ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка заявляет преимущество по предварительной заявке на патент США № 62/832716, поданной 11 апреля 2019 г.; предварительной заявке на патент США № 62/815261, поданной 7 марта 2019 г.; и предварительной заявке на патент США № 62/731252, поданной 14 сентября 2018 г.; при этом полное содержание каждой из них включено в данный документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к области молекулярной биологии растений, более конкретно к воздействию на мужскую фертильность.
ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, ПРЕДОСТАВЛЕННЫЙ В ЭЛЕКТРОННОМ ВИДЕ
Официальная копия перечня последовательностей предоставлена в электронном виде посредством EFS-Web в виде перечня последовательностей в формате ASCII с названием файла 7795WOPCT_seqlisting_ST25.txt, созданном 8 сентября 2019 г. и имеющем размер 97 килобайт, и подана одновременно с описанием. Перечень последовательностей, содержащийся в данном документе в формате ASCII, является частью описания и включен в данный документ посредством ссылки в полном объеме.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Развитие селекции гибридных растений обеспечило возможность значительного прогресса в отношении качественных и количественных свойств получаемых культур. Увеличение урожайности и сочетание необходимых характеристик, таких как устойчивость к заболеваниям и насекомым, выносливость к высоким температурам и засухе, наряду с вариациями состава растений, возможны благодаря процедурам гибридизации. Эти процедуры часто в значительной степени основаны на обеспечении мужского родительского растения, которое передает пыльцу женскому родительскому растению с получением в результате гибрида.
Селекцию полевых культур осуществляют с помощью методик, в которых используется преимущество способа опыления растений. Растение является самоопыляемым, если пыльца с одного цветка переносится на тот же или другой цветок того же растения или генетически идентичного растения. Растение является перекрестно опыляемым, если пыльца происходит из цветка на другом растении.
У определенных видов, таких как Brassica campestris, растение, как правило, является самостерильным и может быть только перекрестно опыляемым. У самоопыляемых видов, таких как соя, хлопчатник и пшеница, мужские и женские растения являются сопоставимыми с анатомической точки зрения. Во время естественного опыления мужские репродуктивные органы данного цветка опыляют женские репродуктивные органы того же цветка. Маис имеет мужские цветки, расположенные на метелке, и женские цветки, расположенные на початке, на одном и том же растении, и его скрещивание можно проводить с помощью методик, предусматривающих как самоопыление, так и перекрестное опыление.
При получении гибридных семян желательно предупреждать самоопыление женского инбредного растения для того, чтобы избежать получения и сбора семян женских инбредных растений, так как они проявляют меньшую мощность, чем гибридные семена. Для того, чтобы повысить товарные количества получаемых в результате гибридных семян, гибридные семена часто получают с применением женских родительских растений с мужской стерильностью и мужских родительских растений.
КРАТКОЕ ОПИСАНИЕ
В данном документе предусмотрены композиции и способы восстановления мужской фертильности у растения с мужской стерильностью. Мужская фертильность может быть восстановлена у растения с мужской стерильностью путем предоставления донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, от других вида или растения, отличающихся от растения с мужской стерильностью. Донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, содержит локус, обеспечивающий восстановление мужской фертильности, который функционально комплементирует фенотип мужской стерильности растения, приведенного в состояние мужской стерильности за счет одной или нескольких гомозиготных рецессивных мутаций в полинуклеотиде, обеспечивающем мужскую фертильность. Локус, обеспечивающий восстановление мужской фертильности, сцеплен с происходящим из растения полинуклеотидом, который обеспечивает фенотипический маркер растения. Также в данном документе предусмотрены растения, растительные клетки и семена, которые имеют одну или несколько гомозиготных мутаций в полинуклеотиде, обеспечивающем мужскую фертильность, и содержат донорный хромосомный компонент, обеспечивающий восстановление фертильности растения. В некоторых примерах в растении донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, замещает хромосому, являющуюся нативной по отношению к растению с мужской стерильностью, вследствие чего у растения образуются эуплоидные семена.
В некоторых вариантах осуществления, раскрытых в данном документе, донорные хромосомные компоненты, обеспечивающие восстановление фертильности растения, модифицированы в отношении одного или нескольких происходящих из растения полинуклеотидов, которые обеспечивают фенотипический маркер растения, и/или одного или нескольких локусов, обеспечивающих восстановление мужской фертильности. Например, модифицированные происходящие из растения полинуклеотиды и/или локус, обеспечивающий восстановление мужской фертильности, могут быть модифицированы в отношении их полинуклеотидной последовательности, числа копий, уровня экспрессии или положения в пределах донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, по сравнению с нативным немодифицированным происходящим из растения полинуклеотидом, который обеспечивает фенотипический маркер растения, или локусом, обеспечивающим восстановление мужской фертильности. Также в данном документе раскрыты растения, растительные клетки и семена, имеющие такие донорные хромосомные компоненты, обеспечивающие восстановление фертильности растения.
Соответственно, в данном документе описаны способы получения и применения таких растений. Растения-закрепители стерильности, имеющие какой-либо из описанный в данном документе донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, можно применять для получения семян путем обеспечения их самоопыления. Семена, растения или их части, содержащие донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, могут быть идентифицированы с применением фенотипического маркера растения, например, при сортировке семян. Отсутствие экспрессии фенотипического маркера растения в семенах указывает на то, что семена не содержат донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, и после высева обеспечат развитие женских растений с мужской стерильностью. И наоборот, наличие фенотипического маркера растения в семенах указывает на то, что семена содержат донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, и после высева обеспечат развитие растений с мужской фертильностью. Такие семена и растения могут применяться для закрепления мужской стерильности, женских инбредных растений с мужской стерильностью для повышения уровня получения гибридов и семян.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На фиг. 1 продемонстрированы примеры фенотипов поколения F3 пшеницы. Часть A) демонстрирует окрашенное в голубой цвет семя пшеницы, из которого сформируются растения пшеницы с мужской фертильностью, и часть B) демонстрирует окрашенное в белый цвет семя пшеницы (не окрашенное в голубой цвет), из которого сформируются женские растения пшеницы с мужской стерильностью.
На фиг. 2 продемонстрирован пример распределения семян из растения пшеницы поколения F3 по цвету семян с частью семян пшеницы, окрашенных в белый цвет, которые обеспечат развитие женских растений пшеницы с мужской стерильностью, частью семян пшеницы, окрашенных в светло-голубой цвет (гетерозиготных по признаку голубого алейрона), которые обеспечат развитие растений пшеницы с мужской фертильностью; и частью семян пшеницы, окрашенных в темно-голубой цвет (гомозиготных по признаку голубого алейрона), которые обеспечат развитие растений пшеницы с мужской фертильностью.
На фиг. 3 проиллюстрирован один вариант осуществления перицентрической инверсии Ms1 для увеличения сцепления с BA (голубым алейроном) на хромосоме.
На фиг. 4 продемонстрирован один вариант осуществления транслокации и замещения хромосомного плеча пшеницы донорным хромосомным компонентом, обеспечивающим восстановление фертильности растения, 4E-Ms45. Часть A демонстрирует хромосомный компонент, обеспечивающий восстановление фертильности растения, телосомной добавочной хромосомы 4EL, которая предоставляет Ms45 и BA. Часть B демонстрирует донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, 4EL, которая предоставляет Ms45 и BA, а также транслоцирует и замещает хромосому 5AL пшеницы.
ПОДРОБНОЕ ОПИСАНИЕ
Все публикации и заявки на патенты, упомянутые в настоящем описании, ориентированы на уровень специалиста в области техники, к которой относится настоящее изобретение. Все публикации и заявки на патенты включены в данный документ посредством ссылки в той же степени, как если бы каждая отдельная публикация или заявка на патент была конкретно и индивидуально указана как включенная посредством ссылки.
В данном документе предусмотрены растения, части растений, клетки растений или семена, имеющие донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, который можно применять для восстановления мужской фертильности у растений с мужской стерильностью и способствования идентификации частей растений, клеток растений или семян, имеющих донорный хромосомный компонент, обеспечивающий восстановление фертильности растения. Используемый в данном документе термин “донорный хромосомный компонент, обеспечивающий восстановление фертильности растения” представляет собой хромосому или ее фрагмент, которые содержат происходящий из растения полинуклеотид, который обеспечивает фенотипический маркер растения, сцепленный с локусом, обеспечивающим восстановление мужской фертильности. Применяемый в данном документе термин "происходящий из растения" указывает на то, что полинуклеотид, который обеспечивает фенотипический маркер растения, получен из растения. В некоторых примерах происходящий из растения полинуклеотид, который обеспечивает фенотипический маркер растения, является эндогенным по отношению к донорному хромосомному компоненту, обеспечивающему восстановление фертильности растения, и обеспечивающему восстановление мужской фертильности локусу. Применяемый в данном документе термин "эндогенный", или “нативный”, или “нативно” означает в норме присутствующий в указанном растении, присутствующий в его нормальном состоянии или положении в хромосоме (немодифицированной), растительной клетке или растении.
В некоторых примерах происходящий из растения полинуклеотид является экзогенным по отношению к донорному хромосомному компоненту, обеспечивающему восстановление фертильности растения, растению или растительной клетке, в которые его вводят. Термин "экзогенный" означает не присутствующий в норме в хромосомном компоненте, растении, растительной клетке; не присутствующий в его нормальном состоянии или положении в хромосомном компоненте, растении или растительной клетке, вводимый в хромосомный компонент, растение или растительную клетку, или происходящий из других хромосомы, типа растения, вида растения, или, в случае происхождения из тех же хромосомы, типа растения, вида растения, находящийся в другом положении, модифицированный по сравнению с его нативной формой в отношении состава и/или геномного локуса путем целенаправленного вмешательства человека. В некоторых примерах донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, предусматривает один или несколько происходящих из растения полинуклеотидов, которые обеспечивают фенотипический маркер растения, включая без ограничения один или несколько нативных, подвергнутых редактированию, перемещению, замене или вставке происходящих из растения полинуклеотидов, которые обеспечивают фенотипический маркер растения, или их комбинации. В вариантах осуществления, где имеется два или более происходящих из растения полинуклеотидов, полинуклеотиды могут быть одинаковыми или отличаться друг от друга, например, в отношении их последовательностей, как, например, в отношении их происхождения, например, типа или вида растения, полинуклеотидной или аминокислотной последовательности или положения в донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения. В некоторых примерах один или несколько происходящих из растения полинуклеотидов, которые обеспечивают фенотипический маркер растения, являются экзогенными по отношению к растению, растительной клетке или донорному хромосомному компоненту, обеспечивающему восстановление фертильности растения, в которые его вводят, или их комбинации.
Экспрессия фенотипического маркера растения в семени позволяет подвергать семя идентификации, отбору и/или отделению при сортировке от семян, которые не содержат донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, т.е. не имеют происходящий из растения полинуклеотид, который обеспечивает фенотипический маркер растения. В некоторых аспектах фенотипический маркер растения является неразрушающим маркером.
Фенотипический маркер растения может относиться к цвету, физиологии или морфологии растения или семени. Примеры фенотипов семени, которые являются подходящими маркерами, включают без ограничения цвет семени, цветовые интенсивность или паттерн семени, форму семени, текстуру поверхности семени, размер семени, включая размер семени по длине и/или ширине, плотность семени или другие характеристики семени. Примеры относящихся к цвету фенотипических маркеров семян включают без ограничения голубой алейрон, ген P, который регулирует синтез флавоноидов у маиса, антоциан, Kala 4 и другие характеристики окраски эндосперма. В некоторых примерах происходящий из растения полинуклеотид представляет собой ThMYC4E и обуславливает фенотип голубого алейрона. См. Li, Na et al. “ThMYC4E, Candidate Blue Aleurone 1 Gene Controlling the Associated Trait in Triticum Aestivum.” Ed. Harsh Raman. PLoS ONE 12.7 (2017): e0181116. PMC. Web. 13 Sept. 2018, включенную в данный документ посредством ссылки во всей своей полноте. См. в опубликованной заявке на патент согласно PCT № WO2019090496, включенной в данный документ посредством ссылки во всей своей полноте, и в относящемся к ней перечне последовательностей. В некоторых вариантах осуществления, где имеется два или более происходящих из растения полинуклеотидов, два происходящих из растения полинуклеотида являются полинуклеотидами, кодирующими относящиеся к цвету маркеры голубого алейрона, которые обуславливают у семени фенотип голубого алейрона и могут применяться для идентификации, отбора и сортировки семян. Относящиеся к цвету маркеры могут быть одинаковыми или отличаться друг от друга, например, в отношении их последовательностей, как, например, в отношении их происхождения, например, типа или вида растения, полинуклеотидной или аминокислотной последовательности или положения в донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения.
При необходимости происходящие из растения полинуклеотиды, которые обеспечивают фенотипический маркер растения, могут быть модифицированы для усиления его экспрессии в растении, например, для усиления экспрессии фенотипического маркера растения в растении, его соответствующей части растения или семени. В некоторых аспектах регуляторный участок происходящего из растения полинуклеотида может быть модифицирован для повышения экспрессии фенотипического маркера растения, например, путем редактирования существующего регуляторного участка с заменой, удалением и/или вставкой нуклеотидов с целью обеспечения улучшенной экспрессии. См., например, публикацию патента согласно PCT № WO2018183878, опубликованного 4 октября 2018 г., включенную в данный документ посредством ссылки во всей своей полноте. Дополнительно или в качестве альтернативы нуклеотиды в фенотипическом маркере растения, находящемся на донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения, могут быть модифицированы с изменением полинуклеотида таким образом, чтобы в нем использовались кодоны, которым отдает предпочтение растение-хозяин.
В некоторых вариантах осуществления, ген голубого алейрона в хромосомном компоненте, обеспечивающем восстановление фертильности растения, характеризуется последовательностью нуклеиновой кислоты, кодирующей аминокислотную последовательность на по меньшей мере 80%, на по меньшей мере 85%, на по меньшей мере 90%, на по меньшей мере 95%, на по меньшей мере 96%, на по меньшей мере 97%, на по меньшей мере 98% или на по меньшей мере 99% идентичную аминокислотной последовательности под SEQ ID NO: 41 или 42 или ее вариантам или фрагментам. Последовательность нуклеиновой кислоты, кодирующая полипептид, который обуславливает фенотип голубого алейрона, может быть синтезирована, выделена или получена из любого числа источников, включающих без ограничения представителей родов Agropyron, Thinopyrum или Triticum, таких как Agropyron elongatum, Agropyron trichophorum, Triticum thaoudar, Triticum Aestivum, Triticum monococcum и Thinopyrum ponticum. Источником голубого алейрона и донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, может являться Sebesta Blue, Blue Sando, Blue Baart, Blue Onas, Blue 1, PBB или Blue Norco или другая добавочная линия, которая содержит полинуклеотид, кодирующий голубой алейрон. Присутствие голубого алейрона в семени может быть обнаружено по появлению голубого окрашивания в алейроновом слое семени, подтверждено с использованием ПЦР или любого другого подходящего анализа. Подразумевается, что включенные в данный документ примеры фенотипических маркеров растений не являются ограничивающими. Любой требуемый фенотипический маркер растения может применяться в способах и композициях, описанных в данном документе.
Семена могут быть подвергнуты сортировке на разные популяции с применением любого подходящего фенотипического маркера семян. Например, отсутствие в семени фенотипического маркера растения, например семя, не имеющее донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, указывает на то, что из семени после высева вырастет женское растение с мужской стерильностью. Растения, полученные из этого семени, могут применяться в качестве женских инбредных растений с мужской стерильностью для повышения уровня получения гибридов и семян. Присутствие фенотипического маркера растения в семени, например семени, имеющем донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, указывает на то, что из семени вырастет растение с мужской фертильностью, которое может быть использовано в качестве закрепителя стерильности для женского растения с мужской стерильностью. Как обсуждалось в других разделах настоящего документа, семена можно подвергать сортировке с применением любого подходящего подхода или инструмента при условии, что он характеризуется достаточной чувствительностью для выявления различий между экспрессирующими и не экспрессирующими фенотипический маркер семенами.
В дополнение к происходящему из растения полинуклеотиду, который обеспечивает фенотипический маркер растения, донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, содержит локус, обеспечивающий восстановление мужской фертильности, который способен функционально комплементировать фенотип с мужской стерильностью растения. Применяемый в данном документе термин “локус, обеспечивающий восстановление мужской фертильности” относится к одному или нескольким полинуклеотидам, обеспечивающим мужскую фертильность, которые в случае их экспрессии в женском растении с мужской стерильностью восстанавливают у растения мужскую фертильность путем комплементации состояния мужской стерильности у женского растения, вызванного одной или несколькими гомозиготными мутациями в полинуклеотиде, обеспечивающем мужскую фертильность. Такие мутации могут являться заменами, удалениями и/или вставками одного или нескольких нуклеотидов, что приводит к приданию растению свойства мужской стерильности. Применяемый в данном документе термин “полинуклеотид, обеспечивающий мужскую фертильность” означает один из полинуклеотидов, критически важных для конкретной стадии микроспорогенеза, причем термин применяется по отношению ко всему процессу образования пыльцы. В некоторых примерах один или несколько полинуклеотидов, обеспечивающих мужскую фертильность, включают без ограничения Ms1, Ms5, Ms9, Ms22, Ms26 или Ms45. В некоторых вариантах осуществления локус, обеспечивающий восстановление мужской фертильности, предусматривает использование без ограничения двух или более полинуклеотидов, обеспечивающих мужскую фертильность, представляющих собой Ms1, Ms5, Ms9, Ms22, Ms26 или Ms45, для обеспечения комплементации состояния мужской стерильности у женского растения, вызванного одной или несколькими гомозиготными мутациями в полинуклеотиде, обеспечивающем мужскую фертильность, представляющем собой Ms1, Ms5, Ms9, Ms22, Ms26 или Ms45 соответственно.
При необходимости локус, обеспечивающий восстановление мужской фертильности, может быть модифицирован для повышения экспрессии полинуклеотида, обеспечивающего мужскую фертильность, в растении, например, для обеспечения комплементации мужской стерильности или восстановления мужской фертильности у растения с мужской стерильностью. В некоторых аспектах регуляторный участок может быть модифицирован для повышения экспрессии локуса, обеспечивающего восстановление мужской фертильности, или полинуклеотида, обеспечивающего мужскую фертильность, например, путем редактирования существующего регуляторного участка с заменой, удалением и/или вставкой нуклеотидов с целью обеспечения улучшенной экспрессии. См., например, публикацию патента согласно PCT № WO2018183878, опубликованного 4 октября 2018 г., включенную в данный документ посредством ссылки во всей своей полноте. Дополнительно или в качестве альтернативы нуклеотиды в обеспечивающем восстановление мужской фертильности локусе или полинуклеотиде, обеспечивающем мужскую фертильность, находящихся на донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения, могут быть модифицированы с изменением полинуклеотида таким образом, чтобы в нем использовались кодоны, которым отдает предпочтение растение-хозяин.
В некоторых примерах локус, обеспечивающий восстановление мужской фертильности, ассоциирован с повышенной выраженностью фенотипа мужской фертильности у растения. Состояние мужской фертильности у растения можно оценить с помощью любой подходящей методики, например, путем наблюдения за развитием мужской ткани у растения, как например фенотипирование пыльников и образующихся семян на отдельных растениях. См., например, пример 1 в данном документе.
В некоторых примерах локус, обеспечивающий восстановление мужской фертильности, предусматривает без ограничения один или несколько нативных, подвергнутых редактированию, перемещению, замене или вставке полинуклеотидов, или их комбинации. В вариантах осуществления где в локусах, обеспечивающих восстановление мужской фертильности, имеется два или более полинуклеотидов, обеспечивающих мужскую фертильность, локусы, обеспечивающие восстановление мужской фертильности, могут являться одинаковыми или отличаться друг от друга, например, в отношении их последовательностей, как, например, в отношении их происхождения, например, типа или вида растения, полинуклеотидной или аминокислотной последовательности или положения в донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения. В некоторых аспектах один или несколько локусов, обеспечивающих восстановление мужской фертильности, являются эндогенными по отношению к растению, растительной клетке или донорному хромосомному компоненту, обеспечивающему восстановление фертильности растения, в которые его вводят, или их комбинации. В некоторых аспектах один или несколько локусов, обеспечивающих восстановление мужской фертильности, являются экзогенными по отношению к растению, растительной клетке или донорному хромосомному компоненту, обеспечивающему восстановление фертильности растения, в которые его вводят, или их комбинации. Восстановление мужской фертильности может предусматривать один или несколько полинуклеотидов, обеспечивающих мужскую фертильность, известных специалистам в данной области, описанных в данном документе, включая гомологи и ортологи всех вышеупомянутых.
В одном примере локус, обеспечивающий восстановление мужской фертильности, функционально комплементирует фенотип мужской стерильности применительно к одной или нескольким гомозиготным мутациям в эндогенном полинуклеотиде, обеспечивающем мужскую фертильность, которые придают мужскую стерильность растению. Это включает без ограничения один или несколько гомозиготных рецессивных аллелей для Ms1, Ms5, Ms9, Ms22, Ms26 или Ms45. Поскольку Ms1 в растениях пшеницы ведет себя как моногенный рецессив, в некоторых вариантах осуществления для придания мужской стерильности растению пшеницы может потребоваться только внесение мутации в полинуклеотид, обеспечивающий мужскую фертильность, представляющий собой ms1, или аллель, расположенный на хромосоме 4BS.
Существует целый ряд известных полинуклеотидов, обеспечивающих мужскую фертильность, и мутантов с мужской фертильностью, относящихся к пшенице и другим видам, включая без ограничения Ms1, Ms5, Ms9, Ms22, Ms26 или Ms45.
В публикации патента согласно PCT № WO2016048891, опубликованном 31 марта 2016 г., описан ген мужской фертильности, обозначаемый как “MS1”, который расположен на хромосоме пшеницы 4BS и кодирует заякоренный гликозилфосфатидилинозитолом (GPI) полипептид nsLTP (LTPG) (также обозначаемый как TaLTPGI), имеющий важное значение для обеспечения мужской фертильности. Примеры последовательностей ДНК и полипептидов для последовательностей Ms1 пшеницы раскрыты в WO2016048891, опубликованном 31 марта 2016 г., и WO2019118342, опубликованном 20 июня 2019 г., каждый из которых включен в данный документ во всей своей полноте, а также в относящихся к ним перечням последовательностей.
Ms5 пшеницы является заякоренным гликозилфосфатидилинозитолом белком-переносчиком липидов, необходимым для нормального развития экзины пыльцы, и данный ген расположен на хромосоме 3A пшеницы. Примеры последовательностей ДНК и полипептидов для Ms5 пшеницы раскрыты в WO 2019118342, опубликованном 20 июня 2019 г., каждый из которых включен в данный документ во всей своей полноте, а также в относящемся к нему перечне последовательностей.
В публикации патента США № US20150191743 A1, опубликованного 9 июля 2015 г., описывается ген мужской фертильности, обозначаемый как “MS9”, который расположен на хромосоме 1 маиса и кодирует транскрипционный фактор myb, являющийся критически важным для обеспечения мужской фертильности. Фенотип Ms9 впервые был идентифицирован у маиса в 1932 г. Beadle, (1932) Genetics 17:413-431. Было обнаружено, что он связан с геном P1 на хромосоме 1. Прекращение развития мужской репродуктивной ткани происходит на очень ранней стадии в период премейоза; тапетальные клетки также могут поражаться. Greyson, et al., (1980) Can. J. Genet. Cytol. 22:153-166. Примеры последовательностей геномной ДНК и полипептида для Ms9 маиса раскрыты в публикации патента США № 20150191743, опубликованного 9 июля 2015 г., включенной в данный документе во всей своей полноте. Ms9 пшеницы расположен на длинном плече хромосомы 4 пшеницы. Примеры последовательностей геномной ДНК и полипептида для Ms9 пшеницы раскрыты в публикации патента США № US20190177722A1, опубликованного 13 июня 2019 г., включенной в данный документе во всей своей полноте, а также в относящемся к ней перечне последовательностей.
В публикации патента США № US20090038026A1, опубликованного 5 февраля 2009 г., описывается ген мужской фертильности, обозначаемый как “Msca1” или “MS22”, который расположен на хромосоме 7 маиса и кодирует белок имеющий критически важное значение для обеспечения мужской фертильности. Мутации, обозначаемые как ms22 или msca1, впервые отметили как обуславливающие фенотипическую мужскую стерильность в случае пыльников, которые не выступали из метелки и не имели спорообразующей ткани. West and Albertsen (1985) Maize Newsletter 59:87; Neuffer et al. (1977) Mutants of maize. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. Мутантный локус изначально был обозначен как ms22, но позже изменен на msca1 или видоизмененный пыльник с мужской стерильностью. См. Chaubal et al. “The transformation of anthers in the msca1 mutant of maize” Planta (2003)216:778-788. Ms22 пшеницы расположен на длинном плече хромосомы 2 пшеницы. Примеры последовательностей геномной ДНК и полипептида для Ms22 пшеницы раскрыты в публикации патента США № US20190177722A1, опубликованного 13 июня 2019 г., включенной в данный документ во всей своей полноте, а также в относящемся к ней перечне последовательностей.
В патенте США № 7517975, выданном 14 апреля 2009 г., описан ген мужской фертильности, обозначаемый как “MS26” (также известный как SB200 или SBMu200), который расположен на хромосоме 1 маиса. Последовательности Ms26 для маиса или риса, например, раскрыты в патентах США № 7919676 или № 8293970. У пшеницы ген Ms26 расположен на хромосоме 4AS пшеницы. Примеры последовательностей геномной ДНК и полипептида для Ms26 пшеницы раскрыты в публикации патента США № US20190177722A1, опубликованного 13 июня 2019 г., включенной в данный документе во всей своей полноте, а также в относящемся к ней перечне последовательностей.
В патенте США № 5478369, выданном 26 декабря 1995 г., описан ген мужской фертильности, обозначаемый как “MS45”, клонированный в составе хромосомы 9 маиса. У пшеницы ген Ms45 расположен на длинном плече хромосомы 4 пшеницы. Примеры последовательностей геномной ДНК и полипептида для Ms45 пшеницы раскрыты в публикации патента США № US20190177722A1, опубликованного 13 июня 2019 г., включенной в данный документе во всей своей полноте, а также в относящемся к ней перечне последовательностей.
Женские растения с мужской стерильностью для применения в способах и композициях, описанных в данном документе, могут быть получены с применением любого числа способов, признанных в данной области, включая без ограничения мутагенез, супрессию и редактирование генома. Мутации, которые вызывают мужскую стерильность у культурных растений, таких как маис, пшеница и рис, были получены посредством ряда различных способов, таких как рентгеновское или УФ-облучение, разновидности химической обработки или вставки мобильных элементов (Chaubal et al. 2000) Am J Bot 87:1193-1201). Обеспечивающие супрессию технологии, такие как антисмысловые молекулы, косупрессия, RNAi, шпилечные структуры, могут применяться для нарушения или предотвращения экспрессии аллелей генов фертильности с целью получения женского растения с мужской стерильностью, например, получения гомозиготных рецессивных аллелей, которые обуславливают мужскую стерильность растения. В некоторых случаях мужская стерильность является результатом использования технологии редактирования генома с введением генетической модификации (мутации) вблизи эндогенного гена мужской фертильности или в него, например, полинуклеотида, обеспечивающего мужскую фертильность, с целью индуцирования стерильности. См. WO2015026883, опубликованный 26 февраля 2015 г., и Singh, M., Kumar, M., Albertsen, M.C. et al. Plant Mol Biol (2018) 97: 371-383. Соответственно, в способах можно применять технологию CRISPR с использованием системы направляющая РНК/эндонуклеаза Cas, где эндонуклеаза Cas направляется направляющей РНК для обеспечения распознавания и необязательного внесения двухнитевого разрыва в специфический целевой сайт в геноме клетки растения. В некоторых примерах геномы пшеницы (A, B и D) содержат гомологичные гены, которые характеризуются сходной генной структурой и функцией, что требует получения тройных мутантов для того, чтобы в результате получить фенотип мужской стерильности, например наличия одной, двух или трех гомозиготных мутаций в полинуклеотиде, обеспечивающем мужскую фертильность. В некоторых вариантах осуществления фенотип мужской стерильности обусловлен внесением генетической модификации (мутации) в целевой сайт, расположенный в одном или нескольких локусах эндогенных генов мужской фертильности Ms1, Ms5, Ms9, Ms22, Ms26 или Ms45 или вблизи них в геноме растительной клетки. См., например, US20190177722A1, опубликованный 13 июня 2019 г., и WO2019118342, опубликованный 20 июня 2019 г., каждый из которых включен в данный документ во всей своей полноте.
Соответственно, известные или созданные отредактированные или мутантные растения c мужской стерильностью ms1/ms1/; ms5/ms5; ms9/ms9/; ms22/ms22/; ms26/ms26/ или ms45/ms45 могут применяться в способах и композициях, описанных в данном документе, например, для применения в качестве женских растений с мужской стерильностью при получении гибридов и гибридных семян. Поскольку Ms1 в растениях пшеницы ведет себя как моногенный рецессив, в некоторых вариантах осуществления для придания мужской стерильности растению пшеницы может потребоваться только внесение мутации в полинуклеотид, обеспечивающий мужскую фертильность, представляющий собой ms1, или аллель, расположенный на хромосоме 4BS.
В некоторых вариантах осуществления в данном документе предусматриваются композиции и способы для комплементации и восстановления мужской фертильности у женских растений пшеницы с мужской стерильностью, содержащих одну или несколько гомозиготных мутаций в полинуклеотиде Ms1, Ms5, Ms9, Ms22, Ms26 или Ms45, обеспечивающем мужскую фертильность, которые вызывают у растения пшеницы состояние мужской стерильности. В некоторых примерах растение с мужской стерильностью содержит одну или несколько гомозиготных мутаций в полинуклеотиде рецессивного спорофитного гена, обеспечивающего мужскую фертильность. В некоторых примерах у таких растений пшеницы с мужской стерильностью можно восстановить мужскую фертильность в случае, когда донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, происходящий из вида, отличного от вида пшеницы, применяют для функциональной комплементации одной или нескольких гомозиготных мутаций Ms1, Ms5, Ms9, Ms22, Ms26 или Ms45, которые вызывают у растения пшеницы состояние мужской стерильности.
В некоторых вариантах осуществления последовательности полинуклеотида Ms1 пшеницы, обеспечивающего мужскую фертильность, включают (a) полинуклеотид, содержащий последовательность, представленную под SEQ ID NO:1, 3 или 5; (b) полинуклеотид, характеризующийся по меньшей мере 85%, 90% или 95% идентичностью последовательности с SEQ ID NO: 1, 3 или 5; (c) полинуклеотид, который кодирует полипептид, характеризующийся по меньшей мере 85%, 90% или 95% идентичностью последовательности с SEQ ID NO:2, 4 или 6; и (d) полинуклеотид, который кодирует полипептид под SEQ ID NO: 2, 4 или 6.
В некоторых вариантах осуществления последовательности полинуклеотида Ms5 пшеницы, обеспечивающего мужскую фертильность, включают (a) полинуклеотид, содержащий последовательность, представленную под SEQ ID NO:7, 9, 12 или 14; (b) полинуклеотид, характеризующийся по меньшей мере 85%, 90% или 95% идентичностью последовательности с SEQ ID NO: 7, 9, 12 или 14; (c) полинуклеотид, который кодирует полипептид, характеризующийся по меньшей мере 85%, 90% или 95% идентичностью последовательности с SEQ ID NO: 8, 10, 11, 13 или 15; и (d) полинуклеотид, который кодирует полипептид под SEQ ID NO: 8, 10, 11, 13 или 15.
В некоторых вариантах осуществления последовательности полинуклеотида Ms9 пшеницы, обеспечивающего мужскую фертильность, включают (a) полинуклеотид, содержащий последовательность, представленную под SEQ ID NO:16, 18 или 20; (b) полинуклеотид, характеризующийся по меньшей мере 85%, 90% или 95% идентичностью последовательности с SEQ ID NO: 16, 18 или 20; (c) полинуклеотид, который кодирует полипептид, характеризующийся по меньшей мере 85%, 90% или 95% идентичностью последовательности с SEQ ID NO:17, 19 или 21; и (d) полинуклеотид, который кодирует полипептид под SEQ ID NO: 17, 19 или 21.
В некоторых вариантах осуществления последовательности полинуклеотида Ms22 пшеницы, обеспечивающего мужскую фертильность, включают (a) полинуклеотид, содержащий последовательность, представленную под SEQ ID NO:22, 24 или 26; (b) полинуклеотид, характеризующийся по меньшей мере 85%, 90% или 95% идентичностью последовательности с SEQ ID NO: 22, 24 или 26; (c) полинуклеотид, который кодирует полипептид, характеризующийся по меньшей мере 85%, 90% или 95% идентичностью последовательности с SEQ ID NO: 23, 25 или 27; и (d) полинуклеотид, который кодирует полипептид под SEQ ID NO: 23, 25 или 27.
В некоторых вариантах осуществления последовательности полинуклеотида Ms26 пшеницы, обеспечивающего мужскую фертильность, включают (a) полинуклеотид, содержащий последовательность, представленную под SEQ ID NO:28, 30 или 32; (b) полинуклеотид, характеризующийся по меньшей мере 85%, 90% или 95% идентичностью последовательности с SEQ ID NO: 28, 30 или 32; (c) полинуклеотид, который кодирует полипептид, характеризующийся по меньшей мере 85%, 90% или 95% идентичностью последовательности с SEQ ID NO: 29, 31 или 33; и (d) полинуклеотид, который кодирует полипептид под SEQ ID NO: 29, 31 или 33.
В некоторых вариантах осуществления последовательности полинуклеотида Ms45 пшеницы, обеспечивающего мужскую фертильность, включают (a) полинуклеотид, содержащий последовательность, представленную под SEQ ID NO:34, 36 или 38; (b) полинуклеотид, характеризующийся по меньшей мере 85%, 90% или 95% идентичностью последовательности с SEQ ID NO: 34, 36 или 38; (c) полинуклеотид, который кодирует полипептид, характеризующийся по меньшей мере 85%, 90% или 95% идентичностью последовательности с SEQ ID NO: 35, 37 или 39; и (d) полинуклеотид, который кодирует полипептид под SEQ ID NO: 35, 37 или 39.
Таблица 1. Краткое описание SEQ ID NO:
В некоторых аспектах происходящий из растения полинуклеотид, который обеспечивает фенотипический маркер растения, и локус, обеспечивающий восстановление мужской фертильности, в нативном состоянии сцеплены друг с другом на донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения. В некоторых аспектах происходящий из растения полинуклеотид, который обеспечивает фенотипический маркер растения, и локус, обеспечивающий восстановление мужской фертильности, в нативном состоянии сцеплены друг с другом и оба расположены по одну сторону от центромеры на донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения, то есть на одном плече хромосомы. В некоторых вариантах осуществления они оба сцеплены друг с другом и располагаются на длинном плече донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, или оба располагаются на коротком плече донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, и не разделены центромерой. Настоящее изобретение, описанное в данном документе, частично основывается на открытии того, что хромосома 4EL из Blue Norco способна комплементировать мутации Tams45-abd, см., например, пример 1 в данном документе. В одном варианте осуществления фенотипический маркер растения представлен голубым алейроном и находится по ту же сторону от центромеры, что и локус, обеспечивающий восстановление мужской фертильности, который функционально комплементирует фенотип мужской стерильности у женского растения пшеницы с мужской стерильностью, которое имеет гомозиготные мутации Ms45.
Донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может происходить из любого растения при условии, что он способен восстанавливать фертильность у растения с мужской стерильностью, вследствие чего растение продуцирует жизнеспособную пыльцу, способную к оплодотворению растения, например, к самооплодотворению. В некоторых аспектах донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, происходит из маиса, риса, сорго, ржи, ячменя, пшеницы, проса, разновидностей овса, сахарного тростника, газонной травы, тритикале, проса прутьевидного, пырея, фонио, сои, канолы, люцерны, подсолнечника, хлопчатника, табака, арахиса, картофеля, табака, Arabidopsis, сафлора, Thinopyrum, Aegilops, Secale, Haynaldia, Elyymus, Hordeum или родственного им вида. В некоторых аспектах донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, предусматривает хромосому 4 растения. В некоторых вариантах осуществления донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, предусматривает хромосому 4E, 4EL или 4H, происходящую из вида рода Triticum, Thinopyrum, Aegilops, Secale, Haynaldia, Elyymus или Hordeum или родственного им вида. Ортологичный аналог донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, у других видов может находиться на той же хромосоме, например 4, или может располагаться на другой хромосоме. В некоторых аспектах фенотипический маркер растения и локус, обеспечивающий восстановление мужской фертильности, сцеплены друг с другом на хромосоме 4, которая может применяться как донорный хромосомный компонент, обеспечивающий восстановление фертильности растения.
В некоторых случаях локус, обеспечивающий восстановление мужской фертильности, и происходящий из растения полинуклеотид, который обеспечивает фенотипический маркер растения, не являются в высокой степени сцепленными или сцепленными в нативном состоянии. Молекулярные и биологические методики, включая технологию редактирования генома, как например CRISPR, Talons, мегануклеазы и т.п., могут использоваться для повышения их генетического сцепления и/или сокращения физической дистанции между ними на донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения.
В некоторых примерах один или несколько происходящих из растения полинуклеотидов, обуславливающих фенотипический маркер растения, подвергают вставке в донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, перемещению в его пределах или перегруппировке на нем с целью обеспечения повышения степени генетического сцепления, сокращения частоты рекомбинации, понижения частоты кроссинговера и/или сокращения физического расстояния между ними или их комбинаций, выполняемым по отношению к одному или нескольким обеспечивающим восстановление мужской фертильности локусам на донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения. Один или несколько происходящих из растения полинуклеотидов, обуславливающих фенотипический маркер растения, могут являться эндогенными или экзогенными по отношению к донорному хромосомному компоненту, обеспечивающему восстановление фертильности растения, и/или одному или нескольким обеспечивающим восстановление мужской фертильности локусам. Один или несколько локусов, обеспечивающих восстановление мужской фертильности, могут являться эндогенными или экзогенными по отношению к донорному хромосомному компоненту, обеспечивающему восстановление фертильности растения.
В некоторых примерах один или несколько локусов, обеспечивающих восстановление мужской фертильности, подвергают вставке в донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, перемещению в его пределах или перегруппировке на нем с целью обеспечения повышения степени генетического сцепления, сокращения частоты рекомбинации, понижения частоты кроссинговера и/или сокращения физического расстояния между ними или их комбинаций, выполняемым по отношению к одному или нескольким происходящим из растения полинуклеотидам, обуславливающим фенотипический маркер растения, на донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения. Один или несколько локусов, обеспечивающих восстановление мужской фертильности, могут являться эндогенными или экзогенными по отношению к донорному хромосомному компоненту, обеспечивающему восстановление фертильности растения, или одному или нескольким происходящим из растения полинуклеотидам, обуславливающим фенотипический маркер растения. Один или несколько происходящих из растения полинуклеотидов, обуславливающих фенотипический маркер растения, могут являться эндогенными или экзогенными по отношению к донорному хромосомному компоненту, обеспечивающему восстановление фертильности растения.
В других вариантах осуществления локус, обеспечивающий восстановление мужской фертильности, и происходящий из растения полинуклеотид, обуславливающий фенотипический маркер растения, были подвергнуты редактированию, вставке, перемещению, или перегруппировке, или их комбинациям, вследствие чего они оба находятся в отличающихся положениях на донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения, по сравнению с их нативными положениями на донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения. В некоторых примерах фенотипический маркер растения и локус, обеспечивающий восстановление мужской фертильности, расположены по одну сторону от центромеры донорного хромосомного компонента, обеспечивающего восстановление фертильности растения. В некоторых вариантах осуществления, где имеется более одного локуса, обеспечивающего восстановление мужской фертильности, и/или происходящего из растения полинуклеотида, обуславливающего фенотипический маркер растения, по меньшей мере один локус, обеспечивающий восстановление мужской фертильности, и по меньшей мере один происходящий из растения полинуклеотид находятся в отличающихся положениях на донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения, по сравнению с их нативными положениями. В некоторых вариантах осуществления происходящий из растения полинуклеотид, обуславливающий фенотипический маркер растения, и один или несколько локусов, обеспечивающих восстановление мужской фертильности, характеризуются меньшим физическим расстоянием между ними по сравнению с физическим расстоянием между исходным происходящим из растения полинуклеотидом и исходным локусом, обеспечивающим восстановление мужской фертильности, в случае их расположения в своих нативных положениях. В некоторых вариантах осуществления происходящий из растения полинуклеотид, обуславливающий фенотипический маркер растения, и один или несколько локусов, обеспечивающих восстановление мужской фертильности, характеризуются повышенным генетическим сцеплением между ними по сравнению с генетическим сцеплением между исходным происходящим из растения полинуклеотидом и исходным локусом, обеспечивающим восстановление мужской фертильности, в случае их расположения в своих нативных положениях. В некоторых аспектах происходящий из растения полинуклеотид, обуславливающий фенотипический маркер растения, и один или несколько локусов, обеспечивающих восстановление мужской фертильности, характеризуются пониженной частотой рекомбинации по сравнению с частотой рекомбинации между исходным происходящим из растения полинуклеотидом и исходным локусом, обеспечивающим восстановление мужской фертильности, в случае их расположения в своих нативных положениях. В некоторых аспектах происходящий из растения полинуклеотид, обуславливающий фенотипический маркер растения, и один или несколько локусов, обеспечивающих восстановление мужской фертильности, характеризуются более низкой частотой кроссинговера по сравнению с частотой кроссинговера между исходным происходящим из растения полинуклеотидом и исходным локусом, обеспечивающим восстановление мужской фертильности, в случае их расположения в своих нативных положениях.
В некоторых вариантах осуществления, где один или несколько локусов, обеспечивающих восстановление мужской фертильности, были вставлены в донорный хромосомный компонент, обеспечивающий восстановление фертильности растения,
нативный или исходный локус, обеспечивающий восстановление мужской фертильности, может быть нарушен таким образом, что он больше не экспрессируется и не может восстанавливать мужскую фертильность у растения с мужской стерильностью. В некоторых аспектах способ может включать нарушение нативного или исходного локуса, обеспечивающего восстановление мужской фертильности, в случае, когда он не находится по ту же сторону от центромеры на донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения, что и происходящий из растения полинуклеотид, обуславливающий фенотипический маркер растения, или не является в высокой степени сцепленными с происходящим из растения полинуклеотидом с генетической точки зрения.
В некоторых вариантах осуществления, где один или несколько происходящих из растения полинуклеотидов, обуславливающих фенотипический маркер растения, были вставлены в донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, нативный или исходный происходящий из растения полинуклеотид, обуславливающий фенотипический маркер растения, может быть нарушен таким образом, что он больше не экспрессируется и не может обуславливать маркерный фенотип у растения или его части. В некоторых аспектах способ может включать нарушение нативного или исходного происходящего из растения полинуклеотида, обуславливающего фенотипический маркер растения, в случае, когда он не находится по ту же сторону от центромеры на донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения, что и локус, обеспечивающий восстановление мужской фертильности, или не является в высокой степени сцепленным с локусом, обеспечивающим восстановление мужской фертильности, с генетической точки зрения.
В некоторых вариантах осуществления донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, содержит один или несколько модифицированных происходящих из растения полинуклеотидов, которые обеспечивают фенотипический маркер растения, сцепленных с одним или несколькими локусами, обеспечивающими восстановление мужской фертильности. Например, модифицированные происходящие из растения полинуклеотиды, которые обеспечивают фенотипический маркер растения, могут быть модифицированы в отношении их полинуклеотидной последовательности, числа копий, уровня экспрессии или положения в пределах донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, по сравнению с полинуклеотидной последовательностью, числом копий, уровнем экспрессии или положением нативного немодифицированного происходящего из растения полинуклеотида, который обеспечивает фенотипический маркер растения.
В некоторых вариантах осуществления донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, содержит один или несколько происходящих из растения полинуклеотидов, которые обеспечивают фенотипический маркер растения, сцепленных с одним или несколькими локусами, обеспечивающими восстановление мужской фертильности. Например, модифицированные локусы, обеспечивающие восстановление мужской фертильности, могут быть модифицированы в отношении их полинуклеотидной последовательности, числа копий, уровня экспрессии или положения в пределах донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, по сравнению с полинуклеотидной последовательностью, числом копий, уровнем экспрессии или положением нативного немодифицированного локуса, обеспечивающего восстановление мужской фертильности.
В некоторых вариантах осуществления донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, содержит один или несколько модифицированных происходящих из растения полинуклеотидов, которые обеспечивают фенотипический маркер растения, сцепленных с одним или несколькими локусами, обеспечивающими восстановление мужской фертильности. Например, модифицированные локусы, обеспечивающие восстановление мужской фертильности, могут быть модифицированы в отношении их полинуклеотидной последовательности, числа копий, уровня экспрессии или положения в пределах донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, по сравнению с полинуклеотидной последовательностью, числом копий, уровнем экспрессии или положением нативного немодифицированного локуса, обеспечивающего восстановление мужской фертильности. Например, модифицированные происходящие из растения полинуклеотиды, которые обеспечивают фенотипический маркер растения, могут быть модифицированы в отношении их полинуклеотидной последовательности, числа копий, уровня экспрессии или положения в пределах донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, по сравнению с полинуклеотидной последовательностью, числом копий, уровнем экспрессии или положением нативного немодифицированного происходящего из растения полинуклеотида, который обеспечивает фенотипический маркер растения.
В некоторых аспектах происходящий из растения полинуклеотид, который обеспечивает фенотипический маркер растения, и локус, обеспечивающий восстановление мужской фертильности, находятся на противоположных плечах хромосомы. Любые подходящие технология и методика могут применяться для того, чтобы обеспечить расположение происходящего из растения полинуклеотида, который обеспечивает фенотипический маркер растения, и локуса, обеспечивающего восстановление мужской фертильности, на одном и том же плече хромосомы, например, транслокация, редактирование генома, перицентрические инверсии или их комбинации.
В одном варианте осуществления локус, обеспечивающий восстановление мужской фертильности, Ms1 и происходящий из растения полинуклеотид, кодирующий фенотипический маркер растения, представленный геном голубого алейрона (Ba1), изначально располагаются на разных плечах хромосомы. В некоторых аспектах локус, обеспечивающий восстановление мужской фертильности, Ms1 находится на хромосоме 4ES из Agropyron elongatum или 4HS из ячменя (Hordeum vulgare), а ген голубого алейрона (Ba1) на 4EL из Blue Norco. В одном варианте осуществления двухнитевые разрывы в гибридном (химерном) донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения, представленным 4HS-4EL, индуцируются, например, одним двухнитевым разрывом между геном ячменя Ms1-H и теломерой 4HS и одним двухнитевым разрывом между геном Ba1 и центромерой 4HS-4EL. Фрагмент хромосомы, содержащий центромеру и Ms1-H, будет снова соединен с теломерными концами в противоположной ориентации, что приведет в результате к перицентрической инверсии, при этом Ms1-H и Ba1 теперь располагаются на одном и том же плече новой хромосомы. См., например, фиг. 3.
Донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может быть введен в растительную клетку, часть растения или растение, например, растение с мужской стерильностью, с применением любой подходящей методики, известной специалистам в данной области. В некоторых подходах донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, вводится в растительную клетку, часть растения или растение с применением редактирования генома, трансформации, культуры зародышей или методик хромосомной транслокации. В некоторых вариантах осуществления растительной клеткой является клетка растения пшеницы, которая имеет одну или несколько гомозиготных мутаций в полинуклеотиде, обеспечивающем мужскую фертильность, например, Ms45, и донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, который функционально комплементирует опосредующие мужскую стерильность гомозиготные мутации, вызывающие у растения пшеницы состояние мужской стерильности.
Донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может быть транслоцирован в геном растения с применением любых подходящих способов. Транслокация может являться Робертсоновской или неробертсоновской транслокацией. В некоторых аспектах донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, подвергается транслокации и замещает существующее плечо хромосомы растения, например, растения пшеницы, ячменя или ржи. В некоторых вариантах осуществления донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может применяться для замены короткого или длинного плеча хромосомы растения, например, пшеницы, ячменя или ржи.
В некоторых вариантах осуществления донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, вводится с применением предусматривающего нацеливание подхода, вследствие чего донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, вводится в геном растения-реципиента таким образом, что любые потенциальные нежелательные эффекты в отношении растения-реципиента минимизируются, избегаются или целевое положение предоставляет потенциальные полезные преимущества. Например, донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может быть подвергнут введению или замещению плеча хромосомы в растении пшеницы, где растение пшеницы с отсутствием данного хромосомного плеча или пары хромосомных плеч (моносомик или дителосомик) характеризуется нормальным фенотипом. В другом примере донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может быть подвергнут введению или замещению плеча хромосомы в растении пшеницы, где отсутствие плеча хромосомы имело бы эффект снижения частоты передачи донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, например, снижения частоты передачи для мужского и женского растения. В одном варианте осуществления донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, замещает длинное плечо хромосомы 5A (5AL) в растении пшеницы, вследствие чего результатом является комплекс донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, и 5AS.
В некоторых примерах наличие транслоцированного и/или замещенного донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, может обеспечивать преимущества по сравнению с независимым (добавочным) донорным хромосомным компонентом, обеспечивающим восстановление фертильности растения. Одним из преимуществ является сниженная передача гаметами независимого (добавочного) донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, что обеспечивает повышение процентной доли семян, которые содержат одну или несколько гомозиготных мутаций в полинуклеотиде, обеспечивающем мужскую фертильность, и не содержат донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, получение которых увеличивается в ходе формирования семян женским растением.
В некоторых вариантах осуществления подвергнутый транслоцированию, замещению донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может быть получен путем Робертсоновской транслокации. См., например, фиг. 4. Анеуплоидный исходный генетический материал может применяться для получения Робертсоновских транслокаций с участием донорного хромосомного компонента в растении пшеницы, осуществляемых направленным образом, путем получения подходящих хромосом донора и пшениц в моносомном состоянии. Подвергнутый транслоцированию, замещению донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может быть интрогрессирован вместе с одной или несколькими гомозиготными мутациями в полинуклеотиде, обеспечивающем мужскую фертильность, или моногенным рецессивом, опосредующим фертильность, в различные элитные женские линии для способствования продуцированию гибридных семян. Поскольку Ms1 в растениях пшеницы ведет себя как моногенный рецессив, в некоторых вариантах осуществления для получения мужской стерильности может потребоваться интрогрессирование только одной мутации в полинуклеотиде Ms1 или аллеле, расположенном на хромосоме 4BS, в женские линии пшеницы.
В одном примере донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, вводят в растение с мужской стерильностью с использованием методик селекции. В некоторых примерах растение с мужской стерильностью скрещивают с растением, содержащим донорный хромосомный компонент, обеспечивающий восстановление фертильности растения. Донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может быть введен в растение, например растение с мужской стерильностью, с применением растения, которое характеризуется таким же базовым числом хромосом, как и растение-реципиент (например, с мужской стерильностью). В некоторых примерах растение с мужской стерильностью является растением пшеницы, например, являющимся диплоидным, тетраплоидным или гексаплоидным. В некоторых примерах донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может происходить из или быть введен при переносе от растения пшеницы, например диплоидной, тетраплоидной или гексаплоидной пшеницы, растения рода Aegilops, Secale, Agropyrin, Haynaldia, Hordeum или Elyymus или любого другого растения, с которым растение с мужской стерильностью является перекрестно-совместимым и которое способно к его оплодотворению.
В некоторых аспектах донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может являться экзогенным по отношению к растению-реципиенту, например растению с мужской стерильностью, или его растению-хозяину, например растению с мужской фертильностью. Например, донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может происходить из растения, которое относится к отличающемуся от растения с мужской стерильностью виду или растению, например, наличие в растении пшеницы донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, происходящего из пырея или ячменя. Соответственно, в некоторых примерах донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, происходит из отличного от пшеницы растения или вида. В некоторых вариантах осуществления донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, не вступает в конъюгацию или рекомбинацию с любой из хромосом женского растения с мужской стерильностью, например с хромосомами пшеницы.
Донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может происходить от любого дикого или культурного растения, включая без ограничения маис, рис, сорго, рожь, ячмень, пшеницу, просо, виды овса, сахарный тростник, газонную траву, тритикале, просо прутьевидное, пырей, Thinopyrum, Aegilops, Secale, Haynaldia, Elyymus, Hordeum, сою, канолу, люцерну, подсолнечник, хлопчатник, маис или родственные им виды. Донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может происходить от любого числа видов растений, включая без ограничения роды Thinopyrum, Aegilops, Secale, Haynaldia, Elyymus или видов рода Hordeum. В некоторых вариантах осуществления донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может происходить от линии пшеницы, имеющей одну или несколько хромосом от маиса, риса, сорго, ржи, ячменя, пшеницы, проса, видов овса, сахарного тростника, газонной травы, тритикале, проса прутьевидного, пырея,Thinopyrum, Aegilops, Secale, Haynaldia, Elyymus или Hordeum, сои, канолы, люцерны, подсолнечника, хлопчатника, маиса или родственных им видов. Донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может быть введен и/или от телосомной синтетической линии, такой как монотелосомная или дителосомная добавочная линия, или от дисомной добавочной линии. В некоторых примерах растение, обладающее донорным хромосомным компонентом, обеспечивающим восстановление фертильности растения, происходит от линии пшеницы, включая без ограничения линии пшеницы Blue Sando, Blue Baart, Blue Onas, Blue 1, PBB или Blue Norco. Неограничивающие примеры включают без ограничения сорт пшеницы Blue Baart, который имеет дисомную добавочную хромосому 4E от Thinopyron ponticum, Blue Norco, который имеет дителосомную добавочную хромосому 4E от Thinopyron ponticum, или линию пшеницы, которая имеет дисомную добавочную хромосому 4H от Hordeum vulgare (ячмень). Blue Norco, анэуплоидные, моносомные, дисомные линии и другие линии пшеницы находятся в открытом доступе и могут быть получены в ряде центров, таких как Национальная коллекция мелких зерновых культур (министерство сельского хозяйства США - служба сельскохозяйственных исследований, Национальная коллекция мелких зерновых культур, Абердин, Id. 83210, США) или ресурсный центр по вопросам генетики пшеницы (университет штата Канзас, Канзас, США).
В некоторых вариантах осуществления донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, происходит от двух, трех или более одинаковых или разных видов, для того, чтобы создать гибридный (химерный) донорный хромосомный компонент, обеспечивающий восстановление фертильности растения. В некоторых вариантах осуществления гибридный (химерный) донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, имеет происходящий из растения полинуклеотид, который обеспечивает фенотипический маркер растения, от одного растения или вида, и локус, обеспечивающий восстановление мужской фертильности, от разных растений или видов. Происходящий из растения полинуклеотид, который обеспечивает фенотипический маркер растения в гибридном (химерном) донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения, может иметь одну или несколько хромосом или хромосомных фрагментов от маиса, риса, сорго, ржи, ячменя, пшеницы, проса, видов овса, сахарного тростника, газонной травы, тритикале, проса прутьевидного, пырея, Thinopyrum, Aegilops, Secale, Haynaldia, Elyymus, Hordeum, сои, канолы, люцерны, подсолнечника, хлопчатника, маиса или родственных им видов. Локус, обеспечивающий восстановление мужской фертильности, в гибридном (химерном) донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения, может иметь одну или несколько хромосом или хромосомных фрагментов от маиса, риса, сорго, ржи, ячменя, пшеницы, проса, видов овса, сахарного тростника, газонной травы, тритикале, проса прутьевидного, пырея, Thinopyrum, Aegilops, Secale, Haynaldia, Elyymus, Hordeum, сои, канолы, люцерны, подсолнечника, хлопчатника, маиса или родственных им видов. В одном примере фенотипический маркер растения происходит от, например, хромосомы 4E Thinopyrum ponticum, и обеспечивающий восстановление фертильности локус от, например, хромосомы 4H Hordeum vulgare (ячмень), для того, чтобы создать гибридный (химерный) донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, 4H-4E. В некоторых вариантах осуществления гибридный (химерный) донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, включает короткое плечо хромосомы 4 от Hordeum vulgare (ячмень) и длинное плечо хромосомы 4E от Thinopyron ponticum, например, от Blue Norco для того, чтобы создать гибридный (химерный) донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, 4HS-4EL. Фенотипический маркер растения и обеспечивающий восстановление фертильности локус могут быть от любых подходящих линий, включая без ограничения дисомные добавочные хромосомы, такие как 4H ячменя, хромосома 4E Thinopyrum ponticum, 4E Agropyron или их комбинации.
Подтверждение того, что растение, часть растения, растительная клетка или семя содержат донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, включающий фенотипический маркер или локус, обеспечивающий восстановление мужской фертильности, или оба, можно осуществить с использованием традиционных и хорошо известных способов. Донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, должен сохранять активность восстановления мужской фертильности в растениях, в частности способность содействовать развитию мужской ткани. Состояние мужской фертильности у растения можно оценить с помощью любой подходящей методики, например, путем наблюдения за развитием мужской ткани у растения, как например фенотипирование пыльников и образующихся семян на отдельных растениях. См., например, пример 1 в данном документе.
Отсутствие донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, например, утраченного в результате кроссинговера или мутации, может подтверждаться отсутствием у растений фенотипа мужской фертильности или отсутствием фенотипа, придаваемого происходящим из растения фенотипическим маркером, у растений, частей растений, клеток растений или семян, по сравнению с контролем. В качестве альтернативы или дополнения подтверждение присутствия или отсутствия донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, может происходить позднее, например, после множественных скрещиваний растений или от последующих поколений.
Семена, полученные от растений или в результате скрещиваний, могут быть собраны вместе и рассортированы на отдельные популяции при необходимости. Например семена, содержащие происходящий из растения фенотипический маркер семян растений, сцепленный с обеспечивающим восстановление фертильности локусом, являются семенами, которые могут быть рассортированы и разделены для применения в качестве закрепителей, и семена без происходящего из растения фенотипического маркера растений, сцепленного с обеспечивающим восстановление фертильности локусом (донорного хромосомного компонента, обеспечивающего восстановление фертильности растения), могут быть выращены и находить применение в качестве женских родительских растений с мужской стерильностью в гибридных скрещиваниях. Семена могут быть рассортированы на такие популяции вручную, механически или оптически. Для того, чтобы способствовать высокой пропускной способности и анализу, при сортировке можно использовать автоматический или полуавтоматический подход. Популяции семян могут быть рассортированы с применением оптических сенсорных технологий, включая мульти- или гиперспектральную визуализацию, спектроскопические системы в УФ, видимом и ближнем ИК спектрах и/или оптическое сканирование. Например, когда фенотипический маркер растения с донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, является цветовым маркером, таким как алейрон, гомозиготное голубое семя будет окрашено более интенсивно, чем гетерозиготное голубое семя, и на этом основании может быть рассортировано, например, семена имеющие разные концентрации или уровни экспрессии голубого алейрона. См., например, фиг. 2. В качестве дополнения или альтернативы семена могут быть оценены на предмет присутствия донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, например, фенотипического маркера растения, с применением любой другой подходящей методики, включая без ограничения проточную цитометрию или qPCR.
В качестве альтернативы, семена могут быть смешанными (несортированными), так что первая часть семян содержит одну или несколько гомозиготных мутаций в полинуклеотиде, обеспечивающем мужскую фертильность (семена, из которых получатся женские растения с мужской стерильностью), и вторая часть семян содержит одну или несколько гомозиготных мутаций в полинуклеотиде, обеспечивающем мужскую фертильность, и донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения, содержащем происходящий из растения полинуклеотид, который обеспечивает фенотипический маркер растения, сцепленный с локусом, обеспечивающим восстановление мужской фертильности (семена, из которых получатся растения с мужской фертильностью). Фенотипический маркер растения, который является маркером цвета семени, может применяться для рассортировывания и разделения семян гибридной пшеницы или семян инбредной пшеницы от семян линии-закрепителя стерильности пшеницы. В некоторых примерах смесь семян высажена совместно для повышения количества производимых женских семян с мужской стерильностью. Смесь семян может быть помещена в мешок или другой подходящий контейнер. Способ может включать высаживание смеси семян мужских и женских родительских растений в один и тот же ряд на поле, а не в отдельные ряды. Выращивают мужские и женские родительские растения и для получения семян мужские родительские растения опыляют женские родительские растения. Получаемые семена будут смесью семян, которые, будучи высажены, обеспечат развитие растений с мужской стерильностью или мужской фертильностью. В некоторых примерах процентная доля полученных семян, которые обеспечат развитие растений с мужской фертильностью, может составлять по меньшей мере приблизительно 20%, 25%, 30%, 35%, 40% или 45% от числа образованных семян. В некоторых примерах процентная доля семян, которые обеспечат развитие женских растений с мужской стерильностью, составляет по меньшей мере, приблизительно 55%, 60%, 65%, 70%, 75%, 80%, 85% или 90% от числа полученных семян. В некоторых аспектах с помощью данного способа повышают число полученных семян, которые обеспечат развитие женских растений с мужской стерильностью, по сравнению с числом женских семян с мужской стерильностью, полученных на поле, где семена с женской стерильностью и семена с мужской стерильностью не выращивают вместе в одном общем ряду, но каждое выращивают в мужских и женских отдельных рядах.
Также в данном документе предусматриваются растительная клетка, растение или семя, полученные или произведенные с применением способов, описанных в данном документе. В некоторых способах женские растения с мужской стерильностью опыляют пыльцой от растений, содержащих донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, который дополняет состояние мужской стерильности у женских растений. С помощью данного способа получают семена, которые могут быть смесью семян, которые будучи высажены, обеспечат развитие растений с мужской стерильностью или мужской фертильностью. Отсутствие фенотипического маркера растения в семенах указывает на то, что в семенах отсутствует донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, и они, будучи высажены, обеспечат развитие женских растений с мужской стерильностью. Растения, полученные из таких семян, могут применяться как женские инбредные растения с мужской стерильностью для получения гибридных семян. Присутствие фенотипического маркера растения в семени указывает на то, что семя содержит донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, и обеспечит развитие растений с мужской фертильностью. Растения, выращенные из данных семян, могут при необходимости применяться в качестве закрепителей. Семена могут быть нерассортированными, смешанными с другими семенами или рассортированными и разделенными на популяции с применением обычных методик и инструментов, как описано в другом месте в данном документе, и известных специалисту в данной области техники.
Поскольку обладающее мужской стерильностью растение, например обладающее мужской стерильностью растение пшеницы, не может быть самоподдерживающимся, в данном документе предусмотрены композиции и способы для поддержания состояния гомозиготной рецессивной мужской стерильности у растения пшеницы, которые включают применение донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, для восстановления мужской фертильности у растения пшеницы. Например, мутация в гене, критически важном для мужской фертильности, может придать фенотип с мужской стерильностью растению пшеницы, когда данный мутантный аллель находится в гомозиготном состоянии, например, в эндогенных полинуклеотидах пшеницы Ms1, Ms5, Ms9, Ms22, Ms26 и Ms45, кодирующих полипептиды пшеницы Ms1, Ms5, Ms9, Ms22, Ms26 и Ms45 соответственно. Поскольку Ms1 в растениях пшеницы ведет себя как моногенный рецессив, в некоторых вариантах осуществления для придания мужской стерильности растению пшеницы может потребоваться только внесение мутации в полинуклеотид, обеспечивающий мужскую фертильность, представляющий собой ms1, или аллель, расположенный на хромосоме 4BS.
Когда локус, обеспечивающий восстановление мужской фертильности, который способен к функциональной комплементации гомозиготного рецессивного аллеля, который обуславливает мужскую стерильность, вводится и экспрессируется в растении с мужской стерильностью, мужская фертильность восстанавливается в растении, так что оно может производить жизнеспособную пыльцу и способно к опылению перекрестно совместимого женского растения.
Поддержание состояния гомозиготной рецессивной мужской стерильности может включать введение в растение пшеницы донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, для создания растения-закрепителя. Донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, будучи введенным в растение, которое имеет одну или несколько гомозиготных мутаций в полинуклеотиде, обеспечивающем мужскую фертильность, восстанавливает в растении мужскую фертильность, так что растение производит жизнеспособную пыльцу, способную к самоопылению или опылению перекрестно совместимого растения. В некоторых вариантах осуществления донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может присутствовать в растении линии-закрепителя стерильности пшеницы в качестве добавочной хромосомы в геноме пшеницы, в качестве транслоцированной хромосомы в геноме пшеницы или в качестве замещенной хромосомы вместо хромосомы пшеницы в геноме пшеницы. Соответственно, в данном документе предусматриваются растение пшеницы или клетка пшеницы, которые имеют одну или несколько их хромосом пшеницы, замещенных донорным хромосомным компонентом, обеспечивающим восстановление фертильности растения, от отличных от пшеницы растения или вида. Например, в некоторых вариантах осуществления, растение пшеницы или клетка пшеницы имеют гомеологичную хромосомную пару из двух хромосом. Первая хромосома является нативной по отношению к растению пшеницы, а вторая хромосома содержит донорный хромосомный компонент, обеспечивающий восстановление фертильности растения. Донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может замещать любую из хромосом пшеницы или быть транслоцированным в пшеницу, при условии, что он не оказывает никакого вредного эффекта на растение пшеницы. В некоторых вариантах осуществления добавление, транслокация или замещение не должны препятствовать одной или нескольким гомозиготным мутациям у растения пшеницы, обуславливающих фенотип с мужской стерильностью. В некоторых вариантах осуществления донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может замещать хромосому 5AL пшеницы или быть траслоцированным на нее.
Семена пшеницы от таких растений, имеющих хромосомные замещения, являются эуплоидными, но имеют одну или несколько хромосом, замещенных на донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, или содержащих его. С помощью любой подходящей методики, такой как геномная in situ гибридизация (GISH) или флуоресцентная in situ гибридизация (FISH), можно подтвердить количество хромосом в растении или клетке пшеницы или определить транслокацию или замещение донорного хромосомного компонента, обеспечивающего восстановления фертильности растения.
Состояние мужской стерильности у растений пшеницы может поддерживаться самоопылением растения линии-закрепителя, содержащего донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, что приведет к созданию смешанной популяции семян. Часть семян будет содержать одну или несколько гомозиготных мутаций мужской стерильности, а часть семян будет содержать одну или несколько гомозиготных мутаций мужской стерильности и донорный хромосомный компонент, обеспечивающий восстановление фертильности растения. Семена, образующиеся в результате самоопыления растений линии-закрепителя, могут быть высажены и семена от таких растений могут быть отобраны и рассортированы для применения в поддержании мужской стерильности или получении гибридных семян. В некоторых примерах в способе отбора используется любой подходящий маркер семени в качестве фенотипического маркера растения для идентификации такого семени. Соответственно, в данном документе предусматриваются способы получения семян от растений пшеницы путем самоопыления растений линий-закрепителей. В других вариантах осуществления обладающее мужской стерильностью растение пшеницы , гомозиготное по одной или нескольким мутациям в полинуклеотиде, обеспечивающем мужскую фертильность, может быть опылено пыльцой от растения с мужской фертильностью, которое является перекрестно совместимым с ним. В некоторых аспектах растение с мужской фертильностью содержит донорный хромосомный компонент, обеспечивающий восстановление фертильности растения.
Также в данном документе включены способы и композиции для восстановления мужской фертильности у растений, имеющих одну или несколько гомозиготных рецессивных мутаций в полинуклеотиде, обеспечивающем мужскую фертильность, который обуславливает мужскую стерильность растений. В некоторых аспектах способ включает введение донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, который функционально комплементирует фенотип мужской стерильности от одной или нескольких гомозиготных мутаций в растении пшеницы с мужской стерильностью таким образом, что растение пшеницы становилось обладающим мужской фертильностью. Донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, может быть введен с применением любого числа подходов, как описано в данном документе, и известных специалисту в данной области техники.
Растения или хромосомный компонент, обеспечивающий восстановление фертильности растения, для применения в способах и композициях, описанных в данном документе, могут быть однодольными или двудольными. Однодольные включают без ограничения маис, рис, сорго, рожь, ячмень, пшеницу, просо, виды овса, тритикале, фонио, сахарный тростник, газонную траву, просо прутьевидное, Thinopyrum, Aegilops, Secale, Haynaldia, Elyymus или Hordeum или родственные им виды. Двудольные включают без ограничения сою, канолу, люцерну, подсолнечник, хлопчатник, табак, арахис, картофель, табак, Arabidopsis или сафлор или родственные им виды.
Дополнительные определения
Применяемые в данном документе и в прилагаемой формуле изобретения формы единственного числа включают ссылки на множественное число, если контекст явно не предусматривает иное. Таким образом, например, ссылка на “растение” включает множество таких растений; ссылка на “клетку” включает одну или несколько клеток и их эквивалентов, известных специалистам в данной области техники, и т. д.
Термин “скрещенные” или “скрещивать” или “скрещивание” в контексте данного изобретения означает слияние гамет посредством опыления для получения потомства (т. е., клеток, семян или растений). Термин охватывает как половое скрещивание (опыление одного растения другим) таки и самооплодотворение (самоопыление, т. е., когда пыльца и семяпочка (или микроспоры и мегаспоры) принадлежат одному растению или генетически идентичным растениям).
Термин “экспрессия” в общем относится к получению функционального продукта. Например, экспрессия фрагмента нуклеиновой кислоты может относиться к транскрипции фрагмента нуклеиновой кислоты (например, транскрипции, приводящей к образованию mRNA или функциональной РНК) и/или трансляции mRNA с образованием белка-предшественника или зрелого белка.
Термин “гамета" относится к репродуктивной клетке, обладающей одинарным набором хромосом 1n (гаплоидная), которая способна к слиянию с другой гаметой противоположного пола в ходе оплодотворения у организмов, осуществляющих половое размножение. Применяемый в данном документе термин гамета в организмах, осуществляющих бесполое размножение, относится к клеткам, имеющим 2n число хромосом (без уменьшения).
Термин “геном” относится ко всей совокупности генетического материала (генам и некодирующим последовательностям), который присутствует в каждой клетке организма, или вирусе, или органелле; и/или полному набору хромосом, унаследованных как набор (гаплоидный) от одного из родительских организмов.
Применяемое в данном документе "обладающее мужской стерильностью растение" является растением, которое не образует мужских гамет, которые являются жизнеспособными или иным образом способными к опылению, и которое представляет собой растение с женской фертильностью.
Термин “растение” относится к целому растению, растительным органам, растительным тканям, семенам, растительным клеткам, семенам и их потомству. Растительные клетки включают без ограничения клетки семян, суспензионных культур, зародышей, меристематических участков, каллюсной ткани, листьев, корней, побегов, гаметофитов, спорофитов, пыльцы и микроспор. Части растения включают дифференцированные и недифференцированные ткани, включая без ограничения, корни, стебли, побеги, листья, пыльцу, семена, опухолевую ткань и различные формы клеток и культуры (например, одиночные клетки, протопласты, зародыши и каллюсную ткань). Ткань растения может быть в растении или в органе растения, тканевой или клеточной культуре. Термин “орган растения” относится к растительной ткани или группе тканей, которые представляют собой морфологически и функционально отличимую часть растения.
“Полинуклеотид”, “последовательность нуклеиновой кислоты”, “нуклеотидная последовательность” или “фрагмент нуклеиновой кислоты” применяются взаимозаменяемо и представляют собой полимер РНК или ДНК, который является одно- или двухнитевым, необязательно содержащим синтетические, не являющиеся природными или измененные нуклеотидные основания.
Применяемый в данном документе термин "полинуклеотид" включает ссылку на дезоксирибополинуклеотид, рибополинуклеотид или их аналоги, которые обладают основными свойствами природного рибонуклеотида в том, что они гибридизуются в жестких условиях гибридизации практически с той же нуклеотидной последовательностью, что и нуклеотиды, встречающиеся в природе, и/или обеспечивают возможность трансляции в ту же аминокислоту(-ы),что и нуклеотид(-ы), встречающийся(-еся) в природе. Полинуклеотид может представлять собой последовательность полной длины или подпоследовательность нативного или гетерологичного структурного или регуляторного гена. Если не указано иное, термин включает ссылку на определенную последовательность, а также последовательность, комплементарную ей. Таким образом, ДНК или РНК с остовами, модифицированными для стабильности, или по иным причинам являются “полинуклеотидами” согласно тому, как этот термин подразумевается в данном документе. Более того, ДНК или РНК, содержащие необычные основания, такие как инозин или модифицированные основания, такие как тритилированные основания, если назвать лишь пару примеров, являются полинуклеотидами согласно тому, как термин применяют в данном документе.
“Полипептид”, “пептид”, “аминокислотная последовательность” и “белок” применяют в данном документе взаимозаменяемо по отношению к полимеру из аминокислотных остатков.
"Потомство" включает любое последующее поколение растения.
"Промотор, функционирующий в растении," является промотором, способным контролировать
транскрипцию в растительной клетке независимо от того, происходит ли он из растительной клетки.
Применяемый в данном документе "промотор" включает ссылку на участок ДНК выше точки начала транскрипции и он задействован в распознавании и связывании РНК-полимеразы и других белков для инициации транскрипции.
"Растительный промотор" представляет собой промотор, способный к инициации транскрипции в клетках растений.
Термины “супрессировать”, “супрессированный”, “супрессия”, “супрессирование” и “сайленсинг” применяются в данном документе взаимозаменяемо и включают понижение, сокращение, падение, уменьшение, ингибирование, устранение или препятствование. "Сайленсинг" или "сайленсинг генов" не определяет механизм и включает без ограничения супрессию, опосредованную антисмысловыми олигонуклеотидами, косупрессию, супрессию, опосредованную вирусами, супрессию, опосредованную шпильками, супрессию, опосредованную структурами типа "стебель-петля", подходы на основе RNAi и подходы на основе малых РНК и т. п.
Применяемый в данном документе термин "пшеница" относится к любому из видов рода Triticum, в том числе его предкам, а также его потомству, полученному посредством скрещивания с другим видом. Пшеница включает "гексаплоидную пшеницу" с организацией генома AABBDD, содержащего 42 хромосомы, и "тетраплоидную пшеницу" с организацией генома AABB, содержащего 28 хромосом. Гексаплоидная пшеница включает T. aestivum, T. spelta, T. mocha, T. compactum, T. sphaerococcum, T. vavilovii и гибриды, полученные при межвидовом скрещивании таковых. Тетраплоидная пшеница включает T. durum (также называемую твердой пшеницей или Triticum turgidum подвида durum), T. dicoccoides, T. dicoccum, T. polonicum и их гибриды, полученные при межвидовом скрещивании таковых. Кроме того, выражение «пшеница» включает возможных предков гексаплоидных или тетраплоидных Triticum sp., таких как T. urartu, T. monococcum или T. boeoticum для генома A, Aegilops speltoides для генома B и T. tauschii (также известный как Aegilops squarrosa или Aegilops tauschii) для генома D. Культурный сорт пшеницы для применения в настоящем изобретении может принадлежать без ограничения к любому из вышеперечисленных видов. Также настоящее изобретение охватывает растения, которые получены с помощью традиционных методик с использованием Triticum sp. в качестве родительского растения при половом скрещивании с видом, отличным от Triticum, такими как рожь (Secale cereale), включая без ограничения тритикале. В некоторых аспектах растение пшеницы, как, например, коммерческие сорта гексаплоидной пшеницы или твердой пшеницы, имеющие подходящие агрономические характеристики, известные специалистам в данной области техники, является подходящим для коммерческого получения зерна.
ПРИМЕРЫ
В следующих примерах, если не указано иное, части и процентные доли приведены по весу и в градусах Цельсия. Необходимо понимать, что данные примеры, хоть и указывают на варианты осуществления настоящего изобретения, приведены лишь для иллюстрации. Исходя из вышеизложенного рассмотрения и данных примеров, специалист в данной области техники может осуществлять различные изменения и модификации настоящего изобретения для адаптации к различным путям применения и условиям. Такие модификации также, как предполагается, находятся в пределах объема прилагаемой формулы изобретения.
ПРИМЕР 1. Монотелесомная хромосома от Thinopyrum ponticum может комплементировать тройные мутации Ms45 и восстанавливать фертильность
Как продемонстрировано в данном документе, хромосома 4EL от Blue Norco способна комплементировать мутации Tams45-abd. Blue Norco, пшеница, характеризующая наличием голубого алейрона, представляет собой дителосомную добавочную линию, которая имеет дополнительный набор 4EL (2n=42” + 2t), которая доступна от USDA-ARS (PI 542465). Растения с Tams45-abd скрещивали с Blue Norco в качестве мужского родительского растения. Высевали семена (голубого цвета), полученные в результате скрещивания, и потомство F1 подвергали скринингу для подтверждения присутствия мутаций TaMS45-A, -B и -D и обеспечивали самоопыление.
Высевали светло-голубые семена F2 (моносомические по 4EL, т. е. одна копия 4EL) и потомство генотипировали в отношении мутантных аллелей TaMs45 и обеспечивали самоопыление и образование семян. Из семян, полученных от таких растений, каждое из 64 светло-голубых семян (с 4EL) и белых семян (без 4EL) высевали с получением растений F3, которые генотипировали в отношении мутантных аллелей TaMs45. Идентифицировали тройные гомозиготные (Tams45-abd) мутантные растения, полученные как из линии белых, так и из линии голубых зерен, и анализировали в отношении мужской фертильности путем фенотипирования пыльников и образовавшихся семян на отдельных растениях. В общем анализировали 12 тройных гомозиготных растений с Tams45-abd, шесть, полученных из белых семян, и шесть, полученных из голубых семян (таблица 2). Пыльники из растений с Tams45-abd, полученных из белых семян, характеризовались сморщенной формой и меньшим размером, при этом пыльники растений с Tams45-abd, полученных из голубых зерен, были подобны растениям дикого типа (фиг. 1). Все шесть растений с Tams45-abd, которые несли хромосому 4EL, характеризовались мужской фертильностью с образованием семян, сопоставимым с растениями дикого типа, при этом шесть растений без хромосомы 4EL продемонстрировали незначительное образование семян (таблица 2; примечание: семя, наблюдаемое у этих растений, скорее всего возникло из-за открытого опыления, поскольку эти колоски не накрывались мешком и такие растения находились в непосредственной близости с фертильными цветущими растениями).
Для испытания рабочей системы поддержания мужской стерильности на основе селекции голубых и белых семян, собирали семена от фертильных растений F3 и 12 визуально отсортированных белых и светло-голубых семян, полученных от трех растений F3 (в общем=36 как белых, так и голубых семян), высевали с получением поколения F4. Все 36 растений, полученные из белых семян, характеризовались мужской стерильностью, в то время как все 36 растений, полученные из голубых семян, характеризовались мужской фертильностью (таблица 2). Эти данные демонстрируют, что моносомная хромосома 4EL может комплементировать и восстанавливать мужскую фертильность и поддерживать тройные гомозиготные мутанты Tams45. Дополнительно, возможной является идентификация из семян растения линии-закрепителя, основанная на цвете, семян, которые обеспечат получение растения с мужской стерильностью или мужской фертильностью.
Таблица 2. Комплементарность тройных гомозиготных мутантов TaMs45 с хромосомой 4EL.
*Примечание: колоски не накрывали мешком и выращивали в непосредственной близости с фертильными растениями на той же стадии.
ПРИМЕР 2. Получение гибридной хромосомы 4HS-4EL
В данном примере гибридная хромосома 4HS-4EL создана с применением Робертсоновской транслокации. Например, анеуплоидный исходный материал может применяться для создания Робертсоновских транслокаций направленным образом путем изготовления подходящего донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, в моносомном состоянии. В данном примере применяли линию пшеницы Chinese Spring, которая характеризуется наличием дисомной добавочной (2n=44) хромосомы 4H от ячменя Betzes. (Исходный генетический материал пшеницы TA#3700, который характеризуется дисомной добавочной (2n=44) хромосомой 4H Hordeum vulgare (ячмень), полученный из Центра генетических ресурсов пшеницы в университете штата Канзас, США.) В данном примере применяли разновидность пшеницы Blue Baart, которая характеризовалась дисомной добавочной (2n=44) хромосомой 4E Thinopyrum ponticum и наличием голубого алейрона. (Burešová V, Kopecký D, Bartoš J, Martinek P, Watanabe N, Vyhnánek T и Doležal J. 2015. Variation in genome composition of blue-aleurone wheat. Theor. Appl. Genet. 128:273-282). Осуществляли скрещивание двух этих линий и полученные семена F1 выращивали и обеспечивали самоопыление растений. Поколения F2 или F3 можно подвергать скринингу в отношении присутствия новой хромосомы 4HS-4EL с применением любого подходящего способа, например, в отношении наличия голубого цвета, указывающего на присутствие 4EL, отсутствие маркеров для 4ES, присутствие маркера 4HS, такого как Mmag 053 и отсутствие маркера 4HL, такого как HVM40 (Molnár I, Linc G, Dulai S, Nagy ED и Molnár-Láng MM. 2007. Ability of chromosome 4H to compensate for 4D in response to drought stress in a newly developed and identified wheat-barley 4H(4D) disomic substitution line. Plant Breeding 126: 369-374). Кандидатные линии могут дополнительно подвергаться скринингу, например, путем применения стандартных цитогенетических методик, таких как GISH (геномная гибридизация in situ), для подтверждения присутствия гибридной хромосомы 4HS-4EL.
ПРИМЕР 3. Применение CRISPR-cas9-индуцированной перицентрической инверсии для расположения Ms1 и Ba1 на одном и том же плече хромосомы
В данном примере перицентрические инверсии применяют в отношении пшеницы для расположения генов Ms1 и Ba1 на одном и том же плече хромосомы в тех случаях, где комплементарный ген Ms1 (гомеолог) и ген Ba1 находятся на разных плечах хромосомы, например, расположены на 4ES или 4HS и 4EL соответственно. Одно преимущество данного подхода заключается в том, что он обеспечит расположение генов Ms1 и Ba1, кодирующих характеристики, на одном и том же плече хромосомы и позволит избежать их разделения, обусловленного разрушением хромосомы при центромере, и возможного ошибочного определения семян как характеризующихся мужской стерильностью, в то время как на самом деле они характеризуются мужской фертильностью.
В данном примере в качестве исходного материала можно применять хромосому 4HS-4EL, описанную в примере 2 в данном документе. Будут индуцированы два DSB, один между геном Ms1-H и теломерой 4HS, и один между геном Ba1 и центромерой 4HS-4EL. Ген Ms1-H ячменя ориентирован так, что 5’-конец находится ближе всего к теломере, и 3’-конец находится ближе всего к центромере. Последовательность 5’ ДНК гена Ms1-H (http://plants.ensembl.org/Hordeum_vulgare/Tools/Blast) будут анализировать в отношении последовательности NGG прилегающего к протоспейсеру мотива (PAM) для cas9. Для каждого найденного сайта вышерасположенная последовательность в 17 п. о. будет оцениваться для обнаружения уникального сайта разрезания, который отсутствует в геноме пшеницы.
Фрагменты маркера, расположенного на длинном плече хромосомы 4J (4Eb) и расположенного между центромерой и геном BaThb, будут клонировать и секвенировать; полученные последовательности будут анализировать в отношении последовательности NGG прилегающего к протоспейсеру мотива (PAM) для cas9. Для каждого сайта вышерасположенная последовательность в 17 п. о. будет оцениваться для обнаружения уникального сайта разрезания, который отсутствует в геноме пшеницы.
Когда подходящий(подходящие) сайт(сайты) и направляющие РНК для двух расположений идентифицированы, будут индуцированы два DSB с помощью CRISPR-cas9 с применением стандартных способов. С определенной частотой фрагмент хромосомы, содержащий центромеру и Ms1-H, будет воссоединен с теломерными концами в противоположной ориентации, что приведет к перицентрической инверсии, при этом Ms1-H и Ba1 теперь расположены на одном и том же плече новой хромосомы. Будут разработаны праймеры для ПЦР, которые обеспечат получение продукта ПЦР только если произошла перицентрическая инверсия.
ПРИМЕР 4. Альтернативная конструкция для системы 4E-ms45
В данном примере описан один вариант осуществления конструкции системы гибридизации с применением 4E-ms45. Вместо маркера цвета семени (голубой алейрон=BA) и функционально доминантного аллеля Ms45, обеспеченных добавочной телоцентрической хромосомой, такой как 4EL, плечо хромосомы 4EL будет транслоцировано в геном пшеницы, заменяя существующее плечо хромосомы пшеницы.
Правильный выбор плеча хромосомы пшеницы, выбранного в качестве цели для замещения с помощью 4EL, может обеспечивать некоторые преимущества по сравнению с независимой (добавочной) 4EL. Одно возможное преимущество представляет собой сниженную передачу добавочной телосомной хромосомы в гаметах, например, сниженную частоту передачи в мужских и/или женских гаметах. Это обеспечит получение повышенной процентной доли семени, отличного от голубого, во время повышения уровня получения женского семени (самоопыление). В данном примере 4EL будет замещена на плечо хромосомы 5AL пшеницы. Гаметы, содержащие хромосому 5AS-4EL, будут характеризоваться недостачей копии 5AL. См., например, фиг. 4.
Даже в случае сниженной частоты передачи в мужских и/или женских гаметах может быть получено некоторое гомозиготное 4EL (голубое) семя. Присутствие таких гомозиготных голубых семян может быть идентифицировано и обнаружено с применением сортировки семени. Например, гомозиготное голубое семя будет более интенсивно окрашено, чем гетерозиготное голубое семя и, основываясь на этом, может быть отсортировано. Полагают, что гомозиготные растения 5AS-4EL будут скорее всего характеризоваться мужской стерильностью, для удаления таких семян не потребуется сортировка или потребуется незначительная сортировка семян из популяции голубых семян.
Хромосома 5AS-4EL может быть создана с применением Робертсоновской транслокации. В данном примере анеуплоидный исходный материал может применяться для создания направленным образом Робертсоновских транслокаций в донорном хромосомном компоненте, обеспечивающем восстановление фертильности растения, пшеницы путем создания подходящих донорных хромосомных компонентов восстановления фертильности растения пшеницы в моносомном состоянии. В данном примере применяли линию анеуплоидной пшеницы Chinese Spring, которая сегрегирует по моносомии хромосомы 5A пшеницы. (Исходный генетический материал TA#3045, который сегрегирует по моносомии хромосомы 5A пшеницы, получали из Центра генетических ресурсов пшеницы в университете штата Канзас, США.) В данном примере применяли разновидность пшеницы Blue Baart, которая имеет дисомную добавочную (2n=44) хромосому 4E Thinopyron ponticum и имеет голубой алейрон (Burešová V, Kopecký D, Bartoš J, Martinek P, Watanabe N, Vyhnánek T и Doležal J. 2015. Variation in genome composition of blue-aleurone wheat. Theor. Appl. Genet. 128:273-282). Выращивали 10 семян линии анеуплоидной пшеницы Chinese Spring, которая сегрегирует по моносомии хромосомы пшеницы 5A, отбирали образцы корневых кончиков и проводили подсчет хромосом с помощью стандартных способов для идентификации моносомных (2n-1=41) растений. Моносомные растения продуцировали ~75% нуллисомных (n-1=20) женских гамет. Идентифицированные моносомные растения в качестве женских особей скрещивали с Blue Baart. Полученные из нуллисомных гамет семена F1 будут характеризоваться композицией из 42 хромосом, включая 1 5A+1 4E, обе получены из мужского родительского растения Blue Baart, и имеют цвет голубого алейрона. Выращивали семена F1 и обеспечивали самоопыление растений.
Можно проводить скрининг в отношении присутствия новой хромосомы 5AS-4EL с помощью любого подходящего способа, например, путем присутствия у семени цвета голубого алейрона, указывающего на присутствие 4EL и отсутствие маркеров 4ES. Это будет указывать на разрушение хромосомы 4E. Кандидатные линии могут дополнительно подвергаться скринингу, например, путем применения стандартных цитогенетических методик, таких как FISH (флуоресцентная гибридизация in situ), для подтверждения присутствия гибридной хромосомы 5AS/4EL.
После создания будет обеспечено самоопыление таких линий для подтверждения мужской стерильности гомозигот 5AS/4EL, а также взаимное скрещивание мужских особей и женских особей для определения частот передачи пыльцы и яйцеклеток. Хромосому 5AS/4E можно интрогрессировать наряду с рецессивными аллелями ms45 на 4A, 4B и 4D в различные элитные женские линии для облегчения получения гибридного семени.
ПРИМЕР 5. Применение добавочных хромосом 4E или 4H для комплементации тройных мутаций Ms9 пшеницы и для восстановления фертильности
В данном примере конструкция CAS9-CRISPR пшеницы была нацелена на ген TA-Ms9 с применением направляющей TA-Ms9-CR2 ggaggtacaccaactacctg (SEQ ID NO:40).
Растения пшеницы будут трансформированы с помощью данной конструкции, и эндогенные гены Ms9 пшеницы будут оценены в отношении мутаций во всех трех геномах пшеницы. Будут осуществляться скрещивания и последующие самоопыления для комбинирования мутаций Ms9 в гомозиготном состоянии в отдельном растении. Гомозиготные в отношении мутаций Ms9 растения пшеницы во всех трех геномах будут оцениваться в отношении мужской стерильности путем изучения пыльников и образования семени на отдельных растениях.
Если мужская стерильность достигнута, то такие растения пшеницы могут поддерживаться путем скрещивания их с линией-закрепителем пшеницы, содержащей добавочную хромосому 4EL, такой как Blue Norco, описанной в примере 1. Генетические карты TA-Ms9 для длинного плеча хромосомы 4 в пшенице, которое характеризуется синтенией с длинным плечом добавочной хромосомы 4EL, что должно обеспечивать дополняющую функцию для мутации TA-ms9, восстанавливающей мужскую фертильность. Линия пшеницы, содержащая хромосому 4 ячменя в качестве добавочной хромосомы (Hordeum vulgare 4H), также может оцениваться в отношении возможного восстановления Ta-ms9. Восстановление мужской фертильности для таких характеризующихся мужской стерильностью растений пшеницы с помощью добавочных хромосом 4E, 4EL или 4H в линии-закрепителе пшеницы будет оцениваться в отношении способности комплементировать мутации ms9 в таких растениях, что определено путем изучения пыльников и образования семени на отдельных растениях.
Как описано в примере 1, добавочная хромосома 4EL также содержит ген голубого алейрона (BA) на длинном плече, который обеспечивает доминантный фенотип с голубым семенем. Следовательно, семя из самоопыленных гомозиготных мутантных по TA-ms9 растений и восстановленных с помощью присутствия добавочной хромосомы 4EL, будут подвергать сегрегации в отношении голубого и отличного от голубого семени. Эти полученные семена можно выращивать и оценивать в отношении мужской фертильности и стерильности. Как продемонстрировано в примере 1, ожидания заключаются в том, что растения, полученные из отличного от голубого семени, будут характеризоваться мужской стерильностью, при этом растения, полученные из голубого семени, будут характеризоваться мужской фертильностью, и последние могут выступать в качестве линии-закрепителя для мутации TA-ms9 в гибридной системе.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Pioneer Hi-Bred International, Inc
<120> ВОССТАНОВЛЕНИЕ ФЕРТИЛЬНОСТИ У РАСТЕНИЙ
<130> 7795WOPCT
<150> 62/832716
<151> 2019-04-11
<150> 62/815261
<151> 2019-03-07
<150> 62/731252
<151> 2018-09-14
<160> 42
<170> PatentIn версия 3.5
<210> 1
<211> 1857
<212> ДНК
<213> Triticum aestivum
<400> 1
atggagagat cccgccgcct gctgctggtg gcgggcctgc tcgccgcgct gctcccggcg 60
gcggcggccg ccttcgggca gcagccgggg gcgccgtgcg agcccacgct gctggcgacg 120
caggtggcgc tcttctgcgc gcccgacatg cccaccgcgc agtgctgcga gcccgtcgtc 180
gccgccgtcg acctcggcgg cggggtcccc tgcctctgcc gcgtcgccgc ggagccgcag 240
ctcgtcatgg cgggcctcaa cgccacccac ctcctcacgc tctacagctc ctgcggcggc 300
ctccgtcccg gcggcgccca cctcgccgcc gcctgcgaag gtacgcgcac gttcaccgcc 360
ctccgtccct ccctctctct gtctacgtgc agattttctg tgctctcttt cctgcttgcc 420
tagtacgtag tgttccatgg cctctcgggc cgctagcgct ccgatttgcg ttggtttcct 480
tgctgttctg ccggatctgt tggcacggcg cgcggcgtcg ggttctcgcc gtctcccgtg 540
gcgagcgacc tgcgcagcgc gcgcggcctg gctagcttca taccgctgta ccttgagata 600
cacggagcga tttagggtct actctgagta tttcgtcatc gtaggatgca tgtgccgctc 660
gcgattgttt catcgatttg agatctgtgc ttgttcccgc gagttaagat ggatctagcg 720
ccgtacgcag atgcagagtc tgttgctcga gttaccttat ctaccgtcgt tcgactatgg 780
tatttgcctg cttccttttg gctgggttta tcgtgcagta gtagtagaca tgtggacgcg 840
ttcttcttat tttgtgccga ccatcgtcga gatacttttc ctgctacagc gtttcatcgc 900
ctgcaccatc ccgttcgtga tagcactttt gtgtcaaacc gcaacgcagc tttgctttct 960
gcggtatctt ctgccttgtt tgtcgccttg cttggtcaaa actgagaact cttgctgttt 1020
gatcgaccga gggcagaggc agagcaagag cctgccgtgc ttttggctct gcagtgcgtc 1080
gtctctgcct cctttgccaa acatttccat gttgatcctc tgggggcact gctttttcgc 1140
atgcggtttc cgtagccttc ctctttcatg aaaaaaggtt tgggtcaaat caaatggatc 1200
gcctattggc agagcagcag cagatagctg gctgtctcac agctttggca gaatcggtct 1260
gttgcctgcc accgtgtctc ttatcttgcc tgccaccgtg tctcttttct tgttgcgcac 1320
gtcgtcacct cctcctactt cttttccagt tttgtttact tttgatgaaa tacggacgaa 1380
cggctggtaa tcattaactt tggttgctgt tgttactgtg gattttggac gcaggacccg 1440
ctcccccggc cgccgtcgtc agcagccccc cgcccccgcc accgtcgacc gcacctcgcc 1500
gcaagcagcc agcgcgtgcg tacctctccc tctcgcccgc atctcgctcc gtattaactg 1560
attgtgtctg catactgacg tgtgctttgg ctttggatct gtttcgcaga cgacgcacca 1620
ccgccgccgc cgccgtccag cgacaagccg tcgtccccgc cgccgtccca ggaacacgac 1680
ggcgccgctc cccacgccaa ggccgccccc gcccaggcgg ctacctcccc gctcgcgccc 1740
gctgctgcca tcgccccgcc gccccaggcg ccacactccg ccgcgcccac ggcgtcatcc 1800
aaggcggcct tcttcttcgt cgccacggcc atgctcggcc tctacatcat cctctga 1857
<210> 2
<211> 222
<212> БЕЛОК
<213> Triticum aestivum
<400> 2
Met Glu Arg Ser Arg Arg Leu Leu Leu Val Ala Gly Leu Leu Ala Ala
1. 5 10 15
Leu Leu Pro Ala Ala Ala Ala Ala Phe Gly Gln Gln Pro Gly Ala Pro
20 25 30
Cys Glu Pro Thr Leu Leu Ala Thr Gln Val Ala Leu Phe Cys Ala Pro
35 40 45
Asp Met Pro Thr Ala Gln Cys Cys Glu Pro Val Val Ala Ala Val Asp
50 55 60
Leu Gly Gly Gly Val Pro Cys Leu Cys Arg Val Ala Ala Glu Pro Gln
65 70 75 80
Leu Val Met Ala Gly Leu Asn Ala Thr His Leu Leu Thr Leu Tyr Ser
85 90 95
Ser Cys Gly Gly Leu Arg Pro Gly Gly Ala His Leu Ala Ala Ala Cys
100 105 110
Glu Gly Pro Ala Pro Pro Ala Ala Val Val Ser Ser Pro Pro Pro Pro
115 120 125
Pro Pro Ser Thr Ala Pro Arg Arg Lys Gln Pro Ala His Asp Ala Pro
130 135 140
Pro Pro Pro Pro Pro Ser Ser Asp Lys Pro Ser Ser Pro Pro Pro Ser
145 150 155 160
Gln Glu His Asp Gly Ala Ala Pro His Ala Lys Ala Ala Pro Ala Gln
165 170 175
Ala Ala Thr Ser Pro Leu Ala Pro Ala Ala Ala Ile Ala Pro Pro Pro
180 185 190
Gln Ala Pro His Ser Ala Ala Pro Thr Ala Ser Ser Lys Ala Ala Phe
195 200 205
Phe Phe Val Ala Thr Ala Met Leu Gly Leu Tyr Ile Ile Leu
210 215 220
<210> 3
<211> 1856
<212> ДНК
<213> Triticum aestivum
<400> 3
atggagagat cccgcgggct gctgctggtg gcggggctgc tggcggcgct gctgccggcg 60
gcggcggcgc agccgggggc gccgtgcgag cccgcgctgc tggcgacgca ggtggcgctc 120
ttctgcgcgc ccgacatgcc gacggcccag tgctgcgagc ccgtcgtcgc cgccgtcgac 180
ctcggcggcg gggtgccctg cctctgccgc gtcgccgccg agccgcagct cgtcatggcg 240
ggcctcaacg ccacccacct cctcacgctc tacagctcct gcggcggcct ccgccccggc 300
ggcgcccacc tcgccgccgc ctgcgaaggt acgttgtccg cctcctcccc tccctccctc 360
cctccctctc tctctacgtg ctcgctttcc tgcttaccta gtagtacgta gtttcccatg 420
ccttcttgac tcgctagaag tgctccggtt tgggtctgtt aatttcctcg ctgtactacc 480
ggatctgtcg tcggcacggc gcgcggcgtc gggtcctcgc cttctcccgt ggcgaccgac 540
ctgcgcagcg cgcgcgcggc ctagctagct tcataccgct gtacctcgac atacacggag 600
cgatctatgg tctactctga gtatttcctc atcgtagaac gcatgcgccg ctcgcgattg 660
tttcgtcgat tctagatccg tgcttgttcc cgcgagttag tatgcatctg cgtgcatatg 720
ccgtacgcac gcagatgcag agtctgttgc tcgagttatc tactgtcgtt cgctcgacca 780
tatttgcctg ttaatttcct gttcatcgtg catgcagtag tagtagccat gtccacgcct 840
tcttgttttg aggcgatcat cgtcgagatc catggctttg ctttctgcac tatcttctgc 900
cttgttttgt tctccgcagt acgtacgtct tgcttggtca aaactgaaaa acgctttgct 960
gtttgtttga tcggcaagag ctggccgtgc ttttggcacc gcagtgcgtc gcctctgccg 1020
cttttgcgaa acatttccat gttgatcctc tggcggaact actttttcgc gtgcggtttg 1080
cgtggccttc ctctctcgtg aaaagaggtc gggtcaaacc aaatggatcg cctcttggca 1140
gagcagcggc agcagatagc tggccgtctc gcagctttgg cagaaccggt ctgtggccat 1200
ctgtcgccgc ctgccaccgt ttccctgatg tttgtttctc tctcgcctgc cactgtttct 1260
tttcttgttg cgcacgtacg tcgtcacctc ctcctacttt tttgccagtt ttgtttactt 1320
ttgatgaaat atacggatga atcggctggt gattaacttt ggctgctgct gttaattact 1380
gtggattttg gatgcaggac ccgctccccc ggccgccgtc gtcagcagcc ccccgccccc 1440
gcctccaccg tccgccgcac ctcgccgcaa gcagccagcg cgtaagaacc tctccctctc 1500
cctctctctc tccctctcgc ctgcatctcg ctatgtttat ccatgtccat atgttgatca 1560
gccttgttta gttactaaca tgtgcaccgg atcgggttct cgcagacgac gcaccaccgc 1620
cgccaccgcc gtcgagcgag aagccgtcgt ccccgccgcc gtcccaggac cacgacggcg 1680
ccgccccccg cgccaaggcc gcgcccgccc aggcggccac ctccacgctc gcgcccgccg 1740
ccgccgccac cgccccgccg ccccaggcgc cgcactccgc cgcgcccacg gcgccgtcca 1800
aggcggcctt cttcttcgtc gccacggcca tgctcggcct ctacatcatc ctctga 1856
<210> 4
<211> 220
<212> БЕЛОК
<213> Triticum aestivum
<400> 4
Met Glu Arg Ser Arg Gly Leu Leu Leu Val Ala Gly Leu Leu Ala Ala
1. 5 10 15
Leu Leu Pro Ala Ala Ala Ala Gln Pro Gly Ala Pro Cys Glu Pro Ala
20 25 30
Leu Leu Ala Thr Gln Val Ala Leu Phe Cys Ala Pro Asp Met Pro Thr
35 40 45
Ala Gln Cys Cys Glu Pro Val Val Ala Ala Val Asp Leu Gly Gly Gly
50 55 60
Val Pro Cys Leu Cys Arg Val Ala Ala Glu Pro Gln Leu Val Met Ala
65 70 75 80
Gly Leu Asn Ala Thr His Leu Leu Thr Leu Tyr Ser Ser Cys Gly Gly
85 90 95
Leu Arg Pro Gly Gly Ala His Leu Ala Ala Ala Cys Glu Gly Pro Ala
100 105 110
Pro Pro Ala Ala Val Val Ser Ser Pro Pro Pro Pro Pro Pro Pro Ser
115 120 125
Ala Ala Pro Arg Arg Lys Gln Pro Ala His Asp Ala Pro Pro Pro Pro
130 135 140
Pro Pro Ser Ser Glu Lys Pro Ser Ser Pro Pro Pro Ser Gln Asp His
145 150 155 160
Asp Gly Ala Ala Pro Arg Ala Lys Ala Ala Pro Ala Gln Ala Ala Thr
165 170 175
Ser Thr Leu Ala Pro Ala Ala Ala Ala Thr Ala Pro Pro Pro Gln Ala
180 185 190
Pro His Ser Ala Ala Pro Thr Ala Pro Ser Lys Ala Ala Phe Phe Phe
195 200 205
Val Ala Thr Ala Met Leu Gly Leu Tyr Ile Ile Leu
210 215 220
<210> 5
<211> 1895
<212> ДНК
<213> Triticum aestivum
<400> 5
atggagagat cccgcggcct gctgctggcg gcgggcctgc tggcggcgct gctgccggcg 60
gcggcggccg cgttcgggca gcagccgggg gcgccgtgcg agcccacgct gctggcgacg 120
caggtggcgc tcttctgcgc gcccgacatg cccacggccc agtgctgcga gcccgtcgtc 180
gccgccgtcg acctcggcgg cggggtgccc tgcctctgcc gcgtcgccgc ggagccgcag 240
ctcgtcatgg cgggcctcaa cgccacccac ctcctcacgc tctacggctc ctgcggcggc 300
ctccgtcccg gcggcgccca cctcgccgcc gcctgcgaag gtacgtcgcg cacgttcacc 360
gcctccctcc ctccctcgct ctctctctct ctctctctct ctctctctac gtgccgattc 420
tctgtgttcg cttccctgct tacctagcac gtagttttcc atggcttctc gactcgctgg 480
tcctccgatt tgggtcggtt aatttcctcg ctgtactacc ggatctgtcg gcacggcgcg 540
cggcgtcggg ttctcgccgt ctcccgtggc gagcgacctg cgcagcgcgc gcgcggccta 600
gctagcttca taccgctgta ccttcagata cacggagcga tttagggtct actctgagta 660
tttcgtcatc gtaggatgca tgtggcagtc gcgattgttt catcgatttt agatctgtgc 720
ttgttcccgc gagttaagat ggatctagcg ccgtacgcag acgcagatgg tcttgctgtc 780
tctgttgctc gagttatctt atctactgtc gttcgagtat atttgcctgc ttccttttga 840
tctgtgttta tcgtgcagta gcagtagcca tgtccacgcc ttcttgtttc gaggcgatca 900
tcgtcgagat agcgctttgt ttcaaaccgc aacgcagcct ttgctttctg cggtatcttc 960
tgccttgttt ttgttctgtg cagtacgtct tgcttggtca aaagtaaaaa ctcttgctgt 1020
tcgatcgacc gaggcctgat gcagagcaag agctggccgt gcttttcgct ctgcagtgca 1080
tcgcctctgc ctctttggcc aaacatttcc atgttgatcc tctggtgtgg tactactttt 1140
ttgcatgcgg tttgcgtagc cttcctcttt cgtgaaaaaa ggtcgggtcg cctattggca 1200
gagcagcagc agcagcaaca gatagctggc tgtctcgcag ctttgacaga accggtctgt 1260
ggccatctgt cgccgcctgc caccgtttcc ctgatgtttg tttctctcgt ctcatctcgc 1320
ctgccactgt ttcttttctt gttgcgcacg tcgtcacctc ctcctacttt tttttccagt 1380
tttgtttact tttgagatac ggacgaacgg ctggtaatta ctaactttgg ttgctgttgt 1440
tactgtggat tttggacgca ggacccgctc ccccggccgc catcgtcagc agccccccgc 1500
ccccgccacc accgtccgcc gcacctcgcc gcaagcagcc agcgcgtacg aacctctccc 1560
tccctctctc tcgcctgcat ctcgctctgt attagctgat tgtgtttact tactgacgtg 1620
tgctttggct ttggatctgt ttcgcagacg acgcaccgcc gccgccgccg ccgtctagcg 1680
agaagccgtc gtccccgccg ccgtcccagg agcacgacgg cgccgccccc cgcgccaagg 1740
ccgcgcccgc ccaggcgacc acctccccgc tcgcgcccgc tgccgccatc gccccgccgc 1800
cccaggcgcc acactccgcg gcgcccacgg cgtcgtccaa ggcggccttc ttcttcgtcg 1860
ccacggccat gctcggcctc tacatcatcc tctga 1895
<210> 6
<211> 223
<212> БЕЛОК
<213> Triticum aestivum
<400> 6
Met Glu Arg Ser Arg Gly Leu Leu Leu Ala Ala Gly Leu Leu Ala Ala
1. 5 10 15
Leu Leu Pro Ala Ala Ala Ala Ala Phe Gly Gln Gln Pro Gly Ala Pro
20 25 30
Cys Glu Pro Thr Leu Leu Ala Thr Gln Val Ala Leu Phe Cys Ala Pro
35 40 45
Asp Met Pro Thr Ala Gln Cys Cys Glu Pro Val Val Ala Ala Val Asp
50 55 60
Leu Gly Gly Gly Val Pro Cys Leu Cys Arg Val Ala Ala Glu Pro Gln
65 70 75 80
Leu Val Met Ala Gly Leu Asn Ala Thr His Leu Leu Thr Leu Tyr Gly
85 90 95
Ser Cys Gly Gly Leu Arg Pro Gly Gly Ala His Leu Ala Ala Ala Cys
100 105 110
Glu Gly Pro Ala Pro Pro Ala Ala Ile Val Ser Ser Pro Pro Pro Pro
115 120 125
Pro Pro Pro Ser Ala Ala Pro Arg Arg Lys Gln Pro Ala His Asp Ala
130 135 140
Pro Pro Pro Pro Pro Pro Ser Ser Glu Lys Pro Ser Ser Pro Pro Pro
145 150 155 160
Ser Gln Glu His Asp Gly Ala Ala Pro Arg Ala Lys Ala Ala Pro Ala
165 170 175
Gln Ala Thr Thr Ser Pro Leu Ala Pro Ala Ala Ala Ile Ala Pro Pro
180 185 190
Pro Gln Ala Pro His Ser Ala Ala Pro Thr Ala Ser Ser Lys Ala Ala
195 200 205
Phe Phe Phe Val Ala Thr Ala Met Leu Gly Leu Tyr Ile Ile Leu
210 215 220
<210> 7
<211> 1694
<212> ДНК
<213> Triticum aestivum
<400> 7
atggccccca ccgccctcct catcgtcgtc ctcgccgtcg ctgccctcca cgcccccgcc 60
gcctccgccg cgttgtccca ggagccacca gcgacgccgt gcgcggcagc catcgtgtcc 120
ttctcgccgt gcctggcgca cgtcgcggtg gtggcgccgc ccgccctgcc ctcgcccgcg 180
cccaccagcg cctgctgcgc ggcgttcctg cgcgccgttt cctccgggga cggcgaaggc 240
ggcggagggg agggatgctt ctgccacctg ctccgcaacc cgctcctcct cggcttcccc 300
gtcgacgccg cgcgcctcgg cactctcctc cccacctgcg cctccgcgaa aacctccgcc 360
gccacggccg ccgaggccga ggccctcttc gccgacaagt gccgaggtga gaaatctgtc 420
ccttcgtgct ccctatttat gctcgtgcaa tatgtatgct tcgatcaatt ctcgtgcgcc 480
atatgcgcgt gctgtcgcgt tcctgttgtt gatcgccgat cgaagcaaat tttactctgc 540
aaagtcctaa ctactgttgt tctcatgatc cttgtacaac tactttcgta atggttggtt 600
gtaatgcgga ttactgacga gctctgaatg ctactaggga tattagcact tgcattactg 660
aactagtgga atgggggaga aacaccgcgg gatttttctt ttgttattaa ggagaagata 720
ctgagtggcc cagacttact tctgcgtttt tgctgcgatt tgtcatgatt accgttgatt 780
taagcagttt gttgggttgc ttgtttgata gtagtagtaa tttcacaaaa tattggcgat 840
atttataaat agctaagggc ttcgttactt ggtggtttct ctcaaagagt tcacaaaagt 900
caattattaa attcaattaa gggggcaagt attagtagta ctggctaact ccatttgcca 960
tttgccaatt gaaacagagc tcaagtcact gcctgagatg cattttacac ctccatcgcc 1020
acctcccgca ccaaaacttt ctccaggtaa atgttctgct gcttgtctaa tgattccata 1080
gcttgttaaa aaaaatgatt ccataaatct gtgcccagta atgctatttc ggatttcggt 1140
tgaatgaacc aattggcatt tgggcagaca tgtcatatgt cctctcctac caaatgaaac 1200
ttgaacttgt tttatcttgt ggtgctccat ccatttcgtg ttctatgcca caactgtaca 1260
ggttcaaatg atagtagaac aactaatttt gcatcggata ctctctgcag cctctgttct 1320
gttcctacta attgcaggac cgagtctgtc tttgtgatac tatctgcttt gcgtgcactg 1380
ctcttttagg ttgcagctgc accttctatt ttgttcacct gagcattcca ttttggtcga 1440
ctgaacaccg caagtcctga atatctttct tgaatcgtcg gtcttacttg gtgtgcacta 1500
gttaaactgc tgtgtatgcc cttgcagctg ccgttacaga accagcgtcc ccgactccga 1560
agatggagga gcattcgacc tcgacgacgc ctgtgtcgga tgatcggtcg ggatccgatg 1620
ccttgtgtgc ctgccgggtc ttccttgtgg ccttggtctt gggagcagca gtcttgatca 1680
cgctgcagtt ctga 1694
<210> 8
<211> 213
<212> БЕЛОК
<213> Triticum aestivum
<400> 8
Met Ala Pro Thr Ala Leu Leu Ile Val Val Leu Ala Val Ala Ala Leu
1. 5 10 15
His Ala Pro Ala Ala Ser Ala Ala Leu Ser Gln Glu Pro Pro Ala Thr
20 25 30
Pro Cys Ala Ala Ala Ile Val Ser Phe Ser Pro Cys Leu Ala His Val
35 40 45
Ala Val Val Ala Pro Pro Ala Leu Pro Ser Pro Ala Pro Thr Ser Ala
50 55 60
Cys Cys Ala Ala Phe Leu Arg Ala Val Ser Ser Gly Asp Gly Glu Gly
65 70 75 80
Gly Gly Gly Glu Gly Cys Phe Cys His Leu Leu Arg Asn Pro Leu Leu
85 90 95
Leu Gly Phe Pro Val Asp Ala Ala Arg Leu Gly Thr Leu Leu Pro Thr
100 105 110
Cys Ala Ser Ala Lys Thr Ser Ala Ala Thr Ala Ala Glu Ala Glu Ala
115 120 125
Leu Phe Ala Asp Lys Cys Arg Glu Leu Lys Ser Leu Pro Glu Met His
130 135 140
Phe Thr Pro Pro Ser Pro Pro Pro Ala Pro Lys Leu Ser Pro Ala Ala
145 150 155 160
Val Thr Glu Pro Ala Ser Pro Thr Pro Lys Met Glu Glu His Ser Thr
165 170 175
Ser Thr Thr Pro Val Ser Asp Asp Arg Ser Gly Ser Asp Ala Leu Cys
180 185 190
Ala Cys Arg Val Phe Leu Val Ala Leu Val Leu Gly Ala Ala Val Leu
195 200 205
Ile Thr Leu Gln Phe
210
<210> 9
<211> 1629
<212> ДНК
<213> Triticum aestivum
<400> 9
atgcccccca ccgccctcct cctcgtcgtc ctcgccgtcg ccgccctcca cgcccctgcc 60
gcctccgccg cgttgtccca ggagccacca gcgacgccgt gcgcggcggc catcgtgtcc 120
ttctcgccgt gcctggcgcc gtcgcggtgg tggcgccgcc cgccctgccc tctcccgcgc 180
ccaccagcgc ctgctgcgcg gcgttcctgc gcgccgtttc cgccggggac ggcgaaggcg 240
gcggagggga gggatgcttc tgccacctgc tccgcgaccc gctcctcctc ggcttccccg 300
tcgacgccgc gcgcctcggc gctctcctcc ccacctgcgc ctccgcgaaa acctccgccg 360
ccacggccgt cgaggctgag gccctcttcg ccgacaagtg ccgaggtgag aaatctgtcc 420
cttcgtgctc cctatttatg ctcgtgcaat atgcatgctt cgatcaattc tcgtgcgcca 480
tatgcgcgag ctgtcgcgtt cctgttgttg atcgccgatc gaaacaaatc ttactctgca 540
aagccctaac tactgttgtt ctcatgatcc ttgtacaact actttcgtaa tggttggttg 600
taatgcggat tactgacgag ctctggatgc tactagggat attagtactt gcatttctga 660
actagttgaa tgggggagaa acaccgcggg atttttcttt tgtggcccag acttacttct 720
gcatttttgc tgcgatttgt catgattacc gttcagttaa gcagtttgtt gggttgcttg 780
tttgatagta gtaatttcac aaaatattgg cgatatttat aaatagctaa aggcttcgtt 840
acttggtggt ttctctcaaa gagttcacaa aagtcaatca ttaaatcaat taagggggca 900
agtattagta gtactggcta actccatttg ccattgaaac agagctcaag tcactgcctg 960
agatgcattt tacacctcca tcgccacctc ccgcaccaaa actttctcca ggtaaacgtt 1020
ctccctcttg tctaatgatt tgataaatct gtgcccagtc tattcgggtt tcggttgaat 1080
aaaccaattg gcatttgggc agcatgttat atggtctctc ctcatcccaa ataaaacttg 1140
aacttgtttt atcttgccgt gctccgtcca ttttgtgctc tatgccgcaa ttgcacaggt 1200
tcaatgatag tagaacacct agttttgcat cgcatactct ctgcagcctc tgttctgttc 1260
ctgctaattg cagtatcgtg tctgtctttg tgatactatc tcctttgcat gcactgcact 1320
tttaggttgc agttgcacgt ttgattctag tttgttcacc tgagcattcc attttggtcg 1380
actgaacacc gcaagtcctg aatatctttc ttgaatcgtc ggtcgtactt cgtgtgcact 1440
agttaaactg ctgtctatgc ccttgcagct gccgttccag aaccagcgtc cccgaagatg 1500
gaggagcatt cgacctcagc gacgcctgtg ccggatgatc ggtcgggatc cgatgccttg 1560
tgtgcctgcc gggtcttcct tgtggccttg gtcttgggag cagcagtctt gatcacgctg 1620
cagttctga 1629
<210> 10
<211> 205
<212> БЕЛОК
<213> Triticum aestivum
<400> 10
Met Pro Pro Thr Ala Leu Leu Leu Val Val Leu Ala Val Ala Ala Leu
1. 5 10 15
His Ala Pro Ala Ala Ser Ala Ala Leu Ser Gln Glu Pro Pro Ala Thr
20 25 30
Pro Cys Ala Ala Ala Ile Val Ser Phe Ser Pro Cys Leu Ala Pro Ser
35 40 45
Arg Trp Trp Arg Arg Pro Pro Cys Pro Leu Pro Arg Pro Pro Ala Pro
50 55 60
Ala Ala Arg Arg Ser Cys Ala Pro Phe Pro Pro Gly Thr Ala Lys Ala
65 70 75 80
Ala Glu Gly Arg Asp Ala Ser Ala Thr Cys Ser Ala Thr Arg Ser Ser
85 90 95
Ser Ala Ser Pro Ser Thr Pro Arg Ala Ser Ala Leu Ser Ser Pro Pro
100 105 110
Ala Pro Pro Arg Lys Pro Pro Pro Pro Arg Pro Ser Arg Leu Arg Pro
115 120 125
Ser Ser Pro Thr Ser Ala Glu Ser Ser Ser His Cys Leu Arg Cys Ile
130 135 140
Leu His Leu His Arg His Leu Pro His Gln Asn Phe Leu Gln Leu Pro
145 150 155 160
Phe Gln Asn Gln Arg Pro Arg Arg Trp Arg Ser Ile Arg Pro Gln Arg
165 170 175
Arg Leu Cys Arg Met Ile Gly Arg Asp Pro Met Pro Cys Val Pro Ala
180 185 190
Gly Ser Ser Leu Trp Pro Trp Ser Trp Glu Gln Gln Ser
195 200 205
<210> 11
<211> 107
<212> БЕЛОК
<213> Triticum aestivum
<400> 11
Met Ala Pro Thr Ala Leu Leu Leu Val Val Leu Ala Val Ala Ala Leu
1. 5 10 15
His Ala Pro Ala Ala Ser Ala Ala Leu Ser Gln Glu Pro Pro Ala Met
20 25 30
Arg Asp Phe Ser Phe Ser Ser Ser His Cys Leu Arg Cys Ile Leu His
35 40 45
Leu His Arg His Leu Pro His Gln Asn Phe Leu Gln Leu Pro Phe Gln
50 55 60
Asn Gln Arg Pro Arg Arg Trp Arg Ser Ile Arg Pro Gln Arg Arg Leu
65 70 75 80
Cys Arg Met Ile Gly Arg Asp Pro Met Pro Cys Val Pro Ala Gly Ser
85 90 95
Ser Leu Trp Pro Trp Ser Trp Glu Gln Gln Ser
100 105
<210> 12
<211> 1665
<212> ДНК
<213> Triticum aestivum
<400> 12
atggccccca ccgccctcct cctcgtcgtc ctcgccgtcg ccgccctcca cgcccccgcc 60
gccaacgccg cgttgtccca ggagccacca gcgaccccgt gcgcggcggc catcgtgtcc 120
ttctcgccgt gcctggcgca cgtcgcggtg gtggcgccgc ccgccctgcc ctcgcccgcg 180
cccaccagcg cctgctgcgc ggcgttcctg cgcgccgttt ccgccgggga cggcgaaggc 240
ggcggagggg agggatgcct ctgccacctg ctccgcgacc cgctcctcct cggcttcccc 300
gtcgacgccg cgcgcctcgg cgctctcctc cccacctgcg cctccgcgaa aacctccgcc 360
gccacggccg tcgaggccga ggccctcttc gccgacaagt gccgaggtga gaaatctgtc 420
ccttcgtgct ccctatttat gctcgtgcaa tatgtatgct ccgatcaatt ctcgtgcgcc 480
atatgcgcgt gctgtcgcgt tcctgttgtt gatcgccgac cgaaacaaat tttactctgc 540
aaagtcctaa ctactgttgt tctcattatc ctcgtacaac tactttcgta atggttggtt 600
gtaatgcgga ttactgacga gctctggatg ctactaggga tattagtact tgcatttctg 660
aactagtgga tgggggagaa agcgggattt ttcttttgtg gcccagactt acttctgcat 720
ttttgctgcg atttgtcatg attaccgttc agttaagcag tttgctgggt tgcctgtttg 780
atagtagtaa tttcacaaaa tattggcgat atttataaat agctaagggc ttcgttactt 840
ggtggtttct ctcaaagagt tcacaaaagt caatcattaa attcaattaa gggggcaagt 900
attagtagta ctggctaact ccatttgcca tttgccaatt gaaacagagc tcaagtcact 960
gcctgagatg cattttacac ctccatcgcc acctcccgca ccaaaacttt ctccaggtaa 1020
atgttctgct gcttgtctaa tgattccata gctcgtcaaa aaaaaaatga ttccataaat 1080
ctgtgcccag taatgctatt tcggatttcg gttgaatgaa ccaattggca tttgggcaga 1140
catgtcatat ttctttcctc ccaaatgaaa cttgaacttg ttttatcttg tggtgctcca 1200
tccatttcgt gctctatgcc gcaactgtac aggttgaaat gatagtagaa catctaattt 1260
tgcatcggat actctctgca gcctctgttc tgtttctact aattgcagga tcgagtctgt 1320
ctttgtgata ctatctgctt tgcgtgcact gctcttttag gttgcagctg caccttctat 1380
tttgttcacc tgagcattcc attttggtcg actgaacacc gcaagtcctg aatatctttc 1440
ttgaatcgtc ggtcgtactt cgtgtgcact agttaaactg ctgtgtatgc ccttgcagct 1500
gccgttccag aaccagcgtc cccgactccg aagatggagg agcattcgac ctcgacgacg 1560
tccatgtcgg atgatcggtc gggatctgat gccttgtgtg cctgccgggt cttccttgtg 1620
gccttggtct tgggagcagc agtcttgatc acgctgcagt tctga 1665
<210> 13
<211> 213
<212> БЕЛОК
<213> Triticum aestivum
<400> 13
Met Ala Pro Thr Ala Leu Leu Leu Val Val Leu Ala Val Ala Ala Leu
1. 5 10 15
His Ala Pro Ala Ala Asn Ala Ala Leu Ser Gln Glu Pro Pro Ala Thr
20 25 30
Pro Cys Ala Ala Ala Ile Val Ser Phe Ser Pro Cys Leu Ala His Val
35 40 45
Ala Val Val Ala Pro Pro Ala Leu Pro Ser Pro Ala Pro Thr Ser Ala
50 55 60
Cys Cys Ala Ala Phe Leu Arg Ala Val Ser Ala Gly Asp Gly Glu Gly
65 70 75 80
Gly Gly Gly Glu Gly Cys Leu Cys His Leu Leu Arg Asp Pro Leu Leu
85 90 95
Leu Gly Phe Pro Val Asp Ala Ala Arg Leu Gly Ala Leu Leu Pro Thr
100 105 110
Cys Ala Ser Ala Lys Thr Ser Ala Ala Thr Ala Val Glu Ala Glu Ala
115 120 125
Leu Phe Ala Asp Lys Cys Arg Glu Leu Lys Ser Leu Pro Glu Met His
130 135 140
Phe Thr Pro Pro Ser Pro Pro Pro Ala Pro Lys Leu Ser Pro Ala Ala
145 150 155 160
Val Pro Glu Pro Ala Ser Pro Thr Pro Lys Met Glu Glu His Ser Thr
165 170 175
Ser Thr Thr Ser Met Ser Asp Asp Arg Ser Gly Ser Asp Ala Leu Cys
180 185 190
Ala Cys Arg Val Phe Leu Val Ala Leu Val Leu Gly Ala Ala Val Leu
195 200 205
Ile Thr Leu Gln Phe
210
<210> 14
<211> 1666
<212> ДНК
<213> Triticum aestivum
<400> 14
atggccccca ccgccctcct cctcgtcgtc ctcgccgtcg ccgccctcca cgcccccgcc 60
gccaacgccg cgttgtcccc aggagccacc agcgaccccg tgcgcggcgg ccatcgtgtc 120
cttctcgccg tgcctggcgc acgtcgcggt ggtggcgccg cccgccctgc cctcgcccgc 180
gcccaccagc gcctgctgcg cggcgttcct gcgcgccgtt tccgccgggg acggcgaagg 240
cggcggaggg gagggatgcc tctgccacct gctccgcgac ccgctcctcc tcggcttccc 300
cgtcgacgcc gcgcgcctcg gcgctctcct ccccacctgc gcctccgcga aaacctccgc 360
cgccacggcc gtcgaggccg aggccctctt cgccgacaag tgccgaggtg agaaatctgt 420
cccttcgtgc tccctattta tgctcgtgca atatgtatgc tccgatcaat tctcgtgcgc 480
catatgcgcg tgctgtcgcg ttcctgttgt tgatcgccga ccgaaacaaa ttttactctg 540
caaagtccta actactgttg ttctcattat cctcgtacaa ctactttcgt aatggttggt 600
tgtaatgcgg attactgacg agctctggat gctactaggg atattagtac ttgcatttct 660
gaactagtgg atgggggaga aagcgggatt tttcttttgt ggcccagact tacttctgca 720
tttttgctgc gatttgtcat gattaccgtt cagttaagca gtttgctggg ttgcctgttt 780
gatagtagta atttcacaaa atattggcga tatttataaa tagctaaggg cttcgttact 840
tggtggtttc tctcaaagag ttcacaaaag tcaatcatta aattcaatta agggggcaag 900
tattagtagt actggctaac tccatttgcc atttgccaat tgaaacagag ctcaagtcac 960
tgcctgagat gcattttaca cctccatcgc cacctcccgc accaaaactt tctccaggta 1020
aatgttctgc tgcttgtcta atgattccat agctcgtcaa aaaaaaaatg attccataaa 1080
tctgtgccca gtaatgctat ttcggatttc ggttgaatga accaattggc atttgggcag 1140
acatgtcata tttctttcct cccaaatgaa acttgaactt gttttatctt gtggtgctcc 1200
atccatttcg tgctctatgc cgcaactgta caggttgaaa tgatagtaga acatctaatt 1260
ttgcatcgga tactctctgc agcctctgtt ctgtttctac taattgcagg atcgagtctg 1320
tctttgtgat actatctgct ttgcgtgcac tgctctttta ggttgcagct gcaccttcta 1380
ttttgttcac ctgagcattc cattttggtc gactgaacac cgcaagtcct gaatatcttt 1440
cttgaatcgt cggtcgtact tcgtgtgcac tagttaaact gctgtgtatg cccttgcagc 1500
tgccgttcca gaaccagcgt ccccgactcc gaagatggag gagcattcga cctcgacgac 1560
gtccatgtcg gatgatcggt cgggatctga tgccttgtgt gcctgccggg tcttccttgt 1620
ggccttggtc ttgggagcag cagtcttgat cacgctgcag ttctga 1666
<210> 15
<211> 141
<212> БЕЛОК
<213> Triticum aestivum
<400> 15
Met Ala Pro Thr Ala Leu Leu Leu Val Val Leu Ala Val Ala Ala Leu
1. 5 10 15
His Ala Pro Ala Ala Asn Ala Ala Leu Ser Pro Gly Ala Thr Ser Asp
20 25 30
Pro Val Arg Gly Gly His Arg Val Leu Leu Ala Val Pro Gly Ala Arg
35 40 45
Arg Gly Gly Gly Ala Ala Arg Pro Ala Leu Ala Arg Ala His Gln Arg
50 55 60
Leu Leu Arg Gly Val Pro Ala Arg Arg Phe Arg Arg Gly Arg Arg Arg
65 70 75 80
Arg Arg Arg Gly Gly Met Pro Leu Pro Pro Ala Pro Arg Pro Ala Pro
85 90 95
Pro Arg Leu Pro Arg Arg Arg Arg Ala Pro Arg Arg Ser Pro Pro His
100 105 110
Leu Arg Leu Arg Glu Asn Leu Arg Arg His Gly Arg Arg Gly Arg Gly
115 120 125
Pro Leu Arg Arg Gln Val Pro Arg Ala Gln Val Thr Ala
130 135 140
<210> 16
<211> 1525
<212> ДНК
<213> Triticum aestivum
<400> 16
atggggcggc cgccgtgctg cgacaaggcc aacgtgaaga aggggccgtg gacggcggag 60
gaggacgcca agctgctggc ctacacctcc aaccatggca ccggcaactg gacctctgtt 120
ccccagaggg caggtcggaa ccctctcccc cggccggccg gatcggcgtc tggcatcgaa 180
atggtgttgg ttttgcatgg tttctggctg atgtgtgttg cgtcatggat gggtgcgtgc 240
tgcagggctg aagcggtgcg ggaagagctg caggctgagg tacaccaact acctgaggcc 300
caacctcaag cacgagaact tcacgcagga ggaggaggag ctcatcgtca ccctccatgc 360
catgctggga agcaggtgca tctctgtaca tgcatgtcat ccatcggtta ctttctcgat 420
tttgtcatca agttttcggt gaatgcgaaa ctttgctatg gacagactac tgtttctgtc 480
atcattcctg ctataaactt tgcatccata tactctagcg tacgttcaga tcttatagat 540
ttatggaagt tctaaaaacc tcacgttggt gtatcattca gacaattgct catacaccgg 600
aaatcacgtc ctacttctat tcaagaattt actttagccg ttcattcata aagtagaaac 660
aaatgtagtt taatccagta accagtctcg tgtctacggt tcttctgaat atattctgtt 720
tcctggtaga atgtagtgaa aagctgaata ttgggtgaag aacagatctg aatagctgtc 780
ttaaattgat agtgtttgac agaaaatact ccacactgag cgtaatataa catgaaagta 840
cgtgtacgtg ttgggtgcat gcaggtggtc tctgatcgcg aaccagctgc cggggcggac 900
ggacaacgac gtcaagaatt actggaacac caagctgagc aagaagctgc ggcagcgggg 960
catcgacccc atcacccatc gccccatcgc cgacctcatg cagagcatcg gcaccctccc 1020
catccgcccg ccgcccagcg ccgcgggtgc ctcctcgtcc tcctacatcc ccgtgaaccc 1080
agcggcggcg ccggggctcc agccgctgca cgacgacgtc aaataccacg cagtcctgaa 1140
ccaccaccag cagcaggtca tcacgctcct cgaccccgac gcgccagggg cggcggcgtc 1200
cccggaccac cagctcaagt ggagcgactt ccttgccgac gacgccgccg ccttcgaggc 1260
ggcgccgcag gtggttcttg gtcagtacca ggaggccgcg gtggctggtg gcggagcagc 1320
gtatggagac actgatagta ttgcagccga tggtgtcggc gggggcgggg aggatagcgc 1380
agcgtcagcg ttcatcgacg cgatgctgga cagcgacaaa aagatgggcg tggaccagct 1440
catcgccgac ctgctcgccg acccggcata ctactacggc ggaggctctt cctcttcgac 1500
gtcggagctg gggcggggcg gttga 1525
<210> 17
<211> 307
<212> БЕЛОК
<213> Triticum aestivum
<400> 17
Met Gly Arg Pro Pro Cys Cys Asp Lys Ala Asn Val Lys Lys Gly Pro
1. 5 10 15
Trp Thr Ala Glu Glu Asp Ala Lys Leu Leu Ala Tyr Thr Ser Asn His
20 25 30
Gly Thr Gly Asn Trp Thr Ser Val Pro Gln Arg Ala Gly Leu Lys Arg
35 40 45
Cys Gly Lys Ser Cys Arg Leu Arg Tyr Thr Asn Tyr Leu Arg Pro Asn
50 55 60
Leu Lys His Glu Asn Phe Thr Gln Glu Glu Glu Glu Leu Ile Val Thr
65 70 75 80
Leu His Ala Met Leu Gly Ser Arg Trp Ser Leu Ile Ala Asn Gln Leu
85 90 95
Pro Gly Arg Thr Asp Asn Asp Val Lys Asn Tyr Trp Asn Thr Lys Leu
100 105 110
Ser Lys Lys Leu Arg Gln Arg Gly Ile Asp Pro Ile Thr His Arg Pro
115 120 125
Ile Ala Asp Leu Met Gln Ser Ile Gly Thr Leu Pro Ile Arg Pro Pro
130 135 140
Pro Ser Ala Ala Gly Ala Ser Ser Ser Ser Tyr Ile Pro Val Asn Pro
145 150 155 160
Ala Ala Ala Pro Gly Leu Gln Pro Leu His Asp Asp Val Lys Tyr His
165 170 175
Ala Val Leu Asn His His Gln Gln Gln Val Ile Thr Leu Leu Asp Pro
180 185 190
Asp Ala Pro Gly Ala Ala Ala Ser Pro Asp His Gln Leu Lys Trp Ser
195 200 205
Asp Phe Leu Ala Asp Asp Ala Ala Ala Phe Glu Ala Ala Pro Gln Val
210 215 220
Val Leu Gly Gln Tyr Gln Glu Ala Ala Val Ala Gly Gly Gly Ala Ala
225 230 235 240
Tyr Gly Asp Thr Asp Ser Ile Ala Ala Asp Gly Val Gly Gly Gly Gly
245 250 255
Glu Asp Ser Ala Ala Ser Ala Phe Ile Asp Ala Met Leu Asp Ser Asp
260 265 270
Lys Lys Met Gly Val Asp Gln Leu Ile Ala Asp Leu Leu Ala Asp Pro
275 280 285
Ala Tyr Tyr Tyr Gly Gly Gly Ser Ser Ser Ser Thr Ser Glu Leu Gly
290 295 300
Arg Gly Gly
305
<210> 18
<211> 1530
<212> ДНК
<213> Triticum aestivum
<400> 18
atggggcggc cgccgtgctg cgacaaggcc aacgtgaaga aggggccgtg gacggcggag 60
gaggacgcca agctgctcgc ctacacctcc aaccatggca ccggcaactg gacctctgtt 120
ccccagaggg caggtcggaa ccctcccccc ggccggccgg atcgacgtcc gtcatggaaa 180
tggtgttggt cttgcatggt ttctggctga tgtgtgttgc atcatggatg cgtgcgtgct 240
gcagggctga agcggtgcgg gaagagctgc aggctgaggt acaccaacta cctgaggccc 300
aacctcaagc acgagaactt cacgcaggag gaggaggagc tcatcgtcac cctccatgcc 360
atgctgggaa gcaggtgcgt ctctctgtgt agtacatgca tgtttctgcc atcagttact 420
ttctcgattt tgtcatcaag ttttcggtga atgcgaaact ttgctatgga ccgactgctg 480
tttctgtcat cattcctgct ataaaccttg catccatata ctctagcgtt cagatcttat 540
agatttatgg aagttctgaa tatctcacgt tggtgtatca gacaattgct catataccgg 600
aaatcacgtc ctacttctat tcaagaattt acttcagtcg ttcgttcaaa aatagaaaga 660
aatgtagttt agtccagtga ccagtctcgt gtctacgtac ggttcttctg aatatatata 720
ttctgtttcc tggtagaatg tagtgaaaag ctgaaaattc ggtgaagaac atatctgaat 780
agctgtctta aattgatagt gtttgacaga aaatactcca cactgagcgt aacgtgaaag 840
tgcgtgtacg tgttgggtgc atgcaggtgg tctctgatcg cgaaccagct gccggggcgg 900
acggacaacg acgtcaagaa ctactggaac accaagctga gcaagaagct gaggcagcgg 960
ggcatcgacc ccatcaccca ccgccccatc gccgatctca tgcagagcat cggcaccctc 1020
gccatccgcc cgccaccgag cgccgcgggt gcctcctcct cctcctacct ccccgtgaac 1080
ccaccggcgg cgccggggct ccagccgctg cacgacgacg tcaaatacca cacagtcctg 1140
aaccagcagc agcagcaggt catcacgctc ctcgaccccg acgcgccagg ggcggcggcg 1200
tccccggagc accagctcaa gtggagcgac ttcctcgcgg acgacgccgc ggccctcgag 1260
gcggcgccgc aggtcgttct tggtcagtac caggaggccg cggtcgctgg tggcggagca 1320
cacgcgtatg gcgacactga cagtactgca gccgatggtg tcggcggggg cggggacgat 1380
agcgcagcgt cagcgttcat cgacgcgatg ctggacagcg acaagaagat gggcgtggac 1440
cagctcatcg ccgacctgct cgccgacccg gcatactact acggcggagg ctcttcctct 1500
tcgacgtcgg agctggggtg gggctgttga 1530
<210> 19
<211> 308
<212> БЕЛОК
<213> Triticum aestivum
<400> 19
Met Gly Arg Pro Pro Cys Cys Asp Lys Ala Asn Val Lys Lys Gly Pro
1. 5 10 15
Trp Thr Ala Glu Glu Asp Ala Lys Leu Leu Ala Tyr Thr Ser Asn His
20 25 30
Gly Thr Gly Asn Trp Thr Ser Val Pro Gln Arg Ala Gly Leu Lys Arg
35 40 45
Cys Gly Lys Ser Cys Arg Leu Arg Tyr Thr Asn Tyr Leu Arg Pro Asn
50 55 60
Leu Lys His Glu Asn Phe Thr Gln Glu Glu Glu Glu Leu Ile Val Thr
65 70 75 80
Leu His Ala Met Leu Gly Ser Arg Trp Ser Leu Ile Ala Asn Gln Leu
85 90 95
Pro Gly Arg Thr Asp Asn Asp Val Lys Asn Tyr Trp Asn Thr Lys Leu
100 105 110
Ser Lys Lys Leu Arg Gln Arg Gly Ile Asp Pro Ile Thr His Arg Pro
115 120 125
Ile Ala Asp Leu Met Gln Ser Ile Gly Thr Leu Ala Ile Arg Pro Pro
130 135 140
Pro Ser Ala Ala Gly Ala Ser Ser Ser Ser Tyr Leu Pro Val Asn Pro
145 150 155 160
Ala Ala Ala Pro Gly Leu Gln Pro Leu His Asp Asp Val Lys Tyr His
165 170 175
Ala Val Leu Asn Gln Gln Gln Gln Gln Val Ile Thr Leu Leu Asp Ala
180 185 190
Asp Ala Pro Gly Ala Ala Ala Ser Pro Asp His Pro Leu Lys Trp Ser
195 200 205
Asp Phe Leu Ala Asp Asp Ala Ala Ala Phe Glu Ala Ala Pro Gln Val
210 215 220
Val Leu Gly Gln Tyr Gln Asp Ala Ala Val Ala Gly Gly Gly Ala His
225 230 235 240
Ala Tyr Gly Asp Thr Asp Ser Thr Ala Ala Asp Gly Val Gly Val Gly
245 250 255
Gly Glu Asp Ser Ala Ala Ser Ala Phe Ile Asp Ala Met Leu Asp Ser
260 265 270
Asp Lys Lys Met Gly Val Asp Gln Leu Ile Ala Asp Leu Leu Ala Asp
275 280 285
Pro Ala Tyr Tyr Tyr Gly Gly Gly Ser Ser Ser Ser Thr Ser Asp Leu
290 295 300
Gly Trp Gly Cys
305
<210> 20
<211> 1530
<212> ДНК
<213> Triticum aestivum
<400> 20
atggggcggc cgccgtgctg cgacaaggcc aacgtgaaga aggggccgtg gacggcggag 60
gaggacgcca agctgctcgc ctacacctcc aaccatggca ccggcaactg gacctctgtt 120
ccccagaggg caggtcggaa ccctcccccc ggccggccgg atcgacgtcc gtcatggaaa 180
tggtgttggt cttgcatggt ttctggctga tgtgtgttgc atcatggatg cgtgcgtgct 240
gcagggctga agcggtgcgg gaagagctgc aggctgaggt acaccaacta cctgaggccc 300
aacctcaagc acgagaactt cacgcaggag gaggaggagc tcatcgtcac cctccatgcc 360
atgctgggaa gcaggtgcgt ctctctgtgt agtacatgca tgtttctgcc atcagttact 420
ttctcgattt tgtcatcaag ttttcggtga atgcgaaact ttgctatgga ccgactgctg 480
tttctgtcat cattcctgct ataaaccttg catccatata ctctagcgtt cagatcttat 540
agatttatgg aagttctgaa tatctcacgt tggtgtatca gacaattgct catataccgg 600
aaatcacgtc ctacttctat tcaagaattt acttcagtcg ttcgttcaaa aatagaaaga 660
aatgtagttt agtccagtga ccagtctcgt gtctacgtac ggttcttctg aatatatata 720
ttctgtttcc tggtagaatg tagtgaaaag ctgaaaattc ggtgaagaac atatctgaat 780
agctgtctta aattgatagt gtttgacaga aaatactcca cactgagcgt aacgtgaaag 840
tgcgtgtacg tgttgggtgc atgcaggtgg tctctgatcg cgaaccagct gccggggcgg 900
acggacaacg acgtcaagaa ctactggaac accaagctga gcaagaagct gaggcagcgg 960
ggcatcgacc ccatcaccca ccgccccatc gccgatctca tgcagagcat cggcaccctc 1020
gccatccgcc cgccaccgag cgccgcgggt gcctcctcct cctcctacct ccccgtgaac 1080
ccaccggcgg cgccggggct ccagccgctg cacgacgacg tcaaatacca cacagtcctg 1140
aaccagcagc agcagcaggt catcacgctc ctcgaccccg acgcgccagg ggcggcggcg 1200
tccccggagc accagctcaa gtggagcgac ttcctcgcgg acgacgccgc ggccctcgag 1260
gcggcgccgc aggtcgttct tggtcagtac caggaggccg cggtcgctgg tggcggagca 1320
cacgcgtatg gcgacactga cagtactgca gccgatggtg tcggcggggg cggggacgat 1380
agcgcagcgt cagcgttcat cgacgcgatg ctggacagcg acaagaagat gggcgtggac 1440
cagctcatcg ccgacctgct cgccgacccg gcatactact acggcggagg ctcttcctct 1500
tcgacgtcgg agctggggtg gggctgttga 1530
<210> 21
<211> 308
<212> БЕЛОК
<213> Triticum aestivum
<400> 21
Met Gly Arg Pro Pro Cys Cys Asp Lys Ala Asn Val Lys Lys Gly Pro
1. 5 10 15
Trp Thr Ala Glu Glu Asp Ala Lys Leu Leu Ala Tyr Thr Ser Asn His
20 25 30
Gly Thr Gly Asn Trp Thr Ser Val Pro Gln Arg Ala Gly Leu Lys Arg
35 40 45
Cys Gly Lys Ser Cys Arg Leu Arg Tyr Thr Asn Tyr Leu Arg Pro Asn
50 55 60
Leu Lys His Glu Asn Phe Thr Gln Glu Glu Glu Glu Leu Ile Val Thr
65 70 75 80
Leu His Ala Met Leu Gly Ser Arg Trp Ser Leu Ile Ala Asn Gln Leu
85 90 95
Pro Gly Arg Thr Asp Asn Asp Val Lys Asn Tyr Trp Asn Thr Lys Leu
100 105 110
Ser Lys Lys Leu Arg Gln Arg Gly Ile Asp Pro Ile Thr His Arg Pro
115 120 125
Ile Ala Asp Leu Met Gln Ser Ile Gly Thr Leu Ala Ile Arg Pro Pro
130 135 140
Pro Ser Ala Ala Gly Ala Ser Ser Ser Ser Tyr Leu Pro Val Asn Pro
145 150 155 160
Pro Ala Ala Pro Gly Leu Gln Pro Leu His Asp Asp Val Lys Tyr His
165 170 175
Thr Val Leu Asn Gln Gln Gln Gln Gln Val Ile Thr Leu Leu Asp Pro
180 185 190
Asp Ala Pro Gly Ala Ala Ala Ser Pro Glu His Gln Leu Lys Trp Ser
195 200 205
Asp Phe Leu Ala Asp Asp Ala Ala Ala Leu Glu Ala Ala Pro Gln Val
210 215 220
Val Leu Gly Gln Tyr Gln Glu Ala Ala Val Ala Gly Gly Gly Ala His
225 230 235 240
Ala Tyr Gly Asp Thr Asp Ser Thr Ala Ala Asp Gly Val Gly Gly Gly
245 250 255
Gly Asp Asp Ser Ala Ala Ser Ala Phe Ile Asp Ala Met Leu Asp Ser
260 265 270
Asp Lys Lys Met Gly Val Asp Gln Leu Ile Ala Asp Leu Leu Ala Asp
275 280 285
Pro Ala Tyr Tyr Tyr Gly Gly Gly Ser Ser Ser Ser Thr Ser Glu Leu
290 295 300
Gly Trp Gly Cys
305
<210> 22
<211> 414
<212> ДНК
<213> Triticum aestivum
<400> 22
atgttgagga tgcagcagca ggtggagggc gtggtgggcg gcggcatcgt ggccgaggcg 60
gaggaggcgg cggtgtacga gcgggtggct cgcatggcca gcggcaacgc cgtggtcgtc 120
ttcagcgcca gcggctgctg catgtgccac gtcgtcaagc gcctcctgct tggcctggga 180
gtcggcccca ccgtctacga gttggaccag atgggcggcg ccgggcggga gatccaggcg 240
gccctggcgc agctgctgcc ccccggaccc ggcgccggcc accaccagca gccgccggtg 300
cccgtggtgt tcgtcggcgg gaggctcctg ggaggcgtgg agaaggtgat ggcgtgccac 360
atcaacggca cgctcgtccc gctcctcaag gacgccggcg cgctctggct ctga 414
<210> 23
<211> 137
<212> БЕЛОК
<213> Triticum aestivum
<400> 23
Met Leu Arg Met Gln Gln Gln Val Glu Gly Val Val Gly Gly Gly Ile
1. 5 10 15
Val Ala Glu Ala Glu Glu Ala Ala Val Tyr Glu Arg Val Ala Arg Met
20 25 30
Ala Ser Gly Asn Ala Val Val Val Phe Ser Ala Ser Gly Cys Cys Met
35 40 45
Cys His Val Val Lys Arg Leu Leu Leu Gly Leu Gly Val Gly Pro Thr
50 55 60
Val Tyr Glu Leu Asp Gln Met Gly Gly Ala Gly Arg Glu Ile Gln Ala
65 70 75 80
Ala Leu Ala Gln Leu Leu Pro Pro Gly Pro Gly Ala Gly His His Gln
85 90 95
Gln Pro Pro Val Pro Val Val Phe Val Gly Gly Arg Leu Leu Gly Gly
100 105 110
Val Glu Lys Val Met Ala Cys His Ile Asn Gly Thr Leu Val Pro Leu
115 120 125
Leu Lys Asp Ala Gly Ala Leu Trp Leu
130 135
<210> 24
<211> 414
<212> ДНК
<213> Triticum aestivum
<400> 24
atgttgagga tgcagcagca ggtggagggc gtggtgggcg gcggcatcgt ggcggaggcg 60
gaggaggcgg cggtgtacga gagggtggct cgcatggcca gcggcaacgc ggtggtcgtc 120
ttcagcgcca gcggctgctg catgtgccac gtcgtcaagc gcctcctgct tggcctggga 180
gtcggcccca ccgtgtacga gttggaccag atgggcggcg ccgggcggga gatccaggcg 240
gccctggcgc agctgctgcc ccccggaccc ggcgccggcc accaccagca gccgccggtg 300
cccgtggtgt tcgttggcgg gaggctcctg ggcggcgtgg agaaggtgat ggcgtgccac 360
atcaacggca cgctcgtccc gctcctcaag gacgccggcg cgctctggct ctga 414
<210> 25
<211> 137
<212> БЕЛОК
<213> Triticum aestivum
<400> 25
Met Leu Arg Met Gln Gln Gln Val Glu Gly Val Val Gly Gly Gly Ile
1. 5 10 15
Val Ala Glu Ala Glu Glu Ala Ala Val Tyr Glu Arg Val Ala Arg Met
20 25 30
Ala Ser Gly Asn Ala Val Val Val Phe Ser Ala Ser Gly Cys Cys Met
35 40 45
Cys His Val Val Lys Arg Leu Leu Leu Gly Leu Gly Val Gly Pro Thr
50 55 60
Val Tyr Glu Leu Asp Gln Met Gly Gly Ala Gly Arg Glu Ile Gln Ala
65 70 75 80
Ala Leu Ala Gln Leu Leu Pro Pro Gly Pro Gly Ala Gly His His Gln
85 90 95
Gln Pro Pro Val Pro Val Val Phe Val Gly Gly Arg Leu Leu Gly Gly
100 105 110
Val Glu Lys Val Met Ala Cys His Ile Asn Gly Thr Leu Val Pro Leu
115 120 125
Leu Lys Asp Ala Gly Ala Leu Trp Leu
130 135
<210> 26
<211> 414
<212> ДНК
<213> Triticum aestivum
<400> 26
atgttgagga tgcagcagca ggtggagggc gtggtgggcg gcggcatcat ggcggaggcg 60
gaggaggcgg cggtgtacga gcgggtggct cgcatggcca gcggcaacgc ggtggtcgtc 120
ttcagcgcca gcggctgctg catgtgccac gtcgtcaagc gcctcctgct tggcctgggg 180
gtcggcccca ccgtctacga gttggaccag atgggcggcg ccgggcgaga gatccaggcg 240
gcgctggcgc agctgctgcc ccccggaccc ggcgccggcc accaccagca gccgccagtg 300
cccgtggtgt tcgtcggcgg gaggctcctg ggcggcgtgg agaaggtgat ggcgtgccac 360
atcaacggca cgctcgtccc gctcctcaag gacgccggcg cgctctggct ctga 414
<210> 27
<211> 137
<212> БЕЛОК
<213> Triticum aestivum
<400> 27
Met Leu Arg Met Gln Gln Gln Val Glu Gly Val Val Gly Gly Gly Ile
1. 5 10 15
Met Ala Glu Ala Glu Glu Ala Ala Val Tyr Glu Arg Val Ala Arg Met
20 25 30
Ala Ser Gly Asn Ala Val Val Val Phe Ser Ala Ser Gly Cys Cys Met
35 40 45
Cys His Val Val Lys Arg Leu Leu Leu Gly Leu Gly Val Gly Pro Thr
50 55 60
Val Tyr Glu Leu Asp Gln Met Gly Gly Ala Gly Arg Glu Ile Gln Ala
65 70 75 80
Ala Leu Ala Gln Leu Leu Pro Pro Gly Pro Gly Ala Gly His His Gln
85 90 95
Gln Pro Pro Val Pro Val Val Phe Val Gly Gly Arg Leu Leu Gly Gly
100 105 110
Val Glu Lys Val Met Ala Cys His Ile Asn Gly Thr Leu Val Pro Leu
115 120 125
Leu Lys Asp Ala Gly Ala Leu Trp Leu
130 135
<210> 28
<211> 1926
<212> ДНК
<213> Triticum aestivum
<400> 28
atgagcagcc ccatggagga agctcaccat ggcatgccgt cgacgacgac ggcgttcttc 60
ccgctggcag ggctccacaa gttcatggcc atcttcctcg tgttcctctc gtggatcttg 120
gtccactggt ggagcctgag gaagcagaag gggccgaggt catggccggt catcggcgcg 180
acgctggagc agctgaggaa ctactaccgg atgcacgact ggctcgtgga gtacctgtcc 240
aagcaccgga cggtcaccgt cgacatgccc ttcacctcct acacctacat cgccgacccg 300
gtgaacgtcg agcatgtgct caagaccaac ttcaacaatt accccaaggt gaaactgaaa 360
gaacccctca gccttgtgaa tttttttgcc aaggttcaga agtttacact gacacaaatg 420
tctgaaattg tacgtgtagg gggaggtgta caggtcctac atggacgtgc tgctcggcga 480
cggcatcttc aacgccgacg gcgagctctg gaggaagcag aggaagacgg cgagcttcga 540
gttcgcttcc aagaacctga gagactttag cacgatcgtg ttcagggagt actccctgaa 600
gctgcgcagc atcctgagcc aggcttgcaa ggccggcaaa gtcgtggaca tgcaggtaac 660
cgaactcagt cccttggtca tctgaacatt gatttcttgg acaaaatttc aagattctga 720
cgcgagcgag cgaattcagg agctgtacat gaggatgacg ctggactcga tctgcaaggt 780
ggggttcggg gtcgagatcg gcacgctgtc gccggagctg ccggagaaca gcttcgcgca 840
ggcgttcgac gccgccaaca tcatcgtgac gctgcggttc atcgacccgc tgtggcgcgt 900
gaagaagttc ctgcacgtcg gctcggaggc gctgctggag cagagcatca agctcgtcga 960
cgagttcacc tacagcgtca tccgccggcg caaggccgag atcgtgcagg cccgggccag 1020
cggcaagcag gagaaggtgc gtacgtgatc gtcgtcgtca agctccggat cgctggtttg 1080
tgtaggtgcc attgatcact gacacactag ctgggtgcgc agatcaagca cgacatactg 1140
tcgcggttca tcgagctggg cgaggccggc ggggacgacg gcggcagcct gttcggggac 1200
gacaagggcc tccgcgacgt ggtgctcaac ttcgtgatcg ccgggcggga caccacggcc 1260
acgacgctct cctggttcac ctacatggcc atgacgcacc cggccgtggc cgagaagctc 1320
cgccgcgagc tggccgcctt cgaggcggac cgcgcccgcg aggatggcgt cgcgctggtc 1380
ccctgcagcg actcagacgg cgacggctcc gacgaggcct tcgccgcccg cgtggcgcag 1440
ttcgcggggc tgctgagcta cgacgggctc gggaagctgg tgtacctcca cgcgtgcgtg 1500
acggagacgc tgcgcctgta cccggcggtg ccgcaggacc ccaagggcat cgcggaggac 1560
gacgtgctcc cggacggcac caaggtgcgc gccggcggga tggtgacgta cgtgccctac 1620
tccatggggc ggatggagta caactggggc cccgacgccg ccagcttccg gccggagcgg 1680
tggatcggcg acgacggcgc gttccgcaac gcgtcgccgt tcaagttcac ggcgttccag 1740
gcggggccgc ggatctgcct cggcaaggac tcggcgtacc tgcagatgaa gatggcgctg 1800
gccatcctgt gcaggttctt caggttcgag ctcgtggagg gccaccccgt caagtaccgc 1860
atgatgacca tcctctccat ggcgcacggc ctcaaggtcc gcgtctccag ggcgccgctc 1920
gcctga 1926
<210> 29
<211> 554
<212> БЕЛОК
<213> Triticum aestivum
<400> 29
Met Ser Ser Pro Met Glu Glu Ala His His Gly Met Pro Ser Thr Thr
1. 5 10 15
Thr Ala Phe Phe Pro Leu Ala Gly Leu His Lys Phe Met Ala Ile Phe
20 25 30
Leu Val Phe Leu Ser Trp Ile Leu Val His Trp Trp Ser Leu Arg Lys
35 40 45
Gln Lys Gly Pro Arg Ser Trp Pro Val Ile Gly Ala Thr Leu Glu Gln
50 55 60
Leu Arg Asn Tyr Tyr Arg Met His Asp Trp Leu Val Glu Tyr Leu Ser
65 70 75 80
Lys His Arg Thr Val Thr Val Asp Met Pro Phe Thr Ser Tyr Thr Tyr
85 90 95
Ile Ala Asp Pro Val Asn Val Glu His Val Leu Lys Thr Asn Phe Asn
100 105 110
Asn Tyr Pro Lys Gly Glu Val Tyr Arg Ser Tyr Met Asp Val Leu Leu
115 120 125
Gly Asp Gly Ile Phe Asn Ala Asp Gly Glu Leu Trp Arg Lys Gln Arg
130 135 140
Lys Thr Ala Ser Phe Glu Phe Ala Ser Lys Asn Leu Arg Asp Phe Ser
145 150 155 160
Thr Ile Val Phe Arg Glu Tyr Ser Leu Lys Leu Arg Ser Ile Leu Ser
165 170 175
Gln Ala Cys Lys Ala Gly Lys Val Val Asp Met Gln Glu Leu Tyr Met
180 185 190
Arg Met Thr Leu Asp Ser Ile Cys Lys Val Gly Phe Gly Val Glu Ile
195 200 205
Gly Thr Leu Ser Pro Glu Leu Pro Glu Asn Ser Phe Ala Gln Ala Phe
210 215 220
Asp Ala Ala Asn Ile Ile Val Thr Leu Arg Phe Ile Asp Pro Leu Trp
225 230 235 240
Arg Val Lys Lys Phe Leu His Val Gly Ser Glu Ala Leu Leu Glu Gln
245 250 255
Ser Ile Lys Leu Val Asp Glu Phe Thr Tyr Ser Val Ile Arg Arg Arg
260 265 270
Lys Ala Glu Ile Val Gln Ala Arg Ala Ser Gly Lys Gln Glu Lys Ile
275 280 285
Lys His Asp Ile Leu Ser Arg Phe Ile Glu Leu Gly Glu Ala Gly Gly
290 295 300
Asp Asp Gly Gly Ser Leu Phe Gly Asp Asp Lys Gly Leu Arg Asp Val
305 310 315 320
Val Leu Asn Phe Val Ile Ala Gly Arg Asp Thr Thr Ala Thr Thr Leu
325 330 335
Ser Trp Phe Thr Tyr Met Ala Met Thr His Pro Ala Val Ala Glu Lys
340 345 350
Leu Arg Arg Glu Leu Ala Ala Phe Glu Ala Asp Arg Ala Arg Glu Asp
355 360 365
Gly Val Ala Leu Val Pro Cys Ser Asp Ser Asp Gly Asp Gly Ser Asp
370 375 380
Glu Ala Phe Ala Ala Arg Val Ala Gln Phe Ala Gly Leu Leu Ser Tyr
385 390 395 400
Asp Gly Leu Gly Lys Leu Val Tyr Leu His Ala Cys Val Thr Glu Thr
405 410 415
Leu Arg Leu Tyr Pro Ala Val Pro Gln Asp Pro Lys Gly Ile Ala Glu
420 425 430
Asp Asp Val Leu Pro Asp Gly Thr Lys Val Arg Ala Gly Gly Met Val
435 440 445
Thr Tyr Val Pro Tyr Ser Met Gly Arg Met Glu Tyr Asn Trp Gly Pro
450 455 460
Asp Ala Ala Ser Phe Arg Pro Glu Arg Trp Ile Gly Asp Asp Gly Ala
465 470 475 480
Phe Arg Asn Ala Ser Pro Phe Lys Phe Thr Ala Phe Gln Ala Gly Pro
485 490 495
Arg Ile Cys Leu Gly Lys Asp Ser Ala Tyr Leu Gln Met Lys Met Ala
500 505 510
Leu Ala Ile Leu Cys Arg Phe Phe Arg Phe Glu Leu Val Glu Gly His
515 520 525
Pro Val Lys Tyr Arg Met Met Thr Ile Leu Ser Met Ala His Gly Leu
530 535 540
Lys Val Arg Val Ser Arg Ala Pro Leu Ala
545 550
<210> 30
<211> 1920
<212> ДНК
<213> Triticum aestivum
<400> 30
atgagcagcc ccatggagga agctcacctt ggcatgccgt cgacgacggc cttcttcccg 60
ctggcagggc tccacaagtt catggccgtc ttcctcgtgt tcctctcgtg gatcctggtc 120
cactggtgga gcctgaggaa gcagaagggg ccacggtcat ggccggtcat cggcgcgacg 180
ctggagcagc tgaggaacta ctaccggatg cacgactggc tcgtggagta cctgtccaag 240
caccggacgg tcaccgtcga catgcccttc acctcctaca cctacatcgc cgacccggtg 300
aacgtcgagc acgtgctcaa gaccaacttc aacaattacc ccaaggtgaa acaatcctcg 360
agatgtcagt caaggttcag tataatcggt actgacagtg ttacaaatgt ctgaaatctg 420
gaattgtgtg tgtaggggga ggtgtacagg tcctacatgg acgtgctgct cggcgacggc 480
atattcaacg ccgacggcga gctctggagg aagcagagga agacggcgag cttcgagttc 540
gcttccaaga acttgagaga cttcagcacg atcgtgttca gggagtactc cctgaagctg 600
tccagcatcc tgagccaggc ttgcaaggca ggcaaagttg tggacatgca ggtaactgaa 660
ctctttccct tggtcatatg aacgttgatt tcttggacaa aatctcaaga ttctgacgcg 720
agcgagccaa ttcaggagct gtacatgagg atgacgctgg actcgatctg caaggtgggg 780
ttcggggtgg agatcggcac gctgtcgccg gagctgccgg agaacagctt cgcgcaggcc 840
ttcgacgccg ccaacatcat cgtgacgctg cggttcatcg acccgctgtg gcgcgtgaag 900
aaattcctgc acgtcggctc ggaggcgctg ctggagcaga gcatcaagct cgtcgacgag 960
ttcacctaca gcgtcatccg ccggcgcaag gccgagatcg tgcaagcccg ggccagcggc 1020
aagcaggaga aggtgcgtac gtggtcatcg tcattcgtca agctcccgat cgctggtttg 1080
tgcagatgcc actgatcact gacacattaa ctgggcgcgc agatcaagca cgacatactg 1140
tcgcggttca tcgagctggg cgaggccggc ggcgacgacg gcggcagcct gttcggggac 1200
gacaagggcc tccgcgacgt ggtgctcaac ttcgtcatcg ccgggcggga cacgacggcc 1260
acgacgctct cctggttcac ctacatggcc atgacgcacc cggccgtggc cgagaagctc 1320
cgccgcgagc tggccgcctt cgagtccgag cgcgcccgcg aggatggcgt cgctctggtc 1380
ccctgcagcg acggcgaggg ctccgacgag gccttcgccg cccgcgtggc gcagttcgcg 1440
ggactcctga gctacgacgg gctcgggaag ctggtgtacc tccacgcgtg cgtgacggag 1500
acgctccgcc tgtacccggc ggtgccgcag gaccccaagg gcatcgcgga ggacgacgtg 1560
ctcccggacg gcaccaaggt gcgcgccggc gggatggtga cgtacgtgcc ctactccatg 1620
gggcggatgg agtacaactg gggccccgac gccgccagct tccggccaga gcggtggatc 1680
ggcgacgacg gcgccttccg caacgcgtcg ccgttcaagt tcacggcgtt ccaggcgggg 1740
ccgcggatct gcctgggcaa ggactcggcg tacctgcaga tgaagatggc gctggccatc 1800
ctgtgcaggt tcttcaggtt cgagctcgtg gagggccacc ccgtcaagta ccgcatgatg 1860
accatcctct ccatggcgca cggcctcaag gtccgcgtct ccagggtgcc gctcgcctga 1920
<210> 31
<211> 551
<212> БЕЛОК
<213> Triticum aestivum
<400> 31
Met Ser Ser Pro Met Glu Glu Ala His Leu Gly Met Pro Ser Thr Thr
1. 5 10 15
Ala Phe Phe Pro Leu Ala Gly Leu His Lys Phe Met Ala Val Phe Leu
20 25 30
Val Phe Leu Ser Trp Ile Leu Val His Trp Trp Ser Leu Arg Lys Gln
35 40 45
Lys Gly Pro Arg Ser Trp Pro Val Ile Gly Ala Thr Leu Glu Gln Leu
50 55 60
Arg Asn Tyr Tyr Arg Met His Asp Trp Leu Val Glu Tyr Leu Ser Lys
65 70 75 80
His Arg Thr Val Thr Val Asp Met Pro Phe Thr Ser Tyr Thr Tyr Ile
85 90 95
Ala Asp Pro Val Asn Val Glu His Val Leu Lys Thr Asn Phe Asn Asn
100 105 110
Tyr Pro Lys Gly Glu Val Tyr Arg Ser Tyr Met Asp Val Leu Leu Gly
115 120 125
Asp Gly Ile Phe Asn Ala Asp Gly Glu Leu Trp Arg Lys Gln Arg Lys
130 135 140
Thr Ala Ser Phe Glu Phe Ala Ser Lys Asn Leu Arg Asp Phe Ser Thr
145 150 155 160
Ile Val Phe Arg Glu Tyr Ser Leu Lys Leu Ser Ser Ile Leu Ser Gln
165 170 175
Ala Cys Lys Ala Gly Lys Val Val Asp Met Gln Glu Leu Tyr Met Arg
180 185 190
Met Thr Leu Asp Ser Ile Cys Lys Val Gly Phe Gly Val Glu Ile Gly
195 200 205
Thr Leu Ser Pro Glu Leu Pro Glu Asn Ser Phe Ala Gln Ala Phe Asp
210 215 220
Ala Ala Asn Ile Ile Val Thr Leu Arg Phe Ile Asp Pro Leu Trp Arg
225 230 235 240
Val Lys Lys Phe Leu His Val Gly Ser Glu Ala Leu Leu Glu Gln Ser
245 250 255
Ile Lys Leu Val Asp Glu Phe Thr Tyr Ser Val Ile Arg Arg Arg Lys
260 265 270
Ala Glu Ile Val Gln Ala Arg Ala Ser Gly Lys Gln Glu Lys Ile Lys
275 280 285
His Asp Ile Leu Ser Arg Phe Ile Glu Leu Gly Glu Ala Gly Gly Asp
290 295 300
Asp Gly Gly Ser Leu Phe Gly Asp Asp Lys Gly Leu Arg Asp Val Val
305 310 315 320
Leu Asn Phe Val Ile Ala Gly Arg Asp Thr Thr Ala Thr Thr Leu Ser
325 330 335
Trp Phe Thr Tyr Met Ala Met Thr His Pro Ala Val Ala Glu Lys Leu
340 345 350
Arg Arg Glu Leu Ala Ala Phe Glu Ser Glu Arg Ala Arg Glu Asp Gly
355 360 365
Val Ala Leu Val Pro Cys Ser Asp Gly Glu Gly Ser Asp Glu Ala Phe
370 375 380
Ala Ala Arg Val Ala Gln Phe Ala Gly Leu Leu Ser Tyr Asp Gly Leu
385 390 395 400
Gly Lys Leu Val Tyr Leu His Ala Cys Val Thr Glu Thr Leu Arg Leu
405 410 415
Tyr Pro Ala Val Pro Gln Asp Pro Lys Gly Ile Ala Glu Asp Asp Val
420 425 430
Leu Pro Asp Gly Thr Lys Val Arg Ala Gly Gly Met Val Thr Tyr Val
435 440 445
Pro Tyr Ser Met Gly Arg Met Glu Tyr Asn Trp Gly Pro Asp Ala Ala
450 455 460
Ser Phe Arg Pro Glu Arg Trp Ile Gly Asp Asp Gly Ala Phe Arg Asn
465 470 475 480
Ala Ser Pro Phe Lys Phe Thr Ala Phe Gln Ala Gly Pro Arg Ile Cys
485 490 495
Leu Gly Lys Asp Ser Ala Tyr Leu Gln Met Lys Met Ala Leu Ala Ile
500 505 510
Leu Cys Arg Phe Phe Arg Phe Glu Leu Val Glu Gly His Pro Val Lys
515 520 525
Tyr Arg Met Met Thr Ile Leu Ser Met Ala His Gly Leu Lys Val Arg
530 535 540
Val Ser Arg Val Pro Leu Ala
545 550
<210> 32
<211> 1920
<212> ДНК
<213> Triticum aestivum
<400> 32
atgagcagcc ccatggagga agctcacggc ggcatgccgt cgacgacggc cttcttcccg 60
ctggcagggc tccacaagtt catggccatc ttcctcgtgt tcctctcgtg gatcttggtc 120
cactggtgga gcctgaggaa gcagaagggg ccgaggtcat ggccggtcat cggcgcgacg 180
ctggagcagc tgaggaacta ctaccggatg cacgactggc tcgtggagta cctgtccaag 240
caccggacgg tgaccgtcga catgcccttc acctcctaca cctacatcgc cgacccggtg 300
aacgtcgagc atgtgctcaa gaccaacttc aacaattacc ccaaggtgaa acaatcctcg 360
agatgtcagt aaaggttcag tataatcggt actgacagtg ttacaaatgt ctgaaatctg 420
aaattgtatg tgtaggggga ggtgtacagg tcctacatgg acgtgctgct cggcgacggc 480
atattcaacg ccgacggcga gctctggagg aagcagagga agacggcgag cttcgagttc 540
gcttccaaga acttgagaga cttcagcacg atcgtgttca gggagtactc cctgaagctg 600
tccagcatac tgagccaggc ttgcaaggcc ggcaaagttg tggacatgca ggtaactgaa 660
ctcattccct tggtcatctg aacgttgatt tcttggacaa aatttcaaga ttctgacgcg 720
agcgagcgaa ttcaggagct gtatatgagg atgacgctgg actcgatctg caaagtgggg 780
ttcggagtcg agatcggcac gctgtcgccg gagctgccgg agaacagctt cgcgcaggcg 840
ttcgacgccg ccaacatcat cgtgacgctg cggttcatcg acccgctgtg gcgcgtgaag 900
aagttcctgc acgtcggctc ggaggcgctg ctggagcaga gcatcaagct cgtcgacgag 960
ttcacctaca gcgtcatccg ccggcgcaag gccgagatcg tgcaggcccg ggccagcggc 1020
aagcaggaga aggtgcgtgc gtggtcatcg tcattcgtca agctcccggt cgctggtttg 1080
tgtagatgcc atggatcact gacacactaa ctgggcgcgc agatcaagca cgacatactg 1140
tcgcggttca tcgagctggg cgaggccggc ggcgacgacg gcggcagtct gttcggggac 1200
gacaagggcc tccgcgacgt ggtgctcaac ttcgtgatcg ccgggcggga caccacggcc 1260
acgacgctgt cctggttcac ctacatggcc atgacgcacc cggacgtggc cgagaagctc 1320
cgccgcgagc tggccgcctt cgaggcggag cgcgcccgcg aggatggcgt cgctctggtc 1380
ccctgcggcg acggcgaggg ctccgacgag gccttcgctg cccgcgtggc gcagttcgcg 1440
gggttcctga gctacgacgg cctcgggaag ctggtgtacc tccacgcgtg cgtgacggag 1500
acgctgcgcc tgtacccggc ggtgccgcag gaccccaagg gcatcgcgga ggacgacgtg 1560
ctcccggacg gcaccaaggt gcgcgccggc gggatggtga cgtacgtgcc ctactccatg 1620
gggcggatgg agtacaactg gggccccgac gccgccagct tccggccgga gcggtggatc 1680
ggcgacgacg gcgccttccg caacgcgtcg ccgttcaagt tcacggcgtt ccaggcgggg 1740
ccgcggattt gcctcggcaa ggactcggcg tacctgcaga tgaagatggc gctggcaatc 1800
ctgtgcaggt tcttcaggtt cgagctcgtg gagggccacc ccgtcaagta ccgcatgatg 1860
accatcctct ccatggcgca cggcctcaag gtccgcgtct ccagggcgcc gctcgcctga 1920
<210> 33
<211> 551
<212> БЕЛОК
<213> Triticum aestivum
<400> 33
Met Ser Ser Pro Met Glu Glu Ala His Gly Gly Met Pro Ser Thr Thr
1. 5 10 15
Ala Phe Phe Pro Leu Ala Gly Leu His Lys Phe Met Ala Ile Phe Leu
20 25 30
Val Phe Leu Ser Trp Ile Leu Val His Trp Trp Ser Leu Arg Lys Gln
35 40 45
Lys Gly Pro Arg Ser Trp Pro Val Ile Gly Ala Thr Leu Glu Gln Leu
50 55 60
Arg Asn Tyr Tyr Arg Met His Asp Trp Leu Val Glu Tyr Leu Ser Lys
65 70 75 80
His Arg Thr Val Thr Val Asp Met Pro Phe Thr Ser Tyr Thr Tyr Ile
85 90 95
Ala Asp Pro Val Asn Val Glu His Val Leu Lys Thr Asn Phe Asn Asn
100 105 110
Tyr Pro Lys Gly Glu Val Tyr Arg Ser Tyr Met Asp Val Leu Leu Gly
115 120 125
Asp Gly Ile Phe Asn Ala Asp Gly Glu Leu Trp Arg Lys Gln Arg Lys
130 135 140
Thr Ala Ser Phe Glu Phe Ala Ser Lys Asn Leu Arg Asp Phe Ser Thr
145 150 155 160
Ile Val Phe Arg Glu Tyr Ser Leu Lys Leu Ser Ser Ile Leu Ser Gln
165 170 175
Ala Cys Lys Ala Gly Lys Val Val Asp Met Gln Glu Leu Tyr Met Arg
180 185 190
Met Thr Leu Asp Ser Ile Cys Lys Val Gly Phe Gly Val Glu Ile Gly
195 200 205
Thr Leu Ser Pro Glu Leu Pro Glu Asn Ser Phe Ala Gln Ala Phe Asp
210 215 220
Ala Ala Asn Ile Ile Val Thr Leu Arg Phe Ile Asp Pro Leu Trp Arg
225 230 235 240
Val Lys Lys Phe Leu His Val Gly Ser Glu Ala Leu Leu Glu Gln Ser
245 250 255
Ile Lys Leu Val Asp Glu Phe Thr Tyr Ser Val Ile Arg Arg Arg Lys
260 265 270
Ala Glu Ile Val Gln Ala Arg Ala Ser Gly Lys Gln Glu Lys Ile Lys
275 280 285
His Asp Ile Leu Ser Arg Phe Ile Glu Leu Gly Glu Ala Gly Gly Asp
290 295 300
Asp Gly Gly Ser Leu Phe Gly Asp Asp Lys Gly Leu Arg Asp Val Val
305 310 315 320
Leu Asn Phe Val Ile Ala Gly Arg Asp Thr Thr Ala Thr Thr Leu Ser
325 330 335
Trp Phe Thr Tyr Met Ala Met Thr His Pro Asp Val Ala Glu Lys Leu
340 345 350
Arg Arg Glu Leu Ala Ala Phe Glu Ala Glu Arg Ala Arg Glu Asp Gly
355 360 365
Val Ala Leu Val Pro Cys Gly Asp Gly Glu Gly Ser Asp Glu Ala Phe
370 375 380
Ala Ala Arg Val Ala Gln Phe Ala Gly Phe Leu Ser Tyr Asp Gly Leu
385 390 395 400
Gly Lys Leu Val Tyr Leu His Ala Cys Val Thr Glu Thr Leu Arg Leu
405 410 415
Tyr Pro Ala Val Pro Gln Asp Pro Lys Gly Ile Ala Glu Asp Asp Val
420 425 430
Leu Pro Asp Gly Thr Lys Val Arg Ala Gly Gly Met Val Thr Tyr Val
435 440 445
Pro Tyr Ser Met Gly Arg Met Glu Tyr Asn Trp Gly Pro Asp Ala Ala
450 455 460
Ser Phe Arg Pro Glu Arg Trp Ile Gly Asp Asp Gly Ala Phe Arg Asn
465 470 475 480
Ala Ser Pro Phe Lys Phe Thr Ala Phe Gln Ala Gly Pro Arg Ile Cys
485 490 495
Leu Gly Lys Asp Ser Ala Tyr Leu Gln Met Lys Met Ala Leu Ala Ile
500 505 510
Leu Cys Arg Phe Phe Arg Phe Glu Leu Val Glu Gly His Pro Val Lys
515 520 525
Tyr Arg Met Met Thr Ile Leu Ser Met Ala His Gly Leu Lys Val Arg
530 535 540
Val Ser Arg Ala Pro Leu Ala
545 550
<210> 34
<211> 1483
<212> ДНК
<213> Triticum aestivum
<400> 34
atggaagaga agaagccgcg gcggcaggga gccgcaggac gcgatggcat cgtgcagtac 60
ccgcacctct tcatcgcggc cctggcgctg gccctggtcc tcatggaccc cttccacctc 120
ggcccgctgg ccgggatcga ctaccggccg gtgaagcacg agctggcgcc gtacagggag 180
gtcatgcagc gctggccgag ggacaacggc agccgcctca ggctcggcag gctcgagttc 240
gtcaacgagg tgttcgggcc agagtccatc gagttcgacc gccagggccg cgggccctac 300
gccgggctcg ccgacggccg cgtcgtgcgg tggatggggg acaaggccgg gtgggagacg 360
ttcgccgtca tgaatcctga ctggtattgg cttactgcag aaaaaccata gcttacctgt 420
gtgtgtgcaa actaaaatag ttttttcgga aaaaaaaagg tcggagaaag tttgtgctaa 480
cggagtggag tcgacgacga agaagcagca cgggaaggag aagtggtgcg gccggcctct 540
cgggctgagg ttccacaggg agaccggcga gctcttcatc gccgacgcgt actatgggct 600
catggccgtt ggcgaaagcg gcggcgtggc gacctccctg gcgagggagg ccggcgggga 660
cccggtccac ttcgccaacg acctcgacat ccacatgaac ggctcgatat tcttcaccga 720
cacgagcacg agatacagca gaaagtgagc ggagtactgc tgccgatctc ctttttctgt 780
tcttgagatt tgtgtttgac aaatgactga tcatgcaggg accatttgaa cattttgctg 840
gaaggagaag gcacggggag gctgctgaga tatgaccgag aaaccggtgc cgttcatgtc 900
gtgctcaacg ggctggtctt cccaaacggc gtgcagatct cacaggacca gcaatttctc 960
ctcttctccg agacaacaaa ctgcaggtga gataaactca ggttttcagt atgatccggc 1020
tcgagagatc caggaactga tgacgccttt attaatcggc tcatgcatgc acactaggat 1080
catgaggtac tggctggaag gtccaagagc gggccaggtg gaggtgttcg cgaacctgcc 1140
ggggttcccc gacaacgtgc gcttgaacag caaggggcag ttctgggtgg cgatcgactg 1200
ctgccggacg ccgacgcagg aggtgttcgc gcggtggccg tggctgcgga ccgcctactt 1260
caagatcccg gtgtcgatga agacgctggg gaagatggtg agcatgaaga tgtacacgct 1320
tctcgcgctc ctcgacggcg aggggaacgt ggtcgaggta ctcgaggacc ggggcggcga 1380
ggtgatgaag ctggtgagcg aggtgaggga ggtggaccgg aggctgtgga tcgggaccgt 1440
tgcgcacaac cacatcgcca cgatccctta tccgttggac tag 1483
<210> 35
<211> 413
<212> БЕЛОК
<213> Triticum aestivum
<400> 35
Met Glu Glu Lys Lys Pro Arg Arg Gln Gly Ala Ala Gly Arg Asp Gly
1. 5 10 15
Ile Val Gln Tyr Pro His Leu Phe Ile Ala Ala Leu Ala Leu Ala Leu
20 25 30
Val Leu Met Asp Pro Phe His Leu Gly Pro Leu Ala Gly Ile Asp Tyr
35 40 45
Arg Pro Val Lys His Glu Leu Ala Pro Tyr Arg Glu Val Met Gln Arg
50 55 60
Trp Pro Arg Asp Asn Gly Ser Arg Leu Arg Leu Gly Arg Leu Glu Phe
65 70 75 80
Val Asn Glu Val Phe Gly Pro Glu Ser Ile Glu Phe Asp Arg Gln Gly
85 90 95
Arg Gly Pro Tyr Ala Gly Leu Ala Asp Gly Arg Val Val Arg Trp Met
100 105 110
Gly Asp Lys Ala Gly Trp Glu Thr Phe Ala Val Met Asn Pro Asp Trp
115 120 125
Ser Glu Lys Val Cys Ala Asn Gly Val Glu Ser Thr Thr Lys Lys Gln
130 135 140
His Gly Lys Glu Lys Trp Cys Gly Arg Pro Leu Gly Leu Arg Phe His
145 150 155 160
Arg Glu Thr Gly Glu Leu Phe Ile Ala Asp Ala Tyr Tyr Gly Leu Met
165 170 175
Ala Val Gly Glu Ser Gly Gly Val Ala Thr Ser Leu Ala Arg Glu Ala
180 185 190
Gly Gly Asp Pro Val His Phe Ala Asn Asp Leu Asp Ile His Met Asn
195 200 205
Gly Ser Ile Phe Phe Thr Asp Thr Ser Thr Arg Tyr Ser Arg Lys Asp
210 215 220
His Leu Asn Ile Leu Leu Glu Gly Glu Gly Thr Gly Arg Leu Leu Arg
225 230 235 240
Tyr Asp Arg Glu Thr Gly Ala Val His Val Val Leu Asn Gly Leu Val
245 250 255
Phe Pro Asn Gly Val Gln Ile Ser Gln Asp Gln Gln Phe Leu Leu Phe
260 265 270
Ser Glu Thr Thr Asn Cys Arg Ile Met Arg Tyr Trp Leu Glu Gly Pro
275 280 285
Arg Ala Gly Gln Val Glu Val Phe Ala Asn Leu Pro Gly Phe Pro Asp
290 295 300
Asn Val Arg Leu Asn Ser Lys Gly Gln Phe Trp Val Ala Ile Asp Cys
305 310 315 320
Cys Arg Thr Pro Thr Gln Glu Val Phe Ala Arg Trp Pro Trp Leu Arg
325 330 335
Thr Ala Tyr Phe Lys Ile Pro Val Ser Met Lys Thr Leu Gly Lys Met
340 345 350
Val Ser Met Lys Met Tyr Thr Leu Leu Ala Leu Leu Asp Gly Glu Gly
355 360 365
Asn Val Val Glu Val Leu Glu Asp Arg Gly Gly Glu Val Met Lys Leu
370 375 380
Val Ser Glu Val Arg Glu Val Asp Arg Arg Leu Trp Ile Gly Thr Val
385 390 395 400
Ala His Asn His Ile Ala Thr Ile Pro Tyr Pro Leu Asp
405 410
<210> 36
<211> 1481
<212> ДНК
<213> Triticum aestivum
<400> 36
atggaagaga agaagccgcg gcggcaggga gccgcagtac gcgatggcat cgtgcagtac 60
ccgcacctct tcatcgcggc cctggcgctg gccctggtcg tcatggaccc cttccacctc 120
ggcccgctgg ctgggatcga ctaccggccg gtgaagcacg agctggcgcc atacagggag 180
gtcatgcagc gctggccgag ggacaacggc agccgcctca ggctcggcag gctcgagttc 240
gtcaacgagg tgttcgggcc ggagtccatc gagttcgaca gccagggccg cgggccctac 300
gccgggctcg ccgacggccg cgtcgtgcgg tggatggggg acaagaccgg gtgggagacg 360
ttcgccgtca tgaatcctga ctggtaattg gcttactgca gataaatcca tagcttacct 420
gtgtgtttgc aaactaaaat gatttcttgg gaaaaaaaaa ggtcggagaa agtttgtgct 480
aacggagtgg agtcaacgac gaagaagcag cacgggaagg agaagtggtg cggccggcct 540
ctcgggctga ggttccacag ggagaccggc gagctcttca tcgccgacgc gtactatggg 600
ctcatggccg tcggcgaaag cggcggcgtg gcgacctccc tggcaaggga ggtcggcggg 660
gacccggtcc acttcgccaa cgacctcgac atccacatga acggctcgat attcttcacc 720
gacacgagca cgagatacag cagaaagtga gcggagtact gtcgctgatc tccatttttg 780
ttcttgagat gttgtgtttg agtgtctgac accatgactg atcatgcagg gatcatttga 840
acattttgct agaaggagaa ggcacgggga ggctgctgag atatgaccga gaaaccggtg 900
ccgttcatgt cgtgctcaac gggctggtct tcccaaacgg cgtgcagatt tcacaggacc 960
agcaatttct cctcttctcc gagacaacca actgcaggtg agataaactc aggttttcag 1020
tatgatccgg ctcgagagat ccaggaactg atgacggatc atgcatgcac gctaggatca 1080
tgaggtactg gctggaaggt ccaagagcgg gccaggtgga ggtgttcgcg aacctgccgg 1140
ggttccccga caacgtgcgc ctgaacagca aggggcagtt ctgggtggcg atcgactgct 1200
gccggacgcc gacgcaggag gtgttcgcga ggtggccgtg gctgcggacc gcctacttca 1260
agatcccggt gtcgatgaag acgctgggga agatggtgag catgaagatg tacacgcttc 1320
tcgcgctcct cgacggcgag gggaacgtgg tggaggtgct cgaggaccgg ggcggcgagg 1380
tgatgaagct ggtgagcgag gtgagggagg tggaccggag gctgtggatc gggaccgttg 1440
cgcacaacca catcgccacg atcccttacc cgctggacta g 1481
<210> 37
<211> 413
<212> БЕЛОК
<213> Triticum aestivum
<400> 37
Met Glu Glu Lys Lys Pro Arg Arg Gln Gly Ala Ala Val Arg Asp Gly
1. 5 10 15
Ile Val Gln Tyr Pro His Leu Phe Ile Ala Ala Leu Ala Leu Ala Leu
20 25 30
Val Val Met Asp Pro Phe His Leu Gly Pro Leu Ala Gly Ile Asp Tyr
35 40 45
Arg Pro Val Lys His Glu Leu Ala Pro Tyr Arg Glu Val Met Gln Arg
50 55 60
Trp Pro Arg Asp Asn Gly Ser Arg Leu Arg Leu Gly Arg Leu Glu Phe
65 70 75 80
Val Asn Glu Val Phe Gly Pro Glu Ser Ile Glu Phe Asp Ser Gln Gly
85 90 95
Arg Gly Pro Tyr Ala Gly Leu Ala Asp Gly Arg Val Val Arg Trp Met
100 105 110
Gly Asp Lys Thr Gly Trp Glu Thr Phe Ala Val Met Asn Pro Asp Trp
115 120 125
Ser Glu Lys Val Cys Ala Asn Gly Val Glu Ser Thr Thr Lys Lys Gln
130 135 140
His Gly Lys Glu Lys Trp Cys Gly Arg Pro Leu Gly Leu Arg Phe His
145 150 155 160
Arg Glu Thr Gly Glu Leu Phe Ile Ala Asp Ala Tyr Tyr Gly Leu Met
165 170 175
Ala Val Gly Glu Ser Gly Gly Val Ala Thr Ser Leu Ala Arg Glu Val
180 185 190
Gly Gly Asp Pro Val His Phe Ala Asn Asp Leu Asp Ile His Met Asn
195 200 205
Gly Ser Ile Phe Phe Thr Asp Thr Ser Thr Arg Tyr Ser Arg Lys Asp
210 215 220
His Leu Asn Ile Leu Leu Glu Gly Glu Gly Thr Gly Arg Leu Leu Arg
225 230 235 240
Tyr Asp Arg Glu Thr Gly Ala Val His Val Val Leu Asn Gly Leu Val
245 250 255
Phe Pro Asn Gly Val Gln Ile Ser Gln Asp Gln Gln Phe Leu Leu Phe
260 265 270
Ser Glu Thr Thr Asn Cys Arg Ile Met Arg Tyr Trp Leu Glu Gly Pro
275 280 285
Arg Ala Gly Gln Val Glu Val Phe Ala Asn Leu Pro Gly Phe Pro Asp
290 295 300
Asn Val Arg Leu Asn Ser Lys Gly Gln Phe Trp Val Ala Ile Asp Cys
305 310 315 320
Cys Arg Thr Pro Thr Gln Glu Val Phe Ala Arg Trp Pro Trp Leu Arg
325 330 335
Thr Ala Tyr Phe Lys Ile Pro Val Ser Met Lys Thr Leu Gly Lys Met
340 345 350
Val Ser Met Lys Met Tyr Thr Leu Leu Ala Leu Leu Asp Gly Glu Gly
355 360 365
Asn Val Val Glu Val Leu Glu Asp Arg Gly Gly Glu Val Met Lys Leu
370 375 380
Val Ser Glu Val Arg Glu Val Asp Arg Arg Leu Trp Ile Gly Thr Val
385 390 395 400
Ala His Asn His Ile Ala Thr Ile Pro Tyr Pro Leu Asp
405 410
<210> 38
<211> 1471
<212> ДНК
<213> Triticum aestivum
<400> 38
atggaagaga agaaaccgcg gcggcaggga gccgcagtac gcgatggcat cgtgcagtac 60
ccgcacctct tcatcgcggc cctggcgctg gccctggtcc tcatggaccc gttccacctc 120
ggcccgctgg ccgggatcga ctaccgaccg gtgaagcacg agctggcgcc gtacagggag 180
gtcatgcagc gctggccgag ggacaacggc agccgcctca ggctcggcag gctcgagttc 240
gtcaacgagg tgttcgggcc ggagtccatc gagttcgacc gccagggccg cgggccttac 300
gccgggctcg ccgacggccg cgtcgtgcgg tggatggggg acaaggccgg gtgggagacg 360
ttcgccgtca tgaatcctga ctggtactgg cttactgcag aaaaacccat agcttacctg 420
tgtgtgtgca gactaaaata gtttctttca taaaaaaaag gtcggagaaa gtttgtgcta 480
acggagtgga gtcgacgacg aagaagcagc acgggaagga gaagtggtgc ggccggcctc 540
tcggcctgag gttccacagg gagaccggcg agctcttcat cgccgacgcg tactatgggc 600
tcatggccgt cggcgaaagg ggcggcgtgg cgacctccct ggcgagggag gccggcgggg 660
acccggtcca cttcgccaac gaccttgaca tccacatgaa cggctcgata ttcttcaccg 720
acacgagcac gagatacagc agaaagtgag cggagtactg ctgccgatct cctttttctg 780
ttcttgagat ttgtgtttga caaatgactg atcatgcagg gaccatttga acattttgct 840
ggaaggagaa ggcacgggga ggctgctgag atatgaccga gaaaccggtg ccgttcatgt 900
cgtgctcaac gggctggtct tcccaaacgg cgtgcagata tcacaggacc agcaatttct 960
cctcttctcc gagacaacaa actgcaggtg agataaactc aggttttcag tatgatccgg 1020
ctcgagagat ccaggaactg atgacggctc atgcatgcac actaggatca tgaggtactg 1080
gctggaaggt ccaagagcgg gccaggtgga ggtgttcgcg aacctgccgg ggttccccga 1140
caatgtgcgc ctgaacagca aggggcagtt ctgggtggcc atcgactgct gccgtacgcc 1200
gacgcaggag gtgttcgcgc ggtggccgtg gctgcggacc gcctacttca agatcccggt 1260
gtcgatgaag acgctgggga agatggtgag catgaagatg tacacgcttc tcgcgctcct 1320
cgacggcgag gggaacgtcg tggaggtgct cgaggaccgg ggcggcgagg tgatgaagct 1380
ggtgagcgag gtgagggagg tggaccggag gctgtggatc gggaccgttg cgcacaacca 1440
catcgccacg atcccttacc cgctggacta g 1471
<210> 39
<211> 413
<212> БЕЛОК
<213> Triticum aestivum
<400> 39
Met Glu Glu Lys Lys Pro Arg Arg Gln Gly Ala Ala Val Arg Asp Gly
1. 5 10 15
Ile Val Gln Tyr Pro His Leu Phe Ile Ala Ala Leu Ala Leu Ala Leu
20 25 30
Val Leu Met Asp Pro Phe His Leu Gly Pro Leu Ala Gly Ile Asp Tyr
35 40 45
Arg Pro Val Lys His Glu Leu Ala Pro Tyr Arg Glu Val Met Gln Arg
50 55 60
Trp Pro Arg Asp Asn Gly Ser Arg Leu Arg Leu Gly Arg Leu Glu Phe
65 70 75 80
Val Asn Glu Val Phe Gly Pro Glu Ser Ile Glu Phe Asp Arg Gln Gly
85 90 95
Arg Gly Pro Tyr Ala Gly Leu Ala Asp Gly Arg Val Val Arg Trp Met
100 105 110
Gly Asp Lys Ala Gly Trp Glu Thr Phe Ala Val Met Asn Pro Asp Trp
115 120 125
Ser Glu Lys Val Cys Ala Asn Gly Val Glu Ser Thr Thr Lys Lys Gln
130 135 140
His Gly Lys Glu Lys Trp Cys Gly Arg Pro Leu Gly Leu Arg Phe His
145 150 155 160
Arg Glu Thr Gly Glu Leu Phe Ile Ala Asp Ala Tyr Tyr Gly Leu Met
165 170 175
Ala Val Gly Glu Arg Gly Gly Val Ala Thr Ser Leu Ala Arg Glu Ala
180 185 190
Gly Gly Asp Pro Val His Phe Ala Asn Asp Leu Asp Ile His Met Asn
195 200 205
Gly Ser Ile Phe Phe Thr Asp Thr Ser Thr Arg Tyr Ser Arg Lys Asp
210 215 220
His Leu Asn Ile Leu Leu Glu Gly Glu Gly Thr Gly Arg Leu Leu Arg
225 230 235 240
Tyr Asp Arg Glu Thr Gly Ala Val His Val Val Leu Asn Gly Leu Val
245 250 255
Phe Pro Asn Gly Val Gln Ile Ser Gln Asp Gln Gln Phe Leu Leu Phe
260 265 270
Ser Glu Thr Thr Asn Cys Arg Ile Met Arg Tyr Trp Leu Glu Gly Pro
275 280 285
Arg Ala Gly Gln Val Glu Val Phe Ala Asn Leu Pro Gly Phe Pro Asp
290 295 300
Asn Val Arg Leu Asn Ser Lys Gly Gln Phe Trp Val Ala Ile Asp Cys
305 310 315 320
Cys Arg Thr Pro Thr Gln Glu Val Phe Ala Arg Trp Pro Trp Leu Arg
325 330 335
Thr Ala Tyr Phe Lys Ile Pro Val Ser Met Lys Thr Leu Gly Lys Met
340 345 350
Val Ser Met Lys Met Tyr Thr Leu Leu Ala Leu Leu Asp Gly Glu Gly
355 360 365
Asn Val Val Glu Val Leu Glu Asp Arg Gly Gly Glu Val Met Lys Leu
370 375 380
Val Ser Glu Val Arg Glu Val Asp Arg Arg Leu Trp Ile Gly Thr Val
385 390 395 400
Ala His Asn His Ile Ala Thr Ile Pro Tyr Pro Leu Asp
405 410
<210> 40
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> синтезировано
<400> 40
ggaggtacac caactacctg 20
<210> 41
<211> 568
<212> БЕЛОК
<213> Thinopyrum ponticum
<400> 41
Met Arg Glu Ile Ala Thr Gln Arg Cys Gly Asn Arg Ser Met Ala Leu
1. 5 10 15
Ser Ala Pro Pro Ser Gln Glu Gln Pro Ser Gly Lys Gln Phe Gly Tyr
20 25 30
Gln Leu Ala Ala Ala Val Arg Ser Ile Asn Trp Thr Tyr Gly Ile Phe
35 40 45
Trp Ser Ile Ser Ala Ser Pro Arg Pro Gly His Ser Ser Val Leu Ala
50 55 60
Trp Lys Asp Gly Phe Tyr Asn Gly Glu Ile Lys Thr Arg Lys Ile Thr
65 70 75 80
Gly Ser Thr Thr Thr Glu Leu Thr Ala Asp Glu Arg Val Met His Arg
85 90 95
Ser Lys Gln Leu Arg Glu Leu Tyr Glu Ser Leu Leu Pro Gly Asn Ser
100 105 110
Asn Asn Arg Ala Arg Arg Pro Thr Ala Ser Leu Ser Pro Glu Asp Leu
115 120 125
Gly Asp Gly Glu Trp Tyr Tyr Thr Ile Ser Met Thr Tyr Thr Phe His
130 135 140
Pro Asn Gln Gly Leu Pro Gly Lys Ser Phe Ala Ser Asn Gln His Val
145 150 155 160
Trp Leu Tyr Asn Ala Gln Tyr Ala Asn Thr Arg Val Phe Pro Arg Ala
165 170 175
Leu Leu Ala Lys Thr Ile Val Cys Ile Pro Phe Met Gly Gly Val Leu
180 185 190
Glu Leu Gly Thr Ser Asp Gln Val Leu Glu Asp Pro Ser Met Val Lys
195 200 205
Arg Ile Ser Thr Ser Phe Trp Glu Leu His Leu Pro Ser Ser Leu Glu
210 215 220
Ser Lys Asp Pro Ser Ser Ser Thr Ser Ala Asn Asp Thr Arg Glu Ala
225 230 235 240
Thr Asp Ile Ile Leu Phe Glu Asp Phe Asp His Asn Asp Thr Val Glu
245 250 255
Gly Val Ile Ser Glu Gln Arg Glu Val Gln Cys Pro Ser Asn Val Asn
260 265 270
Leu Glu Arg Leu Thr Lys Gln Met Asp Glu Phe His Ser Leu Leu Gly
275 280 285
Gly Leu Asp Val His Pro Leu Glu Asp Arg Trp Ile Met Asp Glu Pro
290 295 300
Phe Glu Phe Thr Phe Ser Pro Glu Val Ala Pro Ala Met Asp Met Pro
305 310 315 320
Ser Thr Asp Asp Val Ile Val Thr Leu Ser Arg Ser Glu Gly Ser Arg
325 330 335
Pro Ser Cys Phe Thr Ala Trp Lys Gly Ser Ser Glu Ser Lys Tyr Val
340 345 350
Ala Gly Gln Val Val Gly Glu Ser Gln Lys Leu Leu Asn Lys Val Val
355 360 365
Ala Gly Gly Ala Trp Ala Ser Asn Tyr Gly Gly Arg Thr Met Val Arg
370 375 380
Ala Gln Gly Ile Asn Ser Asn Thr His Val Met Thr Glu Arg Arg Arg
385 390 395 400
Arg Glu Lys Leu Asn Glu Met Phe Leu Val Leu Lys Ser Leu Val Pro
405 410 415
Ser Ile His Lys Val Asp Lys Ala Ser Ile Leu Thr Glu Thr Ile Gly
420 425 430
Tyr Leu Arg Glu Leu Lys Gln Arg Val Asp Gln Leu Glu Ser Ser Arg
435 440 445
Ser Pro Ser His Pro Lys Glu Thr Thr Gly Pro Ser Arg Ser His Val
450 455 460
Val Gly Ala Arg Lys Lys Ile Val Ser Ala Gly Ser Lys Arg Lys Ala
465 470 475 480
Pro Gly Leu Glu Ser Pro Ser Asn Val Val Asn Val Thr Met Leu Asp
485 490 495
Lys Val Val Leu Leu Glu Val Gln Cys Pro Trp Lys Glu Leu Leu Met
500 505 510
Thr Gln Val Phe Asp Ala Ile Lys Ser Leu Cys Leu Asp Val Val Ser
515 520 525
Val Gln Ala Ser Thr Ser Gly Gly Arg Leu Asp Leu Lys Ile Arg Ala
530 535 540
Asn Gln Gln Leu Ala Val Gly Ser Ala Met Val Ala Pro Gly Ala Ile
545 550 555 560
Thr Glu Thr Leu Gln Lys Ala Ile
565
<210> 42
<211> 573
<212> БЕЛОК
<213> Thinopyrum ponticum
<400> 42
Met Arg Glu Ile Ala Thr Gln Arg Cys Gly Asn Arg Ser Met Ala Leu
1. 5 10 15
Ser Ala Pro Pro Ser Gln Glu Gln Pro Ser Gly Lys Gln Phe Gly Tyr
20 25 30
Gln Leu Ala Ala Ala Val Arg Ser Ile Asn Trp Thr Tyr Gly Ile Phe
35 40 45
Trp Ser Ile Ser Ala Ser Pro Arg Pro Gly His Ser Ser Val Leu Ala
50 55 60
Trp Lys Asp Gly Phe Tyr Asn Gly Glu Ile Lys Thr Arg Lys Ile Thr
65 70 75 80
Gly Ser Thr Thr Thr Glu Leu Thr Ala Asp Glu Arg Val Met His Arg
85 90 95
Ser Lys Gln Leu Arg Glu Leu Tyr Glu Ser Leu Leu Pro Gly Asn Ser
100 105 110
Asn Asn Arg Ala Arg Arg Pro Thr Ala Ser Leu Ser Pro Glu Asp Leu
115 120 125
Gly Asp Gly Glu Trp Tyr Tyr Thr Ile Ser Met Thr Tyr Thr Phe His
130 135 140
Pro Asn Gln Gly Leu Pro Gly Lys Ser Phe Ala Ser Asn Gln His Val
145 150 155 160
Trp Leu Tyr Asn Ala Gln Tyr Ala Asn Thr Arg Val Phe Pro Arg Ala
165 170 175
Leu Leu Ala Lys Thr Ala Ser Ile Gln Thr Ile Val Cys Ile Pro Phe
180 185 190
Met Gly Gly Val Leu Glu Leu Gly Thr Ser Asp Gln Val Leu Glu Asp
195 200 205
Pro Ser Met Val Lys Arg Ile Asn Thr Ser Phe Trp Glu Leu His Leu
210 215 220
Pro Ser Ser Leu Glu Ser Lys Asp Pro Ser Ser Ser Thr Ser Ala Asn
225 230 235 240
Asp Thr Arg Glu Ala Thr Asp Ile Ile Leu Phe Glu Asp Phe Asp His
245 250 255
Asn Asp Thr Val Glu Gly Val Ile Ser Glu Gln Arg Glu Val Gln Cys
260 265 270
Thr Ser Asn Val Asn Leu Glu Arg Leu Thr Lys Gln Met Asp Glu Phe
275 280 285
His Ser Leu Leu Gly Gly Leu Asp Val His Pro Leu Lys Asp Arg Trp
290 295 300
Ile Met Asp Glu Pro Phe Glu Phe Thr Phe Ser Pro Glu Val Ala Pro
305 310 315 320
Ala Met Asp Met Pro Ser Thr Asp Asp Val Ile Val Thr Leu Ser Arg
325 330 335
Ser Glu Gly Ser Arg Pro Ser Cys Phe Thr Ala Trp Lys Gly Ser Ser
340 345 350
Glu Ser Lys Tyr Val Ala Gly Gln Val Val Gly Glu Ser Gln Lys Leu
355 360 365
Leu Asn Lys Val Val Ala Gly Gly Ala Trp Ala Ser Asn Tyr Gly Gly
370 375 380
Arg Thr Met Val Arg Ala Gln Gly Ile Asn Ser Asn Thr His Val Met
385 390 395 400
Thr Glu Arg Arg Arg Arg Glu Lys Leu Asn Glu Met Phe Leu Val Leu
405 410 415
Lys Ser Leu Val Pro Ser Ile His Lys Val Asp Lys Ala Ser Ile Leu
420 425 430
Thr Glu Thr Ile Gly Tyr Leu Arg Glu Leu Lys Gln Arg Val Asp Gln
435 440 445
Leu Glu Ser Ser Arg Ser Pro Ser His Pro Lys Glu Thr Thr Gly Pro
450 455 460
Ser Arg Ser His Val Val Gly Ala Arg Lys Lys Ile Val Ser Ala Gly
465 470 475 480
Ser Lys Arg Lys Ala Pro Gly Leu Glu Ser Pro Ser Asn Val Val Asn
485 490 495
Val Thr Met Leu Asp Lys Val Val Leu Leu Glu Val Gln Cys Pro Trp
500 505 510
Lys Glu Leu Leu Met Thr Gln Val Phe Asp Ala Ile Lys Ser Leu Cys
515 520 525
Leu Asp Val Val Ser Val Gln Ala Ser Thr Ser Gly Gly Arg Leu Asp
530 535 540
Leu Lys Ile Arg Ala Asn Gln Gln Leu Ala Val Gly Ser Ala Met Val
545 550 555 560
Ala Pro Gly Ala Ile Thr Glu Thr Leu Gln Lys Ala Ile
565 570
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СВЯЗАННЫЙ С ФЕРТИЛЬНОСТЬЮ ГЕН ТаМS7 ПШЕНИЦЫ И СПОСОБ ЕГО ПРИМЕНЕНИЯ | 2017 |
|
RU2763468C2 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ РЕДУЦИРУЮЩИХ САХАРОВ В РАСТЕНИИ | 2019 |
|
RU2801948C2 |
| СНИЖЕНИЕ ПРЕВРАЩЕНИЯ НИКОТИНА В НОРНИКОТИН В РАСТЕНИЯХ | 2015 |
|
RU2733837C2 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ САХАРОВ И АМИНОКИСЛОТ В РАСТЕНИИ (SULTR3) | 2020 |
|
RU2826107C1 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2015 |
|
RU2741833C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2015 |
|
RU2745322C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ПРОМЫШЛЕННЫХ ПРОДУКТОВ ИЗ РАСТИТЕЛЬНЫХ ЛИПИДОВ | 2015 |
|
RU2743384C2 |
| РАСТЕНИЯ С МОДИФИЦИРОВАННЫМИ ПРИЗНАКАМИ | 2017 |
|
RU2809117C2 |
| ГЕНЫ ПРОТЕАЗ ТАБАКА | 2015 |
|
RU2756102C2 |
| СПОСОБЫ УСОВЕРШЕНСТВОВАНИЯ ОДНОДОЛЬНЫХ РАСТЕНИЙ | 2015 |
|
RU2727428C2 |
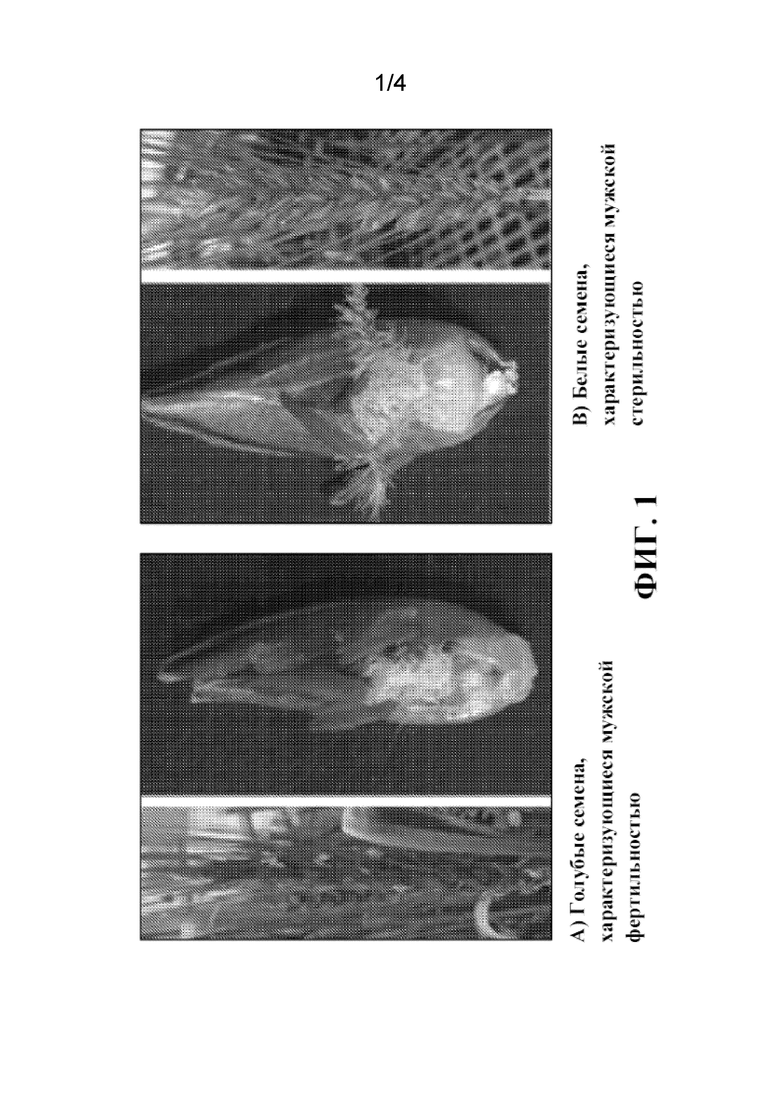
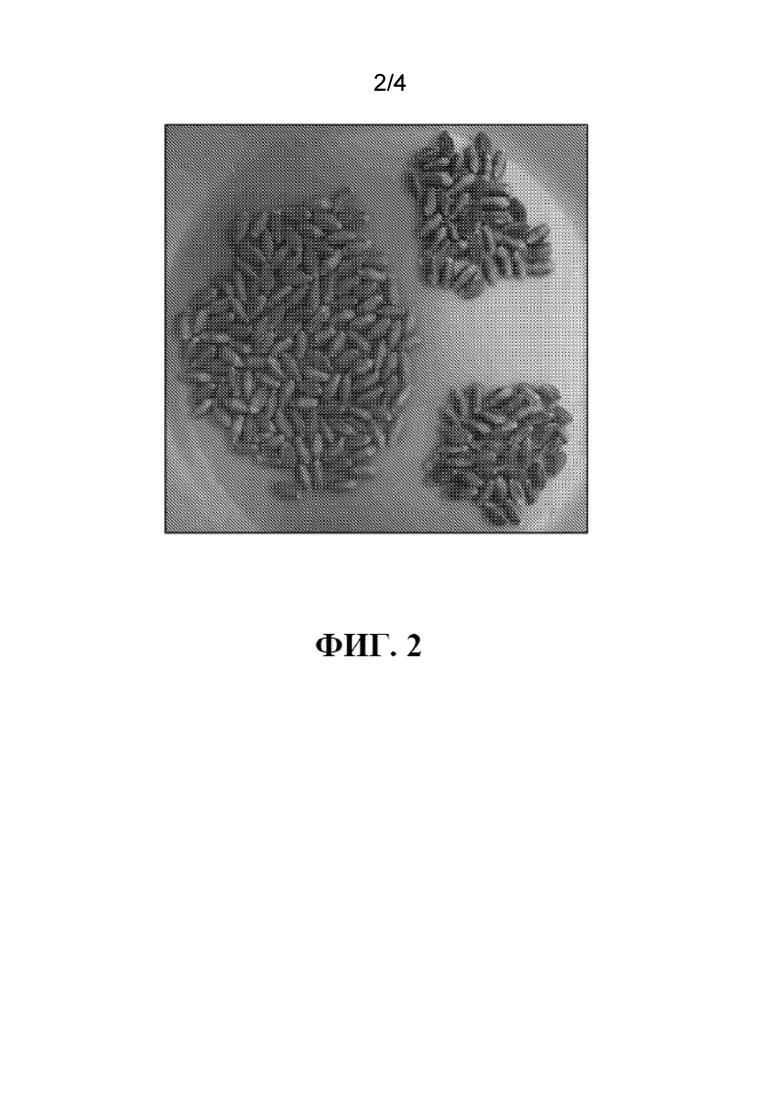
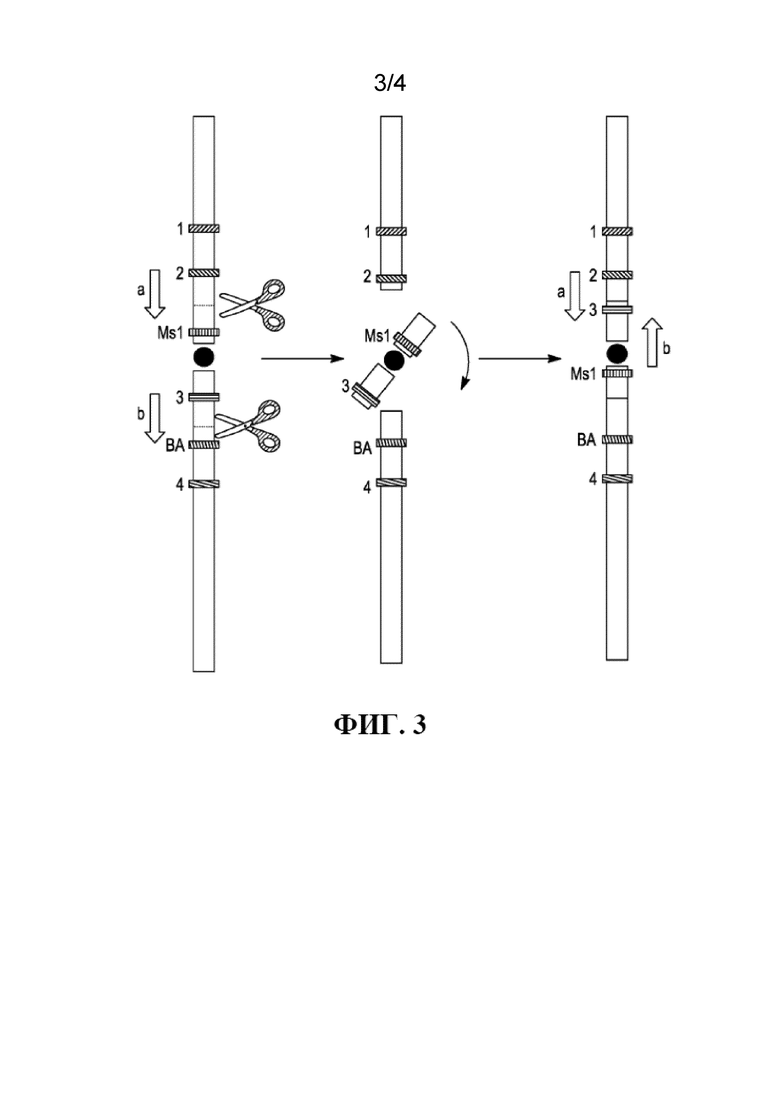
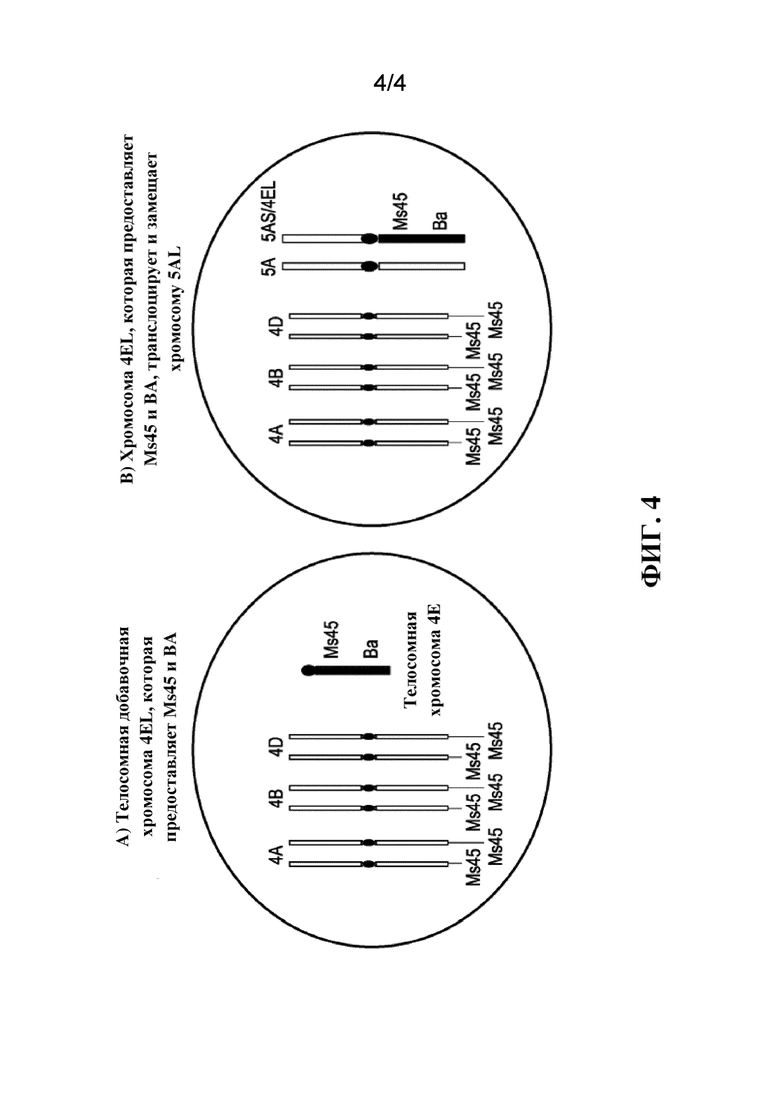
Изобретение относится к области биохимии, в частности к способу восстановления мужской фертильности у растения пшеницы с мужской стерильностью. Изобретение позволяет эффективно восстанавливать мужскую фертильность у растения пшеницы. 9 з.п. ф-лы, 4 ил., 2 табл., 2 пр.
1. Способ восстановления мужской фертильности у растения пшеницы с мужской стерильностью, опосредованной Ms45, включающий:
(a) введение в растение пшеницы с мужской стерильностью, опосредованной Ms45, включающего тройные гомозиготные мутации в полинуклеотиде Ms45, обеспечивающем мужскую фертильность, которые вызывают фенотип мужской стерильности, донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, содержащего хромосомный компонент 4E из Thinopyrum, Agropyron, Aegilops, Secale, Haynaldia или Elymus, или хромосомный компонент 4H из Hordeum, где хромосомный компонент 4E или хромосомный компонент 4H включает:
полинуклеотид растения, который обеспечивает фенотип семени растения, сцепленный с локусом, обеспечивающим восстановление мужской фертильности, опосредованной Ms45; и
(b) восстановление мужской фертильности, опосредованной Ms45, у растения пшеницы с помощью хромосомного компонента 4E или 4H, где экспрессия хромосомного компонента 4E или 4H функционально комплементирует фенотип мужской стерильности по отношению к тройной мутации Ms45 в растении пшеницы с мужской стерильностью, опосредованной Ms45, вследствие чего растение пшеницы с мужской стерильностью, опосредованной Ms45, приобретает мужскую фертильность.
2. Способ по п.1, где экспрессия локуса, обеспечивающего восстановление мужской фертильности, функционально комплементирует фенотип мужской стерильности, опосредованной Ms45, по отношению к тройным мутациям Ms45 в растении пшеницы с мужской стерильностью, опосредованной Ms45.
3. Способ по п.1, где полинуклеотид растения, который обеспечивает фенотип семени растения, расположен на том же хромосомном плече донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, что и локус, обеспечивающий восстановление мужской фертильности, опосредованной Ms45, и они не разделены центромерой.
4. Способ по п.1, предусматривающий введение донорного хромосомного компонента растения путем скрещивания растения пшеницы с мужской стерильностью, опосредованной Ms45, с растением с мужской фертильностью, содержащим донорный хромосомный компонент растения, где донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, происходит из вида, который отличается от вида пшеницы, к которому относится растение пшеницы с мужской стерильностью, опосредованной Ms45, или растение с мужской фертильностью, или оба из них.
5. Способ по п.1, предусматривающий введение донорного хромосомного компонента, обеспечивающего восстановление фертильности растения, путем скрещивания растения пшеницы с мужской стерильностью, опосредованной Ms45, с растением с мужской фертильностью, содержащим донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, где донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, происходит из растения, отличного от растения пшеницы.
6. Способ по п.4, где донорный хромосомный компонент, обеспечивающий восстановление фертильности растения, восстанавливает фертильность у растения с мужской стерильностью, опосредованной Ms45, без рекомбинации с хромосомами пшеницы.
7. Способ по п.4, где растение с мужской фертильностью представляет собой растение пшеницы, ячменя, овса, пырея или ржи или родственного им вида.
8. Способ по п.1, где растением с мужской фертильностью является Sebesta Blue, Blue Sando, Blue Baart, Blue Onas, Blue 1, PBB или Blue Norco, и где донорный хромосомный компонент происходит из вида растения, отличного от вида пшеницы.
9. Способ по п.1, где фенотип семени растения представляет собой маркер, относящийся к цвету, физиологии или морфологии семени.
10. Способ по п.9, где цвет представлен голубым алейроном, геном P, антоцианом или Kala 4.
| US 20180010145, 11.01.2018 | |||
| ГЕН-ВОССТАНОВИТЕЛЬ RF4 ДЛЯ ЦИТОПЛАЗМАТИЧЕСКОЙ МУЖСКОЙ СТЕРИЛЬНОСТИ (CMS) C-ТИПА КУКУРУЗЫ, МОЛЕКУЛЯРНЫЕ МАРКЕРЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2603005C2 |
| WO 2014039815 A9, 13.03.2014. | |||

