ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] По настоящей заявке испрашивается приоритет временной заявки на патент США № 61/847405, поданной 17 июля 2013, полное содержание которой включено в настоящее описание.
ЗАЯВЛЕНИЕ О СПОНСИРУЕМЫХ ПРАВИТЕЛЬСТВОМ НАУЧНЫХ ИССЛЕДОВАНИЯХ И РАЗРАБОТКАХ
[0002] Настоящее изобретение было сделано при поддержке Правительства по Грантам Национального Института Здоровья с номерами PO1DK044935 и 5RO1NS064988. Правительство имеет определенные права на это изобретение.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
[0003] Среди многих вирусных и невирусных генетических векторных систем, векторы на основе вируса простого герпеса (ВПГ) были исследованы для применения в качестве векторов для переноса генов, в том числе для возможного терапевтического применения у пациентов человека. ВПГ представляет собой сложный неинтегративный ДНК-содержащий вирус, способный инфицировать очень широкий спектр клеток человека и животных. Вирусный геном содержит более 80 генов и состоит из двух уникальных сегментов, UL и US, каждый из которых фланкирован инвертированными повторами, которые кодируют критичные диплоидные гены. Важной особенностью репликации ВПГ является экспрессия его генов волнами, называемыми каскадным регулированием (Rajcani, Virus Genes, 28: 293-310 (2004)). Удаление существенных предранних (IE) генов ICP27 и ICP4 делает вирус полностью дефектным и неспособным к экспрессии ранних (Е) генов, участвующих в репликации вирусного генома, и поздних (L) генов, функционирующих при сборке вирионов потомства. Эти дефектные по репликации вирусы можно наращивать на комплементирующих клетках, которые экспрессируют (комплементируют) недостающие продукты генов ICP4 и ICP27, и можно затем применять для инфицирования некомплементирующих клеток, где вирусный геном находится в качестве стабильной ядерной эписомы. При этом векторы, поддерживающие IE-гены ICP0 и ICP22, являются токсичными для клеток, но инактивация или делеция этих генов, в частности, гена ICPO затрудняет экспрессию трансгена.
[0004] Соответственно, сохраняется потребность в векторе на основе ВПГ, способном экспрессировать трансген в любой ткани или клетке in vitro или in vivo без ущерба для клетки или ткани и системы для размножения таких векторов.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0005] Настоящее изобретение относится к открытию в конструировании векторов на основе ВПГ, что дало возможность экспрессировать трансгены в самых разнообразных тканях или клетках (в частности, млекопитающих), in vitro or in vivo, без экспрессии каких-либо вредных вирусных генов. ВПГ-вектор по изобретению не экспрессирует никаких токсичных вирусных генов в некомплементирующих клетках, но несмотря на это способен к мощной, устойчивой (например, в течение по меньшей мере 14 дней, например, по меньшей мере 28 дней и предпочтительно, по меньшей мере 60 дней) трансгенной экспрессии. Как таковой, он заполняет чрезвычайно важную нишу в векторной технологии, поскольку ВПГ является единственным вектором, который сочетает возможность поддержания крупных кассет экспрессии одного или нескольких трансгенов, контролируемых общими или клеточно-специфичными промоторами со свойством высокоэффективной инфекции без интеграции вектора. Вектор по изобретению делает возможной эффективную доставку генов к тканям, таким как печень, для которых эффективные векторы в настоящее время отсутствуют.
[0006] В одном варианте осуществления настоящее изобретение относится к вектору на основе вируса простого герпеса (ВПГ), который не экспрессирует токсичные гены ВПГ в некомплементирующих клетках и который содержит геном, включающий один или несколько трансгенов, причем вектор способен экспрессировать трансген по меньшей мере 28 дней в некомплементирующих клетках. Вектор по изобретению может содержать трансген, встроенный в функциональной связи с одной или несколькими инсуляторными последовательностями в геноме, причем вектор не экспрессируют ICPO, ICP4, ICP22, ICP27 и ICP47 в качестве предранних генов. В зависимости от активности промотора, контролирующего трансген, вектор по изобретению может экспрессировать трансген в любом типе клетки млекопитающего (особенно человека), которую он может инфицировать без цитотоксичности, ассоциированной с экспрессией вирусных генов.
[0007] В другом аспекте, настоящее изобретение относится к комплементирующей клетке для продуцирования вектора по изобретению. Клеточная линия по изобретению получена из клеток U20S, которые были сконструированы, чтобы экспрессировать ICP4 и ICP27 в случае, когда клетка инфицирована ВПГ.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0008] Фиг. 1А схематически показывает экспрессию репортерного гена вектор. Верхняя линия представляет клон KOS ВАС штамма полноразмерного ВПГ-1 дикого типа (Gierash, J. Virol. Meth, 135: 197-206 (2006)), использованный для создания JΔN1-векторов по изобретению. Вторая линия представляет JΔΝΙ5, то есть основу JΔΝΙ7-GFP. Третья линия представляет собой большее увеличение области от LAT до UL4 в JΔΝΙ5. На левой стороне 4-й линии (LAT: CAG-GFP) показано положение кассеты экспрессии репортерного в JΔΝΙ7-GFP. На правой стороне 4-й линии (UL3/4: CAG-GFP) показано положение той же экспрессионной кассеты в JΔΝΙ5-производном контрольном векторе (JΔΝΙ6-CAGGFP). Обозначения на Фиг. 1: TR, концевые повторы; IR, внутренние повторы; UL, уникальная длинная область; US, уникальная краткая область; числа, определяющие расположение различных IE-генов; Δ,делеция; β, ранний промотор; CTRL, инсулятор; LAT Р2, элемент долговременной экспрессии; CAG, кассета CMV/актин/энхансер глобина/промотор/интрон. Фиг. 1B представляет собой набор фотографий, показывающих экспрессию гена-репортера. Обозначения на Фиг. 1B: HDF, фибробласты кожи человека; hpi, часы после инфицирования; dpi, дни после инфицирования; MOI, множественность инфицирования.
[0009] Фиг. 2A-2B представляет собой набор фотографий, сравнивающих экспрессию трансгена в JΔΝΙ6-CAGGFP- и JΔΝΙ7-GFP-инфицированных клетках. Фиг. 2A показывает экспрессию GFP ("EGFP") и mCherry в клетках HDF и U20S, инфицированных с указанной множественностью инфекции. Изображения флуоресцентной микроскопии были получены через 3 дня после инфекции. Фиг. 2B показывает продолжительность трансгенной экспрессии в инфицированной HDF (MOI=0,5). Изображения были получены на 1-14 день после инфицирования.
[0010] Фиг. 3 представляет собой набор фотографий, демонстрирующих размножение вируса JΔΝΙ7-miR302GFP на комплементирующих (U20S-ICP4/ICP27) клетках. Фотографии были сделаны через 3 дня после инфицирования.
[0011] Фиг. 4 представляет собой схематическое изображение конструкции направленной плазмиды для введения тетрациклин-индуцируемого промотора и Gateway рекомбинационной кассеты в локус LAT ВПГ-вектора. Обозначения на Фиг. 4: Zeo, ген устойчивости к зеомицину; Ст, ген устойчивости к хлорамфениколу; ccdB, ген токсина для отрицательной селекции; LATP2, LAT-элемент долговременной экспрессии; CTRL2, граница хроматина/инсуоляторный элемент 2 локуса LAT.
[0012] Фиг. 5 представляет схему конструирования лентивирусной плазмиды дл экспрессии ICP27 в линии клеток U20S-ICP4. Обозначения на Фиг. 5: P, промотор; bla, ген устойчивости к бластицидину.
[0013] Фиг. 6A представляет собой набор фотографий, показывающих иммунофлюоресцентое окрашивание ICP27 клеток U20S-ICP4 и различных клональных клеток U20S-ICP4/ICP27, инфицированных вирусом QOZHG (ICP4-null; ICP27-null). Фиг.6В представляет собой набор фотографий, демонстрирующих рост вируса QOZHG (экспрессия GFP с кассеты промотор HCMV-GFP в геноме вируса) в клетках U20S-ICP4 и U20S-ICP4/ICP27.
[0014] Фиг. 7 представляет собой схематическое изображение в ВПГ-вектора по настоящему изобретению (JΔΝΙ7-GFP, в центре), область LAT JΔΝΙ7-GFP (вверху) и область LAT ВПГ дикого типа (внизу).
[0015] Фиг. 8 представляет собой последовательность области LAT ВПГ-вектора JΔΝΙ7-GFP по настоящему изобретению.
[0016] Фигуры. 9A-9С. структуры векторного генома и комплементирующих клеток для продуцирования вируса. (Фиг. 9) Схема представления генома ВПГ-1 дикого типа KOS в KOS-37 BAC (24) и геномы производных с делецией IE-генов JΔΝΙ2, JΔΝΙ3 и JΔΝΙ5. UL, уникальный длинный сегмент; US, уникальный короткий сегмент. Открытые боксы: терминальные и внутренние инвертированные повторы. Элементы ВАС, в том числе ген устойчивости к хлорамфениколу и кассета экспрессии β-галактозидазы, расположены между участками LoxP в межгенной области UL37-UL38 region (Gierasch et al., J. Virol. Methods 135, 197-206 (2006)). Область US в KOS-37 ВАС и ее производных инвертирована по сравнению со стандартным представлением генома ВПГ. Делеции в конструкциях JΔΝΙ обозначены черными боксами и символом Δ; промотор и кодон инициации трансляции ICP47 делетированы как часть делеции соединения. IE-гены, преобразованные в кинетику ранней экспрессии путем замены промотора (ICPO, ICP27) или делецией TAATGARAT (ICP22), представлены заштрихованными боксами и символом β перед числом ICP. Все JΔΝΙ-рекомбинанты содержат гипер-активационные N/T мутации в гене gB (Uchida et al., J. Virol. 84, 12200-09 (2010)) и кассету промотор убиквитина C (UbCp)-mCherry в локусе ICP4; полиА область SV40 кассеты mCherry представлена небольшим узорным боксом. (Фиг. 9В) Вестерн-блот-анализ комплементирующих клеток. Неинфицированные клетки и клетки, инфицированные вирусом QOZHG (слева) или JΔΝΙ5 (справа) с MOI=1 собирали через 24 hpi, и экстракты подготавливали для гель-электрофореза. Блоты проверяли антителами на ICP4, ICP27 или α-тубулин в качестве контроля нагрузки. (Фиг. 9С) Роста вируса JΔΝΙ2 и JΔΝΙ5 в клетках U20S, U20S-ICP4, U20S-ICP4/27 и Vero-7b. Клетки инфицировали при MOI=0,001, и внеклеточной вирус собирали ежедневно с трех лунок и титровали на клетках U20S-ICP4/27.
[0017] Фиг. 10A-10B. Относительные уровни нуклеарных вирусных ДНК после инфицирования равными gc или БОЕ. HDF инфицировали указанными векторами JΔΝΙ при 5000 gc/клетку (Фиг. 10A) или 1 БОЕ/клетку (Фиг. 10B). Через 2 hpi, ядерную ДНК выделяли, и относительные числа вирусных gc определяли при помощи кПЦР для гена gD, нормированной на гены клеточной 18S рРНК.
[0018] Фиг. 11А-11D. Цитотоксичность JΔΝΙ и экспрессия вирусных генов в некомплементирующих клетках. (Фиг. 11 А) In vitro цитотоксичность. Клетки HDFS и Vero были инфицированы в 25,000 gc/клетку и жизнеспособность клеток в трех лунках была измерена через 5 dpi путем МТТ. Нанесенные значения представляют собой среднее отношение клеток, инфицированных вирусом к имитационно-инфицированных клеткам. Скобки со звездочками указывают статистически значимые различия (р<0,05) между JΔΝΙ2- и JΔΝΙ3-инфицированными клетками Vero и между JΔΝΙ3- и JΔΝΙ5 -инфицированными клетками Vero. (Фиг. 11В). Иммуноблот-анализ продуктов IE-генов в HDF. Клетки инфицировали вирусами KOS, QOZHG или JΔΝΙ с 1 БОЕ/клетку, и экстракты получали через 24 hpi. Блоты зондировали антителами на продукты указанных IE-генов или α-тубулин в качестве контроля нагрузки. (Фиг. 11С) Экспрессия JΔΝΙ IE-гена определяли qRT-PCR. HDF инфицировали указанными вирусами при 1, 000 gc/клетку. мРНК выделяли через 12 hpi и обратно транскрибировали для кПЦР-определения уровней кДНК генов, перечисленных в верхней части. Экспрессию нормировали на уровни рРНК 18S, и она показана по отношению к JΔΝΙ2-инфицированным клеткам. (Фиг. 11D) qRT-PCR-анализ экспрессии ранних (верхние панели) и поздних генов (нижние панели). HDF инфицировали и обрабатывали, как в (Фиг. 11C). ICP6 можно считать задержанным IE-ген и сгруппированным с ранними генами здесь, поскольку его экспрессия, по имеющимся сведениям, в большей степени зависит от ICP0, чем от VP16 или ICP4 (Desai et al., J. Virol 67, 6125-35 (1993); Sze et al., Virus Res. 26, 141-52 (1992); Harkness et al., J. Virol. 55(12) 6847-61 (2014)).
[0019] Фиг. 12A-12D. Экспрессия гена-репортера в JΔΝΙ-инфицированных HDF. (Фиг. 12А) Флуоресценция mCherry. Клетки инфицировали с указанным gc/клетку и фотографировали через 24 hpi. (Фиг. 12В) Относительные уровни мРНК mCherry. HDF заразил инфицировали при 5000 gc/клетку и собирали через 6 hpi для выделения мРНК, обратной транскрипции и кПЦР, как на Фиг. 3C. (Фиг. 12С) Флуоресценция mCherry в клетках U20S. Клетки инфицировали при 1000 gc/клетку и фотографировали через 24 hpi. (Фиг. 12D) Индукция экспрессии mCherry в JΔΝΙ5-инфицированных HDF. Клетки были инфицированы при указанном gc/клетку, супер- инфицировали через 24 ч с QOZHG при 5000 gc/клетку и фотографировали через 24 ч.
[0020] Фиг. 13A-13D. Структуры геномов JΔΝΙ6GFP и JΔΝΙ7GFP и экспрессия гена-репортера. (Фиг. 13 А) JΔΝΙ7GFP содержит экспрессионную кассету промотор-CAG EGFP в 2 т.п.н. интронной области LAT между LATP2 долговременной экспрессии/энхансерной областью и расположенным вниз по течению CTCF-связывающим мотивом (CTRL2) в интроне. LATP2 распространяется от сайта инициации транскрипции LAT до внутренней части 2-т.п.н. интрона. JΔΝΙ6GFP содержит ту же кассету экспрессии промотор CAG-EGFP между генами UL3 и UL4. Поли А область β-глобина кролика кассеты CAGP-EGFP представлена в небольшом узорном боксе. (B-D), Экспрессия EGFP и mCherry в инфицированных HDF. (Фиг. 13B) Клетки инфицировали вирусом JΔΝΙ6GFP или JΔΝΙ7GFP при различных gc/клетку и флуоресценцию визуализировали через 3 dpi. (Фиг. 13С) HDF инфицировали вектором JΔΝΙ6GFP или JΔΝΙ7GFP при 12,500 gc/клетку и собирали через 3 или 5 дней для выделения мРНК и анализа qRT-PCR для 2 генов-репортеров. Экспрессия, нормированная на 18S рРНК, показана в отношении JΔΝΙ6GFP-инфицированных клеток на 3 день. (Фиг. 13D) HDF инфицировали вирусом JΔΝΙ6GFP или JΔΝΙ7GFP при 25000 gc/клетку и EGFP-флуоресценцию фотографировали через 7, 14 и 28 dpi.
[0021] Фиг. 14А-14С. Эффект элементов локуса LAT на EGFP-экспрессию с JΔΝΙ7GFP. (Фиг. 14А) Геномное представление JΔΝΙ7GFP и производных, делетированных по CTRL1 (ACL), CTRL2 (AC2) или LATP2 (ALP2), по отдельности или в комбинации. Делеция положений 8978-9161 в JQ673480 охватывает CTRL1, и делеция положений 5694-5857 в JQ673480 охватывает CTRL2. Эти делеции охватывают несколько пар оснований за пределами CTCF-связывающих мотивов. (Фиг. 14В) Экспрессия гена-гепортера в инфицированных HDF. Клетки инфицировали указанными вирусами при 12500 gc/клетку и флуоресценцию регистрировали через 3 dpi. (Фиг. 14С) Относительные уровни мРНК EGFP в HDF, инфицированных JΔΝΙ7GFP, производные, делетированные по LAT-элементам, или JΔΝΙ6GFP; вирусы идентифицируются сокращенными названиями. Клетки инфицировали при 12500 gc/клетку и процессировали через dpi для qRT-PCR анализа. Уровни экспрессии нормировали на 18S рРНК и представляли по отношению к уровню в JΔΝΙ7GFP-инфицированных клетках.
[0022] Фиг. 15A-15D. Анти-сайленсинговая активность LAT последовательностей, расположенных в других участках вирусного генома. (Фиг. 15 А) Конструирование векторов JΔΝΙ9 и JΔΝΙIO. XhoI-фрагмент, включающий CTRL1, LATP2 и CTRL2, был удален из генома JΔΝΙ5, и GW-рекомбинационную кассету вводили между UL45 и UL46 с образованием JΔΝΙ9GW, или между UL50 и UL51, с образованием JΔΝΙIOGW (верхний). Те же сайты XhoI были использованы, чтобы выделить CAGP-GFP-содержащий LAT-фрагмент из JΔΝΙ7GFP (внизу слева). XhoI-фрагмент клонировали в pENTRlA (внизу справа) и переносили в JΔΝΙ9GW или JΔΝΙIOGW при помощи attL/attR рекомбинации с соответствующими GW-кассетами (LR реакция) для получения JΔΝΙ9LAT-GFP и JΔΝΙ10LAT-GFP, соответственно. В качестве контроля, кассета CAGP-GFP без LAT последовательностей рекомбинировали через pENTRlА интермедиат в GW локус JΔΝΙ9GW или JΔΝΙIOGW, с образоанием JΔΝΙ9GFP и JΔΝΙIOGFP. (Фиг. 15В) Экспрессия гена-репортер в HDF, инфицированных вирусами JΔΝΙ9 или JΔΝΙ10. HDF инфицировали указанными вирусами при 12500 gc/клетку. Флуоресценцию EGFP и mCherry регистрировали через 3 dpi. (Фиг. 15C) Уровни мРНК EGFP в инфицированных HDF определяли при помощи qRT-PCR, как в предыдущих фигурах. Уровни показаны относительно JΔΝΙ9GFP- или JΔΝΙlOGFP-инфицированных клеток. JΔΝΙ10AC12LP2-GFP конструировали передачей XhoI LAT-фрагмента из JΔΝΙ7AC12LP2-GFP в GW сайт JΔΝΙ10GW аналогично конструкции JΔΝΙ10LAT-GFP, описанной выше. (Фиг. 15D) Влияние делеции обеих CTRL и LATP2 из JΔΝΙ10LAT-GFP на трансгенную экспрессию. HDF инфицировали при 12500 gc/клетку и флуоресценцию EGFP и mCherry регистрировали через 3 dpi.
[0023] Фиг. 16A-16D. Экспрессия генов-репортеров с JΔΝΙ-векторов в других некомплементирующих клетках. (Фиг. 16A) Клетки, перечисленные вверху, инфицировали JΔΝΙ6GFP или JΔΝΙ7GFP при gc/клетку, указанной ниже панелей. Флуоресценцию EGFP и mCherry регистрировали через 3 dpi. (Фиг. 16B) Экспрессию гена EGFP в клетках, инфицированных, как в (Фиг. 16 А), определяли через 3 dpi путем анализа qRT-PCR. Результаты, нормированные на 18S рРНК, показаны относительно JΔΝΙ6GFP-инфицированных клеток. (Фиг. 16C) hMDSC были инфицированы вирусом JΔΝΙ6GFP или JΔΝΙ7GFP при 50000 gc/клетку и флуоресценцию EGFP фотографировали на 14 и 28 dpi. (Фиг. 16D) qRT-PCR определение уровня мРНК EGFP в клетках hEK, hPAD and hHEP через 3 дня после инфицирования JΔΝΙ10GFP или JΔΝΙ10:LAT-GFP при 12,500 gc/клетку. Нормированная экспрессия показана относительные JΔΝΙ10GFP-инфицированных клеток.
[0024] Фиг. 17 графически изображает конструирование JΔΝΙ8 от JΔΝΙ5.
[0025] Фиг. 18 приводит данные, касающиеся роста JΔΝΙ8 по отношению к JΔΝΙ5 в комплементирующих клетках (U20S-ICP4/27) после инфицирования на 1 геномной копии (gc)/клетку. На верхней панели представлена экспрессия гена-репортера (mCherry), в то время как нижняя левая панель информирует о выходе вируса в бляшкообразующих единицах (БОЕ), а нижняя правая панель - о выходах вируса в геномных копиях (gc).
[0026] Фиг. 19 приводит данные, показывающие, что вирусные гены экспрессируются раньше с JΔΝΙ8, чем с JΔΝΙ5 в комплементирующих клетках. В каждой панели, нижняя линия представляет данные для JΔΝΙ5, в то время как верхняя линия представляет данные для JΔΝΙ8. Данные были собраны по qRT-PCR и выражены как кратное различие по отношению к данным для JΔΝΙ5 в точке 6 часов после инфекциии (hpi).
[0027] Фиг. 20 графически представляет геномную структуру JDNI7GFP (также известную как JΔΝΙ7GFP) и JDNI8GFP (также известную как JΔΝΙ8GFP).
[0028] Фиг. 21 представляет данные, демонстрирующие, что инфицирование клеток кожных фибробластов человека (HDF) равным количеством JΔΝΙ8 и JΔΝΙ5 (выраженных в gc) приводит равным количествам вирусной ДНК в ядре. Данные представлены для момента два часа после инфицирования (hpi).
[0029] Фиг. 22 приведены данные, сравнивающие жизнеспособность клеток (МТТ) HDF, инфицированных различными ВПГ-векторами при двух M.O.I. Нижняя линия -Х- в каждой панели представляет результаты для JΔΝΙ5, в то время как верхняя линия -Х- в каждом графике представляет результаты для JΔΝΙ8.
[0030] Фиг. 23 приведодит данные, сравнивающие анализ жизнеспособности клеток (МТТ) HDF, инфицированных различными ВПГ-векторами при разных числах вирусных геномных копий на клетку. Для левой панели (12500 gc/клеткe) M.O.I. для JΔΝΙ5 составляла 5 БОЕ/клетку, в то время как M.O.I. для JΔΝΙ8 составляла 18 БОЕ/клетку. Для правой панели (25000 gc/клетки), M.O.I. для JΔΝΙ5 составляла 11, в то время как M.O.I, для JΔΝΙ8 составляла 33. Нижняя линия -Х- в каждой панели представляет результаты для JΔΝΙ5, в то время как верхняя линия -Х- в каждом графике представляет результаты для JΔΝΙ8.
[0031] Фиг. 24 приведодит данные, сравнивающие жизнеспособность клеток, инфицированных KOS, JΔΝΙ5 и JΔΝΙ8 в шести типах клеток (HDF, неонатальных кератиноцитах человека, нервных стволовых клетках человека, Vero, преадипоцитах человека и гепатоцитах человека). МТТ проводили с использованием 25000 gc/клетку, с данными на 5 дней после инфицирования (dpi).
[0032] Фиг. 25 представляет данные доза-ответ через три дня после инфицирования сравнения экспрессии гена-репортера (mCherry или усиленный зеленый флуоресцентный белок (EGFP)) между фибробластов кожи человека (HDF), инфицированных при указанном gc/клетку либо JΔΝΙ7GFP, либо JΔΝΙ8GFP.
[0033] Фиг. 26 представлены данные типа «время-курс», сравнивающие экспрессии гена-репортера (EGFP или mCherry) в фибробластах кожи человека (HDF), инфицированных при 25,000 gc/клетку либо JΔΝΙ7GFP, либо JΔΝΙ8GFP.
[0034] Фиг. 27 представляет результаты экспериментов по сравнению экспрессии гена-репортера (EGFP или mCherry) для неонатальных кератиноцитов человека через три дня после инфицирования либо 12,500 gc/клетку, либо 25,000 gc/клетку либо JΔΝΙ7GFP, либо JΔΝΙ8GFP.
[0035] Фиг. 28 представляет результаты экспериментов по сравнению экспрессии гена-репортера (EGFP или mCherry) для нейронов корней спинного ганглия (DRG) крысы через три дня после инфекции при 6250 gc/клетку либо JΔΝΙ7GFP, либо JΔΝΙ8GFP.
[0036] Фиг. 29 представляет результаты экспериментов по исследованию экспрессии трансгенов (mCherry или EGFP) в нервных клетках, инфицированных JΔΝΙ7GFP. Вверху, отдельные (слева; 40x) или объединенные (центр, справа; 20x) изображения флуоресценции EGFP и mCherry в указанные дни после инфицирования (dpi); снизу, отдельные и объединенные изображения при 15 dpi (10X).
[0037] Фигура 30 представляет схематическую диаграмму pCX4Hyg-Cre.
[0038] Фигура 31 представляет схематическое изображение создания pCX4Hyg-Cre.
[0039] На фиг. 32 приведены данные сравнения уровней мРНК EGFP между JΔΝΙ7GFP- и JΔΝΙ6GFP-инфицированными клетками HDF, определенных количественным обратно-транскрипционным (RT)-PCR.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0040] Следующие патенты и публикации, относящиеся к различным ВПГ-векторным технологиям, включены в данное описание путем ссылки. Патент США № 5658724 относится к штаммам ВПГ, дефектным по ICP4 и ICP27, и способам их получения, роста и применения. Патент США N 5804413 относится к клеточной линии, содержащей ДНК, кодирующую ICP4, ICP27 и ICPO. Патент США № 5849571 относится к промоторам латентносто-активных вирусов герпеса и их использованию. Патент США N 5849572 относится к вектору на основе ВПГ-1, содержащему промотор LAT. Патент США N 5879934 относится к рекомбинантному ВПГ-вектору, содержащему геномные мутации в генах ICP4 и ICP27, таких, что генные продукты ICP4 и ICP27 являются дефектными. Патент США N 5998174 относится к способам получения ВПГ-вектора. Патент США N 6261552 относится к ВПГ-вектору, содержащему геном ВПГ с делецией или мутацией в нативной последовательности TAATGARAT, при этом указанная делеция или мутация вызывает замедление кинетики экспрессии нативного предраннего гена в указанном геноме, когда указанный геном находится внутри клетки, которая содержит генные продукты ВПГ ICP4. Патент США N 7078029 относится к ВПГ, имеющей геном с мутацией последовательности TAATGARAT такой, что в присутствии продукта гена ICP4, нативный предранний ген экспрессируется с генома с задержкой кинетикой. Патент США N 7531167 относится к ВПГ-вектору, содержащему делеции только в гене ICP4, ICP27 и UL55. Публикация заявки на патент США № 2013/0096186 относится к ВПГ-вектору, содержащему мутантный gB и/или мутантный gH гликопротеин. Публикация международной патентной заявки WO 1999/06583 относится к ВПГ, содержащий оболочку, включающую лиганд, отличный от нативного.
[0041] В одном варианте осуществления изобретение относится к вектору на основе вируса простого герпеса (ВПГ), который не экспрессирует токсичные нативные гены ВПГ в некомплементирующих клетках и способен к непрерывной экспрессии трансгена. Например, ВПГ-вектор по изобретению может экспрессировать трансген в течение по меньшей мере 14 дней, например, по меньшей мере 28 дней и предпочтительно по меньшей мере 60 дней, в культивированных клетках дермальных фибробластов человека (HDF). Желательно, чтобы такая экспрессия трансгена с вектора по изобретению происходила в отсутствии детектируемого продукта гена ICPO в таких клетках, например, отсутствие какого-либо продукта гена ICPO в таких клетках. В конкретном варианте осуществления, вектор в соответствии с настоящим изобретением, содержащий трансген (например, кодирующий усиленный зеленый флуоресцентный белок (EGFP) или другой маркер), встроенный в область LAT между элементом LAT P2 и элементом CTRL2, и который имеет как CTRL1, так и CTRL2, может экспрессировать трансген по меньшей мере в 20 (или по меньшей мере около 20) раз превышающий уровень, который дает вектор, имеющий в остальном идентичные генетические мутации, но в котором такой трансген встроен между UL3 и UL4, как определено количественной ОТ-ПЦР через семь дней после инфицирования, и по меньшей мере в 30 (или по меньшей мере около 30) раз превышающий уровень, как определено количественной ОТ-ПЦР, на третий день после инфицирования. Смотри Фиг. 13 и 32. Такой вектор предпочтительно экспрессирует такой трансген на таких уровнях и в течение такого периода времени, при которых трансген находится под функциональным контролем CMV-энхансера/промотора куриного бета-актина/химерного интрона. То, что такие клетки можно культивировать в течение таких периодов, будучи инфицированными ВПГ-вектором по изобретению, также свидетельствует о том, что вектор не является токсичным для таких клеток (то есть, не экспрессирует токсичные нативные ВПГ-гены в клетках). Конечно, некомплементирующая клетка(и) может также представлять собой клетку другого клеточного типа, и может представлять собой клетку in vivo, которая представляет интерес для терапевтического применения.
[0042] Например, ВПГ-вектор по изобретению может включать геном, содержащий трансген, встроенный (а) в генную область инкубационного-ассоциированного транскрипта (LAT), (b) в локус ICP4 (и предпочтительно только в тот, где соединение делетровано, как описано в настоящем документе) и/или (с) в геном вектора в функциональной связи с одной или несколькими инсуляторными последовательностями в геноме. Предпочтительно, вектор не экспрессирует ICPO, ICP4, ICP22, ICP27 и ICP47 в качестве предранних генов (хотя в некоторых вариантах осуществления экспрессия ICP47 может быть желательной). Не желая быть связанными теорией, предполагается, что, в зависимости от активности промотора в трансгене, вектор по изобретению может экспрессировать трансген в клетке млекопитающего любого типа (особенно человека), котору он может инфицировать без цитотоксичности, связанной с вирусной генной экспрессии. Вектор по изобретению может быть представлен как выделенная ДНК, ДНК внутри клетки или упакованный в вирусной оболочке.
[0043] Любой подходящий способ может быть использован для визуализации вектора по изобретению, неспособного экспрессировать ICPO, ICP4, ICP22, ICP27 и ICP47 качестве предранних генов в некомплементирующих клетках. Например, геном вектора по изобретению может быть сконструирован, чтобы содержать инактивирующие мутации (например, делеции) одного из или каждого из этих генов (например, делеции в пределах или всей последовательности, кодирующей один или более из генов ICPO, ICP4, ICP22, ICP27 и ICP47 (предпочтительно содержащий инактивирующие делеции по меньшей мере ICPO, ICP4 и ICP27, более предпочтительно, содержащий инактивирующие делеции ICPO, ICP4, ICP27 и ICP47), или также альтернативно включающий промотор или другие регуляторные последовательности такого гена (в). Альтернативно, один или более из этих ВПГ-генов может быть сконструирован, чтобы экспрессироваться в качестве раннего или позднего гена. Например, геном некоторых вариантов осуществления вектора по изобретению может быть сконструирован, чтобы сохранить последовательность, кодирующую один из более из этих генов, но заменить его промотор с промотором, делающим ген по экспрессии как ранним (бета) или поздним (гамма), но не предранним (альфа) геном. Например, такой ген может быть помещен под контроль промотора, реагирующего на ICP4 (подход предпочтительный по меньшей мере по отношению к ICP22, с тем чтобы экспрессировать ICP22 в качестве раннего гена, а не предраннего гена). Подходящим промотором для экспрессии такого гена с ранней (бета) кинетикой является tk-промотор ВПГ. Промотор ICP22 может быть преобразован в промотор с ранней кинетикой путем укорочения, то есть удаления регуляторных последовательностей, включая TAATGARAT. Можно удалить весь промотор ICP47 и инициирующий кодон. В качестве альтернативы, в некоторых вариантах осуществления, ген ICP47 может экспрессироваться как предранний гена, чтобы защитить инфицированные клетки от иммунного распознавания recognition (Hill er al, Nature 1995, 375(6530): 411-415; Goldsmith et al, J Exp Med. 1998; 187(3): 341-348).
[0044] В дополнение к нарушению экспрессии ICPO, ICP4, ICP22, ICP27 и ICP47 желательно, чтобы вектор по изобретению также не экспрессировал UL41 (т.е. запорный (vhs) ген хозяина). UL41 является РНКазой, которая разрушает многие мРНК хозяина и вирусные мРНК, вызывает быстрое выключение синтеза белка клеткой-хозяином и проникает в клетку в качестве компонента оболочки вириона. Так, например, ген, кодирующий UL41 может быть делетирован из генома вектора по изобретению. Не желая быть связанными теорией, полагают, что такое манипулирование дополнительно усиливает способность вектора по изобретению расти в комплементирующих клетках при помощи сберегания комплементирующих мРНК ICP4 и ICP27, и усиливает экспрессию трансгенов в некомплементирующих клетках при помощи сберегания трансгенных мРНК.
[0045] Следует признать, что геномные последовательности нескольких штаммов ВПГ известны специалистам в данной области техники (например, MacDonald, J. Virol., 86(11): 6371 (2012); McGeoch, J. Gen. Virol., 69: 1531-1574 (1988); инвентарный № GenBank JQ673480; референсная последовательность NCBI: NC_001806.1; NC_001806.1; MacDonald, J. Virol. 86(17): 9540 (2012); инвентарный № GenBank JX 142173, которые включены в настоящий документ в качестве ссылки). Соответственно, манипулирование последовательностью генов и локусов ВПГ находится в пределах компетенции специалистов. Следует также отметить, что эти опубликованные последовательностями являются лишь типичными и что другие штаммы или варианты ВПГ могут использоваться в качестве исходного генома при конструировании вектора по изобретению.
[0046] Кроме того, геном вектора по изобретению может содержать кассету бактериальной искусственной хромосомы (BAC). Включение такого BAC-кассеты облегчает размножение и манипуляцию с геномом вектора по изобретению в бактериях. ВАС-кассета может включать бактериально-экспрессируемые последовательности, которые помогают в использовании бактериальных штаммов, например, селективные гены, такие как гены, придающие бактериям устойчивость к антибиотикам или токсинам (например, предпочтительно хлорамфеникол, но и другие гены устойчивости (например, для тетрациклина, ампициллина, зеоцина и т.д.) также могут быть использованы). ВАС-кассета также может дополнительно включать в себя репортерных генов (например, LacZ (кодирование бета-галактозидазы) или ген, кодирующий флуоресцентный белок (например, GFP (кодирующий зеленый флуоресцентный белок), YFP (кодирующий желтый флуоресцентный белок), RFP (кодирующий красный флуоресцентный белок) и его аналоги (например, кодирующий iRFP, EGFP и тому подобное)) под контролем эукариотического промотора, такого как конститутивный промотор млекопитающего (например, SV40, RSV, CMV, убиквитин C (UbC), CAG или (промотор β-актина, и т.д.).
[0047] ВАС-кассета может быть помещена в геном вектора по изобретению в любом подходящем участке, например, межгенной области UL37-UL38 в векторном генома (например, Gierash, J. Virol. Meth., 135: 197-206 (2006) и Morimoto, Microbiol. Immunol., 53: 155- 161 (2009)). Кроме того, желательно, чтобы ВАС-кассета была фланкирована последовательностями, облегчающими удаление ВАС-кассеты, например, сайтами/консенсусными последовательностями узнавания сайт-специфической рекомбиназой (например, такие, которые узнаются ферментами, такими как ere, dre, flp, KD, B2, B3, R и т.д.). Включение таких сайтов облегчает удаление ВАС-кассеты, если это необходимо, так как ВАС-последовательности, как было показано, ослабляют рост вируса в культуре клеток (например, Gierash, J. Virol. Meth, 135: 197-206 (2006)). Кроме того, удаление ВАС-кассеты может увеличить способность вектора включать один или несколько трансгенов, поскольку BAC-кассеты составляют порядка примерно 11 т.п.н. Следует иметь в виду, что вектор по изобретению также может иметь консенсусную последовательность для фермента рекомбиназы (например, LoxP), особенно не являющуюся нативной для генома ВПГ, например, в результате удаления BAC-кассеты с помощью линии клеток, которые экспрессируют соответствующую сайт-специфическую рекомбиназу для эксцизии ВАС-кассеты, как таковой оставляет одну копию консенсусных последовательностей для фермента рекомбиназы в геноме ВПГ (например, в межгенной области UL37-UL38, если таким было расположение вставки BAC-кассеты).
[0048] Как указано выше, вектор по изобретению может включать в себя по меньшей мере один трансген, встроенный в функциональной связи с одной или несколькими инсуляторными последовательностями в геноме вектора ВПГ. Под "функционально связанный" понимается, что одна или более инсуляторных последовательностей дает возможность трансгену экспрессироваться в клеточной среде, в которой генетические элементы (т.е., "гены"), в противном случае присутствующие в геноме ВПГ, являются транскрипционно молчащими. Не желая быть связанными какой-либо теорией, полагают, что такие инсуляторные последовательности предотвращают образование гетерохроматина в участках таких инсуляторных последовательностей и в пределах примерно от 1 т.п.н. до примерно 5 т.п.н. от этого, что в случае образования заглушает экспрессию генов. Таким образом, инсуляторная последовательность для использования в векторе по изобретению обычно представляет собой последовательность, которая препятствует связыванию или формированию гетерохроматина, который в противном случае заглушает экспрессию трансгена. Неограничивающие примеры подходящих изоляторных последовательностей включают ВПГ-хроматин (CTRL/CTCF-связывающие/инсулятор) связывающие элементы CTRL1 и CTRL2 (которые могут быть нативными для локуса LAT, как описано в настоящем документе, или перемещены в эктопическое положение в геноме), гиперчувствительный сайт 4 инсулятор (cHS4) курицы, повсеместно распространенный в хроматине открывающий элемент (UCOE) HNRА2В1--CBX3 человека и область присоединения к каркасу/матриксу (S/MAR) из гена бета-интерферона человека (IFNB1) (Emery, Hum. Gene Ther. 22, 761- 74 (2011); Antoniou et al, Hum. Gene Ther. 24, 363-74 (2013)).
[0049] Помимо трансгена, встроенного рядом с инсуляторными последовательностями, нативных в геноме ВПГ (такие как последовательности CTRL1 и CTRL2 в пределах области LAT), инсуляторные последовательности могут быть встроены в векторный генома в любом подходящем участке. Эти инсуляторные/граничные элементы могут быть введены в геном вектора по изобретению стандартными способами, и могут быть включены в ту же кассету в качестве трансгена или введены отдельно в геном так, чтобы фланкировать или иным образом находиться в функциональной связи с данной трансгенной кассетой. Таким образом, можно вставить генетическую кассету, включающую, например, одну или более эктопических инсуляторных последовательностей, функционально сходных с таковыми, изначально находящимися в области LAT, фланкирующей, или иным образом функционально связанной с трансгеном. Следует понимать, что вектор по изобретению может включать в себя несколько трансгенов, функционально связанных с несколькими инсуляторными последовательностями, в том числе с участками, эктопическими к LAT. В варианте осуществления, в котором один или более трансгенов встраивается в участок, отличный от LAT и функционально связан с CTRL1 и/или CTRL2, желательно делетировать из LAT или мутировать последовательности CTRL1 и/или CTRL2 в LAT, чтобы свести к минимуму или устранить рекомбинационные события между нативными последовательностями в пределах LAT и теми, сконструированными, чтобы быть в функциональной связи с трансгеном в эктопическом (не LAT) участке вектора по изобретению. Если CTRL1 и CTRL2 остаются в LAT или перемещаются эктопически, предпочтительным участком для встраивания трансгена в геном вектора по изобретению является участок от CTRL1 и CTRL2 (например, в пределах примерно 1-4 т.п.н. от каждого из CTRL1 и CTRL2, причем CTRL1 и CTRL2 фланкируют трансген).
[0050] В пределах вектора по изобретению, одна или более из таких инсуляторных последовательностей функционально связана с трансгеном, так что трансген защищен от генного сайленсинга и экспрессируется. Как правило, трансген и инсуляторная последовательность (и) должны находиться в непосредственной близости в геноме, а именно, разделены менее чем около 5 т.п.н., или меньше, чем примерно 4 т.п.н., или меньше, чем примерно 3 т.п.н., или меньше, чем примерно 2 т.п.н. или меньше, чем примерно 1 т.п.н. Также может быть желательным для экспрессионной кассеты (в том числе трансгена) являться функциональной между двумя инсуляторными последовательностями таким образом, что инсуляторные последовательности фланкируют представляющий интерес трансген (ы) (см, например, Emery, Hum. Gene Ther. 22, 761-74 (2011); Antoniou et al., Hum. Gene Ther. 24, 363- 74 (2013)).
[0051] Один предпочтительный участок для вставки трансгена (например, первого трансгена) в вектор по изобретению находится между инсуляторными последовательностями в пределах генной области LAT векторного генома - специфически встроен между элементами границы хроматина (CTRL/CTCF-связывание/инсулятор), расположенными выше по течению от LAT-промотора LAP1 (CTRL1) и в LAT 2-т.п.н. интроне (CTRL2), соответственно (Amelio et al, J Virol 2006, 80(5): 2358-2368; Bloom, Biochim. Biophys. Acta, 1799: 246-256 (2010)). Эта область называется в настоящем документе как LAT- (ген) область или локус. Поэтому желательно, чтобы геном вектора по изобретению содержал (например, сохранял) CTRL1 и CTRL2 (Фиг.7, вверху). Не желая быть связанными теорией, полагают, что присутствие CTRL1 и CTRL2 защищает регион от образования гетерохроматина и, таким образом, вносит свой вклад в генную область LAT как привилегированный участок для экспрессии трансгенов. Таким образом, вектор экспрессирует трансген, встроенный в генную область LAT в некомплементирующих клетках. В предпочтительном варианте осуществления, вектор содержит множество трансгенных кассет пределах генной области LAT, каждая из которых содержит отдельный промотор и кодирующую область, и при этом каждая может являться моно- или полицистронной.
[0052] Кроме того, предпочтительно, чтобы область LAT вектора по изобретению (который включает по меньшей мере один трансген, как отмечено в настоящем документе) также включала (например, сохраняла) энхансерный элемент LATP2 или LAP2. Опять же, не желая быть связанными теорией, предполагается, что присутствие энхансерного элемента LATP2 или LAP2 способствует способности трансгена в пределах генной области LAT экспрессировать кодирующую последовательность(ти) длительно (Goins, J. Virol., 73: 519-532 (1999); Lilley, J. Virol, 75: 4343-4356 (2001)). В конкретном предпочтительном варианте осуществления трансген (ы) в генной области LAT встроен ниже по течению от энхансерного элемента LATP2 или LAP2. Тем не менее, изобретение предусматривает варианты, в которых трансген(ы) встраиваются выше по течению (по отношению к направлению транскрипции LAT) от энхансерного элемента LATP2 или LAP2. Желательно, чтобы трансген направлялся вне элементов LATP2 или LAP2 в пределах области гена LAT (см, например, Фиг. 7, вверху).
[0053] Другой предпочтительный участок для встраивания трансгена (например, второй трансгенной) в векторе по изобретению находится в локусе гена ICP4. Например, трансген под контролем UBC-промотора, встроенный в этот локус в векторе по изобретению, может продуцировать долговременный сигнал в нейронах гиппокампа и является активным по меньшей мере краткосрочно в DRG крысы.
[0054] Трансгены также могут быть встроены в функциональной связи (например, около (<5 т.п.н.)) с другими инсуляторными последовательностями в геноме вектора по изобретению. Помимо того, что быть встроенными вблизи изоляторных последовательностей, нативных для генома ВПГ, инсуляторные последовательности могут быть встроены в векторный геном в любом подходящем участке. Таким образом, можно вставить генетическую кассету, включающую, например, эктопические инсуляторные последовательности, функционально аналогичные тем, которые изначально присутствуют в LAT-области, фланкирующей трансген. Следует понимать, что вектор по изобретению может включать несколько трансгенов.
[0055] В трансгене (ах), встроенном в вектор по изобретению, присутствует по меньшей мере последовательность промотора и транскрибируемая последовательность, такая, что транскрибируемая последовательность(ти) контролируется промотором. Промотор в трансгене может представлять собой любой необходимый промотор, контролирующий/регулирующий экспрессию транскрибируемой последовательности(тей). Например, промотор может представлять собой клетка-специфический или ткане-специфический промотор (например, EOS, OCT4, Nanog (для ESC/iPSC), SOX2 (для нервных стволовых клеток), aMHC, Brachyury, Tau, GFAP, NSE, синапсина I (для нейронов), Apo A-I, альбумина, ApoE (для печени), МСК, SMC A-актина, тяжелой цепи миозина, легкой цепи миозина (для мышц) и т.д.), такой как промотор, который специфически или предпочтительно экспрессирует гены в определенном типе клеток (например, в клетке печени, клетке легкого, эпителиальной клетке, клетке сердца, нервной клетке, клетке скелетных мышц, эмбриональной, индуцированной плюрипотентной или другой стволовой клетке, раковой клетке и т.д.). Предпочтительные промоторы для использования в сенсорных нейронах включают TRPV1, CGRP и NF200. В других вариантах осуществления промотор в трансгене, встроенном в вектор по изобретению, может представлять собой индуцируемый промотор (например, TRE3G в сочетании с экспрессияй rtTA3G с отдельного промотора в LAT или других индуцируемых промоторов, которые известны в данной области техники). Конечно, промотор в кассете экспрессии трансгена, встроенной в вектор по изобретению, может представлять собой конститутивный промотор млекопитающего, такой, которые известны в данной области (например, SV40, CMV, CAG, EFL КТ, UBC, РСВ, β-актина, PGK и тому подобное).
[0056] В дополнение к промотору (ам) и кодирующей последовательности (ям), трансген (ы), встроенный в геном вектора по изобретению, может также содержать дополнительный регуляторный элемент(ы). Например, трансген(ы) может включать один или несколько сайтов связывания микроРНК. В предпочтительных вариантах трансген (ы) включают тандемных сайты связывания для таких микроРНК, например, 2, 3, 4, 5 или 6 тандемных сайтов (четыре является типичным). Наличие таких сайтов, в частности тандемных участков связывания для таких микроРНК, облегчает подавление экспрессии трансгена в определенных типах клеток. Так, например, вектор, содержащий нужный трансген, для экспрессии в раковой или опухолевой клетке (который может быть токсичным для многих типов клеток) может включать сайты связывания микроРНК в "нормальных" (т.е. не злокачественных) клетках, так что экспрессия трансгена подавляется в незлокачественных клетках.
[0057] Следует отметить, что трансген (ы) в векторе по изобретению может быть моноцистронным (т.е. кодирующим один белок или полипептид) или полицистронным (т.е. кодирующим несколько белков или полипептидов). Кроме того, весь транскрибируемый участок трансгена или его часть может также кодировать нетранслируемую РНК, такую как, например, микроРНК или миРНК. Также вектор по изобретении может содержать несколько отдельных моноцистронных или полицистронных трансгенных единиц (предпочтительно две отдельные трансгенные единицы, но, возможно, больше (например, три, четыре, пять или более отдельных единиц)), каждая со своим собственным соответствующим промотором, транслируемой последовательностью (тями) или нетранслируемой последовательностью(ями) РНК и другими регуляторными элементами.
[0058] Как уже отмечалось, трансген (ы) включает (ют) одну или более транскрибируемых последовательностей (последовательность), которые экспрессируются под контролем промотора и, возможно, другими регуляторными элементами в трансгене (включая функциональную связь с изоляторной последовательностью (ями)). Транскрибируемой последовательность может представлять собой любую последовательностью, желательную для экспрессии в данной клетке, в которую должен быть введен вектор. Неограничивающие примеры транскрибируемых последовательностей, которые могут присутствовать в трансгене в векторе по изобретению включают Oct4, Klf4, Sox2, c-Myc, L- myc, доминантно-негативный p53, Nanog, Glisl, Lin28, TFIID, GATA4, Nkx2.5, Tbx5, Mef2C, Myocd, Hand2, SRF, Mespl, SMARCD3, SERCA2a, РахЗ, MyoD, Lhx2, FoxGl, FoxP2, Isll, Ctip2, Tbrl, Ebfl, Gsx2, Srebp2, фактор VIII, фактор IX, дистрофин, CFTR, GlyRal, энкефалин, GAD67 (или другие изоформы GAD, например, GAD 65), TNFa, IL-4, нейротрофический фактор (например, NGF, BDNF, GDNF, NT-3), Ascl1, Nurr1, Lm×1A, Brn2, Mytll, NeuroDl, FoxA2, Hnf4a, Foxa1, Foxa2 или Foxa3, любую микроРНК или комбинацию микроРНК (например, генный кластер hsa-mir-302/367; hsa-miR200c; hsa-miR369; hsa-mir-124) и/или одну или более других некодирующих РНК ("ncRNA") или ген-репортер для экспрессии в клетках млекопитающих, таких как LacZ (кодирующий бета-галактозидазу), CAT (кодирующий хлорамфениколацетилтрансферазу) или ген, кодирующий флуоресцентный белок (например, GFP, YFP, RFP и их аналоги, такие как iRFP, EGFP и т.п.).
[0059] В дополнение к вышеизложенному, вектор по изобретению дополнительно может также содержать кассету экспрессии, встроенную в участок, отличный от области LAT, или другой, отличный от такового в непосредственной близости от известных инсуляторных последовательностей. Предпочтительным участком для такой кассеты экспрессии является ICP4. Например, если вектор содержит полную или инактивирующую делецию гена ICP4, экспрессионная кассета может быть встроена в участое ICP4-делеции. Желательно, чтобы кодирующая последовательность экспрессионной кассеты, встроенной в участок, отличный от области LAT, контролировалась конститутивным промотором млекопитающего (например, SV40, CMV, CAG, EFLA, UBC, РСВ, β-актин, PGK, и т.п.), однако другие промоторы (как описано в настоящем документе или иным образом известные в данной области техники) могут быть использованы, если это необходимо. Одна типичная экспрессионная кассета включает UbCp, направляющую экспрессию mCherry, сконструированную в делетированном ICP4-локусе. Конечно, такой трансген может также кодировать фактор, представляющий терапевтический интерес.
[0060] В комплементирующе к вышесказанному желательно, чтобы ВПГ-вектор по изобретению также включал делецию области внутреннего повтора (Joint), включающую IRs and IRl. Удаление этой области может внести свой вклад в стабильность векторного генома, и удаление этой последовательности ДНК ВПГ также дает возможность вектору размещать большие трансгены (по меньшей мере, 15 т.п.н.) и, тем не менее, правильно упаковываться в зрелые вирионы. Удаление Joint устраняет одну копию каждого из генов IE ICPO и ICP4, так что остальными копиями можно легко манипулировать. Она также удаляет промотор предраннего гена ICP22 или ICP47. При желании, экспрессия гена ICP47 может быть восстановлена путем вставки предраннего промотора, предпочтительно промотора ICPO или основного IE-промотора HCMV, чтобы свести к минимуму иммунное узнавание инфицированных клеток (Hill er al, Nature 1995, 375(6530): 411-415; Goldsmith et al, J Exp Med. 1998; 187(3): 341-348).
[0061] ВПГ способен инфицировать широкий спектр клеток млекопитающих; таким образом, вектор по изобретению имеет широкую применимость. Тем не менее, для повышения инфекционности, желательно, чтобы оболочка вектора по изобретению могла дополнительно включать мутантный гликопротеин, который усиливает инфекцию и/или латеральное распространение относительно гликопротеина дикого типа. Альтернативно или в комплементирующе, оболочка вектора по изобретению может дополнительно включать в себя мутантный гликопротеин, который направляет вхождение ВПГ в клетки через неканонические рецепторы. Например, такой мутант гликопротеин (ы) может представлять собой gB, gC, gD, gH или gK; конечно, вектор может иметь более одного такого мутантного (с усиленным проникновением или распространением) гликопротеина (например, комбинацию из двух, более или даже всего из этого). Кроме того, известна технология мутирования таких гликопротеинов для усиления ВПГ-инфекции и/или латерального распространения, и любая такая технология может применяться в контексте настоящего изобретения (смотри, например, публикацию заявки на патент США № 2013-0096186 A1, публикация международной заявки на патент WO/1999/006583, Uchida, J. Virol., 84: 12200-12209 (2010), Uchida et al., J. Virol., 87(3). 1430-42 (2013) и Uchida, Mol. Ther., 21: 561-569 (2013), которые включены в настоящий документ в качестве ссылки). Кроме того, геном вектора по изобретению может содержать мутантный ген, кодирующий такой мутантный гликопротеин(ы).
[0062] Типичные векторные "основы", воплощающие изобретение, описаны в настоящем документе как " JΔN15" и "JΔN18" с пониманием, что в качестве "основы" предполагается, что трансгены с посторонними контрольными элементами или без них могут быть встроены в LAT-область этих конкретных векторов (см, например, Фиг.7, вверху).
[0063] Одним из применений векторов по изобретению является перепрограммирование различных типов клеток для продуцирования плюрипотентных стволовых клеток. Стволовые клетки были на переднем крае биомедицинских исследований в последние годы, и содержали в себе много перспектив для понимания развития человека, генетического заболевания и создания новых способов лечения в регенеративной медицине. В 2006 году Яманака и его коллеги обнаружили средства создания эмбрионально-подобных стволовых клеток путем перепрограммирования взрослых фибробластов (Takahashi and Yamanaka, Cell 126: 663-76, 2006). Эти новые клетки, обозначенные как индуцированные плюрипотентные стволовые (iPS) клетки, функционально похожи на ES-клетки ((Wernig et al, Nature 448: 318-24, 2007), и при получении из соматических клеток человека (Takahashi etal, Cell 131: 861-72, 2007; Yu et al, Science 318: 1917-20, 2007) обходят этические проблемы, связанные с использованием ES клеток человека. Четыре перепрограммированных гена были первоначально использованы для продуцирования iPS клеток (Takahashi and Yamanaka, Cell 126: 663-76, 2006; Takahashi et al, Cell 131: 861-72, 2007, однако эффективность перепрограммирования этими или другими генами остается проблематичным из-за неэффективных способов переноса генов. Векторная система по изобретения решает эту проблему путем объединения высокой эффективности трансдукции для многих типов клеток с возможностью экспрессирования нескольких трансгенов одновременно с одного вектора. Например, векторы JΔN17 и JΔN18, описанные в настоящем документе, являются дефектными по репликации и нетоксичными из-за удаления или измененной кинетики экспрессии пяти вирусных генов IE. Так как они не интегрируются в клеточный геном, такие векторы разбавляются во время деления клетки, обеспечивая молниеносную систему доставки генов.
[0064] Для облегчения наращивания, продуцирования и размножения векторов по изобретению и получения их запасов, аспект изобретения предоставляет линию комплементирующих клеток, которая комплементирует ICPO и ICP4, желательно ICPO, ICP4 и ICP27. Предпочтительно, комплементация ICPO достигается без экспрессии ICPO ВПГ, чтобы снизить токсичность в комплементирующей клетке. Таким образом, предпочтительную комплементирующую клетку по настоящему изобретению получают из типа клеток, которые естественным образом комплементируют функцию ICPO ВПГ, такие как клетки U20S (Yao, J. Virol., 69: 6249-6258 (1995), который включен в настоящее описание посредством ссылки). Такие клетки могут быть сконструированы для экспрессии ICP4 или ICP4 и ICP27 способами, известными в данной области (например, путем введения кассет экспрессии ICP4 или ICP4 и ICP27 в клетки таким образом, что они экспрессируют ICP4 и ICP27, соответственно, с генетических конструкций, отличных от ВПГ-генома, таких как клеточные хромосомы). Желательно, чтобы клеточная линия экспрессировала ICP4 и ICP27, соответственно, in trans. Желательно, чтобы введенные кодирующие последовательности ICP4 или ICP4/ICP27 находились под контролем своих родственных вирусных промоторов. Кроме того, одну или обе из кодирующих комплементирующих последовательностей ICP4 и ICP27 можно индуцибельно экспрессировать в таких клеток в ответ на инфекцию ВПГ.
[0065] Как было отмечено выше, варианты осуществления вектора по изобретению включают ВАС, фланкированную последовательностями, обеспечивающими удаление кассеты ВАС путем узнавания сайт-специфической рекомбиназой. Соответственно, комплементирующие клетки по изобретению могут быть сконструированы для дополнительной экспрессии гена, кодирующего сайт-специфическую рекомбиназу, предназначенную для распознавания последовательностей в векторе, продуцируя таким образом белок рекомбиназу. Таким образом, комплементирующая клетка по изобретению может экспрессировать и продуцировать ere, dre, flp, KD, или B2, B3, R и т.п. или мутантное производное этого, в соответствующих случаях. Прохождение через такую клеточную линию, таким образом, восстанавливает место для трансгенов (около 11 т.п.н.) и может улучшать рост вируса более чем на 10 раз, например, более 25 раз или более 50 раз, например, примерно в 100 раз. Улучшение вирусный рост можно оценивать с помощью стандартных процедур (для получения кривых роста) по сравнению с клетками, которые не имеют рекомбиназу. Это включает инфицирование реплицированных лунок клеток при низкой MOI, сбор вирусов в разное время после инфицирования, и титрование выхода при помощи анализ бляшкообразования.
[0066] Кроме того, линия комплементирующих клеток может быть сконструирована для экспрессии гена, кодирующего селективный маркер, такой как маркеры, обычно используемые при конструировании упаковочных клеток или клеток, экспрессирующих любой другой чужеродный ген. Подходящие селективные гены включают такие, которые придают устойчивость к неомицину/G418, гигромицину, пуромицин, бластицидин, зеоцином и тому подобное.
[0067] Следует понимать, что способы конструирования типа исходных клеток (например, клетки U20S) для содержания экспрессирующих конструкций, кодирующих белки ВПГ ICP4 и ICP27, а также другие белки (например, рекомбиназа и/или продукт селективного гена) известны специалистам в данной области техники. Например, представляющий интерес ген с селективным маркером можно субклонировать в лентивирусные векторы, исходную клетку инфицировать лентивирусными векторами, селектируемыми по экспрессии маркера (например, устойчивость к бластицидину), а затем подтвердить экспрессию представляющего интерес трансгена (например, ВПГ ICP27).
[0068] Конечно, комплементирующая клетка по изобретению может быть размножена и клонирована. Таким образом, настоящее изобретение относится к клональной популяции, т.е. линии клеток, включающей или состоящей из или, по существу, из комплементирующей клеточной линии, как описано в настоящем документе.
[0069] ВПГ-вектор по изобретению можно размножать с использованием комплементирующих клеток по изобретению. Таким образом, изобретение предоставляет способ размножения ВПГ-вектора по настоящему изобретению. В соответствии со способом по изобретению, линию комплементирующих клеток трансфицируют векторной ДНК и затем культивируют до образования бляшек. Вирусная ДНК, как уже отмечалось, может содержать BAC, и, если это так, то клетка по изобретению может экспрессировать рекомбиназу, подходящую для вырезания BAC из вирусного генома, если желательно не включать BAC в упакованный вектор. Вирусная популяция амплифицируется путем многократной передачи инфекционных частиц в более крупные, свежие популяции комплементирующих клеток. Для этих многократных передач, множественность инфекции (MOI) может находиться в пределах от примерно 0,001 БОЕ/клетку до примерно 0,03 БОЕ/клетку. В конечном счете, векторы по изобретению (в качестве упакованных вирусов) очищают от клеток при 90% цитопатическом эффекте.
[0070] Как правило, ВПГ-вектор по изобретению наиболее полезен, когда достаточное количество вируса может быть доставлено в клеточную популяцию для обеспечения того, чтобы клетки сталкивались с подходящим количеством вирусов. Таким образом, настоящее изобретение предоставляет сток, предпочтительно гомогенный сток, включающий ВПГ-вектор по изобретению. Получение и анализ стоков ВПГ хорошо известны в данной области техники. Например, вирусный штамм может быть изготовлен в роллер-флаконах, содержащих клетки, трансдуцированные ВПГ-вектором. Вирусный сток затем можно очищать на постоянном градиенте никоденза и разделять на аликвоты и хранить до момента использования. Вирусные стоки значительно различаются в титре, что зависит в значительной степени от генотипа вируса и клеточных линий, используемых для их подготовки. Предпочтительно, такой сток имеет вирусный титр примерно 106 БОЕ/мл или даже более предпочтительно, примерно 107 БОЕ/мл (или по меньшей мере около таких значений). В еще более предпочтительных вариантах, титр может составлять примерно 108 БОЕ/мл, или примерно 109 БОЕ/мл (или по меньшей мере около таких значений), и стоки с высоким титром примерно 1010 БОЕ/мл или примерно 1011 БОЕ/мл или даже примерно 1012 БОЕ/мл (или по меньшей мере около таких значений), являются наиболее предпочтительными. Таким образом, титр ВПГ-стока в соответствии с настоящим изобретением может варьировать от примерно 106 БОЕ/мл до примерно 1012 БОЕ/мл (предпочтительно от примерно 109 до примерно 1011 БОЕ/мл). Числа геномных копий (gc) обеспечивают независимый от клеточной линии показатель количества вирусных частиц, однако включает дефектные частицы. Обычно значения gc для ВПГ-1 дикого типа составляют от нескольких до 20x, до 100х, выше, чем значения БОЕ тех же стоков. Для мутантных вирусов, особенно дефектные вирусы, выращенных на комплементирующих клетках, оно может возрастать до 10,000x выше или даже больше. Значения gc и БОЕ возрастают пропорционально увеличению размера стока.
[0071] Кроме того, изобретение предоставляет композицию, содержащую ВПГ-вектор и носитель, предпочтительно физиологически приемлемый носитель. Носитель композиции может быть любым подходящим носителем для вектора. Носитель в основном бывает жидким, но также может быть твердым, или комбинацией жидких и твердых компонентов. Желательно, чтобы носитель представлял собой фармацевтически приемлемый (например, физиологически или фармакологически приемлемый) носитель (например, наполнитель или разбавитель). Фармацевтически приемлемые носители хорошо известны и довольно доступны. Выбор носителя определяется по меньшей мере частично, конкретным вектором и конкретным способом, используемым для введения композиции. Композиция может дополнительно содержать любые другие подходящие компоненты, особенно для повышения стабильности композиции и/или ее конечного применения. Таким образом, существует большое разнообразие подходящих составов композиции по изобретению. Следующие составы и способы являются лишь иллюстративными и никоим образом не ограничивающими.
[0072] Композиции, пригодные для парентерального введения включают водные и неводные изотонические стерильные растворы для инъекций, которые могут содержать антиоксиданты, буферы, бактериостатические и растворенные вещества, которые придают композиции изотоничность с кровью предполагаемого реципиента, и водные и неводные стерильные суспензии, которые могут включать суспендирующие агенты, солюбилизаторы, загустители, стабилизаторы и консерванты. Составы могут быть представлены в герметичных контейнерах с единичной дозой или несколькими дозами, таких как ампулы и флаконы, и могут храниться в высушенном при замораживании (лиофилизированном) состоянии, требующем только добавления стерильного жидкого наполнителя, например, воды для инъекций, непосредственно перед применением. Инъекционные растворы и суспензии, подготовленные для немедленного приема могут быть подготовлены из стерильных порошков, гранул и таблеток типа описанных ранее.
[0073] Кроме того, композиция может содержать дополнительные терапевтические или биологически активные вещества. Например, могут присутствовать лечебные факторы, используемые при лечении конкретного показания. Факторы, которые контролируют воспаление, такие как ибупрофен или стероиды, могут являться частью композиции, чтобы уменьшить отек и воспаление, связанное с введением вектора in vivo и физиологическим стрессом. Супрессоры иммунной системы могут введиться с помощью композиционного способа для уменьшения любого иммунного ответа на собственно вектор или связанного с расстройством. Кроме того, иммунные усилители могут быть включены в композицию, для активации естественных защитных сил организма против болезни. Могут присутствовать антибиотики, т.е. бактерициды и фунгициды, чтобы уменьшить риск инфекции, связанной с процедурами переноса генов и других расстройств.
[0074] При использовании вектора по изобретению (и стоков и композиций, содержащих вектор) изобретение предоставляет способ экспрессии трансгена в ядросодержащей клетке, особенно в некомплементирующей клетке. В соответствии со способом, вектор по изобретению подвергается действию клетки в условиях, подходящих для инфицирования вектором клетки. После того, как клетка инфицирована, трансген, встроенный в LAT-область вектора будет транскрибироваться (экспрессироваться) в клетке, при условии, что промотор трансгена представляет собой промотор, который активен в клетке и что трансген не подавляется другим регуляторным механизмом (например, микроРНК, обсуждаемые в настоящем документе). Другими словами, векторы по изобретению служат в качестве векторов для переноса генов и экспрессии в клетках млекопитающих.
[0075] Способ по изобретению можно применять для экспрессии трансгена (ов) внутри клеток либо in vivo, либо in vitro, по желанию. Для применения in vivo клетка может представлять собой желаемую клетку любого типа, такую как экзокринные секреторные клетки (например, гландулоциты, такие как клетки слюнных желез, клеток молочной железы, клетки потовых желез, клетки пищеварительной железы и т.д.), секретирующие гормон гландулоциты (например, питуициты, клетки щитовидной железы, клетки паращитовидной железы, клетки надпочечников и т.д.), полученные из эктодермы клетки (например, ороговевающие эпителиальные клетки (например, составляющие кожу и волосы), влажные слоистые барьерные эпителиальные клетки (например, роговицы, языка, полости рта, желудочно-кишечного тракта, мочеиспускательного канала, влагалища и т.д.), клетки нервной системы (например, периферические и центральные нейроны, глия и т.д.)), полученные из мезодермы клетки, клетки многих внутренних органов (например, почек, печени, поджелудочной железы, сердца, легких) клетки костного мозга и раковые клетки, либо внутри опухоли, либо иным образом. Предпочтительные и неограничивающие примеры подходящих клеток для инфицирования вектором по изобретению включают клетки печени, клетки легких, эпителиальные клетки, клетки сердца, мышечные клетки, стволовые клетки и раковые клетки.
[0076] Следует отметить, что при использовании in vivo способом по изобретению можно лечить заболевание или состояние у субъекта, когда трансген в векторе кодирует один или более профилактически или терапевтически активных белков, полипептидов или других факторов (например, некодирующую РНК (нкРНК), такую как миРНК или микроРНК). Таким образом, изобретение предоставляет способ лечения заболевания или состояния у субъекта, включающий введение вектора по настоящему изобретению субъекту, в количестве и в участке, достаточном для инфицирования клеток у субъекта, так, что трансген экспрессируется в клетки субъекта, и где трансген кодирует один или более профилактически или терапевтически активных белков, полипептидов или нкРНК. Например, заболевание или состояние может быть одним из видов рака, причем трансген может кодировать средство, которое усиливает активность уничтожения опухоли (например, TRAIL или фактор некроза опухоли (TNF)). В качестве дополнительного неограничивающего примера, трансген может кодировать агент, пригодный для лечения заболеваний, таких как мышечная дистрофия (подходящий трансген кодирует дистрофин), сердечно-сосудистые заболевания (подходящие трансгены включают, например, SERCA2a, GATA4, Tbx5, Mef2C, Hand2, Myocd и т.д.), нейродегенеративные заболевания (подходящие трансгены включают, например, NGF, BDNF, GDNF, NT-3 и т.д.), хроническая боль (подходящие трансгены кодирует GlyRal, энкефалин или глутаматдекарбоксилазу (например, GAD65, GAD67 или другую изоформу), заболевания легких (например, CFTR) или гемофилия (подходящие трансгены кодируют, например, фактор VIII или фактор IX).
[0077] В других вариантах осуществления способ по изобретению можно использовать in vitro, чтобы вызвать экспрессию трансгена в клетках в культуре. Опять же, любой тип клеток можно инфицировать in vitro способом по изобретению, например, стволовые клетки и фибробласты, например, кожный фибробласт человека (HDF) или фибробласт легких человека (HLF). Другие предпочтительные типы клеток для использования in vitro включают кератиноциты, мононуклеары крови, кроветворные стволовех клетки (CD34+) или мезенхимальных стволовые/клетки-предшественники. В одном варианте осуществления трансген (ы) кодирует один или более факторов, которые могут влиять на дифференцировку клеток. Например, экспрессия одного или более из Oct4, Klf4, Sox2, c-Myc, L-Myc, доминантно-негативного р53, Nanog, Glisl, Lin28, TFIID, mir-302/367 или других микроРНК может привести к тому, что клетка станет индуцировать плюрипотентные стволовые (плюрипотентные) клетки. Смотрите также, Takahashi and Yamanaka, Cell, 126: 663-676 (2006); Takahashi, Cell, 131: 861-872 (2007); Wernig, Nature, 448: 318-324 (2007); и Yu, Science, 318: 1917-1920 (2007), раскрытие которых включено в настоящий документ в качестве ссылки. Кроме того, трансген (ы) в векторе по изобретению может кодировать фактор трансдифференцировки клеток (например, один или более из GATA4, Tbx5, Mef2C, Myocd, Hand2, SRF, Mesp1, SMARCD3 (для кардиомиоцитов), Ascl1, Nurrl, Lmx1A, Brn2, Mytl1, NeuroD1, Foxa2 (для нервных клеток), HNF4, Foxal, Foxa2 или Foxa3 (для печеночных клеток).
[0078] При осуществлении способа по изобретению, включающего инфицирование клетки in vivo или in vitro вектором, композицией или стоком по изобретению, клетка может являться любой ядросодержащей клеткой млекопитающего, в которой желательно экспрессировать трансген. ВПГ имеет широкую инфекционность и, как отмечалось в настоящем документе, вектор по изобретению может быть сконструирован, чтобы изменить свой природный тропизм и повысить инфекционность путем мутирования гликопротеинов вирусной оболочки. Таким образом, вектор можно применять для инфицирования клеток многих видов млекопитающих. Считается, что способы по изобретенныю могут применяться в сельском хозяйстве, например, для экспрессии экзогенных генов или дополнения дефектных генов у животных, таких как крупный рогатый скот, лошади, овцы, козы, свиньи и тому подобное. Аналогичным образом, способ по изобретению можно применять в ветеринарном контексте для домашних животных, таких как кошки, собаки и тому подобное.
[0079] Конечно, способ по изобретению можно также применять in vivo для людей для обеспечения экспрессии профилактически или терапевтически активного средства или фактора, в медицинских учреждениях. Фактор (поставляется посредством экспрессии одного или более трансгенов в векторах по изобретению) может быть экзогенным или таким, который комплементирует генетическую недостаточность.
[0080] Следующие примеры дополнительно иллюстрируют изобретение, но, конечно, не должны быть истолкованы как каким-либо образом ограничивающие его объем.
ПРИМЕР 1
[0081] Этот пример описывает развитие линии комплементирующих клеток для реплицирования и продуцирования ВПГ-векторов по изобретению.
[0082] Некоторые из предранних (IE) генов ВПГ являются существенными для репликации вируса, но эти и другие IE-гены оказывают токсическое воздействие в различных типах клеток. Несмотря на то, что удаление этих генов предотвращает токсичность вектора, существенные продукты должны быть обеспечены для получения инфекционных вирусных частиц. Была сконструирована новая клеточная линия, основанная на U20S-клетках остеосаркомы человека, для условной экспрессии существенных IE-генов ICP4 и ICP27. Эти гены вводили под контролем их родственных промоторов при помощи ретровирус-опосредованной вставки. Как таковые эти гены могут оставаться молчащими до тех пор, пока ВПГ-инфецирование не обеспечит доставку белка оболочки VP ВПГ 16 в ядро, где он стимулирует высокий уровень экспрессии интегрированных генов ICP4 и ICP27 путем активации их промоторов. Поэтому до герпетической инфекции, эти гены могут стабильно поддерживаться в сконструированных U20S-клетках без неподходящей токсичности. Рост ВПГ также зависит от экспрессии ICPO, но этот белок ингибирует репликацию клеток, приводя в результате к остановке клеточного цикла и запрограммированной гибели клетки. Примечательно, что, естественно, U20S клетки комплементируют функции СКП и таким образом введение ICP4 и ICP27 является достаточным для обеспечения сотовой условий для эффективного производства векторов удаленных для всех трех генов IE. Этот новый инженерии клеточной линии стабильно, хорошо растет в культуре и может быть использован для клинического производства нетоксичных векторов ВПГ.
[0083] Клетки U20S-ICP4/27 и клетки 7b (Vero-ICP4/ICP27) были инфицированы вирусами E1G6 (AICP4::HCMVp-EGFP/AICP27/B22/B47) или JDQOZEH1 (AICP4/AICP27/AICP22/AICP0::CMV-EGFP). Через 3 дня вирус в клеточных супернатантах титровали на клетках U20S-ICP4/27. 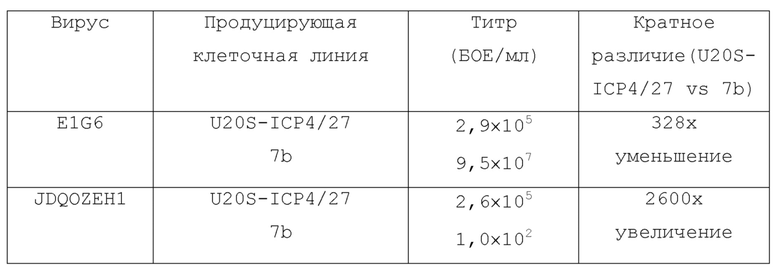
[0084] Эти данные показывают, что E1G6 (делетированы ICP4 и ICP27; экспрессия ICP22 и ICP47 отсутствует) рос на обеих клеточных линиях, тогда как JDQOZEH1 (делетированы ICP0, ICP4, ICP27 и ICP22) был в состоянии расти только на клетках U20S-ICP4/27. Титр JDQOZEH1 в супернатанте 7b, вероятно, представляет остаточное вводимое количество от 7b-инфицирования.
ПРИМЕР 2
[0085] В этом примере описан вариант осуществления ВПГ-вектора, включающий геном, содержащий трансген, встроенный в генную области LAT, причем вектор не экспрессирует ICPO, ICP4, ICP22, ICP27 и ICP47 в качестве предранних генов.
[0086] Векторный геном (Фиг. 1A) содержит Cre-удаляемую кассету бактериальной искусственной хромосомы (ВАС) в межгенной области UL37-UL38, дающую возможность размножения и манипулирования в бактериях. В нем делетирована несущественная область внутренних повторов (Joint, 14 т.п.н.), разделяющая сегменты UL и Us для обеспечения пространства для встраивания трансгена и увеличения стабильности вектора. Вектор является дефектным по репликации из-за делеции генов ICP4 и ICP27, и у него дополнительно делетирован токсичный IE-ген ICPO и промотор и инициирующий кодон IE-гена ICP47; оставшийся токсичный ген, ICP22, контролируется ICP4-зависимым (ранним, β) промотором, чтобы экспрессировать ICP22 во время "ранней", а не "предранней" стадии экспрессии ВПГ-генов в ICP4-комплементирующих клетках. Поскольку U20S-клетки природно комплементируют ICPO, вектор выращивали в вирус-продуцирующей клеточной линии на основе U20S, U20S-ICP4/ICP27, которая комплементирует функции всех этих IE-генов, за исключением таковой несущественного гена ICP47. Векторный геном также содержит пару мутаций в гене gB, которые повышают проникновение вирусов в клетки, и экспрессионную кассету убиквитиновый (UBC) промотор - репортерный гена mCherry в положении делеции ICP4. В некомплементирующих клетках, промоторная область латентность-ассоциированого транскрипта (LAT), которая находится между инсуляторными (CTRL) элементами, которые защищают область от образования гетерохроматина, остается активной. Эта область включает энхансерный элемент, LAT P2 или LAP2, который способствует долговременной генной экспрессии. Кассета экспрессии CAG промотор-GFP была встроена между LATP2 и CTRL2, и сильная экспрессию GFP наблюдалась в инфицированной клетках кожных фибробластов человека (HDF), тогда как минимальная экспрессия наблюдалась с той же GFP-кассеты, встроенной в другие участки генома или с кассеты mCherry (Фиг. IB). Таким образом, при полном отсутствии IE-генной экспрессии LAT-локус является привилегированным местом для экспрессии трансгена.
[0087] Фиг. 1. показывает структуры полного генома ВПГ-1 с последовательностями ВАС в UL (вверху), и основную векторную конструкцию. Области LAT и UL3-UL4 увеличены внизу, и указаны альтернативные положения вставки CAG-GFP. Фиг. IB показывает экспрессию GFP с альтернативных положений в векторном геноме в инфицированных HDF. Ген GFP в этом векторе кодирует EGFP.
ПРИМЕР 3
[0088] В этом примере перечислены структура и свойства различных векторных конструкций ВПГ.
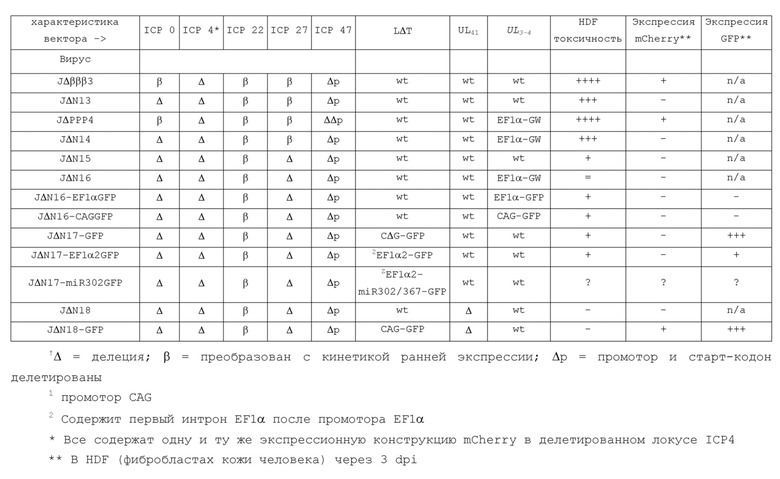
ПРИМЕР 4
[0089] Этот пример демонстрирует экспрессию трансгена в JΔNI7-GFP- и JΔNI6-CAGGFP-инфицированных клетках. Положение CAG-GFP в JΔN17-GFP показана на Фиг. 1 как LAT:CAG-GFP; его положение в JΔN16-CAGGFP показано на Фиг. 1 как UL3/4:CAG-GFP. Кроме того, последовательность области LAT JΔN17-GFP ВПГ изложена на фиг. 8 (SEQ ID NO:1), в том числе последовательности различных генетических элементов в пределах области LAT. Следует отметить, что GFP в этих векторах кодирует EGFP.
[0090] Биологический титр вирусного стока определяли на клетках U20S-ICP4/ICP27, и титр геномных копий (gc) определяли количественным ПЦР в реальном времени для вирусного гена гликопротеина D. Сравнивали отношения частиц (gc) к бляшкообразующим единицам (БОЕ) для JΔN16-CAGGFP и JΔN17-GFP. См Пример 8, Таблица 2.
[0091] Некомплементирующие фибробласты кожи человека (HDF) и ICP0-комплементирующие клетки U20S инфицировали каждым вирусом для сравнения у них экспрессии трансгена. JΔN17-GFP, содержащий кассету CAG-GFP в локусе LAT, показал сильную, вирусную доза-зависимую экспрессию GFP в HDF, в то время как наивысшая доза HDF JΔN16-CAGGFP давала только минимальную экспрессию GFP (Фиг. 2A, левые панели, EGFP). Экспрессия GFP в JΔN17-GFP-инфицированных HDF обнаруживались через 2 недели после инфицирования (Фиг. 3B, EGFP). Однако, слабая экспрессия mCherry или ее отсутствие наблюдалась с любого из вирусов в HDF (Фиг. 2A, 2B, mCherry), что дало основание предполагать, что гены вне локуса LAT являются молчащими в HDF. В отличие от этого, обильная экспрессия GFP и mCherry наблюдалась для обоих вирусов при низком MOI в клетках U20S (Фиг. 2А, правая колонка), что согласуется с интерпретацией, что ICP0-подобная активность этих клеток предотвращает молчание локусов, отличных от LAT, имеющее место в HDF. Взятые вместе, эти результаты дают сильное указание на то, что локус LAT является предпочтительным участком для трансгенной экспрессии с дефектных по репликации, ICP0-дефектных векторов.
ПРИМЕР 5
[0092] Этот пример демонстрирует получение двух изолятов JΔN17-miR302GFP BAC в клетках U20S-ICP4/ICP27.
[0093] Конструкция JΔN17-miR302GFP ВАС несет в локусе LAT кассету экспрессии для кластера miR302s/367 (Anokye-Danso, Cell Stem Cell, 8: 376-388 (2011)) в качестве альтернативы типичному коктейлю Яманака генного перепрограммирования соматических клеток (OKSM:Oct4, Klf4, Sox2, c-Myc) (Takahashi and Yamanaka, Cell, 126: 663-676 (2006); Takahashi, Cell, 131: 861-872 (2007). Генный кластер MiR302s/367 находится в интроне, соединяющим промотор EF1α с кодирующей последовательностью GFP. Два изолята ДНК JΔN17-miR302GFP ВАС очищали и вводили в клетки U20S-ICP4/ICP27 для продуцирования вирусных частиц для исследования роста вируса и экспрессии трансгенов. При наблюдении 90% цитопатического эффекта в культурах, вирус собирали из клеток и надосадочной жидкости и использовали для инфицирования свежих клеток U20S-ICP4/ICP27. Экспрессия трансгена и распространение вируса затем контролировали ежедневно. Как показано на Фиг. 3, через три дня после инфицирования оба ВАС-изолята продуцировали бляшко-образующий вирус, и оба вируса экспрессировали EGFP и mCherry.
ПРИМЕР 6
[0094] В этом примере описывается конструирование направленной плазмиды для введения тетрациклин-индуцируемого промотора и Gateway рекомбинационной кассеты в локус LAT вектора ВПГ.
[0095] Различные стратегии могут быть использованы для предотвращения генетической перестройки и инактивации экспрессионной кассеты OKSM при распространении вируса, когда кассету встраивали в локус LAT JΔNI5 or JΔNI8. Одна из таких стратегий заключается в замене конститутивно активного промотора CAG кассеты OKSM на тетрациклин-индуцируемый промотор. Поскольку тетрациклин-индуцируемый промотор активен только в присутствии и его трансактиватора (rtTA), и тетрациклина/доксициклина, трансгенная экспрессия может жестко регулироваться (репрессироваться) во время распространения вирусов.
[0096] Направленная плазмида для введения тетрациклин-индуцируемого промотора в локус LAT была разработан (Фиг. 4). Лентивирусная конструкция (Plenti-CMVTRE3G-NeoDEST, Addgene), несущая тетрациклин-индуцируемый промотор TRE3G, использовали в качестве исходной конструкции. Промотор TRE3G выделили из Plenti-CMVTRE3G-NeoDEST как фрагмент DraI-SpeI и встроили по сайтам NruI и SpeI плазмиды PCMV-GW, заменив резидентный CMV-промотор. Плазмида pCMV-GW содержала селективный ген зеоцина (Zeo) в Gateway (GW) кассете вместо хлорамфеникол (Cm) селективного гена в GW лентивирусной плазмиды. Кассету экспрессии TRE3G-GW (Zeo) выделили и клонировали между LAT-последовательностями в плазмиду, содержащую часть локуса LAT, для добавления "плеч гомологии" к кассете TRE3G-GW (Zeo) для рекомбинации в ДНК ВАС.
ПРИМЕР 7
[0097] Этот пример демонстрирует иммунофлюоресцентное окрашивание и комплементационный анализ для ICP27.
[0098] Клетки U20S-ICP4 и отдельные клоны ICP27 лентивирус-инфицированных клеток U20S-ICP4, отобранные по приобретению устойчивости к бластицидину, инфицировали вирусом QOZHG (ΔICP4, ΔICP27::HCMV IEp-GFP, β-ICP22, β-ICP47, ΔUL41::ICP0placZ; Chen, J. Virol., 74: 10132-10141 (2000)) с MOI 0,5 для иммунофлуоресцентного окрашивания ICP27 и 0,01 для комплементационного анализа. Для иммунофлуоресцентного окрашивания ICP27 клетки фиксировали и окрашивали через 24 часа после инфицирования (Фиг. 6A). Клоны #1 и #8 показали сильную индукцию экспрессии ICP27 при помощи инфекции ICP27-дефицитными ВПГ (QOZHG). Для комплементационного анализа контролировали роста вируса, и были сделаны фотографии через 24, 48 и 72 часов после инфицирования (Фиг. 6B). Клоны #1 и #8 U20S-ICP4/27 показывали и сильнейшую способность поддерживать рост вируса QOZHG.
ПРИМЕР 8
[0099] Этот пример демонстрирует конструирование и тестирование нескольких ВПГ-векторов.
МАТЕРИАЛЫ И СПОСОБЫ
Клетки
[00100] клетки остеосаркомы U20S человека (АТСС, Manassas, VA, США) выращивали в среде DMEM с 10% FBS и пенициллином-стрептомицином (P/S). Неонатальные кожные фибробласты человека (HDFS) (ATCC, PCS-201-010) и фибробласты крайней плоти BJ человека (ATCC, CRL-2522) выращивали в среде DMEM с 10% подходящей FBS эмбриональных стволовых клеток (Invitrogen) и P/S. Vero, Vero-7b (Krisky et al., Gene Ther. 5, 517-30 (1998)), и клетки 293Т культивировали в DMEM, содержащей 5% FBS и P/S. Гепатоциты человека (hHEP) выделяли и культивировали, как описано (Ueki et al., Hepatology 54, 216-28 (2011); Yoshida et al., Hepatology 58, 163-75 (2013)). Подкожные преадипоциты человека (HPAD) (PT-5020, Lonza) поддерживались на минимальной среде РВМ™-2 (Lonza). Полученные из мышц стволовые/прогениторные клетки человека (hMDSCs) культивировали, как описано (Gao et al., Cell Transplant 22, 2393-408 (2013)). Человеческие неонатальные кератиноциты (hEK) (Invitrogen) культивировали в среде EpiLife®, дополненной ростовыми добавками кератиноцитов человека (обе от Invitrogen). Ганглии спинного корня (rDRG) микро-высекали из 15-дневных эмбрионов крысы, диссоциированные 3 мг/мл коллагеназы тип I (Sigma, St. Louis, МО) в среде Лейбовиц L-15 в течение 30 мин при 37°С при постоянном встряхивании и наносили на покрытые поли-D-лизином (Sigma) покровные стекла примерно 105 клеток/лунку в 24-луночных планшетах в 500 мкл определенной среды Neurobasal с добавлением B27, Glutamax-I, II-Albumax и P/S (Gibco/Invitrogen, Grand Island, NY), дополненной 100 нг/мл 7.0S NGF (Sigma). Через 1-3 г после посева культуры обрабатывали 10 мкΜ уридина и 10 мкΜ фтордезоксиуридина (Sigma) в указанных выше средах в течение 1-2 д для ограничения распространения деления клеток, таких как фибробласты и глия. Затем клетки промывали PBS и инкубировали в среде Neurobasal с добавкой NGF, как описано выше. Вирусные инфекции проводили на 10-15 день после посева.
[00101] Клетки U20S-ICP4 были получены путем инфицирования клеток U20S очищенным лентивирусом ICP4 (смотри ниже), отбором пуромицин-резистентных клонов (2 мг/мл) и иммунофлюоресцентным скринингом на экспрессию ICP4 после инфицирования ICP4-дефектным вирусом (QOZHG) при множественности (MOI) 0,5. Клетки U20S-ICP4/27 получали аналогично инфицированием клеток U20S-ICP4 очищенным лентивирусом ICP27, отбором по устойчивости к пуромицину и бластицидину (10 мг/мл) и скринингом QOZHG-инфицированных клонов по иммунофлуоресценции ICP27 и росту вируса (Фиг. 6).
Лентивирусы
[00102] Лентивирусная ICP4 экспрессионная плазмида pCDH-ICP4-puro была сконструирована путем замены полного HCMV IE промотора плазмиды рCDH-CMV-MCS-EFL-Puro (Systembio) промотором ICP4 и кодирующей областью плазмиды S3, которая состоит из SphI фрагмента pICP4-LOX-PAC pac (Rasty et al., J. Neurovirol. 3, 247-64 (1997)) (GenBank JQ673480.1 HSV-1 KOS положения на карте 131,587-124,379; D. Krisky and J.C.G., неопубликовано), вставленным в сайт SphI в pUC19. Лентивирусная ICP27 экспрессионная плазмида pCDH-ICP27-SV40-bla была сконструирована, во-первых, путем замены кассете пуромицин-устойчивости PCDH-CMV-MCS-EFL-Puro кассетой бластицидин-устойчивости pcDNA6/BioEase™ -DEST (Invitrogen), чтобы создать pCDH-CMV-MCS-SV40-bla (Фиг. 5). Промотор ICP27 и кодирующую область затем выделяли путем расщепления плазмиды PD7, содержащий ген ICP27 и фланкирующие последовательности, между сайтами EcoRV и SacI (GenBank JQ673480.1 положения карты 110,580-115,666, D. Krisky and J.C.G., не опубликовано) с помощью BamHI (Положение на карте 113,244) и SacI, и выделенный фрагмент был использован для замены промотора CMV в pCDH-CMV-MCS-SV40-bla. Полученная плазмида, рCDH-ICP27-SV40-bla, изображена на Фигуре 5.
[00103] Лентивирусы получали с использованием ViraPower™ Lentiviral Packaging Mix (Invitrogen) в соответствии с инструкциями изготовителя. Вкратце, клетки 293Т трансфицировали рCDH-ICP4-Puro или рCDH-ICP27-SV40-bla в смеси ViraPower™. Супернатанты собирали через 2 дня, осветляли, фильтровали через 0,45 мкм фильтр и концентрировали центрифугированием. Лентивирусы, полученные из PCDH-ICP4-Puro, называются в настоящем документе как лентивирус ICP4; лентивирусы, полученные из PCDH-ICP27-SV40-bla, называются лентивирус ICP27.
Конструирование ВПГ-BAC
[00104] Все конструкции ВПГ-BAC, полученные в этом исследовании и превращенные в вирусные частицы, приведены в Таблице 2 и были получены из КОС-37 BAC (24), любезно предоставленной D. Leib (Дартмутская школа медицины, NH). Конструирование всех ВАС выполняли путем бесшрамной Red рекомбинации с pRed/ET (Gene Bridges, Гейдельберг, Германия) и либо плазмиды pBAD-I-sceI (любезно предоставленной Н. Osterrieder, Свободный университет Берлина, Германия), либо штамма E.coli GS1783 (от G. Smith, СеVero-Западный университет, Чикаго, Иллинойс), как описано (Tischer et al., Methods Mol. Biol. 634, 421-30 (2010); Tischer et al., Biotechniques, 40, 191-97 (2006)), или путем in vitro Gateway (GW) рекомбинации в соответствии с Gateway Technology Manual (Invitrogen) (http://tools.lifetechnologies.com/content/sfs/manuals/gatewayman.pdf). Все конструкции были подтверждены с помощью ПЦР-анализа, FIGE-анализа расщеплений ферментами рестрикции и направленным секвенированием ДНК. Направленные плазмиды для Red-рекомбинации сконструировали, как описано (Tischer et al., Biotechniques, 40, 191-97 (2006)). Ген устойчивости к канамицину, фланкированный сайтами рестрикции I-SceI (I-SceI-aphAI фрагмент), амплифицировали с pEPkan-S2 (от N. Osterrieder) (Tischer et al., Biotechniques, 40, 191-97 (2006)) с помощью ПЦР с различными направленными праймерами, указанными ниже и перечисленными в Таблице 1. Части праймеров, ориентированные на вирусный геном, были разработаны на основе последовательности ВПГ-1 штамма-17 (GenBank JN555585.1). Все направленные фрагменты для Red рекомбинации очищали при помощи Qiagen gel extraction kit (Qiagen) или SpinSmart Nucleic Acid Prep & Purification Columns (Denville Scientific). Положения нуклеотидов, приведенные ниже, относятся к последовательности GenBank JQ673480 штамма ВПГ-1 KOS.
[00105] Для построения векторов JΔNI, описанная ранее гиперактивная N/T двойная мутация была введена в ген gB ((Uchida et al., J. Virol., 84, 12200-09 (2010)) KOS-37 BAC, и область внутреннего повтора (joint) была удалена. Фрагмент I-SceI-aphAI был клонирован в сайт SnaBI плазмиды pgBl: D285N/A549T (Uchida et al., J. Virol., 84, 12200-09 (2010)). Полученную в результате плазмиду pgB:N/T-kan использовали в качестве матрицы для амплификации с праймерами 61 и 62 (Таблица 1), и продукт подвергали рекомбинации с нативным геном gB KOS-37 ВАС с последующей I-SceI-усиленной делецией гена aphAI в бактериях, трансформированных плазмиой pBAD-I-sceI. Далее, I-Scel-aphAI амплифицировали при помощи сцепленных прямых праймеров 46, 48 и обратного праймера 47 для Red-опосредованной делеции области joint (положения 117, 080-132, 466 в GenBank JQ673480) вместе с первыми 14 нуклеотидами (nts) из смежного уникального короткого (Us) сегмента генома, с последующим делетированием гена aphAI. Us в KOS-37 ВАС находится в обратной ориентации по отношению к типичному представлению генома ВПГ, с размещением гена Us12 (ICP47) непосредственно рядом с joint и ген Us1 непосредственно рядом с концевым повтором Us. Делеция 14 нуклеотидов за пределами joint (позиции 145,377-145,390 в GenBank JQ673480) при этом удаляла кодон инициации трансляции ICP47.
[00106] Замену локуса ICP4 в joint-делетированной gB:N/T ВАС кассетой экспрессии mCherry получали следующим образом. Плазмида pUbC-mCherry-SV40pA была сконструирована клонированием промотора убиквитина C (UbCp) человека из pBluescriptUB-Flag-mArt (дар H. Nakai, Oregon Health & Science University, Portland, OR), гена mCherry из pEP-miR (Cell Biolabs, Сан-Диего, Калифорния) и области полиаденилирования SV40 (полиА или пА) из рер4-E02SCK2M-EN2L (плазмида Addgene 20924) (Yu et al., Science 324, 797-801 (2009)) в pBluescript Ks + (Stratagene). Фрагмент I-SceI-aphAI затем клонировали в сайт BamHI в pUbC-mCherry-SV40pA на границе между UbCp и mCherry, чтобы получить pUbC-mCherry-SV40pA-KAN. Вставка была ПЦР-амплифицирована с использованием праймеров 51 и 52 (Таблица 1) для Red-опосредованной рекомбинации с локусом-мишенью ICP4. Полученную конструкцию делетировали по ВПГ-1 KOS положениям 146,113-151,581 последовательности GenBank JQ673480, включая мотив TAATGARAT промотора ICP22.
[00107] Замена IE-промоторов ICP0 и ICP27 на ранний (р) ВПГ-1 промотор тимидинкиназы (ТК) для создания JΔΝΙ2 проводили с использованием ПЦР с помощью TK-промотора перед кодирующей областью как ICP0, так и ICP27 в вирусном геноме JΔββ (Craft et al., Stem Cells 26, 3119-29 (2008)) со сцепленными парами праймеров 81/82 и 83/84, соответственно, клонируя продукт каждой реакции в pCRblunt (Invitrogen), с образованием pCRBlunt-P0 и pCRBlunt-P27, соответственно, встройка 1-Scel-aphAI в сайт BglII pCRBlunt-рО и сайт HpaI pCRBlunt-р27, с образованием pCRBlunt-pO-KAN and pCRBlunt-P27-KAN, и ПЦР-амплификация вставок с парами праймеров 85/86 и 87/88, соответственно, для Red-рекомбинации с joint-делетированной UbCp-mCherry gB:N/T ВАС. JΔΝΙ3 затем получили из JΔΝΙ2 путем Red-опосредованной чистой делеции кодирующей последовательности ICP0 с помощью сцепленных направленных прямых праймеров 41, 42 и обратного праймера 43 для амплификации I-SceI-aphAI. JΔΝΙ5 был также получен из JΔΝΙ3 чистым делетированием кодирующей последовательности ICP27 с помощью пары праймеров 44/45 для получения направленного фрагмента I-Scel-aphAI.
[00108] JΔΝΙ6-CAGGFP был создан путем введения экспрессионной кассеты EGFP между генами UL3 и UL4 JΔΝΙ5. Во-первых, плазмида pCAG-GFP была сконструирована путем замены гена gH между промотором CAG (энхансер CMV/промотор бета-актина курицы/химерный интрон) и областью поли А β-глобина кролика плазмиды pPEP100 (дар П. Копье, СеVero-Западный университет) (Pertel et al., Virology 279(1), 313-24 (2001)) геном EGFP из pEGFP-C1 (Clontech). 1-Scel-aphAI затем встраивали в сайт SnaBI pCAG-GFP, чтобы создать плазмиду pCAG-GFPKAN. Отдельно, многоступенчатый ПЦР с использованием ДНК KOS-37 ВАС в качестве исходной матрицы проводили для получения фрагмента, содержащего новые сайты клонирования (MCS) между полиА-областями UL3 и UL4, следующим образом. Во-первых, ПЦР с удлинением проводили для амплификации 3'-UTR из UL3 и UL4 с пар праймеров 57/58 и 59/60, соответственно, что добавило перекрывающуюся MCS область к каждому 3'-UTR фрагменту. 2 ПЦР-продукта были очищены при помощи геля, и 100 нг каждого был использован для перекрывающегося ПЦР с праймерами 57 и 59, чтобы создать непрерывный фрагмент. Этот продукт был клонирован в pCRBlunt (Invitrogen), с образованием плазмиды pCRBluntUL3-41inker. Вставку pCAG-GFPKAN затем клонировали между сайтами AccI и PsiI в MCS pCRBluntUL3-41inker. Полученную плазмиду расщепляли MfeI и PpuMI, и фрагмент UL3-CAG-GFPKAN-UL4 был выделен для рекомбинации с межгенной областью UL3-UL4 из JΔΝΙ5 с последующим удалением гена aphAI.
[00109] Для создания JANHGFP, XhoI-фрагмент (~6,2 т.п.н.), содержащий 2 Ctrl, LAP1 и LATP2 локуса LAT ВПГ-1, был выделен из ДНК от KOS-37 BAC и клонирован в pBluescript KS+. Внутренний KpnI-SalI фрагмент протяженностью от около конца LATP2 до -250 п.н. ниже по течению от CTRL2 выделяли из этой рекомбинантной и клонировали между сайтами KpnI и SalI pSP72 (Promega), с получением pSP72KOS-LAT. Сайт для множественного клонирования затем вводили между 2 сайтами BstXI, расположенных -240 -430 п.н. ниже по течению от сайта KpnI, с получением pSP72KOS-LATlinker. Отдельно, плазмида pCAG-GW была сконструирована путем замены гена gH pPEP100 (Pertel et al., Virology 279(1), 313-24 (2001)) на ПЦР-амплифицированную модифицированную GW рекомбинантную кассету [GW-Zeo; устойчивость к зеоцину вместо устойчивости к хлорамфениколу (Wolfe et al., J. Virol 84, 7360-68 (2010))]. Вставку pCAG-GW затем клонировали в MCS в pSP72KOS-LATlinker с образованием pSP72KOS-LATlinker-GW. Плазмиду переваривали с помощью KpnI и HpaI и выделяли область CAG-GW с фланкирующими последовательностями LAT для Red-опосредованной рекомбинации с локусом LAT JΔΝΙ5 в ccdB-устойчивых HerpesHogs бактериях (Wolfe et al., J. Virol 84, 7360-68 (2010)). Наконец, кассета GW в полученной JΔΝΙ5 рекомбинантной BAC была заменена геном EGFP при помощи Red-опосредованной рекомбинации с фрагментом AatII-PsiI, включая последовательности поли А и CAG, плазмиды pCAG-GFPKAN. Производные JΔΝΙ7GFP с делецией специфических элементов области LAT вне кассеты EGFP (CTRL1, CTRL2, LATP2) были получены с помощью Red-опосредованной рекомбинации ДНК JΔΝΙ7GFP с направленными кассетами 1-Scel-aphAI, производится с помощью ПЦР с соответствующими праймерами F1, F2 и R, перечисленными в таблице 1 (63-65, 66-68 и 69-71, соответственно).
[00110] Для конструирования различных векторов JΔNI9 и JΔNI10 ~6,2 т.п.н. LAT XhoI-фрагмент, описанный выше, делетировали из JΔNI5-генома путем рекомбинации с LAT-направленным I-SceI-aphAI, полученным при помощи ПЦР с праймерами 78, 79 и 80, с образованием JΔNI5ΔL. GW-Zeo затем амплифицировали с направленными праймерами для межгенной области между UL45 и UL46 (74/75) или UL50 и UL51 (76/77), и продукт каждой реакции подвергали рекомбинации с ДНК JΔNI5ΔL ВАС, чтобы создать JΔNI9GW и JΔNI10GW. ДНК JANHGFP ВАС расщепляли XhoI и ~7,2 т.п.н. CAG-GFP-содержащий фрагмент из локуса LAT выделяли и клонировали в pENTR1A (pENTR-LAT-XhoI). Соответствующий ~5,3 т.п.н. XhoI-фрагмент JΔNI7GFPΔC12LP2 также выделяли и клонировали в pENTR1A (pENTR-LATΔ-XhoI) и, наконец, вставку pCAG-GFP (см выше) переносили в pENTR1A (pENTR-CAG-GFP). Реакции LR клоназы (Invitrogen) in vitro затем выполняли для проведения рекомбинации различных pENTR-конструкций с ДНК JΔNI10GW BAC, с образованием JΔNI10LAT-GFP, JΔNI10ΔC12LP2-GFP и JΔNI10GFP, соответственно, и pENTR-LAT-XhoI и pENTR-CAG-GFP рекомбинировали с ДНК JΔNI9GW ВАС для получения JΔNI9LAT-GFP и JΔNI9GFP.
[00111] KNTc конструировали путем введения мутаций gB:N/T в KOS-37 BAC, как описано выше, и кассету UbCp-mCherry в межгенную область между локусами UL3 и UL4 KOS-BAC путем Red рекомбинации с I-SceI-aphAI направляли при помощи амплификации с праймерами 53 и 54.
Вирусы
[00112] ДНК JANI ВАС были преобразованы в инфекционные вирусы путем трансфекции комплементирующих клеток на основе U20S. ДНК в 500 мкл OptiMEM (Invitrogen) инкубировали с 1 мкл Lipofectamine Plus Reagent (Invitrogen) в течение 5 мин при комнатной температуре, добавляли 6,25 мкл Lipofectamine LTX (Invitrogen), полученную смесь инкубировали в течение 30 мин при комнатной температуре и добавляли к клеткам. После инкубации при 37°С в течение 6ч, трансфекционную смесь удаляли и клетки культивировали в течение ночи при 37°С с бессывороточной DMEM, переносили в 33°С инкубатор и контролировали на 100% цитопатический эффект (CPE). Супернатанты титровали и размножали инфицированием последовательно увеличивающихся культур при множественности (MOI) 0,001 КОЕ/клетку. Инфекционный вирус KNTc получали путем трансфекции клеток Vera с использованием 3 мкл Lipofectamine Plus Reagent и 9 мкл Lipofectamine LTX в течение 4 ч при 37°С; множественность инфекции от 0,01 КОЕ/клетку использовали для амплификации вируса KNTc в клетках Vero. Комплементирующие клетки для трансфекции и/или роста вируса были следующими: U20S-ICP4 (JΔΝΙ2, JΔΝΙ3), U20S-ICP4/27 (JΔΝΙ5 и производные), и Vero-7b [вирус QOZHG (AICP4, AICP27::HCMV IEp-GFP-ICP22, -ICP47 и AUL41::ICP0p-LacZ) (Chen et al, J. Virol. 74, 10132-41 (2000))]. Все вирусные стоки титровали на клетках U20S-ICP4/27 (таблица 2). Физические титры [геномные копии (gc)/мл] определяли количественным ПЦР в реальном времени, как описано ниже. Флуоресцентные изображения инфицированных клеток были получены при помощи флуоресцентного микроскопа Nikon Diaphot (Nikon, Melville, PA) при 40-кратном увеличении.
Кривые роста вируса
[00113] Реплицированные лунки клеток Vero-7b, U20S, U20S-ICP4 и U20S-ICP4/27 в 24-луночном планшете инфицировали при множественности (MOI) 0,001 в течение 2 ч, обрабатывали 0,1 М глицина (рН 3,0 ) в течение 1 мин, чтобы инактивировать внеклеточный вирус, и инкубировали при 37°С и 5% CО2. Среду собирали ежедневно и титровали стандартным способом бляшек на клетках U20S-ICP4/27.
Тест на цитотоксичность
[00114] 5×103 клеток HDF и Vero засевали в 96-луночный планшет и инфицировали вирусами KOS, QOZHG или JΔΝΙ с 25000 gc/клетку. Жизнеспособность клеток определяли на 5 дней позже при помощи МТТ теста, как описано (Uchida et al., J. Virol. 87, 1430-42 (2013)).
Вестерн-блоттинг и иммунофлуоресценция
[00115] Подготовку клеточного лизата и Вестерн-блоттинг проводили, как описано (Miyagawa et al., PLoS One 4, e4634 (2009)). Поликлональные антитела анти-ICPO кролика были произведены в нашей лаборатории, анти-ICP27 (10-H44) был из Fitzgerald Industries International (Конкорд, Массачусетс), анти-ICP22 представляло собой дар John Blaho (Mt Sinai School of Medicine, Нью-Йорк), анти-ICP4 (10F1) было от Santa Cruz Biotechnology и анти-α-тубулин (T6793) был от Sigma. Иммунофлуоресценцию с использованием тех же антител ICP4 и ICP27 проводили, по существу, как описано (Uchida et al., J. Virol. 83, 2951-61 (2009)) и исследовали под флуоресцентным микроскопом Nikon.
Количественный обратно-транскрипционный ПЦР (qRT-PCR) и геномный ПЦР
[00116] Для qRT-PCR тотальную РНК экстрагировали при помощи комплекта RNeasy (Qiagen). кДНК синтезировали при помощи комплекта RETROscript® (Ambion). ПЦР в реальном времени проводили с помощью системы ПЦР в реальном времени StepOnePlus (Applied Biosystems). Для небольшого числа клеток, комплект Cells-to-cDNA™ II (Ambion) использовали для лизиса клеток и обратной транскрипции. Результаты для 18S рибосомальной (р) РНК были использованы для нормирования данных. Все qRT-PCR праймеры, использованные в этом исследовании, перечислены в Таблице.1.
[00117] Для определения физических (геномной копии) титров вируса натуральной оспы, 5 мл вируса инкубировали с 300 ед/мл нуклеазы Benzonase (Sigma) в течение 1 ч при 25°С в присутствии 2 мМ MgCl2, и вирусную ДНК экстрагировали при помощи комплекта DNeasy Blood & Tissue (Qiagen). gc-титры определяли количественным ПЦР для гена гликопротеина D (gD) с использованием Gd-праймеров (38/39) и зонда (40), перечисленных в Таблице 1. Количества ядерной вирусной ДНК определяли путем промывки клеток в 2 hpi, выделением ядер, как описано (Dignam et al., Nucleic Acids Res. 11, 1475-89 (1983); Suzuki et al., BMC Res. Notes 3, 294 (2010)), выделения ДНК при помощи комплекта DNeasy Blood & Tissue и кПЦР для гена gD, как указано выше. Клеточные уровни 18S рибосомных ДНК определяли с использованием TaqMan® Ribosomal RNA Control Reagents (Invitrogen) и использовали для нормирования количеств вирусных ДНК.
Статистика
[00118] Все значения представлены как среднее +/- среднеквадратическое отклонение. Различия между парами были проанализированы при помощи t-критерия Стьюдента с использованием Microsoft Excel 14.4.1. Значения P менее 0,05 (P <0,05) считались статистически значимыми.
РЕЗУЛЬТАТЫ
Конструирование векторов и рост вирусов
[00119] Сообщалось об эффективной трансдукции мышиных эмбриональных стволовых клеток (mESC) дефектным ВПГ-вектором, JDββ (Craft et al., Stem Cells 26, 3119-25 (2008)), без видимых повреждений клеток. JDββ делетировали по области внутренних повторов ("joint") ВПГ-генома и по двум IE-генам (ICP4 и ICP22). Кроме того, промоторы двух других IE-генов (ICP0 и ICP27) были заменены копиями вирусной промотора раннего (E или β) гена тимидинкиназы (ТК). JDββ-инфицированные некомплементирующие клетки показали минимальную экспрессию IE-гена, но вектор был способен к эффективной экспрессии трансгенов в mESC без нарушения с формирования эмбриоидного тела или развития программ транскрипции. Для облегчения использования этой векторной основы для разнообразных применений переноса генов, в бактериях проводили Red-опосредованный рекомбиниринг (Tischer et al., Biotechniques 40, 191-97 (2006)), чтобы получить основу, JΔΝΙ2 (Фиг. 9A), из KOS-37 ВАС, бактериальной искусственной хромосомы (ВАС), содержащей полный геном штамма KOS ВПГ-1 (Gierasch et al., Virol. Methods 135:197-206 (2006)). JΔΝΙ2 делетировали по ICP4 и joint, включая промотор ICP47, содержащий те же замены промотора ICPO и ICP27, что и JDββ, и консенсусные VP16-связывающие (TAATGARAT) мотивы в регуляторной области ICP22 делетировали, чтобы изменить кинетику экспрессии ICP22 на таковую для раннего гена. Для визуализации инфекции и отслеживания вирусной транскрипционной активности, кассету экспрессии репортерного гена mCherry вводили в положение удаленного локуса ICP4. Кроме того, ген гликопротеина B был заменен на гиперактивную аллель, gB:N/T (Uchida et al., J. Virol 84:12200-9 (2010)), для повышения проникновение вирусов в клетки. Для устранения потенциального продуцирования на низком уровне токсичных белков ICPO и ICP27 в некомплементирующих клетках, две производные JΔΝΙ2 также были сконструированы путем делеции полных последовательностей, кодирующих для СКП (JΔΝΙ3) или ICPO и ICP27 (JΔΝΙ5) (Фиг. .9A).
[00120] Для преобразования различных векторных конструкций JΔΝΙ в инфекционные вирусные частицы, была создана клеточная линия, способная комплементировать ICPO, ICP4 и ICP27. Клетки остеосаркомы U20S человека по природе комплементируют ICPO (Yao et al., J. Virol. 69, 6249-58 (1995)), что исключает необходимость экспрессировать этот токсичный белок. U20S-клетки трансдуцировали с лентивирусом, несущим ген ICP4 под контролем своих собственных регуляторных последовательностей, и была выделена клональная линия, которые постоянно экспрессирует ICP4 (U20S-ICP4;. Фиг. 9В). Трансдукция этих клеток вторым лентивирусом, несущим ген ICP27 и контрольной областью (Фиг. 6) была затем использована для отбора клеточной линии, U20S-ICP4/27, которая в дополнение экспрессирует ICP27 при инфекции вирусом с делецией ICP4/ICP27 (Фиг. 9В и Фиг. 6). Несмотря на то, что способности клеток U20S активировать промотор ICP4 в отсутствие вирусной белка VP16 и переносить длительную экспрессию ICP4 были неожиданными, неизвестно, относятся ли эти функции к природной ICP0-комплементирующей активности этих клеток. ДНК из BAC конструкций JΔΝΙ2 и JΔΝΙ3 может быть преобразована в инфекционный вирус путем трансфекции клеток U20S-ICP4, тогда как ДНК из конструкции JΔΝΙ5 давала инфекционные частицы только тогда, когда трансфицировали в клетки U20S-ICP4/27 (данные не приведены). Фигура 9С иллюстрирует рост вирусов JΔΝΙ2 и JΔΝΙ5, изначально полученных трансфекцией клеток U20S-ICP4 или U20S-ICP4/27 ДНК ВАС JΔΝΙ2 или JΔΝΙ5, соответственно. На немодифицированных клетках U20S или на клетках Vero-7b, которые комплементируют ICP4 и ICP27, не может расти ни вирус, но и ни ICPO. JΔΝΙ2 был способен расти на клетках U20S-ICP4 без комплементации ICP27, в то время как для роста JΔΝΙ5 требуется эта дополнительная комплементирующая активность. Стабильность JΔΝΙ5-комплементирующих свойств U20S- ICP4/27 клеток проверяли путем анализа бляшек на пассажах различных клеток, и не наблюдали значительного снижения эффективности образования бляшек по меньшей мере в течение 20 пассажей (таблица 3). Эти результаты согласовывались со сконструированными модификациями IE-генов в этих вирусах и позволили сравнить их биологические свойства.
Стандартизация введения вируса
[00121] Оценивали относительные количества каждого вируса, необходимые для доставки равного количества вирусной ДНК в ядро инфицированной клетки. Биологические и физические титры различных вирусных стоков определяли с помощью стандартного анализа образования бляшек на клетках U20S-ICP4/27 и кПЦР для вирусного гена D гликопротеина, соответственно; примерно 3-кратная разница в соотношении геномной копии (gc) к бляшкообразующим единицам (БОЕ) наблюдалась между стоками вируса JΔΝΙ2, JΔΝΙ3 и JΔΝΙ5 (таблица 2). HDF инфицировали 3 вирусами с равной БОЕ или равной gc, и количество ВПГ-геномов в ядрах клеток через 2 часа (h) после инфекции (pi) определяли при помощи кПЦР. При использовании равных gc для инфицирования, число вирусных геномов в ядрах через 2 hpi были сходны у клеток, инфицированных JΔΝΙ2, JΔΝΙ3 и JΔΝΙ5 (фиг. 10А). В противоположность этому, инфицирование с одинаковыми БОЕ привело к более высоким числам геномов JΔΝΙ5 в ядрах, чем геномов JΔΝΙ2 или JΔΝΙ3 (Фиг. 10B). Таким образом, в оставшейся части данного исследования, число gc было использовано для стандартизации введения вируса.
Характеристика векторных свойств в некомплементирующих клетках
[00122] Сначала рассмотрели влияние инфекции JΔΝΙ2, JΔΝΙ3 и JΔΝΙ5 на жизнеспособность некомплементирующих клеток HDF и Vero по сравнению с вирусом дикого типа KOS и предшествующим вектором с делецией гена IE, QOZHG (ICP47-/ICP277-/β-ICP22/β-ICP47; (Chen et al., J. Virol. 74, 10132-41 (2000)). Через 5 дней (д) pi с 25,000 gc/клетку, примерно в 4-5 раз больше жизнеспособных клеток остается в JΔΝΙ-инфицированной HDF-культурах, чем в KOS- или QOZHG-инфицированных культурах, в то время как меньшие различия были отмечены между JΔΝΙ- и QOZHG-инфицированными клетками Vero (Фиг. 11). Среди вирусов JΔΝΙ, JΔΝΙ2 был несколько более токсичным для HDF, чем JΔΝΙ3 (р=0,0017) и 5(р=0,0430), а токсичность для клеток Vero снизилась с JΔΝΙ2 до JΔΝΙ3 (р<0,001) до JΔΝΙ5 (р<0,001). Чтобы соотнести эти данные с экспрессии IE генов, Вестерн-блоты инфицированных HDF при 24 hpi исследовали при помощи антител к 4 из 5 IE-белков (Фиг. 1 IB). Как JΔΝΙ2, так и JΔΝΙ3 показали остаточную экспрессию ICP27, указывая, что ТК-промотор перед геном ICP27 в обоих векторах не является молчащим в этих клетках, но никаких IE-белков не было обнаружено в JΔΝΙ5-инфицированных клетках, несмотря на использование в 2-3 раза более высокое gc/клетку (равное КОЕ/клетку, смотри таблицу 2). Профили KOS и QOZHG согласовывались с описанной понижающей регуляцией экспрессии ICP0 при помощи ICP4 (Douville et al., Virology 207, 107-16 (1995); Resnick et al., J. Virol. 63, 2497-503 (1989)). С использованием количественного обратно-транскрипционного ПЦР (qRT-PCR) анализа была обнаружена мРНК ICP0 в JΔΝΙ2-инфицированных HDF и сниженные уровни мРНК ICP22 и ICP27 среди JΔΝΙ2-, JΔΝΙ3- и JΔΝΙ5-инфицированных клеток через 12 hpi с равным gc/клетку (Фиг. 11С). Эти результаты свидетельствуют о том, что JΔΝΙ2 продуцировал достаточный уровень ICP0 для повышения экспрессии ICP22 и ICP27 и подтвердил, что прогрессивное удаление IE-генов снижает цитотоксичность ВПГ-векторов (Krisky et al., Gene Ther. 5, 1593-603 (1998); Samaniego et al., J. Virol. 72, 3307-20 (1998); Samaniego et al., J. Virol. 71, 4614-25 (1997)).
[00123] qRT-PCR использовали для изучения экспрессии выбранных ранних и поздних вирусных генов (Фиг. 11D). Через 12 hpi, JΔΝΙ2 обычно экспрессирует самые высокие уровни этих генов, а JΔΝΙ5 - самый низкий, по аналогии с наблюдаемой картиной для IE-генов. Эти результаты показали, что замены нативных промоторов ICP0 и ICP27 на ранний промотор было недостаточно, чтобы сделать молчащим вирусный геном в отсутствие ICP4, то время как удаление обоих генов резко понижало остаточную генную экспрессию.
Экспрессия репортертерных генов JΔΝΙ
[00124] Для определения того, была ли подавлена активность экзогенного промотора (р) убиквитина C (UBC) человека, введенного с геном-репортером mCherry в делетированный локус ICP4 векторов JΔΝΙ, также в JΔΝΙ3- и JΔΝΙ5-инфицированных клетках по сравнению с JΔΝΙ2-инфицированными клетками, исследовали экспрессию mCherry в инфицированных HDF. Через 24 hpi с 5000 gc/клетку флуоресценция mCherry легко детектировалась в JΔΝΙ2-инфицированных клетках, но не в JΔΝΙ3- или JΔΝΙ5-инфицированных клетках (Фиг. 12А). Определение уровней мРНК mCherry через 6 hpi по qRT-PCR показал снижение транскрипционной активности с UBC-промотора примерно в 100 раз среди JΔΝΙ2- и JΔΝΙ3-инфицированных клеток, и дополнительное уменьшение приблизительно в 5 раз в JΔΝΙ5-инфицированных клетках (Фиг. 12B); контрольный вирус KNTc, использованный в этом эксперименте, являлся репликацие-компетентным и содержал кассету UbCp-mCherry в межгенной области между UL3 и UL4. Эти данные согласуются с интерпретацией, что остаточное экспрессия ICPO и ICP27 с JΔΝΙ2 (Фиг. 11C) является достаточным для предотвращения глобального транскрипционного сайленсинга вирусного генома, а удаление этих двух генов в существенной степени устраняет активность всех нативных (Фиг. 1 ID), а также экзогенных (Фиг. 12А, 12В) промоторов. В отличие от HDF, стандартные U20S-клетки, инфицированные JΔΝΙ3 или JΔΝΙ5, показали сильную экспрессию mCherry (Фиг. 12С), что позволило предположить, что ICPO-комплементирующая активность U20S-клеток была достаточна, чтобы активизировать эти молчащие в противном случае геномы. Чтобы подтвердить, что вирусный белок ICPO обладал той же активностью, JΔΝΙ5-инфицированные HDF были суперинфицированы вирусом ICP0+ QOZHG (см. Фиг. 1 IB). Как показано на Фиг. 12D, суперинфекции индуцировала экспрессию mCherry, а имитирующая суперинфекция нет. Взятые месте, эти результаты показали, что ICP27 не требуется для дерепрессии молчащего генома JΔΝΙ5 по меньшей мере при наличии обильной экспрессии ICPO или комплементирующей активности, и позволили предположить, что минимального количества ICPO, экспрессируемого в JΔΝΙ2-инфицированных HDF, достаточно для поддержания ограниченной транскрипционной активности на всем протяжении вирусного генома.
Высокий уровень трансгенной экспрессии в молчащем геноме JΔΝΙ5
[00125] При инфицировании нейрональных клеток, ВПГ вступает в латентную стадию, в котором вирусный геном является транскрипционно молчащим, за исключением локуса LAT. Было исследовано, остается ли LAT-локус сходным образом активным в клетках, отличных от нейрональных, в контексте молчащего в ином случае вирусного генома. С этой целью экспрессионную кассету, состоящую из CAG промотора/энхансера и гена EGFP (CAGP-GFP), вводили в LAT 2 т.п.н. интронную область JΔΝΙ5, с образованием векторной конструкции, названной JΔΝΙ7GFP (Фиг. 13 А). В качестве контроля, ту же кассету CAGP-GFP вводили в межгенную область UL3-UL4 JΔΝΙ5, с образованием вектора JΔΝΙ6GFP (Фиг. 13 А); межгенная область UL3-UL4 был использован для безразрывной вставки трансгенных кассет (Baines et al., J. Virol. 65, 938-44 (1991); Menotti et al., J. Virol. 16, 5463-71 (2002)) и близко, но за пределами локуса LAT. В другом месте настоящего описания термин "JΔΝΙ6GFP" упоминается как "JΔΝΙ6-CAGGFP" (смотри также фиг. 1 А). Инфекционные вирусы были получены на клетках U20S-ICP4/27, и соотношения gc: БОЕ новых вирусных стоков были аналогичными JΔΝΙ5 (таблица 2). Как JΔΝΙ6-CAGGFP, так и JΔΝΙ7GFP продуцировали обильную зеленую и красную флуоресценцию в процессе амплификации клеток U20S-ICP4/27, подтверждая целостность их кассет трансгенной экспрессии. Для изучения способности этих вирусов экспрессировать EGFP-трансген в некомплементирующих клетках, HDF инфицировали при увеличенном gc/клетку и EGFP-флуоресценцию регистрировали через 3 dpi (Фиг. 13B). JΔΝΙ7GFP-инфицированные клетки показали обильную, вирусную дозозависимую экспрессию EGFP, в то время как инфекция JΔΝΙ6-CAGGFP продуцировала ограниченную экспрессию даже в высокой дозе. Несмотря на то, что здесь использовали более высокое введение вирус, чем в предыдущих инфекциях JΔΝΙ5, экспрессия mCherry оставалась минимальной. Эти результаты были подтверждены с помощью qRT-PCR определения уровней мРНК EGFP и mCherry через 3 и 5 dpi (Фиг. 13С) и согласуются с предположением, что вирусные геномы JΔΝΙ6-CAGGFP и JΔΝΙ7GFP, как и геном JΔΝΙ5, были молчащими в инфицированных HDF, в то время как локус LAT остался транскрипционно активным. Поскольку уровень мРНК EGFP в JΔΝΙ7GFP-инфицированных клетках был по меньшей мере так же высок через 5 dpi, как и после 3 dpi (Фиг. 13C), было исследовано, возможно ли детектирование экспрессии в последующие моменты времени, когда клетки являются полностью контакт-ингибированными. Результаты показали, что экспрессия может сохраняться в течение не менее 4 недель в некоторых клетках (Фиг. 13D). Взятые вместе, эти наблюдения показали, что локус LAT является привилегированным местом для долговременной экспрессии трансгена с ВПГ-генома, который не является токсичным из-за функционального исключения экспрессия всех IE-генов.
CTRL и LATP2 поддерживают трансгенную экспрессию с локуса LAT
[00126] Было показано, что экспрессия LAT в латентном периоде контролируется латентность-ассоциироваными промоторами (LAP) 1 and 2 (Goins et al., J. Virol. 68, 2239-52 (1994); Zwaagstra et al., Virology 182, 287-97 (1991)). LAP1 расположен выше по течению от сайта инициации транскрипции на ~8,7 т.п.н. нестабильной первичного транскрипта LAT, которые процессируется в стабильный 2 т.п.н. LAT интрон, а LAP2 расположен ниже по течению от LAP1 в первом экзоне и распространяется в 2 т.п.н. интронную область. LAP2 содержащая область, распространяющаяся несколько дальше в интрон, была названа как LATP2. В дополнение к активности в качестве промотора, было показано, что область LAP2/LATP2 может выступать в качестве позиционно-независимого долговременного экспрессионно/энхансерного элемента для трансгенной экспрессии в нейронах (Berthomme et al., J. Virol. 74, 3613-22 (2000); Palmer et al., J. Virol 74, 5604-18 (2000)). Тем не менее, также сообщалось о том, что локус LAT защищен от глобального сайленсинга вирусного генома в латентном периоде при помощи областей, богатых сайтами связывания CTCF, называемыми CTRL, один из которых расположен выше по течению от LAP 1 (CTRL1), а другой в 2 т.п.н. интроне, только вниз по течению от LATP2 (CTRL2) (Blooom et al., Biochim. Biophys. Acta 1799, 246-56 (2010)). Таким образом, потенциальные роли элементов LATP2 и CTRL в позволяя наблюдаемую экспрессия EGFP в JΔΝΙ7GFP-инфицированных HDFS была исследована. Три элемента делетировали из генома JΔΝΙ7GFP по-отдельности и в комбинации (Фиг. 14 А), и исследовали экспрессию гена-репортера в инфицированных HDF (Фиг. 14В, 14С). Делеция либо CTRL1 (ΔC1) или LATP2 (ΔLP2) вызывала заметное снижение уровни зеленой флуоресценции и мРНК EGFP, тогда как делеция CTRL2 (ΔC2) оказывала лишь незначительное влияние. Делеция обеих CTRL (ΔC12) имела тот же эффект, что и делеция CTRL1 только, тогда как делеция всех трех элементов (ΔC12LP2) снижало экспрессию далее до уровня, наблюдаемого в JΔΝΙ6-CAGGFP-инфицированных клетках. Ни в одной из инфицированных клеток флуоресценции mCherry не было обнаружено (фиг. 14B), что позволило предположить, что различные делеции не вызывали дерепрессию других сайтов в вирусном геноме. Эти результаты показывают, что как CTRL1-, так и LATP2-область играет важную роль в защите связанного трансгена от транскрипционного молчания в контексте вирусного генома, который функционально лишен всех IE-генов.
Защита локусом LAT экспрессии трансгена является позиционно-независимой
[00127] Для определения того, являются ли последовательности, ассоциированные с локусом LAT, достаточными для защиты встроенной кассеты экспрессии трансгена от сайленсинга в отсутствие продуктов IE-генов, рестрикционный фрагмент, соответствующий области LAT:CAGp-GFP JΔΝΙ7GFP, в том числе два CTRL, LAP1 и LATP2, был встроен в одно из двух эктопических положений в производном JΔΝΙ5, который был делетирован из той же LAT области, чтобы избежать рекомбинации между исходными и новыми сайтами. Во-первых, Gateway (GW) рекомбинационную кассету вводили в межгенную область между UL45 и 46 (JΔΝΙ9GW) или между UL50 и UL51 (JΔΝΙIOGW) LAT-делетированного генома JΔΝΙ5, а затем в LAT:CAGP-GFP фрагмент или просто CAGp-GFP без LAT-последовательностей вводили рекомбинацией с кассетой GW (Фиг. 15 А). После продуцирования вируса в клетках U20S-ICP4/27 (таблица 2), векторы проверяли на экспрессию гена-репортера в инфицированных HDF. Через 3 dpi векторы, у которых отсутствовали LAT-последовательности, окружающие репортерную кассету (JΔΝΙ9GFP и JΔΝΙIOGFP), показали низкий уровень EGFP флуоресценции (Фиг. 15В) и мРНК (Фиг. 15C) в инфицированных клетках, подобно контрольному вектору JΔΝΙ6-CAGGFP, что позволяет предположить, что два межгенных инсерционных сайта были репрессированы транскрипционно, подобно межгенной области между UL3 и UL4 (Фиг. 13). Однако, когда репортерная кассета была фланкирована с обеих сторон LAT-последовательностями (векторы JΔΝΙ9LAT-GFP и JΔΝΙ10LAT-GFP), экспрессия EGFP увеличивалась до уровня, наблюдаемого в JΔΝΙ7GFP-инфицированных клетках (фиг. 15В), указывая, что анти-сайленсинговая активность LAT-полученных областей является функциональной позиционно-независимым способом. Чтобы подтвердить зависимость этой активности от LATP2 и одного или обоих CTRL, LATP2- и CTRL-делетированная версия LAT:CAGP-GFP фрагмента была введена GW-рекомбинацией в геном JΔΝΙIOGW; делеции были такими же, как таковые в LAT области описанного ранее вектора JΔΝΙ7GFPAC12LP2 (Фиг. 14А). Делеции понижали экспрессию EGFP в инфицированных HDF, хотя и не в полной мере до уровня, наблюдавшегося для не содержащего LAT вектора JΔΝΙ10GFP (Фиг. 15C, 15D); неясно, почему делеции AC12LP2, как оказалось, обладает меньшим эффектом здесь (~3,5 раза), чем в JΔΝΙ7GFP (~в 50 раз, на фиг. 14С). Тем не менее, эти результаты четко показывают, что часть локуса LAT, которая включает 2 CTRL, LAP1 и LATP2 может защитить встроенный кассету экспрессии трансгена независимым от положения образом от глобального сайленсинга вирусного генома при отсутствии IE-генной экспрессии, и показывают, что, по меньшей мере CTRL1 и LATP2 играют определенную роль в этой активности.
Элементы локуса LAT защищают экспрессии трансгена в других типах клеток
[00128] Для оценки применимости этих выводов помимо HDF, экспрессию EGFP и mCherry с выбранных векторов проверяли в других типах клеток. При помощи qRT-PCR, более высокие уровни мРНК EGFP наблюдались в JΔΝΙ7GFP-, чем в JΔΝΙ6-CAGGFP-инфицированных клетках человека через 3 dpi (Фиг. 16В). Наибольшие различия наблюдались в BJ-фибробластах крайней плоти человека (~35 раза), аналогично различию в HDF (~30-кратный, фиг. 13C), и в гепатоцитах человека (hHEP) (~40 раза). Человеческие неонатальные кератиноциты (НЕК) показали наименьший разницу (~4,5 раза), с промежуточными значениями, наблюдаемыми в мышечных стволовых клетках (hMDSC) (~10 раз) и преадипоцитах (HPAD) (~7 раза). Сравнение 2 векторов в нейронах спинных корневых ганглиев плода крысы (rDRG) показали лишь ~2,5-кратное различие. Фигура 16А демонстрирует репрезентативные флуоресцентные изображения через 3 dpi из независимого эксперимента, выполненного при тех же условиях инфекции. За исключением hHEP, которые были от другого донора, чем на Фигуре 16B, результаты в значительной степени согласуются с данными qRT-PCR. Интересно, в то время как ни одна из человеческих клеток не показала значительной флуоресценции mCherry, обильная экспрессия mCherry наблюдалась как в JΔΝΙ6-CAGGFP-, так и в JΔΝΙ7GFP-инфицированных DRG-культурах крысы. Пока еще не известно, является ли это наблюдение уникальным для нейрональных клеток, и может быть функцией промотора перед геном mCherry, расположения кассеты экспрессии в геноме вируса и/или происхождение клеток от крысы. hMDSC поддерживались в течение продолжительного периода времени, что позволяет контролировать экспрессию EGFP с течением времени в JΔΝΙ7GFP-инфицированных клетках. Как показано на фиг 16С, EGFP было легко детектировать по меньшей мере 4 недели после инфицирования в JΔΝΙ7GFP-инфицированных клетках, но не в JΔΝΙ6GFP-инфицированных клетках, подобно наблюдению с JΔΝΙ7GFP-инфицированными HDF (Фиг. 13D).
[00129] Наконец, для некоторых клеток определяли, усиливается ли активность CAGP-GFP в межгенной области UL50/51 фланговыми LAT-последовательностями, как в HDF. Результаты на Фиг. 16D показывают существенное улучшение для всех трех типов клеток, которые могут, по неизвестным причинам, превышать разницу между JΔΝΙ6-CAGGFP и JΔΝΙ7GFP в тех же клетках. В целом, эти результаты показывают, что позиционно-независимая анти-сайленсинговая активность генетических элементов в локусе LAT не ограничивается HDF, но действует в клетках человека различных типов, отличных от нейрональных клеток.
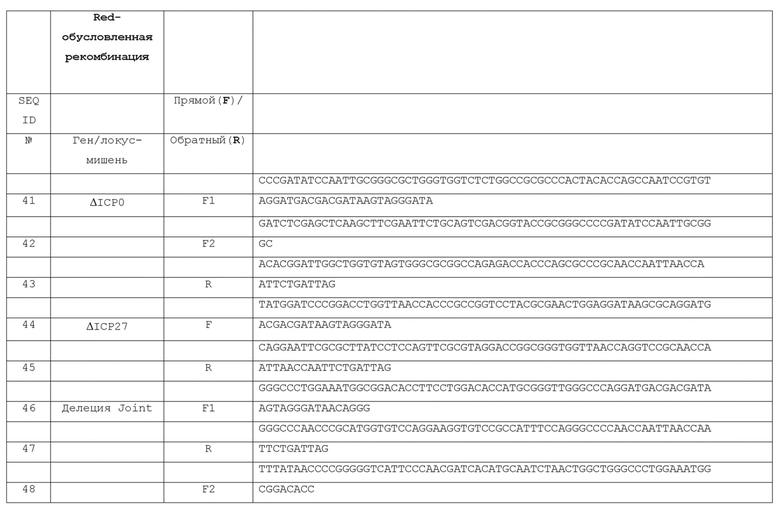
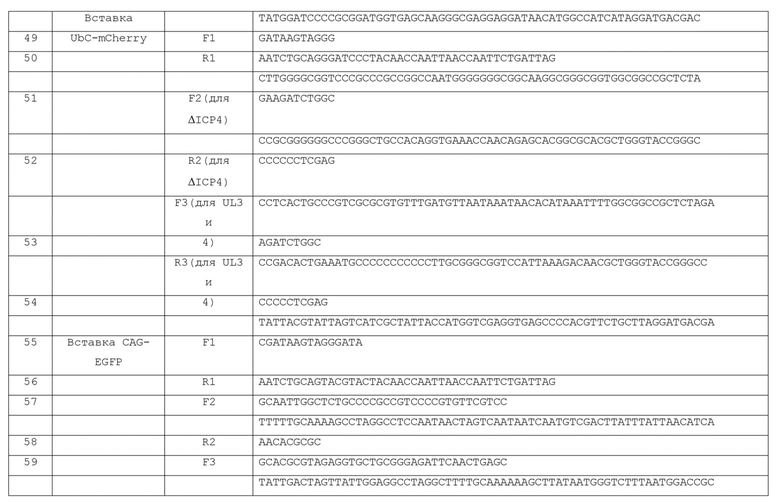
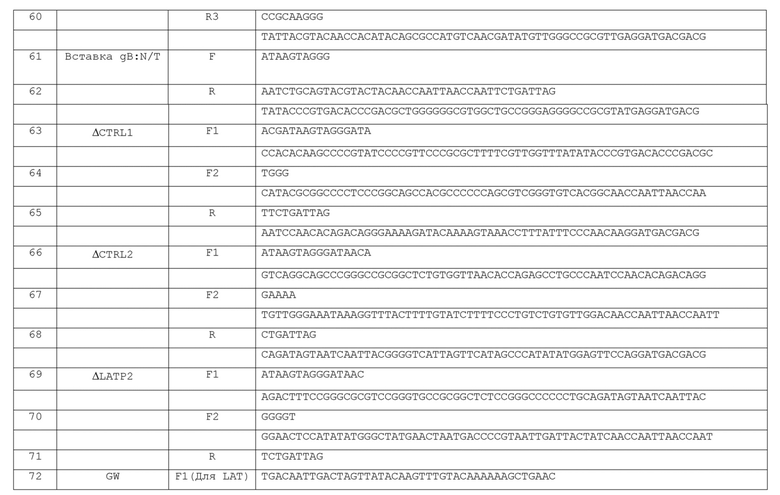
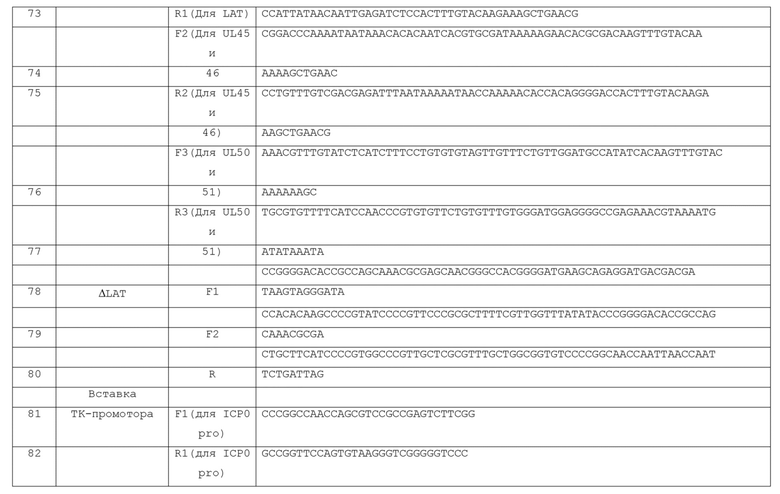

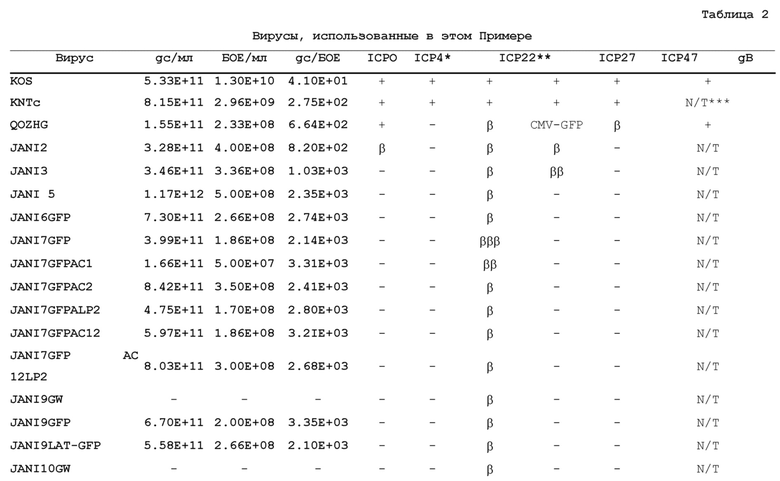

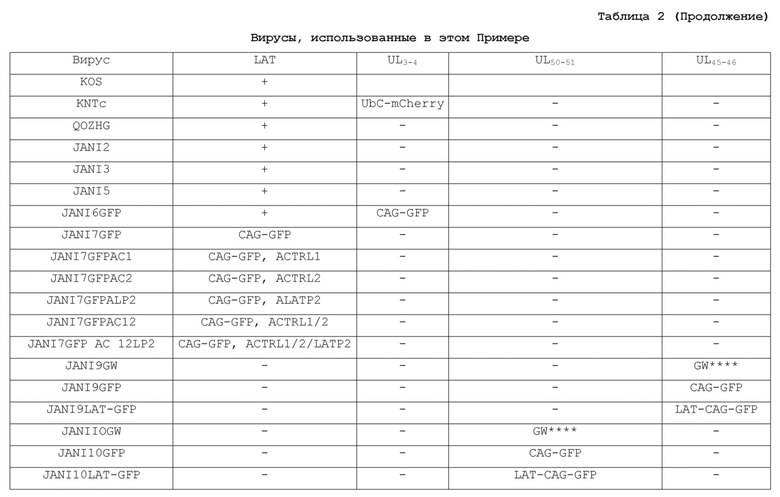


Клетки U20S-ICP4/27 инфицировали вирусом JANI5 при 0,1 gc/клетку, и клетки покрывали метилцеллюлозой через 2 hpi. Бляшки подсчитывали через 4 dpi. Среднее ± стандартное отклонение 3 определений
ПРИМЕР 9
[00130] Этот пример демонстрирует конструирование и свойства JΔΝΙ8.
[00131] JΔΝΙ8 был сконструирован за счет делеции UL41 из вектора JΔΝΙ5, описанного выше. Это графически представлено на Фиг. 17. JΔΝΙ7GFP по сути представляет собой JΔΝΙ5 со вставкой CAGp-EGFP-polyA в локусе LAT между LATP2 и CTRL2. JΔΝΙ8GFP идентичен JΔΝΙ7GFP, за исключением UL41. Смотрите также Фиг. 20.
[00132] Рост JΔΝΙ8 оценивали по отношению к JΔΝΙ5 в комплементирующих клетках, которые, как описано выше, представляют собой клетки U20S, сконструированные для экспрессии ICP4 и ICP27. Комплементирующие клетки были инфицированы соответствующим вектором с 1 копией вирусного генома (gc) на клетку. Как показано на Фигуре 18, как определено по бляшкообразующим единицам (БОЕ), геномной копии (GC) и экспрессии трансгена mCherry, JΔΝΙ8 демонстрирует более высокие темпы роста, чем JΔΝΙ5. Кроме того, нативные гены ВПГ, как наблюдали, экспрессировались ранее в ходе инфекции в комплементирующие клетки с JΔΝΙ8, в отличие от JΔΝΙ5 (Фигура 19, представляющей кратную экспрессию относительно каждого гена JDNI5 через 6 hpi, произвольно присвоено значение 1).
[00133] Как показано в Таблице 9-1, JΔΝΙ8 и JΔΝΙ8GFP продуцируют больше бляшек на геномную копию (gc) (на комплементирующих клетках), чем JΔΝΙ5 и JΔΝΙ7GFP, соответственно. Это показывает, что отношение функциональных частиц к дефектным (мера качества) в препаратах JΔΝΙ8 и JΔΝΙ8GFP выше, чем в препаратах JΔΝΙ5 и JΔΝΙ7GFP.
[00134] Кроме того, было отмечено, что инфекция на равных уровнях gc приводит к равных количествам вирусной ДНК в ядре человеческих кожных фибробластов. Тем не менее, с помощью протокола, обсуждаемого в связи с фиг. 10, это было достигнуто при помощи гораздо более низкого БОЕ для JΔΝΙ8 vs JΔΝΙ5. (Фиг. 21) Эти данные показывают, что инфекции при равных gc должны использоваться для исследований, сравнивающих JΔΝΙ8 с JΔΝΙ5 (или JΔΝΙ7GFP vs JΔΝΙ8GFP). МТТ-анализ жизнеспособности клеток (см. 22) показал, что JΔΝΙ8 (верхняя линия -×- в каждом графике) был заметно менее токсичен (приближающийся или превышающий жизнеспособность неинфицированных контрольных клеток), чем любой из других векторов при 5 или 10 БОЕ/клетку (MOI=5 или = 10). Нижняя линия -×- в каждом графике представляет результаты для JΔΝΙ5. Эти данные также показывают, что векторы, которые имеют интактный ген ICPO, даже под β-промотором (JΔBBB3 (также называется JDBBB3 или JΔβββ3 или JDβββ3)) или без UL41 (QOZHG), являются более токсичными, чем JΔΝΙ5 или JΔΝΙ8. Как и ожидалось, реплицирующийся вирус (KOS) убивает клетки. Дополнительные МТТ эксперименты, использующие HDF, показали, что хотя JΔΝΙ8 демонстрировали незначительную токсичность при более высоких MOI, она была гораздо меньше, чем JΔΝΙ5 при MOI в 3 раза ниже (равный gc/клетку) (Фиг. 23). Другие МТТ-данные показали, что JΔΝΙ8 менее токсичен, чем JΔΝΙ5 в некоторых (HDF, неонатальных кератиноцитах человека и нервных стволовых клеток человека), но не во всех (Vero, преадипоциты человека и гепатоциты человека) типах клеток (см. Фиг. 24).
[00135] Экспрессию репортерного гена с обеих трансгенов (т.е. EGFP и mCherry), присутствующих в JΔΝΙ7GFP и JΔΝΙ8GFP, соответственно, сравнивали в клетках HDF на 3 dpi. Результаты представлены на Фиг. 25 и 26. Было отмечено, что экспрессия GFP в JΔΝΙ8GFP увеличилась с дозой (от 5000 до 50000 gc/клетку, Фигура 25), но выравнивалась для JΔΝΙ7GFP. Не желая быть связанными теорией, полагают, что выравнивание, наблюдаемое для экспрессии в JΔΝΙ7GFP, может быть связано с высокой дозовой токсичностью вектора в клетках HDF, что подразумевает, что такая токсичность ниже в JΔΝΙ8GFP. Кроме того, заметную mCherry наблюдали в клетках HDF, инфицированных JΔΝΙ8GFP, но не в клетках HDF, инфицированных JΔΝΙ7GFP. Аналогичные эксперименты проводили с постоянной дозой либо JΔΝΙ7GFP, либо JΔΝΙ8GFP (25000 gc/клеткe) с уровнем флуоресценции EGFP или mCherry, анализированным в течение долгого времени (2, 4 и 6 дней после инфицирования (Фиг. 26)). Репортерные сигналы в JΔΝΙ8GFP- инфицированных культурах снижались между 4 и 6 dpi, вероятно в результате разбавления вирусной генома за счет неповрежденного деления клеток. Сигналы были более стабильными в JΔΝΙ7GFP-инфицированных культурах, скорее всего, из-за сокращения деления клеток.
[00136] Также исследовали трансгенную (EGFP или mCherry) экспрессию в других клетках, инфицированных либо JΔΝΙ7GFP vs JΔΝΙ8GFP. Как показано на Фигуре 27, на двух измеренных дозах (12500 gc/клетке и 25000 gc/клеткe) JΔΝΙ8GFP экспрессировали больше GFP в неонатальных кератиноцитах человека, чем JΔΝΙ7GFP.
[00137] Кроме того, анализировали трансгенную экспрессию в клетках корней спинного ганглия (DRG) крысы, инфицированных либо JΔΝΙ7GFP, либо JΔΝΙ8GFP (6250 gc/клетку). Как показано на Фигуре 28, экспрессия mCherry (вставляется в ICP4-делеционный локус обоих векторов) с JΔΝΙ7GFP была повышена в DRG-нейронах крысы относительно GFP, по сравнению с клетками, отличными от нейрональных, и относительно больше в JΔΝΙ8GFP-, чем в JΔΝΙ7GFP-инфицированных клетках. Эти результаты показывают, что JΔΝΙ8 является нетоксичным и обеспечивает трансгенную экспрессию в нейронах как с LAT, так и с ICP4 локуса.
ПРИМЕР 10
[00138] Этот пример демонстрирует in vivo экспрессию с вектора JΔΝΙ7-GFP в долговременном эксперименте.
[00139] JDNI7-GFP (8,0×108 геномных копий/2 мкл; 3,7×105 БОЕ) стеротактически вводили в гиппокамп крыс (AP: -1.8; ML: -1.7; P: +3.5) со скоростью 0,1 мл/мин. Животных умерщвляли и перфузировали через 6, 15 или 30 дней после инъекции вектора (dpi), и криосрезы мозга исследовали под флуоресцентным микроскопом. Фигура 29 показывает сильную экспрессию как EGFP, так и mCherry, совместно локализованных в одних и тех же клетках в гиппокампе. На Фигуре 29 сверху, отдельные (слева; 40x) или объединенные (в центре справа; 20x) изображения флуоресценции EGFP и mCherry в указанные дни после инфицирования (dpi); снизу, отдельные и объединенныя изображения представлены через 15 dpi (10X).
ПРИМЕР 11
[00140] Этот пример относится к экспрессии трансгена mCherry с вектора JΔΝΙ7-GFP в инфицированных первичных астроцитах.
[00141] Астроциты мыши инфицировали (M.O.I. = 5) и культивировали. Через 25 дней после инфицирования, клетки фиксировали и подвергали воздействию антитела, специфичного к глиальному фибриллярному кислому белку (GFAP), положительное связывание которого идентифицирует астроциты. Иммунофлуоресцентные изображения показали, что JΔΝΙ7-GFP является нетоксичным и обеспечивает постоянную экспрессию в астроцитах.
ПРИМЕР 12
[00142] Этот пример демонстрирует создание клеточной линии для комплементации ВПГ-векторов по изобретению, и которая также способствует эксцизии из LoxP-фланкированной BAC кассеты во время размножения вирусов.
[00143] Обсуждаемая выше клеточная линия U20S-ICP4/27 была сконструирована, чтобы экспрессировать Cre-рекомбиназу. Полученная в результате клеточная линия U20S-ICP4/27/Cre комплементирует функции каждого из дефектных ВПГ IE-генов в векторах JΔΝΙ5, 7, 8 и их производных, за исключением функции несущественного гена ICP47, и удаляет ВАС кассету во время вирусного роста.
[00144] Клетки U20S-ICP4/27/Cre были получены путем инфицирования клеток U20S-ICP4/27 Cre-экспрессирующего ретровирусного вектора. Для конструирования ретровирусного вектора плазмиду pCX4Hyg (PNAS 2003, 100: 13567-13572; инвентарный номер GenBank AB086387) модифицировали путем вставки Gateway рекомбинантной кассеты в сайт для множественного клонирования, чтобы создать pCX4Hyg-GW. Полную кодирующую последовательность NLS-Cre из плазмиды pTurbo-Cre (инвентарный номер GenBank AF334827.1) встраивали между сайтами attLl и attLl pENTRIA (Invitrogen) и затем переносили в pCX4Hyg-GW путем LR клоназа - опосредованной Gateway рекомбинации, для создания pCX4Hyg-CRE. Эта плазмида и ее конструирование схематически изображены на Фиг. 30 и 31. Ретровирусные частицы получали, как описано в Makino et al. (Exp Cell Res 2009, 315: 2727-2740). Вкратце, pCX4Hyg-Cre совместно трансфицировали с экспрессионными плазмидами Gag-Pol и VSV-G в клетки 293Т, и супернатант собирали через 48 ч, фильтровали через 0,45 микронный фильтр и концентрировали центрифугированием. Подходящие плазмиды Gag-Pol & VSV-G являются коммерчески доступными (например, pCMV-Gag-Pol от Cell Biolabs (Cat. No. RV-111) и pCMV-VSV-G от Addgene).
[00145] Клетки U20S-ICP4/27 инфицировали очищенным Cre-ретровирусом, и клетки отбирали по устойчивости к пуромицину, бластицидину и гигромицину. Устойчивые колонии выделяли, размножали, инфицировали JΔΝΙ7-GFP при 0,001-1 КОЕ/клетку и окрашивали на β-галактозидазную активность через 2 дня после инфицирования (dpi). Клоны, демонстрирующие несколько синих клеток, как было обнаружено, подвергались быстрому цитопатическому эффекту (100% CPE на dpi после инфицирования при MOI=0,01), тогда как родительские клетки U20S-ICP4/27 показали образующие бляшки кластеры голубых клеток и примерно 25% CPE на 9 dpi. Эти результаты показывают, что Cre-опосредованное делетирование ВАС-элементов вместе со связанной экспрессионной кассетой LacZ в JΔΝΙ7-GFP способствует вирусному росту. Точное удаление последовательностей ВАС и LacZ между сайтами LoxP в JΔΝΙ7-GFP был подтвержден ПЦР на вирусной ДНК из отдельных бляшек JΔΝΙ7-GFP на отобранных U20S-ICP4+/27+/Cre+ клонах.
[00146] Таким образом, было замечено, что пассирование через эту клеточную линию U20S-ICP4/27/Cre удаляет примерно 11 т.п.н. последовательности ВАС, восстанавливая место для трансгенов, и заметно ускоряет вирусный рост.
[00147] Все ссылки, включая публикации, патентные заявки и патенты, цитируемые в настоящем документе, включены в качестве ссылок в той же степени, как если бы было индивидуально и конкретно указано, что каждая ссылка включена в качестве ссылки и была изложена во всей ее полноте в данном описании.
[00148] Применение терминов "а" и "an" и "the" и "по меньшей мере, один» и подобные ссылки в контексте описания изобретения (особенно в контексте нижеследующей формулы изобретения) следует истолковывать как охватывающие как единственные, так и множественные числа, если не указано иное в данном документе или явно не противоречит контексту. Использование термина "по меньшей мере, один", за которым следует список из одного или более элементов (например, "по меньшей мере, один из А и В") должно быть истолковано в значении одного элемента, выбранного из перечисленных элементов (А или В) или любого сочетания двух или более из перечисленных элементов (А и В), если не указано иное в данном документе или явно противоречит контексту. Термины "содержащий", "имеющий", "включающий" и "состоящий" следует истолковывать как открытые термины (т.е. означающие "включающий, но не ограничиваясь этим"), если не указано иное. Перечисление диапазонов значений в настоящем документе предназначено только для использования в качестве краткого способа со ссылкой индивидуально на каждое отдельное значение, попадающее в диапазон, если не указано иное в настоящем документе, и каждое отдельное значение включено в описание, как если бы оно было индивидуально указано в настоящем документе. Все описанные здесь способы могут быть выполнены в любом подходящем порядке, если иное не указано в настоящем документе или иным явно не противоречит контексту. Использование любого и всех примеров, или типичной формулировки (например, "такой как"), представленной в настоящем документе, предназначено только для лучшего освещения изобретения и не накладывает ограничения на объем изобретения, если не заявлено иное. Ни одну формулировку в описании не следует толковать как указывающую на какой-либо незаявленный элемент как существенный для практического осуществления изобретения.
[00149] Предпочтительные варианты осуществления настоящего изобретения описаны в данном документе, включая наилучший способ, известный авторам изобретения для осуществления изобретения. Вариации этих предпочтительных вариантов осуществления могут стать очевидными для специалистов в данной области техники после прочтения приведенного выше описания. Авторы изобретения ожидают, что квалифицированные специалисты используют такие вариации соответствующим образом, и авторы изобретения имеют в виду, что изобретение будет осуществлено иначе, чем конкретно описано в настоящем документе. Соответственно, данное изобретение включает все модификации и эквиваленты предмета, изложенного в прилагаемой к этому формуле изобретения в соответствии с действующим законодательством. Кроме того, любая комбинация вышеописанных элементов во всех их возможных вариантах охватывается изобретением, если иное не указано в настоящем документе или явно не противоречит контексту.
| название | год | авторы | номер документа |
|---|---|---|---|
| ОНКОЛИТИЧЕСКИЙ HSV-ВЕКТОР | 2014 |
|
RU2719190C2 |
| РЕКОМБИНАНТНЫЙ ОНКОЛИТИЧЕСКИЙ ВИРУС, СПОСОБ ЕГО ПОЛУЧЕНИЯ, ЕГО ПРИМЕНЕНИЕ И ЛЕКАРСТВЕННЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ | 2020 |
|
RU2795230C1 |
| УСЛОВНО РЕПЛИЦИРУЮЩИЙ АДЕНОВИРУС | 2012 |
|
RU2642293C2 |
| ФЛАВИВИРУС С ДВУХКОМПОНЕНТНЫМ ГЕНОМОМ И ЕГО ИСПОЛЬЗОВАНИЕ | 2008 |
|
RU2527891C2 |
| ВАКЦИНЫ ПРОТИВ ВИРУСА ПРОСТОГО ГЕРПЕСА | 2010 |
|
RU2575067C2 |
| ВИРУСНЫЕ ЧАСТИЦЫ, СОДЕРЖАЩИЕ ВЕКТОР, ПРОИСХОДЯЩИЙ ИЗ АЛЬФАВИРУСА, И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОЙ ВИРУСНОЙ ЧАСТИЦЫ | 2004 |
|
RU2398875C2 |
| ГЕННАЯ ТЕРАПИЯ ОФТАЛЬМОЛОГИЧЕСКИХ НАРУШЕНИЙ | 2016 |
|
RU2762747C2 |
| ОНКОЛИТИЧЕСКИЕ ВИРУСНЫЕ ВЕКТОРЫ И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2771110C2 |
| ДВОЙНОЙ ВЕКТОР ДЛЯ ПОДАВЛЕНИЯ ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА | 2010 |
|
RU2562868C2 |
| СОЗДАНИЕ ГАПЛОИДНЫХ РАСТЕНИЙ И УСОВЕРШЕНСТВОВАНИЕ СЕЛЕКЦИИ РАСТЕНИЙ | 2010 |
|
RU2571927C2 |
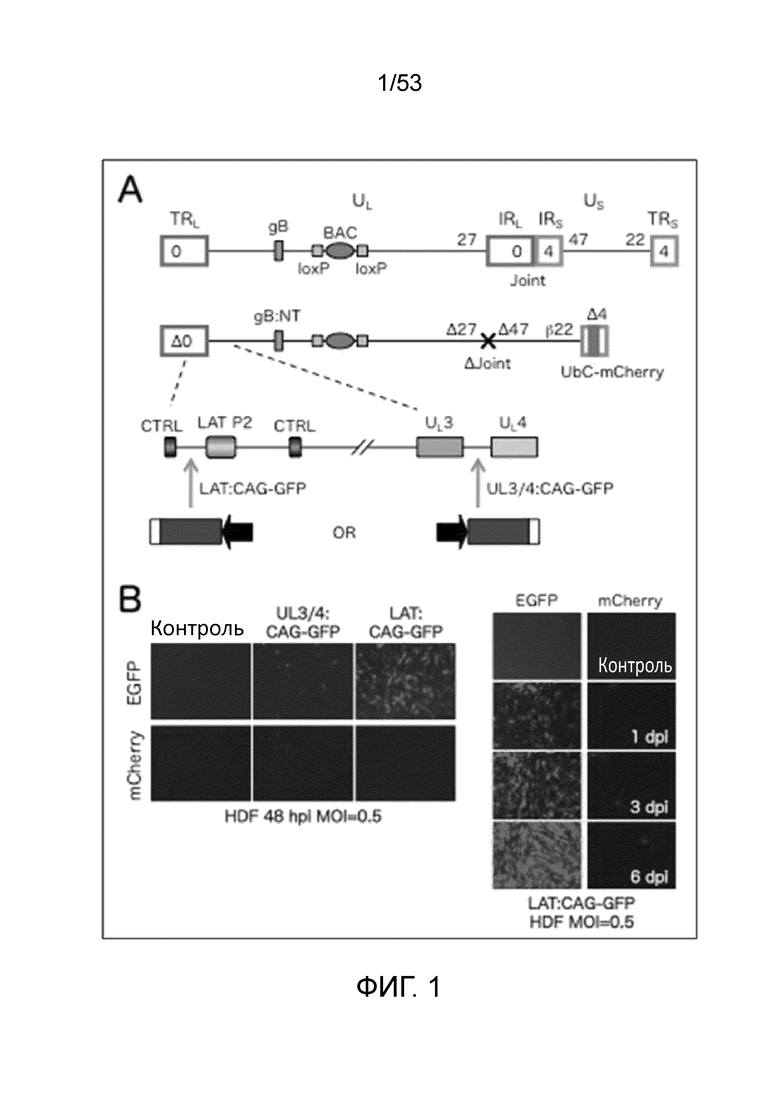


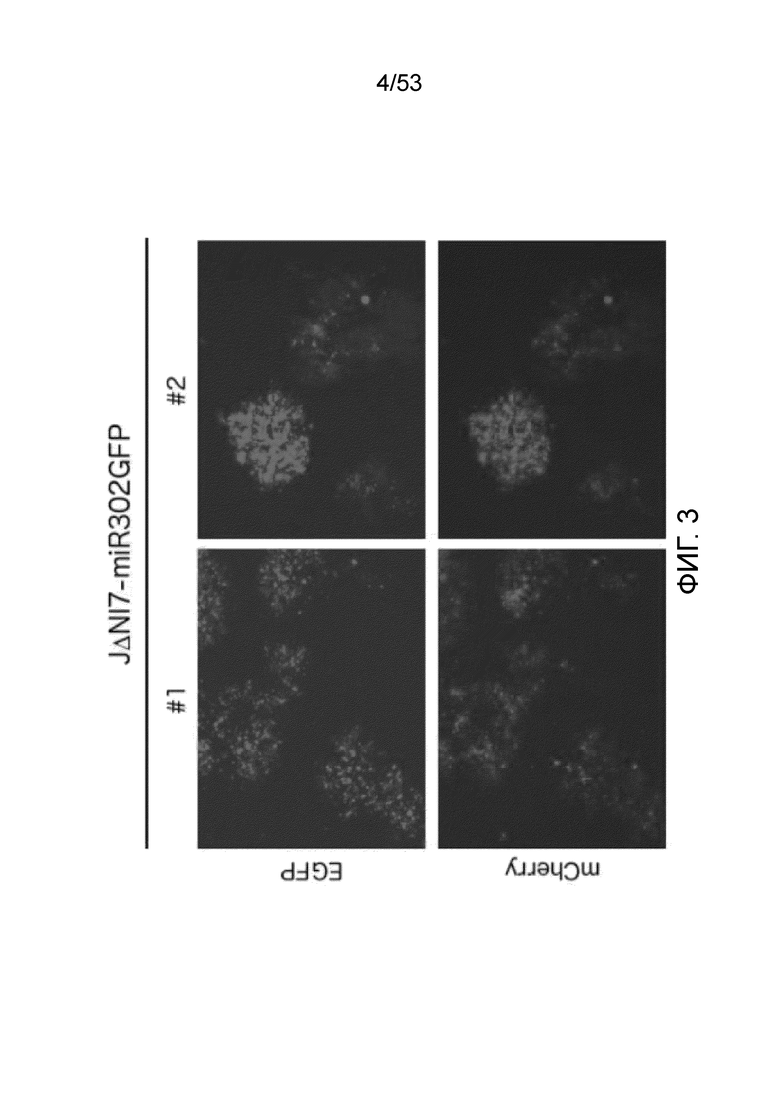
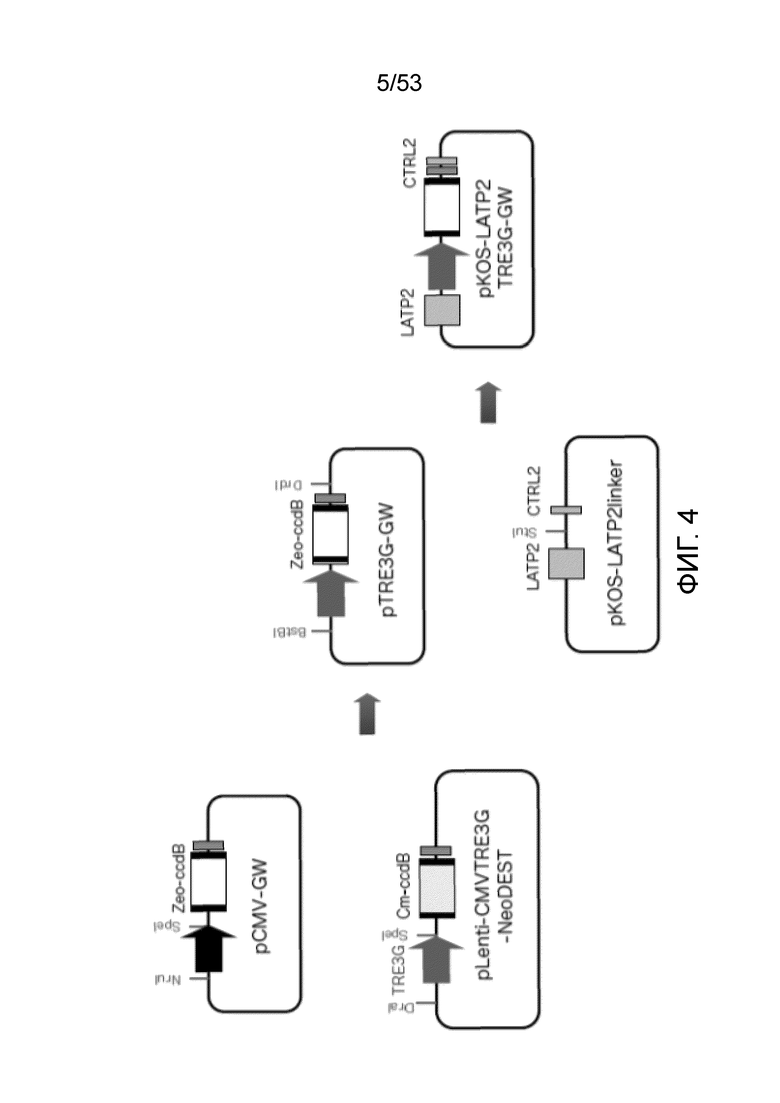
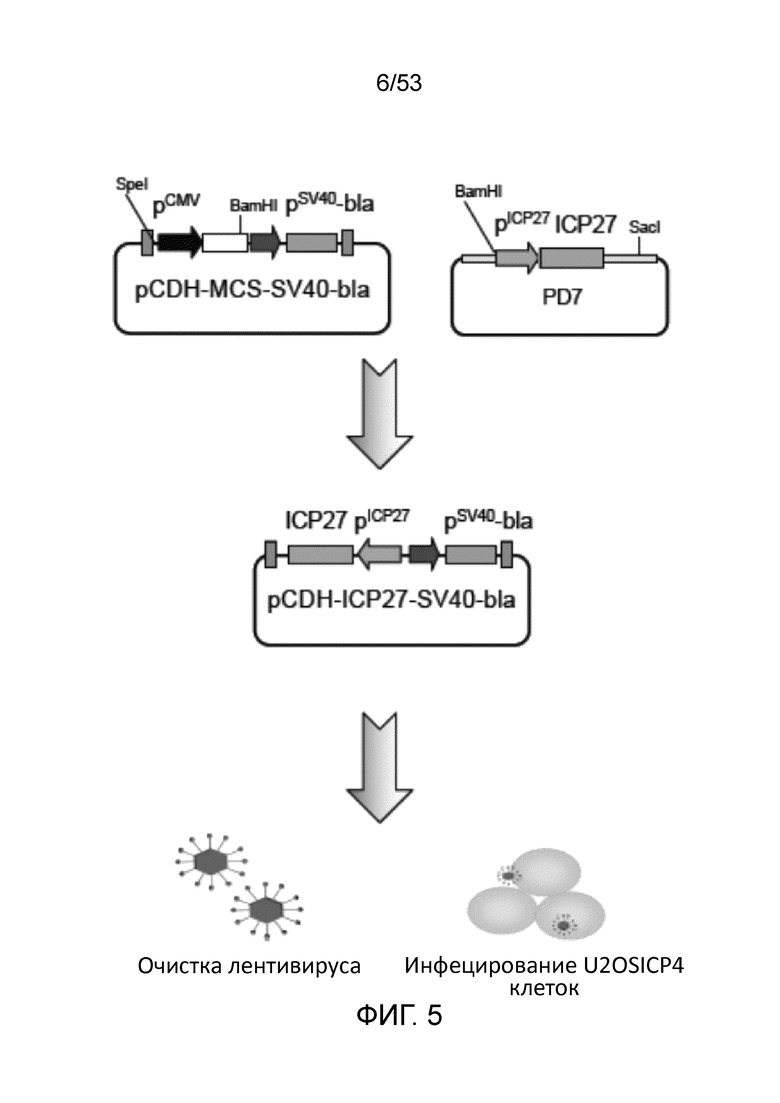
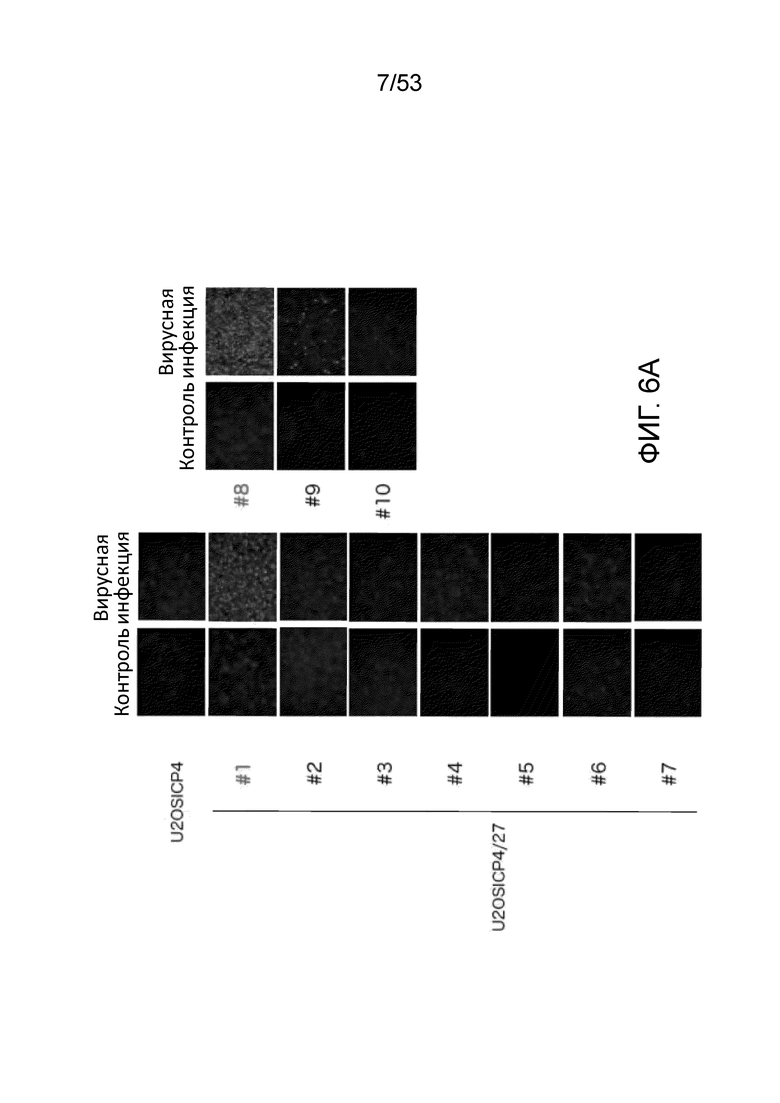
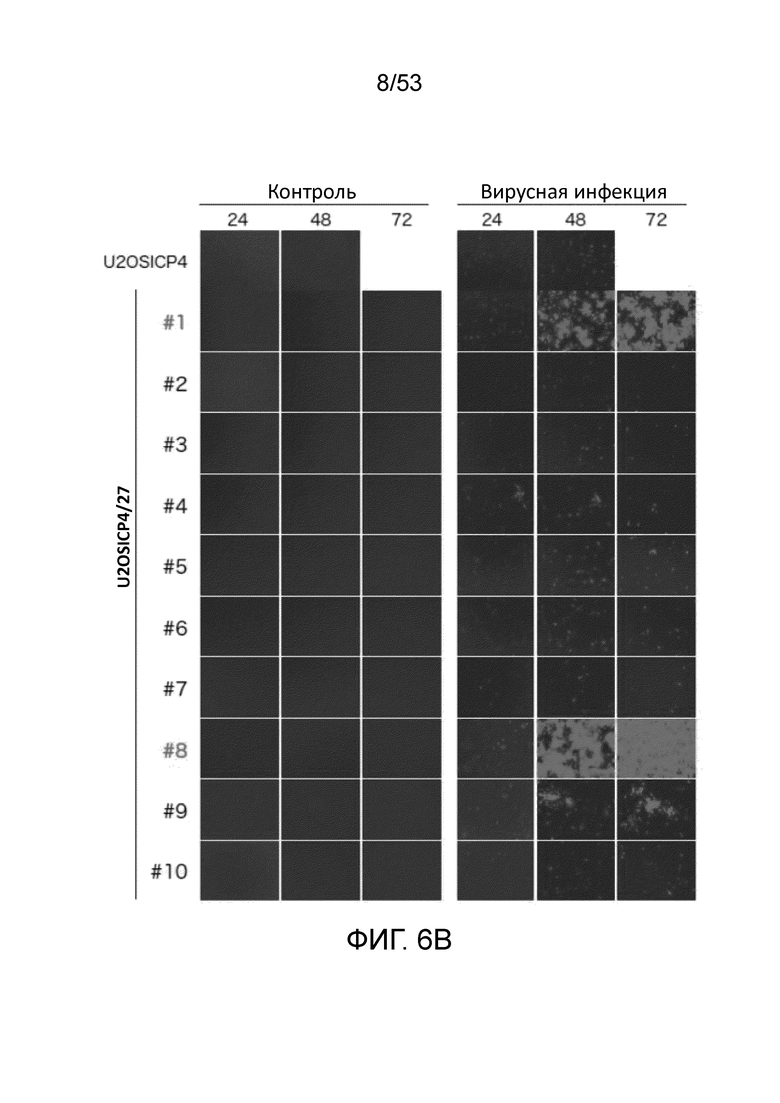
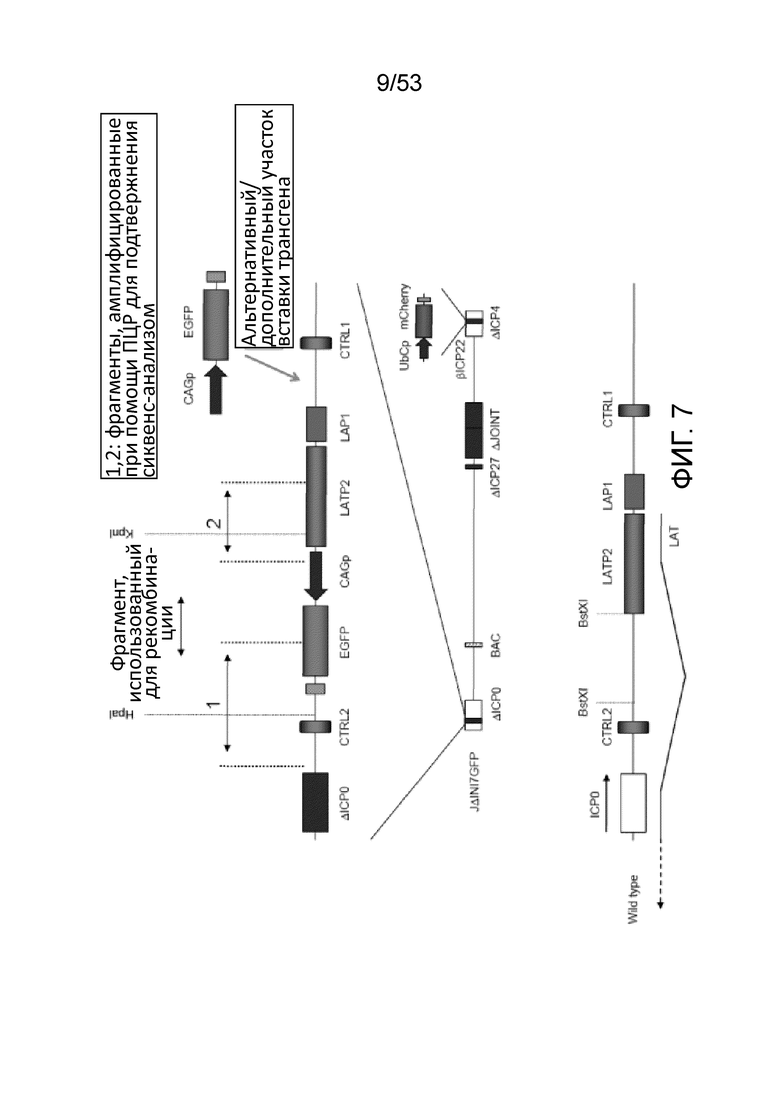





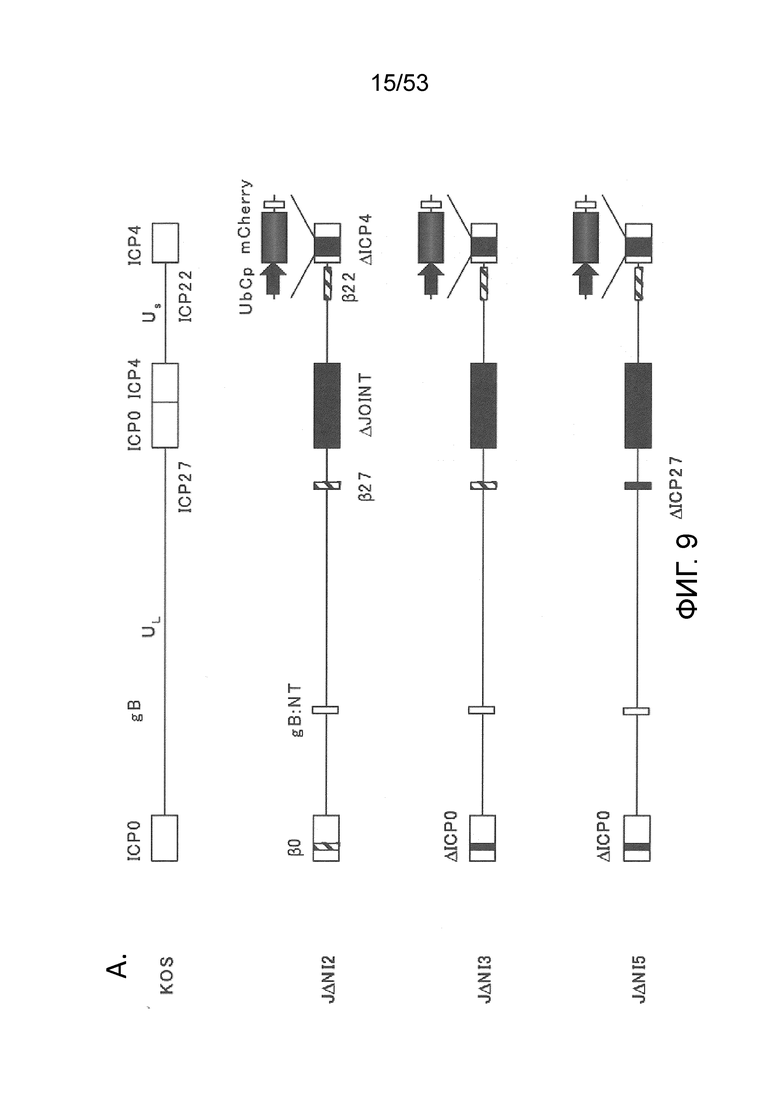
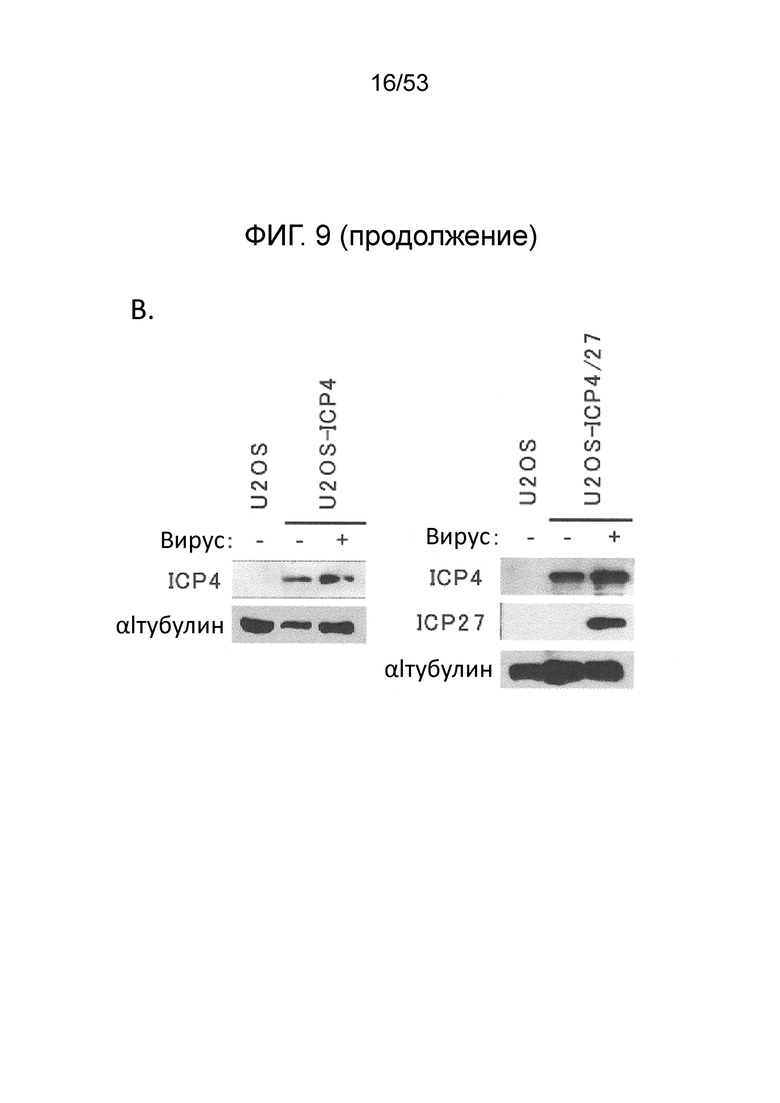
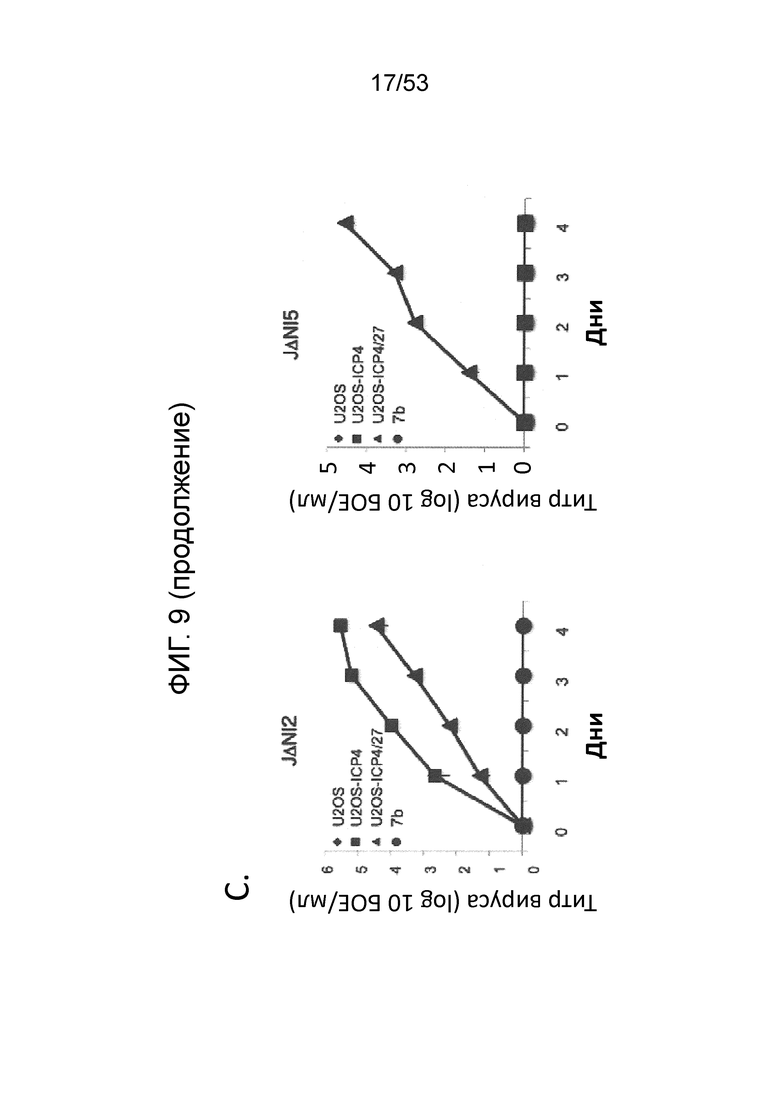
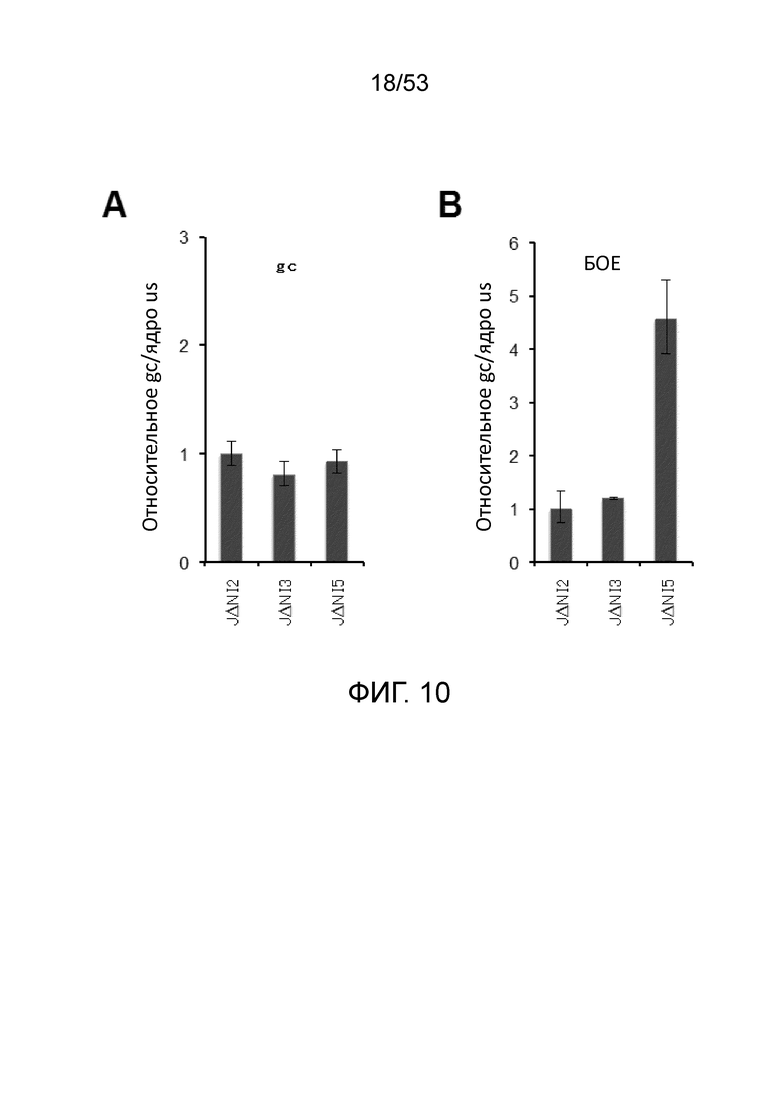
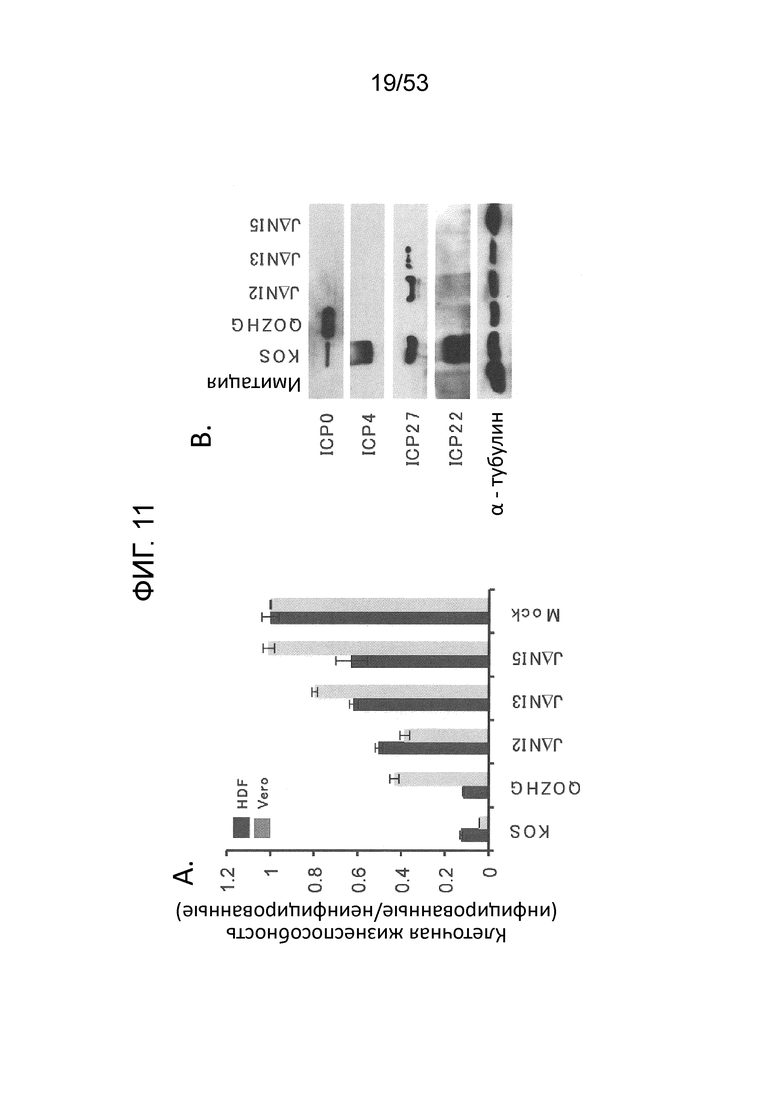
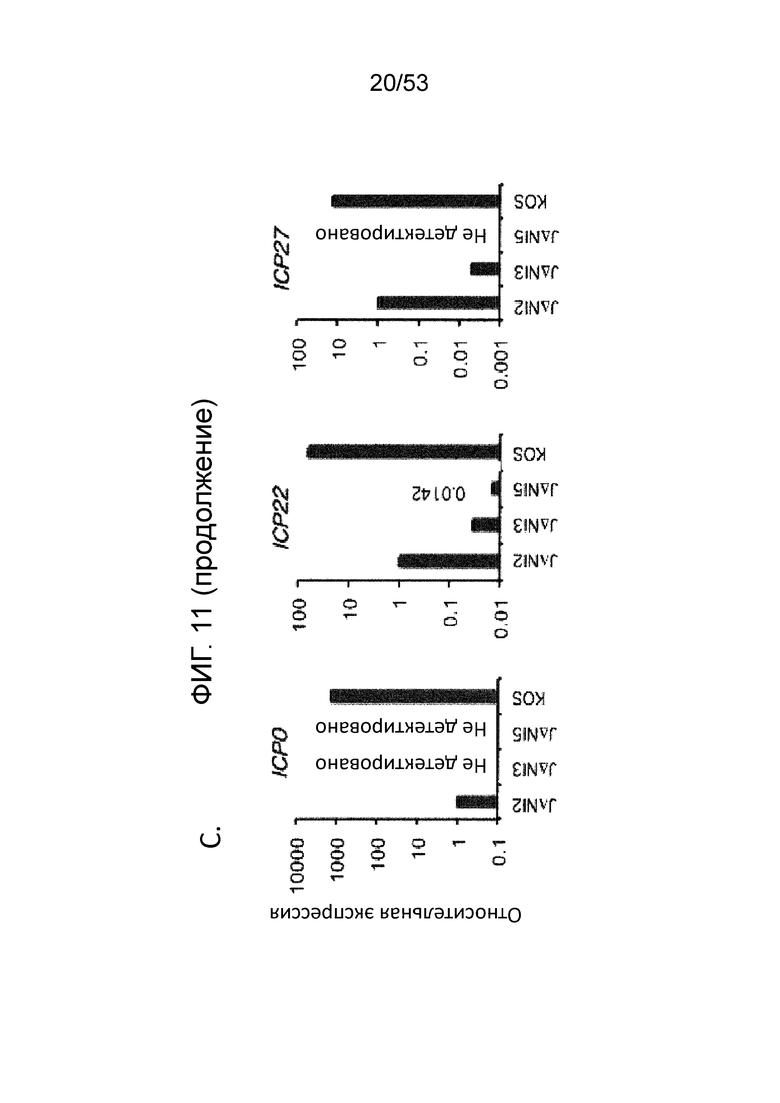
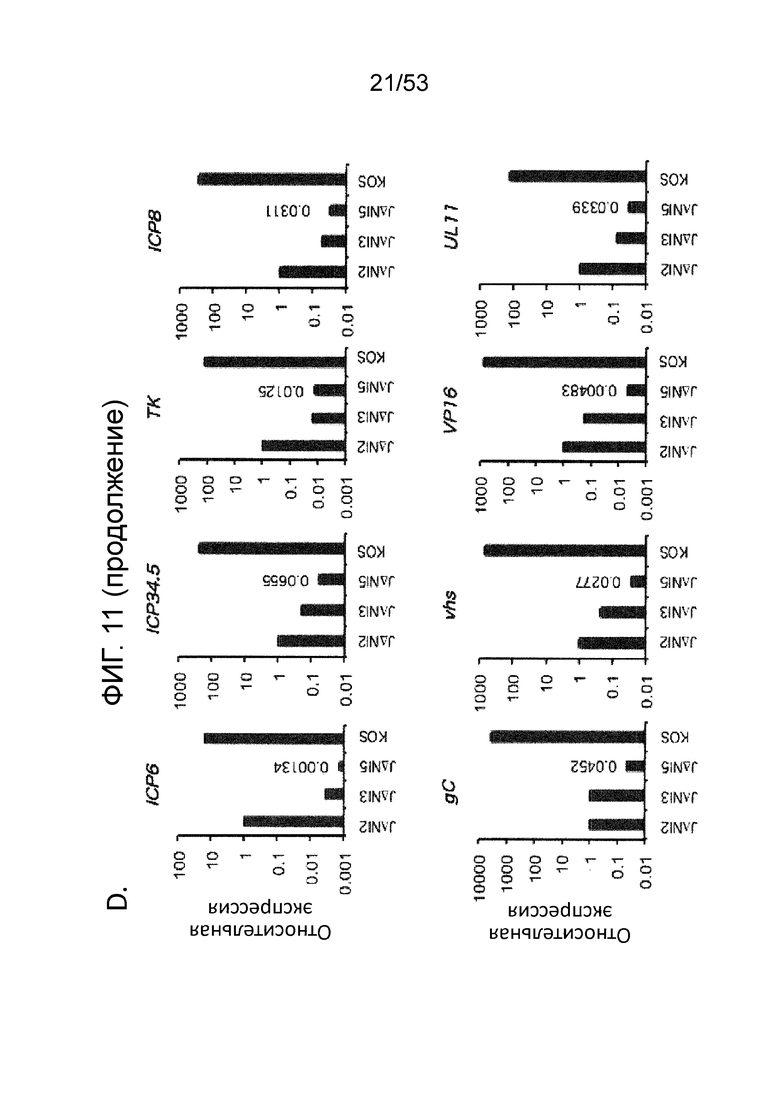
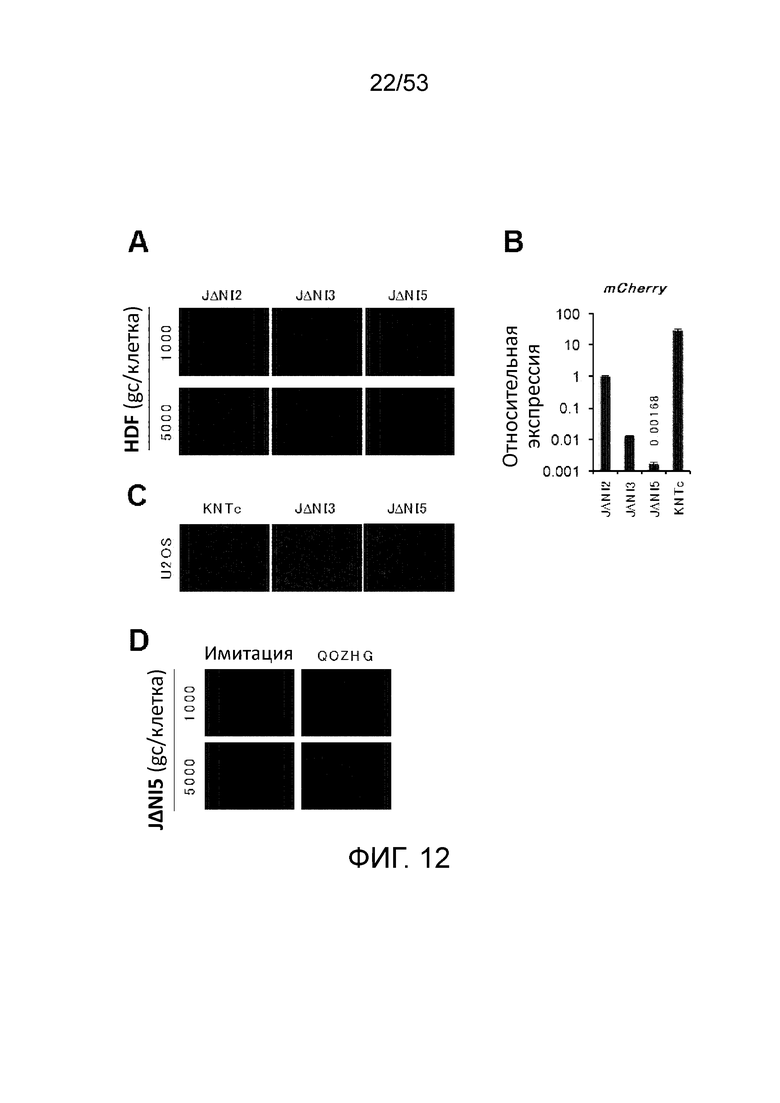
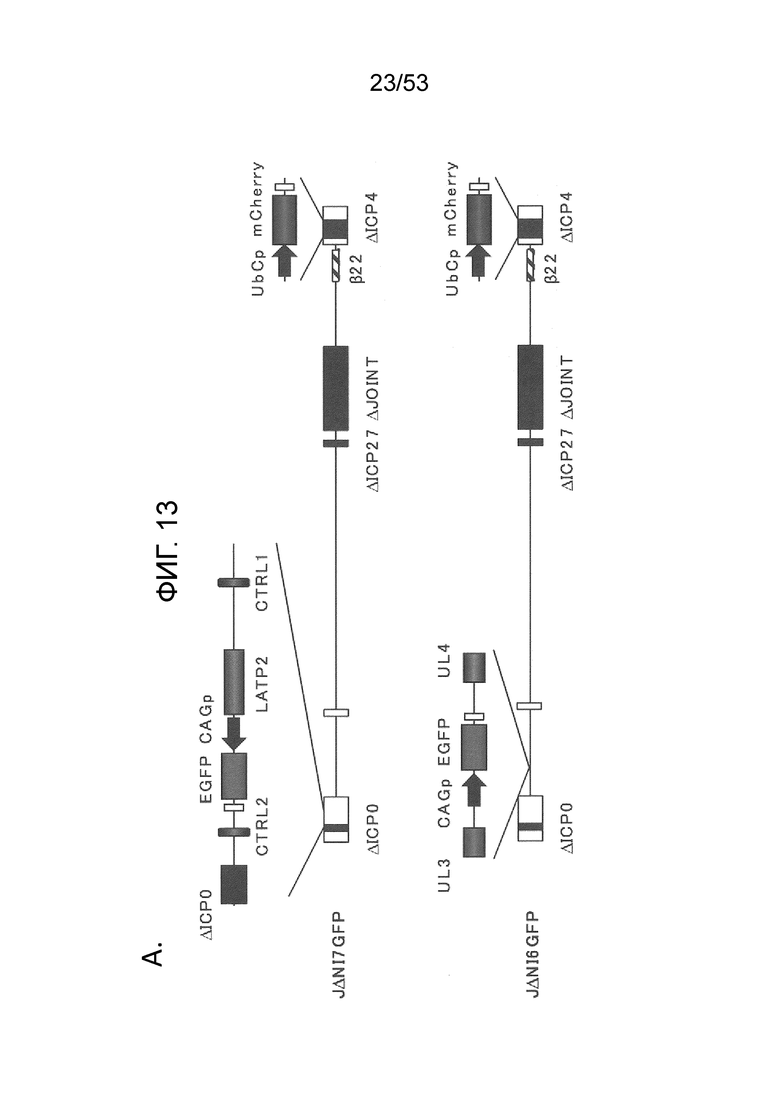
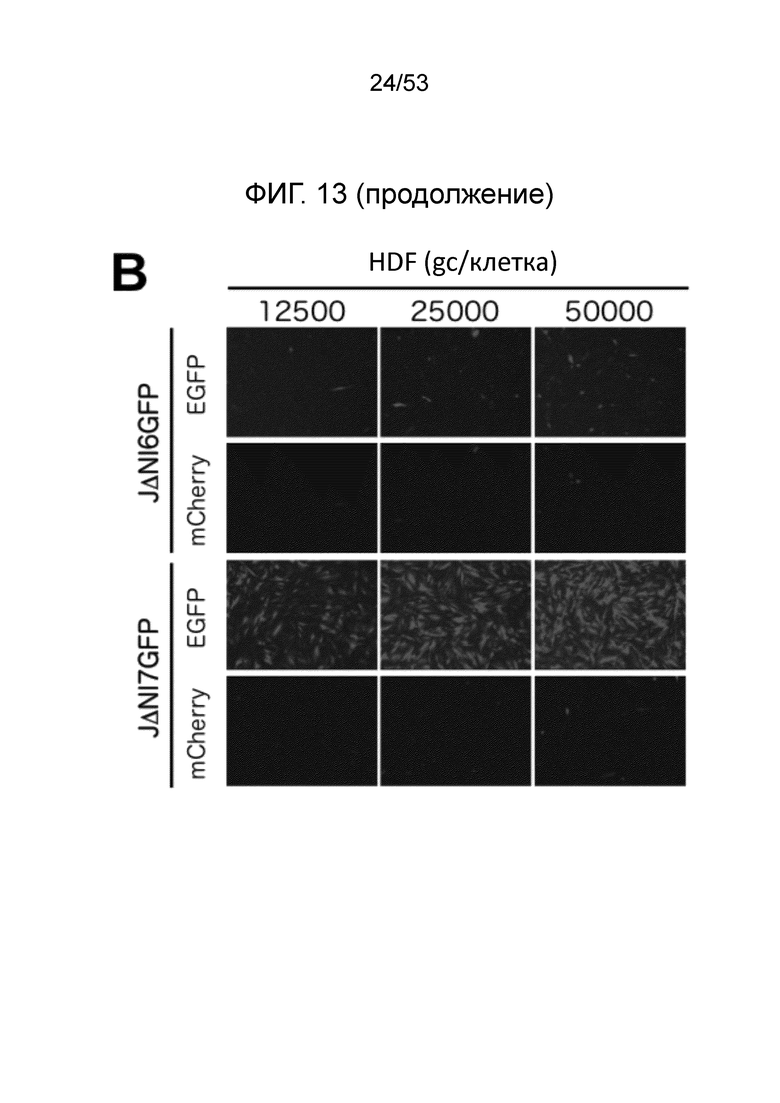
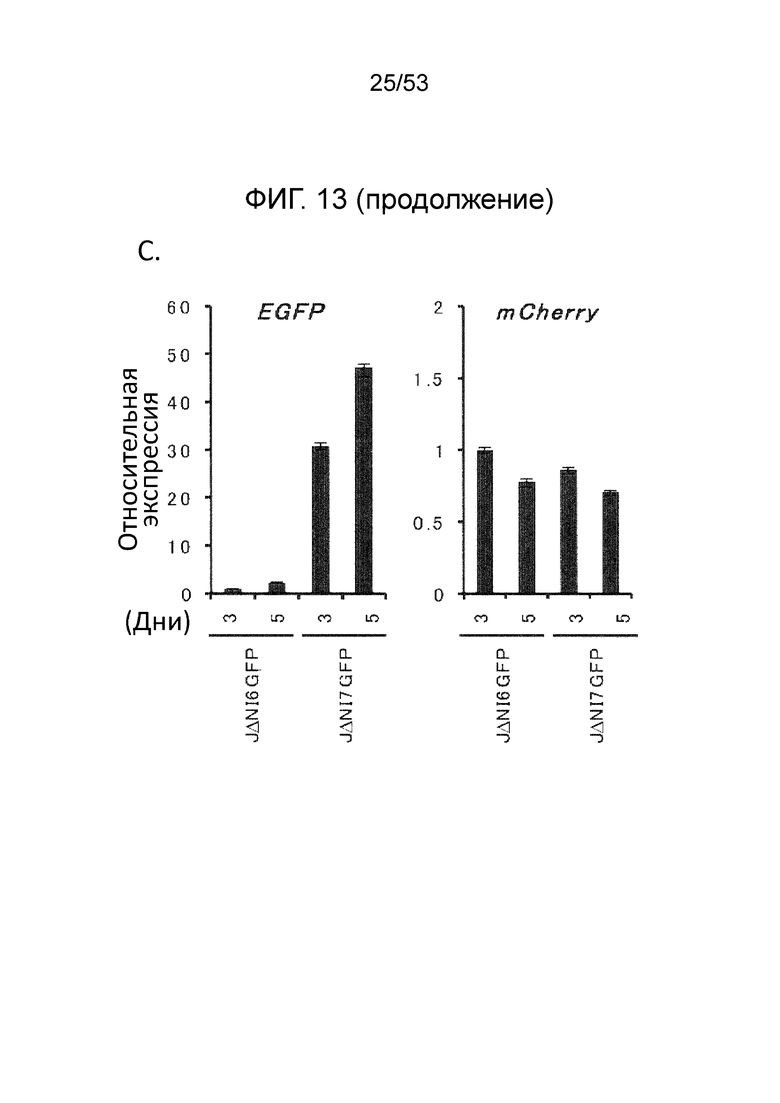
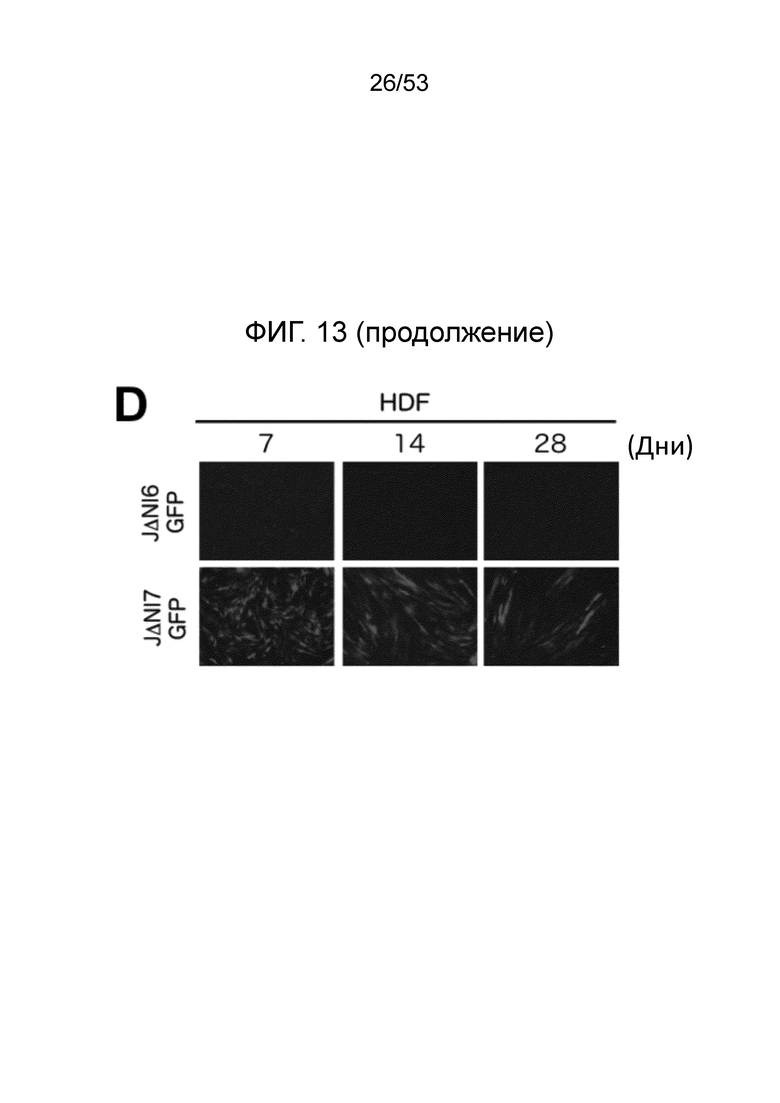
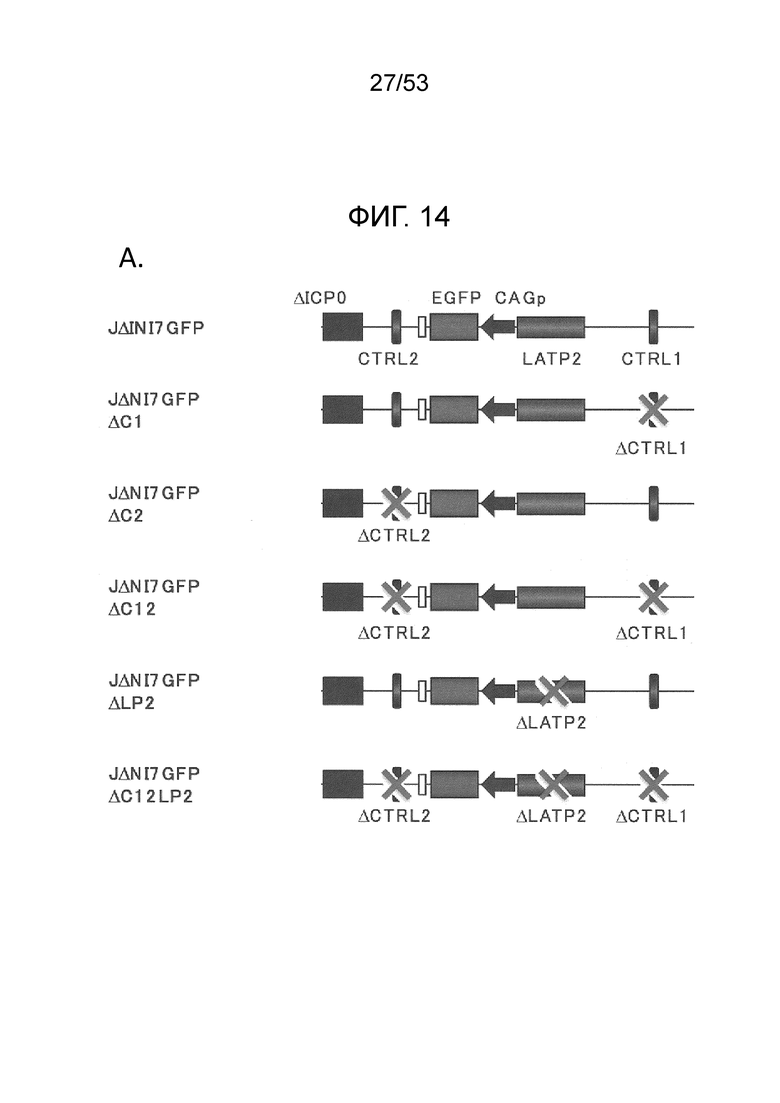

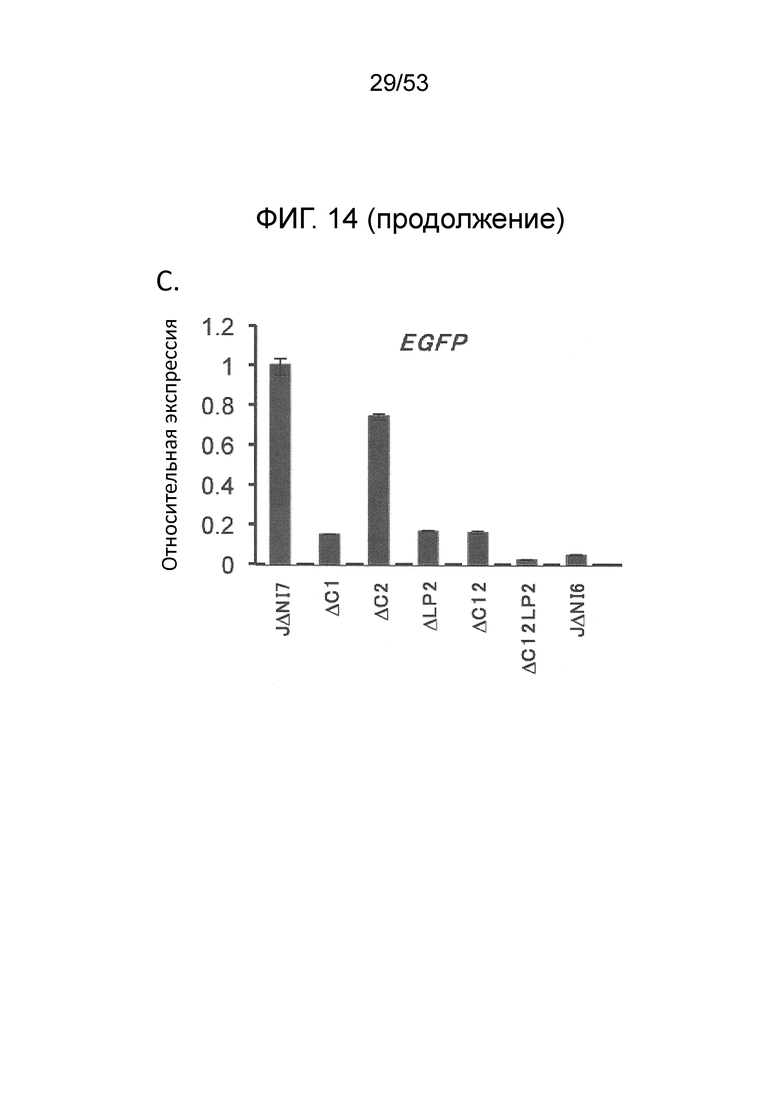
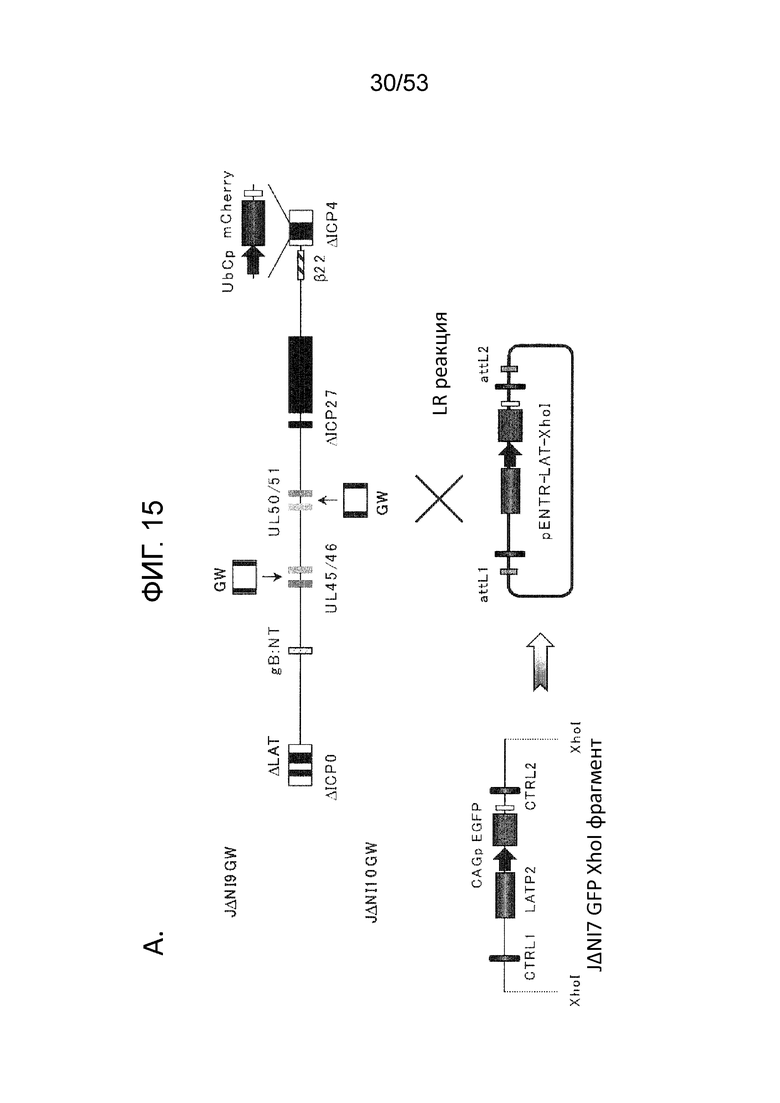
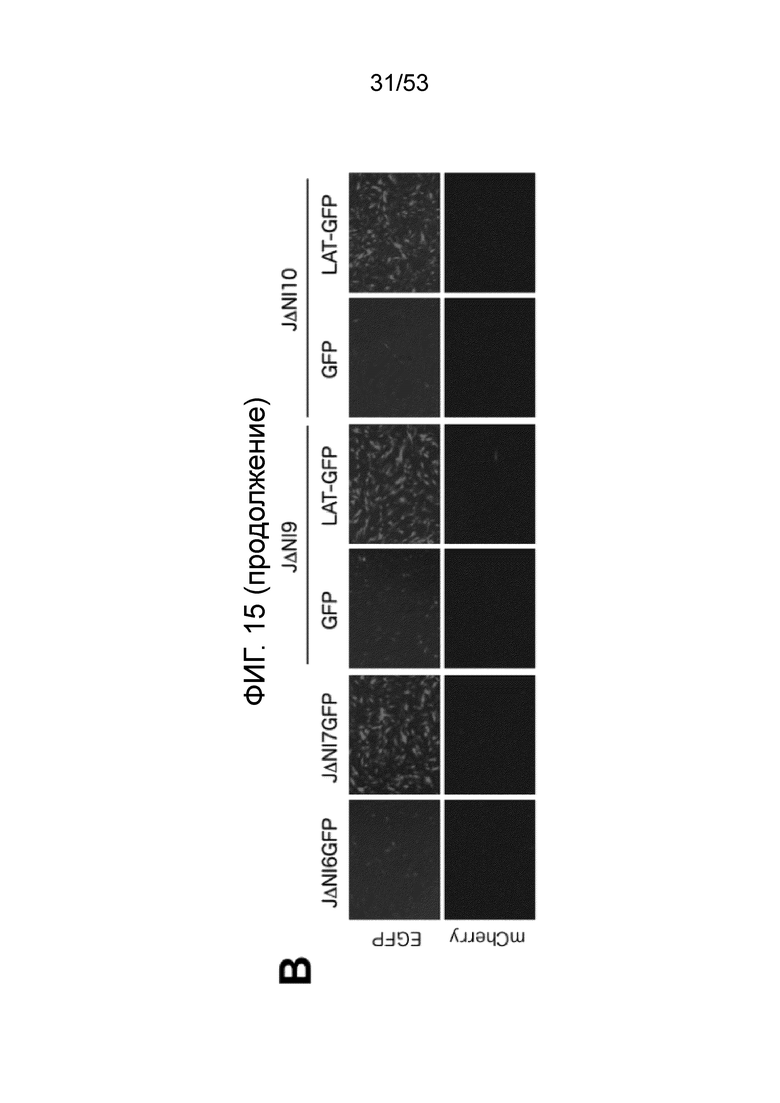
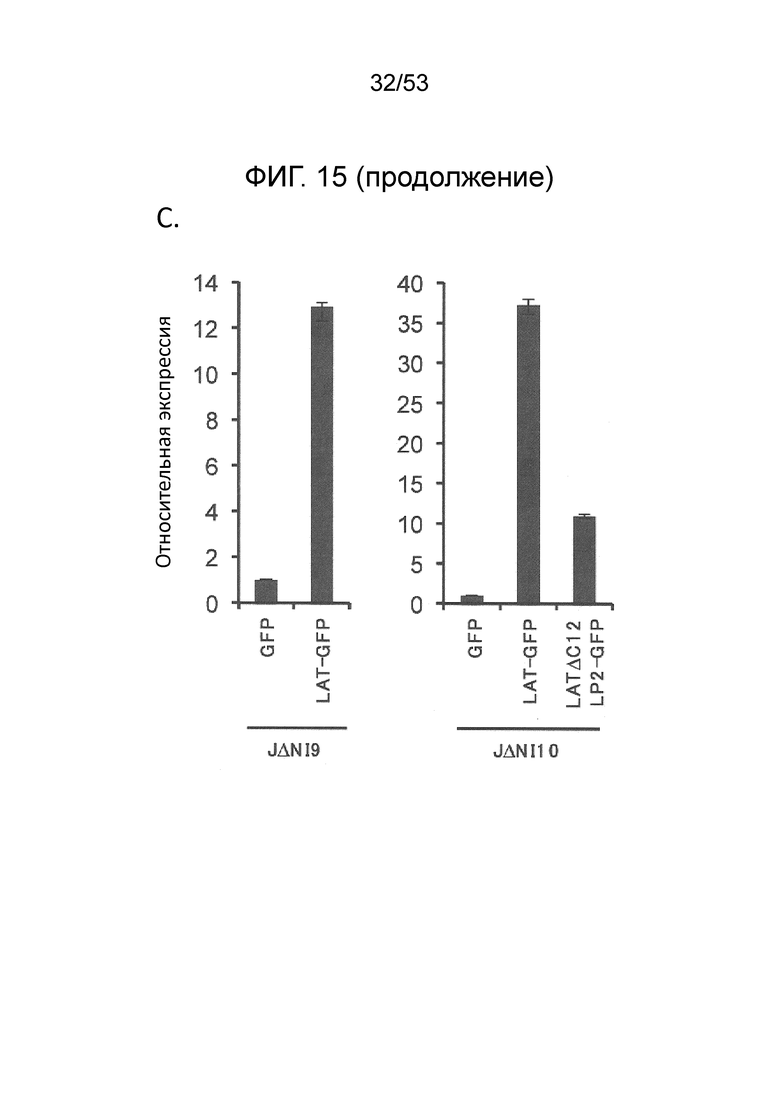
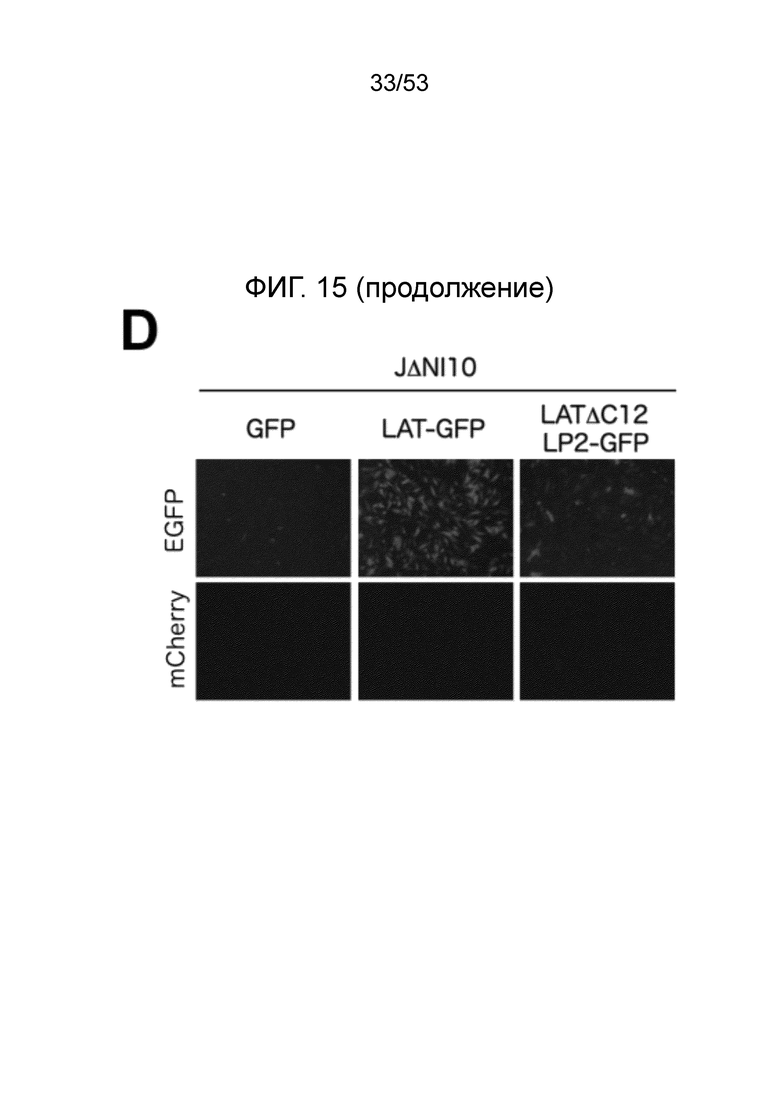
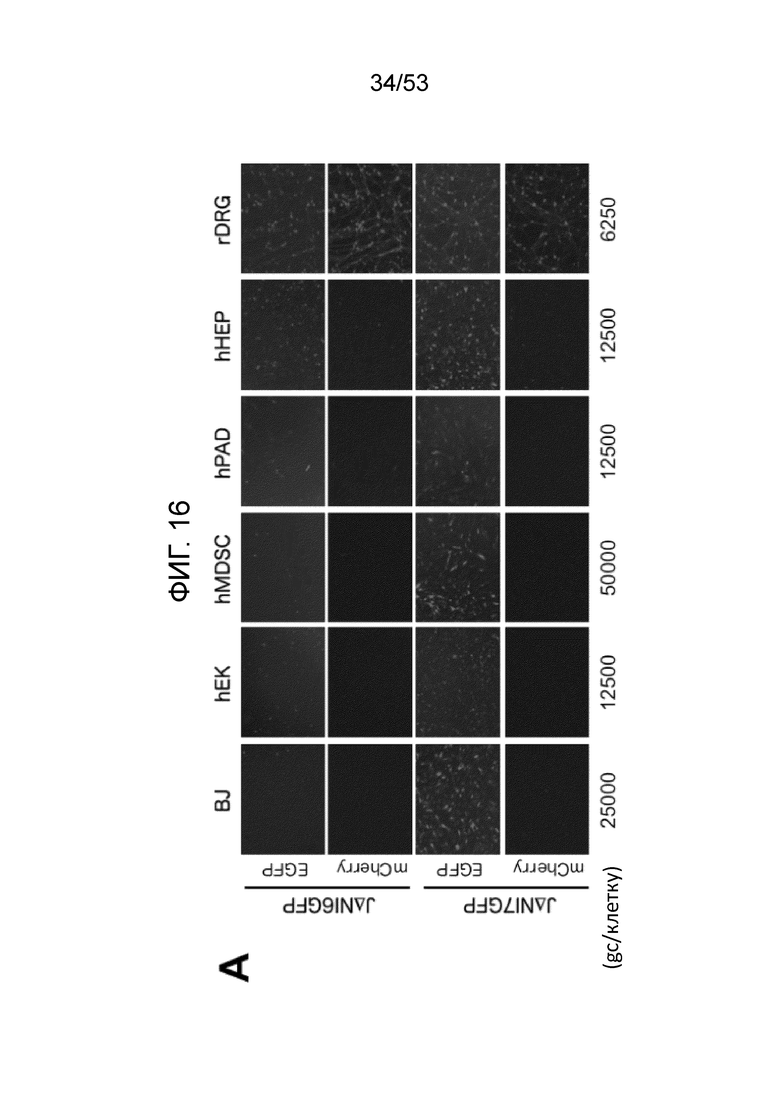
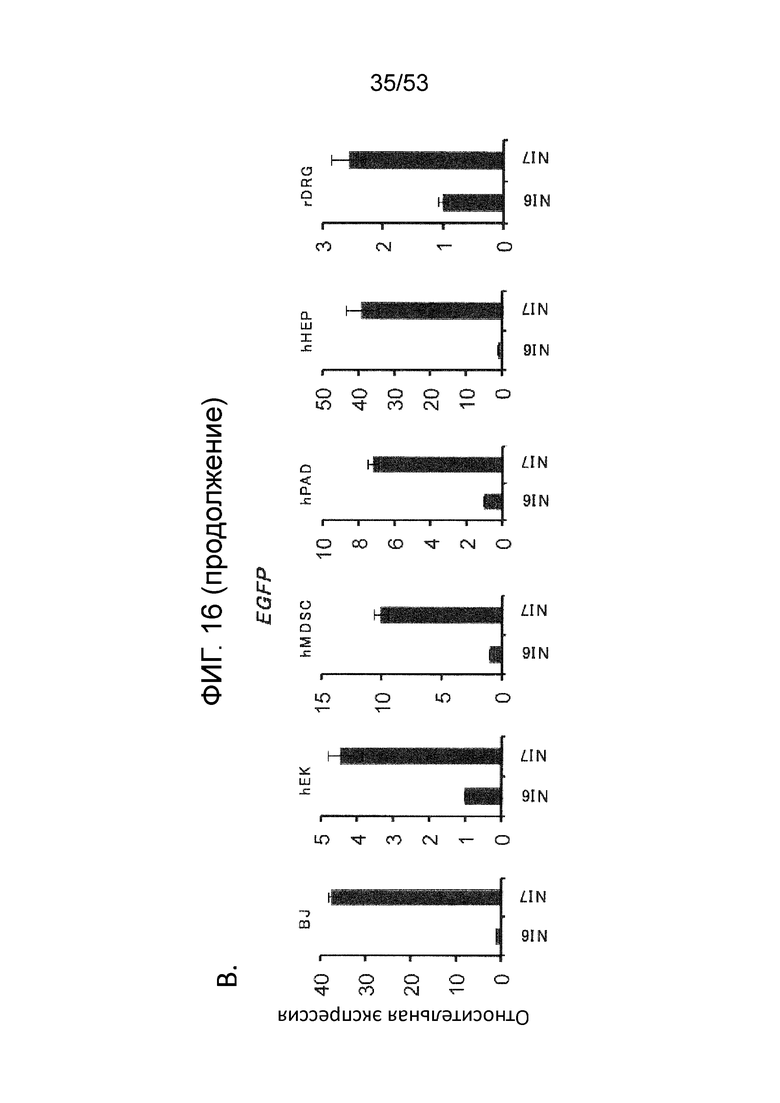
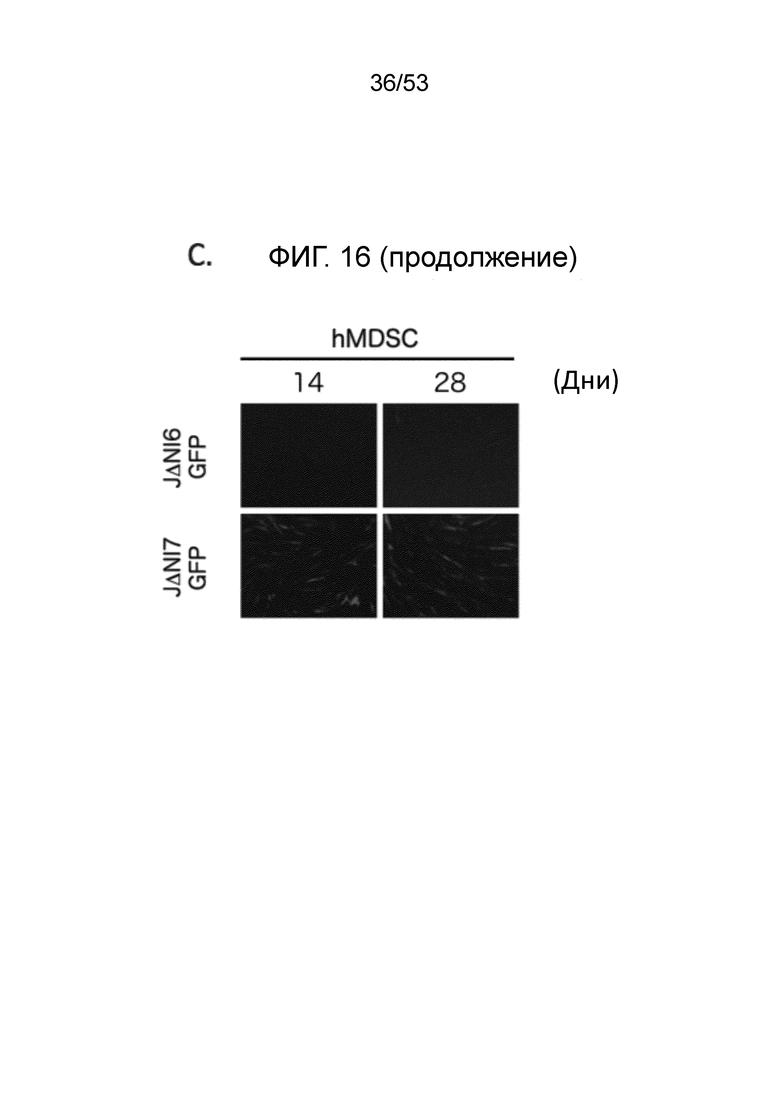
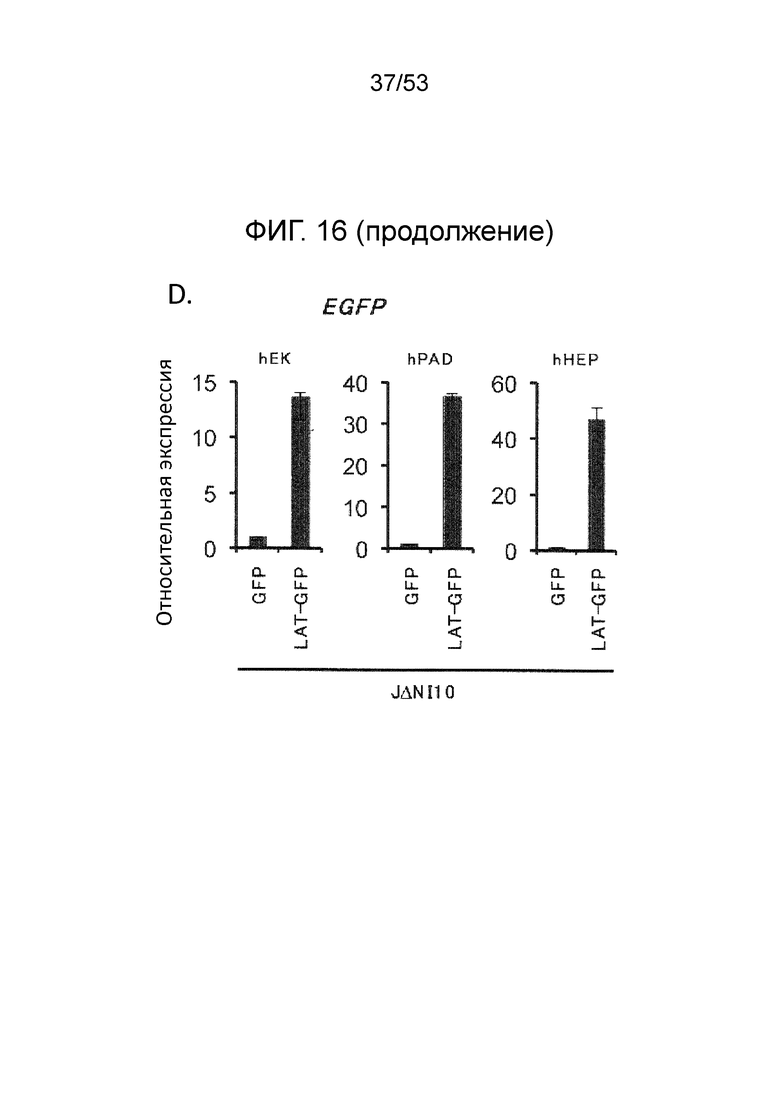
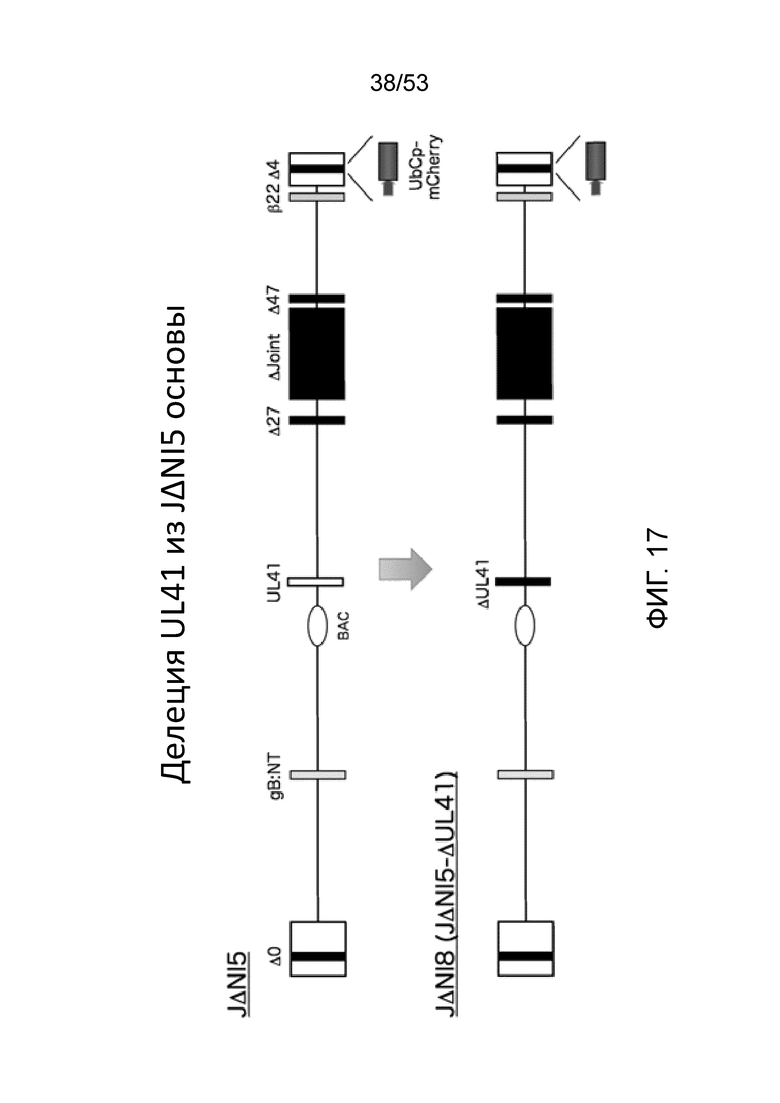
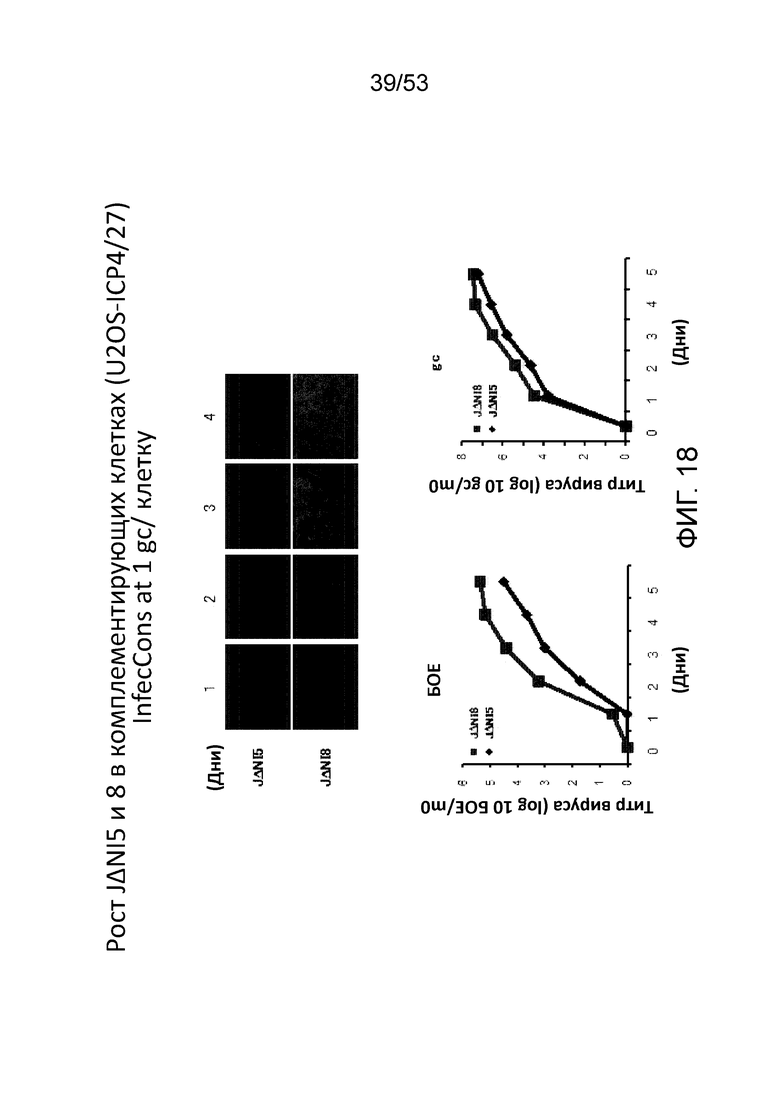
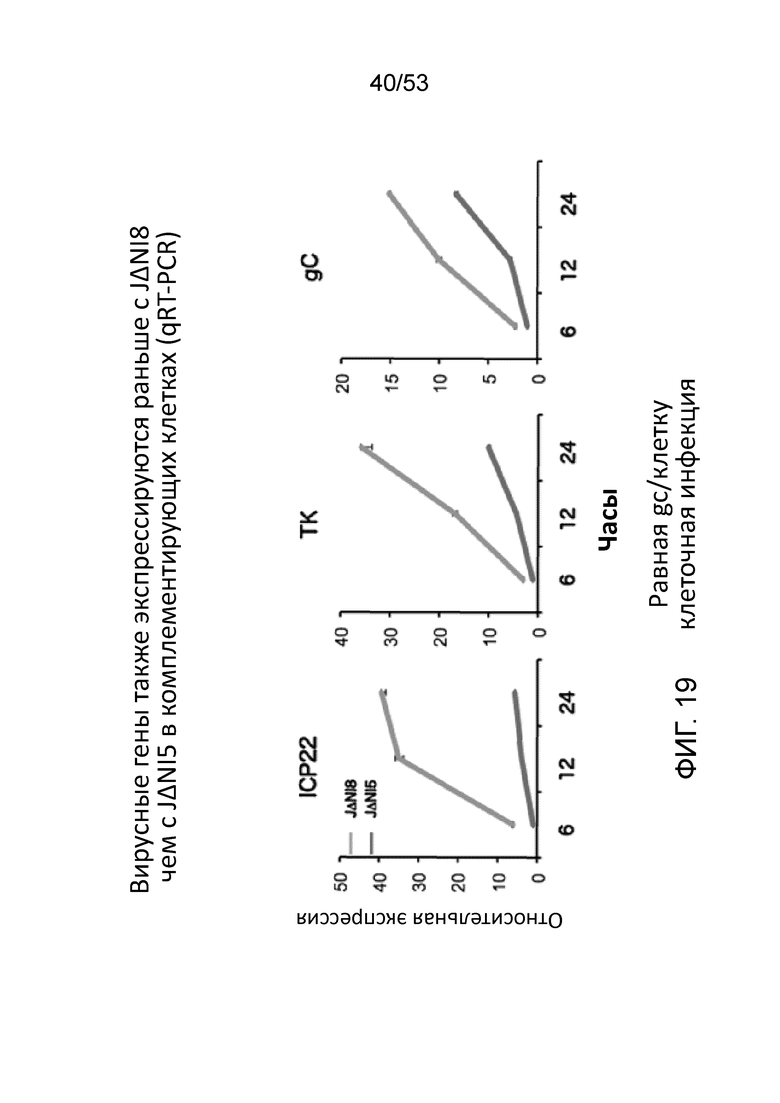
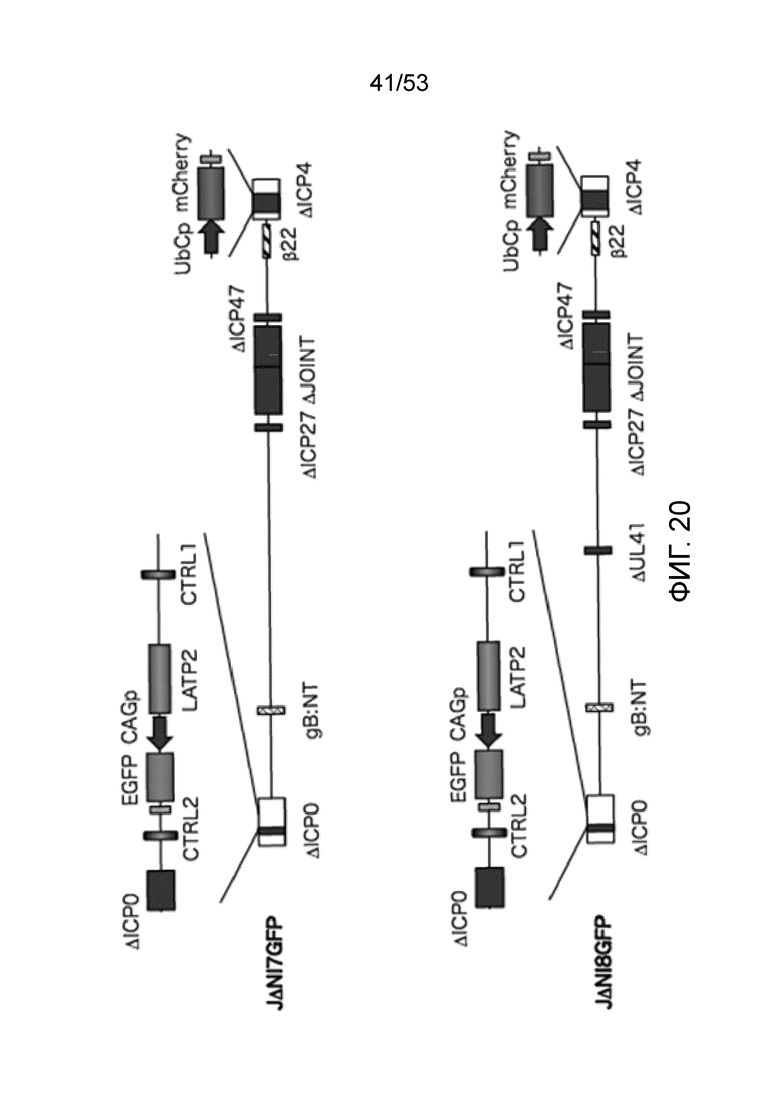
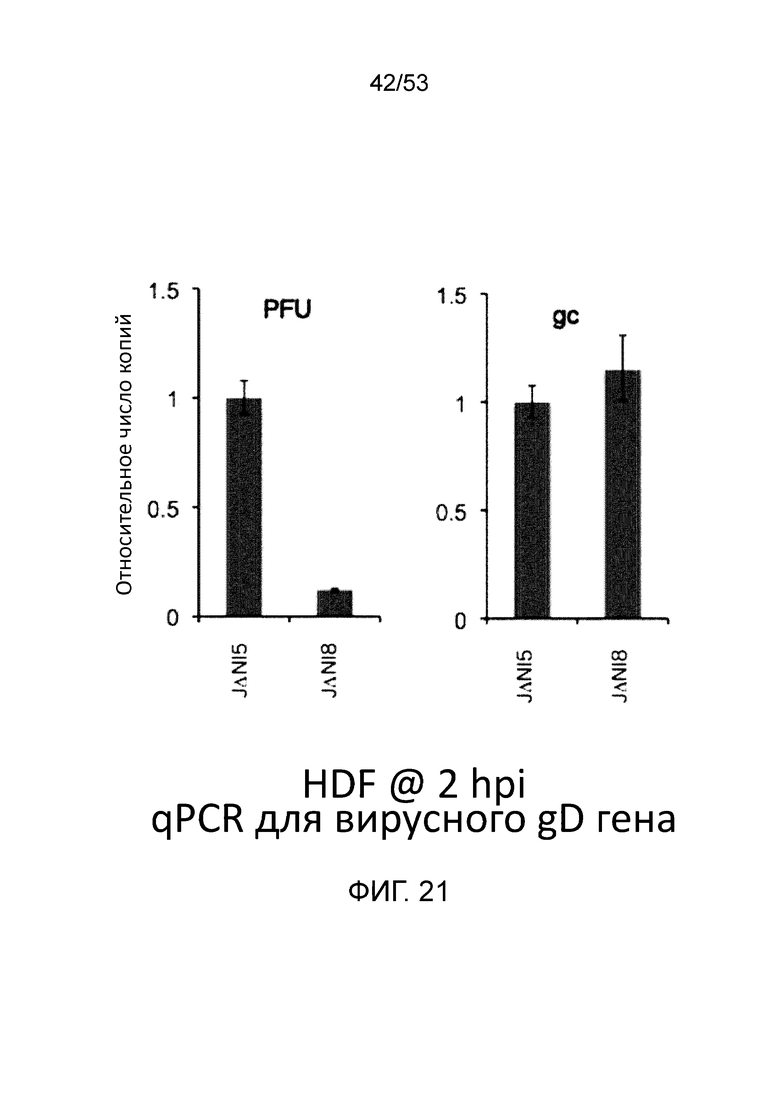
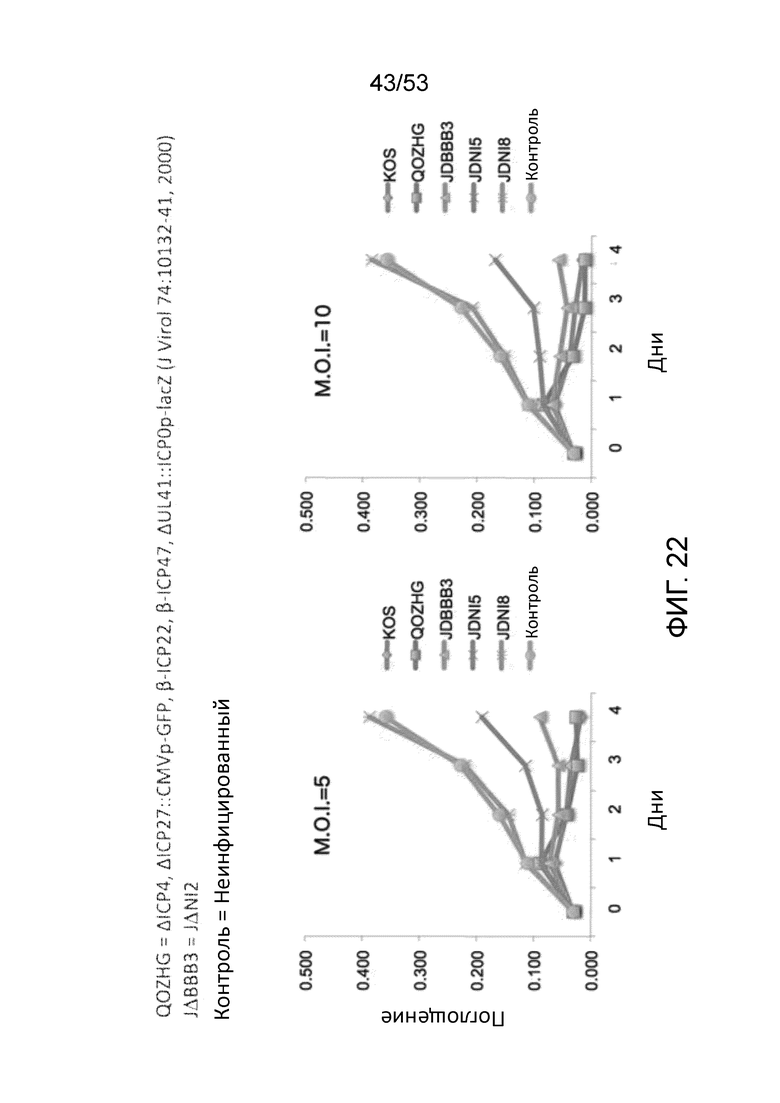
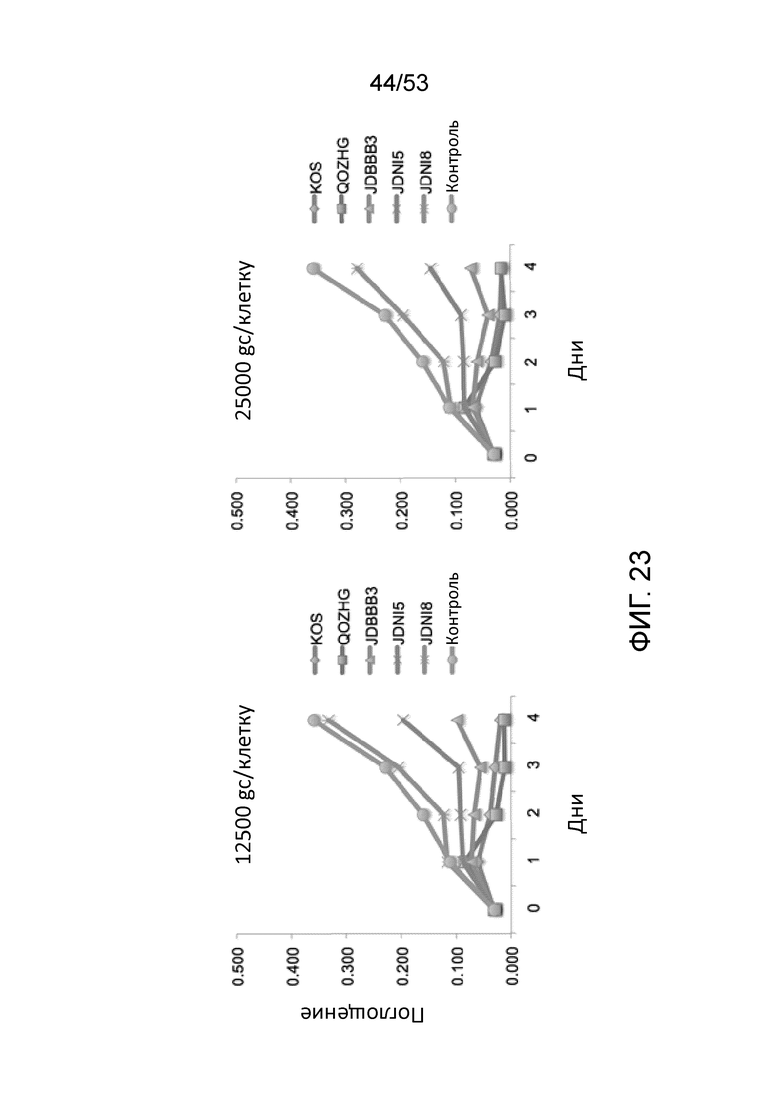
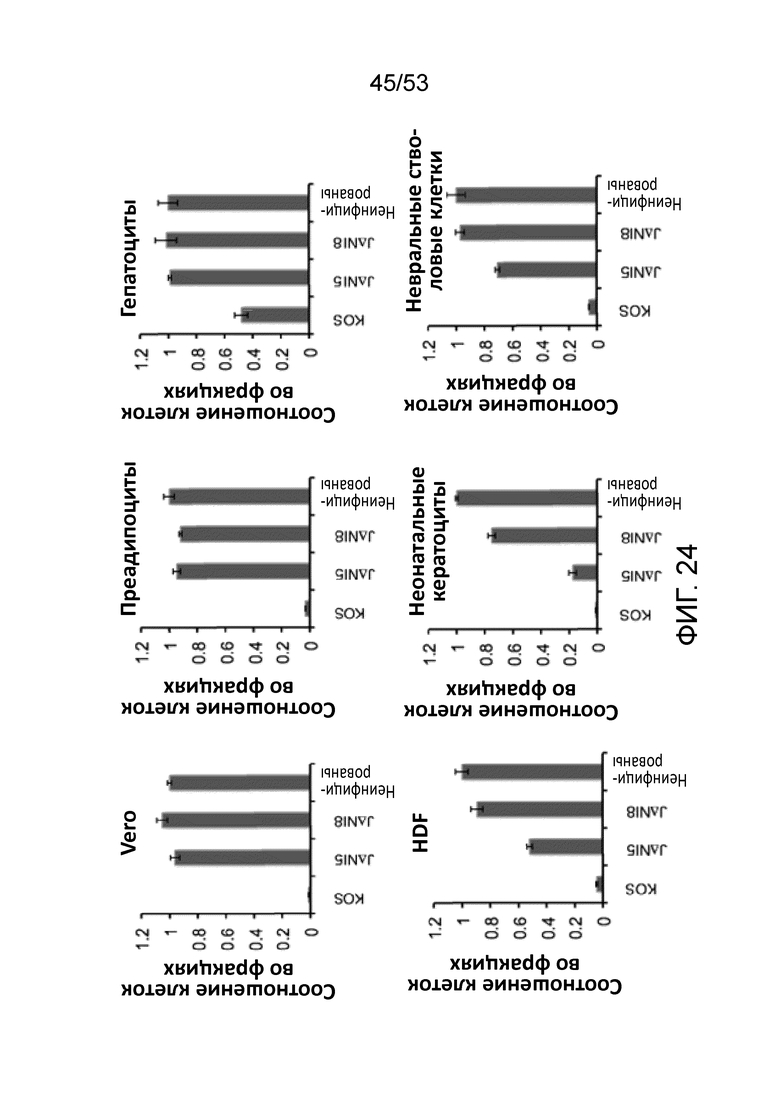
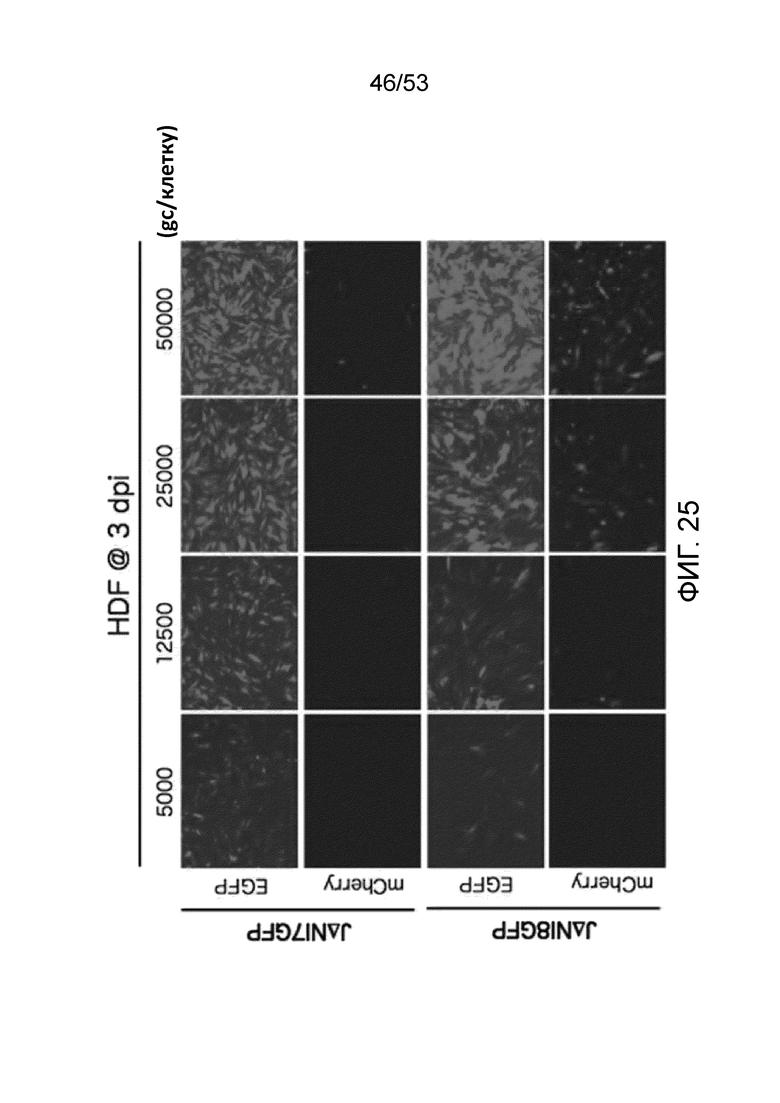
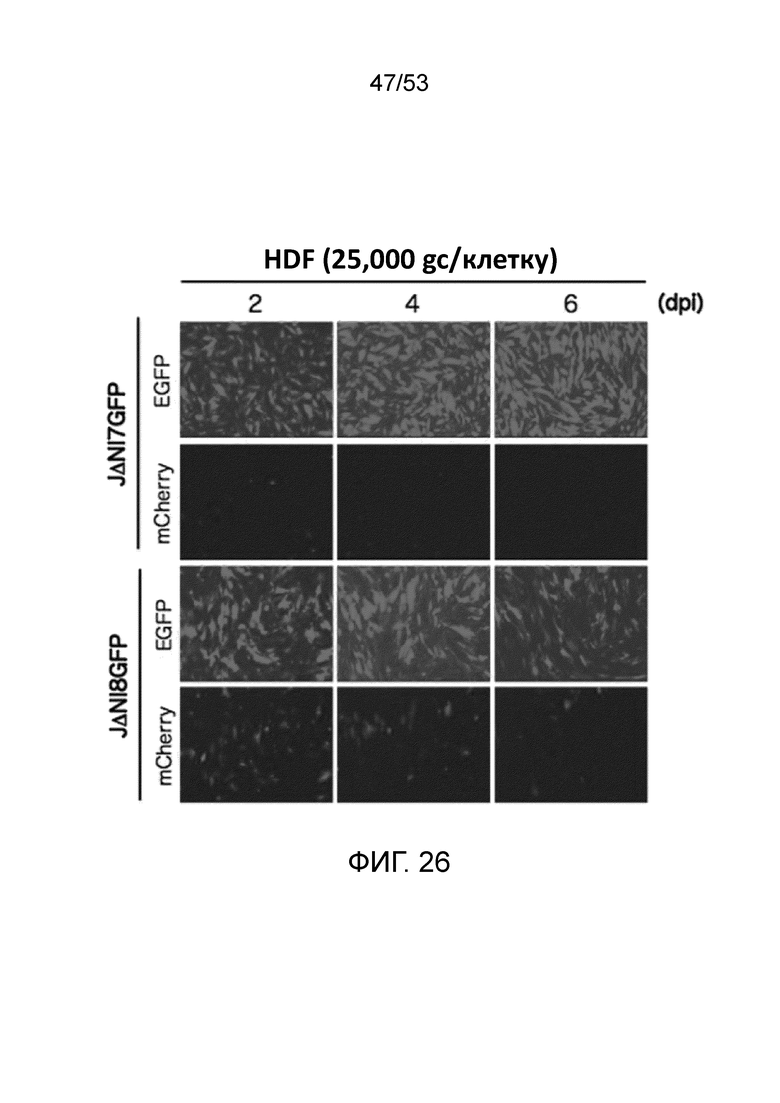
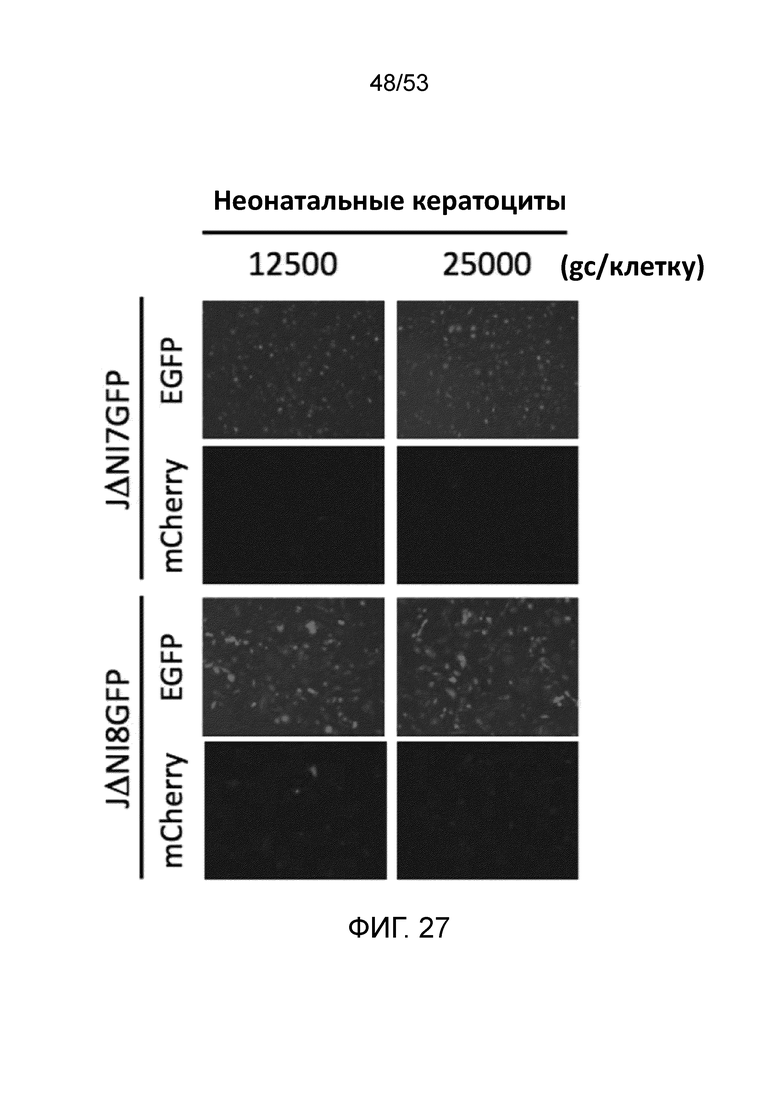
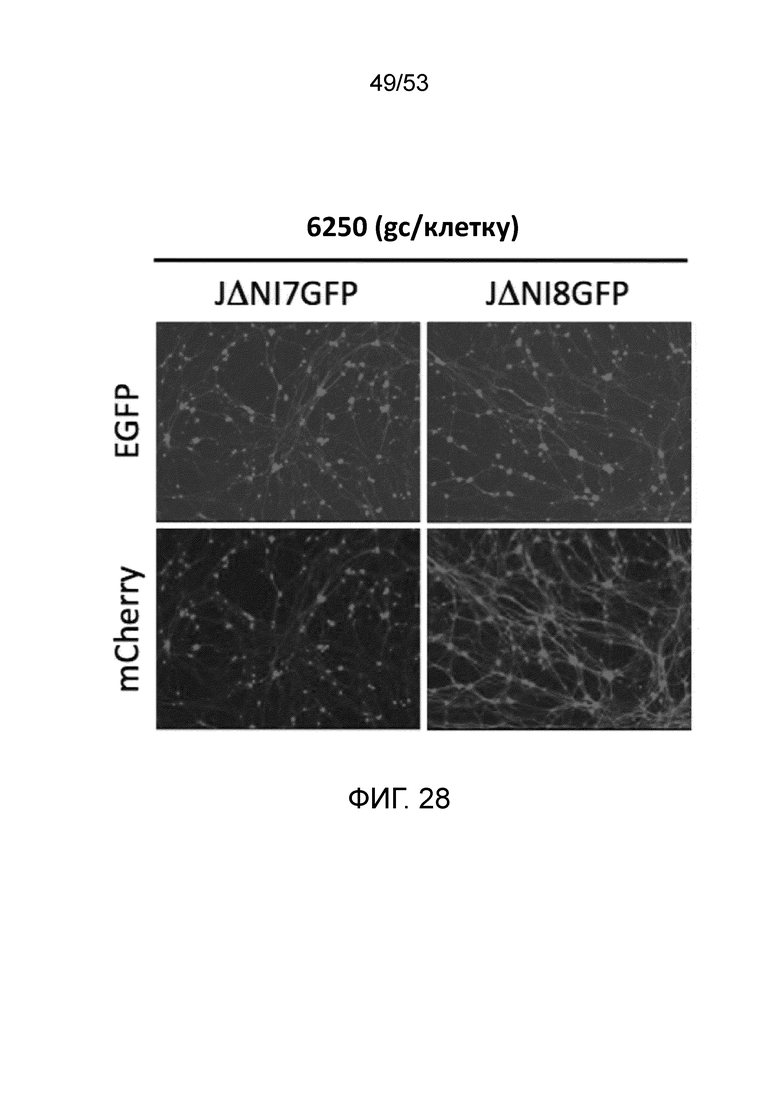
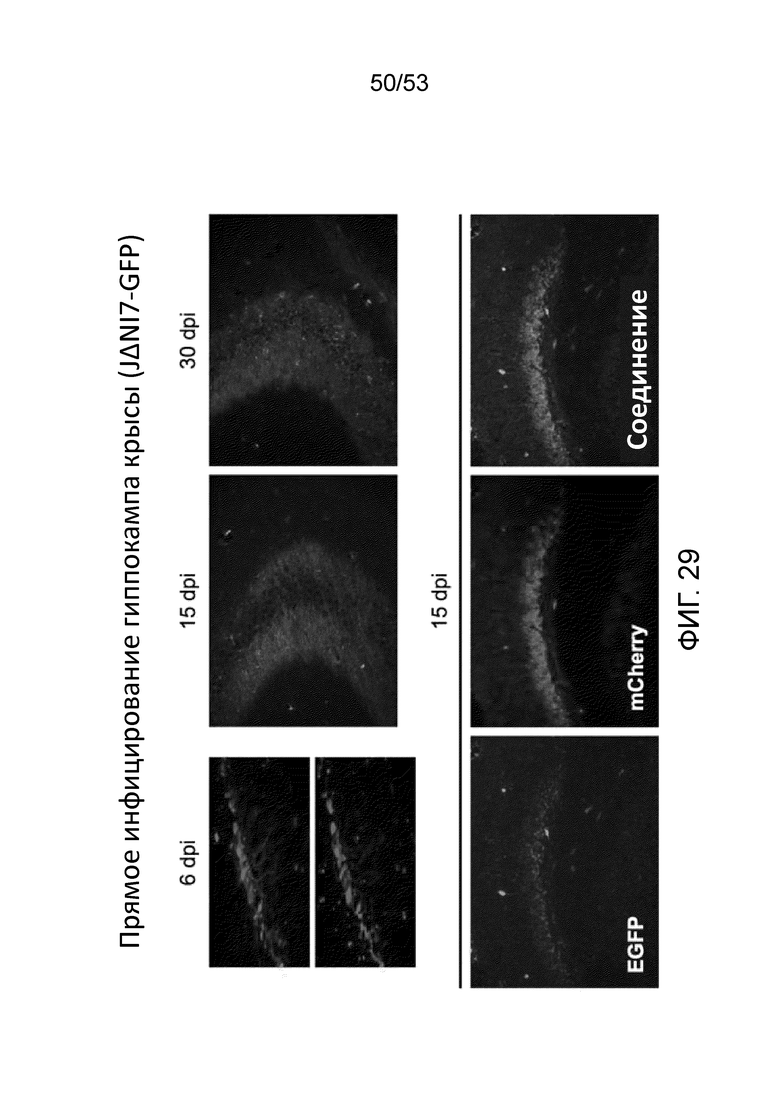
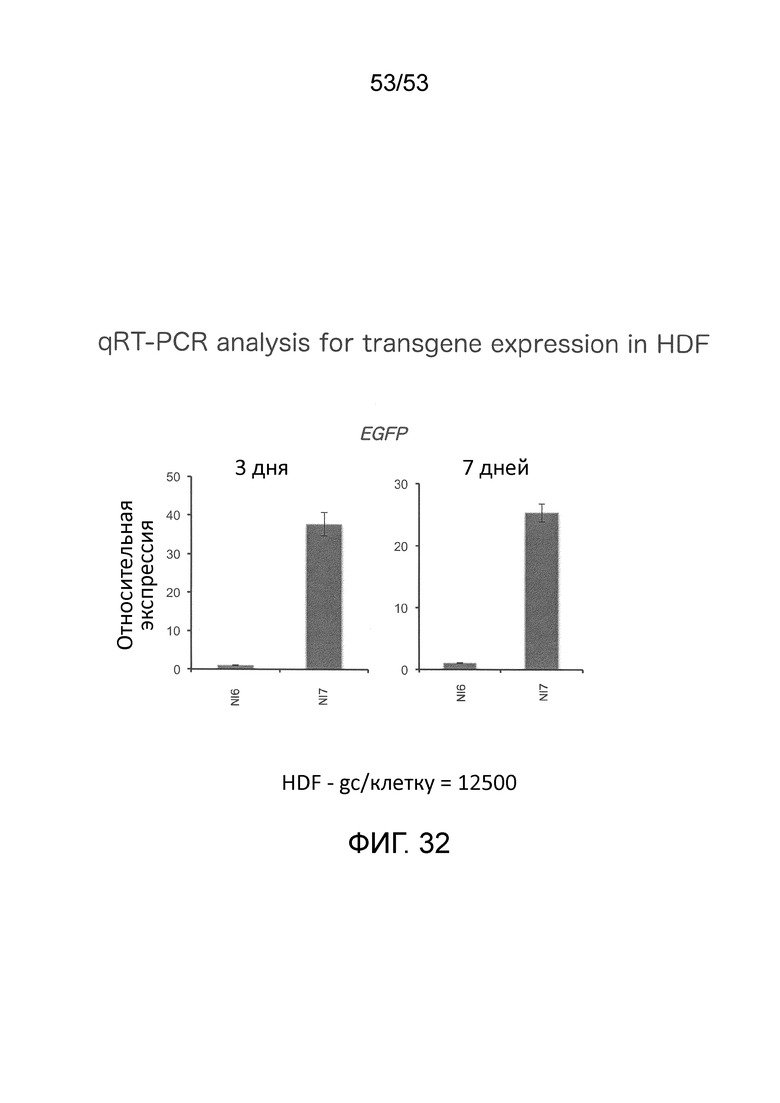
Изобретение относится к области биотехнологии. Предложен вектор на основе вируса простого герпеса (ВПГ), который не экспрессирует токсичные гены ВПГ ICP0, ICP4, ICP22, ICP27 и, в случае необходимости, ICP47 в некомплементирующих клетках. Вектор содержит мутантный геном ВПГ, содержащий генетические перестройки по сравнению к ВПГ дикого типа, а именно инактивирующую делецию гена, кодирующего ICP0, инактивирующую делецию гена, кодирующего ICP4, ген, кодирующий ICP22, который находится под контролем раннего промотора, инактивирующую делецию гена, кодирующего ICP27, и, в случае необходимости, инактивирующую делецию гена, кодирующего ICP47. Вектор способен экспрессировать трансген в течение по меньшей мере 28 дней в некомплементирующих клетках. Изобретение позволяет экспрессировать трансген в любой ткани или клетке in vitro или in vivo без ущерба для клетки или ткани. 33 з.п. ф-лы, 32 ил., 4 табл., 12 пр.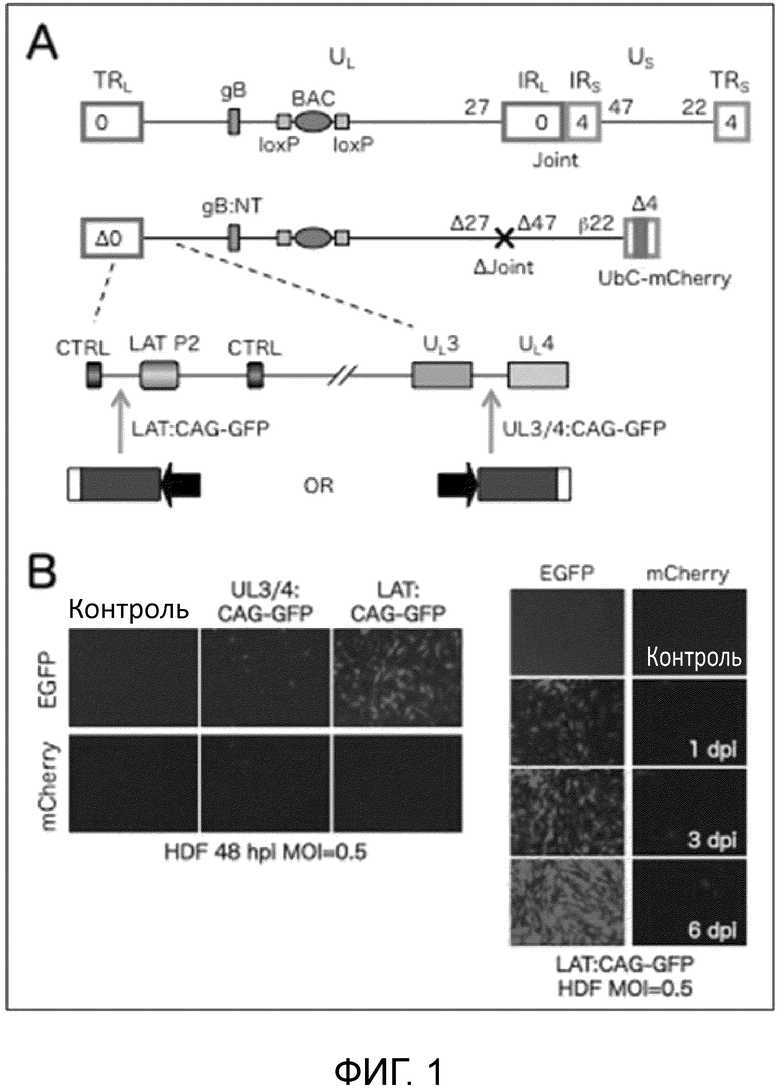
1. Вектор на основе вируса простого герпеса (ВПГ), который не экспрессирует ICP0, ICP4, ICP22, ICP27 и, в случае необходимости, ICP47 в качестве предранних генов в некомплементирующих клетках и который содержит мутантный геном ВПГ, причем мутантный геном ВПГ содержит следующие генетические перестройки по сравнению к ВПГ дикого типа:
(а) инактивирующую делецию гена, кодирующего ICP0,
(b) инактивирующую делецию гена, кодирующего ICP4,
(с) ген, кодирующий ICP22, который находится под контролем раннего промотора,
(d) инактивирующую делецию гена, кодирующего ICP27, и,
(e) в случае необходимости, инактивирующую делецию гена, кодирующего ICP47,
причем цель вектора заключается в способности к экспрессии трансгена, когда он встроен в геном вектора, в течение по меньшей мере 28 дней в некомплементирующих клетках.
2. Вектор на основе ВПГ по п.1, в котором вектор способен к экспрессии трансгена в течение по меньшей мере 28 дней после инфицирования (dpi) в некомплементирующих культивируемых клетках кожных фибробластов человека (HDF).
3. Вектор на основе ВПГ по п.1 или 2, в котором трансген встроен в генную область ассоциированного с латентностью транскрипта (LAT) указанного генома.
4. ВПГ по п.1 или 2, в котором трансген встроен в локус ICP4 указанного генома.
5. Вектор на основе ВПГ по любому из пп.1-4, в котором трансген встроен в указанный геном в функциональной связи с инсуляторной последовательностью.
6. Вектор на основе ВПГ по п.5, в котором инсуляторную последовательность выбирают из группы, состоящей из элемента CTRL1 и CTRL2 граничного хроматина ВПГ, инсулятора гиперчувствительного сайта 4 (cHS4) курицы, повсеместно распространенного элемента открытого хроматина (UCOE) человека HNRPA2B1-CBX3 и области прикрепления каркаса/матрикса (S/MAR) из гена бета-интерферона человека (IFNB1).
7. Вектор на основе ВПГ по п.5, в котором трансген встроен в примерно 5 т.п.н. инсуляторной последовательности в указанном геноме.
8. Вектор по любому из пп.1-7, в котором локус ICP47 указанного генома содержит инактивирующую делецию.
9. Вектор по любому из пп.1-8, в котором по меньшей мере одна из указанных инактивирующих делеций представляет(ют) собой полное удаление кодирующей(их) последовательности(ей) в указанном локусе/локусах.
10. Вектор по любому из пп.1-9, в котором один или более из локусов ICP0, ICP4, ICP27 и ICP47 экспрессируется в качестве раннего или позднего гена.
11. Вектор на основе ВПГ по любому из пп.1-10, в котором трансген фланкирован инсуляторными последовательностями в указанном геноме.
12. Вектор на основе ВПГ по любому из пп.1-11, в котором геном включает граничные элементы хроматина CTRL1 и CTRL2 и в котором трансген находится в примерно 4 т.п.н. обоих из указанных элементов CTRL1 и CTRL2.
13. Вектор по любому из пп.1-12, дополнительно включающий делецию одного или нескольких несущественных генов.
14. Вектор по любому из пп.1-13, в котором вектор не экспрессирует UL41.
15. Вектор по п.14, который содержит инактивирующую делецию UL41 в указанном геноме.
16. Вектор по любому из пп.1-15, который содержит делецию области внутренних повторов (joint) в указанном геноме.
17. Вектор по п.16, который содержит следующие генетические перестройки по сравнению к ВПГ дикого типа:
(а) инактивирующую делецию гена, кодирующего ICP0,
(b) инактивирующую делецию гена, кодирующего ICP4,
(с) делецию joint,
(d) ген, кодирующий ICP22, который находится под контролем раннего промотора,
(e) инактивирующую делецию гена, кодирующего ICP27, и
(f) делецию промотора и инициирующего кодона гена, кодирующего ICP47.
18. Вектор по п.17, который дополнительно содержит инактивирующую делецию UL41.
19. Вектор по п.17, в котором геном содержит генную область LAT, включающую последовательности CTRL1 и CTRL2 и энхансерный элемент LATP2 или LAP2, и в котором первый трансген встроен в указанную генную область LAT между последовательностями CTRL1 и CTRL2.
20. Вектор по п.19, в котором указанный первый трансген встроен в указанную генную область LAT между элементами LATP2 и CTRL1.
21. Вектор по п.19, в котором указанный первый трансген встроен в указанную генную область LAT между элементами LATP2 и CTRL2.
22. Вектор по любому из пп.1-21, содержащий оболочку, которая включает мутантный гликопротеин, который усиливает инфекционность относительно гликопротеина дикого типа или который направляет проникновение ВПГ в клетки через неканонические рецепторы.
23. Вектор по любому из пп.1-22, в котором указанный геном включает ген, кодирующий мутантный гликопротеин, который усиливает инфекционности относительно гликопротеина дикого типа или который направляет проникновение ВПГ в клетки через неканонические рецепторы.
24. Вектор по п.22 или 23, в котором указанный гликопротеин представляет собой gB, gC, gD, gH, gK или их комбинации.
25. Вектор по любому из пп.1-24, в котором указанный геном дополнительно содержит кассету экспрессии, встроенную в участок, отличный от генной области LAT или локуса ICP4.
26. Вектор по любому из пп.1-25, в котором трансген содержит конститутивный промотор млекопитающего.
27. Вектор по любому из пп.1-26, в котором трансген содержит клеточный или ткане-специфический промотор.
28. Вектор по любому из пп.1-27, в котором трансген является полицистронным.
29. Вектор по любому из пп.1-28, в котором трансген содержит один или более сайтов связывания микроРНК.
30. Вектор по любому из пп.1-29, в котором вектор включает множество трансгенных кассет в генной области LAT, каждая из которых содержит отдельный промотор и кодирующую область и при этом каждая из которых является либо моно-, либо полицистронной.
31. Вектор по любому из пп.1-30, в котором трансген кодирует Oct4, Klf4, Sox2, с-Мус, L-Myc, доминантно-негативный р53, Nanog, Glis1, Lin28, TFIID, GATA4, Nkx2.5, Tbx5, MEF2C, Myocd, Hand2, РВСН, Mesp1, SMARCD3, SERCA2a, Pax3, MyoD, Lhx2, FoxGl, FOXP2, Isl1, Ctip2, Tbr1, Ebf1, Gsx2, Srebp2, фактор VIII, фактор IX, дистрофин, CFTR, GlyRα1, энкефалин, изоформу GAD, нейротрофический фактор, Ascl1, Nurr1, Lmx1А, Brn2, Mytl1, NeuroD1, Foxa2, Hnf4a, Foxal, Foxa2 или Foxa3, микроРНК, комбинацию микроРНК либо одну или более некодирующих РНК (нкРНК).
32. Вектор на основе ВПГ по любому из пп.1-31, в котором геном содержит одну или две консенсусные последовательности для рекомбиназного фермента.
33. Вектор на основе ВПГ по любому из пп.1-32, в котором геном содержит кассету бактериальной искусственной хромосомы (BAC), фланкированную консенсусными последовательностями для сайт-специфического рекомбиназного фермента.
34. Вектор на основе ВПГ по п.32 или 33, в котором консенсусная последовательность для сайт-специфического рекомбиназного фермента встроена в межгенную область.
| US 5849572 A1, 15.12.1998 | |||
| US 6261552 B1, 17.07.2001 | |||
| US 5879934 A1, 09.03.1999 | |||
| MARCONI P | |||
| et al | |||
| Replication-defective herpes simplex virus vectors for gene transfer in vivo, PNAS, 1996, Vol.93, pp.11319-11320 | |||
| НАУМОВ Д.А | |||
| Генотерапия злокачественных новообразований | |||
| Состояние проблемы, РМЖ | |||
| ПРИЛОЖЕНИЕ | |||
| ОНКОЛОГИЯ, 2012, N1, с.9-13 | |||
| ЧЕХОНИН В.П |

