ОБЛАСТЬ ТЕХНИКИ
[0001] Описаны генетически модифицированные мыши, клетки, эмбрионы и ткани, содержащие последовательность нуклеиновой кислоты, кодирующую функциональный локус ADAM6. Модификации затрагивают локусы иммуноглобулина человека и/или гуманизированного иммуноглобулина. Описаны мыши, у которых отсутствует функциональный эндогенный ген ADAM6, но присутствует функция ADAM6, в том числе мыши, у которых присутствует эктопическая последовательность нуклеиновой кислоты, кодирующая белок ADAM6. Описаны генетически модифицированные мыши-самцы с модификацией эндогенного локуса иммуноглобулина VH, которая придает мыши неспособность к синтезу функционального белка ADAM6 и приводящей к потере фертильности, а также обеспечивающей функцию ADAM6 у мышей-самцов, в том числе мыши, у которых присутствует эктопическая последовательность нуклеиновой кислоты, восстанавливающая фертильность у указанных самцов мыши.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
[0002] Мыши, у которых присутствует гены антител человека, известны в данной области техники. Фармацевтическое применение антител на протяжении последних двух десятилетий стимулировало значительный объем исследований, касающихся получения антител, подходящих для применения в качестве терапевтических средств для человека. Первые терапевтические средства на основе антител, для которых применялись антитела мыши, не были идеальны в качестве терапевтических средств для человека, так как многократное введение мышиных антител людям приводит к иммуногенности, что может мешать при долгосрочных схемах лечения. Разрабатывались решения, основанные на гуманизации антител мыши для придания им в большей степени качеств антител человека и в меньшей степени - антител мыши. Затем появились способы экспрессирования последовательностей иммуноглобулина человека для применения в антителах, главным образом основанные на in vitro экспрессии фаговых, бактериальных или дрожжевых библиотек иммуноглобулина человека. Наконец, были предприняты попытки получить подходящие антитела человека из лимфоцитов человека in vitro, от мышей, которым были привиты гемопоэтические клетки человека, и от трансхромосомных или трансгенных мышей с инактивированными эндогенными иммуноглобулиновыми локусами. У трансгенных мышей необходимо было инактивировать эндогенные гены иммуноглобулина мыши, чтобы случайным образом встроенные полностью человеческие трансгены функционировали в качестве источника последовательностей иммуноглобулина, экспрессирующихся у таких мышей. Такие мыши способны синтезировать антитела человека, подходящие для применения в качестве терапевтических средств для человека, но у таких мышей наблюдаются значительные проблемы с иммунной системой. Указанные трудности (1) делают использование таких мышей нецелесообразным для получения достаточно разнообразного репертуара антител, (2) требуют применения дорогостоящих реконструктивных доработок, (3) обеспечивают неоптимальный процесс селекции клонов, по-видимому, из-за несовместимости элементов человека и мыши, и (4) делают указанных мышей ненадежным источником больших и разнообразных популяций вариабельных последовательностей человека, которые действительно подходили бы для получения терапевтических средств для человека.
[0003] В данной области техники продолжает существовать потребность в получении улучшенных генетически модифицированных мышей, подходящих для синтеза последовательностей иммуноглобулина, в том числе последовательностей антител человека. Также остается потребность в мышах, способных к реаранжировке сегментов генов иммуноглобулинов с образованием подходящих реаранжированных генов иммуноглобулинов, или способных к синтезу белков из измененных иммуноглобулиновых локусов, в то же время уменьшая или устраняя оказывающие пагубное воздействие изменения, к которым могут приводить генетические модификации.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0004] Согласно одному из аспектов предложены нуклеиновокислые конструкции, клетки, эмбрионы, мыши и способы получения мышей, содержащих модификацию, приводящую к нефункциональности эндогенного мышиного белка ADAM6 или гена ADAM6 (например, к нокауту, или к делеции в эндогенном гене ADAM6), при этом у указанных мышей присутствует последовательность нуклеиновой кислоты, кодирующая белок ADAM6, либо его ортолог, гомолог или фрагмент, функциональный у мышей-самцов.
[0005] Согласно одному из аспектов предложены конструкции нуклеиновых кислот, клетки, эмбрионы, мыши и способы получения мышей, содержащих модификацию эндогенного локуса иммуноглобулина мыши, при этом у указанных мышей присутствует белок ADAM6, либо его ортолог, гомолог или фрагмент, функциональный у мышей-самцов. Согласно одному из вариантов реализации указанный эндогенный локус иммуноглобулина мыши представляет собой локус тяжелой цепи иммуноглобулина, и указанная модификация уменьшает или элиминирует активность ADAM6 в клетке или в ткани у мышей-самцов.
[0006] Согласно одному из аспектов предложены мыши, у которых присутствует эктопическая последовательность нуклеотидов, кодирующая ADAM6 мыши, либо его ортолог, гомолог или функциональный фрагмент; также предложены мыши, у которых присутствует эндогенная последовательность нуклеотидов, кодирующая ADAM6 мыши, либо его ортолог, гомолог или фрагмент, и по меньшей мере одна генетическая модификация локуса тяжелой цепи иммуноглобулина.
[0007] Согласно одному из аспектов предложены способы получения мышей, у которых присутствует модификация эндогенного локуса иммуноглобулина мыши, при этом у указанных мышей присутствует белок ADAM6, либо его ортолог, гомолог или фрагмент, функциональный у мышей-самцов. Мышей согласно настоящему изобретению можно получить, например, с применением описанных в настоящей заявке способов.
[0008] Согласно одному из аспектов предложены способы получения мышей, у которых присутствует генетическая модификация локуса тяжелой цепи иммуноглобулина, при этом применение указанных способов дает мышей-самцов, у которых присутствует модифицированный локус тяжелой цепи иммуноглобулина (или его делеция); указанные мыши-самцы способны производить потомство посредством спаривания. Согласно одному из вариантов реализации указанные мыши-самцы способны продуцировать сперму, способную перемещаться из матки мыши через маточные трубы мыши для оплодотворения мышиной яйцеклетки.
[0009] Согласно одному из аспектов предложены способы получения мышей, у которых присутствует генетическая модификация локуса тяжелой цепи иммуноглобулина, при этом применение указанных способов дает мышей-самцов, у которых присутствует модифицированный локус тяжелой цепи иммуноглобулина (или его делеция); у указанных мышей-самцов наблюдается снижение фертильности; у указанных мышей также имеется генетическая модификация, полностью или частично восстанавливающая сниженную фертильность. Согласно различным вариантам реализации снижение фертильности характеризуется неспособностью спермы мышей-самцов передвигаться из матки мыши по маточным трубам мыши для оплодотворения мышиной яйцеклетки. Согласно различным вариантам реализации снижение фертильности характеризуется проявлением дефекта подвижности у спермы in vivo. Согласно различным вариантам реализации генетическая модификация, полностью или частично восстанавливающая сниженную фертильность, представляет собой последовательность нуклеиновой кислоты, кодирующую ген ADAM6 мыши, либо его ортолог, гомолог или фрагмент, функциональный у мышей-самцов.
[00010] Согласно одному из вариантов реализации указанная генетическая модификация включает замену эндогенных вариабельных локусов тяжелой цепи иммуноглобулина вариабельными локусами тяжелой цепи иммуноглобулина других видов (например, не являющихся мышами). Согласно одному из вариантов реализации указанная генетическая модификация включает вставку ортологичных вариабельных локусов тяжелой цепи иммуноглобулина в эндогенные вариабельные локусы тяжелой цепи иммуноглобулина. Согласно конкретному варианту реализации указанный вид представляет собой человека. Согласно одному из вариантов реализации указанная генетическая модификация включает удаление эндогенного локуса вариабельной области тяжелой цепи иммуноглобулина полностью или частично, при этом указанное удаление приводит к потере функции эндогенного ADAM6. Согласно конкретному варианту реализации указанная потеря функции эндогенного ADAM6 связана со снижением фертильности у мышей-самцов.
[00011] Согласно одному из аспектов предложены мыши, у которых присутствует модификация, которая уменьшает или элиминирует экспрессию ADAM6 мыши из эндогенного аллеля ADAM6, так что у самца мыши, имеющего указанную модификация, наблюдается снижение фертильности (например, значительное снижение способности производить потомство посредством спаривания) или по существу бесплодность из-за снижения или утраты функции эндогенного ADAM6, при этом у указанных мышей также присутствует эктопическая последовательность ADAM6, либо его гомолога, ортолога или функционального фрагмента. Согласно одному из аспектов модификация, которая уменьшает или элиминирует экспрессию ADAM6 мыши, представляет собой модификацию (например, вставку, удаление, замещение, и т.п.) в локусе иммуноглобулина мыши.
[00012] Согласно одному из вариантов реализации снижение или утрата функции ADAM6 включает неспособность или по существу полную неспособность мыши продуцировать сперму, способную передвигаться из матки мыши через маточные трубы мыши для оплодотворения мышиной яйцеклетки. Согласно конкретному варианту реализации по меньшей мере приблизительно 95%, 96%, 97%, 98%, или 99% спермиев в объеме эякулята мыши неспособны проходить через маточные трубы in vivo после копуляции и оплодотворять яйцеклетку мыши.
[00013] Согласно одному из вариантов реализации уменьшение или утрата функции ADAM6 включает неспособность к формированию или по существу полную неспособность к формированию комплекса ADAM2 и/или ADAM3 и/или ADAM6 на поверхности спермия мыши. Согласно одному из вариантов реализации утрата функции ADAM6 включает по существу полную неспособность к оплодотворению мышиной яйцеклетки посредством копуляции с самкой мыши.
[00014] Согласно одному из аспектов предложена мышь, у которой отсутствует функциональный эндогенный ген ADAM6 и присутствует белок (или эктопическая последовательность нуклеотидов, кодирующая белок), обеспечивающий функционал ADAM6 у указанной мыши. Согласно одному из вариантов реализации указанная мышь представляет собой самца мыши, а указанный функционал включает повышение фертильности по сравнению с мышью, у которой отсутствует функциональный эндогенный ген ADAM6.
[00015] Согласно одному из вариантов реализации указанный белок кодируется геномной последовательностью, расположенной в иммуноглобулиновом локусе зародышевой линии мыши. Согласно конкретному варианту реализации указанный иммуноглобулиновый локус представляет собой локус тяжелой цепи. Согласно другому конкретному варианту реализации указанный локус тяжелой цепи содержит по меньшей мере один генный сегмент VH человека, по меньшей мере один генный сегмент DH человека и по меньшей мере один генный сегмент JH человека. Согласно одному из вариантов реализации эктопический белок кодируется геномной последовательностью расположенной в не-иммуноглобулиновом локусе зародышевой линии мыши. Согласно одному из вариантов реализации указанный не-иммуноглобулиновый локус представляет собой транскрипционно активный локус. Согласно конкретному варианту реализации указанный транскрипционно активный локус представляет собой ROSA26 локус. Согласно конкретному варианту реализации указанный транскрипционно активный локус связан с тканеспецифической экспрессией. Согласно одному из вариантов реализации указанная тканеспецифическая экспрессия происходит в репродуктивных тканях. Согласно одному из вариантов реализации указанный белок кодируется геномной последовательностью, случайным образом встраиваемой в зародышевую линию мыши.
[00016] Согласно одному из вариантов реализации у мыши присутствует легкая цепь человека, или гибридная легкая цепь человека/мыши, или гибридная легкая цепь человека/крысы (например, вариабельная область человека, константная область мыши или крысы); и гибридная тяжелая цепь с вариабельной областью человека/константной областью мыши или крысы. Согласно конкретному варианту реализации у мыши присутствует трансген, который содержит ген гибридной легкой цепи с вариабельной областью человека/константной областью крысы или мыши, функционально связанный с транскрипционно активным промотором, например, промотором ROSA26. Согласно еще одному конкретному варианту реализации указанный трансген гибридной легкой цепи человека/мыши или крысы содержит последовательность реаранжированной вариабельной области легкой цепи человека в зародышевой линии мыши.
[00017] Согласно одному из вариантов реализации указанная эктопическая последовательность нуклеотидов расположена в иммуноглобулиновом локусе в зародышевой линии мыши. Согласно конкретному варианту реализации указанный иммуноглобулиновый локус представляет собой локус тяжелой цепи. Согласно одному из вариантов реализации указанный локус тяжелой цепи содержит по меньшей мере один генный сегмент VH человека, по меньшей мере один генный сегмент DH человека и по меньшей мере один генный сегмент JH человека. Согласно одному из вариантов реализации указанная эктопическая последовательность нуклеотидов расположена в не-иммуноглобулиновом локусе в зародышевой линии мыши. Согласно одному из вариантов реализации указанный не-иммуноглобулиновый локус представляет собой транскрипционно активный локус. Согласно конкретному варианту реализации указанный транскрипционно активный локус представляет собой локус ROSA26. Согласно одному из вариантов реализации указанная эктопическая последовательность нуклеотидов случайным образом встроена в зародышевую линию мыши.
[00018] Согласно одному из аспектов предложена мышь, у которой отсутствует функциональный эндогенный ген ADAM6, при этом указанная мышь содержит эктопическую последовательность нуклеотидов, которая восполняет утрату функции ADAM6 мыши. Согласно одному из вариантов реализации эктопическая последовательность нуклеотидов придает указанной мыши способность производить потомство, сопоставимую с соответствующей способностью мыши дикого типа, у которой присутствует функциональный эндогенный ген ADAM6. Согласно одному из вариантов реализации указанная последовательность придает мыши способность к образованию комплекса ADAM2 и/или ADAM3 и/или ADAM6 на поверхности спермия мыши. Согласно одному из вариантов реализации указанная последовательность придает сперме указанной мыши способность перемещаться из матки мыши через маточные трубы мыши к яйцеклетке мыши для оплодотворения указанной яйцеклетки.
[00019] Согласно одному из вариантов реализации мышь, у которой отсутствует функциональный эндогенный ген ADAM6 и присутствует указанная эктопическая последовательность нуклеотидов, производит по меньшей мере приблизительно 50%, 60%, 70%, 80%, или 90% от числа пометов мыши дикого типа того же возраста и линии за период времени, равный шести месяцам.
[00020] Согласно одному из вариантов реализации мышь, у которой отсутствует функциональный эндогенный ген ADAM6 и присутствует указанная эктопическая последовательность нуклеотидов, производит больше потомства по меньшей мере приблизительно в 1,5 раза, приблизительно в 2 раза, приблизительно в 2,5 раза, приблизительно в 3 раза, приблизительно в 4 раза, приблизительно в 6 раз, приблизительно в 7 раз, приблизительно в 8 раз или приблизительно в 10 раз, либо более, при размножении на протяжении равного шести месяцам периода времени, чем мышь того же возраста и той же или сходной линии, у которой отсутствуют функциональный эндогенный ген ADAM6 и указанная эктопическая последовательность нуклеотидов, при размножении на протяжении такого же по существу периода времени и в таких же по существу условиях.
[00021] Согласно одному из вариантов реализации мышь, у которой отсутствует функциональный эндогенный ген ADAM6 и присутствует указанная эктопическая последовательность нуклеотидов, производит в среднем по меньшей мере приблизительно в 2 раза, в 3 раза или в 4 раза больше детенышей на помет на протяжении периода размножения, равного 4 или 6 месяцам, по сравнению с мышью, у которой отсутствует функциональный эндогенный ген ADAM6 и указанная эктопическая нуклеотидная последовательность, размножающейся на протяжении такого же периода времени.
[00022] Согласно одному из вариантов реализации мышь, у которой отсутствует функциональный эндогенный ген ADAM6 и присутствует указанная эктопическая последовательность нуклеотидов, представляет собой самца мыши, и указанный самец мыши продуцирует сперму, которая при извлечении из маточных труб приблизительно через 5-6 часов после копуляции проявляет способность к передвижению по маточным трубам по меньшей мере в 10 раз, по меньшей мере в 20 раз, по меньшей мере в 30 раз, по меньшей мере в 40 раз, по меньшей мере в 50 раз, по меньшей мере в 60 раз, по меньшей мере в 70 раз, по меньшей мере в 80 раз, по меньшей мере в 90 раз, в 100 раз, в 110 раз или в 120 раз, либо более, большую, чем у мыши, у которой отсутствует функциональный эндогенный ген ADAM6 и указанная эктопическая нуклеотидная последовательность.
[00023] Согласно одному из вариантов реализации мышь, у которой отсутствует функциональный эндогенный ген ADAM6 и присутствует эктопическая последовательность нуклеотидов, при копуляции с самкой мыши производит сперму, способную пересечь матку, проникнуть в маточные трубы и переместиться по маточным трубам не более чем приблизительно за 6 часов с эффективностью, приблизительно равной эффективности спермы мыши дикого типа.
[00024] Согласно одному из вариантов реализации мышь, у которой отсутствует функциональный эндогенный ген ADAM6 и присутствует указанная эктопическая последовательность нуклеотидов, производит потомство чаще приблизительно в 1,5 раза, приблизительно в 2 раза, приблизительно в 3 раза, или приблизительно в 4 раза, либо более, на протяжении сопоставимого периода времени по сравнению с мышью, у которой отсутствует функциональный ген ADAM6 и указанная эктопическая нуклеотидная последовательность.
[00025] Согласно одному из аспектов предложена мышь, содержащая в зародышевой линии не происходящую от мыши последовательность нуклеиновой кислоты, кодирующую белок-иммуноглобулин, при этом указанная последовательность не происходящего от мыши иммуноглобулина содержит вставку гена ADAM6 мыши, либо его гомолога, ортолога или функционального фрагмента. Согласно одному из вариантов реализации указанная не происходящего от мыши иммуноглобулина содержит последовательность иммуноглобулина человека. Согласно одному из вариантов реализации указанная последовательность содержит последовательность тяжелой цепи иммуноглобулина человека. Согласно одному из вариантов реализации указанная последовательность содержит последовательность легкой цепи иммуноглобулина человека. Согласно одному из вариантов реализации указанная последовательность содержит один или более генных сегментов V, один или более генных сегментов D и один или более генных сегментов J; согласно одному из вариантов реализации указанная последовательность содержит один или более генных сегментов V и один или более генных сегментов J. Согласно одному из вариантов реализации указанный один или более генных сегментов V, D и J, или один или более генных сегментов V и J, нереаранжирован(ы). Согласно одному из вариантов реализации указанный один или более генных сегментов V, D и J, или один или более генных сегментов V и J, реаранжирован(ы). Согласно одному из вариантов реализации после реаранжировки указанного одного или более генных сегментов V, D и J, или одного или более генных сегментоы V и J, в геноме указанной мыши содержится по меньшей мере одна последовательность нуклеиновой кислоты, кодирующая ген ADAM6 мыши, либо его гомолог, ортолог или функциональный фрагмент. Согласно одному из вариантов реализации после реаранжировки геном указанной мыши содержит по меньшей мере две последовательности нуклеиновой кислоты, кодирующей ген ADAM6 мыши, либо его гомолог, ортолог или функциональный фрагмент. Согласно одному из вариантов реализации после реаранжировки геном указанной мыши содержит по меньшей мере одну последовательность нуклеиновой кислоты, кодирующую ген ADAM6 мыши, либо его гомолог, ортолог или функциональный фрагмент. Согласно одному из вариантов реализации указанная мышь содержит ген ADAM6, либо его гомолог, ортолог или функциональный фрагмент, в B-клетке. Согласно одному из вариантов реализации у мыши присутствует ген ADAM6, либо его гомолог, ортолог или функциональный фрагмент в клетке, не являющейся В-клеткой.
[00026] Согласно одному из аспектов предложены мыши, экспрессирующие вариабельную область тяжелой цепи иммуноглобулина человека или ее функциональный фрагмент из эндогенного локуса тяжелой цепи иммуноглобулина мыши, при этом у указанных мышей присутствует активность ADAM6, функционального у мышей-самцов.
[00027] Согласно одному из вариантов реализации у указанных мышей-самцов присутствует один немодифицированный эндогенный аллель ADAM6 или его ортолог, гомолог или функциональный фрагмент, в эндогенном локусе ADAM6.
[00028] Согласно одному из вариантов реализации у указанных мышей-самцов присутствует эктопическая последовательность ADAM6 мыши, либо ее гомолог, ортолог или функциональный фрагмент, кодирующая(ий) белок, обеспечивающий функцию ADAM6.
[00029] Согласно одному из вариантов реализации у указанных мышей-самцов присутствует последовательность ADAM6, либо ее гомолог, ортолог или функциональный фрагмент, расположение которой в геноме мыши приблизительно соответствует положению эндогенного аллеля ADAM6 мыши, например, в 3′ направлении от последний последовательности генного сегмента V и в 5′ направлении от первого генного сегмента D.
[00030] Согласно одному из вариантов реализации у указанных мышей-самцов присутствует последовательность ADAM6, либо ее гомолог, ортолог или функциональный фрагмент, фланкированная(ый) с 5′-стороны, 3′-стороны, либо с 5′- и 3′-стороны (относительно направления транскрипции указанной последовательности ADAM6) последовательности нуклеиновой кислоты, кодирующей генный сегмент вариабельной области иммуноглобулина. Согласно конкретному варианту реализации указанный вариабельный генный сегмент иммуноглобулина представляет собой генный сегмент человека. Согласно одному из вариантов реализации указанный вариабельный генный сегмент иммуноглобулина представляет собой генный сегмент человека, а указанная последовательность, кодирующая ADAM6 мыши, либо его ортолог, гомолог или фрагмент, функциональный у мыши, расположена между генными сегментами V человека; согласно одному из вариантов реализации указанная мышь содержит два или более генных сегментов V человека, и указанная последовательность находится в положении между последним генным сегментом V и предпоследним генным сегментом V; согласно одному из вариантов реализации указанная последовательность расположена после последнего генного сегмента V и первого генного сегмента D.
[00031] Согласно одному из аспектов предложен самец мыши, в зародышевой линии у которого присутствует нефункциональный эндогенный ген ADAM6 или делеция эндогенного гена ADAM6, при этом спермии указанной мыши способны проходить по маточным трубам самки мыши и оплодотворять яйцеклетку. Согласно одному из вариантов реализации у указанных мышей присутствует экстрахромосомная копия гена ADAM6 мыши, либо его ортолога, гомолога или функционального фрагмента, функционального у мышей-самцов. Согласно одному из вариантов реализации у указанных мышей присутствует эктопический ген ADAM6 мыши, либо его ортолог, гомолог или функциональный фрагмент, функциональный у мышей-самцов.
[00032] Согласно одному из аспектов предложены мыши, у которых присутствует генетическая модификация, снижающая функцию эндогенного ADAM6 мыши, при этом у указанных мышей присутствует по меньшей мере отчасти функционал ADAM6, что обеспечивается либо эндогенным немодифицированным аллелем, функциональным полностью или частично (например, у гетерозигот), либо экспрессией из эктопической последовательности, кодирующей ADAM6 либо его ортолог, гомолог или функциональный фрагмент, функциональный у мышей-самцов.
[00033] Согласно одному из вариантов реализации у указанных мышей присутствует функция ADAM6, достаточная для придания мышам-самцам способности производить потомство посредством спаривания, по сравнению с мышами-самцами, у которых отсутствует функциональный ADAM6. Согласно одному из вариантов реализации функция ADAM6 обеспечивается присутствием эктопической последовательности нуклеотидов, кодирующей ADAM6 мыши, либо его гомолог, ортолог или функциональный фрагмент. Гомологи или ортологи ADAM6, или фрагменты, функциональные у мышей-самцов, включают компенсирующие, полностью или частично, утрату способности производить потомство, наблюдаемую у мышей-самцов, у которых отсутствует достаточная активность эндогенного мышиного ADAM6, например, утрату способности, наблюдаемую у нокаутных по ADAM6 мышей. В этом смысле ADAM6-нокаутные мыши включают мышей, у которых присутствует эндогенный локус или его фрагмент, но не функциональный, т.е. либо совершенно не экспрессирущий ADAM6 (ADAM6a и/или ADAM6b), либо экспрессирущий ADAM6 (ADAM6a и/или ADAM6b) на уровне, недостаточном для поддержания по существу нормальной способности самцов мыши дикого типа производить потомство. Утрата функции может происходить, например, из-за модификации структурного гена указанного локуса {т.е. кодирующей области ADAM6a или ADAM6b), или в регуляторной области локуса (например, в последовательности, расположенной в направлении 5′ от гена ADAM6a, или в направлении 3′ от кодирующей области ADAM6a или ADAM6b, при этом указанная последовательность контролирует, полностью или частично, транскрипцию гена ADAM6, экспрессию РНК ADAM6 или экспрессию белка ADAM6). Согласно различным вариантам реализации ортологи, гомологи или их фрагменты, функциональные у мышей-самцов, представляют собой такие ортологи, гомологи или их фрагменты, которые обеспечивают способность спермы мышей-самцов (или большинству спермиев в эякуляте мышей-самцов) проходить через маточные трубы мыши и оплодотворять яйцеклетку мыши.
[00034] Согласно одному из вариантов реализации у мышей-самцов, экспрессирующих вариабельную область иммуноглобулина человека или ее функциональный фрагмент, присутствует достаточная активность ADAM6 для обеспечения способности указанных мышей-самцов производить потомство посредством спаривания с мышами-самками, и, согласно одному из вариантов реализации у указанных мышей-самцов наблюдается способность производить потомство при спаривании с мышами-самками, составляющая согласно одному из вариантов реализации по меньшей мере 25%, согласно одному из вариантов реализации по меньшей мере 30%, согласно одному из вариантов реализации по меньшей мере 40%, согласно одному из вариантов реализации по меньшей мере 50%, согласно одному из вариантов реализации по меньшей мере 60%, согласно одному из вариантов реализации по меньшей мере 70%, согласно одному из вариантов реализации по меньшей мере 80%, согласно одному из вариантов реализации по меньшей мере 90% от такой способности, а согласно одному из вариантов реализации приблизительно равная такой способности мышей с одним или двумя эндогенными немодифицированными аллелями ADAM6.
[00035] Согласно одному из вариантов реализации мыши-самцы экспрессируют достаточно ADAM6 (или его ортолога, гомолога или функционального фрагмента), чтобы обеспечить способность спермия указанных мышей-самцов проходить через маточные трубы самки мыши и оплодотворять яйцеклетку мыши.
[00036] Согласно одному из вариантов реализации функционал ADAM6 обеспечивается последовательностью нуклеиновой кислоты, смежной с хромосомной последовательностью мыши (например, указанную нуклеиновую кислоту случайным образом встраивают в хромосому мыши; или вводят в конкретный участок, например, направленным встраиванием указанной нуклеиновой кислоты в конкретный участок, например, опосредованным сайт-специфической рекомбиназой (например, опосредованным Cre) встраиванием или с помощью гомологичной рекомбинации). Согласно одному из вариантов реализации указанная последовательность ADAM6 находится на нуклеиновой кислоте, расположенной вне хромосомы мыши (например, указанная последовательность ADAM6 расположена на эписоме, т.е. экстрахромосомно, например, в экспрессионной конструкции, в векторе, на искусственной дрожжевой хромосоме (YAC), в трансхромосоме и т.п.).
[00037] Согласно одному из аспектов предложены генетически модифицированные мыши и клетки, содержащие модификацию эндогенного локуса тяжелой цепи иммуноглобулина, при этом у указанных мышей экспрессируется по меньшей мере часть последовательности тяжелой цепи иммуноглобулина, например, по меньшей мере часть последовательности человека, при этом у указанных мышей присутствует активность ADAM6, функционального у мышей-самцов. Согласно одному из вариантов реализации указанная модификация уменьшает или элиминирует активность ADAM6 мыши. Согласно одному из вариантов реализации указанная мышь модифицирована таким образом, что оба аллеля, кодирующие активный ADAM6, отсутствуют либо экспрессируют ADAM6 с недостаточной для поддержания нормального спаривания у мышей-самцов функциональностью. Согласно одному из вариантов реализации у указанной мыши также присутствует эктопическая последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши, либо его ортолог, гомолог или функциональный фрагмент.
[00038] Согласно одному из аспектов предложены генетически модифицированные мыши и клетки, содержащие модификацию эндогенного локуса тяжелой цепи иммуноглобулина, при этом указанная модификация уменьшает или элиминирует активность ADAM6, экспрессируемого из последовательности ADAM6 указанного локуса, и при этом у указанных мышей присутствует белок ADAM6, либо его ортолог, гомолог или функциональный фрагмент. Согласно различным вариантам реализации указанный белок ADAM6 или его фрагмент кодируется эктопической последовательностью ADAM6. Согласно различным вариантам реализации указанный белок ADAM6 или его фрагмент экспрессируется из эндогенного аллеля ADAM6. Согласно различным вариантам реализации указанная мышь содержит первый аллель тяжелой цепи иммуноглобулина, содержащий первую модификацию, которая уменьшает или элиминирует экспрессию функционального ADAM6 из первого аллеля тяжелой цепи иммуноглобулина; также указанная мышь содержит второй аллель тяжелой цепи иммуноглобулина, который содержит вторую модификацию, существенно не снижающую или не подавляющую полностью экспрессию функционального ADAM6 из второго аллеля тяжелой цепи иммуноглобулина.
[00039] Согласно одному из вариантов реализации вторая модификация расположена в направлении 3′ (относительно направленности транскрипции генного сегмента V мыши) от последнего генного сегмента V мыши и в направлении 5′ (относительно направленности транскрипции последовательности константной области) от гена константной области тяжелой цепи иммуноглобулина мыши (или гибридного гена человека/мыши) или его фрагмента (например, последовательности нуклеиновой кислоты, кодирующей: CH1, и/или шарнир, и/или CH2, и/или CH3 человека и/или мыши).
[00040] Согласно одному из вариантов реализации указанная модификация присутствует в первом аллеле тяжелой цепи иммуноглобулина в первом локусе, кодирующем первый аллель ADAM6, а функция ADAM6 обусловлена экспрессией эндогенного ADAM6 из второго аллеля тяжелой цепи иммуноглобулина во втором локусе, кодирующем функциональный ADAM6, причем указанный второй аллель тяжелой цепи иммуноглобулина содержит по меньшей мере одну модификацию генного сегмента V, D и/или J. Согласно конкретному варианту реализации указанная по меньшей мере одна модификация генного сегмента V, D, и/или J представляет собой удаление, замену на генный сегмент V, D и/или J человека, замену на генный сегмент V, D и/или J верблюдовых, замену на гуманизированный или «камелизированный» генный сегмент V, D и/или J, замену последовательности тяжелой цепи на последовательность легкой цепи, и их комбинации. Согласно одному из вариантов реализации указанная по меньшей мере одна модификация представляет собой удаление одного или более генных сегментов V, D и/или J тяжелой цепи и замене одной или более генных сегментов V и/или J легкой цепи (например, генного сегмента V и/или J легкой цепи человека) в указанном локусе тяжелой цепи.
[00041] Согласно одному из вариантов реализации указанная модификация присутствует в первом аллеле тяжелой цепи иммуноглобулина в первом локусе, и втором аллеле тяжелой цепи иммуноглобулина во втором локусе, и функция ADAM6 обусловлена экспрессией эктопического ADAM6 в не-иммуноглобулиновом локусе в зародышевой линии мыши. Согласно конкретному варианту реализации указанный не-иммуноглобулиновый локус представляет собой локус ROSA26. Согласно конкретному варианту реализации указанный не-иммуноглобулиновый локус транскрипционно активен в репродуктивной ткани.
[00042] Согласно одному из аспектов предложена мышь, гетерозиготная или гомозиготная по нокауту ADAM6. Согласно одному из вариантов реализации у указанной мыши также присутствует модифицированная последовательность иммуноглобулина, представляющая собой последовательность иммуноглобулина человека, или последовательность гуманизированного иммуноглобулина, или последовательность иммуноглобулина верблюдовых, или последовательность «камелизированного» иммуноглобулина человека, или последовательность иммуноглобулина мыши. Согласно одному из вариантов реализации указанная модифицированная последовательность иммуноглобулина находится в эндогенном локусе тяжелой цепи иммуноглобулина мыши. Согласно одному из вариантов реализации указанная модифицированная последовательность иммуноглобулина включает последовательность вариабельных генов тяжелой цепи человека в эндогенном локусе тяжелой цепи иммуноглобулина мыши. Согласно одному из вариантов реализации указанная последовательность вариабельных генов тяжелой цепи человека замещает эндогенную последовательность вариабельных генов тяжелой цепи мыши в эндогенном локусе тяжелой цепи иммуноглобулина мыши.
[00043] Согласно одному из аспектов предложена мышь, неспособная к осуществлению экспрессии функционального эндогенного ADAM6 мыши из эндогенного локуса ADAM6 мыши. Согласно одному из вариантов реализации у мыши присутствует эктопическая последовательность нуклеиновой кислоты, кодирующая ADAM6, или его функциональный фрагмент, функциональный у указанной мыши. Согласно конкретному варианту реализации указанная эктопическая последовательность нуклеиновой кислоты кодирует белок, восстанавливающий утраченную способность производить потомство, наблюдаемую у самца мыши, гомозиготного по нокауту ADAM6. Согласно конкретному варианту реализации указанная эктопическая последовательность нуклеиновой кислоты кодирует мышиный белок ADAM6.
[00044] Согласно одному из аспектов предложена мышь, у которой отсутствует функциональный эндогенный локус ADAM6, и у которой присутствует эктопическая последовательность нуклеиновой кислоты, обеспечивающая функцию ADAM6 мыши. Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты содержит эндогенную последовательность ADAM6 мыши или его функциональный фрагмент. Согласно одному из вариантов реализации указанная эндогенная последовательность ADAM6 мыши содержит ADAM6a- и ADAM6b-кодирующую последовательность, расположенную у мыши дикого типа между крайним с 3′-стороны генным сегментом V тяжелой цепи (VH) иммуноглобулина мыши и крайним с 5′-стороны генным сегментом D тяжелой цепи (DH) иммуноглобулина мыши.
[00045] Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты включает последовательность, кодирующую ADAM6a мыши или его функциональный фрагмент и/или а последовательность, кодирующую ADAM6b мыши или его функциональный фрагмент, при этом указанный(е) ADAM6a и/или ADAM6b или функциональный(е) фрагмент(ы) функционально связан(ы) с промотором. Согласно одному из вариантов реализации указанный промотор представляет собой промотор человека. Согласно одному из вариантов реализации указанный промотор представляет собой промотор ADAM6 мыши. Согласно конкретному варианту реализации указанный промотор ADAM6 содержит последовательность, расположенную между первым кодоном первого гена ADAM6 ближайшим к крайнему с 5′-стороны генному сегменту DH мыши, и последовательностью сигнала рекомбинации крайнего с 5′-стороны DH генного сегмента, где 5′-сторона определяется направлением транскрипции генов иммуноглобулина мыши. Согласно одному из вариантов реализации указанный промотор представляет собой вирусный промотор. Согласно конкретному варианту реализации указанный вирусный промотор представляет собой цитомегаловирусный (CMV) промотор. Согласно одному из вариантов реализации указанный промотор представляет собой убиквитиновый промотор.
[00046] Согласно одному из вариантов реализации указанный промотор представляет собой индуцируемый промотор. Согласно одному из вариантов реализации указанный индуцируемый промотор регулирует экспрессию в не-репродуктивных тканях. Согласно одному из вариантов реализации указанный индуцируемый промотор регулирует экспрессию в репродуктивных тканях. Согласно конкретному варианту реализации экспрессия последовательностей ADAM6a, и/или ADAM6b мыши, или их функциональных фрагментов регулируется в процессе развития указанным индуцируемым промотором в репродуктивных тканях.
[00047] Согласно одному из вариантов реализации указанные ADAM6a и/или ADAM6b мыши выбраны из ADAM6a согласно SEQ ID NO:1 и/или ADAM6b согласно последовательности SEQ ID NO:2. Согласно одному из вариантов реализации промотор ADAM6 мыши представляет собой промотор согласно SEQ ID NO:3. Согласно конкретному варианту реализации указанный промотор ADAM6 мыши включает последовательность нуклеиновой кислоты согласно SEQ ID NO:3, начинающуюся непосредственно после первого кодона ADAM6a в 5′ направлении (относительно направления транскрипции ADAM6a) и продолжающуюся до конца SEQ ID NO:3 в 5′-направлении от кодирующей области ADAM6. Согласно другому конкретному варианту реализации указанный промотор ADAM6 представляет собой фрагмент, начинающийся в пределах приблизительно 5-20 нуклеотидов в 5′-направлении от стартового кодона ADAM6a и продолжающийся на протяжении приблизительно 0,5 т.н., 1 т.н., 2 т.н., или 3 т.н. или более в 5′-направлении от стартового кодона ADAM6a.
[00048] Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты содержит SEQ ID NO:3 или ее фрагмент, введение которой(го) в организм мыши, бесплодной или имеющей низкую фертильность, обусловленную недостатком ADAM6, увеличивает фертильность или восстанавливает фертильность приблизительно до уровня фертильности дикого типа. Согласно одному из вариантов реализации SEQ ID NO:3 или ее фрагмент придает самцу мыши способность произвести спермий, способный пройти через маточные трубы самки мыши для оплодотворения мышиной яйцеклетки.
[00049] Согласно одному из аспектов предложена мышь, у которой удалена эндогенная последовательность нуклеотидов, кодирующая белок ADAM6, заменен эндогенный генный сегмент VH мыши на генный сегмент VH человека, и присутствует эктопическая последовательность нуклеотидов, кодирующая мышиный белок ADAM6, либо его ортолог, гомолог или фрагмент, функциональный у мышей-самцов.
[00050] Согласно одному из вариантов реализации указанная мышь содержит локус тяжелой цепи иммуноглобулина с удаленной нуклеотидной последовательностью локуса эндогенного иммуноглобулина, которая содержит эндогенный ген ADAM6; содержит последовательность нуклеотидов, кодирующую один или более сегментов гена иммуноглобулина человека; и при этом эктопическая последовательность нуклеотидов, кодирующая белок ADAM6 мыши, находится в составе или в непосредственной близости от последовательности нуклеотидов, кодирующей указанные один или более сегментов гена иммуноглобулина человека.
[00051] Согласно одному из вариантов реализации у указанной мыши все или по существу все эндогенные генные сегменты VH заменены последовательностью нуклеотидов, кодирующей один или более генных сегментов VH человека, и эктопическая последовательность нуклеотидов, кодирующая белок ADAM6 мыши, находится в составе или в непосредственной близости от последовательности нуклеотидов, кодирующей указанный один или более генных сегментов VH человека. Согласно одному из вариантов реализации у указанной мыши также заменены один или более эндогенных генных сегментов DH одним или более генными сегментами DH человека в эндогенном локусе гена DH. Согласно одному из вариантов реализации у указанной мыши также заменены один или более эндогенные генные сегменты JH одним или более генными сегментами JH человека в локусе эндогенного JH гена. Согласно одному из вариантов реализации у указанной мыши заменены все или по существу все эндогенные генные сегменты VH, DH и JH; и присутствуют замены на генные сегменты VH, DH и JH человека в эндогенных локусах генов VH, DH, и JH, при этом указанная мышь содержит эктопическая последовательность, кодирующая мышиный белок ADAM6. Согласно конкретному варианту реализации указанная эктопическая последовательность, кодирующая белок ADAM6 мыши, расположена между предпоследним с 3′-конца VH генным сегментом присутствующих генных сегментов VH человека, и последним с 3′-конца VH генным сегментом присутствующих генных сегментов VH человека. Согласно конкретному варианту реализации у указанной мыши удалены все или по существу все генные сегменты VH мыши, и заменены на все или практически все генные сегменты VH человека, и эктопическая последовательность нуклеотидов, кодирующая белок ADAM6 мыши, расположена в 3′-направлении от генного сегмента VH1-2 человека и в 5′-направлении от генного сегмента VH6-1 человека.
[00052] Согласно конкретному варианту реализации у указанной мыши все или по существу все эндогенные генных сегментов VH заменены последовательностью нуклеотидов, кодирующей один или более генных сегментов VH человека, и эктопическая последовательность нуклеотидов, кодирующая белок ADAM6 мыши находится в составе или в непосредственной близости от последовательности нуклеотидов, кодирующей указанный один или более генных сегментов VH человека.
[00053] Согласно одному из вариантов реализации указанная эктопическая последовательность нуклеотидов, кодирующая белок ADAM6 мыши, находится в геноме мыши на трансгене. Согласно одному из вариантов реализации указанная эктопическая последовательность нуклеотидов, кодирующая белок ADAM6 мыши, расположена у указанной мыши экстрахромосомно.
[00054] Согласно одному из аспектов предложена мышь, у которой присутствует модификация эндогенного локуса тяжелой цепи иммуноглобулина, при этом указанная мышь экспрессирует В-клетку, которая содержит реаранжированную последовательность иммуноглобулина, функционально связанную с последовательностью гена константной области тяжелой цепи, и указанная В-клетка содержит в геноме (например, на хромосоме В-клетки) ген, кодирующий ADAM6, либо его ортолог, гомолог или фрагмент, функциональный у мышей-самцов. Согласно одному из вариантов реализации указанная реаранжированная последовательность иммуноглобулина, функционально связанная с последовательностью гена константной области тяжелой цепи, включает последовательность V, D, и/или J тяжелой цепи человека; последовательность V, D, и/или J тяжелой цепи мыши; последовательность легкой цепи V и/или J человека или мыши. Согласно одному из вариантов реализации указанная последовательность гена константной области тяжелой цепи включает последовательность тяжелой цепи человека или мыши, выбранную из группы, состоящей из CH1, шарнира, CH2, CH3 и их комбинации.
[00055] Согласно одному из аспектов предложена генетически модифицированная мышь, отличающаяся тем, что указанная мышь содержит молчащий ген легкой цепи иммуноглобулина, а также заменены один или более эндогенные генные сегменты вариабельной области тяжелой цепи иммуноглобулина одним или более генными сегментами вариабельной области тяжелой цепи иммуноглобулина человека, при этом у указанной мыши отсутствует функциональный эндогенный локус ADAM6, и тем, что указанная мышь содержит эктопическая последовательность нуклеотидов, экспрессирующая белок ADAM6 мыши или его ортолог, гомолог или фрагмент, функциональный у мышей-самцов.
[00056] Согласно одному из аспектов предложена мышь, у которой отсутствует функциональный(ая) эндогенный(ая) локус или последовательность ADAM6 мыши, и присутствует эктопическая последовательность нуклеотидов, кодирующая локус ADAM6 мыши или функциональный фрагмент локуса или последовательности ADAM6 мыши, при этом указанная мышь способна спариваться с мышью противоположного пола с получением потомства, у которого присутствует эктопический(ая) локус или последовательность ADAM6. Согласно одному из вариантов реализации указанная мышь является самцом. Согласно одному из вариантов реализации указанная мышь является самкой.
[00057] Согласно одному из аспектов предложена генетически модифицированная мышь, отличающаяся тем, что у указанной мыши присутствует генный сегмент вариабельной области тяжелой цепи иммуноглобулина человека в эндогенном локусе гена вариабельной области тяжелой цепи иммуноглобулина мыши; отсутствует функциональная последовательность ADAM6 в эндогенном локусе гена вариабельной области тяжелой цепи иммуноглобулина мыши, и при этом указанная мышь содержит эктопическая последовательность нуклеотидов, экспрессирующая белок ADAM6 мыши или его ортолог, гомолог или фрагмент, функциональный у мышей-самцов.
[00058] Согласно одному из вариантов реализации эктопическая последовательность нуклеотидов, экспрессирующая белок ADAM6 мыши, является экстрахромосомной. Согласно одному из вариантов реализации эктопическая последовательность нуклеотидов, экспрессирующая белок ADAM6 мыши, встроена в один или более локусов в геноме мыши. Согласно конкретному варианту реализации указанный один или более локусов включает локус иммуноглобулина.
[00059] Согласно одному из аспектов предложена мышь, экспрессирующая последовательность тяжелой цепи иммуноглобулина из модифицированного эндогенного локуса тяжелой цепи иммуноглобулина мыши, при этом указанную тяжелую цепь получают из генного сегмента V, D генного сегмента, и J генного сегмента человека, при этом у указанной мыши присутствует активность ADAM6, функционального у указанной мыши.
[00060] Согласно одному из вариантов реализации указанная мышь содержит совокупность генных сегментов V человека, совокупность генных сегментов D и совокупность генных сегментов J. Согласно одному из вариантов реализации указанные генные сегменты D представляют собой генные сегменты D человека. Согласно одному из вариантов реализации указанные сегменты J представляют собой генные сегменты J человека. Согласно одному из вариантов реализации у указанной мыши также присутствует гуманизированная последовательность константной области тяжелой цепи, при этом указанная гуманизация включает замену последовательности, выбранной из CH1, шарнира, CH2, CH3 и их комбинации. Согласно конкретному варианту реализации указанную тяжелую цепь получают из генного сегмента V человека, генного сегмента D человека, генного сегмента J человека, последовательности CH1 человека, последовательности шарнира человека или мыши, последовательности CH2 мыши и последовательности CH3 мыши. Согласно другому конкретному варианту реализации у указанной мыши также присутствует последовательность константной области легкой цепи человека.
[00061] Согласно одному из вариантов реализации указанный генный сегмент D фланкирован с 5′ стороны (в отношении направленности транскрипции указанного генного сегмента D) последовательностью, кодирующей активность ADAM6, функционального у указанной мыши.
[00062] Согласно одному из вариантов реализации активность ADAM6, функциональная у указанной мыши, обусловлена экспрессией последовательности нуклеотидов, расположенной в направлении 5′ от крайнего с 5′-стороны D генного сегмента и в направлении 3′ от крайнего с 3′-стороны генного сегмента V (относительно направления транскрипции генного сегмента V) модифицированного эндогенного локуса тяжелой цепи иммуноглобулина мыши.
[00063] Согласно одному из вариантов реализации активность ADAM6, функциональная у указанной мыши, обусловлена экспрессией последовательности нуклеотидов, расположенной между двумя генными сегментами V человека в модифицированном эндогенном локусе тяжелой цепи иммуноглобулина мыши. Согласно одному из вариантов реализации указанные два генных сегмента V человека представляют собой генный сегмент VH1-2 человека и генный сегмент VH6-1.
[00064] Согласно одному из вариантов реализации указанная последовательность нуклеотидов содержит последовательность, выбранную из последовательности ADAM6b мыши или ее функционального фрагмента, последовательности ADAM6a мыши или ее функционального фрагмента, и их комбинации.
[00065] Согласно одному из вариантов реализации указанная последовательность нуклеотидов между двумя генными сегментами V человека ориентирована в направлении, противоположном направлению транскрипции указанных генных сегментов V человека. Согласно конкретному варианту реализации последовательность нуклеотидов кодирует в направлении 5′→3′ относительно направления транскрипции генов ADAM6, и за последовательностью ADAM6a следует последовательность ADAM6b.
[00066] Согласно одному из вариантов реализации у указанной мыши последовательность псевдогена ADAM6 человека между генными сегментами V VH1-2 и VH6-1 человека заменена на последовательность ADAM6 мыши или ее функциональный фрагмент.
[00067] Согласно одному из вариантов реализации последовательность, кодирующая активность ADAM6, функционального у указанной мыши, представляет собой последовательность ADAM6 мыши или его функционального фрагмента.
[00068] Согласно одному из вариантов реализации указанная мышь содержит эндогенный мышиный генный сегмент DFL16.1 (например, у мыши, гетерозиготной по модифицированному локусу тяжелой цепи эндогенного иммуноглобулина мыши) или генный сегмент DH1-1 человека. Согласно одному из вариантов реализации указанный D генный сегмент тяжелой цепи иммуноглобулина, экспрессируемый у мыши, получают из эндогенного мышиного DFL16.1 генного сегмента или генного сегмента DH1-1 человека.
[00069] Согласно одному из аспектов предложена мышь, у которой присутствует последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши (или его гомолог, ортолог или функциональный фрагмент) в ДНК-содержащей клетке нереаранжированной В-клеточной линии, но не присутствует последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши (или его гомолог, ортолог или функциональный фрагмент) в В-клетке, содержащей реаранжированные иммуноглобулиновые локусы; при этом указанная последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши (или его гомолог, ортолог или функциональный фрагмент) находится в геноме в положении, отличном от положения гена ADAM6 мыши у мыши дикого типа. Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши (или его гомолог, ортолог или функциональный фрагмент) присутствует во всех или по существу во всех ДНК-содержащих клетках, не принадлежащих реаранжированной В-клеточной линии; согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты присутствует в клетках зародышевой линии мыши, но не в хромосоме реаранжированной В-клетки.
[00070] Согласно одному из аспектов предложена мышь, у которой присутствует последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши (или его гомолог, ортолог или функциональный фрагмент) во всех или по существу во всех ДНК-содержащих клетках, включая В-клетки, содержащие реаранжированные иммуноглобулиновые локусы; при этом указанная последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши (или его гомолог, ортолог или функциональный фрагмент) находится в геноме в положении, отличном от положения гена ADAM6 мыши у мыши дикого типа. Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши (или его гомолог, ортолог или функциональный фрагмент), расположена на нуклеиновой кислоте, смежной с указанным реаранжированным иммуноглобулиновым локусом. Согласно одному из вариантов реализации указанная нуклеиновая кислота, смежная с указанным реаранжированным иммуноглобулиновым локусом, представляет собой хромосому. Согласно одному из вариантов реализации указанная хромосома представляет собой хромосому, обнаруживаемую у мыши дикого типа, и указанная хромосома содержит модификацию иммуноглобулинового локуса мыши.
[00071] Согласно одному из аспектов предложена генетически модифицированная мышь, отличающаяся тем, что указанная мышь содержит В-клетку, которая содержит в геноме последовательность ADAM6 или ее ортолог или гомолог. Согласно одному из вариантов реализации указанная последовательность ADAM6 или ее ортолог или гомолог располагается в локусе тяжелой цепи иммуноглобулина. Согласно одному из вариантов реализации указанная последовательность ADAM6 или ее ортолог или гомолог располагается в локусе, не являющемся иммуноглобулиновым локусом. Согласно одному из вариантов реализации указанная последовательность ADAM6 располагается на трансгене, управляемом гетерологичным промотором. Согласно конкретному варианту реализации указанный гетерологичный промотор представляет собой не-иммуноглобулиновый промотор. Согласно конкретному варианту реализации В-клетка экспрессирует белок ADAM6 или его ортолог или гомолог.
[00072] Согласно одному из вариантов реализации в 90% или более В-клеток указанных мышей присутствует ген, кодирующий белок ADAM6, или его ортолог, его гомолог или его фрагмент, функциональный у указанной мыши. Согласно конкретному варианту реализации указанная мышь представляет собой самца мыши.
[00073] Согласно одному из вариантов реализации геном В-клетки содержит первый аллель и второй аллель, включающие последовательность ADAM6 или ее ортолог или гомолог. Согласно одному из вариантов реализации геном В-клетки содержит первый аллель, но не второй аллель, содержащий указанную последовательность ADAM6 или ее ортолог или гомолог.
[00074] Согласно одному из аспектов предложена мышь, у которой присутствует модификация в одном или более эндогенных аллелей ADAM6.
[00075] Согласно одному из вариантов реализации указанная модификация придает мыши неспособность к экспрессии функционального белка ADAM6 по меньшей мере из одного из указанных одного или более эндогенных аллелей ADAM6. Согласно конкретному варианту реализации мышь неспособна к экспрессии функционального белка ADAM6 из каждого из эндогенных аллелей ADAM6.
[00076] Согласно одному из вариантов реализации мыши неспособны к экспрессии функционального белка ADAM6 из каждого эндогенного аллеля ADAM6, и у указанных мышей присутствует эктопическая последовательность ADAM6.
[00077] Согласно одному из вариантов реализации мыши неспособны к экспрессии функционального белка ADAM6 из каждого эндогенного аллеля ADAM6, и у указанных мышей присутствует эктопическая последовательность ADAM6, расположенная в пределах 1, 2, 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110 или 120 или более т.н. в 5′ направлении (относительно направления транскрипции локуса тяжелой цепи мыши) от последовательности константной области тяжелой цепи иммуноглобулина мыши. Согласно конкретному варианту реализации указанная эктопическая последовательность ADAM6 находится в эндогенном локусе тяжелой цепи иммуноглобулина (например, в межгенной V-D области, между двумя генными сегментами V, между генными сегментами V и D, между генными сегментами D и J, и т.п.). Согласно конкретному варианту реализации указанная эктопическая последовательность ADAM6 расположена в пределах 90-100 т.н. межгенной последовательности между последним генным сегментом V мыши и первым генным сегментом D мыши. Согласно другому конкретному варианту реализации указанную эндогенную 90-100 т.н. межгенную V-D последовательность удаляют, и эктопическая последовательность ADAM6 располагается между последним V и первым D генными сегментами.
[00078] Согласно одному из аспектов предложен бесплодный самец мыши, отличающийся тем, что указанная мышь содержит делецию двух или более эндогенных аллелей ADAM6. Согласно одному из аспектов предложена самка мыши, представляющая собой носителя признака бесплодности самцов, отличающаяся тем, что в зародышевой линии указанной самки мыши присутствует нефункциональный аллель ADAM6, или нокаутированный эндогенный аллель ADAM6.
[00079] Согласно одному из аспектов предложена мышь, у которой отсутствует эндогенный генный сегмент V, D и J тяжелой цепи иммуноглобулина, при этом большинство В-клеток указанной мыши содержат последовательность ADAM6 или ее ортолог или гомолог.
[00080] Согласно одному из вариантов реализации у указанной мыши отсутствуют генные сегменты тяжелой цепи эндогенного иммуноглобулина, выбранные из двух или более генных сегментов V, двух или более генных сегментов D, двух или более генных сегментов J, и их комбинации. Согласно одному из вариантов реализации у указанной мыши отсутствуют генные сегменты тяжелой цепи иммуноглобулина, выбранные из по меньшей мере 1-89 генных сегментов V, по меньшей мере 1-13 генных сегментов D, по меньшей мере 1-4 генных сегментов J, и их комбинации. Согласно одному из вариантов реализации у указанной мыши отсутствует фрагмент геномной ДНК из хромосомы 12, включающий приблизительно три мегабазы эндогенного локуса тяжелой цепи иммуноглобулина. Согласно конкретному варианту реализации у указанной мыши отсутствуют все функциональные эндогенные генные сегменты V, D и J тяжелой цепи. Согласно конкретному варианту реализации у указанной мыши отсутствуют 89 генных сегмента VH, 13 генных сегментов DH и четыре генных сегмента JH.
[00081] Согласно одному из аспектов предложена мышь, отличающаяся тем, что геном зародышевой линии указанной мыши содержит модификацию локуса тяжелой цепи иммуноглобулина, при этом указанная модификация в локусе тяжелой цепи иммуноглобулина включает замену одной или более последовательностей вариабельной области иммуноглобулина мыши на одну или более последовательностей вариабельной области не происходящего от мыши иммуноглобулина, и при этом указанная мышь содержит последовательность нуклеиновой кислоты, кодирующая белок ADAM6 мыши. Согласно предпочтительному варианту реализации указанные последовательности DH и JH и по меньшей мере 3, по меньшей мере 10, по меньшей мере 20, по меньшей мере 40, по меньшей мере 60, или по меньшей мере 80 последовательностей VH локуса тяжелой цепи иммуноглобулина заменяют последовательностями вариабельной области не происходящего от мыши иммуноглобулина. Согласно еще одному предпочтительному варианту реализации указанные DH, JH, и все VH последовательности локуса тяжелой цепи иммуноглобулина заменяют последовательностями вариабельной области иммуноглобулина, не происходящего от мыши. Указанные последовательности вариабельной области иммуноглобулина, не происходящего от мыши, могут быть переаранжированными. Согласно предпочтительному варианту реализации указанные последовательности вариабельной области иммуноглобулина не происходящего от мыши включают полные переаранжированные DH и JH области и по меньшей мере 3, по меньшей мере 10, по меньшей мере 20, по меньшей мере 40, по меньшей мере 60, или по меньшей мере 80 переаранжированных последовательностей VH указанного не являющегося мышью вида. Согласно еще одному предпочтительному варианту реализации последовательности вариабельной области иммуноглобулина не происходящего от мыши включают вариабельную область полностью, в том числе все VH, DH, и JH области, указанного не являющегося мышью вида. Вид, не являющийся мышью, может представлять собой Homo sapiens, а указанные последовательности вариабельной области иммуноглобулина не происходящего от мыши могут представлять собой последовательности человека.
[00082] Согласно одному из аспектов предложена мышь, экспрессирующая антитело, которое содержит по меньшей мере один вариабельный домен полипептида иммуноглобулина человека/константный домен полипептида иммуноглобулина не происходящий от человека, при этом указанная мышь экспрессирует белок ADAM6 мыши или его ортолог или гомолог из локуса, отличного от иммуноглобулинового локуса.
[00083] Согласно одному из вариантов реализации белок ADAM6 или его ортолог или гомолог экспрессируется в В-клетке мыши, причем указанная В-клетка содержит реаранжированную последовательность иммуноглобулина, которая содержит а вариабельная последовательность человека и константную последовательность не происходящую от человека.
[00084] Согласно одному из вариантов реализации константная последовательность не происходящая от человека представляет собой последовательность грызуна. Согласно одному из вариантов реализации указанный грызун выбран из мыши, крысы и хомяка.
[00085] Согласно одному из аспектов предложен способ получения бесплодного самца-мыши, включающий перевод эндогенного аллеля ADAM6 донорской ЭС-клетки в нефункциональное состояние (или нокаутирование указанного аллеля), введение указанной донорской ЭС-клетки в эмбрион хозяин, вынашивание эмбриона хозяина с помощью суррогатной матери, и обеспечение рождения суррогатной матерью потомства, происходящего полностью или частично из указанной донорской ЭС клетки. Согласно одному из вариантов реализации указанный способ также включает размножение потомства с получением бесплодного самца-мыши.
[00086] Согласно одному из аспектов предложен способ получения мыши с представляющей интерес генетической модификацией, при этом указанная мышь бесплодна; указанный способ включает этапы (а) получения представляющей интерес генетической модификации генома; (b) модифицирование указанного генома с нокаутированием эндогенного аллеля ADAM6, или обеспечение нефункциональности эндогенного аллеля ADAM6; и (с) применение указанного генома для получения мыши. Согласно различным вариантам реализации геном происходит из ЭС-клетки или его используют для экспериментальной ядерной передачи.
[00087] Согласно одному из аспектов предложена мышь, получаемая с применением направленного вектора, нуклеотидной конструкции или клетки согласно настоящему описанию.
[00088] Согласно одному из аспектов предложено потомство мыши согласно настоящему описанию, спаривающейся со второй мышью, представляющей собой мышь дикого типа или генетически модифицированные.
[00089] Согласно одному из аспектов предложен способ поддержания линии мышей, причем у указанной линии мышей последовательность тяжелой цепи иммуноглобулина мыши заменена одной или более гетерологичными последовательностями тяжелой цепи иммуноглобулина. Согласно одному из вариантов реализации указанная одна или более гетерологичных последовательностей тяжелой цепи иммуноглобулина представляют собой последовательность(и) тяжелой цепи иммуноглобулина человека.
[00090] Согласно одному из вариантов реализации у указанной линии мышей удален один или более генных сегментов VH, DH и/или JH мыши. Согласно одному из вариантов реализации указанная мышь дополнительно содержит один или более генных сегментов VH человека, один или более генных сегментов DH человека, и/или один или более генных сегментов JH человека. Согласно одному из вариантов реализации указанная мышь содержит по меньшей мере 3, по меньшей мере 10, по меньшей мере 20, по меньшей мере 40, по меньшей мере 60, или по меньшей мере 80 сегментов VH человека, по меньшей мере 27 генных сегментов DH человека и по меньшей мере шесть генных сегментов JH. Согласно конкретному варианту реализации указанная мышь содержит по меньшей мере 3, по меньшей мере 10, по меньшей мере 20, по меньшей мере 40, по меньшей мере 60, или по меньшей мере 80 сегментов VH человека; указанные по меньшей мере 27 генных сегмента DH человека и указанные по меньшей мере шесть генных сегментов JH функционально связаны с геном константной области. Согласно одному из вариантов реализации указанный ген константной области представляет собой ген константной области мыши. Согласно одному из вариантов реализации указанный ген константной области включает последовательность гена константной области мыши, выбранного из CH1, шарнира, CH2, CH3 и/или CH4, или их комбинации.
[00091] Согласно одному из вариантов реализации указанный способ включает получение самца мыши, гетерозиготного по замене последовательности тяжелой цепи иммуноглобулина мыши, и скрещивание указанного гетерозиготного самца мыши с самкой мыши дикого типа, либо самкой мыши, гомозиготной или гетерозиготной по указанной последовательности тяжелой цепи человека. Согласно одному из вариантов реализации указанный способ включает поддержание линии посредством систематического скрещивания гетерозиготных самцов с самками дикого типа, либо гомозиготных или гетерозиготных по указанной последовательности тяжелой цепи человека.
[00092] Согласно одному из вариантов реализации указанный способ включает получение клеток от самцов или самок мыши, гомозиготных или гетерозиготных по последовательности тяжелой цепи человека, и использование указанных клеток в качестве донорских клеток, либо из ядер в качестве донорских ядер, и применение указанных клеток или ядер для получения генетически модифицированных животных с применением клеток-хозяев и/или вынашивания указанных клеток и/или ядер суррогатными матерями.
[00093] Согласно одному из вариантов реализации только мыши-самцы, гетерозиготные по замене в указанном локусе тяжелой цепи, скрещиваются с мышами-самками. Согласно конкретному варианту реализации указанные мыши-самки гомозиготны, гетерозиготны или относятся к дикому типу в отношении замещенного локуса тяжелой цепи.
[00094] Согласно одному из вариантов реализации у мыши также заменена последовательность вариабельной области λ и/или κ легкой цепи в эндогенном локусе легкой цепи иммуноглобулина гетерологичной последовательностью легкой цепи иммуноглобулина. Согласно одному из вариантов реализации указанные гетерологичные последовательности легкой цепи иммуноглобулина представляют собой последовательности вариабельной области λ и/или κ легкой цепи иммуноглобулина человека.
[00095] Согласно одному из вариантов реализации у мыши также присутствует трансген в локусе, отличном от эндогенного локуса иммуноглобулина, при этом указанный трансген содержит последовательность, кодирующую реаранжированную или нереаранжированную гетерологичную последовательность легкой цепи λ или κ (например, переаранжированный VL и переаранжированный JL, или реаранжированный VJ) функционально связанную (для переаранжированной) или слитую (для реаранжированной) с последовательностью константной области легкой цепи иммуноглобулина. Согласно одному из вариантов реализации указанная гетерологичная последовательность λ или κ легкой цепи является человеческой. Согласно одному из вариантов реализации указанная последовательность константной области выбрана из таковых грызунов, человека и не являющихся человеком приматов. Согласно одному из вариантов реализации указанная последовательность константной области выбрана из таковых мыши, крысы и хомяка. Согласно одному из вариантов реализации указанный трансген содержит не-иммуноглобулиновый промотор, управляющий экспрессией последовательностей легкой цепи. Согласно конкретному варианту реализации указанный промотор представляет собой транскрипционно активный промотор. Согласно конкретному варианту реализации указанный промотор представляет собой промотор ROSA26.
[00096] Согласно одному из аспектов предложена конструкция нуклеиновой кислоты, содержащая 5′-плечо гомологии и 3′-плечо гомологии, при этом указанное 5′-плечо гомологии содержит последовательность, идентичную или по существу идентичную последовательности вариабельной области тяжелой цепи иммуноглобулина человека; указанное 3′-плечо гомологии содержит последовательность, идентичную или по существу идентичную последовательности вариабельной области иммуноглобулина человека или мыши, а между указанными 5′- и 3′-плечами гомологии расположена последовательность, содержащая последовательность нуклеотидов, кодирующая мышиный белок ADAM6. Согласно конкретному варианту реализации последовательность, кодирующая ген ADAM6 мыши, функционально связана с промотором мыши, с которым связан ADAM6 мыши у мышей дикого типа.
[00097] Согласно одному из аспектов предложен направленный вектор, содержащий (а) последовательность нуклеотидов, идентичную или по существу идентичную последовательности нуклеотидов генного сегмента вариабельной области человека; и, (b) последовательность нуклеотидов, кодирующую ADAM6 мыши, либо его ортолог, гомолог или фрагмент, функциональный у мыши.
[00098] Согласно одному из вариантов реализации указанный направленный вектор также содержит промотор, функционально связанный с последовательностью, кодирующей ADAM6 мыши. Согласно конкретному варианту реализации указанный промотор представляет собой промотор ADAM6 мыши.
[00099] Согласно одному из аспектов предложена нуклеотидная конструкция для модифицирования локуса вариабельной области тяжелой цепи иммуноглобулина мыши, при этом указанная конструкция содержит по меньшей мере один участок распознавания сайт-специфической рекомбиназой и последовательность, кодирующую белок ADAM6, либо его ортолог, гомолог или фрагмент, функциональный у мыши.
[000100] Согласно одному из аспектов предложены клетки мыши и эмбрионы мыши, включая, но не ограничиваясь перечисленными, ЭС-клетки, плюрипотентные клетки и индуцированные плюрипотентные клетки, содержащие генетические модификации согласно описанию в настоящей заявке. Предложены ХХ-клетки и XY-клетки. Также предложены клетки, содержащие ядро, где присутствует модификация согласно настоящему описанию, например, модификация, внесенная в клетку посредством пронуклеарной инъекции. Также предложены клетки, эмбрионы и мыши, у которых присутствует введенный с помощью вируса ген ADAM6, например, клетки, эмбрионы и мыши, в/у которых присутствует трансдукционная конструкция, содержащая ген ADAM6, функциональный у мыши.
[000101] Согласно одному из аспектов предложена генетически модифицированная клетка мыши, отличающаяся тем, что в указанной клетке отсутствует функциональный эндогенный локус ADAM6 мыши, и указанная клетка содержит эктопическую последовательность нуклеотидов, кодирующую белок ADAM6 мыши или его функциональный фрагмент. Согласно одному из вариантов реализации указанная клетка также содержит модификацию эндогенной последовательности вариабельных генов тяжелой цепи иммуноглобулина. Согласно конкретному варианту реализации указанная модификация эндогенной последовательности вариабельных генов тяжелой цепи иммуноглобулина включает удаление, выбранное из удаления генного сегмента VH мыши, удаления генного сегмента DH мыши, удаления генного сегмента JH мыши и их комбинации. Согласно конкретному варианту реализации у мыши присутствует замена одной или более последовательностей VH, DH и/или JH иммуноглобулина мыши последовательностями иммуноглобулина человека. Согласно конкретному варианту реализации указанная последовательность иммуноглобулина человека выбрана из VH человека, VL человека, DH человека, JH человека, JL человека и их комбинации.
[000102] Согласно одному из вариантов реализации указанная клетка представляет собой тотипотентную клетку, плюрипотентную клетку или индуцированную плюрипотентную клетку. Согласно конкретному варианту реализации указанная клетка представляет собой ЭС-клетку мыши.
[000103] Согласно одному из аспектов предложена В-клетка мыши, причем указанная мышиная В-клетка содержит реаранжированный ген тяжелой цепи иммуноглобулина, при этом в хромосоме указанной В-клетки присутствует последовательность нуклеиновой кислоты, кодирующая белок ADAM6, либо его ортолог, гомолог или фрагмент, функциональный у мышей-самцов. Согласно одному из вариантов реализации указанная В-клетка мыши содержит два аллеля указанной последовательности нуклеиновой кислоты.
[000104] Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты находится в молекуле нуклеиновой кислоты (например, в хромосоме В-клетки), смежной с указанным локусом реаранжированной тяжелой цепи иммуноглобулина мыши.
[000105] Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты находится в молекуле нуклеиновой кислоты (например, а В-клетка хромосома), отдельной от молекулы нуклеиновой кислоты, которая содержит указанный локус реаранжированной тяжелой цепи иммуноглобулина мыши.
[000106] Согласно одному из вариантов реализации указанная В-клетка мыши содержит реаранжированную последовательность вариабельных генов не происходящего от мыши иммуноглобулина, функционально связанную с геном константной области иммуноглобулина мыши или человека, при этом указанная В-клетка содержит последовательность нуклеиновой кислоты, кодирующую белок ADAM6, либо его ортолог, гомолог или фрагмент, функциональный у мышей-самцов.
[000107] Согласно одному из аспектов предложена соматическая клетка мыши, содержащая хромосому, которая содержит модифицированный локус тяжелой цепи иммуноглобулина и последовательность нуклеиновой кислоты, кодирующую ADAM6 мыши, либо его ортолог, гомолог или фрагмент, функциональный у мышей-самцов. Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты находится в той же хромосоме, что и указанный модифицированный локус тяжелой цепи иммуноглобулина. Согласно одному из вариантов реализации указанная нуклеиновая кислота находится не на той хромосоме, где находится модифицированный локус тяжелой цепи иммуноглобулина. Согласно одному из вариантов реализации указанная соматическая клетка содержит одну копию указанной последовательности нуклеиновой кислоты. Согласно одному из вариантов реализации указанная соматическая клетка содержит по меньшей мере две копии указанной последовательности нуклеиновой кислоты. Согласно конкретному варианту реализации указанная соматическая клетка представляет собой В-клетку. Согласно конкретному варианту реализации указанная клетка представляет собой половую клетку. Согласно конкретному варианту реализации указанная клетка представляет собой стволовую клетку.
[000108] Согласно одному из аспектов предложена половая клетка мыши, содержащая последовательность нуклеиновой кислоты, кодирующую ADAM6 мыши (или его гомолог, ортолог или функциональный фрагмент), на хромосоме указанной половой клетки, при этом указанная последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши (или его гомолог, ортолог или функциональный фрагмент) находится в положении на указанной хромосоме, отличном от положения в хромосоме половой клетки мыши дикого типа. Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты находится в локусе иммуноглобулина мыши. Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты находится на той же хромосоме половой клетки, что и локус иммуноглобулина мыши. Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты находится не на той хромосоме половой клетки, где расположен указанный локус иммуноглобулина мыши. Согласно одному из вариантов реализации в указанном локусе иммуноглобулина мыши по меньшей мере одна последовательность иммуноглобулина мыши заменена по меньшей мере одной последовательностью не происходящего от мыши иммуноглобулина. Согласно конкретному варианту реализации указанная по меньшей мере одна последовательность не происходящего от мыши го иммуноглобулина представляет собой последовательность иммуноглобулина человека.
[000109] Согласно одному из аспектов предложена плюрипотентная, индуцированная плюрипотентная или тотипотентная клетка, происходящая из мыши согласно настоящему описанию. Согласно конкретному варианту реализации указанная клетка представляет собой эмбриональную стволовую (ЭС) клетку мыши.
[000110] Согласно одному из аспектов предложена ткань, происходящая из мыши согласно настоящему описанию. Согласно одному из вариантов реализации указанную ткань получают из селезенки, лимфатического узла или костного мозга мыши согласно описанию в настоящей заявке.
[000111] Согласно одному из аспектов предложено ядро, происходящее из мыши согласно настоящему описанию. Согласно одному из вариантов реализации указанное ядро взят от диплоидной клетки, не являющейся В-клеткой.
[000112] Согласно одному из аспектов предложена последовательность нуклеотидов, кодирующая вариабельную область иммуноглобулина, синтезируемую мышью согласно настоящему описанию.
[000113] Согласно одному из аспектов предложена аминокислотная последовательность вариабельной области тяжелой цепи иммуноглобулина или легкой цепи иммуноглобулина антитела, синтезируемого мышью согласно настоящему описанию.
[000114] Согласно одному из аспектов предложена последовательность нуклеотидов вариабельной области тяжелой цепи иммуноглобулина или легкой цепи иммуноглобулина, кодирующая вариабельную область антитела, синтезируемого мышью согласно настоящему описанию.
[000115] Согласно одному из аспектов предложено антитело или его антигенсвязывающий фрагмент (например, Fab, F(ab)2, scFv), синтезируемое(ый) мышью согласно настоящему описанию. Согласно одному из аспектов предложен способ получения генетически модифицированной мыши, включающий замену одного или более генных сегментоы тяжелой цепи иммуноглобулина в 5′ направлении (в отношении транскрипции указанных генных сегментов тяжелой цепи иммуноглобулина) от эндогенного локуса ADAM6 указанной мыши одним или более генными сегментами тяжелой цепи иммуноглобулина человека, и замену одного или более сегментов гена иммуноглобулина в 3′-направлении (в отношении транскрипции указанных генных сегментов тяжелой цепи иммуноглобулина) указанного локуса ADAM6 указанной мыши одним или более генными сегментами тяжелой цепи или легкой цепи иммуноглобулина человека. Согласно одному из вариантов реализации указанный один или более сегментов гена иммуноглобулина человека, замещающие один или более сегментов эндогенного гена иммуноглобулина в 5′-направлении от эндогенного локуса ADAM6 указанной мыши, включают генные сегменты V. Согласно одному из вариантов реализации указанные сегменты гена иммуноглобулина человека, замещающие один или более сегментов эндогенного гена иммуноглобулина в 5′-направлении от эндогенного локуса ADAM6 указанной мыши, включают генные сегменты V и D. Согласно одному из вариантов реализации указанный один или более сегментов гена иммуноглобулина человека, замещающие один или более сегментов эндогенного гена иммуноглобулина в 3′-направлении от эндогенного локуса ADAM6 указанной мыши, включают генные сегменты J. Согласно одному из вариантов реализации указанные один или более сегментов гена иммуноглобулина человека, замещающие один или более сегментов эндогенного гена иммуноглобулина в 3′-направлении от эндогенного локуса ADAM6 указанной мыши, включают генные сегменты D и J. Согласно одному из вариантов реализации указанные один или более сегментов гена иммуноглобулина человека, замещающие один или более сегментов эндогенного гена иммуноглобулина в 3′-направлении от эндогенного локуса ADAM6 указанной мыши, включают генные сегменты V, D и J.
[000116] Согласно одному из вариантов реализации указанный один или более генных сегментов тяжелой цепи иммуноглобулина, расположенные в 5′ направлении и/или в 3′-направлении от гена ADAM6, заменяют в плюрипотентной, индуцированной плюрипотентной или тотипотентной клетке для формирования генетически модифицированной клетки-предшественника; указанную генетически модифицированную клетку-предшественник вводят хозяину; указанный хозяин, содержащий генетически модифицированную клетку-предшественник, вынашивается с получением мыши, имеющей геном, полученный из указанной генетически модифицированной клетки-предшественника. Согласно одному из вариантов реализации хозяин представляет собой эмбрион. Согласно конкретному варианту реализации указанный хозяин выбран из предморулы мыши (например, на стадии 8 или 4 клеток), тетраплоидного эмбриона, совокупности эмбриональных клеток или бластоцисты.
[000117] Согласно одному из аспектов предложен способ получения генетически модифицированной мыши, включающий замену последовательности нуклеотидов мыши, содержащей генный сегмент иммуноглобулина мыши и последовательность нуклеотидов ADAM6 мыши (или его ортолога, гомолога или фрагмента, функционального у мышей-самцов), на последовательность, содержащую сегмент гена иммуноглобулина человека, с образованием первого гибридного локуса, и последующее встраивание последовательности, содержащей последовательность, кодирующую ADAM6 мыши (или последовательность, кодирующую его ортолог, гомолог или функциональный фрагмент) в указанную последовательность, содержащую сегмент гена иммуноглобулина человека, с образованием второго гибридного локуса.
[000118] Согласно одному из вариантов реализации указанный второй гибридный локус включает вариабельный генный сегмент тяжелой цепи (VH) иммуноглобулина человека. Согласно одному из вариантов реализации указанный второй гибридный локус включает вариабельный генный сегмент легкой цепи (VL) иммуноглобулина человека. Согласно конкретному варианту реализации указанный второй гибридный локус включает генный сегмент VH человека или генный сегмент VL человека, функционально связанный с генным сегментом DH человека и генным сегментом человека JH. Согласно еще одному конкретному варианту реализации указанный второй гибридный локус функционально связан с третьим гибридным локусом, который содержит последовательность CH1 человека, или CH1 человека и последовательность шарнира человека, слитые с последовательностью CH2+CH3 мыши.
[000119] Согласно одному из аспектов предложено применение мыши, у которой присутствует эктопическая последовательность нуклеотидов, содержащая локус или последовательность ADAM6 мыши для получения фертильного самца мыши, при этом указанное применение включает осуществление спаривания мыши, у которой присутствует указанная эктопическая последовательность нуклеотидов, которая содержит локус или последовательность ADAM6 мыши, с мышью, у которой отсутствует функциональный эндогенный локус или последовательность ADAM6 мыши, и получение потомства, представляющего собой самку, способную производить потомство, у которого присутствует указанный(ая) эктопический локус или последовательность ADAM6, либо представляющее собой самца, у которого присутствует указанный(ая) эктопический ADAM6 локус или последовательность, причем у указанного самца наблюдается фертильность, примерно соответствующая фертильности, наблюдаемой у самцов мыши дикого типа.
[000120] Согласно одному из аспектов предложено применение мыши согласно настоящему описанию для получения последовательности нуклеотидов вариабельной области иммуноглобулина.
[000121] Согласно одному из аспектов предложено применение мыши согласно настоящему описанию для получения полностью человеческого Fab или полностью человеческого F(ab)2.
[000122] Согласно одному из аспектов предложено применение мыши согласно настоящему описанию для получения иммортализованной линии клеток.
[000123] Согласно одному из аспектов предложено применение мыши согласно настоящему описанию для получения гибридомы или квадромы.
[000124] Согласно одному из аспектов предложено применение мыши согласно настоящему описанию для получения фаговой библиотеки, содержащей вариабельные области тяжелой цепи человека и вариабельные области легкой цепи человека.
[000125] Согласно одному из аспектов предложено применение мыши согласно настоящему описанию для получения последовательности вариабельной области для получения антитела человека, включающее (а) иммунизацию мыши согласно настоящему описанию представляющим интерес антигеном, (b) выделение лимфоцита из иммунизированной мыши по пункту (а), (с) осуществление контакта указанного лимфоцита с одному или более мечеными антителами, (d) выявление лимфоцита, способного связываться с представляющим интерес антигеном, и (е) амплификация одной или более последовательностей нуклеиновой кислоты вариабельной области указанного лимфоцита с получением таким образом последовательности вариабельной области.
[000126] Согласно одному из вариантов реализации указанный лимфоцит получают из селезенки мыши. Согласно одному из вариантов реализации указанный лимфоцит получают из лимфатического узла мыши. Согласно одному из вариантов реализации указанный лимфоцит получают из костного мозга мыши.
[000127] Согласно одному из вариантов реализации указанное меченое антитело представляет собой конъюгированное с флуорофором антитело. Согласно одному из вариантов реализации указанные одно или более конъюгированные с флуорофором антитела выбраны из IgM, IgG и/или их комбинации.
[000128] Согласно одному из вариантов реализации указанный лимфоцит представляет собой В-клетку.
[000129] Согласно одному из вариантов реализации указанные одна или более последовательности вариабельной области нуклеиновой кислоты включают последовательность вариабельной области тяжелой цепи. Согласно одному из вариантов реализации указанные одна или более последовательности вариабельной области нуклеиновой кислоты включают последовательность вариабельной области легкой цепи. Согласно конкретному варианту реализации указанная последовательность вариабельной области легкой цепи представляет собой последовательность вариабельной области легкой цепи κ (каппа) иммуноглобулина. Согласно одному из вариантов реализации указанные одна или более последовательности вариабельной области нуклеиновой кислоты содержат последовательности вариабельной области тяжелой цепи и легкой цепи κ.
[000130] Согласно одному из вариантов реализации предложено применение мыши согласно настоящему описанию для получения последовательности вариабельной области тяжелой и κ легкой цепи для получения антитела человека, включающее (а) иммунизацию мыши согласно настоящему описанию представляющим интерес антигеном, (b) извлечение селезенки из иммунизированной мыши по пункту (а), (с) осуществление контакта В-лимфоцитов селезенки с одним или более мечеными антителами, (а) выявление В-лимфоцита по пункту (с), способного связываться с представляющим интерес антигеном, и (е) амплификация последовательности нуклеиновой кислоты вариабельной области тяжелой цепи и последовательности нуклеиновой кислоты вариабельной области κ легкой цепи из указанного В-лимфоцита с получением таким образом указанной последовательностей вариабельной области тяжелой цепи и κ легкой цепи.
[000131] Согласно одному из вариантов реализации предложено применение мыши согласно настоящему описанию для синтеза последовательности вариабельной области тяжелой и κ легкой цепи для получения антитела человека, включающее (а) иммунизацию мыши согласно настоящему описанию представляющим интерес антигеном, (b) извлечение одного или более лимфатических узлов из иммунизированной мыши по пункту (а), (с) осуществление контакта В-лимфоцитов из указанных одного или более лимфатических узлов с одним или более мечеными антителами, (d) выявление В-лимфоцита по пункту (с), способного связываться с представляющим интерес антигеном, и (е) амплификация последовательности нуклеиновой кислоты вариабельной области тяжелой цепи и последовательности нуклеиновой кислоты вариабельной области κ легкой цепи из указанного В-лимфоцита с получением таким образом указанных последовательностей вариабельной области тяжелой цепи и κ легкой цепи.
[000132] Согласно одному из вариантов реализации предложено применение мыши согласно настоящему описанию для получения последовательности вариабельной области тяжелой и κ легкой цепи для получения антитела человека, включающее (а) иммунизацию мыши согласно настоящему описанию представляющим интерес антигеном, (b) извлечение костного мозга из иммунизированной мыши по пункту (а), (с) осуществление контакта В-лимфоцитов костного мозга с одним или более мечеными антителами, (d) выявление В-лимфоцита по пункту (с), способного связываться с представляющим интерес антигеном, и (е) амплификация последовательности нуклеиновой кислоты вариабельной области тяжелой цепи и последовательности нуклеиновой кислоты вариабельной области κ легкой цепи из указанного В-лимфоцита с получением таким образом указанных последовательностей вариабельной области тяжелой цепи и κ легкой цепи. Согласно различным вариантам реализации указанные одно или более меченых антител выбраны из IgM, IgG и/или их комбинации.
[000133] Согласно различным вариантам реализации предложено применение мыши согласно настоящему описанию для синтеза последовательности вариабельной области тяжелой и κ легкой цепи для получения антитела человека, включающее также слияние амплифицированных последовательностей вариабельных областей тяжелой и легкой цепей с последовательностями константной области тяжелой и легкой цепей человека, экспрессирование слитых тяжелой и легкой цепи последовательности в клетке и выделение экспрессированных последовательностей тяжелой и легкой цепей с получением таким образом антитела человека.
[000134] Согласно различным вариантам реализации константные области тяжелой цепи человека выбраны из IgM, IgD, IgA, IgE и IgG. Согласно различным конкретным вариантам реализации указанный IgG выбран из IgG1, IgG2, IgG3 и IgG4. Согласно различным вариантам реализации указанная константная область тяжелой цепи человека включает CH1, шарнир, CH2, CH3, CH4 или их комбинации. Согласно различным вариантам реализации константная область легкой цепи представляет собой константную область к иммуноглобулина. Согласно различным вариантам реализации указанная клетка выбрана из клетки HeLa, клетки DU145, клетки Lncap, клетки MCF-7, клетки MDA-МВ-438, клетки РС3, клетки T47D, клетки ТНР-1, клетки U87, клетки SHSY5Y (нейробластомы человека), клетки Saos-2, клетки Vero, клетки СНО, клетки GH3, клетки РС12, клетки сетчатки человека (например, клетки PER.C6™) и клетки МС3Т3. Согласно конкретному варианту реализации указанная клетка представляет собой клетку СНО.
[000135] Согласно одному из аспектов предложен способ получения обратно-гибридных антител грызуна/человека, специфичных к представляющему интерес антигену, включающий этапы иммунизации мыши согласно приведенному в настоящей заявке описанию указанным антигеном, выделение по меньшей мере одной клетки из мыши, продуцирующей обратно-гибридное антитело мыши/человека, специфичное к указанному антигену, культивирование по меньшей мере одной клетки, синтезирующей указанное обратно-гибридное антитело мыши/человека, специфичное к указанному антигену, и получение указанного антитела.
[000136] Согласно одному из вариантов реализации указанное обратно-гибридное антитело мыши/человека содержит вариабельный домен тяжелой цепи человека, слитый с константным геном тяжелой цепи мыши или крысы, и вариабельный домен легкой цепи человека, слитый с константным геном легкой цепи мыши или крысы или человека.
[000137] Согласно одному из вариантов реализации культивирование по меньшей мере одной клетки, синтезирующей указанное обратно-гибридное антитело грызуна/человека, специфичное к указанному антигену, осуществляют на по меньшей мере одной гибридомной клетке, полученной из указанной по меньшей мере одной выделенной из мыши клетки.
[000138] Согласно одному из аспектов предложен способ получения полностью человеческого антитела, специфичного к представляющему интерес антигену, включающий этапы иммунизации мыши согласно приведенному в настоящей заявке описанию указанным антигеном, выделение по меньшей мере одной клетки из мыши, продуцирующей обратно-гибридное антитело грызуна/человека, специфичное к указанному антигену, получение по меньшей мере одной клетки, продуцирующей полностью человеческое антитело, происходящее из указанного обратно-гибридного антитела грызуна/человека, специфичного к указанному антигену, культивирование по меньшей мере одной клетки, синтезирующей указанное полностью человеческое антитело, и получение указанного полностью человеческого антитела.
[000139] Согласно различным вариантам реализации указанная по меньшей мере одна клетка, выделенная из мыши, продуцирующей обратно-гибридное антитело грызуна/человека, специфичное к указанному антигену, представляет собой спленоцит или В-клетку.
[000140] Согласно различным вариантам реализации указанное антитело представляет собой моноклональное антитело.
[000141] Согласно различным вариантам реализации иммунизацию представляющим интерес антителом проводят белком, ДНК, комбинацией ДНК и белка или клетками, экспрессирующими указанный антиген.
[000142] Согласно одному из аспектов предложено применение мыши согласно настоящему описанию для получения последовательности нуклеиновой кислоты, кодирующей вариабельную область иммуноглобулина, или ее фрагмент. Согласно одному из вариантов реализации указанную последовательность нуклеиновой кислоты применяют для получения антитела человека или его антигенсвязывающего фрагмента. Согласно одному из вариантов реализации указанную мышь применяют для получения антигенсвязывающего белка, выбранного из антитела, мультиспецифичного антитела (например, биспецифического антитела), scFv, биспецифического scFv, диатела, триотела, тетратела, V-NAR, VHH, VL, F(ab), F(ab)2, DVD (т.е. антигенсвязывающий белок с двойным вариабельным доменом), SVD (т.е. антигенсвязывающий белок с одним вариабельным доменом) или биспецифического активатора Т-клеток (BiTE).
[000143] Согласно одному из аспектов предложено применение мыши согласно настоящему описанию для введения эктопической последовательности ADAM6 мыши, у которой отсутствует функциональная эндогенная последовательность ADAM6 мыши, при этом указанное применение включает спаривание мыши согласно настоящему описанию с мышью, у которой отсутствует указанная функциональная эндогенная последовательность ADAM6 мыши.
[000144] Согласно одному из аспектов предложено применение генетического материала мыши согласно настоящему описанию для получения мыши, у которой присутствует эктопическая последовательность ADAM6. Согласно одному из вариантов реализации указанное применение включает ядерную передачу с применением ядра клетки мыши согласно описанию в настоящей заявке. Согласно одному из вариантов реализации указанное применение включает клонирование клетки мыши согласно настоящему описанию для получения животного, происходящего из указанной клетки. Согласно одному из вариантов реализации указанное применение включает применение спермы или яйцеклетки мыши согласно настоящему описанию в способе получения мыши, у которой присутствует указанная эктопическая последовательность ADAM6.
[000145] Согласно одному из аспектов предложен способ получения фертильного самца мыши, у которого присутствует модифицированный локус тяжелой цепи иммуноглобулина, включающий оплодотворение первой половой клетки мыши, содержащей модификацию эндогенного локуса тяжелой цепи иммуноглобулина, второй половой клеткой мыши, содержащей ген ADAM6, либо его ортолог, гомолог или фрагмент, функциональный у мышей-самцов; образование оплодотворенной клетки; обеспечение развития указанной оплодотворенной клетки в эмбрион; и суррогатное вынашивание указанного эмбриона для получения мыши.
[000146] Согласно одному из вариантов реализации оплодотворение достигается посредством спаривания самца мыши с самкой мыши. Согласно одному из вариантов реализации у самки мыши присутствует ген ADAM6, либо его ортолог, гомолог или фрагмент. Согласно одному из вариантов реализации у самца мыши присутствует ген ADAM6, либо его ортолог, гомолог или фрагмент.
[000147] Согласно одному из аспектов предложено применение последовательности нуклеиновой кислоты, кодирующей белок ADAM6 мыши, либо его ортолог или гомолог, либо функциональный фрагмент соответствующего белка ADAM6 для восстановления или повышения фертильности мыши, геном которой содержит модификацию локуса тяжелой цепи иммуноглобулина, причем указанная модификация уменьшает или элиминирует функцию эндогенного ADAM6.
[000148] Согласно одному из вариантов реализации указанную последовательность нуклеиновой кислоты внедряют в геном указанной мыши в эктопическом положении. Согласно одному из вариантов реализации указанную последовательность нуклеиновой кислоты внедряют в геном указанной мыши в эндогенный локус иммуноглобулина. Согласно конкретному варианту реализации указанный эндогенный локус иммуноглобулина представляет собой локус тяжелой цепи. Согласно одному из вариантов реализации указанную последовательность нуклеиновой кислоты внедряют в геном указанной мыши в положении, отличном от положения эндогенного локуса иммуноглобулина.
[000149] Согласно одному из аспектов предложено применение мыши согласно настоящему описанию для получения медикамента (например, антигенсвязывающего белка) или для получения последовательности, кодирующей вариабельную последовательность медикамента (например, антигенсвязывающего белка) для лечения заболевания или расстройства человека.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[000150] На Фиг.1А представлена общая иллюстрация, без соблюдения масштаба, прямой геномной замены приблизительно трех мегабаз (Мб) локуса вариабельных генов тяжелой цепи иммуноглобулина мыши (закрашенные символы) приблизительно одной мегабазой (Мб) локуса вариабельных генов тяжелой цепи иммуноглобулина человека (незакрашенные символы).
[000151] На Фиг.1B представлена общая иллюстрация, без соблюдения масштаба, прямой геномной замены приблизительно трех мегабаз (Мб) локуса вариабельных генов κ легкой цепи мышиного иммуноглобулина (закрашенные символы) приблизительно 0,5 мегабазы (Мб) первого, или проксимального, из двух почти идентичных повторов локуса вариабельных генов κ легкой цепи иммуноглобулина человека (незакрашенные символы).
[000152] На Фиг.2А представлена подробная иллюстрация, без соблюдения масштаба, трех первых этапов (А-С) прямой геномной замены локуса вариабельных генов тяжелой цепи иммуноглобулина мыши, которая приводит к удалению всех генных сегментов VH, DH и JH мыши и замене на три генных сегмента VH человека, все генные сегменты DH и JH человека. Показан направленный вектор для первой вставки генных сегментов тяжелой цепи иммуноглобулина человека (3hVH BACvec) с 67 т.н. 5′-плечом гомологии мыши, селективной кассетой (незакрашенный прямоугольник), участком сайт-специфической рекомбинации (незакрашенный треугольник), геномным фрагментом человека размером 145 т.н. и 3′-плечом гомологии мыши размером 8 т.н. Показаны сегменты гена иммуноглобулина человека (незакрашенные символы) и мыши (закрашенные символы), дополнительные селективные кассеты (незакрашенные прямоугольники) и участки сайт-специфической рекомбинации (незакрашенные треугольники), встраиваемые из последующих направленных векторов.
[000153] На Фиг.2B представлена подробная иллюстрация, без соблюдения масштаба, шести дополнительных этапов (D-I) прямой геномной замены локуса вариабельных генов тяжелой цепи иммуноглобулина мыши, которая приводит к вставке 77 дополнительных генных сегментов VH человека и удалению полученной селективной кассеты. Направленный вектор для встраивания дополнительных генных сегментов VH человека (18hVH BACvec) в первую вставку генных сегментов тяжелой цепи человека (3hVH-CRE гибридный аллель) показан с 20 т.н. 5′ мышиным плечом гомологии, селективной кассетой (незакрашенный прямоугольник), 196 т.н. геномным фрагментом человека и 62 т.н. плечом гомологии человека, перекрывающимся с 5′-концом первой вставки генных сегментов тяжелой цепи человека, показанной с участком сайт-специфической рекомбинации (незакрашенный треугольник), расположенной в 5′ направлении от генных сегментов человека. Показаны сегменты гена иммуноглобулина человека (незакрашенные символы) и мыши (закрашенные символы) и дополнительные селективные кассеты (незакрашенные прямоугольники), встраиваемые с помощью последующих направленных векторов.
[000154] На Фиг.2С представлена подробная иллюстрация, без соблюдения масштаба, трех первых этапов (А-С) прямой геномной замены локус вариабельных генов κ легкой цепи мышиного иммуноглобулина, приводящей к удалению всех генных сегментов Vκ и Jκ мыши (Iqκ-cre гибридный аллель). Показаны селективные кассеты (незакрашенные прямоугольники) и участки сайт-специфической рекомбинации (незакрашенные треугольники), встраиваемые из направленных векторов.
[000155] На Фиг.2D представлена подробная иллюстрация, без соблюдения масштаба, пяти дополнительных этапов (D-H) прямой геномной замены локуса вариабельных генов κ легкой цепи иммуноглобулина мыши, приводящей к вставке всех генных сегментов Vκ и Jκ человека проксимального повтора и удалению полученной селективной кассеты (40hVκdHyg гибридный аллель). Показаны сегменты гена иммуноглобулина человека (незакрашенные символы) и мыши (закрашенные символы) и дополнительные селективные кассеты (незакрашенные прямоугольники), встраиваемые с помощью последующих направленных векторов.
[000156] На Фиг.3А представлена общая иллюстрация, без соблюдения масштаба, стратегии скрининга, включая локализацию наборов праймеров/зондов для количественной ПЦР (кПЦР) для детектирования вставки генной последовательности тяжелой цепи человека и удаления генной последовательности тяжелой цепи мыши в эмбриональных стволовых (ЭС) клетках-мишенях. Стратегия отслеживания первой вставки гена тяжелой цепи человека в ЭС-клеток и у мышей показана в виде наборов праймеров/зондов для кПЦР для удаляемой области (зонды «удаления» С и D), встраиваемой области («hIgH» зонды G и Н) и фланкирующих областей (зонды «сохранения» А, В, Е и F) на немодифицированной хромосоме мыши (сверху) и корректно таргетированной хромосоме (снизу).
[000157] На Фиг.3В приведен типовой расчет регистрируемого числа копий зонда в родительских и модифицированных ЭС-клетках для первой вставки генных сегментов тяжелой цепи иммуноглобулина человека. Число регистрируемых копий зондов A-F рассчитывали ка 2/2ΔΔCt. ΔΔCt рассчитывают как ave[ΔCt(проба) - medΔCt(контроль)], где ΔCt представляет собой разность Ct для экспериментальных и эталонных зондов (от 4 до 6 эталонных зондов, в зависимости от анализа). Термин medΔCt(контроль) означает медиану ΔCt для множества (>60) нетаргетированных образцов ДНК из родительских ЭС-клеток. Каждый клон модифицированной ЭС-клетки анализировали в шести повторностях. Для подсчета числа копий IgH-зондов G и Н в родительских ЭС-клетках число копий указанных зондов принимали за равное 1 в модифицированных ЭС-клетках, и использовали максимальное Ct=35, даже если амплификация не наблюдалась.
[000158] На Фиг.3С приведен типовой расчет числа копий для четырех мышей каждого генотипа, рассчитанного с применением только зондов D и Н. Мыши дикого типа: «мыши WT»; Мыши, гетерозиготные по первой вставке сегментов гена иммуноглобулина человека: «мыши НЕТ»; Мыши, гомозиготные по первой вставке сегментов гена иммуноглобулина человека: «мыши Homo».
[000159] На Фиг.4А представлена подробная иллюстрация, без соблюдения масштаба, трех этапов конструирования 3hVH BACvec с применением бактериальной гомологичной рекомбинации (BHR). Показаны сегменты гена иммуноглобулина человека (незакрашенные символы) и мыши (закрашенные символы), селективные кассеты (незакрашенные прямоугольники) и участки сайт-специфической рекомбинации (незакрашенные треугольники), встроенные из направленных векторов.
[000160] На Фиг.4B показан гель-электрофорез в пульсирующем поле (PFGE) трех клонов ВАС (В1, В2 и В3) после расщепления Notl. Маркеры М1, М2 и М3 представляют собой лэддерные PFG-маркеры короткого диапазона, среднего диапазона и фага лямбда, соответственно (New England BioLabs, Ипсвич, Массачусетс).
[000161] На Фиг.5А представлено схематическое изображение, без соблюдения масштаба, последовательных модификаций локуса тяжелой цепи иммуноглобулина мыши увеличивающимся числом генных сегментов тяжелой цепи иммуноглобулина человека. Гомозиготных мышей получали на каждом из трех разных этапов гуманизации тяжелой цепи. Незакрашенные символы обозначают последовательность человека; закрашенные символы обозначают последовательность мыши.
[000162] На Фиг.5В представлено схематическое изображение, без соблюдения масштаба, последовательных модификаций локуса κ легкой цепи иммуноглобулина мыши увеличивающимся числом генных сегментов κ легкой цепи иммуноглобулина человека. Гомозиготных мышей получали на каждом из трех разных этапов гуманизации κ легкой цепи. Незакрашенные символы обозначают последовательность человека; закрашенные символы обозначают последовательность мыши.
[000163] На Фиг.6 приведены полученные на клеточном сортере с активацией флуоресценции (FACS) точечные диаграммы популяций В-клеток у гуманизированных мышей дикого типа и мышей VELOCIMMUNE®. Клетки селезенки (верхний ряд, третий ряд сверху и нижний ряд) или ингвинальных лимфатических узлов (второй ряд сверху) мышей дикого типа (wt), VELOCIMMUNE® 1 (V1), VELOCIMMUNE® 2 (V2) или VELOCIMMUNE® 3 (V3) окрашивали на экспрессированный на поверхности В-клеток IgM (верхний ряд и второй ряд сверху), поверхностный иммуноглобулин, содержащий либо κ, либо λ, легкие цепи (третий ряд сверху) или поверхностный IgM конкретных гаплотипов (нижний ряд), и популяции разделяли с помощью FACS.
[000164] На Фиг.7А приведены типовые CDR3-последовательности тяжелой цепи случайно отобранных антител VELOCIMMUNE® вокруг VH-DH-JH (CDR3) сочленения, демонстрирующие множественность J-сегментов и добавочные нуклеотиды. Последовательности тяжелой цепи CDR3 сгруппированы в соответствии с частотой использования генных сегментов DH, последовательность которого в зародышевой линии приведена над каждой группой и выделена жирным шрифтом. Генные сегменты VH для каждой последовательности тяжелой цепи CDR3 указаны в скобках на 5′-конце каждой последовательности (например, 3-72 представляет собой человека VH3-72). Генные сегменты JH для каждой тяжелой цепи CDR3 указаны в скобках на 3′-конце каждой последовательности (например, 3 представляет собой JH3 человека). Номера (SEQ ID NO) каждой приведенной последовательности следующие, сверху вниз: SEQ ID NO:21; SEQ ID NO:22; SEQ ID NO:23; SEQ ID NO:24; SEQ ID NO:25; SEQ ID NO:26; SEQ ID NO:27; SEQ ID NO:28; SEQ ID NO:29; SEQ ID NO:30; SEQ ID NO:31; SEQ ID NO:32; SEQ ID NO:33; SEQ ID NO:34; SEQ ID NO:35; SEQ ID NO:36; SEQ ID NO:37; SEQ ID NO:38; SEQ ID NO:39.
[000165] На Фиг.7В приведены типовые последовательности легкой цепи CDR3 случайным образом отобранных антител VELOCIMMUNE® вокруг Vκ-Jκ (CDR3) сочленения, демонстрирующие множественность J-сегментов и добавочные нуклеотиды. Генные сегменты Vκ для каждой последовательности легкой цепи CDR3 указаны в скобках на 5′-конце каждой последовательности (например, 1-6 представляет собой Vκ1-6 человека). Генные сегменты Jκ для каждой легкой цепи CDR3 указаны в скобках на 3′-конце каждой последовательности (например, 1 представляет собой человека Jκ1). Номера (SEQ ID NO) каждой показанной последовательности следующие, сверху вниз: SEQ ID NO:40; SEQ ID NO:41; SEQ ID NO:42; SEQ ID NO:43; SEQ ID NO:44; SEQ ID NO:45; SEQ ID NO:46; SEQ ID NO:47; SEQ ID NO:48; SEQ ID NO:49; SEQ ID NO:50; SEQ ID NO:51; SEQ ID NO:52; SEQ ID NO:53; SEQ ID NO:54; SEQ ID NO:55; SEQ ID NO:56; SEQ ID NO:57; SEQ ID NO:58.
[000166] На Фиг.8 приведены частоты соматических сверхмутаций для тяжелой и легкой цепей антител VELOCIMMUNE®, выраженные (после выравнивания с соответствующими последовательностями зародышевой линии) как процент последовательностей, модифицированных по каждому из положений нуклеотида (NT; левый столбец) или аминокислоты (АА; правый столбец) в наборах из 38 (неиммунизированные IgM), 28 (неиммунизированные IgG), 32 (неиммунизированные Igκ из IgG), 36 (иммунизированные IgG) или 36 (иммунизированные Igκ из IgG) последовательностей. Заштрихованными столбцами обозначено расположение последовательностей CDR.
[000167] На Фиг.9А показаны уровни сывороточного иммуноглобулина для изотипов IgM и IgG у мышей дикого типа (незакрашенные столбцы) или мышей VELOCIMMUNE® (закрашенные столбцы).
[000168] На Фиг.9В показаны уровни сывороточного иммуноглобулина для IgA-изотипа у мышей дикого типа (незакрашенные столбцы) или мышей VELOCIMMUNE® (закрашенные столбцы).
[000169] На Фиг.9С показаны уровни сывороточного иммуноглобулина для IgE изотипа у мышей дикого типа (незакрашенные столбцы) или мышей VELOCIMMUNE® (закрашенные столбцы).
[000170] На Фиг.10А приведены титры антиген-специфичного IgG против рецептора интерлейкина-6 (IL-6R) в сыворотке семи мышей VELOCIMMUNE® (VI) и пяти мышей дикого типа (WT) мыши после двух (проба крови 1) или трех (проба крови 2) циклов иммунизации эктодоменом IL-6R.
[000171] На Фиг.10В приведены титры анти-IL-6R-специфичного изотипа IgG для семи мышей VELOCIMMUNE® (VI) и пяти мышей дикого типа (WT).
[000172] На Фиг.11А показано распределение по аффинности моноклональных антител против рецептора интерлейкина-6, синтезируемых мышами VELOCIMMUNE®.
[000173] На Фиг.11В показано антиген-специфическое связывание моноклональных антител против рецептора интерлейкина-6, синтезируемых мышами VELOCIMMUNE® (VI) и мышами дикого типа (WT).
[000174] На Фиг.12 представлено схематическое изображение, без соблюдения масштаба, генов ADAM6a и ADAM6b мыши в локусе тяжелой цепи иммуноглобулина мыши. Показан направленный вектор (направленный вектор mADAM6), применяемый для вставки ADAM6a и ADAM6b мыши в локус гуманизированной эндогенной тяжелой цепи с селективной кассетой (HYG: гигромицин), фланкированной участками сайт-специфической рекомбинации (Frt), включая сконструированные сайты рестрикции на 5′ и 3′-концах.
[000175] На Фиг.13 представлено схематическое изображение, без соблюдения масштаба, псевдогена ADAM6 человека (hADAM6Ψ), расположенного между вариабельными генными сегментами 1-2 (VH1-2) и 6-1 (VH6-1) тяжелой цепи человека. Показан направленный вектор для бактериальной гомологичной рекомбинации (hADAM6Ψ-направленный вектор) для удаления псевдогена ADAM6 человека и встраивания уникальных сайтов рестрикции в локус тяжелой цепи человека с селективной кассетой (NEO: неомицин), фланкированной участками сайт-специфической рекомбинации (loxP), включая сконструированные сайты рестрикции на 5′- и 3′-концах. Приведена иллюстрация, без соблюдения масштаба, полученного таргетированного локуса гуманизированной тяжелой цепи, содержащего геномный фрагмент, кодирующий гены ADAM6a и ADAM6b мыши, включая селективную кассету, фланкированную участками сайт-специфической рекомбинации.
[000176] На Фиг.14А приведены полученные с применением FACS контурные гистограммы лимфоцитов, гейтированных по одиночным параметрам, для экспрессии на поверхности IgM и В220 в костном мозге мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (H+/+κ+/+), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (H+/+A6resκ+/+). Процент незрелых (B220intIgM+) и зрелых (B220highIgM+) В-клеток указан на каждой контурной гистограмме.
[000177] На Фиг.14В приведено общее число незрелых (B220intIgM+) и зрелых (B220highIgM+) В-клеток в костном мозге, выделенном из бедренных костей мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (H+/+κ+/+), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (H+/+A6resκ+/+).
[000178] На Фиг.15А приведены полученные с применением FACS для гейтированных по CD19+ В-клеток контурные гистограммы поверхностной экспрессии c-kit и CD43 в костном мозге мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (H+/+κ+/+), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (H+/+A6resκ+/+). Процент про-В (CD19+CD43+ckit+) и пре-В (CD19+CD43-ckit-) клеток указан в верхнем правом и нижнем левом квадранте, соответственно, каждой контурной гистограммы.
[000179] На Фиг.15В приведено общее число про-B-клеток (CD19+CD43+ckit+) и пре-В-клеток (CD19+CD43-ckit-) в костном мозге, выделенном из бедренных костей мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (H+/+κ+/+), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (H+/+A6resκ+/+).
[000180] На Фиг.16А приведены полученные с применением FACS для гейтированных по одиночному ограничению лимфоцитов контурные гистограммы экспрессии на поверхности CD19 и CD43 в костном мозге мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (Н+/+κ+/+), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (H+/+A6resκ+/+). Процент незрелых В-(CD19+CD43-), пре-В-(CD19+CD43int) и про-В-(CD19+CD43+) клеток указан на каждой контурной гистограмме.
[000181] На Фиг.16В приведены гистограммы для незрелых В (CD19+CD43-) и пре-В (CD19+CD43int) клеток в костном мозге мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (Н+/+κ+/+), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (Н+/+A6resκ+/+).
[000182] На Фиг.17А приведены полученные с применением FACS для гейтированных по одиночному ограничению лимфоцитов контурные гистограммы экспрессии CD19 и CD3 на поверхности спленоцитов у мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (H+/+κ+/+), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (H+/+A6resκ+/+). Процент В-(CD19+CD3-) и Т-(CD19-CD3+) клеток указан на каждой контурной гистограмме.
[000183] На Фиг.17В приведены полученные с применением FACS для гейтированных по CD19+ В-клеток контурные гистограммы поверхностной экспрессии Igλ и Igκ легкой цепи в селезенке мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (Н+/+κ+/+), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (Н+/+A6resκ+/+). Процент Igλ+ (верхний левый квадрант) и Igκ (нижний правый квадрант) В-клеток указан на каждой контурной гистограмме.
[000184] На Фиг.17С приведено общее число CD19+ В-клеток в селезенке мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (Н+/+κ+/+), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (Н+/+A6resκ+/+).
[000185] На Фиг.18А приведены полученные с применением FACS для гейтированных по CD19+ В-клеток контурные гистограммы экспрессии на поверхности IgD и IgM в селезенке мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (Н+/+κ+/+), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (Н+/+A6resκ+/+). Процент зрелых В клеток (CD19+IgDhighIgMint) указан для каждой контурной гистограммы. Стрелка на правой контурной гистограмме иллюстрирует процесс созревания В-клеток в отношении экспрессии на поверхности IgM и IgD.
[000186] На Фиг.18В приведено общее число В-клеток в селезенке мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (Н+/+κ+/+), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (Н+/+A6resκ+/+) во время созревания CD19+IgMhighIgDint до CD19+IgMintIgDhigh.
[000187] На Фиг.19 приведен титр антител в первой и второй пробах крови мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (Н+/+κ+/+; n=5), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (Н+/+A6resκ+/+; n=5), иммунизированных рецептором клеточной поверхности человека (Антиген А).
[000188] На Фиг.20 приведен титр антител в первой и второй пробах крови мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (Н+/+κ+/+; n=5), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (Н+/+A6resκ+/+; n=10), иммунизированных антителом человека, специфичным к рецепторной тирозиновой протеинкиназе человека (Антиген В).
[000189] На Фиг.21 приведен титр антител в первой и второй пробах крови мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (H+/+κ+/+; n=12), и мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (Н+/+A6resκ+/+; n=12), иммунизированных секретируемым белком человека, участвующим в регуляции TGF-β сигнального пути (Антиген С).
[000190] На Фиг.22 приведен титр антител в первой и второй пробах крови мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6 мыши (Н+/+A6resκ+/+; n=12), иммунизированных рецепторной тирозинкиназой человека (Антиген D).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[000191] Настоящее изобретение не ограничивается описанными конкретными способами и экспериментальными условиями, так как такие способы и условия могут варьировать. Кроме того, следует понимать, что используемая в настоящей заявке терминология предназначена исключительно для описания конкретных вариантов реализации, и не предназначена для ограничения, так как объем настоящего изобретения определен формулой изобретения.
[000192] Если не указано иное, значения всех используемых в настоящей заявке терминов и выражений включают значения, которые указанные термины и выражения имеют в данной области техники, кроме случаев, когда иное прямо указано или прямо следует из контекста, в котором использован(о) термин или выражение. Несмотря на то, что любые способы и материалы, сходные или эквивалентные описанным в настоящей заявке, могут применяться для практического применения или тестирования настоящего изобретения, ниже будут описаны конкретные способы и материалы. Все упоминаемые публикации включены в настоящую заявку посредством ссылки.
[000193] Выражение «по существу» или «практически» при использовании в отношении количества генных сегментов (например, «практически все» V генные сегменты) включает как функциональные, так и нефункциональные генные сегменты и включает, согласно различным вариантам реализации, например, 80% или более, 85% или более, 90% или более, 95% или более 96% или более, 97% или более, 98% или более, или 99% или более всех генных сегментов; согласно различным вариантам реализации термин «практически все» генные сегменты включает, например, по меньшей мере 95%, 96%, 97%, 98%, или 99% функциональных (т.е. не являющихся псевдогенами) генных сегментов.
[000194] Термин «замена» включает введение последовательности ДНК в геном клетки таким образом, что последовательность в составе генома замещается гетерологичной последовательностью (например, последовательностью человека у мыши), в указанном локусе геномной последовательности. Размещенная таким образом последовательность ДНК может включать одну или более регуляторную последовательность, являющуюся частью исходной ДНК, использованной для получения размещенной таким образом последовательности (например, промоторы, энхансеры, 5′-или 3′-нетранслируемые области, подходящие сигнальные последовательности рекомбинации и т.п.). Например, согласно различным вариантам реализации замена представляет собой замену эндогенной последовательности на гетерологичную последовательность, что приводит к синтезу генного продукта из размещенной таким образом последовательности ДНК (включая гетерологичную последовательность), но не к экспрессии эндогенной последовательности; замена представляет собой замену эндогенной геномной последовательности последовательностью ДНК, кодирующей белок, имеющий сходную функцию с белком, кодируемым указанной эндогенной геномной последовательностью (например, указанная эндогенная геномная последовательность кодирует ген или домен иммуноглобулина, а указанный фрагмент ДНК кодирует один или более ген или домен иммуноглобулина человека). Согласно различным вариантам реализации эндогенный ген или его фрагмент заменяют соответствующим геном человека или его фрагментом. Соответствующий ген человека или его фрагмент представляет собой ген или фрагмент гена человека, ортологичный, гомологичный или практически идентичный или такой же по структуре и/или функции, что и замещенный эндогенный ген или его фрагмент.
[000195] Мышь как генетическая модель была значительно усовершенствована с помощью трансгенных и нокаутных методик, позволивших изучать эффекты направленной сверхэкспрессии или удаления конкретных генов. Несмотря на все преимущества, применение мышей по-прежнему связано с генетическими сложностями, которые делают их несовершенной моделью для исследования заболеваний человека и несовершенной платформой для тестирования терапевтических средств для человека или их получения. Во-первых, несмотря на то, что приблизительно 99% генов человека имеют мышиные гомологи (Waterston et al. 2002, Initial sequencing and comparative analysis of the mouse genome, Nature 420:520-562), потенциальные терапевтические средства часто оказываются неспособны к перекрестным реакциям с мышиными ортологами предполагаемых мишеней человека, либо перекрестно реагируют неадекватным образом. Чтобы обойти эту проблему, выбранный целевой ген может быть «гуманизирован», то есть мышиный ген может быть устранен и замещен соответствующей ортологичной генной последовательностью человека (например, US 6586251, US 6596541 и US 7105348, включенные в настоящую заявку посредством ссылки). Сначала попытки гуманизации мышиных генов посредством стратегии «нокаут-плюс-трансгенная гуманизация» включали скрещивание мыши, у которой удален (т.е. нокаутирован) эндогенный ген, с мышью, несущей случайным образом встроенный трансген человека (см., например, Bril et al., 2006, Tolerance to factor VIII in a transgenic mouse expressing human factor VIII cDNA carrying an Arg(593) to Cys substitution, Thromb Haemost 95:341-347; Homanics et al., 2006, Production and characterization of murine models of classic and intermediate maple syrup urine disease, BMC Med Genet 7:33; Jamsai et al., 2006, A humanized ВАС transgenic/knockout mouse model for HbE/beta-thalassemia, Genomics 88(3):309-15; Pan et al., 2006, Different role for mouse and human CD3delta/epsilon heterodimer in preT cell receptor (preTCR) function: human CD3delta/epsilon heterodimer restores the defective preTCR function in CD3gamma- and CD3gammadelta-deficient mice, Mol Immuriol 43:1741-1750). Но эти попытки были затруднены по причине ограничений размеров; стандартные методики нокаутирования не позволяли непосредственно заменять большие гены мыши их большими геномными эквивалентами человека. Простой подход с прямым гомологичным замещением, при котором эндогенный мышиный ген непосредственно заменяют эквивалентным геном человека в том же точно определенном положении в гене мыши (т.е. в эндогенном мышином локусе), редко пытались применять ввиду технических трудностей. До настоящего момента попытки прямой замены включали сложные и обременительные процедуры, которые, соответственно, ограничивали протяженность генетического материала, доступного для обработки, и точность, с которой им можно было манипулировать.
[000196] Экзогенно встраиваемые трансгены иммуноглобулина человека реаранжируются в B-клетках-предшественниках у мышей (Alt et al., 1985, Immunoglobulin genes in transgenic mice, Trends Genet 1:231-236). Это открытие было использовано для конструирования мышей с применением подхода "нокаут плюс трансген" для экспрессирования антител человека (Green et al., 1994, Antigen-specific human monoclonal antibodies from mice engineered with human Ig heavy and light chain YACs, Nat Genet 7:13-21; Lonberg et al., 1994, Antigen-specific human antibodies from mice comprising four distinct genetic modifications, Nature 368:856-859; Jakobovits et al., 2007, From XenoMouse technology to panitumumab, the first fully human antibody product from transgenic mice, Nat Biotechnol 25:1134-1143). Локусы тяжелой цепи и κ легкой цепи иммуноглобулина мыши инактивировали у указанных мышей направленным удалением небольших, но важных фрагментов каждого эндогенного локуса, а затем вводили генные локусы иммуноглобулина человека в виде случайным образом встроенных больших трансгенов, согласно приведенному выше описанию, или минихромосомы (Tomizuka et al., 2000, Double trans-chromosomic mice: maintenance of two individual human chromosome fragments containing Ig heavy and kappa loci and expression of fully human antibodies, PNAS USA 97:722-727). Такие мыши представляли собой важный шаг вперед в области генной инженерии; полученные от них полностью человеческие моноклональные антитела обладали многообещающим терапевтическим потенциалом для лечения различных заболеваний человека (Gibson et al., 2006, Randomized phase III trial results of panitumumab, a fully human anti-epidermal growth factor receptor monoclonal antibody, in metastatic colorectal cancer, Clin Colorectal Cancer 6:29-31; Jakobovits et al., 2007; Kim et al., 2007, Clinical efficacy of zanolimumab (HuMax-CD4): two Phase II studies in refractory cutaneous T-cell lymphoma, Blood 109(11):4655-62; Lonberg, 2005, Human antibodies from transgenic animals, Nat Biotechnol 23:1117-1125; Maker et al., 2005, Tumor regression and autoimmunity in patients treated with cytotoxic Т lymphocyte-associated antigen 4 blockade and interleukin 2: a phase I/II study, Ann Surg Oncol 12:1005-1016; McClung et al., 2006, Denosumab in postmenopausal women with low bone mineral density, New Engl J Med 354:821-831). Однако, как говорилось выше, у указанных мышей наблюдаются дефекты развития В-клеток и иммунодефицит по сравнению с мышами дикого типа. Такие проблемы потенциально ограничивают способность указанных мышей поддерживать интенсивный гуморальный ответ и, как следствие, синтезировать полностью человеческие антитела против некоторых антигенов. Дефицит может быть обусловлен: (1) недостаточной функциональностью из-за случайного встраивания трансгенов иммуноглобулина человека и, в результате, некорректной экспрессии, обусловленной недостатком 5′ и 3′ контролирующих элементов (Garrett et al., 2005, Chromatin architecture near a potential 3′ end of the IgH locus involves modular regulation of histone modifications during B-Cell development and in vivo occupancy at CTCF sites, Mol Cell Biol 25:1511-1525; Manis et al., 2003, Elucidation of a downstream boundary of the 3′ IgH regulatory region, Mol Immunol 39:753-760; Pawlitzky et al., 2006, Identification of a candidate regulatory element within the 5′ flanking region of the mouse IgH locus defined by pro-B cell-specific hypersensitivity associated with binding of PU.1, Pax5, and E2A, J Immunol 176:6839-6851); (2) неэффективным межвидовым взаимодействием между константными доменами человека и мышиными компонентами B-клеточного рецепторного сигнального комплекса на поверхности клетки, что может нарушать процессы сигнализации, требуемые для нормального созревания, пролиферации и выживания В-клеток (Hombach et al., 1990, Molecular components of the B-cell antigen receptor complex of the IgM class, Nature 343:760-762); и (3) неэффективными межвидовыми взаимодействиями между растворимыми иммуноглобулинами человека и Fc-рецепторами мыши, что может ослаблять аффинную селекцию (Rao et al., 2002, Differential expression of the inhibitory IgG Fc receptor FcgammaRIIB on germinal center cells: implications for selection of high-affinity В cells, J Immunol 169:1859-1868) и снижать концентрации иммуноглобулина в сыворотке (Brambell et al., 1964, A Theoretical Model of Gamma-Globulin Catabolism, Nature 203:1352-1354; Junghans and Anderson, 1996, The protection receptor for IgG catabolism is the beta2-microglobulin-containing neonatal intestinal transport receptor, PNAS USA 93:5512-5516; Rao et al., 2002; Hjelm et at., 2006, Antibody-mediated regulation of the immune response, Scand J Immunol 64:177-184; Nimmerjahn and Ravetch, 2007, Fc-receptors as regulators of immunity, Adv Immunol 96:179-204). Указанные недостатки могут быть скорректированы in situ гуманизацией только вариабельных областей локусов иммуноглобулина мыши в их естественных положениях в эндогенных локусах тяжелой и легкой цепи. Это должно приводить к успешному получению мышей, синтезирующих «обратно-гибридные» (т.е. содержащие V-области человека/ С-области мыши) антитела, способные нормально взаимодействовать и проходить селекцию у мышей благодаря сохранению константных областей мыши. Кроме того, такие обратно-гибридные антитела могут с легкостью быть преобразованы в полностью человеческие антитела для терапевтических целей.
[000197] Генетически модифицированные животные, у которых в эндогенном локусе тяжелой цепи иммуноглобулина присутствует замена на гетерологичные (например, другого вида) последовательности иммуноглобулина, могут быть получены в сочетании с заменами в эндогенных локусах легких цепей иммуноглобулина или в сочетании с трансгенами легкой цепи иммуноглобулина (например, гибридными трансгенами легкой цепи иммуноглобулина, или полностью человеческими, полностью мышиными, и т.п.). Виды, из которых происходят гетерологичные последовательности тяжелой цепи иммуноглобулина, могут быть различными, как и в случае последовательностей легкой цепи иммуноглобулина, используемых для замены последовательностей легкой цепи иммуноглобулина или в качестве трансгенов легкой цепи иммуноглобулина.
[000198] Последовательности нуклеиновой кислоты вариабельных областей иммуноглобулина, например, V, D и/или J сегменты, получают согласно различным вариантам реализации от человека или от не являющегося человеком животного. Не являющиеся человеком животные, подходящие для получения V, D и/или J сегментов, включают, например костистых рыб, хрящевых рыб, таких как акулы и скаты, амфибий, рептилий, млекопитающих, птиц (например, куры). Не являющиеся человеком животные включают, например, млекопитающих. Указанные млекопитающие включают, например, не являющихся человеком приматов, козьих, баранов, свиней, собак, крупный рогатый скот (например, корову, быка, буйвола), оленьих, верблюдов, куньих и грызунов, и не являющихся человеком приматов (таких как шимпанзе, орангутаны, гориллы, игрунковые, макаки-резусы, бабуины). Подходящие не являющиеся человеком животные выбраны из семейства грызунов, включая крыс, мышей и хомяков. Согласно одному из вариантов реализации указанные не являющиеся человеком животные представляют собой мышей. Как очевидно из контекста, различные не являющиеся человеком животные могут применяться в качестве источника вариабельных доменов или генных сегментов вариабельной области (например, акулы, скаты, млекопитающие (например, верблюды, грызуны, такие как мыши и крысы).
[000199] В соответствии с контекстом не являющихся человеком животных также применяют в качестве источников последовательностей константной области для применения в сочетании с вариабельными последовательностями или сегментами; например, в трансгенах могут быть использованы константные последовательности грызуна, функционально связанные с вариабельными последовательностями человека или не происходящими от человека (например, вариабельные последовательности человека или не являющегося человеком примата функционально связаны, например, с константными последовательностями грызуна, например, мыши, крысы или хомяка). Соответственно, согласно различным вариантам реализации V, D, и/или J сегменты человека функционально связаны с последовательностями генов константной области грызуна(например, мыши, крысы или хомяка). Согласно некоторым вариантам реализации V, D, и/или J сегменты человека (или один или более реаранжированные VDJ или VJ гены) функционально связаны или слиты с последовательностью гена константной области мыши, крысы, или хомяка, например, в трансгене, встроенном в локус, не являющийся эндогенным локусом иммуноглобулина.
[000200] Согласно конкретному варианту реализации предложена мышь, у которой VH, DH и JH сегменты в эндогенном локусе тяжелой цепи иммуноглобулина заменены на один или более сегментовы VH, DH и JH человека, при этом указанный один или более сегментов VH, DH и JH человека функционально связаны с геном тяжелой цепи эндогенного иммуноглобулина; при этом указанная мышь содержит трансген в локусе, отличном от эндогенного локуса иммуноглобулина, при этом указанный трансген содержит переаранжированный или реаранжированный сегмент VL человека и JL человека, функционально связанный с константной областью мыши, крысы или человека.
[000201] Описан способ масштабной in situ генетической замены вариабельных локусов генов иммуноглобулина зародышевой линии мыши на вариабельные локусы генов иммуноглобулина зародышевой линии человека при поддержании способности указанных мышей производить потомство. В частности, описана точная замена шести мегабаз локусов вариабельных генов как тяжелой цепи, так и κ легкой цепи иммуноглобулина мыши их человеческими эквивалентами, при сохранении интактных константных областей мыши. В итоге получены мыши с точной заменой всего репертуара вариабельных иммуноглобулиновых последовательностей зародышевой линии эквивалентными вариабельными иммуноглобулиновыми последовательностями зародышевой линии человека, при сохранении константных областей мыши. Вариабельные области иммуноглобулинов человека соединяют с константными областями иммуноглобулинов мыши с образованием гибридных иммуноглобулиновых локусов человек/мышь, которые реаранжируются и экспрессируются на нормальном физиологическом уровне. Экспрессируемые антитела представляют собой «обратные гибриды», т.е. они содержат последовательности вариабельных областей иммуноглобулинов человека и последовательности константных областей иммуноглобулинов мыши. Таких мышей, у которых имеются гуманизированные вариабельные области иммуноглобулина, экспрессирующих антитела с вариабельными областями человека и константными областями мыши, называют мышами VELCOIMMUNE®.
[000202] У гуманизированных мышей VELOCIMMUNE® наблюдается полностью функциональная гуморальная иммунная система, практически неотличимая от таковой мышей дикого типа. У них наблюдаются нормальные клеточные популяции на всех стадиях развития В-клеток. У них наблюдается нормальная морфология лимфоидных органов. В последовательностях антител мышей VELOCIMMUNE® наблюдается нормальная V(D)J-реаранжировка и нормальная частота соматических сверхмутаций. Популяции антител у указанных мышей отражают распределение изотипов в результате нормального переключения классов (например, нормального cis-переключения изотипов). Иммунизация мыши VELOCIMMUNE® приводит к мощному гуморальному иммунному ответу с генерацией больших разнообразных репертуаров антител, содержащих вариабельные домены иммуноглобулина человека, подходящие для применения в качестве кандидатных терапевтических средств. Эта платформа обеспечивает богатый источник последовательностей вариабельных областей иммуноглобулина человека с естественно созревающей аффинностью для получения фармацевтически приемлемых антител и других антигенсвязывающих белков.
[000203] Получение мышей VELOCIMMUNE® возможно за счет точной замены вариабельных последовательностей иммуноглобулина мыши на вариабельные последовательности иммуноглобулина человека. Однако даже точная замена эндогенных последовательностей иммуноглобулина мыши в локусах тяжелой и легкой цепей на эквивалентные последовательности иммуноглобулина человека, последовательным рекомбинированием очень больших участков последовательностей иммуноглобулина человека, может представлять определенные проблемы в связи с дивергентной эволюции иммуноглобулиновых локусов у мыши и человека. Например, межгенные последовательности, рассеянные по иммуноглобулиновым локусам, не идентичны у мышей и человека и, в определенных обстоятельствах, могут не быть функционально эквивалентными. Различия в иммуноглобулиновых локусах мыши и человека все равно могут приводить к аномалиям у гуманизированных мышей, в частности, при гуманизации или манипулировании определенными фрагментами эндогенных локусов тяжелой цепи иммуноглобулина мыши. Некоторые модификации в локусах тяжелой цепи иммуноглобулина мыши оказывают пагубное воздействие. Оказывающие пагубное воздействие модификации могут включать, например, потерю способности модифицированных мышей к спариванию и производству потомства.
[000204] Осуществляли точную крупномасштабную in situ замену шести мегабаз вариабельных областей тяжелой и легкой цепи иммуноглобулиновых локусов мыши (VH-DH-JH и Vκ-Jκ) соответствующими последовательностями генома человека размером 1,4 мегабазы, оставляя интактными и функциональными фланкирующие мышиные последовательности в гибридных локусах, в том числе все мышиные гены константных цепей и локус-контролирующие области (ФИГ.1А и ФИГ.1В). В частности, генные последовательности VH, DH, JH, Vκ и Jκ человека вводили, поэтапно встраивая 13 гибридных направленных векторов ВАС, несущих перекрывающиеся фрагменты локусов вариабельных областей зародышевой линии человека, в мышиные ЭС-клетки с применением генноинженерной технологии VELOCIGENE® (см., например, патент США №6586251 и Valenzuela et al., 2003, High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nat Biotechnol 21:652-659).
[000205] Гуманизация генов иммуноглобулина мыши представляет собой самую крупную генетическую модификацию генома мыши на сегодняшний день. В то время как предыдущие попытки, задействовавшие случайным образом встроенные трансгены иммуноглобулина человека, увенчались некоторым успехом (см. выше), прямая замена генов иммуноглобулина мыши эквивалентами человека резко увеличивает эффективность синтеза полностью человеческих антител у нормальных во всех прочих отношениях мышей. Далее, у таких мышей наблюдается резкое увеличение разнообразия полностью человеческих антител, чего можно добиться иммунизацией практически любым антигеном, по сравнению с мышами, у которых содержатся деактивированные эндогенные локусы и трансгены полностью человеческих антител. Многочисленные варианты с замещенными гуманизированными локусами демонстрируют полностью нормальные уровни зрелых и незрелых В-клеток, в отличие от мышей со случайным образом встроенными трансгенами человека, у которых наблюдается значительное уменьшение популяций В-клеток на разных стадиях дифференцировки. В то время как попытки увеличить число генных сегментов человека в несущих трансгены человека мышах уменьшили указанные дефекты, расширение репертуара иммуноглобулинов не полностью устранило сокращение популяций В-клеток по сравнению с мышами дикого типа.
[000206] Несмотря на практически дикий тип гуморального иммунитета, наблюдаемый у мышей с замещенными иммуноглобулиновыми локусами (т.е. мышей VELOCIMMUNE®), существуют и другие проблемы, возникающие при использовании прямой замены иммуноглобулина, не свойственные некоторым подходам, при которых используют случайным образом встроенные трансгены. Различия в генетическом составе иммуноглобулиновых локусов мыши и человека привело к открытию последовательностей, благоприятных для размножения мышей с замещенными генными сегментами иммуноглобулина. В частности, оптимально, чтобы гены ADAM мыши, расположенные в эндогенном локусе иммуноглобулина, присутствовали у мышей с замещенными иммуноглобулиновыми локусами, по причине их значения для фертильности.
Положение в геноме и функция ADAM6 мыши
[000207] У мышей-самцов с отсутствием способности к экспрессии любого функционального белка ADAM6 неожиданным образом наблюдается дефект способности к спариванию и производству потомства. У указанных мышей отсутствует способность к экспрессии функционального белка ADAM6 вследствие замены всех или практически всех генных сегментов вариабельных областей иммуноглобулина мыши на генные сегменты вариабельных областей иммуноглобулинов человека. Утрата функции ADAM6 происходит ввиду того, что локус ADAM6 расположен внутри генного локуса вариабельной области тяжелой цепи эндогенного иммуноглобулина мыши, проксимального в отношении 3′-конца локуса VH генного сегмента, расположенного в 5′-направлении от генных сегментов DH. Получение самцов и самок, каждый из которых гомозиготен по указанной замене, и ожидание продуктивного спаривания обычно является трудоемким способом размножения мышей, гомозиготных по замене всех или практически всех эндогенных вариабельных генных сегментов тяжелой цепи мыши на вариабельные генные сегменты тяжелой цепи человека. Успешные пометы редки и немногочисленны. Вместо этого использовали самцов, гетерозиготных по указанной замене, для спаривания с самками, гомозиготными по указанной замене, для получения потомства, гетерозиготного по указанной замене, затем при его размножении получали гомозиготную мышь. Авторы изобретения обнаружили, что вероятной причиной утраты фертильности у мышей-самцов является отсутствие у гомозиготных мышей-самцов функционального белка ADAM6.
[000208] Согласно различным аспектам у мышей-самцов с поврежденным (т.е. нефункциональным или ограниченно функциональным) геном ADAM6 наблюдается снижение или полное подавление фертильности. Так как у мышей (и других грызунов) ген ADAM6 расположен в локусе тяжелой цепи иммуноглобулина, авторы изобретения выяснили, что для размножения мышей, или для создания и поддержания линии мышей с замещенным локусом тяжелой цепи иммуноглобулина, применяются различные модифицированные схемы скрещивания или размножения. Низкая фертильность или бесплодность мышей-самцов, гомозиготных по замене локуса вариабельного гена тяжелой цепи эндогенного иммуноглобулина, усложняет сохранение такой модификации в линии мышей. Согласно различным вариантам реализации поддержание линии включает обход проблем с бесплодностью, наблюдаемой у мышей-самцов, гомозиготных по указанной замене.
[000209] Согласно одному из аспектов предложен способ поддержания линии мышей согласно настоящему описанию. Указанная линия мышей не нуждается в присутствии эктопической последовательности ADAM6, и, согласно различным вариантам реализации, указанная линия мышей гомозиготна или гетерозиготна по нокауту (например, функциональному нокауту) ADAM6.
[000210] У указанной линии мышей присутствует модификация эндогенного локуса тяжелой цепи иммуноглобулина, приводящая к снижению или утрате фертильности у мышей-самцов. Согласно одному из вариантов реализации указанная модификация включает удаление регуляторной области и/или кодирующей области гена ADAM6. Согласно конкретному варианту реализации указанная модификация включает модификацию эндогенного гена ADAM6 (регуляторной и/или кодирующей области), которая уменьшает или элиминирует фертильность мышей-самцов, у которых присутствует указанная модификация; согласно конкретному варианту реализации указанная модификация уменьшает или элиминирует фертильность у мышей-самцов, гомозиготных по указанной модификации.
[000211] Согласно одному из вариантов реализации указанная линия мышей гомозиготна или гетерозиготна по нокауту (например, функциональному нокауту) или делеции гена ADAM6.
[000212] Согласно одному из вариантов реализации указанную линию мышей поддерживают посредством извлечения из мыши, гомозиготной или гетерозиготной по указанной модификации, клетки, введение указанной донорской клетки в эмбрион хозяин, вынашивание указанного эмбриона хозяина с донорской клеткой суррогатной матерью и получение от суррогатной матери потомства, у которого присутствует указанная генетическая модификация. Согласно одному из вариантов реализации донорская клетка представляет собой ЭС-клетку. Согласно одному из вариантов реализации донорская клетка представляет собой плюрипотентную клетку, например, индуцированную плюрипотентную клетку.
[000213] Согласно одному из вариантов реализации линию мышей поддерживают посредством извлечения из мыши, гомозиготной или гетерозиготной по указанной модификации, последовательности нуклеиновой кислоты, содержащей указанную модификацию, и введение указанной последовательности нуклеиновой кислоты в ядро хозяина, и вынашивание клетки, содержащей последовательность нуклеиновой кислоты и ядро хозяина, подходящим животным. Согласно одному из вариантов реализации указанную последовательность нуклеиновой кислоты вводят в ооцит хозяина.
[000214] Согласно одному из вариантов реализации указанную линию мышей поддерживают посредством извлечения из мыши, гомозиготной или гетерозиготной по указанной модификации, ядра, и введения указанного ядра в клетку хозяин, вынашивание указанного ядра и клетки хозяина подходящим животным для получения потомства, гомозиготного или гетерозиготного по указанной модификации.
[000215] Согласно одному из вариантов реализации указанную линию мышей поддерживают с помощью применения in vitro оплодотворения (IVF) самки мыши (дикого типа, гомозиготной по указанной модификации или гетерозиготной по указанной модификации) спермой самца мыши, содержащей указанную генетическую модификацию. Согласно одному из вариантов реализации самец мыши гетерозиготен по указанной генетической модификации. Согласно одному из вариантов реализации самец мыши гомозиготен по указанной генетической модификации.
[000216] Согласно одному из вариантов реализации указанную линию мышей поддерживают с помощью скрещивания самца мыши, гетерозиготного по указанной генетической модификации с самкой мыши для получения потомства, у которого присутствует указанная генетическая модификация, выявление в потомстве самцов и самок, у которых присутствует указанная генетическая модификация, и применение самца, гетерозиготного по указанной генетической модификации для скрещивания с самкой дикого типа, гомозиготной или гетерозиготной по указанной генетической модификации для получения потомства, у которого присутствует указанная генетическая модификация. Согласно одному из вариантов реализации этап скрещивания самца, гетерозиготного по указанной генетической модификации, с самкой дикого типа, самкой, гетерозиготной по указанной генетической модификации, или самкой, гомозиготной по указанной генетической модификации, повторяют с целью сохранения указанной генетической модификации в данной линии мышей.
[000217] Согласно одному из аспектов предложен способ поддержания линии мышей, включающий замену локуса вариабельного гена тяжелой цепи эндогенного иммуноглобулина одной или более последовательностями тяжелой цепи иммуноглобулина человека, включающий разведение указанной линии мышей для получения гетерозиготных мышей-самцов, где указанные гетерозиготные мыши-самцы размножаются для сохранения указанной генетической модификации в данной линии. Согласно конкретному варианту реализации указанную линию не поддерживают путем какого-либо скрещивания гомозиготного самца с самкой дикого типа, или самкой, гомозиготной или гетерозиготной по указанной генетической модификации.
[000218] Белок ADAM6 является членом семейства белков ADAM, где ADAM представляет собой сокращение от «А-дезинтегрин и металлопротеаза». Семейство белков ADAM многочисленно и разнообразно, выполняет различные функции, включая клеточную адгезию. Некоторые члены семейства ADAM задействованы в сперматогенезе и фертилизации. Например, ADAM2 кодирует субъединицу белка фертилина, задействованного во взаимодействии спермия и яйцеклетки. ADAM3, или циритестин, по-видимому, необходим для связывания спермы с zona pellucida. Отсутствие любого из ADAM2 или ADAM3 приводит к бесплодию. Высказывалось предположение, что ADAM2, ADAM3, и ADAM6 образуют комплекс на поверхности спермиев мыши. Ген ADAM6 человека, который в норме находится между генными сегментами VH VH1-2 и VH6-1 человека, по-видимому, является псевдогеном (Фиг.12). У мышей присутствует два гена ADAM6 - ADAM6a и ADAM6b, которые расположены в межгенной области между генными сегментами VH и DH мыши, и у мышей ориентация транскрипции генов ADAM6a и ADAM6b противоположна относительно окружающих сегментов гена иммуноглобулина (ФИГ.12). У мышей функциональный локус ADAM6 с очевидностью необходим для нормальной фертилизации. Функциональный ADAM6 локус или последовательность в этом случае относится к локусу или последовательности ADAM6, способной увеличивать или восстанавливать существенно сниженную фертильность, наблюдаемую у мышей-самцов с отсутствующими или нефункциональными эндогенными локусами ADAM6.
[000219] Положение межгенной последовательности, кодирующей ADAM6a и ADAM6b, у мышей позволяет модифицировать указанную межгенную последовательность при модифицировании эндогенной тяжелой цепи мыши. Если генные сегменты VH удалены или заменены, либо если генные сегменты DH удалены или заменены, имеется высокая вероятность того, что у полученной мыши будет наблюдаться серьезное снижение фертильности. Чтобы компенсировать указанное снижение, мышь модифицируют, вводя последовательность нуклеотидов, кодирующую белок, восстанавливающий активность ADAM6, утраченную в результате модификации эндогенного локуса ADAM6 мыши. Согласно различным вариантам реализации комплементирующая последовательность нуклеотидов представляет собой последовательность, кодирующую ADAM6a мыши, ADAM6b мыши, или их гомолог, ортолог или функциональный фрагмент, восполняющий дефицит фертильности.
[000220] Последовательность нуклеотидов, восстанавливающая фертильность может быть размещена в любом подходящем положении. Она может находиться в межгенной области, или в любом подходящем положении в геноме (т.е. эктопически). Согласно одному из вариантов реализации указанная последовательность нуклеотидов может быть введена в трансген, который случайным образом встраивают в геном мыши. Согласно одному из вариантов реализации указанная последовательность может располагаться эписомно, то есть в нуклеиновой кислоте вне хромосомы мыши. Подходящие положения включают положения, транскрипционно пермиссивные или транскрипционно активные, например, локус ROSA26 (Zambrowicz et al., 1997, PNAS USA 94:3789-3794), a BT-5 locus (Michael et al., 1999, Mech. Dev. 85:35-47), или локус Oct4 (Wallace et al., 2000, Nucleic Acids Res. 28:1455-1464). Нацеливание нуклеотидных последовательностей на транскрипционно активные локусы описаны, например, в US 7473557, включенном в настоящую заявку посредством ссылки.
[000221] Как вариант, последовательность нуклеотидов, восстанавливающая фертильность, может сочетаться с индуцируемым промотором таким образом, чтобы способствовать оптимальной экспрессии, в подходящих клетках и/или тканях, например, репродуктивных тканях. Типовые индуцируемые промоторы включают промоторы, активируемые физическими (например, промотор теплового шока) и/или химическими факторами (например, ИПТГ или тетрациклином).
[000222] Далее, экспрессия указанной последовательности нуклеотидов могут быть связаны с другими генами таким образом, чтобы добиться экспрессии на конкретных стадиях развития или в конкретных тканях. Такой экспрессии можно добиться установлением функциональной связи последовательности нуклеотидов с промотором гена, экспрессируемого на конкретной стадии развития. Например, последовательности иммуноглобулина из одного вида, встроенные в геном вида-хозяина, функционально связывают с последовательностью промотора гена CD19 (гена, специфичного для В-клетки) вида-хозяина. Достигается специфичная для В-клеток экспрессия на точно определенных стадиях развития, когда экспрессируются иммуноглобулины.
[000223] Еще один способ достижения мощной экспрессии встроенной последовательности нуклеотидов состоит в применении конститутивного промотора. Типовые конститутивные промоторы включают SV40, CMV, UBC, EF1A, PGK и CAGG. Сходным образом, требуемая последовательность нуклеотидов функционально связана с выбранным конститутивным промотором, который обеспечивает высокий уровень экспрессии белка(ов), кодируемых указанной нуклеотидной последовательностью.
[000224] Предполагается, что термин «эктопический» включает перемещение, или размещение в положении, в норме не встречающемся в природе (например, размещение последовательности нуклеиновой кислоты в положении, не совпадающем с положением, которое указанная последовательность нуклеиновой кислоты занимает у мыши дикого типа). Указанный термин, согласно различным вариантам реализации, применяют для обозначения объекта, находящегося вне его нормального, или присущего ему, положения. Например, выражение «эктопическая последовательность нуклеотидов, кодирующая…» относится к последовательности нуклеотидов, находящейся не в том положении, в котором она обычно встречается у мыши. Например, в случае эктопической последовательности нуклеотидов, кодирующей белок ADAM6 мыши (или его ортолог, гомолог или фрагмент, оказывающий такое же или сходное благоприятное действие на фертильность мышей-самцов), указанная последовательность может быть размещена в положении в геноме мыши, не совпадающем с положением, в котором она в норме обнаруживается у мыши дикого типа. В таких случаях за счет размещения указанной последовательности в геноме мыши в положении, отличном от положения у мыши дикого типа, образуется новое сочленение последовательности мыши. Функциональный гомолог или ортолог ADAM6 мыши представляет собой последовательность, восстанавливающую утраченную фертильность (например, потерю способности мышей-самцов производить потомство посредством спаривания), наблюдаемую у мыши ADAM6-/-. Функциональные гомологи или ортологи включают белки, идентичные по меньшей мере приблизительно на 89% или более, например, до 99%, аминокислотной последовательности ADAM6a и/или аминокислотной последовательности ADAM6b, и способные улучшать или восстанавливать способность успешно спариваться у мыши, генотип которой включает удаление или нокаут ADAM6a и/или ADAM6b.
[000225] Эктопическим положением может быть любое положение (например, при случайном встраивании трансгена, содержащего последовательность ADAM6 мыши), или, например, положение, приблизительно соответствующее (но не совпадающее) с расположением у мыши дикого типа (например, в модифицированном эндогенном локусе иммуноглобулина мыши, но в 5′ направлении либо в 3′-направлении от естественного положения, например, внутри модифицированного иммуноглобулинового локуса, но между другими генными сегментами, или в другом положении в межгенной последовательности мыши V-D). Одним из примеров эктопического расположения является расположение в локусе тяжелой цепи гуманизированного иммуноглобулина. Например, мышь, у которой один или более эндогенные генные сегменты VH заменены на генные сегменты VH человека, причем в результате указанной замены удаляется эндогенная последовательность ADAM6, может быть сконструирована таким образом, что у нее присутствует последовательность ADAM6 мыши, расположенная внутри последовательности, включающей генные сегменты VH человека. Полученная модификация будет давать (эктопическую) последовательность ADAM6 мыши внутри генной последовательности человека, и указанное (эктопическое) размещение последовательности ADAM6 мыши в генной последовательности человека может приблизительно соответствовать положению псевдогена человека ADAM6 (т.е. между двумя V сегментами), либо может приблизительно соответствовать положению последовательности ADAM6 мыши (т.е. внутри V-D межгенной области). Границы полученной последовательности, образованной при присоединении (эктопической) последовательности ADAM6 мыши к генной последовательности человека (например, последовательности гена иммуноглобулина) или в непосредственной близости от нее в зародышевой линии мыши, будут новыми относительно того же или сходного положения в геноме мыши дикого типа.
[000226] Согласно различным вариантам реализации предложены не являющиеся человеком животные, у которых отсутствует ADAM6 или его ортолог или гомолог, при этом указанное отсутствие приводит к бесплодности указанного не являющегося человеком животного, или существенно уменьшает фертильность указанного не являющегося человеком животного. Согласно различным вариантам реализации отсутствие ADAM6 или его ортолога или гомолога является результатом модификации эндогенного локуса тяжелой цепи иммуноглобулина. Существенное снижение фертильности представляет собой, например, снижение фертильности (например, частоты скрещивания, числа детенышей в помете, количества пометов в год, и т.п.) приблизительно на 50%, 60%, 70%, 80%, 90%, или 95% или более. Согласно различным вариантам реализации указанным не являющимся человеком животным вводят ген ADAM6 мыши, либо его ортолог, гомолог или функциональный фрагмент, функциональный у самцов указанного не являющегося человеком животного, при этом указанный добавленный ген ADAM6 либо его ортолог, гомолог или функциональный фрагмент восстанавливает сниженную фертильность полностью или в значительной степени. Восстановление фертильности в значительной степени представляет собой, например, такое восстановление фертильности, при котором у не являющегося человеком животного наблюдается фертильность, составляющая по меньшей мере 70%, 80% или 90%, или более от фертильности при немодифицированном (т.е. у животного без модификации гена ADAM6, или его ортолога или гомолога) локусе тяжелой цепи.
[000227] Последовательность, придающая генетически модифицированному животному (т.е. животному, у которого отсутствует функциональный ADAM6 или его ортолог или гомолог, например, в результате модификации локуса тяжелой цепи иммуноглобулина) выбрана, согласно различным вариантам реализации, из гена ADAM6 или его ортолога или гомолога. Например, у мыши утрата функции ADAM6 восстанавливается добавлением, согласно одному из вариантов реализации, мышиного гена ADAM6. Согласно одному из вариантов реализации утраченная функция ADAM6 у мыши восстанавливается при добавлении ортолога или гомолога близкородственного для указанной мыши вида, например, грызуна, в частности, мыши другой линии или вида, крысы любого вида, грызуна; при этом указанное добавление ортолога или гомолога мыши восстанавливает фертильность, утраченную в результате утраты функции ADAM6 или утраты гена ADAM6. Ортологи и гомологи других видов согласно различным вариантам реализации выбраны из филогенетически родственных видов и, согласно различным вариантам реализации, демонстрируют процент идентичности эндогенному ADAM6 (или ортологу), составляющий приблизительно 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, или 97% или более; и восстанавливают фертильность, утрата которой связана с ADAM6 или (у не-мышей) с ортологом ADAM6. Например, у генетически модифицированного самца крысы, у которого отсутствует функция ADAM6 (например, крысы, у которой вариабельная область тяжелой цепи эндогенного иммуноглобулина заменена на вариабельную область тяжелой цепи иммуноглобулина человека, или с нокаутом в области тяжелой цепи иммуноглобулина крысы), утраченная фертильность восстанавливается при добавлении ADAM6 крысы, либо, согласно некоторым вариантам реализации, ортолога ADAM6 крысы (например, ортолога ADAM6 другой линии или вида крыс, или, согласно одному из вариантов реализации, мыши).
[000228] Соответственно, согласно различным вариантам реализации у генетически модифицированных животных, у которых наблюдается отсутствие фертильности или снижение фертильности из-за модификации последовательности нуклеиновой кислоты, кодирующей белок ADAM6 (или его ортолог или гомолог) или регуляторной области, функционально связанной с указанной последовательностью нуклеиновой кислоты, присутствует последовательность нуклеиновой кислоты, которая восполняет, или восстанавливает, утрату фертильности, при этом указанная последовательность нуклеиновой кислоты, которая восполняет или восстанавливает утрату фертильности, взята из другой линии того же вида или из филогенетически родственного вида. Согласно различным вариантам реализации указанная комплементирующая последовательность нуклеиновой кислоты представляет собой ортолог, гомолог или функциональный фрагмент ADAM6. Согласно различным вариантам реализации указанный комплементирующий ортолог, гомолог или функциональный фрагмент ADAM6 взят от не являющегося человеком животного, близкородственного генетически модифицированному животному, у которого присутствует дефект фертильности. Например, если генетически модифицированное животное представляет собой мышь конкретной линии, ортолог, гомолог или функциональный фрагмент ADAM6 может быть получен из мыши другой линии или мыши родственного вида. Согласно одному из вариантов реализации в случае, когда указанное генетически модифицированное животное с дефектом фертильности относится к порядку Rodentia, ортолог или гомолог ADAM6 или его функциональный фрагмент взят от другого животного порядка Rodentia. Согласно одному из вариантов реализации указанное генетически модифицированное животное с дефектом фертильности относится к подотряду Myomoropha (например, тушканчики, мышовковые, мышевидные хомяки, хомяки, американские крысы и мыши, полевки, истинные мыши и крысы, песчанки, иглистые мыши, косматые хомяки, лазающие мыши, скальные крысы, белохвостые крысы, мадагаскарские крысы и мыши, колючие сони, кроты, бамбуковые крысы, цокоры), а ортолог или гомолог ADAM6, или его функциональный фрагмент, выбран из таковых от животного порядка Rodentia или подотряда Myomorpha.
[000229] Согласно одному из вариантов реализации указанное генетически модифицированное животное относится к надсемейству Dipodoidea, и ортолог или гомолог ADAM6 или его функциональный фрагмент происходит из надсемейства Muroidea. Согласно одному из вариантов реализации указанное генетически модифицированное животное относится к надсемейству Muroidea, и ортолог или гомолог ADAM6 или его функциональный фрагмент происходит из надсемейства Dipodoidea.
[000230] Согласно одному из вариантов реализации указанное генетически модифицированное животное представляет собой грызуна. Согласно одному из вариантов реализации указанный грызун выбран из надсемейства Muroidea, и ортолог или гомолог ADAM6 взят от другого вида надсемейства Muroidea. Согласно одному из вариантов реализации указанное генетически модифицированное животное относится к семейству, выбранному из Calomyscidae (например, мышевидные хомяки), Cricetidae (например, хомяк, американские крысы и мыши, полевки), Muridae (истинные мыши и крысы, песчанки, иглистые мыши, косматые хомяки), Nesomyidae (лазающие мыши, скальные крысы, белохвостые крысы, мадагаскарские крысы и мыши), Platacanthomyidae (например, колючие сони), и Spalacidae (например, кроты, бамбуковые крысы и цокоры); и ортолог или гомолог ADAM6 выбран из другого вида того же семейства. Согласно конкретному варианту реализации указанный генетически модифицированный грызун выбран из истинной мыши или крысы (семейство Muridae), и ортолог или гомолог ADAM6 взят от вида, выбранного из песчанки, иглистой мыши или косматый хомяк. Согласно одному из вариантов реализации генетически модифицированная мышь является представителем семейства Muridae, и ортолог или гомолог ADAM6 взят от другого вида семейства Muridae. Согласно конкретному варианту реализации генетически модифицированный грызун представляет собой мышь семейства Muridae, и ортолог или гомолог ADAM6 взят от крысы, песчанки, иглистой мыши или косматого хомяка из семейства Muridae.
[000231] Согласно различным вариантам реализации один или более ортологов или гомологов ADAM6 грызуна из какого-либо семейства, или их функциональные фрагменты, восстанавливают фертильность генетически модифицированного грызуна того же семейства, у которого отсутствует ортолог или гомолог ADAM6 (например, Cricetidae (например, хомяки, американские крысы и мыши, полевки); Muridae (например, истинные мыши и крысы, песчанки, иглистые мыши, косматые хомяки).
[000232] Согласно различным вариантам реализации оценивают функциональность ортологов, гомологов ADAM6 и их фрагментов, удостоверяясь, что указанный ортолог, гомолог или фрагмент восстанавливает фертильность генетически модифицированного самца не являющегося человеком животного, у которого отсутствует активность ADAM6 (например, грызуна, в частности, мыши или крысы, у которой присутствует нокаут ADAM6 или его ортолога). Согласно различным вариантам реализации функциональность определяют как способность спермы генетически модифицированного животного, у которого отсутствует эндогенный ADAM6 или его ортолог или гомолог, перемещаться по маточным трубам и оплодотворять яйцеклетку того же вида генетически модифицированного животного.
[000233] Согласно различным аспектам могут быть получены мыши, у которых локус вариабельной области эндогенной тяжелой цепи или его фрагменты удален(ы) или содержи(а)т замены, у которых присутствует эктопическая последовательность нуклеотидов, кодирующую белок, оказывающий благоприятное влияние на фертильность, сходное с таковым ADAM6 мыши (например, его ортолог, гомолог или фрагмент, функциональный у мышей-самцов). Указанная эктопическая последовательность нуклеотидов может включать последовательность нуклеотидов, кодирующую белок, представляющий собой гомолог или ортолог ADAM6 (или его фрагмент) другой линии мышей или другого вида, например, другого вида грызунов, и оказывающую благоприятное действие фертильность, например, увеличение количества пометов за определенный период времени, и/или увеличение количества детенышей в помете, и/или способность спермия мышей-самцов проходить через маточные трубы мыши для оплодотворения мышиной яйцеклетки.
[000234] Согласно одному из вариантов реализации ADAM6 представляет собой гомолог или ортолог, по меньшей мере на 89%-99% идентичный белку ADAM6 мыши (например, по меньшей мере на 89%-99% идентичный ADAM6a мыши или ADAM6b мыши). Согласно одному из вариантов реализации эктопическая последовательность нуклеотидов кодирует один или более белков, независимо выбранные из белка, по меньшей мере на 89% идентичного ADAM6a мыши, белка, по меньшей мере на 89% идентичного ADAM6b мыши, и их комбинации. Согласно одному из вариантов реализации указанный гомолог или ортолог представляет собой белок крысы, хомяка, мыши или морской свинки, который является, либо модифицирован таким образом, чтобы быть приблизительно на 89% или более идентичным ADAM6a мыши и/или ADAM6b мыши. Согласно одному из вариантов реализации идентичность указанного гомолога или ортолога в отношении ADAM6a мыши и/или ADAM6b мыши равна либо составляет по меньшей мере 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, или 99%.
Эктопическая ADAM6 в гуманизированной тяжелой цепи мыши
[000235] Развитие методов направленного воздействия на гены, например, разработка искусственных бактериальных хромосом (ВАС), в настоящее время позволяет рекомбинировать достаточно большие геномные фрагменты. Конструирование ВАС обеспечило возможность удалять и встраивать большие участки в мышиные ЭС-клетки.
[000236] Мыши, синтезирующие антитела человека, существуют уже какое-то время. Несмотря на то, что такие мыши представляют собой важный шаг вперед в разработке терапевтических средств для человека на основе антител, у них наблюдается ряд существенных аномалий, ограничивающих их применимость. Например, у них наблюдается дефекты развития В-клеток. Указанные дефекты развития могут быть результатом разнообразных отличий трансгенных мышей и мышей дикого типа.
[000237] Антитела человека могут не оптимальным образом взаимодействовать с мышиными пре-В-клеточными или В-клеточными рецепторами на поверхности клеток мыши, подающими сигналы для созревания, пролиферации или выживания при селекции клонов. Полностью человеческие антитела могут не оптимальным образом взаимодействовать с Fc-рецепторной системой мыши; мыши экспрессируют Fc-рецепторы, у которых не наблюдается взаимно однозначного соответствия с Fc-рецепторами человека. Наконец, у некоторых мышей, синтезирующих полностью человеческие антитела, имеются не все природные последовательности мыши, например, 3′-энхансерные элементы и другие локус-контролирующие элементы, которые могут требоваться для развития В-клеток дикого типа.
[000238] У мышей, синтезирующих полностью человеческие антитела, как правило, имеются эндогенные иммуноглобулиновые локусы, деактивированные тем или иным образом, и трансгены человека, содержащие вариабельные и константные сегменты гена иммуноглобулина, вводят в случайное положение в геноме мыши. При условии, что указанный эндогенный локус в достаточной степени деактивирован, так что не происходит реаранжировка генных сегментов с образованием функционального гена иммуноглобулина, можно добиться синтеза полностью человеческих антител такой мышью, хоть и наряду с дефектами развития В-клеток.
[000239] Несмотря на успешность принудительного синтеза полностью человеческого антитела из трансгенного локуса человека, синтез антител человека у мышей представляет собой очевидно неблагоприятный процесс. У некоторых мышей указанный процесс настолько неблагоприятен, что приводит к образованию гибридных тяжелых цепей с вариабельными областями иммуноглобулинов человека/константными областями мыши (но не легких цепей) за счет механизма trans-переключения. За счет этого механизма транскрипты, кодирующие полностью человеческие антитела, подвергаются изотипическому trans-переключению от изотипа человека к изотипу мыши. Указанный процесс представляет собой trans-процесс, поскольку полностью человеческий трансген расположен вне эндогенного локуса, содержащего незатронутую копию гена константной области тяжелой цепи мыши. Хотя у таких мышей trans-переключение выражено очевидным образом, указанное явление все же не обеспечивает корректировку развития В-клеток, которое остается выражение дефектным. В любом случае, образованные при trans-переключении антитела, синтезируемые такими мышами, сохраняют полностью человеческие легкие цепи, так как явление trans-переключения, очевидно, не затрагивает легкие цепи; trans-переключение предположительно зависит от последовательностей переключения в эндогенных локусах, используемых (хотя и по-разному) при нормальном cis-переключении изотипов. Соответственно, даже в случае, если у мышей, сконструированных для получения полностью человеческого антитела, при синтезе антител с константными областями мыши выбирается механизм trans-переключения, указанная стратегия не способна обеспечить восстановление нормального развития В-клеток.
[000240] Главной задачей при создании терапевтических средств на основе антител человека является получение достаточно большого разнообразия последовательностей вариабельных областей иммуноглобулина человека для выявления подходящих вариабельных доменов, которые, в частности, распознают конкретные эпитопы и связывают их с требуемым сродством, обычно, но не всегда, с высоким сродством. До получения мышей VELOCIMMUNE® (описанных в настоящей заявке) не было каких-либо данных, указывающих на то, что у мышей, экспрессирующих вариабельные области иммуноглобулинов человека с константными областями мыши будут наблюдаться какие-либо существенные отличия от мышей, синтезировавших антитела человека из трансгена. Однако это предположение было неверным.
[000241] У мышей VELOCIMMUNE® с точной заменой вариабельных областей иммуноглобулина мыши вариабельными областями иммуноглобулина человека в эндогенных локусах мыши наблюдается неожиданное примечательное сходство с мышами дикого типа в отношении развития В-клеток. Неожиданным и удивительным образом, у мышей VELOCIMMUNE® наблюдался практически нормальный ответ дикого типа на иммунизацию, отличавшийся от ответа мышей дикого типа только одним важным качеством - вариабельные области, синтезируемые в ответ на иммунизацию, были полностью человеческими.
[000242] В эндогенных локусах мышей VELOCIMMUNE® проводят точную крупномасштабную замену вариабельных областей зародышевой линии тяжелой цепи иммуноглобулина (IgH) и легкой цепи иммуноглобулина (например, κ легкой цепи, Igκ) мыши на соответствующие вариабельные области иммуноглобулина человека. Всего приблизительно шесть мегабаз локусов мыши заменяют приблизительно 1,5 мегабазами геномной последовательности человека. Указанная точная замена приводит к получению мыши с гибридными иммуноглобулиновыми локусами, синтезирующей тяжелой и легкие цепи, содержащие вариабельные области человека и константную область мыши. При указанной точной замене мышиных VH-DH-JH и Vκ-Jκ сегментов фланкирующие последовательности мыши остаются интактными и функциональными в указанных гибридных иммуноглобулиновых локусах. Гуморальная иммунная система указанных мышей функционирует сходным с системой мыши дикого типа образом. Развитие В-клеток происходит беспрепятственно в любом существенном отношении, и у указанной мыши синтезируется большое разнообразие вариабельных областей иммуноглобулинов человека при стимуляции антигеном.
[000243] Получение мышей VELOCIMMUNE® возможно благодаря тому, что сегменты гена иммуноглобулина тяжелой и κ легкой цепей реаранжируются сходным образом у человека и у мыши, что не означает, что их локусы идентичны или хотя бы близки к этому - определенно это не так. Тем не менее, указанные локусы сходны в достаточной степени для того, чтобы обеспечить возможность гуманизации локуса генов вариабельной области тяжелой цепи посредством замены приблизительно трех миллионов п.о. непрерывной последовательности мыши, содержащей все генные сегменты VH, DH и JH, приблизительно одним миллионом оснований непрерывной геномной последовательности человека, в целом охватывающим эквивалентную последовательность локуса иммуноглобулина человека.
[000244] Согласно некоторым вариантам реализации дальнейшая замена определенных генных последовательностей константной области мыши генными последовательностями человека (например, замена последовательности CH1 мыши последовательностью CH1 человека, и замена последовательности CL мыши последовательностью CL человека) приводит к получению мышей с гибридными иммуноглобулиновыми локусами, синтезирующих антитела, содержащие вариабельные области иммуноглобулинов человека и частично человеческие константные области, подходящие, например, для получения фрагментов полностью человеческого антитела, например, полностью человеческих Fab. У мышей с гибридными иммуноглобулиновыми локусами наблюдается нормальная реаранжировка вариабельных генных сегментов, нормальная частота соматических сверхмутаций и нормальное переключение классов. У указанных мышей наблюдается гуморальная иммунная система, неотличимая мышей дикого типа, и нормальные клеточные популяции на всех стадиях развития В-клеток и нормальная структура лимфоидных органов - даже в тех случаях, когда у мышей отсутствует полный репертуар генных сегментов вариабельных областей человека. Иммунизация таких мышей приводит к мощному гуморальному ответу, при котором наблюдается большое разнообразие использования вариабельных генных сегментов.
[000245] Точная замена генных сегментов вариабельных областей зародышевой линии мыши позволяет получить мышей, у которых имеются частично человеческие иммуноглобулиновые локусы. Так как указанные частично человеческие иммуноглобулиновые локусы проходят нормальные реаранжировку, гипермутацию и переключение классов, указанные частично человеческие иммуноглобулиновые локусы дают у мыши антитела, содержащие вариабельные области иммуноглобулинов человека. Нуклеотидные последовательности, кодирующие указанные вариабельные области, могут быть идентифицированы и клонированы, затем слиты (например, в in vitro системе) с любыми выбранными последовательностями, например, иммуноглобулина любого изотипа, подходящими для конкретного применения, с получением антитела или антигенсвязывающего белка, происходящей полностью из последовательностей человека.
[000246] Для модификации мышиных эмбриональных стволовых (ЭС) клеток с точной заменой до трех мегабаз локуса тяжелой цепи иммуноглобулина мыши, включающих практически все генные сегменты VH, DH и JH мыши, эквивалентными генными сегментами человека с применением до 1 мегабазы геномной последовательности человека, содержащей некоторые или практически все генные сегменты VH, DH и JH человека, применяли крупномасштабную гуманизацию с помощью методов рекомбинационной инженерии. Сегмент генома человека размером до половины мегабазы, содержащий один из двух повторов, кодирующих практически все генные сегменты Vκ и Jκ человека, использовали для замены сегмента локуса κ легкой цепи иммуноглобулина мыши размером три мегабазы, содержащего практически все генные сегменты Vκ и Jκ мыши.
[000247] У мышей с такими замещенными иммуноглобулиновыми локусами может быть разрушен или удален эндогенный локус ADAM6 мыши, в норме обнаруживаемый между крайним с 3′-стороны VH генным сегментом и крайним с 5′-конца DH генным сегментом в указанном локусе тяжелой цепи иммуноглобулина мыши. Разрушение этой области может приводить к снижению или полному подавлению функциональности эндогенного локуса ADAM6 мыши. В случае, если при замене используют крайние с 3′-стороны генные сегменты VH репертуара тяжелых цепей человека, межгенная область, содержащая псевдоген, по-видимому, представляющий собой псевдоген ADAM6 человека, располагается между указанными генными сегментами VH, т.е. между VH1-2 и VH1-6 человека. Как бы то ни было, у мышей-самцов, у которых присутствует указанная межгенная последовательность человека, наблюдается снижение фертильности.
[000248] Описаны мыши, у которых имеются замещенные локусы согласно приведенному выше описанию, а также эктопическая последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши, причем у указанных мышей наблюдается практически нормальная фертильность. Согласно одному из вариантов реализации указанная эктопическая последовательность нуклеиновой кислоты содержит последовательность ADAM6a мыши и/или ADAM6b мыши, либо его/их функциональные фрагменты, расположенные между VH1-2 человека и VH6-1 человека в модифицированном эндогенном локусе тяжелой цепи. Согласно одному из вариантов реализации указанная эктопическая последовательность нуклеиновой кислоты представляет собой последовательность SEQ ID NO:3, расположенную между VH1-2 человека и VH6-1 человека в модифицированном эндогенном локусе тяжелой цепи. Направление транскрипции гена ADAM6 в последовательности SEQ ID NO:3 противоположно направлению транскрипции окружающих генных сегментов VH человека. Хотя в настоящей заявке приведены примеры восстановления фертильности путем размещения эктопической последовательности между указанными генными сегментами VH человека, специалистам будет ясно, что размещение указанной эктопической последовательности в любом подходящем транскрипционно-пермиссивном локусе в геноме мыши (либо даже экстрахромосомно) предположительно будет сходным образом восстанавливать фертильность у мышей-самцов.
[000249] Процесс добавления мыши, у которой отсутствует функциональный ADAM6 локус, эктопической последовательности, которая содержит ген ADAM6 мыши, либо его ортолог, гомолог или функциональный фрагмент, представляет собой стандартный способ, применяемый для компенсации нефункциональности или минимальной функциональности эндогенных локусов ADAM6 у любых мышей. Соответственно, у значительного числа мышей, у которых присутствует разрушающая ADAM6 модификация локуса тяжелой цепи иммуноглобулина, это может компенсировано с помощью применения композиций и способов согласно настоящему изобретению. Соответственно, в настоящем изобретении предложены мыши с широким спектром модификаций локусов тяжелой цепи иммуноглобулина, негативно сказывающихся на функции эндогенного ADAM6. Некоторые (неограничивающие) примеры приведены в настоящем описании. Помимо описываемых мышей VELOCIMMUNE®, композиции и способы, связанные с ADAM6, могут иметь множество применений, например, при модифицировании разнообразными способами локуса тяжелой цепи.
[000250] Согласно одному из аспектов предложена мышь, у которой присутствует эктопическая последовательность ADAM6, кодирующая функциональный белок ADAM6 (или его ортолог, гомолог или функциональный фрагмент); все или практически все генные сегменты VH мыши заменены одним или более генными сегментами VH человека; все или практически все генные сегменты DH и генные сегменты JH мыши заменены на генные сегменты DH человека и JH человека; при этом у указанной мыши отсутствует область CH1 и/или область шарнира. Согласно одному из вариантов реализации указанная мышь продуцирует связывающий белок с одним вариабельным доменом, представляющий собой димер цепей иммуноглобулина, выбранных из: (a) VH человека -CH1 мыши -CH2 мыши -CH3 мыши; (b) VH человека - шарнир мыши -CH2 мыши -CH3 мыши; и (с) VH человека -CH2 мыши -CH3 мыши.
[000251] Согласно одному из аспектов последовательность нуклеотидов, восстанавливающая фертильность, располагают в пределах последовательности вариабельной области тяжелой цепи иммуноглобулина человека (например, между генными сегментами человека VH1-2 и VH1-6) у мыши, у которой присутствует замена одного или более генных сегментов вариабельной области тяжелой цепи иммуноглобулина мыши (mVH, mDH и/или mJH) одним или более генными сегментами вариабельной области тяжелой цепи иммуноглобулина человека (hVH, hDH и/или hJH), и у указанной мыши также присутствует замена одного или более генных сегментов вариабельной области легкой цепи к иммуноглобулина мыши (mVκ и/или mJκ) одним или более генными сегментами вариабельной области легкой цепи к иммуноглобулина человека (hVκ и/или hJκ).
[000252] Согласно одному из вариантов реализации указанные один или более вариабельные генные сегменты тяжелой цепи иммуноглобулина мыши содержат приблизительно три мегабазы локуса тяжелой цепи иммуноглобулина мыши. Согласно одному из вариантов реализации указанные один или более вариабельные генные сегменты тяжелой цепи иммуноглобулина мыши содержат по меньшей мере 89 генных сегментов VH, по меньшей мере 13 генных сегментов DH, по меньшей мере четыре генных сегмента JH или комбинацию указанных сегментов локуса тяжелой цепи иммуноглобулина мыши. Согласно одному из вариантов реализации указанный один или более вариабельные генные сегменты тяжелой цепи иммуноглобулина человека содержат приблизительно одну мегабазу локуса тяжелой цепи иммуноглобулина человека. Согласно одному из вариантов реализации указанные один или более вариабельные генные сегменты тяжелой цепи иммуноглобулина человека содержат по меньшей мере 80 генных сегментов VH, по меньшей мере 27 генных сегментов DH, по меньшей мере шесть генных сегментов JH или комбинацию таковых локуса тяжелой цепи иммуноглобулина человека.
[000253] Согласно одному из вариантов реализации указанные один или более вариабельные генные сегменты легкой цепи к иммуноглобулина мыши содержат приблизительно три мегабазы локуса κ легкой цепи иммуноглобулина мыши. Согласно одному из вариантов реализации указанные один или более вариабельные генные сегменты легкой цепи киммуноглобулина мыши содержат по меньшей мере 137 генных сегментов Vκ, по меньшей мере пять генных сегментов Jκ или комбинацию таковых локуса κ легкой цепи иммуноглобулина мыши. Согласно одному из вариантов реализации указанные один или более вариабельные генные сегменты легкой цепи к иммуноглобулина человека содержат приблизительно 1/2 мегабазы локус легкой цепи к иммуноглобулина человека. Согласно конкретному варианту реализации указанные один или более вариабельные генные сегменты легкой цепи к иммуноглобулина человека содержат проксимальные повторы (относительно константной области κ иммуноглобулина) локуса легкой цепи κ иммуноглобулина человека. Согласно одному из вариантов реализации указанный один или более вариабельный генный сегмент легкой цепи κ иммуноглобулина человека содержит по меньшей мере 40 генных сегментов Vκ, по меньшей мере пять генных сегментов Jκ, или комбинацию указанных сегментов локуса легкой цепи к иммуноглобулина человека.
[000254] Согласно одному из вариантов реализации указанная последовательность нуклеотидов расположена между двумя сегментами гена иммуноглобулина человека. Согласно конкретному варианту реализации указанные два сегмента гена иммуноглобулина человека представляют собой генные сегменты тяжелой цепи. Согласно одному из вариантов реализации указанная последовательность нуклеотидов расположена между генными сегментами VH1-2 человека и генными сегментами VH1-6 человека у мыши VELOCIMMUNE® (US 6596541 и US 7105348, включенные в настоящую заявку посредством ссылки). Согласно одному из вариантов реализации у мыши VELOCIMMUNE®, модифицированной таким образом, вариабельные генные сегменты тяжелой цепи иммуноглобулина мыши заменены на по меньшей мере 80 генных сегментов VH человека, 27 генных сегментов DH человека и шесть генных сегментов JH человека, и вариабельные генные сегменты κ легкой цепи иммуноглобулина мыши заменены на по меньшей мере 40 генных сегментов Vκ человека и пять генных сегментов Jκ человека.
[000255] Согласно одному из аспектов функциональный локус ADAM6 мыши (или его ортолог, гомолог или функциональный фрагмент) расположен среди генных сегментов VH человека, замещающих эндогенные генные сегменты VH мыши. Согласно одному из вариантов реализации по меньшей мере 89 генных сегментов VH мыши удаляют и заменяют одним или более генными сегментами VH человека, и локус ADAM6 мыши непосредственно примыкает к 3′-концу указанных генных сегментов VH человека, либо находится между двумя генными сегментами VH человека. Согласно конкретному варианту реализации локус ADAM6 мыши находится между двумя генными сегментами VH в пределах приблизительно от 20 тыс.нуклеотидов (т.н.) до приблизительно 40 тыс.нуклеотидов (т.н.) от 3′-конца встроенных генных сегментов VH человека. Согласно конкретному варианту реализации локус ADAM6 мыши находится между двумя генными сегментами VH в пределах приблизительно от 29 т.н. до приблизительно 31 т.н. от 3′-конца встроенных генных сегментов VH человека. Согласно конкретному варианту реализации локус ADAM6 мыши находится в пределах приблизительно от 30 т.н. от 3′-конца встроенных человека генных сегментов VH. Согласно конкретному варианту реализации локус ADAM6 мыши находится в пределах приблизительно 30 184 п.о. от 3′-конца встроенных генных сегментов VH человека. Согласно конкретному варианту реализации замена включает генные сегменты VHVH1-2 и VH6-1 человека, и локус ADAM6 мыши находится в 3′-направлении от генного сегмента VH1-2 и в 5′-направлении от генного сегмента VH6-1. Согласно конкретному варианту реализации локус ADAM6 мыши находится между генным сегментом VH1-2 человека и генным сегментом VH6-1 человека, при этом 5′-конец локуса ADAM6 мыши располагается приблизительно на расстоянии 13848 п.о. от 3′-конца генного сегмента VH1-2 человека, и 3′-конец локуса ADAM6 располагается приблизительно на расстоянии 29737 п.о. в 5′ направлении от генного сегмента VH6-1 человека. Согласно конкретному варианту реализации локус ADAM6 мыши содержит SEQ ID NO:3 или ее фрагмент, обеспечивающий функцию ADAM6 в клетках мыши. Согласно конкретному варианту реализации генные сегменты VH человека в этом случае расположены следующим образом (от 5′-стороны до 3′-стороны относительно направления транскрипции генных сегментов человека VH): VH1-2 человека - локус ADAM6 мыши - VH6-1 человека. Согласно конкретному варианту реализации псевдоген ADAM6 между VH1-2 человека и VH6-1 человека заменяют локусом ADAM6 мыши. Согласно одному из вариантов реализации ориентация одного или более из ADAM6a мыши и ADAM6b мыши локуса ADAM6 мыши противоположна в отношении направления транскрипции ориентации генных сегментов VH человека. Как вариант, локус ADAM6 мыши находится в межгенной области между крайним с 3′-стороны генным сегментом VH человека и крайним с 5′-конца генным сегментом DH. В этом случае указанный крайний с 5′-конца DH сегмент может быть либо мышиным, либо человеческим.
[000256] Сходным образом, мышь, модифицированная одним или более генными сегментами VL человека (например, сегментами Vκ или Vλ), замещающими все или практически все эндогенные генные сегменты VH мыши, может быть модифицирована таким образом, что эндогенный локус ADAM6 мыши либо сохраняется согласно приведенному выше описанию, например, с помощью применения направленного вектора с 3′-плечом гомологии, включающим локус ADAM6 мыши или его функциональный фрагмент, или поврежденный локус ADAM6 мыши заменяется эктопической последовательностью, расположенной между двумя генными сегментами VL человека или между генными сегментами VL человека и генным сегментом DH (человека или мыши, например, Vλ+m/hDH), или генным сегментом J (человека или мыши, например, Vκ+JH). Согласно одному из вариантов реализации указанная замена включает два или более генных сегментов VL человека, и локус ADAM6 мыши или его функциональный фрагмент находится между двумя крайними с 3′-стороны генными сегментами VL. Согласно конкретному варианту реализации генные сегменты VL человека в этом случае расположены следующим образом (от 5′ до 3′ относительно направления транскрипции генных сегментов человека): VL3′-1 человека - локус ADAM6 мыши -VL3′ человека. Согласно одному из вариантов реализации ориентация одного или более из ADAM6a мыши и ADAM6b мыши локуса ADAM6 мыши противоположна в отношении направления транскрипции ориентации генных сегментов VL человека. Как вариант, локус ADAM6 мыши находится в межгенной области между крайним с 3′-стороны генным сегментом VL человека и крайним с 5′-конца генным сегментом DH. В этом случае указанный крайний с 5′-конца DH сегмент может представлять собой мышиный или человеческий.
[000257] Согласно одному из аспектов предложена мышь с заменой одного или более эндогенных генных сегментов VH мыши, и содержащая по меньшей мере один эндогенный генный сегмент DH мыши. У такой мыши модификация эндогенных генных сегментов VH мыши может включать модификацию одного или более крайних с 3′-стороны генных сегментов VH, но не крайнего с 5′-конца DH генного сегмента, при этом принимаются меры, направленные на то, чтобы указанная модификация одного или более крайнего с 3′-стороны генного сегмента VH не разрушила или не привела к нефункциональности эндогенного локуса ADAM6 мыши. Например, согласно одному из вариантов реализации у указанной мыши заменены все или практически все эндогенные генные сегменты VH мыши одним или более генными сегментами VH человека, и указанная мышь содержит один или более эндогенные генные сегменты DH и функциональный эндогенный локус ADAM6 мыши.
[000258] Согласно другому варианту реализации у указанной мыши модифицированы эндогенные крайние с 3′-стороны генные сегменты мыши VH, и один или более эндогенные генные сегменты Он мыши, и указанную модификацию осуществляют таким образом, чтобы сохранить целостность указанного эндогенного локуса ADAM6 мыши в степени, достаточной для поддержания функциональности эндогенного локуса ADAM6. Согласно одному из примеров такую модификацию осуществляют за два этапа: (1) заменяют крайние с 3′-стороны эндогенные генные сегменты VH мыши одним или более генными сегментами VH человека с применением направленного вектора с 5′-плечом гомологии и 3′-плечом гомологии, причем указанное 3′-плечо гомологии включает полностью или частично функциональный локус ADAM6 мыши; (2) затем заменяют и эндогенный генный сегмент DH мыши с помощью направленного вектора, содержащего 5′-плечо гомологии, включающее весь локус или функциональную часть локуса ADAM6 мыши.
[000259] Согласно различным аспектам применение мыши, у которой присутствует эктопическая последовательность, кодирующая белок ADAM6 мыши, либо его ортолог, гомолог или функциональный гомолог, целесообразно в тех случаях, когда введение модификаций нарушает функционирование эндогенного ADAM6 мыши. Вероятность разрушения эндогенной функции ADAM6 мыши высока при модификации локусов иммуноглобулина мыши, в частности, при модифицировании последовательностей вариабельных областей тяжелых цепей иммуноглобулина мыши и окружающих областей. Соответственно, применение таких мышей особенно целесообразно при получении мышей с локусами тяжелых цепей иммуноглобулина, удаленными полностью или частично, гуманизированными полностью или частично, или замещенными (например, последовательностями Vκ или Vλ) полностью или частично. Способы получения указанных генетических модификаций у мышей, описываемых ниже, известны специалистам в данной области техники.
[000260] Применение мышей, у которых присутствует эктопическая последовательность, кодирующая мышиный белок ADAM6, или практически идентичный или сходный белок, обладающий благоприятными для фертильности свойствами мышиного белка ADAM6, особенно целесообразно в сочетании с модификациями локуса вариабельных генов тяжелой цепи иммуноглобулина мыши, разрушающими или удаляющими эндогенную последовательность ADAM6 мыши. Несмотря на то, что такие мыши в первую очередь описаны в контексте экспрессирования ими антител с вариабельными областями иммуноглобулинов человека и константными областями иммуноглобулинов мыши, они подходят для применения при любых генетических модификациях, разрушающих эндогенные гены ADAM6 мыши. Специалистам будет ясно, что сюда включен широкий спектр генетически модифицированных мышей, у которых имеются модификации вариабельных генных локусов тяжелой цепи иммуноглобулина мыши. Сюда включены, например, мыши, у которых удалены или заменены полностью или частично генные сегменты тяжелой цепи иммуноглобулина мыши, независимо от других модификаций. Неограничивающие примеры приведены ниже.
[000261] Согласно некоторым аспектам предложены генетически модифицированные мыши, у которых присутствует эктопический ген ADAM6 (или ортолог, гомолог или фрагмент) мыши, грызуна или др., функциональный у мыши, и один или более генных сегментов вариабельной и/или константной области иммуноглобулина человека. Согласно различным вариантам реализации другие ортологи, гомологи или фрагменты гена ADAM6, функциональные у мыши, могут включать последовательности крупного рогатого скота, собачьих, приматов, кролика, или другие последовательности не происходящие от человека.
[000262] Согласно одному из аспектов предложена мышь, у которой присутствует эктопическая последовательность ADAM6, кодирующая функциональный белок ADAM6; все или практически все генные сегменты VH мыши заменены одним или более генными сегментами VH человека; все или практически все генные сегменты DH мыши заменены одним или более генными сегментами DH человека; и все или практически все генные сегменты JH мыши заменены одним или более генными сегментами JH человека.
[000263] Согласно одному из вариантов реализации у указанной мыши также присутствует замена последовательности нуклеотидов CH1 мыши нуклеотидной последовательностью CH1 человека. Согласно одному из вариантов реализации у указанной мыши также присутствует замена последовательности нуклеотидов шарнирной области иммуноглобулина мыши нуклеотидной последовательностью шарнирной области иммуноглобулина человека. Согласно одному из вариантов реализации у указанной мыши также присутствует замена локуса вариабельной области легкой цепи иммуноглобулина (VL и JL) на локус вариабельной области легкой цепи иммуноглобулина человека. Согласно одному из вариантов реализации у указанной мыши также присутствует замена последовательности нуклеотидов константной области легкой цепи иммуноглобулина мыши последовательностью нуклеотидов константной области легкой цепи иммуноглобулина человека. Согласно конкретному варианту реализации указанные VL, JL, и CL представляют собой последовательности κ легкой цепи иммуноглобулина. Согласно конкретному варианту реализации указанная мышь содержит последовательности CH2 мыши и CH3 мыши константной области иммуноглобулина, слитые с последовательностями шарнира человека и CH1 человека, так что локусы иммуноглобулина мыши реаранжируются с образованием гена, кодирующего связывающий белок, включая (а) тяжелую цепь с вариабельной областью человека, CH1 областью человека, шарнирной областью человека, и областью CH2 мыши и CH3 мыши; и (b) ген, кодирующий легкую цепь иммуноглобулина, которая содержит вариабельный домен человека и константную область человека.
[000264] Согласно одному из аспектов предложена мышь, у которой присутствует эктопическая последовательность ADAM6, кодирующая функциональный белок ADAM6; все или практически все генные сегменты VH мыши заменены одним или более генными сегментами человека VL, и все или практически все генные сегменты DH и/или генные сегменты JH необязательно заменены одним или более генными сегментами DH человека и/или генными сегментами JH человека, или все или практически все генные сегменты DH и генные сегменты JH необязательно заменены одним или более генными сегментами JL человека.
[000265] Согласно одному из вариантов реализации у указанной мыши заменены все или практически все генные сегменты VH, DH и JH мыши одним или более генными сегментами VL, одним или более генными сегментами DH, и одним или более генными сегментами J (например, Jκ или Jλ), причем указанные генные сегменты функционально связаны с эндогенной областью шарнира мыши, при этом у указанной мыши образуется реаранжированный ген цепи иммуноглобулина, содержащий, от 5′ к 3′, в направлении транскрипции, VL человека - DH человека или мыши - J человека или мыши - шарнир мыши - CH2 мыши - CH3 мыши. Согласно одному из вариантов реализации указанная область J представляет собой Jκ область иммуноглобулина человека. Согласно одному из вариантов реализации указанная область J представляет собой JH область иммуноглобулина человека. Согласно одному из вариантов реализации указанная область J представляет собой Jλ область иммуноглобулина человека. Согласно одному из вариантов реализации указанная VL область человека выбрана из Vλ области иммуноглобулина человека и Vκ области иммуноглобулина человека.
[000266] Согласно конкретным вариантам реализации мышь экспрессирует антитело с одним вариабельным доменом, содержащее константную область иммуноглобулина мыши или человека и вариабельную область, полученную из Vκ человека, DH человека и Jκ человека; Vκ человека, DH человека, JH человека; Vλ человека, DH человека, Jλ человека; Vλ человека, DH человека, JH человека; Vκ человека, DH человека и Jλ человека; человека Vλ, человека DH и Jκ человека. Согласно конкретному варианту реализации последовательности распознавания рекомбинации модифицируют таким образом, чтобы обеспечить продуктивная реаранжировку между перечисленными генными сегментами V, D и J или между перечисленными генными сегментами V и J.
[000267] Согласно одному из аспектов предложена мышь, у которой присутствует эктопическая последовательность ADAM6, кодирующая функциональный белок ADAM6 (или его ортолог, гомолог или функциональный фрагмент), у которой заменены все или практически все генные сегменты VH мыши одним или более генными сегментами VL человека, заменены все или практически все генные сегменты DH и генные сегменты JH мыши на генные сегменты JL человека; при этом у указанной мыши отсутствует область CH1 и/или шарнирная область.
[000268] Согласно одному из вариантов реализации у указанной мыши отсутствует последовательность, кодирующая домен CH1. Согласно одному из вариантов реализации у указанной мыши отсутствует последовательность, кодирующая область шарнира. Согласно одному из вариантов реализации у указанной мыши отсутствует последовательность, кодирующая CH1 домен и область шарнира.
[000269] Согласно конкретному варианту реализации указанная мышь экспрессирует связывающий белок, который содержит вариабельный домен легкой цепи иммуноглобулина человека (λ или κ), слитый с доменом CH2 мыши, присоединенным к домену CH3 мыши.
[000270] Согласно одному из аспектов предложена мышь, у которой присутствует эктопическая последовательность ADAM6, кодирующая функциональный белок ADAM6 (или его ортолог, гомолог или функциональный фрагмент); все или практически все генные сегменты VH мыши заменены одним или более генными сегментами VL человека; все или практически все генные сегменты DH и JH мыши заменены генными сегментами JL человека.
[000271] Согласно одному из вариантов реализации у указанной мыши удалена последовательность гена константной области тяжелой цепи иммуноглобулина, кодирующая CH1 область, область шарнира, CH1 и шарнирную область, или CH1 область, область шарнира и CH2 область.
[000272] Согласно одному из вариантов реализации указанная мышь продуцирует связывающий белок с одним вариабельным доменом, содержащий гомодимер, выбранный из следующих: (а) VL человека - CH1 мыши - CH2 мыши - CH3 мыши; (b) VL человека - шарнирная область мыши - CH2 мыши - CH3 мыши; (с) VL человека - CH2 мыши - CH3 мыши.
[000273] Согласно одному из аспектов предложена мышь с инактивированным эндогенным локусом тяжелой цепи иммуноглобулина, с инактивированным или удаленным эндогенным локусом ADAM6 мыши, при этом указанная мышь содержит последовательность нуклеиновой кислоты, экспрессирующую гибридное антитело человека, или мыши, или человека/мыши, или др. Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты находится на встроенном трансгене, который случайным образом встроен в геном мыши. Согласно одному из вариантов реализации указанная последовательность нуклеиновой кислоты расположена на эписоме (например, хромосоме), отсутствующей у мыши дикого типа.
[000274] Согласно одному из вариантов реализации у указанной мыши также присутствует деактивированный эндогенный локус легкой цепи иммуноглобулина. Согласно конкретному варианту реализации указанный эндогенный локус легкой цепи иммуноглобулина выбран из локусов каппа- (κ) и лямбда- (λ) легких цепей. Согласно конкретному варианту реализации указанная мышь содержит деактивированный эндогенный локус легкой цепи к и деактивированный локус легкой цепи λ, при этом указанная мышь экспрессирует антитело, которое содержит вариабельный домен тяжелой цепи иммуноглобулина человека и домен легкой цепи иммуноглобулина человека. Согласно одному из вариантов реализации указанный домен легкой цепи иммуноглобулина человека выбран из домена κ легкой цепи человека и домена λ легкой цепи человека.
[000275] Согласно одному из аспектов предложено генетически модифицированное животное, экспрессирующее гибридное антитело и экспрессирующее белок ADAM6 или его ортолог или гомолог, функциональный у указанного генетически модифицированного животного.
[000276] Согласно одному из вариантов реализации указанное генетически модифицированное животное выбрано из мыши и крысы. Согласно одному из вариантов реализации указанное генетически модифицированное животное представляет собой мышь, а белок ADAM6 или его ортолог или гомолог взят от линии мышей, отличной от линии указанного генетически модифицированного животного. Согласно одному из вариантов реализации указанное генетически модифицированное животное представляет собой грызуна семейства Cricetidae (например, хомяка, американскую крысу или мышь, полевку), а указанный ортолог или гомолог белка ADAM6 взят от грызуна семейства Muridae (например, истинной мыши или крысы, песчанки, иглистой мыши, косматого хомяка). Согласно одному из вариантов реализации генетически модифицированное животное представляет собой грызуна семейства Muridae, а ортолог или гомолог белка ADAM6 взят от грызуна семейства Cricetidae.
[000277] Согласно одному из вариантов реализации указанное гибридное антитело содержит вариабельный домен человека и последовательность константной области грызуна. Согласно одному из вариантов реализации указанный грызун выбран из грызуна семейства Cricetidae и грызуна семейства Muridae. Согласно конкретному варианту реализации указанный грызун семейства Cricetidae и семейства Muridae представляет собой мышь. Согласно конкретному варианту реализации указанный грызун семейства Cricetidae и семейства Muridae представляет собой крысу. Согласно одному из вариантов реализации указанное гибридное антитело содержит вариабельный домен человека и константный домен животного, выбранного из мыши или крысы; согласно конкретному варианту реализации указанная мышь или крыса выбрана из семейства Cricetidae и семейства Muridae. Согласно одному из вариантов реализации указанное гибридное антитело содержит вариабельный домен тяжелой цепи человека, вариабельный домен легкой цепи человека и последовательность константной области, полученную от грызуна, выбранного из мыши и крысы, при этом указанные вариабельный домен тяжелой цепи человека и легкая цепь человека когнатны. Согласно конкретному варианту реализации в «когнатные» включены вариабельные домены тяжелой цепи человека и легкой цепи человека, полученные из одной В-клетки, которая экспрессирует указанные вариабельный домен легкой цепи человека и вариабельный домен тяжелой цепи человека вместе и презентирует указанные вариабельные домены вместе на поверхности индивидуальной В-клетки.
[000278] Согласно одному из вариантов реализации гибридное антитело экспрессируется из иммуноглобулинового локуса. Согласно одному из вариантов реализации вариабельный домен тяжелой цепи гибридного антитела экспрессируется из реаранжированного эндогенного локуса тяжелой цепи иммуноглобулина. Согласно одному из вариантов реализации вариабельный домен легкой цепи гибридного антитела экспрессируется из реаранжированного эндогенного локуса легкой цепи иммуноглобулина. Согласно одному из вариантов реализации вариабельный домен тяжелой цепи гибридного антитела и/или вариабельный домен легкой цепи гибридного антитела экспрессируется из реаранжированного трансгена (например, реаранжированной последовательности нуклеиновой кислоты, полученной из переаранжированной последовательности нуклеиновой кислоты, встроенной в геном животного в локусе, отличном от эндогенного локуса иммуноглобулина). Согласно одному из вариантов реализации вариабельный домен легкой цепи гибридного антитела экспрессируется из реаранжированного трансгена (например, реаранжированной последовательности нуклеиновой кислоты, полученной из нереаранжированной последовательности нуклеиновой кислоты, встроенной в геном животного в локусе, отличном от эндогенного локуса иммуноглобулина).
[000279] Согласно конкретному варианту реализации трансген экспрессируется из транскрипционно активного локуса, например, локуса ROSA26, например, локуса ROSA26 мышиных (например, мыши).
[000280] Согласно одному из аспектов предложено не являющееся человеком животное, у которого присутствует локус тяжелой цепи гуманизированного иммуноглобулина, при этом указанный локус тяжелой цепи гуманизированного иммуноглобулина содержит последовательность ADAM6 не происходящая от человека или ее ортолог или гомолог.
[000281] Согласно одному из вариантов реализации указанное не являющееся человеком животное представляет собой грызуна, выбранного из мыши, крысы и хомяка.
[000282] Согласно одному из вариантов реализации указанный ортолог или гомолог ADAM6 не происходящий от человека представляет собой последовательность, ортологичную и/или гомологичную последовательности ADAM6 мыши, при этом указанный ортолог или гомолог функционален у указанного не являющегося человеком животного.
[000283] Согласно одному из вариантов реализации указанное не являющееся человеком животное выбрано из мыши, крысы и хомяка, а указанный ортолог или гомолог ADAM6 взят от не являющегося человеком животного, выбранного из мыши, крысы и хомяка. Согласно конкретному варианту реализации указанное не являющееся человеком животное представляет собой мышь, и ортолог или гомолог ADAM6 взят от животного, выбранного из другого вида мыши, крысы и хомяка. Согласно конкретному варианту реализации указанное не являющееся человеком животное представляет собой крысу, и ортолог или гомолог ADAM6 взят от грызуна, выбранного из другого вида крыс, мыши и хомяка. Согласно конкретному варианту реализации указанное не являющееся человеком животное представляет собой хомяка, и ортолог или гомолог ADAM6 взят от грызуна, выбранного из другого вида хомяка, мыши и крысы.
[000284] Согласно конкретному варианту реализации указанное не являющееся человеком животное относится к подотряду Myomorpha, и последовательность ADAM6 взята от животного, выбранного из грызуна надсемейства Dipodoidea и грызуна надсемейства Muroidea. Согласно конкретному варианту реализации грызун представляет собой мышь надсемейства Muroidea, и ортолог или гомолог ADAM6 взят от мыши, или крысы, или хомяка надсемейства Muroidea.
[000285] Согласно одному из вариантов реализации указанный гуманизированный локус тяжелой цепи содержит один или более генных сегментов VH человека, один или более генных сегментов DH человека и один или более генных сегментов JH человека. Согласно конкретному варианту реализации указанный(е) один или более генных сегментов VH человека, один или более генных сегментов DH человека и один или более генных сегментов JH человека функционально связаны с одним или более генами константной области человека, гибридными генами константной области и/или генами константной области грызуна (например, мыши или крысы). Согласно одному из вариантов реализации указанные гены константной области являются генами мыши. Согласно одному из вариантов реализации указанные гены константной области являются генами крысы. Согласно одному из вариантов реализации указанные гены константной области являются генами хомяка. Согласно одному из вариантов реализации указанные гены константной области включают последовательность, выбранную из шарнира, CH2, CH3 и их комбинации. Согласно конкретному варианту реализации указанные гены константной области включают последовательность шарнира, CH2 и CH3.
[000286] Согласно одному из вариантов реализации указанная последовательность ADAM6 не происходящего от человека смежна с последовательностью тяжелой цепи иммуноглобулина человека. Согласно одному из вариантов реализации указанная последовательность ADAM6 не происходящая от человека находится в последовательности тяжелой цепи иммуноглобулина человека. Согласно конкретному варианту реализации указанная последовательность тяжелой цепи иммуноглобулина человека содержит генный сегмент V, D и/или J.
[000287] Согласно одному из вариантов реализации указанная последовательность ADAM6 не происходящая от человека расположена между двумя генными сегментами V. Согласно одному из вариантов реализации указанная последовательность ADAM6 не происходящая от человека соединяет генные сегменты V и D. Согласно одному из вариантов реализации указанная последовательность ADAM6 мыши расположена между генными сегментами V и J. Согласно одному из вариантов реализации указанная последовательность ADAM6 мыши соединяет генные сегменты D и J.
[000288] Согласно одному из аспектов предложено генетически модифицированное отличное от человека животное, содержащее В-клетку, экспрессирующую домен VH иммуноглобулина человека, когнатный домену VL человека из иммуноглобулинового локуса, при этом указанное не являющееся человеком животное экспрессирует не-иммуноглобулиновый белок не происходящий от человека из иммуноглобулинового локуса. Согласно одному из вариантов реализации указанный не-иммуноглобулиновый белок отличного от человека животного представляет собой белок ADAM. Согласно конкретному варианту реализации белок ADAM представляет собой белок ADAM6, либо его гомолог, ортолог или функциональный фрагмент.
[000289] Согласно одному из вариантов реализации указанное не являющееся человеком животное представляет собой грызуна (например, мышь или крысу). Согласно одному из вариантов реализации указанный грызун принадлежит семейству Muridae. Согласно одному из вариантов реализации указанный грызун принадлежит подсемейству Murinae. Согласно конкретному варианту реализации указанный грызун подсемейства Murinae выбран из мыши и крысы.
[000290] Согласно одному из вариантов реализации указанный отличный от иммуноглобулина белок не происходящий от человека представляет собой белок грызуна. Согласно одному из вариантов реализации указанный грызун принадлежит семейству Muridae. Согласно одному из вариантов реализации указанный грызун принадлежит подсемейству Murinae. Согласно конкретному варианту реализации указанный грызун выбран из мыши, крысы и хомяка.
[000291] Согласно одному из вариантов реализации указанные VH и VL домены человека присоединены непосредственно или с помощью линкера к последовательности константного домена иммуноглобулина. Согласно конкретному варианту реализации указанная последовательность константного домена содержит последовательность, выбранную из шарнира, CH2, CH3 и их комбинации. Согласно конкретному варианту реализации указанный домен VL человека выбран из домена Vκ или Vλ.
[000292] Согласно одному из аспектов предложено генетически модифицированные не являющееся человеком животное, в зародышевой линии которого присутствует последовательность иммуноглобулина человека, при этом сперма самца указанного не являющегося человеком животного характеризуется дефектом подвижности in vivo. Согласно одному из вариантов реализации указанный дефект подвижности in vivo включает неспособность спермы указанного самца не являющегося человеком животного перемещаться из матки через маточные трубы самки не являющегося человеком животного того же вида. Согласно одному из вариантов реализации у указанного не являющегося человеком животного отсутствует последовательность нуклеотидов, кодирующая белок ADAM6 или его функциональный фрагмент. Согласно конкретному варианту реализации указанный белок ADAM6 или его функциональный фрагмент включает белок ADAM6a и/или ADAM6b белок или их функциональный фрагмент. Согласно одному из вариантов реализации указанное не являющееся человеком животное представляет собой грызуна. Согласно конкретному варианту реализации указанный грызун выбран из мыши, крысы и хомяка.
[000293] Согласно одному из аспектов предложено не являющееся человеком животное, у которого присутствует последовательность иммуноглобулина человека, смежная с последовательностью не происходящей от человека, кодирующей белок ADAM6, либо его ортолог, гомолог или функциональный фрагмент. Согласно одному из вариантов реализации указанное не являющееся человеком животное представляет собой грызуна. Согласно конкретному варианту реализации указанный грызун выбран из мыши, крысы и хомяка.
[000294] Согласно одному из вариантов реализации указанная последовательность иммуноглобулина человека представляет собой последовательность тяжелой цепи иммуноглобулина. Согласно одному из вариантов реализации указанная последовательность иммуноглобулина содержит один или более генных сегментов VH. Согласно одному из вариантов реализации указанная последовательность иммуноглобулина человека содержит один или более генных сегментов DH. Согласно одному из вариантов реализации указанная последовательность иммуноглобулина человека содержит один или более генных сегментов JH. Согласно одному из вариантов реализации указанная последовательность иммуноглобулина человека содержит один или более генных сегментов VH, один или более генных сегментов DH и один или более генных сегментов JH.
[000295] Согласно одному из вариантов реализации указанная последовательность иммуноглобулина содержит один или более генных сегментов VH, встречающийся с высокой частотой в природных репертуарах человека. Согласно конкретному варианту реализации указанный один или более генных сегментов VH включают не более чем два генных сегмента VH, не более чем три генных сегмента VH, не более чем четыре генных сегмента VH, не более чем пять генных сегментов VH, не более чем шесть генных сегментов VH, не более чем семь генных сегментов VH, не более чем восемь генных сегментов VH, не более чем девять генных сегментов VH, не более чем 10 генных сегментов VH, не более чем 11 генных сегментов VH, не более чем 12 генных сегментов VH, не более чем 13 генных сегментов VH, не более чем 14 генных сегментов VH, не более чем 15 генных сегментов VH, не более чем 16, генных сегментов VH, не более чем 17 генных сегментов VH, не более чем 18 генных сегментов VH, не более чем 19 генных сегментов VH, не более чем 20 генных сегментов VH, не более чем 21 генный сегмент VH, не более чем 22 генных сегмента VH или не более чем 23 генных сегмента VH.
[000296] Согласно конкретному варианту реализации указанные один или более генных сегментов VH включают пять генных сегментов VH. Согласно конкретному варианту реализации указанные один или более генных сегментов VH включают 10 генных сегментов VH. Согласно конкретному варианту реализации указанные один или более генных сегментов VH включают 15 генных сегментов VH. Согласно конкретному варианту реализации указанные один или более генных сегментов VH включают 20 генных сегментов VH.
[000297] Согласно различным вариантам реализации указанные генные сегменты VH выбраны из VH6-1, VH1-2, VH1-3, VH2-5, VH3-7, VH1-8, VH3-9, VH3-11, VH3-13, VH3-15, VH3-16, VH1-18, VH3-20, VH3-21, VH3-23, VH1-24, VH2-26, VH4-28, VH3-30, VH4-31, VH3-33, VH4-34, VH3-35, VH3-38, VH4-39, VH3-43, VH1-45, VH1-46, VH3-48, VH3-49, VH5-51, VH3-53, VH1-58, VH4-59, VH4-61, VH3-64, VH3-66, VH1-69, VH2-70, VH3-72, VH3-73 и VH3-74.
[000298] Согласно различным вариантам реализации указанные генные сегменты VH выбраны из VH1-2, VH1-8, VH1-18, VH1-46, VH1-69, VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-21, VH3-23, VH3-30, VH3-33, VH3-43, VH3-48, VH4-31, VH4-34, VH4-39, VH4-59, VH5-51 и VH6-1.
[000299] Согласно различным вариантам реализации указанные генные сегменты VH выбраны из VH1-18, VH1-46, VH1-69, VH3-7, VH3-11, VH3-15, VH3-21, VH3-23, VH3-30, VH3-33, VH3-48, VH4-34, VH4-39, VH4-59 и VH5-51.
[000300] Согласно различным вариантам реализации указанные генные сегменты VH выбраны из VH1-18, VH1-69, VH3-7, VH3-11, VH3-15, VH3-21, VH3-23, VH3-30, VH3-43, VH3-48, VH4-39, VH4-59 и VH5-51.
[000301] Согласно различным вариантам реализации указанные генные сегменты VH выбраны из VH1-18, VH3-11, VH3-21, VH3-23, VH3-30, VH4-39 и VH4-59.
[000302] Согласно различным вариантам реализации указанные генные сегменты VH выбраны из VH1-18, VH3-21, VH3-23, VH3-30 и VH4-39.
[000303] Согласно различным вариантам реализации указанные генные сегменты VH выбраны из VH1-18, VH3-23 и VH4-39.
[000304] Согласно различным вариантам реализации указанные генные сегменты VH выбраны из VH3-21, VH3-23 и VH3-30.
[000305] Согласно различным вариантам реализации указанные генные сегменты VH выбраны из VH3-23, VH3-30 и VH4-39.
[000306] Согласно конкретному варианту реализации последовательность иммуноглобулина человека содержит по меньшей мере 18 генных сегментов VH, 27 генных сегментов DH и шесть генных сегментов JH. Согласно конкретному варианту реализации указанная последовательность иммуноглобулина человека содержит по меньшей мере 39 генных сегментов VH, 27 генных сегментов DH и шесть генных сегментов JH. Согласно конкретному варианту реализации указанная последовательность иммуноглобулина человека содержит по меньшей мере 80 генных сегментов VH, 27 генных сегментов DH и шесть генных сегментов JH.
[000307] Согласно одному из вариантов реализации указанное не являющееся человеком животное представляет собой мышь, и у указанной мыши эндогенные генные сегменты VH мыши заменены одним или более генными сегментами VH человека, при этом указанные генные сегменты VH человека функционально связаны с геном CH области мыши, так что у указанной мыши происходит реаранжировка генных сегментов VH человека и экспрессируется обратно-гибридная тяжелая цепь иммуноглобулина, которая содержит VH домен человека и CH мыши. Согласно одному из вариантов реализации 90-100% переаранжированных генных сегментов VH мыши заменяют по меньшей мере одним переаранжированным генным сегментом человека VH. Согласно конкретному варианту реализации все или практически все эндогенные генные сегменты VH мыши заменяют по меньшей мере одним переаранжированным генным сегментом VH человека. Согласно одному из вариантов реализации для замены используют по меньшей мере 19, по меньшей мере 39, или по меньшей мере 80, или 81 переаранжированный генный сегмент VH человека. Согласно одному из вариантов реализации для замены используют по меньшей мере 12 функциональных переаранжированных генных сегментов VH человека, по меньшей мере 25 функциональных переаранжированных генных сегментов VH человека, или по меньшей мере 43 функциональных переаранжированных генных сегментов VH человека. Согласно одному из вариантов реализации у указанной мыши все мышиные сегменты DH и JH заменены по меньшей мере одним переаранжированным сегментом DH иммуноглобулина человека и по меньшей мере одним переаранжированным сегментом JH иммуноглобулина человека. Согласно одному из вариантов реализации указанный по меньшей мере один переаранжированный DH сегмент человека выбран из 1-1, 1-7, 1-26, 2-8, 2-15, 3-3, 3-10, 3-16, 3-22, 5-5, 5-12, 6-6, 6-13, 7-27, и их комбинации. Согласно одному из вариантов реализации указанный по меньшей мере один переаранжированный JH сегмент человека выбран из 1, 2, 3, 4, 5, 6 и их комбинации. Согласно конкретному варианту реализации указанный один или более генных сегментов человека VH выбраны из генных сегментов VH человека 1-2, 1-8, 1-24, 1-69, 2-5, 3-7, 3-9, 3-11, 3-13, 3-15, 3-20, 3-23, 3-30, 3-33, 3-48, 3-53, 4-31, 4-39, 4-59, 5-51, 6-1, и их комбинации.
[000308] Согласно различным вариантам реализации указанная последовательность иммуноглобулина человека функционально связана с константной областью зародышевой линии не являющегося человеком животного (например, грызуна, в частности, мыши, крысы или хомяка). Согласно одному из вариантов реализации указанная константная область представляет собой константную область человека; гибридную человека/мыши; или гибридную человека/крысы; или гибридную человека/хомяка; мыши, крысы или хомяка. Согласно одному из вариантов реализации указанная константная область представляет собой константную область грызуна (например, мыши, крысы или хомяка). Согласно конкретному варианту реализации указанный грызун представляет собой мышь или крысу. Согласно различным вариантам реализации указанная константная область содержит по меньшей мере домен CH2 и домен CH3.
[000309] Согласно одному из вариантов реализации указанная последовательность тяжелой цепи иммуноглобулина человека расположена в локусе тяжелой цепи иммуноглобулина зародышевой линии указанного не являющегося человеком животного (например, грызуна, в частности, мыши, или крысы, или хомяка). Согласно одному из вариантов реализации указанная последовательность тяжелой цепи иммуноглобулина человека расположена в локусе, не являющимся локусом тяжелой цепи иммуноглобулина зародышевой линии указанного не являющегося человеком животного, при этом указанный локус, не являющийся локусом тяжелой цепи, представляет собой транскрипционно активный локус. Согласно конкретному варианту реализации указанный локус, не являющийся локусом тяжелой цепи представляет собой локус ROSA26.
[000310] Согласно различным аспектам у указанного не являющегося человеком животного также присутствует последовательность легкой цепи иммуноглобулина человека (например, одна или более переаранжированные последовательности легкой цепи V и J, или одна или более реаранжированные последовательности VJ) зародышевой линии указанного не являющегося человеком животного. Согласно конкретному варианту реализации указанная последовательность легкой цепи иммуноглобулина представляет собой к последовательность легкой цепи иммуноглобулина. Согласно одному из вариантов реализации указанная последовательность легкой цепи иммуноглобулина человека содержит один или более генных сегментов VL. Согласно одному из вариантов реализации указанная последовательность легкой цепи иммуноглобулина человека содержит один или более генных сегментов JL. Согласно одному из вариантов реализации указанная последовательность легкой цепи иммуноглобулина человека содержит один или более генных сегментов VL и один или более генных сегментов JL. Согласно конкретному варианту реализации указанная последовательность легкой цепи иммуноглобулина человека содержит по меньшей мере 16 генных сегментов Vκ и пять генных сегментов Jκ. Согласно конкретному варианту реализации указанная последовательность легкой цепи иммуноглобулина человека содержит по меньшей мере 30 генных сегментов Vκ и пять генных сегментов Jκ. Согласно конкретному варианту реализации указанная последовательность легкой цепи иммуноглобулина человека содержит по меньшей мере 40 генных сегментов Vκ и пять генных сегментов Jκ. Согласно различным вариантам реализации указанная последовательность легкой цепи иммуноглобулина человека функционально связана с константной областью зародышевой линии указанного не являющегося человеком животного (например, грызуна, например, мыши, или крысы или хомяка). Согласно одному из вариантов реализации указанная константная область представляет собой константную область человека, гибридную человека/грызуна, мыши, крысы или хомяка. Согласно конкретному варианту реализации указанная константная область представляет собой константную область мыши или крысы. Согласно конкретному варианту реализации указанная константная область представляет собой к константную область мыши (mCκ) или к константную область крысы (rCκ).
[000311] Согласно одному из вариантов реализации указанное не являющееся человеком животное представляет собой мышь и у указанной мыши заменены все или практически все генные сегменты Vκ и Jκ по меньшей мере шестью генными сегментами Vκ человека и по меньшей мере один генный сегмент Jκ. Согласно одному из вариантов реализации все или практически все генные сегменты Vκ и Jκ заменяют по меньшей мере 16 генными сегментами человеческой Vκ (Vκ человека) и по меньшей мере одним генным сегментом Jκ. Согласно одному из вариантов реализации все или практически все генные сегменты Vκ и Jκ заменяют по меньшей мере 30 генными сегментами Vκ человека и по меньшей мере одним генным сегментом Jκ. Согласно одному из вариантов реализации все или практически все Vκ и Jκ генные сегменты заменяют по меньшей мере 40 генными сегментами Vκ человека и по меньшей мере один Jκ генный сегмент. Согласно одному из вариантов реализации указанный по меньшей мере один Jκ генный сегмент содержит два, три, четыре или пять генных сегмента Jκ человека.
[000312] Согласно одному из вариантов реализации указанные генные сегменты Vκ человека включают Vκ4-1, Vκ5-2, Vκ7-3, Vκ2-4, Vκ1-5 и Vκ1-6. Согласно одному из вариантов реализации указанные генные сегменты Vκ включают Vκ3-7, Vκ1-8, Vκ1-9, Vκ2-10, Vκ3-11, Vκ1-12, Vκ1-13, Vκ2-14, Vκ3-15 и Vκ1-16. Согласно одному из вариантов реализации указанные генные сегменты Vκ человека включают Vκ1-17, Vκ2-18, Vκ2-19, Vκ3-20, Vκ6-21, Vκ1-22, Vκ1-23, Vκ2-24, Vκ3-25, Vκ2-26, Vκ1-27, Vκ2-28, Vκ2-29 и Vκ2-30. Согласно одному из вариантов реализации указанные генные сегменты Vκ человека включают Vκ3-31, Vκ1-32, Vκ1-33, Vκ3-34, Vκ1-35, Vκ2-36, Vκ1-37, Vκ2-38, Vκ1-39 и Vκ2-40.
[000313] Согласно конкретному варианту реализации генные сегменты Vκ включают непрерывные генные сегменты к иммуноглобулина человека, охватывающие участки локуса κ легкой цепи иммуноглобулина человека от Vκ4-1 до Vκ2-40; а генные сегменты Jκ включают непрерывные генные сегменты, охватывающие участки локуса κ легкой цепи иммуноглобулина человека от Jκ1 до Jκ5.
[000314] Согласно одному из вариантов реализации последовательность легкой цепи иммуноглобулина человека расположена в локусе легкой цепи иммуноглобулина зародышевой линии не являющегося человеком животного. Согласно конкретному варианту реализации указанный локус легкой цепи иммуноглобулина зародышевой линии не являющегося человеком животного представляет собой локус κ легкой цепи иммуноглобулина. Согласно одному из вариантов реализации указанная последовательность легкой цепи иммуноглобулина человека расположена в транскрипционно активном локусе, не являющемся локусом легкой цепи иммуноглобулина зародышевой линии указанного не являющегося человеком животного. Согласно конкретному варианту реализации указанный не-иммуноглобулиновый локус представляет собой локус ROSA26.
[000315] Согласно одному из аспектов предложен способ получения антитела человека, при этом указанное антитело человека содержит вариабельные домены, полученные с применением одной или более последовательностей вариабельной области нуклеиновой кислоты, кодируемой в клетке не являющегося человеком животного согласно описанию в настоящей заявке.
[000316] Согласно одному из аспектов предложена фармацевтическая композиция, содержащая полипептид, который включает антитело или фрагмент антитела, который получен с применением одной или более последовательностей нуклеиновой кислоты вариабельной области, выделенных из не являющегося человеком животного согласно описанию в настоящей заявке. Согласно одному из вариантов реализации указанный полипептид представляет собой антитело. Согласно одному из вариантов реализации указанный полипептид представляет собой антитело, состоящее только из тяжелой цепи. Согласно одному из вариантов реализации указанный полипептид представляет собой одноцепочечный вариабельный фрагмент (например, scFv).
[000317] Согласно одному из аспектов предложено применение отличного от человека животного согласно настоящему описанию для получения антитела. Согласно различным вариантам реализации указанное антитело содержит один или более вариабельных доменов, полученные из одной или более последовательностей нуклеиновой кислоты вариабельной области, выделенной из указанного отличного от человека животного. Согласно конкретному варианту реализации указанные нуклеотидные последовательности вариабельных областей включают генные сегменты тяжелой цепи иммуноглобулина. Согласно конкретному варианту реализации указанные нуклеотидные последовательности вариабельных областей включают генные сегменты легкой цепи иммуноглобулина.
ПРИМЕРЫ
[000318] Следующие примеры приведены для описания получения и применения способов и композиций согласно настоящему изобретению, и не предназначены для ограничения заявленного авторами объема настоящего изобретения. Если не указано иное, температура приводится в градусах Цельсия; давление равно или приблизительно равно атмосферному.
Пример 1
Гуманизация генов иммуноглобулина мыши
[000319] Для конструирования 13 разных направленных векторов ВАС (BACvec) для гуманизации локусов тяжелой цепи и κ легкой цепи иммуноглобулина мыши применяли искусственные бактериальные хромосомы (ВАС) человека и мыши. В Таблицах 1 и 2 приведены описания этапов конструирования всех BACvec, использованных для гуманизации локусов тяжелой цепи и κ легкой цепи иммуноглобулина мыши, соответственно.
[000320] Идентификация ВАС человека и мыши. Мышиные ВАС, включающие 5′ и 3′-концы локусов тяжелой цепи и κ легкой цепи иммуноглобулина, идентифицировали гибридизацией фильтров маркированных с применением библиотек ВАС или с помощью ПЦР-скрининга ДНК-пулов ВАС-библиотек. Фильтры гибридизовали в стандартных условиях с применением зондов, соответствующих представляющим интерес областям. Скрининг пулов библиотек проводили с помощью ПЦР с применением уникальных пар праймеров, фланкирующих нужную целевую область. Дополнительно проводили ПЦР с применением тех же праймеров для деконволюции определенных лунок и выделения соответствующих представляющих интерес ВАС. И ВАС-фильтры, и пулы библиотек получали из мышиных ЭС-клеток 129 SvJ (Incyte Genomics/Invitrogen). ВАС человека, содержащие целиком локусы тяжелой цепи и κ легкой цепи иммуноглобулина, идентифицировали либо гибридизацией фильтров, маркированных с применением библиотеки ВАС, (библиотеки Caltech В, С, или D и библиотека RPCI-11, Research Genetics/Invitrogen) скринингом пулов библиотек ВАС человека (Caltech library, Invitrogen) с применением ПЦР, либо используя базу данных концевых последовательностей ВАС (библиотека Caltech D, TIGR).
[000321] Конструирование BACvec с помощью бактериальной гомологичной рекомбинации и лигирования. Бактериальную гомологичную рекомбинацию (BHR) выполняли согласно описанию (Valenzuela et al., 2003; Zhang et al., 1998, A new logic for DNA engineering using recombination in Escherichia coli, Nat Genet 20:123-128). В большинстве случаев линейные фрагменты синтезировали, лигируя полученные с помощью ПЦР гомологичные фрагменты в клонированные кассеты с последующим выделением из геля продуктов лигирования и внедрением электропорацией в BHR-компетентные бактерии, несущие целевую ВАС. После отбора на содержащих подходящий антибиотик чашках Петри корректно рекомбинированные ВАС идентифицировали с помощью ПЦР в обоих новых сочленениях с последующим рестрикционным анализом гель-электрофорезом в пульсирующем поле (Schwartz and Cantor, 1984, Separation of yeast chromosome-sized DNAs by pulsed field gradient gel electrophoresis, Cell 37:67-75) и выборочная проверка с помощью ПЦР с применением праймеров, распределяемых по последовательностям человека.
[000322] 3hVH BACvec конструировали, проводя три последовательных этапа BHR для начального этапа гуманизации локуса тяжелой цепи иммуноглобулина (ФИГ.4А и Таблица 1). На первом этапе (Этап 1) в родительский ВАС человека в 5′-направлении от генного сегмента VH1-3 человека вводили кассету, содержащую область гомологии с локусом тяжелой цепи иммуноглобулина мыши (НВ1), ген, придающий устойчивость к канамицину у бактерий и устойчивость к G418 у животных клеток (kanR), и участок сайт-специфической рекомбинации (например, loxP). На втором этапе (Этап 2), непосредственно за последним JH сегментом в 3′-направлении вводили вторую кассету, содержащую вторую область гомологии с локусом тяжелой цепи иммуноглобулина мыши (НВ2) и ген, придающий устойчивость к спектиномицину у бактерий (specR). Указанный второй этап включал удаление последовательностей локуса тяжелой цепи иммуноглобулина человека в 3′-направлении от JH6 и гена устойчивости к хлорамфениколу ВАС-вектора (cmR). Затем, на третьем этапе (Этап 3), ВАС человека с двойной модификацией (В1) линеаризовали с применением сайтов I-Ceul, добавленных на протяжении первых двух этапов и встроенных в мышиный ВАС (В2) с помощью BHR по двум областям гомологии (НВ1 и НВ2). Селективный отбор с антибиотиками для первого (cm/kan), второго (spec/kan) и третьего (cm/kan) этапа организовали таким образом, чтобы обеспечить специфичность к нужным продуктам. Модифицированные ВАС-клоны анализировали с применением гель-электрофореза в пульсирующем поле (PFGE) после расщепления рестрикционными ферментами для определения подходящей конструкции (ФИГ.4В).
[000323] Сходным образом были сконструированы 12 дополнительных BACvec для гуманизации локусов тяжелой цепи и κ легкой цепи. В некоторых случаях лигирование ВАС выполняли вместо BHR для соединения двух больших ВАС введением редких сайтов рестрикции в оба родительских BACvec с помощью BHR, наряду с точным размещением селектируемых маркеров. Это позволяло сохранить нужный продукт лигирования при отборе с определенными сочетаниями маркеров лекарственной устойчивости. Рекомбинантные ВАС, полученные лигированием после расщепления редкими рестрикционными ферментами идентифицировали и проводили их скрининг аналогично таковым для ВАС, полученных с помощью BHR (согласно описаниям выше).
[000324] Модификация эмбриональных стволовых (ЭС) клеток и получение мышей. Таргетинг ЭС-клеток (F1H4) осуществляли с применением генноинженерной методики VELOCIGENE® согласно описанию (Valenzuela et al., 2003). Получение мышей из модифицированных ЭС-клеток введением либо в бластоцисту (Valenzuela et al., 2003), либо на стадии 8-клеток (Poueymirou et al., 2007, F0 generation mice fully derived from genetargeted embryonic stem cells allowing immediate phenotypic analyses, Nat Biotechnol 25:91-99) проводили согласно описанию. Успешный таргетинг ЭС-клеток и мышей подтверждали скринингом ДНК ЭС-клеток или мышей с применением уникальных наборов зондов и праймеров для анализа на основе ПЦР (например, ФИГ.3А, 3В и 3С). Все исследования на мышах проводились под надзором и с разрешения Комитета по содержанию и использованию лабораторных животных (IACUC) компании Регенерон (Regeneron′s Institutional Animal Care и Use Committee).
[000325] Анализ кариотипа и флуоресцентная гибридизация in situ (FISH). Анализ кариотипа выполняли с использованием банка клеток института медицинских исследований Кориелла (Coriell Institute for Medical Research, Кэмден, Нью-Джерси). Проводили FISH таргетированных ЭС-клеток согласно описанию (Valenzuela et al., 2003). Зонды, соответствующие ДНК мышиной ВАС или ДНК человеческой ВАС, были помечены с помощью ник-трансляции (Invitrogen) флуоресцентно меченными dUTP нуклеотидами в оранжевом или зеленом спектре (Vysis).
[000326] Вариабельный генный локус тяжелой цепи иммуноглобулина. Гуманизация вариабельной области локуса тяжелой цепи достигалась за девять последовательных этапов прямой заменой приблизительно трех млн. п.о. (Мб) непрерывной геномной последовательности мыши, содержащих все VH, DH и генные сегменты JH, на приблизительно 1 Мб непрерывной геномной последовательности человека, содержащей эквивалентные генные сегменты человека (ФИГ.1А и Таблица 1) с применением VELOCIGENE® генноинженерной методики (см., например, патент США №6586251 и Valenzuela et al., 2003).
[000327] Интрон между генными сегментами JH и генами константной области (J-C интрон) содержит энхансер транскрипции (Neuberger, 1983, Expression and regulation of immunoglobulin heavy chain gene transfected into lymphoid cells, EMBO J 2:1373-1378) с следующей за ним областью простых повторов, требующихся для рекомбинации при изотипическом переключеним (Kataoka et al., 1980, Rearrangement of immunoglobulin gamma 1-chain gene and mechanism for heavy-chain class switch, PNAS USA 77:919-923). На стыке областей VH-DH-JH человека и CH области мыши (проксимальное сочленение) оставляли интронный энхансер тяжелой цепи мыши и домен переключения для сохранения у мыши эффективной экспрессии и переключения классов в гуманизированном локусе тяжелой цепи. Точное расположение нуклеотидов этого и следующих сочленений во всех заменах было возможно за счет применения генноинженерной технологии VELOCIGENE® (выше), где используется бактериальная гомологичная рекомбинация, управляемая синтетическими олигонуклеотидами. Соответственно, проксимальное сочленение размещали на расстоянии приблизительно 200 п.о. в 3′-направлении от последнего JH генного сегмента, а дистальное сочленение размещали на расстоянии в несколько сотен в 5′-направлении от крайнего с 5′-стороны VH генного сегмента локуса человека и на расстоянии приблизительно 9 т.н. в 3′-направлении от генного сегмента VH1-86 мыши, также известного как J558.55. Указанный генный сегмент мыши VH1-86 (J558.55) представляет собой самый дистальный вариабельный генный сегмент тяжелой цепи, который, как сообщалось, является псевдогеном у мышей C57BL/6, однако потенциально активным, хотя и со слабой RSS-последовательностью, в целевом 129 аллеле. Дистальный конец локуса тяжелой цепи мыши, как сообщалось, может содержать контролирующие элементы, регулирующие экспрессию и/или реаранжировку локуса (Pawlitzky et al., 2006).
[000328] Первая вставка последовательности ДНК иммуноглобулина человека мыши достигалась с применением 144 т.н. из проксимального конца локуса тяжелой цепи человека, содержащих 3 генных сегмента VH, все 27 генных сегмента DH и 9 генных сегментов JH человека, встраиваемых в проксимальный конец локуса IgH мыши, с одновременным удалением 16,6 т.н. мышиной геномной последовательности, с применением приблизительно 75 т.н. мышиных плеч гомологии (Этап А, ФИГ.2А; Таблицы 1 и 3, 3hVH). Указанную большую вставку размером 144 т.н. и сопутствующее удаление 16,6 т.н. проводили за один этап (Этап А), частота успешности которого составляла 0,2% (Таблица 3). Корректно таргетированные ЭС-клетки оценивали при помощи анализа утраты природных аллелей (loss-of-native-allele assay, LONA) (Valenzuela et al., 2003) с применением зондов для удаленной последовательности мыши и фланкирующих ее областей и для встроенной последовательности человека; целостность большой вставки человека подтверждали с применением нескольких зондов, охватывающих вставку целиком (ФИГ.3А, 3В и 3С). Так как предполагалось проведение множества раундов последовательного таргетинга ЭС-клеток, клоны таргетированных ЭС-клеток на этом и всех последующих этапах подвергали кариотипическому анализу (выше), и для последующих этапов использовали только клоны с кариотипами, нормальными по меньшей мере в 17 спредах из 20.
[000329] Таргетированные ЭС-клетки согласно Этапу А повторно модифицировали посредством BACvec, производившим удаление 19 т.н. на дистальном конце локуса тяжелой цепи (Этап В, ФИГ.2А). BACvec Этапа В содержал ген устойчивости к гигромицину (hyg), а не ген устойчивости к неомицину (neo), содержащийся на BACvec этапа А. Гены устойчивости из указанных двух BACvec были сконструированы таким образом, что при успешном таргетинге одной и той же хромосомы примерно три Мб локуса вариабельных генов тяжелой цепи мыши, содержащих все генные сегменты VH мыши, отличные от VH1-86, и все генные сегменты DH, отличные от DQ52, а также два указанных гена устойчивости, были фланкированы сайтами loxP; DQ52 и все генные сегменты JH цепи мыши удаляли на Этапе А. Клоны ЭС-клеток, повторно модифицированные по той же хромосоме, идентифицировали, переводя 3hVH проксимальную кассету в гомозиготное состояние при высоком содержании G418 (Mortensen et al., 1992, Production of homozygous mutant ES cells with a single targeting construct, Mol Cell Biol 12:2391-2395), а затем обращались как с дистальной hyg кассетой. Сегменты мыши размером до 4 Мб, модифицированные так, чтобы их фланкировали сайты loxP, были успешно удалены в ЭС-клетках транзиторной экспрессией рекомбиназы CRE с высокой эффективностью (до ≈11%) даже в отсутствие отбора по лекарственной устойчивости (Zheng et al., 2000, Engineering mouse chromosomes with Cre-loxP: range, efficiency, and somatic applications, Mol Cell Biol 20:648-655). Сходным образом авторы изобретения добивались удаления 3 Мб в 8% клонов ЭС-клеток после транзиторной экспрессии CRE (Этап С, ФИГ.2А; Таблица 3). Удаление оценивали с помощью анализа LONA с применением зондов на каждом конце удаленной последовательности мыши, а также по утрате neo и hyg и появлению продукта ПЦР участка удаления, содержащего единственный оставшийся сайт loxP. Далее, удаление подтверждали флуоресцентной гибридизацией in situ (данные не показаны).
[000330] Остаток вариабельной области тяжелой цепи человека добавляли в 3hVH аллель за 5 этапов с применением генноинженерной технологии VELOCIGENE® (Этапы Е-Н, ФИГ.2В), при этом каждый этап включал точно определенную вставку до 210 т.н. генных последовательностей человека. Проксимальный конец каждого нового BACvec для каждого этапа был сконструирован так, чтобы перекрывать расположенные с дистальной стороны последовательности человека предыдущего этапа, а дистальный конец каждого нового BACvec содержал такую же дистальную область мышиной гомологии, что и на Этапе A. BACvec этапов D, F и Н содержали neo-селективные кассеты, тогда как BACvec этапов Е и G содержали hyg-селективные кассеты, соответственно, отбор проводили по G418 либо по гигромицину. Таргетинг на Этапе D оценивали по утрате уникального продукта ПЦР в дистальном сайте loxP 3hVH гибридного аллеля. Таргетинг на Этапах E-I оценивали по утрате предыдущей селективной кассеты. На последнем этапе (Этап I, ФИГ.2В) neo-селективную кассету, фланкированную Frt сайтами (McLeod et al., 1986, Identification of the crossover site during FLP-mediated recombination in the Saccharomyces cerevisiae plasmid 2 microns circle, Mol Cell Biol 6:3357-3367), удаляли транзиторной экспрессией FLPe (Buchholz et al., 1998, Improved properties of FLP recombinase evolved by cycling mutagenesis, Nat Biotechnol 16:657-662). Каждую последовательность человека в BACvec для Этапов D, Е и G получали из двух родительских ВАС человека, тогда как последовательности для Этапов F и Н получали из одиночных ВАС. Сохранность последовательностей человека подтверждали на каждом этапе с применением нескольких зондов, охватывающих встроенные последовательности человека (согласно описаниям выше, например, ФИГ.3А, 3В и 3С). Только клоны с нормальными кариотипом и потенциалом зародышевой линии сохраняли на каждом этапе. ЭС-клетки с последнего этапа были по-прежнему способны вносить вклад в зародышевую линию после девяти последовательных манипуляций (Таблица 3). Мыши, гомозиготные по каждому из аллелей тяжелой цепи, были жизнеспособны, выглядели здоровыми и демонстрировали гуморальную иммунную систему практически дикого типа (см. Пример 3).
[000331] Вариабельный генный локус κ легкой цепи иммуноглобулина. Вариабельную область κ легкой цепи гуманизировали за восемь последовательных этапов прямой заменой приблизительно трех Мб последовательности мыши, содержащих все Vκ и Jκ генные сегменты, приблизительно 0,5 Мб последовательности человека, содержащей проксимальные Vκ и Jκ генные сегменты человека, способом, сходным с использованным для тяжелой цепи (ФИГ.1В; Таблицы 2 и 4).
[000332] Локус вариабельной области κ легкой цепи человека содержит два почти идентичных 400 т.н. повтора, разделенных спейсером размером 800 т.н. (Weichhold et al., 1993, The human immunoglobulin kappa locus consists of two copies that are organized in opposite polarity, Genomics 16:503-511). Так как повторы настолько сходны, вариабельность локуса может почти полностью быть воспроизведена у мышей посредством применения проксимального повтора. Также было описан природный аллель локуса κ легкой цепи человека с отсутствующим дистальным повтором (Schaible et al., 1993, The immunoglobulin kappa locus: polymorphism and haplotypes of Caucasoid and non-Caucasoid individuals, Hum Genet 91:261-267). Авторы изобретения заменяли приблизительно три Мб мышиной последовательности κ легкой цепи вариабельных генов на приблизительно 0,5 Мб последовательности к человека легкой цепи вариабельных генов для эффективной замены всех генных сегментов Vκ и Jκ мыши на проксимальные генные сегменты Vκ человека и все генные сегменты Jκ человека (ФИГ.2С и 2D; Таблицы 2 и 4). В отличие от способа, описанного в Примере 1 для локуса тяжелой цепи, полную генную область Vκ мыши, содержащую все Vκ и Jκ генные сегменты, удаляли с помощью трехэтапного процесса до добавления каких-либо последовательностей человека. Сначала вводили кассету neo в проксимальный конец вариабельной области (Этап А, ФИГ.2С). Затем встраивали кассету hyg в дистальный конец κ-локуса (Этап В, ФИГ.2С). Также в каждой селективной кассете размещали сайты распознавания рекомбиназы (например, loxP), так что обработка CRE индуцировала удаление оставшихся 3 Мб области Vκ мыши, а также обоих генов устойчивости (Этап С, ФИГ.2С).
[000333] Геномный фрагмент человека размером приблизительно 480 т.н., содержащий полную вариабельную область κ легкой цепи иммуноглобулина, встраивали за четыре последовательных этапа (ФИГ.2D; Таблицы 2 и 4), с встраиванием за один этап 150 т.н. κ-последовательности легкой цепи иммуноглобулина человека при помощи способов, сходных с используемыми для тяжелой цепи (см. Пример 1). Полученный ген устойчивости к гигромицину удаляли транзиторной экспрессией FLPe. Как и в случае тяжелой цепи, у таргетированных ЭС-клеточных клонов оценивали целостность полной вставки человека, нормальность кариотипа и потенциал зародышевой линии после выполнения каждого этапа. Получали мышей, гомозиготных по каждой из аллелей κ легкой цепи, и было установлено, что они здоровы и имеют нормальные внешние признаки.
Пример 2
Получение полностью гуманизированных мышей комбинированием нескольких гуманизированных иммуноглобулиновых аллелей
[000334] В разные моменты ЭС-клетки, несущие часть вариабельных наборов тяжелой цепи иммуноглобулина или κ легкой цепи человека согласно описанию в Примере 1, микроинъецировали, и полученных мышей размножали с получением нескольких вариантов мышей VELOCIMMUNE® с прогрессивно увеличивающимися долямми репертуаров иммуноглобулинов зародышевой линии человека (Таблица 5; ФИГ.5А и 5В). У мышей VELOCIMMUNE® 1 (V1) присутствует 18 генных сегментов VH человека и все генные сегменты DH и JH человека, скомбинированные с 16 генными сегментами Vκ человека и всеми генными сегментами Jκ человека. У мышей VELOCIMMUNE® 2 (V2) и VELOCIMMUNE® (V3) имеются расширенные репертуары вариабельных областей, содержащие в общей сложности 39 VH и 30 Vκ, и 80 VH и 40 Vκ, соответственно. Так как геномные области, кодирующие генные сегменты VH, DH и JH, мыши, а также генные сегменты Vκ и Jκ, полностью замещены, антитела, синтезируемые всеми вариантами мышей VELOCIMMUNE®, содержат вариабельные области человека, соединенные с константными областями мыши. Локусы λ легкой цепи мыши остаются интактными согласно различным вариантам реализации мышей VELOCIMMUNE® и служат для сравнения эффективности экспрессии различные локусов κ легкой цепи VELOCIMMUNE®.
[000335] Мышей с двойной гомозиготностью по гуманизации как тяжелой цепи, так и κ легкой цепи иммуноглобулина получали из подгруппы аллелей, описанных в Примере 1. Все генотипы наблюдали на протяжении цикла размножения для получения мышей с двойной гомозиготностью, встречаемость которых приблизительно соответствует менделевским пропорциям. У самцов из потомства, гомозиготных по каждому из аллелей тяжелой цепи человека, наблюдалось снижение фертильности, обусловленное утратой активности мышиного ADAM6. Локус генов вариабельных областей тяжелой цепи мыши содержит два внедренных функциональных гена ADAM6 (ADAM6a и ADAM6b). При гуманизации указанного локуса генов вариабельных областей тяжелой цепи мыши встраиваемая геномная последовательность человека содержала псевдоген ADAM6. Мышиный ADAM6 может быть необходим для фертильности, и таким образом отсутствие генов ADAM6 мыши в гуманизированных локусах генов вариабельных областей тяжелой цепи может приводить к снижению фертильности, несмотря на присутствие псевдогена человека. В Примерах 7-11 описано реконструирование генов ADAM6 мыши в гуманизированном вариабельном генном локусе тяжелой цепи, и восстановление соответствующего дикому типу уровня фертильности у мышей с гуманизированным локусом тяжелой цепи иммуноглобулина.
Пример 3
Популяции лимфоцитов у мышей с гуманизированными генами иммуноглобулина
[000336] Зрелые популяции В-клеток трех разных вариантов мышей VELOCIMMUNE® оценивали с помощью проточной цитометрии.
[000337] Вкратце, суспензии клеток из костного мозга, селезенки и тимуса получали с применением стандартных способов. Клетки ресуспендировали до концентрации 5×105 клеток/мл в FACS-буфере для окрашивания BD Pharmingen, блокировали анти-мышиным CD16/32 (BD Pharmingen), окрашивали подходящей смесью антител и фиксировали в BD CYTOFIX™ в соответствии с инструкциями производителей. Полученную клеточную массу ресуспендировали в 0,5 мл буфера для окрашивания и анализировали с применением программного обеспечения BD FACSCALIBUR™ и BD CELLQUEST PRO™. Все антитела (BD Pharmingen) готовили в разведении и добавляли до конечной концентрации 0,5 мг/105 клеток.
[000338] Для окрашивания костного мозга брали следующие смеси антител (A-D): А: ФИТЦ-меченые против мышиного IgMb, ФЭ-меченые против мышиного IgMa, против мышиного CD45R(B220)-APC; В: ФЭ-меченые против мышиного CD43(S7), против мышиного CD45R(B220)-APC; С: ФЭ-меченые против мышиного CD24(HSA); против мышиного CD45R(B220)-APC; D: ФЭ-меченые против мышиного ВР-1, против мышиного CD45R(B220)-APC.
[000339] Для окрашивания селезенки и ингвинальных лимфатических узлов (Е-Н) брали следующие смеси антител: Е: ФИТЦ-меченые против мышиного IgMb, ФЭ-меченые против мышиного IgMa, против мышиного CD45R(B220)-APC; F: ФИТЦ-меченые против мышиного Ig, λ1, λ2, λ3 легкие цепи, ФЭ-меченые против мышиного Igκ легкие цепи, против мышиного CD45R(B220)-APC; G: ФИТЦ-меченые против мышиного Ly6G/C, ФЭ-меченые против мышиного CD49b(DX5), против мышиного CD11b-APC; Н: ФИТЦ-меченые против мышиного CD4(L3T4), ФЭ-меченые против мышиного CD45R(B220), против мышиного CD8a-APC. Результаты приведены на ФИГ.6.
[000340] Лимфоциты, выделенные из селезенки или лимфатического узла гомозиготных мышей VELOCIMMUNE®, окрашивали на поверхностную экспрессию маркеров В220 и IgM и анализировали с применением проточной цитометрии (ФИГ.6). Размеры зрелых В220+ IgM+ популяции В-клеток у всех протестированных вариантов мышей VELOCIMMUNE® были практически идентичны таковым у мышей дикого типа, независимо от числа имеющихся у них генных сегментов VH. Кроме того, у мышей с гомозиготными гибридными гуманизированными локусами тяжелой цепи иммуноглобулина, даже при присутствии только 3 генных сегмента VH, но с нормальными локусами κ легкой цепи иммуноглобулина мыши, или у мышей с гомозиготными гибридными гуманизированными локусами κ легкой цепи и нормальными локусами тяжелой цепи иммуноглобулина мыши, также содержалось нормальное количество B220+ IgM+ клеток периферических компартментов (не показано). Эти результаты указывают на то, что гибридные локусы с вариабельными генными сегментами человека и константными областями мыши способны полностью обеспечить популяцию зрелых В-клеток. Далее, число вариабельных генных сегментов в локусах как тяжелой цепи, так и κ легкой цепи, и, соответственно, теоретическое разнообразие репертуара антител, не коррелирует со способностью к образованию популяций зрелых В-клеток дикого типа. Напротив, у мышей со случайным образом встроенными полностью человеческими трансгенами иммуноглобулина и инактивированными локусами иммуноглобулина количество В-клеток этих компартментов снижено, при этом тяжесть указанного дефицита зависит от числа вариабельных генных сегментов, включенных в трансген (Green and Jakobovits, 1998, Regulation of B cell development by variable gene complexity in mice reconstituted with human immunoglobulin yeast artificial chromosomes, J Exp Med 188:483-495). Это указывает на то, что стратегия «генетической гуманизации in situ» приводит к принципиально иному функциональному результату по сравнению со случайным образом встраиваемыми трансгенами при подходе «нокаут-плюс-трансген».
[000341] Аллельное исключение и выбор локуса. Способность поддерживать аллельное исключение исследовали у мышей, гетерозиготных по различным вариантам гуманизированного локуса тяжелой цепи иммуноглобулина.
[000342] Гуманизацию иммуноглобулиновых локусов проводили в F1 линии ЭС (F1H4, Valenzuela et al., 2003), полученных из гетерозиготных эмбрионов 129S6/SvEvTac и C57BL/6NTac. Последовательности генов вариабельных областей тяжелой цепи зародышевой линии человека таргетируют в аллель 129S6, несущий гаплотип IgMa, тогда как немодифицированный аллель мыши C576BL/6N несет гаплотип IgMb. Указанные аллельные формы IgM можно различить с помощью проточной цитометрии с применением антител, специфичных к полиморфизмам, обнаруживаемым в аллелях IgMa или IgMb. Как видно из ФИГ.6 (нижний ряд), В-клетки, идентифицированные у мышей, гетерозиготных по каждому варианту гуманизированного локуса тяжелой цепи, экспрессируют исключительно один аллель, либо IgMa (гуманизированный аллель), либо IgMb (аллель дикого типа). Это указывает на то, что механизмы, вовлеченные в аллельное исключение, у мышей VELOCIMMUNE® интактны. Кроме того, относительное число В-клеток, позитивных по гуманизированному аллелю (IgMa), примерно пропорционально числу присутствующих генных сегментов VH. Гуманизированный иммуноглобулиновый локус экспрессируется приблизительно в 30% В-клеток гетерозиготных мышей VELOCIMMUNE® 1, у которых присутствует 18 генных сегментов VH человека, и в 50% В-клеток гетерозиготных мышей VELOCIMMUNE® 2 и 3 (не показано), у которых присутствует 39 и 80 генных сегментов VH человека, соответственно. Примечательно, что доля клеток, экспрессирующих гуманизированный/дикого типа аллель мыши (0,5 для мышей VELOCIMMUNE® 1 и 0,9 для мышей VELOCIMMUNE® 2) выше, чем доля числа вариабельных генных сегментов, содержащихся в указанных гуманизированных локусах/локусах дикого типа (0,2 для мышей VELOCIMMUNE® 1 и 0,4 для мышей VELOCIMMUNE® 2). Это может указывать на то, что вероятность выбора аллеля определяется случайным выбором той или иной хромосомы и случайным выбором любой RSS V-сегмента. Далее, в части В-клеток, но не во всех, один аллель может становиться доступным для рекомбинации, завершать процесс и останавливать рекомбинацию до того, как становится доступен другой аллель. Кроме того, равномерное распределение клеток с поверхностным IgM (sIgM), происходящим либо из гибридного гуманизированного локуса тяжелой цепи, либо из локуса тяжелой цепи мыши дикого типа, представляет собой доказательство того, что гибридный локус функционирует на нормальном уровне. Напротив, случайным образом встроенные трансгены иммуноглобулина человека не в состоянии адекватно конкурировать с локусами иммуноглобулина мышей дикого типа (Bruggemann et al., 1989, A repertoire of monoclonal antibodies with human heavy chains from transgenic mice, PNAS 86:6709-6713; Green et al., 1994; Tuaillon et al., 1993, Human immunoglobulin heavy-chain minilocus recombination in transgenic mice: gene-segment use in mu and gamma transcripts, PNAS USA 90:3720-3724). Это также подтверждает, что иммуноглобулины, синтезируемые у мышей VELOCIMMUNE®, функционально отличны от синтезируемых с помощью случайным образом встраиваемых трансгенов у мышей, полученных с применением методик «нокаут-плюс-трансген».
[000343] Для линий 129S6 или C57BL/6N данные по полиморфизму областей Сκ для изучения аллельного исключения в гуманизированных\негуманизированных локусах κ легкой цепи недоступны. Как бы то ни было, у всех мышей VELOCIMMUNE® присутствуют локусы λ легкой цепи мыши дикого типа, следовательно, есть возможность отследить, может ли реаранжировка и экспрессия гуманизированных локусов κ легкой цепи предотвращать экспрессию λ легкой цепи мыши. Отношение числа клеток, экспрессирующих гуманизированные κ легкую цепь и числа клеток, экспрессирующих λ легкую цепь мыши, оставалось относительно неизменным у мышей VELOCIMMUNE® по сравнению мышами дикого типа, независимо от числа генных сегментов Vκ человека, встроенных в локус κ легкой цепи (ФИГ.6, третий ряд сверху). Кроме того, количество двойных позитивных (κ плюс λ) клеток не возрастало, что указывает на то, что продуктивная рекомбинация в гибридных локусах κ легкой цепи приводит к соответствующему подавлению рекомбинации локусов λ легкой цепи мыши. Напротив, мыши, у которых имеются случайным образом встроенные κ легкой цепи трансгены с инактивированными локусами κ легкой цепи мыши - но локусы λ легкой цепи мыши дикого типа - наблюдается резкое увеличение отношения λ/κ (Jakobovits, 1998), что означает, что встроенные трансгены κ легкой цепи не функционируют у таких мышей правильным образом. Это еще раз подтверждает иной функциональный результат, наблюдаемый для иммуноглобулинов, синтезируемых мышами VELOCIMMUNE® по сравнению с полученными методом «нокаут-плюс-трансген» мышами.
[000344] Развитие В-клеток. Так как популяции зрелых В-клеток у мышей VELOCIMMUNE® напоминают таковые мышей дикого типа (описанные выше), возможно, дефекты ранней дифференцировки В-клеток компенсируются размножением популяций зрелых В-клеток. Различные стадии дифференцировки В-клетки исследовали посредством анализа В-клеточных популяций с применением проточной цитометрии. В Таблице 6 представлено относительное содержание типов клеток в каждой В-клеточной линии, определенные с помощью РАС, с применением специфических маркеров клеточной поверхности, у мышей VELOCIMMUNE® по сравнению с однопометными мышами дикого типа.
[000345] Раннее развитие В-клеток происходит в костном мозге, и разные стадии дифференцировки В-клеток характеризуются изменениями типов и интенсивности экспрессии маркеров клеточной поверхности. Указанные различия в экспрессии на поверхности коррелируют с молекулярными изменениями, происходящими в иммуноглобулиновых локусов внутри клетки. Для перехода от про-В к пре-В клеткам необходимы успешные реаранжировка и экспрессия функционального белка тяжелой цепи, а переход от пре-В к стадии зрелых В-клеток регулируется корректными реаранжировкой и экспрессией κ либо λ легкой цепи. Соответственно, неуспешный переход между стадиями дифференцировки В-клеток можно отследить по изменениям относительного содержания популяций В-клеток на определенной стадии.
CD43hi B220lo
CD24hi B220lo
B220lo IgM+
B220hi IgM+
B220hi IgM+ IgD+
B220hi IgM+
[000346] Значительных дефектов дифференцировки В-клеток у каких-либо мышей VELOCIMMUNE® не наблюдалось. Введение генных сегментов тяжелой цепи человека по-видимому, не влияют на переход от про-В к пре-В, а введение генные сегменты κ легкой цепи человека не влияет на переход от пре-В к В у мышей VELOCIMMUNE®. Это указывает на то, что «обратно-гибридные» молекулы иммуноглобулина, содержащие вариабельные области человека и константные области мыши функционируют нормально в контексте В-клеточной сигнализации и ко-рецепторных молекул, что приводит к надлежащей дифференцировке В-клеток у мыши. И напротив, у мышей со случайным образом встроенными трансгенами иммуноглобулина и инактивированными эндогенными локусами тяжелой цепи или κ легкой цепи баланс между разными популяциями на протяжении дифференцировки В-клеток в той или иной степени нарушен (Green и Jakobovits, 1998).
Пример 4
Репертуар вариабельных генов у мышей с гуманизированным иммуноглобулином
[000347] Частоту использования вариабельных генных сегментов человека в репертуаре гуманизированных антител у мышей VELOCIMMUNE® анализировали при помощи полимеразной цепной реакции с обратной транскриптазой (ОТ-ПЦР) вариабельных областей человека из нескольких источников, включая спленоциты и гибридомные клетки. Оценивали последовательность вариабельной области, частоту использования генных сегментов, соматические сверхмутации и множественность J-сегментов реаранжированных генных сегментов вариабельной области.
[000348] Вкратце, из 1×107 - 2×107 спленоцитов или приблизительно 104-105 гибридомных клеток выделяли тотальную РНК с применением набора TRIZOL™ (Invitrogen) или Qiagen RNEASY™ Mini Kit (Qiagen) и обрабатывали специфическими праймерами константных областей мыши с применением системы SUPERSCRIPT™ III One-Step RT-PCR (Invitrogen). Реакции проводили на 2-5 мкл РНК из каждого образца с применением вышеуказанных 3′-специфических к константным областям праймеров, спаренных со смешанными праймерами лидерных областей, для каждого семейства вариабельных областей человека, для тяжелой цепи и κ легкой цепи, по отдельности. Использовали объемы реагентов и праймеров и условия ОТ-ПЦР/ПЦР, соответствующие инструкциям производителя. Последовательности праймеров основаны на разных источниках (Wang and Stollar, 2000, Human immunoglobulin variable region gene analysis by single cell RT-PCR, J Immunol Methods 244:217-225; Ig-primer sets, Novagen). При необходимости проводили вторичную вложенную ПЦР со смешанными специфическими праймерами каркасных участков семейства и тем же специфическим для константных областей иммуноглобулина мыши 3′-праймером, что и для первичной реакции. Аликвоты (5 мкл) из каждой реакции анализировали электрофорезом в агарозном геле и продукты реакции очищали от агарозы с применением набора MONTAGE™ Gel Extraction Kit (Millipore). Очищенные продукты клонировали с применением системы клонирования ТОРО™ ТА Cloning System (Invitrogen) и трансформировали в DH10β E.coli клетки электропорацией. Индивидуальные клоны отбирали из каждой реакции трансформации и культивировали в 2 мл культурального бульона LB с отбором по устойчивости к антибиотику в течение ночи при 37°С. Плазмидную ДНК выделяли из бактериальных культур с применением наборов (Qiagen).
[000349] Частота использования вариабельных генов иммуноглобулина. Плазмидную ДНК клонов как тяжелой цепи, так и κ легкой цепи секвенировали либо Т7, либо М13 обратными праймерами на анализаторе ABI 3100 Genetic Analyzer (Applied Biosystems). Необработанные данные последовательностей импортировали в SEQUENCHER™ (v4.5, Gene Codes). Каждую последовательность собирали в контиг и выравнивали по последовательностям иммуноглобулина человека с применением поисковой функции IMGT V-Quest (Brochet et al., 2008, IMGT/V-QUEST: the highly customized and integrated system for IG and TR standardized V-J and V-D-J sequence analysis, Nucleic Acids Res 36:W503-508) для определения частоты использования сегментов VH, DH, JH и Vκ, Jκ человека. Последовательности сравнивали с последовательностями зародышевой линии для анализа соматической сверхмутации и рекомбинации сочленений.
[000350] Из ЭС-клеток, содержащих первую модификацию тяжелой цепи (гибридный аллель 3hVH-CRE, внизу на ФИГ.2А), RAG-комплементацией получали мышей (Chen et al., 1993, RAG-2-deficient blastocyst complementation: an assay of gene function in lymphocyte development, PNAS USA 90:4528-4532), и выделяли кДНК из РНК спленоцитов. кДНК амплифицировали с применением наборов специфических праймеров (описанных выше) для предсказанной иРНК гибридной тяжелой цепи, которые должны возникать при V(D)J рекомбинации во встроенных генных сегментах человека с последующим сплайсингом с образованием константных доменов мышиных IgM или IgG. По последовательностям, полученным из указанных кДНК-клонов (не показаны), видно, что произошла полноценная V(D)J рекомбинация последовательностей генов вариабельных областей человека, что реаранжированные V(D)J генные сегменты человека были надлежащим образом сплайсированы внутри рамки считывания в константные домены мыши, и что произошла рекомбинация с переключением классов. Выполняли дальнейший анализ последовательностей иРНК-продуктов следующих гибридных иммуноглобулиновых локусов.
[000351] В сходном эксперименте В-клетки не-иммунизированных мышей дикого типа и мышей VELOCIMMUNE® разделяли проточной цитометрией на основании экспрессии на поверхности В220 и IgM или IgG. Клетки с В220+ IgM+ или поверхностным IgG+ (sIgG+) объединяли и получали VH и Vκ последовательности после ОТ-ПЦР-амплифицирования и клонирования (описано выше). Регистрировали типовые частоты использования генов в наборе ОТ-ПЦР-амплифицированных кДНК из неиммунизированных мышей VELOCIMMUNE® 1 (Таблица 7) и мышей VELOCIMMUNE® 3 (Таблица 8) (*дефектные RSS; † отсутствующий или псевдоген). Звездочка: генные сегменты с дефектными RSS. †: генный сегмент отсутствует или представляет собой псевдоген.
[000352] Как видно из Таблиц 7 и 8, использованы почти все функциональные генные сегменты VH, DH, JH, Vκ и Jκ человека. Некоторые из функциональных вариабельных генных сегментов, описанные, но не обнаруженные у мышей VELOCIMMUNE® в этом эксперименте, как сообщалось, содержат дефектные сигнальные последовательности рекомбинации (RSS) и, соответственно, ожидаемо не должны экспрессироваться (Feeney, 2000, Factors that influence formation of B cell repertoire, Immunol Res 21:195-202). Анализ нескольких других наборов последовательностей иммуноглобулина различных мышей VELOCIMMUNE®, выделенных как из интактных, так и иммунизированных репертуаров, свидетельствовал об использовании указанных генных сегментов, хотя и с более низкими частотами (данные не показаны). Из совокупности данных по частотам использования генов видно, что в различных интактных и иммунизированных репертуарах наблюдались все функциональные генные сегменты VH, DH, JH, Vκ и Jκ человека, присутствующие у мышей VELOCIMMUNE® (данные не показаны). Хотя генный сегмент человека VH7-81 и обнаружен при анализе последовательностей локуса тяжелой цепи человека (Matsuda et al., 1998, The complete nucleotide sequence of the human immunoglobulin heavy chain variable region locus, J Exp Med 188:2151-2162), он отсутствует у мышей VELOCIMMUNE®, что подтверждается повтороным секвенированием полного генома мышей VELOCIMMUNE® 3.
[000353] Известно, что последовательности тяжелых и легких цепей антител проявляют исключительную вариативность, в особенности короткие полипептидные сегменты реаранжированного вариабельного домена. Указанные области, известные как гипервариабельные области или участки, определяющие комплементарность (CDR-области), формируют сайты связывания антигена в структуре молекулы антитела. Промежуточные полипептидные последовательности называются каркасными областями (FR). И тяжелые, и легкие цепи содержат по три CDR (CDR1, CDR2, CDR3) и 4 FR (FR1, FR2, FR3, FR4). Одна из последовательностей CDR, CDR3, уникальна в том отношении, что она образуется в результате рекомбинации как генных сегментов VH, DH и JH, так и генных сегментов Vκ и Jκ, и вносит значительный вклад в разнообразие репертуара еще до контакта с антигеном. Сочленение неточное, что обусловлено как удалением нуклеотидов в результате экзонуклеазной активности, так и нематричного присоединения терминальной дезоксинуклеотидилтрансферазой (TdT) и, соответственно, обеспечивает появление новых последовательностей в результате процесса рекомбинации. Несмотря на то, что и для FR-последовательностей могут наблюдаться существенные уровни соматических мутаций из-за высокой мутабильности вариабельной области в целом, вариабельность, тем не менее, не распределена равномерно по всей вариабельной области. CDR представляют собой сконцентрированные локализованные области высокой вариабельности на поверхности молекулы антитела, обеспечивающие связывание антигена. Последовательности тяжелой цепи и легкой цепи отобранных антител мышей VELOCIMMUNE® вокруг CDR3-сочленения, демонстрирующие множественность J-сегментов, показаны на ФИГ.7А и 7В, соответственно.
[000354] Как видно из ФИГ.7А, в антителах мышей VELOCIMMUNE® нематричное присоединение добавочных нуклеотидов (N-присоединение) наблюдается как в VH-DH, так и в DH-JH сочленениях, указывая на надлежащее функционирование TdT в сегментах человека. Сравнение концов VH, DH и JH сегментов с эквивалентами зародышевой линии указывает также на присутствие экзонуклеазной активности. В отличие от локуса тяжелой цепи, при реаранжировке κ легкой цепи человека наблюдается незначительное число или отсутствие присоединений TdT в последовательности CDR3, образующейся в результате рекомбинации сегментов Vκ и Jκ (ФИГ.7В). Предположительно, это происходит ввиду отсутствия экспрессии TdT у мышей во время реаранжировки легкой цепи при переходе от пре-В к В-клеткам. Наблюдаемое разнообразие CDR3 реаранжированных Vκ областей человека обусловлено главным образом экзонуклеазной активностью на протяжении процесса рекомбинации.
[000355] Соматическая сверхмутация. Дополнительное разнообразие в вариабельные области реаранжированных генов иммуноглобулина во время реакции в зародышевом центре вносит процесс, называемый соматической сверхмутацией. В-клетки, экспрессирующие соматически мутированные вариабельные области, конкурируют с другими В-клетками за доступ к антигену, презентируемому фолликулярными дендритными клетками. В-клетки с более высокой аффинностью к антигену в дальнейшем будут размножаться и перетерпевать переключение классов перед выходом на периферию. Соответственно, В-клетки, экспрессирующие переключенные изотипы, как правило, сталкивались с антигеном, прошли реакции в зародышевом центре, и содержат более число мутаций по сравнению с необученными В-клетками. Далее, последовательности вариабельной области преимущественно необученных sIgM+ В-клеток предположительно должны содержать относительно меньше мутаций по сравнению с вариабельными последовательностями sIgG+ В-клеток, прошедших антигенную селекцию.
[000356] Последовательности произвольно выбранных клонов VH или Vκ из sIgM+ или sIgG+ В-клеток не-иммунизированных мышей VELOCIMMUNE® или sIgG+ В-клеток иммунизированных мышей сравнивали с вариабельными генными сегментами их зародышевых линий и отмечали изменения относительно последовательностей зародышевой линии. Полученные нуклеотидные последовательности транслировали in silico и отмечали также мутации, приводящие к заменам аминокислот. Данные по всем вариабельным областям сопоставляли и рассчитывали процент изменений в определенной позиции (ФИГ.8).
[000357] Как видно из ФИГ.8, в вариабельных областях тяжелой цепи человека, полученных из sIgG+ В-клеток не-иммунизированных мышей VELOCIMMUNE® наблюдается значительно больше нуклетидов относительно sIgM+ В-клеток из того же пула спленоцитов, и даже большее количество изменений наблюдается в вариабельных областях тяжелых цепей, полученных от иммунизированных мышей. Количество изменений увеличивается на определяющих комплементарность участках (CDR) по сравнению с каркасными областями, что указывает на селекцию антигенов. В соответствующих аминокислотных последовательностях вариабельных областей тяжелой цепи человека также обнаруживаются значительно большие количества мутаций в IgG, чем IgM, и еще большие - в иммунизированных IgG. В свою очередь, указанные мутации, по-видимому, чаще встречаются в CDR-последовательностях по сравнению с каркасными последовательностями, что указывает на то, что антитела проходили селекцию in vivo. Сходное увеличение количества нуклеотидных и аминокислотных мутаций обнаруживается в последовательностях Vκ, полученных из IgG+ В-клеток иммунизированных мышей.
[000358] Частота использования генов и частота соматических сверхмутаций, наблюдаемые у мышей VELOCIMMUNE®, показывают, что практически все присутствующие генные сегменты способны к реаранжировке с образованием полностью функциональных обратных гибридных антител у указанных мышей. Далее, антитела VELOCIMMUNE® в полной мере задействованы иммунной системой мышей, претерпевая аффинную селекцию и созревание с получением полностью зрелых антител человека, способных эффективно нейтрализовать целевые антигены. VELOCIMMUNE® мыши способны давать мощный иммунный ответ на разнообразные классы антигенов, что приводит использованию широкого спектра антител человека, и высокоаффинных, и подходящие для применения в качестве терапевтических средств (данные не показаны).
Пример 5
Анализ структуры лимфоидных органов и изотипов сыворотки
[000359] Макроструктуру селезенки, ингвинальных лимфатических узлов, пейеровых бляшек и тимуса из образцов тканей мышей дикого типа или мышей VELOCIMMUNE®, окрашенных гематоксилином-эозином, исследовали с применением оптической микроскопии. Уровни изотипов иммуноглобулина в сыворотке, взятой у мышей дикого типа и мышей VELOCIMMUNE®, анализировали с применением технологии LUMINEX™.
[000360] Структура лимфоидных органов. Структура и функция лимфоидных тканей частично зависят от правильного развития гемопоэтических клеток. Нарушение развития или функции В-клеток может проявляться в виде изменения структуры лимфоидных тканей. Анализ окрашенных тканевых срезов не показал значительных внешних различий вторичных лимфоидных органов мышей дикого типа и мышей VELOCIMMUNE® (данные не показаны).
[000361] Уровни сывороточного иммуноглобулина. Уровни экспрессии каждого изотипа сходны у мышей дикого типа и мышей VELOCIMMUNE® (ФИГ.9А, 9В и 9С). Это указывает на то, что гуманизация вариабельных генных сегментов не оказывает очевидного негативного влияния на переключение классов или экспрессию и секрецию иммуноглобулинов, следовательно, все эндогенные последовательности мыши, необходимые для указанных функций, сохраняются.
Пример 6
Иммунизация и синтез антител у мышей с гуманизированным иммуноглобулином
[000362] Различные варианты мышей VELOCIMMUNE® иммунизировали антигеном для изучения гуморального ответа на стимуляцию чужеродным антигеном.
[000363] Иммунизация и получение гибридом. Мыши VELOCIMMUNE® и мыши дикого типа могут быть иммунизированы антигеном в виде белка, ДНК, комбинации ДНК и белка или клеток, экспрессирующих указанный антиген. Как правило, животные получают бустерные прививки каждые три недели, всего 2-3 раза. После каждой бустерной прививки антигена получают образцы сыворотки от каждого животного и анализируют на антиген-специфический ответ с образованием антител определением титра в сыворотке. Перед слиянием мыши получают последнюю бустерную прививку 5 мкг белка или ДНК, в зависимости от ситуации, в виде внутрибрюшинной или внутривенной инъекции. Спленоциты собирают и сливают с клетками миеломы Ag8.653 в камере для электрослияния в соответствии с предложенным изготовителем протоколом (Cyto Pulse Sciences Inc., Glen Burnie, MD). Через 10 дней культивирования проводят скрининг гибридом на антиген-специфичность с применением ИФА-анализа (Harlow and Lane, 1988, Antibodies: A Laboratory Manual, Cold Spring Harbor Press, New York). Как вариант, антиген-специфические В-клетки выделяют непосредственно из иммунизированных мышей VELOCIMMUNE® и проводят их скрининг с применением стандартных методик, включая описанные в настоящей заявке, для получения специфических антител человека к представляющему интерес антигену (например, см. US 2007/0280945A1, полное содержание которого включено в настоящую заявку посредством ссылки).
[000364] Определение титра в сыворотке. Для отслеживания ответа на антигены в сыворотке животных пробы сыворотки отбирают приблизительно через 10 дней после каждой бустерной прививки, и определяют титры с применением антиген-специфического ИФА. Вкратце, 96-луночные планшеты Nunc MAXISORP™ покрывают 2 мкг/мл антигена, оставляют на ночь при 4°С и блокируют альбумином бычьей сыворотки (Sigma, St. Louis, МО). Образцы сыворотки в серийных 3-кратных разведениях оставляют для связывания в планшетах на протяжении 1 часа при комнатной температуре. Затем планшеты промывают ФСБ, содержащим 0,05% ТВИН-20, и связанный IgG определяют с применением HRP-конъюгированного козьего анти-мышиного Fc (Jackson Immuno Research Laboratories, Inc., West Grove, PA) для получения общего титра IgG, или биотин-меченых изотип-специфичных или специфичных κ легкой цепи поликлональных антител (Southern Biotech Inc.) для получения титров конкретных изотипов, соответственно. При применении меченых биотином антител после промывания планшетов добавляют HRP-конъюгированный стрептавидин (Pierce, Rockford, IL). Во все планшеты добавляют колориметрические субстраты, такие как BD OPTEIA™ (BD Biosciences Pharmingen, Сан-Диего, Калифорния). После того, как реакцию останавливают 1 М фосфорной кислоты, регистрируют оптическое поглощение при 450 нм и анализируют данные с применением программного обеспечения PRISM™ от Graph Pad. Титр определяют как разведение, необходимое для получения сигнала, в два раза превышающего фоновый.
[000365] В одном из экспериментов мышей VELOCIMMUNE® иммунизировали рецептором интерлейкина-6 человека (hIL-6R). Типовые серии титров в сыворотке мышей VELOCIMMUNE® и мышей дикого типа, иммунизированных hIL-6R, приведены на ФИГ.10А и 10В.
[000366] У мышей VELOCIMMUNE® и мышей дикого типа развивался мощный ответ на IL-6R со сходными диапазонами титров (ФИГ.10А). У некоторых мышей из когорт мышей VELOCIMMUNE® и мышей дикого типа максимальный ответ развивался после одной бустерной прививки антигена. Эти результаты указывают на то, что сила и кинетика иммунного ответа на указанный антиген сходны у мышей VELOCIMMUNE® и мышей дикого типа. Затем указанный антиген-специфический ответ с образованием антител анализировали для изучения конкретных изотипов антигенспецифичных антител, обнаруживаемых в сыворотке. И в группах мышей VELOCIMMUNE®, и в группах мышей дикого типа в основном вырабатывался IgG1 (ФИГ.10В), что указывает на то, что переключение классов при гуморальном ответе происходит сходным образом у мышей всех типов.
[000367] Определение аффинности связывания антитела с антигеном в растворе. Как правило, для определения аффинности связывания антитела с антигеном проводят конкурентный анализ в растворе на основе ИФА.
[000368] Вкратце, антитела в кондиционированной среде предварительно смешивают с серийно разведенным в диапазоне от 0 до 10 мг/мл антигенным белком. Затем растворы смесей антитела и антигена инкубируют от двух до четырех часов при комнатной температуре для достижения равновесного связывания. Затем количества свободного антитела в смесях определяют с помощью количественного сэндвич-ИФА. 96-луночные планшеты MAXISORB™ (VWR, Вест Честер, Пенсильвания) покрывают 1 мкг/мл антигенного белка в растворе ФСБ и оставляют на ночь при 4°C с последующей блокировкой неспецифического связывания с применением БСА. Затем растворы смесей антитело-антиген переносят на планшеты и инкубируют в течение 1 часа. Затем планшеты промывают отмывочным буфером, связанные с планшетом антитела выявляют с применением препарата HRP-конъюгированных козьих поликлональных антител против IgG мыши (Jackson Immuno Research Lab) и обрабатывают колориметрическими субстратами, такими как BD OPTEIA™ (BD Biosciences Pharmingen, Сан Диего, Калифорния). После того, как реакцию останавливают 1 М фосфорной кислоты, регистрируют оптическое поглощение при 450 нм и анализируют данные с применением программного обеспечения PRISM™ от Graph Pad. Зависимость сигналов от концентраций антигена в растворе анализируют с применением четырехпараметрической оптимизации и представляют в виде IC50, концентрации антигена, которая требуется для 50%-ного снижения сигнала от образцов антител при отсутствии антигена в растворе.
[000369] В одном из экспериментов мышей VELOCIMMUNE® иммунизировали hIL-6R (согласно описаниям выше). На ФИГ.11А и 11В приведен типовая серия показателей аффинности анти-hILGR антител мышей VELOCIMMUNE® и мышей дикого типа.
[000370] После получения иммунизированными мышами третьей антигенной бустерной прививки, определяют титры в сыворотке с применением анализа ИФА. Спленоциты выделяют из мышей отобранных когорт дикого типа и VELOCIMMUNE®, сливают с клетками миеломы Ag8.653 с образованием гибридом и культивируют в селективных условиях (согласно описаниям выше). Среди полученных 671 анти-IL-6R гибридом обнаружено 236, экспрессирующих антиген-специфичные антитела. Среду, собранную из антиген-положительных лунок, использовали для определения способности антитела к связыванию антигена, с применением конкурентного ИФА-анализа в растворе. Антитела, полученные от мышей VELOCIMMUNE®, демонстрируют широкий диапазон аффинности связывания антигенов в растворе (ФИГ.11А). При этом обнаружено, что 49 из 236 анти-IL-6R гибридом блокируют связывание IL-6 с рецептором в биоанализе in vitro (данные не показаны). Далее, указанные 49 анти-IL-6R блокирующие антитела демонстрировали диапазон высоких значений аффинности в растворе, сходный с диапазоном блокирующих антител, полученных при параллельной иммунизации мышей дикого типа (ФИГ.11В).
Пример 7
Конструирование направленного вектора ADAM6 мыши
[000371] Из-за замены локусов вариабельных генов тяжелой цепи иммуноглобулина мыши локусами вариабельных генов тяжелой цепи иммуноглобулина человека, у первых вариантов мышей VELOCIMMUNE® отсутствовала экспрессия генов ADAM6 мыши. В частности, у мышей-самцов VELOCIMMUNE® наблюдалось снижение фертильности. Соответственно, способность экспрессировать ADAM6 была реконструирована у мышей VELOCIMMUNE® для восстановления нарушенной фертильности.
[000372] Направленный вектор для вставки генов ADAM6a и ADAM6b мыши в гуманизированный локус тяжелой цепи конструировали с применением VELOCIGENE® генноинженерной методики (выше) для модификации искусственной бактериальной хромосомы (ВАС) 929d24, предоставленной д-ром Фредериком Альтом (Dr. Frederick Alt, Гарвардский университет). ДНК ВАС 929d24 была сконструирована таким образом, чтобы включать геномные фрагменты, содержащие гены ADAM6a и ADAM6b мыши и гигромициновую кассету для направленного удаления псевдогена ADAM6 человека (hADAM6Ψ), расположенного между генными сегментами VH1-2 и VH6-1 человека в гуманизированном локусе тяжелой цепи (ФИГ.12).
[000373] Сначала геномный фрагмент, содержащий ген ADAM6b мыши, ~800 п.о. расположенной в обратном направлении (5′) последовательности и ~4800 п.о. расположенной в прямом направлении (3′) последовательности субклонировали из клона ВАС 929d24. Второй геномный фрагмент, содержащий ген ADAM6a мыши, ~300 п.о. расположенной в обратном направлении (5′) последовательности и ~3400 п.о. расположенной в прямом направлении (3′) последовательности, отдельно субклонировали из клона ВАС 929d24. Указанные два геномных фрагмента, содержащие гены ADAM6b и ADAM6a мыши, лигировали в гигромициновую кассету, фланкированную сайтами рекомбинации Frt, с получением направленного вектора (направленный вектор ADAM6 мыши, ФИГ.12; SEQ ID NO:3). Сайты разных рестрикционных ферментов встраивали в 5′-конец направленного вектора после гена ADAM6b мыши и в 3′-конец после гена ADAM6a мыши (снизу на ФИГ.12) для лигирования в гуманизированный локус тяжелой цепи.
[000374] Отдельно модифицировали клон ВАС, содержащий замену локусов вариабельных генов тяжелой цепи мыши локусами вариабельных генов тяжелой цепи человека, в том числе псевдоген ADAM6 человека (hADAM6Ψ), расположенный между генными сегментами VH1-2 и VH6-1 человека в гуманизированном локусе, для последующего лигирования направленного вектора ADAM6 мыши (ФИГ.13).
[000375] Вкратце, неомициновую кассету, фланкированную сайтами рекомбинации loxP, конструировали таким образом, чтобы она содержала плечи гомологии, включающие геномную последовательность человека в положении 3′ от генного сегмента VH1-2 человека (5′ относительно hADAM6Ψ) и 5′ от генного сегмента человека VH6-1 (3′ относительно hADAM6Ψ; см. середину ФИГ.13). Сайт вставки указанной направленной конструкции располагался приблизительно в пределах от 1.3 т.н. в 5′ направлении и до ~350 п.о. в 3′ направлении от псевдогена человека ADAM6. Указанная направленная конструкция включала также те же сайты рестрикции, что и направленный вектор ADAM6 мыши, чтобы позволить последующее лигирование ВАС модифицированного клона ВАС с делецией псевдогена ADAM6 человека и направленного вектора ADAM6 мыши.
[000376] После расщепления ДНК ВАС, полученной из обеих конструкций, геномные фрагменты лигировали совместно для создания сконструированного клона ВАС, содержащего гуманизированный локус тяжелой цепи с эктопически расположенной геномной последовательностью, содержащей нуклеотидные последовательности ADAM6a и ADAM6b мыши. Полученная направленная конструкция для удаления гена ADAM6 человека в гуманизированном локусе тяжелой цепи и вставки последовательностей ADAM6a и ADAM6b мыши в ЭС-клетки включала, начиная с 5′- и до 3′-областей, 5′ геномный фрагмент, содержащий ~13 т.н. геномной последовательности человека 3′ от генного сегмента VH1-2 человека, ~800 п.о. геномной последовательности мыши в 3′-направлении от гена ADAM6b мыши, ген ADAM6b мыши, ~4800 п.о. геномной последовательности в 5′-направлении от гена ADAM6b мыши, 5′ сайт Frt, гигромициновую кассету, 3′сайт Frt, ~300 п.о. геномной последовательности мыши в 3′-направлении от гена ADAM6a мыши, ген ADAM6a мыши, ~3400 п.о. геномной последовательности мыши в 5′-направлении от гена ADAM6a мыши и 3′ геномный фрагмент, содержащий ~30 т.н. геномной последовательности человека в 5′направлении от генного сегмента VH6-1 человека (внизу на ФИГ.13).
[000377] Сконструированный клон ВАС (описанный выше) применяли для электропорации ЭС-клеток мыши, содержащих гуманизированный локус тяжелой цепи, для создания модифицированных ЭС-клеток с эктопически размещенной геномной последовательностью мыши, содержащей последовательности ADAM6a и ADAM6b мыши в гуманизированном локусе тяжелой цепи. Позитивные ЭС-клетки, содержащие указанный эктопический фрагмент генома мыши в гуманизированном локусе тяжелой цепи, идентифицировали с помощью количественной ПЦР с применением зондов TAQMAN™ (Lie and Petropoulos, 1998, Advances in quantitative PCR technology: 5′nuclease assays, Curr Opin Biotechnol 9(1):43-48). 5′- и 3′-области за пределами модифицированной части гуманизированного локуса тяжелой цепи проверяли с помощью ПЦР с применением праймеров и зондов внутри модифицированной области для подтверждения присутствия эктопической геномной последовательности мыши в гуманизированном локусе тяжелой цепи, а также гигромициновой кассеты. Последовательность нуклеотидов 5′-вставки включала следующую последовательность, подтверждающую присутствие геномной последовательности тяжелой цепи человека в 5′-направлении от вставки и сайта рестрикции I-Ceul (приведенного ниже в скобках), прилегающего к геномной последовательности мыши, в месте вставки: (CCAGCTTCAT TAGTAATCGT TCATCTGTGG TAAAAAGGCA GGATTTGAAG CGATGGAAGA TGGGAGTACG GGGCGTTGGA AGACAAAGTG CCACACAGCG CAGCCTTCGT CTAGACCCCC GGGCTAACTA TAACGGTCCT AAGGTAGCGA G) GGGATGACAG ATTCTCTGTT CAGTGCACTC AGGGTCTGCC TCCACGAGAA TCACCATGCC CTTTCTCAAG ACTGTGTTCT GTGCAGTGCC CTGTCAGTGG (SEQ ID NO:4). Последовательность нуклеотидов 3′-вставки на 3′-конце целевой области включала следующую последовательность, подтверждающую присутствие геномной последовательности мыши и сайта рестрикции PI-Scel (приведенного ниже в скобках), прилегающего к геномной последовательности тяжелой цепи человека в 3′ направлении от места вставки: (AGGGGTCGAG GGGGAATTTT ACAAAGAACA AAGAAGCGGG CATCTGCTGA CATGAGGGCC GAAGTCAGGC TCCAGGCAGC GGGAGCTCCA CCGCGGTGGC GCCATTTCAT ТАССТСТТТС TCCGCACCCG ACATAGATAAAGCTT) АТСССССАСС AAGCAAATCC CCCTACCTGG GGCCGAGCTT CCCGTATGTG GGAAAATGAA TCCCTGAGGT CGATTGCTGC ATGCAATGAA ATTCAACTAG (SEQ ID NO:5).
[000378] Таргетированные ЭС-клетки, описанные выше, применяли в качестве донорских ЭС-клеток и вводили в мышиные эмбрионы на стадии 8 клеток с помощью технологии получения мышей VELOCIMOUSE® (VELOCIMOUSE® mouse engineering method) (см., например, патенты США №76598442, №7576259, №7294754). Мышей, несущих гуманизированный локус тяжелой цепи, включающий эктопическую геномную последовательность мыши, содержащую последовательности ADAM6a и ADAM6b мыши, идентифицировали генотипированием с применением модифицированного аллельного анализа (Valenzuela et al., 2003), определяющего наличие генов ADAM6a и ADAM6b мыши в гуманизированном локусе тяжелой цепи.
[000379] Мышей, несущих гуманизированный локус тяжелой цепи, содержащий гены ADAM6a и ADAM6b мыши, скрещивают с мышами линии FLPe deleter (см., например, Rodriguez et al., 2000, High-efficiency deleter mice show that FLPe is an alternative to Cre-loxP. Nature Genetics 25:139-140) для устранения любой Frt-фланкированной гигромициновой кассеты, введенной посредством направленного вектора, не удаленной, например, на стадии ЭС-клеток или эмбриона. Опционально, указанную гигромициновую кассету у мышей сохраняют.
[000380] Детенышей генотипируют, и детеныша, гетерозиготного по гуманизированному локусу тяжелой цепи, содержащему эктопический фрагмент генома мыши с последовательностями ADAM6a и ADAM6b, отбирают для исследования характеристик экспрессии гена ADAM6 мыши и фертильности.
Пример 8
Исследование характеристик мышей с компенсацией функции ADAM6
[000381] Проточная цитометрия. Трех мышей возрастом 25 недель, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепей человека (Н+/+κ+/+) и трех мышей возрастом 18-20 недель, гомозиготных по тяжелой и κ легкой цепям человека, у которых присутствует эктопический фрагмент генома мыши, кодирующий гены ADAM6a и ADAM6b мыши в составе обоих аллелей локуса тяжелой цепи человека (Н+/+A6resκ+/+), умерщвляли для идентификации и анализа популяций клеток лимфоцитов с помощью FACS в системе для проточной цитометрии BD LSR II System (BD Bioscience). Лимфоциты гейтировали по конкретным клеточным линиям и анализировали прогрессию по мере прохождения различных стадий развития В-клеток. Ткани, извлекаемые из животных, включали кровь, селезенку и костный мозг. Кровь отбирали в пробирки BD Microtainer с ЭДТА (BD Biosciences). Костный мозг извлекали из бедренных костей промыванием полной средой RPMI с добавлением эмбриональной телячьей сыворотки, пирувата натрия, HEPES, 2-меркаптоэтанола, заменимых аминокислот и гентамицина. Эритроциты из препаратов крови, селезенки и костного мозга лизировали лизирующим буфером на основе хлорида аммония (например, лизирующим буфером ACK), с последующим промыванием полной средой RPMI.
[000382] Для окрашивания популяций клеток 1×106 клеток из различных тканей инкубировали с анти-мышиными CD16/CD32 (2.4G2, BD Biosciences) на льду в течение 10 минут с последующим мечением одним или комбинацией следующей смесь антител на протяжении 30 минут на льду.
[000383] Костный мозг: ФИТЦ-меченые против мышиного CD43 (1В11, BioLegend), ФЭ-меченые ckit (2B8, BioLegend), PeCy7-IgM (II/41, eBioscience), PerCP-Cy5.5-IgD (11-26c.2a, BioLegend), APC-eFluor780-B220 (RA3-6B2, eBioscience), A700-CD19 (1D3, BD Biosciences).
[000384] Периферическая кровь и селезенка: ФИТЦ-меченые против κ мыши (187.1, BD Biosciences), ФЭ-меченые λ (RML-42, BioLegend), PeCy7-IgM (II/41, eBioscience), PerCP-Cy5.5-IgD (11-26с.2а, BioLegend), APC-CD3 (145-2С11, BD), A700-CD19 (1D3, BD), APC-eFluor780-B220 (RA3-6B2, eBioscience). После инкубирования с мечеными антителами клетки промывали и фиксировали в 2% формальдегиде. Регистрацию данных проводили на проточном цитометре LSRII и анализировали с помощью FlowJo (Treestar, Inc.). Результаты для типовых мышей H+/+κ+/+ и Н+/+A6resκ+/+ показаны на ФИГ.14-18.
[000385] Результаты показывают, что В-клетки мышей Н+/+A6resκ+/+ прогрессируют через стадии развития В-клеток сходным с клетками мышей H+/+κ+/+ образом в костном мозге и периферических областях, и демонстрируют нормальные паттерны созревания, как только попадают на периферию. У мышей Н+/+A6resκ+/+ мыши наблюдалось увеличение популяции CD43intCD19+ клеток по сравнению с мышами Н+/+κ+/+ (ФИГ.16В). Это может свидетельствовать о повышении экспрессии IgM из гуманизированного локуса тяжелой цепи, включающего эктопический фрагмент генома мыши, содержащий последовательности ADAM6a мыши и ADAM6b, у мышей Н+/+A6resκ+/+. На периферии у мышей Н+/+A6resκ+/+ обнаруживаются нормальные сходные с таковыми у мышей Н+/+κ+/+популяции В- и Т-клеток.
[000386] Морфология семенников и характеристики спермы. Для определения того, обусловлена ли бесплодность мышей с гуманизированными вариабельными локусами тяжелой цепи иммуноглобулина дефектами семенников и/или продуцирования спермы, исследовали морфологию семенников и содержание спермы в эпидидимисе.
[000387] Вкратце, семенники двух групп (n=5 на группу; группа 1: мыши, гомозиготные по локусам вариабельных генов тяжелой и κ легкой цепей человека, H+/+κ+/+; группа 2: мыши, гетерозиготные по локусам вариабельных генов тяжелой цепи человека и гомозиготные по локусам вариабельных генов κ легкой цепи человека, Н+/-κ+/+) извлекали, оставляя интактным эпидидимис, и взвешивали. 3атем образцы фиксировали, заливали парафином, изготавливали срезы и окрашивали гематоксилином-эозином (НЕ). Срезы семенников (2 семенника/мышь, всего 20) исследовали на дефекты морфологии и признаки продуцирования спермы, а срезы эпидидимиса исследовали на присутствие спермы.
[000388] В этом эксперименте не обнаружено различий в массе или морфологии семенников мышей Н+/+κ+/+ и Н+/-κ+/+. Сперма обнаружена как в семенниках, так и в эпидидимисе для всех генотипов. Эти результаты доказывают, что отсутствие генов ADAM6a и ADAM6b мыши не приводит к детектируемым изменениям морфологии семенников, и что сперма продуцируется у мышей в присутствии и отсутствие указанных двух генов. Таким образом, дефекты фертильности самцов Н+/+κ+/+, по-видимому, обусловлены низкими уровнями продуцирования спермы.
[000389] Подвижность и перемещение спермы. Мыши, у которых отсутствует другие члены семейства генов ADAM, бесплодны из-за дефектов подвижности или перемещения спермы. Перемещение спермы определяется как способность спермы передвигаться из матки в маточные трубы, и в норме необходимо для оплодотворения у мышей. Для определения, влияет ли на указанный процесс удаление мышиных ADAM6a и ADAM6b, оценивали способность к перемещению и подвижность спермы у мышей Н+/+κ+/+.
[000390] Вкратце, сперму получали из семенников (1) мышей, гетерозиготных по локусам вариабельных генов тяжелой цепи человека и гомозиготных по локусам вариабельных генов κ легкой цепи человека (Н+/-κ+/+); (2) мышей, гомозиготных по локусам вариабельных генов тяжелой цепи человека и гомозиготных по локусам вариабельных генов κ легкой цепи человека (H+/+κ+/+); (3) мышей, гомозиготных по локусам вариабельных генов тяжелой цепи человека и гомозиготных по κ легкой цепи дикого типа (H+/+mκ); и (4) мышей дикого типа С57 BL/6 (WT). При подсчете спермиев или оценке общей подвижности спермы при наблюдении не обнаружено значимых отклонений. У всех мышей наблюдалась дисперсия кумулюса, указывая на то, что каждый образец спермы был способен проникать через клетки кумулюса и связываться с zona pellucida in vitro. Эти результаты доказывают, что сперма мышей Н+/+κ+/+ способна проникать в кумулюс и связываться с zona pellucida.
[000391] Оплодотворение яйцеклетки мыши in vitro (IVF) осуществляли с применением спермы мышей согласно приведенному выше описанию. У мышей Н+/+κ+/+ на следующий день после IVF наблюдалось несколько меньшее число делящихся эмбрионов, а также пониженное количество связавшихся с яйцеклетками спермиев. Эти результаты доказывают, что сперма мышей Н+/+κ+/+ при контакте с яйцеклеткой способна проникать через кумулюс и связываться с zona pellucida.
[000392] В еще одном эксперименте с помощью анализа перемещения спермы определяли способность спермы мышей H+/+κ+/+ перемещаться из матки и через маточные трубы.
[000393] Вкратце, первую группу мышей-самок с суперовуляцией (n=5) размещали с самцами Н+/+κ+/+ (n=5), а вторую группу мышей-самок с суперовуляцией (n=5) размещали с самцами Н+/-κ+/+ (n=5). Наблюдали копуляции у пар, через пять-шесть часов после копуляции извлекали матку с присоединенными маточными трубами из всех самок и делали смывы для анализа. Смывы проверяли на наличие яйцеклеток для подтверждения овуляции и и подсчитывали количество спермы. Перемещение спермы оценивали двумя различными способами. Во-первых, обе маточные трубы извлекали из матки, делали смывы солевым раствором и подсчитывали всю замеченную сперму. Также отмечали присутствие яйцеклеток как подтверждение овуляции. Во-вторых, маточные трубы оставляли прикрепленными к матке и обе ткани фиксировали, заливали парафином, изготавливали срезы и окрашивали (согласно описаниям выше). Срезы исследовали на присутствие спермы, как в матке, так и в обеих маточных трубах.
[000394] У самок, спаривавшихся с пятью самцами Н+/+κ+/+, в смыве из маточных труб обнаруживалась очень незначительное количество спермы. В смывах из маточных труб самок, спаривавшихся с самцами Н+/-κ+/+, обнаружено приблизительно в 25-30 раз больше спермы (avg, n=10 маточные трубы), чем в смывах из маточных труб самок, спаривавшихся с самцами Н+/+κ+/+. Сравнение типового скрещивания мышей Н+/+κ+/+ и Н+/+A6resκ+/+ приведено в Таблице 9.
[000395] Готовили гистологические срезы матки и маточных труб. Срезы исследовали на присутствие спермы в матке и маточных трубах (colliculus tubarius). Исследование гистологических срезов маточных труб и матки показало, что у мышей-самок, спаривавшихся с мышами Н+/+κ+/+, сперма обнаруживается в матке, но не в маточных трубах. Далее, на срезах от самок, спаривавшихся с мышами Н+/+κ+/+, видно, что сперма не обнаруживается в маточно-трубном сочленении (UTJ). На срезах от самок, спаривавшихся с мышами Н+/-κ+/+, сперма обнаружена в UTJ и в маточных трубах.
[000396] Эти результаты доказывают, что мыши, у которых отсутствуют гены ADAM6a и ADAM6b, продуцируют сперму, у которой наблюдается дефект подвижности in vivo. В любом случае, сперма обнаруживалась внутри матки, указывая на то, что копуляция и высвобождение спермы, очевидно, происходят нормальным образом, но в маточных трубах после копуляции обнаруживается небольшое количество спермы или вообще не обнаруживается сперма, что определяют либо подсчетом спермиев, либо при гистологическом исследовании. Эти результаты доказывают, что мыши, у которых отсутствуют гены ADAM6a и ADAM6b, продуцируют сперму, у которой наблюдается неспособность перемещаться из матки в маточные трубы. Указанный дефект, по-видимому, приводит к бесплодию, поскольку сперма не способна проходить через маточно-трубное сочленение в маточные трубы, где оплодотворяются яйцеклетки. Вместе все указанные результаты подтверждают гипотезу, что гены ADAM6 мыши способствуют направленному перемещению спермы с нормальной подвижностью из матки через маточно-трубное сочленение и маточные трубы и, таким образом, достижению яйцеклетки с осуществлением успешного оплодотворения. Механизм, при помощи которого ADAM6 это обеспечивает, может регулироваться одним или обоими белками ADAM6, либо координированной экспрессией других белков, например, других белков ADAM, в спермии, согласно приведенному ниже описанию.
[000397] Экспрессия генов семейства ADAM. Известно, что комплекс белков ADAM присутствует в виде комплекса на поверхности созревающей спермы. Мыши, у которых отсутствует другие члены семейства генов ADAM, утрачивают указанный комплекс по мере созревания спермы, и у них наблюдается уменьшение количества различных белков ADAM в зрелой сперме. Для определения возможного сходного влияния отсутствия генов ADAM6a и ADAM6b на другие белки ADAM, проводили вестерн-блоттинг белковых экстрактов из семенников (незрелая сперма) и эпидидимиса (созревающая сперма) для определения уровней экспрессии других члены семейства генов ADAM.
[000398] В этом эксперименте анализировали белковые экстракты из групп (n=4 на группу) мышей H+/+κ+/+ и Н+/-κ+/+. Результаты показали, что экспрессия ADAM2 и ADAM3 не оказывала влияния на экстракты из семенников. Однако в экстрактах из эпидидимиса количество и ADAM2, и ADAM3 существенно уменьшалось. Это указывает на то, что отсутствие ADAM6a и ADAM6b в сперме мышей Н+/+κ+/+ может оказывать непосредственный эффект на экспрессию и, возможно, функцию других белков ADAM по мере созревания спермы (например, ADAM2 и ADAM3). Это дает основание полагать, что ADAM6a и ADAM6b составляют часть белкового комплекса ADAM на поверхности спермы, что может быть критически важно для нормальной миграции спермы.
Пример 9
Использование вариабельных генов тяжелой цепи человека у мышей с компенсацией функции ADAM6
[000399] У мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепи человека, у которых либо отсутствуют гены ADAM6a и ADAM6b мыши (H+/+κ+/+), либо присутствует эктопический геномный фрагмент, кодирующий гены ADAM6a и ADAM6b мыши (H+/+A6resκ+/+), с помощью количественной ПЦР с применением зондов TAQMAN™ (согласно приведенному выше описанию) определяли частоту использования выбранных вариабельных генов тяжелой цепи человека.
[000400] Вкратце, CD19+ В-клетки выделяли из селезенок мышей Н+/+κ+/+ и H+/+A6resκ+/+ с применением микробус с CD19 мыши (Miltenyi Biotec), и общей РНК очищали с применением набора RNEASY™ Mini kit (Qiagen). Геномную РНК извлекали на колонке с применением не содержащей рибонуклеаз ДНКазы (RNase-Free DNase) (Qiagen). Приблизительно 200 нг иРНК обратно транскрибировали в кДНК с применением набора First Stand cDNA Synthesis kit (Invitrogen) и затем амплифицировали, используя смесь TAQMAN™ Universal PCR Master Mix (Applied Biosystems) с применением системы обнаружения последовательностей ABI 7900 (ABI 7900 Sequence Detection System) (Applied Biosystems). Относительную экспрессию каждого гена нормализовали по экспрессии константной области κ легкой цепи мыши (mCκ). В Таблице 10 приведены комбинации смысловых/антисмысловых/TAQMAN™ MGB зондов, использованных в этом эксперименте.
[000401] В этом эксперименте в проанализированных образцах обнаружена экспрессия всех четырех генов человека VH. Далее, уровни экспрессии были сопоставимы у мышей Н+/+κ+/+ и Н+/+A6resκ+/+. Эти результаты показывают, что VH гены человека, как дистальные относительно участка модификации (VH3-23 и VH1-69), так и проксимальные относительно участка модификации (VH1-2 и VH6-1), способны к рекомбинации с образованием функционально экспрессируемой тяжелой цепи человека. Эти результаты показывают, что эктопический геномный фрагмент, содержащий последовательности ADAM6a и ADAM6b мыши, встроенные в геномную последовательность тяжелой цепи человека, не влияет на V(D)J рекомбинацию генных сегментов тяжелой цепи человека внутри указанного локуса, и указанные мыши способны к рекомбинации генных сегментов тяжелой цепи человека нормальным образом с синтезом функциональных белков тяжелой цепи иммуноглобулина.
Пример 10
Гуморальный иммунный ответ у мышей с компенсацией функции ADAM6
[000402] У мышей, гомозиготных по локусам вариабельных генов тяжелой и κ легкой цепи человека, у которых либо отсутствуют гены ADAM6a и ADAM6b мыши (Н+/+κ+/+), либо присутствует эктопический геномный фрагмент, кодирующий гены ADAM6a и ADAM6b мыши (H+/+A6resκ+/+), определяли гуморальный иммунный ответ с применением мульти-антигенной схемы иммунизации с последующим выделением и характеризацией антител. Результаты сравнивали для определения какого-либо влияния на V(D)J-рекомбинацию, задействующую сегменты генов иммуноглобулина человека, оценки повышения титра в сыворотке, синтеза антител гибридомами и аффинности к антигену.
[000403] Протокол иммунизации. Для сравнительной иммунизации в группах мышей применяли А-рецептор клеточной поверхности человека (Антиген А), специфичное к рецепторной тирозиновой протеинкиназе человека антитело человека (Антиген В), секретируемый белок человека, участвующий в регуляции TGF-β сигнального пути (Антиген С) и рецепторную тирозинкиназу человека (Антиген D). Перед иммунизацией вышеуказанными антигенами у групп мышей брали сыворотку. Каждый антиген (по 2,3 мкг) вводили при начальной примирующей иммунизации в смеси с 10 мкг олигонуклеотида CpG в качестве адъюванта (Invivogen). Иммуноген вводили в подушечки лап в объеме 25 мкл на мышь. После этого мышей бустеризовали введением в подушечки лап 2,3 мкг антигена, а также 10 мкг CpG и 25 мкг Adju-Phos (Brenntag) в качестве адъювантов на 3, 6, 11, 13, 17, и 20 день, в общей сложности шестью бустерными прививками. У мышей брали кровь на 15 и 22 день после четвертой и шестой бустерных прививок, соответственно, и антисыворотки анализировали на титр антител к каждому конкретному антигену.
[000404] Титры антител определяли в сыворотке иммунизированных мышей с применением анализа ИФА. 96-луночные титрационные микропланшеты (Thermo Scientific) покрывали соответствующим антигеном (2 мкг/мл) в солевом растворе с фосфатным буфером (ФСБ, Irvine Scientific) в течение ночи при 4°С. На следующий день планшеты промывали солевым раствором с фосфатным буфером, содержащим 0,05% ТВИН-20 (ФСБ-Т, Sigma-Aldrich) четырехкратно с применением устройства для отмывки планшетов (Molecular Devices). Затем планшеты блокировали 250 мкл 0,5% альбумина бычьей сыворотки (БСА, Sigma-Aldrich) в ФСБ и инкубировали на протяжении 1 часа при комнатной температуре. Затем планшеты промывали четырехкратно ФСБ-Т. Сыворотку от иммунизированных мышей и преиммунную сыворотку серийно разводили три раза в 0,5% БСА-ФСБ, начиная с 1:300 или 1:1000, и добавляли в блокированные планшеты в двух повторностях и инкубировали на протяжении 1 часа при комнатной температуре. Последние две ячейки оставляли пустыми для использования в качестве контроля для вторичных антител. Планшеты повторно промывали четырехкратно ФСБ-Т в устройстве для отмывки планшетов. Козье анти-мышиное IgG-Fc с пероксидазой хрена (HRP, Jackson Immunoresearch) или козье анти-мышиное IgG-каппа-HRP-(Southern Biotech)конъюгированное вторичное антитело в разведении 1:5000/1:10000 добавляли в планшеты и инкубировали на протяжении 1 часа при комнатной температуре. Планшеты повторно промывали восьмикратно ФСБ-Т и обрабатывали с применением ТМВ/Н2О2 в качестве субстрата. Субстрат инкубировали в течение 20 минут и реакцию останавливали 2 N H2SO4 (VWR) или 1 N Н3РО4 (JT Baker). Планшеты считывали на спектрофотометре (Victor, Perkin Elmer) на 450 нм. Титры антител подсчитывали с применением программного обеспечения Graphpad PRISM.
[000405] Титр в сыворотке рассчитывали как разведение сыворотки в пределах экспериментального диапазона титрования при сигнале связывания антигена, эквивалентном превышающему фоновый в два раза. Результаты гуморального иммунного ответа показаны на ФИГ.19 (Антиген А), ФИГ.20 (Антиген В), ФИГ.21 (Антиген С) и ФИГ.22 (Антиген D). Уровни антиген-положительных гибридом, полученных из двух селезенок, извлеченных из мышей каждой группы, иммунизированной выбранным антигеном, приведены в Таблице 11 (Показатель антигена соответствует 2Х/фон).
[000406] Как видно из этого Примера, титры антител, синтезируемых у мышей с компенсацией функции ADAM6 (Н+/+A6resκ+/+) были сопоставимы с таковыми синтезируемых у мышей, у которых отсутствовали ADMA6a и ADAM6b и присутствовали гуманизированная тяжелая цепь (Н+/+κ+/+). Далее, селезенки мышей Н+/+A6resκ+/+ давали антиген-положительные гибридомы для всех протестированных антигенов, включая антитела с высокой аффинностью, на уровнях, сопоставимых с мышами H+/+κ+/+. Соответственно, предполагается, что у мышей с компенсацией функции ADAM6 не нарушена V(D)J-рекомбинация сегментов гена иммуноглобулина человека, учитывая синтез антител с высокой аффинностью, задействующий гены иммуноглобулина человека.
Пример 11
Определение антигенсвязывающей способности
[000407] Аффинность антител, демонстрирующих специфическое связывание с Антигеном В, отслеживали с применением биосенсора на основе поверхностного плазмонного резонанса в реальном времени (BIAcore 2000). Для скрининга BIAcore использовали кондиционированные среды от гибридом, выделенных из мышей двух линий, иммунизированных Антигеном В (H+/+κ+/+ и H+/+A6resκ+/+). Сначала поверхность сенсора BIAcore дериватизировали поликлональным кроличьим анти-мышиным антителом (GE) для захвата анти-Антиген В антител из кондиционированной среды. На протяжении всего процесса скрининга в качестве подвижного буфера применяли HBST (0,01 М HEPES рН 7.4; 0,15М NaCl; 3мМ ЭДТА; 0,005 об.% сурфактанта Р20). Fab фрагмент Антигена В инжектировали на поверхность с захваченным анти-Антиген В антителом со скоростью 50 мкл/мин в концентрации 100 нМ. Связывание антиген-антитело отслеживали на протяжении трех минут, а диссоциацию антигена и захваченного антитела отслеживали на протяжении пяти минут в подвижном буфере HBST. Эксперимент проводили при 25°С. Константы скорости связывания (ka) и скорости диссоциации (kd) определяли с помощью обработки и приведения данных к 1:1 модели связывания с применением программного обеспечения для аппроксимации кривых Scrubber 2.0. Константы равновесия при диссоциации связей (KD) и время полужизни при диссоциации (Т½) вычисляли из констант скорости реакции как: KD (М)=kd/ka; и Т½ (мин) = (In2/(60*kd). Результаты для отобранных анти-Антиген В антител показаны в Таблице 12.
[000408] В сходном эксперименте кинетику связывания различных моноклональных антител, присутствующих в кондиционированных гибридомных средах, с Антигеном А определяли с применением биосенсорного анализа на основе поверхностного плазмонного резонанса в реальном времени (BIAcore 4000). Все клоны гибридом, применяемые для этого анализа, синтезировались мышами H+/+A6resκ+/+.
[000409] Вкратце, для захвата Антиген А-специфичных антител сначала иммобилизировали на сенсорном чипе поликлональное кроличье анти-мышиное антитело (GE Catalog# BR-1008-38). Проводили скрининг BIAcore в двух разных буферах - PBSP с рН7.2 и PBSP с рН6.0. В оба буфера добавляли по 0,1 мг/мл БСА. После захвата анти-Антиген А антител из кондиционированной среды 1 мкМ мономера Антигена А (приготовленного в соответствующем подвижном буфере) инжектировали на поверхность с захваченными антителами в течение 1,5 минут со скоростью 30 мкл/мин и отслеживали диссоциацию связанного мономера Антигена А в течение 1,5 минут в соответствующем подвижном буфере при 25°С. Константы скорости связывания (ka) и диссоциации (kd) определяли обработкой и приведению данных к 1:1 модели связывания с применением программного обеспечения для аппроксимации кривых Scrubber 2.0. Константы равновесия при диссоциации связей (KD) и время полужизни при диссоциации (Т½) подсчитывали из констант скорости реакции как: KD (М) = kd/ka; и Т½ (мин) = (In2/(60*kd). В Таблице 13 приведены параметры кинетики связывания для связывания выбранного анти-Антиген А антитела с мономером Антигена А при рН7.2 и рН6.0. NB: связывания не обнаружено в рассматриваемых экспериментальных условиях.
[000410] Как показано выше, антитела с высокой аффинностью получали и от H+/+A6resκ+/+, и от H+/+κ+/+ мышей аналогичным способом. Из 25 антител, представленных в Таблице 12, у двадцати, синтезированных H+/+A6resκ+/+ мышами, наблюдался диапазон аффинности от 0,5 нМ до 1 мкМ, в то время как у пяти, синтезированных H+/+κ+/+ мышами, наблюдался диапазон аффинности от 10 нМ до 150 нМ. Далее, у 25 антител, приведенных в Таблице 13, наблюдался диапазон аффинности от 20 пМ до 350 нМ в отношении связывания с мономером Антигена А.
[000411] Как видно из этого Примера, повторное встраивание генов Adam6 мыши в локус тяжелой цепи гуманизированного иммуноглобулина не нарушает способность мышей давать мощный иммунный ответ на разнообразные антигены, характеризующийся репертуарами антител человека с различной аффинностью в субнаномолярном диапазоне, полученных из генных сегментов человека, реаранжированных из сконструированной зародышевой линии.
| название | год | авторы | номер документа |
|---|---|---|---|
| МЫШИ ADAM6 | 2012 |
|
RU2722373C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664181C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2761639C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664232C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2787781C2 |
| ГУМАНИЗИРОВАННЫЕ НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕКУ ЖИВОТНЫЕ С ОГРАНИЧЕННЫМИ ЛОКУСАМИ ТЯЖЕЛОЙ ЦЕПИ ИММУНОГЛОБУЛИНА | 2013 |
|
RU2656155C2 |
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ, КОТОРЫЕ ЭКСПРЕССИРУЮТ ТЯЖЕЛЫЕ ЦЕПИ, СОДЕРЖАЩИЕ ДОМЕНЫ VL | 2013 |
|
RU2664185C2 |
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ, КОТОРЫЕ ЭКСПРЕССИРУЮТ ТЯЖЕЛЫЕ ЦЕПИ, СОДЕРЖАЩИЕ ДОМЕНЫ VL | 2013 |
|
RU2770016C2 |
| ЖИВОТНЫЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ, ИМЕЮЩИЕ СКОНСТРУИРОВАННУЮ ЛЕГКУЮ ЦЕПЬ ЛЯМБДА ИММУНОГЛОБУЛИНА, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2778410C2 |
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ ДЛЯ ТЕСТИРОВАНИЯ ТЕРАПЕВТИЧЕСКИХ АГЕНТОВ | 2019 |
|
RU2805212C2 |
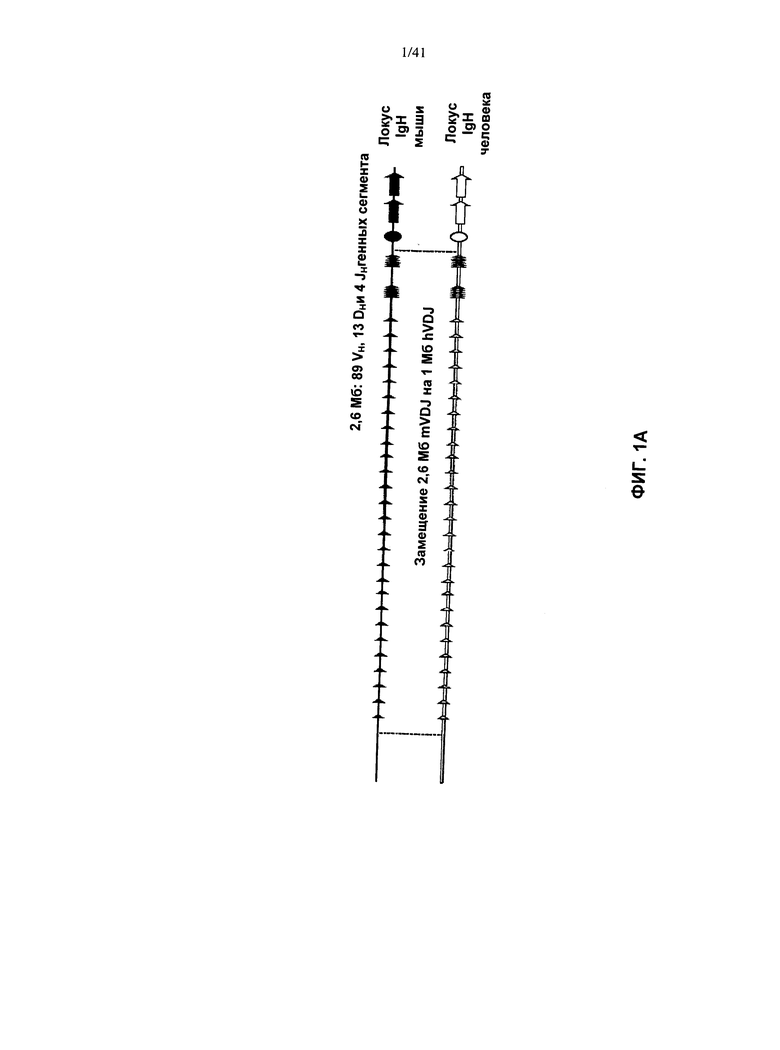
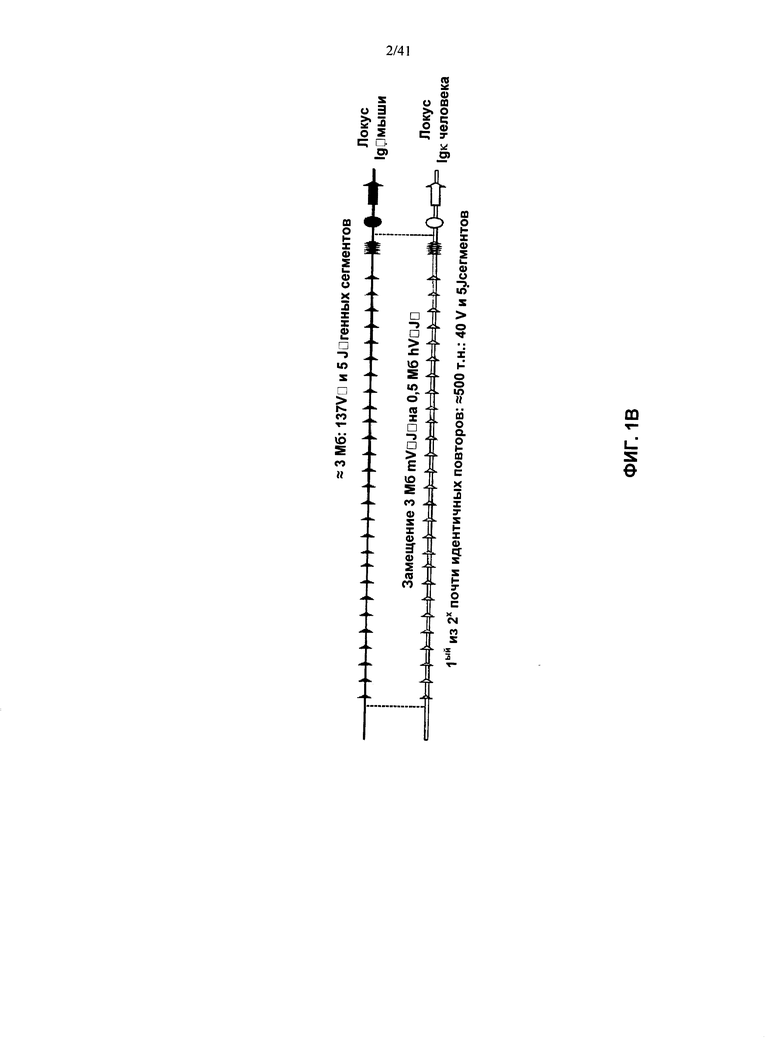


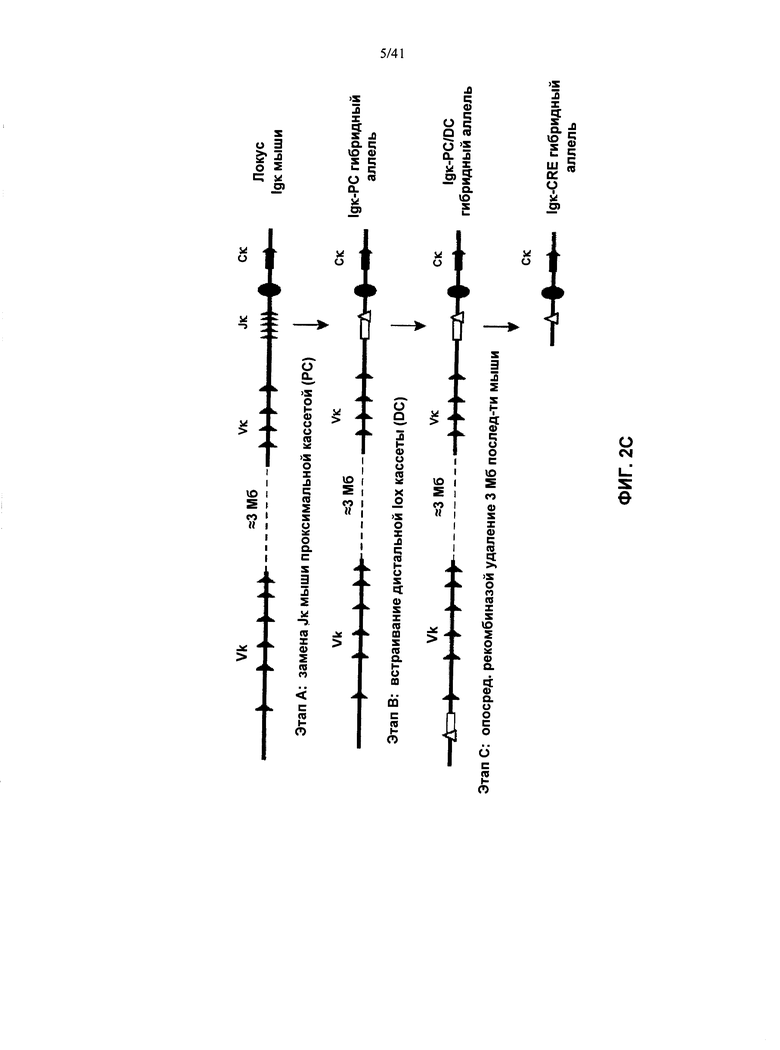
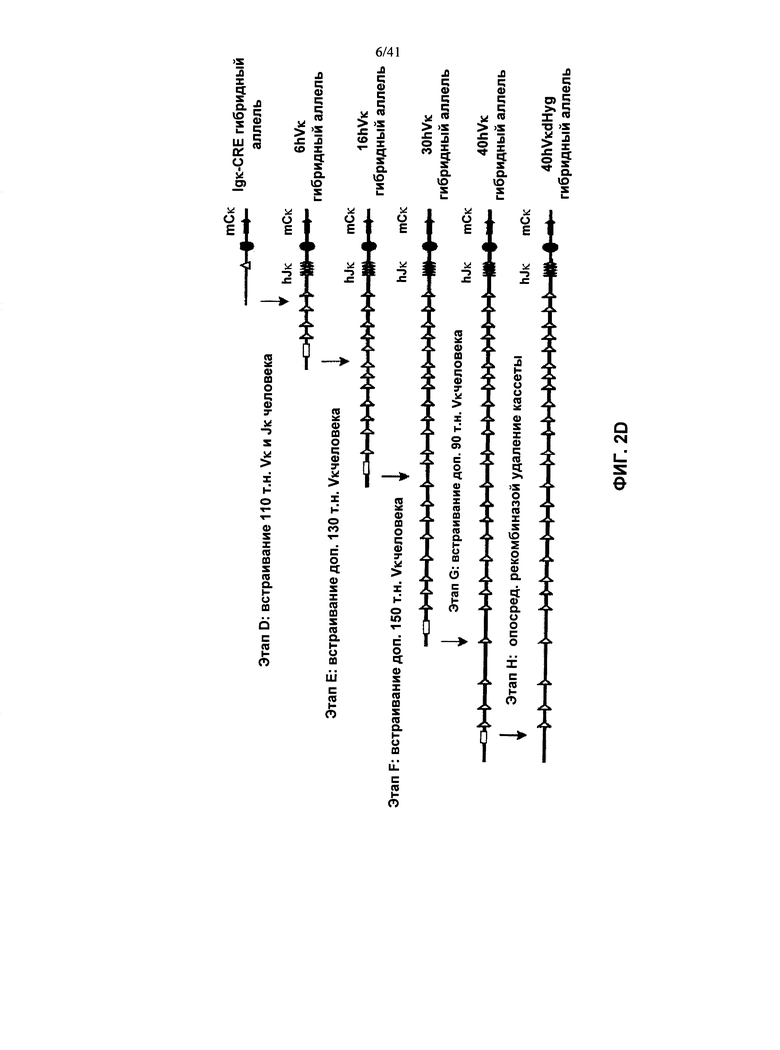
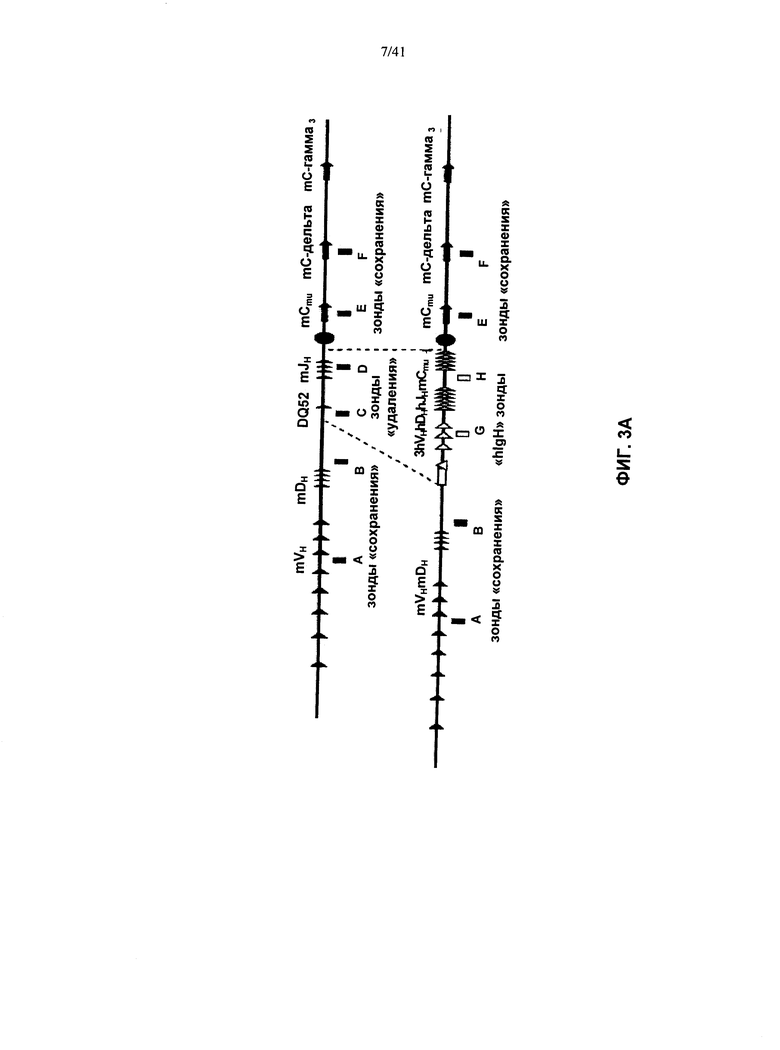
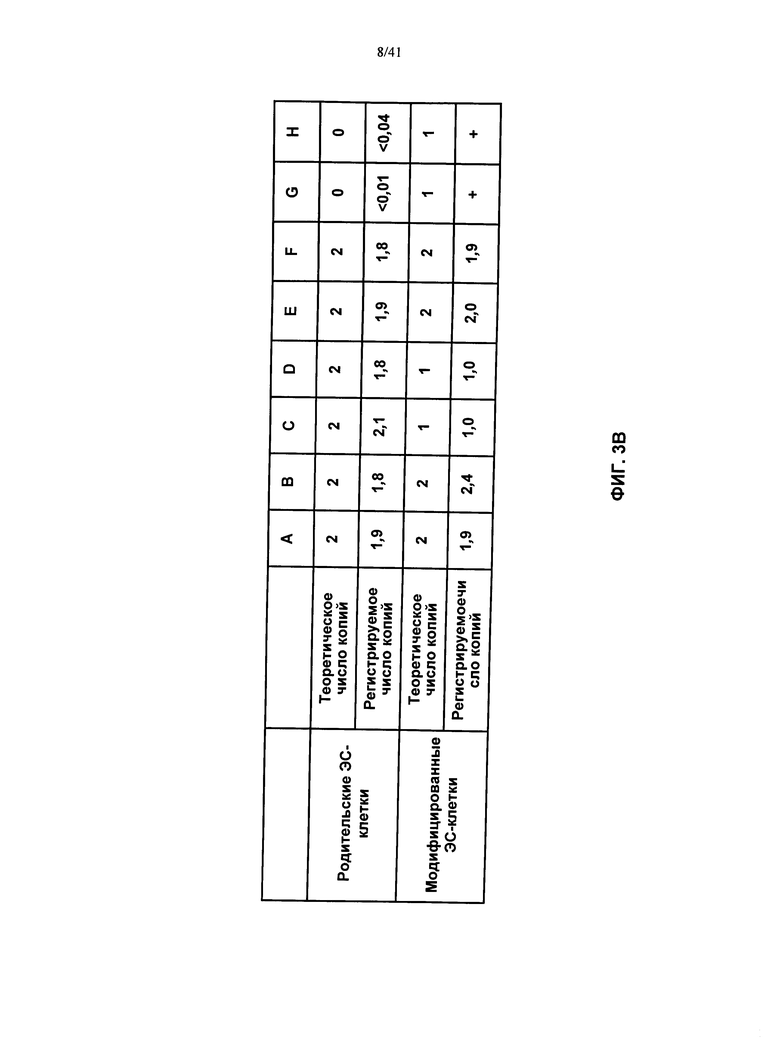
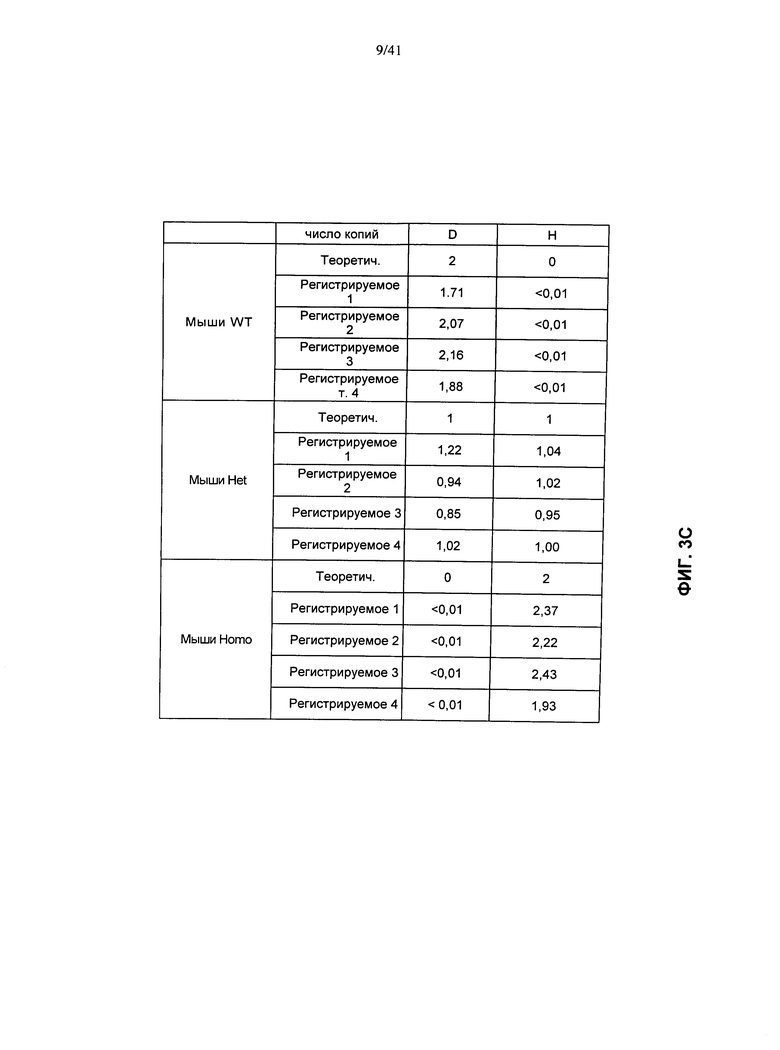

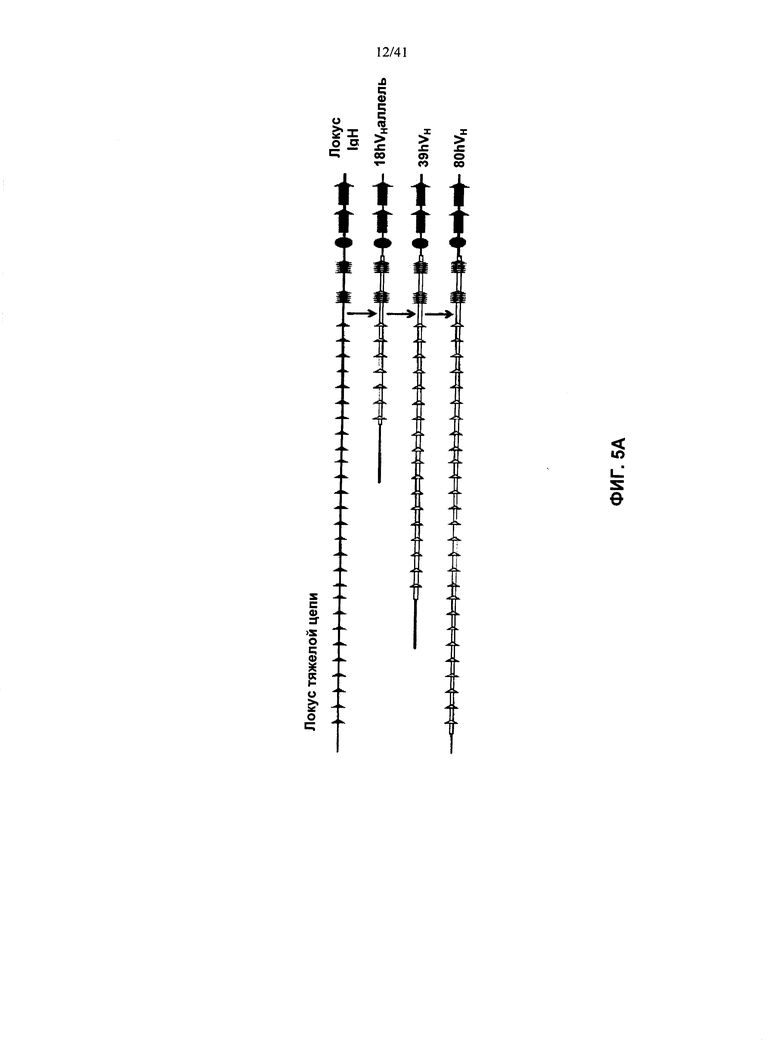
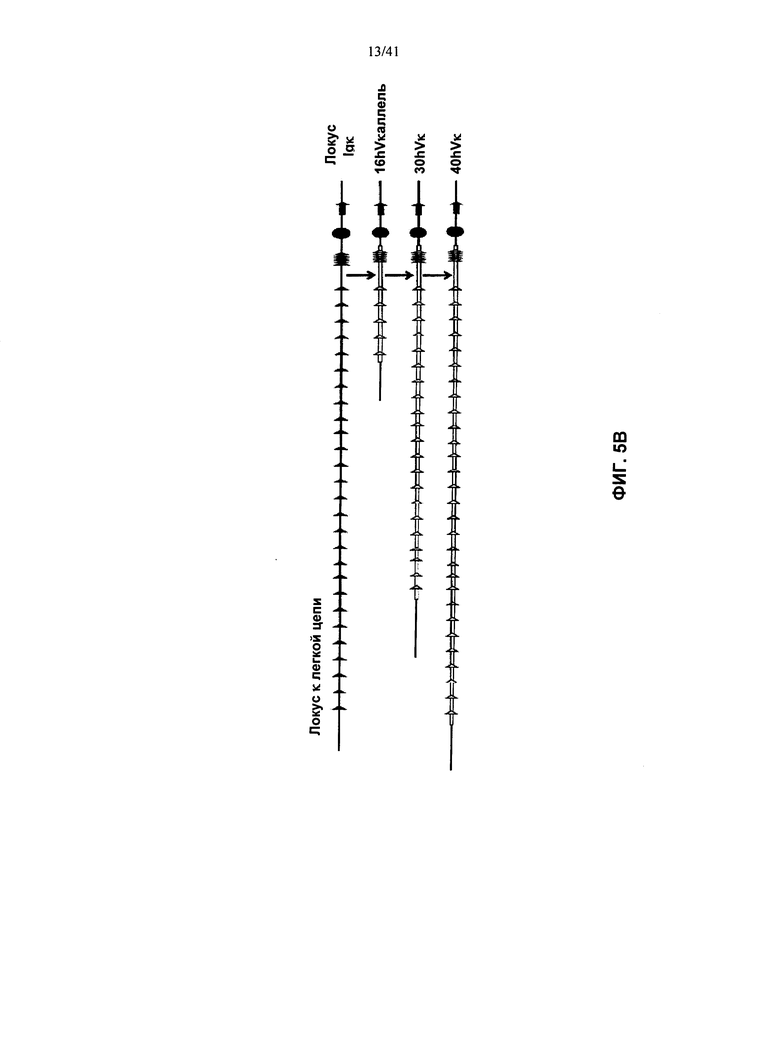
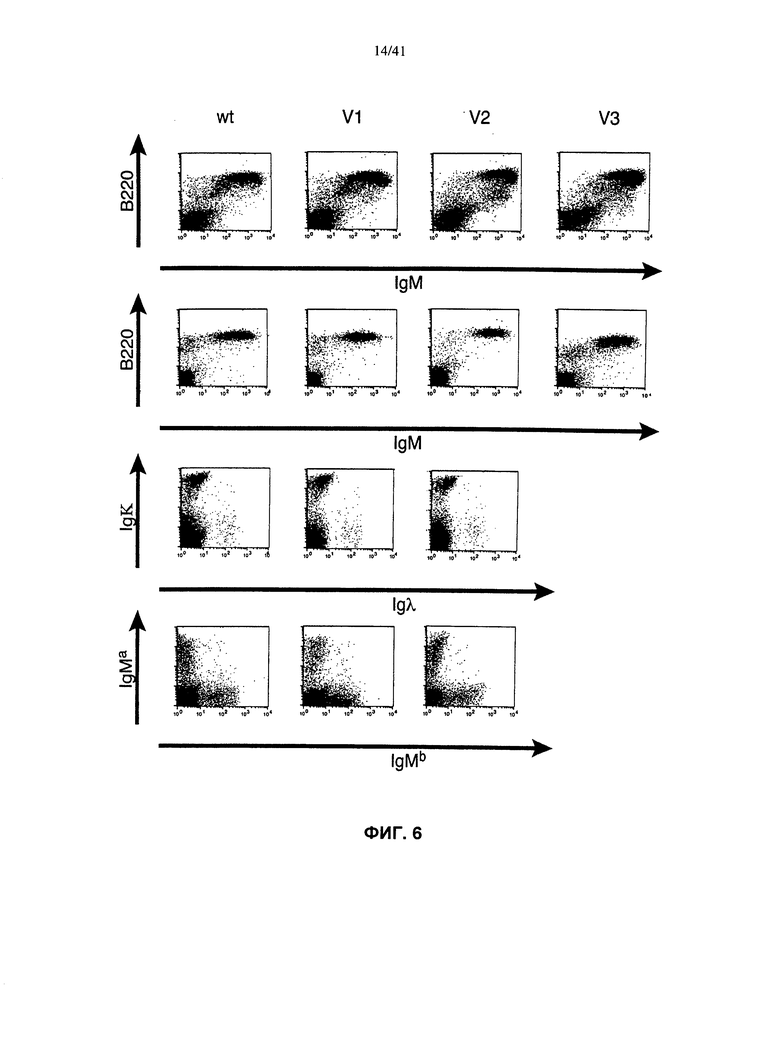
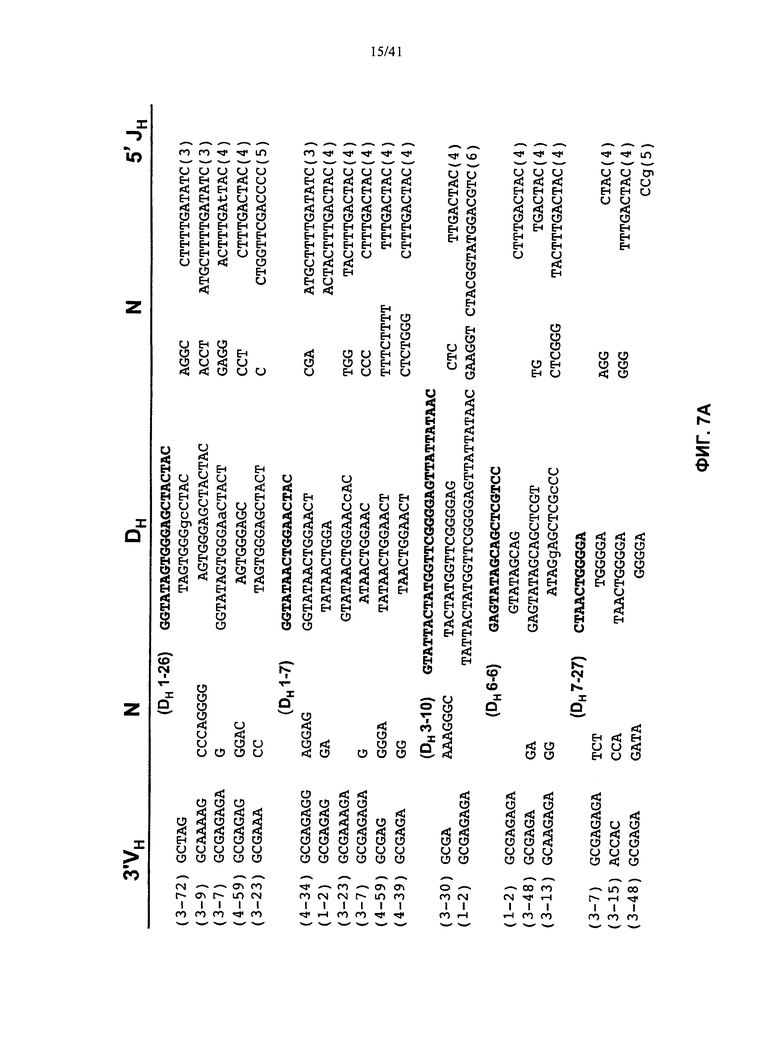

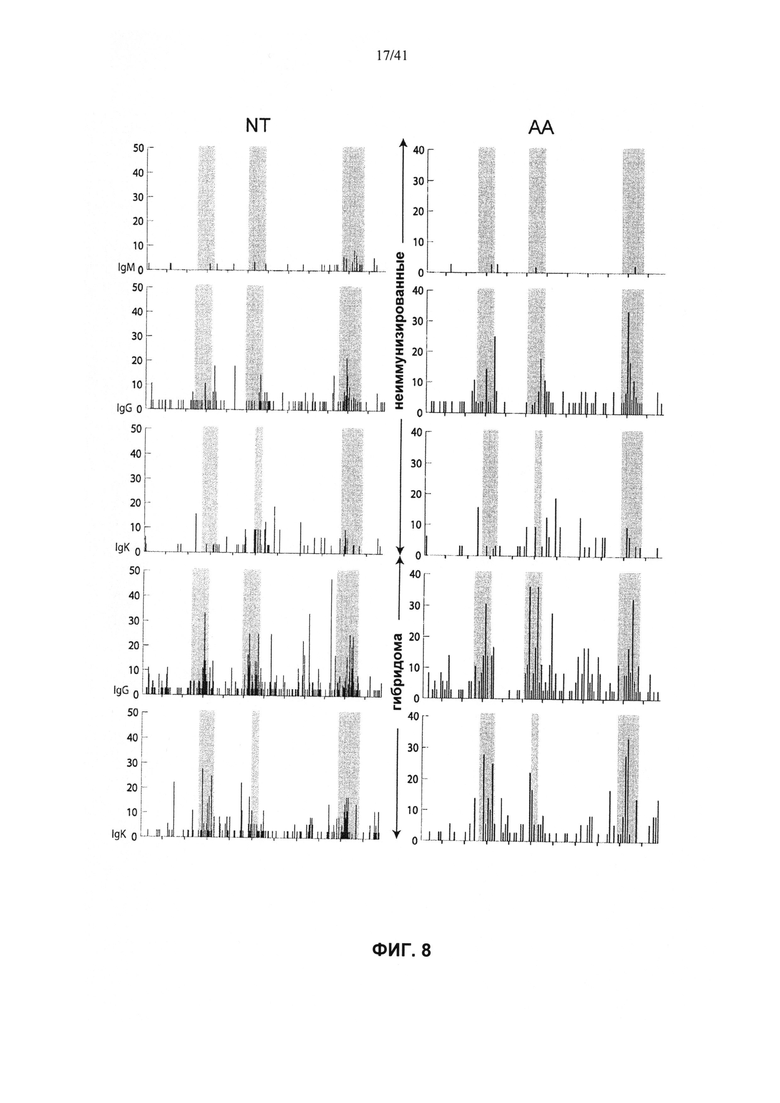
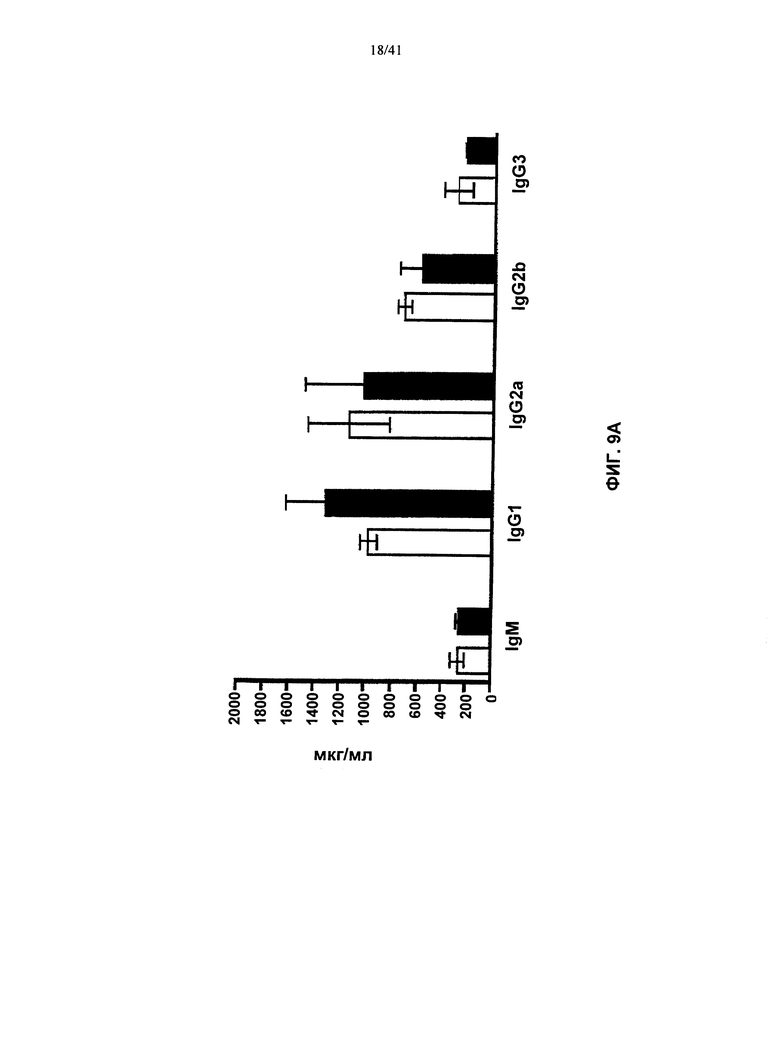
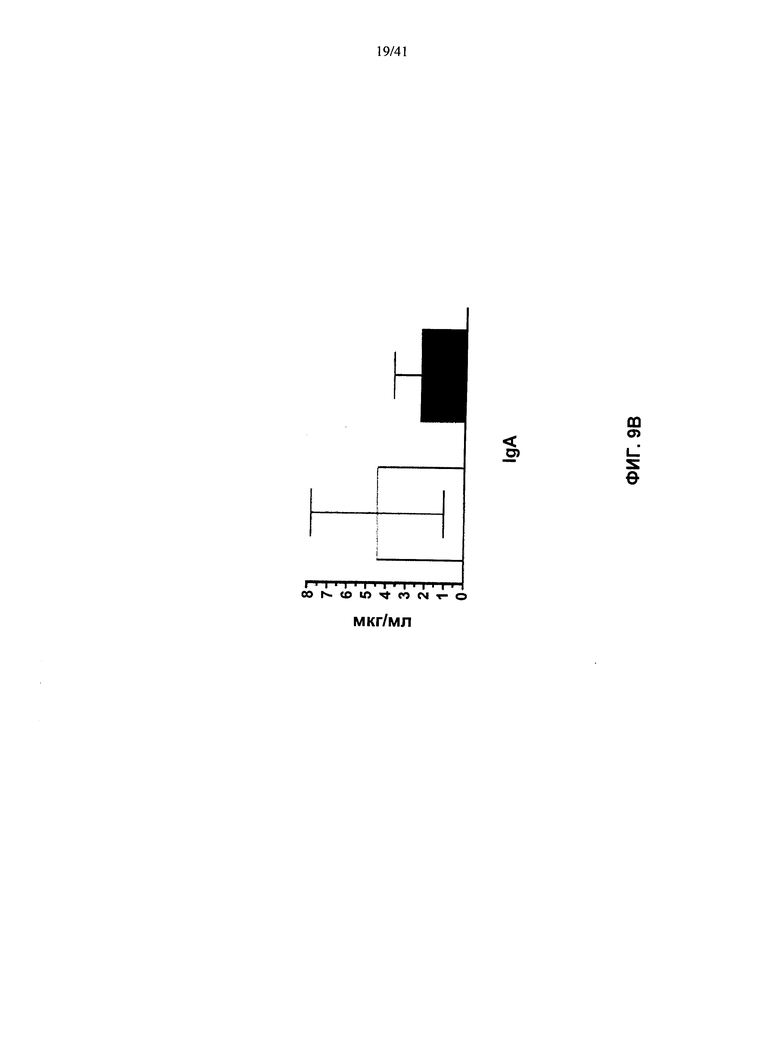
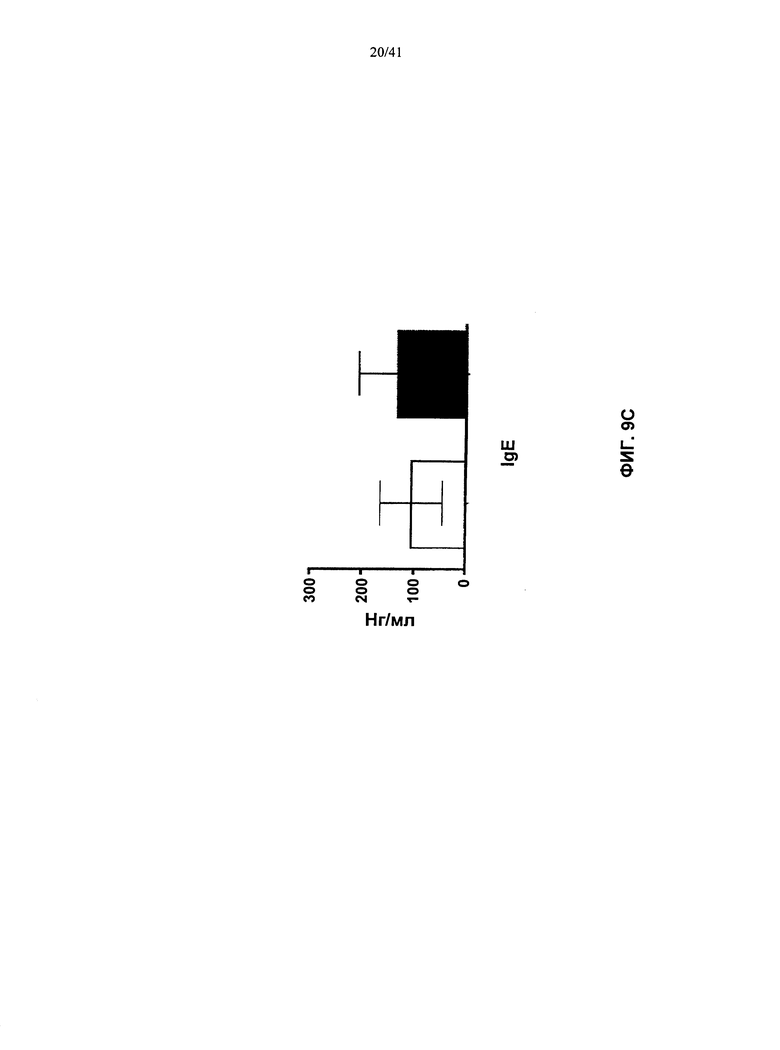
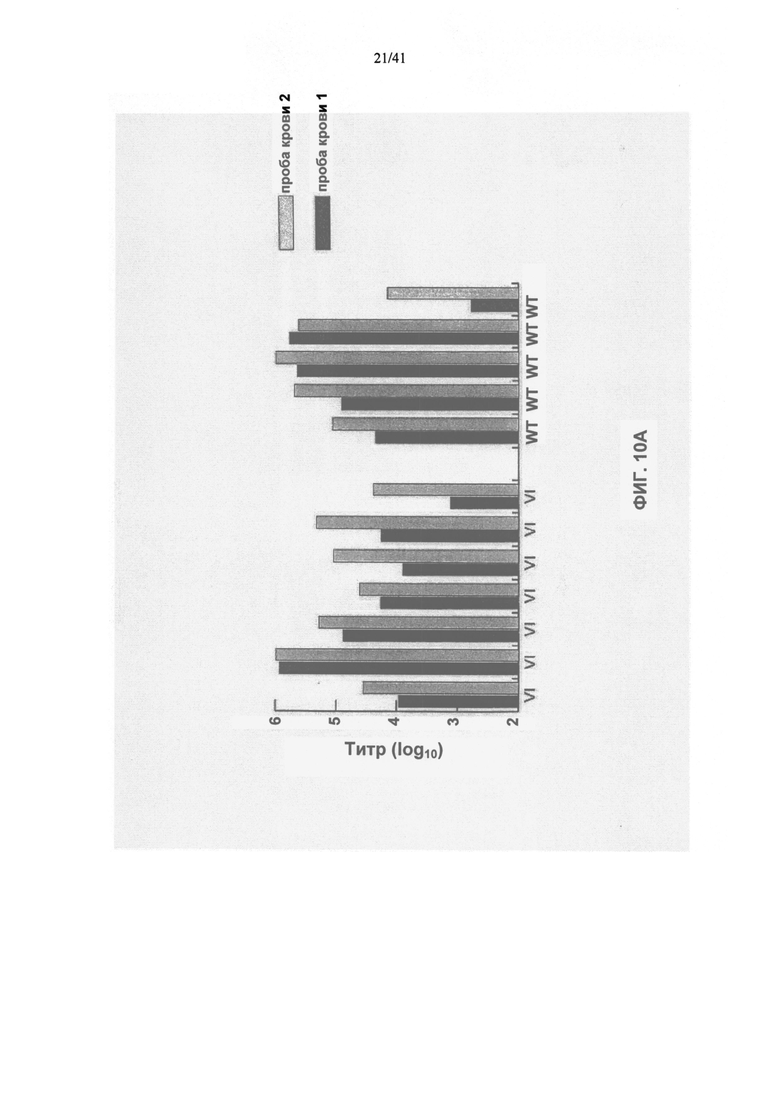

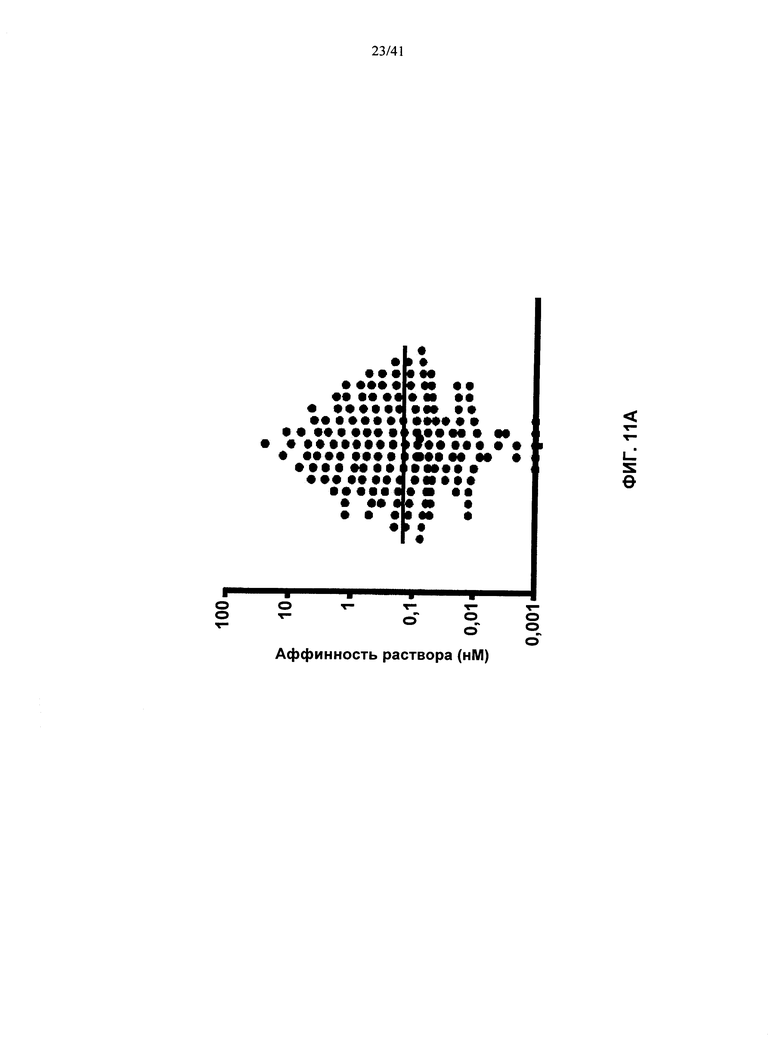
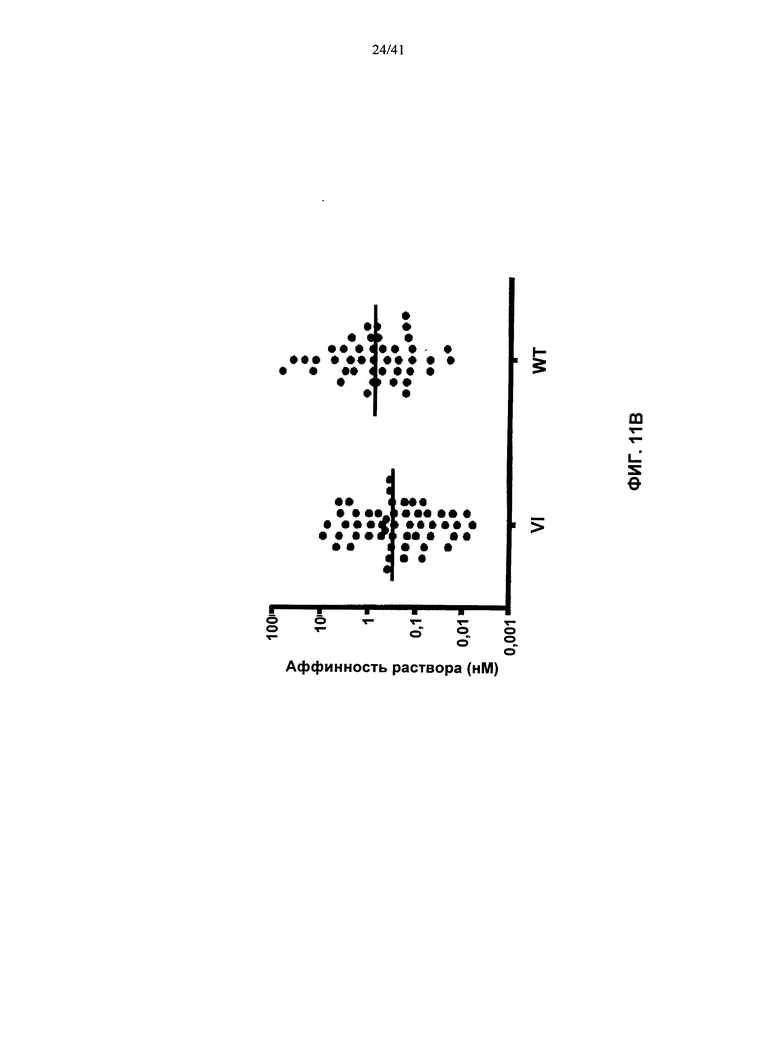
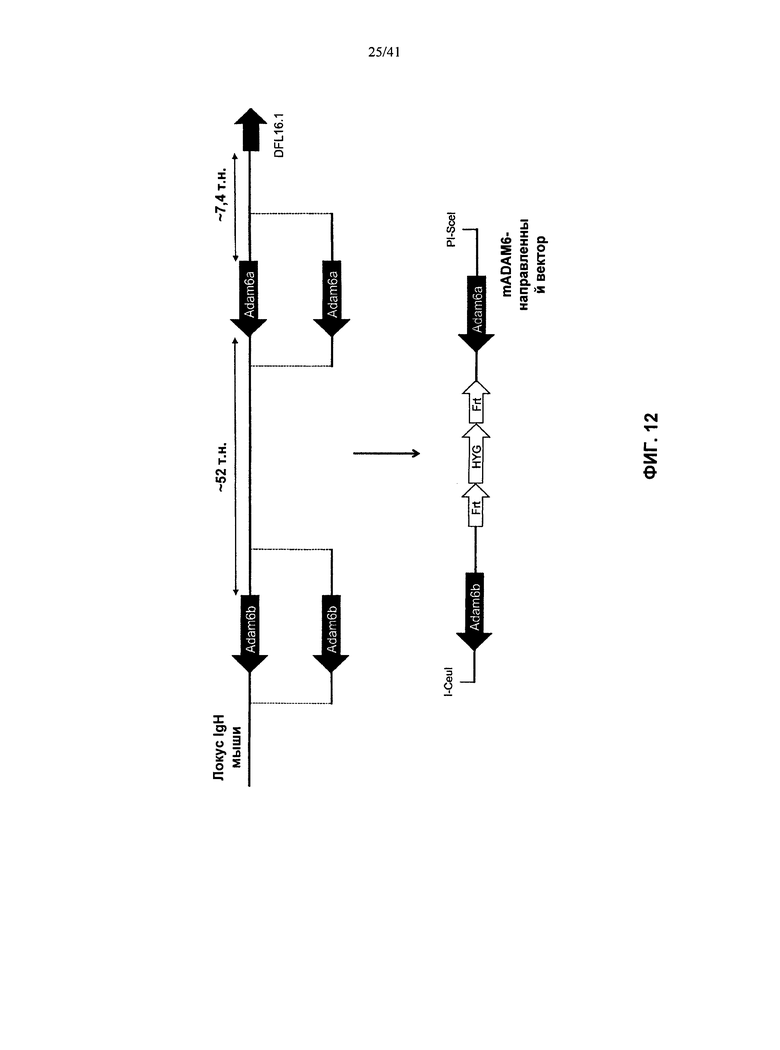
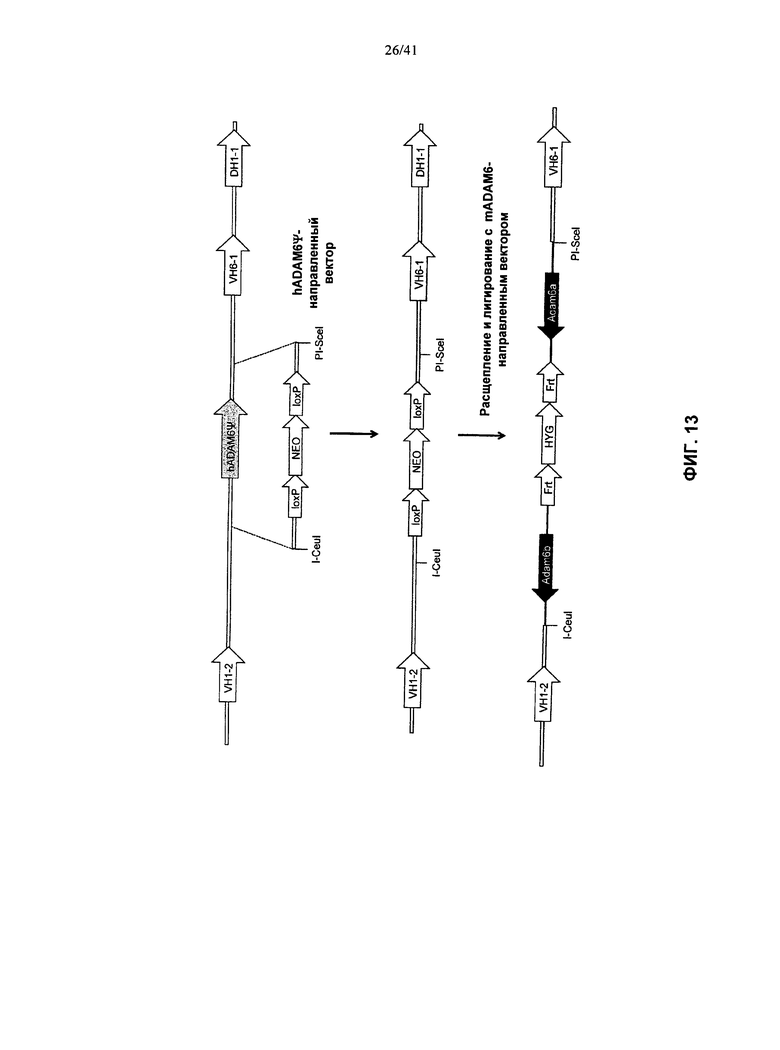
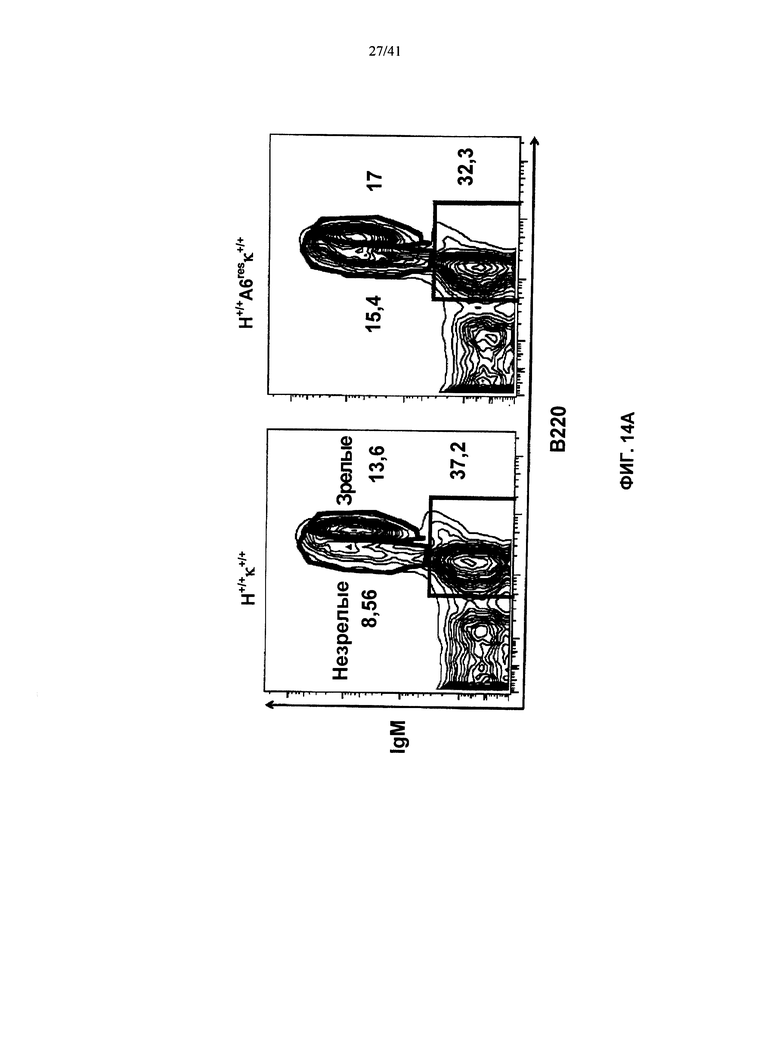
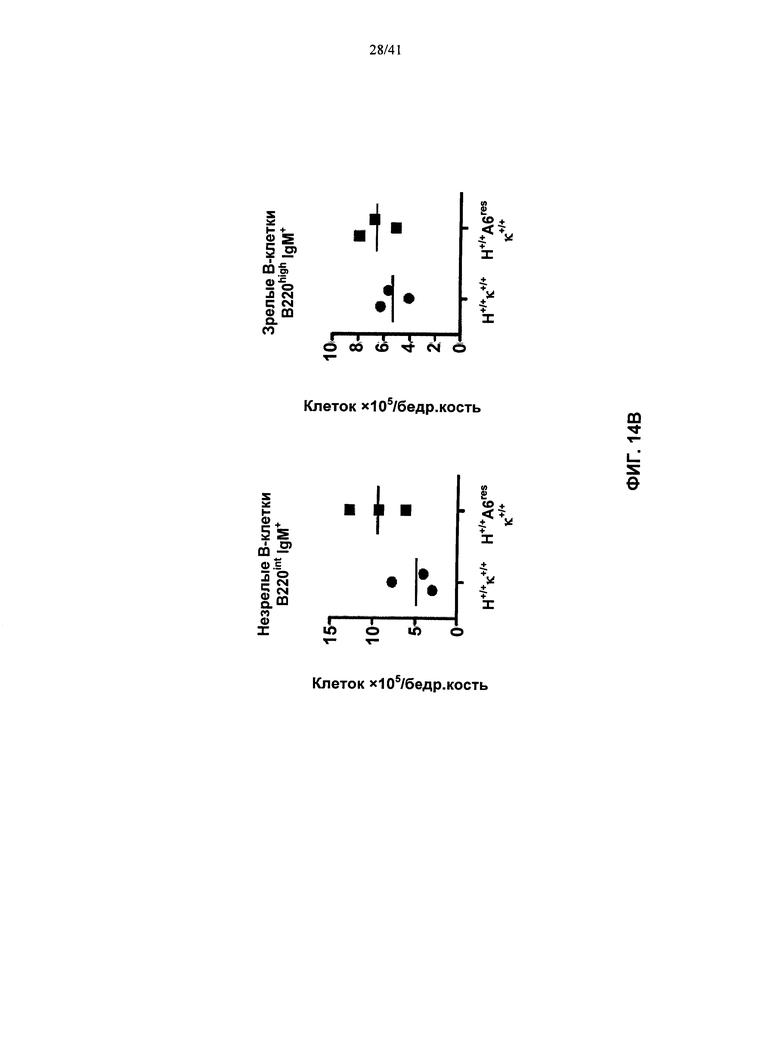
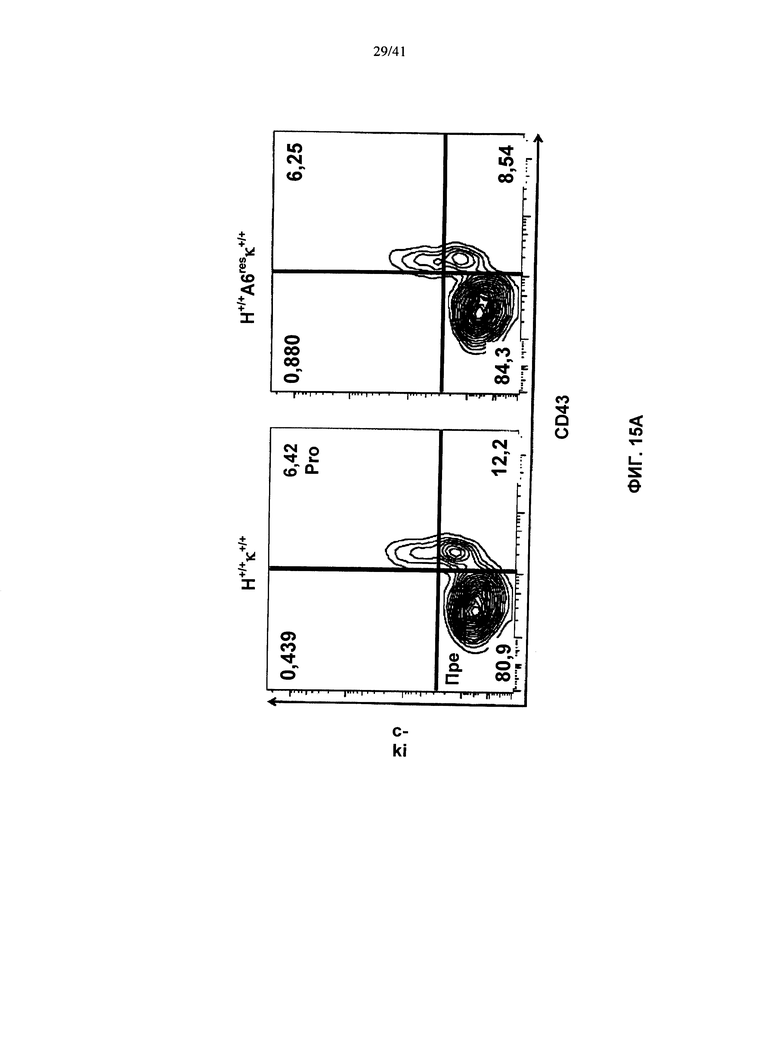
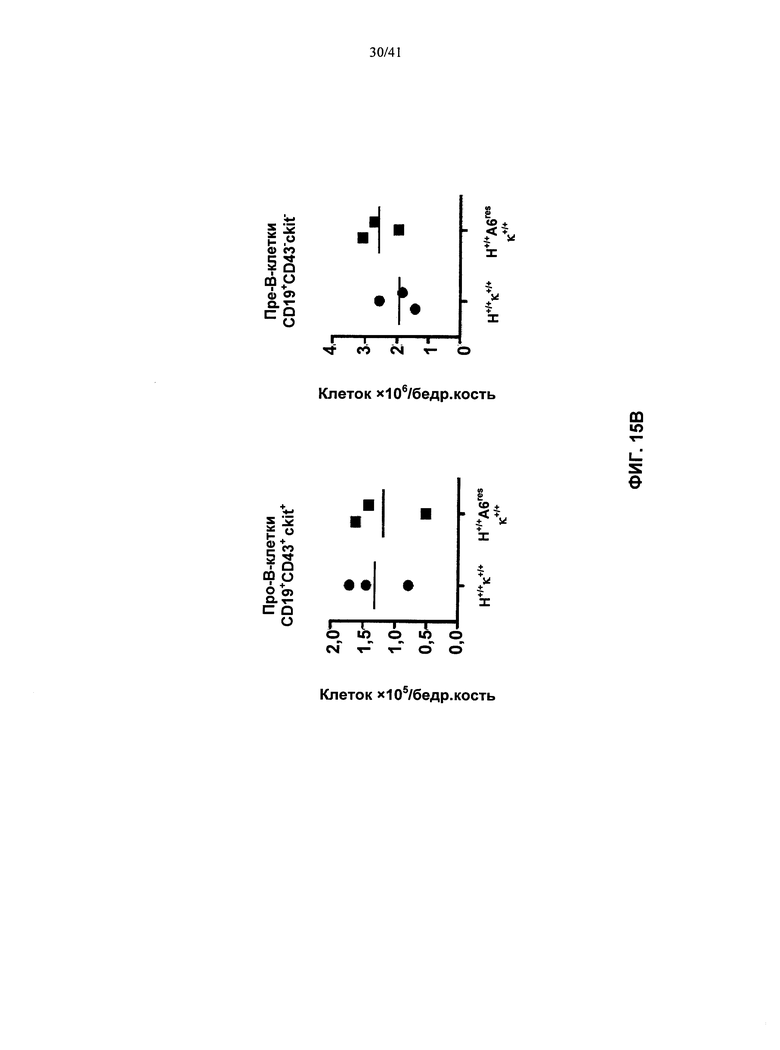

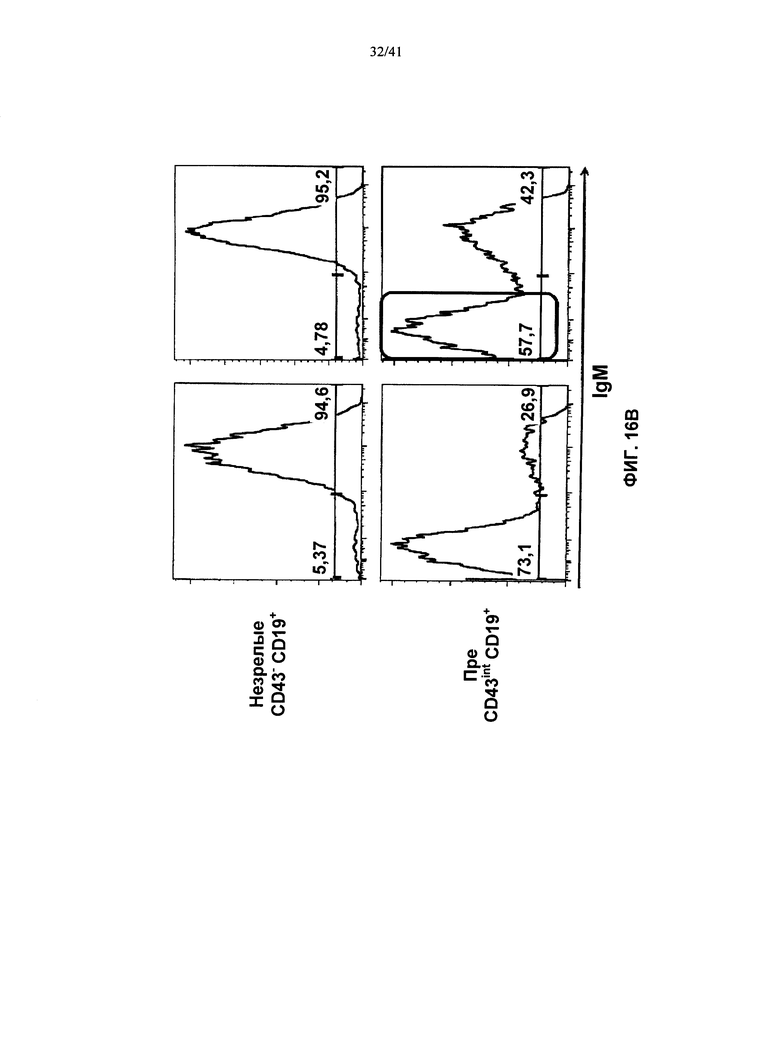
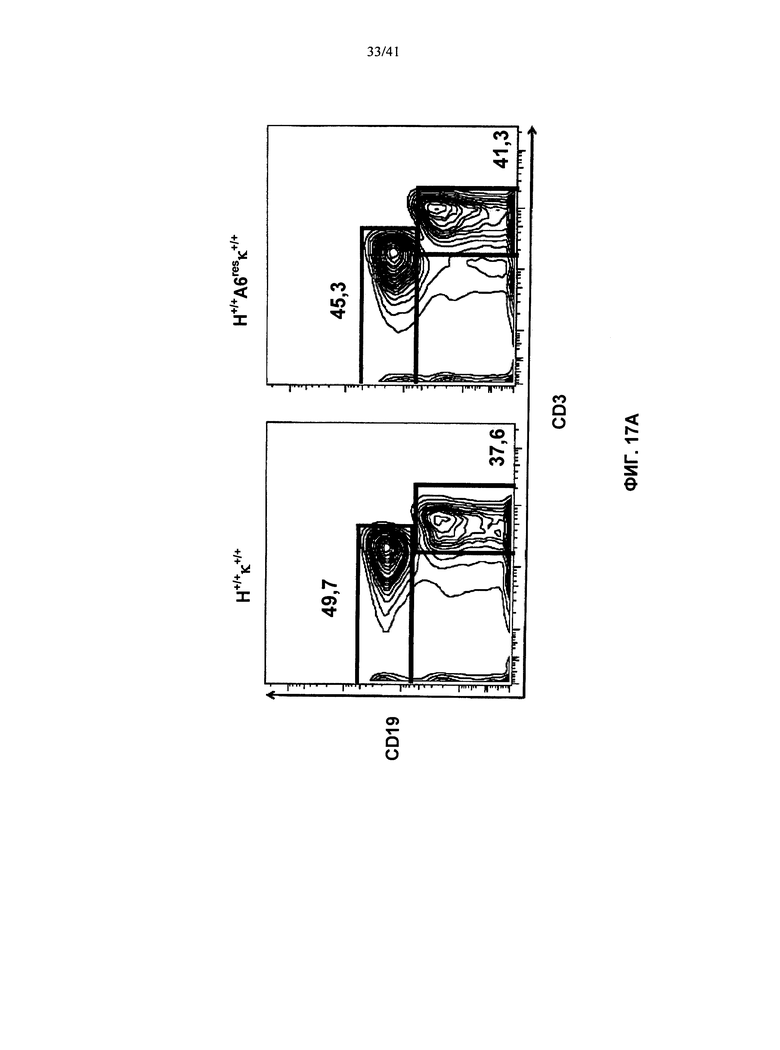
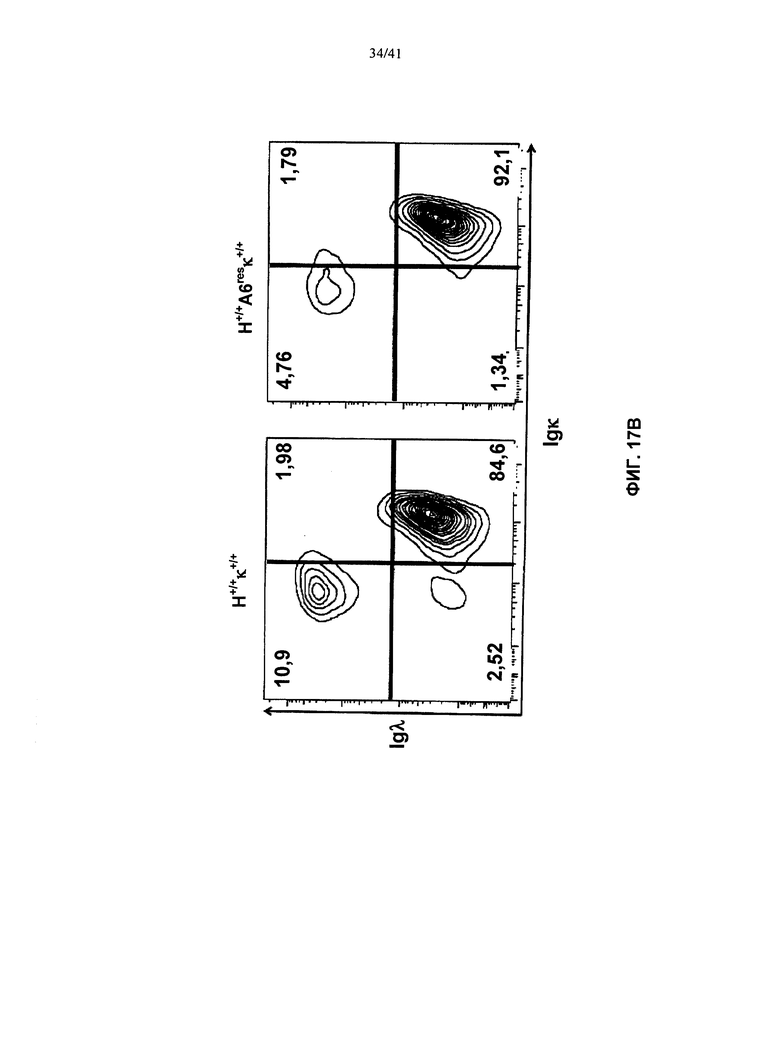
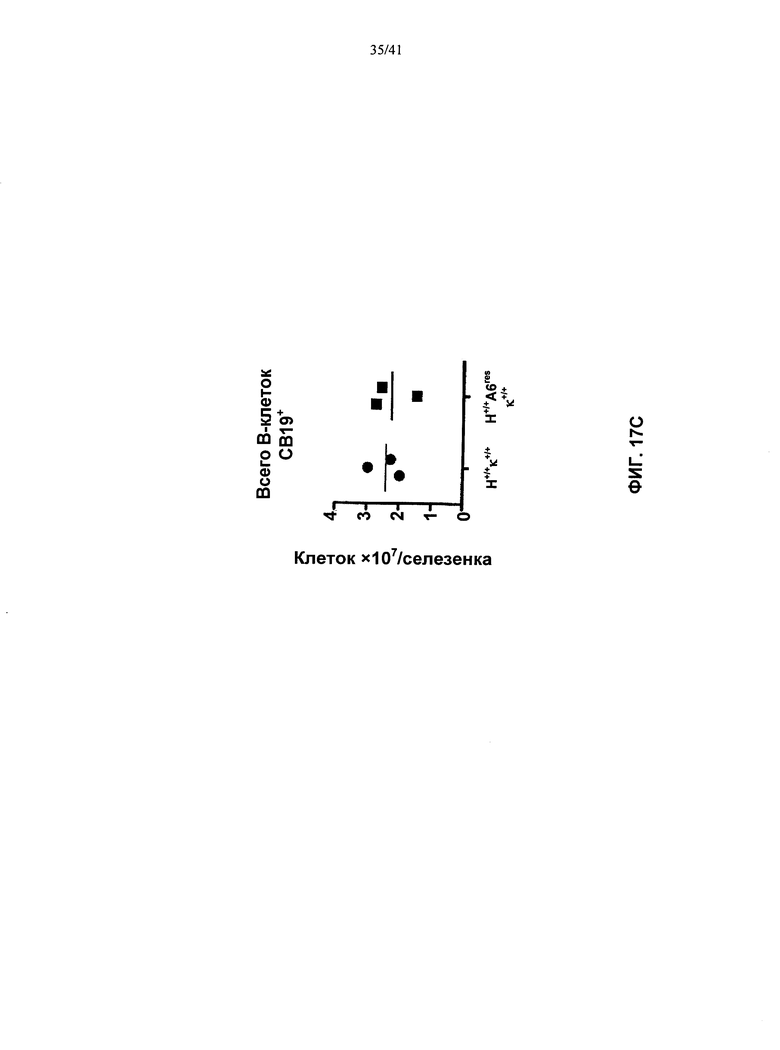
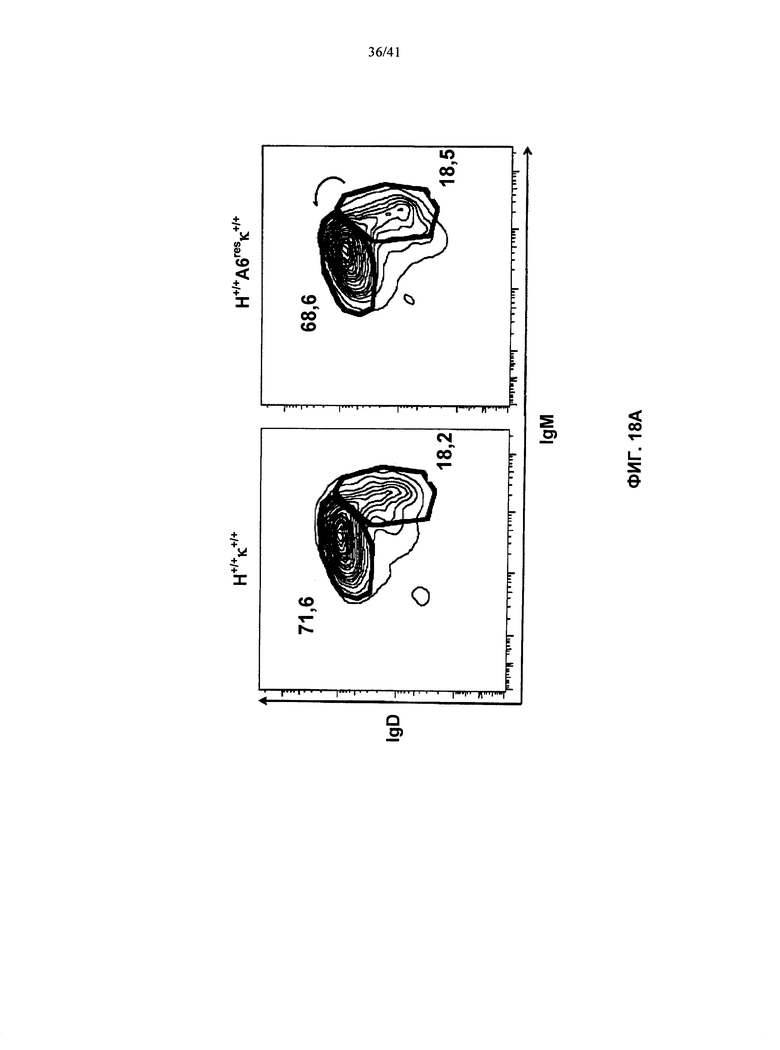
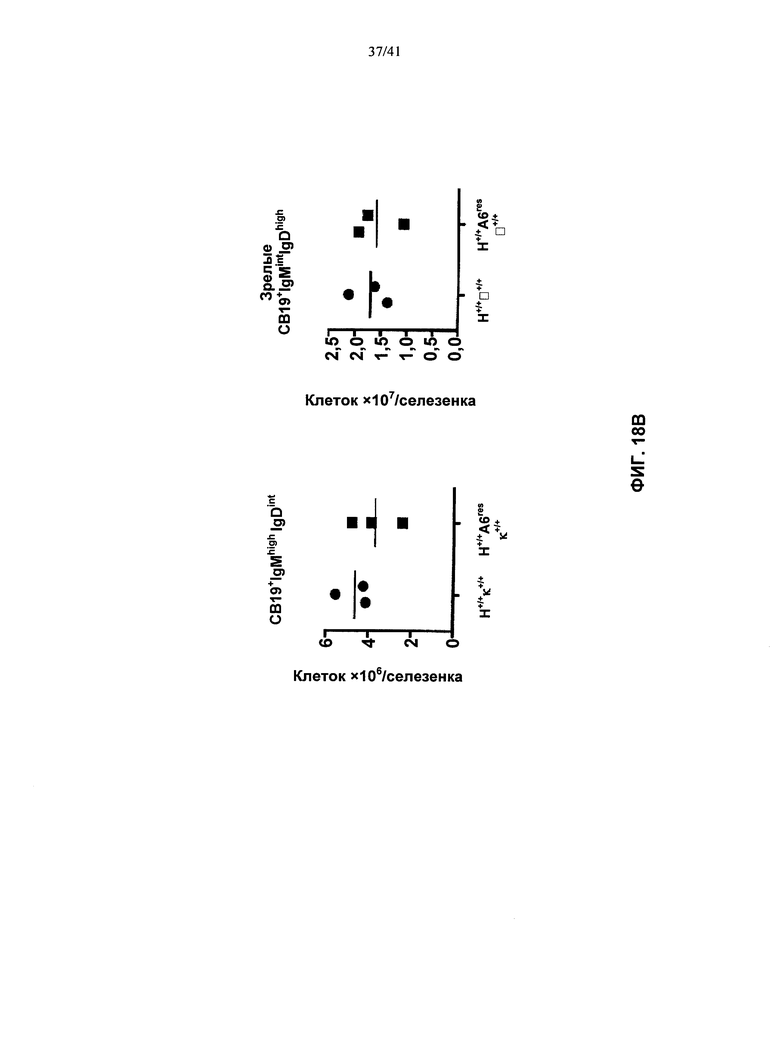

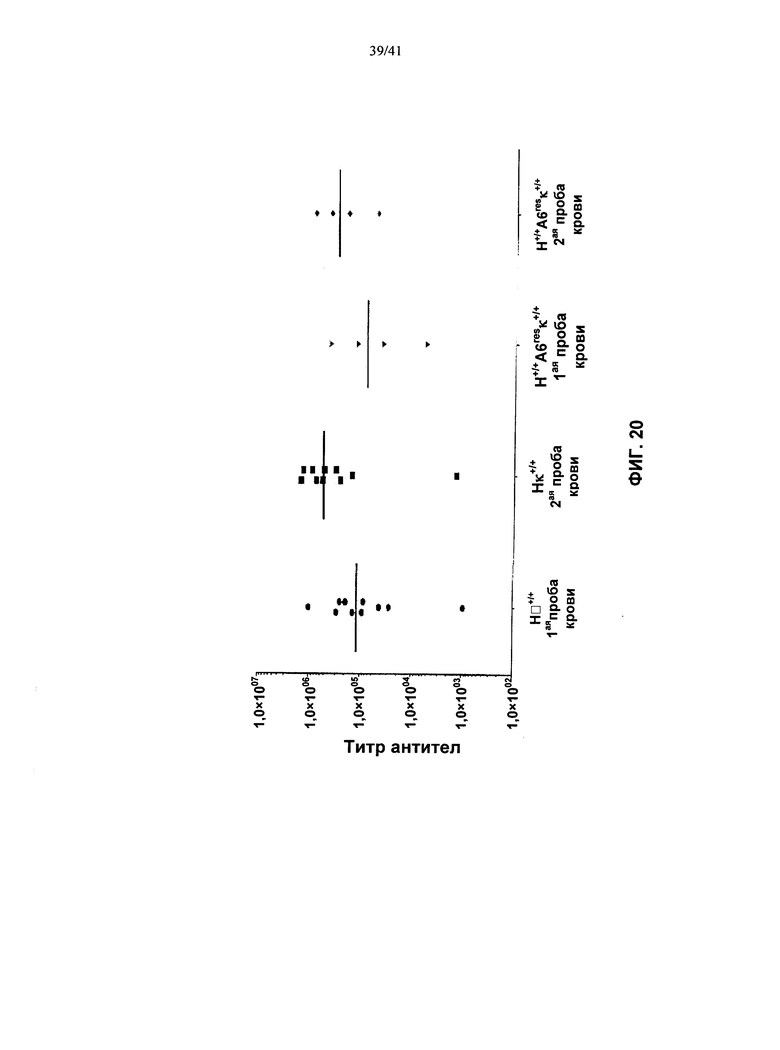

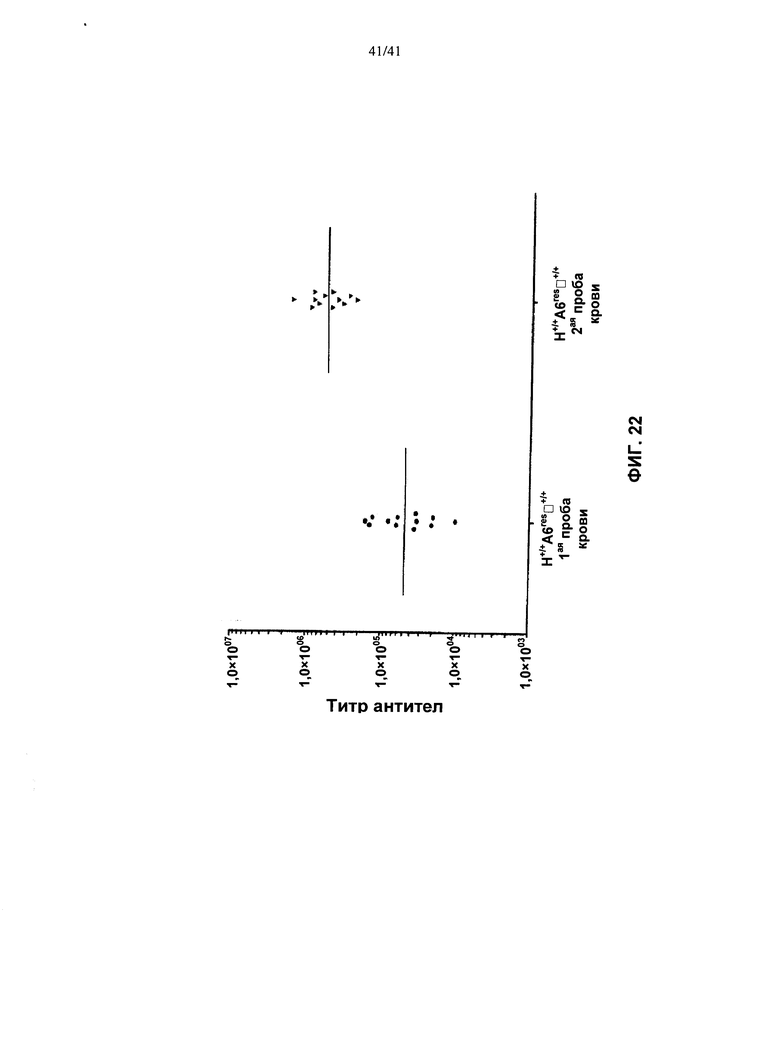
















Изобретение относится к области биотехнологии и генной инженерии. Предложены мыши со сниженной или отсутствующей активностью ADAM6, обеспечиваемой эндогенным локусом ADAM6, либо с отсутствующим эндогенным локусом, кодирующим белок ADAM6 мыши, отличающиеся тем, что у указанных мышей присутствует последовательность, кодирующая ADAM6, либо его ортолог, гомолог или фрагмент, функциональный у мышей-самцов. Согласно одному из вариантов реализации указанная последовательность представляет собой эктопическую последовательность ADAM6 или последовательность, придающую самцам мыши способность производить потомство посредством спаривания. Также предложены мыши и клетки с генетически модифицированными локусами тяжелой цепи иммуноглобулина, содержащими эктопическую последовательность нуклеотидов, кодирующую ADAM6 мыши, либо его функциональный фрагмент, гомолог или ортолог. Предложенное изобретение может быть использовано в иммунологии. 6 н. и 17 з.п. ф-лы, 41 ил., 13 табл., 11 пр.
1. Мышь, генетически модифицированная с образованием последовательностей вариабельной области иммуноглобулина человека и для улучшения или восстановления фертильности указанной мыши и/или ее потомства, генетические модификации указанной мыши обеспечивают:
a) эктопическое расположение гена ADAM6, при этом расположение на хромосоме гена ADAM6 таково, что происходит экспрессия указанного гена, что приводит к улучшению или восстановлению фертильности у мышей-самцов; и
b) модифицированный локус вариабельной области тяжелой цепи иммуноглобулина, при этом указанный модифицированный локус вариабельной области тяжелой цепи иммуноглобулина содержит вставку одного или более сегментов гена VH человека, одного или более сегментов гена DH человека и одного или более сегментов гена JH человека в эндогенный локус тяжелой цепи иммуноглобулина.
2. Мышь по п. 1, отличающаяся тем, что указанная модификация указанного локуса вариабельной области тяжелой цепи иммуноглобулина уменьшает или элиминирует активность ADAM6 в клетке или ткани самца мыши.
3. Мышь по п. 1, отличающаяся тем, что указанный ген ADAM6 присутствует в модифицированном локусе вариабельной области тяжелой цепи.
4. Мышь по п. 1, отличающаяся тем, что указанный ген ADAM6 присутствует в положении, отличном от модифицированного локуса вариабельной области тяжелой цепи.
5. Мышь по п. 1, отличающаяся тем, что указанный ген ADAM6 включает ADAM6a мыши и/или ADAM6b мыши.
6. Мышь по п. 1, отличающаяся тем, что указанные один или более генных сегментов VH человека включают по меньшей мере 18, по меньшей мере 39 или по меньшей мере 80 генных сегментов VH человека.
7. Мышь по п. 1, отличающаяся тем, что указанный модифицированный локус вариабельной области тяжелой цепи иммуноглобулина содержит по меньшей мере 18 генных сегментов VH человека, по меньшей мере один и максимум 27 генных сегментов DH человека и по меньшей мере один и максимум шесть генных сегментов JH человека.
8. Мышь по п. 1, отличающаяся тем, что указанный модифицированный локус вариабельной области тяжелой цепи иммуноглобулина содержит по меньшей мере 39 генных сегментов VH человека, по меньшей мере один и максимум 27 генных сегментов DH человека и по меньшей мере один и максимум шесть генных сегментов JH человека.
9. Мышь по п. 1, отличающаяся тем, что указанный модифицированный локус вариабельной области тяжелой цепи иммуноглобулина содержит 80 генных сегментов VH человека, по меньшей мере один и максимум 27 генных сегментов DH человека и по меньшей мере один и максимум шесть генных сегментов JH человека.
10. Мышь по п. 1, отличающаяся тем, что указанные один или более генные сегменты VH человека, один или более генные сегменты DH человека и один или более генные сегменты JH человека функционально связаны с геном константной области.
11. Мышь по п. 10, отличающаяся тем, что указанный ген константной области представляет собой ген константной области мыши.
12. Мышь по п. 11, отличающаяся тем, что указанный ген константной области мыши содержит константную область, выбранную из CH1, шарнирной области, СН2, СН3 и/или СН4 либо их комбинации.
13. Мышь по п. 1, у которой дополнительно присутствует один или более генных сегментов Vκ человека и один или более генных сегментов Jκ человека.
14. Мышь по п. 13, отличающаяся тем, что указанные один или более генных сегментов Vκ человека и один или более генных сегментов Jκ человека находятся в эндогенном локусе легкой цепи κ иммуноглобулина.
15. Выделенная клетка мыши по п. 1 для получения мыши по любому из пп. 1-14, причем указанная выделенная клетка содержит эктопично расположенный ген ADAM6 согласно этапу а) и указанный модифицированный локус вариабельной области тяжелой цепи иммуноглобулина согласно этапу b).
16. Выделенная клетка по п. 15, отличающаяся тем, что указанная клетка представляет собой эмбриональную стволовую клетку.
17. Выделенная клетка мыши по п. 1 для получения гибридомы, при этом указанная клетка представляет собой В-клетку.
18. Эмбрион мыши для получения мыши по любому из пп. 1-14, при этом указанный эмбрион содержит эмбриональную стволовую клетку по п. 16.
19. Гибридома для получения антитела или его фрагмента, полученная из В-клетки по п. 17.
20. Способ получения антитела, специфичного в отношении представляющего интерес антигена, включающий следующие этапы:
(a) иммунизацию мыши по п. 1 представляющим интерес антигеном;
(b) выделение из указанной мыши по меньшей мере одной клетки, которая синтезирует антитело, специфичное в отношении указанного антигена; и
(c) культивирование указанной по меньшей мере одной клетки, синтезирующей антитело согласно этапу (b) и получение указанного антитела.
21. Способ по п. 20, отличающийся тем, что культивирование на этапе (с) осуществляют в отношении по меньшей мере одной гибридомной клетки, происходящей из по меньшей мере одной клетки, полученной на этапе (b).
22. Способ по п. 20, отличающийся тем, что указанная по меньшей мере одна клетка, выделенная на этапе (b), происходит из селезенки, лимфатического узла или костного мозга мыши согласно этапу (а).
23. Способ по п. 20, отличающийся тем, что иммунизацию представляющим интерес антигеном на этапе (а) проводят белком, ДНК, комбинацией ДНК и белка или клетками, экспрессирующими указанный антиген.
| WO 2000073323 A2, 07.12.2000 | |||
| US 2004018626 A1, 29.01.2004 | |||
| ГУМАНИЗИРОВАННЫЙ ИММУНОГЛОБУЛИН, СПЕЦИФИЧНЫЙ ДЛЯ БЕЛКА L-СЕЛЕКТИНА ЧЕЛОВЕКА | 1993 |
|
RU2151612C1 |
| РЕКОМБИНАНТНОЕ АНТИТЕЛО К ЧЕЛОВЕЧЕСКОМУ ИНТЕРЛЕЙКИНУ 1β | 2001 |
|
RU2264413C2 |
| Воздушная форсунка | 1927 |
|
SU10506A1 |
| Способ очистки хлороформа | 1927 |
|
SU10469A1 |

