Настоящее изобретение относится к семенам гибридного салата (Lactuca sativa), а также к способу получения указанных семян. Изобретение также относится к гибридным растениям Lactuca sativa и к клеткам гибридных растений Lactuca sativa.
К числу причин, обуславливающих производство в широком масштабесемян гибридных растений относят, в частности, гетерозис (или гибридную силу), гомеостаз (стабильность растений в различных условиях окружающей среды), способность к накоплению генов устойчивости к насекомым, грибам, бактериям или вирусам, а также способность адаптироваться к абиотическим стрессам, в том числе, например, устойчивость к экстремальным температурам, т.е. к температурам ниже 5°C или выше 30°C, или устойчивость к низкой освещенности.
Следует отметить, например, что для, салата огородного (Lactuca sativa) невозможно или чрезвычайно трудно обеспечить накопление некоторых генов устойчивости в гомозиготных линиях, поскольку указанные гены находятся в одном и том же локусе (по одному в каждой аллели) или в очень близких локусах.
Поэтому представляет большой интерес получение гибридов F1 салата огородного, поскольку это, могло бы обеспечить, в частности, получение в промышленном масштабе гибридов, в которых накоплено несколько обладающих сельскохозяйственной ценностью кодоминантных или доминантных генов, находящихся в различных аллелях одного локуса или в очень близких локусах.
Род Lactuca насчитывает более 100 видов, среди которых - Lactuca sativa (культурный вид), Lactuca saligna (дикорастущий вид), Lactuca serriola (дикорастущий вид) и Lactuca virosa (дикорастущий вид).
Lactuca sativa представляет собой диплоидный (2n=18) вид с высокой автогамностью; образование его семян на 98% происходит путем самоопыления. Тычинки (мужские органы) сгруппированы и образуют тычиночную трубку, в которой раскрываются пыльники. Пестик (женский орган) состоит из завязи, столбика и раздвоенного рыльца. При раскрытии цветка столбик удлиняется внутрь тычиночной трубки, вследствие чего рыльце покрывается пыльцой и растение подвергается самооплодотворению. Одно растение Lactuca sativa обычно дает от 0,5 до 6 граммов и даже до 10 граммов семян в зависимости от условий возделывания; в каждом грамме обычно насчитывается от 600 до 1000 семян.
Классическая технология получения гибридов растений в промышленном масштабе состоит в возделывании двух сортов, используемых в качестве "родительских форм", на близком расстоянии друг от друга и в применении насекомых для опыления, при этом одна из родительских форм является носителем мужской стерильности, что позволяет исключить контаминацию вследствие самоопыления. Затем собирают семена мужской стерильной родительской формы.
Подобная технология позволяет накапливать целевые гены, присутствующие у каждой из родительских форм в гомозиготной форме, при сохранении однородности делянки, поскольку поколение F1 на стадии сбора урожая (до раскрытия цветков) имеет фенотип, однородный на 100%.
Обнаружено, что получение гибридов латука огородного (Lactuca sativa) в широком масштабе по данной технологии является очень трудным. В самом деле, цветок Lactuca sativa раскрывается только один раз, всего на нескольких часов, например, от одного до четырех часов, во время суток (ранним утром), когда обычные насекомые-опылители, в том числе некоторые виды пчел (Apis mellifera) или шмелей (Bombus spp.) неактивны.
Опыление Lactuca sativa под действием ветра также не происходит.
При этом идентификация насекомого-опылителя, пригодного для применения в получении гибридов салата, также затруднена по многим причинам, среди которых можно назвать, в частности, следующие:
- длительность раскрытия цветка является краткой, кроме того раскрытие происходит утром, что затрудняет идентификацию насекомого, посещающего цветки;
- цветки раскрываются только один раз, вследствие чего для максимального и надежного опыления мужских стерильных цветков выбранное насекомое-опылитель должно представлять собой насекомое, посещающее цветки ежедневно.
Кроме того, получение гибридов F1 требует наличия "мужской" линии и "женской" линии.
"Женская" линия может быть получена путем стерилизации растений вручную. Однако в случае салата, каждый цветок которого раскрывается только один раз и на очень короткое время (несколько часов), данный способ трудно использовать в коммерческом масштабе вследствие того, что он требует очень большого количества рабочей силы.
"Женская" линия также может быть получена путем создания стерильности на генном (или ядерном) уровне или на уровне цитоплазмы, что проявляется в отсутствии пыльников, их пустоте или нежизнеспособности пыльцы. Подобная стерильность передается потомству частично в случае ядерной (или генетической) стерильности и полностью - в случае цитоплазматической стерильности.
В работах Губара и Такасаки (Goubara & Takasaki, Appl. Entomo. Zool. 38(4): 571-581, 2003 [1]) описано проведение исследований на открытом и закрытом грунте с целью идентификации потенциальных насекомых-опылителей, которых можно использовать для получения гибридов салата Lactuca sativa. Проведенные наблюдения для 22 видов насекомых (включая 21 вид пчел и один вид мух, питающихся нектаром, Syrphidae Eristalis tenax), позволили выделить в качестве наилучшего потенциального опылителя пчелу Lagioglossum villosulum trichopse.
Goubara & Takasaki {Appl. Entomo. Zool. 39(1): 163-189, 2004 [2]) также проводили эксперименты по маломасштабной гибридизации между салатом, имеющим ген мужской стерильности, и мужским фертильным салатом на закрытом грунте в присутствии пчелы Lagioglossum villosulum trichopse. Авторы сообщают о получении гибридов F1 салата, однако с очень низким выходом.
На настоящее время других экспериментов по получению гибридов Lactuca sativa с применением насекомых-опылителей не известно.
До сих пор ни один сортовой гибрид F1 салата не производится в коммерческом масштабе.
В настоящем изобретении используются следующие термины в приведенном ниже значении.
"Салат огородный" (латук огородный) обозначает вид Lactuca sativa. Существует пять основных сортогрупп салата огородного (см. Фиг.1): Lactuca sativa var. angustana (спаржевый салат), Lactuca sativa var, capitata (кочанный салат), Lactuca sativa var. crispa (салат Батавия, или Айсберг), Lactuca sativa var. longifolia (салат ромен) и Lactuca sativa var. acephala (листовой салат). Настоящее изобретение охватывает применение каждого из указанных типов салата.
"Опыление" обозначает явление переноса пыльцы с пыльника на рыльце того же или другого цветка. Подобная половая система является преимущественным способом размножения цветковых растений (покрытосеменных и голосеменных). Данная система позволяет зерну пыльцы достичь рыльца, затем при помощи столбика образовать пыльцевую трубку, доходящую до семязачатка, и наконец оплодотворить его.
"Самоопыление" обозначает опыление особи или биотипа собственной пыльцой; при этом образовавшиеся особи называют самоопыленными.
Термин "автогамия" обозначает способность растения к самооплодотворению, при котором обе гаметы происходят от одной и той же особи.
Термин "аллогамия" обозначает явление, при котором цветки особи оплодотворяются пыльцой, происходящей от одной или нескольких других особей.
"Насекомое-опылитель" обозначает насекомых (в том числе пчел, бабочек, двукрылых или некоторых жесткокрылых), которые при посещении цветков (например, в поисках нектара) сталкиваются с тычинками и тем самым собирают несколько зерен пыльцы, которые они затем оставляют на другом цветке.
Термин "локус" обозначает место, занимаемое геном или аллелью в хромосоме.
"Аллель" обозначает внутривидовые варианты одного гена, расположенного в определенном месте хромосомы (локусе), Различные аллели одного гена порождают различные проявления одного признака.
"Кластер" обозначает два гена или более, расположенные поблизости один от другого в одной хромосоме. Кластеры генов устойчивости, найденные у салата, описаны, в частности, Кессели с соавт.(Kesseli et al. [12]).
"Доминантный ген" обозначает ген, сообщающий фенотип вне зависимости от того, имеется ли он в обеих хромосомах пары или только в одной из них.
"Рецессивный ген" обозначает ген, сообщающий фенотип только в том случае, если он присутствует в каждой из двух гомологичных хромосом.
"Кодоминантность" обозначает свойство, при котором в фенотипе совместно проявляются свойства, обуславливаемые и одной и второй аллелями гена. У гетерозиготной особи, имеющей две кодоминантных аллели гена, в фенотипе полностью проявляется генотипическая информация, содержащаяся в данных генах.
"Гетерозиготный" обозначает клетку или особь, имеющую два различных аллельных гена в определенном локусе одной пары хромосом.
"Гомозиготный" обозначает клетку или особь, имеющую два идентичных аллельных гена в определенном локусе одной пары хромосом.
"Гибрид" обозначает результат скрещивания особей с различной генетической конституцией, предпочтительно одного вида..
"Гибрид F1" обозначает первое поколение потомства, полученное в результате скрещивания особей с различной генетической конституцией. Вследствие этого гибриды F1 являются гетерозиготными по меньшей мере по одному гену.
"Возвратное скрещивание" (или обратное скрещивание, или "бэк-кросс») обозначает скрещивание гибрида с одной из его родительских форм. "Культивар" обозначает сорт.
"Генотип" обозначает совокупность генетического материала, имеющегося у особи и составляющего его «наследственный капитал».
Термин "фенотип" обозначает совокупность морфологических или функциональных признаков, характерных для особи и соответствующих как выраженной составляющей генотипа, так и явлениям, определяемым внешней средой.
"Фенотип, обладающий сельскохозяйственной ценностью" обозначает фенотип, полученный, например, при скрещивании двух гомозиготных генотипов и обладающий свойствами, представляющими интерес для сельского хозяйства, в том числе, например, совокупностью признаков устойчивости к различным патогенам или насекомым, гибридной силой (т.е. средним уровнем одного из признаков гибрида, превосходящим средний уровень того же признака у обеих родительских форм), гомеостазом, способностью приспосабливаться к абиотическим стрессам, морфологическими свойствами, в том числе цветом, формой, гибкостью или жесткостью листьев, составом питательных веществ или вкусовыми качествами растения.
"Гетерозис" или "гибридная сила" обозначает явление, при котором гибрид F1 значительно превосходит лучшую из его родительских форм по одному или нескольким признакам, в частности, по силе.
Термин "гомеостаз" обозначает способность растения приспосабливаться к окружающей среде, в том числе к различным характеристикам окружающей среды.
Термин "мужской стерильный" обозначает растение, неспособное к размножению самоопылением вследствие стерильности мужских органов цветков. Например, возможными являются нефункциональность пыльцы или структурные аномалии мужских органов размножения, например, выстилающего слоя (tapetum) пыльников.
"Цитоплазматическая мужская стерильность" обозначает стерильность, равномерно передающуюся от матери в соответствии с цитоплазматическим механизмом наследственности.
"Ядерная (или генная) мужская стерильность" обозначает стерильность, передающуюся по законам Менделя, переносимую ДНК ядра и зависящую либо от рецессивного гена, либо от доминантного гена.
"Моногенная мужская стерильность" обозначает мужскую стерильность, связанную с одним геном.
"Мультигенная мужская стерильность" обозначает мужскую стерильность, связанную с несколькими генами.
"Устойчивость" обозначает способность сорта к ограничению роста и развития определенного патогена или вредителя и/или вызываемых ими повреждений по сравнению с восприимчивыми сортами в сходных условиях окружающей среды и давления данного патогена или данного вредителя. Подобные сорта, однако, могут проявлять некоторые симптомы болезни или слегка повреждаться в случае сильного давления данного патогена или данного вредителя.
"Стандартная, или высокая устойчивость" обозначает способность сорта к значительному ограничению роста и развития определенного патогена или вредителя в условиях нормального давления данного патогена или данного вредителя по сравнению с восприимчивыми сортами. Подобные сорта, однако, могут проявлять симптомы или повреждаться в случае сильного давления данного патогена или данного вредителя.
"Промежуточная, или умеренная устойчивость" обозначает способность сорта к ограничению роста и развития определенного патогена или вредителя при наличии более выраженных симптомов или более сильных повреждений по сравнению с сортами с высокой/стандартной устойчивостью. Сорта с промежуточной устойчивостью проявляют симптомы или повреждаются менее сильно, чем это наблюдается у восприимчивых сортов в сходных условиях окружающей среды и/или силы давления патогена или вредителя.
"Молекулярный маркер" обозначает особый фрагмент ДНК, обнаруживаемый в полном геноме особи, который можно применять для локализации целевого гена или подтверждения наследования особью определенного свойства одного из организмов-родителей. Молекулярный маркер может (не обязательно) представлять собой кодирующую последовательность. При генетическом скрещивании целевой ген обычно сохраняет сцепленность с молекулярным маркером. Обнаружение молекулярного маркера в этом случае позволяет осуществить идентификацию особей, несущих целевой ген, без необходимости в определении последовательности данного гена.
Применение молекулярных маркеров в сельском хозяйстве обеспечивает быстроту исследования растений в ходе селекции и отбор тех из них, которые обладают искомыми характеристиками. В некоторых случаях наличие маркеров, сцепленных с признаком, позволяет селекционеру не проводить некоторые испытания фенотипов или наблюдения над ними. В частности, применение определенных молекулярных маркеров гена мужской стерильности, предпочтительно доминантного гена мужской стерильности, обеспечивает ранний отбор мужских стерильных растений до цветения. Подобный отбор позволяет удалить мужские фертильные растения, возможно, составляющие часть популяции растений, используемых в качестве условно "женских" (мужских стерильных) родительских форм для получения семян гибридов, например, в случае применения доминантного гена мужской стерильности, вследствие чего снижается риск контаминации, связанный с самооплодотворением указанных мужских фертильных растений.
Настоящее изобретение относится к семенам, растениям и клеткам гибридных растений Lactuca sativa, которые обладают генотипом мужской стерильности и являются гетерозиготными по меньшей мере по одному гену, не связанному с мужской стерильностью и сообщающему растению распознаваемый фенотип.
Мужская стерильность, которой обладают семена, растения или клетки растений Lactuca sativa, может иметь ядерную или цитоплазматическую природу. В случае цитоплазматической природы стерильность передается женской родительской формой. Подобная стерильность часто является следствием взаимодействия между митохондриями, находящимися в цитоплазме, и ядерными генами. Цитоплазматическая мужская стерильность отличается появлением потомства, на 100% обладающего мужским стерильным фенотипом, в отсутствие в геноме мужской родительской формы гена, восстанавливающего фертильность. Мужская стерильность потомства может быть восстановлена при наличии в геноме "мужской" родительской формы гена (Rf), восстанавливающего фертильность.
Настоящее изобретение относится, в частности, к случаю, когда генотип мужской стерильности связан по меньшей мере с одним ядерным геном и при котором семя, клетка или растение является гетерозиготным по гену или генам, обуславливающим мужскую стерильность.
Ядерная мужская стерильность обусловлена одним или несколькими генами мужской стерильности, передающимися в ядерной ДНК, при этом данные ген или гены могут быть доминантными или рецессивными.
К числу указанных генов мужской стерильности, в частности, относится доминантный ядерный ген Ms7, описанный Райдером (Ryder) (J.Amer. Soc. Hort. Sci 96(6) 826-828, 1971 [8]). Мужские стерильные растения являются гетерозиготными, Ms7ms7. Фертильные растения являются гомозиготными ms7ms7. Получение доминантных гомозигот Ms7Ms7 остается очень трудным и даже невозможным. К тому же, по Райдеру, доминантный характер упомянутой мужской стерильности является препятствием для применения данного гена при получении гибридов салата. Образец семян Lactuca sativa, полученных способом согласно настоящему изобретению, поступивший в NCIMB (Национальная коллекция промышленных, пищевых и морских микроорганизмов Великобритании, NCIMB Ltd., Ferguson Building, Craibstone Estate, Баксберн, Абердин (Bucksburn, Aberdeen), A21 9YA, Шотландия, Великобритания) 13 февраля 2007 г.под входящим номером NCIMB 41470, является источником гена Ms7. Кроме того, молекулярный маркер RAPD ВА05-675 (SEQ ID NO: 2), разработанный авторами настоящего изобретения, заслуживает особого интереса и помогает производить отбор мужских стерильных растений салата Ms7, поскольку он обеспечивает возможность проведения испытаний на молекулярном уровне на ранней стадии до цветения с целью идентификации мужских стерильных растений, используемых при дальнейшем скрещивании. Обнаружение (детектирование) данного маркера, последовательность которого представлена на Фиг.8, позволяет идентифицировать мужские стерильные растения в среднем в 96% случаев у всех сортотипов салата (Табл.18).
Линдквистом (Lindqvist) также были идентифицированы три рецессивных гена мужской стерильности, ms1, ms2 и ms3 {Hereditas 46: 387-470, 1960 [3]); еще три подобных гена, ms4, ms5 и ms6, были идентифицированы Райдером (Ргос.Am. Soc. Hort. Sci 83: 585-595, 1963 [6], Proc. Am. Soc. Hort. Sci 91: 366-368, 1967 [7]). Однако в своей опубликованной в 1979 г.работе (Leafy Salad Vegetables, p.30 [9]) Райдер утверждает, что, поскольку все гены стерильности ms1, ms2, ms3, ms4, ms5, ms6 и Ms7 являются ядерными, возможность их использования для получения гибридов F1 является маловероятной. Вопреки данному утверждению, авторы настоящего изобретения успешно получили гибридные семена, растения и клетки с применением мужской стерильности ядерной природы.
Ядерная мужская стерильность может быть мультигенной или моногенной. В случае мультигенной мужской стерильности она может быть получена, например, при помощи комплекса рецессивных генов, например, путем накопления генов ms1, ms2 и ms3, указанных выше. Настоящее изобретение предпочтительно относится к случаю моногенной мужской стерильности.
Ядерная мужская стерильность может быть доминантной или рецессивной. Настоящее изобретение, в частности, относится к случаю, когда ген мужской стерильности является единственным и доминантным. Еще более конкретно, настоящее изобретение относится к случаю, когда мужская стерильность связана с доминантным геном Ms7, упомянутым выше.
Геном семян, растений или клеток согласно настоящему изобретению предпочтительно содержит двунитевую последовательность ДНК из 650-700 нуклеотидов, например, из 655-695, 660-690, 665-685, 670-680, 673-677 либо из 674, 675 или 676 нуклеотидов, 5'-концы каждой из двух нитей которой начинаются с последовательности "5' TGCGTTCCAC 3'" (SEQ ID NO:1). Согласно предпочтительному варианту осуществления, геном семян, растений или клеток согласно настоящему изобретению содержит последовательность нуклеотидов, показанную на Фиг.8 (SEQ ID N0:2), или производную от нее последовательность, в которой произведено от 1 до 10, предпочтительно от 1 до 5 или от 1 до 3 заменен, делеций или вставок оснований.
Настоящее изобретение предпочтительно относится к случаю, когда распознаваемый фенотип, обусловленный одним или несколькими гетерозиготными генами, представляет собой фенотип, обладающий сельскохозяйственной ценностью, например, устойчивость к различным патогенам или насекомым, гибридную силу (или гетерозис), гомеостаз (стабильность растения при изменениях окружающей среды), способность адаптироваться к абиотическим стрессам, в том числе, например, к экстремальным температурам или к низкой освещенности, урожайность, превышающую урожайность сортов, от которых происходит гибрид, морфологические характеристики, в том числе цвет, форму, размер, гибкость или жесткость листьев, состав питательных веществ или вкусовые качества растения.
Кроме того, авторами настоящего изобретения установлено, что гибридные растения, выращенные из семян согласно настоящему изобретению, полученных в результате различных операций скрещивания между различными салатами, при возделывании в зимний период в умеренном климате неожиданно отличаются более быстрым ростом и достигают спелости в среднем на 7-10 суток раньше, чем родительские растения, возделываемые в тех же условиях. Также установлено, что равномерность развития гибридов F1 на одной делянке (при численности растений на делянке, примерно равной 30) превосходит равномерность развития линий родительских растений. Другими словами, количество растений с задержкой или опережением в развитии по сравнению во всеми растениями на той же делянке, на делянках с гибридом F1 является существенно меньшей по сравнению с исходными сортами. Вследствие этого подобные свойства гибридов F1 позволяют не только сократить циклы производства салата, но и сосредоточить сбор урожая на более коротком временном отрезке.
Фенотип, обладающий сельскохозяйственной ценностью, в частности, может быть связан с одним или несколькими генами стандартной или промежуточной устойчивости к вирусным или бактериальным инфекциям, насекомым или грибам, в частности, к одному из следующих грибов: Bremia lactucae, Fusarium oxysporum, Sclerotinia minor или sclerotorum, Botrytis cinerea, Rhizictonia solani, Microdochium panattonianum, Verticiulium dahliae, Erysiphe chicocearum или Pithium tracheiphilum, к одному из следующих насекомых: Nasonovia ribisnigri, Myzus persicae, Macrosiphum euphorbia, Nematodes pratylenchus или meloidogyne, минирующим мушкам Liriomyza huidobrensis или Pemphigus busarius; к одной из следующих бактерий: pseudomonas, xanthomonas или rhizomonas, либо к одному из следующих вирусов: LMV (вирусу мозаики салата), TSWV (вирусу бронзовой пятнистости томата), "Big vein" (или разрастанию жилок салата, вызываемому LBW (вирусом разрастания жилок салата) и MILV (вирусом Мирафьори салата)), TBSV (вирусу кустистой карликовости томата), LNSV (вирусу некротической пятнистости салата), TuMV (вирусу мозаики репы), CMV (вирусу огуречной мозаики) или BWYV (вирусу западной желтухи свеклы).
В случае, когда фенотип, обладающий сельскохозяйственной ценностью, обусловлен одним или несколькими генами стандартной или промежуточной устойчивости к вирусным или бактериальным инфекциям, насекомым или грибам, указанные ген или гены стандартной или промежуточной устойчивости, связанные с упомянутым фенотипом, в частности, могут быть выбраны среди генов устойчивости к ложной мучнистой росе Dm10, R17, Dm5, Dm8, R36, R37 (расположенных в кластере 1 Lactuca sativa), Dm1, Dm2, Dm3, Dm6, Dm14, Dm15, Dm16, Dm18 (расположенных в кластере 2 Lactuca sativa), Dm4, Dm7, Dm11, R38 (расположенных в кластере 4 Lactuca sativa), а также гена Tu устойчивости к TuMV, расположенного в кластере 1, гена Nr устойчивости к Nasonovia, расположенного в кластере 2, или генов mol.1 и mol.2 устойчивости к LMV, расположенных в кластере 4. Указанные кластеры 1, 2 и 4, в частности, определены Р.У.Мичелмором (R.W.Michelmore, Plant Pathol., 1997, vol.36, №4: 499-514 [4]; Theor. Appl. Genet., 1993, vol.85, №8: 985-993 [5]).
Фенотип, обладающий сельскохозяйственной ценностью, в частности, может быть обусловлен одним или несколькими генами стандартной или промежуточной устойчивости к одной из основных болезней, поражающих салат -ложной мучнистой росе Bremia lactucae, грибу, вызывающему появление белесого, похожего на пыль пушка на внутренней поверхности листовой пластины. Ложная мучнистая роса дополнительно обладает высокой способностью к адаптации, что приводит к появлению новых разновидностей, способных преодолевать устойчивость сортов, выведенных селекционерами.
Также настоящее изобретение предпочтительно относится к случаю, когда семена, растения или клетки растений Lactuca sativa являются гетерозиготными по меньшей мере по двум генам, не связанным с мужской стерильностью и сообщающих растению распознаваемый фенотип. Два или более генов, сообщающие растению распознаваемый фенотип, еще более предпочтительно расположены в одном кластере, например, в кластере генов устойчивости, в том числе в кластерах 1, 2 или 4 Lactuca sativa.
Настоящее изобретение также относится к совокупности семян гибридного Lactuca sativa, обладающих генотипом мужской стерильности, гетерозиготных по меньшей мере по одному гену, не связанному с мужской стерильностью и сообщающему растениям, выросшим из данных семян, распознаваемый фенотип, и, возможно, обладающих дополнительными характеристиками семян, описанными выше, причем указанная совокупность содержит по меньшей мере 105 семян, предпочтительно по меньшей мере 106 семян, еще более предпочтительно по меньшей мере 107 семян.
В случае, когда семена получены путем скрещивания между двумя растениями Lactuca sativa, одно из которых ("женская" родительская форма) является носителем ядерной мужской стерильности, обусловленной одним или несколькими доминантными генами, и при этом растения, используемые в качестве "женских" родительских форм, сами являются результатом скрещивания, растения, используемые в качестве "женских" родительских форм, обязательно являются гетерозиготными по гену стерильности. В самом деле, для того, чтобы они были гомозиготными по доминантному гену или доминантным генам мужской стерильности, необходимо, чтобы оба растения, от скрещивания которых были получены растения, используемые в качестве "женской" родительской формы, сами являлись носителями гена мужской стерильности, однако в таком случае скрещивание между двумя подобными растениями невозможно.
Вследствие этого совокупность семян согласно настоящему изобретению, полученных в результате скрещивания между двумя разновидностями растения Lactuca sativa, одно из которых ("женская" родительская форма) является носителем ядерной мужской стерильности, сообщаемой одним или несколькими доминантными генами, состоит, по причине явления расщепления хромосом в ходе мейоза, из семян Lactuca sativa, являющихся носителями доминантного гена или доминантных генов мужской стерильности, и семян, не являющихся носителями указанных гена или генов. В случае моногенной доминантной мужской стерильности "женской" родительской формы доля мужских стерильных семян обычно составляет по меньшей мере 40%.
В основу настоящего изобретения, положено, в частности, наблюдение, сделанное его авторами, согласно которому оказалось, что хотя двукрылые не являются обычными опылителями цветков Lactuca sativa, и тем более не известно об их питании нектаром указанных цветков, данные насекомые, в частности, виды Calliphora vomitaria, Calliphora erythrocephala и Lucilia caesar, при помещении их в большом количестве на закрытый грунт выступаютв качестве опылителей Lactuca sativa.
Настоящее изобретение также относится к применению насекомых отряда двукрылых для осуществления опыления в закрытой среде мужских стерильных растений Lactuca sativa мужскими фертильными растениями, в частности, с целью получения гибридных растений Lactuca sativa. Используемые мужские стерильные растения Lactuca sativa предпочтительно обладают мужской стерильностью, носителем которой является один доминантный ген, предпочтительно мужской стерильностью, обусловленной геном Ms7, описанным выше. Используемые мужские фертильные растения предпочтительно представляют собой растения Lactuca sativa, еще более предпочтительно культивар.
Закрытая среда (закрытый грунт) согласно настоящему изобретению может, в частности, представлять собой теплицу либо каркасное или туннельное укрытие, предпочтительно обладающее поверхностью больше 30 м2, еще более предпочтительно больше 300 м2, например, от 30 до 1500 м2 или от 50 до 1000 м2. Высота закрытого грунта обычно составляет от 2 до 4 м, предпочтительно от 2,5 до 3,5 м, например, 3 м. Закрытый грунт может содержать, в частности, средства вентиляции, полива, контроля температуры и освещенности. Указанный закрытый грунт имеет оболочку, непроницаемую для насекомых.
Используемые двукрылые предпочтительно должны присутствовать в количестве по меньшей мере 100 двукрылых на 1 м2, предпочтительно по меньшей мере 250 двукрылых на 1 м2, например, от 100 до 1000 двукрылых на 1 м2. Количество двукрылых также может быть определено как их число на 1 м3, и предпочтительно составляет по меньшей мере 25 двукрылых на 1 м3, более предпочтительно по меньшей мере 50 двукрылых на 1 м3, еще более предпочтительно по меньшей мере 75 двукрылых на 1 м3, например, от 25 до 500 двукрылых на 1 м3 или от 75 до 250 двукрылых на 1 м3.
Объектом настоящего изобретения также является способ получения семян гибридного Lactuca sativa, включающий следующие стадии:
- возделывание в закрытой среде растений Lactuca sativa с фенотипом мужской стерильности, используемых в качестве "женских" родительских форм, и растений Lactuca sativa с фенотипом мужской фертильности, используемых в качестве "мужских" родительских форм на близком расстоянии друг от друга, причем дополнительной характеристикой одной из двух родительских форм является гомозиготность по гену, сообщающему ему распознаваемый фенотип, отличающийся от мужской стерильности, и причем другая родительская форма не имеет указанного гена;
- опыление двукрылыми, помещенными в указанную закрытую среду во время цветения растений в количестве более 100 двукрылых на 1 м2, предпочтительно более 250 двукрылых на 1 м2;
- сбор урожая семян, выросших на мужских стерильных растениях. Вторая родительская форма предпочтительно также является гомозиготной по меньшей мере по одному гену, сообщающему ей распознаваемый фенотип, отличающийся от мужской стерильности, который отсутствует у первой родительской формы. Закрытая среда предпочтительно имеет оболочку, непроницаемую для насекомых.
Наилучшие результаты были достигнуты при большом числе двукрылых; так, их количество предпочтительно составляет более 400 насекомых на 1 м2, более предпочтительно более 500 насекомых на 1 м2.
Двукрылые могут быть введены на стадии яиц, личинок, куколок или взрослых особей. Предпочтительным является их введение в форме куколок.
Используемые двукрылые предпочтительно являются короткоусыми, например, круглошовными короткоусыми, еще более предпочтительно короткоусыми семейств настоящих мух (Muscidae) или падальных мух (Calliphorides), например, Calliphora vomitaria, Calliphora erythrocephala или Lucilia caesar.
Максимальной урожайности обычно достигают в случае, когда введение двукрылых повторяют по меньшей мере один раз в неделю, предпочтительно два раза в неделю, в течение 3-4 недель.
Максимальной урожайности также достигают в случае, когда количество мужских стерильных растений на закрытом грунте превышает количество мужских фертильных растений. Количество мужских стерильных растений в закрытой среде составляет, например, по меньшей мере 2000, а количество мужских фертильных растений - например, по меньшей мере 1000.
Осуществление способа согласно настоящему изобретению требует одновременного цветения обеих родительских форм. Последнее может быть достигнуто путем отбора по данному свойству или при помощи соответствующих агротехнических мероприятий.
Мужская стерильность растений, используемых в качестве женских родительских форм, предпочтительно является моногенной доминантной и ядерной, еще более предпочтительно - обусловленной геном Ms7.
Геном растений, используемых в качестве женских родительских форм, предпочтительно содержит последовательность двунитевой ДНК, состоящую из 650-700 нуклеотидов, например, 655-695, 660-690, 665-685, 670-680, 673-677 нуклеотидов, или из 674, 675 или 676 нуклеотидов, 5'-концы каждой из двух нитей которой начинаются с последовательности "5' TGCGTTCCAC 3'" (SEQ ID N01). Согласно предпочтительному варианту осуществления, геном растений, используемых в качестве женских родительских форм, содержит последовательность нуклеотидов, показанную на Фиг.8 (SEQ ID NO:2), или производную от нее последовательность, содержащую от 1 до 10, предпочтительно от 1 до 5 или от 1 до 3 замен, делеций или вставок нуклеотидов.
Как было объяснено выше, вследствие расщепления хромосом в ходе мейоза получение классическими способами скрещивания гомогенной популяции растений, используемых в качестве "женских" родительских форм и имеющих доминантный ген мужской стерильности, является невозможным. Вследствие этого в случае применения доминантного гена мужской стерильности растения, используемые в качестве женских родительских форм, могут быть получены способом, включающим следующие стадии:
- скрещивание между растениями Lactuca sativa, гетерозиготными по доминантному ядерному гену мужской стерильности, и мужскими фертильными растениями Lactuca sativa, не имеющими генов стерильности;
- возделывание семян, полученных в результате упомянутого скрещивания;
- удаление растений, имеющих фенотип мужской фертильности. Стадия удаления растений, обладающих фенотипом мужской
фертильности, может быть выполнена, например, вручную на основе визуально различимых признаков, позволяющих отличить мужские стерильные растения Lactuca sativa от мужских фертильных растений Lactuca sativa. Например, в случае стерильности, связанной с геном Ms7, отбор может быть осуществлен на основе того, что соцветия мужских стерильных растений остаются раскрытыми дольше, чем соцветия мужских фертильных растений, и того, что мужские стерильные растения не имеют пыльцы.
Согласно альтернативному варианту осуществления стадию удаления растений, обладающих фенотипом мужской фертильности, осуществляют при помощи обнаружения в образце каждого из растений определенного молекулярного маркера доминантного гена мужской стерильности. Подобным маркером может быть, например, либо маркер RAPD (Random Amplification of Polymorphic DNA) длиной примерно 675 пар оснований (например, от 650 до 700, от 655 до 695, от 660 до 680, от 665 до 685, от 670 до 680, от 673 до 677, 674, 675 или 676 пар оснований), 5'-концы каждой из двух нитей которой начинаются с последовательности "5' TGCGTTCCAC 3'" (SEQ ID NO:1), в том числе маркер ВА05-675 (SEQ ID NO: 2), разработанный авторами изобретения, либо маркер SCAR (Sequenced Characterized Amplified Region Marker), либо маркер CAPS (Cleaved Amplified Polymorphic Sequence), либо любой другой из маркеров, обычно называемых STS (Sequence Tagged Site - меченый участок последовательности), разработанных на основе упомянутого маркера RAPD. Согласно предпочтительному варианту осуществления, обнаружение присутствия или отсутствия определенного молекулярного маркера доминантного гена мужской стерильности выполняют до цветения растений, используемых в качестве женских родительских форм, предпочтительно на ранней стадии роста растений, например, на стадии от 1 до 5 листьев, еще более предпочтительно на стадии от 1 до 2 листьев.
Используемый молекулярный маркер предпочтительно обеспечивает обнаружение мужских стерильных растений с чувствительностью, равной по меньшей мере 70%, 75%, 80%, 85%, 95%, 98% или 99%, либо 100%, причем специфичность определяется как отношение количества мужских стерильных растений, в которых молекулярный маркер присутствует (истинный положительный результат) к сумме количества истинных положительных результатов и количества мужских стерильных растений, в которых молекулярный маркер отсутствует (ложный отрицательный результат) (См. Табл.17).
Используемый молекулярный маркер также предпочтительно обеспечивает обнаружение мужских стерильных растений со специфичностью, равной по меньшей мере 70%, 75%, 80%, 85%, 95%, 98% или 99% либо 100%, причем специфичность определяется как отношение между количеством мужских фертильных растений, в которых молекулярный маркер отсутствует (истинный отрицательный результат) и суммой количества истинных отрицательных результатов и количества мужских фертильных растений, в которых молекулярный маркер присутствует (ложный положительный результат) (См. Табл.17).
MMS (масса одной тысячи семян) семян, полученных способом согласно настоящему изобретению, обычно по меньшей мере на 10%, даже по меньшей мере на 20% или по меньшей мере на 30% превосходит MMS семян, полученных при самооплодотворении мужских фертильных растений Lactuca sativa, используемых в качестве "мужских" родительских форм.
Настоящее изобретение также относится к совокупности семян, полученной которой описанным выше способом и которая содержит по меньшей мере 105 семян, предпочтительно по меньшей мере 106 семян, еще более предпочтительно по меньшей мере 107 семян.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1: происхождение Lactuca sativa.
Фиг.2: размещение растений салата, использованных в качестве "мужских" и "женских" родительских форм, в ходе эксперимента №1.
Фиг.3: размещение растений салата, использованных в качестве "мужских" и "женских" родительских форм, в ходе эксперимента №2.
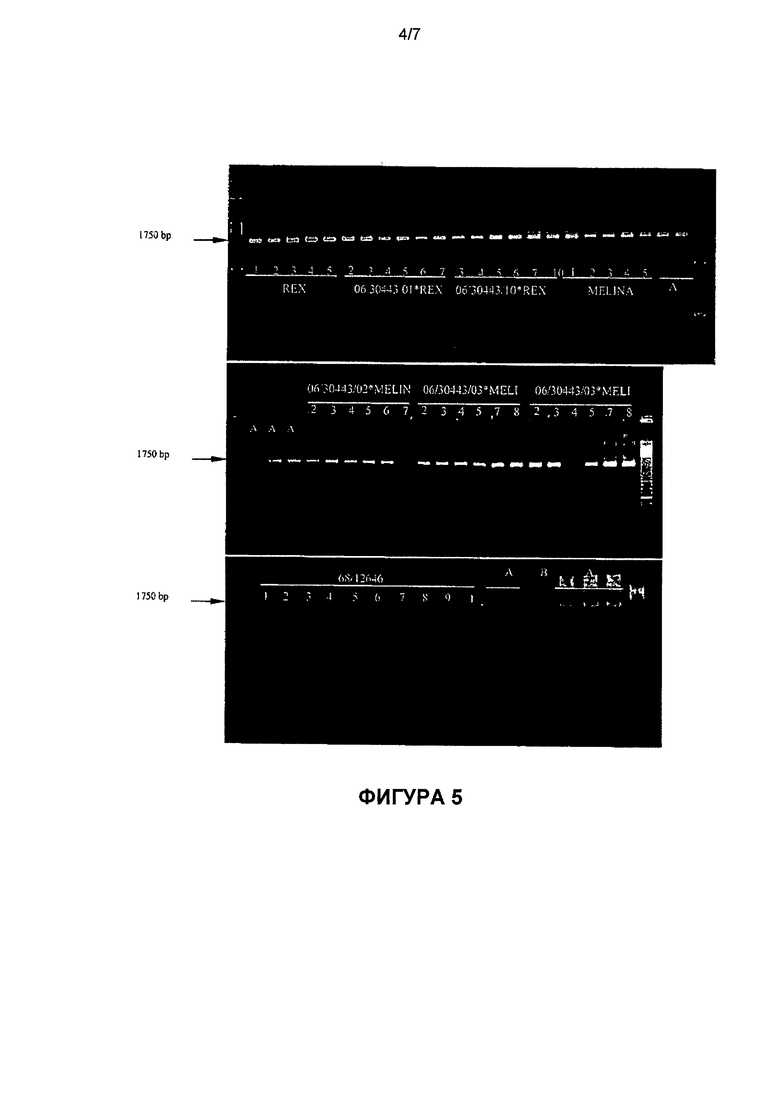
Фиг.4: результат генотипирования 50 гибридных растений в эксперименте №10; профиль амплификации молекулярного маркера SCW09 после расщепления Taql. Маркер SCW09 является маркером генов Dm6 и Dm18 устойчивости к Bremia lactucae. A - Dm18+/Dm18+, B - Dm18-/Dm18-, C - Dm6+, D - не расщепляется Taql.
Фиг.5: результат генотипирования 50 гибридных растений в эксперименте №10; профиль амплификации молекулярного маркера В1, являющегося маркером гена Dm3 устойчивости к Bremia lactucae. A - Dm3+; B - Dm3-.
Фиг.6: электрофоретический профиль праймера ОВРА05 с маркером ВА05-675 (стрелка). Электрофорез осуществляли на агарозном геле при концентрации 1,0% в течение 1 ч 45 мин при напряжении 190 В. M - маркер молекулярной массы 100 п.о. «лестница» (компания Pharmacia Biotech, арт. 27.4001.01).
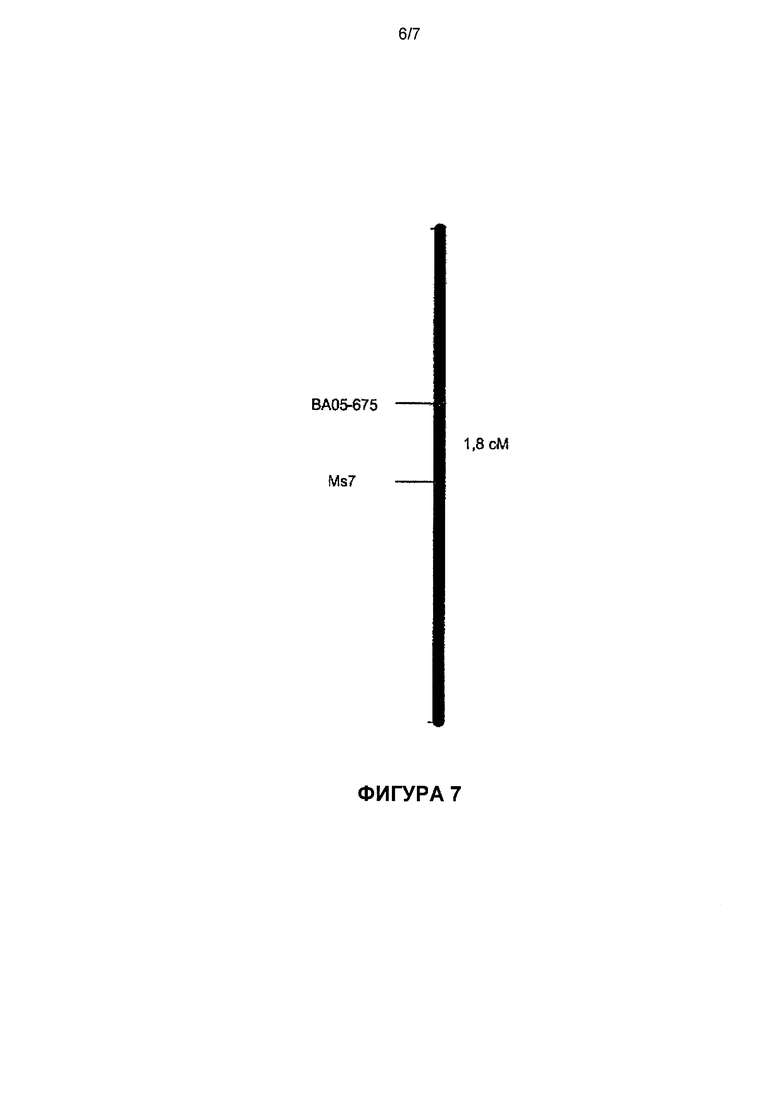
Фиг.7: картограмма молекулярного маркера ВА05-675, сцепленного с локусом Ms7 салата огородного Lactuca sativa (расстояние рассчитано с применением функции Косамби).
Фиг.8: последовательность нуклеотидов 5'-3' маркера ВА05-675, сцепленного с мужской стерильностью МБ7 салата огородного Lactuca sativa (SEQ ID NO: 2).
ПРИМЕРЫ
Пример 1. Получение семян гибридов салата
Мужские стерильные растения Lactuca sativa, использованные в различных экспериментах, описанных ниже, в качестве "женских" родительских форм, являлись носителями гена стерильности Ms7. Указанные растения являлись результатом возвратного скрещивания 5-го поколения (ВС5) кочанного салата для закрытого грунта, являющегося носителем гена Ms7 (X Girelle 94-9538-1) и доступного в различных исследовательских организациях, в том числе в Национальном институте сельскохозяйственных исследований Франции (INRA), Депертаменте сельского хозяйства США (USDA) и др.
Поскольку ген Ms7 является доминантным, скрещивание между двумя растениями-носителями гена Ms7 невозможно. Вследствие этого растения, обладающие фенотипом мужской стерильности, обязательно являются гетерозиготными по гену Ms7 и могут быть получены только путем скрещивания растений Ms7/ms7 (стерильных) с растениями ms7/ms7 (фертильными).
Из этого следует, что для удаления мужских фертильных растений необходима стадия чистки. Последняя оказывается возможной вследствие того, что растения Ms7/ms7 не имеют пыльцы, и вследствие того, что соцветия растений Ms7/ms7 остаются раскрытыми дольше, чем соцветия растений ms7/ms7.
Эксперимент 1
Условия данного эксперимента были следующими:
- В качестве "женских" родительских форм использовали 450 растений кочанного салата Lactuca sativa для закрытого грунта, являющихся результатом возвратного скрещивания 3-го поколения (ВС3) кочанных салатов Ms7 с кочанными салатами культивара Nacre/Cambria Dm18/R38 (ген устойчивости Dm 18, расположенный в кластере 2, и ген устойчивости R38, расположенный в кластере 4, сочетание восприимчиво к Bremia lactucae разновидности 24 (BI24)); примерно половина из указанных растений являлась мужскими стерильными;
- В качестве "мужских" родительских форм использовали 100 растений Lactuca sativa кочанного культивара для закрытого грунта Dm18/R37 (ген устойчивости Dm18, расположенный в кластере 2, и ген устойчивости R37, расположенный в кластере 1, сочетание устойчиво к BI24).
Обе родительские формы обладали превосходной синхронностью цветения, что обеспечивало наилучшее возможное оплодотворение.
Растения размещали в закрытой оболочке площадью 36 м2, непроницаемой для насекомых (голландская теплица с возможностью контроля влажности и температуры, 11×3,30 м), следующим образом (см. Фиг.2):
- растения, используемые в качестве "мужских" родительских форм, размещали в два ряда вдоль стенок закрытой оболочки, причем расстояние между растениями составляло 20 см;
- растения, используемые в качестве "женских" родительских форм, размещали в семь рядов между двумя рядами "мужских" растений на расстоянии от них, равном 50 см, причем расстояние между растениями в ряду составляло 15 см, а между рядами - 25 см.
Посев осуществляли на неделе 13, высадку рассады - на неделе 16.
Вследствие доминантного характера гена Ms7 оказалась необходимой чистка (удаление) растений, используемых в качестве женских родительских форм и обладающих фенотипом мужской фертильности. Данную стадию осуществляли вручную на 26-й и 27-й неделях на основе того, что растения Ms7/ms7 не имеют пыльцы и что соцветия растений Ms7/ms7 (мужских стерильных) остаются раскрытыми дольше, чем соцветия растений ms7/ms7 (мужских фертильных).
Из 450 растений, использованных в качестве женских родительских форм, 217 оказались мужскими стерильными и были эффективно использованы.
В качестве насекомых-опылителей использовали два вида насекомых отряда двукрылых семейства Calliphorides:
- мухи Calliphora vomitaria и Calliphora erythrocephala в виде «личинок» (также называемые синими или мясными мухами), используемые при 15-20°C;
- мухи Lucilia caesar, в виде «червячков» (pinkies) (также называемые зелеными падальными мухами), используемые при температуре 20-25°С.
Ввод насекомых-опылителей осуществляли частотой два раза в неделю в течение недель 28 и 31 в количестве 300 мух на 1 м2 на один ввод.
Сбор урожая мужских стерильных растений ("женских" родительских форм) и мужских фертильных растений ("мужских" родительских форм), также использованных в качестве контроля самооплодотворения, осуществляли на неделе 33.
Растения сушили, молотили, после чего зерна пропускали сквозь колонку с импульсным потоком воздуха с целью окончательной очистки и удаления легких остаточных отходов и калибровали по длине и ширине.
Образец из 2500 семян, полученный в ходе эксперимента 1, 13 февраля 2007 г.поступил в NCIMB (Национальная коллекция промышленных, пищевых и морских микроорганизмов Великобритании, NCIMB Ltd., Ferguson Building, Craibstone Estate, Баксберн, Абердин (Bucksburn, Aberdeen), A21 9YA, Шотландия, Великобритания) под входящим номером NCIMB 41470.
Эксперимент 2
Второй эксперимент, аналогичный первому, осуществили в более широком масштабе в закрытой оболочке площадью 336 м2, непроницаемой для насекомых (длина 56 м, ширина 6 м, высота 3 м).
Условия данного эксперимента были следующими:
- В качестве "женских" родительских форм использовали 3000 растений кочанного салата Lactuca sativa для закрытого грунта, являющихся результатом возвратного скрещивания 4-го поколения (ВС4) кочанных салатов Ms7 с кочанными салатами культивара Nacre/Cambria Dm18/R38 по Bremia (сочетание восприимчиво к BI24); примерно половина из указанных растений являлась мужскими стерильными;
- В качестве "мужских" родительских форм использовали 1500 растений Lactuca sativa кочанного культивара для закрытого грунта (BRA) Dm18/R37, устойчивого к BI24.
Растения размещали в закрытой оболочке следующим образом (См. Фиг.3):
- растения, используемые в качестве "мужских" родительских форм, размещали в четыре ряда: по одному вдоль каждой стенки закрытой оболочки (на расстоянии 80 см) и два в центре (с промежутком 20 см); растения размещали в шахматном порядке с промежутком 15 см;
- растения, используемые в качестве "женских" родительских форм, размещали в восемь рядов между рядами мужских фертильных растений на расстоянии 65 и 70 см от них; расстояние между растениями составляло 15 см, между рядами - 25 см.
Посев осуществляли на неделе 12, высадку рассады - на неделе 16.
Аналогично эксперименту 1, оказалась необходимой чистка (удаление) растений, используемых в качестве женских родительских форм и обладающих фенотипом мужской фертильности. Данную стадию осуществляли вручную на 25-й и 26-й неделях на основе того факта, что растения Ms7/ms7 не имеют пыльцы и что соцветия растений Ms7/ms7 (мужских стерильных) остаются раскрытыми дольше, чем соцветия растений ms7/ms7 (мужских фертильных).
Из 3000 растений, использованных в качестве женских родительских форм, 1250 оказались мужскими стерильными и были эффективно использованы.
В то же время 16 растений ms7/ms7 (мужских фертильных) в ходе чистки сохранили и подвергли самооплодотворению под непроницаемой оболочкой, состоящей из куска сетки или ткани, не пропускающего насекомых, с целью использования для контроля самооплодотворения.
Были использованы те же насекомые-опылители, что и в эксперименте №1.
Ввод насекомых-опылителей осуществляли один раз в неделю в течение недель 25-29 в среднем количестве 1100 мух на 1 м2 на один ввод.
Параллельно 10 мужских фертильных растений (мужских родительских форм) и 16 фертильных растений ms7/ms7, сохраненных на стадии чистки, подвергали самооплодотворению под непроницаемой оболочкой, состоящей из сетки или ткани, не пропускающей насекомых, с целью контроля самооплодотворения.
Сбор урожая мужских стерильных растений ("женских" родительских форм), а также 10 мужских фертильных растений (мужских родительских форм) и 16 фертильных растений ms7/ms7, сохраненных на стадии чистки и использованных для контроля самооплодотворения, осуществляли на неделе 33.
Растения сушили, молотили, затем семена пропускали через колонку с импульсным потоком воздуха с целью окончательной очистки и удаления легких остаточных отходов, и калибровали по длине и ширине.
Высевали образец полученных семян; устойчивость полученных растений к BI24 определяли согласно официальному протоколу GEVES (Группы сортосеменных исследований и сортосеменного контроля). Растения продемонстрировали устойчивость к BI24.
Результаты экспериментов 1 и 2
Результаты экспериментов 1 и 2 представлены в приведенных ниже таблицах.
"женской" родительской формы, опыление мухами)
"женской" родительской формы, опыление мухами)
Обсуждение результатов
В ходе первого эксперимента с 217 мужских стерильных растений Lactuca sativa (женских родительских форм) было получено 730 граммов семян; урожайность составила 3,36 г на растение, что составило 15,30% от контроля, подвергнувшегося самооплодотворению (мужских родительских форм).
В ходе второго эксперимента с 1170 мужских стерильных растений Lactuca sativa (женских родительских форм) тем же способом было получено 7,730 кг семян; урожайность составила 6,60 г на растение, что составило в среднем 50% от контроля, подвергнувшегося самооплодотворению ("мужских" родительских форм и мужских стерильных "женских" родительских форм ms/ms, сохраненных в ходе чистки и оплодотворенных под оболочкой, непроницаемой для насекомых).
В ходе второго эксперимента доля годных семян после калибровки гибридов F1 на 10% превышала соответствующую величину для контроля, подвергшегося самооплодотворению.
В ходе обоих экспериментов MMS (масса одной тысячи семян) гибрида F1 на 20-30% превышала MMS контроля, подвергшегося самооплодотворению.
Растения, выращенные из семян, полученных в ходе второго эксперимента, обладали хорошей устойчивостью к BI24.
Эксперименты 3-8
Параллельно было проведено шесть аналогичных экспериментов с использованием различных сортов кочанного салата и салата батавия, помещенных под непроницаемые для насекомых каркасы размером 3,3×1,1 м, насчитывающие по 8 растений Lactuca sativa, использованных в качестве "женских" родительских форм (на 50% мужских стерильных, полученных возвратным скрещиванием растений Lactuca sativa, являющихся носителями гена Ms7, с фертильными растениями Lactuca sativa), и по 4 мужских фертильных растения Lactuca sativa, использованных в качестве "мужских" родительских форм", и в свою очередь помещенные в вентилируемую теплицу.
Посев осуществляли на неделе 22, высадку рассады - на неделе 27, отбор мужских стерильных и мужских фертильных "женских" растений - в начале цветения на неделе 31 ввод мух - неделях на 32, 33 и 34 (один ввод в неделю в течение трех недель в количестве примерно 100 мух/м2), сбор урожая - на неделе 37.
Результаты экспериментов 3-8
- Каркас 1: CHARLIN (68/12624)×CHARLIN (кочанный) 1 женское растение: 9,20 г
1 мужское растение: 28,30 г
- Каркас 2: CHARLIN (ВС/77)×CHARLIN (кочанный) 1 женское растение: 5,45 г
1 мужское растение: 19,40 г
- Каркас 3: BRA 68/12588 (кочанный салат для закрытого грунта)×BRA 68/12588
6 женских растений
растение 1: 7,00 г
растение 2: 2,00 г
растение 3: 5,20 г
растение 4: 3,40 г
растение 5: 9,60 г
растение 6: 8,40 г
в среднем 5,9 г на одно растение
1 мужское растение: 34,40 г
- Каркас 4: BRA 68/12588×BRA 68/12588
4 женских растения
растение 1: 11,95 г
растение 2: 7,15 г
растение 3: 15,90 г
растение 4: 11,90 г
в среднем 11,25 г на одно растение
1 мужское растение: 29,00 г
- Каркас 5: BVA 68/12553 (салат батавия для закрытого грунта)×BVA 68/12553
5 женских растений
растение 1: 3,50 г
растение 2: 1,30 г
растение 3: 2,20 г
растение 4: 1,70 г
растение 5: 4,20 г
в среднем 3,30 г на одно растение
1 мужское растение: 3,60 г
- Каркас 6: BVA 68/12553×BVA 68/12553
3 женских растения
растение 1: 4,60 г
растение 2: 3,20 г
растение 3: 3,70 г
в среднем 3,80 г на одно растение
1 мужское растение: 23,40 г
Обсуждение результатов
Средняя продуктивность "женских" растений составила от 3,3 до 11,2 г/растение, причем сортотип батавия (BVA 68/12553) обладал существенно более низкой урожайностью (примерно 3,5 г/растение), чем с кочанный сортотип (Charlin, BRA 68/12588), урожайность которого составила примерно 8,5 г/растение, что может дать повод для предположения, что имеет место сильное влияние сортотипа на конечную урожайность.
Эксперимент 9
Еще один эксперимент осуществили под небольшими каркасами диаметром 0,40 м, непроницаемыми для насекомых, помещенными в вентилируемую теплицу, установленную на грунте.
Посев осуществляли на неделе 22, пересадку - на неделе 25, отбор мужских стерильных "женских" растений и мужских фертильных растений - в начале цветения на неделе 30, высадку рассады - на неделе 31, ввод мух - на неделях 32, 33 и 34 неделях (один ввод в неделю в течение трех недель в количестве примерно 100 мух/м2) сбор урожая - на неделе 35.
Результаты
Совокупность полученных результатов ниже приведена в таблицах 4 и 5.
Обсуждение результатов
Результаты являются удовлетворительными с той точки зрения, что средняя продуктивность, достигнутая для салата батавия (BVA 68/12533), составляет примерно 1 г/растение по сравнению с 1,7 г/растение для подвергшегося самооплодотворению контроля, и при этом средняя продуктивность, достигнутая для кочанного салата для закрытого грунта (сорт Charlin), составляет примерно 1 г/на растение и находится на одном уровне с подвергшимся самооплодотворению контролем.
Эксперимент 10. Накопление Dm3/Dm18 в гибридных комбинациях
Способом, описанным в приведенных выше экспериментах, были получены гибриды F1 салата с накоплением двух доминантных генов Dm3 и Dm 18 устойчивости к Bremia lactucae, соответственно разновиности 24 (BI24) и разновидности 23 (BI23), принадлежащих к кластеру 2 и расположенных в одном локусе.
Растения, использованные в качестве "женских" родительских форм (06/30443: MS7/21 NACRExDEVONIAxCAMBRIA), являлись носителями генов устойчивости Dm18 и R38. Растения, использованные в качестве "мужских" родительских форм, представляли собой растения кочанного салата для закрытого грунта, являющиеся носителями гена устойчивости Dm3 (ВС:06/30443 х Rex или ВС:06/30443×Melina).
Выполняли пять следующих операций скрещивания:
- 06/30443/01: (MS7/21NACRExDEVONIAxCAMBRIA-01) Dm18/R38×Rex Dm3;
- 06/30443/10: (MS7/21NACRExDEVONIAxCAMBRIA-10) Dm18/R38×Rex Dm3,
- 06/30443/02: (MS7/21NACRExDEVONIAxCAMBRIA-02) Dm18/R38×Melina Dm3;
- 06/30443/03: (MS7/21NACRExDEVONIAxCAMBRIA-03) Dm18/R38×Melina Dm3;
- 06/30443/04: (MS7/21NACRExDEVONIAxCAMBRIA-04) Dm18/R38×Melina Dm3.
Семена, полученные в результате пяти указанных скрещиваний, были высеяны; на растениях F1, а также на "мужских" (Rex/Melina) и "женских" родительских формах проводили тесты инокуляции листовых пластинок и молекулярного маркирования.
I. Тест инокуляции на листовые пластинки
Материалы. Растительный материал состоял из образцов листьев в виде дисков диаметром 1,5 см в количестве 5 дисков на одно растение и на одну разновидность; использовали 10 растений каждого гибрида и две разновидности Bremia lactucae (BI24, которая поражает Dm18/R38 и к которой придает устойчивость Dm3, и BI23, которая поражает Dm3/R38 и к которой придает устойчивость Dm18). Таким образом, всего 80 растений подвергли тестированию по двум разновидостям.
Методика. Листовые диски инокулировали путем распыления спороцист, разновидностями Bremia BI23 и BI24 раздельно.
1. Сбор листовых дисков. Собирали по пять образцов листьев с растения и помещали их на двойной слой увлажненной фильтровальной бумаги. Каждая пачка содержала 6 слоев испытываемых образцов и один слой восприимчивого контрольного образца.
2. Инокуляция. Количество инокулята, необходимое для инокулирования 6 пачек дисков из листьев, составляло 11 мл. Готовили маточный раствор Tween 80 в количестве 4 капель Tween 80 на 100 мл деминерализованной воды. Количество экстракционного раствора по отношению к объему инокулята составило 5 мл маточного раствора Tween 80 на 100 мл инокулята. Инокулят получали путем отделения спороцист в водопроводной воде с добавлением 5% маточного раствора Tween 80. Спороцисты собирали в количестве 1 растение/1 мл инокулята с целью получения концентрации спороцист, близкой к 108 спор/мл. Выполняли фильтрацию путем пропускания раствора через тампон в стакан объемом 250 мл.
3. Распыление. Инокуляцию выполняли в тот же день, что и сбор образцов листьев. Отмеряли объем раствора инокулята, затем инокулировали разложенные пачки. Инокулят равномерно распыляли тонкими каплями по всей поверхности каждого образца. Пачки герметично закрывали и инкубировали при 15°C 14 ч днем и 10 ч ночью в течение 7 сут.
Результаты. Результаты инокуляции приведены в следующих далее таблицах 6-13 (S - восприимчив, R - устойчив). Гибриды, полученные в результате различных скрещиваний, оказались устойчивыми к обеим разновидностям BI23 и BI24, что указывает на присутствие обоих генов Dm3 и Dm18 кластера 2.
II. Молекулярное маркирование
Накопление двух сильно связанных (расположенных в одном локусе) доминантных генов устойчивости к двум штаммам Bremia lactucae у Lactuca sativa может быть выявлено при помощи двух молекулярных маркеров.
1. Растительный материал. Растительный материал состоял из образцов листьев в виде дисков диаметром 1,5 см в количестве одной пластинки на растение. ДНК из 5-10 растений на один генотип (всего от 50 растений) получали путем экстракции ЦТАБ в хлороформе с последующим суспендированием в растворе TE 0,1Х в концентрации 5 нг/мкл.
2. Генотипирование. Для генотипирования 50 растений были использованы маркеры SCW09 и В1, описанные соответственно Мезонневом и др. (Maisonneuve et al., [10]) и X.Куаном и др. (H.Kuang et al., [11]) (таблица 14). Условия ПЦР описаны в таблице 15. Циклы ПЦР SCW09 и В1 соответственно являлись следующими:
- 94°C/30 с-94°C/1 мин, 60°C/1 мин, 72°C/2 мин; 40 циклов - 72°C/5 мин-4°C;
- 94°C/30 с-94°C/1 мин, 63°C/1 мин, 72°C/2 мин; 40 циклов - 72°C/5 мин-8°C.
Перед стадией миграции маркер SCW09 подвергли ферментативному разложению ферментом Taq I (буфер 1Х, BSA 1Х, Taq I 1U). Электрофорез осуществляли на агарозном геле при концентрации 2% (SCW09) и 1,5% (В1) в буфере ТВЕ 1Х в течение 1 ч (В1) - 1 ч 30 мин (SCW09) при постоянном напряжении, равном 220 В. Проявление электрофоретических профилей осуществляли путем окрашивания этидийбромидом (BET) в свете УФ-лампы.
Результаты
Результаты приведены в таблице 16 и на фиг.4 и 5. Генотипы REX и MELINA имеют профиль Dm18-/Dm18- и Dm3+. Генотип 06/30443 имеет профиль Dm18+/Dm18+и Dm3-. Совокупность скрещиваний с одной или другой 5 родительской формой (REX или MELINA) имеет профиль Dm18+/Dm18- и Dm3+, т.е. является устойчивой по генам Dm18 и Dm3.
Пример 2. Идентификация молекулярного маркера гена Ms7
Способ получения молекулярного маркера, сцепленного с Мэ7-мужской стерильностью Lactuca sativa (салата), описанный далее, приведен для наглядности.
1. Получение популяций для разработки маркера, сцепленного с ядерным геном мужской стерильности Ms7 у салата (Lactuca sativa)
Путем возвратного скрещивания были получены две популяции салата различных сортотипов, обладающие мужской стерильностью Ms7. Полученные подобным образом две рекуррентные популяции четвертого поколения (ВС4), состоящие из 200 растений, затем подвергались фенотипированию по признаку "отсутствие жизнеспособной пыльцы", соответствующему фенотипу мужской стерильности. Были идентифицированы две группы фенотипов, мужская фертильная (F) и мужская стерильная (S), в отношении 1:1.
Величина данного отношения успешно подтверждает моногенную доминантную природу мужской стерильности, пурудающуюся от гетерозиготной по локусу Ms7 родительской формы.
2. Экстракция ДНК растений салата
Экстракцию ДНК осуществляли из свежих молодых листьев по видоизмененной методике с применением ЦТАБ (Томас и др., Tomas et al., 1989 [20]; Дойл и др., Doyle et al., 1990 [15]; Эдварде и др., Edwards et al., 1991 [16]).
Образцы листьев в виде дисков, полученные при помощи пробирки объемом 1,5 мл, измельчали в 500 мкл экстракционного буфера (0,1 М трис-HCI, 0,7 М NaCl, 10 мМ ЭДТА, 1% ЦТАБ, 1% В-меркаптоэтанола). Измельченный материал инкубировали при 65°C в течение 1 ч с 2-3-кратным перемешиваниям путем переворачивания в ходе инкубации.
Затем добавляли 200 мкл раствора изоамилового спирта в хлороформе в отношении 1:24 и перемешивали путем переворачивания. После центрифугирования при 6000 об/мин и 20°C в течение 10 мин отделяли 400 мкл надосадочной жидкости, который смешивали с 400 мкл изопропанола.
После выдерживания при -20°C в течение 1 ч смесь центрифугировали при 6000 об/мин и 4°C в течение 10 мин. Пробирки опорожняли. Осадок ДНК, оставшийся на стенках пробирки, оставляли сушиться на воздухе в течение 12 ч.
ДНК суспендировали в 200 мкл раствора ТЕ 0,1-Х (1 мМ трис-HCI, 0,1 мМ ЭДТА). Конечную концентрацию измеряли путем анализа с фрагментами ДНК фага лямбда / Hind III на агарозном геле (1%).
3. Поиск RAPD-маркера, сцепленного с мужской стерильностью, обусловленной геном Ms7, L.sativa, метод BSA (Bulk Segregant Analysis, анализ объединенных сегрегантов)
В целях идентификации молекулярного маркера, сцепленного с мужской стерильностью Ms7, применяли наборы декамеров RAPD® компании Орегоп Technologies Inc. (Хантсвилл (Huntsville), AL 35805, США) и метода BSA (Мичелмор и др., Michelmore et al., 1991 [17], и Паран и др., Paran et al., 1991 [18]).
Исходя из фенотипирования двух популяций ВС4, описанных выше, для "мужской стерильной" и "мужской фертильной" групп было проведено два отбора образцов смесей 10 растений; способом RAPD, описанным Уильямом и др. (William et al., 1990 [22]) и Уэлшем и др. (Welsh et al., 1990 [21]), на 4 образцах на каждую популяцию испытывали 1200 праймеров компании Орегоп Biotechnologies Inc. (Хантсвилл (Huntsville), AL 35805, США) от ОРА-01 до ОРВН-20. Были отобраны праймеры, обладающие полосой, характерной для "мужских стерильных" образцов, и легко читаемым профилем электрофореза.
ПЦР осуществляли в общем реакционном объеме в 25 мкл, состоящем из буфера для ПЦР 1Х, 3 мМ MgCl2, 200 мкМ dNTP, 400 нМ праймера, 1 ед. ДНК-полимеразы (AmpliTaq компании Perkin-Elmer Cetus). ПЦР-реакция состояла в следующих нескольких циклах: 1 стадия - 30 с при 94°C; 45 циклов при 94°C по 1 мин, затем при 35°C по 1 мин и при 72°C по 2 мин; окончание реакции при 72°C в течение 5 мин и блокирование реакции при 4°C.
Затем продукты амплификации разделяли на агарозном геле при концентрации 2% при следующих условиях электрофореза: миграционный буфер ТВЕ 1-X (Трис-борат-ЭДТА), 190 В, 2 ч 15 мин.
С целью подтверждения эффективности отобранных RAPD-праймеров каждое из 200 растений обеих популяций ВС4 по отдельности тестировали с применением праймера ОВРА05, последовательность которого имеет вид "5' TGCGTTCCAC 3'" (SEQ ID NO: 1) и которой соответствует маркеру ВА05-675 длиной 675 п.о. (SEQ ID NO:2), который отделяется вместе с Ms7. Электрофоретический профиль, полученный для данного маркера, показан на Фиг.6.
Подтверждение эффективности данного маркера на различных сортотипах салата (батавия, кочанного, айсберг, ромен, "дубовый лист" и лолло росса) было проведено примерно на 25 растениях 12 популяций. Условия проведения испытаний были идентичны описанным выше для стадий экстракции, амплификации и электрофореза.
Расчет чувствительности и специфичности маркера RAPD ВА05-675 дал различные значения в зависимости от сортотипа; средние значения составили 96% и 94% соответственно (см. таблицы 17 и 18).
(VN/((VN+(FP)
4. Генное картирование маркера ВА05-675, сцепленного с мужской стерильностью Ms7 у Lactuca sativa
Для 200 растений одной популяции ВС4, описанной выше, вне зависимости от наличия или отсутствия Ms7 при помощи программ JoinMap®4 (Стам и др., Stam et al., 1996 [19]) и Carte Blanche® (Keygene N.P., P.O.Box 216, 6700 AE Вагенинген (Wageningen), Нидерланды) выполнили генное картирование по Уильяму и др. (William et al., 1993 [23]) маркера ВА05-675, сцепленного с Ms7. Статистический критерий X2 позволил подтвердить нулевую гипотезу расщепления по Менделю в отношении 1:1 (критерий является незначимым при p>0,05) для доминантного маркера ВА05-675 (см. таблицу 19). Оценка связи с отношением правдоподобия (или показателем сцепления генов LOD), большим или равным 3,0, позволила провести картирование данного маркера на 1,8 сМ от Ms7 (Фиг.6).
5. Клонирование и секвенирование молекулярного маркера RAPD ВА05-675, сцепленного с мужской стерильностью салата Lactuca sativa На основе электрофоретического профиля на агарозном геле, был выделен фрагмент ДНК длиной 675 нуклеотидов, который суспендировали в буфере ТЕ (10 мМ Tris-Cl, 1 мМ ЭДТА) и затем повторно амплифицировали в условиях ПЦР, описанных выше. Выделенный и повторно амплифицированный фрагмент клонировали при помощи коммерчески доступного набора pCR®4-TOPO® (компания Invitrogen, Карлсбад (Carlsbad), Калифорния, 92008, США). После подтверждения успеха клонирования при помощи набора для очистки на колонке Promega (Мэдисон (Madison), Висконсин, США) была проведена экстракция ДНК (система Midiprep).
Очищенные клоны, сконцентрированные до 75 нг/мкл, секвенировали в компании Cogenics (38944 Мелан (Meylan), Франция). При этом была получена последовательность маркера ВА05-675 (SEQ ID NO:2), показанная на Фиг.8.
6. Результаты
Проведенное авторами настоящего изобретения исследование позволило определить RAPD-маркер ВА05-675 (SEQ ID NO:2), последовательность которого представлена на Фиг.8; данный маркер расположен на 1,8 сМ от Ms7 (Фиг.7) и обладает средними чувствительностью и специфичностью, равными соответственно 94% и 96%, вне зависимости от сортотипа салата (таблица 18). Оценка данного маркера на отдельных растениях двух популяций ВС4 позволила определить предсказательную способность (Олтмен, Altman, 1994а [13]) идентификации мужских стерильных растений, в среднем составляющую около 97%.
СПИСОК ЛИТЕРАТУРЫ
1. Goubara & Takasaki (2003) Flower visitors of lettuce under field and enclosure conditions, Appl. Entomo. Zool. 38(4): 571-581.
2. Goubara & Takasaki (2004) Pollination effects of the sweat bee Lasioglossum vilosulum trichopse on genic-male lettuce, Appl. Entomo. Zool. 39(1): 163-169,2004.
3. Lindqvist, K. (1960) Inheritance studies in lettuce, Heriditas 46: 387-470.
4. Michelmore, R.W. et al. (1987) Genetic analysis of factors for resistance to downey mildew in lettuce, Plant Pathol., vol. 36, №4: 499-514.
5. Michelmore, R.W. et al. (1993) Development of reliable PCR-based markers linked to downey mildew resistance genes in lettuce, Theor. Appl. Genet., vol. 85, №8: 985-993.
6. Ryder, E.J. (1963) An epistatically controlled pollen sterile in lettuce (Lactuca sativa L), Proc. Am. Soc. Hort. Sci. 96: 826-828.
7. Ryder, E.J. (1967) A recessive male sterile gene in lettuce, Proc. Am. Soc. Hort. Sci 91: 366-368.
8. Ryder, E.J. (1971) Genetic Studies in Lettuce {Lactuca sativa L.), J. Amer. Soc. Hort. Sci 96(6) 826-828.
9. Ryder, E.J. (1979) Leafy Salad Vegetables, Avi. Pub. Co., page 30.
10. Maisonneuve, B. et al. (1994) Rapid mapping of two genes for resistance to downy mildew from Lactuca serriola to existing clusters of resistance genes, Theor. Appl. Genet. 89: 96-104.
11. Kuang H., et al. (2004) Multiple genetic processes result in heterogeneous rates of evolution within the major cluster disease resistance genes in lettuce, The plant cell, Vol.16, 2870-2894.
12. Kesseli R.V., et al. (1994) Analysis of a detailed linkage map of Lactuca sativa (lettuce) constructed from RFLP and RAPD markers, Genetics 136: 1435-1446.
13. Altman D.G., Bland J.M., 1994a. Statistics Notes: Diagnostic tests 1: sensitivity and specificity. British Medical Journal, 308:1552.
14. Altman D.G., Bland J.M., 1994b. Statistics Notes: Diagnostic tests 2: predictive values. British Medical Journal, 30:102.
15. Doyle J.J., and Doyle J.L., 1990. Isolation of plant DNA from fresh tissue. Focus 12, 13-15.
16. Edwards К., Johnstone С.and Thompson C. 1991. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acid Res 19: 1349.
17. Michelmore R.W., Paran I., Kesseli R.V., 1991. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations Proc Natl Acad Sci USA 88: 9828-9832.
18. Paran I., Kesseli R., Michelmore R.W., 1991. Identification of restriction fragment length polymorphism and random amplified polymorphic DNA markers linked to downy mildew genes in lettuce, using near-isogenic unes Genome 34: 1021-1027.
19. Stam P., & J.W. van Ooijen, 1996. JoinMap, version 2.0. Software for the calculation of genetic linkage maps. Ed. CPRO-DLO, 60 pp.
20. Tomas H.T., and Tanksley S.D., 1989. A rapid and inexpensive method for isolation of total DNA from dehydrated plant tissue. Plant Mol Biol Rep 12: 106-109.
21. Welsh J, McClelland M., 1990. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res, 18: 7213-7218.
22. Williams J.K.G., Kubelik A.R., Livak K.J., Rafalsky J.A., Tynger S.V., 1990. DNA polymorphisme amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res, 18: 6531-6535.
23. Williams J.K.G., Reiter R.S., Young R.M., Scolnik P.A., 1993. Genetic mapping of mutations using phenotypic pools and mapped RAPD markers. Nucleic Acids Res., 21(11): 2697-2702.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУЧЕНИЕ ГИБРИДНЫХ СЕМЯН ЗЕРНОВЫХ КУЛЬТУР | 2015 |
|
RU2685907C2 |
| СПОСОБ ПОВЫШЕНИЯ СЕМЕННОЙ ПРОДУКТИВНОСТИ ЛЮЦЕРНЫ | 2017 |
|
RU2679837C2 |
| ГЕН-ВОССТАНОВИТЕЛЬ RF4 ДЛЯ ЦИТОПЛАЗМАТИЧЕСКОЙ МУЖСКОЙ СТЕРИЛЬНОСТИ (CMS) C-ТИПА КУКУРУЗЫ, МОЛЕКУЛЯРНЫЕ МАРКЕРЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2603005C2 |
| ФРАГМЕНТ ДНК, ДЕТЕРМИНИРУЮЩИЙ СТЕРИЛЬНОСТЬ OQURA И СООБЩАЮЩИЙ ЦИТОПЛАЗМИТИЧЕСКУЮ МУЖСКУЮ СТЕРИЛЬНОСТЬ ПРИ НАЛИЧИИ ЕГО В МИТОХОНДРИАЛЬНОМ ГЕНОМЕ РАСТЕНИЙ И ПРОБА ДНК ДЛЯ ГИБРИДИЗАЦИИ С ДАННЫМ ФРАГМЕНТОМ | 1991 |
|
RU2117704C1 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЙ С ЦИТОПЛАЗМАТИЧЕСКОЙ МУЖСКОЙ СТЕРИЛЬНОСТЬЮ И СПОСОБ ПОЛУЧЕНИЯ ГИБРИДНЫХ СЕМЯН С МУЖСКОЙ ФЕРТИЛЬНОСТЬЮ | 1995 |
|
RU2139347C1 |
| КУКУРУЗНЫЕ ПРОДУКТЫ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2012 |
|
RU2650764C2 |
| КРАСНЫЙ САЛАТ ЛАТУК | 2006 |
|
RU2460282C2 |
| СПОСОБ СЕЛЕКЦИИ ГАПЛОИДОВ И ДВОЙНЫХ ГАПЛОИДОВ РАСТЕНИЙ | 1994 |
|
RU2158502C2 |
| ВОССТАНОВЛЕНИЕ ФЕРТИЛЬНОСТИ У РАСТЕНИЙ | 2019 |
|
RU2831789C2 |
| КАНОЛА HO/LL С УСТОЙЧИВОСТЬЮ К ЗАБОЛЕВАНИЮ КИЛОЙ КРЕСТОЦВЕТНЫХ | 2012 |
|
RU2618846C2 |





Изобретение относится к области биохимии, в частности к семени гибридного культурного салата (Lactuca sativa), которое обладает генотипом мужской стерильности, обеспечиваемым геном стерильности Ms7, а также к способу его получения, растению гибридного культурного салата, которое обладает гетерозиготным генотипом мужской стерильности, и к клетке растения гибридного культурного салата, которая обладает гетерозиготным генотипом мужской стерильности. Также раскрыто применение насекомых отряда двукрылых, принадлежащих к семейству Calliphorides, в количестве более 100 двукрылых на 1 м2 или более 25 двукрылых на 1 м3 для осуществления опыления в закрытой среде мужских стерильных растений Lactuca sativa и получения гибридного семени Lactuca sativa. Изобретение позволяет эффективно получать растение гибридного культурного салата, которое обладает гетерозиготным генотипом мужской стерильности. 5 н. и 34 з.п. ф-лы, 8 ил., 19 табл., 2 пр.
1. Семя гибридного культурного салата (Lactuca sativa), которое обладает генотипом мужской стерильности, обеспечиваемым геном стерильности Ms7.
2. Семя по п. 1, отличающееся тем, что его геном включает последовательность двунитевой ДНК, которая содержит от 650 до 700 нуклеотидов, и 5′-конец каждой из двух нитей которой начинается с последовательности 5′ TGCGTTCCAC 3′ (SEQ ID NO: 1).
3. Семя по п. 1, отличающееся тем, что указанное семя является гетерозиготным по меньшей мере по еще одному гену, сообщающему растению, выращенному из упомянутого семени, распознаваемый фенотип, обладающий сельскохозяйственной ценностью.
4. Семя по п. 3, отличающееся тем, что упомянутый фенотип обусловлен геном стандартной устойчивости или промежуточной устойчивости к заражению вирусом, бактерией, насекомым или грибом.
5. Семя по п. 4, отличающееся тем, что упомянутый фенотип представляет собой стандартную или промежуточную устойчивость к одному из следующих грибов: Bremia lactucae, Fusarium oxysporum, Sclerotinia minor или sclerotorum, Botrytis cinerea, Rhizictonia solani, Microdochium panattonianum, Verticiulium dahliae, Erysiphe chicocearum или Pithium tracheiphilum, к одному из следующих насекомых: Nasonovia ribisnigri, Myzus persicae, Macrosiphum euphorbia, Nematodes pratylenchus или meloidogyne, минирующим мушкам Liriomyza huidobrensis или Pemphigus busarius; к одной из следующих бактерий: pseudomonas, xanthomonas или rhizomonas, либо к одному из следующих вирусов: LMV, TSWV, "Big vein", TBSV, LNSV, TuMV, CMV или BWYV.
6. Семя по п. 4 или 5, которое обладает вторым геном стандартной или промежуточной устойчивости, отличающимся от первого, в гетерозиготной форме.
7. Семя по п. 4 или 6, отличающееся тем, что указанные гены стандартной или промежуточной устойчивости выбраны из следующих генов устойчивости к ложной мучнистой росе: Dm10, R17, Dm5/Dm8, R36/R37 (кластер 1); Dm1, Dm2, Dm3, Dm6, Dm14, Dm15, Dm16, Dm18 (кластер 2), или Dm4, Dm7, Dm11, R38 (кластер 4), либо гена Tu устойчивости к TuMV (кластер 1), либо гена Nr устойчивости к тле (кластер 2), либо генов mo1.1, mo1.2 устойчивости к LMV (кластер 4).
8. Растение гибридного культурного салата {Lactuca sativa), которое обладает гетерозиготным генотипом мужской стерильности, обеспечиваемым геном стерильности Ms7.
9. Растение по п. 8, отличающееся тем, что геном указанного растения включает последовательность двунитевой ДНК, которая содержит от 650 до 700 нуклеотидов и 5′-конец каждой из двух нитей которой начинается с последовательности 5′ TGCGTTCCAC 3′ (SEQ ID NO: 1).
10. Растение по п. 8 или 9, отличающееся тем, что указанное растение является гетерозиготным по меньшей мере по еще одному другому одному гену, сообщающему растению, выращенному из упомянутого семени, распознаваемый фенотип, обладающий сельскохозяйственной ценностью.
11. Растение по п. 10, отличающееся тем, что упомянутый фенотип обусловлен геном стандартной устойчивости или промежуточной устойчивости к заражению вирусом, бактерией, насекомым или грибом.
12. Растение по п. 11, отличающееся тем, что упомянутый фенотип представляет собой стандартную или промежуточную устойчивость к одному из следующих грибов: Bremia lactucae, Fusarium oxysporum, Sclerotinia minor или sclerotorum, Botrytis cinerea, Rhizictonia solani, Microdochium panattonianum, Verticiulium dahliae, Erysiphe chicocearum или Pithium tracheiphilum, к одному из следующих насекомых: Nasonovia ribisnigri, Myzus persicae, Macrosiphum euphorbia, Nematodes pratylenchus или meloidogyne, минирующим мушкам Liriomyza huidobrensis или Pemphigus busarius; к одной из следующих бактерий: pseudomonas, xanthomonas или rhizomonas, либо к одному из следующих вирусов: LMV, TSWV, "Big vein", TBSV, LNSV, TuMV, CMV или BWYV.
13. Растение по п. 11 или 12, отличающееся тем, что указанный еще один ген стандартной или промежуточной устойчивости находится в гетерозиготной форме.
14. Растение по п. 11 или 12, отличающееся тем, что указанные гены стандартной или промежуточной устойчивости выбраны из следующих генов устойчивости к ложной мучнистой росе: Dm10, R17, Dm5/Dm8, R36/R37 (кластер 1); Dm1, Dm2, Dm3, Dm6, Dm14, Dm15, Dm16, Dm18 (кластер 2), или Dm4, Dm7, Dm11, R38 (кластер 4), либо гена Tu устойчивости к TuMV (кластер 1), либо гена Nr устойчивости к тле (кластер 2), либо генов mo1.1, mo1.2 устойчивости к LMV (кластер 4).
15. Клетка растения гибридного культурного салата (Lactuca sativa), которая обладает гетерозиготным генотипом мужской стерильности, обеспечиваемым геном стерильности Ms7.
16. Клетка по п. 15, отличающаяся тем, что геном указанной клетки включает последовательность двунитевой ДНК, которая содержит от 650 до 700 нуклеотидов, и 5′-конец каждой из двух нитей которой начинается с последовательности 5′ TGCGTTCCAC 3′ (SEQ ID NO: 1).
17. Клетка по п. 16, отличающаяся тем, что указанная клетка является гетерозиготной по меньшей мере по еще другому одному гену, сообщающему ей фенотип, распознаваемый в растении.
18. Клетка по п. 17, отличающаяся тем, что упомянутый фенотип обусловлен геном стандартной устойчивости или промежуточной устойчивости к заражению вирусом, бактерией, насекомым или грибом.
19. Клетка по п. 18, отличающаяся тем, что упомянутый фенотип представляет собой стандартную или промежуточную устойчивость к одному из следующих грибов: Bremia lactucae, Fusarium oxysporum, Sclerotinia minor или sclerotorum, Botrytis cinerea, Rhizictonia solani, Microdochium panattonianum, Verticiulium dahliae, Erysiphe chicocearum или Pithium tracheiphilum, к одному из следующих насекомых: Nasonovia ribisnigri, Myzus persicae, Macrosiphum euphorbia, Nematodes pratylenchus или meloidogyne, минирующим мушкам Liriomyza huidobrensis или Pemphigus busarius; к одной из следующих бактерий: pseudomonas, xanthomonas или rhizomonas, либо к одному из следующих вирусов: LMV, TSWV, "Big vein", TBSV, LNSV, TuMV, CMV или BWYV.
20. Клетка по п. 18 или 19, отличающаяся тем, что указанная клетка содержит второй отличный ген стандартной или промежуточной устойчивости в гетерозиготной форме.
21. Клетка по п. 18 или 19, отличающаяся тем, что указанные гены стандартной или промежуточной устойчивости выбраны из следующих генов устойчивости к ложной мучнистой росе: Dm10, R17, Dm5/Dm8, R36/R37 (кластер 1); Dm1, Dm2, Dm3, Dm6, Dm14, Dm15, Dm16, Dm18 (кластер 2), или Dm4, Dm7, Dm11, R38 (кластер 4), либо гена Tu устойчивости к TuMV (кластер 1), либо гена Nr устойчивости к тле (кластер 2), либо генов mo1.1, mo1.2 устойчивости к LMV (кластер 4).
22. Применение насекомых отряда двукрылых, принадлежащих к семейству Calliphorides, в количестве более 100 двукрылых на 1 м2 или более 25 двукрылых на 1 м3 для осуществления опыления в закрытой среде мужских стерильных растений Lactuca sativa, используемых в качестве "женских" родительских форм мужскими фертильными растениями Lactuca sativa, используемыми в качестве "мужских" родительских форм, и получения гибридного семени Lactuca sativa согласно любому из пп. 1-7, причем мужская стерильность растений, применяемых в качестве женских родительских форм, обеспечивается геном мужской стерильности Ms7.
23. Применение по п. 22, отличающееся тем, что одна из двух родительских форм дополнительно характеризуется гомозиготностью по гену, который придает ей различимый фенотип, отличный от мужской стерильности, и при этом другая родительская форма не обладает этим геном.
24. Применение по п. 22, при котором двукрылые присутствуют в количестве по меньшей мере 250 двукрылых на 1 м2 или по меньшей мере 75 двукрылых на 1 м3.
25. Применение по любому из пп. 22-24 для получения гибридных растений Lactuca sativa.
26. Применение по любому из пп. 22-24, отличающееся тем, что геном указанных растений Lactuca sativa, характеризующихся мужской стерильностью, включает последовательность двунитевой ДНК, которая содержит от 650 до 700 нуклеотидов и 5′-конец каждой из двух нитей которой начинается с последовательности 5′ TGCGTTCCAC 3′ (SEQ ID NO: 1).
27. Способ получения семян гибридного Lactuca sativa, содержащий следующие стадии:
- возделывание в закрытой среде растений Lactuca sativa с фенотипом мужской стерильности, используемых в качестве "женских" родительских форм, и растений Lactuca sativa с фенотипом мужской фертильности, используемых в качестве "мужских" родительских форм, поблизости друг от друга;
- внесение двукрылых, принадлежащих к семейству Calliphorides, в указанную закрытую среду во время цветения растений в количестве более 100 двукрылых на 1 м2 или более 25 двукрылых на 1 м3, предпочтительно по меньшей мере 250 двукрылых на 1 м2 или по меньшей мере 75 двукрылых на 1 м3;
- сбор семян согласно любому из пп. 1-7, выросших на мужских стерильных растениях,
причем мужская стерильность растений, применяемых в качестве женских родительских форм, обеспечивается геном мужской стерильности Ms7.
28. Способ по п. 27, в котором одна из двух родительских форм дополнительно характеризуется гомозиготностью по гену, который придает ей различимый фенотип, отличный от мужской стерильности, и при этом другая родительская форма не обладает этим геном.
29. Способ по п. 28, в котором вторая родительская форма также является гомозиготной по меньшей мере по одному гену, сообщающему ей распознаваемый фенотип, отличающийся от мужской стерильности, которым не обладает другая родительская форма.
30. Способ по любому из пп. 27-29, в котором указанное количество введенных двукрылых составляет более 400 двукрылых на 1 м2, предпочтительно более 50 двукрылых на 1 м2.
31. Способ по любому из пп. 27-29, отличающийся тем, что указанных двукрылых вводят в виде куколок.
32. Способ по любому из пп. 27-29, в котором упомянутая закрытая среда представляет собой оболочку, непроницаемую для насекомых.
33. Способ по любому из пп. 27-29, в котором указанные двукрылые представляют собой Calliphora vomitaria, Calliphora erythrocephalka или Lucilia caesar.
34. Способ по любому из пп. 27-29, в котором количество мужских стерильных растений в закрытой среде превышает количество мужских фертильных растений.
35. Способ по любому из пп. 27-29, в котором количество мужских стерильных растений в закрытой среде составляет по меньшей мере 2000, а количество мужских фертильных растений - по меньшей мере 1000.
36. Способ по любому из пп. 27-29, в котором ввод двукрылых повторяют по меньшей мере один раз в неделю в течение 3-4 недель.
37. Способ по любому из пп. 27-29, в котором растения, используемые в качестве женских родительских форм, получают способом, включающим следующие стадии:
- скрещивание между растениями Lactuca sativa, гетерозиготными по одному доминантному ядерному гену мужской стерильности Ms7, и мужскими фертильными растениями Lactuca sativa, не имеющими генов стерильности;
- возделывание семян, полученных в результате упомянутого скрещивания;
- удаление растений, имеющих фенотип мужской фертильности.
38. Способ по п. 37, в котором стадию удаления растений, имеющих фенотип мужской фертильности, осуществляют путем обнаружения в образце каждого растения отсутствия молекулярного маркера, специфичного для доминантного гена мужской стерильности.
39. Способ по п. 38, в котором геном растений, используемых в качестве женских родительских форм, содержит последовательность двунитевой ДНК, которая содержит от 650 до 700 нуклеотидов и 5′-концы каждой из двух нитей которой начинаются с последовательности TGCGTTCCAC (SEQ ID NO №1).
| US2005144672 A1 (KNERR LARRY D et al), 30.06.2005 (см | |||
| реферта, формулу) | |||
| US2006021077 A1 (VAN SCHIJNDEL JOHANNES T ?[NL]), 26.01.2006 (см | |||
| реферат, формулу) | |||
| GOUBARA M | |||
| et al., Pollination effects of the sweat bee Lasioglossum villosulum trichopse (Hymenoptera: Halictidae) on genic male-sterile lettuce, Appl | |||
| Entomol | |||
| Zool, 2004, Vol.39, |

