ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к способам идентификации соединений-кандидатов для регуляции массы или функции скелетных мышц или для регуляции активности или экспрессии рецептора рилизинг-фактора-2 кортикотропина (CRF2R). Изобретение также относится к способам лечения атрофии скелетных мышц или к способам индуцирования гипертрофии скелетных мышц, в которых CRF2R применяется как мишень для воздействия, и к способам лечения мышечных дистрофий, в которых CRF2R и рецептор рилизинг-фактора-1 кортикотропина (CRF1R) применяются как мишени для воздействия.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
CRFR и лиганды
Известно два рецептора рилизинг-фактора кортикотропина, идентифицированных к настоящему времени (CRF1R и CRF2R), которые относятся к классу рецепторов (GPCR), сопряженных с G-белком. Активация агонистом CRF1R или CRF2R ведет к Gαs-активации аденилатциклазы. Аденилатциклаза катализирует образование цАМФ, который, в свою очередь, обладает многочисленными эффектами, включая активацию протеинкиназы A, высвобождение внутриклеточного кальция и активацию митоген-активированной протеинкиназы (МАР-киназы). Усиление внутриклеточного синтеза инозиттрифосфата после активации агонистом рецепторов CRF, обнаруженное в других исследованиях, означает, что рецепторы CRFR также сопряжены с Gαq.
CRF1R и CRF2R клонировали от человека, крысы, мыши, курицы, коровы, сома, лягушки и овцы. Для каждого из CRF1R и CRF2R есть единственные в своем роде профили распределения. У человека клонировали три изоформы рецептора CRF2R: альфа, бета и гамма. У крысы были идентифицированы гомологи альфа и бета-CRF2R.
Известно несколько лигандов/агонистов рецепторов CRFR. Рилизинг-фактор кортикотропина (или гормон, CRF или CRH) связывается с CRF1R и CRF2R и активирует их. CRF является главным модулятором ответов организмов на стресс. Такой пептид, состоящий из 41 аминокислоты, контролирует защиту нейронных, эндокринных и иммунных процессов как первичный регулятор гормональной оси системы гипоталамус-гипофиз-надпочечники (ось HPA). Кроме того, существует значительная гомологичность последовательностей CRF и пептида саувагина амфибий, также как пептида telostian уротензина, оба из которых действуют как агонисты CRF1R и CRF2R. Указанные три пептида имеют похожие биологические свойства в качестве гипотензивных агентов и средств, усиливающих секрецию АКТГ. Кроме того, для млекопитающих был описан родственный уротензину урокортин.
Рецепторы CRF можно отличить от нерецепторов CRF фармакологически, путем применения селективных агонистов и антагонистов рецепторов. Такие селективные агонисты и антагонисты, наряду с мышами CRFR-нокаут, применялись при определении того, какой из рецепторов CRF опосредует специфичные биологические ответы.
Довольно хорошо установлена роль CRF1R. У мышей с удаленным CRF1R-геном (CRF1R-нокаут) наблюдается ослабленный ответ на стресс и ослабленная реакция тревоги. CRF1R является главным медиатором оси HPA. В частности, рилизинг-фактор кортикотропина, который высвобождается из гипоталамуса и транспортируется в переднюю долю гипофиза через гипоталамогипофизарную воротную систему, взаимодействует с CRF1R, находящимся на клетках, локализованных в передней доле гипофиза. Активация агонистом CRF1R приводит к высвобождению АКТГ из клеток передней доли гипофиза в большой круг кровообращения. Высвобожденный АКТГ связывает рецептор АКТГ, находящийся на клетках, локализованных в коре надпочечников, что приводит к высвобождению гормонов коры надпочечников, включая кортикостероиды. Кортикостероиды опосредуют многие эффекты, включая без ограничения угнетение иммунной системы через механизм, который заключается в атрофии тимуса и селезенки. Таким образом, активация CRF1R косвенно приводит к регуляции иммунной системы по типу отрицательной регуляции через активацию оси HPA.
Роль CRF2R исследована меньше. У мышей с удаленным CRF2R-геном (CRF2R-нокаут) обнаружено снижение замедленного поглощения пищи после стимуляции урокортином, отсутствие вазодилатации, однако нормальный ответ на стресс. Эксперименты с CRF2R показали, что CRF2R является ответственным за гипотензивные/вазодилатационные эффекты агонистов CRFR и за уменьшение поглощения пищи, наблюдаемое после лечения мышей агонистами CRFR.
Атрофия и гипертрофия скелетных мышц
Скелетные мышцы представляют собой пластичную ткань, которая легко адаптируется к изменениям либо в случае физиологической необходимости для работы или метаболической потребности. Гипертрофия связана с увеличением массы скелетных мышц, в то время как атрофия скелетных мышц связана с уменьшением массы скелетных мышц. Острая атрофия скелетных мышц имеет в основе ряд причин, включая без ограничения: бездействие, обусловленное операцией, постельным режимом или переломом костей; повреждение нерва/денервацию, обусловленное травмой спинного мозга, аутоиммунным заболеванием или инфекционным заболеванием; применение глюкокортикоидов при состояниях, не связанных с заместительной гормональной терапией; сепсис, обусловленный инфекцией или другими причинами; ограниченное поступление питательных веществ, обусловленное болезнью или голоданием; и полет в космос. Атрофия скелетных мышц возникает благодаря нормальным биологическим процессам, однако при определенных медицинских состояниях такой нормальный биологический процесс приводит к инвалидизирующему уровню мышечной атрофии. Например, острая атрофия скелетных мышц является значительным ограничением при реабилитации пациентов после неподвижности, включая без ограничения реабилитацию после ортопедической процедуры. В таких случаях период реабилитации, необходимый для обращения атрофии скелетных мышц, часто оказывается гораздо более длительным, чем период, требуемый для устранения первоначального повреждения. Такая острая атрофия, связанная с бездействием, представляет определенную проблему для пожилого человека, который может уже страдать от значительного, связанного с возрастом дефицита мышечной функции и массы, поскольку такая атрофия может привести к постоянной инвалидности и преждевременной смерти.
Атрофия скелетных мышц может также быть результатом хронических состояний, таких как раковая кахексия, хроническое воспаление, кахексия при СПИДе, хроническая обструктивная болезнь легких (ХОБЛ), застойная сердечная недостаточность, генетические нарушения, например мышечные дистрофии, нейродегенеративные заболевания и саркопения (связанная с возрастом потеря мышечной массы). При таких хронических состояниях атрофия скелетных мышц может приводить к преждевременной потере подвижности, таким образом усугубляя связанную с заболеванием болезненность.
О молекулярных процессах, которые управляют атрофией или гипертрофией скелетных мышц, известно немного. Несмотря на различие инициирующих пусковых механизмов атрофии скелетных мышц в случаях, когда атрофия вызвана различными явлениями, в пораженном болезнью волокне скелетных мышц происходит несколько общих биохимических изменений, включая уменьшение синтеза белков и усиление деградации белков, и изменения как сократительных характеристик, так и свойств белковых изозимов метаболических ферментов при переключении с медленных (высокий окислительный метаболизм/изоформы медленного сократительного белка) на быстрые (высокий гликолитический метаболизм/изоформы быстрого сократительного белка) волокна. Дополнительные изменения, происходящие в скелетной мышце, включают сокращение сосудистой сети и перестройку внеклеточного матрикса. Как быстрое, так и медленное подергивание мышцы свидетельствует об атрофии при соответствующих состояниях, связанных с относительной потерей мышечной массы, зависящей от конкретных стимулов, вызывающих атрофию, или состояния. Важно то, что все указанные изменения согласованно отрегулированы и включаются или выключаются в зависимости от изменений, связанных с физиологической и метаболической потребностью.
Процессы, при которых наступает атрофия и гипертрофия, одни и те же для видов млекопитающих. Многократные исследования показали, что во время атрофии как у грызунов, так и у человека имеют место одни и те же основные молекулярные, клеточные и физиологические процессы. Таким образом, модели атрофии скелетных мышц грызуна успешно применяются для понимания и предсказания атрофических ответов человека. Например, атрофия, индуцированная различными способами, как у грызунов, так и у человека приводит к похожим изменениям в анатомии мышцы, площади поперечного сечения, функции, типе переключения волокна, экспрессии сократительного белка и гистологии. Кроме того, было обнаружено несколько агентов, регулирующих атрофию скелетных мышц как у грызунов, так и у человека. Такие агенты включают анаболические стероиды, гормон роста, инсулиноподобный фактор роста I и бета-адренергические агонисты. Взятые вместе, указанные данные свидетельствуют, что атрофия скелетных мышц возникает в результате общих механизмов как у грызунов, так и у человека.
Несмотря на то, что обнаружены некоторые агенты, регулирующие атрофию скелетных мышц, и одобрено их применение для человека по такому показанию, указанные агенты обладают нежелательными побочными эффектами, такими как гипертрофия сердечной мышцы, неоплазия, гирсутизм, андрогенизация женщин, повышенная заболеваемость и смертность, повреждение печени, гипогликемия, мышечно-скелетная боль, повышенный тургор тканей, тахикардия и отеки. В настоящее время не существует высокоэффективного и избирательного лечения ни острой, ни хронической атрофии скелетных мышц. Таким образом, существует потребность в идентификации других лекарственных средств, регулирующих атрофию скелетных мышц.
Мышечные дистрофии
Мышечные дистрофии охватывают группу наследственных, прогрессирующих мышечных расстройств, отличающихся клинически по избирательной локализации слабости скелетных мышц. Двумя наиболее распространенными формами мышечной дистрофии являются дистрофии Дюшенна и Беккера, каждая из которых возникает в результате наследования мутации в гене дистрофина, который локализован в локусе-Xp21. Другие дистрофии включают без ограничения мышечную дистрофию тазового пояса, которая является результатом мутации множественных генетических локусов, включающих локусы кальпаина p94, адхалоина, γ-саркогликана и β-саркогликана; плече-лопаточно-лицевую (Ландузи-Дежирина) мышечную дистрофию, миотоническую дистрофию и мышечную дистрофию Эмери-Драйфуса. Симптомы мышечной дистрофии Дюшенна, которая наблюдается почти исключительно у мужчин, включают походку вперевалку, \ходьбу на носках ног\, лордоз, частые падения, трудность вставания (из положения сидя) и передвижения вверх по лестнице. Симптомы проявляются приблизительно в возрасте 3-7 лет, к 10-12 годам большинство пациентов прикованы к инвалидному креслу и многие умирают приблизительно в возрасте 20 лет из-за осложнений дыхания. Традиционное лечение мышечной дистрофии Дюшена включает введение преднизона (кортикостероидного лекарственного средства), который, несмотря на то, что не является исцеляющим, замедляет упадок мышечной силы и задерживает потерю трудоспособности. Считается, что кортикостероиды, такие как преднизон, действуют как блокаторы активации и инфильтрации иммунных клеток, которые усиливаются при повреждении мышечных волокон в результате заболевания. К сожалению, лечение кортикостероидами также приводит к атрофии скелетных мышц, которая сводит на нет часть потенциальной выгоды от блокирования иммунного ответа у таких пациентов. Таким образом, существует потребность в идентификации лекарственных средств, которые замедляют повреждение мышечного волокна и задерживают наступление инвалидности у больных с мышечными дистрофиями, однако вызывают меньшую степень атрофии скелетных мышц, чем современные терапевтические методики.
Одной из проблем, связанных с идентификацией соединений, применяемых для лечения атрофии скелетных мышц или мышечных дистрофий, было отсутствие хороших способов скрининга для идентификации таких соединений. Теперь авторами установлено, что CRF2R вовлеченыв регуляцию массы или функции скелетных мышц, и что агонисты CRF2R способны блокировать атрофию скелетных мышц и/или индуцировать гипертрофию скелетных мышц. Настоящее изобретение решает проблему идентификации соединений для лечения мышечной атрофии путем обеспечения способов скрининга с применением CRF2R, которые можно применять для идентификации соединений-кандидатов, применимых для лечения мышечной атрофии. Настоящее изобретение также решает проблему идентификации соединений для лечения мышечной дистрофии путем обеспечения способа скрининга для идентификации соединений-кандидатов, которые активируют как CRF1R, так и CRF2R.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к применению CRFR для идентификации соединений-кандидатов, которые потенциально применимы для лечения атрофии скелетных мышц и/или для индуцирования гипертрофии скелетных мышц. В частности, изобретение относится к способам in vitro идентификации соединений-кандидатов для регуляции массы или функции скелетных мышц, предусматривающим взаимодействие тестируемого соединения с клеткой, экспрессирующей CRF2R, или взаимодействие тестируемого соединения с выделенным CRF2R и определение того, соединяется ли тестируемое соединение с CRF2R или активирует ли оно CRF2R. Другой пример осуществления изобретения относится к способу идентификации терапевтических соединений-кандидатов из группы, состоящей из одного или более соединений-кандидатов, для которых обнаружено, что они связываются с CRF2R или активируют CRF2R, включающему введение соединения-кандидата животному, отличному от человека, и определение того, регулирует ли соединение-кандидат массу скелетных мышц или мышечную функцию у подвергнутого лечению животного. Еще один пример осуществления изобретения относится к способу идентификации соединений-кандидатов для регуляции массы или функции скелетных мышц, включающему в любом порядке: (i) взаимодействие тестируемого соединения с клеткой, экспрессирующей функциональный CRF2R, и определение уровня активации CRF2R, вызванного тестируемым соединением; (ii) взаимодействие тестируемого соединения с клеткой, экспрессирующей функциональный CRF1R, и определение уровня активации CRF1R, вызванного тестируемым соединением; последующее (iii) сравнение уровня активации CRF2R и уровня активации CRF1R; и (iv) идентификация тех тестируемых соединений, которые проявляют похожую активность в отношении CRF2R и CRF1R или проявляют селективность в отношении CRF2R, в качестве соединений-кандидатов для регуляции массы или функции скелетных мышц.
Изобретение также относится к способам идентификации соединений-кандидатов, которые пролонгируют или усиливают агонист-индуцированную активацию CRF2R или активацию пути передачи сигнала CRF2R. Данные способы предусматривают в любом порядке или одновременно: (i) взаимодействие тестируемого соединения с клеткой, которая экспрессирует функциональный CRF2R; (ii) обработку клетки агонистом CRF2R в течение достаточного времени и при достаточной концентрации, чтобы вызвать десенсибилизацию CRF2R в контрольных клетках; последующее (iii) определение уровня активации CRF2R и идентификацию тестируемых соединений, которые пролонгируют или усиливают активацию CRFR или активацию пути передачи сигнала CRF2R, в качестве соединений-кандидатов для регуляции массы или функции скелетных мышц. В частности, настоящее изобретение относится к способу идентификации терапевтических соединений-кандидатов из группы, состоящей из одного или более соединений-кандидатов, для которых обнаружено, что они пролонгируют или усиливают активацию CRF2R или активацию пути передачи сигнала CRF2R, включающему: введение соединения-кандидата в сочетании с агонистом CRF2R животному, отличному от человека, и определение, регулирует ли соединение-кандидат массу или функцию скелетных мышц у подвергнутого лечению животного.
Изобретение также относится к способам идентификации соединений-кандидатов, увеличивающих экспрессию CRF2R, предусматривающим взаимодействие тестируемого соединения с клеткой или клеточным лизатом, содержащим репортерный ген, оперативно связанный с регуляторным элементом CRF2R-гена, и обнаружение экспрессии репортерного гена. Тестируемые соединения, увеличивающие экспрессию репортерного гена, идентифицируют в качестве соединений-кандидатов для увеличения экспрессии CRF2R. В частности, настоящее изобретение относится к способу определения, можно ли те соединения-кандидаты, которые увеличивают экспрессию CRF2R, применять для регуляции массы или функции скелетных мышц in vivo путем введения соединения-кандидата животному, отличному от человека, и определения, регулирует ли соединение-кандидат массу или функцию скелетных мышц у подвергнутого лечению животного.
Изобретение также относится к способам идентификации соединений-кандидатов, увеличивающих экспрессию CRF, предусматривающим взаимодействие тестируемого соединения с клеткой или клеточным лизатом, содержащим репортерный ген, оперативно связанный с регуляторным элементом CRF-гена, и обнаружение экспрессии репортерного гена. Тестируемые соединения, которые увеличивают экспрессию репортерного гена, идентифицируют в качестве соединений-кандидатов для увеличения экспрессии CRF. В частности, настоящее изобретение относится к способу определения, можно ли те соединения-кандидаты, которые увеличивают экспрессию CRF, применять для регуляции массы или функции скелетных мышц in vivo путем введения соединения-кандидата животному, отличному от человека, и определения, регулирует ли соединение-кандидат массу или функцию скелетных мышц у подвергнутого лечению животного.
Настоящее изобретение также относится к применению агонистов CRF2R, экспрессирующих векторов, кодирующих функциональный CRF2R, экспрессионных векторов, кодирующих конститутивно активный CRF2R или соединений, увеличивающих экспрессию CRF2R или CRF, для лечения атрофии скелетных мышц. В частности, изобретение обеспечивает способы лечения атрофии скелетных мышц у субъекта, который нуждается в таком лечении, предусматривающие введение субъекту безопасного и эффективного количества агониста CRF2R, экспрессионного вектора, кодирующего функциональный CRF2R, экспрессионного вектора, кодирующего конститутивно активный CRF2R, экспрессионного вектора, кодирующего CRF или аналог CRF, или соединение, увеличивающее экспрессию CRF2R или CRF. В конкретном примере осуществления настоящее изобретение относится к способу лечения атрофии скелетных мышц у субъекта, который нуждается в таком лечении, предусматривающему введение субъекту безопасного и эффективного количества агониста CRF2R в сочетании с безопасным и эффективным количеством соединения, которое пролонгирует или увеличивает индуцированную агонистом активацию CRF2R или активацию пути передачи сигнала CRF2R.
Настоящее изобретение также относится к применению агониста CRF2R для увеличения массы или функции скелетных мышц у субъекта. В частности, изобретение обеспечивает способы увеличения массы или функции скелетных мышц у субъекта, которому требуется такое увеличение, предусматривающие идентификацию субъекта, которому требуется увеличение мышечной массы или функции и введение субъекту безопасного и эффективного количества агониста CRFR.
Изобретение также относится к фармацевтическим композициям, содержащим безопасное и эффективное количество агониста CRF2R и фармацевтически приемлемого носителя. В конкретном примере осуществления изобретения фармацевтическая композиция содержит химерное антитело или антитело человека, специфичное к CRF2R. В другом конкретном примере осуществления фармацевтическая композиция содержит CRF или аналог CRF, предпочтительно урокортин II.
Настоящее изобретение также обеспечивает антитела к CRF2R и, в частности, химерные или человеческие антитела, которые являются агонистами CRF2R.
На всем протяжении данной заявки упоминаются различные публикации. Описания данных публикаций во всей их полноте включены тем самым в данную заявку путем ссылок для того, чтобы более полно описать современное состояние области техники, к которой данное изобретение относится.
ОПИСАНИЕ СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Каждая из нуклеотидных и белковых последовательностей CRFR или белковых последовательностей аналогов CRF, включенная в список последовательностей вместе с соответствующим регистрационным номером(ами) в банках генов (Genbank) или Derwent и видом животного, от которого она клонирована, приведена в таблице 1. Также приведены регистрационные номера родственных нуклеотидных последовательностей, которые кодируют идентичные, или почти идентичные, аминокислотные последовательности в виде последовательности, показанной в списке последовательностей. Такие родственные последовательности отличаются, главным образом, количеством показанных 5′ или 3′ нетранслируемых последовательностей.
β-CRF2R
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 показан антиатрофический эффект агониста CRF1R/CRF2R саувагина (вводимого подкожно 2 раза в сутки) на медиальную головку икроножной мышцы в модели атрофии, вызванной путем денервации седалищного нерва у мыши.
На фиг.2 показан антиатрофический эффект саувагина (вводимого непрерывно при помощи осмотического мининасоса) на переднюю большеберцовую мышцу в модели атрофии, вызванной путем денервации седалищного нерва у мыши.
На фиг.3A и 3B показан антиатрофический эффект саувагина (вводимого непрерывно при помощи осмотического мининасоса) на индуцированную глюкокортикоидами атрофию передней большеберцовой мышцы (фиг.3A) и медиальной головки икроножной мышцы (фиг.3B).
На фиг.4A показан антиатрофический эффект саувагина (вводимого подкожно 2 раза в сутки) на индуцированную наложением гипсовой повязки атрофию передней большеберцовой мышцы и влияние индуцированной гипертрофии на незагипсованную (нормальную) переднюю большеберцовую мышцу. На фиг.4B показан антиатрофийный эффект саувагина на индуцированную наложением гипсовой повязки атрофию медиальной головки икроножной мышцы и гипертрофию, индуцированную влиянием саувагина на незагипсованную (нормальную) медиальную головку икроножной мышцу.
На фиг.5 показаны антиатрофический и гипертрофический эффекты саувагина и урокортина (вводимых непрерывно при помощи осмотического мининасоса) на переднюю большеберцовую мышцу в модели атрофии, индуцированной путем денервации седалищного нерва у мыши.
На фиг.6A и 6B показан антиатрофический эффект урокортина (вводимого подкожно 2 раза в сутки) на индуцированную бездействием атрофию передней большеберцовой мышцы (фиг.6A) и медиальной головки икроножной мышцы (фиг.6B).
На фиг.7 показан антиатрофический эффект саувагина (вводимого подкожно 2 раза в сутки) на индуцированную путем денервации атрофию передней большеберцовой мышцы (фиг.7A), разгибателя большого пальца (EDL) (фиг.7B), камбаловидной мышцы (фиг.7C), медиальной головки икроножной мышцы (фиг.7D) и подошвенной мышцы в модели атрофии, индуцированной путем денервации седалищного нерва у крысы, подвергнутой резекции надпочечника. Кроме того, индуцированная саувагином гипертрофия неденервированной мышцы EDL (фиг.7B).
На фиг.8 показано, что для модели атрофии, вызванной путем денервации седалищного нерва у мыши, саувагин (вводимый непрерывно при помощи осмотического мининасоса) имел антиатрофический эффект на переднюю большеберцовую мышцу у мышей дикого типа, однако не у мышей CRF2R-нокаут.
На фиг.9A и 9B показано, что в модели атрофии, вызванной бездействием загипсованной конечности у мыши, саувагин имел антиатрофический эффект на EDL и камбаловидную мышцу, что определяли по измерению массы (фиг.9A) или мышечной функции (фиг.9B).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
I. Термины и определения
Ниже приводится список определений применяемых здесь терминов.
\Агонист\ означает любое соединение, включая без ограничения антитела, которые активируют рецептор. Например, агонисты CRFR включают без ограничения CRF и аналоги CRF.
\Аллельный вариант\ означает вариантную форму данного гена или генного продукта. Для специалиста в данной области очевидно, что большое количество генов в популяции существует в двух или более аллельных формах, а некоторые гены имеют многочисленные аллели.
Термин \антитело\ в его различных грамматических формах означает молекулы иммуноглобулина и иммунологически активные части молекул иммуноглобулина, то есть молекулы, которые содержат антиген-связывающий сайт, который специфически связывается с антигеном. \Очищенное антитело\ означает антитело, которое было частично или полностью отделено от белков и встречающихся в природе органических молекул, с которыми оно связано в природе. Предпочтительно, препарат содержит, по меньшей мере, 60% антител, более предпочтительно, по меньшей мере, 75% антител, еще более предпочтительно, по меньшей мере, 90% антител и наиболее предпочтительно, по меньшей мере, 99% (по сухому весу) антител.
\Аффинность связывания\ означает склонность лиганда к взаимодействию с рецептором, она обратно пропорционально связана с константой диссоциации реакции взаимодействия определенных CRF и CRFR-лиганда. Константу диссоциации можно измерить непосредственным образом через стандартное насыщение, конкурентный анализ или кинетику связывания, или косвенно фармакологическими методами, включающими функциональный анализ и крайние точки.
\Химерное антитело\ означает антитело, которое содержит структурные элементы от двух или более молекул различных антител, то есть от различных видов животных. Химерные антитела включают без ограничения антитела, известные как \гуманизированные антитела\, которые включают без ограничения химерные антитела, образуемые при помощи известной технологии пересадки области, определяющей комплементарность (гипервариабельного участка).
\CRF\ означает рилизинг-фактор кортикотропина, который представляет то же самое, что рилизинг-гормон кортикотропина (CRH). Примеры пептидов CRF включают r/h CRF и CRF овцы (см. патент США № 4415558) и т.п.
\Аналог CRF\ означает вещества, которые действуют как лиганды CRFR. Соответствующие аналоги CRF можно получить от различных видов позвоночных, они включают без ограничения такие вещества, как саувагин (см., например, патент США № 4605642), уротензин (см., например, патенты США №№ 4908352 и 4533654), мышиный урокортин II (SEQ ID NO:43), пептид, родственный урокортину человека (SEQ ID NO:44) (Reyes T.M. и др., Proc. Natl Acad Sci 98:2843-2848 (2001)), урокортин (см., например, заявку WO 97/00063) и аналоги CRF, описанные в патентах США №№: 4415558; 4489163; 4594329; 4605642; 5109111; 5235036; 5278146; 5439885; 5493006; 5663292; 5824771; 5844074 и 5869450, каждый из которых включен здесь в виде ссылки. Предпочтительные аналоги CRF представляют саувагин, урокортин, пептид, родственный урокортину, урокортин II и уротензин.
\Агонист CRFR\ означает соединение или молекулу, которые способны активировать CRF1R или CRF2R, или обоих. Активацию CRFR можно измерить, как описано дальше.
\CRFR\ означает CRF1R или CRF2R.
\CRF1R\ означает любые изоформы CRF1R любого вида животных. Ранее CRF1R был описан как CRF-RA, PC-CRF, CRF (Perrin M.H. и др. Endocrinology 133:3058-3061 (1993), Chen R. и др., Proc. Natl. Acad. Sci. USA, 90:8967-8971 (1993), Chang C-P. и др., Neuron 11:1187-1195 (1993), Kishimoto T. и др., Proc. Natl. Acad. Sci. USA, 92:1108-1112 (1995) и Vita N. и др., FEBS Lett. 335:1-5 (1993)) или рецептор CRH.
Определение CRF1R включает без ограничения те рецепторы, для которых кДНК или геномная последовательность, кодирующая рецептор, депонирована в базе данных последовательностей. Такие последовательности имеют регистрационные номера: X72304, E11431, L23332, I92584, T37068, T28968, Q81952, L23333, NM_004382, AF180301, T28970, L25438, L24096, I92586, Q81954, AH006791, NM_007762, X72305, AF054582, Y14036, AF229359, AF229361, AB055434 и L41563. Нуклеотидные и белковые последовательности таких рецепторов доступны из банков данных GenBank или Derwent, а для удобства здесь в списке последовательностей приведены типичные последовательности.
\CRF2R\ означает любую изоформу CRF2R любого вида животных. CRF2R также был описан как НМ-CRF, CRF-RB (Kishimoto T. и др., Proc. Natl. Acad. Sci. USA, 92:1108-1112 (1995) и Perrin M. и др., Proc. Natl. Acad. Sci. USA, 92:2969-2973 (1995)).
Определение рецептора CR2FR включает без ограничения те рецепторы, для которых последовательность-ДНК, кодирующая рецептор, размещена в базе данных последовательностей. Такие последовательности имеют регистрационные номера: U34587, E12752, NM_001883, T12247, T66508, AF011406, AF019381, U16253, T12244, T28972, U17858, NM_009953, Y14037 и AF229360. Нуклеотидные и белковые последовательности таких рецепторов доступны из банков данных GenBank или Derwent, а для удобства здесь в списке последовательностей приведены типичные последовательности.
Термин \CRFR\ также означает усеченные и/или мутированные белки, в которых области рецепторной молекулы, не требуемые для связывания лиганда или передачи сигнала, удалены или модифицированы. Например, для специалиста в данной области очевидно, что для настоящего изобретения можно было бы применить CRFR с одной или более консервативными заменами в первичной аминокислотной последовательности. В данной области известно, что замена определенных аминокислот другими аминокислотами с похожей структурой или свойствами (консервативные замены) может приводить к \молчащей\ замене, то есть к замене, которая существенно не изменяет функцию. Консервативные замены хорошо известны в данной области. Например, известно, что GPCR могут допускать замену аминокислотных остатков в трансмембранных альфа-спиралях, ориентированных по направлению к липиду, на другие гидрофобные аминокислоты и оставаться функциональными. Рецепторы CRF1R с последовательностями, отличающимися от существующих в природе тем, что они усечены и/или мутированы, так же, как консервативные замены аминокислот, также включены в определение CRF1R. CRF2R с последовательностью, отличающейся от встречающейся в природе тем, что они усечены и/или мутированы, так же, как консервативные замены аминокислот, также включены в определение CRF2R.
Для специалиста в данной области очевидно, что в настоящем изобретении можно применять рецепторы CRFR других видов, отличающиеся от перечисленных выше, в частности видов млекопитающих. Кроме того, для специалиста в данной области очевидно, что при помощи зондов из видов, последовательности CRFR которых известны, известными способами клонирования можно получить кДНК или геномные последовательности, гомологичные известной последовательности, от тех же самых или альтернативных видов. Такие CRF1R также включены в определение CRF1R, и такие CRF2R также включены в определение CRF2R.
Кроме того, для специалиста в данной области очевидно, что функциональные аллельные варианты или функциональные варианты сплайсинга CRFR могли бы существовать для определенных видов, и что такие варианты могут применяться в настоящем изобретении. Варианты сплайсинга CRFR известны, например, в патентах США №№ 5888811, 5786203 и 5728545, каждый из которых включен здесь в виде ссылки. Такие варианты CRF1R также включены в определение CRF1R, и такие варианты CRF2R также включены в определение CRF2R.
Гибриды полипептида CRF1R или CRF2R или фрагмента полипептида CRF1R или CRF2R с полипептидом, не являющимся CRFR, называются гибридными белками CRFR. Применяя известные способы, специалист в данной области может получить гибридные белки CRF1R или CRF2R, которые, несмотря на то, что отличаются от нативных CRF1R и CRF2R, остаются применимыми для целей настоящего изобретения. Например, полипептид, не являющийся CRFR, может быть сигнальной (или лидерной) полипептидной последовательностью, которая одновременно с трансляцией или после завершения трансляции направляет транспорт белка от его сайта синтеза к другому сайту (например, лидерный α-фактор дрожжей). Или полипептид, не являющийся CRFR, может быть добавлен для облегчения очистки или идентификации CRFR (например, поли-His или Flag-пептид). Гибридные белки CRF1R также включены в определение CRF1R, и гибридные белки CRF2R также включены в определение CRF2R.
\Путь передачи сигнала CRF2R\ означает любой сигнальный путь (например, цАМФ, МАР-киназы) или сочетание сигнальных путей, которые модулируются путем присоединения эндогенных или экзогенных лигандов к CRF2R.
Термин \функциональные CRFR\ относится к CRFR, которые присоединяют CRF или аналог CRF in vivo или in vitro вследствие присоединения лигандов.
\Гибридный ген\ означает две или более кодирующие последовательности-ДНК, оперативно связанные для кодирования одного гибридного белка. \Гибридный белок\ представляет собой белковый продукт гибридного гена.
\Ингибировать\ означает частично или полностью блокировать определенный процесс или активность. Например, соединение ингибирует атрофию скелетных мышц, если оно либо полностью, либо частично предотвращает мышечную атрофию.
Как здесь принято, две последовательности-ДНК считаются \оперативно связанными\, если природа связи между двумя последовательностями-ДНК (1) не приводит к внесению мутации \сдвига рамки\, (2) не мешает способности области промотора управлять транскрипцией кодирующих последовательностей или (3) не мешает способности соответствующего РНК-транскрипта транслироваться в белок. Например, кодирующая последовательность и регуляторные последовательности \оперативно связаны\, когда они связаны ковалентно таким образом, чтобы происходила транскрипция кодирующей последовательности под влиянием или контролем регуляторных последовательностей. Таким образом, область промотора оперативно связана с кодирующей последовательностью, где область промотора способна к проведению транскрипции последовательности ДНК так, что образующийся в результате транскрипт способен к трансляции в требуемый белок или полипептид.
\Степень идентичности\ означает процент нуклеотидов или аминокислот, которые в двух сравниваемых последовательностях являются общими, и рассчитывается следующим образом. Для вычисления степени идентичности определенной последовательности (запрашиваемой) нужную часть запрашиваемой последовательности сравнивают с описанной последовательностью, применяя компьютерную программу сравнения BestFit, Wisconsin Package, версия 10.1, предоставляемую Genetics Computer Group Inc. В такой программе применяется алгоритм Smith и Waterman, Advances in Applied Mathematics, выпуск 2: 482-489 (1981). Степень идентичности рассчитывают со следующими параметрами программы BestFit, выбираемыми по умолчанию: матрицей для расчета является blosum62.cmp, штраф за создание разрыва составляет 8, и штраф за удлинение разрыва составляет 2. При сравнении последовательности с описанной последовательностью нужную часть запрашиваемой последовательности представляет та, которая получена из CRFR-последовательности. Например, когда запрашиваемая последовательность является гибридным белком, помеченным при очистке/CRFR, для расчета степени идентичности выстраивается только полипептидная часть CRFR-последовательности.
\Полипептид\ означает любую цепь аминокислот, независимо от длины или модификации после трансляции (например, фосфорилирования или гликозилирования).
\Промотор\ означает последовательность ДНК, которая контролирует инициацию транскрипции и скорость транскрипции гена или кодирующей области.
\Профилактическое лечение\ означает превентивное лечение субъекта, в настоящее время не имеющего признаков атрофии скелетных мышц, для того, чтобы полностью или частично блокировать явление атрофии скелетных мышц. Для специалиста в данной области очевидно, что некоторые лица входят в группу риска в отношении атрофии скелетных мышц, что обсуждалось здесь в разделе \предшествующий уровень техники\. Кроме того, для специалиста в данной области очевидно, что, если бы биохимические замены, приводящие к атрофии скелетных мышц, соответственно регулировались, явление атрофии было бы предотвращено или уменьшено для лиц, входящих в группу риска. Например, пациенты с мышечной дистрофией, начинающие лечение кортикостероидами, входят в группу риска в отношении развития атрофии скелетных мышц, что указывает на то, что профилактическое лечение таких пациентов должно быть соответствующим.
Термин \регулировать\ во всех его грамматических формах означает \увеличивать\, \уменьшать\ или \сохранять\, например, термин \регулировать массу или функцию скелетных мышц\ означает увеличивать, уменьшать или сохранять уровень массы или функции скелетных мышц.
Термин \регуляция массы или функции скелетных мышц\ предусматривает регуляцию массы скелетных мышц, функции скелетных мышц или регуляцию того и другого.
\Регуляторный элемент\ означает последовательность ДНК, которая способна контролировать уровень транскрипции оперативно связанной последовательности ДНК. В данное определение регуляторного элемента включены промоторы и энхансеры. Например, регуляторный элемент CRFR-гена представляет собой последовательность ДНК, способную контролировать уровень транскрипции от CRFR-гена.
\Репортерный ген\ означает кодирующую последовательность, продукт которой можно обнаружить, предпочтительно количественно, в которой репортерный ген является оперативно связанным с гетерологичным промотором или элементом энхансера, чувствительным к сигналу, который следует измерять. Промотор или элемент энхансера упоминаются в данном контексте как \отвечающий элемент\.
Термин \селективный агонист\ означает, что агонист обладает существенно большей активностью в отношении определенного рецептора(ов) по сравнению с другими рецепторами, но что он не является полностью неактивным в отношении других рецепторов.
\Гипертрофия скелетных мышц\ означает увеличение массы скелетных мышц или функции скелетных мышц или увеличение обеих.
\Атрофия скелетных мышц\ означает то же самое, что \утрата мышцы\, и означает уменьшение массы скелетных мышц или функции скелетных мышц или уменьшение обеих.
\Вариант сплайсинга\ означает мРНК или белок, который образуется в результате использования альтернативного экзона. Для специалиста в данной области очевидно, что, в зависимости от типа клеток или даже в пределах одного типа клеток, мРНК может экспрессироваться в различной форме, как вариант сплайсинга, и таким образом транслируемый белок будет отличаться в зависимости от мРНК, которая экспрессируется.
\Терапевтически эффективное количество\ вещества означает количество, способное давать требуемый, с медицинской точки зрения, результат для подвергаемого лечению пациента, например уменьшение атрофии скелетных мышц, увеличение массы скелетных мышц или увеличение функции скелетных мышц, с приемлемым отношением преимущество:риск для человека или для млекопитающего, отличного от человека.
\Терапевтическое лечение\ означает лечение субъекта, для которого требуется увеличение мышечной массы или мышечной функции. Например, лечение субъекта с признаками атрофии скелетных мышц, проявляющимися в настоящее время, для того, чтобы частично или полностью обратить развившуюся атрофию скелетных мышц либо частично или полностью блокировать дальнейшее развитие атрофии скелетных мышц, будет терапевтическим лечением такого субъекта. Термин \терапевтическое лечение\ также включает, например, лечение субъекта без признаков атрофии скелетных мышц с целью индуцирования гипертрофии скелетных мышц, например обработку домашнего скота для увеличения мышечной массы.
Термин \лечение\ означает профилактическое или терапевтическое лечение.
Если не оговорено иначе, все технические и научные термины, применяемые здесь, имеют то же самое значение, которое обычно понимается специалистом в области химии белков, фармакологии или молекулярной биологии. Способы, материалы и примеры, описанные здесь, не предназначены для ограничения изобретения. Другие способы и материалы, подобные или эквивалентные тем, что описаны здесь, могут применяться на практике или при тестировании согласно настоящему изобретению.
II. Роль рецепторов CRF в регуляции массы скелетных мышц
Для специалиста в данной области очевидно, что применимость настоящего изобретения обусловлена информацией о прототипе и методиках, приводимых ниже. Описанные здесь результаты показывают, что введение агониста рецептора CRF, который активирует как CRF1R, так и CRF2R (неселективный агонист CRFR), блокирует и/или ингибирует атрофию скелетных мышц, индуцированнную путем денервации, бездействием или лечением дексаметазоном, в моделях атрофии скелетных мышц. Кроме того, данные показывают, что агонисты CRFR не проявляют такого антиатрофийного эффекта у мышей с выключенным CRF2R. То же самое у крыс, у которых ось HPA, опосредованная CRF1R, нарушена путем удаления надпочечников (хирургическая резекция надпочечника); при лечении таких животных неселективными агонистами CRFR обнаруживается антиатрофийный эффект, указывающий, что CRF2R опосредует антиатрофийные эффекты. Кроме того, результаты показывают, что при введении неселективного агониста CRFR обнаруживается эффект, индуцирующий гипертрофию. Взятые вместе, такие данные показывают модулирующую роль CRFAR в процессе атрофии скелетных мышц. Специфичную роль CRFR исследовали in vivo, применяя фармакологические агенты саувагин (Bachem Biosciences Inc., King of Prussia, PA) и урокортин (Bachem Biosciences Inc.), которые являются селективными агонистами рецепторов CRF в различных моделях атрофии скелетных мышц, описанных дальше. Такие агенты хорошо охарактеризованы и описаны в научной литературе.
На фигурах 1-7 и 9 приведены результаты экспериментов, демонстрирующие, что введение селективных агонистов CRFR приводит к статистически значимому ингибированию атрофии скелетных мышц. На фигуре 8 показано, что антиатрофийный эффект агониста CRFR саувагина опосредован через CRF2R. Агонисты CRFR вводили два раза в сутки в сочетании с ингибитором фосфодиэстеразы теофиллином, что приводило к ингибированию атрофии скелетных мышц в животных моделях атрофии скелетных мышц. Теофиллин добавляли для увеличения продолжительности и величины воздействия агониста CRFR, что приводило, следовательно, к повышенной эффективности таких соединений. Теофиллин, вводимый отдельно (в единственном числе), в таких моделях атрофии не давал никакого эффекта, указывая на то, что антиатрофийный эффект агониста CRFR в сочетании с теофиллином обусловлен эффектом агониста CRFR. Кроме того, непрерывное дозирование агониста CRFR в отсутствии теофиллина через осмотический мининасос также приводило к ингибированию атрофии скелетных мышц и/или гипертрофии скелетных мышц. Статистическую значимость результатов определяли при помощи ANCOVA (Douglas C. Montgomery, Design and Analysis of Experiments, John Wiley and Sons, Нью-Йорк (2-ая ред. 1984)). Сокращения, применяемые на фигурах 1-9: г означает грамм; SEM означает стандартное отклонение значения.
В частности, на фигуре 1 (фиг.1) показано, что саувагин ингибирует атрофию, индуцированную денервацией медиальной головки икроножной мышцы в модели атрофии, вызванной денервацией седалищного нерва у мышей. Условные обозначения: A означает физиологический раствор (контрольный); B означает саувагин (0,01 мг/кг)+теофиллин; C означает саувагин (0,03 мг/кг)+теофиллин; D означает саувагин (0,1 мг/кг)+теофиллин; E означает саувагин (1,0 мг/кг)+теофиллин; * означает p≤0,05, по сравнению с физиологическим раствором. Вслед за денервацией правого седалищного нерва самцу мыши подкожно два раза в сутки вводили саувагин в межлопаточную область в указанных выше дозах или контрольный носитель (физиологический солевой раствор) в течение девяти дней. Саувагин вводили вместе с 30 мг/кг теофиллина. На девятый день медиальную головку икроножной мышцу удаляли и взвешивали, чтобы определить степень атрофии.
На фигуре 2 (фиг.2) показано, что саувагин ингибирует индуцированную путем денервации атрофию передней большеберцовой мышцы в модели атрофии, вызванной денервацией седалищного нерва у мышей. Условные обозначения: А означает воду (контрольный); B означает саувагин (0,1 мг/кг/сутки); C означает саувагин (0,3 мг/кг/сутки); D означает саувагин (1,0 мг/кг/сутки); * означает p≤0,05 по сравнению с водой. Вслед за денервацией правого седалищного нерва самцу мыши давали определенными дозами либо саувагин, либо контрольный носитель (физиологический солевой раствор) путем непрерывной инфузии 5 мкл/час при помощи осмотического мининасоса (Alzet osmotic minipump) до конца экспериментального периода (без дополнительного теофиллина). Ежедневно подаваемая доза саувагина указана выше. Имплантацию мининасоса проводили во время денервации седалищного нерва. На девятый день переднюю большеберцовую мышцу удаляли и взвешивали, чтобы определить степень атрофии.
На фигуре 3 (фиг.3) показано, что саувагин ингибирует индуцированную глюкокортикоидами мышечную атрофию передней большеберцовой (фиг.3A) и медиальной головки икроножной мышц (фиг.3B) в модели атрофии, индуцированной глюкокортикоидами у мышей. Условные обозначения: А означает только воду без дексаметазона, добавленного в питьевую воду (контрольный, не вызывающий атрофию); B означает воду+дексаметазон (контрольный, вызывающий атрофию); C означает саувагин (0,1 мг/кг/сутки)+дексаметазон; D означает саувагин (0,3 мг/кг/сутки)+дексаметазон; E означает саувагин (1,0 мг/кг/сутки)+дексаметазон; * означает p≤0,05 по сравнению с водой; # означает p≤0,05 по сравнению с водой+дексаметазоном. Вслед за добавлением глюкокортикоида дексаметазона к питьевой воде (1,2 мг/кг/сутки), самцу мыши вводили определенными дозами указанные выше агенты или контрольный носитель (физиологический солевой раствор) путем непрерывной инфузии 5 мкл/час с применением осмотического мининасоса (Alzet osmotic minipump) до конца экспериментального периода (без дополнительного теофиллина). Ежедневно подаваемая доза саувагина указана выше. Имплантацию мининасоса проводили во время инициации действием дексаметазона. Девять дней спустя после начала дозированного введения саувагина медиальную головку икроножной и переднюю большеберцовую мышцы удаляли и взвешивали, чтобы определить степень атрофии.На фигуре 4 (фиг.4) показано, что саувагин ингибирует индуцированную бездействием атрофию передней большеберцовой (фиг.4A) и медиальной головки икроножной (фиг.4B) мышц. Кроме того, при лечении саувагином также наблюдалась статистически значимая гипертрофия медиальной головки икроножной и передней большеберцовой мышцы незагипсованной конечности. Условные обозначения: А означает физиологический солевой раствор (контрольный); B означает теофиллин; C означает саувагин (0,03 мг/кг)+теофиллин; D означает саувагин (0,1 мг/кг)+теофиллин; E означает саувагин (0,3 мг/кг)+теофиллин; * означает p≤ 0,05 по сравнению с солевым раствором. Вслед за наложением гипсовой повязки на правую заднюю конечность самцу мыши два раза в сутки подкожно в межлопаточную область вводили саувагин или контрольный носитель (физиологический солевой раствор) в течение десяти дней при обеспечении указанной дозы. Саувагин вводили два раза в сутки внутрибрюшинно дозами вместе с ингибитором фосфодиэстеразы теофиллином (30 мг/кг). На десятый день удаляли и взвешивали медиальную головку икроножной и переднюю большеберцовую мышцы, чтобы определить степень атрофии.
На фигуре 5 (фиг.5) показано, что как саувагин, так и урокортин ингибируют индуцированную денервацией атрофию передней большеберцовой мышцы в модели атрофии, вызванной денервацией седалищного нерва у мышей. Кроме того, при лечении урокортином наблюдалась гипертрофия неденервированной конечности. Условные обозначения: A означает воду (контрольный); B означает саувагин (1 мг/кг/сутки); C означает урокортин (1,0 мг/кг/сутки); * означает p≤0,05 по сравнению с водой. Вслед за денервацией правого седалищного нерва самцу мыши давали определенными дозами указанные выше агенты или контрольный носитель (физиологический солевой раствор) путем непрерывной инфузии 5 мкл/час при помощи осмотического мининасоса (Alzet osmotic minipump) до конца экспериментального периода (без дополнительного теофиллина). Ежедневно подаваемая доза агентов указана выше. Имплантацию мининасоса проводили в то же время, что денервацию седалищного нерва. На девятый день переднюю большеберцовую мышцу удаляли и взвешивали, чтобы определить степень атрофии.
На фигуре 6 (фиг.6) показано, что урокортин ингибирует индуцированную бездействием атрофию передней большеберцовой (фиг.6A) и медиальной головки икроножной (фиг.6B) мышцы в модели атрофии, вызванной бездействием загипсованной конечности у мыши. Условные обозначения: А означает физиологический солевой раствор (контрольный); B означает урокортин (0,3 мг/кг)+теофиллин; * означает p≤0,05 по сравнению с солевым раствором. Вслед за наложением гипсовой повязки на правую заднюю конечность самцу мыши вводили подкожно в межлопаточную область два раза в сутки урокортин или контрольный носитель (физиологический солевой раствор) в течение десяти дней. Урокортин вводили в дозах, указанных в описании фигур 6A и 6B. Два раза в сутки внутрибрюшинно определенными дозами вводили урокортин вместе с ингибитором фосфодиэстеразы теофиллином (30 мг/кг). На десятый день медиальную головку икроножной и переднюю большеберцовую мышцы удаляли и взвешивали, чтобы определить степень атрофии.
На фигуре 7 (фиг.7) показано, что саувагин ингибирует индуцированную путем денервации атрофию передней большеберцовой мышцы (фиг.7A), EDL (фиг.7B), камбаловидной мышцы (фиг.7C), медиальной головки икроножной мышцы (фиг.7D) и подошвенной мышцы. Кроме того, саувагин вызывает статистически значимую гипертрофию неденервированной EDL мышцы (фиг.7B). Условные обозначения: А означает физиологический солевой раствор (контрольный); B означает саувагин (0,003 мг/кг)+теофиллин; C означает саувагин (0,01 мг/кг)+теофиллин; D означает саувагин (0,03 мг/кг)+теофиллин; # означает p≤0,05 по сравнению с соответствующим контрольным раствором. Вслед за денервацией правого седалищного нерва, самцам крыс, подвергнутых резекции надпочечника (крыс с резекцией надпочечника применяли для удаления влияния на индуцированную атрофию скелетных мышц активации оси HPA через агонистов CRF1R), вводили подкожно в межлопаточную область два раза в сутки либо саувагин, либо контрольный носитель (физиологический солевой раствор) в течение девяти дней в дозах, указанных выше. Саувагин вводили вместе с 30 мг/кг теофиллина. На девятый день переднюю большеберцовую мышцу, разгибатель большого пальца (EDL), камбаловидную мышцу, медиальную головку икроножной и подошвенную мышщу удаляли и взвешивали, чтобы определить степень атрофии.
На фигуре 8 (фиг.8) показано, что саувагин ингибирует атрофию, наблюдаемую у мышей дикого типа, но не у мышей-нокаут CRF2R, в модели атрофии, вызванной денервацией седалищного нерва у мышей. Условные обозначения: A-C означает мышей дикого типа; D-F означает мышей-нокаут CRF2R. А и D означают воду (контрольный); B и E означает саувагин (0,3 мг/кг/сутки); C и F означают саувагин (1,0 мг/кг/сутки); * означает p≤0,05 по сравнению с солевым раствором. Вслед за денервацией правого седалищного нерва самкам мышей дикого типа и мышей-нокаут CRF2R вводили определенными дозами саувагин или контрольный носитель путем непрерывной инфузии 5 мкл/час при помощи осмотического мининасоса (Alzet osmotic minipump) в течение девяти дней, ежедневно обеспечивая дозу, указанную выше. На девятый день переднюю большеберцовую мышцу удаляли и взвешивали, чтобы определить степень атрофии.
На фигуре 9 (фиг.9) показано, что саувагин ингибирует индуцированную бездействием потерю EDL и массы камбаловидной мышцы (фиг.9A) и ингибирует потерю мышечной функции, что оценивали путем измерения абсолютной силы (фиг.9B) для модели атрофии, вызванной бездействием из-за наложения гипсовой повязки на конечность у мышей. Условные обозначения: А означает контрольную незагипсованную мышцу; B означает загипсованную мышцу, контрольный солевой раствор; C означает загипсованную мышцу, саувагин (0,3 мг/кг)+теофиллин (30 мг/кг); * означает p≤0,05 по сравнению с солевым раствором. Вслед за наложением гипсовой повязки на правую заднюю конечность самцу мыши вводили подкожно два раза в сутки в межлопаточную область либо саувагин, либо контрольный носитель (физиологический солевой раствор) в течение десяти дней в дозах, указанных выше. Саувагин вводили вместе с 30 мг/кг теофиллина. На десятый день EDL и камбаловидную мышцу удаляли и проводили измерения массы и абсолютной силы, чтобы определить степень атрофии.
III. Получение рецепторов CRFR, CRF или аналогов CRF, или клеточных линий, экспрессирующих CRFR
CRF1R, CRF2R, CRF и аналоги CRF можно получить для ряда применений, включая без ограничения выработку антител, применяемых в качестве реагентов в скрининг-тестах настоящего изобретения и применяемых в качестве фармацевтических агентов для лечения атрофии скелетных мышц. Для специалиста в данной области очевидно, что для некоторых примеров осуществления изобретения наиболее применимы очищенные полипептиды, в то время как для других примеров осуществления изобретения наиболее применимы клеточные линии, экспрессирующие полипептиды. Например, в ситуациях, когда важно сохранить структурные и функциональные характеристики CRFR, например в способе проведения скрининга для идентификации соединений-кандидатов, которые активируют рецепторы CRFR, желательно применять клетки, которые экспрессируют функциональные CRFR.
Так как CRF и аналоги CRF представляют собой короткие полипептиды, для специалиста очевидно, что такие полипептиды наиболее удобно получать путем прямого синтеза, быстрее, чем рекомбинантным способом, применяя методики, известные в данной области. Кроме того, многие из таких полипептидов имеются в продаже.
Когда источником рецепторов CRFR является клеточная линия, экспрессирующая полипептид, клетки, способные, например, эндогенно экспрессировать CRFR, стимулируют для увеличения эндогенной экспрессии CRFR или конструируют при помощи генной инженерии для того, чтобы экспрессировать CRFR. Способы определения, экспрессирует ли клеточная линия полипептид, представляющий интерес, известны в данной области, например, обнаружение полипептида при помощи соответствующего антитела, применение ДНК-пробы для обнаружения мРНК, кодирующей белок (например, Нозерн-блоттинг или метод ПЦР), или измерение степени связывания агента, селективного для полипептида, представляющего интерес (например, селективный агонист, помеченый радиоактивным изотопом).
В частности, рассмотрено применение технологии рекомбинантной ДНК для получения CRF1R, CRF2R или клеточных линий, экспрессирующих такие полипептиды. Такие рекомбинантные способы известны в данной области. Для экспрессии рекомбинантного CRF1R или CRF2R получают экспрессионный вектор, содержащий нуклеиновую кислоту, которая кодирует полипептид, представляющий интерес, под контролем одного или более регуляторных элементов. Геномные последовательности или последовательности кДНК, кодирующие CRF1R и CRF2R от нескольких видов, описаны и легко доступны для такого применения в базе данных GenBank (расположена на сайте <http://www.ncbi.nlm.nih.gov/>) или в базе данных Derwent (расположена на сайте <http://www.derwent.co.uk/geneseq/index.html>), так же, как в списке последовательностей. Регистрационные номера для последовательностей CRF1R и CRF2R и соответствующие SEQ ID NOS приведены в таблице 1. При использовании такой публично доступной информации о последовательностях один из способов выделения молекулы нуклеиновой кислоты, кодирующей CRF1R или CRF2R, заключается в проведении скрининга геномной библиотеки ДНК или кДНК при помощи природной или искусственно синтезированной ДНК-пробы и с применением хорошо известных в данной области способов, например ПЦР-амплификации последовательности из соответствующей библиотеки. Другой способ заключается в применении олигонуклеотидных праймеров, специфичных для рецептора, представляющего интерес, для ПЦР-амплификации кДНК напрямую из мРНК, выделенной из определенной ткани (такой как скелетная мышца). Такая выделенная мРНК имеется в продаже. Для специалиста в данной области также очевидно, что при применении проб нуклеиновых кислот, соответствующих частям известных последовательностей рецептора CRFR, при помощи известных способов можно получить гомологичные кДНК или геномные последовательности от других видов. В частности, в способах согласно настоящему изобретению применяются рецепторы CRFR от видов, включающих без ограничения человека, мышь, крысу, свинью, обезьяну, шимпанзе, игрунку, собаку, корову, овцу, кошку, цыпленка и индейку. Известными в данной области способами выделенную молекулу нуклеиновой кислоты, кодирующую представляющий интерес CRFR, затем лигируют в подходящий экспрессионный вектор. Экспрессионный вектор, полученный таким образом, экспрессируется в клетке-хозяине, а клетки-хозяева, экспрессирующие рецептор, применяются напрямую в скрининг-тесте, или рецептор выделяют из клеток-хозяев, экспрессирующих рецептор, и выделенный рецептор применяют в скрининг-тесте.
Системы \экспрессионный вектор-хозяин\, которые можно применять для целей изобретения, включают без ограничения микроорганизмы, такие как бактерии (например, E.coli, B.subtilis), трансформированные при помощи рекомбинантной бактериофаговой ДНК, ДНК-плазмиды, или векторы экспрессии космидной ДНК, содержащие нуклеотидные последовательности CRFR; дрожжи (например, Saccharomyces, Pichia), трансформированные при помощи экспрессионных векторов рекомбинантных дрожжей, содержащие нуклеотидные последовательности CRFR; системы клеток насекомых, инфицированные векторами экспрессии рекомбинантных вирусов (например, бакуловируса), содержащие нуклеотидные последовательности CRFR; системы растительных клеток, инфицированные векторами экспрессии рекомбинантных вирусов (например, вирусом мозаики цветной капусты, вирусом табачной мозаики), или трансформированные при помощи экспрессионных векторов рекомбинантных плазмид (например, Ti-плазмиды), содержащие нуклеотидные последовательности CRFR; или системы клеток млекопитающих (например, COS, CHO, HEK293, NIH3T3), предусматривающие рекомбинантные экспрессируемые конструкции, содержащие промоторы, полученные из генома клеток млекопитающих (например, металлотионеиновый промотор) или из вирусов млекопитающих (например, ретровирус LTR), и также содержащие нуклеотидные последовательности CRFR.
Клетка-хозяин применяется для продуцирования полипептидов, представляющих интерес. Поскольку CRFR представляет собой молекулу, связанную с мембраной, его выделяют из мембран клеток-хозяев, или CRFR применяется в тот промежуток времени, когда закрепляется на клеточной мембране, то есть применяются целые клетки или мембранные фракции клеток. Очистку или обогащение CRFR из таких экспрессирующих систем проводят с применением подходящих детергентов и липидных мицелл при помощи способов, известных специалистам в данной области.
В бактериальных системах можно успешно выбрать ряд экспрессионных векторов, в зависимости от предназначения генного продукта, который экспрессируется. Например, когда большое количество такого белка продуцируется для создания антител к рецепторам CRF, требуются векторы, которые направляют экспрессию с высокими уровнями белковых продуктов. Специалист в данной области способен создать такие конструкции векторов и очищать белки при помощи различных методик, включая технологии селективной очистки, такие как селективные колонки для гибридного белка и колонки для антител, и технологии неселективной очистки.
В системах экспресии белков у насекомых в качестве вектора для экспрессии чужеродных генов в клетках S. frugiperda применяется бакуловирус ядерного полиэдроза А. californica (AcNPV). В данном случае нуклеотидные последовательности CRFR клонируются на не-незаменимых областях вируса и размещаются под контролем AcNPV-промотора. Затем рекомбинантные вирусы применяют для инфицирования клеток, в которых встроенный ген экспрессируется, и очищают белок при помощи одной из многочисленных методик, известных специалисту в данной области.
В клетках-хозяевах млекопитающих можно применять ряд экспрессионных систем, основанных на вирусах. Применение таких экспрессионных систем часто требует создания специфичных инициирующих сигналов в векторах для эффективной трансляции встроенной нуклеотидной последовательности. Сказанное особенно важно, если применяется область CRFR-гена, которая не содержит эндогенный инициирующий сигнал. Размещение такого инициирующего сигнала в рамке с кодирующей областью встроенной нуклеотидной последовательности, так же, как добавление элементов, усиливающих транскрипцию и трансляцию, и очистка рекомбинантного белка достигаются при помощи одной из многочисленных методик, известных специалисту в данной области. Для клеток-хозяев млекопитающих также важен отбор соответствующего типа клеток, которые способны к необходимым посттрансляционным модификациям рекомбинантного белка. Такие модификации, например расщепление, фосфорилирование, гликозилирование и т.д. требуют отбора соответствующей клетки-хозяина, которая содержит модифицирующие ферменты. Такие клетки-хозяева включают без ограничения CHO, HEK293, NIH3T3, COS и т.д. и известны специалистам в данной области.
Для высокой экспрессии рекомбинантных белков в течение длительного времени предпочтительна стабильная экспрессия. Например, можно сконструировать клеточные линии, которые стабильно экспрессируют CRFR. Специалист в данной области, следуя известным методикам, таким как электропорация, трансфекция с фосфатом кальция или опосредованная липосомами трансфекция, может создать клеточную линию, которая стабильно экспрессирует рецепторы CRFR. Сказанное обычно выполняется трансфицированными клетками при помощи экспрессионных векторов, которые содержат соответствующие элементы для контроля экспрессии (например, последовательности промоторов, последовательности энхансеров, последовательности завершения транскрипции, сайты полиаденилирования, сайты начала трансляции и т.д.), селектируемый маркер и ген, представляющий интерес. Селектируемый маркер может либо находиться в пределах того же самого вектора, что и ген, представляющий интерес, либо на отдельном векторе, который трансфицируется вместе с последовательностью CRFR, содержащей вектор. Селектируемый маркер в экспрессионном векторе может обеспечивать резистентность к селекции и дать возможность клеткам стабильно интегрировать вектор в их хромосомы и расти для формирования локусов, которые, в свою очередь, могут быть клонированы и размножены в клеточных линиях. С другой стороны, экспрессионный вектор может предоставить возможность для селекции клетки, экспрессирующей селектируемый маркер, используя физически характерный признак маркера, например, экспрессия зеленого флуоресцентного белка (GFP) делает возможным селекцию клеток, экспрессирующих маркерный ген, при помощи клеточного сортера с возбуждением флуоресценции (FACS-анализ).
Специалист в данной области способен выбрать подходящий тип клеток для трансфекции, для того чтобы сделать возможным селекцию клеток, в которых успешно интегрирован ген, представляющий интерес. Например, когда селектируемый маркер представляет собой тимидинкиназу вируса простого герпеса, гипоксантингуанинфосфорибозилтрансферазу или аденинфосфорибозилтрансферазу, подходящим типом клеток могут быть tk-, hgprt- или aprt-клетки, соответственно. Или можно применять нормальные клетки, когда селектируемый маркер представляет собой dhfr, gpt, neo или hygro, которые обеспечивают резистентность к метотрексату, микофенольной кислоте, G-418 или гигромицину, соответственно. Такие линии рекомбинантных клеток применимы для идентификации соединений-кандидатов, которые влияют на активность CRFR.
IV. Получение антител против CRFR
Изобретение также включает антитела, которые селективно распознают один или более эпитопов CRFR. Такие антитела включают, например, поликлональные антитела, моноклональные антитела, химерные антитела, человеческие антитела, одноцепочечные антитела, Fab-фрагменты, F(ab')2-фрагменты, молекулы, продуцируемые при помощи библиотеки экспрессируемых Fab-фрагментов, человеческие антитела (поликлональные или моноклональные), продуцируемые трансгенными мышами, и эпитоп-связывающие фрагменты любого соединения из вышеупомянутых. Для терапевтических применений предпочтительны химерные или человеческие антитела, наиболее предпочтительны человеческие антитела.
Антитела можно применять в сочетании со схемами скрининга соединений, описанными здесь для оценки тестируемых соединений, например, для иммобилизации CRFR-полипептидов, или такие антитела можно применять в сочетании с методиками генной терапии для оценки, например, экспрессии CRFR либо в клетках, либо непосредственно в тканях пациента, которому такие гены вводились. Кроме того, антитела согласно настоящему изобретению применимы для лечения атрофии скелетных мышц. Селективные в отношении CRFR антитела можно подвергнуть скринингу при помощи способов, описанных в настоящем изобретении, чтобы идентифицировать субпопуляцию антител, являющихся агонистами CRFR. Кроме того, антиидиотипические антитела, созданные против антител, специфичных для CRF или аналога CRF, можно применять в качестве агонистов CRFR, а подобные анти-CRFR-антитела можно отобрать по их способности активировать CRFR при помощи способов согласно настоящему изобретению.
Для получения антител можно иммунизировать различных домашних животных при помощи инъекции CRFR, CRF или аналога CRF, анти-CRF-антитела, антитела аналога анти-CRF или их иммуногенных фрагментов при помощи способов, известных в данной области. Иммуноген для получения антиидиотипического антитела представляет собой анти-CRF-антитело или антитело аналога анти-CRF. Получение антиидиотипических антител описано, например, в патенте США № 4699880, включенном здесь в виде ссылки. Подходящие домашние животные включают без ограничения кроликов, мышей, коз, овец и лошадей. Методики иммунизации хорошо известны в данной области. Поликлональные антитела можно выделить из сыворотки иммунизированных животных, или моноклональные антитела можно создать при помощи способов, известных в данной области. Такие методики включают без ограничения известные гибридомные технологии Колера и Мильштейна (Kohler и Milstein), технологии В-клеточных гибридом человека и технологию EBV-гибридом. Моноклональные антитела могут представлять любой класс иммуноглобулинов, включая IgG, IgE, IgM, IgA и IgD, содержащих либо каппа-, либо лямбда-легкие цепи.
Из-за иммуногенности для человека антител, не принадлежащих человеку, для терапевтического лечения пациентов (человека) предпочтительны химерные антитела. Технологии продуцирования и применения химерных антител известны в данной области, и описаны, например, в патентах США №№5807715; 4816397; 4816567; 5530101; 5585089; 5693761; 5693762; 6180370 и 5824307, все из которых включены здесь в виде ссылок.
Для терапевтического лечения пациентов (человека) особенно желательны полностью человеческие антитела, потому что они менее иммуногенны, чем антитела, не принадлежащие человеку, или химерные антитела. Такие антитела можно продуцировать, используя трансгенных мышей, которые, в основном, не способны экспрессировать гены эндогенного иммуноглобулина с тяжелой и легкой цепями, но которые могут экспрессировать гены человека с тяжелой и легкой цепями. Трансгенных мышей иммунизируют обычным образом при помощи выбранного антигена, например при помощи CRF2R или его части. Моноклональные антитела, направленные против антигена, получают с применением общепринятой гибридомной технологии от таких иммунизированных трансгенных мышей. Данная технология подробно описана в патентах США №№ 5874299; 5877397; 5569825; 5661016; 5770429 и 6075181, все из которых включены здесь в виде ссылок. В качестве альтернативы получения иммуноглобулинов человека напрямую из культуры гибридомных клеток гибридомные клетки можно применять как источник реаранжированных локусов тяжелой цепи и легкой цепи для последующей экспрессии или генетической манипуляции. Выделение генов из таких клеток, продуцирующих антитела, правильно, так как дает возможность получить высокие уровни соответствующих мРНК. Восстановленными перестроенными локусами можно манипулировать, как требуется. Например, константную область можно отщепить или обменять, для чего можно соединить различные изотипы или вариабельные области для кодирования Fv-областей одиночной цепи. Такие методики описаны в публикациях WO 96/33735 и WO 96/34096, которые включены здесь в виде ссылок.
V. Отбор тестируемых соединений
Соединения, которые можно подвергнуть скринингу в соответствии с анализом согласно изобретению, включают без ограничения библиотеки известных соединений, включая природные продукты, такие как растительные или животные экстракты, синтетические химические вещества, биологически активные вещества, включая белки, пептиды, такие как растворимые пептиды, включая без ограничения представителей библиотек случайных пептидов и библиотеки молекул, получаемых при помощи комбинаторной химии, аминокислот D- или L-конфигурации, фосфопептиды (включая без ограничения представителей библиотек случайных или частично вырожденных, прямых фосфопептидов), антитела (включая без ограничения поликлональные, моноклональные, химерные, человеческие, антиидиотипические или одноцепочечные антитела, Fab, F(ab′)2, и библиотечные экспрессируемые Fab-фрагменты, и эпитопы, соединяющие фрагменты указанного), органические и неорганические молекулы.
В дополнение к более традиционным источникам тестируемых соединений компьютерное моделирование и технологии поиска разрешают рациональный отбор тестируемых соединений с применением структурной информации о лиганде, связывающимся с сайтом CRFR, или о уже идентифицированных агонистах CRFR. Такой рациональный отбор тестируемых соединений может уменьшать число тестируемых соединений, которые должны быть отобраны для того, чтобы идентифицировать терапевтическое соединение-кандидат. Рецепторы CRF представляют собой рецепторы GPCR, и, таким образом, знание белковой последовательности CRFR дает возможность создавать модель соединения, образующего связь с сайтом, которое можно применять для скрининга потенциальных лигандов. Такой процесс можно осуществить несколькими способами, известными в данной области. Кратко говоря, наиболее надежный подход включает создание последовательности, выровненной в соответствии с CRFR-последовательностью на матрице (полученной из бактерио-родопсина или кристаллических структур родопсина, или другой GPCR-модели), преобразование аминокислотных структур и усовершенствование модели при помощи молекулярной механики и визуальной экспертизы. Если строгое выравнивание последовательности получить нельзя, то можно создать модель путем построения моделей гидрофобных спиралей. Затем спирали подгоняются друг к другу путем вращения и перемещения каждой спирали относительно других, исходя из общей схемы известных структур родопсина. Можно также применять данные о мутациях, которые относятся к контактам остаток-остаток, для установки спиралей относительно друг друга так, чтобы достичь их контактирования. В процессе этого можно также встраивать известные лиганды в полость сайта связывания в спиралях, чтобы помочь сориентировать спирали через взаимодействия, которые стабилизировали бы связывание лиганда. Построение модели можно завершить усовершенствованием, применяя молекулярную механику и выстраивая внутриклеточные и внеклеточные петли при помощи стандартных технологий моделирования гомологии. Общую информацию относительно структуры GPCR и моделирования можно найти в следующих публикациях: Schoneberg T. и др., Molecular and Cellular Endocrinology 151:181-193 (1999), Flower D., Biochimica и Biophysica Acta 1422:207-234 (1999) и Sexton P.M., Current Opinion in Drug Discovery and Development, 2(5):440-448 (1999).
Как только модель закончена, ее можно применять в сочетании с одной из нескольких существующих компьютерных программ, чтобы уменьшить количество соединений, которые нужно отобрать при помощи способов скрининга, соответственно настоящему изобретению. Наиболее общей из них является DOCK-программа (UCSF Molecular Disign Institute, 533 Parnassus Ave, U-64, Box 0446, Сан-Франциско, Калифорния 94143-0446). В некоторых из ее вариантов она может проводить скрининг баз данных коммерческих и/или патентованных соединений для стерической подгонки и обрабатывать начерно электростатическую составляющую для связывающего сайта. Часто обнаруживали, что молекулы, которые хорошо обсчитаны при помощи DOCK-программы, имеют больше шансов стать лигандами. Другая программа, которую можно применять, представляет собой FLEXX (Tripos Inc., 1699 South Hanley Rd., St. Louis, Missouri, 63144-2913 (www.tripos.com)). Данная программа существенно медленнее и обычно ограничивается исследованиями более маленьких баз данных соединений. Схема подсчета в рамках FLEXX более детализирована и обычно дается лучшая оценка способности к связыванию, чем в DOCK-программе. FLEXX лучше применять для подтверждения предложений DOCK-программы или для исследования библиотек соединений, которые созданы комбинаторно из известных лигандов или матриц.
VI. Скрининг-тест для идентификации соединений-кандидатов для регуляции массы или функции скелетных мышц
Открытие, что CRF2R играет роль в регуляции атрофии скелетных мышц, дает возможность различными способами скрининга одного или более тестируемых соединений идентифицировать соединения-кандидаты, которые непременно можно применять для профилактического или терапевтического лечения атрофии скелетных мышц. Данное изобретение обеспечивает способы скрининга тестируемых соединений по их способности образовывать связь с CRF2R, активировать CRF2R, пролонгировать или увеличивать агонист-индуцированную активацию CRF2R или активацию пути передачи сигнала CRF2R, или увеличивать экспрессию CRF2R или CRF-генов.
Поскольку CRF2R и CRF1R являются гомологичными белками, ожидается, что определенная часть агонистов CRF2R будет функционировать так же, как агонисты CRF1R. Как обсуждалось выше, активация CRF1R индуцирует активацию оси HPA и сопутствующую выработку кортикостероидов. В большинстве случаев, когда требуется увеличение мышечной массы или функции, нежелательно активировать ось HPA. Поэтому в дополнение к скринингу тестируемых соединений по их способности активировать CRF2R изобретение также обеспечивает применение CRF2R и CRF1R для отбора селективных агонистов CRF2R. При отборе соединения-кандидата, применимого для лечения острой или хронической мышечной атрофии, которая не связана с мышечной дистрофией, предпочтительно, чтобы соединения-кандидаты были селективными в отношении CRF2R. Предпочтительно, когда соединение-кандидат обладает селективностью в отношении CRF2R, 10-кратно превосходящей селективность в отношении CRF1R (то есть 10-кратно более активное в отношении CRF2R, чем в отношении CRF1R), более предпочтительно 100-кратное превосходство в селективности и наиболее предпочтительно 1000-кратное или более превосходство в селективности. Поскольку в опубликованных исследованиях показано преимущество кортикостероидной терапии для лечения мышечных дистрофий, может быть выгодно, если агонист CRF2R сохраняет некоторый уровень сродства к CRF1R, когда применяется для лечения мышечных дистрофий. Таким образом, для лечения мышечных дистрофий предпочтительно соединение с более низкой селективностью, которое активирует CRF2R так же, как CRF1R в диапазоне похожих концентраций. Предпочтительным является соединение-кандидат с селективностью в отношении CRF2R, 100-кратно превышающей селективность в отношении CRF1R, более предпочтительно 10-кратно превышающей селективность и наиболее предпочтительно соединение, не селективное в отношении CRF2R по сравнению с CRF1R (то есть активность соединения-кандидата является, в основном, похожей для CRF2R и CRF1R). Также в данном случае может быть более предпочтительно, когда соединение является полным агонистом CRF2R и в то же время частичным агонистом CRF1R. Следовательно, для такого соединения-кандидата было бы свойственно ограничивать максимальную степень повышения кортизола и потенциально вызывать мышечную атрофию, в то время как антиатрофический эффект, опосредованный CRF2R, мог бы быть увеличен при увеличении дозы. Для специалиста в данной области легко определить, является ли соединение-кандидат полным или частичным агонистом CRF1R или CRF2R, при помощи способов, известных в данной области.
Для скрининга соединений, которые непременно будут применяться для регуляции массы или функции скелетных мышц при помощи CRF2R у человека, предпочтительно, чтобы начальный скрининг in vitro проводился с применением CRF2R с аминокислотной последовательностью, которая более чем на 80% идентична SEQ ID NO:10, и более предпочтительно, когда идентичность SEQ ID NO:10 составляет более чем 90%. Более предпочтительно, когда тестируемые соединения будут отобраны в отношении CRF2R человека, мыши или крысы, наиболее предпочтительно в отношении CRF2R человека. Для скрининга соединений, которые непременно будут применяться для регуляции массы или функции скелетных мышц с участием CRF2R у видов, не являющихся человеком, предпочтительно применять CRF2R того вида, для которого лечение предназначено.
Для скрининга по определению уровня активности, которым тестируемое соединение или соединение-кандидат обладает в отношении CRF1R, и по определению селективности, если она существует, которой соединение-кандидат обладает в отношении CRF2R по сравнению с CRF1R, предпочтительно, чтобы первоначальный отбор проводился при помощи CRF1R с аминокислотной последовательностью, которая более чем на 80% идентична SEQ ID NO:2, и более предпочтительно, когда идентичность SEQ ID NO:2 составляет более чем 90%. Более предпочтительно, когда тестируемые соединения будут отобраны в отношении CRF1R человека, мыши или крысы, наиболее предпочтительно в отношении CRF1R человека. Для скрининга соединений, которые непременно будут применяться для регуляции массы или функции скелетных мышц у видов, не являющихся человеком, предпочтительно применять CRF1R того вида, для которого лечение предназначено.
Способы согласно настоящему изобретению подходят для прикладных программ с высокой пропускной способностью; однако применение способа для немногих, например для одного тестируемого соединения, также включено в термин \скрининг\. Тестируемые соединения, которые связываются с CRF2R, активируют CRF2R, пролонгируют или увеличивают агонист-индуцированную активацию CRF2R или активацию передачи сигнала CRF2R, или увеличивают экспрессию генов CRF2R или CRF, что установлено при помощи способа согласно настоящему изобретению, упомянуты здесь как \соединения-кандидаты\. Такие соединения-кандидаты можно применять для регуляции массы или функции скелетных мышц. Однако более типично, когда такой первый уровень отбора in vitro обеспечивается способом, при помощи которого выбирают более узкий диапазон соединений, то есть соединений-кандидатов, которые заслуживают дальнейшего изучения на дополнительных уровнях скрининга. Для специалиста очевидно, что применимость настоящего изобретения заключается в идентификации из группы, состоящей из одного или более тестируемых соединений, субпопуляции соединений, которые заслуживают дальнейшего изучения. Для специалиста в данной области также очевидно, что тест настоящего изобретения применим для ранжирования вероятной пригодности определенного соединения-кандидата против других соединений-кандидатов. Например, соединение-кандидат, которое активирует CRF2R при 1000 нМ (но не активирует при 10 нМ), представляет меньший интерес, чем соединение, которое активирует CRF2R при 10 нМ. При помощи такой информации специалист может выбрать субпопуляцию соединений-кандидатов, идентифицированных на первоначальном уровне скрининга, для дальнейшего изучения. Между прочим, только в качестве примера, соединения, которые активируют CRF2R при концентрациях меньших, чем 200 нМ, можно было бы далее тестировать на животной модели атрофии скелетных мышц, принимая во внимание то, что соединения с более высоким порогом не будут далее тестироваться. Специалисту также очевидно, что в зависимости от того, каким образом выбрана группа тестируемых соединениий, и какие положительные стороны выбраны, только некоторая часть тестируемых соединений будет идентифицирована в качестве соединениий-кандидатов, и что такая часть может быть очень мала.
Системы тестов, описанные ниже, можно сделать в виде наборов, включающих CRF2R или клетки, экспрессирующие CRF2R, которые можно упаковать в различные контейнеры, например, пробирки, титрационные луночные микропланшеты, бутылки и т.п. Другие реактивы можно включить в отдельных контейнерах и обеспечить вместе с набором, например, положительные контрольные образцы, отрицательные контрольные образцы, буферы и питательные среды для клеточных культур.
В одном из примеров осуществления изобретение обеспечивает способ скрининга одного или более тестируемых соединений для идентификации соединений-кандидатов, которые связываются с CRF2R. Способы определения связывания соединения с рецептором известны в данной области. Как правило, тесты включают стадии инкубирования источника CRF2R с меченым соединением, о котором известно, что оно связывается с рецептором в присутствии или отсутствии тестируемого соединения, и определение количества связанного меченого соединения. Источником CRF2R могут быть либо клетки, экспрессирующие CRF2R, либо другая форма CRF2R, выделенного как здесь описано. Соединением с меткой может быть CRF или любой аналог CRF, помеченный таким образом, что это можно измерить, предпочтительно количественно (например, 125I-меченое, меченое европием, меченое флуоресцеином, GFP-меченое, меченное 35S-метионином). Такие способы внесения метки известны в данной области. Тестируемые соединения, которые связываются с CRFR, вызывают понижение количества меченого лиганда, связанного с рецептором, таким образом, понижая уровень сигнала, по сравнению с сигналом от контрольных образцов (отсутствие тестируемого соединения). Описаны варианты такой методики, в которых по связыванию рецептора в присутствии и отсутствии агентов, не связывающихся с G-белком, можно отличить агонисты от антагонистов (например, связывание в отсутствии и присутствии аналога нуклеотида гуанина, т.е. GpppNHp). См. Keen M., Radioligand Binding Methods for Membrane Preparations and Intact cell in Receptor Signal Transduction Protocols. R.A.J. Challis, (ed), Humana Press Inc., Totoway N.J. (1997).
Поскольку требуется различить соединения, которые специфично связываются с CRF2R по сравнению с CRF1R, следует провести тесты, описанные выше, применяя клетку или клеточную мембрану, которая экспрессирует только CRF2R, или можно провести тесты с источником рекомбинантного CRF2R. Клетки, экспрессирующие обе формы CRFR, можно модифицировать при помощи гомологичной рекомбинации, чтобы инактивировать или иначе сделать непригодным CRF1R-ген. Альтернативно, если источник CRFR содержит более чем один CRFR-тип, фоновый сигнал, продуцируемый рецептором, который не представляет интерес, следует вычесть из сигнала, полученного в тесте. Фоновый ответ можно определить рядом способов, включая устранение сигнала от CRFR, который не представляет интереса, при помощи антисмысловых, антительных или селективных антагонистов. Известные антагонисты CRFR включают анталармин (CRF1R-селективный), антисаувагин-30 (CRF2R-селективный) и астрессин (неселективный в отношении CRF1R/CRF2R).
В другом примере осуществления изобретение обеспечивает способы скрининга тестируемых соединений для идентификации соединений-кандидатов, которые активируют CRF2R и/или CRF1R. Как правило, тесты основаны на клетках; однако известны тесты в бесклеточных системах, при помощи которых возможно дифференцировать связывание агониста и антагониста, как описано выше. Тесты, основанные на клетках, включают стадии контактирования клеток, которые экспрессируют CRF1R или CRF2R, с тестируемым соединением или контрольным образцом и измерение активации CRFR путем измерения экспрессии или активности компонентов пути передачи сигнала CRFR.
Как описано выше в разделе \предшествующий уровень техники\, рецепторы CRFR, по-видимому, связываются друг с другом посредством различных путей, включая Gαs, Gαq или Gαi, в зависимости от типа клеток. Полагают, что активация агонистом CRFR позволяет рецептору передать сигнал при помощи любого из таких путей, при условии, что необходимые компоненты пути присутствуют в конкретном типе клеток. Таким образом, для скрининга соединений на предмет активации CRFR можно использовать в качестве показателя оценку любых путей передачи сигнала, даже если обрабатываемые in vivo клетки связываются с CRFR, опосредующим атрофию скелетных мышц, другим путем. Для специалиста в данной области очевидно, что скрининг-тест будет эффективен для идентификации приемлемых агонистов CRFR независимо от пути передачи, при помощи которого измеряется активация рецептора. Тесты для измерения активации таких сигнальных путей известны в данной области.
Например, после контактирования с тестируемым соединением можно получить лизаты клеток и провести тест индукции цАМФ. цАМФ индуцируется в ответ на активацию Gαs. Поскольку Gαs активируется другими рецепторами, чем CRFR, и поскольку тестируемое соединение может усиливать его влияние через рецепторы CRF или при помощи другого механизма, уместно провести два контрольных сравнения для определения, повышает ли тестируемое соединение уровни цАМФ через активацию CRFR. В одном контрольном тесте сравнивается уровень цАМФ клеток, контактировавших с тестируемым соединением, с уровнем цАМФ клеток, контактировавших с контрольным соединением (то есть с носителем, в котором тестируемое соединение растворено). Если тестируемое соединение увеличивает уровни цАМФ по сравнению с контрольным соединением, это указывает на то, что тестируемое соединение увеличивает цАМФ при помощи некоторого механизма. В другом контрольном тесте сравниваются уровни цАМФ клеточной линии, экспрессирующей CRFR, и клеточной линии, которая является по существу той же самой, за исключением того, что она не экспрессирует CRFR после обработки обеих клеточных линий тестируемым соединением. Если тестируемое соединение повышает уровни цАМФ в клеточной линии, экспрессирующей CRFR, по сравнению с клеточной линией, которая не экспрессирует CRFR, это указывает на то, что тестируемое соединение повышает цАМФ через активацию рецепторов CRF.
В одном из конкретных примеров осуществления изобретения измеряется индукция цАМФ с применением конструкций ДНК, содержащих элемент, чувствительный к цАМФ, связанный с любым из ряда репортерных генов, который можно ввести в клетки, экспрессирующие CRFR. Такие репортерные гены включают без ограничения хлорамфениколацетилтрансферазу (САТ), люциферазу, глюкуронидсинтетазу, гормон роста, флуоресцентные белки (например, зеленый флуоресцентный протеин) или щелочную фосфатазу. После экспозиции клеток с тестируемым соединением можно количественно определить уровень экспрессии репортерного гена, чтобы определить способность тестируемого соединения(ний) увеличивать уровни цАМФ, и таким образом определить способность тестируемых соединений активировать CRFR.
Клетки, применяемые в таком тесте, являются теми же самыми, что для теста по связыванию CRFR, описанного выше, за исключением того, что клетки, применяемые в тестах активации предпочтительно экспрессируют функциональный рецептор, который дает статистически значимый ответ на CRF или на один, или более аналогов CRF. Помимо применения клеток, экспрессирующих полноразмерные CRFR, можно сконструировать клетки, которые экспрессируют CRFR, содержащие лиганд-связывающий домен рецептора, сопряженного или физически модифицированного для включения в него репортерных элементов или для взаимодействия с сигнальными белками. Например, CRFR дикого типа или фрагмент CRFR можно слить с G-белком, в результате чего происходит активация гибридного G-белка при связывании агониста, с CRFR-частью гибридного белка (Siefert R. и др., Trends Pharmacol. Sci. 20: 383-389 (1999)). Также предпочтительно, чтобы клетки обладали рядом характеристик, в зависимости от показания, для максимизации индуктивного ответа при помощи CRF или аналога CRF, например, для обнаружения сильной индукции репортерного CRE-гена; должны быть выгодны: (a) низкий естественный уровень цАМФ; (b) G-белки, способные взаимодействовать с CRFR; (c) высокий уровень аденилилциклазы; (d) высокий уровень протеинкиназы A; (е) низкий уровень фосфодиэстераз и (f) высокий уровень белка, связывающегося с элементом ответа цАМФ. Для увеличения ответа на CRF или аналог CRF, клетки-хозяева можно сконструировать так, чтобы они экспрессировали большее количество благоприятных факторов или меньшее количество неблагоприятных факторов. Кроме того, альтернативные пути индукции репортерного CRE-гена можно было бы удалить для понижения базовых уровней.
В некоторых примерах ответы рецепторов, сопряженных с G-белком, убывают или становятся десенсибилизированными после продолжительной экспозиции с агонистом. В другом примере осуществления изобретения обеспечиваются способы идентификации соединений, которые пролонгируют или увеличивают агонист-индуцированную активацию CRF2R или активацию пути передачи сигнала CRF2R, по ответу на агонист CRF2R. Такие соединения можно применять, например, для лечения атрофии скелетных мышц в сочетании с агонистом CRF2R. Обычно при таком способе применяется тест, основанный на клетках, и включающий в любом порядке или одновременно (i) взаимодействие клеток с тестируемым соединением; (ii) обработку клеток, экспрессирующих функциональный CRF2R, агонистом CRF2R при концентрации агониста и периоде экспозиции агонист-рецептор, достаточных для десенсибилизации рецептора; последующее (iii) определение уровня активации CRF2R. Для специалиста в данной области очевидно, что некоторые механизмы способствуют десенсибилизации рецептора, включая без ограничения фосфорилирование рецептора, интернализацию рецептора или деградацию и пониженную модуляцию пути передачи сигнала CRFR. Специалист в данной области может определить подходящее время (то есть до, в течение или после обработки агонистом) для контактирования клеток с тестируемыми соединениями, в зависимости от которого механизм десенсибилизации достигает цели. Например, взаимодействие клеток с тестируемыми соединениями после обработки агонистом может обнаружить тестируемые соединения, которые блокируют десенсибилизацию рецептора, происходящую в результате фосфорилирования рецептора.
В другом примере осуществления изобретение обеспечивает способ скрининга одного или более тестируемых соединений для идентификации соединений-кандидатов, которые регулируют транскрипцию CRF2R-гена или регулируют экспрессию CRF2R. Соединения-кандидаты, которые регулируют активность транскрипции CRFR-генов, можно идентифицировать, применяя репортерный ген, оперативно связанный с регуляторной областью CRF2R (репортерная генная конструкция). Такие способы известны в данной области. В одном из таких способов репортерная генная конструкция контактирует с тестируемым соединением в присутствии источника клеточных факторов, и определяется уровень экспрессии репортерного гена. Тестируемое соединение, которое вызывает увеличение уровня экспрессии, сравнимое с контрольным образцом, указывает на соединение-кандидат, которое увеличивает транскрипцию CRF2R-гена. Чтобы обеспечить клеточные факторы, требуемые для транскрипции in vitro или in vivo, подходящие клетки или клеточные экстракты получают из любого типа клеток, которые нормально экспрессируют CRF2R.
Соединения-кандидаты, регулирующие экспрессию CRF2R, можно также идентифицировать при помощи способа, при котором клетка контактирует с тестируемым соединением, и определяется экспрессия CRFR. Уровень экспрессии CRF2R в присутствии тестируемого соединения сравнивают с уровнем экспрессии при отсутствии тестируемого соединения. Тестируемые соединения, которые увеличивают экспрессию CRF2R, идентифицируют в качестве соединений-кандидатов для увеличения мышечной массы или мышечной функции. При помощи такого способа обнаруживают соединения-кандидаты, которые увеличивают транскрипцию или трансляцию CRF2R, или которые увеличивают стабильность мРНК или CRF2R-белка.
В другом примере осуществления данное изобретение обеспечивает способы скрининга одного или более тестируемых соединений для идентификации соединений-кандидатов, которые регулируют экспрессию CRF или аналога CRF. Такие тесты проводили, по существу, как описано выше для тестов по идентификации соединений-кандидатов, которые регулируют экспрессию CRFR с последующими модификациями. Для идентификации соединения-кандидата, которое регулирует транскрипцию от CRF-гена или от аналога CRF-гена, репортерный ген оперативно связывается с регуляторной областью CRF-гена или аналога CRF-гена, представляющего интерес, а источником клеточных факторов должен быть тип клеток, которые экспрессируют ген, представляющий интерес.
VII. Скрининг соединений-кандидатов с применением моделей атрофии скелетных мышц
Соединения-кандидаты, выбранные из одного или более тестируемых соединений в тесте in vitro, как описано выше, можно затем протестировать в отношении их способности регулировать массу или функцию скелетных мышц в модельных системах атрофии скелетных мышц и/или гипертрофии. Такие модели атрофии скелетных мышц или гипертрофии включают как модели атрофии скелетных мышц на клеточных культурах in vitro, так и животные модели in vivo. Такие дополнительные уровни скрининга применяются для дальнейшего сужения диапазона соединений-кандидатов, которые заслуживают дополнительного изучения, например, в клинических испытаниях.
Модели мышечной атрофии на клеточных культурах
В данной области известны модели атрофии скелетных мышц in vitro. Такие модели описаны, например, в публикациях: Vandenburgh H.H., In Vitro 24:609-619(1988), Vandenburgh H.H. и др., J. of Biomechanics, 24 Suppl 1:91-99(1991), Vandenburgh H.H и др., In Vitro Cell. Dev. Biol., 24(3):166-174(1988), Chromiak J.A. и др., In Vitro Cell. Dev. Biol. Anim., 34(9):694-703(1998), Shansky J. и др., In Vitro Cell. Dev. Biol. Anim., 33(9):659-661(1997), Perrone C.E. и др., J. Biol. Chem. 270(5):2099-2106 (1995), Chromiac J.A. и Vandenburgh H.H., J. Cell. Physiol. 159(3):407-414(1994) и Vandenburgh H.H. и Karlisch P., In Vitro Cell. Dev. Biol. 25(7):607-616 (1989). Такие модели применимы, но не являются обязательными, вслед за скринингом in vitro, описанным выше, для того, чтобы еще более сузить диапазон соединений-кандидатов, которые заслуживают тестирования на животных моделях. Модели клеточных культур обрабатывают при помощи соединений-кандидатов и измеряют ответ модели на обработку, оценивая изменения в мышечных показателях, таких как: синтез мышечного белка или деградация, изменения в массе скелетных мышц или сократительная функция. Для тех соединений, которые стимулируют значительные изменения в мышечных показателях, затем, как правило, проводят скрининг на животной модели атрофии скелетных мышц.
Животные модели атрофии скелетных мышц
Соединения-кандидаты вводят животным, которые не являются человеком, и контролируют ответ животных, например, оценивая изменения в показателях атрофии или гипертрофии, таких как: масса скелетных мышц, функция скелетных мышц, площадь поперечного сечения мышцы или миоволокна, содержание сократительных белков, содержание несократительных белков или биохимического или генетического маркера, который коррелирует с изменениями массы или функции скелетных мышц. Соединения-кандидаты, которые индуцируют гипертрофию скелетных мышц или предотвращают любой аспект атрофии скелетных мышц, должны рассматриваться как предполагаемые кандидаты для терапевтического лечения атрофии скелетных мышц у человека, и упоминаются здесь как терапевтические соединения-кандидаты. В дополнение к оценке способности соединения-кандидата регулировать атрофию скелетных мышц при таком скрининге можно также обнаружить нежелательные побочные эффекты, такие как токсичность. Отсутствие недопустимо высоких уровней побочных эффектов можно применять в качестве еще одного критерия для отбора терапевтических соединений-кандидатов.
В данной области известны различные животные модели атрофии скелетных мышц, такие как те, что описаны в следующих публикациях: Herbison G.J. и др., Arch. Phys. Med. Rehabil. 60:401-404 (1979), Appell H-J. Sports Medicine 10:42-58 (1990), Hasselgren P-0. и Fischer J.E. World J. Surg. 22:203-208 (1998), Agbenyega E.T. и Wareham A.C., Comp. Biochem. Physiol. 102A:141-145 (1992), Thomason D.B. и Booth F.W. J. Appl. Physiol. 68:1-12 (1990), Fitts R.H. и др., J. Appl. Physiol 60:1946-1953 (1986), Bramanti P. и др., Int.J. Anat. Embryol. 103:45-64 (1998), Cartee G.D. J. Gerontol. A Biol. Sci. Med. Sci. 50:137-141 (1995), Cork L.C. и др., Prog. Clin. Biol. Res. 229:241-269 (1987), Booth F.W. и Gollnick P.D. Med. Sci. Sports Exerc. 15:415-420 (1983), Bloomfield S.A. Med. Sci. Sports Exerc. 29:197-206 (1997). Предпочтительно животными для таких моделей являются мыши и крысы. Такие модели включают, например, модели атрофии, индуцированной бездействием, такие как наложение гипсовой повязки или фиксирование конечностей другим образом, прекращение работы задней конечности, общее обездвижение животного и ситуации с пониженной силой тяготения. Модели атрофии, индуцированной повреждением нерва, включают, например, уничтожение нерва, удаление частей нервов, которые иннервируют определенные мышцы, применение токсина для нервов и инфицирование нервов при помощи вирусных, бактериальных или эукариотических возбудителей инфекции. Модели атрофии, индуцированной глюкокортикоидами, включают применение индуцирующих атрофию доз экзогенного глюкокортикоида для животных и стимуляцию выработки эндогенных кортикостероидов, например, при помощи применения гормонов, которые активируют ось гипоталамус-гипофиз-надпочечники (ось HPA). Модели атрофии, индуцированной сепсисом, включают, например, инокуляцию организмов, индуцирующих сепсис, таких как бактерии; обработку животного иммуноактивирующими соединениями, такими как экстракт бактериальных клеточных стенок или эндотоксин; и прокол кишечных стенок. Модели атрофии, индуцированной кахексией, включают, например, инокуляцию животного онкогенными клетками, потенциально способными к формированию кахексии, инфицирование животного возбудителями инфекции (такими как вирусы, вызывающие СПИД), которые приводят к кахексии, и обработку (лечение) животного гормонами или цитокинами, такими как CNTF, TNF, IL-6, IL-1 и т.д., которые индуцируют кахексию. Модели атрофии, индуцированной остановкой сердца, включают такое обращение с животным, чтобы произошла остановка сердца с сопутствующей атрофией скелетных мышц. Модели атрофии, индуцированной нейродегенеративными заболеваниями, включают аутоиммунные животные модели, типа тех, которые являются следствием иммунизации животного нейронными компонентами. Модели атрофии, индуцированной мышечной дистрофией, включают естественные или созданные руками человека модели генетически-индуцированной мышечной дистрофии, такой как мутация гена дистрофина, которая имеет место у Mdx-мыши.
Животные модели гипертрофии скелетных мышц включают, например, использование мышц конечности, увеличенных благодаря инактивации другой конечности, повторную нагрузку после явления атрофии, индуцированной бездействием, повторное использование мышцы, которая была атрофирована из-за кратковременного повреждения нерва, использование отдельных мышц, увеличенных благодаря инактивации синергической мышцы (например, компенсационная гипертрофия), использование мышцы, увеличенной при повышенной нагрузке, прикладываемой к ней, и гипертрофию вследствие удаления глюкокортикоида после атрофии, индуцированной глюкокортикоидами. Предпочтительные животные модели атрофии включают модель атрофии при денервации седалищного нерва, модель атрофии, индуцированной глюкокортикоидами, и модель атрофии, вызванной бездействием загипсованной конечности, которые подробно описаны ниже.
Модель атрофии, вызванной денервацией седалищного нерва, включает проведение анестезии животного с последующим хирургическим удалением короткого сегмента либо правого, либо левого седалищного нерва, например, у мышей седалищный нерв выделяют приблизительно в средней точке по длине бедренной кости и удаляют сегмент размером 3-5 мм. Это денервирует мускулатуру голени задней конечности, приводя к атрофии таких мышц. Как правило, иннервация двуглавой мышцы бедра слева не нарушается, чтобы обеспечить удовлетворительную подвижность колена для нормального, по сути, передвижения. Как правило, у не подвергавшихся лечению животных мышечная масса денервированных мышц уменьшается на 30-50% на десятый день после денервации. После денервации вводили тестируемые соединения, например, при помощи инъекции или непрерывной инфузии, например, через имплантированный осмотический мининасос (например, Alzet, Palo Alto, СА), определяли их влияние на атрофию скелетных мышц, индуцированную при денервации. Животных забивали после различных отрезков времени, прошедших после денервации, и быстро отделяли мышцы голени, как от денервированных, так и от неденервированных конечностей; мышцы, очищенные от сухожилий и соединительной ткани, взвешивали. Степень атрофии поврежденных мышц анализировали, например, путем измерения мышечной массы, площади поперечного сечения мышцы, площади поперечного сечения миоволокна, содержания сократительных белков.
Модель атрофии, индуцированной глюкокортикоидами, включает введение глюкокортикоида тестируемому животному, например 1,2 мг/кг/сутки дексаметазона с питьевой водой. Как правило, у животных, не подвергавшихся лечению, масса скелетных мышц уменьшается на 30-50% на десятый день после введения дексаметазона. Одновременно с введением глюкокортикоида или после вводили тестируемые соединения, например, при помощи инъекции или непрерывной инфузии, чтобы определить их влияние на атрофию скелетных мышц, индуцированную глюкокортикоидом. Анализировали степень атрофии в поврежденных мышцах, как описано выше для модели денервации, для различных отрезков времени, прошедших после введения глюкокортикоида.
Модель атрофии, вызванной бездействием загипсованной конечности, включает наложение гипсовой повязки на одну заднюю конечность животного от колена вниз до стопы. Как правило, мышечная масса уменьшается на 20-40% на десятый день после наложения гипсовой повязки. После наложения гипсовой повязки вводили тестируемые соединения при помощи инъекции или непрерывной инфузии через имплантированный осмотический мининасос (например, Alzet, Palo Alto, СА), чтобы определить их влияние на атрофию скелетных мышц, индуцированную при помощи гипсования конечности. Для различных отрезков времени, прошедших после гипсования конечности, анализировали степень атрофии поврежденных мышц, как описано выше для модели денервации.
Для специалиста в данной области очевидно, что поскольку имеются различия между CRF2R человека и CRF2R других видов животных, при скрининге соединений, применяемых для человека, могут иметься некоторые ложно положительные или отрицательные результаты, которые возникают, когда проводят скрининг с применением CRF2R, не принадлежащего человеку. Таким образом, предпочтительно первоначально делать скрининг in vitro, применяя CRF2R человека. При некоторых обстоятельствах идентифицированные соединения-кандидаты могут быть активны только в отношении рецептора человека, а не в отношении рецептора, не принадлежащего человеку. При таких обстоятельствах, тем не менее, может быть желательно определить, способны ли такие соединения-кандидаты регулировать массу или функцию скелетных мышц на втором уровне скрининга. Поскольку такие кандидаты не активируют CRF2R, не принадлежащий человеку, стандартный скрининг in vivo на животном, не являющимся человеком, проводить не рекомендуется. При таких обстоятельствах второй уровень скрининга для таких кандидатов можно выполнить на трансгенных животных, которые экспрессируют CRFR человека.
Животных любого вида, особенно млекопитающих, включая без ограничения мышей, крыс, кроликов, морских свинок, свиней, коз, собак и приматов, не являющихся человеком, можно применять для создания CRFR-трансгенных животных. Предпочтительны мыши и крысы, более всего предпочтительны мыши. В данной области известен ряд методик, которые можно применять для введения CRFR-трансгенов человека животным, чтобы продуцировать базовые линии трансгенных животных. Такие методики включают без ограничения пронуклеарную микроинъекцию, передачу гена посредством ретровируса в линии зародышевых клеток, целевое помещение гена в эмбриональные стволовые клетки, электропорацию эмбрионов и передачу гена посредством сперматозоида.
VIII. Способы генной терапии для лечения атрофии скелетных мышц
Общую активность CRF2R можно увеличить путем сверхэкспрессии гена CRF2R (для увеличения экспрессии CRF2R) или конститутивно активного CRF2R в соответствующей ткани. Уровни CRF можно увеличить in vivo путем подобной сверхэкспрессии CRF-гена. Сверхэкспрессия таких генов увеличит общую клеточную активность CRF2R, таким образом регулируя атрофию скелетных мышц. Ген или гены, представляющие интерес, встраиваются в вектор, подходящий для экспрессии у субъекта. Такие векторы включают без ограничения аденовирус, аденовирус, ассоциированный с вирусом, ретровирус и векторы вируса герпеса в дополнение к другим частицам, которые вводятся при помощи ДНК в клетки (например, липосомы, частицы золота и т.д.), или путем непосредственной инъекции экспрессионного вектора ДНК, содержащего ген, представляющий интерес, в ткань человека (например, мышечную).
IX. Методики приготовления лекарственного средства и способы применения
Соединения-кандидаты или терапевтические соединения-кандидаты, идентифицированные при помощи способов скрининга, описанных здесь, можно вводить пациентам для лечения атрофии скелетных мышц или для индуцирования гипертрофии скелетных мышц. С такой целью настоящее изобретение охватывает способы и композиции для модуляции атрофии скелетных мышц, включая без ограничения атрофию скелетных мышц, индуцированную бездействием, обусловленным операцией, постельным режимом, переломом костей; индуцированную повреждением нерва/денервацией, обусловленным травмой спинного мозга, аутоиммунным заболеванием, инфекционным заболеванием; атрофию, индуцированную применением глюкокортикоидов для состояний, не связанных с гормональным лечением; сепсисом, обусловленным инфекцией или другими причинами; ограничением питательных веществ, обусловленным болезнью или голоданием; атрофию, индуцированную раковой кахексией; хроническим воспалением; кахексией при СПИДе; COPD; застойной сердечной недостаточностью; саркопенией и генетическими нарушениями; например, мышечными дистрофиями, нейродегенеративными заболеваниями. Агонисты CRF2R можно применять для ингибирования атрофии скелетных мышц. Нет необходимости в том, чтобы эффективные соединения обладали абсолютной специфичностью в отношении CRFR. Предполагается, что вместе с эффективным, но неспецифичным агонистом можно вводить специфичный антагонист других пораженных болезнью рецепторов. С другой стороны, такое отсутствие специфичности может быть исправлено при помощи только модулирования дозы или режима дозирования.
Соединения-кандидаты или терапевтические соединения-кандидаты, идентифицированные при помощи способов скрининга согласно настоящему изобретению, можно вводить в сочетании с соединениями, которые пролонгируют или увеличивают активацию CRF2R или активацию пути передачи сигнала CRF2R. Ими могут быть известные соединения, например теофиллин, или такие соединения можно идентифицировать при помощи способов скрининга согласно данному изобретению для пролонгирования или увеличения активации рецептора CRF2R или пути передачи сигнала CRF2R.
Определения дозы
Безопасную и терапевтическую эффективность соединений, которые являются агонистами CRFR, можно определить при помощи стандартных методик, применяя методики либо in vitro, либо in vivo. Соединения, которые обладают большими терапевтическими индексами, предпочтительны, хотя применимы и соединения с более низкими терапевтическими индексами, если приемлем уровень побочных эффектов. Данные, полученные при помощи токсикологических и фармакологических методик in vitro и in vivo, можно применять для определения диапазона доз, которые можно применять для человека. Предпочтительная доза находится в диапазоне, при котором циркулирующая (в крови) концентрация соединения максимальна терапевтически с приемлемым уровнем безопасности. Циркулирующая концентрация соединения может меняться в зависимости от формы дозы, времени после введения дозы, пути введения и т.д. Дозы вне данного диапазона также применимы, если обеспечен приемлемый уровень побочных эффектов. Такие вопросы для обсуждения, как возраст и вес пациента и т.п., можно решать общепринятым способом. Фармакогенетические подходы могут применяться при отборе оптимальных соединений, доз и режима дозирования на клинических популяциях.
Методика приготовления лекарственного средства и применение
Фармацевтические композиции, применимые для модулирования атрофии скелетных мышц в соответствии с настоящим изобретением, можно приготовить при помощи общепринятых методик с применением фармацевтически приемлемых носителей и наполнителей. Композиции в соответствии с данным изобретением предпочтительно обеспечиваются в виде дозированной лекарственной формы. Как здесь принято, \дозированная лекарственная форма\ означает композицию согласно данному изобретению, содержащую агонист CRF2R в количестве, которое подходит для введения животному, предпочтительно млекопитающему, более предпочтительно человеку, в разовой дозе, в соответствии с хорошей медицинской практикой. Фармацевтические композиции можно приготовить для доставки, например, интраназальным способом, подкожно, путем ингаляции, для парэнтерального, кожного, орального или ректального введения. Для орального введения фармацевтической композиции можно придать форму таблеток или капсул, содержащих фармакологически активное соединение и вспомогательные вещества, включая без ограничения связующие агенты, наполнители, смазывающие вещества, дезинтеграторы или смачивающие агенты. Таблетки могут быть с покрытием. Жидкие препараты для орального введения включают без ограничения сиропы, суспензии или сухие продукты, которые повторно суспендируются с жидким носителем перед применением, содержащие фармакологически активное соединение и вспомогательные вещества, включая без ограничения суспендирующие агенты, эмульгаторы, неводные носители, консерванты, буферные соли, агенты, придающие вкус, окраску, подсластители и т.д. Фармацевтические композиции для орального введения можно приготовить для контролируемого высвобождения фармакологически активных соединений либо во рту, либо в желудочно-кишечном тракте.
Для введения путем ингаляции соединения для применения согласно настоящему изобретению, можно выпускать без ограничения в следующих формах: в жидком виде, в виде порошка, геля или в форме аэрозольного спрея, применяя пропелленты либо под давлением, либо при нормальном давлении, либо в предварительно измеренных, либо предварительно неизмеренных дозах. Фармакологически активное соединение можно приготовить с соответствующими наполнителями, носителями, консервантами, буферами и т.д. Для парэнтерального введения фармакологически активное соединение можно приготовить с приемлемыми физиологическими носителями, консервантами и т.д. и приготовить в виде суспензий, растворов, эмульсии, порошков, готовых к приготовлению, и т.д. либо для инъекции ударной дозы вещества, либо для инфузии. Дозы таких соединений можно вводить при помощи различных способов, включая иглы для подкожных инъекций, устройства высокого давления и т.д. Для ректального введения фармакологически активное соединение можно приготовить с приемлемыми физиологическими носителями, консервантами и т.д. для введения в виде свечей, клизм и т.д. Для введения через кожу фармакологически активное соединение можно приготовить с приемлемыми физиологическими носителями, включая лосьоны, смягчающие вещества и т.д., или встроить в приспособления типа кусочков пластыря. Для продолжительного введения фармакологически активное соединение и соответствующие вспомогательные вещества, такие как, но ограниченно, полимеры, гидрофобные материалы, смолы и т.д., можно приготовить в виде базового препарата либо для инъекции, либо для имплантации на многочисленных участках, включая без ограничения внутримышечную и подкожную локализации. Кроме того, фармакологически активное соединение можно вводить при помощи распределяющего устройства.
Мониторинг эффектов в время клинических испытаний
Мониторинг влияния соединений (например, в виде лекарств) на экспрессию или активность CRF2R можно применять не только при основном скрининге лекарственного средства, но также и при клинических испытаниях. Например, эффективность соединения для увеличения активности рецептора CRF2R или для экспрессии рецептора CRF2R, установленную при скрининге, можно оценить при клинических испытаниях на пациентах с атрофией скелетных мышц или относящихся к группе риска. Влияние соединения на пациента при различных интервалах времени, прошедших после введения тестируемого соединения или плацебо, можно определить, например, путем наблюдения изменения массы скелетных мышц, функции скелетных мышц, биохимических маркеров мышечной деградации или критериев качества жизни. Способы измерения массы скелетных мышц у человека известны в данной области и включают, например: измерение периметра конечности; измерение плотности мышцы, например, при помощи компьютерной томографии, MRI или ультразвука; или биопсию мышцы для исследования морфологических и биохимических параметров (например, площади поперечного сечения волокна, диаметра волокна или ферментативной активности). Кроме того, поскольку масса скелетных мышц коррелирует с функцией скелетных мышц, мышечную функцию можно применять в качестве суррогатного маркера массы, а изменения мышечной массы можно оценить путем функциональных измерений, например, мышечной силы, силы группы мышц-синергистов или сократительных характеристик, устанавливаемых при электромиографических записях. Кроме того, потерю мышечного белка в результате атрофии мышцы можно измерить количественно по уровням аминокислот или производных аминокислот, то есть 3-метилгистидина, в моче или крови субъекта. Обзор таких способов приведен в публикации: Appell, Sports Med. 10:42-58(1990). Критерии качества жизни включают без ограничения легкость вставания со стула, число шагов, сделанных до того, как утомиться, или способность подниматься по лестнице.
ПРИМЕРЫ
Пример 1. Конструирование векторов экспрессии рецептора CRF2R человека.
Нашли последовательность ДНК CRF2R человека (hCRF2R)с регистрационным номером E12752 и синтензировали два олигонуклеотида, включая один, содержащий 5′-конец гена, начинающегося в инициирующем кодоне (5′-олигонуклеотид), и один, содержащий 3′-конец гена, обладающего терминирующим кодоном (3′-олигонуклеотид). Данные олигонуклеотиды сконструированы для того, чтобы содержать сайты рестрикции, которых нет в hCRF2R-гене, с одним уникальным сайтом в 5′-олигонуклеотиде, и отличающийся уникальный сайт рестрикции в 3′-олигонуклеотиде. Кроме того, 3′-олигонуклеотид содержит сигнальную последовательность дополнительного полиаденилирования. Двойная цепь кДНК скелетных мышц человека приобретена из универсальной коллекции кДНК QUICK-Clone (Clonetech Inc., Palo Alto, СА, США). Применяя вышеупомянутые 5′- и 3′-олигонуклеотиды, кДНК hCRF2R проводили амплификацию кДНК скелетных мышц человека по ПЦР-методике, используя набор AdvanTaq PCR (Clonetech Inc., Palo Alto, СА, США). Продукт ПЦР hCRF2R-гена очищали от ПЦР-артефактов электрофорезом в агарозном геле, а фрагмент ДНК hCRF2R-гена очищали от агарозного геля, применяя очищающий продукт, такой как NucleoTrap (Clonetech Inc.,Palo Alto,СА, США). Клонирование продукта ПЦР hCRF2R в pIRESneo-вектор (Clonetech Inc., Palo Alto, СА, США) проводили сначала разрезанием продукта ПЦР hCRF2R и pIRESneo-вектора при помощи соответствующих рестрикционных эндонуклеаз так, чтобы 5′- и 3′-сайты рестрикции подготовить к лигированию. ДНК PIRESneo-вектора лигируется с ДНК продукта ПЦР hCRF2R с участием ДНК-лигазы, полученной от AdvantAgeTM PCR Cloning Kit (Clonetech Inc., Palo Alto,СА, США), согласно рекомендациям изготовителя. Затем лигированный вектор и конструкцию (pIRESneo/hCRF2R) применяли для трансформации TOP10F′ в компетентных клетках E.coli (Clonetech Inc.,Palo Alto,СА,США). Трансформированные клетки высевали на агар, содержащий LB/X-gal/IPTG плюс ампициллин. Белые колонии (положительные клоны) отбирали и отдельно культивировали в LB-среде. ДНК-плазмиду выделяли, применяя систему очистки ДНК NucleoBond DNA Purification System (Clonetech Inc.,Palo Alto,СА, США). Вставку, по меньшей мере, от одного клона секвенировали, чтобы гарантировать, что последовательность hCRF2R корректна. В клетки HEK293, содержащие стабильно интегрированную плазмиду Mercury CRE-LUC (Clonetech Inc., Palo Alto, СА, США), трансфицировали очищенную ДНК pIRESneo/hCRF2R, имеющую откорректированную вставку в последовательности, при помощи набора для трансфекции CalPhosТМ Mammalian Transfection Kit (Clonetech Inc., Palo Alto,СА, США). Клетки, стабильно трансфицированные ДНК pIRESneo/hCRF2R отбирали при культивировании клеток в G418. Стабильно трансфицированные клетки (клетки HEK293/CRE-LUC/pIRESneo/hCRF2R) размножали в среде DMEM (Life Technologies, Rockville, MD), содержащей 10% фетальной бычьей сыворотки (Clonetech Inc., Palo Alto, СА, США), раствор пенициллина/стрептомицина (Life Technologies, Rockville, MD), L-глутамин (Life Technologies, Rockville, MD) и не-незаменимую аминокислоту (Life Technologies, Rockville, MD) при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха. Клоны характеризовали как по связыванию CRF, так и по CRE-LUC-активации, после экспозиции с CRF, как описано в примере 2 и примере 3. Затем клетки, экспрессирующие рецептор hCRF2R на соответствующем уровне, и, соответственно, сопряженные с репортерной системой CRE-LUC, применяли для дальнейшего анализа.
Пример 2. Тесты на связывание с рецептором.
Анализ связывания соединений с рецептором проводили на целых клетках при посеве клеток HEK293/CRE-LUC/pIRESneo/hCRF2R из примера 1 в 96-луночные планшеты, покрытые полилизином. Проводили посев клеток в среде DMEM, содержащей 10% фетальной бычьей сыворотки, раствор пенициллина/стрептомицина, L-глутамин и не-незаменимую аминокислоту, при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха, и инкубировали в течение ночи. Культуральную среду удаляли и добавляли соответствующее количество CRF, меченого европием при образовании ковалентной связи (Eu-CRF), в среде MEM (Life Technologies, Rockville, MD)+10% Seablock (Clonetech Inc., Palo Alto, СА, США). Клетки инкубировали с Eu-CRF 90 минут при комнатной температуре, затем промывали 4 раза фосфатным буферным солевым раствором, не содержащим магния и кальция (Life Technologies, Rockville, MD). После заключительного промывания добавляли обогащенный раствор (Wallac Inc., Gaithersburg, MD) и планшет считывали на ридере планшетов Wallac plate reader (Wallac Inc., Gaithersburg, MD) с применением программы BioWorks Europian program. Для анализа насыщенности связывания к клеткам добавляли дозы Eu-CRF, ранжированные в пределах 10-12-10-3 М, и анализировали связывание как при отсутствии, так и при наличии концентрации насыщения непомеченного CRF для оценки неспецифичного связывания. Для конкурентного связывания добавляли Eu-CRF при концентрации, которая наполовину меньше максимальной, при условиях связывания, помимо различных концентраций соединения, представляющего интерес.
Пример 3. Тест активации рецепторов.
Анализ активации рецепторов выполняли путем посева клеток HEK293/CRE-LUC/pIRESneo/hCRF2R из примера 1 в 96-луночные планшеты Packard View Plate-96 (Packard Inc., СА). Клетки высевали в среде DMEM, содержащей 10% фетальной бычьей сыворотки, раствор пенициллина/стрептомицина, L-глутамин и не-незаменимую аминокислоту, при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха, и инкубировали в течение ночи. Затем среду удаляли и заменяли на DMEM (Life Technologies, Rockville, MD), содержащую 0,01% фракции V бычьего альбумина (SIGMA, St.Louis, MO), содержащую соединение, представляющее интерес. Затем клетки инкубировали в течение четырех часов при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха, после чего среду удаляли, а клетки дважды промывали солевым раствором Hanks Balanced Salt Solution (Life Technologies, Rockville, MD). Затем к промытым клеткам добавляли лизирующий реагент (Promega Inc., Madison, WI) и инкубировали клетки 20 минут при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха. Затем клетки помещали на 20 минут при -80°C, после чего 20 минут инкубировали при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха. После такой инкубации к клеточным лизатам добавляли буфер (Luciferase Assay Buffer) и субстрат люциферазы (Luciferase Assay Substrate) (Promega Inc., Madison, WI) и количественно измеряли активность люциферазы при помощи люминометра. Относительную активность соединения оценивали, сравнивая уровень люциферазы после экспозиции с соединением с уровнем люциферазы в HEK-клетках, которые содержат конструкцию CRE-LUC без hCRF2R. Специфичность ответа также проверяли, оценивая люцеферазный ответ hCRF2R/CRE-LUC HEK-клеток на соединение в присутствии и в отсутствии 10-кратного избытка антагониста hCRF2R.
Пример 4. Скрининг для идентификации соединений-кандидатов, которые пролонгируют или увеличивают активацию CRF2R и/или активацию пути передачи сигнала рецептора CRF2R.
Идентификация соединений, которые пролонгируют или увеличивают агонист-индуцированную активацию CRF2R или активацию пути передачи сигнала CRF2R, включает различные тесты активации рецептора, описанные в примере 3. В частности, такой тест проводят путем посева клеток с HEK293/CRE-LUC/pIRESneo/hCRF2R-рецептором в 96-луночные планшеты Packard View Plate-96 (Packard Inc., СА). Клетки высевали в среде DMEM, содержащей 10% фетальной бычьей сыворотки, раствор пенициллина/стрептомицина, L-глутамин, не-незаменимую аминокислоту и количества CRF до насыщения, при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха, и инкубировали в течение 48 часов. Затем среду удаляли и заменяли на DMEM (Life Technologies, Rockville, MD), содержащую 0,01% фракции V бычьего альбумина (SIGMA, St.Louis, MO) и CRF в дополнение к соединению, представляющему интерес. Затем клетки инкубировали в течение четырех часов при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха, после чего среду удаляли, а клетки дважды промывали соляным раствором Hanks Balanced Salt Solution (Life Technologies, Rockville, MD). Затем к промытым клеткам добавляли лизирующий реагент (Promega Inc., Madison, WI) и инкубировали клетки в течение 20 минут при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха. Затем клетки помещали на 20 минут при -80°C, после чего инкубировали 20 минут при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха. После такой инкубации к клеточным лизатам добавляли буфер (Luciferase Assay Buffer) и субстрат люциферазы (Luciferase Assay Substrate) (Promega Inc., Madison, WI), и количественно измеряли активность люциферазы при помощи люминометра. Тестируемые соединения, стимулирующие флуоресценцию существенно выше уровней флуоресценции контрольных необработанных клеток, после коррекции различий в клеточной плотности, рассматривали в качестве соединений-кандидатов для регуляции массы или функции скелетных мышц. К соединениям, представляющим наибольший интерес, относятся те соединения, которые индуцируют относительно высокие уровни флуоресценции.
Пример 5. Скрининг для идентификации соединений-кандидатов, специфичных для CRF2R.
Соединения, активирующие CRF2R, идентифицировали так, как в примере 3. Для того чтобы выбрать те соединения, которые проявляют селективность к CRF2R по сравнению с CRF1R, проводили скрининг таких соединений в отношении CRF1R. Клетки HEK293/CRE-LUC/pIRESneo/hCRF1R создавали, по сути, так, как описано в примере 1, за исключением того, что для начальной ПЦР-амплификации применяли последовательность ДНК CRF1R(hCRF1R) человека с регистрационным номером X72304. Чтобы определить активность соединений в отношении CRF1R, проводили тест по активации, по существу, так, как описано в примере 3, за исключением того, что для посева на планшеты применяли клетки HEK293/CRE-LUC/pIRESneo/hCRF1R. Величину флуоресценции, стимулируемой соединением в клетках, экспрессирующих CRF2R, сравнивали с величиной флуоресценции, стимулируемой соединением в клетках, экспрессирующих CRF1R. Те соединения, которые обладают 10-кратно большим (исходя из молярной концентрации) ответом в клетках, экспрессирующих CRF2R, чем в клетках, экспрессирующих CRF1R, затем проверяли дальше на специфичность ответа, с поправкой на различия, обусловленные клональной изменчивостью. HEK293/CRE-LUC/pIRESneo/hCRF2R-клетки тестировали с соединением в присутствии или отсутствии 10-кратного избытка антагониста CRF2R, антисаувагина-30. Те соединения, которые показывали более чем 10-кратную селективность в отношении CRF2R, и активность которых ингибировалась антисаувагином-30, выбирали в качестве соединений-кандидатов.
Пример 6. Скрининги для идентификации соединений-кандидатов, увеличивающих экспрессию CRF2R.
В базе данных генома человека была найдена последовательность, содержащая область промотора hCRF2R-гена, которая начинается на достаточном расстоянии вверх по течению от сайта инициации транскрипции, чтобы содержать все регуляторные элементы, необходимые для физиологической экспрессии hCRF2R-гена в соответствующей ткани. Были синтезированы два олигонуклеотида, один, содержащий 5′-конец области промотора (5′-олигонуклеотид), и один, содержащий 3′-конец области промотора, включая сайт начала транскрипции (3′-олигонуклеотид). Указанные олигонуклеотиды также содержат сайты рестрикции, которых нет в регуляторной области hCRF2R-гена, один с уникальным сайтом рестрикции в 5′-олигонуклеотиде и другой с уникальным сайтом рестрикции в 3′-олигонуклеотиде. 5′ и 3′-олигонуклеотиды применяли для ПЦР-амплификации регуляторной области hCRF2R-гена ДНК человека (Clonetech Inc., Palo Alto, СА, США) при помощи ПЦР-набора Advantage® Genomic PCR kit (Clonetech Inc., Palo Alto, СА, США). Продукт ПЦР регуляторной области hCRF2R-гена очищали от ПРЦ-артефактов при помощи электрофореза в агарозном геле, а фрагмент ДНК регуляторной области hCRF2R-гена очищали от агарозного геля, применяя продукт для очистки, такой как NucleoTrap (Clonetech Inc., Palo Alto, СА, США). Клонирование продукта ПЦР регуляторной области hCRF2R-гена в вектор pECFP-1 (Clonetech Inc., Palo Alto, СА, США) проводили сначала разрезанием продукта ПЦР регуляторной области hCRF2R-гена и вектора pECFP-1 при помощи соответствующей рестрикционной эндонуклеазы так, чтобы 5′ и 3′-сайты рестрикции подготовить к лигированию. Лигирование ДНК pECFP-1-вектора и ДНК продукта ПЦР регуляторной области hCRF2R-гена проводили при помощи ДНК-лигазы от AdvantAgeTM PCR Cloning Kit (Clonetech Inc., Palo Alto, СА, США), согласно рекомендациям изготовителя. Затем лигированный вектор и инсерционную конструкцию применяли для трансформации TOP10F' в компетентных клетках E. coli (Clonetech Inc., Palo Alto, СА, США). Проводили посев клеток в LB-среду плюс агар, содержащий канамицин, и для дальнейшего анализа выбирали колонии, резистентные к канамицину. Клоны, резистентные к канамицину, культивировали в LB-среде, содержащей канамицин, и c помощью системы очистки NucleoBond DNA Purification System (Clonetech Inc., Palo Alto, СА, США) выделяли ДНК-плазмиду, конструкцию, содержащую регуляторную область hVPAC2-гена, анализировали секвенированием ДНК, чтобы гарантировать правильность и целостность конструкции. Затем очищенную ДНК-плазмиду конструкции, содержащую регуляторную область hCRF2R-гена, трансфицировали в HEK293-клетки, применяя трансфекцию, опосредованную фосфатом кальция, при помощи CalPhosTM Mammalian Transfection Kit (Clonetech Inc., Palo Alto, СА, США). Клоны трансфицированных клеток отбирали при помощи G418, выделяли и размножали в DMEM (Life Technologies, Rockville, MD), содержащей 10% фетальной бычьей сыворотки (Clonetech Inc., Palo Alto, СА, США), раствор пенициллина/стрептомицина (Life Technologies, Rockville, MD), L-глутамин (Life Technologies, Rockville, MD), не-незаменимую аминокислоту (Life Technologies, Rockville, MD) и G418 (Life Technologies, Rockville, MD) при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха. Клоны, резистентные к G418, характеризовали при помощи Саузерн-блоттинга, чтобы гарантировать, что они содержат последовательность промотора hCRF2R-гена; кроме того, анализировали активацию регуляторной области hCRF2R-гена, применяя соответствующий стимулирующий агент. Клетки, экспрессирующие регуляторную ECFP-область hCRF2R-гена на соответствующем уровне, затем применяли в тестах, предназначенных для оценки соединений, которые могут модулировать активность регуляторной области hCRF2R-гена, как следует далее. Анализы активации регуляторной области проводили путем посева HEK293-клеток, содержащих регуляторную ECFP-область hCRF2R-гена, при соответствующей плотности в черные 96-луночные микротитрационные планшеты с прозрачным дном и давали возможность расти в течение ночи. На следующий день среду удаляли, а в свежую питательную среду добавляли тестируемое соединение. Клетки инкубировали в течение 16 часов при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха, после чего измеряли флуоресценцию (возбуждение при 433(453)нм, детектирование эмиссии при 475(501)нм) на флуориметре (bioluminТМ 960, Molecular Dynamics/Amersham Pharmacia Biotech, Piscataway, NJ). Тестируемые соединения, которые стимулируют существенно более высокие уровни флуоресценции по сравнению с уровнями контрольных необработанных клеток, с поправкой на различие клеточной плотности, рассматривали в качестве соединений-кандидатов для регуляции массы или функции скелетных мышц. Соединениями, представляющими наибольший интерес, являются те, которые индуцируют относительно высокие уровни флуоресценции.
Пример 7. Скрининги для идентификации соединений, увеличивающих экспрессию CRF человека.
Способы идентификации соединений, которые увеличивают экспрессию CRF человека (hCRF), по существу, идентичны способам идентификации соединений, которые увеличивают экспрессию hVPAC2-рецептора, за исключением того, что применяется такая же регуляторная область, что для hCRF-гена. В базе данных генома человека была найдена последовательность, содержащая регуляторную область hCRF-гена, начинающуюся на достаточном расстоянии вверх по течению от сайта инициации транскрипции, чтобы содержать все регуляторные элементы, необходимые для физиологической экспрессии hCRF-гена в соответствующей ткани. Синтезировали два олигонуклеотида, один, содержащий 5'-конец регуляторной области (5'-олигонуклеотид), и один, содержащий 3'-конец регуляторной области, включая сайт начала транскрипции (3'-олигонуклеотид). Указанные олигонуклеотиды также содержат сайты рестрикции, которых нет в регуляторной области hCRF-гена, один с уникальным сайтом рестрикции в 5'-олигонуклеотиде и другой уникальный сайт рестрикции в 3'-олигонуклеотиде. 5'- и 3'-олигонуклеотиды применяют для ПЦР-амплификации регуляторной области hCRF-гена ДНК человека (Clonetech Inc., Palo Alto, СА, США) при помощи ПЦР-набора Advantage® Genomic PCR kit (Clonetech Inc., Palo Alto, СА, США). Продукт ПЦР регуляторной области hCRF-гена очищали от ПЦР-артефактов электрофорезом в агарозном геле, а фрагмент ДНК регуляторной области hCRF-гена очищали от агарозного геля при помощи очищающего продукта NucleoTrap (Clonetech Inc., Palo Alto, СА, США). Клонирование продукта ПЦР регуляторной области hCRF-гена в вектор pECFP-1 (Clonetech Inc.,Palo Alto, СА, США) проводили сначала разрезанием продукта ПЦР регуляторной области hCRF-гена и вектора pECFP-1 при помощи соответствующей рестрикционной эндонуклеазы так, чтобы 5'- и 3'-сайты рестрикции подготовить к лигированию. Лигирование ДНК вектора pECFP-1 и ДНК продукта ПЦР регуляторной области hCRF-гена проводили при помощи ДНК-лигазы от AdvantAgeTM PCR Cloning Kit (Clonetech Inc., Palo Alto, СА, США) согласно рекомендациям изготовителя. Затем лигированный вектор и инсерционную конструкцию применяли для трансформации TOP10F' в компетентных клетках E. coli (Clonetech Inc., Palo Alto, СА, США). Проводили посев клеток в LB-среде плюс агар, содержащий канамицин, и для дальнейшего анализа выбирали колонии, резистентные к канамицину. Клоны, резистентные к канамицину, культивировали в LB-среде, содержащей канамицин, и c помощью системы очистки NucleoBond DNA Purification System (Clonetech Inc.,Palo Alto, СА, США) выделяли ДНК-плазмиду и анализировали конструкцию, содержащую регуляторную область hCRF-гена путем секвенирования ДНК, чтобы гарантировать правильность и целостность конструкции. Затем очищенную конструкцию ДНК-плазмиды, содержащую регуляторную область hCRF-гена, трансфицировали в HEK293-клетки, применяя трансфекцию, опосредованную фосфатом кальция, при помощи CalPhosTM Mammalian Transfection Kit (Clonetech Inc., Palo Alto, СА, США). Клоны трансфицированных клеток отбирали при помощи G418, выделяли и размножали в DMEM (Life Technologies, Rockville, MD), содержащей 10% фетальной бычьей сыворотки (Clonetech Inc., Palo Alto, СА, США), раствор пенициллина/стрептомицина (Life Technologies, Rockville, MD), L-глутамин (Life Technologies, Rockville, MD), не-незаменимую аминокислоту (Life Technologies, Rockville, MD) и G418 (Life Technologies, Rockville, MD) при 37°C в атмосфере, состоящей из смеси 5% углекислого газа/95% воздуха. Клоны, резистентные к G418, характеризовали при помощи Саузерн-блоттинга, чтобы гарантировать, что они содержат последовательность регуляторной области hCRF-гена; кроме того, анализировали активацию регуляторной области hCRF-гена, применяя соответствующий стимулирующий агент. Клетки, экспрессирующие регуляторную ECFP-область hCRF-гена на соответствующем уровне, затем применяли в тестах, предназначенных для оценки соединений, которые могут модулировать активность регуляторной области hCRF-гена, как следует далее. Анализы активации регуляторной области проводили, как описано в примере 5, за исключением того, что применяли клоны, содержащие конструкцию регуляторной области hCRF-гена.
Пример 8. Способ создания антител человека, активирующих hCRF2R.
Целиком человеческие моноклональные антитела, активирующие hCRF2R, продуцировали при помощи первоначально созданного рекомбинантного hCRF2R-белка, как следует далее. Для получения продукта ПЦР hCRF2R придерживались методики из примера 1. Затем полученный продукт ПЦР hCRF2R клонировали в pHAT20-вектор (Clonetech Inc., Palo Alto, СА, США) первоначальным разрезанием продукта ПЦР hCRF2R-гена и pHAT20-вектора при помощи соответствующей рестрикционной эндонуклеазы так, чтобы 5' и 3'-сайты рестрикции подготовить к лигированию. Лигирование ДНК pHAT20-вектора и ДНК продукта ПЦР hCRF2R-гена проводили при помощи ДНК-лигазы от AdvantAgeTM PCR Cloning Kit (Clonetech Inc.,Palo Alto, СА, США) согласно рекомендациям изготовителя. Конструкцию лигированный вектор/вставка затем применяли для трансформации TOP10F′ в компетентных клетках E. coli (Clonetech Inc., Palo Alto, СА, США). Трансформированные клетки высеивали на LB-среде плюс агар, содержащий ампициллин, и для дальнейшего анализа отбирали колонии, резистентные к ампициллину. Позитивные клоны культурировали в LB-среде, содержащей ампициллин, и выделяли ДНК-плазмиду, применяя систему очистки NucleoBond DNA Purification System (Clonetech Inc., Palo Alto, СА, США), конструкцию, содержащую hCRF2R-ген, анализировали секвенированием ДНК для гарантии правильности и целостности конструкции. ДНК hCRF2R-pHAT20-вектора затем применяли для дополнительного ПЦР-клонирования с применением 5'-олигонуклеотида, содержащего начало последовательности НАТ, и уникального сайта рестрикции, которого нет в hCRF2R-pHAT20-конструкции, и 3'-олигонуклеотида hCRF2R, полученного предварительно. Затравки олигонуклеотидов применяли для ПЦР-амплификации гибридного НАТ-hCRF2R-гена из hCRF2R-pHAT20-конструкции, а продукт ПЦР очищали, как описано выше. Затем продукт ПЦР гибридного НАТ-hCRF2R-гена применяли для клонирования в pBacPAK8-вектор, применяя систему экспрессии BacPAK Baculovirus Expression System от Clonetech (Clonetech Inc., Palo Alto, СА, США). Лигирование гибридного НАТ-hCRF2R-гена в pBacPAK8-вектор осуществляли, как описано выше. Затем конструкцию hCRF2R/HAT-pBacPAK8 трансфицировали в TOPIO'F компетентные клетки E. coli, отбирали клетки, резистентные к ампициллину, выделяли ДНК-плазмиду и проверяли целостность конструкции, как описано выше. Затем такую конструкцию переносили совместно с выровненной ДНК BacPAK6 в Sf21-клетки-хозяева насекомых, применяя набор для трансфекции CalPhosТМ Mammalian Trasfection Kit (Clonetech Inc., Palo Alto, СА, США). Затем клетки насекомых инкубировали в течение 2-3 дней, с последующим сбором вируса из отдельных прозрачных планшетов. Затем вирус амплифировали в Sf21-клетках, собранный вирус оттитровывали, и оттитрованнный вирус применяли для инфекционирования Sf21-клеток в большом масштабе, применяя среду для клеток насекомых BacPAK Insect Cell Media - все в соответствии с рекомендациями изготовителей (Clonetech Inc.,Palo Alto, СА, США). Гибридный белок рекомбинантного НАТ-CRF2R затем очищали при помощи набора TALON® CellThru Purification Kit от Clonetech (Clonetech Inc., Palo Alto, СА, США) при соблюдении условий, рекомендуемых изготовителем. Кратко, инфицированные Sf21-клетки собирали через 48 часов после инфицирования и разрушали ультразвуком в буфере загрузки/экстракции. Клеточный лизат затем пропускали через колонку TALON® CellThro colum. Колонку промывали два раза буфером загрузки/экстракции, и связанный НАТ-hCRF2R-белок элюировали элюирующим буфером. Элюированный белок анализировали при помощи SDS-PAGE на целостность, а концентрацию белка количественно определяли при помощи системы Bio-Rad SDS-PAGE system и системы количественного определения белка согласно рекомендациям изготовителя (Bio-Rad Laboratories, Hercules, Ca). Очищенный гибридный НАТ-hCRF2R-белок затем применяли для иммунизации животных XenoMouse (Abgenix Inc., Fremont, СА) с целью получения моноклональных антител человека, как описано ниже. Для многократной вакцинации 10 животных XenoMouse в течение восьми недель применяли 10 мкг очищенного гибридного рекомбинантного НАТ-hCRF2R-белка в сочетании с 25 мкг адьювантного монофосфориллипида A (SIGMA, St.Louis, MO). Сыворотку, полученную от привитых животных, применяли в тесте-ELISA по фиксированию антигена, применяя очищенный гибридный НАТ-hCRF2R-белок для обнаружения антител к НАТ-hCRF2R-белку, путем покрытия полистироловых планшетов для теста-ELISA (Corning Glass Works, Corning, NY) гибридным НАТ-hCRF2R-белком, блокированным PBS, содержащим 1% BSA, промывали и инкубировали при 37°C в течение 1 часа с образцами сыворотки в разведении 1:50. После 5-кратного промывания PBS планшеты инкубировали при 37°C в течение 1 часа со щелочной фосфатазой, конъюгированной с козьими антителами к G-иммуноглобулину человека. Затем планшеты 5-кратно промывали PBS и обнаруживали антитела при помощи с p-нитрофенилфосфатного субстрата (SIGMA, St.Louis, MO) в буфере. Измеряли оптическую плотность при 405 нм при помощи ридера планшетов и количественной оценки сигнала. Для формирования гибридом применяли мышей с выраженным высоким продуцированием антител. Гибридомы создавали путем слияния клеток селезенки животных XenoMouse с линией несекретирующих миеломных NSA-bcl2-клеток при отношении клеток селезенки к NSA-bcl2-клеткам 4:1 в присутствии 30% полиэтиленгликоля PEG1450. Гибридные клетки клонировали отдельно при ограниченном разбавлении в 96-луночных планшетах и культивировали в среде RPMI-1640, содержащей 10% фетальной бычьей сыворотки, не-незаменимые аминокислоты, пируват натрия, L-глутамин, 100 ед./мл пенициллин-стрептомицина и гипоксантин-аминоптерин-тимидин (все от Life Technologies, Rockville, MD). Надосадочные жидкости от гибридом, отобранных из гипоксантин-аминоптерин-тимидина, подвергали скринингу для получения антител человека при помощи теста-ELISA, как описано ранее. Гибридомы, которые продуцируют человеческие антитела к гибридному НАТ-hCRF2R-белку, отбирали для получения антител в большом масштабе. Моноклональные антитела очищали хроматографией Protein G-Sepharose. Коротко, надосадочную жидкость от культивированных гибридомных клонов загружали в колонку Protein G-Sepharose (SIGMA, St.Louis, MO) в загрузочном буфере, промывали 3 раза и элюировали IgG элюирующим буфером. Затем такие антитела применяли для скрининга, чтобы оценить потенциал активации (агонизм) hCRF2R. Скрининг выполняли по методике, описанной в примере 3. Те моноклональные антитела человека, которые обладают активностью агониста в отношении hCRF2R, обозначали как соединения-кандидаты.
Пример 9. Экспериментальное измерение абсолютной силы мышцы.
Из загипсованной конечности мыши \сухожилие за сухожилием\ удаляли разгибатель большого пальца (EDL) и камбаловидной мышцы. К каждому сухожилию отделенных мышц привязывали шелковый шовный материал и помещали мышцы в камеру из плексигласа, заполненную раствором Рингера (137 мМ хлорида натрия, 24 мМ бикарбоната натрия, 11 мМ глюкозы, 5 мМ хлорида калия, 1 мМ сульфата магния, 1 мМ фосфата натрия, 0,025 мМ тубокукарина - все при pH 7,4 и оксигенировании смесью 95% кислорода/5% диоксида углерода), который постоянно барботировался смесью 95% кислорода/5% диоксида углерода и поддерживался при 25°C. Мышцы натягивали горизонтально между плечом рычага сервомотора (Model 305B-LR Cambridge Technology Inc., Watertown MA, USA) и крючком из нержавеющей стали преобразователя силы (Model BG-50; Kulite Semiconductor Products Inc., Leonia, NJ, USA) и возбуждали поле, пропуская импульсы между двумя платиновыми электродами, помещенными продольно с обеих сторон мышцы. Импульсы прямоугольных колебаний (продолжительностью 0,2 мс), созданные при помощи персонального компьютера с платой Labview (Model PCI-MIO 16E-4), Labview Inc., Austin, TX, USA), усиливали (Acurus power amplifier model (модель усилителя мощности) A25, Dobbs Ferry, NY, USA), чтобы увеличить тетаническое сокращение. Стимулирующее напряжение и длину мышцы (Lo) регулировали для получения максимальной силы изометрической судороги. Максимальное продуцирование тетанической силы (Ро) определяли по положению плато на кривой зависимости частота-сила.
Пример 10. Терапевтическое лечение атрофии скелетных мышц с применением антитела человека, являющегося агонистом hCRF2R-рецептора.
Мужчину, весом 50 кг, со значительной мышечной атрофией рук и ног, обусловленной длительным постельным режимом, лечили с целью устранения атрофии скелетных мышц. Один раз в неделю в течение 3-х месяцев субъекту путем внутривенной инъекции вводили 15 мл водного раствора с pH 6, содержащего антитело, активирующее рецептор CRF2R. Раствор содержит следующие компоненты:
По окончании лечения у субъекта наблюдалось измеримое увеличение мышечной массы, силы и подвижности рук и ног.
Пример 11. Профилактическое лечение атрофии скелетных мышц с применением антитела человека, являющегося агонистом рецептора hCRF2R.
Женщине, весом 55 кг, была назначена операция по замене тазобедренного сустава в течение месяца. Субъект лечили до и после операции с целью увеличить массу скелетных мышц, чтобы в конечном счете понизить уровень атрофии скелетных мышц, обусловленный бездействием мышцы во время пост-операционного восстановления. В частности, раз в неделю в течение одного месяца до операции и два месяца после операции субъекту внутривенно вводили 18 мл водного раствора с pH 6,0, содержащего антитело, активирующее рецептор CRF2R. Раствор содержит следующие компоненты:
По окончании периода лечения у субъекта наблюдалось измеримое сохранение мышечной массы, силы и подвижности рук и ног по сравнению с ожидаемым состоянием субъекта без терапии с применением антитела.
Пример 12. Профилактическое лечение атрофии скелетных мышц с применением антитела человека, агониста рецептора CRF2R.
Женщине, весом 45 кг, была наложена гипсовая повязка для лечения простого перелома плечевой кости после падения. Лечение субъекта проводили с целью предотвращения атрофии скелетных мышц поврежденных руки и плеча, обусловленной бездействием и ограниченным применением во время заживления перелома. В частности, один раз в неделю, начиная со дня наложения гипсовой повязки, субъекту путем внутривенной инъекции вводили 13 мл раствора с pH 6,0, содержащего рецептор анти-CRF2R. Раствор содержит следующие компоненты:
По окончании периода лечения у субъекта наблюдалось измеримое сохранение мышечной массы, силы и подвижности поврежденной руки и плеча, и требовался пониженный курс физиотерапии, по сравнению с ожидаемым состоянием субъекта и завершающим лечением без терапии при помощи антитела.
Пример 13. Профилактическое лечение атрофии скелетных мышц с применением урокортина-II.
Женщина, весом 60 кг, поступила на госпитализацию в коматозном состоянии. Субъект лечили указанным способом для профилактики атрофии скелетных мышц всего тела, обусловленной бездействием в коматозном состоянии. В частности, один раз в сутки, в то время как субъект находился в коме, ей вводили путем медленной внутривенной инфузии приблизительно 500 мл водного раствора, который готовили добавлением к 500 мл стерильного физиологического раствора 5 мл следующего основного раствора:
В результате лечения у субъекта наблюдалось измеримое сохранение массы и функции скелетных мышц, и требовалась пониженная физиотерапия во время комы и после восстановления сознания, по сравнению с состоянием субъекта без лекарственной терапии.
Пример 14. Терапевтическое лечение пациента с мышечной дистрофией Дюшенна с применением CRF.
Мужчину, весом 40 кг, с установленным диагнозом мышечной дистрофии Дюшенна лечили при помощи соединения, которое проявляет сродство к CRF1-R и CRF2-R, применяя диапазон доз, выше тех доз, при которых они похожи. Субъект лечили при помощи базовой композиции долговременного высвобождения соединения, чтобы улучшить или сохранить мышечную силу и функцию при прогрессировании болезни. В частности, один раз в месяц субъекту вводили путем внутримышечной инъекции 3 мл водного раствора с pH 6,0, содержащего следующие компоненты:
В результате лечения субъект испытывал либо улучшение, либо ослабление хронического прогрессирующего заболевания, связанного с потерей мышечной силы или мышечной функции, на основе определений временной зависимости такой функции, по сравнению с такими же показателями, наблюдаемыми во время естественного развития болезни.
Настоящее изобретение не должно быть ограничено в объеме описанными конкретными примерами осуществления, которые предназначены исключительно для иллюстрации отдельных аспектов изобретения и в качестве функционального эквивалента способов и компонентов, находящихся в пределах объема изобретения. Они включают без ограничения вид тестируемого животного, природу и тип агонистов CRFR, пол животного, модель атрофии, способ активации CRFR, включая методики генной инженерии и т.д. Различные модификации изобретения, в дополнение к тем, что показаны и описаны здесь, будут очевидны специалистам в данной области при ознакомлении с предшествующим описанием и сопроводительными фигурами. Такие модификации предназначены для того, чтобы уложиться в объем приобщенной формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| АГОНИСТЫ РЕЦЕПТОРА КОРТИКОТРОПИН-РИЛИЗИНГ ФАКТОРА 2 (CRFR) ПРИМЕНЕНИЕ | 2003 |
|
RU2294333C2 |
| АГОНИСТЫ РЕЦЕПТОРА КОРТИКОТРОПИН-РИЛИЗИНГ ФАКТОРА 2 | 2003 |
|
RU2286995C2 |
| АГОНИСТЫ РЕЦЕПТОРА КОРТИКОТРОПИН-РИЛИЗИНГ ФАКТОРА 2 И ИХ ПРИМЕНЕНИЕ | 2003 |
|
RU2305110C2 |
| БЕЛКИ УРОКОРТИНА И ИХ ПРИМЕНЕНИЯ | 2001 |
|
RU2278871C2 |
| СОПРЯЖЕННЫЕ С G-БЕЛКОМ РЕЦЕПТОРЫ DROSOPHILA, НУКЛЕИНОВЫЕ КИСЛОТЫ И СПОСОБЫ, ИМЕЮЩИЕ К НИМ ОТНОШЕНИЕ | 2003 |
|
RU2326385C2 |
| КОМПОЗИЦИИ И СПОСОБЫ, СВЯЗАННЫЕ С РАСТВОРИМЫМИ СОПРЯЖЕННЫМИ С G-БЕЛКОМ РЕЦЕПТОРАМИ (SGPCR) | 2006 |
|
RU2423378C2 |
| АНТИТЕЛО ПРОТИВ РЕЦЕПТОРА IGF-I | 2018 |
|
RU2785305C2 |
| ГЕН НОВОЙ СЕРИНОВОЙ ПРОТЕАЗЫ, РОДСТВЕННОЙ DPPIV | 2001 |
|
RU2305133C2 |
| ТКАНЕСПЕЦИФИЧЕСКИЕ БИОМАРКЕРЫ СТАРЕНИЯ | 2010 |
|
RU2557313C2 |
| АНАЛИЗЫ | 2020 |
|
RU2834240C2 |
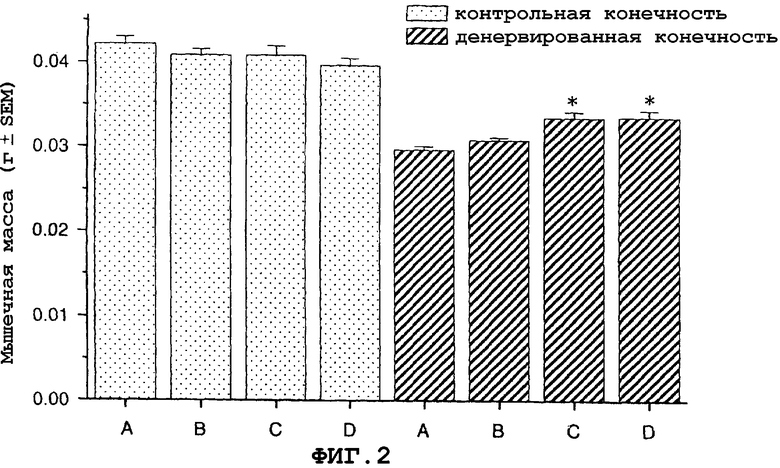

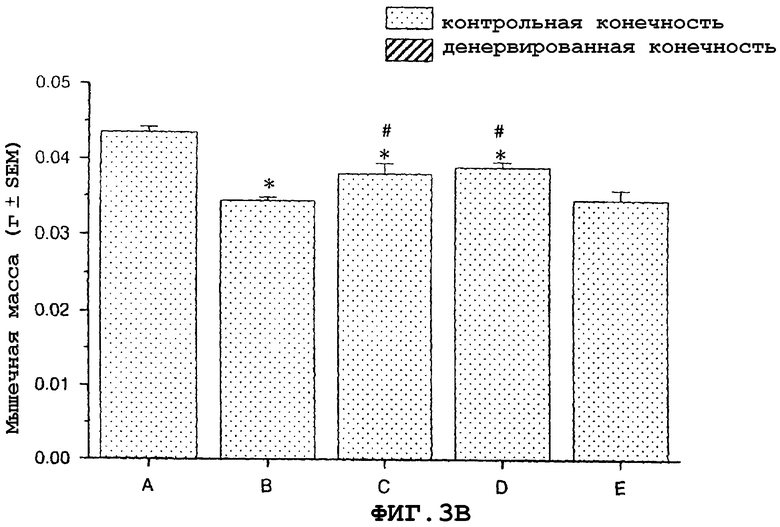
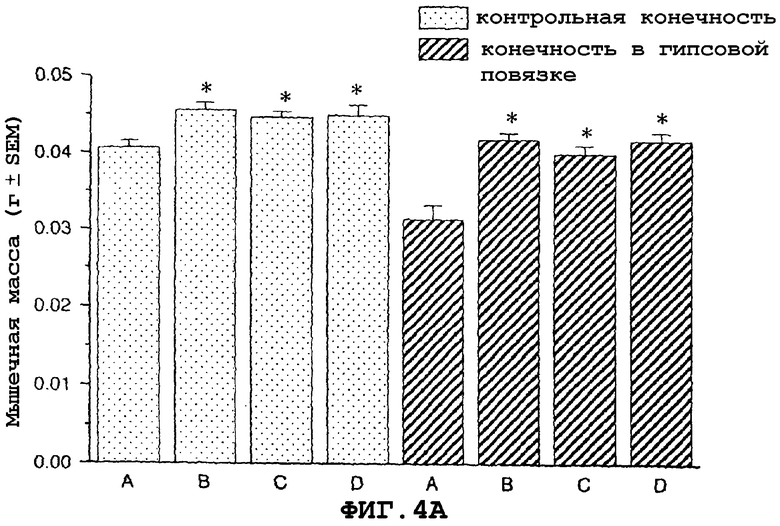
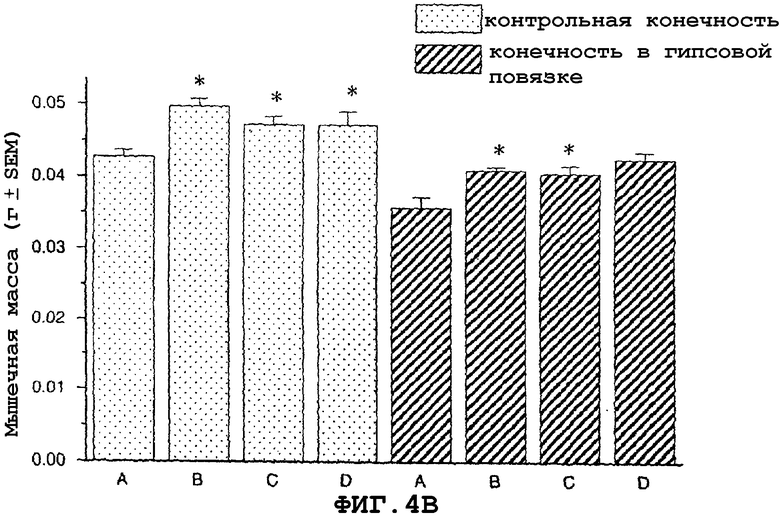
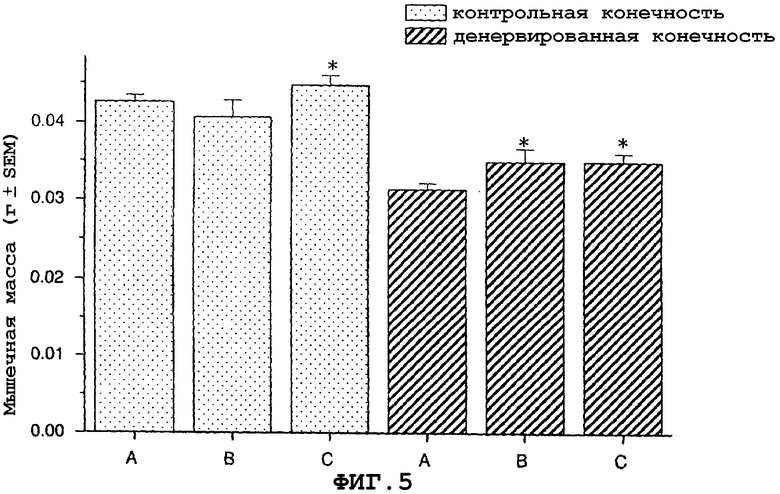
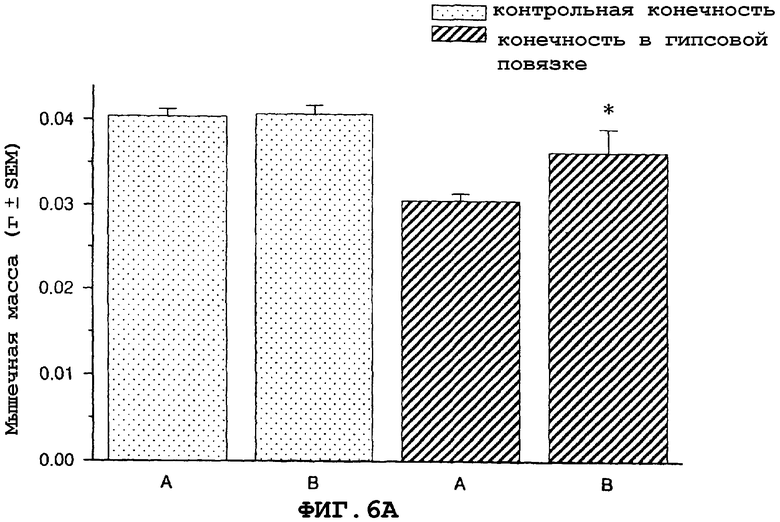
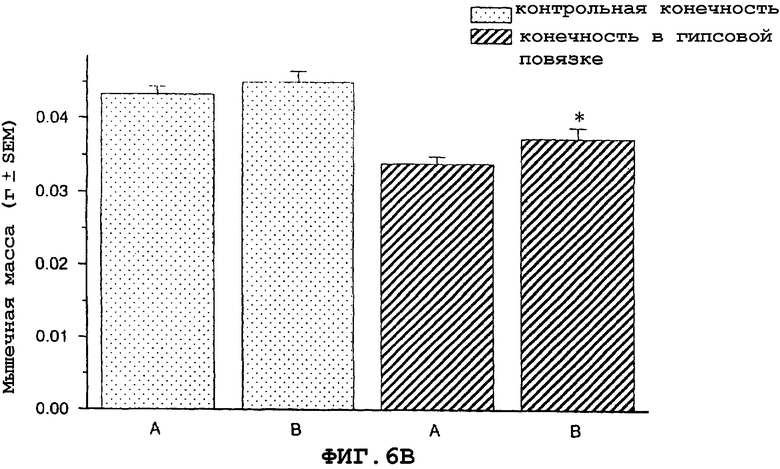
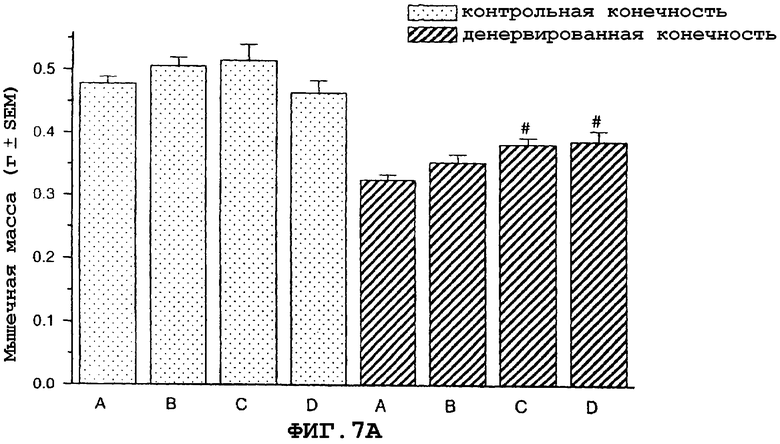
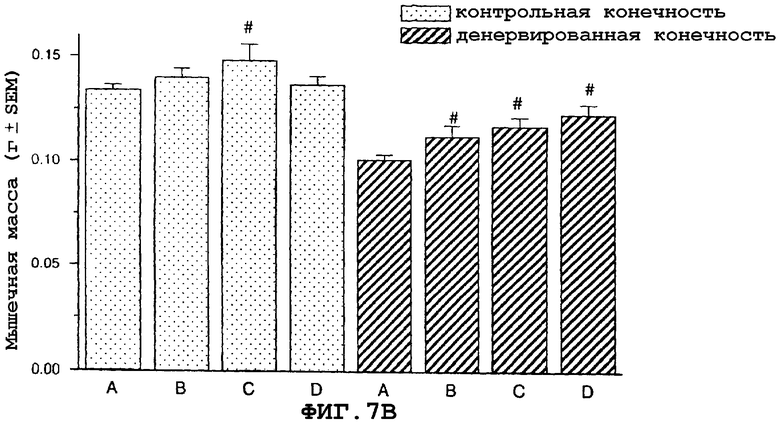
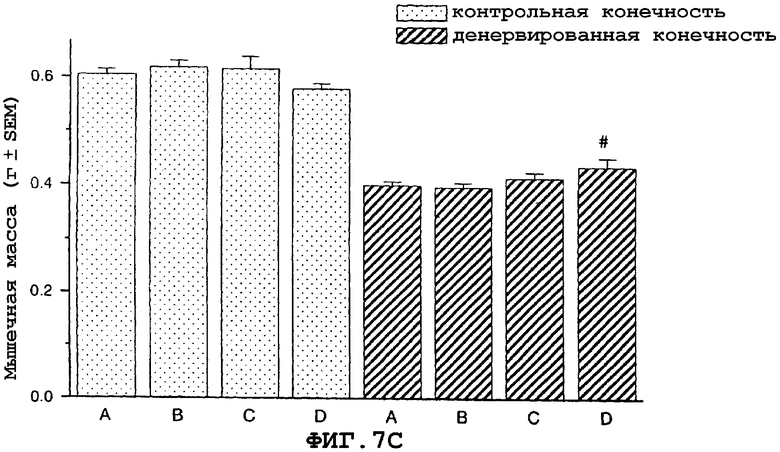
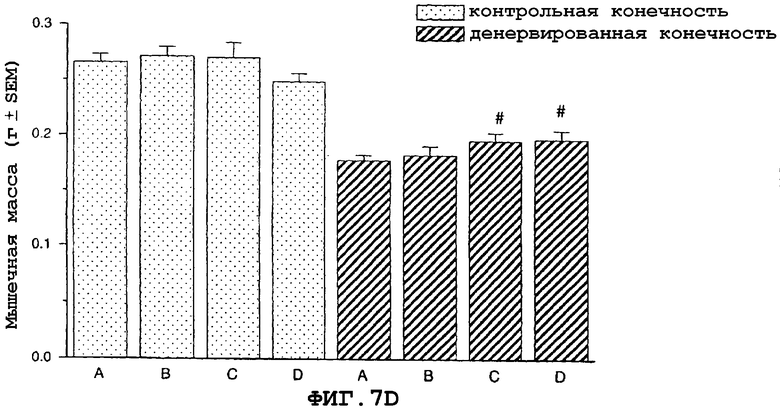
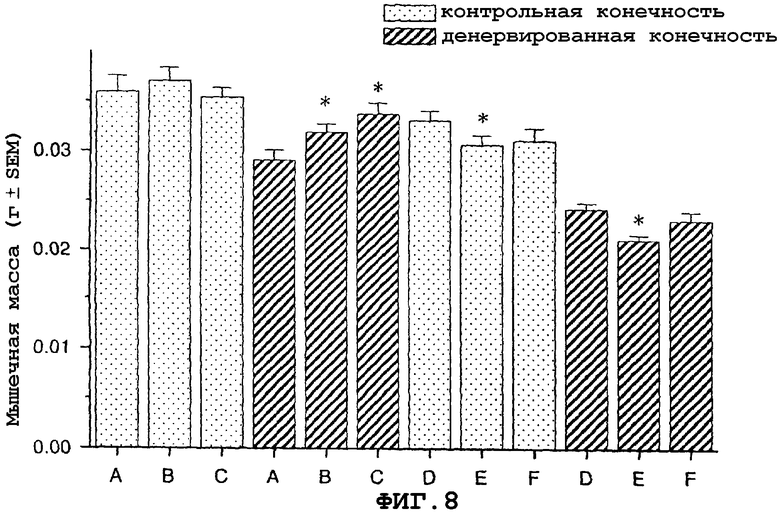
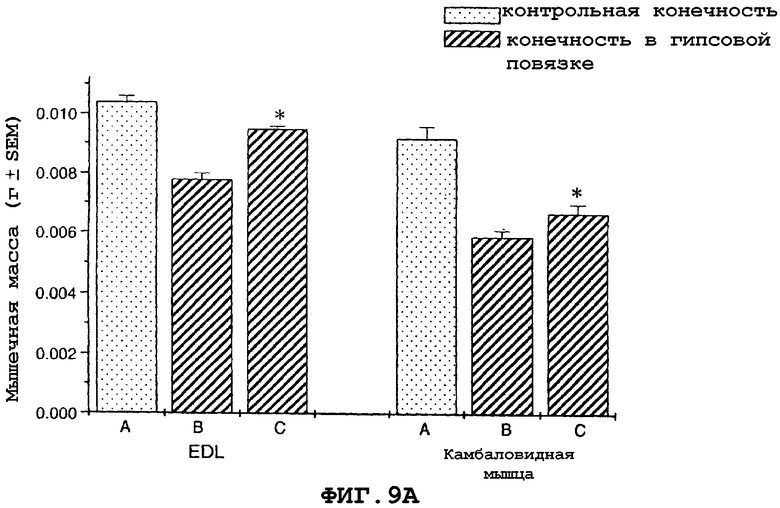
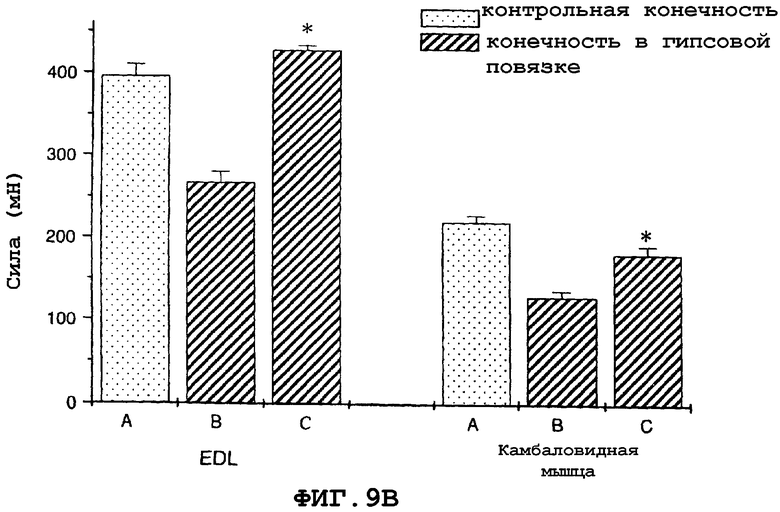
Изобретение относится к области фармакологии. Сущность изобретения состоит в том, что разработаны способы идентификации соединений-кандидатов для регуляции массы или функции скелетных мышц, включающие взаимодействие тестируемого соединения с рецептором рилизинг-фактора 2 кортикотропина. Описаны также фармацевтическая композиция и способ лечения атрофии скелетных мышц. Технический результат - расширение арсенала средств, предназначенных для регуляции массы или функции скелетных мышц. 9 н. и 26 з.п. ф-лы, 1 табл., 9 ил.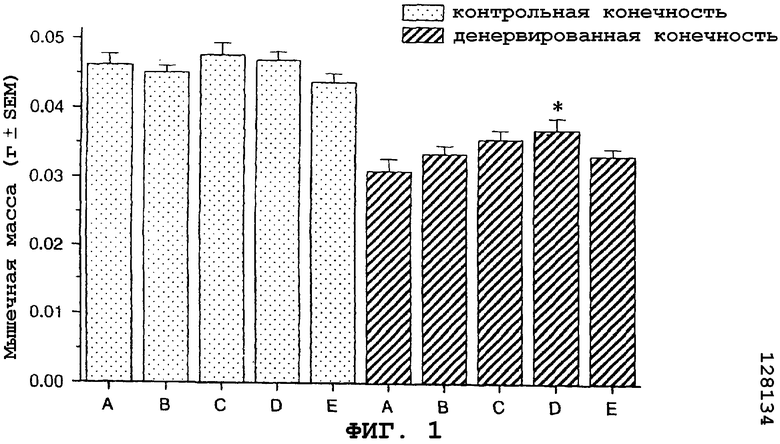
(a) взаимодействие тестируемого соединения с рецептором рилизинг-фактора 2 кортикотропина (CRF2R);
(b) определение, связывается ли тестируемое соединение с CRF2R; и
(c) идентификацию тех тестируемых соединений, которые связываются с CRF2R, в качестве соединений-кандидатов для регуляции массы или функции скелетных мышц.
(a) взаимодействие тестируемого соединения с клеткой, экспрессирующей функциональный CRF2R,
(b) определение, активирует ли тестируемое соединение CRF2R; и
(c) идентификацию тех тестируемых соединений, которые активируют CRF2R, в качестве соединений-кандидатов для регуляции массы или функции скелетных мышц.
(a) взаимодействие тестируемого соединения с клеткой, экспрессирующей функциональный CRF2R, и определение уровня активации CRF2R, возникающего под действием тестируемого соединения;
(b) взаимодействие тестируемого соединения с клеткой, экспрессирующей функциональный рецептор рилизинг-фактора 1 кортикотропина (CRF1R), и определение уровня активации CRF1R, возникающего под действием тестируемого соединения; с последующим
(c) сравнением уровня активации CRF2R и уровня активации CRF1R и
(d) идентификацию тех тестируемых соединений, которые обладают похожей активностью в отношении CRF2R и CRF1R или обладают селективностью в отношении CRF2R, в качестве соединений-кандидатов для регуляции массы или функции скелетных мышц.
(a) введение соединения-кандидата животному, отличному от человека; и
(b) определение, регулирует ли соединение-кандидат массу или функцию скелетных мышц животного, подвергнутого лечению.
(a) взаимодействие тестируемого соединения с клеткой, которая экспрессирует функциональный CRF2R;
(b) обработку клетки агонистом CRF2R в течение достаточного времени и при достаточной концентрации, чтобы имела место десенсибилизация CRF2R в контрольных клетках; с последующим
с) определением уровня активации CRF2R; и
d.) идентификацию тех тестируемых соединений, которые пролонгируют или увеличивают активацию CRF2R или активацию пути передачи сигнала CRF2R, в качестве соединений-кандидатов для регуляции массы или функции скелетных мышц.
a) введение соединения-кандидата в сочетании с агонистом CRF2R животному, отличному от человека; и
b) определение, регулирует ли соединение-кандидат массу или функцию скелетных мышц животного, подвергнутого лечению.
a) взаимодействие тестируемого соединения с клеткой или клеточным лизатом, содержащим репортерный ген, оперативно связанный с регуляторным элементом CRF2R или регуляторным элементом CRF;
b) определение экспрессии репортерного гена и
c) идентификацию тех тестируемых соединений, которые увеличивают экспрессию репортерного гена, в качестве соединений-кандидатов для регуляции массы или функции скелетных мышц.
(а) идентификацию субъекта, для которого желательно увеличение мышечной массы или функции и
b) введение субъекту безопасного и эффективного количества агониста CRF2R.
(a) идентификацию субъекта, которому требуется лечение атрофии скелетных мышц; и
(b) введение субъекту безопасного и эффективного количества соединения, выбранного из группы, состоящей из агониста CRF2R, экспрессирующего вектора, кодирующего функциональный CRF2R, экспрессирующего вектора, кодирующего конститутивно активный CRF2R, экспрессирующего вектора, кодирующего CRF, и соединения, увеличивающего экспрессию CRF2R или CRF.
(a) взаимодействие тестируемого соединения с CRF2R,
(b) определение, активирует ли тестируемое соединение CRF2R; и
(c) идентификацию тех тестируемых соединений, которые активируют CRF2R, в качестве соединений-кандидатов для регуляции массы или функции скелетных мышц.
(a) идентификацию субъекта, которому требуется лечение атрофии скелетных мышц;
(b) введение субъекту безопасного и эффективного количества агониста CRF2R и
(c) введение субъекту безопасного и эффективного количества соединения, которое пролонгирует или увеличивает активацию CRF2R или активацию пути передачи сигнала CRF2R.
a) безопасное и эффективное количество агониста CRF2R и
b) фармацевтически приемлемый носитель.
| US 5786203 | |||
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| Valdenaire O | |||
| et al, \A new functional isoform of the human...\ | |||
| Biochimica et Biophysica Acta | |||
| Электрическое сопротивление для нагревательных приборов и нагревательный элемент для этих приборов | 1922 |
|
SU1997A1 |
| Приспособление к штампу для выделки обувных кольчиков | 1923 |
|
SU1352A1 |
| Palchaudhuri, M.R | |||
| et al, \Isolation a pharmacological char acter: sation of two functional splice variantes...\ | |||
| J | |||
| of Neuroendocrinology, 1999, v.11, p.419-428. | |||

