Область техники
Настоящее изобретение относится к эпитопу, специфичному для вируса гепатита B (здесь и далее именуемого «ВГВ»), и его применению. Поскольку описанный в настоящем изобретении эпитоп представляет собой консервативное положение, в котором вследствие мутации («мутагенеза») не возникают модификации, композиция, содержащая антитела к описанному эпитопу, или композиция вакцины, имеющая в своем составе описанный выше эпитоп, обладают очень низкой вероятностью вызывать снижение терапевтической эффективности вследствие мутаций ВГВ, что делает их очень эффективными в лечении гепатита В.
Настоящее изобретение также относится к способу получения антиген-специфичного антитела к описанному выше эпитопу, и такое антиген-специфичное антитело к эпитопу, полученное в соответствии с настоящим изобретением, проявляет высокую специфичность при введении in vivo.
Предпосылки создания изобретения
Вирус гепатита В представляет собой вирус, обладающий ДНКгеномом, и принадлежит к семейству гепаднавирусов и является причиной развития острого и/или хронического гепатита. Как правило, выделяют восемь генотипов ВГВ, которые различаются между собой не менее чем на 8% по последовательности генов, или девять серологических типов (adw, adr, ayw, ayr и проч.) на основании двух антигенных детерминант (другими словами, эпитопов) (d/y, w/r) поверхностного антигена ВГВ (HBsAg). Во всем мире 350 миллионов человек хронически инфицированы вирусом гепатита В, в частности, от 5 до 8% населения Кореи и Китая хронически инфицировано ВГВ. Заражение вирусом гепатита В является основной причиной заболеваний печени и рака печени в этих регионах. В настоящее время, несмотря на разработку вакцин, которые могут в какой-то степени защитить от вышеописанной инфекции, множество пациентов по-прежнему страдает от хронического гепатита, вызванного ВГВ. Хроническая инфекция, вызванная вирусом гепатита В, может стать причиной развития гепатита, а также цирроза печени и рака печени, вероятность возникновения которого среди хронически инфицированных людей в 300 раз выше по сравнению с неинфицированными людьми. В соответствии с исследованием ВОЗ, хронический гепатит В считается основной причиной развития примерно 80% случаев рака печени.
Препараты для лечения хронического гепатита В, недавно разработанные на основе аналогов нуклеозидов и выпущенные на рынок, включают, например, ламивудин, адефовир дипивоксил и другие. Данные препараты могут взаимодействовать с обратной транскриптазой полимеразы ВГВ, в свою очередь, ингибируя репликацию ДНК вируса гепатита В. Однако в данном случае, когда любой из описанных выше препаратов назначают в течение такого длительного периода, как 3 года, у 75% пациентов появляются устойчивые к препаратам вирусы, что влечет за собой проблему ухудшения терапевтической эффективности. Для предотвращения вертикальной передачи или инфицирования при пересадке печени указанные препараты широко используют в сочетании с иммуноглобулинами против гепатита В (HBIG).
В настоящее время иммуноглобулины против гепатита В производят с использованием ионно-обменной очистки и инактивации вируса в плазме доноров с высоким титром антител к поверхностному антигену ВГВ (HBsAg).
Однако доступные в настоящее время иммуноглобулины против гепатита В не могут считаться идеальным источником получения терапевтических антител вследствие их ограниченной доступности, низкой специфичности и возможного загрязнения инфекционными агентами.
Известно, что антитела, вырабатываемые in vivo вследствие введения вакцин, используемых в настоящее время в области техники, представляют собой антитела к эпитопу ВГВ «а». Однако недавно был представлен устойчивый к действию таких антител мутант G145R, получающийся в результате замены в поверхностном антигене ВГВ остатка глицина в положении 145 на аргинин. В дополнение к тому были обнаружены и другие устойчивые к антителам мутанты; таким образом, у существующих лекарственных средств против ВГВ имеются ограничения в реализации достаточной терапевтической эффективности. Соответственно, существует возрастающая потребность в получении антител для лечения ВГВ и/или вакцин, специфично связанных с эпитопами, соответствующими сайтам, связанными с репликацией ВГВ и необходимыми для выживания ВГВ, и которые не вызывают мутации, а значит не вызывают снижение терапевтической эффективности в результате возникновения мутаций.
Раскрытие сущности изобретения
Техническая проблема
Для решения вышеизложенных проблем настоящее изобретение представляет эпитопы, специфичные для ВГВ, содержащие последовательности RFLWE (номер последовательности 4) или KFLWE (номер последовательности 5), и, в частности, эпитоп, содержащий такие аминокислотные последовательности, как FARFLWEWASVRFSW (номер последовательности 6) или FGKFLWEWASARFSW (номер последовательности 7), которые представляют собой необходимый для выживания вируса сайт, соответствуя, таким образом, области, в которой не возникают мутации.
Другой целью настоящего изобретения является представление способов производства описанного выше эпитопа, композиции вакцины против ВГВ или вакцины, содержащей эпитоп и антитело, способное специфически связываться с эпитопом при добавлении описанного выше эпитопа, а также композиции препарата для лечения ВГВ или лекарственного препарата, содержащего антитело, произведенное, как описано выше.
Другой целью настоящего изобретения является разработка композиции или набора реактивов для обнаружения ВГВ, содержащей описанный выше эпитоп или полинуклеотидную последовательность, кодирующую данный эпитоп.
Техническое решение
Авторами настоящего изобретения было обнаружено, что эпитопы для человеческого антитела, специфично связывающего поверхностный антиген ВГВ, узнаваемые человеческим антителом (см. PCT/KR2010/004445, здесь и далее называемого «патентоспособным антителом»), соответствуют последовательностям, содержащим RFLWE (последовательности номер 4) или KFLWE (последовательности номер 5) и, в частности, последовательностями, полученным из FARFLWEWASVRFSW (последовательности номер 6) или FGKFLWEWASARFSW (последовательности номер 7) или их частям; и такие эпитопные сайты выгодно консервативны, важны для репликации ВГВ и необходимы для выживания ВГВ. Таким образом, настоящее изобретение было реализовано с учетом описанных выше открытий. Среди вышеописанных эпитопов эпитопы, имеющие номера последовательности 4 и 6, относятся к эпитопам подтипа adr (последовательность номер 1) ВГВ, в то время как эпитопы с последовательностями номер 5 и 7 соответствуют эпитопам подтипа ayw (последовательность номер 2) вируса гепатита В.
Эпитоп, специфичный для ВГВ, определенный любой из последовательностей с номерами от 4 до 7, в соответствии с настоящим изобретением может сохранять трехмерную структуру или может быть использован в виде конъюгата с носителем с целью увеличения эффективности при применении в составе композиции, такой как вакцина. Носитель, применяемый в настоящем изобретении, может представлять собой любое биологически доступное вещество, которое обеспечивает эффекты, предусмотренные настоящим изобретением, и может быть выбран из пептидов, сывороточного альбумина, иммуноглобулинов, гемоцианина, полисахаридов и т.п., без особого ограничения ими.
Эпитоп, специфичный для ВГВ, определенный любой из последовательностей с номерами от 4 до 7, в индивидуальном виде или в виде композиции, конъюгированный с носителем, может применяться в композиции вакцины для лечения ВГВ. В этом отношении композиция вакцины может также включать фармацевтически приемлемый адъювант или вспомогательное вещество. Такой адъювант способствует выработке антитела при введении его in vivo и может включать любое вещество, которое соответствует целям настоящего изобретения, конкретнее, по меньшей мере, одну из солей алюминия (Al(OH)3, AlPO4), сквален, сорбитан, полисорбат 80, CpG, липосомы, холестерин, монофосфорил липида А (MPL) и глюкопиранозил липида (GLA), без особого ограничения ими.
Полинуклеотид, кодирующий эпитоп, специфичный для ВГВ, определенный любой из последовательностей с номерами от 4 до 7 и представленный в настоящем изобретении, может быть использован в качестве ДНК-вакцины. В данном случае полинуклеотид может использоваться в виде индивидуального компонента без вектора или же в составе вирусного или невирусного вектора. Вирусный и невирусный векторы, применяемые в настоящем изобретении, могут включать любой вектор, доступный в области техники, к которой относится настоящее изобретение. В качестве вирусных векторов предпочтительно применять аденовирусы, аденоассоциированные вирусы, лентивирусы, ретровирусы и т.п.), в то время как в качестве невирусных векторов можно применять катионные полимеры, неионные полимеры, липосомы, липиды, фосфолипиды, гидрофильные полимеры, гидрофобные полимеры и комбинации с, по меньшей мере, одним из описанных выше компонентов, без особого ограничения ими.
Настоящее изобретение представляет рекомбинантный вектор, содержащий полинуклеотид, кодирующий эпитоп, специфичный для ВГВ, определенный любой из последовательностей с номерами от 4 до 7, в соответствии с настоящим изобретением, хозяйскую клетку с рекомбинантным вектором и способ получения эпитопа, специфичного для ВГВ, определенного любой из последовательностей с номерами от 4 до 7, в соответствии с настоящим изобретением с применением рекомбинантного вектора или хозяйской клетки, описанных выше.
В настоящем изобретении «рекомбинантным вектором» называется экспрессионный вектор, представляющий целевой белок из соответствующей хозяйской клетки, который является генным продуктом, содержащим необходимые регуляторные элементы, функционально связанные с генной вставкой для обеспечения ее экспрессии. В настоящем изобретении термин «функционально связанный» относится к регулирующей экспрессию нуклеотидной последовательности, функционально связанной с нуклеотидной последовательностью, кодирующей целевой белок, с целью обеспечения выполнения общих функций. Функциональная связь с рекомбинантным вектором может обеспечиваться за счет методов рекомбинации, широко известных в области техники, к которой относится настоящее изобретение. Также легко может быть применено сайт-специфичное разрезание и сшивание ДНК с помощью ферментов, широко известных в области техники, к которой относится настоящее изобретение.
Приемлемые экспрессионные векторы, применяемые в настоящем изобретении, могут включать сигнальные последовательности для встраивания в мембрану или секреции, а также такие регуляторные элементы, как промоторы, старт-кодоны, стоп-кодоны, сигналы полиаденилирования, энхансер и т.п. Старт- и стоп-кодоны, как правило, считаются частью нуклеотидной последовательности, кодирующей иммуногенный целевой белок, и при введении генного продукта должны проявлять действие в организме пациента, находясь в одной рамке считывания с кодирующей последовательностью. Главный промотор может быть структурным или индуцибельным. В прокариотических клетках можно применять, например, промоторы lac, tac, T3, Т7, без особого ограничения ими. В эукариотических клеток можно применять, например, промоторы вируса обезьян 40 (SV40), вируса рака молочной железы мышей (MMTV), вируса иммунодефицита человека (ВИЧ) и, в частности, промотор длинного концевого повтора (LTR) ВИЧ, вируса Молоуни, цитомегаловируса (ЦМВ), вируса Эпштейна-Барр, вируса саркомы Рауса, а также промоторы генов β-актина, человеческого гемоглобина, человеческого мышечного креатина, человеческого металлотионеина, без особого ограничения ими.
Экспрессионный вектор может содержать маркеры для селекции хозяйской клетки, содержащей вектор. Селекционный маркер служит для секции клеток, трансформированных в векторы, и могут включать маркеры, дающие такие селектируемые фенотипы, как лекарственная устойчивость, потребность в определенных питательных веществах, устойчивость к цитотоксическому действию, экспрессия поверхностных белков и т.п. Поскольку в селективных условиях выживают только клетки, экспрессирующие селективный маркер, можно скринировать трансформированные клетки. Для обеспечения репликации экспрессионный вектор может содержать точку начала репликации, специфическую нуклеотидную последовательность, с которой начинается репликация. Экспрессионный рекомбинантный вектор может быть представлен плазмидами, вирусами, космидами и т.д. Выбор рекомбинантного вектора не ограничен, если он обеспечивает экспрессию целевого гена и синтез целевого белка прокариотическими или эукариотическими хозяйскими клетками, однако предпочтительно использовать вектор, который обеспечивает синтез чужеродного белка в большом количестве, сравнимом с уровнем естественных белков, и который содержит промотор с высокой активностью, обеспечивающий высокий уровень экспрессии.
В частности, для экспрессии эпитопов, специфичных для ВГВ, определенных любой из последовательностей с номерами от 4 до 7, может быть применен целый ряд комбинаций хозяин-вектор. Экспрессионный вектор для эукариотических клеток может содержать регуляторные элементы, полученные, например, из SV40, бычьего папилломавируса, аденовируса, аденоассоциированного вируса, цитомегаловируса, лентивируса и/или ретровируса, без особого ограничения ими. Экспрессионный вектор для бактериальных клеток может содержать, например, бактериальные плазмиды, полученные из Escherichia coli: pET, pRSET, pBluescript, pGEX2T, pUC вектор, col E1, pCR1, pBR322, pMB9, и их производные; такие плазмиды, как RP4 с широким спектром хозяев; фаговую ДНК, например, такие производные фага лямбда, как λgt10 и λgt11, NM980 и т.д.; другие ДНК-содержащие фаги, например, одноцепочечный филаментный ДНК-фаг, М13 и т.п. Для клеток насекомых можно применять вектор pVL941.
Рекомбинантный вектор вводят в хозяйскую клетку для получения трансформанта, и в качестве хозяйских клеток в настоящем изобретении можно применять, например, прокариотические клетки E. coli, Bacillus subtilis, Streptomyces sp., Pseudomonas sp., Proteus mirabilis или Staphylococcus sp.; грибы Aspergillus sp.; дрожжи Pichia pastoris, Saccharomyces cerevisiae, Schizosaccharomyces sp., Neurospora crassa и т.д.; эукариотические клетки, например, клетки низших эукариот, клетки высших эукариот, в т.ч. клетки насекомых и т.п. Предпочтительно применять хозяйские клетки, полученные из растений и/или млекопитающих, в частности, клетки почки обезьяны 7 (COS7), клетки NSO, SP2/O, клетки яичника китайского хомячка (CHO), W138, клетки почки новорожденного хомячка (BHK), MDCK, линии клеток миеломы, клетки HuT 78 и/или HEK293, без особого ограничения ими. Наиболее предпочтительно применять клетки CHO.
В настоящем изобретении термин «трансформация в хозяйскую клетку» подразумевает любой способ введения нуклеиновой кислоты в органические вещества, клетки, ткани и/или органы, и хорошо известный в области техники стандартный способ, который может быть надлежащим образом подобран для осуществления трансформации в соответствии с типом хозяйских клеток. К числу таких способов относятся электропорация, слияние протоплазм, осаждение фосфатом кальция (CaPO4), осаждение хлоридом кальция (CaCl2), трансформация с применением нитевидных кристаллов карбида кремния, агробактерий, ПЭГ, сульфата декстрана, липофектамина, трансформация способом высушивания/ингибирования, без особого ограничения ими. Инкубируя трансформанты, которые экспрессируют рекомбинантный вектор, в среде для культивирования, можно синтезировать большое количество эпитопа, специфичного для ВГВ, определенного любой из последовательностей с номерами от 4 до 7. Среда для культивирования и условия культивирования могут быть подобраны в соответствии с типом применяемых хозяйских клеток. Во время культивирования можно контролировать такие параметры, как температура, pH среды, время культивирования и т.п., для обеспечения надлежащего уровня роста клеток и синтеза белков. Как описано выше, эпитоп, специфичный для ВГВ, определенный любой из последовательностей с номерами от 4 до 7, можно получить непосредственно из среды культивирования или из лизата клеток с помощью рекомбинации и выделить любым стандартным биохимическим способом выделения (Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press (1989); Deuscher, M., Guide to Protein Purification Methods Enzymology, Vol. 182. Academic Press, Inc., San Diego, CA (1990)). Для этого можно применять разнообразные способы, в том числе электрофорез, центрифугирование, гель-фильтрацию, преципитацию, диализ, хроматографию (ионно-обменную, аффинную, иммуно-адсорбционную, эксклюзионную хроматографию и т.п.), изоэлектрическое фокусирование, различные варианты и комбинации описанных способов, без особого ограничения ими.
Настоящее изобретение представляет способ экспрессии эпитопа, специфичного для ВГВ, определенного любой из последовательностей с номерами с 4 по 7, на поверхности микроорганизмов или вирусов. В этом случае можно применять рекомбинантный вектор, содержащий последовательность, кодирующую индуцибельный промотор или сигнальный белок, а также различные микроорганизмы или вирусы, несущие описанный выше рекомбинантный вектор. В частности, рекомбинантные E. coli, дрожжи и/или бактериофаги представляют собой подходящие микроорганизмы и/или вирусы, без особого ограничения ими. Для экспрессии эпитопа, специфичного для ВГВ, определенного любой из последовательностей с номерами с 4 по 7, на поверхности описанных выше микроорганизмов или вирусов можно использовать методы дисплея, широко известные в области техники, к которой может быть применено настоящее изобретение. В частности, полинуклеотидную последовательность, кодирующую эпитоп, специфичный для ВГВ, определенный любой из последовательностей с номерами с 4 по 7, можно комбинировать (или соединять) с последовательностью, кодирующей промотор или сигнальный белок, который экспрессируется на поверхности микроорганизма и/или вируса, обеспечивая таким образом экспрессию эпитопа, специфичного для ВГВ. Кроме того, после удаления участков гена, кодирующих экспрессирующийся на поверхности белок, в делетированную область можно вставить полинуклеотидную последовательность, кодирующую эпитоп, специфичный для ВГВ, определенный любой из последовательностей с номерами с 4 по 7. Однако настоящее изобретение не ограничивается только вышеизложенными способами. В соответствии с описанными выше способами эпитоп, специфичный для ВГВ, определенный любой из последовательностей с номерами с 4 по 7, который экспрессируется на поверхности микроорганизма или вируса, может быть выделен и очищен для целевого применения в соответствии с целями настоящего изобретения. Кроме того, патентоспособный эпитоп может использоваться для скрининга антитела, специфически связанного с эпитопом, специфичным для ВГВ, определенным любой из последовательностей с номерами с 4 по 7, который экспрессируется на поверхности, и для дальнейшего получения прошедшего скрининг антитела.
Кроме того, настоящее изобретение представляет способ производства антитела, специфически связанного с эпитопом, специфичным для ВГВ, определенным любой из последовательностей с номерами с 4 по 7, или фрагментов антитела, который предполагает применение эпитопа, специфичного для ВГВ, определенного любой из последовательностей с номерами с 4 по 7, композиции, содержащей описанный выше эпитоп, или полинуклеотида, кодирующего описанный выше эпитоп. Такое антитело может быть поликлональным или моноклональным антителом, и поскольку его фрагменты характеризуются способностью связываться с эпитопом, специфичным для ВГВ, определенным любой из последовательностей с номерами с 4 по 7, они тоже включены в предмет настоящего изобретения. Конкретнее, патентоспособное антитело и его фрагменты могут включать, например, одноцепочечные антитела, димерные антитела, тримерные антитела, тетрамерные антитела, Fab-, F(ab)'2-фрагменты, Fd, scFv, доменные антитела, антитела с двойной специфичностью, миниантитела, scap антитела, IgD антитела, IgE антитела, IgM антитела, IgG1 антитела, IgG2 антитела, IgG3 антитела, IgG4 антитела, производные инвариантных областей антител, синтетические антитела на основе белковых остовов, каждое из которых способно связываться с эпитопом, специфичным для ВГВ, определенным любой из последовательностей с номерами с 4 по 7, без особого ограничения ими. При условии сохранения характеристик патентоспособного антитела, антитела с мутациями в вариабельных участках также могут быть включены в предмет настоящего изобретения. Это относится, например, к консервативным аминокислотным заменам в вариабельной области. В настоящем изобретении к такой «консервативной замене» обычно относится замена одного аминокислотного остатка на другой с сохранением свойств исходной аминокислотной последовательности. Например, лизин, аргинин и гистидин содержат основные боковые цепи и проявляют схожие свойства. Вместе с тем, аспарагиновая и глутаминовые кислоты содержат кислые боковые цепи и проявляют схожие друг с другом свойства. Кроме того, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин и триптофан похожи друг на друга, поскольку содержат незаряженные боковые цепи, в то время как аланин, валин, лейцин, треонин, изолейцин, пролин, фенилаланин и метионин похожи друг на друга по наличию неполярных боковых цепей. Далее, тирозин, фенилаланин, триптофан и гистидин похожи друг на друга, поскольку содержат ароматические боковые цепи. Следовательно, для специалистов в данной области техники будет очевидно, что даже если произойдет замена аминокислотных остатков внутри каждой из описанных выше групп с одинаковыми свойствами, это может не оказывать существенного влияния на характеристики. Таким образом, при сохранении специфических свойств патентоспособного антитела способ производства мутированных антител, содержащих консервативные замены в вариабельной области, может быть включен в предмет настоящего изобретения.
Антитело, связывающееся с эпитопом, специфичным для ВГВ, определенным любой из последовательностей с номерами с 4 по 7, может быть получено любым известным способом, известным в области техники, к которой относится настоящее изобретение. Конкретнее, после инокуляции животного эпитопом, специфичным для ВГВ, определенным любой из последовательностей с номерами с 4 по 7, композицией, включающей эпитоп, или полинуклеотидом, кодирующим описанный выше эпитоп, из инокулированного животного получают и скринируют антитело, специфично связывающееся с эпитопом, специфичным для ВГВ, определенным любой из последовательностей с номерами с 4 по 7, которое затем можно выделить.
Животное, примененное в настоящем изобретении, может включать трансгенное животное, в частности, трансгенную мышь, способную производить то же самое антитело как последовательность человеческого происхождения. Так называемое полностью человеческое антитело с пониженной иммуногенностью, получаемое с применением трансгенной мыши, может быть получено в соответствии с одним из способов, описанных в патентах США №№ 5569825, 5633425 и 7501552, и тому подобных. В том случае, если описанное выше животное не было трансформировано надлежащим образом для получения антитела с последовательностью, аналогичной человеческой, можно в дальнейшем провести гуманизацию или деиммунизацию, применяя полученное из животного антитело, в соответствии со способами, описанными в патентах США №№ 5225539, 5859205, 6632927, 5693762, 6054297, 6407213; и в выложенном письменном заключении на патент № 1998/52976, делая, таким образом, антитело пригодным для терапевтического применения in vivo. Конкретнее, такая гуманизация или деиммунизация может включать пересадку гипервариабельного участка, то есть перемещение гипервариабельного участка произведенного в животном антитела в каркасную область человеческого антитела, а затем проведение направленного мутагенеза гипервариабельной области для замены, добавления или удаления, по меньшей мере, одного аминокислотного остатка с целью увеличения аффинности или снижения иммуногенности.
Вместо эпитопа, специфичного для ВГВ, определенного любой из последовательностей с номерами от 4 до 7, композиции, содержащей эпитоп, и/или полинуклеотида, кодирующего эпитоп, если в качестве иммуногена использовался вирус гепатита В целиком, может быть применен способ преимущественного скрининга (часто «пэннинг) антител, имеющих способность связываться с эпитопом, специфичным для ВГВ (иногда сокращенно называемого «связыванием»), и последующий дополнительный пэннинг антител для специфичного узнавания эпитопа, специфичного для ВГВ, определенного любой из последовательностей с номерами от 4 до 7, из антител, прошедших первичный скрининг. Также можно использовать способ скрининга антител, которые не связываются или обладают пониженной способностью связываться с вирусом гепатита В, мутированным в важных положениях эпитопа, специфичного для ВГВ, определенного любой из последовательностей с номерами от 4 до 7, среди прошедших первичный скрининг антител, связывающихся с вирусом гепатита В, при этом способ включает получение мутаций в важных участках эпитопа, специфичного для ВГВ, определенного любой из последовательностей с номерами от 4 до 7.
В то же время, в соответствии со способами дисплея, хорошо известными в области техники, можно получить и скринировать человеческие антитела, связанные с эпитопом, специфичным для ВГВ, определенным любой из последовательностей с номерами от 4 до 7. Такие способы дисплея можно выбрать из фагового дисплея, бактериального дисплея или рибосомного дисплея, без особого ограничения ими. Производство и дисплей библиотек может быть легко осуществлен в соответствии с традиционными способами, описанными, например, в патентах США №№ 5733743, 7063943, 6172197, 6348315, 6589741 и т.п. В частности, библиотеки, применяемые в описанном выше дисплее, могут быть составлены так, чтобы содержать последовательности антител человеческого происхождения. Конкретнее, описанный выше способ может подразумевать скриниг (или пэннинг) антител, специфично связанных с эпитопом, специфичным для ВГВ, определенным любой из последовательностей с номерами от 4 до 7, при применении эпитопа ВГВ, определенного любой из последовательностей с номерами от 4 до 7, или композиции, содержащий данный эпитоп.
Наконец, настоящее изобретение представляет композицию или набор реактивов для обнаружения ВГВ, который содержит эпитоп, определенный любой из последовательностей с номерами от 4 до 7, композицию, включающую эпитоп или полинуклеотид, кодирующий эпитоп. Композиция или набор реактивов для определения ВГВ, в соответствии с настоящим изобретением, может иметь преимущества, позволяющие проводить быструю и точную диагностику заражения вирусом гепатита В, без значительного влияния мутаций ВГВ. Набор реактивов для выявления ВГВ, который включает эпитоп, определенный любой из последовательностей с номерами от 4 до 7, композицию, содержащую эпитоп или полинуклеотид, кодирующий эпитоп, может быть составлен с учетом возможности использования разнообразных способов, включая, например, общий твердофазный иммуноферментный анализ (ELISA), сортинг флуоресцентно-активированных клеток и т.п. Кроме того, при использовании полинуклеотида, кодирующего эпитоп, применяемый в настоящем изобретении, гибридизацию можно выявлять с помощью одного из гибридизационных способов.
Полезные эффекты изобретения
Как становится ясно из подробного описания, эпитоп, специфичный для ВГВ, представленный в настоящем изобретении, по существу является консервативным участком, не подверженным мутагенезу. Таким образом, довольно маловероятно, что терапевтический эффект композиции или композиции вакцины, включающей антитело против описанного выше эпитопа, будет снижаться вследствие мутаций ВГВ, что делает указанные препараты эффективными в лечении ВГВ и/или диагностике.
Краткое описание фигур
Вышеупомянутые и другие цели, особенности и преимущества настоящего изобретения станут очевидными из следующего описания предпочтительных вариантов осуществления, представленного параллельно с сопутствующими фигурами:
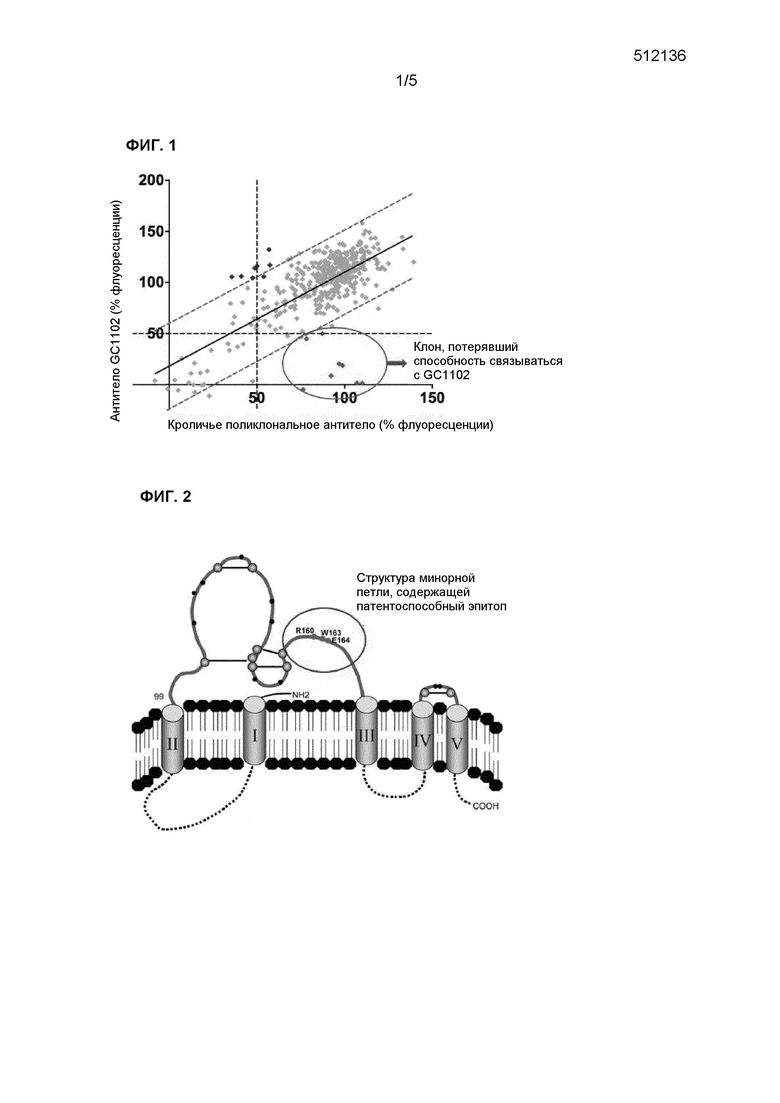
Фиг.1 иллюстрирует результаты анализа разной способности антитела связываться с мутантными белками поверхностного антигена ВГВ, проведенного с целью идентификации эпитопов для патентоспособного антитела;
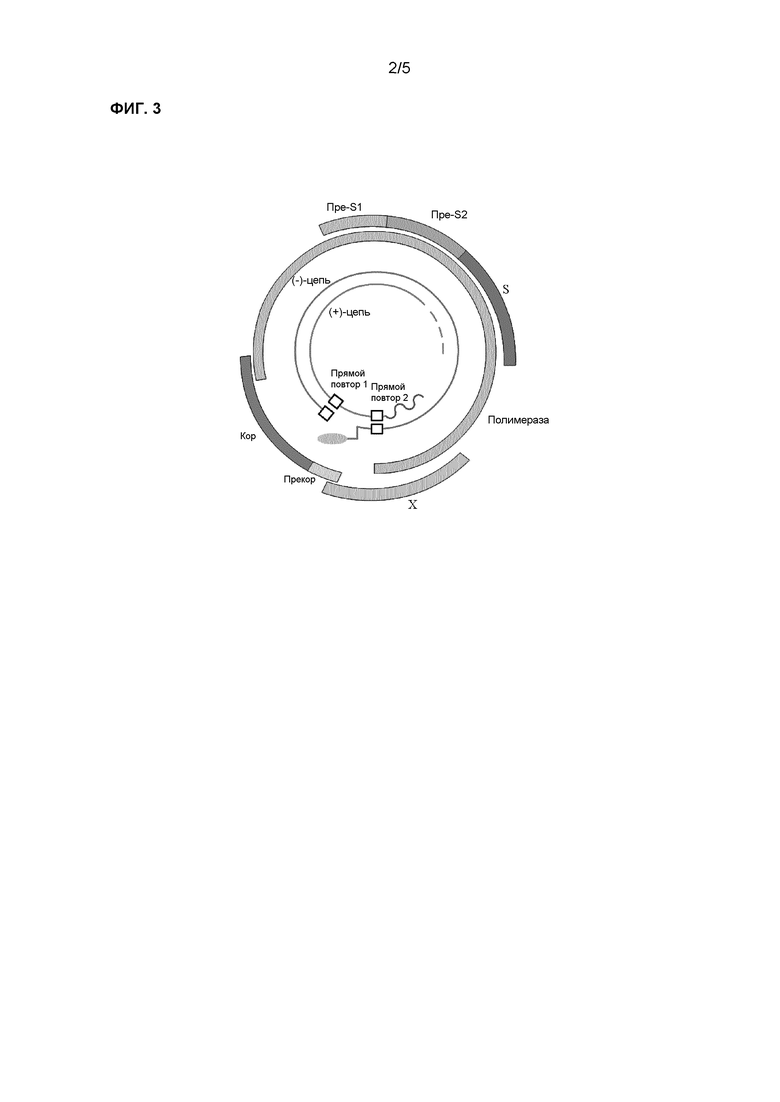
Фиг.2 показывает структуру петли в белке поверхностного антигена ВГВ, содержащей патентоспособный эпитоп;
Фиг.3 иллюстрирует геномную структуру ВГВ, в которой геном открытой рамки считывания S ORF, кодирующей белок поверхностного антигена, частично перекрывается с геномом P ORF, кодирующей полимеразу;
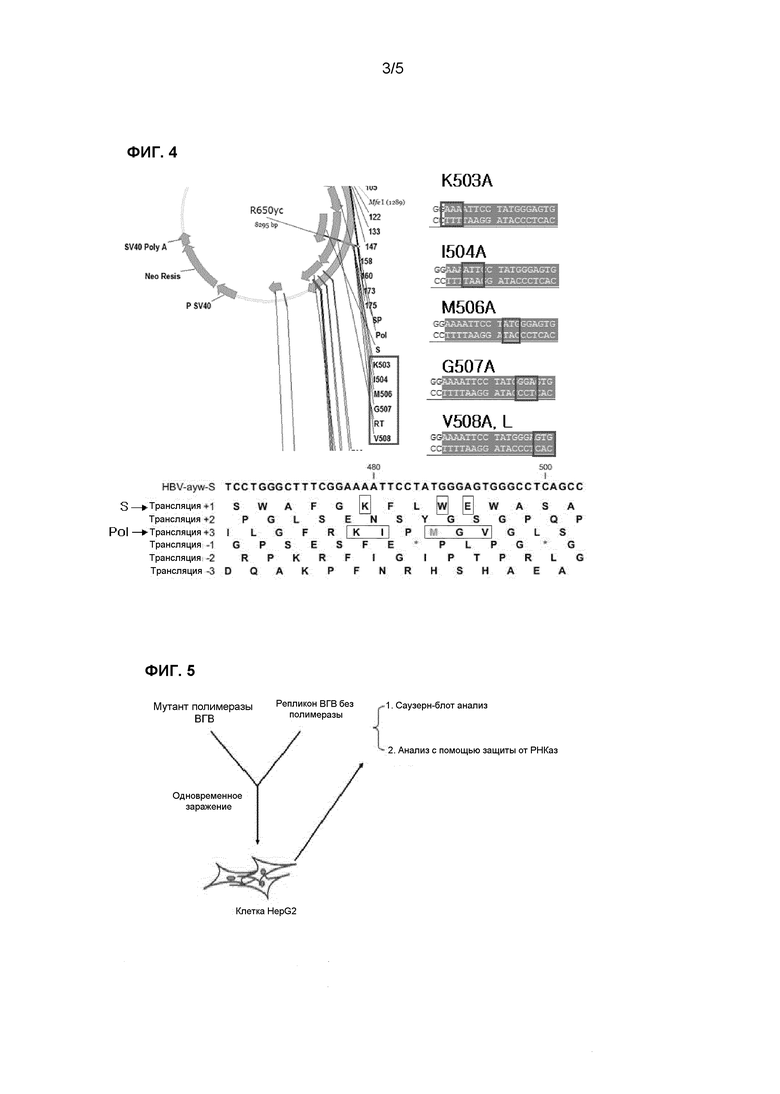
Фиг.4 иллюстрирует способ получения мутантов полимеразы ВГВ;
Фиг.5 иллюстрирует процесс комплементационного теста, проводимого путем одновременного заражения клетки HepG2 репликоном ВГВ без полимеразы и мутантом полимеразы ВГВ;
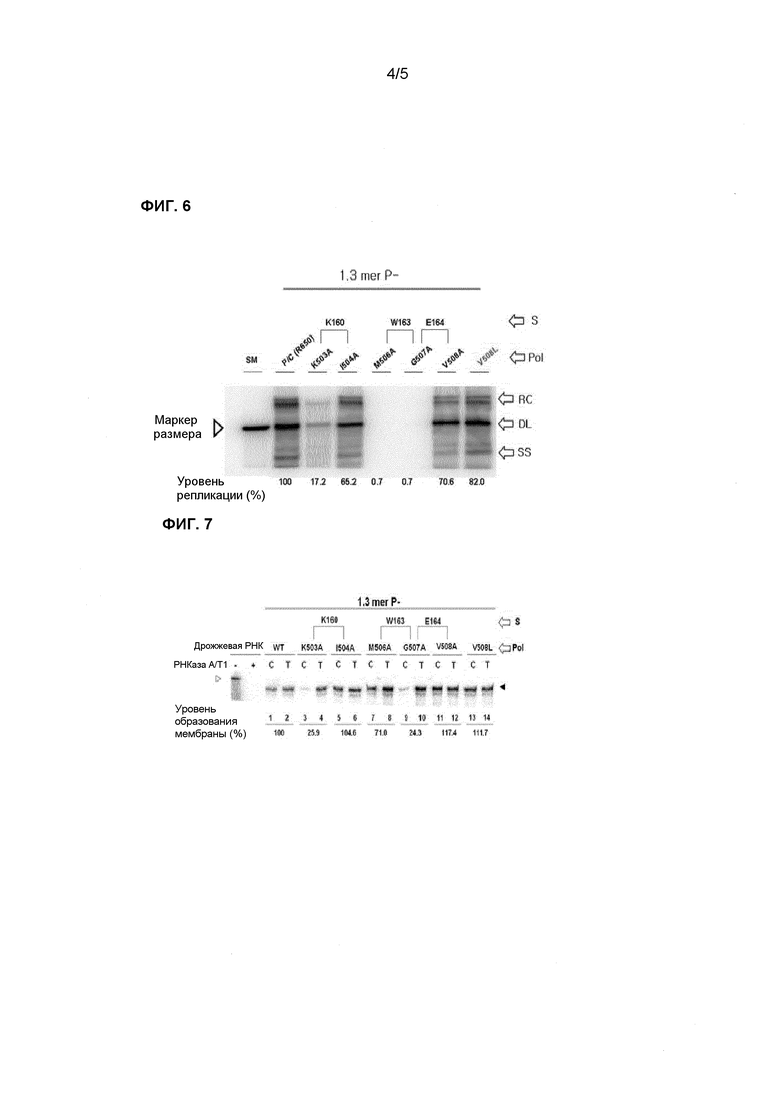
Фиг.6 демонстрирует результаты анализа репликативной способности ВГВ каждого из мутантов полимеразы ВГВ, полученные способом Саузерн-блота (сравнение репликативных интермедиатов ДНК ВГВ, включая релаксированную кольцевую (RC), дендримероподобную (DL), одноцепочечную (SS) ДНК справа на графике);
Фиг.7 демонстрирует результаты анализа влияния соответствующих мутантов полимеразы ВГВ на упаковку прегеномной ДНК, проведенного способом защиты от РНКаз; и
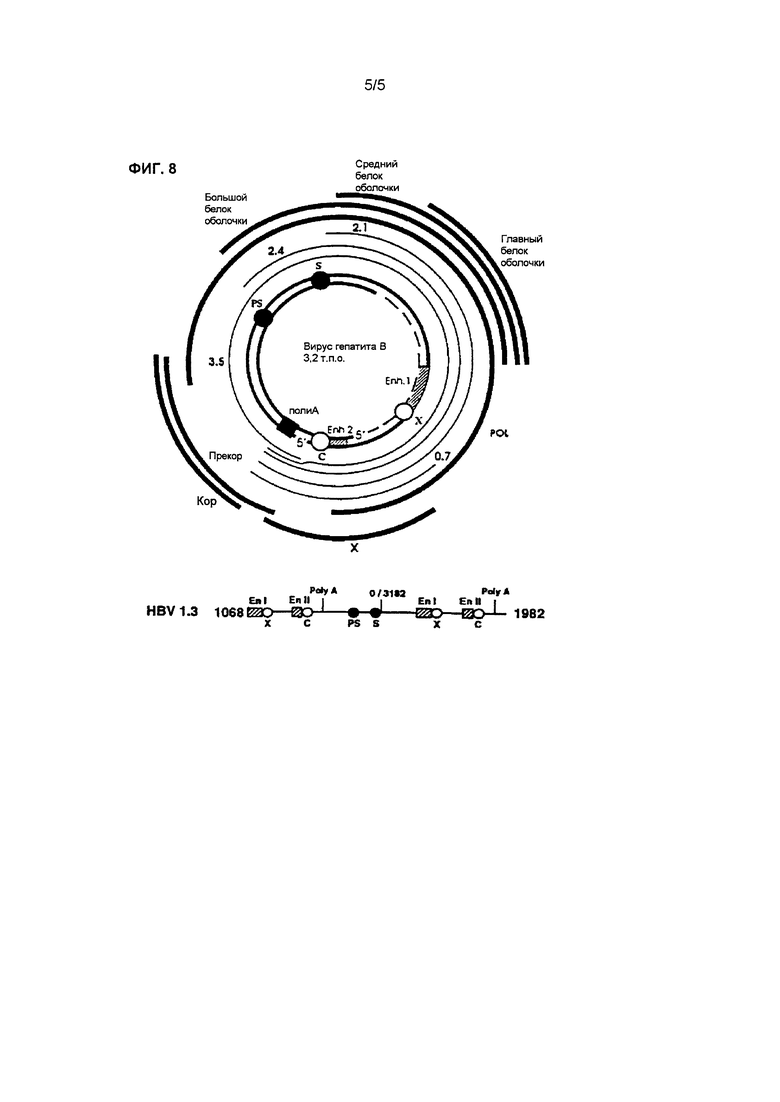
Фиг.8 показывает карту сцепления генов ВГВ в векторе, который применялся для гидродинамической инъекции с целью получения частиц ВГВ в мыши.
Наилучший режим работы
Здесь и далее подробно будут описаны предпочтительные варианты осуществления настоящего изобретения со ссылками на примеры, однако данные примеры приведены с демонстрационной целью, и не предполагаются лимитировать границы возможностей настоящего изобретения.
Пример 1. Идентификация эпитопа для патентоспособного антитела
Для идентификации эпитопа для патентоспособного антитела после проведения случайного мутагенеза в белке поверхностного антигена ВГВ подтипов adr (см. последовательность номер 1) исследовали связывание патентоспособного антитела с соответствующими мутантами. В данном случае получение мутантов и исследование связывания патентоспособного антитела проводили с помощью мутагенеза по способу дробовика, описанного в Integral Molecular Co. (J Am Chem Soc. 2009; 131(20): 6952~6954). Характеристики мутантных библиотек, применяемых для идентификации эпитопа, приведены в приведенной ниже таблице 1. После инъекции клеток HEK-293T клонами, содержащими описанные выше библиотеки, связывание патентоспособного антитела исследовали с помощью иммунофлуоресцентного анализа.
Связывание патентоспособного антитела определяли путем усреднения результатов тестов, повторенных трижды, и нормализации на основании связывания белка поверхностного антигена ВГВ дикого типа. В этом случае, применяя кроличьи поликлональные антитела против белка поверхностного антигена ВГВ, исследовали экспрессию мутантных белков поверхностного антигена и связывание с ними патентоспособного антитела.
Как следует из данных таблицы, патентоспособное антитело теряло способность связываться с восемью (8) клонами, содержащими мутации трех аминокислотных остатков (а.к.) белка поверхностного антигена ВГВ (см. фиг.1). Иными словами, по отношению к восьми клонам, приведенным на фиг.1, было показано, что кроличьи поликлональные антитела проявляли связывающую способность, при этом клоны нормально экспрессировали мутантный белок поверхностного антигена ВГВ, однако патентоспособное антитело не связывалось с ними.
В результате исследования восьми клонов было обнаружено, что каждый клон содержал, по меньшей мере, одну мутацию в положении 160R (160R обозначает аминокислоту R, находящуюся в положении 160; здесь и далее обозначения, как описано выше), 163W и 163E (последовательность номер 1), соответственно. Иными словами, описанная выше последовательность может быть определена как сайт, соответствующий эпитопу патентоспособного антитела. Полученный результат показал, что эпитоп патентоспособного антитела содержит RFLWE (последовательность номер 4), а эпитоп подтипа ayw ВГВ со связывающей способностью содержит KFLWE (последовательность номер 5).
В частности, эпитоп, описанный последовательностями номер 4 и 5, может быть FARFLWEWASVRFSW (последовательность номер 6) или FGKFLWEWASARFSW (последовательность номер 7), соответствуя минорной петле среди двух петель на участке поверхности ВГВ, содержащем описанный выше эпитоп (см. фиг.2).
Пример 2. Идентификация характеристик эпитопа для патентоспособного антитела
(1) Получение мутантов полимеразы ВГВ
Эпитопы для патентоспособного антитела включают остатки 160K, 163W и 164E (последовательность номер 2) в открытой рамке считывания поверхностного антигена (S ORF) ВГВ подтипа ayw, при этом последовательность открытой рамки считывания поверхностного антигена ВГВ, кодирующая эпитопы, перекрывается с рамкой считывания полимеразы ВГВ (P ORF). В частности, остатки 504I, 506M, 507G и 508V (см. последовательность номер 3) полимеразы ВГВ могут соответствовать местам, где эпитоп кодируется генами в открытой рамке считывания, кодирующей эпитоп (см. фиг.3). Вкратце, мутация в описанных выше положениях S ORF ВГВ также ведет к появлению мутации в P ORF ВГВ.
Полимераза ВГВ имеет особенности, отличающие ее от других вирусных полимераз. Во-первых, полимераза ВГВ обладает активностью обратной транскриптазы, которая позволяет синтезировать ДНК по матрице РНК (прегеномная РНК: пгРНК); во-вторых, при инициации обратной транскрипции полимераза ВГВ использует саму себя в качестве белковой затравки; и в-третьих, транслокация затравки и переключение матрицы происходят во время репликации, хотя точный механизм до сих пор остается неизвестным.
Кроме того, как описано выше, открытая рамка считывания («ORF»), кодирующая эпитоп для патентоспособного антитела, нейтрализующего вирус гепатита В, который представляет собой эпитоп для патентоспособного антитела в поверхностном антигене ВГВ, может перекрываться с другой ORF, кодирующей полимеразу ВГВ. Таким образом, для исследования влияния сайта полимеразы ВГВ, закодированного в P ORF ВГВ, перекрывающейся с ORF, кодирующей эпитоп для патентоспособного антитела, на репликацию ВГВ наблюдали за возможностью возникновения мутаций в описанном выше эпитопе.
С этой целью создавали мутант с заменой аминокислоты, которая присутствует в сайте, перекрывающемся с эпитопом для патентоспособного антитела, на аланин и исследовали влияние полученного мутанта на обратно транскриптазную активность полимеразы ВГВ. Сначала получали таких мутантов, как K503A (K503A означает, что аминокислота K в положении 503 мутирована на A; здесь и далее обозначения, как описано выше) I504A, M506A, G507A и V508A, которые получали заменой 503K, 504I, 506M, 507G и 508V полимеразы ВГВ на аланин, а также получали естественный мутант V508L, как показано на фиг.4. Затем с помощью комплементационных тестов исследовали вариации функции репликации генома полимеразы ВГВ при мутации в описанном выше эпитопе. В частности, репликон ВГВ без полимеразы, представляющий собой мутант ВГВ, в котором была получена мутация, приведшая к сдвигу рамки считывания в ORF P, и в котором полимераза ВГВ не была активной, а также плазмида, экспрессирующая полимеразу ВГВ с мутацией, полученной, как описано выше, были применены для инфицирования клеток HepG2 (см. фиг.5). Затем репликацию генома ВГВ исследовали с помощью Саузерн-блота и способа защиты от РНКаз.
(2) Саузерн-блот анализ
Как описано выше, репликоном ВГВ без полимеразы и мутантом, несущим мутацию в полимеразе ВГВ, одновременно инфицировали клетку HepG2, затем через 4 дня выделяли реплицированную вирусную ДНК. Полученный материал использовали для оценки репликации ДНК ВГВ.
В результате, в мутанте K503A репликация вирусной ДНК составляла 17% от уровня репликации дикого типа. Этот результат указывает на то, что сайт 503К в полимеразе ВГВ играет существенную роль в механизме репликации вирусной ДНК. В мутантах M506A и G507A, напротив, редко наблюдалась репликация вирусной ДНК. Этот факт указывает на то, что 506M и 507G являются необходимыми сайтами для репликации вирусной ДНК полимеразой ВГВ. В мутантах I504A, V508A и V508L уровень репликации вирусной ДНК составлял, соответственно, 65%, 70% и 82% от уровня репликации в диком типе. Иными словами, эти мутанты реплицировали вирусную ДНК на уровне, схожем с диким типом. Следовательно, было показано, что описанные мутанты принимают относительно незначительное участие в репликации вирусной ДНК (см. фиг.6).
(3) Результаты анализа с помощью защиты от РНКаз
Инкапсулирование РНК (прегеномной РНК: пгРНК) как стадия, предшествующая репликации ДНК, было исследовано способом защиты от РНКаз (см. Kim et al., 2009, J. Virol. 83: 8032-8040).
Как описано выше, репликоном ВГВ без полимеразы и мутантом, несущим мутацию в полимеразе ВГВ, одновременно инфицировали клетку HepG2, затем через 3 дня выделяли вирусные коры и общую пгРНК. Полученный материал использовали для количественного анализа степени упаковки пгРНК, при котором пгРНК применяется в качестве матрицы для репликации ДНК ВГВ.
Согласно полученным результатам, уровень упаковки пгРНК в мутантах K503A и G507A составлял 25% от уровня дикого типа. Это указывает на то, что K503 и G507 играют существенную роль в механизме упаковки пгРНК в коровые вирусные частицы. Вместе с тем, в мутанте М506А уровень упаковки пгРНК составлял 71% от уровня дикого типа. Иными словами, 506М принимает сравнительно незначительное участие в упаковке пгРНК. В других мутантах, в т.ч. I504A, V508A и V508L, уровень упаковки пгРНК был почти такой же, как в диком типе, следовательно, эти сайты почти не участвуют в упаковке пгРНК (см. фиг.7).
(4) Общий обзор влияния мутантов полимеразы ВГВ на репликацию ВГВ
В мутанте полимеразы ВГВ K503A уровень упаковки пгРНК составлял всего 25% по сравнению с диким типом. По результатам количественного анализа вирусной ДНК как конечного продукта вирусной репликации, было обнаружено, что репликация происходила только в той мере, в какой происходила упаковка пгРНК. В соответствии с этим, можно предположить, что сайт 503К важен в основном для начала упаковки пгРНК (см. таблицу 2). Вместе с тем, в мутанте полимеразы ВГВ М506А уровень упаковки пгРНК составлял 71%, что по существу соответствует уровню в диком типе. Однако результаты количественного анализа вирусной ДНК как конечного продукта вирусной репликации показывают отсутствие репликации. Этот факт означает, что хотя М506 в полимеразе ВГВ никогда не принимает участия в упаковке пгРНК, М506 может играть существенную роль в механизме репликации вирусной ДНК при синтезе (-)-цепей ДНК по матрице пгРНК, т.е. в таком механизме обратной транскрипции, как белковый прайминг и транслокация затравки.
В мутанте полимеразы ВГВ G507A уровень упаковки пгРНК составлял всего 24% по сравнению с диким типом, и репликация вирусной ДНК происходила на очень низком уровне, из чего следует, что сайт М507 выполняет важные функции как в связывании пгРНК, так и в обратной транскрипции полимеразой. Более того, сайт М507 может принимать участие во взаимодействии с таким белком, как Hsp90, в качестве хозяйского фактора и/или с коровым белком ВГВ во время инкапсулирования.
Между тем, в остальных мутантах полимеразы ВГВ I504A, V508A и V508L уровень упаковки пгРНК и/или репликации вирусной ДНК по существу соответствовал уровню в диком типе. В соответствии с этим, среди последовательностей полимеразы ВГВ, закодированных в P ORF ВГВ, перекрывающейся с S ORF ВГВ, которая кодирует сайты 160K, 163W и 164Е поверхностного антигена ВГВ, сайты 160K и 164W тесно связаны с репликацией вируса. В тех случаях, когда мутация происходит в этих сайтах, вирусная репликация может не идти, что указывает на высокую консервативность этих положений. В соответствии с этим, два описанных выше мутанта не существуют, и антитело, специфичное к описанным выше сайтам, может быть эффективным для лечения мутантов, появившихся естественным образом, и/или мутантов, устойчивых к противовирусным препаратам.
Пример 3. Эффекты связывания и нейтрализации патентоспособного антитела с мутантными эпитопами
(1) Получение мутантов
По меньшей мере один из остатков 163W и 164E (последовательность номер 1) в белке поверхностного антигена ВГВ, являющихся эпитопами для патентоспособного антитела, был заменен аланином при получении мутанта. Поскольку при мутации 160К возникают проблемы с серотипами, такие мутанты исключали. Кроме того, поскольку недавно были описаны мутанты, полученные в результате замены 164E на 164D, создавали и исследовали мутант E164D. Поскольку мутанты получали, как описано выше, мутации вводили также в 506M, 507G и 508V (последовательность номер 2) полимеразы ВГВ, закодированные в P ORF ВГВ, перекрывающейся с S ORF ВГВ, которая кодирует описанных выше мутантов. В этом случае даже при мутации одной и той же аминокислоты мутанты полимеразы ВГВ имеют разную аминокислотную последовательность в зависимости от варианта кодона 163W и 164Е белка поверхностного антигена ВГВ (см. таблицу 3).
(2) Анализ и оценка эффективности in vivo с применением мыши с острым гепатитом В
Вводя ДНК ВГВ в мышь C57BL6 с помощью гидродинамической инъекции, добивались развития симптомов острого гепатита В, и использовали обработанную таким образом мышь для исследования связывания патентоспособного антитела с вирусом гепатита В и/или для исследования способности нейтрализовать вирус гепатита В в крови мыши с мутацией эпитопа, полученной, как описано выше. Использовали самку мыши C57BL6 в возрасте 6 недель, весом около 20 г, приобретенную в лаборатории Charles Liver (США). Как показано в таблице 4, всего было исследовано 12 групп, по 5 мышей в каждой группе.
Каждую мышь обрабатывали путем введения 20 мкг вектора pHBV-MBRI (Shin et al., Virus Research 119, 146-153, 2006; см. фигуру 8), содержащего последовательность ДНК ВГВ, вставленную в pcDNA3.1 (Invitrogen, США), в хвостовую вену мыши со скоростью 0,3 мл/мин в количестве 9,5% (объемная доля на вес мыши), вызывая развитие острого гепатита В. Через 48 часов, как показано в таблице 4, в хвостовую вену мышей вводили внутривенно (в/в) 0,2 мл патентоспособного антитела. Перед (за 24 часа и 48 часов) и после (через 72 часа и через 96 часов) инъекции патентоспособного антитела отделяли сыворотку и разводили в 10 раз козьей сывороткой, после чего в крови измеряли концентрацию белка поверхностного антигена ВГВ с применением твердофазного иммуноферментного анализа Genedia HBsAg ELISA 3.0 (Green Cross Corp. MS, Корея). Что кается ДНК ВГВ, до (за 48 часов) и после (через 72 часа) инъекции патентоспособного антитела брали кровь и анализировали с помощью ПЦР в реальном времени для количественной оценки содержания ДНК ВГВ в крови, а затем проводили сравнительный анализ способности патентоспособного антитела нейтрализовать вирус гепатита В.
В результате выявления поверхностного антигена ВГВ в крови с помощью твердофазного иммуноферментного анализа Genedia HBsAg ELISA 3.0 было подтверждено, что при вставке 10 мутантов все поверхностные антигены ВГВ экспрессируются на надлежащем уровне. При анализе 10 вариантов поверхностных антигенов ВГВ на предмет связывания с патентоспособным антителом обнаружили, что вариант, в котором оба положения 163W и 164Е были заменены аланином, не связывался с патентоспособным антителом. Вместе с тем обнаружили, что вариант поверхностного антигена ВГВ с заменой 163W на аланин демонстрировал связывающую способность на уровне 70% или выше по сравнению с диким типом. Кроме того, вариант поверхностного антигена ВГВ с заменой 164Е на аланин демонстрировал связывающую способность на уровне 30% по сравнению с диким типом. Связывающие характеристики варианта E164D были по существу сопоставимы с диким типом (см. таблицу 5).
Мутация в поверхностном антигене ВГВ влияет на последовательность полимеразы ВГВ, как описано выше. Поэтому исследовали эффекты варианта полимеразы, полученного заменой аминокислотных остатков поверхностного антигена ВГВ на аланины, на репликацию ДНК ВГВ. Данные анализа показали, что репликация ДНК ВГВ не происходила при мутации остатков 163W и 164E. В частности, результаты исследования репликации ДНК ВГВ при замене каждого из остатков 163W и 164E на аланин показали, что в варианте 164Е репликация ДНК ВГВ была на уровне от 30 до 70%, в то время как в варианте 163W репликации не было. Таким образом, были установлено, что аминокислотные остатки в полимеразе, соответствующие сайту 163W, очень важны для репликации.
Варианты 164E с экспрессией поверхностного антигена ВГВ и репликацией ДНК ВГВ исследовали на предмет выявления способности патентоспособного антитела нейтрализовать вирус гепатита В. Полученные результаты показали, что способность нейтрализовать вирус гепатита В существенно снижена, так как связывающая способность патентоспособного антитела снижена до 70% по сравнению с диким типом. Однако с вариантом 164D, известным в современной области техники как естественный вариант, патентоспособное антитело демонстрировало такую же связывающую способность, как и с диким типом.
Не определено: верификационное исследование нейтрализующей способности не было проведено (не определено)
Как описано выше, эпитопы для патентоспособного антитела в поверхностном антигене ВГВ включают 160K (ayw) или 160R (adr), 163W и 164Е. В частности, сайт 164Е был идентифицирован в экспериментах с вариантами замены на аланин как наиболее важное положение для связывания патентоспособного антитела. В настоящее время известно, что это положение может быть мутировано в 164D, и патентоспособное антитело обладает нейтрализующей способностью по отношению к варианту 164D. Вместе с тем, несмотря на то, что положение 163W не играет значительной роли в связывании патентоспособного антитела, мутация в этом положении вызывает мутацию в последовательности полимеразы, важную для репликации, что в свою очередь влияет на репликацию ДНК ВГВ. Следовательно, можно предсказать, что описанный выше сайт является высоко консервативным положением, то есть таким положением, в котором мутации происходят крайне редко. Фактически еще не было получено сведений о мутациях в положении 163W. Наконец, 160К (для подтипа ayw) или 160R (для подтипа adr) представляют собой аминокислотные остатки, определяющие серотип. Результаты функционального анализа показали, что они тесно связаны с репликацией ВГВ, таким образом было предсказано, что они представляют собой консервативные положения, в которых мутации происходят крайне редко.
Список последовательностей
Последовательность номер 1 определяет аминокислотную последовательность (подтипа adr) белка поверхностного антигена ВГВ
Последовательность номер 2 определяет аминокислотную последовательность (подтипа ayw) белка поверхностного антигена ВГВ
Последовательность номер 3 определяет аминокислотную последовательность полимеразы ВГВ
Последовательность номер 4 определяет эпитоп (подтипа adr) для патентоспособного антитела, RFLWE
Последовательность номер 5 определяет эпитоп (подтипа ayw) для патентоспособного антитела, KFLWE
Последовательность номер 6 определяет эпитоп (подтипа adr) для патентоспособного антитела, FARFLWEWASVRFSW
Последовательность номер 7 определяет эпитоп (подтипа ayw) для патентоспособного антитела, FGKFLWEWASARFSW






Представлена группа изобретений, касающаяся эпитопа, специфичного для вируса гепатита В (ВГВ), полинуклеотида, кодирующего эпитоп, экспрессионного рекомбинантного векора, рекомбинантного микроорганизма, вируса или клетки млекопитающих, способа получения эпитопа и способа получения антитела, специфично связывающегося с эпитопом. Описанный эпитоп представляет собой консервативное положение ВГВ, которое не подвергается мутагенезу. Представленная группа изобретений может быть использована для получения композиций с высокой терапевтической эффективностью вследствие отсутствия возможности мутаций в используемом эпитопе. 8 н. и 14 з.п. ф-лы, 8 ил., 5 табл., 3 пр.
1. Эпитоп, специфичный для вируса гепатита В (ВГВ), определенный RFLWE (SEQ ID NO: 4) или KFLWE (SEQ ID NO: 5), для получения антиген-специфичного антитела к указанному эпитопу.
2. Эпитоп по п. 1, который содержится в последовательности, представленной FARFLWEWASVRFSW (SEQ ID NO: 6) или FGKFLWEWASARFSW (SEQ ID NO: 7).
3. Полинуклеотид, кодирующий эпитоп, специфичный для ВГВ, по п. 1.
4. Экспрессионный рекомбинантный вектор, включающий полинуклеотид по п. 3.
5. Экспрессионный рекомбинантный вектор по п. 4, дополнительно включающий последовательность, кодирующую промотор или сигнальный белок, обеспечивает экспрессию эпитопа, специфичного для ВГВ, на поверхности клетки микроорганизма или вируса, или клеток млекопитающих.
6. Рекомбинантный микроорганизм для экспрессии эпитопа по п. 1, трансформированный экспрессионным рекомбинантным вектором по п. 4 или 5.
7. Рекомбинантный вирус для экспрессии эпитопа по п. 1, трансформированный экспрессионным рекомбинантным вектором по п. 4 или 5.
8. Рекомбинантная клетка млекопитающих для экспрессии эпитопа по п. 1, трансформированная экспрессионным рекомбинантным вектором по п. 4 или 5.
9. Рекомбинантный микроорганизм по п. 6, в котором трансформированный рекомбинантный микроорганизм представлен рекомбинантной Е. coli или рекомбинантными дрожжами.
10. Рекомбинантный вирус по п. 7, в котором вирус представлен рекомбинантными бактериофагами.
11. Способ получения эпитопа, специфичного для ВГВ, определенного любой из SEQ ID NO: 4-7, включающий культивирование рекомбинантного микроорганизма по п. 6 или 9 или вируса по п. 7 или 10, или рекомбинантных клеток млекопитающих по п. 8 и выделение эпитопа, специфичного для ВГВ.
12. Способ получения антитела, специфично связанного со специфичным для ВГВ эпитопом по п. 1, включающий инокуляцию животного указанным эпитопом, специфичным для ВГВ, композицией, содержащей описанный выше эпитоп, или вектором, содержащим полинуклеотид, который кодирует описанный выше эпитоп, скрининг и получение антитела, специфично связанного с эпитопом, специфичным для ВГВ, из инокулированного животного.
13. Способ по п. 12, в котором антитело является поликлональным антителом или моноклональным антителом.
14. Способ по п. 12, дополнительно включающий процесс гуманизации или деиммунизации.
15. Способ по п. 14, в котором процесс гуманизации включает пересадку гипервариабельной последовательности антитела, полученного из животного, в каркасный участок человеческого антитела.
16. Способ по п. 15, дополнительно включающий процесс замены, вставки или делеции по меньшей мере одного аминокислотного остатка с целью увеличения аффинности и снижения иммуногенности.
17. Способ по п. 12, в котором животное представлено трансгенным животным, способным производить антитело с такой же последовательностью, как в человеческом антителе.
18. Способ по п. 17, в котором трансгенное животное представлено трансгенной мышью.
19. Способ по п. 12, в котором применяется способ дисплея.
20. Способ по п. 19, в котором способ дисплея представлен фаговым дисплеем, бактериальным дисплеем, дрожжевым дисплеем или рибосомным дисплеем.
21. Способ по п. 19 или 20, в котором библиотека, применяемая в способе дисплея, создана таким образом, что содержит последовательности антител человеческого происхождения.
22. Способ по п. 19 или 20, дополнительно включающий применение эпитопа, специфичного для ВГВ, определенного одной из SEQ ID NO: 4-7, композиции, содержащей описанный выше эпитоп, для скрининга (для проведения пэннинга) антитела, специфично связанного с эпитопом, специфичным для ВГВ, определенным одной из SEQ ID NO: 4-7.
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| WO 2004108753 A1, 16.12.2004 | |||
| WO 00/12547 A2, 09.03.2000 | |||
| КОМПОЗИЦИИ НА ОСНОВЕ БЕЛКА ВИРУСА ГЕПАТИТА В И СТРЕССОВОГО БЕЛКА И ИХ ПРИМЕНЕНИЕ | 2002 |
|
RU2295536C2 |

