Перекрестные ссылки на родственную(ые) заявку(и)
По настоящей заявке в соответствии с 35 U.S.C. 119(e) Свода законов США испрашивается приоритет предварительной заявки США 61/238015, поданной 28 августа 2009, которая настоящим включена в качестве ссылки в полном объеме.
Предпосылки изобретения
Область изобретения
Изобретение охватывает антикиновые антитела или антитела, которые связываются с двумя, тремя, четырьмя, пятью или более CC-хемокинами (CC-хемокины также известны как β-хемокины), в частности, те антитела, которые связываются с по меньшей мере двумя хемокинами, выбранными из группы, состоящей из RANTES/CCL5, MIP-1α/CCL3, MIP-1β/CCL4 и MCP-1/CCL2. В отличие от антител, которые связываются только с одним CC-хемокином, антикиновые антитела практически решают проблему функциональной избыточности среди СС-хемокинов путем связывания, выявления и/или нейтрализации более одного СС-хемокина одновременно. Другие аспекты изобретения включают диагностические и терапевтические цели применения антикиновых антител, в том числе лечение состояний, нарушений или заболеваний, опосредованных CC-хемокинами; гибридомные линии клеток, продуцирующие антикиновые антитела, и способы получения гибридом путем последовательной иммунизации; способы гуманизации антикиновых антител; и способы усовершенствования антикиновых антител путем созревания аффинности.
Описание уровня техники
Хемокины являются ключевыми медиаторами воспаления и вовлечены в развитие аутоиммунных заболеваний; Viola & Luster, Ann. Rev. Pharmacol. Toxicol. 48: 171-197 (2008). Они образуются в участках воспаления или инфекции и вызывают миграцию лейкоцитов из кровотока к тканям. Уже давно ведутся поиски простых и эффективных способов регулирования воспаления или иммунологических процессов, опосредованных хемокинами. Эти усилия осложнены избыточностью и перекрыванием функций многих различных хемокинов и их рецепторов. Например, ингибиторы хемокинов были использованы для лечения аутоиммунных состояний в доклинических моделях на животных, но до сих пор в клинике не достигнуты цели по лечению аутоиммунных показаний. Предполагается, что это отсутствие эффекта может быть связано с избыточностью функций хемокинов. Было идентифицировано более 50 различных хемокинов, и каждый обладал различными структурными и функциональными свойствами. Специфичность некоторых хемокинов совпадает, то есть они связываются с одинаковым типом рецепторов или действуют на аналогичные типы клеток; Vergunst, et al., Arthritis Rheum. 58: 1931-1939 (2008). Определенные хемокины могут связываться с более чем одним типом хемокиновых рецепторов, и данный хемокиновый рецептор может связываться с более чем одним типом хемокинов. Таким образом, крайне желательна разработка единого агента, способного связываться с хемокинами, блокировать связывание хемокинов с рецептором или, в других случаях, нейтрализовать активность более чем одного хемокина.
Хемокины, которые получили свое название от хемотаксических цитокинов, представляют собой небольшие секретируемые полипептиды, которые регулируют движение иммунных клеток в тканях; Baggiolini, et al., Adv. Immunol. 55:97-179 (1994); Oppenheim et al., Ann. Rev. Immunol. 9:617-648 (1991).
Все хемокины образуют структуру «греческий ключ», которая стабилизируется путем дисульфидных связей между консервативными остатками цистеина. Однако хемокины подразделяются на четыре различных семейства на основании количества и положения этих консервативных остатков цистеина. Каждый из α- и β-хемокинов содержит четыре консервативных остатка цистеина. Первые два цистеина α-хемокина разделены одной аминокислотой, образуя тем самым характерный аминокислотный мотив CXC. Первые два консервативных цистеина β-хемокина находятся рядом. Поэтому β-хемокины также известны как CC-хемокины. Напротив, лимфотактин является единственным представителем третьего класса XC-хемокинов и содержит только второй и четвертый консервативные остатки цистеина. Четвертый класс хемокинов, в котором фракталкин является единственным представителем, представляет собой класс CXXXC или CX3C, который имеет 3 аминокислоты, разделяющие первые два консервативных цистеина. У людей α-хемокины в основном кодируются генами, сгруппированными на хромосоме 4, а β-хемокины в основном кодируются генами на хромосоме 17. Лимфотактин кодируется на хромосоме 1, а фракталкин - на хромосоме 16.
Хемокины образуют градиенты, которые служат в качестве хемоаттрактантов и сигналов потенциальной пролиферации для иммунных и других клеток, таких как моноциты, макрофаги, базофилы, эозинофилы, T-лимфоциты и фибробласты. CC-хемокины проявляют хемоаттрактантные свойства за счет образования градиентов концентрации, распознаваемых хемотаксическими клетками; CC-хемокины также дают сигнал к размножению определенным типам клеток, включая фибробласты и клетки иммунной системы, такие как моноциты, макрофаги, Т-лимфоциты, базофилы и эозинофилы. Мишени-рецепторы и клетки-мишени хемокинов, включая CC-хемокины, описаны в публикации Viola, et al., Annu. Rev. Pharmacol. Toxicol. 48: 171-197 (2008), см., например, фиг. 1, которая включена в настоящий документ посредством ссылки в отношении хемоаттрактантных и сигнальных функций CC-хемокинов.
Хемокины обладают структурными особенностями, связанными с определенными функциями хемокинов, такими как связывание с хемокиновым рецептором. Общие структуры включают удлиненный N-концевой сегмент (N-концевой домен), который предшествует первому остатку цистеина, N-петлю, 310-спираль, бета-цепи β1, β2 и β3, 30's-, 40's- и 50's-петли; расположение дисульфидных связей и C-концевой α-спиральный сегмент.
Эти и другие структуры CC-хемокинов, в том числе консервативные или гомологичные аминокислотные остатки среди различных CC-хемокинов, а также доступные для растворителя, частично доступные для растворителя и скрытые аминокислотные остатки CC-хемокинов, включены посредством ссылки на Fernandez, et al., Annu. Rev. Pharmacol. Toxicol. 42:469-99 (2002), см., например, фигуры 1 и 2. Скрытые аминокислотные остатки интактных, неденатурированных хемокинов вряд ли образуют эпитопы или антигенные детерминанты, контактирующие с антителами к хемокину. В отличие от этого, растворимые или представленные на поверхности остатки CC-хемокинов являются более доступными для связывания с антителами.
Остатки CC-хемокина, ассоциированные со связыванием с рецептором хемокина CCL3/MIP-1α, включают остатки 11-15 (CCFSY), остатки 17-24 (SRQIPQNF), остатки 34-35 (QC) и остатки 57-67 (EWVQKYVSDLE) SEQ ID NO:71;
остатки, ассоциированные со связыванием с рецептором хемокина CCL4/MIP-1β, включают остатки 11-15 (CCFSY), остатки 17-24 (ARKLPHNF), остатки 34-35 (LC) или остатки 57-67 (SWVQEYVYDLE) SEQ ID NO:72;
остатки, ассоциированные со связыванием CCL5/RANTES, включают остатки 10-14 (CCFAY), остатки 16-23 (ARPLPRAH), остатки 33-34 (KC) или остатки 56-66 (KWVREYINSLE) SEQ ID NO:73;
остатки, ассоциированные со связыванием CCL23/MPIF-1, включают остатки 9-13 (CCISY), остатки 15-22(PRSIPCSL), остатки 32-33 (ЕС) или остатки 55-65 (KQVQVCMRMLK) SEQ ID NO:81; и
остатки, ассоциированные со связыванием CCL15/HCC-2, включают остатки 8-12 (CCTSY), остатки 14-21 (SQSIPCSL), остатки 31-32 (ЕС) или остатки 54-64 (PGVQDCMKKLK) SEQ ID NO:79. Соответствующие аминокислотные остатки других CC-хемокинов изображены, например, на фиг. 1 публикации Fernandez, et al., id. (2002).
Консервативные домены CC-хемокинов раскрыты на сайте http://www.ncbi.nlm.nih.gov/. Эти структурные данные включены посредством ссылки на информацию о базе данных белков и консервативных доменов на указанном выше сайте в последнее обращение 24 августа 2010 года.
Хемокины класса CC-хемокинов взаимодействуют с семью соединенными с трансмембранными G-белками рецепторами, называемыми CC-хемокиновыми рецепторами или CCR, Rossi & Zlotnik, Ann. Rev. Immunol. 18:217-242 (2002). Взаимодействие хемокина со своим рецептором регулирует активацию молекул адгезии и влияет на диапедез и экстравазацию иммунных клеток из кровотока в ткани.
Хемокины вовлечены в развитие и поддержание многочисленных воспалительных и иммунологических состояний, нарушений и заболеваний, которые включают ревматоидный артрит, рассеянный склероз, атеросклероз, псориаз, воспалительные заболевания кишечника (в том числе болезнь Крона, язвенный колит, целиакию), сосудистый рестеноз, волчаночный нефрит, гломерулонефрит, отторжение трансплантата, склеродермию, фиброзные заболевания, астму (и другие воспалительные состояния легких). Например, уровни CC-хемокинов повышены в пораженных тканях у пациентов с ревматоидным артритом, рассеянным склерозом (РС), атеросклерозом и др. Доклинические животные модели этих заболеваний показывают, что ингибирование отдельных хемокинов может, по меньшей мере частично, смягчить симптомы болезни. Например, Kasama et al., J. Clin. Invest. 95: 2868- 2876 (1995) показали, что введение антитела, которое ингибирует MIP-1α/CCL3, может снизить артритную клиническую оценку приблизительно на 50% в модели ревматоидного артрита на грызунах. Аналогичным образом, Ogata et al., J. Pathol. 182: 106-1 14 (1997) показали, что антитело против MCP-1/CCL2 может уменьшить опухание суставов приблизительно на 30%. Рецепторы для MIP-1α/CCL3 (в том числе CCR1 и CCR5) и MCP-1/CCL2 (CCR2) экспрессируются перекрывающимся образом в лейкоцитах, и поэтому не исключено, что ингибитор как MIP-1α/CCL3, так и MCP-1/CCL2 мог бы быть более эффективным, чем отдельные ингибиторы каждого из хемокинов по отдельности. Публикация Viola, et al., Annu. Rev. Pharmacol. Toxicol. 48: 171-197 (2008) включена посредством ссылки для раскрытия информации об определенных классах или типах заболеваний и нарушений, связанных с такими CC-хемокинами или опосредованных ими, см., например, таблицу 1.
Известны природные ингибиторы активности хемокинов и разработаны специфические агенты, такие как антитела или ингибиторы малых молекул, которые связываются с хемокинами или препятствуют активности определенных хемокинов, см. публикацию Fernandez, et al., Annu. Rev. Pharmacol. Toxicol. 42: 469-99 (2002), в которой такие ингибиторы и агенты включены посредством ссылок, см., например, страницы 482-488. Вирус вакцины и родственный поксвирус продуцируют растворимый белок vCCI массой 35 кДа (SEQ ID NO:117), который связывает и ингибирует несколько хемокинов класса CC-хемокинов. CC-класс хемокинов, как правило, действует на лейкоциты, включая T-клетки и моноциты; Smith et al., Virology 236: 316-327 (1997), Burns et al., J. Biol. Chem. 277: 2785-2789 (2002). Было показано, что рекомбинантный vCCI эффективен для уменьшения лейкоцитарной инфильтрации в нескольких моделях хронических воспалительных заболеваний, включая экспериментальный аутоиммунный энцефалит; Jones et al., Cytokine 43: 220-228 (2008) и астму; Dabbagh, et al., J. Immunol. 165: 3418-3422 (2000). Однако применение природных веществ, таких как вирусные белки, подобные vCCI, чужеродные иммунной системе субъекта, поднимает вопросы безопасности. Введение веществ, таких как vCCI, может вызывать нежелательные физиологические или иммунные реакции, и такие вещества могут быть нейтрализованы, удалены или разрушены как чужеродные системой очистки хозяина или защитными механизмами.
С учетом этого, авторы изобретения сосредоточились на разработке способа продуцирования антител, особенно гуманизированных антител, которые могут специфически связываться и нейтрализовать более одного хемокина, но которые не представляют риски, связанные с молекулами, подобными vCCI. Прототипные ингибиторы хемокинов, такие как антитела, связывающиеся с одним CC-хемокином, имеют проблему избыточности рецепторов CC-хемокинов. Например, каждое из CCL3/MIP-1α и CCL5/RANTES связывается с рецепторами хемокинов 1 (CCR1) и 5 (CCR5), см. фиг. 1 в публикации Viola, id. (2008). Антитело, которое ингибирует только связывание CCL3 с этими хемокиновыми рецепторами, не будет предотвращать активацию рецепторов путем связывания с другими CC-хемокинами, такими как CCL5.
Авторы изобретения изначально ориентировались на CC-хемокины MIP-1α/CCL3, MIP-1β/CCL4 и RANTES/CCL5, которые составляют первичные лиганды для хемокиновых рецепторов CCR1 и CCR5. CCL2/MCP-1 также нацелен в качестве лиганда для CCR2. Как показано на фиг. 1 в публикации Viola, et al., id. (2008), эти рецепторы широко экспрессируются на моноцитах и T-клетках, а также в других подмножествах лейкоцитов. Они вовлечены в многочисленные состояния воспалительных заболеваний как в доклинических моделях болезни на животных, так и при болезнях человека.
Краткое описание изобретения
Один аспект изобретения относится к выделенному антикиновому антителу или его антигенсвязывающему фрагменту, которое может связываться с по меньшей мере двумя, тремя, четырьмя, пятью, шестью или семью или более различными CC-хемокинами. Одним из примеров антикинового антитела является антитело, которое может связываться с двумя или более CC-хемокинами, включая по меньшей мере один CC-хемокин, который взаимодействует с хемокиновым рецептором CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9 или CCR10. В другом варианте осуществления антикин связывается с по меньшей мере двумя или тремя из CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES и CCL2/MCP-1, которые взаимодействуют с CCR1, CCR2 и CCR5. Другие примеры антикинового антитела или антигенсвязывающего фрагмента антикинового антитела включают такие, которые связываются с по меньшей мере тремя различными CC-хемокинами, выбранными из группы, состоящей из CCL2/MCP-1, CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES, CCL14/HCC-1, CCL15/HCC-2, CCL18/PARC и CCL23/MPIF-1.
Связывание происходит посредством контактирования антикинового антитела и по меньшей мере одной детерминантой CC-хемокина. Например, связывание может происходить между антителом и детерминантой CCL2/MCP-1, CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES, CCL14/HCC-1, CCL15/HCC-2, CCL18/PARC или CCL23/MPIF-1, которая расположена между остатками CC CC-хемокина и последним остатком C CC-хемокина. Положение смежных остатков CC (цистеин-цистеин) и последнего остатка цистеина в CC-хемокинах известно в данной области и может быть легко идентифицировано в последовательностях CC-хемокинов, приведенных в списке последовательностей.
Антикиновые антитела и их антигенсвязывающие фрагменты могут также связываться с по меньшей мере одной детерминантой CC-хемокина, включая CCL2/MCP-1, CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES, CCL14/HCC-1, CCL15/HCC-2, CCL18/PARC и CCL23/MPIF-1, которая расположена в N-петле, 30's-петле или 40's-петле указанного CC-хемокина.
Антикиновое антитело также можно характеризовать по его способности связываться с некоторыми CC-хемокинами, но не с другими. Например, антикиновое антитело может связываться с CCL3/MIP-1α и CCL4/MIP-1β, но не связываться с CCL5/RANTES, CCL2/MCP-1, CCL8/MCP-2 или CCL7/MCP-3. Другие не будут связываться с некоторыми из хемокинов или со всеми хемокинами и другими биологически активными молекулами, обозначенными на фигурах, включая MIP-1α, MIP-1β, RANTES, MPIF-1, HCC-1, HCC-2, HCC-4, Parc, MCP-2, MCP-3, MCP-4, эотаксин, MDC, ELC, I-309, IL-8, SDF или фракталкин, а также с другими хемокинами. Например, антикины, которые не связываются с по меньшей мере одним из MCP-1, MCP-2 или MCP-3, иллюстрируются в настоящем документе.
Антикин можно также характеризовать по способности связываться с двумя или более CC-хемокинами одного вида, но не с соответствующим CC-хемокином других видов. Например, антикин может связываться с CCL3 человека, но фактически не связываться с CCL3 мыши. Конкретные примеры специфичности связывания антикиновых антител представлены на фигурах 8-12.
Некоторые антикиновые антитела и их антигенсвязывающие фрагменты могут также ингибировать взаимодействие CC-хемокинов с соответствующим рецептором. Такое ингибирование может иметь функциональные эффекты, такие как ингибирующий хемотаксис или другие эффекты, активируемые связыванием хемокинов с рецептором.
Антикиновое антитело может связываться по меньшей мере с одной детерминантой среди остатков связывания СС-хемокина с СС-рецептором. Аминокислотные остатки связывания с рецептором и сегменты CC-хемокинов, которые, как считается или известно, ассоциированы со связыванием с CC-хемокиновым рецептором, подтверждены документально в рассматриваемой области.
Считается, что для семейства CC-хемокинов, как правило, N-конец, N-петля, 30s-петля и остатки, граничащие с дисульфидами и альфа-спиралью, принимают участие в связывании с рецептором. Описание конкретных структурных особенностей хемокинов, которые соотносятся с их различными функциями, включено посредством ссылки на следующие две публикации. Baysal, et al., Proteins 43(2): 150-60 (2001) и Kuloglu, et al., Biochemistry, 40(42): 12486-96 (2001). См. базу данных консервативных доменов NCBI CDD 29111 для предлагаемых функциональных доменов CC-хемокинов.
Авторы изобретения создали и идентифицировали специфические антикиновые моноклональные антитела 3C12F, 7D1G, 7D12A, 18V4F и 18P7E. Эти антикины могут быть использованы для идентификации других антител или веществ, которые конкурентно блокируют связывание этих антикиновых моноклональных антител с одним или несколькими CC-хемокинами, которые они распознают, используя тесты по конкурентному ингибированию, известные в данной области. Конкурентные ингибиторы, такие как антитела, которые ингибируют связывание антикинового антитела, также охватываются изобретением, а также способы выявления таких конкурентных ингибиторов с использованием антикиновых моноклональных антител 3C12F, 7D1G, 7D12A, 18V4F и 18P7E.
Антикиновые антитела могут представлять собой антитела человека, гуманизированные антитела, химерные антитела человека-мыши, антитела мыши или других позвоночных, птиц или млекопитающих, или их антигенсвязывающие фрагменты.
Один тип антикинового антитела по изобретению обладает специфичностью связывания, идентичной или схожей с таковой моноклонального антитела 3C12F, и может связываться с по меньшей мере двумя, тремя, четырьмя или пятью CC-хемокинами, выбранными из группы, состоящей из CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES, CCL15/HCC-2 и CCL23/MPIF-1. Они могут не проявлять или по существу не проявлять никакого связывания с другими хемокинами, включая другие хемокины, показанные на фиг. 8, такие как HCC-1, PARC или MCP-1, 2 или 3. Антитела этого типа могут связываться с доменами, которые могут быть важны для связывания с хемокиновыми рецепторами, в том числе:
детерминанта в N-петле, 30s-петле или 40's-петле CC-хемокина;
по меньшей мере одна антигенная детерминанта CCL3/MIP-1α, расположенная в пределах остатков 11-15 (CCFSY), остатков 17-24 (SRQIPQNF), остатков 34-35 (QC) или остатков 57-67 (EWVQKYVSDLE) SEQ ID NO:71;
по меньшей мере одна антигенная детерминанта CCL4/MIP-1β, расположенная в пределах остатков 11-15 (CCFSY), остатков 17-24 (ARKLPHNF), остатков 34-35 (LC) или остатков 57-67 (SWVQEYVYDLE) SEQ ID NO:72;
по меньшей мере одна антигенная детерминанта CCL5/RANTES, расположенная в пределах остатков 10-14 (CCFAY), остатков 16-23 (ARPLPRAH), остатков 33-34 (KC) или остатков 56-66 (KWVREYINSLE) SEQ ID NO:73;
по меньшей мере одна антигенная детерминанта CCL23/MPIF-1 расположенная в пределах остатков 9-13 (CCISY), остатков 15-22 (PRSIPCSL), остатков 32-33 (ЕС) или остатков 55-65 (KQVQVCMRMLK) SEQ ID NO:81; или
по меньшей мере одна антигенная детерминанта CCL15/HCC-2, расположенная в пределах остатков 8-12 (CCTSY), остатков 14-21 (SQSIPCSL), остатков 31-32 (ЕС) или остатков 54-64 (PGVQDCMKKLK) SEQ ID NO:79.
Этот тип антитела может включать по меньшей мере один определяющий комплементарность участок (CDR) Mab 3C12F, выбранный из группы, состоящей из SEQ ID NO:3, 4, 5, 8, 9 или 10, или SEQ ID NO:53, 54, 55, 58, 59 или 60, или по меньшей мере один CDR антитела, которое конкурентно ингибирует или блокирует связывание Mab 3C12F c CC-хемокинами, с которыми оно связывается, или блокирует связывание RANTES с vCCI (см. фиг. 2). Антикиновое антитело этого типа может содержать 1, 2, 3, 4, 5 или 6 CDR Mab 3C12F или CDR, в которых 1, 2, 3, 4, 5, 6, 7, 8 или более аминокислотных остатков SEQ ID NO:3, 4, 5, 8, 9 и/или 10, или SEQ ID NO:53, 54, 55, 58, 59 и/или 60 были удалены, встроены или замещены. Таким образом, последовательности CDR могут быть идентичны таковым антитела, продуцируемого гибридомной клеточной линией 3C12F либо ее субкультурой; могут соответствовать таковым аналога антикинового антитела 3C12F или соответствовать таковым антикинового моноклонального антитела, которое конкурентно блокирует или ингибирует связывание Mab 3C12F с двумя или более СС-хемокинами, с которыми оно связывается. Такие антитела могут быть в виде антитела человека, гуманизированного антитела, химерного антитела человека-мыши, антитела мыши, птицы или других позвоночных; или их антигенсвязывающего фрагмента.
Второй тип антитела обладает специфичностью связывания, которая сходна или идентична 7D1G, и связывает по меньшей мере два, три, четыре, пять или шесть CC-хемокинов, выбранных из группы, состоящей из CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES, CCL14/HCC-1, CCL23/MPIF-1 и CCL18/PARC. Этот тип антитела может не проявлять или по существу не проявлять никакого связывания с другими хемокинами, такими как другие хемокины, показанные на фиг. 10, такие как HCC-2, эотаксин или MCP-1, 2, 3 или 4.
Антитела этого второго типа могут связываться со структурными детерминантами хемокина или CC-хемокина,
включая детерминанту в N-петле или 40's-петле по меньшей мере одного из CC-хемокинов, связанного с Mab 7D1G;
по меньшей мере одну антигенную детерминанту CCL3/MIP-1α, расположенную в пределах остатков 11-15 (CCFSY), остатков 17-24 (SRQIPQNF), остатков 34-35 (QC) или остатков 57-67 (EWVQKYVSDLE) SEQ ID NO:71;
по меньшей мере одну антигенную детерминанту CCL4/MIP-1β, расположенную в пределах остатков 11-15 (CCFSY), остатков 17-24 (ARKLPHNF), остатков 34-35 (LC) или остатков 57-67 (SWVQEYVYDLE) SEQ ID NO:72;
по меньшей мере одну антигенную детерминанту CCL5/RANTES, расположенную в пределах остатков 10-14 (CCFAY), остатков 16-23 (ARPLPRAH), остатков 33-34 (KC) или остатков 56-66 (KWVREYINSLE) SEQ ID NO:73;
по меньшей мере одну антигенную детерминанту CCL23/MPIF-1, расположенную в пределах остатков 9-13 (CCISY), остатков 15-22 (PRSIPCSL), остатков 32-33 (EC) или остатков 55-65 (KQVQVCMRMLK) SEQ ID NO:81;
по меньшей мере одну антигенную детерминанту CCL14/HCC-1, расположенную в пределах остатков 8-12 (CCFTY), остатков 14-21 (TYKIPRQR), остатков 31-32 (QC) или остатков 54-64 (KWVQDYIKDMK) SEQ ID NO:78; или
по меньшей мере одну антигенную детерминанту CCL18/PARC, расположенную в пределах остатков 10-14 (CCLVY), остатков 16-23 (SWQIPQKF), остатков 33-34 (QC) или остатков 56-66 (KWVQKYISDLK) SEQ ID NO:82.
Структурно эти антитела могут содержать один или несколько CDR Mab 7D1G, выбранных из группы, состоящей из SEQ ID NO:23, 24, 25, 28, 29 или 30, или CDR антитела, которое конкурентно ингибирует или блокирует связывание Mab 7D1G с CC-хемокинами, с которыми оно связывается. Один вид антикинового антитела этого типа будет содержать 1, 2, 3, 4, 5 или 6 CDR Mab 7D1G или CDR, в которых 1, 2, 3, 4, 5, 6, 7, 8 или более аминокислотных остатков SEQ ID NO:23, 24, 25, 28, 29 и/или 30 были удалены, вставлены или заменены. Таким образом, последовательности CDR могут быть идентичны таковым антитела, продуцируемого гибридомной клеточной линией 7D1G либо ее субкультурой; могут соответствовать таковым аналога антикинового антитела 7D1G или соответствовать таковым антикинового моноклонального антитела, которое конкурентно блокирует или ингибирует связывание Mab 7D1G с двумя или более CC-хемокинами, с которыми оно связывается. Такие антитела могут быть в виде антитела человека, гуманизированного антитела, химерного антитела человека-мыши, антитела мыши, птицы или других позвоночных; или их антигенсвязывающего фрагмента.
Третий тип антикинового антитела обладает специфичностью связывания, сходной или идентичной с 7D12A, и связывается с по меньшей мере двумя, тремя или четырьмя CC-хемокинами, выбранными из группы, состоящей из CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES и CCL23/MPIF-1. Они могут не проявлять или по существу не проявлять никакого связывания с другими хемокинами, включая другие хемокины, показанные на фиг. 9, такие как CCL2/MCP-1.
Такой продукт антитела может связываться с детерминантой в N-петле или 40's-петле по меньшей мере одного из указанных CC-хемокинов; может связываться с по меньшей мере одной антигенной детерминантой CCL3/MIP-1α, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (SRQIPQNF), остатков 34-35 (QC) или остатков 57-67 (EWVQKYVSDLE) SEQ ID NO:71;
по меньшей мере одной антигенной детерминантой CCL4/MIP-1β, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (ARKLPHNF), остатков 34-35 (LC) или остатков 57-67 (SWVQEYVYDLE) SEQ ID NO:72;
по меньшей мере одной антигенной детерминантой CCL5/RANTES, расположенной в пределах остатков 10-14 (CCFAY), остатков 16-23 (ARPLPRAH), остатков 33-34 (KC) или остатков 56-66 (KWVREYINSLE) SEQ ID NO:73; или
по меньшей мере одной антигенной детерминантой CCL23/MPIF-1, расположенной в пределах остатков 9-13 (CCISY), остатков 15-22 (PRSIPCSL), остатков 32-33 (EC) или остатков 55-65 (KQVQVCMRMLK) SEQ ID NO:81.
Структурно этот третий тип антител может включать по меньшей мере один CDR Mab 7D12A, выбранный из группы, состоящей из SEQ ID NO:13, 14, 15, 18, 19 или 20, или по меньшей мере один CDR антитела, которое конкурентно ингибирует связывание Mab 7D12A с CC-хемокинами, с которыми оно связывается. Антикиновое антитело этого типа может содержать 1, 2, 3, 4, 5 или 6 CDR Mab 7D12A или CDR, в которых 1, 2, 3, 4, 5, 6, 7, 8 или более аминокислотных остатков SEQ ID NO:13, 14, 15, 18, 19 и/или 20 были удалены, вставлены или заменены. Таким образом, последовательности CDR могут быть идентичны таковым антитела, продуцируемого гибридомной клеточной линией 7D12A или ее субкультурой; могут соответствовать таковым аналога антикинового антитела 7D12A или соответствовать таковым антикинового моноклонального антитела, которое конкурентно блокирует или ингибирует связывание Mab 7D12A с двумя или более CC-хемокинами, с которыми оно связывается. Такие антитела могут быть в виде антитела человека, гуманизированного антитела, химерного антитела человека-мыши, антитела мыши, птицы или других позвоночных; или их антигенсвязывающего фрагмента.
Четвертый тип антикинового антитела обладает специфичностью связывания, сходной или идентичной таковой 18V4F, и связывается с по меньшей мере двумя или тремя CC-хемокинами, выбранными из группы, состоящей из CCL3/MIP-1α, CCL4/MIP-1β и CCL5/RANTES. Такие антитела могут не проявлять или по существу не проявлять никакого связывания с другими хемокинами, включая другие хемокины, показанные на фиг. 11, такие как CCL2/MCP-1. Такой продукт антитела может связываться с детерминантой в N-петле или 40's-петле по меньшей мере одного из указанных CC-хемокинов; может связываться с по меньшей мере одной антигенной детерминантой CCL3/MIP-1α, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (SRQIPQNF), остатков 34-35 (QC) или остатков 57-67 (EWVQKYVSDLE) SEQ ID NO:71;
по меньшей мере одной антигенной детерминантой CCL4/MIP-1β, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (ARKLPHNF), остатков 34-35 (LC) или остатков 57-67 (SWVQEYVYDLE) SEQ ID NO:72;
по меньшей мере одной антигенной детерминантой CCL5/RANTES, расположенной в пределах остатков 10-14 (CCFAY), остатков 16-23 (ARPLPRAH), остатков 33-34 (KC) или остатков 56-66 (KWVREYINSLE) SEQ ID NO:73.
Структурно этот четвертый тип антител может включать по меньшей мере один CDR Mab 18V4F, выбранный из группы, состоящей из SEQ ID NO:33, 34, 35, 38, 39 или 40, или SEQ ID NO:63, 64, 65, 68, 69 или 70, или по меньшей мере один CDR антитела, которое конкурентно ингибирует связывание Mab 18V4F с CC-хемокинами, с которыми оно связывается. Антикиновое антитело этого типа может содержать 1, 2, 3, 4, 5 или 6 CDR Mab 18V4F или CDR, в которых 1, 2, 3, 4, 5, 6, 7, 8 или более аминокислотных остатков SEQ ID NO:33, 34, 35, 38, 39 и/или 40; или SEQ ID NO:63, 64, 65, 68, 69 и/или 70 были удалены, вставлены или заменены другими аминокислотными остатками. Таким образом, последовательности CDR могут быть идентичны таковым антитела, продуцируемого гибридомной клеточной линией 18V4F либо ее субкультурой; могут соответствовать таковым аналога антикинового антитела 18V4F или соответствовать таковым антикинового моноклонального антитела, которое конкурентно блокирует или ингибирует связывание Mab 18V4F с двумя или более CC-хемокинами, с которыми оно связывается. Такие антитела могут быть в виде антитела человека, гуманизированного антитела, химерного антитела человека-мыши, антитела мыши, птицы или других позвоночных; или их антигенсвязывающего фрагмента.
Пятый вид антикинового антитела обладает специфичностью связывания, сходной или идентичной таковой 18P7E, и связывается с по меньшей мере двумя или тремя CC-хемокинами, выбранными из группы, состоящей из CCL3/MIP-1α, CCL4/MIP-1β и CCL5/RANTES. Такие антитела могут не проявлять или по существу не проявлять никакого связывания с другими хемокинами, включая другие хемокины, показанные на фиг. 12, такие как CCL2/MCP-1. Такой продукт антитела может связываться с детерминантой в N-петле или 40's-петле по меньшей мере одного из трех CC-хемокинов, упомянутых выше.
Такой продукт антитела может связываться с детерминантой в N-петле или 40's-петле по меньшей мере одного из указанных CC-хемокинов; может связываться с
по меньшей мере одной антигенной детерминантой CCL3/MIP-1α, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (SRQIPQNF), остатков 34-35 (QC) или остатков 57-67 (EWVQKYVSDLE) SEQ ID NO:71;
по меньшей мере одной антигенной детерминантой CCL4/MIP-1β, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (ARKLPHNF), остатков 34-35 (LC) или остатков 57-67 (SWVQEYVYDLE) SEQ ID NO:72;
по меньшей мере одной антигенной детерминантой CCL5/RANTES, расположенной в пределах остатков 10-14 (CCFAY), остатков 16-23 (ARPLPRAH), остатков 33-34 (KC) или остатков 56-66 (KWVREYINSLE) SEQ ID NO:73.
Структурно этот пятый тип антитела может включать по меньшей мере один CDR Mab 18P7E, выбранный из группы, состоящей из SEQ ID NO:43, 44, 45, 48, 49 или 50, или по меньшей мере один CDR антитела, которое конкурентно ингибирует связывание Mab 18P7E с CC-хемокинами, с которыми оно связывается. Антикиновое антитело этого типа может содержать 1, 2, 3, 4, 5 или 6 CDR Mab 18P7E или CDR, в которых 1, 2, 3, 4, 5, 6, 7, 8 или более аминокислотных остатков SEQ ID NO:43, 44, 45, 48, 49 и/или 50 были удалены, вставлены или заменены другими аминокислотами. Таким образом, последовательности CDR могут быть идентичны таковым антитела, продуцируемого гибридомной клеточной линией 18P7E либо ее субкультурой; могут соответствовать таковым аналога антикинового антитела 18P7E или соответствовать таковым антикинового моноклонального антитела, которое конкурентно блокирует или ингибирует связывание Mab 18P7E с двумя, тремя или более CC-хемокинами, с которыми оно связывается. Такие антитела могут быть в виде антитела человека, гуманизированного антитела, химерного антитела человека-мыши, антитела мыши, птицы или других позвоночных; или их антигенсвязывающего фрагмента.
Последовательности легких и тяжелых вариабельных доменов, включая таковые каждого CDR, 3C12F, 7D1G, 7D12A, 18V4F, 18P7E и их аналогов, или других антикиновых антител, могут быть использованы в качестве основной структуры для лекарственной разработки имитаторов антител как конкурентных ингибиторов, которые препятствуют связыванию CC-хемокинов с рецептором, в качестве лигандов для выделения или идентификации хемокинов или антиидиотипических антител к CC-хемокиновым антителам, или в качестве иммуногенов для индуцирования антиидиотипических антител против антител к CC-хемокинам. Такие пептиды включают модифицированные или стабилизированные пептиды или конформационно ограниченные пептиды, такие как циклический или петлеобразный пептид, содержащий CDR антикинового антитела. Способы конструирования пептидов с использованием CDR антител известны в данной области и включены посредством ссылки на публикации Takahashi, et al., Chem. Eur. J. 6(17):3196-3203 или Feng, et al., Cell. Host. Microb. 98(2): 311-316. Эти CDR включают таковые последовательностей SEQ ID NO:3-5, 8-10, 13-15, 18-20, 23-25, 28-30, 33-35, 38-40, 43-45, 48-50, 53-55, 58-60, 63-65 и 68-70, а также аналоги этих пептидных последовательностей, производимых путем созревания аффинности. Комбинации различных CDR либо в виде сочетаний отдельных пептидов, содержащих различные CDR, либо в виде конъюгата, гибрида или слияния двух или более пептидных последовательностей CDR для образования единого пептидного продукта могут быть использованы для регуляции или ингибирования связывания или активности CC-хемокинов, ингибирования димеризации или мультимеризации хемокинов, или индуцирования полезных физиологических или иммунологических реакций.
Антикиновые антитела по изобретению можно составлять в виде композиции, включающей антикиновое антитело или его антигенсвязывающий фрагмент с носителем, эксципиентом или буфером, как описано более подробно ниже.
Способы получения гибридомных клеточных линий, продуцирующих антикиновые антитела, включают последовательную иммунизацию млекопитающего, такого как мышь, определенным CC-хемокином, с последующей реиммунизацией одним или несколькими различными CC-хемокинами, а затем продуцирование гибридомной клеточной линии от указанного млекопитающего, например, путем слияния клеток его селезенки с миеломной или иммортализованной B-клеточной линией, и выделение гибридомной клеточной линии, которая продуцирует антикиновое антитело, связывающееся с двумя или более хемокинами или CC-хемокинами.
Способы лечения заболевания, нарушения или состояния, опосредованных одним или несколькими CC-хемокинами, особенно таких, которые опосредованы по меньшей мере двумя или тремя CC-хемокинами, распознаваемыми антикиновым антителом, включают введение нуждающемуся в этом субъекту антикинового антитела или его антигенсвязывающего фрагмента. Заболевание, нарушение или состояние могут характеризоваться воспалением или аутоиммунной реакцией.
Другие аспекты изобретения станут наглядными из чертежей и следующего далее подробного описания вариантов осуществления.
Краткое описание чертежей
Фиг. 1 представляет связывание очищенного 3C12F с хемокинами при помощи ELISA.
Фиг. 2 показывает, что 3C12F блокирует связывание vCCI с RANTES/CCL5.
Фиг. 3 демонстрирует титрование ингибирующей активности очищенного 3C12F при хемотаксическом анализе.
Фиг. 4 демонстрирует титрование ингибирующей активности очищенного 7D12A при хемотаксическом анализе.
Фиг. 5 демонстрирует титрование ингибирующей активности очищенного 7D1G при хемотаксическом анализе.
Фиг. 6 демонстрирует титрование ингибирующей активности очищенного 18V4F при хемотаксическом анализе.
Фиг. 7 демонстрирует титрование ингибирующей активности очищенного 18P7E при хемотаксическом анализе.
Фиг. 8 представляет специфичность связывания хемокинов для 3C12F с помощью MSD-платформы.
Фиг. 9 представляет специфичность связывания хемокинов для 7D12A с помощью MSD-платформы.
Фиг. 10 представляет специфичность связывания хемокинов для 7D1G с помощью MSD-платформы.
Фиг. 11 представляет специфичность связывания хемокинов для 18V4F с помощью MSD-платформы.
Фиг. 12 представляет специфичность связывания хемокинов для 18P7E с помощью MSD-платформы.
Фиг. 13 a, b, c и d демонстрируют выравнивание последовательностей хемокинов, распознаваемых пятью антикиновыми антителами.
Подробное описание изобретения
Термин «антитело» следует трактовать расширенно как описывающий единичные моноклональные антитела, композиции антител с полиэпитопной специфичностью, а также фрагменты антител (например, Fab, F(ab')2, scFv и Fv), при условии, что они обладают желательной биологической активностью, такой как способность связываться с определенным антигеном, эпитопом или антигенной детерминантой. Этот термин включает интактные антитела, полноразмерные антитела или неукороченные антитела, а также фрагменты антител и производные, варианты и аналоги антител.
Антитела, в том числе описанные ниже антикиновые антитела, могут иметь различные изотипы - например, IgA, IgD, IgE, IgG, IgM и IgY, а также различные подклассы изотипов, такие как IgG человека подклассов 1, 2, 3 и 4 или IgA человека подклассов 1 и 2. Поливалентные антитела могут быть охарактеризованы по их авидности к поливалентному антигену. Авидность усиливает связывание с антигенами с повторяющимися одинаковыми эпитопами, и некоторые хемокины характеризуются димерными или тетрамерными структурами. Чем больше антигенсвязывающих участков содержит отдельная молекула антитела, тем выше его авидность к антигену. Антитела могут быть получены или могут происходить от различных позвоночных, включая млекопитающих и птиц.
В настоящем документе термин «моноклональное антитело» относится к антителу, полученному из множества по существу однородных антител, т.е. индивидуальные антитела, входящие в популяцию, являются идентичными, за исключением, возможно, природных мутаций, которые могут присутствовать в небольших количествах. Моноклональные антитела являются высоко специфическими, направленными против одного антигенного сайта. Более того, в отличие от обычных (поликлональных) антител препараты, которые обычно включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. В дополнение к своей специфичности моноклональные антитела имеют преимущество в том, что они синтезируются гибридомной культурой, не загрязненной другими иммуноглобулинами. Модификатор «моноклональное» указывает на характер антитела, полученного из по существу однородной популяции антител, и не должен толковаться как требующий получения антител каким-либо конкретным способом. Например, моноклональные антитела, предназначенные для использования в соответствии с настоящим изобретением, можно получить гибридомным способом, впервые описанным Köhler & Milstein, Nature, 256:495 (1975), или можно получить по технологии рекомбинантных ДНК, см., например, Cabilly, et al., патент США № 4816567.
«Химерное антитело» относится к антителам, содержащим последовательности аминокислот из двух разных источников, например, такого, который содержит консервативные сегменты антитела человека, сплайсированные с вариабельными сегментами антитела мыши, для которых известно, что они связываются с конкретным эпитопом или антигеном. Одна часть каждой из аминокислотных последовательностей тяжелых и легких цепей является идентичной или гомологичной последовательностям антител, полученных от определенных видов или принадлежащих к определенному классу, в то время как остальной сегмент цепей гомологичен или идентичен последовательностям от других видов. В одном варианте осуществления изобретение характеризует химерное антитело или антигенсвязывающий фрагмент, в котором вариабельные области как легких, так и тяжелых цепей имитируют вариабельные области антител, полученных от одного вида млекопитающих, а константные части гомологичны последовательностям антител, полученных от другого вида. В другом варианте осуществления изобретения химерные антитела получены пересадкой CDR из антитела мыши в каркасные области антитела человека. Таким образом, моноклональные антитела по изобретению включают «химерные» антитела, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующей последовательности антител, полученных от определенных видов или принадлежащих к определенному классу или подклассу антител, в то время как оставшаяся часть цепи(ей) идентична или гомологична соответствующим последовательностям антител, полученных от других видов или принадлежащих к другим классам или подклассам антител, а также фрагментам таких антител, сохраняющих способность связывания с CC-хемокинами. Особенности химерных антител и способы их получения включены посредством ссылки на публикации Cabilly, et al., патент США № 4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA, 81: 6851-6855 (1984).
«Определяющий комплементарность участок (CDR)» является частью вариабельной области легкой или тяжелой цепи молекулы иммуноглобулина. Эти участки цепи антитела образуют часть участка, который определяет специфичность антитела к определенному эпитопу на антигене, и образуют части молекулы антитела, которые могут непосредственно связываться с эпитопом. CDR3 демонстрирует наибольшую вариабельность среди различных CDR, образующих антитело. CDR опосредуют контакт между антителом и эпитопом, который оно распознает. Выделенные пептиды, включающие или аналогичные последовательностям CDR, могут проявлять дополнительные функциональные активности: Polonelli, et al., PLoS One 3:e2371 (2008).
«Гуманизированные антитела» относятся к антителам, которые включают по меньшей мере одну цепь, содержащую остатки каркаса вариабельной области, в значительной степени, от цепи антитела человека (называемого акцепторным иммуноглобулином или антителом) и по меньшей мере один определяющий комплементарность участок (CDR), в значительно степени, от антитела, отличного от антитела человека (например, мыши). В дополнение к пересадке CDR, гуманизированные антитела обычно подвергаются дополнительным изменениям для усиления аффинности и/или снижения иммуногенности. «Гуманизированное антитело» включает антитела, не полностью являющиеся антителами человека, которые представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина, отличного от иммуноглобулина человека. По большей части, гуманизированные антитела представляют собой иммуноглобулины человека (реципиентное антитело), в котором остатки гипервариабельной области реципиента заменяются остатками гипервариабельной области видов, отличных от человека (антитело-донор), таких как мышь, крыса, кролик или примат, отличный от человека, обладающий желаемой специфичностью, аффинностью и активностью. В некоторых случаях остатки каркасной области (FR) иммуноглобулина человека заменяются соответствующими остатками иммуноглобулина, отличного от иммуноглобулина человека. Кроме того, гуманизированные антитела могут включать остатки, которые не обнаруживаются в реципиентном антителе или в антителе-доноре. Эти модификации производятся для дополнительного увеличения эффективности антитела. Как правило, гуманизированное антитело должно содержать по существу все из по меньшей мере одного и обычно двух вариабельных доменов, в которых все или по существу все из гипервариабельных областей соответствуют таковым иммуноглобулина, отличного от иммуноглобулина человека, и все или по существу все из FR представляют собой таковые последовательности иммуноглобулина человека. Также гуманизированное антитело необязательно будет включать по меньшей мере часть константной области иммуноглобулина (Fc), обычно ту часть иммуноглобулина человека, которая иммуноспецифически связывается с двумя или более CC-хемокинами, которые были изменены путем введения замен, удалений или добавлений аминокислотных остатков (т.е. мутаций). В некоторых вариантах осуществления гуманизированное антитело представляет собой производное. Такое гуманизированное антитело содержит замены, удаления или добавления аминокислотных остатков в одном или нескольких CDR, отличных от CDR человека. Производное гуманизированного антитела может обладать по существу таким же связыванием, лучшим связыванием или худшим связыванием по сравнению с гуманизированным антителом, не являющимся производным. В определенных вариантах осуществления один, два, три, четыре или пять аминокислотных остатков в CDR были заменены, удалены или добавлены (т.е. мутированы). Способы получения гуманизированных антител включены посредством ссылки на европейские патенты №№ EP 239400, EP 592106 и EP 519596; международные патенты N№№ WO 91/09967 и WO 93/17105; патенты США N№№ 5225539, 5530101, 5565332, 5585089, 5766886 и 6407213; и публикации Padlan, Molecular Immunology 28(4/5):489-498 (1991); Studnicka, et al., Protein Engineering 7(6):805-814 (1994); Roguska, et al., Proc. Natl. Acad. Sci. USA 91:969-973 (1994); Tan, et al., J. Immunol. 169:1119-25 (2002); Caldas, et al., Protein Eng. 13:353-60 (2000); Morea, et al., Methods 20:267-79 (2000); Baca, et al., J. Biol. Chem. 272:10678-84 (1997); Roguska, et al., Protein Eng. 9:895-904 (1996); Couto, et al., Cancer Res. 55 (23 Supp):5973s-5977s (1995); Couto, et al., Cancer Res. 55:1717-22 (1995); Sandhu, Gene 150:409-10 (1994); Pedersen, et al., J. Mol. Biol. 235:959-73 (1994); Jones, et al., Nature 321:522-525 (1986); Reichmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992).
«Вариант» или «аналог» антитела относится в настоящем документе к молекуле, которая отличается по аминокислотной последовательности от последовательности аминокислот «родительского» антитела благодаря добавлению, удалению и/или замене одного или нескольких аминокислотных остатков(а) в последовательности родительского антитела. В одном варианте осуществления вариант содержит одну или несколько аминокислотных замен(ы) в одной или нескольких гипервариабельных областях(и) родительского антитела. Например, вариант может содержать по меньшей мере один, например, от примерно одного до примерно десяти, и предпочтительно примерно от двух до примерно пяти замен в одной или нескольких гипервариабельных областей родительского антитела. Обычно этот вариант будет содержать аминокислотную последовательность, обладающую по меньшей мере 75% идентичностью по аминокислотной последовательности с последовательностями вариабельного домена тяжелой и легкой цепи родительского антитела, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95%. Идентичность или гомология по отношению к этой последовательности определяется в настоящем документе как процент аминокислотных остатков в последовательности-кандидате, которые совпадают с остатками родительского антитела, после выравнивания последовательностей и добавления пробелов, если необходимо, для достижения максимального процента идентичности последовательности. Никакие из N-концевых, C-концевых или внутренних расширений, делеций или вставок в последовательности антитела не следует рассматривать как затрагивающие идентичность или гомологию последовательности. Вариант сохраняет способность связывания с рецептором и, предпочтительно, обладает свойствами, превосходящими таковые родительского антитела. Например, вариант может обладать более сильной аффинностью связывания, увеличенной способностью активации рецептора и т.д. Для анализа таких свойств необходимо сравнивать Fab-форму варианта с Fab-формой родительского антитела или полноразмерную форму варианта с полноразмерной формой родительского антитела, например, поскольку было установлено, что формат антитела влияет на его активность при анализе биологической активности, раскрытой в настоящем документе. Представляющее особый интерес в настоящем документе вариантное антитело является антителом, которое обнаруживает по меньшей мере примерно 10-кратное, предпочтительно по меньшей мере примерно 20-кратное и наиболее предпочтительно по меньшей мере примерно 50-кратное увеличение биологической активности по сравнению с родительским антителом.
«Родительское» антитело в настоящем документе представляет собой антитело, которое кодируется аминокислотной последовательностью, используемой для получения варианта. Предпочтительно, родительское антитело имеет каркасную область антитела человека и имеет константную область(и) антитела человека. Например, родительское антитело может представлять собой антитело человека, в которое встроены CDR донорного (мышиного) антитела.
«Выделенное» антитело представляет собой антитело, которое было идентифицировано и выделено и/или извлечено из компонентов окружающей его природной среды. Загрязняющие компоненты окружающей его природной среды представляют собой материалы, которые будут препятствовать диагностическому или терапевтическому использованию антитела и могут включать ферменты, гормоны и другие белковоподобные или отличные от белковоподобных растворы. В некоторых вариантах осуществления антитело должно быть очищено (1) до более чем 95% по массе антитела, определяемых способом Лоури, и наиболее предпочтительно более 99% (по массе), (2) до уровня, достаточного для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с помощью секвенатора с вращающимся стаканом, или (3) до гомогенности путем SDS-PAGE в восстанавливающих и невосстанавливающих условиях, используя окрашивание кумасси синим или, предпочтительно, серебром. Выделенное антитело включает антитело in situ в рекомбинантных клетках, поскольку по меньшей мере один компонент окружающей антитело природной среды не будет присутствовать. Как правило, однако, выделенное антитело следует получать путем по меньшей мере одной стадии очистки.
Выражение «по существу не содержит клеточного материала» подразумевает препараты антитела или фрагмента антитела, в которых антитело или фрагмент антитела отделяется от клеточных компонентов клеток, из которых оно выделено или получено рекомбинантными способами. Таким образом, антитело или фрагмент антитела, которое по существу не содержит клеточного материала, включает препараты антитела или фрагментов антитела, содержащие менее чем примерно 30%, 20%, 10%, или 5% (по сухой массе) гетерологичного белка (также называемого в настоящем документе как «загрязняющий белок»). Когда антитело или фрагмент антитела продуцируется рекомбинантно, оно также предпочтительно по существу не содержит культуральной среды, т.е. культуральная среда составляет менее чем примерно 20%, 10% или 5% от объема белкового препарата. Когда антикиновое антитело или фрагмент этого антитела получают путем химического синтеза, предпочтительно его получают свободным от химических прекурсоров или других химических веществ, т.е. его отделяют от химических прекурсоров или других химических веществ, которые участвуют в синтезе белка. Соответственно, такие препараты антитела или фрагмента антитела содержат менее чем приблизительно 30%, 20%, 10%, 5% (по сухой массе) химических прекурсоров или соединений, отличных от представляющего интерес антитела или фрагмента антитела. В одном варианте осуществления антитела по изобретению или его фрагменты выделяют или очищают.
Термин «антикиновое антитело» относится к антителу, как это определено выше, которое связывается с двумя или более хемокинами, предпочтительно антикиновое антитело связывает три или более CC-хемокинов человека. Антикиновое антитело может представлять собой моноклональное и поликлональное антитело и предпочтительно выделенное или очищенное моноспецифическое или моноклональное антитело. Антикиновое антитело может представлять собой антитело млекопитающего, такое как антитело мыши или человека, или химерное или гуманизированное антитело. Полностью человеческие антитела могут быть получены от людей или от трансгенных животных, или с платформ фагового дисплея, см. публикацию Lonberg, Curr. Opin. Immunol. 20(4):450-9 (2008), в которой методики получения антител человека от трансгенных животных включены посредством ссылки. Антикиновые антитела могут представлять собой синтетические антитела, однодоменные антитела, такие как нанотела (VHH) или камелизированные антитела, одноцепочечные Fvs (csFv), одноцепочечные антитела, Fab-фрагменты, F(ab')-фрагменты, дисульфид-связанные Fvs, внутриклеточные антитела и антиидиотипические (анти-Id) антитела (включая антиидиотипические и анти-антиидиотипические антитела к антикиновым антителам по изобретению, такие как 3C12F и т.д.), биспецифические и фрагменты любого из вышеперечисленных, которые связывают детерминанты или эпитопы CC-хемокина. Различные структурные формы сконструированных антител включены посредством ссылки на публикацию Antibody Engineering: A Practical Approach, edited by McCafferty, et al., Oxford University Press (1996). Термин «антикиновое антитело» охватывает молекулы иммуноглобулина и иммунологически активные фрагменты молекул иммуноглобулина, содержащие по меньшей мере один сайт связывания антигена (ABS). Они могут представлять собой любой изотип, включая IgA, IgD, IgE, IgG, IgM и IgY, и могут быть получены от позвоночных, таких как млекопитающие и птицы, которые продуцируют антитела. Предпочтительно, антикиновые антитела представляют собой антитела человека или гуманизированные антитела, хотя антикиновые антитела могут быть пригодны для введения животным, таким как домашние животные или животные, содержащиеся в коммерческих целях, или дикие животные, или животные, содержащиеся в неволе. Они включают домашних животных, таких как собаки и кошки; домашний скот, такой как крупный рогатый скот, лошади, буйволы, азиатские буйволы, свиньи, козы, овцы, верблюды, ламы и т.д.; и птиц, таких как куры, индюки, гуси, соколы и т.д. Специалистам в рассматриваемой области понятно, как получать и адаптировать антикиновые антитела для применения, отличного от применения на человеке, например, путем индуцирования антител к CC-хемокинам, экспрессируемым определенным типом животного, и/или с помощью способа конструирования антител, аналогичного гуманизации.
Антикиновые антитела или их фрагменты будут связываться со специфическими CC-хемокинами и не связываться неспецифически с другими хемокинами или полипептидами. Антикиновые антитела или их фрагменты, которые иммуноспецифически связываются с CC-хемокином или фрагментом CC-хемокина, могут перекрестно реагировать с другими антигенами. Однако можно выбрать антикиновые антитела или фрагменты, иммуноспецифически связывающиеся с CC-хемокином или его фрагментом, которые перекрестно не реагируют с другими антигенами. Антикиновые антитела или их фрагменты, которые иммуноспецифически связываются с определенными CC-хемокинами, можно идентифицировать, например, с помощью иммуноанализа или других способов, известных специалистам в данной области.
«Фрагмент» описывает часть интактной полипептидной молекулы, такой как CC-хемокин или антикиновый иммуноглобулин. «Фрагмент» может включать пептид или полипептид, содержащий аминокислотную последовательность по меньшей мере из 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 75, 80, 90, 100, 120, 130, 150, 175, 200, или 250 смежных аминокислотных остатков аминокислотной последовательности, такой как последовательность интактного зрелого CC-хемокина или легкая или тяжелая полипептидная цепь антитела. Для CC-хемокина фрагмент представляет собой часть молекулы, которая короче, чем длина зрелого хемокина, например, фрагмент CC-хемокина, включающий аминокислотную последовательность из по меньшей мере 5 смежных аминокислотных остатков, по меньшей мере 10 смежных аминокислотных остатков, по меньшей мере 15 смежных аминокислотных остатков, по меньшей мере 20 смежных аминокислотных остатков, по меньшей мере 25 смежных аминокислотных остатков, по меньшей мере 40 смежных аминокислотных остатков, по меньшей мере 50 смежных аминокислотных остатков, или любое промежуточное значение вплоть до, но не включая, полной аминокислотной последовательности зрелого CC-хемокина. Для антикинового антитела фрагмент включает пептид или полипептид, содержащий аминокислоты из по меньшей мере 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50 смежных аминокислотных остатков, или вплоть до, но не включая, полной длины VH- и/или VL-части антитела, которая специфически связывается с CC-хемокином, и которая связывается с по меньшей мере двумя или тремя CC-хемокинами, связанными интактным антикиновым антителом. Фрагмент антикинового антитела может представлять собой одноцепочечный фрагмент, например, легкую или тяжелую цепь или часть легкой или тяжелой цепи, но также включает фрагменты с несколькими цепями, такие как Fab- или F(ab')2-фрагменты.
«Аффинно-зрелые антитела» представляют собой антитела, которые обладают своей аффинностью связывания и/или биологической активностью, увеличенной путем изменения типа или положения одного или нескольких остатков в вариабельной области. Примером изменения является мутация, которая может находиться либо в CDR, либо в каркасной области. Аффинно-зрелое антитело должно, как правило, обладать аффинностью связывания, увеличенной по сравнению с выделенным или природным антителом или его фрагментом в 2-500 раз. Аффинно-зрелые антитела могут обладать наномолярной или даже пикомолярной аффинностью к рецепторному антигену. Аффинно-зрелые антитела продуцируют с использованием методик, известных в рассматриваемой области. В публикации Marks, J. D. et al., Bio/Technology 10:779-783 (1992), которая включена посредством ссылки, описано созревание аффинности путем перетасовки VH- и VL-доменов. Случайный мутагенез CDR и/или каркасных остатков включен посредством ссылки на публикации Barbas, C. F. et al. Proc Nat. Acad. Sci, USA 91:3809-3813 (1994), Schier, R. et al. Gene 169:147-155 (1995), Yelton, D. E. et al. J. Immunol. 155; 1994-2004 (1995), Jackson, J. R. et al. J. Immunol. 154(7):3310-9 (1995) и Hawkins, R. E. et al., J. Mol. Biol. 226:889-896 (1992).
«Производное антикинового антитела» относится к полипептиду, который содержит аминокислотную последовательность антикинового антитела или фрагмента антитела, связывающего CC-хемокин, которое специфически связывается с по меньшей мере двумя, тремя, четырьмя или более CC-хемокинами, которое было изменено путем введения замен, удалений или добавлений аминокислотных остатков. В настоящем документе термин «производное» также относится к антикиновому антителу или фрагменту антикинового антитела, которое было ковалентно модифицировано, например, путем гликозилирования, ацетилирования, пегилирования, фосфорилирования, амидирования, получения производных по известным защитным/блокирующим группам, протеолитического расщепления, соединения с клеточным лигандом или другим белком и т.д. Такое производное антикинового антитела можно продуцировать путем химических модификаций с использованием способов, известных специалистам в рассматриваемой области, включая, но ими не ограничиваясь, специфическое химическое расщепление, ацетилирование, формилирование, метаболический синтез туникамицина и т.д. Производное антикинового антитела должно сохранять способность специфически связываться с двумя или более CC-хемокинами.
«Аналог антикинового антитела» относится к полипептиду, содержащему по существу аналогичные аминокислотные последовательности, как известное антикиновое антитело, и который сохраняет способность известного антикинового антитела связываться с двумя или более CC-хемокинами. Аналог также может содержать 1, 2, 3, 5 или 10 или более неклассических аминокислот. Полипептид, который имеет аминокислотную последовательность, аналогичную полипептиду антикинового антитела, может быть описан посредством ссылки на идентичность с другим белком или путем ссылки на кодирующую полинуклеотидную последовательность, включенную как:
(i) полипептид, имеющий аминокислотную последовательность, которая по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99% идентична аминокислотной последовательности по меньшей мере одной легкой или тяжелой цепи, или по меньшей мере одному CDR известного антикинового антитела;
(ii) полипептид, кодируемый полинуклеотидной последовательностью, которая гибридизуется в строгих условиях с нуклеотидной последовательностью, кодирующей антикиновое антитело или фрагмент антикинового антитела, описанного в настоящем документе, полипептид, который содержит по меньшей мере 5, 10, 15, 20, 25, 30, 35, 40, 50, 75, 90, 100, 125 или по меньшей мере 150 аминокислотных остатков. Условия температуры и ионной силы определяют «строгость» гибридизации. Условия гибридизации низкой строгости, соответствующие Tm 55°C, включают, например, 5× SSC, 0,1% SDS, 0,25% молоко и без формамида; или 30% формамида, 5× SSC, 0,5% SDS. Условия гибридизации умеренной строгости соответствуют более высокой Tm, например, 40% формамида, с 5× или 6× SSC, и условия гибридизации высокой строгости соответствуют самой высокой Tm, например, 50% формамида, 0,1× SSC; и
(iii) полипептид, кодируемый нуклеотидной последовательностью, которая по меньшей мере на 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или по меньшей мере 99% идентична нуклеотидной последовательности, кодирующей известное антикиновое антитело или фрагмент антикинового антитела. Полипептид со структурой, аналогичной антикиновому антителу или фрагменту антикинового антитела, описанного в настоящем документе, относится к полипептиду, который имеет аналогичные вторичную, третичную и четвертичную структуру с известным антикиновым антителом или его фрагментом. Полипептидную и белковую структуру можно определить способами, известными специалистам в рассматриваемой области, включая, но не ограничиваясь ими, рентгеновскую кристаллографию, ядерный магнитный резонанс и кристаллографическую электронную микроскопию.
Термин «гипервариабельная область» относится к аминокислотным остаткам антитела, которые ответственны за связывание антигена. Гипервариабельная область включает аминокислотные остатки из «определяющего комплементарность участка» или «CDR» в тяжелой цепи и легкой цепи вариабельных доменов; Kabat, et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991) и/или те остатки «гипервариабельной петли» в тяжелой цепи и легкой цепи вариабельных доменов; Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987). Остатки «каркасной области» или «FR» представляют собой те остатки вариабельного домена, которые отличны от остатков гипервариабельной области, как определено в настоящем документе. Структурные особенности антител и способы определения или анализа структуры антител, известные специалистам в рассматриваемой области, включены посредством ссылки на публикации Kabat, id., Chothia, et al., id.; Deret, et al., Comput. Appl. Biosci. 11(4):435-9 (1995); Martin, Protein 25(1):130-3 (1996) и Abhinandan, et al., Mol. Immunol. 45(14):3832-9 (2008); и Abhinandan, et al., J. Mol. Biol. 369(3):852-62 (2007).
«CC-хемокин» относится к полипептиду семейства хемотаксических цитокинов, содержащих четыре консервативных остатка цистеина, первые два из которых находятся рядом, как описано, например, в публикации Van Coillie, et al., Cytokine & Growth Factor Rev. 10:61-86 (1999). CC-хемокины также известны как β-хемокины. По наличию этих соседних остатков цистеин-цистеин (cys-cys, или C-C) β-хемокины известны как «CC»-хемокины, где «CC» означает прилегающие остатки цистеина. Примерами CC-хемокинов являются хемокины, связывающиеся с CCR1 и CCR5, включая CCL3/MIP-1α, CCL4/MIP-1β и CCL5/RANTES. CC-хемокины существуют в организме млекопитающих и птиц. Термины, используемые для вариантов определенных хемокинов человека или мыши, например, CCL3/MIP-1α человека, могут быть использованы для идентификации соответствующей молекулы в других видах, например, «MIP-1α мыши» или «MIP-1α быка», и должны пониматься по отношению к аналогу, структурно сходной молекуле в видах, о которых идет речь. Аналогичные хемокины среди видов млекопитающих и птиц могут иметь по меньшей мере 40%, 50%, 60%, 70%, 80%, 90%, 95% или 99%, или любое промежуточное значение в этом диапазоне, идентичности последовательности и должны демонстрировать те же или аналогичные функциональные иммунологические активности.
Последовательности для CC-хемокинов позвоночных, в том числе млекопитающих и птиц, включены посредством ссылки на базу данных NCBI с конкретными ссылками на те последовательности, которые идентифицированы номерами доступа последнего обновления в этой базе данных (последнее посещение 9 августа 2010 г.), и на публикацию Fernandez & Lolis, Annu. Rev. Pharmacol. Toxicol. 42:469 (2002), которая включена в описание посредством ссылки.
Конкретные номера доступа для полинуклеотидных и полипептидных последовательностей хемокинов человека CCL2/MCP-1, CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES, CCL14/HCC-1, CCL15/HCC-2, CCL18/PARC и CCL23/MPIF-1 и др. приведены в таблице 4. Каждая специально включена посредством ссылки на каталог, поставщика или номер доступа NCBI, указанный в таблице 4. Эти хемокины коммерчески доступны от поставщиков, указанных в таблице 4.
В настоящем документе термин «детерминанта CC-хемокина» относится к частям CC-хемокина, которые контактируют с антигенсвязывающими остатками антитела. Этот термин охватывает как традиционные линейные эпитопы, распознаваемые антикиновым антителом на CC-хемокине, так и конформационные эпитопы, в том числе нелинейные эпитопы. Одна или несколько частей CC-хемокина, связанных с антикиновым антителом или непосредственно находящихся в контакте с его аминокислотными остатками, связывающими антиген, представляют собой его детерминанты. Удаление, замена, нарушение или денатурация антигенной детерминанты CC-хемокина может уменьшать или устранять способность антикинового антитела связываться с CC-хемокином. Способы определения эпитопов связывания антитела известны в рассматриваемой области, как показано в публикации Ladner, Biotechnol. Genet. Eng. Rev. 24:1-30(2007), которая включена посредством ссылки. Детерминанты CC-хемокина включают сегменты или домены, участвующие в связывании с хемокиновым рецептором (например, CCR1 или CCR5), такие как N-петля, цепи β-1, 2 и 3, 30's-, 40's-и 50's-петли и его С-концевой спиральный сегмент, эти детерминанты включены посредством ссылки на публикации Fernandez & Lolis, Annu. Rev. Pharmacol. Toxicol. 42:469 (2002), Viola, et al., Annu. Rev. Pharmacol. Toxicol. 48:171 и Pakianathan, et al., Biochem. 36(32):9642 (1997).
Детерминанты CC-хемокинов также определяют путем связывания белков из патогенов, например, детерминанты CC-хемокинов, связанные с vCCI вируса вакцины, эти детерминанты включены посредством ссылки на публикацию Zhang et al., Proc. Natl. Acad. Sci. 103(38): 13985-13990 (2006).
Термины «связывать» или «связывание» относятся к химическому или физическому контакту или взаимодействию между двумя различными веществами. Они включают соединение между антикиновым антителом и соответствующей детерминантой, например, эпитоп на молекуле CC-хемокина. Такой контакт или взаимодействие может происходить за счет одной или нескольких ионных, неионных, водородных связей, ван-дер-ваальсовых связей, гидрофобных связей или других известных типов межмолекулярного взаимодействия. Связывание может включать непосредственный контакт между двумя молекулами, например, между лигандом и соответствующим ему рецептором, или опосредованно через вмешательство молекулы, например, ковалентное или нековалентное связывание через линкер или би- или поливалентную группу.
«Аффинность связывания» описывается аффинной константой Ka, которая представляет собой соотношение между константами скоростей связывания и диссоциации антитела и антигена. Типичные аффинности для антител IgG составляют 10-5-10-9 моль/л. Аффинность антитела определяют многими различными способами, известными в рассматриваемой области, включая: равновесный диализ, поверхностный плазмонный резонанс (BIAcore®) и анализ кинетического исключения (KinExA®). Соотношение между связанным и свободным антигеном и аффинностью антитела выражается уравнением Скэтчарда, r/c=Kn-Kr, где r = соотношение [связанного антигена] ко [всему антителу], c = [свободный антиген], K = аффинность и n = количество сайтов связывания на молекулу антитела (валентность). Если все антитела имеют одинаковую аффинность к антигену (например, моноклональное антитело), график зависимости r/c от r даст прямую линию с наклоном -K и r-отсечение стремится к n. Если антитело является гетерогенным (например, поликлональным), график зависимости r/c от r даст кривую линию; среднюю аффинность можно определить по наклону кривой, когда заполнена половина сайтов связывания (r=1).
Выражение «специфически связывается с CC-хемокином» описывает превышающее фон взаимодействие антитела или фрагмента антитела с определенным CC-хемокином или набором CC-хемокинов, но не с другими хемокинами. CC-хемокины включают, но не ограничиваются ими, CCL3/MIP-1α, CCL4/MIP-1β и CCL5/RANTES, и примеры сегментов или доменов CC-хемокина включают его остатки, связывающиеся с хемокиновым рецептором, его N-петлю, цепи β-1, 2 и 3, 30's-, 40's- и 50's-петли и его С-концевой спиральный сегмент. Антитело, которое специфически связывается с CC-хемокином или с одним из его доменов или сегментов, может связываться с другими антигенами с более низкой аффинностью, что определяется, например, иммуноанализом, поверхностным плазмонным резонансом (BIAcore®) или другими тестами, известными в рассматриваемой области. Например, антикиновое антитело, которое обладает четырехкратной или более высокой способностью связывания с определенным CC-хемокином по сравнению с другим или контрольным антителом, при сравнении в той же концентрации и в тех же условиях, будет рассматриваться как специфически связывающееся с CC-хемокином. Однако могут быть использованы другие сравнительные критерии связывания, которые устанавливают существенное различие в количестве или аффинности связывания, они будут включать в 2, 3, 4, 5, 10, 20, 50, 100 или 1000 раз большее количество связывания для антикинового антитела или 2, 5, 10, 15, 20, 100 или по меньшей мере 1000-кратное увеличение аффинности связывания.
Необходимо, чтобы антитела или фрагменты, которые специфически связываются с двумя, тремя, четырьмя, пятью или более CC-хемокинами, перекрестно не реагировали с другими нехемокиновыми антигенами или хемокинами, отличными от CC-хемокинов. Антикиновые антитела или фрагменты, которые специфически связываются с CC-хемокинами и их фрагментами, можно идентифицировать с помощью иммуноанализа, BIAcore или других способов, известных специалистам в рассматриваемой области. Антитело или его фрагмент специфически связывается с антигеном CC-хемокина или его фрагментом с большей аффинностью, чем с любым перекрестно реагирующим антигеном, что определено экспериментальными методами, такими как Вестерн-блот-анализ, радиоиммуноанализ (РИА) и ферментный иммуносорбентный анализ (ELISA). См., например, публикацию Paul, ed, Fundamental Immunology, Second Edition, Raven Press, New York (1989) на страницах 332-336 для обсуждения, касающегося специфичности антител.
Термин «ингибирующая концентрация» относится к концентрации антитела, такого как антикиновое антитело или его CC-хемокинсвязывающие фрагменты, которое, по сравнению с контролем, уменьшает активность CC-хемокина, например, на 5%, 10%, 20%, 30%, 50%, 80%, 90%, 95%, 99% или вплоть до 100%. Активность CC-хемокина можно определить in vivo или in vitro и включать анализы активностей CC-хемокина, таких как хемотаксис. Такие анализы включают перенос кальция или активацию интегрина. Модулирующие эффекты антикинового антитела или его CC-хемокинсвязывающих фрагментов, а также контрольных антител, таких как антитела, которые не связываются с CC-хемокинами или которые связываются с одним хемокином, можно также определять in vivo в моделях на животных, включая модели для определения активации CC-хемокинами иммунных явлений, таких как воспаление и аутоиммунные заболевания. Ингибирование активности хемокина относится к вызыванию относительного снижения по меньшей мере одной активности хемокина в присутствии ингибирующего агента по сравнению с активностью в отсутствие агента. Ингибирование может включать антагонизм или нейтрализацию активности хемокина, например, путем связывания антитела с активным участком хемокина, либо путем связывания, которое приводит к эффективному удалению, иммобилизации или инактивации хемокина. Термин «ингибирование хемотаксиса» относится к снижению относительной величины хемотаксической активности клеток в присутствии антитела или его антигенсвязывающего фрагмента по сравнению с хемотаксической активностью, наблюдаемой в отсутствие антитела или его антигенсвязывающего фрагмента. Известные способы определения хемотаксического ингибирования определенных типов клеток, в том числе различных видов лейкоцитов, доступны и включены посредством ссылки на публикацию Chemokine Protocols, Meth. in Mol. Biol. 138 (2000), Humana Press, Eds. AEI Proudfoot, TNC Wells, and CA Power.
«Выделенный» или «очищенный» продукт или компонент по существу не содержит клеточного материала или других загрязняющих белков источника клеток или тканей, из которого получен продукт или компонент, или по существу не содержит химических прекурсоров или других химических веществ в случае химического синтеза. Выделенный или очищенный компонент, молекула или другое вещество (включая пептид, полипептид, антитело, хемокин, полинуклеиновую кислоту или клетку) представляет собой элемент, который был удален из, или синтезирован отдельно от, окружающей его обычной, природной или собственной среды. Выделенный или очищенный продукт или компонент можно также физически или химически удалить или отделить от смеси или ингредиентов, с которыми он связан, включая нежелательные биологические загрязнители, или, в случае синтеза, от субстратов или побочных продуктов, связанных с его синтезом. Очистку можно осуществлять до любой степени, включая удаление 1, 5, 10, 50, 75, 90, 95 или 100% других компонентов. В случае антитела выделение может представлять собой удаление из крови или сыворотки, или для моноклонального антитела - из асцитной жидкости или жидкости культуры ткани.
Термины «нуклеиновые кислоты» и «нуклеотидные последовательности» включают молекулы ДНК (например, кДНК или геномной ДНК), молекулы РНК (например, мРНК), комбинации молекул ДНК и РНК или гибридные ДНК/РНК-молекулы и аналоги молекул ДНК или РНК. Последовательности нуклеиновых кислот по изобретению могут кодировать части аналога или варианта антикинового антитела. Такие аналоги можно создать с помощью, например, нуклеотидных аналогов, которые включают, но не ограничиваются ими, инозин или тритилированные основания. Такие аналоги могут также включать молекулы ДНК или РНК, содержащие модифицированные главные цепи, которые придают полезные свойства молекулам, такие как, например, устойчивость к нуклеазам или усиленная способность пересекать клеточные мембраны. Нуклеиновые кислоты или нуклеотидные последовательности могут быть одноцепочечными, двухцепочечными, могут содержать как одноцепочечные, так и двухцепочечные части, и могут содержать трехцепочечные части, однако предпочтительно являются двухцепочечными ДНК. «Выделенная» молекула нуклеиновой кислоты представляет собой молекулу, которая отделена от других молекул нуклеиновых кислот, которые присутствуют в природном источнике молекулы нуклеиновой кислоты. «Выделенная» молекула нуклеиновой кислоты, такая как молекула кДНК, может по существу не содержать другой клеточный материал, или культуральную среду, в случае продуцирования рекомбинантными способами, или по существу не содержать химических прекурсоров или других химических веществ в случае химического синтеза. В одном варианте осуществления выделяют или очищают молекулы нуклеиновой кислоты, кодирующие антитела по изобретению или их фрагменты.
В настоящем документе термин «клетка-хозяин» относится к клетке определенного субъекта, трансфицированной молекулой нуклеиновой кислоты, и к потомству или потенциальному потомству такой клетки. Потомство такой клетки может не быть идентичным родительской клетке, трансфицированной молекулой нуклеиновой кислоты, из-за мутаций или воздействий окружающей среды, которые могут возникнуть в последующих поколениях, или интеграции молекулы нуклеиновой кислоты в геном клетки-хозяина. Клетка-хозяин может быть использована для рекомбинантной экспрессии антикинового антитела или одного из его компонентов, например, легкой или тяжелой цепи или ее фрагмента.
Чтобы определить «процент идентичности» двух нуклеиновых кислот или аминокислотных последовательностей, последовательности сначала выравнивают в целях оптимального сравнения (например, пробелы могут быть введены в последовательность первой аминокислоты или последовательность нуклеиновой кислоты для оптимального выравнивания с последовательностью второй аминокислоты и последовательностью нуклеиновой кислоты). Аминокислотные остатки или нуклеотиды в положениях соответствующих аминокислот или положениях нуклеотидов затем сравниваются. Если положение в первой последовательности занимается тем же аминокислотным остатком или нуклеотидом, что и в соответствующем положении во второй последовательности, то молекулы идентичны в этом положении. Процент идентичности между двумя последовательностями является функцией от числа идентичных положений, разделяемых последовательностями (т.е. % идентичности = количеству идентичных перекрывающихся положений/общее количество положений, умноженному на 100%). В одном варианте осуществления две последовательности имеют одну и ту же длину. Определение процента идентичности между двумя последовательностями также может быть выполнено с помощью математического алгоритма. Предпочтительный, неограничивающий пример математического алгоритма, который используется для сравнения двух последовательностей, представляет собой алгоритм Karlin and Altschul, Proc. Natl. Acad. Sci. U.S.A. 87:2264-2268 (1990), модифицированный как в публикации Karlin and Altschul, Proc. Natl. Acad. Sci. U.S.A. 90:5873-5877 (1993). Такой алгоритм заложен в программы NBLAST и XBLAST Altschul et al., J. Mol. Biol. 215:403 (1990). Нуклеотидный поиск BLAST может быть выполнен с параметрами установки нуклеотидной программы NBLAST, например, для балльной оценки = 100, длины слова = 12, чтобы получить нуклеотидные последовательности, гомологичные молекуле нуклеиновой кислоты по настоящему изобретению. Поиски белка BLAST можно выполнять с параметрами установки программы XBLAST, например, для балльной оценки = 50, длины слова = 3, чтобы получить аминокислотные последовательности, гомологичные молекуле белка по настоящему изобретению. Для получения выравниваний с пробелами для целей сравнения можно использовать Gapped BLAST, как описано в публикации Altschul et al., Nucleic Acids Res. 25:3389-3402 (1997). Альтернативно, PSI-BLAST можно использовать для выполнения повторного поиска, который обнаруживает отдаленные отношения между молекулами (Id.). При использовании программ BLAST, Gapped BLAST и PSI-Blast параметры соответствующей программы (например, XBLAST и NBLAST) можно применять по умолчанию. Другой предпочтительный, неограничивающий пример математического алгоритма, используемого для сравнения последовательностей, представляет собой алгоритм Myers and Miller, CABIOS 4: 11-17 (1988). Такой алгоритм включен в программу ALIGN (версия 2.0), которая является частью пакета программного обеспечения для выравнивания последовательности GCG. При использовании программы ALIGN для сравнения аминокислотных последовательностей можно использовать таблицу массы остатков PAM-120, штраф длины пробела 12 и штраф пробела 4. Процент идентичности двух последовательностей можно определить с помощью методики, аналогичной методикам, которые описаны выше, с разрешением пробелов или без них. При расчете процента идентичности обычно учитываются только точные совпадения.
«Консервативная замена» относится к изменениям, которые по существу являются нейтральными в конформационном или антигенном отношении, вызывают минимальные изменения в третичной структуре пептидного или полипептидного варианта, или вызывают минимальные изменения в антигенных детерминантах вариантов или аналогичных пептидов или полипептидов, по сравнению с родительским или нативным пептидом или полипептидом. В контексте хемокинов консервативные замены включают аминокислотные замены, которые по существу не влияют на специфичность и/или аффинность полученного в результате варианта или аналога хемокина, например, как определяется по его способности проявлять по меньшей мере одну функцию родительского хемокина, включая такие функции, как индукция хемотаксиса, участие в сети цитокинов или, в ином случае, индуцирование продукции фермента или цитокина, или связывание с рецептором родительского или нативного хемокина. Применительно к вариантам или аналогам антител, фрагментам антител, в том числе CDR-сегментам, консервативная замена относится к аминокислотной замене, дающей продукт антитела, который способен связываться с тем же эпитопом или антигеном, как соответствующий неизмененный продукт антитела. Предсказание того, какие замены аминокислот поддерживают конформационную и антигенную нейтральность молекулы, находятся в компетенции специалистов в рассматриваемой области, как описано в публикациях Berzofsky, Science 229:932-940 (1985) и Bowie, et al., Science 247:1306-1310 (1990). Рекомендации относительно того, какие замены будут наиболее вероятно поддерживать конформационную и антигенную нейтральность, включают: (a) маловероятно, чтобы замена гидрофобных аминокислот влияла на антигенность, поскольку гидрофобные остатки расположены, наиболее вероятно, во внутренней части белка, (b) маловероятно, чтобы замена биохимически похожими аминокислотами влияла на конформацию, поскольку замененная аминокислота структурно имитирует нативную аминокислоту; и (c) изменение эволюционно консервативных последовательностей скорее всего негативно повлияет на конформацию, поскольку такой консерватизм предполагает, что аминокислотные последовательности могут иметь функциональное значение.
«Композиция» или «фармацевтическая или терапевтическая композиция» относится к комбинации носителя, эксципиента или раствора, содержащей антикиновое антитело или его CC-хемокинсвязывающие фрагменты, или иные его фрагменты, которые непосредственно или опосредованно уменьшают тяжесть или облегчают лечение состояний, нарушений или заболеваний, опосредованных CC-хемокинами, или по меньшей мере один их симптом. Термин «фармацевтически приемлемый носитель» включает любые и все носители и эксципиенты, такие как разбавители, растворители, диспергирующие агенты, эмульсии, липидные бислои, липосомы, покрытия, консерванты, включая антибактериальные и противогрибковые средства, изотонические агенты, рН-буферы и вещества, модулирующие абсорбцию, и тому подобное, совместимые с молекулами по настоящему изобретению и подходящие для фармацевтического введения. Применение таких носителей, агентов, вызывающих дезинтеграцию, эксципиентов и средств для введения фармацевтически активных веществ известно в рассматриваемой области, см. публикацию Handbook of Pharmaceutical Excipients, 3rd edition, Am. Pharm. Assoc. (2000), которая включена посредством ссылки. Фармацевтические композиции по изобретению, как правило, составляют в готовую форму для совместимости с намеченным способом введения, такого как парентеральное, пероральное или местное введение.
Терапевтические композиции по изобретению включают по меньшей мере одно антитело или фрагмент антитела по изобретению в фармацевтически приемлемом носителе. «Фармацевтически приемлемый носитель» должен представлять собой по меньшей мере один компонент, традиционно смешанный с активным ингредиентом, биологическим продуктом или лекарственным препаратом и используемый для их введения. Носитель может содержать любой фармацевтический эксципиент, применяемый в рассматриваемой области, и любую форму основы для введения. Композиции могут представлять собой, например, инъекционные растворы, водные суспензии или растворы, неводные суспензии или растворы, аэрозоли, твердые и жидкие пероральные препараты, бальзамы, гели, мази, внутрикожные пластыри, кремы, лосьоны, таблетки, капсулы, составы непрерывного высвобождения и тому подобное. Дополнительные эксципиенты могут включать, например, красители, агенты, маскирующие вкус, вещества, способствующие растворимости, суспендирующие агенты, вещества, способствующие прессованию, энтеросолюбильные покрытия, вещества, способствующие непрерывному высвобождению, и тому подобное. Подходящая лекарственная форма может быть выбрана специалистом в рассматриваемой области из таких форм, которые описаны в U.S. FDA CDER Data Standards Manual C-DRG-00201, Version 08; или перечислены на веб-сайте FDA http://www. fda.gov/Forlndustry/DataStandards/StructuredProductLabeling/
ucm162038.htm (последнее посещение 9 августа 2010 года); которые включены в настоящий документ посредством ссылки.
Вводимые перорально композиции могут включать твердые носители или эксципиенты, или могут быть составлены в виде жидких или гелеобразных препаратов и могут включать съедобный или инертный носитель, и могут быть заключены в капсулы, спрессованы в таблетки или составлены в виде пастилок. Вводимые перорально композиции можно получить в форме с пролонгированным действием или в капсулированной форме для предотвращения деградации в желудке и оптимизирования поглощения молекулы.
Инъекционные композиции могут быть составлены в готовые формы способами, известными в рассматриваемой области, и могут включать стерильные растворы или дисперсии терапевтических молекул. Таковые, как правило, включают стерильный разбавитель, такой как вода, физиологический раствор или другой буфер, совместимый с молекулами по изобретению. Инъекционные композиции могут быть получены в единичных дозировках или в контейнерах с единичными дозами, например, флаконах, ампулах или шприцах.
Могут быть использованы традиционные буферы и изотонические агенты, и pH можно регулировать с помощью известных агентов, таких как HCl или NaOH, или буферов. Могут присутствовать противомикробные или бактериостатические вещества, комплексообразующие агенты, такие как ЭДТА или ЭГТА, и антиоксиданты и консерванты.
Терапевтические композиции по изобретению можно вводить любым приемлемым способом введения, включая местный, на слизистую оболочку, пероральный или энтеральный, или парентеральный. Эти способы включают, но не ограничиваются ими, местный, через слизистую оболочку, пероральный (в том числе трансбуккальный, подъязычный), на слизистую оболочку (конъюнктивальный, назальный, синусальный, уретральный, вагинальный, интестинальный, ректальный), энтеральный, трансдермальный, внутрикожный, подкожный (s.c.), внутримышечный, внутрибрюшинный, внутривенный (i.v.), внутрисердечный, внутрисуставный или внутрикостный, внутриорганный (мозг, спинной мозг, глаз, ухо, печень, селезенка, почки, желчный пузырь, мочевой пузырь), внутрикостный, в хрящ или ткани сустава, ингаляционный (например, интраназальный, интратрахеальный, внутрилегочный или внутрибронхиальный), пероральный, суббукальный. Способы могут быть выбраны специалистами в рассматриваемой области из тех, которые перечислены в the U.S. FDA, CDER, Data Standards Manual "Routes of Administration", CDRG-00301, Version 004; или из таблицы 2, доступной на http://www.fda.gov/Drugs/GuidanceComplianceRegulatoryInformation/DrugRegistrationandListing/ucm084039.htm (последнее посещение 9 августа 2010 года), которые включены в настоящий документ посредством ссылки.
«Терапевтически эффективное количество» относится к такому количеству терапевтического агента, которое достаточно для снижения тяжести или облегчения лечения состояния, нарушения и заболевания, опосредованных CC-хемокинами, для повышения терапевтической эффективности другой терапии состояния, нарушения и заболевания, или для предотвращения повторения или увеличения тяжести состояния, нарушения или заболевания, или по меньшей мере одного из его симптомов. Терапевтически эффективное количество также может относиться к количеству терапевтического агента, достаточному для того, чтобы отсрочить или свести к минимуму начало заболевания. Терапевтически эффективное количество также может относиться к количеству терапевтического агента, которое обеспечивает терапевтическую пользу при лечении или управлении течением заболевания. Кроме того, терапевтически эффективное количество в отношении терапевтического агента по изобретению означает то количество терапевтического агента самого по себе или в сочетании с другими видами терапии, которое обеспечивает терапевтический эффект при лечении или управлении течением заболевания, например, достаточный для повышения терапевтической эффективности терапевтических антител, достаточной для лечения или управления течением заболевания. Используемый в связи с количеством антитела по изобретению, этот термин может охватывать количество, которое улучшает общую терапию, снижает или предотвращает нежелательные эффекты, или аддитивно повышает терапевтическую эффективность другого терапевтического средства, или суммируется с ним.
В настоящем документе термин «субъект» или «пациент» относится к позвоночному, которое экспрессирует CC-хемокины, включая птиц и млекопитающих (например, крупный рогатый скот, лошадей, свиней, коз, овец, собак или кошек) и предпочтительно человека. Субъект или пациент может представлять собой такового, нуждающегося в лечении эффективным количеством антикинового антитела, которое модулирует CC-хемокиновую активность CC-хемокинов, с которыми оно связывается.
«Воспалительное состояние, нарушение или заболевание» относится к явному или поддающемуся количественному определению физиологическому явлению, включая, но не ограничиваясь ими, отек, лихорадку, хемотаксис или миграцию лейкоцитов, пролиферацию кровеносных сосудов, пролиферацию соединительной ткани, покраснение, локализованный жар, экссудацию и другие признаки, как описано в публикации Robbins, The Pathological Basis of Disease, 6th edition, Cotran, et al. (eds.), W. B. Saunders, Co. (1999), especially Chapters 3, 7 and 15. В контексте антикиновых антител к CC-хемокинам этот термин относится к явлениям, связанным с, или прямо или опосредованно обусловленных, CC-хемокином, таким как MIP-1α, MIP-1β, RANTES, MCP-1 или другие хемокины.
«Иммуноаффинная смола» относится к твердому субстрату, с которым связывается по меньшей мере один иммунологический лиганд или рецептор. Например, антикиновое антитело может связывается со смолой через участок, отличный от его сайта соединения с антигеном, таким образом, предоставляя возможность антителу связываться с антигенными детерминантами на СС-хемокинах. Аналогичным образом, СС-хемокин может быть функционально связан со смолой, что позволяет ему связываться с антикиновым антителом. В рассматриваемой области известны многие смолы, пригодные для иммобилизации иммунологических лигандов или рецепторов, и их обычно используют для образования иммуноаффинных смол. Такие смолы полезны при анализе компонентов, связанных определенным лигандом или рецептором, в иммунологических тестах или в методах очистки. Любая такая смола может быть использована для образования иммуноаффинных смол по изобретению.
Авторы изобретения изыскивали средства для решения проблем известного уровня техники, связанных с введением нескольких антител к CC-хемокинам. Удивительно, что они обнаружили антитела, которые могут связываться с более чем одним типом CC-хемокинов: антикиновые антитела. Эти антикиновые антитела обеспечивают улучшения по сравнению с ингибированием одного хемокина или одного хемокинового рецептора при регулировании воспалительного заболевания, поскольку только одно антитело необходимо для блокирования функций нескольких хемокинов. Антикиновые антитела по изобретению также устраняют недостатки введения двух или более антител, обладающих различными фармакокинетическими свойствами, такие как применение затруднительных отдельных правил приема для каждого антитела. Антикины устраняют недостатки биспецифических антител, которые являются менее эффективными при связывании с определенным хемокиновым антигеном и образуют комплексы, которые легче связываются с Fc-рецепторами и, следовательно, поглощаются клетками и деградируют в лизосомах.
Как показано в настоящем документе, авторы изобретения предлагают несколько антикиновых антител, которые могут упростить лечение воспалительных и иммунологических состояний, нарушений и заболеваний человека несколькими хемокинами. Эти антикиновые антитела будут служить в качестве структурных и функциональных прототипов для продуцирования еще более эффективных и безопасных антикиновых антител с помощью процессов, таких как «созревание аффинности» и гуманизация антител.
Антикиновые антитела связываются с двумя или более CC-хемокинами и ингибируют их активности. Путем нацеливания на CC-хемокины, включая MIP-1α, MIP-1β, RANTES и MCP-1, авторы изобретения идентифицировали антитела, полезные для модулирующих хемокинов, активирующих одинаковые или различные хемокиновые рецепторы (включая CCR1, CCR2, CCR3, и CCR5). Например, поскольку каждый из MIP-1α, MIP-1β и RANTES связывается с хемокиновым рецептором CCR5, антикиновое антитело, которое распознает эти три CC-хемокина, может в более полном объеме модулировать активность CCR5, чем антитело к одному CC-хемокину. Активация CCR5 нацеливается на незрелые миелоидные дендритные клетки, моноциты, Th 1, Treg, NK и плазмацитоидные дендритные клетки. Аналогичным образом, каждый из MIP-1α, RANTES, MPIF-1 и HCC-1 связывается с CCR1, активация которого нацеливается на моноциты, T-лимфоциты памяти и NK-клетки. Антикиновое антитело, которое связывается с двумя или более из этих CCR1-связывающих хемокинов, в более полном объеме модулирует активацию рецептора. Антитело, которое может связывать и ингибировать как MIP-1α, так и RANTES, могло бы тогда эффективно блокировать два из первичных лигандов как CCR1, так и CCR5. Таким образом, авторы изобретения идентифицировали антикиновые антитела, которые ингибируют первичные лиганды для различных хемокиновых рецепторов, экспрессируемых на лейкоцитах, включая моноциты и T-лимфоциты, вовлеченные в воспалительные заболевания. Ингибиторы любой из этих комбинаций хемокинов обеспечивают улучшения по сравнению с ингибированием отдельного хемокина (или антител, которые блокируют связывание с одним хемокиновым рецептором) при регулировании воспалительного заболевания, поскольку только одно антитело необходимо для блокирования функций нескольких хемокинов на одном рецепторе или активности хемокина на нескольких рецепторах.
Антикиновые антитела имеют различные особенности и функциональные активности в зависимости от хемокинов, с которыми они связываются. Изобретение предоставляет антикиновые антитела, которые специфически связываются с различными CC-хемокинами, и предпочтительно с по меньшей мере двумя из RANTES, MIP-1α, MIP-1β и/или MCP-1. Изобретение дополнительно предоставляет антитела, которые специфически связываются как с RANTES, так и с MIP-1α; как с RANTES, так и с MIP-1β; как с RANTES, так и с MCP-1, как с MIP-1α, так и с MIP-1β; как с MIP-1α, так и с MCP-1, или как с MIP-1β, так и с MCP-1. Аналогичным образом, изобретение предоставляет антикиновые антитела, которые связываются с тремя или четырьмя из RANTES, MIP-1α, MIP-1β и/или MCP-1, а также с другими тесно связанными CC-хемокинами.
Связывание антитела и антигенсвязывающего фрагмента антитела с хемокином происходит путем контакта между аминокислотными остатками в антигенсвязывающих участках (ABS) антитела и антигенными детерминантами или эпитопами хемокина. Известно, что эпитопы могут представлять собой линейные пептидные последовательности или конформационные эпитопы, включающие несколько антигенных детерминант, даже отдельные аминокислоты и боковые группы аминокислот, которые не образуют часть линейной последовательности смежных аминокислот. Например, остатки, расположенные на одной стороне α-спирали, могут связываться посредством ABS антитела, в то время как остатки на противоположной стороне не могут.
Антитела и антигенсвязывающие фрагменты антитела по изобретению могут связываться с N-концевой или C-концевой половиной зрелого CC-хемокина, который не содержит сигнальный пептид. Например, в зрелой форме RANTES человека отсутствуют остатки 1-23 сигнального пептида. Таким образом, ее N-концевой участок должен находиться в диапазоне остатков 1-35 и ее C-концевой участок должен находиться в диапазоне остатков 36-68 (SEQ ID NO:73).
Антитела и антигенсвязывающие фрагменты антитела по изобретению могут блокировать связывание CC-хемокинов путем связывания с остатками CC-хемокинов, участвующими в связывании хемокина с его рецептором. См. NCBI Conserved Domain Database CDD 29111 [uid] для предлагаемых функциональных доменов CC-хемокинов; http://www.ncbi.nlm.nih.gov/sites/entrez (последнее посещение 9 августа 2010 года). Остатки перспективного сайта связывания указаны для каждого CC-хемокина ниже:
CCL3/MIP-1α остатки 11-15 (CCFSY), остатки 17-24 (SRQIPQNF), остатки 34-35 (QC) или остатки 57-67 (EWVQKYVSDLE) SEQ ID NO:71;
CCL4/MIP-1β остатки 11-15 (CCFSY), остатки 17-24 (ARKLPHNF), остатки 34-35 (LC) или остатки 57-67 (SWVQEYVYDLE) SEQ ID NO:72;
CCL5/RANTES остатки 10-14 (CCFAY), остатки 16-23 (ARPLPRAH), остатки 33-34 (KC) или остатки 56-66 (KWVREYINSLE) SEQ ID NO:73.
CCL14/HCC-1 остатки 8-12 (CCFTY), остатки 14-21 (TYKIPRQR), остатки 31-32 (QC) или остатки 54-64 (KWVQDYIKDMK) SEQ ID NO:78.
CCL15/HCC-2 остатки 8-12 (CCTSY), остатки 14-21 (SQSIPCSL), остатки 31-32 (EC) или остатки 54-64 (PGVQDCMKKLK) SEQ ID NO:79.
CCL23/MPIF-1 остатки 9-13 (CCISY), остатки 15-22 (PRSIPCSL), остатки 32-33 (EC) или остатки 55-65 (KQVQVCMRMLK) SEQ ID NO:81.
CCL18/PARC остатки 10-14 (CCLVY), остатки 16-23 (SWQIPQKF), остатки 33-34 (QC) или остатки 56-66 (KWVQKYISDLK) SEQ ID NO:82.
Такое антитело или антигенсвязывающий фрагмент может связываться с сегментами по меньшей мере двух из MCP-1, MIP-1α, MIP-1β и RANTES, а также других родственных CC-хемокинов, участвующих в связывании указанного хемокина с его рецептором. Антикины, такие как 3C12F, могут связываться с теми же детерминантами, что и хемокинсвязывающие вирусные белки, такие как vCCI, или могут ингибировать связывание таких вирусных белков с хемокинами.
Моноклональные антитела 3C12F, 7D1G, 7D12A, 18V4F и 18P7E, которые связывают CC-хемокины RANTES, MIP-1α и/или MIP-1β, были получены и охарактеризованы авторами изобретения. Эти антитела предоставляют необходимую информацию, включая структурную информацию и гипервариабельных областей и CDR легкой и тяжелой цепи, для разработки гуманизированных антител, которые могут одновременно модулировать активность нескольких CC-хемокинов, и для лечения воспалительных заболеваний, обусловленных CC-хемокинами.
Гипервариабельная и CDR последовательности трех вышеприведенных Mab представлены последовательностями SEQ ID NO:2-5 и 7-10 для 3C12F; SEQ ID NO:52-55 и 57-60 для гуманизированного 3C12F; SEQ ID NO:12-15 и 17-20 для 7D12A; SEQ ID NO:22-25 и 27-30 для 7D1G; SEQ ID NO:32-35 и 37-40 для 18V4F, SEQ ID NO: 62-65 и 67-70 для гуманизированного 18V4F; и SEQ ID NO:42-45 и 47-50 для 18P7E.
Раскрыты пять специфических типов антикиновых антител, характеризующихся специфическими антикиновыми антителами 3C12F, 7D1G, 7D12A, 18V4F и 18P7E. Кроме того, раскрыты гуманизированные варианты 3C12F и 18V4F.
Первый тип антител характеризуется специфичностью связывания моноклональных антител, вырабатываемых гибридомной клеточной линией 3C12F или ее субкультурой. Типичным антителом этого типа моноклонального антитела (Mab) является 3C12F. Таким образом, этот тип включает Mab 3C12F, а также его аналоги и производные, включая антитела, вырабатываемые в процессе созревания аффинности или при гуманизации. Антитела типа 3C12F могут содержать вариабельную область тяжелой цепи, представленную SEQ ID NO:2 или 52, или тяжелую цепь, содержащую по меньшей мере один из CDR1 (SEQ ID NO:3 или 53), CDR2 (SEQ ID NO:4 или 54) и CDR3 (SEQ ID NO:5 или 55) тяжелой цепи 3C12F. Такие антитела могут содержать вариабельную область легкой цепи, включающей SEQ ID NO:7 или 5, или содержащей по меньшей мере один из CDR1 SEQ ID NO:8 или 58, CDR2 как SEQ ID NO:9 или 59 и CDR3 в порядке, определяемом SEQ ID NO:10 или 60. SEQ ID NO:2-5 и 7-10 описывают негуманизированное 3C12F, в то время как SEQ ID NO:52-55 и 57-60 описывают гуманизированное 3C12F. Гуманизированное 3C12F содержит вариабельную область тяжелой цепи, представленной SEQ ID NO:52, или тяжелую цепь, содержащую по меньшей мере один из CDR1 (SEQ ID NO:53), CDR2 (SEQ ID NO:54) и CDR3 (SEQ ID NO:55) тяжелой цепи 3C12F. Такие антитела могут содержать вариабельную область легкой цепи, включающую SEQ ID NO:57, или содержащую по меньшей мере один из CDR1 SEQ ID NO:58, CDR2 как SEQ ID NO:59 и CDR3 в порядке, определяемом SEQ ID NO 60.
Второй тип антитела характеризуется специфичностью связывания моноклональных антител, вырабатываемых гибридомной клеточной линией 7D12A или ее субкультурой. Типичным антителом этого типа моноклонального антитела (Mab) является 7D12A. Таким образом, этот тип включает Mab 7D12, а также его аналоги и производные, включая антитела, вырабатываемые в процессе созревания аффинности или при гуманизации. Антитела типа антитела 7D12A могут содержать вариабельную область тяжелой цепи, представленную SEQ ID NO:12, или тяжелую цепь, содержащую по меньшей мере один из CDR1 (SEQ ID NO:13), CDR2 (SEQ ID NO:14) и CDR3 (SEQ ID NO:15) тяжелой цепи 7D12A. Они могут также содержать вариабельную область легкой цепи, включающую SEQ ID NO:17 или содержащую по меньшей мере один из CDR1 SEQ ID NO:18, CDR2 SEQ ID NO:19 и CDR3, как определено в SEQ ID NO:20.
Третий тип антитела характеризуется специфичностью связывания моноклональных антител, вырабатываемых гибридомной клеточной линией 7D1G или ее субкультурой. Типичным антителом этого типа моноклонального антитела (Mab) является 7D1G. Этот тип включает Mab 7D1G и его аналоги и производные, включая антитела, продуцируемые в процессе созревания аффинности или при гуманизации. Тип 7D1G антитела может содержать вариабельную область тяжелой цепи, представленную SEQ ID NO:22, или тяжелую цепь, содержащую по меньшей мере один из CDR1 (SEQ ID NO:23), CDR2 (SEQ ID NO:24) и CDR3 (SEQ ID NO:25) тяжелой цепи 7D1G. Они могут также содержать вариабельную область легкой цепи, включающую SEQ ID NO:27 или содержащую по меньшей мере один из CDR1 SEQ ID NO:28, CDR2 как SEQ ID NO:29, и CDR3, как определено в SEQ ID NO:30.
Четвертый тип антитела характеризуется специфичностью связывания моноклональных антител, вырабатываемых гибридомной клеточной линией 18V4F или ее субкультурой. Типичным антителом этого типа моноклонального антитела (Mab) является 18V4F. Этот тип включает Mab 18V4F и его аналоги и производные, включая антитела, продуцируемые в процессе созревания аффинности или при гуманизации. Тип 18V4F антитела может содержать вариабельную область тяжелой цепи, представленную SEQ ID NO:32 или 62, или тяжелую цепь, содержащую по меньшей мере один из CDR1 (SEQ ID NO:33 или 63), CDR2 (SEQ ID NO:34 или 64) и CDR3 (SEQ ID NO:35 и 65) тяжелой цепи 18V4F. Они могут также содержать вариабельную область легкой цепи, включающую SEQ ID NO:37 или 67 или содержащую по меньшей мере один из CDR1 SEQ ID NO:38 или 68, CDR2 как SEQ ID NO:39 или 69 и CDR3, как определено в SEQ ID NO:40 или 70. SEQ ID NO:32-35 и 37-40 описывают негуманизированное 18V4F, в то время как SEQ ID NO:62-65 и 67-70 описывают гуманизированное 18V4F. Гуманизированное 18V4F содержит вариабельную область тяжелой цепи, представленную SEQ ID NO:62, или тяжелую цепь, содержащую по меньшей мере один из CDR1 (SEQ ID NO:63), CDR2 (SEQ ID NO:64) и CDR3 (SEQ ID NO:65) тяжелой цепи 18V4F. Такие антитела могут содержать вариабельную область легкой цепи, включающую SEQ ID NO:67 или содержащую по меньшей мере один из CDR1 SEQ ID NO:68, CDR2 как SEQ ID NO:69 и CDR3, как определено в SEQ ID NO:70.
Пятый тип антитела характеризуется специфичностью связывания моноклональных антител, вырабатываемых гибридомной клеточной линией 18P7E или ее субкультурой. Типичным антителом этого типа моноклонального антитела (Mab) является 18P7E. Этот тип включает Mab 18P7E и его аналоги и производные, включая антитела, продуцируемые в процессе созревания аффинности или при гуманизации. Тип 18P7E антитела может содержать вариабельную область тяжелой цепи, представленную SEQ ID NO:42, или тяжелую цепь, содержащую по меньшей мере один из CDR1 (SEQ ID NO:43), CDR2 (SEQ ID NO:44) и CDR3 (SEQ ID NO:45) тяжелой цепи 18P7E. Они могут также содержать вариабельную область легкой цепи, включающую SEQ ID NO:47 или содержащую по меньшей мере один из CDR1 SEQ ID NO:48, CDR2 как SEQ ID NO:49 и CDR3, как определено в SEQ ID NO:50.
В дополнение к специфическим типам антикиновых антител, описанным выше, изобретение охватывает антикиновые антитела, которые связываются с тремя, четырьмя, пятью или более CC-хемокинами, например, антикиновые антитела, такие как Mab 3C12F и другие, описанные выше, которые связываются с MIP-1α, MIP-1β, RANTES и/или другими родственными CC-хемокинами. Предпочтительно, антикиновые антитела могут быть получены на все CC-хемокины, которые связываются с определенным CCR, или могут быть нацелены на те СС-хемокины, которые вовлечены в определенное состояние, нарушение или заболевание.
Антикиновое антитело будет иметь аффинность связывания, достаточную для того, чтобы позволить ему связываться с CC-хемокином. Типичные аффинности связывания включают 1000, 900, 800, 400, 200, 150, 100, 75, 50, 40, 30, 20, 10, 5, 1, 0,1 нМ или меньше СС-хемокинов, с которыми оно связывается. При этом оно может иметь разные аффинности связывания для различных CC-хемокинов, как показано на фигурах.
Выбор антитела, обладающего соответствующей аффинностью связывания, находится в компетенции специалистов в рассматриваемой области. Для многих терапевтических целей антитело с высокой аффинностью является более желательным, чем антитело с низкой аффинностью, например, было продемонстрировано, что пассивно введенные высокоаффинные антитела более эффективно удаляют антиген in vivo, чем низкоаффинные антитела; более высокоаффинные антитела продуцировали более низкие уровни циркулирующих иммунных комплексов и приводили к меньшему ослаблению гломерулярной функции; Steward, Antibodies: Their Structure and Function, Taylor and Francis (1984), см. таблицу 4.5, в которой способы определения аффинности антител, а также характеристики и функции низко- и высокоаффинных антител включены посредством ссылки. С другой стороны, имея в виду, что нейтрализующая сила антитела зависит не только от его аффинности, но также и от его концентрации, валентности и молекулярной конфигурации, а также от участка, в который оно вводится, для некоторых приложений может быть желательна более высокая концентрация низкоаффинного антитела.
Иммуногены, используемые для получения антикиновых антител, могут включать препараты выделенных, нативных CC-хемокинов, рекомбинантно экспрессированных CC-хемокинов или химически синтезированных СС-хемокинов и, необязательно, адъювант. Различные адъюванты, применяемые для увеличения иммунологического ответа, включают, но не ограничиваются ими, адъювант Фрейнда (полный и неполный), минеральные гели (например, гидроксид алюминия), поверхностно-активные вещества (например, лизолецитин, полиолы плюроники, полианионы, пептиды, масляные эмульсии, динитрофенол и т.д.), адъюванты человека, такие как бацилла Кальмета-Герена и Corynebacterium parvum, или аналогичные иммуностимулирующие вещества.
Последовательности белков MIP-1α, MIP-1β, RANTES и MCP-1 и полинуклеотиды, кодирующие такие белки, известны и могут быть найдены, например, в общедоступных базах данных последовательностей, таких как GenBank, или путем ссылки на номер доступа в таблице 4. Если не указано иное, соответствующими версиями этих последовательностей следует рассматривать те, которые помещены в эти базы данных непосредственно перед датой подачи этой заявки. Кроме того, последовательности различных CC-хемокинов, включая MIP-1α, MIP-1β, RANTES и MCP-1, были опубликованы и могут быть найдены, например, в публикациях Furutani, et al., Biochem. Biophys. Res. Commun. 159:249-255 (1989) (MCP-1), Obaru, et al., J. Biochem. 99:885-894 (1986) (MIP-1α), Lipes, et al., Proc. Natl. Acad. Sci. USA 85:9704-9708 (1988) (MIP-1β), Schall, et al., J. Immunol. 141:1018-1025 (1988) (RANTES), раскрытие каждой из которых включено в настоящий документ посредством ссылки в полном объеме. Известно, что аллельные варианты существуют для некоторых или всех этих хемокинов. SEQ ID NO:71-74 представляет последовательности человека, используемые для иммунизации и скрининга антикиновых антител для MIP-1α, MIP-1β, RANTES и MCP-1.
Для получения поликлональных антител можно иммунизировать различных подходящих животных-хозяев (например, кролика, козу, мышь или другое млекопитающее) путем инъекции нативным белком, или его синтетическим вариантом, или производным CC-хемокина, или комбинацией более чем одного CC-хемокина. Например, смесь MIP-1α, MIP-1β, RANTES и МСР-1 можно использовать в качестве иммуногена, чтобы индуцировать иммунный ответ против этих CC-хемокинов.
Если требуется, поликлональные молекулы антитела, направленные против CC-хемокинов, могут быть выделены у млекопитающего (например, из крови) и дополнительно очищены известными способами, такими как хроматография белка A, чтобы получить фракцию иммуноглобулина, и иммуноаффинная очистка для отбора антикиновых антител, связывающихся с двумя или более CC-хемокинами.
Для многих приложений предпочтительно получать антикиновое антитело в виде моноклонального антитела. Можно использовать любой способ, который обеспечивает получение молекул антитела с помощью непрерывной культуры линии клеток. Такие способы включают, но не ограничиваются ими, гибридомный способ, см. Köhler & Milstein, Nature 256:495-497 (1975); триомный способ; способ B-клеточной гибридомы человека, см. Kozbor, et al., Immunol. Today 4:72 (1983) и способ EBV-гибридомы для получения моноклональных антител человека, см. Cole et al., In: Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp. 77-96 (1985). Моноклональные антитела человека можно использовать в практике по настоящему изобретению и можно получать с помощью гибридом человека, как описано в публикации Cote, et al., Proc. Natl. Acad. Sci. USA 80: 2026-2030 (1983), или путем трансформации B-клеток человека вирусом Эпштейна-Барра in vitro, см. Cole, et al. In: Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp. 77-96 (1985). Все документы, упомянутые в этом абзаце, включены посредством ссылки.
Кроме этих традиционных способов получения моноклональных антител, авторы изобретения раскрыли, что последовательные способы иммунизации удивительным образом, учитывая проблемы первичного антигенного стимула (эффект Хоскинса), продуцируют антикиновые антитела, распознающие более одного CC-хемокина. Было установлено, что последовательность иммунизации влияет на специфичность полученных в результате антител. Было обнаружено, что этими способами вырабатываются антитела, которые связываются с определенными CC-хемокинами, и некоторые, которые блокируют связывание белка vCCI вакцинного вируса с СС-хемокинами (RANTES/CCL5).
Последовательная иммунизация позвоночного, предпочтительно мыши, различными хемокинами выполняется, например, путем иммунизации животного первым хемокином в соответствующем адъюванте, бустер-иммунизации спустя несколько недель вторым хемокином и, наконец, бустер-иммунизации во второй раз третьим хемокином. Однако антикиновые антитела можно продуцировать способами, включающими несколько иммунизаций и бустер-иммунизаций, используя, например, 2, 3, 4 или более различных хемокинов в различных комбинациях. Можно проводить как иммунизацию белком хемокина, так и иммунизацию на основе ДНК. ДНК-иммунизация известна в рассматриваемой области и включена посредством ссылки на публикацию Antibodies: A Laboratory Manual (Eds. E. Harlow & D. Lane, 1988). Полинуклеотидные последовательности хемокинов известны в рассматриваемой области и также включены посредством ссылки на публикацию Yoshie, et al., Adv. Immunol. 78: 57-110 (2001). Иммунизации можно проводить с использованием адъювантов или конъюгатов хемокинов и белков-носителей, таких как описано в публикации Antibodies: A Laboratory Manual (Eds. E. Harlow & D. Lane, 1988), которая включена посредством ссылки. Предпочтительно, вводят примерно 500 мкг/кг массы тела или примерно 10 мкг хемокина на мышь, и предпочтительные адъюванты представляют собой полный адъювант Фрейнда для первичной иммунизации и неполный адъювант Фрейнда для последующих бустер-иммунизаций. Типичные CC-хемокины для использования в качестве иммуногенов включают по меньшей мере два из CCL2/MCP-1, CCL3/MIP-1α, CCL4/MIP-1β и CCL5/RANTES.
В одном варианте осуществления последовательной иммунизации мышь сначала иммунизируют MIP-1α и затем проводят бустер-иммунизацию RANTES и/или МСР-1, или любой другой последовательностью из СС-хемокинов RANTES, MIP-1α, MIP-1β и/или МСР-1. Мышиную сыворотку тестируют на реактивность с использованием RANTES, MIP-1α, MIP-1β и МСР-1 методом ELISA. Селезенку мыши с соответствующими сывороточными ответами затем используют для продуцирования гибридом.
Антитела, продуцированные гибридомными клеточными линиями, могут быть проверены на их способность распознавать CC-хемокины методом ELISA, а на блокирующую активность - либо in vitro-тестом на хемотаксис, либо тестом на vCCI/хемокин-связывание. С использованием этой стратегии было идентифицировано несколько уникальных моноклональных антител, которые связывают и ингибируют несколько CC-хемокинов. Эти антитела были названы «антикинами». Продуцированные гибридомами антитела также можно проверять в других функциональных тестах. Гибридомные клеточные линии, продуцирующие антитела с несколькими СС-хемокиновыми реактивностями, затем клонируют и размножают для продуцирования и очистки антител. Скрининг-тесты для определения специфичности связывания антител известны и регулярно практикуются в рассматриваемой области. Для подробного обсуждения таких тестов см. Harlow, et al. (Eds.), Antibodies: A Laboratory Manual; Cold Spring Harbor Laboratory; Cold Spring Harbor, N.Y., Chapter 6 (1988). После своей первоначальной идентификации антикиновое антитело может подвергаться усилению аффинности, например, с помощью способа созревания аффинности или путем конструирования его CDR или каркасной последовательности, или его можно гуманизировать. Для использования в терапии человека, предпочтительно, антикиновое антитело может быть полностью антителом человека или гуманизированным и распознавать два, три, четыре или более CC-хемокинов.
В одном варианте осуществления способы скрининга антител, которые обладают желательной специфичностью, включают, но не ограничиваются ими, ферментный иммуносорбентный анализ (ELISA) и другие иммунологически-опосредованные методы, известные в рассматриваемой области. В конкретном варианте осуществления выбор антител, специфичных для определенного домена CC-хемокинов, облегчается за счет образования гибридом, которые связываются с фрагментом CC-хемокинов, обладающих таким доменом. Антитела, специфичные для одного или нескольких доменов в CC-хемокинах, например, консервативных доменов белков CC-хемокинового семейства, или их производных, фрагментов, аналогов или гомологов, также представлены в настоящем документе.
Антикиновые антитела и их фрагменты по изобретению можно тестировать на специфичность связывания с CC-хемокинами, в частности, MIP-1α, MIP-1β, RANTES, МСР-1 и/или другими хемокинами, в иммуноферментных анализах по конкурентному и неконкурентному связыванию. Известные методики иммуноферментных анализов включены в настоящий документ посредством ссылки на Stites and Terr (Eds.) Basic and Clinical Immunology, 7th ed., (1991); Maggio (Ed.) Enzyme Immunoassay, CRC Press, Boca Raton, Fla. (1980); Tijan, Practice and Theory of Enzyme Immunoassays, Laboratory Techniques in Biochemistry and Molecular Biology, Elsevier Science Publishers B. V., Amsterdam (1985); Harlow and Lane (Eds.) Antibodies, a Laboratory Manual, Cold Spring Harbor, N.Y. (1988); Chan (Ed.), Immunoassay: A Practice Guide, Academic Press, Orlando, Fla. (1987); Price and Newman (Eds.) Principles and Practice of Immunoassays, Stockton Press, N.Y. (1991) и Ngo (Ed.) (1988) Non-isotopic Immunoassays, Plenum Press, N.Y. (1988).
Иммуноферментные анализы для определения связывания антитела могут быть либо конкурентными, либо неконкурентными. Как правило, в контексте антител конкурентный анализ предполагает конкуренцию за связывание с лигандом между двумя антителами. Например, меченый МСР-1 можно использовать для оценки того, может ли одно антитело конкурировать с другим антителом за связывание с меченым МСР-1. Анализ может быть основан на таких стандартных тестах, как, например, ферментный иммуносорбентный анализ (ELISA) или радиоиммуноанализ (RIA).
Альтернативно, антитела и фрагменты антител по изобретению можно тестировать в неконкурентных тестах на связывание с субстратами. Например, можно использовать стандартный ELISA, в котором лиганд (например, МСР-1) иммобилизован на планшете для ELISA. Тестируемое антитело инкубируют с лигандом и дают возможность связаться. Планшет промывают и после этого конъюгированное с ферментом вторичное антитело (например, антитело мыши к Fc-области иммуноглобулина человека) связывается с тестируемым антителом, если тестируемое антитело связано с лигандом. После промывки добавляют субстрат фермента и дают ему возможность взаимодействовать с ферментом. Как правило, изменение цвета указывает на присутствие антитела, которое взаимодействует с лигандом. ELISA можно повторить на различных CC-хемокинах, чтобы определить, какие хемокины распознают тестируемое антитело.
В иммуноферментном анализе часто используют меченые компоненты анализа. Метка может быть в различных формах и может быть связана непосредственно или опосредованно с желательным компонентом анализа в соответствии со способами, известными в рассматриваемой области. Общепринятые метки для компонентов анализа включают радиоактивные изотопы, в том числе 3H, 125I, 35S, 14C и 32P, флуорофоры, хемилюминесцентные вещества и ферменты. Выбор конкретной метки будет зависеть от требуемой чувствительности, простоты конъюгации с соединением, требований к устойчивости и имеющихся приборов, и должен без труда определяться любым специалистом в рассматриваемой области.
Тесты для выявления того, ингибируют ли антитела по изобретению CC-хемокиновую активность, в частности, MIP-1α, MIP-1β, RANTES, МСР-1 и/или других хемокинов, можно без труда выполнить с использованием известных тестов на хемотаксис, увеличение внутриклеточного кальция и тому подобное. Например, но не путем ограничения, анализы хемокинов на хемотаксис могут быть выполнены в 96-луночных пластиковых камерах. Лунки разделены фильтром на два отсека. Фильтр позволяет клеткам проходить из одного отсека в другой в ответ на химические градиенты. Тестируемые клетки помещают в один отсек камеры в культуральной среде, а CC-хемокин, например, помещают в культуральной среде в другой отсек. Клетки, проходящие через фильтр, учитываются. В других лунках CC-хемокин смешивают с тестируемым антителом, чтобы определить, способно ли антитело блокировать миграцию клеток.
Для дальнейшего конструирования антикиновых моноклональных антител, идентифицированных описанной выше методикой скрининга, последовательности ДНК антител были определены, как изложено в примере 7.
Аффинность антикиновых антител по изобретению можно также улучшить с помощью способов, известных в рассматриваемой области, например, тех, которые описаны в публикациях Raipal, et al., Proc. Natl. Acad. Sci. U S A. 102:8466-8471 (2005); Lippow, et al,, Nat. Biotechnol. 25:1171-1176 (2007); Wu, et al., J. Mol. Biol. 368:652-665 (2007); Yang, et al., J. Mol. Biol. 254: 392-403 (1995); и Huse, et al., J. Immunol. 149:3903-3913 (1992), каждая из которых включена посредством ссылки, в качестве обучающих способов созревания аффинности или усовершенствования антител. Антикиновые антитела по изобретению могут иметь аффинности менее 200, 100 нМ (например, первоначально выделенное мышиное антитело 3C12F имеет аффинность 49 нМ для MIP-1α) и могут быть улучшены до менее чем 40, 30, 20, 10, 5, 1, 0,5 или 0,1 нМ в процессе осуществления способов созревания аффинности. Способы, пригодные для созревания аффинности моноклональных антител, известны в рассматриваемой области и включены посредством ссылки на Antibody Engineering (Humana Press, 2004) и Phage Display, T. Clackson and H.B. Lowman, editors (Oxford University Press, 2004). Такие способы можно начинать с химерной формы такого антикинового антитела, которое можно гуманизировать как отдельно, так и одновременно с созреванием аффинности. Вариабельные области антитела могут быть экспрессированы в качестве Fab-фрагмента на поверхности нитевидного фага. Стратегия мутагенеза может быть использована для изменения каждого остатка в шести CDR, чтобы получить комбинаторную библиотеку Fab-фрагментов, экспрессированных на фаге. Их можно скринировать на связывание с хемокиновыми антигенами и анализировать по усилению в аффинности. Впоследствии предпочтительные замены аминокислот могут быть объединены для еще большего улучшения аффинности. В то же время мутации внутри каркасных областей вариабельных доменов могут быть проанализированы для поиска последовательностей с улучшенными свойствами.
«Вариантное» или «аналоговое» антитело, как описано выше, отличается по аминокислотной последовательности от последовательности аминокислот родительского антитела в силу добавления, удаления и/или замены одного или нескольких аминокислотных остатков(ка) в последовательности родительского антитела. В одном варианте осуществления вариант содержит одну или несколько аминокислотных замен(у) в одной или нескольких гипервариабельных областях(ти) родительского антитела.
Аналог антитела можно сконструировать путем замены 1, 2, 3, 4, 5, 6, 7, 8 или более аминокислотных остатков в CDR, гипервариабельной или каркасной области тяжелой и легкой цепи последовательности 3C12F, 7D1G, 7D12A, 18V4F или 18P7E. Аналогичным образом, изотипы такого антитела можно изменить методом переключения изотипа, известным в рассматриваемой области, или с помощью методов генной инженерии, таких как пересадка CDR, образование химерного антитела или гуманизация.
Аналоги могут быть получены в процессе созревания аффинности моноклональных антител типа 3C12F, 7D1G, 7D12A, 18V4F или 18P7E. Обычно аналоги, выбранные по более высокой аффинности связывания или более широкому профилю связывания хемокинов, можно обнаружить в одно и то же время. Аналоги можно легко идентифицировать путем определения того, связываются ли они с теми же хемокинами, с которыми связываются родительские антитела, например, методом ELISA или другими известными тестами. Аналог включает те варианты, в которых тяжелые и легкие цепи обладают примерно 90, 95, 99 и 100% идентичности последовательности с соответствующими последовательностями тяжелой и легкой цепей 3C12F, 7D1G, 7D12A, 18V4F или 18P7E. Можно выбрать варианты созревания аффинности антител, продуцированных гибридомными клеточными линиями 3C12F, 7D1G, 7D12A, 18V4F или 18P7E, или их субкультурами, чтобы иметь аффинности связывания 1000, 800, 400, 200, 100, 75, 50, 40, 30, 20, 10, 5, 1 или 0,1 нМ или меньше. Некоторые аналоги или варианты будут иметь одну, две, три, четыре, пять, шесть, семь, десять или до двадцати модификаций аминокислотной последовательности, таких как делеции, вставки или замены, аналог может иметь от 2 до 10 аминокислотных замен в одной или нескольких гипервариабельных областях или CDR антител 3C12F, 7D1G, 7D12A, 18V4F или 18P7E.
Аналог может представлять собой антитело или его антигенсвязывающий фрагмент, которое связывается с двумя или более из MIP-1α, MIP-1β, RANTES, MCP-I, которое включает по меньшей мере одну из следующих комбинаций CDR: CDR1 и CDR2; CDR1 и CDR3; CDR2 и CDR3; и CDR1, CDR2 и CDR3, 3C12F, 7D1G, 7D12A, 18V4F или 18P7E вариабельных областей тяжелой цепи и/или легкой цепи.
Химерные антитела, в которых антигенсвязывающий вариабельный домен животного соединен с константным доменом человека, известны в рассматриваемой области, и способы их получения включены посредством ссылки на Cabilly et al., патент США № 4816567; Morrison, S. L. et al., Proc. Natl. Acad. Sci. USA 81:6851-6855 (1984); Boulianne, G. L. et al., Nature 312:643-646 (1984); и Neuberger, M. S. et al., Nature 314:268-270 (1985).
Можно выбрать изотип константного домена человека, чтобы адаптировать химерное антитело для участия в антитело-зависимой клеточной цитотоксичности (ADCC) и комплемент-зависимой цитотоксичности (см., например, публикации Bruggemann, M. et al., J. Exp. Med. 166:1351-1361 (1987); Riechmann, L. et al., Nature 332:323-327 (1988); Love et al., Methods in Enzymology 178:515-527 (1989); Bindon, et al., J. Exp. Med. 168:127-142 (1988), которые включены посредством ссылки. Химерные и гуманизированные моноклональные антитела можно продуцировать методами рекомбинантной ДНК, известными в рассматриваемой области, например, с помощью способов, описанных в международной заявке РСТ № PCT/US86/02269; европейской патентной заявке № 184187; европейской патентной заявке № 171496; европейской патентной заявке № 173494; международной публикации РСТ № WO 86/01533; патенте США № 4816567; европейской патентной заявке № 125023; в публикации Better et al., Science 240:1041-1043 (1988).
Антикиновые антитела по изобретению можно гуманизировать с использованием известных методик, включая методики, раскрытые в публикациях Carter, патент США № 6719971, Almagro, et al., Front. Biosci. 13:1619-1633 (2008); Pini, et al., Comb. Chem. High Throughput Screen. 5:503-510 (2002); и Wu, et al., J. Mol. Biol. 294:151-162 (1999), которые включены посредством ссылки. Путем гуманизации продуцируется молекула иммуноглобулина со специфичностью антитела донорного антитела животного (мыши), но с уменьшенной иммуногенностью и лучшими эффекторными функциями в организме человека. Этот процесс предполагает вставку известных последовательностей антикинового CDR в последовательности каркаса вариабельной области легкой и тяжелой цепей антитела человека. Как правило, гуманизированные антитела продуцируются путем замены CDR мыши в каркасе вариабельного домена человека, которая, скорее всего, приведет к сохранению правильной пространственной ориентации CDR, если каркас вариабельного домена человека принимает конформацию, такую же или аналогичную конформации вариабельного каркаса мыши, от которого CDR произошли. Это достигается путем получения вариабельных доменов человека из антител человека, чьи каркасные последовательности показывают высокую степень идентичности последовательности с вариабельными каркасными доменами мыши, от которых CDR произошли. Вариабельные каркасные области тяжелой и легкой цепей могут быть получены из той же или другой последовательности антитела человека. Последовательности антитела человека могут представлять собой последовательности антител человека природного происхождения или могут представлять собой консенсусные последовательности нескольких антител человека. См. Kettleborough, et al., Protein Engineering 4:773-783 (1991); Kolbinger, et al., Protein Engineering 6:971-980 (1993) и Carter, et al., WO 92/22653.
После идентификации определяющих комплементарность участков донорного иммуноглобулина мыши и соответствующих акцепторных иммуноглобулинов человека, следующей стадией является определение того, какие, если вообще существуют, остатки этих компонентов должны быть заменены, чтобы оптимизировать свойства полученного гуманизированного антитела. Как правило, замена аминокислотных остатков человека мышиными должна быть сведена к минимуму, поскольку введение мышиных остатков повышает риск антитела вызывать у людей иммунный ответ в виде продуцирования человеческого антимышиного антитела (НАМА).
Определенные аминокислоты остатков каркаса вариабельной области человека выбирают для замены, основанной на ее возможном влиянии на конформацию CDR и/или связывание с антигеном. Неестественное сочленение CDR-участков мыши с вариабельной каркасной областью человека может привести к неестественным конформационным ограничениям, которые, при отсутствии коррекции путем замены определенных аминокислотных остатков, приводят к потере аффинности связывания.
Выбор аминокислотных остатков для замены определяется, в частности, путем компьютерного моделирования. Как правило, молекулярные модели создают, начиная от разрешенных структур для иммуноглобулиновых цепей или их доменов. Цепи для моделирования сравнивают по сходству аминокислотной последовательности с цепями или доменами разрешенной трехмерной структуры, и цепи или домены, демонстрирующие наибольшее сходство последовательностей, выбирают в качестве отправной точки для конструирования молекулярной модели. Для моделирования выбирают цепи или домены, обладающие по меньшей мере 50% идентичности последовательности, и, предпочтительно, для моделирования выбирают такие цепи или домены, которые обладают по меньшей мере 60%, 70%, 80%, 90%, 95% идентичности последовательности или более. Разрешенные начальные структуры моделируют таким образом, чтобы допустить различия между фактическими аминокислотами в цепях иммуноглобулина или доменах, которые моделируются, и таковыми в начальной структуре. Модифицированные структуры затем компонуют в составной иммуноглобулин. Наконец, модель уточняется по минимизации энергии и по проверке того, что все атомы находятся в пределах соответствующих расстояний один от другого, и что длины связей и углы находятся в химически допустимых пределах.
Выбор аминокислотных остатков для замены также можно определить, в частности, путем изучения характеристик аминокислот в определенных положениях, или эмпирического наблюдения эффектов замены или мутагенеза конкретных аминокислот. Например, когда аминокислота отличается от остатка каркаса вариабельной области мыши и выбранного остатка каркаса вариабельной области человека, тогда аминокислоту каркаса человека, как правило, следует заменить эквивалентной каркасной аминокислотой из антитела мыши в случае, когда есть основания полагать, что аминокислота нековалентно связывает антиген непосредственно, по соседству с CDR-участком, в ином случае, взаимодействует с CDR-участком (например, находится внутри примерно 3-6 Ǻ CDR-участка, как определяется компьютерным моделированием) или принимает участие в VL-VH-интерфейсе.
Остатки, которые «нековалентно связывают антиген непосредственно», включают аминокислоты в положениях в каркасных областях, которые имеют хорошую вероятность непосредственного взаимодействия с аминокислотами на антигене, в соответствии с установленными химическими силами, например, посредством водородных связей, ван-дер-Ваальсовских сил, гидрофобных взаимодействий и тому подобное.
Остатки, которые «прилегают к CDR-участку», включают аминокислотные остатки в положениях, непосредственно примыкающих к одному или нескольким CDR в первичной последовательности цепи гуманизированного иммуноглобулина, например, в положениях, непосредственно примыкающих к CDR, как определено в публикации Kabat-Wu & Kabat, J. Exp. Med. 132:211-250 (1970), или CDR, как определено в публикации Chothia-Chothia & Lesk J. Mol. Biol. 196:901-917 (1987). Эти аминокислоты являются особенно склонными к взаимодействию с аминокислотами в CDR и, если выбраны из акцептора, к искажению донорных CDR и уменьшению аффинности. Кроме того, прилегающие аминокислоты могут непосредственно взаимодействовать с антигеном (публикация Amit, et al., Science 233:747-753 (1986), которая включена посредством ссылки в настоящий документ), и выбор этих аминокислот из донора может быть желательным для сохранения всех контактов антигена, которые обеспечивают аффинность у исходного антитела.
Остатки, которые «иным образом взаимодействуют с CDR-участком», включают остатки, определенные анализом вторичной структуры, которые в пространственной ориентации достаточны для воздействия на CDR-участок. Остатки, которые «иным образом взаимодействуют с CDR-участком», могут быть идентифицированы путем анализа трехмерной модели донорного иммуноглобулина (например, компьютерной модели). Трехмерная модель, как правило, исходного донорного антитела показывает, что определенные аминокислоты вне CDR близки к CDR и обладают хорошей вероятностью взаимодействия с аминокислотами в CDR благодаря водородным связям, ван-дер-Ваальсовским силам, гидрофобным взаимодействиям и т.д. По этим аминокислотным положениям можно выбирать аминокислоту донорного иммуноглобулина, а не аминокислоту акцепторного иммуноглобулина. Аминокислоты, в соответствии с этим критерием, как правило, должны иметь атом боковой цепи в пределах примерно 3Ǻ некоторого атома в CDR и должны содержать атом, который может взаимодействовать с атомами CDR в соответствии с установленными химическими силами, такими как силы, перечисленные выше.
Аминокислоты, которые способны взаимодействовать с аминокислотами в CDR, можно идентифицировать еще одним способом. Область поверхности каждой каркасной аминокислоты, доступная растворителю, рассчитывается двумя способами: (1) в интактном антителе и (2) в гипотетической молекуле, состоящей из антитела с удаленными CDR. Достоверная разница между этими цифрами примерно в 10 Ǻ2 или более показывает, что доступ каркасной аминокислоты к растворителю по меньшей мере частично заблокирован CDR, и поэтому аминокислота создает контакт с CDR. Область поверхности аминокислоты, доступную для растворителя, можно рассчитать на основе трехмерной модели антитела, с помощью алгоритмов, известных в рассматриваемой области, например, из публикаций Connolly, J. Appl. Cryst. 16:548 (1983) и Lee & Richards, J. Mol. Biol. 55:379 (1971), каждая из которых включена в данный документ посредством ссылки. Каркасные аминокислоты могут также иногда взаимодействовать с CDR опосредованно, влияя на конформацию другой каркасной аминокислоты, которая, в свою очередь, контактирует с CDR.
Остатки, которые «принимают участие в VL-VH интерфейсе», или «упаковочные остатки» включают остатки на стыке VL и VH, как определено, например, в публикациях Novotny and Haber, Proc. Natl. Acad. Sci. USA 82:4592-66 (1985) или Chothia & Lesk, см. выше. Как правило, необычные упаковочные остатки должны сохраняться в гуманизированном антителе, если они отличаются от таковых в каркасах человека.
Обычно заменяют одну или несколько аминокислот, удовлетворяющих вышеизложенным критериям. В некоторых вариантах осуществления заменяют все или большинство аминокислот, удовлетворяющих вышеупомянутым критериям. Иногда существует некоторая неопределенность относительно того, соответствует ли конкретная аминокислота указанным выше критериям, и продуцируется альтернативный вариант иммуноглобулинов, один из которых имеет такую конкретную замену, а другой не имеет. Продуцируемые таким образом альтернативные вариантные иммуноглобулины можно тестировать в любом из тестов, описанных в настоящем документе, для желательной активности, и выбирают предпочтительный иммуноглобулин.
Как правило, CDR-участки в гуманизированных антителах являются по существу идентичными и чаще идентичны соответствующим CDR-участкам донорного антитела. Несмотря на то, что это, как правило, нежелательно, иногда можно осуществить одну или несколько консервативных аминокислотных замен CDR-остатков без существенного влияния на аффинность связывания полученного гуманизированного иммуноглобулина. Под консервативными заменами подразумеваются комбинации, такие как gly, ala; val, ile, leu; asp, glu; asn, gln; ser, thr; lys, arg; и phe, tyr.
Дополнительные кандидаты для замены представляют собой аминокислоты акцепторного каркаса человека, которые являются необычными или «редкими» для иммуноглобулина человека в этом положении. Эти аминокислоты могут быть заменены аминокислотами из эквивалентного положения донорных антител мыши или из эквивалентного положения более типичных иммуноглобулинов человека. Например, замена может быть желательна в случае, когда аминокислота в каркасной области акцепторного иммуноглобулина человека является редкой для такого положения и соответствующая аминокислота в донорном иммуноглобулине является общей для этого положения в последовательностях иммуноглобулина человека; или в случае, когда аминокислота в акцепторном иммуноглобулине является редкой для этого положения и соответствующая аминокислота в донорном иммуноглобулине также является редкой, по сравнению с другими последовательностями человека. Эти критерии позволяют гарантировать то, что атипичная аминокислота в каркасе человека не нарушит структуру антитела. Кроме того, путем замены необычной акцепторной аминокислоты человека аминокислотой из донорного антитела, которая оказывается типичной для антител человека, гуманизированное антитело может быть получено менее иммуногенным.
В настоящем документе термин «редкая» означает аминокислоту, встречающуюся в этом положении менее чем примерно в 20%, но, как правило, менее чем примерно в 10% последовательностей в репрезентативной выборке последовательностей, и термин «общий», используемый в настоящем документе, означает аминокислоту, встречающуюся более чем примерно в 25%, но, как правило, более чем примерно в 50% последовательностей в репрезентативной выборке. Например, все последовательности вариабельных областей легких и тяжелых цепей человека, соответственно, сгруппированы в «подгруппы» последовательностей, которые особенно гомологичны друг к другу и имеют одни и те же аминокислоты в определенных критических положениях (Wu & Kabat, выше). При решении вопроса о том, является ли аминокислота в акцепторной последовательности человека «редкой» или «обычной» среди последовательностей человека, часто будет предпочтительнее рассматривать только те последовательности человека, которые находятся в той же подгруппе, что и акцепторная последовательность. Дополнительными кандидатами для замены являются аминокислоты акцепторных каркасов человека, которые были бы идентифицированы как часть CDR-участка под альтернативным определением, предложенным Chothia & Lesk, см. выше.
Другие кандидаты для замены представляют собой остатки акцепторных каркасов, которые соответствуют редкому или необычному остатку донорного каркаса. Редкими и необычными остатками донорного каркаса являются такие остатки, которые являются редкими или необычными (как определено в настоящем документе) в данном положении для антител мыши. Для антител мыши подгруппа может быть определена в соответствии с Kabat и идентифицированы положения остатков, которые отличаются от консенсуса. Эти специфические различия доноров могут указывать на соматические мутации в последовательности мыши, которая повышает активность. Необычные остатки, которые, согласно прогнозам, влияют на связывание, сохраняются, в то время как остатки, согласно прогнозам не являющиеся важными для связывания, могут быть заменены.
Другими кандидатами для замены являются остатки, не относящиеся к гаметическому типу, встречающиеся в акцепторной каркасной области. Например, в случае, когда цепь акцепторного антитела (т.е. цепь антитела человека, обладающая значительной идентичностью последовательности с цепью донорного антитела) выравнивается с цепью антитела гаметического типа (точно так же обладающей значительной идентичностью последовательности с донорной цепью), остатки, не совпадающие между каркасом акцепторной цепи и каркасом цепи гаметического типа, можно заменять соответствующими остатками последовательности гаметического типа.
Помимо специфических аминокислотных замен, которые обсуждались выше, каркасные области гуманизированных иммуноглобулинов, как правило, по существу идентичны и чаще идентичны каркасным областям антител человека, из которых они были получены. Конечно, многие из аминокислот каркасной области вносят незначительный вклад или не вносят никакого вклада в специфичность или аффинность антитела. Таким образом, могут быть допущены многие отдельные консервативные замены остатков каркасной области без заметного изменения специфичности или аффинности полученного в результате гуманизированного иммуноглобулина. Таким образом, в одном варианте осуществления вариабельная каркасная область гуманизированного иммуноглобулина обладает по меньшей мере 85% идентичности последовательности с последовательностью вариабельной каркасной области человека или консенсусом таких последовательностей. В другом варианте осуществления вариабельная каркасная область гуманизированного иммуноглобулина обладает по меньшей мере 90%, предпочтительно 95%, более предпочтительно 96%, 97%, 98% или 99% идентичности последовательности с последовательностью вариабельной каркасной области человека или консенсусом таких последовательностей. Как правило, однако, такие замены являются нежелательными.
В некоторых вариантах осуществления гуманизированные антитела предпочтительно проявляют специфическую аффинность связывания с антигеном, сходную с таковой или превышающую таковую антитела мыши, из которого они были сконструированы. Обычно верхний предел аффинности связывания гуманизированных антител для антигена не превышает трех, четырех или пяти раз таковую донорного иммуноглобулина. Часто нижний предел аффинности связывания также не превышает трех, четырех или пяти раз таковую донорного иммуноглобулина. Альтернативно, аффинность связывания можно сравнить с таковой гуманизированного антитела, не имеющего никаких замен (например, антитело, имеющее донорные CDR и акцепторные каркасные области, но без замен каркасной области). В таких случаях, связывание антитела (с заменами) предпочтительно по меньшей мере в два-три раза превышает, или в три-четыре раза превышает таковое антитела без замен. Для проведения сравнения может быть определена активность различных антител, например, методом BIAcore® (т.е. поверхностным плазмонным резонансом с использованием немеченых реагентов) или тестами по конкурентному связыванию.
Рекомбинантное антитело может быть сконструировано на основе последовательностей моноклональных антител, раскрытых в настоящем документе, с использованием стандартных способов созревания аффинности, таких как способы, описанные в публикации Wu, et al., J. Mol. Biol. 350: 126-144 (2005) (включена посредством ссылки), в целях повышения аффинности антитела к первоначально идентифицированным хемокинам и/или расширения избирательности хемокинов для других CC-хемокинов, которые также могут быть вовлечены в воспалительные заболевания.
Можно выбрать класс или изотип или подкласс (например, IgG1, IgG2, IgG3 или IgG4), конкретный для химерных, гуманизированных или рекомбинантных продуктов антител, для нацеливания или адаптации антитела к конкретной функции, известной для выбранного класса или подкласса. Например, можно выбрать IgG2 человека, чтобы свести к минимуму прохождение продукта антитела через плаценту или свести к минимуму взаимодействие Fc-рецепторов с другими компонентами иммунной системы субъекта, и подкласс IgG4, чтобы свести к минимуму способность продукта антитела активировать комплемент. Можно выбрать изотип IgA для продуцирования секреторного антитела и пентамерного продукта антитела IgM для повышения связывания антитела по сравнению с мономерными антителами. Функции различных классов и подклассов молекул антител включены посредством ссылки на публикацию Kuby, Immunology, WH Freeman (1997).
«Одноцепочечные Fv» или «sFv»-фрагменты антител включают домены VH и VL антикинового антитела, где эти домены представляют одну полипептидную цепь. Как правило, Fv-полипептид дополнительно содержит полипептидный линкер между доменами VH и VL, который позволяет sFv образовывать желательную структуру для связывания антигена; см. публикацию Plückthun in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp. 269-315 (1994), которая включена посредством ссылки. Одноцепочечные антитела, специфичные для CC-хемокинов, можно продуцировать известными способами, такими как способы, раскрытые в патенте США № 4946778, который включен посредством ссылки.
«Диатела» относятся к небольшим фрагментам антител с двумя антигенсвязывающими участками, которые включают вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одной и той же полипептидной цепи (VH и VL). При использовании линкера, который является слишком коротким, чтобы обеспечить объединение двух доменов на одной и той же цепи, домены вынуждены объединяться с комплементарными доменами другой цепи и создавать два антигенсвязывающих участка. Антикиновые антитела по изобретению могут быть составлены в виде диател, которые могут быть получены способами, включенными посредством ссылки на EP 404097; WO 93/11161; и публикацию Hollinger et al., Proc. Natl. Acad. Sci. USA 90:6444-6448 (1993).
«Линейные антитела» относятся к антителам, описанным в публикации Zapata et al. Protein Eng. 8(10): 1057-1062 (1995). Кратко, эти антитела содержат пару тандемных Fd-сегментов (VH-CH 1-VH-CH1), которые образуют пару антигенсвязывающих участков. Линейные антитела могут быть биспецифическими или моноспецифическими. Антикиновые антитела по изобретению могут быть получены в соответствии с методиками, описанными в публикации Zapata, et al., которая включена в настоящий документ посредством ссылки.
Fab-экспрессионные библиотеки могут быть сконструированы с помощью других традиционных способов, таких как способы, раскрытые в публикации Huse et al., Science 246:1275-1281 (1989) и включенные посредством ссылки на нее, чтобы сделать возможным быструю и эффективную идентификацию моноклональных Fab-фрагментов с желательной специфичностью для CC-хемокинов или их производных, фрагментов, аналогов или гомологов.
Уникальные фрагменты антител, проиллюстрированные ниже, такие как их CDR-последовательности и последовательности, содержащие часть CDR, могут быть использованы для продуцирования антиидиотипических антител. Эти антитела распознают один или несколько идиотипов на вариабельном сегменте антикинового антитела и могут быть использованы для идентификации или очистки антикинового антитела или модулировать его активность. Такие антиидиотипические антитела могут быть продуцированы с использованием способов, известных в рассматриваемой области, таких как конъюгация вариабельной пептидной последовательности антитела с иммуногенным носителем и повторная иммунизация. Такие способы также включены посредством ссылки на публикацию Cavenaugh, et al., Pharm. Res. 21:1480-1488 (2004).
Антиидиотипическое антитело представляет собой антитело, которое распознает детерминанты другого антитела (антитело-мишень). Как правило, антиидиотипическое антитело распознает детерминанты антигенсвязывающего участка антитела-мишени. Обычно антитело-мишень является моноклональным антителом. Антиидиотипическое антитело, как правило, получают путем иммунизации животного (в частности, мыши) такого же вида и генетического типа, как у источника моноклонального антитела-мишени, моноклональным антителом-мишенью. Иммунизированное животное усиливает иммунный ответ на идиотипические детерминанты моноклонального антитела-мишени и продуцирует антитела против идиотипических детерминант моноклонального антитела-мишени. Продуцирующие антитело клетки, такие как клетки селезенки иммунизированного животного, могут быть использованы для создания антиидиотипических моноклональных антител. Кроме того, антиидиотипическое антитело также можно использовать для иммунизации животных с целью продуцирования анти-антиидиотипических антител. Этих иммунизированных животных можно использовать для создания анти-антиидиотипических моноклональных антител с помощью стандартных способов. Анти-антиидиотипические антитела могут связываться с тем же эпитопом, что и первоначальное моноклональное антитело-мишень, использованное для получения антиидиотипического антитела. Анти-антиидиотипические антитела представляют другие моноклональные антитела с той же антигенной специфичностью, что и первоначальное моноклональное антитело-мишень.
Если связывание антиидиотипического антитела с антителом-мишенью ингибируется соответствующим антигеном антитела-мишени, и если антиидиотипическое антитело индуцирует образование антитела с той же специфичностью, что и антитело-мишень, то оно имитирует антиген антитела-мишени. Такое антиидиотипическое антитело является «антиидиотипом, несущим внутренний образ антигена», и способно индуцировать образование антитела, как если бы это был исходный антиген, см. публикации Bona and Kohler, Anti-idiotypic Antibodies and Internal Image in Monoclonal and Anti-idiotypic Antibodies: Probes for Receptor Structure and Function, Venter J. C, Frasser, C. M., Lindstrom, J. (Eds.), Alan R. Liss, N. Y., pp 141-149 (1984). Было показано, что вакцины, включающие антиидиотипические антитела, несущие внутренний образ антигена, индуцируют защитные реакции против вирусов, бактерий и паразитов; Kennedy, et al. Science 232:220-223 (1986); McNamara, et al., Science 226:1325-1326 (1985). Также было показано, что антиидиотипические антитела, несущие внутренний образ антигена, индуцируют иммунитет к опухолеспецифическим антигенам; Raychauhuri, et al., J. Immunol. 137:1743-1749 (1986); Raychauhuri et al., J. Immunol. 139:3902-3910 (1987); Bhattacharya-Chatterjee et al., J. Immunol. 139:1354-1360 (1987); Bhattacharya- Chatterjee, et al., J. Immunol. 141:1398-1403 (1988). Изложения документов, упомянутых в этом абзаце, включены посредством ссылки.
Антиидиотипические антитела для CC-хемокинов могут быть получены, например, путем иммунизации животного, такого как мышь, с иммуногенным количеством композиции, включающей CC-хемокины или их иммуногенные части, содержащие по меньшей мере один антигенный эпитоп CC-хемокинов. Композиция также может содержать подходящий адъювант и любой носитель, необходимый для обеспечения иммуногенности. Моноклональные антитела, распознающие CC-хемокины, можно получить из клеток иммунизированного животного, как описано выше. Моноклональное антитело, распознающее общий эпитоп CC-хемокинов, затем отбирают и используют для получения композиции, содержащей иммуногенное количество моноклонального антитела против CC-хемокина. Как правило, бывает достаточно дозы в 25-200 мкг очищенного CC-хемокинового моноклонального антитела в подходящем адъюванте. Животных можно иммунизировать 2-6 раз с интервалом между дозами в 14-30 дней. Обычно животных иммунизируют любым подходящим способом введения, таким как внутрибрюшинный, подкожный, внутривенный или комбинация из них. Продукцию антиидиотипических антител можно контролировать во время периода иммунизации с использованием стандартных способов иммунологического анализа. Животных с подходящими титрами антител, реактивных в отношении моноклональных антител-мишеней, можно повторно иммунизировать моноклональным антителом, используемым в качестве иммуногена, за три дня до сбора клеток, продуцирующих антитело. Предпочтительно используют клетки селезенки, хотя могут быть выбраны и другие клетки, продуцирующие антитело. Продуцирующие антитело клетки собирают и сливают с миеломными клетками для получения гибридом, как описано выше, и отбирают клетки, продуцирующие подходящее антиидиотипическое антитело. Анти-антиидиотипические антитела продуцируются в результате другого раунда иммунизации и получения гибридом с помощью антиидиотипического моноклонального антитела в качестве иммуногена.
Как описано выше, вариабельные сегменты антикиновых антител, такие как CDR-сегменты, или пептидные фрагменты вариабельных сегментов, включая фрагменты с 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более смежными аминокислотными остатками, особенно такие, которые обладают активностью связывания хемокинов, можно использовать для различных приложений, в дополнение к терапевтическому лечению аутоиммунных заболеваний, включая, в качестве фармацевтических препаратов, вакцины на основе пептидов, получение антиидиотипических антител, или в качестве иммуногенов. Другой аспект настоящего изобретения относится к способам индуцирования иммунного ответа у млекопитающего в отношении полипептида по изобретению путем введения млекопитающему количества полипептидного препарата, достаточного для индуцирования иммунного ответа. Количество будет зависеть от вида животного, размера животного и тому подобное, но может быть определено специалистом в рассматриваемой области.
Изобретение также включает антитела или фрагменты антител, полученные из химерных или гуманизированных антикиновых антител посредством способа созревания аффинности. Можно идентифицировать замены аминокислот в CDR, которые существенно повышают аффинность антитела к CC-хемокинам и поэтому включены в настоящий документ. Способы созревания аффинности описаны, например, в публикациях Wu et al., Proc. Natl. Acad. Sci. USA 95:6037-6042 (1998) и Clackson and Loman, Phage Display, Oxford University Press (2004), каждая из которых также включена посредством ссылки.
Антикиновые антитела против CC-хемокинов могут быть использованы в способах, известных в рассматриваемой области, относящихся к локализации и/или количественному определению CC-хемокинов (например, для использования при определении уровней CC-хемокинов в соответствующих физиологических образцах, для использования в диагностических способах, для использования в визуализации белка и тому подобное). В данном варианте осуществления антитела к CC-хемокинам или их производные, фрагменты, аналоги или гомологи, которые содержат полученный из антитела связывающий домен, используются в качестве фармакологически активных соединений, лекарственных препаратов или терапевтических соединений.
Антикиновое антитело против CC-хемокинов (например, моноклональное антитело) можно использовать для выделения CC-хемокинов стандартными способами, такими как аффинная хроматография или иммунопреципитация. Антитело против CC-хемокинов может способствовать очистке природных CC-хемокинов из клеток и рекомбинантно продуцированных CC-хемокинов, экспрессируемых в клетках-хозяевах. Кроме того, пан-антитело против CC-хемокинов можно использовать для обнаружения CC-хемокинов (например, в клеточном лизате или клеточном супернатанте) для того, чтобы оценить относительное содержание и профиль экспрессии CC-хемокинов. Антикиновые антитела против CC-хемокинов могут быть использованы диагностически для контролирования уровней белка в тканях в рамках методики клинического тестирования, для того чтобы, например, определить эффективность данного режима лечения. Детектирование может быть облегчено соединением (т.е. физической связью) антитела с детектируемым веществом. Примеры детектируемых веществ включают различные ферменты, простетические группы, флуоресцентные вещества, люминесцентные вещества, биолюминесцентные вещества и радиоактивные вещества. Примеры подходящих ферментов включают пероксидазу хрена, щелочную фосфатазу, β-галактозидазу или ацетилхолинэстеразу; примеры подходящих комплексов простетических групп включают стрептавидин/биотин и авидин/биотин; примеры подходящих люминесцентных веществ включают умбеллиферон, флуоресцеин, флуоресцеин изотиоцианат, родамин, дихлортриазиниламин флуоресцеин, дансилхлорид или фикоэритрин, пример люминесцентного вещества включает люминол; примеры биолюминесцентных веществ включают люциферазу, люциферин и экворин, и примеры подходящего радиоактивного вещества включают 125I, 131I, 35S и 3H.
Кроме того, антитела по настоящему изобретению можно конъюгировать с токсинами, такими как радиоизотопы, белковые токсины и химические токсины, которые могут быть конъюгированы с антителами. Такие токсины включают, но не ограничиваются ими, свинец-212, висмут-212, астат-211, йод-131, скандий-47, рений-186, рений-188, иттрий-90, йод-123, йод-125, бром-77, индий-111, бор-10, актиниды, рицин, адриамицин, калихеамицины, 5-фторурацил, ауристатины и майтансиноиды.
Химерные, гуманизированные антитела и антитела человека, а также их антигенсвязывающие фрагменты обычно получают путем рекомбинантной экспрессии. Нуклеиновые кислоты, кодирующие гуманизированные вариабельные области легких и тяжелых цепей, необязательно связанных с константными областями, встраивают в векторы экспрессии. Легкие и тяжелые цепи можно клонировать в одном и том же или в разных векторах экспрессии. Сегменты ДНК, кодирующие цепи иммуноглобулина, функционально связаны с контролирующими последовательностями в векторе(ах) экспрессии, которые обеспечивают экспрессию иммуноглобулиновых полипептидов. Контролирующие экспрессию последовательности включают, но не ограничиваются ими, промоторы (например, природно-связанные или гетерологичные промоторы), сигнальные последовательности, энхансерные элементы и последовательности терминации транскрипции. Предпочтительно, контролирующие экспрессию последовательности представляют собой системы эукариотических промоторов в векторах, способных трансформировать или трансфицировать эукариотические клетки-хозяева. После того как вектор был введен в соответствующего хозяина, хозяин поддерживается в условиях, подходящих для обеспечения высокого уровня экспрессии нуклеотидных последовательностей, а также сбора и очистки перекрестно-реагирующих антител.
Эти векторы экспрессии, как правило, способны реплицироваться в организмах хозяев либо в виде эписом, либо в виде составной части хромосомной ДНК хозяина. Обычно векторы экспрессии содержат селективные маркеры (например, устойчивости к ампициллину, устойчивости к гигромицину, устойчивости к тетрациклину или устойчивости к неомицину), чтобы дать возможность обнаружения клеток, трансформированных необходимыми последовательностями ДНК, см., например, Itakura, et al., патент США № 4704362, который включен посредством ссылки. E. coli является одним из прокариотических хозяев, пригодных для клонирования полинуклеотидов (например, последовательностей ДНК) по настоящему изобретению. Другие микробные хозяева, подходящие для использования, включают бациллы, такие, как Bacillus subtilus, и другие кишечные бактерии, такие как Salmonella, Serratia, и различные виды Pseudomonas. В этих прокариотических хозяевах также можно получать векторы экспрессии, которые обычно содержат контролирующие экспрессию последовательности, совместимые с клеткой-хозяином (например, участок начала репликации). Кроме того, будет представлено любое количество из множества известных промоторов, например, система лактозного промотора, система триптофанового (trp) промотора, система бета-лактамазного промотора или система промотора фага лямбда. Промоторы, как правило, контролируют экспрессию, необязательно с операторной последовательностью, и содержат последовательности сайта связывания с рибосомой и т.п. для инициирования и завершения транскрипции и трансляции.
Другие микроорганизмы, такие как дрожжи, также являются пригодными для экспрессии. Saccharomyces является предпочтительным дрожжевым хозяином, с подходящими векторами, имеющими контролирующие экспрессию последовательности (например, промоторы), участок начала репликации, терминирующие последовательности и тому подобное, по желанию. Типичные промоторы включают промоторы генов 3-фосфоглицераткиназы и других гликолитических ферментов. Индуцируемые промоторы дрожжей включают, среди прочего, промоторы генов алкогольдегидрогеназы, изоцитохрома C и ферментов, ответственных за утилизацию мальтозы и галактозы.
В дополнение к микроорганизмам, могут быть также использованы культуры клеток тканей млекопитающих для экспрессии и продуцирования полипептидов по настоящему изобретению (например, полинуклеотиды, кодирующие иммуноглобулины или их фрагменты). См. Winnacker, From Genes to Clones, VCH Publishers, N.Y., N.Y. (1987). Эукариотические клетки фактически являются предпочтительными, поскольку в рассматриваемой области был разработан ряд подходящих линий клеток-хозяев, способных секретировать гетерологичные белки (например, интактные иммуноглобулины), и включают клеточные линии СНО, различные линии Cos-клеток, клетки HeLa, предпочтительно, миеломные линии клеток, или трансформированные B-клетки или гибридомы. Предпочтительно, клетки не являются клетками человека. Векторы экспрессии для этих клеток могут включать контролирующие экспрессию последовательности, такие как участок начала репликации, промотор и энхансер, Queen et al., Immunol. Rev. 89:49-68 (1986), и необходимые информационные сайты для процессинга, такие как сайты связывания с рибосомой, сайты сплайсинга РНК, сайты полиаденилирования и транскрипционные терминаторные последовательности. Предпочтительные контролирующие экспрессию последовательности представляют собой промоторы, полученные из иммуноглобулиновых генов, SV-40, аденовируса, вируса папилломы крупного рогатого скота, цитомегаловируса и тому подобное. См. Co, et al., J. Immunol. 148:1149-1154 (1982), каждый из упомянутых выше документов включен посредством ссылки.
Альтернативно, кодирующие антитело последовательности могут быть включены в трансгены для введения в геном трансгенного животного и последующей экспрессии в молоке трансгенного животного, как описано, например, Deboer, et al., патент США № 5741957, Rosen, патент США № 5304489 и Meade, et al., патент США. № 5849992, каждый из которых включен посредством ссылки. Подходящие трансгены включают кодирующие последовательности для легких и/или тяжелых цепей в функциональной связи с промотором и энхансером определенного гена молочной железы, например, казеин или бета-лактоглобулин.
Векторы, содержащие представляющие интерес полинуклеотидные последовательности (например, последовательности, кодирующие тяжелую и легкую цепи или их фрагменты, и контролирующие экспрессию последовательности), могут быть перенесены в клетку-хозяина известными способами, которые варьируются в зависимости от типа клетки-хозяина. Например, трансфекцию с использованием хлорида кальция обычно используют для прокариотических клеток, в то время как обработку фосфатом кальция, электропорацию, липофекцию, биолистику или трансфекцию на основе вируса можно использовать для других клеток-хозяев. Другие способы, используемые для трансформации клеток млекопитающих, включают использование полибрена, слияния протопластов, липосом, электропорации и микроинъекции (см., как правило, Sambrook, et al., выше). Для получения трансгенных животных трансгены можно микроинъецировать в оплодотворенные яйцеклетки, или можно встроить в геном эмбриональных стволовых клеток, ядер таких клеток, перенесенных в безъядерные ооциты. Все эти способы включены посредством ссылки на публикацию Sambrook, et al., Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Press, 2nd ed. (1989).
При клонировании тяжелых и легких цепей в разных векторах экспрессии векторы совместно трансфицируют для получения экспрессии и образования интактных иммуноглобулинов. Будучи экспрессированы, целые антитела, их димеры, отдельные легкие и тяжелые цепи или другие формы иммуноглобулинов могут быть очищены в соответствии со стандартными методами рассматриваемой области, в том числе путем использования осаждения сульфатом аммония, аффинных колонок, хроматографии на колонках, ВЭЖХ-очистки, гель-электрофореза и других аналогичных методов, включая методы, которые раскрыты в публикации Scopes, Protein Purification, Springer-Verlag, N.Y. (1982), которая включена посредством ссылки. Для применения в фармацевтических целях предпочтительно использовать по существу чистые иммуноглобулины, имеющие степень чистоты и/или гомогенности не менее 90-95% или даже 98-99%.
Для связывания и нейтрализации одного или нескольких CC-хемокинов могут быть использованы фрагменты антикиновых антител или усеченные антикиновые антитела, такие как Fab-фрагмент, Fab'-фрагмент, F(ab')2-фрагмент и Fv-фрагмент, а также цепи одного антитела, включающие CDR отдельной легкой и/или тяжелой цепи, участвующие в связывании хемокинов.
Для обеспечения желаемого эффекта антикиновые антитела можно химически модифицировать или получать их производные.
Конъюгат антитела или конъюгированное антитело можно получить с использованием антикинового антитела или его фрагмента, присоединенного через пептидные связи к гетерологичному белку. В таких вариантах осуществления антигенсвязывающая часть антитела связывается с MIP-1α, MIP-1β, RANTES, МСР-1 и/или другими родственными CC-хемокинами. Антикиновые антитела могут также быть конъюгированы с ферментами, токсинами или цитокинами в качестве эффекторных веществ. Они могут также содержать вспомогательные вещества или быть дополнительно конъюгированными или ковалентно связанными со вспомогательными веществами, такими как химические или радиологические маркеры, токсины, биологически активные или направляющие вещества, или другие вещества, которые увеличивают их биологическое время полужизни или биологическую доступность, например, путем конъюгации с полиэтиленгликолем или альбумином. Антикиновое антитело по изобретению может содержать вспомогательное вещество или быть конъюгированным со вспомогательным веществом, таким как раскрыто в публикации Carter and Senter, Cancer J. 14: 154-169 (2008), которая включена посредством ссылки. Антитела и продукты антител по изобретению могут быть иммобилизованы на твердом субстрате или иммуноаффинной смоле, описанных в публикации Antibodies: A Laboratory Manual; Eds. E. Harlow & D. Lane, (1988), которая включена посредством ссылки.
Пегилирование антител и фрагментов антител по изобретению может быть осуществлено с помощью любой реакции пегилирования, известной в рассматриваемой области, как описано, например, в следующих документах: Focus on Growth Factors 3:4-10 (1992); EP 0154316 и EP 0401384, каждый из которых включен посредством ссылки в настоящий документ в полном объеме. Предпочтительно, пегилирование осуществляют посредством реакции ацилирования или реакции алкилирования с реакционноспособной молекулой полиэтиленгликоля (ПЭГ) (или аналогичным реакционноспособным водорастворимым полимером). Предпочтительный водорастворимый полимер для пегилирования антител и фрагментов антител по изобретению представляет собой ПЭГ. В настоящем документе «полиэтиленгликоль» означает включение любой из форм ПЭГ, которые были использованы для получения производных других белков, таких как моно(Cl-ClO)алкокси- или арилоксиполиэтиленгликоль.
Способы получения пегилированных антител и фрагментов антител по изобретению должны, как правило, включать стадии (a) взаимодействия антитела или фрагмента антитела с ПЭГ, таким как реакционноспособный сложный эфир или альдегидное производное ПЭГ, в условиях, при которых антитело или фрагмент антитела становится присоединенным к одной или нескольким ПЭГ-группам, и (b) получения продуктов реакции. Для любого специалиста в рассматриваемой области не составит труда выбрать оптимальные условия реакции или реакции ацилирования на основе известных параметров и желаемом результате.
Пегилированные антитела и фрагменты антител обычно можно использовать для лечения состояний, которые могут быть смягчены или модулированы путем введения антител и фрагментов антител, описанных в настоящем документе. Как правило, пегилированные антитела и фрагменты антител обладают увеличенным временем полужизни по сравнению с непегилированными антителами и фрагментами антител. Пегилированные антитела и фрагменты антител можно использовать по отдельности, вместе или в комбинации с другими фармацевтическими композициями.
В других вариантах осуществления изобретения антитела или их антигенсвязывающие фрагменты конъюгируют с альбумином, используя способы, признанные в рассматриваемой области.
В другом варианте осуществления изобретения антитела или их фрагменты модифицируют, чтобы уменьшить или устранить потенциальные сайты гликозилирования. Такие модифицированные антитела часто называют «негликозилированными» антителами. В целях улучшения аффинности связывания антитела и его антигенсвязывающего фрагмента сайты гликозилирования антитела могут быть изменены, например, путем мутагенеза (например, сайт-направленного мутагенеза). «Сайты гликозилирования» относятся к аминокислотным остаткам, которые распознаются эукариотической клеткой в виде участка прикрепления остатков сахара. Аминокислоты, к которым прикрепляется углевод, такой как олигосахарид, обычно представляют собой остатки аспарагина (N-связь), серина (O-связь) и треонина (O-связь). В целях идентификации потенциальных сайтов гликозилирования в антителе или антигенсвязывающем фрагменте последовательность антитела анализируют, например, с помощью общедоступных баз данных, таких как веб-сайт, предоставляемый Центром анализа биологических последовательностей (см. http://www.cbs.dtu.dk/services/NetNGlyc/ для предсказания N-связанных сайтов гликозилирования и http://www.cbs.dtu.dk/services/NetOGlyc/ для предсказания O-связанных сайтов гликозилирования). Дополнительные способы изменения сайтов гликозилирования антител описаны в патентах США N№№ 6350861 и 5714350, которые включены посредством ссылки.
В еще одном варианте осуществления изобретения антитела или их фрагменты могут быть изменены путем модифицирования константной области антитела, чтобы ослабить по меньшей мере одну биологическую эффекторную функцию, опосредованную константной областью, по сравнению с немодифицированным антителом. Чтобы модифицировать антитело по изобретению так, чтобы оно проявляло пониженное связывание с Fc-рецептором, сегмент константной области иммуноглобулина антитела может быть мутирован в конкретных областях, необходимых для взаимодействий с Fc-рецептором (FcR), см., например, публикации Canfield, S. M. and S. L. Morrison, J. Exp. Med. 173:1483-1491 (1991); и Lund, J. et al., J. Immunol. 147:2657-266 (1991), каждая из которых включена посредством ссылки. Снижение способности антитела связывать FcR может также ослабить другие эффекторные функции, которые основываются на FcR-взаимодействиях, такие как опсонизация и фагоцитоз и антиген-зависимая клеточная цитотоксичность. Производные константной области с усиленными или ослабленными эффекторными функциями антител и с ослабленной или пролонгированной сывороточной устойчивостью описаны в публикации Carter, P. J., Nat. Rev. Immunol. 6:343-357 (2006), которая включена посредством ссылки.
Антикиновые антитела по изобретению могут быть включены в фармацевтические композиции, такие как описано выше, или в композиции, используемые для хранения, консервации или введения антител субъектам, нуждающимся в лечении.
Наборы для применения в описанных выше терапевтических или диагностических приложениях могут содержать еще одно антикиновое антитело, антитела положительного или отрицательного контроля, CC-хемокины, связанные антикиновыми антителами, контрольные хемокины, не связываемые антикиновым антителом, а также другие фармакологические или диагностические компоненты и письменные инструкции относительно использования набора.
Антитела и антигенсвязывающие фрагменты по изобретению, полезны, например, в терапевтических целях (путем регулирования активности CC-хемокинов), диагностических целях для выявления или количественной оценки CC-хемокинов, и для очистки CC-хемокинов. Таким образом, наборы, содержащие антитело по изобретению, для любых целей, описанных в настоящем документе, также входят в объем изобретения.
Было показано, что CC-хемокины, в частности, MIP-1α, MIP-1β, RANTES и/или MCP-1, играют роль при патологических состояниях, ассоциированных с воспалением. Было показано, что каждый из MIP-1α, MIP-1β, RANTES и/или MCP-1 обладает мощной хемотаксической активностью по отношению к лейкоцитам, особенно моноцитам и T-лимфоцитам. Эти патологические состояния включают состояния, которые описаны в публикации Johnson et al., Trends Immunol. 26:268-274 (2005), которая включена посредством ссылки.
MIP-1α, MIP-1β, RANTES и/или МСР-1 являются молекулами с мощной хемотаксической активностью по отношению к моноцитам и T-лимфоцитам. Учитывая совпадение активности и увеличенной экспрессии каждого из этих четырех хемокинов при заболеваниях человека, блокада двух, трех или четырех молекул CC-хемокина должна оказывать большее положительное действие, чем просто ингибирование одного CC-хемокина отдельно. Соответственно, антитела и фрагменты антител по изобретению пригодны для модуляции активности этих хемокинов и влияют на патологию нарушений, связанных с этими хемокинами. Как таковые, эти антитела и фрагменты полезны в терапевтических композициях для лечения воспалительных заболеваний и патологических состояний, ассоциированных с экспрессией молекул CC-хемокинов. В этих вариантах осуществления пациент идентифицируется как имеющий одно из заболеваний, которое следует лечить, например, по проявлению по меньшей мере одного признака или симптома заболевания или нарушения. По меньшей мере одно антитело или его антигенсвязывающий фрагмент по изобретению, или композиции, содержащие по меньшей мере одно антитело или его антигенсвязывающий фрагмент по изобретению, вводят в достаточном количестве, чтобы смягчить по меньшей мере один симптом заболевания или нарушения, или снизить активность по меньшей мере одного из MIP-1α, MIP-1β, RANTES и/или МСР-1.
Нарушения, поддающиеся профилактике или лечению. В настоящем документе термины «нарушение, при котором активность CC-хемокинов вредна» и «нарушение, связанное с СС-хемокинами» предназначены для включения заболеваний и других нарушений, в которых присутствие СС-хемокинов, в том числе MIP-1α, MIP-1β, RANTES, МСР-1 и других СС-хемокинов, у субъекта, страдающего нарушением, как было показано, является или предполагается либо ответственным за патофизиологию нарушения, либо фактором, который способствует нарушению. Соответственно, нарушение, при котором активность CC-хемокинов вредна, представляет собой нарушение, при котором ингибирование активности CC-хемокинов, как ожидается, предотвратит или облегчит симптомы и/или прогрессирование нарушения. Такие нарушения могут быть подтверждены, например, увеличением концентрации CC-хемокинов в биологической жидкости субъекта, страдающего от нарушения, например, увеличением концентрации RANTES в сыворотке, плазме, синовиальной жидкости, моче и т.д. субъекта, которые можно обнаружить, например, при использовании антитела против RANTES. Имеются многочисленные примеры нарушений, при которых активность CC-хемокинов является вредной. Применение антител и частей антител по изобретению в профилактике или лечении специфических нарушений обсуждаются дополнительно ниже.
Ревматоидный артрит (РА) характеризуется притоком лейкоцитов, в том числе T- и B-лимфоцитов, макрофагов и нейтрофилов, к синовиальной оболочке сустава; см. обзор Koch, Arthritis Rheum. 52: 710-721 (2005). У больных РА MIP-1α в синовиальной жидкости находится на высоком уровне, и повышение уровня коррелирует с тяжестью заболевания. Аналогичным образом, у грызунов, в модели РА на мышах в артритных суставах MIP-1α мыши находится на высоких уровнях. Нейтрализующие антитела снижают артритную балльную оценку в коллаген-индуцированной артритной модели примерно на 50%; Kasama, et al., J. Clin. Invest. 95: 2868-2876 (1995).
Уровни RANTES также увеличены в суставах в адъювант-индуцированной модели артрита на грызунах. Антитела к RANTES ослабляют симптомы в этой модели; Barnes et al., J. Clin. Invest. 101: 2910-2919 (1998).
Также было показано, что МСР-1 продуцируется синовиальными клетками и способствует инфильтрации лейкоцитов у пациентов с РА. Нейтрализующие антитела к МСР-1 также снижают клиническую оценку в модели артрита на грызунах; Ogata, et al., J. Pathol. 182: 106-114 (1997).
Рассеянный склероз (РС) характеризуется разрушением миелиновой оболочки вокруг нервов и притоком лейкоцитов к нервной ткани. Высокие уровни хемокинов, включая MIP-1α, RANTES, и МСР-1, обнаружены в поражениях головного мозга больных РС; Sorensen, et al., J. Clin. Invest. 103: 807-815 (1999).
Экспериментальный аутоиммунный энцефалит (ЕАЕ) представляет собой модель заболевания, которая точно имитирует РС человека. MIP-1α и MCP-1 были вовлечены как в индукцию симптомов заболевания, так и в развитие рецидивов, как показано с нейтрализующими антителами; Kennedy, et al., J. Neuroimmunol. 92: 98-108 (1998).
Фиброзное заболевание включает любое состояние, характеризующееся увеличением интерстициальной фиброзной ткани. CC-хемокины, как известно, связаны с фиброзными состояниями. Например, уровни каждого из МСР-1, MIP-1α и MIP-1β повышены у пациентов с системной склеродермией, и высокие уровни CC-хемокинов коррелировали с развитием фиброза легких у этих больных; Hasegawa et al., Clin. Exp. Immunol. 117: 159-165(1999).
Атеросклероз характеризуется сосудистыми жировыми отложениями с высоким проникновением макрофагов. Мыши с нокаутом МСР-1 демонстрируют заметное снижение макрофагов и отложение липидов в мышиной модели атеросклероза; Gu, et al., Mol. Cell 2: 275-281 (1998).
Больные астмой обладают выраженной инфильтрацией лейкоцитов и повышенными уровнями CC-хемокинов в легких, приводящими к сверхчувствительности дыхательных путей. В модели астмы на грызунах нейтрализующие антитела к MIP-1α, МСР-1 или RANTES уменьшали воспаление и/или сверхреактивность дыхательных путей, характерные для этого заболевания; Lukacs, et al., J. Immunol. 158: 4398-4404 (1997).
Животные модели, описанные в вышеприведенных документах, могут быть использованы для оценки эффективности антикиновых антител при лечении связанных с этим состояний, нарушений или заболеваний, и эти модели и способы их применения включены посредством ссылки на вышеприведенные документы.
Кроме заболеваний, о которых сообщалось выше, ряд других состояний, нарушений и заболеваний были связаны с активностью одного или нескольких хемокинов, они включают, но не ограничиваются ими, следующие: онкогенные заболевания, воспалительные заболевания кишечника, атопический дерматит, псориаз, инсульт, трансплантация органов, ХОБЛ (хроническая обструктивная болезнь легких), гломерулонефрит, волчаночный нефрит, склеродермия, цирроз печени, болезнь Альцгеймера, CHF-ишемия, коронарный рестеноз, диабетическая нефропатия/нейропатия/ретинопатия, остеоартрит, периодонтит, дрожжевые и вирусные инфекции и нарушения регуляции беременности. В случаях, когда CC-хемокины играют роль в этих патологиях, антикиновое антитело может быть введено для уменьшения тяжести этих состояний.
Такие способы, как правило, включают введение количества антикинового антитела обычно в фармацевтически приемлемом носителе субъекту, такому как животное или человек, нуждающемуся в этом. Количество антикинового антитела или его усеченной версии, или его фрагмента выбрано таким образом, чтобы ингибировать активность одного или нескольких хемокинов у субъекта. Аналогичные способы можно выполнять ex vivo на биологических материалах (например, крови, костном мозге, органических тканях), взятых у субъекта, или на биологических материалах, поддерживаемых в условиях in vitro.
Антикиновые антитела можно применять для блокирования хемотаксиса. Такие способы включают смешивание или контактирование антикинового антитела со средой, содержащей CC-хемокины, с которыми антикиновое антитело связывается, или средой, содержащие клетки, продуцирующие эти CC-хемокины.
В диагностических анализах можно использовать антикиновые антитела по изобретению. Такие анализы включают контактирование антикинового антитела, обладающего известной специфичностью к CC-хемокинам, с биологическим образцом, предположительно содержащим по меньшей мере один хемокин, с которым антикиновое антитело связывается, и определение величины связывания, например, путем определения комплексообразования. Антикиновые антитела можно использовать в свободной или в субстрат-связанной форме. Изобретение дополнительно предоставляет in vitro-иммуноанализ для обнаружения CC-хемокинов в образцах.
Антитела и фрагменты антител по изобретению можно использовать для обнаружения CC-хемокинов в образцах с помощью различных широко известных иммунологических тестов. Антитела могут быть использованы, например, в ELISA, Вестерн-блот-анализе, радиоиммуноанализах, способе иммуннопреципитации, иммуноаффинной хроматографии, иммунном окрашивании срезов тканей, детектировании способом иммунного окрашивания золотом в образцах тканей с электронной микроскопией и тому подобное. Протоколы для этих и других анализов известны в рассматриваемой области и находятся в компетенции специалистов.
Иммуноанализ с использованием антител и фрагментов антител по изобретению можно применять для детектирования присутствия и относительного количества CC-хемокинов в образце. Образцы могут включать, но ими не ограничиваясь, гомогенизированную ткань или клетки, гистологические срезы тканей для световой и электронной микроскопии, белковые экстракты тканей или клеток, колониестимулирующий фактор, синовиальную жидкость, кровь, плазму, сыворотку, слизистые выделения, сперму, вагинальную жидкость, слезы, слюну, пот, мочу, фекалии и т.п. Наличие увеличенных количеств CC-хемокина(ов) относительно нормальных образцов, например, может указывать на наличие болезненного состояния, и может быть показано лечение терапевтическим средством по изобретению. В некоторых случаях можно наблюдать снижение количества CC-хемокина(ов) относительно нормальных образцов, и лечение соответствующим(и) CC-хемокином(ми) или антителами внутреннего образа, которые имитируют CC-хемокин(ны), можно применять для стимулирования иммунной функции.
В некоторых вариантах осуществления можно использовать иммуноанализ для содействия очистке CC-хемокинов. Например, можно использовать иммуноаффинную смолу, в которой антитела или фрагменты антител по изобретению иммобилизованы на подложке. Образец, содержащий CC-хемокин(ны), добавляют к иммуноаффинной смоле, и антитела становятся связанными со смолой, тогда как другие компоненты образца остаются в растворе. Смолу промывают и CC-хемокины впоследствии элюируют со смолы, по существу очищенные и выделенные. Предпочтительно, антитела, используемые в иммуноанализе, должны иметь высокую аффинность связывания, как определено в настоящем документе.
ПРИМЕРЫ
Настоящее изобретение дополнительно иллюстрируется следующими примерами, которые не должны быть истолкованы как ограничивающие. Содержание всех ссылок, патентов и опубликованных заявок на патенты, приводимых в настоящем приложении, а также фигуры и перечень последовательностей включены в настоящий документ посредством ссылки.
Пример 1 - Получение гибридом, продуцирующих антикиновые антитела
Мышей последовательно иммунизировали тремя CC-хемокинами, в случайном порядке, из набора, который первоначально включал MIP-1α, RANTES и МСР-1 (PeproTech). Для гибридомного слияния 3, которое продуцировало Mab 3C12F, мышь вначале иммунизировали MIP-1α, после чего следовала бустер-иммунизация RANTES, затем МСР-1, затем снова МСР-1. Для гибридомного слияния 7, которое продуцировало Mab 7D12A и 7D1G, мышь вначале иммунизировали MIP-1α, затем проводили бустер-иммунизацию МСР-1, затем RANTES, затем сочетанием MIP-1α и RANTES. Для гибридомного слияния 18, которое продуцировало Mab 18V4F и 18P7E, мышей иммунизировали вначале MIP-1β, затем проводили бустер-иммунизацию MIP-1α, затем RANTES, потом еще одну бустер-иммунизацию RANTES.
Для каждой иммунизации было использовано 10 мкг белка, в соответствии со стандартными протоколами иммунизации, см. Current Protocols in Immunology, Eds. JE Coligan, et al. (2006), section 2.1. Начальные иммунизации проводили в полном адъюванте Фрейнда, после чего с 3-недельным интервалом следовали бустер-иммунизации двумя различными хемокинами в неполном адъюванте Фрейнда.
Через десять дней после последней бустер-иммунизации сыворотку собирали и тестировали на реактивность с CC-хемокинами MIP-1α, RANTES, МСР-1 и MIP-1α методом ELISA, как описано в публикации Current Protocols in Immunology, 2006, Eds. JE Coligan et al., section 2.4, которая включена посредством ссылки. Для MIP-1α хемокин наносили на 96-луночные планшеты для ELISA (1 мкг/мл) и инкубировали с сывороткой в диапазоне разведений от 1:50 до 1:6400. Для RANTES, МСР-1 и MIP-1β биотинилированные хемокины (0,5 мкг/мл) добавляли к покрытым стрептавидином планшетам (Pierce), а затем инкубировали с разбавленной сывороткой. Антитела, связанные с нанесенными на подложку хемокинами, детектировали с помощью вторичного антитела против мыши, конъюгированного с HRP.
Биотинилирование хемокинов для тестов ELISA выполняли с использованием сульфо-NHS-LC-биотина (Pierce). Хемокин в PBS смешивали примерно с 2-кратным молярным избытком сульфо-NHS-LC-биотина и инкубировали при комнатной температуре в течение 30 мин. Свободный биотиновый реагент удаляли диализом против PBS в течение ночи, используя установку для минидиализа Slide-a-lyzer с отсечением молекулярной массы 3500 (Pierce).
Результаты ELISA на сыворотке из первоначального набора иммунизации с использованием MIP-1α, RANTES и МСР-1 показали разнообразие ответов (см. таблицу 1), в том числе некоторые ответы на MIP-1β, даже если он не был использован в качестве прямого иммуногена. Несколько мышей дали ответы на 3 и даже 4 хемокина. Это были кандидаты на гибридомные слияния с целью обнаружить отдельные антитела, которые взаимодействуют с несколькими хемокинами. Как показано в приведенной ниже таблице 1, иммунизированные мыши продуцировали поликлональные сыворотки, которые взаимодействовали с несколькими СС-хемокинами.
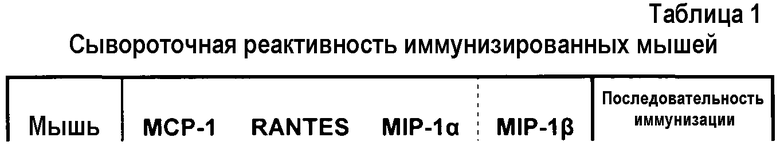
Мышей, которые продемонстрировали значительную сывороточную реактивность по меньшей мере с 3 целевыми хемокинами, отбирали для гибридомных слияний; см. Antibodies: A Laboratory Manual, Eds. E. Harlow & D. Lane (1988), chapter 6.
Выбранную мышь подвергали бустер-иммунизации смесью хемокинов (20 мкг каждого) в PBS в дни -4 и -3 до отбора селезенки для слияния с клетками NS1 с целью создания гибридом. Суспензию отдельных клеток готовили из селезенки с помощью сдавливания между матовыми концами двух стекол и сбора клеток в 10 мл RPMI. Клетки пропускали через 70-мкм нейлоновый фильтр, промывали дополнительными 10 мл RPMI и осаждали при 300×g в течение 5 мин. Клетки промывали и центрифугировали еще 2 раза и затем подсчитывали. Клетки селезенки объединяли с клетками NS1 (ATCC) в соотношении 5:1 селезенка:NS1, а затем осаждали. Один мл 50% ПЭГ добавляли по каплям к осажденным клеткам в течение 1 мин при перемешивании с вращением, затем смесь перемешивали с вращением еще в течение 1 мин. Дополнительно добавляли 1 мл RPMI в течение 1 мин с вращением, затем еще 3 мл RPMI добавляли в течение 1 мин с вращением, затем еще 16 мл RPMI добавляли в течение 2 мин с вращением. Клетки осаждали при 300×g в течение 10 мин и ресуспендировали в 800 мл гибридомной селекционной среды. Клетки оставляли в покое в течение 2 часов и затем распределяли по 40 96-луночным планшетам по 200 мкл/лунку. Планшеты помещали в инкубатор с 37°C и подпитывали каждые 2-3 дня в течение следующих 8-10 дней путем отбора и замены половины объема среды. Альтернативно, слившиеся клетки суспендировали в полутвердой среде на основе метилцеллюлозы CloneMatrix (Genetix), содержащей CloneDetect (Genetix) плюс 4 мкг/мл Alexa488-tagged MIP-1α, и помещали в однолуночный 10 см планшет (Genetix).
Пример 2- Идентификация антикиновых антител
Антитела в гибридомном супернатанте, полученном в примере 1, тестировали на их способность распознавать MIP-1α, RANTES, МСР-1 и MIP-1β методом ELISA (аналогично сывороточным тестам, описанным выше). Результаты ELISA для начальных гибридомных супернатантов приведены ниже в таблице 2. Клетки из лунок со слиянием, которые показали реакционную способность, по меньшей мере в 4 раза превышающую фон с более чем одним хемокином, размножали в 24-луночных планшетах для дальнейшего тестирования. Клетки из лунок, которые взаимодействовали по меньшей мере с 3 хемокинами, клонировали путем ограниченного разведения или путем серийного разведения по меньшей мере два раза. Супернатанты клонов планшета еще раз тестировали методом ELISA для идентификации отдельных клонов, которые продуцируют антитело, реактивное против нескольких хемокинов. Положительные лунки были подтверждены как клональные при визуальном осмотре.
Альтернативно, гибридомы, выращенные как колонии в полутвердой среде, анализировали через 16 дней по росту и связыванию флуоресцентных хемокинов с помощью ClonePixFL (Genetix) и лучшие колонии отбирали в 96-луночные планшеты. Содержащие антитела супернатанты от гибридомных клонов анализировали через 4 дня на реактивность методом ELISA с MIP-1α (как непосредственно покрытым, так и биотинилированным), RANTES и MIP-1β, как описано выше для анализа сывороток. Клоны 18V4F и 18P7E показали значительную реактивность со всеми тремя CC-хемокинами. Они были повторно клонированы путем серийного разведения для подтверждения клональности и снова протестированы методом ELISA для подтверждения продукции антикиновых антител, проявляющих реактивность к нескольким хемокинам.
Сигналы ELISA от лунок первоначального гибридомного слияния для антикиновых моноклональных антител
Пример 3: Характеристика специфичности связывания хемокинов антикиновых антител
Более полный анализ специфичности связывания хемокинов для выделенных антикиновых антител проводили с использованием MSD-анализа (Мезо Scale Discovery). Он очень похож на анализы ELISA, выполненные во время скрининга антитела, однако используется электрохемилюминесцентное детектирование реагентов SULFO-TAG. В этом случае набор хемокинов наносят в концентрации 1 мкг/мл на MSD 96-луночного многослойного планшета (MA2400). Антикиновые антитела добавляют либо в концентрации 3 мкг/мл (очищенные антитела 3C12F, 7D12, 7D1G, 18V4F), либо в виде содержащего антитело супернатанта не прошедшей количественный анализ гибридомы (18P7E) и детектируют SULFO-TAG анти-мышиными антителами, используемыми в концентрации 1 мкг/мл. Планшеты считывали на MSD sector imager 2400. Фоновые уровни, показанные пунктиром на фигурах, основаны на связывании с нереактивным белком, бычьим сывороточным альбумином (BSA).
Пример 4: Изучение функциональных активностей антикиновых антител
Супернатанты от распределенных по лункам слияний, полученных в примере 2, тестировали на возможность блокирования хемотаксиса, опосредованного хемокином-мишенью. Это тестировали в экспериментах in vitro с использованием трансфектантов CCR2 (рецептор для МСР-1) или трансфектантов CCR5 (рецептор для MIP-1α, RANTES и MIP-1β) в 96-луночных планшетах системы Трансвелл (Millipore). В нижней лунке каждой камеры системы Трансвелл 75 мкл раствора хемокина (10 нг/мл в RPMI с 2% FBS) смешивали с 75 мкл гибридомного супернатанта. В верхнюю лунку каждой камеры добавляли 4×105 клеток в 75 мкл RPMI с 2% FBS. Клеткам давали возможность мигрировать в течение 2 часов при 37°C, затем количество клеток в нижней камере количественно оценивали путем подсчета клеток с использованием FACScalibur. Клетки от гибридомных лунок, которые демонстрировали ингибирование по меньшей мере 2 хемокинов, затем также клонировали путем серийных разведений. Очищенные антитела аналогично тестировали на ингибирование хемотаксиса в диапазоне концентраций.
Трансфектанты хемокиновых рецепторов, используемые в тестах на хемотаксис, были созданы с помощью клеток Ba/F3 (полученных из немецкой коллекции микроорганизмов и клеточных культур - DSMZ). Открытые рамки считывания CCR2 и CCR5 человека были амплифицированы с помощью ПЦР (см. Molecular Cloning: A Laboratory Manual, Eds. Sambrook et al.) с кДНК-клонов, приобретенных у Origene. ПЦР-праймеры были сконструированы на основании опубликованных последовательностей. 5'-Праймер перекрывал инициирующий кодон Met и содержал сайт для клонирования XhoI. 3'-Праймер перекрывал терминирующий кодон и содержал сайт для клонирования XbaI. Амплифицированный фрагмент расщепляли XhoI и XbaI и встраивали в эквивалентные сайты экспрессионной плазмиды pNEF38 (Running Deer & Allison, Biotechnol Prog 20, 880-889, 2004). Клетки Ba/F3 трансфицировали экспрессионной плазмидой путем электропорации (Amaxa) и отбирали на G418. Клетки, экспрессирующие функциональный рецептор, отбирали по хемотаксису к лигандам родственных хемокинов и мигрирующие клетки клонировали путем предельных разведений для получения стабильного клона клеток.
Пример 5 - Идентификация и характеристика антикинового антитела
Пять уникальных моноклональных антител, которые распознавали несколько CC-хемокинов, были идентифицированы и обозначены 3C12F, 7D1G, 7D12A, 18V4F и 18P7E. Изотипы антител были определены с использованием IsoStrips (Roche). Было определено, что тяжелая цепь 3C12F является членом подгруппы IgG1 мыши, в то время как легкая цепь 3C12F, как было определено, является членом группы κ мыши. Было определено, что тяжелая цепь 7D1G является членом подгруппы IgG1 мыши, в то время как легкая цепь 7D1G, как было определено, является членом группы κ мыши. Было определено, что тяжелая цепь 7D12A является членом подгруппы IgG1 мыши, в то время как легкая цепь 7D12A, как было определено, является членом группы κ мыши. Было определено, что тяжелая цепь 18V4F является членом подгруппы IgG2a мыши, в то время как легкая цепь 18V4F, как было определено, является членом группы λ мыши. Было определено, что тяжелая цепь 18P7E является членом подгруппы IgG1, а его легкая цепь, как было определено, также является членом группы λ мыши.
Mab 3C12F распознавало MIP-1α, RANTES и MIP-1β, как показано с использованием очищенных антител на фиг. 1 и как первоначально проверено методом ELISA (см. таблицу 2), и блокировало связывание молекулы вируса vCCI с RANTES (фиг. 2). В тестах по хемотаксису 3C12F ингибировало функцию MIP-1α и RANTES, со значением IC50 3-5 мкг/мл, при использовании 5 нг/мл хемокина для индуцирования хемотаксиса (фиг. 3). По BIAcore®, аффинность 3C12F для MIP-1α составляет 49 нМ.
Каждое из антител 7D1G и 7D12A распознавало MIP-1α и MIP-1β, когда гибридомные супернатанты анализировали методом ELISA (таблица 2а). Эти же антитела также блокировали функции обоих хемокинов в тестах на хемотаксис, как показано на фиг. 4 и фиг. 5.
Каждое из антител 18V4F и 18P7E распознавало MIP-1α, MIP-1β и RANTES, когда гибридомные супернатанты анализировали методом ELISA (таблица 2b). Эти же антитела также блокировали функции (по меньшей мере частично) хемокинов MIP-1α, RANTES и MIP-1β в тестах на хемотаксис, как показано на фиг. 6 и фиг. 7.
С помощью MSD-платформы для анализа реактивности очищенных антикиновых антител (или супернатантов антител от клональных гибридом, в случае 18P7E) с матрицей хемокинов было обнаружено, что антикиновое антитело 3C12F связывало MIP-1α, RANTES, MIP-1β, MPIF-1 и HCC-2 по меньшей мере четырехкратно по сравнению с фоном (фиг. 8). Аналогично, антикиновое антитело 7D12A связывало MIP-1α, RANTES, MIP-1β и MPIF-1 (фиг. 9), и антикиновое антитело 7D1G связывало MIP-1α, RANTES, MIP-1β, HCC-1, MPIF-1 и PARC (фиг. 10). Каждое из антикиновых антител 18V4F и 18P7E связывало MIP-1α, RANTES и MIP-1β (фиг. 11 и фиг. 12).
В таблице 3 суммированы данные по специфичности связывания этих пяти антикиновых моноклональных антител.
Специфичности и функциональные активности Mab 3C12F, 7D1G, 7D12A, 18V4F и 18P7E
(MPIF-1,
HCC-2)
(HCC-1,
MPIF-1,
PARC)
(MPIF-1)
Пример 6: Анализ структур, общих для CC-хемокинов, связанных антикиновыми антителами
Проводили выравнивание аминокислотных последовательностей CC-хемокинов, связанных моноклональными антителами 7D1G, 7D12A, 3C12F, 18V4F и 18P7E. Как показано на фиг. 13, было выявлено несколько областей гомологии или имеющих высокую степень структурного сходства, с которыми связываются антикиновые моноклональные антитела по изобретению. Идентификация этих общих или консервативных сегментов CC-хемокинов характеризует связывание антитела и служит в качестве основных структур для получения или скрининга новых антикиновых антител.
Фиг. 13 a-d показывают выравнивание CC-хемокинов, связанных каждым из антикиновых антител 3C12F (фиг. 13a), 7D12A (фиг. 13b), 7D1G (фиг. 13c), 18V4F (фиг. 13d) и 18P7E (фиг. 13d). Идентичные аминокислотные остатки, общие для CC-хемокинов, связанных определенным антикиновым антителом, затенены. Аналогичные аминокислотные остатки были обозначены в соответствии с таблицей 5, например, аминокислотный остаток аланин (А) сходен с глицином, серином или треонином.
Общие идентичные или аналогичные остатки дополнительно были охарактеризованы как недоступные для растворителя, частично доступные для растворителя или полностью доступные для растворителя на основании фиг. 1 и 2 публикаций Fernandez, id. и Czaplewski et al., J. Biol. Chem. 274:16077-84 (1999), каждая из которых включена посредством ссылки.
В дальнейшем анализе сопоставляли общие остатки, доступные для растворителя, выровненных идентичных или идентичных + аналогичных остатков с остатками CC-хемокинов, ассоциированных со связыванием с CC-хемокиновым рецептором. Структурная информация, касающаяся сайтов связывания хемокиновых рецепторов, и другая структурная, химическая или функциональная информация включены посредством ссылки на поступление хемокинов или других биологических молекул, описанных в настоящем документе, в базу данных консервативных доменов NCBI CDD 29111 [uid] и на веб-адреса, указанные ниже, каждый последним посещением 9 августа 2010 года.
Остатки предполагаемого сайта связывания рецептора и другие структурные особенности для MIP-1α/CCL3 (SEQ ID NO:71) описаны на
http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi?INPUT_TYPE=
live&SEQUENCE=NP_00 2974.1.
Остатки предполагаемого сайта связывания рецептора и другие структурные особенности для MIP-1β/CCL4 (SEQ ID NO:72) описаны на
http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi?INPUT_TYPE=
live&SEQUENCE=AAH70 310.1.
Остатки предполагаемого сайта связывания рецептора и другие структурные особенности для RANTES/CCL5 (SEQ ID NO:73) описаны на
http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi?INPUT_TYPE=
live&SEQUENCE=P13501.3.
Остатки предполагаемого сайта связывания рецептора и другие структурные особенности для MPIF-1/CCL23 (SEQ ID NO:81) описаны на
http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi?INPUT_TYPE=
live&SEQUENCE=P55773.2.
Остатки предполагаемого сайта связывания рецептора и другие структурные особенности для НСС-1/CCL14 (SEQ ID NO:78) описаны на
http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi?INPUT_TYPE=
live&SEQUENCE=NP_004 157.1.
Остатки предполагаемого сайта связывания рецептора и другие структурные особенности для НСС-2/CCL15 (SEQ ID NO:79) описаны на
http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi?INPUT_TYPE=
live&SEQUENCE=Q16663.2.
Остатки предполагаемого сайта связывания рецептора и другие структурные особенности для PARC/CCL18 (SEQ ID NO:82) описаны на
http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi?INPUT_TYPE=
live&SEQUENCE=P55774. 1.
Mab 3C12F распознает CC-хемокины, которые обладают идентичными аминокислотными остатками C, C, Y, P, Y, T, C, S, P, V, F, T, C, A, P, V и L в относительных положениях 11, 12, 15, 21, 28, 31, 35, 36, 38, 40, 42, 44, 51, 52, 54, 59 и 66 на фигуре. 13a. Среди этих остатков, остатки 11, 12, 15, 21, 28, 31, 35, 36, 38, 54 и 66 частично или полностью доступны для растворителя на основании фиг. 1 и 2 Fernandez, id and Czaplewski, id.
Кроме того, CC-хемокины, распознаваемые Mab 3C12F, обладают химически схожими аминокислотными остатками в положениях 9, 14, 20, 25, 29, 33, 39, 41, 45, 46, 48 и 63 на фиг. 13а. Из этих остатков, остатки в положениях 14, 20, 29, 33, 39, 45, 46, и 48 частично или полностью доступны для растворителя (сравн. фиг. 13 и фиг. 1 и 2 Fernandez, et al.).
На основе этого структурного анализа идентичные и аналогичные остатки 11, 12, 14, 15, 20, 21, 28, 29, 31, 33, 35, 36, 38, 39, 45, 46, 48, 54 и 66 образуют общие части молекул CC-хемокинов, связываемых Mab 3C12F, пригодных в качестве детерминант для связывания между этим антителом и CC-хемокинами, которые оно распознает.
Из этих общих, доступных для растворителя остатков, остатки 14, 15, 20, 21 и 66 были охарактеризованы как остатки, контактирующие с хемокиновым рецептором в соответствии с базой данных консервативных доменов NCBI CDD 29111. Блокирование или связывание антикиновым антителом контактирования с остатками CC-хемокинового рецептора позволило бы воздействовать на способность CC-хемокина связываться со своими рецепторами или регулировать ее.
Общие, доступные для растворителя остатки CC-хемокинов, связываемых Mab 3C12F, также коррелировали со структурой CC-хемокинов и представлены в N-петле между начальными CC-остатками СС-хемокинов и β1-цепью, а также в 30's-петле CC-хемокинов между цепями β1 и β2.
Mab 7D12A распознает CC-хемокины с аминокислотными остатками C, C, Y, R, P, Y, T, C, S, P, V, F, T, C, A, P, V и L в относительных положениях 11, 12, 15, 18, 21, 28, 31, 35, 36, 38, 40, 42, 44, 51, 52, 54, 59 и 66 на фиг. 13b. Из этих остатков, остатки 11, 12, 15, 18, 21, 28, 31, 35, 36, 38, 54 и 66 частично или полностью доступны для растворителя на основании фиг. 1 и 2 Fernandez, id и Czaplewski, id.
Кроме того, CC-хемокины, распознаваемые Mab 7D12A, обладают подобными аминокислотными остатками в положениях 9, 14, 20, 25, 29, 33, 39, 41, 45, 46, 48 и 63 на фиг. 13b. Из этих остатков, остатки в положениях 14, 20, 29, 33, 39, 45, 46 и 48 частично или полностью доступны для растворителя (сравн. фиг. 13b и фиг.1 и 2 Fernandez, et al.).
На основе этого структурного анализа идентичные и аналогичные остатки 14, 15, 18, 20, 21, 28, 29, 31, 33, 35, 36, 38, 39, 45, 46, 48, 54 и 66 образуют общие части молекул CC-хемокинов, связываемых Mab 7D12A, пригодных в качестве детерминант для связывания между этим антителом и СС-хемокинами, которые оно распознает.
Из этих остатков, остатки 14, 15, 18, 20, 21 и 66 могут быть охарактеризованы как остатки, контактирующие с хемокиновым рецептором в соответствии с базой данных консервативных доменов NCBI CDD 29111. Конкурентное связывание антител, охватывающее эти остатки, могло бы влиять на способность CC-хемокинов связываться со своими рецепторами. Для CC-хемокинов, распознаваемых Mab 7D12A, несколько общих доступных для растворителя остатков представлено в N-петле между начальными CC-остатками CC-хемокина и β1-цепью, в 30's-петле CC-хемокинов между β1- и β2-цепями, а также в 40's-петле между β2- и β3-цепями.
Mab 7D1G распознает CC-хемокины с идентичными аминокислотными остатками C, C, Y, P, Y, T, C, P, T, C, P и V в относительных положениях 11, 12, 15, 21, 28, 31, 35, 38, 44, 51, 54 и 59 на фиг. 13c. Из этих остатков, остатки 11, 12, 15, 21, 28, 31, 35, 38 и 54 частично или полностью доступны для растворителя на основании фиг.1 и 2 Fernandez, id и Czaplewski, id.
Кроме того, CC-хемокины, распознаваемые Mab 7D1G, обладают сходными аминокислотными остатками в положениях 20, 25, 39-41, 45, 46, 63 и 66 на фиг. 13c. Из этих остатков, остатки в положениях 20, 39, 45, 46 и 66 частично или полностью доступны для растворителя (сравн. фиг. 13c и фиг. 1 и 2 Fernandez, et al.).
На основе этого структурного анализа идентичные и аналогичные остатки 11, 12, 15, 20, 21, 28, 31, 35, 38, 39, 45, 46, 54 и 66 образуют общие части молекул CC-хемокинов, связываемых Mab 7D1G, пригодных в качестве детерминант для связывания между этим антителом и CC-хемокинами, которые оно распознает.
Из этих общих остатков, остатки 15, 20, 21, 24 и 66 могут быть охарактеризованы как остатки, контактирующие с хемокиновым рецептором в соответствии с базой данных консервативных доменов NCBI CDD 29111. Конкурентное связывание антител, охватывающее эти остатки, могло бы повлиять на способность CC-хемокина связываться со своим рецептором. Для CC-хемокинов, распознаваемых Mab 7D1G, несколько общих, доступных для растворителя остатков представлены в N-петле между начальными CC-остатками CC-хемокинов и β1-цепью, а также в 40's-петле между β2- и β3-цепями.
Mab 18V4F и Mab 18P7E распознают CC-хемокины с аминокислотными остатками C, C, Y, R, P, Y, T, C, S, P, V, F, T, C, A, P, V и L в относительных положениях 11, 12, 15, 18, 21, 28, 31, 35, 36, 38, 40, 42, 44, 51, 52, 54, 59 и 66 на фиг. 13d. Из этих остатков, остатки 11, 12, 15, 18, 21, 28, 31, 35, 36, 38, 54 и 66 частично или полностью доступны для растворителя на основании фиг. 1 и 2 Fernandez, id и Czaplewski, id.
Кроме того, CC-хемокины, распознаваемые Mab 18V4F и 18P7E, обладают сходными аминокислотными остатками в положениях 9, 14, 20, 25, 29, 33, 39, 41, 45, 46, 48 и 63 на фиг. 13d. Из этих остатков, остатков в положениях 14, 20, 29, 33, 39, 45, 46 и 48 частично или полностью доступны для растворителя (сравн. фиг. 13d и фиг. 1 и 2 Fernandez, et al.).
На основе этого структурного анализа идентичные и аналогичные остатки 14, 15, 18, 20, 21, 28, 29, 31, 33, 35, 36, 38, 39, 45, 46, 48, 54 и 66 образуют общие части молекул СС-хемокинов, связываемых Mab 18V4F и 18P7E, пригодных в качестве детерминант для связывания между этими антителами и CC-хемокинами, которые они распознают.
Из этих остатков, остатки 14, 15, 18, 20, 21 и 66 были охарактеризованы как остатки, контактирующие с хемокиновым рецептором в соответствии с базой данных консервативных доменов NCBI CDD 29111. Конкурентное связывание антител, охватывающее эти остатки, могло бы повлиять на способность CC-хемокина связываться со своим рецептором. Для CC-хемокинов, распознаваемых Mab 18V4F и 18P7E, несколько общих, доступных для растворителя остатков представлены в N-петле между начальными CC-остатками CC-хемокинов и β1-цепью, в 30's-петле CC-хемокинов между β1- и β2-цепями, а также в 40's-петле между β2- и β3-цепями.
Пример 7: Полинуклеотидные последовательности антикиновых антител
Последовательности вариабельных доменов антител были определены после ПЦР-амплификации этих областей с использованием праймеров внутри константных областей. РНК экстрагировали из 107 гибридомных клеток с использованием RNeasy (Qiagen), затем проводили обратную транскрипцию с использованием набора SuperScript III Reverse Transcription от Invitrogen. CDR-участки были амплифицированы с помощью ПЦР с использованием набора праймеров либо для легкой цепи, либо для тяжелой цепи (Jones & Bendig, Biotechnology 9, 88-89, 1991). Полученные в результате фрагменты были встроены в плазмиду pCRII с использованием набора Zero Blunt TOPO PCR cloning от Invitrogen и секвенированы с помощью праймеров M13 и M13rev. Полинуклеотидные последовательности, кодирующие сегменты легкой и тяжелой цепи, такие как CDR, моноклональных антител 3C12F, 7D1G, 7D12A, 18V4F и 18P7E, описаны ниже. Последовательность ДНК, кодирующей вариабельную область тяжелой цепи 3C12F, представлена в SEQ ID NO:1, и аминокислотная последовательность вариабельной области тяжелой цепи 3C12F представлена в SEQ ID NO:2. Последовательность ДНК, кодирующей вариабельную область легкой цепи 3C12F, представлена в SEQ ID NO:6, и аминокислотная последовательность вариабельной области легкой цепи 3C12F представлена в SEQ ID NO:7. Кроме того, CDR антитела 3C12F описаны как SEQ ID NO:3-5 (вариабельная область тяжелой цепи) и SEQ ID NO:8-10 (вариабельная область легкой цепи).
Последовательность ДНК, кодирующей вариабельную область тяжелой цепи 7D12A, представлена в SEQ ID NO:1, и аминокислотная последовательность вариабельной области тяжелой цепи 7D12A, представлена в SEQ ID NO:12. Последовательность ДНК, кодирующей вариабельную область легкой цепи 7D12A, представлена в SEQ ID NO:16, и аминокислотная последовательность вариабельной области легкой цепи 7D12A представлена в SEQ ID NO:17. Кроме того, CDR антитела 7D12A описаны как SEQ ID NO:13-15 (вариабельная область тяжелой цепи) и SEQ ID NO:18-20 (вариабельная область легкой цепи).
Последовательность ДНК, кодирующей вариабельную область тяжелой цепи 7D1G, представлена в SEQ ID NO:21, и аминокислотная последовательность вариабельной области тяжелой цепи 7D1G, представлена в SEQ ID NO:22. Последовательность ДНК, кодирующей вариабельную область легкой цепи 7D1G, представлена в SEQ ID NO:26, и аминокислотная последовательность вариабельной области легкой цепи 7D1G представлена в SEQ ID NO:27. Кроме того, CDR антитела 7D1G описаны как SEQ ID NO:23-25 (вариабельная область тяжелой цепи) и SEQ ID NO:28-30 (вариабельная область легкой цепи).
Последовательность ДНК, кодирующей вариабельную область тяжелой цепи 18V4F, представлена в SEQ ID NO:31, и аминокислотная последовательность вариабельной области тяжелой цепи 18V4F представлена в SEQ ID NO:32. Последовательность ДНК, кодирующей вариабельную область легкой цепи 18V4F, представлена в SEQ ID NO:36, и аминокислотная последовательность вариабельной области легкой цепи 18V4F представлена в SEQ ID NO:37. Кроме того, CDR, как определено Chothia и Lesk, антитела 18V4F описаны как SEQ ID NO:33-35 (вариабельная область тяжелой цепи) и SEQ ID NO:38-40 (вариабельная область легкой цепи).
Последовательность ДНК, кодирующей вариабельную область тяжелой цепи 18P7E, представлена в SEQ ID NO:41, и аминокислотная последовательность вариабельной области тяжелой цепи 18P7E представлена в SEQ ID NO:42. Последовательность ДНК, кодирующей вариабельную область легкой цепи 18P7E, представлена в SEQ ID NO:46, и аминокислотная последовательность вариабельной области легкой цепи 18P7E представлена в SEQ ID NO:47. Кроме того, CDR, как определено Chothia и Lesk, антитело 18P7E описано как SEQ ID NO:43-45 (вариабельная область тяжелой цепи) и SEQ ID NO:48-50 (вариабельная область легкой цепи).
Пример 8: Гуманизация моноклонального антитела 3C12F
Способы гуманизации антитела предназначены для получения молекулы с уменьшенной иммуногенностью у человека. Способы гуманизации антитела известны специалистам в рассматриваемой области и включают способы, описанные в публикации Almagro and Frannson, Frontiers Bioscience 13:1619-1633 (2008). Способ пересадки CDR (Jones et al., Nature 321:522-525, 1986; Riechmann, et al., Nature 332:323-327, 1988) был использован для встраивания последовательностей CDR 3C12F в каркасные последовательности вариабельной области тяжелой и легкой цепи IgG аналогичных генов гаметического типа человека. В этом случае, последовательности CDR грызунов были встроены в каркасные последовательности вариабельной области тяжелой и легкой цепи IgG выбранных генов гаметического типа человека, которые обладают наиболее высокой идентичностью с соответствующими вариабельными последовательностями тяжелой и легкой цепи грызунов. См. публикации Jones et al., Nature 321:522-525 (1986); Riechmann, et al., Nature 332:323-327 (1988), которые включены в настоящий документ посредством ссылки в полном объеме, для подходящих способов.
В гуманизированном 3C12F последовательности гаметического типа человека для вариабельных тяжелой и легкой цепей, которые имеют наибольшее сходство с последовательностью 3C12F мыши, были использованы в качестве каркасных последовательностей человека. В дополнение к общему высокому сходству последовательности с антителом мыши, выбранные последовательности гаметического типа были также найдены в изобилии в выборке перестроенных последовательностей антител. Мышиные остатки, которые находятся в определяющих комплементарность участках (CDR), определяемых по Kabat, были пересажены в каркасные последовательности человека с дополнительными остатками, возвращенными аминокислоте мыши, когда с очевидностью было доказано, что они могут быть важны для связывания. Полинуклеотидные последовательности, кодирующие тяжелую и легкую цепи гуманизированного 3C12F Mab, приводятся в SEQ ID NO:51 и 56. Соответствующие аминокислотные последовательности и CDR определены в SEQ ID NO:52-55 и 57-60.
Пример 9: Гуманизация моноклонального антитела 18V4F
Для гуманизаци 7D12A и 18V4F использовали другую стратегию, отличную от той, которую использовали при гуманизации 3C12F. Вместо пересадки CDR антитела 18V4F мыши в наиболее сходную последовательность гаметического типа человека, CDR пересаживали в консенсусную последовательность, составленную из наиболее наполненного семейства вариабельных тяжелых цепей, VHIII, и наиболее наполненного семейства вариабельных легких лямбда-цепей, VLλIII. Далее, только те остатки в CDR, которые были определены по Chothia и Lesk как структурно значимые для связывания, первоначально были пересажены в консенсусный каркас. Дополнительные положения CDR, определенные более широко по Kabat, а также каркасные остатки, были обратно замещены на аминокислоты мыши, когда c очевидностью было доказано, что они могут быть важны для сохранения связывания. Полинуклеотидные последовательности, кодирующие тяжелую и легкую цепи гуманизированного 18V4F Mab, представлены в SEQ ID NO:61 и 66. Соответствующие аминокислотные последовательности и CDR определены в SEQ ID NO:62-65 и 67-70.
Пример 10: Созревание аффинности гуманизированных Mab 3C12F и Mab 18V4F
Аффинность связывания антикиновых моноклональных антител по изобретению можно дополнительно увеличить, при необходимости, путем созревания аффинности, как описано, например, в публикации Wu et al. (1998), Proc. Natl. Acad. Sci. USA 95: 6037-6042, которая включена посредством ссылки.
Для улучшения характеристик связывания гуманизированного Mab 3C12F и гуманизированного Mab 18V4F с различными хемокинами (например, RANTES/CCL5, MIP-1α/CCL3 и MIP-1β/CCL4) и для образования более сильнодействующих агентов использовали стратегию сайт-направленного мутагенеза в сочетании с фаговым дисплеем. Fab-фрагмент гуманизированного 3C12F или гуманизированного 18V4F представлен на поверхности нитчатого фага М13 в качестве слияния тяжелой цепи с геном III белка фага М13, с коэкспрессией легкой цепи гуманизированного 3C12F или гуманизированного 18V4F, соответственно. Далее, каждое положение всех шести CDR гуманизированного Mab 3C12F или гуманизированного 18V4F методично подвергали мутагенезу, 4-6 смежных CDR-остатков в одно время, используя протокол Sidhu and Weiss (Phage Display, A Practical Approach, Clackson, T. and Lowman, H. B., ed., Oxford University Press, 2004, Chapter 4). В некоторых случаях аминокислоты, окружающие определенные CDR, также подвергали мутагенезу в случае, если они были вовлечены в связывание антигена. В общей сложности для каждого антитела были сконструированы 15 библиотек.
Для каждой библиотеки были получены методом фагового дисплея мутантные Fab-фрагменты гуманизированного 3C12F или гуманизированного 18V4F для селекции связывания. Было проведено до шести раундов фаговой селекции вариантов гуманизированного 3C12F или гуманизированного 18V4F с более высокими аффинностями, используя протоколы созревания аффинности Nielsen and Marks (Clackson and Lowman, ibid, Chapter 14). Кратко, селекцию связывания осуществляют, используя биотинилированные хемокины, и улавливание проводят с помощью покрытых стрептавидином магнитных шариков для идентификации вариантов гуманизированного 3C12F Fab или гуманизированного 18V4F Fab с увеличенной аффинностью. Варианты с более высокой аффинностью сначала идентифицируют, используя способы, известные в рассматриваемой области (см. Clackson и Lowman и ссылки в них), а затем объединяют для достижения более высокой аффинности.
Аффинность антител к их мишеням также можно повысить с помощью различных способов, известных в рассматриваемой области. Аффинность можно увеличить с помощью прямой мутации, фагового дисплея или перестановки цепей в нуклеиновых кислотах, кодирующих молекулы антитела. Отдельные остатки или несколько остатков можно рандомизировать так, чтобы в популяции в ином случае идентичных антигенных участков антитела все двадцать аминокислот или часть из них находились в определенных положениях внутри или рядом с CDR. Полезные для этой цели способы включены посредством ссылки на публикации Yang et al., J. Mol. Biol. 254, 392-403 (1995); Hawkins et al., J. Mol. Biol. 226, 889-896 (1992); или Low et al., J. Mol. Biol. 250, 359-368 (1996). В публикации Marks et al., Bio/Technology, 10:779-783 (1992) описано созревание аффинности путем перестановки VH- и VL-доменов; случайный мутагенез CDR и/или каркасных остатков описан в публикациях Barbas et al., Proc. Nat. Acad. Sci., USA 91:3809-3813 (1994); Schier et al., Gene, 169:147-155, 1995, Yelton et al., J. Immunol., 155:1994-2004 (1995); Jackson et al., J. Immunol. 154(7):3310-3319 (1995); и Hawkins et al., J. Mol. Biol., 226:889-896 (1992), каждая из которых также включена посредством ссылки.
Созревание аффинности иногда осуществляют с использованием случайного мутагенеза. Аминокислотные замены, сделанные в определенных положениях, могут быть случайными или сделаны с использованием упрощенных правил. Например, все остатки могут быть изменены на аланин, что называется аланиновым сканированием, см., например, WO 9523813. Основанные на последовательности способы созревания аффинности также могут быть использованы для увеличения аффинности связывания антител, см. патенты США 2003/022240 A1 и 2002/177170 A1, каждый из которых включен посредством ссылки. Другие способы включают основанный на олигонуклеотидах мутагенез аминокислот в некоторых или всех положениях с помощью некоторых или всех CDR, с последующим скринингом на более высокую аффинность, как описано в публикации Wu et al., Proc. Natl. Acad. Sci. U.S.A., 95, 6037-6042 (1998), или путем селекции на более высокую аффинность с помощью фагового дисплея, как описано в публикации Rajpal, et al., Proc. Natl. Acad. Sci. U.S.A., 102:8466-8471 (1996). Селекцию антител с созревшей аффинностью также можно осуществить путем представления антитела или его фрагмента на поверхности клеток дрожжей, как описано в публикации Methods Mol. Biol., 504:351-83 (2009), или на поверхности клеток млекопитающих, как описано в публикации Ho, et al., Proc. Natl. Acad. Sci. U.S.A., 103:9637-9642 (2006). Каждый из способов, описанных в вышеуказанных документах, которые могут быть использованы для процесса созревания аффинности, включены посредством ссылки на документы, упомянутые в данном примере.
Идентификация последовательностей CC-хемокина и вируса вакцины
Идентичности хемокинов и других продуктов, раскрытых в настоящем документе, включены посредством ссылки на номера доступа в таблице 4.
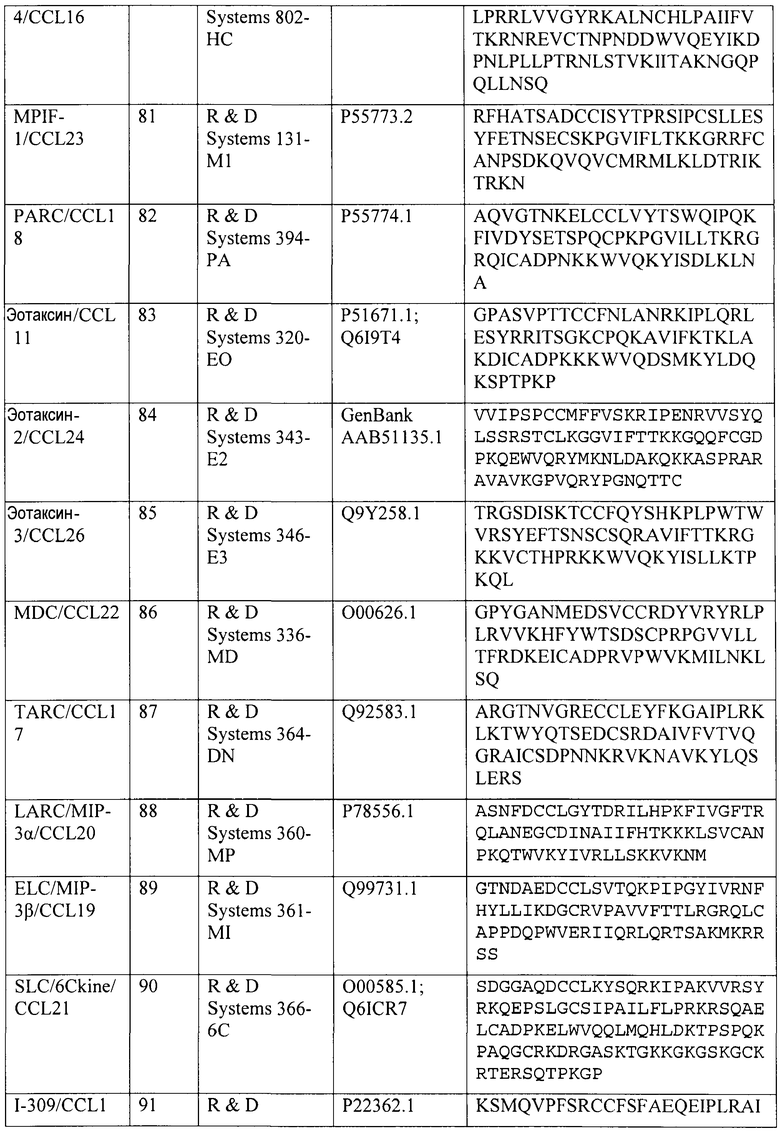
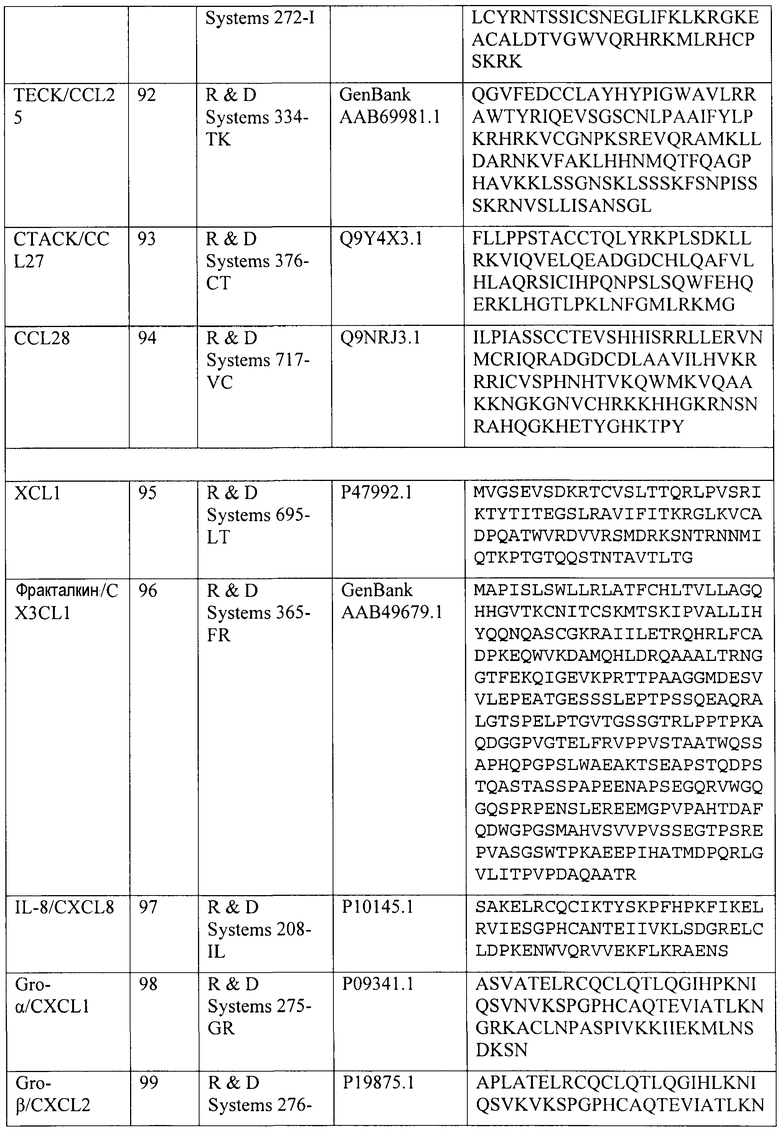
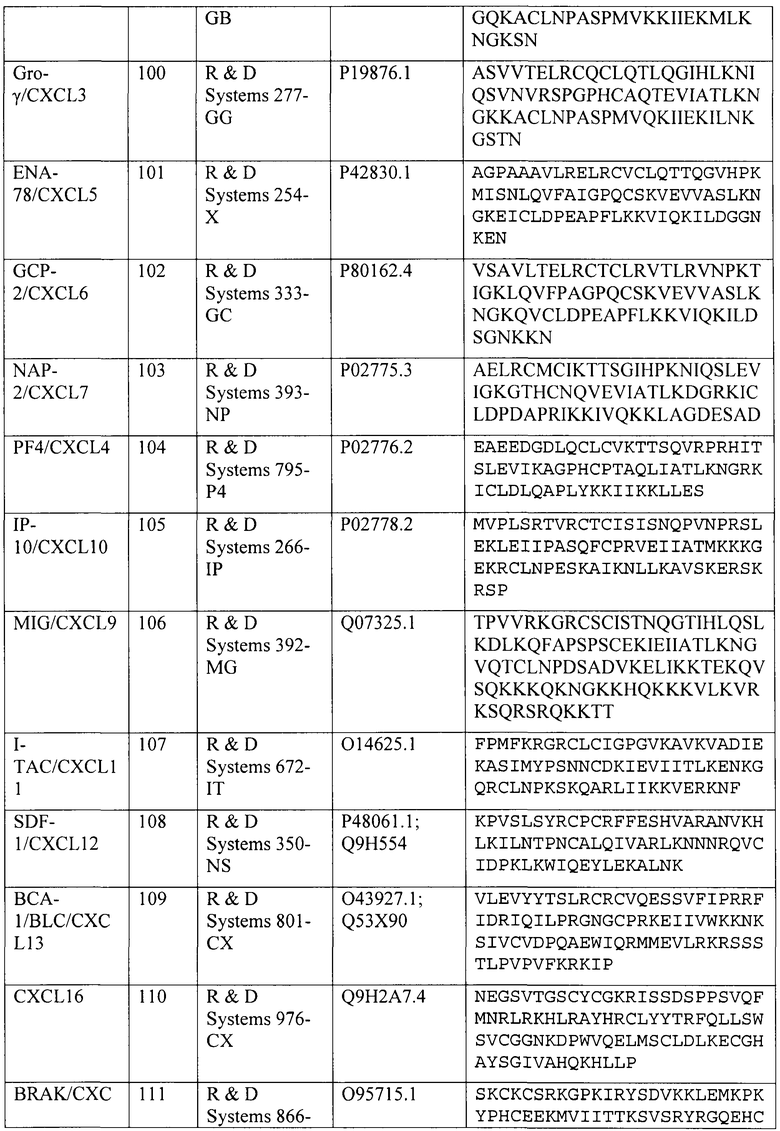
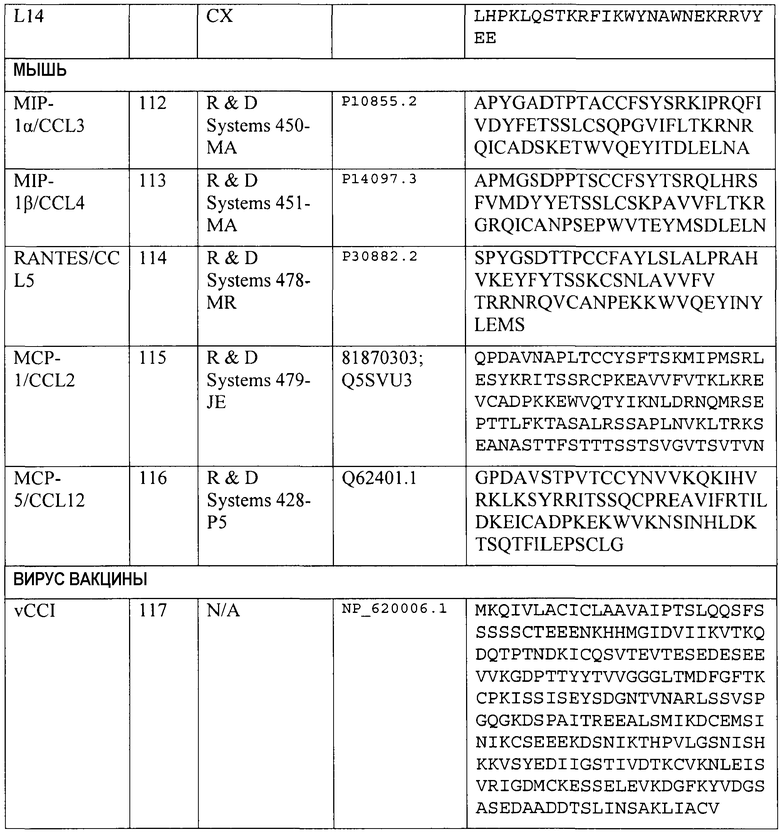
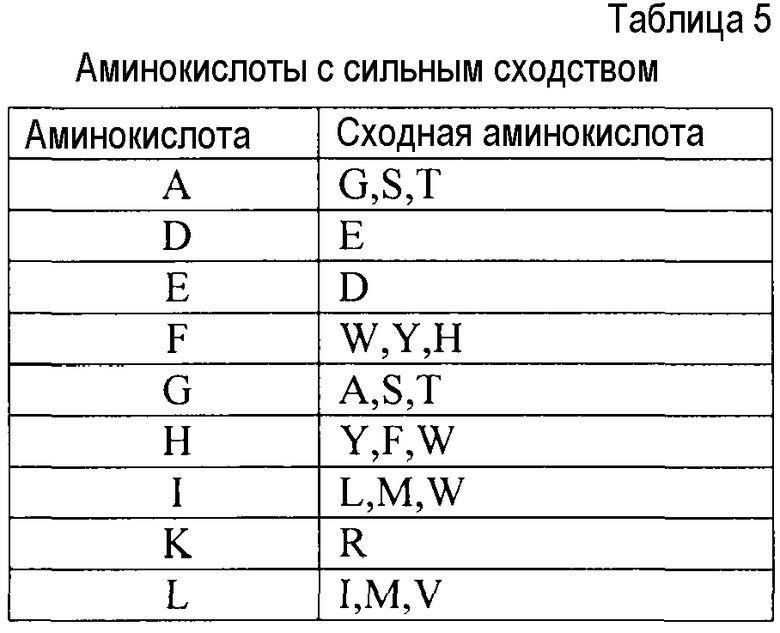
Депозиты гибридом
Гибридомные линии клеток 3C12F, 7D12A, 7D1G, 18V4F и 18P7E были депонированы 12 августа 2010 года в American Type Culture Collection (ATCC), 10801 University Blvd., Manassas, Virginia 201 10-2209, США под следующими номерами доступа:

Модификации и другие варианты осуществления
Модификации и вариации описанных антикиновых антител и композиций и способы их получения и применения, а также концепция изобретения должны быть понятны специалистам в рассматриваемой области без отступления от объема и сущности изобретения. Несмотря на то, что изобретение было описано применительно к конкретным вариантам осуществления, следует понимать, что изобретение, как заявлено, не подразумевает ограничение такими конкретным вариантами осуществления.
Включение путем ссылки
Каждый документ, патент, заявки на патент или патентная публикация, цитируемые или упоминаемые в данном раскрытии, включены посредством ссылки в полном объеме, особенно в отношении конкретного содержания, окружающего цитирование ссылок в тексте. При этом не делается допущение о том, что любая такая ссылка представляет прототип, и сохраняется право оспаривать точность и применимость цитируемых документов. В случае различий между определениями термина, определение термина, указанное в настоящем описании, является определяющим.
| название | год | авторы | номер документа |
|---|---|---|---|
| Антитела против эотаксина 2, которые распознают дополнительные связывающие CCR3 хемокины | 2015 |
|
RU2705255C2 |
| СВЯЗЫВАЮЩИЕ IL-1 БЕЛКИ | 2011 |
|
RU2615173C2 |
| ИММУНОГЛОБУЛИН С ДВОЙНЫМИ ВАРИАБЕЛЬНЫМИ ДОМЕНАМИ И ЕГО ПРИМЕНЕНИЯ | 2006 |
|
RU2515108C2 |
| ЛЕЧЕНИЕ ОСТЕОАРТРИТА И БОЛИ | 2012 |
|
RU2563830C2 |
| АНТИТЕЛО ПРОТИВ CCR5 | 2003 |
|
RU2322454C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ IL-10 ДЛЯ ЛЕЧЕНИЯ СИСТЕМНОЙ КРАСНОЙ ВОЛЧАНКИ (SLE) | 2010 |
|
RU2581812C2 |
| IL-1 АЛЬФА И БЕТА БИСПЕЦИФИЧЕСКИЕ ИММУНОГЛОБУЛИНЫ С ДВОЙНЫМИ ВАРИАБЕЛЬНЫМИ ДОМЕНАМИ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2627171C2 |
| IL-1бета-СВЯЗЫВАЮЩИЕ АНТИТЕЛА И ИХ ФРАГМЕНТЫ | 2006 |
|
RU2518295C2 |
| ИММУНОГЛОБУЛИНЫ K CD52 ЧЕЛОВЕКА | 2010 |
|
RU2603743C2 |
| ИНТЕРЛЕЙКИН-13-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2012 |
|
RU2605327C2 |






















































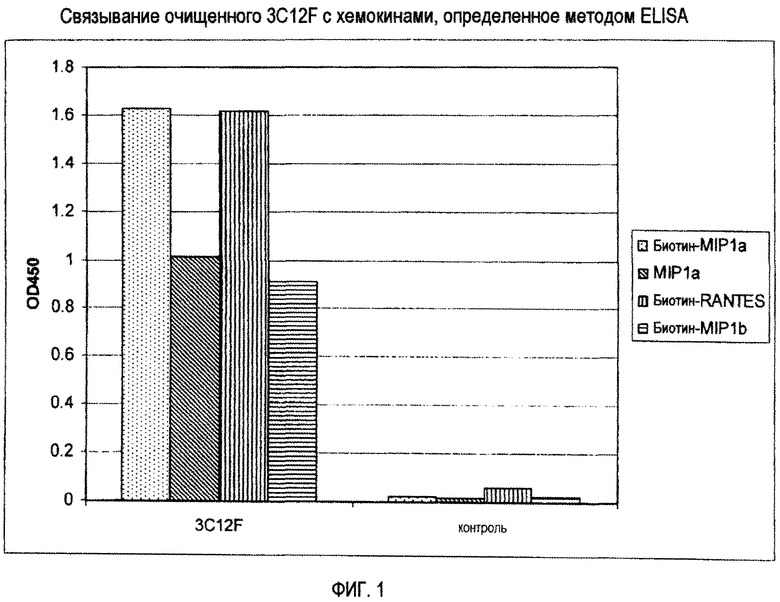
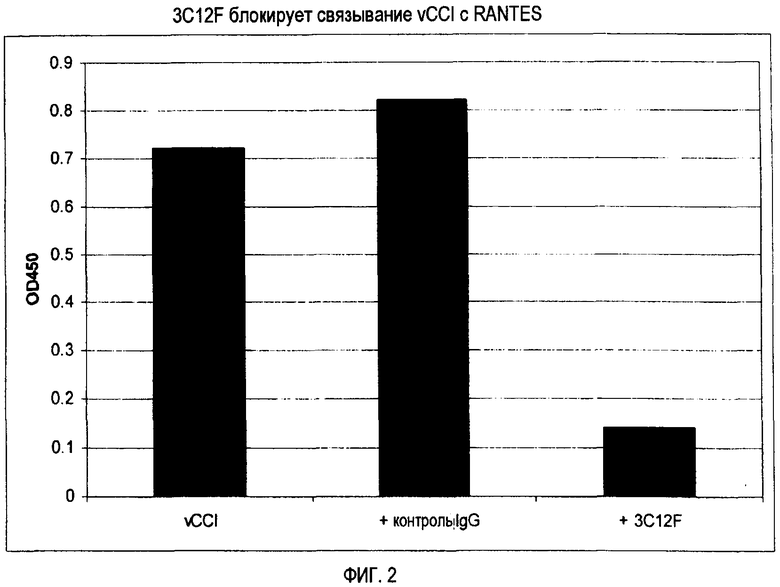
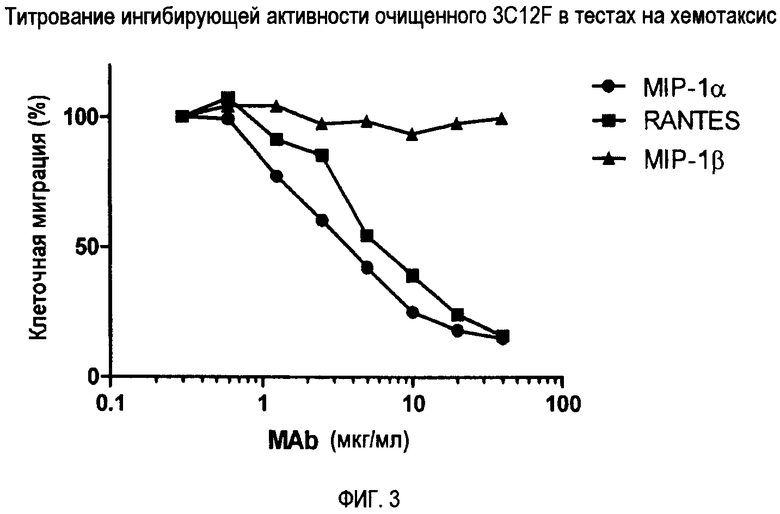
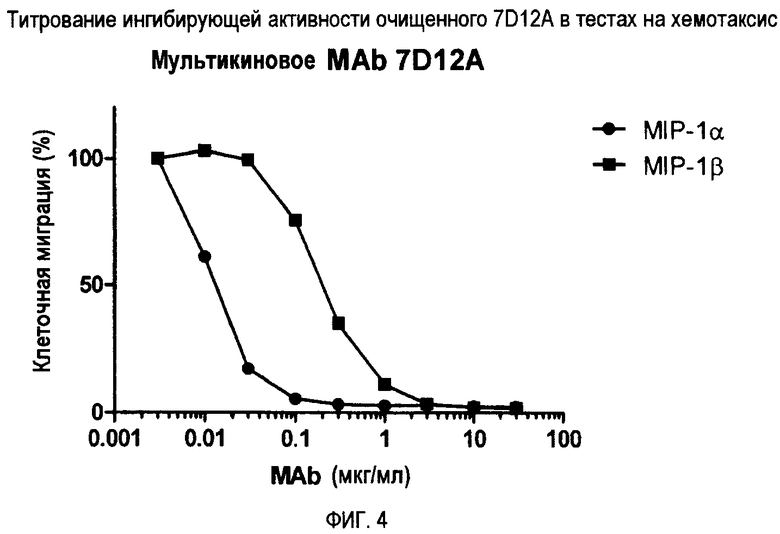
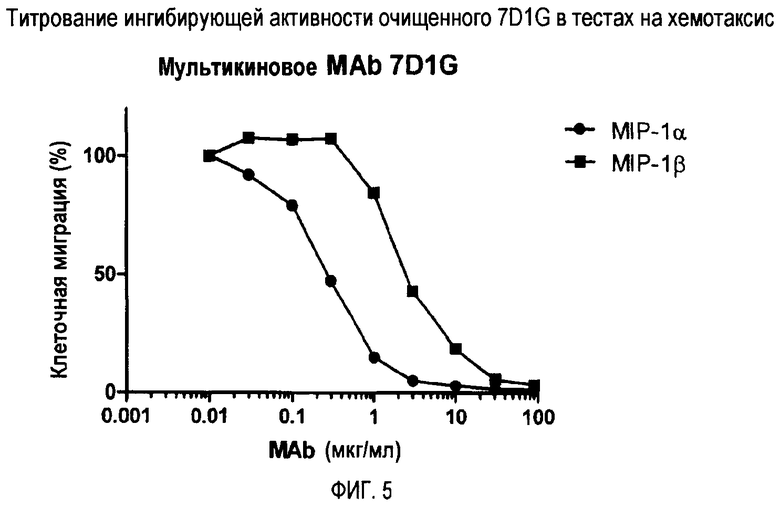
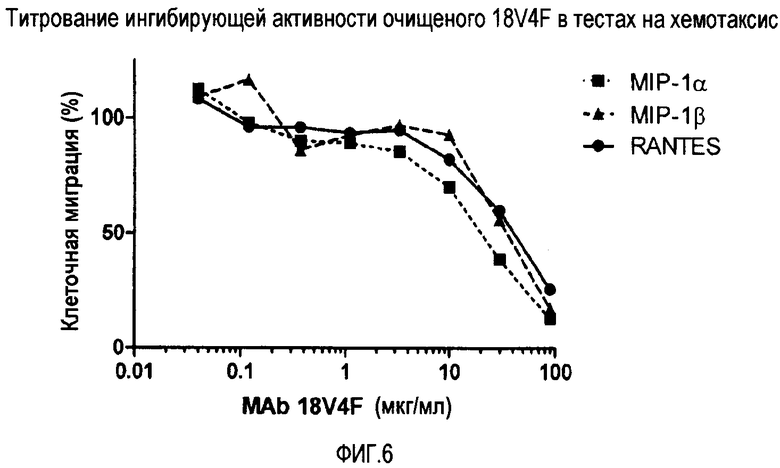
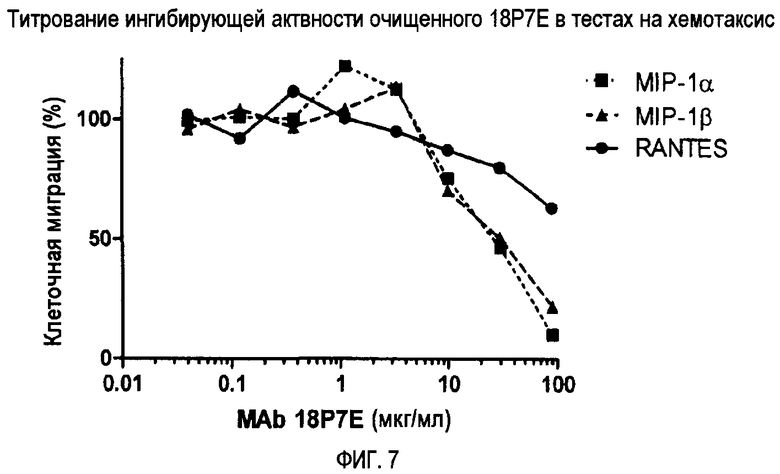
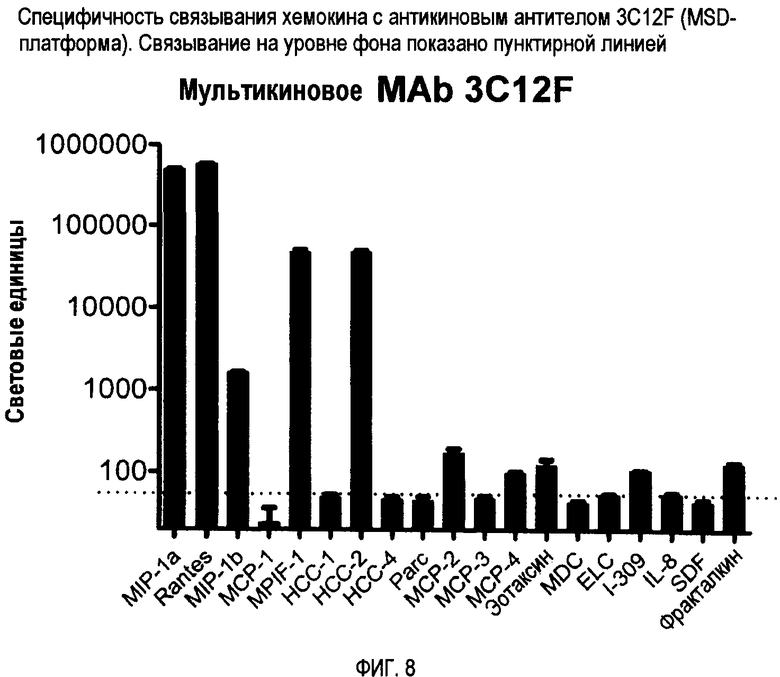
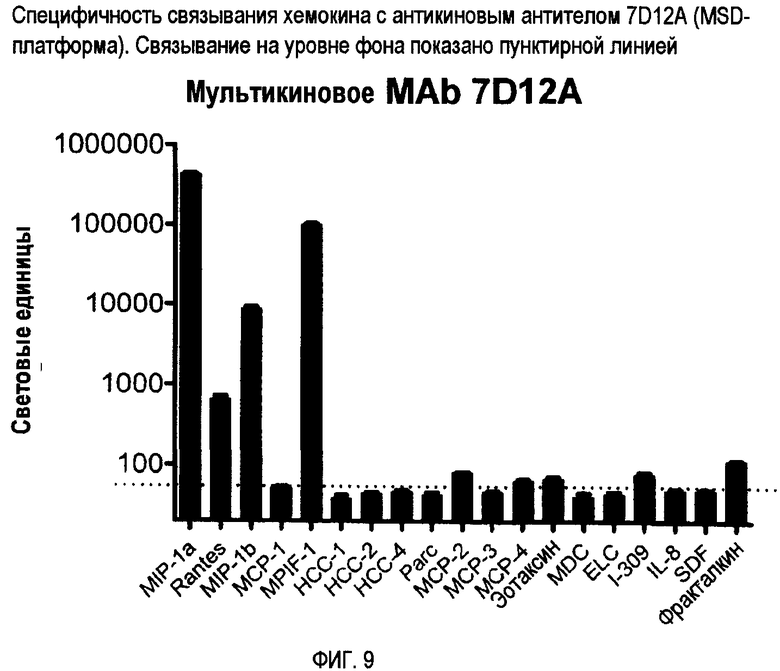
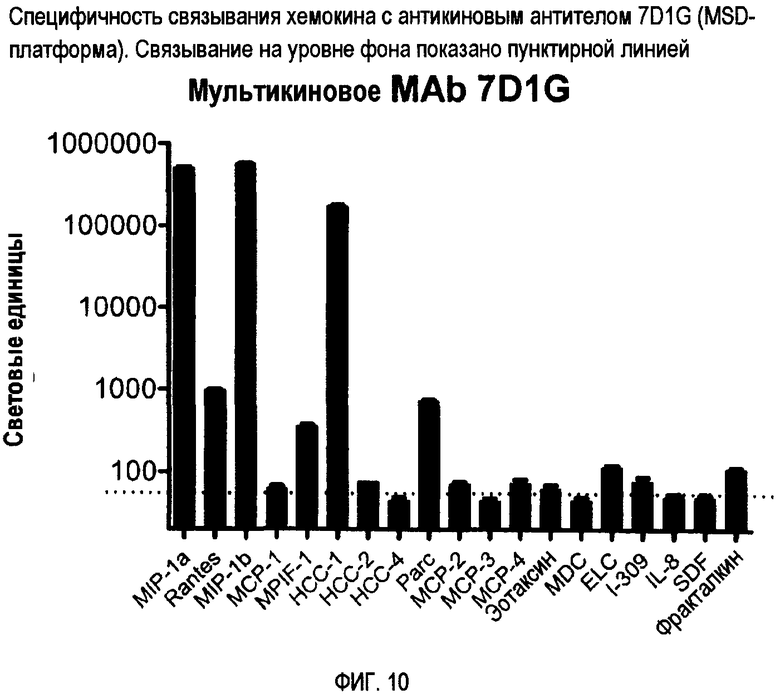
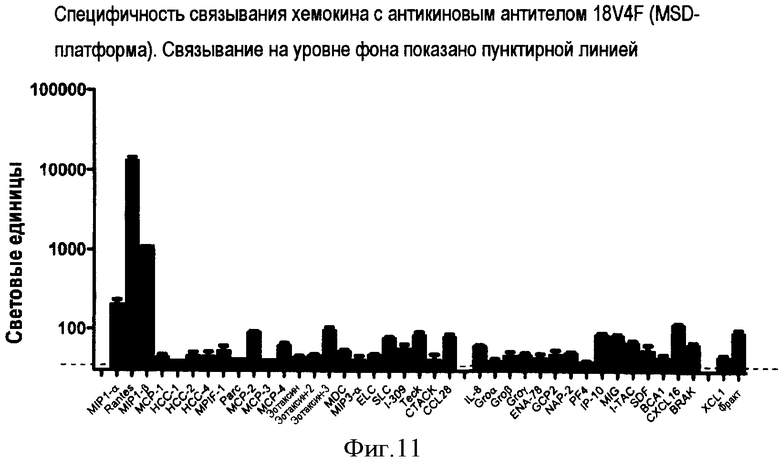
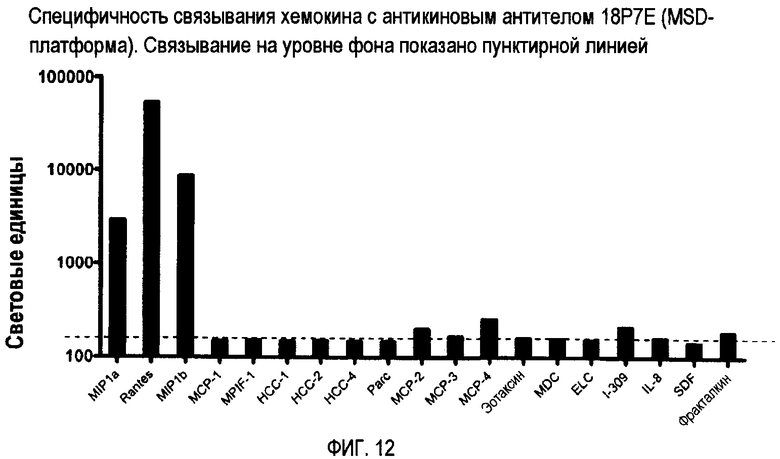
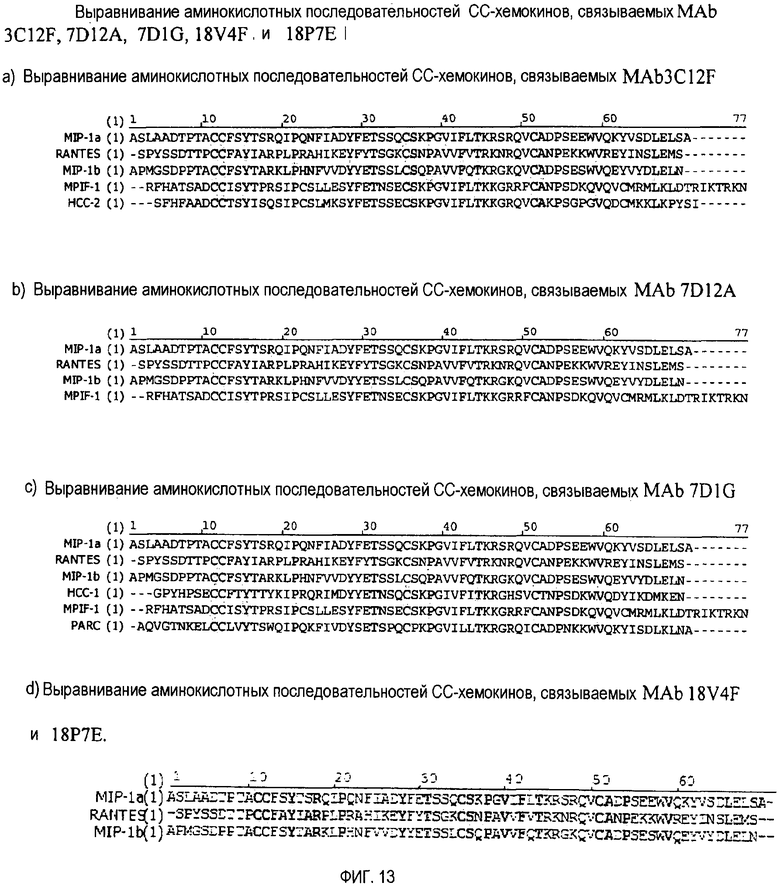
Настоящее изобретение относится к области иммунологии. Предложены моноклональное антитело, которое связывается с по меньшей мере тремя CC-хемокинами, такими как RANTES/CCL5, MIP-1α/CCL3 или MIP-1β/CCL4, и его антигенсвязывающий фрагмент. Также рассмотрены гибридомные линии клеток, продуцирующие антитела по изобретению, композиция, содержащая антитело по изобретению или его антигенсвязывающий фрагмент, и способ лечения заболевания, нарушения или состояния, опосредованного одним или несколькими CC-хемокинами. Данное изобретение может найти дальнейшее применение в регулировании различных воспалительных и аутоиммунных заболеваний. 4 н. и 40 з.п. ф-лы, 13 ил., 5 табл., 10 пр.
1. Моноклональное антитело или его антигенсвязывающий фрагмент, которое связывается с по меньшей мере тремя различными СС-хемокинами, где по меньшей мере один СС-хемокин представляет собой CCL3/MIP-1α, CCL4/MIP-1β или CCL5/RANTES; и
где антитело или антигенсвязывающий фрагмент:
имеет пару аминокислотных последовательностей вариабельных областей легкой и тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 2/7, 12/17, 22/27, 32/37, 42/47, 52/57 и 62/67.
2. Моноклональное антитело по п.1, которое обладает аффинностью связывания, составляющей по меньшей мере 20 нМ в отношении по меньшей мере трех различных СС-хемокинов, где по меньшей мере один СС-хемокин представляет собой CCL3/MIP-1α, CCL4/MIP-1β или CCL5/RANTES.
3. Моноклональное антитело или его антигенсвязывающий фрагмент по п.1, которое связывается с по меньшей мере четырьмя различными СС-хемокинами.
4. Моноклональное антитело или его антигенсвязывающий фрагмент по п.1, которое не связывается с МСР-1, МСР-2 или МСР-3.
5. Моноклональное антитело или его антигенсвязывающий фрагмент по п.1, которое связывается с по меньшей мере тремя различными СС-хемокинами, выбранными из группы, состоящей из CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES, CCL14/HCC-1, CCL15/HCC-2, CCL18/PARC и CCL23/MPIF-1.
6. Моноклональное антитело или антигенсвязывающий фрагмент по п.1, которое связывается с по меньшей мере одной детерминантой из CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES, CCL14/HCC-1, CCL15/HCC-2, CCL18/PARC или CCL23/MPIF-1, которая расположена между остатками СС указанного СС-хемокина и последним остатком С указанного хемокина.
7. Моноклональное антитело или антигенсвязывающий фрагмент по п.1, которое связывается с по меньшей мере одной детерминантой из CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES, CCL14/HCC-1, CCL15/HCC-2, CCL18/PARC или CCL23/MPIF-1, которая расположена в N-петле, 30′s-петле или 40′s-петле указанного СС-хемокина.
8. Моноклональное антитело или антигенсвязывающий фрагмент по п.1, которое связывается с по меньшей мере одной детерминантой СС-хемокина в пределах остатков связывания с СС-рецептором.
9. Моноклональное антитело или антигенсвязывающий фрагмент по п.1, которое нейтрализует хемотаксическую активность СС-хемокина, с которым оно связывается.
10. Моноклональное антитело или антигенсвязывающий фрагмент по п.1, которое продуцируется гибридомной линией клеток 3C12F, 7D1G, 7D12A, 18V4F или 18Р7Е или которое конкурентно блокирует связывание антитела, продуцируемого гибридомной линией клеток 3C12F, 7D1G, 7D12A, 18V4F или 18Р7Е, где
- упомянутая гибридомная линия клеток 3C12F депонирована в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11261,
- упомянутая гибридомная линия клеток 7D1G депонирована в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11257,
- упомянутая гибридомная линия клеток 7D12A депонирована в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11259,
- упомянутая гибридомная линия клеток 18V4F депонирована в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11260, и
- упомянутая гибридомная линия клеток 18Р7Е депонирована в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11258.
11. Моноклональное антитело или его антигенсвязывающий фрагмент по п.1, которое представляет собой гуманизированное антитело, или химерное антитело человека-мыши, или их антигенсвязывающий фрагмент.
12. Моноклональное антитело или его антигенсвязывающий фрагмент по п.1, которое связывается с по меньшей мере пятью СС-хемокинами, выбранными из группы, состоящей из CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES, CCL15/HCC-2 и CCL23/MPIF-1, и которое по существу не связывается с CCL2/MCP-1.
13. Моноклональное антитело или его антигенсвязывающий фрагмент по п.12, которое связывается с детерминантой в N-петле, 30′s-петле или 40′s-петле по меньшей мере одного из указанных СС-хемокинов.
14. Моноклональное антитело или антигенсвязывающий фрагмент по п.12,
которое связывается с по меньшей мере одной антигенной детерминантой CCL3/MIP-1α, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (SRQIPQNF), остатков 34-35 (QC) или остатков 57-67 (EWVQKYVSDLE) SEQ ID NO:71;
которое связывается с по меньшей мере одной антигенной детерминантой CCL4/MIP-1β, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (ARKLPHNF), остатков 34-35 (LC) или остатков 57-67 (SWVQEYVYDLE) SEQ ID NO:72;
которое связывается с по меньшей мере одной антигенной детерминантой CCL5/RANTES, расположенной в пределах остатков 10-14 (CCFAY), остатков 16-23 (ARPLPRAH), остатков 33-34 (КС) или остатков 56-66 (KWVREYINSLE) SEQ ID NO:73.
15. Моноклональное антитело или его антигенсвязывающий фрагмент по п.12, содержащее комбинации CDR последовательностей тяжелых и легких цепей (HCDR1/HCDR2/HCDR3/LCDR1/LCDR2/LCDR3), выбранных из группы, состоящей из SEQ ID NO: 3/4/5/8/9/10 и 53/54/55/58/59/60.
16. Моноклональное антитело или антигенсвязывающий фрагмент по п.12, которое продуцируется гибридомной линией клеток, продуцирующей Mab 3C12F, либо ее субкультурой; где упомянутая гибридомная линия клеток 3C12F депонирована в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11261.
17. Моноклональное антитело или его антигенсвязывающий фрагмент по п.12, которое представляет собой гуманизированное антитело, или химерное антитело человека-мыши, или их антигенсвязывающий фрагмент.
18. Моноклональное антитело или его антигенсвязывающий фрагмент по п.1, которое связывается с по меньшей мере четырьмя СС-хемокинами, выбранными из группы, состоящей из CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES, CCL14/HCC-1 и CCL18/PARC, и которое по существу не связывается с CCL2/MCP-1.
19. Моноклональное антитело или его антигенсвязывающий фрагмент по п.18, которое связывается с детерминантой в N-петле, 30′s-петле или 40′s-петле по меньшей мере одного из указанных СС-хемокинов.
20. Моноклональное антитело или антигенсвязывающий фрагмент по п.18,
которое связывается с по меньшей мере одной антигенной детерминантой CCL3/MIP-1α, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (SRQIPQNF), остатков 34-35 (QC) или остатков 57-67 (EWVQKYVSDLE) SEQ ID NO:71;
которое связывается с по меньшей мере одной антигенной детерминантой CCL4/MIP-1β, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (ARKLPHNF), остатков 34-35 (LC) или остатков 57-67 (SWVQEYVYDLE) SEQ ID NO:72;
которое связывается с по меньшей мере одной антигенной детерминантой CCL5/RANTES, расположенной в пределах остатков 10-14 (CCFAY), остатков 16-23 (ARPLPRAH), остатков 33-34 (КС) или остатков 56-66 (KWVREYINSLE) SEQ ID NO:73.
21. Моноклональное антитело или его антигенсвязывающий фрагмент по п.18, содержащее комбинацию CDR последовательностей тяжелых и легких цепей (HCDR1/HCDR2/HCDR3/LCDR1/LCDR2/LCDR3) SEQ ID NO: 23/24/25/28/29/30.
22. Моноклональное антитело или антигенсвязывающий фрагмент по п.18, которое продуцируется гибридомной линией клеток, продуцирующей Mab 7D1G, либо ее субкультурой; где упомянутая гибридомная линия клеток 7D1G депонирована в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11257.
23. Моноклональное антитело или его антигенсвязывающий фрагмент по п.18, которое представляет собой гуманизированное антитело, или химерное антитело человека-мыши; или их антигенсвязывающий фрагмент.
24. Моноклональное антитело или его антигенсвязывающий фрагмент по п.1, которое связывается с по меньшей мере четырьмя СС-хемокинами, выбранными из группы, состоящей из CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES и CCL23/MPIF-1, и которое по существу не связывается с CCL2/MCP-1.
25. Моноклональное антитело или его антигенсвязывающий фрагмент по п.24, которое связывается с детерминантой в N-петле, 30′s-петле или 40′s-петле по меньшей мере одного из указанных СС-хемокинов.
26. Моноклональное антитело или антигенсвязывающий фрагмент по п.24,
которое связывается с по меньшей мере одной антигенной детерминантой CCL3/MIP-1α, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (SRQIPQNF), остатков 34-35 (QC) или остатков 57-67 (EWVQKYVSDLE) SEQ ID NO:71;
которое связывается с по меньшей мере одной антигенной детерминантой CCL4/MIP-1β, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (ARKLPHNF), остатков 34-35 (LC) или остатков 57-67 (SWVQEYVYDLE) SEQ ID NO:72;
которое связывается с по меньшей мере одной антигенной детерминантой CCL5/RANTES, расположенной в пределах остатков 10-14 (CCFAY), остатков 16-23 (ARPLPRAH), остатков 33-34 (КС) или остатков 56-66 (KWVREYINSLE) SEQ ID NO:73.
27. Моноклональное антитело или его антигенсвязывающий фрагмент по п.24, содержащее комбинацию CDR последовательностей тяжелых и легких цепей (HCDR1/HCDR2/HCDR3/LCDR1/LCDR2/LCDR3) SEQ ID NO: 13/14/15/18/19/20.
28. Моноклональное антитело или антигенсвязывающий фрагмент по п.24, которое продуцируется гибридомной линией клеток, продуцирующей Mab 7D12A, либо ее субкультурой; где упомянутая гибридомная линия клеток 7D12A депонирована в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11259.
29. Моноклональное антитело или его антигенсвязывающий фрагмент по п.24, которое представляет собой гуманизированное антитело, или химерное антитело человека-мыши; или их антигенсвязывающий фрагмент.
30. Моноклональное антитело или его антигенсвязывающий фрагмент по п.1, которое связывается с тремя СС-хемокинами, выбранными из группы, состоящей из CCL3/MIP-1α, CCL4/MIP-1β и CCL5/RANTES, и которое по существу не связывается с CCL2/MCP-1.
31. Моноклональное антитело или его антигенсвязывающий фрагмент по п.30, которое связывается с детерминантой в N-петле, 30′s-петле или 40′s-петле по меньшей мере одного из указанных СС-хемокинов.
32. Моноклональное антитело или антигенсвязывающий фрагмент по п.30,
которое связывается с по меньшей мере одной антигенной детерминантой CCL3/MIP-1α, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (SRQIPQNF), остатков 34-35 (QC) или остатков 57-67 (EWVQKYVSDLE) SEQ ID NO:71;
которое связывается с по меньшей мере одной антигенной детерминантой CCL4/MIP-1β, расположенной в пределах остатков 11-15 (CCFSY), остатков 17-24 (ARKLPHNF), остатков 34-35 (LC) или остатков 57-67 (SWVQEYVYDLE) SEQ ID NO:72;
которое связывается с по меньшей мере одной антигенной детерминантой CCL5/RANTES, расположенной в пределах остатков 10-14 (CCFAY), остатков 16-23 (ARPLPRAH), остатков 33-34 (КС) или остатков 56-66 (KWVREYINSLE) SEQ ID NO:73.
33. Моноклональное антитело или его антигенсвязывающий фрагмент по п.30, содержащее комбинацию CDR последовательностей тяжелых и легких цепей (HCDR1/HCDR2/HCDR3/LCDR1/LCDR2/LCDR3), выбранную из группы, состоящей из SEQ ID NO: 33/34/35/38/39/40 и 63/64/65/68/69/70.
34. Моноклональное антитело или антигенсвязывающий фрагмент по п.30, которое продуцируется гибридомной линией клеток, продуцирующей Mab 18V4F, либо ее субкультурой; где упомянутая гибридомная линия клеток 18V4F депонирована в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11260.
35. Моноклональное антитело или его антигенсвязывающий фрагмент по п.30, которое представляет собой гуманизированное антитело, или химерное антитело человека-мыши, или их антигенсвязывающий фрагмент.
36. Моноклональное антитело или его антигенсвязывающий фрагмент по п.30, содержащее комбинацию CDR последовательностей тяжелых и легких цепей (HCDR1/HCDR2/HCDR3/LCDR1/LCDR2/LCDR3) SEQ ID NO: 43/44/45/48/49/50.
37. Моноклональное антитело или антигенсвязывающий фрагмент по п.30, которое продуцируется гибридомной линией клеток, продуцирующей Mab 18Р7Е, либо ее субкультурой; где упомянутая гибридомная линия клеток 18Р7Е депонирована в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11258.
38. Гибридомная линия клеток, продуцирующая моноклональное антитело по п.1, где гибридомная линия клеток выбрана из группы, состоящей из гибридомной линии клеток 3C12F, депонированной в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11261, гибридомной линии клеток 7D1G, депонированной в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11257, гибридомной линии клеток 7D12A, депонированной в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11259, гибридомной линии клеток 18V4F, депонированной в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11260, и гибридомной линия клеток, 18Р7Е, депонированной в Августе 12, 2010 в Американской коллекции типовых культур (АТСС) под номером доступа РТА-11258.
39. Композиция для лечения заболевания, нарушения или состояния, опосредованного одним или несколькими СС-хемокинами, включающая терапевтически эффективное количество моноклонального антитела или его антигенсвязывающего фрагмента по п.1 в сочетании с носителем, наполнителем или буфером.
40. Композиция по п.39, которая включает более одного моноклонального антитела по п.1.
41. Композиция по п.39, в которой указанное моноклональное антитело обладает аффинностью связывания, составляющей по меньшей мере 20 нМ в отношении по меньшей мере трех различных СС-хемокинов; где по меньшей мере один СС-хемокин представляет собой CCL3/MIP-1α, CCL4/MIP-1β или CCL5/RANTES.
42. Способ лечения заболевания, нарушения или состояния, опосредованного одним или несколькими СС-хемокинами, включающий введение субъекту, нуждающемуся в этом, моноклонального антитела по п.1 или его антигенсвязывающего фрагмента.
43. Способ по п.42, где указанное заболевание, нарушение или состояние представляет собой воспалительное заболевание или заболевание, опосредованное или связанное с воспалением.
44. Способ по п.42, где указанное заболевание, нарушение или состояние представляет собой аутоиммунное заболевание.

