochemistry, 17:4900 (1978)], таких как енолаза, глицеральдегид-3-фосфатдегидрогеназа, гекокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфатизомераза, 3-фосфоглицератмутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкозоизомераза и глюкокиназа.
Другие дрожжевые промоторы, которые являются индуцибельными промоторами, имеющими дополнительное преимущество в отношении транскрипции, контролируемой условиями роста, являются промоторными областями для алкогольдегидрогеназы 2, изоцитохрома C, кислой фосфатазы, ферментов деградации, ассоциированных с метаболизмом азота, металлотионеина, глицеральдегид-3-фосфат-дегидрогеназы, и ферментов, ответственных за утилизацию мальтозы и галактозы. Векторы и промоторы, подходящие для использования при экспрессии в дрожжевых клетках, дополнительно описаны в патенте EP 73657.
Транскрипция из векторов в клетках-хозяевах млекопитающих контролируется, например, промоторами, полученными из геномов вирусов, таких как вирус полиомы, вирус оспы кур (UK 2211504 опубликованный 5 июля 1989), аденовирус (такой как аденовирус 2), вирус папилломы крупного рогатого скота, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита B и вакуолизирующий обезьяний вирус 40 (SV40), из гетерологичных промоторов млекопитающих, например, актинового промотора или иммуноглобулинового промотора, а также из промоторов белков теплового шока, при условии, что такие промоторы являются совместимыми с системами клеток-хозяев.
Транскрипция ДНК, кодирующей химерный полипептид FGF19 у высших эукариот, может быть усилена путем инсерции энхансерной последовательности в такой вектор. Энхансеры являются цис-действующими элементами ДНК, обычно насчитывающими приблизительно от 10 до 300 п.н., которые действуют на промотор, усиливая его транскрипцию. Многие энхансерные последовательности известны в настоящее время из генов млекопитающих (глобинового гена, гена эластазы, альбумина, α-фетопротеина и инсулина). Однако обычно используют энхансер из вируса эукариотических клеток. Примеры включают в себя энхансер SV40 на поздних ориджинах репликации (100-270 п.н.), энхансер раннего промотора цитомегаловируса, энхансер вируса полиомы на поздних ориджинах репликации и аденовирусные энхансеры. Можно осуществить сплайсинг энхансера внутри вектора в положении 5' или 3' по отношению к последовательности, кодирующей химерный полипептид FGF19, но предпочтительно на 5'-участке от промотора.
Экспрессирующие векторы, используемые в эукариотических клетках-хозяевах (дрожжевых, грибковых клетках, клетках насекомых, растительных, животных клетках, клетках человека или содержащих ядро клетках из других многоклеточных организмов), также будут содержать последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности обычно доступны от 5'- и, необязательно, 3'-нетранслируемых областей эукариотических или вирусных ДНК или кДНК. Указанные области содержат нуклеотидные сегменты, транскрибируемые в виде полиаденилированных фрагментов в нетранслируемом участке мРНК, кодирующем химерный полипептид FGF19.
Другие способы, векторы и клетки-хозяева, подходящие для адаптации к синтезу химерного полипептида FGF19 в культурах рекомбинантных клеток позвоночных, описаны в публикациях Gething et al., Nature, 293: 620-625 (1981); Mantei et al., Nature, 281: 40-46 (1979); EP 117060; и EP 117058.
4. Детекция амплификации/экспрессии гена
Амплификацию и/или экспрессию гена можно напрямую измерить в образце, например, путем общеизвестных методов саузерн-блоттинга, назерн-блоттинга, чтобы количественно оценить транскрипцию мРНК [Thomas, Proc. Natl. Acad. Sci. USA, 77: 5201-5205 (1980)], дот-блоттинга (анализ ДНК) или in situ гибридизации, с использованием соответствующим образом меченого зонда, на основе приведенных здесь последовательностей. Альтернативно, могут быть использованы антитела, которые могут распознавать специфические дуплексы, включая ДНК-дуплексы, РНК-дуплексы и гибридные ДНК-РНК-дуплексы или дуплексы ДНК-белок. В свою очередь, могут быть помечены антитела, и анализ может быть осуществлен, когда дуплекс связан с поверхностью, так, чтобы при образовании дуплекса на поверхности можно было детектировать присутствие антитела, связанного с дуплексом.
Альтернативно, экспрессию генов можно измерять иммунологическими методами, такими как иммуногистохимическое окрашивание клеток или срезов тканей, и исследовать клеточную культуру или жидкости организма, чтобы напрямую количественно определить экспрессию продуктов генов. Антитела, применимые для окрашивания и/или анализа образцов жидкостей, могут быть либо моноклональными, либо поликлональными антителами, и они могут быть получены у любого животного. Для удобства могут быть получены антитела против химерного полипептида FGF19 или против синтетического пептида на основе последовательностей представленных здесь ДНК, или против экзогенной последовательности, слитой с ДНК, кодирующей химерный полипептид FGF19 и кодирующей специфический эпитоп антитела.
5. Очистка полипептида
Формы химерного полипептида FGF19 могут быть получены из культуральной среды или из лизатов клеток-хозяев. Если они мембрано-связанные, их можно высвободить из мембраны с помощью раствора соответствующего детергента (например, тритона X-100) или путем ферментативного расщепления. Клетки, используемые для экспрессии химерного полипептида FGF19, могут быть разрушены различными физическими или химическими способами, такими как повторное замораживание-оттаивание, обработка ультразвуком, механическое разрушение или обработка средствами, осуществляющими лизис клеток.
Может потребоваться очистка химерного полипептида FGF19 от белков или полипептидов рекомбинантных клеток. Следующие процедуры представляют собой примеры соответствующих процедур очистки: путем фракционирования на колонке с ионообменником; осаждение этанолом; обращенно-фазовая ВЭЖХ; хроматография на силикагеле или на катионообменной смоле, такой как DEAE; хроматофокусирование; ДСН-ПААГ; осаждение сульфатом аммония; гель-фильтрация, например, с помощью сефадекса G-75; колонки с протеин A-сефарозой для удаления примесей, таких как IgG; и колонки с веществом, вызывающим образование хелатных соединений с металлом, для связывания направленных на эпитоп форм химерного полипептида FGF19. Могут быть использованы разнообразные способы очистки белка, и такие способы являются известными в данной области, и они описаны, например у Deutscher, Methods in Enzymology, 182 (1990); Scopes, Protein Purification: Principles and Practice, Springer-Verlag, New York (1982). Выбранная стадия(стадии) очистки будут зависеть, например, от природы процесса получения и от конкретного продуцируемого химерного полипептида FGF19.
X. Нуклеиновые кислоты, кодирующие химерные полипептиды FGF19, и их применение
В другом аспекте настоящее изобретение включает нуклеотидные последовательности (или комплементарные им последовательности), которые кодируют химерные полипептиды FGF19 (или "химерные нуклеиновые кислоты FGF19") согласно настоящему изобретению. Нуклеиновые кислоты химерных FGF19 согласно настоящему изобретению имеют различные применения в области молекулярной биологии, включая применения в качестве зондов для гибридизации, применения в картировании генов и хромосом, а также в получении антисмысловых РНК и ДНК. Нуклеиновые кислоты химерных FGF19 найдут применение также и в получении химерных полипептидов FGF19 с использованием описанных здесь рекомбинантных технологий.
В некоторых воплощениях химерные полипептиды FGF19 могут включать в себя один или более эпитопов-маркеров. В некоторых воплощениях маркерный эпитоп расположен на N-конце химерного полипептида FGF19. В некоторых воплощениях маркерный эпитоп расположен на C-конце химерного полипептида FGF19. В определенных воплощениях маркерный эпитоп расположен на N-конце химерного полипептида FGF19.
В некоторых воплощениях химерные полипептиды FGF19 могут включать в себя один или более эпитопов-маркеров. В некоторых воплощениях маркерный эпитоп расположен на N-конце химерного полипептида FGF19. В определенных воплощениях маркерный эпитоп расположен на C-конце химерного полипептида FGF19. В определенных воплощениях маркерный эпитоп расположен на N-конце химерного полипептида FGF19. В определенных воплощениях маркерный эпитоп содержит аминокислотную последовательность DYKDDDDK (SEQ ID NO: 279).
В иллюстративном воплощении нуклеиновая кислота химерного FGF19 согласно настоящему изобретению включает следующую последовательность:
CACCCCATCCCTGACTCCAGTCCTCTCCTGCAATTCGGGGGCCAAGTCCGGCAGCGGTACCTCTACACCTCCGGCCCCCACGGGCTCTCCAGCTGCTTCCTGCGCATCCGTGCCGACGGCGTCGTGGACTGCGCGCGGGGCCAGAGCGCGCACAGTTTGCTGGAGATCAAGGCAGTCGCTCTGCGGACCGTGGCCATCAAGGGCGTGCACAGCGTGCGGTACCTCTGCATGGGCGCCGACGGCAAGATGCAGGGGCTGCTTCAGTACTCGGAGGAAGACTGTGCTTTCGAGGAGGAGATCCGCCCAGATGGCTACAATGTGTACCGATCCGAGAAGCACCGCCTCCCGGTCTCCCTGAGCAGTGCCAAACAGCGGCAGCTGTACAAGAACAGAGGCTTTCTTCCACTCTCTCATTTCCTGCCCATGCTGCCCATGGTCCCAGAGGAGCCTGAGGACCTCAGGGGCCACTTGGAATCTGACATGTTCTCTTCGCCCCTGGAGACCGACAGCATGGACCCATTTGGGCTTGTCACCGGACTGGAGGCCGTGAGGAGTCCCAGCTTTGAGAAG (SEQ ID NO: 7). Указанная иллюстративная последовательность нуклеиновой кислоты кодирует полипептид, имеющий аминокислотную последовательность, которая соответствует химерному полипептиду FGF19 cFGF21/19-2, как показано в таблице 3.
В другом иллюстративном воплощении нуклеиновая кислота химерного FGF19 согласно настоящему изобретению включает следующую последовательность:
ATGGACTCGGACGAGACCGGGTTCGAGCACTCAGGgCTGTGGGTTTCTGTGCTGGCTGGTCTTCTGCTGGGAGCCTGCCAGGCACACCCCATCCCTGACTCCAGTCCTCTCCTGCAATTCGGGGGCCAAGTCCGGCAGCGGTACCTCTACACCTCCGGCCCCCACGGGCTCTCCAGCTGCTTCCTGCGCATCCGTGCCGACGGCGTCGTGGACTGCGCGCGGGGCCAGAGCGCGCACAGTTTGCTGGAGATCAAGGCAGTCGCTCTGCGGACCGTGGCCATCAAGGGCGTGCACAGCGTGCGGTACCTCTGCATGGGCGCCGACGGCAAGATGCAGGGGCTGCTTCAGTACTCGGAGGAAGACTGTGCTTTCGAGGAGGAGATCCGCCCAGATGGCTACAATGTGTACCGATCCGAGAAGCACCGCCTCCCGGTCTCCCTGAGCAGTGCCAAACAGCGGCAGCTGTACAAGAACAGAGGCTTTCTTCCACTCTCTCATTTCCTGCCCATGCTGCCCATGGTCCCAGAGGAGCCTGAGGACCTCAGGGGCCACTTGGAATCTGACATGTTCTCTTCGCCCCTGGAGACCGACAGCATGGACCCATTTGGGCTTGTCACCGGACTGGAGGCCGTGAGGAGTCCCAGCTTTGAGAAGGACTACAAAGACGATGACGACAAGTGA (SEQ ID NO: 281). Указанная иллюстративная последовательность нуклеиновой кислоты кодирует полипептид, имеющий аминокислотную последовательность, которая включает в себя последовательность химерного полипептида FGF19 cFGF21/19-2, как показано в таблице 3. Указанный полипептид включает также C-концевой маркерный эпитоп DYKDDDK (SEQ ID NO: 280) и N-концевую нативную сигнальную последовательность полипептида hFGF21 (MDSDETGFEHSGLWVSVLAGLLLGACQA; SEQ ID NO:282).
В другом иллюстративном воплощении нуклеиновая кислота химерного FGF19 согласно настоящему изобретению включает следующую последовательность
ATGCGGAGCGGGTGTGTGGTGGTCCACGTATGGATCCTGGCCGGCCTCTGGCTGGCCGTGGCCGGGCGCCCCCTCGCCTTCTCGGACGCGGGGCCCCACGTGCACTACGGCTGGGGCGACCCCATCCGCCTGCGGCACCTGTACACAGATGATGCCCAGCAGACAGAAGCCCACCTGGAGATCAGGGAGGATGGGACGGTGGGGGGCGCTGCTGACCAGAGCCCCGAAAGTCTCCTGCAGCTGAAAGCCTTGAAGCCGGGAGTTATTCAAATCTTGGGAGTCAAGACATCCAGGTTCCTGTGCCAGCGGCCAGATGGGGCCCTGTATGGATCGCTCCACTTTGACCCTGAGGCCTGCAGCTTCCGGGAGCTGCTTCTTGAGGACGGATACAATGTTTACCAGTCCGAAGCCCACGGCCTCCCGCTGCACCTGCCAGGGAACAAGTCCCCACACCGGGACCCTGCACCCCGAGGACCAGCTCGCTTCCTGCCACTACCAGGCCTGCCCCCCGCACTCCCGGAGCCACCCGGAATCCTGGCCCCCCAGCCCCCCGATGTGGGCTCCTCGGACCCTCTGAGCATGGTGGGACCTTCCCAGGGCCGAAGCCCCAGCTACGCTTCCGACTACAAGGACGACGATGACAAGTGA (SEQ ID NO: 283). Данная иллюстративная последовательность нуклеиновой кислоты кодирует полипептид, имеющий аминокислотную последовательность, которая включает последовательность химерного полипептида FGF19 cFGF19/21-2, как показано в таблице 6. Указанный полипептид включает также C-концевой маркерный эпитоп DYKDDDK (SEQ ID NO: 280) и N-концевую нативную сигнальную последовательность полипептида hFGF19 (MRSGCVVVHVWILAGLWLAVAG; SEQ ID NO: 284).
В другом иллюстративном воплощении нуклеиновая кислота химерного FGF19 согласно настоящему изобретению включает следующую последовательность:
ATGGAcTCGGACGAGACCGGGTTCGAGCACTCAGGGCTGTGGGTTTCTGTGCTGGCTGGTCTTCTGCTGGGAGCCTGCCAGGCACACCCCATCCCTGACTCCAGTCCTCTCCTGCAATTCGGGGGCCAAGTCCGGCAGCGGTACCTCTACACAGATGATGCCCAGCAGACAGAAGCCCACCTGGAGATCAGGGAGGATGGGACGGTGGGGGGCGCTGCTGACCAGAGCCCCGAAAGTCTCCTGCAGCTGAAAGCCTTGAAGCCGGGAGTTATTCAAATCTTGGGAGTCAAGACATCCAGGTTCCTGTGCCAGCGGCCAGATGGGGCCCTGTATGGATCGCTCCACTTTGACCCTGAGGCCTGCAGCTTCCGGGAGCTGCTTCTTGAGGACGGATACAATGTTTACCAGTCCGAAGCCCACGGCCTCCCGCTGCACCTGCCAGGGAACAAGTCCCCACACCGGGACCCTGCACCCCGAGGACCAGCTCGCTTCCTTCCACTCTCTCATTTCCTGCCCATGCTGCCCATGGTCCCAGAGGAGCCTGAGGACCTCAGGGGCCACTTGGAATCTGACATGTTCTCTTCGCCCCTGGAGACCGACAGCATGGACCCATTTGGGCTTGTCACCGGACTGGAGGCCGTGAGGAGTCCCAGCTTTGAGAAGGACTACAAAGACGATGACGACAAGTGA (SEQ ID NO: 285). Данная иллюстративная последовательность нуклеиновой кислоты кодирует полипептид, имеющий аминокислотную последовательность, которая включает последовательность химерного полипептида FGF19 cFGF21/19-13, как показано в таблице 5. Указанный полипептид включает также C-концевой маркерный эпитоп DYKDDDK (SEQ ID NO: 280) и N-концевую нативную сигнальную последовательность полипептида hFGF21 (MDSDETGFEHSGLWVSVLAGLLLGACQA; SEQ ID NO: 282).
Полноразмерная нативная последовательность нуклеиновой кислоты гена hFGF19 (SEQ ID NO: 5), полноразмерная нативная последовательность нуклеиновой кислоты гена FGF21 (SEQ ID NO: 6), полноразмерная нативная последовательность нуклеиновой кислоты химерного полипептида FGF19 согласно настоящему изобретению или любые их вышеуказанные части могут быть использованы в качестве зондов для гибридизации при детектировании или скрининге нуклеиновых кислот, которые кодируют химерные полипептиды FGF19 согласно настоящему изобретению. Длина зондов необязательно будет составлять от приблизительно 20 до приблизительно 50 оснований. В качестве примера, данный способ скрининга будет включать в себя выделение кодирующей области гена FGF19 с помощью известной последовательности ДНК для синтеза выбранного зонда длиной приблизительно в 40 оснований. Зонды для гибридизации могут быть маркированы разными метками, включая радионуклиды, такие как 32P или 35S, или ферментативной меткой, такой как щелочная фосфатаза, соединенная с указанным зондом через системы связывания авидин/биотин. Меченые зонды, имеющие последовательность, комплементарную таковой гена FGF19 согласно настоящему изобретению, могут быть использованы для скрининга библиотек человеческих кДНК, геномной ДНК или мРНК для определения того, какие члены указанных библиотек гибридизуются с указанным зондом. Технологии гибридизации в дальнейших подробностях описаны в приведенных ниже примерах.
Любые EST-последовательности, описанные в настоящей заявке, могут сходным образом быть использованы в качестве зондов с помощью описанных здесь способов.
Другие полезные фрагменты нуклеиновых кислот химерных FGF19 включают в себя антисмысловые или смысловые олигонуклеотиды, содержащие последовательность одноцепочечной нуклеиновой кислоты (либо РНК, либо ДНК), способной связываться с последовательностями мРНК-мишени химерного FGF19 (смысловая) или ДНК-мишени химерного FGF19 (антисмысловая). Антисмысловой или смысловой олигонуклеотиды согласно настоящему изобретению включают в себя фрагмент кодирующей области ДНК химерного FGF19. Такой фрагмент обычно содержит, по меньшей мере, приблизительно 14 нуклеотидов, предпочтительно, приблизительно от 14 до 30 нуклеотидов. Возможность получить антисмысловой или смысловой олигонуклеотид на основе последовательности кДНК, кодирующей данный белок, описана, например, в публикациях Stein and Cohen (Cfncer Res. 48: 2659, 1988) и van der Krol et al. (BioTechniques 6: 958, 1988).
Связывание антисмыслового или смыслового олигонуклеотидов для нацеливания на последовательности нуклеиновой кислоты приводит к образованию дуплексов, которые блокируют транскрипцию или трансляцию последовательности-мишени одним из следующих способов, включая усиленную деградацию дуплексов, преждевременную терминацию транскрипции или трансляции, или другим способом. Таким образом, антисмысловые олигонуклеотиды могут быть использованы для блокирования экспрессии химерных полипептидов FGF19. Антисмысловые или смысловые олигонуклеотиды дополнительно включают в себя олигонуклеотиды, имеющие модифицированные сахаро-фосфодиэфирные остовы (или другие связи сахара, описанные, например, в документе WO 91/06629), и в которых такие связи сахаров являются резистентными к действию эндогенных нуклеаз. Такие олигонуклеотиды с резистентными связями сахара являются стабильными in vivo (то есть они способны сопротивляться ферментативной деградации), но сохраняют специфичность последовательности и могут связываться с нуклеотидными последовательностями-мишенями.
Другие примеры смысловых или антисмысловых олигонуклеотидов включают в себя такие олигонуклеотиды, которые ковалентно связаны с органическими фрагментами, такими, какие описаны в документе WO 90/10048, и с другими фрагментами, которые повышают аффинность олигонуклеотида в отношении последовательности нуклеиновой кислоты-мишени, такими как поли-(L-лизин). Кроме того, к смысловым или антисмысловым олигонуклеотидам могут быть привязаны интеркалирующие вещества, такие как эллиптиоцин, а также алкилирующие вещества или комплексные соединения металла, чтобы модифицировать специфичности связывания антисмыслового или смыслового олигонуклеотида с нуклеотидной последовательностью-мишенью.
Антисмысловые или смысловые олигонуклеотиды могут быть введены в клетку, содержащую последовательность нуклеиновой кислоты-мишени, любым из способов переноса генов, включая, например, CaPO4-опосредованную трансфекцию ДНК; электропорацию, или с использованием векторного переноса генов, например, вируса Эпштейна-Барр. В предпочтительном варианте антисмысловой или смысловой олигонуклеотид встраивают в подходящий ретровирусный вектор. Клетку, содержащую последовательность нуклеиновой кислоты-мишени, приводят в контакт с рекомбинантным ретровирусным вектором, либо in vivo, либо ex vivo. Подходящие ретровирусные векторы включают в себя, но не ограничены перечисленным, векторы, полученные из мышиного ретровируса M-MuLV, N2 (ретровирус, полученный из M-MuLV), или двухкопийные векторы, обозначаемые DCT5A, DCT5B и DCT5C (см. WO 90/13641).
Смысловые или антисмысловые олигонуклеотиды могут также быть встроены в клетку, содержащую нуклеотидную последовательность-мишень, путем образования конъюгата с лиганд-связывающей молекулой, как описано в документе WO 91/04753. Подходящие лиганд-связывающие молекулы включают в себя, но не ограничены перечисленным, рецепторы клеточной поверхности, факторы роста, другие цитокины или другие лиганды, которые связываются с рецепторами клеточной поверхности. Предпочтительно, чтобы конъюгация лиганд-связывающей молекулы по существу не мешала способности лиганд-связывающей молекулы связываться с соответствующей ей молекулой или рецептором или блокировать проникновение смыслового или антисмыслового олигонуклеотида или его конъюгированного варианта в клетку.
Альтернативно, смысловой или антисмысловой олигонуклеотид может быть встроен в клетку, содержащую рассеянный склероз нуклеиновой кислоты-мишени, путем образования олигонуклеотид-липидного комплекса, как описано в международной заявке WO 90/10448. Смысловой или антисмысловой олигонуклеотид-липидный комплекс предпочтительно диссоциирует внутри клетки под действием эндогенной липазы.
Зонды могут быть использованы также в технологии ПЦР для выработки пула последовательностей для идентификации близкородственных последовательностей, кодирующих химерный FGF19.
Нуклеиновая кислота, кодирующая химерный полипептид FGF19, может быть использована также в генной терапии. При использовании в генной терапии гены вводят в клетку с тем, чтобы достичь in vivo синтеза терапевтически эффективного генетического продукта, например, для замены дефективного гена. "Генная терапия" включает как обычно применяемую генную терапию, когда длительный эффект достигается однократным воздействием, так и введение средств генной терапии, при котором предусматривается однократное или многократное введение терапевтически эффективной ДНК или мРНК. Антисмысловые мРНК или смысловые ДНК могут быть использованы в качестве терапевтических средств для блокирования экспрессии определенных генов in vivo. Было уже показано, что короткие антисмысловые олигонуклеотиды могут быть внесены в клетки, где они действуют в качестве ингибиторов, несмотря на их малые внутриклеточные концентрации, обусловленные их ограниченным поглощением клеточной мембраной (Zamecnik et al., Proc. Natl. Acad. Sci. USA 83: 4143-4146 [1986]). Олигонуклеотиды могут быть модифицированы с тем, чтобы повысить их поглощение, например, путем замещения их отрицательно заряженных фосфодиэфирных групп незаряженными группами.
Существует целый ряд доступных технологий для введения нуклеиновых кислот в жизнеспособные клетки. Указанные технологии варьируют, в зависимости от того, намереваются ли вводить нуклеиновую кислоту в культивируемые клетки in vitro или в клетки предполагаемого хозяина in vivo. Технологии, подходящие для переноса нуклеиновой кислоты в клетки млекопитающего in vitro, включают в себя применение липосом, электропорации, микроинъецирования, слияния клеток, DEAE-декстрана, метода кальций-фосфатного осаждения и т.д. В настоящее время предпочтительные технологии переноса гена in vivo включают в себя трансфекцию вирусными (обычно ретровирусными) векторами и трансфекцию, опосредованную вирусными оболочечными белками-липосомами (Dzau et al., Trends in Biotechnology 11, 205-210 [1993]). В некоторых ситуациях бывает необходимо снабдить источник нуклеиновой кислоты средством, которое нацелено на клетки-мишени, таким как антитело, специфичное в отношении мембранного белка клеточной поверхности или клетки-мишени, лиганда рецептора на клетке-мишени, и т.д. Когда используют липосомы, для нацеливания и/или для облегчения поглощения можно использовать белки, которые связываются с мембранным белком клеточной поверхности, ассоциированным с эндоцитозом, например, капсидные белки или их фрагменты, тропные в отношении конкретного типа клеток, антитела против белков, которые периодически подвергаются интернализации, против белков, которые нацелены на внутриклеточно локализованные мишени и увеличивают внутриклеточное время полужизни. Технология рецептор-опосредованного эндоцитоза описана, например, Wu et al., J. Biol. Chem. 262, 4429-4432 (1987); и Wagner et al., Proc. Natl. Acad. Sci. USA 87, 3410-3414 (1990). В качестве обзора в отношении методов маркировки генов и протоколов генной терапии см. Anderson et al., Science 256, 808-813 (1992).
X. Изделия производства
В другом воплощении настоящего изобретения предложено изделие производства, содержащие вещества, используемые для лечения связанных с метаболизмом расстройств, состояний или симптомов, как описано выше. Предпочтительно, чтобы такое изделие производства содержало (a) контейнер, содержащий композицию, включающую в себя описанный здесь химерный полипептид FGF19 и приемлемый фармацевтический носитель или разбавитель внутри указанного контейнера; и (b) вкладыш с инструкциями по введению композиции индивиду, страдающему от признаков связанных с метаболизмом расстройств, состояний или симптомов или проявляющему такие признаки.
В некоторых воплощениях индивид имеет связанные с метаболизмом расстройства, состояния или симптомы. В определенных воплощениях у индивида есть риск развития связанного с метаболизмом расстройства, состояния или симптома. В определенных воплощениях индивид имеет одну или более из характеристик, выбранных из группы, состоящей из (a) обхвата талии, приблизительно составляющего 102 см или более у мужчин и приблизительно 88 см или более у женщин, (b) уровня триглицеридов натощак, составляющего приблизительно 150 мг/дл или более, (c) уровня глюкозы натощак, приблизительно составляющего 95 мг/дл или выше, и (d) высоких уровней окисленных ЛНП. В некоторых воплощениях у индивида дополнительно имеется воспаление, ассоциированное с диабетом. В некоторых воплощениях у индивида обнаруживается уровень глюкозы в крови, приблизительно составляющий 95 мг/дл или выше после голодания в течение ночи. В некоторых воплощениях у индивида обнаруживается уровень глюкозы в крови, приблизительно составляющий 126 мг/дл или выше после голодания в течение ночи. В определенных воплощениях у индивида уровень глюкозы в крови составляет приблизительно 140 мг/дл после двухчасового орального теста на толерантность к глюкозе. В определенных воплощениях у индивида обнаруживается уровень глюкозы в крови, приблизительно составляющий 200 мг/дл после двухчасового орального теста на толерантность к глюкозе. В определенных воплощениях у индивида обнаруживается состояние пре-диабета. В некоторых воплощениях у индивида обнаруживается диабет. В определенных воплощениях диабет выбран из группы, состоящей из диабета I типа, диабета II типа и диабета беременных. В определенных воплощениях диабетом является диабет II типа.
Указанное изделие производства включает в себя контейнер и этикетку или вкладыш на контейнере или ассоциированный с контейнером. Подходящие контейнеры включают в себя, например, бутыли, ампулы, шприцы и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. В контейнере хранится или он содержит композицию, которая эффективна для лечения множественного склероза, и стерильное входное отверстие (например, такой контейнер может быть мешком для внутривенного раствора или пузырьком, имеющим пробку, поддающуюся прокалыванию иглой для подкожного введения). По меньшей мере, одним активным средством в композиции такой является химерный полипептид FGF19. На этикетке или на вкладыше указано, что композиция используется для лечения связанных с метаболизмом расстройств, состояний или симптомов у индивида, страдающего ими, со специальным руководством относительно дозировок и интервалов введения антитела и других предлагаемых лекарственных средств. Указанное изделие производства может дополнительно включать в себя второй контейнер, содержащий фармацевтически приемлемый буфер-разбавитель, такой как бактериостатическая вода для инъекций (BWFI), забуференный фосфатом физиологический раствор, раствор Рингера и раствор декстрозы. Указанное изделие производства может дополнительно включать в себя другие материалы, необходимые с коммерческой и пользовательской точки зрения, включая и другие буферы, разбавители, фильтры, иглы и шприцы.
Указанное здесь изделие производства необязательно может дополнительно включать в себя контейнер, содержащий второе лекарственное средство, отличное от указанного полипептида, для лечения, и дополнительно содержащий инструкции относительно лечения млекопитающего таким средством. В определенных воплощениях указанное второе средство является противовоспалительным средством, средством против диабета и/или понижающим уровень холестерина средством класса "статинов". В некоторых воплощениях вторым активным средством является инсулин. В определенных воплощениях инсулин является инсулином быстрого действия, кратковременного действия, систематического действия, промежуточного действия или пролонгированного действия. В определенных воплощениях инсулин содержит и/или представляет собой хумалог, лиспро, новолог, апидру, хумулин, аспарт, инсулин систематического действия, NPH, инсулин Ленте, ультраленте, лантус, гларжин, левемир или детемир. В некоторых воплощениях вторым активным средством является статин. В определенных воплощениях статин содержит и/или представляет собой аторвастатин (например, липитор или торваст), церивастатин (например, липобай или бейкол), флувастатин (например, лескол), ловастатин (например, мевакор, алтокор или алтопрев) мевастатин, питавастатин (например, ливало или питава), правастатин (например, правахол, селектин или липостат), розувастатин (например, крестор), симвастатин (например, зокор или липекс), виторин, адвикор, безилат, кадуэт или симкор.
"Листовка-вкладыш в упаковке" служит для предоставления инструкций, обычно вкладываемых в коммерческие упаковки терапевтических продуктов, она содержит информацию о показаниях, применении, дозировке, способе введения, противопоказаниях, о других терапевтических продуктах, которые можно сочетать с продуктом, содержащимся в упаковке, и/или о предостережениях, связанных с применением таких терапевтических продуктов, и т.д.
Следующие примеры предложены лишь в иллюстративных целях и ни в коем случае не предназначены для ограничения объема притязаний настоящего изобретения.
Все цитируемые в настоящем описании патенты и литературные ссылки в полном объеме включены в настоящее описание в виде ссылок.
ПРИМЕРЫ
Указанные в примерах коммерчески доступные реагенты, если специально не указано иное, были использованы в соответствии с инструкциями производителей. Источником клеток, идентифицируемых в следующих примерах и в описании настоящего изобретения, с указанием номеров доступа ATCC, является Американская Коллекция Типовых Культур, Manassas, Вирджиния. Клетки культивировали в модифицированной Дульбекко среде Игла (DMEM), обогащенной 10% эмбриональной бычьей сывороткой (ЭБС), при 37°C в атмосфере, содержащей 5% CO2, если специально не указано иное.
Пример 1: KLB-независимая FGFR-связывающая активность химерного и нативного полипептидов FGF
Связывающую активность FGF-рецептора in vitro в отношении химерного полипептида FGF согласно настоящему изобретению измеряли с помощью твердофазного иммуноферментного анализа (ELISA). На фиг. 3A (вверху) представлена схематическая диаграмма ELISA для измерения активности связывания рецептора FGF (FGFR) in vitro и соответствующий ей контроль.
Моноклональные антитела, специфичные в отношении человеческого IgG-фрагмента Fc (Jackson ImmunoResearch, West Grove, PA, USA), иммобилизовывали в лунках 96-луночных плоскодонных планшетов MaxisorpTM (Nunc, Thermo Fisher Scientific, Rochester, New York), инкубируя их в течение ночи со 100 мкл на лунку раствора, содержащего 2 мкг/мл антитела. Затем каждую лунку инкубировали либо с 1 мкг/мл FGFR4-Fc (рекомбинантный полипептид, содержащий внеклеточный домен человеческого FGFR4, слитый с фрагментом Fc человеческого IgG1; каталожный no. 685-FR-050, R&D Systems, Inc., Minneapolis, Minnesota), или 1 мкг/мл FGFR1c-Fc (рекомбинантный полипептид, содержащий внеклеточный домен человеческого GFR1c, слитый с фрагментом Fc человеческого IgG1; каталожный no. 658-FR-050, R&D Systems).
Иммобилизованные на поверхности полипептиды FGFR4-Fc или FGFR1c-Fc инкубировали в течение 1 ч с нативным человеческим полипептидом FGF19-Flag с C-концевым эпитопом-маркером (См. FGF19-Flag в таблице 10, SEQ ID NO: 237) в концентрациях 1 мкг/мл, 0,4 мкг/мл, 0,16 мкг/мл, 0,064 мкг/мл, 0,0256 мкг/мл, 0,004096 мкг/мл или 0,0016384 мкг/мл, каждый с 2 мкг/мл гепарина, для создания возможности связывания полипептида FGF19 с рецепторным доменом. Сходным образом, иммобилизованные на поверхности полипептиды FGFR4-Fc или FGFR1c-Fc инкубировали в течение 1 ч с химерным полипептидом FGF19 с C-концевым эпитопом-маркером (см. cFGF21/19-2/Flag в таблице 10; SEQ ID NO: 242) в концентрациях 1 мкг/мл, 0,4 мкг/мл, 0,16 мкг/мл, 0,064 мкг/мл, 0,0256 мкг/мл, 0,004096 мкг/мл или 0,0016384 мкг/мл, каждый с 2 мкг/мл гепарина, для создания возможности связывания химерного полипептида FGF19 с рецепторным доменом. После инкубации количество нативного или химерного полипептида FGF19, связанного с рецепторным доменом, при данной концентрации полипептида FGF19, определяли с помощью биотинилированного поликлонального антитела против человеческого FGF19 (каталожный no. BAF969, R&D Systems), стрептавидина-пероксидазы хрена (HRP) (каталожный no. RPN1231V, Amersham Biosciences, Pittsburgh, Pennsylvania) и 3,3',5,5'-тетраметилбензидинового субстрата (каталожный no. TMBE-1000, Moss, Inc., Pasadena, Maryland), и путем измерения концентрации HRP-зависимого продукта по его поглощению при 450 нм. Контрольные эксперименты ELISA предпринимали для того, чтобы показать, что нативные и химерные полипептиды FGF19 распознаются поликлональным антителом против человеческого FGF19 с равноценной эффективностью (данные не приведены).
Если обратиться к фиг. 3A, результаты анализа связывания FGFR in vitro с иммобилизованными на поверхности полипептидами FGFR4-Fc или FGFR1c-Fc показали, что нативный человеческий полипептид FGF19 связывается с FGFR4-Fc зависимым от концентрации образом, Klotho-бета-независимым образом, однако существенно не связывается с FGFR1c-Fc. Klotho-бета-независимое связывание химерного полипептида FGFR19 (cFGF21/19-2/Flag; SEQ ID NO:242) с FGFR4-Fc или с FGFR1c-Fc зарегистрировано не было.
На фиг. 3B представлена схематическая диаграмма анализа активации FGFR. В данном анализе транзитно трансфицированные клетки L6 экспрессируют на своей поверхности рецептор FGF, такой как человеческий FGFR1c или человеческий FGFR4. Эффективное связывание лиганда рецептора FGF может приводить к активации пути эндогенной MAP-киназы, что может приводить к фосфорилированию химерного активатора транскрипции, имеющего активационный домен Elk-1 и ДНК-связывающий домен GAL4. Фосфорилированный активатор транскрипции может активировать экспрессию репортерного гена под контролем соответствующей лежащей выше активационной последовательности (UAS), такой как дрожжевая UAS GAL4. Репортерный ген может кодировать фермент, такой как фермент люцифераза, в частности, фермент люциферазу светлячка. Клетки L6 могут быть дополнительно трансфицированы конститутивно экспрессируемой люциферазой Renilla, которая может служить в качестве контроля нормализации индуцибельной люциферазы светлячка.
В данном анализе крысиные миобласты L6 в 96-луночном планшете транзитно трансфицировали экспрессирующим вектором, кодирующим человеческий полипептид FGFR4, экспрессирующим вектором, кодирующим активатор транскрипции GAL4-Elk-1 (каталожный no. 219005, pFA2-Elk1, Stratagene, La Jolla, California), экспрессирующим вектором, кодирующим репортерный ген люциферазы светлячка, под контролем лежащего выше дрожжевого активатора GAL4 (каталожный no. 219050, pFR-luc, Stratagene). Вектор для конститутивной экспрессии люциферазы Renilla (каталожный no. E2231, pRL-SV40, Promega, Madison, Wisconsin) также трансфицировали в указанные клетки. Трансфекции были предприняты с помощью реагента для трансфекции FuGENE HD (каталожный no. 04 709 705 001, Roche Applied Science, Indianapolis, Indiana), в соответствии с инструкциями производителей.
Трансфицированные клетки L6 культивировали в течение ночи в среде DMEM (полученной из Cellgro 50-013-PC, Mediatech, Inc., Manassas, Virginia), содержащей 10% ЭБС (каталожный no. F2442, Sigma-Aldrich, St. Louis, Missouri). Затем клетки отмывали и культивировали в течение дополнительных 6 часов в обогащенной бессывороточной среде, полученной из смеси F12/DME 50:50, содержащей 25 мг/л свиного гепарина и конкретной концентрации полипептида FGF19. Изучаемыми полипептидами FGF19 были нативный полипептид FGF19-Flag (см. FGF19-Flag в таблице 10; SEQ ID NO: 237), нативный полипептид FGF21-His (см. FGF21-His в таблице 10; SEQ ID NO: 238) и химерный полипептид FGF19-Flag (см. cFGF21/19-2/Flag в таблице 10; SEQ ID NO: 242). Клетки инкубировали с полипептидом в концентрациях 10 мкг/мл, 1666,7 нг/мл, 277,8 нг/мл, 46,3 нг/мл, 7,7 нг/мл, 1,3 нг/мл, 0,21 нг/мл, 0,036 нг/мл, 0,0060 нг/мл, 0,00099 нг/мл, 0,00017 нг/мл или 0,000028 нг/мл. Затем клетки лизировали с помощью реагента PLB (каталожный no. E1941, Promega), и в каждой лунке определяли люциферазную активность с помощью системы Dual-Glo Luciferase Assay System (каталожный no. E2940, Promega) и EnVision Multilabel Reader (каталожный no. 2103, PerkinElmer, Waltham, Massachusetts), согласно соответствующим инструкциям производителей. Каждую активность люциферазы светлячка нормировали к совместно экспрессируемой люциферазной активности Renilla, и условия каждого образца воспроизводили в трех повторах.
Полипептидные последовательности
Если еще раз обратиться к фиг. 3B, здесь каждая нормированная люциферазная активность представлена в виде среднего и стандартной ошибки среднего из трех повторов. Результаты показывают, что клетки L6, экспрессирующие FGFR4, но не Klotho-бета, при обработке нативным полипептидом FGF19 проявляют дозо-зависимую активацию люциферазной активности, тогда как ни нативный полипептид FGF21, ни химерный полипептид FGF19 такой активностью не обладали.
Пример 2: KLB-зависимая FGFR4-связывающая активность химерного и нативного полипептидов FGF
В данном анализе крысиные миобласты L6 в 96-луночном планшете транзитно трансфицировали экспрессирующим вектором, кодирующим либо человеческий полипептид FGFR4 (на основе NCBI со ссылочным номером последовательности: NM_002011.3), либо человеческий полипептид FGFR1c (на основе NCBI со ссылочным номером последовательности: NM_015850,3), экспрессирующим вектором, кодирующим полипептид Klotho-бета (KLB) (на основе NCBI со ссылочным номером последовательности: NM_175737,3, слитой с C-концевой последовательностью LEDYKDDDDK эпитопа), экспрессирующим вектором, кодирующим активатор транскрипции GAL4-Elk-1 (pFA2-Elk1, Stratagene), и экспрессирующим вектором, кодирующим репортерный ген люциферазы светлячка, под контролем лежащего выше дрожжевого активатора GAL4 (pFR-luc, Stratagene). Вектор для конститутивной экспрессии люциферазы Renilla (pRL-SV40, Promega) также трансфицировали в указанные клетки. Трансфекции были предприняты с помощью реагента для трансфекции FuGENE HD (Roche Applied Science), в соответствии с инструкциями производителей.
Трансфицированные клетки L6 культивировали в течение ночи в среде DMEM, содержащей 10% ЭБС, как указано выше. Затем клетки отмывали и культивировали в течение дополнительных 6 часов в обогащенной бессывороточной среде, содержащей 25 мг/л свиного гепарина и конкретной концентрации полипептида FGF. Изучаемыми полипептидами FGF были нативный человеческий полипептид FGF19-Flag (см. FGF19-Flag в таблице 11; SEQ ID NO: 237), нативный человеческий полипептид FGF21-His (см. FGF21-His в таблице 11; SEQ ID NO: 238) и химерный полипептид FGF19-Flag (см. cFGF21/19-2/Flag в таблице 11; SEQ ID NO: 242). Клетки инкубировали с полипептидом FGF19 в концентрациях 500 нг/мл, 83,3 нг/мл, 13,9 нг/мл, 2,3 нг/мл, 0,39 нг/мл, 0,064 нг/мл или 0,011 нг/мл. Клетки инкубировали с химерным полипептидом FGF19 в концентрациях 2667 нг/мл, 444,4 нг/мл, 74,1 нг/мл, 12,3 нг/мл, 2,06 нг/мл, 0,34 нг/мл или 0,057 нг/мл. Затем клетки лизировали с помощью реагента PLB (Promega), и в каждой лунке определяли люциферазную активность с помощью системы Dual-Glo Luciferase Assay System (Promega) и EnVision Multilabel Reader (PerkinElmer), согласно соответствующим инструкциям производителей. Каждую люциферазную активность светлячка нормировали к совместно экспрессируемой люциферазной активности Renilla, и условия каждого образца воспроизводили в трех повторах.
Если обратиться к фиг. 4, здесь каждая нормированная люциферазная активность представлена в виде среднего и стандартной ошибки среднего из трех повторов. Результаты показывают, что нативный полипептид FGF19 и химерный полипептид FGF19 вызывают сходную дозо-зависимую активацию люциферазы в присутствии KLB и FGFR1c, с EC50, составляющей 34,3 нг/мл и 22,7 нг/мл, соответственно. В клетках, трансформированных FGFR4 и KLB, дозо-зависимая активация люциферазы под действием химерного полипептида FGF19 была значительно ниже, чем дозо-зависимая активация люциферазы под действием нативного FGF19, с EC50, составляющей 269 нг/мл и 2,6 нг/мл, соответственно. Селективность каждого полипептида FGF19 в отношении FGFR1c и FGFR4 оценивали на основе значений EC50, вычисляемых с помощью программного обеспечения Prism 5 (GraphPad Software, La Jolla, California). В данном примере химерный полипептид FGF19 проявлял более высокую относительную селективность в отношении FGFR1c, которая была выше, чем в отношении FGFR4 (на основе соответствующих вычисленных значений EC50), чем соответствующая относительная селективность нативного полипептида FGF19.
Пример 3: Индукция специфичных для печени генов химерным и нативным полипептидами FGF
В данном примере мыши FVB голодали в течение ночи. Группе выборки (n = 5 или 6) голодавших мышей инъецировали через хвостовую вену нативный человеческий полипептид FGF19-Flag (FGF19-Flag в таблице 11, нативный человеческий полипептид FGF21-His (FGF21-His в таблице 11), химерный полипептид FGF19-Flag (cFGF21/19-2/Flag в таблице 11; SEQ ID NO: 242) или контрольный наполнитель в виде забуференного фосфатом физиологического раствора (PBS). Полипептиды вводили в PBS в дозе 1 мг/кг. Через 4 часа после инъекции из каждой мыши изымали ткань печени и быстро замораживали в жидком азоте. Из собранной ткани печени с использованием реагента Qiazol (каталожный no. 79306, Qiagen, Germantown, Maryland) выделяли тотальную тканевую РНК и использовали в качестве матрицы для синтеза кДНК (набор для обратной транскрипции Quantitect, каталожный no. 205311, Qiagen). Следуя стандартным протоколам для осуществления количественной ПЦР в реальном времени, определяли количество кДНК с помощью зеленого красителя SYBR (каталожный no. 11760500, Invitrogen, Carlsbad, California) и системы 7900HT Fast Real-Time PCR System (Applied BioSystems, Inc., Foster City, California), используя ген 36B4 в качестве стандарта. Если обратиться к фиг. 5, результаты показывают, что уровни мРНК Egr-1 и cFos были наиболее высокими в образце, который был получен после инъекции нативного полипептида FGF19, тогда как уровни мРНК Egr-1 и cFos либо не определялись, либо были значительно ниже в образцах, которые были получены после инъекции нативного полипептида FGF21 или химерного полипептида FGF19. Относительные уровни мРНК БТШ или мРНК Cyp7A1 были сравнимы между образцами, полученными после инъекции указанных полипептидов. Уровни со значениями p < 0,05, < 0,01 и < 0,0005 отмечены, соответственно, как "*", "**" и "***".
Пример 4: Индукция адипоцит-специфичных генов химерным и нативным полипептидами FGF
Данный пример был выполнен так, как описано в примере 3, с бурой жировой тканью (BAT) и белой жировой тканью (WAT), изъятыми через 4 часа после инъекции и быстро замороженными в жидком азоте. Если обратиться к фиг. 6, результаты показывают, что уровни мРНК Egr-1 в случае WAT и мРНК UCP-3 в случае BAT, ни в одной из которых не экспрессировались детектируемые количества FGFR4, сходным образом регулировались используемыми здесь полипептидами FGF. Уровни со значениями p < 0,05, < 0,01 и < 0,001 отмечены, соответственно, как "*", "**" и "***".
Пример 5: Понижение глюкозы в крови у диабетических мышей с ожирением под действием химерного полипептида FGF19 и нативного полипептида FGF21
В данном примере 11-недельным мышам ob/ob (stock#000632, The Jackson Laboratory, Bar Harbor, Maine) подкожно имплантировали осмотический насос (каталожный no. 2001, Alzet, Cupertino, California), содержащий 200 мкл нативного человеческого полипептида FGF21 (FGF21-FlagN в таблице 11; 1 мг/мл в PBS), химерный полипептид FGF19 (cFGF21/19-2/Flag в таблице 11; SEQ ID NO: 242) (1 мг/мл в PBS) или контрольный носитель (PBS). Каждая группа выборки состояла из девяти (9) мышей. Осмотический насос был сконструирован таким образом, чтобы полипептид поступал со скоростью ~ 0,4 мг/кг/день.
Если обратиться к фиг. 7A, здесь измеряли вес тела и уровень глюкозы в крови каждой мыши при беспорядочном кормлении, начиная за три (3) дня до имплантации осмотического насоса и кончая пятым (5) днем после его имплантации, в указанных временных точках. Глюкозу в крови измеряли с помощью системы мониторинга глюкозы в крови One Touch 2 Ultra Dlood Glucose monitoring system (LifeScan, Milpitas, California). На фиг. 7A показан средний вес тела и уровень глюкозы в крови для каждой группы выборки (уровни со значениями p < 0,05, < 0,001 и < 5 × 10-7 отмечены, соответственно, как "*", "**" и "***"). На день 5 оставляли голодать в течение ночи, и на следующее утро натощак определяли глюкозу в крови. На фиг. 7B приведены уровни глюкозы в крови для каждой мыши (голодавшей в течение ночи) на дни 5 и 6 (уровни со значениями p < 0,002, < 0,0005 и < 5 × 10-10 отмечены, соответственно, как "*", "**" и "***"). Полученные результаты показывают, что как нативный человеческий полипептид FGF21, так и химерный полипептид FGF19 способствовали понижению глюкозы в крови у этих мышей до сходных уровней.
Пример 6: Толерантность к вводимой внутрибрюшинно глюкозе у диабетических мышей с ожирением, вызываемая химерным полипептидом FGF19 и нативным полипептидом FGF21
Мышам из примера 5 внутрибрюшинно осуществляли болюсную инъекцию глюкозы в PBS (1 г/кг) с последующим голоданием в течение ночи на день 6, чтобы проверить толерантность к глюкозе. Болюсную инъекцию осуществляли в момент, соответствующий времени = 0, фиг. 8A. После болюсной инъекции у каждой мыши в определенных временных точках измеряли уровни глюкозы в крови, и средние уровни глюкозы в крови для каждой выборки приведены на фиг. 8A. На фиг. 8B отражена площадь под кривой (AUC) во временном интервале от t = 0 до 120 мин в процессе проведения теста на толерантность к глюкозе (GTT, glucose tolerance test) для каждого животного. Значения p для каждой выборки животных, которым инъецировали нативный человеческий полипептид FGF21 или химерный полипептид FGF19, по сравнению с контролем PBS в обоих случаях составляли < 0,001 в соответствии с t-тестом Стьюдента. Полученные результаты показывают, что как нативный человеческий полипептид FGF21, так и химерный полипептид FGF19 вызывали у этих голодавших мышей сходную толерантность к глюкозе.
Пример 7: Активность слитых полипептидов нативного и химерного FGF-Fc
В данном примере кондиционированную среду, содержащую слитый полипептид FGF-Fc, собирали из-под клеток, трансфицированных соответствующим экспрессирующим вектором. Клетки HEK293S транзитно трансфицировали экспрессирующим вектором, кодирующим нативный человеческий полипептид FGF19, слитый с N-концом человеческого фрагмента Fc IgG1 посредством линкера, состоящего из 21 аминокислоты GGGGSGGGGSDYKDDDDKGRAQVT (SEQ ID NO: 286), нативный человеческий полипептид FGF21, слитый с N-концом человеческого фрагмента Fc IgG1 посредством линкера, состоящего из 4-аминокислот GGGG, или химерный человеческий полипептид FGF19 (cFGF21/19-2), слитый с N-концом человеческого фрагмента Fc IgG1 посредством линкера, состоящего из 4-аминокислот GGGS. Ложно-трансфицированные клетки использовали в качестве контроля. Клетки культивировали в течение ночи в среде DMEM, содержащей 10% ЭБС, как указано выше. Затем клетки промывали и в течение двух (2) дней культивировали в обогащенной бессывороточной среде, полученной из смеси F12/DME 50:50. Из каждого образца собирали кондиционированную среду. Равные объемы (6,5 мкл) каждой кондиционированной среды из каждого образца использовали для анализа иммуноблоттинга с использованием антител, специфичных в отношении фрагмента Fc IgG. Результаты иммуноблоттинга представлены на фиг. 9B, на котором видно присутствие фрагмент Fc-содержащего полипептида с ожидаемой молекулярной массой в кондиционированной среде, которую собирали из-под клеток, трансфицированных слиянием FGF19-Fc, FGF21-Fc и cFGF21/19-2-Fc.
В данном примере кондиционированную среду использовали для демонстрации активности слитых полипептидов FGF-Fc. Клетки HEK293S в 96-луночном планшете транзитно трансфицировали экспрессирующим вектором, кодирующим активатор транскрипции GAL4-Elk-1 (pFA2-Elk1, Stratagene), и экспрессирующий вектор, кодирующий репортерный ген люциферазы светлячка под контролем лежащего выше дрожжевого активатора GAL4 (pFR-luc, Stratagene). В некоторых экспериментах клетки трансфицировали также экспрессирующим вектором, кодирующим полипептид Klotho-бета (KLB). Вектор для конститутивной экспрессии люциферазы Renilla (pRL-SV40, Promega) также трансфицировали в указанные клетки. Трансфекции были предприняты с помощью реагента для трансфекции FuGENE HD (Roche Applied Science), в соответствии с инструкциями производителей.
Трансфицированные клетки культивировали в течение ночи в среде DMEM, содержащей 10% ЭБС, как указано выше. Затем клетки промывали и культивировали в течение дополнительных 6 часов в среде, составленной из 1 части кондиционированной среды, разбавленной 3 частями обогащенной бессывороточной среды, полученной из смеси F12/DME 50:50, при этом конечная среда содержала 25 мг/л свиного гепарина. Затем клетки лизировали с помощью реагента PLB (каталожный no. E1 941, Promega), и в каждой лунке определяли люциферазную активность с помощью системы Dual-Glo Luciferase Assay System (Promega) и EnVision Multilabel Reader (PerkinElmer), согласно соответствующим инструкциям производителей. Каждую люциферазную активность светлячка нормировали к совместно экспрессируемой люциферазной активности Renilla, и условия каждого образца воспроизводили в трех повторах.
Если обратиться к фиг. 9A, здесь каждая нормированная люциферазная активность представлена в виде среднего и стандартной ошибки среднего из трех повторов. Результаты показывают, что активность люциферазы трансформированных клеток HEK293S, в присутствии KLB, может быть активирована нативным человеческим полипептидом FGF19-Fc или химерным полипептидом FGF19-Fc способом наподобие того, которым [активировали] соответствующие им не слитые с Fc аналоги. Однако, в отличие от соответствующего не слитого с Fc аналога нативного полипептида FGF21, слитый полипептид FGF21-Fc вызывал существенно более слабую активацию люциферазы светлячка даже в присутствии KLB.
Пример 8: Специфичность нативного и химерного полипептидов FGF в отношении рецептора
В данном примере крысиные миобласты L6 в 48-луночном планшете транзитно трансфицировали экспрессирующим вектором, кодирующим либо человеческий полипептид FGFR4, либо человеческий полипептид FGFR1c, либо контрольным вектором. Клетки каждого образца трансфицировали также экспрессирующим вектором, кодирующим полипептид Klotho-бета (KLB), экспрессирующим вектором, кодирующим активатор транскрипции GAL4-Elk-1 (pFA2-Elk1, Stratagene), экспрессирующим вектором, кодирующим репортерный ген люциферазы светлячка, под контролем расположенного выше дрожжевого активатора GAL4 (pFR-luc, Stratagene), и вектором для конститутивной экспрессии люциферазы Renilla (pRL-SV40, Promega). Трансфекции осуществляли с помощью реагента для трансфекции FuGENE HD (Roche Applied Science), в соответствии с инструкциями производителей.
Трансфицированные клетки L6 культивировали в течение ночи в среде DMEM, содержащей 10% ЭБС, как описано выше. Затем клетки промывали и культивировали в течение дополнительных 6 часов в бессывороточной кондиционированной среде (каждую кондиционированную среду продуцировали и собирали в соответствии с тем, как описано в примере 7, содержащей 25 мг/л свиного гепарина. Кондиционированная среда содержала либо контрольный вектор (группа A на фиг. 10) (pUC-полученный вектор, содержащий промотор CMV); либо нативный человеческий полипептид FGF21-FlagC (FGF21-FlagC в таблице 10; группа B на фиг. 10); либо нативный человеческий полипептид FGF19-Flag (FGF19-Flag в таблице 11; группа C на фиг. 10); либо первый химерный полипептид FGF19, имеющий N-концевую последовательность, полученную из нативного человеческого FGF21 (группа D на фиг. 10) (cFGF21/19-13/Flag в таблице 11);
HPIPDSSPLLQFGGQVRQRYLYTDDAQQTEAHLEIREDGTVGGAADQSPESLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPMLPMVPEEPEDLRGHLESDMFSSPLETDSMDPFGLVTGLEAVRSPSFEKDYKDDDDK; SEQ ID NO: 253); либо второй химерный полипептид FGF19, имеющий N-концевую последовательность, полученную из нативного человеческого FGF21 (группа E на фиг. 10) (cFGF21/19-2/Flag в таблице 11;
HPIPDSSPLLQFGGQVRQRYLYTSGPHGLSSCFLRIRADGVVDCARGQSAHSLLEIKAVALRTVAIKGVHSVRYLCMGADGKMQGLLQYSEEDCAFEEEIRPDGYNVYRSEKHRLPVSLSSAKQRQLYKNRGFLPLSHFLPMLPMVPEEPEDLRGHLESDMFSSPLETDSMDPFGLVTGLEAVRSPSFEKDYKDDDDK; SEQ ID NO: 242), либо химерный полипептид FGF19, имеющий N-концевую последовательность, полученную из нативного человеческого FGF19 (группа F на фиг. 10) (cFGF19/21-2/Flag в таблице 11;
RPLAFSDAGPHVHYGWGDPIRLRHLYTDDAQQTEAHLEIREDGTVGGAADQSPESLLQLKALKPGVIQILGVKTSRFLCQRPDGALYGSLHFDPEACSFRELLLEDGYNVYQSEAHGLPLHLPGNKSPHRDPAPRGPARFLPLPGLPPALPEPPGILAPQPPDVGSSDPLSMVGPSQGRSPSYASDYKDDDDK; SEQ ID NO: 255). Затем клетки лизировали с помощью реагента PLB (Promega), и в каждой лунке определяли люциферазную активность с помощью системы Dual-Glo Luciferase Assay System (Promega) и En Vision Multilabel Reader (PerkinElmer), согласно соответствующим инструкциям производителей. Каждую активность люциферазы светлячка нормировали к совместно экспрессируемой люциферазной активности Renilla, и условия каждого образца воспроизводили в трех повторах.
Если обратиться к фиг. 10, здесь каждая нормированная люциферазная активность представлена в виде кратности индукции по отношению к соответственным образом трансфицированным клеткам в выборке А, которую инкубировали в контрольной кондиционированной среде, полученной из-под клеток, трансфицированных контрольным вектором. Каждая кратность индукции представлена в виде среднего и стандартной ошибки среднего из трех повторов. Результаты показывают, что кратность индукции нормированной активности люциферазы в клетках L6, экспрессирующих FGFR1c, сравнима между неконтрольными образцами. Однако кратность индукции в клетках L6, экспрессирующих FGFR4, была значительно выше в клетках, инкубируемых с нативным FGF19, чем в клетках, инкубируемых либо с нативным полипептидом FGF21, либо с химерным полипептидом FGF19.
Пример 9: Активность химерных полипептидов FGF19 - Часть 1
В данном примере химерные полипептиды FGF19, имеющие N-концевые домены, полученные из нативного человеческого полипептида FGF19, анализировали на предмет их активности. Все исследованные полипептиды содержали также C-концевой эпитоп-маркер Flag. Клетки HEK293S транзитно трансфицировали экспрессирующим вектором, кодирующим полипептид Klotho-бета (KLB), экспрессирующим вектором, кодирующим активатор транскрипции GAL4-Elk-1 (pFA2-Elk1, Stratagene), экспрессирующим вектором, кодирующим репортерный ген люциферазы светлячка, под контролем расположенного выше дрожжевого активатора GAL4 (pFR-luc, Stratagene), и вектором для конститутивной экспрессии люциферазы Renilla (pRL-SV40, Promega). Миобласты крысы L6 транзитно трансфицировали экспрессирующим вектором, кодирующим либо человеческий полипептид FGFR4, либо человеческий полипептид FGFR1c, экспрессирующим вектором, кодирующим полипептид Klotho-бета (KLB), экспрессирующим вектором, кодирующим активатор транскрипции GAL4-Elk-1 (pFA2-Elk1, Stratagene), экспрессирующим вектором, кодирующим репортерный ген люциферазы светлячка, под контролем расположенного выше дрожжевого активатора GAL4 (pFR-luc, Stratagene), и вектором для конститутивной экспрессии люциферазы Renilla (pRL-SV40, Promega). Трансфекции осуществляли с помощью реагента для трансфекции FuGENE HD (Roche Applied Science), в соответствии с инструкциями производителей.
Трансфицированные клетки HEK293S и клетки L6 культивировали в течение ночи в среде DMEM, содержащей 10% ЭБС, как описано выше. Затем клетки промывали и культивировали в течение дополнительных 6 часов в бессывороточной кондиционированной среде (каждую кондиционированную среду продуцировали и собирали в соответствии с тем, как описано в примере 7, разбавляли для использования равным объемом бессывороточной среды), содержащей 25 мг/л свиного гепарина. Кондиционированную среду собирали из-под клеток, трансфицированных нативным человеческим полипептидом FGF21-FlagC (A на фиг. 11), химерным полипептидом FGF19, cFGF19/21-1/Flag (cFGF19/21-1/Flag в таблице 11; B на фиг. 11), химерным полипептидом FGF19, cFGF19/21-2/Flag (cFGF19/21-2/Flag в таблице 11; С на фиг. 11), химерным полипептидом FGF19, cFGF19/21-3/Flag (cFGF19/21-3/Flag в таблице 11; D на фиг. 11), химерным полипептидом FGF19, cFGF19/21-4/Flag (cFGF19/21-4/Flag в таблице 11; E на фиг. 11), химерным полипептидом FGF19, cFGF19/21-5/Flag (cFGF19/21-5/Flag в таблице 11; F на фиг. 11), химерным полипептидом FGF19, cFGF19/21-6/Flag (cFGF19/21-6/Flag в таблице 11; G на фиг. 11), химерным полипептидом FGF19, cFGF19/21-7/Flag (cFGF19/21-7/Flag в таблице 11; H на фиг. 11), химерным полипептидом FGF19, cFGF19/21-8/Flag (cFGF19/21-8/Flag в таблице 11; I на фиг. 11), химерным полипептидом FGF19, cFGF19/21-9/Flag (cFGF19/21-9/Flag в таблице 11; J на фиг. 11), химерным полипептидом FGF19, cFGF19/21/19-29/Flag (cFGF19/21/19-29/Flag в таблице 11; K на фиг. 11), или нативным полипептидом FGF19-Flag (L на фиг. 11).
На фиг. 11 активность люциферазы светлячка для каждого образца нормировали к совместно экспрессируемой люциферазной активности Renilla, и условия каждого образца воспроизводили в трех повторах. Нормированную люциферазную активность сравнивали с активностью нативного человеческого полипептида FGF19-Flag, где "+" означает активность, по существу эквивалентную активности нативного человеческого слитого полипептида FGF19-Fc, "+/-" означает промежуточную активность, и "-" означает отсутствие детектируемой активности. Кондиционированную среду, в которой не выявлялась детектируемая или промежуточная активность в клетках HEK293S, не тестировали в клетках L6.
Пример 10: Активность химерных полипептидов FGF19 - Часть 2
В данном примере химерные полипептиды FGF19, имеющие N-концевые домены, полученные из нативного человеческого полипептида FGF21, анализировали на предмет их активности. Все исследованные полипептиды содержали также C-концевой эпитоп-маркер Flag. Анализ осуществляли так, как описано в примере 9. Кондиционированную среду собирали из-под клеток, трансфицированных нативным человеческим полипептидом FGF19-Flag (FGF19-Flag в таблице 11; A на фиг. 12), химерным полипептидом FGF19, cFGF21/19-1/Flag (cFGF21/19-1/Flag в таблице 11; B на фиг. 12), химерным полипептидом FGF19, cFGF21/19-2/Flag (cFGF21/19-2/Flag в таблице 11; C на фиг. 12), химерным полипептидом FGF19, cFGF21/19-7/Flag (cFGF21/19-7/Flag в таблице 11; D на фиг. 12), химерным полипептидом FGF19, cFGF21/19-8/Flag (cFGF21/19-8/Flag в таблице 11; E на фиг. 12), химерным полипептидом FGF19, cFGF21/19-9/Flag (cFGF21/19-9/Flag в таблице 11; F на фиг. 12), химерным полипептидом FGF19, cFGF21/19-10/Flag (cFGF21/19-10/Flag в таблице 11; G на фиг. 12), химерным полипептидом FGF19, cFGF21/19-11/Flag (cFGF21/19-11/Flag в таблице 11; H на фиг. 12), химерным полипептидом FGF19, cFGF21/19-12/Flag (cFGF21/19-12/Flag в таблице 11; I на фиг. 12), химерным полипептидом FGF19, cFGF21/19-13/Flag (cFGF21/19-13/Flag в таблице 11; J на фиг. 12), или нативным полипептидом FGF21-FlagC (FGF21-FlagC в таблице 10; K на фиг. 12).
Если обратиться к фиг. 12, здесь люциферазную активность светлячка для каждого образца нормировали к совместно экспрессируемой люциферазной активности Renilla, и условия каждого образца воспроизводили в трех повторах. Нормированную люциферазную активность сравнивали с активностью нативного человеческого слитого полипептида FGF19-Fc, где "+" означает активность, по существу эквивалентную активности нативного человеческого слитого полипептида FGF19-Fc, "+/-" означает промежуточную активность, и "-" означает отсутствие детектируемой активности. Кондиционированную среду, в которой не выявлялась детектируемая или промежуточная активность в клетках HEK293S, в клетках L6 не тестировали.
Пример 11: Активность химерных полипептидов FGF19 - Часть 3
В данном примере химерные полипептиды FGF19, имеющие N-концевые домены, полученные из нативного человеческого полипептида FGF21, анализировали на предмет их активности. Все исследованные полипептиды содержали также C-концевой эпитоп-маркер Flag. Анализ осуществляли так, как описано в примере 9, за исключением того, что в данном анализу были использованы только HEK293S и FGFR4-трансфицированные клетки L6. Кондиционированную среду собирали из-под клеток, трансфицированных нативным человеческим полипептидом FGF21-FlagC (FGF21-FlagC в таблице 11; A на фиг. 13), нативным человеческим полипептидом FGF19-Flag (FGF19-Flag в таблице 11; B на фиг. 13), химерным полипептидом FGF19, cFGF21/19-1/Flag (cFGF21/19-1/Flag в таблице 11; C на фиг. 13), химерным полипептидом FGF19, cFGF21/19-2/Flag (cFGF21/19-2/Flag в таблице 11; D на фиг. 13), химерным полипептидом FGF19, cFGF21/19-3/Flag (cFGF21/19-3/Flag в таблице 11; E на фиг. 13), химерным полипептидом FGF19, cFGF21/19-4/Flag (cFGF21/19-4/Flag в таблице 11; F на фиг. 13), химерным полипептидом FGF19, cFGF21/19-5/Flag (cFGF21/19-5/Flag в таблице 11; G на фиг. 13) или химерным полипептидом FGF19, cFGF21/19-6/Flag (cFGF21/19-6/Flag в таблице 11; G на фиг. 13).
Если обратиться к фиг. 13, здесь люциферазную активность светлячка для каждого образца нормировали к совместно экспрессируемой люциферазной активности Renilla, и условия каждого образца воспроизводили в трех повторах. Нормированную люциферазную активность сравнивали с активностью нативного человеческого полипептида FGF19-Flag, где "+" означает активность, по существу эквивалентную активности нативного человеческого полипептида FGF19-Flag, "+/-" означает промежуточную активность, и "-" означает отсутствие детектируемой активности. Кондиционированную среду, в которой не выявлялась детектируемая или промежуточная активность в клетках HEK293S, в клетках L6 не тестировали.
На фиг. 13 дополнительно показано предложенное выравнивание соответствующих аминокислотных последовательностей N-концевых частей изучаемых полипептидов. Выбранные аминокислотные остатки, которые соответствуют консервативным мотивам LYT и LxxIxxG в каждом полипептиде, в приведенном выравнивании обведены рамками.
Пример 12: Активность химерных полипептидов FGF19 - Часть 4
В данном примере химерные полипептиды FGF19, имеющие N- концевой и внутренний домены, полученные из нативного человеческого полипептида FGF21, анализировали на предмет их активности. Все исследованные полипептиды содержали также C-концевой эпитоп-маркер Flag. Анализ осуществляли так, как описано в примере 9. Кондиционированную среду собирали из-под клеток, трансфицированных нативным человеческим полипептидом FGF21-FlagC (как показано на фиг. 14), нативным человеческим полипептидом FGF19-Flag (как показано на фиг. 14), химерным полипептидом FGF19, cFGF21/19-2/Flag (cFGF21/19-2/Flag в таблице 11; A на фиг. 14), химерным полипептидом FGF19, cFGF21/19-3/Flag (cFGF21/19-3/Flag в таблице 11; B на фиг. 14), химерным полипептидом FGF19, cFGF21/19-4/Flag (cFGF21/19-4/Flag в таблице 11; C на фиг. 14), химерным полипептидом FGF19, cFGF21/19-5/Flag (cFGF21/19-5/Flag в таблице 11; D на фиг. 14), химерным полипептидом FGF19, cFGF21/19-6/Flag (cFGF21/19-6/Flag в таблице 11; E на фиг. 14), химерным полипептидом FGF19, cFGF19/21/19-1/Flag (cFGF19/21/19-1/Flag в таблице 11; F на фиг. 14), химерным полипептидом FGF19, cFGF19/21/19-2/Flag (cFGF19/21/19-2/Flag в таблице 11; G на фиг. 14), химерным полипептидом FGF19, cFGF19/21/19-3/Flag (cFGF19/21/19-3/Flag в таблице 11; H на фиг. 14), химерным полипептидом FGF19, cFGF19/21/19-4/Flag (cFGF19/21/19-4/Flag в таблице 11; I на фиг. 14), химерным полипептидом FGF19, cFGF19/21/19-5/Flag (cFGF19/21/19-5/Flag в таблице 11; J на фиг. 14), химерным полипептидом FGF19, cFGF19/21/19-6/Flag (cFGF19/21/19-6/Flag в таблице 11; K на фиг. 14), или химерным полипептидом FGF19, cFGF19/21-1/Flag (cFGF19/21-1/Flag в таблице 11; L на фиг. 14).
Если обратиться к фиг. 14, здесь люциферазную активность светлячка для каждого образца нормировали к совместно экспрессируемой люциферазной активности Renilla, и условия каждого образца воспроизводили в трех повторах. Нормированную люциферазную активность сравнивали с активностью нативного человеческого полипептида FGF19-Flag, где "+" означает активность, по существу эквивалентную активности нативного человеческого полипептида FGF19-Flag, "+/-" означает промежуточную активность, и "-" означает отсутствие детектируемой активности. Кондиционированную среду, в которой не выявлялась детектируемая или промежуточная активность в клетках HEK293S, в клетках L6 не тестировали.
На фиг. 14 дополнительно показано предложенное выравнивание соответствующих аминокислотных последовательностей N-концевых частей изучаемых полипептидов. Выбранные аминокислотные остатки, которые соответствуют консервативным мотивам LYT и LxxIxxG в каждом полипептиде, в приведенном выравнивании обведены рамками.
Пример 13: Пониженное дефосфорилирование STAT5 под действием химерных полипептидов FGF19
В данном примере a химерный полипептид FGF19 согласно настоящему изобретению тестировали на предмет его воздействия на дефосфорилирование STAT5. Пятинедельным самцам мышей C57BL/6J (каждый весом приблизительно от 18 до 19 граммов), в двух повторах, подкожно инъецировали нативный человеческий полипептид FGF21-His (каталожный no. 2539-FG-025/CF, R&D Systems, Inc., Minneapolis, Minnesota), химерный полипептид FGF19-Flag (cFGF21/19-2/Flag в таблице 11; SEQ ID NO: 242) или носитель, забуференный фосфатом физиологический раствор (PBS), в качестве контроля. Полипептид растворяли в PBS и вводили в дозе 1 мг/кг (приблизительно 20 мкг полипептида на инъекцию) дважды в день в течение двух дней. На следующее утро третьего дня мышам внутривенно делали пятую, и последнюю, инъекцию соответствующего полипептида в дозе 1 мг/кг или носителя в контроле, и через 2 часа после этого их забивали. Из каждой мыши изымали печень, и из печени получали ядерный экстракт с использованием набора для выделения ядер, Nuclear Extraction Kit (каталожный no. 10009277, Cayman Chemical, Ann Arbor, Michigan). Для каждого образца ядерного экстракта 22,5 мкг белка разделяли методом ДСН-электрофореза в полиакриламидном геле и анализировали методом иммуноблоттинга с помощью антитела, специфичного в отношении белка Stat5, который фосфорилирован по Tyr694 (каталожный no. 9314, Cell Signaling Technology, Danvers, Massachusetts). Показано, что неспецифическая полоса ("NS"), которую использовали при нанесении контроля, детектируется в каждой полосе приблизительно в таких же количествах. Сходные результаты наблюдались (данные не представлены) в отношении уровней Try694-фосфорилированного Stat5 с другим моноклональным антителом, специфичным в отношении Tyr694- фосфорилированного Stat5 (каталожный no. 9359, Cell Signaling Technology).
Представленные на фиг. 15 результаты показывают, что Tyr694- фосфорилированный белок Stat5 не детектируется у мышей, которым инъецировали нативный человеческий полипептид FGF21. Однако у мышей, которым инъецировали химерный полипептид FGF19, обнаруживаются значительные уровни фосфорилированного белка Stat5.
Пример 14: Пониженная стимуляция «безъякорного» роста клеток под действием химерных полипептидов FGF19
В данном примере химерный полипептид FGF19 согласно настоящему изобретению тестировали на предмет его воздействия на «безъякорный» рост клеток человеческой гепатомы HepG2, которая экспрессирует KLB и FGFR4. 96-луночный планшет заполняли 50 мкл на лунку жидкого основного агара (DMEM, 0,5% агарозы и 10% ЭБС). После того как основной агар застынет, приблизительно 670 клеток HepG2, суспендированных в 50 мкл жидкого раствора верхнего агара (DMEM, 0,35% агарозы и 10% ЭБС), добавляли в каждую лунку к основному агару и давали застыть.
После застывания клеточной суспензии в каждую лунку добавляли по 20 мкл ростовой среды (DMEM и 10% ЭБС) в условленный нулевой день (день 0). Для данного эксперимента ростовая среда в образце дополнительно включала в себя либо нативный человеческий полипептид FGF19-Flag, либо нативный человеческий полипептид FGF21-His (каталожный no. 2539-FG-025/CF, R&D Systems, Inc., Minneapolis, Minnesota), либо химерный FGF19 flag-нацеленный полипептид (cFGF21/19-2/Flag; SEQ ID NO: 242), либо не-FGF-полипептид в качестве контроля. Концентрация полипептида в ростовой среде, которую добавляли в нулевой день, составляла либо 120 нг/мл, либо 1200 нг/мл, так, чтобы концентрация в каждой лунке составила 20 нг/мл или 200 нг/мл, соответственно. В каждый из последующих дней 2, 4, 6 и 8 в каждую лунку добавляли дополнительные 20 мкл ростовой среды, при этом указанная дополнительно добавленная в данную лунку ростовая среда содержала тот же самый полипептид FGF, который и ранее добавлялся в данную лунку, но в концентрации, составляющей одну шестую прежней концентрации полипептида FGF (то есть 20 нг/мл или 200 нг/мл), которая была добавлена в нулевой день. Часть лунок данного образца обрабатывали также ингибитором синтеза белка G418, чтобы получить фоновый флуоресцентный сигнал.
На день 9 в каждую лунку добавляли по 10 мкл реагента AlamarBlue (каталожный no. DAL 1100, Invitrogen) и планшет дополнительно инкубировали в течение 5 часов для анализа суммарной метаболической активности в каждой лунке. Полученную в результате интенсивность флуоресценции измеряли на установке EnVision Multilabel Reader (PerkinElmer). Тестировали по пять (5) параллельных проб каждого образца.
Представленные на фиг. 16 результаты приведены в виде интенсивности флуоресценции над фоном и представляют собой среднее и стандартное отклонение по пяти (5) параллельным пробам. Результаты показывают, что суммарная метаболическая активность, которая, как предполагается, является показателем «безъякорного» клеточного роста [то есть не зависящего от их прикрепления], была стимулирована добавлением нативного человеческого полипептида FGF19, но такая активность была ослаблена добавлением нативного человеческого полипептида FGF21 или химерного полипептида FGF19 (значения p <0,05 по сравнению с ложно-обработанными образцами, согласно t-тесту Стьюдента).
Пример 15: FGF19 регулирует клеточную пролиферацию, метаболизм глюкозы и желчных кислот через FGFR4-зависимый и FGFR4-независимый пути
Чтобы изучить роль FGFR4 в опосредовании активности белка FGF19, использовали Fgfr4-дефицитных мышей, а также вариант белка FGF19, способность которого активировать FGFR4 была специально ослаблена.
Материалы и методы
Экспрессия рекомбинантного белка FGF.
Были созданы аминокислотные последовательности FGF19, FGF21 и химерных полипептидов, и полученные химерные конструкции схематически изображены на фигуре 18B). Приведенные на фигуре 18B конструкции, пронумерованные от 1 до 17, соответствуют конструкциям, содержащим аминокислотные последовательности SEQ ID NO: 1, SEQ ID NO: 270

SEQ ID NO: 74, SEQ ID NO: 5, SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 271, SEQ ID NO: 272, SEQ ID NO: 273, SEQ ID NO: 274, SEQ ID NO: 275, SEQ ID NO: 276, SEQ ID NO: 277, SEQ ID NO: 278, SEQ ID NO: 85 и SEQ ID NO: 2, соответственно. Все приведенные на фигуре 18B конструкции включали в себя также сигнальные последовательности на N-концевом участке (отщепляемые при секреции) и метку flag (DYKDDDDK (SEQ ID NO: 279)) на C-концевом участке.
Если специально не указано иное, в экспериментах были использованы рекомбинантные человеческие FGF21, FGF19 и варианты, полученные в транзитно трансфицированных клетках CHO и очищенные в PBS до состояния гомогенности. В некоторых экспериментах был использован FGF21 (2539-FG/CF, R&D systems), полученный из E. coli. До проведения остальных анализов тестировали активность всех очищенных белков путем анализа GAL-Elk1 на клеточной основе. Для экспериментов, отраженных на фиг. 18B, 18C и 20, белки FGF экспрессировали в транзитно трансфицированных клетках HEK293, и для анализов использовали бессывороточную кондиционированную среду без очистки.
Анализ люциферазы. Все клетки культивировали в модифицированной Дульбекко среде Игла (DMEM), обогащенной 10% эмбриональной бычьей сывороткой (ЭБС), при 37°C в атмосфере, содержащей 5% CO2. Крысиные миобласты L6 в 96-луночном планшете транзитно трансфицировали экспрессирующими векторами, кодирующими люциферазу Renilla (pRL-SV40, Promega), человеческий KLB, подходящий человеческий FGFR, активатор транскрипции GAL4-Elk-1 (pFA2-Elk1, Stratagene) и участки связывания репортерного гена люциферазы светлячка под контролем GAL4 (pFR-luc, Stratagene), с помощью реагента для трансфекции FuGENE HD (Roche Applied Science). На следующий день трансфицированные клетки культивировали в течение дополнительных 6-8 часов в бессывороточной среде, содержащей 25 мг/л свиного гепарина (Sigma) и белок FGF в различных концентрациях. Затем клетки лизировали с помощью реагента PLB (Promega), и в каждой лунке определяли люциферазную активность с помощью системы Dual-Glo Luciferase Assay System (Promega) и установки EnVision Multilabel Reader (PerkinElmer). Люциферазную активность светлячка нормировали к совместно экспрессируемой люциферазной активности Renilla, и результаты были представлены в виде среднего и стандартной ошибки среднего по трем повторам.
Анализ «безъякорной» клеточной пролиферации. 96-луночный планшет заполняли 50 мкл/лунку 0,5% жидкой агарозы в ростовой среде. После того как основной агар застынет, приблизительно 670 клеток HepG2, суспендированных в 50 мкл верхнего раствора жидкой агарозы (0,35% агарозы в ростовой среде), добавляли в каждую лунку к основному агару, и давали застыть. После застывания в каждую лунку добавляли по 20 мкл ростовой среды, содержащей соответствующее количество белка FGF в условленный нулевой день (день 0). В каждый из последующих дней 2, 4, 6 и 8 в каждую лунку добавляли дополнительные 20 мкл ростовой среды с соответствующим количеством белка FGF. Часть лунок данного образца обрабатывали также ингибитором синтеза белка генетицином (invitrogen), чтобы получить фоновый флуоресцентный сигнал. На день 9 в каждую лунку добавляли по 10 мкл реагента AlamarBlue (Invitrogen), и планшет дополнительно инкубировали в течение 5 часов для анализа суммарной метаболической активности в каждой лунке. Полученную интенсивность флуоресценции измеряли на установке EnVision Multilabel Reader (PerkinElmer) и использовали в качестве индикатора суммарной метаболической активности в каждой лунке. Тестировали по пять (5) параллельных проб каждого образца.
Анализ связывания FGFR/лиганд. Активность связывания FGF19 FGF19v с FGFR измеряли, как описано у Desnoyers et al., Oncogene 27(l): 85-97 (2008), с помощью биотинилированного анти-FGF19-антитела (BAF969, R&D systems) в присутствии 2 мкг/мл гепарина. Контрольные эксперименты ELSA осуществляли с помощью анти-FGF19-антитела (BAF969, R&D systems) и биотинилированного анти-FGF19-антитела (BAF969, R&D systems), чтобы подтвердить, что указанное антитело неотличимым образом реагирует с FGF19 и FGF19v.
Исследования на мышах. Мышей содержали в стерильном виварии при температуре 21°C в условиях чередования 12-часового дня и 12-часовой ночи, со свободным доступом к корму (стандартный корм для грызунов (Labdiet 5010, 12,7% калорий из жировой составляющей корма) или диета с высоким содержанием жира, высоким содержанием углеводов (Harlan Teklad TD.03584, 58,4% калорий из жировой составляющей корма) и к воде. Во всех экспериментах использовали самцов мышей. Мышей FGFR4 KO, выведенных на основе линии мышей C57BL/6, которые были описаны ранее Weinstein et al., Development 1998 125(18): 3615-23 (1998), мышей C57BL/6, мышей ob/ob, выведенных на основе линии мышей C57BL/6, и мышей FVB/NJ закупали в компании Jackson Laboratory. Для непрерывной инфузии белка FGF подкожно имплантировали осмотический насос (Alzet 2001). В тесте толерантности к глюкозе уровни глюкозы измеряли с помощью глюкометра One Touch Ultra. Статистический анализ проводили с помощью t-теста Стьюдента. Значения были представлены в виде средних ± SEM (стандартное отклонение). BrdU-окрашивание производили так, как описано {Nicholes, 2002 #79}, и BrdU-положительные гепатоциты подсчитывали, используя автоматическую систему анализа изображений Ariol. Все исследования на животных проводили в соответствии с протоколами, принятыми Комитетом Genentech по биоэтике.
Анализ сыворотки. Общий (суммарный) холестерин, триглицерид, бета-гидроксибутират (BHB), лактат (Thermo DMA) и неэстерифицированные жирные кислоты (Roche) определяли с использованием ферментативных реакций. Уровни инсулина в сыворотке определяли методом ELISA (crystal chem). Состав желчных кислот (BA) определяли путем анализа жидкостной хроматографии-масс-спектрометрии, как было описано ранее Stedman et al., J Biol Chem. 279(12): 11336-43 (2004).
Анализ экспрессии генов. Тканевые РНК выделяли с помощью реагента QIAzol (Qiagen). кДНК синтезировали с использованием набора для обратной транскрипции Quantitect (Qiagen). Для проведения метода количественной ПЦР в режиме реального времени образцы прогоняли в трех повторах в высокопроизводительной системе ABI Prism 7900HT (Applied Biosystems) с использованием системы SYBR green universal mix (Invitrogen) или системы Taqman universal mix (Roche) и нормировали к уровням 36B4. Предварительно созданные праймеры Quantitect для GK, БТШ, Cyp8b1, IGFBP2 и альфа-фетопротеина были получены от компании Qiagen, а все остальные праймеры были сконструированы с использованием программного обеспечения для экспрессии праймеров (Applied Biosystems).
A. FGFR4 регулирует желчные кислоты в сыворотке, но не улучшает толерантность к глюкозе под действием рекомбинантного FGF19
Чтобы определить, какие из метаболических эффектов, выявляемых у FGF19, опосредованы рецептором FGFR4, мышей дикого типа (WT) или мышей FGFR4 KO, которым скармливали корм с высоким содержанием жира, обрабатывали рекомбинантным FGF19 или контрольным носителем, и изучали метаболические фенотипы и экспрессию генов. Для достижения непрерывного контакта с FGF19, 12 - 15-недельных мышам WT и FGFR4 KO, которым в течение 6 недель скармливали корм с высоким содержанием жира, имплантировали осмотический насос для непрерывной инфузии 1 нг/час FGF19. Это позволяло достичь средней сывороточной концентрации FGF19, составляющей 26 нг/мл и определяемой методом ELISA, что приблизительно в 50 - 250 раз выше, чем циркулирующие концентрации FGF19 у человека. На день 6 мышей, голодавших в течение ночи, подвергали тестированию на толерантность к глюкозе, путем в/б инъекции глюкозы в количестве 1 г/кг. Инфузия FGF19 сходным образом улучшала толерантность к глюкозе как у мышей WT, так и у мышей FGFR4 KO (Фиг. 17A), что указывает на то, что FGFR4 не является необходимым для улучшения толерантности к глюкозе у мышей, потребляющих корм с высоким содержанием жира. Непрерывная инфузия FGF19 не вызывала значительной потери веса, и, таким образом, повышение толерантности к глюкозе не зависело от веса тела. К 7 дню под действием FGF19 снижался вес печени и уровень сывороточного инсулина, а также усиливалось образование кетоновых тел (BHB) как у мышей дикого типа, так и у Fgfr4-нокаутных (KO) мышей (Фиг. 17B). Под действием FGF19 понижался также уровень сывороточного лактата и триглицеридов у мышей WT, но не понижался у мышей Fgfr4 KO (Фиг. 17B), несмотря даже на то, что у последних еще до обработки уровни лактата и триглицеридов были ниже.
Для оценки изменений в метаболизме желчных кислот методом жидкостной хроматографии-масс-спектрометрии определяли состав сывороточных желчных кислот (Фиг. 17C). Инфузия FGF19 у мышей дикого типа понижала уровни свободной и конъюгированной с таурином холевой кислоты (CA) и полученной из холевой кислоты вторичной желчной кислоты, дезоксихолевой кислоты (CDCA), у мышей дикого типа, но при этом оказывала минимальное воздействие на CDCA-метаболиты. Это согласуется со смещением синтеза желчных кислот на альтернативный (кислотный) путь обмена, в обход FGF19-подавляемого Cyp7a1 и продолжающийся через Cyp7b1 (Фиг. 17D). Соответственно, утрата Fgfr4 повышала уровни холевой кислоты и ее метаболитов, тогда как уменьшение мурихолевой кислоты (гидроксилированные CDCA-метаболиты) указывало на то, что FGFR4 не только важен в качестве регулятора синтеза желчных кислот, но также играет определяющую роль в соотношении продуцируемых CA к CDCA. Чтобы определить роль FGFR4 в регуляции экспрессии печеночных генов, методом QPCR исследовали ряд печеночных мРНК (Фиг. 17D). Инфузия FGF19 индуцировала экспрессию маркеров клеточной пролиферации, таких как Egr-1, c-Fos и АЛЬФА-ФЕТОПРОТЕИН, и, в отличие от мышей Fgfr4 KO, вызывала торможение экспрессии Cyp7a1 у мышей дикого типа. В отличие от этого, FGF19 вызывал супрессию Cyp8b1 и глюкокиназы (GK) как у мышей дикого типа, так и у мышей Fgfr4 KO, тогда базальные уровни экспрессии Cyp8b1 и Cyp27a1 были значительно выше у мышей Fgfr4 KO по сравнению с мышами дикого типа. Cyp8b1 является необходимым для синтеза холевой кислоты, но не CDCA, таким образом, наблюдаемые изменения в экспрессии Cyp8b1 вносят свой вклад в изменение баланса между метаболитами CA и CDCA (мурихолевые кислоты) у мышей FGFR4 KO (Фиг. 17C и D). В совокупности полученные результаты позволили выявить, что FGFR4 является регулятором синтеза желчных кислот и воздействует на пролиферацию гепатоцитов, но не является необходимым для регуляции утилизации глюкозы, чувствительности к инсулину и образованию кетоновых тел под действием FGF19.
B. Идентификация вариантов FGF19 со специфическим понижением активации FGFR4.
Чтобы количественно оценить специфическую активацию рецепторов FGFR под действием FGF19, FGF-чувствительный репортер люциферазы GAL-Elk1 вводили в крысиные клетки L6. В данном анализе эффективное связывание лиганда FGFR приводило к активации эндогенного пути MAP-киназы, приводя к активации химерного активатора транскрипции, содержащего активационный домен Elk-1 и ДНК-связывающий домен GAL4. Клетки L6 лишены функциональных FGFR или KLB и являются чувствительными только к FGF19 или FGF21, когда осуществляют их совместную трансфекцию узнаваемыми рецепторами. Клетки L6 ко-трансфицировали векторами, экспрессирующими KLB и FGFR (FGFG1c или FGFR4), совместно с GAL-Elk1, SV40-люциферазой renilla и Gal-чувствительным репортером люциферазы светлячка. Перед проведением анализа люциферазы, трансфицированные клетки в течение 6 часов инкубировали со средой, содержащей повышенные концентрации FGF19 или FGF21. Результаты такого анализа показали, что FGF19 и FGF21 со сходной силой и эффективностью активировали FGFR1c, 2c и 3c в присутствии KLB (Фиг. 18A и 19). В отличие от этого, FGF19, но не FGF21, эффективно активировал FGFR4 даже в присутствии KLB (Фиг. 18A). Для картирования сигналов, необходимых для активации FGFR4, был создан целый ряд химерных конструкций с образованием соединений между FGF19 и FGF21 с помощью консервативных остатков (Фиг. 18B). Последовательности обсуждаются выше в разделе «Материалы и методы» под рубрикой «Экспрессия рекомбинантного белка FGF». Каждую конструкцию FGF экспрессировали в транзитно трансфицированных клетках HEK293, и культуральные супернатанты, содержащие секретируемые химерные белки FGF, тестировали на предмет обнаружения активации FGFR1c и/или FGFR4 в KLB-экспрессирующих клетках L6 с помощью GAL-Elk1-репортерного анализа. На основе активности FGFR1c и FGFR4, химерные конструкции были разделены на 4 класса: с высокой активностью FGFR1c и FGFR4 (класс I, FGF19-подобные); с высокой активностью FGFR1c и с низкой, но детектируемой активностью FGFR4 (класс II); с высокой активностью FGFR1c и без детектируемой активности FGFR4 (класс III, FGF21-подобные); и с очень низкой или не детектируемой активностью FGFR1c и FGFR4 в силу слабой экспрессии (класс IV) (I-III, Фиг. 18C и 20; IV не показан). Такое картирование указывало на то, что 39 N-концевых аминокислот FGF19, при их переносе на FGF21, достаточно для придания FGFR4 некоторой активности. Кроме того, 24 N-концевые аминокислоты и 49 C-концевых аминокислот FGF19 являются необходимыми для обеспечения полной активности FGFR4, однако при их переносе на FGF21, их оказывается недостаточно для придания активности рецептору FGFR4. Таким образом, многочисленные сигналы как на N-конце, так и на C-конце FGF19 вносят свой вклад в активацию рецептора FGFR4.
Одна из химерных конструкций, классифицированная как молекула класса II, состоящая из аминокислот 1-20 полипептида FGF21 и аминокислот 25-194 полипептида FGF19 (> 90% идентичная полипептиду FGF19), была выбрана для крупномасштабного синтеза в клетках CHO, и данный вариант был назван "FGF19v". При сравнении с FGF19 с помощью анализа люциферазного репортера, белок FGF19v проявлял сходную дозо-зависимую активность в отношении FGF19 в клетках L6, совместно трансфицированных KLB и FGFR1c (фиг. 18D). Однако активность FGF19v была значительно понижена в клетках L6, совместно трансфицированных либо только FGFR4, либо сочетанием FGFR4 и KLB (фиг. 18D), при этом, как было показано ранее, FGF19 напрямую связывался в FGFR4 даже в отсутствие KLB. FGF19, в отличие от FGF19v, проявлял дозо-зависимую активность связывания с FGFR4 (фиг. 18E и F).
C. FGFR4 опосредует пролиферацию гепатоцитов in vitro и in vivo
Активность FGF19v дополнительно тестировали in vivo по сравнению с активностью FGF19 и FGF21 путем его внутривенного введения голодавшим в течение ночи мышам FVB. Через 4 часа после инъекции у мышей изымали печень, и методом QPCR определяли экспрессию печеночной мРНК. Гены, которые остро индуцировались под действием FGF19, но не под действием FGF21, такие как Egr-1 и c-Fos, эффективно индуцировались под действием FGF19v, что согласуется с пониженной FGFR4-активностью FGF19v (Фиг. 21A). FGF19v проявлял активность, сходную с таковой FGF19 или FGF21, в отношении генов, совместно регулируемых FGF19 и FGF21, таких как GK. На мышах FGFR4 KO показали, что FGFR4 содействует регуляции Egr-1 и c-Fos, но не GK, под действием FGF19 (Фиг. 21B). Неожиданно оказалось, что FGF21 (а также FGF19 и FGF19v) изменяет экспрессию БТШ и Cyp7a1 (Фиг. 21A), которые, как предполагалось, являются основными мишенями FGFR4-зависимой регуляции со стороны FGF19. Изменения в экспрессии БТШ и Cyp7a1 под действием FGF19 и FGF21 наблюдались даже у мышей FGFR4 KO, что указывает на то, что при такой острой обработке оба указанных эндокринных фактора роста FGF могут модулировать экспрессию указанных генов посредством FGFR4-независимого пути (Фиг. 21B).
FGF19 повышал «безъякорную» пролиферацию клеток HepG2 в мягком агаре, и такой эффект был значительно менее выражен для белков FGF19v или FGF21 (Фиг. 21C). Чтобы узнать, проявляет ли FGF19v также ослабленную способность индуцировать пролиферацию гепатоцитов in vivo, мышам посредством осмотического насоса вливали FGF19, FGF19v (1 нг/час) или контрольный носитель. Кроме того, тем же самым мышам в течение 7 дней внутрибрюшинно инъецировали 1 мг/кг/день белка FGF, чтобы добиться высокого пикового воздействия. Чтобы уловить периодические пролиферативные явления, раствор BrdU (30 мг/кг) инъецировали дважды в день, в общей сложности 13 раз. Пролиферацию гепатоцитов определяли путем измерения BrdU-положительных гепатоцитов в печени, изымаемой на день 7. Как было сказано ранее, обработка FGF19 приводила к драматическому повышению включения BrdU; однако такой ответ был существенно ослаблен в случае FGF19v (Фиг. 21D и E). Все печеночные мРНК для Egr-1, c-Fos и для маркера пролиферации гепатоцитов альфа-фетопротеина драматически индуцировались под действием FGF19, и такая индукция под действием FGF19v по большей части отсутствовала, тогда как регуляция GK, Cyp7a1 и Cyp8b1 не отличалась между FGF19 и FGF19v (Фиг. 21F).
D. FGFR4 не является необходимым для улучшения гипергликемии у мышей ob/ob под действием FGF19.
В связи с описанными выше результатами in vitro и in vivo встает вопрос о том, может ли FGF19v, вариант FGF19 с пониженной активностью FGFR4 и пролиферативным потенциалом, облегчить гипергликемию у диабетических животных, как это делает FGF21. FGF21, FGF19v (1 нг/час) или контрольный носитель непрерывно инфузировали мышам ob/ob подкожно с помощью осмотических мини-насосов. Несмотря на то, что инфузия не оказывала значительного влияния на вес тела (Фиг. 22A), тем не менее как FGF21, так и FGF19v драматически понижали уровни глюкозы в крови как у бессистемно питающихся мышей, так и у голодавших мышей (Фиг. 22A и B), понижали уровни циркулирующих свободных жирных кислот (Фиг. 22C) и улучшали толерантность к глюкозе (Фиг. 22D).
Для визуализации пролиферации гепатоцитов животным инъецировали BrdU за 4 часа до умерщвления на день 7. Ни FGF21, ни FGF19v не способствовали повышению включения BrdU (не показано), но при этом значительно уменьшалась суммарная масса печени (Фиг. 22E) и не обнаруживалось никаких существенных изменений на уровне экспрессии мРНК альфа-фетопротеина (Фиг. 22F). Таким образом, FGF19v может улучшать метаболический статус мышей с ожирением без индукции пролиферации гепатоцитов.
Был идентифицирован ряд генов, экспрессия которых, как правило, менялась у мышей ob/ob, обработанных FGF21 и FGF19v. В печени оба этих белка индуцировали IGFBP2 (обнаруженный в настоящее время анти-диабетический белок) и вызывали супрессию стеароил-KoA-десатуразы 1 (SCD-1; липогенный ген) и Cyp8b1 (фактор, определяющий баланс между продуцированием CA и CDCA). Кроме того, как FGF21, так и FGF19v индуцировали UCP-1 (адаптивный термогенез), SCD-1 и среднецепочечную ацил- KoA-дегидрогеназу (MCAD; митохондриальное окисление жирных кислот) в бурой жировой ткани, и SREBP-1c (липогенный фактор транскрипции) в белой жировой ткани (Фиг. 22F). Таким образом, воздействия на уровне различных тканей могут вызывать опосредованные противодиабетические эффекты FGF21 и FGF19v, действующих через FGFR4-независимый механизм.
Путем изучения индивидуальных сывороточных желчных кислот было показано, что рекомбинантный FGF19, действующий через FGFR4, подавляет [ген] Cyp7a1, вызывающий синтез желчных кислот, и далее переходит на Cyp7a1-независимый альтерантивный (кислотный) путь, приводя к продуцированию CDCA за счет CA. Экспрессия Cyp8b1 повышается в несколько раз у мышей с «выбитым» геном FGFR4, и обработка полипептидом FGF19 вызывает супрессию Cyp8b1, необходимой ферментативной стадии синтеза CA. FGFR4 оказался фактором, определяющим баланс между продуцированием CA и CDCA путем отрицательной регуляции как Cyp7a1, так и Cyp8b1. Активация FGFR4 смещает продуцирование желчных кислот в сторону CDCA, тогда как его упразднение приводит к образованию CA. Кроме того, FGF19 повышает экспрессию альфа-фетопротеина печенью FGFR4-зависимым образом. FGF19 улучшает толерантность к глюкозе у мышей FGFR4 KO, которым давали корм с высоким содержанием жира (Фиг. 17), а FGF19v, белок, у которого специфически нарушено связывание с [рецептором] FGFR4 и его активация, ослабляет гипергликемию у мышей ob/ob (Фиг. 18 и 21-22). Помимо влияния на резистентность к инсулину и метаболизм глюкозы, FGF19, подобно FGF21, повышает уровни сывороточного BHB (бета-гидроксибутирата) даже у FGFR4-нокаутных мышей (Фиг. 17). Как FGF19, так и FGF21 могут связываться с FGFR1c, FGFR2c и FGFR3c в присутствии KLB и активировать их. Таким образом FGFR1c, FGFR2c или FGFR3c, в совокупности с KLB, могут опосредованно осуществлять общие метаболические эффекты FGF19 и FGF21.



















































































































































































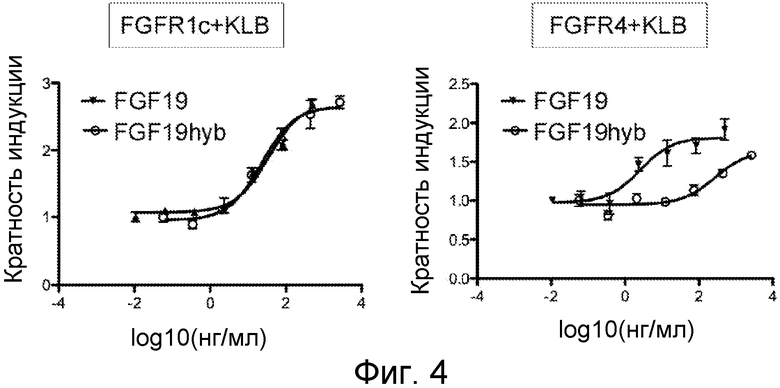














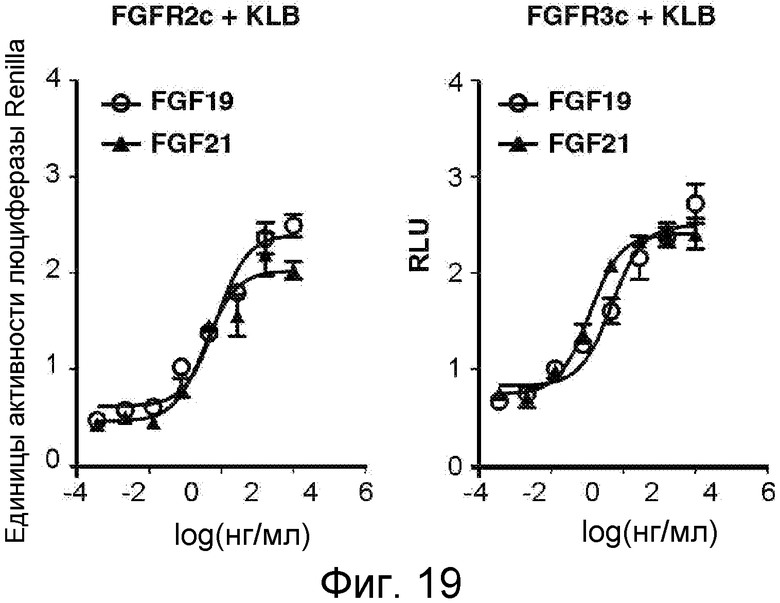
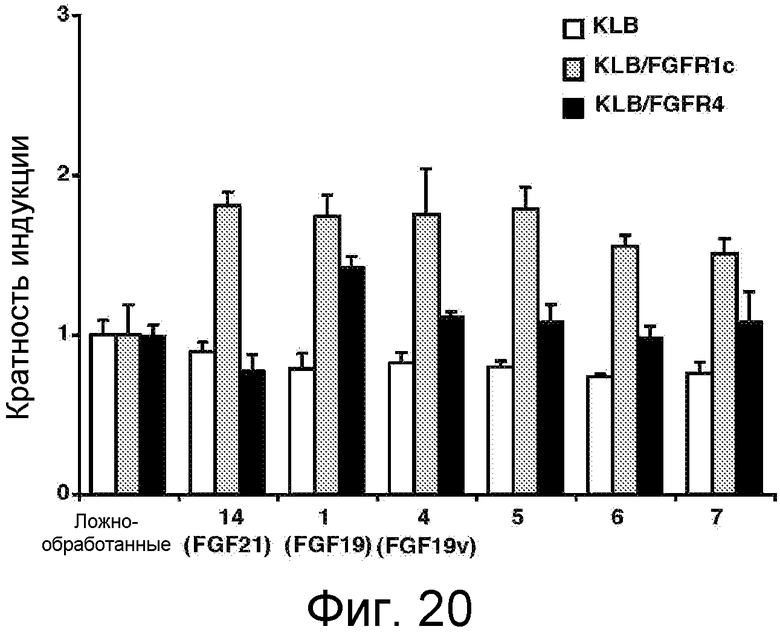

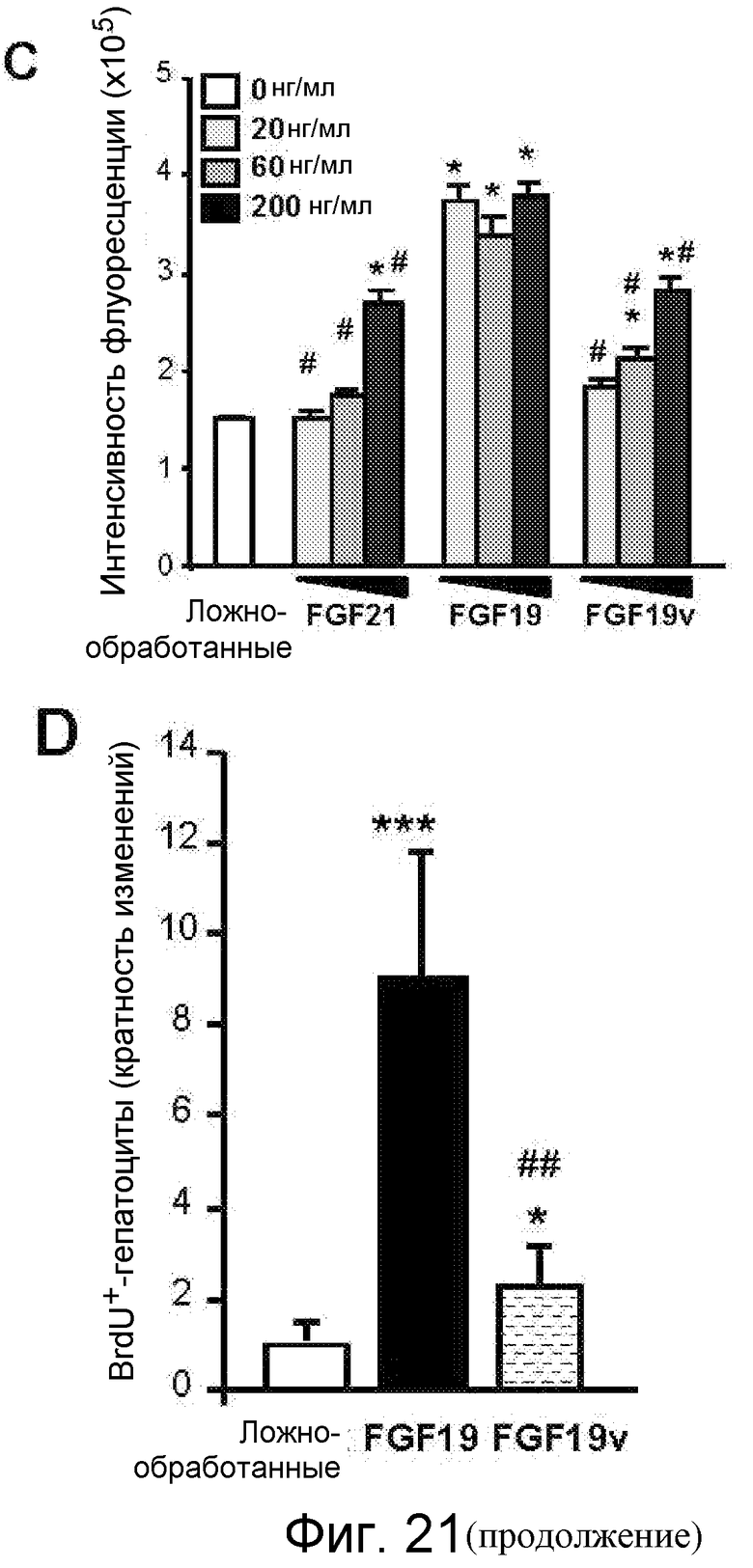
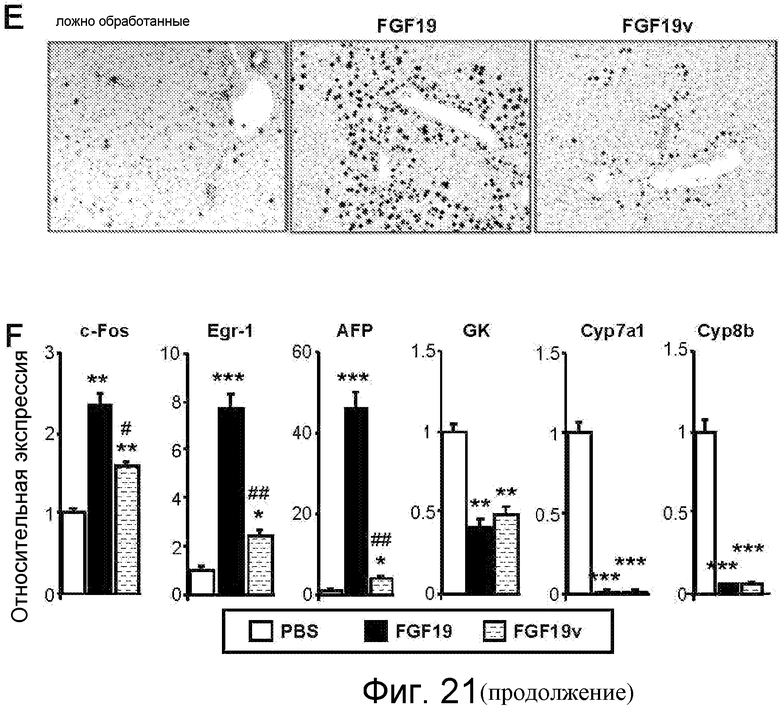
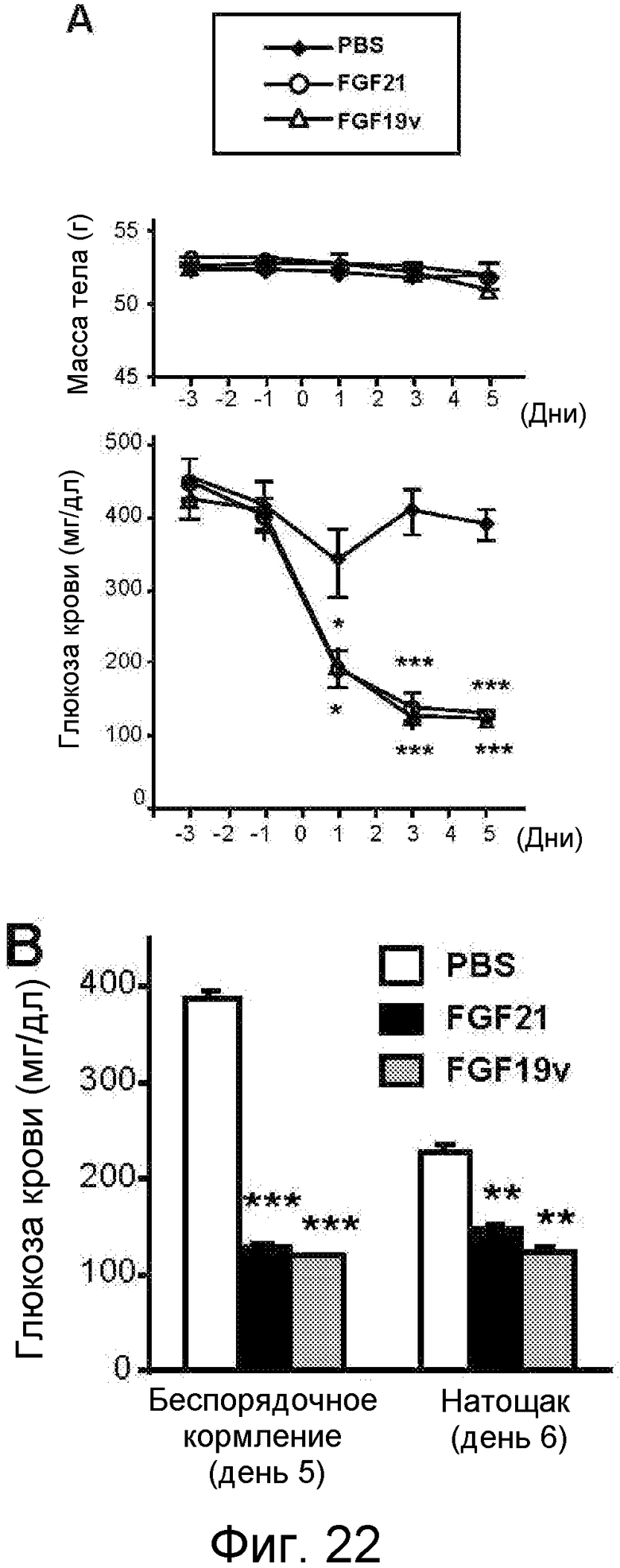

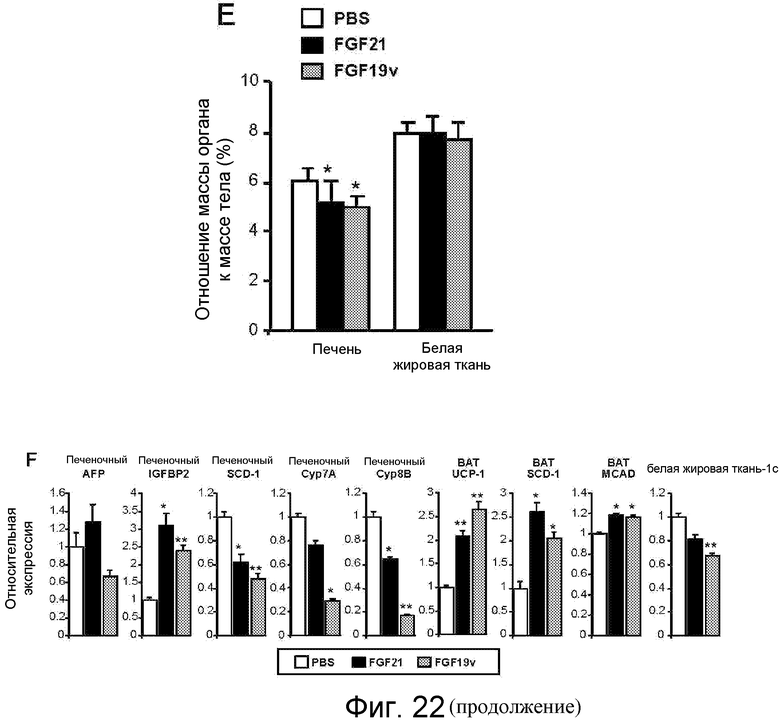
Изобретение относится к области биотехнологии, конкретно к химерным полипептидам фактора роста фибробластов (FGF), и может быть использовано в медицине для лечения ожирения, диабета, высокого содержания глюкозы в крови, метаболического синдрома и гиперхолестеринемии. Получают химерный FGF19, имеющий С-концевую часть последовательности полипептида hFGF19 и N-концевую часть последовательности полипептида hFGF21. Изобретение позволяет получить химерный полипептид FGF19, который активирует FGFRlc Klotho-бета-зависимым образом, но не активирует существенным образом FGFR4 ни Klotho-бета-независимым, ни Klotho-бета-зависимым образом. 12 н. и 6 з.п. ф-лы, 22 ил., 11 табл., 15 пр.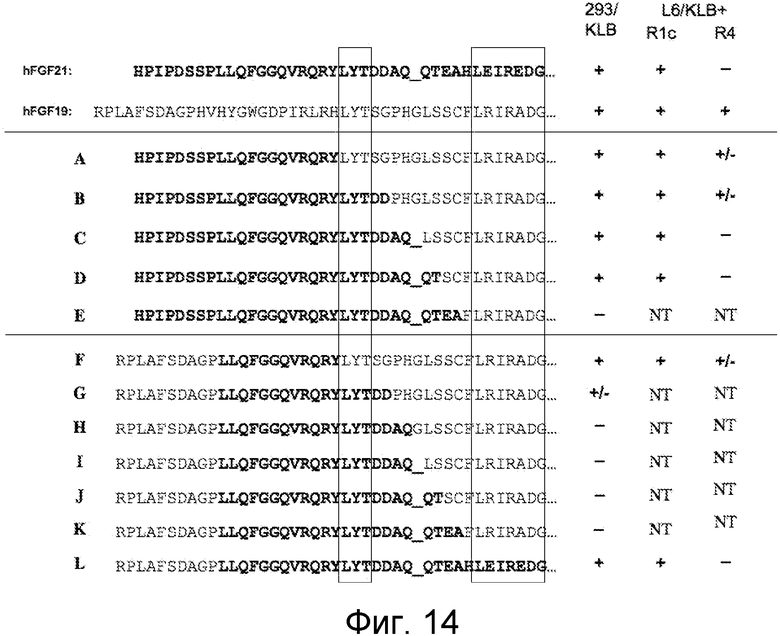
1. Химерный полипептид фактора роста фибробластов 19 (FGF19), причем последовательность указанного полипептида имеет:
С-концевую часть, которая имеет С-концевую часть последовательности полипептида hFGF19; и
N-концевую часть, которая имеет N-концевую часть последовательности полипептида hFGF21, где
С-концевая часть последовательности полипептида hFGF19 имеет длину от 45 до 185 остатков, причем С-концевая часть последовательности полипептида hFGF19 имеет первое положение и последнее положение,
N-концевая часть последовательности полипептида hFGF21 имеет длину от 7 до 140 остатков, причем N-концевая часть последовательности полипептида hFGF21 имеет первое положение и последнее положение,
где первое положение С-концевой части последовательности полипептида hFGF19 соответствует положению в SEQ ID NO: 1, выбранному из группы, состоящей из положений 25, 26, 27, 28, 30, 33 и 35, и последнее положение С-концевой части последовательности полипептида hFGF19 соответствует положению 194 в SEQ ID NO: 1, и где первое положение N-концевой части последовательности полипептида hFGF21 соответствует положению 1 в SEQ ID NO: 2; и последнее положение N-концевой части последовательности полипептида hFGF21 соответствует положению в SEQ ID NO: 2, выбранному из группы, состоящей из положений 20, 21, 22, 23, 25, 27 и 29, и
где химерный полипептид FGF19 не активирует существенным образом FGFR4 ни Klotho-бета-независимым, ни Klotho-бета-зависимым образом; и где химерный полипептид FGF19 активирует FGFRlc Klotho-бета-зависимым образом.
2. Химерный полипептид FGF19 по п. 1, где
первое положение С-концевой части последовательности полипептида hFGF19 соответствует положению 25 в SEQ ID NO: 1, и последнее положение С-концевой части последовательности полипептида hFGF19 соответствует положению 194 в SEQ ID NO: 1; и/или
первое положение N-концевой части последовательности полипептида hFGF21 соответствует положению 1 в SEQ ID NO: 2; и последнее положение N-концевой части последовательности полипептида hFGF21 соответствует положению 20 в SEQ ID NO: 2.
3. Химерный полипептид FGF19, имеющий последовательность SEQ ID NO: 5, где химерный полипептид FGF19 не активирует существенным образом FGFR4 ни Klotho-бета-независимым, ни Klotho-бета-зависимым образом; и где химерный полипептид FGF19 активирует FGFRlc Klotho-бета-зависимым образом.
4. Химерный полипептид фактора роста фибробластов 19 (FGF19), причем последовательность указанного полипептида имеет
С-концевую часть, которая имеет С-концевую часть последовательности полипептида hFGF19; и
N-концевую часть, которая имеет N-концевую часть последовательности полипептида hFGF21, где
С-концевая часть последовательности полипептида hFGF19 имеет длину от 45 до 185 остатков, причем С-концевая часть последовательности полипептида hFGF19 имеет первое положение и последнее положение,
N-концевая часть последовательности полипептида hFGF21 имеет длину от 7 до 140 остатков, причем N-концевая часть последовательности полипептида hFGF21 имеет первое положение и последнее положение,
где первое положение С-концевой части последовательности полипептида hFGF19 соответствует положению в SEQ ID NO: 1, выбранному из группы, состоящей из положений 25, 26, 27, 28, 30, 33 и 35, и последнее положение С-концевой части последовательности полипептида hFGFl9 соответствует положению 194 в SEQ ID NO: 1, и где первое положение N-концевой части последовательности полипептида hFGF21 соответствует положению 1 в SEQ ID NO: 2; и последнее положение N-концевой части последовательности полипептида hFGF21 соответствует положению в SEQ ID NO: 2, выбранному из группы, состоящей из положений 20, 21, 22, 23, 25, 27 и 29,
и где химерный полипептид hFGF19 слит со вторым полипептидом непосредственно или посредством линкера, а второй полипептид выбран из группы, состоящей из участка Fc иммуноглобулина, аналога участка Fc иммуноглобулина и одного или более фрагментов участка Fc иммуноглобулина; и
где химерный полипептид FGF19 не активирует существенным образом FGFR4 ни Klotho-бета-независимым, ни Klotho-бета-зависимым образом; и где химерный полипептид FGF19 активирует FGFRlc Klotho-бета-зависимым образом.
5. Химерный полипептид FGF19 по п. 4, где С-конец химерного полипептида hFGFl9 слит с N-концом второго полипептида посредством линкера, где указанный линкер выбран из группы, состоящей из линкера [Gly]n, линкера [Gly3Ser]m и линкера [Gly4Ser]m, где n является целым числом от 1 до 30, a m является целым числом от 1 до 6.
6. Химерный полипептид FGF19 по п. 4 или 5, где
(i) иммуноглобулин выбран из группы, состоящей из IgG-1, IgG-2, IgG-3, IgG-4, IgA-1, IgA-2, IgE, IgD и IgM; и/или
(ii) участок Fc является человеческим или гуманизированным; и/или
(iii) С-конец химерного полипептида hFGF19 слит с N-концом второго полипептида.
7. Химерный полипептид FGF19 по любому из пп. 1-5, где химерный полипептид hFGF19
(i) при введении его индивиду, не понижает уровень фосфорилированного полипептида STAT5 у индивида, где необязательно индивидуумом является человек;
(ii) при введении его индивиду, понижает количество фосфорилированного полипептида STAT5 у индивида, но указанное количество фосфорилированного полипептида STAT5 превышает количество фосфорилированного полипептида STAT5 при введении индивиду нативного hFGF21, где необязательно индивидуумом является человек;
(iii) при введении его индивиду, понижает количество фосфорилированного полипептида STAT5 до любого из количеств, составляющих от 100% до 5%, от 100% до 10%, от 100% до 20%, от 100% до 30%, от 100% до 40%, от 100% до 50%, от 100% до 60%, от 100% до 70%, от 100% до 80%, от 100% до 90% или от 100% до 95% от количества фосфорилированного полипептида STAT5 у индивида без такого введения, где необязательно индивидуумом является человек;
(iv) при введении химерного полипептида FGF19 индивиду понижение количества фосфорилированного полипептида STAT5 является меньшим, чем понижение количества фосфорилированного полипептида STAT5 при введении нативного hFGF21, где необязательно указанное понижение представляет собой любую из степеней: от 0% до 5%, от 0% до 10%, от 0% до 20%, от 0% до 30%, от 0% до 40%, от 0% до 50%, от 0% до 60%, от 0% до 70%, от 0% до 80%, от 0% до 90% или от 0% до 95% от понижения количества фосфорилированного полипептида STAT5, имеющего место при введении нативного hFGF21, где необязательно индивидуумом является человек;
(v) при введении его индивиду, не вызывает резистентности к гормону роста, где необязательно индивидуумом является человек;
(vi) имеет физиологическое время полужизни in vivo по меньшей мере или приблизительно такое же, что и у полипептида FGF19, или
(vii) имеет физиологическое время полужизни in vivo по меньшей мере или приблизительно такое же, что и у полипептида FGF21.
8. Фармацевтическая композиция, содержащая:
(a) терапевтически эффективное количество химерного полипептида FGF19 по любому из пп. 1-7; и
(b) приемлемый фармацевтический носитель,
где композиция используется для лечения индивида, у которого проявляется один или более из следующих: ожирение, диабет 1 типа, диабет 2 типа, высокое содержание глюкозы в крови, метаболический синдром и гиперхолестеринемия.
9. Способ лечения индивида, у которого проявляется один или более из следующих: ожирение, диабет 1 типа, диабет 2 типа, высокое содержание глюкозы в крови, метаболический синдром и гиперхолестеринемия, где указанный способ включает введение указанному индивиду терапевтически эффективного количества фармацевтической композиции по п. 8, где необязательно индивидуумом является человек.
10. Способ понижения уровня глюкозы в крови индивида, нуждающегося в таком лечении, где указанный способ включает введение индивиду терапевтически эффективного количества фармацевтической композиции по п. 8, где необязательно индивидуумом является человек.
11. Изолированная молекула нуклеиновой кислоты, кодирующая полипептид по п. 1.
12. Изолированная молекула нуклеиновой кислоты по п. 11, где
(i) молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты SEQ ID NO: 7;
(ii) молекула нуклеиновой кислоты содержит молекулу ДНК, кодирующую полипептид, содержащий последовательность аминокислотных остатков от 1 до 190 SEQ ID NO: 5, или (b) комплемента молекулы ДНК подпункта (а);
(iii) молекула нуклеиновой кислоты содержит молекулу ДНК, кодирующую полипептид, аминокислотная последовательность которого, по меньшей мере, на 85% идентична остаткам от 1 до 190 SEQ ID NO: 5, или (b) комплементу молекулы ДНК подпункта (а); или
(iv) молекула нуклеиновой кислоты содержит ДНК, последовательность которая, по меньшей мере, на 90%, по меньшей мере, на 95% или, по меньшей мере, на 99% идентична последовательности SEQ ID NO: 7, где молекула нуклеиновой кислоты используется для получения кодируемого полипептида.
13. Изолированная молекула нуклеиновой кислоты, кодирующая полипептид по п. 4.
14. Изолированная молекула нуклеиновой кислоты по п. 13, где
(i) молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты SEQ ID NO: 7;
(ii) молекула нуклеиновой кислоты содержит молекулу ДНК, кодирующую полипептид, содержащий последовательность аминокислотных остатков от 1 до 190 SEQ ID NO: 5, или (b) комплемента молекулы ДНК подпункта (а);
(iii) молекула нуклеиновой кислоты содержит молекулу ДНК, кодирующую полипептид, аминокислотная последовательность которого, по меньшей мере, на 85% идентична остаткам от 1 до 190 SEQ ID NO: 5, или (b) комплементу молекулы ДНК подпункта (а); или
(iv) молекула нуклеиновой кислоты содержит ДНК, последовательность которой, по меньшей мере, на 90%, по меньшей мере, на 95% или, по меньшей мере, на 99% идентична последовательности SEQ ID NO: 7,
где кодируемый полипептид дополнительно содержит аминокислотные остатки, соответствующие участку Fc иммуноглобулина,
где молекула нуклеиновой кислоты используется для получения кодируемого полипептида.
15. Вектор экспрессии, включающий молекулу нуклеиновой кислоты по любому из пп. 11-14.
16. Клетка-хозяин, содержащая молекулу нуклеиновой кислоты по п. 11 или 13 или вектор экспрессии по п. 15, где клетка используется для получения химерного полипептида FGF19.
17. Способ получения изолированного химерного полипептида FGF19, включающий:
культивирование клетки-хозяина по п. 16 в условиях, подходящих для экспрессии кодируемого полипептида; и
извлечение кодируемого полипептида из клеточной культуры.
18. Изолированный химерный полипептид FGF19, полученный способом по п. 17, где химерный полипептид FGF19 не активирует существенным образом FGFR4 ни Klotho-бета-независимым, ни Klotho-бета-зависимым образом; и где химерный полипептид FGF19 активирует FGFRlc Klotho-бета-зависимым образом.
| US 20090192087 A1, 30.07.2009 | |||
| WU X | |||
| et al., C-terminal Tail of FGF19 Determines Its Specificty toward Klotho Co-receptors, THE JOURNAL OF BIOLOGICAL CHEMISTRY, 2008, v | |||
| ПРИБОР ДЛЯ ОПРЕДЕЛЕНИЯ ОСАДКИ ВАЛОВ ПАРОВЫХ ТУРБИН | 1917 |
|
SU283A1 |
| ПОДСТАВКА ДЛЯ УСТАНОВКИ ГЕОДЕЗИЧЕСКИХ ИНСТРУМЕНТОВ НА ШТАТИВЕ И ПРИВЕДЕНИЯ ИХ В ГОРИЗОНТАЛЬНОЕ ПОЛОЖЕНИЕ | 1932 |
|
SU33304A1 |
| US 20040175703 A1, 09.09.2004 | |||
| EA 200501846 A1, 28.12.2007. | |||

