Область техники, к которой относится изобретение
Настоящее изобретение относится в целом к трансгенным конструкциям, трансгенным животным, отличным от человека, содержащим трансгенные конструкции, способам создания и способам применения трансгенных животных, отличных от человека, содержащих трансгенные конструкции. Вариант осуществления данного изобретения относится к способам количественного анализа GPCR лигандов неинвазивно в интактных животных, тканевых срезах или в нативных клетках, используя трансгенную модель, содержащую систему биолюминесцентного трансгена-репортера, которая является чувствительной к модуляции путей после связывания лиганда с рецепторами GPCR.
Предпосылки создания изобретения
В разработке лекарственных средств коэффициенты отсева являются высокими, и только одно из пяти соединений проходит от разработки до утверждения в Управлении по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) (DiMasi, J.A., et al, J Health Econ 22,151-185, 2003). Более того, несмотря на резко возросший объем инвестирования, уровень внедрения новых лекарственных средств остается относительно постоянным в течение последних 30 лет со всего лишь двумя-тремя продвижениями в классах новых лекарственных средств в год, в конечном итоге доходящих до рынка (Lindsay M.A., Nature Rev. Drug Discovery, 2, 831-838, 2003).
Молекулярная и функциональная визуализация, применяемая на начальных стадиях разработки лекарственного средства, может предоставить данные о биологической активности и подтвердить, что предполагаемое лекарственное средство оказывает влияние на предназначенную для него мишень. Таким образом, существуют большие ожидания, что инвестирование в технологию молекулярной визуализации увеличит разработку лекарственных средств (Rudin M., Progress in Drug Res., vol. 62). Преимущества методов молекулярной визуализации над более традиционными считываниями состоят в том, что они могут быть выполнены в интактном организме с достаточным пространственным и временным разрешением для изучения биологических процессов in vivo. Кроме того, такие методы позволяют проведение повторяющегося, неинвазивного, единообразного и относительно автоматизированного исследования одной и той же биологической модели в разные моменты времени, таким образом, повышая статистическую мощность длительных исследований и, кроме того, уменьшая количество требуемых животных, снижая, таким образом, стоимость разработки лекарственного средства.
Молекулярная визуализация
Молекулярная визуализация относится к сближению подходов из различных дисциплин (клеточной и молекулярной биологии, химии, медицины, фармакологии, физики, биоинформатики и инженерии) для использования и интеграции методов визуализации в оценке конкретных молекулярных процессов на клеточном и субклеточном уровнях в живом организме. (Massoud T.F., Genes Dev. 17:545-580, 2003).
Появление генной инженерии привело к значительным изменениям в прикладной науке, включая, например, процесс разработки при поиске новых лекарственных средств. Таким же образом, разработка и использование методик визуализации на животных предоставляют новые средства для доклинических исследований (Maggie A. and Ciana P., Nat.Rev.Drug Discov. 4, 249-255, 2005). Животные модели традиционно были трудоемкими из-за трудности с количественной оценкой физиологических событий в режиме реального времени. С годами были разработаны новые способы визуализации для преодоления этой трудности, такие как магнитно-резонансная томография (MRI) и позитронно-эмиссионная томография (PET). Совсем недавно для неинвазивного обнаружения была использована биолюминесцентная визуализация на основе in vivo экспрессии люциферазы, светоизлучающего фермента светлячка.
Молекулярная визуализация: биолюминесценция
Биолюминесцентная визуализация (BLI) in vivo является чувствительным инструментом, который основан на регистрации светового излучения от клеток и тканей. Полезность технологии генов-репортеров позволяет анализировать конкретные клеточные и биологические процессы в живом животном посредством способов визуализации in vivo. Биолюминесценция, ферментативное генерирование видимого света живым организмом, является естественным явлением у многих не млекопитающих видов. (Contag, C.H., et al, Mol. Microbiol. 18:593-603, 1995). Люциферазы представляют собой ферменты, которые катализируют окисление субстрата для выделения фотона света (Greer L.F., III, Luminescence 17:43-74, 2002). Биолюминесценция североамериканского светлячка является наиболее широко изученной. Экспрессия гена люциферазы светлячка (luc) производит фермент люциферазу, который преобразует субстрат D-люциферин в неактивный оксилюциферин, давая в результате излучение зеленого света при 562 нм. Поскольку ткани млекопитающих естественно не испускают биолюминесценцию, in vivo BLI имеет большую привлекательность, поскольку изображения могут быть генерированы с очень небольшим фоновым сигналом.
BLI требует генной инженерии клеток или тканей с кассетой экспрессии, состоящей из биолюминесцентного гена-репортера под контролем выбранного промотора гена, конститутивно управляющего световым репортером (фиг.3). Для того чтобы индуцировать производство света, субстраты, такие как люциферин, вводят посредством интрацеребровентрикулярной (icv), внутривенной (iv), внутрибрюшинной (ip) или подкожной (sq) инъекции.
Свет, излучаемый люциферазой, может проникать на глубину от нескольких миллиметров до сантиметров; однако интенсивность фотонов снижается в 10 раз на каждый сантиметр глубины ткани (Contag C.H., et al, Mol. Microbiol. 18:593-603, 1995). Для обнаружения биолюминесценции in vivo должны использоваться чувствительные свето-детекторные инструменты. Детекторы измеряют количество фотонов, испускаемых на единицу площади. Низкие уровни света с длиной волны от 400 до 1000 нм могут быть обнаружены с помощью камер с полупроводниковой светочувствительной матрицей, которые конвертируют световые фотоны, попадающие на кремниевые пластины, в электроны (Spibey C.P. et al electrophoresis 22:829-836, 2001). Программное обеспечение способно конвертировать электронные сигналы в двухмерное изображение. Программное обеспечение также способно количественно оценивать интенсивность испускаемого света (количество испускаемых фотонов, попадающих на детекторы) и конвертировать эти числовые значения в псевдоцветное графическое или полутоновое изображение (фиг.2A и 2B). Фактические данные измеряются в фотонах, но псевдоцветная графика делает возможным быструю визуальную интерпретацию. Количественные измерения в представляющей интерес области могут быть необходимы для обнаружения более тонких различий. Применение охлажденных камер с полупроводниковой светочувствительной матрицей (CCD) снижает тепловой шум, а светонепроницаемый бокс позволяет оптимально визуализировать и количественно оценить производимый люциферазой свет (Contag C.H. and Bachmann, M.H., Annu. Rev. Biomed. Eng. 4:235-260, 2002).
Полезно, когда люциферазное изображение накладывается на изображение другого типа, такое как автограф или рентгенограмма, для анатомического расположения испускаемого сигнала (фиг.2B). Программное обеспечение накладывает изображения для визуализации и интерпретации.
Посредством объединения животной инженерии с методами молекулярной визуализации стало возможным проведение динамических исследований конкретных молекулярных процессов в живых животных. Этот подход способен потенциально повлиять на доклинические протоколы, таким образом, значительно изменяя все аспекты медицины (Maggie A. Trends Pharmacolo. Sci 25, 337-342, 2004).
Рецепторы, сопряженные с G-белком (GPCR) - GPCR в качестве мишени для лекарственных средств
GPCR составляют большое суперсемейство рецепторов клеточной поверхности, которые классифицируются на более чем 100 подсемейств на основе их общей топологической структуры; GPCR также называют семь трансмембранных (7TM) рецепторов. К GPCR наиболее часто обращаются как к мишеням для лекарственных средств в фармацевтической промышленности. Приблизительно 30% из всех продаваемых по рецепту лекарственных средств нацелены на GPCR, что делает это белковое семейство фармацевтически наиболее успешным классом мишеней (Jacoby, E; Chem. Med. Chem., 1: 761-782, 2006).
Взаимодействие между GPCR и их внеклеточными лигандами оказалось привлекательным моментом для вмешательства терапевтических средств. По этой причине фармацевтическая промышленность разработала биохимические количественные анализы для поиска новых лекарственных средств, чтобы исследовать эти взаимодействия лиганд-GPCR. Взаимодействие активированного GPCR с гетеротримерным G-белком катализирует обмен гуанозиндифосфата (GDP) на гуанозинтрифосфат (GTP), позволяя взаимодействие с несколькими нижележащими эффекторами (Cabrera-Vera T.M., Endocr. Rev. 24:765-781, 2003). Нижележащая передача сигнала зависит от G-альфа изоформы, которая предпочитается представляющим интерес GPCR. Белки семейства G-альфаq/11 стимулируют фосфолипазу С (PLC), в то время как представители семейств G-альфаi/0 и G-альфаs главным образом модулируют активность аденилатциклазы (AC). Если представляющий интерес GPCR передает сигнал через PLC, тогда наиболее широко применяемым методом на основе репортера для измерения активации GPCR является анализ высвобождения кальция (Ca+2), либо измеренный во флуоресцентном формате, при использовании Ca+2-чувствительных флуорофор (Sullivan E, Methods Mol. Biol. 114:125-133, 1999), либо в люминесцентном формате, при использовании экворина и хемилюминесцентного субстрата (Dupriez V.J., Receptors Channels 8: 319-330, 2002). Если представляющий интерес GPCR передает сигнал через AC, тогда цитозольное содержание циклического аденозинмонофосфата (цАМФ) может быть определено с использованием различных технологий обнаружения (Gabriel D. Assay Drug Dev. Technol. 1:291-303, 2003).
Количественные анализы на основе GPCR-репортера обширно используются в современных программах поиска новых лекарственных средств. Как правило, GPCR-репортеры вводят в клеточные системы для обеспечения in vitro высокопроизводительного скрининга (HTS) крупных фармацевтических библиотек для идентификации лигандов или соединений, которые активируют или модулируют конкретный GPCR. Вторичные и последующие клеточные анализы подтверждают и уточняют любые «хиты», идентифицированные в HTS в отношении конкретного GPCR; но опять же, эти анализы полагаются на способы рекомбинантного ДНК для введения клонированного GPCR в трансформированный тип клеток. В то время как трансформированные типы клеток обладают отличной пролиферативной способностью для поддержания крупных скрининговых программ, они часто демонстрируют аберрантные генетические и функциональные характеристики и, следовательно, при использовании этой парадигмы встречается значительный отсев предполагаемых «хитов» из HTS.
В течение нескольких лет для измерения функциональной активности GPCR использовались основанные на биолюминесценции анализы генов-репортеров (Hill, S.J. Curr.Opin.Pharmacol.1: 526-532, 2001). Этот формат анализов является очень чувствительным из-за низкого фонового сигнала биолюминесцентного считывания и этапов усиления сигнала между активацией GPCR и совокупной экспрессией гена-репортера.
Ответный элемент цАМФ (CRE) в промоторе гена-репортера делает возможным специфический мониторинг G-белок-зависимой передачи сигнала. Когда лиганд связывается с GPCR, это вызывает конформационное изменение в GPCR, которое позволяет ему активировать связанный G-белок. Фермент аденилатциклаза представляет собой клеточный белок, который может регулироваться G-белками. Активность аденилатциклазы активируется или ингибируется, когда она связывается с субъединицей активированного G-белка. Сигнальная трансдукция зависит от типа G-белка. Аденилатциклаза влияет на увеличение или уменьшение производства цАМФ в клетке. Произведенный цАМФ является вторичным мессенджером в клеточном метаболизме и аллостерическим активатором протеинкиназы А (PKA). Когда цАМФ отсутствует, комплекс PKA является неактивным. Когда цАМФ связывается с регуляторной субъединицей PKA, ее конформация меняется, вызывая диссоциацию регуляторных субъединиц, которые активируют протеинкиназу А и позволяют дальнейшие биологические эффекты. PKA затем фосфорилирует и активирует транскрипционный фактор CREB. CREB связывается с определенными последовательностями ДНК, называемыми чувствительными элементами цАМФ (CRE), и, тем самым, увеличивает или уменьшает транскрипцию, и, таким образом, и экспрессию, определенных генов, таких как ген-репортер люциферазы.
Трансген CreLuc предназначен для анализа активации всех трех главных GPCR либо непосредственно через цАМФ внутриклеточный путь передачи сигнала, либо опосредованно с помощью передачи сигнала посредством PLC. Поскольку любой из типов клеток содержит множество различных типов GPCR на клеточной поверхности (таким образом, любая клетка будет иметь передачу сигнала GPCR через G-альфаq/11, G-альфаi/0 и G-альфаs, происходящую одновременно внутри клетки), общепринятым взглядом будет предположение, что маловероятно, что такой трансген, как CreLuc, будет достаточно специфичен для различения любого конкретного лиганда GPCR. Однако авторы показывают в данном случае, что трансген CreLuc способен различать лиганды GPCR. Авторы прогнозируют, что биолюминесцентный сигнал для люциферазного репортера в клетках, тканевых срезах или интактном животном будет увеличен с форсколином и модулирован лигандами для Gs, Gq или Gi рецепторов. В таблице 1 показан ожидаемый эффект, который активация/ингибирование GPCR будет иметь на системе CreLuc репортера при связывании с лигандом GPCR. Кроме того, авторы показывают данные, что их новая система CreLuc репортера может различать различные классы лигандов GPCR и что такая система репортера применима для выявления новых лигандов GPCR при использовании в клетках, тканевых срезах и интактном животном.
Прогнозируемые изменения биолюминесцентного сигнала от репортера CreLuc при связывании лиганда с определенными GPCR
Трансгенная модель репортера GPCR биовизуализации
Значительный отсев потенциальных лекарств-кандидатов в существующей парадигме поиска новых лекарственных средств встречается на фазе перехода от анализов клеточного репортера к моделям in vivo. Доступны многочисленные in vivo модели, которые повторяют полностью или частично отдельное заболевание человека. Демонстрация активности ведущих соединений в этих моделях является значительной вехой в продвижении новых химических GPCR лекарственных средств. Животные модели заболевания, как правило, требуют большого числа животных и количества времени, позволяющего развитие их фенотипа и точный анализ влияния кандидатных соединений на изменение исхода заболевания. После испытания in vitro следующим уровнем испытания лекарства-кандидата в комплексной системе является использование испытаний in vivo или моделей in vivo болезненных состояний на механистической основе. Неудачи в изменении исходов индуцированных заболеваний плохо понятны, но все еще приводят к большому уровню отсева соединений-кандидатов в процессе разработки лекарственного средства.
Трансгенная модель, содержащая анализ репортера связывания и активации лиганда GPCR, станет значительным улучшением существующей парадигмы поиска новых лекарственных средств для GPCR. Например, вариант осуществления данного изобретения описывает содержащий трансген цАМФ-репортерный анализ, основанный на люциферазном репортере (CreLuc), который объединен с молекулярным визуализированием в интактных животных, тканях или клетках, который смог бы значительно ускорить поиск нового лекарственного средства - лиганда GPCR (Bhaumik, S. and Gambhir, S.S., Proc. Natl. Acad. Sci. USA, 99:377-382 2002; Hasan M.T., et al., Genesis 29:116-122, 2001). Как описано в настоящем описании, варианты осуществления трансгенного животного, отличного от человека, данного изобретения предлагают следующие не ограничивающие преимущества:
1. Анализы на основе тканей или клеток имеют ту же репортерную систему, что и анализ трансгенной in vivo модели, таким образом, уменьшая число неизвестных в комплексных интактных биологических системах.
2. Неинвазивная визуализация позволяет количественный анализ активности лиганда или соединения в анализе временной зависимости у одного и того же животного.
3. Неинвазивная визуализация снижает количество животных в исследовании и приводит к большей статистической мощности на каждое животное, являющееся своим собственным контролем, где контролем будет проанализированное животное в нулевой момент времени.
4. Трансгенное животное будет источником клеток и тканей для обеспечения параллельных анализов, проводимых in vitro или ex vivo.
5. Анализ трансгенного животного обеспечит анализ активности лиганда в нативных типах клеток, что приводит к более реалистичному профилю взаимодействия лиганд:рецептор.
6. Трансгенное животное позволяет одновременную оценку фармакодинамики и фармакокинетики лигандов GPCR.
7. Трансгенное животное позволяет одновременную идентификацию тканевой и клеточной специфичности на уровне органа или интактного животного.
8. Трансгенное животное позволяет скрещивание с другими генетически измененными моделями для выявления новых сигнальных путей и их ответа на конкретные лиганды.
Многие трансгенные животные, сконструированные с различными репортерами, применяются в исследовании молекулярных процессов, таких как метаболизм лекарственных средств (Zhang W., et al., Drug Metab. Dispos. 31:1054-1064, 2003), генотоксичность (Gossen J.A., et al., Proc. Natl. Acad. Sci. USA 86:7971-7975, 1989) и эффекты токсических соединений (Sacco M.G. et al., Nat. Biotechnol. 15:1392-1397, 1997). Для достижения своих целей проектирования, животное-GPCR-репортер, подходящее для исследований молекулярной визуализации, должно включать несколько элементов, приспособленных, чтобы позволить одновременно высокие уровни экспрессии репортера для обеспечения большого окна биолюминесцентного обнаружения, а также экспрессию в каждом типе клеток для обеспечения обширных тонких анализов in vivo биораспределения лиганда или исследуемого соединения.
Сложность и разнообразие механизмов, вовлеченных в экспрессию генов, никогда не позволят исследователям сконструировать гены, способные во всех случаях экспрессироваться в трансгенных животных полностью предсказуемым образом (Pinkert, C. A. (ed.) 1994. Transgenic animal technology: A laboratory handbook. Academic Press, Inc., San Diego, Calif.; Monastersky G. M. and Robl, J. M. (ed.) (1995) Strategies in Transgenic Animal Science. ASM Press. Washington D.C.). Только через обширные пробы и ошибки можно достичь уникальных комбинаций трансгенных структур для выполнения целей проектирования модели, требуемых для биовизуализации GPCR репортеров.
Полезность трансгенного GPCR репортера по сравнению с рекомбинантными клеточными анализами
По мере того, как скрининговая технология приближается к моменту понимания поведения отдельных GPCR, ясно, что скорее, чем быть выключателем, эти рецепторы действуют больше как микропроцессоры информации. Это ввело феномен функциональной селективности, при котором определенные лиганды инициируют только часть сигнального механизма, опосредованного данным рецептором, что открыло новые горизонты для поиска новых лекарственных средств. Необходимость в раскрытии взаимоотношений новых лигандов GPCR и количественной оценки эффекта лекарственного средства на эту комплексную систему для направления медицинской химии ставит значительно более высокие требования к любому фармакологическому репортерному анализу. Эта концепция ведет к возвращению к полным системным анализам от редукционистских рекомбинантных клеточных скрининговых систем. Профилирование лигандной активности с конкретным GPCR или набором GPCR в нативном клеточном окружении, как ожидается, улучшит успешность выявления новых лекарственных средств против ключевого класса фармацевтически важных рецепторов (Kenakin TP, Nat. Rev. Drug Discov. 8,617-625, 2009). Животная модель, содержащая биолюминесцентный GPCR репортерный трансген, является весьма желательной стратегией молекулярной визуализации для определения активности лиганда GPCR в интактной биологической комплексной системе с целью улучшения поиска новых лекарственных средств для борьбы с заболеваниями человека.
Поскольку активация CRE/CREB затрагивает множество различных биологических процессов, существует значительный интерес в исследовании активации CRE с использованием репортерной экспрессирующей системы CRE/CREB. Циклический аденозинмонофосфат (цАМФ) является вторичным мессенджером во внутриклеточной сигнальной трансдукции после активации рецептора и последующей активации протеинкиназы и, таким образом, вовлечен в регуляцию многих биологических процессов. CREB (белок, связывающий чувствительный элемент цАМФ), фосфорилированный киназой, активированной цАМФ, связывается с чувствительным элементом цАМФ (CRE) в промоторной области многих генов и активирует транскрипцию (Shaywitz and Greenberg, Annul. Rev. Biochem., 68:821-861, 1999). Трансгенных мышей, несущих шесть тандемов CRE с минимальным промотором вируса простого герпеса (HSV), управляющим экспрессией бета-галактозидазы, использовали для исследования CRE-опосредованной экспрессии генов в срезах мозга в ответ на хроническую обработку антидепрессантом (Thome J., et al., J. Neurosci. 20:4030-4036, 2000). Аналогичным образом, трансгенные мыши, несущие четыре копии промотора CRE гена крысиного соматостатина, соединенного с промотором тимидинкиназы и геном люциферазы, были использованы для исследования активации CRE в гистологических срезах мозга или гомогенатах (Boer et al., PloS One, May 9; 2(5):e431, 2007). Однако исследования до сих пор затруднены необходимостью сортировки большого количества трансгенных линий для обнаружения подходящей животной модели. Кроме того, после обнаружения подходящей линии относительно низкие уровни экспрессии репортера требуют эвтаназии трансгенного животного для измерения гена-репортера, что требует большого количества животных для использования в одной экспериментальной парадигме.
Вариантом осуществления данного изобретения является разработка трансгена, содержащего инсуляторные элементы, ответные элементы, промоторные элементы, гены-репортеры и функциональные элементы. Трансген может быть быстро введен в животных, не относящихся к человеку, благодаря высокой скорости интеграции и высокого уровня экспрессии гена-репортера, таким образом, трансгенные животные могут быть легко созданы в качестве моделей для исследования активации регуляторного элемента in vivo (т.е. в живом животном), in situ (например, срезы мозга, интактные цельные органы) или in vitro (например, первичные культуры клеток из трансгенного животного, гомогенаты тканей).
Вариантом осуществления данного изобретения является трансген, содержащий CRE Luc репортерную систему, используемую в трансгенных животных, не относящихся к человеку, в качестве моделей для количественной оценки активностей лигандов GPCR через регуляцию уровней внутриклеточного цАМФ in vivo. В качестве неограничивающего примера авторы продемонстрировали изменения в люциферазном репортере с помощью биолюминесценции в выделенных первичных клетках и в интактных животных, используя общепринятые регуляторы цАМФ. В другом варианте осуществления активация репортера количественно проанализирована и подтверждена в тканевых экстрактах при использовании люциферазных количественных анализов ex vivo. Ответ трансгена CRE Luc задокументирован во множестве мышиных линий и показывает одиночные или множественные тканевые активационные профили. Более того, в качестве неограничивающих примеров авторы показывают, что конкретные лиганды GPCR активировали трансген CRE Luc в интактных животных, тканевых срезах и первичных клетках.
Краткое описание изобретения
В целом, данное изобретение предоставляет трансгенные конструкции, трансгенных животных, отличных от человека, содержащих трансгенные конструкции, способы создания и способы применения трансгенных животных, отличных от человека, содержащих трансгенные конструкции. Вариант осуществления данного изобретения предоставляет трансгенную конструкцию, содержащую CRE Luc репортерную систему. Вариантом осуществления данного изобретения является введение трансгенной конструкции, содержащей CRE Luc репортерную систему, в животное, отличное от человека.
Поскольку модуляция цАМФ является одним из ключевых активационных путей для GPCR, данное изобретение служит платформой для количественного определения в интактных животных, тканевых срезах или клетках, причем активация GPCR лигандом или соединением через активацию гена-репортера, где ген-репортер обеспечивает измеряемый биолюминесцентный сигнал, например метаболизм люциферина с помощью люциферазы. Данное изобретение представляет инструменты для улучшения перехода объектов открытия новых лекарственных средств, таких как лиганды или соединения, от клеточных количественных анализов к интактным животным. В варианте осуществления данного изобретения применяют такую же репортерную систему в нативных клетках, которая снижает скорость отсева новых лигандов GPCR, одновременно предоставляя данные биодоступности.
Вариантом осуществления данного изобретения является трансгенное животное, отличное от человека, имеющее геном, содержащий трансген, который содержит первый инсуляторный элемент, ответный элемент, промотор, биолюминесцентный репортер, функциональный элемент и второй инсуляторный элемент.
Вариантом осуществления данного изобретения является трансгенное животное, отличное от человека, где первый инсуляторный элемент выбран из группы, состоящей из участков прикрепления к ядерному матриксу (MAR), ДНКаза I-гиперчувствительного сайта (HS4) и инвертированных концевых повторов (ITR). Дополнительным вариантом осуществления данного изобретения является трансгенное животное, отличное от человека, где второй инсуляторный элемент выбран из группы, состоящей из элемента прикрепления к ядерному матриксу (MAR), HS4 и ITR. Дополнительным вариантом осуществления данного изобретения является трансгенное животное, отличное от человека, где первый инсуляторный элемент является таким же, как второй инсуляторный элемент.
Вариант осуществления данного изобретения охватывает трансгенное животное, отличное от человека, где ответный элемент выбран из группы, состоящей из ответного элемента цАМФ (CRE), активирующего белка 1 (ASP1), ответного элемента глюкокортикоида (GRE), ответного элемента теплового шока (HSE), ответного элемента сыворотки (SRE), ответного элемента тиреоида (TRE) и ответного элемента эстрогена (ERE). Дополнительным вариантом осуществления данного изобретения является трансгенное животное, отличное от человека, где ответный элемент повторяется в тандеме от двух до двадцати четырех раз. Дополнительным вариантом осуществления данного изобретения является трансгенное животное, отличное от человека, где ответный элемент повторяется в тандеме шесть раз. Дополнительным вариантом осуществления данного изобретения является трансгенное животное, отличное от человека, где ответный элемент представляет собой CRE, дополнительно, где ответный элемент CRE может быть единственным элементом или повторяемым от двух до двадцати четырех раз.
Вариантом осуществления данного изобретения является трансгенное животное, отличное от человека, где промотор представляет собой минимальный промотор тимидинкиназы вируса простого герпеса (HSV TK min).
Вариантом осуществления данного изобретения является трансгенное животное, отличное от человека, где биолюминесцентный репортер выбран из группы, состоящей из люциферазы, хлорамфениколацетилтрансферазы (CAT), бета-галактозидазы, секретированной щелочной фосфатазы (SEAP), человеческого гормона роста (HGH) и зеленого флуоресцентного белка (GFP).
Вариантом осуществления данного изобретения является трансгенное животное, отличное от человека, где функциональный элемент представляет собой ген человеческого гормона роста (hGH).
Вариантом осуществления данного изобретения является трансгенное животное, отличное от человека, где трансген содержит SEQ ID NO:18.
Вариантом осуществления данного изобретения является трансгенное животное, отличное от человека, где трансген содержит SEQ ID NO:19.
Вариантом осуществления данного изобретения является клетка, выделенная из трансгенного животного, отличного от человека, или тканевый срез, выделенный из трансгенного животного, отличного от человека, по п.1.
Вариантом осуществления данного изобретения является способ идентификации лиганда рецептора, сопряженного с G-белком (GPCR), включающий (a) измерение количества биолюминесценции в трансгенном животном, отличном от человека, раскрытом в данном описании; (b) введение тестируемого средства трансгенному животному, отличному от человека; (c) измерение количества биолюминесценции трансгенного животного, отличного от человека, в один или более моментов времени после введения тестируемого средства; и (d) сравнение количества биолюминесценции, измеренной в (a), с количеством биолюминесценции, измеренной в (c), где разница в количестве биолюминесценции в (a) по сравнению с (c) идентифицирует тестируемое средство как лиганд GPCR.
Вариантом осуществления данного изобретения является способ идентификации лиганда рецептора, сопряженного с G-белком (GPCR), включающий (a) приготовление тканевого среза из трансгенного животного, отличного от человека, раскрытого в данном описании; (b) измерение количества биолюминесценции в тканевом срезе; (c) введение тестируемого средства в тканевой срез; (d) измерение количества биолюминесценции тканевого среза в один или более моментов времени после введения тестируемого средства; и (e) сравнение количества биолюминесценции, измеренной в (b), с количеством биолюминесценции, измеренной в (d), где разница в количестве биолюминесценции в (b) по сравнению с (d) идентифицирует тестируемое средство как лиганд GPCR.
Вариантом осуществления данного изобретения является способ идентификации лиганда рецептора, сопряженного с G-белком (GPCR), включающий (a) приготовление клетки, выделенной из трансгенного животного, отличного от человека, раскрытого в данном описании; (b) измерение количества биолюминесценции в клетке; (c) введение тестируемого средства в клетку; (d) измерение количества биолюминесценции в клетке в один или более моментов времени после введения тестируемого средства; и (e) сравнение количества биолюминесценции, измеренной в (b), с количеством биолюминесценции, измеренной в (d), где разница в количестве биолюминесценции в (b) по сравнению с (d) идентифицирует тестируемое средство как лиганд GPCR.
Вариантом осуществления данного изобретения является способ мониторинга функции GPCR в животном, отличном от человека, включающий (a) трансгенное модифицирование животного, отличного от человека, для экспрессии трансгена, содержащего первый инсуляторный элемент, ответный элемент, промотор, биолюминесцентный репортер, функциональный элемент и второй инсуляторный элемент; (b) мониторинг биолюминесценции из животного, отличного от человека; и (c) коррелирование указанной биолюминесценции с функцией GPCR.
Вариантом осуществления данного изобретения является способ мониторинга функции GPCR в животном, отличном от человека, включающий (a) трансгенное модифицирование животного, отличного от человека, для экспрессии трансгена, содержащего первый инсуляторный элемент, ответный элемент, промотор, биолюминесцентный репортер, функциональный элемент и второй инсуляторный элемент; (b) мониторинг люциферазы животного, отличного от человека; и (c) коррелирование указанной биолюминесценции с функцией GPCR.
Вариантом осуществления данного изобретения является способ мониторинга функции GPCR в животном, отличном от человека, включающий (a) трансгенное модифицирование животного, отличного от человека, для экспрессии трансгена, содержащего первый инсуляторный элемент, ответный элемент, промотор, биолюминесцентный репортер, функциональный элемент и второй инсуляторный элемент; (b) манипулирование животным, отличным от человека, для имитации аспекта болезненного состояния; (c) мониторинг биолюминесценции от животного, отличного от человека; и (d) коррелирование указанной биолюминесценции с функцией GPCR.
Вариантом осуществления данного изобретения является способ мониторинга функции GPCR в животном, отличном от человека, включающий (a) трансгенное модифицирование животного, отличного от человека, для экспрессии трансгена, содержащего первый инсуляторный элемент, ответный элемент, промотор, биолюминесцентный репортер, функциональный элемент и второй инсуляторный элемент; (b) манипулирование животным, отличным от человека, для имитации аспекта болезненного состояния; (c) мониторинг люциферазы животного, отличного от человека; и (d) коррелирование указанной биолюминесценции с функцией GPCR.
Вариантом осуществления данного изобретения является способ создания трансгенного животного, отличного от человека, для применения в мониторинге функции GPCR, включающий (a) трансгенное модифицирование животного, отличного от человека, для экспрессии трансгена, содержащего первый инсуляторный элемент, ответный элемент, промотор, биолюминесцентный репортер, функциональный элемент и второй инсуляторный элемент; (b) измерение количества биолюминесценции в трансгенном животном, отличном от человека, из (a); (c) введение лиганда GPCR трансгенному животному, отличному от человека; (d) измерение количества биолюминесценции трансгенного животного, отличного от человека, в один или более моментов времени после введения лиганда GPCR; и (e) сравнение количества биолюминесценции, измеренной в (b), с количеством биолюминесценции, измеренной в (d), где разница в количестве биолюминесценции в (b) по сравнению с (d) идентифицирует трансгенное животное, отличное от человека, для применения в мониторинге функции GPCR.
Вариантом осуществления данного изобретения является способ идентификации соединения, которое модулирует рецептор, сопряженный с G-белком (GPCR), включающий (a) приготовление клетки, выделенной из трансгенного животного, отличного от человека, раскрытого в данном описании; (b) измерение количества биолюминесценции в клетке; (c) введение тестируемого средства в клетку; (d) измерение количества биолюминесценции в клетке в один или более моментов времени после введения тестируемого средства; и (e) сравнение количества биолюминесценции, измеренной в (b), с количеством биолюминесценции, измеренной в (d), где разница в количестве биолюминесценции в (b) по сравнению с (d) идентифицирует тестируемое средство как лиганд GPCR.
Вариантом осуществления данного изобретения является способ идентификации соединения, которое модулирует рецептор, сопряженный с G-белком (GPCR), включающий (a) предоставление клеток, выделенных из трансгенного животного, отличного от человека, раскрытого в данном описании, в один или более сосудов; (b) введение контроля в один или более сосудов; (c) введение тестируемого средства в один или более сосудов; и (d) измерение количества люциферазы в сосудах, где разница в количестве люциферазы, измеренной в сосуде(ах), содержащем контроль, по сравнению с количеством люциферазы в сосуде(ах), содержащем тестируемое средство, идентифицирует соединение как модулирующее GPCR.
Вариантом осуществления данного изобретения является способ идентификации соединения, которое модулирует рецептор, сопряженный с G-белком (GPCR), включающий (a) предоставление клеток, выделенных из трансгенного животного, отличного от человека, раскрытого в данном описании, в один или более сосудов; (b) введение неспецифического модулятора цАМФ в один или более сосудов; (c) введение тестируемого средства в один или более сосудов; и (d) измерение количества люциферазы в сосудах, где разница в количестве люциферазы, измеренной в сосуде(ах), содержащем только неспецифический модулятор цАМФ, по сравнению с количеством люциферазы в сосуде(ах), содержащем неспецифический модулятор цАМФ и тестируемое средство, идентифицирует соединение как модулирующее GPCR.
Вариантом осуществления данного изобретения является способ идентификации соединения, которое модулирует рецептор, сопряженный с G-белком (GPCR), включающий (a) предоставление тканевых срезов, выделенных из трансгенного животного, отличного от человека, раскрытого в данном описании, в один или более сосудов; (b) введение контроля в один или более сосудов; (c) введение тестируемого средства в один или более сосудов; и (d) измерение количества люциферазы в сосудах, где разница в количестве люциферазы, измеренной в сосуде(ах), содержащем контроль, по сравнению с количеством люциферазы в сосуде(ах), содержащем тестируемое средство, идентифицирует соединение как модулирующее GPCR.
Вариантом осуществления данного изобретения является способ идентификации соединения, которое модулирует рецептор, сопряженный с G-белком (GPCR), включающий (a) предоставление тканевых срезов, выделенных из трансгенного животного, отличного от человека, раскрытого в данном описании, в один или более сосудов; (b) введение неспецифического модулятора цАМФ в один или более сосудов; (c) введение тестируемого средства в один или более сосудов; и (d) измерение количества люциферазы в сосудах, где разница в количестве люциферазы, измеренной в сосуде(ах), содержащем только неспецифический модулятор цАМФ, по сравнению с количеством люциферазы в сосуде(ах), содержащем неспецифический модулятор цАМФ и тестируемое средство, идентифицирует соединение как модулирующее GPCR.
Вариантом осуществления данного изобретения является способ идентификации соединения, которое модулирует рецептор, сопряженный с G-белком (GPCR), включающий (a) предоставление клеток, выделенных из трансгенного животного, отличного от человека, раскрытого в данном описании, в один или более сосудов; (b) введение клеточного стимулятора в один или более сосудов; (c) введение тестируемого средства в один или более сосудов; и (e) измерение количества люциферазы в сосудах, где разница в количестве люциферазы, измеренной в сосуде(ах), содержащем клеточный стимулятор, по сравнению с количеством люциферазы в сосуде(ах), содержащем тестируемое средство и клеточный стимулятор, идентифицирует соединение как модулирующее GPCR.
Вариантом осуществления данного изобретения является способ идентификации соединения, которое модулирует рецептор, сопряженный с G-белком (GPCR), включающий (a) предоставление клеток, выделенных из трансгенного животного, отличного от человека, раскрытого в данном описании, в один или более сосудов; (b) введение клеточного стимулятора в один или более сосудов; (c) введение неспецифического модулятора цАМФ в один или более сосудов; (d) введение тестируемого средства в один или более сосудов; и (e) измерение количества люциферазы в сосудах, где разница в количестве люциферазы, измеренной в сосуде, содержащем клеточный стимулятор и неспецифический модулятор цАМФ, по сравнению с количеством люциферазы в сосуде(ах), содержащем клеточный стимулятор, неспецифический модулятор цАМФ и тестируемое средство, идентифицирует соединение как модулирующее GPCR.
Вариантом осуществления данного изобретения является способ идентификации соединения, которое модулирует рецептор, сопряженный с G-белком (GPCR), включающий (a) предоставление тканевых срезов, выделенных из трансгенного животного, отличного от человека, раскрытого в данном описании, в один или более сосудов; (b) введение клеточного стимулятора в один или более сосудов; (c) введение тестируемого средства в один или более сосудов; и (d) измерение количества люциферазы в сосудах, где разница в количестве люциферазы, измеренной в сосуде(ах), содержащем клеточный стимулятор, по сравнению с количеством люциферазы в сосуде(ах), содержащем тестируемое средство и клеточный стимулятор, идентифицирует соединение как модулирующее GPCR.
Вариантом осуществления данного изобретения является способ идентификации соединения, которое модулирует рецептор, сопряженный с G-белком (GPCR), включающий (a) предоставление тканевых срезов, выделенных из трансгенного животного, отличного от человека, раскрытого в данном описании, в один или более сосудов; (b) введение клеточного стимулятора в один или более сосудов; (c) введение неспецифического модулятора цАМФ в один или более сосудов; (d) введение тестируемого средства в один или более сосудов; и (e) измерение количества люциферазы в сосудах, где разница в количестве люциферазы, измеренной в сосуде, содержащем клеточный стимулятор и неспецифический модулятор цАМФ, по сравнению с количеством люциферазы в сосуде(ах), содержащем клеточный стимулятор, неспецифический модулятор цАМФ и тестируемое средство, идентифицирует соединение как модулирующее GPCR.
Краткое описание фигур
На фиг.1 показана CreLuc биовизуализирующая мышиная модель. На данной фигуре проиллюстрирована внутриклеточная активация трансгена-репортера CRE Luc всеми тремя типами GPCR (Gi, Gs и Gq), или непосредственно через цАМФ путь, или опосредованно через PLC путь (панель A). Изменение в биолюминесценции люциферазного репортера в ответ на индукцию форсколином проиллюстрированы для трех типов GPCR (панель B). Форсколин будет усиливать передачу сигнала Gs и Gq и, таким образом, CreLuc биолюминесценция будет усиливаться, в то время как индукция Gi будет ослаблять сигнал репортера. Gαs активирует цАМФ зависимый путь непосредственной стимуляцией AC, Gαs ингибирует производство цАМФ, и Gαq стимулирует PLC, приводя к генерированию двух вторичных мессенджеров IP3 и DAG. Сокращения: α,α-субъединица G-белка; β,β-субъединица G-белка; γ,γ-субъединица G-белка; AC, аденилатциклаза; PLC, фосфолипаза C; PKA, протеинкиназа A; PKC, протеинкиназа C; DAG, диацилглицерол; IP3, инозитолтрифосфат; Ca+2, кальций; CaMK, кальций/кальмодулин протеинкиназа; цАМФ, циклический аденозинмонофосфат; CRE, чувствительный элемент цАМФ; CREB, белок, связывающий чувствительный элемент цАМФ.
На фиг.2A показана в режиме реального времени in vivo биолюминесцентная визуализация и описаны преимущества использования системы. IVIS100 (Xenogen) биовизуализирующий прибор с компьютерной автоматизированной системой позволяет в режиме реального времени in vivo визуализацию, используя свет, испускаемый биолюминесцентным геном-репортером, например люциферазой, экспрессированным in vivo. Компьютерная программа обеспечивает количественное определение сигнала неинвазивно и длительно. В режиме реального времени in vivo визуализация имеет множество преимуществ над традиционным in vivo испытанием соединений. Традиционные исследования на животных требуют отдельных мышей в многократные моменты обработки, в то время как исследования, использующие биовизуализирующие модели, позволяют отбирать пробы у одних и тех же животных в многократные моменты обработки и повторно использовать для многократных обработок. Как показано на данной фигуре, период времени 0 часов, 2 часа, 4 часа и 8 часов потребует 24 животных (n=6 на момент времени), используя существующую методологию, тогда как при использовании технологии биовизуализации будут необходимы только 6 животных. Это приводит к нескольким преимуществам, которые включают: более высокую пропускную способность, поскольку необходимо меньше подопытных животных, позволяя тестировать больше соединений на эффективность; большее содержание и качество данных, поскольку временные и пространственные данные могут быть получены от одного и того же животного; и уменьшение статистической ошибки, которое улучшает качество принятых решений относительно отдельных соединений.
На фиг.2B показано типичное визуальное изображение индукции соединения в CreLuc трансгенной мыши. Введение изопротеренола (правая панель) усиливает спинномозговую экспрессию репортера CRE Luc по сравнению с базальными уровнями (левая панель). Биолюминесцентное обнаружение представлено визуально на изображении животного в белом свете как псевдоцветное представление в серой шкале.
На фиг. 3 показано схематическое представление структуры трансгена, содержащего многократные элементы ДНК для увеличения экспрессии. Схематическая структура трансгена содержит следующие элементы: инсуляторный элемент, показанный на этой фигуре как участки прикрепления к ядерному матриксу (MAR), для генерирования независимой от положения экспрессии; чувствительный элемент, представленный CRE-цАМФ, повторенным шесть раз (6X CRE); промоторный элемент, показанный как минимальный промотор тимидинкиназы вируса простого герпеса (HSV TK min); репортерный элемент, который представлен геном люциферазы, оптимизированным для экспрессии у млекопитающих (LUC2); и функциональный элемент, изображенный геном человеческого гормона роста с хвостом поли А (hGH poly A) для повышения экспрессии трансгена.
На фиг.4: In vitro подтверждение трансгенных векторов CreLuc. Гибридные или синтетические (synth) векторы с 30, 100 или 199 нг ДНК трансфицировали в клетки CHO (вместе с вектором положительного контроля люциферазы Renilla) Lipofectamine 2000 (Invitrogen, Карлсбад, Калифорния, кат № 11668-019). Через два дня клетки стимулировали 3 мкмоль/л форсколина (Sigma, Сент-Луис, Миссури, кат. № F6886) в течение 4 часов и затем измеряли люциферазную активность с помощью Dual Glo Luciferase Assay System (Promega, Мэдисон, Висконсин, кат. № E2920). Результаты показывают дозозависимое увеличение люциферазного сигнала обоими векторами. Более высокие уровни индукции достигались гибридным вектором.
На фиг.5A показано воздействие ингибиторов PDE на уровни цАМФ у нормальных мышей (воспроизведенный из Cheng JB, JPET, 280, 621-626). Мышам Balb/c вводили или растворитель, или лекарственные средства, как перечислено на оси Х, кровь собирали через 20 минут и количественно анализировали на цАМФ с помощью радиоиммунологического анализа цАМФ. Как CP-80,633, так и ролипрам значительно повышали уровни цАМФ в плазме при 10 мг/кг.
На фиг.5B показана in vivo стимуляция цАМФ в плазме. Самкам FVB/Tac вводили интраперитонеально или растворитель (1% DMSO), или лекарственное средство, затем через 30 минут образцы крови собирали и количественно анализировали на цАМФ с помощью ELISA (Assay Designs, Анн-Арбор, Мичиган, кат. № 900-163). Использованные лекарственные средства были следующими: 5 мг/кг форсколина (F) (Sigma F6886), 5 мг/кг водорастворимого форсколина (H20F) (Calbiochem 344273), 10 мг/кг ролипрама (R) (Sigma R6520) или комбинации любого форсколина плюс ролипрам (F/R). Статистически значимое увеличение, в 14 раз, наблюдали при обработке водорастворимым форсколином в комбинации с ролипрамом, как определено критерием Стьюдента. Форсколин увеличивает уровни цАМФ путем активации аденилатциклазы, в то время как ингибиторы PDE4, такие как ролипрам, повышают цАМФ в плазме путем предотвращения гидролиза цАМФ. Комбинация ролипрама и водорастворимого форсколина увеличивает уровни цАМФ in vivo в 14 раз. Эту комбинацию использовали для обеспечения большого окна индукции для скрининга основателя посредством биовизуализации; репрезентативное исследование показано на фиг.6.
На фиг.6 показаны результаты начальной индукции на основателя и селекции линии для мышиной модели репортера CreLuc. Многочисленные трансгенные линии отсортировали на индукцию люциферазы форсколином и ролипрамом in vivo, и затем ткани выделяли и количественно анализировали на люциферазные ферменты. Трансгенных мышей биовизуализировали перед введением (базальные уровни экспрессии), затем той же мыши вводили интраперитонеально 10 мг/кг ролипрама и 5 мг/кг водорастворимого форсколина и биовизуализировали через 4 часа после введения (индуцированная экспрессия). A (сублиния № 90): введение форсколин/ролипрама увеличивало базальную экспрессию трансгена-репортера CreLuc в легких и других тканях; B (сублиния № 219): индукция базальной экспрессии главным образом в кишечнике; C (сублиния № 44): не обнаруживаемая базальная экспрессия и репортер индуцирован в головной мозг плюс другие ткани; D (сублиния № 28): необнаруживаемая базальная экспрессия, которая увеличена в тимусе и печени; E (сублиния № 187): необнаруживаемая базальная экспрессия, которая индуцирована в головном и спинном мозге. Как и ожидалось от случайно интегрированного трансгена, были вариации между линиями в базальной экспрессии, тканевом распределении и ответе на индукцию. Двадцать линий были определены как имеющие более чем 5X индукцию в одной или нескольких тканях. Вариации в тканевом профиле показывают, что одиночная ткань (т.е. легкие, печень, головной мозг) позволяет генерирование визуализации, лишенной фонового тканевого ответа, в то время как множественные ткани позволяют генерирование широкого профиля ответа соединения.
На фиг.7 показана общая схема CreLuc скрининговых количественных анализов in vivo или ex vivo.
На фиг.8A показаны эффекты изопротеренола (ISO) и AMN082 (AMN) на люминесценцию в срезах цельного мозга мышей CreLuc (линия 187).
На фиг.8B показаны эффекты форсколина на люминесценцию в срезах цельного мозга мышей CreLuc (линия 44) со временем. Время представлено на оси Х в минутах. Форсколин в количестве 50 мкмоль/л или растворитель (DMSO) добавляли в момент времени = 2880, отмеченный стрелкой на нижней панели.
На фиг.9 показано схематическое представление выделения и обработки соединениями первичных нейронных клеток мышей CreLuc.
На фиг.10 показана модуляция Gs через активацию β-адренергического рецептора (ADβR) и активацию D1 допаминового рецептора (DRD1). Нейроны выделяли из коры эмбрионов E18 линии 187. На третий день в культуру добавляли тестируемые соединения форсколин 5 мкмоль/л (F), ролипрам - 10 мкмоль/л (R), форсколин и ролипрам в комбинации (F/R), изопротеренол - 10 мкмоль/л, изопротеренол и ролипрам в комбинации (I/R); SKF82958 - 10 мкмоль/л и SKF82958 и ролипрам в комбинации (S/R). Данные показаны как число импульсов в секунду (cps).
На фиг.11 показаны эффекты пептида прокинетицин 2 (PROK2) на экспрессию люциферазы в первичных нейронах коры. Первичные нейроны коры собирали из линии 187 (индуцируемая люцифераза в головном и спинном мозге) на E18. Количественный анализ проводили на 3-й день в культуре в течение 4 часов или 8 часов. Пептид PROK2 добавляют в виде водного раствора по 1 нмоль/л и 100 нмоль/л. Данные показаны как число импульсов в секунду (cps).
На фиг.12 показаны эффекты пептида прокинетицин 2 (PROK2) на экспрессию люциферазы в первичных нейронах коры из различных линий CreLuc. Первичные нейроны коры собирали из различных линий CreLuc на E18. Количественные анализы проводили в трех повторах на третий день в культуре с 1 нмоль/л или 100 нмоль/л пептида PROK2 в два момента времени, 4 часа и 24 часа. BrightGlo применяли для количественного анализа и считывания на TopCount. Данные показаны как число импульсов в секунду (cps).
На фиг.13A показаны эффекты агониста mGluR7, AMN082 на экспрессию люциферазы в первичных нейронах коры. Нейроны коры собирали из эмбрионов E18 (линия 187). Количественный анализ проводили на 3-й день в культуре. Форсколин применяли в количестве 10 мкмоль/л. Агонист, AMN082 применяли в комбинации с форсколином в количестве 1 нмоль/л, 10 нмоль/л, 100 нмоль/л и 1 мкмоль/л. Количественный анализ считывали на TopCount с Bright Glo (Promega) через 4 часа и 8 часов. Данные показаны как число импульсов в секунду (cps).
На фиг.13B показаны результаты скрининга неизвестных соединений на возможность модулирования активности Gi в первичных нейронах коры. Нейроны коры собирали из эмбрионов E18 (линия 187). Количественный анализ проводили на 3-й день в культуре. Форсколин применяли в количестве 10 мкмоль/л. AMN082 или неизвестные соединения A, B или C испытывали в комбинации с форсколином при различных концентрациях, и рассчитывали уровни EC50. Количественный анализ считывали на TopCount с Bright Glo (Promega) через 4 часа. Данные показаны как число импульсов в секунду (cps).
На фиг.14 показаны эффекты агониста mGluR7, AMN082 на экспрессию люциферазы в первичных нейронах коры. Первичные нейроны коры выделяли из эмбрионов E18 линии 187. Количественный анализ в трех повторах проводили на 7 день в культуре в течение 6 часов. Концентрационная кривая для AMN082 проводилась в комбинации с 50 мкмоль/л форсколина и 10 мкмоль/л ролипрама. Количественный анализ считывали на люминометре TopCount с субстратом Bright Glo (Promega). Данные показаны как число импульсов в секунду (cps).
На фиг.15A показано Gi модулирование экспрессии люциферазы в первичных нейронах коры из различных линий CreLuc с помощью агониста CB1, CP 55,940. Первичные нейроны коры собирали из четырех различных линий CreLuc на E18. Количественный анализ проводили на третий день в культуре. Агонист CB1 применяли в количестве 10 мкмоль/л, форсколин в количестве 5 мкмоль/л и ролипрам в количестве 10 мкмоль/л. Проводили в два момента времени, четыре часа и двадцать четыре часа. Затем добавляли субстрат Bright Glo для количественного анализа люциферазы, и анализ считывали на люминометре Topcount. Показанные данные являются средним от трех повторов. Данные показаны как число импульсов в секунду (cps).
На фиг.15B показано Gi модулирование люциферазы в первичных нейронах коры из мышей CreLuc агонистом CB1, CP 55,940. Нейроны коры выделяли из эмбрионов E18 (линия 187). Количественный анализ проводили на 3-й день в культуре. Форсколин (F) и ролипрам (R) применяли в количестве 10 мкмоль/л. Агонист добавляли в концентрациях 10 мкмоль/л, 1 мкмоль/л и 100 нмоль/л. Количественный анализ считывали на TopCount с BrightGlo (Promega) через 8 часов. Данные показаны как число импульсов в секунду (cps).
На фиг.16 показана экспрессия люциферазы в стриарных нейронах CreLuc, индуцированную с помощью форсколина и ролипрама, и Gs агонистов DRD1 и ADβR. Стриарные нейроны выделяли из эмбрионов E14 (линия 187). Количественный анализ проводили на 4-й день в культуре. Форсколин (F) применяли в количестве 5 мкмоль/л, ролипрам (R) в количестве 10 мкмоль/л. Gs агонист изопротеренол (iso), допамин (dopa) и SKF82958 (chloro) использовали в количестве 10 мкмоль/л, 3 мкмоль/л и 1 мкмоль/л. Количественный анализ считывали через 5 часов с люминометром TopCount и люциферазным реагентом Bright Glo (Promega). Данные показаны как число импульсов в секунду (cps).
На фиг.17 показаны эффекты неспециализированных индукторов цАМФ, таких как форсколин (F) и ролипрам (R), а также Gs агонистов экспрессии люциферазы в препаратах цельных спленоцитов, выделенных из мышей CreLuc. Линию 64 спленоцитов стимулировали в течение 24 часов с антителом к CD3 (CD), другая половина была необработанной (unstim). В 24 часа соединения добавляли в планшеты еще на 4 часа. Совместная обработка форсколином и ролипрамом (F/R) составила 5 мкмоль/л форсколина и 10 мкмоль/л ролипрама. Использованными агонистами Gs стали: EX00000173A (173A) как агонист EP2, BW245C как агонист DP1 и изопротеренол как агонист ADβR. Все Gs агонисты использовали по 10 мкмоль/л. Количественный анализ проводили в трех повторах. Через 4 часа добавляли 100 мкл BrightGlo, и количественный анализ считывали на люминометре TopCount. Данные показаны как число импульсов в секунду (cps).
На фиг.18 показаны эффекты неспециализированной активации цАМФ ролипрамом и форсколином в T-клетках, выделенных из пяти различных сублиний мышей CreLuc. Клетки стимулировали антителами к CD3 (1 мкг/мл). Через 18 часов добавляли в планшеты 10 мкмоль/л ролипрама и 5 мкмоль/л форсколина еще на 4 часа. Добавляли BrightGlo, и количественный анализ считывали на TopCount. Данные показаны как люминесценция (число импульсов в секунду) на верхней панели, и как кратность увеличения по сравнению со средним в контроле на нижней панели.
На фиг.19 показаны эффекты Gs агонистов на уровни люциферазы в стимулированных к CD3 CD4+ T-клетках, выделенных из мышей CreLuc (линия 64). Клетки, по 1,5×105 на лунку, высевали на 96-луночные белые непрозрачные планшеты и затем стимулировали антителами к CD3 (1 мкг/мл). Через 24 часа соединения добавляли еще на 4 часа. Gs агонисты BW245C, EX00000173A (1734A) и изопротеренол (iso) применяли в количестве 10 мкмоль/л. Форсколин (F) добавляли в количестве 5 мкмоль/л и ролипрам (R) в количестве 10 мкмоль/л. Добавляли BrightGlo, и количественный анализ считывали на TopCount. Данные показаны как число импульсов в секунду (cps).
На фиг.20 показаны эффекты неспециализированной активации цАМФ ролипрамом и форсколином в B-клетках, выделенных из двух различных сублиний мышей CreLuc. Клетки высевали по 2,0×105 на лунку на 96-луночные белые непрозрачные планшеты и стимулировали 10 нг/мл липополисахарида (LPS). Через 18 часов 10 мкмоль/л ролипрама и 5 мкмоль/л форсколина добавляли в планшеты еще на 4 часа. Добавляли BrightGlo, и количественный анализ считывали на TopCount. Данные показаны как люминесценция (число импульсов в секунду) и как кратность увеличения по сравнению со средним в контроле.
На фиг.21 показаны эффекты Gs агонистов на уровни люциферазы в LPS стимулированных B220+ B клетках, выделенных из CreLuc. Клетки высевали по 2,0×105 на лунку на 96-луночные белые непрозрачные планшеты и затем стимулировали 10 нг/мл липополисахарида (LPS). Через 24 часа добавляли соединения еще на 4 часа. Gs-агонисты BW245C, EX00000173A (1734A) и изопротеренол (iso) применяли в количестве 10 мкмоль/л. Форсколин (F) добавляли в количестве 5 мкмоль/л и ролипрам (R) - в количестве 10 мкмоль/л. Добавляли BrightGlo (Promega, Мэдисон, Висконсин, кат. №E2610), и количественный анализ считывали на TopCount. Данные показаны как число импульсов в секунду (cps).
На фиг.22 показана индуцированная экспрессия люциферазы в выделенной микроглии (линия 64) неспецифическими цАМФ активаторами, форсколином (F) и ролипрамом (R), и агонистом рецептора DP, BW245C. Первичную микроглию выделяли из коры P2 мышей и высевали в 96-луночном формате на покрытые поли-D-лизином планшеты. Клетки либо оставляли необработанными, либо стимулировали в течение 2 часов 100 нг/мл LPS. Затем добавляли соединения еще на 4 часа до того, как проводили Bright Glo количественный анализ. Использованными соединениями были 5 мкмоль/л форсколина, 10 мкмоль/л ролипрама или их комбинация, или Gs-агонист рецептора DP1, BW245C в количестве 10 мкмоль/л. Данные показаны как число импульсов в секунду (cps).
На фиг.23 показаны эффекты интратекально введенных форсколина (F) и ролипрама (R) на индукцию экспрессии люциферазы в головном и спинном мозге мышей CreLuc (линия 187). N=3-4 мыши на группу, самцы возрастом 3 месяца. Группа A: DMSO контроль, Группа B: 1 мкг форсколина/10 мкг ролипрама, группа C: 10 мкг форсколина/10 мкг ролипрама, группа D: 40 мкг форсколина/10 мкг ролипрама. Животным вводили дозу посредством интратекальной инъекции в поясничную область объемом 5 мкл на мышь. Их визуализировали через 4 часа после введения дозы. Данные как для спинного, так и головного мозга показаны как среднее пиковой яркости, фотоны в секунду на см2.
На фиг.24 показаны эффекты агониста EP2, EX00000173A на экспрессию люциферазы в головном и спинном мозге мышей CreLuc. Мышам (линия 187) интраперитонеально вводили растворитель (5% DMSO, 0,05% твин 80, PBS) или 10 мг/кг EX00000173A. Животных биовизуализировали через 4 часа после введения. Данные показаны как фотоны в секунду на см2.
На фиг.25 показаны эффекты агониста EP2, EX00000173A на экспрессию люциферазы у мышей CreLuc. Мышам вводили или контрольный растворитель, или различные дозы агониста EP2 EX0000173A. Мышам вводили интратекальную инъекцию (5 мкл на мышь) и биовизуализировали через 4 часа на биовизуализаторе IVIS. Данные показаны как среднее пяти мышей, среднее пиковой яркости, фотоны в секунду на см2.
На фиг.26 показана индукция люциферазы в различных тканях агонистом адренорецептора бета 3 (Adrb3), CL316,243 (1 мг/кг, интраперитонеально) у мышей CRE-Luc. Люциферазный количественный анализ выполняли в гомогенатах тканей.
На фиг.27A показана индукция люциферазы агонистом Adrb3 CL316,243 (1 мг/кг, интраперитонеально) в линиях 11 (n=2) и 115 (n=3) мышей CRE-Luc. BLI были взяты до и через 4-5 часов после обработки. Активности люциферазы в гомогенатах тканей показаны под фотографиями.
На фиг.27B показана индукция люциферазы агонистом Adrb3 CL316,243 (1 мг/кг, интраперитонеально) в линиях 31 (n=2) и 175 (n=3) мышей CRE-Luc. BLI были взяты до и через 4-5 часов после обработки. Активности люциферазы в гомогенатах тканей показаны под фотографиями.
На фиг.28 показана индукция люциферазного репортера агонистом рецептора глюкагоноподобного пептида-1 (GLP-1R), AVE0010, в трех независимых линиях мышей CRE-Luc. Базисные изображения были получены на 1-й день. На 2-й день мышей обрабатывали AVE0010 (0,1 мг/кг, подкожно) и визуализировали через 4 часа. Кратность индукции сверх исходной указана внизу.
На фиг.29 показана индукция люциферазного репортера агонистом рецептора глюкагоноподобного пептида-1 (GLP-1R), AVE0010, в трех независимых линиях мышей CRE-Luc. Мышей обрабатывали AVE0010 (0,1 мг/кг, подкожно) в течение 4 часов. Измеряли активности люциферазы в 8 различных тканях.
На фиг.30 показаны эффекты стрептозотоцина (STZ), токсина бета-клеток, на индукцию CRE-Luc с помощью AVE0010. Самцов мышей CRE-luc (линия 11) визуализировали до («неиндуцированные»; верхняя панель) и после того, как был дан AVE0010 в количестве 0,1 мг/кг, подкожно («индукция AVE0010», средняя панель). Все мыши были чувствительны к AVE0010 (средняя панель). Затем животных обработали растворителем (контроль) или STZ (200 мг/кг, интраперитонеально). Четыре дня спустя их визуализировали снова после AVE0010 обработки (нижняя панель).
На фиг.31 показано, что индукция CRE-Luc с помощью AVE0010, вероятно, является специфичной для бета-клеток. Животных обрабатывали, как описано на фиг.30. Уровни глюкозы крови измеряли путем надреза хвостовой вены не голодавших мышей. Уровни глюкозы считывали на глюкометре Bayer. Уровни глюкозы показаны как мг глюкозы/мл. Кратность индукции является уровнями биовизуализации люциферазы при введении дозы AVE10 относительно базисных сигналов. Уровни глюкозы в крови (BG) были увеличены с помощью STZ (верхняя левая панель). Уровни BG не натощак были снижены посредством AVE0010 (0,1 мг/кг, подкожно). BLI данные, показанные на фиг.30, были определены количественно.
На фиг.32 показано приживление трансплантатов костного мозга CreLuc в NOD scid gamma (NSG) мышах. Клетки костного мозга собирали из линий 44 гетерозигот и линии 64 гомозигот. Затем клетки трансплантировали с помощью инъекций клеток в хвостовую вену облученных NSG мышей по 1 миллиону или 5 миллионов клеток на мышь. Для линии 44: мышь 1 и 2 получала 5 миллионов клеток, мышь 3 и 4 получала 1 миллион клеток; для линии 64: мышь 1 получала 5 миллионов клеток, мышь 2, 3 и 4 получала 1 миллион клеток. (4 NSG мыши на CreLuc линию). Животных биовизуализировали на 4-й неделе (данные не показаны) и затем снова на 8 неделе (данные показаны). До визуализации мыши линии 64 были индуцированы 5 часов 5 мг/кг форсколина и 10 мг/кг ролипрама.
На фиг.33 показаны эффекты форсколина, ролипрама и изопротеренола на экспрессию люциферазы в фибробластах эмбриона мыши. Фибробласты эмбриона мыши культивировали из эмбрионов E12 шести независимых линий CreLuc и высевали по 20000 клеток на лунку. Тестируемые соединения включают 10 мкмоль/л форсколина (F), 5 мкмоль/л ролипрама (R) и 10 мкмоль/л изопротеренола (iso). Данные показаны как число импульсов в секунду (cps).
На фиг.34 показаны эффекты обработки зимозаном на уровни люциферазы у мышей CreLuc (линия 187). Животным в обработанной группе вводили подкожно в обе задние лапы зимозан (zymo) для индукции болевого ответа. Затем животных биовизуализировали ежедневно в течение 4 дней (обозначенных как d1, d2, d3 и d4).
На фиг.35 показаны эффекты форсколина и ролипрама и изопротеренола на уровни люциферазы в кардиомиоцитах. Кардиомиоциты выделяли из детенышей P3 (линия 229). Клетки культивировали в 96-луночном планшете. Тестируемые соединения включали 10 мкмоль/л форсколина (F), 5 мкмоль/л ролипрама (R) и 10 мкмоль/л изопротеренола (iso). Данные показаны как число импульсов в секунду (cps).
Подробное описание изобретения
Если не определено иное, то все технические и научные термины, использованные в настоящем описании, имеют то же значение, как это обычно понимается специалистом в данной области техники, к которой относится данное изобретение.
Каждая публикация, патентная заявка, патент и другие ссылки, цитируемые в настоящем описании, включены посредством ссылки во всей своей полноте, при условии, что это не противоречит настоящему раскрытию.
Следует отметить, что применяемые в настоящем описании и формуле изобретения формы единственного числа включают множественное число, если контекст ясно не диктует иное.
Кроме того, в соответствии с настоящим изобретением могут быть использованы обычные методы молекулярной биологии, микробиологии и рекомбинантной ДНК в пределах компетентности специалиста в данной области. Такие методы в полной мере объясняются в литературе. См., например, Sambrook, Fritsch & Maniatis, Molecular Cloning: A Laboratory Manual, Second Edition (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York (в настоящем описании «Sambrook et al., 1989»); DNA Cloning: A Practical Approach, Volumes I and II (D.N. Glover ed. 1985); Oligonucleotide Synthesis (M.J. Gait ed. 1984); Nucleic Acid Hybridization [B.D. Hames & S.J.Higgins eds. (1985)]; Transcription And Translation [B.D. Hames & S.J. Higgins, eds. (1984)]; Animal Cell Culture [R.I. Freshney, ed. (1986)]; Immobilized Cells And Enzymes [IRL Press, (1986)]; B. Perbal, A Practical Guide To Molecular Cloning (1984); F.M. Ausubel et al. (eds.), Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (1994).
«Тестируемое средство» толкуется широко для включения любого материала, такого как соединение или химические соединения, например, органические химические объекты, неорганические химические объекты, биологические соединения или биологические материалы, например, антитела и антиген-распознающие фрагменты и конструкции, нуклеиновые кислоты, например, иРНК и т.д. Тестируемое средство охватывает одиночное средство или многочисленные средства, примененные совместно.
Как используется в настоящем описании, «трансгенное животное» представляет собой животное, отличное от человека; неограничивающим примером является млекопитающее, у которого одна или более клеток животного включают генетическую модификацию, как определено в настоящем описании. Кроме того, неограничивающие примеры включают грызунов, таких как крыса или мышь. Другие примеры трансгенных животных включают приматов, отличных от человека, овец, собак, коров, коз, кур, амфибий и т.д. Выбор трансгенного животного ограничен только способностью света, генерированного репортером, проникать через ткани и достигать поверхности, где может происходить обнаружение.
Как используется в настоящем описании, «генетическая модификация» представляет собой одно или более изменений в генетических последовательностях животного, отличного от человека. Неограничивающий пример представляет собой вставку трансгена в геном трансгенного животного.
Как используется в настоящем описании, термин «трансген» относится к экзогенной ДНК, содержащей промотор, ген-репортер, полиаденилированный сигнал и другие элементы для увеличения экспрессии (инсуляторы, интроны). Эта экзогенная ДНК интегрируется в геном 1-клеточного эмбриона, из которого развивается трансгенное животное, и трансген остается в геноме взрослого животного. Интегрированная трансгенная ДНК может встречаться в одном или многих местах в геноме яйца или мыши, а также одну или множественные (несколько сотен) тандемные копии трансгена можно интегрировать в каждое местоположение в геноме.
Термин «неспецифический модулятор цАМФ» относится к химическим соединениям, например, органическим химическим объектам, неорганическим химическим объектам, биологическим соединениям или биологическим материалам, например, антителам и антиген-распознающим фрагментам и конструкциям, нуклеиновым кислотам, например, иРНК и т.д., способным увеличивать или поддерживать уровни цАМФ. Неограничивающие примеры включают форсколин и ролипрам. Неспецифический модулятор цАМФ охватывает одиночный модулятор цАМФ или многочисленные модуляторы цАМФ, применяемые совместно.
Термин «клеточный стимулятор» относится к химическим соединениям, например, органическим химическим объектам, неорганическим химическим объектам, биологическим соединениям или биологическим материалам, например, антителам и антиген-распознающим фрагментам и конструкциям, нуклеиновым кислотам, например, иРНК и т.д., способным активировать клетку или побуждать клетку быть в более активированном состоянии. Неограничивающие примеры включают липополисахарид и анти-CD3.
В варианте осуществления настоящего изобретения используют контроль. Контроль является термином в данной области техники, хорошо понятным квалифицированным специалистам. Соответствующий контроль может зависеть от используемых параметров анализа или исследуемого экспериментального вопроса. Как правило, контроль представляет собой контрольный растворитель, в котором контроль представляет собой тот же буфер или растворитель, который использовали для растворения тестируемого средства или соединений. Если для растворения соединения используют забуференный фосфатом физиологический раствор, тогда неограничивающий пример представляет собой контрольный растворитель, которым будет являться забуференный фосфатом физиологический раствор. Подобным образом, если используют DMSO для растворения тестируемых средств, тогда контроль представляет собой DMSO. Часто должен быть использован более чем один контроль на эксперимент или анализ, поскольку для тестируемых соединений используют более чем один растворитель.
Как используется в настоящем описании, «люцифераза» относится не только к ферментативной активности люциферазы, но также к фактическим количествам люциферазного белка.
В соответствии с настоящим изобретением можно использовать обычные методы, известные специалистам в данной области, для генерирования трансгенных животных, отличных от человека. Например, Pinkert, C. A. (ed.) 1994. Transgenic animal technology: A laboratory handbook. Academic Press, Inc., San Diego, Calif.; Monastersky G. M. and Robl, J. M. (ed.) (1995) Strategies in transgenic animal science. ASM Press. Washington D. C. and Nagy A, Gertsenstein, M, Vintersten, K, Behringer R 2003. Manipulating the Mouse Embryo; A laboratory Manual 3rd edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Трансгенные элементы
Вариант осуществления настоящего изобретения относится к трансгену. Трансген может включать инсуляторные элементы, ответные элементы, промоторные элементы, репортерные элементы и функциональные элементы.
Ответные элементы представляют собой палиндромные последовательности ДНК, которые отвечают на клеточные сигналы, такие как гормоны, ферменты или другие ключевые сигнальные белки внутри клетки. Неограничивающие примеры чувствительных элементов включают CRE (ответный элемент цАМФ), ответные элементы эстрогена и другие, перечисленные в таблице 2. Ответные элементы могут быть встроены в трансген как одиночная последовательность ДНК или в тандемных повторах. Например, ответные элементы CRE, повторенные четыре раза или шесть раз, использовали в трансгенной конструкции и подтверждали in vitro (Deutsch P.J., et al., J. Biol. Chem., 263; 18466-18472, 1988; Oetjen E JBC 269; 27036-27044, 1994). Ответные элементы CRE также сравнивали in vivo, и увеличение мультимеров коррелировало с увеличением транскрипционного ответа к активаторам цАМФ пути (Montoliu, L. et al., Proc. Natl. Acad. Sci. USA 92; 4244, 1995; Boer et al., PloS One, May 9; 2(5):e431, 2007). В варианте осуществления настоящего изобретения можно использовать любой известный ответный элемент либо в виде одиночной последовательности, либо в многочисленных тандемных повторах, например, в шести тандемных повторах CRE (6X CRE).
Действующие в цис-положении ответные элементы
Промоторные элементы ДНК представляют собой области ДНК, которые способствуют транскрипции отдельного гена. Промоторы, как правило, расположены рядом с генами, которые они регулируют, на той же нити и выше (по направлению к http://en.wikipedia.org/wiki/5%27" \o "5' области смысловой нити). Промоторы содержат специфические последовательности ДНК и ответные элементы, которые предоставляют сайт связывания для РНК-полимеразы и для белков, называемых транскрипционными факторами, которые рекрутируют РНК-полимеразу. Промоторы ДНК сильно варьируют по своему размеру и внутренним подструктурам, которые способствуют регуляции экспрессии отдельных генов во времени и пространстве. В неограничивающем примере промоторных элементов минимальный промотор тимидинкиназы вируса простого герпеса (HSV TK min) предназначен для того, чтобы предусмотреть экспрессию гена-репортера в каждом типе клеток. Сохраняются только его основные элементы экспрессии, чтобы наделить повсеместной экспрессией либо in vitro, либо in vivo (Park, J. et al., DNA Cell Bio., 12:1147-1149, 1994). В варианте осуществления настоящего изобретения можно использовать любой известный промоторный элемент. В качестве неограничивающего примера можно использовать любой известный промоторный элемент так, что промоторный элемент при комбинации с цис-активирующим ответным элементом CRE позволяет генной экспрессии репортера отвечать на модуляцию цАМФ пути.
Поскольку ответные элементы, такие как CRE, и промоторный элемент HSV TK min имеют малую величину, промотор трансгена, содержащий эти элементы, регулирующие транскрипцию репортерного элемента, будет очень чувствительным к эффектам положения и приводит к слабым экспрессионным ответам на лиганды, особенно при низких концентрациях лиганда in vivo. Таким образом, вариантом осуществления настоящего изобретения является включение дополнительных элементов, которые добавляются к трансгену для достижения высоких уровней экспрессии и широкого распределения, функционирующего трансгена по всем клеточным компартментам. Эти элементы включают инсуляторные элементы и функциональные элементы (Sun F.L and Elgin S.C, Cell 99:459-462, 1999).
В варианте осуществления настоящего изобретения используют функциональные элементы или функциональные энхансерные элементы в пределах трансгена. Неограничивающий пример функционального элемента представляет собой ген гормона роста человека (hGH). Последовательность hGH, использованная в трансгене CreLuc, содержит несколько разработанных элементов, которые способствуют и воздействуют на инсуляторный элемент для достижения целей разработанной модели биовизуализации. Последовательность hGH содержит всю геномную структуру hGH и, таким образом, поставляет несколько важных элементов, но не транскрибируется или транслируется в белок. Критическое влияние последовательности hGH для улучшения продуцирования функционирующего трансгена продемонстрировано с 1990 для нескольких трансгенных моделей Erickson LA, Nature 346: 74-76, 1990. Хотя здесь нет сравнительного анализа ее важности, структура hGH действительно содержит несколько важных и критических элементов ДНК:
a. Интронный сплайсинг: первоначально транскрибированная мРНК содержит одновременно интронные и экзонные последовательности, которые затем экспортируются из ядра и дополнительно обрабатываются для удаления интронной последовательности, приводя в результате к зрелой мРНК, содержащей только экзонные последовательности. Этот процесс транспорта и обрезки соединяет созревающую нить мРНК с дополнительным трансляционным аппаратом для конечного продуцирования белка. Показано, что включение интронов в структуру трансгенной кДНК улучшает уровень экспрессии трансгенной кДНК (Palmiter, R.D. et al., Proc. Natl. Acad. Sci. USA 88:478-482, 1988).
b. Геномная структура с интактным 3'UTR: в варианте осуществления настоящего изобретения последовательность hGH в трансгене содержит интактный 3'UTR, который дает высокий процент стабильности мРНК и, таким образом, более высокие уровни экспрессии трансгена.
c. Геномная структура с поли A (PA+) структурами: последовательность hGH содержит свою нативную PA+ структуру, встроенную в природный 3' UTR. Как правило, PA+ сигналы из вирусных последовательностей (SV40, RSV и т.д.) и представляют собой минимальные структуры, добавленные к концу не связанных 3'UTR. В варианте осуществления настоящего изобретения полная структура 3'UTR с природным PA+ сигналом сохраняется в более широком геномном контексте полного hGH гена.
Инсуляторные элементы представляют собой последовательности ДНК, которые генерируют независимую от положения экспрессию (Giraldo et al., Transgenic Research 12: 751-755, 2003). Инсуляторные последовательности были описаны в 1980-х гг. для локуса глобулина (Sun F.L. and Elgin, S.C., Cell 99;459-462, 1999), и сообщилось, что увеличился шанс получения корректной и ответной трансгенной экспрессии в выбранных тканях, чтобы обеспечить цели разработанной модели для биовизуализации (Pinkert, C. A. (ed.) 1994. Transgenic animal technology: A laboratory handbook. Academic Press, Inc., San Diego, Calif.; Monastersky G. M. and Robl, J. M. (ed.) (1995) Strategies in transgenic animal science. ASM Press. Washington D. C). Инсуляторы представляют собой элементы ДНК, которые создают открытые домены хроматина, пермиссивные к экспрессии гена, и составляют барьер против влияния дистальных сайленсерных/энхансерных последовательностей и против явлений ацетилирования и метилирования. Они должны значительно увеличивать число независимых трансгенных линий-основателей, которые имеют ген-репортер, экспрессированный на обнаруживаемых для биовизуализации уровнях. Показано, что инсуляторные элементы повышают число клонов, экспрессирующих люциферазу, в анализе временной трансфекции от 40 до 70%, таким образом, увеличивая индуцируемость экспрессии люциферазы в трансгене ERE-luci. (Ottobrini L., Mol. Cell. Endo. 246, 69-75). Однако полный обзор применения инсуляторов для экспрессии трансгенов-репортеров приводит к выводу, что в практическом смысле остаются трудности использования инсуляторов. Их механизм действия известен только частично, и их эффект неполностью предсказуем. Неограничивающие примеры инсуляторных элементов включены в таблицу 3.
Примеры известных инсуляторных элементов для улучшения экспрессии трансгенной ДНК
В варианте осуществления трансгена Cre-Luc включение инсуляторных элементов значительно увеличило частоту генерирования линий с функциональным репортером, как было обнаружено с помощью биовизуализации. Для анализа вклада инсуляторных элементов в экспрессию люциферазы в наших линиях CreLuc трансгенных мышей см. раздел VI примеров ниже.
Вариант осуществления настоящего изобретения относится к трансгену, содержащему репортерный элемент или ген. Ген-репортер включает любой ген, который экспрессирует обнаруживаемый генный продукт, который может представлять собой РНК или белок. Многие гены-репортеры известны в данной области техники, включая, но, не ограничиваясь ими, бета-галактозидазу и щелочную фосфатазу. В другом варианте осуществления трансген содержит биолюминесцентный ген-репортер. Многие биолюминесцентные гены-репортеры известны в данной области техники, включая, но без ограничений, люциферазу. Существует много источников люциферазы; неограничивающие примеры включают люциферазу светлячков и люциферазу бактерий. В варианте осуществления настоящего изобретения можно использовать любой известный биолюминесцентный репортер, например, люциферазу. Другие примеры репортерных элементов без ограничения представлены в таблице 5.
Генетические репортерные системы
Другие варианты осуществления настоящего изобретения могут объединять модифицированные версии люциферазного фермента, люциферазный фермент из различных видов или любой другой белок, который может производить свет, способный проходить через животные ткани, или любой фермент, который может излучать свет, способный проходить через животные ткани, при предоставлении подходящего субстрата. Репортерный белок настоящего изобретения ограничен только фактом, что ослабление сигнала зависит от длины волны излучаемого света и свойств ткани, окружающей излучающие клетки. Обычно, сине-зеленый свет (400-590 нм) сильно ослабляется, в то время как красный до ближней части инфракрасного диапазона (590-800 нм) испытывает гораздо меньшее ослабление. Большинство типов люциферазы имеют пиковое излучение от синих до желто-зеленых длин волн, спектр излучения является широким, так что существует значительное излучение на красных длинах волн (>600 нм), которое проникает достаточно глубоко в ткань. Для мелких грызунов, таких как мыши, это позволяет обнаруживать сигналы везде во всем животном.
Пределы обнаружения света in vivo зависят от типа биолюминесцентного репортера, окружающей физиологии животного и глубины источника. Как правило, биолюминесцентные клетки в животном можно обнаружить с глубины 13 см с помощью чувствительных CCD камер в зависимости от количества и местоположения клеток. Рассеяние фотонов, поскольку они распространяются через ткань, ограничивает пространственное разрешение изображений, обнаруженных на поверхности животного. В целом, размер пятна или разрешение на поверхности является приблизительно равным глубине источника под поверхностью. Используя основанные на физике диффузионные модели, приближаясь к миллиметровому уровню, можно достигнуть улучшения пространственного разрешения. Используя охлажденные, высокого уровня классы матрицы CCD, предел обнаружения сигнала определяют считыванием шума, связанного со считыванием CCD пикселей после получения изображения, который составляет порядка нескольких фотонов на пиксель (Honigman et al., Mol. Ther. 4:239-249, 2001). Там может быть дополнительный фоновый свет, поступающий от животного из-за фосфоресценции шерсти, кожи или, возможно, загрязнений животного. Как правило, фоновый свет находится на низком уровне и оказывает только отрицательное воздействие на изображения глубоких низкоуровневых биолюминесцентных источников. Однако фоновый свет можно устранить путем использования подходящего оптического фильтра.
В варианте осуществления настоящего изобретения используют CCD камеры, такие как IVIS (Xenogen Corporation, 860 Atlantic Avenue, Аламеда, Калифорния 94501, США). Система визуализации IVIS.RTM. включает чувствительную CCD камеру, темную камеру визуализации для минимизирования отраженного света и специализированное программное обеспечение для количественной оценки и анализа результатов. IVIS является зарегистрированной торговой маркой Xenogen Corporation. Однако в настоящем изобретении можно применять любую подобную систему биолюминесцентной визуализации.
In vivo визуализация в режиме реального времени предусматривает количественную оценку биолюминесцентного гена-репортера неинвазивно, т.е. без необходимости подвергать животное эвтаназии, и продольно, т.е. измерения можно осуществлять непрерывно или повторять в течение длительного периода действия. В in vivo визуализации в режиме реального времени необходимо меньше подопытных животных (например, из-за того, что одно и то же животное можно использовать в течение определенного периода времени) и меньше времени (например, из-за того, что необходимо обрабатывать меньше животных), чем в обычных протоколах, предусматривающих испытание большего количества соединений на эффективность. In vivo визуализация в режиме реального времени предоставляет более высокое содержание данных и более высокое качество данных по многим причинам. Например, временные и пространственные данные можно получить от одного и того же животного и данные можно получить без необходимости трудоемкой гистологической оценки. Более высокое качество данных снижает статистическую ошибку и улучшает качество оценки тестируемого соединения и принятия решения.
Вариант осуществления настоящего изобретения представляет собой применение способов высокопроизводительного скрининга (HTS). HTS - автоматизированное, одновременное испытание тысяч различных химических соединений в анализах, предназначенных для моделирования биологических механизмов или аспектов болезненных патологий. Более чем одно соединение, например, множество соединений, можно испытывать одновременно, например, в одной серии. В одном варианте осуществления термин способ скрининга HTS относится к анализам, в которых испытывают способность одного соединения или множества соединений влиять на считывание показаний.
Жидкостные системы обработки, аналитическое оборудование, такое как устройства для считывания флуоресценции или сцинтилляционные счетчики и робототехники для клеточной культуры и манипуляции с образцами, хорошо известны в данной области техники. Механические системы, такие как роботизированные руки или устройства «избирательного подхода» доступны квалифицированным специалистам. Коммерческие планшет-ридеры доступны для анализа обычных 96-луночных или 384-луночных планшетов. Доступны устройства считывания одного образца, множества образцов или планшетов с образцами, которые анализируют заранее определенные лунки и генерируют отчеты первичных данных. Первичные данные могут быть трансформированы и представлены различными способами.
Вариант осуществления настоящего изобретения включает совокупность сосудов, которые могут вмещать клетки, тканевые срезы и другие материалы, такие как культуральная среда. Совокупность сосудов может быть любым количеством сосудов, по меньшей мере, от одного до более одного сосуда, подходящего для хранения клеток или тканевых срезов в пределах объема настоящего изобретения. Примеры включают, но без ограничения, колбы, чашки для культивирования, пробирки, такие как 1,5-мл пробирки, 12-луночные планшеты, 96-луночные планшеты, 384-луночные планшеты и титрационные микропланшеты минимального объема, возможно, с 4000 сосудами (заявка на патент США 20050255580). Совокупность сосудов можно поправить путем добавления защитного покрытия, таким образом, предотвращая попадание загрязнений или испарение содержимого.
Дополнительной характеристикой сосудов является то, что сосуд может быть предусмотрен для анализа, неограничивающие примеры которого включают спектрофотометрический анализ, сцинтилляционный подсчет и измерения флуоресценции. Однако это не является ограничением для сосудов, которые можно использовать в пределах объема настоящего изобретения, учитывая, что образцы можно переместить в подходящий контейнер, поддающийся поправке для последующего анализа. Неограничивающий пример представляет собой модификацию способа таким образом, что способ дополнительно включает обеспечение второй совокупности сосудов, где стадия лизиса клеток дополнительно включает отделение надосадочной жидкости от клеточных остатков и следующая стадия дополнительно включает добавление определяемого соединения, способного к интеркаляции во фрагменты ДНК, по меньшей мере, в один сосуд указанной второй совокупности сосудов, содержащий образец указанной отделенной надосадочной жидкости.
Дополнительные аспекты и детали настоящего изобретения будут очевидны из последующих примеров, которые предназначены для иллюстрации и ни в коем случае не предназначены для ограничения объема настоящего изобретения.
Всю экспериментальную работу с участием животных, которую выполняли согласно федеральными инструкциям и протоколам, предварительно рассмотрело и утвердило подразделение Sanofi-Aventis институционального комитета по содержанию и использованию животных (IACUC).
ПРИМЕРЫ
I. Векторная основа для системы клонирования MultiSite Gateway Pro Plus (фиг.3)
Вектор назначения pDest2XMARS разработали и клонировали специально для применения с системой клонирования MultiSite Gateway Pro Plus (Invitrogen, Карлсбад, Калифорния, кат. № 12537-100). Все элементы, вставленные в вектор, клонировали с помощью ПЦР (полимеразная цепная реакция) или искусственно генерировали. Для всех стадий клонирования ПЦР фрагменты амплифицировали из указанного вектора с помощью использования ПЦР SuperMIx Hi Fidelity (Invitrogen, Карлсбад, Калифорния, кат. № 10790-020) и фрагмент-специфических праймеров. Продукты ПЦР затем субклонировали в pCR2.1 векторы с помощью TOPO клонирования. TOPO клонирование представляет собой молекулярно-биологический метод, в котором фрагменты ДНК, амплифицированные с помощью Taq или Pfu полимераз, клонируют в специфические векторы без потребности в ДНК-лигазах.
Во-первых, инсуляторные элементы клонировали с помощью полимеразной цепной реакции (ПЦР) из вектора pCpG-LacZ (InvivoGen Сан-Диего, Калифорния, кат. № pcpg-lacz), последовательность проверяли, и затем субклонировали в сайты рестрикции pNEB193 (New England Biolabs, Ипсвич, Массачусетс, кат. № N3051S). Инсуляторные элементы использовали для снижения вариабельности экспрессии трансгена из-за эффектов положения, зависимых от положения сайта интеграции.
ПЦР праймеры, которые использовали для клонирования фрагментов, представляют собой следующие:
a. Праймерные последовательности человеческого IFN-β MAR (праймеры имеют EcoRV сайты для субклонирования):
SEQ ID NO:1, прямой праймер IFN:
5'-GGGGGATATCAGTCAATATGTTCACCCCA-3'
SEQ ID NO:2, обратный праймер IFN:
5'-GGGGGATATCCTACTGTTTTAATTAAGC-3'
b. Праймерные последовательности человеческого β-глобина MAR:
SEQ ID NO:3, прямой праймер β-глобина:
5'-AAGGATCCTTAATTAAAATTATCTCTAAGGC-3'
SEQ ID NO:4, обратный праймер β-глобина:
5'-GGATCCCTGCAGGAATTCCTTTTAAT-3'
ПЦР фрагмент β-глобина topo клонировали с использованием pCR2.1 (Invitrogen, Карлсбад, Калифорния, кат. № K2030-01). После подтверждения последовательности фрагмент вырезали с BamHI и затем субклонировали в сайт BamH1 из pNEB193 (βgloMAR-pNEB193). ПЦР фрагмент IFN-β также topo клонировали с pCR2.1, секвенировали, и затем вырезали с EcoRv и субклонировали в сайт HincII из βgloMAR-pNEB193 (2XMARS-pNEB193).
Для конечного вектора назначения Gateway линкер, содержащий XbaI концы и внутренний сайт EcoRV субклонировали в сайт XbaI из 2XMAR-pNEB193. Конверсионную кассету Gateway с тупыми концами, RfA, затем вставили в сайт EcoRV из 2XMAR-pNEB193 (2XMARSpDest). Конечный вектор 2XMARSpDest был конечным вектором назначения, который использовали для генерирования трансгенов.
SEQ ID NO:5
TCGCGCGTTTCGGTGATGACGGTGAAAACCTCTGACACATGCAGCTCCCGGAGACGGTCACAGCTTGTCTGTAAGCGGATGCCGGGAGCAGACAAGCCCGTCAGGGCGCGTCAGCGGGTGTTGGCGGGTGTCGGGGCTGGCTTAACTATGCGGCATCAGAGCAGATTGTACTGAGAGTGCACCATATGCGGTGTGAAATACCGCACAGATGCGTAAGGAGAAAATACCGCATCAGGCGCCATTCGCCATTCAGGCTGCGCAACTGTTGGGAAGGGCGATCGGTGCGGGCCTCTTCGCTATTACGCCAGCTGGCGAAAGGGGGATGTGCTGCAAGGCGATTAAGTTGGGTAACGCCAGGGTTTTCCCAGTCACGACGTTGTAAAACGACGGCCAGTGAATTCGAGCTCGGTACCCGGGGGCGCGCCGGGATCCTTAATTAAAATTATCTCTAAGGCATGTGAACTGGCTGTCTTGGTTTTCATCTGTACTTCATCTGCTACCTCTGTGACCTGAAACATATTTATAATTCCATTAAGCTGTGCATATGATAGATTTATCATATGTATTTTCCTTAAAGGATTTTTGTAAGAACTAATTGAATTGATACCTGTAAAGTCTTTATCACACTACCCAATAAATAATAAATCTCTTTGTTCAGCTCTCTGTTTCTATAAATATGTACCAGTTTTATTGTTTTTAGTGGTAGTGATTTTATTCTCTTTCTATATATATACACACACATGTGTGCATTCATAAATATATACAATTTTTATGAATAAAAAATTATTAGCAATCAATATTGAAAACCACTGATTTTTGTTTATGTGAGCAAACAGCAGATTAAAAGGAATTCCTGCAGGATCCTTAATTAAGTTCTAGATCACAAGTTTGTACAAAAAAGCTGAACGAGAAACGTAAAATGATATAAATATCAATATATTAAATTAGATTTTGCATAAAAAACAGACTACATAATACTGTAAAACACAACATATCCAGTCACTATGGCGGCCGCATTAGGCACCCCAGGCTTTACACTTTATGCTTCCGGCTCGTATAATGTGTGGATTTTGAGTTAGGATCCGTCGAGATTTTCAGGAGCTAAGGAAGCTAAAATGGAGAAAAAAATCACTGGATATACCACCGTTGATATATCCCAATGGCATCGTAAAGAACATTTTGAGGCATTTCAGTCAGTTGCTCAATGTACCTATAACCAGACCGTTCAGCTGGATATTACGGCCTTTTTAAAGACCGTAAAGAAAAATAAGCACAAGTTTTATCCGGCCTTTATTCACATTCTTGCCCGCCTGATGAATGCTCATCCGGAATTCCGTATGGCAATGAAAGACGGTGAGCTGGTGATATGGGATAGTGTTCACCCTTGTTACACCGTTTTCCATGAGCAAACTGAAACGTTTTCATCGCTCTGGAGTGAATACCACGACGATTTCCGGCAGTTTCTACACATATATTCGCAAGATGTGGCGTGTTACGGTGAAAACCTGGCCTATTTCCCTAAAGGGTTTATTGAGAATATGTTTTTCGTCTCAGCCAATCCCTGGGTGAGTTTCACCAGTTTTGATTTAAACGTGGCCAATATGGACAACTTCTTCGCCCCCGTTTTCACCATGGGCAAATATTATACGCAAGGCGACAAGGTGCTGATGCCGCTGGCGATTCAGGTTCATCATGCCGTTTGTGATGGCTTCCATGTCGGCAGAATGCTTAATGAATTACAACAGTACTGCGATGAGTGGCAGGGCGGGGCGTAAACGCGTGGATCCGGCTTACTAAAAGCCAGATAACAGTATGCGTATTTGCGCGCTGATTTTTGCGGTATAAGAATATATACTGATATGTATACCCGAAGTATGTCAAAAAGAGGTATGCTATGAAGCAGCGTATTACAGTGACAGTTGACAGCGACAGCTATCAGTTGCTCAAGGCATATATGATGTCAATATCTCCGGTCTGGTAAGCACAACCATGCAGAATGAAGCCCGTCGTCTGCGTGCCGAACGCTGGAAAGCGGAAAATCAGGAAGGGATGGCTGAGGTCGCCCGGTTTATTGAAATGAACGGCTCTTTTGCTGACGAGAACAGGGGCTGGTGAAATGCAGTTTAAGGTTTACACCTATAAAAGAGAGAGCCGTTATCGTCTGTTTGTGGATGTACAGAGTGATATTATTGACACGCCCGGGCGACGGATGGTGATCCCCCTGGCCAGTGCACGTCTGCTGTCAGATAAAGTCTCCCGTGAACTTTACCCGGTGGTGCATATCGGGGATGAAAGCTGGCGCATGATGACCACCGATATGGCCAGTGTGCCGGTCTCCGTTATCGGGGAAGAAGTGGCTGATCTCAGCCACCGCGAAAATGACATCAAAAACGCCATTAACCTGATGTTCTGGGGAATATAAATGTCAGGCTCCCTTATACACAGCCAGTCTGCAGGTCGACCATAGTGACTGGATATGTTGTGTTTTACAGTATTATGTAGTCTGTTTTTTATGCAAAATCTAATTTAATATATTGATATTTATATCATTTTACGTTTCTCGTTCAGCTTTCTTGTACAAAGTGGTGATCTAGACTAGAGTCATCAGTCAATATGTTCACCCCAAAAAAGCTGTTTGTTAACTTGTCAACCTCATTCTAAAATGTATATAGAAGCCCAAAAGACAATAACAAAAATATTCTTGTAGAACAAAATGGGAAAGAATGTTCCACTAAATATCAAGATTTAGAGCAAAGCATGAGATGTGTGGGGATAGACAGTGAGGCTGATAAAATAGAGTAGAGCTCAGAAACAGACCCATTGATATATGTAAGTGACCTATGAAAAAAATATGGCATTTTACAATGGGAAAATGATGATCTTTTTCTTTTTTAGAAAAACAGGGAAATATATTTATATGTAAAAAATAAAAGGGAACCCATATGTCATACCATACACACAAAAAAATTCCAGTGAATTATAAGTCTAAATGGAGAAGGCAAAACTTTAAATCTTTTAGAAAATAATATAGAAGCATGCCATCAAGACTTCAGTGTAGAGAAAAATTTCTTATGACTCAAAGTCCTAACCACAAAGAAAAGATTGTTAATTAGATTGCATGAATATTAAGACTTATTTTTAAAATTAAAAAACCATTAAGAAAAGTCAGGCCATAGAATGACAGAAAATATTTGCAACACCCCAGTAAAGAGAATTGTAATATGCAGATTATAAAAAGAAGTCTTACAAATCAGTAAAAAATAAAACTAGACAAAAATTTGAACAGATGAAAGAGAAACTCTAAATAATCATTACACATGAGAAACTCAATCTCAGAAATCAGAGAACTATCATTGCATATACACTAAATTAGAGAAATATTAAAAGGCTAAGTAACATCTGTGGCTTAATTAAAACAGTAGGATGACTGTTTAAACCTGCAGGCATGCAAGCTTGGCGTAATCATGGTCATAGCTGTTTCCTGTGTGAAATTGTTATCCGCTCACAATTCCACACAACATACGAGCCGGAAGCATAAAGTGTAAAGCCTGGGGTGCCTAATGAGTGAGCTAACTCACATTAATTGCGTTGCGCTCACTGCCCGCTTTCCAGTCGGGAAACCTGTCGTGCCAGCTGCATTAATGAATCGGCCAACGCGCGGGGAGAGGCGGTTTGCGTATTGGGCGCTCTTCCGCTTCCTCGCTCACTGACTCGCTGCGCTCGGTCGTTCGGCTGCGGCGAGCGGTATCAGCTCACTCAAAGGCGGTAATACGGTTATCCACAGAATCAGGGGATAACGCAGGAAAGAACATGTGAGCAAAAGGCCAGCAAAAGGCCAGGAACCGTAAAAAGGCCGCGTTGCTGGCGTTTTTCCATAGGCTCCGCCCCCCTGACGAGCATCACAAAAATCGACGCTCAAGTCAGAGGTGGCGAAACCCGACAGGACTATAAAGATACCAGGCGTTTCCCCCTGGAAGCTCCCTCGTGCGCTCTCCTGTTCCGACCCTGCCGCTTACCGGATACCTGTCCGCCTTTCTCCCTTCGGGAAGCGTGGCGCTTTCTCATAGCTCACGCTGTAGGTATCTCAGTTCGGTGTAGGTCGTTCGCTCCAAGCTGGGCTGTGTGCACGAACCCCCCGTTCAGCCCGACCGCTGCGCCTTATCCGGTAACTATCGTCTTGAGTCCAACCCGGTAAGACACGACTTATCGCCACTGGCAGCAGCCACTGGTAACAGGATTAGCAGAGCGAGGTATGTAGGCGGTGCTACAGAGTTCTTGAAGTGGTGGCCTAACTACGGCTACACTAGAAGAACAGTATTTGGTATCTGCGCTCTGCTGAAGCCAGTTACCTTCGGAAAAAGAGTTGGTAGCTCTTGATCCGGCAAACAAACCACCGCTGGTAGCGGTGGTTTTTTTGTTTGCAAGCAGCAGATTACGCGCAGAAAAAAAGGATCTCAAGAAGATCCTTTGATCTTTTCTACGGGGTCTGACGCTCAGTGGAACGAAAACTCACGTTAAGGGATTTTGGTCATGAGATTATCAAAAAGGATCTTCACCTAGATCCTTTTAAATTAAAAATGAAGTTTTAAATCAATCTAAAGTATATATGAGTAAACTTGGTCTGACAGTTACCAATGCTTAATCAGTGAGGCACCTATCTCAGCGATCTGTCTATTTCGTTCATCCATAGTTGCCTGACTCCCCGTCGTGTAGATAACTACGATACGGGAGGGCTTACCATCTGGCCCCAGTGCTGCAATGATACCGCGAGACCCACGCTCACCGGCTCCAGATTTATCAGCAATAAACCAGCCAGCCGGAAGGGCCGAGCGCAGAAGTGGTCCTGCAACTTTATCCGCCTCCATCCAGTCTATTAATTGTTGCCGGGAAGCTAGAGTAAGTAGTTCGCCAGTTAATAGTTTGCGCAACGTTGTTGCCATTGCTACAGGCATCGTGGTGTCACGCTCGTCGTTTGGTATGGCTTCATTCAGCTCCGGTTCCCAACGATCAAGGCGAGTTACATGATCCCCCATGTTGTGCAAAAAAGCGGTTAGCTCCTTCGGTCCTCCGATCGTTGTCAGAAGTAAGTTGGCCGCAGTGTTATCACTCATGGTTATGGCAGCACTGCATAATTCTCTTACTGTCATGCCATCCGTAAGATGCTTTTCTGTGACTGGTGAGTACTCAACCAAGTCATTCTGAGAATAGTGTATGCGGCGACCGAGTTGCTCTTGCCCGGCGTCAATACGGGATAATACCGCGCCACATAGCAGAACTTTAAAAGTGCTCATCATTGGAAAACGTTCTTCGGGGCGAAAACTCTCAAGGATCTTACCGCTGTTGAGATCCAGTTCGATGTAACCCACTCGTGCACCCAACTGATCTTCAGCATCTTTTACTTTCACCAGCGTTTCTGGGTGAGCAAAAACAGGAAGGCAAAATGCCGCAAAAAAGGGAATAAGGGCGACACGGAAATGTTGAATACTCATACTCTTCCTTTTTCAATATTATTGAAGCATTTATCAGGGTTATTGTCTCATGAGCGGATACATATTTGAATGTATTTAGAAAAATAAACAAATAGGGGTTCCGCGCACATTTCCCCGAAAAGTGCCACCTGACGTCTAAGAAACCATTATTATCATGACATTAACCTATAAAAATAGGCGTATCACGAGGCCCTTTCGTC
II. Трансгенные компоненты
A. CRE:
Два различных 6XCRE элемента использовали в генерировании трансгенов. Первый представляет собой искусственный CRE элемент (отмеченный как «искусственный»), который искусственно сгенерировали на DNA2.0, Menlo Park, Калифорния. Искусственная последовательность имеет attL1 и attR5 сайты для 4-фрагментного Gateway клонирования.
Предприняли многочисленные попытки к ПЦР клону из 6X CRE из различных векторов; однако это было невозможно из-за шпилечной структуры в середине CRE. Этот фрагмент затем клонировали искусственно. Второй 6X CRE элемент, который использовали, представляет собой гибридную версию, сгенерированную с помощью ПЦР клонирования CRE элементов из вектора Clontech (Маунтин-Вью, Калифорния, кат. № PT3336-5). Clontech подтверждает, что в векторе есть 2x CRE, но анализ последовательности показал, что фактически присутствовало три. Для клонирования фрагмента применяли две различные ПЦР реакции. Сайты Att для Gateway клонирования, как и сайт EcoR1, ввели в праймеры для того, чтобы два фрагмента «склеить» вместе и затем рекомбинировать в вектор Invitrogen Gateway pDONR P1-P5r (Карлсбад, Калифорния, кат. № 12537-100).
1. Искусственный элемент CRE:
SEQ ID NO:6:
5'AAATAATGATTTTATTTTGACTGATAGTGACCTGTTCGTTGCAACAAATTGATGAGCAATGCTTTTTTATAATGCCAACTTTGTACAAAAAAGCAGGCTTACTGTCGACAATTGCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGATTGACGTCAATGGGGTGTCTGAAAGACGTCACAGTATGACCCGGGCTCGAGCCTCCTTGGCTGACGTCAGAGAGAGAGGCCGGCCCCTTACGTCAGAGGCGAGAATTCGACAACTTTGTATACAAAAGTTGAACGAGAAACGTAAAATGATATAAATATCAATATATTAAATTAGATTTTGCATAAAAAACAGACTACATAATACTGTAAAACACAACATATCCAGTCACTATG3'
2. Гибридный CRE: стандартное клонирование, ПЦР амплифицированный CRE элемент из Clontech pCreLuc вектора, который содержит 3XCRE, ПЦР праймеры имеют EcoR1 сайт рестрикции на одном конце (CRE3X-B, CRE3X-C) и сайт att, либо attB1 или attB5r для Gateway клонирования на другом конце (CRE3X-A, CRE3X-D). Два фрагмента затем скомбинировали для создания одного фрагмента с 6XCRE. Фрагмент повторно рекомбинировали в вектор Gateway pDONR P1-P5r.
SEQ ID NO:7: прямой праймер CRE A
5'-GGGGACAAGTTTGTACAAAAAAGCAGGCTTAGCACCAGACAGTGA-3'
SEQ ID NO:8: обратный праймер CRE B
5'-GGGAATTCGTTCTCCCATTGACGTCA-3'
SEQ ID NO:9: прямой праймер CRE C
5'-GGGAATTCGCACCAGACAGTGACGTC-3'
SEQ ID NO:10: обратный праймер CRE D
5'-GGGGACAACTTTTGTATACAAAGTTGTGTTCTCCCATTGACGTCA-3'
SEQ ID NO:11: Гибридная CRE последовательность:
GCTTAGCACCAGACAGTGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACGAATTCGCACCAGACAGTGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACA
B. Гормон роста человека с хвостом поли А:
Гормон роста человека с хвостом поли А клонировали ПЦР из вектора pOGH (Nichols Institute Diagnostics (Сан-Хуан-Капистрано, Калифорния, кат. № 40-2205)). Праймерные последовательности имеют сайт attB3 на прямом праймере и сайт attB2 на обратном праймере для рекомбинации в вектор Gateway pDONR P3-P2.
SEQ ID NO:12: прямой праймер hGH:
5'-GGGGACAACTTTGTATAATAAAGTTGGATCCCAAGGCCCAACTCC-3'
SEQ ID NO:13: обратный праймер hGH:
5'-GGGGACCACTTTGTACAAGAAAGCTGGGTACAACAGGCATCTACT-3'
C. HSV TK минимальный промотор:
HSV TK минимальный промотор клонировали ПЦР из синтезированного в лаборатории вектора, названного «661 CreLuc». Праймерные последовательности имеют сайт attB5 на прямом праймере и сайт attB4 на обратном праймере для рекомбинации в вектор Gateway pDONR P5-P4.
SEQ ID NO:14: прямой праймер TK
5'-GGGGACAACTTTGTATACAAAAGTTGTGGAACACGCAGATGCAGT-3'
SEQ ID NO:15: обратный праймер TK
5'-GGGGACAACTTTGTATAGAAAAGTTGGGTGGATCTGCGGCACGCT-3'
D. кДНК люцифераза:
кДНК люциферазу клонировали ПЦР из вектора pGL4.10 (Promega Мэдисон, Висконсин, кат. № E6651). Luc-праймерные последовательности имеют сайт attB4r на прямом праймере и сайт attB3r на обратном праймере для рекомбинации в Gateway вектор pDONR P4r-P3r.
SEQ ID NO:16: прямой праймер luci
5'GGGGACAACTTTTCTATACAAAGTTGATGGAAGATGCCAAAAACA3'
SEQ ID NO:17: обратный праймер luci
5'GGGGACAACTTTATTATACAAAGTTGTTTACACGGCGATCTTGCC3'
III. Конечная векторная конструкция
Четыре компонента трансгена, CRE (синтетическая или гибридная версия), HSVTK min, кДНК люцифераза и hGH с хвостом поли А в векторах pDONR рекомбинировали с pDest2XMAR вектором назначения согласно стандартному протоколу Invitrogen. Два трансгена затем секвенировали и трансфицировали в CHOK1, чтобы протестировать функции. Оба трансгена были функциональными in vitro, как показано на фиг.5. На фиг.5 показан in vitro анализ векторов CreLuc с помощью индукции форсколином.
IV. Генерирование трансгенной мыши
A. Подготовка трансгена:
Качество трансгенных конструкций проверяли и подтверждали разгоном аликвот в агарозном геле. Каких-либо следов деградации не было обнаружено. Наконец, рестрикционный анализ с использованием диагностических XhoI и PvuII и PstI ферментов рестрикции дал ожидаемые профили рестрикции. Затем трансгенные плазмиды расщепляли с помощью Acc65I и PmeI, и фрагменты, содержащие трансгены, выделяли из векторных основ путем разгона гидролизатов на агарозном геле. Затем трансгенные фрагменты вырезали из геля, очищали с помощью набора для экстракции из геля Qiaquick (Qiagen, Валенсия, Калифорния, кат. № 28706) и растворяли перед введением в оплодотворенные яйцеклетки. Чистоту и концентрацию выделенного трансгена проверяли с помощью электрофореза в агарозном геле.
SEQ ID NO:18: искусственный трансген CreLuc (Acc65I/PmeI гидролизат, 5550bp)
5'GTACCCGGGGGCGCGCCGGGATCCTTAATTAAAATTATCTCTAAGGCATGTGAACTGGCTGTCTTGGTTTTCATCTGTACTTCATCTGCTACCTCTGTGACCTGAAACATATTTATAATTCCATTAAGCTGTGCATATGATAGATTTATCATATGTATTTTCCTTAAAGGATTTTTGTAAGAACTAATTGAATTGATACCTGTAAAGTCTTTATCACACTACCCAATAAATAATAAATCTCTTTGTTCAGCTCTCTGTTTCTATAAATATGTACCAGTTTTATTGTTTTTAGTGGTAGTGATTTTATTCTCTTTCTATATATATACACACACATGTGTGCATTCATAAATATATACAATTTTTATGAATAAAAAATTATTAGCAATCAATATTGAAAACCACTGATTTTTGTTTATGTGAGCAAACAGCAGATTAAAAGGAATTCCTGCAGGATCCTTAATTAAGTTCTAGATCCAAGTTTGTACAAAAAAGCAGGCTTACTGTCGACAATTGCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGATTGACGTCAATGGGGTGTCTGAAAGACGTCACAGTATGACCCGGGCTCGAGCCTCCTTGGCTGACGTCAGAGAGAGAGGCCGGCCCCTTACGTCAGAGGCGAGAATTCGACAACTTTGTATACAAAAGTTGTGGAACACGCAGATGCAGTCGGGGCGGCGCGGTCCCAGGTCCACTTCGCATATTAAGGTGACGCGTGTGGCCTCGAACACCGAGCGACCCTGCAGCGACCCGCTTAACAGCGTCAACAGCGTGCCGCAGATCCACCCAACTTTTCTATACAAAGTTGCTATGGAAGATGCCAAAAACATTAAGAAGGGCCCAGCGCCATTCTACCCACTCGAAGACGGGACCGCCGGCGAGCAGCTGCACAAAGCCATGAAGCGCTACGCCCTGGTGCCCGGCACCATCGCCTTTACCGACGCACATATCGAGGTGGACATTACCTACGCCGAGTACTTCGAGATGAGCGTTCGGCTGGCAGAAGCTATGAAGCGCTATGGGCTGAATACAAACCATCGGATCGTGGTGTGCAGCGAGAATAGCTTGCAGTTCTTCATGCCCGTGTTGGGTGCCCTGTTCATCGGTGTGGCTGTGGCCCCAGCTAACGACATCTACAACGAGCGCGAGCTGCTGAACAGCATGGGCATCAGCCAGCCCACCGTCGTATTCGTGAGCAAGAAAGGGCTGCAAAAGATCCTCAACGTGCAAAAGAAGCTACCGATCATACAAAAGATCATCATCATGGATAGCAAGACCGACTACCAGGGCTTCCAAAGCATGTACACCTTCGTGACTTCCCATTTGCCACCCGGCTTCAACGAGTACGACTTCGTGCCCGAGAGCTTCGACCGGGACAAAACCATCGCCCTGATCATGAACAGTAGTGGCAGTACCGGATTGCCCAAGGGCGTAGCCCTACCGCACCGCACCGCTTGTGTCCGATTCAGTCATGCCCGCGACCCCATCTTCGGCAACCAGATCATCCCCGACACCGCTATCCTCAGCGTGGTGCCATTTCACCACGGCTTCGGCATGTTCACCACGCTGGGCTACTTGATCTGCGGCTTTCGGGTCGTGCTCATGTACCGCTTCGAGGAGGAGCTATTCTTGCGCAGCTTGCAAGACTATAAGATTCAATCTGCCCTGCTGGTGCCCACACTATTTAGCTTCTTCGCTAAGAGCACTCTCATCGACAAGTACGACCTAAGCAACTTGCACGAGATCGCCAGCGGCGGGGCGCCGCTCAGCAAGGAGGTAGGTGAGGCCGTGGCCAAACGCTTCCACCTACCAGGCATCCGCCAGGGCTACGGCCTGACAGAAACAACCAGCGCCATTCTGATCACCCCCGAAGGGGACGACAAGCCTGGCGCAGTAGGCAAGGTGGTGCCCTTCTTCGAGGCTAAGGTGGTGGACTTGGACACCGGTAAGACACTGGGTGTGAACCAGCGCGGCGAGCTGTGCGTCCGTGGCCCCATGATCATGAGCGGCTACGTTAACAACCCCGAGGCTACAAACGCTCTCATCGACAAGGACGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGTGGACCGGCTGAAGAGCCTGATCAAATACAAGGGCTACCAGGTAGCCCCAGCCGAACTGGAGAGCATCCTGCTGCAACACCCCAACATCTTCGACGCCGGGGTCGCCGGCCTGCCCGACGACGATGCCGGCGAGCTGCCCGCCGCAGTCGTCGTGCTGGAACACGGTAAAACCATGACCGAGAAGGAGATCGTGGACTATGTGGCCAGCCAGGTTACAACCGCCAAGAAGCTGCGCGGTGGTGTTGTGTTCGTGGACGAGGTGCCTAAAGGACTGACCGGCAAGTTGGACGCCCGCAAGATCCGCGAGATTCTCATTAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAACAACTTTGTATAATAAAGTTGCTGATCCCAAGGCCCAACTCCCCGAACCACTCAGGGTCCTGTGGACAGCTCACCTAGCTGCAATGGCTACAGGTAAGCGCCCCTAAAATCCCTTTGGGCACAATGTGTCCTGAGGGGAGAGGCAGCGACCTGTAGATGGGACGGGGGCACTAACCCTCAGGTTTGGGGCTTCTGAATGTGAGTATCGCCATGTAAGCCCAGTATTTGGCCAATCTCAGAAAGCTCCTGGTCCCTGGAGGGATGGAGAGAGAAAAACAAACAGCTCCTGGAGCAGGGAGAGTGCTGGCCTCTTGCTCTCCGGCTCCCTCTGTTGCCCTCTGGTTTCTCCCCAGGCTCCCGGACGTCCCTGCTCCTGGCTTTTGGCCTGCTCTGCCTGCCCTGGCTTCAAGAGGGCAGTGCCTTCCCAACCATTCCCTTATCCAGGCTTTTTGACAACGCTATGCTCCGCGCCCATCGTCTGCACCAGCTGGCCTTTGACACCTACCAGGAGTTTGTAAGCTCTTGGGGAATGGGTGCGCATCAGGGGTGGCAGGAAGGGGTGACTTTCCCCCGCTGGGAAATAAGAGGAGGAGACTAAGGAGCTCAGGGTTTTTCCCGAAGCGAAAATGCAGGCAGATGAGCACACGCTGAGTGAGGTTCCCAGAAAAGTAACAATGGGAGCTGGTCTCCAGCGTAGACCTTGGTGGGCGGTCCTTCTCCTAGGAAGAAGCCTATATCCCAAAGGAACAGAAGTATTCATTCCTGCAGAACCCCCAGACCTCCCTCTGTTTCTCAGAGTCTATTCCGACACCCTCCAACAGGGAGGAAACACAACAGAAATCCGTGAGTGGATGCCTTCTCCCCAGGCGGGGATGGGGGAGACCTGTAGTCAGAGCCCCCGGGCAGCACAGCCAATGCCCGTCCTTCCCCTGCAGAACCTAGAGCTGCTCCGCATCTCCCTGCTGCTCATCCAGTCGTGGCTGGAGCCCGTGCAGTTCCTCAGGAGTGTCTTCGCCAACAGCCTGGTGTACGGCGCCTCTGACAGCAACGTCTATGACCTCCTAAAGGACCTAGAGGAAGGCATCCAAACGCTGATGGGGGTGAGGGTGGCGCCAGGGGTCCCCAATCCTGGAGCCCCACTGACTTTGAGAGCTGTGTTAGAGAAACACTGCTGCCCTCTTTTTAGCAGTCAGGCCCTGACCCAAGAGAACTCACCTTATTCTTCATTTCCCCTCGTGAATCCTCCAGGCCTTTCTCTACACCCTGAAGGGGAGGGAGGAAAATGAATGAATGAGAAAGGGAGGGAACAGTACCCAAGCGCTTGGCCTCTCCTTCTCTTCCTTCACTTTGCAGAGGCTGGAAGATGGCAGCCCCCGGACTGGGCAGATCTTCAAGCAGACCTACAGCAAGTTCGACACAAACTCACACAACGATGACGCACTACTCAAGAACTACGGGCTGCTCTACTGCTTCAGGAAGGACATGGACAAGGTCGAGACATTCCTGCGCATCGTGCAGTGCCGCTCTGTGGAGGGCAGCTGTGGCTTCTAGCTGCCCGGGTGGCATCCCTGTGACCCCTCCCCAGTGCCTCTCCTGGCCCTGGAAGTTGCCACTCCAGTGCCCACCAGCCTTGTCCTAATAAAATTAAGTTGCATCATTTTGTCTGACTAGGCGTCCTTCTATAATATTATGGGGTGGAGGGGGGTGGTATGGAGCAAGGGGCAAGTTGGGAAGACAACCTGTAGGGCCTGCGGGGTCTATTGGGAACCAAGCTGGAGTGCAGTGGCACAATCTTGGCTCACTGCAATCTCCGCCTCCTGGGTTCAAGCGATTCTCCTGCCTCAGCCTCCCGAGTTGTTGGGATTCCAGGCATGCATGACCAGGCTCAGCTAATTTTTGTTTTTTTGGTAGAGACGGGGTTTCACCATATTGGCCAGGCTGGTCTCCAACTCCTAATCTCAGGTGATCTACCCACCTTGGCCTCCCAAATTGCTGGGATTACAGGCGTGAACCACTGCTCCCTTCCCTGTCCTTCTGATTTTAAAATAACTATACCAGCAGGAGGACGTCCAGACACAGCATAGGCTACCTGGCCATGCCCAACCGGTGGGACATTTGAGTTGTTTGCTTGGCACTGTCCTCTCATGCGTTGGGTCCACTCAGTAGATGCCTGTTGTACCCAGCTTTCTTGTACAAAGTGGGATCTAGACTAGAGTCATCAGTCAATATGTTCACCCCAAAAAAGCTGTTTGTTAACTTGTCAACCTCATTCTAAAATGTATATAGAAGCCCAAAAGACAATAACAAAAATATTCTTGTAGAACAAAATGGGAAAGAATGTTCCACTAAATATCAAGATTTAGAGCAAAGCATGAGATGTGTGGGGATAGACAGTGAGGCTGATAAAATAGAGTAGAGCTCAGAAACAGACCCATTGATATATGTAAGTGACCTATGAAAAAAATATGGCATTTTACAATGGGAAAATGATGATCTTTTTCTTTTTTAGAAAAACAGGGAAATATATTTATATGTAAAAAATAAAAGGGAACCCATATGTCATACCATACACACAAAAAAATTCCAGTGAATTATAAGTCTAAATGGAGAAGGCAAAACTTTAAATCTTTTAGAAAATAATATAGAAGCATGCCATCAAGACTTCAGTGTAGAGAAAAATTTCTTATGACTCAAAGTCCTAACCACAAAGAAAAGATTGTTAATTAGATTGCATGAATATTAAGACTTATTTTTAAAATTAAAAAACCATTAAGAAAAGTCAGGCCATAGAATGACAGAAAATATTTGCAACACCCCAGTAAAGAGAATTGTAATATGCAGATTATAAAAAGAAGTCTTACAAATCAGTAAAAAATAAAACTAGACAAAAATTTGAACAGATGAAAGAGAAACTCTAAATAATCATTACACATGAGAAACTCAATCTCAGAAATCAGAGAACTATCATTGCATATACACTAAATTAGAGAAATATTAAAAGGCTAAGTAACATCTGTGGCTTAATTAAAACAGTAGGATGACTGTTT 3'
SEQ ID NO:19: Гибридная трансгенная последовательность CreLuc (Acc65I/PmeI гидролизат, 5562bp)
5'GTACCCGGGGGCGCGCCGGGATCCTTAATTAAAATTATCTCTAAGGCATGTGAACTGGCTGTCTTGGTTTTCATCTGTACTTCATCTGCTACCTCTGTGACCTGAAACATATTTATAATTCCATTAAGCTGTGCATATGATAGATTTATCATATGTATTTTCCTTAAAGGATTTTTGTAAGAACTAATTGAATTGATACCTGTAAAGTCTTTATCACACTACCCAATAAATAATAAATCTCTTTGTTCAGCTCTCTGTTTCTATAAATATGTACCAGTTTTATTGTTTTTAGTGGTAGTGATTTTATTCTCTTTCTATATATATACACACACATGTGTGCATTCATAAATATATACAATTTTTATGAATAAAAAATTATTAGCAATCAATATTGAAAACCACTGATTTTTGTTTATGTGAGCAAACAGCAGATTAAAAGGAATTCCTGCAGGATCCTTAATTAAGTTCTAGATCCAAGTTTGTACAAAAAAGCAGGCTTAGCACCAGACAGTGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACGAATTCGCACCAGACAGTGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACACAACTTTGTATACAAAAGTTGTGGAACACGCAGATGCAGTCGGGGCGGCGCGGTCCCAGGTCCACTTCGCATATTAAGGTGACGCGTGTGGCCTCGAACACCGAGCGACCCTGCAGCGACCCGCTTAACAGCGTCAACAGCGTGCCGCAGATCCACCCAACTTTTCTATACAAAGTTGCTATGGAAGATGCCAAAAACATTAAGAAGGGCCCAGCGCCATTCTACCCACTCGAAGACGGGACCGCCGGCGAGCAGCTGCACAAAGCCATGAAGCGCTACGCCCTGGTGCCCGGCACCATCGCCTTTACCGACGCACATATCGAGGTGGACATTACCTACGCCGAGTACTTCGAGATGAGCGTTCGGCTGGCAGAAGCTATGAAGCGCTATGGGCTGAATACAAACCATCGGATCGTGGTGTGCAGCGAGAATAGCTTGCAGTTCTTCATGCCCGTGTTGGGTGCCCTGTTCATCGGTGTGGCTGTGGCCCCAGCTAACGACATCTACAACGAGCGCGAGCTGCTGAACAGCATGGGCATCAGCCAGCCCACCGTCGTATTCGTGAGCAAGAAAGGGCTGCAAAAGATCCTCAACGTGCAAAAGAAGCTACCGATCATACAAAAGATCATCATCATGGATAGCAAGACCGACTACCAGGGCTTCCAAAGCATGTACACCTTCGTGACTTCCCATTTGCCACCCGGCTTCAACGAGTACGACTTCGTGCCCGAGAGCTTCGACCGGGACAAAACCATCGCCCTGATCATGAACAGTAGTGGCAGTACCGGATTGCCCAAGGGCGTAGCCCTACCGCACCGCACCGCTTGTGTCCGATTCAGTCATGCCCGCGACCCCATCTTCGGCAACCAGATCATCCCCGACACCGCTATCCTCAGCGTGGTGCCATTTCACCACGGCTTCGGCATGTTCACCACGCTGGGCTACTTGATCTGCGGCTTTCGGGTCGTGCTCATGTACCGCTTCGAGGAGGAGCTATTCTTGCGCAGCTTGCAAGACTATAAGATTCAATCTGCCCTGCTGGTGCCCACACTATTTAGCTTCTTCGCTAAGAGCACTCTCATCGACAAGTACGACCTAAGCAACTTGCACGAGATCGCCAGCGGCGGGGCGCCGCTCAGCAAGGAGGTAGGTGAGGCCGTGGCCAAACGCTTCCACCTACCAGGCATCCGCCAGGGCTACGGCCTGACAGAAACAACCAGCGCCATTCTGATCACCCCCGAAGGGGACGACAAGCCTGGCGCAGTAGGCAAGGTGGTGCCCTTCTTCGAGGCTAAGGTGGTGGACTTGGACACCGGTAAGACACTGGGTGTGAACCAGCGCGGCGAGCTGTGCGTCCGTGGCCCCATGATCATGAGCGGCTACGTTAACAACCCCGAGGCTACAAACGCTCTCATCGACAAGGACGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGTGGACCGGCTGAAGAGCCTGATCAAATACAAGGGCTACCAGGTAGCCCCAGCCGAACTGGAGAGCATCCTGCTGCAACACCCCAACATCTTCGACGCCGGGGTCGCCGGCCTGCCCGACGACGATGCCGGCGAGCTGCCCGCCGCAGTCGTCGTGCTGGAACACGGTAAAACCATGACCGAGAAGGAGATCGTGGACTATGTGGCCAGCCAGGTTACAACCGCCAAGAAGCTGCGCGGTGGTGTTGTGTTCGTGGACGAGGTGCCTAAAGGACTGACCGGCAAGTTGGACGCCCGCAAGATCCGCGAGATTCTCATTAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAACAACTTTGTATAATAAAGTTGCTGATCCCAAGGCCCAACTCCCCGAACCACTCAGGGTCCTGTGGACAGCTCACCTAGCTGCAATGGCTACAGGTAAGCGCCCCTAAAATCCCTTTGGGCACAATGTGTCCTGAGGGGAGAGGCAGCGACCTGTAGATGGGACGGGGGCACTAACCCTCAGGTTTGGGGCTTCTGAATGTGAGTATCGCCATGTAAGCCCAGTATTTGGCCAATCTCAGAAAGCTCCTGGTCCCTGGAGGGATGGAGAGAGAAAAACAAACAGCTCCTGGAGCAGGGAGAGTGCTGGCCTCTTGCTCTCCGGCTCCCTCTGTTGCCCTCTGGTTTCTCCCCAGGCTCCCGGACGTCCCTGCTCCTGGCTTTTGGCCTGCTCTGCCTGCCCTGGCTTCAAGAGGGCAGTGCCTTCCCAACCATTCCCTTATCCAGGCTTTTTGACAACGCTATGCTCCGCGCCCATCGTCTGCACCAGCTGGCCTTTGACACCTACCAGGAGTTTGTAAGCTCTTGGGGAATGGGTGCGCATCAGGGGTGGCAGGAAGGGGTGACTTTCCCCCGCTGGGAAATAAGAGGAGGAGACTAAGGAGCTCAGGGTTTTTCCCGAAGCGAAAATGCAGGCAGATGAGCACACGCTGAGTGAGGTTCCCAGAAAAGTAACAATGGGAGCTGGTCTCCAGCGTAGACCTTGGTGGGCGGTCCTTCTCCTAGGAAGAAGCCTATATCCCAAAGGAACAGAAGTATTCATTCCTGCAGAACCCCCAGACCTCCCTCTGTTTCTCAGAGTCTATTCCGACACCCTCCAACAGGGAGGAAACACAACAGAAATCCGTGAGTGGATGCCTTCTCCCCAGGCGGGGATGGGGGAGACCTGTAGTCAGAGCCCCCGGGCAGCACAGCCAATGCCCGTCCTTCCCCTGCAGAACCTAGAGCTGCTCCGCATCTCCCTGCTGCTCATCCAGTCGTGGCTGGAGCCCGTGCAGTTCCTCAGGAGTGTCTTCGCCAACAGCCTGGTGTACGGCGCCTCTGACAGCAACGTCTATGACCTCCTAAAGGACCTAGAGGAAGGCATCCAAACGCTGATGGGGGTGAGGGTGGCGCCAGGGGTCCCCAATCCTGGAGCCCCACTGACTTTGAGAGCTGTGTTAGAGAAACACTGCTGCCCTCTTTTTAGCAGTCAGGCCCTGACCCAAGAGAACTCACCTTATTCTTCATTTCCCCTCGTGAATCCTCCAGGCCTTTCTCTACACCCTGAAGGGGAGGGAGGAAAATGAATGAATGAGAAAGGGAGGGAACAGTACCCAAGCGCTTGGCCTCTCCTTCTCTTCCTTCACTTTGCAGAGGCTGGAAGATGGCAGCCCCCGGACTGGGCAGATCTTCAAGCAGACCTACAGCAAGTTCGACACAAACTCACACAACGATGACGCACTACTCAAGAACTACGGGCTGCTCTACTGCTTCAGGAAGGACATGGACAAGGTCGAGACATTCCTGCGCATCGTGCAGTGCCGCTCTGTGGAGGGCAGCTGTGGCTTCTAGCTGCCCGGGTGGCATCCCTGTGACCCCTCCCCAGTGCCTCTCCTGGCCCTGGAAGTTGCCACTCCAGTGCCCACCAGCCTTGTCCTAATAAAATTAAGTTGCATCATTTTGTCTGACTAGGCGTCCTTCTATAATATTATGGGGTGGAGGGGGGTGGTATGGAGCAAGGGGCAAGTTGGGAAGACAACCTGTAGGGCCTGCGGGGTCTATTGGGAACCAAGCTGGAGTGCAGTGGCACAATCTTGGCTCACTGCAATCTCCGCCTCCTGGGTTCAAGCGATTCTCCTGCCTCAGCCTCCCGAGTTGTTGGGATTCCAGGCATGCATGACCAGGCTCAGCTAATTTTTGTTTTTTTGGTAGAGACGGGGTTTCACCATATTGGCCAGGCTGGTCTCCAACTCCTAATCTCAGGTGATCTACCCACCTTGGCCTCCCAAATTGCTGGGATTACAGGCGTGAACCACTGCTCCCTTCCCTGTCCTTCTGATTTTAAAATAACTATACCAGCAGGAGGACGTCCAGACACAGCATAGGCTACCTGGCCATGCCCAACCGGTGGGACATTTGAGTTGTTTGCTTGGCACTGTCCTCTCATGCGTTGGGTCCACTCAGTAGATGCCTGTTGTACCCAGCTTTCTTGTACAAAGTGGGATCTAGACTAGAGTCATCAGTCAATATGTTCACCCCAAAAAAGCTGTTTGTTAACTTGTCAACCTCATTCTAAAATGTATATAGAAGCCCAAAAGACAATAACAAAAATATTCTTGTAGAACAAAATGGGAAAGAATGTTCCACTAAATATCAAGATTTAGAGCAAAGCATGAGATGTGTGGGGATAGACAGTGAGGCTGATAAAATAGAGTAGAGCTCAGAAACAGACCCATTGATATATGTAAGTGACCTATGAAAAAAATATGGCATTTTACAATGGGAAAATGATGATCTTTTTCTTTTTTAGAAAAACAGGGAAATATATTTATATGTAAAAAATAAAAGGGAACCCATATGTCATACCATACACACAAAAAAATTCCAGTGAATTATAAGTCTAAATGGAGAAGGCAAAACTTTAAATCTTTTAGAAAATAATATAGAAGCATGCCATCAAGACTTCAGTGTAGAGAAAAATTTCTTATGACTCAAAGTCCTAACCACAAAGAAAAGATTGTTAATTAGATTGCATGAATATTAAGACTTATTTTTAAAATTAAAAAACCATTAAGAAAAGTCAGGCCATAGAATGACAGAAAATATTTGCAACACCCCAGTAAAGAGAATTGTAATATGCAGATTATAAAAAGAAGTCTTACAAATCAGTAAAAAATAAAACTAGACAAAAATTTGAACAGATGAAAGAGAAACTCTAAATAATCATTACACATGAGAAACTCAATCTCAGAAATCAGAGAACTATCATTGCATATACACTAAATTAGAGAAATATTAAAAGGCTAAGTAACATCTGTGGCTTAATTAAAACAGTAGGATGACTGTTT
B. Генерирование основателя:
Трансгены по отдельности микроинъецировали в пронуклеусы одноклеточных эмбрионов FVB/Taconic. Общие стратегии для генерирования трансгенных (Tg) животных хорошо описаны и известны специалистам в данной области.
Применяя стратегию ПЦР генотипирования, трансген-положительных мышей-основателей идентифицировали вскоре после рождения посредством биопсий хвостов и ПЦР на генотип. Выбранная пара праймеров позволяет амплифицировать короткую последовательность ДНК внутри трансгенной последовательности, давая специфичный 405-bp ПЦР продукт. Реакционная смесь для ПЦР представляет собой буфер для ПЦР 1X Qiagen с MgCl2 (Qiagen, Валенсия, Калифорния, кат. № 201205), 0,2 ммоль/л дезоксинуклеозидтрифосфатов, 0,4 мкмоль/л праймеров, 0,5 единицами Taq полимеразы. Условия циклирования следующие: 1 цикл при 94°C; 5 мин, 35 циклов при 94°C; 45 с/57°C; 45 с/72°C; 1 мин, 1 цикл при 72°C; 5 мин.
Генотипирующие праймеры следующие:
SEQ ID NO:20: Luc2-прямой праймер:
5'-GAAGATGCCAAAAACATTAAGAAG-3'
SEQ ID NO:21: Luc2-обратный праймер:
5'-GATCTTTTGCAGCCCTTTCT-3'
Для искусственного трансгена CreLuc получали 39 трансген-положительных основателей (из 181 родившегося) и для гибридного трансгена CreLuc, 73 трансген-положительных (из 244 родившихся). Трансгенные линии мышей обозначали аббревиатурой CreLuc.
V. Анализ экспрессии основателей:
На фиг.5А представлено сравнение эффектов различных ингибиторов фосфодиэстеразы (PDE) на уровни цАМФ в плазме у нормальных мышей (воспроизводили по Cheng et al., JPET, 280(2):621-626, 1997). Группам мышей, по четыре животных в каждой, перорально вводили соединение или растворитель (VEH). Через двадцать минут отбирали кровь и обрабатывали для измерений цАМФ. На основе указанного опубликованного эксперимента ролипрам выбрали в качестве общего индуктора наряду с форсколином для скрининга CreLuc-основателя на его способность и пригодность для повышения уровней цАМФ. Аддитивный эффект двух соединений при совместном дозировании обеспечивает более широкий диапазон индукции при исследовании уровней люциферазы у животных.
На фиг.5В представлен анализ цАМФ относительно уровней плазмы у мышей дикого типа после различной обработки форсколином (F), ролипрамом (R) или комбинацией форсколина и ролипрама (F/R). Самкам FVB/Tac возрастом два месяца интраперитонеально вводили одно из следующих: группа A: контрольный растворитель, 1% диметилсульфоксид (DMSO) в забуференном фосфатом растворе (PBS), группа В: 5 мг/кг форсколина (Sigma, Сент-Луис, Миссури, кат. № F6886), и группа C: 10 мг/кг ролипрама (Sigma, Сент-Луис, Миссури, кат. № R6520), группа D: 5 мг/кг форсколина плюс 10 мг/кг ролипрама, группа E: 5 мг/кг водорастворимого форсколина (Calbiochem Гиббстаун, Нью-Йорк, кат. № 344273) и группа F: 5 мг/кг водорастворимого форсколина плюс 10 мг/кг ролипрама. Группа G обработки не получала. По сравнению с самим растворителем ролипрам и (или) форсколин статистически значимы в их повышении уровней циркуляции цАМФ. Сам форсколин незначительно увеличивал циркуляцию цАМФ по сравнению с контрольным растворителем. Комбинация водорастворимого форсколина (H2OF) и ролипрама давала наибольшее повышение уровней цАМФ в плазме и поэтому указывала на то, что данная обработка будет оптимальной для скрининга CreLuc-основателей для экспрессии трансгена in vivo.
Все основатели (основатели, перечисленные в таблицах 5-8, и репрезентативные основатели, показанные на фиг.6) тестировались на экспрессию трансгена путем интраперитонеального введения комбинации 10 мг/кг ролипрама (Sigma, Сент-Луис, Миссури, кат. № R6520) и 5 мг/кг водорастворимого форсколина (Calbiochem, Гиббстаун, Нью-Йорк, кат. № 344273). Через четыре часа мышей биовизуализировали с использованием системы Xenogen IVIS Lumina. Мышей анестезировали изофураном, и им подкожно вводили по 250 мг/кг люциферина. Через восемь минут после инъекции люциферина мышей визуализировали. Из 39 трансген-положительных на искусственный трансген CreLuc 34, или 87%, имели положительную экспрессию. Для гибридного трансгена 55 из 71, или 78% испытанных мышей, имели положительную экспрессию. Экспрессию считали положительной тогда, когда уровни люциферазы в любой ткани выше фоновых уровней для системы.
На фиг.6 проиллюстрированы различные профили отклика на форсколин/ролипрам, проанализированные путем биовизуализации различных независимых линий мышей CreLuc. В неиндуцированном состоянии наблюдали либо обнаруживаемые базальные сигналы биовизуализации, как проиллюстрировано в линиях 90 и 219, либо отсутствие базальной активности, как проиллюстрировано в линиях 44, 28 и 187. После индукции форсколином/ролипрамом в единичных или множественных тканях возникала картина биовизуализации индукции репортера CreLuc, и она была уникальной в каждой независимой линии CreLuc. Многие из линий CreLuc имели одну или более тканей, экспрессирующих обнаруживаемую люциферазу, что биовизуализируется в терапевтически значимых областях.
Вслед за скринингом биовизуализацией экспрессии CreLuc у основателей, как с базальной индукцией, так и с индукцией форсколином/ролипрамом, линии CreLuc, которые соответствовали полосе пропускания фильтра 5x индукции биовизуализации в одной или нескольких тканях (2x в головном мозге), более подробно анализировали путем анализа индуцированных уровней фермента люциферазы в экстрактах тканей.
Для анализа тканевой экспрессии животных либо оставляли без обработки для базисного сигнала, либо индуцировали в течение 4 часов. Для индукции животным интраперитонеально вводили комбинацию 5 мг/кг форсколина (Calbiochem, Гиббстаун, Нью-Йорк, кат. № 344273) и 10 мг/кг ролипрама (Sigma, Сент-Луис, Миссури, кат. № R6520) с 1% DMSO (Sigma, Сент-Луис, Миссури, кат. № D2650) в PBS Дульбекко (Invitrogen, Карлсбад, Калифорния, кат. № 14040). Через четыре часа животных умерщвляли CO2, извлекали различные органы и замораживали на сухом льде. Для анализа люциферазы применяли систему Luciferase Assay System (Promega, Мэдисон, Висконсин, кат. № E1500). Ткани гомогенизировали в 1 мл лизирующего буфера культуры клеток (Promega, Мэдисон, Висконсин, кат. № E1531), инкубировали на льду в течение 5 минут, и затем вращали на центрифуге в течение 5 минут. Согласно промышленной инструкции в анализе применяли 20 мкл надосадочной жидкости. Концентрации белков в образцах измерили с помощью набора для анализа белков DC (BioRad, Геркулес, Калифорния, кат. № 500-0111). Данные показаны в таблицах 5-8 и представлены как кратность индукции (индуцированный RLU/мкг белка, деленный на базальный RLU/мкг белка). Индуцированный RLU представляет собой RLU/мкг белок в образце индуцированной ткани.
Тканевая экспрессия люциферазы в головном мозге, селезенке и почках
Тканевая экспрессия люциферазы в печени, тимусе и поджелудочной железе
Тканевая экспрессия люциферазы в сердце, легких и
спинном мозге
Тканевая экспрессия люциферазы в коже и кишечнике
Указанный подробный уровень экспрессии CreLuc в каждой линии позволил выбрать оптимальные линии с самыми высокими уровнями экспрессии в целевых тканях, а также выбрать линии с уникальными единичными или множественными картинами экспрессии. Анализ ферментов также гарантировал то, что экспрессия CreLuc безусловно связана с отдельной тканью. В целом, как и было разработано в трансгене, наблюдали экспрессию люциферазы почти во всех тканях через большинство линий. Выбрали самые важные 3-5 линий из ключевых фармацевтически значимых целевых тканей (головной мозг, поджелудочная железа, легкие, селезенка) для дальнейшего анализа профилей ответа референтных соединений с применением лигандов GPCR.
VI. Анализ эффектов инсуляторных элементов на уровни экспрессии
В таблице 9 сопоставляется влияние добавления инсуляторных элементов на 11 различных трансгенов, включая 225 независимых трансгенных линий. Производство функциональных трансгенных линий, как правило, колеблется от 10-30% (ретроспективные данные, полученные в лаборатории авторов изобретения). Как видно из таблицы 9, из-за включения минигена hGH или функционального энхансера, 10 из 11 трансгенов и 190 линий достигли уровней экспрессии по верхней границе указанного интервала (среднее количество функциональных линий составляет 37 или 32,7%). Существенно, что включение инсуляторных элементов более чем вдвое повышает указанную частоту экспрессии, и из 112 линий 78% экспрессируют функциональный репортер. Размеры образцов для обоих сравнений и разнообразие трансгенов достаточно велики для того, чтобы преодолеть нормальные биологические отклонения, вызванные случайным интегрированием трансгенов в геном.
Генерирование линий, трансгенных по репортеру для биовизуализации, представляет собой низкочастотный процесс с высоким уровнем отсева. Большое количество независимых линий основателя (50-100) необходимо для получения линий с желаемым уровнем обнаружения для репортера и для того, чтобы репортер экспрессировался для большого выбора мишеневых тканей, подходящих для скрининга кандидатных лекарственных средств. Как правило, для применений, связанных со скринингом лекарственных средств, достаточно 5% линий или менее, поэтому трансген, содержащий репортер, должен включать несколько элементов для максимизирования экспрессии в каждой из генерируемых трансгенных линий. Инсуляторные элементы, соединенные с другими энхансерами экспрессии (минигеном hGH), как было продемонстрировано, приводят к тому, что большинство трансгенных линий экспрессирует трансген, и они являются ключевыми для обнаружения трансгенных линий с картиной экспрессии, которая применима для открытия новых лекарственных средств (в особенности, для различных сигнальных путей GPCR при регистрации понижения сигнала после связывания лиганда с рецептором).
Сравнение частоты генерирования функциональных трансгенных линий, генерированных с минигеном hGH или инсуляторными элементами
позитивных
tetp
Легенда: CD11 ArtTA, CD11a промотор, экспрессирующий обратный активатор для тетрациклинового трансактиватора; TetEGFP является тетрациклиновым промотором, экспрессирующим EGFP репортер; TetIL13 является тетрациклиновым промотором, экспрессирующим IL13; CC10rtTA; CC10 специфический промотор легких, экспрессирующий обратный активатор для тетрациклинового трансактиватора; hCXCR5- CD11a или b является человеческим CXCR5 промотором, экспрессирующим CD11a или CD11b кодирующую кДНК; MBP luci является промотором основного белка миелина (MBP), экспрессирующим люциферазу (luci); GP1 и PF4- GFP; GP1 и PF4 специфическими промоторами тромбоцитов, экспрессирующими GFP репортер.
Депонирование материала:
Подготовка эмбриональных фибробластов мыши: 12-дневные эмбрионы извлекали из беременных самок. Эмбрионы вырезали из матки в 10-см чашку, содержащую забуференный фосфатом раствор (PBS) (Invitrogen, Карлсбад, Калифорния, кат. № 14040). Затем их переносили на новую чашку с PBS. Эмбрионы препарировали хирургическими щипцами, сердце и печень удаляли. Фрагменты тканей затем инкубировали в 2-3 мл холодного 0,25% трипсина (Invitrogen, Карлсбад, Калифорния, кат. № 25200) в течение ночи при 4°С. На следующий день бόльшую часть трипсина отсасывали и тканевую массу инкубировали в течение 30 мин при 37°С. Добавляли 2-3 мл среды: DMEM (Invitrogen, Карлсбад, Калифорния, кат. № 11965), 5% термоинактивированной FBS (Invitrogen, Карлсбад, Калифорния, кат. № 16140), 1% Penstrep (Invitrogen, Карлсбад, Калифорния, кат. № 15140), и пипетировали вверх и вниз для разрушения остающихся комков ткани. Суспензию клеток затем высевали в одну из колб для тканевой культуры T75. Клетки культивировали в течение 3 пассажей, и затем замораживали во флаконах объемом 1 мл по 1-2 миллиона клеток на флакон в замораживающей среде, 15% термоинактивированной FBS (Invitrogen, Карлсбад, Калифорния, кат. № 16140), 5% DMSO (Sigma, Сент-Луис, Миссури, кат. № D2650), DMEM (Invitrogen, Карлсбад, Калифорния, кат. № 11965).
В Американской коллекции типовых культур (ATCC, 10801 University Blvd., Манассас, Виргиния 20110-2209, США) депонированы следующие материалы:
Депонирование материала
Депозит сделан на условиях Будапештского договора о международном признании депонирования микроорганизмов для целей патентной процедуры, а также нормативных документов на его основе (Будапештский договор). Это гарантирует сохранение жизнеспособной культуры на депозите в течение 30 лет со дня депонирования. Депозит будет доступным через ATCC по условиям Будапештского договора и при условии соглашения между Sanofi-Aventis US Inc. и ATCC, что гарантирует постоянную и неограниченную доступность для общественности потомства культуры, находящейся на депозите, после выдачи имеющего отношение к делу патента США или после выкладки для общественности любой патентной заявки США или иностранной патентной заявки, какая бы из них ни выкладывалась первой, и гарантирует доступность потомства для уполномоченных лиц, определяемых Комиссаром США по патентам и торговым маркам.
Правопреемник настоящей заявки дал согласие на то, чтобы, если культура материалов на депозите погибнет или будет утеряна или уничтожена при культивировании в пригодных условиях, материалы были немедленно заменены по уведомлении другими аналогичными материалами. Доступность депонированных материалов не следует истолковывать как предоставление лицензии на практическое использование изобретения в противоречии с правами, гарантируемыми властью любого правительства в соответствии с его патентным законодательством.
VII. Скрининговый количественный анализ in vivo и ex vivo
На фиг.7 показаны варианты осуществления настоящего изобретения путем представления последовательности операций скрининговых количественных анализов CreLuc или in vivo, или ex vivo. Для количественных анализов in vivo животным вводили только дозу люциферина, и затем их биовизуализировали на биовизуализаторе IVIS для получения базисного сигнала. Затем животным вводили дозу соединения и затем повторно биовизуализировали в заданный момент времени с целью изучения влияния соединения на сигнал люциферазы. Данные могут анализироваться как кратность индукции для индуцированного сигнала относительно базисного сигнала. Для количественных анализов ex vivo мышам вводили либо дозу соединения, либо дозу растворителя, и затем в заданный момент времени ткани собирали и замораживали. Затем ткани могут быть гомогенизированы, и на экстрактах тканей может быть проведен количественный анализ люциферазы, такой как, например, система количественного анализа Promega Luciferase (Promega, кат. № E1500), и количественный анализ может быть считан на люминометре, таком как, например, Topcount. Данные могут быть проанализированы в относительных световых единицах на мкг белка путем сравнения групп растворителя и соединения.
Дополнительные аспекты и подробности настоящего изобретения станут ясны из нижеследующих примеров, которые предназначены для иллюстрации и ни в коем случае не подразумеваются как ограничивающие объем настоящего изобретения.
A. Эксперименты на срезах цельного мозга
Для того чтобы показать влияние соединений на люминесценцию у мышей CreLuc (фиг.8А, 8В), использовали срезы цельного мозга. В целом, мозг мышей CreLuc может быть собран, а затем разрезан на срезы на микротоме, или из него также могут быть вырезаны различные подобласти, а затем могут быть сделаны срезы на микротоме. Срезы могут затем инкубироваться с различными соединениями, такими как форсколин и ролипрам, или с GPCR специфичными соединениями. Уровни люциферазы затем могут быть измерены биовизуализатором IVIS или фотоумножителем с карусельным магазином с целью получения количественных измерений с течением времени.
Срезы мозга из гиппокампальной области мышей CreLuc (линия № 187) инкубировали с 1 мкмоль/л изопротеренола (Sigma, кат. № 5627) в течение 2 дней. Изопротеренол (ISO) является Gs-агонистом, который активирует цАМФ путь (см. фиг.1 со схемой пути) и индуцирует уровни экспрессии люциферазы у мышей CreLuc (фиг.8А, нижняя панель). Поэтому изопротеренол используется для увеличения ширины окна сигнала люциферазы, что позволяет выполнять скрининг таких GPCR специфичных соединений, как Gs-агонисты. AMN082 (AMN, Tocris Bioscience, кат. № 2385) является mGluR7-агонистом, и mGluR7 является рецептором, который сопряжен с Gi. При обработке срезов мозга 1 мкмоль/л ISO в сочетании с 1 мкмоль/л AMN082 сигнал люциферазы ослаблялся по сравнению с обработкой только ISO.
Также в жизнеспособном срезе мозга может количественно оцениваться активность соединений с течением времени, и экспрессия люциферазы может визуализироваться при помощи биовизуализатора. На фиг.8В срезы головного мозга мышей CreLuc (линия 44) были чувствительны к форсколину. Форсколин является неспецифичным активатором цАМФ. Обработка среза головного мозга с 50 мкмоль/л форсколина (стрелка отмечает время добавления форсколина к срезу) вызывала индукцию зависящим от времени образом по сравнению с растворителем.
B. Эксперименты на клеточных культурах in vitro
1. Первичные кортикальные нейроны
Общая схема последовательности операций для выделения и обработки соединениями первичных нейронных клеток из модели CreLuc изображена на фиг.9. Кору головного мозга или другие подобласти извлекают из эмбрионов, обычно на эмбриональный (Е) день 17 или 18, однако также и таких ранних, как Е14. Индивидуальные нейроны затем выделяют и высевают на 96-луночные аналитические планшеты. Количественные анализы выполняют или в трех, или в четырех повторах. Количественные анализы выполняют, начиная с 3-го дня в культуре до 7-го или 10-го дня в культуре, в зависимости от уровней экспрессии GPCR-мишени. Затем в установленный момент времени, в планшеты добавляют соединения, также в планшеты добавляют Bright Glo (Promega, кат. № E2610) и затем выполняют количественный анализ на TopCount или другом люминометре.
a) Gs-модуляция посредством β-адренергических рецепторов (ADβR) и D1-дофаминовых рецепторов (DRD1)
Первичные клеточные культуры мышей CreLuc могут использоваться для скрининга соединений, которые способны модулировать Gs-сопряженные рецепторы. Нейроны были выделены из коры головного мозга эмбрионов E18 линии 187. Линия 187, как было показано, имеет индуцируемые уровни люциферазы как в головном, так и в спинном мозге путем количественных анализов люциферазы во всей ткани (см. текст ниже; Фигура 24). Количественный анализ выполняли в трех повторах на третий день в культуре. Форсколин применяли в концентрации 5 мкмоль/л, ролипрам - 10 мкмоль/л, и Gs-агонисты, ADβR - изопротеренол, и дофамин - SKF82958 (MFG, кат. № C130) - 10 мкмоль/л. Через 4 часа добавляли Bright Glo, и количественные анализы выполняли на Topcount. Форсколин представляет собой неспецифический активатор цАМФ. Ролипрам не активирует цАМФ, однако вместо этого ролипрам блокирует разрыв цАМФ и таким образом стабилизирует уровни цАМФ. Форсколин и ролипрам действуют синергично, увеличивая экспрессию люциферазы. При комбинации форсколина и ролипрама наблюдалась более чем 100-кратная индукция. Увеличенные по сравнению с контролем DMSO уровни luci наблюдались для ADβR-агониста изопротеренола (11-кратное повышение). Несмотря на то, что агонист D1 DR, SKF82958, незначительно повышал уровни люциферазы, при введении SKF82958 в сочетании с ролипрамом наблюдалась 2-кратная индукция в сравнении с одним ролипрамом. Таким образом, линия 187 является полезной моделью для скрининга соединений с целью определения, является ли соединение способным модулировать Gs-сопряженные рецепторы.
b) Gq-модуляция с помощью рецептора прокинетицина 2 (PROK2R)
Первичные клеточные культуры мышей CreLuc могут использоваться для скрининга соединений, которые способны модулировать Gs-сопряженные рецепторы. PROK2R является Gq-сопряженным, поэтому прокинецитин 2 (PROK2), который является агонистом для PROK2R, использовали для того, чтобы наблюдать, будут ли мыши CreLuc чувствительными к Gq-модуляции. Gq обходит цАМФ, однако использует PLC путь для активации CREB (см. схему пути на фиг.1). Первичные кортикальные нейроны собирали из линии 187 (которая, как показано, содержит индуцируемую люциферазу в головном и спинном мозге, см. текст ниже и фиг.24) на Е18. Количественный анализ выполняли на 3-й день в культуре в течение 4 или 8 часов. Белок PROK2 (Peprotech, кат. № 100-46) добавляли в виде водного раствора с концентрацией 1 нмоль/л и 100 нмоль/л. Для сигнала люциферазы использовали Bright Glo, и количественный анализ выполняли на люминометре TopCount. Для обоих моментов времени и для обеих концентраций PROK2 наблюдали высокозначимое увеличение уровней люциферазы относительно контроля, содержащего только среду. Таким образом, мыши CreLuc могут использоваться для скрининга соединений, которые модулируют Gq-сопряженные рецепторы.
Были изучены эффекты белка PROK2 на экспрессию люциферазы в первичных кортикальных нейронах различных линий CreLuc. Первичные кортикальные нейроны собирали из четырех различных линий CreLuc на E18. Линии выбирали на основе ранее проведенных экспериментов, которые определяли индуцируемые уровни люциферазы в экстрактах всего мозга (данные не показаны). Количественные анализы выполняли в трех повторах на третий день в культуре либо с 1 нмоль/л, либо со 100 нмоль/л белка PROK2 в два момента времени, в 4 часа и в 24 часа. Для количественного анализа и считывания на TopCount использовали BrightGlo. Статистически значимые увеличения уровней люциферазы наблюдали в оба момента времени и при обеих концентрациях для линий 219 и 175. И хотя все линии в некоторой степени отвечают на лиганд PROK2, различия в уровнях экспрессии люциферазы могут быть связаны с различиями в интеграции трансгена.
c) Gi-модуляция с помощью рецепторов mGluR7
Первичные клеточные культуры мышей CreLuc могут использоваться для скрининга соединений, которые способны модулировать Gi-сопряженные рецепторы. На фиг.13А показаны эффекты mGluR7-агониста, AMN082, на экспрессию люциферазы в первичных кортикальных нейронах. Кортикальные нейроны собирали из эмбрионов Е18 (линия 187). Количественный анализ выполняли на 3-й день в культуре. Для получения более широкого окна, используемого при регистрации Gi-активности, форсколин применяли в концентрации 10 мкмоль/л. Агонист mGluR7, AMN082, применяли в комбинации с форсколином в концентрациях 1 нмоль/л, 10 нмоль/л, 100 нмоль/л и 1 мкмоль/л. Количественный анализ считывали на TopCount с Bright Glo (Promega) через 4 часа и 8 часов. Значимые снижения уровней люциферазы по отношению к одному только форсколину наблюдались при 100 нмоль/л и 1 мкмоль/л AMN082 для обоих моментов времени. Мыши CreLuc могут использоваться для скрининга соединений, которые модулируют Gq-сопряженные рецепторы. Например, «неизвестные» соединения, обозначенные как «A», «B» и «C», подвергали скринингу в культурах первичных кортикальных нейронов с использованием AMN082 в качестве внутреннего контроля с целью определения, проявляют ли какие-либо из соединений способность модулировать Gi (фиг.13В). Соединение А можно считать активным соединением с вычисленной EC50=1,529×10-6 в сравнении с известным mGluR7-агонистом, AMN082, который имеет EC50=1,735×10-7, в то время как соединение В следует считать неспецифичным соединением, и соединение С неактивно.
Скрининг соединений на способность модулировать Gi-сопряженные рецепторы с использованием культуры первичных клеток мышей CreLuc, как продемонстрировано выше, может выполняться в присутствии стимулятора цАМФ, такого как форсколин. Однако для увеличения ширины окна сигнала еще шире используется ролипрам (блокирующий разрыв цАМФ) в комбинации с форсколином. Первичные кортикальные нейроны выделяли из эмбрионов E18 линии 187, которые, как было определено, обладают индуцируемой экспрессией в головном и спинном мозге. Количественный анализ в трех повторах выполняли на 7 день в культуре в течение 6 часов. Построение дозовой кривой для AMN082 в концентрации от 10 мкмоль/л до 1 нмоль/л выполняли в комбинации с 50 мкмоль/л форсколина и 10 мкмоль/л ролипрама. Количественный анализ считывали на люминометре TopCount с субстратом Bright Glo (Promega). Статистически значимые (p<0,0005) снижения уровней люциферазы (59-61% по отношению к контрольным группам форсколина и ролипрама) наблюдаются при 1 мкмоль/л, 100 нмоль/л и 1 нмоль/л.
Gi-модуляция экспрессии люциферазы в первичных кортикальных нейронах из различных линий CreLuc CB1-агонистом, CP 55,940, выполняли в присутствии форсколина и ролипрама (фиг.15). Первичные кортикальные нейроны собирали из линий 69, 187, 175 и 219 на E18. Все используемые линии, как было определено предварительно, обладают индуцируемыми уровнями экспрессии люциферазы в экстрактах тканей всего мозга (данные не показаны). Количественные анализы выполняли на третий день в культуре. CB1-агонист использовали в концентрации 10 мкмоль/л, форсколин - 5 мкмоль/л и ролипрам - 10 мкмоль/л. Выполняли анализы для двух моментов времени, четыре часа и двадцать четыре часа. Затем добавляли субстрат Bright Glo для количественного анализа люциферазы, и количественный анализ считывали на люминометре Topcount. Количественный анализ выполняли в трех повторах. Приведенные данные представляют среднее для трех повторов. Данные показаны как число импульсов в секунду (cps). При добавлении СВ1-агониста наблюдаются сниженные сигналы люциферазы во всех четырех линиях в оба момента времени. Незначительные различия в уровнях ответа могут быть вызваны интеграцией трансгена, однако все линии были чувствительны к Gi-модуляции и могут быть улучшены для скрининга соединений на Gi-активность.
Gi-модуляция люциферазы в первичных кортикальных нейронах мышей CreLuc СВ1-агонистом, CP 55,940, происходила дозозависимым образом (фиг.15В). Кортикальные нейроны выделяли из эмбрионов E18. Количественный анализ выполняли на 3-й день в культуре. Форсколин и ролипрам применяли в концентрации 10 мкмоль/л. Агонист добавляли в концентрациях 10 мкмоль/л, 1 мкмоль/л и 100 нмоль/л. Количественный анализ считывали на TopCount с BrightGlo (Promega) через 8 часов. Значимые снижения уровней люциферазы (агонист плюс форсколин и ролипрам против форсколина и ролипрама по отдельности) наблюдались для всех 3 концентраций.
2. Первичные стриарные нейроны
Любые клеточные культуры, полученные из мышей CreLuc, могут применяться для скрининга соединений на способность модулировать GPCR. Экспрессию люциферазы индуцировали в стриарных нейронах CreLuc посредством форсколина и ролипрама и Gs-агонистов для DRD1 и ADβR (фиг.16). Стриарные нейроны были выделены из эмбрионов Е14 (линия 187). Количественные анализы выполняли на 4-й день в культуре. Форсколин применяли в концентрации 5 мкмоль/л, ролипрам - 10 мкмоль/л. Gs-агонисты (изопротеренол является агонистом у ADRβ, дофамин и SKF82958 являются агонистами у D1DR) применяли в концентрациях 10 мкмоль/л, 3 мкмоль/л и 1 мкмоль/л. Анализ считывали через 5 часов люминометром TopCount и реактивом на люциферазу Bright Glo (Promega). Высокозначимое 28-кратное повышение наблюдалось в клетках, обработанных комбинацией форсколина и ролипрама. Значимые повышения, 2,7-кратные, также наблюдалось для 10 мкмоль/л дофамина и для всех трех концентраций изопротеренола. Таким образом, трансген является функциональным в стриарных, а также кортикальных нейронах (см. фиг.10).
3. Цельные спленоциты
Препараты цельных спленоцитов, выделенных из мышей CreLuc (линия 64), использовали для демонстрации эффектов неспецифических индукторов цАМФ, таких как форсколин и ролипрам, а также Gs-агонистов на экспрессию люциферазы (фиг.16). Селезенки собирали из животных в 1X HBSS (Invitrogen, Карлсбад, Калифорния, кат. № 14025). Затем клетки выделяли путем механического разрушения капсулы селезенки с использованием кончика шприца объемом 5 мл в 5 мл D-PBS (Invitrogen, Карлсбад, Калифорния, кат. № 14190). Суспензию клеток затем пропускали через 70 мкм фильтр в коническую пробирку объемом 50 мл. Затем клетки вращали со скоростью 800 об./мин, повторно суспендировали и инкубировали в течение 6 минут при комнатной температуре в 5 мл раствора 1X Pharm Lyse (BD BioSciences, кат. № 555899). Затем клетки промывали путем добавления 28 мл среды, состоящей из RPMI 1640 (Invitrogen, Карлсбад, Калифорния, кат. № 11875), 10% FCS (Invitrogen, Карлсбад, Калифорния, кат. № 16000), 1% Penstrep (Invitrogen, Карлсбад, Калифорния, кат. № 15070) и 0,1% β-меркаптоэтанола (Invitrogen, Карлсбад, Калифорния, кат. № 21985). Клетки разбавляли до 2×106/мл и высевали по 100 мкл клеток на каждую лунку белого непрозрачного 96-луночного планшета. Половину клеток стимулировали анти-CD3 антителом (BD Pharmingen, кат. № 553058) в течение 24 часов; вторую половину не обрабатывали. Через 24 часа в планшеты добавили соединения еще на 4 часа. Неспецифические индукторы включали ролипрам (Sigma R6520) и форсколин (Sigma F6886). Используемыми Gs-агонистами являлись: EX00000173A (173A; синтезирован в лаборатории), представляющий собой агонист Gs-сопряженного рецептора простагландина E2 (EP2); BW245C (Sigma B9305), представляющий собой агонист Gs-сопряженного рецептора 1 простагландина D2 (DP1); и изопротеренол (Sigma I5627), представляющий собой агонист Gs-сопряженного β-адренергического рецептора (ADβR). Количественный анализ выполняли в трех повторах. Через 4 часа добавляли 100 мкл BrightGlo (Promega, Мэдисон, Висконсин, кат. № E2610), и количественный анализ считывали на люминометре TopCount. В клетках, стимулированных CD3, в присутствии ролипрама и форсколина наблюдается 14-кратное увеличение относительно DMSO. Статистически значимые увеличения (согласно проверке по t-критерию Стьюдента) также наблюдались для всех трех Gs-агонистов по отношению к контрольной группе, содержащей только DMSO. Данный эксперимент показывает, что трансген является функциональным в популяциях цельных спленоцитов. Препарат, использованный в данном эксперименте, являлся препаратом цельных спленоцитов, который представляет собой смешанную популяцию клеток. Описанные ниже эксперименты рассматривают уровни экспрессии люциферазы в таких субпопуляциях, как Т-клетки и В-клетки.
4. Выделенные Т-клетки
Изучали эффекты неспецифической активации цАМФ посредством ролипрама и форсколина в Т-клетках, выделенных из различных сублиний мышей CreLuc (фиг.18). Популяции цельных спленоцитов были, как описано выше, получены путем механического разрушения. Затем с использованием магнитного разделения MACS с колонкой позитивного отбора Positive Selection Column (Miltenyi Biotec, кат. № 130-049-201) были выделены клетки CD4+. Затем клетки высевали на 96-луночные белые непрозрачные планшеты по 1,5×105 на лунку и стимулировали анти-CD3 антителами (BD Pharmingen кат. № 553058). Через 18 часов в планшеты добавляли 10 мкмоль/л ролипрама (Sigma R6520) и 5 мкмоль/л форсколина (Sigma F6886) еще на 4 часа. Добавляли BrightGlo (Promega, Мэдисон, Висконсин, кат. № E2610), и количественный анализ считывали на TopCount. Данные показаны как люминесценция (число импульсов в секунду) и как кратность увеличения относительно контрольных групп, содержащих только среду. Увеличения экспрессии люциферазы наблюдали для всех испытанных линий, где линия 64 дает наибольшие уровни индукции, демонстрируя, что цАМФ путь активирован неспецифическими модуляторами у мышей CreLuc.
Изучали эффекты различных Gs-агонистов на уровни люциферазы в выделенных из мышей CreLuc (линия 64) анти-CD3 стимулированных CD4+ Т-клетках (фиг.19). Популяции цельных спленоцитов были получены путем механического разрушения. Клетки CD4+ затем выделяли путем магнитного разделения MACS с позитивным отбором (Miltenyi Biotec кат. № 130-049-201). Затем клетки высевали на 96-луночные белые непрозрачные планшеты по 1,5×105 на лунку и стимулировали анти-CD3 антителами (BD Pharmingen кат. № 553058). Через 24 часа добавляли соединения еще на 4 часа. Все Gs-агонисты для DP (BW245C), EP2 (EX00000173A) и ADβR (изопротеренол) использовали в концентрациях 10 мкмоль/л. Форсколин 5 мкмоль/л, и ролипрам 10 мкмоль/л. Добавляли BrightGlo (Promega, Мэдисон, Висконсин, кат. № E2610) и считывали количественный анализ на TopCount. В клетках, обработанных форсколином и ролипрамом, наблюдалось 25-кратное увеличение. Высокозначимые увеличения наблюдались для всех трех Gs-агонистов: 3-кратное для BW245C, 2-кратное для 173A и 5-кратное для изопротеренола. Трансген был чувствителен к специфическим Gs-агонистам, а также к неспецифическим модуляторам цАМФ пути.
5. Выделенные В-клетки
Изучали эффекты неспецифической активации цАМФ посредством ролипрама и форсколина в В-клетках, выделенных из двух различных сублиний мышей CreLuc (фиг.20). Популяции цельных спленоцитов были, как описано выше, получены путем механического разрушения. Затем с использованием магнитного разделения MACS с колонкой позитивного отбора Positive Selection Column (Miltenyi Biotec, кат. № 130-049-501) были выделены клетки B220+. Затем клетки высевали на 96-луночные белые непрозрачные планшеты по 2,0×105 на лунку и стимулировали 10 нг/мл липополисахарида (LPS) (Sigma L-2630). Через 18 часов в планшеты добавили 10 мкмоль/л ролипрама (Sigma R6520) и 5 мкмоль/л форсколина (Sigma F6886) еще на 4 часа. Добавили BrightGlo (Promega, Мэдисон, Висконсин, кат. № E2610) и считывали количественный анализ на TopCount. Данные показаны как люминесценция (число импульсов в секунду) и как кратность увеличения относительно контрольных групп, содержащих только среду. Увеличение экспрессии люциферазы наблюдалось в линии 64, но не в линии 229.
6. Микроглия
Изучали индукцию экспрессии люциферазы в микроглии, выделенной из мышей CreLuc, посредством неспецифических активаторов цАМФ, форсколина и ролипрама, и агониста DP-рецептора, BW245C (фиг.22). Первичную микроглию выделяли из коры головного мозга мышей Р2 (линия 64) в среде, состоящей из DMEM (GIBCO кат. № 11995), 10% FBS (GIBCO кат. № 16140) и 1% пенициллина-стрептомицина 100X (GIBCO кат. № 15140). Клетки высевали в планшеты 96-луночного формата, покрытые поли-D-лизином. Клетки либо оставляли необработанными, либо стимулировали в течение 2 часов 100 нг/мл LPS. Затем соединения добавляли еще на 4 часа перед выполнением количественного анализа Bright Glo. Применяли следующие соединения: 5 мкмоль/л форсколина, 10 мкмоль/л ролипрама или их комбинацию, или Gs-агонист для DP1-рецептора, BW245C, в концентрации 10 мкмоль/л. В нестимулируемых условиях микроглия является нечувствительной к неспецифическим модуляторам цАМФ и специфическому Gs-агонисту для DP-рецептора. Однако если микроглия стимулирована LPS, клетки становятся чувствительными к форсколину и BW245C.
7. Эмбриональные фибробласты мыши
Изучали эффекты форсколина, ролипрама и изопротеренола на экспрессию люциферазы в эмбриональных фибробластах мыши (фиг.33). Эмбриональные фибробласты мыши культивировали из эмбрионов E12 из шести независимых линий CreLuc. Клетки высевали в количестве 20000 клеток на лунку. Тестируемые соединения включали 10 мкмоль/л форсколина, 5 мкмоль/л и 10 мкмоль/л изопротеренола (ADβR-агониста). Значимые увеличения наблюдались для всех линий в ответ на комбинацию форсколина и ролипрама. Значимые увеличения также наблюдались в трех линиях в ответ на изопротеренол.
8. Кардиомиоциты
Изучали эффекты форсколина и ролипрама, а также изопротеренола на уровни люциферазы в кардиомиоцитах (фиг.35). Кардиомиоциты выделяли из детенышей Р3 линии 229. Клетки культивировали на 96-луночном планшете. Тестируемые соединения включали 10 мкмоль/л форсколина, 5 мкмоль/л и 10 мкмоль/л изопротеренола (ADβR-агониста). Значимое увеличение (25-кратное) наблюдалось для комбинации форсколина и ролипрама, а также значимое увеличение (2-кратное) наблюдалось для агониста изопротеренола.
B. Эксперименты in vivo и ex vivo
1. Влияние неспецифических модуляторов цАМФ
Эффекты интратекально введенных форсколина и ролипрама на индукцию экспрессии люциферазы изучали в головном и спинном мозге мышей CreLuc линии 187 (фиг.23). Выбранная линия мышей, как было определено предварительно, обладает экспрессией трансгена как в головном, так и в спинном мозге (данные не показаны). Линия мышей, выбранная для данного количественного анализа (линия 187), обладает индуцируемыми уровнями люциферазы как в головном, так и в спинном мозге. N=3-4 мыши на группу, четыре обрабатываемые группы, самцы возрастом 3 месяца. Группа А: контроль DMSO, группа В: 1 мкг форсколина/10 мкг ролипрама, группа C: 10 мкг форсколина/10 мкг ролипрама, группа D: 40 мкг форсколина/10 мкг ролипрама. Животным вводили дозы посредством интратекальной инъекции в поясничную область объемом 5 мкл на мышь. Их визуализировали через 4 часа после введения дозы. Данные для спинного и головного мозга показаны как среднее пиковой яркости, фотон в секунду на см2. Статистически значимое увеличение наблюдается в спинном мозге, значимое увеличение сигнала люциферазы в головном мозге наблюдается только при самой высокой концентрации форсколина. Если неспецифические модуляторы или цАМФ вводят в спинной мозг, происходит локальное увеличение экспрессии трансгена в спинном мозге, и менее высокий ответ наблюдается в головном мозге.
2. Эффекты Gs-агонистов
Изучали эффекты EP2-агониста, EX00000173A, на экспрессию люциферазы в головном и спинном мозге мышей CreLuc (фиг.24). Самцам мышей возрастом 5 месяцев из линии 187, n=5, интраперитонеально вводили или растворитель (5% DMSO, 0,05% твин 80, PBS), или 10 мг/кг EX00000173A (синтезирован в лаборатории). Животных биовизуализировали через 4 часа после введения дозы. Данные показаны как среднее для 5 мышей и для головного, и для спинного мозга, как фотон в секунду на см2. Статистически значимые увеличения наблюдаются при обработке агонистом и в головном, и в спинном мозге. Таким образом, Gs-агонист, вводимый интраперитонеально, активировал трансген.
Кроме того, эффекты EP2-агониста, EX00000173A, на экспрессию люциферазы у мышей CreLuc являются дозозависимыми (фиг.25). Была выбрана линия 187, поскольку эта линия обладает высокой индуцируемой экспрессией и в головном, и в спинном мозге. Количественный анализ состоял из пяти групп мышей возрастом шесть недель с n=5. Мышам вводили дозы или контрольного растворителя (D-PBS; забуференный фосфатом раствор Дульбекко, Invitrogen, кат. № 14040) или варьируемые дозы EP2-агониста EX0000173A (синтезирован в лаборатории): 1,5 нмоль, 5 нмоль, 15 нмоль и 50 нмоль. Мышам вводили дозу путем интраперитонеальной инъекции (5 мкл на мышь) и биовизуализировали через 4 часа на биовизуализаторе IVIS. Данные и для головного, и для спинного мозга показаны как среднее для пяти мышей, средняя пиковая яркость, фотон в секунду на см2. Статистически значимые (согласно проверке по t-критерию Стьюдента) увеличения при обработке агонистом наблюдались при трех концентрациях в спинном мозге, но не в головном мозге. Увеличение (незначительное) в головном мозге наблюдалось только при самой высокой концентрации, как и следовало ожидать для интратекальной инъекции.
На фиг.26 показана индукция люциферазы агонистом адренорецептора бета 3 (Adrb3), CL316,243 (1 мг/кг, интраперитонеально), в различных тканях мышей CRE-Luc. Количественный анализ люциферазы выполняли в гомогенатах тканей. Из 12 подвергнутых скринингу независимых трансгенных линий 7 линий показали более чем 10-кратную индукцию в жировой ткани и легких. 4 линии показали индукцию, видимую посредством биовизуализации (см. фиг.27А и 27В).
3. Специфические эффекты Gs-агониста AVE0010
Активация трансгена локализуется совместно с активацией тканеспецифического рецептора, как показано на примере индукции репортера люциферазы агонистом рецептора глюкагоноподобного пептида 1 (GLP-1R), AVE0010, который изучали в трех независимых линиях мышей CRE-Luc (фиг.28). GLP-1R является Gs-сопряженным. Базисные изображения получали в 1 день. На 2 день мышей обрабатывали AVE0010 (0,1 мг/кг, подкожно) и через 4 часа биовизуализировали. Кратность индукции относительно базисной указана снизу. Как и следовало ожидать, поскольку рецептор GLP-1R экспрессируется, главным образом, в ткани поджелудочной железы, в поджелудочной железе наблюдалась сильная индукция активации трансгена. Активности люциферазы в 8 различных тканях измерялись для линий 11, 16 и 90 (фиг.29) после обработки AVE0010 (0,1 мг/кг, подкожно) в течение 4 часов. Активность люциферазы в гомогенатах тканей подтверждала специфичную для поджелудочной железы индукцию люциферазы. Активность AVE0010 ограничена поджелудочной железой, несмотря на то, что GLP-1R обнаруживается в различных типах тканей.
Индукция Cre-Luc посредством AVE0010, наиболее вероятно, опосредована бета-клетками. Исследовали эффекты токсина бета-клеток стрептозотоцина (STZ) на индукцию CRE-Luc посредством AVE0010 (фиг.30). Самцов мышей CRE-luc (линия 11) визуализировали до и после обработки AVE0010 (0,1 мг/кг, подкожно). Данные указывают на то, что все мыши чувствительны к AVE0010 (см. среднюю панель на фиг.30). Затем животных обрабатывали растворителем или STZ (200 мг/кг, интраперитонеально). Через четыре дня их снова визуализировали после обработки AVE0010. В сравнении с группой растворителя группа STZ имеет сниженную индукцию люциферазы.
Индукция CRE-Luc посредством AVE0010, вероятно, является специфичной для бета-клеток (фиг.31). Животных обрабатывали, как описано на фиг.30. Уровни глюкозы в крови измеряли путем надреза хвостовой вены у не голодавших мышей. Уровни глюкозы считывали на глюкометре Bayer. Уровни глюкозы показаны как мг глюкозы/мл. Кратность индукции является уровнями биовизуализации люциферазы при введении дозы AVE10 относительно базисных сигналов. Уровни глюкозы в крови (BG) были увеличены с помощью STZ (верхняя левая панель). Уровни BG не натощак были снижены посредством AVE0010 (0,1 мг/кг, подкожно). Данные BLI, показанные на фиг.30, были определены количественно.
4. Трансплантации костного мозга
Трансплантации костного мозга CreLuc выполняли с использованием мышей NOD scid gamma (NSG) (фиг.32). Мыши NSD являются мышами с ослабленным иммунитетом и недостатком зрелых Т- и В-клеток, функциональных клеток природных киллеров, и имеют дефицит передачи цитокиновых сигналов, допускающий трансплантацию гемопоэтических клеток. Клетки костного мозга собирали из гетерозигот линии 44 (имеющей высокие базальные уровни люциферазы) и гетерозигот линии 64 (имеющей индуцируемые уровни люциферазы). Затем клетки трансплантировали посредством инъекций клеток в хвостовую вену облученным мышам NSG в количествах 1 миллион или 5 миллионов. Для линии 44 мыши 1 и 2 получали по 5 миллионов клеток, в то время как мыши 3 и 4 получали по 1 миллиону клеток. Для линии 64 мышь 1 получала 5 миллионов клеток, и мыши 2, 3 и 4 получали по 1 миллиону клеток (по 4 мыши NSG на линию CreLuc). Животных биовизуализировали через 4 недели (данные не показаны), и затем еще через 8 недель. Перед визуализацией мышей линии 64 в течение 5 часов индуцировали с 5 мг/кг форсколина и 10 мг/кг ролипрама. Изображения биовизуализации показаны для момента времени 8 недель. Уровни люциферазы у мышей NSG, которым трансплантировали клетки костного мозга линии 44, имитируют изображение, наблюдаемое у CreLuc линии 44 (данные не показаны), с экспрессией, наблюдаемой в суставах, спинном мозге, голове и грудине. Индуцируемая экспрессия люциферазы наблюдается в селезенках мышей NSG, которым трансплантировали клетки костного мозга линии 64.
5. Животные модели
Мыши CreLuc могут быть использованы для исследования животных моделей заболеваний или аспектов болезненных состояний. Кроме того, мыши CreLuc могут быть использованы для скрининга соединений, которые способны модулировать заболевание или аспекты заболевания, которые индуцированы у мышей CreLuc. Например, изучали эффекты обработки зимозаном на уровни люциферазы в CreLuc линии 187 (фиг.34). Мышам CreLuc (линия 187) в обрабатываемой группе в обе задние лапы подкожно вводили зимозан с целью индукции болевого ответа. Затем мышей биовизуализировали ежедневно в течение 4 дней. Статистически значимые увеличения экспрессии люциферазы в лапах животных в ответ на зимозан наблюдались во все моменты времени. Зимозан представляет собой компонент клеточной оболочки дрожжевых клеток, который сильно активирует воспалительную реакцию. Таким образом, мыши CreLuc представляют инструмент, при помощи которого можно вести наблюдение за воспалительной реакцией во времени на одном и том же животном. Кроме того, обработанные зимозаном животные могут быть использованы в качестве инструмента скрининга для оценки способностей тестируемых соединений для того, чтобы обращать или обострять индуцированное зимозаном воспаление.

















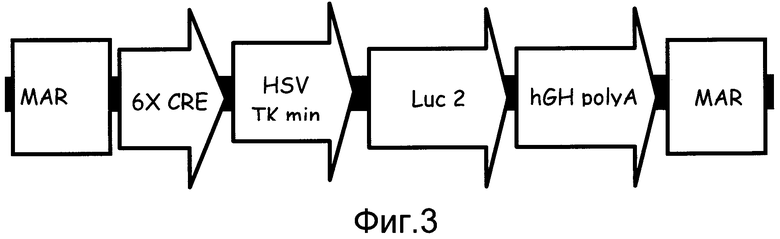

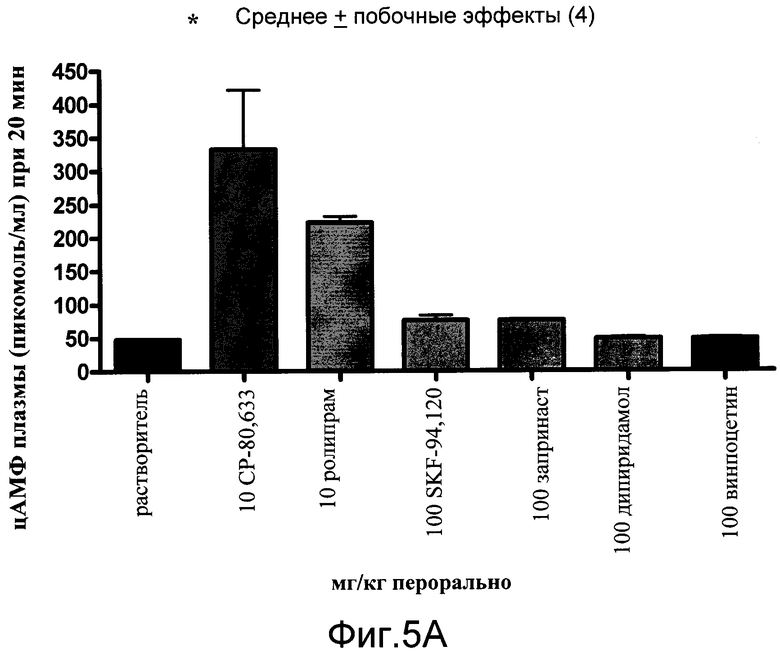

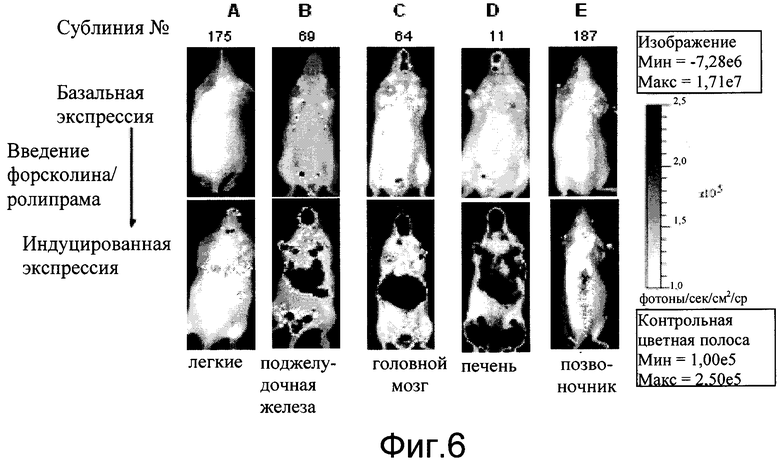
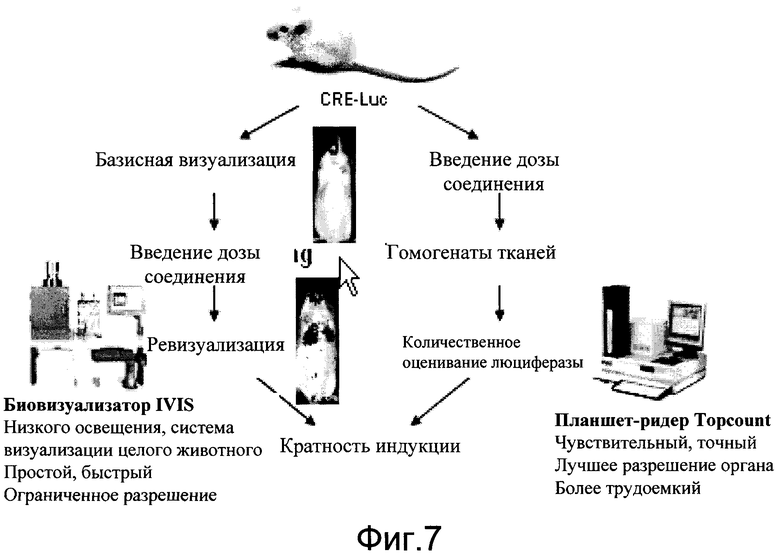

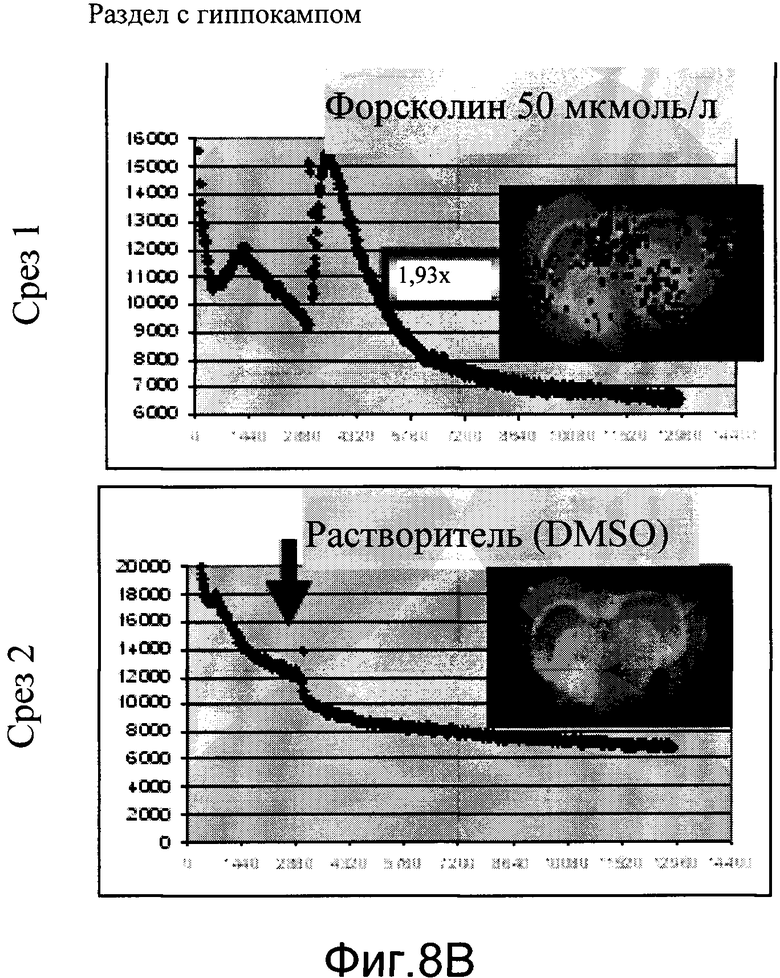
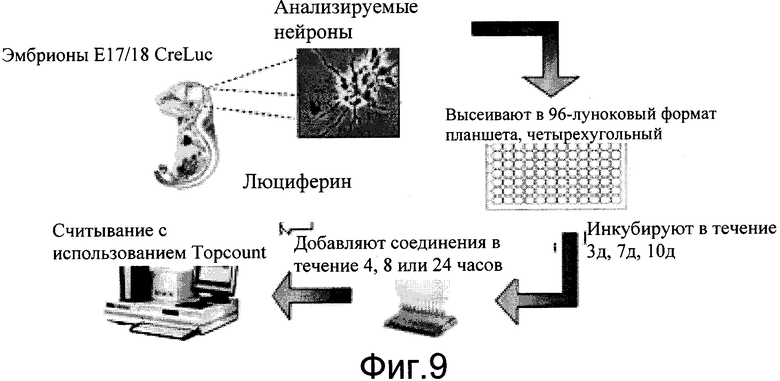

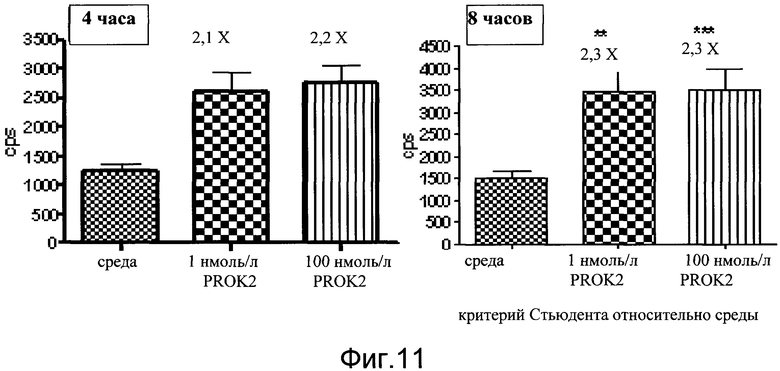

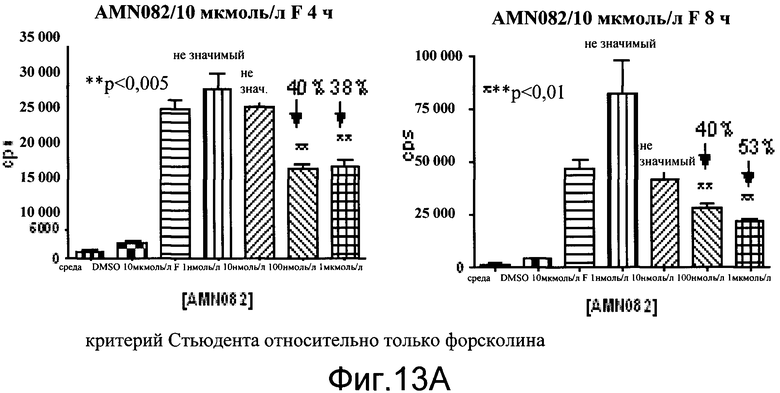
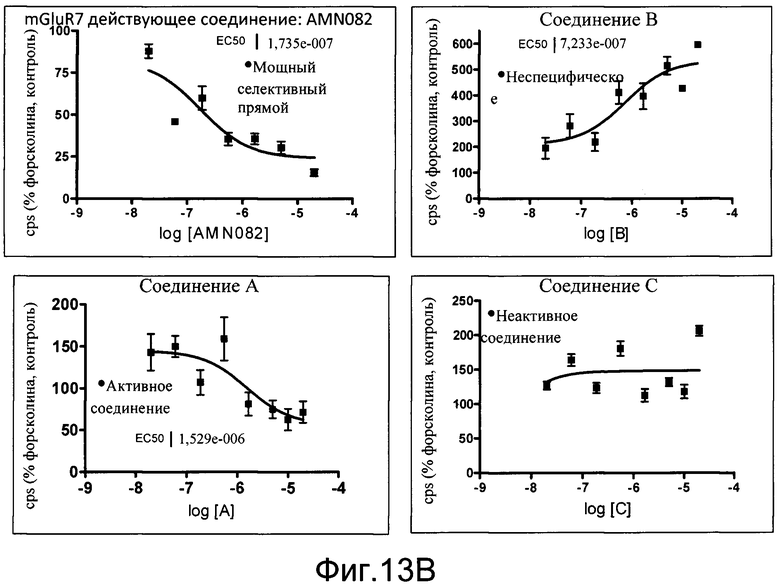
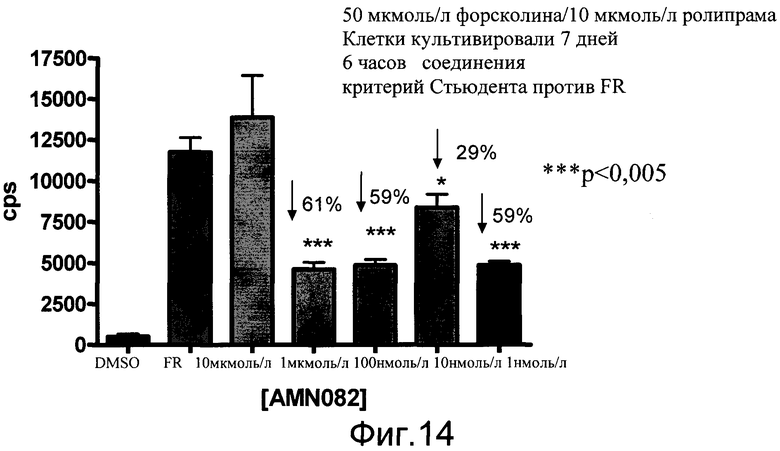
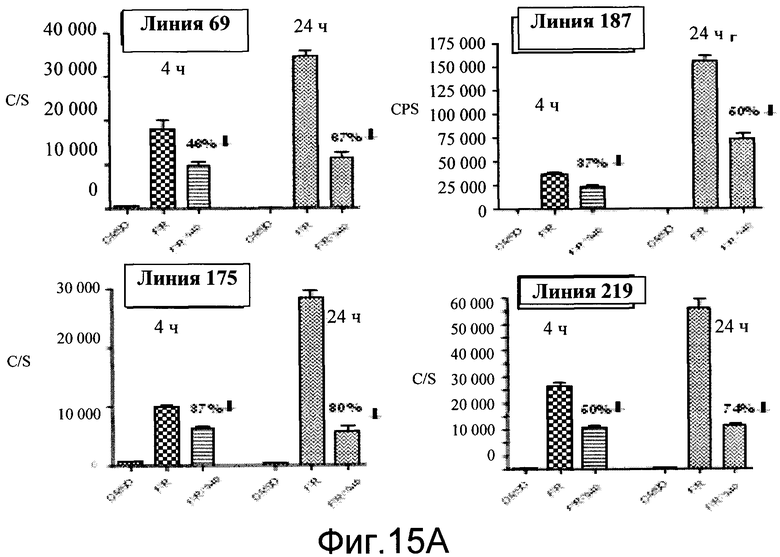
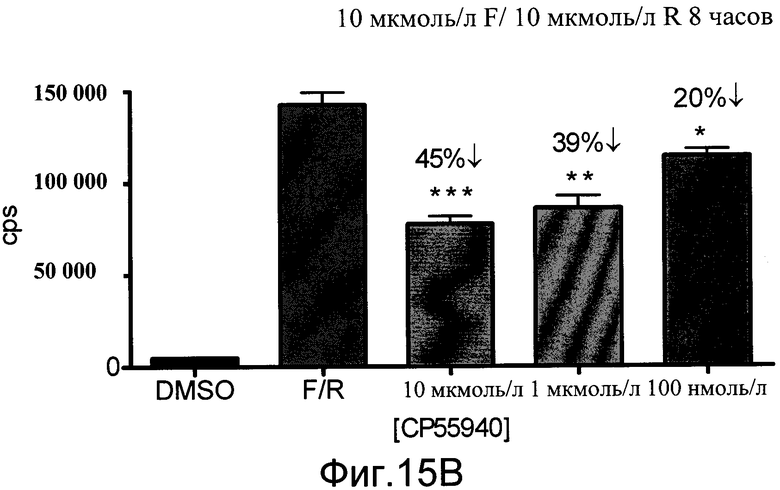
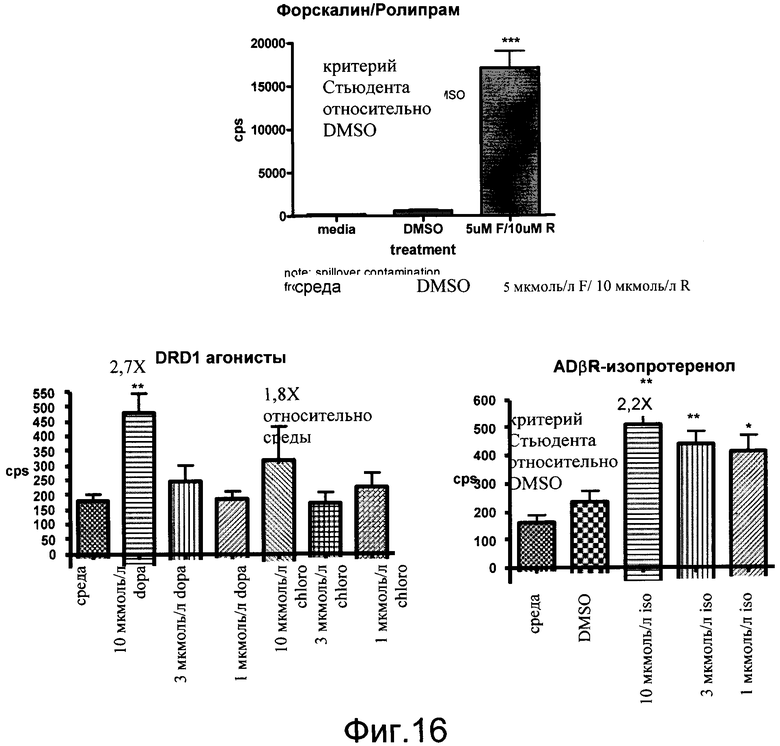
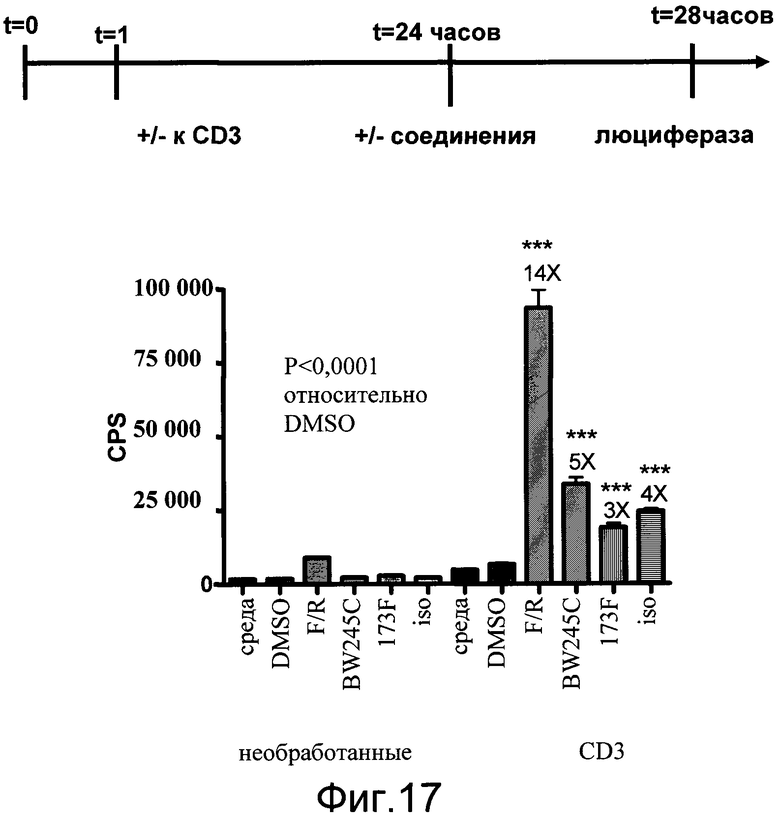
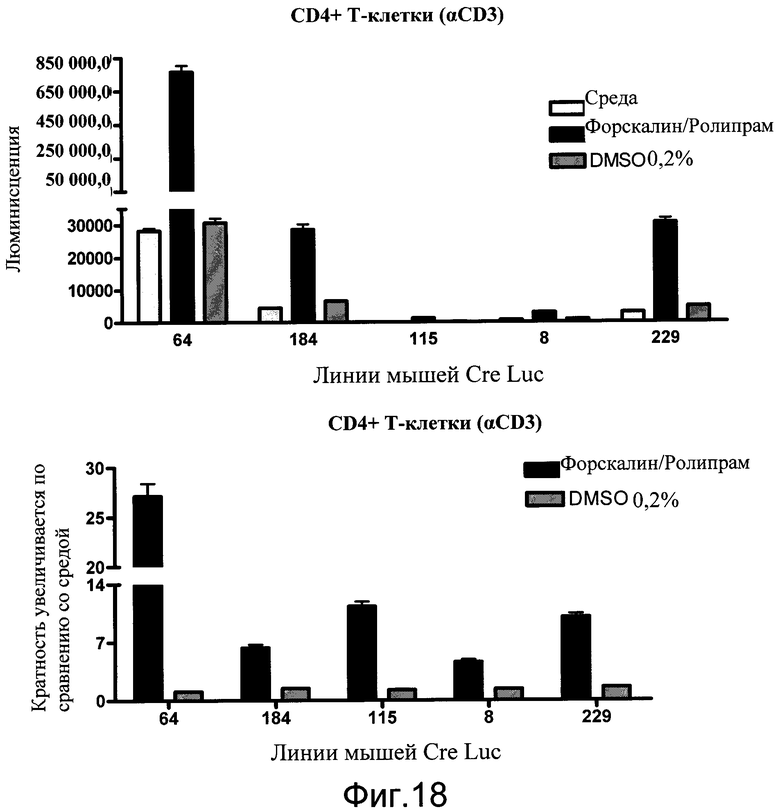

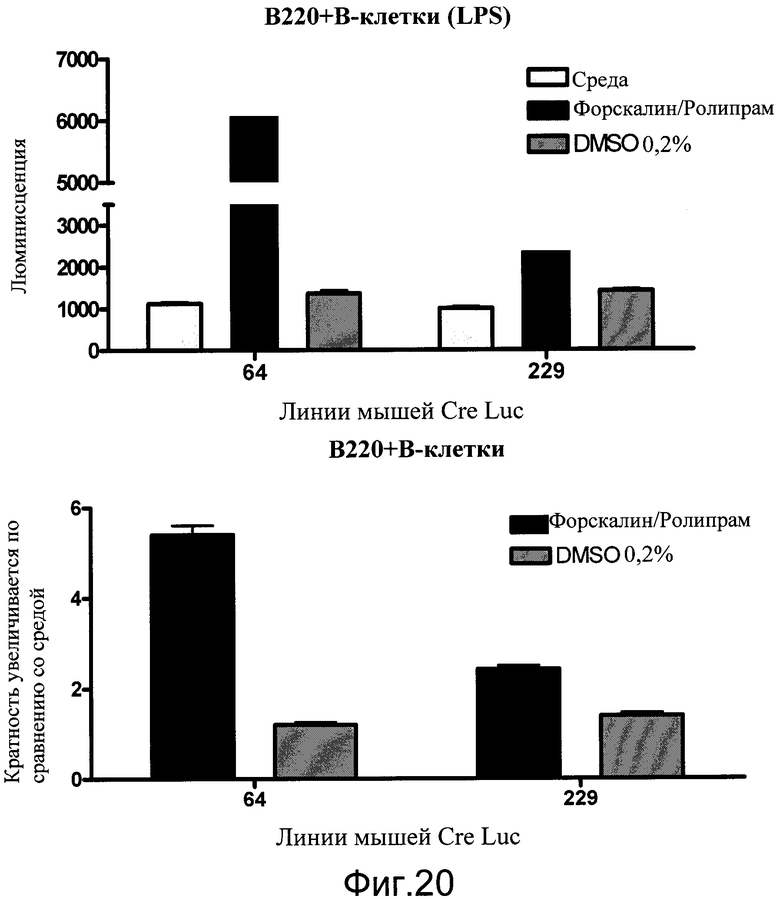
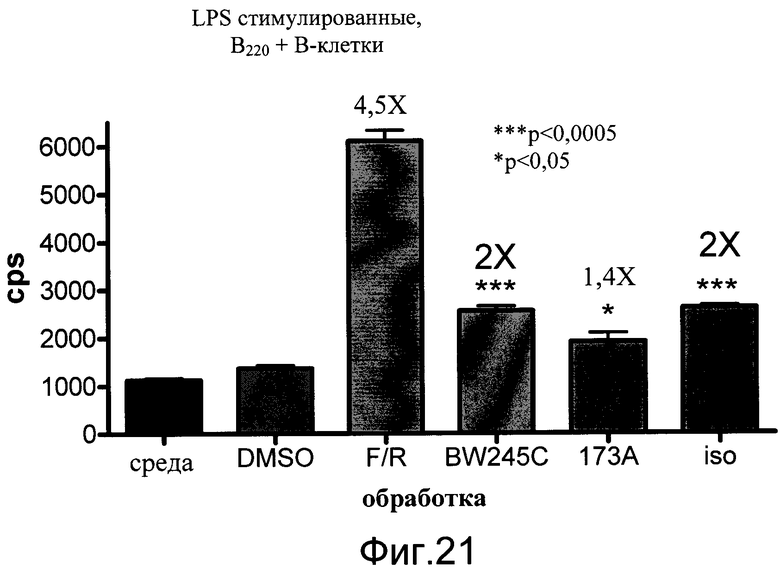
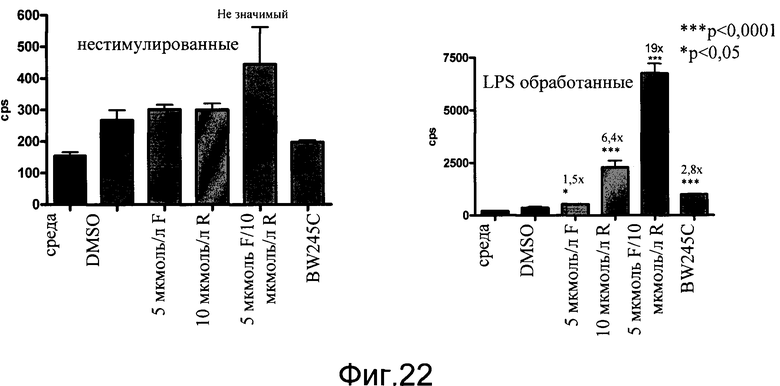
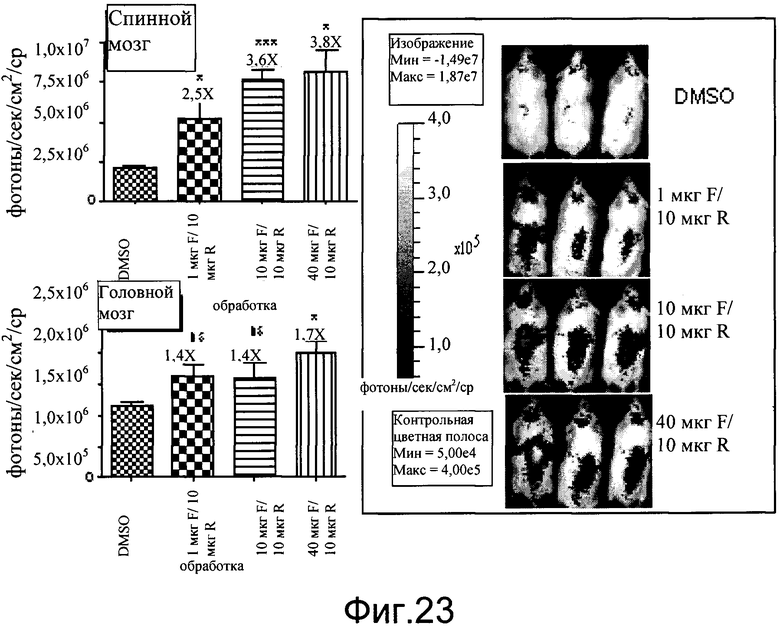
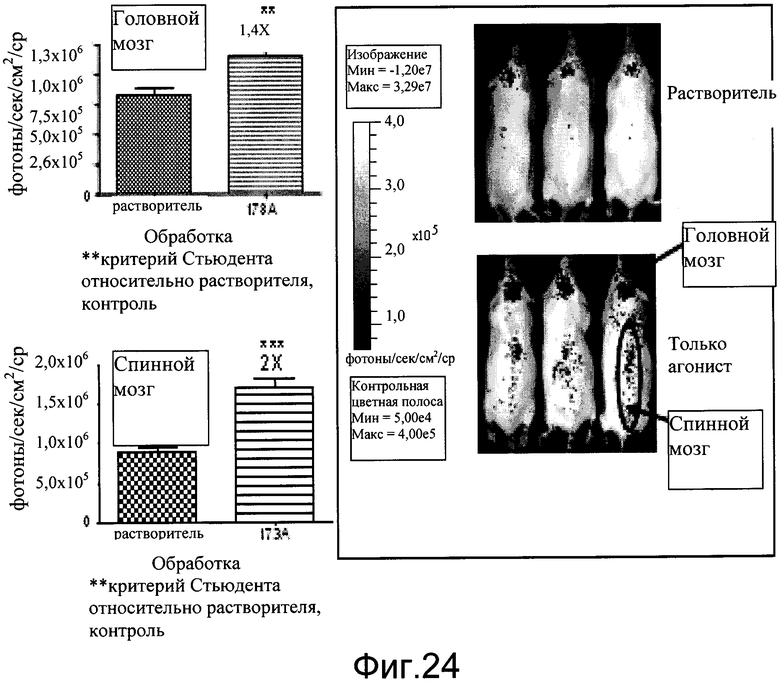




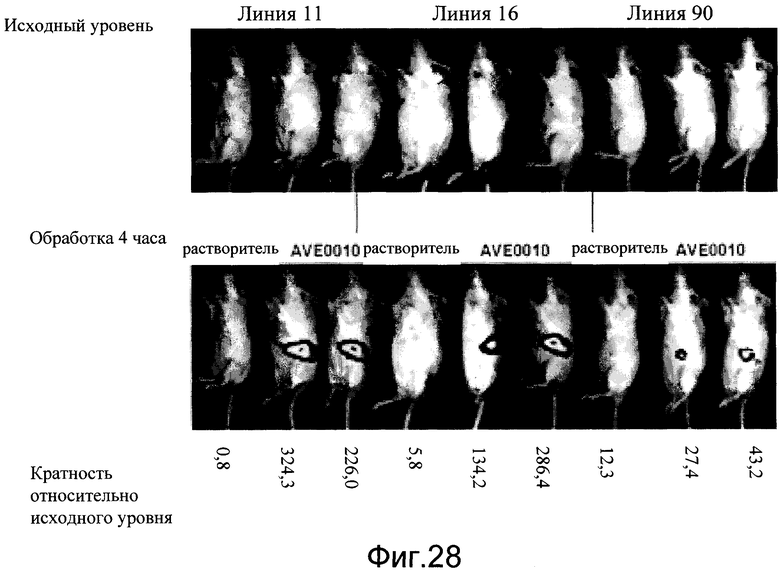
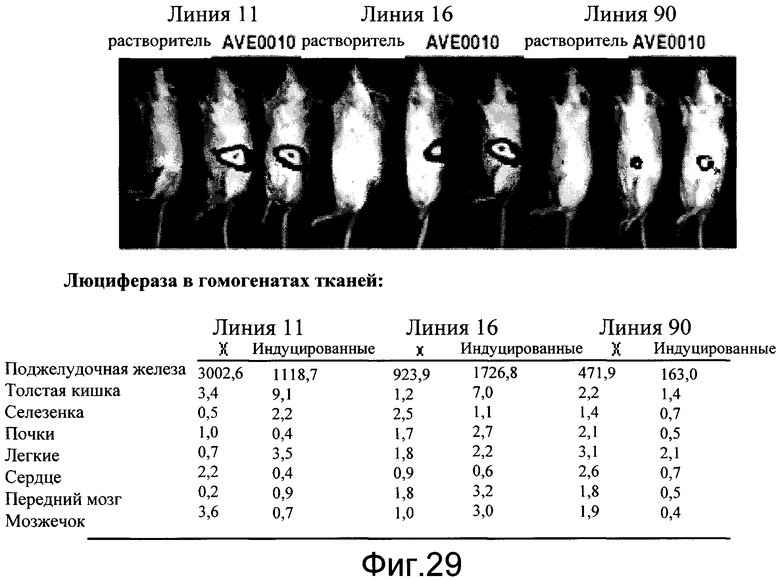
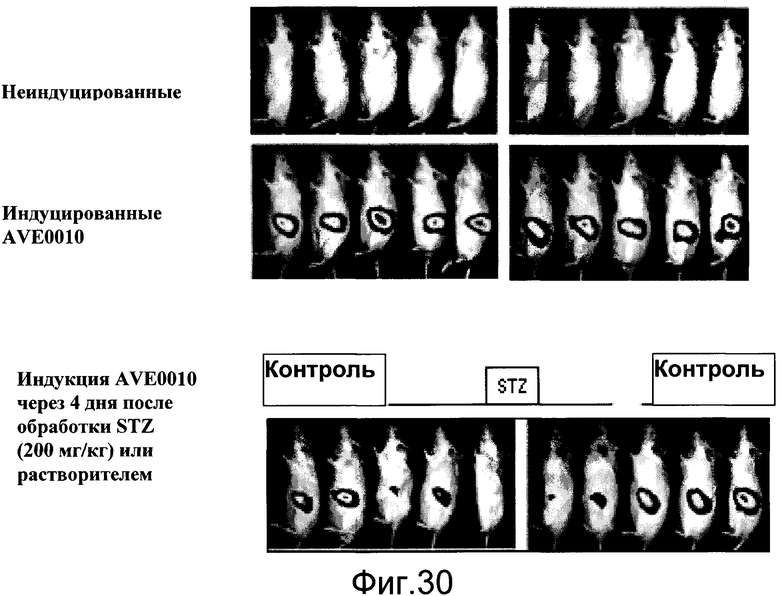
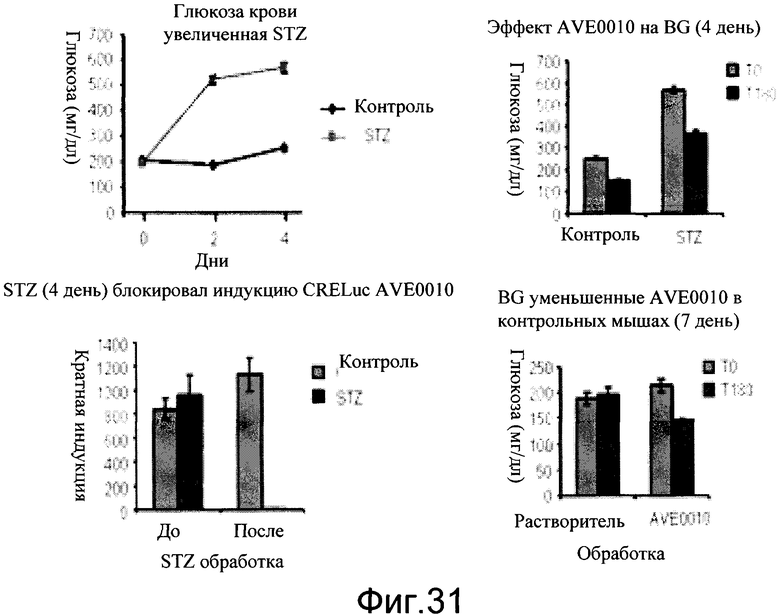

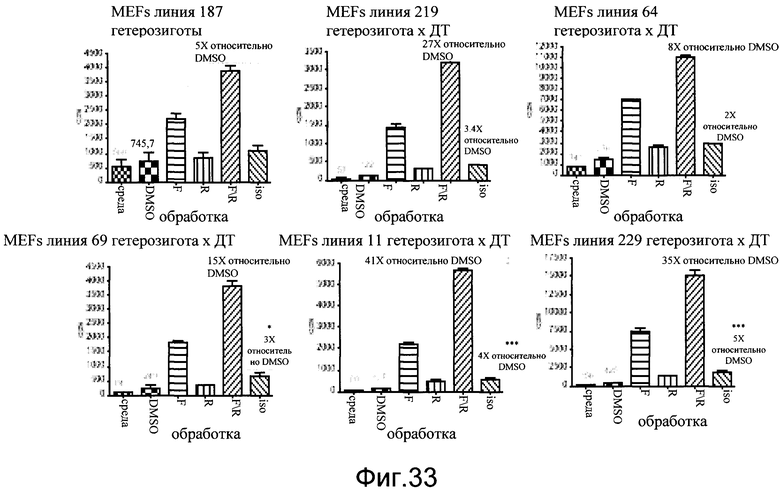
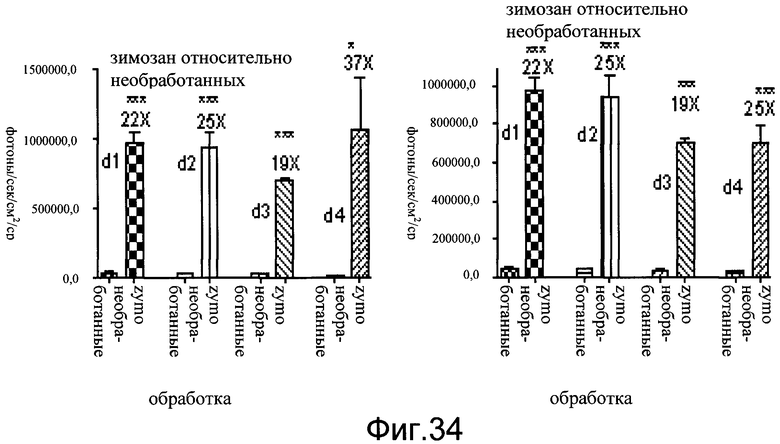
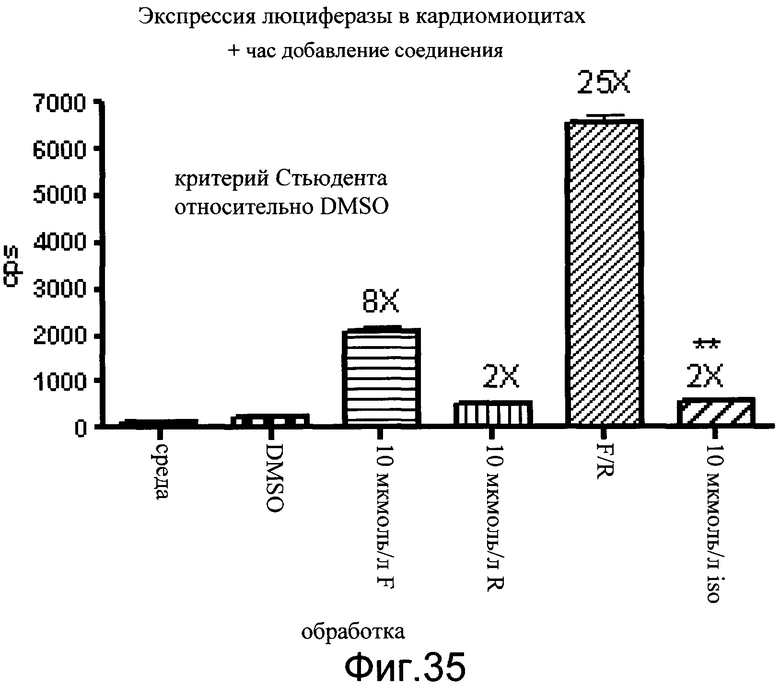
Группа изобретений относится к области генной инженерии, в целом к трансгенным конструкциям, трансгенным животным, отличным от человека, способам количественного анализа активации GPCR лигандов неинвазивно в интактных животных, тканевых срезах или в нативных клетках, используя трансгенную модель, содержащую систему биолюминесцентного трансгена-репортера, которая является чувствительной к модуляции путей после связывания лиганда с рецепторами GPCR. Трансгенное животное, отличное от человека, для оценки функции рецептора, сопряженного с G-белком (GPCR), имеющее геном, содержащий индуцируемый циклическим АМФ (цАМФ), трансген, содержащий, в направлении от 5′-конца к 3′-концу, первый инсуляторный элемент, ответный элемент цАМФ, промотор, биолюминесцентный репортер, миниген человеческого гормона роста (hGH) и второй инсуляторный элемент. Использование трансгенной конструкции приводит к повышению мышиных линий, продуцирующих мРНК и белок, а также к увеличению уровня экспрессии репортерного гена. 9 н. и 11 з.п. ф-лы, 41 ил., 10 табл.
1. Трансгенное животное, отличное от человека, для оценки функции рецептора, сопряженного с G-белком (GPCR), имеющее геном, содержащий индуцируемый циклическим АМФ (цАМФ) трансген, содержащий, в направлении от 5′-конца к 3′-концу, первый инсуляторный элемент, ответный элемент цАМФ, промотор, биолюминесцентный репортер, миниген человеческого гормона роста (hGH) и второй инсуляторный элемент.
2. Трансгенное животное, отличное от человека, по п. 1, где первый инсуляторный элемент выбран из группы, состоящей из участков прикрепления к ядерному матриксу (MAR), ДНКаза I-гиперчувствительного сайта (HS4) и инвертированных концевых повторов (ITR).
3. Трансгенное животное, отличное от человека, по п. 1, где второй инсуляторный элемент выбран из группы, состоящей из элемента прикрепления к ядерному матриксу (MAR), HS4 и ITR.
4. Трансгенное животное, отличное от человека, по п. 1, где первый инсуляторный элемент является таким же, как второй инсуляторный элемент.
5. Трансгенное животное, отличное от человека, по п. 1, где ответный элемент повторяется в тандеме от двух до двадцати четырех раз.
6. Трансгенное животное, отличное от человека, по п. 5, где ответный элемент повторяется в тандеме шесть раз.
7. Трансгенное животное, отличное от человека, по п. 1, где промотор представляет собой минимальный промотор тимидинкиназы вируса простого герпеса (HSV ТК min).
8. Трансгенное животное, отличное от человека, по п. 1, где биолюминесцентный репортер выбран из группы, состоящей из люциферазы, хлорамфениколацетилтрансферазы (CAT), бета-галактозидазы, секретированной щелочной фосфатазы (SEAP), человеческого гормона роста (HGH) и зеленого флуоресцентного белка (GFP).
9. Трансгенное животное, отличное от человека, по п. 1, где трансген содержит SEQ ID NO: 18.
10. Трансгенное животное, отличное от человека, по п. 1, где трансген содержит SEQ ID NO: 19.
11. Трансгенное животное, отличное от человека, по п. 1, где указанные первый и второй инсуляторные элементы фланкируют миниген hGH и где указанное трансгенное животное, отличное от человека, производит больше трансгенных животных-основателей, проявляющих индуцируемую цАМФ экспрессию трансгена, чем трансгенное животное, отличное от человека, экспрессирующее содержащий CRE трансген, включающий функциональный элемент hGH без фланкирующих инсуляторных элементов.
12. Клетка для идентификации лиганда рецептора, сопряженного с G-белком (GPCR), выделенная из трансгенного животного, отличного от человека, по п. 1.
13. Тканевой срез для идентификации лиганда рецептора, сопряженного с G-белком (GPCR), выделенный из трансгенного животного, отличного от человека, по п. 1.
14. Способ идентификации лиганда рецептора, сопряженного с G-белком (GPCR), включающий:
(а) измерение количества биолюминесценции в трансгенном животном, отличном от человека, по п. 1;
(b) введение тестируемого средства трансгенному животному, отличному от человека;
(c) измерение количества биолюминесценции трансгенного животного, отличного от человека, в один или более моментов времени после введения тестируемого средства; и
(d) сравнение количества биолюминесценции, измеренной в (а), с количеством биолюминесценции, измеренной в (с),
где увеличение количества биолюминесценции в (а) по сравнению с (с) идентифицирует тестируемое средство как лиганд GPCR.
15. Способ идентификации лиганда рецептора, сопряженного с G-белком (GPCR), включающий:
(a) приготовление тканевого среза из трансгенного животного, отличного от человека, по п. 1;
(b) измерение количества биолюминесценции в тканевом срезе;
(c) введение тестируемого средства в тканевый срез;
(d) измерение количества биолюминесценции тканевого среза в один или более моментов времени после введения тестируемого средства; и
(e) сравнение количества биолюминесценции, измеренной в (b), с количеством биолюминесценции, измеренной в (d),
где увеличение количества биолюминесценции в (b) по сравнению с (d) идентифицирует тестируемое средство как лиганд GPCR.
16. Способ идентификации лиганда рецептора, сопряженного с G-белком (GPCR), включающий:
(a) приготовление клетки, выделенной из трансгенного животного, отличного от человека, по п. 1;
(b) измерение количества биолюминесценции в клетке;
(c) введение тестируемого средства в клетку;
(d) измерение количества биолюминесценции в клетке в один или более моментов времени после введения тестируемого средства; и
(e) сравнение количества биолюминесценции, измеренной в (b), с количеством биолюминесценции, измеренной в (d),
где увеличение количества биолюминесценции в (b) по сравнению с (d) идентифицирует тестируемое средство как лиганд GPCR.
17. Способ мониторинга функции GPCR в животном, отличном от человека, включающий:
(a) трансгенное модифицирование животного, отличного от человека, для экспрессии индуцируемого цАМФ трансгена, содержащего первый инсуляторный элемент, ответный элемент цАМФ, промотор, биолюминесцентный репортер, миниген человеческого гормона роста (hGH) и второй инсуляторный элемент;
(b) мониторинг биолюминесценции из животного, отличного от человека; и
(c) коррелирование увеличения указанной биолюминесценции с функцией GPCR.
18. Способ создания трансгенного животного, отличного от человека, для применения в мониторинге функции GPCR, включающий:
(а) трансгенное модифицирование животного, отличного от человека, для экспрессии индуцируемого цАМФ трансгена,
содержащего первый инсуляторный элемент, ответный элемент цАМФ, промотор, биолюминесцентный репортер, миниген человеческого гормона роста (hGH) и второй инсуляторный элемент;
(b) измерение количества биолюминесценции в трансгенном животном, отличном от человека, из (а);
(c) введение лиганда GPCR трансгенному животному, отличному от человека;
(d) измерение количества биолюминесценции трансгенного животного, отличного от человека, в один или более моментов времени после введения лиганда GPCR; и
(e) сравнение количества биолюминесценции, измеренной в (b), с количеством биолюминесценции, измеренной в (d),
где увеличение количества биолюминесценции в (b) по сравнению с (d) идентифицирует трансгенное животное, отличное от человека, для применения в мониторинге функции GPCR.
19. Трансгенное животное, отличное от человека, для оценки функции рецептора, сопряженного с G-белком (GPCR), имеющее геном, содержащий индуцируемую циклическим АМФ (цАМФ) трансгенную конструкцию, содержащую, в направлении от 5′-конца к 3′-концу, первый инсуляторный элемент, ответный элемент, промотор, биолюминесцентный репортер, миниген человеческого гормона роста (hGH) и второй инсуляторный элемент, где указанные первый инсуляторный элемент и второй инсуляторный элемент различаются и где указанный ответный элемент представляет собой ответный элемент цАМФ (CRE), содержащий последовательность нуклеиновой кислоты SEQ ID NO: 6 или 11.
20. Трансгенное животное, отличное от человека, по п.1, где ответный элемент цАМФ содержит последовательность нуклеиновой кислоты SEQ ID NO: 6 или 11.
| OTTOBRINI L et al, Molecular imaging: A new way to study molecular processes in vivo, Molecular and cellular endocrinology, vol | |||
| Котел | 1921 |
|
SU246A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| CIANA PAOLO et al, Engineering of a mouse for the in vivo profiling of estrogen receptor activity, Molecular endocrinology, vol.15, no.7, 07.2001, pages | |||

