Настоящее изобретение относится к растению красного салата латука, к кочанному салату, который можно получать из него, и к комплексу генов, который отвечает за красную окраску салата латука.
Сменяются образы жизни и продолжают возникать запросы со стороны ресторанов и предприятий общественного питания о красочном и интересном гарнире и даже от домохозяек о готовых к употреблению салатах. Как результат, компании, занимающиеся выращиванием, находятся в поиске сортов с резко выделяющейся окраской, лучшими вкусовыми свойствами и широким диапазоном текстуры. Рынок салата латука можно подразделить на три группы, а именно цельные кочаны, предварительно нарезанные цельные кочаны и листья проростков.
В настоящее время привлекательный красный цвет в предварительно нарезанных салатных смесях латука часто обеспечивают за счет доступных в настоящее время «красного» салата латука (Latuca sativa), красной разновидности цикория (Cichorium intybus), краснокочанной капусты или листовой свеклы. Не все листья известного до настоящего времени «красного» салата латука являются красными. В частности, разновидности, образующие кочаны, или имеют красную окраску вдоль краев листа, или испещрены красными пятнами. Они никогда не бывают красными в центральной части кочана. Это вызвано тем, что красный цвет выражен только в тех частях кочана, которые подвергаются воздействию дневного света, более конкретно УФ-излучения. Листья так называемых «красных» салатов латуков, таким образом, представляют собой по большей части зеленые и их красная окраска вряд ли может вносить вклад в впечатление красного цвета предварительно нарезанной салатной смеси.
Часто переходная стадия между красной частью и зеленой частью листа окрашена в коричневый цвет. Такая коричневая окраска считается непривлекательной с точки зрения внешнего вида. Салат латук с красными пятнами обычно ассоциируют с болезнями растений или пятнами крови. В настоящее время селекционеры проводят отбор, направленный против коричневого цвета, а также красной окраски пятнами. Дополнительно, также было обнаружено, что покрытые пятнами или коричневые листья салата латука часто выбрасывают, как при производстве упакованного салата латука, так и потребитель.
Недостаток использования листьев овощей, отличающихся от салата латука, состоит в том, что различный вкус указанных других овощей часто ощущается как нежелательный. Красная разновидность цикория представляет собой, например, Cichorium intybus, который обладает горьким вкусом. Текстура краснокочанной капусты совершенно отличается от в значительной степени более мягкой текстуры салата латука.
Синтез антоцианина в салате латуке вызывается под действием УФ-облучения (смотри, например, Voipio and Autio, 1995, Responses of red-leaved lettuce to light intensity, UV-A radiation and root zone temperature. In: Acta Horticulturae 399. Greenhouse environmental control and automation. Eds. BJ Bailey, T Takakura. Kyoto, Japan, p 183-187; Benoit, et al., 1998, Effect of a photoselective greenhouse film on a few vegetable crops in the Belgian North Sea climate. In: 14th International congress on plastics in agriculture, Tel Aviv, Israel, March 1997. Laser Pages Publishing, Jerusalem, Israel, p 81-92; Krizek et al., 1998, Inhibitory effects of ambient levels of solar UV-A and UV-B radiation on growth of cv. New Red Fire lettuce. Physiologia Plantarum 103(1), p 1-7; Kleinhenz et al., 2003, Variety, shading, and growth stage effects on pigment concentrations in lettuce grown under contrasting temperature regimens. Hortechnology 13(4), p 677-683). Развитие красной окраски у многих видов растений зависит от образования антоцианина.
Множество различных разновидностей салата латука обычно группируют в три группы. Наиболее обычным является кочанный салат латук, среди которого хрустящий кочанный салат (или айсберг) и кочанный салат. Салат-ромэн (или летний эндивий) образует рыхлые прямостоячие кочаны. Салат латук «листового» типа не образует кочанов и легко разбирается на листья. В салате латуке, образующем кочаны, центральная часть кочана более или менее закрыта и не может подвергаться действию света. Таким образом, не представляется возможным индуцировать образование антоцианина в закрытых кочанах, таких как салат айсберг (кочанный салат) и кочанный салат латук, или в салате-ромэн, который имеет рыхло сомкнутые кочаны.
Сходная проблема состоит в недостатке красного окрашивания красного салата латука, выращенного в теплицах, теплицах туннельного типа из полимерной пленки и закрытых контейнерах. Из-за недостатка УФ-излучения, которое отражается стеклом или пластиковым покрытием или которого не хватает в искусственном ассимилируемом спектре света, экспрессия антоцианина проходит в значительно меньшей степени, чем при условиях открытого грунта. Это уменьшает возможности получения красного салата латука в условиях закрытого грунта и, исходя из этого, получения красного салата латука, защищенного от неблагоприятных погодных условий, таких как холод, жара и любых типов осадков.
Настоящее изобретение в настоящее время обеспечивает красный салат латук, имеющий красные листья во всем кочане, включая центральную часть кочана. Листья центральной части кочана красного салата латука по изобретению представляют собой в значительной степени полностью красные, что означает, что они содержат исключительно высокое соотношение антоцианин/хлорофилл. Это является неожиданным, поскольку свет, более конкретно УФ-свет, который, как полагалось, необходимым для синтеза антоцианина, не способен проникнуть в центральную часть кочана. Это показывает, что в салате латуке по настоящему изобретению за синтез антоцианина отвечает независимый от света механизм.
Такая независимость от света также позволяет культивировать красный салат латук, кочанный или не образующий кочанов, под стеклом, пластиком или в закрытых контейнерах. В последнем случае используется ассимиляция искусственного света, что обычно вызывает проблемы развития красной окраски. Это связано с тем, что УФ-длины волн (280-400 нм), которые обычно необходимы для синтеза антоцианина, в большей или меньшей степени отсутствуют в спектре света.
Под стеклом или пластиком сходное отсутствие УФ-излучения обычно связано с сильным поглощением УФ-излучения покрытием. Однако, благодаря своей УФ независимой экспрессии антоцианина, красный салат латук по настоящему изобретению пригоден для культивирования в теплице, теплице с пленочным покрытием и закрытом контейнере. Это связано с независящим от света механизмом по настоящему изобретению, который также будет обозначаться как «УФ независимая экспрессия антоцианина» и «УФ независимая красная окраска».
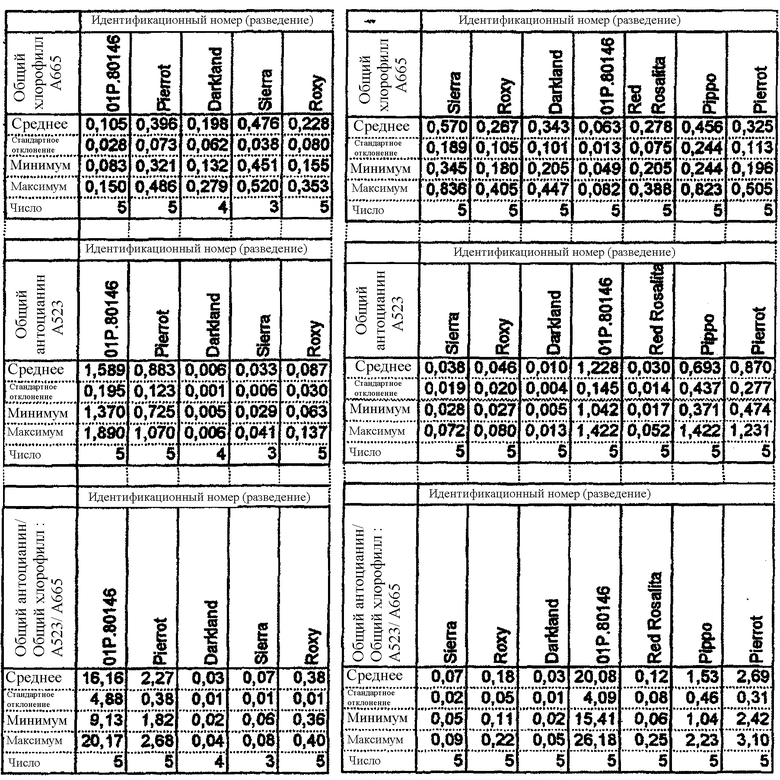
В конкретном варианте осуществления изобретения соотношение коэффициентов поглощения А523/А665 антоцианина и хлорофилла находятся в диапазоне между 4 и 50, предпочтительно между 9 и 27.
Не совсем понятно, является ли экспрессия антоцианина по настоящему изобретению полностью независимой от УФ-излучения или она запускается при в значительной степени меньших уровнях УФ-излучения, чем обычно требуется для экспрессии антоцианина у нормальных растений красного салата латука. Однако указанное различие не влияет на практическую применимость настоящего изобретения.
Хотя настоящее изобретение в особенности пригодно для кочанных разновидностей салата латука, независимую от света красную окраску можно использовать в других разновидностях салата латука, таких как листовой салат латук и при получении листьев проростков, что представляет собой получение молодых растений салата латука для сбора мелких незрелых листьев.
Происхождение красного салата латука по изобретению представлено в Примере 1.
Изобретение относится ко всем потомкам оригинальных родителей, имеющих красные листья в центральной части кочана и которые удовлетворяют одному или нескольким из других требований по изобретению, таким как соотношение между хлорофиллом и антоцианином в листьях центральной части.
Дополнительно, растения салата латука, не образующие кочана, их свободные листья и проростки, которые имеют независящую от УФ красную окраску по изобретению, также рассматривают в качестве потомков полностью красных образующих кочаны растений салата латука и, таким образом, как часть настоящего изобретения.
Отбор растений салата латука, которые уже представляют собой или могут привести к получению растений салата латука по изобретению, может основываться на визуальном отборе молодых растений, имеющих красную окраску (стадия 1). При выращивании растений под стеклом или пластиком можно впоследствии отличить УФ зависимую экспрессию антоцианина, т.е. слабую красную окраску, и УФ независимую экспрессию антоцианина, т.е. относительно сильную красную окраску. Эти последние растения сажают и выращивают до образования кочанов. Затем проводят визуальный выбор растений с красными листьями в центральной части кочана при помощи отрезания верхней части каждого растения. Затем растения с красными листьями в центральной части кочана отбирают для получения дочерних семян.
Гибриды выбирают для увеличения степени красной окраски и/или для получения требуемого уровня образования кочанов у взрослого растения.
Как используется здесь, растение салата латука определяют как кочанное, если оно имеет «открытый кочан» или «закрытый кочан», согласно рекомендациям Международного совета по защите новых сортов растений (International Union for the Protection of New Varieties of Plants (UPOV)) для характеристик «формирования кочана» («TG/13/9. Lettuce (Lactuca sativa L.) Guidelines for the conduct of tests for distinctness, uniformity and stability.» International Union for the Protection of New Varieties of Plants. Geneva. 2004).
Центральные части кочана салата латука по настоящему изобретению имеют соотношение коэффициентов поглощения А523/А665 антоцианина и хлорофилла, которое предпочтительно составляет более 9, более предпочтительно составляет 13 и более. В нормально красном салате латуке такое соотношение никогда не отмечалось выше чем 3. Концентрации хлорофилла и антоцианина, используемые для определения соотношения по изобретению, определяют при использовании спектрофотометрии. Приготовление образца и анализ описаны в Примере 2.
Следует отметить, что такие высокие концентрации антоцианина, как в листьях центральной части кочана по настоящему изобретению, могут быть обнаружены в других сортах салата латука, не образующих кочанов. Некоторые примеры таких не образующих кочанов сортов представляют собой 40-0203103-B (Knerr LD, 2005. Lettuce cultivar 40-0203103-B. US Patent application US 2005/0144672 Al), Галактика (Galactic), Новый Красный Пожар (New Red Fire), Ролина (Rolina) (Kleinhenz et al., 2003. Variety, shading and growth stage effects on pigment concentrations in lettuce grown under contrasting temperature regimens. Horttechnology 13(4) p.677-683), Красная салатница (Red Salad Bowl) и Сезам (Sesam) (Voipio I. and Autio J., 1995. Responses of red-leaved lettuce to light intensity, UV-A radiation and root zone temperature. Acta horticulturae 399. p.183-187).
Однако указанные сорта имеют листья в центральной части кочана с чрезвычайно высокими концентрациями хлорофилла по сравнению с листьями в центральной части кочана по настоящему изобретению, в результате сильного воздействия света, из-за того, что они имеют характеристики некочанного салата. С другой стороны, у разновидностей, образующих кочаны, могут быть обнаружены такие же низкие уровни хлорофилла в листьях центральной части кочана, как и в листьях центральной части кочана по настоящему изобретению. Однако листья таких кочанных разновидностей не демонстрируют высоких концентраций антоцианина по сравнению с листьями центральной части кочана по настоящему изобретению. Из-за низкого воздействия света на листья центральной части кочана, синтез антоцианина обычно индуцируется в малой степени. В настоящем изобретении экспрессия антоцианина представляет собой Уф независимую. Это объясняет, почему соотношение антоцианин/хлорофилл по настоящему изобретению находится на беспрецедентно высоком уровне.
Эта новая форма красной окраски, которая представляет собой УФ независимую, обнаружена и разработана в кочанных растениях салата латука и отличается от красной окраски, известной в предшествующем уровне техники. Однако изобретение также относится к не образующим кочанов растениям салата латука, таким как растения листового салата или проросткам салата латука, или растениям салата латука, которые выращивают в теплицах или под пленкой, где количество УФ понижено, которые обладают характеристиками «независимой от УФ красной окраски» по изобретению.
Красную окраску листьев центральной части кочана также оценивали на основе цветовой таблицы RHS (The Royal Horticultural Society, London, UK). Цвет листовых пластинок центральной части кочана, в особенности в верхней части листа по настоящему изобретению, оценивали как 183А, 184А или 187B, все из которых относятся к группе серовато-пурпурных. Цвет остальной части листовой пластинки оценивали как 180В, 180С, 180D или 181С, все из которых относятся к группе серовато-красных.
Также возможно оценивать красную окраску и экспрессию антоцианина в растениях, выращенных под стеклом или пленкой, т.е. при условиях пониженного уровня УФ-излучения по сравнению со светом в условиях открытого грунта. В этом случае также растения по настоящему изобретению могут показывать более высокое содержание антоцианина и более темную красную окраску по сравнению с обычным красным салатом латуком. В особенности новообразующиеся листья растений по настоящему изобретению представляют собой в значительной степени более красные, по сравнению с новообразующимися листьями обычных растений красного салата латука.
Генетический анализ красного салата латука по изобретению проводили как описано в Примере 2. Было обнаружено, что по крайней мере три гена участвуют в красном окрашивании центральной части кочана.
Настоящее изобретение относится, таким образом, к растениям салата латука, которые способны экспрессировать антоцианин и имеют по крайней мере три гена, которые участвуют в красном окрашивании центральной части кочана. Предпочтительно растения по изобретению включают полный комплекс генов, описанный в Примере 2.
Также возможно оценить наличие локусов, участвующих в красном окрашивании по настоящему изобретению при помощи сцепленных маркеров ДНК, таких как AFLP, RFLP, RAPD, SCAR, CAPS, SSR или SNP. Например, выявление генетической связи маркер-признак в описанной выше популяции и ее потомках может обеспечить такие сцепленные ДНК маркеры.
Наличие в растении салата латука сходной генетической конституции красной окраски внутренних листьев как в настоящем изобретении, т.е. оценка является ли растение салата латука растением по настоящему изобретению, может быть легко оценено при сравнении фенотипов такого потенциального растения по изобретению с фенотипом известного растения по настоящему изобретению. Фенотип можно оценить по, например, красной окраске внутренних листьев, соотношению антоцианин/хлорофилл в листьях средней части кочана и/или красной окраске растений, выращенных под стеклом или пластиком.
Другой путь оценки сходства генетической конституции между потенциальным растением по изобретению и известным растением по изобретению состоит в сравнении маркера генотипа потенциального растения с маркером генотипа известного растения по настоящему изобретению. Маркер генотипа определяется на основе набора маркеров на основе ДНК, таких как AFLP, RFLP, RAPD, SCAR, CAPS, SSR или SNP, которые тесно связаны с локусами, которые участвуют в экспрессии красного цвета по изобретению.
Другой путь оценки сходства генетической конституции состоит в сравнении генотипа потенциального растения по изобретению и генотипа известного растения по изобретению. Это сравнение генотипа проводится на F2-популяции, полученной самооплодотворением F1-растения от скрещивания потенциального растения и известного растения по изобретению. F2-популяцию можно исследовать на отсутствие расщепления фенотипа, т.е. красной окраски внутренних листьев. Во всех сравнениях фенотипы можно также оценивать, например, по соотношению антоцианин/хлорофилл в листьях средней части кочана или красной окраске растений, выращенных под стеклом или пластиком.
Семена растений красного салата латука по изобретению были депонированы в NCIMB 18 июля 2005 под входящими номерами NCIMB 41337, NCIMB 41338 и NCIMB 41339.
Изобретение также относится к потомству этих семян, а также к растениям, которые получили генетическую конституцию или генетический комплекс растений по изобретению, которые относятся к красной окраске по изобретению, за счет гибридизации или при использовании способов молекулярной биологии. Изобретение также относится к потомству этих растений, которые сохранили или приобрели признак красной окраски по изобретению.
Для переноса генов, которые отвечают за красную окраску к другому растению, можно использовать обратное скрещивание. В этих целях родительское растение, с которым гибрид вновь скрещивают, представляет собой желаемый гомозиготный сорт или инбредное растение. Источник переносимого признака называют родителем-донором. Ожидают, что полученное растение будет иметь характеристики родительской формы, с которой гибрид скрещивается (например, сорта), и желаемый признак переносится от родителя-донора. После изначального скрещивания выбирают индивидуальные растения, обладающие фенотипом родителя-донора (красная окраска листьев центральной части кочана), и повторно скрещивают (обратное скрещивание) с родительской формой. Ожидается, что полученное растение будет иметь характеристики родительской формы (например, сорта), и желаемый признак переносится от родителя-донора. В том случае, если наследование красной окраски представляет собой более сложное по сравнению с наследованием других желаемых признаков или комбинации признаков, то растение с красной краской листьев в центре кочана может быть использовано в качестве родительской формы, с которой проводят повторное скрещивание гибрида, и растение с другим желаемым признаком или комбинацией признаков может быть использовано в качестве родителя-донора. Описания других способов селекции, которые обычно используют для различных признаков и культур можно найти в одном или нескольких справочниках (например, "Principles of Plant Breeding" John Wiley and Son, pp.115-161, 1960; Allard, 1960; Simmonds, 1979; Sneep et al., 1979; Fehr, 1987).
Настоящее изобретение дополнительно проиллюстрировано в последующих примерах, которые представлены исключительно в целях иллюстрации и ни каким образом не предназначены для ограничения изобретения.
На Фиг. 1 показано распределение в пределах линии расщепления красной окраски внешних и внутренних листьев для популяции 212 F3-линий, полученных от гибридизации '99P.30637' х сv. “Sharp Shooter”. Для популяции использовали три классификации красной окраски: ”красные внешние”, т.е. частота растений внутри линии с красными внешними листьями вне зависимости от окраски внутренних листьев; “ красные внешние и красные внутренние”, т.е. частота растений внутри линии с красными наружными и красными внешними листьями; “красные внешние/ красные внутренние”, т.е. частота растений внутри линии с красными внутренними листьями за вычетом общего числа растений с красными внешними листьями. О = наблюдаемые, Е = ожидаемые согласно генетической модели.
На Фиг. 2 показана генетическая модель для экспрессии антоцианина на основе расщепления в гибриде '99P.30637' х сv. “Sharp Shooter”.
На Фиг. 3А показано растение по изобретению. Это молодое растение представляет собой растение из семени, полученного от линии 03Р82421, которое получили после трех генераций самоопыления 25 растений 99P38154. На фиг. 3B, 3C и 3D показаны молодые растения от родителей Pierrot, Pippo и Gringo соответственно. На фиг. 3E и 3F показаны молодые растения сравниваемых сортов Apache and Bijou соответственно. Все эти молодые растения выращивали в торфяных горшочках в ростовой камере при 14 ч света при 16°С и 10 ч темноты при 12°С. Свет обеспечивали за счет Philips TLD 36 W 840 REFLEX ламп, с 1 лампой на 0,24 квадратных метра при расстоянии над растением 0,6 м.
На Фиг. 4 показан молодой лист растения по изобретению, обозначенный как «НОВЫЙ» в сравнении с молодыми листьями коммерческих сортов Pippo, Pierrot и Gringo. Указанные молодые растения выращивали в торфяных горшочках в ростовой камере при 14 ч света при 16°С и 10 ч темноты при 12°С. Свет обеспечивали за счет Philips TLD 36 W 840 REFLEX ламп, с 1 лампой на 0,24 квадратных метра при расстоянии над растением 0,6 м.
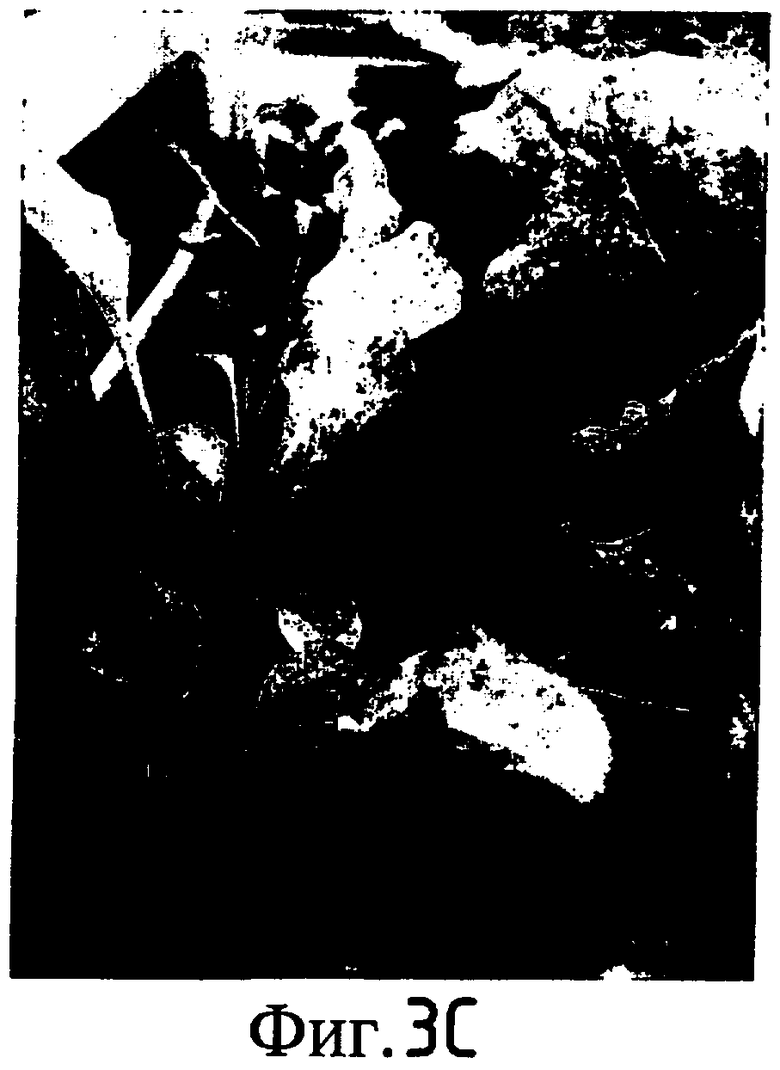
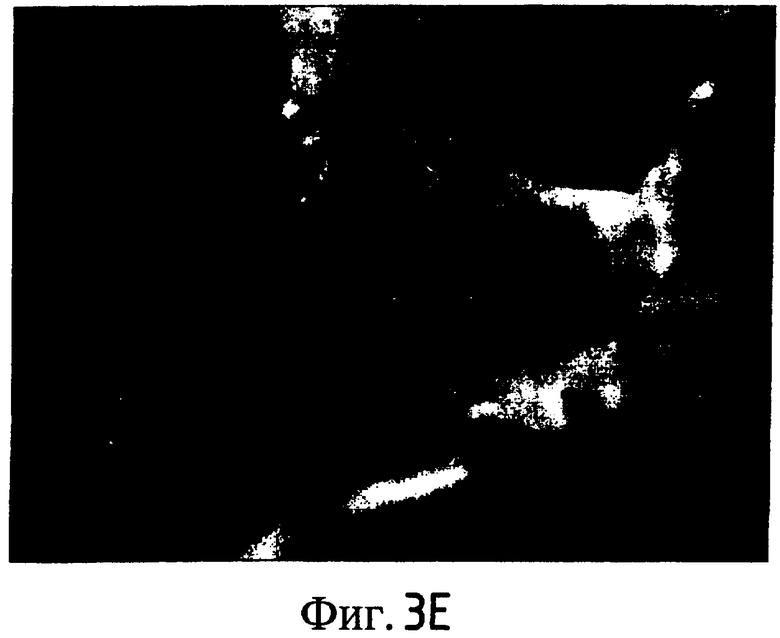
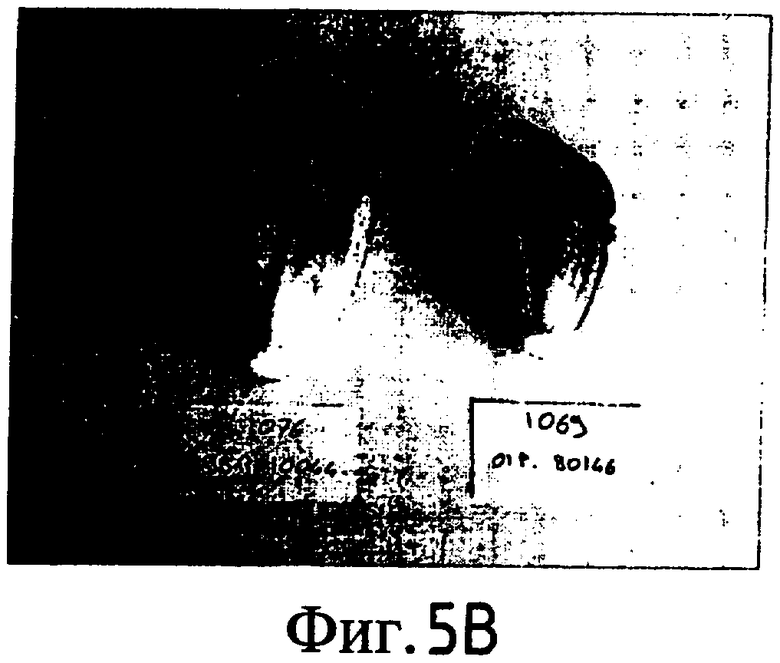
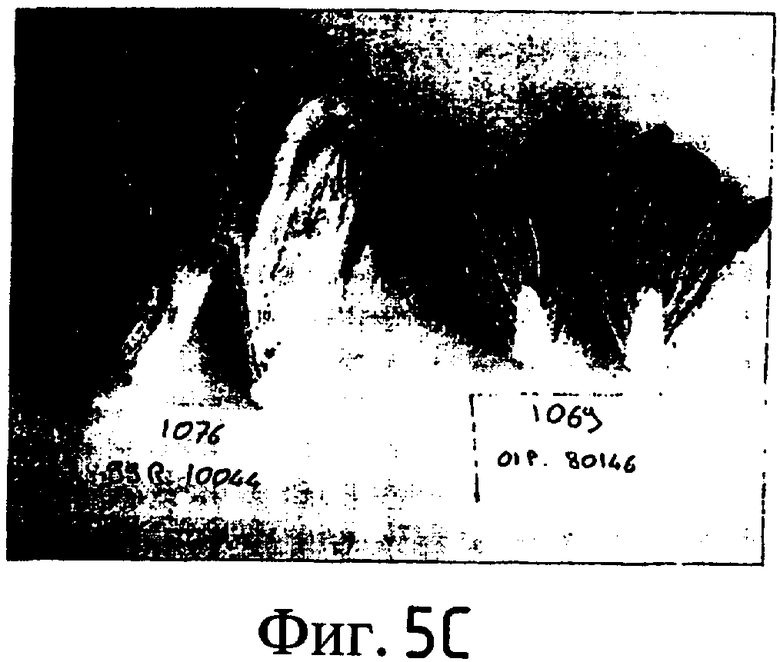
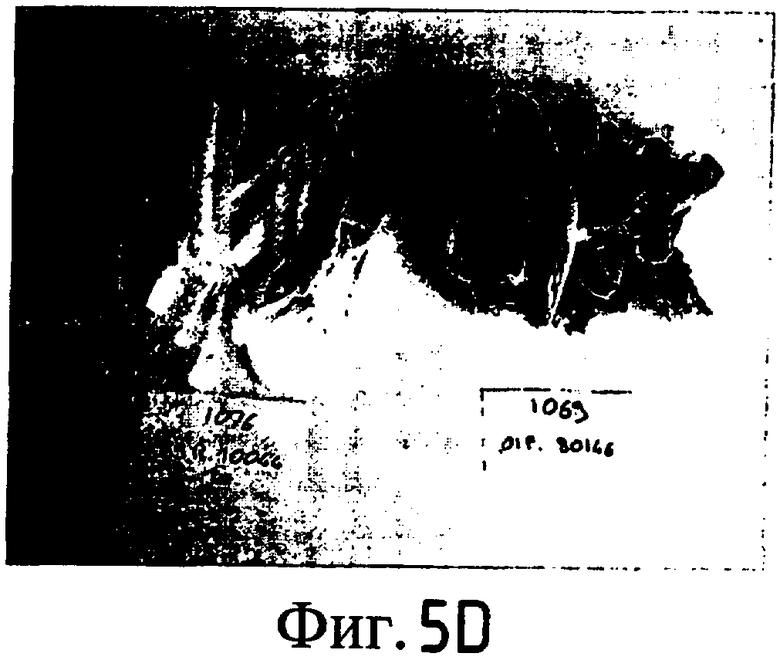
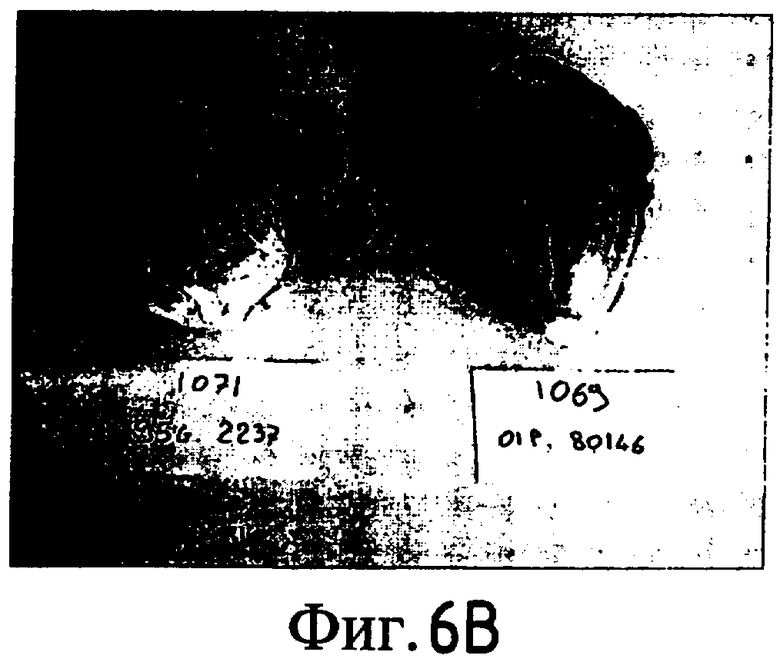
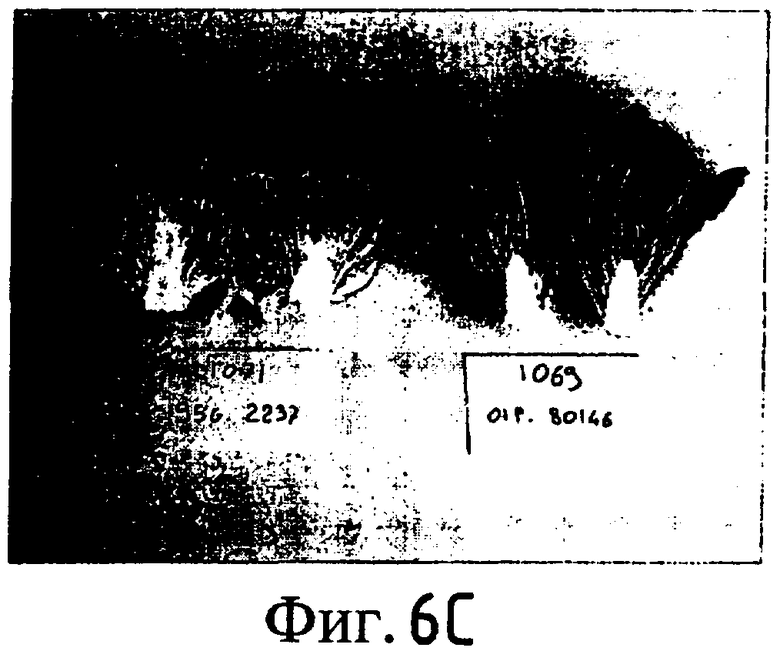
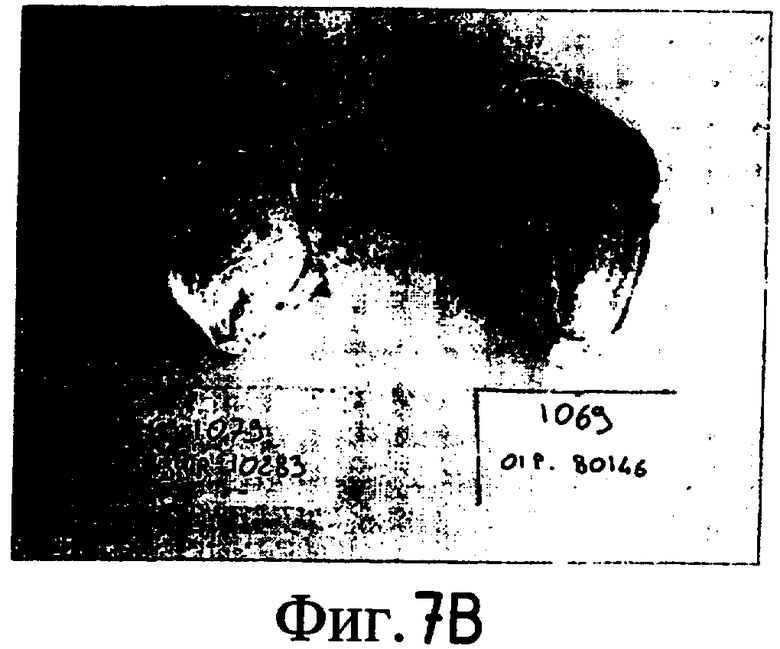
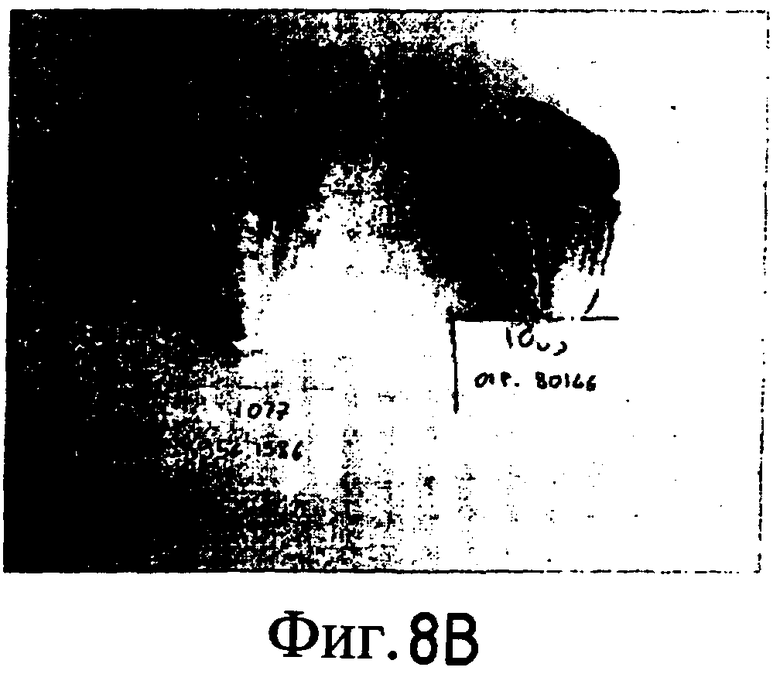
На Фиг. 5-8 показано сравнение между растением по изобретению (01P80146) и коммерческими сортами Darkland (обозначенный как '99R10044'; фиг. 5А-5D), Pierrot (обозначенный как '95G2237'; фиг. 6А-6D), Roxy (обозначенный как '99R10283'; фиг. 7А-7D) и Sierra (обозначенный как '95G1986'; фиг. 8А-8D) соответственно. На А-фиг. (5А, 6А, 7А и 8А) показаны цельные собранные кочаны. На B-фиг. (5B, 6B, 7B и 8B) показана центральная часть кочана без внешних листьев. На С-фиг. (5С, 6С, 7С и 8С) показано продольное сечение центральной части кочана. На D-фиг. (5D, 6D, 7D и 8D) показаны нарезанные листья центральной части кочана. Растения салата латука по изобретению представляют собой полностью красные в центральной части кочана, тогда как другие растения салата латука не являются красными в центральной части кочана. Все растения выращивали в Арамоне (Aramon), Франция в 2002: посеяны 10 января 2002, пересажены на собственное открытое поле 15 февраля 2002, собраны 25 апреля 2002.
ПРИМЕРЫ
ПРИМЕР 1
Селекция красного салата латука по изобретению
Салат латук по изобретению был получен согласно следующей селекции (происхождению): в 1986 был получен гибрид между растением cv. Pippo (Rijk Zwaan; красный) и растением cv. Blonde Maraichere (Caillard; зеленый).
В 1988 красное растение потомок из указанного скрещивания использовали в качестве опылителя при гибридизации с растением cv. Gringo (Rijk Zwaan; красный). В том же году был получен гибрид между растением cv. Pierrot (Rijk Zwaan; красный) и растением cv. Roxette (Rijk Zwaan; зеленый).
В 1989 красное растение потомок из указанного скрещивания использовали в качестве опылителя при гибридизации с растением cv. Krizet (Rijk Zwaan; зеленый).
В 1992 красное F3-растение потомок из указанного скрещивания было выбрано из указанного выше гибрида Krizet х (Pierrot x Roxette) и его использовали в качестве опыляемого растения при гибридизации с опылителем, который представлял собой выбранное красное растение потомок из упомянутого выше скрещивания Gringo х (Pippo x Blonde Maraichere). Красное F4-растение из указанного вновь полученного гибрида 1992 отбирали в 1995 году и использовали в качестве опыляемого растения при гибридизации с cv. Roxette (Rijk Zwaan; зеленый).
Отбор по типу, кочану и цвету, который проводили на F2-, F3- и F4-поколениях от указанного скрещивания привел к получению красного F4-растения (98Р.31582) в 1998. F5-поколение демонстрировало расщепление по цвету (красный и зеленый), но отбор в следующем поколении привел к получению трех кочанных F3-растений с полностью красными листьями во внутренней части кочана (99Р.38152; 99Р.38154; 99Р.30637), которые размножали в течение нескольких лет после этого.
Потомки не показывали выщипления зеленых растений и семена депонировали под номерами 02R.2413, 01R.1439, 02R.2418. Указанные F5-растения и их потомки служили в качестве источника родительских растений для дальнейшей селекции (breeding) салата латука с независимой от света красной окраской.
В заключении можно отметить, что ни один из используемых родительских сортов, т.е. Pippo, Blonde Maraichere, Gringo, Pierrot, Roxette и Krizet, не имел характеристик красных листьев в центральной части кочана. Изобретение включает уникальную и новую комбинацию генов из этих красных и зеленых родительских сортов, которая обеспечивает полностью красные листья в центральной части кочана.
ПРИМЕР 2
Генетический анализ красного салата латука по изобретению
Как используется здесь, термин локус (множественное локусы) определяется как специфический участок хромосомы, в котором локализован ген (Griffiths AJF, Miller JH, Suzuki, DT, Lewontin RC, Gelbart, WM. 'An introduction to genetic analysis.' 6th edition. 1996. WH Freeman and Company, New York). Число локусов, ответственных за красную окраску в представленном растении, можно установить при использовании генетического анализа потомков гибридизации указанного растения с имеющим зеленую окраску cv. 'Sharp Shooter'.
Генетический анализ красного салата латука по изобретению проводили следующим образом: получали гибрид между растением потомком из F5-растения 99P.30637, т.е. растения по настоящему изобретению, и неярко зеленым растением салата латука айсберг cv. Sharp Shooter (SVS, Waycott et al., 1999: US-patent nr 5,973,232). От потомка этой гибридизации 212 случайно отобранных F2-растения разводили с получением F3-линий.
Указанные 212 F3-линии оценивали в собственном испытании в условиях открытого грунта в Fijnaart, Нидерланды. Семена высевали в торфяные горшочки 17 июня 2002, молодые растения выращивали в теплице и пересаживали в открытый грунт 5 июля 2002. Дата сбора урожая (время наблюдения) составляло с 19 августа по 23 августа 2002. Размер делянки составлял 24 растения на F3-линию, т.е. 4 ряда по 6 растений. Линии не повторялись поскольку экспрессия признаков окраски представляет собой чрезвычайно стабильную внутри одного испытания. Наблюдения проводили по F3-растению.
Наблюдаемые признаки были: а) цвет и если красный, его интенсивность и характер экспрессии на внешних листьях; b) цвет и если красный, его интенсивность и характер экспрессии на внутренних листьях; с) степень образования кочана; d) в случае зеленых внешних листьев: классификация на два класса яркие и не яркие.
Интенсивность и характер экспрессии красной окраски классифицировали по нарастающей шкале, как: 1) имеющая оттенок или красноватая, т.е. легкая красная окраска по внешнему или внутреннему краю листа или внутренней стороне основания листа, 2) с красными пятнами, 3) с зеленым пятнами, т.е. зеленые пятна на красной поверхности листа, 4) полностью красные, т.е. с интенсивной красной окраской без пятен на внешних частях листа, в той части, на которую воздействует свет и, в случае внутренних листьев, по всему внутреннему листу.
Кочаны оценивали как 1) слегка открытые кочаны, типа салат-ромэн или летний эндивий, 2) плотные кочаны с явно перекрывающимися листьями, как салат латук айсберг.
Некоторые растения в испытании погибли до сбора. Из планируемых 5088 растений, т.е. 212×24, всего 5007 F3-растений достигли стадии, пригодной для уборки и были оценены (см. Таблицу 1).
Все растения показывали степень образования кочана, которая была по крайней мере сравнима с салатом-ромэн или летним эндивием. Полученные данные показали расщепление 55 полностью зеленых линий из общего числа 212. Из 157 линий, в которых имелось по крайней мере одно растение с красной окраской, 28 линий включали растения, которые все демонстрировали экспрессию антоцианина на внешних листьях. Из указанных 28 линий, 8 линий включали растения, которые все демонстрировали экспрессию антоцианина на листьях центральной части кочана. В 33 линиях из 157 линий растения с красной окраской демонстрировали экспрессию антоцианина только во внешних листьях, но не в листьях центральной части кочана. В 33 других линиях из 157 линий растения с красной окраской всегда демонстрировали экспрессию антоцианина одновременно во внешних листьях, а также в листьях центральной части кочана. Растения с красной окраской оставшейся 91 линии все демонстрировали экспрессию антоцианина во внешних листьях. Однако указанные растения демонстрировали расщепление внутри линии экспрессии антоцианина в листьях центральной части кочана. Ни одно из растений из 5007 растений не демонстрировало экспрессию антоцианина в центральной части кочана в сочетании с внешними листьями зеленой окраски, в которых не происходило экспрессии антоцианина.
Таким образом, сделано заключение, что для экспрессии антоцианина в центральной части кочана требуются один или несколько генов, которые приводят к экспрессии антоцианина во внешних листьях. В случае расщепления между красной и зеленой окраской соотношение расщепления между красной и зеленой окраской изменялось в диапазоне от 23:1 до 1:22 для окраски внешних листьев. Для внутренних листьев соотношение расщепления между красной и зеленой окраской изменялось в диапазоне от 18:1 до 1:23, исключая все растения с зелеными внешними листьями. В таблице 1 показано распределение внутри линий, растений с красной и зеленой окраской внешних и внутренних листьев, для популяции из 212 случайным образом полученных F3-линий от гибрида '99P.30637' x cv. Sharp Shooter.
Указанные данные по расщеплению использовали для построения генетической модели при допущении соотношений менделеевского независимого расщепления. Было обнаружено, что по крайней мере три локуса вовлечены в получение УФ независимой экспрессии антоцианина во внутренних листьях. Первый локус, далее обозначаемый как А-а, также обнаружен в известном красном салате латуке, и во всех случаях требуется доминантный аллель А для получения экспрессии антоцианина. Возможно указанный локус представляет собой С или G-локус (Robinson et al., 1983, The genes of lettuce and closely related species. In: Plant Breeding Reviews 1. Ed. J Janick. p 267-293).
Второй локус обнаружен подгонкой модели, далее обозначается как В-b, где присутствие в гомозиготном состоянии рецессивного аллеля b приводит к экспрессии антоцианина во внешних листьях в комбинации с А. Было обнаружено, что еще два локуса, далее обозначаемых как С-с и D-d, также вовлечены в экспрессию антоцианина во внешних листьях. Присутствие по крайней мере одного доминантного аллеля С или присутствие в гомозиготном состоянии рецессивного аллеля d приводит к экспрессии антоцианина во внешних листьях, но только в том случае, если присутствует один аллель А.
Были обнаружены еще три локуса, далее обозначаемые как Е-е, F-f и G-g. Для получения УФ независимой экспрессии антоцианина во внутренних листьях требуется присутствие по крайней мере одного А-аллеля и двух b-аллелей, в комбинации с или присутствием одной копии доминантного Е-аллеля, или присутствием f-аллеля в гомозиготном состоянии, или присутствием g-аллеля в гомозиготном состоянии. Таким образом, для УФ независимой экспрессии антоцианина во внутренних листьях необходимо присутствие аллелей красной окраски в, по крайней мере, трех локусах, а именно А-а, B-b и или E-e, или F-f, или G-g. Более того, считается, что по крайней мере три, но возможно и все семь упомянутых выше локусов определяют интенсивность красной окраски по настоящему изобретению.
Пример 3
Определение цвета, антоцианинов и хлорофиллов в салате латуке
1. Получение образца и анализы
Использовали спектрофотометр UltrospecIII (Pharmacia) со следующей спецификацией:
- монохроматор: Czerny Turner с голографической дифракционной решеткой (1200 линий/мм)
- точность длины волны: ±1 нм
- воспроизводимость длины волы: ±0,5 нм
- тип детектора: монокристаллический кремниевый фотодиод
ширина полосы: 5 нм
2. Принцип
Красный цвет (антоцианины) и зеленый цвет (хлорофиллы) определяют на основе биохимического способа. Приготовляли два экстракта, один для измерения поглощения при 523 нм, которое является мерой общего содержания антоцианинов, и второй для поглощения при 665 нм, которое является мерой общего содержания хлорофиллов (хлорофилл а и b).
3. Получение образца
Сформировавшееся кочанное растение салата латука собирали и удаляли внешние листья, пока не оставалась центральная часть кочана. Центральную часть кочана салата латука использовали для анализа. Листья центральной части кочана не должны были подвергаться прямому солнечному свету до сбора, за исключением небольшого участка верхушки листа (максимально 10%) в верхней части растения. Центральная часть кочана содержит по крайней мере 10 листьев с длиной 1 см или длиннее. Самые старые листья кочана должны быть изогнутыми.
Центральные части кочанов помещают в пластиковые пакеты и замораживают при <-70°С. После по крайней мере нескольких дней при <-70°С замороженные центральные части кочана измельчали кувалдой. Для получения порошка измельченный образец размалывают в Grindomix (GM 200, Retsch, 5 сек при 3000 об/мин и затем 5 сек при 5000 об/мин) при использовании плавающей крышки в присутствие жидкого азота (образец необходимо поддерживать замороженным).
Порошок затем помещают в пробирку и пробирку с порошком охлаждают в жидком азоте и необязательно хранят при <-70°С до анализа.
4. Анализ
Три грамма порошка взвешивали в четыре пробирки по 50 мл. Две пробирки использовали для анализа антоцианинов и две пробирки для анализа хлорофиллов.
Для измерения антоцианинов в две пробирки немедленно добавляли 1,0 М HCl в 50% метаноле. Использовали 5-10 мл/г образца в зависимости от цвета образца и экстракта. Регистрировали использованный объем (мл). Раствор образца смешивают вручную и помещают на лед. Часть раствора помещают в пробирку типа еппендорф 1,5 мл и пробирку центрифугируют при 4°С 13000 об/мин в течение 4 мин.
Спектр измеряют при использовании спектрофотометра с шириной полосы 5 нм. Использовали кювету 1 см.
Измеряют спектр от 360 до 900 нм и поглощение при 523 нм (если необходимо после разбавления жидкостью для экстракции) и определяют максимальную длину волны (λmax). λmax должна быть близка к 523 нм.
Хлорофиллы измеряли путем немедленного добавления в две пробирки 100% метанола. Использовали 5-10 мл метанола на 1 г образца в зависимости от цвета образца и экстракта. Регистрировали использованный объем (мл). Раствор образца смешивают вручную и подвергают обработке ультразвуком в ультразвуковой бане в течение 5 мин в положении «удаление газа». Часть раствора помещают в пробирку типа еппендорф 1,5 мл и пробирку центрифугируют при 4°С, 13000 об/мин в течение 4 мин.
Измеряют спектр от 360 до 900 нм и поглощение при 665 нм (если необходимо после разбавления жидкостью для экстракции) и определяют максимальную длину волны (λmax). λmax должна быть близка к 665 нм.
А523 и А665 корректируют таким образом, чтобы представлять поглощение раствора 1 г сырой массы в 10 мл экстракционного объема. Корректированные А523 и А665 рассчитывают с поправкой на массу образца (в г), экстракционный объем (в мл) и, если необходимо, разбавление при использование следующей формулы:
Поправка:
Рассчитывают отношение А523/А665.
Таблица 2а показывает поглощение хлорофилла и антоцианина и отношение хлорофилл/антоцианин, наблюдаемые в салате латуке в Арамоне, Франция (высеян 10 января 2002, пересажен 15 февраля 2002, собран 25 апреля 2002). Линия 01Р.80146 представляет собой линию потомков, полученную от растения 99Р.38154 после двух поколений самоопыления. Pierrot (Rijk Zwaan), Darkland (Central Valley), Sierra (Vilmorin) и Roxy (Enza) представляют собой известные сорта салата латука.
Таблица 2b показывает поглощение хлорофилла и антоцианина и отношение хлорофилл/антоцианин, наблюдаемые в салате латуке, выращенном в Fijnaart, Нидерланды (высеян 21 мая 2002, пересажен 10 июня 2002, собран 20 августа 2002). Линия 01Р.80146 представляет собой линию потомков, полученную от растения 99Р.38154 после двух поколений самоопыления. Pierrot (Rijk Zwaan), Pippo (Rijk Zwaan), Red Rosalita (Johnny's Selected & Orsetti), Darkland (Central Valley), Sierra (Vilmorin) и Roxy (Enza) представляют собой известные сорта салата латука.
Информация о депонировании
F5-растения 01R.1439, 02R.2413 и 02R.2418 были депонированы 18 июля 2005 в NCIMB, Ferguson Building, Craibstone Estate, Bucksburn, Aberdeen AB21 9YA, United Kingdom под входящими номерами NCIMB 41337, NCIMB 41338 и NCIMB 41339 соответственно.
| название | год | авторы | номер документа |
|---|---|---|---|
| РАСТЕНИЯ САЛАТ-ЛАТУК, ИМЕЮЩИЕ РЕЗИСТЕНТНОСТЬ ПРОТИВ NASONOVIA RIBISNIGRI БИОТИПА 1 | 2015 |
|
RU2731639C2 |
| УСТОЙЧИВОСТЬ К ФИЗИОЛОГИЧЕСКИМ НАРУШЕНИЯМ В САЛАТЕ | 2007 |
|
RU2480983C2 |
| ПОЛУЧЕНИЕ СЕМЯН ГИБРИДА Lactuca sativa | 2008 |
|
RU2593940C2 |
| РАСТЕНИЕ, УСТОЙЧИВОЕ К НАСЕКОМЫМ | 2008 |
|
RU2522477C2 |
| РАСТЕНИЕ, УСТОЙЧИВОЕ К НАСЕКОМЫМ | 2014 |
|
RU2617955C2 |
| СПОСОБ СКРИНИНГА ДЛЯ ОТБОРА РАСТЕНИЙ, ОБНАРУЖИВАЮЩИХ ПОНИЖЕННОЕ НАРУШЕНИЕ ОКРАСКИ ПОВЕРХНОСТИ, ВЫЗЫВАЕМОЕ ПОВРЕЖДЕНИЕМ, И РАСТЕНИЕ И ЧАСТИ РАСТЕНИЯ, ПОЛУЧАЕМЫЕ ТАКИМ ОБРАЗОМ | 2007 |
|
RU2433585C2 |
| КУКУРУЗНЫЕ ПРОДУКТЫ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2012 |
|
RU2650764C2 |
| РАСТЕНИЯ КАПУСТЫ BRASSICA OLERACEA, УСТОЙЧИВЫЕ К ЗАБОЛЕВАНИЮ КИЛОЙ | 2003 |
|
RU2369090C2 |
| РАСТЕНИЯ ОГУРЦА, УСТОЙЧИВЫЕ К ЗАБОЛЕВАНИЯМ | 2006 |
|
RU2418405C2 |
| РАСТЕНИЕ ТОМАТА, ОБРАЗУЮЩЕЕ ПЛОДЫ С МОДИФИЦИРОВАННЫМ СОДЕРЖАНИЕМ САХАРОВ | 2019 |
|
RU2817600C2 |














Изобретение относится к образующему кочан растению салата латука вида Lactuca sativa, имеющему красные листья по всему кочану, включая центральную часть кочана. Красные листья в центральной части кочана представляют собой красные даже в отсутствие излучения с длинами волн короче чем 400 нм, где отсутствие излучения с длинами волн короче чем 400 нм наблюдается при условиях выращивания в ходе всего периода от высевания до наблюдения. Соотношение между антоцианином и хлорофиллом находится в диапазоне от 4 до 50, предпочтительно от 9 до 27. Описан генетический комплекс, придающий описанные свойства указанному растению, включающий по крайней мере один доминантный А-аллель для экспрессии антоцианина; рецессивный аллель b в гомозиготном состоянии; необязательно, по меньшей мере один доминантный С-аллель или рецессивный аллель d в гомозиготном состоянии; и по крайней мере один доминантный Е-аллель, рецессивный аллель f в гомозиготном состоянии или рецессивный аллель g в гомозиготном состоянии. Также описано применение растений, содержащих описанный комплекс, для селекции. Изобретение позволяет получать растения салата латука вида Lactuca sativa, имеющие красные листья по всему кочану, включая центральную часть кочана. 3 н. и 3 з.п. ф-лы, 25 ил., 2 табл., 3 пр.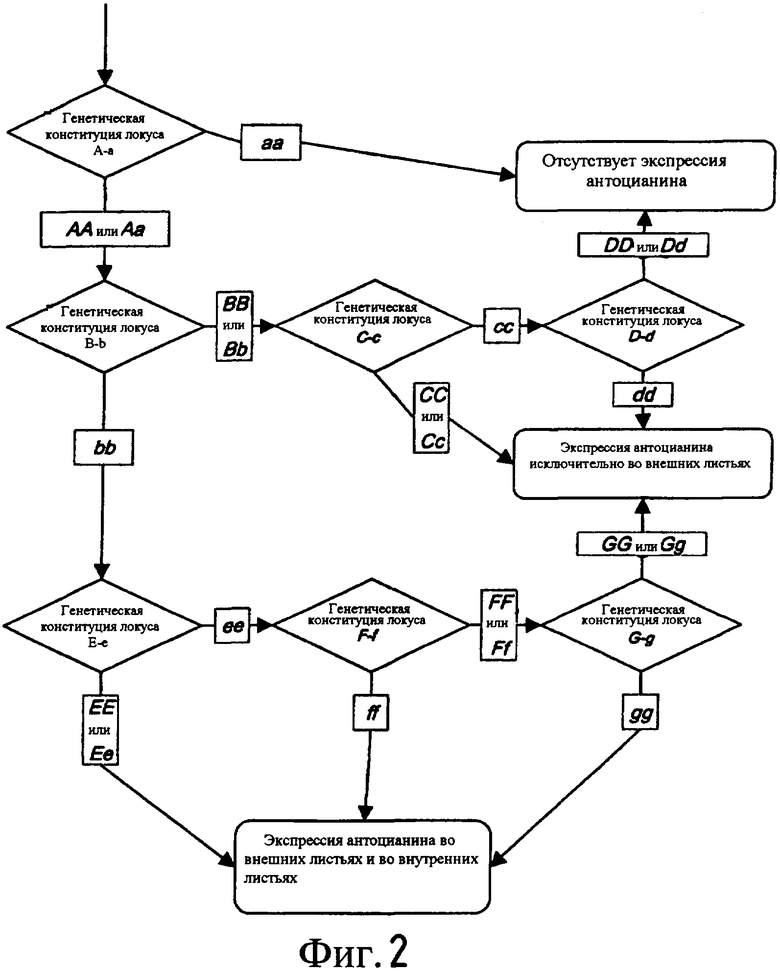
1. Генетический комплекс, который в растении салата латука вида Lactuca sativa приводит к независимой от ультрафиолетового облучения красной окраске листьев всего кочана, включая центральную часть кочана, где генетический комплекс включает:
по крайней мере один доминантный А-аллель для экспрессии антоцианина;
рецессивный аллель b в гомозиготном состоянии;
необязательно, по меньшей мере один доминантный С-аллель или рецессивный аллель d в гомозиготном состоянии; и
по крайней мере один доминантный Е-аллель, рецессивный аллель f в гомозиготном состоянии или рецессивный аллель g в гомозиготном состоянии.
2. Генетический комплекс по п.1, включающий аллели А-а, или A-A, b-b и Е-е, или Е-Е, f-f, или g-g.
3. Генетический комплекс по п.1, включающий аллели А-а, или А-А, b-b, C-с, или С-С, d-d, Е-е, или Е-Е, f-f, или g-g.
4. Генетический комплекс по любому из пп.1-3, такой как присутствует в семенах, депонированных 18 июля 2005 в NCIMB под номерами NCIMB 41337, NCIMB 41338 и NCIMB 41339.
5. Применение салата латука вида Lactuca sativa, имеющего красную окраску листьев всего кочана, включая центральную часть кочана, репрезентативные семена которого депонированы 18 июля 2005 в NCIMB 41337, NCIMB 41338 и NCIMB 41339 для селекции.
6. Применение салата латука вида Lactuca sativa, имеющего красную окраску листьев всего кочана, включая центральную часть кочана, содержащее в своем геноме генетический комплекс по любому из пп.1-4, для селекции.
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ BRASSICA OLERACEA | 1997 |
|
RU2142013C1 |

