Изобретение относится к области биотехнологии, а именно генной инженерии, и касается рекомбинантной вакцины для профилактики вирусного гепатита В, которая содержит в своем составе в качестве активного компонента рекомбинантный поверхностный антиген вируса гепатита В (HBsAg), имеющий мутацию G145R, и может содержать рекомбинантный HBsAg дикого типа серотипа ayw и/или adw.
Гепатит В - одна из самых распространенных вирусных инфекций, которая ведет к тяжелым последствиям таким, как хронический гепатит, цирроз печени и гепатокарцинома. Успешное применение вакцин против гепатита В существенно улучшило эпидемиологическую обстановку по гепатиту В. В тоже время в последние годы было установлено, что имеет место распространение мутантных форм вируса гепатита В (ВГВ), которые не выявляются тестами иммунодетекции HBsAg и «ускользают» от защитного действия поствакцинального иммунитета. Такие мутантные формы ВГВ, отличительной чертой которых является экспрессия HBsAg с атипичными серологическими свойствами, были названы «ускользающими» («эскейп», «escape») мутантами.
Впервые существование таких мутантов ВГВ зарегистрировано в Италии. Позднее было показано (Carman W.F. et al., Vaccine-induced escape mutant of hepatitis В virus, Lancet, 1990, v. 336, p. 325-329), что из 1590 лиц, вакцинированных против гепатита В, 32 (2%) оказались инфицированными ВГВ. У всех инфицированных определялось одновременное присутствие в крови и HBsAg, и антител к HBsAg. Вирус, выделенный из крови одного из инфицированных, оказался «ускользающим» мутантом ВГВ, отличающимся по последовательности нуклеотидов в геноме от дикого типа.
Изменение нуклеотидной последовательности в S-регионе генома ВГВ и последующие аминокислотные замены приводят к конформационным изменениям вирусных белков оболочки. Эти изменения способны вызвать пространственные затруднения во взаимодействии антигенных эпитопов с нейтрализующими антителами, таким образом позволяя ускользать мутантным штаммам из-под вакцинного и иммунного надзора. Особенно важны для ускользания вируса от протективного действия вакцин мутации «а» детерминанты, которая локализована между 124-147 аминокислотными остатками в пределах главной гидрофильной петли (100-170 ао) (Weber В., Genetic variability of the S gene of hepatitis В virus: clinical and diagnostic impact, J.Clin. Virol., 2005, v. 32, p. 102-112).
Наиболее важной среди серологически значимых аминокислотных замен является замена глицина в положении 145 на аргинин (G145R), вызванная точечной мутацией в позиции 587 нуклеотидной последовательности HBsAg дикого типа. Этот мутант стабилен на протяжении долгого времени и может сохранять способность к репликации несколько лет. Он обладает способностью к горизонтальной передаче другим людям. Факты внутрисемейной передачи были отмечены у 3-х из 10-ти носителей мутанта G145R, несмотря на присутствие в высоких концентрациях антител к HBsAg в крови всех заразившихся (Oon C.J. et al., Intra-familial evidence of horizontal transmission of hepatitis В virus surface antigen mutant G145R, J.Infect., 2000, v.41, p. 260-26400). Частота мутаций G145R может колебаться от 0,4 до 5%. При динамическом наблюдении и определении мутантных форм среди новорожденных исследователи Тайваня выявили увеличение числа детей, инфицированных мутантными штаммами ВГВ. Так, в 1984 г. оно составило 7,8%, в 1989 г. - 19,6%, в 1994 г. - 28,1% (Hsu Ну et al., Hepatology, 1999, v. 30(5), p. 1312-7).
На территории РФ, где широко распространены генотип D и серотип «ау», также выявлена циркуляция мутантных штаммов HBV, экспрессирующих HBsAg разных серотипов («ау» и «ad») с заменами G145R. Распространенность серологически значимой мутации G145R в московской области среди хронических носителей HBV составила 0,12%. (А.И. Баженов и др., Эпидемиология и вакцинопрофилактика, 2011, №5, с. 49-53). Учитывая высокий процент носительства HBV в человеческой популяции, выявленное количество мутантов HBV представляется значительным.
Таким образом, «ускользающие» мутанты ВГВ G145R, представляя собой реплицирующие инфекционные вирусные мутанты, избегающие нейтрализующий иммунитет при вакцинации, основанной на нормальном HBsAg, могут быть причиной заболевания ВГ. Существование и распространение «ускользающих» мутантов создает серьезную проблему как для донорства, так и для эффективности современной стратегии вакцинации. В связи с этим существует необходимость в создании вакцины нового поколения против вируса гепатита В, которая обладает максимально широким, приближающимся к естественно индуцируемому спектром эпитопов, способных стимулировать развитие протективного ответа антителообразования как против отдельных субтипов ВГВ - ау и ad, так и против эпитопов мутанта G145R.
Затруднения, препятствующие созданию эффективных вакцин для профилактики гепатита В, вызываемого мутантным ВГВ G145R, связаны с разработкой оптимальных экспрессионных систем для продукции рекомбинантного HBsAg, содержащего мутацию G145R, и обладающего высокими иммуногенными свойствами.
Известно получение полипептида HBsAg, имеющего аминокислотную последовательность с заменой в позиции 145 глицина на аргинин рекомбинантным путем (US 7,038,035, 02.05.2006). Из крови пациента (ребенок из Китая) был изолирован мутантный вирус гепатита В, несущий мутацию в последовательности HBsAg, которая была обусловлена заменой в нуклеотидной последовательности в позиции 587-589. Геном выделенного мутантного вируса относился к «adw» субтипу. Чтобы получить мутантный белок HBsAg был сконструирован экспрессионный вектор, содержащий нуклеотидную последовательность с AGA в позиции 587-589, соответствующую последовательности изолированного штамма вируса. Сконструированный вектор интродуцировали в подходящие клетки-хозяева, в частности клетки млекопитающих. Полученный путем культивирования рекомбинантных клеток млекопитающих мутантный HBsAg имеет кажущуюся молекулярную массу 23 kDa в отличие от 25 kDa HBsAg дикого типа и обладает более высокой иммуногенностью. На основе мутантного HBsAg создана вакцина, содержащая эффективное количество антигена, которое определяется размером и формой полипептида, биоактивностью, способностью к биодеградации.
Thomas H.С. et al. в ряде патентов охарактеризовали вариант белка HBsAg с модифицированной детерминантой «а», в которой аминокислота глицин в позиции 145 заменена на аргинин (US 5,639,637, 1997), последовательность ДНК, кодирующую данный вариант белка (US 5,854,024, 1998), дрожжевой экспрессирующий вектор, включающий указанную последовательность (US 5,851,823, 1998), набор для диагностики антител против варианта HBsAg (US 5,989,865, 1999), антитело против варианта HBsAg (US 6,099,840, 2000).
Известна рекомбинантная вакцина (RU 2238105, 2004) для профилактики вирусного гепатита В, содержащая эффективное количество HBsAg серотипов «adw» и/или «ayw», полученных из штаммов дрожжей Pichia augusta VKPM Y-2412 и VKM Y-2924D, соответственно, адъювант и физиологически приемлемый разбавитель. При вакцинации HBsAg у здоровых людей индуцируется выработка специфических антител (анти-HBs), которые и обеспечивают иммунитет. Вакцину отличает высокая иммуногенность, нетоксичность, отсутствие побочных эффектов. Вакцина внедрена в практику здравоохранения и внесена в Государственный реестр лекарственных средств РФ. Недостатком вакцины, как и других вакцин, основанных на использовании рекомбинантного HBsAg, является их относительно узкая по сравнению с природными антигенами антигенная специфичность, поскольку у лиц, переболевших гепатитом В, в отличие от вакцинированных, индуцировались антитела, взаимодействующие с максимально серологически модифицированным мутантом G145R (одновременное инфицирование различными вирусами гепатита В). Таким образом, вакцина имеет более ограниченный спектр эпитопов, против которых индуцируются нейтрализующие антитела.
Ближайшим аналогом является патент US 5,639,637, 1997, в котором описано получение варианта HBsAg, имеющего модифицированную детерминанту «а» с заменой глицина в положении 145 на аргинин, путем экспрессии кодирующего его гена в клетках дрожжей Saccharomyces cerevisiae. Первоначально получали последовательность ДНК, кодирующую HBsAG серотипа «ау», которая содержала точечную мутацию, приводящую к замене кодона GGA, кодирующего глицин на кодон AGA, кодирующий аргинин, в позиции 145 полипептида HBsAg. Затем конструировали плазмиду pRIT13557, содержащую полученную последовательность, и этой плазмидой трансформировали дрожжевые клетки S. cerevisiae штамма DC5-cir0 (АТСС20820). После культивирования рекомбинантных дрожжевых клеток (Y1648) полученный вариант HBsAg выделяли, очищали и анализировали радиоиммунным и электронномикроскопическим методами. Было установлено присутствие сферических частиц, типичных для HBsAg, антигенность которых отличалась от таковой HBsAg дикого типа. Предложенная вакцина содержит вариант HBsAg (5-40 мкг/мл), гидроокись алюминия (адъювант), мертиолят натрия (консервант) в натрий-фосфатном буфере с рН 6,8, содержащим хлористый натрий. Вакцина может содержать HBsAg дикого типа.
К недостаткам использования системы экспрессии на основе Saccharomyces cerevisiae можно отнести нестабильность рекомбинантных клеток, связанную с относительно высокой частотой потерь автономной плазмиды при делении клеток, что требует постоянного контроля процесса культивирования и подтверждения стабильности используемой плазмиды. При культивировании штамма в неселективных для плазмиды условиях это приводит к накоплению клеток, не содержащих плазмиды и неспособных к синтезу целевого белка. Это существенно осложняет культивирование рекомбинантного штамма и увеличивает себестоимость целевого продукта. Для индукции экспрессии рекомбинантных белков используется достаточно дорогой реагент - галактоза, что увеличивает себестоимость целевого продукта. Данная вакцина в практическом здравоохранении не зарегистрирована, промыщленно не воспроизведена.
Задачей изобретения является создание высокоиммуногенной вакцины против гепатита В, обладающей широким, приближающимся к естественному, спектром эпитопов, способной стимулировать развитие протективного ответа антителообразования как против отдельных субтипов ВГВ - «ау» и «ad», так против эпитопов мутанта G145R.
Задача решена путем создания вакцины, содержащей комбинацию рекомбинантного поверхностного антигена вируса гепатита В (HBsAg), имеющего мутацию G145R (ESC-Ag), и рекомбинантного HBsAg дикого типа серотипа «ayw» и/или «adw». В другом варианте вакцина в качестве активного компонента содержит только поверхностный антиген вируса гепатита В (HBsAg), имеющий мутацию G145R.
Согласно изобретению, для получения рекомбинантного поверхностного антигена вируса гепатита В, имеющего мутацию G145R (ESC-Ag), были созданы четыре различных рекомбинантных штамма дрожжей H. Polymorpha - KBT11/pEMmulti-7, КБТ12/pEMmono-4, КБТ14/pEMmalt-8, КБТ-14/рЕМmono2, каждый из которых представляет собой стабильный эффективный продуцент ESC-Ag, обладающего необходимыми биологическими свойствами (правильная конформация, иммуногенность, чистота) и низкой себесоимостью.
Для получения штамма-продуцента поверхностного антигена вируса гепатита В, имеющего мутацию G145R (ESC-Ag), плазмиду, содержащую фрагмент ДНК с рекомбинантным геном, кодирующим ESC-Ag, под контролем промотора гена метанолоксидазы {МОХ H. polymorpha) и селективным маркером (ген LEU2 S. cerevisiae) интегрируют в геном реципиентного штамма Н. polymorpha DL1-L (Sohn J.H. et. al., J Bacteriol. 1996 Aug; 178(15):4420-8. PMID:8755868). Для получения клонов, содержащих в геноме несколько копий плазмиды, применена процедура селекции множественной интеграции. По результатам проверки клонов, полученных в результате этой процедуры, на способность синтезировать белок ESC-Ag, был выделен трансформант с наибольшей продуктивностью. Т.к. искусственный ген интегрирован в геном дрожжей, т.е. находится в составе одной из хромосом, это обеспечивает высокую митотическую стабильность штамма, давая возможность оптимизировать условия его культивирования без учета риска накопления клеток, потерявших способность синтезировать чужеродный белок. Полученный рекомбинантный штамм, содержащий несколько копий целевого гена под контролем промотора МОХ, позволяет добиваться более высоких уровней синтеза рекомбинантного белка ESC-Ag при более простых способах получения культур высокой плотности. Штамм депонирован 20.05.2014 в коллекции микроорганизмов ЗАО НПК «Комбиотех» под номером КБТН/pEMmulti-7. Штамм является новым и ни в патентной, ни в научно-технической литературе не описан.
Согласно изобретению, второй рекомбинантный штамм-продуцент поверхностного антигена вируса гепатита В, имеющего мутацию G145R (ESC-Ag), получали интеграцией в геном штамма DL1-L (H. polymorpha) одной копии синтетической последовательности ДНК, кодирующей ESC-Ag, под контролем промотора гена МОХ. Для этого в качестве реципиента был использован штамм DLT2, полученный в результате инактивации в штамме DL1-L генов МОХ и TRP3 интеграцией селективного маркера, гена LEU2 S. cerevisiae, аналогично тому, как это было сделано ранее (Bogdanova АЛ., Agaphonov M.О. et. al., Yeast, 1995 Apr 15; 11 (4):343-53, PMID:7785335). Это давало возможность с высокой эффективностью отбирать трансформанты, в которых интеграция синтетической последовательности происходила по механизму гомологичной рекомбинации с последовательностями локуса гена МОХ, в результате чего происходила замена кодирующей области этого гена на последовательность, кодирующую ESC-Ag. Уровень экспрессии ESC-Ag, который достигается у полученного таким образом штамма, подходит для увеличения доли антигена во фракции растворимых клеточных белков. Поскольку такой штамм содержит только одну копию рекомбинантного гена, интегрированную в строго определенный локус генома, реплика этого гена может быть получена при помощи полимеразной цепной реакции для контроля отсутствия в нем мутаций, которые могли образоваться в процессе его введения в геном дрожжей или при культивировании штамма-продуцента. Штамм депонирован 20.05.2014 в коллекции микроорганизмов ЗАО НПК «Комбиотех» под номером КБТ12/pEMmono-4. Штамм является новым и ни в патентной, ни в научно-технической литературе не описан.
Согласно изобретению, для получения еще одного рекомбинантного штамма-продуцента поверхностного антигена вируса гепатита В, имеющего мутацию G145R (ESC-Ag), плазмиду, содержащую фрагмент ДНК с рекомбинантным геном, кодирующим ESC-Ag, под контролем промотора гена мальтазы (MALI H. polymorpha) и селективным маркером (ген LEU2 S. cerevisiae) интегрируют в геном реципиентного штамма H. polymorpha DL1-L. Для получения клонов, содержащих в геноме несколько копий плазмиды, применена процедура селекции множественной интеграции. По результатам проверки полученных в результате этой процедуры клонов на способность синтезировать белок поверхностного антигена вируса гепатита В, имеющего мутацию G145R, был выделен трансформант, наиболее отвечающий требуемым свойствам. Полученный рекомбинантный штамм, содержащий несколько копий целевого гена под контролем промотора MAL1, депонирован 20.05.2014 в коллекции микроорганизмов ЗАО НПК «Комбиотех» под номером KBT14/pEMmalt-8. Штамм является новым и ни в патентной, ни в научно-технической литературе не описан.
Согласно изобретению, для получения 4-го рекомбинантного штамма-продуцента поверхностного антигена вируса гепатита В, имеющего мутацию G145R (ESC-Ag), в геном штамма DL1-L H. polymorpha последовательно интегрируют две экспрессионные кассеты, содержащие по одной копии нужного гена. Первая кассета содержит фрагмент ДНК с рекомбинантным геном, кодирующим ESC-Ag, под контролем промотора гена МОХ (ген метанолоксидазы). Интеграция этой кассеты проходила по механизму гомологичной интеграции в локус гена МОХ. Вторая экспрессионная кассета содержит аналогичный ген ESC-Ag, но под контролем промотора гена DAK (ген кодирует дигидроксиацетон киназу) дрожжей Н. polymorpha. Интеграция этой кассеты проходила по механизму гомологичной интеграции в локус гена LEU2. В результате трансформации получают рекомбинантный штамм, содержащий одну копию целевого гена ESC-Ag под контролем промотора DAK и одну копию под контролем промотора МОХ. При этом искусственные гены интегрированы непосредственно в геном штамма-продуцента, т.е. находятся в составе хромосом дрожжей Н. polymorpha. Это обеспечивает высокую митотическую стабильность штамма, давая возможность оптимизировать условия его культивирования без учета риска накопления клеток, потерявших способность синтезировать чужеродный белок. Кроме того, наличие в каждом из локусов интеграции только одной копии рекомбинантного гена с уникальной промоторной и терминаторной областью позволяет контролировать отсутствие мутаций этого гена в процессе его введения в геном и при культивировании штамма. Контроль за отсутствием мутаций осуществляют путем анализа нуклеотидной последовательности продуктов полимеразной цепной реакции с использованием праймеров, комплементарных специфическим последовательностям, фланкирующим кодирующую область рекомбинантного гена в каждом из локусов интеграции. Использование экспрессионных кассет с двумя разными промоторами {МОХ и DAK), максимальная активность которых достигается при разных условиях культивирования, обеспечивает большую равномерность синтеза продукта в процессе культивирования штамма. При этом не превышается уровень синтеза белка, который может привести к возможному нарушению эффективности укладки и, как следствие, снижению продуктивности штамма. В результате отбора клонов, способных синтезировать белок ESC-Ag, был выделен трансформант, наиболее отвечающий требуемым свойствам. Штамм депонирован 20.05.2014 в коллекции ЗАО НПК «Комбиотех» под номером КБТ-14/рЕМmono2. Штамм является новым и ни в патентной, ни в научно-технической литературе не описан.
Полученные рекомбинантные штаммы-продуценты КБТ11/pEMmulti-7, КБТ12/pEMmono-4, KBT14/pEMmalt-8 и КБТ-14/рЕМmono2 могут быть использованы для получения с высоким выходом белка ESC-Ag в виде вирусоподобных частиц, обладающего антигенными свойствами, необходимыми для его использования в качестве основного компонента вакцины для защиты от мутантного варианта Escape вируса гепатита В человека. Образование вирусоподобных частиц подтверждали с использованием метода электронной микроскопии и гель-фильтрационной хроматографии.
После выделения и очистки рекомбинантные антигены использовали в составе вакцины для профилактики вирусного гепатита В. При этом получены вакцины в нескольких вариантах. В одном варианте рекомбинантная вакцина содержит поверхностный антиген вируса гепатита В, имеющий мутацию G145R (ESC-Ag), полученный путем культивирования рекомбинантного штамма дрожжей H. Polymorpha KBT-11/pEMmulti-7 или Н. Polymorpha КБТ-12/pEMmono-4, или Н. Polymorpha КБТ-14/ pEMmalt-8, или Н. Polymorpha КБТ-14/рЕМmono2 в эффективном количестве, адьювант, физиологически приемлемый разбавитель и может содержать мертиолят в качестве консерванта.
В другом варианте рекомбинантная вакцина содержит также рекомбинантный HBsAg серотипа «ayw» и/или «adw». HBsAg серотипа ayw получали путем культивирования штамма дрожжей Pichia augusta VKM Y-2924D (RU2238105). HBsAg серотипа «adw» получали путем культивирования штамма дрожжей Pichia augusta VKPMY-2412 (RU2238105). Выделенные и очищенные рекомбинантные HBsAg не имеют мутаций, имеют чистоту не менее 95%, обладают антигенной активностью и иммунной специфичностью. В составе вакцины может быть использован коммерческий препарат HBsAg.
Под эффективным количеством понимается такое количество активного начала, которое достаточно для защиты от вируса гепатита В. Точное количество будет зависеть от конкретных обстоятельств и может быть оценено специалистом в данной области с использованием известных методик.
Технический результат предложенного изобретения заключается в создании вакцины, обладающей улучшенным серологическим соответствием природному мутанту за счет широкого, приближающегося к естественному, спектра эпитопов, и способной стимулировать развитие протективного ответа антителообразования как против отдельных субтипов ВГВ - «ау» и «ad», так против эпитопов мутанта G145R, снижении себестоимости.
Вакцина согласно изобретению позволяет наиболее полно удовлетворять потребности практического здравоохранения, снижая затраты на реализацию программ вакцинопрофил актики.
Изобретение может быть проиллюстрировано следующими примерами.
В приводимых примерах все генно-инженерные операции производили согласно стандартным методикам и инструкциям компаний производителей ферментов и наборов для манипуляций с ДНК in vitro. Трансформацию клеток Escherichia coli и Hansenula polymorpha осуществляли согласно ранее описанным методам (Inoue H. et. al., Gene, 1990, 96:23-28. и Bogdanova A.I. et. al., Yeast, 1995, 11(4):343-53, соответственно). Для полимеразной цепной реакции (ПЦР) использовали полимеразу Pwo (Roche). Синтез олигонуклеотидов, получение фрагмента ДНК, содержащего последовательность, кодирующую ESC-Ag (SEQ ID NO: 1), а также определение последовательности нуклеотидов производились ЗАО "Евроген" г. Москва.
Пример 1. Получение штамма Н. polymorpha, содержащего несколько копий рекомбинантного гена, кодирующего полипептид ESC-Ag, под контролем промотора МОХ.
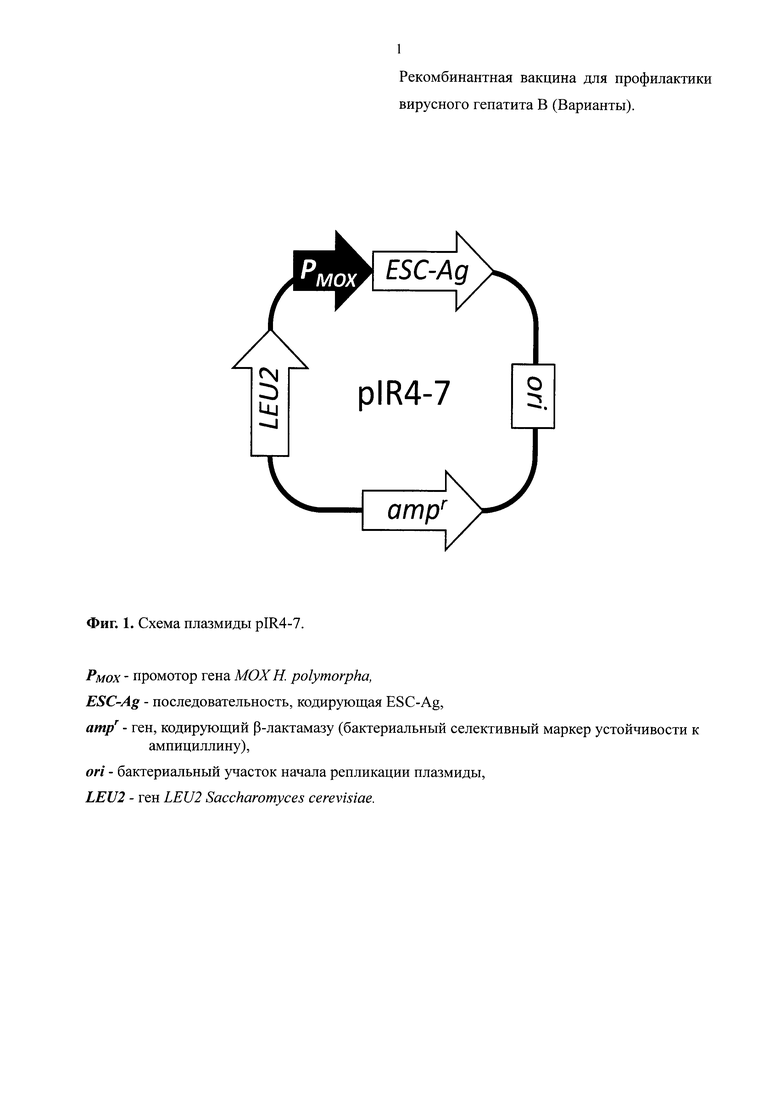
Получали плазмиду pIR4-7 (Фиг. 1; SEQ ID NO: 2), которая содержала рекомбинантный ген, кодирующий полипептид ESC-Ag. Этот ген включал промоторную область гена МОХ Н. polymorpha (SEQ ID NO: 2, нуклеотидные позиции 4338-4862) и синтетическую последовательность (SEQ ID NO: 2, нуклеотидные позиции 4863-5554), содержащую открытую рамку считывания ESC-Ag (SEQ ID NO: 2, нуклеотидные позиции 4874-5551), последовательность которой совпадала с последовательностью открытой рамки считывания гена HBsAg "ayw2" дикого типа за исключением позиции 433, в которой вместо остатка гуанина находился остаток аденина (SEQ ID NO: 2, нуклеотидная позиция 5306). Помимо этого гена, плазмида pIR4-7 состояла из: (а) фрагмента плазмиды рАМ619 (Agaphonov and Alexandrov, FEMS Yeast Res., 2014, 14(7):1048-54), включающего бактериальный селективный маркер устойчивости к ампициллину и дрожжевой селективный маркер - модифицированный ген LEU2 S. cerevisiae (SEQ ID NO: 2, нуклеотидные позиции 895-4337), (б) фрагмента плазмиды AMIpSL1 (Agaphonov et al., Yeast. 1999, 15(7):541-51), несущего терминатор транскрипции и автономно-реплицирующуюся последовательность Н. polymorpha, (SEQ ID NO: 2, нуклеотидные позиции 7-894) и (в) линкерной последовательности (SEQ ID NO: 2, нуклеотидные позиции 1-6).
Штамм DL1-L трансформировали плазмидой pIR4-7. Трансформантов отбирали на среде, не содержащей лейцин. Несколько выросших трансформантов посеяли на твердую среду без лейцина истощающим штрихом, чтобы получить отдельные колонии. Среди выросших колоний отобрали самые крупные и снова посеяли на твердую среду без лейцина истощающим штрихом. Эту процедуру повторяли до тех пор, пока вырастающие при рассеве колонии не стали одинаковыми по скорости роста. По одному такому субклону нескольких независимых трансформантов отобрали для анализа продукции белка ESC-Ag. Один из таких трансформантов был обозначен КБТ-11/pEMmulti-7.
Пример 2. Получение штамма Н. polymorpha, содержащего одну копию рекомбинантного гена, кодирующего полипептид ESC-Ag, под контролем промотора МОХ.
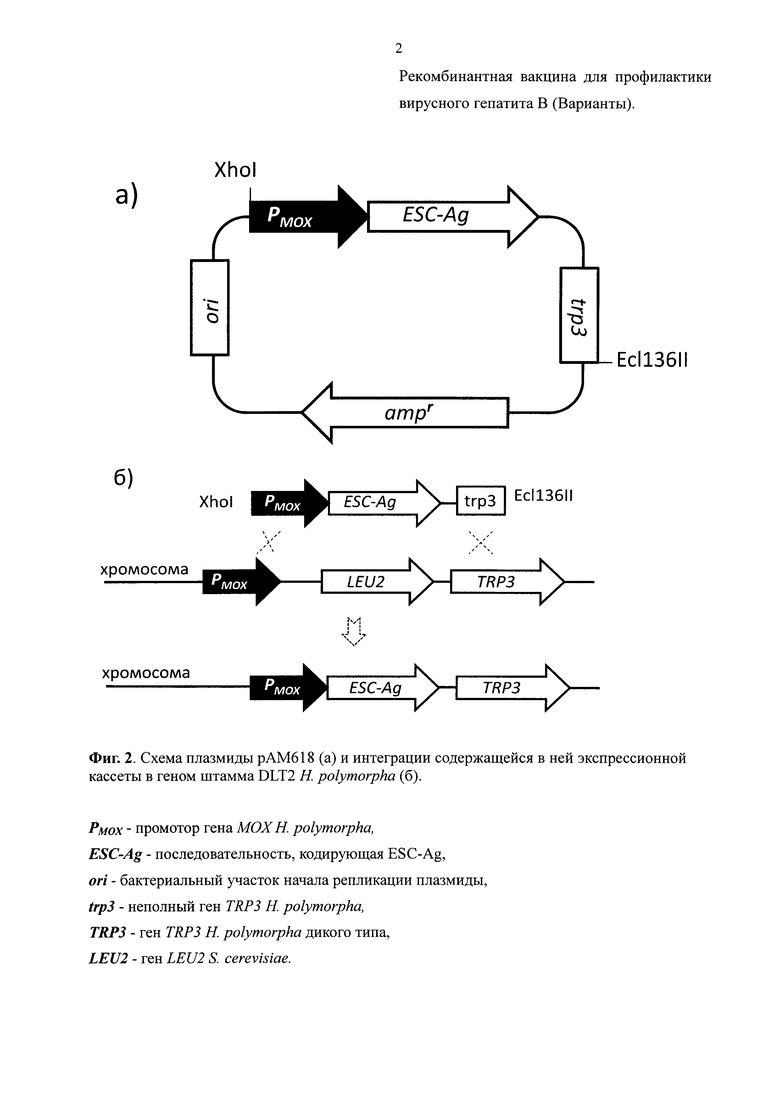
Получали плазмиду рАМ618 (Фиг. 2А; SEQ ID NO: 3), которая содержала рекомбинантный ген, кодирующий полипептид ESC-Ag, под контролем промотора МОХ (SEQ ID NO: 3, нуклеотидные позиции 4188-5715). После терминирующего кодона этого гена находился фрагмент локуса гена МОХ, включающий его терминаторную область и часть гена TRP3 (SEQ ID NO: 3, нуклеотидные позиции 1-1358). Помимо этих последовательностей плазмида рАМ618 содержала векторную последовательность для ее поддержания в клетках Е. coli, состоящую из фрагмента Pcil-Stul плазмиды pUK21 (Vieira J. et. al., Gene. 1991, 100:189-94.) (SEQ ID NO: 3, нуклеотидные позиции 3818-4187) и фрагмента Ecl136II-PciI плазмиды pBlueScriptKS+ (Stratagen, USA) (SEQ ID NO: 3, нуклеотидные позиции 1359-3817).
Штамм DLT2 трансформировали смесью фрагментов плазмиды рАМ618, гидролизованной рестриктазами ЕсL136II и XhoI. Трансформантов отбирали на среде, содержащей лейцин и не содержащей триптофан. В локусе МОХ штамма DLT2 интегрирован селективный маркер, ген LEU2 S. cerevisiae, обеспечивающий штамму способность расти на среде без лейцина. При интеграции рекомбинантного гена в локус МОХ путем двойного кроссинговера, ген LEU2 должен замещаться (Фиг. 2Б). Поэтому полученные трансформанты проверяли на способность расти на среде, не содержащей лейцин. Неспособность трансформантов расти на среде без лейцина свидетельствовала об интеграции рекомбинантного гена в локус МОХ путем двойного кроссинговера. Один из таких трансформантов был обозначен КБТ-12/pEMmono-4.
Пример 3. Получение штамма H. polymorpha, содержащего несколько копий рекомбинантного гена, кодирующего полипептид ESC-Ag, под контролем промотора MAL1.
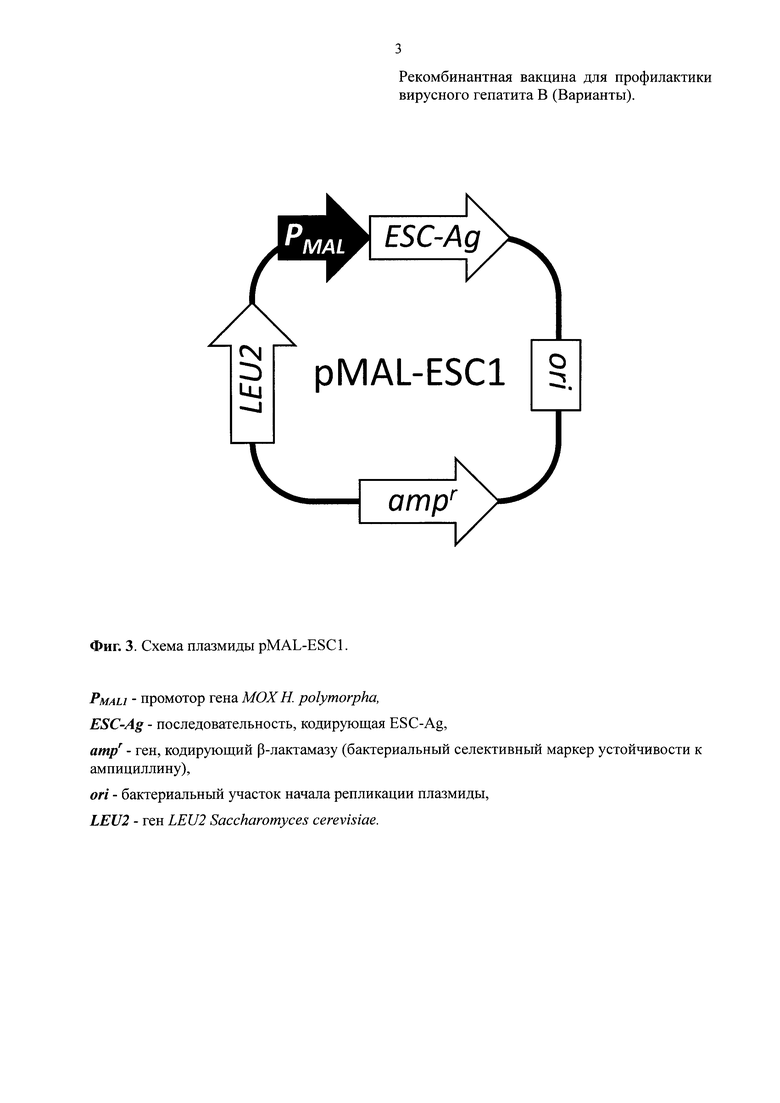
Получали плазмиду pMAL-ESC1 (Фиг. 3; SEQ ID NO: 4), которая отличалась от плазмиды pIR4-7 тем, что фрагмент, несущий промотор МОХ (SEQ ID NO: 2, нуклеотидные позиции 4390-4862) был заменен на фрагмент, несущий промотор гена MAL1 H. polymorpha DL1 (SEQ ID NO: 4, нуклеотидные позиции 1-1326).
Штамм DL1-L трансформировали плазмидой pMAL-ESC1. Трансформантов отбирали на среде, не содержащей лейцин. Несколько выросших трансформантов рассевали истощающим штрихом на твердую среду без лейцина, чтобы получить отдельные колонии. Среди выросших колоний отбирали самые крупные и снова сеяли на твердую среду без лейцина истощающим штрихом. Эту процедуру повторяли до тех пор, пока вырастающие при рассеве колонии не стали одинаковыми по скорости роста. По одному такому субклону нескольких независимых трансформантов отбирали для анализа продукции белка ESC-Ag. Один из таких трансформантов был обозначен KBT-14/pEMmalt-8.
Пример 4. Получение штамма Н. polymorpha, содержащего одну копию рекомбинантного гена, кодирующего полипептид ESC-Ag, под контролем промотора МОХ и одну копию под контролем промотора DAK.
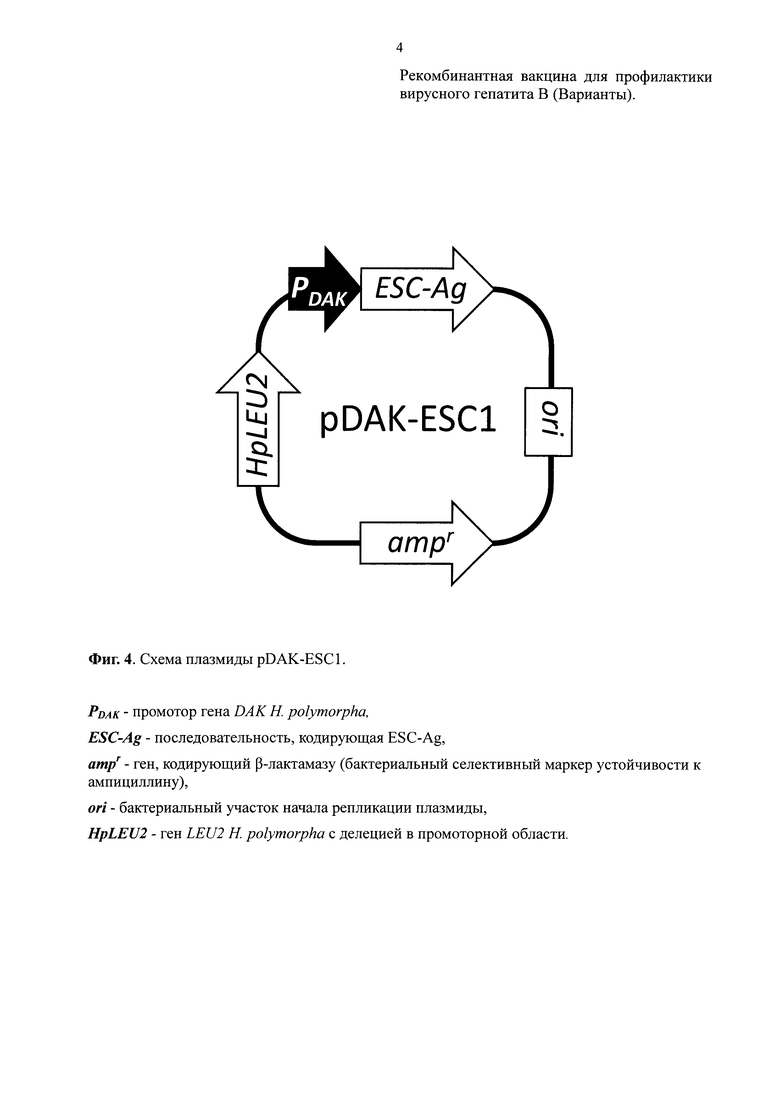
Получали плазмиду pDAK-ESC1 (Фиг. 4; SEQ ID NO: 5), которая содержала рекомбинантный ген, кодирующий полипептид ESC-Ag, под контролем промотора DAK Н. polymorpha DL-1 (SEQ ID NO: 5, нуклеотидные позиции 4404-5727) и последовательность вектора AMIpLD1 (Agaphonov et al., Yeas,. 1999, 15(7):541-51) (SEQ ID NO: 5, нуклеотидные позиции 43-4403) включая дрожжевой селективный маркер ген LEU2 Н. polymorpha с делецией в промоторной области, а также линкерную последовательность (SEQ ID NO: 5, нуклеотидные позиции 1-42).
Штамм КБТ-12/pEMmono-4 (см. Пример 2) трансформировали плазмидой pDAK-ESC1, гидролизованной рестриктазой BstEII. Трансформантов отбирали на среде, не содержащей лейцин. У быстро растущих трансформантов проверяли продукцию белка ESC-Ag. Один из таких трансформантов был обозначен КБТ-14/pEMmono-2.
Пример 5. Культивирование штаммов H. polymorpha КБТ 11/pEMmulti-7, КБТ 12/pEMmono-4 и КБТ-14/рЕМmono2.
Ферментацию штаммов-продуцентов дрожжей H. polymorpha осуществляли в два этапа. На первом этапе культивирования в режиме фед-бэтч при температуре 30°С наращивали биомассу в культуральной среде, содержащей 4% дрожжевого экстракта, 2% бакто-пептона и 4% глицерина. Процесс вели до истощения источника углерода в питательной среде и прекращения роста биомассы. На втором этапе культивирование проводили с подпиткой культуры путем непрерывной подачи 30%-ного раствора дрожжевого экстракта. Одновременно с подпиткой добавляли индуктор до концентрации 0.5-0.8% и поддерживали на этом уровне в течение 48-72 часов. Продолжительность всего процесса составляла около 96 часов.
Пример 6. Культивирование штамма H. polymorpha KBT-14/pEMmalt-8, содержащего рекомбинантный ген ESC-Ag под контролем промотора MALI.
Ферментацию штамма-продуцента KBT-14/pEMmalt-8 проводили в тех же условиях, что и штаммы с промоторами МОХ и DAK (Пример 5). За исключением того, что на втором этапе культивирования в качестве индуктора использовали 40% (вес/объем) раствор сахарозы, который добавляли порциями по 0.5% (вес сахарозы / объем культуральной жидкости) через каждые два часа в течение 48-72 часов.
Пример 7. Выделение и очистка рекомбинантного белка ESC-Ag из клеток H. polymorpha.
После ферментации белок ESC-Ag (аминокислотная последовательность - SEQ ID NO: 6) выделяли согласно опубликованной методике (Lunsdorf H. et al., Microbial Cell Factories, 2011, 10:48) с небольшими изменениями. Клетки из культуральной жидкости осаждали центрифугированием при 4000g в течение 30 минут при 4°С. Осажденную биомассу ресуспендировали до исходного объема в карбонатно-солевом буферном растворе для экстракции (50mM NaHCO3, 150 тМ NaCl, рН 8.0) и снова осаждали центрифугированием при 4000g в течение 30 минут при 4°С. Полученную таким образом отмытую биомассу ресуспендировали в буферном растворе для экстракции с добавлением 1.7mM EDTA и 2 mM PMSF до концентрации 400 г влажных клеток на литр суспензии. Далее клетки разрушали в проточном дезинтеграторе Dyno-Mill типа KDL при температуре 5-10°С, используя стеклянные шары диаметром 0.5-0.7 мм. Для удаления клеточного дебриса полученный гомогенизат центрифугировали при 4000g в течение 60 минут при 4°С.
Полученный осветленный гомогенизат подвергали ультрафильтрации в тангенциальном потоке на системе Sartocon через мембрану с порогом отсечения 300 kDa (Sartorius, Германия), одновременно концентрировали и переводили в фосфатно-солевой буферный раствор (10mM NaH2PO4, 150mM NaCl, 1.7mM EDTA, pH 6.8). При этом отделяется большая часть примесных низкомолекулярных белков.
Дальнейшую очистку проводили методом адсорбционной хроматографии на колонке с макропористым стеклом (МПС) с использованием технологии Streamline (хроматография в восходящем потоке). Концентрированный раствор ESC-Ag, полученный после ультрафильтрации, наносили на колонку при скорости 1 мл/мин, промывали 3-4 объемами фосфатно-солевого буферного раствора (рН 6.8) и элюировали карбонатным буфером (50mM NaHCO3, 150 mM NaCl, 1.7 mM EDTA, рН 9.5).
Объединенные фракции, содержащие ESC антиген (определяли по иммуноблоту и SDS-PAGE), концентрировали ультрафильтрацией в тангенциальном потоке на мембране 300 KDa и разделяли при помощи зонального ультрацентрифугирования (Beckman Coulter, USA) в градиенте плотности сахарозы (20-50% вес/вес). Фракции, содержащие целевой белок, объединяли, а затем очищали гельфильтрацией на сорбенте Toyopearl HW-65 (ToyoSoda Corp., Japan) в фосфатном буферном растворе (10mM NaH2РО4 0.03% Tween-20, рН 7.2).
Чистоту полученного таким образом рекомбинантного белка ESC-Ag определяли методом SDS-PAGE электрофореза. Если чистота белка была недостаточна (менее 95%) производили дополнительную очистку при помощи анионообменной хроматографии на сорбенте Toyopearl DEAE-650M.
В результате этих операций удается выделить около 30% антигена, присутствующего в осветленном клеточном лизате. Выход очищенного ESC-Ag составлял не менее 50 мг/л культуральной жидкости.
Пример 8. Анализ рекомбинантного ESC-Ag.
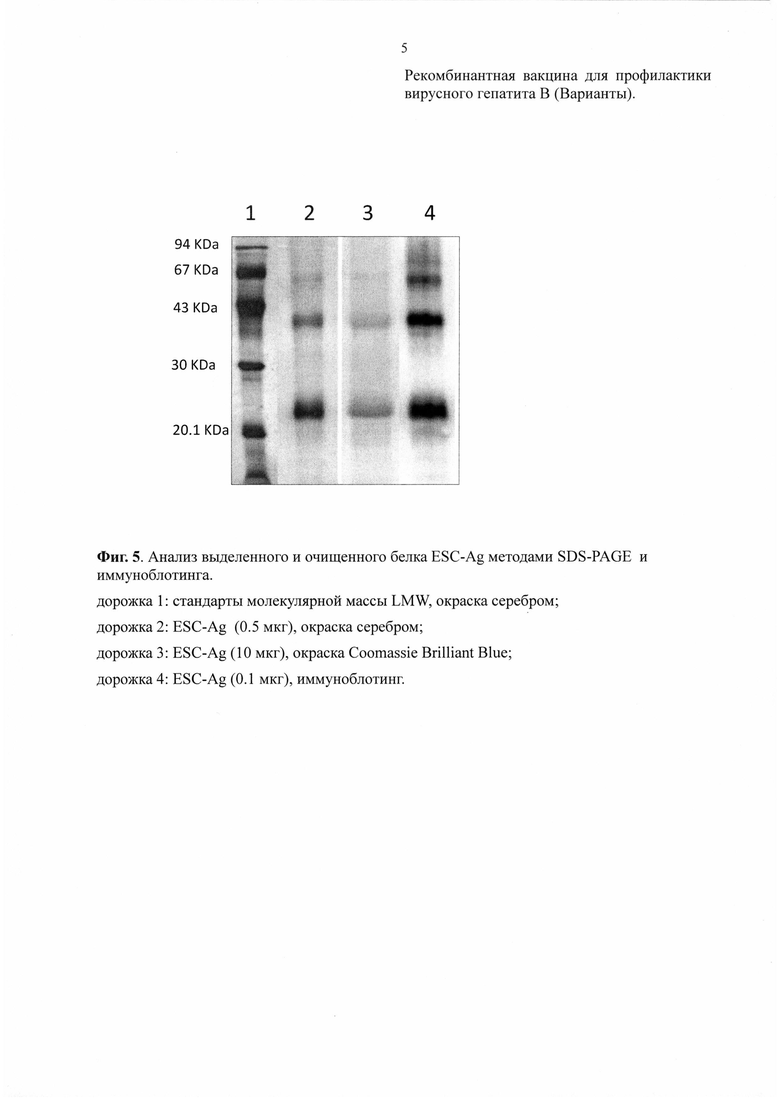
1) Чистота рекомбинантного ESC-Ag определяется электрофорезом в полиакриламидном геле в восстанавливающих условиях (SDS-PAGE) путем окрашивания с Coomassie Brilliant Blue R-250 и/или серебром. Анализ интенсивности полос отсканированных гелей проводится посредством компьютерной программы NIH Image (Фиг. 5).
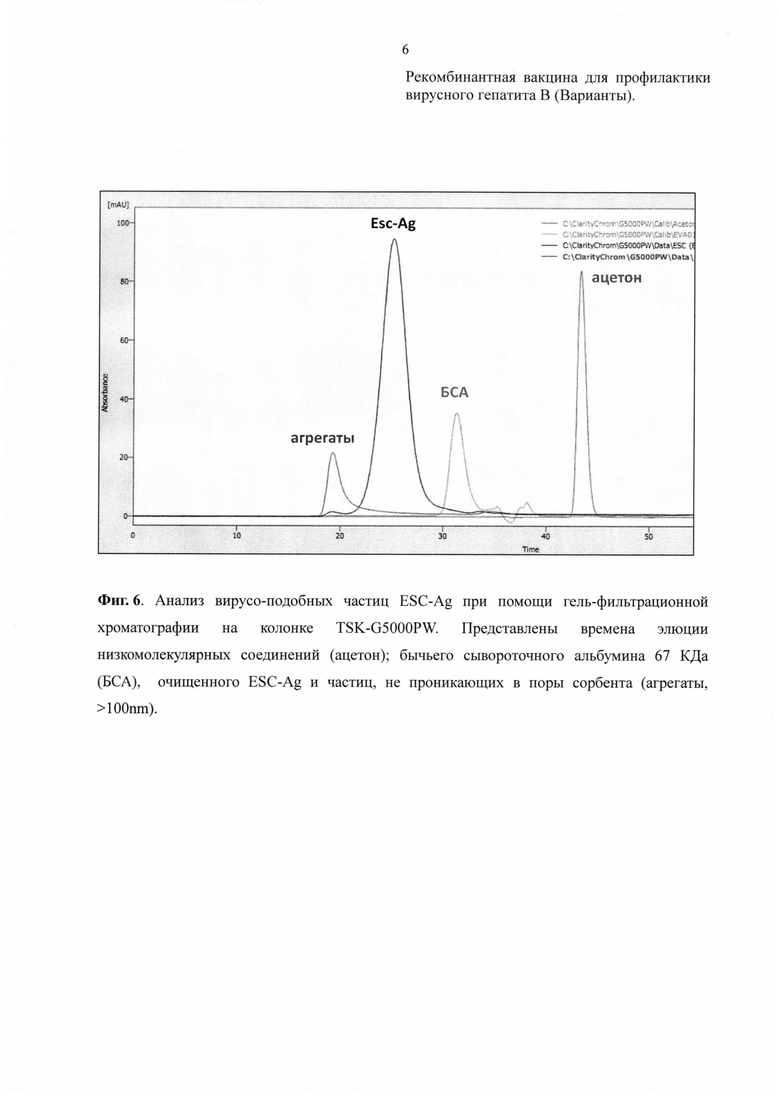
2) Образование вирусоподобных частиц (ВПЧ) рекомбинантного белка ESC-Ag подтверждали методом гель-фильтрационной хроматографии на полимерной колонке TSK G5000PW диаметром 7.5 мм и длиной 60 см, соединенной с предколонкой PrePW диаметром 7.5 мм и длиной 7.5 см (Tosoh Bioscience, Japan). Разделение проводили в фосфатном буферном растворе (10mM КН2РО4, (рН 7.2), 0.03% Tween-20) при скорости потока 0.6 мл/мин. Мониторинг процесса осуществляли по поглощению на 280 нм. Размер пор сорбента 1000Å позволяет отделить правильно собранные ВПЧ от мономеров и агрегатов Esc-антигена. Время элюции очищенного препарата ESC-Ag соответствует времени элюции вирусоподобных частиц 23-26 мин (Фиг. 6).
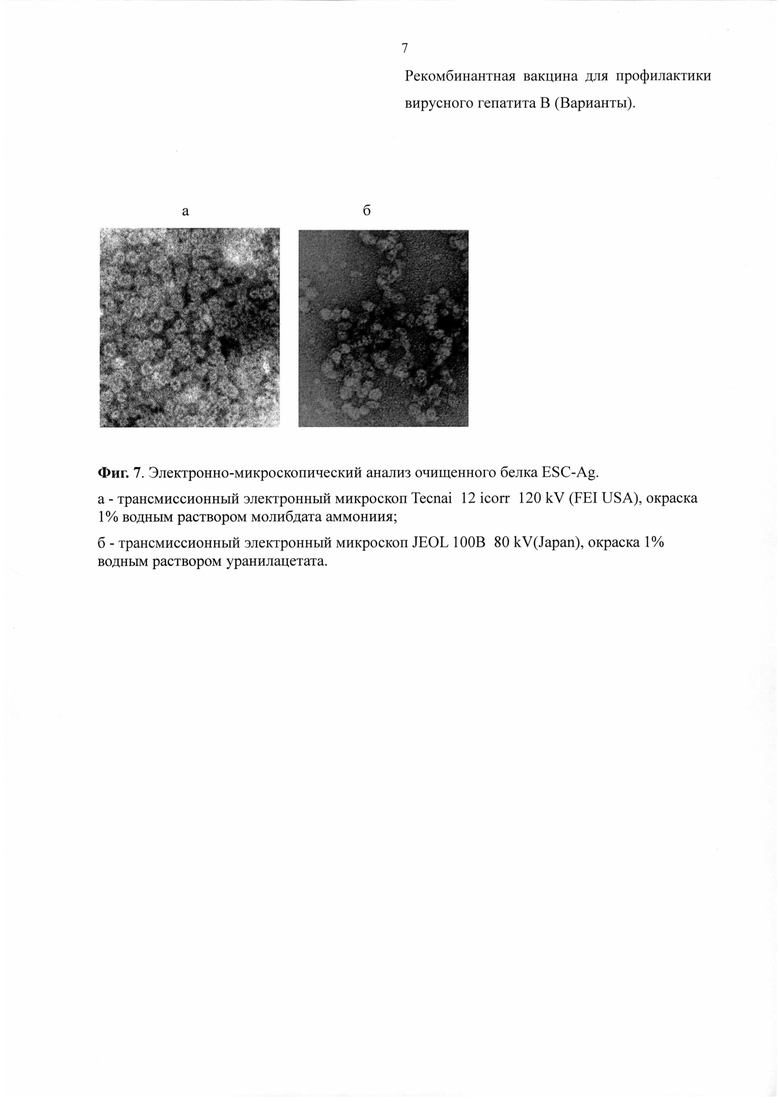
Электронно-микроскопический анализ полученного антигена показал наличие в образце частиц размером 15-25 нм. округлой и овальной формы. Ободок частиц имел низкую электронную плотность и относительно более темный центр при окраске 1% раствором молибдата аммония и уранилацетата (Фиг. 7). Эти данные подтверждают то, что полученный Esc-антиген собирается в вирусоподобные частицы.
3) Иммуноспецифичность рекомбинантного ESC антигена определяют методом иммуноблотинга. Первичные антитела для этой цели получали иммунизацией кроликов рекомбинантным HBsAg (ЗАО НПК «Комбиотех»), восстановленным в присутствии 2% SDS и 5% β-меркаптоэтанола. Из полученных высокоиммунных сывороток специфические поликлональные антитела выделяли на афинной колонке с коньюгированным рекомбинантным HBsAg (Крымский М.А. et al., Биофармацевтический Журнал, 2010, 2(5):8-15) Этими антителами окрашивали иммуноблот с последующей визуализацией при помощи специфических антител к иммуноглобулинам кроликов, коньюгированными с пероксидазой хрена, по методике улучшенной хемилюминисценции ECL (Amersham, UK). Рекомбинантный антиген ESC-Ag представлен в виде полосы мономера белка размером 22-25 кДа и слабых полос димера и тримера (Фиг. 5).
4) Серологический портрет рекомбинантных Esc антигенов проводили по методике, разработанной в лаборатории медиаторов и эффекторов иммунитета ГУ НИИЭМ им. Н.Ф. Гамалеи РАМН, с применением репрезентативной панели конъюгатов моноклональных антител к HBsAg. Поликлональные кроличьи (КАТ) и моноклональные мышиные (11F3, Н10, НВ4, 10D10, 5Н7, 4F5, Н2) антитела к HBsAg, а также их конъюгаты с пероксидазой хрена были получены в ГУ НИИЭМ им. Н.Ф. Гамалеи РАМН. Также использовали коммерческие моноклональные конъюгаты NF5, NE2 («Сорбент», Россия) и Х7 («Фармакс», Россия). Реактивность антигенных образцов с 11-ю пероксидазными конъюгатами антител исследовали методом ИФА. Оптимальные концентрации для каждого конъюгата подбирались в предварительных опытах с помощью внутреннего производственного стандарта, оттитрованного по отраслевому стандартному образцу. Относительную реактивность образцов оценивали по сравнению с соответствующей реактивностью внутреннего производственного стандарта (ВПС) HBsAg. За относительную реактивность каждого образца (обр) с исследованными конъюгатами принимали отношение Собр/Свпс×103. Реактивность образца с данным конъюгатом считали дефектной, если отношение Собр/Свпс для этого конъюгата не превышало 100. Полученные результаты представлены в таблице 1.
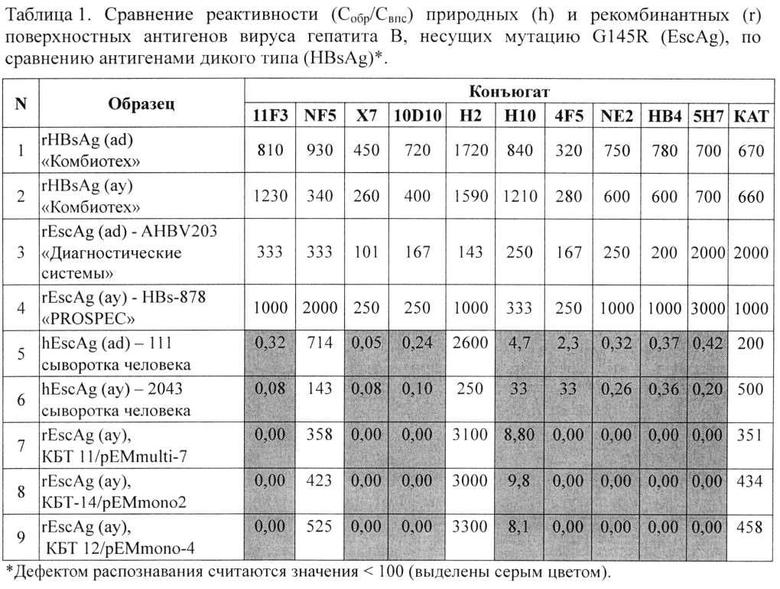
Из таблицы видно, что рекомбинантные антигены HBsAg субтипов «ау» и «ad» - стандартные компоненты вакцины, производимой ЗАО НПК «Комбиотех», распознаются всей панелью конъюгатов. Это означает, что их серологические и конформационные характеристики полностью соответствуют HBsAg дикого типа. Серологическое портретирование антигенов с эскейп-мутацией G145R показало, что все три исследуемых рекомбинантных препарата, рассматриваемые в данном изобретении, (Таблица 1, строки 7, 8, 9) имеют многочисленные дефекты в реактивности с большинством конъюгатов (Табл 1, выделение серым фоном). Точно такую же картину дефектов (или серологический портрет) показывают нативные Esc-антигены, выделенные из человеческих сывороток, несущих природную мутацию G145R (Таблица 1, строки 5, 6), В то же время рекомбинантные образцы AHBV203 (НПО «Диагностические системы», Нижний Новгород) и HBs-878 («PROSPEC», Израиль), тоже содержащие мутацию G145R, не имели ни одного дефекта распознавания моноклональными конъюгатами, т.е. конформация этих рекомбинантных молекул полностью соответствовала HBsAg дикого типа, а не мутантного антигена, и они не могли быть использованы в качестве претендентов для создания искомого вакцинного препарата.
Таким образом, серологическое портретирование показывает полное конформационное соответствие рекомбинантных Esc антигенов, полученных из штаммов, описанных в данном изобретении, с нативными мутантами G145R, в отличие от Esc-Ag других производителей.
Культурально-морфологические особенности штаммов: клетки округлой формы, небольшие по размеру, на агаризованной среде YPD образуют крупные круглые колонии с выраженной выпуклой серединой. Хранение штаммов - при -70°С в виде суспензии клеток в стерильном 30-50%-ном растворе глицерина.
Генетические особенности: Штаммы не являются зоопатогенными или фитопатогенными.
Способ, условия и состав сред для размножения штаммов: инкубирование прокачиванием при 30°С в питательной среде состава: 2% пептона, 1% дрожжевого экстракта, 2% глюкозы.
Условия и состав среды для ферментации: прокачивание при 30°С и рН 5.5-6.0 в питательной среде, содержащей до 4% глицерина.
Выделенный и очищенный ESC-Ag используют для создания рекомбинантной вакцины для профилактики вирусного гепатита В.
Полученные вакцинные композиции представлены в следующих вариантах:
1) Вакцина содержит поверхностный антиген вируса гепатита В, имеющий мутацию G145R (ESC-Ag), полученный путем культивирования рекомбинантного штамма дрожжей H. Polymorpha КБТ-11 /pEMmulti-7 или H. Ρolymorpha КБТ-12/pEMmono-4, или H. Polymorpha KBT-14/pEMmalt-8, или H. Polymorpha КБТ-14/рЕМmono2, адъювант и физиологически приемлемый разбавитель;
2) Вакцина содержит в равных долях смесь поверхностного антигена вируса гепатита В, имеющего мутацию G145R (ESC-Ag), полученного путем культивирования рекомбинантного штамма дрожжей Н. Polymorpha КБТ-11/ pEMmulti-7 или Н. Polymorpha КБТ-12/pEMmono-4, или Н. Polymorpha KBT-14/pEMmalt-8, или H. Polymorpha КБТ-14/рЕМmono2, и рекомбинантного HBsAg серотипа «ayw», адъювант и физиологически приемлемый разбавитель;
3) Вакцина содержит в равных долях смесь поверхностного антигена вируса гепатита В, имеющего мутацию G145R (ESC-Ag), полученного путем культивирования рекомбинантного штамма дрожжей H. Polymorpha КБТ-11/ pEMmulti-7 или Н. Polymorpha КБТ-12/pEMmono-4, или H. Polymorpha КБТ-14/ pEMmalt-8, или Н. Polymorpha КБТ-14/рЕМmono2, и рекомбинантного HBsAg серотипа «adw», адъювант и физиологически приемлемый разбавитель;
4) Вакцина содержит в равных долях смесь поверхностного антигена вируса гепатита В, имеющего мутацию G145R (ESC-Ag), полученного путем культивирования рекомбинантного штамма дрожжей H. Polymorpha КБТ-11/pEMmulti-7 или Н. Polymorpha КБТ-12/pEMmono-4, или H. Polymorpha KBT-14/pEMmalt-8, или H. Polymorpha КБТ-14рЕМmono2, и рекомбинантного HBsAg серотипов «ayw» и «adw», адъювант и физиологически приемлемый разбавитель.
5) Вакцина вариантов 1, 2, 3, 4 дополнительно содержит консервант - мертиолят.
Конкретные примеры композиций вакцин для профилактики вирусного гепатита В:
1. Вакцина для профилактики вирусного гепатита В содержит:
20 мкг ESC-Ag, полученного путем культивирования рекомбинантного штамма дрожжей H. Polymorpha KBT-11/pEMmulti-7 или H. Polymorpha КБТ-12/pEMmono-4, или H. Polymorpha KBT-14/pEMmalt-8, или H. Polymorpha КБТ-14/pEMmono2.
0.5 мг геля гидроокиси алюминия,
до 1.0 мл фосфатно-солевого буфера (ФСБ), представленного 50 mM Na-фосфатного буфера, рН 6.8 и 0.13 M NaCl.
2. Вакцина для профилактики вирусного гепатита В содержит:
10 мкг ESC-Ag, полученного путем культивирования рекомбинантного штамма дрожжей H. Polymorpha KBT-11/pEMmulti-7 или H. Polymorpha КБТ-12/pEMmono-4, или H. Polymorpha KBT-14/pEMmalt-8, или H. Polymorpha КБТ-14/pEMmono2.
10 мкг HBsAg серотипа ayw,
0.5 мг геля гидроокиси алюминия,
до 1.0 мл фосфатно-солевого буфера (ФСБ), представленного 50 mM Na-фосфатного буфера, рН 6.8 и 0.13 M NaCl.
3. Вакцина для профилактики вирусного гепатита В содержит:
10 мкг ESC-Ag, полученный путем культивирования рекомбинантного штамма дрожжей H. Polymorpha КБТ-11/ pEMmulti-7 или H. Polymorpha КБТ-12/pEMmono-4, или H. Polymorpha KBT-14/pEMmalt-8, или H. Polymorpha КБТ-14/pEMmono2,
10 мкг HBsAg серотипа adw,
0.5 мг геля гидроокиси алюминия,
до 1.0 мл фосфатно-солевого буфера (ФСБ), представленного 50 тМ Na-фосфатного буфера, рН 6.8 и 0.13 M NaCl.
4. Вакцина для профилактики вирусного гепатита В содержит:
8 мкг ESC-Ag, полученного путем культивирования рекомбинантного штамма дрожжей H. Polymorpha KBT-11/pEMmulti-7 или H. Polymorpha КБТ-12/pEMmono-4, или H. Polymorpha КБТ-14/pEMmalt-8, или H. Polymorpha КБТ-14/рЕМmono2,
8 мкг HBsAg серотипа ayw,
8 мкг HBsAg серотипа adw,
60 мкг мертиолята,
0,5 мг геля гидроокиси алюминия,
до 1,0 мл фосфатно-солевого буфера (ФСБ), представленного 50 mM Na-фосфатного буфера, рН 6.8 и 0.13 M NaCl.
Согласно изобретению вакцины для профилактики вирусного гепатита В можно получать как с введением в их состав консерванта - мертиолята, так и без него. Отсутствие в вакцине консерванта не сказывается на ее качестве.
Для детей готовят вакцины в прививочной дозе 0.5 мл.
Иммуногенность рекомбинантных вакцин, определенная в тесте на мышах линии Balb/c с массой 12-14 г. в расчете на ED50 (доза антигена, вызывающая сероконверсию у 50% мышей), составляет 0.05-0.10 мкг.
Токсичность вакцины определяли путем введения белым мышам 0.5 мл (60 мкг) вакцины внутрибрюшинно и 0.5 мл (60 мкг) вакцины морским свинкам подкожно. Наблюдение в течение 7 суток показало, что вакцина не токсична.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ МУТАНТНОГО ПОВЕРХНОСТНОГО АНТИГЕНА ВИРУСА ГЕПАТИТА В (ВАРИАНТЫ) | 2015 |
|
RU2586513C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ПОВЕРХНОСТНОГО АНТИГЕНА ВИРУСА ГЕПАТИТА В СЕРОТИПА "ayw" | 2015 |
|
RU2586511C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА В (ВАРИАНТЫ) | 2003 |
|
RU2238105C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ГЛАВНОГО КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 16 | 2014 |
|
RU2546241C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА Е У ЖИВОТНЫХ | 2012 |
|
RU2501568C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО КАПСИДНОГО БЕЛКА ВИРУСА ГЕПАТИТА Е И РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА Е | 2012 |
|
RU2501809C1 |
| Способ получения рекомбинантной вакцины для профилактики папилломавирусной инфекции человека, рекомбинантная вакцина | 2018 |
|
RU2681174C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ГЛАВНОГО КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 56 | 2014 |
|
RU2546240C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ HANSENULA POLYMORPHA - ПРОДУЦЕНТ КАПСИДНОГО БЕЛКА ВИРУСА ГЕПАТИТА Е | 2012 |
|
RU2493249C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ГЛАВНОГО КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 18 | 2014 |
|
RU2546242C1 |




Изобретение относится к биотехнологии, а именно генной инженерии. Предложена рекомбинантная вакцина для профилактики вирусного гепатита В. Вакцина создана на основе поверхностного антигена вируса гепатита В, имеющего мутацию G145R (ESC-Ag), полученного путем культивирования рекомбинантного штамма дрожжей H. polymorpha KBT-11/pEMmulti-7 или H. polymorpha КБТ-12/pEMmono-4, или H. polymorpha KBT-14/pEMmalt-8, или H. polymorpha КБТ-14/рЕМmono2. Вакцина может содержать смесь в равных долях ESC-Ag, HBsAg серотипа «ayw» и/или «adw». Вакцина может содержать только антиген вируса гепатита В, имеющий мутацию G145R (ESC-Ag). Полученные вакцины высокоиммуногенны, нетоксичны, не обладают побочными эффектами. 2 н. и 2 з.п.ф-лы, 7 ил., 1 табл., 8 пр.
1. Рекомбинантная вакцина для профилактики вирусного гепатита В, содержащая эффективное количество поверхностного антигена вируса гепатита, адъювант и физиологически приемлемый разбавитель, отличающаяся тем, что в качестве антигена вируса гепатита В рекомбинантная вакцина содержит в равных долях смесь поверхностного антигена вируса гепатита В, имеющего мутацию G145R (ESC-Ag), полученного путем культивирования рекомбинантного штамма дрожжей Н. Polymorpha КБТ-11/ pEMmulti-7 или Н. Polymorpha КБТ-12/pEMmono-4, или Н. Polymorpha КБТ-14/pEMmalt-8, или Н. Polymorpha КБТ-14/рЕМmono2, и HBsAg серотипа ayw и/или HBsAg серотипа adw.
2. Рекомбинантная вакцина для профилактики вирусного гепатита В по п. 1, отличающаяся тем, что дополнительно содержит мертиолят.
3. Рекомбинантная вакцина для профилактики вирусного гепатита В, содержащая эффективное количество поверхностного антигена вируса гепатита, адъювант и физиологически приемлемый разбавитель, отличающаяся тем, что в качестве антигена вируса гепатита В рекомбинантная вакцина содержит поверхностный антиген вируса гепатита В, имеющий мутацию G145R (ESC-Ag), полученный путем культивирования рекомбинантного штамма дрожжей Н. Polymorpha КБТ-11/pEMmulti-7 или Н. Polymorpha КБТ-12/pEMmono-4, или Н. Polymorpha КБТ-14/ pEMmalt-8, или Н. Polymorpha КБТ-14/рЕМmono2.
4. Рекомбинантная вакцина для профилактики вирусного гепатита В по п. 3, отличающаяся тем, что дополнительно содержит мертиолят.
| US 5639637 A, 17.06.1997 | |||
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА В (ВАРИАНТЫ) | 2003 |
|
RU2238105C1 |
| US 7038035 B1, 02.05.2006 | |||
| US 6099840 A, 08.08.2000.. | |||

