ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0001] Настоящее изобретение относится к сконструированным пестицидным белкам и молекулам нуклеиновых кислот, которые их кодируют, а также к композициям и способам контроля вредителей растений.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0002] Bacillus thuringiensis (Bt) является грамположительной спорообразующей почвенной бактерией, которая отличается своей способностью продуцировать кристаллические включения, которые особенно токсичны для определенных отрядов и видов вредителей растений, в том числе насекомых, но безвредны для растений и других нецелевых организмов. По этой причине композиции, содержащие штаммы Bacillus thuringiensis или их инсектицидные белки, могут применяться в качестве приемлемых с точки зрения охраны окружающей среды инсектицидов для контроля насекомых-вредителей, важных с точки зрения сельского хозяйства, или насекомых-переносчиков различных заболеваний человека или животных.
[0003] Кристаллические (Cry) белки из Bacillus thuringiensis обладают сильной инсектицидной активностью в отношении преимущественно чешуекрылых, двукрылых и жесткокрылых насекомых-вредителей. Эти белки также продемонстрировали активность в отношении вредителей из отрядов Hymenoptera, Homoptera, Phthiraptera, Mallophaga и вредителей из отрядов подкласса Acari, а также других групп беспозвоночных, таких KaKNemathelminthes, Platyhelminthes и Sarcomastigorphora (Feitelson, J. 1993. The Bacillus Thuringiensis family tree. In Advanced Engineered Pesticides. Marcel Dekker, Inc., New York, N.Y.). Эти белки изначально разделили на классы Cry1 - CryVI, исходя преимущественно из их инсектицидной активности. Основными классами были специфические для Lepidoptera (I), специфические для Lepidoptera и Diptera (II), специфические для Coleoptera (III), специфические для Diptera (IV) и специфические для нематод (V) и (VI). Далее белки разделили на подсемейства, белкам с более высокой степенью родства в пределах каждого семейства присвоили буквы для обозначения раздела, как, например: Cry1A, Cry1B, Cry1C и т.д. Еще более близкородственным белкам в пределах каждого раздела присвоили названия, как, например: Cry1C(a), Cry1C(b) и т.д.
[0004] Выражения "токсин Cry" и "дельта-эндотоксин" применялись взаимозаменяемо с выражением "белок Cry". Современная номенклатура белков и генов Cry основана на гомологии аминокислотных последовательностей, а не на специфичности в отношении целевых насекомых (Crickmore et al. (1998) Microbiol. Mol. Biol. Rev. 62:807-813). В этой более распространенной классификации каждому токсину присваивается уникальное название, включающее первичный ранг (арабская цифра), вторичный ранг (заглавная буква), третичный ранг (строчная буква) и четвертичный ранг (еще одна арабская цифра). В современной классификации в первичном ранге римские цифры были заменены на арабские цифры. Например, "CryIA(а)" согласно старой номенклатуре в настоящее время имеет название "Cry1Aa" согласно современной номенклатуре. Согласно публикации Ibrahim et al. (2010, Bioeng. Bugs, 1:31-50) токсины Cry были разделены на шесть основных классов в соответствии с их специфичностью в отношении насекомого-хозяина и они включают: группу 1 чешуекрылые (например, Cry1, Cry9 и Cry15); группу 2 - чешуекрылые и двукрылые (например, Cry2); группу 3 - жесткокрылые (Cry3, Cry7 и Cry8); группу 4 - двукрылые (Cry4, Cry10, Cry11, Cry16, Cry17, Cry19 и Cry20); группу 5 - чешуекрылые и жесткокрылые (Cry1I); и группу 6 - нематоды (Cry6). Токсины Cry1I, Cry2, Cry3, Cry10 и Cry11 (73-82 кДа) являются уникальными, поскольку, по-видимому, они являются результатом естественных усечений белков большего размера Cry1 и Cry4 (130-140 кДа).
[0005] Белки Cry представляют собой молекулы глобулярного белка, которые накапливаются в виде протоксинов в кристаллической форме в ходе стадии спорообразования Bt. После поедания вредителем кристаллы, как правило, растворяются с высвобождением протоксинов, размер которых может варьировать, например от 130-140 кДа в случае белков Cry, активных в отношении многих чешуекрылых, таких как Cry1 и Cry9, и 60-80 кДа в случае белков Cry3, активных в отношении жесткокрылых, и белков Cry2, активных в отношении чешуекрылых/двукрылых. После растворения кристаллов в организме восприимчивого насекомого высвободившиеся протоксины подвергаются обработке протеазами в пищеварительном канале насекомого, например трипсином и химотрипсином, с образованием протеазаустойчивой сердцевины белкового токсина Cry. Данный протеолитический процессинг включает удаление аминокислот из разных участков различных протоксинов Cry. Например, протоксины Cry размером 130-140 кДа, как правило, активируются посредством протеолитического удаления N-концевого пептида из 25-30 аминокислот и примерно половины остального белка с С-конца, результатом чего является образование зрелого токсина Cry размером примерно 60-70 кДа. Протоксины размером 60-80 кДа, например Cry2 и Cry3, также подвергаются процессингу, но не в той же степени, как протоксины более крупного размера. Протоксины меньшего размера, как правило, характеризуются тем, что с N-конца удаляется такое же или большее количество аминокислот, чем у протоксинов более крупного размера, но на С-конце удаляется меньшее количество аминокислот. Например, протеолитическая активация представителей семейства Cry2, как правило, включает удаление примерно 40-50 N-концевых аминокислот. Многие белки Cry являются достаточно токсичными для определенных целевых насекомых, но многие обладают узкими спектрами активности.
[0006] Части токсина белков Cry обычно содержат пять блоков с консервативной последовательностью и три домена с консервативной структурой (см., например, de Maagd et al. (2001) Trends Genetics 17:193-199). Первый домен с консервативной структурой, называемый домен I, как правило, состоит из семи альфа-спиралей и вовлечен во вставку в мембрану и образовании пор. Домен II, как правило, состоит из трех бета-складчатых слоев, расположенных в конфигурации типа греческий ключ, а домен III состоит из двух антипараллельных бета-складчатых слоев в виде структуры 'jelly-roll' (de Maagd et al., 2001, выше). Домены II и III вовлечены в распознавание и связывание рецепторов, и поэтому считаются детерминантами специфичности токсина.
[0007] Пять блоков с консервативной последовательностью пронумерованы от СВ1 до СВ5 с N-конца до С-конца белка Cry (Hofte & Whitely, 1989, Microbiol. Rev. 53:242-255). Консервативный блок 1 (СВ1) содержит приблизительно 29 аминокислот, консервативный блок 2 (СВ2) содержит приблизительно 67 аминокислот, консервативный блок 3 (СВ3) содержит приблизительно 48 аминокислот, консервативный блок 4 (СВ4) содержит приблизительно 10 аминокислот и консервативный блок 5 (СВ5) содержит приблизительно 12 аминокислот. Последовательности до и после этих пяти консервативных блоков высоковариабельны и, таким образом, обозначены как "вариабельные области" V1-V6. Домен I дельта-эндотоксина Bt обычно содержит вариабельную область 1, консервативный блок 1, вариабельную область 2 и 52 N-концевые аминокислоты консервативного блока 2. Домен II обычно содержит приблизительно 15 С-концевых аминокислот консервативного блока 2, вариабельную область 3 и приблизительно 10 N-концевых аминокислот консервативного блока 3. Домен III обычно содержит примерно 38 С-концевых аминокислот консервативного блока 3, вариабельную область 4, консервативный блок 4, вариабельную область 5 и консервативный блок 5. Среди других дельта-эндотоксинов активные в отношении чешуекрылых токсины Cry1 имеют вариабельную область 6 с приблизительно 1-3 аминокислотами, лежащими в домене III.
[0008] Многочисленные коммерчески ценные растения, в том числе широко распространенные сельскохозяйственные культуры, восприимчивы к нападению вредителей растений, в том числе насекомых-вредителей и нематод-вредителей, что приводит к существенным показателям снижения урожайности и качества сельскохозяйственных культур. Например, вредители растений являются главным фактором потери урожая важных мировых сельскохозяйственных культур. Из-за повреждений, вызываемых беспозвоночными вредителями, в том числе насекомыми, ежегодный убыток в США составляет приблизительно 8 миллиардов долларов. Насекомые-вредители также становятся бременем для овощеводов и плодоводов, производителей декоративных цветов, а также владельцев приусадебных хозяйств.
[0009] Насекомых-вредителей контролируют главным образом путем интенсивного применения химических пестицидов, которые действуют путем подавления роста насекомых, препятствования кормлению или размножению, или вызывая гибель. Средства для биологического контроля вредителей, например, штаммы Bacillus thuringiensis, экспрессирующие пестицидные токсины, такие как белки Cry, также применялись в отношении культурных растений с удовлетворительными результатами, что является альтернативой или дополнением к химическим пестицидам. Были выделены гены, кодирующие некоторые из этих белков Cry, и было показано, что их экспрессия в гетерологичных хозяевах, таких как трансгенные растения, обеспечивает другой инструмент для контроля экономически важных насекомых-вредителей.
[0010] Таким образом, можно достичь хорошего контроля насекомых, но продолжительное применение определенных способов химического и биологического контроля повышает вероятность развития у насекомых-вредителей устойчивости к таким мерам контроля. Это было частично ослаблено посредством различных практик контроля устойчивости, но осталась необходимость в развитии новых и эффективных средств для контроля вредителей. Особенно необходимыми являются средства для контроля, которые могут целенаправленно воздействовать на более широкий спектр экономически важных насекомых-вредителей и/или которые обеспечивают эффективный контроль линий насекомых, которые могут стать устойчивыми к существующим средствам для контроля насекомых или являются таковыми.
КРАТКОЕ ОПИСАНИЕ
[0011] В настоящем изобретении предусмотрены нуклеиновые кислоты, полипептиды, композиции и способы придания пестицидной активности (например, инсектицидной активности) бактериям, растениям, растительным клеткам, тканям и семенам. В частности, в настоящем изобретении предусмотрены новые химерные пестицидные белки (например, химерные инсектицидные белки), необязательно с измененной или повышенной активностью по сравнению с исходной молекулой.
[0012] В вариантах осуществления химерные белки по настоящему изобретению являются токсичными для экономически важных насекомых-вредителей (например, путем подавления способности насекомых-вредителей выживать, расти и/или размножаться), особенно насекомых-вредителей, которые заражают растения. Например, в вариантах осуществления химерные инсектицидные белки по настоящему изобретению можно применять для контроля одного или нескольких экономически важных вредителей, относящихся к чешуекрылым, включая без ограничения такие как совка-ипсилон (Agrotis ipsilon), кукурузный мотылек (Ostrinia nubilalis), совка травяная (Spodoptera frugiperda), американская кукурузная совка (Helicoverpa zea), огневка тростниковая (Diatraea saccharalis), гусеница совки бархатных бобов (Anticarsia gemmatalis), соевая совка (Chrysodeixis includes), огневка кукурузная юго-западная (Diatraea grandiosella), западная бобовая совка (Richia albicosta), табачная совка (Heliothis virescens), восточный кукурузный мотылек (Ostrinia furnacalis), хлопковая совка (Helicoverpa armigera), огневка желтая рисовая (Chilo suppressalis), розовая стеблевая совка (Sesamia calamistis), огневка рисовая (Cnaphalocrocis medinalis) и т.п. В вариантах осуществления химерный инсектицидный белок обладает активностью в отношении насекомого-вредителя совка травяная, которая устойчива к белку Vip3A и/или белку Cry1F.
[0013] Соответственно, в качестве одного аспекта в настоящем изобретении предусмотрен химерный инсектицидный белок, который токсичен для насекомого-вредителя, относящегося к чешуекрылым, содержащий в направлении от N-конца к С-концу: (а) N-концевую область первого белка Cry1, которая необязательно является N-концевой областью белка ВТ-0029 под SEQ ID NO: 2 или аминокислотной последовательностью, которая по существу идентична ей, слитую с (b) С-концевой областью другого белка Cry1; где положение перехода между первым белком Cry1 и другим белком Cry1 находится в консервативном блоке 3. В иллюстративных вариантах осуществления другой белок Cry1 представляет собой белок Cry1F (например, Cry1Fa), Cry1G, Cry1I (например, Cry1Ia или Cry1If, такой какВТ-0022), Cry1C (например, Cry1Ca) или Cry1K.
[0014] В вариантах осуществления химерный инсектицидный белок в соответствии с настоящим изобретением обладает инсектицидной активностью в отношении насекомого-вредителя Spodoptera frugiperda или популяции насекомых-вредителей с устойчивостью к белку Vip3A и/или белку Cry1F.
[0015] В качестве дополнительного аспекта в настоящем изобретении предусмотрена нуклеотидная последовательность, кодирующая химерные инсектицидные белки по настоящему изобретению, и кассеты экспрессии и векторы, содержащие ее. В вариантах осуществления полинуклеотид является кодон-оптимизированным для экспрессии в растении (например, однодольном растении, таком как маис, или двудольном растении, таком как соя).
[0016] В качестве дополнительного аспекта в настоящем изобретении предусмотрена трансгенная клетка (например, трансгенная растительная клетка, такая как клетка двудольного или клетка однодольного, или трансгенная бактериальная клетка), трансгенная часть растения, трансгенная культура растения и трансгенное семя растения, которые содержат нуклеотидную последовательность, кассету экспрессии, вектор и/или химерный инсектицидный белок по настоящему изобретению.
[0017] В качестве еще одного аспекта в настоящем изобретении охвачены трансгенные растения, содержащие растительную клетку, часть растения, нуклеотидную последовательность, кассету экспрессии, вектор и/или химерный инсектицидный белок по настоящему изобретению.
[0018] В качестве дополнительного аспекта выступают семена, из которых получают трансгенные растения по настоящему изобретению, и семена, полученные с помощью трансгенных растений по настоящему изобретению.
[0019] Также предусмотрены собранные продукты, полученные из трансгенных растений по настоящему изобретению, где собранный продукт необязательно содержит нуклеотидную последовательность, кассету экспрессии, вектор и/или химерный инсектицидный белок по настоящему изобретению. Дополнительно предусмотрены обработанные продукты, полученные из собранных продуктов по настоящему изобретению, где собранный продукт необязательно содержит нуклеотидную последовательность, кассету экспрессии, вектор и/или химерный инсектицидный белок по настоящему изобретению. В вариантах осуществления собранный продукт или обработанный продукт содержит химерный инсектицидный белок по настоящему изобретению и обладает повышенной устойчивостью к насекомому-вредителю (например, насекомому-вредителю, относящемуся к чешуекрылым).
[0020] В качестве еще одного аспекта в настоящем изобретении предусмотрена инсектицидная композиция, содержащая химерный инсектицидный белок по настоящему изобретению и приемлемый с точки зрения сельского хозяйства носитель.
[0021] Кроме того, в настоящем изобретении предусмотрен в качестве дополнительного аспекта способ получения трансгенного растения с повышенной устойчивостью к насекомому-вредителю (например, насекомому-вредителю, относящемуся к чешуекрылым). В вариантах осуществления способ включает введение в растение полинуклеотида, кассеты экспрессии или вектора по настоящему изобретению, где химерный инсектицидный белок экспрессируется в растении с получением таким образом растения с повышенной устойчивостью к насекомому-вредителю. Стадия введения необязательно включает: (i) трансформацию растительной клетки с помощью полинуклеотида, кассеты экспрессии или вектора и регенерацию трансгенного растения; или (ii) скрещивание первого растения, содержащего полинуклеотид, кассету экспрессии или вектор, со вторым растением. В вариантах осуществления способ дополнительно включает получение семени из трансгенного растения. В вариантах осуществления способ дополнительно включает получение потомства растения из трансгенного растения, где потомство растения содержит полинуклеотид, кассету экспрессии или вектор, экспрессирует химерный инсектицидный белок и обладает повышенной устойчивостью к насекомому-вредителю.
[0022] В качестве еще одного аспекта в настоящем изобретении предусмотрен способ получения трансгенного растения с повышенной устойчивостью к насекомому-вредителю (например, насекомому-вредителю, относящемуся к чешуекрылым), при этом способ включает: (а) посев семени, содержащего полинуклеотид, кассету экспрессии или вектор по настоящему изобретению; и (b) выращивание трансгенного растения из семени, где трансгенное растение содержит полинуклеотид, кассету экспрессии или вектор, и продуцирует химерный инсектицидный белок, и обладает повышенной устойчивостью к насекомому-вредителю. В вариантах осуществления способ дополнительно включает: (с) сбор семени от трансгенного растения из (b), где собранное семя содержит полинуклеотид, кассету экспрессии, вектор и/или химерный инсектицидный белок. Необязательно семя обладает повышенной устойчивостью к насекомому-вредителю (например, насекомому-вредителю, относящемуся к чешуекрылым).
[0023] Кроме того, в качестве другого аспекта в настоящем изобретении предусмотрен способ получения семени. В вариантах осуществления способ включает: (а) получение трансгенного растения, которое содержит полинуклеотид, кассету экспрессии или вектор по настоящему изобретению; и (b) сбор семени от трансгенного растения из (а), где собранное семя содержит полинуклеотид, кассету экспрессии, или вектор, и/или химерный инсектицидный белок по настоящему изобретению. Необязательно семя обладает повышенной устойчивостью к насекомому-вредителю (например, насекомому-вредителю, относящемуся к чешуекрылым).
[0024] В настоящем изобретении дополнительно рассматривается способ получения гибридного семени растения. В иллюстративных вариантах осуществления способ включает: (а) скрещивание первого инбредного растения, которое представляет собой трансгенное растение, содержащее полинуклеотид, кассету экспрессии или вектор по настоящему изобретению, с другим инбредным растением, которое может содержать или не содержать полинуклеотид, кассету экспрессии или вектор по настоящему изобретению; и (b) обеспечение возможности образования гибридного семени. В вариантах осуществления гибридное семя содержит полинуклеотид, кассету экспрессии, или вектор, и/или химерный инсектицидный белок по настоящему изобретению. Необязательно семя обладает повышенной устойчивостью к насекомому-вредителю (например, насекомому-вредителю, относящемуся к чешуекрылым).
[0025] В качестве другого аспекта в настоящем изобретении предусмотрен способ контроля насекомого-вредителя (например, насекомого-вредителя, относящегося к чешуекрылым, такого как совка травяная), при этом способ включает доставку в организм насекомого-вредителя или в среду его обитания композиции, содержащей эффективное количество химерного инсектицидного белка или инсектицидной композиции по настоящему изобретению. В вариантах осуществления данный способ представляет собой способ контроля насекомого-вредителя, относящегося к чешуекрылым (например, совки травяной), который устойчив к белку Vip3A и/или белку Cry1F.
[0026] Соответственно, в качестве другого аспекта в настоящем изобретении предусмотрен способ уменьшения развития устойчивости к белку Vip3A и/или белку Cry1F в популяции целевого насекомого-вредителя, относящегося к чешуекрылым (например, совки травяной). В вариантах осуществления способ включает доставку к целевой популяции или в среду ее обитания трансгенного растения, содержащего (i) полинуклеотид, кассету экспрессии или вектор по настоящему изобретению и (ii) полинуклеотид, содержащий нуклеотидную последовательность, кодирующую белок Vip3A, и/или нуклеотидную последовательность, кодирующую белок Cry1F; где химерный инсектицидный белок и белок Vip3A и/или белок Cry1F продуцируются в трансгенном растении.
[0027] Настоящее изобретение также направлено на способы применения полинуклеотидов по настоящему изобретению, например, в ДНК-конструкциях, или кассетах экспрессии, или векторах для трансформации и экспрессии в организмах, в том числе растениях и микроорганизмах, таких как бактерии. Нуклеотидные или аминокислотные последовательности могут представлять собой нативные или синтетические последовательности, которые были сконструированы для экспрессии в организме, таком как растение или бактерии. Кроме того, настоящее изобретение направлено на способы получения инсектицидных белков по настоящему изобретению и на способы применения полинуклеотидных последовательностей и инсектицидных белков, например, в микроорганизмах, для контроля насекомых, или в трансгенных растениях для обеспечения защиты от повреждения насекомыми.
[0028] Другой аспект по настоящему изобретению включает инсектицидные композиции и составы, содержащие химерные инсектицидные белки по настоящему изобретению, и способы применения композиций или составов для контроля популяций насекомых, например, путем применения композиций или составов в отношении пораженных насекомыми площадей, или для профилактической обработки восприимчивых к насекомым площадей или растений для обеспечения защиты против насекомых-вредителей. Необязательно композиции или составы по настоящему изобретению могут, в дополнение к химерному инсектицидному белку по настоящему изобретению, содержать другие пестицидные средства, такие как химические пестициды, другие пестицидные белки или dsRNA, например, для увеличения или усиления способности композиции или состава обеспечивать контроль насекомых и/или для контроля устойчивости насекомых к пестицидам.
[0029] Композиции и способы по настоящему изобретению являются применимыми для контроля насекомых-вредителей, которые поражают растения, в частности, культурные растения. Композиции по настоящему изобретению также являются применимыми для обнаружения присутствия химерного инсектицидного белка или нуклеиновой кислоты, кодирующей его, в коммерческих продуктах или трансгенных организмах.
[0030] В настоящем изобретении также предусмотрены пути применения химерных инсектицидных белков, нуклеиновых кислот, трансгенных растений, частей растений, семян и инсектицидных композиций по настоящему изобретению, например, для контроля насекомого-вредителя, такого как вредитель, относящийся к чешуекрылым.
[0031] В вариантах осуществления в настоящем изобретении предусмотрен способ применения полинуклеотида, кассеты экспрессии, вектора или клетки-хозяина по настоящему изобретению для получения инсектицидной композиции для контроля насекомого-вредителя (например, насекомого-вредителя, относящегося к чешуекрылым).
[0032] В вариантах осуществления в настоящем изобретении предусмотрен способ применения полинуклеотида, кассеты экспрессии или вектора по настоящему изобретению для получения трансгенного семени, где из трансгенного семени вырастает трансгенное растение с повышенной устойчивостью к насекомому-вредителю.
[0033] В качестве другого аспекта в настоящем изобретении также рассматривается применение трансгенного растения по настоящему изобретению для получения трансгенного семени, которое необязательно является гибридным семенем.
[0034] В вариантах осуществления в настоящем изобретении предусмотрен способ применения химерного инсектицидного белка, полинуклеотида, кассеты экспрессии, вектора, трансгенного растения или инсектицидной композиции по настоящему изобретению для предотвращения развития устойчивости в популяции целевого насекомого-вредителя, относящегося к чешуекрылым, к белку Vip3A и/или белку Cry1F.
[0035] Эти и другие признаки, аспекты и преимущества настоящего изобретения станут более понятны со ссылкой на следующее подробное описание и формулу изобретения.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0036] На фигурах 1А и 1В показано выравнивание аминокислотных последовательностей полноразмерного ВТ-0029 (SEQ ID NO: 2), ВТ-0022 (SEQ ID NO: 1), Cry1Fa (SEQ ID NO: 8) и Cry1Ka (SEQ ID NO: 12). Указаны сердцевина домена III и консервативный блок 3 (СВ3). Идентичные аминокислоты в выровненных последовательностях заштрихованы.
[0037] На фигурах 2А и 2В показано выравнивание аминокислотных
последовательностей полноразмерного ВТ-0029 (SEQ ID NO: 2) с иллюстративными химерами ВТ-0029: Bt29-Bt22 (SEQ ID NO: 3), Bt29-1Fa (SEQ ID NO: 9) и Bt29-1Ka (SEQ ID NO: 13). Указаны сердцевина домена III и консервативный блок 3 (СВ3). Сердцевина домена III, указанная на фигуре, получена из второго белка Cry.
Идентичные аминокислоты в выровненных последовательностях заштрихованы.
[0038] На фигурах 3А, 3В, 3С и 3D показана полноразмерная химера Bt29-Bt22 (SEQ ID NO: 3), выровненная с серией химер Bt29-Bt22, Bt29-Bt22Tr1 (SEQ ID NO: 20), Bt29-Bt22Tr2 (SEQ ID NO: 21), Bt29-Bt22Tr3 (SEQ ID NO: 22), Bt29-Bt22Tr4 (SEQ ID NO: 23), Bt29-Bt22Tr5 (SEQ ID NO: 24) и Bt29-Bt22Tr6 (SEQ ID NO: 25), с C-концевыми усечениями в пределах хвоста протоксина ВТ-0029.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ В ПЕРЕЧНЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0039] SEQ ID NO: 1 представляет собой аминокислотную последовательность белка ВТ-0022.
[0040] SEQ ID NO: 2 представляет собой аминокислотную последовательность белка ВТ-0029.
[0041] SEQ ID NO: 3 представляет собой аминокислотную последовательность химеры Bt29-Bt22. Последовательность ВТ-0022 представляет собой аминокислоты 459-597.
[0042] SEQ ID NO: 4 представляет собой нуклеотидную последовательность, кодирующую химеру Bt29-Bt22 под SEQ ID NO: 3.
[0043] SEQ ID NO: 5 представляет собой иллюстративную, оптимизированную для маиса последовательность, кодирующую химеру Bt29-Bt22 под SEQ ID NO: 3.
[0044] SEQ ID NO: 6 представляет собой иллюстративную, оптимизированную для маиса последовательность, кодирующую химеру Bt29-Bt22 под SEQ ID NO: 3.
[0045] SEQ ID NO: 7 представляет собой иллюстративную, оптимизированную для маиса последовательность, кодирующую химеру Bt29-Bt22 под SEQ ID NO: 3.
[0046] SEQ ID NO: 8 представляет собой аминокислотную последовательность полноразмерного Cry1Fa.
[0047] SEQ ID NO: 9 представляет собой аминокислотную последовательность химеры Bt29-1Fa. Последовательность Cry1Fa представляет собой аминокислоты 459-597.
[0048] SEQ ID NO: 10 представляет собой нуклеотидную последовательность, кодирующую химеру Bt29-1Fa под SEQ ID NO: 9.
[0049] SEQ ID NO: 11 представляет собой иллюстративную, оптимизированную для маиса последовательность, кодирующую химеру Bt29-1Fa под SEQ ID NO: 9.
[0050] SEQ ID NO: 12 представляет собой аминокислотную последовательность полноразмерного Cry1Ka.
[0051] SEQ ID NO: 13 представляет собой аминокислотную последовательность химеры Bt29-1Ka. Последовательность Cry1Ka представляет собой аминокислоты 459-597.
[0052] SEQ ID NO: 14 представляет собой нуклеотидную последовательность, кодирующую химеру Bt29-1Ka под SEQ ID NO: 13.
[0053] SEQ ID NO: 15 представляет собой аминокислотную последовательность химеры Bt29-1Kav2.
[0054] SEQ ID NO: 16 представляет собой нуклеотидную последовательность, кодирующую химеру Bt29-1Ka под SEQ ID NO: 15.
[0055] SEQ ID NO: 17 представляет собой аминокислотную последовательность полноразмерного Cry1Ca.
[0056] SEQ ID NO: 18 представляет собой аминокислотную последовательность химеры Bt29-1Ca. Последовательность Cry1Ca представляет собой аминокислоты 459-597.
[0057] SEQ ID NO: 19 представляет собой нуклеотидную последовательность, кодирующую химеру Bt29-1Ca под SEQ ID NO: 18.
[0058] SEQ ID NO: 20 представляет собой аминокислотную последовательность белка Bt29-Bt22Tr1.
[0059] SEQ ID NO: 21 представляет собой аминокислотную последовательность белка Bt29-Bt22Tr2.
[0060] SEQ ID NO: 22 представляет собой аминокислотную последовательность белка Bt29-Bt22Tr3.
[0061] SEQ ID NO: 23 представляет собой аминокислотную последовательность белка Bt29-Bt22Tr4.
[0062] SEQ ID NO: 24 представляет собой аминокислотную последовательность белка Bt29-Bt22Tr5.
[0063] SEQ ID NO: 25 представляет собой аминокислотную последовательность белка Bt29-Bt22Tr6.
[0064] SEQ ID NO: 26 представляет собой аминокислотную последовательность белка BT29BT22-TL22v1.
[0065] SEQ ID NO: 27 представляет собой аминокислотную последовательность белка BT29BT22-TL22v2.
[0066] SEQ ID NO: 28 представляет собой аминокислотную последовательность белка BT29BT22-TL22v3.
[0067] SEQ ID NO: 29 представляет собой аминокислотную последовательность белка BT29BT22-TL22v4.
[0068] SEQ ID NO: 30 представляет собой аминокислотную последовательность белка BT29BT22-TL22v5.
[0069] SEQ ID NO: 31 представляет собой аминокислотную последовательность белка BT29BT22-TL22v6.
[0070] SEQ ID NO: 32 представляет собой аминокислотную последовательность белка BT291FaTr1.
[0071] SEQ ID NO: 33 представляет собой аминокислотную последовательность белка BT291FaTr2.
[0072] SEQ ID NO: 34 представляет собой аминокислотную последовательность белка BT291FaTr3.
[0073] SEQ ID NO: 35 представляет собой аминокислотную последовательность белка BT291FaTr4.
ПОДРОБНОЕ ОПИСАНИЕ
[0074] Не подразумевается, что данное описание является подробным перечнем всех различных способов, с помощью которых может быть реализовано настоящее изобретение, или всех признаков, которые можно добавить к настоящему изобретению. Например, признаки, проиллюстрированные в отношении одного варианта осуществления, могут быть включены в другие варианты осуществления, а признаки, проиллюстрированные в отношении конкретного варианта осуществления, могут быть удалены из этого варианта осуществления. Таким образом, настоящим изобретением предполагается, что в некоторых вариантах осуществления настоящего изобретения можно исключить или опустить любой признак или комбинацию признаков, изложенных в данном документе. Кроме того, многочисленные вариации и дополнения к различным вариантам осуществления, предлагаемым в данном документе, будут очевидны для специалистов в данной области в свете настоящего раскрытия, которое не отступает от сути настоящего изобретения. Следовательно, следующие описания предназначены для иллюстрации некоторых конкретных вариантов осуществления настоящего изобретения, а не исчерпывающего определения всех их преобразований, комбинаций и вариаций.
[0075] Если не указано иное, все технические и научные термины, применяемые в данном документе, имеют то же значение, которое обычно понятно специалисту в области техники, к которой относится настоящее изобретение. Терминология, применяемая в данном документе при описании настоящего изобретения, предназначена только для описания конкретных вариантов осуществления и не предназначена для ограничения настоящего изобретения.
[0076] Все публикации, заявки на патенты, патенты и другие ссылки, цитируемые в данном документе, включены с помощью ссылки во всей своей полноте для объяснения идей, относящихся к предложению и/или абзацу, в котором приведена данная ссылка.
[0077] Нуклеотидные последовательности, обеспечиваемые в данном документе, представлены в направлении 5'-3' слева направо и представлены с применением стандартного кода для представления нуклеиновых оснований, как изложено в разделе 37 CFR, §§1.821-1.825 и стандарте ST. 25. Всемирной организации интеллектуальной собственности (WIPO), например: аденин (А), цитозин (С), тимин (Т) и гуанин (G).
[0078] Аналогичным образом, аминокислоты обозначены с применением стандарта ST.25 WIPO, например: аланин (Ala; А), аргинин (Arg; R), аспарагин (Asn; N), аспарагиновая кислота (Asp; D), цистеин (Cys; С), глутамин (Gln; Q), глутаминовая кислота (Glu; Е), глицин (Gly; G), гистидин (His; Н), изолейцин (Ile; 1), лейцин (Leu; L), лизин (Lys; K), метионин (Met; М), фенилаланин (Phe; F), пролин (Pro; Р), серии (Ser; S), треонин (Thr; Т), триптофан (Trp; W), тирозин (Tyr; Y) и валин (Val; V).
[0079] Если в контексте не указано иное, специально подразумевается, что различные признаки настоящего изобретения, описанные в данном документе, могут использоваться в любой комбинации. Более того, в настоящем изобретении также предполагается, что в некоторых вариантах осуществления настоящего изобретения любой признак или комбинацию признаков, изложенных в данном документе, можно исключить или опустить. С целью иллюстрации, если в данном описании утверждается, что композиция содержит компоненты А, В и С, то это, в частности, предполагает, что любое из А, В или С или их комбинацию можно опустить и отклонить по отдельности или в любой комбинации.
Определения
[0080] Как предполагается, используемые в описании настоящего изобретения и прилагаемой формуле изобретения формы единственного числа также включают формы множественного числа, если контекст явно не указывает иное.
[0081] Используемое в данном документе выражение "и/или" относится к любой и всем возможным комбинациям одного или нескольких соответствующих перечисленных элементов и охватывает их, а также к отсутствию комбинаций при интерпретации в качестве альтернативы ("или").
[0082] Выражение "приблизительно", используемое в данном документе при упоминании измеряемой величины, такой как дозировка или период времени и т.п., предназначено для охвата изменений определенного количества на ±20%, ±10%, ±5%, ±1%, ±0,5% или даже ±0,1%. Используемые в данном документе фразы, такие как "от приблизительно X до Y", означают "от приблизительно X до приблизительно Y", и фразы, такие как "от приблизительно X до Y", означают "от приблизительно X до приблизительно Y".
[0083] Используемые в данном документе фразы, такие как "от Х до Y", следует интерпретировать как включающие X и Y, если контекст не указывает иное.
[0084] Под "активностью" инсектицидного белка по настоящему изобретению подразумевается, что инсектицидный белок функционирует в качестве перорально активного средства для контроля насекомых, оказывает токсическое действие, например, путем подавления способности насекомого-вредителя к выживанию, росту и/или размножению (например, вызывая заболеваемость и/или смертность), и/или способен нарушать и/или сдерживать кормление насекомых, что может вызывать или не вызывать гибель насекомого. Таким образом, когда инсектицидный белок по настоящему изобретению доставлен в организм насекомого, результатом, как правило, является заболеваемость и/или смертность насекомого, и/или насекомое уменьшает или прекращает кормление на источнике, который делает инсектицидный белок доступным для насекомого.
[0085] В контексте настоящего изобретения "химерный" белок представляет собой белок, созданный путем слияния всего или части из по меньшей мере двух разных белков. В вариантах осуществления настоящего изобретения химерный белок представляет собой химерный белок Cry, содержащий все или часть из двух разных белков Cry, слитых вместе в один полипептид. "Химерный инсектицидный белок" представляет собой химерный белок, который обладает инсектицидной активностью (как описано в данном документе).
[0086] "Кодирующая последовательность" представляет собой последовательность нуклеиновой кислоты, которая транскрибируется в РНК, такую как mRNA, rRNA, tRNA, snRNA, смысловая РНК или антисмысловая РНК. В вариантах осуществления РНК затем транслируется с получением белка.
[0087] Используемое в данном документе выражение "кодон-оптимизированная" нуклеотидная последовательность означает нуклеотидную последовательность рекомбинантного, трансгенного или синтетического полинуклеотида, в которой кодоны выбраны так, чтобы отражать склонность к определенным кодонам, которая может наблюдаться в клетке- или организме-хозяине. Как правило, это выполняется таким образом, чтобы сохранить аминокислотную последовательность полипептида, кодируемого кодон-оптимизированной нуклеотидной последовательностью. В определенных вариантах осуществления нуклеотидная последовательность является кодон-оптимизированной для клетки (например, клетки животного, растения, гриба или бактерии), в которой конструкция будет экспрессироваться. Например, в конструкции, которая будет экспрессироваться в растительной клетке, могут быть кодон-оптимизированы вся последовательность или ее части для экспрессии в растении. См., например, патент США №6121014. В вариантах осуществления полинуклеотиды по настоящему изобретению кодон-оптимизированы для экспрессии в растительной клетке (например, клетке двудольного или клетке однодольного) или бактериальной клетке.
[0088] "Контроль" насекомых-вредителей означает подавление посредством токсического действия способности насекомых-вредителей к выживанию, росту, питанию и/или размножению, и/или ограничение связанного с насекомыми повреждения или гибели культурного растения, вызванной насекомым-вредителем, и/или защиту максимального потенциального урожая сельскохозяйственной культуры, вызванного вредителем, при выращивании в присутствии насекомого-вредителя. "Контроль" насекомого-вредителя может означать или может не означать уничтожение насекомого, хотя в вариантах осуществления настоящего изобретения "контроль" насекомого означает уничтожение насекомого.
[0089] Термин "содержание", "содержит" или "содержащий" при использовании в данном описании указывает на присутствие изложенных признаков, целых чисел, стадий, операций, элементов или компонентов, но не исключает присутствия или добавления одного или нескольких других признаков, целых чисел, стадий, операций, элементов, компонентов и/или их групп.
[0090] Используемая в данном документе переходная фраза "по сути состоящий из" (и ее грамматические варианты) означает, что объем пункта формулы изобретения следует интерпретировать как охватывающий указанные материалы или этапы, перечисленные в пункте формулы изобретения, "и таковые, которые существенно не изменяют основную и новую характеристику (характеристики)" заявленного изобретения. Таким образом, термин "по сути состоящий из" в случае, если он используется в пункте формулы настоящего изобретения, не предназначен для интерпретации как эквивалент термина "содержащий".
[0091] В контексте настоящего изобретения "соответствующий" или "соответствует" означает, что когда аминокислотные последовательности модифицированных или гомологичных белков выравнивают друг с другом, аминокислоты, которые "соответствуют" определенным перечисленным положениям в модифицированном или гомологичном белке, являются такими, которые выравниваются с этими положениями в эталонном белке, но не обязательно находятся именно в этих числовых положениях относительно конкретной эталонной аминокислотной последовательности по настоящему изобретению. Например, если SEQ ID NO: 2 (ВТ-0029) представляет собой эталонную последовательность и выровнена с SEQ ID NO: 1 (ВТ-0022), как на фигуре 1, последовательность TLEAVT, следующая сразу за доменом III в SEQ ID NO: 1 (ВТ-0022), "соответствует" последовательности TFEAEY, следующей сразу за доменом III в SEQ ID NO: 2 (ВТ-0029).
[0092] Применяемый в данном документе термин "белок Cry" означает инсектицидный белок типа кристаллического дельта-эндотоксина из Bacillus thuringiensis. Термин "белок Cry" может относиться к форме протоксина или любому его инсектицидно активному фрагменту или токсину, включая частично подвергнутую процессингу форму и форму зрелого токсина (например, без N-концевого пептидильного фрагмента и/или С-концевого хвоста протоксина).
[0093] Используемое в данном документе выражение "доставка" или "доставлять" (и грамматические варианты) композицию или инсектицидный белок означает, что композиция или инсектицидный белок вступает в контакт с насекомым, способствуя пероральному поеданию композиции или инсектицидного белка, что приводит к токсическому действию и контролю насекомого. Данную композицию или инсектицидный белок можно доставлять множеством известных путей, в том числе без ограничения посредством экспрессии в трансгенном растении, составленной(составленных) белковой(белковых) композиции(композиций), распыляемой(распыляемых) белковой(белковых) композиции(композиций), матрицы с приманкой или с помощью любой другой известной в данной области системы доставки белка.
[0094] Термин "домен" относится к набору аминокислот, консервативных в определенных положениях по длине выравнивания последовательностей эволюционно родственных белков. В то время как аминокислоты в других положениях гомологов могут отличаться, аминокислоты, которые являются высококонсервативными в определенных положениях, указывают на аминокислоты, которые, вероятно, являются необходимыми для структуры, стабильности или функции белка. Идентифицированные по их высокой степени консервативности в выравненных последовательностях семейства гомологов белков, они могут применяться в качестве идентификаторов для определения того, принадлежит ли любой рассматриваемый полипептид к ранее идентифицированной группе полипептидов.
[0095] "Эффективное для контроля насекомых количество" означает концентрацию инсектицидного белка, которая обеспечивает подавление посредством токсического действия способность насекомых к выживанию, росту, питанию и/или размножению или ограничивает связанные с насекомыми повреждение или потерю урожая сельскохозяйственной культуры. "Эффективное для контроля насекомых количество" может означать или может не означать уничтожение насекомого, хотя в вариантах осуществления оно означает уничтожение насекомого.
[0096] Используемый в данном документе термин "кассета экспрессии" означает молекулу нуклеиновой кислоты, способную управлять экспрессией по меньшей мере одного представляющего интерес полинуклеотида, такого как полинуклеотид, кодирующий инсектицидный белок по настоящему изобретению, в соответствующей клетке-хозяине, содержащую промотор, функционально связанный с представляющим интерес полинуклеотидом, который функционально связан с сигналом терминации. "Кассета экспрессии" также, как правило, содержит дополнительные полинуклеотиды для облегчения надлежащей трансляции представляющего интерес полинуклеотида. Кассета экспрессии также может содержать другие полинуклеотиды, которые не связаны с экспрессией представляющего интерес полинуклеотида, но которые присутствует из-за соответствующих сайтов рестрикции для извлечения кассеты из вектора экспрессии. В вариантах осуществления по меньшей мере один из компонентов в кассете экспрессии может быть гетерологичным (т.е. чужеродным) по отношению к по меньшей мере одному из других компонентов (например, гетерологичный промотор, функционально связанный с представляющим интерес полинуклеотидом). Кассета экспрессии может также представлять собой последовательность, которая встречается в природе, но была получена в рекомбинантной форме, пригодной для гетерологичной экспрессии. Тем не менее, как правило, кассета экспрессии является гетерологичной по отношению к хозяину, т.е. кассета экспрессии (или даже представляющий интерес полинуклеотид) не встречается в природе в клетке-хозяине и была введена в клетку-хозяина или ее клетку-предшественника с помощью процесса трансформации или процесса скрещивания. Экспрессия представляющего интерес полинуклеотида(полинуклеотидов) в кассете экспрессии обычно находится под контролем промотора. В случае многоклеточного организма, такого как растение, промотор может также быть специфическим или предпочтительным в отношении конкретной ткани, или органа, или стадии развития (как описано более подробно в данном документе). После трансформации в растение кассета экспрессии или ее фрагмент также могут называться "вставленным полинуклеотидом" или "полинуклеотидом вставки".
[0097] "Ген" определяется в данном документе как единица наследственности, содержащая один или несколько полинуклеотидов, которая занимает определенное местоположение на хромосоме или плазмиде, и которая содержит генетическую инструкцию для определенной характеристики или признака, свойственных организму.
[0098] Используемый в данном документе термин "протеаза пищеварительного канала" относится к протеазе, которая встречается в пищеварительном тракте насекомого в природе. Данная протеаза, как правило, участвует в переваривании поглощенных белков. Примеры протеаз пищеварительного канала включают трипсин, который, как правило, расщепляет пептиды с С-концевой части остатков лизина (К) или аргинина (R), и химотрипсин, который, как правило, расщепляет пептиды с C-концевой части фенилаланина (F), триптофана (W) или тирозина (Y).
[0099] Используемое в данном документе выражение "гетерологичный" означает чужеродный, экзогенный, ненативный и/или не встречающийся в естественных условиях. В вариантах осуществления "гетерологичный" полинуклеотид или полипептид представляет собой полинуклеотид или полипептид, в естественных условиях не связанный с клеткой-хозяином, в которую его вводят, в том числе не встречающиеся в естественных условиях множественные копии встречающейся в естественных условиях нуклеотидной последовательности или полипептида. В вариантах осуществления нуклеотидная последовательность является гетерологичной по отношению к другой последовательности, с которой она функционально связана, например, промотор может быть гетерологичным (т.е. чужеродным) по отношению к функционально связанной кодирующей последовательности.
[00100] Как используется в данном документе, "гомологичный" означает нативный. Например, гомологичная нуклеотидная последовательность или аминокислотная последовательность означает нуклеотидную последовательность или аминокислотную последовательность, которая в естественных условиях связана с клеткой-хозяином, в которую она встроена, гомологичная промоторная последовательность представляет собой промоторную последовательность, которая в естественных условиях связана с кодирующей последовательностью и т.п.
[00101] Термины "увеличивать", "увеличение", "увеличенный", "повышать", "повышенный", "повышение" и "улучшение" (и их грамматические варианты) и аналогичные термины, используемые в данном документе, описывают повышение контроля вредителя растений, например, путем приведения растения в контакт с полипептидом по настоящему изобретению (таким как, например, путем трансгенной экспрессии или способами местного применения). Это увеличение контроля может быть относительно уровня контроля вредителя растений в отсутствие полипептида по настоящему изобретению (например, растение, которое не экспрессирует трансгенно полипептид или не подвергается местной обработке с помощью полипептида). Таким образом, в вариантах осуществления термины "увеличивать", "увеличение", "увеличенный", "повышать", "повышенный", "повышение" и "улучшение" (и их грамматические варианты) и подобные термины могут указывать на повышение на по меньшей мере приблизительно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 100%, 125%, 150%, 200%, 300%, 400%, 500% или более по сравнению с подходящим контролем (например, растением, частью растения, растительной клеткой, которая не находится в контакте с полипептидом по настоящему изобретению).
[00102] Термин "инсектицидный", используемый в данном документе, определяется как токсическая биологическая активность, способная обеспечивать контроль насекомого-вредителя, необязательно, но предпочтительно путем его уничтожения.
[00103] Последовательность нуклеиновой кислоты является "изокодонной" по отношению к эталонной последовательности нуклеиновой кислоты в том случае, когда последовательность нуклеиновой кислоты кодирует полипептид, имеющий такую же аминокислотную последовательность, что и полипептид, кодируемый эталонной последовательностью нуклеиновой кислоты.
[00104] В иллюстративных вариантах осуществления молекулы нуклеиновых кислот, полинуклеотиды или полипептиды по настоящему изобретению являются "выделенными". "Выделенная" молекула нуклеиновой кислоты, полинуклеотид или белок и т.п. представляет собой молекулу нуклеиновой кислоты, полинуклеотид или белок и т.п., которые больше не находятся в своей природной среде. Выделенная молекула нуклеиновой кислоты, полинуклеотид или белок по настоящему изобретению могут находиться в очищенной форме или могут находиться в рекомбинантном хозяине, в таком как трансгенная бактерия или трансгенное растение. В вариантах осуществления выделенная молекула нуклеиновой кислоты, нуклеотидная последовательность или полипептид находятся в очищенной форме, то есть по меньшей мере частично отделены от по меньшей мере некоторых других компонентов встречающегося в естественных условиях организма или вируса, например, структурных компонентов клетки или вируса или других полипептидов или нуклеиновых кислот, обычно обнаруживаемых связанными с полинуклеотидом. В других вариантах осуществления "выделенная" молекула нуклеиновой кислоты, нуклеотидная последовательность или полипептид могут существовать в среде, не являющейся природной, например, такой как рекомбинантная клетка-хозяин. Таким образом, например, в отношении нуклеотидных последовательностей, термин "выделенная" может означать то, что нуклеотидная последовательность отделена от хромосомы и/или клетки, в которой она встречается в естественных условиях. Полинуклеотид также является выделенным, если он отделен от хромосомы и/или клетки, в которой он встречается в естественных условиях, и затем встроен в генетическое окружение, хромосому и/или клетку, в которых он в естественных условиях не встречается (например, другую клетку-хозяина, другие регуляторные последовательности и/или другое положение в геноме по сравнению с таковыми, обнаруживаемыми в природе). Соответственно, рекомбинантные молекулы нуклеиновой кислоты, нуклеотидные последовательности и кодируемые ими полипептиды являются "выделенными", поскольку они посредством вмешательства человека существуют отдельно от их естественной среды и, следовательно, не являются природными продуктами, однако в некоторых вариантах осуществления они могут быть введены в рекомбинантную клетку-хозяина и существовать в ней. В иллюстративных вариантах осуществления выделенная молекула нуклеиновой кислоты, выделенная нуклеотидная последовательность и/или выделенный полипептид являются чистыми на по меньшей мере приблизительно 1%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, или более.
[00105] Термин "мотив", или "консенсусная последовательность", или "сигнатура" относится к короткой консервативной области в последовательности эволюционно родственных белков. Мотивы часто являются высоко консервативными частями доменов, но могут также включать только часть домена или располагаться вне консервативного домена (если все аминокислоты мотива выходят за пределы определенного домена).
[00106] "Нативная" или "дикого типа" нуклеиновая кислота, нуклеотидная последовательность, полипептид или аминокислотная последовательность относится к встречающейся в естественных условиях или эндогенной нуклеиновой кислоте, нуклеотидной последовательности, полипептиду или аминокислотной последовательности. Таким образом, например, "mRNA дикого типа" представляет собой mRNA, которая встречается в естественных условиях в организме или является эндогенной по отношению к нему.
[00107] Термины "нуклеиновая кислота", "молекула нуклеиновой кислоты", "нуклеотидная последовательность", "олигонуклеотид" и "полинуклеотид" используются в данном документе взаимозаменяемо, если в контексте не указано иное, и относятся к гетерополимеру нуклеотидов. Эти термины включают без ограничения молекулы ДНК и РНК, включая cDNA, геномную ДНК, синтетическую (например, химически синтезированную) ДНК и РНК, плазмидную ДНК, mRNA, антисмысловую РНК и гибриды РНК/ДНК, любые из которых могут быть линейными или разветвленными, однонитевыми или двухнитевыми, или их комбинации. В случае если dsRNA получают синтетическим путем, менее распространенные основания, такие как инозин, 5-метилцитозин, 6-метиладенин, гипоксантин и другие, также можно применять для спаривания антисмысловых последовательностей, dsRNA и рибозимов. Например, полинуклеотиды, которые содержат С-5-пропиновые аналоги уридина и цитидина, как было показано, связывают РНК с высокой степенью сродства и являются сильными антисмысловыми ингибиторами экспрессии генов. Также можно производить другие модификации, например, модификацию фосфодиэфирного остова или 2'-гидрокси группы сахара рибозы РНК. В вариантах осуществления выражения "нуклеиновая кислота", "молекула нуклеиновой кислоты", "нуклеотидная последовательность", "олигонуклеотид" или "полинуклеотид" относятся к ДНК.
[00108] Под используемыми в данном документе выражениями "имеющий функциональную связь" или "функционально связанный" подразумевают, что элементы являются функционально связанными друг с другом, а также обычно являются физически связанными. Таким образом, используемые в данном документе выражения "имеющий функциональную связь" или "функционально связанный" относятся к нуклеотидным последовательностям в одной молекуле нуклеиновой кислоты, которые связаны функционально. Таким образом, первая нуклеотидная последовательность, имеющая функциональную связь со второй нуклеотидной последовательностью, означает ситуацию, когда первая нуклеотидная последовательность находится в функциональной взаимосвязи со второй нуклеотидной последовательностью. К примеру, промотор функционально связан с нуклеотидной последовательностью, если промотор влияет на транскрипцию или экспрессию указанной нуклеотидной последовательности. Специалистам в данной области будет понятно, что регуляторные последовательности (например, промотор) не обязательно должны быть смежными с той нуклеотидной последовательностью, с которой они функционально связаны, пока регуляторные последовательности выполняют функцию управления ее экспрессией. Таким образом, например, промежуточные нетранслируемые, но при этом транскрибируемые последовательности могут присутствовать между промотором и нуклеотидной последовательностью, и промотор может все еще считаться "имеющим функциональную связь" или "функционально связанным" с этой нуклеотидной последовательностью.
[00109] Используемый в данном документе термин "растение" относится к любому растению на любой стадии развития.
[00110] Любое растение (или группирования растений, например, в род или более высокий порядок классификации) можно использовать в осуществлении настоящего изобретения, включая покрытосемянные или голосемянные, однодольные или двудольные.
[00111] Типичные растения включают без ограничения кукурузу (Zea mays), канолу (Brassica napus, Brassica rapa ssp.), люцерну (Medicago saliva), рис (Oryza sativa, в том числе без ограничения разновидности Indica и/или Japonica), рапс (Brassica napus), рожь (Secale cereale), сорго (Sorghum bicolor, Sorghum vulgare), подсолнечник (Helianthus annus), пшеницу (Triticum aestivum), сою (Glycine max), табак (Nicotiana tobacum), картофель (Solanum tuberosum), арахис культурный (Arachis hypogaea), хлопчатник (Gossypium hirsutum), сладкий картофель (Ipomoea batatus), маниоку (Manihot esculenta), кофейное дерево (Cofea spp.), кокососвую пальму (Cocos nucifera), ананас (Ananas comosus), цитрусовые деревья (Citrus spp.), дерево какао (Theobroma cacao), чайное растение (Camellia sinensis), банан (Musa spp.), авокадо (Persea americana), фиговое дерево (Ficus casica), гуаву (Psidium guajava), манго (Mangifera indica), оливу (Olea europaea), дынное дерево (Carica papaya), анакардию западную (Anacardium occidentale), макадамию (Macadamia integrifolia), миндаль (Prunus amygdalus), виды сахарной свеклы (Beta vulgaris), яблоню (Malus pumila), ежевику (Rubus), клубнику (Fragaria), грецкий орех (Juglans regia), виноград (Vitis vinifera), абрикос (Prunus armeniaca), вишню (Prunus), персик (Prunus persica), сливу (Prunus domestica), грушевое дерево (Pyrus communis), арбуз (Citrullus vulgaris), ряску (Lemna spp.), виды овса (Avena sativa), ячмень (Hordium vulgare), овощи, декоративные растения, хвойные, и виды газонной травы (например, для декоративных целей, для зоны отдыха или для кормовых целей), и виды трав, образующих биомассу (например, просо прутьевидное и мискантус).
[00112] Овощи включают без ограничения виды пасленовых (например, томаты; Lycopersicon esculentum), салат-латук (например, Lactuca sativa), виды моркови (Caucus carota), цветную капусту (Brassica oleracea), сельдерей (Apium graveolens), баклажан (Solarium melongena), спаржу (Asparagus officinalis), бамию (Abelmoschus esculentus), зеленую фасоль (Phaseolus vulgaris), лимскую фасоль (Phaseolus limensis), виды гороха (Lathyrus spp.), представителей рода Cucurbita, таких как тыква Хаббарда (С. hubbard), тыква мускатная (С. moschata), тыква обыкновенная (С. реро), тыква-горлянка (С. crookneck), С. argyrosperma, С. argyrosperma ssp sororia, С. digitata, С. ecuadorensis, С. foetidissima, С. lundelliana и С. martinezii, и представителей рода Cucumis, таких как огурец (Cucumis sativus), канталупа (С. Cantalupensis) и дыня мускусная (С. melo).
[00113] Декоративные растения включают азалию (Rhododendron spp.), гортензию (Macrophylla hydrangea), гибискус (Hibiscus rosasanensis), виды розы (Rosa spp.), виды тюльпана (Tulipa spp.), виды нарцисса (Narcissus spp.), виды петунии (Petunia hybrida), гвоздику (Dianthus caryophyllus), пуансетию (Euphorbia pulcherima) и хризантему.
[00114] Хвойные, которые можно использовать в осуществлении настоящего изобретения, включают, например, сосны, такие как сосна ладанная (Pinus taeda), сосна Эллиота (Pinus elliotii), сосна желтая (Pinus ponderosa), сосна скрученная широкохвойная (Pinus contorta) и сосна лучистая (Pinus radiata); псевдотсуга Мензиса (Pseudotsuga menziesii); тсуга канадская (Tsuga canadensis); ель сизая (Picea glauca); секвойя вечнозеленая (Sequoia sempervirens); пихты, такие как пихта миловидная (Abies amabilis) и пихта бальзамическая (Abies balsamea); и кедры, такие как туя (Thuja plicata) и кипарис нутканский (Chamaecyparis nootkatensis).
[00115] Газонная трава включает без ограничения представителей рода Зойсия, представители рода Agrostis, виды овсяницы, виды мятлика, представителей рода Stenotaphrum, представителей рода Свинорой, виды бизоновой травы, виды плевела и виды ежи сборной.
[00116] Также включают растения, которые служат преимущественно в качестве лабораторных моделей, например, Arabidopsis.
[00117] "Растительная клетка" представляет собой структурную и физиологическую единицу растения, содержащую протопласт и клеточную стенку. Растительная клетка может находиться в форме выделенной одиночной клетки или культивируемой клетки или в качестве части более высокоорганизованной единицы, такой как, например, растительная ткань, орган растения или целое растение.
[00118] "Культура растительных клеток" означает культуру единиц растения, таких как, например, протопласты, клетки в клеточной культуре, клетки в растительных тканях, пыльца, пыльцевые трубки, семязачатки, зародышевые мешки, зиготы и зародыши на различных стадиях развития.
[00119] "Растительный материал" относится к листьям, стеблям, корням, цветкам или частям цветков, плодам, пыльце, яйцеклеткам, зиготам, семенам, черенкам, клеточным или тканевым культурам, или к любым другим частям или продуктам растения.
[00120] "Орган растения" представляет собой отдельную и визуально структурированную и дифференцированную часть растения, такую как корень, стебель, лист, цветочная почка или зародыш.
[00121] Используемое в настоящем документе выражение "часть растения" включает без ограничения зародыши, пыльцу, семяпочки, семена, листья, цветки, ветки, плоды, стебли, корни, кончики корней, пыльники и/или растительные клетки, включая те растительные клетки, которые являются интактными в растениях и/или частях растений, растительные протопласты, растительные ткани, культуры растительных тканей, растительные каллусы, скопления растительного материала и т.п.
[00122] Выражение "растительная ткань", применяемое в данном документе, означает группу растительных клеток, организованных в структурную и функциональную единицу. Предусматривается любая ткань растения in planta или в культуре. Данный термин включает без ограничения целые растения, органы растений, семена растений, тканевую культуру и любые группы растительных клеток, организованных в структурные или функциональные единицы. Применение данного термина в сочетании с любым специфическим типом растительной ткани, приведенным выше или иным образом охваченным данным определением, или без такового не предназначено для исключения любого другого типа растительной ткани.
[00123] "Представляющий интерес полинуклеотид" относится к любому полинуклеотиду, который при переносе в организм, например, растение, придает организму требуемую характеристику, такую как устойчивость к насекомым, устойчивость к болезням, толерантность к гербицидам, устойчивость к антибиотикам, улучшенная питательная ценность, улучшенные показатели в производственном процессе, продукция коммерчески ценных ферментов или метаболитов или измененная репродуктивная способность и т.п.
[00124] Под "частью" или "фрагментом" полипептида по настоящему изобретению следует понимать аминокислотную последовательность уменьшенной длины относительно эталонной аминокислотной последовательности полипептида по настоящему изобретению. Такая часть или фрагмент согласно настоящему изобретению может быть, в случае когда это является подходящим, включен в больший полипептид, частью которого он является (например, меченый или слитый белок). В вариантах осуществления "часть" или "фрагмент", по существу, сохраняет инсектицидную активность (например, по меньшей мере 40%, 50%, 60%, 70%, 80%, 85%, 90%, 95% или даже 100% активности полноразмерного белка или даже имеет большую инсектицидную активность, чем полноразмерный белок).
[00125] Термины "белок", "пептид" и "полипептид" используются в данном документе взаимозаменяемо.
[00126] Термин "промотор" относится к полинуклеотиду, как правило, расположенному выше (5') от кодирующего полинуклеотида, который осуществляет контроль экспрессии кодирующего полинуклеотида, обеспечивая узнавание РНК-полимеразой и другими факторами, необходимыми для правильной транскрипции.
[00127] Используемый в данном документе термин "протопласт" относится к выделенной растительной клетке без клеточной стенки или только с частями клеточной стенки.
[00128] Используемый в данном документе термин "рекомбинантный" относится к форме нуклеиновой кислоты (например, ДНК или РНК) или белка, или организма, которая обычно не будет встречаться в природе, и которая как таковая была создана посредством вмешательства человека. Используемая в данном документе "молекула рекомбинантной нуклеиновой кислоты" (и аналогичные термины) представляет собой молекулу нуклеиновой кислоты, содержащую комбинацию полинуклеотидов, которые в природе не встречаются вместе и являются результатом вмешательства человека, например, молекулу нуклеиновой кислоты, которая состоит из комбинации по меньшей мере двух полинуклеотидов, гетерологичных по отношению друг к другу, или молекулу нуклеиновой кислоты, синтезированную искусственно и содержащую полинуклеотид, который отличается от полинуклеотида, который будет в норме существовать в природе, или молекулу нуклеиновой кислоты, которая содержит трансген, искусственно введенный в геномную ДНК клетки-хозяина, и ассоциированную фланкирующую ДНК генома клетки-хозяина. Примером молекулы рекомбинантной нуклеиновой кислоты является молекула ДНК, полученная в результате вставки трансгена в геномную ДНК растения, что в конечном итоге может приводить к экспрессии молекулы рекомбинантной РНК или белка в данном организме. В вариантах осуществления "рекомбинантный" белок представляет собой белок, который обычно не существует в природе или присутствует в не встречающемся в естественных условиях окружении и экспрессируется из молекулы рекомбинантной нуклеиновой кислоты. Используемое в данном документе выражение "рекомбинантное растение" представляет собой растение, которое обычно не будет существовать в природе, и оно является результатом вмешательства человека и содержит рекомбинантный полинуклеотид (например, трансген или гетерологичную молекулу нуклеиновой кислоты, встроенную в его геном). В результате такого изменения генома рекомбинантное растение явно отличается от родственного растения дикого типа.
[00129] Термины "снижать", "сниженный", "снижение", "сокращение", "уменьшать" и "подавлять" (и их грамматические варианты) и аналогичные термины, используемые в данном документе, относятся к уменьшению выживаемости, роста и/или размножения вредителя растений, например, путем приведения растения в контакт с полипептидом по настоящему изобретению (таким как, например, путем трансгенной экспрессии или способами местного применения). Это уменьшение выживаемости, роста и/или размножения может быть относительно уровня, наблюдаемого в отсутствие полипептида по настоящему изобретению (например, растение, которое не экспрессирует трансгенно полипептид или не подвергается местной обработке с помощью полипептида). Таким образом, в вариантах осуществления термины "снижать", "сниженный", "снижение", "сокращение", "уменьшать" и "подавлять" (и их грамматические варианты) и аналогичные термины означают уменьшение на по меньшей мере приблизительно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более по сравнению с растением, которое не вступает в контакт с полипептидом по настоящему изобретению (например, растение, которое не экспрессирует трансгенно полипептид или не подвергается местной обработке с помощью полипептида). В иллюстративных вариантах осуществления снижение приводит к отсутствию или по существу отсутствию (т.е. незначительному количеству, например, менее чем приблизительно 10%, менее чем приблизительно 5% или даже менее чем приблизительно 1%) выявляемой выживаемости, роста и/или размножения вредителя растений.
[00130] "Регуляторный элемент" относится к нуклеотидной последовательности, участвующей в контроле экспрессии полинуклеотида. Примеры регуляторных элементов включают промоторы, сигналы терминации и нуклеотидные последовательности, которые облегчают правильную трансляцию полинуклеотида.
[00131] Используемый в данном документе термин "селектируемый маркер" означает нуклеотидную последовательность, которая при экспрессии приводит к отличительному фенотипу растения, части растения и/или растительной клетки, экспрессирующих маркер, и, таким образом, обеспечивает возможность отличать такие трансформированные растения, части растений и/или растительные клетки от тех, которые не имеют маркера. Такая нуклеотидная последовательность может кодировать либо селектируемый, либо подвергаемый скринингу маркер в зависимости от того, придает ли маркер признак, по которому можно провести отбор с помощью химических средств, например, путем применения селективного средства (например, антибиотика, гербицида и т.п.), или от того, является ли маркер просто признаком, который можно идентифицировать посредством наблюдения или тестирования, например, путем скрининга (например, признаком, определяемым в R-локусе).
[00132] Используемая в данном документе "специфическая активность" относится к количеству белка, необходимому для получения инсектицидного эффекта. Следовательно, если первый белок обладает более высокой специфической активностью, чем второй белок, это означает, что для получения инсектицидного эффекта в отношении такой же процентной доли насекомых необходимо меньшее количество первого белка по сравнению с количеством второго белка.
[00133] Фраза "по существу идентичный" в контексте двух последовательностей нуклеиновых кислот или двух аминокислотных последовательностей относится к двум или более последовательностям или подпоследовательностям, которые на по меньшей мере приблизительно 50% идентичны по нуклеотидам или аминокислотным остаткам при сравнении и выравнивании для максимального соответствия, что определяют при помощи одного из следующих алгоритмов сравнения последовательностей или путем визуальной проверки. В определенных вариантах осуществления по существу идентичные последовательности характеризуются по меньшей мере приблизительно 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или большей идентичностью по нуклеотидам или аминокислотным остаткам. В определенных вариантах осуществления значительная идентичность существует в пределах области последовательностей, длина которой составляет по меньшей мере приблизительно 50 остатков, 100 остатков, 150 остатков, 200 остатков, 250 остатков, 300 остатков, 350 остатков, 400 остатков или более. В дополнительных вариантах осуществления последовательности являются по существу идентичными, когда они идентичны на протяжении всей длины кодирующих областей.
[00134] "Идентичность" или "процент идентичности" относится к степени идентичности между двумя последовательностями нуклеиновых кислот или аминокислотными последовательностями. При сравнении последовательностей одна последовательность обычно выступает в качестве эталонной последовательности, с которой сравнивают тестируемые последовательности. При использовании алгоритма сравнения последовательностей тестируемую и эталонную последовательности вводят в компьютер, при необходимости задают координаты подпоследовательности, и задают программные параметры алгоритма сравнения последовательностей. Затем с помощью алгоритма сравнения последовательностей на основе заданных программных параметров вычисляют процентную идентичность последовательностей для тестируемой(тестируемых) последовательности(последовательностей) относительно эталонной последовательности.
[00135] При сравнении последовательностей одна последовательность обычно выступает в качестве эталонной последовательности, с которой сравнивают тестируемые последовательности. При использовании алгоритма сравнения последовательностей тестируемую и эталонную последовательности вводят в компьютер, при необходимости задают координаты подпоследовательности, и задают программные параметры алгоритма сравнения последовательностей. Затем с помощью алгоритма сравнения последовательностей на основе заданных программных параметров вычисляют процентную идентичность последовательностей для тестируемой(тестируемых) последовательности(последовательностей) относительно эталонной последовательности.
[00136] Оптимальное выравнивание последовательностей для сравнения можно проводить, например, с помощью алгоритма поиска локальной гомологии по Smith & Waterman, Adv. Appl. Math. 2: 482 (1981), с помощью алгоритма гомологичного выравнивания Нидлмана-Вунша, J. Mol. Biol. 48: 443 (1970), с помощью способа поиска сходства по Pearson & Lipman, Proc. Nat'l. Acad. Sci. USA 85: 2444 (1988), с помощью программной реализации данных алгоритмов (GAP, BESTFIT, FASTA и TFASTA из Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Мадисон, Висконсин) или с помощью визуального осмотра (см. в общем Ausubel et al., ниже).
[00137] Одним примером алгоритма, который подходит для определения процентной идентичности последовательностей и сходства последовательностей, является алгоритм BLAST, который описан в Altschul et al., J. Mol. Biol. 215: 403-410 (1990). Программное обеспечение для осуществления анализов BLAST находится в открытом доступе благодаря Национальному центру биотехнологической информации (http://www.ncbi.nlm.nih.gov/). Данный алгоритм предусматривает изначальную идентификацию пар последовательностей с наибольшим сходством (HSP) путем идентификации коротких "слов" длиной W в запрашиваемой последовательности, которые либо совпадают, либо удовлетворяют некоторому положительному пороговому показателю Т при выравнивании со "словом" такой же длины в последовательности из базы данных. Т называется пороговым показателем соседнего "слова" (Altschul et al., 1990). Эти исходные совпадения соседних "слов" выступают в качестве затравки для начала поисков с целью обнаружения более длинных HSP, содержащих их. Совпадения "слов" затем продлеваются в обоих направлениях вдоль каждой последовательности до тех пор, пока может увеличиваться совокупный показатель выравнивания. В случае нуклеотидных последовательностей совокупные показатели рассчитывают с применением параметров М (балл-вознаграждение, начисляемый за пару совпадающих остатков; всегда >0) и N (штрафной балл, начисляемый за несовпадающие остатки; всегда <0). В случае аминокислотных последовательностей, для расчета совокупного показателя применяют матрицу замен. Продление совпадений "слов" в каждом направлении прекращается, когда совокупный показатель выравнивания снижается на величину X от его максимального достигнутого значения, при этом совокупный показатель падает до нуля или ниже вследствие накопления одного или нескольких выравниваний остатков с отрицательными показателями, либо в случае достижения конца одной из последовательностей. Параметры W, Т и X алгоритма BLAST определяют чувствительность и скорость выравнивания. В программе BLASTN (для нуклеотидных последовательностей) по умолчанию используется длина "слова" (W), равная 11, ожидаемое значение (Е), равное 10, пороговое значение, равное 100, М=5, N=-4 и сравнение обеих нитей. В случае аминокислотных последовательностей программа BLASTP использует по умолчанию длину "слова" (W), составляющую 3, ожидаемое значение (Е), составляющее 10, а также матрицу замен BLOSUM62 (см. Henikoff & Henikoff, Proc. Natl. Acad. Sci. USA 89: 10915 (1989)).
[00138] В дополнение к расчету процентной идентичности последовательностей алгоритм BLAST также выполняет статистический анализ сходства между двумя последовательностями (см., например, Karlin & Altschul, Proc. Natl. Acad. Sci. USA 90: 5873-5787 (1993)). Одной мерой сходства, предоставляемой алгоритмом BLAST, является наименьшая суммарная вероятность (P(N)), являющаяся показателем вероятности, согласно которому совпадения между двумя нуклеотидными или аминокислотными последовательностями будут наблюдаться случайным образом. Например, тестируемая последовательность нуклеиновой кислоты считается подобной эталонной последовательности, если наименьшая суммарная вероятность при сравнении тестируемой последовательности нуклеиновой кислоты с эталонной последовательностью нуклеиновой кислоты составляет менее приблизительно 0,1, более предпочтительно менее приблизительно 0,01 и наиболее предпочтительно менее приблизительно 0,001.
[00139] Другой широко применяемой и признанной компьютерной программой для осуществления выравниваний последовательностей является CLUSTALW v1.6 (Thompson, et al. Nuc. Acids Res., 22: 4673-4680, 1994). Число совпадающих оснований или аминокислот делят на общее число оснований или аминокислот и умножают на 100 с получением процента идентичности. Например, если бы две последовательности из 580 пар оснований имели 145 совпавших оснований, то они были бы идентичны на 25 процентов. Если две сравниваемые последовательности имеют разную длину, то число совпадений делят на более короткую из двух длин. Например, если в белках из 200 и 400 аминокислот было 100 совпадавших аминокислот, то они идентичны на 50 процентов с учетом более короткой последовательности. Если длина более короткой последовательности составляет менее 150 оснований или 50 аминокислот, то число совпадений делят на 150 (в случае нуклеиновых оснований) или 50 (в случае аминокислот) и умножают на 100 с получением процента идентичности.
[00140] Две нуклеотидные последовательности также могут считаться по существу идентичными, если две последовательности гибридизируются друг с другом при жестких условиях. В иллюстративных вариантах осуществления две нуклеотидные последовательности, которые считаются по существу идентичными, гибридизируются друг с другом при очень жестких условиях.
[00141] Термины "жесткие условия" или "жесткие условия гибридизации" включают ссылку на условия, при которых нуклеиновая кислота будет избирательно гибридизироваться с целевой последовательностью до определяемой более высокой степени, чем с другими последовательностями (например, по меньшей мере в 2 раза более высокой по сравнению с нецелевой последовательностью), и необязательно могут, по существу, исключать связывание с нецелевыми последовательностями. Жесткие условия являются зависимыми от последовательностей и будут различаться при разных обстоятельствах. При помощи контроля жесткости условий гибридизации и/или отмывки можно идентифицировать целевые последовательности, которые могут быть до 100% комплементарными эталонной нуклеотидной последовательности. Альтернативно можно применять условия умеренной или даже низкой жесткости для обеспечения некоторого несовпадения последовательностей, чтобы определять меньшие уровни сходства последовательностей. Например, специалистам в данной области техники будет понятно, что для функционирования в качестве праймера или зонда последовательность нуклеиновой кислоты должна быть лишь достаточно комплементарной целевой последовательности для, по существу, связывания с ней с образованием таким образом стабильной двухнитевой структуры в применяемых условиях. Таким образом, праймеры или зонды можно применять при условиях высокой, умеренной или даже низкой жесткости. Аналогично условия низкой или умеренной жесткости могут быть предпочтительными для обнаружения гомологичных, ортологичных и/или паралогичных последовательностей, характеризующихся более низкими степенями идентичности последовательностей, чем те, которые были бы идентифицированы при очень жестких условиях.
[00142] Как используется в данном документе, термины "комплементарный" или "комплементарность" (и подобные термины) относятся к естественному связыванию полинуклеотидов в условиях содержания солей и температуры, допускающих это, путем спаривания оснований. Например, последовательность "A-G-T" связывается с комплементарной последовательностью "Т-С-А". Комплементарность между двумя однонитевыми молекулами может быть частичной, при которой связываются только некоторые из нуклеотидов, или она может быть полной, когда имеет место абсолютная комплементарность между одноцепочечными молекулами. Степень комплементарности между нитями нуклеиновой кислоты значительно влияет на эффективность и силу гибридизации между молекулами. Как используется в данном документе, термин "по существу комплементарный" (и подобные термины) означает, что две последовательности нуклеиновой кислоты комплементарны на по меньшей мере приблизительно 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или более. Альтернативно термин "по существу комплементарный" (и подобные термины) может означать, что две последовательности нуклеиновой кислоты могут гибридизироваться вместе в условиях высокой жесткости (как описано в данном документе).
[00143] Как используется в данном документе, "специфически" или "селективно" гибридизируется (и подобные термины) относится к связыванию, образованию дуплекса или гибридизации молекулы с конкретной целевой последовательностью нуклеиновой кислоты в жестких условиях, когда эта последовательность присутствует в сложной смеси (например, общей клеточной ДНК или РНК) с, по сути, исключением нецелевых нуклеиновых кислот или даже без выявляемого связывания, образования дуплекса или гибридизации с нецелевыми последовательностями. Специфически или селективно гибридизирующиеся последовательности обычно являются на по меньшей мере приблизительно 40% комплементарными и необязательно по существу комплементарными или даже полностью комплементарными (т.е. идентичными на 100%).
[00144] Для гибридов ДНК-ДНК Tm приблизительно вычислить из уравнения Meinkoth и Wahl, Anal. Biochem., 138:267-84 (1984): Tm=81,5°C+16,6 (log M)+0,41 (% GC>0,61 (% формамида) - 500/л; где M представляет собой молярность раствора одновалентных катионов, % GC представляет собой процентную долю гуанозиновых и цитозиновых нуклеотидов в ДНК, % формамида представляет собой процентную долю формамида в растворе для гибридизации, а L представляет собой длину гибрида в парах оснований. Tm представляет собой температуру (при заданных ионной силе и рН), при которой 50% комплементарной целевой последовательности гибридизируется с идеально совпадающим зондом. Tm снижают приблизительно на 1°С для каждого 1% несовпадения; таким образом, Tm, условия гибридизации и/или отмывки можно скорректировать для гибридизации с последовательностями с требуемой степенью идентичности. Например, если ищут последовательности с идентичностью >90%, Tm можно снизить на 10°С. В целом, жесткие условия выбирают такими, чтобы они были приблизительно на 5°С ниже, чем температура плавления (Tm) для специфической последовательности и ее комплементарной последовательности при определенных ионной силе и рН. Однако при очень жестких условиях можно использовать гибридизацию и/или отмывку при температуре плавления (Tm) или на 1, 2, 3 или 4°С ниже температуры плавления (Tm); при условиях умеренной жесткости можно использовать гибридизацию и/или отмывку при температуре на 6, 7, 8, 9 или 10°С ниже температуры плавления (Tm); при условиях низкой жесткости можно применять гибридизацию и/или отмывку при температуре на 11, 12, 13, 14, 15 или 20°C ниже температуры плавления (Tm). Если необходимая степень несовпадения обуславливает Tm ниже 45°С (водный раствор) или 32°С (раствор формамида), то необязательно концентрацию SSC можно повысить для того, чтобы можно было использовать более высокую температуру. Исчерпывающее руководство по гибридизации нуклеиновых кислот можно найти в Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, часть I, глава 2, "Overview of principles of hybridization and the strategy of nucleic acid probe assays," Elsevier, New York (1993); Current Protocols in Molecular Biology, глава 2, под редакцией Ausubel, et al. Greene Publishing and Wiley-Interscience, New York (1995); и Green & Sambrook в Molecular Cloning, A Laboratory Manual, 4-е издание, Cold Spring Harbor Press, Cold Spring Harbor, N.Y. (2012).
[00145] Как правило, жесткие условия являются такими, при которых концентрация соли составляет менее приблизительно 1,5 М ионов Na, как правило, концентрация ионов Na (или других солей) составляет приблизительно 0,01-1,0 М при приблизительно рН 7,0 - рН 8,3, а температура составляет по меньшей мере приблизительно 30°С для коротких зондов (например, 10-50 нуклеотидов) и по меньшей мере приблизительно 60°С для более длинных зондов (например, более 50 нуклеотидов). Жестких условий можно также достичь при добавлении дестабилизирующих средств, таких как формамид или раствор Денхардта (5 г Ficoll, 5 г поливинилпирролидона, 5 г бычьего сывороточного альбумина в 500 мл воды). Иллюстративные условия низкой жесткости включают гибридизацию с использованием буферного раствора 30%-35% формамида, 1 М NaCl, 1% SDS (додецилсульфата натрия) при 37°С и отмывку в 1Х-2Х SSC (20Х SSC=3,0 М NaCl/0,3 М цитрата тринатрия) при 50°С-55°С. Иллюстративные условия умеренной жесткости включают гибридизацию в 40-45% формамиде, 1 М NaCl, 1% SDS при 37°С и отмывку в 0,5Х-1Х SSC при 55°С-60°С. Иллюстративные условия высокой жесткости включают гибридизацию в 50% формамиде, 1 М NaCl, 1% SDS при 37°С и отмывку в 0,1X SSC при 60°С-65°С. Дополнительный неограничивающий пример условий высокой жесткости включает гибридизацию в 4Х SSC, 5Х растворе Денхардта, 0,1 мг/мл кипяченой ДНК из молок лососевых, 25 мМ фосфата Na при 65°С и отмывку в 0,1Х SSC, 0,1% SDS при 65°С. Другой пример условий гибридизации высокой жесткости включает гибридизацию в 7% SDS, 0,5 М NaPO4, 1 мМ EDTA при 50°C с отмывкой в 2Х SSC, 0,1% SDS при 50°С, альтернативно, с отмывкой в IX SSC, 0,1% SDS при 50°С, альтернативно, с отмывкой в 0,5Х SSC, 0,1% SDS при 50°С или, альтернативно, с отмывкой в 0,1Х SSC, 0,1% SDS при 50°С, или еще с отмывкой в 0,1Х SSC, 0,1% SDS при 65°С. Специалистам в данной области будет понятно, что специфичность, как правило, зависит от отмывок после гибридизации, при этом важными факторами являются ионная сила и температура конечного раствора для отмывки.
[00146] При этом нуклеиновые кислоты, которые не гибридизируются друг с другом при жестких условиях, являются по существу идентичными, если белки, которые они кодируют, являются по существу идентичными (например, вследствие вырожденности генетического кода).
[00147] Еще одним показателем того, что две нуклеиновые кислоты или белка являются по существу идентичными, является то, что белок, кодируемый первой нуклеиновой кислотой, характеризуется иммунологической перекрестной реактивностью с белком, кодируемым второй нуклеиновой кислотой. Таким образом, белок, как правило, является по существу идентичным второму белку, например, если два белка отличаются только консервативными заменами.
[00148] Как используется в данном документе, если модифицированный полипептид или фрагмент (и тому подобное) "по существу, сохраняет" инсектицидную активность, это означает, что модифицированный полипептид или фрагмент сохраняет по меньшей мере приблизительно 40%, 50%, 60%, 70%, 80%, 85%, 90%, 95% или даже 100% инсектицидной активности эталонного белка или характеризуется еще большей инсектицидной активностью.
[00149] "Синтетический" относится к нуклеотидной последовательности, содержащей основания или структурный(-е) признак(и), отсутствующие в природной последовательности. Например, синтетической считается искусственная последовательность, кодирующая белок по настоящему изобретению, которая по содержанию G+C и нормальному распределению кодонов больше похожа на гены двудольных или однодольных растений.
[00150] Используемый в данном документе белок, который является "токсичным" для насекомого-вредителя, представляет собой активное при пероральном применении средство контроля насекомых, которое убивает насекомое-вредителя, вызывает уменьшение роста и/или размножения насекомого-вредителя и/или способно нарушать или сдерживать кормление насекомого, последние два из которых могут вызвать или не вызвать гибель насекомого. Когда белок по настоящему изобретению доставляется в организм насекомого или насекомое вступает в контакт с белком, как правило, результатом является гибель насекомого, рост и/или размножение насекомого замедляется, и/или насекомое уменьшает или прекращает кормление на источнике, который делает токсичный белок доступным для насекомого.
[00151] Термины "токсиновый фрагмент" и "токсиновая часть" используются в данном документе взаимозаменяемо для обозначения фрагмента или части более длинного (например, полноразмерного) химерного инсектицидного белка по настоящему изобретению, где "токсиновый фрагмент" или "токсиновая часть" сохраняет инсектицидную активность. Например, в данной области техники известно, что нативные белки Cry экспрессируются в виде протоксинов, которые подвергаются процессингу с N-конца и С-конца с получением зрелого токсина. В вариантах осуществления "токсиновый фрагмент" или "токсиновая часть" химерного инсектицидного белка по настоящему изобретению усекается с N-конца и/или С-конца. В вариантах осуществления "токсиновый фрагмент" или "токсиновая часть" усекается с N-конца с удалением части или всего N-концевого пептидильного фрагмента и необязательно содержит по меньшей мере приблизительно 400, 425, 450, 475, 500, 510 520, 530, 540, 550, 560, 570, 580 или 590 смежных аминокислот химерного инсектицидного белка, конкретно описанного в данном документе, или аминокислотную последовательность, которая по существу идентична ему. Таким образом, в вариантах осуществления "токсиновый фрагмент" или "токсиновая часть" химерного инсектицидного белка усекается с N-конца (например, чтобы исключить часть или весь пептидильный фрагмент), например, N-концевое усечение одной аминокислоты или более одной аминокислоты, например, до 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60 или более аминокислот. В вариантах осуществления "токсиновый фрагмент" или "токсиновая часть" химерного инсектицидного белка усекается с С-конца (например, чтобы исключить часть или весь хвост протоксина), например, C-концевое усечение одной аминокислоты или более чем одной аминокислоты, например, до 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 70, 80, 90, 100, 125, 150, 175, 200, 225, 250, 275, 300, 325, 350, 375, 400, 425, 450, 475, 500, 525, 550, 560 или более аминокислот. В вариантах осуществления "токсиновый фрагмент" или "токсиновая часть" включает домены 1 и 2 и сердцевинный домен III (например, как показано на фигуре 2). В вариантах осуществления "токсиновый фрагмент" или "токсиновая часть" представляет собой зрелый (т.е. подвергнутый процессингу) токсин (например, токсин Cry).
[00152] "Трансформация" представляет собой процесс введения гетер о логичной нуклеиновой кислоты в клетку- или организм-хозяин. В конкретных вариантах осуществления "трансформация" означает стабильную интеграцию молекулы ДНК в геном представляющего интерес организма (например, растительную клетку).
[00153] Используемые в данном документе термины "трансформированный" и "трансгенный" относятся к организму-хозяину, такому как бактерия или растение, в который ввели гетерологичную молекулу нуклеиновой кислоты. Молекулу нуклеиновой кислоты можно стабильно интегрировать в геном хозяина, или же молекула нуклеиновой кислоты также может присутствовать в виде внехромосомной молекулы. Такая внехромосомная молекула может быть автореплицирующейся. Подразумевается, что "трансформированные" или "трансгенные" клетки, ткани или растения охватывают не только конечный продукт процесса трансформации, но также и их потомство, содержащее гетерологичную молекулу нуклеиновой кислоты. "Нетрансформированный" или "нетрансгенный" хозяин относится к организму дикого типа, например, бактерии или растению, который не содержит гетерологичную молекулу нуклеиновой кислоты.
[00154] Выражение "вектор" относится к композиции для передачи, доставки или введения нуклеиновой кислоты (или нуклеиновых кислот) в клетку. Вектор содержит молекулу нуклеиновой кислоты, содержащую нуклеотидную(нуклеотидные) последовательность(последовательности), подлежащую(подлежащие) переносу, доставке или введению.
Химерные инсектицидные белки
[00155] В настоящем изобретении предусмотрены новые химерные инсектицидные белки, содержащие по меньшей мере одну область из первого белка Cry (например, белка ВТ-0029 [SEQ ID NO: 2] и его по существу идентичных вариантов). В вариантах осуществления в настоящем изобретении предусмотрен химерный инсектицидный белок, содержащий область из двух или больше различных белков Cry. В вариантах осуществления полученный химерный инсектицидный белок характеризуется повышенной активностью в отношении одного или нескольких насекомых-вредителей (например, усиленной активностью или активностью в отношении нового целевого вредителя) и/или другим способом действия в отношении одного или нескольких насекомых-вредителей по сравнению с одним или более (или даже всеми) из исходных белков. В иллюстративных вариантах осуществления химерный инсектицидный белок представляет собой химеру, содержащую области от двух разных белков Cry (например, Cry1), и химера характеризуется повышенной инсектицидной активностью в отношении одного или нескольких насекомых-вредителей по сравнению с обоими исходными белками.
[00156] В вариантах осуществления химерные инсектицидные белки по настоящему изобретению могут обеспечивать новые способы действия в отношении одного или нескольких целевых насекомых-вредителей. Например, химерный инсектицидный белок может обладать инсектицидной активностью в отношении насекомых-вредителей или колоний, которые обычно устойчивы к инсектицидной активности другого средства (например, инсектицидного белка, включая без ограничения белок Bt, такой как белок Cry или белок Vip). В вариантах осуществления исходные белки сами по себе не являются инсектицидно активными или только слабо активны в отношении устойчивых насекомых-вредителей или колоний, что может указывать на то, что химера обладает новым способом действия по сравнению с исходными белками. Например, если исходные белки активны в отношении целевого насекомого-вредителя ("восприимчивого"), но не в отношении устойчивого аналога, а химерный инсектицидный белок активен в отношении устойчивого насекомого-вредителя, это указывает на то, что химерный инсектицидный белок является токсичным в отношении устойчивого вредителя благодаря новому способу действия.
[00157] Соответственно, в вариантах осуществления в настоящем изобретении предусмотрен химерный инсектицидный белок, который токсичен для насекомого-вредителя (например, насекомого-вредителя, относящегося к чешуекрылым), при этом химерный инсектицидный белок содержит область из белка ВТ-0029 (SEQ ID NO: 2) или полипептид, который по существу идентичен области из белка ВТ-0029. В вариантах осуществления химерный инсектицидный белок содержит N-концевую область первого белка Cry1, который необязательно является белком ВТ-0029 (SEQ ID NO: 2), или полипептид, содержащий аминокислотную последовательность, которая по существу идентична N-концевой области белка ВТ-0029. В вариантах осуществления N-концевая область первого белка Cry слита с С-концевой областью из другого белка Cry (например, другого белка Cry1) с образованием химерного инсектицидного белка (например, химерного инсектицидного белка Cry).
[00158] В иллюстративных вариантах осуществления С-концевая область из другого белка Cry может представлять собой С-концевую область другого белка Cry1 или полипептид, содержащий аминокислотную последовательность, которая по существу идентична С-концевой области из другого белка Cry1.
[00159] Таким образом, в иллюстративном варианте осуществления химерный инсектицидный белок по настоящему изобретению содержит, по сути состоит или состоит, в направлении OTN-конца к С-концу, из следующего: (а) N-концевая область первого белка Cry (например, белка Cry1), которая необязательно представляет собой N-концевую областью белка ВТ-0029 или аминокислотную последовательность, которая по существу идентична N-концевой области белка ВТ-0029, слитая с (b) С-концевой областью другого белка Cry (например, другого белка Cry1).
[00160] В иллюстративных вариантах осуществления другой белок Cry (например, белок Cry1) выбран на основании выравнивания С-концевой области из другого белка Cry и соответствующей области из первого белка Cry (например, белка Cry1), например, см. иллюстративные выравнивания на фигурах 1А и 1 В. В вариантах осуществления другой белок Cry выбран таким образом, что аминокислотная последовательность C-концевой области другого белка Cry по существу идентична соответствующей области первого белка Cry. В иллюстративных вариантах осуществления аминокислотная последовательность С-концевой области другого белка Cry (например, белка Cry1) на по меньшей мере приблизительно 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности соответствующей С-концевой области из первого белка Cry. Способы выравнивания и определения идентичности аминокислотных последовательностей в выровненной области хорошо известны специалистам в данной области техники.
[00161] В вариантах осуществления другой белок Cry1 включает в себя без ограничения белок Cry1F (например, белок Cry1Fa или белок Cry1Fb), белок Cry1G (например, белок Cry1Ga, Cry1Gb или Cry1Gc), белок Cry1I (например, белок Cry1Ia, белок Cry1Ib, белок Cry1Ic, белок Cry1Id, белок Cry1Ie, белок Cry1If или белок Cry1Ig), Cry1K (например, белок Cry1Ka) или Cry1C (например, белок Cry1Ca). В вариантах осуществления другой белок Cry1 представляет собой белок Cry1 If, необязательно белок ВТ-0022 (SEQ ID NO: 1). В вариантах осуществления другой белок Cry1 представляет собой белок Cry1Fa (SEQ ID NO: 8). В вариантах осуществления другой белок Cry1 представляет собой белок Cry1K (SEQ ID NO: 12). В вариантах осуществления другой белок Cry1 представляет собой белок Cry1Ca (SEQ ID NO: 17).
[00162] Термины "N-концевая область" и "С-концевая область" не обязательно указывают на то, что большинство N-концевых или С-концевых аминокислот (например, N-конец или С-конец) соответственно из полноразмерного белка включены в область. Например, специалистам в данной области техники хорошо известно, что протоксины Cry подвергаются процессингу как с N-конца, так и с С-конца с получением зрелого (т.е. подвергнутого процессингу) токсина. Таким образом, в вариантах осуществления "N-концевая область" и/или "С-концевая область" исключает некоторые или все из подвергнутых процессингу частей протоксина, так что химерный инсектицидный белок содержит зрелый белок токсина (например, домены I, II и III белка Cry) без части или всего N-концевого пептидильного фрагмента и/или С-концевого хвоста протоксина, или полипептид, который по существу идентичен зрелому белку токсина. В вариантах осуществления химерный инсектицидный белок содержит пептидильный фрагмент и/или хвост протоксина. В вариантах осуществления химерный инсектицидный белок не содержит пептидильный фрагмент или хвост протоксина, т.е. соответствует зрелому подвергнутому процессингу токсину.
[00163] В вариантах осуществления N-концевая область первого белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39 или 40 до приблизительно положения 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499 или 500 SEQ ID NO: 2 (полноразмерный ВТ-0029) и любой комбинации более низких и более высоких положений, как если бы каждая такая комбинация была конкретно изложена в данном документе.
[00164] В вариантах осуществления N-концевая область первого белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 1 до приблизительно положения 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490 или 491 SEQ ID NO: 2.
[00165] В иллюстративном варианте осуществления N-концевая область первого белка Cry1 содержит, по сути состоит или состоит из аминокислотной последовательности, соответствующей аминокислотной последовательности от приблизительно положения 1 до приблизительно аминокислотного положения 458 SEQ ID NO: 2.
[00166] В иллюстративных вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499 или 500 до приблизительно положения 580, 581, 582, 583, 584, 585, 586, 587, 588, 589, 590, 591, 592, 593, 594, 595, 596, 587, 598, 599, 600, 601, 602, 603, 604, 605, 606, 607, 608, 609, 610, 611, 612, 613, 614, 615, 616, 617, 618, 619 или 620 SEQ ID NO: 8 (полноразмерный Cry 1 Fa) и любой комбинации более низких и более высоких положений, как если бы каждая такая комбинация была конкретно изложена в данном документе.
[00167] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 464 до приблизительно положения 580, 581, 582, 583, 584, 585, 586, 587, 588, 589, 590, 591, 592, 593, 594, 595, 596, 587, 598, 599, 600, 601, 602, 603, 604, 605, 606, 607, 608, 609, 610, 611, 612, 613, 614, 615, 616, 617, 618, 619 или 620 SEQ ID NO: 8.
[00168] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499 или 500 до приблизительно положения 602 SEQ ID NO: 8.
[00169] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 464 до приблизительно положения 602 SEQ ID NO: 8.
[00170] В иллюстративных вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499, 500, 501, 502, 503, 504, 505, 506, 507, 508, 509, 510, 511, 512, 513, 514, 515, 516, 517, 518, 519, 520, 521, 522, 523, 524, 525, 526, 527, 528, 529, 530, 531, 532, 533, 534, или 535 до приблизительно положения 615, 616, 617, 618, 619, 620, 621, 622, 623, 624, 625, 626, 627, 628, 629, 630, 631, 632, 633, 634, 635, 636, 637, 638, 639, 640, 641, 642, 643, 644, 645, 646, 647, 648, 649, 650, 651, 652, 653, 654 или 655 SEQ ID NO: 1 (полноразмерный ВТ-0022) и любой комбинации более низких и более высоких положений, как если бы каждая такая комбинация была конкретно изложена в данном документе.
[00171] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 498 до приблизительно положения 615, 616, 617, 618, 619, 620, 621, 622, 623, 624, 625, 626, 627, 628, 629, 630, 631, 632, 633, 634, 635, 636, 637, 638, 639, 640, 641, 642, 643, 644, 645, 646, 647, 648, 649, 650, 651, 652, 653, 654 или 655 SEQ ID NO: 1.
[00172] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499, 500, 501, 502, 503, 504, 505, 506, 507, 508, 509, 510, 511, 512, 513, 514, 515, 516, 517, 518, 519, 520, 521, 522, 523, 524, 525, 526, 527, 528, 529, 530, 531, 532, 533, 534, или 535 до приблизительно положения 636 SEQ ID NO: 1.
[00173] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 498 до приблизительно положения 636 SEQ ID NO: 1.
[00174] В иллюстративных вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499, 500, 501, 502, 503, 504, 505, 506, 507, 508, 509, 510, 511, 512, 513, 514, 515, 516, 517, 518, 519, 520, 521, 522, 523, 524, 525, 526, 527, 528, 529, 530, 531, 532, 533, 534, 535, 536 или 537 до приблизительно положения 590, 591, 592, 593, 594, 595, 596, 597, 598, 599, 600, 601, 602, 603, 604, 605, 606, 607, 608, 609, 610, 611, 612, 613, 614, 615, 616, 617, 618, 619, 620, 621, 622, 623, 624, 625, 626, 627, 628, 629, 630, 631, 632, 633, 634, 635, 636, 637, 638, 639, 640, 641, 642, 643, 644, 645, 646, 647, 648, 649, 650, 651, 652, 653, 654, 655, 656 или 657 SEQ ID NO: 12 (полноразмерный Cry1Ka) и любой комбинации более низких и более высоких положений, как если бы каждая такая комбинация была конкретно изложена в данном документе.
[00175] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 500 до приблизительно положения 590, 591, 592, 593, 594, 595, 596, 597, 598, 599, 600, 601, 602, 603, 604, 605, 606, 607, 608, 609, 610, 611, 612, 613, 614, 615, 616, 617, 618, 619, 620, 621, 622, 623, 624, 625, 626, 627, 628, 629, 630, 631, 632, 633, 634, 635, 636, 637, 638, 639, 640, 641, 642, 643, 644, 645, 646, 647, 648, 649, 650, 651, 652, 653, 654, 655, 656 или 657 SEQ ID NO: 12.
[00176] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499, 500, 501, 502, 503, 504, 505, 506, 507, 508, 509, 510, 511, 512, 513, 514, 515, 516, 517, 518, 519, 520, 521, 522, 523, 524, 525, 526, 527, 528, 529, 530, 531, 532, 533, 534, 535, 536, 537 до приблизительно положения 597 SEQ ID NO: 12.
[00177] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499, 500, 501, 502, 503, 504, 505, 506, 507, 508, 509, 510, 511, 512, 513, 514, 515, 516, 517, 518, 519, 520, 521, 522, 523, 524, 525, 526, 527, 528, 529, 530, 531, 532, 533, 534, 535, 536, 537 до приблизительно положения 610 SEQ ID NO: 12.
[00178] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 500 до приблизительно положения 597 SEQ ID NO: 12.
[00179] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 500 до приблизительно положения 610 SEQ ID NO: 12.
[00180] В иллюстративных вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466 или 467 до приблизительно положения 580, 581, 582, 583, 584, 585, 586, 587, 588, 589, 590, 591, 592, 593, 594, 595, 596, 597, 598, 599, 600, 601, 602, 603, 604, 605, 606, 607, 608, 609, 610, 611, 612, 613, 614, 615, 616 или 617 из SEQ ID NO: 17 (полноразмерный Cry1Ca) и любой комбинации более низких и более высоких положений, как если бы каждая такая комбинация была конкретно изложена в данном документе.
[00181] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 467 до приблизительно положения 580, 581, 582, 583, 584, 585, 586, 587, 588, 589, 590, 591, 592, 593, 594, 595, 596, 597, 598, 599, 600, 601, 602, 603, 604, 605, 606, 607, 608, 609, 610, 611, 612, 613, 614, 615, 616 или 617 SEQ ID NO: 17.
[00182] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466 или 467 до приблизительно положения 617 SEQ ID NO: 17.
[00183] В вариантах осуществления С-концевая область второго белка Cry1 содержит, по сути состоит или состоит из полипептида, соответствующего аминокислотной последовательности от приблизительно положения 467 до приблизительно положения 617 SEQ ID NO: 17.
[00184] Как хорошо известно в данной области, белки Cry из Bt имеют 5 доменов с консервативной последовательностью (консервативный блок [СВ] 1-5]), разделенных более вариабельными областями (Hofte & Whitely, 1989, Microbiol. Rev. 53:242-255), и три домена с консервативной структурой (домены I, II и III) (de Maagd et al., 2001, Trends Genetics 17:193-199). На фигурах 1A и 1B показано выравнивание белков Cry ВТ-0029, BT-0022, Cry1Fa и Cry1Ka с отмеченными СВ3 и доменом III. Специалисты в данной области могут использовать широко известную информацию о последовательности и структуре, доступную для белков Cry, для создания химерного инсектицидного белка Cry в соответствии с настоящим изобретением, например, для выбора подходящей(подходящих) области(областей) перехода между двумя белками Cry. В вариантах осуществления область перехода расположена в консервативном блоке, например, в СВ3. Термин "в консервативном блоке" включает в себя положения на каждом конце консервативного блока. Чтобы проиллюстрировать это, термин "в СВ3" включает положение непосредственно перед (например, между остатками метионина и фенилаланина в пределах последовательности VPMFSW в последовательности ВТ-0029 на фигуре 2А) и сразу после (например, между остатками треонина и аспарагина в пределах последовательности RRTNVG в последовательности ВТ-0029 на фигуре 2А) консервативного блока 3. На фигурах 2А и 2 В показаны иллюстративные химерные инсектицидные белки в соответствии с настоящим изобретением с областью перехода в СВ3.
[00185] Со ссылкой на фигуру 2А в вариантах осуществления переход располагается в положении, соответствующем положению непосредственно перед аминокислотным остатком 1 или сразу после аминокислотного остатка 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50 или 51, или любой их комбинации в СВ3. В вариантах осуществления переход располагается в пределах области (включая остатки на каждом конце области), соответствующей участку от аминокислотного остатка 2 до аминокислотного остатка 7, 8, 9 ли 10 СВ3, в пределах области, соответствующей участку от аминокислотного остатка 11 до аминокислотного остатка 14, 15, 16, 17 или 18 СВ3, в пределах области, соответствующей участку от аминокислотного остатка 19 до аминокислотного остатка 27, 28, 29, 30, 31, 32, 33, 34, 35 или 36 СВ3, и/или в пределах области, соответствующей участку от аминокислотного остатка 37, 38 или 39 до аминокислотного остатка 51, и любой комбинации таких областей. В иллюстративных вариантах осуществления переход располагается в положении в пределах области, соответствующей участку от аминокислотного остатка 19 до аминокислотного остатка 51, от аминокислотного остатка 19 до аминокислотного остатка 33, от аминокислотного остатка 19 до аминокислотного остатка 28, от аминокислотного остатка 19 до аминокислотного остатка 27 или от аминокислотного остатка 19 до аминокислотного остатка 26 в СВ3. В вариантах осуществления положение перехода располагается в специфическом сайте между аминокислотным остатком 18 и аминокислотным остатком 19 в СВ3 (например, конкретно между валином и изолейцином в последовательности DPDVITQ в ВТ-0029, SEQIDNO: 2).
[00186] Химерные инсектицидные белки также можно определить в отношении структурных доменов, полученных из каждой из исходных молекул. Например, в вариантах осуществления N-концевая область первого белка Cry содержит домен I первого белка Cry и весь или, по сути, весь домен П. В вариантах осуществления С-концевая область другого белка Cry содержит весь или, по сути, весь домен III другого белка Cry. Специалисты в данной области понимают, что существует некоторая вариабельность в определении точного местоположения концов структурных доменов белка Cry и линкерных областей между доменами, например, расположения конца линкерной области между доменами II и III и начала домена III, хотя сердцевина домена III легко идентифицируется специалистами в данной области (например, в соответствии с расположением основного домена III, как показано на фигуре 1А и фигуре 2А). В вариантах осуществления переход между N-концевой областью первого белка Cry и С-концевой областью, полученной из другого белка Cry, располагается в СВ3.
[00187] В иллюстративных вариантах осуществления С-концевую область другого белка Cry (например, белка Cry1) выбирают на основании выравнивания домена III (например, сердцевинного домена III, как показано на фигуре 1А) из другого белка Cry и соответствующей последовательности домена III из первого белка Cry (например, такого белка Cry1, как ВТ-0029), например, см. иллюстративные выравнивания на фигурах 1А и 1В с конкретно отмеченным доменом III. В вариантах осуществления другой белок Cry выбран таким образом, что аминокислотная последовательность домена III другого белка Cry по существу идентична соответствующей области домена III первого белка Cry (например, ВТ-0029). В иллюстративных вариантах осуществления аминокислотная последовательность области домена III другого белка Cry (например, белка Cry1) на по меньшей мере приблизительно 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична аминокислотной последовательности соответствующей области домена III из первого белка Cry. Способы выравнивания и определения идентичности аминокислотных последовательностей в выровненной области хорошо известны специалистам в данной области техники.
[00188] В конкретных вариантах осуществления химерный инсектицидный белок по настоящему изобретению содержит, по сути состоит или состоит из (а) аминокислотной последовательности из любой из аминокислот 1-597 из SEQ ID NO: 3, SEQ ID NO: 9 или SEQ ID NO: 13, или аминокислот 1-610 из SEQ ID NO: 15, или аминокислот 1-617 из SEQ ID NO: 18, или токсинового фрагмента из любой из SEQ ID NO: 3, 9, 13, 15 или 18; или (b) аминокислотной последовательности, которая по существу идентична аминокислотной последовательности из (а).
[00189] В конкретных вариантах осуществления химерный инсектицидный белок по настоящему изобретению содержит, по сути состоит или состоит из (а) аминокислотной последовательности из любой из аминокислот 1-603 из SEQ ID NO: 3, SEQ ID NO: 9 или SEQ ID NO: 13, или аминокислот 1-615 из SEQ ID NO: 15 или SEQ ID NO: 18, или их токсинового фрагмента; или (b) аминокислотной последовательности, которая по существу идентична аминокислотной последовательности из (а).
[00190] В конкретных вариантах осуществления химерный инсектицидный белок по настоящему изобретению содержит, по сути состоит или состоит из (а) аминокислотной последовательности из любой из аминокислот 1-607 из SEQ ID NO: 3, SEQ ID NO: 9 или SEQ ID NO: 13, или аминокислот 1-620 из SEQ ID NO: 15 или SEQ ID NO: 18, или их токсинового фрагмента; или (b) аминокислотной последовательности, которая по существу идентична аминокислотной последовательности из (а).
[00191] В конкретных вариантах осуществления химерный инсектицидный белок по настоящему изобретению содержит, по сути состоит или состоит из (а) аминокислотной последовательности из любой из аминокислот 1-610 из SEQ ID NO: 3, SEQ ID NO: 9 или SEQ ID NO: 13, или аминокислот 1-620 из SEQ ID NO: 15 или SEQ ID NO: 18, или их токсинового фрагмента; или (b) аминокислотной последовательности, которая по существу идентична аминокислотной последовательности из (а).
[00192] Как понятно специалистам в данной области техники, нативные токсины Cry экспрессируются в виде протоксинов, которые подвергаются процессингу с получением зрелого токсина, подвергнутого процессингу путем расщепления N-концевого пептидильного фрагмента и С-концевого хвоста протоксина. N-концевой пептидильный фрагмент и/или С-концевой хвост протоксина могут функционировать для повышения стабильности и/или инсектицидной активности токсина Cry. В вариантах осуществления настоящего изобретения химерный инсектицидный белок содержит весь N-концевой пептидильный фрагмент и/или хвост протоксина или их часть. В вариантах осуществления химерный инсектицидный белок не содержит полный N-концевой пептидильный фрагмент и/или полный хвост протоксина. В вариантах осуществления химерный инсектицидный белок не содержит N-концевой пептидильный фрагмент и/или хвост протоксина, т.е. соответствует зрелому подвергнутому процессингу токсину.
[00193] В вариантах осуществления N-концевой пептидильный фрагмент получен из белка Cry (например, из первого белка Cry, который необязательно является белком Cry1). В других вариантах осуществления N-концевой пептидильный фрагмент гетерологичен первому белку Cry, например, не получен из белка Cry и/или является частично или полностью синтетическим. В вариантах осуществления пептидильный фрагмент содержит по меньшей мере приблизительно 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60 или более аминокислот, необязательно из первого белка Cry, включая без ограничения первый белок Cry1 (например, ВТ-0029, SEQ ID NO: 2). В вариантах осуществления пептидильный фрагмент содержит аминокислоты от приблизительно аминокислоты 1 до приблизительно аминокислоты 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59 или 60 первого белка Cry, например, белка Cry1, такого как ВТ-0029.
[00194] В вариантах осуществления химерный инсектицидный белок по настоящему изобретению содержит на С-конце область хвоста протоксина белка Cry Bt, включая модификации хвостов протоксина нативного белка Cry, которые по существу идентичны хвосту протоксина нативного белка Cry. В вариантах осуществления хвост протоксина получен из активного в отношении чешуекрылых белка Cry. В вариантах осуществления хвост протоксина не получен из белка Cry и/или является частично или полностью синтетическим. В вариантах осуществления белок Cry гетерологичен первому белку Cry и/или другому белку Cry. В вариантах осуществления хвост протоксина получен из белка Cry1, например, ВТ-0029, ВТ-0022, Cry1F (например, Cry1Fa), Cry1l (например, Cry1la или Cry1lf), Cry1K (например, Cry1Ka) или Cry1C (например Cry1Ca), или он представляет собой полипептид, который по существу идентичен хвосту протоксина (или его фрагменту) любого из вышеперечисленных. Область хвоста протоксина может содержать целый хвост протоксина белка Cry или любую его часть. В вариантах осуществления область хвоста протоксина содержит по меньшей мере приблизительно 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 38, 40, 45, 50 или более смежных аминокислот из хвоста протоксина белка Cry (например, хвоста протоксина Cry1, такого как ВТ-0029), например, как показано для различных белков Cry на фигуре 1А. В вариантах осуществления хвост протоксина содержит по меньшей мере приблизительно 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 38, 40, 45, 50 или более смежных аминокислот, начиная с аминокислотного положения 598 из SEQ ID NO: 2 (ВТ-0029) или соответствующей области из другого белка Cry, такого как белок Cry1 (например, белок ВТ-0022, Cry1I, Cry1F, Cry1K или Cry1C). В вариантах осуществления хвост протоксина содержит аминокислоты 598-1169, или 598-652, или 598-636, или 598-622, или 598-610, или 598-607, или 598-603, или 598-600 из SEQ ID NO: 2 (ВТ-0029) или соответствующей области из другого белка Cry (см., например, выравнивание ВТ-0029 с ВТ-0022, Cry1Fa и Cry1Ka на фигурах 1А и 1В). В некоторых вариантах осуществления химерный инсектицидный белок содержит любую из SEQ ID NO: 20-25 или SEQ ID NO: 32-35. В других вариантах осуществления хвост протоксина содержит аминокислоты 637-715, или 637-691, или 637-675, или 637-661, или 637-649, или 637-646, или 637-642 из SEQ ID NO: 1 (ВТ-0022). В еще других вариантах осуществления химерный инсектицидный белок по настоящему изобретению содержит, по сути состоит или состоит из любой из SEQ ID NO: 26-31.
[00195] Соответственно, в вариантах осуществления химерный инсектицидный белок по настоящему изобретению содержит, по сути состоит или состоит из (а) аминокислотной последовательности любой из SEQ ID NO: 3, SEQ ID NO: 9, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 18, SEQ ID NO: 20-35, или их токсинового фрагмента; или (b) аминокислотной последовательности, которая по существу идентична аминокислотной последовательности из (а). В необязательных вариантах осуществления химерный инсектицидный белок по настоящему изобретению содержит, по сути состоит или состоит из аминокислотной последовательности из любой из аминокислот из SEQ ID NO: 3, SEQ ID NO: 9, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 18 или SEQ ID NO: 20-35.
[00196] Специалистам в данной области будет понятно, что химерные инсектицидные белки по настоящему изобретению могут дополнительно содержать другие функциональные домены и/или пептидные метки, например, пептидную метку на N-конце и/или С-конце. Например, может быть полезно экспрессировать химерный инсектицидный белок с пептидной меткой, которая может распознаваться коммерчески доступным антителом (например, мотив FLAG), или с пептидной меткой, которая облегчает очистку (например, с помощью добавления метки поли-His) и/или обнаружение. Альтернативно эпитоп можно вводить в химерный белок для облегчения образования антител, которые специфически распознают модифицированный химерный белок, чтобы отличить модифицированный химерный белок от немодифицированной химеры и/или исходного(исходных) белка(белков). Например, одну или несколько аминокислот можно заменить в антигенной петле нативной последовательности для создания нового эпитопа. В одном варианте осуществления антигенная петля расположена в неконсервативной области за пределами домена I нативного белка Cry. В вариантах осуществления антигенная петля не является петлей, вовлеченной в распознавание рецептора в пищеварительном канале насекомого белком Cry, и/или не вовлечена в активацию белка Cry под действием протеазы. В других вариантах осуществления химерный белок можно модифицировать для повышения его стабильности, например, путем слияния связывающего мальтозу белка (МБР) или глутатион-8-трансферазы с полипептидом. В качестве другой альтернативы слитый белок может содержать репортерную группу.
[00197] Химерные инсектицидные белки, которые модифицированы путем введения или удаления сайта процессинга под действием протеазы в подходящем положении (положениях) для обеспечения или устранения протеолитического расщепления протеазой насекомых, растений и/или микроорганизмов, также входят в объем настоящего изобретения. В вариантах осуществления модифицированный химерный инсектицидный белок по существу сохраняет инсектицидную активность. В вариантах осуществления стабильность и/или инсектицидная активность таких модифицированных химерных белков повышается по сравнению с химерным инсектицидным белком, который не содержит такой модификации с введением/устранением сайта расщепления протеазой.
[00198] Таким образом, настоящее изобретение охватывает полипептиды, имеющие аминокислотные последовательности, которые по существу идентичны последовательностям, которые конкретно раскрыты в данном документе, и их фрагментам токсина. Будет понятно, что химерные инсектицидные белки, конкретно раскрытые в данном документе, будут, как правило, переносить модификации в аминокислотной последовательности и по существу сохранять биологическую активность (например, инсектицидную активность). Такие модификации включают вставки, делеции (включая усечения на любом конце) и замены одной или нескольких аминокислот, включая до приблизительно 2, приблизительно 3, приблизительно 4, приблизительно 5, приблизительно 6, приблизительно 7, приблизительно 8, приблизительно 9, приблизительно 10, приблизительно 15, приблизительно 20, приблизительно 25, приблизительно 30, приблизительно 35, приблизительно 40, приблизительно 45, приблизительно 50, приблизительно 55, приблизительно 60, приблизительно 65, приблизительно 70, приблизительно 75, приблизительно 80, приблизительно 85, приблизительно 90, приблизительно 100, приблизительно 105, приблизительно 110, приблизительно 115, приблизительно 120, приблизительно 125, приблизительно 130, приблизительно 135, приблизительно 140, приблизительно 145, приблизительно 150, приблизительно 155 или больше аминокислотных замен, делеций и/или вставок.
[00199] Для выявления полипептидов, по существу идентичных химерным инсектицидным белкам, конкретно раскрытым в данном документе, аминокислотные замены могут основываться на любой характеристике, известной в данной области техники, в том числе относительном сходстве или различиях заместителей боковых цепей аминокислот, например, их гидрофобности, гидрофильности, заряда, размера и т.п.
[00200] Например, при выявлении аминокислотных последовательностей, кодирующих инсектицидные полипептиды, кроме тех, которые конкретно раскрыты в данном документе, можно учитывать индекс гидрофобности аминокислот. Важность индексов гидрофобности аминокислот в обеспечении согласованной биологической функции белка в целом известна в данной области техники (см. Kyte and Doolittle, (1982) J. Mol. Biol. 157:105; включенную в данный документе помощью ссылки во всей своей полноте). Предполагается, что относительная гидрофобная характеристика аминокислоты вносит вклад во вторичную структуру получаемого в результате белка, что, в свою очередь, определяет взаимодействие белка с другими молекулами, например, ферментами, субстратами, рецепторами, ДНК, антителами, антигенами и т.п.
[00201] Каждой аминокислоте был присвоен индекс гидрофобности на основе характеристик ее гидрофобности и заряда (Kyte and Doolittle, Id.), это: изолейцин (+4,5); валин (+4,2); лейцин (+3,8); фенилаланин (+2,8); цистеин/цистин (+2,5); метионин (+1,9); аланин (+1,8); глицин (-0,4); треонин (-0,7); серии (-0,8); триптофан (-0,9); тирозин (-1,3); пролин (-1,6); гистидин (-3,2); глутамат (-3,5); глутамин (-3,5); аспартат (-3,5); аспарагин (-3,5); лизин (-3,9) и аргинин (-4,5).
[00202] Соответственно, индекс гидрофобности аминокислоты (или аминокислотной последовательности) можно учитывать при модификации химерных полипептидов, конкретно раскрытых в данном документе.
[00203] Также из уровня техники известно, что замену аминокислот можно проводить на основании гидрофильности. В патенте США №4554101 указано, что наибольшая средняя локальная гидрофильность белка, которая определяется гидрофильностью его смежных кислот, коррелирует с биологическим свойством белка.
[00204] Как подробно описано в патенте США №4554101, следующие значения гидрофильности были присвоены аминокислотным остаткам: аргинин (+3,0); лизин (.+-.3,0); аспартат (+3,0.+-.1); глутамат (+3,0.+-.1); серии (+0,3); аспарагин (+0,2); глутамин (+0,2); глицин (0); треонин (-0,4); пролин (-0,5+1); аланин (-0,5); гистидин (-0,5); цистеин (-1,0); метионин (-1,3); валин (-1,5); лейцин (-1,8); изолейцин (-1,8); тирозин (-2,3); фенилаланин (-2,5); триптофан (-3,4).
[00205] Таким образом, гидрофильность аминокислоты (или аминокислотной последовательности) можно учитывать при выявлении дополнительных инсектицидных полипептидов помимо тех, которые конкретно раскрыты в данном документе.
[00206] Химерные инсектицидные белки по настоящему изобретению, включая модификации и фрагменты токсинов химерного полипептида, конкретно раскрытые в данном документе, можно получать любым подходящим способом, известным в данной области техники, обычно путем модификации кодирующих последовательностей нуклеиновых кислот. Способы манипулирования и модификации нуклеиновых кислот для достижения желаемой модификации хорошо известны в данной области. Кроме того, методики редактирования генов также можно применять для получения химерного инсектицидного белка по настоящему изобретению или для внесения в него дополнительных модификаций.
[00207] В качестве другого подхода модифицируемый полипептид можно экспрессировать в клетке-хозяине, которая демонстрирует высокую степень ошибочного включения оснований в ходе репликации ДНК, такой как XL-1 Red (Stratagene, Ла-Хойя, Калифорния). После размножения в таких штаммах можно выделять ДНК (например, путем получения плазмидной ДНК или путем амплификации с помощью ПЦР и клонирования полученного ПЦР-фрагмента в вектор), экспрессировать мутантные последовательности белка в штамме, не приводящем к образованию мутаций, и выявлять мутированные гены с инсектицидной активностью, например, путем осуществления анализа для тестирования инсектицидной активности. В иллюстративных способах белок смешивают и применяют в анализах кормления. См., например, Marrone et al. (1985) J. of Economic Entomology 78:290-293. Такие анализы могут включать приведение растений в контакт с одним или несколькими вредителями и определение способности растений к выживанию или способности вызывать гибель вредителей. Примеры мутаций, которые приводят к повышению токсичности, можно найти в Schnepf et al. (1998) Microbiol. Mol. Biol. Rev. 62:775-806.
[00208] В вариантах осуществления химерный инсектицидный белок (включая по существу сходные полипептиды и фрагменты токсинов) по настоящему изобретению является выделенным. В вариантах осуществления химерный инсектицидный белок (включая по существу сходные полипептиды и фрагменты токсинов) по настоящему изобретению является рекомбинантный белком.
[00209] Химерные инсектицидные белки по настоящему изобретению характеризуются инсектицидной активностью в отношении вредителя, относящегося к чешуекрылым. В вариантах осуществления химерный инсектицидный белок характеризуется активностью в отношении одного или нескольких из следующих неограничивающих примеров вредителя, относящегося к чешуекрылым: Ostrinia spp., такой как О. nubilalis (кукурузный мотылек) и/или О. furnacalis (восточный кукурузный мотылек); Plutella spp., такой как P. xylostella (капустная моль); Spodoptera spp., такой как S. frugiperda (совка травяная), S. littoralis (совка хлопчатника египетского), S. ornithogalli (желтополосая совка), S. praefwa (западная желтополосая совка), S. eridania (южная совка) и/или S. exigua (совка малая); Agrotis spp., такой как A ipsilon (совка-ипсилон), A. segetum (озимая совка обыкновенная), A. gladiaria (озимая совка рода Agrotis) и/или A. orthogonia (совка прямоугольная); Striacosta spp., такой как S. albicosta (западная бобовая совка); Helicoverpa spp., такой как H. zea (совка кукурузная), Н. punctigera (совка хлопковая австралийская) и/или Н. armigera (совка хлопковая); Heliothis spp., такой как H. virescens (табачная листовертка); Diatraea spp., такой как D. grandiosella (огневка кукурузная юго-западная) и/или D. saccharalis (точильщик стеблей сахарного тростника); Trichoplusia spp., такой как Т. ni (совка капустная); Sesamia spp., такой как S. nonagroides (мотылек кукурузный средиземноморский) и/или S. calamistis (розовая стеблевая совка); Pectinophora spp., такой как Р. gossypiella (розовый коробочный червь); Cochylis spp., такой как С. hospes (полосатая подсолнечная моль); Manduca spp., такой как М. sexta (табачный бражник) и/или М. quinquemaculata (бражник пятиточечный); Elasmopalpus spp., такой как Е. lignosellus (точильщик стеблей кукурузы малый); Pseudoplusia spp., такой как P. includens (соевая совка); Anticarsia spp., такой как A. gemmatalis (совка бархатных бобов); Plathypena spp., такой как Р. scabra (совка клеверная); Pieris spp., такой как P. brassicae (белянка капустная), Papaipema spp., такой как Р. nebris (точильщик Papaipema nebris); Pseudaletia spp., такой как Р. unipuncta (совка луговая обыкновенная); Peridroma spp., такой как Р. saucia (совка маргаритковая); Keiferia spp., такой как K. lycopersicella (томатная острица); Artogeia spp., такой как A. rapae (белянка репная); Phthorimaea spp., такой как P. operculella (картофельная моль); Chrysodeixis spp., такой как С. includes (соевая совка); Feltia spp., такой как F. ducens (гусеница совки Feltia jaculifera); Chilo spp., такой как С. suppressalis (огневка желтая рисовая), Cnaphalocrocis spp., такой как С. medinalis (огневка рисовая), или любой комбинации вышеуказанных.
[00210] Необязательно химерный инсектицидный белок обладает повышенной активностью в отношении одного или нескольких вредителей, относящихся к чешуекрылым, по сравнению с одной или несколькими исходными молекулами (например, первым белком Cry и другим белком Cry). В вариантах осуществления химерный инсектицидный белок обладает повышенной активностью в отношении одного или нескольких вредителей, относящихся к чешуекрылым, по сравнению с ВТ-0029. В вариантах осуществления химерный инсектицидный белок обладает повышенной активностью в отношении одного или нескольких вредителей, относящихся к чешуекрылым, по сравнению с ВТ-0022, Cry1Fa, Cry1Ia, Cry1If, Cry1Ka или Cry1C.
[00211] В вариантах осуществления химерный инсектицидный белок обладает повышенной активностью в отношении совки травяной (Spodoptera frugiperda) по сравнению с одной или несколькими исходными молекулами (например, первым белком Cry и другим белком Cry). В вариантах осуществления химерный инсектицидный белок обладает повышенной активностью в отношении совки травяной по сравнению с ВТ-0029. В вариантах осуществления химерный инсектицидный белок обладает повышенной активностью в отношении совки травяной по сравнению с ВТ-0022, Cry1Fa, Cry1Ia, Cry1If, Cry1Ka и/или Cry1Ca. В соответствии с вышеизложенными вариантами осуществления химерный инсектицидный белок может необязательно обладать инсектицидной активностью в отношении насекомого-вредителя или колонии совки травяной, которая характеризуется устойчивостью к другому инсектицидному средству, в том числе другому инсектицидному белку (такому как, например, белок Bt). В вариантах осуществления химерный инсектицидный белок обладает повышенной активностью в отношении колонии совки травяной, которая устойчива к белку Vip3A (например, Vip3Aa, включая без ограничения трансформант маиса MIR162) или белку Cry1F (например, Cry1Fa, включая без ограничения трансформант маиса ТС1507). В вариантах осуществления химерный инсектицидный белок обладает повышенной активностью в отношении устойчивой колонии совки травяной по сравнению с одной или несколькими исходными молекулами, например, ВТ-0029, ВТ-0022, Cry1Fa, Cry1Ia, Cry1If, Cry1Ka и/или Cry1Ca.
[00212] Настоящее изобретение также охватывает антитела, которые специфически связываются с химерным инсектицидным белком по настоящему изобретению. Антитело необязательно может быть моноклональным антителом или поликлональной антисывороткой. В вариантах осуществления антитело является селективным по отношению к химерному белку и не связывается с одной или несколькими исходными молекулами (например, ВТ-0029, ВТ-0022, Cry1Fa и т.п.) и может использоваться для различения химерного белка и исходного белка. Такие антитела могут быть получены с применением стандартных иммунологических методик получения поликлональной антисыворотки и при необходимости иммортализации антителопродуцирующих клеток иммунизированного организма-хозяина для получения источников моноклональных антител. Методики получения антител к любому представляющему интерес веществу хорошо известны, например, как описано в Harlow and Lane (1988. Antibodies a laboratory manual, pp.726. Cold Spring Harbor Laboratory) и в Goding (Monoclonal Antibodies: Principles & practice. 1986. Academic Press, Inc., Orlando, FL). Настоящее изобретение также охватывает инсектицидный белок, который перекрестно реагирует с антителом, в частности, моноклональным антителом, вырабатываемым против одного или нескольких химерных инсектицидных белков по настоящему изобретению.
[00213] Антитела согласно настоящему изобретению применимы, например, в иммунологических анализах для определения количества или присутствия химерного инсектицидного белка по настоящему изобретению или антигенно родственного полипептида, например, в биологическом образце. Такие анализы также применимы в получении композиций, содержащих один или несколько химерных инсектицидных белков по настоящему изобретению или антигенно родственного полипептида, с обеспечением контроля качества. Кроме того, антитела можно применять для оценки эффективности рекомбинантного получения одного или нескольких химерных инсектицидных белков по настоящему изобретению или антигенно родственного полипептида, а также для скрининга библиотек экспрессии на предмет наличия нуклеотидной последовательности, кодирующей один или несколько химерных инсектицидных белков по настоящему изобретению или антигенно родственный полипептид. Антитела, кроме того, применимы в качестве аффинных лигандов для очистки или выделения любого одного или нескольких белков по настоящему изобретению или антигенно родственного полипептида.
Нуклеиновые кислоты, кассеты экспрессии и векторы
[00214] В качестве дополнительного аспекта в настоящем изобретении предусмотрены нуклеиновые кислоты, кодирующие полипептиды по настоящему изобретению, включая модифицированные полипептиды и фрагменты токсинов, как описано в данном документе.
[00215] Согласно некоторым вариантам осуществления в настоящем изобретении предусмотрена молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, которая содержит, по сути состоит или состоит из следующего: (а) нуклеотидной последовательности, кодирующей аминокислотную последовательность под SEQ ID NO: 3, SEQ ID NO: 9, SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 18, SEQ ID NO: 20-35, или ее токсинового фрагмента; (b) нуклеотидной последовательности, кодирующей аминокислотную последовательность, которая по существу идентична аминокислотной последовательности из (а); (с) нуклеотидной последовательности, которая гибридизируется при жестких условиях гибридизации с нуклеотидной последовательностью из (а) или (b); или (d) нуклеотидной последовательности, которая отличается от нуклеотидных последовательностей из (а), (b) или (с) вследствие вырожденности генетического кода.
[00216] В вариантах осуществления молекула нуклеиновой кислоты содержит нуклеотидную последовательность, которая содержит, по сути состоит или состоит из следующего: (а) нуклеотидной последовательности под SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 14 или SEQ ID NO: 16 или ее кодирующего токсин фрагмента; (b) нуклеотидной последовательности, которая по существу идентична нуклеотидной последовательности из (а); (с) нуклеотидной последовательности, которая гибридизируется при жестких условиях гибридизации с нуклеотидной последовательностью из (а) или (b); или (d) нуклеотидной последовательности, которая отличается от нуклеотидных последовательностей из (а), (b) или (с) вследствие вырожденности генетического кода. Необязательно нуклеотидная последовательность содержит, по сути состоит или состоит из нуклеотидной последовательности под SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 14 или SEQ ID NO: 16.
[00217] В вариантах осуществления нуклеотидная последовательность является частично или полностью синтетической последовательностью, например, которая была кодон-оптимизирована для экспрессии в организме-хозяине, например, в бактерии-хозяине или растении-хозяине (например, трансгенном однодольном растении-хозяине или трансгенном двудольном растении-хозяине). Неограничивающие примеры нуклеотидных последовательностей, которые кодон-оптимизированы для экспрессии в растении маиса, включают SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7 и SEQ ID NO: 11.
[00218] В иллюстративных вариантах осуществления для экспрессии в трансгенных растениях нуклеотидные последовательности по настоящему изобретению модифицируют и/или оптимизируют. Например, хотя во многих случаях гены микроорганизмов могут экспрессироваться в растениях при высоких уровнях и без модификации, низкий уровень экспрессии в трансгенных растениях может быть обусловлен нуклеотидными последовательностями микроорганизмов, имеющими кодоны, которые не являются предпочтительными для растений. Как известно из уровня техники, для живых организмов характерны определенные предпочтения в отношении частоты использования кодонов, и поэтому кодоны нуклеотидных последовательностей, описанных в данном изобретении, могут быть изменены в соответствии с характерными для растений предпочтениями, при этом с сохранением аминокислот, кодируемых ими. Кроме того, в данной области техники известно, что высокий уровень экспрессии в растениях, например, растениях кукурузы, лучше всего достигается в случае кодирующих последовательностей, которые характеризуются по меньшей мере приблизительно 35% содержанием GC, или по меньшей мере приблизительно 45%, или по меньшей мере приблизительно 50% или по меньшей мере приблизительно 60%. Микробные последовательности нуклеотидов, которые имеют низкое содержание GC, могут плохо экспрессироваться в растениях. Хотя определенные нуклеотидные последовательности могут надлежащим образом экспрессироваться как в видах однодольных, так и двудольных растений, последовательности можно модифицировать с учетом предпочтений в отношении кодонов и предпочтений в отношении содержания GC для однодольных растений или двудольных растений, поскольку было показано, что эти предпочтения отличаются (Murray et al. Nucl. Acids Res. 17:477-498 (1989)). Кроме того, в вариантах осуществления нуклеотидную последовательность модифицируют для удаления незаконных сайтов сплайсинга, которые могут вызывать усечение транскрипта. Такие модификации нуклеотидных последовательностей можно выполнить с применением хорошо известных методик сайт-направленного мутагенеза, ПЦР и конструирования синтетических генов с помощью способов, описанных, например, в патентах США №№5625136; 5500365 и 6013523.
[00219] В некоторых вариантах осуществления настоящего изобретения предусмотрены синтетические кодирующие последовательности или полинуклеотид, полученные согласно процедуре, раскрытой в патенте США №5625136. В этой процедуре применяют предпочтительные для маиса кодоны, т.е. один кодон, который чаще всего кодирует данную аминокислоту в маисе. Предпочтительный для маиса кодон, кодирующий конкретную аминокислоту, может быть определен, например, на основании известных последовательностей генов маиса. Например, данные о частоте использования кодонов у маиса для 28 генов из растений маиса находятся в Murray et al., Nucleic Acids Research 17:477-498 (1989). Известно, что кодоны, оптимизированные для экспрессии в растении одного вида, также будут функционировать в растениях других видов, но, вероятно, не в той же степени, как в виде растения, для которого кодоны были оптимизированы. Подобным образом нуклеотидные последовательности могут быть оптимизированы для экспрессии в любом растении. Следует понимать, что вся нуклеотидная последовательность или любая ее часть могут быть оптимизированными или синтетическими. То есть полинуклеотид может содержать нуклеотидную последовательность, которая частично является нативной последовательностью и частично является последовательностью с оптимизированными кодонами.
[00220] В иллюстративных вариантах осуществления полинуклеотид по настоящему изобретению представляет собой выделенный полинуклеотид. В вариантах осуществления полинуклеотид по настоящему изобретению представляет собой рекомбинантный полинуклеотид.
[00221] В вариантах осуществления в настоящем изобретении дополнительно предусмотрена молекула нуклеиновой кислоты, содержащая полинуклеотид, функционально связанный с промотором (например, гетерологичным промотором). Промоторы могут включать, например, конституитивные, индуцируемые, временно регулируемые, регулируемые развитием, химически регулируемые, тканепредпочтительные и/или тканеспецифические промоторы. В конкретных аспектах промотор, применимый в настоящем изобретении, представляет собой промотор, способный инициировать транскрипцию нуклеотидной последовательности в растительной клетке, например, в клетке однодольного (например, маис или рис) или двудольного (например, соя, хлопчатник) растения.
[00222] В вариантах осуществления гетерологичный промотор является промотором, обеспечивающим экспрессию в растении (например, экспрессируемым в однодольном или экспрессируемым в двудольном). Например, без ограничения промотор, обеспечивающий экспрессию в растении, можно выбрать из промоторов: убиквитина, вируса, поражающего цеструм оранжевый, TrpA кукурузы, OsMADS 6, гистона Н3 маиса, 5'-UTR гена 9 бактериофага Т3, сахарозосинтетазы 1 кукурузы, алкогольдегидрогеназы 1 кукурузы, светособирающего комплекса кукурузы, белка теплового шока кукурузы, mtl маиса, малой субъединицы RuBP карбоксилазы гороха, актина риса, циклофилина риса, маннопинсинтазы Ti-плазмиды, нопалинсинтазы Ti-плазмиды, халкон-изомеразы петунии, богатого глицином белка 1 бобов, пататина картофеля, лектина, 35S CaMV и малой субъединицы S-E9 RuBP карбоксилазы.
[00223] Хотя, как было показано, многие промоторы из двудольных растений функционируют у однодольных и наоборот, в вариантах осуществления для экспрессии в двудольных растениях выбирают промоторы двудольных, а для экспрессии в однодольных растениях выбирают промоторы однодольных. Однако не существует каких-либо ограничений в отношении происхождения выбранных промоторов; при этом достаточно, чтобы они являлись функциональными в отношении управления экспрессией нуклеотидных последовательностей в требуемой клетке.
[00224] Выбор промотора может меняться в зависимости от временных и пространственных требований для экспрессии, а также в зависимости от клетки-хозяина, подлежащей трансформации. Таким образом, например, экспрессия нуклеотидных последовательностей по настоящему изобретению может осуществляться в любом растении и/или части растения, (например, в листьях, в цветоножках или стеблях, в колосьях, в соцветиях (например, колосьях, метелках, початках и т.д.), в корнях, семенах и/или проростках и т.п.). Например, когда желательна экспрессия в конкретной ткани или органе, можно применять тканеспецифический или тканепредпочтительный промотор (например, корнеспецифический/-предпочтительный промотор). В отличие от этого, если желательной является экспрессия в ответ на стимул, можно применять промотор, индуцируемый определенными стимулами или химическими веществами. Если желательной является постоянная экспрессия на относительно стабильном уровне во всех клетках растения, может быть выбран конститутивный промотор.
[00225] Промоторы, применимые в соответствии с настоящим изобретением, включают без ограничения промоторы, которые управляют экспрессией нуклеотидной последовательности конститутивно, промоторы, которые управляют экспрессией при индукции, и промоторы, которые управляют экспрессией тканеспецифический образом или специфическим образом по отношению к стадии развития. Эти различные типы промоторов известны из уровня техники.
[00226] Подходящие конститутивные промоторы включают, например, промотор 35S CaMV (Odell et al., Nature 313:810-812, 1985); промотор At6669 Arabidopsis (см. публикацию согласно PCT № W004081173A2); промотор Ubi 1 маиса (Christensen et al., Plant Mol. Biol. 18:675-689, 1992); актина риса (McElroy et al., Plant Cell 2:163-171, 1990); pEMU (Last et al., Theor. Appl. Genet. 81:581-588, 1991); 19S CaMV (Nilsson et al., Physiol. Plant 100:456-462, 1997); GOS2 (de Pater et al., Plant J November; 2(6):837-44, 1992); убиквитина (Christensen et al., Plant Mol. Biol. 18: 675-689, 1992); циклофилина риса (Bucholz et al., Plant Mol Biol. 25(5):837-43, 1994); гистона НЗ маиса (Lepetit et al., Mol. Gen. Genet. 231: 276-285, 1992); актина 2 (An et al., Plant J. 10(1); 107-121, 1996), конститутивный промотор корневого кончика СТ2 (SEQ ID NO: 1535; см. также заявку согласно PCT №IL/2005/000627) и синтетический Super MAS (Ni et al., The Plant Journal 7: 661-76, 1995). Другие конститутивные промоторы включают описанные в патентах США №№5659026, 5608149; 5608144; 5604121; 5569597: 5466785; 5399680; 5268463 и 5608142.
[00227] Тканеспецифическими или тканепредпочтительными промоторами, применимыми для экспрессии полипептидов по настоящему изобретению в растениях, необязательно в маисе, включают те, которые управляют экспрессией в корне, сердцевине, листе или пыльце. Подходящие тканеспецифические промоторы включают без ограничения специфические для листьев промоторы [такие, которые описаны, например, в Yamamoto et al., Plant J. 12:255-265, 1997; Kwon et al., Plant Physiol. 105:357-67, 1994; Yamamoto et al., Plant Cell Physiol. 35:773-778, 1994; Gotor et al., Plant J. 3:509-18, 1993; Orozco et al., Plant Mol. Biol. 23:1129-1138, 1993; и Matsuoka et al., Proc. Natl. Acad. Sci. USA 90:9586-9590, 1993], предпочтительные для семени промоторы (например, из семяспецифических генов (Simon, et al., Plant Mol. Biol. 5. 191, 1985; Scofield, et al., J. Biol. Chem. 262: 12202, 1987; Baszczynski, et al., Plant Mol. Biol. 14: 633, 1990), промоторы альбумина из бразильского ореха (Pearson et al., Plant Mol. Biol. 18: 235-245, 1992), легумина (Ellis, et al. Plant Mol. Biol. 10: 203-214, 1988), глютелина (Takaiwa, et al., Mol. Gen. Genet. 208: 15-22, 1986; Takaiwa, et al., FEBS Letts. 221: 43-47, 1987), зеина (Matzke et al., Plant Mol Biol, 143).323-32 1990), napA (Stalberg, et al., Planta 199: 515-519, 1996), SPA пшеницы (Albanietal, Plant Cell, 9: 171-184, 1997), олеозина подсолнечника (Cummins, et al., Plant Mol. Biol. 19: 873-876, 1992)], специфические для эндосперма промоторы (например, LMW и HMW пшеницы, глютенина-1 (Mol Gen Genet 216:81-90, 1989; NAR 17:461-2), глиадинов a, b и g пшеницы (ЕМВОЗ:1409-15, 1984), промотор ltrl ячменя, гордеина В1, С, D ячменя (Theor Appl Gen 98:1253-62, 1999; Plant J 4:343-55, 1993; Mol Gen Genet 250:750-60, 1996), DOF ячменя (Mena et al., The Plant Journal, 116(1): 53-62, 1998), Biz2 (EP99106056.7), синтетический промотор (Vicente-Carbajosa et al., Plant J. 13: 629-640, 1998), промотор проламина NRP33 риса, глобулина Glb-1 риса (Wu et al., Plant Cell Physiology 39(8) 885-889, 1998), альфа-глобулина REB/OHP-1 риса (Nakase et al. Plant Mol. Biol. 33: 513-S22, 1997), PP ADP-глюкозы риса (Trans Res 6:157-68, 1997), промотор гена семейства ESR маиса (Plant J 12:235-46, 1997), гамма-кафирина сорго (Plant Mol. Biol 32:1029-35, 1996)], специфические для зародышей промоторы (например, OSH1 риса; Sato et al., Proc. Nati. Acad. Sci. USA, 93: 8117-8122), KNOX (Postma-Haarsma of al, Plant Mol. Biol. 39:257-71, 1999), олеозина риса (Wu et at, J. Biochem., 123:386, 1998), специфические для цветков промоторы, например, AtPRP4, халкон-синтазы (chsA) (Van der Meer, et al., Plant Mol. Biol. 15, 95-109, 1990), LAT52 (Twell et al., Mol. Gen Genet. 217:240-245; 1989), apetala-3 и промоторы, специфические для репродуктивных тканей растений (например, промоторы OsMADS; публикация патента США №2007/0006344).
[00228] Примеры промоторов, подходящих для предпочтительной экспрессии в зеленой ткани, включают в себя множество промоторов, которые осуществляют регуляцию генов, участвующих в фотосинтезе, и многие из них были клонированы как из однодольных, так и из двудольных. Одним из таких промоторов является промотор РЕРС гена фосфоенолкарбоксилазы маиса (Hudspeth & Grula, Plant Molec. Biol. 12:579-589 (1989)). Другим промотором специфической для корня экспрессии является промотор, описанный de Framond (FEBS 290:103-106 (1991) или в патенте США №5466785). Другой промотор, применимый в настоящем изобретении, представляет собой специфический для стебля промотор, описанный в патенте США №5625136, который в естественных условиях управляет экспрессией гена trpA маиса.
[00229] Кроме того, могут применяться промоторы, функционирующие в пластидах. Неограничивающие примеры таких промоторов включают промотор 5'-UTR гена 9 бактериофага Т3 и другие промоторы, раскрытые в патенте США №7579516. Другие промоторы, применимые в соответствии с настоящим изобретением, включают без ограничения промотор малой субъединицы S-E9 RuBP-карбоксилазы и промотор гена ингибитора трипсина Кунитца (Kti3).
[00230] В некоторых вариантах осуществления настоящего изобретения могут применяться индуцируемые промоторы. Таким образом, например, промоторы, регулируемые химическими веществами, можно применять для модуляции экспрессии гена в растении путем применения экзогенного химического регулятора. Регуляция экспрессии нуклеотидных последовательностей по настоящему изобретению посредством промоторов, регулируемых химическими веществами, обеспечивает синтез полипептидов по настоящему изобретению только тогда, когда культурные растения обрабатывают индуцирующими химическими веществами. В зависимости от цели промотор может представлять собой промотор, индуцируемый химическим веществом, где применение химического вещества индуцирует экспрессию гена, или промотор, подавляемый химическим веществом, где применение химического вещества подавляет экспрессию гена. Примеры такой технологии химической индукции экспрессии генов подробно описаны в публикации заявки ЕР 0332104 и патенте США №5614395.
[00231] Промоторы, индуцируемые химическими веществами, известны из уровня техники и включают в себя без ограничения промотор 1п2-1 маиса, который активируется антидотами бензолсульфонамидных гербицидов, промотор GST маиса, который активируется гидрофобными электрофильными соединениями, применяемыми в качестве предвсходовых гербицидов, и промотор о PR-la табака, который активируется салициловой кислотой (например, системы PRla), стероидзависимые промоторы (см., например, промотор, индуцируемый глюкокортикоидами, в Schena et al. (1991) Proc. Natl. Acad. Sci. USA 88, 10421-10425 и McNellis et al. (1998) Plant J. 14, 247-257) и промоторы, индуцируемые тетрациклинами и подавляемые тетрациклинами (см., например, Gatz et al. (1991) Mol. Gen. Genet. 221, 229-237, и патенты США №№5814618 и 5789156), промоторы системы Lac-репрессора, промоторы системы, индуцируемой медью, промоторы системы, индуцируемой салицилатом (например, системы PR1a), промоторы, индуцируемые глюкокортикоидами (Aoyama et al. (1997) Plant J. 11:605-612), и промоторы системы, индуцируемой экдизоном.
[00232] Другие неограничивающие примеры индуцируемых промоторов включают промоторы, индуцируемые ABA и тургорным давлением, промотор гена белка, связывающегося с ауксином (Schwob et al. (1993) Plant J. 4:423-432), промотор UDP-глюкозофлавоноидгликозилтрансферазы (Ralston et al. (1988) Genetics 119:185-197), промотор ингибитора протеиназы MPI (Cordero et al. (1994) Plant J. 6:141-150) и промотор глицеральдегид-3-фосфатдегидрогеназы (Kohler et al. (1995) Plant Mol. Biol. 29:1293-1298; Martinez et al. (1989) J. Mol. Biol. 208:551-565 и Quigley et al. (1989) J. Mol. Evol. 29:412-421). Также включены системы, индуцируемые бензолсульфонамидом (патент США №5364780) и индуцируемые спиртами (публикации Международных патентных заявок №№ WO 97/06269 и WO 97/06268), и промоторы гена глутатион-8-трансферазы. Аналогично можно применять любой из индуцируемых промоторов, описанных в Gatz (1996) Current Opinion Biotechnol. 7:168-172 и Gatz (1997) Annu. Rev. Plant Physiol. Plant Mol. Biol. 48:89-108. Другие химическими индуцируемые промоторы, применимые для управления экспрессией нуклеотидных последовательностей по настоящему изобретению в растениях, раскрыты в патенте США 5614395. Химическая индукция экспрессии генов также подробно изложена в ЕР 0332104 (от Ciba-Geigy) и патенте США №5614395.
[00233] Другая категория промоторов, применимых в настоящем изобретении, представляет собой промоторы, индуцируемые ранением. Были описаны многочисленные промоторы, которые экспрессируются в участках ранения, а также в участках инфицирования фитопатогеном. В идеальном случае такой промотор должен активироваться локально в участках инвазии насекомых и, таким образом, инсектицидные белки накапливаются исключительно в клетках, где необходим синтез инсектицидных белков для уничтожения нападающего насекомого-вредителя. Примеры промоторов этого типа включают описанные Stanford et al. Mol. Gen. Genet. 215:200-208 (1989), Xu et al. Plant Molec. Biol. 22:573-588 (1993), Logemann et al. Plant Cell 1:151-158 (1989), Rohrmeier & Lehle, Plant Molec. Biol. 22:783-792 (1993), Firek et al. Plant Molec. Biol. 22:129-142 (1993) и Warner et al. Plant J. 3:191-201 (1993).
[00234] В вариантах осуществления нуклеиновая кислота по настоящему изобретению может содержать, по сути состоять или состоять из кассеты экспрессии, или может содержаться в кассете экспрессии.
[00235] Кассета экспрессии, содержащая представляющий интерес полинуклеотид, может быть химерной, что означает, что по меньшей мере один из ее компонентов является гетерологичный по отношению по меньшей мере к одному из ее других компонентов. Кассета экспрессии также может представлять собой последовательность, которая встречается в природе, но которая была получена в рекомбинантной форме, применимой для гетерологичной экспрессии. Однако, как правило, кассета экспрессии является гетерологичной по отношению к хозяину, т.е. конкретная последовательность нуклеиновой кислоты в кассете экспрессии не встречается в клетке-хозяине в природе, и ее необходимо было ввести в клетку-хозяина или предка клетки-хозяина с помощью события трансформации.
[00236] В дополнение к промоторам, функционально связанным с нуклеотидными последовательностями по настоящему изобретению, кассета экспрессии по настоящему изобретению может также содержать другие регуляторные элементы. Регуляторные элементы включают без ограничения энхансеры, нитроны, лидерные последовательности, регулирующие трансляцию, сигналы терминации и сигнальные последовательности полиаденилирования. Примеры подходящих сигналов терминации транскрипции доступны и известны в данной области техники (например, tml из CaMV, Е9 из rbcS). В контексте настоящего изобретения можно применять любой доступный терминатор, о котором известно, что он функционирует в растениях.
[00237] В кассеты экспрессии, описанные в настоящем изобретении, можно вводить множество других последовательностей. Они предусматривают последовательности, которые, как показано, усиливают экспрессию, как например, интронные последовательности (например, из Adhl и bronzel) и вирусные лидерные последовательности (например, из TMV, MCMV и AMV).
[00238] Для более эффективной инициации трансляции можно модифицировать
последовательности, прилегающие к инициирующему метионину. Например, их можно модифицировать путем включения последовательностей, которые, как известно, являются эффективными в растениях. Joshi предложил подходящую консенсусную последовательность для растений (NAR 15:6643-6653 (1987)), a Clonetech предлагает дополнительную консенсусную последовательность, являющуюся инициатором трансляции (каталог 1993/1994, стр. 210). Эти консенсусные последовательности подходят для использования с нуклеотидными последовательностями по настоящему изобретению. Последовательности встраивают в конструкции, содержащие нуклеотидные последовательности, выше и включительно ATG (при этом вторая аминокислота остается немодифицированной) или в качестве альтернативы выше и включительно GTC, расположенного после ATG (с возможностью модифицирования второй аминокислоты трансгена).
[00239] В вариантах осуществления предпочтительной может быть направленная экспрессия полипептидов по настоящему изобретению в конкретном клеточном местоположении в растительной клетке. В некоторых случаях может быть желательной локализация в цитозоле, тогда как в других случаях предпочтительной может быть локализация в определенной субклеточной органелле. Для осуществления настоящего изобретения на практике можно применять любой механизм нацеливания генных продуктов, например в растениях, и, как известно, такие механизмы существуют в растениях, и последовательности, регулирующие функционирование таких механизмов, были описаны довольно подробно. Были охарактеризованы последовательности, которые обеспечивают нацеливание генных продуктов в другие компартменты клетки. Например, аминоконцевые последовательности могут обусловливать нацеливание представляющего интерес белка в такой компартмент клетки, как вакуоль, митохондрия, пероксисома, белковые тельца, эндоплазматический ретикулум, хлоропласт, крахмальное зерно, амилопласт, апопласт или клеточная стенка растительной клетки (например, Unger et. al. Plant Molec. Biol. 13: 411-418 (1989); Rogers et. al. (1985) Proc. Natl. Acad. Sci. USA 82: 6512-651; патенты США №7102057; WO 2005/096704. Необязательно сигнальная последовательность может представлять собой N-концевую сигнальную последовательность из waxy, N-концевую сигнальную последовательность из гамма-зеина, домен, связывающий крахмал, С-концевой домен, связывающий крахмал, последовательность, нацеливающую в хлоропласт, которая обеспечивает доставку зрелого белка в хлоропласт (Comai et. al. (1988) J. Biol. Chem. 263: 15104-15109; van den Broeck, et. al. (1985) Nature 313: 358-363; патент США №5639949) или сигнальную последовательность секреции из клеток алейронового слоя (Koehler & Но, Plant Cell 2: 769-783 (1990)). Кроме того, аминоконцевые последовательности вместе с карбоксиконцевыми последовательностями обусловливают нацеливание генных продуктов в вакуоль и могут применяться в настоящем изобретении (Shinshi et. al. (1990) Plant Molec. Biol. 14: 357-368). В одном варианте осуществления выбранная сигнальная последовательность включает известный сайт расщепления, и при конструировании слияния учитываются любые аминокислоты после сайта(сайтов) расщепления, необходимые для расщепления. В некоторых случаях это требование может быть выполнено путем добавления небольшого числа аминокислот между сайтом расщепления и ATG трансгена или, в качестве альтернативы, путем замещения некоторых аминокислот в пределах последовательности трансгена. Эти методики конструирования хорошо известны из уровня техники и одинаково применимы к любому клеточному компартменту.
[00240] Будет понятно, что описанные выше механизмы нацеливания в клетке можно использовать не только с сочетании когнатными для них промоторами, но также в сочетании с гетерологичными промоторами, так что специфическая задача нацеливания в клетке осуществляется под транскрипционным контролем промотора, характеризующегося паттерном экспрессии, отличный от такового для промотора, из которого получен нацеливающий сигнал.
[00241] Кассета экспрессии по настоящему изобретению также может включать нуклеотидную последовательность для селектируемого маркера, который можно применять для отбора трансформированного растения, части растения или растительной клетки. Много примеров подходящих селектируемых маркеров известны из уровня техники и могут применяться в кассетах экспрессии, описанных в данном документе.
[00242] Примеры селектируемых маркеров включают без ограничения нуклеотидную последовательность, кодирующую neo или nptII, которые придают устойчивость к канамицину, G418 и т.п. (Potrykus et al. (1985) Mol. Gen. Genet. 199:183-188); нуклеотидную последовательность, кодирующую bar, который придает устойчивость к фосфинотрицину; нуклеотидную последовательность, кодирующую измененную 5-енолпирувилшикимат-3-фосфатсинтазу (EPSP), которая придает устойчивость к глифосату (Hinchee et al. (1988) Biotech. 6:915-922); нуклеотидную последовательность, кодирующую нитрилазу, такую как bxn от Klebsiella ozaenae, которая придает устойчивость к бромоксинилу (Stalker et al. (1988) Science 242:419-423); нуклеотидную последовательность, кодирующую измененную ацетолактатсинтазу (ALS), которая придает устойчивость к имидазолинону, сульфонилмочевине или другим ALS-ингибирующим химическим веществам (европейская патентная заявка №154204); нуклеотидную последовательность, кодирующую устойчивую к метотрексату дигидрофолатредуктазу (DHFR) (Thillet et al. (1988) J. Biol. Chem. 263:12500-12508); нуклеотидную последовательность, кодирующую далапондегалогеназу, которая придает устойчивость к далапону; нуклеотидную последовательность, кодирующую маннозо-6-фосфатизомеразу (также называемую фосфоманнозоизомеразой (PMI)), которая придает способность к метаболизму маннозы (патенты США №5767378 и №5994629); нуклеотидную последовательность, кодирующую измененную антранилатсинтазу, которая придает устойчивость к 5-метилтриптофану; или нуклеотидную последовательность, кодирующую hph, который придает устойчивость к гигромицину. Специалист в данной области способен выбрать подходящий селектируемый маркер для применения в кассете экспрессии по настоящему изобретению.
[00243] Дополнительные селектируемые маркеры включают без ограничения нуклеотидную последовательность, кодирующую β-глюкуронидазу или uidA (GUS), который кодирует фермент, для которого известны различные хромогенные субстраты; нуклеотидную последовательность R-локуса, которая кодирует продукт, регулирующий продуцирование антоцианиновых пигментов (красного цвета) в растительных тканях (Dellaporta et al., "Molecular cloning of the maize R-nj allele by transposon-tagging with Ac," pp. 263-282, в Chromosome Structure and Function: Impact of New Concepts, 18th Stadler Genetics Symposium (Gustafson & Appels eds., Plenum Press 1988)); нуклеотидную последовательность, кодирующую β-лактамазу, фермент, для которого известны различные хромогенные субстраты (например, PAD АС, хромогенный цефалоспорин) (Sutcliffe (1978) Proc. Natl. Acad. Sci. USA 75:3737-3741); нуклеотидную последовательность, кодирующую xylE, который кодирует катехолдиокситеназу (Zukowsky et al. (1983) Proc. Natl. Acad. Sci. USA 80:1101-1105); нуклеотидную последовательность, кодирующую тирозиназу, фермент, способный окислять тирозин до DOPA и допахинона, который, в свою очередь, конденсируется с образованием меланина (Katz et al. (1983) J. Gen. Microbiol. 129:2703-2714); нуклеотидную последовательность, кодирующую β-галактозидазу, фермент, для которого существуют хромогенные субстраты; нуклеотидную последовательность, кодирующую люциферазу (lux), которая обеспечивает выявление с помощью биолюминесценции (Ow et al. (1986) Science 234:856-859); нуклеотидную последовательность, кодирующую экворин, который может быть использован в обнаружении чувствительной к кальцию биолюминесценции (Prasher et al. (1985) Biochem. Biophys. Res. Comm. 126:1259-1268); или нуклеотидную последовательность, кодирующую зеленый флуоресцентный белок (Niedz et al. (1995) Plant Cell Reports 14:403-406). Специалист в данной области может выбрать подходящий селектируемый маркер для применения в кассете экспрессии по настоящему изобретению.
[00244] В некоторых вариантах осуществления кассета экспрессии по настоящему изобретению также может содержать полинуклеотиды, которые кодируют другие требуемые признаки, помимо химерных инсектицидных белков по настоящему изобретению. Примеры таких других полинуклеотидов включают в себя полинуклеотиды, которые кодируют полипептид или dsRNA для другого представляющего(-их) интерес необходимого(-ых) признака (признаков). Такие кассеты экспрессии, содержащие "пакетированные" признаки, можно применять, например, для получения растений, частей растений или растительных клеток, обладающих требуемым фенотипом с пакетированными признаками (т.е. молекулярное пакетирование). Такие пакетированные комбинации в растениях также можно создавать с помощью других способов, включая без ограничения кроссбридинг растений с помощью любой традиционной методики (т.е. селекционный пакет). При "пакетировании" путем генетической трансформации растений представляющие интерес нуклеотидные последовательности можно комбинировать в любой момент времени и в любом порядке. Например, трансгенное растение, содержащее один или несколько требуемых признаков, можно применять в качестве мишени для введения дополнительных признаков путем последующей трансформации. Дополнительные нуклеотидные последовательности можно вводить согласно протоколу котрансформации одновременно с нуклеотидной последовательностью, молекулой нуклеиновой кислоты, конструкцией нуклеиновой кислоты или композицией по настоящему изобретению, обеспечиваемыми любой комбинацией кассет экспрессии. Например, если будут вводить две нуклеотидные последовательности, то их можно встроить в отдельные кассеты (транс), или их можно встроить в одну кассету (цис). Экспрессия полинуклеотидов может управляться одним и тем же промотором или различными промоторами. Кроме того, известно, что полинуклеотиды можно "пакетировать" в требуемом местоположении в геноме при помощи системы сайт-специфической рекомбинации. См., например, публикации международных заявок на патенты №№ WO 99/25821, WO 99/25854, WO 99/25840, WO 99/25855 и WO 99/25853.
[00245] В иллюстративных вариантах осуществления кассета экспрессии также может содержать дополнительную последовательность, кодирующую один или несколько полипептидов или молекул двухнитевой РНК (dsRNA), обеспечивающих представляющий интерес агрономический признак (например, агрономический признак, который преимущественно приносит пользу компании-производителю семян, сельхозпроизводителю или переработчику зерна). Представляющий интерес полипептид может быть любым полипептидом, кодируемым представляющей интерес нуклеотидной последовательностью. Неограничивающие примеры представляющих интерес полипептидов, которые подходят для продуцирования в растениях, включают такие полипептиды, которые обуславливают возникновение агрономически важных признаков, таких как устойчивость к гербицидам (также иногда называемая "толерантностью к гербицидам"), устойчивость к вирусам, устойчивость к патогенным бактериям, устойчивость к насекомым, устойчивость к нематодам или устойчивость к грибам. См., например, патенты США №№5569823; 5304730; 5495071; 6329504 и 6337431. В вариантах осуществления полипептид, представляющий интерес, также может являться таким, который увеличивает мощность или урожайность растений (включая признаки, которые дают возможность растению произрастать при различных температурах, почвенных условиях и уровнях солнечного освещения и атмосферных осадков), или таким, который дает возможность идентифицировать растение, проявляющее признак, представляющий интерес (например, селектируемый маркер, цвет семенной оболочки и т.д.). Разнообразные представляющие интерес полипептиды, а также способы введения этих полипептидов в растение описаны, например, в патентах США №№4761373; 4769061; 4810648; 4940835; 4975374; 5013659; 5162602; 5276268; 5304730; 5495071; 5554798; 5561236; 5569823; 5767366; 5879903, 5928937; 6084155; 6329504 и 6337431; а также в публикации заявки на патент США №2001/0016956. См. также во всемирной сети Интернет по адресу lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/.
[00246] В некоторых вариантах осуществления настоящего изобретения также могут применяться полинуклеотиды, придающие устойчивость/толерантность к гербициду, ингибирующему конус нарастания или меристему, такому как имидазолинон или сульфонилмочевина. Иллюстративные полинуклеотиды из этой категории кодируют мутантные ферменты ALS и AHAS, как описано, например, в патентах США №№5767366 и 5928937. Патенты США №№4761373 и 5013659 относятся к растениям, устойчивым к различным имидазолиноновым или сульфонамидным гербицидам. Патент США №4975374 относится к растительным клеткам и растениям, содержащим нуклеиновую кислоту, кодирующую мутантную глутаминсинтетазу (GS), устойчивую к ингибированию гербицидами, которые, как известно, ингибируют GS, например, фосфинотрицин и метионинсульфоксимин. В патенте США №5162602 раскрыты растения, устойчивые к ингибированию гербицидами на основе циклогександиона и арилоксифеноксипропановой кислоты. Устойчивость придает измененная ацетил-коэнзим А-карбоксилаза (АССаза).
[00247] Полипептиды, кодируемые нуклеотидными последовательностями, придающими устойчивость к глифосату, также подходят для настоящего изобретения. См., например, патент США №4940835 и патент США №4769061. В патенте США №5554798 раскрыты трансгенные растения маиса, устойчивые к глифосату, устойчивость которым придает ген измененной 5-енолпирувил-3-фосфошикиматсинтазы (EPSP).
[00248] Также подходят полинуклеотиды, кодирующие устойчивость к фосфоновым соединениям, таким как глюфосинат аммония или фосфинотрицин, а также пиридинокси- или феноксипропионовым кислотам и циклогексанонам. См. заявку на европейский патент №0242246. См. также патенты США №№5879903, 5276268 и 5561236.
[00249] Другие подходящие полинуклеотиды включают таковые, кодирующие устойчивость к гербицидам, подавляющим фотосинтез, таким как триазин и бензонитрил (ген нитрилазы). См. патент США №4810648. Дополнительные подходящие полинуклеотиды, кодирующие устойчивость к гербицидам, включают таковые, кодирующие устойчивость к 2,2-дихлорпропионовой кислоте, сетоксидиму, галоксифопу, имидазолиноновым гербицидам, сульфонилмочевинным гербицидам, триазолопиримидиновым гербицидам, s-триазиновым гербицидам и бромоксинилу. Также подходят полинуклеотиды, придающие устойчивость к ингибиторам фермента protox, или которые обеспечивают повышенную устойчивость к заболеваниям растений; повышенную толерантность к неблагоприятным условиям окружающей среды (видам абиотического стресса), в том числе без ограничения к засухе, чрезмерному охлаждению, чрезмерному нагреву, или чрезмерной засоленности почвы, или экстремальной кислотности или щелочности; и изменения строения или развития растений, в том числе изменения сроков развития. См., например, публикацию заявки на патент США №2001/0016956 и патент США №6084155.
[00250] Дополнительные подходящие полинуклеотиды включают таковые, кодирующие пестицидные (например, инсектицидные) полипептиды. Эти полипептиды могут быть получены в количествах, достаточных для контроля, например, насекомых-вредителей (т.е. в количествах, обеспечивающих контроль насекомых). В вариантах осуществления полипептид представляет собой полипептид, активный в отношении чешуекрылых, активный в отношении жесткокрылых, активный в отношении полужесткокрылых и/или активный в отношении двукрылых, или любую их комбинацию. Считается, что количество продуцируемого в растении пестицидного полипептида, необходимое для контроля насекомых или других вредителей, может варьировать в зависимости от сорта, типа вредителя, факторов окружающей среды и т.п. Полинуклеотиды, применимые для придания дополнительной устойчивости к насекомым или вредителям, включают, например, таковые, кодирующие токсины, идентифицированные в организмах Bacillus. Полинуклеотиды, содержащие нуклеотидные последовательности, кодирующие белки Cry Bacillus thuringiensis (Bt) из нескольких подвидов, были клонированы и было обнаружено, что рекомбинантные клоны токсичны для личинок чешуекрылых, двукрылых и жесткокрылых насекомых. Примеры таких инсектицидных белков Bt включают белки Cry, такие как Cry1Aa, Cry1Ab, Cry1Ac, Cry1B, Cry1C, Cry1D, Cry1Ea, Cry1Fa, Cry3A, Cry9A, Cry9B, Cry9C и т.п., а также вегетативные инсектицидные белки, такие как Vip1, Vip2, Vip3 и т.п., и любые комбинации вышеуказанных инсектицидных белков Bt. Полный перечень белков Bt можно найти во всемирной сети Интернет в базе данных номенклатуры токсинов Bacillus thuringiensis, поддерживаемой университетом Сассекса (см. также Crickmore et al. (1998) Microbiol. Mol. Biol. Rev. 62:807-813).
[00251] В вариантах осуществления дополнительный полипептид представляет собой инсектицидный полипептид, полученный из источника, отличного orBt, включая без ограничения альфа-амилазу, пероксидазу, холестеролоксидазу, пататин, протеазу, ингибитор протеазы, уреазу, ингибитор альфа-амилазы, порообразующий белок, хитиназу, лектин, сконструированное антитело или фрагмент антитела, инсектицидный белок Bacillus cereus, инсектицидный белок Xenorhabdus spp. (такого как Х. nematophila или Х. bovienii), инсектицидный белок Photorhabdus spp. (такого как P. luminescens или P. asymobiotica), инсектицидный белок Brevibacillus spp. (такого как В. laterosporous), инсектицидный белок Lysinibacillus spp. (такого как L., sphearicus), инсектицидный белок Chromobacterium spp. (такого как С. subtsugae или С. piscinae), инсектицидный белок Yersinia spp. (такого как Y. entomophaga), инсектицидный белок Paenibacillus spp. (такого как P. propylaea), инсектицидный белок Clostridium spp. (такого как С. bifermentans), Pseudomonas spp. (такого как P. fluorescens) и лигнин.
[00252] Полипептиды, подходящие для продуцирования в растениях, дополнительно включают такие полипептиды, которые улучшают превращение собранных растений или частей растений в коммерчески применимый продукт или иным образом содействующие ему, в том числе, например, обеспечивающие повышенное или измененное содержание или распределение углеводов, улучшенные свойства сбраживаемости, повышенное содержание масла, повышенное содержание белка, улучшенную усвояемость или повышенное содержание нутрицевтиков, например, повышенное содержание фитостерола, повышенное содержание токоферола, повышенное содержание станола или повышенное содержание витаминов. Представляющие интерес полипептиды также включают, например, таковые, обуславливающие снижение содержания нежелательного компонента в собранном урожае, например, фитиновой кислоты или ферментов, расщепляющих сахара, или способствующие этому. Под "обуславливающим" или "способствующим" подразумевается, что представляющий интерес полипептид может прямо или косвенно способствовать наличию представляющего интерес признака (например, путем увеличения расщепления целлюлозы с помощью гетерологичного фермента целлюлазы).
[00253] В некоторых вариантах осуществления полипептид способствует улучшению усвояемости еды или корма. Ксиланазы представляют собой ферменты, расщепляющие гемицеллюлозу, которые усиливают разрушение клеточных стенок растений, что приводит к лучшему использованию питательных веществ растения животным. Это приводит к увеличению темпов роста и конверсии корма. Также можно снижать вязкость кормов, содержащих ксилан. Гетерологичное продуцирование ксиланаз в растительных клетках также может содействовать превращению лигноцеллюлозы в сбраживаемые сахара в ходе промышленной переработки.
[00254] Многочисленные ксиланазы из микроорганизмов, являющихся грибами и бактериями, были идентифицированы и охарактеризованы (см., например, патент США №5437992; Coughlin et al. (1993) "Proceedings of the Second TRICEL Symposium on Trichoderma reesei Cellulases and Other Hydrolases" Espoo; Souminen and Reinikainen, eds. (1993) Foundation for Biotechnical and Industrial Fermentation Research 8:125-135; публикацию патента США №2005/0208178; и публикацию согласно PCT WO 03/16654). В частности, три специфичные ксиланазы (XYL-I, XYL-II и XYL-III) были идентифицированы у T. reesei (Tenkanen et al. (1992) EnzymeMicrob. Technol. 14:566; Torronen et al. (1992) Bio/Technology 10:1461; и Xu et al. (199%) Appl. Microbiol. Biotechnol. 49:718).
[00255] В других вариантах осуществления полипептид, применимый для настоящего изобретения, может представлять собой фермент, расщепляющий полисахариды. Растения по настоящему изобретению, вырабатывающие такой фермент, могут быть применимы для получения, например, сбраживаемого сырья для биологической переработки. В некоторых вариантах осуществления ферменты, применимые для способа сбраживания, включают альфа-амилазы, протеазы, пуллуланазы, изоамилазы, целлюлазы, гемицеллюлазы, ксиланазы, циклодекстрингликозилтрансферазы, липазы, фитазы, лакказы, оксидазы, эстеразы, кутиназы, фермент, гидролизующий гранулированный крахмал, и другие глюкоамилазы.
[00256] Ферменты, расщепляющие полисахариды, включают ферменты, расщепляющие крахмал, такие как альфа-амилазы (ЕС 3.2.1.1), глюкуронидазы (Е.С.3.2.1.131); экзо-1,4-альфа-D-глюканазы, такие как амилоглюкозидазы и глюкоамилаза (ЕС 3.2.1.3), бета-амилазы (ЕС 3.2.1.2), альфа-глюкозидазы (ЕС 3.2.1.20) и другие экзо-амилазы; ферменты, расщепляющие крахмал, такие как а) изоамилаза (ЕС 3.2.1.68), пуллуланаза (ЕС 3.2.1.41) и т.п.; b) целлюлазы, такие как экзо-1,4-3-целлобиогидролаза (ЕС 3.2.1.91), экзо-1,3-бета-D-глюканаза (ЕС 3.2.1.39), бета-глюкозидаза (ЕС 3.2.1.21); с) L-арабиназы, такие как эндо-1,5-альфа-L-арабиназа (ЕС 3.2.1.99), альфа-арабинозидазы (ЕС 3.2.1.55) и т.п.; d) галактаназы, такие как эндо-1,4-бета-D-галактаназа (ЕС 3.2.1.89), эндо-1,3-бета-D-галактаназа (ЕС 3.2.1.90), альфа-галактозидаза (ЕС 3.2.1.22), бета-галактозидаза (ЕС 3.2.1.23) и т.п.; е) маннаназы, такие как эндо-1,4-бета-D-маннаназа (ЕС 3.2.1.78), бета-маннозидаза (ЕС 3.2.1.25), альфа-маннозидаза (ЕС 3.2.1.24) и т.п.; f) ксиланазы, такие как эндо-1,4-бета-ксиланаза (ЕС 3.2.1.8), бета-D-ксилозидаза (ЕС 3.2.1.37), 1,3-бета-D-ксиланаза и т.п.; и g) другие ферменты, такие как альфа-L-фукозидаза (ЕС 3.2.1.51), альфа-L-рамнозидаза (ЕС 3.2.1.40), леваназа (ЕС 3.2.1.65), инуланаза (ЕС 3.2.1.7) и т.п. В одном варианте осуществления альфа-амилаза представляет собой синтетическую альфа-амилазу, Amy797E, описанную в патенте США №8093453.
[00257] Дополнительные ферменты, которые можно применять в соответствии с настоящим изобретением, включают протеазы, такие как протеазы грибов и бактерий. Протеазы грибов включают без ограничений таковые, полученные из Aspergillus, Trichoderma, Mucor и Rhizopus, как, например, A. niger, A. awamori, A. oryzae и М. miehei. В некоторых вариантах осуществления полипептиды по настоящему изобретению могут представлять собой ферменты целлобиогидролазы (СВН) (ЕС 3.2.1.91). В одном варианте осуществления фермент целлобиогидролаза может представлять собой СВН1 или СВН2.
[00258] Другие ферменты, применимые в соответствии с настоящим изобретением, включают без ограничения гемицеллюлазы, такие как манназы и арабинофуранозидазы (ЕС 3.2.1.55); лигниназы; липазы (например, Е.С.3.1.1.3), глюкозооксидазы, пектиназы, ксиланазы, трансглюкозидазы, альфа-1,6-глюкозидазы (например, Е.С.3.2.1.20); эстеразы, такие как естераза феруловой кислоты (ЕС 3.1.1.73) и ацетилксиланэстеразы (ЕС 3.1.1.72); и кутиназы (например, Е.С.3.1.1.74).
[00259] Молекулы двухнитевой РНК (dsRNA), применимые в настоящем изобретении, включают без ограничения такие, которые подавляют экспрессию генов у целевого вредителя (например, насекомого). В вариантах осуществления dsRNA нацеливается на ген насекомого-вредителя, относящегося к чешуекрылым, жесткокрылым, полужесткокрылым или двукрылым или любой комбинации вышеуказанного. Предполагается, что применяемое в данном документе словосочетание "супрессия гена" в совокупности относится к любым хорошо известным способам снижения уровней белка, продуцируемого в результате транскрипции гена в mRNA и дальнейшей трансляции mRNA. Также подразумевается, что супрессия гена означает снижение экспрессии белка с гена или кодирующей последовательности, в том числе посттранскрипционную супрессию гена и транскрипционную супрессию. Посттранскрипционная супрессия гена обусловлена гомологией всей или части mRNA, транскрибируемой с гена или кодирующей последовательности, являющихся мишенями для супрессии, и соответствующей двухцепочечной РНК, используемой для супрессии, и означает значимое и измеряемое снижение количества доступной mRNA, имеющейся в клетке для связывания рибосомами. Транскрибируемая РНК может находиться в смысловой ориентации для обеспечения эффекта, называемого косупрессией, в антисмысловой ориентации для обеспечения эффекта, называемого антисмысловой супрессией, или в обеих ориентациях с получением dsRNA для обеспечения эффекта, называемого РНК-интерференцией (RNAi). Транскрипционная супрессия обусловлена присутствием в клетке dsRNA, средства, обеспечивающего супрессию гена, характеризующейся значительной идентичностью последовательности с промоторной последовательностью ДНК или комплементарной ей нитью, для обеспечения эффекта, который называется супрессией промотора в транс-положении. Супрессия гена может быть эффективной в отношении нативного гена растения, ассоциированного с признаком, например, для обеспечения в растениях сниженных уровней белка, кодируемого нативным геном, или повышенными уровнями измененного метаболита. Супрессия гена также может быть эффективной в отношении генов-мишеней во вредителях растений, которые могут проглатывать или контактировать с растительным материалом, содержащим средства для обеспечения супрессии гена, специально сконструированного для подавления или супрессии одного или нескольких гомологичных или комплементарных последовательностей в клетках вредителя. Такие гены-мишени для супрессии могут кодировать незаменимый белок, основная функция которого выбрана из группы, состоящей из формирования мышц, образования ювенильного гормона, регуляции ювенильного гормона, регуляции и транспорта ионов, синтеза пищеварительных ферментов, поддержания мембранного потенциала клетки, биосинтеза аминокислот, деградации аминокислот, образования спермы, синтеза феромонов, восприятия феромонов, формирования антенн, формирования крыльев, формирования конечностей, развития и дифференциации, формирования яиц, созревания личинок, образования пищеварительных ферментов, синтеза гемолимфы, поддержания постоянства состава гемолимфы, передачи нервных импульсов, клеточного деления, энергетического обмена, развития и дифференциации, дыхания и апоптоза.
[00260] В вариантах осуществления нуклеиновые кислоты по настоящему изобретению могут дополнительно содержать, по сути состоять или состоять из вектора. В вариантах осуществления полинуклеотиды и кассеты экспрессии по настоящему изобретению содержатся в векторе. Векторы для применения в трансформации растений и других организмов хорошо известны из уровня техники. Неограничивающие примеры общих классов векторов включают плазмиду, фаговый вектор, фагмидный вектор, космидный вектор, фосмиду, бактериофаг, искусственную хромосому или вирусный вектор. В вариантах осуществления вектор представляет собой растительный вектор, например, для применения при трансформации растений. В вариантах осуществления вектор представляет собой бактериальный вектор, например, для применения при трансформации бактерий. Подходящие векторы для растений, бактерий и других организмов известны в данной области техники.
Трансгенные растения, части растений, растительные клетки, семена
[00261] Настоящее изобретение также охватывает трансгенную клетку-хозяина, отличную от клетки человека, которая содержит полинуклеотид, молекулу нуклеиновой кислоты, кассету экспрессии, вектор или полипептид по настоящему изобретению. Трансгенная клетка-хозяин, отличная от клетки человека, может включать без ограничения растительную клетку (в том числе клетку однодольного и/или клетку двудольного), дрожжевую клетку, бактериальную клетку или клетку насекомого. Следовательно, в некоторых вариантах осуществления настоящего изобретения предусмотрена бактериальная клетка, выбранная из рода Bacillus, Brevibacillus, Clostridium, Xenorhabdus, Photorhabdus, Pasteuria, Escherichia, Pseudomonas, Erwinia, Serratia, Klebsiella, Salmonella, Pasteurella, Xanthomonas, Streptomyces, Rhizobium, Rhodopseudomonas, Methylophilius, Agrobacterium, Acetobacter, Lactobacillus, Arthrobacter, Azotobacter, Leuconostoc или Alcaligenes. Таким образом, например, в качестве средств для биологического контроля насекомых можно получать химерные инсектицидные белки по настоящему изобретению путем экспрессии полинуклеотида, кодирующего их, в бактериальной клетке. Например, в некоторых вариантах осуществления предусмотрена клетка Bacillus thuringiensis, содержащая полинуклеотид, кодирующий химерный инсектицидный белок по настоящему изобретению.
[00262] В вариантах осуществления трансгенная растительная клетка представляет собой клетку однодольного растения или клетку двудольного растения. В дополнительных вариантах осуществления клетка двудольного растения представляет собой клетку сои, клетку подсолнечника, клетку томата, клетку культурной разновидности капусты, клетку хлопчатника, клетку сахарной свеклы и клетку табака. В дополнительных вариантах осуществления клетка однодольного растения представляет собой клетку ячменя, клетку маиса, клетку овса, клетку риса, клетку сорго, клетку сахарного тростника и клетку пшеницы. В вариантах осуществления настоящего изобретения предусмотрена совокупность клеток двудольных растений или клеток однодольных растений, содержащих полинуклеотид, экспрессирующий химерный инсектицидный белок по настоящему изобретению. В вариантах осуществления клетки в совокупности расположены рядом с образованием апопласта, и их выращивают при естественном солнечном свете. В вариантах осуществления трансгенная растительная клетка не может регенерироваться в целое растение.
[00263] В вариантах осуществления настоящего изобретения инсектицидный белок по настоящему изобретению экспрессируется в высшем организме, например, растении. В этом случае трансгенные растения, экспрессирующие эффективные количества инсектицидного белка, защищают себя от вредителей растений, таких как насекомые-вредители. Когда насекомое начинает кормиться на таком трансгенном растении, оно также поглощает экспрессируемый инсектицидный белок. Это может удерживать насекомое от дальнейшего вгрызания в растительную ткань или даже может причинять вред насекомому или уничтожать его. В вариантах осуществления полинуклеотид по настоящему изобретению вставляют в кассету экспрессии, которая затем стабильно интегрируется в геном растения. В других вариантах осуществления полинуклеотид включен в непатогенный самореплицирующийся вирус.
[00264] В некоторых вариантах осуществления настоящего изобретения трансгенная растительная клетка, содержащая молекулу нуклеиновой кислоты или полипептид по настоящему изобретению, представляет собой клетку части растения, органа растения или культуры растения (каждая, как описано в данном документе), включая без ограничения корень, лист, семя, цветок, плод, клетку пыльцы, орган или культуру растения и т.п., или каллусную клетку или культуру.
[00265] Трансгенное растение или растительная клетка в соответствии с настоящим изобретением может представлять собой однодольное или двудольное растение или растительную клетку и включает без ограничения кукурузу (маис), сою, рис, пшеницу, ячмень, рожь, овес, сорго, просо, подсолнечник, сафлор, сахарную свеклу, хлопчатник, сахарный тростник, масличный рапс, люцерну, табак, арахис, овощные культуры (в том числе батат, фасоль, горох, цикорий, латук, кочанную капусту, цветную капусту, брокколи, репу, морковь, баклажан, огурец, редьку, шпинат, картофель, томат, спаржу, лук, чеснок, дыню, перец, сельдерей, тыкву крупноплодную, тыкву мускатную, тыкву обыкновенную и т.п.), плодовую культуру (включая яблоню, грушу, айву, сливу, вишню, персик, нектарин, абрикос, землянику, виноград, малину, ежевику, ананас, авокадо, папайю, манго, банан и т.п.) и специализированное растение или растительная клетка (такое как Arabidopsis), или древесное растение или растительная клетка (такое как хвойные и/или лиственные деревья). В вариантах осуществления растение или растительная клетка по настоящему изобретению представляет собой культурное растение или растительную клетку, такое как растение или растительная клетка маиса, сорго, пшеницы, подсолнечника, томата, крестоцветного, перца, картофеля, хлопчатника, риса, сои, сахарной свеклы, сахарного тростника, табака, ячменя, масличного рапса и т.п.
[00266] В настоящем изобретении дополнительно предусмотрена часть трансгенного растения по настоящему изобретению. Необязательно часть растения содержит химерный инсектицидный белок по настоящему изобретению и/или нуклеиновую кислоту, кодирующую его.
[00267] В настоящем изобретении дополнительно предусмотрено семя трансгенного растения по настоящему изобретению или семя, из которого получают трансгенное растение по настоящему изобретению. Необязательно семя содержит химерный инсектицидный белок по настоящему изобретению и/или нуклеиновую кислоту, кодирующую его.
[00268] Дополнительные варианты осуществления настоящего изобретения включают собранные продукты, полученные из трансгенных растений, частей растений или семян по настоящему изобретению, а также переработанный продукт, полученный из собранного продукта. Собранный продукт может представлять собой целое растение или любую часть растения, как раскрыто в данном документе. Таким образом, в некоторых вариантах осуществления неограничивающие примеры собранного продукта включают семя, плод, цветок или его часть (например, пыльник, рыльце и т.п.), лист, стебель и т.п. В других вариантах осуществления обработанный продукт включает без ограничения муку, муку грубого помола, масло, крахмал, зерно и т.п., полученные из собранного семени или другой части растения по настоящему изобретению. Необязательно собранный продукт или обработанный продукт содержит химерный инсектицидный белок по настоящему изобретению и/или нуклеиновую кислоту, кодирующую его.
[00269] В других вариантах осуществления настоящего изобретения предусмотрен экстракт из трансгенного растения или части растения по настоящему изобретению, где необязательно экстракт содержит химерный инсектицидный белок по настоящему изобретению и/или нуклеиновую кислоту, кодирующую его. Экстракты из растений или частей растений можно получить согласно процедурам, хорошо известным в данной области (см. de la Torre et al., Food, Agric. Environ. 2(l):84-89 (2004); Guidet, Nucleic Acids Res. 22(9): 1772-1773 (1994); Lipton et al., Food Agric. Immun. 12:153-164 (2000)).
[00270] Химерный инсектицидный белок может функционировать в части растения, растительной клетке, органе растения, семени, собранном продукте, обработанном продукте или экстракте и т.п. в качестве средства для контроля насекомых. Другими словами, химерный инсектицидный белок может продолжать выполнять инсектицидную функцию, которой он обладал в трансгенном растении. Нуклеиновая кислота может функционировать для экспрессии химерного инсектицидного белка. В качестве альтернативы кодированию инсектицидного белка по настоящему изобретению нуклеиновая кислота может функционировать для выявления трансгенных части растения, растительной клетки, органа растения, семени, собранного продукта, обработанного продукта или экстракта по настоящему изобретению.
[00271] В вариантах осуществления трансгенные растение, часть растения, растительная клетка, орган растения или семя по настоящему изобретению являются гемизиготными в отношении полинуклеотида или кассеты экспрессии по настоящему изобретению. В вариантах осуществления трансгенные растение, часть растения, растительная клетка, орган растения или семя по настоящему изобретению являются гомозиготными в отношении полинуклеотида или кассеты экспрессии по настоящему изобретению.
[00272] В вариантах осуществления трансгенные растение, часть растения, растительная клетка, орган растения, семя, собранный продукт, обработанный продукт или экстракт имеют повышенную устойчивость к одному или нескольким насекомым-вредителям (например, вредителю, относящемуся к чешуекрылым, такому как совка травяная) по сравнению с подходящим контролем, который не содержит нуклеиновую кислоту, кодирующую инсектицидный белок по настоящему изобретению.
Трансформация растений
[00273] Процедуры трансформации растений хорошо известны и общеприняты в данной области и описаны в литературе во всех отношениях. Неограничивающие примеры способов трансформации растений включают трансформацию с помощью доставки нуклеиновых кислот, опосредованной бактериями (например, с помощью Agrobacterium), доставки нуклеиновых кислот, опосредованной вирусами, доставки нуклеиновых кислот, опосредованной карбидом кремния или микроиглами с нуклеиновыми кислотами, доставки нуклеиновых кислот, опосредованной липосомами, микроинъекцию, бомбардировку микрочастицами, трансформацию, опосредованную фосфатом кальция, трансформацию, опосредованную циклодекстринами, электропорацию, трансформацию, опосредованную наночастицами, обработку ультразвуком, инфильтрацию, поглощение нуклеиновых кислот, опосредованное PEG, а также любой другой электрический, химический, физический (механический) или биологический механизм, который приводит к введению нуклеиновой кислоты в растительную клетку, включая любую их комбинацию. Общие указания по разнообразным способам трансформации растений, известным в данной области техники, включают Miki et al. ("Procedures for Introducing Foreign DNA into Plants" in Methods in Plant Molecular Biology and Biotechnology, Glick, B.R. and Thompson, J.E., Eds. (CRC Press, Inc., Boca Raton, 1993), страницы 67-88) и Rakowoczy-Trojanowska (Cell. Mol. Biol. Lett. 7:849-858 (2002)).
[00274] Для трансформации, опосредованной Agrobacterium, обычно подходят бинарные векторы или векторы, несущие по меньшей мере одну граничную последовательность T-DNA, тогда как для прямого переноса генов (например, с помощью бомбардировки частицами и т.п.) подходит любой вектор, при этом можно применять линейную ДНК, содержащую только представляющую интерес конструкцию. В случае прямого переноса генов можно применять трансформацию при помощи одного вида ДНК или котрансформацию (Schocher et al., Biotechnology 4:1093-1096 (1986)). В случае как прямого переноса генов, так и переноса, опосредованного Agrobacterium, трансформацию обычно (но не обязательно) выполняют с селектируемым маркером, который может представлять собой средство для положительного отбора (например, фосфоманнозоизомераза), которое обеспечивает устойчивость к антибиотику (например, канамицину, гигромицину или метотрексату) или гербициду (например, глифосату или глюфосинату). Однако выбор селектируемого маркера не является критически важным для настоящего изобретения.
[00275] Трансформация, опосредованная Agrobacterium, представляет собой способ, широко применяемый для трансформации растений в связи с высокой эффективностью трансформации и в связи с широкой применимостью в отношении множества различных видов. Трансформация, опосредованная Agrobacterium, как правило, предполагает перенос бинарного вектора, несущего чужеродную ДНК, представляющую интерес, в соответствующий штамм Agrobacterium, что может зависеть от набора генов vir штамма-хозяина Agrobacterium, расположенного либо в корезидентной Ti-плазмиде, либо в хромосоме (Uknes et al. (1993) Plant Cell 5:159-169). Перенос рекомбинантного бинарного вектора в Agrobacterium можно выполнять с помощью процедуры трехродительского скрещивания с применением Escherichia coli, несущей рекомбинантный бинарный вектор, хелперного штамма Е. coli, несущего плазмиду, которая способна мобилизовать рекомбинантный бинарный вектор в целевом штамме Agrobacterium. В качестве альтернативы, рекомбинантный бинарный вектор можно переносить в Agrobacterium путем трансформации нуклеиновой кислоты (Hofgen & Willmitzer (1988) Nucleic Acids Res. 16:9877).
[00276] Двудольные, а также однодольные можно трансформировать с использованием Agrobacterium. Способы опосредованной Agrobacterium трансформации риса включают хорошо известные способы трансформации риса, такие как описанные в любом из следующего: Европейская патентная заявка ЕР 1198985 Al, Aldemita and Hodges (Planta 199: 612-617, 1996); Chan et al. (Plant Mol Biol 22 (3): 491-506, 1993), Hiei et al. (Plant J 6 (2): 271-282, 1994), раскрытия которых включены в данный документ посредством ссылки, как если бы они были полностью изложены. В случае трансформации кукурузы предпочтительным способом является описанный или в Ishida et al. (Nat. Biotechnol 14(6): 745-50, 1996), или Frame et al. (Plant Physiol 129(1): 13-22, 2002), раскрытия которых включены в данный документ посредством ссылки, как если бы они были полностью изложены. Указанные способы также описаны в качестве примера в в. Jenes et al., Techniques for Gene Transfer, in: Transgenic Plants, Vol.1, Engineering and Utilization, eds. S. D. Kung and R. Wu, Academic Press (1993) 128-143 и в Potrykus Annu. Rev. Plant Physiol. Plant Molec. Biol. 42 (1991) 205-225). Нуклеиновые кислоты или конструкция, подлежащие экспрессии, предпочтительно клонируют в вектор, который является подходящим для трансформации Agrobacterium tumefaciens, например pBinl9 (Bevan et al., Nucl. Acids Res. 12 (1984) 8711). Agrobacteria, трансформированные таким вектором, затем можно применять известным способом для трансформации растений, таких как растения, применяемые в качестве модели, такие как Arabidopsis, или сельскохозяйственные культуры, такие как, в качестве примера, растения табака, например, посредством погружения истолченных листьев или нарубленных листьев в раствор Agrobacteria и затем культивирования их в подходящей среде. Трансформация растений посредством Agrobacterium tumefaciens описана, например, Hagen and Willmitzer в Nucl. Acid Res. (1988) 16, 9877 или известна, в частности, из F.F. White, Vectors for Gene Transfer in Higher Plants; в Transgenic Plants, Vol.1, Engineering and Utilization, eds. S. D. Kung andR. Wu, Academic Press, 1993, pp. 15-38.
[00277] Трансформация растения с помощью рекомбинантной Agrobacterium обычно включает совместное культивирование Agrobacterium с эксплантатами растения, и ее проводят в соответствии со способами, хорошо известными из уровня техники. Трансформированную ткань регенерируют на селективной среде, содержащей маркер устойчивости к антибиотикам или гербицидам между граничными последовательностями T-DNA бинарной плазмиды.
[00278] Как обсуждалось ранее, другой способ трансформации растений, частей растений и растительных клеток включает внедрение инертных или биологически активных частиц в растительные ткани и клетки. См, например, патенты США №№4945050, 5036006 и 5100792. В общем случае этот способ включает внедрение в растительные клетки инертных или биологически активных частиц в условиях, эффективных для проникновения через наружную поверхность клетки и возможности встраивания в ее внутреннюю часть. При использовании инертных частиц вектор можно вводить в клетку путем покрытия частиц вектором, содержащим представляющую интерес нуклеиновую кислоту. В качестве альтернативы, клетка или клетки могут быть окружены вектором так, чтобы вектор переносился в клетку вслед за частицей. Биологически активные частицы (например, высушенная дрожжевая клетка, высушенная бактерия или бактериофаг, каждая(каждый) из которых содержит одну или несколько нуклеиновых кислот, подлежащих введению) также можно внедрять в растительную ткань.
[00279] В других вариантах осуществления полинуклеотид по настоящему изобретению можно напрямую вводить в геном пластид путем трансформации. Основное преимущество трансформации пластид состоит в том, что пластиды обычно способны экспрессировать бактериальные гены без существенной модификации, при этом пластиды способны экспрессировать несколько открытых рамок считывания под контролем одного промотора. Технология трансформации пластид подробно описана в патентах США №№5451513, 5545817 и 5545818, в заявке согласно PCT № WO 95/16783 и в McBride et al. (1994) Proc. Nati. Acad. Sci. USA 91, 7301-7305. Основная методика трансформации хлоропластов включает введение участков клонированной пластидной ДНК, фланкирующих селектируемый маркер, вместе с представляющим интерес геном в подходящую целевую ткань, например, с применением биолистики или трансформации протопластов (например, трансформации, опосредованной хлоридом кальция или PEG). Фланкирующие участки размером 1-1,5, т.о. называемые нацеливающими последовательностями, содействуют гомологичной рекомбинации с геномом пластид и, таким образом, обеспечивают замещение или модификацию специфических участков пластома. Сначала в качестве селектируемого маркера трансформации можно использовать точечные мутации в генах 16S рРНК и rpsl2 хлоропластов, придающие устойчивость к спектиномицину или стрептомицину (Svab, Z., Hajdukiewicz, P., and Maliga, P. (1990) Proc. Natl. Acad. Sci. USA 87, 8526-8530; Staub, J.M., and Maliga, P. (1992) Plant Cell 4, 39-45). Наличие сайтов клонирования между этими маркерами позволяет создать вектор, нацеленный на пластиды, для введения чужеродных генов (Staub, J.М., and Maliga, P. (1993) EMBO J. 12, 601-606). Существенного повышения частоты трансформации достигали путем замещения рецессивных генов rRNA или r-белка, обеспечивающих устойчивость к антибиотикам, на доминантный селектируемый маркер, бактериальный ген aadA, кодирующий фермент аминогликозид-3'-аденилтрансферазу, обезвреживающий спектиномицин (Svab, Z., and Maliga, P. (1993) Proc. Natl. Acad. Sci. USA 90, 913-917). Ранее этот маркер успешно применяли для трансформации генома пластид зеленой водоросли Chlamydomonas reinhardtii с высокой частотой (Goldschmidt-Clermont, М. (1991) Nucl. Acids Res. 19:4083-4089). Другие селектируемые маркеры, применимые для трансформации пластид, известны из уровня техники и включены в объем настоящего изобретения. Как правило, для достижения состояния, при котором все пластиды являются одинаковыми, требуется около 15-20 циклов клеточного деления после трансформации. При экспрессии в пластидах, при которой гены вставлены с помощью гомологической рекомбинации во все несколько тысяч копий кольцевого пластидного генома, присутствующего в каждой растительной клетке, используют преимущество огромного числа копий по сравнению с генами, экспрессируемыми в ядре, что допускает уровни экспрессии, которые легко могут превышать 10% общего количества растворимых растительных белков. В одном варианте осуществления полинуклеотид по настоящему изобретению может быть вставлен в вектор, нацеленный на пластиды, и введен в геном пластид требуемого растения-хозяина путем трансформации. Таким образом, можно получить растения, гомопластические в отношении геномов пластид, содержащих нуклеотидную последовательность по настоящему изобретению, способные экспрессировать полинуклеотид на высоком уровне.
[00280] Способы отбора трансформированных трансгенных растений, растительных клеток или культур растительных тканей являются общепринятыми в данной области техники и могут использоваться в способах настоящего изобретения, предусмотренных в данном документе. Например, рекомбинантный вектор по настоящему изобретению также может включать кассету экспрессии, содержащую нуклеотидную последовательность селектируемого маркера, который можно применять для отбора трансформированного растения, части растения или растительной клетки.
[00281] Дополнительно, как хорошо известно из уровня техники, целые трансгенные растения можно регенерировать из трансформированных растительных клеток, культур растительных тканей или культивируемых протопластов с помощью любой из множества известных методик. Регенерация растений из растительных клеток, культуры растительных тканей или культивируемых протопластов описана, например, в Evans et al. (Handbook of Plant Cell Cultures, Vol. 1, MacMilan Publishing Co. New York (1983)); и Vasil I.R. (ed.) (Cell Culture and Somatic Cell Genetics of Plants, Acad. Press, Orlando, Vol. I (1984), and Vol. II (1986)).
[00282] Кроме того, описанные выше свойства генов, обеспеченные с помощью методик генной инженерии в трансгенных семенах и растениях, частях растения или растительных клетках по настоящему изобретению, могут передаваться путем полового размножения или вегетативного роста и, следовательно, могут поддерживаться и передаваться по наследству растениям-потомкам. Как правило, при поддержании и передаче по наследству применяют известные сельскохозяйственные способы, разработанные для соответствия конкретным целям, таким как уборка урожая, посев или возделывание.
[00283] Следовательно, полинуклеотид можно вводить в растение, часть растения или растительную клетку с помощью любого из целого ряда способов, хорошо известных из уровня техники, как описано выше. Следовательно, не придерживаются какого-либо конкретного способа введения одного или нескольких полинуклеотидов в растение, а наоборот, можно применять любой способ, обеспечивающий стабильную интеграцию одного или нескольких полинуклеотидов в геном растения. Если требуется ввести более одного полинуклеотида, то соответствующие полинуклеотиды можно собрать как части одной молекулы нуклеиновой кислоты или как отдельные молекулы нуклеиновых кислот и можно расположить в пределах одной и той же или различных молекул нуклеиновых кислот. Соответственно, введение полинуклеотидов в представляющую интерес клетку можно осуществлять в ходе одного события трансформации, в ходе отдельных событий трансформации или, например, в растения, в виде части протокола скрещивания.
[00284] После того как требуемый полинуклеотид введен в конкретный вид растения путем трансформации, при помощи традиционных методик размножения он может передаваться в пределах данного вида или переходить в другие разновидности того же вида, в частности, включающие коммерческие разновидности.
Инсектицидные композиции
[00285] В некоторых вариантах осуществления настоящего изобретения предусмотрена инсектицидная композиция, содержащая химерный инсектицидный белок по настоящему изобретению в приемлемом с точки зрения сельского хозяйства носителе. Используемое в данном документе выражение "приемлемый с точки зрения сельского хозяйства носитель" может включать природный или синтетический, органический или неорганический материал, который объединен с активным белком для облегчения его применения на растении или его части или в растении или его части. Примеры приемлемых с точки зрения сельского хозяйства носителей включают без ограничения порошки, дусты, пеллеты, гранулы, аэрозоли, эмульсии, коллоиды и растворы. Приемлемые с точки зрения сельского хозяйства носители дополнительно включают без ограничения инертные компоненты, диспергирующие вещества, поверхностно-активные вещества, вспомогательные вещества, придающие липкость вещества, клейкие вещества, связующие вещества или их комбинации, которые можно применять в составах, используемых в сельском хозяйстве. Такие композиции можно применять любым способом, посредством которого пестицидные белки или другие средства для контроля вредителей приводят в контакт с вредителями. Соответственно, композиции можно наносить на поверхности растений или частей растений, в том числе на семена, листья, цветки, стебли, клубни, корни и т.п. В других вариантах осуществления растение, продуцирующее инсектицидный белок по настоящему изобретению in planta, представляет собой приемлемый с точки зрения сельского хозяйства носитель для экспрессируемого инсектицидного белка. В вариантах осуществления композиции и приемлемые с точки зрения сельского хозяйства носители по настоящему изобретению исключают трансгенные растения.
[00286] В дополнительных вариантах осуществления инсектицидная композиция содержит бактериальную клетку или трансгенную бактериальную клетку по настоящему изобретению, при этом бактериальная клетка или трансгенная бактериальная клетка продуцирует инсектицидный белок по настоящему изобретению. Такую инсектицидную композицию можно получить путем высушивания, лиофилизации, гомогенизации, экстракции, фильтрации, центрифугирования, осаждения или концентрирования культуры клеток Bacillus thuringiensis (Bt), в том числе трансгенной культуры Bt. В вариантах осуществления композиция по настоящему изобретению может содержать по меньшей мере приблизительно 1%, по меньшей мере приблизительно 5%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 25%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 35%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 60%, по меньшей мере 70%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 97% или по меньшей мере 99% по весу полипептида по настоящему изобретению. В дополнительных вариантах осуществления композиция содержит от приблизительно 1% до приблизительно 99% по весу инсектицидного белка по настоящему изобретению.
[00287] Инсектицидные белки по настоящему изобретению можно применять в комбинации с другими средствами для контроля вредителей для увеличения спектра целевых вредителей и/или для предупреждения или контроля устойчивости насекомых. Кроме того, применение инсектицидных белков по настоящему изобретению в комбинации с инсектицидным средством, которое характеризуется другим механизмом действия или целенаправленно воздействует на другой рецептор в пищеварительном канале насекомого, характеризуется особой полезностью в отношении предупреждения появления устойчивости у насекомых и/или ее контроля.
[00288] Следовательно, в некоторых вариантах осуществления настоящего изобретения предусмотрена композиция, которая обеспечивает контроль одного или нескольких вредителей растений (например, насекомого-вредителя, такого как насекомое-вредитель, относящееся к чешуекрылым, насекомое-вредитель, относящееся к жесткокрылым, насекомое-вредитель, относящееся к полужесткокрылым и/или насекомое-вредитель, относящееся к двукрылым), где композиция содержит первое средство для контроля вредителей, которое представляет собой химерный инсектицидный белок по настоящему изобретению, и по меньшей мере второе средство для контроля вредителей, которое отличается от первого средства для контроля вредителей. В других вариантах осуществления композиция представляет собой состав для местного нанесения на растение. В еще одних вариантах осуществления композиция представляет собой трансгенное растение. В дополнительных вариантах осуществления композиция представляет собой комбинацию, предусматривающую состав и трансгенное растение, на которое он наносится. В некоторых вариантах осуществления состав содержит первое средство для контроля вредителей, которое представляет собой химерный инсектицидный белок по настоящему изобретению, если трансгенное растение содержит второе средство для контроля вредителей. В других вариантах осуществления состав содержит второе средство для контроля вредителей, если трансгенное растение содержит первое средство для контроля вредителей, которое представляет собой химерный инсектицидный белок по настоящему изобретению.
[00289] В некоторых вариантах осуществления второе средство для контроля вредителей может представлять собой одно или несколько из химического пестицида, такого как инсектицид, инсектицидный белок Bacillus thuringiensis (Bt) и/или инсектицидное средство из источника, отличного от Bt, включая без ограничения инсектицидный белок Xenorhabdus, инсектицидный белок Photorhabdus, инсектицидный белок Brevibacillus laterosporus, инсектицидный белок Bacillus sphaericus, ингибитор протеаз (как сериновых, так и цистеиновых типов), лектин, альфа-амилаза, пероксидаза, холестериноксидаза или молекула двухнитевой РНК (dsRNA).
[00290] В других вариантах осуществления второе средство для контроля вредителей представляет собой один или несколько химических пестицидов, которые необязательно представляют собой покрытие для семян. Неограничивающие примеры химических пестицидов включают пиретроиды, карбаматы, неоникотиноиды, блокаторы нейрональных натриевых каналов, инсектицидные макроциклические лактоны, антагонисты гамма-аминомасляной кислоты (GABA), инсектицидные мочевины и имитаторы ювенильных гормонов. В других вариантах осуществления химический пестицид представляет собой одно или несколько из абамектина, ацефата, ацетамиприда, амидофлумета (S-1955), авермектина, азадирахтина, азинфос-метила, бифентрина, бинфеназата, бупрофезина, карбофурана, хлорфенапира, хлорфлуазурона, хлорпирифоса, хлорпирифос-метила, хромафенозида, клотианидина, цифлутрина, бета-цифлутрина, цигалотрина, лямбда-цигалотрина, циперметрина, циромазина, дельтаметрина, диафентиурона, диазинона, дифлубензурона, диметоата, диофенолана, эмамектина, эндосульфана, эсфенвалерата, этипрола, фенотикарба, феноксикарба, фенпропатрина, фенпроксимата, фенвалерата, фипронила, флоникамида, флуцитрината, тау-флювалината, флуфенерима (UR-50701), флуфеноксурона, фонофоса, галофенозида, гексафлумурона, имидаклоприда, индоксакарба, изофенфоса, луфенурона, малатиона, метальдегида, метамидофоса, метидатиона, метомила, метопрена, метоксихлора, монокротофоса, метоксифенозида, нитиазина, новалурона, новифлумурона (XDE-007), оксамила, паратиона, паратион-метила, перметрина, фората, фозалона, фосмета, фосфамидона, пиримикарба, профенофоса, пиметрозина, пиридалила, пирипроксифена, ротенона, спиносада, спиромезифена (BSN 2060), сульпрофоса, тебуфенозида, тефлубензурона, тефлутрина, тербуфоса, тетрахлорвинфоса, тиаклоприда, тиаметоксама, тиодикарба, тиосултап-натрия, тралометрина, трихлорфона и трифлумурона, алдикарба, оксамила, фенамифоса, амитраза, хинометионата, хлоробензилата, цигексатина, дикофола, диенохлора, этоксазола, феназаквина, оксида фенбутатина, фенпропатрина, фенпироксимата, гекситиазокса, пропаргита, пиридабена и тебуфенпирада. В еще одних вариантах осуществления химический пестицид выбран из одного или нескольких из циперметрина, цигалотрина, цифлутрина и бета-цифлутрина, эсфенвалерата, фенвалерата, тралометрина, фенотикарба, метомила, оксамила, тиодикарба, клотианидина, имидаклоприда, тиаклоприда, индоксакарба, спиносада, абамектина, авермектина, эмамектина, эндосульфана, этипрола, фипронила, флуфеноксурона, трифлумурона, диофенолана, пирипроксифена, пиметрозина и амитраза.
[00291] В дополнительных вариантах осуществления второе средство для контроля вредителей может представлять собой один или несколько из любого числа инсектицидных белков Bacillus thuringiensis, в том числе без ограничения белок Cry, вегетативный инсектицидный белок (VIP) и инсектицидные химерные варианты любого из перечисленных выше инсектицидных белков. В других вариантах осуществления второе средство для контроля вредителей представляет собой белок Cry, выбранный из: Cry1Aa, Cry1Ab, Cry1Ac, Cry1Ad, Cry1Ae, Cry1Af, Cry1Ag, Cry1Ah, Cry1Ai, Cry1Aj, Cry1Ba, Cry1Bb, Cry1Bc, Cry1Bd, Cry1Be, Cry1Bf, Cry1Bg, Cry1Bh, Cry1Bi, Cry1Ca, Cry1Cb, Cry1Da, Cry1Db, Cry1Dc, Cry1Dd, Cry1Ea, Cry1Eb, Cry1Fa, Cry1Fb, Cry1Ga, Cry1Gb, Cry1Gc, Cry1Ha, Cry1Hb, Cry1Hc, Cry1Ia, Cry1Ib, Cry1Ic, Cry1Id, Cry1Ie, Cry1If, Cry1Ig, Cry1Ja, Cry1Jb, Cry1Jc, Cry1Jd, Cry1Ka, Cry1La, Cry1Ma, Cry1Na, Cry1Nb, Cry2Aa, Cry2Ab, Cry2Ac, Cry2Ad, Cry2Ae, Cry2Af, Cry2Ag, Cry2Ah, Cry2Ai, Cry2Aj, Cry2Ak, Cry2Al, Cry2Ba, Cry3Aa, Cry3Ba, Cry3Bb, Cry3Ca, Cry4Aa, Cry4Ba, Cry4Ca, Cry4Cb, Cry4Cc, Cry5Aa, Cry5Ab, Cry5Ac, Cry5Ad, Cry5Ba, Cry5Ca, Cry5Da, Cry5Ea, Cry6Aa, Cry6Ba, Cry7Aa, Cry7Ab, Cry7Ac, Cry7Ba, Cry7Bb, Cry7Ca, Cry7Cb, Cry7Da, Cry7Ea, Cry7Fa, Cry7Fb, Cry7Ga, Cry7Gb, Cry7Gc, Cry7Gd, Cry7Ha, Cry7Ia, Cry7Ja, Cry7Ka, Cry7Kb, Cry7La, Cry8Aa, Cry8Ab, Cry8Ac, Cry8Ad, Cry8Ba, Cry8Bb, Cry8Bc, Cry8Ca, Cry8Da, Cry8Db, Cry8Ea, Cry8Fa, Cry8Ga, Cry8Ha, Cry8Ia, Cry8Ib, Cry8Ja, Cry8Ka, Cry8Kb, Cry8La, Cry8Ma, Cry8Na, Cry8Pa, Cry8Qa, Cry8Ra, Cry8Sa, Cry8Ta, Cry9Aa, Cry9Ba, Cry9Bb, Cry9Ca, Cry9Da, Cry9Db, Cry9Dc, Cry9Ea, Cry9Eb, Cry9Ec, Cry9Ed, Cry9Ee, Cry9Fa, Cry9Ga, Cry10Aa, Cry11Aa, Cry11Ba, Cry11Bb, Cry12Aa, Cry13Aa, Cry14Aa, Cry14Ab, Cry15Aa, Cry16Aa, Cry17Aa, Cry18Aa, Cry18Ba, Cry18Ca, Cry19Aa, Cry19Ba, Cry19Ca, Cry20Aa, Cry20Ba, Cry21Aa, Cry21Ba, Cry21Ca, Cry21Da, Cry21Ea, Cry21Fa, Cry21Ga, Cry21Ha, Cry22Aa, Cry22Ab, Cry22Ba, Cry22Bb, Cry23Aa, Cry24Aa, Cry24Ba, Cry24Ca, Cry25Aa, Cry26Aa, Cry27Aa, Cry28Aa, Cry29Aa, Cry29Ba, Cry30Aa, Cry30Ba, Cry30Ca, Cry30Da, Cry30Db, Cry30Ea, Cry30Fa, Cry30Ga, Cry31Aa, Cry31Ab, Cry31Ac, Cry31Ad, Cry32Aa, Cry32Ab, Cry32Ba, Cry32Ca, Cry32Cb, Cry32Da, Cry32Ea, Cry32Eb, Cry32Fa, Cry32Ga, Cry32Ha, Cry32Hb, Cry32Ia, Cry32Ja, Cry32Ka, Cry32La, Cry32Ma, Cry32Mb, Cry32Na, Cry32Oa, Cry32Pa, Cry32Qa, Cry32Ra, Cry32Sa, Cry32Ta, Cry32Ua, Cry33Aa, Cry34Aa, Cry34Ab, Cry34Ac, Cry34Ba, Cry35Aa, Cry35Ab, Cry35Ac, Cry35Ba, Cry36Aa, Cry37Aa, Cry38Aa, Cry39Aa, Cry40Aa, Cry40Ba, Cry40Ca, Cry40Da, Cry41Aa, Cry41Ab, Cry41Ba, Cry42Aa, Cry43Aa, Cry43Ba, Cry43Ca, Cry43Cb, Cry43Cc, Cry44Aa, Cry45Aa, Cry46Aa Cry46Ab, Cry47Aa, Cry48Aa, Cry48Ab, Cry49Aa, Cry49Ab, Cry50Aa, Cry50Ba, Cry51Aa, Cry52Aa, Cry52Ba, Cry53Aa, Cry53Ab, Cry54Aa, Cry54Ab, Cry54Ba, Cry55Aa, Cry56Aa, Cry57Aa, Cry57Ab, Cry58Aa, Cry59Aa, Cry59Ba, Cry60Aa, Cry60Ba, Cry61Aa, Cry62Aa, Cry63Aa, Cry64Aa, Cry65Aa, Cry66Aa, Cry67Aa, Cry68Aa, Cry69Aa, Cry69Ab, Cry70Aa, Cry70Ba, Cry70Bb, Cry71Aa, Cry72Aa, Cry73Aa, или любой комбинации вышеуказанного. В вариантах осуществления белок Cry представляет собой Cry1Fa, например, как представлено в трансформанте маиса ТС1507.
[00292] В дополнительных вариантах осуществления второе средство для контроля вредителей представляет собой один или несколько вегетативных инсектицидных белков Vip3, выбранных из Vip3Aa1, Vip3Aa2, Vip3Aa3, Vip3Aa4, Vip3Aa5, Vip3Aa6, Vip3Aa7, Vip3Aa8, Vip3Aa9, Vip3Aa10, Vip3Aa11, Vip3Aa12, Vip3Aa13, Vip3Aal4, Vip3Aa15, Vip3Aa16, Vip3Aa17, Vip3Aa18, Vip3Aa19, Vip3Aa20, Vip3Aa21, Vip3Aa22, Vip3Aa2, Vip3Aa24, Vip3Aa25, Vip3Aa26, Vip3Aa27, Vip3Aa28, Vip3Aa29, Vip3Aa30, Vip3Aa31, Vip3Aa32, Vip3Aa33, Vip3Aa34, Vip3Aa35, Vip3Aa36, Vip3Aa37, Vip3Aa38, Vip3Aa39, Vip3Aa40, Vip3Aa41, Vip3Aa42, Vip3Aa43, Vip3Aa44, Vip3Ab1, Vip3Ab2, Vip3Ac1, Vip3Ad1, Vip3Ad2, Vip3Ae1, Vip3Af1, Vip3Af2, Vip3Af3, Vip3Ag1, Vip3Ag2, Vip3Ag3 HM117633, Vip3Ag4, Vip3Ag5, Vip3Ah1, Vip3Ba1, Vip3Ba2, Vip3Bb1, Vip3Bb2, Vip3Bb3, или любой комбинации вышеуказанного. В вариантах осуществления белок Vip3 представляет собой Vip3Aa (патент США №6137033), например, как представлено в трансформанте кукурузы MIR162 (патент США №8232456; патент США №8455720 и патент США №8618272).
[00293] В вариантах осуществления второе средство для контроля вредителей может быть получено из источников, отличных от B. thuringiensis. Например, второе средство для контроля вредителей может представлять собой альфа-амилазу, пероксидазу, холестеролоксидазу, пататин, протеазу, ингибитор протеазы, уреазу, ингибитор альфа-амилазы, порообразующий белок, хитиназу, лектин, сконструированное антитело или фрагмент антитела, инсектицидный белок Bacillus cereus, инсектицидный белок Xenorhabdus spp. (такого как Х. nematophila или X. bovienii), инсектицидный белок Photorhabdus spp. (такого как P. luminescens или P. asymobiotica), инсектицидный белок Brevibacillus spp. (такого как В. laterosporous), инсектицидный белок Lysinibacillus spp. (такого как L. sphearicus), инсектицидный белок Chromobacterium spp. (такого как С. subtsugae или С. piscinae), инсектицидный белок Yersinia spp. (такого как Y. entomophaga), инсектицидный белок Paenibacillus spp. (такого как P. propylaea), инсектицидный белок Clostridium spp. (такого как С. bifermentans), Pseudomonas spp. (такого как Р. fluorescens) и лигнин. В других вариантах осуществления второе средство может представлять собой по меньшей мере один инсектицидный белок, полученный из инсектицидного токсинового комплекса (Тс) из Photorhabdus, Xenorhabus, Serratia или Yersinia. В других вариантах осуществления инсектицидный белок может представлять собой АДФ-рибозилтрансферазу, полученную из инсектицидной бактерии, такой как Photorhabdus ssp. В других вариантах осуществления инсектицидный белок может представлять собой белок VIP, такой как VIP1 или VIP2 из В. cereus. В еще одних вариантах осуществления инсектицидный белок может представлять собой бинарный токсин, полученный из инсектицидных бактерий, такой как ISP1A и ISP2A из В. laterosporous или BinA и BinB из L. sphaericus. В еще одних вариантах осуществления инсектицидный белок может быть сконструированным или может быть гибридом или химерой из любых предыдущих инсектицидных белков.
[00294] В некоторых вариантах осуществления второе пестицидное средство может быть небелковым, например, молекула интерферирующей РНК, такая как dsRNA, которая может экспрессироваться трансгенно или применяться как часть композиции (например, с применением способов местного нанесения). Как правило, интерферирующая РНК содержит по меньшей мере фрагмент РНК, соответствующий целевому гену, спейсерную последовательность и второй фрагмент РНК, который комплементарен первому, за счет чего может образовываться двухнитевая структура РНК. РНК-интерференция (RNAi) происходит, когда организм распознает молекулы двухнитевой РНК (dsRNA) и гидролизует их. Полученные продукты гидролиза представляют собой небольшие фрагменты РНК длиной приблизительно 19-24 нуклеотидов, называемые малыми интерферирующими РНК (siRNA). Затем siRNA распространяются или разносятся по всему организму, в том числе через клеточные мембраны, где они гибридизируются с mRNA (или другими РНК) и вызывают гидролиз РНК. Интерферирующие РНК распознаются комплексом сайленсинга РНК-интерференции (RISC), в который вводится эффекторная нить (или "направляющая нить") РНК. Данная направляющая цепь действует как матрица для распознавания и разрушения дуплексных последовательностей. Данный процесс повторяется каждый раз, когда siRNA гибридизируется с комплементарной ей целевой РНК, эффективно предотвращая трансляцию таких mRNA, и таким образом "сайленсингу" подвергается экспрессия специфических генов, с которых mRNA были транскрибированы. Интерферирующие РНК известны из уровня техники как применяемые для контроля насекомых (см., например, публикацию WO 2013/192256, включенную в данный документ посредством ссылки). Интерферирующая РНК, предназначенная для применения в контроле насекомых, обеспечивает получение не встречающейся в естественных условиях двухнитевой РНК, которая пользуется нативными путями RNAi у насекомого для запуска подавления целевых генов, что может приводить к прекращению кормления и/или роста и может приводить к гибели насекомого вредителя. Молекула интерферирующей РНК может придавать устойчивость к насекомым в отношении того же целевого вредителя, что и белок по настоящему изобретению, или может целенаправленно воздействовать на другого вредителя. Целевое насекомое-вредитель растения может кормиться путем жевания, сосания или прокалывания. Интерферирующие РНК известны из уровня техники как применимые для контроля насекомых. В вариантах осуществления dsRNA, пригодная для контроля насекомых, описана в предварительных заявках на патент США №№62/371259, 62/371261 или 62/371262, поданных 5 августа 2016 г. В вариантах осуществления dsRNA, пригодная для контроля насекомых, описана в патентах США №№92388223, 9340797 или 8946510. В вариантах осуществления dsRNA, пригодная для контроля насекомых, описана в заявках на патент США №№12/868994, 13/831230, 14/207313 или 14/207318. В других вариантах осуществления интерферирующая РНК может придавать устойчивость в отношении вредителя растений, отличного от насекомого, такого как нематода-вредитель или вирус-вредитель.
[00295] В еще дополнительных вариантах осуществления первое средство для контроля насекомых, которое представляет собой химерный инсектицидный белок по настоящему изобретению, и второе средство для контроля вредителей коэкспрессируются в трансгенном растении. Такой коэкспрессии более чем одного пестицидного компонента в одном и том же трансгенном растении можно достичь путем получения растения, содержащего и экспрессирующего последовательности нуклеиновых кислот, кодирующие средства для контроля насекомых, с помощью методов генной инженерии. Например, коэкспрессии более чем одного пестицидного средства в одном и том же трансгенном растении можно достичь путем получения одиночного рекомбинантного вектора, содержащего кодирующие последовательности более чем одного пестицидного средства в так называемом "молекулярном пакете", и генетического конструирования растения, которое бы содержало и экспрессировало все эти пестицидные средства в трансгенном растении. Такие молекулярные пакеты также можно получить с применением мини-хромосом, которые описаны, например, в патенте США №7235716. В качестве альтернативы растение, родитель 1, может быть получено с помощью методов генной инженерии для экспрессии химерного инсектицидного белка по настоящему изобретению. Второе растение, родитель 2, может быть получено с помощью методов генной инженерии для экспрессии второго средства для контроля вредителей. Путем скрещивания родителя 1 с родителем 2 получают растения-потомки, которые экспрессируют оба средства для контроля насекомых от родителей 1 и 2.
[00296] В других вариантах осуществления настоящего изобретения предусматривается трансгенное растение с пакетированными признаками, устойчивое к заражению вредителями растений, содержащее последовательность нуклеиновой кислоты (например, ДНК), кодирующую dsRNA для супрессии основного гена в организме целевого вредителя, и последовательность нуклеиновой кислоты (например, ДНК), кодирующую химерный инсектицидный белок по настоящему изобретению, проявляющий инсектицидную активность в отношении целевого вредителя. Ранее сообщалось о том, что dsRNA являются неэффективными против определенных вредителей, относящихся к чешуекрылым (Rajagopol et al. 2002. J. Biol. Chem. 277:468-494) вероятно из-за высокого показателя рН в средней кишке, который нарушает устойчивость dsRNA. Следовательно, в некоторых вариантах осуществления, в которых целевой вредитель является вредителем, относящимся к чешуекрылым, химерный инсектицидный белок по настоящему изобретению действует для временного снижения показателя рН в средней кишке, который обеспечивает стабилизацию одновременно поглощаемой dsRNA, делая dsRNA эффективной в сайленсинге генов-мишеней.
[00297] Трансгенные растения или семена, содержащие и/или экспрессирующие инсектицидный белок по настоящему изобретению, также можно обработать инсектицидом или инсектицидным покрытием для семян, которые описаны в патентах США №№5849320 и 5876739. В вариантах осуществления, где как инсектицид или инсектицидное покрытие для семян, так и трансгенное растение или семя по настоящему изобретению являются активными в отношении одного и того же целевого насекомого, например, вредителя, относящегося к чешуекрылым (например, совка травяная), данная комбинация является пригодной (i) в способе дополнительного усиления активности композиции по настоящему изобретению в отношении целевого насекомого и/или (ii) в способе предупреждения развития устойчивости к композиции по настоящему изобретению путем обеспечения еще одного механизма действия в отношении целевого насекомого. Таким образом, в вариантах осуществления настоящего изобретения предусматривается способ повышения контроля популяции насекомых, относящихся к чешуекрылым, предусматривающий обеспечение трансгенного растения или семени по настоящему изобретению и применение по отношению к растению или семени инсектицида или инсектицидного покрытия для семян, предназначенного для трансгенного растения или семени по настоящему изобретению.
[00298] Даже в случае, если инсектицид или инсектицидное покрытие для семян являются активными в отношении другого насекомого, инсектицид или инсектицидное покрытие для семян являются пригодными для расширения диапазона контроля насекомых, например, посредством добавления инсектицида или инсектицидного покрытия для семян, которые характеризуются активностью в отношении насекомых, относящихся к жесткокрылым, к трансгенному семени по настоящему изобретению, которое в некоторых вариантах осуществления характеризуется активностью в отношении насекомых, относящихся к чешуекрылым, при этом полученное покрытое трансгенное семя обеспечивает контроль насекомых-вредителей, относящихся как к чешуекрылым, так и жесткокрылым.
Способы получения и применения химерных инсектицидных белков, нуклеиновых кислот и трансгенных растений.
[00299] В настоящем изобретении также охвачены способы получения трансгенного растения, устойчивого к насекомым (например, устойчивого к насекомым, относящимся к чешуекрылым). В иллюстративных вариантах осуществления способ включает введение в растение полинуклеотида, кассеты экспрессии или вектора по настоящему изобретению, содержащего нуклеотидную последовательность, которые кодируют химерный инсектицидный белок по настоящему изобретению (включая фрагменты токсина и модифицированные формы, которые по существу идентичны полипептидам, конкретно раскрытым в данном документе), где нуклеотидная последовательность экспрессируется в растении с получением химерного инсектицидного белка по настоящему изобретению, тем самым обеспечивая устойчивость растения к насекомому-вредителю и обеспечивая получение устойчивого к насекомым трансгенного растения (например, по сравнению с подходящим контрольным растением, таким как растение, которое не содержит полинуклеотид, кассету экспрессии или вектор по настоящему изобретению и/или не экспрессирует полипептид по настоящему изобретению).
[00300] В вариантах осуществления способ введения полинуклеотида, кассеты экспрессии или вектора по настоящему изобретению в растение включает в себя сначала трансформацию растительной клетки с использованием полинуклеотида, кассеты экспрессии или вектора и регенерацию трансгенного растения из нее, где трансгенное растение содержит полинуклеотид, кассету экспрессии или вектор и экспрессирует химерный инсектицидный белок по настоящему изобретению.
[00301] Альтернативно или дополнительно стадия введения может включать скрещивание первого растения, содержащего полинуклеотид, кассету экспрессии или вектор, со вторым растением (например, растением, отличным от первого растения, например, растением, которое не содержит полинуклеотид, кассету экспрессии или вектор) и необязательно получение растения-потомка, которое содержит полинуклеотид, кассету экспрессии или вектор и экспрессирует химерный инсектицидный белок по настоящему изобретению, с получением тем самым в результате повышенной устойчивости к по меньшей мере одному насекомому-вредителю. Таким образом, трансгенное растение по настоящему изобретению охватывает растение, которое является непосредственным результатом события трансформации, и его потомка (любого поколения), которое содержит полинуклеотид, кассету экспрессии или вектор и необязательно экспрессирует химерный инсектицидный белок, приводящий к повышенной устойчивости к по меньшей мере одному насекомому-вредителю.
[00302] В настоящем изобретении дополнительно предусмотрен способ определения трансгенного растения по настоящему изобретению, включающий обнаружение наличия полинуклеотида, кассеты экспрессии, вектора или химерного инсектицидного белка по настоящему изобретению в растении (или растительной клетке, части растения и т.п., полученных из него) и тем самым определение растения как трансгенного растения по настоящему изобретению на основании наличия полинуклеотида, кассеты экспрессии, вектора или химерного инсектицидного белка по настоящему изобретению.
[00303] В настоящем изобретении дополнительно предусмотрен способ получения трансгенного растения с повышенной устойчивостью по меньшей мере к одному насекомому-вредителю (например, по меньшей мере к одному вредителю, относящемуся к чешуекрылым), включающий: посадку семени, содержащего полинуклеотид, кассету экспрессии или вектор по настоящему изобретению, и выращивание трансгенного растения из этого семени, где трансгенное растение содержит полинуклеотид, кассету экспрессии или вектор и продуцирует химерный инсектицидный белок.
[00304] В вариантах осуществления трансгенные растения, полученные с помощью способов по настоящему изобретению, содержат полинуклеотид, кассету экспрессии или вектор по настоящему изобретению. В вариантах осуществления трансгенное растение, полученное с помощью способов по настоящему изобретению, содержит химерный инсектицидный белок по настоящему изобретению и необязательно характеризуется повышенной устойчивостью к по меньшей мере одному насекомому-вредителю.
[00305] Способы получения трансгенного растения, описанные в данном документе, необязательно включают дополнительную стадию сбора семени от трансгенного растения, где семя содержит полинуклеотид, кассету экспрессии или вектор и продуцирует химерный инсектицидный белок. Необязательно это семя обеспечивает получение дополнительного трансгенного растения, которое содержит полинуклеотид, кассету экспрессии или вектор и продуцирует химерный инсектицидный белок и, таким образом, характеризуется повышенной устойчивостью к по меньшей мере одному насекомому-вредителю.
[00306] В настоящем изобретении дополнительно предусмотрены части растения, растительные клетки, органы растения, растительные культуры, семена, растительные экстракты, собранные продукты и переработанные продукты трансгенных растений, полученных с помощью способов по настоящему изобретению.
[00307] В качестве дополнительного аспекта в настоящем изобретении также предусмотрен способ получения семени, при этом способ включает получение трансгенного растения, которое содержит полинуклеотид, кассету экспрессии или вектор по настоящему изобретению, и сбор семени от трансгенного растения, где семя содержит полинуклеотид, кассету экспрессии, вектор и продуцирует химерный инсектицидный белок. Необязательно это семя обеспечивает получение дополнительного трансгенного растения, которое содержит полинуклеотид, кассету экспрессии или вектор и продуцирует химерный инсектицидный белок и, таким образом, характеризуется повышенной устойчивостью к по меньшей мере одному насекомому-вредителю. В иллюстративных вариантах осуществления стадия получения трансгенного растения включает посев семени, которое обеспечивает получение трансгенного растения.
[00308] В настоящем изобретении дополнительно предусмотрен способ получения гибридного семени растения, при этом способ включает скрещивание первого инбредного растения, которое представляет собой трансгенное растение, содержащее полинуклеотид, кассету экспрессии или вектор по настоящему изобретению и необязательно экспрессирующее химерный инсектицидный белок по настоящему изобретению, с другим инбредным растением (например, инбредным растением, которое не содержит полинуклеотид, кассету экспрессии или вектор по настоящему изобретению) и обеспечение возможности образования гибридного семени. Необязательно способ дополнительно включает сбор гибридного семени. В вариантах осуществления гибридное семя содержит полинуклеотид, кассету экспрессии или вектор по настоящему изобретению и, в вариантах осуществления, может дополнительно содержать химерный инсектицидный белок по настоящему изобретению и обладать повышенной устойчивостью к насекомому-вредителю. В вариантах осуществления гибридное семя обеспечивает получение трансгенного растения, которое содержит полинуклеотид, кассету экспрессии или вектор по настоящему изобретению, экспрессирует химерный инсектицидный белок по настоящему изобретению и обладает повышенной устойчивостью к по меньшей мере одному насекомому-вредителю.
[00309] В некоторых вариантах осуществления трансгенное растение по настоящему изобретению является устойчивым к по меньшей мере одному насекомому-вредителю, относящемуся к чешуекрылым (как описано в данном документе). В вариантах осуществления трансгенное растение обеспечивает контроль насекомого-вредителя или колонии совки травяной, устойчивой к белку Vip3A (например, белку Vip3Aa, например, который экспрессируется в трансформанте маиса MIR162) и/или Cry1F (например, белку Cry1Fa, который экспрессируется в трансформанте маиса ТС 1507).
[00310] В дополнительных вариантах осуществления способ контроля по меньшей мере одного насекомого-вредителя (например, по меньшей мере одного насекомого-вредителя, относящегося к чешуекрылым, такого как совка травяная) включает обеспечение химерного инсектицидного белка по настоящему изобретению. В вариантах осуществления способ включает доставку (например, пероральную доставку) насекомому-вредителю или в среду его обитания эффективного количества химерного инсектицидного белка по настоящему изобретению. Обычно в целях эффективности полипептид поглощается насекомым пероральным путем. Однако химерный инсектицидный белок можно доставлять насекомому с помощью многих известных способов. Способы доставки белка в организм насекомого пероральным путем предусматривают без ограничения обеспечение белка (1) в трансгенном растении, при этом насекомое съедает (поглощает) одну или несколько частей трансгенного растения, поглощая тем самым полипептид, экспрессируемый в трансгенном растении; (2) в составленной белковой(белковых) композиции(композициях), которые можно наносить, например, на питательную среду насекомых или включать в ее состав; (3) в белковой(белковых) композиции(композициях), которые можно наносить на поверхность, например, путем опрыскивания поверхности части растения, и которая затем заглатывается насекомым в силу того, что насекомое съедает одну или несколько подвергнутых опрыскиванию частей растения; (4) в матрице приманки или (5) с помощью любой другой известной в данной области системы доставки белков. Таким образом, для доставки токсичных белков по настоящему изобретению можно применять любой способ пероральной доставки в организм насекомого. В некоторых конкретных вариантах осуществления химерный инсектицидный белок по настоящему изобретению доставляют в организм насекомого пероральным путем, например, когда насекомое поедает одну или несколько частей трансгенного растения по настоящему изобретению.
[00311] В других вариантах осуществления инсектицидный белок по настоящему изобретению доставляют в организм насекомого пероральным путем, при этом насекомое поедает одну или несколько частей растения, подвергнутых опрыскиванию композицией, содержащей инсектицидный белок по настоящему изобретению. Доставку композиции по настоящему изобретению на поверхность растения можно осуществлять с помощью любого способа нанесения соединений, композиций, составов и т.п. на поверхности растений, известного специалистам в данной области техники. Некоторые неограничивающие примеры доставки на растение или его часть или приведения в контакт с ними включают опрыскивание, опыливание, посыпание, распыление, орошение туманом, мелкодисперсное разбрызгивание, разбрасывание, пропитывание, впрыскивание в почву, введение в почву, смачивание (например, обработку корней, почвы), погружение, полив, нанесение покрытия, впитывание в листья или стебли, внесение в междурядья или обработку семян и т.п. и их комбинации. Эти и другие процедуры приведения растения или его части в контакт с соединением(соединениями), композицией(композициями) или составом(составами) хорошо известны специалистам в данной области.
[00312] В дополнительных вариантах осуществления настоящего изобретения предусмотрен способ контроля насекомого-вредителя, относящегося к чешуекрылым, который устойчив к белку Vip3A (например, белку Vip3Aa, например, который экспрессируется трансформантом маиса MFR162) и/или белку Cry1F (например белку Cry1Fa, например, который экспрессируется трансформантом маиса ТС1507), включающий доставку насекомому, относящемуся к чешуекрылым, или в среду его обитания эффективного количества химерного инсектицидного белка или композиции по настоящему изобретению. В иллюстративных вариантах осуществления устойчивое насекомое-вредитель представляет собой устойчивое насекомое-вредителя или колонию совки травяной.
[00313] В других вариантах осуществления настоящего изобретения предусмотрен способ предупреждения развития устойчивости в популяции целевого насекомого-вредителя, относящегося к чешуекрылым, к белку Vip3A (например, белку Vip3Aa, например, который экспрессируется трансформантом маиса MTR162) и/или Cry1F (например белку Cry1Fa, например, который экспрессируется трансформантом маиса ТС1507), экспрессируемому в трансгенном растении, при этом способ включает доставку в целевую популяцию трансгенного растения, содержащего полинуклеотид, который содержит нуклеотидную последовательность, кодирующую белок Vip3A, и/или нуклеотидную последовательность, кодирующую белок Cry1F; и полинуклеотид, кассету экспрессии или вектор по настоящему изобретению, экспрессирующие химерный инсектицидный белок по настоящему изобретению. В некоторых вариантах осуществления целевое насекомое-вредитель, относящееся к чешуекрылым, представляет собой совку травяную. В соответствии с вышеупомянутыми вариантами осуществления трансгенное растение может содержать селекционный пакет из двух или более инсектицидных признаков, молекулярный пакет из двух или более инсектицидных признаков или их комбинацию.
[00314] В некоторых вариантах осуществления настоящее изобретение охватывает способ предоставления фермеру средств для контроля насекомого-вредителя (например, насекомого-вредителя, относящегося к чешуекрылым, такого как совка травяная), при этом способ включает поставку или продажу фермеру растительного материала, такого как семя, при этом растительный материал содержит полинуклеотид, кассету экспрессии, вектор, способные экспрессировать химерный инсектицидный белок по настоящему изобретению. В вариантах осуществления растительный материал содержит химерный инсектицидный белок по настоящему изобретению и необязательно характеризуется повышенной устойчивостью к по меньшей мере одному насекомому-вредителю. В вариантах осуществления растительный материал представляет собой семя, и растение, выращенное из семени, содержит полинуклеотид, кассету экспрессии или вектор по настоящему изобретению, экспрессирует химерный инсектицидный белок по настоящему изобретению и характеризуется повышенной устойчивостью к по меньшей мере одному насекомому-вредителю.
[00315] Помимо обеспечения композиций, в настоящем изобретении предусмотрены способы продуцирования химерного инсектицидного белка, токсичного для вредителя, относящегося к чешуекрылым. Такой способ включает культивирование трансгенной клетки-хозяина, отличной от клетки человека, которая содержит полинуклеотид, кассету экспрессии или вектор по настоящему изобретению, которая экспрессирует химерный инсектицидный белок по настоящему изобретению, в условиях, при которых клетка-хозяин продуцирует химерный инсектицидный белок, токсичный для вредителя, относящегося к чешуекрылым. В некоторых вариантах осуществления трансгенная клетка-хозяин, отличная от клетки человека, представляет собой растительную клетку. В некоторых других вариантах осуществления растительная клетка представляет собой клетку маиса. В других вариантах осуществления условия, при которых выращивают растительную клетку или клетку маиса, включают естественный солнечный свет. В других вариантах осуществления трансгенная клетка-хозяин, отличная от клетки человека, представляет собой бактериальную клетку. В еще одних вариантах осуществления трансгенная клетка-хозяин, отличная от клетки человека, представляет собой дрожжевую клетку.
[00316] В некоторых вариантах осуществления способы по настоящему изобретению обеспечивают контроль по меньшей мере одного насекомого-вредителя, относящегося к чешуекрылым, включая без ограничения одного или несколько из следующего: Ostrinia spp., такой как О. nubilalis (кукурузный мотылек) и/или О. furnacalis (восточный кукурузный мотылек); Plutella spp., такой как P. xylostella (капустная моль); Spodoptera spp., такой как S. frugiperda (совка травяная), S. littoralis (совка хлопчатника египетского), S. ornithogalli (желтополосая совка), S. praefica (западная желтополосая совка), S. eridania (южная совка) и/или S. exigua (совка малая); Agrotis spp., такой как A. ipsilon (совка-ипсилон), A. segetum (озимая совка обыкновенная), A. gladiaria (озимая совка рода Agrotis) и/или A. orthogonia (совка прямоугольная); Striacosta spp., такой как S. albicosta (западная бобовая совка); Helicoverpa spp., такой как Н. zea (совка кукурузная), Н. punctigera (совка хлопковая австралийская) и/или Н. armigera (совкахлопковая); Heliothis spp., такой как H. virescens (табачная листовертка); Diatraea spp., такой как D. grandiosella (огневка кукурузная юго-западная) и/или D. saccharalis (точильщик стеблей сахарного тростника); Trichoplusia spp., такой как Т. ni (совка капустная); Sesamia spp., такой как S. nonagroides (мотылек кукурузный средиземноморский) и/или S. calamistis (розовая стеблевая совка); Pectinophora spp., такой как P. gossypiella (розовый коробочный червь); Cochylis spp., такой как С. hospes (полосатая подсолнечная моль); Manduca spp., такой как М. sexta (табачный бражник) и/или М. quinquemaculata (бражник пятиточечный); Elasmopalpus spp., такой как P. lignosellus (точильщик стеблей кукурузы малый); Pseudoplusia spp., такой как Р. includens (соевая совка); Anticarsia spp., такой как A gemmatalis (совка бархатных бобов); Plathypena spp., такой как P. scabra (совка клеверная); Pieris spp., такой как Р. brassicae (белянка капустная), Papaipema spp., такой какР. nebris (точильщик Papaipema nebris); Pseudaletia spp., такой как P. unipuncta (совка луговая обыкновенная); Peridroma spp., такой как P. saucia (совка маргаритковая); Keiferia spp., такой как K. lycopersicella (томатная острица); Artogeia spp., такой как A. rapae (белянка репная); Phthorimaea spp., такой как P. operculella (картофельная моль); Chrysodeixis spp., такой как С. includes (соевая совка); Feltia spp., такой как F. ducens (гусеница совки Feltia jaculifera); Chilo spp., такой как С. suppressalis (огневка желтая рисовая), Cnaphalocrocis spp., такой как С. medinalis (огневка рисовая), или любую комбинацию вышеуказанных. В вариантах осуществления способы по настоящему изобретению обеспечивают контроль насекомого-вредителя или колонии совки травяной, которая устойчива к белку Vip3A (например, белку Vip3Aa, например, который экспрессируется в трансформанте маиса MIR162) и/или Cry1F (например, белку Cry1Fa, например, который экспрессируется в трансформанте маиса ТС1507).
[00317] В настоящем изобретении также предусмотрено применение химерных инсектицидных белков, нуклеиновых кислот, трансгенных растений, частей растений, семян и инсектицидных композиций по настоящему изобретению, например, для контроля насекомого-вредителя, такого как вредитель, относящийся к чешуекрылым (как описано в данном документе).
[00318] В вариантах осуществления в настоящем изобретении предусмотрен способ применения полинуклеотида, кассеты экспрессии, вектора или клетки-хозяина по настоящему изобретению для получения инсектицидной композиции для контроля насекомого-вредителя (например, насекомого-вредителя, относящегося к чешуекрылым).
[00319] В вариантах осуществления в настоящем изобретении предусмотрен способ применения полинуклеотида, кассеты экспрессии или вектора по настоящему изобретению для получения трансгенного семени, где из трансгенного семени вырастает трансгенное растение с повышенной устойчивостью к насекомому-вредителю.
[00320] В качестве другого аспекта в настоящем изобретении также рассматривается применение трансгенного растения по настоящему изобретению для получения трансгенного семени, которое необязательно является гибридным семенем.
[00321] В вариантах осуществления в настоящем изобретении предусмотрен способ применения химерного инсектицидного белка, полинуклеотида, кассеты экспрессии, вектора, трансгенного растения или инсектицидной композиции по настоящему изобретению для предотвращения развития устойчивости в популяции целевого насекомого-вредителя, относящегося к чешуекрылым, к белку Vip3A и/или белку Cry1F (каждый, как описано в данном документе).
[00322] Заявления по настоящему изобретению включают в себя без ограничения следующее:
[00323] 1. Химерный инсектицидный белок, который является токсичным для насекомого-вредителя, относящегося к чешуекрылым, содержащий в направлении от N-конца к С-концу: (а) N-концевую область первого белка Cry1, которая является N-концевой областью белка ВТ-0029 под SEQ ID NO: 2 или аминокислотной последовательностью, которая на по меньшей мере 90% идентична ей, слитую с (b) С-концевой областью другого белка Cry1; где положение перехода между первым белком Cry1 и другим белком Cry1 находится в консервативном блоке 3.
[00324] 2. Химерный инсектицидный белок по п. 1, где другой белок Cry1 представляет собой белок Cry1F, Cry1G, Cry1I или Cry1K.
[00325] 3. Химерный инсектицидный белок по п. 1, где другой белок Cry1 представляет собой белок Cry1Fa.
[00326] 4. Химерный инсектицидный белок по п. 1, где другой белок Cry1 представляет собой белок Cry1Ia или Cry1If.
[00327] 5. Химерный инсектицидный белок по п. 1, где другой белок Cry1 представляет собой белок ВТ-0022 под SEQ ID NO: 1.
[00328] 6. Химерный инсектицидный белок по п. 1, где С-концевая область другого белка Cry1 содержит аминокислотную последовательность, соответствующую аминокислотной последовательности от положения 464 до 602 SEQ ID NO: 8 или аминокислотной последовательности, которая на по меньшей мере 80% идентична ей.
[00329] 7. Химерный инсектицидный белок по п. 1, где С-концевая область другого белка Cry1 содержит аминокислотную последовательность, соответствующую аминокислотной последовательности от положения 498 до 636 SEQ ID NO: 1 или аминокислотной последовательности, которая на по меньшей мере 80% идентична ей.
[00330] 8. Химерный инсектицидный белок по любому из пп. 1-7, где N-концевая область первого белка Cry1 содержит аминокислотную последовательность, соответствующую аминокислотной последовательности от положения 1 до 458 SEQ ID NO: 2 или аминокислотной последовательности, которая на по меньшей мере 90% идентична ей.
[00331] 9. Химерный инсектицидный белок по любому из пп. 1-8, где химерный инсектицидный белок дополнительно содержит на N-конце пептидильный фрагмент, который отщепляется от протоксина после поглощения насекомым-вредителем, относящимся к чешуекрылым.
[00332] 10. Химерный инсектицидный белок по любому из пп. 1-9, где химерный инсектицидный белок дополнительно содержит на С-конце хвост протоксина из белка Cry, который отщепляется от протоксина после поглощения насекомым-вредителем, относящимся к чешуекрылым.
[00333] 11. Химерный инсектицидный белок по любому из пп. 1-10, где химерный инсектицидный белок содержит (а) аминокислотную последовательность от положения 1 до 597 SEQ ID NO: 3 или ее токсиновый фрагмент, или (b) аминокислотную последовательность, которая на по меньшей мере 80% идентична аминокислотной последовательности из (а).
[00334] 12. Химерный инсектицидный белок по любому из пп. 1-11, где химерный инсектицидный белок содержит (а) аминокислотную последовательность под SEQ ID NO: 3 или ее токсиновый фрагмент, или (b) аминокислотную последовательность, которая на по меньшей мере 80% идентична аминокислотной последовательности из (а).
[00335] 13. Химерный инсектицидный белок по любому из пп. 1-10, где химерный инсектицидный белок содержит (а) аминокислотную последовательность от положения 1 до 597 SEQ ID NO: 9 или ее токсиновый фрагмент, или (b) аминокислотную последовательность, которая на по меньшей мере 80% идентична аминокислотной последовательности из (а).
[00336] 14. Химерный инсектицидный белок по любому из пп. 1-10 или п. 13, где химерный инсектицидный белок содержит (а) аминокислотную последовательность под SEQ ID NO: 9 или ее токсиновый фрагмент, или (b) аминокислотную последовательность, которая на по меньшей мере 80% идентична аминокислотной последовательности из (а).
[00337] 15. Химерный инсектицидный белок по любому из пп. 1-14, где химерный инсектицидный белок содержит аминокислотную последовательность под SEQ ID NO: 3 или SEQ ID NO: 9.
[00338] 16. Химерный инсектицидный белок по любому из пп. 1-15, где химерный инсектицидный белок характеризуется инсектицидной активностью в отношении одного или нескольких из Spodoptera frugiperda (совка травяная), Chrysodeixis includes (совка соевая), Diatraea saccharalis (точильщик стеблей сахарного тростника), Diatraea grandiosella (огневка кукурузная юго-западная) к Agrotis ipsilon (совка-ипсилон).
[00339] 17. Химерный инсектицидный белок по любому из пп. 1-16, где химерный инсектицидный белок характеризуется инсектицидной активностью в отношении насекомого-вредителя или колонии Spodoptera frugiperda с устойчивостью к белку Vip3A и/или белку Cry1F.
[00340] 18. Полинуклеотид, содержащий нуклеотидную последовательность, кодирующую химерный инсектицидный белок по любому из пп. 1-17.
[00341] 19. Полинуклеотид по п. 18, где полинуклеотид является кодон-оптимизированным для экспрессии в растении.
[00342] 20. Полинуклеотид по п. 18 или п. 19, где полинуклеотид содержит нуклеотидную последовательность, которая содержит (а) нуклеотидную последовательность под SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 10, SEQ ID NO: 11 или SEQ ID NO: 14 или ее токсин-кодирующий фрагмент; (b) нуклеотидную последовательность, которая по существу идентична нуклеотидной последовательности из (а); (с) нуклеотидную последовательность, которая гибридизируется при жестких условиях гибридизации с нуклеотидной последовательностью из (а) или (b); или (d) нуклеотидную последовательность, которая отличается от нуклеотидной последовательности из (а), (b) или (с) вследствие вырожденности генетического кода.
[00343] 21. Полинуклеотид по п. 18 или п. 19, где нуклеотидная последовательность содержит нуклеотидную последовательность под SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 10, SEQ ID NO: 11 или SEQ ID NO: 14.
[00344] 22. Молекула нуклеиновой кислоты, содержащая полинуклеотид по любому из пп. 18-21, функционально связанный с гетерологичным промотором.
[00345] 23. Молекула нуклеиновой кислоты по п. 22, где гетерологичный промотор представляет собой промотор, обеспечивающий экспрессию в растении.
[00346] 24. Вектор, содержащий полинуклеотид по любому из пп. 18-21 или молекулу нуклеиновой кислоты по п. 22 или п. 23.
[00347] 25. Трансгенная клетка, содержащая полинуклеотид по любому из пп. 18-21, молекулу нуклеиновой кислоты по п. 22 или п. 23 или вектор по п. 24.
[00348] 26. Трансгенная клетка по п. 25, где клетка представляет собой транс генную бактериальную клетку.
[00349] 27. Трансгенная клетка по п. 25, где клетка представляет собой транс генную растительную клетку.
[00350] 28. Трансгенная растительная клетка по п. 27, где растительная клетка представляет собой: (i) клетку однодольного, необязательно клетку ячменя, клетку маиса, клетку овса, клетку риса, клетку сорго, клетку сахарного тростника или клетку пшеницы; или (ii) клетку двудольного, необязательно клетку сои, клетку подсолнечника, клетку томата, клетку культурной разновидности капусты, клетку хлопчатника, клетку сахарной свеклы или клетку табака.
[00351] 29. Трансгенное растение, содержащее трансгенную растительную клетку по п. 27 или п. 28.
[00352] 30. Трансгенное растение по п. 29, где растение представляет собой: (i) однодольное растение, необязательно растение ячменя, растение маиса, растение овса, растение риса, растение сорго, растение сахарного тростника или растение пшеницы; или (ii) двудольное растение, необязательно растение сои, растение подсолнечника, растение томата, растение культурной разновидности капусты, растение хлопчатника, растение сахарной свеклы или растение табака.
[00353] 31. Трансгенное растение по п. 28 или п. 29, где трансгенное растение содержит дополнительную нуклеотидную последовательность, кодирующую второе средство для контроля насекомых.
[00354] 32. Трансгенная часть растения из трансгенного растения по любому из пп. 29-31, где трансгенная часть растения содержит химерный инсектицидный белок.
[00355] 33. Трансгенное семя трансгенного растения по любому из пп. 29-31.
[00356] 34. Собранный продукт, полученный из трансгенного растения по любому из пп. 29-31, трансгенной части растения по п. 32 или трансгенного семени по п. 33, где собранный продукт содержит химерный инсектицидный белок.
[00357] 35. Переработанный продукт, полученный из собранного продукта по п. 34, где переработанный продукт представляет собой муку, муку грубого помола, масло, крахмал или продукт, полученный из любого из вышеперечисленного.
[00358] 36. Инсектицидная композиция, содержащая химерный инсектицидный белок по любому из пп. 1-17 и приемлемый с точки зрения сельского хозяйства носитель.
[00359] 37. Инсектицидная композиция по п. 36, где приемлемый с точки зрения сельского хозяйства носитель представляет собой порошок, дуст, пеллет, гранулу, аэрозоль, эмульсию, коллоид или раствор.
[00360] 38. Инсектицидная композиция по п. 36 или п. 37, где композиция представляет собой высушенную, лиофилизированную, гомогенизированную, экстрагированную, фильтрованную, центрифугированную и/или осажденную композицию и/или представляет собой концентрат бактериальной культуры.
[00361] 39. Инсектицидная композиция по любому из пп. 36-38, где композиция содержит трансгенную бактериальную клетку, которая продуцирует химерный инсектицидный белок.
[00362] 40. Инсектицидная композиция по любому из пп. 36-39, где композиция содержит от приблизительно 1% до приблизительно 99% по весу химерного инсектицидного белка.
[00363] 41. Инсектицидная композиция по любому из пп. 36-40, где композиция содержит второе средство для контроля насекомых.
[00364] 42. Способ получения трансгенного растения с повышенной устойчивостью к насекомому-вредителю, относящемуся к чешуекрылым, при этом способ включает введение в растение полинуклеотида по любому из пп. 18-21, молекулы нуклеиновой кислоты по п. 22 или п. 23 или вектора по п. 24, где химерный инсектицидный белок экспрессируется в растении с получением таким образом трансгенного растения с повышенной устойчивостью к насекомому-вредителю.
[00365] 43. Способ по п. 42, где стадия введения включает (i) трансформацию растительной клетки полинуклеотидом, молекулой нуклеиновой кислоты или вектором и регенерацию трансгенного растения или (ii) скрещивание первого растения, содержащего полинуклеотид, молекулу нуклеиновой кислоты или вектор, со вторым растением.
[00366] 44. Способ по п. 42 или п. 43, где способ дополнительно включает получение растения-потомка из трансгенного растения, где растение-потомок содержит полинуклеотид, молекулу нуклеиновой кислоты или вектор и характеризуется повышенной устойчивостью к насекомому-вредителю.
[00367] 45. Способ получения трансгенного растения с повышенной устойчивостью к насекомому-вредителю, относящемуся к чешуекрылым, при этом способ включает: (а) посев семени, содержащего полинуклеотид по любому из пп. 18-21, молекулу нуклеиновой кислоты по п. 22 или п. 23 или вектор по п. 24 и (b) выращивание трансгенного растения из семени, где трансгенное растение содержит полинуклеотид, молекулу нуклеиновой кислоты или вектор и продуцирует химерный инсектицидный белок.
[00368] 46. Способ по п. 45, где способ дополнительно включает (с) сбор семени от трансгенного растения из (b), где собранное семя содержит химерный инсектицидный белок.
[00369] 47. Способ получения семени, при этом способ включает (а) получение трансгенного растения, которое содержит полинуклеотид по любому из пп. 18-21, молекулу нуклеиновой кислоты по п. 22 или п. 23 или вектор по п. 24 и (b) сбор семени от трансгенного растения из (а), где собранное семя содержит химерный инсектицидный белок.
[00370] 48. Способ получения гибридного семени растения, при этом способ включает: (а) скрещивание первого инбредного растения, которое представляет собой трансгенное растение, содержащее полинуклеотид по любому из пп. 18-21, молекулу нуклеиновой кислоты по п. 22 или п. 23 или вектор по п. 24, с другим инбредным растением и (b) обеспечение возможности образования гибридного семени.
[00371] 49. Способ контроля насекомого-вредителя, относящегося к чешуекрылым, при этом способ включает доставку в организм насекомого-вредителя или в среду его обитания композиции, содержащей эффективное количество химерного инсектицидного белка по любому из пп. 1-17 или инсектицидной композиции по любому из пп. 36-41.
[00372] 50. Способ контроля насекомого-вредителя, относящегося к чешуекрылым, устойчивого к белку Vip3A и/или белку Cry1F, при этом способ включает доставку в организм устойчивого насекомого-вредителя, относящегося к чешуекрылым, или в среду его обитания инсектицидного белка по любому из пп. 1-17 или инсектицидной композиции по любому из пп. 36-41.
[00373] 51. Способ уменьшения развития устойчивости к белку Vip3A и/или белку Cry1F в популяции целевого насекомого-вредителя, относящегося к чешуекрылым, при этом способ включает доставку в целевую популяцию или в среду ее обитания трансгенного растения, содержащего (i) полинуклеотид по любому из пп. 18-21, молекулу нуклеиновой кислоты по п. 22 или п. 23 или вектор по п. 24, и (ii) полинуклеотид, содержащий нуклеотидную последовательность, кодирующую белок Vip3A, и/или нуклеотидную последовательность, кодирующую белок Cry1F; где химерный инсектицидный белок и белок Vip3 А и/или белок Cry1F продуцируются в трансгенном растении.
[00374] 52. Способ по любому из пп. 42-51, где насекомое-вредитель, относящееся к чешуекрылым, включает одно или несколько из Spodoptera frugiperda (совка травяная), Chrysodeixis includes (совка соевая), Diatraea saccharalis (точильщик стеблей сахарного тростника), Diatraea grandiosella (огневка кукурузная юго-западная) и Agrotis ipsilon (совка-ипсилон).
[00375] 53. Способ по любому из пп. 42-52, где насекомое-вредитель, относящееся к чешуекрылым, включает Spodoptera frugiperda (совка травяная), которая устойчива к белку Vip3A и/или белку Cry1F.
[00376] 54. Способ по любому из пп. 42-53, где растение представляет собой: (i) однодольное растение, необязательно растение ячменя, растение маиса, растение овса, растение риса, растение сорго, растение сахарного тростника или растение пшеницы; или (ii) двудольное растение, необязательно растение сои, растение подсолнечника, растение томата, растение культурной разновидности капусты, растение хлопчатника, растение сахарной свеклы или растение табака.
[00377] 55. Способ по любому из пп. 42-54, где растение представляет собой растение маиса.
[00378] Далее настоящее изобретение будет описано со ссылкой на следующие примеры. Специалистам в данной области будет понятно, что эти примеры не ограничивают объем формулы изобретения настоящего изобретения, а скорее предназначены быть иллюстративными для некоторых вариантов осуществления. Другие варианты осуществления настоящего изобретения могут быть осуществлены на практике без отклонения от сущности и объема настоящего изобретения, объем которого определяется настоящим раскрытием и прилагаемой формулой изобретения.
ПРИМЕРЫ
Пример 1. Обнаружение химерного белка ВТ-0029, Bt29-Bt22, с инсектицидной активностью в отношении совки травяной
[00379] Два активных в отношении чешуекрылых белка из Bacillus thuringiensis (Bt), ВТ-0022 (SEQ ID NO: 1; ближайший известный представитель семейства Cry - Cry1If) и ВТ-0029 (SEQ ID NO: 2; ближайший известный представитель семейства Cry-Cry1Gb), были раскрыты в PCT/US16/038947.
[00380] Как показано в таблице 1 ниже, данные в отношении спектра насекомых для ВТ-0022 и ВТ-0029 указывают на то, что ВТ-0022 не обладает активностью в отношении совки травяной (FAW, Spodoptera frugiperda), а ВТ-0029 обладает только слабой активностью в отношении FAW. Другими протестированными насекомыми-вредителями, относящимися к чешуекрылым, являлись следующие: кукурузный мотылек (ЕСВ; Ostrinia nubilalis), совка-ипсилон (BCW; Agrotis ipsilon), совка кукурузная (CEW; Helicoverpa zed), точильщик стеблей сахарного тростника (SCB; Diatraea saccharalis), огневка кукурузная юго-западная (SWCB; Diatraea grandiosella), западная бобовая совка (WBCW; Striacosta albicosta), соевая совка (SBL; Pseudoplusia includens), совка бархатных бобов (VBC; Anticarsia gemmatalis) и табачная листовертка (TBW; Heliothis virescens).
[00381] Применяли подход инженерии белков в попытке повысить активность ВТ-0029 в отношении FAW. С использованием ВТ-0029 в качестве матрицы создали шесть сконструированных белков путем замены домена III (DIII) из ВТ-0029 доменом Ш из другого белка Cry. В таблице 2 проиллюстрированы шесть химерных белков с информацией об их доменном составе. В таблице 3 предоставлена информация о последовательности для полноразмерных белков.
[00382] Кодирующие сконструированные белки cDNA синтезировали в Genewiz (Саут-Плейнфилд, Нью-Джерси) и клонировали в вектор экспрессии Bacillus thuringiensis (Bt), содержащий промотор Cry1Ac и не содержащий терминатора. Плазмидные ДНК вводили в штамм Bt АВ227 с помощью опосредованной электропорацией процедуры трансформации. Все шесть сконструированных белков получали в виде кристаллических белков в Bt. Кристаллические белки очищали от культур и растворяли в буфере, содержащем 50 мМ Na2CO3, рН 11,0, 2 мМ DTT. Затем каждый растворимый белок оценивали в биологическом анализе с применением насекомых.
[00383] Коротко, инсектицидную активность химерных белков исследовали с помощью биологического анализа с применением насекомых, основанного на искусственном рационе, в котором растворенные кристаллические белки клали на поверхность искусственного рациона для насекомых с конечной концентрацией 2-3,2 мкг/см2. Буфер (50 мМ Na2CO3, рН 11,0, 2 мМ DTT), применяемый для растворения кристаллических белков Bt, служил в качестве отрицательного контроля. Полноразмерный белок Cry1Fa (SEQ ID NO: 8) использовали в качестве положительного контроля активности в отношении FAW. Каждый химерный белок протестировали в двух повторностях. Смертность насекомых оценивали на 7-й день.
[00384] Результаты показаны в таблице 4. Когда химерные белки тестировали в биологическом анализе с применением насекомых, химера Bt29-Bt22 (SEQ ID NO: 3) продемонстрировала сильную инсектицидную активность в отношении FAW, тогда как два других химерных белка (Bt29-Bt67 и Bt29-3A) показали лишь незначительную активность в отношении вредителя. Химера Bt29-1Ab не продемонстрировала какой-либо выявляемой активности в отношении FAW.
[00385] В этом тесте было получено недостаточно белка химер Bt29-Bt51 и Bt29-8D для оценки инсектицидной активности.
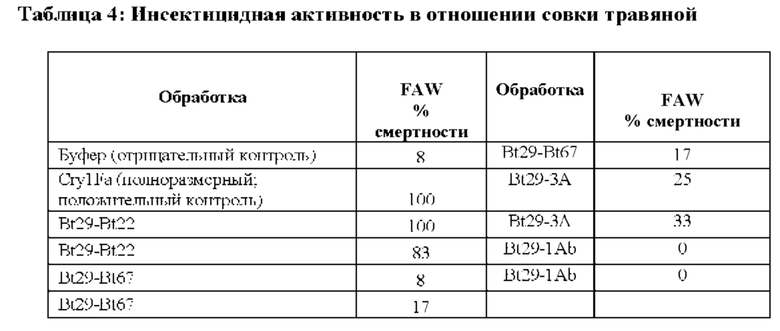
Пример 2. Инсектицидная активность химерных белков ВТ-0029 против различных вредителей, относящихся к чешуекрылым
[00386] Четыре химерных белка, описанных в примере 1 (Bt29-3A, Bt29-Bt22, Bt29-1Ab и Bt29-Bt67), исследовали в отношении их видов инсектицидной активности против нижеперечисленных вредителей, относящихся к чешуекрылым, с применением известных в данной области способов биологических анализов с искусственным рационом: точильщик стеблей сахарного тростника (SCB; Diatraea saccharlis), огневка кукурузная юго-западная (SWCB; Diatraea grandiosella), соевая совка (SBL; Pseudoplusia includens), совка кукурузная (CEW; Helicoverpa zed), табачная листовертка (TBW; Heliothis virescens), совка-ипсилон (BCW; Agrotis ipsilon) и кукурузный мотылек (ЕСВ; Ostrinia nubilalis).
[00387] Как показано в таблице 5, все четыре протестированных химерных белка сохранили сильную активность нативного ВТ-0029 в отношении SBL. Bt29-Bt22 и Bt29-Bt67 обладали слабой активностью в отношении SCB, сравнимой с нативным ВТ-0029, тогда как Bt29-3A и Bt29-1Ab по сути теряли всю активность в отношении SCB. Немного повышенную активность в отношении SWCB наблюдали у Bt29-Bt67 по сравнению с полноразмерным ВТ-0029. Достаточно согласуясь с результатами в таблице 1 для нативного ВТ-0029, только слабая активность в отношении BCW наблюдалась для Bt29-3A, Bt29-Bt22 и Bt29-Bt67, и ни одна из химер не обладала активностью в отношении CEW, TBW или ЕСВ.
Пример 3. Bt29-1F обладает инсектицидной активностью в отношении совки травяной
[00388] Для исследования дополнительных химерных белков разработали и протестировали второй набор химерных белков ВТ-0029 (таблица 6). Bt29-1Fa представляет собой химерный белок, в котором домен III из ВТ-0029 заменили доменом III из Cry1Fa. Полноразмерная аминокислотная последовательность Cry1Fa представлена в виде SEQ ID NO: 8 (см. также номер доступа в Genbank ААВ00376.1). Аминокислотная последовательность химеры Bt29-1Fa представлена в виде SEQ ID NO: 9, где последовательность Cry1Fa представляет собой аминокислоты 459-597.
[00389] Bt29-1Ka представляет собой химерный белок, в котором домен III из ВТ-0029 заменили доменом III из Cry1Ka. Полноразмерная аминокислотная последовательность Cry1Ka представлена в виде SEQ ID NO: 12 (см. также номер доступа в Genbank ААВ00376.1). Аминокислотная последовательность химеры Bt29-1Ka также представлена в виде SEQ ID NO: 13, где последовательность Cry1Ka представляет собой аминокислоты 459-597. Второй вариант химерного белка Bt29-1Ka (обозначенный в данном документе как Bt29-1Kav2) получали путем перемещения места перехода между последовательностью Cry1Ka и последовательностью ВТ-0029 в С-конце домена III с аминокислоты 598 (переход в первой химере Bt 29-1Ka) к аминокислоте 610. Аминокислотная последовательность химеры Bt29-1Kav2 представлена в виде SEQ ID NO: 15, где последовательность Cry1Ka представляет собой аминокислоты 459-609.
[00390] Bt29-1Ca представляет собой химерный белок, в котором домен III из Bt-0029 заменили доменом III из Cry1Ca. Полноразмерная аминокислотная последовательность Cry1Ca представлена в виде SEQ ID NO: 17 (см. также номер доступа в Genbank AF362020.1). Аминокислотная последовательность химеры Bt29-1Са представлена в виде SEQ ID NO: 18, где последовательность Cry1Ca представляет собой аминокислоты 467-617.

[00391] Инсектицидную активность химер Bt29-1Fa (SEQ ID NO: 9), Bt29-1Ka (SEQ ID NO: 13, Bt29-1Kav2 (SEQ ID NO: 16) и Bt29-1Ca (SEQ ID NO: 18) исследовали с помощью биологического анализа с применением насекомых, основанного на искусственном рационе, в котором растворенные белки клали на поверхность искусственного рациона для насекомых с конечной концентрацией 2 мкг/см2. Буфер (50 мМ Na2CO3, рН 10,5, 2 мМ DTT), применяемый для растворения кристаллических белков Bt, служил в качестве отрицательного контроля. Полноразмерный белок Cry1Fa (SEQ ID NO: 8) использовали в качестве положительного контроля активности в отношении FAW. Каждый химерный белок протестировали в трех повторностях. Инсектицидную активность оценивали как эффективную смертность на 7-й день (личинки, которые имеют задержку роста и умирают, оцениваются как эффективно погибшие).
[00392] Как показано в таблице 7, химерный белок, обозначенный Bt29-1Fa, демонстрировал сильную активность в отношении FAW.
[00393] В этом тесте было получено недостаточно белка Bt29-1Ka для оценки в биологическом анализе. Удивительно, но полученный из химеры Bt29-1Kav2 белок был активен в отношении совки травяной (см. таблицу 7).
Пример 4. Bt29-Bt22 и Bt29-1Fa обладают инсектицидной активностью в отношении совки травяной, устойчивой к Cry1F
[00394] Чтобы определить, зависит ли токсичность химерных белков ВТ-0029 в отношении FAW от способа действия (МОА), отличного от Cry1Fa, два химерных белка, Bt29-Bt22 и Bt29-1Fa, получали, как описано в примере 1, и кристаллические белки выделяли и очищали. Очищенные кристаллы растворяли в буфере 1 (50 мМ Na2CO3/NaHCO3, рН 11,2 мМ DTT) и чистоту препарата растворенного белка контролировали с применением системы Bio-Rad Experion (BioRad, Геркулес, Калифорния).
[00395] Очищенные белки тестировали на эффективность в отношении штамма FAW, который устойчив к инсектицидному токсину Cry1Fa. Анализ с покрытием рациона проводили с различными дозами каждого очищенного токсина (125, 500 и 2000 нг/см2), главным образом, как описано в примере 1. Белок Vip3A (положительный контроль) растворяли в PBS, а другие белки в буфере 1 (50 мМ Na2CO3/NaHCO3, рН 11, 2 мМ DTT). Две обработки отрицательного контроля представляли собой PBS и буфер 1. Также тестировали популяцию FAW, которая восприимчива к Cry1Fa или Vip3A, т.е. восприимчивую линию FAW. Каждый белок протестировали в двух повторностях. Инсектицидную активность оценивали как эффективную смертность на 7-й день (личинки, которые имеют задержку роста и умирают, по дочитываются как эффективно погибшие).
[00396] Как показано в таблице 8, личинки восприимчивых FAW подвергались контролю с помощью Cry1Fa, Vip3A, Bt29-F3t22 и Bt29-1Fa даже при самой низкой концентрации. В отличие от этого, устойчивая к Cry1F линия FAW не подвергалась контролю с помощью какой-либо тестируемой дозы Cry1Fa с демонстрацией того, что линия устойчива к этому токсину (таблица 9). Удивительно, но как Bt29-Bt22, так и Bt29-1Fa были очень эффективными при контроле устойчивых к Cry1F FAW даже при очень низкой дозе, составляющей 125 нг/см2, как показано в таблице 9, указывая на то, что способ действия этих двух химерных белков в отношении устойчивых FAW отличается от способа действия белка Cry1Fa.

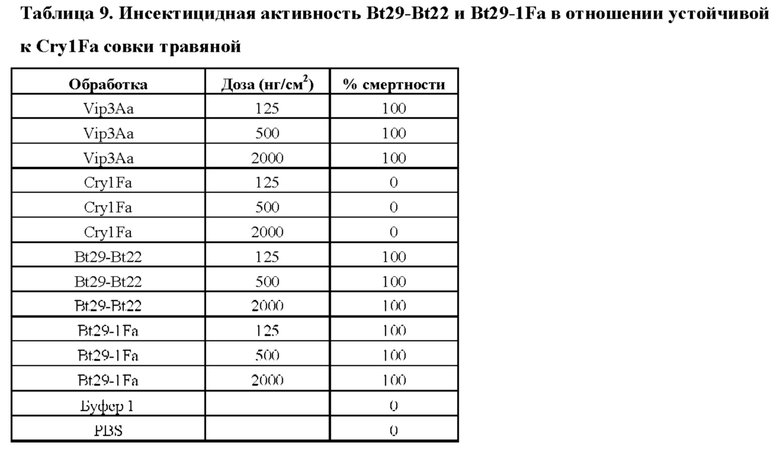
Пример 5. Bt29-Bt22 и Bt29-1Fa обладают инсектицидной активностью в отношении совки травяной, устойчивой к Vip3A
[00397] Чтобы определить, зависит ли токсичность химерных белков ВТ-0029 от МОА, отличного от белка Vip3A, получали белки Bt29-Bt22 и Bt29-1Fa, как описано в примере 1, и кристаллические белки выделяли и очищали. Очищенные кристаллы растворяли в буфере 1 (50 мМ Na2CO3/NaHCO3, рН 11, 2 мМ DTT) и чистоту препарата растворенного белка контролировали с использованием системы Bio-Rad Experion (BioRad, Геркулес, Калифорния).
[00398] Очищенные белки тестировали на эффективность в отношении штамма FAW, который устойчив к инсектицидному токсину Vip3A. Анализ с покрытием рациона проводили с различными дозами каждого очищенного токсина (125, 500 и 2000 нг/см2), главным образом, как описано в примере 1. Белок Vip3 А растворяли в PBS, а другие белки в буфере 1 (50 мМ Na2CO3/NaHCO3, рН 11, 2 мМ DTT). Две обработки отрицательного контроля представляли собой PBS и буфер 1. Белки Cry1Fa использовали в качестве положительного контроля для устойчивой к Vip3A линии FAW. Каждый белок протестировали в двух повторностях. Инсектицидную активность оценивали как эффективную смертность на 7-й день (личинки, которые имеют задержку роста и умирают, оцениваются как эффективно погибшие).
[00399] Устойчивая к Vip3A линия FAW не подвергалась контролю с помощью какой-либо тестируемой дозы Vip3A с демонстрацией того, что линия устойчива к этому токсину (таблица 10). В отличие от этого, как Bt29-Bt22, так и Bt29-1Fa продемонстрировали высокую степень эффективности в отношении устойчивых к Vip3A FAW, что указывает на то, что способ действия этих двух химерных белков в отношении устойчивых FAW отличается от способа действия белка Vip3A.
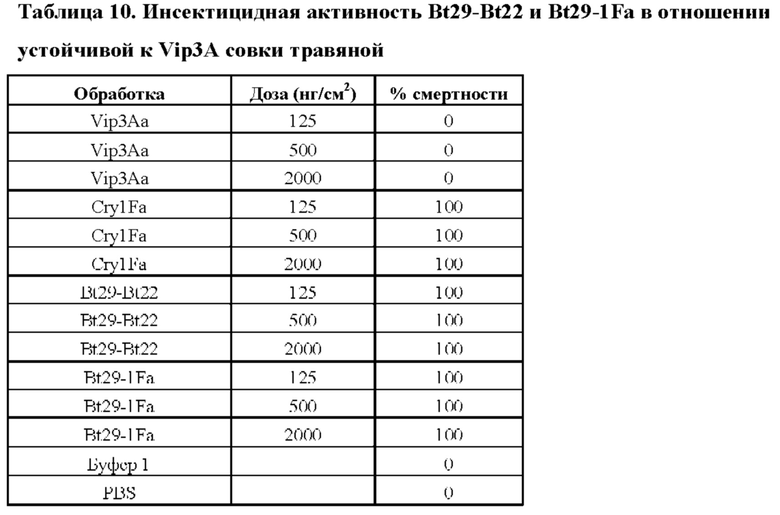
Пример 6. Усечения в области хвоста протоксина
[00400] Хорошо известно, что форма протоксина белков Cry Bt подвергается процессингу как на N-, так и на С-конце с получением зрелого токсина. Шесть усеченных с С-конца форм химеры Bt29-Bt22 (SEQ ID NO: 3), которая содержит область хвоста протоксина ВТ-0029, получали, как показано на фигуре 3. Во всех усеченных вариантах, обозначенных в данном документе как Bt29-Bt22Tr1 (SEQ ID NO: 20), Bt29-Bt22Tr2 (SEQ ID NO: 21), Bt29-Bt22Tr3 (SEQ ID NO: 22), Bt29-Bt22Tr4 (SEQ ID NO: 23), Bt29-Bt22Tr5 (SEQ ID NO: 24), Bt29-Bt22Tr6 (SEQ ID NO: 25), домен III из ВТ-0022 (оканчивающийся PVTA) и по меньшей мере первые 6 аминокислот хвоста протоксина ВТ-0029 (TFEAEY) сохраняли. Специалистам в данной области будет понятно, что конец домена III и начало хвоста протоксина не были точно определены для ВТ-0029 или химер, описанных в данном документе. Сердцевинный домен III, показанный на фигурах 1А и 1В, основан на выравнивании с другими белками Cry, в которых была определена область домена III.
[00401] Все усеченные формы Bt29-Bt22Tr1, -Tr2, -Tr4 и -Tr5 демонстрировали активность в отношении FAW, оцениваемую как подавление роста (не смертность) в биологических анализах с применением насекомых при тестировании в виде лизатов Е. coli. Формы Bt29-Bt22Tr1 и -Tr4 были наиболее перспективными, демонстрируя наибольшую активность в данном анализе (подавление роста на 67% и 75% соответственно).
[00402] Получали вторую полноразмерную химеру Bt29-Bt22, в которой область хвоста протоксина получена из ВТ-0022 (Cry1I) вместо ВТ-0029 (Cry1G). Получали шесть усеченных с С-конца форм этой полноразмерной химеры, представленных в данном документе под SEQ ID NO: 26-31, и тестировали в отношении совки травяной и соевой совки. Cry1Fa использовали в качестве положительного контроля. Результаты показаны в таблице 11.
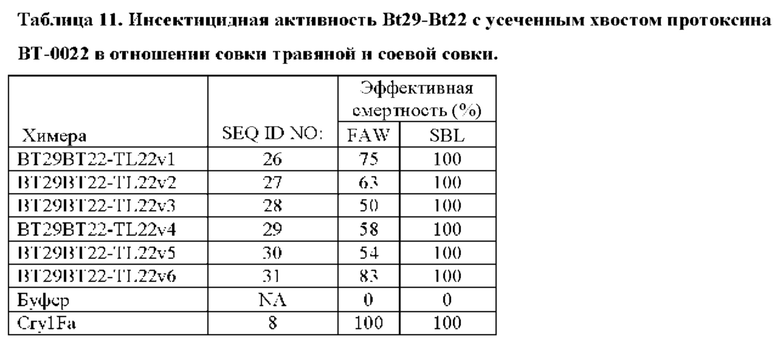
[00403] Четыре усеченные формы Bt29-1Fa (SEQ ID NO: 9), которые содержат область хвоста протоксина ВТ-0029, представленные в данном документе как Bt29-1FaTr1 (SEQ ID NO: 32), Bt29-1FaTr2 (SEQ ID NO: 33), Bt29-1FaTr3 (SEQ ID NO: 34) и Bt29-1FaTr4 (SEQ ID NO: 35), получали в системе экспрессии Е. coli, и лизаты тестировали в отношении совки травяной. Результаты показаны в таблице 12.
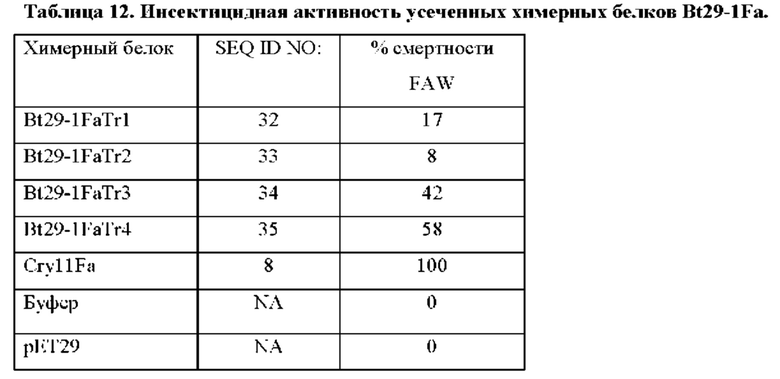
Пример 7. Экспрессия и активность химерных белков Bt29-Bt22 и Bt29-Cry1Fa в растениях маиса
[00404] Перед экспрессией в растениях синтетические полинуклеотиды, содержащие нуклеотидную последовательность, кодирующую химеру Bt29-Bt22 и Bt29-1Fa, синтезировали с использованием платформы для автоматизированного синтеза генов (Genscript, Inc., Пискатауэй, Нью-Джерси). В этом примере получали первую и вторую кассету экспрессии, содержащие убиквитиновый промотор маиса (Ubi1), функционально связанный с химерной последовательностью, кодирующей Bt29-Bt22 или Bt29-1Fa, которая функционально связана с убиквитиновым терминатором маиса, и получали третью кассету экспрессии, содержащую промотор Ubi1, функционально связанный с последовательностью, кодирующей фосфоманнозоизомеразу (PMI), которая функционально связана с терминатором Ubi. Экспрессия PMI позволяет осуществлять положительную селекцию трансгенных растений на маннозе. Для трансформации растений первую и третью кассеты экспрессии и вторую и третью кассеты экспрессии клонировали в подходящий вектор для трансформации маиса, опосредованной Agrobacterium.
[00405] Трансформацию незрелых зародышей маиса выполняли, главным образом, как описано в Negrotto et al., 2000, Plant Cell Reports 19: 798 803. Коротко, штамм Agrobacterium LBA4404 (pSB1), содержащий вектор экспрессии, экспрессирующий Bt29-Bt22 или Bt29-Cry1Fa, выращивали на твердой среде YEP (дрожжевой экстракт (5 г/л), пептон (10 г/л), NaCl (5 г/л), 15 г/л агара, рН 6,8) в течение 2-4 дней при 28°С. Примерно 0,8Х 109 клеток Agrobacterium суспендировали в среде LS-inf, дополненной 100 мкМ As. Бактерий предварительно индуцировали в данной среде в течение примерно 30-60 минут.
[00406] Незрелые зародыши инбредной линии маиса вырезали из початков возрастом 8-12 дней с переносом в жидкую среду LS-inf + 100 мкМ As. Зародыши однократно ополаскивали свежей средой для инфицирования. Затем добавляли раствор Agrobacterium, и зародыши перемешивали на вихревой мешалке в течение 30 секунд и оставляли осесть с бактериями в течение 5 минут. Затем зародыши переносили стороной со щитком зародыша кверху на среду LSAs и культивировали в темноте в течение двух-трех дней. Впоследствии примерно 20-25 зародышей на чашку Петри переносили на среду LSDc, дополненную цефотаксимом (250 мг/л) и нитратом серебра (1,6 мг/л), и культивировали в темноте при примерно 28°С в течение 10 дней.
[00407] Незрелые зародыши, образующие эмбриогенный каллюс, переносили в среду LSD1M0.5S. Селекцию культур на этой среде осуществляли в течение примерно 6 недель, при этом через приблизительно 3 недели проводили стадию субкультивирования. Выжившие каллюсы переносили на среду Reg1, дополненную маннозой. После культивирования на свету (в режиме 16 часов света/8 часов темноты) зеленые ткани затем переносили на среду Reg2 без регуляторов роста и инкубировали в течение приблизительно 1-2 недель. Проростки переносили в контейнеры Magenta GA-7 (Magenta Corp, Чикаго, Иллинойс), содержащие среду Reg3, и выращивали на свету. Спустя приблизительно 2-3 недели растения тестировали с помощью ПНР в отношении наличия генов PMI, а также химерного гена Bt cry. Растения, показавшие положительные результаты в ПЦР-анализе, переносили в теплицу для дальнейшей оценки.
[00408] Трансгенные растения оценивали в отношении числа копий (определяли с помощью анализа Taqman), уровня экспрессии белка (определяли с помощью ELISA) и эффективности в отношении представляющих интерес видов насекомых в биологических анализах с вырезанием листа. В частности, ткань растения (лист или рыльца) вырезали из однокопийных трансформантов (стадии V3 - V4) и заражали новорожденными личинками целевого вредителя, затем инкубировали при комнатной температуре в течение 5 дней. Листовые диски от трансгенных растений, экспрессирующих каждый химерный белок Bt, тестировали в отношении совки травяной (Spodoptera frugiperda, FAW).
[00409] Результаты подтверждают, что трансгенные растения экспрессируют химерные белки по настоящему изобретению и активны в отношении насекомых-вредителей. Экспрессия белка в трансгенных трансформантах для химеры Bt29-Bt22 составляла приблизительно 25-125 нг/мг общего растворимого белка (TSP), а для химеры Bt29-1Fa составляла приблизительно 25-290 нг/мг TSP. Приблизительно 90% и 92% трансгенных транс формантов, экспрессирующих химеры Bt29-Bt22 и Bt29-1Fa соответственно, приводили к 100%-ной смертности и подавлению роста личинок совки травяной.
[00410] Примеры, представленные выше, явным образом иллюстрируют преимущества настоящего изобретения. Несмотря на то, что настоящее изобретение было описано на основании конкретных деталей некоторых его вариантов осуществления, не подразумевается, что такие детали следует считать ограничениями объема заявленного изобретения, за исключением и до такой степени, с которой они включены в прилагаемую формулу данного изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЗИНГЕНТА ПАРТИСИПЕЙШНС АГ
<120> СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ
ВРЕДИТЕЛЕЙ РАСТЕНИЙ
<130> 81090-WO-REG-ORG-P-1
<150> US 62/432,909
<151> 2016-12-12
<160> 35
<170> PatentIn версия 3.5
<210> 1
<211> 715
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность BT-0022, собранная на основе анализа данных
в отношении генома Bacillus.
<400> 1
Met Lys Ser Lys Asn Gln Asn Met His Gln Ser Leu Ser Asn Asn Ala
1 5 10 15
Thr Val Asp Lys Asn Phe Thr Gly Ser Leu Glu Asn Asn Thr Asn Thr
20 25 30
Glu Leu Gln Asn Phe Asn His Glu Gly Ile Glu Pro Phe Val Ser Val
35 40 45
Ser Thr Ile Gln Thr Gly Ile Gly Ile Ala Gly Lys Ile Leu Gly Asn
50 55 60
Leu Gly Val Pro Phe Ala Gly Gln Val Ala Ser Leu Tyr Ser Phe Ile
65 70 75 80
Leu Gly Glu Leu Trp Pro Lys Gly Lys Ser Gln Trp Glu Ile Phe Met
85 90 95
Glu His Val Glu Glu Leu Ile Asn Gln Lys Ile Ser Thr Tyr Ala Arg
100 105 110
Asn Lys Ala Leu Ala Asp Leu Lys Gly Leu Gly Asp Ala Leu Ala Val
115 120 125
Tyr His Glu Ser Leu Glu Ser Trp Ile Lys Asn Arg Asn Asn Thr Arg
130 135 140
Thr Arg Ser Val Val Lys Ser Gln Tyr Ile Thr Leu Glu Leu Met Phe
145 150 155 160
Val Gln Ser Leu Pro Ser Phe Ala Val Ser Gly Glu Glu Val Pro Leu
165 170 175
Leu Pro Ile Tyr Ala Gln Ala Ala Asn Leu His Leu Leu Leu Leu Arg
180 185 190
Asp Ala Ser Ile Phe Gly Lys Glu Trp Gly Leu Ser Asp Ser Glu Ile
195 200 205
Ser Thr Phe Tyr Asn Arg Gln Val Glu Arg Thr Ser Asp Tyr Ser Asp
210 215 220
His Cys Thr Lys Trp Phe Asp Thr Gly Leu Asn Arg Leu Lys Gly Ser
225 230 235 240
Asn Ala Glu Ile Trp Val Lys Tyr Asn Gln Phe Arg Arg Asp Met Thr
245 250 255
Leu Met Val Leu Asp Leu Val Ala Leu Phe Gln Ser Tyr Asp Thr His
260 265 270
Met Tyr Pro Ile Lys Thr Thr Ala Gln Leu Thr Arg Glu Val Tyr Thr
275 280 285
Asn Ala Ile Gly Thr Val His Pro His Pro Ser Phe Thr Ser Thr Thr
290 295 300
Trp Tyr Asn Asn Asn Ala Pro Ser Phe Ser Ala Ile Glu Ala Ala Val
305 310 315 320
Ile Arg Ser Pro His Leu Leu Asp Phe Leu Glu Gln Val Thr Ile Tyr
325 330 335
Ser Leu Leu Ser Arg Trp Ser Asn Thr Gln Tyr Met Asn Met Trp Gly
340 345 350
Gly His Lys Leu Glu Phe Arg Thr Ile Gly Gly Thr Leu Asn Thr Ser
355 360 365
Thr Gln Gly Ser Thr Asn Thr Ser Ile Asn Pro Val Thr Leu Pro Phe
370 375 380
Thr Ser Arg Asp Ile Tyr Arg Thr Glu Ser Leu Ala Gly Leu Asn Leu
385 390 395 400
Phe Leu Thr Gln Pro Val Asn Gly Val Pro Arg Val Asp Phe His Trp
405 410 415
Lys Phe Val Thr His Pro Ile Ala Ser Asp Asn Phe Tyr Tyr Pro Gly
420 425 430
Tyr Ala Gly Ile Gly Thr Gln Leu Gln Asp Ser Glu Asn Glu Leu Pro
435 440 445
Pro Glu Thr Thr Gly Gln Pro Asn Tyr Glu Ser Tyr Ser His Arg Leu
450 455 460
Ser His Ile Gly Leu Ile Ser Ala Ser His Val Lys Ala Leu Val Tyr
465 470 475 480
Ser Trp Thr His Arg Ser Ala Asp Arg Thr Asn Thr Ile His Ser Asp
485 490 495
Ser Ile Thr Gln Ile Pro Leu Val Lys Ala His Thr Leu Gln Ser Gly
500 505 510
Thr Thr Val Val Lys Gly Pro Gly Phe Thr Gly Gly Asp Ile Leu Arg
515 520 525
Arg Thr Ser Gly Gly Pro Phe Ala Phe Ser Asn Val Asn Leu Asp Trp
530 535 540
Asn Leu Ser Gln Arg Tyr Arg Ala Arg Ile Arg Tyr Ala Ser Thr Thr
545 550 555 560
Asn Leu Arg Met Tyr Val Thr Ile Ala Gly Glu Arg Ile Phe Ala Gly
565 570 575
Gln Phe Asn Lys Thr Met Asn Thr Gly Asp Pro Leu Thr Phe Gln Ser
580 585 590
Phe Ser Tyr Ala Thr Ile Asp Thr Ala Phe Thr Phe Pro Thr Lys Ala
595 600 605
Ser Ser Leu Thr Val Gly Ala Asp Thr Phe Ser Ser Gly Asn Glu Val
610 615 620
Tyr Val Asp Arg Phe Glu Leu Ile Pro Val Thr Ala Thr Leu Glu Ala
625 630 635 640
Val Thr Asp Leu Glu Arg Ala Gln Lys Ala Val His Glu Leu Phe Thr
645 650 655
Ser Thr Asn Pro Gly Gly Leu Lys Thr Asp Val Ala Lys Asp His Tyr
660 665 670
Thr Asn Thr Ile Ser Lys Ser Val Gln Ser Val Phe Arg Cys Arg Cys
675 680 685
Ser Glu Arg Thr Arg Ile Tyr Arg Trp Gly Tyr Pro Ser Lys Lys Glu
690 695 700
Tyr Trp Tyr Ile Trp Gly Tyr Thr Ser Lys Tyr
705 710 715
<210> 2
<211> 1169
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность BT-0029, собранная на основе анализа данных
в отношении генома Bacillus.
<400> 2
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala Phe Asn Leu His Ser Gly Ala Thr Ile Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Asn Val Gly Ser Phe
485 490 495
Gly Asp Met Arg Val Asn Ile Thr Ala Pro Leu Ser Gln Arg Tyr Arg
500 505 510
Val Arg Ile Arg Tyr Ala Ser Thr Thr Asp Leu Gln Phe Tyr Thr Asn
515 520 525
Ile Asn Gly Thr Thr Ile Asn Ile Gly Asn Phe Ser Ser Thr Met Asp
530 535 540
Ser Gly Asp Asp Leu Gln Tyr Gly Arg Phe Arg Val Ala Gly Phe Thr
545 550 555 560
Thr Pro Phe Thr Phe Ser Asp Ala Met Ser Thr Phe Thr Ile Gly Ala
565 570 575
Phe Ser Phe Ser Ser Asn Asn Glu Val Tyr Ile Asp Arg Ile Glu Phe
580 585 590
Val Pro Ala Glu Val Thr Phe Glu Ala Glu Tyr Asp Leu Glu Lys Ala
595 600 605
Gln Lys Ala Val Asn Ala Leu Phe Thr Ser Ser Asn Gln Ile Gly Leu
610 615 620
Lys Thr Asp Val Thr Asp Tyr His Ile Asp Lys Val Ser Asn Leu Val
625 630 635 640
Glu Cys Leu Ser Asp Glu Phe Cys Leu Asp Glu Lys Arg Glu Leu Ser
645 650 655
Glu Lys Val Lys His Ala Lys Arg Leu Cys Asp Glu Arg Asn Leu Leu
660 665 670
Gln Asp Pro Asn Phe Arg Gly Ile Asn Arg Gln Pro Asp Arg Gly Trp
675 680 685
Arg Gly Ser Thr Asp Ile Thr Ile Gln Gly Gly Asp Asp Val Phe Lys
690 695 700
Glu Asn Tyr Val Thr Leu Pro Gly Thr Phe Asp Glu Cys Tyr Pro Thr
705 710 715 720
Tyr Leu Tyr Gln Lys Ile Asp Glu Ser Lys Leu Lys Ala Tyr Thr Arg
725 730 735
Tyr Glu Leu Arg Gly Tyr Ile Glu Asp Ser Gln Asp Leu Glu Ile Tyr
740 745 750
Leu Ile Arg Tyr Asn Ala Lys His Glu Thr Val Asn Val Pro Gly Thr
755 760 765
Gly Ser Leu Trp Pro Leu Ser Ala Gln Ser Pro Ile Gly Lys Cys Gly
770 775 780
Glu Pro Asn Arg Cys Ala Thr His Leu Glu Trp Asn Pro Asp Leu Asp
785 790 795 800
Cys Ser Cys Arg Asp Gly Glu Lys Cys Ala His His Ser His His Phe
805 810 815
Ser Leu Asp Ile Asp Val Gly Cys Thr Asp Leu Asn Glu Asp Leu Gly
820 825 830
Val Trp Val Ile Phe Lys Ile Lys Thr Gln Asp Gly His Ala Arg Leu
835 840 845
Gly Asn Leu Glu Phe Leu Glu Glu Lys Pro Leu Val Gly Glu Ala Leu
850 855 860
Ala Arg Val Lys Arg Ala Glu Lys Lys Trp Arg Asp Lys Arg Glu Lys
865 870 875 880
Leu Glu Leu Glu Thr Asn Ile Val Tyr Lys Glu Ala Lys Lys Ser Val
885 890 895
Asp Ala Leu Phe Val Asn Ser Gln Tyr Asp Arg Leu Gln Ala Asp Thr
900 905 910
Asn Ile Ala Ile Ile His Ala Ala Asp Lys Arg Val His Ser Ile Arg
915 920 925
Glu Ala Tyr Leu Pro Glu Leu Ser Val Ile Pro Gly Val Asn Ala Ala
930 935 940
Ile Phe Glu Glu Leu Glu Gly Arg Ile Phe Thr Ala Tyr Ser Leu Tyr
945 950 955 960
Asp Ala Arg Asn Val Ile Lys Asn Gly Asp Phe Asn Asn Gly Leu Ser
965 970 975
Cys Trp Asn Val Lys Gly His Val Asp Val Glu Glu Gln Asn Asn His
980 985 990
Arg Ser Val Leu Val Val Pro Glu Trp Glu Ala Glu Val Ser Gln Glu
995 1000 1005
Val Arg Val Cys Pro Gly Arg Gly Tyr Ile Leu Arg Val Thr Ala
1010 1015 1020
Tyr Lys Glu Gly Tyr Gly Glu Gly Cys Val Thr Ile His Glu Ile
1025 1030 1035
Glu Asp Asn Thr Asp Glu Leu Lys Phe Ser Asn Cys Val Glu Glu
1040 1045 1050
Glu Ile Tyr Pro Asn Asn Thr Val Thr Cys Asn Asp Tyr Thr Ala
1055 1060 1065
Thr Gln Glu Glu Tyr Glu Gly Thr Tyr Thr Ser Arg Asn Arg Gly
1070 1075 1080
Tyr Asp Gly Ala Tyr Glu Ser Asn Ser Ser Val Pro Ala Asp Tyr
1085 1090 1095
Ala Ser Ala Tyr Glu Glu Lys Ala Tyr Thr Asp Gly Arg Arg Asp
1100 1105 1110
Asn Thr Cys Glu Ser Asn Arg Gly Tyr Gly Asp Tyr Thr Pro Leu
1115 1120 1125
Pro Ala Gly Tyr Val Thr Lys Glu Leu Glu Tyr Phe Pro Glu Thr
1130 1135 1140
Asp Lys Val Trp Ile Glu Ile Gly Glu Thr Glu Gly Thr Phe Ile
1145 1150 1155
Val Asp Ser Val Glu Leu Leu Leu Met Glu Glu
1160 1165
<210> 3
<211> 1169
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Химерный белок Bt29-Bt22.
<400> 3
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Phe Glu Ala Glu Tyr Asp Leu Glu Lys Ala
595 600 605
Gln Lys Ala Val Asn Ala Leu Phe Thr Ser Ser Asn Gln Ile Gly Leu
610 615 620
Lys Thr Asp Val Thr Asp Tyr His Ile Asp Lys Val Ser Asn Leu Val
625 630 635 640
Glu Cys Leu Ser Asp Glu Phe Cys Leu Asp Glu Lys Arg Glu Leu Ser
645 650 655
Glu Lys Val Lys His Ala Lys Arg Leu Cys Asp Glu Arg Asn Leu Leu
660 665 670
Gln Asp Pro Asn Phe Arg Gly Ile Asn Arg Gln Pro Asp Arg Gly Trp
675 680 685
Arg Gly Ser Thr Asp Ile Thr Ile Gln Gly Gly Asp Asp Val Phe Lys
690 695 700
Glu Asn Tyr Val Thr Leu Pro Gly Thr Phe Asp Glu Cys Tyr Pro Thr
705 710 715 720
Tyr Leu Tyr Gln Lys Ile Asp Glu Ser Lys Leu Lys Ala Tyr Thr Arg
725 730 735
Tyr Glu Leu Arg Gly Tyr Ile Glu Asp Ser Gln Asp Leu Glu Ile Tyr
740 745 750
Leu Ile Arg Tyr Asn Ala Lys His Glu Thr Val Asn Val Pro Gly Thr
755 760 765
Gly Ser Leu Trp Pro Leu Ser Ala Gln Ser Pro Ile Gly Lys Cys Gly
770 775 780
Glu Pro Asn Arg Cys Ala Thr His Leu Glu Trp Asn Pro Asp Leu Asp
785 790 795 800
Cys Ser Cys Arg Asp Gly Glu Lys Cys Ala His His Ser His His Phe
805 810 815
Ser Leu Asp Ile Asp Val Gly Cys Thr Asp Leu Asn Glu Asp Leu Gly
820 825 830
Val Trp Val Ile Phe Lys Ile Lys Thr Gln Asp Gly His Ala Arg Leu
835 840 845
Gly Asn Leu Glu Phe Leu Glu Glu Lys Pro Leu Val Gly Glu Ala Leu
850 855 860
Ala Arg Val Lys Arg Ala Glu Lys Lys Trp Arg Asp Lys Arg Glu Lys
865 870 875 880
Leu Glu Leu Glu Thr Asn Ile Val Tyr Lys Glu Ala Lys Lys Ser Val
885 890 895
Asp Ala Leu Phe Val Asn Ser Gln Tyr Asp Arg Leu Gln Ala Asp Thr
900 905 910
Asn Ile Ala Ile Ile His Ala Ala Asp Lys Arg Val His Ser Ile Arg
915 920 925
Glu Ala Tyr Leu Pro Glu Leu Ser Val Ile Pro Gly Val Asn Ala Ala
930 935 940
Ile Phe Glu Glu Leu Glu Gly Arg Ile Phe Thr Ala Tyr Ser Leu Tyr
945 950 955 960
Asp Ala Arg Asn Val Ile Lys Asn Gly Asp Phe Asn Asn Gly Leu Ser
965 970 975
Cys Trp Asn Val Lys Gly His Val Asp Val Glu Glu Gln Asn Asn His
980 985 990
Arg Ser Val Leu Val Val Pro Glu Trp Glu Ala Glu Val Ser Gln Glu
995 1000 1005
Val Arg Val Cys Pro Gly Arg Gly Tyr Ile Leu Arg Val Thr Ala
1010 1015 1020
Tyr Lys Glu Gly Tyr Gly Glu Gly Cys Val Thr Ile His Glu Ile
1025 1030 1035
Glu Asp Asn Thr Asp Glu Leu Lys Phe Ser Asn Cys Val Glu Glu
1040 1045 1050
Glu Ile Tyr Pro Asn Asn Thr Val Thr Cys Asn Asp Tyr Thr Ala
1055 1060 1065
Thr Gln Glu Glu Tyr Glu Gly Thr Tyr Thr Ser Arg Asn Arg Gly
1070 1075 1080
Tyr Asp Gly Ala Tyr Glu Ser Asn Ser Ser Val Pro Ala Asp Tyr
1085 1090 1095
Ala Ser Ala Tyr Glu Glu Lys Ala Tyr Thr Asp Gly Arg Arg Asp
1100 1105 1110
Asn Thr Cys Glu Ser Asn Arg Gly Tyr Gly Asp Tyr Thr Pro Leu
1115 1120 1125
Pro Ala Gly Tyr Val Thr Lys Glu Leu Glu Tyr Phe Pro Glu Thr
1130 1135 1140
Asp Lys Val Trp Ile Glu Ile Gly Glu Thr Glu Gly Thr Phe Ile
1145 1150 1155
Val Asp Ser Val Glu Leu Leu Leu Met Glu Glu
1160 1165
<210> 4
<211> 3510
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодирующие последовательности для химерного белка Bt29-Bt22.
<400> 4
atggagatta ataatcagaa ccaatgtgtc ccttataatt gtttgaataa tcctgaaagc
60
gagatattaa acgttgcaat ttttagtagc gaacaggtag cagaaattca cttaaagatc
120
acgcgcttaa ttttagagaa ttttttacca ggtgggagtt ttgcattcgg cttatttgat
180
ttaatatggg ggatttttaa tgaagatcaa tggagcgcat ttcttcggca ggtagaagaa
240
ttaattaatc aaaggataac ggaattcgca agagggcaag caattcagag actagtaggg
300
tttggaagga gttatgatga atatatttta gcactaaaag aatgggaaaa cgatcctgat
360
aacccagctt caaaggaaag agtgcgcact cgatttcgga caactgatga tgccttgcta
420
accggtgttc ctcttatggc aattccaggt tttgaattag ctactttatc tgtttatgct
480
caatcagcca atctacattt agccctatta agagatgctg tattttttgg ggagagatgg
540
ggattgacac aaacaaatat aaatgattta tatagtagat taaaaaactc cattcgtgat
600
tatacaaatc attgtgttcg tttttataat ataggtttag ggaatttaaa tgttataaga
660
ccagagtatt accgtttcca aagagaatta acaatatctg tcttagatct tgtagctctt
720
tttccaaatt acgatatccg aacatatcca ataccaacta aaagtcaatt aacaagagaa
780
atttatacag atccgattat ttcacctggt gcacaggcag gttatactct tcaagatgtt
840
ttgagagaac cacaccttat ggacttttta aaccgactta ttatttatac tggtgagtat
900
cgcggaattc gtcactgggc aggacatgaa gtagaatcta gtagaacagg tatgatgact
960
aatataagat ttcctttgta tggaacagcc gcaacagcag aaccaacacg atttataact
1020
cctagtactt ttcctggtct taatttattt tatagaacat tatcagctcc tatttttaga
1080
gatgaaccgg gagctaatat tattattaga tatagaacga gtttggtgga aggagtagga
1140
tttattcaac caaataacgg tgaacagctt tacagagtga gaggaacatt agattctctt
1200
gatcaattac cacttgaggg tgagagtagt ctaactgaat atagtcatcg attatgccat
1260
gttagatttg cgcaatcatt gaggaatgca gaacctttag attatgcaag ggttccgatg
1320
ttttcttgga cacatcgtag tgcaacccct acaaatacaa ttgatccaga tgtcatcacc
1380
caaataccgt tagtaaaagc acataccctt cagtcaggta ctactgttgt aaaagggcca
1440
gggtttacag gtggagatat cctccgacga actagtggag gaccatttgc ttttagtaat
1500
gttaatttag actggaactt gtcacaaaga tatcgtgcta gaatacgcta tgcttctact
1560
actaatctaa gaatgtacgt aacgattgca ggggaacgaa tttttgctgg tcaatttaat
1620
aaaacaatga atactggtga tccattaaca ttccaatctt ttagttacgc aactattgat
1680
acagcattta cattcccaac gaaagcgagc agcttgactg taggtgctga tacttttagc
1740
tcaggtaatg aagtttatgt agatagattt gaattgatcc cagttactgc aacatttgag
1800
gcagaatatg atttagagaa agctcagaaa gcggtgaatg cgctgtttac ttcttccaat
1860
caaatcgggt taaaaacaga tgtgacggac tatcatattg ataaagtatc caatctagtt
1920
gagtgtttat cagatgaatt ttgtctagat gaaaagcgag aattgtccga gaaagtcaaa
1980
catgcgaagc gactctgtga tgagcggaat ttacttcaag atccaaactt cagaggcatc
2040
aatagacaac cagaccgtgg ttggagagga agtacggata ttaccatcca aggaggagat
2100
gacgtattca aagagaatta cgttacgcta ccgggtacct ttgatgagtg ctatccaacg
2160
tatttatatc aaaaaataga tgagtcgaaa ttaaaagcct atacccgcta tgaattaaga
2220
gggtatatcg aggatagtca agacttagaa atctatttaa ttcgctacaa tgcaaaacat
2280
gaaacagtaa atgtgccagg tacgggttcc ttatggccgc tttcagccca aagtccaatc
2340
ggaaagtgtg gagaaccgaa tcgatgtgcg acacaccttg aatggaatcc tgacttagat
2400
tgttcgtgta gggatggaga aaagtgtgcc catcattcgc atcatttctc cttagacatt
2460
gatgtaggat gtacagacct aaatgaggac ctaggtgtat gggtgatctt taagattaag
2520
acgcaagatg gtcatgcgag actaggaaat ctagaatttc tcgaagagaa accattagta
2580
ggagaagcgc tagctcgtgt gaagagagcg gagaaaaaat ggagagacaa acgcgaaaaa
2640
ttggaattgg aaacaaatat tgtttataaa gaggcaaaaa aatctgtaga tgctttattt
2700
gtgaactctc aatatgatag attacaagcg gatacgaata tcgcgataat tcatgcggca
2760
gataaacgcg ttcatagcat tcgagaagca tatcttccag agttgtctgt aattccgggt
2820
gtaaatgcag ctatttttga agaattagag ggacgtattt tcacagccta ctctctatat
2880
gatgcgagaa atgtcattaa aaatggcgat ttcaataatg gcttatcatg ctggaacgtg
2940
aaagggcatg tagatgtaga agaacagaac aaccatcgtt cggtccttgt tgttccagaa
3000
tgggaagcag aagtgtcaca agaggttcgt gtctgtccag gtcgtggcta tatccttcgt
3060
gttacagcgt acaaagaggg atatggagag ggctgtgtaa cgattcatga gatcgaagac
3120
aatacagacg aactgaaatt cagcaactgt gtagaagagg aaatatatcc aaacaacacg
3180
gtaacgtgta atgattatac tgcgactcaa gaagaatatg agggtacgta cacttctcgt
3240
aatcgaggat atgacggagc ctatgaaagc aattcttctg taccagctga ttatgcatca
3300
gcctatgaag aaaaagcgta tacagatgga agaagagaca atacttgtga atctaacaga
3360
ggatatgggg attacacacc actaccagct ggctatgtga caaaagaatt agagtacttc
3420
ccagaaaccg ataaggtatg gattgagatt ggagaaacgg aaggaacatt tatcgtggac
3480
agcgtggaat tactccttat ggaggaatag
3510
<210> 5
<211> 3510
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная кодирующая последовательность маиса для
химерного белка Bt29-Bt22.
<400> 5
atggaaatca acaaccagaa ccagtgcgtg ccgtacaact gcctcaacaa ccccgagtcc
60
gagatcctga acgtggccat cttctccagc gagcaggtcg cggagatcca cctcaagatc
120
acgcgcctga tcctcgagaa cttcctgccg ggcggctcct tcgctttcgg cctgttcgac
180
ctcatctggg gcatcttcaa cgaggaccag tggagcgcgt tcctcaggca ggtggaggag
240
ctgatcaacc agcgcatcac ggagttcgcc aggggccagg ctatccagcg gctggtgggc
300
ttcggcaggt cctacgacga atacatcctg gccctcaagg agtgggagaa cgaccccgac
360
aacccggcca gcaaggagcg cgtgaggacc cgcttcagga ccaccgacga cgctctcctg
420
acgggcgtcc ccctcatggc tatcccgggc ttcgagctgg ccaccctctc ggtgtacgct
480
cagtcggcca acctgcacct cgccctcctg cgggacgctg tgttcttcgg cgagaggtgg
540
ggcctgaccc aaaccaacat caacgacctc tactccaggc tgaagaacag catccgcgac
600
tacacgaacc actgcgtgcg cttctacaac atcggcctgg gcaacctcaa cgtcatcagg
660
ccggaatact accgcttcca gagggagctg accatcagcg tgctggacct cgtcgccctg
720
ttccccaact acgacatccg cacgtacccg atccccacca agtcccagct cacgagggag
780
atctacaccg acccgatcat ctcgccgggc gcccaggccg gctacaccct gcaggacgtc
840
ctgagggagc cccacctgat ggacttcctg aacaggctca tcatctacac cggcgagtac
900
aggggcatca ggcactgggc gggccacgag gtggagtcca gcaggacggg catgatgacc
960
aacatccgct tcccgctcta cggcaccgcg gccacggccg agccaacccg cttcatcacg
1020
ccgtccacct tccccggcct gaacctcttc tacaggaccc tgtcggctcc catcttccgc
1080
gacgagccgg gcgcgaacat catcatccgc tacaggacct ccctcgtgga gggcgtcggc
1140
ttcatccagc cgaacaacgg cgagcagctg taccgcgtga ggggcacgct ggacagcctg
1200
gaccagctcc cactggaggg cgagtccagc ctcaccgagt actcgcacag gctgtgccac
1260
gtcaggttcg cccagagcct caggaacgcg gagcccctgg actacgccag ggtgcccatg
1320
ttcagctgga cccacaggtc ggctaccccc accaacacca tcgacccaga cgtgatcacg
1380
cagatcccgc tcgtcaaggc ccacaccctg cagtcgggca ccaccgtggt caagggcccc
1440
ggcttcacgg gcggcgacat cctgaggagg acctccggcg gcccattcgc cttcagcaac
1500
gtcaacctcg actggaacct gtcccagcgc tacagggcgc gcatcaggta cgccagcacc
1560
acgaacctgc gcatgtatgt gaccatcgcg ggcgagagga tcttcgccgg ccagttcaac
1620
aagacgatga acaccggcga cccgctcacc ttccagtcct tcagctacgc gacgatcgac
1680
accgccttca cgttccccac gaaggcctcc agcctgaccg tgggcgccga caccttctcc
1740
agcggcaacg aggtctacgt ggaccgcttc gagctgatcc cggtgacggc gaccttcgag
1800
gccgagtacg acctggagaa ggcccagaag gcggtcaacg ccctcttcac ctccagcaac
1860
cagatcggcc tgaagacgga cgtgaccgac taccacatcg acaaggtgtc caacctcgtc
1920
gagtgcctga gcgacgagtt ctgcctcgac gagaagaggg agctgtccga gaaggtcaag
1980
cacgccaagc gcctctgcga cgagaggaac ctcctgcagg acccgaactt caggggcatc
2040
aaccgccagc cggacagggg ctggaggggc agcaccgaca tcaccatcca gggcggcgac
2100
gacgtgttca aggagaacta cgtcacgctc ccgggcacct tcgacgagtg ctaccccacg
2160
tacctgtacc agaagatcga cgagtccaag ctcaaggcct acacccgcta cgagctgagg
2220
ggctacatcg aggacagcca ggacctcgag atctacctga tccgctacaa cgcgaagcac
2280
gagacggtga acgtccccgg cacgggctcc ctgtggcccc tctcggctca gtcgccgatc
2340
ggcaagtgcg gcgagcccaa caggtgcgcc acccacctcg agtggaaccc ggacctggac
2400
tgctcctgcc gggacggcga gaagtgcgct caccactccc accacttcag cctggacatc
2460
gacgtgggct gcacggacct caacgaggac ctgggcgtgt gggtcatctt caagatcaag
2520
acgcaggacg gccacgctag gctgggcaac ctcgagttcc tggaggagaa gccgctggtg
2580
ggcgaggctc tggccagggt caagagggcg gagaagaagt ggcgcgacaa gagggagaag
2640
ctggagctgg agacgaacat cgtctacaag gaggccaaga agtccgtgga cgcgctcttc
2700
gtcaacagcc agtacgacag gctgcaggcg gacaccaaca tcgccatcat ccacgccgcg
2760
gacaagcgcg tgcactccat cagggaggcc tacctccccg agctgagcgt gatcccgggc
2820
gtcaacgctg ccatcttcga ggagctggag ggccgcatct tcaccgccta ctccctgtac
2880
gacgcgagga acgtcatcaa gaacggcgac ttcaacaacg gcctcagctg ctggaacgtg
2940
aagggccacg tggacgtcga ggagcagaac aaccaccgct cggtgctggt ggtccccgag
3000
tgggaggctg aggtcagcca ggaggtgcgc gtctgcccgg gcaggggcta catcctccgc
3060
gtgaccgcgt acaaggaggg ctacggcgag ggctgcgtca cgatccacga gatcgaggac
3120
aacaccgacg agctgaagtt ctccaactgc gtggaggagg agatctaccc gaacaacacg
3180
gtcacctgca acgactacac ggccacccag gaggagtacg agggcacgta cacgtcgagg
3240
aacaggggct acgacggcgc ttacgagtcc aacagctcgg tgccggccga ctacgctagc
3300
gcgtacgagg agaaggccta cacggacggc cgcagggaca acacctgcga gtcgaacagg
3360
ggctacggcg actacacgcc gctcccggcc ggctacgtga ccaaggagct ggagtacttc
3420
ccggagacgg acaaggtctg gatcgagatc ggcgagacgg agggcacctt catcgtggac
3480
agcgtcgagc tgctgctcat ggaggagtag
3510
<210> 6
<211> 3510
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная кодирующая последовательность маиса для
химерного белка BT29-BT22.
<400> 6
atggaaatca acaaccagaa ccagtgcgtg ccgtacaact gccttaacaa ccccgagtcc
60
gagatcctga acgtggccat cttctccagc gagcaggtcg cggagatcca cctcaagatc
120
acgcgcctga tcctcgagaa cttcctgccg ggcggctcct tcgctttcgg cctgttcgac
180
ctcatctggg gcatcttcaa cgaggaccag tggagcgcgt tcctcaggca ggtggaggag
240
ctgatcaacc agcgcatcac ggagttcgcc aggggccagg ctatccagcg gctggtgggc
300
ttcggcaggt cctacgacga atacatcctg gccctcaagg agtgggagaa cgaccccgac
360
aacccggcca gcaaggagcg cgtgaggacc cgcttcagga ccaccgacga cgctctcctg
420
acgggcgtcc ccctcatggc tatcccgggc ttcgagctgg ccaccctctc ggtgtacgct
480
cagtcggcca acctgcacct cgccctcctg cgggacgctg tgttcttcgg cgagaggtgg
540
ggcctgaccc aaaccaacat caacgacctc tactccaggc tgaagaacag catccgcgac
600
tacacgaacc actgcgtgcg cttctacaac atcggcctgg gcaacctcaa cgtcatcagg
660
ccggaatact accgcttcca gagggagctg accatcagcg tgctggacct cgtcgccctg
720
ttccccaact acgacatccg cacgtacccg atccccacca agtcccagct cacgagggag
780
atctacaccg acccgatcat ctcgccgggc gcccaggccg gctacaccct gcaggacgtc
840
ctgagggagc cccacctgat ggacttcctg aacaggctca tcatctacac cggcgagtac
900
aggggcatca ggcactgggc gggccacgag gtggagtcca gcaggacggg catgatgacc
960
aacatccgct tcccgctcta cggcaccgcg gccacggccg agccaacccg cttcatcacg
1020
ccgtccacct tccccggcct gaacctcttc tacaggaccc tgtcggctcc catcttccgc
1080
gacgagccgg gcgcgaacat catcatccgc tacaggacct ccctcgtgga gggcgtcggc
1140
ttcatccagc cgaacaacgg cgagcagctg taccgcgtga ggggcacgct ggacagcctg
1200
gaccagctcc cactggaggg cgagtccagc ctcaccgagt actcgcacag gctgtgccac
1260
gtcaggttcg cccagagcct caggaacgcg gagcccctgg actacgccag ggtgcccatg
1320
ttcagctgga cccacaggtc ggctaccccc accaacacca tcgacccaga cgtgatcacg
1380
cagatcccgc tcgtcaaggc ccacaccctg cagtcgggca ccaccgtggt caagggcccc
1440
ggcttcacgg gcggcgacat cctgaggagg acctccggcg gcccattcgc cttcagcaac
1500
gtcaacctcg actggaacct gtcccagcgc tacagggcgc gcatcaggta cgccagcacc
1560
acgaacctgc gcatgtatgt gaccatcgcg ggcgagagga tcttcgccgg ccagttcaac
1620
aagacgatga acaccggcga cccgctcacc ttccagtcct tcagctacgc gacgatcgac
1680
accgccttca cgttccccac gaaggcctcc agcctgaccg tgggcgccga caccttctcc
1740
agcggcaacg aggtctacgt ggaccgcttc gagctgatcc cggtgacggc gaccttcgag
1800
gccgagtacg acctggagaa ggcccagaag gcggtcaacg ccctcttcac ctccagcaac
1860
cagatcggcc tgaagacgga cgtgaccgac taccacatcg acaaggtgtc caacctcgtc
1920
gagtgcctga gcgacgagtt ctgcctcgac gagaagaggg agctgtccga gaaggtcaag
1980
cacgccaagc gcctctgcga cgagaggaac ctcctgcagg acccgaactt caggggcatc
2040
aaccgccagc cggacagggg ctggaggggc agcaccgaca tcaccatcca gggcggcgac
2100
gacgtgttca aggagaacta cgtcacgctc ccgggcacct tcgacgagtg ctaccccacg
2160
tacctgtacc agaagatcga cgagtccaag ctcaaggcct acacccgcta cgagctgagg
2220
ggctacatcg aggacagcca ggacctcgag atctacctga tccgctacaa cgcgaagcac
2280
gagacggtga acgtccccgg cacgggctcc ctgtggcccc tctcggctca gtcgccgatc
2340
ggcaagtgcg gcgagcccaa caggtgcgcc acccacctcg agtggaaccc ggacctggac
2400
tgctcctgcc gggacggcga gaagtgcgct caccactccc accacttcag cctggacatc
2460
gacgtgggct gcacggacct caacgaggac ctgggcgtgt gggtcatctt caagatcaag
2520
acgcaggacg gccacgctag gctgggcaac ctcgagttcc tggaggagaa gccgctggtg
2580
ggcgaggctc tggccagggt caagagggcg gagaagaagt ggcgcgacaa gagggagaag
2640
ctggagctgg agacgaacat cgtctacaag gaggccaaga agtccgtgga cgcgctcttc
2700
gtcaacagcc agtacgacag gctgcaggcg gacaccaaca tcgccatcat ccacgccgcg
2760
gacaagcgcg tgcactccat cagggaggcc tacctccccg agctgagcgt gatcccgggc
2820
gtcaacgctg ccatcttcga ggagctggag ggccgcatct tcaccgccta ctccctgtac
2880
gacgcgagga acgtcatcaa gaacggcgac ttcaacaacg gcctcagctg ctggaacgtg
2940
aagggccacg tggacgtcga ggagcagaac aaccaccgct cggtgctggt ggtccccgag
3000
tgggaggctg aggtcagcca ggaggtgcgc gtctgcccgg gcaggggcta catcctccgc
3060
gtgaccgcgt acaaggaggg ctacggcgag ggctgcgtca cgatccacga gatcgaggac
3120
aacaccgacg agctgaagtt ctccaactgc gtggaggagg agatctaccc gaacaacacg
3180
gtcacctgca acgactacac ggccacccag gaggagtacg agggcacgta cacgtcgagg
3240
aacaggggct acgacggcgc ttacgagtcc aacagctcgg tgccggccga ctacgctagc
3300
gcgtacgagg agaaggccta cacggacggc cgcagggaca acacctgcga gtcgaacagg
3360
ggctacggcg actacacgcc gctcccggcc ggctacgtga ccaaggagct ggagtacttc
3420
ccggagacgg acaaggtctg gatcgagatc ggcgagacgg agggcacctt catcgtggac
3480
tcagtcgagc tgctgctcat ggaggagtag
3510
<210> 7
<211> 3510
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная кодирующая последовательность маиса для
химерного белка Bt29-Bt22.
<400> 7
atggagatca acaaccagaa ccagtgcgtg ccgtacaact gccttaacaa ccccgagtcc
60
gagatcctga acgtggccat cttctccagc gagcaggtcg cggagatcca cctcaagatc
120
acgcgcctga tcctcgagaa cttcctgccg ggcggctcct tcgctttcgg cctgttcgac
180
ctcatctggg gcatcttcaa cgaggaccag tggagcgcgt tcctcaggca ggtggaggag
240
ctgatcaacc agcgcatcac ggagttcgcc aggggccagg ctatccagcg gctggtgggc
300
ttcggcaggt cctacgacga gtacatcctg gccctcaagg agtgggagaa cgaccccgac
360
aacccggcca gcaaggagcg cgtgaggacc cgcttcagga ccaccgacga cgctctcctg
420
acgggcgtcc ccctcatggc tatcccgggc ttcgagctgg ccaccctctc ggtgtacgct
480
cagtcggcca acctgcacct cgccctcctg cgggacgctg tgttcttcgg cgagaggtgg
540
ggcctgaccc agacgaacat caacgacctc tactccaggc tgaagaacag catccgcgac
600
tacacgaacc actgcgtgcg cttctacaac atcggcctgg gcaacctcaa cgtcatcagg
660
ccggagtact accgcttcca gagggagctg accatcagcg tgctggacct cgtcgccctg
720
ttccccaact acgacatccg cacgtacccg atcccaacca agtcccagct cacgagggag
780
atctacaccg acccgatcat ctcgccgggc gcccaggccg gctacaccct gcaggacgtc
840
ctgagggagc cccacctgat ggacttcctg aacaggctca tcatctacac cggcgagtac
900
aggggcatca ggcactgggc gggccacgag gtggagtcca gcaggacggg catgatgacc
960
aacatccgct tcccgctcta cggcaccgcg gccacggccg agccaacccg cttcatcacg
1020
ccgtccacct tccccggcct gaacctcttc tacaggaccc tgtcggctcc catcttccgc
1080
gacgagccgg gcgcgaacat catcatccgc tacaggacct ccctcgtgga gggcgtcggc
1140
ttcatccagc cgaacaacgg cgagcagctg taccgcgtga ggggcacgct ggacagcctg
1200
gaccagctcc cactggaggg cgagtccagc ctcaccgagt actcgcacag gctgtgccac
1260
gtcaggttcg cccagagcct caggaacgcg gagcccctgg actacgccag ggtgcccatg
1320
ttcagctgga cccacaggtc ggctaccccc accaacacca tcgacccaga cgtgatcacg
1380
cagatcccgc tcgtcaaggc ccacaccctg cagtcgggca ccaccgtggt caagggcccc
1440
ggcttcacgg gcggcgacat cctgaggagg acctccggcg gcccattcgc cttcagcaac
1500
gtcaacctcg actggaacct gtcccagcgc tacagggcgc gcatcaggta cgccagcacc
1560
acgaacctgc gcatgtatgt gaccatcgcg ggcgagagga tcttcgccgg ccagttcaac
1620
aagacgatga acaccggcga cccgctcacc ttccagtcct tcagctacgc gacgatcgac
1680
accgccttca cgttccccac gaaggcctcc agcctgaccg tgggcgccga caccttctcc
1740
agcggcaacg aggtctacgt ggaccgcttc gagctgatcc cggtgacggc gaccttcgag
1800
gccgagtacg acctggagaa ggcccagaag gcggtcaacg ccctcttcac ctccagcaac
1860
cagatcggcc tgaagacgga cgtgaccgac taccacatcg acaaggtgtc caacctcgtc
1920
gagtgcctga gcgacgagtt ctgcctcgac gagaagaggg agctgtccga gaaggtcaag
1980
cacgccaagc gcctctgcga cgagaggaac ctcctgcagg acccgaactt caggggaatc
2040
aaccgccagc cggacagggg ctggaggggc agcaccgaca tcaccatcca gggcggcgac
2100
gacgtgttca aggagaacta cgtcacgctc ccgggcacct tcgacgagtg ctaccccacg
2160
tacctgtacc agaagatcga cgagtccaag ctcaaggcct acacccgcta cgagctgagg
2220
ggatacatcg aggacagcca ggacctcgag atctacctga tccgctacaa cgcgaagcac
2280
gagacggtga acgtccccgg cacgggctcc ctgtggcccc tctcggctca gtcgccgatc
2340
ggcaagtgcg gcgagcccaa caggtgcgcc acccacctcg agtggaaccc ggacctggac
2400
tgctcctgcc gggacggcga gaagtgcgct caccactccc accacttcag cctggacatc
2460
gacgtgggct gcacggacct caacgaggac ctgggcgtgt gggtcatctt caaaatcaag
2520
acgcaggacg gccacgctag gctgggcaac ctcgagttcc tggaggagaa gccgctggtg
2580
ggcgaggctc tggccagggt caagagggcg gagaagaagt ggcgcgacaa gagggagaag
2640
ctggagctgg agacgaacat cgtctacaag gaggccaaga agtccgtgga cgcgctcttc
2700
gtcaacagcc agtacgacag gctgcaggcg gacaccaaca tcgccatcat ccacgccgcg
2760
gacaagcgcg tgcactccat cagggaggcc tacctccccg agctgagcgt gatcccgggc
2820
gtcaacgctg ccatcttcga ggagctggag ggccgcatct tcaccgccta ctccctgtac
2880
gacgcgagga acgtcatcaa gaacggcgac ttcaacaacg gcctcagctg ctggaacgtg
2940
aagggccacg tggacgtcga ggagcagaac aaccaccgct cggtgctggt ggtccccgag
3000
tgggaggctg aggtcagcca ggaggtgcgc gtctgcccgg gcaggggata catcctccgc
3060
gtgaccgcgt acaaggaggg ctacggcgag ggctgcgtca cgatccacga gatcgaggac
3120
aacaccgacg agctgaagtt ctccaactgc gtggaggagg agatctaccc gaacaacacg
3180
gtcacctgca acgactacac ggccacccag gaggagtacg agggcacgta cacgtcgagg
3240
aacaggggct acgacggcgc ttacgagtcc aacagctcgg tgccggccga ctacgctagc
3300
gcgtacgagg agaaggccta cacggacggc cgcagggaca acacctgcga gtcgaacagg
3360
ggctacggcg actacacgcc gctcccggcc ggctacgtga ccaaggagct ggagtacttc
3420
ccggagacgg acaaggtctg gatcgagatc ggcgagacgg agggcacctt catcgtggac
3480
tcagtcgagc tgctgctcat ggaggagtag
3510
<210> 8
<211> 1174
<212> БЕЛОК
<213> Bacillus thuringiensis
<400> 8
Met Glu Asn Asn Ile Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Val Glu Ile Leu Asn Glu Glu Arg Ser Thr Gly Arg Leu
20 25 30
Pro Leu Asp Ile Ser Leu Ser Leu Thr Arg Phe Leu Leu Ser Glu Phe
35 40 45
Val Pro Gly Val Gly Val Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Phe Ile Thr Pro Ser Asp Trp Ser Leu Phe Leu Leu Gln Ile Glu Gln
65 70 75 80
Leu Ile Glu Gln Arg Ile Glu Thr Leu Glu Arg Asn Arg Ala Ile Thr
85 90 95
Thr Leu Arg Gly Leu Ala Asp Ser Tyr Glu Ile Tyr Ile Glu Ala Leu
100 105 110
Arg Glu Trp Glu Ala Asn Pro Asn Asn Ala Gln Leu Arg Glu Asp Val
115 120 125
Arg Ile Arg Phe Ala Asn Thr Asp Asp Ala Leu Ile Thr Ala Ile Asn
130 135 140
Asn Phe Thr Leu Thr Ser Phe Glu Ile Pro Leu Leu Ser Val Tyr Val
145 150 155 160
Gln Ala Ala Asn Leu His Leu Ser Leu Leu Arg Asp Ala Val Ser Phe
165 170 175
Gly Gln Gly Trp Gly Leu Asp Ile Ala Thr Val Asn Asn His Tyr Asn
180 185 190
Arg Leu Ile Asn Leu Ile His Arg Tyr Thr Lys His Cys Leu Asp Thr
195 200 205
Tyr Asn Gln Gly Leu Glu Asn Leu Arg Gly Thr Asn Thr Arg Gln Trp
210 215 220
Ala Arg Phe Asn Gln Phe Arg Arg Asp Leu Thr Leu Thr Val Leu Asp
225 230 235 240
Ile Val Ala Leu Phe Pro Asn Tyr Asp Val Arg Thr Tyr Pro Ile Gln
245 250 255
Thr Ser Ser Gln Leu Thr Arg Glu Ile Tyr Thr Ser Ser Val Ile Glu
260 265 270
Asp Ser Pro Val Ser Ala Asn Ile Pro Asn Gly Phe Asn Arg Ala Glu
275 280 285
Phe Gly Val Arg Pro Pro His Leu Met Asp Phe Met Asn Ser Leu Phe
290 295 300
Val Thr Ala Glu Thr Val Arg Ser Gln Thr Val Trp Gly Gly His Leu
305 310 315 320
Val Ser Ser Arg Asn Thr Ala Gly Asn Arg Ile Asn Phe Pro Ser Tyr
325 330 335
Gly Val Phe Asn Pro Gly Gly Ala Ile Trp Ile Ala Asp Glu Asp Pro
340 345 350
Arg Pro Phe Tyr Arg Thr Leu Ser Asp Pro Val Phe Val Arg Gly Gly
355 360 365
Phe Gly Asn Pro His Tyr Val Leu Gly Leu Arg Gly Val Ala Phe Gln
370 375 380
Gln Thr Gly Thr Asn His Thr Arg Thr Phe Arg Asn Ser Gly Thr Ile
385 390 395 400
Asp Ser Leu Asp Glu Ile Pro Pro Gln Asp Asn Ser Gly Ala Pro Trp
405 410 415
Asn Asp Tyr Ser His Val Leu Asn His Val Thr Phe Val Arg Trp Pro
420 425 430
Gly Glu Ile Ser Gly Ser Asp Ser Trp Arg Ala Pro Met Phe Ser Trp
435 440 445
Thr His Arg Ser Ala Thr Pro Thr Asn Thr Ile Asp Pro Glu Arg Ile
450 455 460
Thr Gln Ile Pro Leu Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr
465 470 475 480
Val Val Arg Gly Pro Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr
485 490 495
Ser Gly Gly Pro Phe Ala Tyr Thr Ile Val Asn Ile Asn Gly Gln Leu
500 505 510
Pro Gln Arg Tyr Arg Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu
515 520 525
Arg Ile Tyr Val Thr Val Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe
530 535 540
Asn Lys Thr Met Asp Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser
545 550 555 560
Tyr Ala Thr Ile Asn Thr Ala Phe Thr Phe Pro Met Ser Gln Ser Ser
565 570 575
Phe Thr Val Gly Ala Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Ile
580 585 590
Asp Arg Phe Glu Leu Ile Pro Val Thr Ala Thr Phe Glu Ala Glu Tyr
595 600 605
Asp Leu Glu Arg Ala Gln Lys Ala Val Asn Ala Leu Phe Thr Ser Ile
610 615 620
Asn Gln Ile Gly Ile Lys Thr Asp Val Thr Asp Tyr His Ile Asp Gln
625 630 635 640
Val Ser Asn Leu Val Asp Cys Leu Ser Asp Glu Phe Cys Leu Asp Glu
645 650 655
Lys Arg Glu Leu Ser Glu Lys Val Lys His Ala Lys Arg Leu Ser Asp
660 665 670
Glu Arg Asn Leu Leu Gln Asp Pro Asn Phe Lys Gly Ile Asn Arg Gln
675 680 685
Leu Asp Arg Gly Trp Arg Gly Ser Thr Asp Ile Thr Ile Gln Arg Gly
690 695 700
Asp Asp Val Phe Lys Glu Asn Tyr Val Thr Leu Pro Gly Thr Phe Asp
705 710 715 720
Glu Cys Tyr Pro Thr Tyr Leu Tyr Gln Lys Ile Asp Glu Ser Lys Leu
725 730 735
Lys Pro Tyr Thr Arg Tyr Gln Leu Arg Gly Tyr Ile Glu Asp Ser Gln
740 745 750
Asp Leu Glu Ile Tyr Leu Ile Arg Tyr Asn Ala Lys His Glu Thr Val
755 760 765
Asn Val Leu Gly Thr Gly Ser Leu Trp Pro Leu Ser Val Gln Ser Pro
770 775 780
Ile Arg Lys Cys Gly Glu Pro Asn Arg Cys Ala Pro His Leu Glu Trp
785 790 795 800
Asn Pro Asp Leu Asp Cys Ser Cys Arg Asp Gly Glu Lys Cys Ala His
805 810 815
His Ser His His Phe Ser Leu Asp Ile Asp Val Gly Cys Thr Asp Leu
820 825 830
Asn Glu Asp Leu Asp Val Trp Val Ile Phe Lys Ile Lys Thr Gln Asp
835 840 845
Gly His Ala Arg Leu Gly Asn Leu Glu Phe Leu Glu Glu Lys Pro Leu
850 855 860
Val Gly Glu Ala Leu Ala Arg Val Lys Arg Ala Glu Lys Lys Trp Arg
865 870 875 880
Asp Lys Arg Glu Lys Leu Glu Leu Glu Thr Asn Ile Val Tyr Lys Glu
885 890 895
Ala Lys Glu Ser Val Asp Ala Leu Phe Val Asn Ser Gln Tyr Asp Gln
900 905 910
Leu Gln Ala Asp Thr Asn Ile Ala Met Ile His Ala Ala Asp Lys Arg
915 920 925
Val His Arg Ile Arg Glu Ala Tyr Leu Pro Glu Leu Ser Val Ile Pro
930 935 940
Gly Val Asn Val Asp Ile Phe Glu Glu Leu Lys Gly Arg Ile Phe Thr
945 950 955 960
Ala Phe Phe Leu Tyr Asp Ala Arg Asn Val Ile Lys Asn Gly Asp Phe
965 970 975
Asn Asn Gly Leu Ser Cys Trp Asn Val Lys Gly His Val Asp Val Glu
980 985 990
Glu Gln Asn Asn His Arg Ser Val Leu Val Val Pro Glu Trp Glu Ala
995 1000 1005
Glu Val Ser Gln Glu Val Arg Val Cys Pro Gly Arg Gly Tyr Ile
1010 1015 1020
Leu Arg Val Thr Ala Tyr Lys Glu Gly Tyr Gly Glu Gly Cys Val
1025 1030 1035
Thr Ile His Glu Ile Glu Asn Asn Thr Asp Glu Leu Lys Phe Ser
1040 1045 1050
Asn Cys Val Glu Glu Glu Val Tyr Pro Asn Asn Thr Val Thr Cys
1055 1060 1065
Asn Asp Tyr Thr Ala Asn Gln Glu Glu Tyr Gly Gly Ala Tyr Thr
1070 1075 1080
Ser Arg Asn Arg Gly Tyr Asp Glu Thr Tyr Gly Ser Asn Ser Ser
1085 1090 1095
Val Pro Ala Asp Tyr Ala Ser Val Tyr Glu Glu Lys Ser Tyr Thr
1100 1105 1110
Asp Gly Arg Arg Asp Asn Pro Cys Glu Ser Asn Arg Gly Tyr Gly
1115 1120 1125
Asp Tyr Thr Pro Leu Pro Ala Gly Tyr Val Thr Lys Glu Leu Glu
1130 1135 1140
Tyr Phe Pro Glu Thr Asp Lys Val Trp Ile Glu Ile Gly Glu Thr
1145 1150 1155
Glu Gly Thr Phe Ile Val Asp Ser Val Glu Leu Leu Leu Met Glu
1160 1165 1170
Glu
<210> 9
<211> 1169
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сконструированный химерный белок BT29-Cry1Fa1.
<400> 9
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Arg Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Tyr Thr Ile Val Asn Ile Asn Gly Gln Leu Pro Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Ile Tyr Val Thr
515 520 525
Val Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asp
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asn
545 550 555 560
Thr Ala Phe Thr Phe Pro Met Ser Gln Ser Ser Phe Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Ile Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Phe Glu Ala Glu Tyr Asp Leu Glu Lys Ala
595 600 605
Gln Lys Ala Val Asn Ala Leu Phe Thr Ser Ser Asn Gln Ile Gly Leu
610 615 620
Lys Thr Asp Val Thr Asp Tyr His Ile Asp Lys Val Ser Asn Leu Val
625 630 635 640
Glu Cys Leu Ser Asp Glu Phe Cys Leu Asp Glu Lys Arg Glu Leu Ser
645 650 655
Glu Lys Val Lys His Ala Lys Arg Leu Cys Asp Glu Arg Asn Leu Leu
660 665 670
Gln Asp Pro Asn Phe Arg Gly Ile Asn Arg Gln Pro Asp Arg Gly Trp
675 680 685
Arg Gly Ser Thr Asp Ile Thr Ile Gln Gly Gly Asp Asp Val Phe Lys
690 695 700
Glu Asn Tyr Val Thr Leu Pro Gly Thr Phe Asp Glu Cys Tyr Pro Thr
705 710 715 720
Tyr Leu Tyr Gln Lys Ile Asp Glu Ser Lys Leu Lys Ala Tyr Thr Arg
725 730 735
Tyr Glu Leu Arg Gly Tyr Ile Glu Asp Ser Gln Asp Leu Glu Ile Tyr
740 745 750
Leu Ile Arg Tyr Asn Ala Lys His Glu Thr Val Asn Val Pro Gly Thr
755 760 765
Gly Ser Leu Trp Pro Leu Ser Ala Gln Ser Pro Ile Gly Lys Cys Gly
770 775 780
Glu Pro Asn Arg Cys Ala Thr His Leu Glu Trp Asn Pro Asp Leu Asp
785 790 795 800
Cys Ser Cys Arg Asp Gly Glu Lys Cys Ala His His Ser His His Phe
805 810 815
Ser Leu Asp Ile Asp Val Gly Cys Thr Asp Leu Asn Glu Asp Leu Gly
820 825 830
Val Trp Val Ile Phe Lys Ile Lys Thr Gln Asp Gly His Ala Arg Leu
835 840 845
Gly Asn Leu Glu Phe Leu Glu Glu Lys Pro Leu Val Gly Glu Ala Leu
850 855 860
Ala Arg Val Lys Arg Ala Glu Lys Lys Trp Arg Asp Lys Arg Glu Lys
865 870 875 880
Leu Glu Leu Glu Thr Asn Ile Val Tyr Lys Glu Ala Lys Lys Ser Val
885 890 895
Asp Ala Leu Phe Val Asn Ser Gln Tyr Asp Arg Leu Gln Ala Asp Thr
900 905 910
Asn Ile Ala Ile Ile His Ala Ala Asp Lys Arg Val His Ser Ile Arg
915 920 925
Glu Ala Tyr Leu Pro Glu Leu Ser Val Ile Pro Gly Val Asn Ala Ala
930 935 940
Ile Phe Glu Glu Leu Glu Gly Arg Ile Phe Thr Ala Tyr Ser Leu Tyr
945 950 955 960
Asp Ala Arg Asn Val Ile Lys Asn Gly Asp Phe Asn Asn Gly Leu Ser
965 970 975
Cys Trp Asn Val Lys Gly His Val Asp Val Glu Glu Gln Asn Asn His
980 985 990
Arg Ser Val Leu Val Val Pro Glu Trp Glu Ala Glu Val Ser Gln Glu
995 1000 1005
Val Arg Val Cys Pro Gly Arg Gly Tyr Ile Leu Arg Val Thr Ala
1010 1015 1020
Tyr Lys Glu Gly Tyr Gly Glu Gly Cys Val Thr Ile His Glu Ile
1025 1030 1035
Glu Asp Asn Thr Asp Glu Leu Lys Phe Ser Asn Cys Val Glu Glu
1040 1045 1050
Glu Ile Tyr Pro Asn Asn Thr Val Thr Cys Asn Asp Tyr Thr Ala
1055 1060 1065
Thr Gln Glu Glu Tyr Glu Gly Thr Tyr Thr Ser Arg Asn Arg Gly
1070 1075 1080
Tyr Asp Gly Ala Tyr Glu Ser Asn Ser Ser Val Pro Ala Asp Tyr
1085 1090 1095
Ala Ser Ala Tyr Glu Glu Lys Ala Tyr Thr Asp Gly Arg Arg Asp
1100 1105 1110
Asn Thr Cys Glu Ser Asn Arg Gly Tyr Gly Asp Tyr Thr Pro Leu
1115 1120 1125
Pro Ala Gly Tyr Val Thr Lys Glu Leu Glu Tyr Phe Pro Glu Thr
1130 1135 1140
Asp Lys Val Trp Ile Glu Ile Gly Glu Thr Glu Gly Thr Phe Ile
1145 1150 1155
Val Asp Ser Val Glu Leu Leu Leu Met Glu Glu
1160 1165
<210> 10
<211> 3510
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодирующая последовательность BT29-Cry1Fa1.
<400> 10
atggagatta ataatcagaa ccaatgtgtc ccttataatt gtttgaataa tcctgaaagc
60
gagatattaa acgttgcaat ttttagtagc gaacaggtag cagaaattca cttaaagatc
120
acgcgcttaa ttttagagaa ttttttacca ggtgggagtt ttgcattcgg cttatttgat
180
ttaatatggg ggatttttaa tgaagatcaa tggagcgcat ttcttcggca ggtagaagaa
240
ttaattaatc aaaggataac ggaattcgca agagggcaag caattcagag actagtaggg
300
tttggaagga gttatgatga atatatttta gcactaaaag aatgggaaaa cgatcctgat
360
aacccagctt caaaggaaag agtgcgcact cgatttcgga caactgatga tgccttgcta
420
accggtgttc ctcttatggc aattccaggt tttgaattag ctactttatc tgtttatgct
480
caatcagcca atctacattt agccctatta agagatgctg tattttttgg ggagagatgg
540
ggattgacac aaacaaatat aaatgattta tatagtagat taaaaaactc cattcgtgat
600
tatacaaatc attgtgttcg tttttataat ataggtttag ggaatttaaa tgttataaga
660
ccagagtatt accgtttcca aagagaatta acaatatctg tcttagatct tgtagctctt
720
tttccaaatt acgatatccg aacatatcca ataccaacta aaagtcaatt aacaagagaa
780
atttatacag atccgattat ttcacctggt gcacaggcag gttatactct tcaagatgtt
840
ttgagagaac cacaccttat ggacttttta aaccgactta ttatttatac tggtgagtat
900
cgcggaattc gtcactgggc aggacatgaa gtagaatcta gtagaacagg tatgatgact
960
aatataagat ttcctttgta tggaacagcc gcaacagcag aaccaacacg atttataact
1020
cctagtactt ttcctggtct taatttattt tatagaacat tatcagctcc tatttttaga
1080
gatgaaccgg gagctaatat tattattaga tatagaacga gtttggtgga aggagtagga
1140
tttattcaac caaataacgg tgaacagctt tacagagtga gaggaacatt agattctctt
1200
gatcaattac cacttgaggg tgagagtagt ctaactgaat atagtcatcg attatgccat
1260
gttagatttg cgcaatcatt gaggaatgca gaacctttag attatgcaag ggttccgatg
1320
ttttcttgga cacatcgtag tgcaacccct acaaatacaa ttgatccaga tgtcatcacc
1380
caaataccgt tagtaaaagc acatacactt cagtcaggta ctactgttgt aagagggccc
1440
gggtttacgg gaggagatat tcttcgacga acaagtggag gaccatttgc ttatactatt
1500
gttaatataa atgggcaatt accccaaagg tatcgtgcaa gaatacgcta tgcctctact
1560
acaaatctaa gaatttacgt aacggttgca ggtgaacgga tttttgctgg tcaatttaac
1620
aaaacaatgg ataccggtga cccattaaca ttccaatctt ttagttacgc aactattaat
1680
acagctttta cattcccaat gagccagagt agtttcacag taggtgctga tacttttagt
1740
tcagggaatg aagtttatat agacagattt gaattgattc cagttactgc aacatttgag
1800
gcagaatatg atttagagaa agctcagaaa gcggtgaatg cgctgtttac ttcttccaat
1860
caaatcgggt taaaaacaga tgtgacggac tatcatattg ataaagtatc caatctagtt
1920
gagtgtttat cagatgaatt ttgtctagat gaaaagcgag aattgtccga gaaagtcaaa
1980
catgcgaagc gactctgtga tgagcggaat ttacttcaag atccaaactt cagaggcatc
2040
aatagacaac cagaccgtgg ttggagagga agtacggata ttaccatcca aggaggagat
2100
gacgtattca aagagaatta cgttacgcta ccgggtacct ttgatgagtg ctatccaacg
2160
tatttatatc aaaaaataga tgagtcgaaa ttaaaagcct atacccgcta tgaattaaga
2220
gggtatatcg aggatagtca agacttagaa atctatttaa ttcgctacaa tgcaaaacat
2280
gaaacagtaa atgtgccagg tacgggttcc ttatggccgc tttcagccca aagtccaatc
2340
ggaaagtgtg gagaaccgaa tcgatgtgcg acacaccttg aatggaatcc tgacttagat
2400
tgttcgtgta gggatggaga aaagtgtgcc catcattcgc atcatttctc cttagacatt
2460
gatgtaggat gtacagacct aaatgaggac ctaggtgtat gggtgatctt taagattaag
2520
acgcaagatg gtcatgcgag actaggaaat ctagaatttc tcgaagagaa accattagta
2580
ggagaagcgc tagctcgtgt gaagagagcg gagaaaaaat ggagagacaa acgcgaaaaa
2640
ttggaattgg aaacaaatat tgtttataaa gaggcaaaaa aatctgtaga tgctttattt
2700
gtgaactctc aatatgatag attacaagcg gatacgaata tcgcgataat tcatgcggca
2760
gataaacgcg ttcatagcat tcgagaagca tatcttccag agttgtctgt aattccgggt
2820
gtaaatgcag ctatttttga agaattagag ggacgtattt tcacagccta ctctctatat
2880
gatgcgagaa atgtcattaa aaatggcgat ttcaataatg gcttatcatg ctggaacgtg
2940
aaagggcatg tagatgtaga agaacagaac aaccatcgtt cggtccttgt tgttccagaa
3000
tgggaagcag aagtgtcaca agaggttcgt gtctgtccag gtcgtggcta tatccttcgt
3060
gttacagcgt acaaagaggg atatggagag ggctgtgtaa cgattcatga gatcgaagac
3120
aatacagacg aactgaaatt cagcaactgt gtagaagagg aaatatatcc aaacaacacg
3180
gtaacgtgta atgattatac tgcgactcaa gaagaatatg agggtacgta cacttctcgt
3240
aatcgaggat atgacggagc ctatgaaagc aattcttctg taccagctga ttatgcatca
3300
gcctatgaag aaaaagcgta tacagatgga agaagagaca atacttgtga atctaacaga
3360
ggatatgggg attacacacc actaccagct ggctatgtga caaaagaatt agagtacttc
3420
ccagaaaccg ataaggtatg gattgagatt ggagaaacgg aaggaacatt tatcgtggac
3480
agcgtggaat tactccttat ggaggaatag
3510
<210> 11
<211> 3510
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Оптимизированная кодирующая последовательность маиса BT29-
Cry1Fa1.
<400> 11
atggaaatca acaaccagaa ccagtgcgtg ccgtacaact gcctcaacaa ccccgagtcc
gagatcctga acgtggccat cttctccagc gagcaggtcg cggagatcca cctcaagatc
120
acgcgcctga tcctcgagaa cttcctgccg ggcggctcct tcgctttcgg cctgttcgac
180
ctcatctggg gcatcttcaa cgaggaccag tggagcgcgt tcctcaggca ggtggaggag
240
ctgatcaacc agcgcatcac ggagttcgcc aggggccagg ctatccagcg gctggtgggc
300
ttcggcaggt cctacgacga atacatcctg gccctcaagg agtgggagaa cgaccccgac
360
aacccggcca gcaaggagcg cgtgaggacc cgcttcagga ccaccgacga cgctctcctg
420
acgggcgtcc ccctcatggc tatcccgggc ttcgagctgg ccaccctctc ggtgtacgct
480
cagtcggcca acctgcacct cgccctcctg cgggacgctg tgttcttcgg cgagaggtgg
540
ggcctgaccc aaaccaacat caacgacctc tactccaggc tgaagaacag catccgcgac
600
tacacgaacc actgcgtgcg cttctacaac atcggcctgg gcaacctcaa cgtcatcagg
660
ccggaatact accgcttcca gagggagctg accatcagcg tgctggacct cgtcgccctg
720
ttccccaact acgacatccg cacgtacccg atccccacca agtcccagct cacgagggag
780
atctacaccg acccgatcat ctcgccgggc gcccaggccg gctacaccct gcaggacgtc
840
ctgagggagc cccacctgat ggacttcctg aacaggctca tcatctacac cggcgagtac
900
aggggcatca ggcactgggc gggccacgag gtggagtcca gcaggacggg catgatgacc
960
aacatccgct tcccgctcta cggcaccgcg gccacggccg agccaacccg cttcatcacg
1020
ccgtccacct tccccggcct gaacctcttc tacaggaccc tgtcggctcc catcttccgc
1080
gacgagccgg gcgcgaacat catcatccgc tacaggacct ccctcgtgga gggcgtcggc
1140
ttcatccagc cgaacaacgg cgagcagctg taccgcgtga ggggcacgct ggacagcctg
1200
gaccagctcc cactggaggg cgagtccagc ctcaccgagt actcgcacag gctgtgccac
1260
gtcaggttcg cccagagcct caggaacgcg gagcccctgg actacgccag ggtgcccatg
1320
ttcagctgga cccacaggtc ggctaccccc accaacacca tcgacccaga cgtgatcacg
1380
cagatcccgc tcgtcaaggc ccacaccctg cagtcgggca ccaccgtggt ccgcggccca
1440
ggcttcaccg gcggcgacat cctgaggagg acgagcggcg gccccttcgc ttacaccatc
1500
gtcaacatca acggccagct gccacagagg tacagggcgc gcatcaggta cgcctccacc
1560
acgaacctgc gcatctacgt gaccgtggcg ggcgagagga tcttcgccgg ccagttcaac
1620
aagacgatgg acaccggcga cccgctgacc ttccagtcct tcagctacgc gacgatcaac
1680
accgccttca cgttccccat gagccagtcc agcttcaccg tgggcgccga cacgttctcc
1740
agcggcaacg aggtgtacat cgaccgcttc gagctgatcc cggtgacggc gaccttcgag
1800
gccgagtacg acctggagaa ggcccagaag gcggtcaacg ccctcttcac ctccagcaac
1860
cagatcggcc tgaagacgga cgtgaccgac taccacatcg acaaggtgtc caacctcgtc
1920
gagtgcctga gcgacgagtt ctgcctcgac gagaagaggg agctgtccga gaaggtcaag
1980
cacgccaagc gcctctgcga cgagaggaac ctcctgcagg acccgaactt caggggcatc
2040
aaccgccagc cggacagggg ctggaggggc agcaccgaca tcaccatcca gggcggcgac
2100
gacgtgttca aggagaacta cgtcacgctc ccgggcacct tcgacgagtg ctaccccacg
2160
tacctgtacc agaagatcga cgagtccaag ctcaaggcct acacccgcta cgagctgagg
2220
ggctacatcg aggacagcca ggacctcgag atctacctga tccgctacaa cgcgaagcac
2280
gagacggtga acgtccccgg cacgggctcc ctgtggcccc tctcggctca gtcgccgatc
2340
ggcaagtgcg gcgagcccaa caggtgcgcc acccacctcg agtggaaccc ggacctggac
2400
tgctcctgcc gggacggcga gaagtgcgct caccactccc accacttcag cctggacatc
2460
gacgtgggct gcacggacct caacgaggac ctgggcgtgt gggtcatctt caagatcaag
2520
acgcaggacg gccacgctag gctgggcaac ctcgagttcc tggaggagaa gccgctggtg
2580
ggcgaggctc tggccagggt caagagggcg gagaagaagt ggcgcgacaa gagggagaag
2640
ctggagctgg agacgaacat cgtctacaag gaggccaaga agtccgtgga cgcgctcttc
2700
gtcaacagcc agtacgacag gctgcaggcg gacaccaaca tcgccatcat ccacgccgcg
2760
gacaagcgcg tgcactccat cagggaggcc tacctccccg agctgagcgt gatcccgggc
2820
gtcaacgctg ccatcttcga ggagctggag ggccgcatct tcaccgccta ctccctgtac
2880
gacgcgagga acgtcatcaa gaacggcgac ttcaacaacg gcctcagctg ctggaacgtg
2940
aagggccacg tggacgtcga ggagcagaac aaccaccgct cggtgctggt ggtccccgag
3000
tgggaggctg aggtcagcca ggaggtgcgc gtctgcccgg gcaggggcta catcctccgc
3060
gtgaccgcgt acaaggaggg ctacggcgag ggctgcgtca cgatccacga gatcgaggac
3120
aacaccgacg agctgaagtt ctccaactgc gtggaggagg agatctaccc gaacaacacg
3180
gtcacctgca acgactacac ggccacccag gaggagtacg agggcacgta cacgtcgagg
3240
aacaggggct acgacggcgc ttacgagtcc aacagctcgg tgccggccga ctacgctagc
3300
gcgtacgagg agaaggccta cacggacggc cgcagggaca acacctgcga gtcgaacagg
3360
ggctacggcg actacacgcc gctcccggcc ggctacgtga ccaaggagct ggagtacttc
3420
ccggagacgg acaaggtctg gatcgagatc ggcgagacgg agggcacctt catcgtggac
3480
agcgtcgagc tgctgctcat ggaggagtag
3510
<210> 12
<211> 1215
<212> БЕЛОК
<213> Bacillus thuringiensis
<400> 12
Met Asn Ser Asn Arg Lys Asn Glu Asn Glu Ile Ile Asn Ala Leu Ser
1 5 10 15
Ile Pro Ala Val Ser Asn His Ser Ala Gln Met Asp Leu Ser Pro Asp
20 25 30
Ala Arg Ile Glu Asp Ser Leu Cys Val Ala Glu Gly Asn Asn Ile Asp
35 40 45
Pro Phe Val Ser Ala Ser Thr Val Gln Thr Gly Ile Ser Ile Ala Gly
50 55 60
Arg Ile Leu Gly Val Leu Gly Val Pro Phe Ala Gly Gln Leu Ala Ser
65 70 75 80
Phe Tyr Ser Phe Leu Val Gly Glu Leu Trp Pro Ser Gly Arg Asp Pro
85 90 95
Trp Glu Ile Phe Met Glu His Val Glu Gln Ile Val Arg Gln Gln Gln
100 105 110
Ile Thr Asp Ser Val Arg Asp Thr Ala Ile Ala Arg Leu Glu Gly Leu
115 120 125
Gly Arg Gly Tyr Arg Ser Tyr Gln Gln Ala Leu Glu Thr Trp Leu Asp
130 135 140
Asn Arg Asn Asp Ala Arg Ser Arg Ser Ile Ile Arg Glu Arg Tyr Ile
145 150 155 160
Ala Leu Glu Leu Asp Ile Thr Thr Ala Ile Pro Leu Phe Ser Ile Arg
165 170 175
Asn Glu Glu Val Pro Leu Leu Met Val Tyr Ala Gln Ala Ala Asn Leu
180 185 190
His Leu Leu Leu Leu Arg Asp Ala Ser Leu Phe Gly Ser Glu Trp Gly
195 200 205
Met Ser Ser Ala Asp Val Asn Gln Tyr Tyr Gln Glu Gln Ile Arg Tyr
210 215 220
Thr Glu Glu Tyr Ser Asn His Cys Val Gln Trp Tyr Asn Thr Gly Leu
225 230 235 240
Asn Arg Leu Arg Gly Thr Thr Ala Glu Thr Trp Val Arg Tyr Asn Gln
245 250 255
Phe Arg Arg Asp Leu Thr Leu Gly Val Leu Asp Leu Val Ala Leu Phe
260 265 270
Pro Ser Tyr Asp Thr Arg Thr Tyr Pro Ile Pro Thr Thr Ala Gln Leu
275 280 285
Thr Arg Glu Val Tyr Thr Asp Pro Asn Gly Val Val Ala Gly Pro Asn
290 295 300
Asn Ser Trp Phe Arg Asn Gly Ala Ser Phe Ser Ala Ile Glu Asn Ala
305 310 315 320
Ile Ile Arg Gln Pro His Leu Tyr Asp Phe Leu Thr Asn Leu Thr Ile
325 330 335
Tyr Thr Arg Arg Ser Gln Val Gly Thr Thr Ile Met Asn Leu Trp Ala
340 345 350
Gly His Arg Ile Thr Phe Asn Arg Ile Gln Gly Gly Ser Thr Ser Glu
355 360 365
Met Val Tyr Gly Ala Ile Thr Asn Pro Val Ser Val Ser Asp Ile Pro
370 375 380
Phe Val Asn Arg Asp Val Tyr Arg Thr Val Ser Leu Ala Gly Gly Leu
385 390 395 400
Gly Ser Leu Ser Gly Ile Arg Tyr Gly Leu Thr Arg Val Asp Phe Asp
405 410 415
Met Ile Phe Arg Asn His Pro Asp Ile Val Thr Gly Leu Phe Tyr His
420 425 430
Pro Gly His Ala Gly Ile Ala Thr Gln Val Lys Asp Ser Asp Thr Glu
435 440 445
Leu Pro Pro Glu Thr Thr Glu Gln Pro Asn Tyr Arg Ala Phe Ser His
450 455 460
Leu Leu Ser His Ile Ser Met Gly Pro Thr Thr Gln Asp Val Pro Pro
465 470 475 480
Val Tyr Ser Trp Thr His Gln Ser Ala Asp Arg Thr Asn Thr Ile Asn
485 490 495
Ser Asp Arg Ile Thr Gln Ile Pro Leu Val Lys Ala His Thr Leu Gln
500 505 510
Ser Gly Thr Thr Val Val Lys Gly Pro Gly Phe Thr Gly Gly Asp Ile
515 520 525
Leu Arg Arg Thr Ser Gly Gly Pro Phe Ala Phe Ser Asn Val Asn Leu
530 535 540
Asp Phe Asn Leu Ser Gln Arg Tyr Arg Ala Arg Ile Arg Tyr Ala Ser
545 550 555 560
Thr Thr Asn Leu Arg Ile Tyr Val Thr Val Ala Gly Glu Arg Ile Phe
565 570 575
Ala Gly Gln Phe Asp Lys Thr Met Asp Ala Gly Ala Pro Leu Thr Phe
580 585 590
Gln Ser Phe Ser Tyr Ala Thr Ile Asn Thr Ala Phe Thr Phe Pro Glu
595 600 605
Arg Ser Ser Ser Leu Thr Ile Gly Ala Asp Thr Phe Ser Ser Gly Asn
610 615 620
Glu Val Tyr Val Asp Arg Phe Glu Leu Ile Gln Val Thr Ala Thr Phe
625 630 635 640
Glu Ala Glu Ser Asp Leu Glu Arg Ala Arg Lys Ala Val Asn Ala Leu
645 650 655
Phe Thr Ser Thr Asn Pro Arg Gly Leu Lys Thr Asp Val Thr Asp Tyr
660 665 670
His Ile Asp Gln Val Ser Asn Leu Val Glu Cys Leu Ser Asp Glu Phe
675 680 685
Cys Leu Asp Lys Lys Arg Glu Leu Leu Glu Glu Val Lys Tyr Ala Lys
690 695 700
Arg Leu Ser Asp Glu Arg Asn Leu Leu Gln Asp Pro Thr Phe Thr Ser
705 710 715 720
Ile Ser Gly Gln Thr Asp Arg Gly Trp Ile Gly Ser Thr Gly Ile Ser
725 730 735
Ile Gln Gly Gly Asp Asp Ile Phe Lys Glu Asn Tyr Val Arg Leu Pro
740 745 750
Gly Thr Val Asp Glu Cys Tyr Pro Thr Tyr Leu Tyr Gln Lys Ile Asp
755 760 765
Glu Ser Gln Leu Lys Ser Tyr Thr Arg Tyr Gln Leu Arg Gly Tyr Ile
770 775 780
Glu Asp Ser Gln Asp Leu Glu Ile Tyr Leu Ile Arg Tyr Asn Ala Lys
785 790 795 800
His Glu Thr Leu Ser Val Pro Gly Thr Glu Ser Pro Trp Pro Ser Ser
805 810 815
Gly Val Tyr Pro Ser Gly Arg Cys Gly Glu Pro Asn Arg Cys Ala Pro
820 825 830
Arg Ile Glu Trp Asn Pro Asp Leu Asp Cys Ser Cys Arg Tyr Gly Glu
835 840 845
Lys Cys Val His His Ser His His Phe Ser Leu Asp Ile Asp Val Gly
850 855 860
Cys Thr Asp Leu Asn Glu Asp Leu Gly Val Trp Val Ile Phe Lys Ile
865 870 875 880
Lys Thr Gln Asp Gly His Ala Lys Leu Gly Asn Leu Glu Phe Ile Glu
885 890 895
Glu Lys Pro Leu Leu Gly Lys Ala Leu Ser Arg Val Lys Arg Ala Glu
900 905 910
Lys Lys Trp Arg Asp Lys Tyr Glu Lys Leu Gln Leu Glu Thr Lys Arg
915 920 925
Val Tyr Thr Glu Ala Lys Glu Ser Val Asp Ala Leu Phe Val Asp Ser
930 935 940
Gln Tyr Asp Lys Leu Gln Ala Asn Thr Asn Ile Gly Ile Ile His Gly
945 950 955 960
Ala Asp Lys Gln Val His Arg Ile Arg Glu Pro Tyr Leu Ser Glu Leu
965 970 975
Pro Val Ile Pro Ser Ile Asn Ala Ala Ile Phe Glu Glu Leu Glu Gly
980 985 990
His Ile Phe Lys Ala Tyr Ser Leu Tyr Asp Ala Arg Asn Val Ile Lys
995 1000 1005
Asn Gly Asp Phe Asn Asn Gly Leu Ser Cys Trp Asn Val Lys Gly
1010 1015 1020
His Val Asp Val Gln Gln Asn His His Arg Ser Val Leu Val Leu
1025 1030 1035
Ser Glu Trp Glu Ala Glu Val Ser Gln Lys Val Arg Val Cys Pro
1040 1045 1050
Asp Arg Gly Tyr Ile Leu Arg Val Thr Ala Tyr Lys Glu Gly Tyr
1055 1060 1065
Gly Glu Gly Cys Val Thr Ile His Glu Phe Glu Asp Asn Thr Asp
1070 1075 1080
Val Leu Lys Phe Arg Asn Phe Val Glu Glu Glu Val Tyr Pro Asn
1085 1090 1095
Asn Thr Val Thr Cys Asn Asp Tyr Thr Thr Asn Gln Ser Ala Glu
1100 1105 1110
Gly Ser Thr Asp Ala Cys Asn Ser Tyr Asn Arg Gly Tyr Glu Asp
1115 1120 1125
Gly Tyr Glu Asn Arg Tyr Glu Pro Asn Pro Ser Ala Pro Val Asn
1130 1135 1140
Tyr Thr Pro Thr Tyr Glu Glu Gly Met Tyr Thr Asp Thr Gln Gly
1145 1150 1155
Tyr Asn His Cys Val Ser Asp Arg Gly Tyr Arg Asn His Thr Pro
1160 1165 1170
Leu Pro Ala Gly Tyr Val Thr Leu Glu Leu Glu Tyr Phe Pro Glu
1175 1180 1185
Thr Glu Gln Val Trp Ile Glu Ile Gly Glu Thr Glu Gly Thr Phe
1190 1195 1200
Ile Val Gly Ser Val Glu Leu Leu Leu Met Glu Glu
1205 1210 1215
<210> 13
<211> 1169
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сконструированный химерный белок BT29-Cry1Ka1.
<400> 13
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Phe Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Ile Tyr Val Thr
515 520 525
Val Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asp Lys Thr Met Asp
530 535 540
Ala Gly Ala Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asn
545 550 555 560
Thr Ala Phe Thr Phe Pro Glu Arg Ser Ser Ser Leu Thr Ile Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Gln Val Thr Ala Thr Phe Glu Ala Glu Tyr Asp Leu Glu Lys Ala
595 600 605
Gln Lys Ala Val Asn Ala Leu Phe Thr Ser Ser Asn Gln Ile Gly Leu
610 615 620
Lys Thr Asp Val Thr Asp Tyr His Ile Asp Lys Val Ser Asn Leu Val
625 630 635 640
Glu Cys Leu Ser Asp Glu Phe Cys Leu Asp Glu Lys Arg Glu Leu Ser
645 650 655
Glu Lys Val Lys His Ala Lys Arg Leu Cys Asp Glu Arg Asn Leu Leu
660 665 670
Gln Asp Pro Asn Phe Arg Gly Ile Asn Arg Gln Pro Asp Arg Gly Trp
675 680 685
Arg Gly Ser Thr Asp Ile Thr Ile Gln Gly Gly Asp Asp Val Phe Lys
690 695 700
Glu Asn Tyr Val Thr Leu Pro Gly Thr Phe Asp Glu Cys Tyr Pro Thr
705 710 715 720
Tyr Leu Tyr Gln Lys Ile Asp Glu Ser Lys Leu Lys Ala Tyr Thr Arg
725 730 735
Tyr Glu Leu Arg Gly Tyr Ile Glu Asp Ser Gln Asp Leu Glu Ile Tyr
740 745 750
Leu Ile Arg Tyr Asn Ala Lys His Glu Thr Val Asn Val Pro Gly Thr
755 760 765
Gly Ser Leu Trp Pro Leu Ser Ala Gln Ser Pro Ile Gly Lys Cys Gly
770 775 780
Glu Pro Asn Arg Cys Ala Thr His Leu Glu Trp Asn Pro Asp Leu Asp
785 790 795 800
Cys Ser Cys Arg Asp Gly Glu Lys Cys Ala His His Ser His His Phe
805 810 815
Ser Leu Asp Ile Asp Val Gly Cys Thr Asp Leu Asn Glu Asp Leu Gly
820 825 830
Val Trp Val Ile Phe Lys Ile Lys Thr Gln Asp Gly His Ala Arg Leu
835 840 845
Gly Asn Leu Glu Phe Leu Glu Glu Lys Pro Leu Val Gly Glu Ala Leu
850 855 860
Ala Arg Val Lys Arg Ala Glu Lys Lys Trp Arg Asp Lys Arg Glu Lys
865 870 875 880
Leu Glu Leu Glu Thr Asn Ile Val Tyr Lys Glu Ala Lys Lys Ser Val
885 890 895
Asp Ala Leu Phe Val Asn Ser Gln Tyr Asp Arg Leu Gln Ala Asp Thr
900 905 910
Asn Ile Ala Ile Ile His Ala Ala Asp Lys Arg Val His Ser Ile Arg
915 920 925
Glu Ala Tyr Leu Pro Glu Leu Ser Val Ile Pro Gly Val Asn Ala Ala
930 935 940
Ile Phe Glu Glu Leu Glu Gly Arg Ile Phe Thr Ala Tyr Ser Leu Tyr
945 950 955 960
Asp Ala Arg Asn Val Ile Lys Asn Gly Asp Phe Asn Asn Gly Leu Ser
965 970 975
Cys Trp Asn Val Lys Gly His Val Asp Val Glu Glu Gln Asn Asn His
980 985 990
Arg Ser Val Leu Val Val Pro Glu Trp Glu Ala Glu Val Ser Gln Glu
995 1000 1005
Val Arg Val Cys Pro Gly Arg Gly Tyr Ile Leu Arg Val Thr Ala
1010 1015 1020
Tyr Lys Glu Gly Tyr Gly Glu Gly Cys Val Thr Ile His Glu Ile
1025 1030 1035
Glu Asp Asn Thr Asp Glu Leu Lys Phe Ser Asn Cys Val Glu Glu
1040 1045 1050
Glu Ile Tyr Pro Asn Asn Thr Val Thr Cys Asn Asp Tyr Thr Ala
1055 1060 1065
Thr Gln Glu Glu Tyr Glu Gly Thr Tyr Thr Ser Arg Asn Arg Gly
1070 1075 1080
Tyr Asp Gly Ala Tyr Glu Ser Asn Ser Ser Val Pro Ala Asp Tyr
1085 1090 1095
Ala Ser Ala Tyr Glu Glu Lys Ala Tyr Thr Asp Gly Arg Arg Asp
1100 1105 1110
Asn Thr Cys Glu Ser Asn Arg Gly Tyr Gly Asp Tyr Thr Pro Leu
1115 1120 1125
Pro Ala Gly Tyr Val Thr Lys Glu Leu Glu Tyr Phe Pro Glu Thr
1130 1135 1140
Asp Lys Val Trp Ile Glu Ile Gly Glu Thr Glu Gly Thr Phe Ile
1145 1150 1155
Val Asp Ser Val Glu Leu Leu Leu Met Glu Glu
1160 1165
<210> 14
<211> 3510
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодирующая последовательность BT29-Cry1Ka1.
<400> 14
atggagatta ataatcagaa ccaatgtgtc ccttataatt gtttgaataa tcctgaaagc
60
gagatattaa acgttgcaat ttttagtagc gaacaggtag cagaaattca cttaaagatc
120
acgcgcttaa ttttagagaa ttttttacca ggtgggagtt ttgcattcgg cttatttgat
180
ttaatatggg ggatttttaa tgaagatcaa tggagcgcat ttcttcggca ggtagaagaa
240
ttaattaatc aaaggataac ggaattcgca agagggcaag caattcagag actagtaggg
300
tttggaagga gttatgatga atatatttta gcactaaaag aatgggaaaa cgatcctgat
360
aacccagctt caaaggaaag agtgcgcact cgatttcgga caactgatga tgccttgcta
420
accggtgttc ctcttatggc aattccaggt tttgaattag ctactttatc tgtttatgct
480
caatcagcca atctacattt agccctatta agagatgctg tattttttgg ggagagatgg
540
ggattgacac aaacaaatat aaatgattta tatagtagat taaaaaactc cattcgtgat
600
tatacaaatc attgtgttcg tttttataat ataggtttag ggaatttaaa tgttataaga
660
ccagagtatt accgtttcca aagagaatta acaatatctg tcttagatct tgtagctctt
720
tttccaaatt acgatatccg aacatatcca ataccaacta aaagtcaatt aacaagagaa
780
atttatacag atccgattat ttcacctggt gcacaggcag gttatactct tcaagatgtt
840
ttgagagaac cacaccttat ggacttttta aaccgactta ttatttatac tggtgagtat
900
cgcggaattc gtcactgggc aggacatgaa gtagaatcta gtagaacagg tatgatgact
960
aatataagat ttcctttgta tggaacagcc gcaacagcag aaccaacacg atttataact
1020
cctagtactt ttcctggtct taatttattt tatagaacat tatcagctcc tatttttaga
1080
gatgaaccgg gagctaatat tattattaga tatagaacga gtttggtgga aggagtagga
1140
tttattcaac caaataacgg tgaacagctt tacagagtga gaggaacatt agattctctt
1200
gatcaattac cacttgaggg tgagagtagt ctaactgaat atagtcatcg attatgccat
1260
gttagatttg cgcaatcatt gaggaatgca gaacctttag attatgcaag ggttccgatg
1320
ttttcttgga cacatcgtag tgcaacccct acaaatacaa ttgatccaga tgtcatcacc
1380
caaataccgt tagtaaaggc gcataccctc caatcgggta caactgtagt aaaagggcca
1440
gggtttacag gaggggatat cctccgtcga acaagtggag gaccatttgc ttttagtaat
1500
gttaatctag attttaactt gtcacaaagg tatcgtgcta gaattcgtta tgcctctact
1560
actaacctaa gaatttacgt aacggttgca ggtgaacgaa tttttgctgg tcaatttgac
1620
aaaacgatgg atgctggtgc cccattaaca ttccaatctt ttagttacgc aactattaat
1680
acagctttta cattcccaga aagatcgagc agcttgacta taggtgccga tacgtttagt
1740
tcaggtaatg aagtttatgt agatagattt gaattaatcc aagttactgc aacatttgag
1800
gcagaatatg atttagagaa agctcagaaa gcggtgaatg cgctgtttac ttcttccaat
1860
caaatcgggt taaaaacaga tgtgacggac tatcatattg ataaagtatc caatctagtt
1920
gagtgtttat cagatgaatt ttgtctagat gaaaagcgag aattgtccga gaaagtcaaa
1980
catgcgaagc gactctgtga tgagcggaat ttacttcaag atccaaactt cagaggcatc
2040
aatagacaac cagaccgtgg ttggagagga agtacggata ttaccatcca aggaggagat
2100
gacgtattca aagagaatta cgttacgcta ccgggtacct ttgatgagtg ctatccaacg
2160
tatttatatc aaaaaataga tgagtcgaaa ttaaaagcct atacccgcta tgaattaaga
2220
gggtatatcg aggatagtca agacttagaa atctatttaa ttcgctacaa tgcaaaacat
2280
gaaacagtaa atgtgccagg tacgggttcc ttatggccgc tttcagccca aagtccaatc
2340
ggaaagtgtg gagaaccgaa tcgatgtgcg acacaccttg aatggaatcc tgacttagat
2400
tgttcgtgta gggatggaga aaagtgtgcc catcattcgc atcatttctc cttagacatt
2460
gatgtaggat gtacagacct aaatgaggac ctaggtgtat gggtgatctt taagattaag
2520
acgcaagatg gtcatgcgag actaggaaat ctagaatttc tcgaagagaa accattagta
2580
ggagaagcgc tagctcgtgt gaagagagcg gagaaaaaat ggagagacaa acgcgaaaaa
2640
ttggaattgg aaacaaatat tgtttataaa gaggcaaaaa aatctgtaga tgctttattt
2700
gtgaactctc aatatgatag attacaagcg gatacgaata tcgcgataat tcatgcggca
2760
gataaacgcg ttcatagcat tcgagaagca tatcttccag agttgtctgt aattccgggt
2820
gtaaatgcag ctatttttga agaattagag ggacgtattt tcacagccta ctctctatat
2880
gatgcgagaa atgtcattaa aaatggcgat ttcaataatg gcttatcatg ctggaacgtg
2940
aaagggcatg tagatgtaga agaacagaac aaccatcgtt cggtccttgt tgttccagaa
3000
tgggaagcag aagtgtcaca agaggttcgt gtctgtccag gtcgtggcta tatccttcgt
3060
gttacagcgt acaaagaggg atatggagag ggctgtgtaa cgattcatga gatcgaagac
3120
aatacagacg aactgaaatt cagcaactgt gtagaagagg aaatatatcc aaacaacacg
3180
gtaacgtgta atgattatac tgcgactcaa gaagaatatg agggtacgta cacttctcgt
3240
aatcgaggat atgacggagc ctatgaaagc aattcttctg taccagctga ttatgcatca
3300
gcctatgaag aaaaagcgta tacagatgga agaagagaca atacttgtga atctaacaga
3360
ggatatgggg attacacacc actaccagct ggctatgtga caaaagaatt agagtacttc
3420
ccagaaaccg ataaggtatg gattgagatt ggagaaacgg aaggaacatt tatcgtggac
3480
agcgtggaat tactccttat ggaggaatag
3510
<210> 15
<211> 1169
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Химерный белок Bt29-1Kav2
<400> 15
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Phe Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Ile Tyr Val Thr
515 520 525
Val Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asp Lys Thr Met Asp
530 535 540
Ala Gly Ala Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asn
545 550 555 560
Thr Ala Phe Thr Phe Pro Glu Arg Ser Ser Ser Leu Thr Ile Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Gln Val Thr Ala Thr Phe Glu Ala Glu Ser Asp Leu Glu Arg Ala
595 600 605
Arg Lys Ala Val Asn Ala Leu Phe Thr Ser Ser Asn Gln Ile Gly Leu
610 615 620
Lys Thr Asp Val Thr Asp Tyr His Ile Asp Lys Val Ser Asn Leu Val
625 630 635 640
Glu Cys Leu Ser Asp Glu Phe Cys Leu Asp Glu Lys Arg Glu Leu Ser
645 650 655
Glu Lys Val Lys His Ala Lys Arg Leu Cys Asp Glu Arg Asn Leu Leu
660 665 670
Gln Asp Pro Asn Phe Arg Gly Ile Asn Arg Gln Pro Asp Arg Gly Trp
675 680 685
Arg Gly Ser Thr Asp Ile Thr Ile Gln Gly Gly Asp Asp Val Phe Lys
690 695 700
Glu Asn Tyr Val Thr Leu Pro Gly Thr Phe Asp Glu Cys Tyr Pro Thr
705 710 715 720
Tyr Leu Tyr Gln Lys Ile Asp Glu Ser Lys Leu Lys Ala Tyr Thr Arg
725 730 735
Tyr Glu Leu Arg Gly Tyr Ile Glu Asp Ser Gln Asp Leu Glu Ile Tyr
740 745 750
Leu Ile Arg Tyr Asn Ala Lys His Glu Thr Val Asn Val Pro Gly Thr
755 760 765
Gly Ser Leu Trp Pro Leu Ser Ala Gln Ser Pro Ile Gly Lys Cys Gly
770 775 780
Glu Pro Asn Arg Cys Ala Thr His Leu Glu Trp Asn Pro Asp Leu Asp
785 790 795 800
Cys Ser Cys Arg Asp Gly Glu Lys Cys Ala His His Ser His His Phe
805 810 815
Ser Leu Asp Ile Asp Val Gly Cys Thr Asp Leu Asn Glu Asp Leu Gly
820 825 830
Val Trp Val Ile Phe Lys Ile Lys Thr Gln Asp Gly His Ala Arg Leu
835 840 845
Gly Asn Leu Glu Phe Leu Glu Glu Lys Pro Leu Val Gly Glu Ala Leu
850 855 860
Ala Arg Val Lys Arg Ala Glu Lys Lys Trp Arg Asp Lys Arg Glu Lys
865 870 875 880
Leu Glu Leu Glu Thr Asn Ile Val Tyr Lys Glu Ala Lys Lys Ser Val
885 890 895
Asp Ala Leu Phe Val Asn Ser Gln Tyr Asp Arg Leu Gln Ala Asp Thr
900 905 910
Asn Ile Ala Ile Ile His Ala Ala Asp Lys Arg Val His Ser Ile Arg
915 920 925
Glu Ala Tyr Leu Pro Glu Leu Ser Val Ile Pro Gly Val Asn Ala Ala
930 935 940
Ile Phe Glu Glu Leu Glu Gly Arg Ile Phe Thr Ala Tyr Ser Leu Tyr
945 950 955 960
Asp Ala Arg Asn Val Ile Lys Asn Gly Asp Phe Asn Asn Gly Leu Ser
965 970 975
Cys Trp Asn Val Lys Gly His Val Asp Val Glu Glu Gln Asn Asn His
980 985 990
Arg Ser Val Leu Val Val Pro Glu Trp Glu Ala Glu Val Ser Gln Glu
995 1000 1005
Val Arg Val Cys Pro Gly Arg Gly Tyr Ile Leu Arg Val Thr Ala
1010 1015 1020
Tyr Lys Glu Gly Tyr Gly Glu Gly Cys Val Thr Ile His Glu Ile
1025 1030 1035
Glu Asp Asn Thr Asp Glu Leu Lys Phe Ser Asn Cys Val Glu Glu
1040 1045 1050
Glu Ile Tyr Pro Asn Asn Thr Val Thr Cys Asn Asp Tyr Thr Ala
1055 1060 1065
Thr Gln Glu Glu Tyr Glu Gly Thr Tyr Thr Ser Arg Asn Arg Gly
1070 1075 1080
Tyr Asp Gly Ala Tyr Glu Ser Asn Ser Ser Val Pro Ala Asp Tyr
1085 1090 1095
Ala Ser Ala Tyr Glu Glu Lys Ala Tyr Thr Asp Gly Arg Arg Asp
1100 1105 1110
Asn Thr Cys Glu Ser Asn Arg Gly Tyr Gly Asp Tyr Thr Pro Leu
1115 1120 1125
Pro Ala Gly Tyr Val Thr Lys Glu Leu Glu Tyr Phe Pro Glu Thr
1130 1135 1140
Asp Lys Val Trp Ile Glu Ile Gly Glu Thr Glu Gly Thr Phe Ile
1145 1150 1155
Val Asp Ser Val Glu Leu Leu Leu Met Glu Glu
1160 1165
<210> 16
<211> 3510
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Химера Bt29-1Ka
<400> 16
atggagatta ataatcagaa ccaatgtgtc ccttataatt gtttgaataa tcctgaaagc
60
gagatattaa acgttgcaat ttttagtagc gaacaggtag cagaaattca cttaaagatc
120
acgcgcttaa ttttagagaa ttttttacca ggtgggagtt ttgcattcgg cttatttgat
180
ttaatatggg ggatttttaa tgaagatcaa tggagcgcat ttcttcggca ggtagaagaa
240
ttaattaatc aaaggataac ggaattcgca agagggcaag caattcagag actagtaggg
300
tttggaagga gttatgatga atatatttta gcactaaaag aatgggaaaa cgatcctgat
360
aacccagctt caaaggaaag agtgcgcact cgatttcgga caactgatga tgccttgcta
420
accggtgttc ctcttatggc aattccaggt tttgaattag ctactttatc tgtttatgct
480
caatcagcca atctacattt agccctatta agagatgctg tattttttgg ggagagatgg
540
ggattgacac aaacaaatat aaatgattta tatagtagat taaaaaactc cattcgtgat
600
tatacaaatc attgtgttcg tttttataat ataggtttag ggaatttaaa tgttataaga
660
ccagagtatt accgtttcca aagagaatta acaatatctg tcttagatct tgtagctctt
720
tttccaaatt acgatatccg aacatatcca ataccaacta aaagtcaatt aacaagagaa
780
atttatacag atccgattat ttcacctggt gcacaggcag gttatactct tcaagatgtt
840
ttgagagaac cacaccttat ggacttttta aaccgactta ttatttatac tggtgagtat
900
cgcggaattc gtcactgggc aggacatgaa gtagaatcta gtagaacagg tatgatgact
960
aatataagat ttcctttgta tggaacagcc gcaacagcag aaccaacacg atttataact
1020
cctagtactt ttcctggtct taatttattt tatagaacat tatcagctcc tatttttaga
1080
gatgaaccgg gagctaatat tattattaga tatagaacga gtttggtgga aggagtagga
1140
tttattcaac caaataacgg tgaacagctt tacagagtga gaggaacatt agattctctt
1200
gatcaattac cacttgaggg tgagagtagt ctaactgaat atagtcatcg attatgccat
1260
gttagatttg cgcaatcatt gaggaatgca gaacctttag attatgcaag ggttccgatg
1320
ttttcttgga cacatcgtag tgcaacccct acaaatacaa ttgatccaga tgtcatcacc
1380
caaataccgt tagtaaaagc acataccctt cagtcaggta ctactgttgt aaaagggcca
1440
gggtttacag gtggagatat cctccgacga actagtggag gaccatttgc ttttagtaat
1500
gttaatttag actttaactt gtcacaaaga tatcgtgcta gaatacgcta tgcttctact
1560
actaatctaa gaatttacgt aacggtagca ggggaacgaa tttttgctgg tcaatttgat
1620
aaaacaatgg atgcaggtgc accattaaca ttccaatctt ttagttacgc aactattaat
1680
acagcattta cattcccaga aagaagtagc agcttgacta ttggtgctga tacttttagc
1740
tcaggtaatg aagtttatgt agatagattt gaattgatcc aggttactgc aacatttgag
1800
gcagaatcag atttagagag agcacgaaaa gcggtgaatg cgctgtttac ttcttccaat
1860
caaatcgggt taaaaacaga tgtgacggac tatcatattg ataaagtatc caatctagtt
1920
gagtgtttat cagatgaatt ttgtctagat gaaaagcgag aattgtccga gaaagtcaaa
1980
catgcgaagc gactctgtga tgagcggaat ttacttcaag atccaaactt cagaggcatc
2040
aatagacaac cagaccgtgg ttggagagga agtacggata ttaccatcca aggaggagat
2100
gacgtattca aagagaatta cgttacgcta ccgggtacct ttgatgagtg ctatccaacg
2160
tatttatatc aaaaaataga tgagtcgaaa ttaaaagcct atacccgcta tgaattaaga
2220
gggtatatcg aggatagtca agacttagaa atctatttaa ttcgctacaa tgcaaaacat
2280
gaaacagtaa atgtgccagg tacgggttcc ttatggccgc tttcagccca aagtccaatc
2340
ggaaagtgtg gagaaccgaa tcgatgtgcg acacaccttg aatggaatcc tgacttagat
2400
tgttcgtgta gggatggaga aaagtgtgcc catcattcgc atcatttctc cttagacatt
2460
gatgtaggat gtacagacct aaatgaggac ctaggtgtat gggtgatctt taagattaag
2520
acgcaagatg gtcatgcgag actaggaaat ctagaatttc tcgaagagaa accattagta
2580
ggagaagcgc tagctcgtgt gaagagagcg gagaaaaaat ggagagacaa acgcgaaaaa
2640
ttggaattgg aaacaaatat tgtttataaa gaggcaaaaa aatctgtaga tgctttattt
2700
gtgaactctc aatatgatag attacaagcg gatacgaata tcgcgataat tcatgcggca
2760
gataaacgcg ttcatagcat tcgagaagca tatcttccag agttgtctgt aattccgggt
2820
gtaaatgcag ctatttttga agaattagag ggacgtattt tcacagccta ctctctatat
2880
gatgcgagaa atgtcattaa aaatggcgat ttcaataatg gcttatcatg ctggaacgtg
2940
aaagggcatg tagatgtaga agaacagaac aaccatcgtt cggtccttgt tgttccagaa
3000
tgggaagcag aagtgtcaca agaggttcgt gtctgtccag gtcgtggcta tatccttcgt
3060
gttacagcgt acaaagaggg atatggagag ggctgtgtaa cgattcatga gatcgaagac
3120
aatacagacg aactgaaatt cagcaactgt gtagaagagg aaatatatcc aaacaacacg
3180
gtaacgtgta atgattatac tgcgactcaa gaagaatatg agggtacgta cacttctcgt
3240
aatcgaggat atgacggagc ctatgaaagc aattcttctg taccagctga ttatgcatca
3300
gcctatgaag aaaaagcgta tacagatgga agaagagaca atacttgtga atctaacaga
3360
ggatatgggg attacacacc actaccagct ggctatgtga caaaagaatt agagtacttc
3420
ccagaaaccg ataaggtatg gattgagatt ggagaaacgg aaggaacatt tatcgtggac
3480
agcgtggaat tactccttat ggaggaatag
3510
<210> 17
<211> 1189
<212> БЕЛОК
<213> Bacillus thuringiensis
<400> 17
Met Glu Glu Asn Asn Gln Asn Gln Cys Ile Pro Tyr Asn Cys Leu Ser
1 5 10 15
Asn Pro Glu Glu Val Leu Leu Asp Gly Glu Arg Ile Ser Thr Gly Asn
20 25 30
Ser Ser Ile Asp Ile Ser Leu Ser Leu Val Gln Phe Leu Val Ser Asn
35 40 45
Phe Val Pro Gly Gly Gly Phe Leu Val Gly Leu Ile Asp Phe Val Trp
50 55 60
Gly Ile Val Gly Pro Ser Gln Trp Asp Ala Phe Leu Val Gln Ile Glu
65 70 75 80
Gln Leu Ile Asn Glu Arg Ile Ala Glu Phe Ala Arg Asn Ala Ala Ile
85 90 95
Ala Asn Leu Glu Gly Leu Gly Asn Asn Phe Asn Ile Tyr Val Glu Ala
100 105 110
Phe Lys Glu Trp Glu Glu Asp Pro Asn Asn Pro Ala Thr Arg Thr Arg
115 120 125
Val Ile Asp Arg Phe Arg Ile Leu Asp Gly Leu Leu Glu Arg Asp Ile
130 135 140
Pro Ser Phe Arg Ile Ser Gly Phe Glu Val Pro Leu Leu Ser Val Tyr
145 150 155 160
Ala Gln Ala Ala Asn Leu His Leu Ala Ile Leu Arg Asp Ser Val Ile
165 170 175
Phe Gly Glu Arg Trp Gly Leu Thr Thr Ile Asn Val Asn Glu Asn Tyr
180 185 190
Asn Arg Leu Ile Arg His Ile Asp Glu Tyr Ala Asp His Cys Ala Asn
195 200 205
Thr Tyr Asn Arg Gly Leu Asn Asn Leu Pro Lys Ser Thr Tyr Gln Asp
210 215 220
Trp Ile Thr Tyr Asn Arg Leu Arg Arg Asp Leu Thr Leu Thr Val Leu
225 230 235 240
Asp Ile Ala Ala Phe Phe Pro Asn Tyr Asp Asn Arg Arg Tyr Pro Ile
245 250 255
Gln Pro Val Gly Gln Leu Thr Arg Glu Val Tyr Thr Asp Pro Leu Ile
260 265 270
Asn Phe Asn Pro Gln Leu Gln Ser Val Ala Gln Leu Pro Thr Phe Asn
275 280 285
Val Met Glu Ser Ser Ala Ile Arg Asn Pro His Leu Phe Asp Ile Leu
290 295 300
Asn Asn Leu Thr Ile Phe Thr Asp Trp Phe Ser Val Gly Arg Asn Phe
305 310 315 320
Tyr Trp Gly Gly His Arg Val Ile Ser Ser Leu Ile Gly Gly Gly Asn
325 330 335
Ile Thr Ser Pro Ile Tyr Gly Arg Glu Ala Asn Gln Glu Pro Pro Arg
340 345 350
Ser Phe Thr Phe Asn Gly Pro Val Phe Arg Thr Leu Ser Asn Pro Thr
355 360 365
Leu Arg Leu Leu Gln Gln Pro Trp Pro Ala Pro Pro Phe Asn Leu Arg
370 375 380
Gly Val Glu Gly Val Glu Phe Ser Thr Pro Thr Asn Ser Phe Thr Tyr
385 390 395 400
Arg Gly Arg Gly Thr Val Asp Ser Leu Thr Glu Leu Pro Pro Glu Asp
405 410 415
Asn Ser Val Pro Pro Arg Glu Gly Tyr Ser His Arg Leu Cys His Ala
420 425 430
Thr Phe Val Gln Arg Ser Gly Thr Pro Phe Leu Thr Thr Gly Val Val
435 440 445
Phe Ser Trp Thr His Arg Ser Ala Thr Leu Thr Asn Thr Ile Asp Pro
450 455 460
Glu Arg Ile Asn Gln Ile Pro Leu Val Lys Gly Phe Arg Val Trp Gly
465 470 475 480
Gly Thr Ser Val Ile Thr Gly Pro Gly Phe Thr Gly Gly Asp Ile Leu
485 490 495
Arg Arg Asn Thr Phe Gly Asp Phe Val Ser Leu Gln Val Asn Ile Asn
500 505 510
Ser Pro Ile Thr Gln Arg Tyr Arg Leu Arg Phe Arg Tyr Ala Ser Ser
515 520 525
Arg Asp Ala Arg Val Ile Val Leu Thr Gly Ala Ala Ser Thr Gly Val
530 535 540
Gly Gly Gln Val Ser Val Asn Met Pro Leu Gln Lys Thr Met Glu Ile
545 550 555 560
Gly Glu Asn Leu Thr Ser Arg Thr Phe Arg Tyr Thr Asp Phe Ser Asn
565 570 575
Pro Phe Ser Phe Arg Ala Asn Pro Asp Ile Ile Gly Ile Ser Glu Gln
580 585 590
Pro Leu Phe Gly Ala Gly Ser Ile Ser Ser Gly Glu Leu Tyr Ile Asp
595 600 605
Lys Ile Glu Ile Ile Leu Ala Asp Ala Thr Phe Glu Ala Glu Ser Asp
610 615 620
Leu Glu Arg Ala Gln Lys Ala Val Asn Ala Leu Phe Thr Ser Ser Asn
625 630 635 640
Gln Ile Gly Leu Lys Thr Asp Val Thr Asp Tyr His Ile Asp Gln Val
645 650 655
Ser Asn Leu Val Asp Cys Leu Ser Asp Glu Phe Cys Leu Asp Glu Lys
660 665 670
Arg Glu Leu Ser Glu Lys Val Lys His Ala Lys Arg Leu Ser Asp Glu
675 680 685
Arg Asn Leu Leu Gln Asp Pro Asn Phe Arg Gly Ile Asn Arg Gln Pro
690 695 700
Asp Arg Gly Trp Arg Gly Ser Thr Asp Ile Thr Ile Gln Gly Gly Asp
705 710 715 720
Asp Val Phe Lys Glu Asn Tyr Val Thr Leu Pro Gly Thr Val Asp Glu
725 730 735
Cys Tyr Pro Thr Tyr Leu Tyr Gln Lys Ile Asp Glu Ser Lys Leu Lys
740 745 750
Ala Tyr Thr Arg Tyr Glu Leu Arg Gly Tyr Ile Glu Asp Ser Gln Asp
755 760 765
Leu Glu Ile Tyr Leu Ile Arg Tyr Asn Ala Lys His Glu Ile Val Asn
770 775 780
Val Pro Gly Thr Gly Ser Leu Trp Pro Leu Ser Ala Gln Ser Pro Ile
785 790 795 800
Gly Lys Cys Gly Glu Pro Asn Arg Cys Ala Pro His Leu Glu Trp Asn
805 810 815
Pro Asp Leu Asp Cys Ser Cys Arg Asp Gly Glu Lys Cys Ala His His
820 825 830
Ser His His Phe Thr Leu Asp Ile Asp Val Gly Cys Thr Asp Leu Asn
835 840 845
Glu Asp Leu Gly Val Trp Val Ile Phe Lys Ile Lys Thr Gln Asp Gly
850 855 860
His Ala Arg Leu Gly Asn Leu Glu Phe Leu Glu Glu Lys Pro Leu Leu
865 870 875 880
Gly Glu Ala Leu Ala Arg Val Lys Arg Ala Glu Lys Lys Trp Arg Asp
885 890 895
Lys Arg Glu Lys Leu Gln Leu Glu Thr Asn Ile Val Tyr Lys Glu Ala
900 905 910
Lys Glu Ser Val Asp Ala Leu Phe Val Asn Ser Gln Tyr Asp Arg Leu
915 920 925
Gln Val Asp Thr Asn Ile Ala Met Ile His Ala Ala Asp Lys Arg Val
930 935 940
His Arg Ile Arg Glu Ala Tyr Leu Pro Glu Leu Ser Val Ile Pro Gly
945 950 955 960
Val Asn Ala Ala Ile Phe Glu Glu Leu Glu Gly Arg Ile Phe Thr Ala
965 970 975
Tyr Ser Leu Tyr Asp Ala Arg Asn Val Ile Lys Asn Gly Asp Phe Asn
980 985 990
Asn Gly Leu Leu Cys Trp Asn Val Lys Gly His Val Asp Val Glu Glu
995 1000 1005
Gln Asn Asn His Arg Ser Val Leu Val Ile Pro Glu Trp Glu Ala
1010 1015 1020
Glu Val Ser Gln Glu Val Arg Val Cys Pro Gly Arg Gly Tyr Ile
1025 1030 1035
Leu Arg Val Thr Ala Tyr Lys Glu Gly Tyr Gly Glu Gly Cys Val
1040 1045 1050
Thr Ile His Glu Ile Glu Asp Asn Thr Asp Glu Leu Lys Phe Ser
1055 1060 1065
Asn Cys Val Glu Glu Glu Val Tyr Pro Asn Asn Thr Val Thr Cys
1070 1075 1080
Asn Asn Tyr Thr Gly Thr Gln Glu Glu Tyr Glu Gly Thr Tyr Thr
1085 1090 1095
Ser Arg Asn Gln Gly Tyr Asp Glu Ala Tyr Gly Asn Asn Pro Ser
1100 1105 1110
Val Pro Ala Asp Tyr Ala Ser Val Tyr Glu Glu Lys Ser Tyr Thr
1115 1120 1125
Asp Gly Arg Arg Glu Asn Pro Cys Glu Ser Asn Arg Gly Tyr Gly
1130 1135 1140
Asp Tyr Thr Pro Leu Pro Ala Gly Tyr Val Thr Lys Asp Leu Glu
1145 1150 1155
Tyr Phe Pro Glu Thr Asp Lys Val Trp Ile Glu Ile Gly Glu Thr
1160 1165 1170
Glu Gly Thr Phe Ile Val Asp Ser Val Glu Leu Leu Leu Met Glu
1175 1180 1185
Glu
<210> 18
<211> 1181
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сконструированный химерный белок Bt29-1Ca.
<400> 18
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Gly Phe Arg Val Trp Gly Gly Thr Ser Val Ile Thr Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Asn Thr Phe Gly Asp Phe
485 490 495
Val Ser Leu Gln Val Asn Ile Asn Ser Pro Ile Thr Gln Arg Tyr Arg
500 505 510
Leu Arg Phe Arg Tyr Ala Ser Ser Arg Asp Ala Arg Val Ile Val Leu
515 520 525
Thr Gly Ala Ala Ser Thr Gly Val Gly Gly Gln Val Ser Val Asn Met
530 535 540
Pro Leu Gln Lys Thr Met Glu Ile Gly Glu Asn Leu Thr Ser Arg Thr
545 550 555 560
Phe Arg Tyr Thr Asp Phe Ser Asn Pro Phe Ser Phe Arg Ala Asn Pro
565 570 575
Asp Ile Ile Gly Ile Ser Glu Gln Pro Leu Phe Gly Ala Gly Ser Ile
580 585 590
Ser Ser Gly Glu Leu Tyr Ile Asp Lys Ile Glu Ile Ile Leu Ala Asp
595 600 605
Ala Thr Phe Glu Ala Glu Tyr Asp Leu Glu Lys Ala Gln Lys Ala Val
610 615 620
Asn Ala Leu Phe Thr Ser Ser Asn Gln Ile Gly Leu Lys Thr Asp Val
625 630 635 640
Thr Asp Tyr His Ile Asp Lys Val Ser Asn Leu Val Glu Cys Leu Ser
645 650 655
Asp Glu Phe Cys Leu Asp Glu Lys Arg Glu Leu Ser Glu Lys Val Lys
660 665 670
His Ala Lys Arg Leu Cys Asp Glu Arg Asn Leu Leu Gln Asp Pro Asn
675 680 685
Phe Arg Gly Ile Asn Arg Gln Pro Asp Arg Gly Trp Arg Gly Ser Thr
690 695 700
Asp Ile Thr Ile Gln Gly Gly Asp Asp Val Phe Lys Glu Asn Tyr Val
705 710 715 720
Thr Leu Pro Gly Thr Phe Asp Glu Cys Tyr Pro Thr Tyr Leu Tyr Gln
725 730 735
Lys Ile Asp Glu Ser Lys Leu Lys Ala Tyr Thr Arg Tyr Glu Leu Arg
740 745 750
Gly Tyr Ile Glu Asp Ser Gln Asp Leu Glu Ile Tyr Leu Ile Arg Tyr
755 760 765
Asn Ala Lys His Glu Thr Val Asn Val Pro Gly Thr Gly Ser Leu Trp
770 775 780
Pro Leu Ser Ala Gln Ser Pro Ile Gly Lys Cys Gly Glu Pro Asn Arg
785 790 795 800
Cys Ala Thr His Leu Glu Trp Asn Pro Asp Leu Asp Cys Ser Cys Arg
805 810 815
Asp Gly Glu Lys Cys Ala His His Ser His His Phe Ser Leu Asp Ile
820 825 830
Asp Val Gly Cys Thr Asp Leu Asn Glu Asp Leu Gly Val Trp Val Ile
835 840 845
Phe Lys Ile Lys Thr Gln Asp Gly His Ala Arg Leu Gly Asn Leu Glu
850 855 860
Phe Leu Glu Glu Lys Pro Leu Val Gly Glu Ala Leu Ala Arg Val Lys
865 870 875 880
Arg Ala Glu Lys Lys Trp Arg Asp Lys Arg Glu Lys Leu Glu Leu Glu
885 890 895
Thr Asn Ile Val Tyr Lys Glu Ala Lys Lys Ser Val Asp Ala Leu Phe
900 905 910
Val Asn Ser Gln Tyr Asp Arg Leu Gln Ala Asp Thr Asn Ile Ala Ile
915 920 925
Ile His Ala Ala Asp Lys Arg Val His Ser Ile Arg Glu Ala Tyr Leu
930 935 940
Pro Glu Leu Ser Val Ile Pro Gly Val Asn Ala Ala Ile Phe Glu Glu
945 950 955 960
Leu Glu Gly Arg Ile Phe Thr Ala Tyr Ser Leu Tyr Asp Ala Arg Asn
965 970 975
Val Ile Lys Asn Gly Asp Phe Asn Asn Gly Leu Ser Cys Trp Asn Val
980 985 990
Lys Gly His Val Asp Val Glu Glu Gln Asn Asn His Arg Ser Val Leu
995 1000 1005
Val Val Pro Glu Trp Glu Ala Glu Val Ser Gln Glu Val Arg Val
1010 1015 1020
Cys Pro Gly Arg Gly Tyr Ile Leu Arg Val Thr Ala Tyr Lys Glu
1025 1030 1035
Gly Tyr Gly Glu Gly Cys Val Thr Ile His Glu Ile Glu Asp Asn
1040 1045 1050
Thr Asp Glu Leu Lys Phe Ser Asn Cys Val Glu Glu Glu Ile Tyr
1055 1060 1065
Pro Asn Asn Thr Val Thr Cys Asn Asp Tyr Thr Ala Thr Gln Glu
1070 1075 1080
Glu Tyr Glu Gly Thr Tyr Thr Ser Arg Asn Arg Gly Tyr Asp Gly
1085 1090 1095
Ala Tyr Glu Ser Asn Ser Ser Val Pro Ala Asp Tyr Ala Ser Ala
1100 1105 1110
Tyr Glu Glu Lys Ala Tyr Thr Asp Gly Arg Arg Asp Asn Thr Cys
1115 1120 1125
Glu Ser Asn Arg Gly Tyr Gly Asp Tyr Thr Pro Leu Pro Ala Gly
1130 1135 1140
Tyr Val Thr Lys Glu Leu Glu Tyr Phe Pro Glu Thr Asp Lys Val
1145 1150 1155
Trp Ile Glu Ile Gly Glu Thr Glu Gly Thr Phe Ile Val Asp Ser
1160 1165 1170
Val Glu Leu Leu Leu Met Glu Glu
1175 1180
<210> 19
<211> 3546
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Химера Bt29-1Ca
<400> 19
atggagatta ataatcagaa ccaatgtgtc ccttataatt gtttgaataa tcctgaaagc
60
gagatattaa acgttgcaat ttttagtagc gaacaggtag cagaaattca cttaaagatc
120
acgcgcttaa ttttagagaa ttttttacca ggtgggagtt ttgcattcgg cttatttgat
180
ttaatatggg ggatttttaa tgaagatcaa tggagcgcat ttcttcggca ggtagaagaa
240
ttaattaatc aaaggataac ggaattcgca agagggcaag caattcagag actagtaggg
300
tttggaagga gttatgatga atatatttta gcactaaaag aatgggaaaa cgatcctgat
360
aacccagctt caaaggaaag agtgcgcact cgatttcgga caactgatga tgccttgcta
420
accggtgttc ctcttatggc aattccaggt tttgaattag ctactttatc tgtttatgct
480
caatcagcca atctacattt agccctatta agagatgctg tattttttgg ggagagatgg
540
ggattgacac aaacaaatat aaatgattta tatagtagat taaaaaactc cattcgtgat
600
tatacaaatc attgtgttcg tttttataat ataggtttag ggaatttaaa tgttataaga
660
ccagagtatt accgtttcca aagagaatta acaatatctg tcttagatct tgtagctctt
720
tttccaaatt acgatatccg aacatatcca ataccaacta aaagtcaatt aacaagagaa
780
atttatacag atccgattat ttcacctggt gcacaggcag gttatactct tcaagatgtt
840
ttgagagaac cacaccttat ggacttttta aaccgactta ttatttatac tggtgagtat
900
cgcggaattc gtcactgggc aggacatgaa gtagaatcta gtagaacagg tatgatgact
960
aatataagat ttcctttgta tggaacagcc gcaacagcag aaccaacacg atttataact
1020
cctagtactt ttcctggtct taatttattt tatagaacat tatcagctcc tatttttaga
1080
gatgaaccgg gagctaatat tattattaga tatagaacga gtttggtgga aggagtagga
1140
tttattcaac caaataacgg tgaacagctt tacagagtga gaggaacatt agattctctt
1200
gatcaattac cacttgaggg tgagagtagt ctaactgaat atagtcatcg attatgccat
1260
gttagatttg cgcaatcatt gaggaatgca gaacctttag attatgcaag ggttccgatg
1320
ttttcttgga cacatcgtag tgcaacccct acaaatacaa ttgatccaga tgtcatcacc
1380
caaataccgc tggtgaaagg ctttcgcgtg tggggcggca ccagcgtgat taccggcccg
1440
ggctttaccg gcggcgatat tctgcgccgc aacacctttg gcgattttgt gagcctgcag
1500
gtgaacatta acagcccgat tacccagcgc tatcgcctgc gctttcgcta tgcgagcagc
1560
cgcgatgcgc gcgtgattgt gctgaccggc gcggcgagca ccggcgtggg cggccaggtg
1620
agcgtgaaca tgccgctgca gaaaaccatg gaaattggcg aaaacctgac cagccgcacc
1680
tttcgctata ccgattttag caacccgttt agctttcgcg cgaacccgga tattattggc
1740
attagcgaac agccgctgtt tggcgcgggc agcattagca gcggcgaact gtatattgat
1800
aaaattgaaa ttattctggc ggatgcgaca tttgaggcag aatatgattt agagaaagct
1860
cagaaagcgg tgaatgcgct gtttacttct tccaatcaaa tcgggttaaa aacagatgtg
1920
acggactatc atattgataa agtatccaat ctagttgagt gtttatcaga tgaattttgt
1980
ctagatgaaa agcgagaatt gtccgagaaa gtcaaacatg cgaagcgact ctgtgatgag
2040
cggaatttac ttcaagatcc aaacttcaga ggcatcaata gacaaccaga ccgtggttgg
2100
agaggaagta cggatattac catccaagga ggagatgacg tattcaaaga gaattacgtt
2160
acgctaccgg gtacctttga tgagtgctat ccaacgtatt tatatcaaaa aatagatgag
2220
tcgaaattaa aagcctatac ccgctatgaa ttaagagggt atatcgagga tagtcaagac
2280
ttagaaatct atttaattcg ctacaatgca aaacatgaaa cagtaaatgt gccaggtacg
2340
ggttccttat ggccgctttc agcccaaagt ccaatcggaa agtgtggaga accgaatcga
2400
tgtgcgacac accttgaatg gaatcctgac ttagattgtt cgtgtaggga tggagaaaag
2460
tgtgcccatc attcgcatca tttctcctta gacattgatg taggatgtac agacctaaat
2520
gaggacctag gtgtatgggt gatctttaag attaagacgc aagatggtca tgcgagacta
2580
ggaaatctag aatttctcga agagaaacca ttagtaggag aagcgctagc tcgtgtgaag
2640
agagcggaga aaaaatggag agacaaacgc gaaaaattgg aattggaaac aaatattgtt
2700
tataaagagg caaaaaaatc tgtagatgct ttatttgtga actctcaata tgatagatta
2760
caagcggata cgaatatcgc gataattcat gcggcagata aacgcgttca tagcattcga
2820
gaagcatatc ttccagagtt gtctgtaatt ccgggtgtaa atgcagctat ttttgaagaa
2880
ttagagggac gtattttcac agcctactct ctatatgatg cgagaaatgt cattaaaaat
2940
ggcgatttca ataatggctt atcatgctgg aacgtgaaag ggcatgtaga tgtagaagaa
3000
cagaacaacc atcgttcggt ccttgttgtt ccagaatggg aagcagaagt gtcacaagag
3060
gttcgtgtct gtccaggtcg tggctatatc cttcgtgtta cagcgtacaa agagggatat
3120
ggagagggct gtgtaacgat tcatgagatc gaagacaata cagacgaact gaaattcagc
3180
aactgtgtag aagaggaaat atatccaaac aacacggtaa cgtgtaatga ttatactgcg
3240
actcaagaag aatatgaggg tacgtacact tctcgtaatc gaggatatga cggagcctat
3300
gaaagcaatt cttctgtacc agctgattat gcatcagcct atgaagaaaa agcgtataca
3360
gatggaagaa gagacaatac ttgtgaatct aacagaggat atggggatta cacaccacta
3420
ccagctggct atgtgacaaa agaattagag tacttcccag aaaccgataa ggtatggatt
3480
gagattggag aaacggaagg aacatttatc gtggacagcg tggaattact ccttatggag
3540
gaatag
3546
<210> 20
<211> 652
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Bt29-Bt22Tr1
<400> 20
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Phe Glu Ala Glu Tyr Asp Leu Glu Lys Ala
595 600 605
Gln Lys Ala Val Asn Ala Leu Phe Thr Ser Ser Asn Gln Ile Gly Leu
610 615 620
Lys Thr Asp Val Thr Asp Tyr His Ile Asp Lys Val Ser Asn Leu Val
625 630 635 640
Glu Cys Leu Ser Asp Glu Phe Cys Leu Asp Glu Lys
645 650
<210> 21
<211> 636
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Bt29-Bt22Tr2
<400> 21
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Phe Glu Ala Glu Tyr Asp Leu Glu Lys Ala
595 600 605
Gln Lys Ala Val Asn Ala Leu Phe Thr Ser Ser Asn Gln Ile Gly Leu
610 615 620
Lys Thr Asp Val Thr Asp Tyr His Ile Asp Lys Val
625 630 635
<210> 22
<211> 622
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Bt29-Bt22Tr3
<400> 22
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Phe Glu Ala Glu Tyr Asp Leu Glu Lys Ala
595 600 605
Gln Lys Ala Val Asn Ala Leu Phe Thr Ser Ser Asn Gln Ile
610 615 620
<210> 23
<211> 610
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Bt29-Bt22Tr4
<400> 23
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Phe Glu Ala Glu Tyr Asp Leu Glu Lys Ala
595 600 605
Gln Lys
610
<210> 24
<211> 607
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Bt29-Bt22Tr5
<400> 24
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Phe Glu Ala Glu Tyr Asp Leu Glu Lys
595 600 605
<210> 25
<211> 603
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Bt29-Bt22Tr6
<400> 25
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Phe Glu Ala Glu Tyr
595 600
<210> 26
<211> 676
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> BT29BT22-TL22v1
<400> 26
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
116
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Leu Glu Ala Val Thr Asp Leu Glu Arg Ala
595 600 605
Gln Lys Ala Val His Glu Leu Phe Thr Ser Thr Asn Pro Gly Gly Leu
610 615 620
Lys Thr Asp Val Ala Lys Asp His Tyr Thr Asn Thr Ile Ser Lys Ser
625 630 635 640
Val Gln Ser Val Phe Arg Cys Arg Cys Ser Glu Arg Thr Arg Ile Tyr
645 650 655
Arg Trp Gly Tyr Pro Ser Lys Lys Glu Tyr Trp Tyr Ile Trp Gly Tyr
660 665 670
Thr Ser Lys Tyr
675
<210> 27
<211> 644
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> BT29BT22-TL22v2
<400> 27
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Leu Glu Ala Val Thr Asp Leu Glu Arg Ala
595 600 605
Gln Lys Ala Val His Glu Leu Phe Thr Ser Thr Asn Pro Gly Gly Leu
610 615 620
Lys Thr Asp Val Ala Lys Asp His Tyr Thr Asn Thr Ile Ser Lys Ser
625 630 635 640
Val Gln Ser Val
<210> 28
<211> 634
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> BT29BT22-TL22v3
<400> 28
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Leu Glu Ala Val Thr Asp Leu Glu Arg Ala
595 600 605
Gln Lys Ala Val His Glu Leu Phe Thr Ser Thr Asn Pro Gly Gly Leu
610 615 620
Lys Thr Asp Val Ala Lys Asp His Tyr Thr
625 630
<210> 29
<211> 622
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> BT29BT22-TL22v4
<400> 29
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Leu Glu Ala Val Thr Asp Leu Glu Arg Ala
595 600 605
Gln Lys Ala Val His Glu Leu Phe Thr Ser Thr Asn Pro Gly
610 615 620
<210> 30
<211> 610
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> BT29BT22-TL22v5
<400> 30
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Leu Glu Ala Val Thr Asp Leu Glu Arg Ala
595 600 605
Gln Lys
610
<210> 31
<211> 600
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> BT29BT22-TL22v6
<400> 31
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Lys Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Phe Ser Asn Val Asn Leu Asp Trp Asn Leu Ser Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Met Tyr Val Thr
515 520 525
Ile Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asn
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asp
545 550 555 560
Thr Ala Phe Thr Phe Pro Thr Lys Ala Ser Ser Leu Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Val Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Leu Glu
595 600
<210> 32
<211> 610
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Bt29-1FaTr1
<400> 32
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Arg Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Tyr Thr Ile Val Asn Ile Asn Gly Gln Leu Pro Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Ile Tyr Val Thr
515 520 525
Val Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asp
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asn
545 550 555 560
Thr Ala Phe Thr Phe Pro Met Ser Gln Ser Ser Phe Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Ile Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Phe Glu Ala Glu Tyr Asp Leu Glu Arg Ala
595 600 605
Gln Lys
610
<210> 33
<211> 607
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Bt29-1FaTr2
<400> 33
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Arg Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Tyr Thr Ile Val Asn Ile Asn Gly Gln Leu Pro Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Ile Tyr Val Thr
515 520 525
Val Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asp
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asn
545 550 555 560
Thr Ala Phe Thr Phe Pro Met Ser Gln Ser Ser Phe Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Ile Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Phe Glu Ala Glu Tyr Asp Leu Glu Arg
595 600 605
<210> 34
<211> 603
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Bt29-1FaTr3
<400> 34
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Arg Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Tyr Thr Ile Val Asn Ile Asn Gly Gln Leu Pro Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Ile Tyr Val Thr
515 520 525
Val Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asp
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asn
545 550 555 560
Thr Ala Phe Thr Phe Pro Met Ser Gln Ser Ser Phe Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Ile Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Phe Glu Ala Glu Tyr
595 600
<210> 35
<211> 600
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Bt29-1FaTr4
<400> 35
Met Glu Ile Asn Asn Gln Asn Gln Cys Val Pro Tyr Asn Cys Leu Asn
1 5 10 15
Asn Pro Glu Ser Glu Ile Leu Asn Val Ala Ile Phe Ser Ser Glu Gln
20 25 30
Val Ala Glu Ile His Leu Lys Ile Thr Arg Leu Ile Leu Glu Asn Phe
35 40 45
Leu Pro Gly Gly Ser Phe Ala Phe Gly Leu Phe Asp Leu Ile Trp Gly
50 55 60
Ile Phe Asn Glu Asp Gln Trp Ser Ala Phe Leu Arg Gln Val Glu Glu
65 70 75 80
Leu Ile Asn Gln Arg Ile Thr Glu Phe Ala Arg Gly Gln Ala Ile Gln
85 90 95
Arg Leu Val Gly Phe Gly Arg Ser Tyr Asp Glu Tyr Ile Leu Ala Leu
100 105 110
Lys Glu Trp Glu Asn Asp Pro Asp Asn Pro Ala Ser Lys Glu Arg Val
115 120 125
Arg Thr Arg Phe Arg Thr Thr Asp Asp Ala Leu Leu Thr Gly Val Pro
130 135 140
Leu Met Ala Ile Pro Gly Phe Glu Leu Ala Thr Leu Ser Val Tyr Ala
145 150 155 160
Gln Ser Ala Asn Leu His Leu Ala Leu Leu Arg Asp Ala Val Phe Phe
165 170 175
Gly Glu Arg Trp Gly Leu Thr Gln Thr Asn Ile Asn Asp Leu Tyr Ser
180 185 190
Arg Leu Lys Asn Ser Ile Arg Asp Tyr Thr Asn His Cys Val Arg Phe
195 200 205
Tyr Asn Ile Gly Leu Gly Asn Leu Asn Val Ile Arg Pro Glu Tyr Tyr
210 215 220
Arg Phe Gln Arg Glu Leu Thr Ile Ser Val Leu Asp Leu Val Ala Leu
225 230 235 240
Phe Pro Asn Tyr Asp Ile Arg Thr Tyr Pro Ile Pro Thr Lys Ser Gln
245 250 255
Leu Thr Arg Glu Ile Tyr Thr Asp Pro Ile Ile Ser Pro Gly Ala Gln
260 265 270
Ala Gly Tyr Thr Leu Gln Asp Val Leu Arg Glu Pro His Leu Met Asp
275 280 285
Phe Leu Asn Arg Leu Ile Ile Tyr Thr Gly Glu Tyr Arg Gly Ile Arg
290 295 300
His Trp Ala Gly His Glu Val Glu Ser Ser Arg Thr Gly Met Met Thr
305 310 315 320
Asn Ile Arg Phe Pro Leu Tyr Gly Thr Ala Ala Thr Ala Glu Pro Thr
325 330 335
Arg Phe Ile Thr Pro Ser Thr Phe Pro Gly Leu Asn Leu Phe Tyr Arg
340 345 350
Thr Leu Ser Ala Pro Ile Phe Arg Asp Glu Pro Gly Ala Asn Ile Ile
355 360 365
Ile Arg Tyr Arg Thr Ser Leu Val Glu Gly Val Gly Phe Ile Gln Pro
370 375 380
Asn Asn Gly Glu Gln Leu Tyr Arg Val Arg Gly Thr Leu Asp Ser Leu
385 390 395 400
Asp Gln Leu Pro Leu Glu Gly Glu Ser Ser Leu Thr Glu Tyr Ser His
405 410 415
Arg Leu Cys His Val Arg Phe Ala Gln Ser Leu Arg Asn Ala Glu Pro
420 425 430
Leu Asp Tyr Ala Arg Val Pro Met Phe Ser Trp Thr His Arg Ser Ala
435 440 445
Thr Pro Thr Asn Thr Ile Asp Pro Asp Val Ile Thr Gln Ile Pro Leu
450 455 460
Val Lys Ala His Thr Leu Gln Ser Gly Thr Thr Val Val Arg Gly Pro
465 470 475 480
Gly Phe Thr Gly Gly Asp Ile Leu Arg Arg Thr Ser Gly Gly Pro Phe
485 490 495
Ala Tyr Thr Ile Val Asn Ile Asn Gly Gln Leu Pro Gln Arg Tyr Arg
500 505 510
Ala Arg Ile Arg Tyr Ala Ser Thr Thr Asn Leu Arg Ile Tyr Val Thr
515 520 525
Val Ala Gly Glu Arg Ile Phe Ala Gly Gln Phe Asn Lys Thr Met Asp
530 535 540
Thr Gly Asp Pro Leu Thr Phe Gln Ser Phe Ser Tyr Ala Thr Ile Asn
545 550 555 560
Thr Ala Phe Thr Phe Pro Met Ser Gln Ser Ser Phe Thr Val Gly Ala
565 570 575
Asp Thr Phe Ser Ser Gly Asn Glu Val Tyr Ile Asp Arg Phe Glu Leu
580 585 590
Ile Pro Val Thr Ala Thr Phe Glu
595 600
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2816526C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2015 |
|
RU2741833C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2015 |
|
RU2745322C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2018 |
|
RU2761666C2 |
| НОВЫЕ БЕЛКИ, ИМЕЮЩИЕ ИНГИБИРУЮЩЕЕ ДЕЙСТВИЕ В ОТНОШЕНИИ НАСЕКОМЫХ | 2017 |
|
RU2781075C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2740312C2 |
| ГЕНЫ ТОКСИНОВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2825305C2 |
| ГЕНЫ ТОКСИНОВ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2723717C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2017 |
|
RU2765304C2 |
| КОНТРОЛЬ УСТОЙЧИВОСТИ НАСЕКОМЫХ С ПОМОЩЬЮ КОМБИНАЦИИ БЕЛКОВ Cry1Be И Cry1F | 2010 |
|
RU2575611C2 |


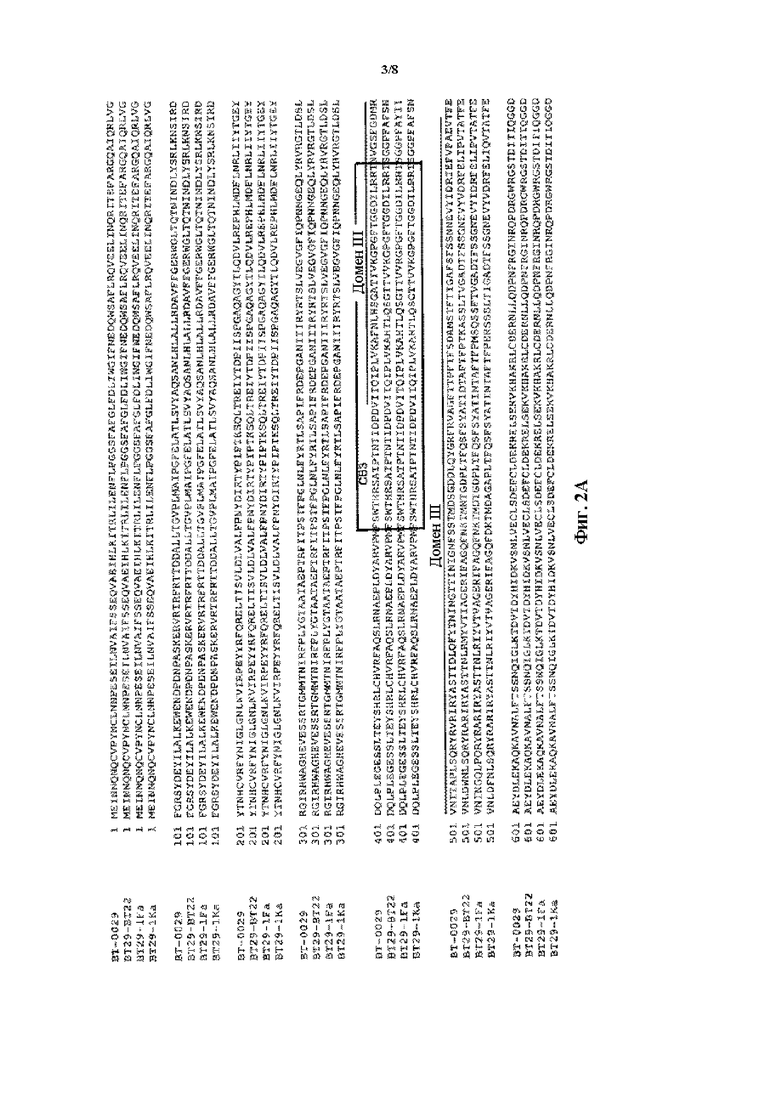





Изобретение относится к области биохимии, в частности к химерному инсектицидному белку, токсичному для насекомого-вредителя, относящегося к чешуекрылым. Раскрыт полинуклеотид, кодирующий указанный белок; кассета экспрессии и клетка, содержащие указанный полинуклеотид; вектор экспрессии, содержащий указанную кассету экспрессии; трансгенное растение, его часть или трансгенное семя, содержащее указанную клетку. Изобретение позволяет эффективно бороться с насекомым-вредителем, относящимся к чешуекрылым. 8 н. и 5 з.п. ф-лы, 3 ил., 12 табл., 7 пр.
1. Химерный инсектицидный белок, токсичный для насекомого-вредителя, относящегося к чешуекрылым, содержащий аминокислотную последовательность SEQ ID NO:18.
2. Химерный инсектицидный белок по п. 1, где химерный инсектицидный белок содержит SEQ ID NO:18.
3. Химерный инсектицидный белок по п. 1 или 2, где химерный инсектицидный белок характеризуется инсектицидной активностью в отношении насекомого-вредителя Spodoptera frugiperda.
4. Полинуклеотид, содержащий нуклеотидную последовательность, кодирующую химерный инсектицидный белок по любому из пп. 1-3.
5. Полинуклеотид по п. 4, где полинуклеотид содержит нуклеотидную последовательность SEQ ID NO: 19.
6. Кассета экспрессии, обеспечивающая экспрессию химерного инсектицидного белка по любому из пп. 1-3, содержащая полинуклеотид по п. 4 или 5, функционально связанный с гетерологичным промотором.
7. Вектор экспрессии химерного инсектицидного белка по любому из пп. 1-3, содержащий кассету экспрессии по п. 6.
8. Трансгенная клетка, экспрессирующая белок по п. 1 и содержащая полинуклеотид по п. 4, где трансгенная клетка представляет собой бактериальную клетку или растительную клетку.
9. Трансгенная клетка по п. 8, которая является растительной клеткой и представляет собой:
(a) клетку однодольного растения, необязательно клетку ячменя, клетку маиса, клетку овса, клетку риса, клетку сорго, клетку сахарного тростника или клетку пшеницы; или
(b) клетку двудольного растения, необязательно клетку сои, клетку подсолнечника, клетку томата, клетку культурной разновидности капусты, клетку хлопчатника, клетку сахарной свеклы или клетку табака.
10. Трансгенное растение, токсичное для насекомого-вредителя, относящегося к чешуекрылым, содержащее трансгенную растительную клетку по п. 9, где растение представляет собой:
(a) однодольное растение, необязательно растение ячменя, растение маиса, растение овса, растение риса, растение сорго, растение сахарного тростника или растение пшеницы; или
(b) двудольное растение, необязательно растение сои, растение подсолнечника, растение томата, растение культурной разновидности капусты, растение хлопчатника, растение сахарной свеклы или растение табака.
11. Трансгенное растение по п. 10, которое дополнительно содержит нуклеотидную последовательность, кодирующую второе средство для борьбы с насекомыми.
12. Трансгенная часть растения, токсичная для насекомого-вредителя, относящегося к чешуекрылым, из трансгенного растения по п. 10, где трансгенная часть растения содержит химерный инсектицидный белок по любому из пп. 1-3.
13. Трансгенное семя, токсичное для насекомого-вредителя, относящегося к чешуекрылым, трансгенного растения по п. 10, где семя содержит химерный инсектицидный белок по любому из пп. 1-3.
| US 8735560 B1, 27.05.2014 | |||
| RU 2007102339, 14.06.2005 | |||
| US 20150148288 A1, 28.05.2015. |
