Область техники, к которой относится настоящее изобретение
Предусмотрены генетически модифицированные отличные от человека фертильные животные, которые экспрессируют иммуноглобулиноподобные связывающие белки человека, содержащие константную область тяжелой цепи иммуноглобулина, слитую с вариабельным доменом легкой цепи иммуноглобулина, а также связывающие белки, содержащие вариабельный домен легкой цепи иммуноглобулина, слитый с константным доменом легкой цепи, и вариабельный домен легкой цепи иммуноглобулина, слитый с константным доменом тяжелой цепи. Описаны генетически модифицированные мыши, клетки, зародыши и ткани, которые содержат последовательность нуклеиновой кислоты, которая кодирует функциональный белок ADAM6, причем мыши, клетки, зародыши и ткани содержат генные сегменты легкой цепи иммуноглобулина человека, функционально связанные с одним или несколькими не относящимися к человеку константными генами тяжелой цепи иммуноглобулина. Модификации включают в себя относящиеся к человеку и/или гуманизированные локусы иммуноглобулина. Описаны мыши, которые содержат функцию ADAM6, включая в себя мышей, которые содержат эктопическую последовательность нуклеиновой кислоты, которая кодирует белок ADAM6. Описаны генетически модифицированные самцы мышей, которые содержат генетическую модификацию эндогенного локуса области VH иммуноглобулина мыши, и которые дополнительно содержат активность ADAM6, включая в себя мышей, которые содержат эктопическую последовательность нуклеиновой кислоты, которая восстанавливает или поддерживает фертильность у самца мыши. Иллюстративная фертильность представляет собой фертильность, которая сопоставима с фертильностью мышей дикого типа.
Предусмотрены генетически модифицированные не относящиеся к человеку фертильные животные, которые содержат делецию или модификацию эндогенного гена ADAM6 или его гомолога или ортолога, и которые содержат генетическую модификацию, которая полностью или частично восстанавливает функцию ADAM6 (или его гомолога или ортолога), причем отличные от человека животные экспрессируют вариабельную последовательность легкой цепи иммуноглобулина человека, ассоциированную с константной последовательностью тяжелой цепи. Также предусмотрены клетки, экспрессирующие такие связывающие белки, грызуны (например, мыши), которые их производят, и относящиеся к этому способы и композиции.
Описаны генетически сконструированные животные, которые экспрессируют антитела, содержащие вариабельные области легкой цепи, слитые с константными областями тяжелой цепи, причем отличные от человека животные не содержат функциональный эндогенный ген ADAM6, но сохраняют функцию ADAM6, включая в себя грызунов (например, мышей), которые содержат модификацию эндогенного локуса вариабельной области тяжелой цепи (VH) иммуноглобулина, который делает мышь неспособной производить функциональный белок ADAM6 и приводит к потере фертильности. Описаны генетически модифицированные мыши, которые содержат локус VH иммуноглобулина, характеризующийся множеством генных сегментов VL, JL и необязательно DH человека или их комбинацией, и которые дополнительно содержат функцию ADAM6, включая в себя мышей, которые содержат эктопическую последовательность нуклеиновой кислоты, которая восстанавливает фертильность у самца мыши. Генетически модифицированные мыши экспрессируют антитела, которые не содержат вариабельные домены тяжелой цепи и вместо этого содержат вариабельные домены, содержащие реаранжированные генные сегменты легкой цепи.
Описаны генетически модифицированные грызуны (например, мыши), клетки, зародыши и ткани, которые содержат последовательность нуклеиновой кислоты, кодирующую функциональный локус ADAM6, причем мыши, клетки, зародыши и ткани экспрессируют тяжелую цепь иммуноглобулина, содержащую вариабельный домен легкой цепи человека. Кроме того, мыши, клетки, зародыши и ткани не содержат функциональный эндогенный ген ADAM6, но сохраняют функцию ADAM6, характеризующуюся присутствием эктопической последовательности нуклеиновой кислоты, которая кодирует белок ADAM6. Предусмотрены способы получения последовательностей антител у фертильных отличных от человека животных, которые применимы для связывания антигенов.
Предшествующий уровень техники настоящего изобретения
За последние два десятилетия фармацевтические применения антител активизировали большое количество исследований в области получения антител, подходящих для применений в качестве терапевтических средств для людей. Ранее существующие терапевтические средства на основе антител, основанные на мышиных антителах, не были оптимальными в качестве терапевтических средств для людей, поскольку повторное введение мышиных антител людям приводит к проблемам с иммуногенностью, которые могут нарушать схемы длительного лечения. Разрабатывали решения, основанные на гуманизации мышиных антител, чтобы сделать их более похожими на антитела человека и менее похожими на антитела мыши. Придерживались способов экспрессии последовательностей иммуноглобулина человека для применения в антителах, преимущественно основанных на in vitro экспрессии библиотек иммуноглобулина человека в фаге, бактериях или дрожжах. В конце концов, были предприняты попытки создания применимых антител человека из лимфоцитов человека in vitro, в организмах мышей, которым пересадили гематопоэтические клетки человека, и в организмах трансхромосомных или трансгенных мышей с недееспособными эндогенными локусами иммуноглобулина.
Для создания таких мышей, было необходимо инактивировать эндогенные гены иммуноглобулина мыши так, чтобы случайным образом интегрированные полностью человеческие трансгены могли бы функционировать в качестве экспрессированного репертуара иммуноглобулинов в организме мыши. Такие мыши могут создавать антитела человека, подходящие для применения в качестве терапевтических средств для людей, но эти мыши демонстрируют существенные проблемы в отношении их иммунных систем. Эти проблемы приводят к некоторым экспериментальным препятствиям, например, мыши становятся неспособными производить в достаточной степени разнообразные репертуары антител, требуют применения решений для широкомасштабного реконструирования, обеспечивают субоптимальный процесс селекции клонов, вероятно, вследствие несовместимости между элементами человека и мыши, и представляют собой ненадежный источник больших и разнообразных популяций вариабельных последовательностей человека, которые должны были быть действительно применимыми для получения терапевтических средств для людей.
Трансгенные мыши, которые содержат трансгены полностью человеческих антител, содержат случайным образом вставленные трансгены, которые содержат нереаранжированные вариабельные последовательности тяжелой цепи иммуноглобулина человека (последовательности V, D и J), соединенные с константными последовательностями тяжелой цепи человека, и нереаранжированные вариабельные последовательности легкой цепи иммуноглобулина человека (V и J), соединенные с константными последовательностями легкой цепи человека. Мыши, следовательно, создают реаранжированные гены антитела из локусов, отличных от эндогенных локусов, причем реаранжированные гены антитела являются полностью человеческими. Как правило, мыши содержат последовательности тяжелой цепи человека и последовательности легкой цепи человека, хотя также сообщалось о мышах по меньшей мере с несколькими последовательностями λ человека. Трансгенные мыши, как правило, содержат поврежденные и нефункциональные эндогенные локусы иммуноглобулина или нокауты эндогенных локусов иммуноглобулина, так чтобы мыши были неспособны к реаранжировке последовательностей антитела человека на эндогенном локусе иммуноглобулина. Изменения у таких трансгенных мышей делают их менее чем оптимальными для создания достаточно разнообразного репертуара антител человека в организме мышей, вероятно, по меньшей мере частично, вследствие субоптимального процесса селекции клонов, который обеспечивает контакт молекул полностью человеческих антител в эндогенной системе селекции, и неблагоприятных эффектов от изменений на эндогенный генетический состав таких мышей.
В настоящей области техники остается потребность в получении улучшенных генетически модифицированных отличные от человека животные, которые применимы в создании последовательностей иммуноглобулина, включая в себя последовательности антител человека, и которые применимы в создании разнообразного репертуара иммуноглобулиноподобных молекул, которые проявляют разнообразие, отличающееся от традиционных молекул антител, при этом в то же время снижая или устраняя неблагоприятные изменения, к которым могут привести генетические модификации. Кроме того, остается необходимость в мышах, которые способны к реаранжировке генных сегментов иммуноглобулина для образования применимых реаранжированных генов иммуноглобулина, включая в себя вариабельные домены легкой цепи иммуноглобулина человека, ассоциированные с константными доменами тяжелой цепи, которые являются когнатными вариабельным доменам легкой цепи иммуноглобулина человека, ассоциированными с константными доменами легкой цепи, или которые способны к созданию белков из измененных локусов иммуноглобулина, включая в себя локусы, которые содержат достаточно разнообразное собрание вариабельных генных сегментов легкой цепи человека. Остается потребность в отличные от человека животные, которые могут производить иммуноглобулиноподобные связывающие белки, причем связывающие белки содержат вариабельные домены легкой цепи иммуноглобулина человека, связанные с константными доменами тяжелой цепи.
Краткое раскрытие настоящего изобретения
Предусмотрены генетически модифицированные отличные от человека животные, содержащие локусы иммуноглобулина, причем локусы иммуноглобулина содержат множество вариабельных генных сегментов легкой цепи (VL) человека, функционально связанных с одной или несколькими не относящимися к человеку константными областями, например, Vκ и Jκ человека или Vλ и Jλ человека, и согласно различным вариантам осуществления локусы не содержат последовательность, которая кодирует эндогенный функциональный белок ADAM6. Отличные от человека животные включают в себя грызунов, например, мышей и крыс.
Предусмотрены локусы, которые способны к реаранжировке и образованию гена, кодирующего вариабельный домен легкой цепи, который происходит из реаранжировки, включающей в себя генный сегмент Vκ или Vλ легкой цепи человека и генный сегмент Jκ или Jλ человека и согласно различным вариантам осуществления дополнительно генный сегмент DH, причем согласно различным вариантам осуществления локусы не содержат эндогенный функциональный ген ADAM6 или его функциональный фрагмент.
Предусмотрены модифицированные локусы иммуноглобулина, которые включают в себя локусы, которые не содержат эндогенный функциональный ген ADAM6 и содержат последовательности иммуноглобулина человека, например, сегмент VL человека, функционально связанный с относящейся человеку или (или относящейся человеку/не относящейся к человеку химерной) не относящейся к человеку константной последовательностью иммуноглобулина (и в функциональной связи, например, с сегментом V и/или J). Предусмотрены модифицированные локусы, которые содержат множественные генные сегменты VL и эктопическую нуклеотидную последовательность, которая кодирует белок ADAM6 или его фрагмент, который является функциональным у отличного от человека животного. Предусмотрены модифицированные локусы, которые содержат множественные генные сегменты VL, функционально связанные с одним или нескольким сегментами DH и/или одним или несколькими сегментами JL или JH, функционально связанными с не относящейся к человеку константной последовательностью иммуноглобулина, например, последовательностью грызуна (например, мыши или крысы) или человека. Также предусмотрены отличные от человека животные, содержащие такие гуманизированные локусы, причем отличные от человека животные проявляют фертильность дикого типа.
Предусмотрены отличные от человека животные, которые содержат вариабельный локус тяжелой цепи иммуноглобулина (например, на трансгене или в качестве вставки или замещения на эндогенном вариабельном локусе тяжелой цепи отличного от человека животного), который содержит множество генных сегментов VL человека, функционально связанных с генным сегментом D человека и/или J человека. Согласно различным вариантам осуществления множеств генных сегментов VL человека функционально связаны с одним или несколькими генными сегментами D человека и/или одним или несколькими генными сегментами J человека на эндогенном вариабельном генном локусе тяжелой цепи иммуноглобулина отличного от человека животного. Согласно различным вариантам осуществления отличные от человека животные дополнительно содержат эктопическую нуклеотидную последовательность, которая кодирует белок ADAM6 или его гомолог или ортолог, который является функциональным у самца отличного от человека животного, который содержит модифицированный локус тяжелой цепи. Согласно различным вариантам осуществления эктопическая нуклеотидная последовательность является смежной по меньшей мере с одним сегментом VL человека, генным сегментом DH или генным сегментом JL. Согласно различным вариантам осуществления эктопическая нуклеотидная последовательность является смежной с не относящейся к иммуноглобулину последовательностью в геноме отличного от человека животного. Согласно одному варианту осуществления эктопическая нуклеотидная последовательность находится на той же хромосоме, что и модифицированный локус тяжелой цепи. Согласно одному варианту осуществления эктопическая нуклеотидная последовательность находится на другой хромосоме, чем модифицированный локус тяжелой цепи.
Предусмотрены отличные от человека животные, которые модифицированы на их локусах вариабельной области тяжелой цепи иммуноглобулина для делеции всех или по существу всех (например, всех функциональных сегментов или почти всех функциональных сегментов) эндогенных сегментов VH иммуноглобулина и которые содержат множество генных сегментов VL человека, функционально связанных с сегментом DH и J или генным сегментом JL на эндогенном локусе вариабельной области тяжелой цепи иммуноглобулина отличного от человека животного. Также предусмотрены отличные от человека животные, содержащие такие локусы и которые не содержат эндогенный ген ADAM6.
Предусмотрены способы получения последовательностей иммуноглобулина человека в отличные от человека животные. Согласно различным вариантам осуществления последовательности иммуноглобулина человека происходят из репертуара последовательностей тяжелой цепи иммуноглобулина, которые содержат генные сегменты VL человека, реаранжированные и находящиеся в функциональной связи с константными областями тяжелой цепи иммуноглобулина, например, VL, и один или несколько сегментов DH и J или один или несколько сегментов JL. Предусмотрены способы получения последовательности иммуноглобулина человека в относящихся к человеку животных, тканях и клетках, причем последовательности иммуноглобулина человека связывают представляющий интерес антиген.
Согласно одному аспекту предусмотрены конструкты нуклеиновых кислот, клетки, зародыши, грызуны (например, мыши) и способы получения грызунов (например, мышей), которые содержат модификацию, которая дает в результате нефункциональный эндогенный белок ADAM6 или ген ADAM6 грызуна (например, мыши) (например, нокаут или делецию в эндогенном гене ADAM6), причем грызуны (например, мыши) содержат последовательность нуклеиновой кислоты, которая кодирует белок ADAM6 или его ортолог или гомолог или фрагмент, который является функциональным у самца грызуна того же вида (например, мыши). Согласно одному варианту осуществления мыши содержат эктопическую нуклеотидную последовательность, кодирующую белок ADAM6 грызуна или его ортолог или гомолог или функциональный фрагмент; согласно конкретному варианту осуществления белок ADAM6 грызуна представляет собой белок ADAM6 мыши. Согласно одному варианту осуществления мыши содержат эктопическую нуклеотидную последовательность, которая кодирует один или несколько белков ADAM6 грызуна, причем один или несколько белков содержат SEQ ID NO: 1 или SEQ ID NO: 2 или ее фрагмент, который является функциональным у мыши.
Согласно различным аспектам последовательность, которая кодирует активность ADAM6, является смежной с последовательностью иммуноглобулина человека. Согласно различным аспектам последовательность, которая кодирует активность ADAM6, является смежной с не относящейся к человеку последовательностью иммуноглобулина. Согласно различным аспектам последовательность присутствует на той же хромосоме, что и эндогенный не относящийся к человеку локус тяжелой цепи иммуноглобулина отличного от человека животного. Согласно различным аспектам последовательность присутствует на другой хромосоме, чем локус тяжелой цепи иммуноглобулина отличного от человека животного.
Описаны генетически модифицированные отличные от человека животные, которые содержат модификацию, которая поддерживает активность гена ADAM6 или его гомолога или ортолога, причем модификация включает в себя вставку одного или нескольких генных сегментов легкой цепи иммуноглобулина человека выше не относящейся к человеку константной области тяжелой цепи иммуноглобулина, и отличные от человека животные дополнительно содержат модификации, которые позволяют им экспрессировать вариабельные области легкой цепи иммуноглобулина человека, когнатные вариабельным областям легкой цепи иммуноглобулина человека. Согласно различным аспектам вариабельные области легкой цепи иммуноглобулина человека экспрессируются в ассоциации с константными областями легкой и тяжелой цепи.
Согласно различным аспектам вставку одного или нескольких генных сегментов легкой цепи иммуноглобулина человека проводят 3’ или ниже по ходу транскрипции гена ADAM6 отличного от человека животного. Согласно различным аспектам вставку одного или нескольких генных сегментов легкой цепи иммуноглобулина человека проводят таким способом, чтобы ген(ы) ADAM6 отличного от человека животного не был(и) разрушен(ы), удален(ы) и/или функционально выключен(ы) так, чтобы активность ADAM6 отличного от человека животного находилась на таком же или сопоставимом уровне, как у отличного от человека животного, которое не содержит такой вставки. Иллюстративные разрывы, делеции и/или модификации функционального сайленсинга включают в себя любую модификацию, которая приводит к снижению, устранению и/или потере активности белка(ов) ADAM6, кодируемого(ых) геном(ами) ADAM6 отличного от человека животного.
Согласно одному аспекту предусмотрены конструкты нуклеиновой кислоты, клетки, зародыши, мыши и способы получения мышей, которые содержат модификацию эндогенного локуса иммуноглобулина, причем мыши содержат белок ADAM6 или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши. Согласно одному варианту осуществления эндогенный локус иммуноглобулина представляет собой локус тяжелой цепи иммуноглобулина, и модификация снижает или устраняет активность ADAM6 клетки или ткани самца мыши.
Согласно одному аспекту предусмотрены мыши, которые содержат эктопическую нуклеотидную последовательность, кодирующую ADAM6 мыши или его ортолог или гомолог или функциональный фрагмент; также предусмотрены мыши, которые содержат эндогенную нуклеотидную последовательность, кодирующую ADAM6 мыши или его ортолог или гомолог или фрагмент, и по меньшей мере одну генетическую модификацию локуса тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления эндогенная нуклеотидная последовательность, кодирующая ADAM6 мыши или его ортолог или гомолог или функциональный фрагмент, расположена в эктопическом положении по сравнению с эндогенным геном ADAM6 мыши дикого типа.
Согласно одному аспекту предусмотрены способы получения мышей, которые содержат модификацию эндогенного локуса иммуноглобулина, причем мыши содержат белок ADAM6 или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши. Согласно различным вариантам осуществления модификация предусматривает вставку одного или нескольких генных сегментов VL человека в эндогенном локусе иммуноглобулина.
Согласно одному аспекту предусмотрены способы получения мышей, которые содержат генетическую модификацию локуса тяжелой цепи иммуноглобулина, причем применение способов дает в результате самцов мышей, которые содержат модифицированный локус тяжелой цепи иммуноглобулина (или его делецию), и самцы мышей способны производить потомство путем спаривания. Согласно одному варианту осуществления самцы мышей способны производить сперму, которая может проходить от матки мыши через яйцевод мыши для оплодотворения яйцеклетки мыши.
Согласно одному аспекту предусмотрены способы получения мышей, которые содержат генетическую модификацию локуса тяжелой цепи иммуноглобулина и локуса легкой цепи иммуноглобулина, причем применение способов модификации локуса тяжелой цепи дает в результате самцов мышей, которые проявляют снижение фертильности, и мыши содержат генетическую модификацию, которая полностью или частично восстанавливает снижение фертильности. Согласно различным вариантам осуществления снижение фертильности характеризуется неспособностью спермы самцов мышей мигрировать от матки мыши через яйцевод мыши для оплодотворения яйцеклетки мыши. Согласно различным вариантам осуществления снижение фертильности характеризуется наличием спермы, которая проявляет in vivo нарушение миграции. Согласно различным вариантам осуществления генетическая модификация, которая полностью или частично восстанавливает снижение фертильности, представляет собой последовательность нуклеиновой кислоты, кодирующую ген ADAM6 мыши или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши.
Согласно одному варианту осуществления генетическая модификация предусматривает замещение эндогенных вариабельных локусов легкой цепи иммуноглобулина вариабельными локусами тяжелой цепи иммуноглобулина другого вида (например, не относящегося к мыши вида). Согласно одному варианту осуществления генетическая модификация предусматривает вставку вариабельных локусов легкой цепи иммуноглобулина другого вида (например, не относящегося к мыши вида) в эндогенные вариабельные локусы тяжелой цепи иммуноглобулина. Согласно конкретному варианту осуществления вид представляет собой человека. Согласно одному варианту осуществления генетическая модификация предусматривает делецию эндогенного вариабельного локуса тяжелой цепи иммуноглобулина полностью или частично, причем делеция приводит к потере эндогенной функции ADAM6. Согласно конкретному варианту осуществления потеря эндогенной функции ADAM6 связана со снижением фертильности у самцов мышей.
Согласно одному варианту осуществления генетическая модификация предусматривает инактивацию эндогенного не относящегося к человеку вариабельного локуса тяжелой цепи иммуноглобулина полностью или частично, причем инактивация не приводит к потере эндогенной функции ADAM6. Инактивация может включать в себя замещение или делецию одного или нескольких эндогенных не относящихся к человеку генных сегментов, что дает в результате эндогенный не относящийся к человеку локус тяжелой цепи иммуноглобулина, который по существу является неспособным к реаранжировке для кодирования тяжелой цепи антитела, которое содержит эндогенные не относящиеся к человеку генные сегменты. Инактивация может включать в себя другие модификации, которые делают эндогенный локус тяжелой цепи иммуноглобулина неспособным к реаранжировке для кодирования тяжелой цепи антитела, причем модификация не включает в себя замещение или делецию эндогенных генных сегментов. Иллюстративные модификации включают в себя хромосомные вставки и/или транслокации, опосредованные молекулярными техниками, например, с использованием точного размещения сайтов сайт-специфической рекомбинации (например, технологии Cre-lox). Другие иллюстративные модификации включают в себя блокирование функциональной связи между не относящимися к человеку вариабельными генными сегментами иммуноглобулина и не относящимися к человеку константными областями иммуноглобулина.
Согласно одному варианту осуществления генетическая модификация предусматривает вставку в геном отличного от человека животного фрагмента ДНК другого вида (например, не относящегося к мыши вида), содержащего один или несколько генных сегментов VL человека и один или несколько генных сегментов JL человека и необязательно один или несколько относящихся к человеку генных сегментов DH, функционально связанных с одной или несколькими последовательностями константной области (например, геном IgM и/или IgG). Согласно одному варианту осуществления фрагмент ДНК способен подвергаться реаранжировке в геноме отличного от человека животного для образования последовательности, которая кодирует вариабельный домен легкой цепи, функционально связанный с константной областью тяжелой цепи. Согласно одному варианту осуществления вид представляет собой человека. Согласно одному варианту осуществления генетическая модификация предусматривает вставку одного или нескольких генных сегментов легкой цепи иммуноглобулина человека ниже или 3’ по отношению к эндогенному гену ADAM6 отличного от человека животного так, чтобы активность ADAM6 (например, экспрессия и/или функция кодируемого белка) являлась одинаковой или сопоставимой с не относящимся к человеку животным, которое не содержит вставку.
Согласно одному аспекту предусмотрены способы получения мышей, которые содержат генетическую модификацию локуса тяжелой цепи иммуноглобулина, причем применение способов дает в результате самцов мышей, которые содержат модифицированный локус тяжелой цепи иммуноглобулина (или его делецию), и самцы мышей проявляют снижение фертильности, и мыши содержат генетическую модификацию, которая восстанавливает полностью или частично снижение фертильности. Согласно различным вариантам осуществления снижение фертильности характеризуется неспособностью спермы самцов мышей мигрировать от матка мыши через яйцевод мыши для оплодотворения яйцеклетки мыши. Согласно различным вариантам осуществления снижение фертильности характеризуется наличием сперматозоидов, которые проявляют нарушение миграции in vivo. Согласно различным вариантам осуществления генетическая модификация, которая полностью или частично восстанавливает снижение фертильности, представляет собой последовательность нуклеиновой кислоты, кодирующую ген ADAM6 мыши или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши.
Согласно одному варианту осуществления генетическая модификация предусматривает замещение эндогенных вариабельных локусов тяжелой цепи иммуноглобулина вариабельными локусами легкой цепи иммуноглобулина, например, одним или несколькими вариабельными генными сегментами легкой цепи (VL), одним или нескольким дополнительными генными сегментами тяжелой цепи (DH) и одним или несколькими соединительными генными сегментами (J) или одним или несколькими соединительными генными сегментами легкой цепи (JL) другого вида (например, не относящегося к мыши вида). Согласно одному варианту осуществления генетическая модификация предусматривает вставку одного ортологичного генного сегмента вариабельных локусов легкой цепи VL иммуноглобулина, по меньшей мере одного генного сегмента DH и по меньшей мере одного генного сегмента J или по меньшей мере одного генного сегмента JL в эндогенные вариабельные локусы тяжелой цепи иммуноглобулина. Согласно конкретному варианту осуществления вид представляет собой человека. Согласно одному варианту осуществления генетическая модификация предусматривает делецию эндогенного вариабельного локуса тяжелой цепи иммуноглобулина полностью или частично, причем делеция приводит к потере эндогенной функции ADAM6. Согласно конкретному варианту осуществления потеря эндогенной функции ADAM6 ассоциирована со снижением фертильности у самцов мышей. Согласно одному варианту осуществления генетическая модификация предусматривает инактивацию эндогенного вариабельного локуса тяжелой цепи иммуноглобулина полностью или частично, причем инактивация не приводит к потере эндогенной функции ADAM6. Инактивация может включать в себя замещение или делецию одного или нескольких эндогенных генных сегментов, которые дают в результате эндогенный локус тяжелой цепи иммуноглобулина, который по существу является неспособным к реаранжировке для кодирования тяжелую цепь антитела, которое содержит эндогенные генные сегменты. Инактивация может включать в себя другие модификации, которые делают эндогенный локус тяжелой цепи иммуноглобулина неспособным к реаранжировке для кодирования тяжелой цепи антитела, причем модификация не включает в себя замещение или делецию эндогенных генных сегментов. Иллюстративные модификации включают в себя хромосомные инверсии и/или изменения, которые дают в результате локус тяжелой цепи, который не находится в функциональной связи с одной или несколькими эндогенными константными областями.
Согласно одному варианту осуществления генетическая модификация предусматривает вставку в геном мыши фрагмента ДНК, содержащего один или несколько генных сегментов VL человека, один или несколько генных сегментов J и необязательно один или несколько генных сегментов D другого вида (например, не относящегося к мыши вида), функционально связанных с одной или несколькими последовательностями константной области (например, геном IgM и/или IgG). Согласно различным вариантам осуществления генные сегменты J включают в себя генные сегменты JH или JL. Согласно одному варианту осуществления фрагмент ДНК способен подвергаться реаранжировке для образования последовательности, которая кодирует тяжелую цепь антитела, причем тяжелая цепь содержит реаранжированный вариабельный генный сегмент легкой цепи человек, слитый с константной областью тяжелой цепи. Согласно одному варианту осуществления генетическая модификация предусматривает вставку по меньшей мере шести, по меньшей мере 16, по меньшей мере 30 или по меньшей мере 40 или больше генных сегментов VL человека и по меньшей мере одного или по меньшей мере 5 генных сегментов JL человека в геном мыши. Согласно конкретному варианту осуществления вид представляет собой человека, и генные сегменты представляют собой генные сегменты κ легкой цепи человека. Согласно одному варианту осуществления генетическая модификация предусматривает делецию эндогенного вариабельного локуса тяжелой цепи иммуноглобулина полностью или частично, чтобы сделать эндогенный локус тяжелой цепи иммуноглобулина нефункциональным, причем делеция дополнительно приводит к потере эндогенной функции ADAM6. Согласно конкретному варианту осуществления потеря эндогенной функции ADAM6 ассоциирована со снижением фертильности у самцов мышей.
Согласно одному аспекту предусмотрены мыши, которые содержат модификацию, которая снижает или устраняет экспрессию ADAM6 мыши из эндогенного аллеля ADAM6 так, что самец мыши с модификацией проявляет сниженную фертильность (например, сильно сниженную способность производить потомство путем спаривания), или является по существу стерильным вследствие снижения или устранения эндогенной функции ADAM6, причем мыши дополнительно содержат эктопическую последовательность ADAM6 или ее гомолог или ортолог или функциональный фрагмент. Согласно одному аспекту модификация, которая снижает или устраняет экспрессию ADAM6 мыши, представляет собой модификацию (например, вставку, делецию, замещение и т.д.) в локусе иммуноглобулина мыши. Согласно одному варианту осуществления локус иммуноглобулина представляет собой локус тяжелой цепи иммуноглобулина.
Согласно одному варианту осуществления снижение или потеря функции ADAM6 предусматривает неспособность или существенную неспособность мыши производить сперму, которая может проходить от матки мыши через яйцевод мыши для оплодотворения яйцеклетки мыши. Согласно конкретному варианту осуществления по меньшей мере приблизительно 95%, 96%, 97%, 98% или 99% сперматозоидов, произведенных в объеме эякулята мыши, являются неспособными пройти через яйцевод in vivo после копуляции и оплодотворить яйцеклетку мыши.
Согласно одному варианту осуществления снижение или потеря функции ADAM6 предусматривает неспособность образования или существенную неспособность образования комплекса ADAM2 и/или ADAM3 и/или ADAM6 на поверхности сперматозоида мыши. Согласно одному варианту осуществления потеря функции ADAM6 предусматривает существенную неспособность оплодотворить яйцеклетку мыши путем копуляции с самкой мыши.
Согласно одному аспекту предусмотрена мышь, которая не содержит функциональный эндогенный ген ADAM6 и содержит белок (или эктопическую нуклеотидную последовательность, которая кодирует белок), который предоставляет мыши функциональность ADAM6. Согласно одному варианту осуществления мышь представляет собой самца мыши, и функциональность включает в себя усиленную фертильность по сравнению с мышью, которая не содержит функциональный эндогенный ген ADAM6.
Согласно одному варианту осуществления белок кодируется геномной последовательностью, расположенной в пределах локуса иммуноглобулина в зародышевой линии мыши. Согласно конкретному варианту осуществления локус иммуноглобулина представляет собой локус тяжелой цепи. Согласно другому конкретному варианту осуществления локус тяжелой цепи содержит по меньшей мере один генный сегмент VH человека, по меньшей мере один генный сегмент DH человека и по меньшей мере один генный сегмент JH человека. Согласно другому конкретному варианту осуществления локус тяжелой цепи содержит по меньшей мере один генный сегмент VL человека и по меньшей мере один генный сегмент JL человека. Согласно другому конкретному варианту осуществления локус тяжелой цепи содержит по меньшей мере один VL человека, по меньшей мере один DH человека и по меньшей мере один JL человека. Согласно другому конкретному варианту осуществления локус тяжелой цепи содержит по меньшей мере один генный сегмент VL человека, по меньшей мере один генный сегмент DH человека и по меньшей мере один генный сегмент JH человека. Согласно другому конкретному варианту осуществления локус тяжелой цепи содержит по меньшей мере один генный сегмент VL человека и по меньшей мере один генный сегмент JL человека. Согласно другому конкретному варианту осуществления локус тяжелой цепи содержит по меньшей мере один генный сегмент VL человека и по меньшей мере один генный сегмент JH человека. Согласно другому конкретному варианту осуществления локус тяжелой цепи содержит шесть генных сегментов Vκ человека и пять генных сегментов Jκ человека. Согласно другому конкретному варианту осуществления локус тяжелой цепи содержит 16 генных сегментов Vκ человека и пять генных сегментов Jκ человека. Согласно другому конкретному варианту осуществления локус тяжелой цепи содержит 30 генных сегментов Vκ человека и пять генных сегментов Jκ человека. Согласно другому конкретному варианту осуществления локус тяжелой цепи содержит 40 генных сегментов Vκ человека и пять генных сегментов Jκ человека.
Согласно одному варианту осуществления эктопический белок кодируется геномной последовательностью, расположенной в пределах не относящегося к иммуноглобулину локуса в зародышевой линии мыши. Согласно одному варианту осуществления не относящийся к иммуноглобулину локус представляет собой транскрипционно активный локус. Согласно конкретному варианту осуществления транскрипционно активный локус представляет собой локус ROSA. Согласно конкретному варианту осуществления транскрипционно активный локус ассоциирован с тканеспецифической экспрессией. Согласно одному варианту осуществления тканеспецифическая экспрессия присутствует в репродуктивных тканях. Согласно одному варианту осуществления белок кодируется геномной последовательностью, случайным образом вставленной в зародышевую линию мыши.
Согласно одному варианту осуществления мышь содержит относящуюся к человеку или химерную относящуюся к человеку/мыши или химерную относящуюся к человеку/крысе (например, относящуюся к человеку вариабельную, относящуюся к мыши или крысе константную) легкую цепь и химерную относящуюся к человеку вариабельную/относящуюся к мыши или крысе константную тяжелую цепь. Согласно конкретному варианту осуществления мышь содержит трансген, который содержит химерный ген относящейся к человеку вариабельной/относящейся к крысе или мыши константной легкой цепи, функционально связанный с транскрипционно активным промотором, например, промотором ROSA26. Согласно дополнительному конкретному варианту осуществления химерный трансген относящейся к человеку/мыши или крысы легкой цепи содержит реаранжированную последовательность вариабельной области легкой цепи человека в зародышевой линии мыши.
Согласно одному варианту осуществления эктопическая нуклеотидная последовательность расположена в пределах локуса иммуноглобулина в зародышевой линии мыши. Согласно конкретному варианту осуществления локус иммуноглобулина представляет собой локус тяжелой цепи. Согласно одному варианту осуществления локус тяжелой цепи содержит по меньшей мере один генный сегмент VL человека и по меньшей мере один генный сегмент JL человека. Согласно конкретному варианту осуществления локус тяжелой цепи содержит по меньшей мере шесть и до 40 генных сегментов Vκ человека и пять генных сегментов Jκ человека. Согласно одному варианту осуществления эктопическая нуклеотидная последовательность расположена в пределах не относящегося к иммуноглобулину локуса в зародышевой линии мыши. Согласно одному варианту осуществления не относящийся к иммуноглобулину локус представляет собой транскрипционно активный локус. Согласно конкретному варианту осуществления транскрипционно активный локус представляет собой локус ROSA26. Согласно одному варианту осуществления эктопическая нуклеотидная последовательность размещена в виде вставленной случайным образом в зародышевую линию мыши.
Согласно одному аспекту предусмотрена мышь, которая не содержит функциональный эндогенный ген ADAM6, причем мышь содержит эктопическую нуклеотидную последовательность, которая восполняет потерю функции ADAM6 мыши. Согласно одному варианту осуществления эктопическая нуклеотидная последовательность предоставляет мыши способность производить потомство, которая является сопоставимой с таковой у соответствующей мыши дикого типа, которая содержит функциональный эндогенный ген ADAM6. Согласно одному варианту осуществления последовательность предоставляет мыши способность образовывать комплекс ADAM2 и/или ADAM3 и/или ADAM6 на поверхности сперматозоида мыши. Согласно одному варианту осуществления последовательность предоставляет сперматозоиду мыши способность проходить от матки мыши через яйцевод мыши к яйцеклетке мыши для оплодотворения яйцеклетки.
Согласно одному варианту осуществления мышь, не содержащая функциональный эндогенный ген ADAM6 и содержащая эктопическую нуклеотидную последовательность, производит по меньшей мере приблизительно 50%, 60%, 70%, 80% или 90% от числа пометов, которые производит мышь дикого типа мышь того же возраста и линии за шестимесячный период времени.
Согласно одному варианту осуществления мышь, не содержащая функциональный эндогенный ген ADAM6 и содержащая эктопическую нуклеотидную последовательность, производит больше по меньшей мере приблизительно в 1,5 раза, приблизительно в 2 раза, приблизительно в 2,5 раза, приблизительно в 3 раза, приблизительно в 4 раза, приблизительно в 6 раз, приблизительно в 7 раз, приблизительно в 8 раз или приблизительно в 10 раз или более потомства при разведении в течение шестимесячного периода времени, чем мышь такого же возраста и такой же или аналогичной линии, которая не содержит функциональный эндогенный ген ADAM6 и которая не содержит эктопическую нуклеотидную последовательность, которую разводят в течение по существу такого же периода времени и по существу при таких же условиях.
Согласно одному варианту осуществления мышь, не содержащая функциональный эндогенный ген ADAM6 и содержащая эктопическую нуклеотидную последовательность, производит в среднем по меньшей мере приблизительно в 2 раза, в 3 раза или в 4 раза большее число детенышей на помет за четырех- или шестимесячный период разведения, чем мышь, которые не содержит функциональный эндогенный ген ADAM6 и которая не содержит эктопическую нуклеотидную последовательность, и которую разводят в течение такого же периода времени.
Согласно одному варианту осуществления мышь, не содержащая функциональный эндогенный ген ADAM6 и содержащая эктопическую нуклеотидную последовательность, представляет собой самца мыши, и самец мыши производит сперму, которая будучи извлеченной из яйцеводов приблизительно через 5-6 часов после копуляции, отражает миграцию в яйцеводе, которая больше по меньшей мере в 10 раз, по меньшей мере в 20 раз, по меньшей мере в 30 раз, по меньшей мере в 40 раз, по меньшей мере в 50 раз, по меньшей мере в 60 раз, по меньшей мере в 70 раз, по меньшей мере в 80 раз, по меньшей мере в 90 раз, в 100 раз, в 110 раз или в 120 раз или более, чем у мыши, которая не содержит функциональный эндогенный ген ADAM6 и которая не содержит эктопическую нуклеотидную последовательность.
Согласно одному варианту осуществления мышь, не содержащая функциональный эндогенный ген ADAM6 и содержащая эктопическую нуклеотидную последовательность, при копуляции с самкой мыши образует сперму, которая способна к прохождению матки и попаданию в яйцевод и его прохождению в течение приблизительно шести часов при результативности, которая приблизительно равна сперме мыши дикого типа.
Согласно одному варианту осуществления мышь, не содержащая функциональный эндогенный ген ADAM6 и содержащая эктопическую нуклеотидную последовательность, производит больше приблизительно в 1,5 раза, приблизительно в 2 раза, приблизительно в 3 раза или приблизительно в 4 раза или более пометов в сопоставимый период времени, чем мышь, которая не содержит функциональный ген ADAM6 и которая не содержит эктопическую нуклеотидную последовательность.
Согласно одному аспекту предусмотрена мышь, содержащая в своей зародышевой линии не относящуюся к мыши последовательность нуклеиновой кислоты, которая кодирует белок иммуноглобулина, причем не относящаяся к мыши последовательность иммуноглобулина содержит вставку гена ADAM6 мыши или его гомолога или ортолога или функционального фрагмента. Согласно одному варианту осуществления не относящаяся к мыши последовательность иммуноглобулина содержит последовательность иммуноглобулина человека. Согласно одному варианту осуществления последовательность содержит последовательность тяжелой цепи иммуноглобулина человека. Согласно одному варианту осуществления последовательность содержит последовательность легкой цепи иммуноглобулина человека. Согласно одному варианту осуществления последовательность содержит последовательность тяжелой цепи человека, смежную с последовательностью легкой цепи человека. Согласно одному варианту осуществления последовательность содержит один или несколько генных сегментов V, один или несколько генных сегментов D и один или несколько генных сегментов J; согласно одному варианту осуществления последовательность содержит один или несколько генных сегментов V и один или несколько генных сегментов J. Согласно одному варианту осуществления один или несколько генных сегментов V, D и J или один или несколько генных сегментов V и J являются нереаранжированными. Согласно одному варианту осуществления один или несколько генных сегментов V, D и J или один или несколько генных сегментов V и J являются реаранжированными. Согласно одному варианту осуществления после реаранжировки одного или нескольких генных сегментов V, D и J или одного или нескольких генных сегментов V и J мышь содержит в своем геноме по меньшей мере одну последовательность нуклеиновой кислоты, кодирующую ген ADAM6 мыши или его гомолог или ортолог или функциональный фрагмент. Согласно одному варианту осуществления после реаранжировки мышь содержит в своем геноме по меньшей мере две последовательности нуклеиновой кислоты, кодирующие ген ADAM6 мыши или его гомолог или ортолог или функциональный фрагмент. Согласно одному варианту осуществления после реаранжировки мышь содержит в своем геноме по меньшей мере одну последовательность нуклеиновой кислоты, кодирующую ген ADAM6 мыши или его гомолог или ортолог или функциональный фрагмент. Согласно одному варианту осуществления мышь содержит ген ADAM6 или его гомолог или ортолог или функциональный фрагмент в B-клетке. Согласно одному варианту осуществления мышь содержит ген ADAM6 или его гомолог или ортолог или функциональный фрагмент в клетке, не относящейся к B-клетке.
Согласно одному аспекту предусмотрены мыши, которые экспрессируют вариабельную область тяжелой цепи иммуноглобулина человека или ее функциональный фрагмент из эндогенного локуса тяжелой цепи иммуноглобулина, причем мыши содержат активность ADAM6, которая является функциональной у самца мыши. Согласно одному варианту осуществления локус тяжелой цепи содержит один или несколько генных сегментов VL человека и один или несколько генных сегментов JL человека и необязательно один или несколько генных сегментов DH человека. Согласно одному варианту осуществления локус тяжелой цепи содержит по меньшей мере шесть генных сегментов Vκ человека и пять генных сегментов Jκ человека. Согласно одному варианту осуществления локус тяжелой цепи содержит по меньшей мере 16 генных сегментов Vκ человека и пять генных сегментов Jκ человека. Согласно одному варианту осуществления локус тяжелой цепи содержит по меньшей мере 30 генных сегментов Vκ человека и пять генных сегментов Jκ человека. Согласно одному варианту осуществления локус тяжелой цепи содержит по меньшей мере 40 генных сегментов Vκ человека и пять генных сегментов Jκ человека.
Согласно одному аспекту предусмотрены мыши, которые экспрессируют вариабельную область легкой цепи иммуноглобулина человека или ее функциональный фрагмент из эндогенного локуса тяжелой цепи иммуноглобулина, причем мыши содержат активность ADAM6, которая является функциональной у самца мыши.
Согласно одному варианту осуществления самцы мышей содержат один немодифицированный эндогенный аллель ADAM6 или его ортолог или гомолог или функциональный фрагмент на эндогенном локусе ADAM6.
Согласно одному варианту осуществления самцы мышей содержат эктопическую последовательность ADAM6 мыши или ее гомолог или ортолог или функциональный фрагмент, который кодирует белок, которые предоставляет функцию ADAM6.
Согласно одному варианту осуществления самцы мышей содержат последовательность ADAM6 или ее гомолог или ортолог или функциональный фрагмент в положении в геноме мыши, которое приблизительно соответствует положению эндогенного аллеля ADAM6, например, 3’ по отношению к последовательности последнего генного сегмента V и 5’ по отношению к первому генному сегменту J.
Согласно одному варианту осуществления самцы мышей содержат последовательность ADAM6 или ее гомолог или ортолог или функциональный фрагмент в положении в мышином геноме, которое отличается от положения эндогенного аллеля ADAM6, например, 5’ по отношению к расположенной наиболее 5’ последовательности генного сегмента V генного локуса V.
Согласно одному варианту осуществления самцы мышей содержат последовательность ADAM6 или ее гомолог или ортолог или функциональный фрагмент, фланкированный выше, ниже или выше и ниже (относительно направления транскрипции последовательности ADAM6) последовательностью нуклеиновой кислоты, кодирующей генный сегмент V иммуноглобулина и/или генный сегмент J иммуноглобулина. Согласно конкретному варианту осуществления генные сегменты V и J иммуноглобулина представляют собой генные сегменты человека. Согласно одному варианту осуществления генные сегменты V и J иммуноглобулина представляют собой генные сегменты человека, и последовательность, кодирующая ADAM6 мыши или его ортолог или гомолог или фрагмент, функциональный у мыши, находится между генными сегментами V и J человека; согласно одному варианту осуществления мышь содержит два или больше генных сегментов V человека, и последовательность находится в положении 5’ по отношению к наиболее 5’ генному сегменту V человека; согласно одному варианту осуществления мышь содержит два или больше генных сегментов V человека, и последовательность находится в положении между последним генным сегментом V и предпоследним генным сегментом V; согласно одному варианту осуществления мышь содержит множество генных сегментов V человека, и последовательность находится в положении выше по отношению к наиболее 5’ генному сегменту V человека; согласно одному варианту осуществления мышь дополнительно содержит генный сегмент D, и последовательность находится в положении, следующем за наиболее 3’ генным сегментом V и наиболее 5’ генным сегментом D; согласно одному варианту осуществления последовательность находится в положении между генным сегментом V и генным сегментом J.
Согласно одному варианту осуществления генные сегменты V человека представляют собой генные сегменты V легкой цепи. Согласно конкретному варианту осуществления генные сегменты V легкой цепи представляют собой генные сегменты Vκ. Согласно другому конкретному варианту осуществления генные сегменты V легкой цепи представляют собой генные сегменты Vλ. Согласно одному варианту осуществления генный сегмент J выбран из генного сегмента JH и JL. Согласно конкретному варианту осуществления генный сегмент JL представляет собой генный сегмент Jκ. Согласно другому конкретному варианту осуществления генный сегмент JL представляет собой генный сегмент Jλ.
Согласно одному варианту осуществления самцы мышей содержат последовательность ADAM6 или ее гомолог или ортолог или функциональный фрагмент, который расположен в положении в эндогенном локусе иммуноглобулина, которое является таким же или по существу таким же, как и у самца мыши дикого типа. Согласно конкретному варианту осуществления эндогенный локус является неспособным кодировать вариабельную область тяжелой цепи антитела, причем вариабельная область содержит или происходит из эндогенного не относящегося к человеку генного сегмента. Согласно конкретному варианту осуществления эндогенный локус размещен в положении в геноме самца мыши, которое делает генные сегменты тяжелой цепи локуса неспособными кодировать вариабельную область тяжелой цепи антитела. Согласно различным вариантам осуществления самцы мышей содержат последовательность ADAM6, расположенную на той же хромосоме, что и генные сегменты иммуноглобулина человека, и последовательность ADAM6 кодирует функциональный белок ADAM6.
Согласно одному аспекту предусмотрен самец мыши, который содержит нефункциональный эндогенный ген ADAM6 или делецию эндогенного гена ADAM6 в своей зародышевой линии; причем сперматозоиды мыши способны проходить яйцевод самки мыши и оплодотворять яйцеклетку. Согласно одному варианту осуществления мыши содержат экстрахромосомную копию гена ADAM6 мыши или его ортолога или гомолога или функционального фрагмента, который является функциональным у самца мыши. Согласно одному варианту осуществления мыши содержат эктопический ген ADAM6 мыши или его ортолог или гомолог или функциональный фрагмент, который является функциональным у самца мыши.
Согласно одному аспекту предусмотрен самец мыши, который содержит функциональный эндогенный ген ADAM6 и модификацию эндогенного локуса иммуноглобулина тяжелой цепи. Согласно одному варианту осуществления модификацию производят ниже, или 3’, по отношению к эндогенному гену или локусу ADAM6. Согласно одному варианту осуществления модификация представляет собой замещение одного или нескольких эндогенных генных сегментов тяжелой цепи иммуноглобулина одним или несколькими генными сегментами легкой цепи иммуноглобулина человека. Согласно одному варианту осуществления модификация представляет собой вставку одного или нескольких генных сегментов легкой цепи иммуноглобулина человека выше по отношению к эндогенному гену константной области тяжелой цепи иммуноглобулина.
Согласно одному аспекту предусмотрены мыши, которые содержат генетическую модификацию, которая снижает эндогенную функцию ADAM6, причем мышь содержит по меньшей мере некоторую функциональность ADAM6, обеспеченную либо эндогенным немодифицированным аллелем, который является полностью или частично функциональным (например, гетерозигота), либо экспрессией из эктопической последовательности, которая кодирует ADAM6 или его ортолог или гомолог или функциональный фрагмент, который является функциональным у самца мыши. Согласно различным вариантам осуществления ADAM6 или его ортолог или гомолог или функциональный фрагмент содержит последовательность нуклеиновой кислоты, которая кодирует белок ADAM6, представленный в SEQ ID NO: 1, SEQ ID NO: 2 или их комбинации.
Согласно одному варианту осуществления мыши содержат функцию ADAM6, достаточную для предоставления самцам мышей способности производить потомство путем спаривания по сравнению с самцами мышей, у которых отсутствует функциональный ADAM6. Согласно одному варианту осуществления функция ADAM6 обеспечивается путем присутствия эктопической нуклеотидной последовательности, которая кодирует ADAM6 мыши или его гомолог или ортолог или функциональный фрагмент. Согласно одному варианту осуществления функция ADAM6 обеспечивается эндогенным геном ADAM6, присутствующим в эндогенном локусе иммуноглобулина, причем мышь является неспособной экспрессировать антитело, содержащее эндогенный генный сегмент тяжелой цепи иммуноглобулина. Гомологи ADAM6 или его ортологи или фрагменты, которые являются функциональными у самца мыши, включают в себя те, которые восстанавливают, полностью или частично, потерю способности производить потомство, наблюдаемую у самца мыши, у которого отсутствует достаточная эндогенная активность ADAM6, например, потерю способности, наблюдаемую у нокаутной в отношении ADAM6 мыши. В этом смысле нокаутные в отношении ADAM6 мыши включают в себя мышей, которые содержат эндогенный локус или его фрагмент, но который не является функциональным, т.е. который вообще не экспрессирует ADAM6 (ADAM6a и/или ADAM6b) или который экспрессирует ADAM6 (ADAM6a и/или ADAM6b) на уровне, недостаточном для поддержания по существу нормальной способности самца мыши дикого типа производить потомство. Потеря функции может быть обусловлена, например, модификацией в структурном гене локуса (т.е. в кодирующей области ADAM6a или ADAM6b) или в регуляторной области локуса (например, в последовательности 5’ по отношению к гену ADAM6a или 3’ по отношению к кодирующей области ADAM6a или ADAM6b, причем последовательность контролирует, полностью или частично, транскрипцию гена ADAM6, экспрессию РНК ADAM6 или экспрессию белка ADAM6). Согласно различным вариантам осуществления его ортологи или гомологи или фрагменты, которые являются функциональными у самца мыши, являются такими, которые позволяют сперме самца мыши (или большинству сперматозоидов в эякуляте самца мыши) проходить яйцевод мыши и оплодотворять яйцеклетку мыши.
Согласно одному варианту осуществления самцы мышей, которые экспрессируют вариабельную область иммуноглобулина человека или ее функциональный фрагмент, содержат достаточную активность ADAM6 для предоставления самцам мышей способности производить потомство путем спаривания с самками мышей и согласно одному варианту осуществления самцы мышей проявляют способность производить потомство при спаривании с самками мышей, которое согласно одному варианту осуществления составляет по меньшей мере 25%, согласно одному варианту осуществления по меньшей мере 30%, согласно одному варианту осуществления по меньшей мере 40%, согласно одному варианту осуществления по меньшей мере 50%, согласно одному варианту осуществления по меньшей мере 60%, согласно одному варианту осуществления по меньшей мере 70%, согласно одному варианту осуществления по меньшей мере 80%, согласно одному варианту осуществления по меньшей мере 90% и согласно одному варианту осуществления является приблизительно одинаковым по сравнению с потомством мышей с одним или двумя эндогенными немодифицированными аллелями ADAM6.
Согласно одному варианту осуществления самцы мышей экспрессируют достаточный ADAM6 (или его ортолог или гомолог или функциональный фрагмент) для предоставления сперматозоиду от самцов мышей возможности проходить яйцевод самки мыши и оплодотворять яйцеклетку мыши.
Согласно одному варианту осуществления функциональность ADAM6 обеспечивается последовательностью нуклеиновой кислоты, которая является смежной с хромосомной последовательностью мыши (например, нуклеиновая кислота случайным образом интегрирована в хромосому мыши; или помещена в конкретное положение, например, путем нацеливания нуклеиновой кислоты в конкретное положение, например, с помощью опосредованной сайт-специфической рекомбиназой (например, опосредованной Cre) вставки или гомологичной рекомбинации). Согласно одному варианту осуществления последовательность ADAM6 присутствует на нуклеиновой кислоте, которая расположена отдельно от хромосомы мыши (например, последовательность ADAM6 присутствует на эписоме, т.е. экстрахромосомно, например, в конструкте экспрессии, векторе, YAC, трансхромосоме и т.д.).
Согласно одному аспекту предусмотрены генетически модифицированные мыши и клетки, которые содержат модификацию эндогенного локуса тяжелой цепи иммуноглобулина, причем мыши экспрессируют по меньшей мере часть последовательности легкой цепи иммуноглобулина, например, по меньшей мере часть последовательности человека, причем мыши содержат активность ADAM6, которая является функциональной у самца мыши. Согласно одному варианту осуществления модификация снижает или устраняет активность ADAM6 мыши. Согласно одному варианту осуществления мышь модифицирована так, что оба аллеля, которые кодируют активность ADAM6, либо отсутствуют, либо экспрессируют ADAM6, который по существу не функционирует для поддержания нормального спаривания у самца мыши. Согласно одному варианту осуществления мышь дополнительно содержит эктопическую последовательность нуклеиновой кислоты, кодирующую ADAM6 мыши или его ортолог или гомолог или функциональный фрагмент. Согласно одному варианту осуществления модификация поддерживает активность ADAM6 мыши и делает эндогенный локус тяжелой цепи иммуноглобулина неспособным кодировать тяжелую цепь антитела. Согласно конкретному варианту осуществления модификация включает в себя хромосомные вставки и/или транслокации, которые делают эндогенный локусе тяжелой цепи иммуноглобулина неспособными к реаранжировке для кодирования вариабельной области тяжелой цепи антитела.
Согласно одному аспекту предусмотрены генетически модифицированные мыши и клетки, которые содержат модификацию эндогенного локуса тяжелой цепи иммуноглобулина, причем модификация снижает или устраняет активность ADAM6, экспрессированного из последовательности локуса ADAM6, и причем мыши содержат белок ADAM6 или его ортолог или гомолог или функциональный фрагмент. Согласно различным вариантам осуществления белок ADAM6 или его фрагмент кодируется эктопической последовательностью ADAM6. Согласно различным вариантам осуществления белок ADAM6 или его фрагмент экспрессируется из эндогенного аллеля ADAM6. Согласно различным вариантам осуществления мышь содержит первый аллель тяжелой цепи, который содержит первую модификацию, которая снижает или устраняет экспрессию функционального ADAM6 из первого аллеля тяжелой цепи, и мышь содержит второй аллель тяжелой цепи, который содержит вторую модификацию, которая по существу не снижает или не устраняет экспрессию функционального ADAM6 из второго аллеля тяжелой цепи.
Согласно различным вариантам осуществления модификация представляет собой вставку одного или нескольких генных сегментов легкой цепи иммуноглобулина человека выше, или 5’, по отношению к эндогенному гену константной области тяжелой цепи иммуноглобулина. Согласно различным вариантам осуществления модификация поддерживает эндогенный ген ADAM6, расположенный на эндогенном локусе тяжелой цепи иммуноглобулина.
Согласно одному варианту осуществления вторая модификация расположена 3’ (по отношению к направлению транскрипции генного сегмента V мыши) последнего генного сегмента V мыши и расположена 5’ (по отношению к направлению транскрипции константной последовательности) относящегося к мыши (или химерного относящегося к человеку/мыши) константного гена тяжелой цепи иммуноглобулина или его фрагмента (например, последовательности нуклеиновой кислоты, кодирующей относящийся к человеку и/или относящийся к мыши: CH1 и/или шарнир и/или CH2 и/или CH3).
Согласно одному варианту осуществления модификация находится на первом аллеле тяжелой цепи иммуноглобулина на первом локусе, который кодирует первый аллель ADAM6, и функция ADAM6 является результатом экспрессии эндогенного ADAM6 на втором аллеле тяжелой цепи иммуноглобулина на втором локусе, который кодирует функциональный ADAM6, причем второй аллель тяжелой цепи иммуноглобулина содержит по меньшей мере одну модификацию генного сегмента V, D и/или J. Согласно конкретному варианту осуществления по меньшей мере одна модификация генного сегмента V, D и/или J представляет собой делецию, замещение генным сегментом V, D и/или J человека, замещение генным сегментом V, D и/или J верблюда, замещение гуманизированным или камелизированным генным сегментом V, D и/или J, замещение последовательности тяжелой цепи последовательностью легкой цепи и их комбинацию. Согласно одному варианту осуществления по меньшей мере одна модификация представляет собой делецию одного или нескольких генных сегментов V, D и/или J тяжелой цепи и замещение одного или нескольких генных сегментов V и/или J легкой цепи (например, генного сегмента V и/или J легкой цепи человека) на локусе тяжелой цепи.
Согласно одному варианту осуществления модификация находится на первом аллеле тяжелой цепи иммуноглобулина на первом локусе и втором аллеле тяжелой цепи иммуноглобулина на втором локусе, и функция ADAM6 является результатом экспрессии эктопического ADAM6 на не относящемся к иммуноглобулину локусе в зародышевой линии мыши. Согласно конкретному варианту осуществления не относящийся к иммуноглобулину локус представляет собой локус ROSA26. Согласно конкретному варианту осуществления не относящийся к иммуноглобулину локус является транскрипционно активным в репродуктивной ткани.
Согласно одному варианту осуществления модификация находится на первом аллеле тяжелой цепи иммуноглобулина на первом локусе и втором аллеле тяжелой цепи иммуноглобулина на втором локусе, и функция ADAM6 является результатом эндогенного гена ADAM6 в зародышевой линии мыши. Согласно конкретному варианту осуществления эндогенный ген ADAM6 находится бок о бок с генными сегментами тяжелой цепи иммуноглобулина мыши.
Согласно одному варианту осуществления модификация находится на первом аллеле тяжелой цепи иммуноглобулина на первом локусе и на втором аллеле тяжелой цепи иммуноглобулина на втором локусе, и функция ADAM6 является результатом экспрессии эктопической последовательности ADAM6 на первом аллеле тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления модификация находится на первом аллеле тяжелой цепи иммуноглобулина на первом локусе и на втором аллеле тяжелой цепи иммуноглобулина на втором локусе, и функция или активность ADAM6 является результатом экспрессии эктопического ADAM6 на втором аллеле тяжелой цепи иммуноглобулина.
Согласно одному аспекту предусмотрена мышь, содержащая гетерозиготный или гомозиготный нокаут ADAM6. Согласно одному варианту осуществления мышь дополнительно содержит модифицированную последовательность иммуноглобулина, которая представляет собой относящуюся к человеку или гуманизированную последовательность иммуноглобулина, или относящуюся к верблюду или камелизированную последовательность иммуноглобулина человека или мыши. Согласно одному варианту осуществления модифицированная последовательность иммуноглобулина присутствует на эндогенном локусе тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления модифицированная последовательность иммуноглобулина содержит последовательность вариабельной области легкой цепи человека на эндогенном локусе тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления последовательность вариабельной области легкой цепи человека замещает эндогенную вариабельную последовательность тяжелой цепи на эндогенном локусе тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления модифицированная последовательность иммуноглобулина содержит последовательность вариабельной области κ легкой цепи человека на эндогенном локусе тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления модифицированная последовательность иммуноглобулина содержит последовательность вариабельной области λ легкой цепи человека на эндогенном локусе тяжелой цепи иммуноглобулина.
Согласно одному аспекту предусмотрена мышь, неспособная к экспрессии функционального эндогенного ADAM6 из эндогенного локуса ADAM6. Согласно одному варианту осуществления мышь содержит эктопическую последовательность нуклеиновой кислоты, которая кодирует ADAM6 или его функциональный фрагмент, который является функциональным у мыши. Согласно конкретному варианту осуществления эктопическая последовательность нуклеиновой кислоты кодирует белок, который восстанавливает потерю способности производить потомство, проявляемую самцом мыши, который является гомозиготным в отношении нокаута ADAM6. Согласно конкретному варианту осуществления эктопическая последовательность нуклеиновой кислоты кодирует белок ADAM6 мыши.
Согласно одному аспекту предусмотрена мышь, которая не содержит функциональный эндогенный локус ADAM6, и которая содержит эктопическую последовательность нуклеиновой кислоты, которая предоставляет мыши функцию ADAM6. Согласно одному варианту осуществления последовательность нуклеиновой кислоты содержит эндогенную последовательность ADAM6 или ее функциональный фрагмент. Согласно одному варианту осуществления эндогенная последовательность ADAM6 содержит кодирующую ADAM6a и ADAM6b последовательность, расположенную у мыши дикого типа между наиболее 3’ генным сегментом V тяжелой цепи (VH) иммуноглобулина мыши и наиболее 5’ генным сегментом D тяжелой цепи (DH) иммуноглобулина мыши.
Согласно одному варианту осуществления последовательность нуклеиновой кислоты содержит последовательность, кодирующую ADAM6а мыши или ее функциональный фрагмент и/или последовательность, кодирующую ADAM6b мыши или ее функциональный фрагмент, причем ADAM6a и/или ADAM6b или их функциональный(е) фрагмент(ы) является(ются) функционально связанным(и) с промотором. Согласно одному варианту осуществления промотор представляет собой промотор человека. Согласно одному варианту осуществления промотор представляет собой промотор ADAM6 мыши. Согласно конкретному варианту осуществления промотор ADAM6 содержит последовательность, расположенную между первым кодоном первого гена ADAM6, ближайшего к наиболее 5’ генному сегменту DH мыши, и сигнальной последовательностью рекомбинации наиболее 5’ генного сегмента DH, причем направление 5’ указано по отношению к направлению транскрипции генов иммуноглобулина мыши. Согласно одному варианту осуществления промотор представляет собой вирусный промотор. Согласно конкретному варианту осуществления вирусный промотор представляет собой промотор цитомегаловируса (CMV). Согласно одному варианту осуществления промотор представляет собой промотор убиквитина.
Согласно одному варианту осуществления промотор представляет собой индуцируемый промотор. Согласно одному варианту осуществления индуцируемый промотор регулирует экспрессию в нерепродуктивных тканях. Согласно одному варианту осуществления индуцируемый промотор регулирует экспрессию в репродуктивных тканях. Согласно конкретному варианту осуществления экспрессия последовательностей ADAM6а мыши и/или ADAM6b или их функционального(ых) фрагмента(ов) регулируется в зависимости от стадии развития индуцируемым промотором в репродуктивных тканях.
Согласно одному варианту осуществления ADAM6а мыши и/или ADAM6b выбраны из ADAM6a согласно SEQ ID NO:1 и/или ADAM6b последовательности SEQ ID NO:2.
Согласно одному варианту осуществления промотор ADAM6 мыши представляет собой промотор согласно SEQ ID NO:3. Согласно конкретному варианту осуществления промотор ADAM6 мыши содержит последовательность нуклеиновой кислоты согласно SEQ ID NO:3 непосредственно выше (относительно направления транскрипции ADAM6a) первого кодона ADAM6a и продолжаясь до конца SEQ ID NO:3 выше кодирующей области ADAM6. Согласно другому конкретному варианту осуществления промотор ADAM6 представляет собой фрагмент, продолжающийся от в пределах приблизительно 5 - приблизительно 20 нуклеотидов выше старт-кодона ADAM6a до приблизительно 0,5 т.п.н., 1 т.п.н., 2 т.п.н. или 3 т.п.н. или более выше старт-кодона ADAM6a.
Согласно одному варианту осуществления последовательность нуклеиновой кислоты содержит SEQ ID NO:3 или ее фрагмент, который, будучи помещенным в мышь, которая является стерильной или которая характеризуется низкой фертильностью вследствие отсутствия ADAM6, улучшает фертильность или восстанавливает фертильность приблизительно до фертильности дикого типа. Согласно одному варианту осуществления SEQ ID NO:3 или ее фрагмент предоставляет самцу мыши способность производить сперматозоид, который способен проходить яйцевод самки мыши для оплодотворения яйцеклетки мыши.
Согласно одному варианту осуществления промотор ADAM6 мыши представляет собой промотор согласно SEQ ID NO: 4. Согласно конкретному варианту осуществления промотор ADAM6 мыши содержит последовательность нуклеиновой кислоты SEQ ID NO: 4 непосредственно выше (относительно направления транскрипции ADAM6a) первого кодона ADAM6a и продолжаясь до конца SEQ ID NO: 4 выше кодирующей области ADAM6. Согласно другому конкретному варианту осуществления промотор ADAM6 представляет собой фрагмент, продолжающийся от в пределах приблизительно 5 - приблизительно 20 нуклеотидов выше старт-кодона ADAM6a до приблизительно 0,5 т.п.н., 1 т.п.н., 2 т.п.н. или 3 т.п.н. или более выше старт-кодона ADAM6a.
Согласно одному варианту осуществления последовательность нуклеиновой кислоты содержит SEQ ID NO: 4 или ее фрагмент, который, будучи помещенным в мышь, которая является стерильной или которая характеризуется низкой фертильностью вследствие отсутствия ADAM6, улучшает фертильность или восстанавливает фертильность приблизительно до фертильности дикого типа. Согласно одному варианту осуществления SEQ ID NO: 4 или ее фрагмент предоставляет самцу мыши способность производить сперматозоид, который способен проходить яйцевод самки мыши для оплодотворения яйцеклетки мыши.
Согласно одному варианту осуществления промотор ADAM6 мыши представляет собой промотор согласно SEQ ID NO: 5. Согласно конкретному варианту осуществления промотор ADAM6 мыши содержит последовательность нуклеиновой кислоты согласно SEQ ID NO: 5 непосредственно выше (относительно направления транскрипции ADAM6a) первого кодона ADAM6a и продолжаясь до конца SEQ ID NO: 5 выше кодирующей области ADAM6. Согласно другому конкретному варианту осуществления промотор ADAM6 представляет собой фрагмент, продолжающийся от в пределах приблизительно 5 - приблизительно 20 нуклеотидов выше старт-кодона ADAM6a до приблизительно 0,5 т.п.н., 1 т.п.н., 2 т.п.н. или 3 т.п.н. или более выше старт-кодона ADAM6a.
Согласно одному варианту осуществления последовательность нуклеиновой кислоты содержит SEQ ID NO: 5 или ее фрагмент, который, будучи помещенным в мышь, которая является стерильной или которая характеризуется низкой фертильностью вследствие отсутствия ADAM6, улучшает фертильность или восстанавливает фертильность приблизительно до фертильности дикого типа. Согласно одному варианту осуществления SEQ ID NO: 5 или ее фрагмент предоставляет самцу мыши способность производить сперматозоид, который способен проходить яйцевод самки мыши для оплодотворения яйцеклетки мыши.
Согласно различным вариантам осуществления эктопическая последовательность нуклеиновой кислоты, которая предоставляет мыши функцию ADAM6, кодирует один или несколько белков ADAM6, причем один или несколько белков ADAM6 содержат SEQ ID NO: 1, SEQ ID NO: 2 или их комбинацию.
Согласно различным вариантам осуществления эктопическая последовательность нуклеиновой кислоты содержит последовательность, выбранную из SEQ ID NO: 3, SEQ ID NO: 4 и SEQ ID NO: 5, причем эктопическая последовательность нуклеиновой кислоты предоставляет мыши функцию ADAM6 посредством одного или нескольких белков ADAM6, кодируемых эктопической последовательностью нуклеиновой кислоты.
Согласно одному варианту осуществления последовательность нуклеиновой кислоты представляет собой любую последовательность, кодирующую ген ADAM6 или его гомолог или ортолог или функциональный фрагмент, который при введении и сохранении у мыши, дает уровень фертильности, который является таким же или сопоставимым с фертильностью мыши дикого типа. Иллюстративный уровень фертильности может быть продемонстрирован способностью самца мыши производить сперматозоид, который способен проходить яйцевод самки мыши для оплодотворения яйцеклетки мыши.
Согласно одному аспекту предусмотрена мышь, которая содержит делецию эндогенной нуклеотидной последовательности, которая кодирует белок ADAM6, замещение эндогенного генного сегмента VH генным сегментом VH человека и эктопическую нуклеотидную последовательность, которая кодирует белок ADAM6 мыши или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши.
Согласно одному аспекту предусмотрена мышь, которая содержит делецию эндогенной нуклеотидной последовательности, которая кодирует белок ADAM6, замещение эндогенного генного сегмента VH генным сегментом VL человека и эктопическую нуклеотидную последовательность, которая кодирует белок ADAM6 мыши или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши. Согласно одному варианту осуществления генный сегмент VL человека представляет собой генный сегмент Vκ. Согласно одному варианту осуществления генный сегмент VL представляет собой генный сегмент Vλ.
Согласно одному варианту осуществления мышь дополнительно содержит генный сегмент JL человека, и эктопическая нуклеотидная последовательность, которая кодирует белок ADAM6 мыши или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши, расположена между генным сегментом VL человека и генным сегментом человека JL. Согласно одному варианту осуществления мышь содержит один или несколько генных сегментов VL человека и один или несколько генных сегментов VL человека и эктопическую нуклеотидную последовательность, которая кодирует белок ADAM6 мыши или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши, расположена выше (или 5’) по отношению к одному или нескольким генным сегментам VL человека. Согласно конкретному варианту осуществления генные сегменты VL и JL человека представляют собой генные сегменты Vκ и Jκ.
Согласно одному варианту осуществления мышь содержит локус тяжелой цепи иммуноглобулина, который содержит делецию эндогенной нуклеотидной последовательности локуса иммуноглобулина, которая содержит эндогенный ген ADAM6, содержит нуклеотидную последовательность, кодирующую один или несколько генных сегментов иммуноглобулина человека, и причем эктопическая нуклеотидная последовательность, кодирующая белок ADAM6 мыши, находится в пределах или непосредственно прилегает к нуклеотидной последовательности, кодирующей один или несколько генных сегментов иммуноглобулина человека.
Согласно одному варианту осуществления мышь содержит замещение всех или по существу всех эндогенных генных сегментов VH нуклеотидной последовательностью, кодирующей один или несколько генных сегментов VL человека, и эктопическая нуклеотидная последовательность, кодирующая белок ADAM6 мыши, находится в пределах или непосредственно прилегает к нуклеотидной последовательности, кодирующей один или несколько генных сегментов VL человека. Согласно одному варианту осуществления мышь дополнительно содержит замещение одного или нескольких эндогенных генных сегментов DL одним или несколькими генными сегментами VL человека и/или JL человека на эндогенном генном локусе DH. Согласно одному варианту осуществления мышь дополнительно содержит замещение одного или нескольких эндогенных генных сегментов JH одним или несколькими генными сегментами JL человека на эндогенном генном локусе JH. Согласно одному варианту осуществления мышь содержит замещение всех или по существу всех эндогенных генных сегментов VH, DH и JH и замещение на эндогенных генных локусах VH, DH и JH генными сегментами VL и JL человека, причем мышь содержит эктопическую последовательность, кодирующую белок ADAM6 мыши. Согласно одному варианту осуществления мышь содержит вставку одного или нескольких генных сегментов VL и JL человека на эндогенном локусе тяжелой цепи иммуноглобулина, причем мышь содержит ген ADAM6, который является функциональным у мыши. Согласно конкретному варианту осуществления генные сегменты VL и JL человека представляют собой генные сегменты Vκ и Jκ. Согласно конкретному варианту осуществления эктопическая последовательность, кодирующая мышь белок ADAM6, расположена между предпоследним наиболее 3’ генным сегментом VL из присутствующих генных сегментов VL человека и последним наиболее 5’ генным сегментом JL из присутствующих генных сегментов JL человека. Согласно конкретному варианту осуществления эктопическая последовательность, кодирующая белок ADAM6 мыши, расположена выше (или 5’) наиболее 5’ генного сегмента VL из присутствующих генных сегментов VL человека. Согласно конкретному варианту осуществления мышь содержит делецию всех или по существу всех генных сегментов VH мыши и замещение по меньшей мере 40 генными сегментами VL человека, и эктопическая нуклеотидная последовательность, кодирующая белок ADAM6 мыши, расположена ниже генного сегмента Vκ4-1 человека и выше генного сегмента Jκ1 человека. Согласно конкретному варианту осуществления мышь содержит делецию всех или по существу всех генных сегментов VH мыши и замещение по меньшей мере 40 генными сегментами VL человека, и эктопическая нуклеотидная последовательность, кодирующая белок ADAM6 мыши, расположена выше генного сегмента Vκ2-40 человека.
Согласно конкретному варианту осуществления мышь содержит замещение всех или по существу всех эндогенных генных сегментов VL нуклеотидной последовательностью, кодирующей один или несколько генных сегментов VH человека, и эктопическая нуклеотидная последовательность, кодирующая белок ADAM6 мыши, находится в пределах или непосредственно прилегает к нуклеотидной последовательности, кодирующей один или несколько генных сегментов VL человека.
Согласно одному варианту осуществления генные сегменты VL представляют собой генные сегменты Vκ. Согласно одному варианту осуществления генные сегменты VL представляют собой генные сегменты Vλ.
Согласно одному варианту осуществления эктопическая нуклеотидная последовательность, которая кодирует мышь белок ADAM6, присутствует на трансгене в геноме мыши. Согласно одному варианту осуществления эктопическая нуклеотидная последовательность, которая кодирует белок ADAM6 мыши, присутствует экстрахромосомно у мыши.
Согласно одному аспекту предусмотрена мышь, которая содержит модификацию эндогенного локуса тяжелой цепи иммуноглобулина, причем мышь экспрессирует B-клетку, которая содержит реаранжированную последовательность иммуноглобулина, функционально связанную с последовательностью гена константной области тяжелой цепи, и B-клетка содержит в своем геноме (например, на хромосоме B-клетки) ген, кодирующий ADAM6 или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши. Согласно одному варианту осуществления реаранжированная последовательность иммуноглобулина, функционально связанная с генной последовательностью константной области тяжелой цепи, содержит генную последовательность V, J легкой цепи человека и необязательно генную последовательность D; последовательность V, D и/или J тяжелой цепи мыши; последовательность V и/или J легкой цепи человека или мыши. Согласно одному варианту осуществления генная последовательность константной области тяжелой цепи содержит последовательность тяжелой цепи человека или мыши, выбранную из группы, состоящей из CH1, шарнира, CH2, CH3 и их комбинации.
Согласно одному варианту осуществления последовательность V и/или J легкой цепи человека выбрана из последовательности Vκ, Vλ, Jκ и Jλ человека.
Согласно одному аспекту предусмотрена мышь, которая содержит функционально выключенный эндогенный локус тяжелой цепи иммуноглобулина, причем у мыши сохраняется функция ADAM6, и дополнительно содержит вставку одного или нескольких генных сегментов иммуноглобулина человека, причем один или несколько генных сегментов иммуноглобулина человека включают в себя генные сегменты VL и JL человека и необязательно генные сегменты DH человека. Согласно одному варианту осуществления один или несколько генных сегментов иммуноглобулина человека включают в себя генные сегменты Vκ, Vλ, Jκ и Jλ человека.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, причем мышь содержит функционально выключенный ген легкой цепи иммуноглобулина, и дополнительно содержит замещение одного или нескольких эндогенных генных сегментов вариабельной области тяжелой цепи иммуноглобулина одним или несколькими генными сегментами вариабельной области легкой цепи иммуноглобулина человека, причем мышь не содержит функциональный эндогенный локус ADAM6, и причем мышь содержит эктопическую нуклеотидную последовательность, которая экспрессирует белок ADAM6 мыши или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, причем мышь содержит функционально выключенный генный локус легкой цепи иммуноглобулина и дополнительно содержит замещение одного или нескольких эндогенных вариабельных генных сегментов легкой цепи иммуноглобулина одним или несколькими вариабельными генными сегментами легкой цепи иммуноглобулина человека, причем мышь не содержит функциональный эндогенный локус ADAM6 и причем мышь содержит эктопическую нуклеотидную последовательность, которая кодирует белок ADAM6 мыши или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши.
Согласно одному варианту осуществления один или несколько вариабельных генных сегментов легкой цепи иммуноглобулина человека являются смежными с эктопической нуклеотидной последовательностью.
Согласно одному аспекту предусмотрена мышь, которая не содержит функциональный эндогенный локус или последовательность ADAM6 и которая содержит эктопическую нуклеотидную последовательность, кодирующую локус ADAM6 мыши или функциональный фрагмент локуса или последовательности ADAM6 мыши, причем мышь способна спариваться с мышью противоположного пола, чтобы производить потомство, которое содержит эктопический локус или последовательность ADAM6. Согласно одному варианту осуществления мышь представляет собой самца. Согласно одному варианту осуществления мышь представляет собой самку.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, причем мышь содержит генный сегмент вариабельной области легкой цепи иммуноглобулина человека на эндогенном генном локусе вариабельной области тяжелой цепи иммуноглобулина, мышь не содержит эндогенную функциональную последовательность ADAM6 на эндогенном генном локусе вариабельной области тяжелой цепи иммуноглобулина, и причем мышь содержит эктопическую нуклеотидную последовательность, которая экспрессирует белок ADAM6 мыши или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, причем мышь содержит вариабельный генный сегмент легкой цепи иммуноглобулина человека на эндогенном генном локусе вариабельной области тяжелой цепи иммуноглобулина, мышь не содержит эндогенную функциональную последовательность ADAM6 на эндогенном вариабельном генном локусе тяжелой цепи иммуноглобулина, и причем мышь содержит эктопическую нуклеотидную последовательность, которая экспрессирует белок ADAM6 мыши или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши.
Согласно одному варианту осуществления эктопическая нуклеотидная последовательность, которая экспрессирует белок ADAM6 мыши, является экстрахромосомной. Согласно одному варианту осуществления эктопическая нуклеотидная последовательность, которая экспрессирует белок ADAM6 мыши, интегрирована в один или несколько локусов в геноме мыши. Согласно конкретному варианту осуществления один или несколько локусов включают в себя локус иммуноглобулина.
Согласно одному аспекту предусмотрена мышь, которая экспрессирует последовательность легкой цепи иммуноглобулина из модифицированного эндогенного локуса тяжелой цепи иммуноглобулина, причем тяжелая цепь происходит из генного сегмента VL человека, генного сегмента J человека и необязательно генного сегмента DH человека, причем мышь содержит активность ADAM6, которая является функциональной у мыши. Согласно одному варианту осуществления генный сегмент VL человека выбран из генного сегмента Vκ человека и генного сегмента Vλ человека. Согласно различным вариантам осуществления генный сегмент J представляет собой генный сегмент JH, Jκ или Jλ или их комбинацию.
Согласно одному варианту осуществления мышь содержит множество генных сегментов VL человека и множество генных сегментов J. Согласно конкретному варианту осуществления генные сегменты J представляют собой генные сегменты JL.
Согласно одному аспекту предусмотрена мышь, которая экспрессирует последовательность легкой цепи иммуноглобулина из модифицированного эндогенного локуса тяжелой цепи иммуноглобулина, причем тяжелая цепь происходит из относящегося к человеку генного сегмента VL и генного сегмента JL, причем мышь содержит активность ADAM6, которая является функциональной у мыши.
Согласно одному варианту осуществления мышь содержит множество относящихся к человеку генных сегментов V, множество генных сегментов J и необязательно множество генных сегментов D. Согласно одному варианту осуществления генные сегменты D представляют собой генные сегменты D человека. Согласно одному варианту осуществления генные сегменты J представляют собой генные сегменты J человека. Согласно одному варианту осуществления мышь дополнительно содержит гуманизированную последовательность константной области тяжелой цепи, причем гуманизация предусматривает замещение последовательности, выбранной из CH1, шарнира, CH2, CH3 и их комбинации. Согласно конкретному варианту осуществления тяжелая цепь происходит из генного сегмента V человека, генного сегмента J человека, последовательности CH1 человека, шарнирной последовательности человека или мыши, последовательности CH2 мыши и последовательности CH3 мыши. Согласно другому конкретному варианту осуществления мышь дополнительно содержит константную последовательность легкой цепи человека. Согласно одному варианту осуществления мышь содержит ген ADAM6, который фланкирован 5’ и 3’ эндогенными генными сегментами тяжелой цепи иммуноглобулина. Согласно конкретному варианту осуществления эндогенные вариабельная генные сегменты тяжелой цепи иммуноглобулина являются неспособными кодировать вариабельную область тяжелой цепи антитела. Согласно конкретному варианту осуществления ген ADAM6 мыши находится в положении, которое является таким же, как и у мыши дикого типа, и эндогенные вариабельные генные локусы тяжелой цепи иммуноглобулина мыши являются неспособными к реаранжировке для кодирования тяжелой цепи антитела.
Согласно одному варианту осуществления множество генных сегментов V человека фланкированы 5’ (относительно направления транскрипции генных сегментов V) последовательностью, кодирующей активность ADAM6, которая является функциональной у мыши. Согласно конкретному варианту осуществления множество генных сегментов V человека включают в себя генные сегменты Vκ человека Vκ4-1, Vκ5-2, Vκ7-3, Vκ2-4, Vκ1-5, Vκ1-6, Vκ3-7, Vκ1-8, Vκ1-9, Vκ2-10, Vκ3-11, Vκ1-12, Vκ1-13, Vκ2-14, Vκ3-15, Vκ1-16, Vκ1-17, Vκ2-18, Vκ2-19, Vκ3-20, Vκ6-21, Vκ1-22, Vκ1-23, Vκ2-24, Vκ3-25, Vκ2-26, Vκ1-27, Vκ2-28, Vκ2-29, Vκ2-30, Vκ3-31, Vκ1-32, Vκ1-33, Vκ3-34, Vκ1-35, Vκ2-36, Vκ1-37, Vκ2-38, Vκ1-39 и Vκ2-40, и генный сегмент Vκ2-40 человека фланкирован 5’ (относительно направления транскрипции генного сегмента Vκ2-40 человека) последовательностью, кодирующей активность ADAM6, которая является функциональной у мыши. Согласно конкретному варианту осуществления последовательность, кодирующая активность ADAM6, которая является функциональной у мыши, расположена в аналогичной ориентации транскрипции по отношению к генным сегментам Vκ человека. Согласно конкретному варианту осуществления последовательность, кодирующая активность ADAM6, которая является функциональной у мыши, расположена в противоположной ориентации транскрипции по отношению к генным сегментам Vκ человека.
Согласно одному варианту осуществления генный сегмент V фланкирован 3’ (относительно направления транскрипции генного сегмента V) последовательностью, кодирующей активность ADAM6, которая является функциональной у мыши.
Согласно одному варианту осуществления генный сегмент D фланкирован 5’ (относительно направления транскрипции генного сегмента D) последовательностью, кодирующей активность ADAM6, которая является функциональной у мыши.
Согласно одному варианту осуществления генный сегмент J фланкирован 5’ (относительно направления транскрипции генного сегмента J) последовательностью, кодирующей активность ADAM6, которая является функциональной у мыши.
Согласно одному варианту осуществления активность ADAM6, которая является функциональной у мыши, является результатом экспрессии нуклеотидной последовательности, расположенной 5’ относительно наиболее 5’ генного сегмента D и 3’ относительно наиболее 3’ генного сегмента V (относительно направления транскрипции генного сегмента V) модифицированного эндогенного локуса тяжелой цепи иммуноглобулина.
Согласно одному варианту осуществления активность ADAM6, которая является функциональной у мыши, является результатом экспрессии нуклеотидной последовательности, расположенной 5’ относительно наиболее 5’ генного сегмента J и 3’ относительно наиболее 3’ генного сегмента V (относительно направления транскрипции генного сегмента V) модифицированного эндогенного локуса тяжелой цепи иммуноглобулина.
Согласно одному варианту осуществления активность ADAM6, которая является функциональной у мыши, является результатом экспрессии нуклеотидной последовательности, расположенной между двумя генными сегментами V человека в модифицированном эндогенном локусе тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления два генных сегмента V человека представляют собой генный сегмент Vκ5-2 человека и генный сегмент Vκ4-1 человека.
Согласно одному варианту осуществления активность ADAM6, которая является функциональной у мыши, является результатом экспрессии нуклеотидной последовательности, расположенной между генным сегментом V человека и генным сегментом J человека в модифицированном эндогенном локусе тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления генный сегмент V человека представляет собой генный сегмент Vκ4-1 человека и сегмент J человека представляет собой генный сегмент Jκ1.
Согласно одному варианту осуществления нуклеотидная последовательность содержит последовательность, выбранную из последовательности ADAM6b мыши или ее функционального фрагмента, последовательности ADAM6a мыши или ее функционального фрагмента и их комбинации.
Согласно различным вариантам осуществления последовательность, кодирующая активность ADAM6, которая является функциональной у мыши, кодирует белок ADAM6b, представленный в SEQ ID NO: 2 и/или кодирует белок ADAM6a представленный в SEQ ID NO: 1.
Согласно одному варианту осуществления нуклеотидная последовательность между двумя генными сегментами V человека расположена в противоположной ориентации транскрипции по отношению к генным сегментам V человека. Согласно конкретному варианту осуществления нуклеотидная последовательность кодирует, в направлении 5’ - 3’ относительно направления транскрипции генов ADAM6, последовательность ADAM6a, за которой следует последовательность ADAM6b.
Согласно одному варианту осуществления нуклеотидная последовательность между генным сегментом V человека и генным сегментом J человека расположена в противоположной ориентации транскрипции по отношению к генным сегментам V и J человека. Согласно конкретному варианту осуществления нуклеотидная последовательность кодирует, в направлении 5’ - 3’ относительно направления транскрипции генов ADAM6, последовательность ADAM6a, за которой следует последовательность ADAM6b.
Согласно одному варианту осуществления мышь содержит гибридную последовательность иммуноглобулина, причем гибридная последовательность иммуноглобулина содержит последовательность κ легкой цепи иммуноглобулина человека, смежную с не относящейся к человеку последовательностью ADAM6.
Согласно одному варианту осуществления мышь содержит последовательность человека, смежную с последовательностью мыши на эндогенном локусе тяжелой цепи иммуноглобулина, причем смежная последовательность содержит по меньшей мере один генный сегмент VL человека, последовательность ADAM6 мыши или ее ортолог или гомолог или функциональный фрагмент и генный сегмент JL человека. Согласно конкретному варианту осуществления последовательность ADAM6 мыши или ее ортолог или гомолог или функциональный фрагмент расположен непосредственно примыкая по меньшей мере к одному генному сегменту VL человека. Согласно одному варианту осуществления генный сегмент VL человека представляет собой генный сегмент Vκ человека. Согласно одному варианту осуществления последовательность ADAM6 мыши или ее ортолог или гомолог или функциональный фрагмент расположена непосредственно примыкая и 3’ к по меньшей мере одному генному сегменту VL человека и непосредственно примыкая и 5’ к генному сегменту JL человека. Согласно конкретному варианту осуществления генный сегмент VL человека представляет собой генный сегмент Vκ человека и генный сегмент JL человека представляет собой генный сегмент Jκ человека.
Согласно одному варианту осуществления последовательность, кодирующая активность ADAM6, которая является функциональной у мыши, представляет собой последовательность ADAM6 мыши или ее функциональный фрагмент.
Согласно одному варианту осуществления мышь содержит эндогенный генный сегмент DFL16.1 мыши (например, у мыши, гетерозиготной в отношении модифицированного эндогенного локуса тяжелой цепи иммуноглобулина мыши) или генный сегмент DH1-1 человека. Согласно одному варианту осуществления генный сегмент D тяжелой цепи иммуноглобулина, экспрессированной мышью, происходит из эндогенного генного сегмента DFL16.1 мыши или генного сегмента DH1-1 человека.
Согласно одному аспекту предусмотрена мышь, которая содержит последовательность нуклеиновой кислоты, кодирующую ADAM6 мыши (или его гомолог или ортолог или функциональный фрагмент) в содержащей ДНК клетке нереаранжированной B-клеточной линии, но не содержит последовательность нуклеиновой кислоты, кодирующую ADAM6 мыши (или его гомолог или ортолог или функциональный фрагмент) в B-клетке, которая содержат реаранжированные локусы иммуноглобулина, причем последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши (или его гомолог или ортолог или функциональный фрагмент) находится в геноме в положении, которое отличается от положения, в котором ген ADAM6 мыши встречается у мыши дикого типа. Согласно одному варианту осуществления последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши (или его гомолог или ортолог или функциональный фрагмент), присутствует во всех или по существу всех содержащих ДНК клетках, которые не происходят из реаранжированной B-клеточной линии; согласно одному варианту осуществления последовательность нуклеиновой кислоты присутствует в зародышевых клетках мыши, но не в хромосоме реаранжированной B-клетки.
Согласно одному аспекту предусмотрена мышь, которая содержит последовательность нуклеиновой кислоты, кодирующую ADAM6 мыши (или его гомолог или ортолог или функциональный фрагмент), во всех или по существу всех содержащих ДНК клетках, включая в себя B-клетки, которые содержат реаранжированные локусы иммуноглобулина, причем последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши (или его гомолог или ортолог или функциональный фрагмент) находится в геноме в положении, которое отличается от положения, в котором ген ADAM6 мыши встречается у мыши дикого типа. Согласно одному варианту осуществления последовательность нуклеиновой кислоты, кодирующая ADAM6 мыши (или его гомолог или ортолог или функциональный фрагмент), находится на нуклеиновой кислоте, которая является смежной с реаранжированным локусом иммуноглобулина. Согласно одному варианту осуществления нуклеиновая кислота, которая является смежной с реаранжированным локусом иммуноглобулина, представляет собой хромосому. Согласно одному варианту осуществления хромосома представляет собой хромосому, которая встречается у мыши дикого типа, и хромосома содержит модификацию локуса иммуноглобулина мыши.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, причем мышь содержит B-клетку, которая содержит в своем геноме последовательность ADAM6 или ее ортолог или гомолог. Согласно одному варианту осуществления последовательность ADAM6 или ее ортолог или гомолог находится на локусе тяжелой цепи иммуноглобулина. Согласно конкретному варианту осуществления локус тяжелой цепи содержит эндогенные генные сегменты тяжелой цепи иммуноглобулина, которые являются неспособными к реаранжировке для кодирования тяжелой цепи антитела. Согласно одному варианту осуществления последовательность ADAM6 или ее ортолог или гомолог находится на локусе, который не является локусом иммуноглобулина. Согласно одному варианту осуществления последовательность ADAM6 находится на трансгене, которым управляет гетерологичный промотор. Согласно конкретному варианту осуществления гетерологичный промотор представляет собой не относящийся к иммуноглобулину промотор. Согласно конкретному варианту осуществления B-клетка экспрессирует белок ADAM6 или его ортолог или гомолог.
Согласно одному варианту осуществления 90% или более B-клеток мыши содержат ген, кодирующий белок ADAM6 или его ортолог или его гомолог или его фрагмент, который является функциональным у мыши. Согласно конкретному варианту осуществления мышь представляет собой самца мыши.
Согласно одному варианту осуществления геном B-клетки содержит первый аллель и второй аллель, содержащий последовательность ADAM6 или ее ортолог или гомолог. Согласно одному варианту осуществления геном B-клетки содержит первый аллель, но не второй аллель, содержащий последовательность ADAM6 или ее ортолог или гомолог.
Согласно одному аспекту предусмотрена мышь, которая содержит модификацию на одном или нескольких эндогенных аллелях тяжелой цепи иммуноглобулина, причем модификация сохраняет один или несколько эндогенных аллелей ADAM6.
Согласно одному варианту осуществления модификация делает мышь неспособной экспрессировать функциональную тяжелую цепь, которая содержит реаранжированные эндогенные генные сегменты тяжелой цепи по меньшей мере из одного аллеля тяжелой цепи и сохраняет эндогенный аллель ADAM6, расположенный в пределах по меньшей мере одного эндогенного аллеля тяжелой цепи иммуноглобулина.
Согласно одному варианту осуществления мыши являются неспособными экспрессировать функциональную тяжелую цепь, которая содержит реаранжированные эндогенные генные сегменты тяжелой цепи по меньшей мере из одного из эндогенных аллелей тяжелой цепи иммуноглобулина, и мыши экспрессируют белок ADAM6 из эндогенного аллеля ADAM6. Согласно конкретному варианту осуществления мыши являются неспособными экспрессировать функциональную тяжелую цепь, которая содержит реаранжированные эндогенные генные сегменты тяжелой цепи из двух эндогенных аллелей тяжелой цепи иммуноглобулина, и мыши экспрессируют белок ADAM6 из одного или нескольких эндогенных аллелей ADAM6.
Согласно одному варианту осуществления мыши являются неспособными экспрессировать функциональную тяжелую цепь из каждого эндогенного аллеля тяжелой цепи, и мыши содержат функциональный аллель ADAM6, расположенный в пределах 1, 2, 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110 или 120 или более миллионов пар нуклеотидов выше (относительно направления транскрипции локуса тяжелой цепи иммуноглобулина мыши) последовательности константной области тяжелой цепи иммуноглобулина мыши. Согласно конкретному варианту осуществления функциональный аллель ADAM6 находится на эндогенном локусе тяжелой цепи (например, в межгенной V-D области, между двумя генными сегментами V, между генным сегментом V и D, между генным сегментом D и J и т.д.). Согласно конкретному варианту осуществления функциональный аллель ADAM6 расположен в пределах 90 - 100 т.п.н. межгенной последовательности между последним генным сегментом V мыши и первым генным сегментом D мыши. Согласно другому конкретному варианту осуществления эндогенную межгенную последовательность V-D размером 90 - 100 т.п.н. удаляют, и эктопическую последовательность ADAM6 помещают между последним генным сегментом V и первым генным сегментом D.
Согласно одному аспекту предусмотрена мышь, которая содержит модификацию на одном или нескольких эндогенных аллелях ADAM6.
Согласно одному варианту осуществления модификация делает мышь неспособной экспрессировать функциональный белок ADAM6 по меньшей мере из одного или нескольких эндогенных аллелей ADAM6. Согласно конкретному варианту осуществления мышь является неспособной экспрессировать функциональный белок ADAM6 из каждого из эндогенных аллелей ADAM6.
Согласно одному варианту осуществления мышь является неспособной экспрессировать функциональный белок ADAM6 из каждого эндогенного аллеля ADAM6, и мыши содержат эктопическую последовательность ADAM6.
Согласно одному варианту осуществления мышь является неспособной экспрессировать функциональный белок ADAM6 из каждого эндогенного аллеля ADAM6, и мышь содержит эктопическую последовательность ADAM6, расположенную в пределах 1, 2, 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110 или 120 или более т.п.н. выше (относительно направления транскрипции локуса тяжелой цепи мыши) последовательности константной области тяжелой цепи иммуноглобулина мыши. Согласно конкретному варианту осуществления эктопическая последовательность ADAM6 находится на эндогенном локусе тяжелой цепи иммуноглобулина (например, в межгенной области V-D, между двумя генными сегментами V, между генным сегментом V и J, между генным сегментом D и J и т.д.). Согласно конкретному варианту осуществления эктопическая последовательность ADAM6 расположена в пределах 90 - 100 т.п.н. межгенной последовательности между последним генным сегментом V мыши и первым генным сегментом D мыши. Согласно другому конкретному варианту осуществления эндогенную 90 - 100 т.п.н. межгенную последовательность V-D удаляют, и эктопическую последовательность ADAM6 размещают между генным сегментом Vκ человека и первым генным сегментом Jκ человека. Согласно другому конкретному варианту осуществления эндогенную межгенную последовательность V-D размером 90 - 100 т.п.н. удаляют, и эктопическую последовательность ADAM6 размещают 5’ или выше относительно генного сегмента Vκ человека, причем генный сегмент Vκ человека выбран из генного сегмента Vκ4-1 или Vκ2-40 человека.
Согласно одному варианту осуществления мышь способна экспрессировать функциональный белок ADAM6 из одного или нескольких эндогенных аллелей ADAM6, и модификация включает в себя вставку последовательности человека, которая кодирует вариабельный домен иммуноглобулина. Согласно одному варианту осуществления последовательность человека содержит нереаранжированные генные сегменты иммуноглобулина. Согласно конкретному варианту осуществления последовательность человека содержит генный сегмент V и генный сегмент J. Согласно другому конкретному варианту осуществления последовательность человека содержит генный сегмент V, генный сегмент J и генный сегмент D.
Согласно одному аспекту предусмотрен стерильный самец мыши, причем мышь содержит делецию двух или более эндогенных аллелей ADAM6. Согласно одному аспекту предусмотрена самка мыши, которая представляет собой носителя признака мужской стерильности, причем самка мыши содержит в своей зародышевой линии нефункциональный аллель ADAM6 или нокаут эндогенного аллеля ADAM6.
Согласно одному аспекту предусмотрена мышь, содержащая эндогенный генный сегмент V, D и/или J тяжелой цепи иммуноглобулина, который является неспособным к реаранжировке для кодирования тяжелой цепи антитела, причем большинство B-клеток мыши содержат функциональный ген ADAM6. Согласно различным вариантам осуществления большинство B-клеток дополнительно содержат один или несколько генных сегментов VL и JL человека выше константной области тяжелой цепи иммуноглобулина мыши. Согласно одному варианту осуществления генные сегменты VL и JL человека представляют собой генные сегменты Vκ и Jκ.
Согласно одному варианту осуществления мышь содержит интактные эндогенные генные сегменты V, D и J тяжелой цепи иммуноглобулина, которые являются неспособными к реаранжировке, чтобы кодировать функциональную тяжелую цепь антитела. Согласно одному варианту осуществления мышь содержит по меньшей мере один и до 89 генных сегментов V, по меньшей мере один и до 13 генных сегментов D, по меньшей мере один и до четырех генных сегментов J и их комбинацию; причем по меньшей мере один и до 89 генных сегментов V, по меньшей мере один и до 13 генных сегментов D, по меньшей мере один и до четырех генных сегментов J являются неспособными к реаранжировке, чтобы кодировать вариабельную область тяжелой цепи антитела. Согласно конкретному варианту осуществления мышь содержит функциональный ген ADAM6, расположенный в пределах интактных эндогенных генных сегментов V, D и J тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления мышь содержит эндогенный локус тяжелой цепи, который включает в себя эндогенный локус ADAM6, причем эндогенный локус тяжелой цепи содержит 89 генных сегментов V, 13 генных сегментов D и четыре генных сегмента J, причем эндогенные генные сегменты тяжелой цепи являются неспособными к реаранжировке, чтобы кодировать вариабельную область тяжелой цепи антитела, и локус ADAM6 кодирует белок ADAM6, который является функциональным у мыши.
Согласно одному аспекту предусмотрен способ получения стерильного самца мыши, предусматривающий приведение эндогенного аллеля ADAM6 донорной ES клетки в нефункциональное состояние (или нокаут указанного аллеля), введение донорной ES клетки в зародыш - хозяин, вынашивание зародыша - хозяина в суррогатной матери и предоставление возможности суррогатной матери родить потомство, происходящее полностью или частично из донорной ES клетки. Согласно одному варианту осуществления способ дополнительно предусматривает скрещивание потомства для получения стерильного самца мыши.
Согласно одному аспекту предусмотрен способ получения мыши с представляющей интерес генетической модификацией, причем мышь является стерильной, причем способ предусматривает стадии (a) осуществления представляющей интерес генетической модификации в геноме; (b) модификации генома для нокаута эндогенного аллеля ADAM6 или приведения эндогенного аллеля ADAM6 в нефункциональное состояние; и (c) использования генома в получении мыши. Согласно различным вариантам осуществления геном происходит из ES клетки или используется в эксперименте ядерного транспорта.
Согласно одному аспекту предусмотрена мышь, которая не содержит эндогенный генный сегмент V, D и J тяжелой цепи иммуноглобулина, причем большинство B-клеток мыши содержат последовательность ADAM6 или ее ортолог или гомолог.
Согласно одному варианту осуществления мышь не содержит эндогенные генные сегменты тяжелой цепи иммуноглобулина, выбранные из двух или более генных сегментов V, двух или более генных сегментов D, двух или более генных сегментов J и их комбинации. Согласно одному варианту осуществления мышь не содержит генные сегменты тяжелой цепи иммуноглобулина, выбранные по меньшей мере из одного и до 89 генных сегментов V, по меньшей мере одного и до 13 генных сегментов D, по меньшей мере одного и до четырех генных сегментов J и их комбинации. Согласно одному варианту осуществления мышь не содержит геномный фрагмент ДНК из хромосомы 12, содержащей приблизительно три миллиона пар нуклеотидов эндогенного локуса тяжелой цепи иммуноглобулина. Согласно конкретному варианту осуществления мышь не содержит все функциональные эндогенные генные сегменты V, D и J тяжелой цепи. Согласно конкретному варианту осуществления мышь не содержит 89 генных сегментов VH, 13 генных сегментов DH и четыре генных сегмента JH.
Согласно одному аспекту предусмотрена мышь, причем мышь содержит геном в зародышевой линии, содержащий модификацию локуса тяжелой цепи иммуноглобулина, причем модификация локуса тяжелой цепи иммуноглобулина включает в себя замещение одной или нескольких последовательностей вариабельной области иммуноглобулина мыши одной или несколькими не относящимися к мыши последовательностями вариабельной области иммуноглобулина, и причем мышь содержит последовательность нуклеиновой кислоты, кодирующую белок ADAM6 мыши. Согласно одному варианту осуществления последовательности DH и JH и по меньшей мере 3, по меньшей мере 10, по меньшей мере 20, по меньшей мере 40, по меньшей мере 60, или по меньшей мере 80 последовательностей VH эндогенного локуса тяжелой цепи иммуноглобулина замещают не относящимися к мыши последовательностями легкой цепи иммуноглобулина. Согласно одному варианту осуществления последовательности DH, JH и все последовательности VH эндогенного локуса тяжелой цепи иммуноглобулина замещают множеством не относящихся к мыши генных сегментов VL иммуноглобулина, одной или нескольким последовательностями генного сегмента JL и необязательно одной или нескольким последовательностями генного сегмента D. Не относящиеся к мыши последовательности иммуноглобулина могут быть нереаранжированными. Согласно одному варианту осуществления не относящиеся к мыши последовательности иммуноглобулина содержат целые нереаранжированные области VL и JL не относящегося к мыши вида. Согласно одному варианту осуществления не относящиеся к мыши последовательности иммуноглобулина способны формировать целую вариабельную область, т.е. реаранжированную вариабельную область, содержащую генные сегменты VL и JL, соединенные вместе для образования последовательности, которая кодирует вариабельную область легкой цепи не относящегося к мыши вида, функционально связанную с одной или несколькими эндогенными константными областями. Не относящийся к мыши вид может представлять собой Homo sapiens, и не относящиеся к мыши последовательности иммуноглобулина могут представлять собой последовательности человека.
Согласно одному аспекту генетически предусмотрена модифицированная мышь, содержащая нуклеотидную последовательность, кодирующую белок ADAM6 или его функциональный фрагмент, которая является смежной с вариабельным генным сегментом легкой цепи иммуноглобулина человека.
Согласно одному варианту осуществления мышь не содержит эндогенную немодифицированную генную последовательность ADAM6. Согласно одному варианту осуществления мышь не содержит функциональную эндогенную генную последовательность ADAM6.
Согласно одному варианту осуществления вариабельный генный сегмент легкой цепи иммуноглобулина человека представляет собой вариабельный генный сегмент κ легкой цепи иммуноглобулина. Согласно одному варианту осуществления вариабельный генный сегмент легкой цепи иммуноглобулина человека представляет собой вариабельный генный сегмент λ легкой цепи иммуноглобулина. Согласно одному варианту осуществления вариабельный генный сегмент легкой цепи иммуноглобулина человека является функционально связанным с константной генной последовательностью тяжелой цепи иммуноглобулина.
Согласно одному варианту осуществления константная генная последовательность тяжелой цепи иммуноглобулина представляет собой генную последовательность тяжелой цепи мыши или крысы или человека. Согласно одному варианту осуществления константная генная последовательность тяжелой цепи содержит CH1 и/или шарнирную область.
Согласно одному варианту осуществления мышь содержит делецию или замещение одной или нескольких эндогенных генных последовательностей тяжелой цепи иммуноглобулина.
Согласно одному варианту осуществления мышь дополнительно содержит нереаранжированный генный сегмент Vκ человека или нереаранжированный генный сегмент Vλ человека, функционально связанный с последовательностью константной области легкой цепи человека или мыши или крысы. Согласно одному варианту осуществления мышь содержит множество нереаранжированных генных сегментов Vκ человека (например, два или больше сегментов Vκ человека и один или несколько сегментов Jκ человека) или множество нереаранжированных генных сегментов Vλ человека (например, два или больше сегментов Vλ человека и один или несколько сегментов Jλ человека). Согласно одному варианту осуществления нереаранжированные генные сегменты Vκ человека или нереаранжированные генные сегменты Vλ человека функционально связаны с последовательностью константной области на эндогенном локусе легкой цепи иммуноглобулина.
Согласно одному варианту осуществления мышь дополнительно содержит модификацию, которая делает эндогенный локус κ легкой цепи и/или эндогенный локус λ легкой цепи нефункциональным. Согласно одному варианту осуществления мышь содержит нокаут или делецию эндогенного локуса κ легкой цепи мыши и/или эндогенного локуса λ легкой цепи мыши.
Согласно одному аспекту предусмотрен способ поддержания линии мыши, причем линия мыши содержит замещение последовательности тяжелой цепи иммуноглобулина мыши одной или несколькими последовательностями легкой цепи иммуноглобулина человека. Согласно одному варианту осуществления одна или несколько последовательности легкой цепи иммуноглобулина человека представляют собой генные сегменты Vκ и/или Jκ иммуноглобулина человека.
Согласно одному варианту осуществления линия мыши содержит делецию одного или нескольких генных сегментов VH, DH и/или JH мыши. Согласно одному варианту осуществления мышь дополнительно содержит один или несколько генных сегментов VL человека и один или несколько генных сегментов JL человека. Согласно одному варианту осуществления мышь содержит по меньшей мере 6, по меньшей мере 16, по меньшей мере 30 или по меньшей мере 40 генных сегментов Vκ человека и по меньшей мере пять генных сегментов Jκ. Согласно конкретному варианту осуществления человека генные сегменты легкой цепи функционально связаны с геном константной области. Согласно одному варианту осуществления ген константной области представляет собой ген константной области мыши. Согласно одному варианту осуществления ген константной области содержит последовательность гена константной области мыши, выбранную из CH1, шарнира, CH2, CH3 и/или CH4 или их комбинации.
Согласно одному варианту осуществления способ предусматривает получение самца мыши, гетерозиготного в отношении замещения последовательности тяжелой цепи иммуноглобулина мыши, и скрещивание гетерозиготного самца мыши с самкой мыши дикого типа или самкой мыши, которая является гомозиготной или гетерозиготной в отношении последовательности тяжелой цепи человека. Согласно одному варианту осуществления способ предусматривает сохранение линии путем повторного скрещивания гетерозиготных самцов с самками, которые являются самками дикого типа или гомозиготными или гетерозиготными в отношении последовательности тяжелой цепи человека.
Согласно одному варианту осуществления способ предусматривает получение клеток от самцов или самок мышей, гомозиготных или гетерозиготных в отношении последовательности тяжелой цепи человека, и использование этих клеток в качестве донорных клеток или ядер из них в качестве донорных ядер и использование клеток или ядер для получения генетически модифицированных животных с использованием клеток-хозяев и/или вынашивания клеток и/или ядер в организмах суррогатных матерей.
Согласно одному варианту осуществления только самцов мышей, которые являются гетерозиготными в отношении замещения на локусе тяжелой цепи, скрещивают с самками мышей. Согласно конкретному варианту осуществления самки мышей являются гомозиготными, гетерозиготными или дикого типа по отношению к замещенному локусу тяжелой цепи.
Согласно одному варианту осуществления мышь дополнительно содержит замещение вариабельных последовательностей λ и/или κ легкой цепи на эндогенном локусе легкой цепи иммуноглобулина гетерологичными последовательностями легкой цепи иммуноглобулина. Согласно одному варианту осуществления гетерологичные последовательности легкой цепи иммуноглобулина представляют собой вариабельные последовательности λ и/или κ легкой цепи иммуноглобулина человека.
Согласно одному варианту осуществления мышь дополнительно содержит трансген в локусе, отличном от эндогенного локуса иммуноглобулина, причем трансген содержит последовательность, кодирующую реаранжированную или нереаранжированную гетерологичную последовательность λ или κ легкой цепи (например, нереаранжированную VL и нереаранжированную JL или реаранжированную VJ), функционально связанную (для нереаранжированной) или слитую (для реаранжированной) с последовательностью константной области легкой цепи иммуноглобулина. Согласно одному варианту осуществления гетерологичная последовательность λ или κ легкой цепи относится к человеку. Согласно одному варианту осуществления последовательность константной области выбрана из грызуна, человека и не относящегося к человеку примата. Согласно одному варианту осуществления последовательность константной области выбрана из мыши, крысы и хомяка. Согласно одному варианту осуществления трансген содержит не относящийся к иммуноглобулину промотор, который управляет экспрессией последовательностей легкой цепи. Согласно конкретному варианту осуществления промотор представляет собой транскрипционно активный промотор. Согласно конкретному варианту осуществления промотор представляет собой промотор ROSA26.
Согласно одному аспекту предусмотрена фертильная мышь, которая содержит модификацию эндогенного гена ADAM6, причем мышь содержит эктопическую последовательность, которая предоставляет функцию ADAM6 мыши, и причем мышь содержит в своей зародышевой линии нереаранжированный генный сегмент легкой цепи иммуноглобулина человека, функционально связанный с последовательностью нуклеиновой кислоты, кодирующей последовательность тяжелой цепи иммуноглобулина.
Согласно одному аспекту предусмотрена фертильная мышь, которая содержит модификацию эндогенного локуса ADAM6, причем модификация делает локус ADAM6 нефункциональным, и причем мышь экспрессирует связывающий белок иммуноглобулина, содержащий вариабельный домен легкой цепи иммуноглобулина человека, смежный с константной последовательностью тяжелой цепи.
Согласно одному варианту осуществления связывающий белок иммуноглобулина дополнительно содержит когнатный вариабельный домен легкой цепи иммуноглобулина человека, слитый с константной последовательностью легкой цепи.
Согласно одному варианту осуществления константная последовательность тяжелой цепи и константная последовательность легкой цепи являются не относящимися к человеку.
Согласно одному аспекту предусмотрена мышь, содержащая локус тяжелой цепи иммуноглобулина, содержащий замещение одного или нескольких генных сегментов вариабельной области тяжелой цепи (VH) иммуноглобулина, дополнительных генных сегментов тяжелой цепи (DH) и соединительных генных сегментов тяжелой цепи (JH) на эндогенном локусе тяжелой цепи иммуноглобулина одним или несколькими генными сегментами вариабельной области легкой цепи (VL) и одним или несколькими генными сегментами соединительной области легкой цепи (JL), причем мышь способна экспрессировать белок ADAM6.
Согласно одному аспекту предусмотрена мышь, содержащая локус тяжелой цепи иммуноглобулина, который содержит замещение всех или по существу всех генных сегментов VH, DH и JH одним или несколькими генными сегментами VL и одним или несколькими генными сегментами JL для образования последовательности генного сегмента VL на эндогенном локусе тяжелой цепи (локусе VLH), причем локус VLH способен к рекомбинации с эндогенным геном CH для образования реаранжированного гена, который происходит из генного сегмента VL, генного сегмента JL и эндогенного гена CH.
Согласно одному варианту осуществления сегменты VL представляют собой VL человека. Согласно одному варианту осуществления сегменты JL представляют собой JL человека. Согласно конкретному варианту осуществления сегменты VL и JL представляют собой сегменты VL человека и сегменты JL человека.
Согласно одному варианту осуществления все или по существу все генные сегменты VH, DH и JH замещают по меньшей мере шестью человека генными сегментами Vκ и по меньшей мере одним генным сегментом Jκ. Согласно одному варианту осуществления все или по существу все генные сегменты VH, DH и JH замещают по меньшей мере 16 генными сегментами Vκ человека (Vκ человека) и по меньшей мере одним генным сегментом Jκ. Согласно одному варианту осуществления все или по существу все генные сегменты VH, DH и JH замещают по меньшей мере 30 генными сегментами Vκ человека и по меньшей мере одним генным сегментом Jκ. Согласно одному варианту осуществления все или по существу все генные сегменты VH, DH и JH замещают по меньшей мере 40 генными сегментами Vκ человека и по меньшей мере одним генным сегментом Jκ. Согласно одному варианту осуществления по меньшей мере один генный сегмент Jκ содержит два, три, четыре или пять генных сегментов Jκ человека.
Согласно одному варианту осуществления сегменты VL представляют собой сегменты Vκ человека. Согласно одному варианту осуществления сегменты Vκ человека включают в себя 4-1, 5-2, 7-3, 2-4, 1-5 и 1-6. Согласно одному варианту осуществления сегменты Vκ человека включают в себя 3-7, 1-8, 1-9, 2-10, 3-11, 1-12, 1-13, 2-14, 3-15 и 1-16. Согласно одному варианту осуществления сегменты Vκ человека включают в себя 1-17, 2-18, 2-19, 3-20, 6-21, 1-22, 1-23, 2-24, 3-25, 2-26, 1-27, 2-28, 2-29 и 2-30. Согласно одному варианту осуществления сегменты Vκ человека включают в себя 3-31, 1-32, 1-33, 3-34, 1-35, 2-36, 1-37, 2-38, 1-39 и 2-40.
Согласно одному варианту осуществления сегменты VL представляют собой сегменты Vκ человека и включают в себя 4-1, 5-2, 7-3, 2-4, 1-5, 1-6, 3-7, 1-8, 1-9, 2-10, 3-11, 1-12, 1-13, 2-14, 3-15 и 1-16. Согласно одному варианту осуществления сегменты Vκ дополнительно включают в себя 1-17, 2-18, 2-19, 3-20, 6-21, 1-22, 1-23, 2-24, 3-25, 2-26, 1-27, 2-28, 2-29 и 2-30. Согласно одному варианту осуществления сегменты Vκ дополнительно включают в себя 3-31, 1-32, 1-33, 3-34, 1-35, 2-36, 1-37, 2-38, 1-39 и 2-40.
Согласно одному варианту осуществления сегменты VL представляют собой сегменты Vλ человека и содержат фрагмент кластера A локуса λ легкой цепи человека. Согласно конкретному варианту осуществления фрагмент кластера A локуса λ легкой цепи человека продолжается от hVλ3-27 до hλΛ3-1.
Согласно одному варианту осуществления сегменты VL содержат фрагмент кластер B локуса λ легкой цепи человека. Согласно конкретному варианту осуществления фрагмент кластера B локуса λ легкой цепи человека продолжается от hVλ5-52 до hVλ1-40.
Согласно одному варианту осуществления сегменты VL содержат последовательность вариабельной области λ легкой цепи человека, которая содержит геномный фрагмент кластера A и геномный фрагмент кластера B. Согласно одному варианту осуществления последовательность вариабельной области λ легкой цепи человека содержит по меньшей мере один генный сегмент кластера A и по меньшей мере один генный сегмент кластера B.
Согласно одному варианту осуществления сегменты VL содержат по меньшей мере один генный сегмент кластера B и по меньшей мере один генный сегмент кластера C.
Согласно одному варианту осуществления сегменты VL включают в себя hVλ3-1, 4-3, 2-8, 3-9, 3-10, 2-11 и 3-12. Согласно конкретному варианту осуществления сегменты VL содержат смежную последовательность локуса λ легкой цепи человека, которая охватывает от Vλ3-12 до Vλ3-1. Согласно одному варианту осуществления смежная последовательность содержит по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 hVλ. Согласно конкретному варианту осуществления hVλ включают в себя 3-1, 4-3, 2-8, 3-9, 3-10, 2-11 и 3-12. Согласно конкретному варианту осуществления hVλ содержат смежную последовательность λ локуса человека, которая охватывает от Vλ3-12 до Vλ3-1.
Согласно одному варианту осуществления hVλ содержат 13 - 28 или больше hVλ. Согласно конкретному варианту осуществления hVλ включают в себя 2-14, 3-16, 2-18, 3-19, 3-21, 3-22, 2-23, 3-25 и 3-27. Согласно конкретному варианту осуществления hVλ содержат смежную последовательность λ локуса человека, которая охватывает от Vλ3-27 до Vλ3-1.
Согласно одному варианту осуществления сегменты VL содержат 29 - 40 hVλ. Согласно конкретному варианту осуществления сегменты VL содержат смежную последовательность λ локуса человека, которая охватывает от Vλ3-29 до Vλ3-1, и a смежную последовательность λ локуса человека, которая охватывает от Vλ5-52 до Vλ1-40. Согласно конкретному варианту осуществления вся или по существу вся последовательность между hVλ1-40 и hVλ3-29 у генетически модифицированной мыши состоит по существу из последовательности λ человека, составляющей приблизительно 959 п.н., встречающейся в природе (например, в человеческой популяции) ниже генного сегмента hVλ1-40 (ниже нетранслируемой части 3’), сайт фермента рестрикции (например, PI-SceI), за которой следует последовательность λ человека, составляющая приблизительно 3431 п.н. выше генного сегмента hVΛ3-29, встречающего в природе.
Согласно одному варианту осуществления Jκ представляет собой Jκ человека и выбран из группы, состоящей из Jκ1, Jκ2, Jκ3, Jκ4, Jκ5 и их комбинации. Согласно конкретному варианту осуществления Jκ содержит Jκ1-Jκ5.
Согласно одному варианту осуществления сегменты VL представляют собой сегменты Vλ человека, и генный сегмент Jκ содержит RSS с 12-мерным спейсером, причем RSS расположена бок о бок на вышележащем конце генного сегмента Jκ. Согласно одному варианту осуществления генные сегменты VL представляют собой Vλ человека, и локус VLH содержит два или больше генных сегментов Jκ, каждый из которых содержит RSS с 12-мерным спейсером, причем RSS расположена бок о бок на вышележащем конце каждого генного сегмента Jκ.
Согласно конкретному варианту осуществления сегменты VL содержат смежные κ генные сегменты человека, охватывающие κ локус человека от Vκ4-1 до Vκ2-40, и сегменты JL содержат смежные генные сегменты, охватывающие κ локус человека от Jκ 1 до Jκ5.
Согласно одному варианту осуществления, если сегменты VL представляют собой сегменты Vλ и ни один сегмент DH не присутствует между сегментами VL и сегментами J, то сегменты VL фланкированы по ходу транскрипции (т.е. расположены бок о бок на стороне по ходу транскрипции с) 23-мерной RSS, и сегменты Jκ, если они присутствуют, или сегменты Jλ, если они присутствуют, фланкированы против хода транскрипции (т.е. расположены бок о бок на стороне против хода транскрипции с) 12-мерной RSS.
Согласно одному варианту осуществления, если генные сегменты V представляют собой генные сегменты Vκ и ни один сегмент DH не присутствует между генными сегментами и генными сегментами J, каждый из генных сегментов Vκ расположен бок о бок на стороне по ходу транскрипции с 12-мерной RSS, и каждый из сегментов Jκ, если они присутствуют, или сегментов Jλ, если они присутствуют, расположен бок о бок на стороне против хода транскрипции с 23-мерной RSS.
Согласно одному варианту осуществления мышь содержит реаранжированный ген, который происходит из генного сегмента VL, генного сегмента JL и эндогенного гена CH. Согласно одному варианту осуществления реаранжированный ген является соматически мутированным. Согласно одному варианту осуществления реаранжированный ген содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или больше N-присоединений. Согласно одному варианту осуществления N-присоединения и/или соматические мутации, наблюдаемые в реаранжированном гене, происходящие из сегмента VL и сегмента JL, в 1,5 раз, 2 раза, 2,5 раза, 3 раза, 3,5 раза, 4 раза, 4,5 раза или по меньшей мере в 5 раз превосходят количество N-присоединений и/или соматических мутаций, наблюдаемых в реаранжированном вариабельном домене легкой цепи (происходящем из того же генного сегмента VL и того же генного сегмента JL), который является реаранжированным на эндогенном локусе легкой цепи. Согласно одному варианту осуществления реаранжированный ген находится в B-клетке, которая специфически связывает представляющий интерес антиген, причем B-клетка связывает представляющий интерес антиген с KD в наномолярном диапазоне или ниже (например, KD, составляющей 10 наномоль или ниже). Согласно конкретному варианту осуществления сегмент VL, сегмент JL или оба сегмента представляют собой генные сегменты человека. Согласно конкретному варианту осуществления сегменты VL и JL представляют собой κ генные сегменты человека. Согласно одному варианту осуществления ген CH мыши выбран из IgM, IgD, IgG, IgA и IgE. Согласно конкретному варианту осуществления IgG мыши выбран из IgG1, IgG2A, IgG2B, IgG2C и IgG3. Согласно другому конкретному варианту осуществления IgG мыши представляет собой IgG1.
Согласно одному варианту осуществления мышь содержит B-клетку, причем B-клетка экспрессирует из локуса на хромосоме B-клетки связывающий белок, состоящий по существу из четырех полипептидных цепей, причем четыре полипептидных цепи состоят по существу из (a) двух идентичных полипептидов, которые содержат эндогенную область CH, слитую с VL; и (b) двух идентичных полипептидов, которые содержат эндогенную область CL, слитую с областью VL, которая является когнатной по отношению к области VL, которая слита с областью CH мыши, и, согласно одному варианту осуществления представляет собой VL область человека (например, κ человека). Согласно одному варианту осуществления область VL, слитая с эндогенной областью CH, представляет собой область VL человека. Согласно конкретному варианту осуществления область VL человека, слитая с областью CH мыши, представляет собой область Vκ. Согласно конкретному варианту осуществления область VL человека, слитая с областью CH мыши, идентична области V, кодируемой реаранжированной зародышевой нуклеотидной последовательностью легкой цепи человека. Согласно конкретному варианту осуществления область VL человека, слитая с областью CH мыши, содержит две, три, четыре, пять, шесть или больше соматических гипермутаций. Согласно одному варианту осуществления область VL человека, слитая с областью CH мыши, кодируется реаранжированным геном, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или больше N-присоединений.
Согласно одному варианту осуществления по меньшей мере 50% из всех молекул IgG, экспрессируемых мышью, содержат полипептид, который содержит область CH и область VL изотипа IgG, причем длина указанного полипептида не превышает 535, 530, 525, 520 или 515 аминокислот. Согласно одному варианту осуществления по меньшей мере 75% из всех молекул IgG содержат полипептид, указанный в настоящем абзаце. Согласно одному варианту осуществления по меньшей мере 80%, 85%, 90%, или 95% из всех молекул IgG содержат полипептид, указанный в настоящем абзаце. Согласно конкретному варианту осуществления все молекулы IgG, экспрессируемые мышью, содержат полипептид, который не превышает в длину полипептид, указанный в настоящем абзаце.
Согласно одному варианту осуществления мышь экспрессирует связывающий белок, содержащий первый полипептид, который содержит эндогенную область CH, слитую с вариабельным доменом, кодируемым реаранжированным генным сегментом V человека и генным сегментом J, но не генным сегментом DH, и второй полипептид, который содержит эндогенную область CL, слитую с доменом V, кодируемым реаранжированным генным сегментом V и генным сегментом J человека, но не генным сегментом DH, и связывающий белок специфически связывает антиген с аффинностью в микромолярном, наномолярном или пикомолярном диапазоне. Согласно одному варианту осуществления сегмент J представляет собой сегмент J человека (например, κ генный сегмент человека). Согласно одному варианту осуществления сегмент V человека представляет собой сегмент Vκ человека. Согласно одному варианту осуществления вариабельный домен, который слит с эндогенной областью CH, содержит большее количество соматических гипермутаций, чем вариабельная область, которая слита с эндогенной областью CL; согласно конкретному варианту осуществления вариабельная область, слитая с эндогенной областью CH, содержит приблизительно в 1,5, 2, 3, 4 или 5 раз или более больше соматических гипермутаций, чем область V, слитая с эндогенной областью CL; согласно конкретному варианту осуществления область V, слитая с областью CH мыши, содержит по меньшей мере на 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 или больше соматических гипермутаций, чем область V, слитая с областью CL мыши. Согласно одному варианту осуществления область V, слитая с областью CH мыши, кодируется реаранжированным геном, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, или 10 или больше N-присоединений.
Согласно одному варианту осуществления мышь экспрессирует связывающий белок, содержащий первый вариабельный домен легкой цепи (VL1), слитый с последовательностью константной области тяжелой цепи иммуноглобулина, и второй вариабельный домен легкой цепи (VL2), слитый с константной областью легкой цепи иммуноглобулина, причем VL1 содержит количество соматических гипермутаций, которое приблизительно в 1,5- приблизительно в 5 раз или более выше, чем количество соматических гипермутаций, присутствующих в VL2. Согласно одному варианту осуществления количество соматических гипермутаций в VL1 приблизительно в 2 - приблизительно в 4 раза выше, чем в VL2. Согласно одному варианту осуществления количество соматических гипермутаций в VL1 приблизительно в 2 - приблизительно в 3 раза выше, чем в VL2. Согласно одному варианту осуществления VL1 кодируется последовательностью, которая содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, или 10 или больше N-присоединений.
Согласно одному аспекту предусмотрена генетически модифицированная мышь, которая экспрессирует иммуноглобулин, который состоит по существу из следующих полипептидов: первых двух идентичных полипептидов, каждый из которых состоит по существу из области CH, слитой с вариабельным доменом, который происходит из генных сегментов, которые состоят по существу из генного сегмента VL и генного сегмента JL, и вторых двух идентичных полипептидов, каждый из которых состоит по существу из области CL, слитой с вариабельным доменом, который происходит из генных сегментов, которые состоят по существу из сегмента VL и сегмента JL.
Согласно конкретному варианту осуществления два идентичных полипептида, которые содержат область CH, содержат область CH мыши.
Согласно конкретному варианту осуществления два идентичных полипептида, которые содержат область CL, содержат область CL мыши.
Согласно одному варианту осуществления вариабельный домен, слитый с областью CL, представляет собой вариабельный домен, который является когнатным вариабельному домену, слитому с областью CH.
Согласно одному варианту осуществления вариабельный домен, который слит с эндогенной областью CH, содержит большее количество соматических гипермутаций, чем вариабельный домен, который слит с эндогенной областью CL; согласно конкретному варианту осуществления вариабельный домен, слитый с эндогенной областью CH, содержит приблизительно в 1,5 раза, 2 раза, 2,5 раза, 3 раза, 3,5 раза, 4 раза, 4,5 раза или 5 раз или более больше соматических гипермутаций, чем вариабельный домен, слитый с эндогенной областью CL. Согласно одному варианту осуществления вариабельный домен, слитый с эндогенной областью CL, кодируется геном, который содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или больше N-присоединений.
Согласно одному варианту осуществления один или несколько сегментов V и сегментов J представляют собой генные сегменты человека. Согласно конкретному варианту осуществления как сегменты V, так и сегменты J представляют собой генные сегменты κ человека. Согласно другому конкретному варианту осуществления как сегменты V, так и сегменты J представляют собой генные сегменты λ человека. Согласно одному варианту осуществления сегменты V сегменты и J независимо выбраны из генных сегментов κ человека и генных сегментов λ человека. Согласно конкретному варианту осуществления сегменты V представляют собой сегменты Vκ, и сегменты J представляют собой сегменты Jλ. Согласно другому конкретному варианту осуществления сегменты V представляют собой сегменты Vλ, и сегменты J представляют собой сегменты Jκ.
Согласно одному варианту осуществления один или несколько вариабельных доменов, слитых с областью CL, и вариабельных доменов, слитых с областью CH, представляют собой вариабельные домены человека. Согласно конкретному варианту осуществления вариабельные домены человека представляют собой домены Vκ человека. Согласно другому конкретному варианту осуществления вариабельные домены человека представляют собой домены Vλ. Согласно одному варианту осуществления человека домены независимо выбраны из доменов Vκ человека и доменов Vλ человека. Согласно конкретному варианту осуществления вариабельный домен человека, слитый с областью CL, представляет собой домен Vλ человека, и вариабельный домен человека, слитый с областью CH, представляет собой домен Vκ человека. Согласно другому варианту осуществления вариабельный домен человека, слитый с областью CL, представляет собой домен Vκ человека, и вариабельный домен человека, слитый с CH, представляет собой домен Vλ человека.
Согласно одному варианту осуществления генный сегмент VL первых двух идентичных полипептидов выбран из сегмента Vλ человека и сегмента Vκ человека. Согласно одному варианту осуществления сегмент VL вторых двух идентичных полипептидов выбран из сегмента Vλ человека и сегмента Vκ человека. Согласно конкретному варианту осуществления сегмент VL первых двух идентичных полипептидов представляет собой сегмент Vκ человека, и сегмент VL вторых двух идентичных полипептидов выбран из сегмента Vκ человека и сегмента Vλ человека. Согласно конкретному варианту осуществления сегмент VL первых двух идентичных полипептидов представляет собой сегмент Vλ человека, и сегмент VL вторых двух идентичных полипептидов выбран из сегмента Vλ человека и сегмента Vκ человека. Согласно конкретному варианту осуществления сегмент VL человека первых двух идентичных полипептидов представляет собой сегмент Vκ человека, и сегмент VL человека вторых двух идентичных полипептидов представляет собой сегмент Vκ человека.
Согласно одному варианту осуществления IgG мышь содержит связывающий белок, произведенный в ответ на антиген, причем связывающий белок содержит полипептид, который состоит по существу из вариабельного домена и области CH, причем вариабельный домен кодируется нуклеотидной последовательностью, которая состоит по существу из реаранжированного сегмента VL и реаранжированного сегмента J, и причем связывающий белок специфически связывает эпитоп антигена с KD в микромолярном, наномолярном или пикомолярном диапазоне.
Согласно одному аспекту предусмотрена мышь, причем весь или по существу весь IgG, произведенный мышью в ответ на антиген, содержит тяжелую цепь, которая содержит вариабельный домен, причем вариабельный домен кодируется реаранжированным геном, происходящим из генных сегментов, которые состоят по существу из генного сегмента V и генного сегмента J. Согласно одному варианту осуществления реаранжированный ген содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или больше N-присоединений.
Согласно одному варианту осуществления сегмент V представляет собой сегмент V легкой цепи. Согласно одному варианту осуществления легкая цепь выбрана из κ легкой цепи и λ легкой цепи. Согласно конкретному варианту осуществления легкая цепь представляет собой κ легкую цепь. Согласно конкретному варианту осуществления сегмент V представляет собой сегмент V человека. Согласно конкретному варианту осуществления сегмент V представляет собой сегмент Vκ человека, и сегмент J представляет собой сегмент Jκ человека.
Согласно одному варианту осуществления сегмент J представляет собой сегмент J легкой цепи. Согласно одному варианту осуществления легкая цепь выбрана из κ легкой цепи и λ легкой цепи. Согласно конкретному варианту осуществления легкая цепь представляет собой κ легкую цепь. Согласно конкретному варианту осуществления сегмент J представляет собой сегмент J человека. Согласно другому варианту осуществления, сегмент J представляет собой сегмент J тяжелой цепи (т.е. JH). Согласно конкретному варианту осуществления тяжелая цепь происходит от мыши. Согласно другому конкретному варианту осуществления тяжелая цепь происходит от человека.
Согласно одному варианту осуществления вариабельный домен тяжелой цепи, который происходит не более чем из сегмента V и сегмента J, представляет собой соматически мутированный вариабельный домен.
Согласно одному варианту осуществления вариабельный домен тяжелой цепи, который происходит не более чем из сегмента V и сегмента J, слит с областью CH мыши.
Согласно конкретному варианту осуществления все или по существу все IgG, произведенные мышью в ответ на антиген, содержат вариабельный домен, который происходит не более чем из одного сегмента V человека и не более чем одного сегмента J человека, и вариабельный домен слит с константной областью IgG мыши, и IgG дополнительно содержит легкую цепь, которая содержит домен VL человека, слитый с областью CL мыши. Согласно конкретному варианту осуществления домен VL, слитый с областью CL мыши, происходит из сегмента Vκ человека и сегмента Jκ человека. Согласно конкретному варианту осуществления домен VL, слитый с областью CL мыши, происходит из сегмента Vλ человека и сегмента Jλ человека.
Согласно одному аспекту предусмотрена мышь, которая производит IgG, содержащий первую CDR3 на полипептиде, содержащем область CH, и вторую CDR3 на полипептиде, содержащем область CL, причем каждая из первой CDR3 и второй CDR3 независимо происходит не более чем из двух генных сегментов, причем два генных сегмента состоят по существу из генного сегмента VL и генного сегмента JL. Согласно одному варианту осуществления CDR3 на полипептиде, содержащем область CH, содержит последовательность, которая происходит из нуклеотидной последовательности CDR3, которая содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или больше N-присоединений.
Согласно одному варианту осуществления сегмент VL и сегмент JL представляют собой генные сегменты человека. Согласно одному варианту осуществления сегмент VL и сегмент JL представляют собой κ генные сегменты. Согласно одному варианту осуществления сегмент VL и сегмент JL представляют собой λ генные сегменты.
Согласно одному аспекту предусмотрена мышь, которая производит IgG, содержащий первую CDR3 на первом полипептиде, содержащем область CH, и вторую CDR3 на втором полипептиде, содержащем область CL, причем каждая из первой CDR3 и второй CDR3, содержит последовательность аминокислот, причем больше чем 75% аминокислот происходят из генного сегмента V. Согласно одному варианту осуществления CDR3 на полипептиде, содержащем область CH, содержит последовательность, которая происходит из нуклеотидной последовательности CDR3, которая содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или больше N-присоединений.
Согласно одному варианту осуществления больше чем 80%, больше чем 90%, или больше чем 95% аминокислот первой CDR3, и больше чем 80%, больше чем 90%, или больше чем 95% аминокислот второй CDR3 происходят из сегмента V легкой цепи.
Согласно одному варианту осуществления не более чем две аминокислоты первой CDR3 происходят из генного сегмента, отличного от сегмента V легкой цепи. Согласно одному варианту осуществления не более чем две аминокислоты второй CDR3 происходят из генного сегмента, отличного от сегмента V легкой цепи. Согласно конкретному варианту осуществления не более чем две аминокислоты первой CDR3 и не более чем две аминокислоты второй CDR3 происходят из генного сегмента, отличного от сегмента V легкой цепи. Согласно одному варианту осуществления ни одна из CDR3 IgG не содержит аминокислотную последовательность, происходящую из генного сегмента D. Согласно одному варианту осуществления CDR3 первого полипептида не содержит последовательность, происходящую из сегмента D.
Согласно одному варианту осуществления сегмент V представляет собой генный сегмент V человека. Согласно конкретному варианту осуществления сегмент V представляет собой генный сегмент Vκ человека.
Согласно одному варианту осуществления первая и/или вторая CDR3 содержат по меньшей мере одну, две, три, четыре, пять или шесть соматических гипермутаций. Согласно одному варианту осуществления первая CDR3 кодируется последовательностью нуклеиновой кислоты, которая содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или больше N-присоединений.
Согласно одному варианту осуществления первая CDR3 состоит по существу из аминокислот, происходящих из генного сегмента V легкой цепи человека и генного сегмента J легкой цепи человека, и вторая CDR3 состоит по существу из аминокислот, происходящих из генного сегмента V легкой цепи человека и генного сегмента J легкой цепи человека. Согласно одному варианту осуществления первая CDR3 происходит из последовательности нуклеиновой кислоты, которая содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или больше N-присоединений. Согласно одному варианту осуществления первая CDR3 происходит не более чем из двух генных сегментов, причем не более чем два генных сегмента представляют собой генный сегмент Vκ человека и генный сегмент Jκ человека; и вторая CDR3 происходит не более чем из двух генных сегментов, причем не более чем два генных сегмента представляют собой генный сегмент Vκ человека и генный сегмент J, выбранный из сегмента Jκ человека, сегмента Jλ человека и сегмента JH человека. Согласно одному варианту осуществления первая CDR3 происходит не более чем из двух генных сегментов, причем не более чем два генных сегмента представляют собой сегмент Vλ человека и сегмент J, выбранный из сегмента Jκ человека, сегмента Jλ человека и сегмента JH человека.
Согласно одному аспекту предусмотрена мышь, которая производит IgG, который не содержит аминокислотную последовательность, происходящую из генного сегмента DH, причем IgG содержит первый полипептид, содержащий первый домен VL, слитый с областью CL мыши, и второй полипептид, содержащий второй домен VL, слитый с областью CH мыши, причем первый домен VL и второй домен VL не являются идентичными. Согласно одному варианту осуществления первый и второй домены VL происходят из различных сегментов V. Согласно другому варианту осуществления первый и второй домены VL происходят из различных сегментов J. Согласно одному варианту осуществления первый и второй домены VL происходят из идентичных сегментов V и J, причем второй домен VL содержит большее количество соматических гипермутаций по сравнению с первым доменом VL.
Согласно одному варианту осуществления первый и второй домены VL независимо выбраны из доменов VL человека и мыши. Согласно одному варианту осуществления первый и второй домены VL независимо выбраны из доменов Vκ и Vλ. Согласно конкретному варианту осуществления первый домен VL выбран из домена Vκ и домена Vλ, и второй домен VL представляет собой домен Vκ. Согласно другому конкретному варианту осуществления домен Vκ представляет собой домен Vκ человека.
Согласно одному аспекту предусмотрена мышь, причем весь или по существу весь IgG, произведенный мышью, состоит по существу из легкой цепи, содержащей первый домен VL человека, слитый с доменом CL мыши, и тяжелой цепи, содержащей второй домен VL человека, слитый с доменом CH мыши.
Согласно одному варианту осуществления домен VL человека, слитый с доменом CH мыши, представляет собой домен Vκ человека.
Согласно одному варианту осуществления первый и второй домены VL человека не являются идентичными.
Согласно одному аспекту предусмотрена мышь, причем по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или приблизительно 100% иммуноглобулина G, произведенного мышью, состоит по существу из димера (a) первого полипептида, который состоит по существу из домена VL иммуноглобулина и области CL иммуноглобулина; и, (b) второго полипептида, составляющего не более чем 535 аминокислот в длину, причем второй полипептид состоит по существу из области CH и домена V, который не содержит последовательность, происходящую из генного сегмента DH.
Согласно одному варианту осуществления второй полипептид составляет приблизительно 435-535 аминокислот в длину. Согласно конкретному варианту осуществления второй полипептид составляет приблизительно 435-530 аминокислот в длину. Согласно конкретному варианту осуществления второй полипептид составляет приблизительно 435-525 аминокислот в длину. Согласно конкретному варианту осуществления второй полипептид составляет приблизительно 435-520 аминокислот в длину. Согласно конкретному варианту осуществления второй полипептид составляет приблизительно 435-515 аминокислот в длину.
Согласно одному варианту осуществления приблизительно у 90% или больше IgG, произведенного мышью, второй полипептид составляет не более чем приблизительно 535 аминокислот в длину.
Согласно одному варианту осуществления приблизительно у 50% или больше IgG, произведенного мышью, второй полипептид составляет не более чем приблизительно 535 аминокислот в длину. Согласно одному варианту осуществления приблизительно в 50% или больше иммуноглобулина G, произведенного мышью, второй полипептид составляет не более чем приблизительно 530, 525, 520, 515, 510, 505, 500, 495, 490, 485, 480, 475, 470, 465, 460, 455 или 450 аминокислот в длину. Согласно одному варианту осуществления приблизительно 60%, 70%, 80%, 90% или 95% или больше IgG, произведенного мышью, характеризуется указанной длиной. Согласно конкретному варианту осуществления весь или по существу весь IgG, произведенный мышью, характеризуется указанной длиной.
Согласно одному варианту осуществления домен V второго полипептида представляет собой домен VL. Согласно конкретному варианту осуществления домен V второго полипептида выбран из домена Vκ и Vλ. Согласно конкретному варианту осуществления домен V второго полипептида представляет собой домен Vκ или Vλ человека.
Согласно одному аспекту предусмотрена мышь, которая экспрессирует из нуклеотидной последовательности в своей зародышевой линии полипептид, который содержит вариабельную последовательность легкой цепи (например, последовательность V и/или J), последовательность DH и константную область тяжелой цепи.
Согласно одному варианту осуществления мышь экспрессирует полипептид из эндогенного локуса тяжелой цепи, который содержит замещение всех или по существу всех функциональных эндогенных генных сегментов вариабельного локуса тяжелой цепи множеством генных сегментов человека на эндогенном локусе тяжелой цепи.
Согласно одному варианту осуществления полипептид содержит последовательность VL, происходящую из генного сегмента Vλ или Vκ, полипептид содержит CDR3, происходящую из генного сегмента DH, и полипептид содержит последовательность, происходящую из генного сегмента JH или Jλ или Jκ.
Согласно одному варианту осуществления мышь содержит эндогенный локус тяжелой цепи иммуноглобулина, содержащий замещение всех функциональных генных сегментов VH одним или нескольким генными сегментами Vλ легкой цепи человека, причем каждый из одного или нескольких сегментов Vλ человека был помещен бок о бок на стороне по ходу транскрипции с содержащей 23-мерный спейсер последовательностью сигнала рекомбинации (RSS), причем сегменты Vλ функционально связаны с сегментом DH человека или мыши, который был помещен бок о бок против и по ходу транскрипции с содержащей 12-мерный спейсер RSS; генный сегмент DH функционально связан с сегментом J, расположенным бок о бок против хода транскрипции с содержащей 23-мерный спейсер RSS, которая является подходящей для рекомбинации с содержащей 12-мерный спейсер RSS, соединяющей генный сегмент DH; причем сегменты V, DH и J функционально связаны с последовательностью нуклеиновой кислоты, кодирующей константную область тяжелой цепи.
Согласно одному варианту осуществления мышь содержит эндогенный локус тяжелой цепи иммуноглобулина, содержащий замещение всех функциональных генных сегментов VH одним или несколькими генными сегментами Vκ человека, каждый из которых расположен бок о бок на стороне по ходу транскрипции с содержащей 12-мерный спейсер последовательностью сигнала рекомбинации (RSS), причем сегменты V функционально связаны с сегментом DH человека или мыши, который расположен бок о бок как против, так и по ходу транскрипции с содержащей 23-мерный спейсер RSS; сегмент DH функционально связан с сегментом J, расположенным бок о бок на стороне против хода транскрипции с содержащей 12-мерный спейсер RSS, которая является подходящей для рекомбинации с содержащей 23-мерный спейсер RSS, соединяющей сегмент DH; причем генные сегменты V, DH и J функционально связаны с последовательностью нуклеиновой кислоты, кодирующей константную область тяжелой цепи.
Согласно одному варианту осуществления константная область тяжелой цепи представляет собой эндогенную константную область тяжелой цепи. Согласно одному варианту осуществления последовательность нуклеиновой кислоты кодирует последовательность, выбранную из CH1, шарнира, CH2, CH3 и их комбинации. Согласно одному варианту осуществления один или несколько из CH1, шарнира, CH2, и CH3 происходят от человека.
Согласно одному варианту осуществления мышь содержит эндогенный локус тяжелой цепи иммуноглобулина, содержащий замещение всех функциональных генных сегментов VH множеством генных сегментов Vλ или Vκ человека, каждый их которых расположен бок о бок по ходу транскрипции с содержащей 23-мерный спейсер RSS, множеством сегментов DH человека, расположенных бок о бок как против, так и по ходу транскрипции с содержащей 12-мерный спейсер RSS, множеством сегментов J человека (JH или Jλ или Jκ), расположенных бок о бок как против, так и по ходу транскрипции с содержащей 23-мерный спейсер RSS, причем локус содержит эндогенную последовательность константной области, выбранную из CH1, шарнира, CH2, CH3 и их комбинации. Согласно конкретному варианту осуществления мышь содержит все или по существу все функциональные сегменты Vλ или Vκ человека, все или по существу все функциональные сегменты DH человека и все или по существу все сегменты JH или Jλ или Jκ.
Согласно одному варианту осуществления мышь экспрессирует антигенсвязывающий белок, содержащий (a) полипептид, который содержит последовательность легкой цепи человека, связанную с константной последовательностью тяжелой цепи, содержащей последовательность мыши; и (b) полипептид, который содержит вариабельную область легкой цепи человека, связанную с константной последовательностью легкой цепи человека или мыши. Согласно конкретному варианту осуществления последовательность легкой цепи представляет собой последовательность легкой цепи человека, и при воздействии протеазы, которая способна расщеплять антитело на Fc и Fab, образуется полностью человеческий Fab, который содержит по меньшей мере четыре CDR легкой цепи, причем по меньшей мере четыре CDR легкой цепи выбраны из λ последовательностей, κ последовательностей и их комбинации. Согласно одному варианту осуществления Fab содержит по меньшей мере пять CDR легкой цепи. Согласно одному варианту осуществления Fab содержит шесть CDR легкой цепи. Согласно одному варианту осуществления по меньшей мере одна CDR Fab содержит последовательность, происходящую из сегмента Vλ или сегмента Vκ, и по меньшей мере одна CDR дополнительно содержит последовательность, происходящую из сегмента D. Согласно одному варианту осуществления по меньшей мере одна CDR представляет собой CDR3, и CDR происходит из сегмента Vκ человека, сегмента D человека и сегмента Jκ человека.
Согласно одному варианту осуществления полипептид содержит вариабельную область, происходящую из генного сегмента Vλ или Vκ человека, генного сегмента DH человека и генного сегмента JH или Jλ или Jκ человека. Согласно конкретному варианту осуществления константная последовательность тяжелой цепи происходит из последовательности CH1 человека и CH2 мыши и CH3 мыши.
Согласно одному аспекту предусмотрена мышь, которая содержит в своей зародышевой линии нереаранжированный генный сегмент Vκ или Vλ человека, функционально связанный с генным сегментом J человека и последовательностью константной области тяжелой цепи, причем мышь экспрессирует связывающий белок VL, который содержит домен Vκ человека, слитый с константной областью тяжелой цепи, и домен VL человека, слитый с константным доменом легкой цепи. Согласно одному варианту осуществления домен VL человека содержит реаранжированный генный сегмент VL человека, выбранный из генного сегмента Vκ человека и Vλ человека. Согласно конкретному варианту осуществления домен VL человека дополнительно содержит реаранжированный генный сегмент JL человека, выбранный из генного сегмента Jκ человека и Jλ человека.
Согласно одному аспекту предусмотрена мышь, которая экспрессирует белок иммуноглобулина из модифицированного эндогенного локуса тяжелой цепи в своей зародышевой линии, причем модифицированный эндогенный локус тяжелой цепи не содержит функциональный генный сегмент V тяжелой цепи мыши и локус содержит нереаранжированные генные сегменты V легкой цепи и нереаранжированные генные сегменты J, причем нереаранжированные генные сегменты V легкой цепи и нереаранжированные генные сегменты J функционально связаны с последовательностью константной области тяжелой цепи; причем белок иммуноглобулина состоит по существу из первого полипептида и второго полипептида, причем первый полипептид содержит последовательность легкой цепи иммуноглобулина и константную последовательность тяжелой цепи иммуноглобулина, и второй полипептид содержит вариабельный домен легкой цепи иммуноглобулина и константную область легкой цепи.
Согласно одному аспекту предусмотрена мышь, которая экспрессирует белок иммуноглобулина, причем белок иммуноглобулина не содержит вариабельный домен тяжелой цепи иммуноглобулина, и белок иммуноглобулина содержит первый вариабельный домен, происходящий из гена легкой цепи, и второй вариабельный домен, происходящий из гена легкой цепи, причем первый вариабельный домен и второй вариабельный домен являются когнатными относительно друг друга, причем первый и второй вариабельные домены легкой цепи не являются идентичными, и причем первый и второй вариабельные домены легкой цепи ассоциируются и, будучи ассоциированными, специфически связывают представляющий интерес антиген.
Согласно одному аспекту предусмотрена мышь, которая экспрессирует из нереаранжированных генных сегментов в своей зародышевой линии белок иммуноглобулина, содержащий вариабельные области, которые полностью происходят из генных сегментов, которые состоят по существу из нереаранжированных генных сегментов человека, причем белок иммуноглобулина содержит константную последовательность легкой цепи иммуноглобулина и константную последовательность тяжелой цепи иммуноглобулина, выбранную из группы, состоящей из CH1, шарнира, CH2, CH3 и их комбинации.
Согласно одному аспекту предусмотрена мышь, которая экспрессирует из нереаранжированных генных сегментов в своей зародышевой линии белок иммуноглобулина, содержащий вариабельные области, причем все CDR3 всех вариабельных областей полностью образованы из генных сегментов V и J легкой цепи, и необязательно одной или нескольких соматических гипермутаций, например, одного или нескольких N-присоединений.
Согласно одному аспекту предусмотрена мышь, которая экспрессирует соматические мутированный белок иммуноглобулина, происходящий из нереаранжированных генных сегментов вариабельной области легкой цепи иммуноглобулина человека в зародышевой линии мыши, причем белок иммуноглобулина не содержит CDR, которая содержит последовательность, происходящую из генного сегмента D, причем белок иммуноглобулина содержит первую CDR3 на вариабельном домене легкой цепи, слитом с константной областью легкой цепи, содержит вторую CDR3 на вариабельном домене легкой цепи, слитом с константной областью тяжелой цепи, и причем вторая CDR3 происходит из реаранжированной последовательности вариабельной области легкой цепи, которая содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или больше N-присоединений.
Согласно одному аспекту описанная предусмотрена в настоящем документе мышь, причем мышь содержит функционально выключенный локус легкой цепи, выбранный из λ локуса, κ локуса и их комбинации. Согласно одному варианту осуществления мышь содержит такую делецию λ и/или κ локуса, полностью или частично, что λ и/или κ локус является нефункциональным.
Согласно одному аспекту предусмотрен зародыш мыши, содержащий клетку, которая содержит описанный в настоящем документе модифицированный локус иммуноглобулина. Согласно одному варианту осуществления мышь представляет собой химеру, и по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 95% клеток зародыша содержат описанный в настоящем документе модифицированный локус иммуноглобулина. Согласно одному варианту осуществления по меньшей мере 96%. 97%, 98%, 99% или 99,8% клеток зародыша содержат описанный в настоящем документе модифицированный локус иммуноглобулина. Согласно одному варианту осуществления зародыш содержит клетку - хозяин и клетку, происходящую из донорной ES клетки, причем клетка, происходящая из донорной ES клетки, содержит описанный в настоящем документе модифицированный локус иммуноглобулина. Согласно одному варианту осуществления зародыш представляет собой зародыш - хозяин на стадии 2, 4, 8, 16, 32, или 64 клеток, или бластоцисту, и дополнительно содержит донорную ES клетку, содержащую описанный в настоящем документе модифицированный локус иммуноглобулина.
Согласно одному аспекту предусмотрена мышь или клетка, полученная с использованием описанного в настоящем документе конструкта нуклеиновой кислоты.
Согласно одному аспекту предусмотрена мышь, полученная с использованием описанной в настоящем документе клетки. Согласно одному варианту осуществления клетка представляет собой ES клетку мыши.
Согласно одному аспекту предусмотрены клетки мыши и зародыши мыши, включая в себя без ограничения ES клетки, плюрипотентные клетки и индуцированные плюрипотентные клетки, которые содержат описанные в настоящем документе генетические модификации. Предусмотрены клетки, которые представляют собой XX, и клетки, которые представляют собой XY. Также предусмотрены клетки, которые содержат ядро, содержащее модификацию, описанную в настоящем документе, например, модификацию, введенную в клетку путем пронуклеарной инъекции. Также предусмотрены клетки, зародыши и мыши, которые содержат введенный с помощью вируса ген ADAM6, например, клетки, зародыши и мыши, содержащие конструкт трансдукции, содержащий ген ADAM6, который является функциональным у мыши.
Согласно одному аспекту предусмотрена клетка или ткань, полученная от описанной в настоящем документе мыши. Согласно одному варианту осуществления клетка или ткань получена из селезенки, лимфатического узла или костного мозга мыши, описанной в настоящем документе. Согласно одному варианту осуществления клетка представляет собой B-клетку. Согласно одному варианту осуществления клетка представляет собой эмбриональную стволовую клетку. Согласно одному варианту осуществления клетка представляет собой половую клетку.
Согласно одному варианту осуществления ткань выбрана из соединительной, мышечной, нервной и эпителиальной ткани. Согласно конкретному варианту осуществления ткань представляет собой репродуктивную ткань.
Согласно одному варианту осуществления клетку и/или ткань, полученную от мыши, описанной в настоящем документе, выделяют для применения в одном или нескольких анализах ex vivo. Согласно различным вариантам осуществления один или несколько анализов ex vivo включают в себя измерения физических, термических, электрических, механических или оптических свойств, хирургическую процедуру, измерения взаимодействий различных типов ткани, разработку техник получения изображений или их комбинацию.
Согласно одному аспекту предусмотрено применение клетки или ткани, полученной от описанной в настоящем документе мыши для получения антитела. Согласно одному аспекту предусмотрено применение клетки или ткани, полученной от описанной в настоящем документе мыши для получения гибридомы или квадромы.
Согласно одному аспекту предусмотрена не относящаяся к человеку клетка, содержащая хромосому или ее фрагмент описанного в настоящем документе отличного от человека животного. Согласно одному варианту осуществления не относящаяся к человеку клетка содержит ядро описанного в настоящем документе отличного от человека животного. Согласно одному варианту осуществления не относящаяся к человеку клетка содержит хромосому или ее фрагмент как результат ядерного транспорта.
Согласно одному аспекту предусмотрено ядро, полученное из мыши, описанной в настоящем документе. Согласно одному варианту осуществления ядро происходит из диплоидной клетки, которая не является B-клеткой.
Согласно одному аспекту предусмотрена генетически модифицированная клетка мыши, причем клетка не способна экспрессировать тяжелую цепь, содержащую реаранжированные эндогенные генные сегменты тяжелой цепи иммуноглобулина, и клетка содержит функциональный ген ADAM6, который кодирует белок ADAM6 мыши или его функциональный фрагмент. Согласно одному варианту осуществления клетка дополнительно содержит вставку генных сегментов легкой цепи иммуноглобулина человека. Согласно конкретному варианту осуществления генные сегменты легкой цепи иммуноглобулина человека представляют собой генные сегменты Vκ и/или Jκ, которые функционально связаны с константными областями тяжелой цепи мышь так, чтобы при реаранжировке кодировать функциональный вариабельный домен легкой цепи, слитый с константным доменом тяжелой цепи мыши.
Согласно одному аспекту предусмотрена генетически модифицированная клетка мыши; причем клетка не содержит функциональный эндогенный локус ADAM6, и клетка содержит эктопическую нуклеотидную последовательность, которая кодирует ADAM6 мыши. Согласно одному варианту осуществления клетка дополнительно содержит модификацию эндогенной последовательности вариабельной области тяжелой цепи иммуноглобулина. Согласно конкретному варианту осуществления модификация эндогенной последовательности вариабельной области тяжелой цепи иммуноглобулина содержит делецию, выбранную из делеции генного сегмента VH мыши, делеции генного сегмента DH мыши, делеции генного сегмента JH мыши и их комбинации. Согласно конкретному варианту осуществления мышь содержит замещение одной или нескольких последовательностей VH, DH, и/или JH иммуноглобулина мыши последовательностью иммуноглобулина человека. Согласно конкретному варианту осуществления последовательность иммуноглобулина человека выбрана из VH человека, VL человека, DH человека, JH человека, JL человека и их комбинации.
Согласно одному варианту осуществления клетка представляет собой тотипотентную клетку, плюрипотентную клетку или индуцированную плюрипотентную клетку. Согласно конкретному варианту осуществления клетка представляет собой ES клетку мыши.
Согласно одному аспекту предусмотрена B-клетка мыши, причем B-клетка мыши содержит реаранжированный ген иммуноглобулина, причем B-клетка содержит на хромосоме B-клетки последовательность нуклеиновой кислоты, кодирующую белок ADAM6 или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши. Согласно одному варианту осуществления B-клетка мыши содержит два аллеля последовательности нуклеиновой кислоты. Согласно одному варианту осуществления реаранжированный ген иммуноглобулина содержит реаранжированную последовательность легкой цепи иммуноглобулина, смежную с константной последовательностью тяжелой цепи. Согласно одному варианту осуществления последовательность легкой цепи представляет собой κ последовательность; согласно одному варианту осуществления последовательность легкой цепи представляет собой λ последовательность.
Согласно одному варианту осуществления последовательность нуклеиновой кислоты находится на молекуле нуклеиновой кислоты (например, хромосоме B-клетки), которая является смежной с реаранжированным локусом тяжелой цепи иммуноглобулина мыши.
Согласно одному варианту осуществления последовательность нуклеиновой кислоты находится на молекуле нуклеиновой кислоты (например, хромосоме B-клетки), которая находится на расстоянии от молекулы нуклеиновой кислоты, которая содержит реаранжированный локус тяжелой цепи иммуноглобулина мыши.
Согласно одному варианту осуществления B-клетка мыши содержит реаранжированную не относящуюся к мыши последовательность вариабельной области легкой цепи иммуноглобулина, функционально связанную с геном константной области тяжелой цепи иммуноглобулина мыши или человека, причем B-клетка содержит последовательность нуклеиновой кислоты, которая кодирует белок ADAM6 или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши.
Согласно одному аспекту предусмотрена соматическая клетка мыши, содержащая хромосому, которая содержит модифицированный локус тяжелой цепи иммуноглобулина, и последовательность нуклеиновой кислоты, кодирующую ADAM6 мыши или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши. Согласно одному варианту осуществления последовательность нуклеиновой кислоты находится на той же хромосоме, что и модифицированный локус тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления нуклеиновая кислота находится на другой хромосоме, чем модифицированный локус тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления соматическая клетка содержит одну копию последовательности нуклеиновой кислоты. Согласно одному варианту осуществления соматическая клетка содержит по меньшей мере две копии последовательности нуклеиновой кислоты. Согласно конкретному варианту осуществления соматическая клетка представляет собой B-клетку. Согласно одному варианту осуществления модифицированный локус тяжелой цепи иммуноглобулина содержит нереаранжированный генный сегмент легкой цепи иммуноглобулина, функционально связанный с последовательностью константной области тяжелой цепи.
Согласно одному аспекту предусмотрена половая клетка мыши, содержащая последовательность нуклеиновой кислоты, кодирующую ADAM6 мыши (или его гомолог или ортолог или функциональный фрагмент) на хромосоме половой клетки, причем последовательность нуклеиновой кислоты, кодирующая мышь ADAM6 (или его гомолог или ортолог или функциональный фрагмент) находится в положении в хромосоме, которое отличается от положения в хромосоме половой клетки мыши дикого типа, причем мышь дополнительно содержит модификацию, содержащую нереаранжированный генный сегмент легкой цепи иммуноглобулина (Vκ и/или Vλ и/или Vκ и Jκ и/или Vλ и Jλ), функционально связанный с последовательностью константной области тяжелой цепи. Согласно одному варианту осуществления последовательность нуклеиновой кислоты находится на локусе иммуноглобулина мыши. Согласно одному варианту осуществления последовательность нуклеиновой кислоты находится на той же хромосоме половой клетки, что и локус иммуноглобулина мыши. Согласно одному варианту осуществления последовательность нуклеиновой кислоты находится на другой хромосоме половой клетки, чем локус иммуноглобулина мыши. Согласно одному варианту осуществления локус иммуноглобулина мыши содержит замещение по меньшей мере одной последовательности иммуноглобулина мыши по меньшей мере одной не относящейся к мыши последовательностью иммуноглобулина. Согласно конкретному варианту осуществления по меньшей мере одна не относящаяся к мыши последовательность иммуноглобулина представляет собой последовательность иммуноглобулина человека.
Согласно одному аспекту предусмотрена плюрипотентная, индуцированная плюрипотентная или тотипотентная клетка, полученная от описанной в настоящем документе мыши. Согласно конкретному варианту осуществления клетка представляет собой эмбриональную стволовую (ES) клетку мыши.
Согласно одному аспекту предусмотрена клетка, содержащая описанный в настоящем документе модифицированный локус иммуноглобулина. Согласно одному варианту осуществления клетка выбрана из тотипотентной клетки, плюрипотентной клетки, индуцированной плюрипотентной стволовой клетки (iPS) и ES клетки. Согласно конкретному варианту осуществления клетка представляет собой клетку мыши, например, ES клетку мыши. Согласно одному варианту осуществления клетка является гомозиготной в отношении модифицированного локуса иммуноглобулина.
Согласно одному аспекту предусмотрена клетка, содержащая последовательность нуклеиновой кислоты, кодирующую первый полипептид, который содержит первую соматически мутированную последовательность Vκ или Vλ человека, слитую с последовательностью константной области тяжелой цепи человека.
Согласно одному варианту осуществления клетка дополнительно содержит вторую полипептидную цепь, которая содержит вторую соматически мутированную последовательность Vκ или Vλ человека, слитую с последовательностью константной области легкой цепи человека.
Согласно одному варианту осуществления последовательность Vκ или Vλ человека первого полипептида является когнатной последовательности Vκ или Vλ человека второго полипептида.
Согласно одному варианту осуществления Vκ или Vλ первого полипептида и Vκ или Vλ человека второго полипептида, будучи ассоциированными, специфически связывают представляющий интерес антиген. Согласно конкретному варианту осуществления первый полипептид содержит вариабельный домен, состоящий по существу из Vκ человека, и второй полипептид содержит вариабельный домен, состоящий из Vκ человека, который является когнатным Vκ человека первого полипептида, и последовательность константной области человека представляет собой последовательность IgG.
Согласно одному варианту осуществления клетка выбрана из клетки CHO, клетки COS, клетки 293, клетки HeLa и ретинальной клетки человека, экспрессирующей вирусную последовательность нуклеиновой кислоты (например, клетки PERC.6™).
Согласно одному аспекту предусмотрена соматическая клетка грызуна (например, мыши), содержащая хромосому, которая содержит описанную в настоящем документе генетическую модификацию.
Согласно одному аспекту предусмотрена половая клетка грызуна (например, мыши), содержащая последовательность нуклеиновой кислоты, которая содержит описанную в настоящем документе генетическую модификацию.
Согласно одному аспекту предусмотрена плюрипотентная, индуцированная плюрипотентная или тотипотентная клетка, полученная от описанного в настоящем документе грызуна (например, мыши). Согласно конкретному варианту осуществления клетка представляет собой эмбриональную стволовую (ES) клетку мыши.
Согласно одному аспекту предусмотрено применение описанной в настоящем документе клетки для получения грызуна (например, мыши), клетки или терапевтического белка (например, антитела или другого антигенсвязывающего белка). Согласно одному варианту осуществления предусмотрено применение описанной в настоящем документе клетки для получения терапевтического белка, причем терапевтический белок содержит вариабельный домен человека. Согласно конкретному варианту осуществления вариабельный домен человека содержит реаранжированные генные сегменты Vκ и Jκ.
Согласно одному аспекту предусмотрен грызун (например, мышь), полученный с использованием описанного в настоящем документе нацеливающего вектора, нуклеотидного конструкта или клетки.
Согласно одному аспекту предусмотрено применение описанного в настоящем документе нацеливающего вектора для получения грызуна (например, мыши) или клетки (например, ES клетки мыши, фибробласта мыши и т.д.). Согласно одному варианту осуществления нацеливающий вектор содержит геномный фрагмент человека, который содержит нереаранжированные генные сегменты иммуноглобулина. Согласно конкретному варианту осуществления нереаранжированные генные сегменты иммуноглобулина включают в себя генные сегменты V и J. Согласно конкретному варианту осуществления нереаранжированные генные сегменты иммуноглобулина включают в себя генные сегменты V, D и J.
Согласно одному аспекту предусмотрен конструкт нуклеиновой кислоты, содержащий вышележащее гомологичное плечо и нижележащее гомологичное плечо, причем вышележащее гомологичное плечо содержит последовательность, которая идентична или по существу идентична последовательности вариабельной области тяжелой цепи иммуноглобулина человека, нижележащее гомологичное плечо содержит последовательность, которая идентична или по существу идентична последовательности вариабельной области иммуноглобулина человека или мыши, и между вышележащим и нижележащим гомологичными плечами расположена последовательность, которая содержит нуклеотидную последовательность, кодирующую белок ADAM6 мыши. Согласно конкретному варианту осуществления последовательность, кодирующая ген ADAM6 мыши, функционально связана с промотором мыши, с которым ADAM6 мыши связан у мыши дикого типа.
Согласно одному аспекту предусмотрен нацеливающий вектор, содержащий (a) нуклеотидную последовательность, которая идентична или по существу идентична нуклеотидной последовательности генного сегмента вариабельной области человека; и (b) нуклеотидную последовательность, кодирующую ADAM6 мыши или его ортолог или гомолог или фрагмент, который является функциональным у мыши.
Согласно одному варианту осуществления нацеливающий вектор дополнительно содержит промотор, функционально связанный с последовательностью, кодирующей ADAM6 мыши. Согласно конкретному варианту осуществления промотор представляет собой промотор ADAM6 мыши.
Согласно одному аспекту предусмотрен нуклеотидный конструкт для модификации вариабельного локуса тяжелой цепи иммуноглобулина мыши, причем конструкт содержит по меньшей мере один сайт распознавания сайт-специфической рекомбиназы и последовательность, кодирующую белок ADAM6 или его ортолог или гомолог или фрагмент, который является функциональным у мыши.
Согласно одному аспекту предусмотрен конструкт нуклеиновой кислоты, который содержит генный сегмент DH человека, расположенный бок о бок против и по ходу транскрипции с содержащей 23-мерный спейсер RSS. Согласно конкретному варианту осуществления конструкт нуклеиновой кислоты содержит гомологичное плечо, которое является гомологичным геномной последовательности человека, содержащей генные сегменты Vκ человека. Согласно одному варианту осуществления нацеливающий конструкт содержит все или по существу все генные сегменты DH человека, каждый из которых расположенный бок о бок против и по ходу транскрипции с содержащей 23-мерный спейсер RSS.
Согласно одному аспекту предусмотрен конструкт нуклеиновой кислоты, который содержит генный сегмент Jκ человека, расположенный бок о бок против хода транскрипции с содержащей 12-мерный спейсер RSS. Согласно конкретному варианту осуществления конструкт нуклеиновой кислоты содержит первое гомологичное плечо, которое содержит гомологию с геномной генной последовательностью DH человека, которая расположена бок о бок против и по ходу транскрипции с содержащей 23-мерный спейсер RSS. Согласно одному варианту осуществления конструкт нуклеиновой кислоты содержит второе гомологичное плечо, которое содержит гомологию с геномной генной последовательностью J человека или которое содержит гомологию с последовательностью константной области тяжелой цепи мыши или которое содержит гомологию с межгенной последовательностью J-C выше относительно последовательности константной области тяжелая цепь мыши.
Согласно одному аспекту предусмотрен конструкт нуклеиновой кислоты, который содержит сегмент Vλ человека, расположенный бок о бок по ходу транскрипции с содержащей 23-мерный спейсер RSS, сегмент DH человека, расположенный бок о бок против и по ходу транскрипции с содержащей 12-мерный спейсер RSS, и сегмент J человека, выбранный из сегмента Jκ, расположенного бок о бок против хода транскрипции с содержащей 23-мерный спейсер RSS, сегмента Jλ человека, расположенного бок о бок против хода транскрипции с содержащей 23-мерный спейсер RSS, и сегмента JH человека, расположенного бок о бок против хода транскрипции с содержащей 23-мерный спейсер RSS. Согласно одному варианту осуществления конструкт содержит гомологичное плечо, которое содержит гомологию с последовательностью константной области мыши, межгенной последовательностью J-C мыши и/или последовательностью Vλ человека.
Согласно одному варианту осуществления конструкт нуклеиновой кислоты содержит последовательность вариабельной области λ легкой цепи человека, которая содержит фрагмент кластера A локуса λ легкой цепи человека. Согласно конкретному варианту осуществления фрагмент кластера A локуса λ легкой цепи человека продолжается от hVλ3-27 до hVλ3-1.
Согласно одному варианту осуществления конструкт нуклеиновой кислоты содержит последовательность вариабельной области λ легкой цепи человека, которая содержит фрагмент кластера B локуса λ легкой цепи человека. Согласно конкретному варианту осуществления фрагмент кластера B локуса λ легкой цепи человека продолжается от hVλ5-52 до hVλ1-40.
Согласно одному варианту осуществления конструкт нуклеиновой кислоты содержит последовательность вариабельной области λ легкой цепи человека, которая содержит геномный фрагмент кластера A и геномный фрагмент кластера B. Согласно одному варианту осуществления последовательность вариабельной области λ легкой цепи человека содержит по меньшей мере один генный сегмент кластера A и по меньшей мере один генный сегмент кластера B.
Согласно одному варианту осуществления последовательность вариабельной области λ легкой цепи человека содержит по меньшей мере один генный сегмент кластера B и по меньшей мере один генный сегмент кластера C.
Согласно одному аспекту предусмотрен конструкт нуклеиновой кислоты, содержащий сегмент DH человека, расположенный бок о бок против и по ходу транскрипции с содержащей 23-мерный спейсер RSS, в норме встречающейся в природе, фланкирующей либо сегмент Jκ, JH, Vλ, либо сегмент VH. Согласно одному варианту осуществления конструкт нуклеиновой кислоты содержит первое гомологичное плечо, гомологичное межгенной области V-J человека или гомологичное геномной последовательности человека, содержащей генный сегмент V человека. Согласно одному варианту осуществления конструкт нуклеиновой кислоты содержит второе гомологичное плечо, гомологичное последовательности константной области тяжелой цепи человека или мыши. Согласно конкретному варианту осуществления последовательность константной области тяжелой цепи человека или мыши выбрана из CH1, шарнира, CH2, CH3 и их комбинации. Согласно одному варианту осуществления конструкт нуклеиновой кислоты содержит генный сегмент J человека, фланкированный против хода транскрипции 12-мерной RSS. Согласно одному варианту осуществления конструкт нуклеиновой кислоты содержит второе гомологичное плечо, которое содержит гомологию с генным сегментом J, фланкированным против хода транскрипции 12-мерной RSS. Согласно одному варианту осуществления генный сегмент J выбран из Jκ человека, Jλ человека и JH человека.
Согласно одному аспекту предусмотрен конструкт нуклеиновой кислоты, который содержит сегмент DH человека, расположенный бок о бок против и по ходу транскрипции с содержащей 23-мерный спейсер RSS, и последовательность распознавания сайт-специфической рекомбиназы, например, такую последовательность, распознаваемую сайт-специфической рекомбиназой, как белок Cre, Flp или Dre.
Согласно одному аспекту предусмотрен конструкт нуклеиновой кислоты, который содержит сегмент Vλ человека или сегмент Vκ человека, сегмент DH, расположенный бок о бок против и по ходу транскрипции с содержащей 12-мерный или 23-мерный спейсер RSS, и сегмент J человека с содержащей 12-мерный или 23-мерный спейсер RSS, причем содержащая 12-мерный или 23-мерный спейсер RSS расположена непосредственно 5’ по отношению к сегменту J человека (т.е. относительно направления транскрипции). Согласно одному варианту осуществления конструкт содержит Vλ человека, расположенный бок о бок с 3’ содержащей 23-мерный спейсер RSS, сегмент DH человека, расположенный бок о бок против и по ходу транскрипции с содержащей 12-мерный спейсер RSS, и сегмент Jκ человека, расположенный бок о бок с 5’ содержащей 23-мерный спейсер RSS. Согласно одному варианту осуществления конструкт содержит Vκ человека, расположенный бок о бок с 3’ содержащей 12-мерный спейсер RSS, сегмент DH человека, расположенный бок о бок против и по ходу транскрипции с содержащей 23-мерный спейсер RSS, и сегмент Jλ человека, расположенный бок о бок с 5’ содержащей 12-мерный спейсер RSS.
Согласно одному аспекту предусмотрен нацеливающий вектор, содержащий (a) первое нацеливающее плечо и второе нацеливающее плечо, причем первое и второе нацеливающие плечи независимо выбраны из нацеливающих плеч человека и мыши, причем нацеливающие плечи направляют вектор на эндогенный или модифицированный генный локус области V иммуноглобулина; и, (b) смежную последовательность генных сегментов VL человека или смежную последовательность генных сегментов VL человека и по меньшей мере одного генного сегмента Jκ человека, причем смежная последовательность выбрана из группы, состоящей из (i) hVκ4-1 - hVκ1-6 и Jκ 1, (ii) hVκ4-1 - hVκ1-6 и Jκ1 - Jκ 2, (iii) hVκ4-1 - hVκ1-6 и Jκ1 - Jκ3, (iv) hVκ4-1 - hVκ1-6 и Jκ 1 - Jκ 4, (v) hVκ4-1 - hVκ1-6 и Jκ1 - Jκ5, (vi) hVκ3-7 - hVκ1-16, (vii) hVκ1-17 - hVκ2-30, (viii) hVκ3-31 - hVκ2-40, и (ix) их комбинации.
Согласно одному варианту осуществления нацеливающие плечи, которые направляют вектор на эндогенный или модифицированный локус иммуноглобулина, являются идентичными или по существу идентичными последовательности на эндогенном или модифицированном локусе иммуноглобулина.
Согласно одному аспекту предусмотрен способ получения генетически модифицированной мыши, предусматривающий замещение одного или нескольких генных сегментов тяжелой цепи иммуноглобулина выше (относительно направления транскрипции генных сегментов тяжелой цепи иммуноглобулина) эндогенного локуса ADAM6 мыши одним или несколькими генными сегментами легкой цепи и/или тяжелой цепи иммуноглобулина человека, и замещение одного или нескольких генных сегментов иммуноглобулина ниже (относительно направления транскрипции генных сегментов тяжелой цепи иммуноглобулина) локуса ADAM6 мыши одним или несколькими генными сегментами тяжелой цепи или легкой цепи иммуноглобулина человека. Согласно одному варианту осуществления один или несколько генных сегментов иммуноглобулина человека, замещающие один или несколько эндогенных генных сегментов иммуноглобулина выше эндогенного локуса ADAM6 мыши, включают в себя генные сегменты V. Согласно одному варианту осуществления генные сегменты иммуноглобулина человека, замещающие один или несколько эндогенных генных сегментов иммуноглобулина выше эндогенного локуса ADAM6 мыши, включают в себя генные сегменты V и D. Согласно одному варианту осуществления один или несколько генных сегментов иммуноглобулина человека, замещающие один или несколько эндогенных генных сегментов иммуноглобулина ниже эндогенного локуса ADAM6 мыши, включают в себя генные сегменты J. Согласно одному варианту осуществления один или несколько генных сегментов иммуноглобулина человека, замещающие один или несколько эндогенных генных сегментов иммуноглобулина ниже эндогенного локуса ADAM6 мыши, включают в себя генные сегменты D и J. Согласно одному варианту осуществления один или несколько генных сегментов иммуноглобулина человека, замещающие один или несколько эндогенных генных сегментов иммуноглобулина ниже эндогенного локуса ADAM6 мыши, включают в себя генные сегменты V, D и J.
Согласно одному аспекту предусмотрен способ получения генетически модифицированной мыши, предусматривающий замещение одного или нескольких генных сегментов тяжелой цепи иммуноглобулина выше (относительно направления транскрипции генных сегментов тяжелой цепи иммуноглобулина) эндогенного локуса ADAM6 мыши одним или нескольким генными сегментами легкой цепи иммуноглобулина человека, и замещение одного или нескольких генных сегментов иммуноглобулина ниже (относительно направления транскрипции генных сегментов тяжелой цепи иммуноглобулина) локуса ADAM6 мыши одним или несколькими сегментами легкой цепи иммуноглобулина человека. Согласно одному варианту осуществления один или несколько генных сегментов иммуноглобулина человека, замещающие один или несколько эндогенных генных сегментов иммуноглобулина выше эндогенного локуса ADAM6 мыши, включают в себя генные сегменты V. Согласно одному варианту осуществления генные сегменты иммуноглобулина человека, замещающие один или несколько эндогенных генных сегментов иммуноглобулина выше эндогенного локуса ADAM6 мыши, включают в себя генные сегменты V и J. Согласно одному варианту осуществления один или несколько генных сегментов иммуноглобулина человека, замещающие один или несколько эндогенных генных сегментов иммуноглобулина ниже эндогенного локуса ADAM6 мыши, включают в себя генные сегменты J. Согласно одному варианту осуществления один или несколько генных сегментов иммуноглобулина человека, замещающие один или несколько эндогенных генных сегментов иммуноглобулина ниже эндогенного локуса ADAM6 мыши, включают в себя генные сегменты V и J. Согласно одному варианту осуществления один или несколько генных сегментов иммуноглобулина человека, замещающие один или несколько эндогенных генных сегментов иммуноглобулина ниже эндогенного локуса ADAM6 мыши, включают в себя генные сегменты V.
Согласно конкретному варианту осуществления генные сегменты V представляют собой генные сегменты VL. Согласно другому конкретному варианту осуществления генные сегменты J представляют собой генные сегменты JL.
Согласно одному варианту осуществления один или несколько генных сегментов тяжелой цепи иммуноглобулина выше и/или ниже гена ADAM6 замещают в плюрипотентной, индуцированной плюрипотентной или тотипотентной клетке для получения генетически модифицированной клетки - предшественника; генетически модифицированную клетку - предшественник вводят хозяину; и осуществляют вынашивание хозяина, содержащего генетически модифицированную клетку - предшественник, для получения мыши, содержащей геном, полученный из генетически модифицированной клетки - предшественника. Согласно одному варианту осуществления хозяин представляет собой зародыш. Согласно конкретному варианту осуществления хозяин выбран из пре-морулы мыши (например, 8- или 4-клеточная стадия), тетраплоидного зародыша, агрегата эмбриональных клеток или бластоцисты.
Согласно одному аспекту предусмотрен способ получения генетически модифицированной мыши, предусматривающий замещение нуклеотидной последовательности мыши, которая содержит нуклеотидную последовательность генного сегмента иммуноглобулина мыши и ADAM6 мыши (или его ортолога или гомолога или фрагмента, функционального у самца мыши) последовательностью, содержащей генный сегмент иммуноглобулина человека для образования первого химерного локуса, затем вставку последовательности, содержащей кодирующую ADAM6 мыши последовательность (или последовательность, кодирующую его ортолог или гомолог или функциональный фрагмент), в последовательность, содержащую генный сегмент иммуноглобулина человека для образования второго химерного локуса.
Согласно одному варианту осуществления второй химерный локус содержит вариабельный генный сегмент тяжелой цепи иммуноглобулина человека (VH). Согласно одному варианту осуществления второй химерный локус содержит вариабельный генный сегмент легкой цепи иммуноглобулина человека (VL). Согласно конкретному варианту осуществления второй химерный локус содержит генный сегмент VH человека или генный сегмент VL человека, функционально связанный с генным сегментом DH человека и генным сегментом JH человека. Согласно конкретному варианту осуществления второй химерный локус содержит генный сегмент VL человека, функционально связанный с генным сегментом JH человека или генным сегментом JL человека. Согласно дополнительному конкретному варианту осуществления второй химерный локус функционально связан с третьим химерным локусом, который содержит последовательность CH1 человека или CH1 человека и шарнирную последовательность человека, слитые с последовательностью CH2 + CH3 мыши.
Согласно одному аспекту предусмотрен способ модификации локуса тяжелой цепи иммуноглобулина мыши, предусматривающий: (a) осуществление первой модификации локуса тяжелой цепи иммуноглобулина мыши, которая приводит к снижению или устранению эндогенной активности ADAM6 мыши у самца мыши; и, (b) осуществление второй модификации для добавления последовательности нуклеиновой кислоты, которая предоставляет мыши активность ADAM6, которая является функциональной у самца мыши.
Согласно одному варианту осуществления первая модификация предусматривает добавление последовательности иммуноглобулина человека или замещение последовательности иммуноглобулина мыши последовательностью иммуноглобулина человека.
Согласно одному варианту осуществления последовательность иммуноглобулина человека представляет собой последовательность тяжелой цепи. Согласно одному варианту осуществления последовательность иммуноглобулина человека представляет собой последовательность легкой цепи.
Согласно одному варианту осуществления первую и вторую модификацию производят в отдельной ES клетке, и отдельную ES клетку вводят в зародыш - хозяин для получения мыши.
Согласно одному аспекту предусмотрено потомство скрещивания описанной в настоящем документе мыши со второй мышью, которая представляет собой мышь дикого типа или генетически модифицированную мышь.
Согласно одному аспекту предусмотрено применение мыши, которая содержит эктопическую нуклеотидную последовательность, содержащую локус или последовательность ADAM6 мыши для получения фертильного самца мыши, причем применение предусматривает спаривание мыши, содержащей эктопическую нуклеотидную последовательность, которая содержит локус или последовательность ADAM6 мыши, с мышью, которая не содержит функциональный эндогенный локус или последовательность ADAM6, и получение потомства, которое представляет собой самку, способную производить потомство, содержащее эктопический локус или последовательность ADAM6, или которое представляет собой самца, который содержит эктопический локус или последовательность ADAM6, и самец проявляет фертильность, которая является приблизительно такой же, как и фертильность, проявляемая самцом мыши дикого типа.
Согласно одному аспекту предусмотрено применение описанной в настоящем документе мыши для введения эктопической последовательности ADAM6 в мышь, которая не содержит функциональную эндогенную последовательность ADAM6, причем применение предусматривает скрещивание описанной в настоящем документе мыши с мышью, которая не содержит функциональную эндогенную последовательность ADAM6.
Согласно одному аспекту предусмотрено применение генетического материала от описанной в настоящем документе мыши для получения мыши, содержащей эктопическую последовательность ADAM6. Согласно одному варианту осуществления применение предусматривает ядерный транспорт с использованием ядра клетки описанной в настоящем документе мыши. Согласно одному варианту осуществления применение предусматривает клонирование клетки описанной в настоящем документе мыши для получения животного, полученного из клетки. Согласно одному варианту осуществления применение предусматривает использование спермы или яйцеклетки описанной в настоящем документе мыши в способе получения мыши, содержащей эктопическую последовательность ADAM6.
Согласно одному аспекту предусмотрен способ получения фертильного самца мыши, содержащего модифицированный локус тяжелой цепи иммуноглобулина, предусматривающий оплодотворение первой половой клетки мыши, которая содержит модификацию эндогенного локуса тяжелой цепи иммуноглобулина, второй половой клеткой мыши, которая содержит ген ADAM6 или его ортолог или гомолог или фрагмент, который является функциональным у самца мыши; образование оплодотворенной клетки; предоставление возможности оплодотворенной клетке развиться в зародыш; и вынашивание зародыша в суррогатной матери для получения мыши.
Согласно одному варианту осуществления оплодотворение достигается путем спаривания самца мыши и самки мыши. Согласно одному варианту осуществления самка мыши содержит ген ADAM6 или его ортолог или гомолог или фрагмент. Согласно одному варианту осуществления самец мыши содержит ген ADAM6 или его ортолог или гомолог или фрагмент.
Согласно одному аспекту предусмотрено применение последовательности нуклеиновой кислоты, кодирующей белок ADAM6 мыши или его ортолог или гомолог или функциональный фрагмент соответствующего белка ADAM6, для восстановления или усиления фертильности мыши с геномом, содержащим модификацию локуса тяжелой цепи иммуноглобулина, причем модификация снижает или устраняет эндогенную функцию ADAM6.
Согласно одному варианту осуществления последовательность нуклеиновой кислоты интегрирована в геном мыши в эктопическом положении. Согласно одному варианту осуществления последовательность нуклеиновой кислоты интегрирована в геном мыши на эндогенном локусе иммуноглобулина. Согласно конкретному варианту осуществления эндогенный локус иммуноглобулина представляет собой локус тяжелой цепи. Согласно одному варианту осуществления последовательность нуклеиновой кислоты интегрирована в геном мыши в положении, отличном от эндогенного локуса иммуноглобулина.
Согласно одному аспекту предусмотрен способ получения генетически модифицированной мыши, предусматривающий замещение на эндогенном локусе тяжелой цепи одного или нескольких генных сегментов тяжелой цепи иммуноглобулина мыши одним или несколькими генными сегментами легкой цепи иммуноглобулина человека. Согласно одному варианту осуществления происходит замещение всех или по существу всех функциональных сегментов тяжелой цепи иммуноглобулина мыши (т.е. сегментов VH, DH и JH) одним или несколькими функциональными сегментами легкой цепи человека (т.е. сегментами VL и JL). Согласно одному варианту осуществления происходит замещение всех или по существу всех функциональных сегменты VH, DH и JH тяжелой цепи мыши всеми или по существу всеми сегментами Vλ или Vκ человека и по меньшей мере одним сегментом Jλ или Jκ. Согласно конкретному варианту осуществления замещение включает в себя все или по существу все функциональные сегменты Jλ или Jκ человека.
Согласно одному аспекту предусмотрен способ получения мыши, которая экспрессирует полипептид, который содержит последовательность, происходящую из сегмента Vλ или Vκ и/или Jλ или Jκ иммуноглобулина человека, слитого с константной областью тяжелой цепи мыши, содержащей замещение эндогенных вариабельных сегментов тяжелой цепи иммуноглобулина (VH, DH и JH) по меньшей мере одним сегментом Vλ или Vκ человека и по меньшей мере одним сегментом Jλ или Jκ человека, причем замещение происходит в плюрипотентной, индуцированной плюрипотентной или тотипотентной клетке мыши для образования генетически модифицированной клетке - предшественнике мыши; генетически модифицированную клетку - предшественник мыши вводят в мышь - хозяин; и, мышь - хозяина, содержащую генетически модифицированную клетку - предшественник, вынашивают для получения мыши, содержащей геном, полученный из генетически модифицированной клетки - предшественника. Согласно одному варианту осуществления хозяин представляет собой зародыш. Согласно конкретному варианту осуществления хозяин выбран из пре-морулы мыши (например, 8- или 4-клеточная стадия), тетраплоидного зародыша, агрегата эмбриональных клеток или бластоцисты.
Согласно одному аспекту предусмотрен способ получения описанной в настоящем документе генетически модифицированной мыши, предусматривающий введение путем ядерного транспорта нуклеиновой кислоты, содержащей описанную в настоящем документе модификацию, в клетку, и содержание клетки в подходящих условиях (например, включающих в себя культивирование клетки и вынашивание зародыша, содержащего клетку в организме суррогатной матери) для развития в описанную в настоящем документе мышь.
Согласно одному аспекту предусмотрен способ получения модифицированной мыши, предусматривающий описанную в настоящем документе модификацию ES клетки мыши или плюрипотентной или тотипотентной или индуцированной плюрипотентной клетки мыши для введения одного или нескольких нереаранжированных вариабельных генных сегментов легкой цепи иммуноглобулина, функционально связанных с константной последовательностью тяжелой цепи иммуноглобулина, культивирование ES клетки, введение культивированной ES клетки в зародыш - хозяин для образования химерного зародыша, и введение химерного зародыша в подходящую мышь - хозяин для развития в модифицированную мышь. Согласно одному варианту осуществления один или несколько нереаранжированных генных сегментов вариабельной области легкой цепи иммуноглобулина представляют собой λ генные сегменты человека или κ генные сегменты человека. Согласно одному варианту осуществления один или несколько нереаранжированные генных сегментов вариабельной области легкой цепи иммуноглобулина содержат сегменты Vλ человека или сегменты Vκ человека и один или несколько сегментов Jλ, Jκ или JH. Согласно одному варианту осуществления константная генная последовательность тяжелой цепи представляет собой последовательность человека, выбранную из CH1, шарнира, CH2, CH3 и их комбинации. Согласно одному варианту осуществления один или несколько нереаранжированных вариабельных генных сегментов легкой цепи иммуноглобулина замещают все или по существу все функциональные эндогенные генные сегменты вариабельной области тяжелой цепи на эндогенном локусе тяжелой цепи, и константная последовательность тяжелой цепи представляет собой последовательность мыши, содержащую CH1, шарнир, CH2 и CH3.
Согласно одному аспекту предусмотрены конструкты нуклеиновых кислот, клетки, зародыши, мыши и способы получения белков, которые содержат один или несколько последовательностей вариабельной области κ и/или λ легкой цепи иммуноглобулина и последовательность константной области тяжелой цепи иммуноглобулина, включая в себя белки, которые содержат вариабельный домен λ или κ легкой цепи человека и последовательность константной области тяжелой цепи человека или мыши.
Согласно одному аспекту предусмотрена нуклеотидная последовательность, кодирующая вариабельную область иммуноглобулина, полученной у описанный в настоящем документе мыши.
Согласно одному аспекту предусмотрена аминокислотная последовательность вариабельной области тяжелой цепи или легкой цепи антитела, образованного у описанной в настоящем документе мыши.
Согласно одному аспекту предусмотрена нуклеотидная последовательность вариабельной области тяжелой цепи иммуноглобулина или легкой цепи иммуноглобулина, кодирующая вариабельную область антитела, образованного у описанной в настоящем документе мыши.
Согласно одному аспекту предусмотрено антитело или его антигенсвязывающий фрагмент (например, Fab, F(ab)2, scFv), образованный у описанной в настоящем документе мыши.
Согласно одному аспекту описаны связывающие белки, которые содержат вариабельные домены иммуноглобулина, которые происходят из вариабельных доменов легкой цепи (т.е. каппа (κ) и/или лямбда ((λ)) иммуноглобулина, но не из полноразмерных вариабельных доменов тяжелой цепи иммуноглобулина. Также предусмотрены способы и композиции для получения связывающих белков, включая в себя генетически модифицированных мышей.
Согласно одному аспекту предусмотрен способ получения антигенсвязывающего белка, который не содержит вариабельный домен тяжелой цепи иммуноглобулина, предусматривающий стадию иммунизации описанной в настоящем документе мыши представляющим интерес антигеном, и содержание мыши в условиях, которые позволяют ей произвести антигенсвязывающий белок, который специфически связывает представляющий интерес антиген.
Согласно одному аспекту предусмотрен способ получения антигенсвязывающего белка, который содержит первый вариабельный домен легкой цепи иммуноглобулина, смежный с последовательностью константной области тяжелой цепи, и второй вариабельный домен легкой цепи иммуноглобулина, который является смежным с константной последовательностью легкой цепи, предусматривающий стадию иммунизации описанной в настоящем документе мыши представляющим интерес антигеном, и содержание мыши в условиях, которые позволяют ей производить антигенсвязывающий белок, который специфически связывает представляющий интерес антиген.
Согласно одному аспекту предусмотрен способ получения антигенсвязывающего белка, который содержит вариабельный домен легкой цепи иммуноглобулина, смежный с последовательностью константной области тяжелой цепи, причем антигенсвязывающий белок не содержит легкую цепь иммуноглобулина, предусматривающий стадию иммунизации описанной в настоящем документе мыши, которая содержит нокаут эндогенного κ и λ локуса (или которая содержит нефункциональный эндогенный κ и/или λ локус) представляющим интерес антигеном, и содержание мыши в условиях, которые позволяют ей производить антигенсвязывающий белок, который специфически связывает представляющий интерес антиген.
Согласно одному аспекту предусмотрено применение описанной в настоящем документе мыши для получения нуклеотидной последовательности вариабельной области иммуноглобулина. Согласно одному варианту осуществления нуклеотидная последовательность вариабельной области содержит реаранжированные генные сегменты VL, DH и JL. Согласно одному варианту осуществления нуклеотидная последовательность вариабельной области содержит реаранжированные генные сегменты VL, DH и JH. Согласно одному варианту осуществления нуклеотидная последовательность вариабельной области содержит реаранжированные генные сегменты Vκ и Jκ. Согласно одному варианту осуществления нуклеотидная последовательность вариабельной области содержит реаранжированные генные сегменты Vλ и Jλ.
Согласно одному аспекту предусмотрено применение описанной в настоящем документе мыши для получения полностью человеческого Fab или полностью человеческого F(ab)2. Согласно одному варианту осуществления полностью человеческий Fab или полностью человеческий F(ab)2 содержит реаранжированные генные сегменты VL, DH и JL. Согласно одному варианту осуществления полностью человеческий Fab или полностью человеческий F(ab)2 содержит реаранжированные генные сегменты VL, DH и JH. Согласно одному варианту осуществления полностью человеческий Fab или полностью человеческий F(ab)2 содержит реаранжированные генные сегменты Vκ и Jκ. Согласно одному варианту осуществления полностью человеческий Fab или полностью человеческий F(ab)2 содержит реаранжированные генные сегменты Vλ и Jλ.
Согласно одному аспекту предусмотрено применение описанной в настоящем документе мыши для получения полностью человеческого Fab (содержащего первый VL человека, слитый с константной областью легкой цепи человека, и второй VL человека, слитый с последовательностью константной области тяжелой цепи человека) или полностью человеческого F(ab)2.
Согласно одному аспекту предусмотрено применение описанной в настоящем документе мыши для получения иммортализованной клеточная линия. Согласно одному варианту осуществления иммортализованная клеточная линия содержит последовательность нуклеиновой кислоты, кодирующую домен Vλ или Vκ человека, функционально связанный с последовательностью нуклеиновой кислоты, которая содержит последовательность нуклеиновой кислоты константной области мыши. Согласно одному варианту осуществления иммортализованная клеточная линия экспрессирует антитело, которое содержит вариабельный домен человека. Согласно конкретному варианту осуществления вариабельный домен человека представляет собой вариабельный домен легкой цепи.
Согласно одному аспекту предусмотрено применение описанной в настоящем документе мыши для получения гибридомы или квадромы. Согласно одному варианту осуществления гибридома или квадрома экспрессирует полипептид, содержащий вариабельный домен человека, который связывает представляющий интерес антиген.
Согласно одному аспекту предусмотрено применение описанной в настоящем документе мыши для получения фаговой библиотеки, содержащей вариабельные области легкой цепи человека. Согласно одному варианту осуществления вариабельные области легкой цепи представляют собой области Vκ человека.
Согласно одному аспекту предусмотрено применение описанной в настоящем документе мыши для создания последовательности вариабельной области для получения антигенсвязывающего белка человека, предусматривающее (a) иммунизацию описанной в настоящем документе мыши представляющим интерес антигеном, (b) выделение лимфоцита из иммунизированной мыши согласно (a), (c) воздействие на лимфоцит одного или нескольких меченых антител, (d) идентификацию лимфоцита, который способен связываться с представляющим интерес антигеном, и (e) амплификацию одной или нескольких последовательностей нуклеиновой кислоты вариабельной области легкой цепи человека из лимфоцита, тем самым создавая последовательность вариабельной области.
Согласно одному варианту осуществления лимфоцит получен из селезенки мыши. Согласно одному варианту осуществления лимфоцит получен из лимфатического узла мыши. Согласно одному варианту осуществления лимфоцит получен из костного мозга мыши.
Согласно одному варианту осуществления меченое антитело представляет собой конъюгированное с флуорофором антитело. Согласно одному варианту осуществления одно или несколько конъюгированных с флуорофором антител выбраны из IgM, IgG и/или их комбинации.
Согласно одному варианту осуществления лимфоцит представляет собой B-клетку.
Согласно одному варианту осуществления одна или несколько последовательностей нуклеиновой кислоты вариабельной области содержат последовательность вариабельной области легкой цепи. Согласно конкретному варианту осуществления последовательность вариабельной области легкой цепи представляет собой последовательность вариабельной области κ легкой цепи иммуноглобулина. Согласно одному варианту осуществления одна или несколько последовательностей вариабельной области нуклеиновой кислоты представляют собой последовательность вариабельной области λ легкой цепи.
Согласно одному варианту осуществления предусмотрено применение описанной в настоящем документе мыши для создания одной или нескольких последовательностей вариабельной области κ легкой цепи для получения антигенсвязывающего белка человека, предусматривающее (a) иммунизацию описанной в настоящем документе мыши представляющим интерес антигеном, (b) выделение селезенки из иммунизированной мыши согласно (a), (c) воздействие на B-лимфоциты из селезенки одного или нескольких меченых антител, (d) идентификацию B-лимфоцита согласно (c), который способен связываться с представляющим интерес антигеном, и (e) амплификацию последовательности нуклеиновой кислоты вариабельной области легкой цепи из B-лимфоцита, тем самым создавая последовательность вариабельной области κ легкой цепи.
Согласно одному варианту осуществления предусмотрено применение описанной в настоящем документе мыши для создания последовательности вариабельной области κ легкой цепи для получения антигенсвязывающего белка человека, предусматривающее (a) иммунизацию описанной в настоящем документе мыши представляющим интерес антигеном, (b) выделение одного или нескольких лимфатических узлов из иммунизированной мыши согласно (a), (c) воздействие на B-лимфоциты из одного или нескольких лимфатических узлов одного или нескольких меченых антител, (d) идентификацию В-лимфоцита согласно (c), который способен связываться с представляющим интерес антигеном, и (e) амплификацию последовательности нуклеиновой кислоты вариабельной области κ легкой цепи из B-лимфоцита, тем самым создавая последовательность вариабельной области κ легкой цепи.
Согласно одному варианту осуществления предусмотрено применение описанной в настоящем документе мыши для создания последовательности вариабельной области κ легкой цепи для получения антигенсвязывающего белка человека, предусматривающее (a) иммунизацию описанной в настоящем документе мыши представляющим интерес антигеном, (b) выделение костного мозга из иммунизированной мыши согласно (a), (c) воздействие на B-лимфоциты из костного мозга одного или нескольких меченых антител, (d) идентификацию B-лимфоцита согласно (c), который способен связываться с представляющим интерес антигеном, и (e) амплификацию последовательности нуклеиновой кислоты вариабельной области κ легкой цепи из B-лимфоцита, тем самым создавая последовательность вариабельной области κ легкой цепи. Согласно различным вариантам осуществления одно или несколько меченых антител выбраны из IgM, IgG и/или их комбинации.
Согласно различным вариантам осуществления предусмотрено применение описанной в настоящем документе мыши для создания последовательности вариабельной области κ легкой цепи для получения антигенсвязывающего белка человека, дополнительно предусматривающее слияние амплифицированной последовательности вариабельной области легкой цепи с последовательностями константной области тяжелой или легкой цепи человека и необязательно последовательностью вариабельной области тяжелой цепи человека, экспрессию слитых последовательностей в клетке и выделение экспрессированных последовательностей, тем самым создавая антигенсвязывающий белок человека.
Согласно различным вариантам осуществления константные области тяжелой цепи человека выбраны из IgM, IgD, IgA, IgE и IgG. Согласно различным конкретным вариантам осуществления IgG выбран из IgG1, IgG2, IgG3 и IgG4. Согласно различным вариантам осуществления константная область тяжелой цепи человека содержит CH1, шарнир, CH2, CH3, CH4 или их комбинацию. Согласно различным вариантам осуществления константная область легкой цепи представляет собой κ константную область иммуноглобулина. Согласно различным вариантам осуществления клетка выбрана из клетки HeLa, клетки DU145, клетки Lncap, клетки MCF-7, клетки MDA-MB-438, клетки PC3, клетки T47D, клетки THP-1, клетки U87, клетки SHSY5Y (нейробластома человека), клетки Saos-2, клетки Vero, клетки CHO, клетки GH3, клетки PC12, ретинальной клетки человека (например, клетки PER.C6™) и клетки MC3T3. Согласно конкретному варианту осуществления клетка представляет собой клетку CHO.
Согласно одному аспекту предусмотрен способ создания вариабельной области легкой цепи человека, специфической к представляющему интерес антигену, предусматривающий стадии иммунизации описанной в настоящем документе мыши антигеном, выделение по меньшей мере одной клетки из мыши, производящей вариабельную область легкой цепи человека, специфическую к антигену, создание по меньшей мере одной клетки, производящей антигенсвязывающий белок человека, содержащий вариабельную область легкой цепи, специфическую к антигену, и культивирование по меньшей мере одной клетки, производящей антигенсвязывающий белок человека, и получение указанного антигенсвязывающего белка человека. Согласно одному варианту осуществления вариабельная область легкой цепи человека представляет собой область Vκ человека.
Согласно различным вариантам осуществления по меньшей мере одна выделенная из мыши клетка, производящая вариабельной области легкой цепи человека, специфическую к антигену, представляет собой спленоцит или B-клетку.
Согласно различным вариантам осуществления антигенсвязывающий белок представляет собой антитело.
Согласно различным вариантам осуществления иммунизацию представляющим интерес антигеном проводят с помощью белка, ДНК, комбинации ДНК и белка или клеток, экспрессирущих антиген.
Согласно одному аспекту предусмотрено применение описанной в настоящем документе мыши для получения последовательности нуклеиновой кислоты, кодирующей вариабельную область иммуноглобулина или ее фрагмент. Согласно одному варианту осуществления последовательность нуклеиновой кислоты применяют для получения антитела человека или его антигенсвязывающего фрагмента. Согласно одному варианту осуществления мышь применяют для получения антигенсвязывающего белка, выбранного из антитела, мультиспецифического антитела (например, биспецифического антитела), scFv, биспецифического scFv, диатела, триатела, тетратела, V-NAR, VHH, VL, F(ab), F(ab)2, DVD (т.е. антигенсвязывающего белка с двойным вариабельным доменом), SVD (т.е. антигенсвязывающего белка с одним вариабельным доменом) или биспецифического проводника T-клеток (BiTE).
Согласно одному аспекту предусмотрено применение описанной в настоящем документе мыши для получения лекарственного средства (например, антигенсвязывающего белка) или для получения последовательности, кодирующей вариабельную последовательность лекарственного средства (например, антигенсвязывающего белка), для лечения заболевания или нарушения человека.
Согласно одному аспекту предусмотрено описанной в настоящем документе применение мыши для получения последовательности нуклеиновой кислоты, кодирующей первую вариабельную последовательность легкой цепи иммуноглобулина человека (VL1), которая является когнатной второй вариабельной последовательности легкой цепи иммуноглобулина человека (VL2), причем VL1, слитая с константной областью легкой цепи иммуноглобулина человека (полипептид 1) экспрессируется с VL2, слитой с константной областью тяжелой цепи иммуноглобулина человека (полипептид 2), в виде димера полипептид 1/полипептид 2, для образования антитела VL1-VL2.
Согласно одному аспекту предусмотрено применение описанной в настоящем документе мыши для получения последовательности нуклеиновой кислоты, кодирующей вариабельную последовательность легкой цепи иммуноглобулина человека, которая слита с последовательностью тяжелой цепи иммуноглобулина человека, причем последовательность нуклеиновой кислоты кодирует полипептид VL-CH человека, причем полипептид VL-CH человека экспрессируется в виде димера, и причем димер экспрессируется при отсутствии легкой цепи иммуноглобулина (например, при отсутствии λ легкой цепи человека или κ легкой цепи человека). Согласно одному варианту осуществления димер VL-CH специфически связывает представляющий интерес антиген при отсутствии λ легкой цепи и при отсутствии κ легкой цепи.
Согласно одному аспекту предусмотрено применение описанной в настоящем документе мыши для получения последовательности нуклеиновой кислоты, кодирующей весь или часть вариабельного домена иммуноглобулина. Согласно одному варианту осуществления вариабельный домен иммуноглобулина представляет собой домен Vλ человека или домен Vκ человека.
Согласно одному аспекту предусмотрено применение описанного в настоящем документе конструкта нуклеиновой кислоты для получения мыши, клетки или терапевтического белка (например, антитела или другого антигенсвязывающего белка).
Согласно одному аспекту предусмотрено применение последовательности нуклеиновой кислоты от описанной в настоящем документе мыши для получения клеточной линии для получения терапевтического средства для человека. Согласно одному варианту осуществления терапевтическое средство для человека представляет собой связывающий белок, содержащий вариабельную последовательность легкой цепи человека (например, происходящую из сегмента Vλ человека или сегмента Vκ человека), слитую с константной последовательностью тяжелой цепи человека. Согласно одному варианту осуществления терапевтическое средство для человека содержит первый полипептид, который представляет собой λ или κ легкую цепь иммуноглобулина человека, и второй полипептид, который содержит вариабельная последовательность Vλ или Vκ человека, слитую с константной последовательностью тяжелой цепи человека.
Согласно одному аспекту предусмотрена система экспрессии, содержащая клетку млекопитающего, трансфектированную с помощью конструкта ДНК, который кодирует полипептид, который содержит соматически мутированный домен VL человека, слитый с доменом CH человека.
Согласно одному варианту осуществления система экспрессии дополнительно содержит нуклеотидную последовательность, которая кодирует домен VL иммуноглобулина, слитый с доменом CL человека, причем домен VL, слитый с доменом CL человека, представляет собой когнатную легкую цепь с доменом VL, слитым с доменом CH человека.
Согласно одному варианту осуществления клетка млекопитающего выбрана из клетки CHO, клетки COS, клетки Vero, клетки 293 и ретинальной клетки, которая экспрессирует вирусный ген (например, клетки PER.C6™).
Согласно одному аспекту предусмотрен способ получения связывающего белка, предусматривающий получение нуклеотидной последовательности, кодирующей домен VL из гена, кодирующего область VL, слитую с областью CH, из клетки описанной в настоящем документе мыши, и клонирование нуклеотидной последовательности, кодирующей последовательность области VL в одной рамке считыванию с геном, кодирующим область CH человека, для образования последовательности связывающего белка человека, экспрессирующей последовательность связывающего белка человека в подходящей клетке.
Согласно одному варианту осуществления мышь была иммунизирована с помощью представляющего интерес антигена, и область VL, слитая с областью CH, специфически связывает (например, с KD в микромолярном, наномолярном или пикомолярном диапазоне) эпитоп представляющего интерес антигена. Согласно одному варианту осуществления нуклеотидная последовательность, кодирующая область VL, слитую с областью CH, является соматически мутированной у мыши.
Согласно одному варианту осуществления подходящая клетка выбрана из B-клетки, гибридомы, квадромы, клетки CHO, клетки COS, клетки 293, клетки HeLa и ретинальной клетки человека, экспрессирующей вирусную последовательность нуклеиновой кислоты (например, клетки PERC.6™).
Согласно одному варианту осуществления область CH содержит изотип IgG человека. Согласно конкретному варианту осуществления IgG человека выбран из IgG1, IgG2 и IgG4. Согласно другому конкретному варианту осуществления IgG человека представляет собой IgG1. Согласно другому конкретному варианту осуществления IgG человека представляет собой IgG4. Согласно другому конкретному варианту осуществления IgG4 человека представляет собой модифицированный IgG4. Согласно одному варианту осуществления модифицированный IgG4 содержит замещение в шарнирной области. Согласно конкретному варианту осуществления модифицированный IgG4 содержит замещение на аминокислотном остатке 228 относительно относящегося к человеку IgG4 дикого типа, пронумерованного согласно индексу нумерации EU по Kabat. Согласно конкретному варианту осуществления замещение на аминокислотном остатке 228 представляет собой S228P замещение, пронумерованного согласно индексу нумерации EU по Kabat.
Согласно одному варианту осуществления клетка дополнительно содержит нуклеотидную последовательность, кодирующую домен VL из легкой цепи, которая является когнатной домену VL, слитому с областью CH, и способ дополнительно предусматривает экспрессию нуклеотидной последовательности, кодирующей когнатный домен VL, слитый с доменом Cκ или человека Cλ.
Согласно одному аспекту предусмотрен способ получения биспецифического антигенсвязывающего белка, предусматривающий воздействие на первую описанную в настоящем документе мышь первого представляющего интерес антигена и идентификация последовательности первого домена VL человека, который специфически связывает первый представляющий интерес антиген; воздействие на вторую описанную в настоящем документе мышь второго представляющего интерес антигена и идентификацию последовательности второго домена VL человека, который специфически связывает второй представляющий интерес антиген; причем первый домен VL человека не связывает второй представляющий интерес антиген, и второй домен VL человека не связывает первый представляющий интерес антиген; и слияние последовательности первого домена VL человека с первой константной последовательностью тяжелой цепи для образования первого антигенсвязывающего полипептида, и слияние последовательности второго домена VL человека со второй константной последовательностью тяжелой цепи для образования второго антигенсвязывающего полипептида; и использование первого и второго антигенсвязывающих полипептидов в биспецифическом связывающем белке.
Согласно одному варианту осуществления антигенсвязывающий белок дополнительно содержит первую легкую цепь иммуноглобулина, которая содержит κ или λ вариабельный домен человека, который является когнатным первому домену VL человека, и вторую легкую цепь иммуноглобулина, которая содержит κ или λ вариабельный домен человека, который является когнатным второму домену VL человека.
Согласно одному варианту осуществления первая константная последовательность тяжелой цепи идентична второй константной последовательности тяжелой цепи. Согласно одному варианту осуществления первый константная последовательность тяжелой цепи содержит модификацию, которая снижает или устраняет связывание первой константной области тяжелой цепи с белком A, и вторая константная последовательность тяжелой цепи связывает белок A.
Согласно одному варианту осуществления первый и второй домены VL человека содержат последовательность, происходящую из генного сегмента Vκ человека и генного сегмента Jκ человека. Согласно одному варианту осуществления первый и второй домены VL человека содержат последовательность, происходящую из генного сегмента Vλ человека и генного сегмента Jλ человека. Согласно одному варианту осуществления первый и второй домены VL человека содержат последовательность, происходящую из генного сегмента DH человека. Согласно одному варианту осуществления первый и второй домены VL человека содержат последовательность, происходящую из генного сегмента JH человека. Согласно конкретному варианту осуществления первый и второй домены VL человека содержат последовательность, происходящую из генного сегмента DH человека и генного сегмента JH человека. Согласно конкретному варианту осуществления первый и второй домены VL человека содержат последовательность, происходящую из генного сегмента DH человека и генного сегмента Jκ человека.
Согласно одному варианту осуществления первый домен VL человека содержит последовательность, происходящую из генного сегмента Vκ человека и генного сегмента Jκ человека, и второй VL домен человека содержит последовательность, происходящую из генного сегмента Vλ человека и генного сегмента Jλ человека. Согласно одному варианту осуществления первый VL домен человека содержит последовательность, происходящую из генного сегмента Vλ человека и генного сегмента Jλ человека, и второй домен VL человека содержит последовательность, происходящую из генного сегмента Vκ человека и генного сегмента Jκ человека.
Согласно одному аспекту предусмотрена вариабельная область иммуноглобулина (VR) (например, содержащая последовательность VL человека, слитую с JL, или JH, или DH и JH, или DH и JL человека), образованная у описанной в настоящем документе мыши. Согласно конкретному варианту осуществления VR иммуноглобулина происходит из зародышевого генного сегмента человека, выбранного из сегмента Vκ и сегмента Vλ, причем VR кодируется реаранжированной последовательностью от мыши, причем реаранжированная последовательность является соматически гипермутированной. Согласно одному варианту осуществления реаранжированная последовательность содержит 1 - 5 соматических гипермутаций. Согласно одному варианту осуществления реаранжированная последовательность содержит по меньшей мере 6, 7, 8, 9 или 10 соматических гипермутаций. Согласно одному варианту осуществления реаранжированная последовательность содержит больше 10 соматических гипермутаций. Согласно одному варианту осуществления реаранжированная последовательность слита с одной или несколькими последовательностями константной области тяжелой цепи человека или мыши (например, выбранными из относящегося к человеку или мыши CH1, шарнира, CH2, CH3 и их комбинации).
Согласно одному аспекту предусмотрена аминокислотная последовательность вариабельного домена иммуноглобулина связывающего белка, образованного у описанной в настоящем документе мыши. Согласно одному варианту осуществления VR слита с одной или несколькими человека или мыши последовательностями константной области тяжелой цепи (например, выбранными из относящегося к человеку или мыши CH1, шарнира, CH2, CH3 и их комбинации).
Согласно одному аспекту предусмотрен вариабельный домен легкой цепи, кодируемый последовательностью нуклеиновой кислоты, происходящей от описанной в настоящем документе мыши.
Согласно одному аспекту предусмотрено антитело или его антигенсвязывающий фрагмент (например, Fab, F(ab)2, scFv), образованный у описанной в настоящем документе мыши, или происходящий из последовательности, образованной у описанной в настоящем документе мыши.
Краткое описание графических материалов
На фиг. 1 показано схематическое изображение (без соблюдения масштаба) локуса тяжелой цепи мыши (вверху) и локуса κ легкой цепи человека (внизу). Локус тяжелой цепи мыши составляет приблизительно 3 миллиона пар нуклеотидов (Mb) в длину и содержит приблизительно 200 вариабельных генных сегментов тяжелой цепи (VH), 13 дополнительных генных сегментов тяжелой цепи (DH) и 4 соединительных генных сегментов тяжелой цепи (JH), а также энхансеры (Enh) и константные области тяжелой цепи (CH). Локус κ легкой цепи человека дуплицируется в дистальные и проксимальные контиги противоположной полярности, охватывающие приблизительно 440 т.п.н. и 600 т.п.н., соответственно. Между двумя контигами находится ДНК, составляющая приблизительно 800 т.п.н., которая, как полагают, не содержит генные сегменты Vκ. Локус κ легкой цепи человека содержит приблизительно 76 генных сегментов Vκ, 5 генных сегментов Jκ, интронный энхансер (Enh) и одну константную область (Cκ).
На фиг. 2 показана иллюстративная стратегия нацеливания для последовательной вставки 40 генных сегментов Vκ человека и пяти генных сегментов Jκ человека в локус тяжелой цепи мыши, дающая в результате модифицированный локус тяжелой цепи иммуноглобулина мыши, содержащий генные сегменты Vκ и Jκ человека, функционально связанные с константными областями тяжелой цепи иммуноглобулина мыши. Гигромициновые (hyg) и неомициновые (neo) кассеты селекции показаны с сайтами распознавания рекомбиназы (R1, R2 и т.д.).
На фиг. 3 показана иллюстративная стратегия нацеливания для последовательной вставки множества генных сегментов Vλ человека и одного генного сегмента Jλ человека в локус тяжелой цепи мыши. Гигромициновые (hyg) и неомициновые (neo) кассеты селекции показаны с сайтами распознавания рекомбиназы (R1, R2 и т.д.).
На фиг. 4 показана иллюстративная стратегия нацеливания для последовательной вставки множества генных сегментов Vλ человека и четырех генных сегментов Jλ человека в локус тяжелой цепи мыши. Гигромициновые (hyg) и неомициновые (neo) кассеты селекции показаны с сайтами распознавания рекомбиназы (R1, R2 и т.д.).
На фиг. 5 показана иллюстративная стратегия нацеливания для последовательной вставки генных сегментов Vλ человека, генных сегментов DH человека и генных сегментов JH человека в локус тяжелой цепи мыши. Гигромициновые (hyg) и неомициновые (neo) кассеты селекции показаны с сайтами распознавания рекомбиназы (R1, R2 и т.д.).
На фиг. 6 показана иллюстративная стратегия нацеливания для последовательной вставки генных сегментов Vλ человека, генных сегментов DH человека и генных сегментов Jκ человека в локус тяжелой цепи мыши. Гигромициновые (hyg) и неомициновые (neo) кассеты селекции показаны с сайтами распознавания рекомбиназы (R1, R2 и т.д.).
На фиг. 7 показаны стадии клонирования геномного фрагмента, кодирующего гены ADAM6 мыши из межгенной области V-D тяжелой цепи иммуноглобулина мыши, и стадии констуирования для модификации геномного фрагмента для вставки в модифицированный локус тяжелой цепи иммуноглобулина.
На фиг. 8 показана стратегия нацеливания для вставки геномного фрагмента, кодирующего гены ADAM6 мыши, в Vκ-Jκ межгенную область модифицированного локуса тяжелой цепи иммуноглобулина мыши, содержащего генные сегменты Vκ и Jκ человека, функционально связанные с константными областями тяжелой цепи иммуноглобулина мыши.
На фиг. 9 показана стратегия нацеливания для вставки геномного фрагмента, кодирующего гены ADAM6 мыши выше (5’) по отношению к генным сегментам Vκ человека (т.е. hVκ2-40) модифицированного локуса тяжелой цепи иммуноглобулина мыши, содержащего генные сегменты Vκ и Jκ человека, функционально связанные с константными областями тяжелой цепи иммуноглобулина мыши.
Подробное раскрытие настоящего изобретения
Настоящее изобретение не ограничивается конкретными способами и описанными экспериментальными условиями, поскольку такие способы и условия могут варьировать. Кроме того, следует понимать, что используемая в настоящем документе терминология предназначена только для цели описания конкретных вариантов осуществления и не предусматривается для ограничения, поскольку объем настоящего изобретения определяется его формулой.
Если не указано иное, все используемые в настоящем документе термины и фразы, включают в себя те значения, которые подразумеваются под терминами и фразами в настоящей области техники, если иное прямо не указано или явно не следует из контекста, в котором используется термин или фраза. Хотя любые способы и материалы, подобные или эквивалентные описанным в настоящем документе, могут быть использованы на практике или при испытании настоящего изобретения, теперь будут описаны конкретные способы и материалы. Все упомянутые публикации включены в настоящий документ посредством ссылки.
Фраза "существенный" или "по существу" при использовании для обозначения количества генных сегментов (например, "по существу все" генные сегменты V) включает в себя как функциональные, так и нефункциональные генные сегменты и включает в себя, согласно различным вариантам осуществления, например, 80% или более, 85% или более, 90% или более, 95% или более 96% или более, 97% или более, 98% или более, или 99% или более всех генных сегментов; согласно различным вариантам осуществления фраза "по существу все" генные сегменты включает в себя, например, по меньшей мере 95%, 96%, 97%, 98% или 99% функциональных (т.е. не относящихся к псевдогенам) генных сегментов.
Термин "замещение" включает в себя помещение последовательности ДНК в геном клетки таким образом, чтобы заменить последовательность в пределах генома гетерологичной последовательностью (например, последовательностью человека у мыши) на локусе геномной последовательности. Помещенная таким образом последовательность ДНК может включать в себя одну или несколько регуляторных последовательностей, которые представляют собой часть исходной ДНК, используемой для получения помещенной таким образом последовательности (например, промоторы, энхансеры, 5’- или 3’- нетранслируемые области, соответствующие последовательности сигналов рекомбинации и т.д.). Например, согласно различным вариантам осуществления замещение представляет собой замену эндогенной последовательности гетерологичной последовательностью, что приводит к производству генного продукта из помещенной таким образом последовательности ДНК (содержащей гетерологичную последовательность), но не экспрессии эндогенной последовательности; замещение эндогенной геномной последовательности последовательностью ДНК, которая кодирует белок, который характеризуется аналогичной функцией, что и белок, кодируемый эндогенной геномной последовательностью (например, эндогенная геномная последовательность кодирует ген или домен иммуноглобулина, и фрагмент ДНК кодирует один или несколько генов или доменов иммуноглобулина человека). Согласно различным вариантам осуществления эндогенный ген или его фрагмент заменяют соответствующим геном человека или его фрагментом. Соответствующий ген человека или его фрагмент представляет собой ген человека или фрагмент, который представляет собой ортолог, гомолог или по существу идентичный или такой же по структуре и/или функции, как эндогенный ген или его фрагмент, который заменен.
Термин "смежный" включает в себя ссылку на расположение на одной молекуле нуклеиновой кислоты, например, две последовательности нуклеиновой кислоты являются "смежными", если они располагаются на одной молекуле нуклеиновой кислоты, но прерываются другой последовательностью нуклеиновой кислоты. Например, реаранжированная последовательность V(D)J является "смежной" с генной последовательностью константной области, хотя за последним кодоном последовательности V(D)J не сразу следует первый кодон последовательности константной области. Согласно другому пример, две последовательности генного сегмента V являются "смежными", если они находятся на одном геномном фрагменте, хотя они могут быть разделены последовательностью, которая не кодирует кодон области V, например, они могут быть разделены регуляторной последовательностью, например, промотором или другой некодирующей последовательностью. Согласно одному варианту осуществления смежная последовательность включает в себя геномный фрагмент, который содержит геномные последовательности, расположенные так, как они встречаются в геноме дикого типа.
Фраза "происходящий из" при использовании в отношении вариабельной области, "происходящей из" указанного гена или генного сегмента, включает в себя способность установить для последовательность конкретный нереаранжированный генный сегмент или генные сегменты, которые были реаранжированы для образования гена, который экспрессирует вариабельный домен (учитывая при необходимости отличия в отношении сплайсинга и соматические мутации).
Фраза "функциональный" при использовании в отношении генного сегмента вариабельной области или соединяющего генного сегмента относится в частоте использования в экспрессируемом репертуаре антитела; например, у людей генные сегменты Vλ 3-1, 4-3, 2-8 и т.д. являются функциональными, тогда как генные сегменты Vλ 3-2, 3-4, 2-5 и т.д. являются нефункциональными.
"Локус тяжелой цепи" включает в себя положение на хромосоме, например, хромосоме мыши, в котором у мыши дикого типа встречаются последовательности ДНК вариабельной области тяжелой цепи (VH), дополнительной области тяжелой цепи (DH), соединяющей области тяжелой цепи (JH) и константной области тяжелой цепи (CH).
Фраза "биспецифический связывающий белок" включает в себя связывающий белок, способный селективно связывать два или больше эпитопов. Биспецифические связывающие белки содержат два различных полипептида, которые содержат первый вариабельный домен легкой цепи (VL1), слитый с первой областью CH, и второй вариабельный домен легкой цепи (VL2), слитый со второй областью CH. Как правило, первая и вторая области CH являются идентичными, или они отличаются на одно или несколько аминокислотных замещений (например, описанных в настоящем документе). VL1 и VL2 специфически связывают различные эпитопы - или на двух различных молекулах (например, антигенах), или на одной и той же молекуле (например, на одном и том же антигене). Если биспецифический связывающий белок селективно связывает два различных эпитопа (первый эпитоп и второй эпитоп), аффинность VL1 для первого эпитопа будет, как правило, по меньшей мере на одну - два или три или четыре порядка величин ниже, чем аффинность VL1 для второго эпитопа, и наоборот в отношении VL2. Эпитопы, распознаваемые биспецифическим связывающим белком, могут находиться на одной и той же или отличной мишени (например, на одном и том же или отличном антигене). Биспецифические связывающие белки могут быть образованы, например, путем комбинации VL1 и VL2, которые распознают различные эпитопы одного и того же антигена. Например, последовательности нуклеиновой кислоты, кодирующие последовательности VL, которые распознают различные эпитопы одного и того же антигена, могут быть слиты с последовательностями нуклеиновой кислоты, кодирующими различные области CH, и такие последовательности могут экспрессироваться в клетке, которая экспрессирует легкую цепь иммуноглобулина, или могут экспрессироваться в клетке, которая не экспрессирует легкую цепь иммуноглобулина. Типичный биспецифический связывающий белок содержит две тяжелых цепи, каждая из которых содержит три CDR легкой цепи, за которыми следует (с N-конца к C-концу) домен CH1, шарнир, домен CH2 и домен CH3, и легкая цепь иммуноглобулина, которая либо не предоставляет антигенсвязывающую специфичность, но которая может ассоциироваться с каждой тяжелой цепью, либо может ассоциироваться с каждой тяжелой цепью и которая может связывать один или несколько эпитопов, связанных VL1 и/или VL2, или которая может ассоциироваться с каждой тяжелой цепью и обеспечивать связывание или содействовать связыванию одной или обеих тяжелых цепей с одним или обоими эпитопами.
Следовательно, два основных типа биспецифических связывающих белков представляют собой (1) VL1-CH (димер) и (2) VL1-CH:легкая цепь + VL2-CH:легкая цепь, причем легкая цепь является одинаковой или различной. В любом случае CH (т.е. константная область тяжелой цепи) может быть дифференциально модифицированной (например, чтобы дифференциально связывать белок A, увеличивать период полужизни в сыворотке и т.д.), как описано в настоящем документе, или может быть одинаковой.
Термин "клетка" при использовании в отношении экспрессии последовательности включает в себя любую клетку, которая является подходящей для экспрессии рекомбинантной последовательности нуклеиновой кислоты. Клетки включают в себя клетки прокариот и эукариот (одноклеточных или многоклеточных), бактериальные клетки (например, штаммы E. coli, Bacillus spp., Streptomyces spp. и т.д.), клетки микобактерий, клетки грибов, дрожжевые клетки (например, S. cerevisiae, S. pombe, P. pastoris, P. methanolica и т.д.), растительные клетки, клетки насекомых (например, SF-9, SF-21, инфицированные бакуловирусом клетки насекомых, Trichoplusia ni и т.д.), клетки отличные от человека животные, клетки человека или такие слияния клеток, как, например, гибридомы или квадромы. Согласно некоторым вариантам осуществления клетка представляет собой клетку человека, мартышки, человекообразной обезьяны, хомяка, крысы или мыши. Согласно некоторым вариантам осуществления клетка является эукариотической и выбрана из следующих клеток: CHO (например, CHO K1, DXB-11 CHO, Veggie-CHO), COS (например, COS-7), ретинальная клетка, Vero, CV1, почечная клетка (например, HEK293, 293 EBNA, MSR 293, MDCK, HaK, BHK), HeLa, HepG2, VVI38, MRC 5, Colo205, HB 8065, HL-60, (например, BHK21), Jurkat, Daudi, A431 (эпидермальная), CV-1, U937, 3T3, клетка L, клетка C127, SP2/0, NS-O, MMT 060562, клетка Сертоли, клетка BRL 3A, клетка HT1080, клетка миеломы, опухолевая клетка и клеточная линия, происходящая из вышеупомянутой клетки. Согласно некоторым вариантам осуществления клетка содержит один или несколько вирусных генов, например, ретинальная клетка, которая экспрессирует вирусный ген (например, клетка PER.C6™).
Термин "когнатный", используемый в смысле "когнатный по отношению к", например, первому домену VL, который "является когнатным" второму домену VL, подразумевает включение в себя ссылки на отношение между двумя доменами VL из одного связывающего белка, образованного мышью в соответствии с настоящим изобретением. Например, мышь, которая является генетически модифицированной согласно варианту осуществления настоящего изобретения, например, мышь, содержащая локус тяжелой цепи, в котором области VH, DH и JH замещены областями VL и JL, производит антителоподобные связывающие белки, которые содержат две идентичных полипептидных цепи, образованные одной и той же областью CH мыши (например, изотипа IgG), слитой с первым доменом VL человека, и две идентичных полипептидных цепи, образованных одной и той же областью CL мыши, слитой со вторым доменом VL человека. В ходе клональной селекции у мыши, первый и второй домены VL человека отбирали с помощью процесса клональной селекции, чтобы оказаться вместе в контексте одного антителоподобного связывающего белка. Таким образом, первый и второй домены VL, которые оказываются вместе в результате процесса клональной селекции в одной антителоподобной молекуле, называются "когнатными". Напротив, домен VL, который оказался в первой антителоподобной молекуле, и домен VL, который оказался во второй антителоподобной молекуле, являются не когнатными, за исключением случая, когда первая и вторая антителоподобные молекулы содержат идентичные тяжелые цепи (т.е. за исключением случая, когда домен VL, слитый с первой областью тяжелой цепи человека, и домен VL, слитый со второй областью тяжелой цепи человека, являются идентичными).
Фраза "определяющая комплементарность область", или термин "CDR", включает в себя аминокислотную последовательность, кодируемую последовательностью нуклеиновой кислоты генов иммуноглобулина организма, которая в норме (т.е., у животного дикого типа) находится между двумя каркасными областями в вариабельной области легкой или тяжелой цепи молекулы иммуноглобулина (например, антитела или T-клеточного рецептора). CDR может кодироваться, например, зародышевой последовательностью или реаранжированной или нереаранжированной последовательностью, и, например, наивной или зрелой B-клеткой или T-клеткой. В некоторых обстоятельствах (например, для CDR3), CDR могут кодироваться двумя или более последовательностями (например, зародышевыми последовательностями), которые не являются смежными (например, в нереаранжированной последовательности нуклеиновой кислоты), но являются смежными в последовательности нуклеиновой кислоты B-клетки, например, в результате сплайсинга или соединения последовательностей (например, V-D-J рекомбинации для образования CDR3 тяжелой цепи).
Фраза "генный сегмент" или "сегмент" включает в себя ссылку на генный сегмент иммуноглобулина V (легкий или тяжелый) или D или J (легкий или тяжелый), который включает в себя нереаранжированные последовательности на локусах иммуноглобулина (например, у людей и мышей), которые могут участвовать в реаранжировке (опосредованной, например, эндогенными рекомбиназами) для образования реаранжированной последовательности V/J или V/D/J. Если не указано иное, сегменты V, D и J содержат сигнальный последовательности рекомбинации (RSS), которые обеспечивают рекомбинацию V/J или рекомбинацию V/D/J согласно правилу 12/23. Если не указано иное, сегменты дополнительно содержат последовательности, с которыми они связаны в природе, или их функциональные эквиваленты (например, для V сегментов промотор(ы) и лидер(ы)).
Фраза "тяжелая цепь" или "тяжелая цепь иммуноглобулина" включает в себя последовательность константной области тяжелой цепи иммуноглобулина из любого организма, и если иное не указано, включает в себя вариабельный домен тяжелой цепи (VH). Домены VH включают в себя три CDR тяжелой цепи и четыре каркасных (FR) области, если не указано иное. Фрагменты тяжелых цепей включают в себя CDR, CDR и FR и их комбинации. Типичная тяжелая цепь состоит по существу из следующего: вариабельный домен (от N-конца к C-концу), домен CH1, шарнир, домен CH2, домен CH3 и необязательно домен СH4 (например, в случае IgM или IgE) и трансмембранный (M) домен (например, в случае мембраносвязанного иммуноглобулина на лимфоцитах). Константная область тяжелой цепи представляет собой область тяжелой цепи, которая продолжается (от N-конца к C-концу) от наружной FR4 до C-конца тяжелой цепи. Константные области тяжелой цепи с минорными девиациями, например, усечениями одного, двух, трех или нескольких аминокислот с C-конца, будут охватываться фразой "константная область тяжелой цепи", а также константные области тяжелой цепи с модификациями последовательности, например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотными замещениями. Аминокислотные замещения могут быть произведены на одном или нескольких положениях, выбранных, например, из (в соответствии с нумерацией EU константной области иммуноглобулина, например, константной области IgG человека), 228, 233, 234, 235, 236, 237, 238, 239, 241, 248, 249, 250, 252, 254, 255, 256, 258, 265, 267, 268, 269, 270, 272, 276, 278, 280, 283, 285, 286, 289, 290, 292, 293, 294, 295, 296, 297, 298, 301, 303, 305, 307, 308, 309, 311, 312, 315, 318, 320, 322, 324, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 337, 338, 339, 340, 342, 344, 356, 358, 359, 360, 361, 362, 373, 375, 376, 378, 380, 382, 383, 384, 386, 388, 389, 398, 414, 416, 419, 428, 430, 433, 434, 435, 437, 438 и 439.
В качестве примера, а не в качестве ограничения, константная область тяжелой цепи может быть модифицирована, чтобы проявлять усиленный период полужизни в сыворотке (по сравнению с аналогичной константной областью тяжелой цепи без указанной(ых) модификации(й)) и может содержать модификацию в положении 250 (например, E или Q); 250 и 428 (например, L или F); 252 (например, L/Y/F/W или T), 254 (например, S или T) и 256 (например, S/R/Q/E/D или T); или модификацию в положении 428 и/или 433 (например, L/R/SI/P/Q или K) и/или 434 (например, H/F или Y); или модификацию в положении 250 и/или 428; или модификацию в положении 307 или 308 (например, 308F, V308F) и 434. Согласно другому примеру модификация может включать в себя модификацию 428L (например, M428L) и 434S (например, N434S); модификацию 428L, 259I (например, V259I) и 308F (например, V308F); модификацию 433K (например, H433K) и 434 (например, 434Y); модификацию 252, 254 и 256 (например, 252Y, 254T и 256E); модификацию 250Q и 428L (например, T250Q и M428L); модификацию 307 и/или 308 (например, 308F или 308P).
Фраза "легкая цепь" включает в себя последовательность константной области легкой цепи (CL) иммуноглобулина из любого организма, и если не указано иное, включает в себя человека κ и λ легкие цепи. Вариабельные домены легкой цепи (VL), как правило, включают в себя три CDR легкой цепи и четыре каркасных (FR) области, если не указано иное. Как правило, полноразмерная легкая цепь (VL + CL) включает в себя, от амино-конца к карбокси-концу, домен VL, который включает в себя FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, и область CL. Легкие цепи (VL + CL), которые могут использоваться согласно настоящему изобретению, включают в себя легкие цепи, например, которые не связывают селективно или первый, или второй (в случае биспецифических связывающих белков) эпитоп, селективно связанный связывающим белком (например, эпитоп(ы), селективно связанный(е) доменом VL, слитым с доменом CH). Домены VL, которые не связывают селективно эпитоп(ы), связанный(е) VL, который слит с доменом CH, включают в себя те, которые могут быть идентифицированы путем скрининга в отношении наиболее часто используемых легких цепей в существующих библиотеках (жидкостных библиотеках или in silico), причем легкие цепи по существу не оказывают неблагоприятного влияния на аффинность и/или селективность связывающих эпитоп доменов связывающих белков. Подходящие легкие цепи включают в себя легкие цепи, которые могут связывать (отдельно или в комбинации с ее когнатным VL, слитым с областью CH) эпитоп, который специфически связывается VL, слитым с областью CH.
Фраза "микромолярный диапазон" подразумевает 1-999 микромоль; фраза "наномолярный диапазон" подразумевает 1-999 наномоль; фраза "пикомолярный диапазон" подразумевает 1-999 пикомоль.
Подразумевается, что термин "отличные от человека животные" включает в себя любые отличные от человека животные, такие как круглоротые, костные рыбы, такие хрящевые рыбы, как акулы и ромбовые скаты, амфибии, рептилии, млекопитающие и птицы. Подходящие отличные от человека животные включают в себя млекопитающих. Подходящие млекопитающие включают в себя не относящихся к человеку приматов, коз, овец, свиней, собак, коров и грызунов. Подходящие отличные от человека животные выбраны из семейства грызунов, включающего в себя крысу и мышь. Согласно одному варианту осуществления отличные от человека животные представляют собой мышей.
Мышь в качестве генетической модели была значительно усовершенствована с помощью трансгенных технологий и технологий нокаута, которые предоставили возможность для изучения эффектов направленной избыточной экспрессии или делеции специфических генов. Несмотря на все свои преимущества, мышь все еще демонстрирует генетические затруднения, которые делают ее несовершенной моделью для заболеваний человека и несовершенной платформой для испытания или получения терапевтических средств для людей. Во-первых, несмотря на то, что приблизительно 99% генов человека имеют мышиный гомолог (Waterston et. al. (2002). Initial sequencing and comparative analysis of the mouse genome, Nature 420, 520-562), потенциальные терапевтические средства зачастую не дают перекрестной реакции или дают неадекватную перекрестную реакцию с мышиными ортологами предполагаемых мишеней человека. Для устранения этой проблемы выбранные целевые гены могут быть "гуманизированы", то есть ген мыши может быть устранен и замещен соответствующей ортологичной генной последовательностью человека (например, патенты США №№ 6586251, 6596541 и 7105348, включенные в настоящий документ посредством ссылки). Изначально, попытки гуманизировать мышиные гены с помощью стратегии "генный нокаут вместе с трансгенной гуманизацией" предусматривали скрещивание мыши, несущей делецию (т.е., нокаут) эндогенного гена, с мышью, несущей интегрированный в произвольном порядке трансген человека (смотрите, например, Bril, et al. (2006), Tolerance to factor VIII in a transgenic mouse expressing human factor VIII cDNA carrying an Arg(593) to Cys substitution. Thromb Haemost 95, 341-347; Homanics et al. (2006), Production and characterization of murine models of classic and intermediate maple syrup urine disease. BMC Med Genet 7, 33; Jamsai, et al. (2006). A humanized BAC transgenic/knockout mouse model for HbE/beta-thalassemia. Genomics 88(3):309-15; Pan et al. (2006), Different role for mouse and human CD3delta/epsilon heterodimer in preT cell receptor (preTCR) function: human CD3delta/epsilon heterodimer restores the defective preTCR function in CD3gamma- and CD3gammadelta-deficient mice, Mol Immunol 43:1741-1750). Но эти попытки затруднялись ограничениями в размере; традиционные технологии генного нокаута были недостаточными для прямого замещения больших мышиных генов их большими геномными эквивалентами человека. Простой подход прямого гомологичного замещения, при котором эндогенный мышиный ген напрямую замещается эквивалентным геном человека в том же точном генетическом положении гена меши (т.е., на эндогенном мышином локусе), редко предпринимался вследствие технических сложностей. До настоящего времени попытки прямого замещения предусматривали усложненные и трудные процедуры, таким образом, ограничивая длину генетического материала, с который могли быть произведены манипуляции, и точность, с которой эти манипуляции могли быть произведены.
Экзогенно введенные трансгены иммуноглобулина человека перестраиваются в предшественниках B-клеток у мышей (Alt et al. (1985), Immunoglobulin genes in transgenic mice, Trends Genet 1:231-236). Это открытие использовалось в конструировании мышей с использованием генного нокаута вместе с трансгенным подходом для экспрессии антител человека (Green et al. (1994), Antigen-specific human monoclonal antibodies from mice engineered with human Ig heavy and light chain YACs, Nat Genet 7:13-21; Lonberg, N. (2005), Human antibodies from transgenic animals. Nat Biotechnol 23:1117-1125; Lonberg et al. (1994), Antigen-specific human antibodies from mice comprising four distinct genetic modifications, Nature 368:856-859; Jakobovits et al. (2007), From XenoMouse technology to panitumumab, the first fully human antibody product from transgenic mice, Nat Biotechnol 25:1134-1143). Эндогенные локусы тяжелой цепи и κ легкой цепи иммуноглобулина мыши инактивировали у этих мышей путем направленной делеции небольших, но критически важных частей каждого эндогенного локуса с последующим введением генных локусов иммуноглобулина человека в виде описанных выше случайным образом интегрированных больших трансгенов или минихромосом (Tomizuka et al. (2000), Double trans-chromosomic mice: maintenance of two individual human chromosome fragments containing Ig heavy and kappa loci and expression of fully human antibodies, PNAS USA 97:722-727). Такие мыши представляли важное преимущество в генной инженерии; выделенные из них полностью человеческие моноклональные антитела давали в результате перспективные терапевтические средства для лечения различных заболеваний человека (Gibson et al. (2006), Randomized phase III trial results of panitumumab, a fully human anti-epidermal growth factor receptor monoclonal antibody, in metastatic colorectal cancer, Clin Colorectal Cancer 6:29-31; Jakobovits et al., 2007; Kim et al. (2007), Clinical efficacy of zanolimumab (HuMax-CD4): two Phase II studies in refractory cutaneous T-cell lymphoma, Blood 109(11):4655-62; Lonberg, 2005; Maker et al. (2005), Tumor regression and autoimmunity in patients treated with cytotoxic T lymphocyte-associated antigen 4 blockade and interleukin 2: a phase I/II study, Ann Surg Oncol 12:1005-1016; McClung et al. (2006), Denosumab in postmenopausal women with low bone mineral density, N Engl J Med 354:821-831). Но, как обсуждалось выше, эти мыши проявляют нарушенное развитие B-клеток и иммунологические дефициты по сравнению с мышами дикого типа. Такие проблемы потенциально ограничивают способность мышей обеспечивать сильный гуморальный ответ и, следовательно, образовывать полностью человеческие антитела к некоторым антигенам. Недостаточности могут быть обусловлены следующим: (1) неэффективная функциональность вследствие введения случайным образом трансгенов иммуноглобулина человека и полученная в результате неправильная экспрессия вследствие отсутствия контролирующих элементов против хода транскрипции и по ее ходу (Garrett et al. (2005), Chromatin architecture near a potential 3' end of the Igh locus involves modular regulation of histone modifications during B-Cell development and in vivo occupancy at CTCF sites, Mol Cell Biol 25:1511-1525; Manis et al. (2003), Elucidation of a downstream boundary of the 3' IgH regulatory region, Mol Immunol 39:753-760; Pawlitzky et al. (2006), Identification of a candidate regulatory element within the 5' flanking region of the mouse Igh locus defined by pro-B cell-specific hypersensitivity associated with binding of PU.1, Pax5, and E2A, J Immunol 176I6839-6851); (2) неэффективные межвидовые взаимодействия между константными доменами человека и мышиными компонентами B-клеточного рецепторного сигнального комплекса на клеточной поверхности, которые могут ослаблять сигнальные процессы, необходимые для нормального созревания, пролиферации и выживания B-клеток (Hombach et al. (1990), Molecular components of the B-cell antigen receptor complex of the IgM class, Nature 343:760-762); и (3) неэффективные межвидовые взаимодействия между растворимыми иммуноглобулинами человека и мышиными Fc рецепторами, которые могут снижать аффинную селекцию (Rao et al. (2002). Differential expression of the inhibitory IgG Fc receptor FcgammaRIIB on germinal center cells: implications for selection of high-affinity B cells, J Immunol 169:1859-1868) и концентрации иммуноглобулинов в сыворотке (Brambell et al. (1964), A Theoretical Model of Gamma-Globulin Catabolism, Nature 203:1352-1354; Junghans and Anderson, (1996), The protection receptor for IgG catabolism is the beta2-microglobulin-containing neonatal intestinal transport receptor, PNAS USA93:5512-5516; Rao et al., 2002; Hjelm et al. (2006), Antibody-mediated regulation of the immune response, Scand J Immunol 64:177-184; Nimmerjahn and Ravetch, (2007), Fc-receptors as regulators of immunity, Adv Immunol 96:179-204). Эти дефициты можно корректировать с помощью гуманизации in situ только вариабельных областей локусов иммуноглобулина мыши в пределах их естественных положений на эндогенных локусах тяжелых и легких цепей. Это будет действительно давать в результате мышей, которые образуют обратные химерные антитела (т.е., V человека: C мыши), которые будут способны к нормальным взаимодействиям и селекции в мышином окружении на основании сохранения константных областей мыши. Более того, такие обратные химерные антитела легко реформатируются в полностью человеческие антитела для терапевтических целей.
Генетически модифицированные животные, которые содержат вставку или замещение на эндогенном локусе тяжелой цепи иммуноглобулина гетерологичными (например, от другого вида) последовательностями иммуноглобулина, могут быть получены в сочетании со вставками или замещениями на эндогенных локусах легкой цепи иммуноглобулина или в сочетании с трансгенами легкой цепи иммуноглобулина (например, химерными трансгенами легкой цепи иммуноглобулина или полностью человеческими полностью мышиными и т.д.). Вид, из которого происходят гетерологичные последовательности иммуноглобулина, может изменяться в широком диапазоне. Иллюстративная гетерологичные последовательности иммуноглобулина включают в себя последовательности человека.
Последовательности нуклеиновой кислоты вариабельной области иммуноглобулина, например, сегменты V, D и/или J согласно различным вариантам осуществления получены от человека или отличного от человека животного. Отличные от человека животные, подходящие для предоставления сегментов V, D и/или J, включают в себя, например, костных рыб, таких хрящевых рыб, как акулы и ромбовидные скаты, амфибий, рептилий, млекопитающих, птиц (например, куриц). Отличные от человека животные включают в себя, например, млекопитающих. Млекопитающие включают в себя, например, отличных от человека приматов, коз, овец, свиней, собак, крупный рогатый скот (например, корову, быка, буйвола), оленя, верблюдов, хорьков и грызунов и не относящихся к человеку приматов (например, шимпанзе, орангутанов, горилл, игрунок, макак-резусов, бабуинов). Подходящие отличные от человека животные выбраны из семейства грызунов, включающего в себя крыс, мышей и хомяков. Согласно одному варианту осуществления отличные от человека животные представляют собой мышей. Из контекста ясно, что различные отличные от человека животные могут быть использованы в качестве источника вариабельных доменов или генных сегментов вариабельной области (например, акулы, ромбовидные скаты, млекопитающие, например, верблюды, такие грызуны, как мыши и крысы).
Согласно контексту отличные от человека животные также используются в качестве источника последовательностей константной области для применения вместе с вариабельными последовательностями или сегментами, например, константные последовательности грызуна могут использоваться в трансгенах, функционально связанных с относящимися к человеку или не относящимися к человеку вариабельными последовательностями (например, вариабельные последовательности человека или не относящегося к человеку примата, функционально связанные, например, с константными последовательностями грызуна, например, мыши или крысы или хомяка). Таким образом, согласно различным вариантам осуществления сегменты V, D и/или J человека функционально связаны с генными последовательностями константной области грызуна (например, мыши или крысы или хомяка). Согласно некоторым вариантам осуществления сегменты V, D и/или J человека (или одного или нескольких реаранжированных генов VDJ или VJ) функционально связаны или слиты с генной последовательностью константной области мыши, крыса или хомяка, например, в трансгене, интегрированном в локус, который не является эндогенным локусом иммуноглобулина.
Согласно конкретному варианту осуществления предусмотрена мышь, которая содержит замещение генных сегментов VH, DH, и JH на эндогенном локусе тяжелой цепи иммуноглобулина одним или несколькими генными сегментами VL человека и одним или несколькими генными сегментами JL человека, причем один или несколько генных сегментов VL человека и один или несколько генных сегментов JL функционально связаны с эндогенным геном тяжелой цепи иммуноглобулина; причем мышь содержит трансген на локусе, отличном от эндогенного локуса иммуноглобулина, причем трансген содержит нереаранжированный или реаранжированный генный сегмент VL человека и JL человека, функционально связанный с константной областью мыши или крысы или человека. Согласно различным вариантам осуществления один или несколько генных сегментов VL человека включают в себя генные сегменты Vκ человека или генные сегменты Vλ человека. Согласно одному варианту осуществления один или несколько генных сегментов JL человека включают в себя генные сегменты Jκ человека или генные сегменты Jλ человека.
Описан способ крупномасштабного in situ генетического замещения зародышевых вариабельных генов тяжелой цепи иммуноглобулина мыши зародышевыми вариабельными генами легкой цепи иммуноглобулина человека при подержании способности мышей к производству потомства. В частности, описано точное замещение вариабельных генных локусов тяжелой цепи мыши вариабельными генными локусами легкой цепи человека, оставляя интактными константные области мыши. В результате были получены мыши, которые экспрессируют иммуноглобулиноподобные связывающие белки, ассоциированные с эндогенными константными областями. Вариабельные области легкой цепи человека связаны с константными областями тяжелой цепи мыши для образования химерных относящихся к человеку-мыши локусов иммуноглобулина, которые реаранжируются и экспрессируют уникальные иммуноглобулиноподобные молекулы. Экспрессированные иммуноглобулиноподобные молекулы представляют собой “обратные химеры”, т.е. они содержат последовательности вариабельной области человека и последовательности константной области мыши.
Конструирование последовательностей иммуноглобулина человека в геном мыши, даже в точных положениях, например, на эндогенных локусах иммуноглобулина мыши, может представлять определенные проблемы вследствие дивергентной эволюции локусов иммуноглобулина между мышью и человеком. Например, межгенные последовательности, разбросанные в пределах локусов иммуноглобулина, не идентичны между мышами и людьми и в некоторых обстоятельствах могут не быть функционально эквивалентными. Различия между мышами и людьми в их локусах иммуноглобулина может все еще приводить к аномалиям у гуманизированных мышей, в частности, при гуманизации или манипуляции с определенными частями эндогенных локусов тяжелой цепи иммуноглобулина мыши. Некоторые модификации на локусах тяжелой цепи иммуноглобулина мыши являются вредными. Вредные модификации могут включать в себя, например, потерю способности модифицированных мышей спариваться и производить потомство. Согласно различным вариантам осуществления конструирование последовательностей иммуноглобулина человека в геноме мыши включает в себя способы, которые сохраняют эндогенные последовательности, которые при отсутствии у модифицированных линий мышей являются вредными. Иллюстративные неблагоприятные эффекты могут включать в себя неспособность воспроизводить модифицированные линии, потерю функции жизненно важных генов, неспособность экспрессировать полипептиды и т.д. Такие неблагоприятные эффекты могут быть напрямую или косвенно связаны с модификацией, разработанной в геноме мыши.
Несмотря на то, что близкая к дикому типу гуморальная иммунная функция, наблюдаемая у мышей с замещенными локусами иммуноглобулина, существуют другие обнаруживаемые при использовании прямого замещения иммуноглобулина сложности, которые не встречаются в некоторых подходах, которые используют интегрированные случайным образом трансгены. Различия в генетическом составе иммуноглобулиновых локусов между мышами и людьми привело к открытию последовательностей, преимущественных для размножения мышей с замещенными иммуноглобулиновыми генными сегментами. В частности, гены ADAM мыши, расположенные в пределах эндогенного локуса тяжелой цепи иммуноглобулина, оптимально присутствуют у мышей с замещенными иммуноглобулиновыми локусами вследствие их роли в фертильности.
Проводили точное, крупномасштабное замещение in situ шести миллионов пар нуклеотидов вариабельных областей локусов тяжелой цепи иммуноглобулина мыши (VH-DH-JH) вариабельными генными локусами легкой цепи иммуноглобулина человека (VL-JL) при сохранении фланкирующих последовательностей мыши интактными и функциональными в пределах гибридных локусов, включающих в себя все гены константной цепи мыши и области транскрипционного контроля локуса (фиг. 2 - фиг. 6). Проводили стадии конструирования для сохранения последовательностей мыши, которые предоставляют мыши способность спариваться и производить потомство сопоставимым с мышью дикого типа образом (фиг. 7 - фиг. 9). В частности, приблизительно полмиллиона пар нуклеотидов локуса κ легкой цепи иммуноглобулина человека, содержащего проксимальное плечо (т.е. 40 функциональных генных сегментов Vκ человека и пять генных сегментов Jκ человека) и гены ADAM6 мыши вводили посредством химерных основанных на BAC нацеливающих векторов в ES клетки мыши с применением технологии генной инженерии VELOCIGENE® (смотрите, например, патент США № 6586251 и Valenzuela et al., 2003, High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nat Biotechnol 21:652-659).
Положение в геноме и функция ADAM6 мыши
Самцы мышей, у которых отсутствует способность экспрессировать какой-либо функциональный белок ADAM6, проявляют тяжелый дефект в способности мышей спариваться и давать потомство. Мыши утрачивают способность экспрессировать функциональный белок ADAM6 посредством замещения всех или по существу всех вариабельных генных сегментов тяжелой цепи иммуноглобулина мыши вариабельными генными сегментами легкой цепи человека. Потеря функции ADAM6 происходит, поскольку локус ADAM6 расположен в пределах области эндогенного генного локуса вариабельной области тяжелой цепи иммуноглобулина, проксимального к 3’ концу локуса генного сегмента VH, который находится выше генных сегментов DH. Разведение мышей, являющихся гомозиготными в отношении замещения всех или по существу всех эндогенных вариабельных генных сегментов тяжелой цепи генными сегментами вариабельной области легкой цепи человека, как правило, представляет собой трудоемкий подход для спаривания самцов и самок, каждый из которых являются гомозиготными в отношении замещения, и ожидания продуктивного спаривания. Частота успешных пометов и средний размер помета являются низкими. Вместо этого, самцы, гетерозиготные в отношении замещения, использовались для спаривания с самками, гомозиготными в отношении замещения, для получения потомства, которое является гетерозиготным в отношении замещения, затем выводя из него гомозиготную мышь. Авторы настоящего изобретения показали, что вероятной причиной потери фертильности у самцов мышей является отсутствие у гомозиготных самцов мышей функционального белка ADAM6.
Согласно различным аспектам самцы мышей, которые содержат поврежденный (т.е. нефункциональный или незначительно функциональный) ген ADAM6, проявляют снижение или устранение фертильности. Поскольку у мышей (и других грызунов) ген ADAM6 расположен в локусе тяжелой цепи иммуноглобулина, авторы настоящего изобретения определили, что для того чтобы воспроизвести мышей или создать и поддержать линию мышей, которые содержат модификации в эндогенном локусе тяжелой цепи иммуноглобулина, использовали различные модифицированные схемы скрещивания или размножения. Низкая фертильность, или стерильность, самцов мышей, гомозиготных в отношении замещения эндогенного вариабельного генного локуса тяжелой цепи иммуноглобулина затрудняет поддержание такой модификации в линии мыши. Согласно различным вариантам осуществления сохранение линии предусматривает устранение проблем со стерильностью, которые проявляют самцы мышей, гомозиготные в отношении замещения.
Согласно одному аспекту предусмотрен способ поддержания линии описанной в настоящем документе мыши. Линия мыши может не содержать эктопическую последовательность ADAM6, и согласно различным вариантам осуществления линия мыши является гомозиготным или гетерозиготным в отношении нокаута (например, функционального нокаута) ADAM6.
Линия мыши содержит модификацию эндогенного локуса тяжелой цепи иммуноглобулина, которая приводит к снижению или потере фертильности у самца мыши. Согласно одному варианту осуществления модификация предусматривает делецию регуляторной области и/или кодирующей области гена ADAM6. Согласно конкретному варианту осуществления модификация предусматривает модификацию эндогенного гена ADAM6 (регуляторной и/или кодирующей области), которая снижает или устраняет фертильность самца мыши, который содержит модификацию; согласно конкретному варианту осуществления модификация снижает или устраняет фертильность самца мыши, который является гомозиготным в отношении модификации.
Согласно одному варианту осуществления линия мыши является гомозиготным или гетерозиготным в отношении нокаута (например, функционального нокаута) или делеции гена ADAM6.
Согласно одному варианту осуществления линию мыши поддерживают путем выделения из мыши, которая является гомозиготной или гетерозиготной в отношении модификации, клетки, и использование донорной клетки в зародыше-хозяине и вынашивание зародыша-хозяина и донорной клетки в суррогатной матери и получение от суррогатной матери потомства, которое содержит генетическую модификацию. Согласно одному варианту осуществления донорная клетка представляет собой ES клетку. Согласно одному варианту осуществления донорная клетка представляет собой плюрипотентную клетку, например, индуцированную плюрипотентную клетку.
Согласно одному варианту осуществления линию мыши поддерживают путем выделения из мыши, которая является гомозиготной или гетерозиготной в отношении модификации, последовательности нуклеиновой кислоты, содержащей модификацию, и введение последовательности нуклеиновой кислоты в ядро-хозяина и вынашивание клетки, содержащей последовательность нуклеиновой кислоты и ядро-хозяин в подходящем животном. Согласно одному варианту осуществления последовательность нуклеиновой кислоты вводят в зародыш из ооцита-хозяина.
Согласно одному варианту осуществления линию мыши поддерживают путем выделения из мыши, которая является гомозиготной или гетерозиготной в отношении модификации, ядра и введение ядра в клетку-хозяин, и вынашивание ядра и клетки-хозяина в подходящем животном для получения потомства, которое является гомозиготным или гетерозиготным в отношении модификации.
Согласно одному варианту осуществления линию мыши поддерживают путем использования оплодотворения in vitro (IVF) самки мыши (дикого типа, гомозиготной в отношении модификации или гетерозиготной в отношении модификации), используя сперму от самца мыши, содержащего генетическую модификацию. Согласно одному варианту осуществления самец мыши является гетерозиготным в отношении генетической модификации. Согласно одному варианту осуществления самец мыши является гомозиготным в отношении генетической модификации.
Согласно одному варианту осуществления линию мыши поддерживают путем скрещивания самца мыши, который является гетерозиготным в отношении генетической модификации, с самкой мыши для получения потомства, которое содержит генетическую модификацию, идентификации мужского и женского потомства, содержащего генетическую модификацию, и использования самца, который является гетерозиготным в отношении генетической модификации, в скрещивании с самкой, которая относится к дикому типу, является гомозиготной или гетерозиготной в отношении генетической модификации, для получения потомства, содержащего генетическую модификацию. Согласно одному варианту осуществления стадию скрещивания самца, гетерозиготного в отношении генетической модификации, с самкой дикого типа, самкой, гетерозиготной в отношении генетической модификации, или самкой, гомозиготной в отношении генетической модификации, повторяют для поддержания генетической модификации в линии мыши.
Согласно одному аспекту предусмотрен способ поддержания линии мыши, который предусматривает замещение эндогенного вариабельного генного локуса тяжелой цепи иммуноглобулина одним или несколькими последовательностями легкой цепи иммуноглобулина человека и необязательно одним или несколькими генными сегментами DH человека, предусматривая скрещивание линии мыши так, чтобы создать гетерозиготных самцов мышей, причем гетерозиготных самцов мышей скрещивают для поддержания генетической модификации в линии. Согласно конкретному варианту осуществления линию не поддерживают путем какого-либо скрещивания гомозиготного самца с самкой дикого типа или самкой, гомозиготной или гетерозиготной в отношении генетической модификации.
Белок ADAM6 представляет собой представителя семейства белков дизинтегринов и металлопротеаз (ADAM), которое представляет собой большое семейство с разнообразными функциями, включающими в себя клеточную адгезию. Некоторые представители семейства ADAM вовлечены в сперматогенез и оплодотворение. Например, ADAM2 кодирует субъединицу белка фертилина, который вовлечен во взаимодействия сперматозоидов и яйцеклетки. ADAM3, или циритестин, оказывается необходимым для связывания сперматозоида с вителлиновым слоем. Отсутствие или ADAM2, или ADAM3 приводит к стерильности. Предположили, что ADAM2, ADAM3 и ADAM6 формируют комплекс на поверхности мышиных сперматозоидов.
У людей, ген ADAM6, который по имеющимся данным является псевдогеном, расположен между VH генными сегментами VH1-2 и VH6-1 человека. У мышей существует два гена ADAM6 - ADAM6a и ADAM6b - которые расположены в межгенной области между генными сегментами VH и DH мыши и ориентированы в транскрипционной ориентации, противоположной ориентации транскрипции окружающих иммуноглобулиновых генных сегментов. У мышей функциональный локус ADAM6 очевидно необходим для нормального оплодотворения. Функциональный локус или последовательность ADAM6, затем относится к локусу или последовательности ADAM6, которая может дополнить или восстановить резко сниженное оплодотворение, демонстрируемое самцами мышей с утраченными или поврежденными эндогенными локусами ADAM6.
Положение межгенной последовательности у мышей, которая кодирует ADAM6a и ADAM6b, делает межгенную последовательность подверженной модификации при модификации эндогенной тяжелой цепи. Когда генные сегменты VH подвергаются делеции или замещаются или когда генные сегменты DH подвергаются делеции или замещаются, существует высокая вероятность того, что полученная мышь будет проявлять тяжелую недостаточность фертильности. Для компенсации этой недостаточности мышь модифицируют, чтобы включить в нее нуклеотидную последовательность, которая кодирует белок, который будет дополнять потерю активности ADAM6 вследствие модификации эндогенного локуса ADAM6. Согласно различным вариантам осуществления дополняющая нуклеотидная последовательность представляет собой такую, которая кодирует мышиный ADAM6a, мышиный ADAM6b или его гомолог или ортолог или функциональный фрагмент, который восстанавливает недостаточность фертильности. Согласно различным вариантам осуществления дополняющая нуклеотидная последовательность кодирует белок ADAM6a мыши, представленный в SEQ ID NO: 1, и/или кодирует белок ADAM6b мыши, представленный в SEQ ID NO: 2. Альтернативно, могут использоваться подходящие способы для сохранения эндогенного локуса ADAM6, при это делая эндогенные последовательности тяжелой цепи иммуноглобулина, фланкирующие локус ADAM6 мыши, неспособными к реаранжировке, чтобы кодировать функциональную эндогенную вариабельную область тяжелой цепи. Иллюстративные альтернативные способы включают в себя манипуляцию с большими частями хромосом мыши, которые располагают эндогенные локусы вариабельной области тяжелой цепи иммуноглобулина таким образом, что они являются неспособными к реаранжировке, чтобы кодировать функциональную вариабельную область тяжелой цепи, которая функционально связана с эндогенным константным геном тяжелой цепи. Согласно различным вариантам осуществления способы предусматривают вставки и/или транслокации хромосомных фрагментов мыши, содержащих эндогенные генные сегменты тяжелой цепи иммуноглобулина.
Нуклеотидная последовательность, которая восстанавливает фертильность, может быть расположена в любом подходящем положении. Она может быть расположена в межгенной области (например, между генными сегментами V и J или выше генных сегментов V), или в любом подходящем положении в геноме (т.е., эктопически). Согласно одному варианту осуществления нуклеотидная последовательность может быть введена в трансген, который случайным образом интегрирован в геном мыши. Согласно одному варианту осуществления последовательность может храниться эписомно, то есть на отдельной нуклеиновой кислоте, а не на хромосоме мыши. Подходящие положения предусматривают положения, которые являются транскрипционно пермиссивными или активными, например, локус ROSA26 (Zambrowicz et al., 1997, PNAS USA 94:3789-3794), локус BT-5 (Michael et al., 1999, Mech. Dev. 85:35-47), или локус Oct4 (Wallace et al., 2000, Nucleic Acids Res. 28:1455-1464). Нацеливание нуклеотидных последовательностей на транскрипционно активные локусы описаны, например, в патенте США № 7473557, включенный посредством ссылки в настоящий документ.
Альтернативно нуклеотидная последовательность, которая восстанавливает фертильность, может быть соединена с индуцируемым промотором так, чтобы облегчать оптимальную экспрессию в соответствующих клетках и/или тканях, например, репродуктивных тканях. Иллюстративные индуцируемые промоторы включают в себя промоторы, активируемые физическими (например, промотор теплового шока) и/или химическими средствами (например, IPTG или тетрациклином).
Кроме того, экспрессия нуклеотидной последовательности может быть ассоциирована с другими генами так, чтобы достичь экспрессии на конкретной стадии развития или в специфических тканях. Такая экспрессия может быть достигнута путем помещения нуклеотидной последовательности в функциональной связи с промотором гена, экспрессируемого на конкретной стадии развития. Например, последовательности иммуноглобулина от одного вида, сконструированные в геноме вида-хозяина, помещают в функциональной связи с промоторной последовательностью гена CD19 (специфический для B-клеток ген) от вида-хозяина. Достигается специфическая для B-клеток экспрессия на точно определенных стадиях развития при экспрессии иммуноглобулинов.
Еще один способ достижения устойчивой экспрессии вставленной нуклеотидной последовательности предусматривает использование конститутивного промотора. Иллюстративные конститутивные промоторы включают в себя SV40, CV, UBC, EF1A, PGK и CAGG. Аналогичным образом, требуемая нуклеотидная последовательность располагают в функциональной связи с выбранным конститутивным промотором, что обеспечивает высокий уровень экспрессии белка(ов), кодируемого(ых) нуклеотидной последовательностью.
Предусматривается, что термин "эктопический" включает в себя перемещение или размещение в положении, которое в норме не встречается в природе (например, размещение последовательности нуклеиновой кислоты в положении, которое не является аналогичным положением, в котором последовательность нуклеиновой кислоты встречается у мыши дикого типа). Термин согласно различным вариантам осуществления используется в том смысле, что его объект находится за пределами его нормального, или правильного, положения. Например, фраза "эктопическая нуклеотидная последовательность, кодирующая…" относится к нуклеотидной последовательности, которая находится в положении, в котором она не встречается в норме у мыши. Например, в случае эктопической нуклеотидной последовательности, кодирующей белок ADAM6 мыши (или ее ортолога или гомолога или фрагмента, который обеспечивает такое же или сходное улучшение фертильности у самцов мышей), последовательность может быть расположена в положении в геноме мыши, которое отличается от встречающегося в норме у мыши дикого типа. В таких случаях, новые участки соединения последовательности для последовательности мыши будут создаваться путем помещения последовательности в положение в геноме мыши, отличное от такового у мыши дикого типа. Функциональный гомолог или ортолог ADAM6 мыши представляет собой последовательность, которая обеспечивает восстановление потери фертильности (например, потери способности самца мыши давать потомство путем спаривания), которая наблюдается у мыши ADAM6-/-. Функциональные гомологи или ортологи предусматривают белки, которые характеризуются по меньшей мере приблизительно 89% идентичности или более, например, до 99% идентичности, с аминокислотной последовательностью ADAM6a и/или с аминокислотной последовательностью ADAM6b, и которые могут дополнять или восстанавливать способность успешно спариваться у мыши, характеризующейся фенотипом, который содержит делецию или нокаут ADAM6a и/или ADAM6b.
Эктопическое положение может находиться где-либо (например, как в случае случайной вставки трансгена, содержащего последовательность ADAM6 мыши), или может находиться, например, в положении, которое приблизительно соответствует (но не является точно таким же как) его расположение у мыши дикого типа (например, в модифицированном эндогенном локусе иммуноглобулина, но или выше, или ниже в отношении хода транскрипции от его естественного положения, например, в пределах модифицированного локуса иммуноглобулина, но между различными генными сегментами, или в другом положении в V-D межгенной последовательности мыши). Один пример эктопического размещения представляет собой поддержание положения, встречающегося в норме у мышей дикого типа в пределах эндогенного локуса тяжелой цепи иммуноглобулина, при этом делая окружающие эндогенные генные сегменты тяжелой цепи неспособными к реаранжировке, чтобы кодировать функциональную тяжелую цепь, содержащую эндогенную константную область тяжелой цепи. Согласно описанному примеру, это можно осуществить путем инверсии хромосомного фрагмента, содержащего эндогенные вариабельные локусы тяжелой цепи иммуноглобулина, например с использованием сконструированных сайтов сайт-специфической рекомбинации, помещенных в положениях, фланкирующих локус вариабельной области. Таким образом, при рекомбинации эндогенные локусы вариабельной области тяжелой цепи помещаются на большом расстоянии от эндогенных генов константной области тяжелой цепи, тем самым предотвращая реаранжировку для кодирования функциональной тяжелой цепи, содержащей эндогенную константную область тяжелой цепи. Другие иллюстративные способы для достижения функционального сайленсинга эндогенного вариабельного генного локуса тяжелой цепи иммуноглобулина при сохранении функционального локуса ADAM6 будут очевидными специалистам в настоящей области техники при прочтении настоящего раскрытия и/или в комбинации со способами, известными в настоящей области техники. При таком размещении эндогенного локуса тяжелой цепи эндогенные гены ADAM6 сохраняются, и эндогенный локус тяжелой цепи иммуноглобулина является функционально выключенным.
Другой пример эктопического размещения представляет собой размещение в пределах модифицированного локуса тяжелой цепи иммуноглобулина. Например, мышь, содержащая замещение одного или нескольких эндогенных генных сегментов VH генными сегментами VL человека, причем замещение удаляет эндогенную последовательность ADAM6, может быть сконструирована так, чтобы содержать последовательность ADAM6 мыши, расположенную в пределах последовательности, которая содержит генные сегменты VL человека. Полученная модификация будет образовывать (эктопическую) последовательность ADAM6 мыши в пределах генной последовательности человека, и (эктопическое) размещение последовательности ADAM6 мыши в пределах генной последовательности человека может приблизительно соответствовать положению псевдогена ADAM6 человека (т.е., между двумя сегментами V) или может приблизительно соответствовать положению последовательности ADAM6 мыши (т.е., в пределах V-D межгенной области). Полученные участки соединения последовательности, созданные соединением (эктопической) последовательности ADAM6 мыши в пределах или рядом с генной последовательностью человека (например, генной последовательностью легкой цепи иммуноглобулина) в зародышевой линии мыши, будут новыми по сравнению с таким же или аналогичным положением в геноме мыши дикого типа.
Согласно различным вариантам осуществления предусмотрены отличные от человека животные, у которых отсутствует ADAM6 или его ортолог или гомолог, причем отсутствие делает отличное от человека животное стерильным или по существу снижает фертильность отличного от человека животного. Согласно различным вариантам осуществления отсутствие ADAM6 или его ортолога или гомолога обусловлено модификацией эндогенного локуса тяжелой цепи иммуноглобулина. Существенное снижение фертильности, например, представляет собой снижение фертильности (например, частоты скрещивания, количества детенышей на помет, количества пометов в год и т.д.) приблизительно на 50%, 60%, 70%, 80%, 90% или 95% или больше. Согласно различным вариантам осуществления отличным от человека животным добавляют ген ADAM6 мыши или его ортолог или гомолог или функциональный фрагмент, который является функциональным у самца отличного от человека животного, причем добавленный ген ADAM6 или его ортолог или гомолог или функциональный фрагмент восстанавливает снижение фертильности полностью или существенную ее часть. Восстановление существенной части фертильности представляет собой, например, такое восстановление фертильности, что отличное от человека животное проявляет фертильность, которая составляет по меньшей мере 70%, 80% или 90% или больше по сравнению с немодифицированным (т.е. животное без модификации гена ADAM6 или его ортолога или гомолога) локусом тяжелой цепи.
Последовательность, которая обеспечивает генетически модифицированное животное (т.е. животное, которое не содержит функциональный ADAM6 или его ортолог или гомолог, вследствие, например, модификации локуса тяжелой цепи иммуноглобулина), согласно различным вариантам осуществления выбрана из гена ADAM6 или его ортолога или гомолога. Например, у мыши потеря функции ADAM6 восстанавливается путем добавления согласно одному варианту осуществления гена ADAM6 мыши. Согласно одному варианту осуществления потеря функции ADAM6 у мыши восстанавливается путем добавления ортолога или гомолога близкородственных видов по отношению к мыши, например, грызуна, например, мыши другой линии или вида, крысы любого вида, грызуна; причем добавление мыши ортолога или гомолога восстанавливает потерю фертильности вследствие потери функции ADAM6 или потери гена ADAM6. Ортологи и гомологи из других видов согласно различным вариантам осуществления выбраны из филогенетически родственного вида и согласно различным вариантам осуществления проявляют процентную идентичность по отношению к эндогенному ADAM6 (или ортологу), которая составляет приблизительно 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, или 97% или более; и которые восстанавливают связанную с ADAM6 или (у не относящихся к мыши) связанную с ортологом ADAM6 потерю фертильности. Например, у генетически модифицированного самца крысы, у которого отсутствует функция ADAM6 (например, крыса с эндогенной вариабельной областью тяжелой цепи иммуноглобулина, замещенной вариабельной областью тяжелой цепи иммуноглобулина человека, или нокаут у крысы области тяжелой цепи иммуноглобулина), потеря фертильности у крысы восстанавливается путем добавления ADAM6 крысы или согласно некоторым вариантам осуществления ортолога ADAM6 крысы (например, ортолога ADAM6 от другой линии или вида крысы или согласно одному варианту осуществления от мыши).
Таким образом, согласно различным вариантам осуществления генетически модифицированные животные, которые не проявляют фертильности или снижение фертильности вследствие модификации последовательности нуклеиновой кислоты, кодирующей белок ADAM6 (или его ортолог или гомолог) или регуляторной области, функционально связанной с последовательностью нуклеиновой кислоты, содержат последовательность нуклеиновой кислоты, которая взаимодополняет или восстанавливает потерю фертильности, если последовательность нуклеиновой кислоты, которая взаимодополняет или восстанавливает потерю фертильности, происходит от другой линии того же вида или из филогенетически родственного вида. Согласно различным вариантам осуществления взаимодополняющая последовательность нуклеиновой кислоты представляет собой ADAM6, его ортолог или гомолог или функциональный фрагмент. Согласно различным вариантам осуществления взаимодополняющий ортолог ADAM6 или его гомолог или функциональный фрагмент происходит от отличного от человека животного, которое является близкородственным генетически модифицированному животному с нарушением фертильности. Например, если генетически модифицированное животное представляет собой мышь конкретной линии, ортолог ADAM6 или его гомолог или функциональный фрагмент может быть получен от мыши другой линии или мыши родственного вида. Согласно одному варианту осуществления если генетически модифицированное животное, содержащее нарушение фертильности, происходит из отряда Rodentia, то ортолог ADAM6 или его гомолог или функциональный фрагмент происходит от другого животного отряда Rodentia. Согласно одному варианту осуществления генетически модифицированное животное, содержащее нарушение фертильности, происходит из подотряда Myomoropha (например, тушканчики, полутушканчики, мышевидные хомяки, хомяки, крысы и мыши Нового Света, полевки, истинные мыши и крысы, карликовые песчанки, иглистые мыши, косматые хомяки, рипидомисы, скалистые хомячки, белохвостые крысы, мадагаскарские крысы и мыши, колючие соневидные хомяки, слепыши, бамбуковые крысы, цокоры), и ортолог ADAM6 или его гомолог или функциональный фрагмент выбран из животного отряда Rodentia или подотряда Myomorpha.
Согласно одному варианту осуществления генетически модифицированное животное происходит из надсемейство Dipodoidea, и ортолог ADAM6 или его гомолог или функциональный фрагмент происходит из надсемейства Muroidea. Согласно одному варианту осуществления генетически модифицированное животное происходит из надсемейство Muroidea, и ортолог ADAM6 или его гомолог или функциональный фрагмент происходит из надсемейства Dipodoidea.
Согласно одному варианту осуществления генетически модифицированное животное представляет собой грызуна. Согласно одному варианту осуществления грызун выбран из надсемейства Muroidea, и ортолог или гомолог ADAM6 происходит от другого вида в пределах надсемейства Muroidea. Согласно одному варианту осуществления генетически модифицированное животное происходит из семейства, выбранного из Calomyscidae (например, мышевидные хомяки), Cricetidae (например, хомяк, крысы и мыши Нового Света, полевки), Muridae (настоящие мыши и крысы, карликовые песчанки, иглистые мыши, косматые хомяки), Nesomyidae (рипидомисы, скалистые хомячки, белохвостые крысы, мадагаскарские крысы и мыши), Platacanthomyidae (например, колючие соневидные хомяки), и Spalacidae (например, слепыши, бамбуковые крысы и цокоры); и ортолог или гомолог ADAM6 выбран из другого вида того же семейства. Согласно конкретному варианту осуществления генетически модифицированный грызун выбран из настоящей мыши или крысы (семейство Muridae), и ортолог или гомолог ADAM6 происходит от вида, выбранного из карликовой песчанки, иглистой мыши, косматого хомяка. Согласно одному варианту осуществления генетически модифицированная мышь происходит из представителя семейства Muridae, и ортолог или гомолог ADAM6 происходит от другого вида семейства Muridae. Согласно конкретному варианту осуществления генетически модифицированный грызун представляет собой мышь семейства Muridae, и ортолог или гомолог ADAM6 происходит от крысы, карликовой песчанки, иглистой мыши, косматого хомяка семейства Muridae.
Согласно различным вариантам осуществления один или несколько ортологов ADAM6 грызуна или их гомологов или функциональных фрагментов грызуна в семействе восстанавливает фертильность генетически модифицированного грызуна того же семейства, который не содержит ортолог или гомолог ADAM6 (например, Cricetidae (например, хомяки, крысы и мыши Нового Света, полевки); Muridae (например, настоящие мыши и крысы, карликовые песчанки, иглистые мыши, косматые хомяки)).
Согласно различным вариантам осуществления ортологи ADAM6, их гомологи и фрагменты оценивают в отношении функциональности путем определения того, восстанавливает ли ортолог, гомолог или фрагмент фертильность генетически модифицированного самца отличного от человека животного, у которого отсутствует активность ADAM6 (например, грызуна, например, мыши или крысы, которая содержит нокаут ADAM6 или его ортолога). Согласно различным вариантам осуществления функциональность определяют как способность спермы генетически модифицированного животного, у которого отсутствует эндогенный ADAM6 или его ортолог или гомолог, проходить яйцевод и оплодотворять яйцеклетку того же вида генетически модифицированного животного.
Согласно различным аспектам могут быть получены мыши, которые содержат делеции или замещения эндогенного локуса вариабельной области тяжелой цепи или его частей, которые содержат эктопическую нуклеотидную последовательность, кодирующую белок, обеспечивающий сходные улучшения фертильности ADAM6 мыши (например, его ортолог или гомолог или фрагмент, который является функциональным у самца мыши). Эктопическая нуклеотидная последовательность может содержать нуклеотидную последовательность, кодирующую белок, который представляет собой гомолог или ортолог ADAM6 (или их фрагмент) другой линии или другого вида, например, другого вида грызунов, и которая обеспечивает улучшение фертильности, например, увеличенное количество пометов в течение определенного периода времени и/или увеличенное количество детенышей на помет и/или способность сперматозоида самца мыши проходить через яйцевод мыши для оплодотворения яйцеклетки мыши.
Согласно одному варианту осуществления ADAM6 представляет собой гомолог или ортолог, которые по меньшей мере на 89% - 99% идентичен белку ADAM6 мыши (например, по меньшей мере на 89% - 99% идентичен ADAM6а мыши или ADAM6b мыши). Согласно одному варианту осуществления эктопическая нуклеотидная последовательность кодирует один или несколько белков, независимо выбранных из белка, по меньшей мере на 89% идентичного ADAM6а мыши, белка, по меньшей мере на 89% идентичного ADAM6b мыши, и их комбинации. Согласно одному варианту осуществления гомолог или ортолог представляет собой белок крысы, хомяка, мыши или морской свинки, который является или модифицирован, чтобы быть приблизительно на 89% или более идентичным ADAM6а мыши и/или ADAM6b мыши. Согласно одному варианту осуществления гомолог или ортолог является или по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичен ADAM6а мыши и/или ADAM6b мыши. Согласно конкретному варианту осуществления мышь ADAM6a содержит SEQ ID NO: 1 или ее функциональный фрагмент, и мышь ADAM6b содержит SEQ ID NO: 2 или ее функциональный фрагмент.
Согласно одному аспекту предусмотрены отличные от человека животные, причем отличные от человека животные содержат (a) вставку одного или нескольких генных сегментов VL и JL человека выше не относящейся к человеку константной области тяжелой цепи иммуноглобулина, (b) вставку одного или нескольких генных сегментов VL и JL человека выше не относящейся к человеку константной области легкой цепи иммуноглобулина и (c) нуклеотидную последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент. Согласно одному варианту осуществления не относящиеся к человеку константные области тяжелой и/или легкой цепи представляют собой константные области грызуна (например, выбранные из константных областей мыши, крысы или хомяка). Согласно одному варианту осуществления не относящаяся к человеку константная область легкой цепи представляет собой константную область грызуна. Согласно конкретному варианту осуществления константная область легкой цепи представляет собой область Cκ мыши или Cκ крысы. Согласно конкретному варианту осуществления константная область легкой цепи представляет собой область Cλ мыши или область Cκ крысы. Согласно одному варианту осуществления генные сегменты VL и JL человека представляют собой генные сегменты Vκ и Jκ. Согласно одному варианту осуществления генные сегменты VL и JL человека представляют собой генные сегменты Vλ и Jλ. Согласно одному варианту осуществления отличное от человека животное дополнительно содержит один или несколько генных сегментов DH человека, присутствующих между человека генными сегментами VL и JL. Подходящие отличные от человека животные включают в себя грызунов, например, мышей, крыс и хомяков. Согласно одному варианту осуществления грызун представляет собой мышь или крысу.
Согласно одному варианту осуществления отличное от человека животное содержит по меньшей мере шесть - по меньшей мере 40 генных сегментов Vκ человека и по меньшей мере один - по меньшей мере пять генных сегментов Jκ человека. Согласно конкретному варианту осуществления отличное от человека животное содержит шесть генных сегментов Vκ человека и пять генных сегментов Jκ человека. Согласно конкретному варианту осуществления отличное от человека животное содержит 16 генных сегментов Vκ человека и пять генных сегментов Jκ человека. Согласно конкретному варианту осуществления отличное от человека животное содержит 30 генных сегментов Vκ человека и пять генных сегментов Jκ человека. Согласно конкретному варианту осуществления отличное от человека животное содержит 40 генных сегментов Vκ человека и пять генных сегментов Jκ человека. Согласно различным вариантам осуществления генные сегменты Jκ человека выбраны из Jκ1, Jκ2, Jκ3, Jκ4, Jκ5 и их комбинации.
Согласно одному варианту осуществления нуклеотидная последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент, является эктопической у отличного от человека животного. Согласно одному варианту осуществления нуклеотидная последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент (который является функциональным у отличного от человека животного), присутствует в том же положении по сравнению с не относящимся к человеку локусом ADAM6 дикого типа. Согласно одному варианту осуществления отличное от человека животное представляет собой мышь, и нуклеотидная последовательность кодирует белок ADAM6 мыши или его функциональный фрагмент и присутствует в эктопическом положении в геноме отличного от человека животного. Согласно одному варианту осуществления отличное от человека животное представляет собой мышь, и нуклеотидная последовательность кодирует белок ADAM6 мыши или его функциональный фрагмент и присутствует в пределах генных сегментов иммуноглобулина. Согласно конкретному варианту осуществления генные сегменты иммуноглобулина представляют собой генные сегменты тяжелой цепи отличного от человека животного. Согласно конкретному варианту осуществления генные сегменты иммуноглобулин представляют собой генные сегменты легкой цепи другого вида. Согласно одному варианту осуществления генные сегменты легкой цепи представляют собой генные сегменты κ легкой цепи человека. Согласно одному варианту осуществления мышь содержит эктопическую смежную последовательность, содержащую один или несколько эндогенных нереаранжированных генных сегментов тяжелой цепи, и последовательность ADAM6 находится в пределах эктопической смежной последовательности.
Согласно одному варианту осуществления отличное от человека животное не содержит эндогенный генный сегмент VL и/или JL иммуноглобулина на эндогенном локусе легкой цепи иммуноглобулина. Согласно одному варианту осуществления отличное от человека животное содержит эндогенные генные сегменты VL и/или JL иммуноглобулина, которые являются неспособными к реаранжировке для образования домена VL иммуноглобулина у отличного от человека животного. Согласно одному варианту осуществления все или по существу все эндогенные генные сегменты Vκ и Jκ иммуноглобулина замещают одним или несколькими генными сегментами Vκ и Jκ человека. Согласно одному варианту осуществления все или по существу все эндогенные генные сегменты Vλ и Jλ иммуноглобулина удаляют полностью или частично. Согласно одному варианту осуществления все или по существу все эндогенные генные сегменты VL и JL иммуноглобулина являются интактными у отличного от человека животного, и отличное от человека животное содержит один или несколько генных сегментов Vκ человека и один или несколько генных сегментов Jκ человека, вставленных между эндогенными генными сегментами VL и/или JL иммуноглобулина и эндогенной константной областью легкой цепи иммуноглобулина. Согласно конкретному варианту осуществления интактные эндогенные генные сегменты VL и JL иммуноглобулина делают неспособными к реаранжировке для образования домена VL антитела у отличного от человека животного. Согласно одному варианту осуществления эндогенный локус легкой цепи иммуноглобулина отличного от человека животного представляет собой локус κ легкой цепи иммуноглобулина. Согласно одному варианту осуществления эндогенные генные сегменты VL и JL иммуноглобулина представляют собой генные сегменты Vκ и Jκ.
Согласно одному аспекту предусмотрены клетки и/или ткани, происходящий из не относящихся к человеку животных, описанных в настоящем документе, причем клетки и/или ткани содержат (a) вставку одного или нескольких генных сегментов Vκ и Jκ человека выше не относящейся к человеку константной области легкой цепи иммуноглобулина, (b) вставку одного или нескольких генных сегментов Vκ и Jκ человека выше не относящейся к человеку константной области тяжелой цепи иммуноглобулина, и (c) нуклеотидную последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент. Согласно одному варианту осуществления не относящиеся к человеку константные области тяжелой и/или легкой цепи представляют собой константный области мыши. Согласно одному варианту осуществления не относящиеся к человеку константные области тяжелой и/или легкой цепи представляют собой константные области крысы. Согласно одному варианту осуществления не относящиеся к человеку константные области тяжелой и/или легкой цепи представляют собой константные области хомяка.
Согласно одному варианту осуществления нуклеотидная последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент, является эктопической в клетке и/или ткани. Согласно одному варианту осуществления нуклеотидная последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент, присутствует в таком же положение по сравнению с не относящимся к человеку локусом ADAM6 дикого типа. Согласно одному варианту осуществления не относящаяся к человеку клетка и/или ткань происходит от мыши, и нуклеотидная последовательность кодирует белок ADAM6 мыши или его функциональный фрагмент и присутствует в эктопическом положении. Согласно одному варианту осуществления не относящаяся к человеку клетка и/или ткань происходит от мыши, и нуклеотидная последовательность кодирует белок ADAM6 мыши или его функциональный фрагмент и присутствует в пределах генных сегментов иммуноглобулина. Согласно конкретному варианту осуществления генные сегменты иммуноглобулина представляют собой генные сегменты тяжелой цепи. Согласно конкретному варианту осуществления генные сегменты иммуноглобулина представляют собой генные сегменты генные сегменты легкой цепи. Согласно одному варианту осуществления смежная последовательность эндогенных генных сегментов тяжелой цепи размещена эктопически у отличного от человека животного, причем смежная последовательность эктопически размещенных эндогенных генных сегментов тяжелой цепи содержит ген ADAM6, который является функциональным у мыши (например, у самца мыши).
Согласно одному аспекту предусмотрено применение описанного в настоящем документе отличного от человека животного для получения антигенсвязывающего белка, причем отличное от человека животное экспрессирует (a) антитело, которое содержит (i) легкую цепь иммуноглобулина, которая содержит домен Vκ человека и не относящуюся к человеку константную область легкой цепи и (ii) тяжелую цепь иммуноглобулина, которая содержит домен Vκ человека и не относящуюся к человеку константную область; и (b) белок ADAM6 или его функциональный фрагмент. Согласно одному варианту осуществления антигенсвязывающий белок является относящимся к человеку. Согласно одному варианту осуществления отличное от человека животное представляет собой грызуна, и не относящиеся к человеку константные области представляют собой константные области грызуна. Согласно конкретному варианту осуществления грызун представляет собой мышь.
Согласно одному аспекту предусмотрена не относящаяся к человеку клетка или ткань, происходящая от описанного в настоящем документе отличного от человека животного. Согласно одному варианту осуществления не относящаяся к человеку клетка или ткань содержит один или несколько генных сегментов Vκ иммуноглобулина человека и по меньшей мере один генный сегмент Jκ иммуноглобулина человека, смежный с не относящимся к человеку геном константной области легкой цепи иммуноглобулина, и один или несколько генных сегментов Vκ человека, человека и один или несколько генных сегментов Jκ человека, смежных с не относящимся к человеку геном константной области тяжелой цепи иммуноглобулина, причем клетка или ткань экспрессирует белок ADAM6 или его функциональный фрагмент. Согласно одному варианту осуществления не относящийся к человеку ген константной области легкой цепи представляет собой Сκ мыши.
Согласно одному варианту осуществления нуклеотидная последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент, является эктопической. Согласно одному варианту осуществления нуклеотидная последовательность, которая кодирует ADAM6 белок или ее функциональный фрагмент, расположена в положении, которое является таким же, как и в не относящейся к человеку клетке дикого типа. Согласно различным вариантам осуществления не относящаяся к человеку клетка представляет собой B-клетку мыши. Согласно различным вариантам осуществления не относящаяся к человеку клетка представляет собой эмбриональную стволовую клетку.
Согласно одному варианту осуществления ткань происходит из селезенки, костного мозга или лимфатического узла отличного от человека животного.
Согласно одному аспекту предусмотрено применение клетки или ткани, происходящей от описанного в настоящем документе отличного от человека животного для получения гибридомы или квадромы.
Согласно одному аспекту предусмотрена не относящаяся к человеку клетка, содержащая описанный в настоящем документе модифицированный геном, причем не относящаяся к человеку клетка представляет собой ооцит, зародыш-хозяин или слияние клетки от описанного в настоящем документе отличного от человека животного и клетки от другого отличного от человека животного.
Согласно одному аспекту предусмотрено применение клетки или ткани, полученной от описанного в настоящем документе отличного от человека животного для получения антигенсвязывающего белка человека. Согласно одному варианту осуществления антигенсвязывающий белок человека содержит домен Vκ человека выделенный из описанного в настоящем документе отличного от человека животного.
Согласно одному аспекту предусмотрен способ получения антигенсвязывающего белка, который связывается с представляющим интерес антигеном, причем способ предусматривает (a) воздействие на описанное в настоящем документе отличное от человека животное представляющего интерес антигена, (b) выделение одного или нескольких B-лимфоцитов отличного от человека животного, причем один или несколько B-лимфоцитов экспрессируют связывающий белок VL, который связывает представляющий интерес антиген, и (c) идентификацию последовательности нуклеиновой кислоты, которая кодирует домен VL связывающего белка VL, который связывает представляющий интерес антиген, причем связывающий белок VL содержит домен Vκ человека и не относящийся к человеку константный домен легкой цепи и домен Vκ человека и не относящийся к человеку константный домен тяжелой цепи, и (d) использование последовательности нуклеиновой кислоты согласно (c) с последовательностью нуклеиновой кислоты константной области иммуноглобулина человека для получения антигенсвязывающего белка человека, который связывает представляющий интерес антиген.
Согласно одному варианту осуществления не относящиеся к человеку константный домен легкой цепи связывающего белка VL представляет собой Cκ мыши. Согласно одному варианту осуществления не относящиеся к человеку константный домен тяжелой цепи связывающего белка VL представляет собой Cγ мыши. Согласно одному варианту осуществления отличное от человека животное представляет собой мышь.
Согласно одному аспекту предусмотрен фертильный самец мыши, содержащий модификацию на локусе тяжелой цепи иммуноглобулина, причем фертильный самец мыши содержит эктопическую последовательность ADAM6, которая является функциональной у самца мыши.
Эктопический ADAM6 в модифицированных локусах тяжелой цепи иммуноглобулина
Разработки в области нацеленного воздействия на гены, например, разработка бактериальных искусственных хромосом (BAC), в настоящее время обеспечивают возможность рекомбинации относительно больших геномных фрагментов. Конструирование BAC предоставила возможность получения больших делеций и больших вставок в ES клетки мыши.
Мыши, которые производят человеческие антитела (т.е. вариабельные области человека), были доступны уже в течение определенного времени. Хотя они представляют собой важный прогресс в разработке человеческих терапевтических антител, эти мыши проявляют ряд значительных нарушений, которые ограничивают их применимость. Например, они проявляют нарушенное развитие B-клеток. Нарушенное развитие может быть обусловлено разнообразными различиями между трансгенными мышами и мышами дикого типа.
Существует вероятность, что человеческие антитела не взаимодействуют оптимально с рецепторами пре-B-клеток или B-клеток мыши на поверхности клеток мыши, которые передают сигнал к созреванию, пролиферации или выживанию в ходе клональной селекции. Полностью человеческие антитела могут не взаимодействовать оптимально с Fc рецепторной системой мыши; мыши экспрессируют Fc рецепторы, которые не проявляют соответствия один к одному с Fc рецепторами человека. В конце концов, различные мыши, которые производят полностью человеческие антитела, не включают в себя все истинные мышиные последовательности, например, нижележащие энхансерные элементы и другие контролирующие элементы локуса, которые могут быть необходимы для развития B-клеток дикого типа.
Мыши, которые производят полностью человеческие антитела, как правило, содержат эндогенные локусы иммуноглобулина, которые являются некоторым образом недееспособными, и человеческие трансгены, которые содержат вариабельные и константные генные сегменты иммуноглобулина, вводят в случайное положение в геноме мыши. При условии, что эндогенный локус является достаточно недееспособным, так чтобы не реаранжировать генные сегменты для образования функционального гена иммуноглобулина, цель получения полностью человеческих антител в такой мыши может быть достигнута, хотя и с нарушенным развитием B-клеток.
Несмотря на принуждение производить полностью человеческие антитела из человеческого трансгенного локуса, образование человеческих антител у мыши является очевидно затрудненным процессом. У некоторых мышей процесс является настолько затрудненным, что приводит к формированию химерных человеческих вариабельных/мышиных константных тяжелых цепей (но не легких цепей) посредством механизма транс-переключения. С помощью этого механизм, транскрипты, которые кодируют полностью человеческие антитела, подвергаются переключению изотипа в транс-конфигурации из изотипа человека на изотип мыши. Процесс происходит в транс-конфигурации, поскольку полностью человеческий трансген расположен в отдалении от эндогенного локуса, который сохраняет неповрежденную копию генной константной области тяжелой цепи мыши. Хотя у таких мышей транс-переключение очевидно выражено, явление остается недостаточным для восстановления развития B-клеток, которое остается фактически нарушенным. В любом случае, транс-переключенные антитела, произведенные у таких мышей, сохраняют полностью человеческие легкие цепи, поскольку явление транс-переключения, очевидно, не происходит в отношении легких цепей; транс-переключение, вероятно, основано на переключении последовательностей в используемых эндогенных локусах (хотя и по-другому) в переключении нормального изотипа в цис-конфигурации. Таким образом, даже когда мыши, сконструированные для получения полностью человеческих антител, выбирают механизм транс-переключения для образования антител с константными областями мыши, стратегия остается все еще недостаточной для восстановления нормального развития B-клеток.
Первоочередной задачей в получении основанных на антителах терапевтических средств для людей состоит в получении достаточно большого разнообразия последовательностей вариабельной области иммуноглобулина человека для выявления применимых вариабельных доменов, которые специфически распознают конкретные эпитопы и связывают их с необходимой аффинностью, как правило, но не всегда, с высокой аффинностью. Перед разработкой мышей VELOCIMMUNE® (описанных в настоящем документе), не существовало ни одного указания на то, что мыши, экспрессирующие вариабельные области человека с константными областями мыши, будут проявлять какие-либо значительные отличия от мышей, которые образуют человеческие антитела из трансгена. Это предположение, тем не менее, было неправильным.
Гуманизированные мыши VELOCIMMUNE®, которые содержат точное замещение вариабельных областей иммуноглобулина мыши вариабельными областями иммуноглобулина человека на эндогенных локусах, проявляют удивительное и существенное сходство с мышами дикого типа по отношению к развитию B-клеток. В удивительном и замечательном развитии мыши VELOCIMMUNE® проявили по существу нормальный ответ дикого типа на иммунизацию, который отличался только в одном существенном аспекте от мышей дикого типа мыши - вариабельные области, образованные в ответ на иммунизацию, являются полностью человеческими.
Мыши VELOCIMMUNE® содержат точное, крупномасштабное замещение зародышевых вариабельных областей тяжелой цепи иммуноглобулина мыши (IgH) и легкой цепи иммуноглобулина (например, κ легкой цепи, Igκ) соответствующими вариабельными областями иммуноглобулина человека на эндогенных локусах. В общем, приблизительно шесть миллионов пар нуклеотидов мышиных локусов замещают приблизительно 1,5 миллиона пар нуклеотидов геномной последовательности человека. Это точное замещение дает в результате мышь с гибридными локусами иммуноглобулина, которая образует тяжелые и легкие цепи, которые содержатся вариабельные области человека и константную область мыши. Точное замещение сегментов VH-DH-JH и Vκ-Jκ мыши оставляет фланкирующие мышиные последовательности интактными и функциональными на гибридных локусах иммуноглобулина. Гуморальная иммунная система мыши функционирует как система у мыши дикого типа. Развитие B-клеток не затруднено каким-либо существенным образом, и у мыши при антигенной сенсибилизации образуется богатое разнообразие вариабельных областей человека.
Мыши VELOCIMMUNE® являются возможными, поскольку иммуноглобулиновые генные сегменты тяжелых и κ легких цепей реаранжируются подобным образом у людей и мышей, что не означает, что их локусы являются одинаковыми или даже приблизительно таковы - ясно, что они не такие. Тем не менее, локусы являются достаточно сходными, чтобы гуманизация вариабельного генного локуса тяжелой цепи могла осуществиться путем замещения приблизительно трех миллионов пар нуклеотидов смежной мышиной последовательности, которая содержит все генные сегменты VH, DH и JH, приблизительно одним миллионом оснований смежной геномной последовательности человека, охватывающей, в сущности, эквивалентную последовательность из локуса иммуноглобулина человека.
Согласно некоторым вариантам осуществления дополнительное замещение определенных генных последовательностей константной области мыши генными последовательностями человека (например, замещение последовательности CH1 мыши последовательностью CH1 человека и замещение последовательность CL мыши последовательностью CL человека) дает в результате мышей с гибридными локусами иммуноглобулина, которые образуют антитела, содержащие вариабельные области человека и частично константные области человека, подходящие, например, для образования полностью человеческих фрагментов антител, например, полностью человеческих Fab. Мыши с гибридными локусами иммуноглобулина проявляют нормальную реаранжировку вариабельных генных сегментов, нормальную соматическую гипермутацию и нормальное переключение класса. Эти мыши проявляют гуморальную иммунную систему, которая неотличима от мышей дикого типа, и демонстрируют нормальные клеточные популяции на всех стадиях развития B-клеток и нормальные структуры лимфомных органов - даже когда если у мышей отсутствует полный репертуар вариабельных генных сегментов человека. Иммунизация указанных мышей приводит к устойчивым гуморальным ответам, которые проявляют широкое разнообразие использования вариабельных генных сегментов.
Точное замещение зародышевых генных сегментов вариабельной области мыши обеспечивают получение мышей, которые содержат частично человеческие локусы иммуноглобулина. Поскольку частично человеческие локусы иммуноглобулина нормально реаранжируются, гипермутируют и подвергаются переключению класса, частично человеческие локусы иммуноглобулина образуют у мыши антитела, которые содержат вариабельные области человека. Нуклеотидные последовательности, которые кодируют вариабельные области, могут быть установлены и клонированы, затем слиты (например, в системе in vitro) с любыми предпочтительными последовательностями, например, любым изотипом иммуноглобулина, подходящим для конкретного применения, давая в результате антитело или антигенсвязывающий белок, происходящий полностью из последовательностей человека.
Крупномасштабную гуманизацию с помощью способов рекомбинационный инженерии использовали для модификации мышиных эмбриональных стволовых (ES) клеток для создания уникального локуса тяжелой цепи иммуноглобулина путем точного замещения до трех миллионов пар нуклеотидов локуса тяжелой цепи иммуноглобулина мыши, включая в себя по существу все генные сегменты VH, DH и JH мыши, с помощью сегмента генома человека длиной до 0,5 миллионов пар нуклеотидов, содержащего один из двух повторов, кодирующих по существу все генные сегменты Vκ и Jκ человека. Кроме того, до 0,5 миллиона пар нуклеотидов сегмент генома человека, содержащий один из двух повторов, кодирующих по существу все генные сегменты Vκ и Jκ человека использовали для замещения сегмента локуса κ легкой цепи иммуноглобулина мыши размером 3 миллиона пар нуклеотидов, содержащего по существу все генные сегменты Vκ и Jκ мыши. Мыши с такими замещенными иммуноглобулиновыми локусами могут содержать разрыв или делецию эндогенного локуса ADAM6 мыши, который в норме встречается между расположенным наиболее 3’ генным сегментом VH и наиболее 5’ генным сегментом DH на локусе тяжелой цепи иммуноглобулина мыши. Разрыв в этой области может привести к снижению или подавлению функциональности локуса ADAM6 мыши.
Описываются мыши, которые содержат описанные выше замещенные локусы, и которые также содержат эктопическую последовательность нуклеиновой кислоты, кодирующую мышиный ADAM6, причем мыши проявляют по существу нормальную фертильность. Согласно одному варианту осуществления эктопическая последовательность нуклеиновой кислоты расположена между генным сегментом VL человека и генным сегментом JL человека или выше расположенного наиболее 5’ генного сегмента VL человека на модифицированном эндогенном локусе тяжелой цепи. Направление транскрипции генов ADAM6 может быть противоположным (фиг. 7) или аналогичным (фиг. 8) по отношению к направлению транскрипции окружающих генных сегментов VL человека. Хотя примеры в настоящем документе демонстрируют восстановление фертильности путем размещения эктопической последовательности между указанными генными сегментами VL и JL человека или выше наиболее 5’ генного сегмента VL человека, специалистам в настоящей области будет понятно, что размещение эктопической последовательности на любом подходящем транскрипционно пермиссивном локусе в мышином геноме (или даже экстрахромосомно), как предполагается, приведет к подобному восстановлению фертильности у самца мыши. Согласно различным вариантам осуществления эктопическая последовательность нуклеиновой кислоты выбрана из SEQ ID NO: 3, SEQ ID NO: 4 и SEQ ID NO: 5, причем эктопическая последовательность кодирует один или несколько белков ADAM6, причем один или несколько белков ADAM6 содержат SEQ ID NO: 1, SEQ ID NO: 2 или их комбинацию.
Явление дополнения у мыши, у которой отсутствует функциональный локус ADAM6, с помощью эктопической последовательности, которая содержит ген ADAM6 мыши или его ортолог или гомолог или функциональный фрагмент, представляет собой общий способ, который является применимым для восстановления любых мышей с нефункциональными или минимально функциональными эндогенными локусами ADAM6. Таким образом, фертильность большого количества мышей, которые содержат разрушающую ADAM6 модификацию локуса тяжелой цепи иммуноглобулина, может быть восстановлена с помощью композиций и способов по настоящему изобретению. Соответственно, настоящее изобретение предусматривает мышей с широким разнообразием модификаций локусов тяжелой цепи иммуноглобулина, которые нарушают функцию эндогенного ADAM6. Некоторые (не ограничивающие) примеры представлены в настоящем описании. В дополнение к описанным мышам относящиеся к ADAM6 композиции и способы могут использоваться в многочисленных применениях, например, при модификации локуса тяжелой цепи разнообразными путями.
Согласно одному аспекту предусматривается мышь, которая содержит эктопическую последовательность ADAM6, которая кодирует функциональный белок ADAM6 (или его ортолог или гомолог или функциональный фрагмент), замещение всех или по существу всех VH генных сегментов мыши одним или несколькими генными сегментами VL человека, замещение всех или по существу всех генных сегментов DH и генных сегментов JH мыши генными сегментами JL человека; причем мышь не содержит CH1 и/или шарнирную область. Согласно одному варианту осуществления мышь производит один связывающий белок вариабельного домена, который представляет собой димер цепей иммуноглобулина, выбранных из: (a) VL человека -CH1 мыши -CH2 мыши -CH3 мыши; (b) VL человека -шарнир мыши -CH2 мыши -CH3 мыши; и, (c) VL человека -CH2 мыши -CH3 мыши.
Согласно одному аспекту нуклеотидная последовательность, которая восстанавливает фертильность, расположена в пределах последовательности вариабельной области легкой цепи иммуноглобулина человека (например, между генными сегментами Vκ4-1 и Jκ1 человека) у мыши, которая содержит замещение всех или по существу всех вариабельных генных сегментов тяжелой цепи иммуноглобулина мыши (mVH, mDH и mJH) одним или нескольким вариабельными генными сегментами κ легкой цепи иммуноглобулина человека (hVκ и hJκ), и мышь дополнительно содержит замещение всех или по существу всех вариабельных генных сегментов κ легкой цепи иммуноглобулина мыши (mVκ, mJκ) одним или несколькими вариабельными генными сегментами κ легкой цепи иммуноглобулина человека (hVκ и hJκ).
Согласно одному аспекту функциональный локус ADAM6 мыши (или его ортолог или гомолог или функциональный фрагмент) может быть помещен среди генных сегментов VL человека или выше наиболее 5’ генного сегмента VL человека, причем генные сегменты VL человека замещают эндогенные генные сегменты VH. Согласно одному варианту осуществления все или по существу все генные сегменты VH мыши удаляют и замещают одним или несколькими генными сегментами VL человека, и локус ADAM6 мыши расположен, непосредственно примыкая к 5’ концу наиболее 5’ генных сегментов VL человека, или между двумя генными сегментами VL человека. Согласно конкретному варианту осуществления локус ADAM6 мыши расположен между двумя генными сегментами VL вблизи 3’ конца вставленных генных сегментов VL человека. Согласно конкретному варианту осуществления после этого расположение генных сегментов VL человека является следующим (сверху вниз относительно направления транскрипции генных сегментов VL человека): Vκ5-2 человека - локус ADAM6 мыши -Vκ4-1 человека. Согласно конкретному варианту осуществления расположение генных сегментов VL человека после этого является следующим (сверху вниз относительно направления транскрипции генных сегментов VL человека): локус ADAM6 мыши -Vκ2-40 человека, причем Vκ2-40 человека является наиболее 5’ генным сегментом VL человека на модифицированном локусе тяжелой цепи иммуноглобулина. Согласно одному варианту осуществления ориентация одного или нескольких ADAM6a мыши и ADAM6b мыши локуса ADAM6 мыши является противоположной по отношению к направлению транскрипции по сравнению с ориентацией генных сегментов VL человека. Согласно одному варианту осуществления ориентация одного или нескольких ADAM6a мыши и ADAM6b мыши локуса ADAM6 мыши является аналогичной относительно направления транскрипции по сравнению с ориентацией генных сегментов VL человека.
Согласно одному аспекту функциональный локус ADAM6 мыши (или его ортолог или гомолог или функциональный фрагмент) может быть расположен между генным сегментом VL человека и генным сегментом JL человека (т.е. в межгенной области между наиболее 3’ генным сегментом VL человека и наиболее 5’ генным сегментом JL), причем генные сегменты VL и JL человека замещают эндогенные генные сегменты VH. Согласно одному варианту осуществления все или по существу все генные сегменты VH мыши удаляют и замещают одним или нескольким генными сегментами VL человека и одним или несколькими генными сегментами JL человека, и локус ADAM6 мыши расположен, непосредственно примыкая к 3’ концу наиболее 3’ генного сегмента VL человека и непосредственно примыкая к 5’ концу наиболее 5’ генного сегмента JL человека. Согласно конкретному варианту осуществления один или несколько генных сегментов VL человека и один или несколько генных сегментов JL человека представляют собой генные сегменты Vκ и Jκ. Согласно конкретному варианту осуществления расположение генных сегментов VL человека после этого является следующим (сверху вниз относительно направления транскрипции генных сегментов VL человека): Vκ4-1 человека - локус ADAM6 мыши - Jκ1 человека. Согласно одному варианту осуществления ориентация одного или нескольких ADAM6a мыши и ADAM6b мыши локуса ADAM6 мыши является противоположной относительно направления транскрипции по сравнению с ориентацией генных сегментов VL человека. Согласно одному варианту осуществления ориентация одного или нескольких ADAM6a мыши и ADAM6b мыши локуса ADAM6 мыши является аналогичной относительно направления транскрипции по сравнению с ориентацией генных сегментов VL человека.
Мышь, модифицированная с помощью одного или нескольких генных сегментов VL человека (например, сегментов Vκ или Vλ), замещающих все или по существу все эндогенные VH генные сегменты, может быть модифицирована так, чтобы или сохранять эндогенный локус ADAM6, как описано выше, например, путем использования нацеливающего вектора, характеризующегося расположенным ниже гомологичным плечом, которое содержит локус ADAM6 мыши или его функциональный фрагмент, или замещать поврежденный локус ADAM6 мыши эктопической последовательностью, расположенной между двумя генными сегментами VL человека или между генными сегментами VL человека и генным сегментом DH (либо человека, либо мыши, например, Vλ + m/hDH), или генным сегментом J (либо человека, либо мыши, например, Vκ + JH). Согласно одному варианту осуществления замещение предусматривает два или более генных сегментов VL человека, и локус ADAM6 мыши или его функциональный фрагмент расположен между двумя наиболее 3’ генными сегментами VL. Согласно конкретному варианту осуществления расположение генных сегментов VL человека после этого является следующим (сверху вниз относительно направления транскрипции генных сегментов человека): VL3’-1 человека - локус ADAM6 мыши -VL3’ человека. Согласно одному варианту осуществления ориентация одного или нескольких из мышиного ADAM6a и мышиного ADAM6b локуса ADAM6 мыши является противоположной относительно направления транскрипции по сравнению с ориентацией VL генных сегментов человека. Альтернативно, локус ADAM6 мыши может быть помещен в межгенную область между наиболее 3’ генным сегментом VL человека и наиболее 5’ генным сегментом JL.
Согласно одному аспекту предусматривается мышь с замещением одного или нескольких эндогенных генных сегментов VH, и которая содержит по меньшей мере один эндогенный генный сегмент DH. У такой мыши модификация эндогенных генных сегментов VH мыши может содержать модификацию одного или нескольких из наиболее 3’ генных сегментов VH, но не наиболее 5’ генный сегмент DH, причем нужно быть внимательным, чтобы модификация одного или нескольких наиболее 3’ VH генных сегментов не разрушила эндогенный локус ADAM6 или не сделала его нефункциональным. Например, согласно одному варианту осуществления мышь содержит замещение всех или по существу всех эндогенных VH генных сегментов одним или несколькими генными сегментами VL человека, и мышь содержит один или несколько эндогенных генных сегментов DH и функциональный эндогенный локус ADAM6.
Согласно другому варианту осуществления мышь содержит модификацию эндогенных наиболее 3’ генных сегментов VH, и модификацию одного или нескольких эндогенных генных сегментов DH, и модификацию проводят так, чтобы сохранить целостность эндогенного локуса ADAM6 до такой степени, чтобы эндогенный ADAM6 локус оставался функциональным. Согласно одному примеру такую модификацию выполняют в две стадии: (1) замещение наиболее 3’ эндогенных генных сегментов VH одним или несколькими генными сегментами VL человека с использованием нацеливающего вектора с вышележащим гомологичным плечом и нижележащим гомологичным плечом, причем нижележащее гомологичное плечо содержит весь или часть функционального локуса ADAM6 мыши; (2) затем замещение и эндогенного генного сегмента DH с помощью нацеливающего вектора, содержащего вышележащее гомологичное плечо, которое содержит весь или функциональную часть локуса ADAM6 мыши.
Согласно различным аспектам использование мышей, которые содержат эктопическую последовательность, которая кодирует белок ADAM6 мыши или его ортолог или гомолог или функциональные гомолог, является применимым, если модификации нарушают функцию эндогенного ADAM6 мыши. Вероятность разрушения функции эндогенного ADAM6 мыши является высокой при осуществлении модификаций в отношении локусов иммуноглобулина мыши, в частности, при модификации вариабельных областей тяжелой цепи иммуноглобулина мыши и окружающих последовательностей. Следовательно, такие мыши представляют конкретное преимущество при получении мышей с локусами тяжелой цепи иммуноглобулина, которые подвергнуты делеции полностью или частично, которые являются гуманизированными полностью или частично или замещенными (например, Vκ или Vλ последовательностями) полностью или частично. Способы получения генетических модификаций, описанных для мышей, описанных ниже, известны специалистам в настоящей области техники.
Мыши, содержащие эктопическую последовательность, кодирующую белок ADAM6 мыши или по существу идентичный или сходный белок, который обеспечивает улучшения фертильности белка ADAM6 мыши, являются особенно применимыми в связи с модификациями относительно вариабельной генного локуса тяжелой цепи иммуноглобулина мыши, которые разрушают или подвергают делеции эндогенную последовательность ADAM6. Несмотря на то, что изначально описаны в связи с мышами, которые экспрессируют антитела с вариабельными областями человека и константными областями мыши, такие мыши применимы в отношении любых генетических модификаций, которые нарушают эндогенные гены ADAM6. Специалистам в настоящей области понятно, что это охватывает широкое разнообразие генетически модифицированных мышей, которые содержат модификации вариабельных генных локусов тяжелой цепи иммуноглобулина мыши. Они предусматривают, например, мышей с делецией или замещением всех или части генных сегментов тяжелой цепи иммуноглобулина мыши, независимо от других модификаций. Не ограничивающие примеры описаны ниже.
Согласно некоторым аспектам предусматриваются генетически модифицированные мыши, которые содержат эктопический ген ADAM6 мыши, грызуна или другого животного (или ортолог или гомолог или фрагмент), функциональный у мыши, и один или несколько генных сегментов вариабельной и/или константной области иммуноглобулина человека. Согласно различным вариантам осуществления другие ортологи или гомологи или фрагменты гена ADAM6, функциональные у мыши, могут включать в себя последовательности от коров, собак, примата, кролика или другие не относящиеся к человеку последовательности.
Согласно одному аспекту предусматривается мышь, которая содержит эктопическую последовательность ADAM6, которая кодирует функциональный белок ADAM6, замещение всех или по существу всех генных сегментов VH мыши одним или несколькими VL генными сегментами человека; замещение всех или по существу всех генных сегментов DH и JH мыши одним или несколькими генными сегментами JL человека.
Согласно одному варианту осуществления мышь дополнительно содержит замещение нуклеотидной последовательности CH1 мыши нуклеотидной последовательностью CH1 человека. Согласно одному варианту осуществления мышь дополнительно содержит замещение шарнирной нуклеотидной последовательности мыши шарнирной нуклеотидной последовательностью человека. Согласно одному варианту осуществления мышь дополнительно содержит замещение вариабельного локуса легкой цепи иммуноглобулина (VL и JL) вариабельным локусом легкой цепи иммуноглобулина человека. Согласно одному варианту осуществления мышь дополнительно содержит замещение нуклеотидной последовательности константной области легкой цепи иммуноглобулина мыши нуклеотидной последовательностью константной области легкой цепи иммуноглобулина человека. Согласно конкретному варианту осуществления VL, JL и CL представляют собой последовательности κ легкой цепи иммуноглобулина. Согласно конкретному варианту осуществления мышь содержит последовательность константной области иммуноглобулина CH2 мыши и CH3 мыши, слитую с шарнирной последовательностью человека и последовательностью CH1 человека, так что локусы иммуноглобулина мыши реаранжируются для образования гена, который кодирует связывающий белок, содержащий (a) тяжелую цепь, которая содержит вариабельную область человека, область CH1 человека, шарнирную область человека и область CH2 мыши и область CH3 мыши; и (b) ген, который кодирует легкую цепь иммуноглобулина, которая содержит вариабельный домен человека и константную область человека.
Согласно одному аспекту предусматривается мышь, которая содержит эктопическую последовательность ADAM6, которая кодирует функциональный белок ADAM6, замещение всех или по существу всех генных сегментов VH мыши одним или несколькими генными сегментами VL человека, и необязательно замещение всех или по существу всех генных сегментов DH и/или генных сегментов JH одним или несколькими генными сегментами DH человека и/или генными сегментами JH человека, или необязательно замещение всех или по существу всех генных сегментов DH и генных сегментов JH одним или несколькими генными сегментами JL человека.
Согласно одному варианту осуществления мышь содержит замещение всех или по существу всех генных сегментов VH, DH и JH мыши одним или несколькими VL, одним или несколькими DH и одним или несколькими генными сегментами J (например, Jκ или Jλ), причем генные сегменты функционально связаны с шарнирной областью мыши, причем мышь образует реаранжированный ген цепи иммуноглобулина, который содержит, от 5’ к 3’ в направлении транскрипции, VL человека -DH человека или мыши -J человека или мыши -шарнир мыши -CH2 мыши -CH3 мыши. Согласно одному варианту осуществления J область представляет собой Jκ область человека. Согласно одному варианту осуществления J область представляет собой JH область человека. Согласно одному варианту осуществления J область представляет собой Jλ область человека. Согласно одному варианту осуществления VL область человека выбрана из Vλ области человека и Vκ области человека.
Согласно конкретным вариантам осуществления мышь экспрессирует антитело с одним вариабельным доменом, содержащее константную область мыши или человека и вариабельную область, происходящую из Vκ человека, DH человека и Jκ человека; Vκ человека, DH человека и JH человека; Vλ человека, DH человека и Jλ человека; Vλ человека, DH человека и JH человека; Vκ человека, DH человека и Jλ человека; Vλ человека, DH человека и Jκ человека. Согласно конкретному варианту осуществления, последовательности распознавания рекомбинации модифицируют так, чтобы обеспечить осуществление продуктивных реаранжировок между указанными генными сегментами V, D и J или между указанными генными сегментами V и J.
Согласно одному аспекту предусматривается мышь, которая содержит эктопическую последовательность ADAM6, которая кодирует функциональный белок ADAM6 (или его ортолог или гомолог или функциональный фрагмент), замещение всех или по существу всех генных сегментов VH мыши одним или несколькими VL генными сегментами человека, замещение всех или по существу всех генных сегментов DH и генных сегментов JH мыши генными сегментами JL человека; причем у мыши отсутствует CH1 и/или шарнирная область.
Согласно одному варианту осуществления у мыши отсутствует последовательность, кодирующая домен CH1. Согласно одному варианту осуществления у мыши отсутствует последовательность, кодирующая шарнирную область. Согласно одному варианту осуществления у мыши отсутствует последовательность, кодирующая домен CH1 и шарнирную область.
Согласно конкретному варианту осуществления мышь экспрессирует связывающий белок, который содержит вариабельный домен легкой цепи (λ или κ) иммуноглобулина человека, слитый с доменом CH2 мыши, который прикреплен к домену CH3 мыши.
Согласно одному аспекту предусматривается мышь, которая содержит эктопическую последовательность ADAM6, которая кодирует функциональный белок ADAM6 (или его ортолог или гомолог или функциональный фрагмент), замещение всех или по существу всех генных сегментов VH мыши одним или несколькими генными сегментами VL человека, замещение всех или по существу всех генных сегментов DH и JH мыши генными сегментами JL человека.
Согласно одному варианту осуществления мышь содержит делецию генной последовательности константной области тяжелой цепи иммуноглобулина, кодирующей область CH1, шарнирную область, CH1 и шарнирную область или область CH1 и шарнирную область и область CH2.
Согласно одному варианту осуществления мышь образует связывающий один белок вариабельного домена, содержащий гомодимер, выбранный из следующего: (a) VL человека - CH1 мыши - CH2 мыши - CH3 мыши; (b) VL человека - шарнир мыши - CH2 мыши - CH3 мыши; (c) VL человека - CH2 мыши - CH3 мыши.
Согласно одному аспекту предусмотрено отличное от человека животное, содержащее модифицированный локус тяжелой цепи иммуноглобулина, причем модифицированный локус тяжелой цепи иммуноглобулина содержит не относящуюся к человеку последовательность ADAM6 или ее ортолог или гомолог.
Согласно одному варианту осуществления отличное от человека животное представляет собой грызуна, выбранного из мыши, крысы и хомяка.
Согласно одному варианту осуществления не относящийся к человеку ортолог или гомолог ADAM6 представляет собой последовательность, которая является ортологичной и/или гомологичной последовательности ADAM6 мыши, причем ортолог или гомолог является функциональным у отличного от человека животного.
Согласно одному варианту осуществления отличное от человека животное выбрано из мыши, крысы и хомяка, и ортолог или гомолог ADAM6 происходит от отличного от человека животного, выбранного из мыши, крысы и хомяка. Согласно конкретному варианту осуществления отличное от человека животное представляет собой мышь, и ортолог или гомолог ADAM6 происходит от животного, которое выбрано из другого вида мыши, крысы и хомяка. Согласно конкретному варианту осуществления отличное от человека животное представляет собой крысу, и ортолог или гомолог ADAM6 происходит от грызуна, который выбран из другого вида крысы, мыши и хомяка. Согласно конкретному варианту осуществления отличное от человека животное представляет собой хомяка, и ортолог или гомолог ADAM6 происходит от грызуна, который выбран из другого вида хомяка, мыши и крысы.
Согласно конкретному варианту осуществления отличное от человека животное происходит из подотряда Myomorpha, и последовательность ADAM6 происходит от животного, выбранного из грызуна надсемейства Dipodoidea и грызуна надсемейства Muroidea. Согласно конкретному варианту осуществления грызун представляет собой мышь надсемейства Muroidea, и ортолог или гомолог ADAM6 происходит от мыши или крысы или хомяка надсемейства Muroidea.
Согласно одному варианту осуществления модифицированный локус тяжелой цепи иммуноглобулина содержит один или несколько генных сегментов VL человека и один или несколько генных сегментов JL человека. Согласно конкретному варианту осуществления один или несколько генных сегментов VL человека и один или несколько генных сегментов Jλ человека функционально связаны с одним или несколькими генами константной области человека, являющихся химерными и/или относящихся к грызуну (например, мыши или крысе). Согласно одному варианту осуществления гены константной области являются относящимися к мыши. Согласно одному варианту осуществления гены константной области являются относящимися к крысе. Согласно одному варианту осуществления гены константной области являются относящимися к хомяку. Согласно одному варианту осуществления гены константной области содержат последовательность, выбранную из шарнира, CH2, CH3 и их комбинации. Согласно конкретному варианту осуществления гены константной области содержат шарнир, последовательность CH2 и CH3. Согласно одному варианту осуществления генные сегменты VL и JL человека представляют собой генные сегменты Vκ и Jκ человека.
Согласно одному варианту осуществления не относящаяся к человеку последовательность ADAM6 является смежной с последовательностью легкой цепи иммуноглобулина человека. Согласно одному варианту осуществления не относящаяся к человеку последовательность ADAM6 расположена в пределах последовательности легкой цепи иммуноглобулина человека. Согласно конкретному варианту осуществления последовательность легкой цепи иммуноглобулина человека содержит генный сегмент V и/или J.
Согласно одному варианту осуществления не относящаяся к человеку последовательность ADAM6 расположена бок о бок с генным сегментом V. Согласно одному варианту осуществления не относящаяся к человеку последовательность ADAM6 расположена между двумя генными сегментами V. Согласно одному варианту осуществления не относящаяся к человеку последовательность ADAM6 расположена бок о бок между генным сегментом V и J. Согласно одному варианту осуществления последовательность ADAM6 мыши расположена бок о бок между двумя генными сегментами J.
Согласно одному аспекту предусмотрено генетически модифицированное отличное от человека животное, содержащее B-клетку, которая экспрессирует домен VL человека, когнатный домену VL человека из локуса иммуноглобулина, причем отличное от человека животное экспрессирует не относящийся к иммуноглобулину не относящийся к человеку белок из локуса иммуноглобулина. Согласно одному варианту осуществления не относящийся к иммуноглобулину не относящийся к человеку белок представляет собой белок ADAM. Согласно конкретному варианту осуществления белок ADAM представляет собой белок ADAM6 или его гомолог или ортолог или функциональный фрагмент.
Согласно одному варианту осуществления отличное от человека животное представляет собой грызуна (например, мышь или крысу). Согласно одному варианту осуществления грызун происходит из семейства Muridae. Согласно одному варианту осуществления грызун происходит из подсемейства Murinae. Согласно конкретному варианту осуществления грызун подсемейства Murinae выбран из мыши и крысы.
Согласно одному варианту осуществления не относящийся к иммуноглобулину не относящийся к человеку белок представляет собой белок грызуна. Согласно одному варианту осуществления грызун происходит из семейства Muridae. Согласно одному варианту осуществления грызун происходит из подсемейства Murinae. Согласно конкретному варианту осуществления грызун выбран из мыши, крысы и хомяка.
Согласно одному варианту осуществления домены VL человека прикреплены напрямую или через линкер к последовательности константного домена иммуноглобулина. Согласно конкретному варианту осуществления последовательность константного домена содержит последовательность, выбранную из шарнира, CH2, CH3 и их комбинации. Согласно конкретному варианту осуществления домен VL человека выбран из домена Vκ или Vλ.
Согласно различным вариантам осуществления домены VL человека представляют собой домены Vκ человека.
Согласно одному аспекту предусмотрено генетически модифицированное отличное от человека животное, содержащее в своей зародышевой линии последовательность иммуноглобулина человека, причем сперма самца отличного от человека животного характеризуется нарушением миграции in vivo. Согласно одному варианту осуществления нарушение миграции in vivo предусматривает неспособность спермы самца отличного от человека животного мигрировать из матки через яйцевод самки отличного от человека животного того же вида. Согласно одному варианту осуществления отличное от человека животное не содержит нуклеотидную последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент. Согласно конкретному варианту осуществления белок ADAM6 или его функциональный фрагмент включает в себя белок ADAM6a и/или ADAM6b или его функциональный фрагменты. Согласно одному варианту осуществления отличное от человека животное представляет собой грызуна. Согласно конкретному варианту осуществления грызун выбран из мыши, крысы и хомяка.
Согласно одному аспекту предусмотрено отличное от человека животное, содержащее последовательность иммуноглобулина человека, смежную с не относящейся к человеку последовательностью, которая кодирует белок ADAM6 или его ортолог или гомолог или функциональный фрагмент. Согласно одному варианту осуществления отличное от человека животное представляет собой грызуна. Согласно конкретному варианту осуществления грызун выбран из мыши, крысы и хомяка.
Согласно одному варианту осуществления последовательность иммуноглобулина человека представляет собой последовательность легкой цепи иммуноглобулина. Согласно одному варианту осуществления последовательность иммуноглобулина содержит один или несколько генных сегментов VL. Согласно одному варианту осуществления последовательность иммуноглобулина человека содержит один или несколько генных сегментов JL. Согласно одному варианту осуществления последовательность иммуноглобулина человека содержит один или несколько генных сегментов VL и один или несколько генных сегментов JL. Согласно различным вариантам осуществления генные сегменты VL и JL человека представляют собой генные сегменты Vκ и Jκ человека.
Согласно одному аспекту предусмотрена мышь с недееспособным эндогенным локусом тяжелой цепи иммуноглобулина, содержащим недееспособный или удаленный эндогенный локус ADAM6, причем мышь содержит последовательность нуклеиновой кислоты, которая экспрессирует относящееся к человеку или мыши или относящееся к человеку/мыши или другое химерное антитело. Согласно одному варианту осуществления последовательность нуклеиновой кислоты присутствует на интегрированном трансгене, который случайным образом интегрирован в мышиный геном. Согласно одному варианту осуществления последовательность нуклеиновой кислоты находится на эписоме (например, хромосоме), не встречающейся у мыши дикого типа.
Согласно одному аспекту предусмотрена мышь с недееспособным эндогенным локусом тяжелой цепи иммуноглобулина, содержащим функциональный эндогенный локус ADAM6, причем мышь содержит последовательность нуклеиновой кислоты, которая экспрессирует относящееся к человеку или мыши или относящееся к человеку/мыши или другое химерное антитело. Согласно одному варианту осуществления последовательность нуклеиновой кислоты присутствует на эндогенном локусе тяжелой цепи иммуноглобулина в положении выше относительно одного или нескольких эндогенных генов константной области тяжелой цепи. Согласно одному варианту осуществления последовательность нуклеиновой кислоты присутствует на интегрированном трансгене, который случайным образом интегрирован в мышиный геном. Согласно одному варианту осуществления последовательность нуклеиновой кислоты находится на эписоме (например, хромосоме), не встречающейся у мыши дикого типа.
Биспецифические связывающие белки
Описанные в настоящем документе связывающие белки и кодирующие их нуклеотидные последовательности могут использоваться для получения мультиспецифических связывающих белков, например, биспецифических связывающих белков. Согласно настоящему аспекту первый полипептид, состоящий по существу из первого домена VL, слитого с областью CH, может ассоциироваться со вторым полипептидом, состоящим по существу из второго домена VL, слитого с областью CH. Если первый домен VL и второй домен VL специфически связывают различный эпитоп, то биспецифическая связывающая молекула может быть образована с использованием двух доменов VL. Область CH может быть одинаковой или различной. Согласно одному варианту осуществления, например, одна из областей CH может быть модифицирована так, чтобы удалять связывающую белок A детерминанту, тогда как другая константная область тяжелой цепи не модифицирована таким образом. Это конкретное расположение упрощает выделение биспецифическогой связывающего белка, например, из смеси гомодимеров (например, гомодимеров первого или второго полипептидов).
Согласно одному аспекту описанные в настоящем документе способы и композиции используются для получения биспецифических связывающих белков. Согласно настоящему аспекту каждый из первого VL, который слит с областью CH, и второго VL, который слит с областью CH, независимо клонируют в одной рамке считывания с последовательностью IgG человека аналогичного изотипа (например, IgG1, IgG2, IgG3 или IgG4 человека). Первый VL специфически связывает первый эпитоп, и второй VL специфически связывает второй эпитоп. Первый и второй эпитопы могут находиться на различных антигенах или на одном и том же антигене.
Согласно одному варианту осуществления изотип IgG области CH, слитой с первым VL, и изотип IgG области CH, слитой со вторым VL, представляют собой один и тот же изотип, но отличающийся в том, что один изотип IgG содержит по меньшей мере одно аминокислотное замещение. Согласно одному варианту осуществления по меньшей мере одно аминокислотное замещение делает тяжелую цепь, содержащую замещение, неспособной или по существу неспособной связывать белок A по сравнению с тяжелой цепью, которая не содержит замещение.
Согласно одному варианту осуществления первая область CH содержит первый домен CH3 IgG человека, выбранного из IgG1, IgG2 и IgG4; и вторая область CH содержит второй домен CH3 IgG человека, выбранного из IgG1, IgG2 и IgG4, причем второй домен CH3 содержит модификацию, которая снижает или устраняет связывание второго домена CH3 с белком A.
Согласно одному варианту осуществления второй домен CH3 содержит модификацию 435R, пронумерованную согласно индексу EU по Kabat. Согласно другому варианту осуществления второй домен CH3 дополнительно содержит модификацию 436F, пронумерованную согласно индексу EU по Kabat.
Согласно одному варианту осуществления второй домен CH3 представляет собой домен CH3 IgG1 человека, который содержит модификацию, выбранную из группы, состоящей из D356E, L358M, N384S, K392N, V397M и V422I, пронумерованных согласно индексу EU по Kabat.
Согласно одному варианту осуществления второй домен CH3 представляет собой домен CH3 IgG2 человека, который содержит модификацию, выбранную из группы, состоящей из N384S, K392N, и V422I, пронумерованных согласно индексу EU по Kabat.
Согласно одному варианту осуществления второй домен CH3 представляет собой домен CH3 IgG4 человека, который содержит модификацию, выбранную из группы, состоящей из Q355R, N384S, K392N, V397M, R409K, E419Q и V422I, пронумерованных согласно индексу EU по Kabat.
Согласно одному варианту осуществления связывающий белок содержит области CH с одной или несколькими модификациями, указанными в настоящем документе, причем константная область связывающего белка является неиммуногенной или по существу неиммуногенной у человека. Согласно конкретному варианту осуществления области CH содержат аминокислотные последовательности, которые не представляют иммуногенный эпитоп у человека. Согласно другому конкретному варианту осуществления связывающий белок содержит область CH, которая не встречается в относящейся к человеку тяжелой цепи дикого типа, и область CH не содержит последовательность, которая образует T-клеточный эпитоп.
ПРИМЕРЫ
Следующие примеры предусмотрены для описания получения и применения способов и композиций согласно настоящему изобретению, и не предусмотрены для ограничения объема того, что авторы настоящего изобретения рассматривают как свое изобретение. Если не указано иное, температура указана в градусах Цельсия и давление равно или близко атмосферному.
Пример 1. Введение генных сегментов легкой цепи человека в локус тяжелой цепи
Получали различные нацеливающие конструкты с использованием технологии генной инженерии VELOCIGENE® (см., например, патент США № 6586251 и Valenzuela et al. (2003), High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nat Biotechnol 21:652-659) для модификации геномных библиотек мышиных бактериальных искусственных хромосом (BAC). BAC ДНК мыши модифицировали путем гомологичной рекомбинацией для инактивации эндогенного локуса тяжелой цепи посредством направленной делеции генных сегментов VH, DH и JH для обеспечения вставки нереаранжированных зародышевых генных последовательностей κ легкой цепи человека (например, смотрите верхнюю часть фиг. 2).
Кратко, локус тяжелой цепи мыши удаляли за два последовательных нацеливающих события с использованием опосредованной рекомбиназой рекомбинации. Первое нацеливающее событие предусматривало нацеливание на 5’ конец локуса тяжелой цепи мыши с использованием нацеливающего вектора, содержащего в направлении 5’ - 3’ 5’ гомологичное плечо мыши, сайт распознавания рекомбиназы, неомициновую кассету и 3’ гомологичное плечо. 5’ и 3’ гомологичные плечи содержали последовательность 5’ относительно локуса тяжелой цепи мыши. Второй нацеливающее событие предусматривало нацеливание на 3’ конец локуса тяжелой цепи мыши в области генных сегментов JH с использованием второго нацеливающего вектора, который содержал в направлении 5’ - 3’ 5’ гомологичное плечо мыши, 5’ сайт распознавания рекомбиназы, второй сайт распознавания рекомбиназы, гигромициновую кассету, третий сайт распознавания рекомбиназы и 3’ гомологичное плечо мыши. 5’ и 3’ гомологичные плечи содержали последовательность, фланкирующую генные сегменты JH мыши и 5’ относительно интронного энхансера и константных областей. Положительные ES клетки, содержащие модифицированный локус тяжелой цепи, нацеленные с помощью обоих нацеливающих векторов (описанных выше), подтверждали с помощью способов кариотипирования. ДНК затем выделяли из дважды нацеленных ES клеток и подвергали обработке рекомбиназой, тем самым опосредуя делецию геномной ДНК локуса тяжелой цепи мыши между 5’ сайтом распознавания рекомбиназы в первом нацеливающем векторе и 5’ сайтом распознавания рекомбиназы во втором нацеливающем векторе, оставляя один сайт распознавания рекомбиназы и гигромициновую кассету, фланкированную двумя сайтами распознавания рекомбиназы (верхняя часть фиг. 2). Таким образом, создавали модифицированный локус тяжелой цепи мыши, содержащий интактные гены CH, для последовательной вставки зародышевых κ генных сегментов человека точным способом с использованием нацеливающих векторов, описанных ниже.
Четыре отдельных нацеливающих вектора конструировали для последовательной вставки 40 генных сегментов Vκ человека и пяти генных сегментов человека Jκ в инактивированный локус тяжелой цепи мыши (описанный выше) с использованием стандартных молекулярных техник, принятых в настоящей области техники (фиг. 2). κ генные сегменты человека, используемые для конструирования четырех нацеливающих конструктов, встречаются в природе в проксимальном контиге зародышевого локуса κ легкой цепи человека (нижняя часть фиг. 1 и таблица 1).
Конструировали геномный фрагмент человека, составляющий ~110499 п.н., содержащий первые шесть генных сегментов Vκ человека и пять генных сегментов Jκ человека, чтобы он содержал сайт PI-SceI на 431 п.н. ниже (3’) относительно генного сегмента Jκ5 человека. Другой сайт PI-SceI конструировали на 5’ конце ~7852 п.н. геномного фрагмента, содержащего интронный энхансер тяжелой цепи мыши, область переключения IgM (Sµ) и ген IgM локуса тяжелой цепи мыши. Этот фрагмент мыши использовали в качестве 3’ гомологичного плеча путем лигирования с ~110,5 т.п.н. фрагментом человека, что создавало 3’ участок соединения, содержащий, в направлении 5’ - 3’, ~110,5 т.п.н. геномной последовательности локуса κ легкой цепи человека, содержащей первые шесть последовательных генных сегментов Vκ и пять генных сегментов Jκ, сайт PI-SceI, ~7,852 п.н. последовательности тяжелой цепи мыши, содержащей интронный энхансер мыши, Sμ и константный ген IgM мыши. Выше (5’) относительно генного сегмента Vκ1-6 человека находилось добавление 3710 п.н. κ последовательности человека перед началом 5’ гомологичного плеча мыши, которое содержало 19752 п.н. геномной ДНК мыши, соответствующих последовательности 5’ локуса тяжелой цепи мыши. Между 5’ гомологичным плечом и началом κ последовательности человека находилась неомициновая кассета, фланкированная тремя сайтами распознавания рекомбиназы (смотрите нацеливающий вектор 1, фиг. 2). Конечный нацеливающий вектор для первой вставки κ последовательности человека в направлении 5’ - 3’ включал в себя 5’ гомологичное плечо, содержащее ~20 т.п.н. геномной последовательности мыши 5’ относительно локуса тяжелой цепи, первый сайт распознавания рекомбиназы (R1), неомициновую кассету, второй сайт распознавания рекомбиназы (R2), третий сайт распознавания рекомбиназы (R3), ~110,5 т.п.н. геномной κ последовательности человека, содержащей первые шесть последовательных генных сегментов Vκ человека и пять генных сегментов Jκ человека, сайт PI-SceI и 3’ гомологичное плечо, содержащее ~8 т.п.н. геномной последовательности мыши, включая в себя интронный энхансер, Sμ и константный ген IgM мыши (фиг. 2, нацеливающий вектор 1). Гомологичная рекомбинация с помощью этого нацеливающего вектора создавала модифицированный локус тяжелой цепи мыши, содержащий шесть генных сегментов Vκ человека и пять генных сегментов Jκ человека, функционально связанных с эндогенными константными генами тяжелой цепи, который при рекомбинации приводит к образованию гибридной тяжелой цепи (т.е. домен Vκ человека и область CH мыши).
вектор
κ последовательности человека
1-12, 1-13, 2-14, 3-15, 1-16
1-22, 1-23, 2-24, 3-25, 2-26,
1-27, 2-28, 2-29, 2-30
2-36, 1-37, 2-38, 1-39, 2-40
Введение десяти дополнительных генных сегментов Vκ человека в гибридный локус тяжелой цепи. Второй нацеливающий вектор конструировали для введения 10 дополнительных генных сегментов Vκ человека в описанный выше модифицированный локус тяжелой цепи мыши (смотрите фиг. 2, нацеливающий вектор 2). Составляющий 140058 п.н. геномный фрагмент человека, содержащий 12 последовательных генных сегментов Vκ человека из локуса κ легкой цепи человека, конструировали с 5’ гомологичным плечом, содержащим геномную последовательность мыши 5’ относительно локуса тяжелой цепи мыши и 3’ гомологичным плечом, содержащим геномную κ последовательность человека. Выше (5’) относительно генного сегмента Vκ1-16 человека находились дополнительные 10170 п.н. κ последовательности человека перед началом 5’ гомологичного плеча мыши, которое было аналогичным 5’ гомологичным плечом, используемым для конструирования нацеливающего вектора 1 (фиг. 2). Между 5’ гомологичным плечом и началом κ последовательности человека находилась гигромициновая кассета, фланкированная сайтами распознавания рекомбиназы. 3’ гомологичное плечо включало в себя составляющее 31165 п.н. перекрывание геномной κ последовательности человека, соответствующее эквивалентному 5’ концу ~110,5 т.п.н. фрагменту геномной κ последовательности человека нацеливающего вектора 1 (фиг. 2). Конечный нацеливающий вектор для вставки 10 дополнительных генных сегментов Vκ человека в направлении 5’ - 3’ включал в себя 5’ гомологичное плечо, содержащее ~20 т.п.н. геномной последовательности мыши 5’ относительно локуса тяжелой цепи, первый сайт распознавания рекомбиназы (R1), гигромициновую кассету, второй сайт распознавания рекомбиназы (R2) и ~140 т.п.н. геномной κ последовательности человека, содержащих 12 последовательных генных сегментов Vλ человека, ~31 т.п.н. которых перекрывается с 5’ концом κ последовательности человека нацеливающего вектора 1 и служит в качестве 3’ гомологичного плеча для указанного нацеливающего конструкта. Гомологичная рекомбинация с помощью указанного нацеливающего вектора создавала модифицированный локус тяжелой цепи мыши, содержащий 16 генных сегментов Vκ человека и пять генных сегментов Jκ человека, функционально связанных с константными генами тяжелой цепи мыши, который при рекомбинации приводит к образованию гибридной тяжелой цепи.
Введение четырнадцати дополнительных генных сегментов Vκ человека в гибридный локус тяжелой цепи. Третий нацеливающий вектор конструировали для введения 14 дополнительных генных сегментов Vκ человека в описанный выше модифицированный локус тяжелой цепи мыши (фиг. 2, нацеливающий вектор 3). Составляющий 160579 п.н. геномный фрагмент человека, содержащий 15 последовательных генных сегментов Vκ человека, конструировали с 5’ гомологичным плечом, содержащим геномную последовательность мыши 5’ относительно локуса тяжелой цепи мыши и 3’ гомологичным плечом, содержащим геномную κ последовательность человека. Выше (5’) относительно генного сегмента Vκ2-30 человека находились дополнительные 14687 п.н. κ последовательности человека перед началом 5’ гомологичного плеча мыши, которое являлось таким же 5’ гомологичным плечом, как и используемое для предыдущих двух нацеливающих векторов (описанных выше, смотрите также фиг. 2). Между 5’ гомологичным плечом и началом κ последовательности человека находилась неомициновая кассета, фланкированная сайтами распознавания рекомбиназы. 3’ гомологичное плечо включало в себя 21275 п.н. перекрывание геномной κ последовательности человека, соответствующее эквивалентному 5’ концу ~140 т.п.н. фрагмента геномной κ последовательности человека нацеливающего вектора 2 (фиг. 20). Конечный нацеливающий вектор для вставки 14 дополнительных генных сегментов Vκ человека в направлении 5’ - 3’ включал в себя 5’ гомологичное плечо, содержащее ~20 т.п.н. геномной последовательности мыши 5’ относительно локуса тяжелой цепи мыши, первый сайт распознавания рекомбиназы (R1), неомициновую кассету, второй сайт распознавания рекомбиназы (R2) и ~161 т.п.н. геномной κ последовательности человека, содержащие 15 генных сегментов Vκ человека, ~21 т.п.н. из которых перекрывается с 5’ концом κ последовательности человека нацеливающего вектора 2 и служит в качестве 3’ гомологичного плеча для указанного нацеливающего конструкта. Гомологичная рекомбинация с помощью указанного нацеливающего вектора создавала модифицированный локус тяжелой цепи мыши, содержащий 30 генных сегментов Vκ человека и пять генных сегментов Jκ человека, функционально связанных с константными генами тяжелой цепи мыши, который при рекомбинация приводит к образованию химерной κ тяжелой цепи.
Введение десяти дополнительных генных сегментов Vκ человека в гибридный локус тяжелой цепи. Четвертый нацеливающий вектор конструировали для введения 10 дополнительных генных сегментов Vκ человека в описанный выше модифицированный локус тяжелой цепи мыши (фиг. 2, нацеливающий вектор 4). Составляющий 90398 п.н. геномный фрагмент человека, содержащий 16 последовательных генных сегментов Vκ человека, конструировали с 5’ гомологичным плечом, содержащим геномную последовательность мыши 5’ относительно локуса тяжелой цепи мыши, и 3’ гомологичным плечом, содержащим геномную κ последовательность человека. Выше (5’) относительно генного сегмента Vκ2-40 человека находились дополнительные 8484 п.н. κ последовательности человека перед началом 5’ гомологичного плеча мыши, которое являлось таким же 5’ гомологичным плечом, как и в предыдущих нацеливающих векторах (описанных выше, фиг. 2). Между 5’ гомологичным плечом и началом κ последовательности человека находилась гигромициновая кассета, фланкированная сайтами распознавания рекомбиназы. 3’ гомологичное плечо включало в себя 61615 п.н. перекрывание геномной κ последовательности человека, соответствующее эквивалентному 5’ концу ~160 т.п.н. фрагмента геномной κ последовательности человека нацеливающего вектора 3 (фиг. 2). Конечный нацеливающий вектор для вставки 10 дополнительных генных сегментов Vκ человека в направлении 5’ - 3’ включал в себя 5’ гомологичное плечо, содержащее ~20 т.п.н. геномной последовательности мыши 5’ относительно локуса тяжелой цепи мыши, первый сайт распознавания рекомбиназы (R1), гигромициновую кассету, второй сайт распознавания рекомбиназы (R2) и ~90 т.п.н. геномной κ последовательности человека, содержащие 16 генных сегментов Vκ человека, ~62 т.п.н. которых перекрывается с 5’ концом κ последовательности человека нацеливающего вектора 3 и служит в качестве 3’ гомологичного плеча для указанного нацеливающего конструкта Гомологичная рекомбинация с указанным нацеливающим вектором создавала модифицированный локус тяжелой цепи мыши, содержащий 40 генных сегментов Vκ человека и пять генных сегментов Jκ человека, функционально связанных с константными генами тяжелой цепи мыши, который при рекомбинации приводит к образованию химерной κ тяжелой цепи (нижняя часть фиг. 2).
С использованием подхода, аналогичного описанному выше, конструировали другие комбинации вариабельных доменов легкой цепи человека, ассоциированных с константными областями тяжелой цепи мыши. Дополнительные вариабельные домены легкой цепи могут происходить из генных сегментов Vλ и Jλ человека (фиг. 3 и фиг. 4).
Локус λ легкой цепи человека охватывает свыше 1000 т.п.н. и содержит свыше 80 генов, которые кодируют вариабельные (V) или соединительные (J) сегменты. Среди 70 генных сегментов Vλ локуса λ легкой цепи человека, в пределах 30-38 оказываются функциональными генными сегментами согласно опубликованным данным. 70 последовательностей Vλ организованы в три кластера, все из которых содержат различных представителей отдельных групп V генного семейства (кластеры A, B и C). В пределах локуса λ легкой цепи человека свыше половины всех наблюдаемых доменов Vλ кодируются генными сегментами 1-40, 1-44, 2-8, 2-14 и 3-21. Существуют семь генных сегментов Jλ, только четыре из которых рассматриваются в качестве в основном функциональных генных сегментов Jλ:Jλ1, Jλ2, Jλ3 и Jλ7. В некоторых аллелях пятая пара генных сегментов Jλ -Cλ представляет собой, по имеющимся данным, псевдоген (Cλ6). Встраивание множественных генных сегментов Jλ человека в гибридный локус тяжелой цепи, описанный в настоящем документе, осуществляют путем синтеза de novo. Таким образом, геномный фрагмент, содержащий множественные генные сегменты Jλ человека в зародышевой конфигурации, конструируют со множественными генными сегментами Vλ человека и обеспечивают нормальную рекомбинацию V-J в контексте константной области тяжелой цепи.
Объединение вариабельных доменов легкой цепи с константными областями тяжелой цепи представляет собой потенциально богатый источник разнообразия для создания уникальных связывающих белков VL с областями VL человека у отличные от человека животные. Использование указанного разнообразия локуса λ легкая цепь человека (или описанного выше κ локуса человека) у мышей приводит к конструированию уникальных гибридных тяжелых цепей и приводит к появлению другого измерения связывающих белков к иммунному репертуару генетически модифицированных животных и их последующему применению в качестве платформы следующего поколения для создания терапевтических средств.
Кроме того, генные сегменты DH и JH (или Jκ ) человека могут быть встроены либо с генными сегментами Vκ, либо Vλ человека для конструирования новых гибридных локусов, которые будут приводить к образованию, при рекомбинации, новых сконструированных вариабельных доменов (фиг. 5 и 6). В этом последнем случае, конструирование комбинаций генных сегментов, которые в природе не содержаться в одном локусе, будет требовать особого внимания к последовательностям сигнала рекомбинации (RSS), которые ассоциированы с соответствующими генными сегментами так, чтобы нормальная рекомбинация могла осуществиться, когда их комбинируют в один локус. Например, известно, что V(D)J рекомбинацией руководит консервативные некодирующие последовательности ДНК, известные как гептамерные и нонамерные последовательности, которые встречаются расположенными смежно с каждым генным сегментом в точном положении, в котором происходит рекомбинация. Между этими некодирующими последовательностями ДНК расположены неконсервативные спейсерные области, которые составляют или 12, или 23 пар нуклеотидов (п.н.) в длину. Как правило, рекомбинация происходит только на генных сегментах, расположенных на одной хромосоме, и указанные генные сегменты, фланкированные составляющим 12 п.н. спейсером, могут быть соединены с генным сегментом, фланкированным составляющим 23 п.н. спейсером, т.е. согласно правилу 12/23, ходя соединение двух генных сегментов DH (каждый из которых фланкирован составляющими 12 п.н. спейсерами) наблюдалось у небольшой пропорции антител. Чтобы обеспечить возможность рекомбинации между генными сегментами, которые в норме не содержат совместимые спейсеры (например, Vκ и DH или DH и Jλ), уникальные, совместимые спейсеры синтезируют в смежных положениях с требуемыми генными сегментами для конструирования уникальных гибридных тяжелых цепей, которые обеспечивают успешную рекомбинацию для образования уникальных тяжелых цепей, содержащих вариабельные области легкой цепи.
Таким образом, использование изложенной выше стратегии для встраивания генных сегментов κ легкой цепи человека в эндогенный локус тяжелой цепи обеспечивает возможность применения других комбинаций генных сегментов λ легкой цепи человека, а также конкретных генных сегментов тяжелой цепи человека (например, DH и JH) и их комбинаций.
Пример 2. Идентификация нацеленных ES клеток и получение генетически модифицированных мышей, содержащих генные сегменты легкой цепи человека на эндогенном локусе тяжелой цепи
Нацеленную BAC ДНК, полученную в предыдущих примерах, используют для электропорации мышиных ES клеток для создания модифицированных ES клеток для получения химерных мышей, которые экспрессируют связывающие белки VL (т.е. κ генные сегменты легкой цепи человека, функционально связанные с константными областями тяжелой цепи мыши). Нацеленные ES клетки, содержащие вставку нереаранжированных генных сегментов κ легкой цепи человека, идентифицируют с помощью анализа количественной ПЦР, Taqman® (Lie, Y.S., and Petropoulos, C.J. (1998) Advances in quantitative PCR technology: 5’ nuclease assays. Curr Opin Biotechnol 9(1): 43-48). Специфические наборы праймеров и зонды разрабатывают для обнаружения вставки κ последовательностей человека и ассоциированных кассет селекции, потери последовательностей тяжелой цепи мыши и удержания последовательностей мыши, фланкирующих эндогенный локус тяжелой цепи.
ES клетки, содержащие генные сегменты κ легкой цепи человека, могут трансфектировать с помощью конструкта, который экспрессирует рекомбиназу для удаления любых нежелательных кассет селекции, введенных путем вставки нацеливающего конструкта, содержащего κ генные сегменты человека. Необязательно, мышей, содержащих сконструированный локус тяжелой цепи, содержащий генные сегменты κ легкой цепи человека, могут скрещивать с линией делеторных мышей FLPe (смотрите, например, Rodriguez, C.I. et al. (2000) High-efficiency deletor mice show that FLPe is an alternative to Cre-loxP. Nature Genetics 25: 139-140; патент США № 6774279) для удаления любой фланкированной Frt кассеты, введенной нацеливающим вектором, который не удаляют, например, на стадии ES клетки или в зародыше. Необязательно, кассету селекции оставляют у мышей.
Описанный выше нацеленные ES клетки используют в качестве донорных ES клеток и вводят в зародыш мыши на стадии 8 клеток с помощью способ VELOCIMOUSE® (выше). Мышей, содержащие модифицированный локус тяжелой цепи, содержащий генные сегменты Vκ и Jκ человека, функционально связанные с генами константной области тяжелой цепи иммуноглобулина мыши, идентифицируют путем генотипирования с использованием модификации аллельного анализа (Valenzuela et al., выше), который обнаруживал присутствие и/или отсутствие последовательностей кассеты, генных сегментов Vκ и Jκ человека и эндогенных последовательностей тяжелой цепи.
Детенышей генотипируют и детеныша, гетерозиготного в отношении модифицированного локуса тяжелой цепи, содержащего эктопический геномный фрагмент мыши, который содержит последовательности ADAM6a и ADAM6b мыши, выбирают для определения характеристик экспрессии и фертильности гена ADAM6 мыши.
Пример 3. Размножение мышей, экспрессирующих связывающие белки VL
Для создания нового поколения VL связывающих белков, мышей, содержащих нереаранжированные κ генные сегменты человека, могут скрещивать с другой мышью, содержащей делецию противоположного или ненацеленного эндогенного аллеля тяжелой цепи (т.е. с мышью, гетерозиготной в отношении модификации). Таким образом, полученное потомство будет экспрессировать только гибридные тяжелые цепи, как описано в примере 1. Скрещивание проводят согласно стандартным техникам, принятым в настоящей области техники, и альтернативно с помощью коммерческих компаний, например, The Jackson Laboratory. Линии мышей, содержащие модифицированный локус тяжелой цепи, подвергают скринингу в отношении присутствия уникальных тяжелых цепей, содержащих вариабельные домены легкой цепи человека.
Альтернативно, мышей, содержащих нереаранжированные κ генные сегменты человека на локусе тяжелой цепи мыши, могут оптимизировать путем скрещивания с другими мышами, содержащими одну или несколько делеций в локусах легкой цепи мыши (κ и λ). Таким образом, полученное потомство будет экспрессировать только антитела с уникальной κ тяжелой цепью человека, как описано в примере 1. Скрещивание аналогично проводят согласно стандартным техникам, принятым в настоящей области техники, и альтернативно с помощью коммерческих компаний, например, The Jackson Laboratory. Линии мышей, содержащие модифицированный локус тяжелой цепи и одну или несколько делеций локусов легкой цепи мыши, подвергают скринингу в отношении присутствия уникальных тяжелых цепей, содержащих домены Vκ человека и константные домены тяжелой цепи мыши, и отсутствия эндогенных легких цепей.
Мышей, содержащих модифицированный локус тяжелой цепи (описанный выше), также скрещивают с мышами, которые содержат замещение эндогенного вариабельного генного локуса κ легкой цепи вариабельным генным локусом κ легкой цепи человека (смотрите патент США № 6596541, Regeneron Pharmaceuticals, The VELOCIMMUNE® Humanized Mouse Technology). Гуманизированная мышь VELOCIMMUNE® включает в себя, частично, геном, содержащий человека вариабельные области κ легкой цепи, функционально связанные с эндогенными вариабельными локусами константной области κ легкой цепи так, что мышь производит антитела, содержащие вариабельный домен κ легкой цепи человека и константный домен тяжелой цепи мыши в ответ на антигенную стимуляцию. ДНК, кодирующая вариабельные области легкой цепи антител, могут выделять и функционально связывать с ДНК, кодирующей константные области легкой цепи человека. Затем ДНК может экспрессироваться в клетке, способной экспрессировать полностью человеческую легкую цепь антитела. При подходящей схеме скрещивания получают мышей, содержащих замещение эндогенной κ легкой цепи локусом κ легкой цепи человека и модифицированный локус тяжелой цепи согласно примеру 1. Уникальные связывающие белки VL, содержащие соматически мутированные домены Vκ человека, могут выделять при иммунизации с помощью представляющего интерес антигена.
Пример 4. Реконструирование генов ADAM в модифицированный локус тяжелой цепи
Мыши с модифицированными локусами тяжелой цепи иммуноглобулина, в которых эндогенные генные сегменты вариабельной области (т.е. VDJ) были замещены и/или удалены, не содержат экспрессии эндогенных генов ADAM6. В частности, самцы мышей, содержащие такие модификации локусов тяжелой цепи иммуноглобулина, демонстрируют снижение фертильности. В настоящем примере показаны два способа реконструирования способности экспрессировать ADAM6 у мышей с модифицированными локусами тяжелой цепи согласно примеру 1, таким образом, обеспечивая возможность для поддержания модифицированных линий мышей с использованием способов нормального скрещивания.
Реконструирование генов ADAM6 в пределах генных сегментов легкой цепи человека. Модифицированный локус тяжелой цепи иммуноглобулина, содержащий генные сегменты Vκ и Jκ человека, реконструировали так, чтобы он содержал геномный фрагмент, кодирующий мышиный ADAM6a и ADAM6b путем гомологичной рекомбинации с использованием BAC ДНК. Это осуществляли с помощью технологии генной инженерии VELOCIGENE® (выше) в серии из шести стадий, которые включали в себя модификацию BAC ДНК, содержащей последовательности мыши и человека, которая давала в результате конечный нацеливающий вектор, содержащий генные сегменты Vκ и Jκ человека, смежные с генами ADAM6 мыши и константными областями тяжелой цепи мыши.
Клон BAC мыши (VI149) содержащий в направлении 5’ - 3’ уникальный сайт рестрикции (I-CeuI), гены Adam6a и Adam6b, регуляторный элемент IGCR1 (Guo et al., 2011), генные сегменты DH и JH иммуноглобулина, энхансер Eµ и ген константной области IgM, использовали в качестве исходного материала для реконструирования генов ADAM6 в модифицированный локус тяжелой цепи, содержащий генные сегменты VL и JL (фиг. 7). VI149 модифицировали с помощью бактериальной гомологичной рекомбинации (BHR) для удаления все генных сегментов DH и JH и гена IgM от приблизительно 53 п.н. 5’ относительно наиболее дистального D сегмента (DFL16.1) до 3’ конца BAC. Эту область замещали кассетой устойчивости к спектиномицину (pSVi0029), которая содержали уникальный сайт AscI на своем 5’ конце для получения клона BAC VI413.
Дополнительные модификации BHR проводили для создания клонов BAC, содержащих гены Adam6a и Adam6b мыши, а также элемент IGCR1. Первый клон BAC создавали путем замещения 47199 п.н. области между Adam6a и Adam6b фланкированной Frt кассетой устойчивости к неомицину с уникальными сайтами рестрикции I-CeuI (5’) и AscI (3’) (pLMa0294). Эта делеция охватывала область от 4779 п.н. 3’ относительно Adam6b CDS до 290 п.н. 5’ относительно Adam6b CDS. Полученный клон BAC назвали VI421. Второй клон BAC создавали путем вставки такой же фланкированной Frt кассеты между Adam6a и Adam6b в положении 4782 п.н. 3’ относительно Adam6a CDS в VI413 для получения VI422.
Клон BAC VI421 содержал, в направлении 5’ - 3’, уникальный сайт I-CeuI, Adam6a, включая в себя 751 п.н. 5’ и 4779 п.н. 3’ относительно CDS, фланкированную Frt кассету устойчивости к неомицину, Adam6b, включая в себя 290 п.н. 5’ и 7320 п.н. 3’ относительно CDS, IGCR1 и уникальный сайт AscI (SEQ ID NO: 3).
Клон BAC VI422 содержал, в направлении 5’ - 3’, уникальный сайт I-CeuI, Adam6a, включая в себя 751 п.н. 5’ и 4779 п.н. 3’ относительно CDS, фланкированную Frt кассету устойчивости к гигромицину, Adam6b, включая в себя 47490 п.н. 5’ и 7320 п.н. 3’ относительно CDS, IGCR1 и уникальный сайт AscI (SEQ ID NO: 4).
Реконструирование генов ADAM6 осуществляли путем вставки VI421 и VI422 в межгенную область модифицированной версии нацеливающего вектора 1 (фиг. 2), описанного в примере 1. Нацеливающий вектор 1 модифицировали в две стадии BHR для вставки фрагментов ADAM6 мыши из VI421 и VI422. В первой стадии BHR неомициновую кассету из нацеливающего вектора 1 удаляли с помощью гигромициновой кассеты (pLMa0100). Полученный клон BAC назвали VI425, который содержал, в направлении 5’ - 3’, кассету устойчивости к гигромицину, четыре наиболее проксимальных сегмента Vκ человека, 23552 п.н. межгенную область Vκ-Jκ, и все пять сегментов Jκ человека, которые функционально связаны с 8 т.п.н. 3’ гомологичного плеча мыши, содержащего энхансер Eμ мыши и ген константной области IgM. Для второй стадии BHR VI425 модифицировали для удаления 740 п.н. в пределах межгенной области Vκ-Jκ с кассетой устойчивости к хлорамфениколу, фланкированной уникальными сайтами рестрикции I-CeuI и AscI (pDBa0049; фиг. 8). Положение 740 п.н. делеции находилось от 16858 до 17597 п.н. 3’ относительно наиболее проксимального генного сегмента Vκ человека (Vκ4-1). Полученный от обеих BHR клон BAC назвали VI426 (фиг. 8).
Фрагмент ДНК, содержащий гены ADAM6 мыши от VI421 и VI422, независимо использовали для замещения кассеты устойчивости к хлорамфениколу VI426 с помощью обработки I-CeuI/AscI и повторным лигированием совместимых концов. На фиг. 8 показаны конечные нацеливающие векторы, названные VI429 и VI428, соответственно. Каждый использовали для электропорации в ES клетки, предварительно модифицированные с помощью нацеливающего вектора 4 (описанного в примере 1, смотрите фиг. 2) для обеспечения возможности рекомбинации с уникальным локусом тяжелой цепи, модифицированным согласно примеру 1, и вставки фрагмента ДНК, кодирующего гены ADAM6 мыши. Положительные колонии подвергали селекции с помощью неомицина.
Реконструирование генов ADAM6, фланкирующих генные сегменты легкой цепи человека. Модифицированный локус тяжелой цепи иммуноглобулина, содержащий генные сегменты Vκ и Jκ человека, расположенные выше всех эндогенных константных областей тяжелой цепи, реконструировали так, чтобы он содержал геномный фрагмент, кодирующий ADAM6a и ADAM6b мыши с помощью гомологичной рекомбинации с использованием BAC ДНК. Это осуществляли с помощью технологии генной инженерии VELOCIGENE® (выше) в серии стадий, которые включали в себя модификацию BAC ДНК, содержащую последовательности мыши и человека, которая давала в результате конечный нацеливающий вектор, содержащий генные сегменты Vκ и Jκ человека, смежные с генами ADAM6 мыши и константными областями тяжелой цепи мыши.
Нацеливающий вектор 4, полученный согласно примеру 1 (смотрите фиг. 2 и верхнюю часть фиг. 9), модифицировали с помощью BHR для замещения фланкированной Frt гигромициновой кассеты кассетой устойчивости к хлорамфениколу, содержащей уникальные сайты рестрикции AscI (5’) и I-CeuI (3’) (pLMa0231; фиг. 9). Нацеливающий вектор 4 содержал, в направлении 5’ - 3’, ~20 т.п.н. дистальное гомологичное плечо IgH мыши, фланкированную Frt кассету устойчивости к гигромицину и генные сегменты Vκ2-40 человека - Vκ3-25 человека.
Далее клон BAC, названный VI444, использовали для вставки фрагмента ДНК, кодирующего гены ADAM6 мыши в положении 5’ относительно генных сегментов Vκ человека клона BAC VI477 путем обработки AscI/I-CeuI и повторного легирования совместимых концов. Клон VI444 содержал, в направлении 5’ - 3’, уникальный сайт I-CeuI, ген Adam6a, включая в себя 751 п.н. 5’ и 4779 п.н. 3’ относительно CDS, фланкированную Frt кассету устойчивости к неомицину, ген Adam6b, включая в себя 290 п.н. 5’ и 1633 п.н. 3’ относительно CDS и уникальный сайт AscI (SEQ ID NO: 5). Полученный клон BAC, используемый в качестве нацеливающего вектора для вставки генов ADAM6 мыши выше генных сегментов Vκ человека, назвали VI478, который, в отличие от VI421 и VI422, располагал гены ADAM6 мыши в VI478 в обратной ориентации (т.е. таком же направлении транскрипции относительно генных сегментов Vκ человека; фиг. 9). Конечный нацеливающий вектор для вставки генов ADAM6 мыши на дистальном конце генных сегментов Vκ человека, содержал, в направлении 5’ - 3’, ~20 т.п.н. дистальное гомологичное плечо IgH мыши, уникальный сайт AscI, Adam6b мыши, фланкированную Frt кассету устойчивости к неомицину, Adam6a мыши, a уникальный I-CeuI сайт и генные сегменты Vκ2-40 человека - Vκ3-25 человека. Указанный нацеливающий вектор использовали для электропорации в ES клетки, предварительно модифицированные с помощью нацеливающего вектора 4 (фиг. 2) для обеспечения возможности рекомбинации с уникальным локусом тяжелой цепи, модифицированным согласно примеру 1, и вставки фрагмента ДНК, кодирующего гены ADAM6 мыши. Положительные колонии подвергали селекции с помощью неомицина.
Селекция и подтверждение нацеленных ES клеток. Каждый из конечных нацеливающих векторов (описанных выше) использовали для электропорации мышиных ES клеток для создания модифицированных ES клеток, содержащих эктопически размещенную геномную последовательность мыши, которая содержит последовательности ADAM6a и ADAM6b мыши в пределах модифицированного локуса тяжелой цепи, содержащего генные сегменты Vκ и Jκ человека. Положительные ES клетки, содержащие эктопический геномный фрагмент мыши в пределах модифицированного локуса тяжелой цепи, идентифицировали с помощью анализа количественной ПЦР с использованием зондов TAQMAN™ (Lie and Petropoulos (1998), выше).
Описанные выше нацеленные ES клетки использовали в качестве донорных ES клеток и вводили в мышиный зародыш на стадии 8 клеток с помощью способа генной инженерии мышей VELOCIMOUSE® (смотрите, например, патенты США №№ 76598442; 7576259 и 7294754). Мышей, содержащих модифицированный локус тяжелой цепи, содержащий генные сегменты κ легкой цепи человека и эктопическую геномную последовательность мыши, содержащую последовательности ADAM6a и ADAM6b мыши, идентифицировали путем генотипирования с использованием модификации аллельного анализа (Valenzuela et al., 2003), который обнаруживал присутствие генов ADAM6a и ADAM6b мыши в пределах модифицированного локуса тяжелой цепи, а также последовательности κ легкой цепи человека.
Детенышей генотипировали и детеныша, гетерозиготного в отношении модифицированного локуса тяжелой цепи, содержащего эктопический геномный фрагмент мыши, который содержит последовательности ADAM6a и ADAM6b мыши, выбирали для определения характеристик экспрессии гена ADAM6 мыши и фертильности.
В частных воплощениях настоящее изобретение может относиться к следующим вариантам:
Вариант 1. Отличное от человека животное, содержащее
(a) вставку одного или нескольких генных сегментов VL человека и одного или нескольких генных сегментов JL человека выше не относящейся к человеку константной области легкой цепи иммуноглобулина,
(b) вставку одного или нескольких генных сегментов VL человека и одного или нескольких генных сегментов JL человека выше не относящейся к человеку константной области тяжелой цепи иммуноглобулина, и
(c) нуклеотидную последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент, причем белок ADAM6 экспрессируется из эктопической последовательности нуклеиновой кислоты ADAM6.
Вариант 2. Отличное от человека животное согласно варианту 1, у которого не относящиеся к человеку константные области тяжелой и/или легкой цепи представляют собой константные области грызуна.
Вариант 3. Отличное от человека животное согласно варианту 1 или 2, у которого константная область легкой цепи представляет собой Cκ мыши.
Вариант 4. Отличное от человека животное согласно любому из предыдущих вариантов, у которого генные сегменты VL и JL человека выше не относящейся к человеку константной области легкой цепи иммуноглобулина представляют собой генные сегменты Vκ и Jκ человека.
Вариант 5. Отличное от человека животное согласно любому из предыдущих вариантов, у которого генные сегменты VL и JL человека выше не относящейся к человеку константной области тяжелой цепи иммуноглобулина представляют собой генные сегменты Vκ и Jκ человека.
Вариант 6. Отличное от человека животное согласно любому из предыдущих вариантов, у которого нуклеотидная последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент, присутствует в таком же положении по сравнению с не относящимся к человеку локусом ADAM6 дикого типа.
Вариант 7. Отличное от человека животное согласно любому из предыдущих вариантов, у которого нуклеотидная последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент, присутствует в пределах генных сегментов иммуноглобулина.
Вариант 8. Отличное от человека животное согласно варианту 7, у которого генные сегменты иммуноглобулина представляют собой генные сегменты κ легкой цепи человека.
Вариант 9. Отличное от человека животное согласно варианту 7 или 8, у которого генные сегменты иммуноглобулина представляют собой эндогенные генные сегменты тяжелой цепи отличного от человека животного.
Вариант 10. Отличное от человека животное согласно любому из предыдущих вариантов, причем отличное от человека животное содержит эндогенные генные сегменты VL и/или JL иммуноглобулина, которые являются неспособными к реаранжировке для образования домена VL иммуноглобулина у отличного от человека животного.
Вариант 11. Генетически модифицированное отличное от человека животное, которое содержит:
(a) один или несколько нереаранжированных генных сегментов Vκ человека и один или несколько нереаранжированных генных сегментов Jκ человека на эндогенном локусе тяжелой цепи иммуноглобулина указанного отличного от человека животного,
(b) один или несколько нереаранжированных генных сегментов Vκ человека и один или несколько нереаранжированных генных сегментов Jκ человека на эндогенном локусе легкой цепи иммуноглобулина указанного отличного от человека животного, причем указанное отличное от человека животное способно экспрессировать белок ADAM6 или его функциональный фрагмент.
Вариант 12. Клетка, полученная от отличного от человека животного согласно варианту 1 или 11.
Вариант 13. Клетка согласно варианту 12, причем клетка представляет собой B-клетку.
Вариант 14. Гибридома, образованная из B-клетки согласно варианту 13.
Вариант 15. Клетка согласно варианту 12, причем отличное от человека животное представляет собой грызуна.
Вариант 16. Клетка согласно варианту 15, причем грызун выбран из мыши и крысы.
Вариант 17. Способ получения антигенсвязывающего белка, который связывается с представляющим интерес антигеном, причем способ предусматривает
(a) воздействие на отличное от человека животное согласно варианту 1 или 11 представляющего интерес антигена,
(b) выделение одного или нескольких B-лимфоцитов отличного от человека животного, причем один или несколько B-лимфоцитов экспрессируют связывающий белок VL, который связывает представляющий интерес антиген, и
(c) идентификацию последовательности нуклеиновой кислоты, которая кодирует домен VL связывающего белка VL, который связывает указанный представляющий интерес антиген, причем связывающий белок VL содержит домен Vκ человека и не относящийся к человеку константный домен легкой цепи и домен Vκ человека и не относящийся к человеку константный домен тяжелой цепи, и
(d) использование последовательности нуклеиновой кислоты согласно (c) с последовательностью нуклеиновой кислоты константной области иммуноглобулина человека для получения антигенсвязывающего белка человека, который связывает представляющий интерес антиген.
Вариант 18. Способ согласно варианту 17, при котором не относящаяся к человеку константная область легкой цепи связывающего белка VL представляет собой Cκ мыши, и не относящаяся к человеку константная область тяжелой цепи представляет собой константную область тяжелой цепи мыши.
Вариант 19. Способ согласно варианту 17 или 18, при котором отличное от человека животное представляет собой мышь.
Вариант 20. Грызун, содержащий один или несколько генных сегментов легкой цепи иммуноглобулина человека, функционально связанных с не относящимся к человеку геном константной области тяжелой цепи иммуноглобулина, причем грызун экспрессирует один или несколько белков ADAM6.
Вариант 21. Грызун согласно варианту 20, у которого один или несколько генных сегментов легкой цепи иммуноглобулина человека представляют собой генные сегменты Vκ и Jκ человека.
Вариант 22. Грызун согласно варианту 20 или 21, у которого не относящийся к человеку ген константной области тяжелой цепи иммуноглобулина представляет собой ген константной области тяжелой цепи мыши или крысы.
Вариант 23. Грызун согласно любому из вариантов 20 - 22, у которого ген константной области тяжелой цепи иммуноглобулина содержит CH1 и/или шарнирную область.
Вариант 24. Грызун согласно любому из вариантов 20 - 23, причем грызун содержит делецию или замещение одной или нескольких эндогенных генных последовательностей тяжелой цепи иммуноглобулина.
Вариант 25. Грызун согласно любому из вариантов 20 - 24, у которого эндогенные генные сегменты VH, DH и JH являются неспособными к реаранжировке для образования реаранжированной последовательности V/D/J.
Вариант 26. Грызун согласно любому из вариантов 20 - 25, причем грызун содержит делецию эндогенного гена ADAM6 и дополнительно содержит эктопический ген ADAM6 мыши.
Вариант 27. Грызун согласно любому из вариантов 20 - 26, причем грызун выбран из мыши или крысы.
Вариант 28. Способ модификации локуса тяжелой цепи иммуноглобулина грызуна, предусматривающий:
(a) осуществление первой модификации локуса тяжелой цепи иммуноглобулина грызуна, которая приводит к снижению или устранению эндогенной активности ADAM6 у самца грызуна, причем первая модификация включает в себя вставку генного сегмента, выбранного из одного или нескольких генных сегментов VL человека, одного или нескольких генных сегментов JL человека и их комбинации; и,
(b) осуществление второй модификации для добавления последовательности нуклеиновой кислоты в организм грызуна, которая предоставляет грызуну активность ADAM6, которая является функциональной у самца грызуна.
Вариант 29. Способ согласно варианту 28, при котором первая модификация дополнительно предусматривает вставку одного или нескольких генных сегментов DH человека, которые способны к реаранжировке с одним или несколькими генными сегментами VL и одним или несколькими генными сегментами JL.
Вариант 30. Способ согласно варианту 28, при котором один или несколько генных сегментов VL человека и один или несколько генных сегментов JL человека представляют собой генные сегменты Vκ и Jκ или генные сегменты Vλ и Jλ.
Вариант 31. Способ согласно любому из вариантов 28 - 30, при котором последовательность нуклеиновой кислоты, которая предоставляет грызуну активность ADAM6, которая является функциональной у самца грызуна, является смежной с одним или несколькими генными сегментами VL человека и/или одним или несколькими генными сегментами JL человека.
Вариант 32. Способ согласно любому из вариантов 28 - 31, при котором первую и вторую модификацию осуществляют в отдельной ES клетке, и отдельную ES клетку вводят в зародыш - хозяин для получения грызуна.
--->
Перечень последовательностей
<110> Regeneron Pharmaceuticals, Inc.
<120> Humanized Rodents That Express Heavy Chains Containing VL Domains
<130> 7350A-WO
<140> To be assigned
<141> Filed herewith
<150> 61/593,463
<151> 2012-02-01
<150> 61/677,538
<151> 2012-07-31
<160> 5
<170> FastSEQ for Windows Version 4.0
<210> 1
<211> 754
<212> PRT
<213> Mus musculus
<400> 1
Met Leu Ser Leu Thr Trp Gly Met Arg Leu Val Glu Arg Pro Val Val
1 5 10 15
Pro Arg Val Leu Leu Leu Leu Phe Ala Leu Trp Leu Leu Leu Leu Val
20 25 30
Pro Val Trp Cys Ser Gln Gly His Pro Thr Trp Arg Tyr Ile Ser Ser
35 40 45
Glu Val Val Ile Pro Arg Lys Glu Ile Tyr His Thr Lys Gly Leu Gln
50 55 60
Ala Gln Arg Leu Leu Ser Tyr Ser Leu Arg Phe Arg Gly Gln Arg His
65 70 75 80
Ile Ile His Leu Arg Arg Lys Thr Leu Ile Trp Pro Arg His Leu Leu
85 90 95
Leu Thr Thr Gln Asp Asp Gln Gly Ala Leu Gln Met Glu Tyr Pro Phe
100 105 110
Phe Pro Val Asp Cys Tyr Tyr Ile Gly Tyr Leu Glu Gly Ile Leu Gln
115 120 125
Ser Met Val Thr Val Asp Thr Cys Tyr Gly Gly Leu Ser Gly Val Ile
130 135 140
Lys Leu Asp Asn Leu Thr Tyr Glu Ile Lys Pro Leu Asn Asp Ser Gln
145 150 155 160
Ser Phe Glu His Leu Val Ser Gln Ile Val Ser Glu Ser Asp Asp Thr
165 170 175
Gly Pro Met Asn Ala Trp Lys His Trp Ser His Asn Thr Gly Ser Pro
180 185 190
Ser Ser Arg Leu Glu Tyr Ala Asp Gly Ala Pro Arg Leu Ser Ser Lys
195 200 205
Asn Tyr Ala Thr His Pro Ala Ala Ile Lys Gly His Phe Gln Ala Thr
210 215 220
His Ser Val Tyr Ser Ala Ser Gly Gly Asp Lys Leu Ser Ser Thr Val
225 230 235 240
Glu Tyr Leu Phe Lys Val Ile Ser Leu Met Asp Thr Tyr Leu Thr Asn
245 250 255
Leu His Met Arg Tyr Tyr Val Phe Leu Met Thr Val Tyr Thr Glu Ala
260 265 270
Asp Pro Phe Ser Gln Asp Phe Arg Val Pro Gly Gly Gln Ala His Thr
275 280 285
Phe Tyr Glu Arg Val Phe Tyr Ala His Phe Arg Pro Asp Ala Gly Ala
290 295 300
Ile Ile Asn Lys Asn Ser Pro Gly Asp Asp Ala Val Asn Pro Ala Glu
305 310 315 320
Arg Ser Ile Cys Ser Pro Ser Ala Leu Ile Cys Leu Gly Gln His Gly
325 330 335
Arg Asn Pro Leu Phe Leu Ser Ile Ile Ile Thr Asn Arg Val Gly Arg
340 345 350
Ser Leu Gly Leu Lys His Asp Glu Gly Tyr Cys Ile Cys Gln Arg Arg
355 360 365
Asn Thr Cys Ile Met Phe Lys Asn Pro Gln Leu Thr Asp Ala Phe Ser
370 375 380
Asn Cys Ser Leu Ala Glu Ile Ser Asn Ile Leu Asn Thr Pro Asp Leu
385 390 395 400
Met Pro Cys Leu Phe Tyr Asp Arg His Val Tyr Tyr Asn Thr Ser Leu
405 410 415
Thr Tyr Lys Phe Cys Gly Asn Phe Lys Val Asp Asn Asn Glu Gln Cys
420 425 430
Asp Cys Gly Ser Gln Lys Ala Cys Tyr Ser Asp Pro Cys Cys Gly Asn
435 440 445
Asp Cys Arg Leu Thr Pro Gly Ser Ile Cys Asp Lys Glu Leu Cys Cys
450 455 460
Ala Asn Cys Thr Tyr Ser Pro Ser Gly Thr Leu Cys Arg Pro Ile Gln
465 470 475 480
Asn Ile Cys Asp Leu Pro Glu Tyr Cys Ser Gly Ser Lys Phe Ile Cys
485 490 495
Pro Asp Asp Thr Tyr Leu Gln Asp Gly Thr Pro Cys Ser Glu Glu Gly
500 505 510
Tyr Cys Tyr Lys Gly Asn Cys Thr Asp Arg Asn Ile Gln Cys Met Glu
515 520 525
Ile Phe Gly Val Ser Ala Lys Asn Ala Asn Ile Lys Cys Tyr Asp Ile
530 535 540
Asn Lys Gln Arg Phe Arg Phe Gly His Cys Thr Arg Ala Glu Glu Ser
545 550 555 560
Leu Thr Phe Asn Ala Cys Ala Asp Gln Asp Lys Leu Cys Gly Arg Leu
565 570 575
Gln Cys Thr Asn Val Thr Asn Leu Pro Phe Leu Gln Glu His Val Ser
580 585 590
Phe His Gln Ser Val Ile Ser Gly Val Thr Cys Phe Gly Leu Asp Glu
595 600 605
His Arg Gly Thr Glu Thr Ala Asp Ala Gly Leu Val Arg His Gly Thr
610 615 620
Pro Cys Ser Arg Gly Lys Phe Cys Asp Arg Gly Ala Cys Asn Gly Ser
625 630 635 640
Leu Ser Arg Leu Gly Tyr Asp Cys Thr Pro Glu Lys Cys Asn Phe Arg
645 650 655
Gly Val Cys Asn Asn Arg Arg Asn Cys His Cys His Phe Gly Trp Ser
660 665 670
Pro Pro Lys Cys Lys Glu Glu Gly His Ser Gly Ser Ile Asp Ser Gly
675 680 685
Ser Pro Pro Val Gln Arg Arg Ile Ile Lys Gln Asn Leu Glu Pro Val
690 695 700
Val Tyr Leu Arg Ile Leu Phe Gly Arg Ile Tyr Phe Leu Phe Val Ala
705 710 715 720
Leu Leu Phe Gly Ile Ala Thr Arg Val Gly Val Thr Lys Ile Phe Arg
725 730 735
Phe Glu Asp Leu Gln Ala Ala Leu Arg Ser Trp Gln Glu Gln Ala Lys
740 745 750
Asp Lys
<210> 2
<211> 756
<212> PRT
<213> Mus musculus
<400> 2
Met Leu Ser Leu Thr Trp Gly Met Arg Leu Val Glu Arg Pro Val Val
1 5 10 15
Pro Arg Val Leu Leu Leu Leu Phe Ala Leu Trp Leu Leu Leu Leu Val
20 25 30
Pro Val Trp Cys Ser Gln Gly His Pro Thr Trp Arg Tyr Ile Ser Ser
35 40 45
Glu Val Val Ile Pro Arg Lys Glu Ile Tyr His Thr Lys Gly Leu Gln
50 55 60
Ala Gln Arg Leu Leu Ser Tyr Ser Leu His Phe Arg Gly Gln Arg His
65 70 75 80
Ile Ile His Leu Arg Arg Lys Thr Leu Ile Trp Pro Arg His Leu Leu
85 90 95
Leu Thr Thr Gln Asp Asp Gln Gly Ala Leu Gln Met Asp Tyr Pro Phe
100 105 110
Phe Pro Val Asp Cys Tyr Tyr Ile Gly Tyr Leu Glu Gly Ile Pro Gln
115 120 125
Ser Met Val Thr Val Asp Thr Cys Tyr Gly Gly Leu Ser Gly Val Met
130 135 140
Lys Leu Asp Asp Leu Thr Tyr Glu Ile Lys Pro Leu Asn Asp Ser Gln
145 150 155 160
Ser Phe Glu His Leu Val Ser Gln Ile Val Ser Glu Ser Asp Asp Thr
165 170 175
Gly Pro Met Asn Ala Trp Lys His Trp Ser His Asn Thr Gly Ser Pro
180 185 190
Ser Ser Arg Leu Glu Tyr Ala Asp Gly Ala Pro Arg Ile Ser Ser Lys
195 200 205
Asn Tyr Ala Thr His Pro Ala Ala Ile Lys Gly His Phe Gln Ala Thr
210 215 220
Asn Ser Val Tyr Asn Ser Ala Ala Gly Asp Lys Leu Ser Ser Thr Val
225 230 235 240
Gly Tyr Leu Phe Gln Val Ile Ser Leu Met Asp Thr Tyr Leu Thr Asn
245 250 255
Leu His Met Arg Tyr Tyr Val Phe Leu Met Thr Val Tyr Thr Asn Ser
260 265 270
Asp Pro Phe Arg Leu Glu Phe Ala Val Pro Gly Gly Ser Ala Tyr Asn
275 280 285
Tyr Tyr Val Ser Val Phe Tyr Asn Lys Phe Lys Pro Asp Ala Gly Val
290 295 300
Leu Leu Asn Lys Tyr Gly Pro Gln Asp Asn Gln Val Asn Pro Ala Glu
305 310 315 320
Arg Ser Ile Cys Ser Ser Leu Ala Leu Ile Cys Ile Gly Lys Tyr Asp
325 330 335
Arg Asn Pro Leu Phe Leu Ser Pro Ile Ile Thr Asn Arg Val Gly Arg
340 345 350
Ser Leu Gly Leu Lys Tyr Asp Glu Gly Tyr Cys Val Cys Gln Arg Arg
355 360 365
Asn Thr Cys Ile Met Phe Arg His Pro Gln Leu Thr Asp Ala Phe Ser
370 375 380
Asn Cys Ser Leu Ala Glu Ile Ser Asn Ile Leu Asn Thr Pro Gly Leu
385 390 395 400
Met Pro Cys Leu Phe Tyr Asp Arg His Val Tyr Tyr Asn Thr Ser Leu
405 410 415
Thr Tyr Lys Phe Cys Gly Asn Phe Lys Val Asp Asn Asp Glu Gln Cys
420 425 430
Asp Cys Gly Ser Gln Lys Ala Cys Tyr Ser Asp Pro Cys Cys Gly Asn
435 440 445
Asp Cys Arg Leu Thr Pro Gly Ser Ile Cys Asp Lys Glu Leu Cys Cys
450 455 460
Ala Asn Cys Thr Tyr Ser Pro Ser Gly Thr Leu Cys Arg Pro Ile Gln
465 470 475 480
Asn Ile Cys Asp Leu Pro Glu Tyr Cys Asn Gly Thr Lys Tyr Ile Cys
485 490 495
Pro Asp Asp Thr Tyr Leu Gln Asp Gly Thr Pro Cys Ser Glu Asp Gly
500 505 510
Tyr Cys Tyr Lys Gly Asn Cys Thr Asp Arg Asn Ile Gln Cys Met Glu
515 520 525
Ile Phe Gly Val Ser Ala Lys Asn Ala Asn Ile Lys Cys Tyr Asp Ile
530 535 540
Asn Lys Gln Arg Phe Arg Phe Gly His Cys Thr Arg Ala Glu Glu Ser
545 550 555 560
Leu Thr Phe Asn Ala Cys Ala Asp Gln Asp Lys Leu Cys Gly Arg Leu
565 570 575
Gln Cys Thr Asn Val Thr Asn Leu Pro Tyr Leu Gln Glu His Val Ser
580 585 590
Phe His Gln Ser Ile Ile Ser Gly Phe Thr Cys Phe Gly Leu Asp Glu
595 600 605
His Arg Gly Thr Glu Thr Thr Asp Ala Gly Met Val Arg His Gly Thr
610 615 620
Pro Cys Ser Lys Ser Lys Phe Cys Asp Gln Gly Ala Cys Ser Gly Ser
625 630 635 640
Leu Ser His Leu Gly Tyr Asp Cys Thr Pro Glu Lys Cys Ser Phe Arg
645 650 655
Gly Val Cys Asn Asn His Arg Asn Cys His Cys His Phe Gly Trp Lys
660 665 670
Pro Pro Glu Cys Lys Glu Glu Gly Leu Ser Gly Ser Ile Asp Ser Gly
675 680 685
Ser Pro Pro Val Gln Arg His Thr Ile Lys Gln Lys Gln Glu Pro Val
690 695 700
Val Tyr Leu Arg Ile Leu Phe Gly Arg Ile Tyr Phe Leu Phe Val Ala
705 710 715 720
Leu Leu Phe Gly Ile Ala Thr Arg Val Gly Val Thr Lys Ile Phe Arg
725 730 735
Phe Glu Asp Leu Gln Ala Thr Leu Arg Ser Gly Gln Gly Pro Ala Arg
740 745 750
Asp Lys Pro Lys
755
<210> 3
<211> 20468
<212> DNA
<213> Artificial Sequence
<220>
<223> synthetic
<400> 3
taactataac ggtcctaagg tagcgaggga tgacagattc tctgttcagt gcactcaggg 60
tctgcctcca cgagaatcac catgtccttt ctcaagactg tgttctgtgc agtgccctgt 120
cagtggaaat ctggagagca tgcttccatg agcttgtgag tagtatatct agtaagccat 180
ggctttgtgt taatggtgat gttctacata tcagttctct ggcttaataa tgaggtgatg 240
attctatgtt cctgtaacgc ttcctcaact gggtcctaag tctttcttca ctccatctat 300
tcctctaagg aatgatcctg aaaatcccat cacaaactat aggagatggg aaccatcaaa 360
aaacacagtg acaaagaggt gggaacgcat cagggttcag gaaccatatt ttaaaaagat 420
atcgtaaata acttcttaaa agagatatag acaaatctcc attaatacgg agaccagagg 480
cctaaggcta agaaccaatg gtggctcaag gtctcctgct acccgaggag caaacgtaga 540
gcagtttcta atgatttatt taaaatatag aatcaaaagt accagtttgc aattttgaaa 600
gatttatttc agcaatgcaa caacatcagg tggtgccgag tccaacacgt cttatgtccc 660
atgatataaa caaaggccat ccagaactgt ggactggagt tctaccttgt cccctaatga 720
cattcagatt ttttttccat tctctttatc ttagaggaga cagggggcta actcatttta 780
cttgtccttt gcttgttctt gccaagaacg taaagcagct tgcaagtctt caaacctaaa 840
tatcttagta actcctacac gagtggcaat gccaaagagc agtgcaacaa agaggaagta 900
aatacgacca aagagtattc ttaaatacac tactggctct aggttctgtt ttattatgcg 960
cctttgaacc ggaggggacc cactgtctat gctcccactg tgtccctctt ctttgcactt 1020
tggagggctc caaccaaaat ggcaatggca attccgacga ttgttacaca ctcctctgaa 1080
attgcatttt tctggggtgc agtcataacc caaacgagat aaacttccat tgcaagctcc 1140
tcgatcacag aacttacccc ttgaacacgg ggtaccatgt ctcaccaatc cagcatctgc 1200
tgtttctgtc ccacgatgtt catcaagccc aaagcaggta accccagaga taaccgattg 1260
atggaatgaa acatgttctt gcaaaaatgg aagattggtg acattggtac actgcaacct 1320
tccacacagc ttgtcctgat cagcacaagc attgaatgtg aggctttctt ctgctctagt 1380
acaatgccca aatcgaaacc gttgtttgtt gatgtcatag cacttaatat tagcattctt 1440
agcacttaca ccaaagattt ccatgcattg tatgttgcga tcagtgcagt tacctttata 1500
gcagtaaccc tcttctgagc atggtgtccc atcttgcaga taagtgtcat ctgggcaaat 1560
gaacttagag ccactacagt actctggaag atcacatatg ttctggatag gtctgcagag 1620
tgtcccagaa ggactgtaag tgcaatttgc acagcataat tctttatcac aaatgctacc 1680
aggtgttaac ctgcaatcat ttccacagca gggatctgaa taacatgcct tttgggagcc 1740
acagtcacac tgctcattgt tatctacttt gaagtttcca caaaacttat aagtcaatga 1800
tgtattataa taaacatgac ggtcatagaa aagacatggc atcagatcag gagtattaag 1860
tatgttgctt atctctgcaa gggaacaatt gctgaaagca tctgttaatt gaggattttt 1920
gaacatgatg caggtgttcc ttctctggca gatacagtac ccctcatcat gttttaggcc 1980
taaactcctt ccaacacgat tggttattat aatagataaa aataaaggat ttcgaccatg 2040
ttgaccaaga caaattaggg ctgagggaga acatatactc ctctcagctg gattaacagc 2100
atcatctcct ggcgaattct tgttaattat agctcctgca tcaggcctaa aatgagcata 2160
aaatactctc tcatagaaag tatgagcctg ccctcctgga actcgaaaat cttgtgaaaa 2220
tggatcagcc tcggtataca cagtcatgag aaagacatag taccgcatat gaagattggt 2280
cagataggtg tccattaaac taatgacttt aaacaaatac tcaacagtag atgaaagttt 2340
gtcacctcca gaagcactat atacagaatg ggttgcttga aagtggcctt ttatagcagc 2400
tggatgtgta gcgtaattct tactagatag tctgggagct ccatctgcat attccaatct 2460
ggaggaggga gaacctgtat tatggctcca gtgcttccat gcattcatag gccctgtgtc 2520
atcagactca gatactatct gagaaacaag gtgttcaaag ctctgtgaat cattgagggg 2580
tttgatttca taggtaaggt tatccaactt tatgacccct gacaggcccc cataacaagt 2640
atccacagtg accatggatt gcaggatccc ctccaggtag ccaatatagt aacaatctac 2700
aggaaaaaag gggtactcca tctgtaaggc tccttggtca tcttgagttg tcagcaacaa 2760
gtgtctgggc caaatgagtg tctttctccg caggtggatg atatgtctct ggccccgaaa 2820
acgcaagcta tacgagagca gtctttgtgc ttgaagtcct ttggtatggt agatctcctt 2880
ccgaggaata accacctccg atgagatgta acgccaagtg ggatggcctt gagaacacca 2940
gactggaacc aggaggagca gccagagtgc aaatagcaag aggaggaccc tggggaccac 3000
aggtctttcc actagcctca tgccccaggt cagagataac atcctgggtg gagctaactc 3060
cctctgctgt ggccactgcc tggtctagaa aatactgaca gaggactaaa aacctcctca 3120
ggctcccaac ctaagtggtt acccagacaa ctggagttag gtaacagtca ctgggtgtgg 3180
caggaattga gtctgaatgt gttagctgag gttgaggtta aatattgtca aaagggatgt 3240
ctataaatgt gcctggacaa gaaaagtcag aagcagcaag gagtgtctct gacaggctca 3300
atcctttctt ttcttttttt gaagttcaaa atatcatttc cacgtgaatg tatttggttc 3360
ccagtgtgac tctgggtctc tttctaggag tcaatatttc tttatatctt ggctcatgtt 3420
tttcacagtt gttctaactt cttgttttgt tttgtttgtt tgtttgtttg aaagttagaa 3480
gtaaatactg tctatattag ccttttagct ataaatgatt gtttttattt cttctaatca 3540
tgttttgttt gagttttggt taaactattt acaaatgagt tttttttttc cttttgggtg 3600
ttgctcgaaa gtttggagct ttctgttaat attgtgttgt tgtttctcca atattattag 3660
acctgagaat tctacctggg tacctgtgaa ctccagaatt tttaaaaatt ccatctcttg 3720
ggaacattat ctctgacccc gtctgaggcc gaagtggctg tccccctcca acctttagta 3780
tctttctttc ctgactattg ggatttcttc aagcaatcag gctgatgggt tctcagcagt 3840
gagaccagta gactgtcggt atgaacgtcg aagagtctgc cacacactcc gggttcatca 3900
acagtgcttt cgcgtctctt acttttgtag aaggaaatgc agcctctgag ttttctccaa 3960
gaaatcattg atgaaagggt gaaaagatgg gtatcacccg gagttcatga caagccctgg 4020
ctcagacacg tgagcaaggt ctacagcccc aaagataggc tgccctgcaa catgtattta 4080
taagatagga gaaaaaaatg ggtagttgga gggttgatca acttacttcc tctcaaacat 4140
atatatctca tctaagtgtg caggggaaaa ctctgtagaa ctactgggat acctgctcac 4200
ccccaggagc ctcatgaata agtctctgct tctgccttgt agccatgagc attactgcac 4260
ctgatacccc tgcagcttcc tagggaagag ggaggaagtg acttggcccc tgtctggtta 4320
aggtaagagg agataaatcc cttctcattg attagggtga gaggggtcat gtgctctatc 4380
attggtgacc cagttgggac atgggtttat accaaagtca tcactctgag gttctgtgta 4440
ccaccaggct gaactcccat atcctacatg gacataggac aacaccaagc agaaggaggt 4500
tttaggacta aactgaagga cagagatgcg gtttctaaac aactagggag tgccagggcc 4560
agcctctcta accactatag gacactgtgg agtctggtta caaagagaga ttactcaagg 4620
tccttagcac tgattacaga gcatatctca gatgccttct gctgaccaga tgtatctttg 4680
cataatctgc ctatccagat tcagaaaatt gatgccacat agccaagtgg actttcagga 4740
acagacgatt taaaaacagg cagagagatg tgagagaaag gagaaggaga gagagaaggg 4800
agagggagag aagagagagg gagacggaga aggaaagagg gagaaggaga aggagagaag 4860
gggcatggac agagggaggg acagaaggag agaggagata gagaggggga taaggaagaa 4920
gggagggagg gagagagaga gaaggctaag tctttccata cctgggtccc aatacctctt 4980
ataacccaag cacatggttt cacatatcac aatgcggttg ggatatagat aactgtaaat 5040
acttgtgaaa ataatggggc tgagatctgg ggttttcatg atagtttcaa agtcaccgta 5100
ctgactaaaa ccttccactg gcccatctcc agcttcctaa tctgagggta tcaaatttcc 5160
cactaagtgt gtttagaaag atctccacct ttttgccctt gtcttccagt gccccaccta 5220
cgttctggtc tcccacatct gatgtcttct cagtgattct ggccctgcct gctccacagc 5280
tacaaacccc ttcctataat gagctctgtg ctgagccatc atcctgaatc aatccacctt 5340
aagcagatgt tttgcttatt tttcctgtgt ccatactaca gaggaaaggt aggcatgtag 5400
aagctgaagc atctcacctc attccaagca ccctcagtct ctaaatgtgc ccccttgttt 5460
ccagaagtgc aacctcaagc atcttttatt cattcatctt agagggccac atgtgctgta 5520
gtgttataag atgaaattta aagcattaat tattcctaac aagccaatta aacaagccaa 5580
aaacattcat cagtcattcc catggaacct ctgaagcatc ttcctgctct aaccttgggt 5640
tttccagggc tgctctggga tcacaggagc tgtcctgtct accagccata taaaggcaga 5700
cctatcagaa ttacaccaga cttctcacca tagactataa aagccagaat atcctggaca 5760
gatgttatac agaaactaag agaacacaaa tgccagccca ggctactata cccagcaaaa 5820
ctctcaatta ccatcgatga agaaaccaag atattccatt acaagtccaa atttacacaa 5880
tatctttcca taaatccagc cctacaaagg atagcagatg gaaaactcca acacaggtag 5940
gaaaactaca ccctagaaag agcactaaag taatcatctt tcaacacact caaaagaaga 6000
taaccacaca aacataattc cacctctaac aacaaaaata aagtaggcaa caatcactat 6060
tccttaatat ctcttttaac atcaatggac tcaattctcc aataaaaaga catagactaa 6120
cagactgaat acataaacag gacacagcat tttgctgcat aaagcaaaca cagcgttact 6180
tttttttttc taaatgacat tttttattag atattgtctt tattgacatt tcaaatgtta 6240
tcccctttcc tggtttaccc tctgaaatcc cctatctcct ccccctcccc ctgctcacca 6300
atccacccac tcccacttcc aggccctggc aatcccctat atttgggcat agagccttca 6360
caggaccaag gtactctcct tgcattgatg accaactagt ccattctctg ctacaaatgc 6420
agctagatct atgagtccca ccatgttttc ttttgttggt ggtttcatgc cagggagctc 6480
ttggagtact gattggttca tattgttgtt ctccctatgg ggttacaaaa cccttcaact 6540
tcttgggtcc tttctctggc tgcctcattg gggaccttgt gcgaagtcca atggatgact 6600
gtgagcatcc acttctgtat ttgccaggca ctggcagagc ctctcagaag acagctatat 6660
caagatcctg gcagcaagct cttgttggta tccacaaaag tgtctggtgg ttgtctatgg 6720
gatggatccc caaaggggca gtctctggat ggtcattcct tcagtctctg ttccacactt 6780
tgtctcttta actccttcca tgactatttt attcctccct ctaagaagga ccgaagtatt 6840
catactttgg tcttccttct tgaaattcat gtgttttgtg aattgtatct ttgatattcc 6900
gaacttctgg gctaatatcc acttatcagt gagtgaatat catgtgtgtt cttatgtgat 6960
tgagttacct cactcaggat gatatcctcc agaaccatcc atttgtctaa gaatttaatg 7020
aattcattgt ttttaatagc tgaggagtac tccattgtgt aaatgtacca cattttctgt 7080
acccattgtt ctcttgaggg acatctgggt tctttaaagc ttctggacat taaatataag 7140
gctgctatgg aaatagtgga gaatgtgtcc ttattacatg ttggagcatc ttctgggtat 7200
atgcccagga gtgctattgc tggatcctct gatagtacta tgtccaattt tctgaggaac 7260
tgccaaactg atttacagag tggttgtacc agcttgcaat tccaccagca atggagaaat 7320
gttccccttc ctccacatcc tcaccaacat ctgctgtcac ctcaatttgt tcttagtgat 7380
tcagacaggt gtgaggtgga atatcagggt tgtttggcat ttccctgatg actagtgata 7440
ttgaaaaaaa ttttaagtgt ttctcagcca ttcagtattc ttcagttgag aattcactgt 7500
ttagctctgt actcaggttt ttttaatagg gttatttggt tttctggagt ctaacgtctt 7560
gaattctttc tatatattgg atattagccc tctgtcatat ttaggattgg taaagatctt 7620
tcccaatatg ttggctgcct ttttgtgtcc tttgccttac agaacctttt taattttatg 7680
aggtcccatt tgctaattct tcattttaca gcacaagcca ttggtgttct gttcaaaaat 7740
ctttccccct gaaccctatc ttcgaggatc ttccccactt tctcctctat aagtttcagt 7800
gtctctatta ttgtgctgag ggcgatcgca ccggtgaaac cctaggaagt tcctatactt 7860
tctagagaat aggaacttcg gaataggaac ttctctaggg gctgcaggtc gaggtctgat 7920
ggaattagaa cttggcaaaa caatactgag aatgaagtgt atgtggaaca gaggctgctg 7980
atctcgttct tcaggctatg aaactgacac atttggaaac cacagtactt agaaccacaa 8040
agtgggaatc aagagaaaaa caatgatccc acgagagatc tatagatcta tagatcatga 8100
gtgggaggaa tgagctggcc cttaatttgg ttttgcttgt ttaaattatg atatccaact 8160
atgaaacatt atcataaagc aatagtaaag agccttcagt aaagagcagg catttatcta 8220
atcccacccc acccccaccc ccgtagctcc aatccttcca ttcaaaatgt aggtactctg 8280
ttctcaccct tcttaacaaa gtatgacagg aaaaacttcc attttagtgg acatctttat 8340
tgtttaatag atcatcaatt tctgcagact tacagcggat cttcagaaga actcgtcaag 8400
aaggcgatag aaggcgatgc gctgcgaatc gggagcggcg ataccgtaaa gcacgaggaa 8460
gcggtcagcc cattcgccgc caagctcttc agcaatatca cgggtagcca acgctatgtc 8520
ctgatagcgg tccgccacac ccagccggcc acagtcgatg aatccagaaa agcggccatt 8580
ttccaccatg atattcggca agcaggcatc gccatgggtc acgacgagat cctcgccgtc 8640
gggcatgcgc gccttgagcc tggcgaacag ttcggctggc gcgagcccct gatgctcttc 8700
gtccagatca tcctgatcga caagaccggc ttccatccga gtacgtgctc gctcgatgcg 8760
atgtttcgct tggtggtcga atgggcaggt agccggatca agcgtatgca gccgccgcat 8820
tgcatcagcc atgatggata ctttctcggc aggagcaagg tgagatgaca ggagatcctg 8880
ccccggcact tcgcccaata gcagccagtc ccttcccgct tcagtgacaa cgtcgagcac 8940
agctgcgcaa ggaacgcccg tcgtggccag ccacgatagc cgcgctgcct cgtcctgcag 9000
ttcattcagg gcaccggaca ggtcggtctt gacaaaaaga accgggcgcc cctgcgctga 9060
cagccggaac acggcggcat cagagcagcc gattgtctgt tgtgcccagt catagccgaa 9120
tagcctctcc acccaagcgg ccggagaacc tgcgtgcaat ccatcttgtt caatcatggt 9180
ggatggttta gttcctcacc ttgtcgtatt atactatgcc gatatactat gccgatgatt 9240
aattgtcaac aattcgagag acccgggttc ccaatctaga gctcgacgtc taacaaaaaa 9300
gccaaaaacg gccagaattt agcggacaat ttactagtct aacactgaaa attacatatt 9360
gacccaaatg attacatttc aaaaggtgcc taaaaaactt cacaaaacac actcgccaac 9420
cccgagcgca tagttcaaaa ccggagcttc agctacttaa gaagataggt acataaaacc 9480
gaccaaagaa actgacgcct cacttatccc tcccctcacc agaggtccgg cgcctgtcga 9540
ttcaggagag cctaccctag gcccgaaccc tgcgtcctgc gacggagaaa agcctaccgc 9600
acacctaccg gcaggtggcc ccaccctgca ttataagcca acagaacggg tgacgtcacg 9660
acacgacgag ggcgcgcgct cccaaaggta cgggtgcact gcccaacggc accgccataa 9720
ctgccgcccc cgcaacagac gacaaaccga gttctccagt cagtgacaaa cttcacgtca 9780
gggtccccag atggtgcccc agcccatctc acccgaataa gagctttccc gcattagcga 9840
aggcctcaag accttgggtt cttgccgccc accatgcccc ccaccttgtt tcaacgacct 9900
cacagcccgc ctcacaagcg tcttccattc aagactcggg aacagccgcc attttgctgc 9960
gctcccccca acccccagtt cagggcaacc ttgctcgcgg acccagacta cagcccttgg 10020
cggtctctcc acacgcttcc gtcccaccga gcggcccggc ggccacgaaa gccccggcca 10080
gcccagcagc ccgctactca ccaagtgacg atcacagcga tccacaaaca agaaccgcga 10140
cccaaatccc ggctgcgacg gaactagctg tgccacaccc ggcgcgtcct tatataatca 10200
tcggcgttca ccgccccacg gagatccctc cgcagaatcg ccgagaaggg actacttttc 10260
ctcgcctgtt ccgctctctg gaaagaaaac cagtgcccta gagtcaccca agtcccgtcc 10320
taaaatgtcc ttctgctgat actggggttc taaggccgag tcttatgagc agcgggccgc 10380
tgtcctgagc gtccgggcgg aaggatcagg acgctcgctg cgcccttcgt ctgacgtggc 10440
agcgctcgcc gtgaggaggg gggcgcccgc gggaggcgcc aaaacccggc gcggaggcct 10500
cggaggccag acgcgcgaga agttcctata ctttctagag aataggaact tcggaatagg 10560
aacttcggta ccgcgatcgc gatgtgggaa cgcttcagtg ttcaggaacc atatgattta 10620
tttaaaatat agaatcaaaa gtaccaattt gcagttttga aagatttatt ccagtgtaag 10680
cattagcaat gcaccaacat caggtgattt ctgaatccaa cacgtcttat gtcctcatga 10740
tattaaaaaa aaaaaaaggc catccagaac tgtgaacttg agttctacct tgttccctac 10800
tgacattcag attttctttt ttgcattctc tttatcttac aggagacagg aggggagggc 10860
taactcattt tactttggct tgtcccttgc tggtccttgc ccagaacgta aagtagcttg 10920
caagtcttca aatctaaaaa tcttagtaac tcctacacga gtggcaatgc caaagagcag 10980
tgcaacaaag aggaagtaaa tacgaccaaa gagtattctt aaatacacca ctggctcttg 11040
tttttgtttt attgtgtgcc tttgaactgg aggggaccca ctgtctatgc tcccacttag 11100
tccctcttct ttgcactctg gaggcttcca accaaaatga caatggcaat tccgatgatt 11160
gttacacact cctctaaaac tgcatttttc tggggtgcag tcataaccca aatgagataa 11220
acttccactg caagctcctt gatcacagaa cttacttttg gagcaggggg taccatgtct 11280
caccattcca gcatctgttg tttctgtccc acgatgttca tcaagcccaa agcaggtaaa 11340
cccagagata atcgattgat ggaatgaaac atgttcttgc aaatatggaa gattggtgac 11400
attggtacac tgcaaccttc cacacagctt gtcctgatca gcacaagcat tgaatgtgag 11460
gctttcttct gctctagtac aatgcccaaa tcgaaaccgt tgtttgttga tgtcatagca 11520
cttaatatta gcattcttag cacttacacc aaagatttcc atgcattgta tgttgcgatc 11580
agtgcagtta cctttatagc agtaaccatc ttctgagcat ggtgtcccat cttgcagata 11640
agtgtcatct gggcaaatgt atttagtccc attacagtac tctggaagat cacatatgtt 11700
ctggataggt ctgcagagtg tcccagaagg actgtaagtg caatttgcac agcataattc 11760
tttatcacaa atgctaccag gtgttaacct gcaatcattt ccacagcagg gatctgaata 11820
acatgccttt tgggagccac agtcacactg ctcatcgtta tctactttga agtttccaca 11880
aaacttataa gtcaatgatg tattataata aacatgacgg tcatagaaaa gacatggcat 11940
cagaccagga gtattaagta tgttgcttat ctctgcaagg gaacaattgc tgaaagcatc 12000
tgttaattga ggatgtctga acataatgca ggtgttcctt ctctggcaga cacagtaccc 12060
ctcatcatat tttaagccta aactccttcc aacacgattg gttattatag gagataaaaa 12120
taaaggattt cgatcatatt taccaataca aattagggct aaggaagaac atatactcct 12180
ctcagctgga ttaacctggt tatcttgtgg cccatactta ttaagtaaaa ctcctgcatc 12240
aggcttaaat ttattataaa agactgacac atagtaatta taagccgacc ctcctggaac 12300
tgcaaactca agtcgaaatg gatcagaatt ggtgtacaca gtcatgagaa agacatagta 12360
ccgcatatga agattggtca gataggtgtc cattaaacta atgacttgaa acaaataccc 12420
aacagtagat gaaagtttgt cacctgcagc agaattatat acagaattgg ttgcttgaaa 12480
gtggcctttt atagcagctg gatgtgtagc gtagttctta ctagatattc tgggagctcc 12540
atctgcatat tccaatctgg aggagggaga acctgtatta tggctccagt gcttccatgc 12600
attcataggc cctgtgtcat cagactcaga tactatctga gaaacaaggt gttcaaagct 12660
ctgtgaatca ttgaggggtt tgatttcata ggtaaggtca tctaacttca tgacccctga 12720
caggccccca taacaagtat ccacagtgac catggattgt gggatcccct ccaggtagcc 12780
aatatagtaa caatctacag gaaaaaaggg gtaatccatc tgtaaggctc cttggtcatc 12840
ttgagttgtc agcaacaagt gtctgggcca aatgagtgtc tttctccgca ggtggatgat 12900
atgtctctgg ccccgaaaat gcaagctata tgagagcagt ctttgtgctt gaagtccttt 12960
ggtatggtag atctccttcc gaggaataac cacctccgat gagatgtaac gccaagtagg 13020
atggccttga gaacaccaga ctggaaccag gaggagcagc cagagtgcaa atagcaagag 13080
gaggaccctg gggaccacag gtctttccac tagcctcatg ccccaggtca gagataacat 13140
cctgggtgga gctaaatccc tctgctgtgg ccactgcctg gtctagaaaa tactgacaga 13200
ggactaaaaa cctcctcagg ctcccaacct aagtggttac ccagacaact ggagttaggt 13260
aacagtcact gggtgtggca ggaattgagt ctgaatgtgt tagctgaggt tgaggttaaa 13320
tattgtcaaa agggatgtct ataaatgtgc ctggacaaga aaagtcagaa gcagcaagga 13380
gtgtctctga caggctcaat cctttctttt ctttttttga agttcaaaat atcatttcca 13440
cgtgaatgta tttggttccc agtgtgactc tgggtctctt tctaggagtc aatatttctt 13500
tatatcttgg ctcatgtttc tcacagttgt tctaatttct tgttttgttt tgtttgtttg 13560
tttgaacgtt agtagtaaat actgtctata ttagcctttt agctataaat gattgttttt 13620
atttcttcta atcatatttt gtttgagttt tggttaaact atttacaaat gagttttttt 13680
tttttccttt tgggtgttgc tcgaaagttt ggagctttct gttaatattg tgttgttatt 13740
tttccaatat tattagacct gagaattcta tctgggtacc tgtgaactct agaattttta 13800
aaaattccat ctcttgggaa cattacctct gaccccgtct gaggccgaag tggctgtccc 13860
cctccaacct ttagtatctt tctttcctga ctattgggat ttcttcaagc aatcaggctg 13920
atgggttctc agcagtgaga ccagtagact gccggtatga acgtcgaaga gactgccaca 13980
cactccaggt tcatcaacag tgctttcgcg tctcttactt ttgtagaagg aaaagcagcc 14040
tctgagttat ctccaagaaa tcattaatga aagagttaaa agatgggtat cacccggagt 14100
tcatgacaag ccctggctca gacacgtgag caaggtctac agccccaaag ataggctgcc 14160
ctgcaacatg tatttataag atagaagaaa aaaatgggtg gttggagggt tgatcaactt 14220
acttcctctc aaacatatat atctcatcta agtgtgcagg ggaaaactct gtaggactac 14280
tgggattgtt attatcatta ttattattat tattattatt attattatta ttattattat 14340
taacttaagg cattttatta gatattttct tcatttagtt ttcaaatgtt atccccggaa 14400
cctcctatac tctctccctg ccctgctccc caacccaccc actcctacat cctggccctg 14460
gcattcccct atactgtggc agatgatctt cgtaagacca agagcctttc ctcccattga 14520
tggcctacta ggctatcctc ttttacatat gcaactagag tcacagctct ggggaggtat 14580
tgcttagttc atattgtttt tcctcctata gggttgcaga tccctttagc tccttgggta 14640
ctttctctag ctcctccatt gggggccctg tgttccatcc aatagatgac tgtgagcatc 14700
cacttctgta tttgccaggt attggcatgg atcttactgc accttctgaa ctctctaagc 14760
agctttcctg gtcacctcca ggagcctcat gaataagtct ctgcttcccc cttgtggcta 14820
tgagcattac tgcacctgat acaccctgca gcttcctagg gaagagggag gaagtggctt 14880
ggcccctgtc tggttaaggt aagaggagat aaatcccttc tcatgaatta gggtgagaag 14940
ggtcatgtgc tctatcattg gtgaccaact tggggacatg ggcttataca gtcatcactc 15000
tgaggctctg tgtaccacca gactgaactc ccatatccta catgcacata ggacaacacc 15060
aagtagaagg aggttttagg actaaactga aggacagaga tggggtttct aaacaactag 15120
ggagtgccag ggccagcctc tctaaccact ataggacact atggagtctg gttacaaaga 15180
gagattactc aaggtcctta gcactgatta cagagcatat ctcagatgcc ttctgctgac 15240
cagatgtatc tttgcataat ctgcctatcc agattcagaa aattgatgcc acatagccaa 15300
gtggactttc aggaacagac gatttaaaaa caggcagaga gatgtgagag aaaggagaag 15360
gagagagaga agggagaggg agagaagaga gagggagacg gagaaggaaa gagggagaag 15420
gagaaggaga gaaggggcat ggacagaggg agggacagaa ggagagagga gatagagagg 15480
gggataagga agaaaggagg gagggagaga gagagaaggc taagtctttc catacctggg 15540
tcccaatacc tcttataacc caagcacatg gtttcagata tcacaatgcg gttgggatat 15600
agataactgt aaatacttgt gaaaataatg gggctgagat ctggggtttt catgatagtt 15660
tcaaagtcac tgtactgact aaaaccttcc actggcccat ctccagcttg ttaatctgag 15720
ggtatcaaat ttcccactaa gtgtgtttag aaagatctcc acctttttgc cctagtcttc 15780
cagtgcccca cctacgttct ggtctcccac atctgatgtc ttctcagtga ttctggccct 15840
gcctgctcca cagctacaaa ccccttccta taatgagctc tgtgctgagc catcatcctg 15900
aatcaatcca ccttaagcag atgttttgct tatttttcct gtgtccatac tacagaggaa 15960
gggtaggcat gtagaagctg aggcatctca tctcactcta agcaccctca gtctctaaat 16020
gtgccccttt gtttccagca gttcagcctc aagcatcttt tattcactcg tcttagaggg 16080
acacatgtgc tgtagtgtta taagatgaaa tttaaagcat tagttattcc caacaagcca 16140
attaaacaag ccaaaaacat tcatcagtca ttcccatgga acctctgaag catcttcctg 16200
ctctaacctt gagtttccta gggctgctgt gggatcacag gagctgtcct gtttaccagc 16260
ctatcctgtc ccacgggatt cagttattag tgggtgcgag ggggaccgca aacctggaag 16320
aaaatgggat tggaagagaa aagagaaacg aagaccaagt agatcttttc ctatcaaggt 16380
cttcgtttat taggctgagg tgcctggtgt aaagcatgca tcgcggggaa taggaagggg 16440
tcgaggggga attttacaaa gaacaaagaa gcgggcatct gctgacatga gggccgaagt 16500
caggctccag gcagcggaca ctctgcatct tatctctgga acatagatcc tccttgacag 16560
ccttggggtg tcaggccagg ctcaggtgta actcatgtcc ttggatggca tgggagttca 16620
ggaagagata gggaagaggg gactataatt cagcttttac agcctcaggt gccaggaagg 16680
caacagggag gagggagtga ctacaggctc ctagcacgag gccatttggc ctgtcagggt 16740
gggagattgt gaagggctca ctttctcatg gtatggtctc tgacaccagc cagaaaaaaa 16800
aaaaatctcc agtcaatcta cagaaaggca gaaatatgga gaaccttcta gaagaacagc 16860
aacccttgac tgactctgca gggcagtcta agcacacagg ttcctctgag cccagtccct 16920
ggctgtattc actcacacag ggcccagcag tctggagatg cctgcagcag gaaggttctc 16980
actgtacctc agtctcacag ctcagtcact ttgtagtctc cccacagcaa atcacaacca 17040
atgcagttgt cacccttgag gccattcctc agtgacgcct gggaaacact gtccatttgc 17100
cctgctcata tgctgtctca gctcacataa attccctcca agctccttta cagaaacaac 17160
caattgacat acaaattaaa gtaatatgaa gctctgtcct gtaccaattc ctagaacacc 17220
tcagctgcta aaatctaact gtgcaataca gagaactacc tagataatac ctaactccaa 17280
tcttgaggta tctgttgaag aggctttcag ctatgaatta ccaagaggga tgcctgtgtc 17340
tcctacatct taaggcatct ttctgaagct cacactaccc ttcacagcgc cacagcagca 17400
gggagatctg tatctcttcc tctccacaag agggcagaat tcgggtgtca actgtgacaa 17460
ccctccagca ttaatttcta ttagtagata ttttctttac ttacatttca aatgttatcc 17520
ccttccccag tttttcctcc ctgaaacccc ctatcccatc acccatctcc ctgctcacca 17580
acccacccat tccataagaa ccaatcagta ccccaccccc ccagagctcc cagggactaa 17640
accaggaacc aaagagtaca catggaagga cccatagctc cagctgcata ggtagcacta 17700
ctttgaaaag ggggaagatg atgaaatcat tactgtgggg gaaatgcaag gaagattcca 17760
acacatctag gcatctatga agattttaag tcttcaaaat ccaaaaccac cacaaaattt 17820
aaaaaaaaaa aaagtagatt ctaaatgcag tcacctgcac caggtgcctg gggaatcact 17880
cagcagccct agactgagaa agcttggaga aagtagaaat agagaaagtg tacagccagt 17940
atcctctagc tactcacatc caaacagggc ctcctgactg ctctgagcct gtcctaagaa 18000
cagcaatgat gccacagaaa tttttagagt gagccctgaa ggaacttgag gctgatatga 18060
gcaagccagt cccagaggaa aggaaaaccc atagagagaa aacaggtgag ttagtgcatt 18120
aaaggggctg agcagggagt tctcatcgct ccccagcacc agaaataaga gcctctccgg 18180
agctgctggg acatggaatg cagatgattc ggaccatcag ccccacagag acctttccca 18240
ctctggctca gaaagaggca ctggaccaca gttggagagg agaatcgaaa gctgatatct 18300
ctgtattcac ttagcctgtt acccacccat gcacccaagt ccaaggtggg agaaacactg 18360
agggtctaaa cacagcccca gagcaactgc cagtattaaa taccagattt cagaagatgg 18420
gaaatcacct ctctggtcat ttttgggaca tgtaaactgt aacaggaaac ataggaccaa 18480
tttaagatgg agcagtccta tatccctaac ccagttgtaa ataaaacatt caagagtgcc 18540
atcagacacc acagtggtac aggagagatg agtgtaccta gtgcatcaag agttccctca 18600
ctagataaac caagatgtag ccccaggacc acccaggcac ctaccaggac tcccctccag 18660
aggtctgagc cagttagctc tagttcattc tatgtttcag accaaaacat cagaaacaac 18720
agcatctcca ctgcagatga acccctaagc catacagtgt acccaaaggc agcaccacag 18780
atggggaatg tgggggcgaa gaagcttgta tcacattaag agtgttgccc aatgactatt 18840
tgctatcttc acagtaagat gggcactgat cccacccacc aaaacatagt aggacaagga 18900
cccttaaata acatctgtca aggggagctg tcaaatagcc actgagctga gatggctcat 18960
acggggtgat atagaaaaac aggccaaaga acctcctgtg ttgcaagcac aaatgggaag 19020
ctgtgaatct ccactacctt acaagaaaac tgtaccaatc tgcaagaaca ggggatacac 19080
agctttccac actgtgtagg aaggcattgg actctcagat catccaagga ctagggctaa 19140
agtggccatg tgtgggaaat acatccactt tataaaccta ccttgtatcc acatcacaca 19200
tggagcatct gctcctggtc cctacctcat gtgggtgctg cggtgctgta cattgagtct 19260
gggggcatga gtgtgcccgg taaattcctt atcactcaga tgaatttcca gtccacactc 19320
atcaccttgg agtaggaatt ttaaaagtta gtgtagatat aagtaaagag agtcaggagg 19380
gggggctgaa aggcagacct gcagaaattc tccagagggt agagctcaga agcagcaagc 19440
aaaacttggg ctaacagaag agcaaacaca gagctcaggc agaactacct ggcaatgcga 19500
ctgggcacac tgaaagcact gggcatcagc actgagcccc aaatatgcac tcaggatcct 19560
ctgcataata atgtaacata acaggaaggt tagaacaggc caaaagagga aacagaacaa 19620
atgcccctaa ccaaagaagt ataaacaaat tgggaagagt aaagaaggat tgtaaggatt 19680
gagtaccaca cagaacatgc tcttaatggc ctcaatgctg aagctaggaa gaactaagtt 19740
aaaagaaaca tgttcaacgg gattccctgt cactggactt cacaacaagc aaaattcaat 19800
ctttctgtta aggagatgag aagagaatat ctgaaccttg tgttgacagt gccccacccc 19860
gactgtcagg ctgtgggaaa tgccagagca atcactagga acacacaagg atgagggaga 19920
cgagggttag gacacaacca tcatgatatc ccacaagtat ggaagagcaa gaacttgtag 19980
agagcagaga atggcagaca aagcagcata tacataagta gatggccaga ctatacagga 20040
aagatctaca tagcctgtga ggctttctga cagaaaaggg caggcatgtc tcaaagcaca 20100
atgcctggct tgggacactg tctgccctgg atcctctcac tccacatgta aggagctcac 20160
agcaaaacca cacagccttc cacaagagga gaagaaaagg tagcttgtca gtgaggaagt 20220
cccccagaga cagaccattc cagtagttct tatcattcct cccaaagcag ccaccatcca 20280
ggcactgaga gaccaaaggc tgttgggagg tcagcctaga ggcaggctca gacctttctt 20340
gctccctagg accttcctga aaagatagaa gcacagaagt gaataatctg gacctcaact 20400
caggatgaca actgaaactc aaccgtgctg cctgggcccc caatgctctc tacacctgca 20460
ggcgcgcc 20468
<210> 4
<211> 67667
<212> DNA
<213> Artificial Sequence
<220>
<223> synthetic
<400> 4
taactataac ggtcctaagg tagcgaggga tgacagattc tctgttcagt gcactcaggg 60
tctgcctcca cgagaatcac catgtccttt ctcaagactg tgttctgtgc agtgccctgt 120
cagtggaaat ctggagagca tgcttccatg agcttgtgag tagtatatct agtaagccat 180
ggctttgtgt taatggtgat gttctacata tcagttctct ggcttaataa tgaggtgatg 240
attctatgtt cctgtaacgc ttcctcaact gggtcctaag tctttcttca ctccatctat 300
tcctctaagg aatgatcctg aaaatcccat cacaaactat aggagatggg aaccatcaaa 360
aaacacagtg acaaagaggt gggaacgcat cagggttcag gaaccatatt ttaaaaagat 420
atcgtaaata acttcttaaa agagatatag acaaatctcc attaatacgg agaccagagg 480
cctaaggcta agaaccaatg gtggctcaag gtctcctgct acccgaggag caaacgtaga 540
gcagtttcta atgatttatt taaaatatag aatcaaaagt accagtttgc aattttgaaa 600
gatttatttc agcaatgcaa caacatcagg tggtgccgag tccaacacgt cttatgtccc 660
atgatataaa caaaggccat ccagaactgt ggactggagt tctaccttgt cccctaatga 720
cattcagatt ttttttccat tctctttatc ttagaggaga cagggggcta actcatttta 780
cttgtccttt gcttgttctt gccaagaacg taaagcagct tgcaagtctt caaacctaaa 840
tatcttagta actcctacac gagtggcaat gccaaagagc agtgcaacaa agaggaagta 900
aatacgacca aagagtattc ttaaatacac tactggctct aggttctgtt ttattatgcg 960
cctttgaacc ggaggggacc cactgtctat gctcccactg tgtccctctt ctttgcactt 1020
tggagggctc caaccaaaat ggcaatggca attccgacga ttgttacaca ctcctctgaa 1080
attgcatttt tctggggtgc agtcataacc caaacgagat aaacttccat tgcaagctcc 1140
tcgatcacag aacttacccc ttgaacacgg ggtaccatgt ctcaccaatc cagcatctgc 1200
tgtttctgtc ccacgatgtt catcaagccc aaagcaggta accccagaga taaccgattg 1260
atggaatgaa acatgttctt gcaaaaatgg aagattggtg acattggtac actgcaacct 1320
tccacacagc ttgtcctgat cagcacaagc attgaatgtg aggctttctt ctgctctagt 1380
acaatgccca aatcgaaacc gttgtttgtt gatgtcatag cacttaatat tagcattctt 1440
agcacttaca ccaaagattt ccatgcattg tatgttgcga tcagtgcagt tacctttata 1500
gcagtaaccc tcttctgagc atggtgtccc atcttgcaga taagtgtcat ctgggcaaat 1560
gaacttagag ccactacagt actctggaag atcacatatg ttctggatag gtctgcagag 1620
tgtcccagaa ggactgtaag tgcaatttgc acagcataat tctttatcac aaatgctacc 1680
aggtgttaac ctgcaatcat ttccacagca gggatctgaa taacatgcct tttgggagcc 1740
acagtcacac tgctcattgt tatctacttt gaagtttcca caaaacttat aagtcaatga 1800
tgtattataa taaacatgac ggtcatagaa aagacatggc atcagatcag gagtattaag 1860
tatgttgctt atctctgcaa gggaacaatt gctgaaagca tctgttaatt gaggattttt 1920
gaacatgatg caggtgttcc ttctctggca gatacagtac ccctcatcat gttttaggcc 1980
taaactcctt ccaacacgat tggttattat aatagataaa aataaaggat ttcgaccatg 2040
ttgaccaaga caaattaggg ctgagggaga acatatactc ctctcagctg gattaacagc 2100
atcatctcct ggcgaattct tgttaattat agctcctgca tcaggcctaa aatgagcata 2160
aaatactctc tcatagaaag tatgagcctg ccctcctgga actcgaaaat cttgtgaaaa 2220
tggatcagcc tcggtataca cagtcatgag aaagacatag taccgcatat gaagattggt 2280
cagataggtg tccattaaac taatgacttt aaacaaatac tcaacagtag atgaaagttt 2340
gtcacctcca gaagcactat atacagaatg ggttgcttga aagtggcctt ttatagcagc 2400
tggatgtgta gcgtaattct tactagatag tctgggagct ccatctgcat attccaatct 2460
ggaggaggga gaacctgtat tatggctcca gtgcttccat gcattcatag gccctgtgtc 2520
atcagactca gatactatct gagaaacaag gtgttcaaag ctctgtgaat cattgagggg 2580
tttgatttca taggtaaggt tatccaactt tatgacccct gacaggcccc cataacaagt 2640
atccacagtg accatggatt gcaggatccc ctccaggtag ccaatatagt aacaatctac 2700
aggaaaaaag gggtactcca tctgtaaggc tccttggtca tcttgagttg tcagcaacaa 2760
gtgtctgggc caaatgagtg tctttctccg caggtggatg atatgtctct ggccccgaaa 2820
acgcaagcta tacgagagca gtctttgtgc ttgaagtcct ttggtatggt agatctcctt 2880
ccgaggaata accacctccg atgagatgta acgccaagtg ggatggcctt gagaacacca 2940
gactggaacc aggaggagca gccagagtgc aaatagcaag aggaggaccc tggggaccac 3000
aggtctttcc actagcctca tgccccaggt cagagataac atcctgggtg gagctaactc 3060
cctctgctgt ggccactgcc tggtctagaa aatactgaca gaggactaaa aacctcctca 3120
ggctcccaac ctaagtggtt acccagacaa ctggagttag gtaacagtca ctgggtgtgg 3180
caggaattga gtctgaatgt gttagctgag gttgaggtta aatattgtca aaagggatgt 3240
ctataaatgt gcctggacaa gaaaagtcag aagcagcaag gagtgtctct gacaggctca 3300
atcctttctt ttcttttttt gaagttcaaa atatcatttc cacgtgaatg tatttggttc 3360
ccagtgtgac tctgggtctc tttctaggag tcaatatttc tttatatctt ggctcatgtt 3420
tttcacagtt gttctaactt cttgttttgt tttgtttgtt tgtttgtttg aaagttagaa 3480
gtaaatactg tctatattag ccttttagct ataaatgatt gtttttattt cttctaatca 3540
tgttttgttt gagttttggt taaactattt acaaatgagt tttttttttc cttttgggtg 3600
ttgctcgaaa gtttggagct ttctgttaat attgtgttgt tgtttctcca atattattag 3660
acctgagaat tctacctggg tacctgtgaa ctccagaatt tttaaaaatt ccatctcttg 3720
ggaacattat ctctgacccc gtctgaggcc gaagtggctg tccccctcca acctttagta 3780
tctttctttc ctgactattg ggatttcttc aagcaatcag gctgatgggt tctcagcagt 3840
gagaccagta gactgtcggt atgaacgtcg aagagtctgc cacacactcc gggttcatca 3900
acagtgcttt cgcgtctctt acttttgtag aaggaaatgc agcctctgag ttttctccaa 3960
gaaatcattg atgaaagggt gaaaagatgg gtatcacccg gagttcatga caagccctgg 4020
ctcagacacg tgagcaaggt ctacagcccc aaagataggc tgccctgcaa catgtattta 4080
taagatagga gaaaaaaatg ggtagttgga gggttgatca acttacttcc tctcaaacat 4140
atatatctca tctaagtgtg caggggaaaa ctctgtagaa ctactgggat acctgctcac 4200
ccccaggagc ctcatgaata agtctctgct tctgccttgt agccatgagc attactgcac 4260
ctgatacccc tgcagcttcc tagggaagag ggaggaagtg acttggcccc tgtctggtta 4320
aggtaagagg agataaatcc cttctcattg attagggtga gaggggtcat gtgctctatc 4380
attggtgacc cagttgggac atgggtttat accaaagtca tcactctgag gttctgtgta 4440
ccaccaggct gaactcccat atcctacatg gacataggac aacaccaagc agaaggaggt 4500
tttaggacta aactgaagga cagagatgcg gtttctaaac aactagggag tgccagggcc 4560
agcctctcta accactatag gacactgtgg agtctggtta caaagagaga ttactcaagg 4620
tccttagcac tgattacaga gcatatctca gatgccttct gctgaccaga tgtatctttg 4680
cataatctgc ctatccagat tcagaaaatt gatgccacat agccaagtgg actttcagga 4740
acagacgatt taaaaacagg cagagagatg tgagagaaag gagaaggaga gagagaaggg 4800
agagggagag aagagagagg gagacggaga aggaaagagg gagaaggaga aggagagaag 4860
gggcatggac agagggaggg acagaaggag agaggagata gagaggggga taaggaagaa 4920
gggagggagg gagagagaga gaaggctaag tctttccata cctgggtccc aatacctctt 4980
ataacccaag cacatggttt cacatatcac aatgcggttg ggatatagat aactgtaaat 5040
acttgtgaaa ataatggggc tgagatctgg ggttttcatg atagtttcaa agtcaccgta 5100
ctgactaaaa ccttccactg gcccatctcc agcttcctaa tctgagggta tcaaatttcc 5160
cactaagtgt gtttagaaag atctccacct ttttgccctt gtcttccagt gccccaccta 5220
cgttctggtc tcccacatct gatgtcttct cagtgattct ggccctgcct gctccacagc 5280
tacaaacccc ttcctataat gagctctgtg ctgagccatc atcctgaatc aatccacctt 5340
aagcagatgt tttgcttatt tttcctgtgt ccatactaca gaggaaaggt aggcatgtag 5400
aagctgaagc atctcacctc attccaagca ccctcagtct ctaaatgtgc ccccttgttt 5460
ccagaagtgc aacctcaagc atcttttatt cattcatctt agagggccac atgtgctgta 5520
gtgttataag atgaaattta aagcattaat tattcctaac aagccaatta aacaagccaa 5580
aaacattcat cagtcattcc catggaacct ctgaagcatc ttcctgctct aaccttgggt 5640
tttccagggc tgctctggga tcacaggagc tgtcctgtct accagccata taaaggcaga 5700
cctatcagaa ttacaccaga cttctcacca tagactataa aagccagaat atcctggaca 5760
gatgttatac agaaactaag agaacacaaa tgccagccca ggctactata cccagcaaaa 5820
ctctcaatta ccatcgatga agaaaccaag atattccatt acaagtccaa atttacacaa 5880
tatctttcca taaatccagc cctacaaagg atagcagatg gaaaactcca acacaggtag 5940
gaaaactaca ccctagaaag agcactaaag taatcatctt tcaacacact caaaagaaga 6000
taaccacaca aacataattc cacctctaac aacaaaaata aagtaggcaa caatcactat 6060
tccttaatat ctcttttaac atcaatggac tcaattctcc aataaaaaga catagactaa 6120
cagactgaat acataaacag gacacagcat tttgctgcat aaagcaaaca cagcgttact 6180
tttttttttc taaatgacat tttttattag atattgtctt tattgacatt tcaaatgtta 6240
tcccctttcc tggtttaccc tctgaaatcc cctatctcct ccccctcccc ctgctcacca 6300
atccacccac tcccacttcc aggccctggc aatcccctat atttgggcat agagccttca 6360
caggaccaag gtactctcct tgcattgatg accaactagt ccattctctg ctacaaatgc 6420
agctagatct atgagtccca ccatgttttc ttttgttggt ggtttcatgc cagggagctc 6480
ttggagtact gattggttca tattgttgtt ctccctatgg ggttacaaaa cccttcaact 6540
tcttgggtcc tttctctggc tgcctcattg gggaccttgt gcgaagtcca atggatgact 6600
gtgagcatcc acttctgtat ttgccaggca ctggcagagc ctctcagaag acagctatat 6660
caagatcctg gcagcaagct cttgttggta tccacaaaag tgtctggtgg ttgtctatgg 6720
gatggatccc caaaggggca gtctctggat ggtcattcct tcagtctctg ttccacactt 6780
tgtctcttta actccttcca tgactatttt attcctccct ctaagaagga ccgaagtatt 6840
catactttgg tcttccttct tgaaattcat gtgttttgtg aattgtatct ttgatattcc 6900
gaacttctgg gctaatatcc acttatcagt gagtgaatat catgtgtgtt cttatgtgat 6960
tgagttacct cactcaggat gatatcctcc agaaccatcc atttgtctaa gaatttaatg 7020
aattcattgt ttttaatagc tgaggagtac tccattgtgt aaatgtacca cattttctgt 7080
acccattgtt ctcttgaggg acatctgggt tctttaaagc ttctggacat taaatataag 7140
gctgctatgg aaatagtgga gaatgtgtcc ttattacatg ttggagcatc ttctgggtat 7200
atgcccagga gtgctattgc tggatcctct gatagtacta tgtccaattt tctgaggaac 7260
tgccaaactg atttacagag tggttgtacc agcttgcaat tccaccagca atggagaaat 7320
gttccccttc ctccacatcc tcaccaacat ctgctgtcac ctcaatttgt tcttagtgat 7380
tcagacaggt gtgaggtgga atatcagggt tgtttggcat ttccctgatg actagtgata 7440
ttgaaaaaaa ttttaagtgt ttctcagcca ttcagtattc ttcagttgag aattcactgt 7500
ttagctctgt actcaggttt ttttaatagg gttatttggt tttctggagt ctaacgtctt 7560
gaattctttc tatatattgg atattagccc tctgtcatat ttaggattgg taaagatctt 7620
tcccaatatg ttggctgcct ttttgtgtcc tttgccttac agaacctttt taattttatg 7680
aggtcccatt tgctaattct tcattttaca gcacaagcca ttggtgttct gttcaaaaat 7740
ctttccccct gaaccctatc ttcgaggatc ttccccactt tctcctctat aagtttcagt 7800
gtctctatta ttgtgctgag gtcgcgatcg caccggtgaa accctaggaa gttcctatac 7860
tttctagaga ataggaactt cggaatagga acttctctag gggctgcagg tcgaggtctg 7920
atggaattag aacttggcaa aacaatactg agaatgaagt gtatgtggaa cagaggctgc 7980
tgatctcgtt cttcaggcta tgaaactgac acatttggaa accacagtac ttagaaccac 8040
aaagtgggaa tcaagagaaa aacaatgatc ccacgagaga tctatagatc tatagatcat 8100
gagtgggagg aatgagctgg cccttaattt ggttttgctt gtttaaatta tgatatccaa 8160
ctatgaaaca ttatcataaa gcaatagtaa agagccttca gtaaagagca ggcatttatc 8220
taatcccacc ccacccccac ccccgtagct ccaatccttc cattcaaaat gtaggtactc 8280
tgttctcacc cttcttaaca aagtatgaca ggaaaaactt ccattttagt ggacatcttt 8340
attgtttaat agatcatcaa tttctgcaga cttacagcgg atcttcagaa gaactcgtca 8400
agaaggcgat agaaggcgat gcgctgcgaa tcgggagcgg cgataccgta aagcacgagg 8460
aagcggtcag cccattcgcc gccaagctct tcagcaatat cacgggtagc caacgctatg 8520
tcctgatagc ggtccgccac acccagccgg ccacagtcga tgaatccaga aaagcggcca 8580
ttttccacca tgatattcgg caagcaggca tcgccatggg tcacgacgag atcctcgccg 8640
tcgggcatgc gcgccttgag cctggcgaac agttcggctg gcgcgagccc ctgatgctct 8700
tcgtccagat catcctgatc gacaagaccg gcttccatcc gagtacgtgc tcgctcgatg 8760
cgatgtttcg cttggtggtc gaatgggcag gtagccggat caagcgtatg cagccgccgc 8820
attgcatcag ccatgatgga tactttctcg gcaggagcaa ggtgagatga caggagatcc 8880
tgccccggca cttcgcccaa tagcagccag tcccttcccg cttcagtgac aacgtcgagc 8940
acagctgcgc aaggaacgcc cgtcgtggcc agccacgata gccgcgctgc ctcgtcctgc 9000
agttcattca gggcaccgga caggtcggtc ttgacaaaaa gaaccgggcg cccctgcgct 9060
gacagccgga acacggcggc atcagagcag ccgattgtct gttgtgccca gtcatagccg 9120
aatagcctct ccacccaagc ggccggagaa cctgcgtgca atccatcttg ttcaatcatg 9180
gtggatggtt tagttcctca ccttgtcgta ttatactatg ccgatatact atgccgatga 9240
ttaattgtca acaattcgag agacccgggt tcccaatcta gagctcgacg tctaacaaaa 9300
aagccaaaaa cggccagaat ttagcggaca atttactagt ctaacactga aaattacata 9360
ttgacccaaa tgattacatt tcaaaaggtg cctaaaaaac ttcacaaaac acactcgcca 9420
accccgagcg catagttcaa aaccggagct tcagctactt aagaagatag gtacataaaa 9480
ccgaccaaag aaactgacgc ctcacttatc cctcccctca ccagaggtcc ggcgcctgtc 9540
gattcaggag agcctaccct aggcccgaac cctgcgtcct gcgacggaga aaagcctacc 9600
gcacacctac cggcaggtgg ccccaccctg cattataagc caacagaacg ggtgacgtca 9660
cgacacgacg agggcgcgcg ctcccaaagg tacgggtgca ctgcccaacg gcaccgccat 9720
aactgccgcc cccgcaacag acgacaaacc gagttctcca gtcagtgaca aacttcacgt 9780
cagggtcccc agatggtgcc ccagcccatc tcacccgaat aagagctttc ccgcattagc 9840
gaaggcctca agaccttggg ttcttgccgc ccaccatgcc ccccaccttg tttcaacgac 9900
ctcacagccc gcctcacaag cgtcttccat tcaagactcg ggaacagccg ccattttgct 9960
gcgctccccc caacccccag ttcagggcaa ccttgctcgc ggacccagac tacagccctt 10020
ggcggtctct ccacacgctt ccgtcccacc gagcggcccg gcggccacga aagccccggc 10080
cagcccagca gcccgctact caccaagtga cgatcacagc gatccacaaa caagaaccgc 10140
gacccaaatc ccggctgcga cggaactagc tgtgccacac ccggcgcgtc cttatataat 10200
catcggcgtt caccgcccca cggagatccc tccgcagaat cgccgagaag ggactacttt 10260
tcctcgcctg ttccgctctc tggaaagaaa accagtgccc tagagtcacc caagtcccgt 10320
cctaaaatgt ccttctgctg atactggggt tctaaggccg agtcttatga gcagcgggcc 10380
gctgtcctga gcgtccgggc ggaaggatca ggacgctcgc tgcgcccttc gtctgacgtg 10440
gcagcgctcg ccgtgaggag gggggcgccc gcgggaggcg ccaaaacccg gcgcggaggc 10500
ctcggaggcc agacgcgcga gaagttccta tactttctag agaataggaa cttcggaata 10560
ggaacttcgg taccgcgatc gcccttgatc cacttgaact tgagcattgt tcaaggagat 10620
aagaatggat caattcgaat tcttctacat gataacagcc agttgagcca gcaccatttg 10680
ttgaaaattc tcttttttgc actggatggt tttagcactt ttgtcaaaga tcaagtgact 10740
atggctcttc aactatggct cattccattg atcaacttgt ctgtcactgt acaagcacca 10800
tgcaattttt attgcaattg cttagtatta caccttgagg tcaaggatgg tcattccacc 10860
agaggttctt ttatggttga gaagagtttt tgctatccta ggtttttgtt attccagatg 10920
aatttgcaaa tggccctttc taactcagtg aagaattgag gtggaatttt gatgggaatt 10980
ttattgaatc tgtagattgc attcaacaag atagccattt ataatacatt aatcctgcca 11040
gtccatgagc atgggagatc tttccatctt ccgagatctt cttcgatttc tttcttcaga 11100
gacttgaagt ttttatcata cagatctttc acttccttag ttagagtcac accaaggtat 11160
tttatattat ttgtgactac tgtgaaggtt gttgtttccc taatttcttt ctcagcctgt 11220
tcatcctttg tgtagagaaa ggccactgat ttatttgagt taatattgta tccagctaat 11280
tcactgaagt tgtttatcag gtttaggagt tctcttgtgg aatttttgga atcacatgtg 11340
tatactatta tatcatctgc aattagtgat attttgactt cttctttccc aaattgtatc 11400
cctttgatct ccttttgttg tctaattgcc cacactagga ctcgggcagc cttagtgcct 11460
agtccctgat tttagtgtga tttgttcaag tttctctcca cttagtcgga tgttggctac 11520
tgatttgctg tatattgctt ttattatgtt taggtatggg ccttgaattc ctgatctttc 11580
caatactttt atcatgaatg ggtgttgaat tttgtcaaat gctttctcaa catctacaaa 11640
gatgatcatg tagattttgt ctttcagttt gattatatag tgtattatgt tgatggattt 11700
ccatatatta aaccatccct gcatccctgg gatgaagcct acttggtcat gatagacgat 11760
tgttttgatg tgttcttgga ttcagttagt gagaaatata ttgagtattt ttacatcgat 11820
attcataagg gaaattggtc tgaagttctc tttctttgtt gggtctttat gtggtttagt 11880
tatcagagtc attgtagctt catagaacaa attgagtaga gtaccttctg tctctatttt 11940
gtggtatagt ttgaggagat ttggaaatat gtcttcttgg aacgtctgag agaattctgc 12000
actaaaccca tctgatcctg ggcttctttg gggggggggg actattaatg actgcttcta 12060
tttctttagg ggaaatggga ctgtttagat tgttaatatg atcctgaata gaaatctgat 12120
ctgatctaga aaattgtcca ttttattcag gttttccagt tttgttgagt attgcctttt 12180
gtggtaggat ctgatgatgt tttggatttc cttaggttct gttgttatgt cttctttttc 12240
atttctcatt ttgttaatta ggatactgtc cctgtgtcct ctagttactc tggctaagcg 12300
tttatctatc ttattgattt tctcaaagaa ccagctcctg gtttggttga ttctttgtat 12360
agttcttttt gtttccactt gattgatttc tgccctaagt ttgattgttt cctgctgtct 12420
actcctcttg ggtgaatttg cttccttttg ttctagagct tttaggtgtg ctgtcaagct 12480
gatagggtat gctctctcta gtttcttttt ggcggcactc atagctagga gttttcctct 12540
tagcagtgct ttcattacgt cctgtaagtt tgggtatgtt gtggcttcat ttgcattaaa 12600
ttctaataag tctttaatct ctttccttct ttcttccttg accgagttat cattgactag 12660
agtgttcatc agcttccaca tcaatgttgg ctttttatta tttatgtttt tattgaagat 12720
cagcctttgt cggtggtgat cttctaggat gcatgggaaa ttttcaatat ttttgtatct 12780
attgaggcct gttttgtgac caattatacg gtcaattttg gagaaagtac cgtgaggtac 12840
tgagaagatg gtatatcttt ttgttttagg ataaaatgtt ctgtagatat ctgttaaatc 12900
catttgtttc ataacttctg ttagtttcac tgtgtctctg tttagtttct gattccagaa 12960
tctgtccaat gataagagta gggtattaaa ttctcccact actattgtgt gaggtacaat 13020
gtgtggtttg agctttaaaa gagtttcctt aatgaatgtg gatggccttg catttggagc 13080
atagttattc agaattgaga gttcctcttg gaagatttta cctttgatga gtataaaatg 13140
cccctccttg tcttttttga tacctttggg ttagaagtgg attttattcg atattagaat 13200
ggctaatcca tcttgtttct ttgagatgtt tgcttggaaa attattttcc tgccctttac 13260
tcggtggtag tgtctgtctt agtccctgag gtgggtttcc tgtatacagc aaaatgttgg 13320
gtcctggtta tgtagccagt ctgttagtct gtctttttat caggtaattg agtccattga 13380
tattaagagc tattaaggaa aagtaattgg tgcttcctgt tatttttgtt gttagacttg 13440
ggattctgtt cttgtggcta tcttctttta ggtttgttga aggattactt tcttgctttt 13500
tttagggtgt aatttccctc tttgtgttgg agttttctct ttattatcct ttgaagggct 13560
ggattcatgg aaagatgttg ggtgaatttg gttttgtcat ggaattcttt ggtttctcca 13620
tctataattg agagttttgc tgggtatagt agcctaagct ggcatttgtg ctctcttagt 13680
gtctataaca tctgtccagg atcttctggc tttcataatc tctggtgaga agtctggtgt 13740
aattctgata ggcctgcctt tatatgttac ttgacctttt tcccttactg ctttaaatat 13800
tctatcttca tttagtgcat tttttttctg attttttatg tgtcaggagg aatttctttt 13860
ctgctccagt ctatttggat tctgtaggct acttctatgt tcatgggcat ctccttcttt 13920
aggttacgga cgttttcttc tataattttg ttgaagatat ttactggccc tttaagttga 13980
aaatctccat tctcatctat acctattatc tttaggtttg gtcttctcat tgtgtcctgg 14040
atttcctgga tgttttgagt taggatcttt tttgcatttt gcattttttt ttattgttgt 14100
gcccatgttt tctacggaat cttatgcacc tgagactctc tcttctacct cttgtattct 14160
attggctgat gcttccacct atgtttctcg atttctttcc taggatttct atccccagag 14220
ttgtctccct ttgtgatttc tttattgttt ctacttccat ttttagattt tgaatggttc 14280
tgttcgattc catcgcctgt tgggttgtgt ttttctgtat ttctttgagg gatttttgtg 14340
cttcatcttt aaggtcttct acctgtttag gagtgttttc ctataattct ttgagggatt 14400
tttgtgtttt ctctttaagg gcttctagca atttagcagt gttctcctgt atttctttaa 14460
gtgagttatt aatgcccttc ttaaaatcct ctaccaacat cattagatat gattttaaat 14520
ccgaatcttg cttttcaggt gtgttggggt atccaggact cactgtgggg ggagtactgg 14580
gttctgatga tgaaaactgg tcttggtttt tattagtaag attcctactt ttgccttcca 14640
ccatctgata atatctgttg ttagatattc tagctgtctc tggctggagc ttgttcctcc 14700
tgtgattctg tcagcctctg tcagcactcc tgggagtaca actcttttct gagtcccaat 14760
gttcagagca ttctctgcag gcaagctctc ctctggcagg taaggtgccc agagctcttg 14820
agctcagctc cacctcctga ctgcagatga agacccaaag ggaccctgtc caataagctc 14880
tgttgcttct gccacccaca tgctctcctg tgcgaactgg tctctgagag acccgggata 14940
caagatggta ctctcacctg aatcccaggg tcaaagccct ccctggaggc tgactctcct 15000
cttgtgggaa ggtgcacaga ggtctggagc tcagctctgc ctcctggctg aagatgaagg 15060
cccgaaggga ccctgtccaa gaagctttgt tgcttctggg acccacatgc tctcctacat 15120
ggactggtct ctgagagacc agggattcaa gatggtgctc tcacctgagt cccagggtca 15180
gagccctctc tggaggccaa ctctcctcag tgatcctaag atcctgggta tgctagggtg 15240
cctatggcat ggagagtctt ctgaggaatg tgggactgtc tgctgagttt ccacccaagg 15300
tggtgctggg ctggctccag tcagaatgaa cccagactct ggttgggcag gtttcctgtc 15360
ctgctggccc aagcccctct gggttgtttt agaacagatg ttgctttcca ctcaccagtg 15420
atcccaagat cctgggtgtg ctagggtgcc tgctatgtgg agagtccact ggggacctta 15480
ggagcataca tcaagttcac acccatggtg gcaaggagct ggtgcctacc agaacaaacc 15540
ccgggcactt ttactgaccc tttaagttga aaatcttcat tctcatctat acctattatc 15600
cttaggtttg gtcttctcat tgtgtcctgg atttcctgga tgttttgact taagatcttt 15660
ttgcattttg catttttttg attgttgtgt ccatgttctc tctggaatct tctacacctg 15720
agattctctc ctctgtatct tgtattctgt tggtgatgct tgcatctatt gctcctgatc 15780
tctttcctag ggtttctatc tccagagttt tctccctttg tgatttcttt attatttcta 15840
cttccatttt tagatcctag atgattttgt taaattcctt cacctgtttg gttgtgtgtt 15900
cctgtaattc tttaagggat ttttgtgttt cctctttaag ggcttctacc tgtttagctg 15960
tgttctactg tatttcttta aaggacttat gaatgtcctt cttaaaaacc tctaccagca 16020
tcatgagatg tgatcttaaa tgtcaatctt ccctttctgg tgtgttgggg tatccaggac 16080
ttgctgtggt tggagttctg ggttctggta aacctgcctt agagggtcac cacagagtaa 16140
tgatagcact acttttaaac aggggaagat gatgaaataa ttgctgtggg aaaatgcaag 16200
gaagactcca acacatgtag gcatctatga aggtctcaaa tcttcaaaat ccaaaaccac 16260
caagaaagaa agaaagaaag aaagaaagaa agaaagaaag aaagaaagaa agaaagaaag 16320
aaagaaagaa agaaaggaag gaaggaagga aggaaggaag gaaggaagga aggaaggaag 16380
gaaggaagga aggaaggaag gaaggaagat tctaaaagta gtcacctgca ccaggtgcct 16440
ggggagtcac tcagcagccc tagactgaga aagcttgaag aaagtagaaa tagagaaagt 16500
gtacagccag tatcctctag ctactcacat ccaaacaggg cctcctgact gctctgagcc 16560
tgtcctaaga acagcaatga tgccacagaa atttttagag tgaaccctga aggaacttga 16620
ggccgatatg agcaagccag tcccagagga aaggaaaacc catagagaga aaacaggtga 16680
gttagtgcat taaaggggct gagcagggag ttctcatcgc tccccagcac cagaaataag 16740
agtctctcca gagctgctgg gacatggaat gcagatgatt tggaccatca gccccacaga 16800
gacctttccc actctggatc agaaagaggc aagcatgtaa ccatctggac cacagttgga 16860
gaggagaatt gaaagctgat atctctgcat tcacttagcc tgttacccac ccatgcaccc 16920
aagtccaagg tgggagaaat actgagggtc taaacacagc cccagatcaa ctgccagtat 16980
taaataccag atttcagaag atgggaaatc gactctctgg tcagttttgg gacatgtaaa 17040
ctgaaagaga aaacatagat ggaccaaatt aacatggagc ctaattaaaa tggagcagtc 17100
ctatatccct aacccagttg taaataaaac attcaagagt gccatcagac accagagtgg 17160
tacaggagag atgagtgtac ctagtacatc aagagttccc tcactagata aaccaagagt 17220
agccccagga ccacccaggc acctaccagg agtcccctcc agaggtctga gccagttagc 17280
tctggttcat tcagttccag accaaaatat cagaaacaat agcatctcca ctgcaggtga 17340
atccctgagc aatacagtgt acccaaagaa aacaccatgg atggaggaca agggaggcag 17400
aggagtctgt atcatactaa gagtattaca caatgagttt tttctctttt cacagtgacg 17460
aagcactgat tccccctgtc ttagttaggg ttctactgct gtgaacagac attataacca 17520
agacaggact tataaaggac aaaatttaat tggggctggc ttacaggttc agagtgtcag 17580
tccattatca tcaatgcaag agcatggcaa catccaggca ggtatggtac agtaaaagct 17640
tagaggtcta catcttcatc tgaaggctgc tagcagaata ctgacttcca ggcatctagg 17700
atgaggggtt tttcaacaac atccacagtg acacacctac ttcaacaggg ccacatcttt 17760
taactgtgcc aatacctagg cagagcatat acaaaccatg acatcacact atttggctcc 17820
tgtaggctgg ttcaaacaca tgagtcaatg gtggccacat ctggtcatag tgtaattaaa 17880
aaaaagtaca gttagtccaa cttccaacgt ccccatagtg tatagcaatc tcaacaatgt 17940
taaaagtcaa aagttcaaag tctcttctaa aatccattca attacttaac tgtaacccca 18000
aatcaagaca ggaaaccatc tgagcaaact ccacactctg catctccaag tctgacgtca 18060
aagcaatttt cagatctcca atctatcttt tcatattgtg actacaacaa acttcttttc 18120
ctgggatggt cctactccct gttagcagca ttcctcagaa gatactctgt ggctttgata 18180
tttccaaaac cttgggtctt caaggcagat tcaatgttac agcttgtttc agtatctggg 18240
atccacacat gattttctag gctcctccaa agggctggca ccacttcttt agctctgccc 18300
tctgtggcat tctaagctca ggttgatcca ctccactgct gctgctgctg ctgttcttgg 18360
tgaccatcca ccccatggta ttggcatctc cactatgatg aggacttctg ctgcaactag 18420
gcttcaccaa tagcctctct taggctttct tcataatgac aagggtcaaa tcctttacat 18480
gaccccttca gttctgggcc accaactgca actggggttg caccttcccc agtggtggtc 18540
catagcctct cactccaagc ctcagatgtt cttcatgacc ccttcatgcc ttcaaaacca 18600
gtaccacctg ggtgactctt atacattacc aagtatagtt gcagcacaag gtacaacctt 18660
ggctatcact ggaacacaac ttctttgtgc tttcagaaaa caatttccag aagatttcac 18720
ctttcttaat cactactaat ttcttagctc cagctaacca gtattaattg tcccagtatc 18780
aattgtctcc ttctcctctt gactctaaag ccagaatcac atggacaaag gtgctgagtt 18840
ctgctgcttg caggatctag atcatggctg ccttgttcta ttacattatc attagcttcc 18900
tctttttcaa ctcctttact gcctaagctt ggctgtcctg gatcttgctc tatagattga 18960
cctttagctc agagatctgc atgaatatct cctgggacta aaggtgtgtg ccaccatgcc 19020
tggatttaag cttttcttca cctagaactt gctctgtctc aggctagcct tgaactcaaa 19080
gatctgcttg gctttatctc ctgggattaa aagtgtgtag caccatgcct gtatctaaat 19140
ttagctgggt aggatcttac cccaacatcc caccccctta attcaattta atatccttga 19200
acataggatt cagctccatt tcacttcatg ttgcccattt aatactcgaa ccatacattt 19260
tgtagttccc ttttctcagc gtgctgtgct tgtttgaaat gttcttcatg agaattaacc 19320
agacaaaaag tctatgatgg atatttctga gacttccttt gtcagtgcaa ttaatctgag 19380
tctctttacc ttagcctcag gcagattcct cagaaaagag caaaaaaaaa aaaaaagtag 19440
ccactttctt caccaatata ccacaaaaac agtctctagg ccacatattg aaattctcct 19500
ctgaaatctt gtgggccggg tccatacagt tcaaattgct ctcagtaaca aagacttcca 19560
tagtcctact aggataaccc atcaagcacc atttaaatta ccccactgct ttccaaatcc 19620
aaagtcccca aatccacatt ctcccaaaca aaaatgcatg gtcagaccta tcacagcaat 19680
accccactga gtttactgtt gtgaacagac accatgactg aggcaagtct tatgaaggac 19740
aacatttaac tgaggcaggc tcaaaggttc acagtctcag tccattatca tcaaggcagg 19800
aacatggcag catccaagca ggcacggtgc aggaggagct gagagttctc catcttcatc 19860
tgaaggctgt tagcagaatg ctgacgtcca gacagatagg atgagggtct tttcgacaac 19920
atccacagtg acacacctac tccaacaggg caacatcttc taatcatgcc agtctttagg 19980
caagcatata caaaccatta cacacacaca cacacacaca ccctcaaaaa aaagaaaaaa 20040
gaaaacataa cagaacaagg acccttacat aacatctgac gaggggagct gtcaaaatag 20100
ccactgagct taggtgtctc atacaggtta atatagaatg acaggccaat gaaactcctg 20160
tgtggcaacc acaaagggga agctgtggat ctccatcacc ttacaagcaa actgtgccaa 20220
tcagcaagga ctagggaagc agagacacac caagactttc acactgtgtg taaatggtgt 20280
gggactctgc aggtcatcca aggactgtgg gcccagatga acatgtgtgg gaaatacata 20340
cactttacat gcctaccttg tatttataac atatatgtaa caccttcttc tattttttat 20400
cttctatgac tgctaaagtg ctgcacattg agcactgagg catgtccatg ccaggtaaat 20460
ttcttaccac tctgatgcat ttccattccc aactcatcat cttgggaaag gaattttcaa 20520
agttaatgta gatataagta aagagagtca ggagggtggg ctgaaaggca gacctgcaga 20580
aattctccag agggtagagc tcagaagcag caagcagaac ttgggctaac agaagagcaa 20640
acacagagct cagacagaac tacctggcaa tgcgactggg cacactggaa gcactgtgca 20700
tcagcactga gccccaaata tgcccccaga atcctctgca taacaatgta acataacaga 20760
aaggctggaa taggccaaaa gaggaaacaa aaaaagacac ccccccaaga aaaaataaac 20820
aaattgggaa gagtaaaaaa gaattgtaag tgtaccacac agaacctgct cataatggct 20880
tgcatgcata gaataggaag gaccacgtta aaagaaaaat gttcagcagg attccttctc 20940
cctcaaactc ccaacaagca aaattcactc tcattaggag atgagaagag aaagtctgag 21000
ccctgtgttg acagtgccag gaaccactgc cagcctgtgg gaaatgtctg gctctcctga 21060
cagagcctgc tgcttctgcc ctgaatcctc tcactccacc tataaggggc tcatggcatc 21120
accacacagc ctccccaaag aggggggaaa agggcagctt gtcattgagt aagtcaccca 21180
gagacatgcc tttctagtag ttcttataat cgccctactc cagcagcctc catccaggca 21240
ttgagagacc aaaggatatt tggaagtcag ccttatggta gccccagacc ctgcttgttg 21300
cctatgatct tcctgaagag atagaagccc aggagaaaaa agttgtcccc caactcaaga 21360
tggcccctga gactctgcac tgatgtctgt gcccccacta ggccaaaggt agtcacccta 21420
tgcaaaccaa ccaatgcatg aaagggttac atgctacacc aattagaata agccatctag 21480
ccctgttgcc taaatctgcc ccttatgagg tataaaaagt gtgttctttc tttgttcagg 21540
gtctctcctc ttgcctgcat gctgaggggg attccccaac atattggatt aataataata 21600
ataataataa taataataat aaatcttgtg gtttgcatca atttcagtct ctgttgtctt 21660
tgtgagtgag ggtcttttgg tgaactccaa cacctactcc tgccaaacca gagctcttac 21720
tttgcagggt ttttggagaa aggctctact actgtgtata taactaaagt ggtagctcac 21780
acaatggtgt atctagtagc aaaagtttgt tgtgcccagc agattcttca gctcaaaaca 21840
gtaattctag agtagctgac atctaagggt ctgtatctta gtcttgacca ctgaggctta 21900
attaactgat attaccacac acacacagca attctgactg accacagaga catcaaagca 21960
ccctaaagtc agatcccaat agccccttac ccagagtttg taaggccagc agagagcctc 22020
tcagctctcc acctggacca atggctcctt ttagtaggaa gagacctaga gaactttccc 22080
tcagaaaaac atgggaaaca ctccaagctg agctcagaca gccttccaca gccacttgga 22140
aaatactgat ttggctatct ttacctggga ccaaaaagac attactacct tagtcacagc 22200
ctaaacatgt tctttcagtc acttggtggt ggtccacaag gagcttatgg taggttctac 22260
caagatcttt tgaagactac caggagcata ttgctaaatc agagtctaac ttcagctacc 22320
taggagctcc ctgtatcatg gtggagcacc caaagaatgg aagcatctcc caatctactg 22380
cggacctgag aaaggttttt ggttttgtga agttttttgg attttatgac ccatttattg 22440
gaacatgatg gatctgttaa gggacagaat gccagtggta accatcctgc tgtcacaaat 22500
ccaaagacct tcccatagcc tactctagaa gaccagatac aggcctacac aacttccctt 22560
ctgattaatg ttggggacaa attttaacat ttggcttcta tatgtggatc tctgtcttct 22620
ctgtcctctc tttgtcttac atactgcaga gactagggga agataactgt ggacatgaaa 22680
catctgtgtc atgtagtgtt ggctgtgtca ctaggaagct acagtgtgtt tctcactatg 22740
gaacagatac cagaaggtcc tgtgtctttg atgaactgac ccttagggcc catcccataa 22800
ttcctgcaag tctggaccag gcatggccac atgacatcag actgtctcat atttcatctc 22860
tgctagagcc atggtacaga tattgacata gatcaggtac agaaatagca gagtgctcct 22920
gaagagggtt cttctccatc tgcaatcagc atggcaggaa gaccacagga gcagaattca 22980
gtatctgcct ctctcactgt ggagtaccta ccacagtgat acagagcatc catggaagaa 23040
cgacccctca tctacagtgg ctagccagtg aggataagtg ggacatgaag tttgacctgg 23100
tgtcacagct ttgagtctag agaggcccag tggagttagg attgagagtc ccaagttttg 23160
gtctagccat actgacacaa tctctataac agtatctgtg gtggcttcag gaagctggct 23220
tctgaagtgc ccccacccaa cattccatct gcactggaac caaaaagttt cagatctcag 23280
cagataaata cttggattaa aagaacacac acacacatac acacacacac acacacacac 23340
acacacacac acacactaca aggtcaaata tgctccaggg gacacaaacc accatggctt 23400
gtgaaaagac cacaagccaa ggagtttctg ttacaggttg ggttactcag taccatggta 23460
acactcaatg atacagaaac acatccagga caattctaga agatgatcca ggctgttcag 23520
atgaggagat aggtcctgga gagatcaatt cctcactctc cattctaaga tgactgaaac 23580
aacccaacta gggcatgagc cagaattaca cacacttaca catgaactca cacacacata 23640
ctcatatgca cgaacatgtg tgcatacaca tacacttgca ttcacacata catgcacaag 23700
tgggcgtaca cacactcata tacattcata cacatatcca tgcacataag tgcacacaca 23760
cattcatgta cataggcact ggtacacaca catatacaca tgcatgcaca catgcccttg 23820
tatacacaca tacacacaca tgctcacata cacacacaca tacacaatgt cataagcaga 23880
aagtacccaa cagtctttcc cactcaaacc agcccatcaa gagtgaaggg atgaagactg 23940
atagtatcaa tgttacaggg ggccctgtta taatggggcc tggaagtaga cacacaatgg 24000
agaggaatct gtgccagtct caggcactgt gcctacctat actagtggat ggggccacat 24060
agtcatacgt acactatagc tgggcatggt ggcacatgca tttaatcaga gggctcagaa 24120
tgcaaaatca agtagatctc taagttggac tccagcacag tcagggctac atgaagaaaa 24180
ccaatatttt aaaaagaaac aaagaaataa agaaaagaaa aggaaaaaag aaaaaggaaa 24240
agaaaaggaa aaaaggaaaa ggaaaaagaa aaggaaaaga aaaagaaaaa gaaaaagaaa 24300
aagaaaaaga aaaagaagaa aaagaaaaag aaaaagaaag agaaaaagaa aaggcttgcc 24360
ctgaaaccac tcagtgctca cacagctctg cccacagggg aaacctaccc atgcctgtcc 24420
ccactcagca ggaaggctgt gaagtctctg acaggatttt gaacaagtta ctgtcacagt 24480
gggcacagct cgggctacca ctgtaagaaa agctcaaacc aaaactgccc tccaaaactc 24540
atccccagac ctattccctt tctggctttg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 24600
tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgta catgtgcatg tatatgtgta 24660
tacataccca accagcaagg tgctcctgaa tatcaagtcc tttataactt ggagtaggcc 24720
atccttaccc tttcatcttg gctcaggact ctccaacaaa gctgaccctt gagggagaag 24780
ctacaggcag ataggatcag tcttctctta ccaccatgaa tacaggcagt taactgcagg 24840
tatgaacctg tgacatccag taatttctgc aaaagtaaca gacaccatac cacccctgga 24900
gcaccatgac accaaggccc taaactgtca tcttctccat gtttccagca acgaatgatc 24960
ttcctgttct gtcctccgag aagggttgga aaggagcttt gcatgaatag gcatgagaag 25020
caacactgtg gtgtcaatgt caaccacaaa cccctcagaa aggctacaca agcagctgtt 25080
cagacaatgg cactcagagt gaatggacac acagacatca gattggtgcc tggcacagag 25140
gcccaggcca aagcactgga ttaaaaatac ctgtgcccct ttggacaatg gcagccagcc 25200
atggtcatcc ccatggcaag gtcctccatt atgttcagga gttgaaactc agaatgacac 25260
tcactgacct ccagtgatgg gtcccttgac cagcactggt cccagacaga cagacagaca 25320
gtaacagaaa gacagagaga gaaagacaga gggagggaga ggagaggagg gagggagaga 25380
gaaagggagg gagggaggga gggagggagg gagggaggga gggagggagg gagcaggaaa 25440
gtcaaaacta gaagtagtta attttaacaa caggaaaggg gtttagaatc actcaccaaa 25500
gtgttaagag cctgttttgg ataccaataa gatggctatt ctggctgact cttctctctt 25560
tgctttgcta aggcaggcag tgttcaaaca gtgtcatcat aatcttgtgg ctgctccact 25620
aggccaccct gaacgtgatc ctcccatagt cttttctgtg gtggttctcc actcctgtga 25680
tgcaacacac ttccttcctc cctccctccc tccctccctc ccttccttcc ttccttcctt 25740
ccttccttcc ttccttcctt ccttccttcc ttccttcctc ccttcctccc ttccttcctt 25800
ccctccaggc aagacccgga gatccctaat ctgtaccaac agctagttat gccaaagtta 25860
ctttccgaac ttcagattcc tatgctgtgc ccagccatgc ttctacatgg gtccctttca 25920
cactttcatt cagagctccc taggctgtac ccagacctgc atgtgcctag gtacctttca 25980
aaccttcact cagaggtccc tatgctgtac ctagccatgt ctataccaag gaaacattca 26040
gactttcact cagagctctc tatgctgtat ccagcactgc ttgtgcctag gtccccgaaa 26100
gaccttcact cagagctcac tatgaagtac ccagccctgc ttgtgccaag gtaattttca 26160
taccttccct cagaaatccc tatgctgtac ccacacctga ttgtggctag atacctttca 26220
gaccttcgct caaaactccc tatgctgtac ccagccctgc ttgtgccaag gtacatttca 26280
gaccttcaat cagagctcct tatggtgtac caagcctggc ttgtgccaag gtaacttttt 26340
tttccttttt ttttttataa tgtagcttac aagcacaatg cagtgacttt atataatttt 26400
gttgtctatg gtataaatga ccatgatgtg cacttggtta tcagctgcac caccattaaa 26460
gagcccatag aatattcaaa tccacacaca catacacaca catacacaca aacacacatt 26520
tttatgagtt tcagacctag ttgtgaattg ttctgtatac agagtataag catcttcaaa 26580
ttgacaacat aatttgtgtt ggttttccac cgtttctact tctgttaaaa acagcatatt 26640
ttaactgcgg ccccgtctgc ctggttattt ctttgcaaac attgtgagtc tgatacagtc 26700
cgatatgctc ggtgagtgga tggactccct ttctcaaccc agaacttcta caagaatgtc 26760
tttctttttg ttttgttttt tttttctttt tttaatatct cttttctttt caaattttta 26820
ttagatattt tcttcattta catttcaaat gctatcccta aagtccccta taccctcccc 26880
ccggcctgct cccataccca cccactccca cttcatggtc ctgacattcc cctgtactgg 26940
ggcatataaa gtgtgcaaga ccaagagacc tctcttccca atgatggccg actaggccat 27000
cttctgctac atatgcatct agagacacgg gctctggggg tactggttag ttcatattgt 27060
tgttccactt acagggctgc agaccccttc agctccttgg gtactttttc tagctcctcc 27120
attgggagcc ctaggttcta tccaagagat gactgtgagc atccacttct gtagttgcca 27180
ggcagtggca tagcctcaga agagaaagct atatcagggt cctttcagca aaatcttagt 27240
ggcatatgca atagtgtctg tgtttggtgg ctgattatgg gatggatccc tggttggagc 27300
agtctctgga tgttccatcc tttcatttca gctccaaaat tttctctgta atgccttcca 27360
cgggtgtttt gttccctgtt ctaagaagga atgaactatc cacaaattgg tcgtccttct 27420
tgattttctt ggtttttgtt ttgttttgtt ttgttttgtt ttttgcaaat tgtatcttgg 27480
gtattctaag tttctgggct aatattcact tatcagtgag tgcatatcaa ctgagttctt 27540
ttgtgattgg gttatcccct agatacatcc atttgcctaa gaatttcata aatttattgt 27600
ttttaatagc tgagtagtac tccattgtgt aaatgtacca cattttctgt atacattcct 27660
atgttgaggg acatctgggt tctttccagt ttgtggctat tataaataag gctgctatga 27720
acatagtgaa gcacgtgtcc ttattaccag ttggaacatc ttctgggtta tgcccaggag 27780
aggtatttcg ggatcctcca gtagtactat gtccactttt ctgaggaacc accagactga 27840
tttccagagt agttgtacaa gcttgcaatc ccaccagcaa tggaggagtg ttcctttttc 27900
tccacatcct cgccagcatc tgctgttacc tgagtttttg atcttagcca ttctgactgg 27960
tatgaggtgg aatctcaggg ttgttttgat ttgcatttct ctgaagatta aggatgttga 28020
atattttttc aggtgcttct cagtcattca gtattcctca gttgagaatt ctttagctct 28080
gtaccccatt tttaatacga ttatttcaat ttctggagtc cagcttctta agctatttct 28140
atatattgga tattagtccc ctatcatatt tagcattggt aaaaaatcct ttcccactct 28200
gttggtggcc tttttgtctt atttacagta tcgtttgcct tacagaagct ttgcaatttt 28260
atgaagcctc atttgtcaat ttttgatctt aaggtacaag ccattggtgt tctgttcaaa 28320
aatctttccc tctgtaccct aacttcgagg atcttcccca ctttctcctc tataaatttc 28380
agtgtctctg gttttatgtg gagttctatg atccacttag acttgagctt tatacaagga 28440
gataagaatg gatcaattct cattcatcta catgataact gccagttgtg ccagcaccat 28500
ttgttgaaaa tgctgtcatt tttccactcg acggttttag ctcccttgtc aaagatcaag 28560
taccccttga taggctgtag gataggtatc tgggttcatt tctggatctt cagttctatt 28620
ccattgatct acctgtctgt cactgtacca gtaccatgca gtttttatca caattgctct 28680
gtagtatagc ttgaggtcag gcatggtgat tccaccagag gttcttttat tgttgagaac 28740
agtttttact atcctagggt ttttgttatt ccagatgaat ttgcaaattg gcctttctaa 28800
ctcagtgaag aattgaattg gaattttgat gaggattgca ttgaatctgt agatagcttt 28860
cagtaggata gccattttga ctatattaat cctgccaatc catgagcatg ggagatcttt 28920
ccaacttctg agatcttcaa tttctttctt cagagacttg aagttcttat catacagatc 28980
tttcacttcc ttagttaggg tcacaccaag atattttata ttatttgtga ctattgtgaa 29040
gggtgttgtt tccctaattt ctttctcagc ctgtttatcc tttgtgtaaa gaagggccac 29100
tgatttgttt gagttaattt tatatccaac tactgcactg aagctgttta tcagctttag 29160
gagttctcta gtggaatttt tagggtcact tatatatact atcatatcat ctgcaaatag 29220
tgatgttttg acttcttcct ttccaatttg tatccccttg atctcctttt tttgtcgaat 29280
tgctctggct aggatttcaa gtactaaatt gaatagatag ggagaaagtg ggtagccttg 29340
cctagtccct gattttagta ggattgcttc aagtttttct ccacttagtt tgatgttggc 29400
tactgatttt ctgtatatta cttttattat gtttagttat gggccttcaa ttcctgatat 29460
ttccattact tttatcatga aggggtgttg gatattgtca aatgctttct cggcatctaa 29520
cgagatgatc atgtgatttt tgtctttgag tttgtttata tagtggatta tgtcgataga 29580
tttctgtata ttaaaccatc cctgcttccc tgggatggag ccaacttggt catcatggat 29640
gatcatttcg atatgttctt ggattcggtt tgcgaggatt ttattgagta tttttgcagt 29700
gatattcata agggaaactg gtctgaagtt ctctttcttt gttggatctt tgtgtagttt 29760
aagtatcaga gtaattgtag cttcatagat tgaattgggt agagtacctt ctgtttctat 29820
tttgtggaat agtttgagga gtgttggaat taggtcttct ttgaaggtct gatagaactc 29880
tacactaaac ccatctggtc ctgggctttt ttttggttgg gagactatta atgactgctt 29940
ctatttcttt aggggaagtg ggactgttta gattgtgtta atctgatcct gatttaactt 30000
tggtacctgg tatctttcta gaaaattgtt catttcatcc aggttttcca gttttgttga 30060
gtattgcctt ttgtggtagg atctgatgat gttttggatt tcctcaggtt ctgttgttat 30120
gtctcctttt tcatttctca ttttgttaat taggatactg tccctgtgtc ctctagttag 30180
tctggctaag ggtttatcta tcatagattt tctcaaagaa ccagctcctg gtttggttga 30240
ttctttgaac agttcttttt gttccaactt ggttgatttc agccctaagt ttgattattt 30300
cctgctgtct actcctcttt ggtgaatttg cttcctttcg ttctagagct tctaggtgtg 30360
ctgtcaggct gctagtgtat gctctatcta gcttcttttt ggaggcactc ggggctatga 30420
attttcctct taggactgcc ttcattgtgt cccataagtt tgggtatttt gtggctacat 30480
tttcatttaa attctaaaaa gtcttgaatt tctttctttg tttctgcctt gaccaaggta 30540
tgattgagaa gagtgttgtt cagtttccat gtgaatgttg gctttctatt atttatgctg 30600
ttaatgaaca tcagccttag tctgtggtga tctgatagga tgcatgggat aatttcaata 30660
tttttgtatc tgtttaggcc agttttttga ctgattatat ggtcagtttt ggaggatgta 30720
ccatgaggtg ctgagaagaa gttatagcct tttgttttag gataaaatgt tctagagata 30780
tctgttaaat ccatttcttt cataacttct gttagtttca acgtgtctct gtttagtttc 30840
tgtttccagg atctgtccat tggtgagagt ggagtgttga agtcccccac tatttttgtg 30900
tgaggtgcaa tgtgtgcttt gagctttact aaagtgtcat taatgaatgt ggctgccctt 30960
gcatttggag catagatatt cagaattgag agttcctctt ggagtatttt acctttgatg 31020
agtatgaagt gtccctcttt gtcttctttg ataactttgg gttggaagtc aattttattc 31080
gatattagaa tggctactcc agcttgtttc ttcagaccat ttgtttggaa aattgttttc 31140
cagcctttca ctctgaggta gtgtctgtct ttttccctga gatgggtttc ctgtaagcag 31200
cagaatgttg ggtcctgttt gtactgtacc cagccctgct tgtgctaagg tacctttcag 31260
gccttcagtc agagctccct atgctgtacc cagacttact tgtgcctacg tccacttcag 31320
acgtttgctc agaaccctct atgctgtatc cagccctgct tgtttgtgcc tagatcccct 31380
tgggtgcttt gctcacacta ctcaccatta ccaacaaatg aatggtttcc agatgttcca 31440
gaaatttccc tgattcccaa aagaaaatga tacagaacag tcaactatat gcatgttttt 31500
ttttaactac caaacttcaa aattagcaaa gtagggtcct aaatgagcaa gagcctccat 31560
ctcagagctc tttgaaccat gcaccttttt gtcaacttga tatcagctgg aatcatctga 31620
gaagcaggaa ccacagttta gaaaatgcct ccatagaaga ttggtccata gacaatctct 31680
gggggtatcg tcttggttag tgtttgatgt gggagctagg cacacatcgt tgtgagtggt 31740
accacccatg ggtaggtagt cctcaatgcc atttaaaaaa aatcaggtgg agaaagtcct 31800
gaagaagaac ccttgttttt ccatgtctta tgttcaacca caacctctaa tttcctgccg 31860
acttccctca gtgacagtgt gtgacctaga gttgtaagat gaaataaacc attttctctc 31920
cattttgctt ttggtcatgg tgtttgtcac agcaatagaa acctaaatta gacatctagt 31980
gtgtcaacac ctattcaacc ttattggaaa gaagacttga tctaataatc aataacccta 32040
gattcagata tatttttcac atgtacagtc ttgaggattt ttttttgctt gaacccaacc 32100
tcactgaata agatttacat gaatacacaa ttctatttat aaacttacat gcatcttttt 32160
tcatgtaaaa taatatttag tgttgtgtat gatatagaat attgacaaac aaagttacct 32220
acatgaatgt gaatatgttt tatttatgtg aatatgttta ctggataaaa atacaaaagt 32280
gagcaaggaa gacaaaacca cctgaagtaa atgtcagcac aatgaggcag agatcaggaa 32340
cccccacagt ggtaacagcg ggtttgtgtg tgtgagtgaa ctcactgggc aactctgtcc 32400
ccagaaatac taggcaggat cagagggttg agatgttcag ccttccacct ccagtgaaga 32460
aagggtctag aagctgaatt ggtcattaat ggccaacaga ttccatcctg cttatatgca 32520
aggctgccat caaagccaaa aatatggtgt taggagaact cccacataac tggacacaga 32580
ggagactgtg gggaggtaac ccagaggcac agaagccctt cacaggctgt cagagttcat 32640
tcctcaccct gtgtgcctct tcatctctct gcttcctcct atcctttgta atccagcaat 32700
catggtgggt gtccaactgc tccagtaaat ttgtctaaag agtatgtggc aggatcctta 32760
accaaaagct ggtgagtcag cacaggctaa aacctgggat acacaaccag ctggcaatgt 32820
agagagactt gtagtgctga agcgtcaaac cctgaggtct gtgctgtccc atgatggtta 32880
gcatcagaaa cccctttccc agaaggttct gctgtcacta gtcagctaag tgaaggttct 32940
tttgctctac ctaggtagac ctcatgggtc agcaggaagc tttgacctct ggtagccttc 33000
ttctcacctg ccaggggcca gccctggtgt ggggttcttc tttttgtcag ttcttcttga 33060
gccagtggaa ggggaagaat gtcagtggag ggtaagactt ggtcttacga aattcccagg 33120
taagactttg tcttacatga tatttagggt tgctgtataa taaaaagttt acttatctaa 33180
gtctaagtta ctatgctatt atagctgacc tgccttctgt gttcctctga ggccagaaac 33240
tgcagtctca ggcactttag cacctcatcc taaaacagcc ggagattaag taaacaatta 33300
attgctagag ccttttccct agtgttcaga tgcctcttgc ttgtcaggct agggggatgt 33360
ttgtagcaat aaacttctac tgaacccaaa gagaataaca cttgcagaag gtcaaacttt 33420
tacataatcc aatatgcata atctcacata tgtacataaa atgattcatg tatgctcatt 33480
tatacatttt cattcaaacc cagttgggcc cacatgcatg cattgtcaca attacatact 33540
tacacacaca aatgcataca ttctcactgt gatggtttgt atattcttgg accagggaat 33600
gctaccatct ggagatgtgg ctttgttgga ataggtgtga cctggttgga ataggtgtgt 33660
cactgtgggt gtggacttaa gactctcacc ccagttgcct ggaagtcagt cttccactag 33720
cagcctttgg atgaagacat agaactctca gctctgcctg caccatgcct tcctgaatac 33780
tgccatgctc ccaccttgat gataatggac tgaacctgta agcctgcccc aattaaatgt 33840
tgtttttata agacttccct tggtcatggt gtctgttcac agcagtaaaa ctctaactaa 33900
taccgaagtt agtaccaggg actggggtat tgttgtgata ggcctgacca tgcttttatt 33960
tgaaagaatg tgaattttgg gactttggat ttagaaagca gtggaattgg ggcttaatgg 34020
gtcatcctag taggaatatg gaagactttg ttgctgggag taatttgaac tgtgttgaac 34080
ctggcccaag agatttcaaa ggaaaagaat ttcagaatgt ggcataaaga ctgttttgtg 34140
gtattttggt gaagaatgtg gctacttttt gcccctgtct gaaaagtctg cctgagactt 34200
aggtgaagag actcaaatta attgcattgg caaatgaagt tttaaaaatg cccagcagag 34260
actttgttct ctggttaagt cttatgaaga gaagtttgaa caagcattgc aaacttagaa 34320
aggaaaaata taacatatat gatttgagta ttaaaggggt tgtgacatgg ttgaattcaa 34380
agatgaagaa tgtgtaaaga aagcattaga aactatgaac aaatatgacc ttagtggaag 34440
gcctttgaat attaaagagg atcccgatag agaaaatgct cgcaggacac tgcagcaaac 34500
aggaacatca tttcaaggat catgtgcctc cgatgtggga tctgggttgc taaatttacc 34560
accttccatt ctcaatagtc taaacattcc tcctgaggtt atcagcaatt ttcaggctgc 34620
tagacttggt tccacaattt ttgttgctaa tcttgacttc aaagttggtt ggaagaaatt 34680
aaaggaagtg ttcagcatag ctggaactgt gaagcgtcta ttagtgccag ccagctgaac 34740
ataactggta taataagaaa tttgggtcca agtgttggtg agtttgcata cgtggtactc 34800
tttaaggaag tagaaggaaa atcaagggat catgcagagc agctgaggat cggcactgtg 34860
agaggccatg gaaggccatt ggtgaaggtg cagcctcagt tgcaattgat ggcccaggac 34920
agaaggggtc atgcaaagga gatgaggctt ggcaccatga agaaagccta tgagaggctg 34980
ttggtgaagc ctagttacag tggaagacag cagcattttg gagatgccag taccatgaga 35040
tgaccaccaa gaacagcagc agtagttgag tacaggcagc tgaagcctag aagacaacat 35100
gtgtgctaca aaggacaggg ctagagaagt gacccaagcc cttggaggag cccagaagat 35160
cgtgagttgg ataccagaca ttggacagtt ggaatttaat ttttgttttt gattgtgaca 35220
gtgccctgat atttttccct cttgaaggaa gaaaatattt tagtgaatcc cacagttaag 35280
agacttttaa ttgtaaaaag actttggatt tcaaaagata ttgaatattt ttaaaggatt 35340
gaacttttaa tatgtaaaga gtgtaggact tttaaagtta tttaagatcg tggggatgaa 35400
tacgaactaa gggttgaggc ttactagtgt tgtgtttgtg tgtcaagttg acaatgggtc 35460
aattgtactg gctagttttg tgtcaacctg acacaggctg gagttatcac agagaaagga 35520
gcttcagttg gggaaatgcc tccatgagat ccagctgcaa ggcattttct caattagtga 35580
tcaagggggg aagtcctctt gtgggtggtg ccatctctgg gctggtagtc ttgggttcta 35640
taagagagcg gactgagcaa tccaggggag gcaagccagt aaagaacatc cttccatgtc 35700
ctctgcatca gctcctgctt tctgacctgc ttgagttcca gtcttgactg ccttggtgat 35760
gaacagcagt atagaagtgt aagctgaata aaccctttcc tccccaattt gttccgtggt 35820
catgatgttt gtgcaggagt agaaacactg actaagacac tcacaattac atacttatac 35880
acacacccat acttacatat gcatatggag caaaagacca agcactggtg cagtaagaac 35940
ttactgattc tagatgaaaa ataagctcta atctttattg catagagaaa caaagggctt 36000
ccaccctttt actgttaaat gaattaaacc catgtttgta agaaaaaaaa agctttgttc 36060
cttttaataa gactaagcct gccttgttat taagaaaaga cctgttctgt cccacggtaa 36120
ggagaaggct gcctgctcct tctgctctgc agtgtcttct tttttcccct ttccctcttc 36180
attctatacc ttgctctctt aatattttat cttacatata gtttctacgt tattccactc 36240
tgcattctcc ttctaattgt tttctctcct cctgaagtga tgtcacaata atggtcttac 36300
tctcaatctc ttttctttaa aagctatacc tatgcttgaa actttacttg tctagggatg 36360
ggcctcccct tttctttata aagcatgttt gctgacatta aaattgaacc ttgatcagaa 36420
tgactgtctt ggtttcatct ttctttcatc tgcctagctc cctcttctct tccagattcc 36480
aagatacctt tccagactag aacccagaca agtgagccgc tggctggcca caacaggtgt 36540
acttttgcta agcctaccct gagactttaa ctcttatcca gtaaaatact gtaagaaagc 36600
atgcaagaca ctccacaata ttgcagggac aaatctgggg cattctagct atgggaatgc 36660
caaggtccag gaggctaagt ttccttgaaa ctctttgcct tgtgactgct tccaggcttt 36720
taggcctacc aagtgaatca ctactggagt ggatgtagca tttggttgct tccagtcacc 36780
ccagtcatct ggcatgagga agaggcagtt gctagtcttt ccacacattg atcaatcatt 36840
cataaaaact catacccaga ttctcaaaga gaattttgtt accccatttc agcctggctt 36900
atatgtgcaa tggtgtcaac agtttcccat tggtgcaaga ccttggagtt atcaatatgg 36960
taatgcctat atcaacaata cacagtcact aaggacttca cactctgtcc ttctccattc 37020
cctatacatt tgtcttctga ctgcagacat aatgtgacta gctacttcaa gctcctgctg 37080
ccttgacttc cccaccatta aagataatac tttgaactgc tagcagaaat aagcctatgc 37140
tttcttaggc tgcttttgtt aaagcatttt gtcagagcaa caaaagagta atggagacag 37200
acattggtac caagaattgg gcgtcattgc agagatgaac ctgactgaca ttcctagggc 37260
actgaaaatg tttctgtggg aggaatttgg agcttccagt ttgaaaagac ttagaattct 37320
gtaagccaaa tgtaatcatt cattctgctg gaagctttgg aagacaaaaa taattaaaga 37380
aatgaggaca atggtccccc agctcatgaa gttacataga tgaataagaa gtccaacagg 37440
acctggggtt gggaccattt gtgtgatctt taagccaaga gttgggcttc attctgctat 37500
gtcctgagaa cttgtgtgat actaaattta aatctcatag ttgaatttat gtagcagaag 37560
aaatttgaag ccaggagagt attcagactg gtgccaggaa aacaactaca cctgttaaag 37620
agataatgtc attgaaaaga agcctcctta agaagggtgt gctgagcttc ttaagaagat 37680
gacaggtgac agaagaagaa cttcttattg tcatccacat ggtagtggtt ttggaattat 37740
ggaagatgca agactgggga gagtcactgt ggcttgctcc atgtgttcaa agagctgctc 37800
agggcaaaca atgtgtggca cagttagagt ccattcaaag agatcctgaa aggccactgt 37860
ttgagtatgt gaagaggaag tctttccact gctgttctct tatctttccc tcttcctcca 37920
gagttgctga tgctcattcc tttgctgaca gtagatctag catttgcaag ctttggtcat 37980
ggactaaaga ccagctgcta tccaggaact tcaagggctc cagctttaga tttctgaagc 38040
attctttcct taaaaatctg gaactcttgt agcaaccagg atctgcagaa tcaagaagtt 38100
acagaggaga agaacgtatt agtaagaata tattaatcat atattcttga gatattttgg 38160
caaagaatat ggtttccctt tgccattgtc caaaaaaatc tgcctgaggc tacattaaag 38220
agttttggat tgcccagtcc ctccaagaga gagcaagtct cccaaggagt acaaagatca 38280
caggtgagat gaccactcct gcccaaatac aaggtccaca tgggacctgc ccagagccca 38340
caagacccag gaacaacaga gcagcctggt acaagatact tctggtttcc atctgaatcc 38400
agggctgacc ctgtgccaca gctatctaga ccccagtccc acccccaccc aagtcctgcc 38460
caaagaaagg aggtctcaca ggagtgctca caacactctt ctaacaacaa aaataacagg 38520
aagcaatgat cactggtctt taatatctct taacatcaat ggactcagtc ccccaattaa 38580
aagattaaca gactggatat gtaaagagga cccagcattt tgctgcatac aggaaacata 38640
cctcagtgac aaaaacaaac actatctcaa agtaaaaggc tggaaaacaa ttttccaagc 38700
aaatggtccc aagaaacaat ctggagtcgc cattttaata tcaaataaaa tcgactttca 38760
aacaaaagtt atcaaaaaaa gatgaggaag acatttcata cttaaaaaaa gaaaaattct 38820
gtcaaaatga actctcaatt ctgaacattt ttgctgcaaa tgttagggta cccacattca 38880
taaaagaaaa actgataaag ctcaaagcat acattgcacc ttactcaata ataatgggag 38940
actttgccac cccactctca tcaatggaca gatcatggaa acacaaagta aacagagaca 39000
cagtgaaacg aacataagtt atgaatcaaa gagactgaac agatatatct agaacatttc 39060
atcctaaaat gaaagaatat atcttctcag cacctcatgg taccttctcc aaaactgaca 39120
atagaattgg tcacaaaaaa aaaagcctcc acagatacaa gaaatctgat atatcccata 39180
tatcctataa gatcaccatg gactaaagct ggtcttcaat aacaaaaaca caaaagaaag 39240
tccacatact aatggaagac gaacagttct ctactcaatg aaaattgggt cagggaagaa 39300
acaaagaatg aaattaaaga ctttttagaa tttaatgaaa atgaaggcac aatattccca 39360
aacttatggg acacaatgaa agcactgcta agagaaaact cataacactg agtgccttca 39420
aaagaaacta aagagagcat acacttgcac attgaaagca cacctgaaag ctctagagca 39480
aaagaagcaa atacacccaa gaggagtaga aagcaggaaa taatcaaact cgttactgaa 39540
atcaaccaag tagaaaccaa aagaactata caaggaatca acaaaaccag gagatggttc 39600
tttgagaaaa tcaacaagat agataaaccc ttagccagac taaccagata ctgaacagag 39660
acagtatcct aattaacaaa accagaaatg aaaaggaaga catagcaaca aaaaccaaag 39720
aaattaaata tatatatgat atatatatat atgatatata tatatatcat atcctactat 39780
aaaccctata ctcaacaaaa ctggaaaatc tggatgaaat ggacaatttt ctagaaaggt 39840
accaggtacc aaagttaaat cagggacaga taaataatct aaacggtccc ataaccccta 39900
cagaaataga agccttcatt aaaagaaaga acactaccta attcattcta tgaagccaca 39960
gttatgctta tacctaaacc aaacaaaaac ccaacaaaga cagagaacct caaaccaatt 40020
tcacttatga atattgatgc aaaaatactg actaaaatgc tcacaaactg aatccaaaaa 40080
gacattgaaa tgatcattca ccatgatcaa gtaggcttca tcctaaacat gaagggatgg 40140
ttccatatat ggaaatccat caatgtaata cactatataa acaaactcaa agaaaaaaaa 40200
gcaaatgatc atctccttag atgctaaaaa aagaatttga caaaattcaa ctccccttaa 40260
tgttaaaagt cgtggaaaga tcaggaattc aaagcctata cctaaacata gtaaaagcgg 40320
tatatggcaa accaggagcc atcatcaaac taaatgcaga gaaacaaagt aatcccacta 40380
aaatccattc ttatcttcat gtacaaatct caagtcaaaa tgaaacaagc acctccataa 40440
gaaaccagat acactgaatc taatagaaga gatagtgggt aaaattctca aacacataag 40500
acaggaaaaa gatactgaaa agaacaccaa tggcttatgc tctacgacca aggactgaca 40560
aatgggacct cataaaattg aaaagcttat ataaggcaaa ggacactgtc agtaagacaa 40620
aacgggaacc aacagattgg gaaaagatat ttaccattcc ttcatcttat agagggttaa 40680
tatccaatat gaacaaagaa ctcaagaagt tagaccccat agaaccaaaa aggtctttca 40740
aaaaatgggg aacagagcta aacagagaat tctaaaccaa ggaagttcaa atggctgaga 40800
agcacctaaa aaatgttcaa catccttagt ctttagggaa acacaaatca aaaccactct 40860
gagattccac ctcataccag tcagaatggc taagatgaaa aactcaggtg acagcagatg 40920
ctggtgaggc tgtagagaaa gagaaacact cctccattgc tggtgggatt gcaagttggt 40980
acaaccactc tggaaatcag tgtggtgttt tctcagaaaa ctgaacatag tactacctaa 41040
ggacccagct atatcactcc tgggcatata cccagaagat gttccaactt gtaataagga 41100
cacatgctcc actatgttca taaaagcctt atttataata gctagaagtt tgaaagaacc 41160
cagatgtccc tcaacatagg aatggataca gaaaatgtgg tacatttaca caatggaata 41220
attagtagta actaaactat taaaagcaat gacttcatga atttcccagg caaatggatg 41280
gaacttgaga atatcatcct gagtgaggtc actcagtcac ctaggaaaac aaacagtatg 41340
tactcacaga taagtggata ttagcccaag agttcagaat acccaagatt caattcaaag 41400
accatgtaaa acgtaagaag aaggaagatg aaaatgtgga tgtttcagta ctttttagaa 41460
gggttcctat ttgttctgtt tgtcccatat ctagtccatt aaagaagccc actgtattaa 41520
gcccttgcaa gttatggttt ccaattgtaa ttaaaaagaa gttcctgttc ttagatatga 41580
tataaactgc aaaacatatt ttcaacctac actaaacttg aaacctcatt aatgtgatgt 41640
atattcttct atttgttctt tctctttcat tgtatttttc ttttttagca tttttatgat 41700
attttcttca tttacatttc aaatgctatc ctgaaggtcc cctataccct ccccctgccc 41760
tgctccccta cccaaccact cccacttctt ggccctggca ttcccctgta ctggggcata 41820
taaagtttgc aagaccaagg ggcctctttt cccaatgatg gctgactagg ccatcttctg 41880
gtacatatgc agctagagac acttgctctg gagttgatgg ttagttcata ttgttgttcc 41940
acctataggg ttgcagaccc cttcagctct tgggtacttt ctctacatcc tccactgggg 42000
gcctgtgctc catccaatac atgactgtga gcatctactt ctgtatattc caggcactgg 42060
catagcctta cactagacag ctatatcaga gtcctttcag caaaatcttg ctggcatgtg 42120
caatattatc tgcgtttggt ggctgattat gggatggatc cccagttgtg gcagtctctg 42180
gatggtctca actcaaaact ttgtctctgt aactccttcc atggaaattt tgttccctat 42240
tataaagagg aatgaagtat ccacacattg gtcttccttc ttctggattt tcttgtgttt 42300
tgcaaatttc agaatggcta agatcacaaa ttcaggtgac agcagatact agcgaggatg 42360
tggagaaaga gggacactcc tccattgatg gtgggactgc aagcttgtaa agccattctg 42420
gaaatcagtc tggcagttcc tcagaaaatt gaacatagta ttaccagaag atccagcaat 42480
acctctcctg ggcatttacc cagaagatgt tccaactggt gataagaaca catgctccgc 42540
tatgttcata gcagccttat ttataatagc cagaaactgg aaagaaccca gaggtccctc 42600
aacagaggaa tgaatacaga aaatatgata catttacaca atggagtact actcagctat 42660
taaaaacaat taatttataa aattcttagg caaatggatg aatctggagg atatcatcct 42720
gagggaggta acccgctcac aaaagaactc acttgatatg cactaactga taagtggaca 42780
ttagcccaga aatttagaat actcgtattt ttctaacaat agctaacagc cagcaatctc 42840
acccttgtaa gcacttcctc cctcccataa aagggacttt aagaagcctt ctcaggctgt 42900
tctctaaaac cctgacttag agggatacag ccaaatggca cccaaaacta aagcctgctt 42960
cattcagaca gttgtggact tggtcttctc ctcacaattc taatgtttaa catacagtga 43020
catccctagt aagatgtacc tggactgcca gctgagcaag tcagaggtaa caaggtggta 43080
agtggcattc ttctattatc tctgcttcaa attctgcctt catttcctac cttgactctc 43140
ctcaaacaca gatgatgacc tgtaagccag ataagccctt tcttcctcca gttgctttaa 43200
aggcaggctg tttatcacag taacagaaaa tcaaactaca tgttgggccc agtcttctcc 43260
atgctctcat ttcctctaca ctatggatca aaatttacct acctatttat accttcctga 43320
gtgccagtct gctgtaagga acatctgagc cacactggta ctgtctttat aagagcagct 43380
atgcctactt gtgaaggacc tttccctcct aacagaactg tatggagaag attattagaa 43440
cctcacctga gcttccagcc actactccaa gccatttatg tgtagttttt ccataattac 43500
aattagcatg aggagtctaa ggaagtgcta taatagcagc ttggagaggg taaatgacag 43560
ggacaacacc tatgtggcca tgcttagtac agacacacca gcatgtctgc tgtaggatcg 43620
ctggtattct tacccataca ttccattcat gttctgtaag gaagcataat aaactcactg 43680
attctcccat ttttttttgg tgggttcaaa cttggactgg tcattgggtt ttgggagaca 43740
tgccaatcca aggcttttcc tggaactcta atgtatgaag gaggaacata cttgacaaag 43800
ctagtaatag tttagtgtct ggaccttggg ggaacttttt cctggggttc tgaccccagg 43860
tatagacaag aaatgtattc cttctctaat accaggaata tgcttagcaa ggccagtaat 43920
ggcctggggc tagggcttta aagaagttgt gatatttgga tcttaaaaac ccacctcttt 43980
cccttttgat tggagccagc accgggtcac cttggacgca gagtcggcag atacccccaa 44040
gttccctaga ggactctcca tgtgatctta ggacctctgg tgagtggaac acaacttctg 44100
ttctaatcca attgtgcagg acctgagaac attaattagt gaagcagaaa acccagcctc 44160
accagggtca caagtccctt ctggtcggtg ccacaccagg tcacattggg catggagtta 44220
gcagacaccc ccaaggtgtc caaaagactc ttcaagtgat cttaggacct ctgctgagtg 44280
gaacacaact tctgcaggaa gcaggttcga acccaagata tctgggcatc ttccctgcaa 44340
gagaagagct tgcctgcaga gagtgctctg accactgaaa ctcaggagag aactagtctc 44400
ccaggtctgc tgatagaggc taacagaatc acgagaggaa caagctctaa ccagagacaa 44460
ctataacaac taactgaaga gaataccaga tggagaaagg caaacgtaag aatattacta 44520
acagaaacca agactattca ccatcatcag aacccagcac tcccacctca cccagtcctg 44580
ggcaccccaa aacacccgaa aagctagacc tggatttaaa agcatatctc aggatgatgg 44640
tagagaacat aaagaaggac attaataact cacttaaaga aatacaggag aacactgcta 44700
aagagttaca agtcctaaaa gaaaaacagg aaaacacaac caaaccggta gaagttcttg 44760
aagaaaaaca ggaaaacaca accagacatg tgatggaaat gaacaaaaac atactaagaa 44820
gggaagtaga cacaatagag aaaacccaaa gtgaggcaat gctgaagaca gaaaccctag 44880
gaaagaaatc tggaaccata gaggcgagca tcagcaacag aatacaagag atggaagaga 44940
gaatctcagg tgcagaagat ttcatagaga acataagcac aacaatcaaa gaaaatgcaa 45000
aatgcaaaag gatcctaact caaaacatcc aggaaatcca ggacacaatg agaagaccaa 45060
acctacagat aataggagta gatgggaatg aagattttca acttaaaggg ccagcaaata 45120
tcttcaacaa aattttagaa gaaaacttcc caaacctaaa gaaagagatg cccatgaaca 45180
tacaagaggc ccacagaact ccaaatagac tggaccagaa aagaaattcc tcccgacaca 45240
taataatcaa aacaacaaat gcactaaata aagatagaat attaaaagca gcaagggaaa 45300
aaggtcaggt aagagatata ggcaggccta tcagaattac accagatttt tcaccagaga 45360
ctatgaaggc cagaagatcc tgaacagatg ttatacagac aaaaagaaca caaatgccag 45420
ccaagcctac tatacccagc caaactctca attacaaaac caaattcaca cattaacttt 45480
caaggaatcc agcccttcaa aggataataa caggaaaaaa aacaaacaaa aacaaaaaca 45540
aaaaaaaaca aggacggaaa tcacacccta gaagaagcaa gaaagtaatc cctcaacaaa 45600
ccaaaaagaa gacagccaca agaacagaat gccaactcta acaacaaaaa taataggaag 45660
caacaattac ttttccttaa tatctcttaa tatcaatgga ctcaattccc caataagaag 45720
acatagacta acagactgac tacacaaaca agacccaaca ttctgctgct tacaggaaac 45780
ccatctcagg gaaaaagaca gacactacct cagagtgaaa ggctggaaaa caattttcca 45840
agcaaatggc ctgaagaaac aagctggaat agccattcta atatcgaata aaatcgactt 45900
ccaacccaaa gttatcaaaa aagacaagga gggacacttc atactcatca aaggtaaaat 45960
cttccaggag taactctcaa ttctgtatat gtatgctcca aatgcaaggg cagccacata 46020
cattaaagaa actttagtaa agctcaaagc acacattgca cctcccacaa taatagtggg 46080
agacttcaac acaccacttt catcaatgga cagatcgtgg aagcagaaac taaacaggaa 46140
cacagtgaaa ctaacagaag ttatgaaaca aatggacctg acagatatct acagaacatt 46200
ttatcctaaa acaaaaggat ataccttctt ctgaacacct catggtacct tctccaaaat 46260
tgagcatata attcatcaca aaacaggcct caacagatac aaaaatattg aaattgtccc 46320
atgcatccta tcagactacc atggactaag gctgatcttc aataaaaaca taaataatag 46380
aaatccaaca ttcacgtgga aactgaacaa cacacttctc aatgatacct tggtcaagga 46440
aagaataagg aaagaaatta aagacttttt agagttcaat gaaaatgaaa ccacaacata 46500
cccaaactta tgggacaaaa tgaaagcatt tctaagaggg aaacacatag ctctgagtgc 46560
ctccaaaaag aaactagaga gagcacacac tagcggcttg acaacacaac taaaagctct 46620
agaaataaaa aaggaagcaa attcactcaa gaggagtata cggcaggaaa taatcaaact 46680
cagtggtgaa atcaaccaag tggaaataag aagaactatt caaagaatca accaaaggag 46740
gagctggttc tttgataaaa tcaacaagat agataaaccc ttagccagac tcactagagg 46800
gcacagggac agcatcctac ttaacaaaat cagaaatgaa aagggagaca taacaacaga 46860
tcctgaagaa atccaaaaca ccatcagatc cttctacaaa aggctatact caacaaaact 46920
ggaaaacctg gaagaaatgg acaaatttct agacagatac caggtacaat agttaaatca 46980
ggatcaggtt aaggatctaa acagtcctat atcccctaaa gaaatagaag cagtcattaa 47040
tagtctccca accaaaaaca gtccaggacc acatgggttt agggcagagc tctatcagac 47100
cttcaaagaa gatctaattc cagttctgca caaactattc cacaaaatag aagcacaagg 47160
tacgctaccc aattcattct atgaagccac aattactctg atacctaaat cacagacaga 47220
tccaacaaag atagagaact ttcagaccaa tttcccttat gaatattgat gcaaaaatcc 47280
tcaataaaat tctcgctaac cgaatgcaag aacccattaa aacaatcatc catcctgacc 47340
aagtaggttt tattccaggg atgcagtaat ggtttaatat gcgaaaatcc atcaatgtaa 47400
tccattatat aaacaaactc aaagacaaaa ccacatgatc atctcatttg agaaaatcca 47460
acacccattc atgataaaag tcttggaaag atcaaaaatt caaggcccat acctaaacat 47520
gacaaaagca atctatggca aaccagtagc caacatcaaa gtaagtggtg agaagctgga 47580
agcaatccca ctaaaatcag ggactagaca aggctgccca ctttctccct acctattcaa 47640
cattgtactt gaagtcctag tcagagcaat tcgacaaaaa aaaggagatc aaggggatac 47700
aaattggaaa agaagcagtc aaaatatcac tttttgcaga tgatatgaca gtatatataa 47760
gtgagcctaa aaattccacc agagaactcc taaacctgat aaatagcttc aatgaagtag 47820
ctggatataa agtttactca aacaagtcaa tggcctttct ctacaccaag aataaacagg 47880
ctgagaaaga aattagggaa acaacaccct tctcaatagt cacaaataat ataaaatatc 47940
ttggcgtgac tctaactaag gaagtgaaag atctgtatga taaaaacttc aagtctctga 48000
agaaagaaat taaagaagat ctcagaagat ggaaagatct cccatgctca tggattggca 48060
gtatcaacac tgtaaaaatg gctatcttgc caaaagcaat ctacagattc agtgcaatcc 48120
ccatcaaatt ccaactcaat tcttcaagga attataaaga gcaatctgca aattcacctg 48180
gaataacaaa aaacctagga tagcaaaaac tcttctcaag gataaaagaa cctctggtgg 48240
aatcaacatg cctgacctaa agctgtacta cagagcaatt gtgataaaaa ctgcatggta 48300
ctggtatagc aacagacaag tagaccaatg gactagaatt gaagacccag aaatgaaacc 48360
acatacctat ggtcacttga tctttgacat gggagctaaa accatccagt gaaaaaatga 48420
cagcattttc aaaaaatggt gctcacacaa ctggctgtta tcatgtaaaa gaatgcaaat 48480
tgatccattc ctatctcctt gtactaaggt caaaactaag tggatcaagg aacttcacat 48540
aaaaccagag acactgaaac ttatagagga gaaagtgggg aaaaccttga agatatgggc 48600
aaagggggaa aattcctgaa tagaacagca atggcttgtc ctgtaagatc aagaattgac 48660
aaatggaacc tcatgaaact gcaaagcttt tgtaaggcaa aaggcaccgt caataagaca 48720
aaaaggccac caacagactg ggaaaggatc tttacctatc ctaaatcaga taggagacta 48780
atatccaata tgtagagagc acaagaacat ggactccaga aaatcaaata accccattaa 48840
aaaatggtgc tcagggctaa aaaaagaatt ctcacctaag gaatactgaa tggctgagaa 48900
gcacctgaaa acatgttcag catccttaat catcagggaa atgaaaatca gaacaacact 48960
gagattcttc ttcacaccag tcagaatggc taagaaaaaa aattcaggtg acagcagatg 49020
ctggcaagga tgtggagaaa gagaacactc ctccattgtt ggtgagattg aagcttatac 49080
aaccactctg gaaatcagtc tggcagttcc tcagaaaact ggacatagta ctaccagagg 49140
atcccacaat acctctcctg ggcatatatc cagaagatgt tccaactggt aagaaagaca 49200
catgctccac tatgtccata gcagccttat ttataatagc cagaagctgg aaagaaccca 49260
gatgcccctc agcagaggaa tgggtacaga aaatgtggta catttacata atggaatact 49320
agtcagctat taaaaagaat gaatttatga aatttctagg caaatggatg gacctggagg 49380
gcatcatcct gagtgaggta acacaaacac aaaagaactc acatgatatg tacacactga 49440
taagtggata ttagcccaga aacttagaat acccaagata taagatacaa tttgtgaaac 49500
acatgaaact caagaagaat gaagatcaaa gtgtgggcac tttgcccctt cttagaattg 49560
ggagcaaaac acccatggaa ggagttacaa agacaaagtt tggagctgag acaaaaggat 49620
ggaccatcta gagactgtca tatctgggga tccatcccat aatcagactc caaacactga 49680
caccattgca tacactagca agattttgct aaaaggaccc agatatagct gtctcttttg 49740
agactatgcc ggcgcctagc aagcacataa gtggatgctc acaatcagct attggatgga 49800
tcacagggcc cccaatggag tagctagaga aagtacccaa ggagctaaag ggatatgcaa 49860
ccctttaagt gaaacatcaa tataaactaa ccagtacccc actcccaccc cagagctcgt 49920
atctatagct gcatatttag cagaagatgg cctagtcggc catcactgga aagagaggcc 49980
cattggtcgt gcaaacttta tatgcctcag tacaggggaa cgccagggcc aagaattggg 50040
agtgggtagg taggggagtg tgggggaggg tatgggggac ttttgggata acattagaaa 50100
tgtaaatgag gaaaatacct aataaaaatg ttaaaaaaca aacaaacaaa caaacaactc 50160
tcctgccact gaagggtttt agttatcagt cccaaggtca ctatgtgctt agtctttgag 50220
tgcttgagat atcctctgtt ccctatgtgc aacatttctt gattataact ttgtctcatg 50280
tatgatggat ttctgctgat tgacacattt ttccttgcaa actttatata atacacttta 50340
attttttaat attttttatt acgtattttc ctcaattaca tttccaatgc tctcccaaaa 50400
atcccccata ccctcccccc cacttcccta accacccatt cccatttttg gccctggcgt 50460
tcccctgtac tggggcatat aaagtttgtg tgtccaatgg gcctctcttt ccagtgatgg 50520
ccaactaggc catcttttga tacatatgca actatagtca ggagctgttt gtttatataa 50580
tggattacat tgatggattt ccatagatta aaccatccct gcatccctgg aataaaacct 50640
acttggtcag gattgatgat tgttttaaag tgttcttgga ttcggttagt gagaatttta 50700
ttgagtattt ttttaattag gtattttcct cctttatatt ttcaaacgct gacaccattg 50760
catacactag caagattttg ctgaaaggtc tcagatatag ctgtctcttg tgagactagg 50820
ccgggcctag caaacacaga agtggatgct cacagtcagc tattggatgg atcacagggc 50880
tccctatgga ggagctagag aaagtacaca aagagctaaa gggatctgca accctgtagg 50940
tgcaacatta tgaactaacc agtaccaccg gagctcatgt ctctagctgc atatgtatca 51000
gaagatggcc tagtcggcca tcactggaaa gagaggccca ttggacacac aaactttata 51060
tgccccaata caggggaacg ccagggccaa aaaaatggga atgggtgggt cggagattgg 51120
gggggtgggt ataagggacc tttgcgatag cattgaaaat gtaaactttg tctctgtaac 51180
tcattccatg ggtgtttgtt cccaattcta aagaggggca tagagtccac acttcagtct 51240
tcattcttga gtttcatgtg ttttgcaaat tgtatcttat atcttgggca tcctaagttt 51300
ctgggctaat atccacttat cagtgagtac atattgtgtg agttcctttg tgattgggtt 51360
acctcactca ggatgatgct ctccaggtcc aaccatttgc ctaggaattt cataaactca 51420
ttctttttaa tagctgacta gtacttcatt gtgtaaatgt accacatttt ctgtatccat 51480
tcctctgtta aggggcatct gggttctttc cagcttctgg ctattataaa taaggcttct 51540
atgaacatag tggaacatgt gtccttctta acggttggga catcttctgg atatatgccc 51600
aggagaggta ttgctggatc ttccggtagt attatgtcca attttctgag gaaccaccag 51660
actgatttcc agagtggttg tacaagcttg caatcccatc tacagtagag gagtgttcct 51720
ctttctccac atcctggcca gcatctgctt tcacctgaat ttttggtctt agccattctg 51780
actggtgtga ggtggaatct cagggttgtt ttgatttgca tttccctgat gattaaggat 51840
gctgaacatt ttttcaaatg cttctcttcc attcggtatt cctcaggtga gaattctttg 51900
ttcaattctg agccccattt tttaatgggg ttatttgaaa ttctggagtc caccttcttg 51960
ggttctttat atatattgga tattagtccc ctatctgatt taagataggt aaagatcctt 52020
tcccaatctg ttggtggtct ttttgtctta ttgacggtgt cttttgcctt gcagaagctt 52080
tgcagtttca tgaggtccca tttgtcaatt ctcaatctta ctgcacaagc cattgctgtt 52140
ctattcagta attttcccct ttgcccatgt cttcaaggct ttcccccact ttctcctcta 52200
taagtttcag tgtctggttt tatgtgaagt tccttgatcc acttagattt gaccttagta 52260
caaggagata ggaatggatc aattcgcatt cttctacatg ataacaacca gttgtgccaa 52320
caccatttgt tgaatatgct gtctttcttc cactggatgg ttttagctcc cttgtcgaat 52380
atcaagtgac cataggtgtg tgggttcatt tctgggtctt caattctagt ccattggtct 52440
acttgtctgt ctctatacca ctaccatgca gtttttatca caatctagat gcatgctcga 52500
ggccatttca ttacctcttt ctccgcaccc gacatagata aagcttatcg ataccgtcga 52560
cctcgagggg gggcccttgc tctgtagtaa agctttagat caggcatggt gattccacca 52620
gaggttcttt tatccttgag aagagttttt gctatcctag gttttttgtt attccacatg 52680
aatttgcaga ttgctctttc taattccttg aagaattgag ttggaatttg atggggattg 52740
cattaaatct gtagattcct tttggcaaga cagccatttt tacaatgtta atcctgccaa 52800
tccatgagca tgggagatct ttccatcttc tgagatcttc tttaatttct ttcttcagag 52860
acttgaagtt tttatcatac agatctttca cttccttagt tagagtcacg ccaagatatt 52920
ttatattatt tgtgactatt gagaagggtg ttgtttccct aatttctttc tcagcctgtt 52980
tattcttggt gtagagaaag gccattgact tgtttgagtt agttttatat ccagctactt 53040
cactgaagct gtttatcagg tttaggagtt ctctggtgga attttagggt cacttacata 53100
tactatcaca tcctctgcaa aaagtgatat tttgacttca tcttttccaa tttgtatccc 53160
cttgatctcg ttttgttgtc aaattgctct ggctaggact tcaagtataa tgttgaatag 53220
gtatggagag agtggacagc cttgtctagt tcctgatttt agtgggattg cttcaagctt 53280
ctcaacattt actttgatgt tggctactgg tttgctgtag attgctttta taatgtttag 53340
gtatgggcct tgaattcctg atctttccaa gacttttatc atgaatgggt gttggatttg 53400
tcgaatgctt tttccgcatc taacgagatg atcatgtggt ttttgtcttt gagtttgttt 53460
atataatgga ttacgttgat ggatttccat atattaaacc atccctgcat ccctgaaatg 53520
aaacctactt ggtcgggatg gatgattgct ttaatgtgtt cttggatttg gttagcgaga 53580
attttattga ggatttttgc atctatattc ataagggaaa ttggtctgaa attctctatc 53640
tttgttggat ctttctgtgg tttaggtatc agagtaatag tggcttcata gaatgagtat 53700
ggtagagttc cttctacttc tattttgtgg aataatttgt gcagaactgg aattagatcg 53760
tctttgaagg tctggtagaa ctctgcacta aaccctctga tccagggctt tttttggctg 53820
ggagactatt aatgactgct tctatttctt taggggatat gggactgttt agatcattaa 53880
cttgatcctg acttaacttt ggtaccttgt ttctgtctag aaatttgtcc atttcttcca 53940
ggttttccag ttttgttgag tatagccttt tgtagaagga tctgatggtg ttttggattt 54000
cttcaggatc tgttgttatg tctccctttt catttctgat tttgttaatt aggatgctgt 54060
ccctttgccc tctagtgagt ctaggtaagg gtttatctat cttgttgatt ttatcaaaga 54120
accaactcct cctttggtta attctttgaa tagttcttct tgtttccact tggttgattt 54180
cacccctgag tttgattatt tcctgccgtc tactcctctt gggtgaattt tcttcctttt 54240
tttctagggc ttttagatgt gttgtcaggc tgctagtgtg tgctctctcc agttactttt 54300
tggaggcact cagagctatg agtttccctc ttagaaatgc tttcattctg tcccataggt 54360
ttgggtatgt tgtggattca ttttcattaa gctctaaaga gtctttaatt tttttcttta 54420
ttccttcatt gaccaagtta tcattgagaa gagtgatgtt cagtttccac gtgaatgttg 54480
gctttccttt atttatgatg ttattgaaga tcagccttag gccatgatgg tctgatagga 54540
tgcatgggac aatttcaata tttttgtatc tgttgaggcc tgttttgtga ccaattatat 54600
ggtcaatttt ggagaaggtc ccgtgaggtg ctgtgaagaa ggtatatcct tttgttttag 54660
aataaaatga tctgtagata tctgttaagt ccatttgttt catcacttct gttagtttca 54720
ctgtgtcctt gtttagtttc tgtttccacg atctgtccat tgatgaaagt ggtgtgttga 54780
agtctcccac tattattgtg tgaggtgcaa tgtgtgcttt gagctttact aaagtgtctt 54840
taatgaatgt ggctgccctt gtatttggag catacatatt cagaattgag agttcctctt 54900
ggaggatttt acctttgatg agtatgaagt gtccctcctt gtcttttttg ataactttgg 54960
gttggaagtc gattttatcc gatattagaa tggctattcc agcttgttta ttcaggctat 55020
ttgcttggaa aattgttttc cagcctttca ctctgaggta gtgtctgtct ttttccctga 55080
gatgggtttc ctgtaagcag cagaatgttg ggtcttgttt gtgtagccag tctgttagtc 55140
taagtctttt tattggggaa ttgagtctat tgatattaag agatattaag gaaaagtaat 55200
tgttgcttcc ttttcttttt gttgttagat ttggcattct gttcttgtgg ctgtcttctt 55260
tttggtttgt tgagggatta ctttctggct ttatctaggg tgtaatttct gtccttgtat 55320
tggttttttt tctgttatta tcctttgaaa ggctggattc gtggaaagat aatgtgtgaa 55380
tttggttttg tcgtggaata ctttggtttc tccatctatg gtgattgaga gtttggctgg 55440
gtatcatagc ctggactggc atttgtgttc tcttagtgtc tgtataacat ctgtccagaa 55500
tcttctggct ttcatagtct ctggtgaaaa gtctggtgta attctgatag gtctgccttt 55560
atatgttact tgacctttct cccttactgc ttttaatatt ctctctttat ttagtgcatt 55620
tgttgttctg attattatgt gtcgggagga atttcttttc tggtccagtc tatttggagt 55680
tctgtaggtt tcttatatgt tcatgggcat gtcattcttt aggtttggga agttttcttc 55740
tataattttg ttgaagatat ttgcaggccc tttaagttga aaatcttcat tctcatcaac 55800
tcctattatc cgtaggtttg gtcttctcat tgtgtcctgg atttcctgga tgttttgagt 55860
taggatcctt ttgcattttg tattttcttt gattgttgtg ccgatgttct ctatggaatc 55920
ttctgcacct gagattctct cttccatctc ttgtattctg ttgctgatgc taacatctat 55980
ggttccagat ttctttccta gggtttctat ctccagcatt gcctcacttt gggttttctt 56040
tattgtgtct acttcccttt ttaggtctag tatggttttg ttcatttcca tcacctgttt 56100
ggatgtgttt tcctgttttt ctataaggac ttctacctgt ttggttgagt tttcctgctt 56160
ttctttaagg acttgtaact ctttagcagt gttctcctgt atttctttaa gtgagttatt 56220
aaagttcttc ttgatgtcct ctaccatcat catgagatat gcttttaaat ctgggtctag 56280
cttttcagtt gtgttggggt gcccaggacc aggtggggtg ggagtgctgc gttctgatgt 56340
tggtgagtgg tcttgatttc tgttagtagg attcttgtgt ttgcctttcg ccatctggta 56400
atctctgggg cctgtgcaga atacactcgg ttgggtcccg gaaccaagat ttctgctgct 56460
gatgctcagg caaagcgctc cctctctgct tggaccccct tcctttggcc gggaaggtgg 56520
ccagatgtct ggctacacta actttcttaa taaacttcct atttataagg aatcagctta 56580
tgaaagatgt cacatcaaaa tgttgatact gccagactgt caggtgctgt ttaaacagag 56640
aagctgaccc tgcacctgga atttgttcca ttccacttta aacatagtgt ttggttcaaa 56700
aggtcctcac ctgggtctta atacctaaag agatgattgt tctactccag aacacacttt 56760
tcccattgga caattaccct ccctttattt aaaaaatagc atgtctctct gcctttatta 56820
cacacacaca cacacacaca cacacacaca cacacacaca cactttcctg tcttcattat 56880
tttctcattt tagtccttct acttaggacc ctgtcaataa aaaaaaaaaa atggcctgat 56940
ggtccagatc aattggaact ttataaaaaa ggagaggagg gaataaatgt tgttcatgac 57000
tccctcaggg aaaaattcag tcgttgcaca tctgtcacag cttttgtagc tcttgttagc 57060
tacagtatgc tagatgaagt cagccttgca tacataccct gcacacctac acatgcaagg 57120
aataaaagag gctaaggtga accttagatc tggtttaaac accagttagt gggcttcagg 57180
gcctccagag ggatgaactg cagactctca agatgttctg ggagctccac attcacacaa 57240
aacaagtcaa tccctcttct ctgcttgctt ggtgacccag tggtagcttt tgtgttgcca 57300
gaactttctt tgcatcggtt aagaattatt aggaagtgca ggatcaaggg caacagtaac 57360
tatgatgtgt cccaaattct gccagtagag ggatgtgaat tatctcttgt caagctagag 57420
aaaacctgag gatgaacagc tgagaaagga gtaaggcaga ctgcagtcag cagctacaga 57480
cactgtacat catttttcag ttgactaata tacatcgatg cagctaagaa ggcagtgatc 57540
ccaaggaagg ttattggcgt tccgaaaaat atatgagcac attaacagaa cagcccttac 57600
cccaaactcc ttcagaataa actagtttca aacacagttg tctagcatgt aatcccatca 57660
gtctcgtgga ggaagactct gagtgcatgg aacagagaat ctgtcagcag tgtaccctga 57720
atctgaatat gaccttctgc tataggagaa aggaaccatc aaaaaagcac agtgataatg 57780
atgtgggaac gcttcagtgt tcaggaacca tatgatttat ttaaaatata gaatcaaaag 57840
taccaatttg cagttttgaa agatttattc cagtgtaagc attagcaatg caccaacatc 57900
aggtgatttc tgaatccaac acgtcttatg tcctcatgat attaaaaaaa aaaaaaggcc 57960
atccagaact gtgaacttga gttctacctt gttccctact gacattcaga ttttcttttt 58020
tgcattctct ttatcttaca ggagacagga ggggagggct aactcatttt actttggctt 58080
gtcccttgct ggtccttgcc cagaacgtaa agtagcttgc aagtcttcaa atctaaaaat 58140
cttagtaact cctacacgag tggcaatgcc aaagagcagt gcaacaaaga ggaagtaaat 58200
acgaccaaag agtattctta aatacaccac tggctcttgt ttttgtttta ttgtgtgcct 58260
ttgaactgga ggggacccac tgtctatgct cccacttagt ccctcttctt tgcactctgg 58320
aggcttccaa ccaaaatgac aatggcaatt ccgatgattg ttacacactc ctctaaaact 58380
gcatttttct ggggtgcagt cataacccaa atgagataaa cttccactgc aagctccttg 58440
atcacagaac ttacttttgg agcagggggt accatgtctc accattccag catctgttgt 58500
ttctgtccca cgatgttcat caagcccaaa gcaggtaaac ccagagataa tcgattgatg 58560
gaatgaaaca tgttcttgca aatatggaag attggtgaca ttggtacact gcaaccttcc 58620
acacagcttg tcctgatcag cacaagcatt gaatgtgagg ctttcttctg ctctagtaca 58680
atgcccaaat cgaaaccgtt gtttgttgat gtcatagcac ttaatattag cattcttagc 58740
acttacacca aagatttcca tgcattgtat gttgcgatca gtgcagttac ctttatagca 58800
gtaaccatct tctgagcatg gtgtcccatc ttgcagataa gtgtcatctg ggcaaatgta 58860
tttagtccca ttacagtact ctggaagatc acatatgttc tggataggtc tgcagagtgt 58920
cccagaagga ctgtaagtgc aatttgcaca gcataattct ttatcacaaa tgctaccagg 58980
tgttaacctg caatcatttc cacagcaggg atctgaataa catgcctttt gggagccaca 59040
gtcacactgc tcatcgttat ctactttgaa gtttccacaa aacttataag tcaatgatgt 59100
attataataa acatgacggt catagaaaag acatggcatc agaccaggag tattaagtat 59160
gttgcttatc tctgcaaggg aacaattgct gaaagcatct gttaattgag gatgtctgaa 59220
cataatgcag gtgttccttc tctggcagac acagtacccc tcatcatatt ttaagcctaa 59280
actccttcca acacgattgg ttattatagg agataaaaat aaaggatttc gatcatattt 59340
accaatacaa attagggcta aggaagaaca tatactcctc tcagctggat taacctggtt 59400
atcttgtggc ccatacttat taagtaaaac tcctgcatca ggcttaaatt tattataaaa 59460
gactgacaca tagtaattat aagccgaccc tcctggaact gcaaactcaa gtcgaaatgg 59520
atcagaattg gtgtacacag tcatgagaaa gacatagtac cgcatatgaa gattggtcag 59580
ataggtgtcc attaaactaa tgacttgaaa caaataccca acagtagatg aaagtttgtc 59640
acctgcagca gaattatata cagaattggt tgcttgaaag tggcctttta tagcagctgg 59700
atgtgtagcg tagttcttac tagatattct gggagctcca tctgcatatt ccaatctgga 59760
ggagggagaa cctgtattat ggctccagtg cttccatgca ttcataggcc ctgtgtcatc 59820
agactcagat actatctgag aaacaaggtg ttcaaagctc tgtgaatcat tgaggggttt 59880
gatttcatag gtaaggtcat ctaacttcat gacccctgac aggcccccat aacaagtatc 59940
cacagtgacc atggattgtg ggatcccctc caggtagcca atatagtaac aatctacagg 60000
aaaaaagggg taatccatct gtaaggctcc ttggtcatct tgagttgtca gcaacaagtg 60060
tctgggccaa atgagtgtct ttctccgcag gtggatgata tgtctctggc cccgaaaatg 60120
caagctatat gagagcagtc tttgtgcttg aagtcctttg gtatggtaga tctccttccg 60180
aggaataacc acctccgatg agatgtaacg ccaagtagga tggccttgag aacaccagac 60240
tggaaccagg aggagcagcc agagtgcaaa tagcaagagg aggaccctgg ggaccacagg 60300
tctttccact agcctcatgc cccaggtcag agataacatc ctgggtggag ctaaatccct 60360
ctgctgtggc cactgcctgg tctagaaaat actgacagag gactaaaaac ctcctcaggc 60420
tcccaaccta agtggttacc cagacaactg gagttaggta acagtcactg ggtgtggcag 60480
gaattgagtc tgaatgtgtt agctgaggtt gaggttaaat attgtcaaaa gggatgtcta 60540
taaatgtgcc tggacaagaa aagtcagaag cagcaaggag tgtctctgac aggctcaatc 60600
ctttcttttc tttttttgaa gttcaaaata tcatttccac gtgaatgtat ttggttccca 60660
gtgtgactct gggtctcttt ctaggagtca atatttcttt atatcttggc tcatgtttct 60720
cacagttgtt ctaatttctt gttttgtttt gtttgtttgt ttgaacgtta gtagtaaata 60780
ctgtctatat tagcctttta gctataaatg attgttttta tttcttctaa tcatattttg 60840
tttgagtttt ggttaaacta tttacaaatg agtttttttt ttttcctttt gggtgttgct 60900
cgaaagtttg gagctttctg ttaatattgt gttgttattt ttccaatatt attagacctg 60960
agaattctat ctgggtacct gtgaactcta gaatttttaa aaattccatc tcttgggaac 61020
attacctctg accccgtctg aggccgaagt ggctgtcccc ctccaacctt tagtatcttt 61080
ctttcctgac tattgggatt tcttcaagca atcaggctga tgggttctca gcagtgagac 61140
cagtagactg ccggtatgaa cgtcgaagag actgccacac actccaggtt catcaacagt 61200
gctttcgcgt ctcttacttt tgtagaagga aaagcagcct ctgagttatc tccaagaaat 61260
cattaatgaa agagttaaaa gatgggtatc acccggagtt catgacaagc cctggctcag 61320
acacgtgagc aaggtctaca gccccaaaga taggctgccc tgcaacatgt atttataaga 61380
tagaagaaaa aaatgggtgg ttggagggtt gatcaactta cttcctctca aacatatata 61440
tctcatctaa gtgtgcaggg gaaaactctg taggactact gggattgtta ttatcattat 61500
tattattatt attattatta ttattattat tattattatt aacttaaggc attttattag 61560
atattttctt catttagttt tcaaatgtta tccccggaac ctcctatact ctctccctgc 61620
cctgctcccc aacccaccca ctcctacatc ctggccctgg cattccccta tactgtggca 61680
gatgatcttc gtaagaccaa gagcctttcc tcccattgat ggcctactag gctatcctct 61740
tttacatatg caactagagt cacagctctg gggaggtatt gcttagttca tattgttttt 61800
cctcctatag ggttgcagat ccctttagct ccttgggtac tttctctagc tcctccattg 61860
ggggccctgt gttccatcca atagatgact gtgagcatcc acttctgtat ttgccaggta 61920
ttggcatgga tcttactgca ccttctgaac tctctaagca gctttcctgg tcacctccag 61980
gagcctcatg aataagtctc tgcttccccc ttgtggctat gagcattact gcacctgata 62040
caccctgcag cttcctaggg aagagggagg aagtggcttg gcccctgtct ggttaaggta 62100
agaggagata aatcccttct catgaattag ggtgagaagg gtcatgtgct ctatcattgg 62160
tgaccaactt ggggacatgg gcttatacag tcatcactct gaggctctgt gtaccaccag 62220
actgaactcc catatcctac atgcacatag gacaacacca agtagaagga ggttttagga 62280
ctaaactgaa ggacagagat ggggtttcta aacaactagg gagtgccagg gccagcctct 62340
ctaaccacta taggacacta tggagtctgg ttacaaagag agattactca aggtccttag 62400
cactgattac agagcatatc tcagatgcct tctgctgacc agatgtatct ttgcataatc 62460
tgcctatcca gattcagaaa attgatgcca catagccaag tggactttca ggaacagacg 62520
atttaaaaac aggcagagag atgtgagaga aaggagaagg agagagagaa gggagaggga 62580
gagaagagag agggagacgg agaaggaaag agggagaagg agaaggagag aaggggcatg 62640
gacagaggga gggacagaag gagagaggag atagagaggg ggataaggaa gaaaggaggg 62700
agggagagag agagaaggct aagtctttcc atacctgggt cccaatacct cttataaccc 62760
aagcacatgg tttcagatat cacaatgcgg ttgggatata gataactgta aatacttgtg 62820
aaaataatgg ggctgagatc tggggttttc atgatagttt caaagtcact gtactgacta 62880
aaaccttcca ctggcccatc tccagcttgt taatctgagg gtatcaaatt tcccactaag 62940
tgtgtttaga aagatctcca cctttttgcc ctagtcttcc agtgccccac ctacgttctg 63000
gtctcccaca tctgatgtct tctcagtgat tctggccctg cctgctccac agctacaaac 63060
cccttcctat aatgagctct gtgctgagcc atcatcctga atcaatccac cttaagcaga 63120
tgttttgctt atttttcctg tgtccatact acagaggaag ggtaggcatg tagaagctga 63180
ggcatctcat ctcactctaa gcaccctcag tctctaaatg tgcccctttg tttccagcag 63240
ttcagcctca agcatctttt attcactcgt cttagaggga cacatgtgct gtagtgttat 63300
aagatgaaat ttaaagcatt agttattccc aacaagccaa ttaaacaagc caaaaacatt 63360
catcagtcat tcccatggaa cctctgaagc atcttcctgc tctaaccttg agtttcctag 63420
ggctgctgtg ggatcacagg agctgtcctg tttaccagcc tatcctgtcc cacgggattc 63480
agttattagt gggtgcgagg gggaccgcaa acctggaaga aaatgggatt ggaagagaaa 63540
agagaaacga agaccaagta gatcttttcc tatcaaggtc ttcgtttatt aggctgaggt 63600
gcctggtgta aagcatgcat cgcggggaat aggaaggggt cgagggggaa ttttacaaag 63660
aacaaagaag cgggcatctg ctgacatgag ggccgaagtc aggctccagg cagcggacac 63720
tctgcatctt atctctggaa catagatcct ccttgacagc cttggggtgt caggccaggc 63780
tcaggtgtaa ctcatgtcct tggatggcat gggagttcag gaagagatag ggaagagggg 63840
actataattc agcttttaca gcctcaggtg ccaggaaggc aacagggagg agggagtgac 63900
tacaggctcc tagcacgagg ccatttggcc tgtcagggtg ggagattgtg aagggctcac 63960
tttctcatgg tatggtctct gacaccagcc agaaaaaaaa aaaatctcca gtcaatctac 64020
agaaaggcag aaatatggag aaccttctag aagaacagca acccttgact gactctgcag 64080
ggcagtctaa gcacacaggt tcctctgagc ccagtccctg gctgtattca ctcacacagg 64140
gcccagcagt ctggagatgc ctgcagcagg aaggttctca ctgtacctca gtctcacagc 64200
tcagtcactt tgtagtctcc ccacagcaaa tcacaaccaa tgcagttgtc acccttgagg 64260
ccattcctca gtgacgcctg ggaaacactg tccatttgcc ctgctcatat gctgtctcag 64320
ctcacataaa ttccctccaa gctcctttac agaaacaacc aattgacata caaattaaag 64380
taatatgaag ctctgtcctg taccaattcc tagaacacct cagctgctaa aatctaactg 64440
tgcaatacag agaactacct agataatacc taactccaat cttgaggtat ctgttgaaga 64500
ggctttcagc tatgaattac caagagggat gcctgtgtct cctacatctt aaggcatctt 64560
tctgaagctc acactaccct tcacagcgcc acagcagcag ggagatctgt atctcttcct 64620
ctccacaaga gggcagaatt cgggtgtcaa ctgtgacaac cctccagcat taatttctat 64680
tagtagatat tttctttact tacatttcaa atgttatccc cttccccagt ttttcctccc 64740
tgaaaccccc tatcccatca cccatctccc tgctcaccaa cccacccatt ccataagaac 64800
caatcagtac cccacccccc cagagctccc agggactaaa ccaggaacca aagagtacac 64860
atggaaggac ccatagctcc agctgcatag gtagcactac tttgaaaagg gggaagatga 64920
tgaaatcatt actgtggggg aaatgcaagg aagattccaa cacatctagg catctatgaa 64980
gattttaagt cttcaaaatc caaaaccacc acaaaattta aaaaaaaaaa aagtagattc 65040
taaatgcagt cacctgcacc aggtgcctgg ggaatcactc agcagcccta gactgagaaa 65100
gcttggagaa agtagaaata gagaaagtgt acagccagta tcctctagct actcacatcc 65160
aaacagggcc tcctgactgc tctgagcctg tcctaagaac agcaatgatg ccacagaaat 65220
ttttagagtg agccctgaag gaacttgagg ctgatatgag caagccagtc ccagaggaaa 65280
ggaaaaccca tagagagaaa acaggtgagt tagtgcatta aaggggctga gcagggagtt 65340
ctcatcgctc cccagcacca gaaataagag cctctccgga gctgctggga catggaatgc 65400
agatgattcg gaccatcagc cccacagaga cctttcccac tctggctcag aaagaggcac 65460
tggaccacag ttggagagga gaatcgaaag ctgatatctc tgtattcact tagcctgtta 65520
cccacccatg cacccaagtc caaggtggga gaaacactga gggtctaaac acagccccag 65580
agcaactgcc agtattaaat accagatttc agaagatggg aaatcacctc tctggtcatt 65640
tttgggacat gtaaactgta acaggaaaca taggaccaat ttaagatgga gcagtcctat 65700
atccctaacc cagttgtaaa taaaacattc aagagtgcca tcagacacca cagtggtaca 65760
ggagagatga gtgtacctag tgcatcaaga gttccctcac tagataaacc aagatgtagc 65820
cccaggacca cccaggcacc taccaggact cccctccaga ggtctgagcc agttagctct 65880
agttcattct atgtttcaga ccaaaacatc agaaacaaca gcatctccac tgcagatgaa 65940
cccctaagcc atacagtgta cccaaaggca gcaccacaga tggggaatgt gggggcgaag 66000
aagcttgtat cacattaaga gtgttgccca atgactattt gctatcttca cagtaagatg 66060
ggcactgatc ccacccacca aaacatagta ggacaaggac ccttaaataa catctgtcaa 66120
ggggagctgt caaatagcca ctgagctgag atggctcata cggggtgata tagaaaaaca 66180
ggccaaagaa cctcctgtgt tgcaagcaca aatgggaagc tgtgaatctc cactacctta 66240
caagaaaact gtaccaatct gcaagaacag gggatacaca gctttccaca ctgtgtagga 66300
aggcattgga ctctcagatc atccaaggac tagggctaaa gtggccatgt gtgggaaata 66360
catccacttt ataaacctac cttgtatcca catcacacat ggagcatctg ctcctggtcc 66420
ctacctcatg tgggtgctgc ggtgctgtac attgagtctg ggggcatgag tgtgcccggt 66480
aaattcctta tcactcagat gaatttccag tccacactca tcaccttgga gtaggaattt 66540
taaaagttag tgtagatata agtaaagaga gtcaggaggg ggggctgaaa ggcagacctg 66600
cagaaattct ccagagggta gagctcagaa gcagcaagca aaacttgggc taacagaaga 66660
gcaaacacag agctcaggca gaactacctg gcaatgcgac tgggcacact gaaagcactg 66720
ggcatcagca ctgagcccca aatatgcact caggatcctc tgcataataa tgtaacataa 66780
caggaaggtt agaacaggcc aaaagaggaa acagaacaaa tgcccctaac caaagaagta 66840
taaacaaatt gggaagagta aagaaggatt gtaaggattg agtaccacac agaacatgct 66900
cttaatggcc tcaatgctga agctaggaag aactaagtta aaagaaacat gttcaacggg 66960
attccctgtc actggacttc acaacaagca aaattcaatc tttctgttaa ggagatgaga 67020
agagaatatc tgaaccttgt gttgacagtg ccccaccccg actgtcaggc tgtgggaaat 67080
gccagagcaa tcactaggaa cacacaagga tgagggagac gagggttagg acacaaccat 67140
catgatatcc cacaagtatg gaagagcaag aacttgtaga gagcagagaa tggcagacaa 67200
agcagcatat acataagtag atggccagac tatacaggaa agatctacat agcctgtgag 67260
gctttctgac agaaaagggc aggcatgtct caaagcacaa tgcctggctt gggacactgt 67320
ctgccctgga tcctctcact ccacatgtaa ggagctcaca gcaaaaccac acagccttcc 67380
acaagaggag aagaaaaggt agcttgtcag tgaggaagtc ccccagagac agaccattcc 67440
agtagttctt atcattcctc ccaaagcagc caccatccag gcactgagag accaaaggct 67500
gttgggaggt cagcctagag gcaggctcag acctttcttg ctccctagga ccttcctgaa 67560
aagatagaag cacagaagtg aataatctgg acctcaactc aggatgacaa ctgaaactca 67620
accgtgctgc ctgggccccc aatgctctct acacctgcag gcgcgcc 67667
<210> 5
<211> 14781
<212> DNA
<213> Artificial Sequence
<220>
<223> synthetic
<400> 5
ggcgcgccga ccaggaaagc tgcttagaga gttcagaagg tgcagtaaga tccatgccaa 60
tacctggcaa atacagaagt ggatgctcac agtcatctat tggatggaac acagggcccc 120
caatggagga gctagagaaa gtacccaagg agctaaaggg atctgcaacc ctataggagg 180
aaaaacaata tgaactaagc aatacctccc cagagctgtg actctagttg catatgtaaa 240
agaggatagc ctagtaggcc atcaatggga ggaaaggctc ttggtcttac gaagatcatc 300
tgccacagta taggggaatg ccagggccag gatgtaggag tgggtgggtt ggggagcagg 360
gcagggagag agtataggag gttccgggga taacatttga aaactaaatg aagaaaatat 420
ctaataaaat gccttaagtt aataataata ataataataa taataataat aataataata 480
ataatgataa taacaatccc agtagtccta cagagttttc ccctgcacac ttagatgaga 540
tatatatgtt tgagaggaag taagttgatc aaccctccaa ccacccattt ttttcttcta 600
tcttataaat acatgttgca gggcagccta tctttggggc tgtagacctt gctcacgtgt 660
ctgagccagg gcttgtcatg aactccgggt gatacccatc ttttaactct ttcattaatg 720
atttcttgga gataactcag aggctgcttt tccttctaca aaagtaagag acgcgaaagc 780
actgttgatg aacctggagt gtgtggcagt ctcttcgacg ttcataccgg cagtctactg 840
gtctcactgc tgagaaccca tcagcctgat tgcttgaaga aatcccaata gtcaggaaag 900
aaagatacta aaggttggag ggggacagcc acttcggcct cagacggggt cagaggtaat 960
gttcccaaga gatggaattt ttaaaaattc tagagttcac aggtacccag atagaattct 1020
caggtctaat aatattggaa aaataacaac acaatattaa cagaaagctc caaactttcg 1080
agcaacaccc aaaaggaaaa aaaaaaaact catttgtaaa tagtttaacc aaaactcaaa 1140
caaaatatga ttagaagaaa taaaaacaat catttatagc taaaaggcta atatagacag 1200
tatttactac taacgttcaa acaaacaaac aaaacaaaac aagaaattag aacaactgtg 1260
agaaacatga gccaagatat aaagaaatat tgactcctag aaagagaccc agagtcacac 1320
tgggaaccaa atacattcac gtggaaatga tattttgaac ttcaaaaaaa gaaaagaaag 1380
gattgagcct gtcagagaca ctccttgctg cttctgactt ttcttgtcca ggcacattta 1440
tagacatccc ttttgacaat atttaacctc aacctcagct aacacattca gactcaattc 1500
ctgccacacc cagtgactgt tacctaactc cagttgtctg ggtaaccact taggttggga 1560
gcctgaggag gtttttagtc ctctgtcagt attttctaga ccaggcagtg gccacagcag 1620
agggatttag ctccacccag gatgttatct ctgacctggg gcatgaggct agtggaaaga 1680
cctgtggtcc ccagggtcct cctcttgcta tttgcactct ggctgctcct cctggttcca 1740
gtctggtgtt ctcaaggcca tcctacttgg cgttacatct catcggaggt ggttattcct 1800
cggaaggaga tctaccatac caaaggactt caagcacaaa gactgctctc atatagcttg 1860
cattttcggg gccagagaca tatcatccac ctgcggagaa agacactcat ttggcccaga 1920
cacttgttgc tgacaactca agatgaccaa ggagccttac agatggatta cccctttttt 1980
cctgtagatt gttactatat tggctacctg gaggggatcc cacaatccat ggtcactgtg 2040
gatacttgtt atgggggcct gtcaggggtc atgaagttag atgaccttac ctatgaaatc 2100
aaacccctca atgattcaca gagctttgaa caccttgttt ctcagatagt atctgagtct 2160
gatgacacag ggcctatgaa tgcatggaag cactggagcc ataatacagg ttctccctcc 2220
tccagattgg aatatgcaga tggagctccc agaatatcta gtaagaacta cgctacacat 2280
ccagctgcta taaaaggcca ctttcaagca accaattctg tatataattc tgctgcaggt 2340
gacaaacttt catctactgt tgggtatttg tttcaagtca ttagtttaat ggacacctat 2400
ctgaccaatc ttcatatgcg gtactatgtc tttctcatga ctgtgtacac caattctgat 2460
ccatttcgac ttgagtttgc agttccagga gggtcggctt ataattacta tgtgtcagtc 2520
ttttataata aatttaagcc tgatgcagga gttttactta ataagtatgg gccacaagat 2580
aaccaggtta atccagctga gaggagtata tgttcttcct tagccctaat ttgtattggt 2640
aaatatgatc gaaatccttt atttttatct cctataataa ccaatcgtgt tggaaggagt 2700
ttaggcttaa aatatgatga ggggtactgt gtctgccaga gaaggaacac ctgcattatg 2760
ttcagacatc ctcaattaac agatgctttc agcaattgtt cccttgcaga gataagcaac 2820
atacttaata ctcctggtct gatgccatgt cttttctatg accgtcatgt ttattataat 2880
acatcattga cttataagtt ttgtggaaac ttcaaagtag ataacgatga gcagtgtgac 2940
tgtggctccc aaaaggcatg ttattcagat ccctgctgtg gaaatgattg caggttaaca 3000
cctggtagca tttgtgataa agaattatgc tgtgcaaatt gcacttacag tccttctggg 3060
acactctgca gacctatcca gaacatatgt gatcttccag agtactgtaa tgggactaaa 3120
tacatttgcc cagatgacac ttatctgcaa gatgggacac catgctcaga agatggttac 3180
tgctataaag gtaactgcac tgatcgcaac atacaatgca tggaaatctt tggtgtaagt 3240
gctaagaatg ctaatattaa gtgctatgac atcaacaaac aacggtttcg atttgggcat 3300
tgtactagag cagaagaaag cctcacattc aatgcttgtg ctgatcagga caagctgtgt 3360
ggaaggttgc agtgtaccaa tgtcaccaat cttccatatt tgcaagaaca tgtttcattc 3420
catcaatcga ttatctctgg gtttacctgc tttgggcttg atgaacatcg tgggacagaa 3480
acaacagatg ctggaatggt gagacatggt accccctgct ccaaaagtaa gttctgtgat 3540
caaggagctt gcagtggaag tttatctcat ttgggttatg actgcacccc agaaaaatgc 3600
agttttagag gagtgtgtaa caatcatcgg aattgccatt gtcattttgg ttggaagcct 3660
ccagagtgca aagaagaggg actaagtggg agcatagaca gtgggtcccc tccagttcaa 3720
aggcacacaa taaaacaaaa acaagagcca gtggtgtatt taagaatact ctttggtcgt 3780
atttacttcc tctttgttgc actgctcttt ggcattgcca ctcgtgtagg agttactaag 3840
atttttagat ttgaagactt gcaagctact ttacgttctg ggcaaggacc agcaagggac 3900
aagccaaagt aaaatgagtt agccctcccc tcctgtctcc tgtaagataa agagaatgca 3960
aaaaagaaaa tctgaatgtc agtagggaac aaggtagaac tcaagttcac agttctggat 4020
ggcctttttt tttttttaat atcatgagga cataagacgt gttggattca gaaatcacct 4080
gatgttggtg cattgctaat gcttacactg gaataaatct ttcaaaactg caaattggta 4140
cttttgattc tatattttaa ataaatcata tggttcctga acactgaagc gttcccacat 4200
cgcgatcgcg gtaccgaagt tcctattccg aagttcctat tctctagaaa gtataggaac 4260
ttctcgcgcg tctggcctcc gaggcctccg cgccgggttt tggcgcctcc cgcgggcgcc 4320
cccctcctca cggcgagcgc tgccacgtca gacgaagggc gcagcgagcg tcctgatcct 4380
tccgcccgga cgctcaggac agcggcccgc tgctcataag actcggcctt agaaccccag 4440
tatcagcaga aggacatttt aggacgggac ttgggtgact ctagggcact ggttttcttt 4500
ccagagagcg gaacaggcga ggaaaagtag tcccttctcg gcgattctgc ggagggatct 4560
ccgtggggcg gtgaacgccg atgattatat aaggacgcgc cgggtgtggc acagctagtt 4620
ccgtcgcagc cgggatttgg gtcgcggttc ttgtttgtgg atcgctgtga tcgtcacttg 4680
gtgagtagcg ggctgctggg ctggccgggg ctttcgtggc cgccgggccg ctcggtggga 4740
cggaagcgtg tggagagacc gccaagggct gtagtctggg tccgcgagca aggttgccct 4800
gaactggggg ttggggggag cgcagcaaaa tggcggctgt tcccgagtct tgaatggaag 4860
acgcttgtga ggcgggctgt gaggtcgttg aaacaaggtg gggggcatgg tgggcggcaa 4920
gaacccaagg tcttgaggcc ttcgctaatg cgggaaagct cttattcggg tgagatgggc 4980
tggggcacca tctggggacc ctgacgtgaa gtttgtcact gactggagaa ctcggtttgt 5040
cgtctgttgc gggggcggca gttatggcgg tgccgttggg cagtgcaccc gtacctttgg 5100
gagcgcgcgc cctcgtcgtg tcgtgacgtc acccgttctg ttggcttata atgcagggtg 5160
gggccacctg ccggtaggtg tgcggtaggc ttttctccgt cgcaggacgc agggttcggg 5220
cctagggtag gctctcctga atcgacaggc gccggacctc tggtgagggg agggataagt 5280
gaggcgtcag tttctttggt cggttttatg tacctatctt cttaagtagc tgaagctccg 5340
gttttgaact atgcgctcgg ggttggcgag tgtgttttgt gaagtttttt aggcaccttt 5400
tgaaatgtaa tcatttgggt caatatgtaa ttttcagtgt tagactagta aattgtccgc 5460
taaattctgg ccgtttttgg cttttttgtt agacgtcgag ctctagattg ggaacccggg 5520
tctctcgaat tgttgacaat taatcatcgg catagtatat cggcatagta taatacgaca 5580
aggtgaggaa ctaaaccatc caccatgatt gaacaagatg gattgcacgc aggttctccg 5640
gccgcttggg tggagaggct attcggctat gactgggcac aacagacaat cggctgctct 5700
gatgccgccg tgttccggct gtcagcgcag gggcgcccgg ttctttttgt caagaccgac 5760
ctgtccggtg ccctgaatga actgcaggac gaggcagcgc ggctatcgtg gctggccacg 5820
acgggcgttc cttgcgcagc tgtgctcgac gttgtcactg aagcgggaag ggactggctg 5880
ctattgggcg aagtgccggg gcaggatctc ctgtcatctc accttgctcc tgccgagaaa 5940
gtatccatca tggctgatgc aatgcggcgg ctgcatacgc ttgatccggc tacctgccca 6000
ttcgaccacc aagcgaaaca tcgcatcgag cgagcacgta ctcggatgga agccggtctt 6060
gtcgatcagg atgatctgga cgaagagcat caggggctcg cgccagccga actgttcgcc 6120
aggctcaagg cgcgcatgcc cgacggcgag gatctcgtcg tgacccatgg cgatgcctgc 6180
ttgccgaata tcatggtgga aaatggccgc ttttctggat tcatcgactg tggccggctg 6240
ggtgtggcgg accgctatca ggacatagcg ttggctaccc gtgatattgc tgaagagctt 6300
ggcggcgaat gggctgaccg cttcctcgtg ctttacggta tcgccgctcc cgattcgcag 6360
cgcatcgcct tctatcgcct tcttgacgag ttcttctgaa gatccgctgt aagtctgcag 6420
aaattgatga tctattaaac aataaagatg tccactaaaa tggaagtttt tcctgtcata 6480
ctttgttaag aagggtgaga acagagtacc tacattttga atggaaggat tggagctacg 6540
ggggtggggg tggggtggga ttagataaat gcctgctctt tactgaaggc tctttactat 6600
tgctttatga taatgtttca tagttggata tcataattta aacaagcaaa accaaattaa 6660
gggccagctc attcctccca ctcatgatct atagatctat agatctctcg tgggatcatt 6720
gtttttctct tgattcccac tttgtggttc taagtactgt ggtttccaaa tgtgtcagtt 6780
tcatagcctg aagaacgaga tcagcagcct ctgttccaca tacacttcat tctcagtatt 6840
gttttgccaa gttctaattc catcagacct cgacctgcag cccctagaga agttcctatt 6900
ccgaagttcc tattctctag aaagtatagg aacttcctag ggtttcaccg gtgcgatcgc 6960
cctcagcaca ataatagaga cactgaaact tatagaggag aaagtgggga agatcctcga 7020
agatagggtt cagggggaaa gatttttgaa cagaacacca atggcttgtg ctgtaaaatg 7080
aagaattagc aaatgggacc tcataaaatt aaaaaggttc tgtaaggcaa aggacacaaa 7140
aaggcagcca acatattggg aaagatcttt accaatccta aatatgacag agggctaata 7200
tccaatatat agaaagaatt caagacgtta gactccagaa aaccaaataa ccctattaaa 7260
aaaacctgag tacagagcta aacagtgaat tctcaactga agaatactga atggctgaga 7320
aacacttaaa atttttttca atatcactag tcatcaggga aatgccaaac aaccctgata 7380
ttccacctca cacctgtctg aatcactaag aacaaattga ggtgacagca gatgttggtg 7440
aggatgtgga ggaaggggaa catttctcca ttgctggtgg aattgcaagc tggtacaacc 7500
actctgtaaa tcagtttggc agttcctcag aaaattggac atagtactat cagaggatcc 7560
agcaatagca ctcctgggca tatacccaga agatgctcca acatgtaata aggacacatt 7620
ctccactatt tccatagcag ccttatattt aatgtccaga agctttaaag aacccagatg 7680
tccctcaaga gaacaatggg tacagaaaat gtggtacatt tacacaatgg agtactcctc 7740
agctattaaa aacaatgaat tcattaaatt cttagacaaa tggatggttc tggaggatat 7800
catcctgagt gaggtaactc aatcacataa gaacacacat gatattcact cactgataag 7860
tggatattag cccagaagtt cggaatatca aagatacaat tcacaaaaca catgaatttc 7920
aagaaggaag accaaagtat gaatacttcg gtccttctta gagggaggaa taaaatagtc 7980
atggaaggag ttaaagagac aaagtgtgga acagagactg aaggaatgac catccagaga 8040
ctgccccttt ggggatccat cccatagaca accaccagac acttttgtgg ataccaacaa 8100
gagcttgctg ccaggatctt gatatagctg tcttctgaga ggctctgcca gtgcctggca 8160
aatacagaag tggatgctca cagtcatcca ttggacttcg cacaaggtcc ccaatgaggc 8220
agccagagaa aggacccaag aagttgaagg gttttgtaac cccataggga gaacaacaat 8280
atgaaccaat cagtactcca agagctccct ggcatgaaac caccaacaaa agaaaacatg 8340
gtgggactca tagatctagc tgcatttgta gcagagaatg gactagttgg tcatcaatgc 8400
aaggagagta ccttggtcct gtgaaggctc tatgcccaaa tataggggat tgccagggcc 8460
tggaagtggg agtgggtgga ttggtgagca gggggagggg gaggagatag gggatttcag 8520
agggtaaacc aggaaagggg ataacatttg aaatgtcaat aaagacaata tctaataaaa 8580
aatgtcattt agaaaaaaaa aagtaacgct gtgtttgctt tatgcagcaa aatgctgtgt 8640
cctgtttatg tattcagtct gttagtctat gtctttttat tggagaattg agtccattga 8700
tgttaaaaga gatattaagg aatagtgatt gttgcctact ttatttttgt tgttagaggt 8760
ggaattatgt ttgtgtggtt atcttctttt gagtgtgttg aaagatgatt actttagtgc 8820
tctttctagg gtgtagtttt cctacctgtg ttggagtttt ccatctgcta tcctttgtag 8880
ggctggattt atggaaagat attgtgtaaa tttggacttg taatggaata tcttggtttc 8940
ttcatcgatg gtaattgaga gttttgctgg gtatagtagc ctgggctggc atttgtgttc 9000
tcttagtttc tgtataacat ctgtccagga tattctggct tttatagtct atggtgagaa 9060
gtctggtgta attctgatag gtctgccttt atatggctgg tagacaggac agctcctgtg 9120
atcccagagc agccctggaa aacccaaggt tagagcagga agatgcttca gaggttccat 9180
gggaatgact gatgaatgtt tttggcttgt ttaattggct tgttaggaat aattaatgct 9240
ttaaatttca tcttataaca ctacagcaca tgtggccctc taagatgaat gaataaaaga 9300
tgcttgaggt tgcacttctg gaaacaaggg ggcacattta gagactgagg gtgcttggaa 9360
tgaggtgaga tgcttcagct tctacatgcc tacctttcct ctgtagtatg gacacaggaa 9420
aaataagcaa aacatctgct taaggtggat tgattcagga tgatggctca gcacagagct 9480
cattatagga aggggtttgt agctgtggag caggcagggc cagaatcact gagaagacat 9540
cagatgtggg agaccagaac gtaggtgggg cactggaaga caagggcaaa aaggtggaga 9600
tctttctaaa cacacttagt gggaaatttg ataccctcag attaggaagc tggagatggg 9660
ccagtggaag gttttagtca gtacggtgac tttgaaacta tcatgaaaac cccagatctc 9720
agccccatta ttttcacaag tatttacagt tatctatatc ccaaccgcat tgtgatatgt 9780
gaaaccatgt gcttgggtta taagaggtat tgggacccag gtatggaaag acttagcctt 9840
ctctctctct ccctccctcc cttcttcctt atccccctct ctatctcctc tctccttctg 9900
tccctccctc tgtccatgcc ccttctctcc ttctccttct ccctctttcc ttctccgtct 9960
ccctctctct tctctccctc tcccttctct ctctccttct cctttctctc acatctctct 10020
gcctgttttt aaatcgtctg ttcctgaaag tccacttggc tatgtggcat caattttctg 10080
aatctggata ggcagattat gcaaagatac atctggtcag cagaaggcat ctgagatatg 10140
ctctgtaatc agtgctaagg accttgagta atctctcttt gtaaccagac tccacagtgt 10200
cctatagtgg ttagagaggc tggccctggc actccctagt tgtttagaaa ccgcatctct 10260
gtccttcagt ttagtcctaa aacctccttc tgcttggtgt tgtcctatgt ccatgtagga 10320
tatgggagtt cagcctggtg gtacacagaa cctcagagtg atgactttgg tataaaccca 10380
tgtcccaact gggtcaccaa tgatagagca catgacccct ctcaccctaa tcaatgagaa 10440
gggatttatc tcctcttacc ttaaccagac aggggccaag tcacttcctc cctcttccct 10500
aggaagctgc aggggtatca ggtgcagtaa tgctcatggc tacaaggcag aagcagagac 10560
ttattcatga ggctcctggg ggtgagcagg tatcccagta gttctacaga gttttcccct 10620
gcacacttag atgagatata tatgtttgag aggaagtaag ttgatcaacc ctccaactac 10680
ccattttttt ctcctatctt ataaatacat gttgcagggc agcctatctt tggggctgta 10740
gaccttgctc acgtgtctga gccagggctt gtcatgaact ccgggtgata cccatctttt 10800
caccctttca tcaatgattt cttggagaaa actcagaggc tgcatttcct tctacaaaag 10860
taagagacgc gaaagcactg ttgatgaacc cggagtgtgt ggcagactct tcgacgttca 10920
taccgacagt ctactggtct cactgctgag aacccatcag cctgattgct tgaagaaatc 10980
ccaatagtca ggaaagaaag atactaaagg ttggaggggg acagccactt cggcctcaga 11040
cggggtcaga gataatgttc ccaagagatg gaatttttaa aaattctgga gttcacaggt 11100
acccaggtag aattctcagg tctaataata ttggagaaac aacaacacaa tattaacaga 11160
aagctccaaa ctttcgagca acacccaaaa ggaaaaaaaa aactcatttg taaatagttt 11220
aaccaaaact caaacaaaac atgattagaa gaaataaaaa caatcattta tagctaaaag 11280
gctaatatag acagtattta cttctaactt tcaaacaaac aaacaaacaa aacaaaacaa 11340
gaagttagaa caactgtgaa aaacatgagc caagatataa agaaatattg actcctagaa 11400
agagacccag agtcacactg ggaaccaaat acattcacgt ggaaatgata ttttgaactt 11460
caaaaaaaga aaagaaagga ttgagcctgt cagagacact ccttgctgct tctgactttt 11520
cttgtccagg cacatttata gacatccctt ttgacaatat ttaacctcaa cctcagctaa 11580
cacattcaga ctcaattcct gccacaccca gtgactgtta cctaactcca gttgtctggg 11640
taaccactta ggttgggagc ctgaggaggt ttttagtcct ctgtcagtat tttctagacc 11700
aggcagtggc cacagcagag ggagttagct ccacccagga tgttatctct gacctggggc 11760
atgaggctag tggaaagacc tgtggtcccc agggtcctcc tcttgctatt tgcactctgg 11820
ctgctcctcc tggttccagt ctggtgttct caaggccatc ccacttggcg ttacatctca 11880
tcggaggtgg ttattcctcg gaaggagatc taccatacca aaggacttca agcacaaaga 11940
ctgctctcgt atagcttgcg ttttcggggc cagagacata tcatccacct gcggagaaag 12000
acactcattt ggcccagaca cttgttgctg acaactcaag atgaccaagg agccttacag 12060
atggagtacc ccttttttcc tgtagattgt tactatattg gctacctgga ggggatcctg 12120
caatccatgg tcactgtgga tacttgttat gggggcctgt caggggtcat aaagttggat 12180
aaccttacct atgaaatcaa acccctcaat gattcacaga gctttgaaca ccttgtttct 12240
cagatagtat ctgagtctga tgacacaggg cctatgaatg catggaagca ctggagccat 12300
aatacaggtt ctccctcctc cagattggaa tatgcagatg gagctcccag actatctagt 12360
aagaattacg ctacacatcc agctgctata aaaggccact ttcaagcaac ccattctgta 12420
tatagtgctt ctggaggtga caaactttca tctactgttg agtatttgtt taaagtcatt 12480
agtttaatgg acacctatct gaccaatctt catatgcggt actatgtctt tctcatgact 12540
gtgtataccg aggctgatcc attttcacaa gattttcgag ttccaggagg gcaggctcat 12600
actttctatg agagagtatt ttatgctcat tttaggcctg atgcaggagc tataattaac 12660
aagaattcgc caggagatga tgctgttaat ccagctgaga ggagtatatg ttctccctca 12720
gccctaattt gtcttggtca acatggtcga aatcctttat ttttatctat tataataacc 12780
aatcgtgttg gaaggagttt aggcctaaaa catgatgagg ggtactgtat ctgccagaga 12840
aggaacacct gcatcatgtt caaaaatcct caattaacag atgctttcag caattgttcc 12900
cttgcagaga taagcaacat acttaatact cctgatctga tgccatgtct tttctatgac 12960
cgtcatgttt attataatac atcattgact tataagtttt gtggaaactt caaagtagat 13020
aacaatgagc agtgtgactg tggctcccaa aaggcatgtt attcagatcc ctgctgtgga 13080
aatgattgca ggttaacacc tggtagcatt tgtgataaag aattatgctg tgcaaattgc 13140
acttacagtc cttctgggac actctgcaga cctatccaga acatatgtga tcttccagag 13200
tactgtagtg gctctaagtt catttgccca gatgacactt atctgcaaga tgggacacca 13260
tgctcagaag agggttactg ctataaaggt aactgcactg atcgcaacat acaatgcatg 13320
gaaatctttg gtgtaagtgc taagaatgct aatattaagt gctatgacat caacaaacaa 13380
cggtttcgat ttgggcattg tactagagca gaagaaagcc tcacattcaa tgcttgtgct 13440
gatcaggaca agctgtgtgg aaggttgcag tgtaccaatg tcaccaatct tccatttttg 13500
caagaacatg tttcattcca tcaatcggtt atctctgggg ttacctgctt tgggcttgat 13560
gaacatcgtg ggacagaaac agcagatgct ggattggtga gacatggtac cccgtgttca 13620
aggggtaagt tctgtgatcg aggagcttgc aatggaagtt tatctcgttt gggttatgac 13680
tgcaccccag aaaaatgcaa tttcagagga gtgtgtaaca atcgtcggaa ttgccattgc 13740
cattttggtt ggagccctcc aaagtgcaaa gaagagggac acagtgggag catagacagt 13800
gggtcccctc cggttcaaag gcgcataata aaacagaacc tagagccagt agtgtattta 13860
agaatactct ttggtcgtat ttacttcctc tttgttgcac tgctctttgg cattgccact 13920
cgtgtaggag ttactaagat atttaggttt gaagacttgc aagctgcttt acgttcttgg 13980
caagaacaag caaaggacaa gtaaaatgag ttagccccct gtctcctcta agataaagag 14040
aatggaaaaa aaatctgaat gtcattaggg gacaaggtag aactccagtc cacagttctg 14100
gatggccttt gtttatatca tgggacataa gacgtgttgg actcggcacc acctgatgtt 14160
gttgcattgc tgaaataaat ctttcaaaat tgcaaactgg tacttttgat tctatatttt 14220
aaataaatca ttagaaactg ctctacgttt gctcctcggg tagcaggaga ccttgagcca 14280
ccattggttc ttagccttag gcctctggtc tccgtattaa tggagatttg tctatatctc 14340
ttttaagaag ttatttacga tatcttttta aaatatggtt cctgaaccct gatgcgttcc 14400
cacctctttg tcactgtgtt ttttgatggt tcccatctcc tatagtttgt gatgggattt 14460
tcaggatcat tccttagagg aatagatgga gtgaagaaag acttaggacc cagttgagga 14520
agcgttacag gaacatagaa tcatcacctc attattaagc cagagaactg atatgtagaa 14580
catcaccatt aacacaaagc catggcttac tagatatact actcacaagc tcatggaagc 14640
atgctctcca gatttccact gacagggcac tgcacagaac acagtcttga gaaaggacat 14700
ggtgattctc gtggaggcag accctgagtg cactgaacag agaatctgtc atccctcgct 14760
accttaggac cgttatagtt a 14781
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ, КОТОРЫЕ ЭКСПРЕССИРУЮТ ТЯЖЕЛЫЕ ЦЕПИ, СОДЕРЖАЩИЕ ДОМЕНЫ VL | 2013 |
|
RU2664185C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664181C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2761639C2 |
| ГУМАНИЗИРОВАННЫЕ НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕКУ ЖИВОТНЫЕ С ОГРАНИЧЕННЫМИ ЛОКУСАМИ ТЯЖЕЛОЙ ЦЕПИ ИММУНОГЛОБУЛИНА | 2013 |
|
RU2656155C2 |
| МЫШИ ADAM6 | 2012 |
|
RU2582261C2 |
| МЫШИ ADAM6 | 2012 |
|
RU2722373C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664232C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2787781C2 |
| СПОСОБЫ ПРЕОДОЛЕНИЯ ИММУНОЛОГИЧЕСКОЙ ТОЛЕРАНТНОСТИ С ИСПОЛЬЗОВАНИЕМ МНОЖЕСТВА НАПРАВЛЯЮЩИХ РНК | 2017 |
|
RU2745563C2 |
| ЖИВОТНЫЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ, ИМЕЮЩИЕ СКОНСТРУИРОВАННУЮ ЛЕГКУЮ ЦЕПЬ ЛЯМБДА ИММУНОГЛОБУЛИНА, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2778410C2 |
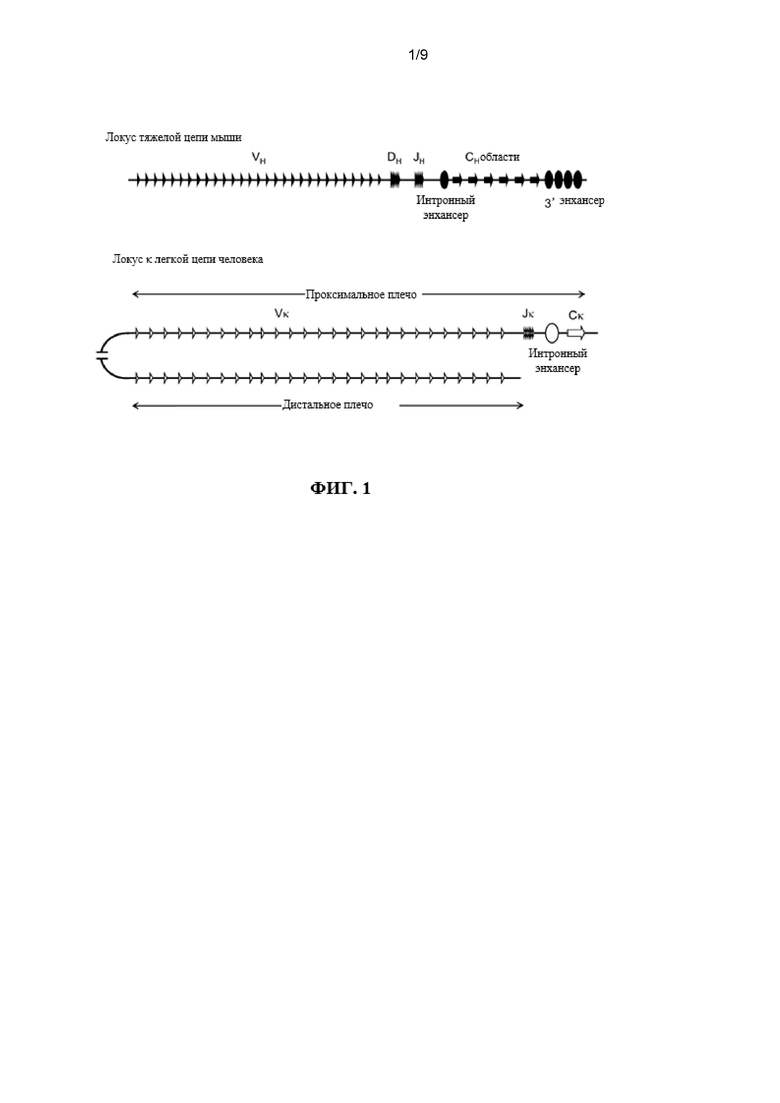

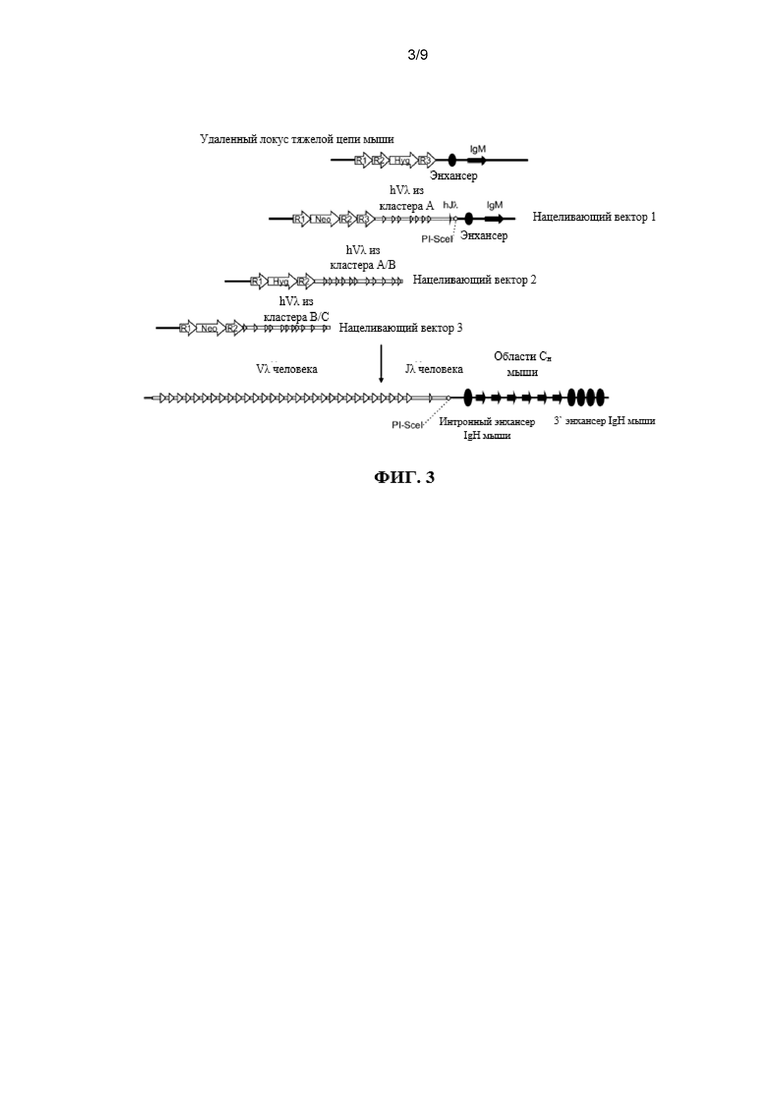
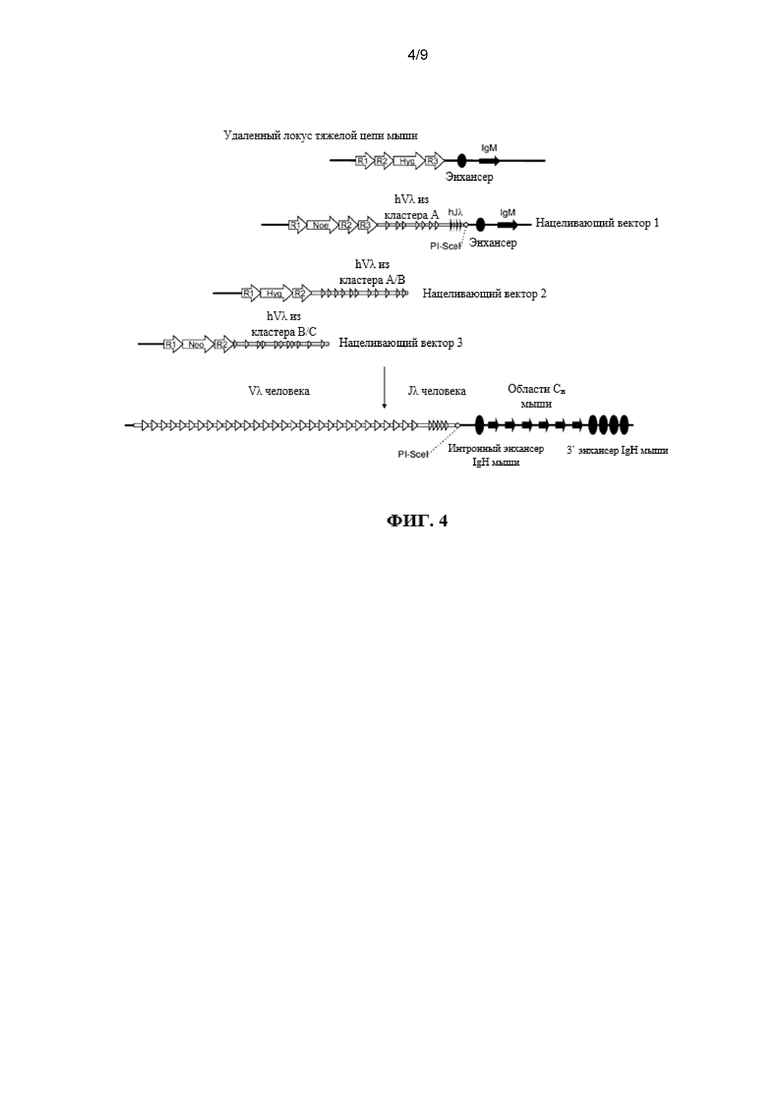
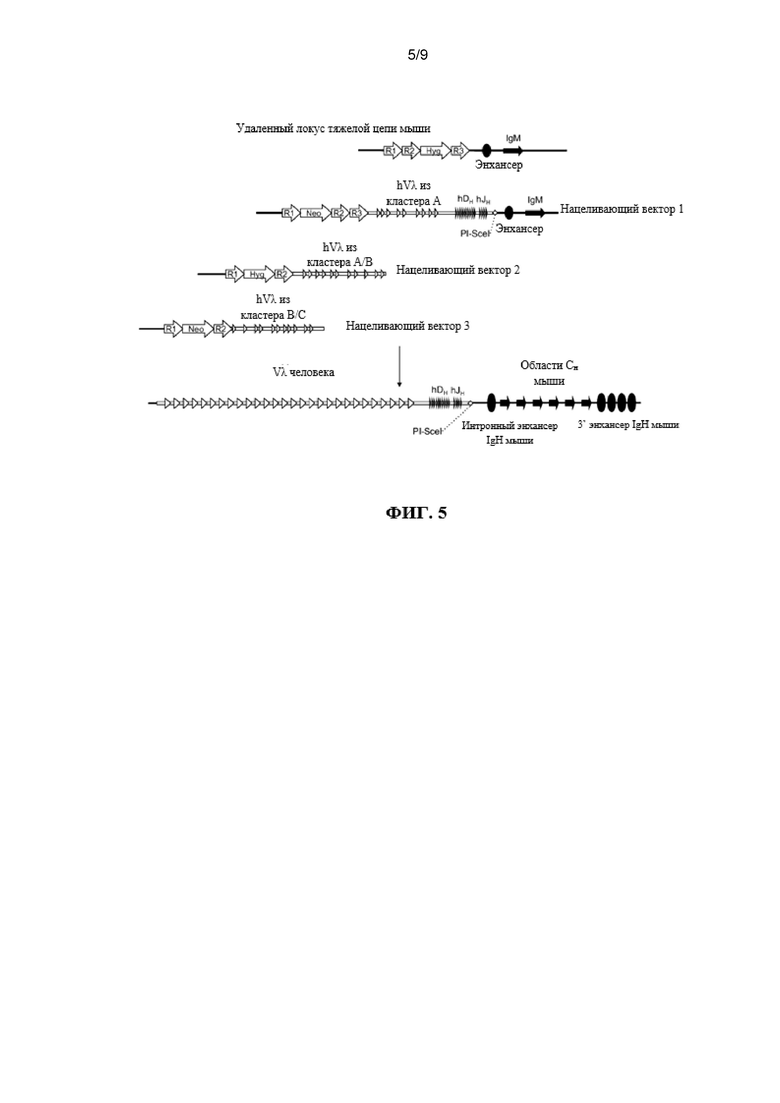
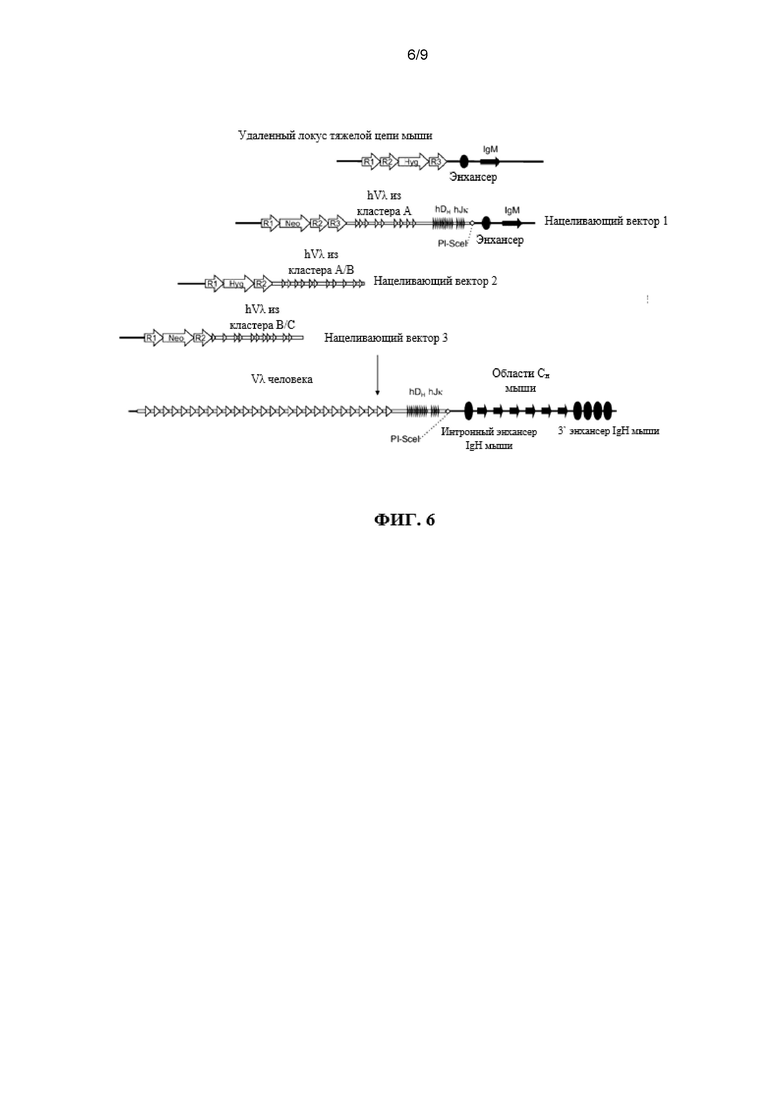


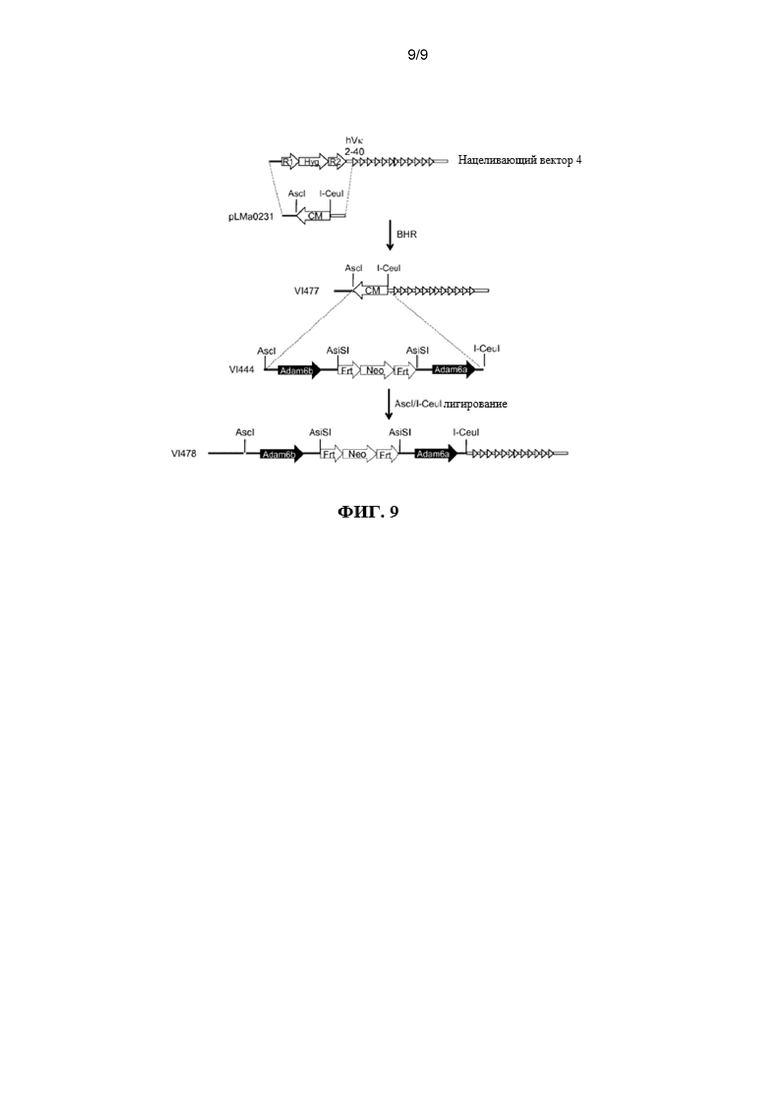
Изобретение относится к области биотехнологии, в частности к способу получения антитела, которое связывается с представляющим интерес антигеном. Способ включает получение клетки млекопитающего, сконструированной для экспрессии антитела, которое является специфическим в отношении представляющего интерес антигена, вставку одного или нескольких генных сегментов VL человека и одного или нескольких генных сегментов JL человека и вставку одного или нескольких генных сегментов VL человека и одного или нескольких генных сегментов JL человека и эктопическую нуклеотидную последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент. Изобретение эффективно для получения антитела, применимого для получения терапевтических средств для людей. 18 з.п. ф-лы, 9 ил., 1 табл., 4 пр.
1. Способ получения антитела, которое связывается с представляющим интерес антигеном, причем способ предусматривает:
(а) получение клетки млекопитающего, сконструированной для экспрессии антитела, которое является специфическим в отношении представляющего интерес антигена, причем антитело содержит две тяжелые цепи иммуноглобулина и две легкие цепи иммуноглобулина, причем каждая из тяжелых цепей иммуноглобулина включает вариабельный домен легкой цепи иммуноглобулина человека и константный домен тяжелой цепи иммуноглобулина, причем каждый из вариабельных доменов легкой цепи иммуноглобулина человека в пределах тяжелых цепей иммуноглобулина содержит аминокислотную последовательность, идентичную аминокислотной последовательности вариабельного домена легкой цепи иммуноглобулина человека тяжелой цепи иммуноглобулина антитела, экспрессированного в В-клетке генетически модифицированной мыши, которая содержит в своем геноме:
(i) вставку одного или нескольких генных сегментов VL человека и одного или нескольких генных сегментов JL человека выше мышиной константной области легкой цепи иммуноглобулина,
(ii) вставку одного или нескольких генных сегментов VL человека и одного или нескольких генных сегментов JL человека выше мышиной константной области тяжелой цепи иммуноглобулина и
(iii) эктопическую нуклеотидную последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент, причем белок ADAM6 или его функциональный фрагмент экспрессируется из эктопической последовательности нуклеиновой кислоты ADAM6.
2. Способ по п. 1, при котором константный домен тяжелой цепи иммуноглобулина антитела согласно (а) представляет собой константный домен тяжелой цепи иммуноглобулина человека.
3. Способ по п. 1, при котором константный домен тяжелой цепи иммуноглобулина антитела согласно (а) представляет собой константный домен тяжелой цепи IgG1, IgG2, IgG3 или IgG4 человека.
4. Способ по п. 1, при котором константный домен тяжелой цепи иммуноглобулина человека согласно (а) включает CH1, шарнир, CH2, CH3, CH4 или их комбинацию.
5. Способ по п. 1, при котором один или несколько генных сегментов VL человека и один или несколько генных сегментов JL человека согласно (ii) представляют собой один или несколько генных сегментов Vλ человека и один или несколько генных сегментов Jλ человека.
6. Способ по п. 1, при котором один или несколько генных сегментов VL человека и один или несколько генных сегментов JL человека согласно (ii) представляют собой один или несколько генных сегментов Vκ человека и один или несколько генных сегментов Jκ человека.
7. Способ по п. 1, при котором каждая из иммуноглобулиновых легких цепей антитела согласно (а) включает вариабельный домен легкой цепи иммуноглобулина человека и константный домен легкой цепи иммуноглобулина.
8. Способ по п. 7, при котором константный домен легкой цепи иммуноглобулина представляет собой константный домен легкой цепи иммуноглобулина человека.
9. Способ по п. 1, при котором один или несколько генных сегментов VL человека и один или несколько генных сегментов JL человека согласно (i) представляют собой один или несколько генных сегментов Vλ человека и один или несколько генных сегментов Jλ человека.
10. Способ по п. 1, при котором один или несколько генных сегментов VL человека и один или несколько генных сегментов JL человека согласно (i) представляют собой один или несколько генных сегментов Vκ человека и один или несколько генных сегментов Jκ человека.
11. Способ по п. 1, при котором мышиная константная область легкой цепи иммуноглобулина согласно (i) представляет собой Сκ мыши.
12. Способ по п. 1, при котором эктопическая нуклеотидная последовательность, которая кодирует белок ADAM6 или его функциональный фрагмент согласно (iii), присутствует между генными сегментами иммуноглобулина.
13. Способ по п. 1, при котором генетически модифицированная мышь содержит один или несколько эндогенных генных сегментов VL иммуноглобулина и/или один или несколько эндогенных генных сегментов JL иммуноглобулина, которые являются неспособными к реаранжировке для образования домена VL иммуноглобулина у генетически модифицированной мыши.
14. Способ по п. 1, при котором генетически модифицированная мышь содержит делецию или замещение одного или нескольких эндогенных генных сегментов тяжелой цепи иммуноглобулина.
15. Способ по п. 1, при котором генетически модифицированная мышь содержит один или несколько эндогенных генных сегментов VH, один или несколько эндогенных генных сегментов DH и один или несколько эндогенных генных сегментов JH, причем эндогенные генные сегменты VH, генные сегменты DH и генные сегменты JH являются неспособными к реаранжировке для образования реаранжированного домена VH иммуноглобулина у генетически модифицированной мыши.
16. Способ по п. 1, при котором генетически модифицированная мышь содержит делецию эндогенного гена ADAM6.
17. Способ по п. 1, при котором клетку млекопитающего выбирают из группы, состоящей из клетки HeLa, клетки DU145, клетки Lncap, клетки MCF-7, клетки MDA-MB-438, клетки РС3, клетки T47D, клетки ТНР-1, клетки U87, клетки SHSY5Y, клетки Saos-2, клетки Vero, клетки СНО, клетки GH3, клетки PC12, ретинальной клетки человека и клетки МС3Т3.
18. Способ по п. 1, причем способ дополнительно предусматривает:
(a) воздействие на генетически модифицированную мышь представляющим интерес антигеном;
(b) выделение по меньшей мере одного В-лимфоцита генетически модифицированной мыши, причем по меньшей мере один В-лимфоцит экспрессирует антитело, которое связывается с представляющим интерес антигеном, причем экспрессированное антитело включает вариабельный домен легкой цепи иммуноглобулина человека, функционально связанный с мышиным константным доменом тяжелой цепи иммуноглобулина;
(c) идентификацию последовательности нуклеиновой кислоты, которая кодирует вариабельный домен легкой цепи иммуноглобулина человека;
(d) слияние последовательности нуклеиновой кислоты, которая кодирует вариабельный домен легкой цепи иммуноглобулина человека, с последовательностью нуклеиновой кислоты, которая кодирует константный домен тяжелой цепи иммуноглобулина человека; и
(e) введение слитой нуклеиновой кислоты в клетку млекопитающего, тем самым создавая клетку млекопитающего, сконструированную для экспрессии антитела, которое является специфическим в отношении представляющего интерес антигена.
19. Способ по п. 1 или 18, причем способ дополнительно предусматривает выделение экспрессированных тяжелых цепей иммуноглобулина и легких цепей иммуноглобулина из клетки млекопитающего.
| WO 2011158009 A1, 22.12.2011 | |||
| WO 2009143472 A2, 26.11.2009 | |||
| HAN C | |||
| et al., Comprehensive analysis of reproductive ADAMs: relationship of ADAM4 and ADAM6 with an ADAM complex required for fertilization in mice, Biol Reprod., 2009, Vol.80(5), pp.1001-1008 | |||
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ МЫШЕЙ | 2009 |
|
RU2425880C2 |

