Уровень изобретения
Люди выращивают кукурузу для использования в качестве пищи и источника энергии. Насекомые поедают и повреждают сельскохозяйственные растения и таким образом подрывают усилия человека.
В настоящее время применяют имплантируемый трансгенный контроль этих вредителей посредством экспрессии растением гена дельта-эндотоксина Crystal (Cry), кодирующего белок Cry1Fa из Bacillus thuringiensis. Белок Cry1Fa представляет собой современный токсин марки Herculex™ от производителя трансгенных семян сельскохозяйственных культур Dow AgroSciences (Herculex, Herculex-Extra и Herculex-RW), которые обладают устойчивостью к насекомым-вредителям, а именно, к кукурузной листовой совке (FAW, Spodoptera frugiperda) и европейскому кукурузному мотыльку (ECB, Ostrinia nubilalis). Указанный белок действует путем связывания со специфичным рецептором (рецепторами), расположенным в средней кишке насекомых, и образует поры в клетках кишечника. Образование этих пор препятствует регуляции осмотического равновесия у насекомых, что вызывает их гибель.
Вместе с тем, существует проблема, связанная с возможным развитием у насекомых устойчивости к действию Cry1Fa посредством генетических изменений связывающихся с Cry1Fa кишечных рецепторов насекомых. Насекомые, имеющие рецепторы с уменьшенной способностью связываться с Cry1Fa, могут быть устойчивыми к действию Cry1Fa, и таким образом, выживать на растениях, которые экспрессируют этот белок.
В период роста у растения постоянно присутствует единственный токсин Cry, и в этой связи возникает проблема возможности развития у насекомых устойчивости к действию упомянутого белка посредством генетических изменений рецептора, который связывается с токсином Cry1Fa в кишечнике насекомого. Снижение связывания с токсином, обусловленное такими изменениями в рецепторе, будет вызывать уменьшение токсичности Cry1Fa, что может приводить к вероятному снижению эффективности белка, которое экспрессирует сельскохозяйственное растение. См., например, патент США 2009 0313717, который рассматривает белок Cry2 плюс Vip3Aa, Cry1F или Cry1A для контроля Helicoverpa zea или armigera. Патент WO 2009 132850 относится к белкам Cry1F или Cry1A и Vip3Aa для контроля Spodoptera frugiperda. Патент США 2008 0311096 относится к Cry1Ab для контроля ECB с устойчивостью к Cry1F.
Дополнительные токсины Cry перечислены на вебсайте комитета по официальной номенклатуре B.t. (Crickmore et al.; lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/). См. Приложение A к указанному источнику. В настоящее время существует около 60 основных групп токсинов "Cry" (Cry1-Cry59), с дополнительными токсинами Cyt, токсинами VIP и подобными токсинами. Многие из этих многочисленных групп имеют подгруппы, обозначаемые прописными буквами, и в подгруппах с прописными буквами выделяют подподгруппы, обозначаемые строчными буквами (например, Cry1 имеет подгруппы A-L, и в подгруппе Cry1A выделяют подподгруппы а-i).
В публикации van Frankenhuyzen (2009) (J. Invert. Pathol. 101:1-16), например, показано, что существует множество целевых вредителей и большое количество токсинов, которые потенциально можно выбирать для контроля целевых вредителей. См., например, фигуру 3, van Frankenhuyzen. В число многих вредителей, которые могут быть мишенью, включен один из них, Ostrinia nubilalis, и для указанного насекомого показано 17 токсинов, обладающих активностью против ECB, и один токсин с вероятной активностью (фигура 3, van Frankenhuyzen). Этот список вариантов не является исчерпывающим.
На фигуре 3 (van Frankenhuyzen) также показано, что каждый белок Cry имеет уникальный спектр действия, то есть указанные белки обладают активностью против одних насекомых и не активны против других. Белки Cry обычно связываются с рецепторами на клетках кишечника насекомого, и это представляет собой один из факторов, который влияет на спектр действия. Рецепторы для одного белка Cry могут присутствовать у одних насекомых, и отсутствовать у других; у конкретного насекомого могут быть рецепторы для одного или больше белков Cry, и отсутствовать рецепторы для других белков Cry.
Учитывая, что множество насекомых может быть мишенью и что множество белков Cry могут активно действовать против любого данного насекомого, сложность проблемы управления устойчивостью насекомых демонстрируется даже одними цифрами. Принимая во внимание, что только в исследованиях van Frankenhuyzen были идентифицированы 18 белков в качестве активных или возможно активных против ECB, существует возможность тестирования комбинаций из сотен возможных пар токсинов.
Дополнительно, нелегкой задачей является исследование конкурентного/неконкурентного связывания. Такие исследования могут охватывать радиоактивные метки и анализ перемещения радиомеченых белков. Такие анализы представляют собой сложные методики.
Также сложной задачей является попытка непосредственного использования устойчивых насекомых. Необходимо выводить штаммы насекомых, устойчивые к заданному белку. В публикации Siqueira (июнь 2004г., J. Econ. Entomol, 97 (3): 1049-1057) утверждается (реферативно), что "...тесты на перекрестную устойчивость среди различных токсинов лимитированы нехваткой устойчивых колоний". Это является примером проблемы, связанной с получением устойчивых штаммов насекомых в целях оценки потенциала белков по управлению устойчивостью. Для скрининга развития устойчивости насекомых можно использовать любой белок, задействуя пару белков.
В упомянутой публикации Siqueira также утверждает, что селекция с Cry1Ab (то есть, выведение колоний ECB, обладающих устойчивостью к Cry1Ab), "... вызывала снижение восприимчивости ко многим другим токсинам...". Это является проявлением феномена перекрестной устойчивости. Cry1Ab-устойчивые ECB обладают перекрестной устойчивостью ко "многим другим токсинам".
Таким образом, исходным аспектом анализа для решения проблемы устойчивости является просто селекция двух белков, обладающих активностью против одного и того же насекомого (не проявляющего устойчивость). Другой фактор представляет собой уровень активности против неустойчивых насекомых. Фигура 11 (van Frankenhuyzen) показывает, что даже среди группы из 12 токсинов, отобранных для тестирования против ECB (не проявляющих устойчивость), другие белки Cry (такие как Cry1Ac, Cry1Bb и Cry2Aa) могли быть более активными, чем белки, указанные в формуле изобретения для контроля ECB.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение частично относится к комплексу белков Cry1Be с белками Cry1Fa, который создает продукты, более стабильные и менее подверженные развитию устойчивости насекомых к любому из отдельных упомянутых белков.
Как более подробно описано в изобретении, рассматриваемая пара белков представляет собой комбинацию с особым преимуществом, поскольку никакая другая известная пара белков не обеспечивает высокий уровень контроля и действие против насекомых Spodoptera frugiperda (FAW) и против Ostrinia nubilalis (ECB) без перекрестной устойчивости.
Это двойное действие без перекрестной устойчивости также имеет преимущество, поскольку с помощью многоцелевых белков без перекрестной устойчивости можно сократить количество белков/генов, необходимое для воздействия на упомянутых насекомых. При этом можно уменьшить необходимую для посевов «убежищ» площадь земли в акрах, или устранить необходимость в площадях под «убежища». Соответственно, настоящее изобретение также относится в целом к использованию четырех генов для получения трех белков для контроля FAW без перекрестной устойчивости, и трех белков для контроля ECB без перекрестной устойчивости.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКОГО МАТЕРИАЛА
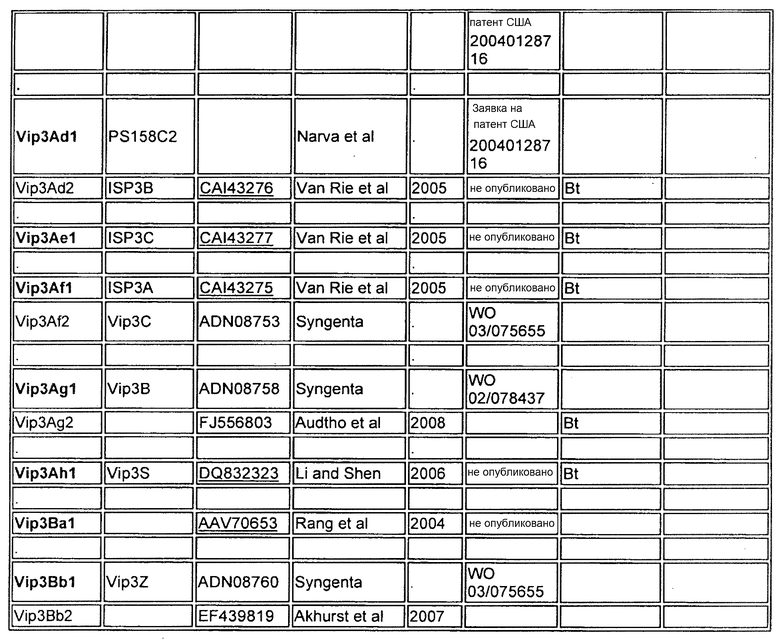
Фиг.1 показывает конкурентное связывание меченого 125I Cry1Fa по сравнению со связыванием Cry1Fa или Cry1Be с мембранными везикулами щеточной каймы, полученными от Spodoptera frugiperda (кукурузная листовая совка, FAW).
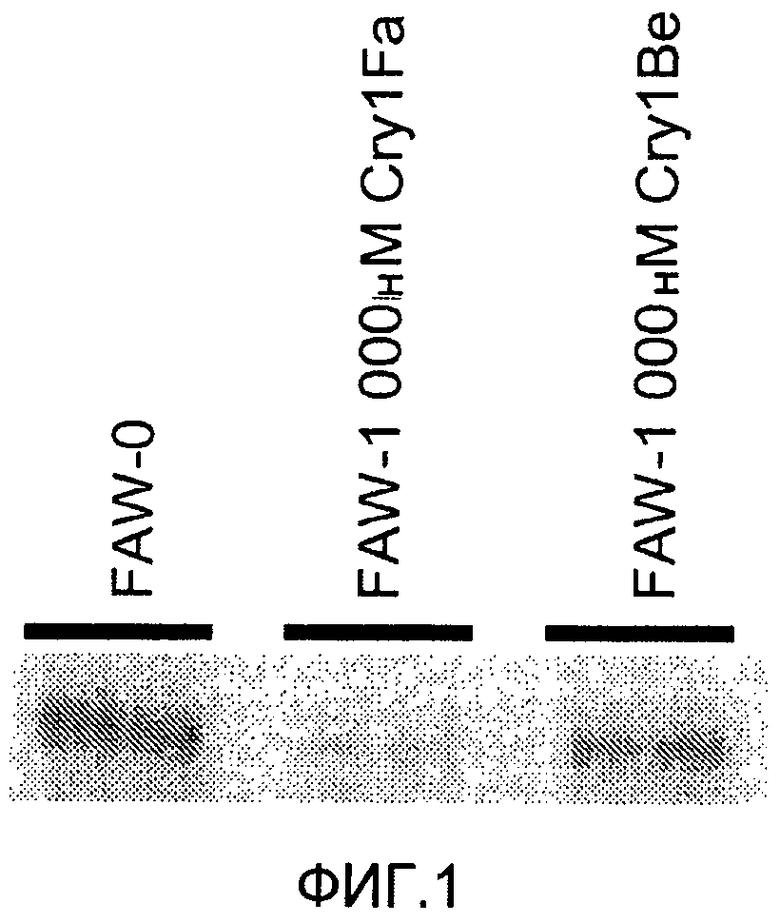
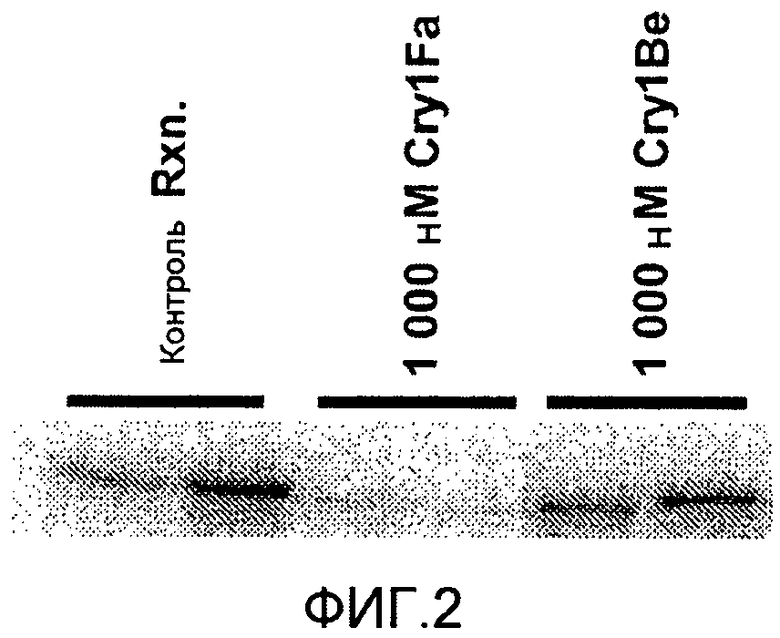
Фиг.2 показывает конкурентное связывание меченого 125I Cry1Fa по сравнению со связыванием Cry1Fa или Cry1Be с мембранными везикулами щеточной каймы, полученными из Ostrinia nubilalis (европейский кукурузный мотылек, ЕСВ).
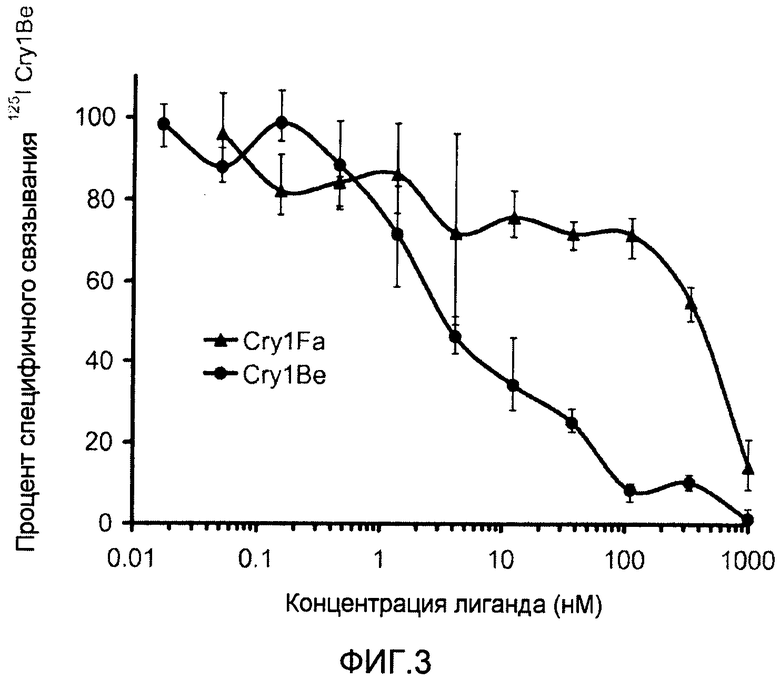
Фиг.3 показывает конкурентное смещение связывания меченого 125I Cry1Be с мембранными везикулами щеточной каймы, полученными из Spodoptera frugiperda Cry1Fa (▲) и Cry1Be (•).
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение включает в себя применение белков Cry1Be с белками Cry1Fa в спаренном виде. Настоящее изобретение также частично относится к тройным комплексам или "пирамидам" из трех (или больше) токсинов с белками Cry1Fa и Cry1Be в качестве основной пары. Рассматриваемая основная пара белков представляет два белка, обеспечивающие действие без перекрестной устойчивости против двух насекомых, а именно кукурузной листовой совки (FAW; Spodoptera frugiperda) и европейского кукурузного мотылька (ECB; Ostrinia nubilalis). Это делает рассматриваемую пару белков комбинацией с особыми преимуществами, поскольку ни одна из других известных пар белков не обеспечивает высокий уровень контроля и действие против двух указанных насекомых без перекрестной устойчивости.
В некоторых предпочтительных вариантах изготовления пирамиды к основной паре настоящего изобретения можно добавлять другой белок, чтобы получать третий белок, обладающий активностью против ECB. Некоторые из этих предпочтительных пирамидных комбинаций представляют собой белок Cry1Fa плюс белок Cry1Be плюс другой токсин/ген, выбираемый из группы, состоящей из белков Cry1Ab, Cry2Aa, Cry1I и DIG-3.
В некоторых предпочтительных вариантах осуществления и в свете действия обоих белков Cry1F и Cry1Be как против ECB, так и против FAW, настоящее изобретение дает возможность применять четыре белка, при этом три из этих четырех белков обладают действием против ECB без перекрестной устойчивости, и три из этих четырех белков обладают неконкурентным действием против FAW. Предпочтительные комплексы из четырех партнеров представляют собой Cry1Fa плюс Cry1Be плюс: Cry1C, Cry1D, Cry1E или Vip3 (предназначенные для FAW), плюс Cry1Ab, Cry2A, Cry1I или DIG-3 (предназначенные для ECB).
Одновременно зарегистрированная заявка под названием "Использование Vip3Ab для контроля устойчивых насекомых" ("Use of Vip3Ab for management of resistant insects") представляет данные, демонстрирующие полезность Vip3Ab вместе с Cry1F для управления устойчивостью FAW к инсектицидным белкам, и показывает, что Vip3Ab и Cry1F конкурентно не связываются с препаратами мембран FAW.
В заявке US 61/284281 (подана 16 декабря 2009 г.) показано, что Cry1C является активным против Cry1F-устойчивых FAW, и в заявке US 61/284252 (подана 16 декабря 2009 г.) показано, что Cry1D является активным против Cry1F-устойчивых FAW. В этих двух заявках также указано, что Cry1C не конкурирует с Cry1F за связывание в препаратах мембран FAW и что Cry1D не конкурирует с Cry1F за связывание в препаратах мембран FAW.
В заявке US 61/284278 (подана 16 декабря 2009 г.) приведены сведения об активности Cry2A против Cry1F-устойчивых ECB.
В патенте США 2008 0311096 раскрыта полезность Cry1Ab для контроля Cry1F-устойчивых ECB.
В патенте США 2010 0269223 раскрыт белок DIG-3.
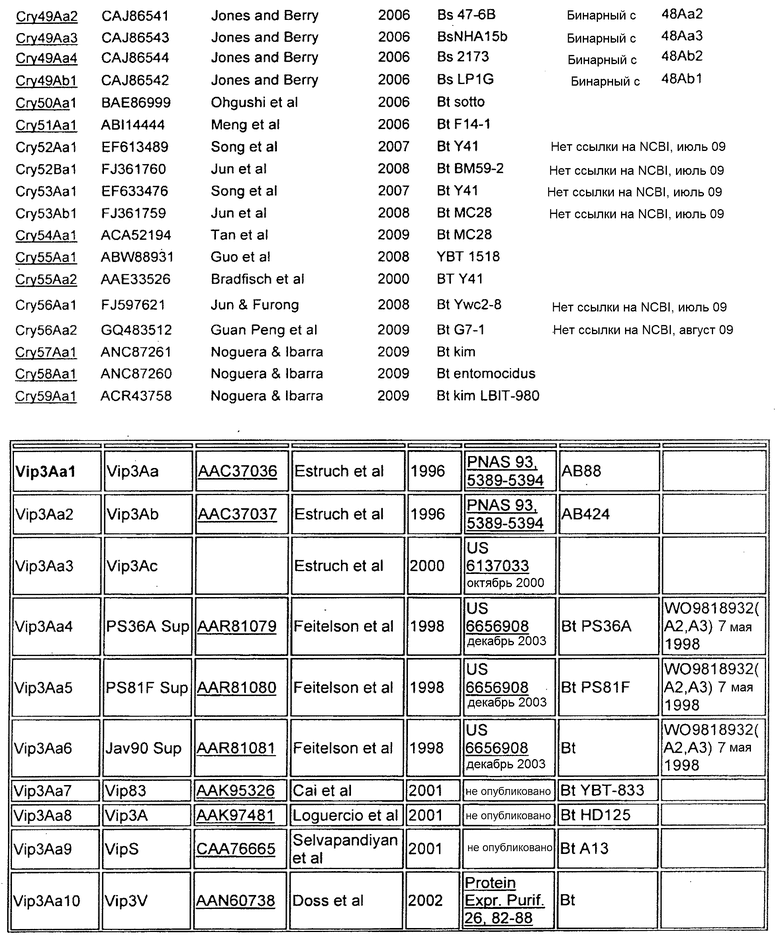
В Приложении A настоящего изобретения перечислены, например, токсины Vip3 (включающие в себя в некоторых предпочтительных вариантах осуществления Vip3Ab). Также перечислены белки Cry. Упомянутые номера GENBANK также можно использовать для получения последовательности любого из генов и белков, раскрытых или упомянутых в изобретении.
Настоящее изобретение также относится в целом к применению трех инсектицидных белков (белков Cry в некоторых предпочтительных вариантах осуществления), которые не вызывают перекрестную устойчивость друг с другом против единственного целевого вредителя. Настоящее изобретение в целом также относится к использованию четырех инсектицидных белков (белки Cry и Vip в некоторых предпочтительных вариантах осуществления), комбинация которых обеспечивает высокий уровень контроля и действие против двух целевых насекомых без перекрестной устойчивости.
В объем настоящего изобретения включены растения (и площадь земли в акрах под такими растениями), которые продуцируют комбинации рассматриваемых белков. Также можно добавлять дополнительные токсины/гены, но предпочтительные комплексы белок/ген (из трех и четырех партнеров), согласно настоящему изобретению, неожиданно будут иметь преимущество для получения трех белков с неконкурентным действием против FAW и/или ECB. Это может способствовать уменьшению или устранению необходимости для выделения земли (в акрах) под площади для «убежища» (например, площадь «убежища» будет менее 40%, менее 20%, менее 10%, менее 5% или даже будет составлять 0%). Таким образом, установленная площадь поля более 10 акров входит в объем настоящего изобретения.
Полинуклеотид (полинуклеотиды) по настоящему изобретению, предпочтительно, находятся в генетической конструкции под контролем промотора, не происходящего из Bacillus thuringiensis (функционально связанного с ним/содержащими его). Рассматриваемые полинуклеотиды могут содержать кодон, используемый для усиления экспрессии в растении.
Для борьбы со способностью насекомых развивать устойчивость к белку Cry1Fa авторы определили токсины Cry, которые неконкурентно (по отношению к Cry1Fa) связываются с рецепторами белка. Белок Cry1Fa не перемещает связывание Cry1Be с рецепторами, расположенными в кишечнике насекомых FAW и личинок ECB. Авторы обнаружили, что Cry-белки Cry1Be или взаимодействуют с абсолютно разными рецепторами, или их действие только частично накладывается на взаимодействие с рецепторами по сравнению с Cry1Fa. Способность упомянутых токсинов Cry1Be быть токсичными для личинок FAW и ECB, при неполном взаимодействии с теми же участками рецептора, что и белки Cry1Fa, показывает, что на их токсичность не будут влиять развивающиеся у насекомых генетические изменения их рецептора Cry1Fa, что представляет собой механизм развития устойчивости к токсичности Cry1Fa. Таким образом, насекомые, у которых развивается устойчивость к Cry1Fa посредством снижения способности рецепторов кишечника этих насекомых к связыванию с Cry1Fa, тем не менее, будут восприимчивы к токсичности белков Cry1Be, которые связываются с альтернативными участками рецептора. В подтверждение этого авторы получили биохимические данные.
Таким образом, наличие комбинаций упомянутых белков, экспрессируемых трансгенными растениями, будет полезным и ценным механизмом для уменьшения вероятности развития устойчивости насекомых в поле, что приведет к сокращению требуемых посевов под «убежища». Белки Cry1Be изучались в отношении их активности против других основных насекомых-вредителей как восприимчивых, так и устойчивых к Cry1Fa (rFAW и rECB); в таблице 1 показано, что Cry1Be обладает активностью и против устойчивых и против восприимчивых личинок ECB. Эти данные показывают, что токсин Cry1Be взаимодействует на отдельном целевом участке(участках) в кишечнике насекомого, по сравнению с Cry1Fa, и таким образом создаются превосходные партнеры по комплексу.
Создание комплексов белков Cry1Fa, которые экспрессируются сельскохозяйственными культурами, с одним или больше дополнительными генами Cry, например, с генами, которые экспрессируют белки-токсины Cry1Be, даст в результате эффективную управляющую стратегию по предотвращению способности насекомых развивать переносимость к действию трансгенных растений, экспрессирующих эти белки-токсины. Авторы показали, что белки Cry1Be и белки Cry1Fa взаимодействуют на разных участках, и если устойчивость должна возникать посредством изменения аффинности рецепторов в кишечнике насекомого, которые связываются с токсинами Cry, изменение должно возникать по меньшей мере одновременно в двух разных рецепторах, и это позволит насекомым выживать на растениях, экспрессирующих многоцелевые белки. Вероятность возникновения таких изменений чрезвычайно мала, таким образом, повышается устойчивость трансгенного продукта к отдельной популяции насекомых, которые способны развивать переносимость упомянутых белков.
Авторы готовили меченые радиоактивным йодом усеченные трипсином формы белков-токсинов Cry1Be и использовали технологии анализа радиорецепторного связывания для измерения их взаимодействия в связывании с предлагаемыми рецепторными белками, расположенными в мембранах кишечника насекомого. Препараты кишечных мембран изготовляли из мембранных везикул щеточной каймы (BBMV) по способу Wolfersberger. Йодирование токсинов осуществляли или с помощью Iodo-beads, или в пробирках путем обработки Iodogen, от фирмы Pierce Chemicals. Специфичная активность радиомеченого токсина составляла примерно 1-4 мкКи/мг белка. Исследования связывания проводили в основном согласно методикам Liang.
Далее в разделе примеров также представлены дополнительные данные по конкурентному связыванию с использованием меченого Cry1Fa. Эти данные также показывают действие без перекрестной устойчивости белков Cry1Fa и Cry1Be как против ECB, так и против FAW.
Данные, представленные в изобретении, указывают на взаимодействие белков Cry1Be на отдельном целевом участке в кишечнике насекомого по сравнению с Cry1Fa. Таким образом, эти два белка являются превосходными партнерами по комплексу.
Гены и токсины, полезные согласно настоящему изобретению, включают в себя не только полноразмерные раскрытые в изобретении последовательности, но также и фрагменты этих последовательностей, варианты, мутанты и слитые белки, которые сохраняют свойства пестицидной активности токсинов, конкретно описанных в изобретении в качестве примеров. Используемые в изобретении термины "варианты" или "изменения" генов относятся к нуклеотидным последовательностям, которые кодируют одинаковые токсины, или которые кодируют эквивалентные токсины, обладающие пестицидным действием. Используемый в изобретении термин "эквивалентные токсины" относится к токсинам, имеющим одинаковое или по существу одинаковое биологическое действие против целевых вредителей, как и токсины, указанные в формуле изобретения.
Используемые в настоящем изобретении пределы идентичности последовательностей составляют около 95% (белки Cry1Fa и 1Be), 78% (Cry1F и Cry1B) и 45% (Cry1), см. публикации "Revision of the Nomenclature for the Bacillus thuringiensis Pesticidal Crystal Proteins", N. Crickmore, D.R. Zeigler, J. Feitelson, E. Schnepf, J. Van Rie, D. Lereclus, J. Baum, и D.H. Dean. Microbiology and Molecular Biology Reviews (1998) Vol 62: 807-813. Эти пределы также можно применять только для коровых токсинов (например, для токсинов Cry1Fa и CryBe).
Фрагменты и эквиваленты, которые сохраняют пестицидное действие рассматриваемых токсинов, входят в объем настоящего изобретения. Кроме того, благодаря избыточности генного кода множество разных последовательностей ДНК может кодировать раскрытые в изобретении аминокислотные последовательности. Специалистам в данной области техники будут очевидны способы создания таких альтернативных последовательностей ДНК, кодирующих одинаковые или по существу аналогичные токсины. Эти вариантные последовательности ДНК входят в объем настоящего изобретения. Используемое в изобретении понятие "по существу аналогичная" последовательность относится к последовательностям, имеющим аминокислотные замены, делеции, добавления или инсерции, которые фактически не влияют на пестицидное действие. В это определение также включены фрагменты генов, которые кодируют белки, сохраняющие пестицидное действие.
Использование олигонуклеотидных зондов является дополнительным способом идентификации генов, кодирующих токсины, и генных участков, полезных согласно настоящему изобретению. Эти зонды представляют собой обнаружимые нуклеотидные последовательности. Детекцию таких последовательностей можно проводить с помощью подходящей метки или можно задействовать их флуоресцентные свойства, как описано в международной патентной заявке WO 93/16094. Как известно в данной области техники, если при гибридизации молекулы зонда и образца нуклеиновой кислоты образуется прочная связь между этими двумя молекулами, то можно обоснованно предполагать наличие значительной гомологии зонда и образца. Предпочтительно, проводить гибридизацию при жестких условиях способами, известными в данной области техники, которые описаны, например, авторами Keller, G. H., M. M. Manak (1987) DNA Probes, Stockton Press, New York, N.Y., pp. 169-170. Некоторые примеры комбинаций показателей концентраций солей и температуры представляют собой (в порядке увеличения степени жесткости): 2 X SSPE или SSC (раствор хлорида и цитрата натрия) при комнатной температуре; 1 X SSPE или SSC при 42°C; 0,1 X SSPE или SSC при 42°C; 0,1 X SSPE или SSC при 65°C. Детекция зонда представляет собой общепринятую методику определения факта проведения гибридизации. Такой анализ зондов обеспечивает быстрый способ идентификации кодирующих токсины генов настоящего изобретения. Нуклеотидные сегменты, которые используются в качестве зондов согласно изобретению, можно синтезировать с помощью синтезатора ДНК и стандартных методик. Указанные нуклеотидные последовательности можно также использовать в качестве ПЦР-праймеров для амплификации генов настоящего изобретения.
Конкретные белки настоящего изобретения приведены в изобретении в качестве конкретных примеров. Поскольку упомянутые белки являются просто примерами белков настоящего изобретения, будет очевидно, что настоящее изобретение содержит варианты или эквиваленты белков (и нуклеотидных последовательностей, кодирующих эквивалентные белки), обладающие пестицидным действием, которое одинаково или сходно с действием рассматриваемого белка. Эквивалентные белки и белок по изобретению имеют аминокислотную гомологию. Эта аминокислотная гомология обычно превышает 75%, предпочтительно, составляет более 90% и, наиболее предпочтительно, более 95%. Наиболее выраженную аминокислотную гомологию выявляют в самых важных участках белка, которые обуславливают биологическое действие или вовлечены в предопределение трехмерной конфигурации, которая в конечном счете отвечает за биологическое действие. В этом отношении подходят конкретные замены аминокислоты, и можно предполагать, что эти замены находятся в областях, не имеющих решающего значения для действия, или они представляют собой консервативные аминокислотные замены, которые не затрагивают трехмерную конфигурацию молекулы. Например, аминокислоты могут относиться к следующим классам: неполярные, незаряженные полярные, кислотные и основные. Консервативные замены, посредством которых аминокислоту одного класса заменяют на другую аминокислоту того же типа, входят в объем настоящего изобретения, если в результате такой замены не происходит значительного изменения биологической активности соединения. Далее приведен перечень примеров аминокислот, относящихся к каждому классу. В некоторых случаях можно делать консервативные замены. Решающим фактором является то, что такая замена в значительной степени не изменяет биологическую активность белка.
Трансформация растений. Предпочтительным рекомбинантным хозяином, продуцирующим инсектицидные белки настоящего изобретения, является трансформированное растение. Гены, кодирующие Bt белки-токсины, раскрытые в изобретении, можно вставлять в клетки растения, используя множество способов, известных в данной области техники. Например, для подготовки к вставке чужих генов в высшие растения доступно большое количество векторов клонирования, содержащих систему репликации в Escherichia coli и маркер, позволяющий проводить селекцию трансформированных клеток. Векторы содержат, например, среди прочего, pBR322, серии pUC, серии M13mp, pACYC184. Соответственно, фрагмент ДНК, несущий последовательность, кодирующую белок-токсин Bt, можно вставлять в вектор в подходящий сайт рестрикции. Полученную плазмиду используют для трансформации в E. coli. Клетки E. coli выращивают в подходящей питательной среде, затем собирают и лизируют. Восстанавливают плазмиду. В качестве способов анализа обычно проводят анализ последовательностей, анализ рестрикции, электрофорез и другие биохимические и молекулярно-биологические тесты. После каждой манипуляции используемую последовательность ДНК можно расщеплять и соединять со следующей последовательностью ДНК. Каждую последовательность плазмиды можно клонировать в той же плазмиде или в других плазмидах. В зависимости от способа вставки желательных генов в растение могут потребоваться другие последовательности ДНК. Например, если для трансформации растительной клетки используют плазмиду Ti или Ri, то по меньшей мере правую границу, но часто и правую, и левую границу Ti или Ri плазмиды T-ДНК необходимо соединять в качестве фланкирующей области генов, предназначенных для вставки. Проведены интенсивные исследования использования T-ДНК для трансформации растительных клеток, что в достаточном объеме описано в патенте EP 120516, Lee and Gelvin (2008), Hoekema (1985), Fraley et al., (1986) и An et al., (1985), и хорошо известно в данной области техники.
После включения в геном растения вставленная ДНК является относительно стабильной. Вектор трансформации обычно содержит селектируемый маркер, который придает трансформированным растительным клеткам резистентность к биоциду или антибиотику, такому как биалафос, канамицин, G418, блеомицин или гигромицин, среди прочего. Конкретный используемый маркер должен соответственно позволять отбор трансформированных клеток, а не клеток, не содержащих ДНК-вставку.
Для вставки ДНК в растительную клетку-хозяин доступно большое количество способов. Эти способы включают в себя трансформацию с T-ДНК, с использованием в качестве агента трансформации Agrobacterium tumefaciens или Agrobacterium rhizogenes, слияние, инъекцию, биолистику (бомбардировку микрочастицами) или электропорацию, а также другие возможные способы. Если для трансформации используют Agrobacteria, вставляемую ДНК необходимо клонировать в специальные плазмиды, а именно или в промежуточный вектор, или в бинарный вектор. Промежуточные векторы можно интегрировать в плазмиды Ti или Ri путем гомологичной рекомбинации, обусловленной последовательностями, которые являются гомологичными последовательностям в T-ДНК. Плазмиды Ti или Ri также содержат vir-область, необходимую для переноса T-ДНК. Промежуточные векторы не могут самостоятельно реплицироваться в Agrobacteria. Промежуточный вектор можно переносить в Agrobacterium tumefaciens посредством хелперной плазмиды (конъюгация). Бинарные векторы могут реплицироваться и в E. coli, и в Agrobacteria. Они содержат ген маркера селекции и линкер или полилинкер, которые ограничены правыми и левыми пограничными областями T-ДНК. Они могут быть трансформированы непосредственно в Agrobacteria (Holsters et al., 1978). Используемая в качестве клетки-хозяина Agrobacterium, должна содержать плазмиду, несущую vir-область. Эта vir-область необходима для переноса T-ДНК в клетку растения. Может присутствовать дополнительная T-ДНК. Трансформированная указанным путем бактерия используется для трансформации клеток растения. Растительные экспланты, предпочтительно, можно культивировать с Agrobacterium tumefaciens или Agrobacterium rhizogenes для переноса ДНК в клетку растения. Затем можно восстанавливать целые растения из инфицированного растительного материала (например, из частей листьев, сегментов стеблей, корней, а также из протопластов или суспензии культивированных клеток) в подходящей среде, которая может содержать антибиотики или биоциды для селекции. Полученные указанным путем растения затем можно тестировать на присутствие вставки ДНК. Специальных требований к плазмидам в случае инъекции и электропорации не существует. Можно использовать обычные плазмиды, например, такие как производные pUC.
Трансформированные клетки растут внутри растений обычным путем. Они могут формировать зародышевые клетки и передавать трансформированный признак (признаки) растениям-потомкам. Такие растения можно выращивать обычным путем и скрещивать с растениями, которые имеют те же самые трансформированные наследственные факторы или другие наследственные факторы. Получаемые гибридные субъекты имеют соответствующие фенотипические свойства.
В предпочтительном варианте осуществления настоящего изобретения растения трансформируют генами, в которых используемый кодон был оптимизирован для растений. См., например, патент США № 5380831, включенный в изобретение путем ссылки. Некоторые усеченные токсины рассмотрены в изобретении в качестве примера, вместе с тем в области техники Bt-белков известно, что токсины 130 кДа-типа (полноразмерные) имеют N-концевую группу, которая представляет собой коровый токсин, и C-концевой остаток, который является "хвостом" протоксина. Таким образом, соответствующие "хвосты" можно использовать с усеченными/коровыми токсинами настоящего изобретения. См., например, патент США № 6218188 и патент США № 6673990. Дополнительно, в данной области техники известны способы создания синтетических Bt-генов для использования в растениях (Stewart and Burgin, 2007). Одним из неограничивающих примеров предпочтительного трансформированного растения является фертильное растение кукурузы, содержащее растительный экспрессируемый ген, кодирующий белок Cry1Fa, и дополнительно содержащее второй растительный экспрессируемый ген, кодирующий белок Cry1Ca.
Перенос (или вставку) признака(признаков) Cry1Fa и Cry1Ca в инбредные линии кукурузы можно осуществлять размножением путем рекуррентной селекцией, например, с помощью обратного скрещивания. В этом случае, желательный рекуррентный родитель вначале скрещивают с инбредным донором (нерекуррентным родителем), который несет соответствующий ген(гены) для признаков Cry1F и Cry1C. Потомство от этого скрещивания затем обратно спаривают с рекуррентным родителем, с последующей селекцией получаемого потомства на желательный признак(признаки), переносимый от нерекуррентного родителя. Через три, предпочтительно, четыре, более предпочтительно, через пять или больше поколений обратных скрещиваний с рекуррентным родителем с селекцией на желательный признак(признаки), потомство будет гетерозиготным по локусам, отвечающим за переносимый признак(признаки), но будет похожим на рекуррентного родителя по большинству или почти по всем остальным генам (см., например, Poehlman & Sleper (1995) Breeding Field Crops, 4th Ed., 172-175; Fehr (1987) Principles of Cultivar Development, Vol. 1: Theory and Technique, 360-376).
Стратегии управления устойчивостью насекомых (IRM). Авторы Roush et al., например, выделяют стратегию двух токсинов, также называемый "пирамидинг" или "комплекс", для управления инсектицидными трансгенными сельскохозяйственными культурами (The Royal Society. Phil. Trans. R. Soc. Lond. B. (1998) 353, 1777-1786).
На вебсайте Управления по охране окружающей среды США (United States Environmental Protection Agency: epa.gov/oppbppdl/biopesticides/pips/bt_corn_refuge_2006.htm) опубликованы следующие требования для обеспечения посевов нетрансгенных (то есть, не-Bt) «убежищ» (блок не-Bt сельскохозяйственных культур/кукурузы), высеваемый вместе с трансгенными зерновыми культурами, продуцирующими единственный Bt белок, активный против целевых вредителей.
Существуют следующие конкретные структурированные требования для кукурузной продукции, Bt-защищенной от кукурузного мотылька (Cry1Ab или Cry1F):
Структурированные «убежища»:
20% площади в кукурузной зоне отводят под «убежище» для Bt-незащищенной от чешуекрылых вредителей кукурузы
50% площади в хлопковой зоне отводят для «убежища» культур, Bt-незащищенных от чешуекрылых вредителей
Блоки
1. Внутренние (то есть в пределах Bt-поля)
2. Внешние (то есть отдельные поля в пределах ½ мили (¼ мили, по возможности) от Bt-поля, для максимального увеличения частоты случайного спаривания)
Полосы внутри поля
Ширина полос должна составлять по меньшей мере 4 ряда (предпочтительно, 6 рядов), для уменьшения эффекта перемещения личинок.
Дополнительно, на вебсайте Национальной ассоциации производителей кукурузы (ncga.com/insect-resistance-management-fact-sheet-bt-com) также опубликовано подобное руководство в отношении требований. Например:
Требования IRM по кукурузному мотыльку:
- По меньшей мере 20% площадей отводят под выращивание «убежищ» гибридной кукурузы;
- В зонах выращивания хлопка «убежища» должны составлять 50%;
- Выращивание должно быть на расстоянии в пределах 1/2 мили от гибридов «убежища»;
- «Убежища» можно выращивать в виде полос внутри Bt-поля; полосы «убежища» должны иметь ширину по меньшей мере 4 ряда;
- «Убежище» можно обрабатывать обычными пестицидами только в случае достижения экономического порога для целевого насекомого;
- В «убежище» кукурузы нельзя применять распыляемые инсектициды на основе Bt;
- Соответствующее «убежище» должно выращиваться на каждой ферме с Bt-кукурузой.
Авторы Roush et al. утверждают (например, правая колонка, стр. 1780 и 1784), что комплекс или «пирамидинг» из двух разных белков, каждый из которых эффективен против целевых вредителей, и не имеет какой-либо перекрестной устойчивости или слабую перекрестную устойчивость, может дать возможность применять убежища меньшего размера. Roush предполагает, что в случае успешного комплекса размер убежища менее 10% убежища может дать контроль устойчивости, сопоставимый с уровнем 50% убежища для единственного (не-пирамидного) признака. Для доступных в настоящее время пирамидных Bt-кукурузных продуктов согласно требованиям Управления по охране окружающей среды США необходимо структурированное убежище не-Bt кукурузы значительно меньшей площади (обычно 5%), чем для продуктов с единственным признаком (обычно 20%).
Существуют разные способы обеспечения эффектов IRM в отношении убежищ, включающие в себя выращивание в полях с различной геометрической планировкой (как упомянуто выше), и упаковки с семенной смесью, как дополнительно рассмотрено Roush и, например, в патенте США № 6551962.
Вышеупомянутые процентные показатели или сходные соотношения % убежищ можно применять для рассматриваемых двойных или тройных комплексов или пирамид. Целью для тройных комплексов с тремя механизмами действия против единственного целевого вредителя является нулевая площадь убежища (или, например, площадь убежища менее 5%). Это особенно справедливо для площади коммерческих посевов в акрах, например, больше 10 акров.
Все патенты, патентные заявки, предварительные заявки и публикации, упомянутые или цитируемые в изобретении, включены в изобретение путем ссылки во всей полноте при условии, что они не противоречат идее настоящей заявки. Если конкретно не обозначено или подразумевается, используемые в изобретении термины в единственном числе обозначают "по меньшей мере один".
Ссылки

ПРИМЕРЫ
Пример 1
Биологическая активность
Результаты биотестов рассматриваемых белков Cry, действующих на FAW, ECB, и устойчивых к Cry1Fa насекомых FAW и ECB, показаны в таблице 1. Оба белка обладают высокой активностью против личинок FAW (обсуждение указанного вредителя можно найти, например, Tabashnik, PNAS (2008), vol 105 No 49, 19029-19030). Белок Cry1Fa имеет намного меньшую активность против FAW, которые устойчивы к токсичности Cry1Fa (rFAW), по сравнению с восприимчивыми FAW. Белок Cry1Be обладает такой же активностью или более активен против rFAW по сравнению с восприимчивыми FAW.
Таблица 1. Биологическое действие белков Cry против четырех разных типов насекомых, плюс против FAW с устойчивостью к Cry1Fa и против личинок ECB. Неподчеркнутые значения в зеленом цвете представляют собой значения LC-50, выраженными как диапазоны значений, полученных от множественных измерений. Подчеркнутые значения отражают показатели GI-50, в случаях, когда белок против конкретного насекомого не вызывает смертность. Значения приведены в нг/см2.
Пример 2
Исследования связывания
Фигура 1 показывает конкурентное связывание меченого 125I Cry1Fa по сравнению со связыванием Cry1Fa или Cry1Be с мембранными везикулами щеточной каймы, полученными от Spodoptera frugiperda (кукурузная листовая совка, FAW).
Исследование проводили дважды, применяя способ с понижением. FAW-0 демонстрирует связывание 125ICry1Fa с рецепторами в отсутствии какого-либо конкурентного лиганда (контроль). FAW-1000 нМ Cry1Fa демонстрирует значительное уменьшение уровня связывания, возникающее в присутствии гомологичного немеченого Cry1Fa, который смещает связывание радиомеченого Cry1Fa из его рецептора. FAW-1000 нМ Cry1Be демонстрирует связывание, происходящее в присутствии немеченого Cry1Be, который не способен перемещать связывание радиомеченого Cry1Fa из его рецептора.
Фигура 2 показывает конкурентное связывание меченого 125I Cry1Fa по сравнению со связыванием Cry1Fa или Cry1Be с мембранными везикулами щеточной каймы, полученными из Ostrinia nubilalis (европейский кукурузный мотылек, ECB). Исследование проводили дважды, применяя способ с понижением. "Контроль Rxn" демонстрирует связывание 125I Cry1Fa с рецепторами в отсутствии какого-либо конкурентного лиганда. 1000 нМ Cry1Fa демонстрирует значительно сниженный уровень связывания, возникающий в присутствии гомологичного немеченого Cry1Fa, который смещает связывание радиомеченого Cry1Fa из его рецептора. 1000 нМ Cry1Be демонстрирует связывание, происходящее в присутствии немеченого Cry1Be, который не способен перемещать связывание радиомеченого Cry1Fa из его рецептора.
Фигура 3 показывает конкурентное смещение связывания меченого 125I Cry1Be с мембранными везикулами щеточной каймы, полученными из Spodoptera frugiperda Cry1Fa (▲) и Cry1Be (●). Белок Cry1Fa эффективно смещает связывание 0,5 нМ 125I Cry1Be только при концентрациях, превышающих 100 нМ (в 200 раз от концентрации радиомеченого Cry1Be, используемого в исследовании). Белок Cry1Be проявляет намного больший эффект в смещении по сравнению с Cry1Fa, даже притом, что Cry1Fa обладает большей активностью, чем Cry1Be, против указанного вредителя.
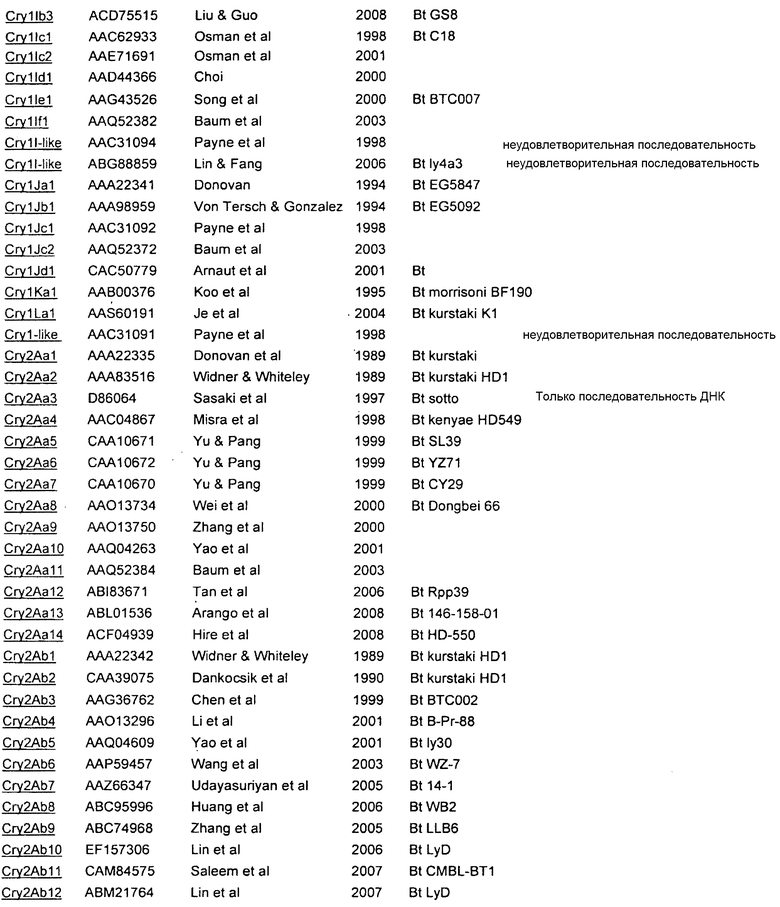
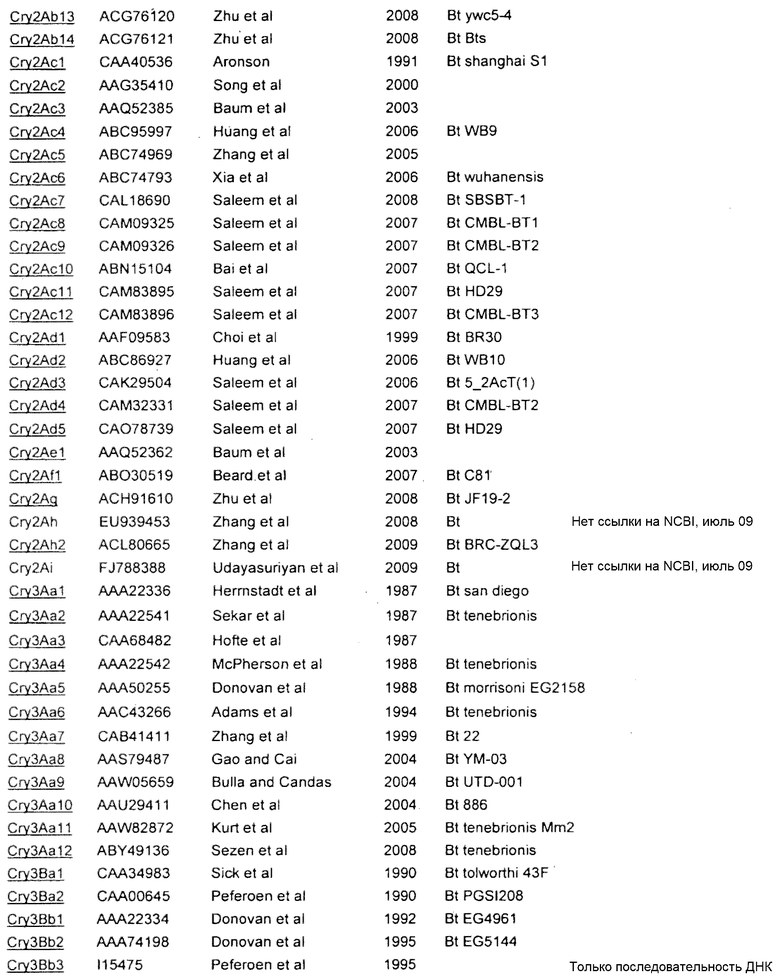
Приложение А
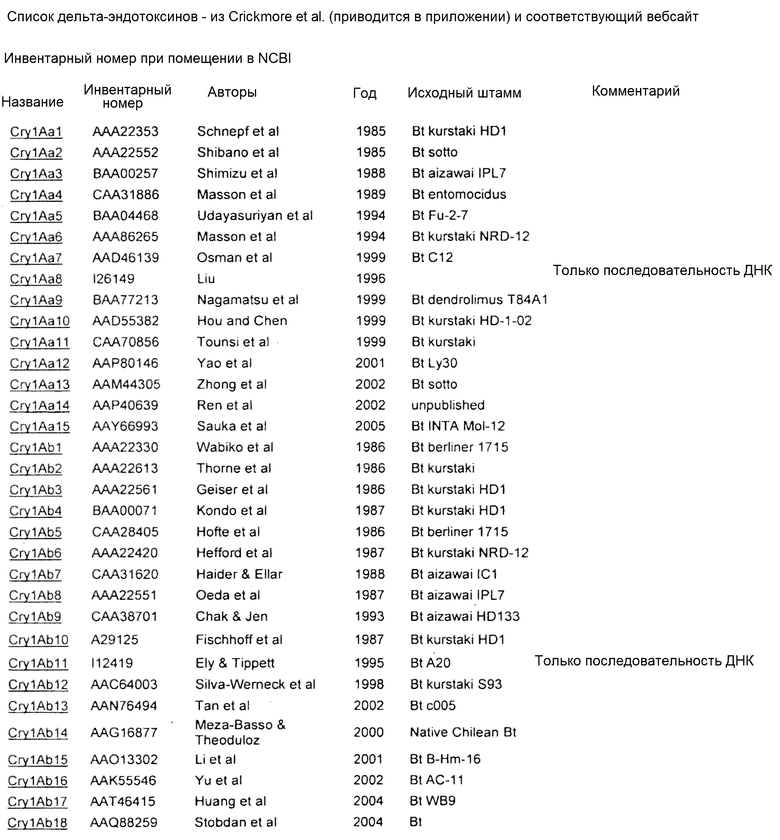
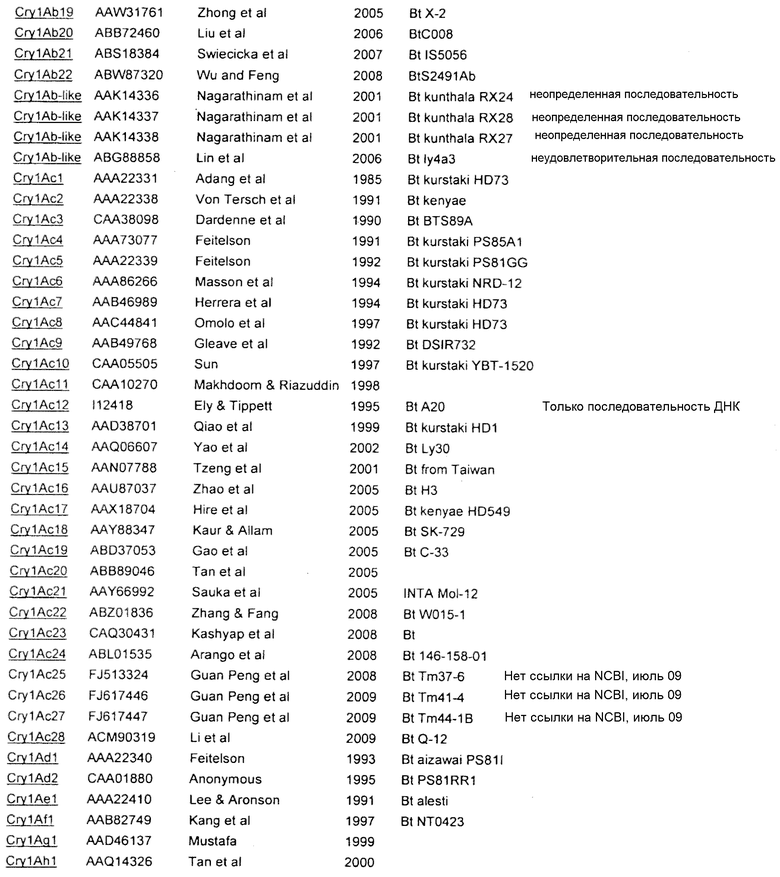
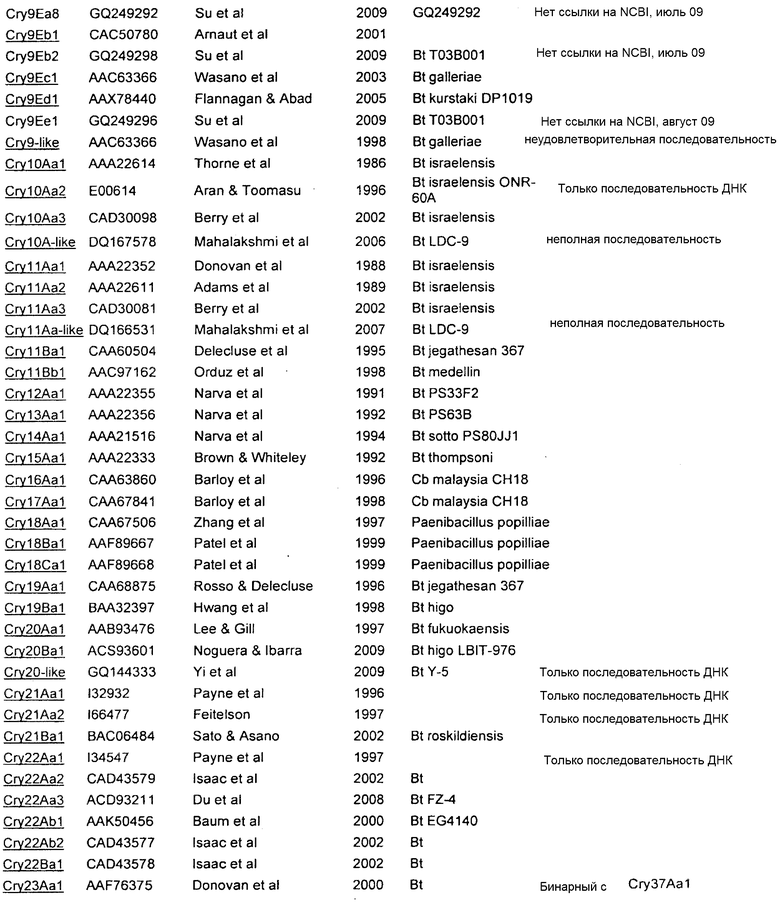
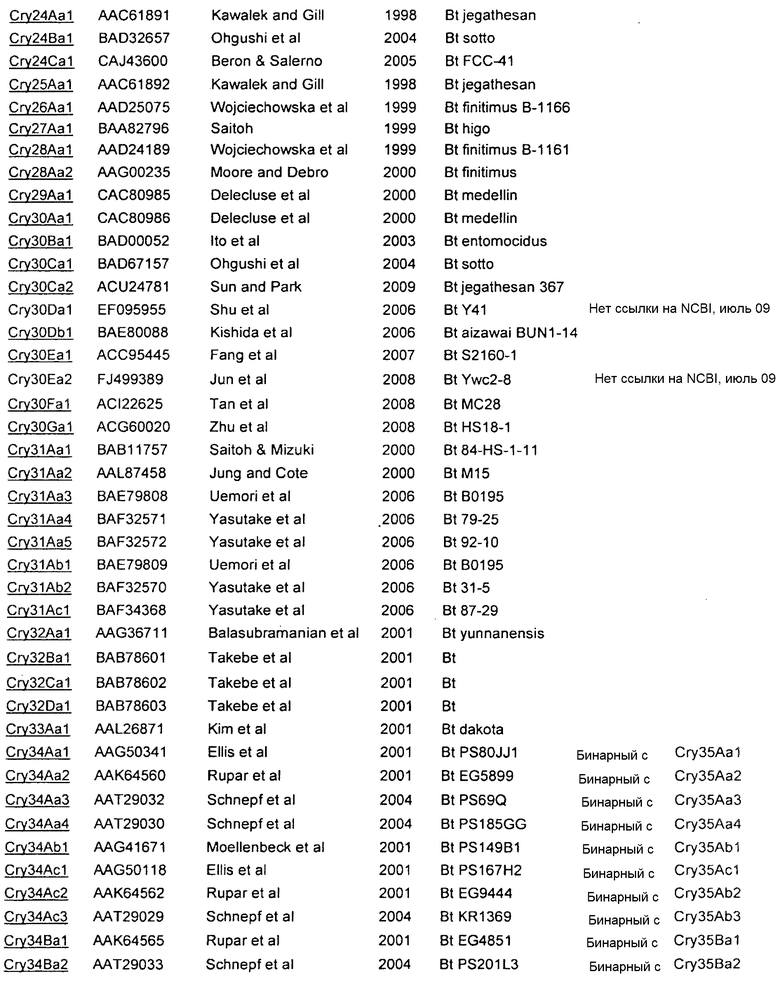
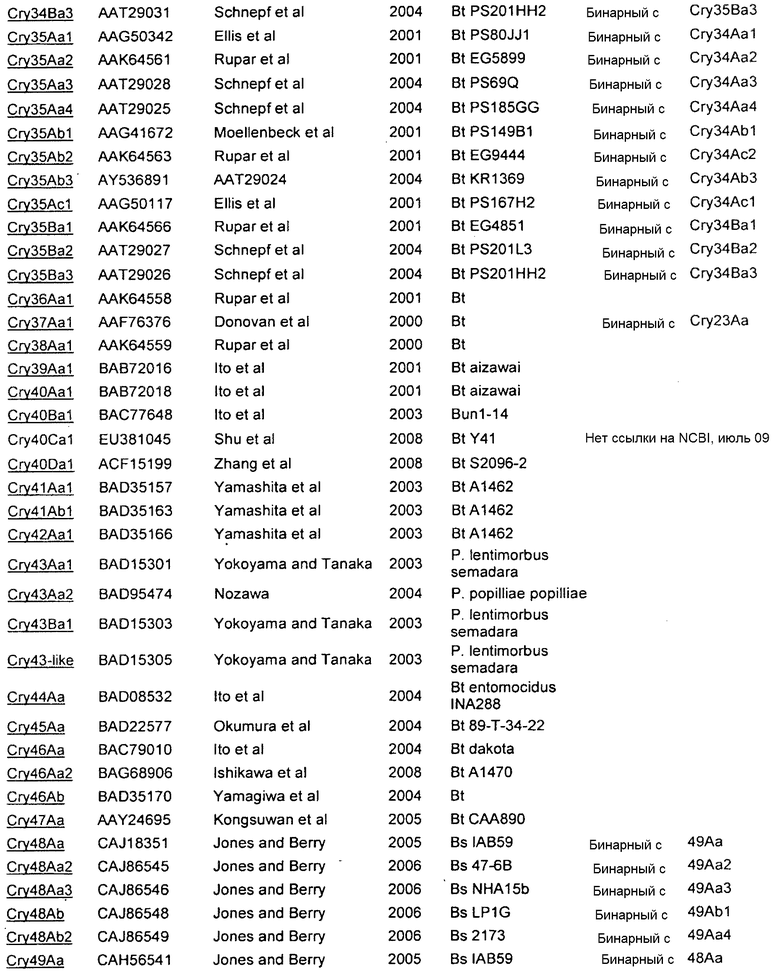
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Dow AGROSCIENCES LLC
<120> КОНТРОЛЬ УСТОЙЧИВОСТИ НАСЕКОМЫХ С ПОМОЩЬЮ КОМБИНАЦИИ БЕЛКОВ CRY1BE И CRY1F
<130> DAS-P0162-US-05
<160> 2
<170> PatentIn version 3.5
<210> 1
<211> 605
<212> Белок
<213> Искусственная последовательность
<220>
<223> Cry1Fa
<400> 1
Met Glu Asn Asn Ile Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Val Glu Ile Leu Asn Glu Glu Arg Ser Thr Gly Arg Leu
20 25 30
Pro Leu Asp Ile Ser Leu Ser Leu Thr Arg Phe Leu Leu Ser Glu Phe
35 40 45
Val Pro Gly Val Gly Val Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Phe Ile Thr Pro Ser Asp Trp Ser Leu Phe Leu Leu Gln Ile Glu Gln
65 70 75 80
Leu Ile Glu Gln Arg Ile Glu Thr Leu Glu Arg Asn Arg Ala Ile Thr
85 90 95
Thr Leu Arg Gly Leu Ala Asp Ser Tyr Glu Ile Tyr Ile Glu Ala Leu
100 105 110
Arg Glu Trp Glu Ala Asn Pro Asn Asn Ala Gln Leu Arg Glu Asp Val
115 120 125
Arg Ile Arg Phe Ala Asn Thr Asp Asp Ala Leu Ile Thr Ala Ile Asn
130 135 140
Asn Phe Thr Leu Thr Ser Phe Glu Ile Pro Leu Leu Ser Val Tyr Val
145 150 155 160
Gln Ala Ala Asn Leu His Leu Ser Leu Leu Arg Asp Ala Val Ser Phe
165 170 175
Gly Gln Gly Trp Gly Leu Asp Ile Ala Thr Val Asn Asn His Tyr Asn
180 185 190
Arg Leu Ile Asn Leu Ile His Arg Tyr Thr Lys His Cys Leu Asp Thr
195 200 205
Tyr Asn Gln Gly Leu Glu Asn Leu Arg Gly Thr Asn Thr Arg Gln Trp
210 215 220
Ala Arg Phe Asn Gln Phe Arg Arg Asp Leu Thr Leu Thr Val Leu Asp
225 230 235 240
Ile Val Ala Leu Phe Pro Asn Tyr Asp Val Arg Thr Tyr Pro Ile Gln
245 250 255
Thr Ser Ser Gln Leu Thr Arg Glu Ile Tyr Thr Ser Ser Val Ile Glu
260 265 270
Asp Ser Pro Val Ser Ala Asn Ile Pro Asn Gly Phe Asn Arg Ala Glu
275 280 285
Phe Gly Val Arg Pro Pro His Leu Met Asp Phe Met Asn Ser Leu Phe
290 295 300
Val Thr Ala Glu Thr Val Arg Ser Gln Thr Val Trp Gly Gly His Leu
305 310 315 320
Val Ser Ser Arg Asn Thr Ala Gly Asn Arg Ile Asn Phe Pro Ser Tyr
325 330 335
Gly Val Phe Asn Pro Gly Gly Ala Ile Trp Ile Ala Asp Glu Asp Pro
340 345 350
Arg Pro Phe Tyr Arg Thr Leu Ser Asp Pro Val Phe Val Arg Gly Gly
355 360 365
Phe Gly Asn Pro His Tyr Val Leu Gly Leu Arg Gly Val Ala Phe Gln
370 375 380
Gln Thr Gly Thr Asn His Thr Arg Thr Phe Arg Asn Ser Gly Thr Ile
385 390 395 400
Asp Ser Leu Asp Glu Ile Pro Pro Gln Asp Asn Ser Gly Ala Pro Trp
405 410 415
Asn Asp Tyr Ser His Val Leu Asn His Val Thr Phe Val Arg Trp Pro
420 425 430
Gly Glu Ile Ser Gly Ser Asp Ser Trp Arg Ala Pro Met Phe Ser Trp
435 440 445
Thr His Arg Ser Ala Thr Pro Thr Asn Thr Ile Asp Pro Glu Arg Ile
450 455 460
Thr Gln Ile Pro Leu Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr
465 470 475 480
Val Val Arg Gly Pro Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr
485 490 495
Ser Gly Gly Pro Phe Ala Tyr Thr Ile Val Asn Ile Asn Gly Gln Leu
500 505 510
Pro Gln Arg Tyr Arg Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu
515 520 525
Arg Ile Tyr Val Thr Val Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe
530 535 540
Asn Lys Thr Met Asp Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser
545 550 555 560
Tyr Ala Thr Ile Asn Thr Ala Phe Thr Phe Pro Met Ser Gln Ser Ser
565 570 575
Phe Thr Val Gly Ala Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Ile
580 585 590
Asp Arg Phe Glu Leu Ile Pro Val Thr Ala Thr Leu Glu
595 600 605
<210> 2
<211> 641
<212> Белок
<213> Искусственная последовательность
<220>
<223> Cry1Be
<400> 2
Met Thr Ser Asn Arg Lys Asn Glu Asn Glu Ile Ile Asn Ala Leu Ser
1 5 10 15
Ile Pro Ala Val Ser Asn His Ser Ala Gln Met Asn Leu Ser Thr Asp
20 25 30
Ala Arg Ile Glu Asp Ser Leu Cys Ile Ala Glu Gly Asn Asn Ile Asp
35 40 45
Pro Phe Val Ser Ala Ser Thr Val Gln Thr Gly Ile Asn Ile Ala Gly
50 55 60
Arg Ile Leu Gly Val Leu Gly Val Pro Phe Ala Gly Gln Ile Ala Ser
65 70 75 80
Phe Tyr Ser Phe Leu Val Gly Glu Leu Trp Pro Arg Gly Arg Asp Pro
85 90 95
Trp Glu Ile Phe Leu Glu His Val Glu Gln Leu Ile Arg Gln Gln Val
100 105 110
Thr Glu Asn Thr Arg Asp Thr Ala Leu Ala Arg Leu Gln Gly Leu Gly
115 120 125
Asn Ser Phe Arg Ala Tyr Gln Gln Ser Leu Glu Asp Trp Leu Glu Asn
130 135 140
Arg Asp Asp Ala Arg Thr Arg Ser Val Leu Tyr Thr Gln Tyr Ile Ala
145 150 155 160
Leu Glu Leu Asp Phe Leu Asn Ala Met Pro Leu Phe Ala Ile Arg Asn
165 170 175
Gln Glu Val Pro Leu Leu Met Val Tyr Ala Gln Ala Ala Asn Leu His
180 185 190
Leu Leu Leu Leu Arg Asp Ala Ser Leu Phe Gly Ser Glu Phe Gly Leu
195 200 205
Thr Ser Gln Glu Ile Gln Arg Tyr Tyr Glu Arg Gln Val Glu Lys Thr
210 215 220
Arg Glu Tyr Ser Asp Tyr Cys Ala Arg Trp Tyr Asn Thr Gly Leu Asn
225 230 235 240
Asn Leu Arg Gly Thr Asn Ala Glu Ser Trp Leu Arg Tyr Asn Gln Phe
245 250 255
Arg Arg Asp Leu Thr Leu Gly Val Leu Asp Leu Val Ala Leu Phe Pro
260 265 270
Ser Tyr Asp Thr Arg Val Tyr Pro Met Asn Thr Ser Ala Gln Leu Thr
275 280 285
Arg Glu Ile Tyr Thr Asp Pro Ile Gly Arg Thr Asn Ala Pro Ser Gly
290 295 300
Phe Ala Ser Thr Asn Trp Phe Asn Asn Asn Ala Pro Ser Phe Ser Ala
305 310 315 320
Ile Glu Ala Ala Val Ile Arg Pro Pro His Leu Leu Asp Phe Pro Glu
325 330 335
Gln Leu Thr Ile Phe Ser Val Leu Ser Arg Trp Ser Asn Thr Gln Tyr
340 345 350
Met Asn Tyr Trp Val Gly His Arg Leu Glu Ser Arg Thr Ile Arg Gly
355 360 365
Ser Leu Ser Thr Ser Thr His Gly Asn Thr Asn Thr Ser Ile Asn Pro
370 375 380
Val Thr Leu Gln Phe Thr Ser Arg Asp Val Tyr Arg Thr Glu Ser Phe
385 390 395 400
Ala Gly Ile Asn Ile Leu Leu Thr Thr Pro Val Asn Gly Val Pro Trp
405 410 415
Ala Arg Phe Asn Trp Arg Asn Pro Leu Asn Ser Leu Arg Gly Ser Leu
420 425 430
Leu Tyr Thr Ile Gly Tyr Thr Gly Val Gly Thr Gln Leu Phe Asp Ser
435 440 445
Glu Thr Glu Leu Pro Pro Glu Thr Thr Glu Arg Pro Asn Tyr Glu Ser
450 455 460
Tyr Ser His Arg Leu Ser Asn Ile Arg Leu Ile Ser Gly Asn Thr Leu
465 470 475 480
Arg Ala Pro Val Tyr Ser Trp Thr His Arg Ser Ala Asp Arg Thr Asn
485 490 495
Thr Ile Ser Ser Asp Ser Ile Thr Gln Ile Pro Leu Val Lys Ser Phe
500 505 510
Asn Leu Asn Ser Gly Thr Ser Val Val Ser Gly Pro Gly Phe Thr Gly
515 520 525
Gly Asp Ile Ile Arg Thr Asn Val Asn Gly Ser Val Leu Ser Met Gly
530 535 540
Leu Asn Phe Asn Asn Thr Ser Leu Gln Arg Tyr Arg Val Arg Val Arg
545 550 555 560
Tyr Ala Ala Ser Gln Thr Met Val Leu Arg Val Thr Val Gly Gly Ser
565 570 575
Thr Thr Phe Asp Gln Gly Phe Pro Ser Thr Met Ser Ala Asn Glu Ser
580 585 590
Leu Thr Ser Gln Ser Phe Arg Phe Ala Glu Phe Pro Val Gly Ile Ser
595 600 605
Ala Ser Gly Ser Gln Thr Ala Gly Ile Ser Ile Ser Asn Asn Ala Gly
610 615 620
Arg Gln Thr Phe His Phe Asp Lys Ile Glu Phe Ile Pro Ile Thr Ala
625 630 635 640
Thr
| название | год | авторы | номер документа |
|---|---|---|---|
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2816526C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2817591C2 |
| НОВЫЕ БЕЛКИ, ИМЕЮЩИЕ ИНГИБИРУЮЩЕЕ ДЕЙСТВИЕ В ОТНОШЕНИИ НАСЕКОМЫХ | 2017 |
|
RU2781075C2 |
| НОВЫЕ БЕЛКИ, ОБЛАДАЮЩИЕ ИНГИБИРУЮЩЕЙ АКТИВНОСТЬЮ В ОТНОШЕНИИ НАСЕКОМЫХ | 2015 |
|
RU2740313C2 |
| ИНСЕКТИЦИДНЫЕ ПОЛИПЕПТИДЫ С ШИРОКИМ СПЕКТРОМ АКТИВНОСТИ И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2750459C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2015 |
|
RU2745322C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2015 |
|
RU2741833C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2018 |
|
RU2761666C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2740312C2 |
| ГЕНЫ ТОКСИНОВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2723717C2 |






Изобретение относится к области биохимии, в частности к трансгенному растению, которое устойчиво к насекомым-вредителям: кукурузной листовой совке и европейскому кукурузному мотыльку, и содержит ДНК, кодирующую инсектицидный белок Cry1Be, и ДНК, кодирующую инсектицидный белок Cry1Fa, а также к его семени. Раскрыто множество трансгенных растений на поле растений, включающее растения, не экспрессирующие инсектицидные белки, а также раскрыта смесь семян, включающая семена, которые не экспрессируют трансгенные инсектицидные белки и множество вышеуказанных трансгенных семян. Изобретение также относится к композиции для контроля или профилактики Cry-устойчивых чешуекрылых вредителей, содержащая клетки, которые экспрессируют эффективное количество белка Cry1Fa и белка Cry1Be, а также к способу контроля или профилактики Cry-устойчивых чешуекрылых вредителей с ее использованием. Изобретение позволяет эффективно бороться с насекомыми-вредителями: кукурузной листовой совкой и европейским кукурузным мотыльком. 8 н. и 16 з.п. ф-лы, 3 ил., 1 табл., 2 пр.
1. Трансгенное растение, которое устойчиво к насекомым-вредителям кукурузной листовой совке (FAW: Spodoptera frugiperda) и европейскому кукурузному мотыльку (ЕСВ, ostrinia nubilalis), и содержит ДНК, кодирующую инсектицидный белок Cry1Be с SEQ ID NO: 2, и ДНК, кодирующую инсектицидный белок Cry1Fa с SEQ ID NO: 1.
2. Семя растения по п. 1, где указанное семя содержит ДНК, кодирующую инсектицидный белок Cry1Be с SEQ ID NO: 2, и ДНК, которая кодирует инсектицидный белок Cry1Fa с SEQ ID NO: 1, где указанное семя устойчиво к насекомым кукурузной листовой совке (FAW: Spodoptera frugiperda) и европейскому кукурузному мотыльку (ЕСВ, ostrinia nubilalis).
3. Трансгенное растение по п. 1, где ДНК, кодирующая инсектицидный белок Cry1Be с SEQ ID NO: 2, и ДНК, кодирующая инсектицидный белок Cry1Fa с SEQ ID NO: 1, были внедрены в указанное растение.
4. Семя растения по п. 3, содержащее ДНК, которая кодирует инсектицидный белок Cry1Be с SEQ ID NO: 2, и ДНК, которая кодирует инсектицидный белок Cry1Fa с SEQ ID NO: 1, где указанное семя засеивают для задерживания развития устойчивости у кукурузной листовой совки и европейского кукурузного мотылька к белкам Cry1Be и Cry1Fa.
5. Множество растений на поле, содержащее не-Bacillus thuringiensis (не-Bt) растения, которые не экспрессируют трансгенные инсектицидные белки, и множество трансгенных растений по п. 1, которые обладают устойчивостью к насекомым-вредителям кукурузной листовой совке и европейскому кукурузному мотыльку, где указанные трансгенные растения содержат ДНК, которая кодирует инсектицидный белок Cry1Be с SEQ ID NO: 2, и ДНК, которая кодирует инсектицидный белок Cry1Fa с SEQ ID NO: 1, где указанные не-Bt растения составляют 5-40% всех сельскохозяйственных растений в указанном множестве растений, где указанное множество трансгенных растений задерживает развитие устойчивости к белку Cry насекомыми кукурузной листовой совкой и европейским кукурузным мотыльком.
6. Множество трансгенных растений на поле по п. 5, где указанные не-Bt растения составляют 5-30% всех сельскохозяйственных растений в указанном множестве растений.
7. Множество трансгенных растений на поле по п. 5, где указанные не-Bt растения составляют 5-20% всех сельскохозяйственных растений в указанном множестве растений.
8. Множество трансгенных растений на поле по п. 5, где указанные не-Bt растения составляют 5-10% всех сельскохозяйственных растений в указанном множестве растений.
9. Множество трансгенных растений на поле по любому из пп. 5-8, где указанные не-Bt растения высажены блоками или полосами.
10. Смесь семян, содержащая семена от не-Bt растений, которые не экспрессируют трансгенные инсектицидные белки, и множество семян по п. 4, где указанные семена от не-Bt растений составляют 5-40% всех семян в смеси, где указанную смесь семян выращивают с целью контроля развития устойчивости кукурузной листовой совки и европейского кукурузного мотылька к инсектицидным белкам.
11. Смесь семян по п. 10, где указанные семена от не-Bt растений составляют 5-30% всех семян в смеси.
12. Смесь семян по п. 10, где указанные семена от не-Bt растений составляют 5-20% всех семян в смеси.
13. Смесь семян по п. 10, где указанные семена от не-Bt растений составляют 5-10% всех семян в смеси.
14. Трансгенное растение по п. 1, где указанное растение дополнительно содержит ДНК, кодирующую коровый Cry1Ab токсин-содержащий белок.
15. Множество растений на поле, содержащее не-Bt растения, которые не экспрессируют трансгенные инсектицидные белки, и множество трансгенных растений по п. 14, где указанные не-Bt растения составляют 5-20% всех сельскохозяйственных растений на указанном множестве растений, где указанное множество растений задерживает развитие устойчивости к белкам Cry1Be и Cry1Fa насекомыми кукурузной листовой совкой и европейским кукурузным мотыльком.
16. Множество растений на поле растений, включающее не-Bt растения, которые не экспрессируют трансгенные инсектицидные белки, и множество трансгенных растений по п. 14, где указанное множество растений содержит 5-10% не-Bt растений, где указанное множество растений задерживает развитие устойчивости к белкам Cry1Be и Cry1Fa насекомыми кукурузной листовой совкой и европейским кукурузным мотыльком.
17. Композиция для контроля или профилактики Cry-устойчивых чешуекрылых вредителей, содержащая клетки, которые экспрессируют эффективное количество и корового токсин-содержащего белка Cry1Fa с SEQ ID NO: 1 и корового токсин-содержащего белка Cry1Be с SEQ ID NO: 2, где указанные чешуекрылые вредители выбраны из группы, состоящей из европейского кукурузного мотылька и кукурузной листовой совки.
18. Композиция по п. 17, содержащая хозяина, трансформированного для экспрессии и корового токсин-содержащего белка Cry1Fa с SEQ ID NO: 1 и корового токсин-содержащего белка Cry1Be с SEQ ID NO: 2, где указанный хозяин представляет собой клетку микроорганизма или растения.
19. Способ контроля или профилактики Cry-устойчивых чешуекрылых вредителей, включающий представление указанным вредителям или среде обитания указанных вредителей эффективного количества композиции по п. 17, где указанные чешуекрылые вредители выбраны из группы, состоящей из европейского кукурузного мотылька и кукурузной листовой совки.
20. Множество растений на поле по любому из пп. 5 и 15, где указанные растения занимают площадь более 10 акров.
21. Растение по любому из пп. 1 и 14, где указанное растение выбирают из группы, состоящей из кукурузы, сои и хлопка.
22. Растение по любому из пп. 1 и 14, где указанным растением является растение кукурузы.
23. Растение по любому из пп. 1 и 14, где указанный инсектицидный белок Cry1Fa содержит SEQ ID NO: 1 и указанный инсектицидный белок Cry1Be содержит SEQ ID NO: 2.
24. Растение по любому из пп. 1 и 14, где указанный инсектицидный белок Cry1Be содержит SEQ ID NO: 2 и указанный инсектицидный белок Cry1Fa содержит SEQ ID NO: 1.

